o zooplâncton da plataforma continental de santa catarina ...siaibib01.univali.br/pdf/renata...
TRANSCRIPT

UNIVERSIDADE DO VALE DO ITAJAÍ
MESTRADO EM CIÊNCIA E TECNOLOGIA AMBIENTAL
O zooplâncton da plataforma continental de
Santa Catarina. Ênfase em Copepoda e
Cladocera e suas relações com os processos
físicos costeiros
Renata Domingos Nunes
Itajaí, 29 de outubro de 2010

ii
UNIVERSIDADE DO VALE DO ITAJAÍ
MESTRADO EM CIÊNCIA E TECNOLOGIA AMBIENTAL
O zooplâncton da plataforma continental de
Santa Catarina. Ênfase em Copepoda e
Cladocera e suas relações com os processos
físicos costeiros
Renata Domingos Nunes
Dissertação apresentada à Universidade do Vale do Itajaí, como parte dos requisitos para obtenção do grau de Mestre em Ciência e Tecnologia Ambiental.
Orientador: Dr. Charrid Resgalla Junior
Itajaí, 29 de outubro de 2010

iii
DEDICATÓRIA
Dedico à todos que acreditaram em mim e me encheram de esperanças para a conclusão desta nova etapa.

iv
AGRADECIMENTOS
Agradeço,
À Deus, esta força superior que me manteve em paz em muitos momentos difíceis, e
conseguir, enfim, alcançar um dos meus objetivos.
Aos meus pais pela amizade, amor incondicional, por seus exemplos e lições de vida,
essenciais para a formação do meu caráter. Aos meus irmãos por serem minha âncora e meu
sinônimo de equilíbrio. Por mais que eu percorra caminhos, vocês fazem parte de todos eles.
Sempre serão o outro lado da balança.
Ao meu orientador, Dr. Charrid, que é muito mais que um ótimo professor e
cientista, é um amigo admirável. Desde o início de minha graduação, sempre disposto a
ensinar, agradeço pela paciência nas minhas faltas. Desejo ser uma profissional com o mesmo
caráter dele.
Às minhas queridas amigas, as FF’s, agradeço que pela amizade e tudo o que esta
palavra significa. São todas especiais para mim e dedico a elas, esta nova vitória. Os amigos
se amam...os verdadeiros. E quem me conhece sabe o quanto eu valorizo e faço questão de
manter minhas amizades bem floridas!!
Precisamos permitir que pessoas especiais possam entrar em nossa vida e nos olhar
com amor...
E eu conheci muitas pessoas especiais na graduação e que continuaram presentes no
mestrado, são estas: Thais (que de promoter e sinônimo de organização e agilidade agora é a
mãe da Lari. Simples assim!), Lud (um furacão em pessoa...sou muito orgulhosa por ser tua
discípula), a Fê e o Rafa (casal Letox que faz muiiiiiiita falta lá), a Dani e a Jéssica (novas
aquisições do Letox e em pouco tempo nasceu uma amizade, Obrigada por gostarem tanto de
mim, pelo menos é o que parece!!!!), Vah (minha irmã “preta”), o Martin (obrigada pela
paciência em me ensinar a ser fera no ArcGis), tem a Cris (sem ela o mestrado não existe!), a
Telma (sempre muito alto astral!), toda a galera do LETOX. Outras amigas também ficarão
pra sempre comigo, a Ana e sua gargalhada mais engraçada do que qualquer piada, a Débora
que sempre ia roubar um café no LETOX, a Benta e a Magda que sempre atendem o pessoal
com carinho e atenção.

v
À todos aqueles que passaram pela minha vida, à todas as pessoas importantes para
mim que já partiram, não é possível citar todo mundo aqui, porém saibam que são especiais.
E não podia deixar de agradecer ao Jaison que me concedeu a sorte de um amor
tranqüilo e por acreditar tanto no meu potencial que me convence a cada dia que posso mais.
Preenche um lugar especial no meu coração.

vi
SUMÁRIO
LISTA DE FIGURAS .......................................................................................................................... viii
LISTA DE TABELAS ............................................................................................................................ x
RESUMO ............................................................................................................................................... xi
1.0 INTRODUÇÃO .............................................................................................................................. 13
1.1 Copepoda e Cladocera................................................................................................................. 14
1.2 Massas de Água........................................................................................................................... 15
1.3 Processos Oceanográficos na Região de Estudo ......................................................................... 16
2.0 OBJETIVOS.................................................................................................................................... 18
2.1 Objetivo Geral ............................................................................................................................. 18
2.2 Objetivos Específicos .................................................................................................................. 18
3.0 MATERIAL E MÉTODOS............................................................................................................. 19
3.1 Área de Estudo ............................................................................................................................ 19
3.2 Amostras...................................................................................................................................... 19
4.0 RESULTADOS............................................................................................................................... 23
4.1 Condições oceanográficas durante o período de estudo.............................................................. 23
4.2 Comunidade zooplanctônica ....................................................................................................... 25
4.2.1 Densidade total do zooplâncton............................................................................................ 25
4.2.2 Grandes grupos do zooplâncton ........................................................................................... 26
4.3 MOLLUSCA MEROPLANCTÔNICO (LARVAS)................................................................... 27
4.4 DECAPODA ............................................................................................................................... 29
4.5 CHAETOGNATHA.................................................................................................................... 30
4.6 SALPIDAE.................................................................................................................................. 31
4.7 APENDICULARIA..................................................................................................................... 32
4.8 PTEROPODA ............................................................................................................................. 33
4.9 COPEPODA................................................................................................................................ 34
4.9.1 Índices de Riqueza, Diversidade e Equitabilidade para Copepoda ...................................... 42

vii
4.10 CLADOCERA .......................................................................................................................... 45
4.10.1 Índices de Riqueza, Diversidade e Equitabilidade para Cladocera .................................... 48
4.11 Variáveis ambientais e comunidade zooplanctônica ................................................................. 50
5.0 DISCUSSÃO................................................................................................................................... 54
6.0 CONCLUSÕES............................................................................................................................... 62
REFERÊNCIAS .................................................................................................................................... 63

viii
LISTA DE FIGURAS
Figura 1 – Mapa de localização do litoral do estado de Santa Catarina com as amostras oceanográficas
realizadas em Primavera de 2005 e Outono de 2006 dentro do Projeto PROSAR. .............................. 20
Figura 2 – Diagramas TS e classificação das massas de água referentes aos cruzeiros de Primavera de
2005 (A) e Outono de 2006 (B) do projeto PROSAR (modificado de Hille et al., 2007) com valores
médios da coluna de água. Sendo AC= Água Costeira, ASP= Água Subtropical de Plataforma,
ACAS= Água Central do Atlântico Sul, AT= Água tropical e APP= Água da Pluma do Prata. .......... 25
Figura 3 – Distribuição espacial dos valores de densidade para o zooplâncton total (Org. m-3) em (A)
Primavera de 2005 e (B) Outono de 2006. ............................................................................................ 26
Figura 10 – Distribuição espacial da densidade (Org. m-3) de larvas de Mollusca bentônicos nos
cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006. ................................................................. 28
Figura 7 – Distribuição espacial da densidade (Org. m-3) de Larvas de Decapoda nos cruzeiros de (A)
Primavera de 2005 e (B) Outono de 2006. ............................................................................................ 29
Figura 6 – Distribuição espacial da densidade (Org. m-3) de Chaetognatha nos cruzeiros de (A)
Primavera de 2005 e (B) Outono de 2006. ............................................................................................ 30
Figura 5 – Distribuição espacial da densidade (Org. m-3) de Salpidae nos cruzeiros de (A) Primavera
de 2005 e (B) Outono de 2006. ............................................................................................................. 31
Figura 9 – Distribuição espacial da densidade (Org. m-3) de Apendicularia nos cruzeiros de (A)
Primavera de 2005 e (B) Outono de 2006. ............................................................................................ 32
Figura 8 – Distribuição espacial da densidade (Org. m-3) de Pteropoda nos cruzeiros de (A) Primavera
de 2005 e (B) Outono de 2006. ............................................................................................................. 33
Figura 11 - Distribuição espacial da densidade (Org. m3) de Copepoda nos cruzeiros de (A) Primavera
de 2005 e (B) Outono de 2006. ............................................................................................................. 34
Figura 12 - Distribuição espacial da densidade (Org. m-3) das espécies de Copepoda: (A) Acartia
lilljeborgi, (B) Centropages velificatus, (C) Oithona Plumifera, (D) Oncaea clevei, (E) Temora
stylifera, (F) Temora turbinata, referente ao cruzeiro de Primavera de 2005....................................... 37
Figura 13 - Distribuição espacial da densidade (Org. m-3) da espécie de Copepoda Undinula vulgaris,
referente ao cruzeiro de Primavera de 2005. ......................................................................................... 38
Figura 14 – Distribuição espacial da densidade (Org. m-3) das espécies de Copepoda: (A) Acartia
lilljeborgi¸ (B) Clausocalanus furcatus, (C) Temora stylifera, (D) Temora turbinata, (E) Oncaea
clevei, (F) Oithona plumifera, referente ao cruzeiro de Outono de 2006.............................................. 39
Figura 15 - Distribuição espacial da densidade (Org. m-3) das espécies de Copepoda: (A) Temora
turbinata, (B) Subeucalanus pileatus, (C) Corycaeus speciosus, (D) Nannocalanus minor, (E)
Euchaeta marina, (F) Macrosetella gracilis referente ao cruzeiro Outono de 2006............................. 41

ix
Figura 16 - Distribuição espacial da densidade (Org. m-3) das espécies de Copepoda (A) Neocalanus
gracilis e (B) Undinula vulgaris, referente ao cruzeiro de Outono de 2006. ........................................ 42
Figura 17 - Distribuição espacial da Riqueza de espécies (Nits) para o grupo de Copepoda nos
cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006. ................................................................. 43
Figura 18 - Distribuição espacial da Diversidade (Nits) para o grupo de Copepoda nos cruzeiros de (A)
Primavera de 2005 e (B) Outono de 2006. ............................................................................................ 43
Figura 19 - Distribuição espacial da Equitabilidade (Nits) para o grupo de Copepoda nos cruzeiros de
(A) Primavera de 2005 e (B) Outono de 2006. ..................................................................................... 44
Figura 20 - Distribuição espacial da densidade (Org. m3) de Cladocera nos cruzeiros de (A) Primavera
de 2005 e (B) Outono de 2006. ............................................................................................................. 45
Figura 21 - Distribuição espacial da densidade (Org. m--3) das espécies de Cladocera: (A) Penilia
avirostris, (B) Pleopis schmackeri, (C) Evadne spinifera, (D) Pseudoevadne tergestina, referente ao
cruzeiro de Primavera de 2005.............................................................................................................. 46
Figura 22 – Distribuição espacial da densidade (Org. m-3) das espécies de Cladocera: (A) Penilia
avirostris, (B) Pleopis schmackeri, (C) Pseudoevadne spinifera, (D) Pseudoevadne tergestina
referente ao cruzeiro de Outono de 2006. ............................................................................................. 47
Figura 23 – Distribuição espacial da Riqueza de espécies (Nits) para Cladocera nos cruzeiros de
Primavera de 2005 e Outono de 2006. .................................................................................................. 48
Figura 24 – Distribuição espacial da Diversidade (Nits) para Cladocera nos cruzeiros de Primavera de
2005 e Outono de 2006. ........................................................................................................................ 49
Figura 25 – Distribuição espacial da Equitabilidade (Nits) para Cladocera nos cruzeiros de Primavera
de 2005 e Outono de 2006..................................................................................................................... 49
Figura 26 - Resultado da ACP para a comunidade zooplanctônica e os parâmetros ambientais referente
ao cruzeiro de Primavera de 2005. ........................................................................................................ 51
Figura 27 - Resultado da ACP para a comunidade zooplanctônica e os parâmetros ambientais referente
ao cruzeiro de Outono de 2006.............................................................................................................. 53

x
LISTA DE TABELAS
Tabela 1 - Valores mínimos, médios e máximos de Salinidade e temperatura (ºC) referentes aos
cruzeiros de Primavera de 2005 e Outono de 2006 do projeto PROSAR modificado de Hille et al.
(2008). ................................................................................................................................................... 24
Tabela 2 - Abundância relativa média (%) dos grandes grupos ocorrentes para o cruzeiro de (A)
Primavera de 2005 e (B) Outono de 2006. ............................................................................................ 27
Tabela 3 - Frequência de Ocorrência (%) dos grandes grupos do zooplâncton para o cruzeiro de
Primavera de 2005 e Outono de 2006. .................................................................................................. 27
Tabela 4 - Frequência de Ocorrência (FO) e Abundância Relativa (AR) para as espécies de Copepoda
observados nos cruzeiros de Primavera de 2005 e Outono de 2006. .................................................... 35
Tabela 5 - Frequência de Ocorrência (FO) e Abundância Relativa (AR) para as espécies de Cladocera
referente aos cruzeiros de Primavera de 2005 e Outono de 2006. ........................................................ 45
Tabela 6 – Peso das variáveis para ACP da comunidade zooplanctônica e parâmetros ambientais
referente ao cruzeiro de Primavera de 2005. ......................................................................................... 50
Tabela 7 – Peso das variáveis para ACP da comunidade zooplanctônica e parâmetros ambientais
referente ao cruzeiro de Outono de 2006. ............................................................................................. 52

xi
RESUMO
Estudos exclusivamente destinados a investigação da comunidade zooplanctônica para a
plataforma continental do Estado de Santa Catarina ainda não haviam sido realizados apesar
da importância econômica e ecológica da área. Esta região costeira do sul do Brasil apresenta
processos oceanográficos de alta relevância como forte influência de aportes continentais,
ressurgência no Cabo de Santa Marta Grande e influência da Frente do Prata ao sul. Devido a
esta complexidade hidrológica foram realizados dois cruzeiros em Primavera de 2005 e
Outono de 2006 com o objetivo de estudar as variações temporais destes processos e suas
influências sobre a biota. Para este estudo foram analisados amostras de zooplâncton obtidas
por arrastos oblíquos com rede Bongo em 33 pontos amostrais dispostos em perfis
perpendiculares a costa para cada cruzeiro. Os grupos dominantes nas amostras foram
Copepoda, Cladocera, Salpas e Chaetognatha, apresentando maiores densidades nos pontos
mais costeiros. Para o cruzeiro de Primavera de 2005 os gradientes de salinidade e
temperatura perpendiculares a costa, promovidos pelos aportes continentais ao norte e pela
ressurgência ao sul da área, determinaram os limites de distribuição de Acartia lilljeborgi e
Penilia avirostris. No cruzeiro de Outono de 2006, devido a menor variação da temperatura e
salinidade, os gradientes longitudinais a costa não explicaram a variação da distribuição das
espécies, indicando que forçantes bióticos poderiam estar atuando na ecologia do sistema.

xii
ABSTRACT
Studies intended exclusively for the investigation of the zooplankton community from the
continental shelf of Santa Catarina State had not yet been achieved despite the economic and
ecological importance of the area. This coastal region of southern Brazil presents high
relevant oceanographic processes as a strong influence of continental inputs, upwelling at
Cape Santa Marta Grande and the influence of the Plate Front from the south. Due to this
complexity hydrology two cruises were made, December 2005 and May 2006, aiming to
study the temporal variations of these processes and their influences on biota. For this study,
samples of zooplankton collected by oblique hauls with Bongo net at 33 sampling points
arranged in profiles perpendicular to the coast for each cruise were analyzed. The dominant
groups in the samples were Copepoda, Cladocera, Chaetognatha and salps, with highest
densities in coastal points. For the cruise of December 2005 the temperature and salinity
gradients perpendicular to the coast, driven by continental inputs on north and upwelling on
south of the area, determined the distribution limits of Acartia lilljeborgi and Penilia
avirostris. In May 2006 cruise, due to less variation in temperature and salinity, the
longitudinal gradients to coast could not explain the variation of species distribution,
indicating that biotic forcings could be acting in the ecology of the system.

