obtenÇÃo de biocatalisador para...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
Programa de Pós-Graduação de Tecnologia de Processos Químicos e Bioquímicos
Escola de Química
Tese de Doutorado
OBTENÇÃO DE BIOCATALISADOR PARA RESOLUÇÃO
CINÉTICA DE DERIVADOS DE MIO-INOSITOL
Evelin Andrade Manoel
RIO DE JANEIRO
MAIO/2014

OBTENÇÃO DE BIOCATALISADOR PARA RESOLUÇÃO
CINÉTICA DE DERIVADOS DE MIO-INOSITOL
Evelin Andrade Manoel
TESE SUBMETIDA AO CORPO DOCENTE DO CURSO DE PÓS-GRADUAÇÃO EM
TECNOLOGIA DE PROCESSOS QUÍMICOS E BIOQUÍMICOS DA ESCOLA DE
QUÍMICA DA UNIVERSIDADE FEDERAL DO RIO DE JANEIRO, COMO PARTE DOS
REQUISITOS NECESSÁRIOS PARA A OBTENÇÃO DO GRAU DE DOUTOR EM
CIENCIAS.
Orientadores:
Maria Alice Zarur Coelho, D.Sc.
Denise Maria Guimarães Freire, D.Sc.
RIO DE JANEIRO
MAIO/2014

OBTENÇÃO DE BIOCATALISADOR PARA RESOLUÇÃO
CINÉTICA DE DERIVADOS DE MIO-INOSITOL
Evelin Andrade Manoel
TESE SUBMETIDA AO CORPO DOCENTE DO CURSO DE PÓS-GRADUAÇÃO EM
TECNOLOGIA DE PROCESSOS QUÍMICOS E BIOQUÍMICOS DA ESCOLA DE
QUÍMICA DA UNIVERSIDADE FEDERAL DO RIO DE JANEIRO, COMO PARTE DOS
REQUISITOS NECESSÁRIOS PARA A OBTENÇÃO DO GRAU DE DOUTOR EM
CIÊNCIAS.
Aprovada por:
Orientador (es)
Banca Examinadora:
RIO DE JANEIRO
MAIO/2014

MANOEL, EVELIN ANDRADE
OBTENÇÃO DE BIOCATALISADOR PARA RESOLUÇÃO
CINÉTICA DE DERIVADOS DE MIO-INOSITOL
[Rio de Janeiro] – 2014.
180p. 29,7 cm
Tese (Doutorado) -Universidade Federal do Rio de Janeiro - UFRJ,
Programa de Pós-Graduação em Tecnologia de Processos Químicos e
Bioquímicos – EQ, 2014.
Orientador: Profa. Dra. Maria Alice Zarur Coelho
Profa. Dra. Denise Maria Guimarães Freire
1.Biocatálise. 2. myo-inositol. 3. Lipase. 4. Biorreator. 5. Substâncias
bioativas–I. Título
II. Universidade Federal do Rio de Janeiro-EQ.
III- Série

Dedicatória,
Aos meus queridos pais, Jorge e Rosa, por sempre estarem do meu lado.

AGRADECIMENTOS
Primeiramente gostaria de agradecer a Deus, por ter me concedido saúde e perseverança para realizar o trabalho.
Aos meus pais, Jorge e Rosa, irmã, Erika, cunhado Alex e ao meu sobrinho lindo Isaac, que chegou as nossas
vidas para mostrar que o mundo pode ser muito melhor. Obrigada pelo amor, carinho, compreensão, apoio e
principalmente paciência incondicional, uma vez que me ausentei muito nesta trajetória de tese.
Às minha queridas orientadoras, Profa. Maria Alice Zarur Coelho e Profa. Denise Maria Guimarães Freire, pela
orientação dedicada, pelas oportunidades, idéias e estímulo que me proporcionaram. Pelos apoios e sugestões
não só na vida profissional, como na pessoal, que me atentei com todo carinho. Por tudo que aprendi, e ainda,
pelo que vou aprender sou muito grata a vocês.
Ao Prof. Alessandro Bolis Costa Simas (NPPN) pela colaboração neste trabalho, pela livre entrada e
permanência em seu laboratório e, sobretudo, pela livre utilização de seus reagentes e colunas para o HPLC. A
sua contribuição foi muito além do que uma simples colaboração.
Ao prof. Rodrigo Octávio Sousa, pela colaboração e pela livre entrada e permanência em seu laboratório. Muito
obrigada!
À Suema Branco, minha querida irmã, obrigada pela grandiosa amizade e carinho todos estes 10 anos. Obrigada
pela compreensão e paciência.
A Jaqueline Nascimento, pela amizade e companheirismo, não somente na bancada, mas na vida. Obrigada por
sempre estar a disposição, inclusive em um dos momentos mais importantes da minha vida, durante meu período
de concurso público me dando caronas, cafés, almoços etc, rsrs. Nunca vou esquecer isso. Você faz parte desta
vitória!
Aos amigos mais que queridos da família LaBiM, Laboratório de Biotecnologia Microbiana, por permitir que eu
fizesse parte desta grande família.
Aos amigos do Laboratório de Microbiologia Molecular e de Proteínas, o LaMMP, que além da colaboração,
fizeram do ambiente de trabalho um lugar mais divertido e familiar.
Aos meus queridos companheiros Marcelo, em especial pela paciência, pelo carinho, pela alegria nos tempos
vagos; Jaqueline Nascimento, pela eterna amizade, ombro e apoio de sempre; Joab Sampaio, pela amizade e
ombro sempre que necessário; Erika Aguilheiras, Mateus Godoy, Carolina Guimarães e aos meninos Arthur e
Bruno, pelo enorme carinho, sou grata pela amizade de vocês. À querida amiga Lívia, a ―Liv‖, como eu gosto de
chamar. Obrigada pela amizade e consideração em todos os momentos. Ao Bruno César, Roberta Branco e a
eterna Valéria Soares, saudades Valzinha!!. Val, muito obrigada pela mão amiga, pelos conselhos profissionais e
pessoais, pelas risadas na bancadas.
Aos meus queridos amigos do BOSS group. Muito obrigada pelo carinho de vocês!
À profa. Melissa Gutarra, a eterna Mel, amiga e profissional admirável. Agradeço pelo enorme carinho e
amizade de sempre.
À Aline Fernandes, pelas ajudas na concessão de materiais e por sempre estar disposta a ajudar.
Aos professores do laboratório LaMMP, Rodrigo, Bianca e Marcia pela livre entrada e permanência em seu
laboratório durante todo o período do curso.
Aos amigos do laboratório BIOSE (EQ-UFRJ), em especial ao amigo Bernardo, pelas dicas no ramo
profissional, as amigas Roberta e Tatiana por serem tão queridas.
Aos verdadeiros amigos que estiveram presentes nos bons e maus momentos em toda a minha vida e, como não
poderia deixar de ser, também nestes últimos três anos. A Secretaria da Escola de Química, principalmente
Roselee, por sempre estar alegre e pronta a ajudar a nós, alunos da pós-graduação e coordenação do curso de
pós-graduação.
Ao CNPq pela bolsa de doutorado concedida nos dois primeiros anos de estudo, a qual foi muito importante para
realização de trabalhos, idas e vindas a congressos e cursos importantes para a minha carreira.
À FAPERJ, pela bolsa nota 10 concedida posteriormente.
A todas as demais pessoas que de forma direta ou indireta contribuíram para a realização deste trabalho.
Muito Obrigada!!!

E não há outro lugar onde eu gostaria de estar, amar o ciência só me
faz sonhar... Manoel, E.A.

Resumo da Tese de doutorado apresentada a Escola de Química da Universidade Federal do
Rio de Janeiro (EQ/UFRJ) como parte dos requisitos necessários para a obtenção do título de
Doutor em Ciências (D.Sc.)
Manoel, Evelin Andrade. OBTENÇÃO DE BIOCATALISADOR PARA RESOLUÇÃO
CINÉTICA DE DERIVADOS DE MIO-INOSITOL. Rio de Janeiro, 2014. Programa de
Pós-Graduação em Tecnologia de Processos Químicos e Bioquímicos, Escola de Química,
Universidade Federal do Rio de Janeiro.
Nos últimos anos, grandes esforços têm sido feitos para o desenvolvimento de processos
eficientes na elaboração de compostos enantiomericamente puros, principalmente para as
indústrias farmacêuticas. O interesse na obtenção de derivados de mio-inositol
enantiomericamente puros, deve-se, principalmente, à síntese de novas drogas e à importância
da sinalização celular como segundo mensageiro. Lipases vem sendo aplicadas nas resoluções
enantiosseletivas por serem altamente regio-, quimio-, diastero- e enantiosseletivas. Com isso,
esta tese teve como objetivo principal o estudo de um novo biocatalisador home-made tão
eficiente quanto as suas respectivas versões comerciais para resolução de diferentes derivados
farmacológicos de mio-inositol. Para isso, lipases produzidas pelos laboratórios de pesquisa
da UFRJ (LaBiM e BIOSE) e lipases liofilizadas comerciais foram imobilizadas em diferentes
suportes home-made (tipo casca-núcleo: PS-co-DVB/PS-co-DVB e PMMA-co-DVB/
PMMA-co-DVB) e comercial (para fins comparativos: Accurel MP 1000). Posteriormente, os
biocatalisadores imobilizados foram utilizados na resolução cinética de diferentes derivados
farmacológicos previamente sintetizados no Lab. Roderick Barnes, NPPN-UFRJ, tais como:
(±)-1,4,5,6-tetra-O-benzil-mio-inositol (rac-1); (±)-1,4,5,6-tetra-O-alil-mio-inositol (rac-2);
(±)-1,3,4-tri-O-benzil-mio-inositol (rac-3) e (±)-3,6-O-dibenzil-1,2-O-isopropilideno-mio-
inositol. As influências do solvente, agente acilante, água, utilização de diferentes suportes de
imobilização e reatores foram estudadas no desempenho do biocatalisador. Dentre as lipases
utilizadas, destacou-se a lipase B de Candida antarctica na resolução dos derivados rac-3 e
rac-4. Planejamentos experimentais foram realizados com a Novozym 435 utilizando acetato
de vinila em hexano em modo de batelada simples. Através do modelo matemático
construído, as condições ótimas para a reação de acetilação de rac-3 foram estabelecidas para
o biocatalisador comercial imobilizado. O produto L-1-O-acetil-2,3-O-ciclohexilideno-mio-
inositol (L-5) foi obtido com 49,7 ±0,2% de conversão e eep maior que 99%. Este sistema
permitiu o aumento da produtividade em 15 vezes, comparando-se com as condições não
otimizadas. Por outro lado, a abordagem utilizando o sistema de fluxo contínuo mostrou ser
muito eficiente para Novozym 435 na resolução de rac-3, levando um aumento em 531 vezes
(3.188mgproduto/mgenzima.h) na produtividade em relação ao sistema BSTR
(0.006mgproduto/mgenzima.h). Testes em maiores volumes foram feitos com a condição
otimizada obtendo alta conversão (50±1%) e alta pureza óptica (eep maior que 99%). O reuso
do biocatalisador foi feito em até 7 ciclos sem que houvesse queda na conversão (49% ± 0.7)
e eep. Após estudar as condições reacionais ótimas de Novozym 435 na resolução cinética de
rac-3 e rac-4, as versões home-made e comercial (LIPB e CALB, respectivamente) foram
imobilizadas nos suportes supramencionados e comparadas com Novozym 435. LIPB
imobilizada mostrou melhor desempenho quando comparadas com CALB comercial após
imobilizada nos mesmos suportes e desempenho similar a Novozym 435, uma das lipases
mais utilizadas no setor industrial. O processo permitiu a produção de L-5 em diferentes
escalas de produção, importante intermediário farmacológico no combate a doenças de
Chagas.

Abstract of doctor thesis presented to Escola de Química of Universidade Federal do Rio de
Janeiro (EQ/UFRJ) as partial fulfillment of the requiriments for the degree of Doctor of
Science (D.sc.)
Manoel, Evelin Andrade. ATTAINMENT OF BIOCATALYSTS TO KINETIC
RESOLUTION OF MIO-INOSITOL DERIVATIVES. Rio de Janeiro, 2014. Programa de
Pós-Graduação em Tecnologia de Processos Químicos e Bioquímicos, Escola de Química,
Universidade Federal do Rio de Janeiro.
In recent years, great efforts have been made to develop efficient processes in the preparation
of enantiomerically pure compounds, particularly for the pharmaceutical industries. There are
great interest in obtaining derivatives of myo-inositol enantiomerically pure mainly in the
synthesis of new drugs due to importance of cellular signaling as a second messenger. Lipases
has been used due to the regio-, chemo-, diastereo- and enantioselectivity. Thus, this thesis
aimed the study of a new home-made biocatalyst as efficient as its respective commercial
form to the kinetic resolution of myo-inositol derivatives. For this, lipases produced by
differents research groups at UFRJ (LaBIM and BIOSE) and commercial lyophilized lipases
were immobilized on different supports home-made (core-shell particles: PS-co-DVB/PS-co-
DVB and PMMA-co-DVB / PMMA-co-DVB) and commercial (for comparative purposes:
Accurel MP 1000). Subsequently, the immobilized biocatalysts were used for kinetic
resolution of different myo-inositol derivatives previously synthesized in Lab Roderick
Barnes, NPPN-UFRJ such as (±) -1,4,5,6-tetra-O-benzyl-myo-inositol (rac-1); (±) -1,4,5,6-
tetra-O-allyl-myo-inositol (rac-2); (±) -1,3,4-tri-O-benzyl-myo-inositol (rac-3) and (±) -3,6-O-
dibenzyl-1 ,2-O-isopropylidene-myo-inositol. The influence of solvent, acyl donor, water,
immobilization on different supports and reactors will be studied in the performance of the
biocatalyst. Among the lipases used, the best performance was the lipase B from Candida
antarctica in the kinetic resolution of rac-3 and rac-4. Response surface method to the
optimization of kinetic resolution of (±)-1,3,6-tri- O-benzyl-myo-inositol by Novozym 435
(immobilized lipase B from Candida antarctica) with vinyl acetate in hexane was applied on
batch mode. Through the constructed mathematical model, optimum condition for this
enzymatic condition was established. L-1-O-acetyl-2,3- tri-O-benzyl-myo-inositol was
obtained with a 49,7 ±0,2% conversion and eep higher than 99%. This system allowed the
productivity increase 15 times compared with non-optimized conditions. Moreover, the
approach using continuous flow system proved to be very effective for the resolution of
Novozym 435 rac-3, with productivity of 531 times higher (3.188mgproduct/mgenzyme h)
than of the optimized condition for the corresponding batch process
(0.006mgproduct/mgenzyme h). The feasibility of enzyme recycle was demonstrated without
loss of conversion and eep values up to the seventh recycle. Tests were made in larger
volumes with the optimized conditions obtaining high conversion (50 ± 1%) and high optical
purity (greater than 99% eep). The reuse of the biocatalyst was made within 7 cycles without
any decrease in conversion (49% ± 0.7) and eep. After the study of the optimal reaction
conditions of Novozym 435 in the kinetic resolution of rac-3 and rac-4, the lipases home-
made and commercial (LipB and CALB, respectively) were immobilized on different
supports and compared with Novozym 435. LIPB immobilized showed better performance
when compared with comercial CALB after immobilization process on the same support and
similar performance with Novozym 435, the one most used lipase in industrial sector. The
process showed the production of L-5 at different scales of production. This compound is an
important pharmacological intermediate against Chagas' Disease.

ÍNDICE GERAL
1. INTRODUÇÃO ............................................................................................................................................ 3
1.1 JUSTIFICATIVA .......................................................................................................................................... 5
2. OBJETIVOS ................................................................................................................................................. 9
2.1- OBJETIVOS ESPECÍFICOS ....................................................................................................................... 9
3. REVISÃO BIBLIOGRÁFICA ....................................................................................................................... 10
3.1- O MIO-INOSITOL..................................................................................................................................... 10 3.1.1 O papel do mio-inositol nas atividades biológicas ............................................................................... 12
3.1.2 Mio-Inositol no tratamento da doença de chagas ............................................................................... 16 3.2 ESTEREOSSELETIVIDADE: CONCEITO, ROTAS PARA OBTENÇÃO DE COMPOSTOS
ENANTIOMERICAMENTE PUROS E CINÉTICA DOS ENANTIÔMEROS ............................................... 16 3.2.1 Rotas para a obtenção de compostos enantiomericamente puros ..................................................... 17
3.2.2 Cinética dos enantiômeros ................................................................................................................... 21 3.3 LIPASES ..................................................................................................................................................... 26
3.3.1 Características das lipases ................................................................................................................... 26
3.3.2 Principais micro-organismos utilizados para lipases ........................................................................... 29
3.3.3 Estrutura proteica de lipases ............................................................................................................... 30
3.3.4 Sítios de ligação do substrato em lipases ............................................................................................ 32
3.3.5 Fenômeno da ativação interfacial em lipases ...................................................................................... 33
3.3.6 Mecanismo cinético ............................................................................................................................. 36
3.3.7 Aplicação das lipases ........................................................................................................................... 39
3.3.8 Aplicação de lipases na síntese de fármacos ....................................................................................... 42
3.3.9 Lipases promissoras para o estudo dos derivados de mio-inositol ...................................................... 47
3.3.10 Fatores que podem influenciar a estereosseletividade das lipases ................................................... 50
Quanto ao Biocatalisador: Tecnologia da Imobilização de Enzimas............................................................. 51
Quanto ao substrato: Influência dos grupamentos substituintes dos substratos......................................... 64
Quanto ao design do processo ...................................................................................................................... 64
4. MATERIAIS E MÉTODOS ........................................................................................................................... 77
4.1 MATERIAIS ............................................................................................................................................... 77 4.1.1 Biocatalisadores utilizados .................................................................................................................. 77
4.1.2 Substratos utilizados ............................................................................................................................ 77
4.1.3 Suportes utilizados na imobilização por adsorção ............................................................................... 78 4.2. MÉTODOS ................................................................................................................................................ 80
4.2.1 Seleção de lipases e substratos por meio da reação de acetilação realizados em sistema descontínuo
(ou modo de batelada) ................................................................................................................................. 80
4.2.2 Efeito do agente acilante na conversão e na enantiosseletividade ..................................................... 82
4.2.3 Efeito do solvente na conversão e na enantiosseletividade ................................................................. 82
4.2.4 Estudo da resolução cinética dos substratos/lipases selecionadas utilizando o sistema de fluxo
contínuo ........................................................................................................................................................ 83
4.2.6 Teste de estabilidade de estocagem das lipases imobilizadas ............................................................. 96
4.2.7 Desenho Experimental e Análise Estatística para resolução cinética do rac-3 .................................... 96
4.2.8 Testes prévios com aumento de volume .............................................................................................. 98
4.2.9 Estudo do reuso das reações de acetilação dos substratos/lipases selecionadas utilizando o sistema
de batelada simples ...................................................................................................................................... 98

5. RESULTADOS E DISCUSSÃO..................................................................................................................... 99
5.1 SELEÇÃO DE SUBSTRATOS E LIPASES HOME-MADE ...................................................................... 99 5.2 PLANEJAMENTO EXPERIMENTAL DA REAÇÃO DE ACETILAÇÃO DE RAC-3 UTILIZANDO A
LIPASE NOVOZYM 435 EM SISTEMA DE BATELADA .......................................................................... 106 5.2.1- Análise estatística dos dados ............................................................................................................ 107
5.2.3 PLANEJAMENTO EXPERIMENTAL DA REAÇÃO DE ACETILAÇÃO DE RAC-3 UTILIZANDO A LIPASE
NOVOZYM 435 EM SISTEMA DE FLUXO CONTÍNUO ................................................................................... 124
5.2.5- Imobilização da lipase B de Candida antarctica expressa em Pichia pastoris (LIPB) por adsorção
hidrofóbica em diferentes suportes ............................................................................................................ 131
5.2.6- Estabilidade dos biocatalisadores ao longo do tempo ..................................................................... 135
5.2.7- Imobilização da lipase CALB em Accurel MP 1000 e comparações com LIPB ................................... 136
5.2.8- Resolução cinética de rac-3 utilizando lipase B de Candida antarctica expressa em Pichia pastoris,
imobilizada sobre diferentes suportes ........................................................................................................ 137
5.2.9- Resolução cinética de rac-4 utilizando lipase B de Candida antarctica expressa em Pichia pastoris,
imobilizada em diferentes suportes ............................................................................................................ 138
6. CONCLUSÕES ............................................................................................................................................. 141
7. SUGESTÕES PARA TRABALHOS FUTUROS ....................................................................................... 142
8. REFERÊNCIAS BIBLIOGRÁFICAS......................................................................................................... 143

ÍNDICE DE TABELAS
TABELA 1. LIPASES SELECIONADAS PRODUZIDAS PELO LABIM/UFRJ E PELO BIOSE/UFRJ ......................................................... 8
TABELA 2. PRINCIPAIS ÁREAS APLICAÇÕES, PRODUTOS E TIPOS DE REAÇÕES DESEMPENHADAS PELAS LIPASES ................................ 40
TABELA 3. RELAÇÃO DOS TRABALHOS ATUAIS QUE UTILIZAM MÉTODOS E SUPORTES DISTINTOS DURANTE O PROCESSO DE
IMOBILIZAÇÃO. ................................................................................................................................................. 59
TABELA 4. VALOR DE LOG P DE SOLVENTES ORGÂNICOS (ADLERCREUTZ, 2000). ................................................................ 72
TABELA 5. SOLUÇÕES SALINAS SATURADAS ADEQUADAS AO CONTROLE DA ATIVIDADE DE ÁGUA (ADLERCREUTZ, 2000). ............ 74
TABELA 6. DADOS DE CARACTERIZAÇÃO DOS SUPORTES POLIMÉRICOS SELECIONADOS PARA ESTA TESE (BESTETI, ET AL., 2009, 2010)
..................................................................................................................................................................... 79
TABELA 7. VALORES REAIS E CODIFICADOS DAS VARIÁVEIS EMPREGADAS NO PLANEJAMENTO EXPERIMENTAL FRACIONÁRIO. ............ 96
TABELA 8. VALORES REAIS E CODIFICADOS DAS VARIÁVEIS EMPREGADAS NO PLANEJAMENTO EXPERIMENTAL COMPOSTO CENTRAL. . 97
TABELA 9. SELEÇÃO INICIAL DE LIPASES POR CCF PRODUZIDAS PELOS LABORATÓRIOS LABIM E BIOSE (UFRJ) NA RESOLUÇÃO CINÉTICA
DE DERIVADOS DE MIO-INOSITOL. ....................................................................................................................... 101
TABELA 10. RESULTADOS DO SCREENING REALIZADO COM AS DIFERENTES LIPASES NA RESOLUÇÃO CINÉTICA DE RAC-3 UTILIZANDO
ACETATO DE VINILA COMO SOLVENTE E AGENTE ACILANTE. ...................................................................................... 102
TABELA 11. RESULTADOS DO SCREENING REALIZADO COM AS DIFERENTES LIPASES NA RESOLUÇÃO CINÉTICA DE RAC-4 UTILIZANDO
ACETATO DE VINILA COMO SOLVENTE E AGENTE ACILANTE. ...................................................................................... 103
TABELA 12. INFLUÊNCIA DA VARIÁVEL TEMPERATURA NA CONVERSÃO EM 48H DE TEMPO REACIONALA ..................................... 109
TABELA 13. MATRIZ DO PLANEJAMENTO FATORIAL FRACIONÁRIO COM OS VALORES REAIS E CODIFICADOS PARA AS VARIÁVEIS
INDEPENDENTES E PARA A VARIÁVEL DE RESPOSTA CONVERSÃO (X%). ....................................................................... 110
TABELA 14. MATRIZ DO PLANEJAMENTO COMPOSTO CENTRAL ROTACIONAL COM OS VALORES REAIS E CODIFICADOS PARA AS
VARIÁVEIS INDEPENDENTES E PARA A VARIÁVEL DE RESPOSTA CONVERSÃO (X%), TEMPO REACIONAL DE 24HS. .................. 112
TABELA 15. EFEITOS ESTIMADOS PARA A ANÁLISE DA VARIÁVEL DE RESPOSTA CONVERSÃO NA RESOLUÇÃO CINÉTICA DE RAC-3 (APENAS
COM AS VARIÁVEIS QUE APRESENTARAM EFEITO SIGNIFICATIVO). .............................................................................. 113
TABELA 16. ANÁLISE DE VARIÂNCIA PARA DESENVOLVIMENTO EXPERIMENTAL DA RESOLUÇÃO CINÉTICA DE RAC-3.A ..................... 114
TABELA 17. RESULTADOS NA CONDIÇÃO ÓTIMA DE RAC-3 (45 °C, 4 MG/ML DA CONCENTRAÇÃO DE SUBSTRATO E 222,8 U DE
ATIVIDADE ENZIMÁTICA) EM SISTEMA DE BATELADA UTILIZANDO ESCALA DE MG E UMA ESCALA MAIOR (100X). ................. 122
TABELA 18. RESOLUÇÃO CINÉTICA DE RAC-3 EM ACETATE DE ETILA COMO SOLVENTE DA REAÇÃO EM SISTEMA DE FLUXO CONTÍNUO.
................................................................................................................................................................... 125
TABELA 19. RESOLUÇÃO CINÉTICA DE RAC-3 EM TBME COMO SOLVENTE DA REAÇÃO EM SISTEMA DE FLUXO CONTÍNUO. ............. 127
TABELA 20. RESOLUÇÃO CINÉTICA DE RAC-3/ACETATO DE VINILA (1:10) EM TBME CATALISADO PELA NOVOZYM 435 A 50OC EM
CONDIÇÕES DE FLUXO CONTÍNUO (TEMPO DE RESIDÊNCIA DE 3MIN) USANDO DIFERENTES AGENTES ACILANTES. ................. 129
TABELA 21. VALORES DE ATIVIDADE HIDROLÍTICA E PARÂMETROS DE IMOBILIZAÇÃO DE LIPB SOBRE DIFERENTES SUPORTES
POLIMÉRICOS. ................................................................................................................................................ 132
TABELA 22. ATIVIDADE DE ESTERIFICAÇÃO DA LIPB IMOBILIZADA SOBRE DIFERENTES SUPORTES POLIMÉRICOS. ........................... 135
TABELA 23. VALORES DE ATIVIDADE HIDROLÍTICA E PARÂMETROS DE IMOBILIZAÇÃO DE CALB E LIPB EM ACCUREL MP 1000. ...... 137
TABELA 24. RESULTADOS DA RESOLUÇÃO ENANTIOSSELETIVA DE RAC-3 PARA AS DIFERENTES VERSÕES DA LIPASE B DE CANDIDA
ANTARCTICA. .................................................................................................................................................. 138
TABELA 25. RESULTADOS DA RESOLUÇÃO ENANTIOSSELETIVA DE RAC-4 PARA AS DIFERENTES VERSÕES DA LIPASE B DE CANDIDA
ANTARCTICA. .................................................................................................................................................. 139

INDICE DE FIGURAS
FIGURA 1. ESTEREOISÔMEROS DO INOSITOL (REPRODUZIDO DE ALMEIDA ET AL., 2003). ....................................................... 10
FIGURA 2. PLANO DE SIMETRIA DO MIO-INOSITOL ILUSTRANDO COMO AS NOMENCLATURAS SÃO FEITAS PARA ESTA MOLÉCULA
(REPRODUZIDO DE POTTER E LAMPE, 1995). ..................................................................................................... 11
FIGURA 3. MECANISMO DE TRANSDUÇÃO CELULAR (REPRODUZIDO POR ALMEIDA ET AL., 2003). ........................................... 12
FIGURA 4. CASCATA DOS FOSFOINOSITÍDEOS (REPRODUZIDO POR ALMEIDA ET AL., 2003). .................................................... 14
FIGURA 5. O MIO-INOSITOL E SEU ANÁLOGO BIOATIVO 6FINS ............................................................................................. 16
FIGURA 6. MÉTODOS PARA OBTENÇÃO DE COMPOSTOS ENANTIOMERICAMENTE PUROS (REPRODUZIDO DE GHANEM ET AL., 2004).
..................................................................................................................................................................... 18
FIGURA 7. OS DOIS TIPOS DE CONVERSÃO ASSIMÉTRICA: A) QUANDO OCORRE EM ESTRUTURAS PROQUIRAL COM DUPLA LIGAÇÃO; B)
DISTINÇÃO ENTRE ÁTOMOS OU GRUPOS ENANTIOTÓPICOS. ....................................................................................... 18
FIGURA 8. RESOLUÇÃO CINÉTICA DINÂMICA DE Α-HIDROXI-ÉSTERES. SUPONDO QUE A CONVERSÃO DO ISÔMERO-R SEJA O MAIS
RÁPIDO, A REAÇÃO FICARÁ MAS LENTA PARA O OUTRO ISÔMERO (S). (MODIFICADO DE GHANEM ET AL., 2004). ............. 19
FIGURA 9. ESQUEMA CONSIDERADO PARA SISTEMAS DO TIPO MICHAELIANO, ASSUMINDO QUE A REAÇÃO É IRREVERSÍVEL E QUE OS
PRODUTOS GERADOS NÃO INTERFEREM NA REAÇÃO. ................................................................................................ 22
FIGURA 10. EFEITO DA ENANTIOSSELETIVIDADE ENZIMÁTICA DEMONSTRADA COMO A RELAÇÃO ENTRE A RAZÃO ENANTIOMÉRICA (E) E
O EXCESSO ENANTIOMÉRICO DO PRODUTO (EEP) (REPRODUZIDO POR STRAATHOF E JONGEJAN, 1997). ............................. 24
FIGURA 11. REAÇÃO DE HIDRÓLISE CATALISADA POR LIPASES. UM TRIACILGLICEROL PODE SER HIDROLISADO PARA FORMAR GLICEROL E
ÁCIDOS GRAXOS OU, NA DIREÇÃO REVERSA, EM AMBIENTES COM BAIXO TEOR DE ÁGUA, PODE SER FORMADO O ÉSTER (REAÇÃO
DE SÍNTESE). .................................................................................................................................................... 26
FIGURA 12. REAÇÕES CATALISADAS POR LIPASE. ............................................................................................................... 28
FIGURA 13. MODELO ESTRUTURAL DE Α/Β HIDROLASES. OXIÂNION: RESÍDUOS QUE ESTABILIZAM O OXIÂNION, NU: RESÍDUO
NUCLEOFILICO QUE É A SERINA PARA LIPASES, PROTEASES E ESTERASES. Α -HÉLICES ESTÃO REPRESENTADAS POR RETÂNGULOS, Β-
HÉLICES POR SETAS. (REPRODUZIDO DE BORNSCHEUER E KAZLAUSKAS, 2006B). ................................................... 31
FIGURA 14. MODELO PROPOSTO PARA OS SÍTIOS DE LIGAÇÃO DO SUBSTRATO À TRÊS LIPASES SINTETICAMENTE ÚTEIS. O RESÍDUO
CATALÍTICO SER ENCONTRA-SE NA PARTE INFERIOR DA CAVIDADE COM A SUA CATALÍTICA HIS À ESQUERDA. EMBORA OS
DETALHES DA CAVIDADE SE DIFEREM PARA CADA LIPASE, CADA CAVIDADE CONTÉM UMA GRANDE BOLSA HIDROFÓBICA
(CINZENTO CLARO) E UM MENOR BOLSO (CINZA MÉDIO), CONHECIDOS COMO "BOLSOS ESTEREOSELETIVOS”. AS REGIÕES QUE SE
LIGAM AO GRUPAMENTO ACILA DO ÉSTER DIFERE SIGNIFICATIVAMENTE ENTRE AS TRÊS ESTRUTURAS. PARA CRL, O
GRUPAMENTO ACILA LIGA-SE NUM TÚNEL (REPRESENTADO POR CINZA ESCURO ONDE ESTÁ ESCRITO TUNNEL MOUTH). NA CALB
E PCL, O GRUPAMENTO SE LIGA NO ENORME BOLSO HIDROFÓBICO DA CAVIDADE (REPRODUZIDO DE BORNSCHEUER E
KASLAUSKAS, 2006B). ................................................................................................................................... 32
FIGURA 15. CONFORMAÇÕES TAMPA FECHADA (AZUL) E TAMPA ABERTA (AMARELO) EM VÁRIAS LIPASES. REPRODUZIDO DE ALOULOU
ET AL., (2006). ................................................................................................................................................ 34
FIGURA 16. ESQUEMA ILUSTRANDO O FUNCIONAMENTO DA CATAPULTA ELETROSTÁTICA. O SÍTIO ATIVO COM POTENCIAL NEGATIVO
FORÇA, ATRAVÉS DE REPULSÃO ELETROSTÁTICA, A SAÍDA DO ÁCIDO CARBOXÍLICO FORMADO, QUE TAMBÉM POSSUI CARGA
NEGATIVA. MODIFICADO DE PETERSEN ET AL., (2001) POR ALMEIDA, (2005). ............................................................ 35
FIGURA 17 – MECANISMO CINÉTICO (PING PONG BI BI) DE REAÇÕES COM MÚLTIPLOS PRODUTOS E SUBSTRATOS CATALISADAS POR
LIPASES (E: ENZIMA; ES: ÉSTER; AL: ÁLCOOL; AC: ÁCIDO; W: ÁGUA; I = 1,2,...,I; J=1,2,...,J) (REPRODUZIDO DE PAIVA,
BALCÃO E MALCATA, 2000). ......................................................................................................................... 37
FIGURA 18. ESQUEMA DA REAÇÃO DE HIDRÓLISE DE LIGAÇÕES ÉSTER CATALISADA POR ESTERASES E LIPASES. ONDE TD1 E TD2
REPRESENTAM O PRIMEIRO E O SEGUNDO INTERMEDIÁRIOS TETRAÉDRICOS. (MODIFICADO A PARTIR DE BORNSCHEUER E
KAZSLAUSKAS, 2006B E SIMAS ET AL., 2011). ................................................................................................. 38
FIGURA 19. MERCADO MUNDIAL DE TECNOLOGIA QUIRAL POR SEGMENTOS: SÍNTESE QUIRAL, ANÁLISE QUIRAL E RESOLUÇÃO QUIRAL
(REPRODUZIDO DE DEWAN, 2012). ................................................................................................................... 43
FIGURA 20. REAÇÃO ENANTIOSSELETIVA DE (±)-9 CATALISADA PELA LIPASE DE PSEUDOMONAS FLUORESCENS, UTILIZANDO ACETATO DE
VINILA COMO AGENTE ACILANTE EM 75H A TEMPERATURA AMBIENTE (MODIFICADO DE EVANS ET AL., 1992). ................. 45

FIGURA 21. RESOLUÇÃO CINÉTICA DE (±)-2,3,4,9-TETRAHIDRO-1H-CARBAZOL-3-OL CATALISADO POR CALB UTILIZANDO ACETATO DE
VINILA EM THF A 250RPM (REPRODUZIDO POR BUSTO ET AL., 2012). ..................................................................... 45
FIGURA 22. RESOLUÇÃO RACÊMICA DE (1) CATALISADO POR CALB APÓS 120H DE REAÇÃO (REPRODUZIDO DE PARKER ET AL.,
2012). ........................................................................................................................................................... 46
FIGURA 23. SÍNTESE DE (R)-1 E APERFEIÇOAMENTO DA ENANTIOSSELETIVIDADE UTILIZANDO LIPASE DE PSEUDOMONAS CEPACIA
(MODIFICADO POR KAWANO ET AL., 2012). ....................................................................................................... 47
FIGURA 24. ESTRUTURA TRIDIMENSIONAL DA LIPASE B DE CANDIDA ANTARCTICA (CALB) EM SUA FORMA ABERTA (A) E FECHADA (B).
REGIÃO DA TAMPA HIDROFÓBICA (EM AZUL) REPRESENTA OS DOBRAMENTOS DA HÉLICE (Α5, COMPARTIMENTO SUPERIOR EM
(A) E Α10, COMPARTIMENTO INFERIOR EM (A) ). (REPRODUZIDO POR GANJALIKHANY ET AL., 2012). .......................... 48
FIGURA 25. MÉTODOS GERAIS DE IMOBILIZAÇÃO DE BIOCATALISADOR: 1-ADSORÇÃO FÍSICA; 2-LIGAÇÃO COVALENTE: A- DERIVADOS
MONOMÉRICOS MULTIPONTUAIS, B- DERIVADO MONOMÉRICO UNIPONTUAL, C- DERIVADO BIMOLECULAR MULTIPONTUAL (O
CÍRCULO PREENCHIDO DE AMARELO REPRESENTA O SÍTIO ATIVO); 3-CROSS-LINKING; 4- ENTRELAÇAMENTO EM MATRIX
POLIMÉRICA; 5- ENCAPSULAMENTO EM GEL; 6- LIGAÇÃO METÁLICA, 7-LIGAÇÃO POR PONTES DISSULFETO, 8-9: REATOR DE
MEMBRANA- 8- ENZIMA IMOBILIZADA NA CAMADA EXTERNA DA MEMBRANA (ITEM 8-MODIFICADO A PARTIR DE LAU ET AL.
2010); 9- MEMBRANA CONTÉM BIOCATALISADORES IMOBILIZADOS EM SEU INTERIOR (ITEM 9-MODIFICADO A PARTIR DE
NAGY, 2012). ................................................................................................................................................ 58
FIGURA 26. HIPERATIVAÇÃO DE LIPASES SOBRE SUPERFÍCIES ALTAMENTE HIDROFÓBICAS (MODIFICADO DE PALOMO ET AL., 2002).
..................................................................................................................................................................... 61
FIGURA 27. ENANTIOMERO RÁPIDO (A) E ENANTIOMERO LENTO (B)- MODELO DE SÍTIO ATIVO POR LIPASES (REPRODUZIDO DE
KAZLAUSKAS ET AL., 1991). ............................................................................................................................ 64
FIGURA 28. RESOLUÇÃO CINÉTICA DE AMINAS RACÊMICAS RAC -1A-C POR ACETILAÇÃO DO ACETATO DE ETILA CATALISADO POR CALB
SOB VARIADOS MÉTODOS DE IMOBILIZAÇÃO (MODIFICADO DE BOROS ET AL., 2013). .................................................. 66
FIGURA 29. RESOLUÇÃO CINÉTICA GLOBAL E PROCESSO DE PURIFICAÇÃO DO ENANTIÔMERO S EM SISTEMA DE FLUXO CONTÍNUO
(MODIFICADO DE TAMBORINI ET AL., 2012). ..................................................................................................... 67
FIGURA 30. EXEMPLOS DE AGENTES ACILANTES PARA ACILAÇÕES IRREVERSÍVEIS A PARTIR DE ÁLCOOIS (A-B-D:REPRODUZIDO DE
BORNSCHEUER E KAZLAUSKAS, 2006B; C- REPRODUZIDO DE SIMON ET AL., 2012). ........................................... 69
FIGURA 31. SÍNTESE DE OLIGOSSACARÍDEOS ARTIFICIAIS UTILIZANDO A LIPASE PS-30 E ACETATO DE VINILA DURANTE 11H DE TEMPO
REACIONAL (MODIFICADO DE BERKOWITZ ET AL., [1994] POR BORNSCHEUER E KAZLAUSKAS, 2006B). ................ 69
FIGURA 32. EFEITO DA ATIVIDADE DE ÁGUA NA ATIVIDADE ENZIMÁTICA DE LIPASES DE ORIGENS DIVERSAS: RHIZOPUS ARRHIZUS ( )
; CANDIDA RUGOSA ( ); PSEUDOMONAS SP. ( ) (REPRODUZIDO DE WEHTJE E ADLERCREUTZ, 1997). ............... 75
FIGURA 33. DERIVADOS DE MIO-INOSITOL UTILIZADOS NA RESOLUÇÃO CINÉTICA POR LIPASES. ................................................... 77
FIGURA 34. SÍNTESE DE POTENCIAIS SUBSTRATOS PARA LIPASES ........................................................................................... 78
FIGURA 35- MICROGRAFIA ÓPTICA E MICROGRAFIA ELETRÔNICA DE VARREDURA DOS SUPORTES PMMA-CO-DVB/ PMMA-CO-DVB
25% E PS-CO-DVB/PS-CO-DVB 25%. (REPRODUZIDO DE BESTETI ET AL., 2011). .................................................... 80
FIGURA 36. FOTO DA APARELHAGEM UTILIZADA NO SISTEMA OPERACIONAL EMPREGADO NAS REAÇÕES DE ACETILAÇÃO DOS DERIVADOS
RACÊMICOS. ..................................................................................................................................................... 81
FIGURA 37. REATOR ASIA SYSTEM, SISTEMA EM FLUXO CONTÍNUO DA EMPRESA SYRRIS MOSTRANDO A COLUNA UTILIZADA PARA AS
REAÇÕES: A) DETALHE DA BOMBA DO EQUIPAMENTO; B) COLUNA TERMOESTATIZADA. .................................................. 83
FIGURA 38. COLUNA DE LEITO FIXO UTILIZADA NO SISTEMA DE FLUXO CONTÍNUO ADQUIRIDO PELA EMPRESA SYRRIS, ONDE A) DETALHE
DO LÚMEN DA COLUNA ONDE O BIOCATALISADOR É COLOCADO; B) SETAS REPRESENTANDO A ALTURA QUE O BIOCATALISADOR
PREENCHE A COLUNA, EM CENTÍMETROS (CM), PARA CÁLCULO DO VOLUME DA COLUNA. ................................................ 84
FIGURA 39. CROMATOGRAMA REFERENTE AO PADRÃO DO SUBSTRATO (RAC-3) POR CLAE EM FASE REVERSA COM TR DE 8,178MIN
(REPRODUZIDO DE MANOEL, 2011). ................................................................................................................. 87
FIGURA 40 CROMATOGRAMA REFERENTE A MISTURA DO PADRÃO RACÊMICO (RAC-3) DO PRODUTO ANALISADOS POR CLAE EM FASE
REVERSA. O TR DE 12,456MIN É REFERENTE AO PRIMEIRO MONOACETILADO, O TR =13,296MIN É REFERENTE AO SEGUNDO. O
TR =21,797MIN É REFERENTE AO DIACETILADO E O TR = 38,745MIN AO TRIACETILADO. O TR DE 8,325MIN É REFERENTE AO
SUBSTRATO (REPRODUZIDO DE MANOEL, 2011). ................................................................................................. 88
FIGURA 41. CURVA DE CALIBRAÇÃO CALCULADA A PARTIR DA CURVA PADRÃO PARA O RAC-3 (REPRODUZIDO DE MANOEL, 2011). 88

FIGURA 42. CURVA DE CALIBRAÇÃO CALCULADA A PARTIR DA CURVA PADRÃO PARA O RAC-4 (REPRODUZIDO DE MANOEL, 2011)..
..................................................................................................................................................................... 89
FIGURA 43. CROMATOGRAMA EM SISTEMA DE FASE REVERSA (SHIMADZU C-18) DO PADRÃO DIOL. ........................................... 89
FIGURA 44. CROMATOGRAMA EM SISTEMA DE FASE REVERSA (SHIMADZU C-18) DA MISTURA SINTÉTICA DOS PADRÕES: DIOL; OS DOIS
POSSÍVEIS MONOÉSTERES E O DIÉSTER, VISUALIZADOS DA ESQUERDA PARA A DIREITA (MODIFICADO DE CUNHA 2011). ...... 90
FIGURA 45 – CROMATOGRAMA EM SISTEMA QUIRAL (CHIRACEL OD-H) DO PADRÃO DO PRODUTO (D)-ISÔMERO E (L)-ISÔMERO DO
(±)- 1,3,4-TRI-O-BENZIL-MIO-INOSITOL (RAC-3). REPRODUZIDO DE MANOEL, 2011. ................................................... 91
FIGURA 46. CROMATOGRAMA EM SISTEMA DE QUIRAL (CHIRACEL-OD-H) DO (D)-ISÔMERO E (L)-ISÔMERO DO RAC-4.
(REPRODUZIDO DE CUNHA, 2011). .................................................................................................................... 92
FIGURA 47- DERIVADOS DE MIO-INOSITOL UTILIZADOS NA RESOLUÇÃO CINÉTICA POR LIPASES. ................................................ 100
FIGURA 48. RESOLUÇÃO CINÉTICA ENZIMÁTICA DO RACEMATO RAC-3 CATALISADA POR DIFERENTES LIPASES HOME-MADE EM ACETATO
DE VINILA. ..................................................................................................................................................... 102
FIGURA 49. RESOLUÇÃO CINÉTICA ENZIMÁTICA DO RACEMATO RAC-4 CATALISADA POR DIFERENTES LIPASES HOME-MADE EM ACETATO
DE VINILA. ..................................................................................................................................................... 103
FIGURA 50. CROMATOGRAMA EM SISTEMA DE CLAE EM FASE REVERSA (SHIMADZU C-18) DA REAÇÃO DE ACETILAÇÃO DO RAC-3
CATALISADO PELA LIPB EM SUA FORMA LIOFILIZADA EM ACETATO DE VINILA APÓS 24 HORAS DE REAÇÃO. OS PICOS NOS
TEMPOS DE 8,102 E 12,04 SÃO SUBSTRATO E PRODUTO FORMADO, RESPECTIVAMENTE............................................... 105
FIGURA 51. CROMATOGRAMA EM SISTEMA DE CLAE EM FASE REVERSA (SHIMADZU C-18) DA REAÇÃO DE ACETILAÇÃO DO RAC-4
CATALISADO PELA LIPB EM SUA FORMA LIOFILIZADA EM ACETATO DE VINILA APÓS 24 HORAS DE REAÇÃO. OS PICOS NOS
TEMPOS DE 9,323 E 15,425 SÃO SUBSTRATO E PRODUTO FORMADO, RESPECTIVAMENTE............................................. 105
FIGURA 52. GRÁFICO DE PARETO APÓS O PLANEJAMENTO FATORIAL FRACIONÁRIO MOSTRANDO AS VARIÁVEIS SIGNIFICATIVAS PARA A
VARIÁVEL CONVERSÃO (X%), APÓS O TRACEJADO VERMELHO (* P-LEVEL < 0,1). ......................................................... 108
FIGURA 53. GRÁFICO DA PROBABILIDADE NORMAL DOS RESÍDUOS GERADO APÓS O DCCR DA ANÁLISE DA RESOLUÇÃO CINÉTICA DE
RAC-3. .......................................................................................................................................................... 115
FIGURA 54. GRÁFICO DA HOMOGENEIDADE DA VARIÂNCIA-VALORES OBSERVADOS X RESÍDUOS GERADO APÓS O DCCR DA ANÁLISE DA
RESOLUÇÃO CINÉTICA DE RAC-3. ........................................................................................................................ 115
FIGURA 55. GRÁFICO DE SHAPIRO E KOMOGOROV COM P= 0,9844 E P>0,20, RESPECTIVAMENTE. ......................................... 116
FIGURA 56. SUPERFÍCIE DE RESPOSTA (ESQUERDA) E GRÁFICO DE CONTORNO (DIREITA) NA INTERAÇÃO ENTRE AS VARIÁVEIS
TEMPERATURA E ATIVIDADE ENZIMÁTICA PARA A VARIÁVEL DE RESPOSTA CONVERSÃO (X%). O VALOR DA CONCENTRAÇÃO DO
SUBSTRATO FOI FIXADA EM 4MG/ML. ................................................................................................................. 117
FIGURA 57. SUPERFÍCIE DE RESPOSTA (ESQUERDA) E GRÁFICO DE CONTORNO (DIREITA) NA INTERAÇÃO ENTRE AS VARIÁVEIS
CONCENTRAÇÃO DE SUBSTRATO E ATIVIDADE ENZIMÁTICA, PARA A VARIÁVEL DE RESPOSTA CONVERSÃO (X%). O VALOR DA
TEMPERATURA FOI FIXADA EM 40º C. ................................................................................................................. 118
FIGURA 58. CROMATOGRAMA DA REAÇÃO DE ACETILAÇÃO DO RAC-3 COM A NOVOZYM 435 EM ACETATO DE VINILA E HEXANO. ... 119
FIGURA 59. CROMATOGRAMA DO PRODUTO L-(-)-2 OBTIDO NA REAÇÃO DE ACETILAÇÃO DO RAC-3 COM A NOVOZYM 435 EM
HEXANO. ....................................................................................................................................................... 119
FIGURA 60. CURVA DE PROGRESSO DA RESOLUÇÃO CINÉTICA DE RAC-3 EM HEXANO CATALISADA PELA NOVOZYM 435 NAS CONDIÇÕES
ÓTIMAS ENCONTRADAS NO PLANEJAMENTO EXPERIMENTAL: 45 °C, 4 MG/ML DE SUBSTRATO E 222,8U DE ATIVIDADE
ENZIMÁTICA. .................................................................................................................................................. 119
FIGURA 61. GRÁFICO DE CONTORNO PARA A VARIÁVEL DE RESPOSTA CONVERSÃO (X%) USANDO 220U DA LIPASE NOVOZYM 435.
VALORES DE CONCENTRAÇÃO DE SUBSTRATO FORAM FIXADAS (A) 4 MG/ML, (B) 8,5 MG/ML, AND (C) 14 MG/ML. .......... 120
FIGURA 62. PRODUTIVIDADE E EVOLUÇÃO DA CONVERSÃO NA RESOLUÇÃO CINÉTICA DE RAC-3 NAS CONDIÇÕES ÓTIMAS NÃO
OTIMIZADAS (MANOEL, 2011) E NAS CONDIÇÕES OTIMIZADAS (45 °C, 4 MG/ML DA CONCENTRAÇÃO DE SUBSTRATO E 222,8
U DE ATIVIDADE ENZIMÁTICA). .......................................................................................................................... 121
FIGURA 63. ESTABILIDADE OPERACIONAL NA RESOLUÇÃO CINÉTICA DE RAC-3 POR NOVOZYM 435 SOB AS CONDIÇÕES CONCEBIDAS
PARA: 4 MG / ML DE SUBSTRATO, 222,8 U DE ENZIMA, A 45 ° C DE TEMPERATURA EM SISTEMA DE BATELADA. ............... 123
FIGURA 64. RESOLUÇÃO CINÉTICA DE RAC-3 UTILIZANDO A LIPASE NOVOZYM 435: (A) SISTEMA EM BATELADA (B) SISTEMA DE FLUXO
CONTÍNUO EM LEITO FIXO. ................................................................................................................................ 124
FIGURA 65. RESOLUÇÃO CINÉTICA DE RAC-3 UTILIZANDO A LIPASE NOVOZYM 435, ACETATO DE VINILA COMO AGENTE ACILANTE E
ACETATO DE ETILA COMO SOLVENTE. ................................................................................................................... 125

FIGURA 66. RESOLUÇÃO CINÉTICA DE RAC-3 UTILIZANDO A LIPASE NOVOZYM 435, ACETATO DE VINILA COMO AGENTE ACILANTE
ACETATO DE ETILA COMO SOLVENTE. ................................................................................................................... 126
FIGURA 67. INTEGRIDADE DO REACTOR DE LEITO FIXO DEPOIS DE NOVE CICLOS NA RESOLUÇÃO CINÉTICA DE RAC-3 UTILIZANDO A
NOVOZYM 435 COMO BIOCATALISADOR, NAS MELHORES CONDIÇÕES EXPERIMENTAIS: TBME COMO SOLVENTE, ACETATO DE
VINILA, COMO DOADOR DE ACILA (PROPORÇÃO 1:10), A 50◦C. TEMPO DE RESIDÊNCIA: 3MIN. ...................................... 130
FIGURA 68. CINÉTICA DE IMOBILIZAÇÃO DA LIPB SOBRE OS SUPORTES ACCUREL MP 1000, PS-CO-DVB/PS-CO-DVB 25%, PMMA-
CO-DVB/PMMA-CO-DVB 25%. ATIVIDADE INICIAL DE 350U. ............................................................................. 134
FIGURA 69. CURVA DE ESTABILIDADE DA LIPB IMOBILIZADA SOBRE OS SUPORTES ACCUREL MP 1000, PS-CO-DVB/PS-CO-DVB
25% E PMMA-CO-DVB/ PMMA-CO-DVB 25%, ESTOCADA A 4 ºC. ..................................................................... 136
FIGURA 70. RESOLUÇÃO ENANTIOSSELTICA DE RAC-3 CATALISADA POR CALB COMERCIAL E LIPB (HOME-MADE) IMOBILIZADAS EM
DIFERENTES SUPORTES. .................................................................................................................................... 137
FIGURA 71. RESOLUÇÃO ENANTIOSSELTICA DE RAC-4 CATALISADA POR CALB COMERCIAL E LIPB (HOME-MADE) IMOBILIZADAS EM
DIFERENTES SUPORTES. .................................................................................................................................... 139

ÍNDICE DE ABREVIAÇÕES
CALB – lipase comercial fração B de Candida antarctica
LIPB- lipase B de Candida antarctica expressa em Pichia pastoris
E - razão enantiomérica.
eep - excesso enantiomérico do produto.
ees - excesso enantiomérico do substrato.
p-NFL – p-nitro fenil laurato.
rac-1 - 1,4,5,6-tetra-O-benzil-mio-inositol
rac-2 - 1,4,5,6-tetra-O-alil-mio-inositol
rac-3 - 1,3,4-tri-O-benzil-mio-inositol
rac-4 - (±)-3,6-O-dibenzil-1,2-O-isopropilideno-mio-inositol
Ins – Nomenclatura utilizada para identificação do mio-inositol de configuração D.
Px- Números de fosforilações presentes na estrutura do mio-inositol
g- grama

3
Capítulo I – INTRODUÇÃO
1. INTRODUÇÃO
O desenvolvimento de processos para a fabricação de compostos ativos e/ou
intermediários farmacêuticos muitas vezes envolve sínteses químicas complexas que podem
resultar em problemas, como a instabilidade da molécula nas condições de reação e/ou
formação de misturas racêmicas, quando a molécula de interesse é um dos enantiômeros. O
emprego de enzimas em etapas intermediárias da síntese de substâncias bioativas tem sido
utilizado para contornar estes problemas (JAEGER e REETZ, 1998; SCHULZE e
WUBBOLTZ, 1999; BEVILÁQUA et al., 2005).
Com a intensificação das ações dos órgãos de vigilância sobre as substâncias
racêmicas bioativas, principalmente as rigorosas inspeções praticadas pelo FDA ("Food and
Drug Administration") a partir de 1992, a maioria das companhias farmacêuticas foi obrigada
a desenvolver enantiômeros na sua forma estereoisomericamente pura (FABER e
FRANSSEN, 1993). O mercado de enantiômeros puros ocupa mais de dois terços do total e
vem crescendo em uma média anual de aproximadamente 7%, enquanto as substâncias
racêmicas vêm perdendo mercado numa média de 30% ao ano.
Durante os últimos anos, grandes esforços têm sido feitos para estabelecer rotas
enzimáticas na preparação de compostos enantiomericamente puros devido à sua importância
nas indústrias alimentícia, agrícola e principalmente, para a indústria farmacêutica. A maioria
das substâncias biologicamente ativas (fármacos, herbicidas, fungicidas, inseticidas,
aromatizantes e fragrâncias) são moléculas quirais (ZWINGENBERGER e WENDT, 1996;
NAIK et al., 2010). Em geral, apenas um dos enantiômeros possui a atividade biológica
desejada. O antípoda ótico pode não apresentar atividade alguma ou mesmo produzir efeitos
indesejáveis. O caso de maior repercussão foi o da talidomida, substância amplamente
receitada nos anos 60 contra os enjôos em gestantes e que causou o nascimento de milhares de
bebês com deformações congênitas. Posteriormente, descobriu-se que o efeito teratogênico da
talidomida era proveniente do emprego do enantiômero de configuração absoluta (S),
enquanto seu antípoda era desprovido de ação teratogênica em humanos
(ZWINGENBERGER e WENDT, 1997; LIMA, 2001).
A síntese de moléculas biologicamente ativas se tornou um desafio maior para o
mercado farmacêutico devido ao aumento da exigência de segurança e qualidade nos
produtos, a procura por processos industriais ambientalmente corretos e também devido a
enorme demanda por estes compostos obtidos em estratégias economicamente atrativas
(NESTL, NEBEL e HAUER, 2011; SOLANO, et al., 2012). Em 2002, por exemplo, a venda

4
Capítulo I – INTRODUÇÃO
de um determinado isômero em sua forma pura atingiu cerca de U$159 bilhões (ROUHI,
2003; GARCIA-URDIALES et al., 2011).
Dentre os muitos substratos que carecem de resolução quiral, encontra-se o mio-
inositol, um ciclocitol aquiral. A literatura mostra o interesse considerável na síntese de
derivados de mio-inositol (POTTER e LAMPE, 1995; ALMEIDA et al., 2003). Um dos
obstáculos para trabalhos de síntese orgânica e química medicinal relacionados aos inositóis é
a dificuldade de síntese destas substâncias (POTTER e LAMPE, 1995). Devido à sua
disponibilidade, boa parte dos estudos realizados nesta área fez uso do próprio mio-inositol
para a construção de derivados quirais desta substância. As rotas baseadas no uso do mio-
inositol têm como vantagem o uso de reações simples e disponibilidade de protocolos
estabelecidos. Entretanto, existe a necessidade de resolução óptica dos intermediários
racêmicos sintetizados quimicamente (POTTER e LAMPE, 1995; SURESHAN et al.,
2003a,b).
Lipases têm sido utilizadas com sucesso em reações que incluem resolução cinética de
álcoois racêmicos, ácidos, ésteres ou aminas, assim como dessimetrização de compostos pro-
quirais (ADER et al., 1992; CHEN e FANG, 1997; BERNSMANN et al., 2000; GOTOR-
FERNANDEZ, BUSTO e GOTOR, 2006; NAIK et al., 2010). Estes biocatalisadores
constituem uma classe de enzimas de grande interesse na pesquisa da biocatálise aplicada à
química orgânica devido às suas propriedades de especificidade e seletividade química,
posicional e estérica. A boa estabilidade das lipases na presença de solventes orgânicos
propicia sua utilização em etapas intermediárias de processos químicos convencionais e na
catálise de reações químicas envolvendo substratos insolúveis em meio aquoso (GOTOR et
al., 2006).
A variedade de sistemas reacionais e enzimas disponíveis para biocatálise e
biotransformações aumentou quase exponencialmente nos últimos 10 anos. Novas estratégias
como evolução dirigida, mutagênese (WATSON, 2012), engenharia metabólica, engenharia
genética, imobilização de enzimas etc. estão em desenvolvimento principalmente para tornar
o biocatalisador mais enantiosseletivo e a tecnologia mais econômica (MAGNUSSON et al.,
2005a; MAGNUSSON et al., 2005b; MARTON et al., 2010; BARBOSA et al., 2011; DÍAZ-
RODRÍGUEZ e DAVIS, 2011; BRUSTAD e ARNOLD, 2011; BOMMARIUS, BLUM e
ABRAHAMSON, 2011). Além destas estratégias, o estudo da variação de doadores de acila e
de solvente em reações de transesterificação também são empregados na otimização de
reações mostrando bons resultados na literatura (REETZ et al., 2002; LIU et al., 2009),
principalmente por serem técnicas que apresentam menor custo, maior flexibilidade e

5
Capítulo I – INTRODUÇÃO
facilidade na execução. Outras tecnologias vem sendo destacadas em biocatálise como a
irradiação de micro-ondas e estratégias que visam a eliminação parcial ou total de solventes
tóxicos ao ser humano e ao meio ambiente, por meio dos solventes não clássicos, como o uso
dos líquidos iônicos e fluidos supercríticos, em diferentes processos reacionais (LAU et al.,
2000; BERGLUND, 2001; TURNER, 2003; BORNSCHEUER e KZLAUSKAS, 2006a;
JAIN et al., 2005; ZHAO et al., 2009, 2010).
Dentro desta abordagem, o emprego de lipases em etapas intermediárias da síntese de
derivados de mio-inositol pode ser considerado uma alternativa atrativa para a obtenção de
derivados enantiomericamente puros ou enriquecidos.
1.1 JUSTIFICATIVA
O interesse na obtenção de derivados de mio-inositois enantiomericamente puros se
justifica pela sua grande versatilidade em síntese química, principalmente na síntese de
substancias bioativas e desempenhando papéis importantes como segundo mensageiro em
sistemas de contração muscular, metabolismo, secreção, entre outros (ALMEIDA et al.,
2003).
Estudos preliminares demonstraram a necessidade de uma maior pesquisa na área uma
vez que os inositóis polifosforilados são biologicamente importantes, mas poucos disponíveis
na natureza. Os conhecimentos sobre o sistema de fosfoinositídeos continuam a progredir
rapidamente; entretanto, muitos fatores ainda permanecem obscuros, como por exemplo, a
importância fisiológica da transformação do Ins(1,4,5)P3 (mio-inositol-1,4,5-trifosfato) em
diversos compostos fosforilados e a função destes metabólitos intermediários. Poucos
análogos dos inositóis polifosforilados sintetizados têm apresentado efeito significativo na
cascata dos fosfoinositídeos. Portanto, a síntese destes compostos é de fundamental
importância para que se possa conhecer melhor a cascata dos fosfoinositídeos e, talvez,
possibilitar a descoberta de algum composto com propriedades farmacológicas a partir destes
conhecimentos.
O mio-inositol é um componente essencial para o crescimento do agente etiológico da
doença de chagas, o Tripanossoma cruzi, participando da formação de lipídeos complexos
derivados do fosfatidil-inositol. O 1-D-6-desóxi-6-flúor-mio-inositol (Figura 5, pag. 10), um
análogo fluoretado de mio-inositol, é um forte inibidor da síntese de mio-inositol e,
conseqüentemente, inibe o crescimento do T. cruzi. Além disso, esta substância é altamente

6
Capítulo I – INTRODUÇÃO
seletiva contra o parasita, não interferindo sobre o crescimento de células de mamíferos
(EINICKER-LAMAS et al., 1999; OLIVEIRA et al., 2000).
Um dos racematos selecionados para a tese, (±)-1,3,4-tri- O-benzil-mio-inositol (rac-
3), é um substrato intermediário da síntese de D-2,4,5 tri-O-benzil-mio-inositol, uma estrutura
análoga ao D-1,4,5 tri-benzil-mio-inositol. Este composto é bem conhecido e descrito na
literatura por desempenhar um papel fundamental na mobilização de cálcio intracelular.
Como muitas células de mamíferos possuem receptores para D-1,4,5 tri-benzil-mio-inositol, o
estudo destes compostos, derivados de mio-inositol, são de extrema importância para um
melhor entendimento dos processos de sinalização celular (DESAI et al., 1994; MILLS e
POTTER, 2002).
Nos últimos anos, inúmeros artigos podem ser encontrados na literatura envolvendo a
síntese e/ou os estudos biológicos, bioquímicos, farmacológicos e imunológicos. dos inositóis
fosfatos, evidenciando que a pesquisa científica nesta área continua sendo promissora e de
grande relevância. Cunha et al., (2010) e Simas et al., (2011) testaram novos derivados de
mio-inositol utilizados pela primeira vez na resolução cinética catalisadas por lipases.
Recentemente, outros trabalhos de seleção de lipases, modelagem molecular e cinética de
resolução com derivados de mio-inositol foram publicados por nosso grupo de pesquisa e
relatam a obtenção bem sucedida de enantiômeros puros (CUNHA et al., 2011; MANOEL,
2011; MANOEL et al., 2012a; CUNHA et al., 2012). Entretanto ainda persiste a necessidade
do emprego de outros derivados como materiais de partida em resoluções cinéticas, os quais
precisam ser explorados para um maior compreensão do estudo em resolução de mio-
inositóis.
Dentro deste contexto, pretendeu-se dar continuidade ao conhecimento da resolução
cinética de diferentes racematos derivados de mio-inositol produzidos previamente por síntese
química, utilizando-os como substratos. Portanto, o trabalho aqui apresentado vem contribuir
principalmente para os estudos da biocatálise, no que diz respeito ao aumento da
enantiosseletividade de produtos de interesse farmacêutico a partir do estudo de fatores que
podem influenciar o processo reacional. Fatores como aqueles relacionados com o substrato
(tipo de grupo substituinte), com a enzima (liofilizada ou imobilizada) e condições reacionais
(tipo de solvente, agente acilante, temperatura e o uso de diferentes reatores). O efeito das
variáveis no processo foi determinado por meio de planejamentos experimentais, criando a
perspectiva de realização de um processo otimizado para futuro escalonamento.
No ponto de vista dos biocatalisadores, a literatura demonstra um grande potencial
biotecnológico para as lipases de Candida antarctica, Pyrococcus furiosus, Rhizomucor

7
Capítulo I – INTRODUÇÃO
miehei e de Yarrowia lipolytica (LARSEN et al., 2008, ROTTICCI-MULDER et al., 2001;
MARCHETTI, MIGUEL e ERRAZU, 2007; HA et al., 2007; RODRIGUEZ e
FERNANDEZ-LAFUENTE, 2010). Todavia, versões destas lipases produzidas pelos grupos
LaBiM/UFRJ, LaMMP/UFRJ e pelo BIOSE/UFRJ (via micro-organismos isolados ou de
forma heteróloga), têm se tornado promissoras. Estas são lipase de Candida antarctica
expressa em Pichia pastoris, Pyrococcus furiosus expressa em E.coli, Rhizomucor miehei e
lipase de Yarrowia lipolytica. Tais biocatalisadores são excelentes na síntese de inúmeras
moléculas com importâncias biotecnológicas distintas (Tabela 1), justificando o uso destes
neste trabalho.
Assim, nesta tese, investigou também a eficácia das lipases próprias (Tabela 1)
comparativamente às suas versões comerciais nas reações de hidrólise e esterificação de
derivados de mio-inositol.
O estudo ainda tem a finalidade de possibilitar o desenvolvimento de bioprodutos
nacionais em substituição aos atualmente importados e de elevado custo, viabilizando e
ampliando a utilização destes em diversos segmentos industriais.