13
1.0 INTRODUÇÃO
A comunidade zooplanctônica tem uma posição relevante na cadeia alimentar
pelágica marinha, uma vez que transfere a energia produzida pelas algas unicelulares
fotossintetizantes aos níveis tróficos superiores (LENZ, 2000). Por essa razão, o
conhecimento de sua abundância, distribuição espacial e temporal são necessários para o
entendimento dos ciclos biogeoquímicos e da dinâmica das comunidades marinhas
(WISNHER et al., 1998). Além disto, na comunidade zooplanctônica, estão incluídas muitas
larvas de importantes recursos pesqueiros e de interesse comercial, como larvas de crustáceos,
moluscos e de peixes. De forma geral, a estrutura e a distribuição do zooplâncton são
influenciadas por predação, competição, bem como por processos físicos que atuam em
diferentes escalas temporais e espaciais (ASHJIAN, 1993; WIAFE & FRID, 1996).
O estudo do zooplâncton da plataforma continental brasileira conta com diversos
trabalhos que abordam a sistemática (BJÖRNBERG, 1967; CAMPANER, 1978. ALVAREZ,
1986; VEGA-PÉREZ & BOWMAN, 1992; BERSANO & BOXSHALL, 1994), aspectos
ecológicos (MOREIRA, 1973; BJÖRNBERG, 1980; CAMPANER, 1985; LIANG & VEGA-
PÉREZ, 1994; 1995; VEGA- PEREZ & HERNANDEZ, 1997), fisiológicos (YAMASHITA,
1977; YAMASHITA & MOREIRA, 1981) e biogeográficos (e.g. PALÁCIO, 1982) de
diferentes grupos taxonômicos.
Na costa de Santa Catarina existe uma carência de informações sobre a comunidade
zooplanctônica, o que vem sendo destacado por trabalhos sobre a revisão do tema
apresentados por Valentin et al. (1994) e por Brandini et al. (1997). Entretanto, as
características biogeográficas do zooplâncton da costa catarinense em comparação com outras
áreas mais exaustivamente investigadas ao norte e ao sul do estado, tais como São Paulo,
Paraná e Rio Grande do Sul foram apresentadas por Resgalla Jr. (no prelo).
Dentre os grupos do zooplâncton marinho, merecem destaque os Copepoda e
Cladocera como os mais importantes representantes do holopâncton costeiro (OMORI &
IKEDA, 1984). Por apresentarem altas freqüências de ocorrência e abundância, estes grupos
foram estudados ao nível de espécie com o intuito de relacionar as suas ocorrências com as
massas de água típicas da plataforma continental do estado de Santa Catarina, sul do Brasil,
em dois períodos e com processos oceanográficos distintos.

14
1.1 Copepoda e Cladocera
A Classe Maxillopoda, Subclasse Copepoda constitui os organismos do
mesozooplâncton mais representativos nas amostras do zooplâncton marinho. Na costa
brasileira, estudos de Björnberg (1981); Bersano (1994); Bradford-Grieve et al. (1999);
Cavalcanti & Larrazabál (2004); Sant’anna & Björnberg (2006); Sterza & Fernandez (2006);
Cornils et al. (2007) e Nunes (2007) apontam o grupo Copepoda como o responsável pela
maior diversidade de riqueza de espécies. Sua importância é tal que praticamente toda a
ecologia pelágica é dependente direta ou indiretamente destes organismos. É um grupo que
tem sido utilizado como indicador biológico ao longo da história da oceanografia brasileira.
Para a costa brasileira, este grupo tem sido utilizado como indicadores de
ressurgências (VALENTIN et al., 1976), de impacto ambiental (RESGALLA Jr., 2001) e de
produção biológica em ambientes costeiros (MONTÚ, 1980).
Estes organismos são, historicamente, bons indicadores de massas de água
(BJÖRNBERG, 1981 e BAKER et al.,1990). Em um estudo de abrangência da presente área
de estudo, Muxagata (1999) relacionou agrupamentos de espécies de Copepoda com as
massas de água, estabelecendo assembléias zooplanctônicas da Plataforma Continental
Sudeste Brasileira, durante o inverno de 1995.
Para a zona de arrebentação da Praia de Tramandaí, RS, Ávila et al. (2009) definiu a
variação temporal do zooplâncton dando ênfase aos Copepoda por apresentar melhor
descrição e maior conhecimento no Oceano Atlântico Sul (BOLTOVSKOY, 1999).
A antiga ordem Cladocera (Ctenopoda e Onychopoda) agrega representantes
marinhos de distribuição preferencialmente costeiros e com poucas espécies registradas no
mundo mas igualmente importantes na ecologia pelágica marinha.
Apesar de sua baixa diversidade, é um grupo abundante e que reflete diretamente a
produção primária de uma área (RAMIREZ, 1981) sendo excelentes indicadores de massas de
água e de processos oceanográficas ocorrentes nas regiões costeiras (RESGALLA Jr. &
MONTÚ, 1993). Além disto, este grupo pode ser significativo em águas costeiras e próximas
a estuários, servindo como um elo importante nas transferências energéticas da cadeia
alimentar (ONBÉ, 1977).

15
Posta (2006) estudou a influência de fatores ambientais sobre a distribuição dos
Cladocera marinho em um estuário em Ubatuba (SP). Trabalhos abrangendo a plataforma sul
do Brasil, como os de Resgalla Jr. & Montú (1993) e Muxagata & Montú (2006), são
importantes referências já que existem poucos estudos com este grupo quando comparado aos
ambientes de água doce.
1.2 Massas de Água
A circulação na PCSE, segundo Castro (1996), é determinada pelos regimes de
ventos e ação da Corrente do Brasil (CB) na região. De uma maneira geral, durante o verão,
os ventos mais frequentes apontam de NE para SW causando um transporte superficial de
água da costa para o mar. A divergência superficial costeira, possibilita a penetração da
ACAS (ressurgência) sobre a plataforma e a diminuição do nível do mar junto a costa,
levando, consequentemente, um empilhamento de água em regiões de quebra da plataforma.
Segundo Castro et al. (1987) esses movimentos opostos podem gerar uma estratificação em
duas camadas, uma acima e outra abaixo da termoclina permanente.
Já durante o inverno a situação se reverte e os ventos apontam de SW para NE,
também com componentes paralelos a costa, causando assim um transporte de água
superficial do mar aberto para a costa. A convergência superficial costeira, assim causada,
condiciona a elevação do nível do mar junto à costa e o afastamento da ACAS.
A energia fornecida pelo sol, na forma de radiação solar, é o fator fundamental para a
circulação oceânica e a distribuição e estratificação de massas de água. Estes processos físicos
determinam a formação de uma termoclina sazonal e a conseqüente estratificação vertical em
duas camadas durante o verão, seguida do desaparecimento da termoclina sazonal e
relaxamento da estratificação vertical durante o inverno, fazendo com que a parte interna da
plataforma continental seja ocupada por águas com características físicas mais homogêneas
durante este período (CASTRO et al, 1987; CASTRO, 1996).
A identificação das diversas massas de água da costa até a borda da plataforma pode
seguir as características apresentadas por Emílsson (1961), Miranda (1982) e Castro &
Miranda (1998), acrescidas de algumas informações sobre nutrientes (CASTRO, B.M.;
LORENZZETTI, J.A.; SILVEIRA, I.C.A. & MIRANDA, L.B. de, 2006).

16
Água Tropical (AT) – Água quente com reduzido teor de nutrientes. Temperatura
superior a 20ºC e salinidade maior que 36,4. A AT é transportada em direção sudoeste pela
camada superficial (0-200 m) da Corrente do Brasil (CB), próximo à borda da plataforma.
Água Central do Atlântico Sul (ACAS) – Água fria e rica em nutrientes. Temperaturas
inferiores a 20ºC e salinidade entre 35 e 36,4. Esse índice termohalino é o mais indicado para
definir a ACAS entre 22ºS e 25ºS (MIRANDA, 1982). A ACAS apresenta as mesmas
propriedades da massa de água formada na zona de confluência entre as correntes do Brasil
(CB) e das Malvinas, fluindo ao longo do talude continental pela camada inferior da CB (200-
500 m), próximo à borda da plataforma. Altos teores de nutrientes caracterizam essas águas,
não raro evidenciando afloramentos de águas em regiões sobre a plataforma.
Água Costeira (AC) – É uma mistura de água doce oriunda da drenagem continental
com as águas da plataforma continental. Temperatura com variação sazonal e salinidade
inferior a 35. Apresenta salinidade baixa devido à influencia de vários estuários de pequeno e
médio porte presentes na costa sudeste-sul do Brasil, sendo caracterizada por alto teor de
silicato e teores significativos de nitrato e fosfato, segundo a maior ou menor influência
continental.
Água de Plataforma (AP) – Origina-se da mistura entre duas massas de água: a AT e a
AC (SILVA et al., 2004; SILVA et al., 2005). A temperatura é variável e a salinidade entre
35 e 36. Apresenta teores variáveis em nutrientes. Esta AP pode ser definida como Água
Subtropical de Plataforma (ASP), formada pela mistura da Água da Pluma do Prata com a
Água Tropical (MÖLLER JR. et al., 2008 apud HILLE, 2009).
Água da Pluma do Prata (APP)- É uma água de baixa salinidade influenciada pela
vazão do Rio da Prata. As características físico-quimicas desta massa de água sofre variações
sazonais, principalmente pelo regime de ventos (MÖLLER JR. et al., 2008 apud HILLE, 20).
1.3 Processos Oceanográficos na Região de Estudo
Na Plataforma de Santa Catarina ocorre diversos processos oceanográficos, como a
Ressurgência que é o movimento de ascensão de águas de subsuperfície, mais frias, até a
superfície, resultante da divergência positiva do transporte horizontal induzido pelo vento na
camada superior do oceano (RODRIGUES, 1997). A divergência positiva do transporte na
camada superior pode ocorrer próxima a costa, sendo por isso denominada ressurgência
costeira. A topografia de fundo, nestes casos, é um dos fatores principais no desencadeamento

17
desse fenômeno. As ressurgências trazem para a zona fótica importante quantidade de
nutrientes das camadas mais profundas, promovendo o aumento do potencial trófico local.
Há também os aportes de águas continentais representam a entrada de água
proveniente da drenagem continental e de estuários nos sistemas costeiros, contribuindo para
a diminuição da salinidade e a alteração na composição química da água, promovendo
alterações na constância das proporções entre os elementos maiores da água do mar. Isto
causa enriquecimento em nutrientes, sobretudo em silicato, que caracteriza esses aportes.
Influencias antropogênicas também podem ser evidenciadas nos aportes continentais via
sinais de poluição, como efluentes e lixos domésticos.
A influência da Frente do Prata, embora o estuário do Prata esteja a mais de 1.000
km ao sul, pode ser detectada durante o período de inverno quando as condições
oceanográficas das correntes do Brasil e das Malvinas favorecem o deslocamento de águas de
baixa salinidade para o norte. Embora seja um processo ainda pouco explorado, esta “frente”
pode chegar até o litoral do Rio de Janeiro (PIOLA et al., 2005). Borzone et al. (1999)
observaram águas de salinidade inferior a 32 em toda a coluna de água em uma região de 40
m de profundidade numa região cerca de 100 km ao norte da área de estudo, o que reforça o
efeito do rio da Prata na região durante os meses de inverno (SCHETINNI, 2005).

18
2.0 OBJETIVOS
2.1 Objetivo Geral
Este trabalho tem como objetivo analisar a comunidade zooplanctônica da
plataforma de Santa Catarina em dois períodos distintos, Primavera de 2005 e Outono de
2006.
2.2 Objetivos Específicos
• Diagnosticar os grupos zooplanctônicos mais representativos nas amostras da
plataforma de Santa Catarina para Primavera de 2005 e Outono de 2006.
• Descrever a composição e distribuição espacial das espécies de Copepoda e Cladocera
ocorrentes na plataforma de Santa Catarina.
• Relacionar a ocorrência das espécies mais abundantes e freqüentes de Copepoda e
Cladocera com as massas de águas encontradas.
• Comparar as espécies de Copepoda e Cladocera a outras áreas da costa brasileira, ao
sul e ao norte da área de estudo.