8
Capítulo I – INTRODUÇÃO
Tabela 1. Lipases selecionadas produzidas pelo LaBiM/UFRJ e pelo BIOSE/UFRJ
Lipases Reação catalisada Aplicações
biotecnológicas Citação
LIPB-Lipase B de Candida
antarctica expressa em
Pichia pastoris
Hidrólise Biodiesel
Farmacêutica
GUTARRA et al., 2011.
CASTRO et al., 2009
(Pedido de Patente ao INPI:
PI0905122-8
Depositada 17/12/2009).
Lipase de Pyrococcus
furiosus expressa em E.coli ------------- -------------
BRANCO et al., 2010;
BRANCO, 2012.
Lipase de Rhizomucor
miehei (fermentado em
torta de babaçu)
------------- -------------
Ainda não há referencia.
Lipase vegetal de Jatropha
curcas L.
Acidólise e
hidrólise
Industria alimentícia e
produção de biodiesel
SOUZA, et al., 2009.
SOUZA, et al., 2010.
Lipase de Yarrowia
lypolitica
hidrólise de
triacilgliceróis,
esterificação,
transesterificação e
interesterificação
Produção de Biodiesel;
Biorremediação,
produção de
biossurfactante, na
indústria de alimentos;
detergentes; Cosméticos
e Oleoquímica; indústria
farmacêutica gerando
compostos com alta
pureza optica
MARTINS, 2012; FONTES,
2012; OSÓRIO et al., 2001;
ISO et al.,, 2001; GÜVENÇ
et al., 2002; AKIN et al.,
2003; UNDURRAGA et
al., 2001; OKAZAKI et
al.,1997; MARCHETTI et
al., 2007; HA et al., 2007;
SHARMA et al., 2001.

9
Capítulo II – OBJETIVOS
2. OBJETIVOS
Esta tese tem como foco principal o estudo de um novo biocatalisador com potencial
tecnológico na resolução cinética de derivados de mio-inositol. A pesquisa visa a obtenção de
produtos-alvo de relevância à indústria farmacêutica.
2.1- OBJETIVOS ESPECÍFICOS
Selecionar biocatalisadores produzidos pelo nosso grupo de pesquisa (Home-made), a
fim de se encontrar aqueles capazes de catalisar a reação com substratos selecionados.
Selecionar biocatalisadores comerciais que sejam produzidos pelos mesmos micro-
organismos, daqueles produzidos pelo nosso grupo de pesquisa, através da otimização
das condições de reação, por meio de planejamento de experimentos, a fim de
posterior comparação com as lipases home-made.
Otimização das condições operacionais em reatores de batelada simples e em reatores
de coluna de Leito Fixo com o melhor par lipase imobilizada/substrato.
Obtenção de produtos-alvo de relevância industrial que será dada pela imobilização da
enzima selecionada sobre suportes comerciais Accurel MP1000 e sobre os suportes
produzidos pelo LMSCP (COPPE/UFRJ) - PS-DVB/PS-DVB e PMMA-
DVB/PMMA-DVB visando obter um biocatalisador eficiente e inédito, tanto do ponto
de vista da enzima quanto do suporte.
Aplicação do biocatalisador selecionado na resolução cinética dos
intermediários/substratos do 6FIns, buscando o melhor par enzima
imobilizada/substrato.
Verificar o potencial de reuso do biocatalisador em cada condição de reação em
pequena e grande escala (escala de g). Análise da perda da atividade esterásica (ou
atividade hidrolítica) ao longo das reações de reuso e/ ou perda de proteína que pode
ser dessorvida do suporte.

10
Capítulo II – REVISÃO BIBLIOGRÁFICA
3. REVISÃO BIBLIOGRÁFICA
3.1- O MIO-INOSITOL
Inositóis são ciclitóis ou ciclo-hexanóis e, portanto, moléculas com esqueleto ciclo-
hexano, onde cada carbono encontra-se ligado a um grupo hidroxila. A orientação espacial
diferenciada das hidroxilas gera estereoisômeros, sendo nove os isômeros conhecidos. Os
isômeros mio-, neo-, D-quiro-, L-quiro- e scilo-inositol são de ocorrência natural, enquanto
cis-, epi-, allo- e muco-inositol são isômeros sintéticos não-naturais (Figura 1) (POTTER e
LAMPE, 1995; ALMEIDA et al., 2003).
Figura 1. Estereoisômeros do inositol (Reproduzido de ALMEIDA et al., 2003).

11
Capítulo II – REVISÃO BIBLIOGRÁFICA
Dos estereoisômeros do inositol, o mio-inositol é o mais abundante na natureza, sendo
produzido a partir da glicose. A molécula pode ser encontrada na sua forma livre e
combinada, principalmente, como inositóis fosfato. O mio-inositol possui somente uma
hidroxila na posição axial em C2 e um plano de simetria passando por C2 e C5, sendo
considerado um composto meso (estereoisômero que tem um número igual de grupos
identicamente ligados, sendo aquiral).
A incorporação de um substituinte em C1 (enantiotópico para C3) e/ou C4
(enantiotópico para C6) levam a um par de enantiômeros (Figura 1). Logo apesar desta
configuração (meso), seus derivados normalmente apresentam atividade óptica. Em
contrapartida a incorporação de um substituinte em C2 ou C5 resulta em um composto
aquiral, logo opticamente inativo (retenção do plano de simetria) (POTTER e LAMPE, 1995).
Quanto às regras de nomenclatura, os seis carbonos do anel de mio-inositol são
numerados da seguinte forma: conta-se a partir da 5ª posição substituída vizinha ao átomo de
carbono ligado ao grupo hidroxila axial. Caso a numeração se dê no sentido anti-horário, o
enantiômero obtido é o D-mio-inositol (por exemplo, D-1,4-IP2) (Figura 2), sendo L-mio-
inositol (por exemplo, L-1,4-IP2) aquele obtido pela numeração no sentido horário (IUPAC-
IUB, 1975 e 1989). O símbolo Ins é utilizado para o mio-inositol com configuração D
(ALMEIDA et al, 2003). Caso seja de configuração L, esta deve ser previamente mencionada.
A terminação Px em itálico indica o número de fosforilações presentes no inositol.
Figura 2. Plano de simetria do mio-inositol ilustrando como as nomenclaturas são feitas para esta molécula
(Reproduzido de POTTER e LAMPE, 1995).

12
Capítulo II – REVISÃO BIBLIOGRÁFICA
3.1.1 O papel do mio-inositol nas atividades biológicas
Os Inositóis apresentam participação na cascata dos fosfoinositídeos. O entendimento
desta cascata é indispensável a compreensão de como se dá a atividade biológica dos
compostos derivados dos inositóis.
As comunicações entre células nos organismos superiores são necessárias para o
controle do desenvolvimento celular, de suas organizações em tecidos e órgãos, de seus
crescimentos e de suas multiplicações necessárias também à coordenação de suas atividades.
O mecanismo de transdução de sinal é responsável por esta transmissão de
informações que hormônios e outros sinalizadores químicos, por exemplo, podem transportar
da superfície celular ao seu interior.
No sistema de comunicação celular, a primeira transmissão de sinal é dada por um
―primeiro mensageiro‖ (hormônio, fator de crescimento, etc.). Este é secretado e circula no
meio extracelular. O primeiro mensageiro é captado na superfície da célula por receptores que
lhe são específicos. A ocupação dos receptores, inseridos na membrana celular, dá início a
eventos complexos na membrana plasmática e no interior da célula até a liberação do segundo
mensageiro, ocasionando diversas respostas em nível celular (ALMEIDA et al, 2003). O
mecanismo de transdução celular está apresentado na Figura 3.
Figura 3. Mecanismo de transdução celular (Reproduzido por ALMEIDA et al., 2003).

13
Capítulo II – REVISÃO BIBLIOGRÁFICA
O primeiro contato do primeiro mensageiro ao receptor ativa a proteína G. Esta
constitui uma família de proteínas que está ligada ao nucleotídeo guanina, sendo acopladas
aos canais iônicos, ou a outras enzimas, controlando a liberação de novos mensageiros
intracelulares. Os novos mensageiros intracelulares são conhecidos como ―segundo
mensageiros‖ (mensageiros secundários) que constituem a última ligação da cadeia de
comunicação intracelular antes da resposta fisiológica sendo ponto de grande interesse para a
compreensão dos mecanismos de transdução do sinal.
A partir de uma estimulação induzida por um agonista, ao nível do receptor na
membrana, uma proteína G ativa a fosfolipase C (PLC), a qual catalisa a hidrólise da ligação
fosfodiéster do PI(4,5) P2, liberando no meio intracelular dois novos segundos mensageiros: o
mio-inositol 1,4,5-trifosfato [Ins(1,4,5)P3] e o sn-1,2-diacilglicerol. Este fica na membrana
plasmática e age ativando a proteína quinase C- PKC, responsável pelo estímulo de
numerosas proteínas intracelulares (NISHIZUKA, 1988; WOOTEN e WRENN 1984). A
liberação dos dois mensageiros supradescritos dispara a cascata de fosfoinositídeos, uma série
de reações seqüenciais (Figura 4).
O mio-inositol-1,4,5-trifosfato (Ins(1,4,5)P3) é um mediador na liberação de Ca2+
intracelular do retículo endoplasmático para o citoplasma. Ele ativa um receptor situado na
membrana externa do retículo endoplasmático ligado ao canal de cálcio. Esta ativação
provoca a abertura do canal de cálcio e, assim, a liberação de Ca2+
para o citoplasma. Sabe-se
que a capacidade dos inositóis fosfatos de provocar a liberação do cálcio depende do número
e da posição dos grupos fosfatos na molécula, e que a presença de um grupo fosfato na
posição C1 é indispensável para fazer a ligação com o receptor. Estudos de relação estrutura-
atividade indicam que a presença de grupos fosfatos vicinais nas posições C4 e C5 do mio-
inositol se mostra essencial para a liberação de Ca2+
(IRVINE et al., 1984; KOZIKOWSKI et
al., 1990).
Sabe-se também que a posição C2 no mio-inositol (hidroxila axial) tem um papel
particular, pois esta posição é importante para o reconhecimento dos inositóis fosfatos por
diferentes enzimas (BAKER et al., 1989).
Ins(1,4,5)P3, mediador na liberação Ca2+
, é amplamente encontrado em células
eucarióticas (KOZIKOWSKI et al., 1990) e responsável pela regulação de numerosos
processos celulares, como a secreção, o metabolismo e a proliferação (ALMEIDA et al.,,
2003). Entretanto, o seu papel continua a ser elucidado (KOZIKOWSKI et al., 1990). Além
disso, não se sabe qual é a importância fisiológica da transformação do Ins(1,4,5)P3 em

14
Capítulo II – REVISÃO BIBLIOGRÁFICA
diversos compostos fosforilados. Especula-se que eles tanto podem ser apenas metabólitos
intermediários, quanto também possuir atividade biológica própria (ALMEIDA et al., 2003).
Atualmente, sabe-se que existem duas vias para o metabolismo do Ins(1,4,5)P3. A
primeira via se inicia com uma desfosforilação específica, resultando em inositol 1,4-
bifosfato. Este bifosfato é seqüencialmente desfosforilado, principalmente via inositol 4-
fosfato até o inositol livre. Esta via, provavelmente, serve para finalizar o sinal do
Ins(1,4,5)P3. A segunda via começa com uma fosforilação específica do D-mio-inositol 1,4,5-
trifosfato, gerando o inositol 1,3,4,5-tetraquisfosfato. Algumas pesquisas sugerem que esse
inositol 1,3,4,5-tetraquisfosfato pode ter uma função própria como segundo mensageiro,
afetando o influxo de cálcio na célula do meio extracelular. O tetraquisfosfato é metabolizado
a um segundo trifosfato, o inositol 1,3,4-trifosfato, e os dois bifosfatos posteriores, os
inositóis 1,3- e 3,4-bifosfatos, são degradados a inositóis monofosfatos. Os monofosfatos são
convertidos em inositol livre, como no início do ciclo (Figura 4) (POTTER e LAMPE, 1995;
ALMEIDA et al., 2003).
Figura 4. Cascata dos fosfoinositídeos (Reproduzido por ALMEIDA et al., 2003).
A participação em processos celulares por parte de derivados do mio-inositol,
efetivamente como segundos mensageiros, despertou grande interesse no sentido de se obter
sinteticamente essas moléculas e de estudar a melhor forma para obtenção das mesmas.

15
Capítulo II – REVISÃO BIBLIOGRÁFICA
Todavia, como o enantiômero D é o mensageiro natural, os derivados de mio-inositol
sintetizados devem ser eficientemente resolvidos ou obtidos a partir de materiais de partida
quirais (POTTER e LAMPE, 1995).
Os análogos do mio-inositol 1,4,5-trifosfato constituem o objetivo principal dos
pesquisadores. O mio-inositol 1,4,5-tritiofosfato é um análogo fosforotioato agonista do
Ins(1,4,5)P3 pela mobilização do Ca2+
intracelular em uma grande variedade de sistemas. Este
derivado é resistente à desfosforilação catalisada pela 5-fosfatase, sendo o mais poderoso
inibidor já descrito desta enzima. Outros derivados halogenados e os mono- e ditiofosfatos
foram sintetizados e testados com o objetivo de melhorar a interação com as enzimas da
cascata, mas eles não trouxeram vantagens suplementares (ALMEIDA et al., 2003).
Estudos têm demonstrado que o mio-inositol, o isômero mais abundante no cérebro, e
os inositóis fosfatos podem representar uma via para o tratamento de diversas patologias
como Alzheimer e Síndrome de Down (SHIMOHAMA et al., 1998, HUANG et al., 1999,
apud ALMEIDA et al., 2003; MCLAURIN et al., 2000). Além disso, os análogos de D-3-
desoxi-mio-inositol são inibidores do crescimento de células v-sis-transformada NIH 3T3. Os
compostos mais ativos são D-3-desoxi-3-cloro-mio-inositol, D-3-desoxi-3-azido-mio-inositol
e D-3-desoxi-3-flúor-mio-inositol. O 5-desoxi e o 5-desoxi-5-flúor-D-mio-inositol associam-
se, respectivamente, 4 e 5 vezes mais facilmente a PI-sintetase que o mio-inositol natural, mas
não são transformados pela PI-sintetase no fosfatidil-inositol correspondente (ALMEIDA et
al., 2003).
Um dos obstáculos para trabalhos de Síntese Orgânica/Química Medicinal voltados
para inositóis é a dificuldade de síntese destas substâncias. Processos de etapas múltiplas
estão envolvidos (SURESHAN et al., 2003a,b; POTTER e LAMPE, 1995). A elaboração de
inositóis quirais não racêmicos pode ser realizada ou partir do abundante mio-inositol ou pelo
emprego de substâncias naturais quirais (e enantiopuras) (POTTER e LAMPE, 1995).
Aparentemente, a não ser em casos muito específicos, estas duas estratégias mostram-se
equiparáveis quanto à praticidade. As rotas baseadas no uso do mio-inositol têm como
vantagem o uso de reações mais simples e maior disponibilidade de protocolos estabelecidos
(SURESHAN et al., 2003a,b). Há, porém, nesta opção, a necessidade de resolução óptica de
intermediários racêmicos (POTTER e LAMPE, 1995).

16
Capítulo II – REVISÃO BIBLIOGRÁFICA
3.1.2 Mio-Inositol no tratamento da doença de chagas
Avanços no conhecimento das vias metabólicas do Trypanosoma cruzi têm permitido
identificar moléculas alvo para o desenvolvimento de fármacos mais eficientes e seguros para
o tratamento da doença de Chagas.
O mio-inositol é um componente essencial para o crescimento do T. cruzi,
participando da formação de lipídeos complexos derivados do fosfatidil-inositol. Estas
moléculas contendo mio-inositol estão presentes na superfície das células destes protozoários
flagelados em todos os estágios de sua vida e sua rota biossintética tem sido considerada um
potencial alvo terapêutico (EINICKER-LAMAS et al., 1999; OLIVEIRA et al., 2000). O 1-D-
6-desóxi-6-flúor-mio-inositol (6-FIns) (Figura 5), um análogo fluoretado de mio-inositol, é
um forte inibidor da síntese de mio-inositol e, conseqüentemente, inibe o crescimento do
Trypanosoma cruzi (OLIVEIRA et al., 2000). Além disto, esta substância é altamente seletiva
contra o parasita, não interferindo sobre o crescimento de células de mamíferos.
Figura 5. O mio-inositol e seu análogo bioativo 6Fins
3.2 ESTEREOSSELETIVIDADE: CONCEITO, ROTAS PARA
OBTENÇÃO DE COMPOSTOS ENANTIOMERICAMENTE PUROS E
CINÉTICA DOS ENANTIÔMEROS
A quiralidade é uma propriedade geométrica e diz se uma molécula que não pode ser
sobreposta à sua imagem especular é quiral, enquanto uma molécula aquiral é aquela em que
sua imagem especular pode ser sobreposta à original sem a quebra e formação de ligações.
Assim, compostos que apresentam centro assimétrico irão apresentar-se na forma de

17
Capítulo II – REVISÃO BIBLIOGRÁFICA
estereoisômeros. Este elemento de simetria, frequentemente, é o carbono tetraédrico no qual
estão ligados quatro diferentes grupamentos, ou seja, carbono quiral.
Os estereoisômeros são divididos em diastereoisômeros, cujas moléculas possuem
diferentes arranjos espaciais de seus ligantes, porém não são imagens especulares
sobreponíveis umas das outras, e enantiômeros, nos quais a molécula apresenta a geometria e
a disposição espacial de seus átomos igual à imagem especular do seu par complementar,
sendo que somente compostos quirais apresentam esta característica (MARCH, 1992;
CAREY e SUNDBERG, 2007).
Os enantiômeros têm propriedades físicas e químicas idênticas, além da mesma
solubilidade. Entretanto exibem diferentes comportamentos quirais fisiológicos e
bioquímicos. Frequentemente os enantiômeros causam respostas fisiológicas diferentes uma
vez que os receptores biológicos são quirais. Por exemplo, o odor dos enantiomeros R e S da
carvona, enquanto o primeiro isômero tem o odor de hortelã, o segundo tem o odor de óleo da
semente de cominho. A resposta fisiológica e a possibilidade de distintas funções na
farmacodinâmica, farmacocinética e atividades toxicológicas fazem o estudo dos
enantiômeros ser de suma importância (ANOVÁ e HUTTA, 2003; CAREY e SUNDBERG,
2007).
3.2.1 Rotas para a obtenção de compostos enantiomericamente puros
A preparação de compostos enantiomericamente puros para processos farmacológicos,
agroquímicos, entre outros, requer um número robusto de métodos disponíveis para a
preparação de um composto. Os métodos utilizados para a obtenção de compostos
enantiomericamente puros podem ser divididos em três categorias (GHANEM et al., 2004;
ANTHONSEN, 2005; AHMED et al., 2012):
i. Transformação química de produtos enantiomericamente puros (―Chiral pool‖
synthesis).
ii. Síntese assimétrica (a partir de substratos proquirais)
iii. Resolução de racematos
A escolha do método a ser utilizado vai depender do tipo do material de partida utilizado
(Figura 6).

18
Capítulo II – REVISÃO BIBLIOGRÁFICA
Figura 6. Métodos para obtenção de compostos enantiomericamente puros (Reproduzido de GHANEM et al.,
2004).
O método por meio da transformação química de substâncias enantiopuras de
interesse, conhecido como ―chiral pool‖, está relacionado a estratégia de se obter o aumento
da eficiência da síntese quiral a partir de substratos enantiomericamente puros. O termo
―chiral pool‖ refere a uma rota sintética de produtos naturais enantiomericamente puros (e
seus derivados) de forma econômica. Substratos tais como aminoácidos e acido láctico podem
ser transformados em produtos sintéticos quirais por rotas de síntese química. Por exemplo,
no processo de produção de L-acido ascórbico (vitamina C) a partir da D-glicose, o isômero L
da vitamina C é obtido, contudo a conversão ocorre com seis etapas (AGER, 2006).
Outra categoria seria aquela que inclui o método realizado com os substratos
proquirais. Nesta síntese, dois tipos podem ser distinguidos, como apresentados na Figura 7.
Figura 7. Os dois tipos de conversão assimétrica: a) quando ocorre em estruturas proquiral com dupla ligação; b)
Distinção entre átomos ou grupos enantiotópicos.

19
Capítulo II – REVISÃO BIBLIOGRÁFICA
Este método visa à obtenção de compostos quirais por síntese química ou por meio de
biocatalisadores. Desta maneira, a conversão é chamada de síntese assimétrica e se pode obter
até 100% de conversão do produto (diferente da resolução cinética que veremos a seguir).
Pelo fato de somente a metade do material de partida ser convertido à produto (50%) na
resolução cinética faz com que esta técnica seja considerada, por alguns autores, desvantajosa,
frente a síntese assimétrica (FABER, 2001).
Uma alternativa para o aumento da conversão em reações enantiosseletivas é a
racemização simultânea do material de partida, conhecida como resolução cinética dinâmica
(WARD, 1995; STECHER e FABER, 1997; FABER, 2001; REETZ et al., 2002;
PELLISSIER, 2003; SCHNELL et al., 2003). A combinação da acilação enantiosseletiva,
catalisada por lipase, com a racemização do substrato catalisada por rutênio (Ru) resulta numa
resolução cinética dinâmica de α-hidroxi-ésteres com rendimento teórico de 100% e alto
excesso enantiomérico (Figura 8) (GHANEM et al. 2004).
Figura 8. Resolução cinética dinâmica de α-hidroxi-ésteres. Supondo que a conversão do isômero-R seja o mais
rápido, a reação ficará mas lenta para o outro isômero (S). (Modificado de GHANEM et al., 2004).
A síntese assimétrica é geralmente catalisada por oxidoredutases e liases, enquanto a
resolução cinética é feita exclusivamente pelas hidrolases (STRAATHOF et al., 2002).
Na terceira categoria, encontra-se a fonte dos mais importantes racematos e será
melhor discutida na seção abaixo.
Resolução de racematos
Apesar do progresso impressionante na síntese assimétrica, o método de produção
dominante para se obter um único enantiómero na síntese industrial consiste na resolução de
racematos. Diferente da síntese assimétrica que se inicia com um substrato aquiral, a
resolução dos racematos se inicia com uma mistura de dois enantiômeros, onde apenas um
deles deve ser convertido, preferencialmente, por um (bio)catalisador (no caso o
biocatalisador pode ser enzima ou o próprio micro-organismo), sendo este considerado então

20
Capítulo II – REVISÃO BIBLIOGRÁFICA
enantiosseletivo (máxima conversão teórica do produto em 50%) (FABER, 2000,
STRAATHOF et al., 2002 e FABER, 2011).
Nos processos em batelada, quando há reações em equilíbrio, a reação precisa ser
interrompida antes que o enantiômero mais lento comece a ser convertido. A finalidade é
manter o enantiômero desejado na sua forma enantiomericamente pura. (CHEN et al., 1987).
A resolução dos enantiômeros pode ser dividida em três técnicas de separação, que
consistem em a) cristalização: cristalização preferencial direta e cristalização de sais
diastereoméricos; b) cromatografia e c) resolução cinética (Figura 6, pag. 12). Apesar da
técnica clássica ser a cristalização, a técnica mais utilizada, até o presente momento, é a
resolução cinética, catalisada por enzimas (FABER, 2001; SARIC et al., 2011). O sucesso do
método é dependente do fato de os dois enantiômeros reagirem com velocidades
diferenciadas. Então um dos enantiomeros é transformado em produto de forma mais rápida
que o outro (FABER, 2001; GHANEM et al., 2004).
Resolução de racematos derivados de mio-inositol, exemplos de cristalização e
cromatografia
As resoluções ópticas dos intermediários racêmicos normalmente são realizadas
empregando agentes de resolução quiral, que dependem da conversão destes intermediários
em diastereoisômeros, que são separados por cristalização ou cromatografia como já discutido
anteriormente. Um dos primeiros métodos disponíveis foi o procedimento de Shvets
(STEPANOV e SHVETS, 1979), que empregava ortoésteres de D-manose na resolução dos
derivados. Garegg e colaboradores (1985) separaram os enantiômeros de (±)-4-O-benzil-mio-
inositol utilizando o (S)-ácido O-acetil-mandélico como agente de resolução quiral, o
emprego desse auxiliar de quiralidade funciona bem para derivados de mio-inositol,
originando ésteres diastereoisoméricos facilmente separados por cromatografia.
Wewers e colaboradores (2005) conseguiram obter, por meio de duas etapas de
cristalização, os dois enantiômeros do derivado tetra-O-benzil-mio-inositol utilizando o
isômero D da cânfora. Ozaki e colaboradores (1986, 1987) foram os primeiros a utilizar como
agente de quiralidade derivados do L-mentol. Os autores sintetizaram o derivado D-mio-
inositol 1,4,5-tris(fosfato) opticamente ativo usando o cloreto do ácido mentoxiacético na
resolução. Entretanto, as desvantagens da utilização de agentes de quiralidade são:
possibilidade de migração do grupo na molécula; hidrólise do grupo na etapa de cristalização;

21
Capítulo II – REVISÃO BIBLIOGRÁFICA
número maior de etapas, com perda do rendimento total; e pouca regiosseletividade quando
realizada em dióis e polióis (WHITESELL e REYNOLDS, 1983).
Resolução de racematos: Derivados de mio-inositol catalisados por lipases
O uso de lipases na resolução cinética de derivados quirais de mio-inositol tem sido
um método muito atraente e empregado por diversos autores. Os intermediários racêmicos
podem ser hidrolisados ou esterificados de forma enantiosseletiva e separados por
cristalização ou cromatografia.
Ling e Ozaki (1994) utilizaram a lipase de Pseudomonas sp. (lipase CES) na resolução
cinética de 2,3-O-ciclohexilideno-mio-inositol em 1,4-dioxano utilizando anidrido acético
como agente acilante. A reação foi regiosseletiva e enantiosseletiva acetilando apenas o grupo
hidroxila da posição C-1 do L-isômero produzindo o L-1-Oacetil- 2,3-O-ciclohexilideno-mio-
inositol com 49% de conversão e 98% de excesso enantiomérico.
Chung e colaboradores (1998) obtiveram o produto monoacetilado L-4-O-acetil-
1,2:5,6-O-diisopropilideno-mio-inositol com 48% de conversão e 84% de excesso
enantiomérico, a partir da resolução cinética do diol 1,2:5,6-O-diisopropilideno-mio-inositol
catalisada pela lipase de Candida rugosa com anidrido acético em éter dietílico.
As principais vantagens do uso deste método são: alta especificidade e seletividade
química, posicional e estéreo apresentada pelas lipases, diminuição do número de etapas
necessárias na resolução, alto rendimento e condições suaves das reações evitando a formação
de subprodutos.
3.2.2 Cinética dos enantiômeros
Com o objetivo do reconhecimento quiral por enzimas, alguns modelos descrevem o
mecanismo de enantiosseletividade da reação enzimática pela capacidade da enzima de reagir
com diferentes taxas, com os dois enantiômeros, que se constituem em ―substratos
alternativos‖, tendo formação preferencial de um enantiômero do produto em relação ao outro
(FABER, 2000; STRAATHOF et al., 2002; FABER, 2011). Estes modelos usados no
tratamento quantitativo dos dados bioquímicos e acompanhados pela química sintética,
podem gerar predições úteis.
Chen e colaboradores (1982) foram os primeiros a encontrar relações úteis entre a
extensão de conversão do substrato racêmico (c), a pureza óptica expressa como o excesso
enantiomérico (ee) do produto ou do substrato remanescente, e a razão enantiomérica no

22
Capítulo II – REVISÃO BIBLIOGRÁFICA
estudo de resoluções de racematos (E). O parâmetro razão enantiomérica, como proposto
pelos autores tem um significado mais amplo do que a simples aplicação de misturas de
enantiômeros. Este parâmetro pode ser aplicado a qualquer reação enzimática com substratos
alternativos, enantiômeros ou não (OSTREICHER et al., 2001).
Para que o processo de resolução seja eficiente, é necessário que as constantes de
especificidade das reações para os enantiômeros sejam diferentes. Quanto maior a diferença,
maior o poder da resolução. Diversos processos como farmacêuticos, geoquímicos e
alimentícios requerem uma alta pureza óptica (eep maior que 98%) (STRAATHOF e
JONGEJAN, 1997). Logo esta abordagem é de grande interesse para estes segmentos.
Admitindo que A é o enantiômero que reage mais rapidamente e B é o enantiômero
que reage mais lentamente, e que ambos competem pelo mesmo sitio ativo da enzima, o
seguinte esquema (Figura 9) pode ser considerado para sistemas do tipo Michaeliano,
assumindo que a reação é virtualmente irreversível e que os produtos gerados não interferem
na reação:
Figura 9. Esquema considerado para sistemas do tipo Michaeliano, assumindo que a reação é irreversível e que
os produtos gerados não interferem na reação.
No estado estacionário, a razão entre as duas equações de velocidade é dada por:
B
A
K
K
V
V
v
v
A
B
B
A
B
A (Eq. 1)
onde VA, KA e VB, KB são velocidade máxima e constante de Michaelis dos substratos
A e B, respectivamente. A equação 1 pode ser integrada em função do tempo:
EKV
KV
BB
AA
BB
AA 0
0
ln
ln (Eq. 2)
onde, Ao, A, Bo, B, são as áreas dos cromatogramas dos substratos no início e no final
da reação, respectivamente.
A razão enantiomérica (E), parâmetro cinético que informa a especificidade da enzima
para os substratos alternativos, é a relação entre as duas eficiências catalíticas, Vmax/KM. Esta

23
Capítulo II – REVISÃO BIBLIOGRÁFICA
equação pode ser desenvolvida em termos de conversão (c ou X) e excesso enantiomérico do
substrato (ee(S)). Uma vez que a conversão do racemato é a soma das conversões dos
enantiômeros:
00
1BA
BAc
(Eq. 3)
E o excesso enantiomérico é a diferença das frações molares dos enantiômeros da
mistura com relação ao substrato, dado por:
BA
ABSee
(Eq. 4)
Logo, torna-se possível calcularmos a razão enantiomérica, substituindo as equações 3
e 4 na equação 2:
EKV
KV
Seec
Seec
BB
AA
11ln
11ln (Eq. 5)
Sendo assim, conclui-se que o parâmetro E, um parâmetro qualitativo e quantitativo
(COSTA e AMORIM, 1999), é dependente da razão das constantes de especificidade
(Vmax/KM) e é independente da concentração dos substratos. Este parâmetro nos mostra
quantas vezes um enantiômero é melhor substrato que o outro para a enzima utilizada
(STRAATHOF e JONGEJAN, 1997).
Quando o catalisador é altamente seletivo, 50% de conversão do produto desejado será
obtido enquanto 50% do substrato não reagido, na sua forma enantiopura, permanecerá até o
fim da reação.
Em experimentos de resolução cinética usando enzimas hidrolíticas, pode-se ainda
relacionar a extensão da conversão (c ou X) e do excesso enantiomérico do produto (ee(P)) ao
valor da razão enantiomérica (E). Esta relação pode ser obtida convertendo a equação 2 na
equação 6:
EKV
KV
Peec
Peec
BB
AA
11ln
11ln (Eq. 6)
onde o excesso enantiomérico com relação ao produto é dado por:
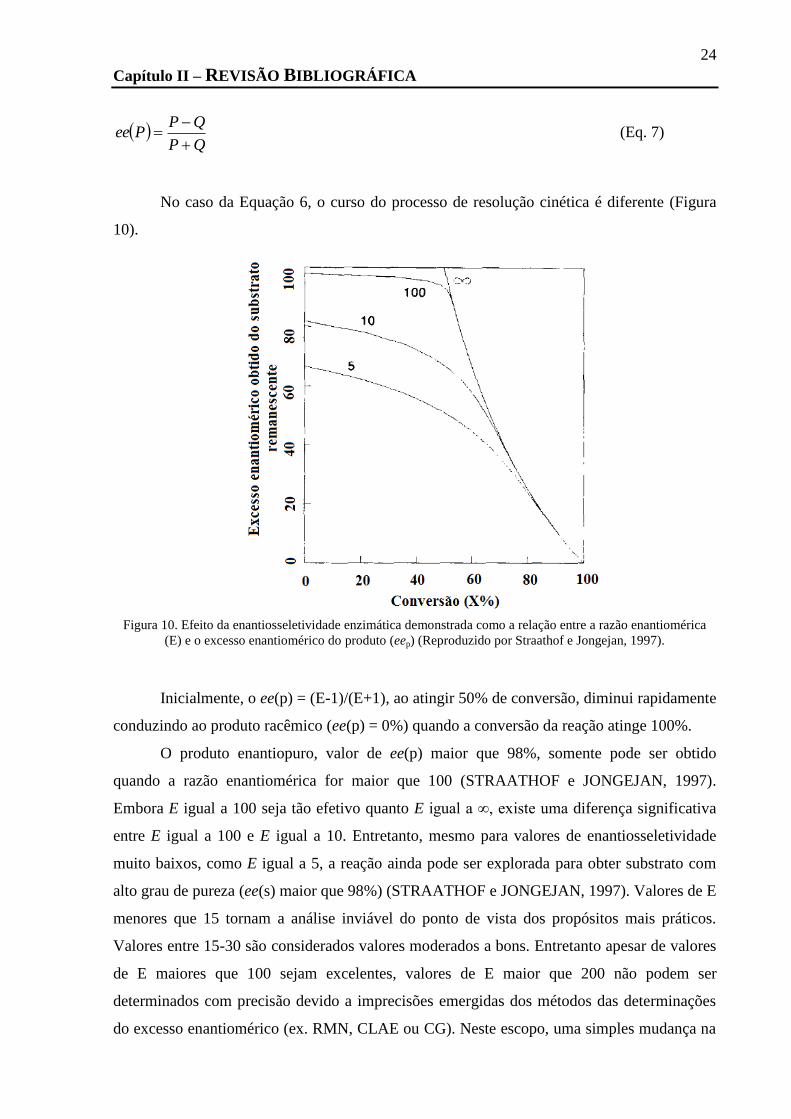
24
Capítulo II – REVISÃO BIBLIOGRÁFICA
QP
QPPee
(Eq. 7)
No caso da Equação 6, o curso do processo de resolução cinética é diferente (Figura
10).
Figura 10. Efeito da enantiosseletividade enzimática demonstrada como a relação entre a razão enantiomérica
(E) e o excesso enantiomérico do produto (eep) (Reproduzido por Straathof e Jongejan, 1997).
Inicialmente, o ee(p) = (E-1)/(E+1), ao atingir 50% de conversão, diminui rapidamente
conduzindo ao produto racêmico (ee(p) = 0%) quando a conversão da reação atinge 100%.
O produto enantiopuro, valor de ee(p) maior que 98%, somente pode ser obtido
quando a razão enantiomérica for maior que 100 (STRAATHOF e JONGEJAN, 1997).
Embora E igual a 100 seja tão efetivo quanto E igual a ∞, existe uma diferença significativa
entre E igual a 100 e E igual a 10. Entretanto, mesmo para valores de enantiosseletividade
muito baixos, como E igual a 5, a reação ainda pode ser explorada para obter substrato com
alto grau de pureza (ee(s) maior que 98%) (STRAATHOF e JONGEJAN, 1997). Valores de E
menores que 15 tornam a análise inviável do ponto de vista dos propósitos mais práticos.
Valores entre 15-30 são considerados valores moderados a bons. Entretanto apesar de valores
de E maiores que 100 sejam excelentes, valores de E maior que 200 não podem ser
determinados com precisão devido a imprecisões emergidas dos métodos das determinações
do excesso enantiomérico (ex. RMN, CLAE ou CG). Neste escopo, uma simples mudança na

25
Capítulo II – REVISÃO BIBLIOGRÁFICA
variação do excesso enatiomérico causa uma mudança significativa no valor numérico de E
(SIH et al., 1982; FABER, 1997).
Embora a qualidade do produto da resolução de um racemato seja caracterizada pelo
excesso enantiomérico, a razão enantiomérica (E) é um parâmetro de grande importância,
uma vez que a enzima apresentará uma eficiência catalítica própria para cada substrato. Um
valor elevado de E, para um determinado par enzima-substrato, é essencial para a constatação
do sucesso da resolução enantiomérica (E).
Freqüentemente, quando não existe uma enzima apropriada para uma dada reação,
então a reação deve ser explorada para se obter o substrato remanescente enantiopuro ao invés
do produto enantiopuro (STRAATHOF e JONGEJAN, 1997).

26
Capítulo II – REVISÃO BIBLIOGRÁFICA
3.3 LIPASES
3.3.1 Características das lipases
As lipases (glicerol éster hidrolases - E.C. 3.1.1.3) compreendem um grupo de
enzimas hidrolíticas que atuam, geralmente, na interface água/lipídeo catalisando a hidrólise
de ligações éster carboxílicas presentes em triacilgliceróis (TG) resultando na formação de
diacilgliceróis (DG), monoacilgliceróis (MG), ácidos graxos (AG) e glicerol (Figura 13)
(JAEGER et al., 1994; SHARMA et al., 2001). Entretanto, as lipases também podem catalisar
a reação de síntese de mono, di ou triacilglicerol a partir de ácidos graxos e glicerol em
condições em que a água é restrita no meio reacional (SAXENA et al., 1999).
Figura 11. Reação de hidrólise catalisada por lipases. Um triacilglicerol pode ser hidrolisado para formar glicerol
e ácidos graxos ou, na direção reversa, em ambientes com baixo teor de água, pode ser formado o éster (reação
de síntese).
Os principais substratos das lipases são os triglicerídeos de cadeia longa (com mais de
dez átomos de carbono), enquanto as enzimas que hidrolisam triacilgliceróis compostos de
ácidos graxos de cadeia curta (menos de dez átomos de carbono) são reconhecidas como as
esterases-carboxil éster hidrolases (E.C. 3.1.1.1) (JAEGER, DIJKSTRA e REETZ, 1999;

27
Capítulo II – REVISÃO BIBLIOGRÁFICA
NINI et al., 2001; SHARMA et al., 2001; BORNSCHEUER, 2002; CASTRO-OCHOA et al.,
2005). Como produto da reação das hidrolases, há a liberação de ácidos graxos e glicerol
(Figura 11) (JAEGER, RANSAC e DIJKSTRA, 1994; JAEGER; DIJKSTRA; REETZ, 1999).
Além dos substratos citados, as lipases também catalisam uma vasta quantidade de
ésteres não lipídicos (substratos não naturais), sendo ativas não apenas em meio aquoso mas
também em solventes orgânicos, permitindo a transformação em compostos não solúveis em
água enquanto mantêm sua alta regio-, quimio e enantiosseletividade, além de não
requisitarem cofatores para realização da catálise enzimática. Esta é uma das características
que tornam as lipases catalisadores ideais para a síntese orgânica (BORNSCHEUER e
KAZLAUSKAS 2006; PRIEGO et al., 2009; de CARVALHO et al., 2011).
A facilidade com que estas enzimas aceitam uma gama de substratos não naturais e de
tamanhos diversos sugere que a espinha dorsal polipeptídica seja flexível, podendo adotar
diferentes conformações. Como consequência, a baixa barreira de energia que é necessário
para que ocorram as mudanças conformacionais dificulta a modelagem e a previsão de
interações estereoquímicas para este grupo de biocatalisadores (COSTA e AMORIN, 1999).
As lipases são amplamente encontradas na natureza, principalmente devido à sua
importante função no metabolismo de lipídeos. Estas podem ser obtidas a partir de fontes
animais (lipase pancreática, por exemplo), vegetais (extraída de soja, do centeio e do algodão,
por exemplo) e microbiana. Esta última, do ponto de vista econômico e industrial, é a fonte
mais utilizada para obtenção destas enzimas. Isso devido à relativa facilidade de produção
(beneficiada pelo rápida taxa de crescimento), maior variedade das espécies microbianas
disponíveis para sintetizá-las, alto rendimento de conversão de substrato em produto, grande
versatilidade e maior simplicidade na manipulação ambiental e genética, além da utilização de
meios de baixo custo e mais estáveis que suas correspondentes de plantas e animais
(BALCÃO et al., 1996; PANDEY et al., 1999a,b; HASAN et al., 2006; ILLANES, 1994;
FREIRE e CASTILHO, 2010). Dos micro-organismos utilizados, os fungos são
especialmente valorizados porque as enzimas por eles produzidas normalmente são
extracelulares, o que facilita a sua recuperação do meio de fermentação e também porque a
maioria dos fungos não é nociva à saúde humana, (GRAS - Generally Regarded as Save),
JAEGER, RANSAC e DIJKSTRA, 1994).
Os processos fermentativos vêm sendo os principais meios utilizados na produção de
lipases. Isso porque estes apresentam maior facilidade de controle, maior capacidade
produtiva e custo de obtenção reduzido (BALCÃO et al., 1996; PANDEY et al., 1999a).
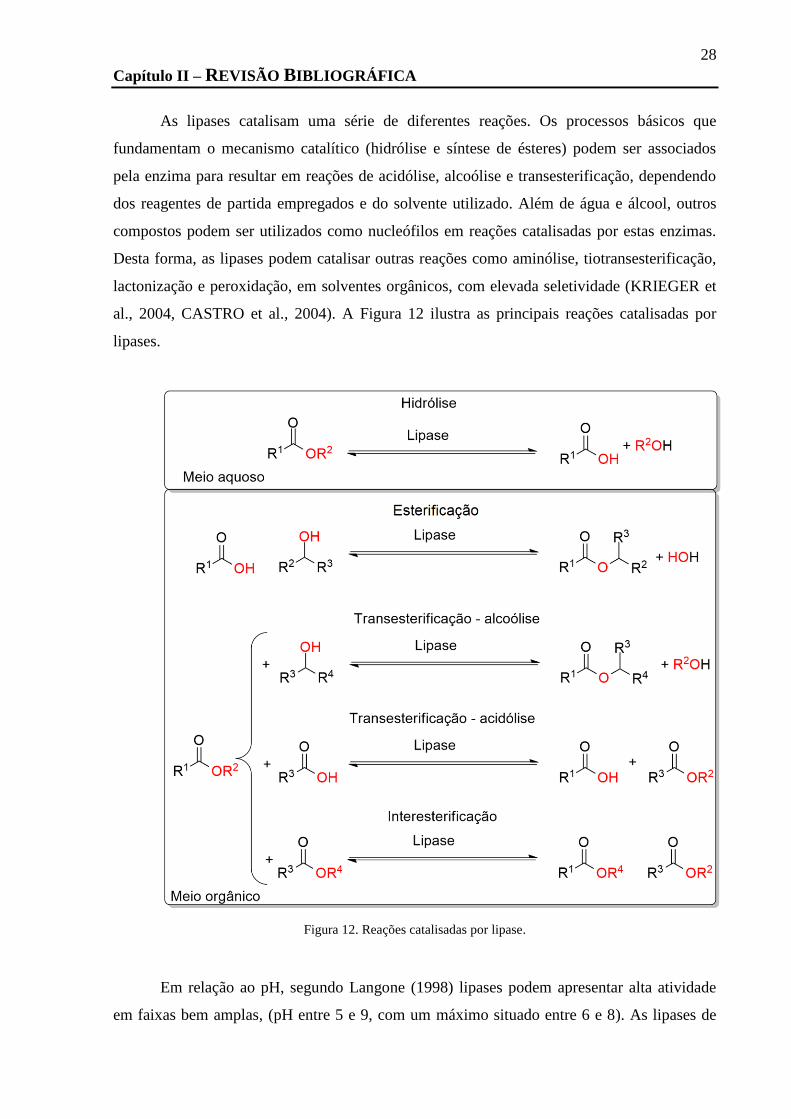
28
Capítulo II – REVISÃO BIBLIOGRÁFICA
As lipases catalisam uma série de diferentes reações. Os processos básicos que
fundamentam o mecanismo catalítico (hidrólise e síntese de ésteres) podem ser associados
pela enzima para resultar em reações de acidólise, alcoólise e transesterificação, dependendo
dos reagentes de partida empregados e do solvente utilizado. Além de água e álcool, outros
compostos podem ser utilizados como nucleófilos em reações catalisadas por estas enzimas.
Desta forma, as lipases podem catalisar outras reações como aminólise, tiotransesterificação,
lactonização e peroxidação, em solventes orgânicos, com elevada seletividade (KRIEGER et
al., 2004, CASTRO et al., 2004). A Figura 12 ilustra as principais reações catalisadas por
lipases.
Figura 12. Reações catalisadas por lipase.
Em relação ao pH, segundo Langone (1998) lipases podem apresentar alta atividade
em faixas bem amplas, (pH entre 5 e 9, com um máximo situado entre 6 e 8). As lipases de

29
Capítulo II – REVISÃO BIBLIOGRÁFICA
cepas do gênero Pseudomonas apresentam atividade máxima em pH entre 6,0 e 9,0 (HAKI e
RAKSHIT, 2003). Conforme encontrado em muitos trabalhos acadêmicos, a maioria das
lipases bacterianas mostra uma tendência de maior estabilidade em pH básico. Todavia, esse
comportamento pode ser diferente conforme se altere a espécie microbiana. São exemplos de
lipases estáveis em ampla faixa de pH a lipase de Bacillus megaterium, estável em pH de 4 a
12, e a lipase de Staphylococcus epidermidis, estável entre pH entre 2 e 10 (SIMONS et al.,
1998; RUIZ et al., 2002).
Na área industrial existem muitos processos que ocorrem em temperaturas em torno de
50º C. Uma vez que a termostabilidade é uma característica requerida para as enzimas
utilizadas no ramo industrial, faz-se necessário o seu estudo. A estabilidade pode variar
dependendo da espécie, gênero e até entre isoformas produzidas por uma mesma cepa. A
maioria das lipases apresenta temperatura de atividade ótima na faixa entre 30 e 40o
C, outras
apresentam atividade ótima em temperaturas mais elevadas, entre 40 e 70°C. Algumas lipases
apresentam atividade acima de 70oC, como as de Bacillus thermoleovorans e Bacillus
thermocatenulatus (CHO, YOO e KIM, 2000; QUYEN, SCHMIDT-DANNERT e SCHIMD,
2003). Rathi et al., (2000) mencionam ainda a atividade ótima para a lipase de Pseudomonas
sp. de 90 oC.
3.3.2 Principais micro-organismos utilizados para lipases
Fungos de diversos gêneros demonstraram ser bons produtores de lipases e as suas
enzimas têm sido estudadas sob o ponto de vista acadêmico e industrial. Por exemplo, lipases
de Aspergillus niger, A. oryzae, Mucor javanicus, Rhizopus niveus, R. oryzae, Penicillium
camembertii, P. roqueforti e da levedura Candida rugosa são comercializadas atualmente pela
Amano (Amano Europe Enzyme Ltd. UK) para processamento de óleos, gorduras e queijos,
para a determinação de triglicerídeos, como aditivos em preparações digestivas e para síntese
quiral (http://www.amano-enzyme.co.jp/).
Dentre as bactérias produtoras de lipases comercialmente estão disponíveis as enzimas
de Pseudomonas sp., P. fluorescens, Bulkholderia (Pseudomonas) cepacia e Arthrobacter sp.
para a aplicação em síntese quiral. O rápido crescimento celular, em relação aos fungos é uma
das vantagens citadas das fontes bacterianas como produtoras destas enzimas (JAEGER,
DIJKSTRA e REETZ, 1999). Apesar de lipases produzidas por vegetais terem sido
purificadas e estudadas quanto às suas características bioquímicas e quanto o seu potencial em
biotransformação de lipídeos, estas são pouco exploradas em escala industrial. Dentre as
enzimas lipolíticas vegetais estudadas estão as triacilglicerol lipases, as acil-hidrolases não

30
Capítulo II – REVISÃO BIBLIOGRÁFICA
específicas como as fosfolipases A1, A2 e B, as glicolipases, as sulfolipases e
monoacilglicerol lipases, além das fosfolipases C e D (MUKHERJEE, 1994). Entre as
espécies estudadas estão Ricinus communis, Vernonia, Pinus, Brassica napus, Cuphea
racemosa (HELLYER, CHANDLER e BOSLEY, 1999), Euphorbia characias e E. wulfenii
(PALOCCI, FIORILLO e MONACHE, 2003; CAVALCANTI et al., 2007).
Lipases humanas têm sido largamente estudadas devido às implicações que sua
estimulação ou inibição têm sobre a saúde humana, como problemas associados com a
obesidade. São elas lipases gástrica, pancreática, a estimulada por sais biliares que ajudam na
digestão e assimilação de lipídeos da dieta, e as lipases hepática e endotelial, as quais atuam
no metabolismo de lipoproteínas. (MILED et al., 2000, ROUSSEL et al., 2002, VAGANAY
et al., 1998).
O interesse na medicina está principalmente na inativação da atividade lipolítica para
controlar a infecção de micro-organismos, na inibição da lipase digestiva para controle da
obesidade, na estimulação da lipase metabólica para diminuir a lipidemia ou no uso de lipase
pancreática suplementar para controle da fibrose cística (MUKHERJEE, 2003). Dentre outras
lipases animais estudadas, pode-se destacar a lipase pancreática canina (STEINER e
WILLIAMS, 2002); as lipases ácidas lisossomais humana e de rato (GROENER et al., 2000);
a fosfolipase pancreática de peru (BEN SALAH et al., 2003) e as pancreáticas de esquilo
(Spermophilus tridecemlineatus) (SQUIRE et al., 2003), gato (Felis catus) (STEINER,
WILSON e WILLIAMS, 2003) e bovina (YAQOOB, NABI e MASSOM-YASINZAI, 1999).
3.3.3 Estrutura proteica de lipases
Até o momento, as lipases microbianas caracterizadas apresentam massa molar entre
19 e 60 kDa e apresentam uma estrutura terciária comum com um dobramento de α/β
hidrolase (Figura 13). Embora lipases difiram significativamente nas suas sequências de
aminoácidos, 11 lipases, cujas estruturas foram resolvidas, mostraram semelhantes estruturas
3-D (CAMBILLAU e TILBEURGH, 1993; CYGLER, SCHRAG e ERGAN, 1992;
DEREWENDA et al., 1994a,b; DODSON, LAWSON e WINKLER, 1992; RANSAC et al.,
1996; BORNSCHEUER e KAZLAUSKAS, 2006b). O dobramento α/β hidrolase consiste em
uma estrutura que possui um núcleo formado por uma folha β central, consistindo de oito
diferentes fitas β (β1-β8), conectadas com seis α hélices (A-F) (POUDEROYEN et al., 2001).
As fitas β têm orientação para a esquerda.

31
Capítulo II – REVISÃO BIBLIOGRÁFICA
Apesar das lipases apresentarem baixa homologia na seqüência de aminoácidos
(estrutura primária), estas enzimas apresentam uma alta similaridade em sua arquitetura
tridimensional (estruturas secundária e terciária). Assim como as outras hidrolases, as lipases
também possuem uma tríade catalítica, em seu sítio ativo formada por três resíduos: um
nucleófilo (serina), um ácido (aspartato ou glutamato) e um aceptor/doador de próton
(histidina) que ocorrem, necessariamente, nesta ordem na sequência primária (JAEGER,
DIJKSTRA e REETZ, 1999).
O nucleófilo catalítico, serina é responsável pela catálise e está unido por ligações de
hidrogênio a um resíduo de histidina; o resíduo carboxilado ligado ao mesmo resíduo de
histidina poderá ser um aspartato ou glutamato (EGGERT et al., 2001; JAEGER e REETZ,
1998; SCHRAG et al., 1991; EGLOFF et al., 1995; JAEGER, RANSAC e DIKSTRA, 1994;
JAEGER, DIJKSTRA e REETZ, 1999). A serina catalítica está localizada no C-terminal da
fita β5, faz parte de um pentapeptídeo altamente conservado GXSXG, onde G= glicina; S=
serina; X1= histidina e X2= ácido glutâmico ou aspártico (JAEGER e REETZ, 1998).
Figura 13. Modelo estrutural de α/β hidrolases. Oxiânion: resíduos que estabilizam o oxiânion, Nu: resíduo
nucleofilico que é a serina para lipases, proteases e esterases. α -hélices estão representadas por retângulos, β-
hélices por setas. (Reproduzido de BORNSCHEUER e KAZLAUSKAS, 2006b).
Além da região que compõe o sítio ativo, para a ligação do substrato, existe uma
estrutura anfipática móvel que cobre o sítio ativo catalítico na estrutura da maioria das lipases,
chamada região de tampa hidrofóbica ou lid, que está envolvida na seletividade de lipases
(SECUNDO et al., 2006). Em lipases, esta estrutura está presente na posição β8, que
prolonga-se sobre o conjunto central de fitas β e sobre o sítio catalítico, sendo chamada de
tampa hidrofóbica. Segundo Ghanem (2007), a provável função da tampa hidrofóbica é na
interação com a interface lipídeo-água, no fenômeno de ativação interfacial, onde esta

32
Capítulo II – REVISÃO BIBLIOGRÁFICA
estrutura sofre uma mudança conformacional, expondo o sítio ativo e favorecendo a ligação
com o substrato.
3.3.4 Sítios de ligação do substrato em lipases
Estudos utilizando as técnicas de cristalografia de raios-X mostraram que estruturas
em estado de transição análogas, ligadas ao sítio ativo de lipases, têm identificado distintos
sítios de ligação para a porção álcool e para a porção ácido (oriundos de esteres). O sítio de
ligação da porção álcool é similar para todas as lipases. O sítio ativo é representado por duas
cavidades de diferentes tamanhos, uma grande bolsa hidrofóbica, que está aberta para o
solvente, e uma pequena bolsa que está em direção a base da cavidade. Sabe-se que a forma
desta bolsa define a estereosseletividade de lipases em relação a álcoois secundários. O sítio
de ligação para a porção ácido varia consideravelmente entre as lipases. Para CRL (lipase de
Candida rugosa) o grupo acila se liga em um túnel de comprimento suficiente para acomodar
uma cadeia de pelo menos 18 átomos de carbono. RML (lipase de Rhizomucor miehei) e
CALB (lipase de Candida antarctica), possuem uma região com um espaço menor. Em todas
as três estruturas o carbono α da cadeia acila se liga imediatamente abaixo da grande região
hidrofóbica do sítio de ligação de álcool (Figura 14) (BORNSCHEUER e KAZLAUSKAS,
2006b).
Figura 14. Modelo proposto para os sítios de ligação do substrato à três lipases sinteticamente úteis. O resíduo
catalítico Ser encontra-se na parte inferior da cavidade com a sua catalítica His à esquerda. Embora os detalhes
da cavidade se diferem para cada lipase, cada cavidade contém uma grande bolsa hidrofóbica (cinzento claro) e
um menor bolso (cinza médio), conhecidos como "bolsos estereoseletivos‖. As regiões que se ligam ao
grupamento acila do éster difere significativamente entre as três estruturas. Para CRL, o grupamento acila liga-se
num túnel (representado por cinza escuro onde está escrito tunnel mouth). Na CALB e PCL, o grupamento se
liga no enorme bolso hidrofóbico da cavidade (Reproduzido de BORNSCHEUER e KASLAUSKAS, 2006b).

33
Capítulo II – REVISÃO BIBLIOGRÁFICA
3.3.5 Fenômeno da ativação interfacial em lipases
Em 1958, Sarda e Desnuelle propuseram definir lipases a partir de suas características
cinéticas, utilizando como critério a propriedade de ativação na presença de substratos
insolúveis em água e emulsionados, ou seja, na presença de uma interface lipídeo/água. Os
autores estudaram as diferenças na atividade das esterases e das lipases. Estes observaram que
a lipase pancreática de porco sofria um aumento da atividade enzimática quando o limite de
solubilidade dos substratos era ultrapassado (esse aumento não foi observado para as
esterases). A atividade das esterases era função da concentração do substrato de acordo com o
modelo clássico de Michaelis e Menten, no qual a velocidade máxima ocorria antes do limite
de solubilidade do substrato ser atingido e da formação de agregados e/ou interfaces.
Enquanto a atividade lipásica se caracterizou por manter-se constante até que a concentração
micelar crítica (CMC) do substrato no sistema fosse alcançado, e aumentando a partir desse
ponto. A este fenômeno de aumento de atividade enzimática quando há formação de
interfaces, chamou-se de ativação interfacial (COSTA e AMORIM, 1999; SARDA e
DESNUELLE, 1958).
Os aspectos básicos sobre os mecanismos de catálise, incluindo o processo de ativação
interfacial foram elucidados a partir da resolução da estrutura tridimensional de várias
enzimas. Métodos de purificação, seqüenciamento, expressão genética, clonagem,
cristalização e cristalografia de raio-X foram empregados permitindo obter a resolução da
estrutura tridimensional de algumas lipases como Rhizomucor miehei, Pseudomonas glumae,
Pseudomonas cepacia, lipase de pâncreas humano, lipase gástrica humana e de cão, Candida
rugosa e Geotrichum candidum em sua forma inativa e ativada interfacialmente utilizando
inibidor para a formação de complexo com o sítio ativo (BRZOZOWSKI et al., 1991;
BRZOZOWSKI et al., 2000; GROCHULSKI et al., 1994 e ROUSSEL et al., 2002) (Figura
15). Na forma inativa, o sítio ativo está totalmente oculto sob um curto segmento helicóide,
formado por um ou mais loops-tampa lid ou aba flap. Na forma ativada, a tampa é deslocada
para fora do sítio ativo, deixando-o totalmente acessível ao solvente e substrato. Nesse
movimento, o lado hidrofóbico fica totalmente exposto, expandindo consideravelmente a
superfície não polar do sítio ativo (COSTA e AMORIM, 1999).