19
3.0 MATERIAL E MÉTODOS
3.1 Área de Estudo
A região do presente estudo abrange a Plataforma Continental do estado de Santa
Catarina, desde a fronteira com o Rio Grande do Sul até a fronteira com o Paraná, e entre as
isóbatas de 10 até 150 metros (Fig. 1).
A maior parte da plataforma continental catarinense está inserida na Plataforma
Continental Sudeste do Brasil (PCSE), ao norte do Cabo de Santa Marta, e sua porção mais
austral está na Plataforma Continental Sul do Brasil (PCS), que se estende do Cabo de Santa
Marta até o Arroio Chuí. No trecho da PCSE, o alinhamento da costa é aproximadamente
norte-sul, e no trecho da PCS o alinhamento é nordeste-sudoeste.
A quebra de plataforma se dá a uma profundidade que varia de 120 à 180 metros, e a
largura varia entre 100 à 220 quilômetros, no Cabo de Santa Marta e na Baía da Babitonga,
respectivamente. As principais massas de água presentes neste trecho do litoral brasileiro são:
Água Tropical - AT, (ii) Água Central do Atlântico Sul – ACAS, (iii) Água de Plataforma –
AP, (iv) Água Costeira – AC, e Água da Pluma do Prata – APP. Estas massas de água foram
descritas anteriormente para a região por Castro Filho (1977); Miranda (1982); Lentini
(1997); Carvalho et al. (1998); Mémery et al. (2000); Piola et al. (2000 e 2005); Silva et al.
(2004); Schettini et al. (2005) e Möller Jr. et al. (2008).
3.2 Amostras
As amostras utilizadas neste estudo originaram do projeto “Prospecção de ovos e
larvas da Sardinha-Verdadeira (Sardinella brasiliensis) na costa catarinense” (PROSAR)
executado pela Universidade do Vale do Itajaí em parceria com o Sindicato dos Trabalhadores
da Indústria de Pesca de Itajaí (SITRAPESCA) e o Centro de Pesquisa e Gestão de Recursos
Pesqueiros do Litoral Sudeste e Sul (CEPSUL/IBAMA).
Neste projeto foram realizados cruzeiros oceanográficos entre os anos de 2005 e
2006 na costa de Santa Catarina, limitadas pelas fronteiras com os estados do Paraná (ao

20
norte) e do Rio Grande do Sul (ao sul) (Lat. 26° 00` S à 29° 30` S e Long. 49° 41` W à 47°
05` W) e entre as isóbatas de 10 a 150 m.
Os cruzeiros foram realizados pelo Navio de Pesquisa do IBAMA Soloncy Moura
em duas campanhas em Primavera de 2005 e Outono de 2006. O plano amostral constou em
estações amostrais distribuídas em 10 perfis perpendiculares a costa totalizando 33 amostras
por cruzeiro (Fig. 1).
Os dados de temperatura e salinidade foram obtidos em cada cruzeiro mediante o uso
de uma sonda tipo CTD, marca SAIV S/A modelo SD204 em perfis verticais em cada estação
oceanográfica (HILLE et al., 2008).
Figura 1 – Mapa de localização do litoral do estado de Santa Catarina com as amostras oceanográficas realizadas em Primavera de 2005 e Outono de 2006 dentro do Projeto PROSAR.
As amostras de zooplâncton foram obtidas mediante arrastos oblíquos integrando da
superfície ao fundo utilizando uma rede tipo Bongo de 330 µm de tamanho de malha, 60 cm
de diâmetro de boca equipada com fluxômetro calibrado. Após as coletas, as amostras foram
fixadas em solução de formaldeido a 4% e estocadas no Laboratório de Oceanografia
Biológica do CTTMar/UNIVALI.
Em laboratório, as análises qualitativas e quantitativas foram realizadas em câmaras
do tipo Bogorov sob microscópio estereoscópico segundo as recomendações de Boltovskoy
(1981), que constou na retirada de alíquotas que variaram de 3 a 10% da amostra total, através

21
do sub-amostrador do tipo pistão. Foram quantificados todos os organismos zooplanctônicos
presentes nas amostras e a classificação ao menor nível taxonômico possível com o auxílio
dos trabalhos de Ramirez (1981) e Onbé (1999) para Cladocera e Björnberg (1981), Bradford-
Grieve et al. (1999) e Hernandez & Morales (1994) para Copepoda.
Dados de densidade foram obtidos a partir das estimativas de densidade total da
amostra que foram expressos pelo número de organismos por metro cúbico (Org. m-3) de água
filtrada pela rede. Além disto, foram calculados os índices de riqueza de Margalef (OMORI &
IKEDA, 1984), de diversidade de Shannon-Weaver (LUDWIG & REYNOLDS, 1988) e de
equitabilidade de Pielou (PIELOU, 1966).
Índice de Riqueza de Margalef (D)
LnN
SD
1−= (1)
Sendo:
S = número de espécies
N = número total de indivíduos
Índice de Diversidade de Shannon-Weaver (H’)
∑=
×−=
S
i NnLn
NnH ii
1
' (2)
Sendo:
S = número total de espécies amostradas
N = número total de indivíduos
ni = número de indivíduos amostrados para a i-ésima espécie
Índice de Equitabilidade de Pielou (J’)
max
''
H
HJ = (3)
Sendo:
Hmax = ln(S)

22
O tratamento dos dados constou na caracterização das associações de espécies de
Copepoda e Cladocera e sua associação com as massas de água ocorrentes apresentadas por
Hille et al. (2008). Para isto foi aplicado a análise de ordenação (Análises de Componentes
Principais – ACP) (PIELOU, 1984) para identificação dos grupos de espécies de similares
exigências ecológicas e identificação de forçantes ambientais que poderiam influenciar a
formação dos grupos. Para este caso, foram utilizadas as espécies que apresentaram uma
freqüência de ocorrência superior a 20%, além de dados de temperatura e salinidade dos
pontos intermediários a coluna de água amostrada nos arrastos oblíquos (Fig. 1). Com o
intuito de aproximar a distribuição dos dados a uma distribuição normal, os valores das
variáveis bióticas e abióticas foram previamente transformados em Log (x+1).
As comparações faunísticas com outras áreas geográficas, tanto ao norte como ao sul
do estado de Santa Catarina, foram realizadas a partir de comparações, entre outros, com os
trabalhos de Sartori & Lopes (2000), Muxagata (1999), Oliveira (1999), Resgalla Jr. & Montú
(1993), Vega-Perez (1993), Fernández Aráoz (1991) e Rocha (1977). Para a identificação dos
indicadores de ressurgências foram utilizados os trabalhos de Valentin (1988) e Valentin et al.
(1976).

23
4.0 RESULTADOS
4.1 Condições oceanográficas durante o período de estudo
Para o período de estudo deste trabalho, as condições oceanográficas foram
estudadas por Hille et al. (2008) que identificou as massas de água ocorrentes na região. De
uma maneira geral, para Primavera de 2005, as águas foram caracterizadas por baixas
salinidades e altas temperaturas, principalmente entre São Francisco do Sul e a Península de
Porto Belo, indicando forte influência de aportes continentais e a formação da Água Costeira
disposta localmente pelo aporte de água doce proveniente dos principais rios da região como
o Itajaí-Açú, Itapocu e Tijucas. Essas Regiões de Influência Fluvial (RIFs) destacam-se por
serem ricas em nutrientes e, conseqüentemente, apresentam alto valor em termos de recursos
pesqueiros (HILLE et al., 2008; SCHETTINI, et al., 2005). Já ao Sul, ocorreram as águas de
altas salinidades e baixas temperaturas. As águas de menores temperaturas foram observadas
nos pontos amostrais próximos ao Cabo de Santa Marta. Para a salinidade, os maiores valores
(>34) estiveram concentrados nos pontos ao sul da Ilha de Santa Catarina, sugerindo a
intrusão da ACAS na região. Externamente a isóbata de 100 m a Água Tropical (AT)
dominou ao longo da plataforma. Este cruzeiro foi o que apresentou maior variação de
temperatura entre os pontos amostrais, com valor mínimo de 14,0 e máximo de 24,6ºC
(Tabela 1).
Já no cruzeiro de Outono de 2006, as águas que se concentram ao norte foram
essencialmente de altas temperaturas e salinidades (AT), enquanto ao sul, baixas temperaturas
e salinidades foram observadas. Esta mancha de baixa salinidade penetrou na plataforma
proveniente do sul do Estado devido a influencia da Frente do Prata (mistura da Corrente das
Malvinas com a descarga do Rio da Prata e, em menor escala, da Lagoa dos Patos). Uma faixa
com valores de salinidade mais elevados foi observada em quase toda área de estudo
localizada externamente a isóbata de 100 m indicando a presença das AT.
Devido a técnica de amostragem empregada para as coletas dos organismos
zooplanctônicos, que envolveu o uso de rede Bongo integrando da superfície ao fundo da
coluna de água (máximo de 130 m de profundidade), foi estimado os valores médios da

24
temperatura e salinidade e caracterizado as massas de água dominantes em grupos de pontos
amostrais para cada cruzeiro (Fig. 2). Desta forma, águas típicas descritas por Hille et al.
(2008) foram redefinidas como: Água Costeira (AC) na região de RIFs para o cruzeiro de
Primavera de 2005, ainda para este cruzeiro foram definidas a Água Subtropical de
Plataforma (ASP) na plataforma externa e Água Central do Atlântico Sul (ACAS) na
ressurgência do Cabo de Santa Marta. Já o cruzeiro de Outono de 2006 foi caracterizado pela
presença de ASP na região dos RIFs, Água Tropical (AT) na região central da plataforma e
Água da Pluma do Prata (APP) na altura do Cabo de Santa Marta.
Tabela 1 - Valores mínimos, médios e máximos de Salinidade e temperatura (ºC) referentes aos cruzeiros de Primavera de 2005 e Outono de 2006 do projeto PROSAR modificado de Hille et al. (2008).
Salinidade Temperatura
Mínima Média Máxima Mínima Média Máxima Primavera 2005 10,99 35,02 36,79 14,00ºC 20,07ºC 24,61ºC Outono 2006 9,92 34,66 36,86 18,16ºC 21,58ºC 25,29ºC

25
Figura 2 – Diagramas TS e classificação das massas de água referentes aos cruzeiros de Primavera de 2005 (A) e Outono de 2006 (B) do projeto PROSAR (modificado de Hille et al., 2007) com valores médios da coluna de água. Sendo AC= Água Costeira, ASP= Água Subtropical de Plataforma, ACAS= Água Central do Atlântico Sul, AT= Água tropical e APP= Água da Pluma do Prata.
4.2 Comunidade zooplanctônica
4.2.1 Densidade total do zooplâncton
Em termos de densidade da comunidade zooplanctônica total na plataforma
catarinense não foi observado um padrão claro de variações sazonais e espaciais. Observou-
se, entretanto, tendências dos pontos amostrais mais costeiros apresentarem as maiores
concentrações de organismos zooplanctônicos. Entre os cruzeiros, os valores médios de
densidade observados foram de 482 Org. m-3 para Primavera de 2005 e 593 Org. m-3 para
Outono de 2006 (Fig. 3).

26
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
Figura 3 – Distribuição espacial dos valores de densidade para o zooplâncton total (Org. m-3) em (A) Primavera de 2005 e (B) Outono de 2006.
4.2.2 Grandes grupos do zooplâncton
A comunidade zooplanctônica da plataforma de Santa Catarina foi composta por 14
grandes grupos zooplanctônicos. Em Primavera de 2005 o grupo mais abundante foi o de
Copepoda (51,7%), seguido por Cladocera (15,7%) e Salpas (13,6%), enquanto que em
Outono de 2006, Copepoda também foi o dominante (74,6%), mas houve diminuição da
representatividade dos demais grupos como Salpas (7,3%), Chaetognatha (5,69%) e
Cladocera (3,7%) (Tabela 2). Grupos menos representativos envolveram Siphonophorae,
Hydromedusae, Polychaeta, larvas de Mollusca, Ostracoda, larvas de Cirripedia, Amphipoda,
Euphausiacea e Pteropoda (Tabela 3).
A B

27
Tabela 2 - Frequência de Ocorrência (%) dos grandes grupos do zooplâncton para o cruzeiro de Primavera de 2005 e Outono de 2006.
FO (%) Grupos Primavera
2005 Outono
2006 Siphonoforae 27,27 33,33
Hydromedusae 21,21 12,12
Polychaeta 0,00 3,03
Mollusca 48,48 69,7
Cladocera 100 90,91
Ostracoda 0,00 9,09
Copepoda 100 100
Cirripedia 0,00 30,3
Amphipoda 39,39 0,00
Euphausiae 6,06 3,03
Decapoda 100 100
Chaetognatha 100 93,94
Salpidae 69,7 57,58
Appendicularia 57,58 57,58
Pteropoda 66,67 51,52
Tabela 3 - Abundância relativa média (%) dos grandes grupos ocorrentes para o cruzeiro de (A) Primavera de 2005 e (B) Outono de 2006.
Grupos Primavera 2005 Outono 2006
Copepoda 60,38 71,94
Cladocera 12,46 3,98
Decapoda 4,14 5,12
Chaetognatha 3,40 8,18
Salpidae 14,08 5,22
Appendicularia 2,45 0,68
Outros 1,80 3,92

28
4.3 MOLLUSCA MEROPLANCTÔNICO (LARVAS)
Para o grupo de larvas de Mollusca, foram observados formas véligers de Gastropoda
e Bivalvia ao longo da plataforma, mas sem um padrão espacial bem definido (Fig. 10). Os
valores médios de densidade foram de 1 Org. m-3 em Primavera de 2005 e 7 Org. m-3 em
Outono de 2006.
Figura 4 – Distribuição espacial da densidade (Org. m-3) de larvas de Mollusca bentônicos nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
A B
SC

29
4.4 DECAPODA
As larvas de Decapoda apresentaram uma ampla distribuição e ocorreram em todos
os pontos amostrais nos dois cruzeiros, mas com altos valores na região costeira. Foram
registradas 4 estágios de desenvolvimento (protozoea, mysis, zoea e decapodito) e 1 espécie
holoplanctônica Lucifer faxoni (Fig. 7). Os valores médios de densidade foram de 12 Org. m-3
em Primavera de 2005 e 11 Org. m-3 em Outono de 2006.
Figura 5 – Distribuição espacial da densidade (Org. m-3) de Larvas de Decapoda nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
A B
SC