34
Capítulo II – REVISÃO BIBLIOGRÁFICA
Figura 15. Conformações tampa fechada (azul) e tampa aberta (amarelo) em várias lipases. Reproduzido de
Aloulou et al., (2006).
A diferença estrutural entre as duas conformações de lipase, tampa fechada e tampa
aberta, varia de um simples movimento para trás da tampa até um modelo mais complexo de
múltiplas alças com uma profunda mudança na estrutura secundária e terciária da lipase. As
lipases de Rhizomucor miehei e Thermomyces lanuginosa apresentam uma única tampa
helicoidal. Nas lipases de pâncreas humano e de Candida rugosa foi encontrado um modelo
mais complexo de múltiplas alças (SVENDSEN, 2000; ALOULOU et al., 2006).
Do ponto de vista estrutural, existem evidências que sugerem que a lid pode propiciar
a atividade catalítica e determina seletividade de algumas lipases (SECUNDO et al., 2006).
As lipases de Geotrichum candidum (SCHRAG et al., 1991), Thermomyces (Humicola)
lanuginosa e R. miehei (BRADY et al., 1990; OLLIS et al., 1992; CAJAL et al., 2000 a,b) são
exemplos de lipases que apresentam a tampa em suas estruturas e sofrem ativação interfacial.
Entretanto, em outros casos, a existência da lid não implica necessariamente em ativação
interfacial. Este é o caso das lipases de P. aeruginosa, Burkholderia glumae e C. antarctica B
(JAEGER e REETZ, 1998a, JAEGER, RANSAC e DIJKSTRA, 1994), que possuem a lid
mas não sofrem necessariamente ativação interfacial. Por outro lado, as cutinases, que são as
menores lipases de estrutura conhecida (19 kDa), não apresentam a tampa catalítica e não
precisam da interface para exercer sua atividade hidrolítica (CYGLER e SCHRAG, 1997;
YAO e KOLLER, 1994). Portanto, nem o critério de ativação interfacial, nem o da existência
da tampa são suficientes para a caracterização de uma lipase. Por isto, a caracterização de
lipases com base no conceito de ativação interfacial foi substituída pela definição de que
lipases são carboxilesterases capazes de catalisar a hidrólise de acilgliceróis de cadeia longa
(VERGER, 1997; FREIRE e CASTILHO, 2010). O fato de a maioria das lipases conhecidas

35
Capítulo II – REVISÃO BIBLIOGRÁFICA
ser ativada na interface orgânico-aquosa proporcionou o desenvolvimento de diversas
metodologias de estudo da ativação interfacial (FREIRE e CASTILHO, 2010).
O fenômeno de ativação interfacial ocorre quando a tampa hidrofóbica encontra outra
molécula hidrofóbica na interface. Esta então se move, sofrendo uma mudança
conformacional, expondo o seu sítio ativo e permitindo a catálise da reação (ALOULOU et
al., 2006). A mudança conformacional durante o processo de ativação pode ser alterada de
acordo com a lipase utilizada. Por exemplo, nas lipases de Rhizomucor miehei a espinha
dorsal da enzima é deslocada 7Å expandindo 750Å a área hidrofóbica da superfície da enzima
(DEREWENDA et al., 1992). Entretanto outras lipases sofrem alterações conformacionais
mais complexas, como o caso da lipase do pâncreas humano, onde a mudança ocorre na
estrutura secundária e terciária.
Petersen e colaboradores (2001) demonstraram por meio de um estudo que a superfície
das lipases apresenta diversos resíduos apolares. Este caráter apolar aumenta ainda mais
quando estas sofrem ativação interfacial. Os autores descreveram ainda que os resíduos em
volta do sítio ativo das lipases apresentam caráter eletrostático negativo nos valores de pH
ótimos, que são tipicamente próximos de 8.
O potencial negativo caracterizado no sitio ativo das lipases e esterases também pode
atuar repelindo o ácido carboxílico formado após a reação de hidrólise de um éster, forçando
então o produto para fora do sítio ativo da enzima, deixando-o livre para outras reações. Este
mecanismo foi descrito por Petersen e colaboradores (2001) e foi chamado de mecanismo da
catapulta eletrostática. O esquema se encontra na Figura 16.
Figura 16. Esquema ilustrando o funcionamento da catapulta eletrostática. O sítio ativo com potencial negativo
força, através de repulsão eletrostática, a saída do ácido carboxílico formado, que também possui carga negativa.
Modificado de Petersen et al., (2001) por Almeida, (2005).

36
Capítulo II – REVISÃO BIBLIOGRÁFICA
Em algumas lipases, a própria estrutura do sítio ativo é afetada pela mudança
conformacional na presença de seu substrato. Em lipase pancreática humana, por exemplo,
ambos o lid e a fita β5 se adaptaram totalmente para a diferente conformação na presença de
fosfolipídios e sais biliares. O movimento de abertura da tampa cria uma região eletrofílica
próxima do resíduo serina do sítio ativo (cavidade do oxiânion), que é responsável pela
estabilização das cargas negativas do intermediário tetraédrico do estado de transição formado
durante a hidrólise do éster. As lipases que não possuem uma tampa helicoidal, como por
exemplo, a lipase de Pseudomonas aeruginosa, apresentam a cavidade do oxiânion pré-
formado. A abertura da tampa helicoidal gera uma grande superfície hidrofóbica em torno do
sítio ativo. A face interna da tampa da lipase tem sido a principal responsável pelo
reconhecimento e interação da lipase com a interface água-lipídeo (ALOULOU et al., 2006).
Dependendo da lipase, a localização topológica da tampa hidrofóbica se encontra de
maneira variada. Além disso, o tamanho e complexidade geralmente aumentam com o
aumento da molécula protéica (JAEGER, DIJKSTRA e REETZ 1999).
3.3.6 Mecanismo cinético
O mecanismo cinético aceito para descrever a ação das lipases é o Ping Pong Bi Bi,
independente do tipo de reação catalisada (hidrólise, esterificação ou interesterificação). Este
mecanismo consiste de duas etapas principais:
Entrada do primeiro substrato e o ataque nucleofílico sobre a ligação éster do
substrato pelo átomo de oxigênio do grupo hidroxil do resíduo serina do sítio
ativo após a abertura da tampa (resultando na formação de complexo
enzimático acilado e desprendimento de espécie álcool do substrato original-
Intermediário tetraédrico)
Entrada do segundo substrato formando o segundo intermediário tetraédrico;
hidrólise do complexo enzimático acilado, resultando na formação do produto
e regeneração da enzima (JAEGER, DIJKSTRA e REETZ 1999; PAIVA,
BALCÃO e MALCATA, 2000).

37
Capítulo II – REVISÃO BIBLIOGRÁFICA
O mecanismo Ping Pong Bi Bi envolve substratos e produtos múltiplos. Usando a
notação de Cleland, este mecanismo pode ser representado de forma esquemática como na
Figura 17.
Figura 17 – Mecanismo cinético (Ping Pong Bi Bi) de reações com múltiplos produtos e substratos catalisadas
por lipases (E: enzima; Es: éster; Al: álcool; Ac: ácido; W: água; i = 1,2,...,I; j=1,2,...,J) (Reproduzido de
PAIVA, BALCÃO e MALCATA, 2000).
Segundo o mecanismo Ping Pong Bi Bi, a taxa de hidrólise (r) de um éster gerando um
ácido e um álcool, considerando a inibição pelo substrato e pelo produto, pode ser
genericamente descrita através da Equação 8 (PAIVA, BALCÃO e MALCATA, 2000).
AcAlK
vAcW
K
vKAlEs
KK
vKWEsv
AcK
vKAl
K
vKWKvEsKv
K
AcAlWEsvv
r
eq
f
Acii
rEsm
Esiieq
fAcm
r
eq
fAlm
eq
fAcm
EsmrWmr
eq
rf
max,
,
max,,
,
max,,
max,
max,,max,,
,max,,max,
max,max,
(Eq.8)
onde, Ac: ácido; Al: álcool; Es: éster; W: água; os subscritos f e r denotam o sentido
direto (hidrólise) e reverso (síntese) das reações, respectivamente; e max,f; max,r; Keq; Km,Ac;
Km,Es; Ki,Es; Km,W; Km,Al e Ki,Ac são parâmetros globais relativos às taxas máximas, constante
de equilíbrio, constantes de Michaelis-Menten e constantes de inibição, respectivamente, que
podem ser ajustados independentemente a partir de dados de concentração de produtos e
reagentes.
Quando água está presente em grande excesso no meio reacional (concentração de
água permanece constante), a equação genérica que descreve o mecanismo Ping Pong Bi Bi
pode ser simplificada em uma expressão bem mais simples, semelhante a expressão de taxa de
reação de Michaelis-Menten, representada pela Equação 9 (PAIVA, BALCÃO e MALCATA,
2000):
Es Al W Ac E E.Es F.Al F F.W E.Ac E
K1 K-1 K2 K-2 K3 K-3 K4 K-4

38
Capítulo II – REVISÃO BIBLIOGRÁFICA
EsK
Esvr
aparentem
aparente
,
max,
(Eq.9)
onde, max,aparente definida como max,f[W]/(Km,W + [W]) denota a taxa máxima
aparente da reação no sentido direto (hidrólise) e Km, aparente definido como Km,Es[W]/(Km,W +
[W]) denota uma constante de Michaëlis-Menten aparente.
A Figura 18 apresenta de forma esquemática as etapas de hidrólise catalisada por
lipases e os estados transientes formados (Modificado a partir de BORNSCHEUER e
KAZSLAUSKAS, 2006b e SIMAS et al., 2011).
Figura 18. Esquema da reação de hidrólise de ligações éster catalisada por esterases e lipases. Onde Td1 e Td2
representam o primeiro e o segundo intermediários tetraédricos. (Modificado a partir de BORNSCHEUER e
KAZSLAUSKAS, 2006b e SIMAS et al., 2011).
A participação dos resíduos de aminoácidos da tríade catalítica da lipase é de extrema
importância para que uma determinada reação ocorra. A primeira etapa que precisa ocorrer é a
ativação do grupo hidroxila do resíduo de aminoácido serina a partir da ligação hidrogênio
com a histidina vizinha. Como consequência da reação, o resíduo de aminoácido serina libera
um próton, que é aceito no anel imidazólico da histidina (2ª etapa). Após este processo ocorre

39
Capítulo II – REVISÃO BIBLIOGRÁFICA
a interação do terceiro resíduo de aminoácido, o aspartato (ou glutamato), o qual estabiliza a
carga positiva formada na histidina. Com isso, haverá um ataque nucleofílico ao átomo de
carbono da carbonila do primeiro substrato. Se considerarmos uma reação de hidrólise, por
exemplo, o ataque nucleofílico será do carbono da carbonila de um éster, quebrando a ligação
π do grupo C=O, resultando na formação do complexo do primeiro intermediário tetraédrico.
O complexo do primeiro intermediário tetraédrico formado é estabilizado por ligações
hidrogênio com os grupos amida dos resíduos da cavidade do oxiânion. O intermediário
tetraédrico é desfeito, pelo retorno da ligação π (C=O) e conseqüente quebra da ligação éster.
O primeiro produto, álcool, produzido na reação de clivagem é liberado.
Após a liberação do primeiro produto ocorrerá a formação do intermediário covalente
(enzima acilada), onde o componente ácido do substrato encontra-se esterificado com o
resíduo serina da enzima. O segundo substrato, molécula de água ou álcool, que se aproxima é
ativada pelo resíduo histidina vizinho e o íon hidroxila resultante promove o ataque
nucleofílico ao átomo de carbono da carbonila formando o complexo do segundo
intermediário tetraédrico. O retorno da ligação π (C=O), desfaz o complexo do segundo
intermediário tetraédrico e finalmente ocorre a liberação do segundo produto, ácido
carboxílico ou éster, e da enzima livre.
3.3.7 Aplicação das lipases
Lipases são as enzimas comumente mais empregadas em biotecnologia e representa
atualmente cerca de 5% do mercado mundial de enzimas. Certamente, este percentual tende a
crescer de forma constante devido às suas aplicações em diversas áreas industriais, incluindo
aplicações em síntese orgânica, resolução de compostos racêmicos, energia e alimentos. Fato
no qual torna cada vez mais proeminente o interesse industrial por esses biocatalisadores
(CHOWDARY, RAMESH e PRAPULLA, 2001; LEAL et al., 2002; BARON, 2003;
CAMAROTA e FREIRE, 2006; TAN et al., 2010; RIBEIRO et al. 2011; YENIAD, NAIK e
HEISE, 2011; KAPPOR e GUPTA, 2012; SOLANO et al., 2012; GOSWAMI, BASU e DE,
2013; SOUMANOU, PÉRIGNON e VILLENEUVE, 2013, CUNHA et al., 2014).
Há inúmeras razões para o interesse nessas enzimas, como por exemplo, a capacidade de
atuarem em meios heterogêneos e poderem catalisar reações diversas como hidrólise,
esterificação (CHOWDARY, RAMESH e PRAPULLA, 2001), transesterificação, acilação
regioseletiva de mentóis e pentóis, síntese de peptídeos (ZHANG et al. 2001) etc. Além disso
podem ser empregadas sob condições suaves de reação apresentando alta químiosseletidade,

40
Capítulo II – REVISÃO BIBLIOGRÁFICA
regiosseletividade e enantiosseletividade, capacidade de aceitarem um grande número de
substratos não naturais (SNELLMAN, SULLIVAN e COLWELL, 2002; KAPPOR e
GUPTA, 2012), não necessitarem de cofatores (BORNSCHEUER e KAZLAUSKAS, 2006b)
etc.
As condições mais sustentáveis oferecidas pelo uso de tecnologias biocatalíticas frente aos
processos químicos (como o reuso do biocatalisador, uso de materiais biodegradáveis, de alta
economia (economia de átomos, alta seletividade etc.), condições brandas (em relação à
temperatura, pressão e pH) fazem com que estes biocatalisadores sejam uma alternativa
interessante. Devido a estes e a outros fatores, estas enzimas possuem um alto potencial de
utilização sendo utilizadas em diferentes setores industriais. Algumas áreas deste setor estão
citadas na Tabela 2.
Tabela 2. Principais áreas aplicações, produtos e tipos de reações desempenhadas pelas lipases
Tipos de reações Áreas de
aplicação
Aplicações Produtos Referências
Hidrólise
Alimentos
(latícinios)
Hidrólise da
gordura
do leite
Agentes
flavorizantes
para
queijos e
derivados.
VILLENEUVE et
al., 2000;
HASAN et al.,,
2006;
BARROS et al.,
2010.
Química
(Processamento
do Óleo)
Hidrólise de óleos
e gorduras
Ácidos graxos,
diglicerídeos e
monoglicerídeos
(emulsificantes,
reagentes para
análise de
lipídios)
OSÓRIO et al.,
2001;
UNDURRAGA,
MARKOVITS e
ERAZO,
2001; HASAN et
al., 2006;
VILLENEUVE et
al., 2000; SILVA
et al., 2003.
Química
(Detergente)
Remover
manchas
de óleo
Detergentes para
lavanderias e
uso
SHARMA,
CHISTI e
BANERJEE,
2001; KIRK,
BORCHERT e
FUGLSANG,
2002a; HASAN
41
Capítulo II – REVISÃO BIBLIOGRÁFICA
doméstico SHAH e
HAMEED, 2006.
Tratamento de
efluentes
Hidrólise de
lipídeos,
proteases,
celulases,
matérias
orgânicas etc.
Ácidos graxos,
triglicerídeos
CAMMAROTA e
FREIRE, 2006;
DAMASCENO,
et al. 2012;
SILVA, et al.
2013;
CAMMAROTA,
et al. 2013.
Esterificação
Química
Fina
Síntese de ésteres Intermediários
quirais
ésteres,
emulsificantes
BUCALÁ et al.,
2006;
FERNANDES et
al., 2004;
ROMERO et al.,
2005;
TRUBIANO,
BORIO
e FERREIRA,
2004;
FERNANDES et
al.,
2006; FORESTI e
FERREIRA,
2006;
HASAN, SHAH e
HAMEED, 2006;
YENIAD, NAIK
e HEISE, 2011;
MANOEL et al.
2012a;
CUNHA et al.
2014.
Química
Alimentos
Esterificação ou
transesterificação
Óleos ou
gorduras.
Flavorizantes e
aromatizantes.
HASAN, SHAH e
HAMEED, 2006
42
Capítulo II – REVISÃO BIBLIOGRÁFICA
O alto custo na preparação destas enzimas pode ser considerado como uma das
principais desvantagens desta tecnologia. Contudo, com os avanços de outras tecnologias,
como a da engenharia de imobilização de enzimas, surgem alternativas que permitem reciclos
cada vez eficientes destes biocatalisadores e aumentam a atividade enzimática, o que resulta
em melhor relação custo-benefício para um determinado processo (DÍAZ-RODRÍGUEZ e
DAVIS, 2011; BRUSTAD e ALNOLD, 2011; BOMMARIUS, BLUM e ABRAHAMSON,
2011).
3.3.8 Aplicação de lipases na síntese de fármacos
A importância da quiralidade tem aumentado nas últimas décadas e é hoje o foco
central das indústrias farmacêutica e de química fina, especialmente no desenvolvimento de
novas moléculas bioativas. O valor de mercado para tecnologia quiral atingiu cerca de 2,7
Química
Farmacêutica
Síntese de
intermediários de
medicamentos
preparação
de intermediários
homoquirais
substâncias
antiinflamatórias
como
naproxeno,
ibuprofeno,
cetoprofen,
suprofen.
Intermediários
para substâncias
anti- asmáticas.
WILKINSON e
BACHMANN,
2006;
JAEGER e
EGGERT,
2002; ZHANG et
al.,
2002;
BEVILAQUA et
al., 2004 e 2005
Transesterificação
Biocombustíveis Transesterificação
de óleos vegetais
Biodiesel ABIGOR et al.,
2000; ISO et al.,
2001;
SHIMADA et al.,
2002; ZHANG et
al., 2002;
SALIS et al.,
2003;
HSU et al., 2004;
SOUZA et al.,
2010
CAVALCANTI-
OLIVEIRA et al.,
2011.

43
Capítulo II – REVISÃO BIBLIOGRÁFICA
bilhões de dólares nos EUA em 2007, com uma taxa de crescimento anual de 10,8%
(GHANEM et al., 2010). Em 2008 este valor foi para 4,3 bilhões de dólares, com um aumento
de 3,3% de crescimento no ano seguinte (4,5 bilhões de dólares em 2009). Já em 2011, o
mercado global de tecnologia quiral foi de quase US $ 5,3 bilhões. Estima-se que o mercado
deverá aumentar a uma taxa CAGR de 6,5% de 2011-2016 e alcançará 7,2 bilhões dólares até
o final do período de previsão (Figura 19) (AHMED et al., 2012; DEWAN, 2012).
Figura 19. Mercado mundial de tecnologia quiral por segmentos: Síntese quiral, análise quiral e resolução quiral
(Reproduzido de DEWAN, 2012).
Impulsionado pela importância da quiralidade na eficácia de inúmeros produtos
agroquímicos e farmacêuticos, estabelecida pelas legislações brasileira e de outros países, o
interesse pela produção de um único enantiômero a partir de intermediários quirais tem
aumentado significativamente (SKORIDOU et al., 2004). As exigências são em relação à
separação dos enantiômeros em um racemato, ao estudo dos efeitos biológicos dos
enantiômeros e a comprovação dos efeitos tóxicos e adversos dos enatiômeros e do racemato,
dependendo do composto quiral. A FDA (U.S. Food and Drug Administration, RockVille,
EUA) determina para um dado produto se ele pode ser comercializado como racemato ou
como enantiopuro, baseado em protocolos estabelecidos para a classe ou utilização deste
composto quiral (FDA, 2009). No Brasil, a comercialização de medicamentos quirais é
regulamentada pela ANVISA seguindo normas da RDC (Resolução da Diretoria Colegiada)
que dispõem sobre comercialização e uso de medicamentos (ANVISA, 2012).

44
Capítulo II – REVISÃO BIBLIOGRÁFICA
Ao analisar potenciais fármacos, a pureza enantiomérica faz-se necessária, uma vez
que o antípoda ótico pode não apresentar atividade ou mesmo produzir efeitos indesejáveis no
processo biológico, como o caso discutido anteriormente sobre a Talidomida (vide pag. 19).
Dentre as inúmeras serino-hidrolases que catalisam as biotransformações, as lipases
são as mais empregadas na síntese orgânica de processos industriais de substâncias quirais
pelas grandes companhias farmacêuticas, tais como Pfizer, Schering-Plough, GlaxoSmithkline
Pharmaceuticals, Bristol-Myers Squibb, DSM, Tanabe Pharmaceutical, Celltech Group etc.
As lipases mais empregadas são produzidas por: Candida rugosa (também conhecida como
Candida cylindracea); Candida antarctica , uma das enzimas de maior versatilidade no
campo da biotransformação com destaque em duas diferentes isoformas, CALA e CALB;
Pseudomonas fluorescens, uma das mais empregadas na resolução de compostos racêmicos;
lipase pancreática (suína) provavelmente a mais econômica do mercado; Pseudomonas
cepacia, biocatalisador que tem se mostrado bem seletivo; Serratia marcescens, menos
utilizada e conhecida, porém igualmente utilizada por algumas companhias na preparação de
intermediários quiral (SOLANO et al., 2012).
A lipase de Candida rugosa foi utilizada como catalisador na síntese dos seguintes
anti-flamatórios não esteroidais (AINEs ou no inglês NSAIDs): ácido (S)-2-arilpropionico e o
(S)-ibuprofeno (ácido(S)-2-4-isobutilfenil propanóico), isômeros ativos produzidos. A
companhia americana Pfizer desenvolveu um processo industrial do (S)-ibuprofeno a partir da
hidrólise enantiosseletiva do correspondente racêmico metoxi-metil ester pela lipase
mencionada em sua forma imobilizada em reator de membrana. O isômero ativo é produzido
em escala de kg com 90% de rendimento e eep igual a 99% (SHELDON, 1993).
A empresa americana Celltech Group utilizou a lipase de Pseudomonas fluorescens
(SOLANO et al., 2012) na síntese de (1S, 4R)-1-acetoxi-4-(trifenilmetoximetil)-ciclopent-2-
en- (composto (+)-3 da Figura 20), um intermediário necessário na síntese do nucleosideo
similar ao Carbovir, um antiviral usado no tratamento da AIDS (EVANS et al., 1992). Os
autores utilizaram acetato de vinila como agente acilante na reação de alcoólise por 75h, em
temperatura ambiente (22% de conversão e eep maior que 95%).

45
Capítulo II – REVISÃO BIBLIOGRÁFICA
Figura 20. Reação enantiosseletiva de (±)-9 catalisada pela lipase de Pseudomonas fluorescens, utilizando
acetato de vinila como agente acilante em 75h a temperatura ambiente (Modificado de EVANS et al., 1992).
Busto et al., (2012) utilizaram as lipases CALB, CAL-A e PSC (Pseudomonas cepacia
atual Burkholderia cepacia) na resolução cinética de (±)-2,3,4,9-tetrahidro-1H-carbazol-3-ol,
um intermediário da síntese de Ramatroban, substância ativa com potencial farmacológico,
que é comercializado no Japão para tratamento de rinite alérgica e asma. CALB, mostrou-se
ser a lipase mais promissora catalisando a conversão de 50% do racemato supra-citado com
alto excesso enantiomérco e razão enantiomérica (eep de 99% e E maior que 200). Os autores
obtiveram em 8 horas o isômero R, que foi posteriormente usado na síntese do Ramatroban,
como ilustrado na Figura 21.
Figura 21. Resolução cinética de (±)-2,3,4,9-tetrahidro-1H-carbazol-3-ol catalisado por CALB utilizando acetato
de vinila em THF a 250rpm (Reproduzido por BUSTO et al., 2012).

46
Capítulo II – REVISÃO BIBLIOGRÁFICA
CALB tem sido empregada na desimetrização do composto proquiral dietil3-(3’, 4’-
diclorofenil) glutarato para obter (S)-3-(3,4-diclorofenil)-5-etoxi-5- acido oxopentanoico, um
intermediário quiral na síntese do antagonista dos receptores de taquicinina, NK 1 e NK2,
compostos utilizados no tratamento contra asma, artrite e enxaqueca (HOMANN et al., 2001).
Os autores obtiveram 80% de conversão e alto excesso enantiomérico (maior que 99%). A
empresa Schering-Plough tem realizado este processo com um aumento de escala (produção
de 200 kg) obtendo os mesmos rendimentos que os autores (SOLANO et al., 2012).
A resolução racêmica de 1-Ciclopropil-2-metoxietanamina (1) foi estudada por
PARKER et al., (2012) igualmente utilizando a CALB (Figura 22). Os autores utilizaram
caprato de etila (2) como agente acilante e TBME como solvente da reação. Após 120h de
reação, o isômero (S)-1- Ciclopropil -2- metoxietanamina (3) foi obtido com 35% (91,3% de
eep). Este intermediário é necessário para a síntese de um factor de libertação do hormônio
corticotropina-1, receptor antagonista, mediador de resposta ao stress celular (BORSODY e
WEISS, 1996).
Figura 22. Resolução racêmica de (1) catalisado por CALB após 120h de reação (Reproduzido de PARKER et
al., 2012).
β-hidroxinitrilas quirais são comumente utilizados como intermediários utilizados em
síntese orgânica. O isômero (R)-3-hidroxipentanitrila, por exemplo, é um importante
intermediário na síntese do imunossupressivo inosina 5´monofosfao dehidrogenase, no qual
um excesso enantiomérico maior que 99% é requerido. A lipase de Pseudomonas cepacia tem
sido empregada no melhoramento da atividade óptica do isômero ativo de interesse. Kawano
et al., (2012) realizaram a síntese de (R)-3-hidroxipentanitrila através de uma redutase todavia
um eep de 81,5% foi obtido. Posteriormente a lipase de Pseudomonas cepacia foi utilizada na
hidrólise enantiosseletiva (Figura 23) resultando na máxima enaniosseletividade requerida
para o processo (99% de eep).

47
Capítulo II – REVISÃO BIBLIOGRÁFICA
Figura 23. Síntese de (R)-1 e aperfeiçoamento da enantiosseletividade utilizando lipase de Pseudomonas cepacia
(Modificado por KAWANO et al., 2012).
Recentemente, Fernandez-Alvaro et al. (2014) utilizaram CALB na hidrólise
enantiosseletiva de compostos tetrahydropyrazolo[1,5-α]pirimidina (THPPs). Os produtos
enantiomericamente puro são importantes agentes terapêuticos contra tuberculose, hepatite C
e osteoporose. Os autores obtiveram altos valores de conversão e de excesso enantiomérico
para os derivados utilizados (51% e >93%, respectivamente).
3.3.9 Lipases promissoras para o estudo dos derivados de mio-inositol
Lipases de Candida antarctica
Nos últimos anos, a lipase comercial de Candida antarctica fração B demonstrou ser o
melhor biocatalisador para as reações de acetilação para alguns derivados de mio-inositol
(SIMAS et al., 2011; CUNHA et al., 2011; CUNHA et al., 2012). Por este motivo, este
biocatalisador será melhor descrito.
A lipase B de Candida antarctica (CALB) é dentre diversas enzimas, uma das mais
utilizadas em biocatálise (KIRK e WURTZ, 2002; BORNSCHEUER e KAZLAUSKAS,
2006b; DUNN, WELLS e WILLIAMS, 2010; BOROS et al. 2013). A CALB é altamente
regiosseletiva na síntese de monoésteres de hidratos de carbono e enantiosseletiva na
resolução de inúmeros compostos como aminas, ácidos e álcoois (KOTIK et al., 2005).
CALB apresenta diversas aplicações como a síntese de ésteres constituintes de flavors
e fragrâncias (LARIOS et al., 2004), resolução de compostos nitrogenados utilizados na
produção de fármacos e manufaturados no setor industrial (GOTOR-FERNANDEZ et al.,

48
Capítulo II – REVISÃO BIBLIOGRÁFICA
2006b). C. antarctica é considerada uma levedura extremófila, pois está adaptada a
temperaturas baixas, e inúmeros pesquisadores buscam o aumento da estabilidade térmica de
sua lipase (SIDDIQUI e CAVICCHIOLI, 2005). CALB se encontra disponível no mercado na
forma livre ou imobilizada (imobilização por adsorção). A lipase comercial de C. antarctica,
denominada Novozym 435, corresponde a CALB imobilizada em resina acrílica macroporosa
(DU et al., 2004).
CALB tem uma massa molecular de 33 kDa e a enzima disponível comercialmente
contém 16-51% em peso de proteína. É uma lipase constituída por 317 aminoácidos, três
pontes dissulfeto e pela tríade catalítica Ser 105- His 224- Asp 187. Sua estrutura terciária
com dobramento de α/β hidrolase apresenta uma tampa hidrofóbica (lid) curta e flexível que é
responsável pelas conformações aberta e fechada. Simulação por dinâmica molecular vem
mostrando um potencial para a região do sítio catalítico devido a flexibilidade e a poucas
alterações conformacionais da posição α5, presente na lid. Apesar de CALB ser conhecida por
não apresentar ativação interfacial, alguns autores relatam a possibilidade da lipase sofrer
algum tipo de ativação. Este fenômeno pode ser explicado pela conformação da enzima:
atuação da posição α5 da lid (141-147: AGPLDAL)) e a posição α10 (280-
288:PAAAAIVAG), que funciona como um elemento de ativação (Figura 24). Estes dois
dobramentos da lid constroem um estreito sítio ativo hidrofóbico com os clássicos resíduos de
aminoácidos supracitados (UPPENBERG et al., 1994; WEBER et al., 1995b; UPPENBERG
et al., 1995; SKJOT et al., 2009, FERRARIO, et al., 2011; GANJALIKHANY et al., 2012).
Figura 24. Estrutura tridimensional da lipase B de Candida antarctica (CALB) em sua forma aberta (A) e
fechada (B). Região da tampa hidrofóbica (em azul) representa os dobramentos da hélice (α5, compartimento
superior em (A) e α10, compartimento inferior em (A) ). (Reproduzido por GANJALIKHANY et al., 2012).

49
Capítulo II – REVISÃO BIBLIOGRÁFICA
A lipase de CALB é uma proteína recombinante expressa em fungo do gênero
Aspergillus (HOEGH et al., 1995). A lipase possui alta atividade e elevada
enantiosselectividade para uma vasta gama de álcoois. Sua enantioseletividade geralmente é
baixa para ácidos carboxílicos (ANDERSON et al., 1998).
Lipase B de Candida antarctica expressa em Pichia pastoris (LIPB)
Os Laboratórios de Biologia Molecular (CEL-UnB) e Laboratório de Biotecnologia
Microbiana (IQ-UFRJ), coordenados pelos professores Fernando Araripe Torres e Denise
Maria Guimarães Freire, recentemente clonaram, expressaram e produziram com sucesso o
gene LIPB, referente a lipase B de Candida antarctica em P. pastoris. Apesar da lipase
comercial CALB (Candida antarctica fração B) ser glicosilada, esta glicosilação não é
necessária para manter a atividade hidrolítica da lipase (BLANK et al., 2006). A glicosilação
é uma das mais comuns modificações pós-traducionais de proteínas em sistemas biológicos,
afetando não somente as propriedades das proteínas em seus vários níveis, mas também a
interação entre a proteína com o sistema biológico (SOLÁ et al., 2007). Assim como a CALB,
a lipase LIPB, produzida pelo grupo, possui uma proteína glicosilada no resíduo Asn 74
(GUTARRA et al., 2011). O vetor de expressão utilizado foi o pPGKΔ3_PRO_LIPB, que
utiliza o promotor constitutivo da enzima fosfogliceratoquinase de P. pastoris, da via
glicolitica. Assim a expressão não precisa de indutor e a produção da enzima é associada ao
crescimento celular (ARRUDA, 2008).
A levedura Pichia pastoris foi selecionada como plataforma de expressão, pois sendo
um micro-organismo eucarioto, é capaz de realizar modificações pós traducionais na enzima,
as quais não são possíveis em sistemas de expressão bacterianos. Além disso, a levedura
Pichia pastoris possui uma alta capacidade de secreção de proteínas (GELLISSEN et al.,
2000). A proteína secretada por P. pastoris pode ser purificada a partir do sobrenadante do
meio de cultura e então caracterizada.
Os resultados obtidos, com a caracterização preliminar deste biocatalisador,
principalmente por sua seletividade e termoestabilidade, foram patenteados em cooperação
com a PETROBRÁS e apontam para a potencialidade de sua aplicação em diversos processos
biotecnológicos (CASTRO et al., 2009). Contudo, por se tratar de um produto com alta
densidade tecnológica e valor agregado, possivelmente sua melhor utilização (relação
custo/benefício) se dará em processos envolvendo produtos finais, igualmente com elevado
valor agregado, como intermediários de fármacos e/ou química fina.

50
Capítulo II – REVISÃO BIBLIOGRÁFICA
3.3.10 Fatores que podem influenciar a estereosseletividade das lipases
O processo da biocatálise depende de diversos fatores que podem influenciar a
enantiosseletividade da reação. Para que ocorra a biotransformação com eficiência, alguns
fatores devem ser avaliados (ANTHONSEN et al., 2005; BORNSCHEUER e
KAZLAUSKAS, 2006b):
1) Biocatalisador: enzima livre ou imobilizada, ou até mesmo a utilização da própria
célula do micro-organismo (whole cell). Novos biocatalisadores têm sido estudados
por meio da engenharia genética com o objetivo do aumento da enantiosseletividade
para um determinado processo específico. A purificação de enzimas a partir de um
meio bruto também é utilizada para esta finalidade (ANTHONSEN et al., 2005;
BORNSCHEUER e KAZLAUSKAS, 2006b; LUETZ, GIVER e LALONDE, 2008;
BOTTCHER e BORNSCHEUER, 2010; KOURIST, BRUNDIEK e
BORNSCHEUER, 2010; BOMMARIUS, BLUM, e ABRAHAMSON, 2011).
Geralmente, como primeira etapa quando não se tem a idéia do biocatalisador a ser
empregado, uma seleção (screening) dos catalisadores é realizada (QUARTEY et al.,
1996), com uma posterior escolha do mais enantiosseletivo e produtivo.
2) Substrato: O procedimento mais utilizado na química orgânica é a utilização de
substratos ligeiramente modificados com grupos de proteção, que são facilamente
removidos ou manipulados no meio da reação.
3) Meio reacional: A literatura apresenta muitos trabalhos que mostram o estudo do
aumento da enantiosseletividade com a alteração deste fator. O estudo de diferentes
meios reacionais envolve, dentre inúmeros fatores, otimização de solventes, análises
da atividade de água e diferentes agentes acilantes utilizados nas reações de
transesterificação e alcoólise (CHEN e SIH, 1989; FABER, OTTOLINA e RIVA,
1993, MANOEL et al., 2012a).
O sucesso da reação enzimática dependerá dos estudos destes e de outros fatores. Por isso
alguns pontos relevantes serão mencionados sobre os tópicos supramencionados.

51
Capítulo II – REVISÃO BIBLIOGRÁFICA
Quanto ao Biocatalisador: Tecnologia da Imobilização de Enzimas
Breve Histórico
O processo de imobilização tem sido realizado no início do século passado quando
Nelson e Griffin (1916) observaram pela primeira vez o fenômeno da imobilização por meio
do aprisionamento da invertase em carvão ativo. A base para as tecnologias atuais teve início
em 1960s com um aumento considerado no número de publicações (TOSA et al., 1966).
Entretanto somente em 1971, motivado pela importância nos processos industriais, que o
conceito de enzima imobilizada foi estabilizado como sendo ―enzimas ligados a suportes
insolúveis confinados a espaços físicos definidos‖ -na primeira conferência de Engenharia
Enzimática em Henniker (USA) por Katschalski-Katzir (HARTMEIER, 1988 apud CASTRO
et al. 2008; GUISÁN, 2006)-na qual são mantidas suas atividades catalíticas em processos de
operação contínuo ou descontínuo, com a possibilidade de reutilização das mesmas
(FREEMAN e LILLY 1998).
Importância Industrial
Os setores acadêmicos e industriais têm se dedicado ardualmente à obtenção de
melhores biocatalisadores como alta enantiosseletividade e estabilidade frente a diferentes
condições reacionais, quer seja a aplicação, pequena ou em larga escala (HARTMANN e
JUNG 2010).
Uma enzima pode ser considerada útil para uma determinada reação em sua forma
livre (em suspensão ou liofilizada), todavia a instabilidade do biocatalisador muitas vezes é
prejudicada pelas condições de processos aplicados (como efeito de solvente que podem
desnaturar a enzima, efeitos mecânicos, custo da produção, difícil recuperação do
biocatalisador etc.). A tecnologia de imobilização, ou ―aprisionamento‖ do biocatalisador, é
empregada como ferramenta para melhorar as propriedades da enzima, como o aumento da
atividade catalítica e/ou enantiosseletividade (BAGI e SIMON 1999; BORNSCHEUER e
KAZLAUSKAS, 2006b; GUISÁN, J. M. 2006; BAI et al. 2006b, DESAI et al. 2006;
MATEO et al., 2007; PALOMO et al. 2007; KOSZELEWSKI et al. 2007; PAULA et al.
2007; CABALLERO et al. 2009a; LEE et al. 2009; HARTMANN e JUNG 2010; LU et al.
2010; ZOU et al. 2010; RUFINO et al. 2010; HE et al. 2010; XU et al. 2011; WANG et al.
2011; CALGAROTO et al. 2011; HERNÁNDEZ et al. 2011; RODRIGUES et al. 2011;
MENDES et al. 2012).
Novos métodos de imobilização surgem constantemente sendo o principal objetivo o
reuso do biocatalisador e a possibilidade da realização em processos contínuos. A viabilidade

52
Capítulo II – REVISÃO BIBLIOGRÁFICA
do biocatalisador para sua aplicação industrial dependerá do seu custo, do suporte e do
método de imobilização a ser empregado. Como resultado, a enzima imobilizada apresenta
uma série de vantagens como a recuperação do biocatalisador no meio reacional para sua
possível reutilização, a possibilidade do aumento da atividade enzimática, do aumento da
enantiosseletividade e estabilidade térmica (GUISÁN, J. M. 2006; LU et al. 2010, ZOU et al.
2010, RUFINO et al. 2010, HE et al. 2010; XU et al. 2011; WANG et al. 2011;
CALGAROTO et al. 2011; HERNÁNDEZ et al. 2011; RODRIGUES et al. 2011; MENDES
et al. 2012).
Atualmente, diferentes estratégias como técnicas da engenharia genética, evolução
sítio dirigida, modificação química sítio-dirigida de proteínas, uso de líquidos iônicos como
suporte de imobilização e como solvente, nanotecnologia (através de inovadores
nanomateriais para imobilização de enzimas) etc. tem sido descritas e utilizadas por diferentes
autores para aumentar a performance dos derivados enzimáticos imobilizados (WANG et al.
2009; XU et al. 2009; GODOY et al. 2010; LEE et al. 2012; BUCKO et al. 2012; LIMA et al.
2013).
Vantagens da técnica de imobilização
Como já relatado anteriormente, o uso de enzimas imobilizadas tem sido utilizado e
estudado extensivamente por facilitar o desenvolvimento de processos, principalmente em
escala comercial (TISCHER e KASCHE, 1999, HARTMANN e JUNG 2010). Este processo
oferece vantagens importantes nos setores industriais. As principais vantagens apontadas por
diversos autores (FREIRE, 1988; KATCHALSKI-KATZIR, 1993; BENJAMIN e PANDEY,
1998; VILLENEUVE et al., 2000; MATEO et al., 2007) são:
Facilidade de recuperação do biocatalisador e separação de produtos. As técnicas
empregadas na separação do biocatalisador imobilizado do meio reacional são
operações unitárias simples, como filtração, centrifugação ou decantação, facilitando a
aplicação de enzimas imobilizadas em biocatálise, principalmente quando o
biocatalisador apresentar alto custo, pois estes poderão ser reutilizados.
Biocatalisadores imobilizados podem ainda ser indicados quando um produto final
requerer elevado grau de pureza, pois o procedimento de separação entre a mistura
reacional e o biocatalisador são operações unitárias simples, como mencionado
anteriormente.

53
Capítulo II – REVISÃO BIBLIOGRÁFICA
Processo pode ser operado continuamente. A facilidade em se reter o biocatalisador
permite que processos contínuos sejam implementados, utilizando reatores variados,
como os reatores tubulares (Plug Flow Reactor - PFR) ou reatores tanques de agitação
(Continuous Stirred-Tank Reactor –CSTR) e o controle do processo é facilitado.
Previne a formação de agregados em meio orgânico. A imobilização de enzimas
surge como uma alternativa às enzimas livres, uma vez que estas, quando suspensas
em um solvente orgânico, tendem a se agregar e aderir às paredes do reator,
principalmente se for adicionado água ao sistema.
Manutenção de microambiente com alta atividade de água. O processo de
imobilização permite a manutenção de uma camada de solvatação no microambiente
em torno da enzima capaz de manter uma dinâmica molecular, mínima, essencial à
atividade enzimática em ambientes orgânicos.
Estabilização da enzima. A imobilização da enzima sobre diferentes suportes pode
aumentar a rigidez da estrutura molecular minimizando a ocorrência de desnaturação
por temperatura, pH ou solvente.
Propriedades enzimáticas podem ser alteradas favoravelmente. A interação entre a
enzima e o suporte pode ocasionar mudanças conformacionais na enzima, podendo
gerar alterações desejadas nas propriedades catalíticas, por exemplo, o aumento da
atividade e estereoespecificidade da enzima.
Custos com manejo de materiais são minimizados. O aumento da estabilidade da
enzima imobilizada facilita a sua estocagem e diminui as exigências para sua
manipulação.
Efeitos causados pelos processos de imobilização
O processo de imobilização normalmente modifica as propriedades físico-químicas da
enzima. A interação enzima-suporte pode causar alteração na estabilidade e nas propriedades
cinéticas da enzima imobilizada, seus principais aspectos serão apresentados a seguir
(FREIRE, 1988):

54
Capítulo II – REVISÃO BIBLIOGRÁFICA
Efeitos conformacionais. Efeitos associados às modificações na conformação da
proteína devido às interações entre a enzima e suporte durante o processo de
imobilização covalente.
Efeitos estereoquímicos. A imobilização pode tornar certas regiões da enzima pouco
acessíveis ao substrato, causando um efeito de exclusão. Algumas propriedades do
suporte, tais como: natureza hidrofóbica ou hidrofílica, constante dielétrica e presença
de cargas fixas no suporte podem interferir na performance do biocatalisador. A
minimização desses efeitos pode ser realizada pela utilização de espaçadores capazes
de manter os sítios ativos da enzima mais distantes dos efeitos de exclusão do suporte.
Efeitos de partição. São os efeitos causados pela desigualdade na distribuição das
espécies químicas entre o microambiente e o seio do meio reacional. Esses efeitos
resultam de interações hidrofóbicas, hidrofílicas e eletrostáticas entre o suporte e o
substrato.
Limitações difusionais. Assim como ocorre com a catálise heterogênea, a ação
catalítica de enzimas imobilizadas pode ser dividida em pelo menos cinco etapas:
Difusão externa de substratos. Difusão do substrato do seio da fase
líquida até a superfície do suporte;
Difusão interna de substratos. Transporte do substrato da superfície
do suporte para o domínio da enzima;
Catálise. A catálise propriamente dita da reação química;
Difusão interna de produtos. Transporte do produto do domínio da
enzima para a superfície do suporte;
Difusão externa de produtos. Difusão do produto da superfície do
suporte para o seio do meio reacional.
As interações entre os efeitos supra-citados podem causar alterações das propriedades
enzimáticas. Dependendo do efeito que controla a reação, os parâmetros cinéticos podem ser
diferenciados. Esses parâmetros podem ser classificados em três categorias principais
(FREIRE, 1988):
Parâmetros cinéticos intrínsecos. Parâmetros que refletem o comportamento cinético
verdadeiro da enzima imobilizada. Esses parâmetros só podem ser determinados no

55
Capítulo II – REVISÃO BIBLIOGRÁFICA
macro-ambiente, caso a concentração de substrato e produto seja a mesma do
microambiente. Devido à existência de efeitos conformacionais e estereoquímicos, os
parâmetros cinéticos intrínsecos da enzima imobilizada geralmente são diferentes dos
parâmetros da enzima solúvel.
Parâmetros cinéticos inerentes. Esses parâmetros refletem o comportamento cinético
da enzima na ausência de limitações difusionais e só podem ser determinados quando
o transporte de substratos e produtos ocorrer de forma infinitamente rápida. Essa
condição pode ser alcançada na prática, pelo uso de membranas finas, enzimas com
baixa atividade e agitação suficiente do meio reacional. Os parâmetros cinéticos
inerentes e intrínsecos diferem entre si devido a interação da matriz sólida com os
substratos e os produtos solúveis.
Parâmetros cinéticos efetivos. Parâmetros globais que refletem o comportamento da
enzima imobilizada na presença de efeitos difusionais, de partição, conformacionais e
estereoquímicos. Esses parâmetros são determinados a partir da velocidade da reação
nas condições experimentais normalmente empregadas.
Como relatado, a imobilização de uma enzima pode produzir distorções na estrutura
terciária da enzima com alteração da configuração do sítio ativo e redução da flexibilidade da
proteína, aprisionando-a em sua nova configuração. As enzimas que sofrem grandes
mudanças conformacionais durante a catálise podem ter estas mudanças distorcidas quando
imobilizadas, produzindo enzimas com propriedades catalíticas inteiramente diferentes
(MATEO et al., 2007). Portanto, para que seja possível utilizar a distorção da enzima como
ferramenta, é necessário obter uma grande quantidade de suportes e de protocolos de
imobilização que permitam investigar os diferentes tipos de imobilização, com o objetivo de
se ter o controle das modificações da estrutura terciária da enzima. Assim, a proteína pode ser
imobilizada a partir de diferentes regiões, conferindo diferença em sua rigidez e distorcendo a
enzima de maneiras muito diferentes. Os diferentes microambientes gerados em torno da
enzima durante o processo de imobilização podem alterar as propriedades físicas da enzima.
Desta forma, torna-se possível conseguir uma grande biblioteca de enzimas imobilizadas com
propriedades muito distintas (MATEO et al., 2007).

56
Capítulo II – REVISÃO BIBLIOGRÁFICA
Protocolos de imobilização
A imobilização de enzimas tem sido feita em diferentes suportes inertes. Este processo
vem sendo utilizado para melhorar o desempenho do biocatalisador em relação a: facilidade
de recuperação e purificação de enzimas (via adsorção em suportes hidrofóbicos); ao aumento
da estabilidade térmica e pH; a redução da inativação do biocatalisador etc. Apesar de não
existir um método padrão para ser utilizado, a escolha do método pode ser feita com base em
critérios como a máxima atividade de enzima imobilizada, estabilidade operacional, custo do
suporte, toxidade do reagente de imobilização etc. (MALCATA et al., 1990, BRENA e
BATISTA-VIEIRA, 2006).
O suporte a ser utilizado no processo pode ser orgânico (natural: colágeno, celulose
etc. ou não natural: poliestireno) ou inorgânico (e.g.: sílica) e deve ter uma grande área
superficial, alta capacidade de adsorção enzimática, resistência física e resistência ao ataque
de micro-organismos, caráter hidrofóbico/hidrofílico, baixo custo além da característica
desejável de não apresentar limites difusionais no transporte de substrato e produto
(VILLENEUVE et al. 2000; NASRATUN et al., 2009; MING-MING ZHENG et al., 2013).
Não é possível definir um suporte ideal para todos biocatalisadores. Apesar de muitos
parecerem promissores na literatura, estes requerem ainda mais estudos e otimizações em seus
respectivos processos (CUNHA et al., 2014).
Existe uma grande variedade de materiais disponíveis no mercado para o processo de
imobilização, como por exemplo, SepabeadsR, SpherezymeTM, EupergitR etc. A interação
entre a enzima e o suporte fornecerá um biocatalizador imobilizado com diferentes
propriedades específicas: químicas, bioquímicas, mecânicas e cinética (MALCATA et al.,
1990, NASRATUM et al., 2009, ZHENG et al., 2013, GODOY et al., 2010,
BORNSCHEUER et al., 1994). As características físicas do suporte, como o tamanho do poro
e da partícula, estão relacionados com a capacidade de ligação da enzima ao mesmo. Apesar
de suportes não-porosos apresentarem baixo efeito difusional após a imobilização
(característica desejável), estes apresentam pouca capacidade de imobilização-carga de
enzima. Entretanto suportes porosos são preferíveis por apresentarem alta área superficial,
permitindo que ocorra uma grande quantidade de carga de enzima imobilizada, além do
processo de imobilização garantir a proteção da proteína do ambiente externo (BRENA e
BATISTA-VIEIRA, 2006).
A alteração das propriedades enzimáticas, que irão resultar na melhor performance,
está relacionada com os processos de imobilização que precisarão ser selecionados para tornar
a imobilização mais adequada para um determinado biocatalisador. Em alguns casos, o uso de

57
Capítulo II – REVISÃO BIBLIOGRÁFICA
mais de um protocolo de imobilização pode ser a melhor solução para o aumento da eficiência
do biocatalisador em uma determinada reação (GARCIA-GALAN et al., 2011; SHELDON,
2011; COWAN e FERNANDEZ-LAFUENTE, 2011; RODRIGUES et al., 2011;
HERNANDEZ e FERNANDEZ- LAFUENTE, 2011).
Os métodos de imobilização vem sendo classificados de diversas formas segundo
critérios estipulados por diferentes pesquisadores (BRENA e BATISTA-VIEIRA; 2006;
KRAJEWSKA 2009; MILETIC et al., 2012; BUCKO et al. 2012, MENDES et al. 2012).
Basicamente, os métodos de imobilização podem ser classificados em imobilização por
ligação a superfície, como adsorção física, iônica e metálica; ligação covalente e
confinamento (Incorporação em redes poliméricas; em membranas ou inclusão em
membranas) que podem ser visualizados na Figura 25. Estes diferentes métodos vem sendo
empregados constantemente na literatura.

58
Capítulo II – REVISÃO BIBLIOGRÁFICA
Figura 25. Métodos gerais de imobilização de biocatalisador: 1-Adsorção física; 2-Ligação covalente: A- Derivados monoméricos multipontuais, B- Derivado monomérico
unipontual, C- Derivado bimolecular multipontual (o círculo preenchido de amarelo representa o sítio ativo); 3-Cross-Linking; 4- Entrelaçamento em matrix polimérica; 5-
Encapsulamento em gel; 6- Ligação metálica, 7-Ligação por pontes dissulfeto, 8-9: Reator de membrana- 8- Enzima imobilizada na camada externa da membrana (item 8-
modificado a partir de LAU et al. 2010); 9- Membrana contém biocatalisadores imobilizados em seu interior (item 9-modificado a partir de NAGY, 2012).
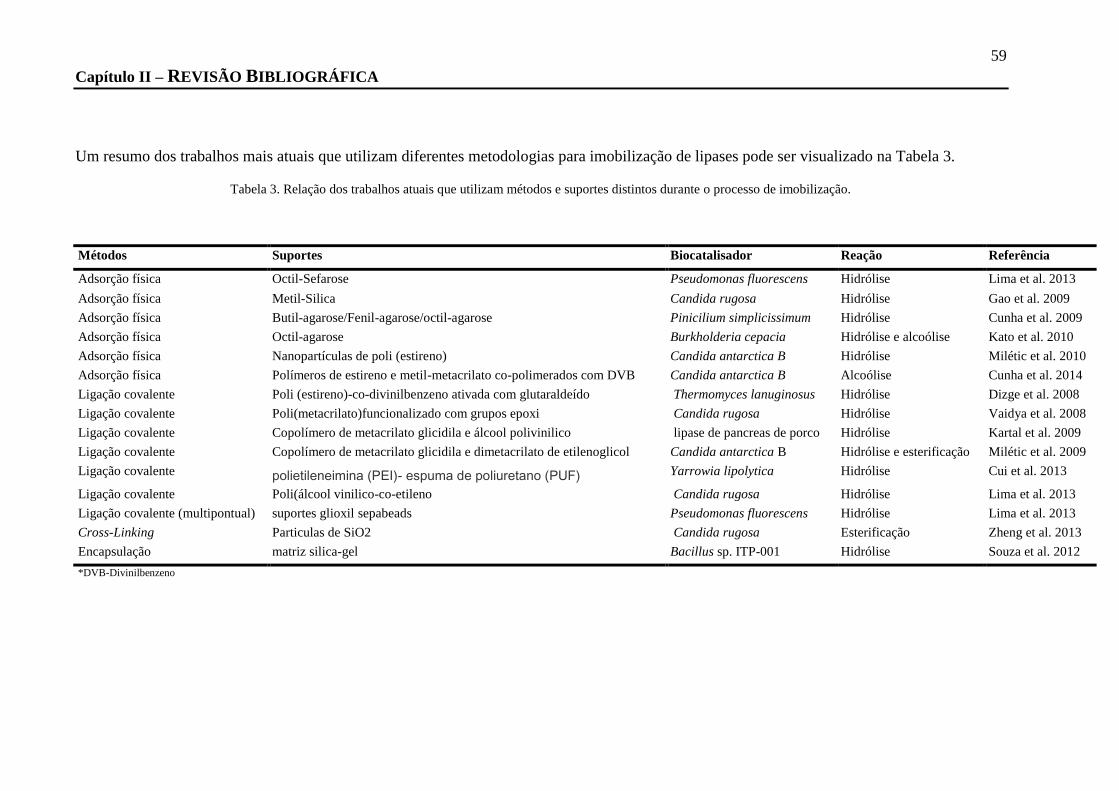
59
Capítulo II – REVISÃO BIBLIOGRÁFICA
Um resumo dos trabalhos mais atuais que utilizam diferentes metodologias para imobilização de lipases pode ser visualizado na Tabela 3.
Tabela 3. Relação dos trabalhos atuais que utilizam métodos e suportes distintos durante o processo de imobilização.
Métodos Suportes Biocatalisador Reação Referência
Adsorção física Octil-Sefarose Pseudomonas fluorescens Hidrólise Lima et al. 2013
Adsorção física Metil-Silica Candida rugosa Hidrólise Gao et al. 2009
Adsorção física Butil-agarose/Fenil-agarose/octil-agarose Pinicilium simplicissimum Hidrólise Cunha et al. 2009
Adsorção física Octil-agarose Burkholderia cepacia Hidrólise e alcoólise Kato et al. 2010
Adsorção física Nanopartículas de poli (estireno) Candida antarctica B Hidrólise Milétic et al. 2010
Adsorção física Polímeros de estireno e metil-metacrilato co-polimerados com DVB Candida antarctica B Alcoólise Cunha et al. 2014
Ligação covalente Poli (estireno)-co-divinilbenzeno ativada com glutaraldeído Thermomyces lanuginosus Hidrólise Dizge et al. 2008
Ligação covalente Poli(metacrilato)funcionalizado com grupos epoxi Candida rugosa Hidrólise Vaidya et al. 2008
Ligação covalente Copolímero de metacrilato glicidila e álcool polivinilico lipase de pancreas de porco Hidrólise Kartal et al. 2009
Ligação covalente Copolímero de metacrilato glicidila e dimetacrilato de etilenoglicol Candida antarctica B Hidrólise e esterificação Milétic et al. 2009
Ligação covalente polietileneimina (PEI)- espuma de poliuretano (PUF) Yarrowia lipolytica Hidrólise Cui et al. 2013
Ligação covalente Poli(álcool vinilico-co-etileno Candida rugosa Hidrólise Lima et al. 2013
Ligação covalente (multipontual) suportes glioxil sepabeads Pseudomonas fluorescens Hidrólise Lima et al. 2013
Cross-Linking Particulas de SiO2 Candida rugosa Esterificação Zheng et al. 2013
Encapsulação matriz silica-gel Bacillus sp. ITP-001 Hidrólise Souza et al. 2012
*DVB-Divinilbenzeno

60
Capítulo III – REVISÃO BIBLIOGRÁFICA
Como visualizado na Figura 25, as imobilizações podem ser reversíveis ou
irreversíveis. A imobilização reversível pode ser por simples adsorção física, ligações iônicas,
metálicas ou por ligações que envolvam pontes dissulfeto. Nesta tese será focado a
imobilização por adsorção física, como parte dos objetivos da mesma.
Imobilização da enzima por adsorção física
O procedimento de adsorção física é um método que se baseia na imobilização de uma
proteína sobre um suporte sólido por ligações de baixa energia, tais como interações de Van
der Waals, iônicas e ligações hidrogênio. Este é um dos métodos mais utilizados por ser
considerado relativamente simples, econômico e rápido. A técnica consiste na incubação do
suporte em uma solução enzimática ou precipitação da lipase sobre a superfície do suporte
pelo uso de solventes (VILLENEUVE et al. 2000).
Suportes de naturezas distintas vem sendo utilizados para esta proposta, a saber,
polietileno, polipropileno, celite, resina sintética, celulose, octil-agarose, estireno, divinil-
benzeno, sefadex e outros (VILLENEUVE et al., 2000; PALOMO et al., 2002a,b;
AYBASTIER e DEMIR, 2010; HERNANDEZ, GARCIA-GALAN e FERNANDEZ-
LAFUENTE, 2011).
O sucesso e a eficiência da adsorção de uma proteína sobre um determinado suporte
dependerá de diversos parâmetros, como a área superficial do adsorvente, tamanho da
proteína a ser adsorvida e principalmente o tamanho e número dos poros. Portanto o uso de
suportes porosos pode ser considerado vantajoso para a técnica já que a enzima é adsorvida no
interior dos mesmos. A concentração da enzima (em termos de unidade de atividade) também
é outro parâmetro importante na eficiência da adsorção. Isso porque a quantidade de enzima
adsorvida por quantidade do suporte aumenta com a concentração da enzima livre até atingir a
um patamar de saturação (DALLA-VECCHIA et al., 2004).
A adsorção hidrofóbica de lipases sobre suportes porosos, ocorre via ativação
interfacial. Basicamente, a ativação interfacial ocorre quando as enzimas reconhecem estes
suportes como sendo uma interface hidrofóbica/hidrofílica, sofrendo ativação interfacial. Com
isso, tem-se a adsorção da proteína sobre os suportes por meio dos grandes bolsos
hidrofóbicos (formados pela face interna da tampa que recobre o sítio ativo) e da área
hidrofóbica próxima ao sítio da enzima, estabilizando a forma aberta e ativa da lipase (Figura
26) (BASTIDA et al., 1998; PAIVA et al., 2000; PALOMO et al., 2002). A imobilização
permite a fixação da lipase em sua conformação aberta, ou seja, o sitio ativo fica exposto pelo
deslocamento da lid, permitindo a acessibilidade de distintos substratos hidrofóbicos e

61
Capítulo III – REVISÃO BIBLIOGRÁFICA
solúveis (FERNANDEZ-LORENTE et al. 2008; BYZOZOWSKI et al.1991; MATEO et al.,
2007).
Figura 26. Hiperativação de lipases sobre superfícies altamente hidrofóbicas (Modificado de PALOMO et al.,
2002).
Entretanto, se o substrato for muito grande ou hidrofílico, pode ocorrer impedimento
estereoquímico devido à proximidade da superfície do suporte hidrofóbico em relação ao sítio
ativo da lipase, reduzindo a atividade da enzima (MATEO et al., 2007). Geralmente a
adsorção hidrofóbica de lipases sobre suportes porosos ocorre na superfície. Os suportes
porosos possuem superfície interna inerte recoberta por espessas camadas bem definidas e
altamente hidrofóbicas a baixa força iônica (BATISDA et al., 1998; PAIVA et al., 2000;
PALOMO et al., 2002).
Como já supra-mencionado, o mecanismo de ativação interfacial das lipases pode ser
explorada no desenvolvimento de métodos de purificação. Lipases com pequenas diferenças
estruturais podem reconhecer interfaces hidrofóbicas de maneiras diferentes, modificando a
taxa e a força de adsorção. Esta diferença na afinidade por superfícies com graus de
hidrofobicidade distintos pode ser explorada para separar ou enriquecer diferentes isoformas
por adsorção seletiva, variando as condições experimentais como tempo de equilíbrio, pH,
força iônica e temperatura de equilíbrio da solução. Com isso, pode-se obter enzimas
purificadas e imobilizadas de forma eficiente (SABUQUILLO et al., 1998, FERNÁNDEZ-
LORENTE et al., 2001).
Pela técnica de imobilização por adsorção envolver interações fracas, torna-se possível
a dessorção do catalisador sem atividade e, consequentemente, a recuperação e reciclagem do
suporte com o uso de substâncias como detergente, ureia, guanidina, variação do pH, solução
salina, etc. A possibilidade da dessorção da proteína pode ser considerado uma vantagem
desta técnica de imobilização na recuperação de suportes de alto custo. Em contra-partida, a
possibilidade de dessorção pode ser considerado uma das principais desvantagens para a
técnica quando o biocatalisador está sendo utilizado no meio reacional, em sistemas aquosos.

62
Capítulo III – REVISÃO BIBLIOGRÁFICA
Todavia essa dessorção pode ser minimizada em meio orgânico, uma vez que há baixa
afinidade entre enzimas e solventes orgânicos (principalmente os mais apolares) (PALOMO
et al., 2002).
A técnica de adsorção em suportes hidrofóbicos tem sido a mais empregada para
imobilização de enzimas em reatores contínuos e do tipo tanques agitados (SOARES, et al.
2007; SOUZA et al., 2012), a mais adequada para imobilização de lipases para uso em
solventes orgânicos e empregada em diferentes preparações industriais como na preparação da
CALB- Novozym 435 (CALB recombinante expressa em Aspergillus niger e imobilizada em
resina acrílica), comercializada pela Novozymes.
A técnica de adsorção de lipases em suportes hidrofóbicos tem sido bem relatada na
literatura como um fator chave para o melhoramento da enantiosseletividade
(BYZOZOWSKI et al.1991; MATEO et al., 2007; FERNANDEZ-LORENTE et al. 2008;
LIMA et al., 2013), sendo considerada ainda uma estratégia para modificação de
enantiopreferencia de muitas lipases. Palomo e colaboradores (2004) observaram que a
mesma lipase imobilizada sobre diferentes suportes, ou utilizando diferentes protocolos de
imobilização, pode exibir enantiosseletividade muito diferente e, em alguns casos, pode
ocorrer a modificação do enantiômero catalisado pela lipase sob as mesmas condições
experimentais.
Zhang et al. (2012) estudaram a influência das características das superfícies
hidrofóbica/hidrofílica na composição do suporte de imobilização e na atividade de CRL.
Diferentes proporções dos monômeros acetato de vinila (monômero hidrofóbico) e acrilamida
(monômero hidrofílico) foram utilizados na polimerização formando microesferas de Poli
(acetato de vinila–acrilamida). Quando as características das superfícies
hidrofóbica/hidrofílica das microesferas foram modificadas pela mudança na proporção de
acetato de vinila-acrilamida de 50:50 para 86:24, a eficiência de imobilização mudou de 45%
para 92% e a atividade da enzima imobilizada aumentou de 202,5 para 598,0 U/g. O processo
de imobilização resultou ainda em uma maior atividade específica (4.65 U/mg lipase) se
comparado com a enzima livre (3.00 U/mg lipase).
Zheng et al. (2013) confeccionaram um suporte por cross linking (Hyper-Cross-
Linked Polymer- Coated Silica com posterior imobilizacao de Candida rugosa (CRL) por
adsorção para síntese biocatalítica de ésteres fitoesteróis (composto ativo natural com
propriedades anti-inflamatórias, antioxidantes, anti-cancerígenas etc.). A lipase imobilizada
(CRL-HPCS) mostrou uma estabilidade térmica e condições de reutilização do biocatalisador
superiores do que aquelas apresentadas pela enzima na sua forma livre (CRL). Alem disso, foi

63
Capítulo III – REVISÃO BIBLIOGRÁFICA
possivel observar um alto valor de conversao do produto linolenato de fitostanila (95.6%) se
comparado com a lipase imobilizada em polímero de sílica (46,5%) (MING-MING ZHENG
et al. 2013).
A influência de lipases imobilizadas em suportes distintos submetidos a diferentes
processos de imobilização, tais como adsorção e ligação covalente, tem sido estudada na
hidrólise enantiosseletiva de (R,S) -2-hidroxi-4-fenilbutirato ester etílico [HPBET] (LIMA et
al. 2013, FERNANDEZ-LORENTE et al. 2008, PALOMO et al. 2005). O comportamento
frente a diferentes suportes, mesmo utilizando o mesmo método de imobilização, pode alterar
a especificidade, atividade e estabilidade do biocatalisador. Lipase B de Candida Antarctica
(CALB), lipase from Thermomyces lanuginose and lipase de Bacillus thermocatenulatus,
imobilizadas por ativação interfacial em quatro diferentes suportes hidrofóbicos (hexil- e
butil-toyopearl [resina hidrofílica] e ainda, butil- e octil-agarose, apresentaram diferentes
resultados de estabilidade, atividade recuperada (casos com hiperativação e outros com quase
a inativação do biocatalisador), especificidade e força de adsorção (FERNANDEZ-
LORENTE et al. 2008).
Lima et al. (2013) obtiveram maior atividade específica (1,2 IU/mg) da lipase de
Pseudomonas fluorescens quando imobilizada em octil-Sefarose (adsorção) com alta
enantiosseletividade (E>100) na hidrolise enantiosseletiva de HPBET.
No processo de imobilização, se objetiva ter o máximo de retenção de atividade.
Quando este valor ultrapassa os 100%, diz-se que ocorreu a hiperativação. O fenômeno da
hiperativação de lipases em suportes hidrofóbicos tem sido relatado por muitos autores.
Bastida et al., (1998) mostraram a hiperativação de seis diferentes lipases (dentre elas lipase
C. antarctica A, Rhizomucor miehei e Humicola lanuginosa com 200, 700 e 2000% da
atividade relativa, respectivamente). Palomo et al., (2002, 2004) obtiveram a hiperativação de
Mucor miehei (500%) e Thermus thermophilus (400%) quando imobilizadas em octil-
sepabeads. Cunha et al., (2009) imobilizaram Penicilium simplicissimum sobre diferentes
suportes com hidrofobicidades distintas (butil-, fenil- e octil-agarose) observando a maior
atividade relativa (%) com o suporte mais hidrofóbico (octil-agarose/ 1.133%) (BATISDA et
al., 1998; PALOMO et al., 2002; PALOMO et al., 2004; CUNHA et al., 2009). Vale ressaltar
que o fenômeno de hiperativação é observado principalmente em baixas concentrações de
enzima (em unidades de atividade). Por esse motivo, nem sempre um suporte que mostra
hiperativação se apresenta como boa alternativa biocatalítica, uma vez que muitas vezes, para
que a reação ocorra, faz-se necessário uma grande quantidade de biocatalisador por grama de
suporte (CUNHA et al., 2014).

64
Capítulo III – REVISÃO BIBLIOGRÁFICA
Quanto ao substrato: Influência dos grupamentos substituintes dos substratos
O mecanismo de enantiosseletividade em uma reação enzimática pode ser descrito por
alguns modelos que preveem a enantiopreferência pelo substrato. Fitzpatrick e Klibanov
(1991) tentaram explicar a enantiosseletividade da enzima a partir do tamanho dos
grupamentos substituintes dos substratos. Kazlauskas e colaboradores (1991) predisseram a
enantiosseletividade de lipases de Pseudomonas cepacia e Candida rugosa frente a álcoois
secundários com base no tamanho e na forma das moléculas quando estas estão no sítio ativo
da enzima. A estrutura leva a uma enantiopreferência por um determinado enantiômero. A
Figura 27 ilustra o modelo de sítio ativo de lipases que consiste de duas regiões de diferentes
tamanhos, uma grande e outra pequena.
Figura 27. Enantiomero rápido (a) e enantiomero lento (b)- modelo de sítio ativo por lipases (Reproduzido de
KAZLAUSKAS et al., 1991).
Quanto ao design do processo
Utilização de diferentes reatores na síntese de compostos bioativos em sistemas
multifásicos
A seleção do reator mais apropriado para um dado bioprocesso dependerá de alguns
aspectos chaves como controle do pH, riscos de contaminação, conhecimento da cinética do
processo e do biocatalisador, em sua forma livre (recomendável o uso de micro-organismos
seguros - GRAS- Generally Regarded As Safe) ou imobilizado etc (PALOMARES et al.,
2009; TUFVESSON et al., 2010). Estes vão determinar o modo de condução do processo, as
características do fluxo e se estes estão diretamente relacionadas com a estabilidade e re-uso
da enzima, pontos fundamentais a serem avaliados na obtenção de moléculas bioativas
visando a diminuição de custos na catálise enzimática em escala comercial.
O tipo de configuração do biorreator para preparações enzimáticas livres ou
imobilizadas deverão levar em consideração o mecanismo de reaproveitamento do

65
Capítulo III – REVISÃO BIBLIOGRÁFICA
biocatalisador da corrente de produtos. No caso de utilização de biocatalisadores livres,
unidades de ultra- filtração ou centrifugação poderão ser acopladas ao sistema. Entretanto,
maiores opções de processo e bioreatores são possíveis se o biocatalizador estiver na forma
imobilizada (FREIRE et al., 2011).
As características da bioconversão incluem as propriedades bioquímicas dos
substratos/produtos (pontos de fusão e ebulição, solubilidade em água, estabilidade ao pH e
temperatura); além das características reacionais (pH, temperatura, controle da atividade da
água). As características do biocatalisador incluem a utilização de enzimas (desintegração
celular) ou células íntegras (necessidade de co-fatores), livres ou imobilizadas, a localização
da atividade enzimática (normalmente na fase aquosa, exceto para as lipases que são ativadas
por interfaces), e por fim a quantidade de água no sistema para manutenção da atividade
catalítica do biocatalisador.
É importante considerar ainda as interações entre o biocatalisador, substratos e
produtos. Em tanques agitados, os suportes dos biocatalisadores imobilizados nem sempre
possuem resistência ao estresse mecânico. Portanto reatores descontínuos podem não ser a
melhor escolha uma vez que com a danificação do suporte utilizado, as proteínas serão
lixiviadas e o reuso ficará comprometido (HARTMANN e JUNG, 2010).
Dentre os reatores usados em sistemas multifásicos destacam-se, os reatores agitados
mecanicamente (descontínuos ou contínuos), os reatores de leito fixo e os reatores agitados
por uma fase líquida (AIRES-BARROS, 2002). Nesta tese, os reatores agitados
mecanicamente (descontínuos ou batelada simples) e reatores de leito fixo em fluxo contínuo
serão destacados.
A maior parte dos trabalhos da literatura relata a utilização de reatores descontínuos do
tipo tanque agitado operando em modo de batelada simples em escala reduzida. Entretanto,
durante os últimos anos, houve um aumento considerável no número de trabalhos que surgiu
na literatura na área de biocatálise utilizando o processo de fluxo contínuo. Esta tecnologia
pode melhorar a eficiência de reações enzimáticas e ainda, estudos utilizando ambos sistemas,
descontínuo e contínuo, na produção de diversos produtos com aplicações biotecnológicas,
como produção de biodiesel (HASSAN, et al., 1995; HASSAN et al., 1996; KEE et al., 1999;
RAO et al., 2009; NIE et al., 2006; CHEN et al., 2010; LEE et al., 2010; TONGBORIBOON
et al., 2010; BRASK et al., 2011; KAWAKAMI et al., 2011; ITABAIANA Jr., et al. 2011,
2012a,b).
Watson (2012) realizou um estudo de investigação do interesse da tecnologia em
sistema de fluxo contínuo pelas empresas farmacêuticas. O resultado mostrou que cerca de

66
Capítulo III – REVISÃO BIBLIOGRÁFICA
48% de todas as empresas entrevistadas manifestaram alto interesse em processos contínuos,
com algumas destas empresas já realizando processos com este sistema. A maior parte deste
percentual de empresas estudadas são as maiores companhias farmacêuticas, que possuem
fontes disponíveis para investimentos em tecnologias novas e/ou alternativas.
Cunha et al., (2010, 2012) estudaram a resolução cinética de derivados de mio-inositol
catalisados por lipase comercial imobilizada de Candida antartica (Novozym 435). Os
autores utilizaram os reatores do tipo tanque agitado operando em modo batelada, em escala
reduzida. Os autores obtiveram elevada pureza e conversão.
Boros et al. (2013) investigaram diversos métodos de imobilização e suportes
utilizando a lipase CALB (Figura 28). Os autores avaliaram a resolução cinética de aminas
secundárias em dois tipos de reatores, em sistema de batelada simples e de fluxo contínuo e
encontraram maiores produtividades para as lipases submetidas em sistema de fluxo contínuo.
Figura 28. Resolução cinética de aminas racêmicas rac -1a-c por acetilação do acetato de etila catalisado por
CalB sob variados métodos de imobilização (Modificado de BOROS et al., 2013).
A literatura reporta diversos trabalhos que apresentam o estudo de derivados do
ibuprofeno em diferentes reatores. A resolução cinética de flurbiprofeno tem sido feita em
sistema de batelada simples e fluxo contínuo. Apesar do vasto campo de aplicação do
flurbiprofeno, substância antiflamatória não-esteroidal, recentemente tem sido realizados
ensaios clínicos para tratamento de metástase contra o câncer de próstata, no qual o isômero
(R)-fluorbiprofeno é o que apresenta o efeito ativo para o tratamento (TAMBORINI et al.,
2012). O esquema da reação de esterificação em sistema de fluxo contínuo para a obtenção

67
Capítulo III – REVISÃO BIBLIOGRÁFICA
dos isômeros (S) e (R)- flurbiprofeno, em suas formas enantiopuras (eep maior que 98%),
pode ser visualizado na Figura 29.
Figura 29. Resolução cinética global e processo de purificação do enantiômero S em sistema
de fluxo contínuo (Modificado de TAMBORINI et al., 2012).
A diminuição do tempo reacional e o aumento da produtividade são claramente
observados em determinados processos quando o sistema em batelada é substituído pelo
sistema de fluxo contínuo. Tamborini et al. (2012) conseguiram a redução de até 10 vezes no
tempo reacional e o aumento de 10 vezes na produtividade quando alteraram o tipo de
processo para fluxo contínuo.
Resoluções cinéticas catalisadas por lipases em sistemas de fluxo contínuo foram
abordados por diferentes autores. No entanto, a aplicação desta tecnologia para a síntese de
inositóis não foi previamente estudada.
Uma outra alternativa interessante aos reatores supracitados seria a utilização de
bioreatores acoplados a membranas para separação simultânea de produto e recuperação do
biocatalizador que pode estar livre ou imobilizado na membrana. A escolha da membrana
mais adequada será função do tipo de biocatalizador empregado (LAU et al., 2010; LAU et
al., 2011).
Para os reatores relatados, o desenvolvimento de produtos, em nível industrial ainda é
limitado devido ao seu alto custo e baixa estabilidade (GUMÍ et al., 2008). A maioria dos
autores operam os reatores em escala reduzida ou laboratorial.

68
Capítulo III – REVISÃO BIBLIOGRÁFICA
Influência do agente acilante
Os primeiros relatos do uso de doadores de acila em catalise enantiosseletiva por
lipase ocorreram em 1986-1987 (DEGUEIL-CASTAING et al., 1987; SWEERS e WONG,
1986). Um ano depois, as reações de acetilação enantioselectiva foram mais descritas na
literatura (LAUMEN et al., 1988; TERAO et al., 1988; WANG et al., 1988; WANG e
WONG, 1988). A partir deste momento, inúmeros pesquisadores vêm desenvolvendo estudos
da influencia dos agentes acilantes nas reações enzimáticas.
A presença do agente acilante (ou doador de acila) é de suma importância em certas
reações como reações de transesterificação e alcoólise, devido à necessidade de um composto
éster no início da resolução racêmica ou na síntese assimétrica (WEBER et al., 1995a;
CARREA et al., 1995).
Ésteres ativados são mais adequados como doadores de acila para reações
enantioseletivas catalisadas por lipase, uma vez que estas moléculas evitam a reversibilidade
do processo (WANG et al., 1988; GOTOR-FERNÁNDEZ, BRIEVA e GOTOR, 2006;
PARAVIDINO e HANEFELD, 2011).
Acetato de vinila é doador de acila mais comum dos ésteres enólicos que são usados
em reações enantiosseletivas (PARAVIDINO e HANEFELD, 2011; SIMON et al., 2012).
Apesar do acetato de vinila ser mais reativo, acetato de isopropenila também pode ser usado
nestas reações. Estes ésteres podem ser empregados não apenas como agente acilante, mas
também como solvente da reação. O produto álcool tautomeriza a um composto carbonila,
impulsionando assim a reação e eliminando a inibição da reação para obtenção do produto
requerido. (PARAVIDINO e HANEFELD, 2011). Novos doadores de acila como dietil
malonato, também são citados na literatura (SIMON et al., 2012).
Um doador de acila ideal seria aquele com baixo custo, capaz de acilar de forma
rápida e irreversível, além de ser completamente inerte a lipase e ao meio ambiente.
Infelizmente nenhum dos doares de acila cumpre todos estes requisitos (BORNSCHEUER e
KAZLAUSKAS, 2006b; PARAVIDINO e HANEFELD, 2011). Diferentes agentes acilantes
são citados na literatura e utilizados em reações enantiosseletivas. Estes estão ilustrados na
Figura 30.

69
Capítulo III – REVISÃO BIBLIOGRÁFICA
Figura 30. Exemplos de agentes acilantes para acilações irreversíveis a partir de álcoois (A-B-D:Reproduzido de
BORNSCHEUER e KAZLAUSKAS, 2006b; C- Reproduzido de SIMON et al., 2012).
Lipases que catalisam acilações com estes ésteres ativados (Figura 30-A) permitem
uma catálise de até duas vezes mais rápidas do que com os ácidos ou ésteres simples. Além
disso, estes ésteres deslocam a constante de equilíbrio a favor da acilação (BORNSCHEUER
e KAZLAUSKAS, 2006b).
No caso dos enol-ésteres e anidridos, a acilação é praticamente irreversível. Como
reações irreversíveis são requeridas para resolução cinética de álcoois, estes são
frequentemente utilizados (BORNSCHEUER e KAZLAUSKAS, 2006b; CUNHA et al.,
2011; MANOEL et al., 2012a). Em reações do tipo reversíveis, ocorre a diminuição da pureza
enantiomérica do substrato não reagido (LUNDH et al., 1995; BORNSCHEUER e
KAZLAUSKAS, 2006b).
BERKOWITZ et al. (1992) desenvolveram de maneira eficiente uma rota para
oligossacarídeos artificiais. Os autores obtiveram 47% de conversão em 11h de tempo
reacional (Figura 31):
Figura 31. Síntese de oligossacarídeos artificiais utilizando a lipase PS-30 e acetato de vinila durante 11h de
tempo reacional (Modificado de BERKOWITZ et al., [1994] por BORNSCHEUER e KAZLAUSKAS, 2006b).
A literatura tem mostrado que muitas lipases toleram o acetaldeído liberado pela
tautomerização do acetato de vinila, com exceção das lipases CRL (lipase de C. rugosa) e
GCL (lipase de Geotrichum candidum). A inativação da lipase pelo acetaldeído se dá
A B
C D

70
Capítulo III – REVISÃO BIBLIOGRÁFICA
provavelmente devido a formação da base de Schiff com o resíduo de aminoácido Lisina que
inativa lentamente a enzima. Por outro lado, quando o acetato de isopropenila é utilizado
como agente acilante, tem-se a liberação da acetona (WEBER et al., 1995). Diferente do
aldeído, a acetona é menos reativa. Outra alternativa é utilizar o acetato 1-etoxivinila, que
libera acetato de etila.
Anidridos cíclicos (anidrido succínico, por exemplo) são mais frequentemente
utilizados para a resolução cinética de álcoois do que seu equivalente acíclico, principalmente
escala industrial. O uso dos anidridos acíclicos é feito de maneira limitada uma vez que estes
são agentes irritantes, desativam algumas enzimas ao acilá-las, liberam ácido em suas reações
podendo prejudicar o sistema (PARAVIDINO e HANEFELD, 2011).
O doador de acila ideal que mais se adeque aos 12 princípios da química verde ainda é
desconhecido. Em reações de grande escala, o ácido carboxílico aparece como sendo o doador
de acila mais indicado na perspectiva da química verde. Apesar de muitos esforços,
pesquisadores enfatizam a importância de estudos na busca de alternativas mais sustentáveis,
uma vez que os agentes acilantes mais utilizados na literatura (enol-ésteres), não se
enquadram nos 12 princípios da química verde, portanto a redução da utilização dos mesmos
se faz necessário (PARAVIDINO e HANEFELD, 2011).
Influência do solvente
O emprego de diferentes solventes, agentes acilantes e quantidade da água no meio
reacional pode afetar drasticamente a reação de biotransformação. Diversos fatores afetam
uma reação de resolução enantiomérica desde o sistema de solvente empregado, que tem o
poder de alterar a especificidade, quimiosseletividade, regiosseletividade e
enantiosseletividade das lipases, até o teor de água presente na reação.
No início de um trabalho, há a necessidade da realização de experimentos
preliminares, os quais identificarão o melhor sistema de solvente, as enzimas que serão
capazes de catalisar a reação e quais agentes acilantes poderão ser empregados.
Apesar da grande influência da natureza do solvente sobre a enantio-, régio-e
quimiosseletividade da enzima, ainda não foi estabelecido um consenso sobre qual parâmetro
melhor descreve a influência do solvente sobre a reação enzimática (COSTA e AMORIM,
1999; LIU et al., 2009).
Uma vez que o conceito de biotransformação está associado ao uso de
biocatalisadores, onde a água é o solvente preponderante, é fácil compreender que a mesma

71
Capítulo III – REVISÃO BIBLIOGRÁFICA
função tenha sido desempenhada por este solvente em inúmeros estudos da cinética
enzimática. Contudo, o uso exclusivo da água como solvente restringia a gama de aplicações
da biocatálise, bem como limitava a produtividade de diversos processos que envolvessem
solventes hidrofóbicos. Por outro lado, a constatação de que muitas enzimas funcionam in
vivo em ambientes ricos em lipídeos hidrofóbicos levou a conclusão de que os meios não
aquosos são igualmente adequados à atividade biocatalítica. A convergência dos dois aspectos
conduziu a incorporação do meio reacional de solventes orgânicos, fluídos supercríticos, fases
gasosas ou sólidas, conhecidos hoje como meios não convencionais (AIRES, 2002).
Solventes orgânicos têm sido utilizados como meios reacionais para
biotransformações enzimáticas em inúmeros processos de síntese orgânica. Muitos estudos
vêm empregando diferentes sistemas de solventes na resolução enantiosseletiva (COSTA e
AMORIM, 1999; LIU et al., 2009). A seletividade das enzimas, incluindo enantio, regio e
quimiosseletividades, é alterada em diferentes solventes, o que normalmente é atribuído às
modificações das conformações da enzima (alteração de estrutura e flexibilidade) causadas
pelo estabelecimento de diferentes tipos de interações enzima-solvente (SHEN et al., 2008).
Em biocatálise, as propriedades dos solventes como polaridade (1/ε), hidrofobicidade
(geralmente determinada como log P1), geometria molecular e habilidade de solvatação são
fatores significativos. Apesar de ainda não ter sido estabelecido um parâmetro que melhor
descreva a influência do solvente sobre a reação enzimática, dois parâmetros vêm sendo
utilizados por diferentes autores para caracterizar as propriedades físico-químicas dos
solventes, log P e constante dielétrica (ε). Dentre eles, o log P é o mais utilizado para a
escolha do solvente. Primeiro, porque este parâmetro é uma medida direta da hidrofobicidade;
segundo, porque seu valor pode ser facilmente determinado por método padrão ou calculado
com base nas constantes hidrofóbicas tabeladas; e terceiro, devido à sua sensibilidade a
pequenas diferenças de polaridade das moléculas (CHUA e SARMIDI, 2006).
Apesar das numerosas exceções, o parâmetro log P é o que apresenta melhor correlação
com a atividade enzimática e, geralmente, as enzimas são mais estáveis em solventes com
caráter hidrofóbico (log P>2) (YANG e RUSSEL, 1996). Contudo o efeito do solvente é
distinto para cada enzima.
A Tabela 4, a seguir, lista alguns solventes de uso comum e os valores de log P relativos
a eles.

72
Capítulo III – REVISÃO BIBLIOGRÁFICA
Tabela 4. Valor de log P de solventes orgânicos (ADLERCREUTZ, 2000).
A forma como os solventes afetam a atividade enzimática e a enantiosseletividade ainda
não é bem compreendida e as hipóteses apresentadas para explicar esse fenômeno possuem
discrepâncias entre si (COSTA e AMORIM, 1999). Pode-se observar nos trabalhos que
exploram esse tema, a preocupação em salientar que os resultados alcançados são restritos aos
sistemas estudados, evitando-se generalizações.
O fato das enzimas manterem a atividade catalítica em solventes orgânicos não possui
explicação simples. A hipótese mais aceita é de que, quando a enzima é colocada em solvente
orgânico anidro, esta é cineticamente ―congelada‖ no seu estado nativo. Isso ocorre em parte
devido à baixa constante dielétrica do meio, que produz uma maior efetividade nas forças
eletrostáticas dos resíduos carregados responsáveis pela integridade estrutural da proteína,
(responsáveis pela manutenção da estrutura enzimática) embora possam tornar as enzimas
menos ativas (ZAKS e KLIBANOV, 1988; ZACKS et al., 1996).
Diversos autores têm relatado a influência da hidrofobicidade do solvente na
enantiosseletividade de enzima. De maneira geral, o efeito tem sido analisado em termos de
partição, de moléculas de água presentes no sítio ativo da enzima (SAKURAI et al., 1988) ou
de grupos funcionais dos substratos (TAWAKI e KLIBANOV, 1992).
Sakurai et al., (1988) observaram uma correlação linear entre a diferença de energia
livre de ativação (G) para a relação de transesterificação entre o éster N-acetil-2-cloroetil-
alanina e propanol e os valores de log P dos 13 diferentes solventes orgânicos empregados,
utilizando subtilisina como catalisador da reação. Os autores encontraram uma relação inversa
entre o valor G e da hidrofobicidade, expressa em valores de log P, como consequência, uma
relação inversa a enantiosseletividade. A dependência entre estes fatores ocorreu
Solvente log P Solvente log P
Metanol -0,76 TBME 1,5
Acetonitrila -0,33 Clorofórmio 2,0
Etanol -0,24 Tolueno 2,5
Acetato de etila 0,68 Hexano 3,5

73
Capítulo III – REVISÃO BIBLIOGRÁFICA
principalmente pela dificuldade no deslocamento das moléculas de água presente na cavidade
hidrofóbica do sítio de ligação da enzima durante a ligação enzima-substrato. O deslocamento
destas moléculas de água é termodinamicamente menos favorecido em solventes hidrofóbicos
do que naqueles hidrofílicos.
Os solventes hidrofóbicos não retiram a água de solvatação da estrutura protéica,
prevenindo a desnaturação e mantendo a estrutura conformacional da enzima. Moléculas do
solvente podem ainda interagir com a estrutura protéica e exercer efeitos ativadores ou
estabilizadores, efeitos esses que também podem estar relacionados com a presença do
suporte quando se trabalha com enzimas imobilizadas (PENCREACH e BARATTI, 1997).
Nesta interação com a estrutura protéica, ainda deve ser considerado que solventes apolares
podem manter a enzima em sua conformação aberta, ou seja, na presença destes solventes, a
tampa hidrofóbica não estaria cobrindo o sítio ativo, o que poderia manter ou até aumentar a
atividade enzimática (RÚA et al., 1993).
Nakamura et al. (1991) estabeleceram um modelo empírico para a escolha do solvente
adequado em reações catalisadas por lipases. Segundo os autores, solventes cíclicos são os
mais adequados para encontrar uma maior enantiosseletividade. Os autores estudaram a
influência da estrutura do solvente na reação entre 1-nitro-propanol e acetato de vinila,
utilizando lipase de Pseudomonas sp. como biocatalisador. Nakamura e colaboradores
observaram que em solventes lineares obtinham maior valor de enantiosseletividade do que
quando os respectivos solventes ramificados eram usados. Constataram ainda que em série
homóloga aos cicloalcanos a enantiosseletividade diminuía conforme o aumento de carbonos
na estrutura dos solventes cíclicos. Isso foi explicado pela facilidade dos solventes lineares e
cíclicos (com um menor número de carbono) se incorporarem melhor na cavidade menor do
sítio ativo da enzima.
Influência da atividade da água
A camada de solvatação presente nas enzimas tem um papel fundamental atuando
como componente primário do microambiente enzimático. Este ambiente é capaz de manter
uma dinâmica molecular mínima, essencial à atividade enzimática em ambientes orgânicos
(ZAKS e KLIBANOV, 1988).
De fato a água presente no meio reacional pode estar dissolvida no solvente, ligada à
enzima ou ao suporte, logo a quantidade de água do meio não indica o grau de hidratação da
enzima. O conceito termodinâmico da atividade de água (aw) é o melhor parâmetro para se

74
Capítulo III – REVISÃO BIBLIOGRÁFICA
referir à quantidade de água disponível em um sistema orgânico (ADLERCREUTZ, 1996;
HALLING, 2003;). A atividade de água (aw), é definida como a razão entre a pressão parcial
da água no sistema (pw) e a pressão parcial da água pura (p
0
w), como mostra a Equação 10.
(Eq. 10)
O parâmetro aw representa a umidade relativa do sistema considerado, situando-se
entre 0 e 1, sendo que quando a atividade de água iguala-se a 1, já há formação de uma fase
aquosa distinta (apud BARON, 2003).
A Tabela 5 apresenta uma lista com os sais mais adequados ao controle da atividade
de água e as atividades de suas respectivas soluções.
Tabela 5. Soluções salinas saturadas adequadas ao controle da atividade de água (ADLERCREUTZ, 2000).
Sal Atividade de água
(25oC)
LiCl 0,113
Acetato de potássio 0,225
MgCl2 0,328
K2CO3 0,432
Mg(NO3)2 0,529
SrCl2 0,708
KCl 0,843
KNO3 0,936
K2SO4 0,973
Como relatado anteriormente, solventes hidrofílicos (aqueles com maior afinidade pela
água) tendem a remover a camada de água da superfície proteica em maior quantidade,
podendo até inativá-la (GORMAN e DORDICK, 1992). Logo a quantidade ótima de água
necessária à reação está relacionada também a polaridade do solvente. Metanol (log P = –
0,76) foi capaz de promover a dessorção de até 60% da água ligada a enzimas (quimiotripsina,
subtilisina e peroxidase) enquanto hexano (log P = 3,5) promoveu a dessorção de apenas 0,5%
da água ligada às essas mesmas enzimas (YANG e RUSSEL, 1996).

75
Capítulo III – REVISÃO BIBLIOGRÁFICA
A quantidade de água presente no meio reacional, assim como qualquer outro
substrato ou produto, afeta também o equilíbrio termodinâmico entre as reações catalisadas
pelas enzimas, alterando a conversão de equilíbrio, embora não modifique a constante de
equilíbrio (PAIVA et al., 2000).
A influência da atividade de água nos sistemas reacionais tem sido estudada por
diferentes autores (SVENSSON et al., 1994; VALIVETY et al., 1992a; MANOEL et al.,
2011; CUNHA et al., 2012). Os resultados obtidos pelos mesmos mostraram que lipases
respondem diferentemente para um aumento da atividade de água. Enquanto algumas lipases
possuem a atividade ótima em baixa atividade de água (Rhizopus arrhizus), outras possuem o
aumento da atividade com o aumento da atividade de água (Pseudomonas sp.). Ainda há
aquelas que possuem perfis intermediários (Candida rugosa) (WEHTJE e ADLERCREUTZ,
1997). A Figura 32 mostra a variação no perfil de atividade de lipases de Rhizopus arrhizus,
Candida rugosa e Pseudomonas sp. em um sistema empregando como reação modelo a
esterificação entre ácido decanóico e dodecanol (WEHTJE e ADLERCREUTZ, 1997).
Figura 32. Efeito da atividade de água na atividade enzimática de lipases de origens diversas: Rhizopus arrhizus
( ) ; Candida rugosa ( ); Pseudomonas sp. ( ) (Reproduzido de WEHTJE e ADLERCREUTZ, 1997).
A quantidade máxima de água requerida para maximizar a atividade enzimática pode
ser determinada para a maioria das lipases e dependerá de cada sistema de solvente orgânico
empregado, de cada biocatalisador e do suporte, quando a lipase estiver em sua forma
imobilizada. Cunha et al., (2012) e Manoel et al. (2011) estudaram por meio de planejamento
experimental a quantidade de água como uma variável independente na reação de alcoólise de
diferentes derivados de mio-inositol utilizando Novozym 435 (C. antarctica tipo B
imobilizada em resina acrílica e PS-IM (P. cepacia imobilizada em terra de diatomácea). Para

76
Capítulo III – REVISÃO BIBLIOGRÁFICA
ambos os casos a variável independente (quantidade de água) mostrou um efeito significativo
negativo para o aumento da conversão enzimática.

77
Capítulo IV – MATERIAL E MÉTODOS
4. MATERIAIS E MÉTODOS
4.1 MATERIAIS
4.1.1 Biocatalisadores utilizados
As lipases comerciais de Rhizomucor miehei (Lipozyme RM IM) e de Candida
antarctica tipo B (CALB, Novozym 435) foram adquiridas da empresa Novozymes.
A lipase B de Candida antarctica expressa em Pichia pastoris - extrato liofilizado;
lipase de Pyrococcus furiosus expressa em E.coli (imobilizada em diferentes suportes); lipase
de Rhizomucor miehei (produzida por Fermentação em estado solido utilizando torta de
babaçu); lipase vegetal de Jatropha curcas L. foram fornecidas pelo laboratório de
Biotecnologia Microbiana (LaBim-IQ/UFRJ). Lipase liofilizada de Yarrowia lipolytica foi
obtida pelo Grupo Biose, da Escola de Química (EQ-UFRJ). Os solventes e demais reagentes
empregados neste trabalho para a resolução cinética dos derivados de mio-inositol e
imobilização foram de grau analítico.
4.1.2 Substratos utilizados
Os substratos foram sintetizados pelo laboratório Roderick Barnes, especificamente
pela química Karla Ceodaro e o farmacêutico Angelo Amaro, ambos alunos de doutorado,
orientados pelo Professor Dr. Alessandro B. C. Simas, NPPN (Núcleo de Pesquisas de
Produtos Naturais)/UFRJ. Os racematos (±)-1,4,5,6-tetra-O-benzil-mio-inositol (rac-1); (±)-
1,4,5,6-tetra-O-alil-mio-inositol (rac-2); (±)-1,3,4-tri-O-benzil-mio-inositol (rac-3) e (±)-3,6-
di-O-benzil-1,2-O-isopropilideno-mio-inositol (rac-4) carecem de resolução óptica e estão
apresentados na Figura 33.
Figura 33. Derivados de mio-inositol utilizados na resolução cinética por lipases.
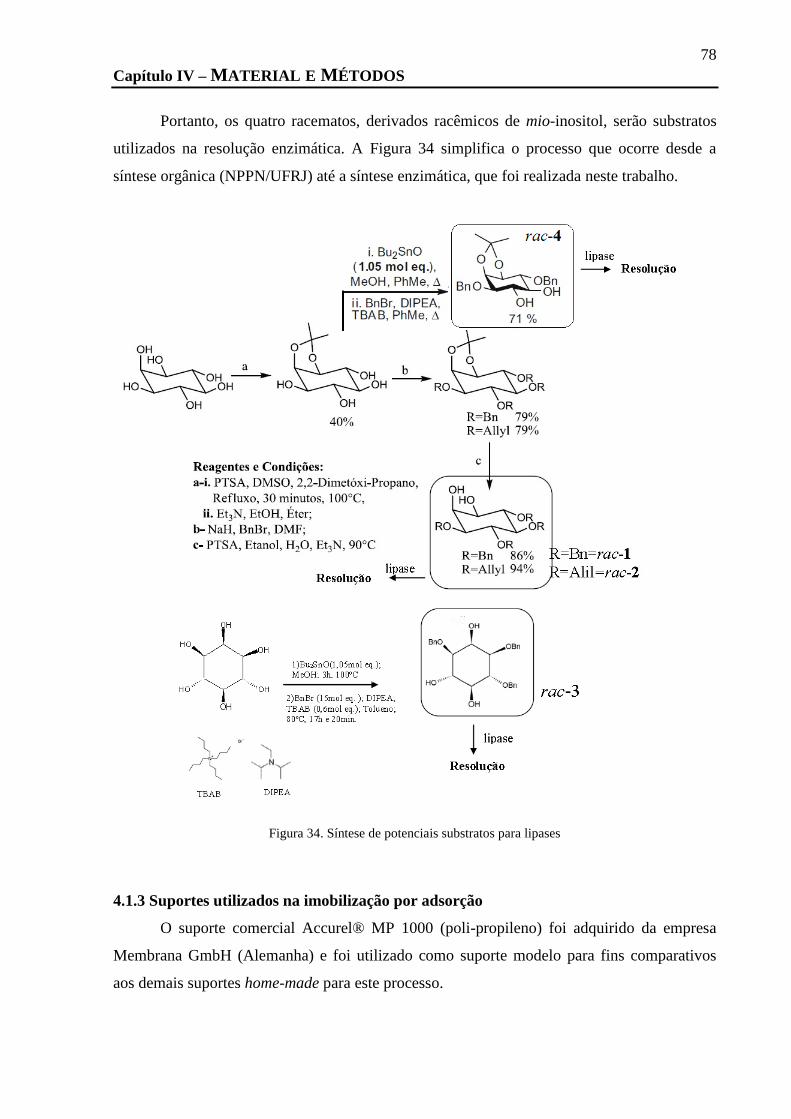
78
Capítulo IV – MATERIAL E MÉTODOS
Portanto, os quatro racematos, derivados racêmicos de mio-inositol, serão substratos
utilizados na resolução enzimática. A Figura 34 simplifica o processo que ocorre desde a
síntese orgânica (NPPN/UFRJ) até a síntese enzimática, que foi realizada neste trabalho.
Figura 34. Síntese de potenciais substratos para lipases
4.1.3 Suportes utilizados na imobilização por adsorção
O suporte comercial Accurel® MP 1000 (poli-propileno) foi adquirido da empresa
Membrana GmbH (Alemanha) e foi utilizado como suporte modelo para fins comparativos
aos demais suportes home-made para este processo.

79
Capítulo IV – MATERIAL E MÉTODOS
Os suportes do tipo casca-núcleo utilizados para imobilização neste trabalho foram
cedidos pelo Laboratório de Modelagem, Simulação e Controle de Processos-
PEQ/COPPE/UFRJ segundo a metodologia de Lenzi et al. (2003), Pinto et al. (2006) e Besteti
(2009). A metodologia de produção dos suportes, realizada pelo laboratório supra-citado,
combina processos em suspensão e em emulsão. Primeiramente, é feita uma polimerização
em suspensão, gerando um núcleo não poroso. Em seguida, promove-se, no mesmo reator,
sem qualquer separação de componentes, uma polimerização em emulsão. Em função da alta
viscosidade e caráter pegajoso, o núcleo atua como um centro de aglomeração das partículas
que são geradas pela emulsão, originando polímeros com morfologia casca-núcleo.
A eficiência de recobrimento do núcleo está diretamente relacionada ao instante em
que a emulsão tem início (ou seja, a duração da etapa de suspensão). Os monômeros
utilizados foram estireno (PS) e metil metacrilato (PMMA), ambos foram reticulados com
divinilbenzeno. Os suportes selecionados foram aqueles que apresentaram maior atividade de
esterificação após o processo de imobilização, além de boas estabilidades frente à diferentes
solventes orgânicos (Figura 35). Estes tiveram por base os resultados de Cunha (2011) e
Cunha et al. (2014). Algumas propriedades das superfícies poliméricas, como a área
específica (m2/g) e o diâmetro de poro (Å) também foram analisadas (Tabela 6):
1. PS-co-DVB/PS-co-DVB 25% (2 horas de suspensão) - núcleo de poli(estireno-co-
divinilbenzeno) e casca de poli(estireno-co-divinilbenzeno) contendo 25% de
divinilbenzeno.
2. PMMA-co-DVB/PMMA-co-DVB 25% (2 horas de suspensão) - núcleo de
poli(metacrilato de metila-co-divinilbenzeno) e casca de poli(metacrilato de metila-co-
divinilbenzeno) contendo 25% de divinilbenzeno.
Tabela 6. Dados de caracterização dos suportes poliméricos selecionados para esta tese (BESTETI, et al., 2009,
2010)
Suportes Área específica (m2/g) Diâmetro do poro (Å)
Accurel MP 1000 39,0 230,0
PS-co-DVB/PS-co-DVB 25% 29,0 190,4
PMMA-co-DVB/PMMA-co-DVB 25% 12,6 193,4

80
Capítulo IV – MATERIAL E MÉTODOS
Figura 35- Micrografia óptica e micrografia eletrônica de varredura dos suportes PMMA-co-DVB/ PMMA-co-
DVB 25% e PS-co-DVB/PS-co-DVB 25%. (Reproduzido de BESTETI et al., 2011).
4.2. MÉTODOS
4.2.1 Seleção de lipases e substratos por meio da reação de acetilação realizados em
sistema descontínuo (ou modo de batelada)
Com o objetivo de selecionar o melhor par lipase-substrato, as reações de acetilação
foram realizadas com as lipases home-made em suas formas liofilizadas e imobilizadas, e com
os racematos mencionados (descritos no item 4.1.1. e 4.1.2, pag. 77), conduzidas em frascos
fechados termostatizados e agitados magneticamente (Figura 36).

81
Capítulo IV – MATERIAL E MÉTODOS
Figura 36. Foto da aparelhagem utilizada no sistema operacional empregado nas reações de acetilação dos
derivados racêmicos.
A seleção inicial foi feita em sistema de batelada simples em reatores (encamisados),
realizada com 5 mg de cada racemato, dissolvidos em 2,5 mL de acetato de vinila utilizado
como solvente e agente acilante, 50 U de enzima, a 30º C, 40º C e 50º C de temperatura por
48 horas. As reações foram realizadas utilizando, a princípio, acetato de vinila como agente
acilante e solvente da reação, uma vez que este agente acilante tem se mostrado o mais
adequado nas reações com mio-inositol (SIMAS et al., 2011) e para a literatura (KALER et
al., 2011; HERBS et al., 2012), além dos benefícios supracitados (pag.68).
Após o período citado (48 horas), as reações foram interrompidas pela remoção do
biocatalisador (quer seja em sua forma livre ou imobilizado) do meio reacional da seguinte
forma: a amostra foi centrifugada com o objetivo de separar o biocatalisador do produto. O
sobrenadante contendo o produto foi levado ao concentrador a vácuo de velocidade (Speed
vac concentrator-Savant SPD 1010-Thermo scientific). O biocatalisador foi lavado três vezes
com 1mL de acetato de etila afim de se retirar todo o produto contido no frasco. O
biocatalisador, já separado, foi levado ao Concentrador a vácuo de velocidade (Speed vac
concentrator) para a evaporação do solvente residual para posterior reutilização do mesmo.
As reações foram acompanhadas por cromatografia em camada fina (CCF), conforme descrito
no item 4.2.5.

82
Capítulo IV – MATERIAL E MÉTODOS
4.2.2 Efeito do agente acilante na conversão e na enantiosseletividade
Para a reação onde a acetilação resultou em formação de produto para um determinado
par substrato-lipase home-made utilizada, ou ainda, substrato lipase comercial imobilizada
utilizada, seguiu-se com a análise do efeito do agente acilante. Agentes acilantes distintos
foram usados com o objetivo de se estudar a influência dos mesmos na conversão e na
enantiosseletividade, em sistema livre de solvente.
As reações enzimáticas foram realizadas em frascos fechados termostatizados
contendo 5 mg de substrato dissolvidos em 2,5 mL de agente acilante (acetato de etila, acetato
de vinila ou acetato de isopropenila) em sistema livre de solvente e 50 U de enzima, a 30° C.
Alíquotas de 100 µL foram retiradas nos intervalos de 0,5, 6, 24, 48, 72, 96 e 120 h. As
reações foram paralisadas pela remoção do biocatalisador do meio reacional e o solvente foi
evaporado no concentrador a vácuo de velocidade (Speed Vac Concentrator- Savant SPD
1010-Thermo scientific). As amostras foram resolubilizadas com 1 mL de fase móvel
acetronitrila:H2O (60:40) para a análise por Cromatografia Líquida de Alta Eficiência em fase
reversa e em coluna quiral, conforme descrito no item 4.2.5.
4.2.3 Efeito do solvente na conversão e na enantiosseletividade
Após a seleção do melhor agente acilante, seguiu-se com a análise do efeito do
solvente na conversão e enantiosseletividade. De fato, sabe-se que a influência do solvente na
enantiosseletividade é relevante (SHEN et al., 2008). Diferentes solventes orgânicos (grau
analítico, Tedia®) foram avaliados, em triplicata, para a reação de resolução cinética dos
derivados do mio-inositol catalisada pelas diferentes lipases: terc-metil butil éter, hexano,
acetato de etila, acetato de vinila e acetato de isopropenila. As reações enzimáticas foram
realizadas em frascos fechados termostatizados contendo 5 mg do substrato selecionado
dissolvidos em 2,0 mL de solvente e agente acilante selecionado (1,5:1), e 136,4 U de enzima
(condição já otimizada), a 30°C. Alíquotas de 100 µL foram retiradas nos intervalos de 0,5, 6,
24, 48, 72, 96 e 120h. As reações foram feitas para cada solvente citado.
As reações foram interrompidas pela remoção do biocatalisador do meio reacional. As
amostras foram ressolubilizadas com 1mL de fase móvel acetronitrila: água (60:40) para a
análise por Cromatografia Líquida de Alta Eficiência (CLAE) em fase reversa e em coluna
quiral. O volume injetado foi de 20µL na coluna em fase reversa e 10 µL na coluna quiral.

83
Capítulo IV – MATERIAL E MÉTODOS
4.2.4 Estudo da resolução cinética dos substratos/lipases selecionadas utilizando o
sistema de fluxo contínuo
O equipamento do sistema em fluxo contínuo
As reações enzimáticas realizadas em condições de fluxo contínuo foram conduzidas
usando o sistema de fluxo Asia system da empresa Syrris (Figura 37).
Figura 37. Reator Asia system, sistema em fluxo contínuo da empresa Syrris mostrando a coluna utilizada para as
reações: A) Detalhe da bomba do equipamento; B) Coluna termoestatizada.
Basicamente o sistema em reatores Asia system funciona da seguinte maneira: A
solução é impulsionada pela bomba (Figura 37.A) ao interior da coluna termostatizada pelo
equipamento (Figura 37.B) contendo o biocatalisador da reação, em uma coluna de leito fixo.
Posteriormente, a solução é recolhida em sua parte superior e preparada para a análise por
CLAE. A taxa do fluxo e a temperatura foram variáveis monitoradas durante todo o processo.
A coluna utilizada foi do tipo ajustável, modelo Omnfit 600psi fornecido igualmente
pela empresa Syrris, como ilustrado na Figura 38.
A
B

84
Capítulo IV – MATERIAL E MÉTODOS
Figura 38. Coluna de leito fixo utilizada no sistema de fluxo contínuo adquirido pela empresa Syrris, onde A)
detalhe do lúmen da coluna onde o biocatalisador é colocado; B) Setas representando a altura que o
biocatalisador preenche a coluna, em centímetros (cm), para cálculo do volume da coluna.
A massa do biocatalisador (de valor conhecido) foi inserida na coluna. A empresa
fornece informações prévias para os cálculos de volume para cada coluna, como pode ser
visualizado na Figura 40.A. Neste caso, 0,7854 x BED (cm) será o valor do volume utilizado.
O valor (em cm) dependerá da altura após preenchimento com o biocatalisador.
O tempo de residência (tr) foi calculado da seguinte forma (CABRAL e TRAMPER,
2005):
tr= V/F
Onde V (mL) é o volume ocupado pela coluna e F é o fluxo utilizado (mL/min).
Porosidade do leito
A fração de vazios ou porosidade média é definida como a razão entre o volume vazio
em relação ao volume total do leito. Esta é uma propriedade estrutural global do leito, sendo
utilizado para caracterizar o espaço total disponível para o fluxo. Uma vez que a porosidade
pode influenciar os perfis de velocidade, podendo causar perda de carga ao longo do leito
(FREUND, 2003; FOLTIN, 2013), este valor foi calculado quando a enzima Novozym 435
foi utilizada:
A
B

85
Capítulo IV – MATERIAL E MÉTODOS
Onde Ɛ é adimensional:
Ɛ = 1 –[(1,60mL)/(2,1)]
Ɛ= 0,238.
Condições reacionais no sistema em fluxo contínuo
O estudo em condições de fluxo contínuo foi realizado com a lipase Novozym 435
utilizando como ponto de partida TBME (Terc-Butil-Metil-Eter) e Hexano como solvente,
uma vez que estes foram os melhores solventes encontrados anteriormente (MANOEL et al.,
2012b). Os agentes acilantes testados foram acetato de vinila, acetato de etila, acetato de
isopropenila e anidrido acético.
Uma solução contendo 4,86 mM de solvente foi usada em condições de fluxo
contínuo. A coluna Omnifit com capacidade de 12,3 mL foi usada preenchida com 527 mg de
enzima imobilizada (atividade de esterificação de Novozym 435 por meio da reação de síntese
do oleato de etila foi de 2728+ 339 U/gsuporte). A coluna de leito fixo preenchida com 527mg
de biocatalisador foi usada para conduzir todas as reações neste tipo de sistema.
Diferentes proporções de substrato/agente acilante foram usados (1:1; 1:5: 1:10 e 1:15
mmol, respectivamente). Os parâmetros reacionais selecionados foram temperatura (30–60º
C) e taxa de fluxo (0,5; 1 e 1,5mL/min). Alíquotas de 0,2 mL e 1,5 mL foram coletadas por
5min para análises posteriores de conversão e excesso enantiomérico em CLAE.
Diversos ciclos de operação em sistema de fluxo contínuo foram feitos utilizando-se a
mesma coluna inicial (preenchida com 527 mg do biocatalisador) durante todo este estudo.
Após o término de cada corrida, a coluna foi lavada com solvente puro (acetato de etila)
utilizando um fluxo de 0,5 mL/min (durante 15 min) para a retirada de possíveis produtos, a
fim de preparar a coluna para uma próxima utilização. No começo e no final de cada dia de
trabalho no equipamento, 1,5 mL foi coletado para a dosagem de proteínas (método Bradford)
e para atividade hidrolítica/esterásica, com o objetivo de analisar perda protéica (dessorção do
catalisador do suporte) ou perda da atividade enzimática.
O mesmo procedimento foi feito utilizando como substrato o rac- 4.
4.2.5 Métodos analíticos

86
Capítulo IV – MATERIAL E MÉTODOS
Análise por CCF (Cromatografia de Camada Fina/delgada ou do inglês, TLC – Thin
Layer Chromatography)
Análises qualitativas por Cromatografia em Camada Fina (CCF) foram feitas
previamente à análise por CLAE (no inglês HPLC).
A cromatografia em camada delgada foi realizada aplicando uma pequena amostra do
meio reacional da fase orgânica. A aplicação foi feita com auxílio de tubo capilar. Utilizou-se
uma solução de acetato de etila: hexano (3:7) como fase móvel e a sílica, da placa de CCF,
como a fase estacionária. Como revelador KMnO4 foi utilizado seguido da secagem da placa
de CCF em placa de aquecimento. O fator de retenção (Rf) foi utilizado como critério de
distinção entre o Rf do substrato e o do produto formado pela Equação 7:
Rf = deslocamento da substância
Deslocamento do eluente (Eq. 7)
O termo Rf expressa a migração de uma substância em relação ao deslocamento total do
solvente (Front do solvente) em um sistema cromatográfico, sendo constante e característico
para cada substância.
A análise qualitativa do grau de conversão da reação por CCF foi avaliada por um
critério baseado na visualização das marcas do substrato e produto, comparado com seus
respectivos padrões, assim como a intensidade dessas marcas quando a placa foi revelada
como já mencionado.
Para as análises em CCF foram utilizadas folhas de alumínio KieselGel 60 F254 e
como fase móvel hexano:acetato de etila (7:3). A visualização das substâncias
cromatografadas foi realizada por revelação de KMnO4 (Permanganato de Potássio).
KMnO4 é um agente oxidante de coloração violeta e foi preparado da seguinte forma:
1º Adiciona-se 150mL de água e 10g de K2CO3; 2º Adiciona-se 2,5mL de NaOH (5% aq.) e
homogeneizar o sistema; 3º Adicionar KMnO4 (1,5g) aos poucos.
Análise das reações de acetilação por Cromatografia Líquida de Alta Eficiência (CLAE)
Análise da conversão
A análise da conversão dos substratos: rac-1, rac-2, rac-3 e rac-4, além dos seus
respectivos compostos acetilados foi feita em CLAE em fase reversa, com coluna Kromasil-
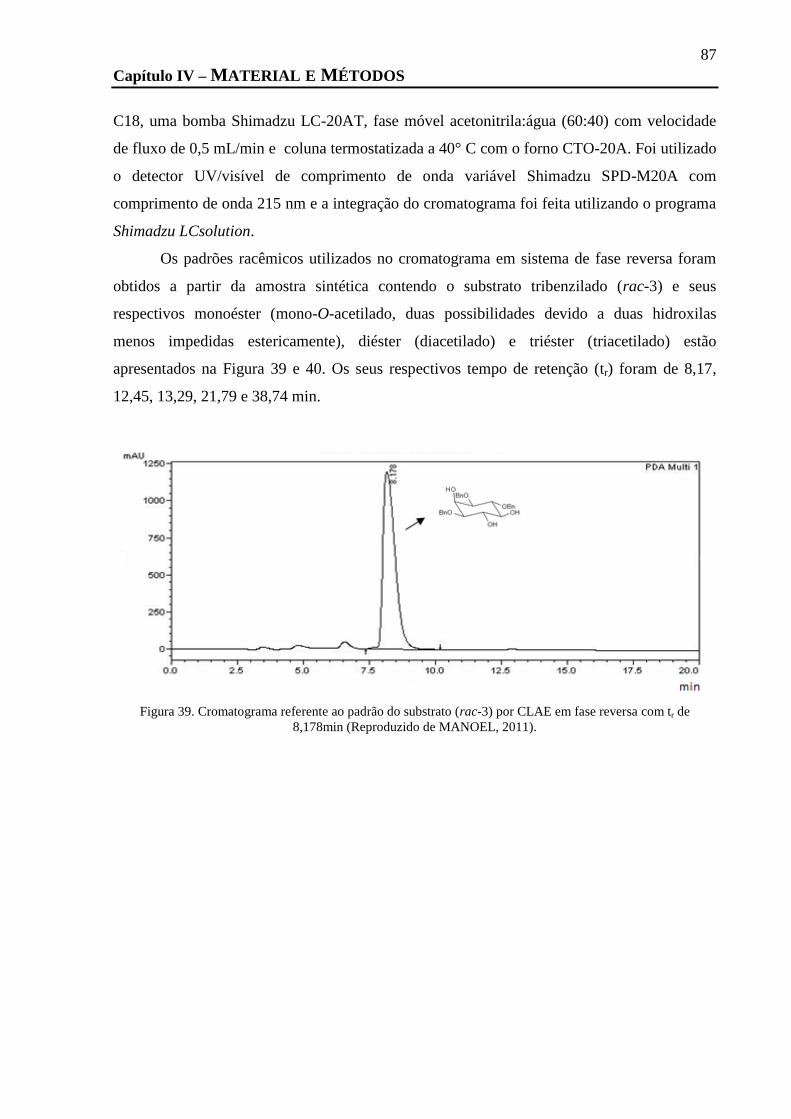
87
Capítulo IV – MATERIAL E MÉTODOS
C18, uma bomba Shimadzu LC-20AT, fase móvel acetonitrila:água (60:40) com velocidade
de fluxo de 0,5 mL/min e coluna termostatizada a 40° C com o forno CTO-20A. Foi utilizado
o detector UV/visível de comprimento de onda variável Shimadzu SPD-M20A com
comprimento de onda 215 nm e a integração do cromatograma foi feita utilizando o programa
Shimadzu LCsolution.
Os padrões racêmicos utilizados no cromatograma em sistema de fase reversa foram
obtidos a partir da amostra sintética contendo o substrato tribenzilado (rac-3) e seus
respectivos monoéster (mono-O-acetilado, duas possibilidades devido a duas hidroxilas
menos impedidas estericamente), diéster (diacetilado) e triéster (triacetilado) estão
apresentados na Figura 39 e 40. Os seus respectivos tempo de retenção (tr) foram de 8,17,
12,45, 13,29, 21,79 e 38,74 min.
Figura 39. Cromatograma referente ao padrão do substrato (rac-3) por CLAE em fase reversa com tr de
8,178min (Reproduzido de MANOEL, 2011).

88
Capítulo IV – MATERIAL E MÉTODOS
Figura 40 Cromatograma referente a mistura do padrão racêmico (rac-3) do produto analisados por CLAE em
fase reversa. O tr de 12,456min é referente ao primeiro monoacetilado, o tr =13,296min é referente ao segundo.
O tr =21,797min é referente ao diacetilado e o tr = 38,745min ao triacetilado. O tr de 8,325min é referente ao
substrato (Reproduzido de MANOEL, 2011).
A Figura 40 representa o padrão sintético do rac-3. A Figura 40 representa o padrão
sintético dos derivados acetilados de rac-3 obtido como componente de uma mistura contendo
dois possíveis mono-acetilados, nos tr= 12,456 e tr=13,296 (acetila nas posições C5 e C6,
respectivamente), diacetilado e uma pequena quantidade do substrato não acetilado (tr=
8,325). A mistura de padrões foi importante para o acompanhamento inicial das reações
enzimáticas. A quantificação da conversão, foi feita segundo Chen et al. (1982) (eq. 3, pag.
23) após a análise por CLAE.
A concentração do substrato rac-3 foi comparada com os valores das áreas da curva de
calibração (Figura 41) (SHAH e MARYANOFF, 2001).
Figura 41. Curva de calibração calculada a partir da curva padrão para o rac-3 (Reproduzido de MANOEL,
2011).

89
Capítulo IV – MATERIAL E MÉTODOS
A concentração do substrato rac-4 foi comparada com os valores das áreas da curva de
calibração (Figura 42) (SHAH e MARYANOFF, 2001).
Figura 42. Curva de calibração calculada a partir da curva padrão para o rac-4 (Reproduzido de MANOEL,
2011)..
Os padrões para o rac-4 foram analisados no cromatograma em sistema de fase
reversa e obtidos a partir da amostra sintética contendo o diol (rac-4) (Figura 43), dois
monoésteres e o diéster (CUNHA et al., 2010) (Figura 44). Os tempos de retenção para o diol
(rac-4), monoéster e diéster foram 9,3, 15,4 e 26,8 min, respectivamente.
Figura 43. Cromatograma em sistema de fase reversa (Shimadzu C-18) do padrão diol.
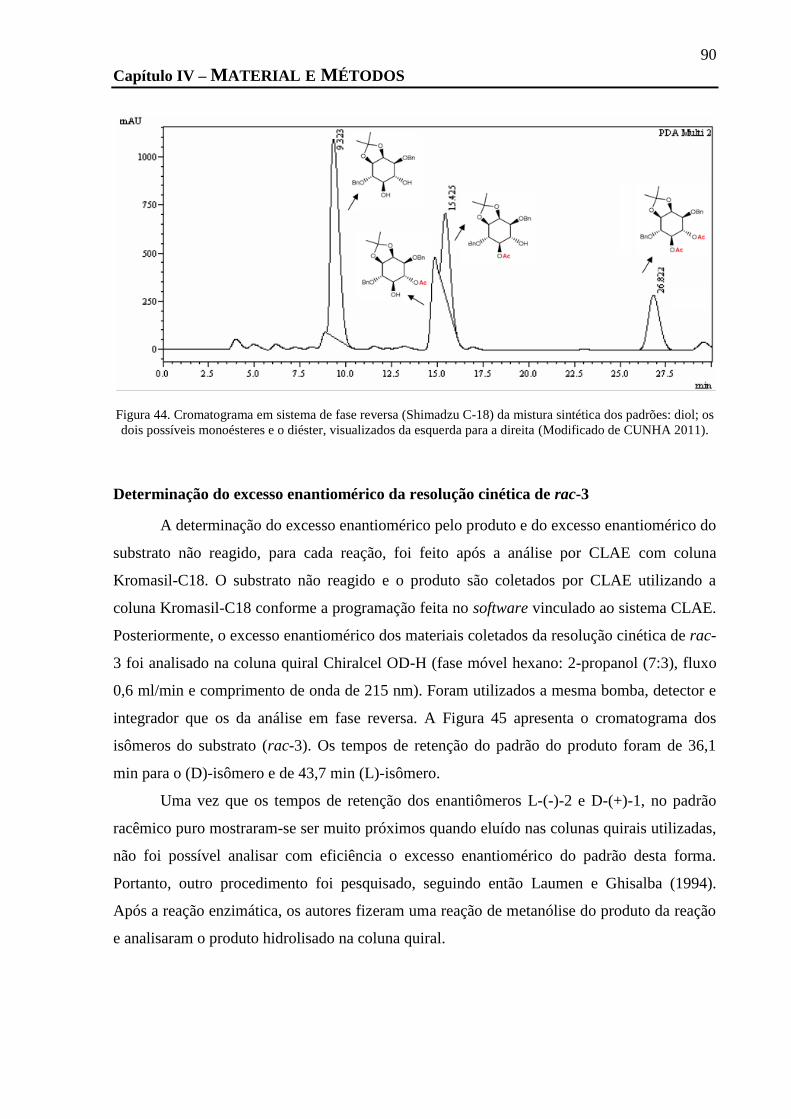
90
Capítulo IV – MATERIAL E MÉTODOS
Figura 44. Cromatograma em sistema de fase reversa (Shimadzu C-18) da mistura sintética dos padrões: diol; os
dois possíveis monoésteres e o diéster, visualizados da esquerda para a direita (Modificado de CUNHA 2011).
Determinação do excesso enantiomérico da resolução cinética de rac-3
A determinação do excesso enantiomérico pelo produto e do excesso enantiomérico do
substrato não reagido, para cada reação, foi feito após a análise por CLAE com coluna
Kromasil-C18. O substrato não reagido e o produto são coletados por CLAE utilizando a
coluna Kromasil-C18 conforme a programação feita no software vinculado ao sistema CLAE.
Posteriormente, o excesso enantiomérico dos materiais coletados da resolução cinética de rac-
3 foi analisado na coluna quiral Chiralcel OD-H (fase móvel hexano: 2-propanol (7:3), fluxo
0,6 ml/min e comprimento de onda de 215 nm). Foram utilizados a mesma bomba, detector e
integrador que os da análise em fase reversa. A Figura 45 apresenta o cromatograma dos
isômeros do substrato (rac-3). Os tempos de retenção do padrão do produto foram de 36,1
min para o (D)-isômero e de 43,7 min (L)-isômero.
Uma vez que os tempos de retenção dos enantiômeros L-(-)-2 e D-(+)-1, no padrão
racêmico puro mostraram-se ser muito próximos quando eluído nas colunas quirais utilizadas,
não foi possível analisar com eficiência o excesso enantiomérico do padrão desta forma.
Portanto, outro procedimento foi pesquisado, seguindo então Laumen e Ghisalba (1994).
Após a reação enzimática, os autores fizeram uma reação de metanólise do produto da reação
e analisaram o produto hidrolisado na coluna quiral.