30
4.5 CHAETOGNATHA
Este grupo apresentou alta freqüência de ocorrência, e uma tendência de maiores
valores de densidade nos pontos mais costeiros. Esta evidência ficou mais clara no cruzeiro de
Outono de 2006 (Fig. 6). Os valores médios de densidade foram de 7 Org. m-3 em Primavera
de 2005 e 15 Org. m-3 em Outono de 2006.
Figura 6 – Distribuição espacial da densidade (Org. m-3) de Chaetognatha nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
A B
31
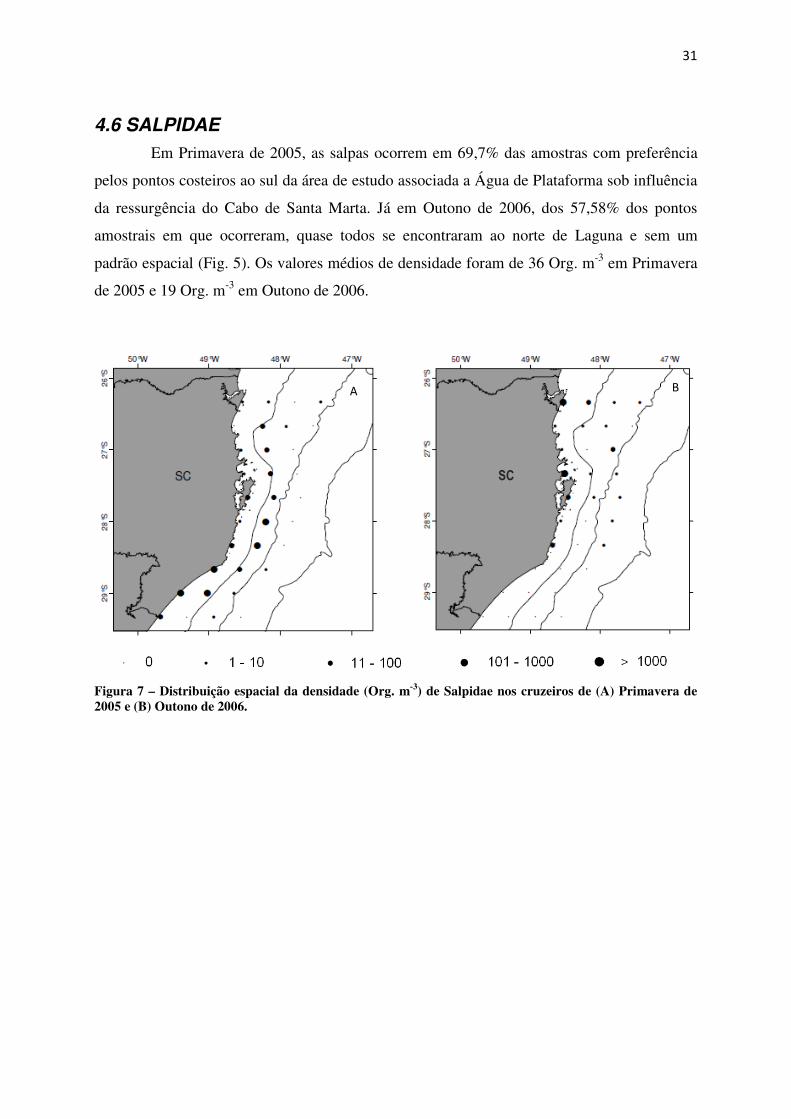
4.6 SALPIDAE Em Primavera de 2005, as salpas ocorrem em 69,7% das amostras com preferência
pelos pontos costeiros ao sul da área de estudo associada a Água de Plataforma sob influência
da ressurgência do Cabo de Santa Marta. Já em Outono de 2006, dos 57,58% dos pontos
amostrais em que ocorreram, quase todos se encontraram ao norte de Laguna e sem um
padrão espacial (Fig. 5). Os valores médios de densidade foram de 36 Org. m-3 em Primavera
de 2005 e 19 Org. m-3 em Outono de 2006.
Figura 7 – Distribuição espacial da densidade (Org. m-3) de Salpidae nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
A B
SC
32
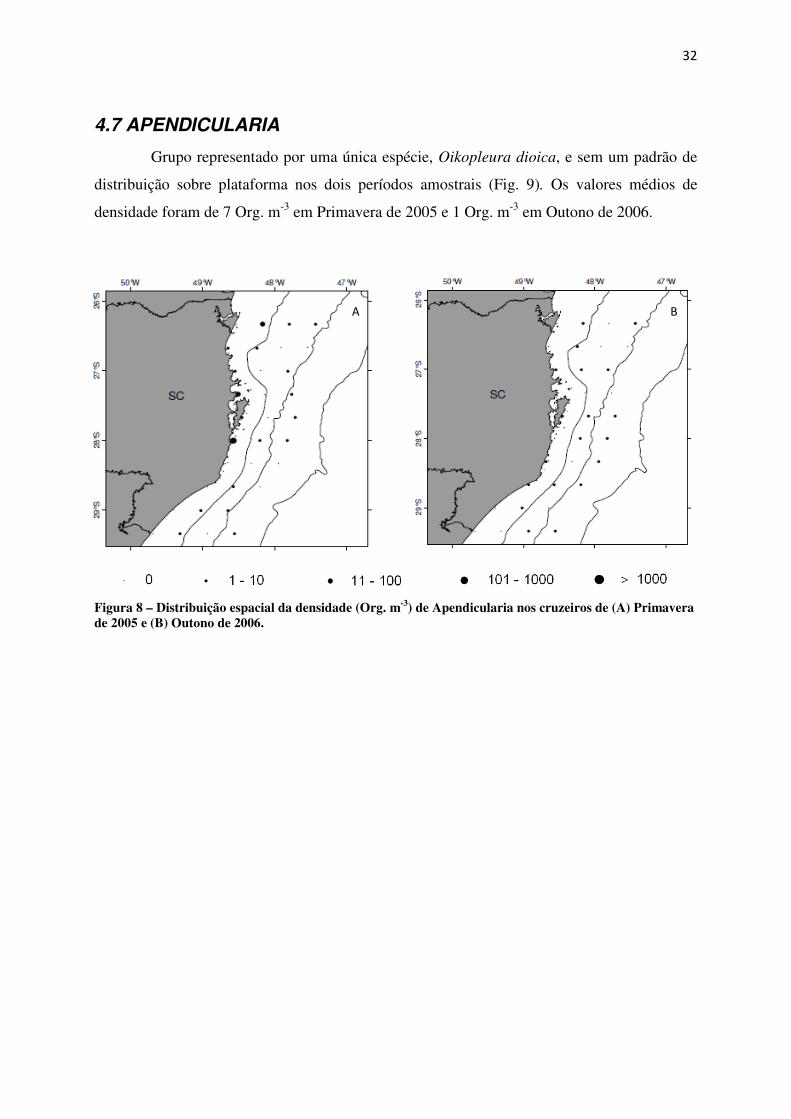
4.7 APENDICULARIA
Grupo representado por uma única espécie, Oikopleura dioica, e sem um padrão de
distribuição sobre plataforma nos dois períodos amostrais (Fig. 9). Os valores médios de
densidade foram de 7 Org. m-3 em Primavera de 2005 e 1 Org. m-3 em Outono de 2006.
Figura 8 – Distribuição espacial da densidade (Org. m-3) de Apendicularia nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
A B

33
4.8 PTEROPODA
A espécie Cresseis sp foi a única representante deste grupo e em formas juvenis,
ocorrendo com maior freqüência e densidade no cruzeiro de Primavera de 2005. No geral,
este grupo apresentou distribuição em baixas densidades e evitando pontos amostrais
costeiros (Fig. 8). Os valores médios de densidade foram de 3 Org. m-3 em Primavera de 2005
e 1 Org. m-3 em Outono de 2006.
Figura 9 – Distribuição espacial da densidade (Org. m-3) de Pteropoda nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
A B
SC

34
4.9 COPEPODA
Em termos de distribuição espacial, Copepoda apresentou os maiores valores de
densidade nos pontos amostrais na costa e na plataforma interna em ambos os cruzeiros (Fig.
11).
Figura 10 - Distribuição espacial da densidade (Org. m3) de Copepoda nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
Foram registrados, nos dois cruzeiros, 43 taxa de Copepoda, entre ordem, família,
gênero e espécies. Os médios de densidade foram de 153 Org. m-3 em Primavera de 2005 e
196 Org. m-3 em Outono de 2006.
Para o cruzeiro de Primavera de 2005, as espécies de maiores densidades foram
Temora stylifera (média de 40 Org. m-3), Oithona plumifera (média de 18 Org. m-3), Oncaea
clevei (média de 17 Org. m-3), Temora turbinata (média de 16 Org. m-3), Centropages
velificatus (média de 10 Org. m-3) e Acartia lilljeborgi (média de 4 Org. m-3) (Tabela 4).
A B

35
Tabela 4 - Frequência de Ocorrência (FO) e Abundância Relativa (AR) para as espécies de Copepoda observados nos cruzeiros de Primavera de 2005 e Outono de 2006.
FO (%) AR (%) Taxa
Primavera 2005 Outono 2006 Primavera 2005 Outono 2006 COPEPODA 100,00 100,00 60,38 71,95
Copepodito 100,00 84,21 14,69 10,54
Naupliu 24,24 21,05 0,64 0,34
Acartia sp 6,06 0,00 0,04 0,00
Acartia lilljeborgi Giesbrecht, 1889 36,36 39,47 1,78 10,96
Aetideopsis carinata Bradford, 1969 3,03 2,63 0,01 0,02
Calanoides carinatus (Krøyer, 1849) 3,03 2,63 0,08 0,00
Calocalanus pavo (Dana, 1852) 3,03 23,68 0,08 0,38
Candacia curta (Dana, 1849) 6,06 26,32 0,08 0,28
Candacia pachydactila (Dana, 1849) 3,03 21,05 0,02 0,25
Centropages velificatus (Oliveira, 1947) 57,58 60,53 6,39 1,74
Clausocalanus furcatus (Brady, 1883) 0,00 57,89 0,00 15,87
Copilia mirabilis Dana, 1852 0,00 10,53 0,00 0,12
Corycaues giesbrecthi (F. Dahl, 1894) 6,06 63,16 0,27 4,37
Corycaeus speciosus Dana, 1849 72,73 57,89 0,03 3,45 Corycaeus sp 3,03 7,89 5,38 0,83
Eucalanus sp 54,55 28,95 6,26 1,79
Euchaeta marina (Prestandrea, 1833) 3,03 36,84 0,04 0,54
Euchaeta media Giesbrecht, 1888 3,03 2,63 0,03 0,01
Farranula gracilis (Dana, 1849) 3,03 7,89 0,04 0,22
Labidocera fluviatilis Dahl, 1894 12,12 13,16 0,13 0,14
Lucicutia flavicornis (Claus, 1863) 0,00 5,26 0,00 0,02
Macrosetella gracilis (Dana, 1847) 0,00 26,32 0,00 0,17
Microsetella rosea (Dana, 1849) 15,15 2,63 0,32 0,04
Miracia efferata Dana, 1849 0,00 13,16 0,00 0,13
Nannocalanus minor (Claus,1863) 0,00 50,00 0,00 2,36
Neocalanus gracilis (Dana, 1849) 6,06 18,42 0,21 0,33
Oithona plumifera Baird, 1843 87,88 76,32 12,81 7,64
Oithona setigera (Dana, 1849) 0,00 5,26 0,00 0,03
Oncaea sp 21,21 5,26 1,85 0,71
Oncaea Antarctica Heron, 1977 0,00 10,53 0,00 0,07
Oncaea clevei Früchtl, 1923 63,64 84,21 11,98 12,54
Paracalanus sp 63,64 0,00 1,28 0,00
Paracalanus quasimodo Bowman, 1971 9,09 2,63 0,46 0,01
Paracandacia simplex (Giesbrecht, 1889) 0,00 10,53 0,00 0,07
Pareucalanus sewelli (Fleminger, 1973) 3,03 10,52 0,03 12,55
Temora stylifera (Dana, 1849) 96,97 81,58 25,75 6,66
Temora turbinata (Dana, 1849) 78,79 55,26 7,30 0,04
Sapphirina sp 3,03 7,89 0,26 0,01
Scolecithrix bradyi Giesbrecht, 1888 0,00 5,26 0,00 0,23
Scolecithrix danae (Lubbock, 1856) 3,03 21,05 0,02 0,34
Subeucalanus crassus (Giesbrecht, 1888) 0,00 10,52 0,00 2,76
Subeucalanus pielatus (Giesbrecht, 1888) 9,09 47,26 0,95 1,20
Undinula vulgaris (Dana, 1849) 24,24 39,47 0,78 0,05

36
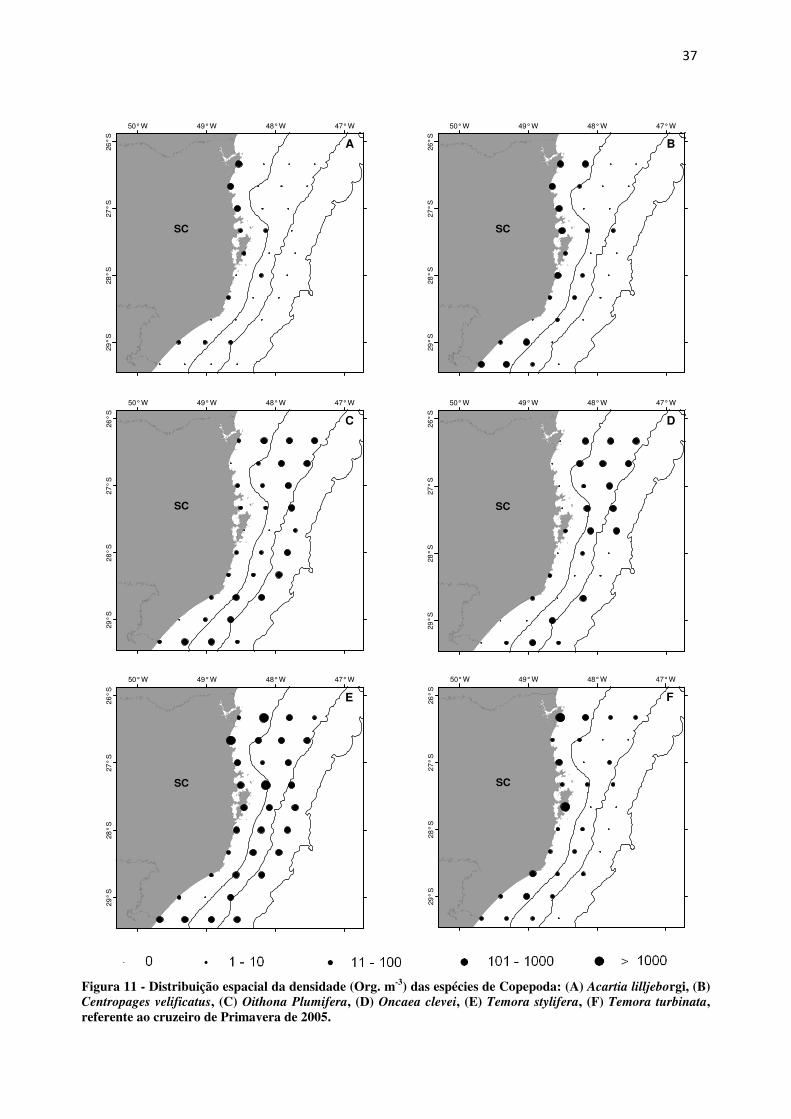
A espécie Acartia lilljeborgi ocorreu nos pontos bem próximos da costa, assim como
Centropages velificatus e Temora turbinata. Contrariamente, Oithona plumifera e Oncaea
clevei aumentam em densidade em direção aos pontos da plataforma externa. Já Temora
stylifera apresentou-se bem distribuída em toda a plataforma catarinense (Fig. 12).
37
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
A
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
B
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
C
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
D
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
E
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
F
Figura 11 - Distribuição espacial da densidade (Org. m-3) das espécies de Copepoda: (A) Acartia lilljeborgi, (B) Centropages velificatus, (C) Oithona Plumifera, (D) Oncaea clevei, (E) Temora stylifera, (F) Temora turbinata, referente ao cruzeiro de Primavera de 2005.