91
Capítulo IV – MATERIAL E MÉTODOS
Figura 45 – Cromatograma em sistema quiral (Chiracel OD-H) do padrão do produto (D)-isômero e (L)-isômero
do (±)- 1,3,4-tri-O-benzil-mio-inositol (rac-3). Reproduzido de Manoel, 2011.
Adicionou-se ao produto de reação previamente separado do substrato não reagido
(separado por CLAE) o carbonato de potássio (K2CO3 -0,4g, 2,9mmol) e metanol (MeOH -
100ml). A reação de hidrólise ocorreu durante um período de 2h a temperatura ambiente. Os
frascos foram agitados magneticamente para maior interação do sistema. Após a reação,
adicionou-se no sistema 1mL do solvente orgânico diclorometano e 1mL de água, para uma
extração líquido-líquido do sistema (para maior purificação do produto hidrolisado). O
mesmo foi posteriormente seco no concentrador a vácuo de alta velocidade. Em seguida, a
fase móvel foi então adicionada para a análise na coluna quiral Chiracel OD-H, pelo qual os
valores dos excessos enantioméricos foram obtidos eficientementes. O produto da reação uma
vez isolado foi analisado por Ressonância Magnética Nuclear (RMN) por alunos do
laboratório Roderick Barnes, no Núcleo de Pesquisas de Produtos Naturais que confirmaram
o isômero obtido da resolução cinética.
Determinação do excesso enantiomérico da resolução cinética de rac-4
A análise do excesso enantiomérico pelo produto e do excesso enantiomérico do
substrato não reagido da resolução cinética de rac-4 foi feito pelo mesmo processo que por
rac-3, salvo algumas exceções: Após a análise por CLAE com coluna Kromasil-C18, o
substrato não reagido e o produto foram coletados por CLAE utilizando a coluna Kromasil-
C18 conforme a programação feita no software vinculado ao sistema CLAE. Posteriormente,
o excesso enantiomérico dos materiais coletados da resolução cinética de rac-4 foi analisado
na coluna quiral Chiralcel OD-H (fase móvel hexano: 2-propanol (9:1), fluxo 0,8 ml/min e
comprimento de onda de 215 nm), conforme descrito por Cunha (2011). Foram utilizados a

92
Capítulo IV – MATERIAL E MÉTODOS
mesma bomba, detector e integrador que os da análise em fase reversa. A Figura 46 apresenta
o cromatograma dos isômeros do substrato (rac-4). Os tempos de retenção do padrão do
produto foram de 15,7 min para o (D)-isômero e de 22,3 min para o (L)-isômero.
Figura 46. Cromatograma em sistema de quiral (Chiracel-OD-H) do (D)-isômero e (L)-isômero do rac-4.
(Reproduzido de CUNHA, 2011).
A conversão e o excesso enantiomérico do substrato e do produto foram determinados
pelas equações 3, 4 e 7 respectivamente (pags.23 e 24). A razão enantiomérica (E) do produto
formado foi determinada pela equação 6 (pag. 23) de acordo com Chen e colaboradores
(1982).
Dosagem de atividade hidrolítica de lipase método espectrofotométrico
As atividades hidrolíticas das enzimas na forma solúvel foram determinadas a partir da
reação de hidrólise do p-nitrofenil laurato catalisada pela lipase com a formação do produto
cromóforo p-nitro fenol. 2,5 mL de solução contendo 2,5 mM de p-nitrofenil laurato (p-NFL)
em tampão fosfato de sódio (25 mM e pH 7,0) foi termostatizada a 30°C. A reação foi
iniciada pela adição de 0,05 mL de solução enzimática liofilizada (enzima liofilizada diluída
no mesmo tampão supracitado). O progresso da reação foi acompanhado por
espectrofotômetro com leituras de absorbância a 410 nm. No caso de preparações com
enzimas imobilizadas, utilizou-se o seguinte procedimento experimental: 1 mL de solução
contendo 2,5 mM de p-nitrofenil laurato (p-NFL) em 9 mL de tampão fosfato de sódio (25
mM e pH 7,0), termostatizada a 30°C. A reação foi iniciada pela adição de 3mg da enzima
imobilizada. A reação se inicia pela adição da enzima imobilizada ao meio reacional, em um
reator aberto, provido de agitação magnética e conectado a um banho termostático. Alíquotas

93
Capítulo IV – MATERIAL E MÉTODOS
de 1mL foram retiradas do meio reacional em triplicata, e inseridas na cubeta para leituras no
tempo zero e em tempos de reação determinados. O progresso da reação foi acompanhado por
espectrofotômetro com leituras de absorbância a 410 nm. Como se trata de enzima
imobilizada, o cálculo da atividade foi realizado utilizando a equação (8), porém com o valor
da massa de enzima (Menz) pesada no denominador da equação.
Uma unidade internacional (UI) foi definida como a quantidade de enzima necessária
para formar 1,0 µmol de p-nitrofenol por minuto nas condições de ensaio.
O cálculo da atividade foi realizado utilizando-se a equação (8).
(Eq. 8)
onde A é atividade lipásica (U/mL) ou (U/g de suporte); Vfinal é volume final de meio
reacional (mL); f é fator obtido na curva de calibração de p-nitro fenol (µmol/mL)
(0,1223±0,0168), fdil é o fator de diluição, caso seja feita diluição da solução para a dosagem;
Xenz é o Venz, volume da solução enzimática (mL), no caso de dosagem de atividade
hidrolítica de enzimas liofilizadas ou Menz, massa de enzima (g), para as versões imobilizadas.
Dosagem da atividade de esterificação por método titulométrico
A atividade de esterificação da enzima imobilizada foi quantificada pelo consumo de
ácido oléico na reação de esterificação com etanol na razão molar de ácido: álcool de 1:1. A
reação foi conduzida à temperatura de 40°C utilizando-se 3% (m/m) de enzima imobilizada
sob agitação constante (CUNHA, 2011). A reação foi iniciada pela adição da enzima ao meio
reacional, em um reator aberto, provido de agitação magnética e conectado a um banho
termostático. Alíquotas de 200 µL foram retiradas do meio reacional (volume reacional de 5
mL) em triplicata, no tempo zero e em tempos de reação determinados, até um limite máximo
de 10%, afim de garantir que a reação estivesse em velocidade inicial. A cada amostra foram
adicionados 20 mL de uma solução de acetona-etanol (1:1) (v/v). A quantidade de ácido
consumido foi determinada por titulação com NaOH 0,02 M. Uma unidade de atividade
enzimática de esterificação foi definida como a quantidade de enzima que esterifica 1,0 μmol
de ácido graxo por minuto, nas condições do ensaio. A atividade enzimática foi calculada
segundo a Equação 9:

94
Capítulo IV – MATERIAL E MÉTODOS
(Eq. 9)
onde A é a atividade de esterificação (μmol/ min. gsuporte ou UE/gsuporte); Va é o volume
de NaOH gasto na titulação da amostra retirada no tempo zero (mL); Vb é o volume de NaOH
gasto na titulação da amostra retirada após o tempo t (mL); M é a molaridade da solução de
NaOH (mmol/mL); Vfinal é o volume final de meio reacional (mL); m é a massa da enzima
imobilizada utilizada na reação (g); Ve é o volume da alíquota do meio reacional retirada para
titulação (mL) e t é o tempo (min).
Imobilização da lipase por adsorção hidrofóbica
Os suportes hidrofóbicos descritos no item 4.1.3 foram primeiramente lavados com
etanol e posteriormente com água destilada. Este tratamento é feito com o objetivo da retirada
de ar de dentro dos poros do suporte. O processo ocorreu da seguinte forma: 1º- Lavagem do
suporte com etanol (100%) durante 30 min, 2º- Lavagem com etanol:água destilada (50:50)
por 10min, 3º- Lavagem com água destilada (100%) até a retirada total do etanol do meio com
os suportes. Uma vez que os suportes já se encontravam hidratados, os mesmos foram pré-
equilibrados com a solução tampão utilizada na imobilização.
O experimento padrão de imobilização consiste em 1 g de suporte hidratado em 10 mL
de solução lipásica selecionada. Utilizou-se diferentes quantidades em unidades enzimáticas
(U) a saber: 50U, 200U, 350U, 700U para cada suporte, afim de se estudar qual a capacidade
máxima de carga/g de suporte. A suspensão foi mantida sob agitação (250 rpm) à temperatura
ambiente. O processo de imobilização foi acompanhado pela determinação da atividade
hidrolítica com p-NFL da suspensão e do sobrenadante em intervalos de tempo de 0, 30, 60,
90, 120 e 180 min. Após estabelecido o melhor tempo, este foi selecionado para dar
continuidade ao processo. Ao término da imobilização, a enzima imobilizada foi removida
por filtração simples, lavada com água destilada e armazenada a 4ºC.
Cálculo dos parâmetros de imobilização
O processo de imobilização foi acompanhado pela determinação da atividade
hidrolítica da solução com p-NFL e da suspensão em tempos determinados. Os parâmetros de
imobilização foram calculados de acordo com as fórmulas a seguir:

95
Capítulo IV – MATERIAL E MÉTODOS
A eficiência de imobilização (Eimo) foi calculada pela equação (10):
(Eq. 10)
O rendimento de imobilização em atividade (P) foi calculado pela equação (11):
(Eq. 11)
A retenção de atividade (Ra) foi calculada pela equação (12):
(Eq. 12)
Onde Uimo são as unidades de enzima medida após a imobilização (U/g de suporte);
UC são as unidades totais de enzima que são colocadas na etapa de imobilização, no
sobrenadante (U/g de suporte); US são as unidades de enzima do sobrenadante, que não
reagiram com o suporte e que saíram nas primeiras lavagens (U/g de suporte); e UTeo são as
unidades de enzima teoricamente imobilizada (UC-US)/gsuporte.
Dosagem da concentração de proteína
A concentração de proteínas foi determinada pelo método de Bradford, utilizando a
albumina de soro bovino (BSA) como padrão (BRADFORD, 1976). Para o preparo da
solução de Bradford, 10 mg de Coomassie Brilliant Blue G 250 foram solubilizados em 5 mL
de etanol 95%. Após completa homogeneização, adicionaram-se 10 mL de ácido fosfórico
85%, e o volume foi completado com água ultrapura até 100 mL. Posteriormente, esta solução
foi filtrada em papel Whatmam n°1 e estocada ao abrigo da luz. Para a dosagem de proteínas
foram utilizados 1000 μL da solução de Bradford mais 50 μL da amostra diluída, sendo a
leitura feita em espectrofotômetro a 595 nm. Todas as análises de proteína foram realizadas
com triplicatas.
c
imo
U
U 100.(%)
Teo
imoa
U
UR
100.(%)
c
Teoimo
U
UE
100.(%)

96
Capítulo IV – MATERIAL E MÉTODOS
4.2.6 Teste de estabilidade de estocagem das lipases imobilizadas
A lipase imobilizada sobre os suportes poliméricos foi estocada à temperatura média
de 4º C. A estabilidade da enzima foi testada pela determinação da atividade hidrolítica em
tempos determinados (a cada 10 dias).
4.2.7 Desenho Experimental e Análise Estatística para resolução cinética do rac-3
Para determinação das condições experimentais que maximizassem a resolução
cinética do rac-3, selecionado nos experimentos anteriores, realizou-se previamente um
Planejamento Fracionário 24-1
para a escolha das variáveis que fossem significativas para a
reação de acetilação. Tomou-se por base os conhecimentos prévios para as escolhas das
variáveis independentes (MANOEL, 2011; MANOEL et al., 2012a; CUNHA et al., 2013).
As reações enzimáticas foram realizadas em frascos agitados e termostatizados
contendo diferentes concentrações do substrato, acetato de vinila e Novozym 435. A
quantidade de Novozym 435 utilizada foi calculada a partir da atividade de esterificação da
lipase (estão expressas em U-unidade de atividade da enzima). A atividade de esterificação da
Novozym 435 (conforme descrito na pag. 93) obtida por meio da reação de síntese do oleato
de etila foi de 2728 ± 400 U/gsuporte. O solvente utilizado nas reações foi hexano, uma vez que
foi o melhor solvente encontrado após o estudo da influência de solvente para esta reação
(MANOEL et al., 2012a). O volume final da reação foi de 2,0 mL.
As variáveis selecionadas (com seus valores reais e codificados), para o planejamento
fracionário, estão descritas na Tabela 7.
Tabela 7. Valores reais e codificados das variáveis empregadas no Planejamento experimental Fracionário.
*p-level <0,1
Variáveis independentes Siglas
Intervalos de estudo
-1 0 1
Concentração de substrato (mg/mL) S1 2 8 14
Concentração de agente acilante (mg/mL) S2 186,8 373,6 560,4
Quantidade de Enzima (U) E 54,56 81,84 136,4
Temperatura (oC) T 30 40 50

97
Capítulo IV – MATERIAL E MÉTODOS
O tempo de reação selecionado foi de 24 horas (alíquotas foram previamente retiradas
nos intervalos 0, 12, 24 e 36h. O tempo de 24h foi considerado o melhor tempo para o
planejamento experimental). Ao final da reação, o biocatalisador foi removido do meio
reacional e o solvente evaporado a vácuo. Os produtos das reações foram analisados em
sistema de CLAE em fase reversa, para a determinar o grau de conversão da reação, e em
coluna quiral, para determinar o excesso enantiomérico do produto.
Após a seleção das variáveis que fossem significativas para o processo, o
Planejamento Composto Central Rotacional (ou DCCR) foi realizado. No presente trabalho, o
planejamento foi feito para 3 variáveis: (23) mais 6 ensaios axiais com 4 repetições no ponto
central totalizando 18 ensaios. Em cada caso a conversão e o excesso enantiomérico foram
verificados. As variáveis que apresentassem valores menores que 0,05% (i.e. 95% de
intervalo de confiança) eram consideradas de valor significativo e portanto, de influência
significativa na maximização da resolução do racemato selecionado (rac-3). As variáveis e
suas faixas foram escolhidas com base nos experimentos do planejamento experimental
fracionário.
As variáveis selecionadas para o DCCR (com seus valores reais e codificados) para o
estudo da resolução cinética de rac-3 catalisada pela Novozym 435 estão descritas na Tabela
8.
Tabela 8. Valores reais e codificados das variáveis empregadas no Planejamento experimental Composto
Central.
A equação do polinômio de segunda ordem para as variáveis foi a seguinte:
𝑌 = 𝑎0 + 𝑎𝑖𝑥𝑖
𝑛
𝑖=0
+ 𝑎𝑖𝑗 𝑥𝑖𝑥𝑗
𝑛
𝑗=𝑖+1
𝑛−1
𝑖=1
(Eq. 13)
Variáveis independentes Siglas
Intervalos de estudo
-1,68 -1 0 1 1,68
Concentração de substrato (mg/mL) S1 2 5 8,5 11,5 15
Quantidade de Enzima (U) E 80,5 136,4 218,2 300,1 355,8
Temperatura (oC) T 23 30 40 50 57

98
Capítulo IV – MATERIAL E MÉTODOS
onde Y é a variável de resposta, ao é o termo independente, ai o coeficiente do efeito linear, aij
o coeficiente do efeito de interação, xi e xj, são as variáveis independentes.
Os dados de ambos planejamentos experimentais foram tratados com o software
STATISTICA 7.0.
4.2.8 Testes prévios com aumento de volume
Com o objetivo de estudar o comportamento da reação selecionada em um maior
volume, realizou-se um teste para a reação enzimática em uma escala em massa de 100 vezes
maior (escala em grama, antes apenas realizada em miligrama) utilizando as respectivas
proporções de reagentes encontrados no planejamento experimental (para Novozym 435) e na
melhor condição encontrada com o mesmo, em sistema de batelada.
4.2.9 Estudo do reuso das reações de acetilação dos substratos/lipases selecionadas
utilizando o sistema de batelada simples
Após cada reação de 24h de tempo reacional, o experimento foi interrompido e o meio
reacional foi centrifugado. A fase líquida foi recolhida para análise cromatográfica de
conversão e excesso enantiomérico, enquanto o biocatalisador foi separado e cuidadosamente
lavado com acetato de etila para possível retirada do produto e substrato remanescentes da
reação anterior. Posteriormente a lipase foi conduzida para a próxima reação, nas condições
ótimas encontradas. Em cada corrida uma amostra de 100 μL foi retirada para análise da
atividade hidrolítica/esterásica. O reuso do biocatalisador foi feito em sistema de batelada até
que houvesse queda na conversão e/ou perda da atividade enzimática.

99
Capítulo V- RESULTADOS E DISCUSSÃO
5. RESULTADOS E DISCUSSÃO
Neste capítulo serão apresentados os resultados com as lipases comerciais e àquelas
produzidas na UFRJ, abordando as diferenças no desempenho nas reações enantiosseletivas.
5.1 SELEÇÃO DE SUBSTRATOS E LIPASES HOME-MADE
Os substratos selecionados para este trabalho foram pesquisados com base na literatura
(DESAI et al., 1994; EINICKER-LAMAS et al., 1999; OLIVEIRA et al., 2000; MILLS e
POTTER, 2002; SIMAS et al., 2011) a fim de escolher não apenas moléculas inéditas para a
aplicação farmacológica, mas também aqueles substratos pouco estudados e/ou com rotas
dispendiosas do ponto de vista da química orgânica.
Com isso foram realizados experimentos preliminares que tinham como objetivo
identificar: os solventes capazes de solubilizar o substrato; as enzimas que seriam capazes de
catalisar a reação de resolução cinética de derivados de mio-inositol (enzimas provenientes da
coleção de culturas do LABIM/UFRJ e BIOSE/UFRJ) e os agentes acilantes que poderiam ser
empregados.
Deste modo, para a realização deste trabalho, foram escolhidas cinco lipases
promissoras do ponto de vista biotecnológico: LIPB (Lipase B de Candida antarctica
expressa em Pichia pastoris - extrato liofilizado); Lipase de Pyrococcus furiosus expressa em
E.coli (imobilizada em diferentes suportes-lipase proveniente da coleção do LABIM/UFRJ);
Lipase vegetal de Jatropha curcas L. e Lipase de Yarrowia lypolitica (extrato liofilizado).
Algumas são citadas na literatura (SOUZA et al., 2009, 2010; GUTARRA et al., 2011;
MARTINS, 2012; FONTES, 2012), outras estão sendo apresentadas pela primeira vez neste
trabalho como biocatalisadores para diversos substratos intermediários farmacológicos.
Os substratos (±)-1,4,5,6-tetra-O-benzil-mio-inositol (rac-1), (±)-1,4,5,6-tetra-O-alil-
mio-inositol (rac-2), (±)-1,3,4-tri-O-benzil-mio-inositol (rac-3) e (±)-3,6-di-O-benzil-1,2-O-
isopropilideno-mio-inositol (rac-4) foram os substratos propostos para investigar a viabilidade
do emprego de lipases na resolução cinética enzimática de derivados de mio-inositol por
reações de alcoólise, em reações em modo de batelada (Figura 47).

100
Capítulo V- RESULTADOS E DISCUSSÃO
Bn= Benzil=
Alil=
Figura 47- Derivados de mio-inositol utilizados na resolução cinética por lipases.
Nesta primeira etapa, teve-se como objetivo apenas a realização do screening inicial
para a seleção do biocatalisador mais eficiente para estas reações. O efeito do solvente foi
estudado posteriormente.
A seleção inicial (screening) realizada em sistema de batelada e o resultado dos
produtos das resoluções cinéticas (analisados por CCF) dos racematos podem ser visualizados
na Tabela 9, juntamente com a atividade esterásica determinada para cada biocatalisador.
Como se pode notar, não foi observado formação de produto nas reações de acetilação dos
rac-1 e rac-2 com acetato de vinila (em reação livre de solvente) com os biocatalisadores
testados. A catálise do rac-2 pode não ter ocorrido pela não reatividade do substrato com os
biocatalisadores. Possivelmente a catálise do rac-1 não ocorreu devido ao impedimento
estérico dado pelos substituintes benzila ao lado do grupamento hidroxila, presente neste
substrato.
Outros autores também observaram a não formação de produtos com outros derivados
de mio-inositol benzilados e igualmente atribuíram este fato ao efeito do impedimento
estérico do substrato volumoso no sítio ativo da enzima (LING e OZAKI, 1995; SIMAS et
al., 2011). Ling e Ozaki (1995) notificaram a não formação do produto no estudo da resolução
do 3,4,5,6-O-tetrabenzil-mio-inositol por meio da esterificação enzimática dos grupos
hidroxila (C-1 e C-2) e da hidrólise enzimática do éster correspondente (1,2-O-diacetil-
3,4,5,6-O-tetrabenzil-mio-inositol). Os biocatalisadores utilizados pelos autores foram as
Lipases P, PS e CES (Pseudomonas sp., Amano), KWI-56 (Pseudomonas sp.), Lipase AY
(Candida rugosa) e SP-435 (lipase B de Candida antarctica).

101
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 9. Seleção inicial de lipases por CCF produzidas pelos laboratórios LaBiM e Biose (UFRJ) na resolução cinética de derivados de mio-inositol.
Enzimas
Reação de alcoólise
30º C
Reação de alcoólise
40º C
Reação de alcoólise
50º C
Atividade esterásica
(U/mL ou U/g)*
rac-1 rac-2 rac-3 rac-4 rac-1 rac-2 rac-3 rac-4 rac-1 rac-2 rac-3 rac-4
Lipase B de Candida antarctica expressa
em Pichia pastoris (LIPB) - - + + - - + + - - + + 33,2 U/mL
Lipase de Pyrococcus furiosus expressa em
E.coli imobilizada em Octilagarose - - - - - - - - - - + + 1,7 U/g
Lipase de Pyrococcus furiosus expressa em
E.coli imobilizada em Glioxil - - - - - - - - - - - + 2,1 U/g
Lipase de Pyrococcus furiosus expressa em
E.coli imobilizada em Accurel - - - - - - - - - - - + 2 U/g
Lipase vegetal de Jatropha curcas L. - - - - - - - + - - - - ---
Lipase de Yarrowia lypolitica liofilizada - - - - - - - - - - - - 1,1 U/mL
Condições das reações em batelada simples: Tempo reacional: 48h; Solvente e agente acilante: Acetato de vinila; Enzima: 220 U; substrato: 5 mg.
Atividade hidrolítica/esterásica pelo método espectrofotométrico utilizando pNFL como substrato. (unidade U/mL refere a unidade das enzimas liofilizadas e em U/g para as imobilizadas.
Símbolos (-) não houve reação; (+) houve reação.

102
Capítulo V- RESULTADOS E DISCUSSÃO
A resolução cinética dos racematos e das lipases supra-citadas foi feita utilizando a
princípio acetato de vinila (como agente acilante e solvente da reação), uma vez que este
agente acilante tem se mostrado o mais adequado nas reações com mio-inositol como já
relatado pelo grupo (SIMAS et al. 2011) gerando uma maior conversão.
No estudo com os demais substratos (rac-3 e rac-4) foi possível visualizar a formação
de produto pela análise em CCF. Das cinco lipases utilizadas, três mostraram a capacidade de
catálise nas condições estudadas: lipase B de C. antarctica expressa em P. pastoris, em sua
forma liofilizada; lipase de P. furiosus expressa em E. coli, imobilizada nos três suportes e
lipase vegetal de J. curcas L. (Tabela 10). Posteriormente às análises por CCF, as amostras
foram analisadas por CLAE e o valor da conversão foi calculado. O esquema da possível
reação de acetilação enantiosseletiva de rac-3 pode ser visualizado na Figura 48. Os
resultados da acetilação enantiosseletiva de rac-3, após a realização do screening, podem ser
visualizados na Tabela 10.
Figura 48. Resolução cinética enzimática do racemato rac-3 catalisada por diferentes lipases home-made em
acetato de vinila.
Tabela 10. Resultados do screening realizado com as diferentes lipases na resolução cinética de rac-3 utilizando
acetato de vinila como solvente e agente acilante.
Lipase Conversão
(X%) eep
b E
c
Tempo reacional/
Temperatura
LIPB- Lipase B de
Candida antarctica
expressa em Pichia
pastoris
3 n.d n.d 24h/ 30º C
Lipase de Pyrococcus
furiosus expressa em
E.coli imobilizada em
Octilagarose
2,8 n.d. n.d. 48h/ 50º C
Lipase de Pyrococcus
furiosus expressa em
E.coli imobilizada em
2,5 n.d. n.d. 48h/ 50º C

103
Capítulo V- RESULTADOS E DISCUSSÃO
Glioxil
Lipase vegetal de
Jatropha curcas L.
2 n.d. n.d. 24h/ 40º C
O esquema da possível reação de acetilação enantiosseletiva para o rac-4 pode ser
visualizado na Figura 49. Os resultados da acetilação enantiosseletiva para o rac-4, após a
realização do screening, pode ser visualizado na Tabela11.
Figura 49. Resolução cinética enzimática do racemato rac-4 catalisada por diferentes lipases home-made em
acetato de vinila.
Tabela 11. Resultados do screening realizado com as diferentes lipases na resolução cinética de rac-4 utilizando
acetato de vinila como solvente e agente acilante.
Lipase Conversão*
(X%) eep
b E
c
Tempo reacional /
Temperatura
LIPB- Lipase B de
Candida antarctica
expressa em Pichia
pastoris
18 99 >200 24h/ 30º C
Lipase de Pyrococcus
furiosus expressa em
E.coli imobilizada em
Octilagarose
2,5 n.d. n.d. 48h/ 50º C
Lipase de Pyrococcus
furiosus expressa em
E.coli imobilizada em
Glioxil
1,5 n.d. n.d. 48h/ 50º C
Condições das reações em batelada simples: Solvente e agente acilante: Acetato de vinila; Enzima: 220 U; substrato:
5 mg. *n.d.=não detectado

104
Capítulo V- RESULTADOS E DISCUSSÃO
Lipase de Pyrococcus
furiosus expressa em
E.coli imobilizada em
Accurel
2 n.d. n.d. 48h/ 50º C
Lipase vegetal de
Jatropha curcas L.
6 n.d. n.d. 24h/ 40º C
*Condições das reações em batelada simples: Solvente e agente acilante: Acetato de vinila; Enzima: 200 U;
substrato: 5 mg.
Como se pode observar nas tabelas 10 e 11, a reação de alcoólise mostrou maior
conversão quando a LIPB, em sua forma liofilizada, foi utilizada como biocatalisador para o
substrato rac-4. Apesar da baixa conversão na resolução cinética de rac-3 (3%), como esta
apresentou melhor resultado em comparação com as demais, este substrato foi selecionado
juntamente com o rac-4 (18%) para os estudos de imobilização, que serão apresentados mais
adiante nesta tese. Por esta razão, selecionou-se esta lipase para os estudos da otimização da
resolução cinética para ambos os substratos com a finalidade de, ao final do estudo,
compararmos os resultados deste biocatalisador com o comercial: Novozym 435 (lipase de C.
antarctica B imobilizada em resina acrílica), CALB em sua forma liofilizada e CalB
imobilizada com os suportes inéditos preparados na UFRJ.
As Figuras 50 e 51 apresentam os cromatogramas das reações de acetilação da LIPB
com o rac-3 e rac-4 mostrando os produtos formados: L-(-)-2 e L-5. A reação foi
regiosseletiva formando apenas um dos produtos mono-O-acetilados possíveis. Não houve a
formação do produto diacetilado (os cromatogramas das triplicatas não apresentaram pico
referente ao mesmo, como observado na Figura 40 e 44, respectivamente).
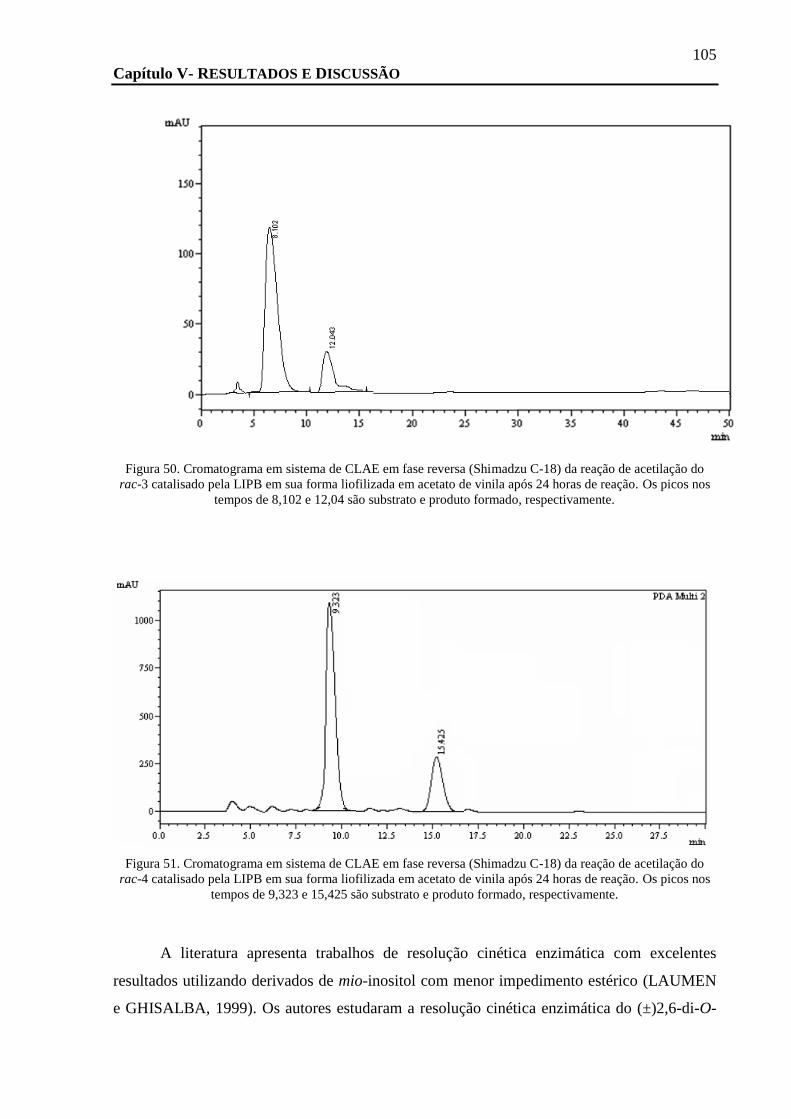
105
Capítulo V- RESULTADOS E DISCUSSÃO
Figura 50. Cromatograma em sistema de CLAE em fase reversa (Shimadzu C-18) da reação de acetilação do
rac-3 catalisado pela LIPB em sua forma liofilizada em acetato de vinila após 24 horas de reação. Os picos nos
tempos de 8,102 e 12,04 são substrato e produto formado, respectivamente.
Figura 51. Cromatograma em sistema de CLAE em fase reversa (Shimadzu C-18) da reação de acetilação do
rac-4 catalisado pela LIPB em sua forma liofilizada em acetato de vinila após 24 horas de reação. Os picos nos
tempos de 9,323 e 15,425 são substrato e produto formado, respectivamente.
A literatura apresenta trabalhos de resolução cinética enzimática com excelentes
resultados utilizando derivados de mio-inositol com menor impedimento estérico (LAUMEN
e GHISALBA, 1999). Os autores estudaram a resolução cinética enzimática do (±)2,6-di-O-

106
Capítulo V- RESULTADOS E DISCUSSÃO
benzil-mio-inositol utilizando a lipase B de Candida antarctica (Novozym 435) como
biocatalisador em acetato de vinila. A reação foi regiosseletiva e enantiosseletiva
esterificando apenas o grupo hidroxila do C-5 do isômero L, o produto monoacetilado L-5-O-
acetil-2,6-di-O-benzil-mio-inositol foi obtido com 49% de conversão e 99% de excesso
enantiomérico. A posição do grupo hidroxila quiral do mio-inositol reconhecido pela enzima
foi dependente das substituições existentes no substrato.
A investigação do melhoramento da resolução cinética de rac-4 com a Novozym 435
foi estudada recentemente pelo nosso grupo de estudo. O estudo da otimização de solventes
mostrou que o uso do acetato de vinila como solvente e agente acilante resulta em 49,9% de
conversão do isômero L-5 (L-(-)-5-O-acetyl-3,6-di-O-benzil-1,2-O-isopropilideno-mio-
inositol), com alto enantiosseletividade (>99%) e razão enantiomérica (E>100) em 48h de
tempo reacional (CUNHA et al., 2010). Por outro lado, quando TBME foi usado como
solvente (acetato de vinila como agente acilante), em apenas 24h foi possível obter 48% de
conversão (eep>99% e E>100). Com base nestes resultados, os autores realizaram um
planejamento experimental em batelada simples e acetato de vinila como agente acilante
(meio livre de solvente). As variáveis independentes utilizadas foram temperatura, percentual
de água, quantidade de enzima e quantidade de substrato. A condição que maximizou a
resolução cinética de rac-4 (49,2% de conversão; eep>99%; E > 100) foi 45º C de
temperatura, 5mg/mL de substrato, 71 U de enzima e 0% de água. Com o planejamento
experimental foi possível reduzir o tempo reacional para 24h (antes 48h). Os autores
aumentaram ainda em 14 vezes a produtividade da reação (0,0145mgproduto/mgenzima.h
comparado com a condição não otimizada [0,001mgproduto/mgenzima.h]) (CUNHA et al., 2012).
Uma vez tendo as condições otimizadas para o racemato 4 (rac-4), em batelada
simples, utilizando a lipase comercial Novozym 435 (CUNHA et al., 2013), resolveu-se
estudar o comportamento deste biocatalisador para o rac-3, no mesmo sistema reacional.
5.2 PLANEJAMENTO EXPERIMENTAL DA REAÇÃO DE
ACETILAÇÃO DE RAC-3 UTILIZANDO A LIPASE NOVOZYM 435 EM
SISTEMA DE BATELADA
O estudo da otimização da resolução cinética de rac-3 catalisada pela enzima
Novozym 435 foi feito por meio de métodos estatísticos: Planejamento Fatorial Fracionário e
Planejamento Composto Central Rotacional. O objetivo principal da otimização do processo
foi encontrar os parâmetros que resultassem na máxima formação do produto

107
Capítulo V- RESULTADOS E DISCUSSÃO
enantiomericamente puro, em um menor intervalo de tempo, aumentando a produtividade e
reduzindo os custos da produção.
Primeiramente, para determinar quais seriam as variáveis de efeito significativo na
resolução cinética, um Planejamento Fatorial Fracionado foi realizado. As variáveis
selecionadas foram temperatura, concentração de enzima, concentração de substrato e
concentração de agente acilante. A concentração de água foi de 0 mg/mL, uma vez que em
trabalhos anteriores, pôde-se notar um melhor desempenho da lipase Novozym 435 com os
derivados de mio-inositol na ausência de água (CUNHA et al., 2013). Logo, foi realizado um
planejamento com 24-1
ensaios e três pontos centrais. As faixas das variáveis estudadas foram
escolhidas com base em experimentos prévios de reação de resolução cinética e testes de
solubilidade.
Recentemente, estudou-se o efeito de diferentes solventes e agentes acilantes na
resolução cinética de rac-3 com a enzima Novozym 435 (MANOEL et al., 2012a). Neste
trabalho, obteve-se 48,3% de conversão (97% eep e E>200) quando hexano foi utilizado como
solvente e acetato de vinila como agente acilante (1,5:1), em 112h de tempo reacional. Por
este motivo este sistema de solventes foi selecionado para as reações dos planejamentos
experimentais. Após os ensaios preliminares o tempo reacional selecionado foi de 24h.
5.2.1- Análise estatística dos dados
Seleção das variáveis pelo Planejamento Fatorial Fracionário
O primeiro tratamento estatístico realizado, Planejamento Fatorial Fracionário, com os
valores reais e codificados para as variáveis independentes e para a variável de resposta
conversão (X%), está apresentado na Tabela 6 (pag.110). Para todos os casos, o eep foi de
98% (± 0,8).
Os resultados do planejamento Fatorial Fracionário revelaram que as concentrações de
enzima e de substrato são as variáveis mais relevantes para a variável de resposta conversão.
O gráfico de pareto apresenta de forma rápida e clara estes fatores estatisticamente
importantes (Figura 52).
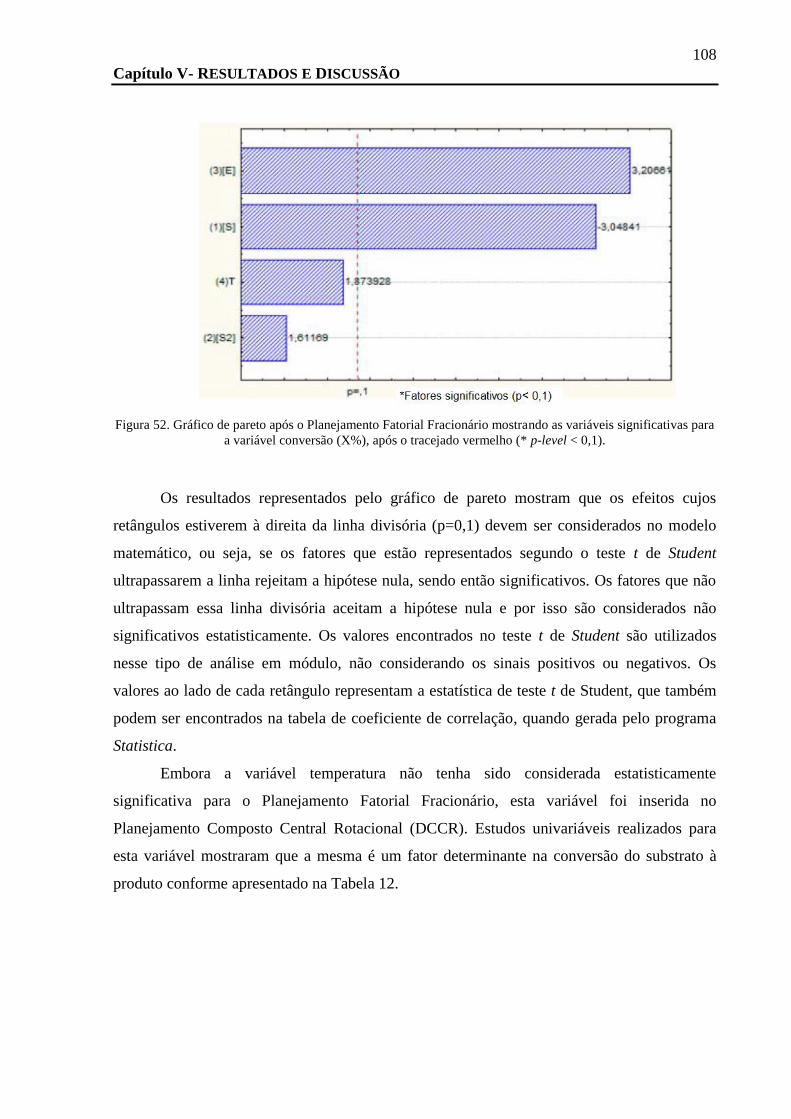
108
Capítulo V- RESULTADOS E DISCUSSÃO
Figura 52. Gráfico de pareto após o Planejamento Fatorial Fracionário mostrando as variáveis significativas para
a variável conversão (X%), após o tracejado vermelho (* p-level < 0,1).
Os resultados representados pelo gráfico de pareto mostram que os efeitos cujos
retângulos estiverem à direita da linha divisória (p=0,1) devem ser considerados no modelo
matemático, ou seja, se os fatores que estão representados segundo o teste t de Student
ultrapassarem a linha rejeitam a hipótese nula, sendo então significativos. Os fatores que não
ultrapassam essa linha divisória aceitam a hipótese nula e por isso são considerados não
significativos estatisticamente. Os valores encontrados no teste t de Student são utilizados
nesse tipo de análise em módulo, não considerando os sinais positivos ou negativos. Os
valores ao lado de cada retângulo representam a estatística de teste t de Student, que também
podem ser encontrados na tabela de coeficiente de correlação, quando gerada pelo programa
Statistica.
Embora a variável temperatura não tenha sido considerada estatisticamente
significativa para o Planejamento Fatorial Fracionário, esta variável foi inserida no
Planejamento Composto Central Rotacional (DCCR). Estudos univariáveis realizados para
esta variável mostraram que a mesma é um fator determinante na conversão do substrato à
produto conforme apresentado na Tabela 12.
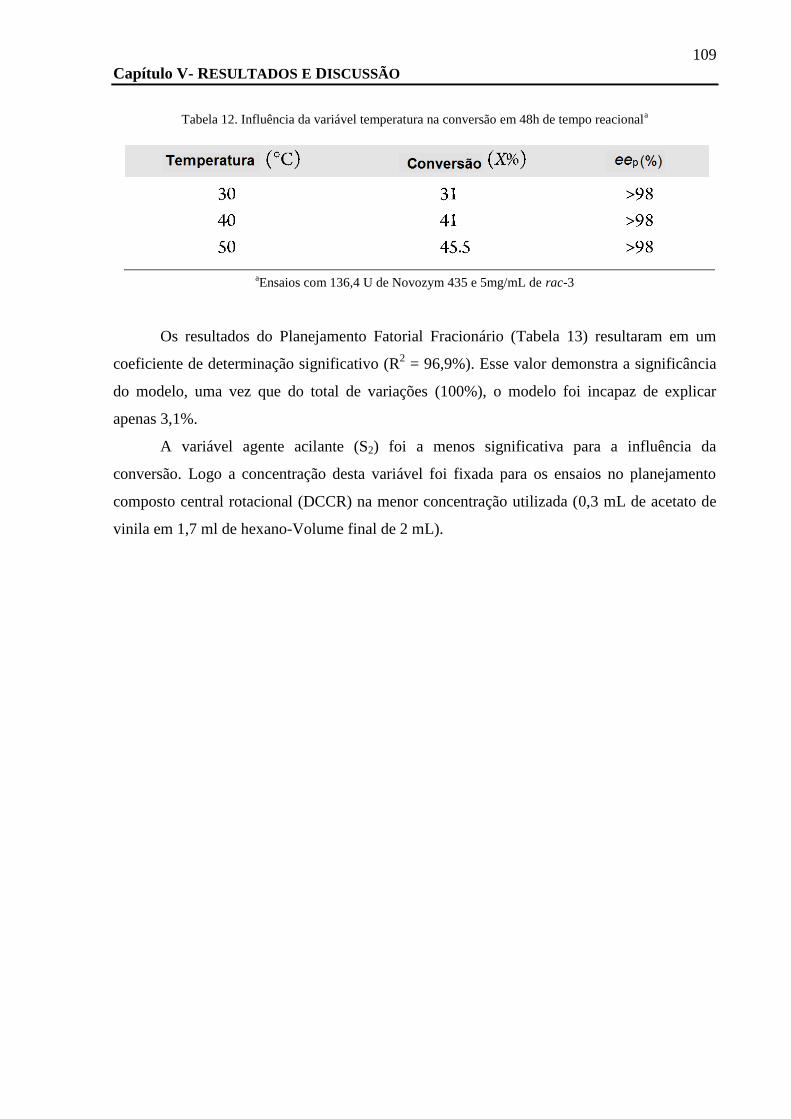
109
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 12. Influência da variável temperatura na conversão em 48h de tempo reacionala
aEnsaios com 136,4 U de Novozym 435 e 5mg/mL de rac-3
Os resultados do Planejamento Fatorial Fracionário (Tabela 13) resultaram em um
coeficiente de determinação significativo (R2 = 96,9%). Esse valor demonstra a significância
do modelo, uma vez que do total de variações (100%), o modelo foi incapaz de explicar
apenas 3,1%.
A variável agente acilante (S2) foi a menos significativa para a influência da
conversão. Logo a concentração desta variável foi fixada para os ensaios no planejamento
composto central rotacional (DCCR) na menor concentração utilizada (0,3 mL de acetato de
vinila em 1,7 ml de hexano-Volume final de 2 mL).
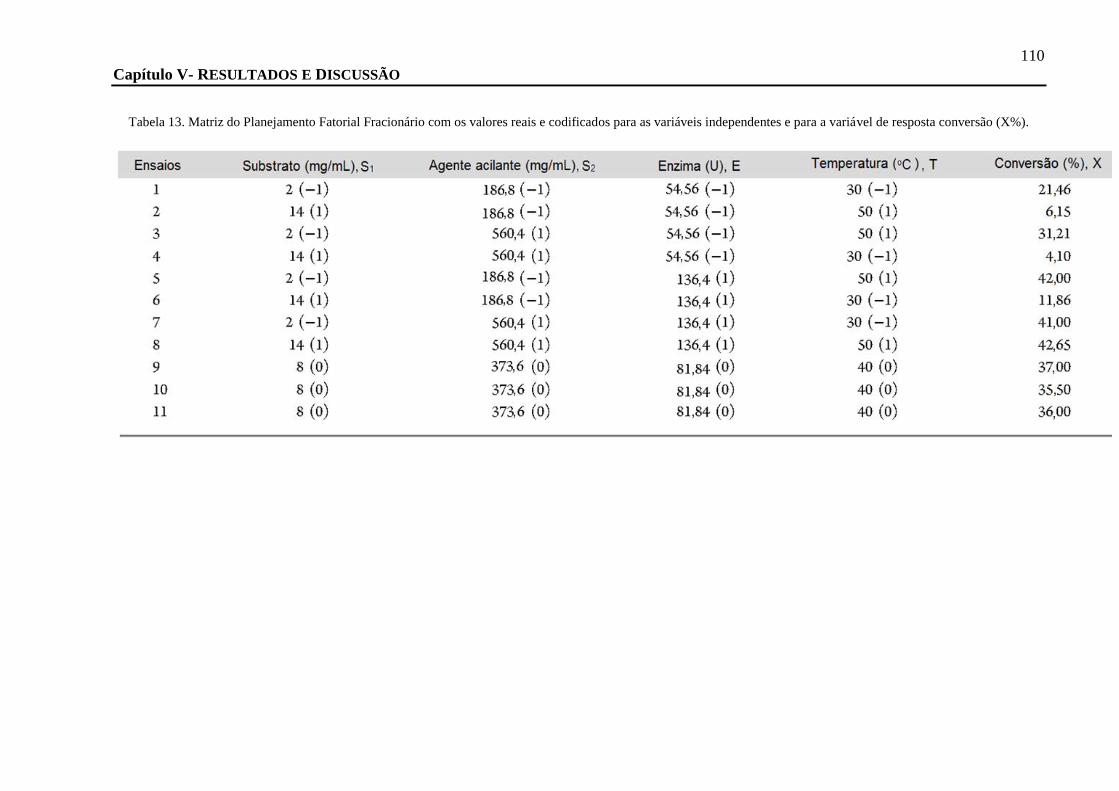
110
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 13. Matriz do Planejamento Fatorial Fracionário com os valores reais e codificados para as variáveis independentes e para a variável de resposta conversão (X%).

111
Capítulo V- RESULTADOS E DISCUSSÃO
Delineamento Composto Central Rotacional (DCCR)
Com base nos resultados do modelo fracionário, o Planejamento Composto Central
Rotacional (DCCR) com 23 ensaios, incluindo quatro réplicas no ponto central e seis pontos
axiais, foi empregado para obter o modelo de segunda ordem a fim de avaliar a influência das
variáveis do processo (variáveis independentes) no rendimento final de L-(-)-2. As seguintes
variáveis foram selecionadas para o DCCR: atividade enzimática (U ou µmol/min),
concentração do substrato (mg/mL) e temperatura (oC). Os resultados do tratamento
estatístico estão apresentados na Tabela 14. Para todos os casos, eep permaneceu maior que
98%.
O maior valor de conversão obtido foi de 51,66% no ensaio 2. Todas as amostras
apresentaram um baixo percentual de erro. Os valores das conversões foram similares aos
seus respectivos valores preditos com um máximo de desvio de -4,7% (ensaio 10).
A análise do DCCR revelou que a interação entre os termos lineares da concentração
do substrato e da temperatura (S.T) e a interação entre os termos lineares da temperatura e da
atividade enzimática (E.T) não mostraram significância estatística.
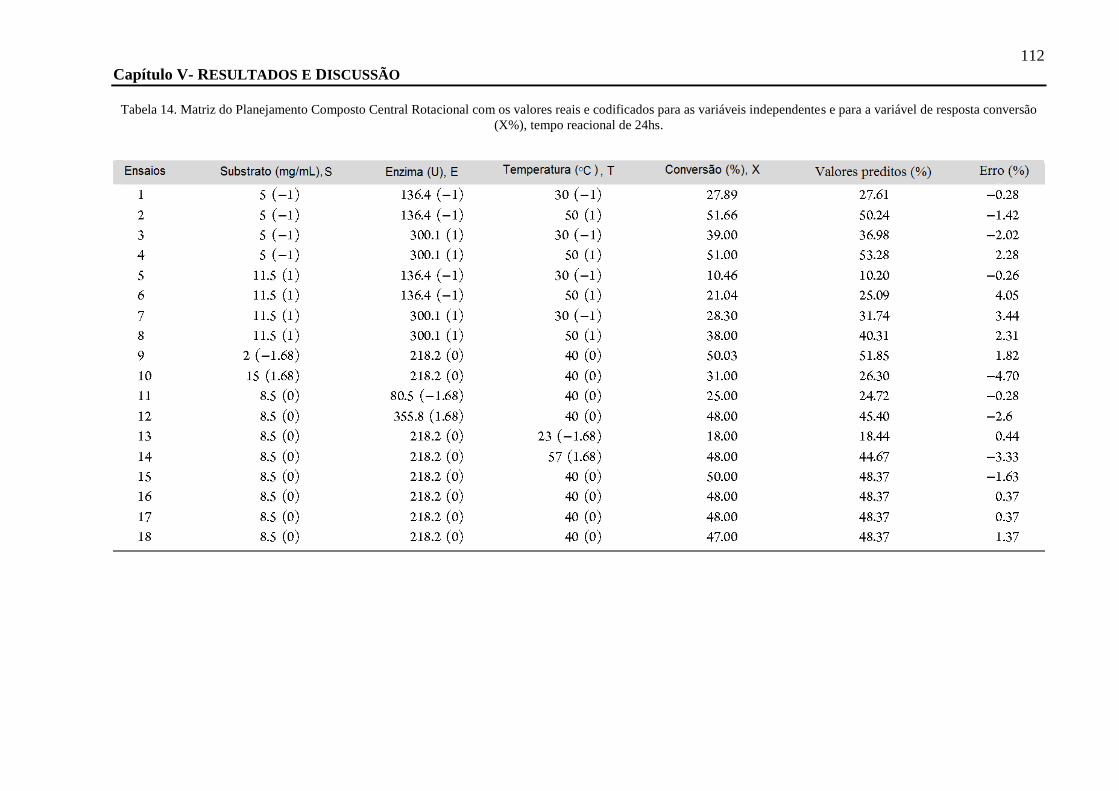
112
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 14. Matriz do Planejamento Composto Central Rotacional com os valores reais e codificados para as variáveis independentes e para a variável de resposta conversão
(X%), tempo reacional de 24hs.

113
Capítulo V- RESULTADOS E DISCUSSÃO
Os efeitos estimados (em termos das variáveis escalonadas) gerados a partir do DCCR
podem ser visualizados na Tabela 15 (apenas as variáveis significativas estão apresentadas na
Tabela). Igualmente o valor do test t está apresentado na tabela, logo a significância estatística
dos parâmetros pode ser analisada. A análise da significância dos coeficientes pode ser
encontrada na Tabela dos efeitos estimados (coeficientes das variáveis escalonadas, uma vez
que as variáveis originais apresentam valores com diferentes ordens de grandeza).
Tabela 15. Efeitos estimados para a análise da variável de resposta conversão na resolução cinética de rac-3
(apenas com as variáveis que apresentaram efeito significativo).
aFatores significativos p<0,05.
Os termos lineares e quadráticos da variável concentração de substrato e os termos
quadráticos da quantidade de enzima adicionada e da temperatura mostraram um efeito
negativo para a variável de resposta conversão na faixa estudada. Por outro lado, os termos
lineares da temperatura e da atividade enzimática mostraram um efeito significativo na faixa
estudada. Como esperado, o aumento da conversão foi observado. De fato, a forte
dependência da temperatura e na quantidade de enzima adicionada na conversão foi
estabelecida.
Além disso, o ajuste do modelo estatístico para os dados experimentais foi confirmado
pela Tabela da ANOVA (Tabela 16).

114
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 16. Análise de variância para desenvolvimento experimental da resolução cinética de rac-3.a
aCoeficiente de determinação (R
2) = 0,9687.
bF0,05; 9; 8 (Ftabelado) = 3,50.
O valor do teste F (27,52) mostrou ser significativo, uma vez que o F-calculado foi
maior que o valor do teste Ftabelado (3,50), levando então a rejeição da hipótese nula. Portanto,
uma vez que o valor do teste F foi maior aproximadamente 8 vezes ao test F tabelado, com
alta significancia (p = 0,000044), o modelo pode ser considerado predito. Além disso, o
coeficiente de determinação foi altamente significativo (R2 = 96.87%), o que significa que o
modelo não explica somente 3,13% do total de variação.
Diante do exposto, o modelo de segunda ordem que considera os parâmetros
estatisticamente significativos (p <0,05) e que descreve o efeito do aumento da conversão na
resolução cinética de rac-3, foi gerado a partir das variáveis escalonadas, originando a
equação 14:
X%= 48,37 – 7,60.S – 3,29.S2 + 6,14. E – 4,70. E
2 + 7,80.T – 5,94. T
2 + 3,04. S.E
Eq. (14)
onde S, E e T representam a concentração de substrato, quantidade de enzima na reação (U) e
temperatura, respectivamente. As siglas S2, E
2 e T
2 representam os termos quadráticos para a
concentração de substrato, quantidade de enzima adicionada e temperatura. As interações
entre os termos lineares concentração de substrato e enzima foram representados por ―S.E‖.
Análise dos resíduos
A verificação da adequação do modelo pode ainda ser estudada pela análise dos
resíduos. Para isso é preciso realizamos a análise da variância. Este é um teste exato de
hipóteses de nenhuma diferença nas médias dos tratamentos. Dessa forma, é primordial
verificar que essas suposições sejam válidas, por meio de testes dos resíduos.
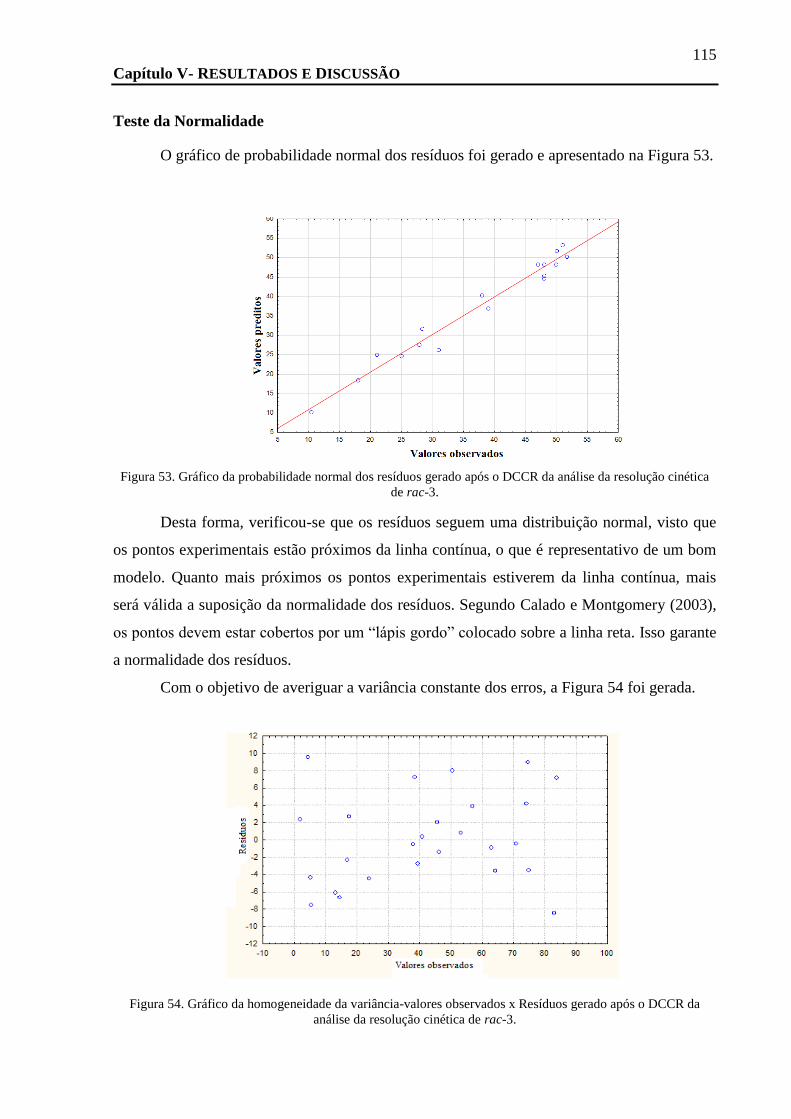
115
Capítulo V- RESULTADOS E DISCUSSÃO
Teste da Normalidade
O gráfico de probabilidade normal dos resíduos foi gerado e apresentado na Figura 53.
Figura 53. Gráfico da probabilidade normal dos resíduos gerado após o DCCR da análise da resolução cinética
de rac-3.
Desta forma, verificou-se que os resíduos seguem uma distribuição normal, visto que
os pontos experimentais estão próximos da linha contínua, o que é representativo de um bom
modelo. Quanto mais próximos os pontos experimentais estiverem da linha contínua, mais
será válida a suposição da normalidade dos resíduos. Segundo Calado e Montgomery (2003),
os pontos devem estar cobertos por um ―lápis gordo‖ colocado sobre a linha reta. Isso garante
a normalidade dos resíduos.
Com o objetivo de averiguar a variância constante dos erros, a Figura 54 foi gerada.
Figura 54. Gráfico da homogeneidade da variância-valores observados x Resíduos gerado após o DCCR da
análise da resolução cinética de rac-3.
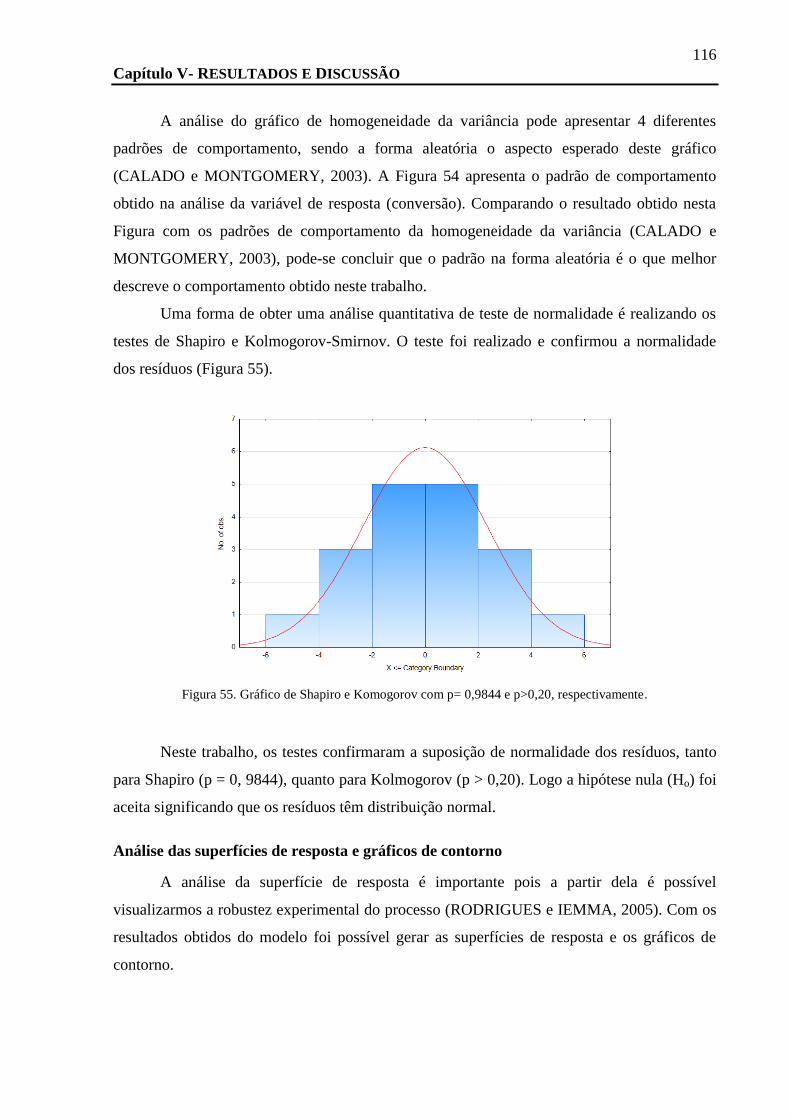
116
Capítulo V- RESULTADOS E DISCUSSÃO
A análise do gráfico de homogeneidade da variância pode apresentar 4 diferentes
padrões de comportamento, sendo a forma aleatória o aspecto esperado deste gráfico
(CALADO e MONTGOMERY, 2003). A Figura 54 apresenta o padrão de comportamento
obtido na análise da variável de resposta (conversão). Comparando o resultado obtido nesta
Figura com os padrões de comportamento da homogeneidade da variância (CALADO e
MONTGOMERY, 2003), pode-se concluir que o padrão na forma aleatória é o que melhor
descreve o comportamento obtido neste trabalho.
Uma forma de obter uma análise quantitativa de teste de normalidade é realizando os
testes de Shapiro e Kolmogorov-Smirnov. O teste foi realizado e confirmou a normalidade
dos resíduos (Figura 55).
Figura 55. Gráfico de Shapiro e Komogorov com p= 0,9844 e p>0,20, respectivamente.
Neste trabalho, os testes confirmaram a suposição de normalidade dos resíduos, tanto
para Shapiro (p = 0, 9844), quanto para Kolmogorov (p > 0,20). Logo a hipótese nula (Ho) foi
aceita significando que os resíduos têm distribuição normal.
Análise das superfícies de resposta e gráficos de contorno
A análise da superfície de resposta é importante pois a partir dela é possível
visualizarmos a robustez experimental do processo (RODRIGUES e IEMMA, 2005). Com os
resultados obtidos do modelo foi possível gerar as superfícies de resposta e os gráficos de
contorno.