38
A espécie Undinula vulgaris ocorreu em pontos esparsos, porém limitados a região
norte da área de estudo (Fig. 13).
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
G
Figura 12 - Distribuição espacial da densidade (Org. m-3) da espécie de Copepoda Undinula
vulgaris, referente ao cruzeiro de Primavera de 2005.
Para a amostragem de Outono de 2006 as espécies mais abundantes foram Acartia
lilljeborgi (média de 71 Org. m-3)¸ Clausocalanus furcatus (média de 23 Org. m-3), Temora
stylifera (média de 18 Org. m-3), Temora turbinata (média de 18 Org. m-3), Oncaea clevei
(média de 16 Org. m-3), Oithona plumifera (média de 8 Org. m-3), Corycaeus giesbrechti
(média de 6 Org. m-3) e Subeucalanus pileatus (média de 5 Org. m-3) (Tabela 3).
Em relação a distribuição espacial, Acartia lilljeborgi ocorreu em toda a plataforma,
mas com os maiores valores de densidade observados ao norte da Ilha de Santa Catarina. A
espécie Clausocalanus furcatus Corycaeus giesbrecthi, Oithona plumifera, Oncaea clevei e
Temora stylifera não apresentaram um padrão definido de distribuição, ocorrendo em toda a
plataforma (Fig. 14). Temora turbinata apresentou preferência por ocorrências em pontos
amostrais mais costeiros e Subeucalanus pileatus mostrou maiores densidades ao sul da área
de estudo (Fig. 15).

39
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
A
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
F
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
O
47° W48° W49° W50° W26
° S
27°
S28
° S
29°
S
SC
P
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
N
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
M
Figura 13 – Distribuição espacial da densidade (Org. m-3) das espécies de Copepoda: (A) Acartia
lilljeborgi¸ (B) Clausocalanus furcatus, (C) Temora stylifera, (D) Temora turbinata, (E) Oncaea clevei, (F) Oithona plumifera, referente ao cruzeiro de Outono de 2006.
A B
C D
E F

40
A distribuição de outros Copepoda, como Corycaeus speciosus e Nannocalanus
minor mostram-se sem padrão definido na distribuição espacial. Para estas espécies, sugere-se
que os maiores valores de densidade estiveram próximos da influencia dos RIFs. Já as
espécies Euchaeta marina, Macrosetella gracilis e Neocalanus gracilis mostraram-se
presentes em pontos afastados da costa, porém em baixa densidade entre 1 e 10 Org. m-3 (Fig.
15 e 16).

41
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
P
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
R
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
H
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
K
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
I
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
J
Figura 14 - Distribuição espacial da densidade (Org. m-3) das espécies de Copepoda: (A) Temora turbinata, (B) Subeucalanus pileatus, (C) Corycaeus speciosus, (D) Nannocalanus minor, (E) Euchaeta
marina, (F) Macrosetella gracilis referente ao cruzeiro Outono de 2006.
A B
C D
E F

42
O Copepoda Undinula vulgaris também não apresenta padrão em relação a
distribuição, mas foi possível visualizar maiores densidades ao sul da Ilha de Santa Catarina
(Fig. 16).
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
L
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
S
Figura 15 - Distribuição espacial da densidade (Org. m-3) das espécies de Copepoda (A) Neocalanus
gracilis e (B) Undinula vulgaris, referente ao cruzeiro de Outono de 2006.
4.9.1 Índices de Riqueza, Diversidade e Equitabilidade para Copepoda
Para os Copepoda, os maiores índices de riqueza ocorreram em Outono de 2006
(0,92 para Primavera e 1,90 para Outono), assim como há uma tendência dos pontos mais
externos da plataforma serem os mais ricos (Fig. 17), enquanto que para o cruzeiro de
Primavera de 2005 não foi observado um padrão claro de distribuição espacial.
A B

43
Figura 16 - Distribuição espacial da Riqueza de espécies (Nits) para o grupo de Copepoda nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
De maneira geral, a diversidade, nos pontos mais costeiros, se manteve baixa para os
dois cruzeiros, contudo, um gradiente em direção a plataforma externa só foi observado em
Outono de 2006 (Fig. 18). A diversidade média para Copepoda foi de 0,98 Nits para o
cruzeiro de Primavera de 2005 e de 1,67 Nits para Outono de 2006.
Figura 17 - Distribuição espacial da Diversidade (Nits) para o grupo de Copepoda nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
A B
A B
SC
SC

44
Para a equitabilidade, foi possível observar um gradiente perpendicular a costa em
Primavera de 2005, com menores índices próximo a costa. No cruzeiro de Outono de 2006
não há um padrão na distribuição (Fig. 19). Os valores médios de Equitabilidade foram de
0,59 Nits para Primavera de 2005 e 0,71 para Outono de 2006.
Figura 18 - Distribuição espacial da Equitabilidade (Nits) para o grupo de Copepoda nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
A B

45
4.10 CLADOCERA
Este grupo apresentou distribuição espacial preferencialmente costeiro, com
ocorrências ao longo da costa para o cruzeiros de Primavera de 2005 e limitada a região norte
da área de estudo e associada a RIFs em Outono de 2006 (Fig. 20). Os valores médios de
densidade para Cladocera foram de 42 Org. m-3 para Primavera de 2005 e 10 Org. m-3 para
Outono de 2006.
Figura 19 - Distribuição espacial da densidade (Org. m3) de Cladocera nos cruzeiros de (A) Primavera de 2005 e (B) Outono de 2006.
Foram observados 4 taxa de Cladocera em ambos os cruzeiros. A espécie Penilia
avirostris foi a mais abundante e freqüente das espécies, seguido por Pseudoevadne tergestina
(Tabela 5).
Tabela 5 - Frequência de Ocorrência (FO) e Abundância Relativa (AR) para as espécies de Cladocera referente aos cruzeiros de Primavera de 2005 e Outono de 2006.
FO (%) AR (%) Taxa
Primavera 2005 Outono 2006 Primavera 2005 Outono 2006 CLADOCERA 100,00 91,00 12,47 3,99
Penilia avirostris Dana, 1852 93,94 51,52 63,37 33,77
Pleopsis schmackeri (Poppe, 1889) 18,18 27,27 3,51 4,61
Evadne spinifera Müller, 1868 33,33 36,36 11,14 12,23
Pseudevadne tergestina (Claus, 1877) 72,73 75,76 21,98 40,30
SC SC
A B
46
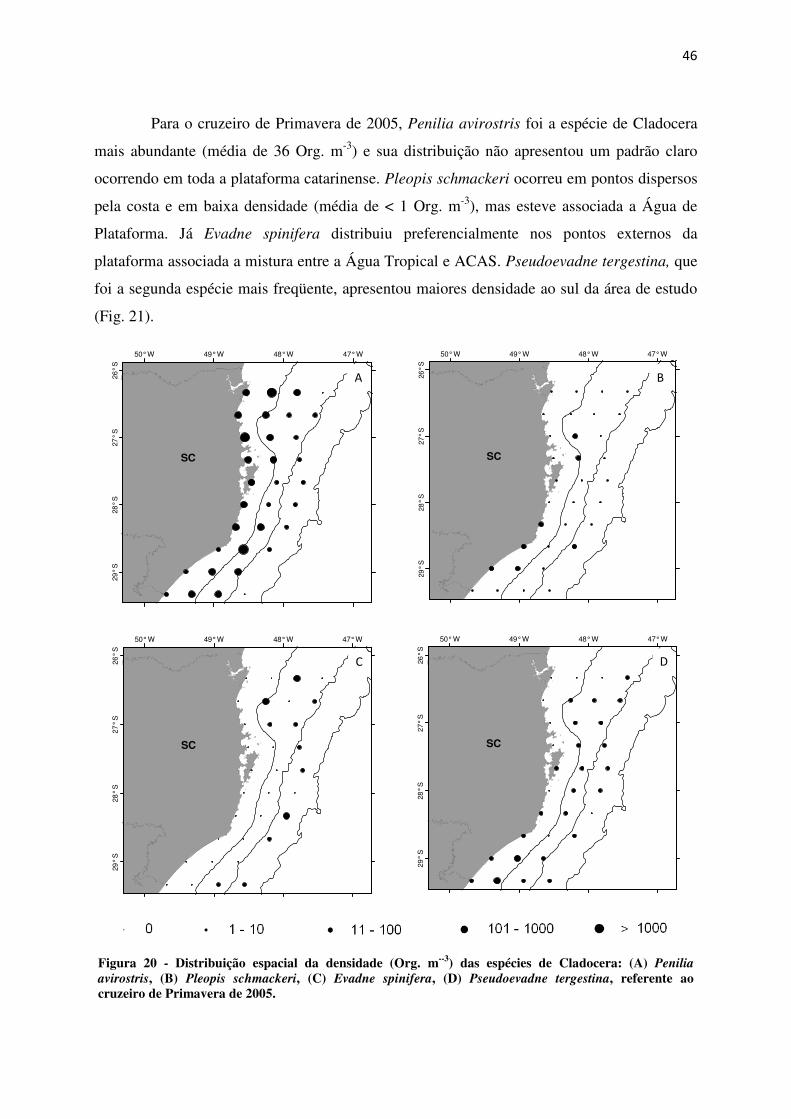
Para o cruzeiro de Primavera de 2005, Penilia avirostris foi a espécie de Cladocera
mais abundante (média de 36 Org. m-3) e sua distribuição não apresentou um padrão claro
ocorrendo em toda a plataforma catarinense. Pleopis schmackeri ocorreu em pontos dispersos
pela costa e em baixa densidade (média de < 1 Org. m-3), mas esteve associada a Água de
Plataforma. Já Evadne spinifera distribuiu preferencialmente nos pontos externos da
plataforma associada a mistura entre a Água Tropical e ACAS. Pseudoevadne tergestina, que
foi a segunda espécie mais freqüente, apresentou maiores densidade ao sul da área de estudo
(Fig. 21).
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
H
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
I
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
J
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
K
Figura 20 - Distribuição espacial da densidade (Org. m--3) das espécies de Cladocera: (A) Penilia
avirostris, (B) Pleopis schmackeri, (C) Evadne spinifera, (D) Pseudoevadne tergestina, referente ao cruzeiro de Primavera de 2005.
A B
C D
C

47
Em Outono de 2006, Penilia avirostris apresentou uma distribuição mais específica,
ocorrendo principalmente no norte da área de estudo, com os maiores valores na região dos
RIFs (Fig. 22). Pleopis schmackeri apresentou distribuição próximo a costa associada a Água
de Plataforma e Tropical, enquanto Pseudoevadne spinifera mostrou preferências por pontos
na plataforma externa. Pseudoevadne tergestina foi a espécie de Cladocera mais freqüente, e
apresentou uma ampla distribuição, mas evitando a região das RIFs ao norte da área de
estudo.
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
T
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
U
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
V
47° W48° W49° W50° W
26°
S27
° S
28°
S29
° S
SC
X
Figura 21 – Distribuição espacial da densidade (Org. m-3) das espécies de Cladocera: (A) Penilia
avirostris, (B) Pleopis schmackeri, (C) Pseudoevadne spinifera, (D) Pseudoevadne tergestina referente ao cruzeiro de Outono de 2006.
A
C
B
C
C
C
D
C

48
4.10.1 Índices de Riqueza, Diversidade e Equitabilidade para Cladocera
Os valores de riqueza para o grupo Cladocera foram mais baixos do que os
Copepoda, porém o padrão de distribuição se mantém semelhante, de modo que os pontos
mais costeiros apresentam baixa riqueza (Fig. 23). Os valores médios de Riqueza para
Cladocera foram de 0,44 Nits para Primavera de 2005 e de 0,42 Nits para Outono de 2006.
Figura 22 – Distribuição espacial da Riqueza de espécies (Nits) para Cladocera nos cruzeiros de Primavera de 2005 e Outono de 2006.
Em relação a diversidade, alguns pontos com valores maiores ocorrem,
esparsamente, ao longo da costa, principalmente em Primavera de 2005, porém não
mostraram um padrão espacial em ambos os cruzeiros (Fig. 24). Os valores médios de
diversidade para Cladocera foram de 0,52 Nits para Primavera de 2005 e de 0,39 Nits para
Outono de 2006.
A B
SC