117
Capítulo V- RESULTADOS E DISCUSSÃO
A Figura 56 representa a superfície de resposta (esquerda) e o gráfico de contorno
(direita) na interação entre a temperatura e a quantidade enzimática (em termos de unidades
de enzima adicionada-U) na conversão.
Figura 56. Superfície de resposta (esquerda) e gráfico de contorno (direita) na interação entre
as variáveis temperatura e atividade enzimática para a variável de resposta conversão (X%). O
valor da concentração do substrato foi fixada em 4mg/mL.
Como pode ser observado na Figura 56, a interação da temperatura e da atividade
enzimática contribui para o aumento da conversão, indicando o efeito benéfico de ambas as
variáveis (efeito significativamente positivo). A superfície indica que o melhor valor de
conversão encontrado foi obtido no ponto médio do eixo da atividade enzimática e da
temperatura, passando em um ponto ótimo. Valores acima de 55º C no entanto, mostraram
uma queda da conversão. Esse resultado já era esperado, uma vez que a utilização de
temperaturas muito altas pode diminuir a estabilidade da enzima, diminuindo a sua resposta
sobre a conversão. Além disso, as altas temperaturas podem degradar o substrato ou os
produtos lábeis e aumentar o custo do processo (CUNHA, 2011).
Yadav e Sivakumar (2004) igualmente observaram o aumento da conversão com a
temperatura durante o estudo de hidrólise de R,S-(+)-mandelato de metila catalisada pela
Novozym 435 em meio orgânico, na faixa de temperatura de 30-60°C. Quando a temperatura
passou de 30 para 50°C, houve um aumento da conversão. Entretanto para temperaturas
superiores a 60°C, ocorreu a diminuição da concentração. Os autores atribuíram este fato à
desnaturação da proteína a altas temperaturas, causando a redução da atividade enzimática.

118
Capítulo V- RESULTADOS E DISCUSSÃO
Por outro lado, na Figura 57, no gráfico de superfície de resposta e de contorno para a
interação entre o substrato e a atividade enzimática, os efeitos entre estas variáveis foram
diferentes para se obter a máxima conversão.
Figura 57. Superfície de resposta (esquerda) e gráfico de contorno (direita) na interação entre as variáveis
concentração de substrato e atividade enzimática, para a variável de resposta conversão (X%). O valor da
temperatura foi fixada em 40º C.
Quanto menor a concentração do substrato e maior atividade enzimática, em termos de
unidades total adicionada, maior a conversão. Outros resultados foram obtidos por Cunha
(2011). Neste trabalho, a autora observou que a concentração do derivado de mio-inositol
utilizado não influenciava o aumento ou a diminuição da conversão. Possivelmente, os
distintos grupos constituintes dos derivados de mio-inositol podem estar influenciando a
regiosseletividade da enzima, alterando os valores de conversão.
Com isso, o modelo para a resolução cinética de rac-3 utilizando Novozym 435 com
acetato de vinila em hexano resultou nas seguintes condições ótimas dentro das faixas
estudadas: 45º C, 4mg/mL de substrato e 222,8 U de enzima (valores obtidos pela função
desirability do Statistic). A acurácia do modelo foi validada com três replicas nas condições
ótimas supramencionadas. Os resultados da conversão após a validação do modelo foram de
49,7% ± 0,2% depois de 24h de reação contra o valor predito de 52% de conversão (Figura
58).
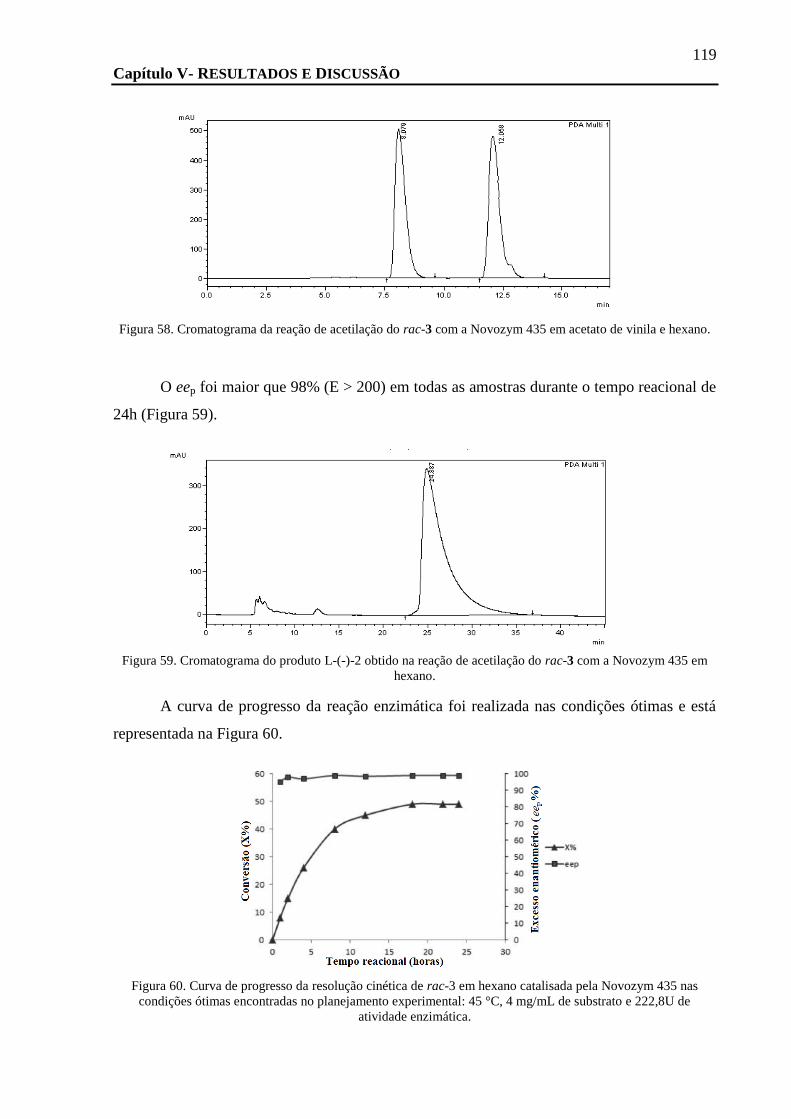
119
Capítulo V- RESULTADOS E DISCUSSÃO
Figura 58. Cromatograma da reação de acetilação do rac-3 com a Novozym 435 em acetato de vinila e hexano.
O eep foi maior que 98% (E > 200) em todas as amostras durante o tempo reacional de
24h (Figura 59).
Figura 59. Cromatograma do produto L-(-)-2 obtido na reação de acetilação do rac-3 com a Novozym 435 em
hexano.
A curva de progresso da reação enzimática foi realizada nas condições ótimas e está
representada na Figura 60.
Figura 60. Curva de progresso da resolução cinética de rac-3 em hexano catalisada pela Novozym 435 nas
condições ótimas encontradas no planejamento experimental: 45 °C, 4 mg/mL de substrato e 222,8U de
atividade enzimática.

120
Capítulo V- RESULTADOS E DISCUSSÃO
Embora o melhor valor da concentração do substrato no planejamento experimental
para a resolução cinética de rac-3, tenha sido de 4mg/mL, o desejável seria um aumento da
concentração de substrato por uma questão de economicidade e viabilidade do processo
(maior concentração de substrato com a mesma quantidade de enzima). Por isso testes foram
realizados na tentativa de aumentar esta concentração, mantendo as outras variáveis, de forma
que não houvesse queda na conversão. Os resultados podem ser analisados na Figura 61.
Figura 61. Gráfico de contorno para a variável de resposta conversão (X%) usando 220U da lipase Novozym
435. Valores de concentração de substrato foram fixadas (a) 4 mg/mL, (b) 8,5 mg/mL, and (c) 14 mg/mL.
Os resultados mostraram que nas concentrações de substrato dentro do intervalo de 4 a
8,5mg/mL e utilizando 220U de lipase, ainda foi possível obter altos valores de conversão
(Figura 61a,b). Um aumento adicional da concentração de substrato de até 11 mg/mL não
mostrou perdas significativas na conversão. O uso de maiores valores de concentração de
substrato (14 mg/mL -Figura 61c) exigiria maior carga de enzima (280U) para obter o mesmo
nível de conversão. Dependendo do processo, estes valores poderão tornar o processo menos
vantajoso do ponto de vista econômico. Por outro lado, dependendo do processo, maiores
valores de carga enzimática podem gerar maiores quantidades de produto formado, logo esse
valor precisa ser estudado.
Claramente, o planejamento experimental reduziu o tempo reacional, uma vez que
antes das condições otimizadas eram necessárias 112h para obter a máxima conversão de
48,3% na resolução cinética do rac-3. De fato, o efeito da otimização na produtividade
específica da reação foi significativa (Figura 62).
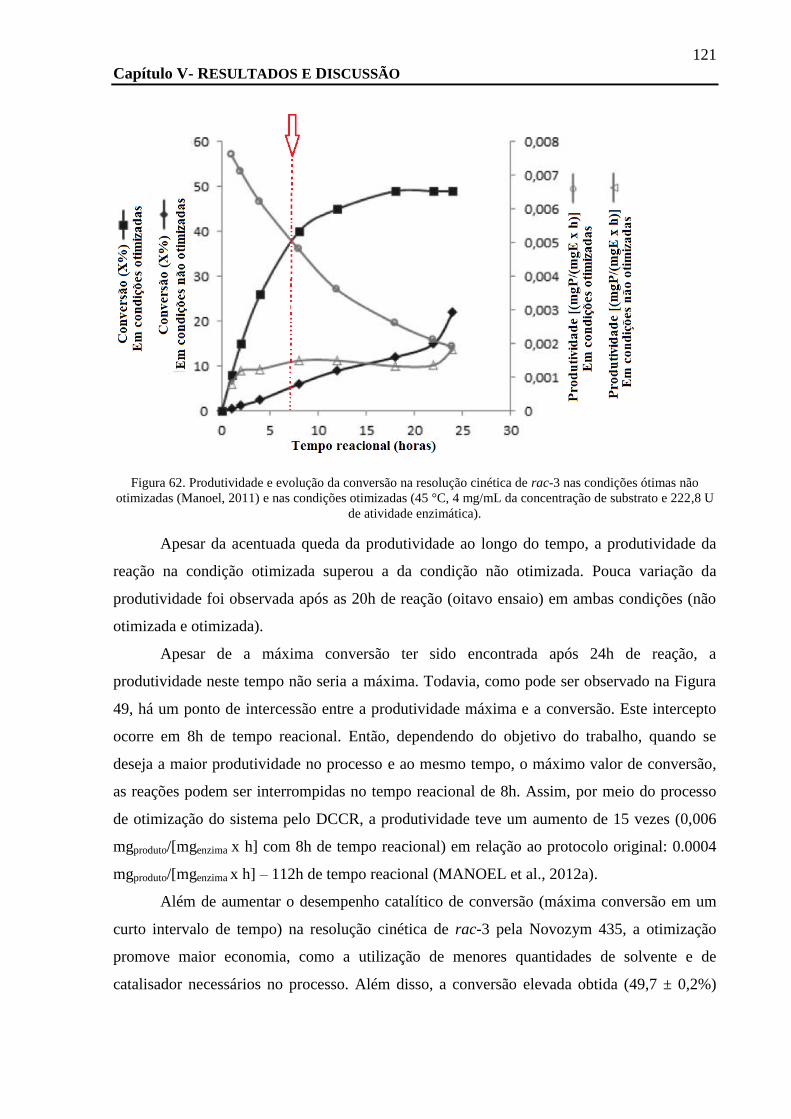
121
Capítulo V- RESULTADOS E DISCUSSÃO
Figura 62. Produtividade e evolução da conversão na resolução cinética de rac-3 nas condições ótimas não
otimizadas (Manoel, 2011) e nas condições otimizadas (45 °C, 4 mg/mL da concentração de substrato e 222,8 U
de atividade enzimática).
Apesar da acentuada queda da produtividade ao longo do tempo, a produtividade da
reação na condição otimizada superou a da condição não otimizada. Pouca variação da
produtividade foi observada após as 20h de reação (oitavo ensaio) em ambas condições (não
otimizada e otimizada).
Apesar de a máxima conversão ter sido encontrada após 24h de reação, a
produtividade neste tempo não seria a máxima. Todavia, como pode ser observado na Figura
49, há um ponto de intercessão entre a produtividade máxima e a conversão. Este intercepto
ocorre em 8h de tempo reacional. Então, dependendo do objetivo do trabalho, quando se
deseja a maior produtividade no processo e ao mesmo tempo, o máximo valor de conversão,
as reações podem ser interrompidas no tempo reacional de 8h. Assim, por meio do processo
de otimização do sistema pelo DCCR, a produtividade teve um aumento de 15 vezes (0,006
mgproduto/[mgenzima x h] com 8h de tempo reacional) em relação ao protocolo original: 0.0004
mgproduto/[mgenzima x h] – 112h de tempo reacional (MANOEL et al., 2012a).
Além de aumentar o desempenho catalítico de conversão (máxima conversão em um
curto intervalo de tempo) na resolução cinética de rac-3 pela Novozym 435, a otimização
promove maior economia, como a utilização de menores quantidades de solvente e de
catalisador necessários no processo. Além disso, a conversão elevada obtida (49,7 ± 0,2%)

122
Capítulo V- RESULTADOS E DISCUSSÃO
permitiu a síntese tanto do produto acilado (ee%> 99) quanto do substrato restante (aquele
que não reagiu, com ee igual a 95%), ambos em sua forma enantiomericamente pura.
Neste trabalho, todo o produto formado (intermediário farmacológico L-(-)-2) foi
posteriormente utilizado para a síntese final do fármaco de interesse. Por este motivo o
objetivo nesta etapa do trabalho foi obter a maior conversão deste produto, por intermédio da
catálise enzimática, para então utilizá-la nas etapas finais na síntese orgânica. Deste modo, as
reações posteriores foram interrompidas com tempo de reação de 24h, onde foi obtido o
máximo valor de conversão.
Reações em escala maior
Com o objetivo de avaliar se a condição ótima em escala de bancada resultaria na
mesma conversão quando as reações fossem submetidas a outras escalas de trabalho e
sabendo a importância do ajuste do processo na escala laboratorial para a escala de planta
piloto ou em escala comercial, um estudo de aumento de volume reacional foi realizado em
reatores do tipo batelada (manteve-se as mesmas relações geométricas dos reatores).
A condição estipulada após o DCCR pôde ser reproduzida em escala maior (aumento
de 100x), em escala de grama (g). O processo pôde ser reproduzido e os resultados
encontrados mostraram alta conversão (50% ± 1), seletividades (eep maior que 99%; E maior
que 100) e ainda valores de produtividade idênticos aos obtidos nas reações com menor
volume. (Tabela 17).
Tabela 17. Resultados na condição ótima de rac-3 (45 °C, 4 mg/mL da concentração de substrato e 222,8 U de
atividade enzimática) em sistema de batelada utilizando escala de mg e uma escala maior (100x).
Escala X(%) eep(%) E Produtividade
(mgproduto/[mgenzima x h)
1x 49,7± 0,20 >99 >100 0,006
100x 50± 1 >99 >100 0.006
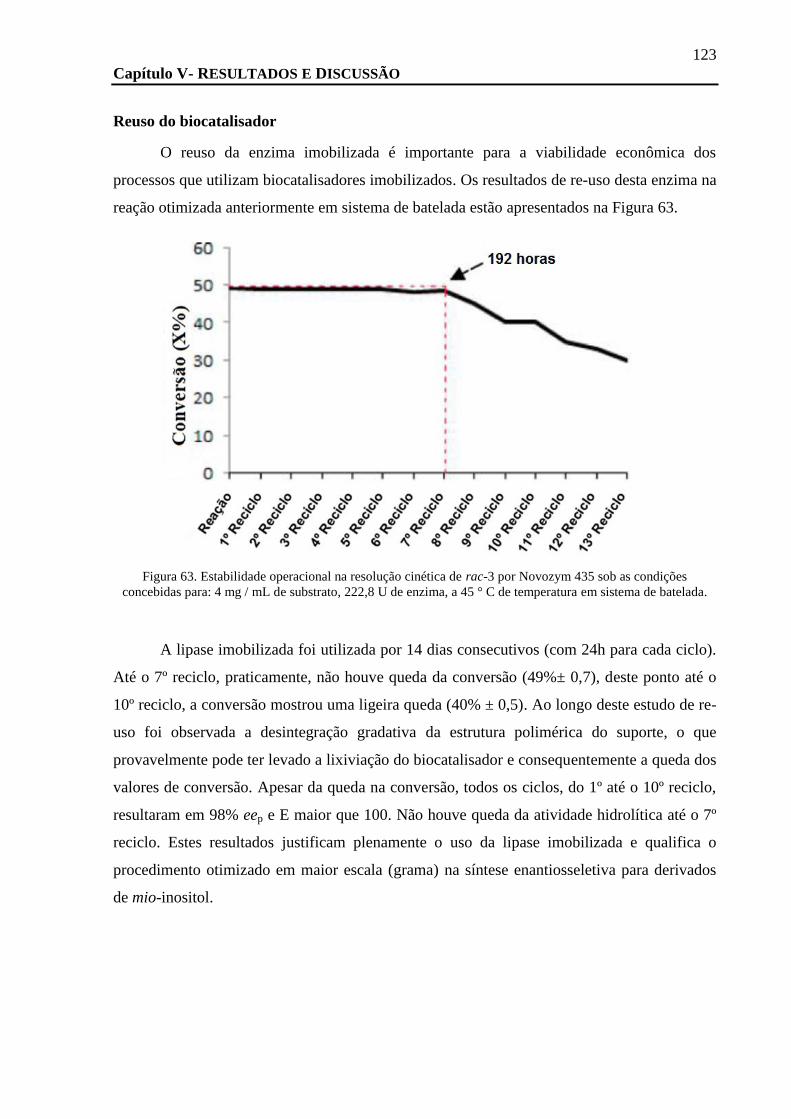
123
Capítulo V- RESULTADOS E DISCUSSÃO
Reuso do biocatalisador
O reuso da enzima imobilizada é importante para a viabilidade econômica dos
processos que utilizam biocatalisadores imobilizados. Os resultados de re-uso desta enzima na
reação otimizada anteriormente em sistema de batelada estão apresentados na Figura 63.
Figura 63. Estabilidade operacional na resolução cinética de rac-3 por Novozym 435 sob as condições
concebidas para: 4 mg / mL de substrato, 222,8 U de enzima, a 45 ° C de temperatura em sistema de batelada.
A lipase imobilizada foi utilizada por 14 dias consecutivos (com 24h para cada ciclo).
Até o 7º reciclo, praticamente, não houve queda da conversão (49%± 0,7), deste ponto até o
10º reciclo, a conversão mostrou uma ligeira queda (40% ± 0,5). Ao longo deste estudo de re-
uso foi observada a desintegração gradativa da estrutura polimérica do suporte, o que
provavelmente pode ter levado a lixiviação do biocatalisador e consequentemente a queda dos
valores de conversão. Apesar da queda na conversão, todos os ciclos, do 1º até o 10º reciclo,
resultaram em 98% eep e E maior que 100. Não houve queda da atividade hidrolítica até o 7º
reciclo. Estes resultados justificam plenamente o uso da lipase imobilizada e qualifica o
procedimento otimizado em maior escala (grama) na síntese enantiosseletiva para derivados
de mio-inositol.

124
Capítulo V- RESULTADOS E DISCUSSÃO
5.2.3 PLANEJAMENTO EXPERIMENTAL DA REAÇÃO DE ACETILAÇÃO DE
RAC-3 UTILIZANDO A LIPASE NOVOZYM 435 EM SISTEMA DE FLUXO
CONTÍNUO
Apesar do sucesso das reações catalisadas pela Novozym 435 em reatores
descontínuos, o uso do sistema em fluxo contínuo foi realizado para a possível redução do
tempo reacional e do biocatalisador.
A Figura 64 representa o esquema da reação em sistema de fluxo contínuo.
Figura 64. Resolução cinética de rac-3 utilizando a lipase Novozym 435: (A) sistema em batelada (B) sistema de
fluxo contínuo em leito fixo.
Mesmo conhecendo a melhor condição de reação para o rac-3 em sistema de batelada,
reator do tipo BSTR, esta condição não necessariamente seria a mesma para o sistema de
fluxo contínuo em leito fixo, pois inúmeras são as variáveis que distinguem um sistema do
outro. A possibilidade de melhorar ainda mais a eficiência reacional com o aumento da
conversão, estereoseletividade e produtividade em um novo sistema, levou ao estudo em
reator de fluxo contínuo.
Escolha do melhor sistema de solvente-agente acilante para o sistema de fluxo contínuo
Tomou-se como ponto de partida, a investigação dos solventes e agentes acilantes que
têm sido considerados na resolução de rac-3 quando Novozym 435 foi usada (MANOEL et
al., 2012a). Assim, uma solução contendo o rac-3, acetato de vinila e acetato de etila (como
solvente e agente acilante, respectivamente) foi bombeada através do reator de leito fixo
carregado com 527 mg de Novozym 435 (2728 U/gsuporte). Utilizaram-se diferentes

125
Capítulo V- RESULTADOS E DISCUSSÃO
temperaturas, tempos de residência e proporções de rac-3 e acetato de vinila nesta primeira
investigação. O esquema da reação está apresentado na Figura 65.
Figura 65. Resolução cinética de rac-3 utilizando a lipase Novozym 435, acetato de vinila como agente acilante
e acetato de etila como solvente.
Os resultados da conversão em função da temperatura para diferentes tempos de
residência, no reator de fluxo contínuo, estão apresentados na Tabela 18.
Tabela 18. Resolução cinética de rac-3 em acetate de etila como solvente da reação em sistema de fluxo
contínuo.
Os dados apresentados na Tabela 18 (Ensaios 1 ao 4) mostram que baixos valores de
conversão foram encontrados na resolução cinética de rac-3 no sistema de fluxo contínuo. O
Ensaio
rac-3 /
acetato de
vinila
Temperatura
Tempo de
Residência
Conversão
(X%)
1 1:5 30ºC
3 min 0,1
1,5 min 0,1
2 1:5 60ºC
3 min 5,8
1,5 min 2,4
3 1:15 30ºC
3 min 2
1,5 min 1,3
4 1:15 60ºC
3 min 9
1,5 min 3

126
Capítulo V- RESULTADOS E DISCUSSÃO
aumento da temperatura e do tempo de residência aumentaram os valores de conversão,
entretanto estes valores ainda permaneceram muito baixos. Outros valores de temperatura
foram igualmente estudados (40 e 50º C), todavia resultados semelhantes foram obtidos.
Os resultados apresentados na Tabela 18 são diferentes daqueles apresentados por
Manoel et al., (2012a) quando estudaram diferentes solventes na resolução de rac-3, em
sistema de batelada. Neste trabalho, os autores observaram que o uso do acetato de etila
revelou uma elevada conversão quando foi utilizado como solvente do meio de reação (com
29,4% de conversão em 112h de tempo de reação). Isso revela o quão diferentes sistemas
reacionais podem resultar em diferentes comportamentos.
Outra possível explicação para o baixo rendimento observado na Tabela 18, para as
reações em fluxo contínuo, seria a possibilidade de uma reação de transesterificação
preferencial entre o acetato de etila e o acetato de vinila, que conduz ao consumo do agente de
acetilação mais reativo.
Recentemente, foi obtida a melhora significativa no rendimento na biotransformação
de rac-3 quando hexano foi utilizado como solvente, seguido do TBME (MANOEL et al.,
2012a, TREVISAN et al., 2013). Entretanto, devido à baixa solubilidade do substrato em
hexano, decidiu-se realizar as reações em TBME (Tabela 19). Com exceção do solvente, os
ensaios foram feitos sob as mesmas condições anteriores (Figura 66).
Figura 66. Resolução cinética de rac-3 utilizando a lipase Novozym 435, acetato de vinila como agente acilante
acetato de etila como solvente.
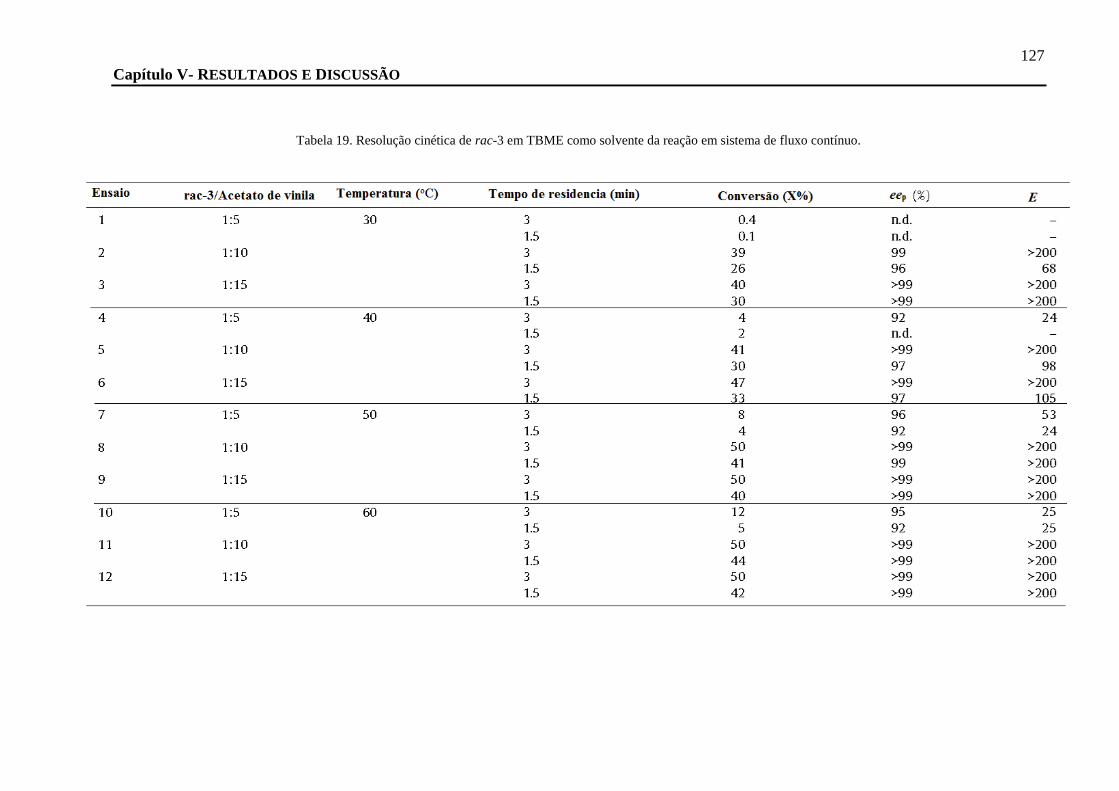
127
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 19. Resolução cinética de rac-3 em TBME como solvente da reação em sistema de fluxo contínuo.

128
Capítulo V- RESULTADOS E DISCUSSÃO
Como mostrado na Tabela 19, os resultados obtidos pela utilização de TBME como
solvente são muito melhores comparativamente às reações onde acetato de etila foi avaliado
como solvente (Tabela 18). Em geral, as proporções de 1:10 e 1:15 (substrato / acetato de
vinila) resultaram nos melhores valores de conversão / seletividade. A utilização de um menor
excesso de acetato de vinila (1:5) gerou baixos valores de conversão e E inferior (ensaios 1, 4,
7 e 10, Tabela 19).
O sistema de fluxo contínuo com TBME como solvente, além de resultar em maiores
produtividades, permitiu o uso de menores quantidades de acetato de vinila, em comparação
ao processo de batelada (1:300 proporção de substrato/acetato de vinila), o que torna o
processo mais interessante do ponto de vista econômico.
Em relação a temperatura, os melhores rendimentos foram alcançados no intervalo
entre 50 e 60 °C (Tabela 19). Os resultados obtidos a 30 e 40 °C podem ter conversões
melhoradas com o uso de uma proporção maior de agente de acetilação (Tabela 19, ensaios 1
ao 3 (30º C) e 4 ao 6 (40º C)) ou por tempos de residência mais longos, com a consequente
diminuição de produtividade. Independentemente da temperatura escolhida, um tempo de
permanência de apenas 3 minutos gerou a mais alta conversão para o produto (L-(-)-2) com
elevado excesso enantiomérico (eep) e razões enantioméricas (E). Por esta razão, as melhores
condições obtidas pela resolução cinética de rac-3, com o acetato de vinila sob condições de
fluxo contínuo (1:10, 50°C e 3 minutos de tempo de residência) resultaram em uma
produtividade 531 vezes maior (3,188 mgproduto/mgenzima. h) do que no processo em batelada
(0,006 mgproduto/mgenzima. h).
A condição de reação que resultou no melhor desempenho para o rac-3, quando
submetidas em sistema de fluxo, foi aplicada a outros agentes de acetilação a fim de
confirmar o uso do acetato de vinila como a melhor opção para o sistema de fluxo contínuo.
Os resultados obtidos são mostrados na Tabela 20.
129
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 20. Resolução cinética de rac-3/acetato de vinila (1:10) em TBME catalisado pela Novozym 435 a 50oC
em condições de fluxo contínuo (tempo de residência de 3min) usando diferentes agentes acilantes.
Os resultados obtidos por meio da utilização de acetato de isopropenila e anidrido
acético em TBME (ensaios 1 e 2, Tabela 20) foram satisfatórios. Contudo, quando
comparados com a sequência anterior, utilizando acetato de vinila (ensaio 8, Tabela 19),
percebe-se uma pequena diminuição na conversão e na seletividade. TBME, como solvente, e
acetato de etila, como agente de acilação, conduziram a baixa conversão e seletividade
moderada (ensaio 3, Tabela 20). Uma vez que o TBME é considerado um bom solvente para
esta reação (MANOEL et al., 2012a), uma probabilidade para a diminuição da seletividade e
conversão seria pelo uso do acetato de etila, menos reativo que o acetato de vinila.
A reutilização do biocatalisador empacotado na coluna foi avaliada por vários dias,
efetuando a resolução cinética de (±) -1,3,6-tri-O-benzil-mio-inositol (rac-3), nas melhores
condições, e os resultados são mostrados na Figura 67.

130
Capítulo V- RESULTADOS E DISCUSSÃO
Figura 67. Integridade do reactor de leito fixo depois de nove ciclos na resolução cinética de rac-3 utilizando a
Novozym 435 como biocatalisador, nas melhores condições experimentais: TBME como solvente, acetato de
vinila, como doador de acila (proporção 1:10), a 50◦C. Tempo de residência: 3min.
A integridade do biocatalisador foi investigada em todas as reações. A coluna
empacotada com 527 mg de Novozym 435 foi reutilizada 9 vezes sem que houvesse perda de
conversão. Além disso, durante as reações não foi observado lixiviação de proteína e nem
queda da atividade enzimática. Este fato foi comprovado por meio da dosagem de proteína e
de atividade na solução (rac-3 solubilizado em TBME e acetato de vinila) a todo tempo. A
lixiviação de proteínas precisa ser investigada uma vez que esta pode jusificar uma possível
queda na conversão, uma vez que há menos catalisador disponível para a reação
enantiosseletiva.
O sistema em fluxo contínuo tem sido cada vez mais utilizado em síntese orgânica e é
uma ferramenta importante para o desenvolvimento de novas metodologias reacionais (RAO,
et al., 2009). Todavia, apesar de numerosos trabalhos estarem utilizando este sistema, o reuso
do biocatalisador não tem sido muito explorado nos trabalhos recentes de catálise enzimática
neste tipo de sistema (ITABAIANA Jr. et al., 2011).

131
Capítulo V- RESULTADOS E DISCUSSÃO
5.2.5- Imobilização da lipase B de Candida antarctica expressa em Pichia pastoris (LIPB)
por adsorção hidrofóbica em diferentes suportes
Existe uma constante busca pela síntese de novos suportes que resulte em
biocatalisadores mais eficientes. O Laboratório de Modelagem, Simulação e Controle de
Processos-PEQ-UFRJ (LMSCP) vem se empenhado nessa busca com a síntese novos suportes
poliméricos por meio de reações de polimerização simultâneas em suspensão e emulsão,
formando partículas do tipo casca-núcleo (PINTO et al., 2006; BESTETI, 2009, BESTETI,
FREIRE e PINTO, 2011; BESTETI et al., 2014). A escolha dos melhores suportes para este
estudo foi realizada com base na literatura (CUNHA, 2011); com base nos parâmetros de
imobilização, como atividade hidrolitica, atividade de esterificação, retenção da atividade e
rendimento de imobilização além dos parâmetros relacionados com a estrutura do suporte.
Neste tópico, procurou-se avaliar diferentes suportes na imobilização de LIPB e posterior
comparações com as versões comerciais, CalB, versão liofilizada e Novozym 435.
Os suportes estudados para a imobilização por adsorção hidrofóbica foram PMMA-co-
DVB/PMMA-co-DVB 25% e PS-co-DVB/PS-co-DVB 25%. Os dois últimos suportes foram
sintetizados com adição de agentes ―cross-linking‖ divinil-benzeno com o objetivo de
aumento das propriedades de porosidade do suporte, a resitencia mecânica e a solventes
químicos. O terceiro suporte estudado foi o Accurel MP 1000, suporte comercial, que foi
utilizado como modelo de comparação.
A adsorção hidrofóbica de LIPB sobre os suportes, citados anteriormente, foi
conduzida utilizando diferentes quantidades de biocatalisador obtidas por atividade hidrolítica
(em unidades de atividade, 50, 200, 350, 700U). Este procedimento foi feito com o objetivo
de avaliar qual a capacidade máxima de enzima, na solução de imobilização, que resultasse na
obtenção de um biocatalisador com elevada atividade (Tabela 21).
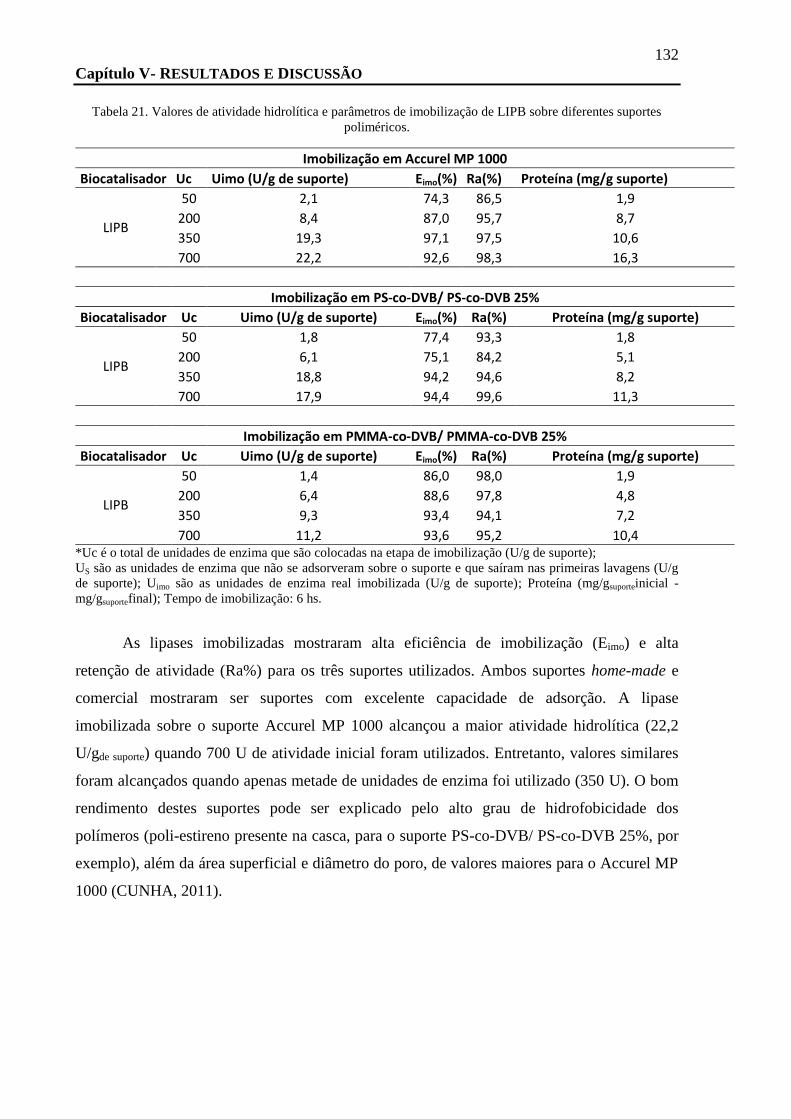
132
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 21. Valores de atividade hidrolítica e parâmetros de imobilização de LIPB sobre diferentes suportes
poliméricos.
Imobilização em Accurel MP 1000
Biocatalisador Uc Uimo (U/g de suporte) Eimo(%) Ra(%) Proteína (mg/g suporte)
LIPB
50 2,1 74,3 86,5 1,9
200 8,4 87,0 95,7 8,7
350 19,3 97,1 97,5 10,6
700 22,2 92,6 98,3 16,3
Imobilização em PS-co-DVB/ PS-co-DVB 25%
Biocatalisador Uc Uimo (U/g de suporte) Eimo(%) Ra(%) Proteína (mg/g suporte)
LIPB
50 1,8 77,4 93,3 1,8
200 6,1 75,1 84,2 5,1
350 18,8 94,2 94,6 8,2
700 17,9 94,4 99,6 11,3
Imobilização em PMMA-co-DVB/ PMMA-co-DVB 25%
Biocatalisador Uc Uimo (U/g de suporte) Eimo(%) Ra(%) Proteína (mg/g suporte)
LIPB
50 1,4 86,0 98,0 1,9
200 6,4 88,6 97,8 4,8
350 9,3 93,4 94,1 7,2
700 11,2 93,6 95,2 10,4 *Uc é o total de unidades de enzima que são colocadas na etapa de imobilização (U/g de suporte);
US são as unidades de enzima que não se adsorveram sobre o suporte e que saíram nas primeiras lavagens (U/g
de suporte); Uimo são as unidades de enzima real imobilizada (U/g de suporte); Proteína (mg/gsuporteinicial -
mg/gsuportefinal); Tempo de imobilização: 6 hs.
As lipases imobilizadas mostraram alta eficiência de imobilização (Eimo) e alta
retenção de atividade (Ra%) para os três suportes utilizados. Ambos suportes home-made e
comercial mostraram ser suportes com excelente capacidade de adsorção. A lipase
imobilizada sobre o suporte Accurel MP 1000 alcançou a maior atividade hidrolítica (22,2
U/gde suporte) quando 700 U de atividade inicial foram utilizados. Entretanto, valores similares
foram alcançados quando apenas metade de unidades de enzima foi utilizado (350 U). O bom
rendimento destes suportes pode ser explicado pelo alto grau de hidrofobicidade dos
polímeros (poli-estireno presente na casca, para o suporte PS-co-DVB/ PS-co-DVB 25%, por
exemplo), além da área superficial e diâmetro do poro, de valores maiores para o Accurel MP
1000 (CUNHA, 2011).

133
Capítulo V- RESULTADOS E DISCUSSÃO
O maior rendimento de imobilização foi obtido quando Accurel MP 1000 foi utilizado,
seguido de PS-co-DVB/ PS-co-DVB 25% e PMMA-co-DVB/ PMMA-co-DVB 25%. Os três
suportes mostraram alta capacidade de adsorção da enzima. Para os três suportes, o
rendimento de imobilização permaneceu alto quando houve aumento de 350U para 700U de
atividade inicial.
Nos experimentos realizados, o suporte Accurel MP 1000 foi aquele que adsorveu a
maior quantidade de proteína (16,3 mg/gsuporte) seguido de PS-co-DVB/ PS-co-DVB 25%
(11,3 mg/gsuporte). Cabrera et al. (2009) realizaram a imobilização de CalB em suportes
poliméricos hidrofóbicos distintos (octil-agarose, butil-agarose e octadecil-sepabeads)
obtendo capacidade de adsorção em termos de proteína entre 20 a 23 mg/gsuporte.
Besteti et al., (2014) imobilizaram CALB comercial em diferentes suportes do tipo
casca-núcleo. Os autores utilizaram uma quantidade menor de enzima em termos de unidade
de atividade durante o processo de imobilização. Estes obtiveram atividade hidrolítica de
1,4U/gsuporte de CALB comercial imobilizada (atividade inicial de 41,3U) quando o suporte
PMMA/PS foi utilizado. Estes valores foram similares aos desta tese quando LIPB foi
imobilizada em PMMA-co-DVB (casca e núcleo-Tabela 21). Entretanto, os autores
encontraram uma baixa retenção de atividade durante todo o processo de imobilização
(máxima de 34,3%), diferente dos resultados aqui encontrados.
Cunha et al., (2014) relatou sobre o aumento da atividade hidrolítica e dos parâmetros
de imobilização quando divinil-benzeno foi incorporado na casca dos polímeros. Os autores
imobilizaram CalB comercial em diversos suportes do tipo casca-núcleo e obteve baixos
rendimentos de imobilização quando utilizou PS-PS e PMMA/PMMA, na ausência do
divinilbenzeno. O aumento da área supercial e do diâmetro do poro promovidos pelo
divinilbenzeno levou, consequentemente, ao aumento dos valores dos parâmetros de
imobilização. A imobilização de CALB comercial em PS-co-DVB/ PS-co-DVB 25% foi
realizada pelos autores. Estes apresentaram resultados de atividades enzimáticas do
biocatalisador imobilizado similares aos desta tese, quando LIPB foi imobilizada no mesmo
suporte (ambos com 350U de atividade inicial).
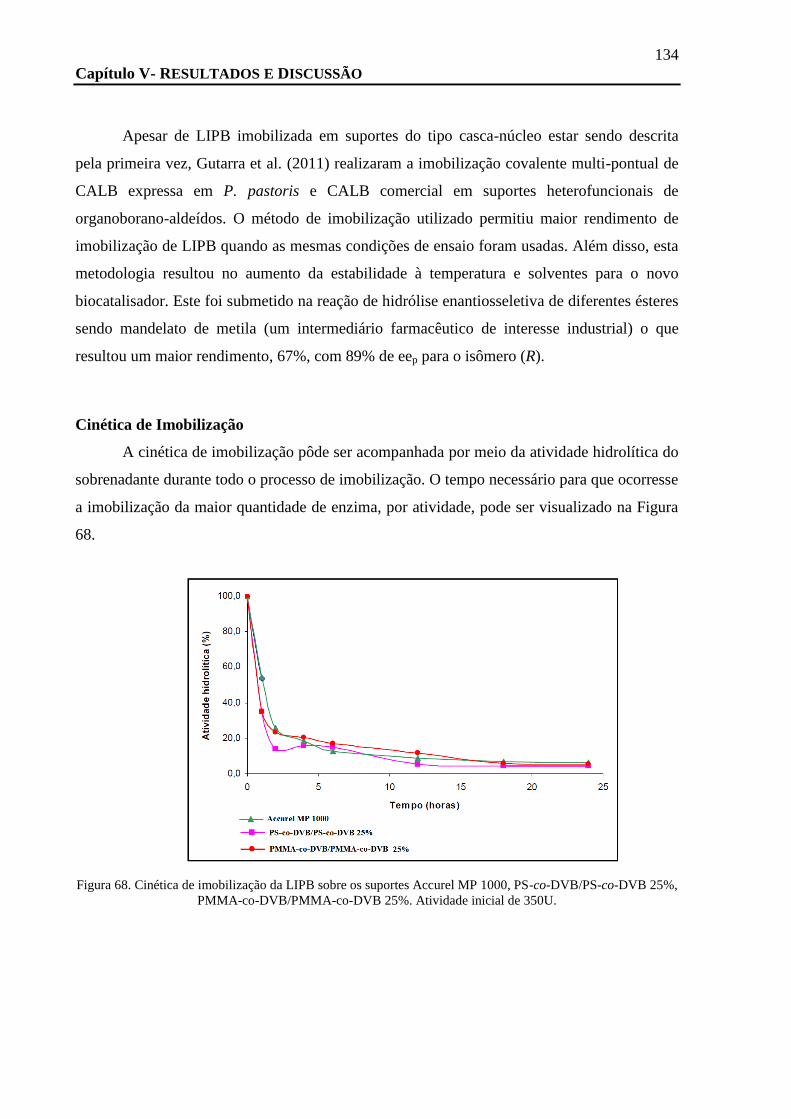
134
Capítulo V- RESULTADOS E DISCUSSÃO
Apesar de LIPB imobilizada em suportes do tipo casca-núcleo estar sendo descrita
pela primeira vez, Gutarra et al. (2011) realizaram a imobilização covalente multi-pontual de
CALB expressa em P. pastoris e CALB comercial em suportes heterofuncionais de
organoborano-aldeídos. O método de imobilização utilizado permitiu maior rendimento de
imobilização de LIPB quando as mesmas condições de ensaio foram usadas. Além disso, esta
metodologia resultou no aumento da estabilidade à temperatura e solventes para o novo
biocatalisador. Este foi submetido na reação de hidrólise enantiosseletiva de diferentes ésteres
sendo mandelato de metila (um intermediário farmacêutico de interesse industrial) o que
resultou um maior rendimento, 67%, com 89% de eep para o isômero (R).
Cinética de Imobilização
A cinética de imobilização pôde ser acompanhada por meio da atividade hidrolítica do
sobrenadante durante todo o processo de imobilização. O tempo necessário para que ocorresse
a imobilização da maior quantidade de enzima, por atividade, pode ser visualizado na Figura
68.
Figura 68. Cinética de imobilização da LIPB sobre os suportes Accurel MP 1000, PS-co-DVB/PS-co-DVB 25%,
PMMA-co-DVB/PMMA-co-DVB 25%. Atividade inicial de 350U.

135
Capítulo V- RESULTADOS E DISCUSSÃO
Apesar do comportamento de adsorção enzimática ser relativamente similar entre os
suportes, PS-co-DVB/PS-co-DVB 25% mostrou ter uma adsorção inicial mais rápida que os
demais, apresentando cerca de 83% de atividade hidrolítica após 2 horas de imobilização de
LIPB.
Os biocatalisadores produzidos (LIPB imobilizada em Accurel MP 1000, PS-co-
DVB/PS-co-DVB 25% e PMMA-co-DVB/PMMA-co-DVB 25%) ainda foram avaliados
quanto ao seu comportamento em meio orgânico. Para tal os mesmos foram utilizados na
reação de síntese do oleato de etila. A Tabela 22 apresenta os resultados da atividade de
esterificação.
Tabela 22. Atividade de esterificação da LIPB imobilizada sobre diferentes suportes poliméricos.
Suportes Atividade de esterificação (U/gsuporte)
Accurel MP 1000 1.970,00 + 220
PS-co-DVB/PS-co-DVB 25% 1.958,00 + 109
PMMA-co-DVB/ PMMA-co-DVB 25% 1.755,00 + 170
Novozym 435 2.728,00 + 339
Dos biocatalisadores imobilizados neste trabalho, os maiores valores de atividade de
esterificação foram observados com LIPB imobilizada em Accurel MP 1000 e LIPB
imobilizada em PS-co-DVB/PS-co-DVB 25% com atividade de esterificação de 1.970,00 +
220 e 1.958,00+ 109, respectivamente. A maior atividade de esterificação pode ser explicada
pela dependência da área específica e do diâmetro do poro do suporte, com valores superiores
para o Accurel MP 1000, o que já foi observado em outros estudos (CUNHA, 2011).
5.2.6- Estabilidade dos biocatalisadores ao longo do tempo
A fim de estudar a estabilidade dos biocatalisadores ao longo do tempo, os
biocatalisadores foram estocados a 30º C e suas atividades hidrolíticas determinadas
periodicamente, utilizando como substrato o p-NFL (Figura 69). O estudo da estabilidade de
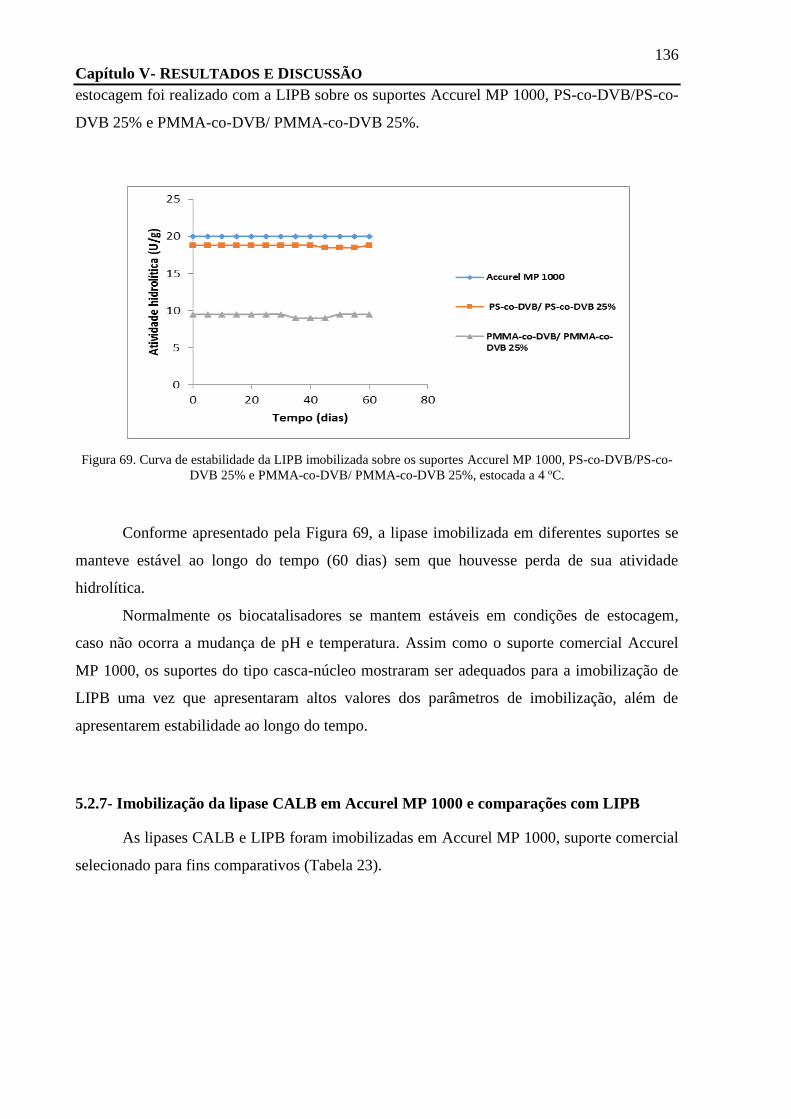
136
Capítulo V- RESULTADOS E DISCUSSÃO
estocagem foi realizado com a LIPB sobre os suportes Accurel MP 1000, PS-co-DVB/PS-co-
DVB 25% e PMMA-co-DVB/ PMMA-co-DVB 25%.
Figura 69. Curva de estabilidade da LIPB imobilizada sobre os suportes Accurel MP 1000, PS-co-DVB/PS-co-
DVB 25% e PMMA-co-DVB/ PMMA-co-DVB 25%, estocada a 4 ºC.
Conforme apresentado pela Figura 69, a lipase imobilizada em diferentes suportes se
manteve estável ao longo do tempo (60 dias) sem que houvesse perda de sua atividade
hidrolítica.
Normalmente os biocatalisadores se mantem estáveis em condições de estocagem,
caso não ocorra a mudança de pH e temperatura. Assim como o suporte comercial Accurel
MP 1000, os suportes do tipo casca-núcleo mostraram ser adequados para a imobilização de
LIPB uma vez que apresentaram altos valores dos parâmetros de imobilização, além de
apresentarem estabilidade ao longo do tempo.
5.2.7- Imobilização da lipase CALB em Accurel MP 1000 e comparações com LIPB
As lipases CALB e LIPB foram imobilizadas em Accurel MP 1000, suporte comercial
selecionado para fins comparativos (Tabela 23).
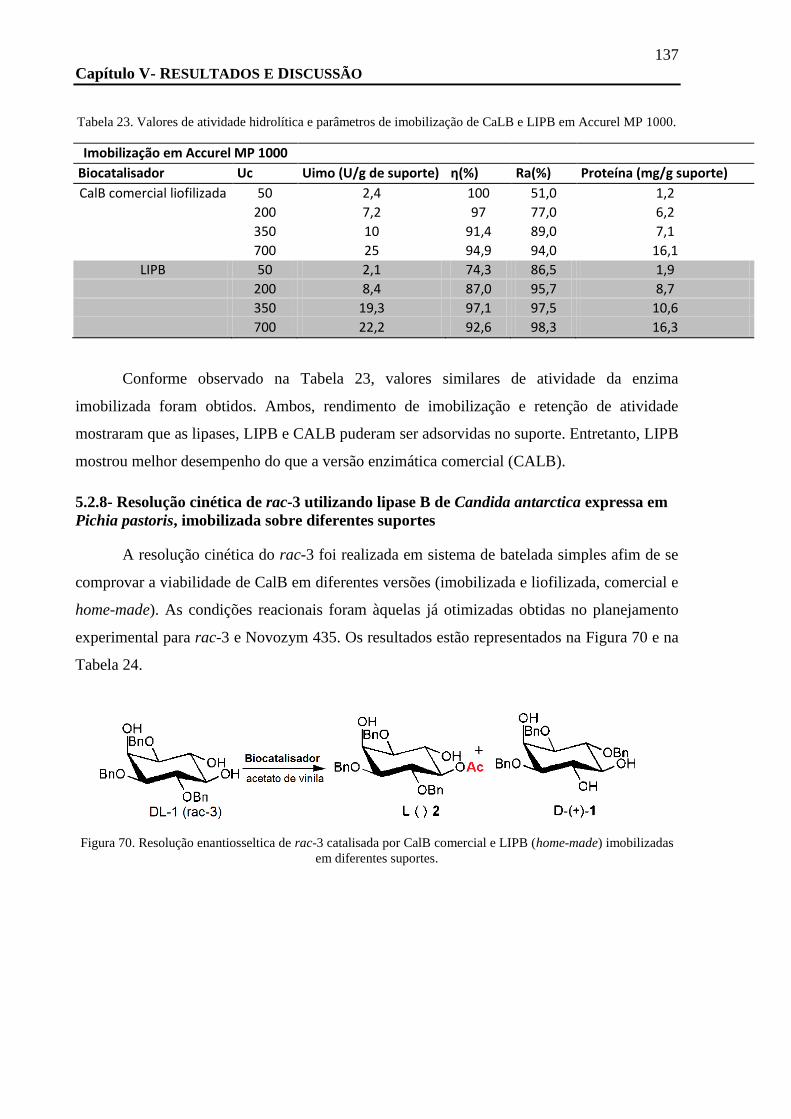
137
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 23. Valores de atividade hidrolítica e parâmetros de imobilização de CaLB e LIPB em Accurel MP 1000.
Imobilização em Accurel MP 1000
Biocatalisador Uc Uimo (U/g de suporte) η(%) Ra(%) Proteína (mg/g suporte)
CalB comercial liofilizada 50 2,4 100 51,0 1,2
200 7,2 97 77,0 6,2
350 10 91,4 89,0 7,1
700 25 94,9 94,0 16,1
LIPB 50 2,1 74,3 86,5 1,9
200 8,4 87,0 95,7 8,7
350 19,3 97,1 97,5 10,6
700 22,2 92,6 98,3 16,3
Conforme observado na Tabela 23, valores similares de atividade da enzima
imobilizada foram obtidos. Ambos, rendimento de imobilização e retenção de atividade
mostraram que as lipases, LIPB e CALB puderam ser adsorvidas no suporte. Entretanto, LIPB
mostrou melhor desempenho do que a versão enzimática comercial (CALB).
5.2.8- Resolução cinética de rac-3 utilizando lipase B de Candida antarctica expressa em
Pichia pastoris, imobilizada sobre diferentes suportes
A resolução cinética do rac-3 foi realizada em sistema de batelada simples afim de se
comprovar a viabilidade de CalB em diferentes versões (imobilizada e liofilizada, comercial e
home-made). As condições reacionais foram àquelas já otimizadas obtidas no planejamento
experimental para rac-3 e Novozym 435. Os resultados estão representados na Figura 70 e na
Tabela 24.
Figura 70. Resolução enantiosseltica de rac-3 catalisada por CalB comercial e LIPB (home-made) imobilizadas
em diferentes suportes.
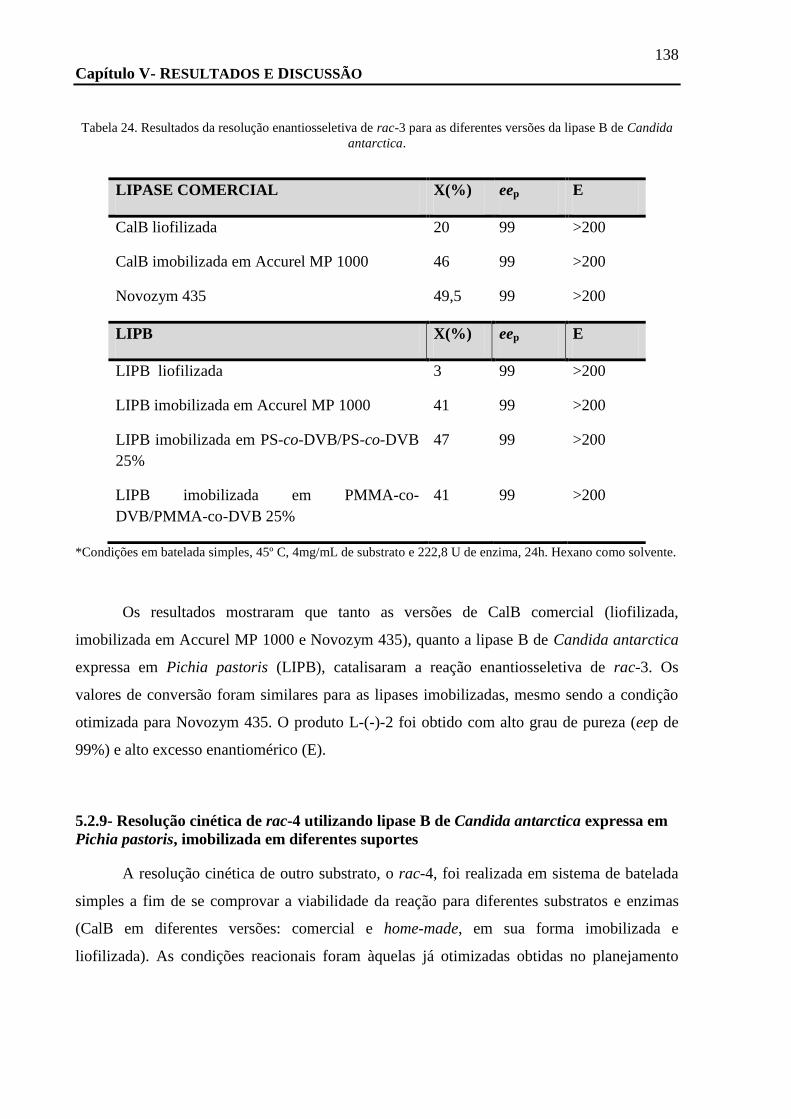
138
Capítulo V- RESULTADOS E DISCUSSÃO
Tabela 24. Resultados da resolução enantiosseletiva de rac-3 para as diferentes versões da lipase B de Candida
antarctica.
*Condições em batelada simples, 45º C, 4mg/mL de substrato e 222,8 U de enzima, 24h. Hexano como solvente.
Os resultados mostraram que tanto as versões de CalB comercial (liofilizada,
imobilizada em Accurel MP 1000 e Novozym 435), quanto a lipase B de Candida antarctica
expressa em Pichia pastoris (LIPB), catalisaram a reação enantiosseletiva de rac-3. Os
valores de conversão foram similares para as lipases imobilizadas, mesmo sendo a condição
otimizada para Novozym 435. O produto L-(-)-2 foi obtido com alto grau de pureza (eep de
99%) e alto excesso enantiomérico (E).
5.2.9- Resolução cinética de rac-4 utilizando lipase B de Candida antarctica expressa em
Pichia pastoris, imobilizada em diferentes suportes
A resolução cinética de outro substrato, o rac-4, foi realizada em sistema de batelada
simples a fim de se comprovar a viabilidade da reação para diferentes substratos e enzimas
(CalB em diferentes versões: comercial e home-made, em sua forma imobilizada e
liofilizada). As condições reacionais foram àquelas já otimizadas obtidas no planejamento
LIPASE COMERCIAL X(%) eep E
CalB liofilizada 20 99 >200
CalB imobilizada em Accurel MP 1000 46 99 >200
Novozym 435 49,5 99 >200
LIPB X(%) eep E
LIPB liofilizada 3 99 >200
LIPB imobilizada em Accurel MP 1000 41 99 >200
LIPB imobilizada em PS-co-DVB/PS-co-DVB
25%
47 99 >200
LIPB imobilizada em PMMA-co-
DVB/PMMA-co-DVB 25%
41 99 >200

139
Capítulo V- RESULTADOS E DISCUSSÃO
experimental para rac-4 e Novozym 435 (CUNHA et al. 2013). Os resultados estão
representados na Figura 71 e na Tabela 25.
Figura 71. Resolução enantiosseltica de rac-4 catalisada por CalB comercial e LIPB (home-made) imobilizadas
em diferentes suportes.
Tabela 25. Resultados da resolução enantiosseletiva de rac-4 para as diferentes versões da lipase B de Candida
antarctica.
rac-4
LIPASE COMERCIAL X(%) eep E
CalB liofilizada 28 99 >200
CalB imobilizada em Accurel MP 1000 40,8 99 >200
Novozym 435 49 99 >200
LIPB X(%) eep E
LIPB liofilizada 18 99 >200
LIPB imobilizada em Accurel MP 1000 48 99 >200
LIPB imobilizada em PS-co-DVB/PS-co-DVB 25% 49 99 >200
LIPB imobilizada em PMMA-co-DVB/PMMA-co-
DVB 25%
49 99 >200
*Condições em batelada simples, 45º C, 14mg/mL de substrato e 218 U de enzima, tempo reacional de 24h,
reação sem solvente, acetato de vinila como agente acilante e solvente.
Os resultados mostraram que tanto as versões de CalB comercial (versão liofilizada,
imobilizada em Accurel MP 1000 e Novozym 435), quanto a lipase B de Candida antarctica
expressa em Pichia pastoris (LIPB), catalisaram a reação enantiosseletiva de rac-4. Os

140
Capítulo V- RESULTADOS E DISCUSSÃO
valores de conversão foram similares para as lipases imobilizadas, mesmo as reações sendo
feitas na condição otimizada de Novozym 435. O produto L-5 foi obtido com alto grau de
pureza (eep de 99%) e alto excesso enantiomérico (E).
Os biocatalisadores imobilizados home-made mostraram o mesmo comportamento que
a lipase comercial Novozym 435, nas mesmas condições reacionais. Para a lipase imobilizada
em Accurel MP 1000, o valor de conversão foi ligeiramente menor (48%).
Cunha (2011) observou conversões de 47% (eep de 99%) quando CalB comercial foi
imobilizada em PS-co-DVB/PS-co-DVB 25%, PMMA-co-DVB/PMMA-co-DVB 25% e em
Accurel MP 1000.
Os três suportes, o comercial e os produzidos por laboratórios de pesquisa (PS-co-
DVB/PS-co-DVB 25% e PMMA-co-DVB/PMMA-co-DVB 25%), mostraram ser adequados
não apenas para a imobilização de LIPB, mantendo sua atividade catalítica e enantiosseletiva,
mas também para as reações cinéticas de rac-3 e rac-4. Este trabalho mostrou que LIPB,
apesar de estar sendo utilizada nas condições previamente otimizadas para o biocatalisador
comercial (Novozym 435), apresentou grande eficiência no processo reacional. Este resultou
em valores de conversão similares àqueles encontrados para a versão comercial.