49
Figura 23 – Distribuição espacial da Diversidade (Nits) para Cladocera nos cruzeiros de Primavera de 2005 e Outono de 2006.
O índice de equitabilidade se distribuiu sem um padrão definido nos dois cruzeiros
(Fig. 25). Os valores médios de Equitabilidade para Cladocera foram de 0,75 Nits para
Primavera de 2005 e de 0,62 Nits para Outono de 2006.
Figura 24 – Distribuição espacial da Equitabilidade (Nits) para Cladocera nos cruzeiros de Primavera de 2005 e Outono de 2006.
A B
SC
A B

50
4.11 Variáveis ambientais e comunidade zooplanctônica
O resultado das Análises de Componentes Principais (ACP), para o cruzeiro de
Primavera de 2005 apresentou uma percentagem acumulada de explicabilidade de 56,01%
para os dois primeiros eixos, mostrando a temperatura e a salinidade com alto peso sobre o
eixo 1, sendo a temperatura positivamente correlacionada e a salinidade negativamente
correlacionada. Estas variáveis correlacionaram um grupo de espécies constituído por
Centropages velificatus, Acartia lilljeborgi, Temora turbinata e Penilia avirostris como
espécies de maiores temperaturas e baixas salinidades (Tabela 6 e Fig. 26).
Tabela 6 – Peso das variáveis para ACP da comunidade zooplanctônica e parâmetros ambientais referente ao cruzeiro de Primavera de 2005.
Primavera 2005 Axis 1 Axis 2
Porcentagem acumulada de explicabilidade
38,55 56,01
Acartia lilljeborgi 0,374 0,101
Centropages velificatus 0,395 -0,017
Oithona plumifera -0,285 0,247
Oncaea clevei -0,29 0,307
Temora stylifera 0,053 0,492
Temora turbinata 0,302 -0,128
Undinula vulgaris 0,05 0,387
Penilia avirostris 0,307 0,101
Pleopsis schmackeri -0,016 -0,412
Evadne spinifera -0,211 0,203
Pseudoevadne tergestina -0,175 -0,42
Salinidade -0,338 -0,167
Temperatura 0,397 -0,006

51
Figura 25 - Resultado da ACP para a comunidade zooplanctônica e os parâmetros ambientais referente ao cruzeiro de Primavera de 2005.
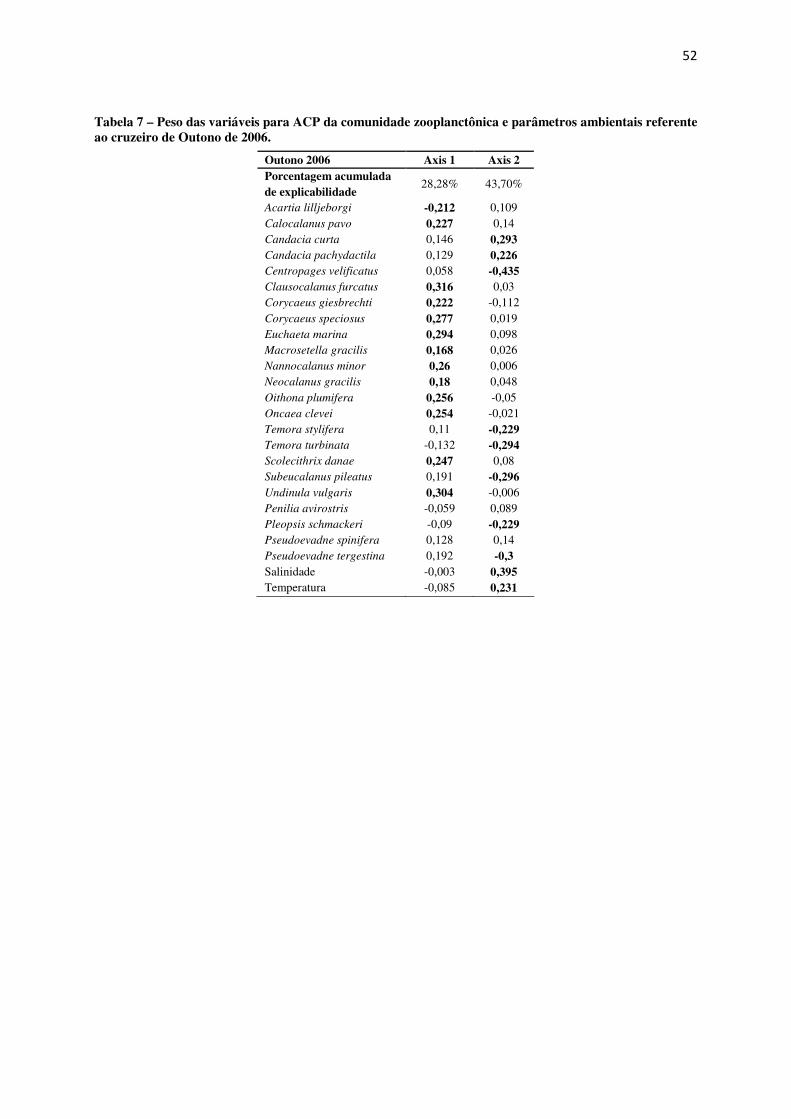
A ACP das amostragens de Outono de 2006 não apresentou um alto peso das
variáveis abióticas (temperatura e salinidade) com o eixo 1, e uma menor explicabilidade dos
dois primeiros eixos (43,7%). Observou-se, em base do ordenamento do eixo 1 dois grupos de
espécies, um positivamente correlacionado constituído por Calocalanus pavo, Clausocalanus
furcatus, Corycaeus giesbrechti, Corycaeus speciosus, Euchaeta marina, Macrosetella
gracilis, Nannocalanus minor, Neocalanus gracilis, Oithona plumifera, Oncaea clevei,
Scolecithrix danae e Undinula vulgaris e outro negativamente correlacionado constituído
somente por Acartia lilljeborgi (Tabela 7 e Fig. 27).
Para o eixo 2, Centropages velificatus e Pleopsis schmackeri apresentaram relação
inversa com a temperatura e salinidade (Fig. 27).
52
Tabela 7 – Peso das variáveis para ACP da comunidade zooplanctônica e parâmetros ambientais referente ao cruzeiro de Outono de 2006.
Outono 2006 Axis 1 Axis 2 Porcentagem acumulada de explicabilidade
28,28% 43,70%
Acartia lilljeborgi -0,212 0,109 Calocalanus pavo 0,227 0,14 Candacia curta 0,146 0,293 Candacia pachydactila 0,129 0,226 Centropages velificatus 0,058 -0,435 Clausocalanus furcatus 0,316 0,03 Corycaeus giesbrechti 0,222 -0,112 Corycaeus speciosus 0,277 0,019 Euchaeta marina 0,294 0,098 Macrosetella gracilis 0,168 0,026 Nannocalanus minor 0,26 0,006 Neocalanus gracilis 0,18 0,048 Oithona plumifera 0,256 -0,05 Oncaea clevei 0,254 -0,021 Temora stylifera 0,11 -0,229 Temora turbinata -0,132 -0,294 Scolecithrix danae 0,247 0,08 Subeucalanus pileatus 0,191 -0,296 Undinula vulgaris 0,304 -0,006 Penilia avirostris -0,059 0,089 Pleopsis schmackeri -0,09 -0,229 Pseudoevadne spinifera 0,128 0,14 Pseudoevadne tergestina 0,192 -0,3 Salinidade -0,003 0,395 Temperatura -0,085 0,231

53
Figura 26 - Resultado da ACP para a comunidade zooplanctônica e os parâmetros ambientais referente ao cruzeiro de Outono de 2006.

54
5.0 DISCUSSÃO
A Plataforma Continental Catarinense tem sido amplamente estudada devido a
ocorrência de uma alta diversidade de fenômenos oceanográficos. Por estar inserida, ao norte
do Cabo de Santa Marta na Plataforma Continental Sudeste do Brasil (PCSE) e ao sul na
Plataforma Continental Sul do Brasil (PCS), apresentam processos físicos como a advecção
para sul e meandramento da Corrente do Brasil (AT), a presença da ACAS em sub-superfície
e sua ressurgência no Cabo de Santa Marta, e a advecção para norte da Frente do Prata, além
das contribuições fluviais locais (HILLE, et al., 2008; SCHETTINI, et al., 2005). Esta
complexidade hidrológica determina também uma complexidade faunística que muitas vezes
restringem o uso dos organismos zooplanctônicos como indicadores hidrológicos quando
existem limitações do amostrador. No presente estudo, o emprego de um amostrador que
integra a coluna de água prejudica um refinamento na ocorrência e distribuição dos
organismos indicadores.
A composição da comunidade zooplanctônica foi caracterizada por 14 grandes
grupos e a distribuição espacial encontrada, para os dois cruzeiros, foi semelhante a relatada
por Vega-Pérez (1993), Muxagata (1999) e Sant’anna & Björnberg (2006) em estudos da
costa sul e sudeste brasileira, ou seja, as maiores abundâncias foram observadas na plataforma
interna.
Dos grupos holoplanctônicos observados nas amostras, Salpas, Chaetogntha,
Copepoda e Cladocera sempre foram os grupos mais representativos em águas costeiras do
estado de Santa Catarina (RESGALLA Jr., no prelo).
Pelo padrão de distribuição do grupo Salpidae, em Primavera de 2005, verificou-se
uma maior concentração na região do Cabo de Santa Marta, indicando associação com a
intrusão da ACAS como destacado por Hille et al. (2008). Muxagata (1999) encontrou num
ponto situado ao sudoeste da Baía de Guanabara/RJ a maior concentração de Salpidae
associada a presença de AT e ACAS. No trabalho de Resgalla Jr. (no prelo) é destacado que
este organismo pode estar relacionado com processos de ressurgência de curta duração devido
ao domínio de ventos nordeste nesta região. Nestas intrusões da água de maior profundidade
para a zona eufótica ocorre o aumento da biomassa do fitoplâncton e que não é consumido
pelo zooplâncton devido a intermitência do processo, ficando um excedente de produção

55
primária que entra na alça microbiana, o que favorece o desenvolvimento de organismos que
se alimentam por muco (RESGALLA et. al., 2001).
O grupo Chaetognatha ocorreu preferencialmente nos pontos costeiros da área de
estudo. A distribuição deste grupo tem sido relacionada com a ocorrência de Copepoda, seu
item alimentar preferencial (BOLTOVSKOY, 1981b). Liang & Vega-Pérez (1994) também
sugeriram esta relação entre os grupos para a região de Ubatuba, relatando que as maiores
densidades de Chaetognatha foram encontradas em águas costeiras, também mais ricas em
Copepoda. Muxagata (1999) encontrou as maiores concentrações destes organismos na área
ao norte de Cabo Frio/RJ, ao sul de São Sebastião/SP e ao norte de Paranaguá/PR e sugeriu
uma ocorrência prévia de ressurgência na região de Cabo Frio. Segundo Parsons et al. (1984),
valores elevados de predadores são costumeiros após os primeiros dias de ressurgência,
quando já existe uma elevada densidade de organismos herbívoros. Apesar da ampla
distribuição de Copepoda e Chaetognatha observados no presente estudo, pouca relação pôde
ser observada entre estes grupos.
As larvas Decapoda ocorreram em toda a área de estudo, com maiores densidades
nos pontos da plataforma interna. A ocorrência deste grupo e sua importância já tem sido
destacada em trabalhos anteriores na costa de Santa Catarina (RESGALLA JR. et al., 2008).
Também Muxagata (1999) caracterizou as larvas de Decapoda como sendo abundantes nas
regiões costeiras de Santa Catarina, Paraná e São Paulo. Além do interesse comercial, este
grupo é importante nos processos ecológicos dos ambientes aquáticos, pois atuam em
diferentes níveis da cadeia trófica, quer como herbívoros, predadores, necrófagos ou presas de
outros grupos (MAGALHÃES, 2000).
O Pteropoda Cresseis sp é um indicador da corrente do Brasil (VAN DER SPOEL &
BOLTOVSKOY, 1981). As altas densidades deste Mollusco planctônico também foram
observadas nos meses quentes entre a primavera e outono dos anos de 1997 e 2003 em
Garopaba e Governador Celso Ramos em acumulações massivas de conchas nas praias
(RESGALLA JR., no prelo). A ocorrência destes organismos apresenta similaridade com as
Salpas e podem estar associadas a processos de ressurgências, fato não confirmado neste
estudo.
A espécie Oikopleura dioica, única representante neste estudo do grupo
Appendicularia, obteve ampla distribuição, porém com baixas densidades em águas de alta
salinidade. A ocorrência desta espécie foi relatada por Campos (2004) entre a Belomonte/BA

56
e São Tomé/RJ, em Arraial do Cabo/RJ e entre Cabo Frio/RJ e Cabo de Santa Marta
Grande/SC, sendo que no primeiro e último trecho estavam sob influência da ACAS.
Os Mollusca meroplanctônicos, representados pelas larvas de Gastropoda e Bivalvia,
ocorreram em toda a área de estudo sendo que a ocorrência conjunta destas formas larvais
podem ser explicadas pela proximidade do fundo e da presença de estoques reprodutores de
adultos bentônicos (VAN DER SPOEL & BOLTOVSKOY, 1981).
5.1 COPEPODA
Entre os grupos, Copepoda foi o dominante em densidade e riqueza, grupo
reconhecidamente importante no zooplâncton marinho (OMORI & IKEDA, 1984). Foram 43
taxa de Copepoda, entre ordem, família, gênero e espécie, onde as espécies de maiores
abundâncias em Primavera de 2005 foram Acartia lilljeborgi, Centropages velificatus,
Oithona plumifera, Oncaea clevei, Temora turbinata e Temora stylifera. Em Outono de 2006,
as espécies de maior abundância foram: Acartia lilljeborgi, Clausocalanus furcatus,
Corycaeus giesbrecthi, Oithona plumifera, Oncaea clevei, Temora stylifera, Temora
turbinata e Subeucalanus pileatus. Todas estas espécies são consideradas típicas da costa
brasileira segundo Björnberg (1981).
O número de taxa foi maior para o cruzeiro de Outono de 2006, apresentando 11
espécies exclusivas. Este fato sugere que as condições oceanográficas observadas para este
período indicam menores gradientes abióticos, fato confirmado pelos diagramas TS
apresentado na figura 1.
Os gradientes perpendiculares a costa registrados para Primavera de 2005 e os
longitudinais para Outono de 2006 também refletiram na distribuição espacial das espécies de
Copepoda. Exemplos são: A. lilljeborgi foi costeira em Primavera e limitou-se a região norte
em Outono. Da mesma forma, O. plumifera e O. clevei ocorreram na plataforma externa em
Primavera passando a ter ampla distribuição em Outono. Somado a isto, a grande maioria das
espécies exclusivas do cruzeiro de Outono de 2006 como Clausocalanus furcatus e
Corycaeus giesbrechti apresentaram ampla distribuição sobre a plataforma continental. Este
padrão de distribuição, formado por gradientes abióticos, são frequentemente citados nos
estudos de indicadores biológicos aplicados ao zooplâncton (BOLTOVSKOY, 1979 e
RESGALLA Jr., 2008).

57
Em termos de importância quanto a densidade e frequência de ocorrência, podem ser
descritas as seguintes espécies de Copepoda para uso na caracterização ambiental da
plataforma do Estado de Santa Catarina:
Acartia lilljeborgi é uma espécie freqüente em estudos na costa catarinense, como na
enseada do Saco dos Limões (Florianópolis) e na enseada da Armação do Itapocoroi (Penha)
(VEADO, 2002; VEADO & RESGALLA Jr, 2005; NUNES, 2007). Esteve relacionada com
AT e AC, indicando gradientes abióticos nos dois cruzeiros estudados.
Clausocalanus furcatus e Corycaeus giesbrecthi também são espécies comumente
encontradas na plataforma catarinense, registradas em todos os pontos amostrais no trabalho
de Muxagata (1999) na plataforma continental sudeste do Brasil. Esteve presente somente no
cruzeiro de Outono de 2006 e são indicadoras da Corrente do Brasil sobre a plataforma sul do
Brasil (BJORNBERG, 1981).
Oithona plumifera tem ampla distribuição em águas tropicais e subtropicais nos
oceanos Atlântico, Pacífico e Índico (BJORNBERG,1981; MONTÚ & GLOEDEN, 1986;
BRADFORD-GRIEVE, et al.,1999; DIAS & ARAUJO, 2006). É abundante no nordeste
brasileiro (CAVALCANTI & LARRAZABÁL, 2004). É uma espécie típica da costa de Santa
Catarina e indicadora de águas quentes (RESGALLA Jr., no prelo), Neste trabalho respondeu
aos gradientes abióticos perpendiculares a costa.
Oncaea clevei apresentou ampla distribuição. Para a PCSE, Muxagata (1999)
encontrou a espécie junto com AT, ACAS e água costeira fria. Já Bjornberg (1981) associou
esta espécie com Água subantártica e subtropical. Neste estudo ocorreu em ambos os
cruzeiros e respondeu aos gradientes abióticos, similar a Oithona plumifera.
Temora stylifera, abundante e frequente em águas quentes costeiras, água de
plataforma na superfície e no fundo e em águas tropicais. Para este estudo, teve maior
densidade no cruzeiro de Primavera de 2005, onde a temperatura média foi maior. Lopes et al.
(1999) relacionou T. stylifera com massas de água de temperatura mais elevada em frente a
Cabo Frio/RJ. Lindley & Daykin (2005) observaram que a abundância da espécie no norte do
Oceano Atlântico responde positivamente com a temperatura.
Temora turbinata apresentou distribuição mais costeira em relação a T. stylifera e
mais tolerante a água de menores salinidades (RESGALLA Jr. et al., 2008). É considerada
espécie invasora na costa brasileira por Lopes (2004).

58
Centropages velificatus é uma espécie tipicamente costeira, ocorre nas costas norte,
nordeste, central e sul do Brasil (BJORNBERG, 1981; BRADFORD-GRIEVE, et al.,1999;
DIAS & ARAUJO, 2006). Foi uma espécie dominante também na enseada da Armação do
Itapocoroi (Penha, SC) (NUNES, 2007).
Subeucalanus pileatus tem sido descrita por Ávila et al. (2009) como típica da costa
do Rio Grande do Sul, sendo tolerante ao deslocamento para água de maior profundidade. Pra
este estudo, sua maior densidade esteve relacionada com a água de plataforma sob influência
da ressurgência da ACAS em Primavera de 2005.
A diversidade de Copepoda mostrou diferenças entre os cruzeiros sendo determinada
pela equitabilidade em Primavera de 2005 e pela riqueza em Outono de 2006. Estes resultados
indicam forçantes bióticos para o último cruzeiro.
5.2 CLADOCERA
O grupo Cladocera foi freqüente e abundante nos dois cruzeiros analisados. É um
grupo tipicamente costeiro e indicador para áreas de estudos próximos a costa (RAMIREZ,
1981). A presença deste grupo esteve normalmente associada a águas de baixas salinidades.
As espécies encontradas, Evadne spinifera, Penilia avirostris, Pleopis schmackeri, e
Pseudoevadne tergestina, estão de acordo com as encontradas por Muxagata (1999) e Rocha
(1985) na costa de São Paulo e típicas de águas quentes costeiras e de plataforma.
Evadne spinifera mostrou preferência por pontos amostrais na plataforma externa
para os dois cruzeiros. Segundo Raymont (1983) é a menos costeira dos Cladocera, sendo que
aparentemente evita água de baixas salinidades e de menor qualidade.
Penilia avirostris, típica de AC, foi a única espécie a ser encontrada em toda a
plataforma. Em Outono de 2006, apresentou preferência pelo norte da área de estudo, onde a
temperatura da água foi mais alta. Este comportamento em relação a baixa temperatura é
facilmente explicada devido a natureza termófila desta espécie (RAMIREZ, 1981;
RAYMONT, 1983; ROCHA, 1985). Das espécies de Cladocera, foi a que apresentou resposta
aos gradientes abióticos perpendiculares e longitudinais a costa.
Pleopsis schmackeri ocorreu em pontos mais costeiros associada a AP. Considerada
uma espécie invasora, mas já está estabelecida nas águas costeiras do Brasil e com registro
desde a Baía da Guanabara até o Rio Grande do Sul (LOPES, 2004 e CASTRO, 2008). Indica
águas quentes e junto com a ocorrência de P. avirostris tem sido relacionada com a

59
dominância da Corrente do Brasil, caracterizada por apresentar água limpa e com alta
salinidade, na região costeira do Rio Grande do Sul (RESGALLA Jr. & MONTÚ, 1993 e
RESGALLA Jr., 2008) e na região costeira do Rio de Janeiro (MARAZZO; VALENTIN,
2003).
Pseudoevadne tergestina indica águas quentes costeiras e oceânicas e pode estar
presente em águas temperadas e tropicais em ambos hemisférios (RAMIREZ, 1981; ONBÉ,
1999). Ocorreu em toda a plataforma sem nenhum padrão de distribuição, porém com maior
densidade em Outono de 2006. Posta (2006) observou altas densidades da espécie durante o
inverno na região de Ubatuba/SP.
Como é um grupo representado por poucas espécies, os índices de diversidade não
demonstraram variações importantes que possam determinar alguma informação ecológica da
plataforma em estudo.
5.3 ASSOCIAÇÕES E INDICADORES HIDROLÓGICOS
Segundo a classificação das massas de água apresentada na figura 1, destaca-se que
os forçantes abióticos de temperatura e salinidade foram mais evidentes para o cruzeiro de
Primavera de 2005, envolvendo neste caso, gradientes perpendiculares a costa. Para o cruzeiro
de Outono de 2006, somente a salinidade apresentou gradientes longitudinais a costa.
Estas diferenças foram evidentes para as espécies de Copepoda e Cladocera mais
abundantes, demonstrando para Acartia lilljeborgi distribuição ao longo dos pontos amostrais
da plataforma interna em Primavera de 2005 e limitada ao norte da área de estudo em Outono
de 2006. O mesmo comportamento foi observado para Penilia avirostris, indicando, que estas
espécies típicas de águas costeiras quentes, as estratificações horizontais na área de estudo.
Para a Análise de Componentes Principais (ACP) de Primavera de 2005, as variáveis
ambientais apresentaram maior peso sobre o primeiro eixo indicando que estes forçantes
podem explicar a distribuição das espécies zooplanctônicas. Nas amostragens de Outono de
2006, pelo contrário, a ausência de altos pesos das variáveis abióticas (temperatura e
salinidade) com o eixo 1 e ao menor percentual de explicabilidade dos eixos sugerem que
forçantes bióticos podem apresentar maior peso sobre a distribuição das espécies ocorrentes
na área de estudo. Segundo Fager (1963) a medida que forçantes abióticos diminuem a
variação, maior é a influência dos forçantes bióticos. Desta forma, as análises de ACP
confirmam alguns padrões de distribuição das espécies, ou seja, distinção entre espécies

60
costeiras e de plataforma externa para o cruzeiro de Primavera de 2005 e ampla distribuição
para grande maioria no cruzeiro de Outono de 2006.
Entretanto, os processos físicos de ressurgência e da entrada da frente do Prata na
área de estudo não demonstraram associações típicas segundo estudos realizados por outros
autores. Calanoides carinatus, observada em ambos os períodos, é indicadora de processos de
ressurgências, mas somente associada a ocorrência de Ctenocalanus vanus (não observada
neste estudo), como destacado por VALENTIN et al. (1976); CAMPANER (1985);
VALENTIN (1988) e MONTEIRO-RIBAS (1988) na região de Cabo Frio (RJ). Desta forma,
estes processos de entrada de águas de menores temperaturas, destacados pelos parâmetros
físicos-químicos, não foram confirmados pelos indicadores biológicos, como por exemplo a
ausência das espécies de Cladocera Pleopis polyphemoides e Podon intermedius já citados em
estudos na costa de Santa Catarina. O tipo de amostrador utilizado (limitação de
profundidade) e o período de amostragem realizada (estágio inicial de desenvolvimento do
processo físico) podem ter limitado o uso dos indicadores hidrológicos.
Por outro lado, as análises de ACP indicaram espécies típicas das RIFs, ou seja,
espécies inversamente correlacionadas com as altas salinidades como Acartia lilljeborgi,
Temora turbina, Centropages velificatus e Penilia avirostris para o cruzeiro de Primavera de
2005. Para Outono de 2006, os forçantes bióticos, que normalmente são de difícil explicação
como competições, predações e outros (RESGALLA Jr., 2008), separaram Acartia lilljeborgi
do grupo de Calocalanus pavo, Clausocalanus furcatus, Corycaeus giesbrechti, Corycaeus
speciosus, Euchaeta marina, Macrosetella gracilis, Nannocalanus minor, Neocalanus
gracilis, Oithona plumifera, Oncaea clevei, Scolecithrix danae e Undinula vulgaris.
5.4 COMPARAÇÃO DAS ESPÉCIES DE COPEPODA E CLADOCERA COM OUTRAS ÁREAS DA COSTA BRASILEIRA
A comunidade zooplanctônica da Plataforma Continental Catarinense, caracterizada
neste estudo, foi semelhante a relatada em outros trabalhos na costa brasileira, como em
Vega-Pérez (1993) e Muxagata (1999), com maiores densidades nas regiões costeiras e
predomínio de Copepoda em todos os pontos amostrais.
As espécies Acartia lilljeborgi, Oithona plumifera. Penilia avirostris, Pleopis
schmackeri, ocorrentes nos dois cruzeiros do presente estudo, são comuns na costa brasileira e
podem ser encontradas desde o estado do Pará até Rio Grande do Sul. Em Santa Catarina, a

61
ocorrência destas espécies foram registradas em estudos na Baia do Itapocoroi, Estuário do
Rio Itajaí-Açu, Praia Navegantes, Reserva Biológica Marinha do Arvoredo e Ilha de SC.
Outras espécies ocorrentes, como U. vulgaris, E. marina, C. speciosus e T. stylifera,
são consideradas, segundo Omori & Ikeda (1984), “espécies-chave”, pois ocorrem durante
todo o ano em abundância na região nordeste do Brasil (MAFALDA Jr. et al., 2009). Este
padrão pode ser observado neste trabalho por ocorrerem em dois períodos distintos.
A não ocorrência de algumas espécies, como Pleopis polyphemoides, Evadne
nordmanni, que são representantes de água fria é relevante visto que P. polyphemoides é
comum em estudos na PCSE e E. nordmanni foi registrada em uma praia do Rio Grande do
Sul. Além disto, os diferentes processos que ocorrem na plataforma catarinense influenciam
diretamente na diversidade de espécies.
Resgalla Jr. (no prelo) sugere que a costa é uma área de transição entre as faunas
tropical, subtropical e temperado entre as regiões sul e sudeste do Brasil. A mistura ou a
sobreposição vertical das massas de água podem gerar áreas de transição faunística,
caracterizadas pela coexistência de espécies que integram as diferentes associações (LOPES,
et al., 2006). Por ser um dos poucos trabalhos que abrange toda a plataforma catarinense, a
comparação com outros períodos ficou prejudicada, além disto, há diversos processos
oceanográficos que tornam o ambiente instável para estabelecimento de grupos bem definidos
de espécies, porém, foi possível verificar uma fauna similar a encontrada nas regiões sudeste,
central e nordeste do Brasil, onde prevalecem águas costeiras quentes.