141
Capítulo VI- CONCLUSÕES
6. CONCLUSÕES
Com a otimização realizada em sistema de batelada para a resolução cinética de rac-3,
catalisada por Novozym 435, foi possível obter o valor máximo de conversão (50%) com alta
pureza óptica (> 99%) para o produto e para o substrato remanescente. De fato, a condição
otimizada, por meio do DCCR, reduziu o tempo reacional, para 24h (tempo reacional no
protocolo original, antes da otimização era de 120h) levando o aumento considerado da
produtividade (3,188mgproduto/mgenzima.h, aumento de 15 vezes sob o protocolo original).
Em contrapartida, a abordagem desenvolvida utilizando o sistema de fluxo contínuo
em leito fixo também mostrou ser muito eficiente. Os máximos valores de conversão do rac-3
foram obtidos em apenas 3 min de tempo de residência. Com isso foi garantido o aumento da
produtividade em 531 vezes em relação ao sistema BSTR.
Em ambos os sistemas, contínuo e descontínuo, a reutilização do biocatalisador foi realizada
com êxito durante vários reciclos, o que pode contribuir com a economicidade do processo.
A tentativa do aumento da escala utilizada no processo descontínuo (BSTR), gerou
altos valores de conversão (50% ± 1) e seletividades (eep igual a 99%; E maior que 100).
Após o processo de seleção de biocatalisadores home-made se estabeleceu que LIPB
foi o mais promissor. Os suportes home-made utilizados no processo de imobilização de LIPB
mostraram alta capacidade de adsorção, atividade hidrolítica e esterásica, similar a
imobilização feita no suporte comercial.
LIPB em suportes home-made PS-co-DVB/PS-co-DVB e PMMA-co-DVB/ PMMA-
co-DVB mostrou grande desempenho na resolução cinética dos substratos utilizados em
sistema BSTR, com destaque para o rac-4, mostrando valores de conversão (entre 47% e
49%, para rac-3 e rac-4, respectivamente) e eep (>99%) muito parecidos com a versão
imobilizada comercial, Novozym 435.
Uma vez otimizada a produção dos derivados racêmicos (rac- 3 e rac-4), seus
respectivos produtos e substratos não reagidos podem então ser inseridos na rota sintética para
a produção de fármacos opticamente puros.
A otimização realizada neste trabalho permitiu utilizarmos pela primeira vez suportes
e biocatalisadores, ambos home-made, no processo de imobilização, aplicados na área
farmacêutica. Além disso, a rota enzimática na produção do fármaco permitiu a redução do
número de etapas, comparando-se com a literatura.

142
Capítulo VII- SUGESTÕES PARA TRABALHOS FUTUROS
7. SUGESTÕES PARA TRABALHOS FUTUROS
Os trabalhos desenvolvidos nesta tese fortaleceram a linha de pesquisa da biocatálise
para produção de fármacos, abrindo possibilidades para novas investigações sobre o tema
estudado.
As sugestões para os trabalhos futuros podem ser mencionados:
- Produção de novos biocatalisadores para outros derivados farmacológicos. Realizar
ainda a caracterização bioquímica dos novos biocatalisadores produzidos.
- Empregar a catálise enzimática proporcionada por lipases em reações de resolução
cinética e dessimetrizações para novas moléculas com potencial ativo.
- Aprofundar os estudos na imobilização de LIPB em suportes hidrofóbicos e testar os
mesmos nas reações mencionadas anteriormente.
- Desenvolver novos biocatalisadores com base na produção de lipase por diferentes
fungos e bactérias da cepa dos laboratórios deste grupo de pesquisa.
- As reações empregadas nesta tese ainda fazem uso de solventes orgânicos. Uma das
metas para trabalhos futuros é desenvolver reações com o uso dos Líquidos Iônicos,
contribuindo mais eficientemente para os princípios da química verde.
- Escalonamento do processo, em nível industrial, na produção do fármaco de interesse.

143
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
8. REFERÊNCIAS BIBLIOGRÁFICAS
ABIGOR, R.D., UADIA, P.O., FOGLIA, T.A., HAAS, M.J., JONES, K.C., OKPEFA, E., OBIBUZOR, J.U.e
BAFOR, M.E. Lipase-catalysed production of biodiesel fuel from some Nigerian lauric oils. Biochem. Soc.
Transactions., v. 28, p. 979-981, 2000.
ABIGOR, R.D., UADIA, P.O., FOGLIA, T.A., HAAS, M.J.; SCOTT, K.; SAVARY, B.J. Partial purification
and properties of lipase from germinating seeds of Jatropha curcas L., JAOCS, v. 79 (11), p. 1123-1126.
ADLERCREUTZ, P. Enzyme-catalyzed lipid modification. Biotechnol. Genet. Eng. Rev., v. 12, p.231-254,
1994.
ADLERCREUTZ, P. Modes of using enzymes in organic media‖ In: Enzymatic reactions in organic media, Eds:
Koskinen e Klibanov, Ed.Blackie academic & professional, Great Britain. 1996.
ADLERCREUTZ, P. Biocatalysis in non-conventional media In: Applied biocatalysis, Eds.: Straathof e
Adlercreutz, Ed. Harwood academic publishers Netherlands. 2000.
ADER, U.; ANDERSCH, P.; BERGER, M.; GOERGENS, U.; SEEMAYER, R. e SCHNEIDER, M., Pure Appl.
Chem. v. 64, p.1165–1170, 1992.
AGER, D.A. (Ed.), Handbook of Chiral Chemicals, Taylor & Francis, 2 edition. USA, 2006, 626p.
AHMED, M., Kelly, T., GHANEM, A. Applications of enzymatic and non-enzymatic methods to access
enantiomerically pure compounds using kinetic resolution and racemization, Tetrahedron, v. 68, p. 6781-6802,
2012.
AIRES-BARROS M.R. Biocatálise em solventes orgânicos. Boletim Biotecnologia, n. 72, p.2-12, 2002.
AKIN, N., AYDEMIR, S., KÇAK, C. e YILDIZ, M.A., Changes of free fatty acid contents and sensory
properties of white pickled cheese during ripening, Food Chemistry, v.80, p. 77–83, 2003.
ALMEIDA, M.V., SILVA, D.S., SOUZA, M.V.N. e BENÍCIO, A.A.A. A Cascata dos fosfoinositídeos. Quim.
Nova, v.26, n.1, p.105-111, 2003.
ALMEIDA, R.V., Clonagem, Expressão, Caracterização e Modelagem estrutural de uma esterase termoestável
de Pyrococcus furiosus. Tese doutorado. Engenharia Química, COPPE/UFRJ, Rio de janeiro, RJ – Brasil nov.
2005. 109p.
ALOULOU, A., RODRIGUEZ, J. A., FERNANDEZ, S., OSTERHOUT, D., PUCCINELLI, D., CARRIÈRE,
F., ―Exploring the specific features of interfacial enzymology based on lipase studies‖, Biochimica et Biophysica
Acta, v.1761, p. 995–1013, 2006.
AMUD, A.E.; da SILVA, R.P.; TARDIOLI, P.W.; SOARES, C.M.F.; MORAES, F.F. ZANIN, G.M. Methods
ans support for immobilization of cyclomaltodextrin glucanotransferase from Thermoanaerobacter, App.
Biochem. Biotechnol., v.146, p.189-201, 2008.
ANDERSON, E. M., LARSSON, K. M., KIRK, O., One biocatalyst – many applications: The use of Candida
antarctica B-lipase in organic synthesis, Biocatal. Biotransform. v.16, p.181-204, 1998.
ANOVÁ, C.M. e HUTTA, M. Role of biological matrices during the analysis of chiral drugs by Liquid
Chromatography. J. Chromatog. B, Amsterdam, v. 797, p. 91-109, 2003.
ANTHONSEN, T. Reactions Catalysed by enzymes. In: Straathof, A., Adlercreutz, P. (Eds.) Applied
Biocatalysis, second edition. Harwood academic publishers. Netherlands. 2005. 545p.
ANVISA. Disponível em: http://www.anvisa.gov.br/Legis/index.htm. Acesso em 04 jan. 2012.
ARRUDA, A. Utilização do promotor do gene PGK1 de Pichia pastoris para expressão heteróloga. 2008.
Dissertação de mestrado apresentado ao corpo docente de Biologia Celular do Instituto de Ciências da
Universidade de Brasília. Brasília. 2008, 90p.
AYBASTIER T.; DEMIR C.; J. Mol. Catal. B: Enzym. 63 (2010) 170–178.
BAGI K., SIMON L. M., Biotechnol. Tech. 13 (1999) 309–312.
BAI, Y. X., YAN, F. L., YANG., Y., YI, L. X., Covalent immobilization of triacylglycerol lipase onto
functionalized novel mesoporous silica supports, Journal of Biotechnology, v.125, n.4, p. 574-582, 2006.

144
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
BAI Y. X., Li Y. F., YANG Y., Yi L. X., Process Biochem. 41 (2006b) 770–777.
BAIA, Z. W., ZHOU, Y. K., A novel enzyme support derived from aminated silica gel and polysuccinimide:
preparation and application for the immobilization of porcine pancreatic lipase, Reactive & Functional Polymers,
v. 59, p. 93–98, 2004.
BAKER, R.; KULAGOWSKI, J.K.; BILLINGTON, D.C.; LEESON, P.D.; LENNOM, J.C.; LIVERTON, N. J.;
Journal Chem. Soc. Chem. Communication, p. 1383, 1989.
BAKER, G.A., BAKER, S., PANDEY, S., BRIGHT, F.V., Analyst, v.130, p. 800–808, 2005.
BALCÃO, V. M., PAIVA, A. L., MALCATA, F. X. Bioreactors with immobilized lipases, Enzyme and
Microbial Technology, v. 18, p. 392-416, 1996.
BARBOSA, O., ORTIZ, C., TORRES, R., FERNANDEZ-LAFUENTE, R., Effect of the immobilization
protocol on the properties of lipase B from Candida antarctica in organic media: Enantiospecifc production of
atenolol acetate, Journal of Molecular Catalysis B: Enzymatic, v.71, p. 24–132, 2011.
BARON, A.M. Biocatálise em ambientes aquo-restrictos:comparação de diferentes sistemas reacionais‖, tese de
mestrado, Universidade federal do Paraná, Curitiba. 2003.
BARROS M., FLEURI L.F. E MACEDO, G.A., Seed lipases: sources, applications and properties- a review.
Brazilian Journal of Chemical Engineering, v. 27, n. 1, p.15-29, 2010.
BASTIDA, A., SABUQUILLO, P., ARMISEN, P., FERNÁNDEZ-LAFUENTE, R., HUGUET, J., GUISÁN, J.
M., A single step purification, immobilization, and hyperactivation of lipases via interfacial adsorptions on
strongly hydrophobic supports, Biotechnology and Bioengineering, v. 58, n.5, p. 486-493, 1998.
BEHRENS, G.A., HUMMEL, A., PADHI, S.K., SCHÄTZLE, S., BORNSCHEUER, U.T., Discovery and
protein engineering of biocatalysts for organic synthesis. Adv. Synth. Catal., v.353, n.13, p.2191–2215, 2011.
BELLEZZA, F., CIPICIANI, A., COSTANTINO, U., Esterase activity of biocomposites constituted by lipases
adsorbed on layered zirconium phosphate and phosphonates: selective adsorption of different enzyme isoformas,
Journal of Molecular Catalysis B: Enzymatic, v. 26, p. 47–56, 2003.
BENJAMIN, S., PANDEY, A. Yeast, v. 14, p.69–87, 1998.
BEN SALAH, R., ZOUARI, N., REINBOLT, J. e MEJDOUB, H., 2003. Purification of turkey pancreatic
phospholipase A2. Biosci. Biotechnol. Biochem. 67, p.2139–2144.
BERGLUND P. Controlling lipase enantioselectivity for organic synthesis. Biomol Eng., v.18, p.13–22, 2001.
BERKOWITZ, D. B., DANISHEFSKY, S. J., SCHULTE, G. K. A route to artificial glycoconjugates and
oligosaccharides via enzymatically resolved glycals: Dramatic effects of the handedness of the sugar domain
upon the properties of an anthracycline drug, J. Am. Chem. Soc., v.114, n.12, p.4518- 4529, 1992.
BERNSMANN, H; GRUNER, M. e METZ, P., Tetrahedron Lett., v. 41, p. 7629–7633, 2000.
BESTETI, M. D. Dissertação de M.Sc., COPPE/UFRJ, Rio de Janeiro, RJ, Brasil, 2009.
BESTETI, M. D.; CUNHA, A. G.; FREIRE, D.M.G. e PINTO, J.C. Core/Shell Polymer Particles by Semibatch
Combined Suspension/Emulsion Polymerizations for Enzyme Immobilization. Macromol. Mater. Eng., 299,
135–143, 2014.
BESTETI, M.D., CUNHA, A. G., FREIRE, D. M. G., LIMA, E. L., PINTO, J. C. CBPol.- 10º CONGRESSO
BRASILEIRO DE POLÍMEROS, Foz do Iguaçu, Brasil, 2009.
BESTETI, M.D., SANTOS, D.P., CUNHA, A.G., ALMEIDA, R.V., FREIRE, D.M.G., PINTO, J. C. COBEQ
2010 – XVIII CONGRESSO BRASILEIRO DE ENGENHARIA QUÍMICA, Foz do Iguaçu, Brasil, 2010.
BESTETI, M.D., Tese de Doutorado em Engenharia Química, EQ/UFRJ, Rio de Janeiro, 2011.
BESTETI, M.D.; FREIRE, D.M.G.; PINTO, J.C. Production of Core–shell Particles by Combined Semibatch
Emulsion/Suspension Polymerizations. Macromol. React. Eng. 2011, 5, 518–532.
BESTETI, M.D.; CUNHA, A.G.; FREIRE, D.M.G.; PINTO, J.C. Core/Shell Polymer Particles by
Semibatch Combined Suspension/Emulsion Polymerizations for Enzyme Immobilization. Macromolecular
Materials and Engineering Volume 299, Issue 2, Article first published online: 22 OCT 2013.

145
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
BEVILAQUA, J.V., PINTO, J.C., LIMA, L.M., BARREIRO, E.J., ALVES, T.L.M., FREIRE, D.M.G.
Enzymatic hydrolysis by immobilized lipase applied to a new prototype anti-asthma drug, Biochemical
Engineering Journal, v. 21, p. 103–110, 2004.
BEVILAQUA, J.V., CUNHA, A.G., LIMA, L.M., ALVES, T.L.M., BARREIRO, E.J., PAIVA, L.M.C.,
FREIRE, D.M.G., Applied Biochemistry and Biotechnology, v. 121, p. 117-128, 2005.
BLANCO, R. M., TERREROS, P., FERNÁNDEZ-PÉREZ, M., OTERO, C., DIAZ-GONZÁLEZ G.,
Functionalization of mesoporous silica for lipase immobilization characterization of the support and the
catalysts, Journal of Molecular Catalysis B: Enzymatic, v. 30, p. 83–93, 2004.
BLANK, K., MOFILL, J., GUMPP, H., GAUB, H.E. Functional expression of Candida antarctica lipase B in
Escherichia coli, J. Biotechnol. v.125, p.474–483, 2006.
BOMMARIUS, S. ; BLUM, J. K. e ABRAHAMSON, M. J. Curr. Opin. Chem. Biol., 2011, 15, 194–200.
BORNSCHEUER, U.T. Microbial carboxyl esterases: classification, properties and application in biocatalysis.
FEMS Microbiol. Review., v. 733, p. 1-9, 2002.
BORNSCHEUER, U.T., Angewandte Chemie International Edition, v. 42, p.3336, 2003.
BORNSCHEUER, U. T. e KAZLAUSKAS, R.J. 2006. Ionic Liquids. In: BORNSCHEUER, U. T. e
KAZLAUSKAS, R.J. Hydrolases in Organic Synthesis. Regio- and Stereoselective Biotransformations. 2nd
edition. WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim. 2006a, 355p.
BORNSCHEUER, U. T. e KAZLAUSKAS, R.J. 2006. Lipases and esterases. In: BORNSCHEUER, U. T. e
KAZLAUSKAS, R.J. Hydrolases in Organic Synthesis. Regio- and Stereoselective Biotransformations. 2nd
edition. WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim. 2006b, pp.61-183 (Chapter 5).
BORNSCHEUER; U.T., HULT, K. Expression of Candida antarctica lipase B in Pichia pastoris and various
Escherichia coli systems Marianne Wittrup Larsen; Protein Expression and Purification, v.62, p. 90–97, 2008.
BOROS, Z.; FALUS, P.; MÁRKUS, M.; WEISER, D.; OLÁH, M.; HORNYÁNSZKY, G.; NAGY, J. e
POPPEA, L. How the mode of Candida antarctica lipase B immobilization affects the continuous-flow kinetic
resolution of racemic amines at various temperatures Journal of Molecular Catalysis B: Enzymatic, v. 85–86,
p.119–125, 2013.
BORSODY, M.K. e WEISS, J.M., Influence of corticotropin-releasing hormone on electrophysiological
activity of locus coeruleus neurons Brain Research, v. 724, p. 149-168, 1996.
BÖTTCHER D. e BORNSCHEUER U. T., Curr. Opin. Microbiol., 2010, 13, 274–282.
BRADFORD, M.M. Analytical Biochemistry, 1976, 72, 248-254.
BRADFORD, M.M., Analytical Biochemistry, v. 72, p. 248-554, 1976.
BRADY, L., BROZOZOWSKI, A. M., DEREWENDA, Z. S., DODSON, E., DODSON, G., TOLLEY, S.,
TURKENBURG, J. P., CHRISTIANXEN, L., JUGE-JENSEN, B., ORSKOV, L. e MENGE, U. A serine
protease triad forms the catalytic center of a triacylglycerol lipase. Nature, v. 343, p. 767-770, 1990.
BRANCO, R.V.; GUTARRA, M.L.E.; FREIRE, D.M.G.; ALMEIDA, R.V. Immobilization and Characterization
of a Recombinant Thermostable lipase (Pf2001) from Pyrococcus furiosus on supports with different degrees of
hydrophobicity, Enzyme Research, v.2010, 8p., 2010. Doi:10.4061/2010/180418.
BRANCO, R.V. Estudo de diferentes métodos de imobilização da lipase termoestável recombinante de
Pyrococcus furiosus: Estabilidade, enantiosseletividade e engenharia de proteínas. Tese submetida ao corpo
docente do programa de pós-graduação em Bioquímica da Universidade Federal do Rio de Janeiro, UFRJ, Rio
de Janeiro, 2012.109p.
BRASK, J., DAMSTRUP, M.L., NIELSEN, P.M., HOLM, H.C., MAES, J., GREYT, W. Applied Biochemistry
and Biotechnology, v.163, p. 918, 2011.
BRENA B.B. e BATISTA-VIEIRA F., Immobilization of enzymes, In: J. M. Guisan, Immobilized of Enzyme
and Cells. Methods in Biotechnology, 2nd Edition, Humana Press, Totowa, New Jersey, 2006, ch. 2, pp. 15-30.
BRUSTAD E. M. e ARNOLD, F. H. Curr. Opin. Chem. Biol., 2011, 15, 201–210.
BRZOZOWSKI, A.M., DEREWENDA, U., DEREWENDA, Z. S., DODSON, G. G., LAWSON, D. M.,
TURKENBURG, J. P., BJORKLING, F., HUGE-JENSEN, B., PATKAR, S. A., THIM, L., A model for

146
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
interfacial activation in lipases from the structure of a fungal lipase–inhibitor complex, Nature, v.351, p. 491–
494, 1991.
BRZOZOWSKI, A.M., SAVAGE, H., VERMA, C.S., TURKENBURG, J.P., LAWSON, D.M., SVENDSEN,
A., PATKAR, S., Structural origins of the interfacial activation in Thermomyces (Humicola) lanuginosa lipase,
Biochemistry, v. 39, p. 5071–15082, 2000.
BUCALÁ, V., FORESTI, M.L., TRUBIANO, G., FERREIRA, M.L., BRIOZZO, M. e OTTINI, S. Analysis of
solvent-free ethyl oleate enzymatic synthesis at equilibrium conditions. Enzyme Microbial Technol., v. 38, p.
914-920. 2006.
BUČKO M., MISLOVIČOVÁ D., NAHÁLKA J., VIKARTOVSKÁ A., ŠEFČOVIČOVÁ J., KATRLÍK J.,
TKÁČ J., GEMEINER P., LACÍK I., ŠTEFUCA V.; POLAKOVIČ M., ROSENBERG M., REBROŠ M.,
ŠMOGROVIČOVÁ D. e ŠVITEL J. Immobilization in biotechnology and biorecognition: from macro- to
nanoscale systems, Chemical Papers 66 (11) 983–998 (2012) DOI: 10.2478/s11696-012-0226-3
BUSTO, E.; GOTOR-FERNÁNDEZ e GOTOR, V. Asymmetric Chemoenzymatic Synthesis of Ramatroban
Using Lipases and Oxidoreductases. J. Org. Chem. v. 77, p.4842-4848, 2012.
BYZOZOWSKI, A. M., DEREWENDA, U., DEREWENDA, Z. S., DODSON, G. G., LAWSON, D. M.,
TURKEMBURG, J. P., BJORKLING, F., HUGE-JENSEN, B., PATKAR, S. S., THIM, L. 1991. A model for
interfacial activation in lipases from the structure of a fungal lipase–inhibitor complex. Nature 351: 491–494
CAO, L., van RANTWIJK, F., SHELDON, R. A., Cross-linked enzyme aggregates: a simple and effective
method for the immobilization of penicillin acylase, Org. Lett., v.2, p. 1361-1364, 2000.
CAO, L., van LANGEN, L. M., van RANTWIJK, F., SHELDON, R. A., Cross-linked aggregates of penicillin
acylase: robust catalysts for the synthesis of β-lactam antibiotics, J. Mol. Catal. B: Enzym. v.11, p. 665-670,
2001.
CABALLERO V., BAUTISTA F. M., CAMPELO J. M., LUNA D., MARINAS J. M., ROMERO A. A.,
HIDALGO J. M., LUQUE R., MACARIO A., GIORDANO G., Process Biochem. 44 (2009a) 334–342.
CABALLERO, E.; WILSON, L.; AROCA, G. New Biotechnol. 2009b, 25, S138.
CABRAL, J. M. S. e TRAMPER, J. Bioreactor Design in: STRAATHOF, A. J. J. e ADLERCREUTZ, P. (Eds.)
Applied Biocatalysis, second edition, hardwood academic publishers, Amsterdam, Netherlands, 2005, pags.392-
440.
CABRERA Z., FERNANDEZ-LORENTE G., FERNANDEZ-LAFUENTE R., PALOMO, J.M., GUISÁN, J.M.;
Novozym 345 displays very different selectivity compared to lipase from Candida Antarctica B adsorved on
other hydrophobic supports, Journal of Molecular Catalysis B: Enzymatic, v. 57, p. 171-176, 2009.
CAJAL, Y., SVENDSEN, A., GIRONA, V., PATKAR, S. A. and ALSINA, M. A. Interfacial control of lid
opening in Thermomyces lanuginosa lipase. Biochemistry, v.39, p.413–423, 2000.
CAJAL, Y., SVENDSEN, A., DE BOLÓS, J., PATLAR, S.A. e ALSINA, M.A. Effect of the lipid interface on
the catalytic activity and spectroscopic of a fungal lipase. Biochimie., v.82, p.1053-1061. 2000b.
CALADO, V. e MONTGOMERY, D. C. Planejamento de experimentos usando o Statistica. Rio de Janeiro: E-
papers Serviços editoriais, 260 p., 2003.
CALGAROTO C., SCHERER R. P., CALGAROTO S., OLIVEIRA J. V., de OLIVEIRA D., PERGHER S. B.
C., Appl. Catal. A 394 (2011) 101–104.
CAMBILLAU, G., TILBEURGH, H. VAN., Structure of hidrolases- lipases and celulases, Curr. Opin. Struct.
Biol., v.3, p.885-895, 1993.
CAMMAROTA, M. C.; ROSA, D. R.; DUARTE, T. C. R.; SAAVEDRA, N. K.; VARESCHE, M. B. A.;
ZAIAT, M.; FREIRE, D. M. G. The effect of enzymatic pre-hydrolysis of dairy wastewater on the granular and
immobilized microbial community in anaerobic bioreactors. Environmental Technology, v. 34, 4, pages 417-
428, 2013.
CAMMAROTA, M. C. e FREIRE, D. M. G. A review on hydrolytic enzymes in the treatment of wastewater
with high oil and grease content. Bioresource Technology, v.97, n.2, p.195-210, 2006.
CAMMAROTA, M. C., TEIXEIRA, G. A. e FREIRE, D. M. G. Enzymatic prehydrolysis and anaerobic
degradation of wastewater with high fat content. Biotechnology Letters, v.71, p.44-50, 2001.

147
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
CAREY, F.A. e SUNDBERG, R.J. Stereochemistry, Conformation, and Stereoselectivity In: Advanced Organic
Chemistry Part A:Structure and Mechanisms. 2ª edition, Springer Science+Business Media, New York, NY,
USA, 2007, 1199p.
de CARVALHO. C.C.C.R., Enzymatic and whole cell catalysis: Finding new strategies for old processes,
Biotechnology Advances, v.29, p. 75–83, 2011.
CASTRO, H.F., MENDES, A.A., SANTOS, J.C. e AGUIAR, C.L. Modificação de Óleos e Gorduras por
Biotransformação. Quimica Nova, v. 27, p. 146-156, 2004.
CASTRO-OCHOA, L. D., RODRÍGUEZ-GÓMEZ, C., VALERIO-ALFARO, G. e ROS, R.O. Screening,
purification and characterization of the thermoalkalophilic lipase produced by Bacillus thermoleovorans CCR11.
Enzyme and Microbial Technology, v. 37, n.6, p.648-654, 2005.
CASTRO H. F., ZANIN G. M., de MORAES F. F., SÁ-PEREIRA P. Imobilização de enzimas e sua
estabilização. In: E. P. S. [et al] BON, M. A. FERRARA, and M. L. CORVO, Enzimas em biotecnologia:
produção, aplicações e mercado, Interciência, 2008, ch 6, pp.123-151.
CASTRO, A.M., BEVILAQUA, J.V., FREIRE, D. M. G., TORRES, F. A. G., SANTAANNA, L. M. M.,
GUTARRA, M. L. E., BARBOSA, C. A., V. ALMEIDA, R., MENEZES, R. R. e CUNHA, A. G. Processo de
produção de lipases por meio de modificação genética de levedura. Pedido de Patente ao INPI: Privilégio e
Inovação no. PI0905122-8 depositada em 17/12/2009, 2009.
CAVALCANTI, E.D.C., MACIEL, F.M., VILLENEUVE, P., LAGO, R. C. A., MACHADO, O. L. e FREIRE,
D.M.G. Acetone Powder from Dormant Seeds of Ricinus communis L.: Lipase Activity and Presence of Toxic
and Allergenic Compounds. Applied Biochemistry and Biotechnology, v.136, p.57 – 65, 2007.
CAVALCANTI-OLIVEIRA, E.A., SILVA, P.R., RAMOS, A.P., DONATO, D.A.G. e FREIRE, D.M.G. Study
of Soybean Oil Hydrolysis Catalyzed by Thermomyces lanuginosus. Lipase and Its Application to Biodiesel
Production via Hydroesterification. Enzyme Research, 2011, doi:10.4061/2011/618692
CHAE, M.-H., PARK, H.-K., KWON, K.-I., KIM, J.-W., HONG, S.I., KIM, Y., KIM, B.H.,
CHARTRAIN, M., GREASHAM, R., MOORE, J., REIDER, P., ROBINSON, D., BUCKLAND, B., Journal of
Molecular Catalysis B: Enzymatic, v.11, p. 503, 2001.
CHEN, C. S.; FUJIMOTO, Y.; GIRDAUKAS, G.; SIH, C. J. J. Amerian Chem. Society, v.104, p.7294–7299,
1982.
CHEN, C.-S. e SIH, C.J., General Aspects and Optimization of Enantioselective Biocatalysis in Organic
Solvents: The Use of Lipases. Angew. Chem. Int. Ed. Engl., v. 28, p. 695–707, 1989.
CHEN, C.S.; FUJIMOTO, Y.; GIRDAUKAS, G.; SIH, C.J., Quantitative analyses of biochemical kinetic
resolutions of enatiomers, Journal American Chemical Society, v. 104, p. 7294–7299, 1982.
CHEN, S.T. e FANG, J.M. J. Org. Chem., v. 62, p. 4349–4357, 1997.
CHEN, Y.-H., HUANG, Y.-H., LIN, R.-H., SHANG, N.-C. Bioresource Technology, v. 101, p.668, 2010.
CHEN., B., MILLER, M.E., GROSS, R.A., Langmuir, v. 23, p. 6467-6474, 2007.
CHIOU, S.H., WU, W.T., Immobilization of Candida rugosa lipase on chitosan with activation of the hydroxil
groups, Biomaterials, v. 25, p. 197-204, 2004.
CHOWDARY GV, RAMESH MN, PRAPULLA SG. Enzymic synthesis of isoamyl isovalerate using
immobilized lipase from Rhizomucor miehei: multivariate analysis. Process Biochem 2001;36:331–9.
CHUA, L. e SARMIDI, M. Effect of solvent and initial water content on (R, S)-1- phenylethanol resolution.
Enzyme and Microbial Technology, v. 38, p. 551–556, 2006.
CHO, A. R., YOO, S.K., KIM, E. J. FEMS Microbiology Letters, v. 186, p. 235-238, 2000.
CHUNG, S.K., CHANG, Y.T., LEE E. J., SHIN, B.G., KWON, Y.U., KIM K.C., LEE, D.H., KIM, M.J.,
Syntheses of two enantiomeric pairs of myo-inositol(1,2,4,5,6) and (1,2,3,4,5) pentakisphosphate, Bioorganic &
Medicinal Chemistry Letters, v. 8, p. 1503-1506, 1998.
CLAIR, N. L., NAVIA, M. A., Cross-linked enzyme crystals as robust biocatalysts, J. Am.Chem. Soc. v.114,
n.18, p. 7314-7316. 1992.
CORZO, G. e REVAH, S. (1999). ―Production and characteristics of the lipase from Yarrowia lipolytica 681‖,
Bioresource Technol., v.70, pp.173-180

148
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
COSTA, V.E.U., AMORIN, H.L., O emprego de lipases como agentes de resolução cinética de enantiomeros em
síntese orgânica: aspectos gerais sobre a influencia do solvente, Química Nova, v. 22, n. 6, p. 863-873, 1999.
COWAN D. A. e FERNANDEZ-LAFUENTE R., Enhancing the functional properties of thermophilic enzymes
by chemical modification and immobilization. Enzyme Microb. Technol., 2011, 49, 326–346. DOI:
10.1016/j.enzmictec.2011.06. 023.
CUNHA, A.G., FERNÁNDEZ-LORENTE, G., BEVILAQUA, J.V., DESTAIN, J., PAIVA, L.M.C., FREIRE,
D.M.G., FERNÁNDEZ-LAFUENTE, R., GUISÁN, J.M., AppliedBiochemistry and Biotechnology, v.146, p. 49-
56, 2008.
CUNHA, A.G.; FERNÁNDEZ-LORENTE, G.; GUTARRA, M.L.E.; BEVILAQUA, J.V.; ALMEIDA, R.V.;
PAIVA, L.M.C.; FERNÁNDEZ-LAFUENTE, R.; GUISÁN, J.M.; FREIRE, D.M.G.. Appl Biochem Biotechnol,
v.156, p. 563-575, 2009.
CUNHA, A.G.; da SILVA, A.A.T.; da SILVA, A.J.; TINOCO, L.W.; ALMEIDA, R.V.; de ALENCASTRO,
R.B.; SIMAS, A.B.C. e Freire, D.M.G., Efficient kinetic resolution of (±)-1,2-O-isopropylidene-3,6-di-O-
benzyl-myo-inositol with the lipase B of Candida Antarctica. Tetrahedron: Asymmetry, v. 21, p.2899-2903,
2010.
CUNHA, A.G. Resolução cinética de derivados de mio-inositol catalisada por lipases. Tese de Doutorado,
IQ/UFRJ, Rio de Janeiro. 140p. 2011.
CUNHA, A. G., da SILVA, A. A. T., GODOY, M. G., ALMEIDA, R. V., SIMAS, A. B. C. e FREIRE, D. M.
G., Experimental design of the kinetic resolution of a key precursor of high-value bioactive myo-inositols by an
immobilized lipase. J. Chem. Technol. Biotechnol. doi: 10.1002/jctb.3806. 2012.
CUNHA, A.G.; da SILVA, A.A.T.; GODOY, M.G.; ALMEIDA, R.V.; SIMAS, A.B.C. e Freire, D.M.G.
Experimental design of the kinetic resolution of a key precursor of high-value bioactive myo-inositols by an
immobilized lipase. J Chem Technol Biotechnol. v. 88, p. 205–211, 2013.
CUNHA, A.G.; BESTETI, M.D.; MANOEL, E.A.; da SILVA, A.A.T.; ALMEIDA, R.V.; SIMAS, A.B.C.;
FERNANDEZ-LAFUENTE, R.; PINTO, J.C.; FREIRE, D.M.G. Preparation of core–shell polymer supports to
immobilize lipase B from Candida antarctica. Effect of the support nature on catalytic properties. J. Mol.
Catalysis B: Enzymatic., v. 100, p. 59-67, 2014.
CYGLER, M., SCHRAG, J. D., ERGAN, F., Advances in structural understanding of lipases, Biotechnol. Genet.
Eng. Rev. v.10, p.143-184, 1992.
CYGLER, M. e SCHRAG, J. D. 1997. Structure as basis for understanding interfacial properties of lipases.
Methods Enzymol, 284, part A, p. 3-27.
DAMASCENO, F.S.C.; CAMMAROTA, M.C.; FREIRE, D.M.G. The combined use of a biosurfactant and an
enzyme preparation to treat an effluent with a high fat contente. Colloids and Surfaces B: Biointerfaces, v. 95,
n.15, p. 241–246, 2012.
DALLA-VECCHIA, R., NASCIMENTO, M.G., SOLDI, V. Química Nova, v. 27, n. 4, p. 1-16, 2004.
DEGUEIL-CASTAING, M., de JESO, B., DROUILLARD, S. e MAILLARD, B. Enzymatic reactions in organic
synthesis: 2-Ester interchange of vinyl esters, Tetrahedron Lett. v. 28, n.9, p. 953-954, 1987.
DEREWENDA, U., BRZOZOWSKI, A.M.; LAWSON, D.M. e DEREWENDA, Z. S., Biochemistry, v.31,
p.532-1541, 1992.
DEREWENDA, U., SWENSON, L., GREEN, R., WEI, Y., DODSON, G. G., YAMAGUCHI, S., HAAS, M. J.
e DEREWENDA, Z. S., An unsusal buried polar cluster in a family of fungal lipases, Nature Struct. Biol., v.1,
p.36-47, 1994a.
DEREWENDA, U., SWENSON, L., GREEN, R., WEI, Y., YAMAGUCHI, S., JOERGER, R., HAAS, M. J.,
DEREWENDA, Z. S., Current progress in crystallographic studies of new lipases from filamentous fungi,
Protein Engineer, v.7, p.551-557, 1994b.
DESAI, T.; GIGG, J.; GIGG, R.; MARTÍN-ZAMORA, E. SCHNETZ, N.; Carbohydr. Res., v. 258, p. 135-144,
1994.
DESAI, T., GIGG, J., GIGG, R. e MARTIN-ZAMORA, E., The preparation of racemic nd enantiomerically
pure myo-inositol derivatives as intermediates for the synthesis of phosphatidylinositol3-, 3,4-bis-and 3,4,5-tris-
phosphates and for the synthesis of analogues of lD-myo-inositol 1,3,4,5tetrakisphosphate. Carbohydrate
Research, v. 296, p.97-133, 1996.

149
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
DESAI P.D., DAVE A.M., DEVI S., Food Chem. 95 (2006) 193–199.
DEWAN, S. S. BCC Research; BCC Research: 2012. Disponível em http://bccresearch.com/report/chiral-
technology-bio012e.html. Acesso em 10. Jan. 2013.
DÍAZ-RODRÍGUEZ e B. G. DAVIS, Curr. Opin. Chem. Biol., 2011, 15, 211–219.
DIJKSTRA, Z. J.; MERCHANT, R.; KEURENTJES, J. T. F. J. Supercritic. Fluids 2007, 41, 102-108.
DIZGE, N., KESKINLER, B., TANRISEVEN, A., Colloids and Surfaces B: Biointerfaces, v. 66, p. 34–38,
2008.
DODSON, G. G., LAWSON, D. M. e WINKLER, F. K., Structural and evolutionary relationships in lipase
mechanism and activation, Faraday Discuss. p.95-105. 1992.
DU, W. J. Mol. Cat. B: Enzym., 2004, 30, 125–129.
DUNN P.J., WELLS A.S., WILLIAMS M.T. (Eds.), Green Chemistry in the Pharmaceutica, Industry, Wiley-
VCH, Weinheim, 2010, pp. 269–287 (Chapter 13).
EINICKER-LAMAS, M., MALAQUIAS, A.T., DE CASTRO, S.L., SMITH, G.A., OLIVEIRA, M.M.,
Parasitology Research, v. 85, p. 232–238, 1999.
EGGERT, T., VAN POUDEROYEN G., DIJKSTRA, B.W., JAEGER, K.E. Lipolytic Enzymes LipA and LipB
from Bacillus subtilis differ in regulation of gene expression, biochemical properties, and three-dimensional
structure. FEBS Lett 502:89–92, 2001.
EGLOFF, P., RANSAC, S., MARGUET, F., ROGALSKA, E., VAN TILBEURGH, H., BUONO, G.,
CAMBILLAU, C. e BERGER, R. Les lipases: cinétiques, spécificités et aspects structuraux. In MALCATA,
F.X. (Ed). Engineering of/with lipases. Dordrecht: Kluwer Academic Publishers, 1995.
EVANS, C.T., ROBERTS, S.M., SHOBERU, K.A., SUTHERLAND, A.G., Potential use of carbocylic
nucleosides for the treatment of AIDS: chemo-enzymatic syntheses of the enantiomers of carbovir. J. Chem. Soc.
Perkin Trans. v.1, p.589–592, 1992.
FABER, K. Biotransformations in Organic Chemistry, Sringer-Verlag New York INC.: New York, 1997.
FABER, K. Biotransformations in Organic Chemistry, 4 ed. Springer: Berlin, 2000.
FABER, K., Non-sequential processes for the transformation of a racemate into a single stereoisomeric product:
proposal for stereochemical classification, Chem. Eur. J., v.7, p.5004-5010, 2001.
FABER, K. Biocatalytic Applications In: Biotransformations in organic chemistry, 2011. 423p.
FABER, K., FRANSSEN, M.C.R., Prospectsor the increase application of biocatayst in organic transformations,
Trends in Biotechnol., v. 11, p.461-470, 1993.
FABER, K., OTTOLINA, G. e RIVA, S., Selectivity-Enhancement of Hydrolase Reactions. Biocatalysis , v. 8,
p. 91–132, 1993.
FERNANDES, M.L.M., KRIEGER, N., BARON, A.O, ZAMORA, P.P., RAMOS, L.P. e MITCHELL, D.A.
Hydrolysis and synthesis reactions catalysed by Thermomyces lanuginosa lipase in OT/Isooctane reversed
micellar system. J. Mol. Catalysis B: Enzymatic., v. 30, p. 43-49, 2004.
FERNANDES, M.L.M., SAAD, E.B., MEIRA, J.A., RAMOS, L.P., MITCHELL, D.A. e KRIEGER, N.
Esterification and transesterification reactions catalysed by addition of fermented solids to organic reaction
media. J. Mol. Catalysis B: Enzymatic., v. 263, p. 8-13, 2006.
FERNANDEZ-ÁLVARO, E.; ESQUIVIAS, J.; PÉREZ-SÁNCHEZ, M.; DE MARIA, P.D.; REMUINÁN-
BLANCO, M.J. Assessing biocatalysis for the synthesis of optically activetetrahydropyrazolo[1,5- _]pyrimidines
(THPPs) as noveltherapeutic agentes. Journal of Molecular Catalysis B: Enzymatic 100 (2014) 1– 6.
FERNANDEZ-LAFUENTE, R., ARMISÉN, P., SABUPUILLO, P., FERNÁNDEZ-LORENTE, G., GUISÁN,
J. M., ―Immobilization of lipases by selective adsorption on hydrophobic supports‖, Chemistry and Physics of
Lipids, v. 93, pp. 185-197, 1998.
FERNÁNDEZ-LORENTE, G., TERRENI, M., MATEO, C., BASTIDA, A, FERNÁNDEZ-LAFUENTE, R.,
DALMASES, P., HUGUET, J., GUISÁN, J.M., Modulation of lipase properties in macro-aqueous systems by
controlled enzyme immobilization: enantioseletive hydrolysis of a chiral ester by immobilized Pseudomonas
lipase, Enz. Microbial Technol.,, v. 28, p. 389-396, 2001.

150
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
FERNÁNDEZ-LORENTE, G.; PALOMO J.M.; FUENTES M., MATEO, C.; GUISÁN, J.M.; FERNANDEZ-
LAFUENTE, R. Self-assembly of Pseudomonas fluorescens lipase into bimolecular aggregates dramatically
affects functional properties. Biotechnol Bioeng 2003;82:232–7.
FERNÁNDEZ-LORENTE G, CABRERA Z, GODOY C, FERNANDEZ-LAFUENTE R, PALOMO JM,
GUISAN JM. Interfacially activated lipases against hydrophobic supports: effect of the support nature on the
biocatalytic properties. Process Biochem 2008;43:1061–7.
FERRARIO V.; EBERT, C.; KNAPIC, L.; FATTOR, D.; BASSO, A.; et al. Conformational changes of lipases
in aqueous media: A comparative computacional study and experimental implications. Adv. Synth Catal. 353:
2466-2480, 2011.
FITZPATRICK, P.A., KLIBANOV, A.M., Journal American Chemical Society, v.113, p. 3166, 1991.
FOLTIN, J.P. Avaliação da perda de carga em leito fixo de partículas irregulars utilizando xisto betuminoso,
analisando a modelagem matemárica através do efeito de parede e porosidade. Dissertação apresentada a curso
de mestrado em engenharia química da Universidade Federal do Paraná, Curitiba, Brasil.2013,104p.
FONTES, G.C.; RAMOS, N. M.; AMARAL, P. F. F.; NELE, M.; COELHO, M. A. Z., Renewable resources for
biosurfactant production by Yarrowia lipolytica. Brazilian Journal of Chemical Engineering (Impresso), v. 29, p.
483-494, 2012.
FORDE, J.; VAKUROV, A.; GIBSON, T. D.; MILLNER, P.; WHELEHAN, M.; MARISON, I. W.;
Ó'FÁGÁIN, C. J.Mol. Catal. B: Enzym. 2010, 66, 203-209.
FORESTI, M.L, FERREIRA, M.L., Chitosan-immobilized lipases for the catalysis of fatty acid esterifications,
Enzyme and Microbial Technology, v. 40, n.4, p. 769-777, 2006.
FREEMAN A. e LILLY M. D., Enzyme Microb. Technol., 1998, 23, 335–345.
FREIRE, D.M.G. Imobilização de amiloglicosidase em quitina – caracterização e testes em reatores contínuos de
leito expandido, Dissertação de Mestrado, EQ/UFRJ, Rio de Janeiro, 1988.
FREIRE, D.M.G. e CASTILHO L. R. 2010. Lipases em Biocatálise. In: Elba Pinto da Silva BON;Maria
Antonieta Ferrara;Maria Luisa Corvo. (Org.). Enzimas em Biotecnologia. Produção, Aplicações e Mercado..
1ed.Rio de Janeiro: Editora Interciência, 2008, v. 1, p. 369.
FREIRE, D.M.G.; de SOUSA, J.S.; CAVALCANTI E.D.C. Biotechnological Methods to Produce Biodiesel.In:
PANDEY, A.; LARROCHE, C.; RICKE, S.C.; DUSSAP, C.G. e GNANSOUNOU, E. (Eds.). Biofuels,
Burlington: Academic Press, 2011, p. 315-337. ISBN: 978-0-12-385099-7.
FREUND, H. et al. Numerical simulations of single phase reacting flows in randomly packed fixed-bed reactors
and experimental validation, Chemical Engineering Science, Netherlands, v. 58, p. 903-910, 2003.
FUENTES, I.E., VISERAS, C.A., UBIALI, D., TERRENI, ALCÂNTARA, A.R., Different phyllosilicates as
supports for lipase immobilisation, Journal of Molecular Catalysis B: Enzymatic, v. 11, p.657–663, 2001.
GANJALIKHANY,M.R.; RANJBAR, B.; TAGHAVI, A.H.; MOGHADAM, T.T.. 2012. Functional motions of
Candida Antarctica lipase B: A survey through open-close conformations. Plosone, v. 7, issue 7, p. 1-11.
GAO, S., WANG, Y., WANG, T., LUO, G., DAÍ, Y., Bioresource Technology v. 100, p.996–999, 2009.
GARCIA-GALAN, C.; BERENGUER-MURCIA, A.; FERNANDEZ-LAFUENTE R.; RODRIGUES, R. C.
Potential of different enzyme immobilization strategies to improve enzyme performance. Adv. Synth. Catal.,
2011, 353, 2885–2904. DOI: 10.1002/adsc.201100534.
GARCÍA-URDIALES, E., AFONSO, I. E GOTOR, V., Enantioselective Enzymatic Desymmetrizations in
Organic Synthesis, Chem. Rev., v.111, p.110-180, 2011.
GAREGG, P.; LINDBERG, B.; KVARNSTRÖM, I.; C.T. SVENSSON, S. Carbohydrate Research, v. 139, p.
209-215, 1985.
GELLISSEN G. Heterologous protein production in methylotrophic yeasts. Appl Microbiol Biotechnol, v.54,
p.741–750, 2000.
GHANEM, A.; ABOUL-ENEIN, H. Y. Tetrahedron: Asymmetry, v. 15, p. 3331-3351, 2004.
GHANEM, A. 2007. Trends in lipase-catalyzed asymmetric access to enantiomerically pureenriched
compounds. Tetrahedron, v. 63, p.1721–1754, 2007.

151
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
GHANEM, A., ABOUL-ENEIN, M.N., EL-AZZOUNY, A. e EL-BEHAIRY, M.F. Lipase-mediated
enantioselective kinetic resolution of racemic acidic drugs in non-standard organic solvents: Direct chiral liquid
chromatography monitoring and accurate determinationof the enantiomeric excesses. J. Chromatogr. A 1217,
p.1063–1074, 2010.
GONG, P.-F. e XU, J.H. Bio-resolution of a chiral epoxide using whole cells of Bacillus megaterium ECU1001
in a biphasic system. Enzyme and Microbial Technology, v.36, p.252–257, 2005.
GONG, P.-F. e XU, J.-H., Bio-resolution of a chiral epoxide using whole cells of Bacillus megaterium ECU1001
in a biphasic system. Enzyme and Microbial Technology, v. 36, p. 252–257, 2005.
GONZALEZ-SABÍN, J., LAVANDERA, I., REBOLLEDO, F., GOTOR, V., Redesigning the mechanism of the
lipase-catalysed aminolysis of esters, Tetrahedron: Asymmetry, v. 17, n.8, p. 1264–1274, 2006.
GORMAN, L. A. e DORDICK, J. S. Biotechnol. Bioeng. v. 39, p.392, 1992.
GOSWAMI D.; BASU J.K.; DE D.; Crit. Rev. Biotechnol. 33 (2013) 81–96.
GOTOR, V. F., BRIEVA, R., GOTOR, V., Lipases: Useful biocatalysts for the preparation of pharmaceuticals,
Journal of Molecular Catalysis B: enzymatic, v. 40, p. 111-129, 2006.
GOTOR-FERNANDEZ, V., BUSTO E., GOTOR, V., Candida antarctica Lipase B: An Ideal Biocatalyst for the
Preparation of Nitrogenated Organic Compounds, Adv. Synth. Catal., v. 348, p.797–812, 2006.
GROCHULSKI, P., BOUTHILLIER, F., KAZLAUSKAS, R.J., SERREQI, A.N., SCHRAG, J.D., ZIOMEK, E.,
CYGLER, M., ―Analogs of reaction intermediates identify a unique substrate binding site in Candida rugosa
lipase‖, Biochemistry, v. 33, pp. 3494–3500, 1994.
GROENER, J.E.M., BAX, W., STUANI C. e PAGANI, F. Biochimica et Biophysica Acta Molecular and Cell
Biology of Lipids, v. 27, p. 155-162. 2000.
GU, Q.; LIN, Q.; HU, C.; YANG, B. J. Appl. Polym. Sci., 2005, 95, 404-412.
GUISÁN, J.M., ALVARO, G., FERNANDEZ-LAFUENTE, R., ROSELL, C.M., GARCIALOPEZ, J.L.,
TAGLIATI, A., Biotechnology Bioeng., v. 42, p. 455–464, 1993.
GUISÁN, J. M., SABUQUILLO, P., FERNANDEZ-LAFUENTE, R., FERNANDEZ-LORENTE, G.,
Preparation of new lipases derivatives with high activity-stability in anhydrous media: adsorption on
hydrophobic supports plus hydrophilization with polyethylemine, Journal of molecular Catalysis B: enzymatic,
v. 11, p. 817-824, 2001.
GUISAN J.M. Immobilization of Enzymes as the 21st Century Begins In: Guisan, J.M. Immobilized of Enzyme
and Cells. Methods in Biotechnology, second Edition, Humana Press, Totowa, New Jersey, 2006. 465p.
GUMÍ, T., ALBACETE, J.F.D., PAOLUCCI-JEANJEAN, D., BELLEVILLE, M.P., RIOS, G.M. Study of the
influence of the hydrodynamic parameters on the performance of an enzymatic membrane reactor, Journal of
Membrane Science, v.311, p.147–152, 2008.
GUTARRA, M.L.E; MATEO, C.; FREIRE, D.M.G; TORRES, F.A.G.; CASTRO, A.M., GUISAN, J.M. e
PALOMO, J.M., Oriented irreversible immobilization of a glycosylated Candida antarctica B lipase on
heterofunctional organoborane-aldehyde support., Catal. Sci. Technol., v.1, p.260–266, 2011.
GÜVENÇ, A., KAPUCU, N. e MEHMETOGÙLU, U., The production of isoamyl acetate using immobilized
lipases in a solvent-free system‖, Proc. Biochem., v.38, p.379-386, 2002.
HA, S.H., LAN, M.N., LEE, S.H., HWANG, S.M. e KOO, Y.M. Lipase catalyzed biodiesel production from
soybean oil in ionic liquids, Enzyme and Microbial Technology, v. 41, p. 480-483, 2007.
HAKI, G.D., RAKSHIT, S.K., Bioresource Technology, v. 89, p. 17–34, 2003.
HALLING, P.J. Protein Structure and Molecular. Biochimica et Biophysica Acta. Enzymology, v. 1040, n.2, p.
225-228, 2003.
HANEFELD, U., GARDOSSI, L., MAGNER, E., Understanding enzyme immobilisation. Chem. Soc. Rev., v.
38, n.2, p.453–468, 2009.
HASAN, F., SHAH, A. A. e HAMEED, A. Industrial applications of microbial lipases, Enzyme and Microbial
Technology, v. 39, p. 235-251, 2006.
HARA, P.; HANEFELD, U.; KANERVA, L. T. J. Mol. Catal. B: Enzym. 2008, 50, 80-86.

152
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
HARTMANN, M., JUNG, D., Journal of Materials Chemistry, v. 20, p. 844, 2010.
HASSAN, M.M., ATIQULLAH, M., BEG, S.A., CHOWDHURY, M.H.M. Chemical Engineering Journal and
the Biochemical Engineering Journal, v.58, p.275, 1995.
HASSAN, M.M., ATIQULLAH, M., BEG, S.A., CHOWDHURY, M.H.M., Journal of Chemical Technology
and Biotechnology, v. 66, p.41, 1996.
HE F., WANG C.F., JIANG T., HAN B., ZHUO R.X., Biomacromolecules 11 (2010) 3028–3035.
HERBS, D.; PEPER, S.; NIEMEYER, B. Enzyme catalysis in organic solvents: influence of water content,
solvent composition and temperature on Candida rugosa lipase catalyzed transesterification, Journal of
Biotechnology, 162, 398-403p., 2012.
HELLYER, S. A., CHANDLER, I. C. e BOSLEY, J. A., Can the Fatty Acid Selectivity of Plant Lipases be
Predicted from the Composition of the Seed Triglyceride? Biochemica et Biophysica Acta, v.1440, n. 2-3, p.215,
1999.
HERNÁNDEZ K., FERNÁNDEZ-LAFUENTE R., Enzyme Microb. Technol. 48 (2011) 107–122.
HERNÁNDEZ K., GARCIA-GALAN, C.; FERNÁNDEZ-LAFUENTE R., Enzyme Microb. Technol. 49 (2011)
72–78.
HIDE, W., CHAN, L. e LI, W. J. Lipid. Res., v. 33, p. 167-178, 1992.
HOEGH, I., PATKAR, S., HALKIER, T., HANSEN, M. T., Two lipases from Candida antarctica - cloning and
expression in Aspergillus oryzae, Can. J. Botan. v. 73, p. 869-875, 1995.
HOMANN, M.J., VAIL, R., MORGAN, B., SABESAN, V., LEVY, C., DODDS, D.R., ZAKS, A., 2001.
Enzymatic hydrolysis of a prochiral 3-substituted glutarate ester, na intermediate in the synthesis of an
NK1/NK2 dual antagonist. Adv. Synth. Catal. 343 (6–7), 744–749.
HSU, AN-FEI, JONES, K.C., FOGLIA, T.A. e MARMER, W.N. Transesterification activity of lipase
immobilized in a phyllosilicate sol-gel matrix. Biotechnol. Letters, v. 26, p.917-921, 2004.
HUANG, W.; ALEXANDER, G. E.; DALY, E. M.; SHETTY, H. U.; KRASUSKI, J. S.; RAPOPORT, S. I.;
SCHAPIRO, M. B.; Am. J. Psychiatry, 1999, 156, 1879-1999.
HUANG, D.F., HAN, S.Y., HAN, Z.L., LIN, Y., Biodiesel production catalyzed by Rhizomucor miehei lipase-
displaying Pichia pastoris whole cells in an isooctane system. Biochem. Eng. J., v. 63, p.10–14, 2010.
ILLANES, A. Biotecnologia de enzimas, Ediciones Universitarias de Valparaíso, Chile, 1994.
IRVINE, R.F., BROWN, K.D., BERRIDGE, M.J., Biochemistry Journal, v. 222, p. 269,1984.
ISO, M., CHEN, B., EGUCHI, M., KUDO T. e SHRESTHA, S. Production of biodiesel fuel from triglycerides
and alcohol using immobilized lipase, Journal of Molecular Catalysis B: Enzymatic, v.16, p.53–58, 2001.
ITABAIANA Jr., F.K. SUTILI, F.K., LEITE, S.G.F., MIRANDA, L.S.M., LEAL, I.C.R. e SOUZA, R.O.M.A.,
Journal of Molecular Catalysis B-Enzymatic, v.72, p.313, 2011.
ITABAIANA Jr., I. FLORES, M.C., SUTILI, F.K., LEITE, S.G.F., MIRANDA, L.S.M., LEAL, I.C.R. e
SOUZA, R.O.M.A. Journal of Molecular Catalysis B-Enzymatic v.77, p. 53, 2012.
ITABAIANA Jr., I., MIRANDA, L.S.M., de SOUZA, R.O.M.A, Towards a continuous flow environment for
lipase-catalyzed reactions. Journal of Molecular Catalysis B: Enzymatic, v. 85– 86, p.1– 9, 2013.
ITABAIANA Jr., I., FLORES, M.C.; SUTILI, F.K., LEITE, S.G.F., MIRANDA, L.S.M., LEAL, I.C.R. e
SOUZA R.O.M.A., Lipase-Catalyzed Monostearin Synthesis under Continuous Flow Conditions Org. Process
Res. Dev., v.16, p.1098–1101, 2012.
JAEGER, K., RANSAC, S., DIJKSTRA, B.W., FEMS Microbiology Reviews, v. 15, p. 29-63, 1994.
JAEGER, K.E.; REETZ, M.T. 1998. Microbial Lipases form versatile tools for biotechnology. TIBTECH, 16,
396-403.
JAEGER, K.E., DIJKSTRA, B.W., REETZ, M.T., Bacterial biocatalysts: molecular biology, three-dimensional
structures, and biotechnological applications of lipases, Annual Review of Microbiology, v.53, p. 315-351, 1999.
JAEGER, K. E. e EGGERT, T. 2002. Lipases for biotechnology. Curr. Opin. Biotechnol., 13,390-397.

153
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
JAEGER, K-E. e EGGERT, T. Enantioselective biocatalysis optimized by directed evolution. Curr. Opin.
Biotechnol. v.15, p.305–313, 2004.
JAIN, N., KUMAR, A., CHAUHAN, S., CHAUHAN, S.M.S. Tetrahedron, v.61, p. 1015, 2005.
JIN, Z., NTWALI, J., HAN, S.Y., ZHENG, S.P., LIN, Y., Production of flavor esters catalyzed by CALB-
displaying Pichia pastoris whole-cells in a batch reactor. J. Biotechnol., v.159, p.108–114, 2012.
JIN, Z., HAN, S.Y., ZHANG, L., ZHENG, S.P., WANG, Y., LIN, Y., Combined utilization of lipase-displaying
Pichia pastoris whole-cell biocatalysts to improve biodiesel production in co-solvent media, Bioresource
Technology, v.130, p.102–109, 2013.
KADEMI, A.; AÏT-ABDELKADER, N.; FAKHREDDINE, L.; BARATTI, J.C., Journal of Molecular Catalysis
B: enzymatic, v. 10, p. 395–401, 2000.
KALER, A.; MEENA, V.S.; SING, M.; PUJALA, B.; CHAKRABORTI, A.K., BANERJEE, U.C. Lipase-
mediated kinetic resolution of (RS)-1-bromo-3-[4-(2-methoxy-ethyl)-phenoxy]-propan-2-ol to ®-1-bromo-3-(4-
(2-methoxyethyk)ohenoxy) propan-2-yl acetate. Tetrahedron Letters, v. 52, issue 41, 5355-5358p., 2011.
KAPPOR, M. e GUPTA, M.N. Lipase promiscuity and its biochemical applications. V. 47, Issue 4, 2012, p.
555–569 Process Biochemistry.
KATO, K., SEELAN, S., Journal of Bioscience and Bioengineering, v. 109, n. 6, p. 615–617, 2010.
KATCHALSKI-KATZIR E., Trends Biotechnol. 11 (1993) 471–478.
KAUR, J. e SHARMA, R. Directed evolution: an approach to engineer enzymes. Crit. Rev. Biotechnol., v. 26,
p.165–199, 2006.
KAWAKAMI, K., ODA, Y., TAKAHASHI, R. Biotechnology for Biofuels, 2011.
KAWANO, S. HASEGAWA J., YASOHARA Y., Efficient Preparation of (R)-3-Hydroxypentanenitrile with
High Enantiomeric Excess by Enzymatic Reduction with subsequent enhancement of the optical purity by
lipase-catalysed ester hydrolysis. Biosci. Biotechnol. Biochem., v. 76, n. 9, p. 1796-1798, 2012.
KAZLAUSKAS, R.J., WEISSFLOCH, A.N.E., RAPPAPORT, A.T., CUCCIA, L.A., Journal Organic
Chemical, v. 56, p. 2656–2665, 1991.
KEE, C.Y.G., HASSAN, M., RAMACHANDRAN, K.B., Artificial Cells Blood Substitutes and Immobilization
Biotechnology, v.27, p. 393, 1999.
KIRK, O, BORCHERT T.V. e FUGLSANG C.C. Industrial Enzymes applications. Current Op. Biotechnol., v.
13, p.345–351, 2002a.
KIRK O., WURTZ M., Org. Process Res. Dev. 6 (2002b) 446–451.
KISS, V., EGRI, G, BÁLINT, J., LING, I., BARKÓCZ, J., FOGASSY, E., ―Kinetic and chemical resolution of
different 1-phenyl-2-propanol derivatives‖, Tetrahedron: Asymmetry, v. 17, p. 2220–2234, 2006.
KITA, Y., TAKEBE, Y., MURATA, K., NAKA, T., AKAI, S. 1-Ethoxyvinyl acetate as a novel, highly reactive,
and reliable acyl donor for enzymatic resolution of alcohols, Tetrahedron Lett. v.37, p.7369-7372, 1996.
KNEZEVIC, Z., MILOSAVIC, N., BEZBRADICA, D., JAKOVLJEVIC, Z., PRODANOVIC, R.,
Immobilization of lipase from Candida rugosa on Eupergit® C supports by covalent attachment, Biochemical
Engineering Journal, v. 30, p. 269–278, 2006.
KOTIK M., BRICHAC J. e KYSLÍK P. 2005. Novel microbial epoxide hydrolases for biohydrolysis of glycidyl
derivatives. J Biotechnol., v. 120, p. 364–375, 2005.
KOURIST R., BRUNDIEK H., e BORNSCHEUER U. T., Eur. J. Lipid Sci. Technol., 2010, 112, 64–74.
KOSZELEWSKI D., REDZEJ A., OSTASZEWSKI R., J. Mol. Catal. B: Enzym. 47 (2007)51–57.
KOZIKOVSKI, A. P., FAUQ, A. H., POWIS, G. e MELDER, D. C. J. Am. Chem. Soc., v. 112, p. 4528-4531,
1990.
KRAJEWSKA B. Ureases. II. Properties and customizing by enzyme immobilizations: A review.
J. Mol. Catal. B: Enzym., 2009, 59, 22-40. doi: 10.1016/j.molcatb.2009.01.004.

154
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
KRIEGER, N., BHATNAGAR, T., BARATTI, J.C., BARON, A.M., LIMA, V.M.G. e MITCHELL, D. Non-
Aqueous Biocatalysis in Heterogeneous Solvent Systems. Food Technol. Biotechnol., v. 42, n.4, p.279–286,
2004.
LANGONE, M. Síntese de triglicerídeos utilizando lipase, Tese de Doutorado, COPPE/UFRJ, Rio de Janeiro,
1998.
LARIOS, A. Appl. Microbiol.Biotechnol., v.65, n.4, p.373-376, 2004.
LARSEN, M.W., BORNSCHEUER, U. e HULT, K. Expression of Candida antarctica lipase B in Pichia
pastoris and various Escherichia coli, Protein Expression and Purification, v.62, p.90–97, 2008.
LAU, R.M., VAN RANTWIJK, F., SEDDON, K.R., SHELDON, R.A. Organic Letters v. 2, p. 4189, 2000.
LAU, S.Y., UZIR, M.H., KAMARUDDIN, A.H., BHATIA, S., Lipase-catalyzed dynamic kinetic resolution of
racemic ibuprofen ester via hollow fiber membrane reactor: Modeling and simulation, Journal of Membrane
Science, v.357, p.109–121, 2010.
LAU, S.Y., GONAWAN, F.N., BHATIA, S., KAMARUDDIN, A.H. e UZIR, M.H. Conceptual design and
simulation of a plant for the production of high purity (S)-ibuprofen acid using innovative enzymatic membrane
technology, Chemical Engineering Journal, v.166, p. 726–737, 2011.
LAUMEN, K., BREITGOFF, D., SCHNEIDER, M. P. Enzymic preparation of enantiomerically pure secondary
alcohols. Ester synthesis by irreversible acyl transfer using a highly selective ester hydrolase from Pseudomonas
sp.; an attractive alternative to ester hydrolysis, J. Chem. Soc., Chem. Commun. p.1459-1461, 1988a.
LAUMEN, K., SCHNEIDER, M. P. A highly selective ester hydrolase from Pseudomonas sp. for the enzymic
preparation of enantiomerically pure secondary alcohols chiral auxiliaries in organic synthesis, J. Chem. Soc.,
Chem. Commun. p.598-600, 1988b.
LAUMEN, K., GHISALBA, O., Preparative scale Chemo-enzymatic Synthesis of Optically Pure D-myo-
Inositol-1-phosphate, Bioscience Biotechnology and Biochemistry, v. 58 (11), p. 2046-2049, 1994.
LAUMEN, K., GHISALBA, O., ―Chemo-enzymatic synthesis of both enantiomers of myo-inositol 1,3,4,5-
tetrakisphosphate‖, Bioscience Biotechnology and Biochemistry, v. 63, p. 1374-1377, 1999.
LEAL, M.C.C.R., CAMMAROTA, M.C., FREIRE, D.M.G. e SANT’ANNA Jr., G.L. 2002. Hydrolytic enzymes
as coadjuvants in the anaerobic treatment of dairy wastewaters. Brazilian J. Chemical Eng., v. 19, n.2, 2002.
LEE, D.G., PONVEL, K.M., KIM, M., HWANG, S., AHN, I.S., LEE, C.H. Journal of Molecular Catalysis B:
Enzymatic, 2008.
D.G. Lee, K.M. Ponvel, M. Kim, S. Hwang, I.-S. Ahn, C.-H. Lee, J. Mol. Catal. B: Enzym. 57 (2009) 62–66.
LEE, J.H., KIM, S.B., PARK, C., TAE, B., HAN, S.O., KIM, S.W. Applied Biochemistry and Biotechnology,
v.161, p.365, 2010.
LENZI, M. K.; SILVA, F. M.; LIMA, E.L.; PINTO, J. C. J. Appl. Polym. Sci., v.89, p.3021, 2003.
LENZI, M.K., Modelagem da Polimerização Simultânea de Estireno em Suspensão e Emulsão, D.Sc. Thesis,
COPPE/Universidade Federal do Rio de Janeiro, 2002.
LIMA, L.M., Planejamento, síntese e avaliação farmacológica de novos candidatos a fármacos de agentes
antiinflamatórios e antiasmáticos, Tese de Doutorado, IQ/UFRJ, Rio de Janeiro, 2001.
de LIMA L. N.; ARAGON C. C.; MATEO C.; PALOMO J. M.; GIORDANO R. L. C.; TARDIOLI P. W.;
GUISAN J. M., FERNANDEZ-LORENTE G. Immobilization and stabilization of a bimolecular aggregate of the
lipase from Pseudomonas fluorescens by multipoint covalent attachment, Process Biochem., 2013, 48, 118–123.
doi:10.1016/j.procbio.2012.11.008.
LING, L., OZAKI, S., A chemoenzymatic synthesis of D-myo-inositol 1,4,5-triphosphate, Carbohydrate
Research, v. 256, p. 49-58, 1994.
LING, L., OZAKI, S., Enzymatic resolution of the sterically hindered myo-inositol derivative, Bull Chemical
Society Jpn., v. 68, p. 1200-1205, 1995.
LIU Y, WANG F, TAN T. Cyclic resolution of racemic ibuprofen via coupled eficiente lipase and acid-base
catalysis. Chirality, v. 21, n.3, p.349–353, 2009.
LUETZ S., GIVER L., e LALONDE J., Biotechnol. Bioeng., 2008, 101, 647–653.

155
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
LUNDH, H., NORDIN, O., HEDENSTRÖM, E., HÖGBERG, H. E. Enzyme catalysed irreversible
transesterifications with vinyl acetate - are they really irreversible? Tetrahedron: Asymmetry, v. 6, p.2237-2244,
1995.
MACHADO, A.C.O., da SILVA, A.A.T.T., BORGES, C.P., SIMAS, A.B.C., FREIRE, D.M.G., Kinetic
resolution of (R,S)-1,2-isopropylidene glycerol (solketal) ester derivatives by lipases, Journal of Molecular
Catalysis B: Enzymatic, v.69, p.42–46, 2011.
MACHADO, A.C.O.; da SILVA, A.A.T.; BORGES, C.P.; SIMAS, A.B.C.; FREIRE, D.M.G., Kinetic resolution
of (R,S)-1,2-isopropylidene glycerol (solketal) ester derivatives by lipases. Journal of Molecular Catalysis B:
Enzymatic v.69, p. 42–46, 2011.
MAGNUSSON, A.; TAKWA, M.; HAMBERG, M.A. e HULT, K., Angew. Chem. Int. Ed. 44, p.4582–4585,
2005a.
MAGNUSSON, A.O.; ROTTICCI-MULDER, J.C; SANTAGOSTINO, A. e HULT, K., Chem.Bio.Chem. v.6,
p.1051–1056, 2005b.
MALCATA, F.X. H.R. Reyes, H.S. Garcia, C.G. Hill Jr., C.H. Amundson, J. Am. Oil Chem. Soc. 67 (1990)
890–910.
MANOEL, E.A. Resolução de derivados racêmicos do mio-inositol catalisada por lipases. Dissertação de
mestrado. Escola de Química. Rio de Janeiro, RJ. 2011, 117p.
MANOEL, E.A.; PAES, K.C.; CUNHA, A.G.; COELHO, M.A.Z.; FREIRE, D.M.G. e SIMAS, A.B.C., On the
kinetic resolution of sterically hindered myo-inositol derivatives in organic media by lipase. Tetrahedron:
Asymmetry, v. 23, p. 47, 2012a.
MANOEL, E.A.; PAES, K.C.; CUNHA, A.G.; SIMAS, A.B.C.; COELHO, M.A.Z. e FREIRE, D.M.G., Kinetic
Resolution of 1,3,6-Tri‑O‑benzyl-myo-Inositol by Novozym 435: Optimization and Enzyme Reuse, Org. Process
Res. Dev., v. 16, p.1378-1384. 2012b.
MANOEL, E.A., PAIS, K.C., FLORES, M.C., MIRANDA, L.S.M., COELHO, M.A.Z., SIMAS, A.B.C.,
FREIRE, D.M.G. e SOUZA, R.O.M.A. Kinetic resolution of a precursor for myo-inositol phosphates under
continuous flow conditions, Journal of Molecular Catalysis B: Enzymatic, v. 87, p.139–143, 2013.
MARCH, J. 1992. Advanced Organic Chemistry: reactions, mechanisms and structure. 4. ed. John Wiley &
Sons: New York
MARCHETTI, J.M., MIGUEL, V.U. e ERRAZU, A.F., Possible methods for biodiesel production, Renewable
and Sustainable Energy Reviews, v. 11, p. 1300–1311, 2007.
MARGOLIN, A. L. Novel crystalline catalysts, Trends Biotechnol. v.14, p. 223-230, 1996.
MARGOLIN, A. L., Enzym. Microbial Technol., v.15, p.266-280, 1993.
MARTINS, F.F., FERREIRA, T.F., AZEVEDO, D., COELHO, M.A.Z., Evaluation of crude oil degradation by
Yarrowia lipolytica. Chemical Engineering Transactions, v. 27, p. 223-228, 2012.
MARTON, Z.; LÉONARD-NEVERS, V.; SYRÉN, P.; BAUER, C.; LAMARE, S.; HULT, K.K., J. Mol. Catal.
B: Enzym. v. 65, p. 11–17, 2010.
MATEO, C.; ABIAN, O.; BERNEDO, M.; CUENCA, E.; FUENTES, M.; FERNÁNDEZ- LORENTE, G.;
PALOMO, J. M.; GRAZU, V.; PESSELA, B.C.C.; GIACOMINI, C.; IRAZOQUI, G.; VILLARINO, A.;
OVSEJEVI, K.; BATISTA-VIERA, F.; FERNÁNDEZ -LAFUENTE, R. e GUISAN, J. M., Improvement of
enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol., v.36, p. 447–
454. 2005
MATEO, C., PALOMO, J.M., FUENTES, M., BETANCOUR, L., GRAZU, V., LÓPEZ-GALEGO, F.,
PESSELA, B.C.C., HIDALGO, A., FERNANDEZ-LORENTE, G., FERNANDEZ-LAFUENTE, R., GUISÁN,
J.M., Glyoxyl Agarose: a fully inert and hydrophilic support for immobilization and high stabilization of
proteins, Enzyme and Microbial Technology, v. 39, p. 274-280, 2006.
MATEO, C., PALOMO, J.M., FERNANDEZ-LORENTE, G., GUISÁN, J.M., FERNANDEZLAFUENTE, R,
Enzyme and Microbial Technology, v. 40, p. 1451–1463, 2007.
MATSUSHIMA, A., KODERA, Y., HIROTO, M., Bioconjugates of proteins and polyethylene glycol: potent
tools in biotechnological processes, Journal of Molecular Catalysis B: enzymatic, v. 2, p. 1-17, 1996.

156
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
MAY, O. Green Chemistry with Biocatalysis for Production of Pharmaceuticals 305-321p. In: TAO, J.; LIN,
G.Q.; LIESE A. Biocatalysis for the pharmaceutical industry: Discovery, Development, and Manufacturing.
John Wiley & Sons (Asia) Pte Ltd. Singapura.2009.324p.
MCCLEAN, K.H. Biocatalyst Identification and Scale-up: Molecular Biology for Chemists. In: John Whittall &
Peter Sutton. Practical Methods for Biocatalysis and Biotransformations. John Wiley and Sons, Ltd. 2010. Reino
Unido. 402p.
MENDES A. A., OLIVEIRA P. C., de CASTRO H. F.; Properties and biotechnological applications of porcine
pancreatic lipase Journal of Molecular Catalysis B: Enzymatic 78 (2012) 119– 134
MENSAH, P., GAINER, J.L., CARTA, G. Biotechnology and Bioengineering, v. 60, p.445, 1998.
MCLAURIN, J., GOLOMB, R., JUREWICZ, A., ANTEL, J. P., FRASER, P. E., Journal of Biological
Chemistry, v. 275, p.18495, 2000.
MILÉTIC, N., VUKOVIC, Z., NASTASOVIC, A., LOOS, K., Macroporous poly(glycidyl methacrylate-co-
ethylene glycol dimethacrylate) resins—Versatile immobilization supports for biocatalysts, Journal of Molecular
Catalysis B: Enzymatic, v. 56, p. 196-201, 2009.
MILÉTIC, N., ABETZ, V., EBERT, K., LOOS, K., Macromolecular Rapid communications, v. 31, no. 1, p. 71–
74, 2010.
MILETÍC N., NASTASOVIC A. LOOS K. Immobilization of biocatalysts for enzymatic polymerizations:
Possibilities, advantages, applications, Bioresour. Technol., 2012, 115, 126–135,
doi:10.1016/j.biortech.2011.11.054.
MILED, N., CANAAN, S., DUPUIS, L., ROUSSEL, A., RIVIÈRE, M., CARRIÈRE, F., DE CARO, A.,
CAMBILLAU, C. e VERGER, R. Digestive lipases: From three-imensional structure to physiology. Review
article. Biochimie., v. 82, p. 973-986, 2000.
MILLS S.J. e POTTER B.V.L. Synthesis of D- and L-myo-Inositol 1,4,6- Trisphosphate, Regioisomers of a
Ubiquitous Second Messenger. J. Org. Chem. v. 61, n. 25, p.8980-8987, 1996.
MILLS, S. J., LIU, C. e POTTER, B. V. L. Synthesis of D- and L-myo-inositol 2,4,5-triphosphate and
triphosphorothioate: structural analogues of D-myo-inositol 1,4,5- triphosphate. Carbohydr. Res. v. 337, p.1795-
1801, 2002.
MILNER, S. E. e MAGUIRE, A. R. Arkivoc, 2012, 321–382.
MING-MING ZHENG, YONG LU, FENG-HONG HUANG, LIAN WANG, PING-MEI GUO, YU-QI FENG, e
QIAN-CHUN DENG Lipase Immobilization on Hyper-Cross-Linked Polymer-Coated Silica for Biocatalytic
Synthesis of Phytosterol Esters with Controllable Fatty Acid Composition dx.doi.org/10.1021/jf3042962 J.
Agric. Food Chem. 2013, 61, 231−237
MONTGOMERY, D.C. e RUNGER, G.C. Estatística Aplicada e probabilidade para engenheiros.Segunda
edição. LTC editora. S.A. 2003, 463p.
MONTGOMERY, D.C. Design and Analysis of Experiments. 5aedição. John Wiley & Sons, I.N.C. New York
Chechester Weinheim Brisbane Toronto Singapura. 2001, 680p.
MUKHERJEE, K. D. Plant lipases and their application in lipid biotransformations. Prog. Lipid Res., v. 33, n.1-
2, p.165-174, 1994.
MUKHERJEE, M. Human digestive and metabolic lipases—a brief review J. Mol.Catalysis B: Enzymatic., v.22,
p. 369–376. 2003.
NAIK, S., BASU, A., SAIKIA, R., MADAN, B., PAUL, P., CHATERJEE, R., BRASK, J., SVENDSEN, A.,
Lipases for use in industrial biocatalysis: Specificity of selected structural groups of lipases, Journal of
Molecular Catalysis B: Enzymatic, v. 65, p.18–23, 2010.
NAGY E., Basic equations of the mass transport through a membrane layer, Elsevier, Amsterdam ; Boston, 1st
ed., 2012, ch. 9, pp. 213-247
NAKAMURA, K., TAKEBE, Y., KITAYAMA, T. e OHNO. A. Effect of solvente structure on
enantioselectivity of lipase-catalyzed transesterfication. Tetrahedron, 32, 4941-4944, 1991.
NASRATUN, M.; SAID, H. A.; NORAZIAH, A.; ALLA, A. N. Immobilization of lipase from Candida rugosa
on chitosan beads for transesterification reaction. Am. J. Appl. Sci. 2009, 6 (9), 1653−1657.

157
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
NAUSHADA, M., ALOTHMANA, Z.A., KHANB, A.B., ALI, M., Effect of ionic liquid on activity, stability,
and structure of enzymes: A review, International Journal of Biological Macromolecules, v. 51, p.555– 560,
2012.
NELSON J. M. e GRIFFIN, E. G. J. Am. Chem. Soc., 1916, 38, 1109–1115.
NESTL, B.M., NEBEL, B.A., HAUER, B., Recent progress in industrial biocatalysis. Curr. Opin. Chem. Biol. v.
15, n.2, p. 187–193, 2011.
NIE, K., XIE, F., WANG, F., TAN, T. Journal of Molecular Catalysis B-Enzymatic, v.43, p. 142, 2006.
NINI, L., SARDA, L., LOUIS-CLAUDE, C., BOITARD, E., JEAN-PAUL, DD. E CHAHINIAN. H., Lipase-
catalysed hydrolysis of short-chain substrates in solution and in emulsion: a Kinetic study. Biochim. Biophys.
Acta., v.534, p.34-44, 2001.
NISHIO, T., KAMIMURA, M., MURATA, M., TERAO, Y., ACHIWA, K. 1989. Production of optically active
esters and alcohols from racemic alcohols by lipase-catalyzed stereoselective transesterification in nonaqueous
reaction system, J. Biochem. v.105, p.510-512.
NISHIZUKA, Y. Nature , v. 334, p. 356, 1988.
OKAZAKI, S., KAMIYA, N., GOTO, M. e NAKASHIO, F., Purification and characterization of a novel
lipolytic enzyme from Aspergillus oryzae, J. Ferment. Bioeng., v.78, n.6, p.413-419, 1997.
OLIVEIRA, D., FEIHRMANN, A. C., DARIVA, C., CUNHA, A. G., BEVILAQUA, J. V., DESTAIN, J.,
OLIVEIRA, J. V., FREIRE, D. M. G., Influence of compressed fluids treatment on the activity of Yarrowia
lipolytica lipase, Journal of Molecular Catalysis B: Enzymatic, v. 39, p. 117-123, 2006.
OLIVEIRA, D.; FEIHRMANN, A.C.; DARIVA, C.; CUNHA, A.G.; BEVILAQUA, J.V.; DESTAIN, J.;
OLIVEIRA, J.V.; FREIRE, D.M.G. J. Mol. Cat. B: enzym., v. 39, p.117-123. 2006.
OLIVEIRA, M.M.; EINICKER-LAMAS, M. Anais da Academia Brasileira de Ciências, Brasil, v.72, p.413-419,
2000.
ONG, A.L.; KAMARUDDIN, A.H., BHATIA, S.W.; LONG W.S.; LIM S.T.; KUMARI R. Enzyme Microb
Technol., v.39, p.924–929. 2006.
OSÓRIO, N.M., FERREIRA-DIAS, S., GUSMÃO, J.H. e FONSECA, M.M.R., Response surface modelling of
the production of v-3 polyunsaturated fatty acidsenriched fats by a commercial immobilized lipase, Journal of
Molecular Catalysis B: Enzymatic, v.11, p.677–686, 2001.
OSTREICHER, E.G., PINTO, G.F. e ANTUNES, O.A.C. 2001. Cadernos didáticos de pós graduação. Catálise
Assimétrica. Editor Angelo C. Pinto. Setor Científico Cultural Intituto de Química-UFRJ. v. 2, 1-114p.
OTERO C., BALLESTERO, A., GUISAN, J. M., Immobilization/stabilization of lipase from Candida rugosa,
Applied Biochemistry and Biotechnology, v. 19, p. 163-175, 1991.
OTTEN, L.G., HOLLMANN, F. e ARENDS, I. W.C.E.; Enzyme engineering for enantioselectivity: from trial-
and-error to rational design? Trends in Biotechnology, v.28, n.1, p. 46-54, 2009.
OZAKI, S.; WATANABE, Y.; OGASAWARA, T.; KONDO, Y.; SHIOTANI, N.; NISHII, H.;
MATSUKI, T. Tetrahedron Letters, v. 27, p. 3157-3160, 1986.
OZAKI, S.; KONDO, Y.; NAKAHIRA, H.; YAMAOKA, S.; WATANABE, Y. Tetrahedron Letters, v. 28, p.
4691-4694, 1987.
OZYILMAZ, G., The effect of spacer arm on hydrolytic and synthetic activity of Candida rugosa lipase
immobilized on silica gel, Journal of Molecular Catalysis B: Enzymatic, v. 56, p. 231–236, 2009.
PAIVA, A. L., BALCÃO, V. M., MALCATA, F. X., Kinetics and mechanics of reactions catalyzed by
immobilized lipases, Enzyme and Microbial Technology, v. 27, p. 187-204, 2000.
PALOCCI, C., FIORILLO, F. e MONACHE, D. Lipolytic isoenzymes from Euphorbia latex. Plant Sci., v. 165,
p. 577– 582, 2003.
PALOMO, J. M.; FERNÁNDEZ- LORENTE, G.; MATEO, C.; FERNÁNDEZ -LAFUENTE, R. e GUISAN, J.
M. Tetrahedron: Asymmetry, v. 13, n.21, p.2375–2381, 2002a.
PALOMO, J. M., MUNOZ, G., FERNANDEZ-LORENTE, G., MATEO, C., FERNANDEZ-LAFUENTE, R.,
GUISAN, J.M., Interfacial adsorption of lipases on very hydrophobic support (octadecyl-Sepabeads):

158
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
immobilization, hyper activation and stabilization of the open form of lipases, Journal of Molecular Catalysis B:
enzymatic, v. 19, p. 279-286, 2002b.
PALOMO, J. M.; FUENTES M.; FERNANDEZ-LORENTE, G.; MATEO, C, GUISAN, J.M.; FERNANDEZ-
LAFUENTE, R. General trend of lipase to auto-assemble giving bi-molecular aggregates greatly modifies the
enzyme functionality. Biomacromolecules 2003;4:1–6.
PALOMO, J. M., SEGURA, R. L., FERNANDEZ-LORENTE, G., PERNA, M., RUA, M. L., GUISAN, J.M,
FERNANDEZ-LAFUENTE, R., Purification, immobilization and stabilization of a lipase from Bacillus
thermocatenulatus by adsorption on hydrofobic supports, Biotechnology Program, v. 20, p. 630-635, 2004.
PALOMO, J. M., SEGURA, R. L., MATEO, C., TERRENI, M., FERNANDEZ-LAFUENTE, R., GUISAN,
J.M., Synthesis of enantiomerically pure glycidol via a fully enantioselective lipase-catalyzed resolution,
Tetrahedron: Asymmetry, v. 16, p. 869–874, 2005a.
PALOMO, J. M.; ORTIZ C.; FERNANDEZ-LORENTE, G.; FUENTES M.; GUISAN, J.M.; FERNANDEZ-
LAFUENTE, R. Lipase–lipase interactions as a new tool to immobilize and modulate the lipase properties.
Enzyme Microb. Technol., 2005b, 36, 447–454. Doi: 10.1016/j.enzmictec.2004.09.013
PALOMO, J. M., SEGURA, R. L., FERNANDEZ-LORENTE, G., FERNANDEZ-LAFUENTE, R., GUISAN,
J.M., Glutaraldehyde modification of lipases adsorbed on aminated supports: A simple way to improve their
behaviour as enantioselective biocatalyst, Enzyme and Microbial Technology, v. 40, n. 4, p. 704-707, 2007.
PANDEY, A., BENJAMIN, S., SOCCOL, C.R., NIGAM, P., KRIEGER, N., SOCCOL, V. T., The realm of
microbial lipases in biotechnology, Applied Biotechnology Biochemistry, v. 29, p. 119-131, 1999.
PANDEY, A.; BENJAMIN, S.; SOCCOL, C.R. Biotechnol. Appl. Biochem., v.29, p.119-131, 1999.
PANZAVOLTA, F., SORO, S., D’AMATO, R., PALOCCI, C., CERNIA, E., RUSSO, M. V., Acetylenic
polymers as new immobilization matrices for lipolytic enzymes, Journal of Molecular Catalysis B: Enzymatic, v.
32, p. 67–76, 2005.
PARAVIDINO, M. e HANEFELD, U. Enzymatic acylation: assessing the greenness of diferente acyl donors,
Green Chem., v.13, p.2651-2657, 2011.
PARK, H.J., JOO, J.C., PARK, K., KIMD, Y.H., YOO, Y.J., Prediction of the solvent affecting site and the
computational design of stable Candida antarctica lipase B in a hydrophilic organic solvent, Journal of
Biotechnology, v. 163, p.346–352, 2013.
PARKER, W.L.; HANSON, R.L.; GOLDBERG, S.L.; TULLY, T.P. e GOSWAMI, A. Preparation of (S)-1-
Cyclopropyl-2-methoxyethanamine by a Chemoenzymatic route using leucine dehydrogenase. Org. Process Res.
Dev. v. 16, p. 464−469, 2012.
PAULA A.V., URIOSTE D., SANTOS J.C., de CASTRO H.F., J. Chem. Technol. Biotechnol. 82 (2007) 281–
288.
PENCREACH, G. e BARATTI, J. C. Activity of Pseudomonas cepacia lipase in organic media is greatly
enhanced after immobilization on a polypropylene support. Applied Microbiology and Biotechnology, v.47, p.
630-635, 1997.
PELLISSIER, H., Dynamic kinetic resolution, Tetrahedron, v.59, p.8291-8327, 2003.
PETERSEN, M.T..N.; FOJAN, P.; PETERSEN, S.B., How do lipases and esterases work: the electrostatic
contribution. Journal of Biotechnology v. 85, p. 115–147, 2001.
PETKAR, M., LALI, A., CAIMI, P., DAMINATI, M., Immobilization of lipases for non-aqueous synthesis,
Journal of Molecular Catalysis B: Enzymatic, v. 39, p. 83–90, 2006.
PINTO, J.C.; LOPEZ, G.E.; NIEMEYER, M.A.L.; SILVA, F.M.; NELE, M.; MELO, P.A.; Process for the
Synthesis of Poly(Vinyl Alcohol) and/or Poly(Vinyl Acetate) With Spherical Morphology ans Shell and Nucleus
Structure and Its Use in Vascular Embolization, Pedido de patente internacional PCT/WO2006/050591 A2,
2006.
POTTER, B.V.L e LAMPE, D. Angew. Chem. Int. Ed. Engl. v.34, p.1993-1972, 1995.
POUDEROYEN, G.V., THORSTEN, E., JAEGER, K.E. e DIJKSTRA, B.W. The Crystal Structure of Bacillus
subtilis Lipase: A Minimal a/b Hydrolase Fold Enzyme‖, J. Mol. Biol., v. 309, p. 215-226, 2001.
PRIEGO, J., ORTÍZ-NAVA, C.; CARRILLO-MORALES, M.; LÓPEZ-MUNGUÍA, A.; ESCALANTE, J.;
CASTILLO, E. Tetrahedron v.65, p.536-539, 2009.

159
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
PUSCH, J.; VAN HERK, A. M. Macromolecules, v.38, p.909-6914, 2005.
QUARTEY, E.G.K., HUSTAD, J.A., FABER, K. e ANTHONSEN, T., Selectivity enhancement of PPL-
catalysed resolution by enzyme-fractionation and mediumengineering: Synthesis of both enantiomers of
tetrahydropyran-2-methanol. Enzyme.Microb. Technol., v.19, p. 361–366, 1996.
QUYEN, D.T.; SCHMIDT-DANNERT, C.; SCHMID, R.D. Protein Expr. Purification., v. 28, p. 102-110, 2003.
RANSAC, S., CARRIÉRE, F., ROGALSKA, E., VERGER, R., MARGUET, F., BUONO, G., MELO, E. P.,
CABRAL, J. M. S., EGLOFF, M. P. E., van TILBEURGH, H., CAMBILLAU, C., The kinetics specificities and
structural features of lipases, In: Molecular Dynamics of Biomembranes (de Kamp, J. A. F.; Ed.), Berlin:
Springer. v. 96, p. 254-304. 1996.
RAO, N.N., LÜTZ, S., WÜRGES, K., MINÖR, D., Organic Process Research & Development, v.13, p. 607,
2009.
RATHI, P., SAPNA, B., SEXENA, R., GUPTA, R. Biotechnology Letters, v. 22, p. 495– 498, 2000.
REETZ, M. T.; Current Opin. Chem. Biol., v. 6, p. 145-150, 2002.
REETZ, M.T. et al. Learning from directed evolution: further lessons from theoretical investigations into
cooperative mutations in lipase enantioselectivity. ChemBioChem., v. 8, p.106–112, 2007.
RIBEIRO B.D., de CASTRO A.M., COELHO M.A.Z., FREIRE D.M.G. Production and use of lipases in
bioenergy: a review from the feedstocks to biodiesel production. Enzyme Res 2011, doi:10.4061/2011/615803
ROBERTS S.M., TURNER N.J., WILLETTS A.J., TURNER M.K., Introduction to biocatalysis using enzymes
and microorganisms. 1st ed. Cambridge: Cambridge University Press; 1995.
RODRIGUES, M. I. e IEMMA, A. F. Planejamento de Experimentos e Otimização de Processos. Editora Casa
do Pão, Campinas. 2005, 325p.
RODRIGUEZ, R.C., FERNANDEZ-LAFUENTE, R., Lipase from Rhizomucor miehei as an industrial
biocatalyst in chemical process., Journal of Molecular Catalysis B: Enzymatic, v. 64, p.1–22, 2010.
RODRIGUES R.C., BERENGUER-MURCIA Á., FERNÁNDEZ-LAFUENTE R., Adv. Synth.Catal. 353 (2011)
2885–2904.
ROMERO, M.D., CALVO, L., ALBA, C., HABULIN, M., PRIMOZIC, M. e KENS, Z. Enzymatic synthesis
115 of isoamyl acetate with immobilized Candida antarctica lipase in supercritical carbon dioxide.
J.Supercritical Fluids., v. 33, p. 77-84, 2005.
ROTTICCI-MULDER, J.C. GUSTAVSSON, M. HOLMQUIST, M. HULT, K. MARTINELLE, M. Expression
in Pichia pastoris of Candida antarctica lipase B and lipase B fused to a cellulose-binding domain, Protein
Expr. Purif., v.1, p. 386–392, 2001.
ROUHI, A. M., Chem. Eng. News, v. 81, n.18, p.45. 2003.
ROUSSEL, A., MILED, N., BERTI-DUPUIS, L., RIVIERE, M., SPINELLI, S., BERNA, P., GRUBER, V.,
VERGER, R., CAMBILLAU, C., Crystal structure of the open form of dog gastric lipase in complex with a
phosphonate inhibitor, Journal of Biological Chemistry, v. 277, p. 2266–2274, 2002.
RÚA, M.L., DIAZMAURINO, T., FERNANDEZ, V.M., OTERO, C. e BALLESTEROS, A.
Purification na characterization of 2 distinct lipases from Candida cylindracea. Biochimica and Biophysica Acta,
v. 1156, p. 181-189, 1993.
RUFINO A.R., BIAGGIO F.C., SANTOS J.C., de CASTRO H.F., Int. J. Biol. Macromol. 47 (2010) 5–9.
RUIZ, C., BLANCO, A., PASTOR, J., DIAZ, P. FEMS Microbiology Letters, v. 217, p. 263-
267, 2002.
SABBANI, S., HEDENSTRÖM, H., NORDIN, O., The enantioselectivity of Candida rugosa lipase is
influenced by the particle size of the immobilising support material Accurel, Journal of Molecular Catalysis B:
Enzymatic, v. 42, p. 1-9, 2006.
SABUQUILLO, P., REINA, J., FERNANDEZ-LORENTE, G., GUISAN, J. M., FERNANDEZ-LAFUENTE,
R. Interfacial affinity chromatography’ of lipases: separation of different fractions by selective adsorption on
supports actived with hydrophobic groups, Biochimica et Biophysica Acta, v. 1388, p. 337-348, 1998.

160
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
SAKURAI, T., MARGOLIN, A. L., RUSSEL, A. J. e KLIBANOV, A. M. Control of enzyme enatioselectivity
by the reaction medium. J. Am. Chem. Soc., 110, 7236-7237, 1988.
SARDA, L., DESNUELE, P., Action de la lipase pancréatique sur les esters en émulsion, Biochimica et
Biophysica Acta, v. 58, p. 513-521, 1958.
SARDONINI, C.A., and DIBIASIO, D. (1992) An investigation of the diffusion-limited growth of animal cells
around single hollow fibers. Biotechnol. Bioeng. 40, 1233_1242.
SARIC, M.,VAN DER WIELEN, L.A.M.; STRAATHOF, A.J.J., Theoretical performance of countercurrent
reactors for production of enantiopure compounds, Chemical Engineering Science, v. 66, n.3, p. 510–518, 2011.
SAXENA, R. K., GHOSH, P. K., GUPTA, R., Microbial lipases: Potential biocatalysts for the future industry,
Current Science, v. 77, p. 101-115, 1999.
SCHRAG, J. D.; LI, Y.; WU, S.; CYGLER, M. Ser-His-Glul triad form the catalytic site of the lipase from the
catalytic site of the lipase from Geotrichum candidum. Nature, v.351, p.761-764, 1991.
SCHNELL, B., FABER, K., KROUTIL, W., Enzymatic racemization and its application to synthetic
biotransformations, Adv. Synth. Catal. v.345, p.653-666, 2003.
SCHUDOK, M., KRETZSCHMAR, G. 1997. Enzyme catalyzed resolution of alcohols using ethoxyvinyl
acetate, Tetrahedron Lett. v.38, p.387-388.
SCHULZE, B. e WUBBOLTS. M.G., Biocatalysis for industrial production of fine chemicals, Current Opinion
in Biotechnology, v.10, n.6, p.609-615, 1999.
SECUNDO, F.; CARREA, G.; TARABIONO, C.; GATTI-LAFRANCONI, P.; BROCCA, S.; LOTTI, M.;
JAEGER, K. E.; PULS, M.; EGGERT, T. The lid is a structural and funcional determinant of lipase activity and
selectivity. Journal of Molecular Catalysis B: Enzymatic, Amsterdam, v. 39, p. 166-170, 2006.
SHAH, R.D. e MARYANOFF, C.A. Reversed phase HPLC in: Swadesh J. K..(eds). HPLC Practical and
Industrial Applications. 2nd
edition.142-189p.ch4.2001.
SHARMA, R., CHISTI, Y. e BANERJEE, U.C., Research review paper production, purification,
characterization, and applications of lipases, Biotechnology Advances, v.19, p.627–662, 2001.
SHELDON, R.A. Chirotechnology. Marcel Dekker Inc., New York. 1993.
SHELDON, R.A. Characteristic features and biotechnological applications of cross-linked enzyme aggregates
(CLEAs), Appl. Microbiol. Biotechnol., 2011, 92, 467–477. DOI:10.1007/s00253-011-3554-2.
SHEN, L., WANG, F., MUN, H., SUH, M. e JEONG, J. Solvent-dependent reactivity in porcine pancreatic
lipase (PPL)-catalyzed hydrolysis. Tetrahedron: Asymmetry, v. 19, p. 1647–1653, 2008.
SHIBATANI, T.; OMORI, K.; AKATSUKA, H.; KAWAI, E. e MATSUMAE, H., J. Mol. Catal. B. Enzym.
v.10, p.141–149, 2000.
SHIMADA, Y., WATANABE, H., SUGIHARA, A. e TOMINAGA, Y., Enzymatic alcoholysis for biodiesel
fuel production and application of the reaction to oil processing. J.Mol. Catal. B: Enzym. v. 17, p. 133–142,
2002.
SHIMOHAMA, S.; TANINO, H.; SUMIDA, Y.; TSUDA, J.; FUJIMOTO, S.; Neuroscience Lett., v.245, p.159,
1998.
SIDDIQUI, K.S. e CAVICCHIOLI, R. Extremophiles, v.9, p.471-476, 2005.
SIH, C.J., GIRDAUKAS, G., FUJIMOTO, Y. e CHEN, C.S. Quantitative analyses of biochemical Kinetic
resolutions of enantiomers. J. Am. Chem. Soc. v. 104, p. 7294, 1982.
SIH, C.J. e WU, S.H. Resolution of enatiomers via biocatalysis. Topics Sterochem., v.19, p. 63-125, 1989a.
SIH, C.J. e CHEN, C.S. Aspects and optimization of enantioselective biocatalysis in organic solvents: the use of
lipases. Angew. Chem. Int. Ed. Engl., v. 28, p. 695-707, 1989b.
SILVA, M.A.M., MEDEIROS, V.C., LANGONE, PEREIRA, M.A. e FREIRE, D.M.G. Synthesis of
Monocaprin Catalyzed by Lipase. Applied Biochemistry and Biotechnology. v.105, p.757 – 767, 2003.
SILVA A.A.T., Dissertação de MSc., NPPN/Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brasil,
2009.

161
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
SILVA, J. do N.; GUTARRA, M.L.E.; FREIRE, D.M.G.; CAMAROTA, M.C. Application of Home-Made
Enzyme and Biosurfactant in the Anaerobic Treatment of Effluent with High Fat Content. J. Bioprocess Biotech
3, 139, 2013.
SIMAS, A.B.C.; PAIS, K.C.; DA SILVA, A.A.T. J. Org. Chem., v.68, p.5426-5428, 2003.
SIMAS, A.B.C.; DA SILVA, A.A.T.; DOS SANTOS FILHO, T.J., BARROSO, P.T.W. Tetrahedron Letters,
v.50, p.2744-2746, 2009.
SIMAS, A.B.C.; DA SILVA, A.A.T.; CUNHA, A.G., ASSUMPÇÃO, R.S., HOELZ, L.V.B., NEVES, B.C.,
GALVÃO, T.C., ALMEIDA, R.V., ALBUQUERQUE, M.G., FREIRE, D.M.G. e ALENCASTRO, R.B.,
Kinetic resolution of (±)-1,2-O-isopropylidene-3,6-di-O-benzyl-myo-inositol by lipases: An experimental and
theoretical study on the reaction of a key precursor of chiral inositols, Journal of Molecular Catalysis B:
Enzymatic., v. 70, p. 32–40, 2011.
SIMONS.; OßWALD S.; ROOS J. e GRÖGER H., Z. Naturforsch. B, 2012, 67, 1123–1126.
SIMONS, J.W.F.A.; GÖTZ, F.; EGMOND, M.R.; VERHEIJ, H.M. Chem. Phys of Lipids, v. 93, p. 27-37, 1998.
SNELLMAN E.A., SULLIVAN E.R., COLWELL R.R., Purification and properties of the extracellular lipase,
LipA, of Acinetobacter sp. RAG-1. FEBS J; v. 269, p.5771–9. 2002.
SKORIDOU, V., CHRYSINA, E., STAMATIS, H., OIKONOMAKOS, N. e KOLISIS, F. Kinetic and
modelling studies on the lipase catalysed enantioselective esterification of (±)- perillyl alcohol. Journal of
Molecular Catalysis B: Enzymatic, v. 29, p. 9-12, 2004.
SKJOT, M.; DE MARIA, L.; CHATTERJEE, R.; SVENDSEN A.; PATKAR, S.A. et al. Undestanding the
plasticity of the α/β hydrolase fold: lid swapping on the Candida Antarctica lipase B results in Chimeras with
interesting biocatalytic Properties. ChemBioChem, 10:520-527, 2009.
SOARES, M.F. FORESTI M.L., FERREIRA M.L; Colloid Surface A 294 (2007) 147–155.
SOLÁ, R., RODRÍGUEZ-MARTÍNEZ, J., GRIEBENOW, K. Modulation of protein biophysical properties by
chemical glycosylation: biochemical insights and biomedical implications, Cell. Mol. Life Sci., v.64, p. 2133–
2152, 2007.
SOLANO, D.M.; HOYOS, P.; HERNAIZ, M.J.; ALCANTARA, A.R. e SANCHEZ-MONTERO, J.M.,
Industrial biotransformations in the synthesis of building blocks leading to enantiopure drugs Bioresource
Technology v. 115, p.196–207, 2012.
SOUSA, J. S.; CAVALCANTI E.D.C. ; ARANDA D.A.G. ; FREIRE D.M.G. . Application of Vegetable Lipase
from Physic nut (Jatropha curcas Lin.) in a New Hybrid Process (Enzymatic/Quemical) of Hidresterification ion
Biofuel Production. In: 9th International Symposium on Biocatalysis, 2009, Berne, CH. Book of Abstracts
BIOTRANS 2009, 2009. p. 73-73.
SOUSA, J. S., CAVALCANTI E.D.C., ARANDA D.A.G., FREIRE D.M.G., Application of lipase from the
physic nut (Jatropha curcas L.) to a new hybrid (enzyme/chemical) hydroesterification process for biodiesel
production. Journal of Molecular Catalysis B: Enzymatic, v.65, p.133–137, 2010.
SOUMANOU M.M.; PÉRIGNON M.; VILLENEUVE P.; Eur. Lipid J. Technol. Sci. 115 (2013) 270–285.
SOUZA R. L., RESENDE W. C., BARAO C. E., ZANIN G. M., de CASTRO H. F., SANTOS O. A. A.,
FRICKS A. T., FIGUEIREDO R. T., CLEIDE A. S. L., Influence of the use of Aliquat 336 in the
immobilization procedure in sol–gel of lipase from Bacillus sp. ITP-001, J. Mol. Catal. B: Enzym., 2012, 84,
152– 159. doi: 10.1016/j.molcatb.2012.05.013
SQUIRE, T.L.; LOWE, M.E.; BAUER, V.W. e ANDREWS, M.T. Pancreatic triacylglycerol lipase in a
hibernating mamal. II Cold adapted function and differential expression. Physiological Genomics., v.16, p. 131-
140, 2003.
STAEBLER A, CRUZ A, VAN DER GOOT W, PINHEIRO HM, CABRAL JMS, FERNANDES P.
Optimization of androstenedione production in an organic–aqueous two-liquid phase system. J. Mol. Catal. B.
Enzym., v. 29, p.19–23, 2004.
STAUBMANN, R..; NCUBE, I; GUBITZ, G.M.; STEINER, W.; READ, J.S. Esterase and lipase activity in
Jatropha curcas L. Seeds. Journal of Biotechnology, 75 (1999), 117-126.
STECHER, H., FABER, K., Biocatalytic deracemization techniques: Dynamic Resolutions and Stereoinversions,
Synthesis v.1, p. 1-16, 1997.

162
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
STEINER, J.M. e WILLIAMS, D.A. Purification of classical pancreatic lipase from dog pancreas. Biochimie., v.
84, p. 1245-1253, 2002.
STEINER, J.M., WILSON, B.G. e WILLIAMS, D.A. Purification and partial characterization of feline classical
pancreatic lipase. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology, v.134, p.
151-159, 2003.
STEPANOV, A., SHVETS, V. Chemistry and Physics of Lipids, v. 25, p. 247-263, 1979.
STRAATHOF, A.J.J.; PANKE, S. e SCHMID, A., The production of fine chemicals by biotransformations,
Current Opinion in Biotechnology, v. 13, p.548–556, 2002.
STRAATHOF, A.J.J. e ADLERCREUTZ, P. (Eds.) Taylor & Francis e-Library. Netherlands. Amsterdam.,
2005, 545p.
STRAATHOF, A.J.J., JONGEJAN, J.A., Enzyme and Microbial Technology, v. 21, p. 559-571, 1997.
SUGIHARA, A.; UESHIMA, M.; SHIMADA, Y.; TSUNASAWA, S; TOMINAGA, Y. Journal of
Biochemistry, v. 112, p. 598 – 603, 1992.
SURESHAN, K.M. MYSORE S. SHASHIDHAR, M.S., PRAVEEN, T. e DAS, T., Regioselective Protection
and Deprotection of Inositol Hydroxyl. Chem. Rev., v. 103, p.4477-4503, 2003a.
SURESHA K.M.; YAMASAKI, T.; HAYASHI, M.; WATANABE, Y. A simple and practical resolution of
1,2:4,5-di-O-isopropylidene-myo-inositol, Tetrahedron: Asymmm., v.14, n.13, p.1771-1774, 2003b.
SURESHKUMAR, M., LEE, C.-K. Biocatalytic reactions in hydrophobic ionic liquids. Journal of Molecular
Catalysis B: Enzymatic, v. 60, p. 1–12, 2009.
SVENDSEN, A., Biochimica et Biophysica Acta, v. 1543, p. 223-238, 2000.
SVENSSON, I., WEHTJE, E., ADLERCREUTZ, P. e MATTIASSON, B. Effects of water activity on reaction
rates and equilibrium positions in enzymatic esterifications. Biotechnology and Bioengineering, v. 44, n.5, p.
549-556, 2004.
SWEERS, H. M., WONG, C.-H. Enzyme-catalyzed regioselective deacylation of protected sugars in
carbohydrate synthesis, J. Am. Chem. Soc., v.108, p.6421-6422, 1986.
TAMBORINI, L., ROMANO, D., PINTO, A., BERTOLANI, A. MOLINARI, F. e CONTI, P. An efficient
method for the lipase-catalysed resolution and in-line purification of racemic flurbiprofen in a continuous-flow
reactor. Journal of Molecular Catalysis B: Enzymatic, v.84, p.78– 82, 2012.
TAN T.; LU J.; NIE K.; DENG L.; WANG F.; Biotechnol. Adv. 28 (2010) 628–634.
TAO, J., KAZLAUKAS, R.J., 2011. Biocatalysis For Green Chemistry and Chemical Process Development.
John Wiley & Sons Ltd., Hoboken, New Jersey.
TAO. J. e XU, J.H. Enzymes and their synthetic applications: an overview-1-19p. In: Tao, J.; Lin, G.Q.; Liese A.
Biocatalysis for the pharmaceutical industry: Discovery, Development, and Manufacturing. John Wiley & Sons
(Asia) Pte Ltd. Singapura. 2009, 324p.
TARDIOLI, P.W; VIEIRA, M.F.; VIEIRA, A.M.S.; ZANIN, G.M.; BETANCOR, L., MATEO, C.;
FERNANDEZ-LORENTE, G.; GUISÁN, J.M. Immobilization-stabilization of glucoamylase: Chemical
modification of the enzyme followed by covalent attachment on highly activated glyoxyl-agarose support,
Process Biochemisry, v. 46, p.409-412, 2011.
TAWAKI, S. e KLIBANOV, A. M. Inversion of enzyme enantioselectivity mediated by the solvent. J. Am.
Chem. Soc., v. 114, p. 1882-1884, 1992.
TERAO, Y., MURATA, M., ACHIWA, K. Highly-efficient lipase-catalyzed asymmetric synthesis of chiral
glycerol derivatives leading to practical synthesis of (S)-propranolol, Tetrahedron Lett. v.29, p.5173-5176, 1988.
TISCHER, W., KASCHE, V., "Immobilized enzymes: crystal or carriers?", Trends in Biotechnology, v. 17, p.
326-335, 1999.
TONGBORIBOON, K., CHEIRSILP, B., KITTIKUN, A. H., Journal of Molecular Catalysis B: Enzymatic,
v.67, p. 52, 2010.
TORRES, R., ORTIZ, C. PESSELA, B.C.C., PALOMO, J.M., MATEO, C., GUISAN, J. M., FERNANDEZ-
LAFUENTE R., ―Improvement of the enentioselectivity of lipase (fraction B) from Candida antarctica via

163
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
adsorption on polyethynimine-agarose under different experimental conditions‖, Enzyme and Microbial
Tecnology, 39, pp. 167-171, 2006.
TOSA T., MORI T., FUSE N. CHIBATA I. Enzymologia, 1966, 31, 214–224.
TREVISAN, E.; LEAL NETO, M.; MORO, P.; HOSHINO, S.O.; ARROYO, P.A. Avaliação dos solventes
hexano e clorofórmio em reações de transesterificação in situ de Chlorella vulgaris. VIII Encontro Internacional
de Produção Científica, 1-4p., 22-25/ out, 2013.
TRUBIANO, G. D., BORIO, e FERREIRA, M.L. Ethyl Oleate Synthesis Using Candida rugosa Lipase in a
Solvent-Free System. Role of Hydrophobic Interactions Biomacromolecules, v. 5, p. 1832-1840, 2004.
TUFVESSON, P., Fu, W., JENSEN, J.S., WOODLEY, J.M., Process considerations for the scale-up and
implementation of biocatalysis, Food Bioprod. Process, v.88, n. 1, p.3–11, 2010.
TURNER, N.J. Controlling chirality. Cur Opin Biotechnol, v.14, p.401–406, 2003.
UNDURRAGA, D., MARKOVITS, A. e ERAZO, S., Cocoa butter equivalente through enzymic
interesterification of palm oil midfraction, Proc. Biochem., v.36, p.933–939, 2001.
UPPENBERG, J., HANSEN, M.T., PATKAR, S., JONES, T.A., The sequence, crystal sctruture determination
and refinement of two crystal form of lipase B from Candida Antarctica. Structure, v. 2, p. 293–308, 1994.
Crystallographic and molecular-modeling studies of lipase B from Candida Antarctica reveal a stereospecificity
pocket for secondary alcohols. Biochemistry 34, 16838-16851, 1995.
UPPENBERG, J.; OEHRNER, N.; NORIN, M.; HULT, K.; KLEYWEGT, G.J.; et al.
VAGANAY, S., JOLIFF, G., BERTAUX, O., TOSELLI, E., DEVIGNES, M.D. e BÉNICOURT, C. The
complete cDNA sequence encoding dog gastric lipase, v. 8, n.4, p.257-262, 1998.
VAIDYA, B.K., INGAVLE, G.C., PONRATHNA, S., KULKARNI, B.D., NENE, S.N., Bioresourch
Technology, v.99, p. 3623-3629, 2008.
VALIVETY, R. H., HALLING, P. J. e MACRAE, A. R. Reaction rate with lipase catalyst shows similar
dependence on water activity in different organic solvents, Biochim. Biophys. Acta, v.1118, p. 218–222, 1992a.
VALIVETY, R. H., HALLING, P. J., PEILOW, A. D. e MACRAE, A. R. Lipases from different sources vary
widely in dependence of catalytic activity on water activity, Biochim. Biophys. Acta 1122, p. 143–146, 1992b.
VALIVETY, R. H., HALLING, P. J. e MACRAE, A. R. Rhizomucor miehei lipase remains highly active at
water activity below 0. 0001, FEBS Lett. v.301, p. 258–260, 1992c.
VERGER, R. Interfacial activation of lipases: facts and artifacts. Trens Biotechnol., v. 15, 32-38, 1997.
VILLENEUVE, P., MUDERHWA, J.M., GRAILLE, J., HAAS, M.J., Customizing lipases for biocatalysis: a
survey of chemical, physical and molecular biological approches, J. Mol. Cat. B: enzym., v. 9, p. 113-148, 2000.
WAKABAYASHI, H., WAKABAYASHI, M., EISENREICH,W. e ENGEL, K.H. Stereoselectivity of the
Generation of 3-Mercaptohexanal and 3-Mercaptohexanol by Lipase- Catalyzed Hydrolysis of 3-
Acetylthioesters J. Agric. Food Chem., v. 51, n.15, p 4349–4355, 2003.
WANDREY, C., LIESE, A., KIHUMBU, D., Organic Process Research & Development, v. 4, p.286, 2000.
WANG, Y. F., LALONDE, J. J., MOMONGAN, M., BERGBREITER, D. E., WONG, C.-H. Lipasecatalyzed
irreversible transesterifications using enol esters as acylating reagents: preparative enantio- and regioselective
syntheses of alcohols glycerol derivatives sugars and organometallics, J. Am. Chem. Soc. v.110, p.7200-7205,
1988.
WANG, Y. F., WONG, C. H. Lipase -catalyzed irreversible transesterification for preparative synthesis of chiral
glycerol derivatives, J. Org. Chem. v.53, p.3127-3129, 1988.
WANG, Y.F., LALONDE, J.J., MOMONGAN, M., BERGBREITER, D.E., WONG, C.H., Lipase catalyzed
irreversible transesterifications using enol esters as acylating reagents: preparative enantio- and regioselective
syntheses of alcohols, glycerol derivatives, sugars, and organometallics, Journal American Chemical Society, v.
110, p. 7200–7205, 1988.
WANG X., ZHOU G., ZHANG H., DU S., XU Y., WANG C., J. Non-Cryst. Solids 357 (2011) 3027–3032.
WARD, R. S., Dynamic kinetic resolution. Tetrahedron: Asymmetry, v. 6, p. 1475-1490, 1995.
WASSERSCEID, P., WELTON, T. (Eds.), Ionic Liquids in Synthesis, Wiley-VCH, 2003.

164
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
WATSON. W.J.W., How do the fine chemical, pharmaceutical, and related industries approach green chemistry
and sustainability? Green Chem. v. 14, p.251, 2012.
WEBER, H. K., STECHER, H., FABER, K. Sensitivity of microbial lipases to acetaldehyde formed by acyl-
transfer reactions from vinyl esters, Biotechnol. Lett. v.17, p.803-808, 1995a.
WEBER, H. K., STECHER, H. e FABER, K., Some properties of commercially available crude lipase
preparations, In: Preparative Biotransformations (Roberts, S. M.; Eds.), New York: Wiley. 1995b.
WEHTJE, E., COSTES, D. e ADLERCREUTZ, P. Enantioselectivity of lipases: effects of water activity.
Journal of Molecular Catalysis B: Enzymatic, v. 3, p. 22 l-230, 1997.
WEWERS, W.; GILLANDT, H.; TRAUB, H. Tetrahedron: Asymmetry, v. 16, p. 1723-1728,2005.
WHITESELL, J. K.; REYNOLDS, D. The Journal Organic Chemistry, v. 48, p. 3548-3551, 1983.
WILKINSON B. e BACHMANN, B.O. Biocatalysis in pharmaceutical preparation and alteration. Curr Opin
Chem Biol v. 10, p. 169-176, 2006.
WILSON, L., PALOMO, J. M., FERNÁNDEZ-LORENTE, G., ILLANES, A., GUISÁN, J. M., FERNÁNDEZ-
LAFUENTE R., ―Improvement of the properties of a thermostable lipase from Alcaligenes sp. via strong
adsorption on hydrophobic supports‖, Enzyme and Microbial Tecnology, v. 38, p. 975-980, 2006.
WOOTEN, M. W.; WRENN, R. W.; FEBS Lett., v. 171, p.183, 1984.
XIE, X. and TANG, Y., Efficient synthesis of simvastatin by use of whole-cell biocatalysis. Appl. Environ.
Microbiol., v.73, p.2054–2060, 2007.
XU Y.Q., ZHOU G.W,. WU C.C,. LI T.D, SONG H.B., Solid State Sci. 13 (2011) 867–874.
YADAV, G. D. e SIVAKUMAR, P. Biochemical Engineering Journal, v. 19, p.101-107, 2004.
YANG, RUSSEL, Fundamentals of non-aqueous enzymology, Enzymatic reactions in organic media, Eds:
Koskinen e Klibanov, Ed.Blackie academic & professional, Great Britain, 1996.
YAO, C. e KOËLLER, W. Diversity of cutinases from plant pathogenic fungi: Cloning and sequence analysis of
a cutinase gene from Alternaria brassicicola. Physiol. Mol. Plant Pathol. v. 44, p. 81-92, 1994.
YAQOOB, M., NABI, A. e MASSOM-YASINZAI, M. Bovine pancreatic lipase: Purification and applications
in immobilized form for transesterification reactions. J. Chem. Society of Pakistan., v.21, p. 114-119, 1999.
YENIAD B., NAIK H., HEISE A., Adv. Biochem. Eng. Biotechnol. 125 (2011) 69–95.
ZAKS, A. e KLIBANOV, A.M. Enzymic catalysis in non-aqueous solvents. Journal of Biological Chemistry, v.
263, p. 3194-3201, 1988.
ZAKS, A. ―New enzymatic properties in organic media‖ In: Enzymatic reactions in organic media, Eds:
Koskinen e Klibanov, Ed.Blackie academic & professional, Great Britain, 1996.
ZHANG LQ, ZHANG YD, XU L, LI XL, YANG XC, XU GL, WU XX, GAO HY, DU WB, ZHANG XT,
ZHANG XZ. Lipasecatalyzed synthesis of RGD diamide in aqueous water-miscible organic solvents. Enzyme
Microb Technol. 2001;29:129–35.
ZHANG, J., HOU, Z., YAO, C. e YU, Y. Purification and properties of a lipase from a Bacillus strain for
catalytic resolution of (R)-Naproxen. J. Mol. Catalalysis B: Enzymatic., v.18, p. 205-214, 2002.
ZHANG D. H., YUWEN L. X., Li C., Li Y. Q., Effect of poly (vinyl acetate–acrylamide) microspheres
properties and steric hindrance on the immobilization of Candida rugosa lipase, Bioresour. Technol., 2012, 124,
233-236. doi:10.1016/j.biortech.2012.08.083.
ZHAO, H., BAKER, G.A., SONG, Z., OLUBAJO, O., ZANDERS, L., CAMPBELL, S.M., Journal of
Molecular Catalysis B: Enzymatic, v. 57, p.149, 2009.
ZHAO, H., Journal of Chemical Technology & Biotechnology, v. 85, p.891, 2010.
ZHENG, C.Z., WANG, J.L., Li, X., LIU, B.K., WU, Q. e LIN, X.F., Regioselective synthesis of amphiphilic
metoprolol-saccharide conjugates by enzymatic strategy in organic media. v. 46, n. 1, p.123-127, 2011.
ZHENG M. M., LU Y., HUANG F. H., WANG L., GUO P. M., FENG Y. Q. e DENG Q. C., Lipase
Immobilization on Hyper-Cross-Linked Polymer-Coated Silica for Biocatalytic Synthesis of Phytosterol Esters
with Controllable Fatty Acid Composition, J. Agric. Food Chem., 2013, 61, 231−237. doi: 10.1021/jf3042962.

165
Capítulo VIII- REFERÊNCIAS BIBLIOGRÁFICAS
ZHOU, R., XU, J.H. Biochemical Engineering Journal, v. 23, p. 11–15, 2005.
ZOU B., HU Y., YU D., XIA J., TANG, S., LIU W., HUANG H., Biochem. Eng. J. 53 (2010) 150–153.
ZWINGENBERGER K.,WENDT S., Immunomodulation by thalidomide: systematic review of literature and of
unpublished observations. J Inflamm, v.46, p.177–211, 1996.