62
6.0 CONCLUSÕES
1. A comunidade zooplanctônica da costa de Santa Catarina apresenta grupos típicos do
holoplâncton marinho, onde 14 grupos, como Copepoda, Cladocera, Salpas e
Chaetognatha, apresentaram as maiores abundâncias em pontos amostrais próximos a
costa.
2. A metodologia aplicada no estudo para coleta dos dados biológicos foi elaborada para
coleta do ictioplâncton, sendo portanto um obstáculo na caracterização mais completa
do padrão de distribuição dos organismos zooplanctônicos indicadores.
3. A ocorrência de Salpas na região do Cabo de Santa Marta indicou associação com a
intrusão da ACAS.
4. As espécies de Copepoda e Cladocera registradas indicam grande similaridade com a
fauna da região sudeste e nordeste do Brasil, típicas de águas quentes costeiras e de
plataforma, confirmado pela abundância de Acartia lilljeborgi, Oithona plumifera,
Penilia avirostris e Pleopis schmackeri, sendo encontradas desde o estado do Pará até
Rio Grande do Sul.
5. Os gradientes abióticos perpendiculares e longitudinais a costa proporcionados por
processos de aportes continentais, ressurgências e a frente do Prata podem determinar
a distribuição das espécies mais abundantes como Acartia lilljeborgi e Penilia
avirostris, sendo os gradientes perpendiculares (aportes continentais e ressurgências)
mais determinantes que os longitudinais (frente do Prata). Este fato pode explicar o
maior número de espécies exclusivas em Outono de 2006, onde a maioria destas,
como Clausocalanus furcatus e Corycaeus giesbrechti apresentaram ampla
distribuição sobre a plataforma continental.
6. Os diferentes processos que ocorrem na plataforma catarinense influenciam
diretamente na diversidade de espécies e indicaram, para o período de estudo, uma
fauna tropical com uma grande similaridade também com o nordeste do Brasil.

63
REFERÊNCIAS
ALVAREZ, M.P.J. New Calanoid Copepods (Aetideidae) of the Genera Comantenna,
Mesocomantenna, New Genus, and Paracomantenna off the Brazilian Coast. Journal of
Crustacean Biology, 6(4): 858-877,1986.
ASHJIAN, C. J. & WISHNER, K. F. Temporal consistency of copepod species groups in the
Gulf Stream. Deep-Sea Research, 40: 483-516, 1993.
ÁVILA, T.R.; PEDROZO, C.S. & BERSANO, J.G.F. Variação temporal do zooplâncton da
Praia de Tramandaí, Rio Grande do Sul, com ênfase em Copepoda. Iheringia, Sér. Zool.
99(1):18-26, 2009.
BAKER, A.C.; BODEN, B.P. & BRINTON, E. 1990. A practical Guide to the Euphausiids of
the World. Publ. British Museum (Natural History), London. 96 p.
BERSANO, J. G. F. 1994. Zooplâncton da zona de arrebentação de praias arenosas, situadas
ao sul de Rio Grande, RS. Primavera de 1990, Verão de 1991. 163p. Dissertação
(Mestrado). Fundação Universidade Federal do Rio Grande, Rio Grande. BERSANO, J.G.F. & BOXSHALL, G.A. Planktonic copepods of the genus Oncaea Philippi
(Poecilostomatoida: Oncaeidae) from the waters off southern Brazil. Nauplius 2: 29-41,
1994.
BJÖRNBERG, T.K.S. The larva and young form of two Eucalanus (Copepoda) rom Tropical
Atlantic Waters. Crustaceana, Holanda, v. 12, n. 1, p. 59-73, 1967.
BJÖRNBERG, T.K.S. Revisão da distribuição dos gêneros Paracalanus, Clausocalanus e
Ctenocalanus (Copepoda, Crustacea) ao largo do Brasil. Boletim do Instituto
Oceanográfico, v. 29, n. 2, p. 65-68, 1980.
BJÖRNBERG, T.K.S. Copepoda. In: Atlas del Zooplancton del Atlántico Sudoccidental y
métodos de trabajo con el zooplancton marino, Boltovskoy, D.(ed.).Pub. Esp. INIDEP.
Mar del Plata, Argentina. 587-679, 1981.
BOLTOVSKOY, D. Zooplankton of the south-western. Atlantic. South Afr. J. Sci., 75: 541-
544, 1979.
BOLTOVSKOY, D. Submuestreo. In: Atlas del Zooplancton del Atlántico Sudoccidental y
métodos de trabajo con el zooplancton marino, Boltovskoy, D.(ed.).Pub. Esp. INIDEP.
Mar del Plata, Argentina. 143-146, 1981a.

64
BOLTOVSKOY, D. 1981b. Chaetognatha. In: Atlas del Zooplancton del Atlántico
Sudoccidental y métodos de trabajo con el zooplancton marino, Boltovskoy, D.(ed.).Pub.
Esp. INIDEP. Mar del Plata, Argentina. 759-791.
BOLTOVSKOY, D. South Atlantic Zooplancton. Leiden: Backhuys Publishers, 1706p. 1999.
BORZONE, C.A.; PEZZUTO, P.R. & MARONE, E. 1999. Oceanographic characteristics of
a multi-specific fishing ground of the Central South Brazil Bight.Marine Ecology,
20(2):131-146.
BRADFORD-GRIEVE, J. M.; MARKHASEVA, E. L.; ROCHA, C. E. F. & ABIAHY, B.
1999. Copepoda. In: BOLTOVSKOY, D. ed. South Atlantic Zooplankton. Leiden,
Backhuys. v.2. p.869-1098.
BRANDINI, F.P., LOPES, R.M., GUTSEIT, K.S., SPACH, H.L., SASSI, R. 1997.
Planctonologia na plataforma continental do Brasil - Diagnose e revisão bibliográfica.
MOD Com. Visual. Rio de Janeiro. 196 p.
CAMPANER, A.F. 1985. Occurrence and distribution of copepods (Crustacea) in the
epipelagial off southern Brazil. Bolm. Inst. oceanogr. 33(1): 5-27.
CAMPANER, A.F., HONDA, S. 1987. Distribution and co-occurrence of Calanoides
carinatus and larvae of Sardinella brasiliensis and Engraulis anchoita over the southern
brazilian continental shelf. Bolm. Inst. oceanogr. 35(1): 7-16.
CARVALHO JLB, SCHETTINI CAF AND RIBAS TM. 1998. Estrutura termohalina do
litoral centro-norte catarinense. Notas Téc. FACIMAR 2: 181-197.
CASTRO, M.C.T. 2008. O Porto do Rio de Janeiro sob o enfoque da água de lastro.
Dissertação de mestrado, Universidade do estado do Rio de Janeiro, 160 p.
CASTRO, B.M.; LORENZZETTI, J.A.; SILVEIRA, I.C.A. & MIRANDA, L.B. de. Estrutura
termohalina e circulação na região entre o Cabo de São Tomé (RJ) e o Chuí (RS). In:
ROSSI-WONGTSCHOWSKI, C.L.Del B. & MADUREIRA, L.S-P. O Ambiente
Oceanográfico da Plataforma Continental e do Talude na Região Sudeste-Sul do Brasil.
São Paulo: EdUSP, cap.1, p. 11-120, 2006.
CASTRO, B.M. & MIRANDA, L.B.de. Physical Oceanography of the Western Atlantic
Continental Shelf located between 4ºN and 34ºS. The Sea. v.11, c.8, p. 209-251, 1998.
CASTRO BM de. 1996. Correntes e Massas de Água da Plataforma Continental Norte de São
Paulo. Tese de Livre-Docência, Instituto Oceanográfico, Universidade de São Paulo, 248
p.

65
CASTRO FILHO BM. 1977. Análise termohalina de massas de água da região oeste do
Oceano Atlântico Sul Tropical. Dissertação de mestrado, Universidade de São Paulo, 104
p.
CAVALCANTI, E. A. H. & LARRAZÁBAL, M. E. L. 2004. Macrozooplâncton da Zona
Econômica Exclusiva do Nordeste do Brasil (Segunda Expedição Oceanográfica –
REVIZEE/NE II) com ênfase em Copepoda (Crustacea). Revista Brasileira de Zoologia
21(3):467-475.
CORNILS, A.; SCHNACK-SCHIEL, S. B.; AL-NAJJAR, T.; BADRAN, C. M. I.;
RASHEED, M.; MANASREH, R.; RICHTER, C. 2007. The seasonal cycle of the
epipelagic mesozooplankton in the northern Gulf of Aqaba (Red Sea). Journal of Marine
Systems 68:278-292. DIAS, C. O. & ARAUJO, A. V. 2006. Copepoda. In: Atlas de zooplâncton da região central
da Zona Econômica Exclusiva brasileira / editor Sérgio Luiz Costa Bonecker. – Rio de
Janeiro: Museu Nacional, 21-100.
EMILSON, I. The Shelf and Coastal Waters off Southern Brazil. Boletim do Instituto
Oceanográfico. v.11, n.2, p. 101-112, 1961.
FAGER, E.W. 1963. Communities of organisms. In: The sea, Hill, M.N.(ed.).John Wiley &
Sons. New York. 415-437.
OLIVEIRA, M. R. 1999. Zooplâncton da plataforma continental sudeste do Brasil, entre
Mongaguá-SP (24º 05'S) e o Cabo de Santa Marta Grande-SC (28º 40'S), com especial
referência aos copépodos Eucalanidae. Unpublished Ph.D. Dissertation, Universidade de
São Paulo, São Paulo. 147p.
GUSMÃO, L.M.O.; S. NEUMANN-LEITÃO; D.A. NASCIMENTO-VIEIRA; T.A. SILVA;
A.P. SILVA; F.F. PORTO-NETO & M.C.O. MOURA. 1997. Zooplâncton oceânico
entre os Estados do Ceará e Pernambuco-Brasil. Trabalhos Oceanográficos da
Universidade Federal de Pernambuco, Recife, 25: 17-30.
HERNÁNDEZ, A.P., MORALES, E.S. 1994. Copépodos pelágicos del Golfo de México y
Mar Caribe - I. Biologia y sistemática. Centro de Investigaciones de Quintana Roo
(CIQRO). México.360 p.
HILLE, E. 2009. Hidrografia da Plataforma Continental Catarinense. Trabalho de Conclusão
de Curso (Graduação em Oceanografia) – Centro de Ciências Tecnológicas da Terra e do
Mar, Universidade do Vale do Itajaí, Itajaí. 68 p.

66
HILLE, E. SCHETTINI, C.A.F. AND RIBEIRO, M.R. 2008. Estrutura termohalina no litoral
de Santa Catarina nos anos de 2005 e 2006. In: BRAGA ES (Ed). Oceanografia e
mudanças globais. São Paulo: Edusp. p. 371-381.
LARRAZÁBAL, M.E., CAVALCANTI, E.A.H., VIEIRA, D.A.N., OLIVEIRA-KOBLITZ,
V.S., ARAÚJO, E.M., BARRETO, T.M.S.P., NUNES, T.R.S. Macrozooplâncton na
Zona Econômica Exclusiva do Nordeste do Brasil. In: Programa Revizee - Score
Nordeste. v. 2, p. 48-102, 2009.
LENTINI, C.A.D. 1997. Estudo das variabilidades da temperatura da superfície do mar na
Plataforma Continental Sudeste da América do Sul. Tese de Mestrado. Universidade de
São Paulo – USP, São Paulo. 133 p.
LENZ, J. 2000. Introduction. In: ICES Zooplankton Methodology Manual. R.P et al. (eds.).
Academic Press. San Diego (California).
LIANG, T.H., VEGA-PEREZ, L.A. 1994. Studies on chaetognaths off Ubatuba region,Brazil.
I. distribution and abundance. Bolm. Inst. oceanogr. 42(1/2): 73-84.
LIANG, T. H. & VEJA-PEREZ, L.A. 1995. Studies on chaetognaths off Ubatuba region,
Brazil. II. Feeding habits. Bolm Inst. oceanogr., S Paulo, 43(1):27-40.
LINDLEY, J.A. & DAYKIN, S. 2005. Variations in the distributions of the Centropages
chierchiae and Temora stylifera (Copepoda: Calanoida) in the north-eastern Atlantic
Ocean and western European shelf waters, ICES J. Mar. Sci. 62 (5), pp. 869–877.
LOPES, R.M.; BRANDINI, F.; GAETA, S.A. Distribution patterns of patterns of epipelagic
copepods off Rio de Janeiro (SE Brazil) in summer 1991-1992 and winter 1992.
Hydrobiology, v. 411, p. 161-174, 1999.
LOPES R.M. 2004. Bioinvasões aquáticas por organismos zooplanctônicos: uma breve
revisão In: J.S.V. Silva; R.C.C.L. Souza. (Org.). Água de Lastro e Bioinvasão. Rio de
Janeiro: Interciência, p. 113-131.
LOPES RM. 2007. Marine zooplankton studies in Brazil – A brief evaluation and
perspectives. An. Acad. Bras. Cienc. 79(3): 369-379.
LUDWIG, J.A., and REYNOLDS, J.F., 1988, Statistical ecology; a primer in methods and
computing: New York, John Wiley & Sons, 368 p.
MAGALHÃES C. 2000. Diversity and abundance of decapod crustaceans in the Rio Negro
basin, Pantanal, Mato Grosso do Sul, Brazil. In: Chernoff, B.; Alonso, L. E.;
Montambault, J. R. & Lourival, R. (Eds). A biological assessment of the aquatic

67
ecossystems of the Pantanal, Mato Grosso do Sul, Brazil. RAP Bulletin of Biological
Assessment 18. conservation International, Washington, USA, p.56-62
MARAZZO, A.; VALENTIN, J. L. Population dynamics of Penilia avirostris (Dana,1852)
(CLADOCERA) in a tropical bay. Crustaceana, v. 76, n. 7, p. 803-817, 2003.
MÉMERY, L.; ARHAN, M.; ALVAREZ-SALGADO, X.A.; MESSIAS, M.J.; MERCIER,
H.; CASTRO, C.G. & RIOS, A.F. 2000. The water masses along the western boundary of
the south and equatorial Atlantic. Progress in Oceanography, v. 47, p. 69-98.
MIRANDA, L.B. 1982. Análise de massas de água da plataforma continental e da região
oceânica adjacente: Cabo de São Tomé‚ (RJ) a Ilha de São Sebastião (SP). Tese de Livre
Docência. Instituto Oceanográfico da Universidade de São Paulo – IOUSP, São Paulo.
123 p.
MÖLLER Jr., O.O.; PIOLA, A.R.; FREITAS, A.C. & CAMPOS, E.J.D. 2008. The effects of
river discharge and seasonal winds on the shelf off southeastern South America.
Continental Shelf Research, v. 28, p.1607-1624.
MONTEIRO-RIBAS, W.M. 1988. Os copépodes na região de Cabo Frio: variação qualitativa
e quantitativa, durante um ciclo de ressurgência. Rio de Janeiro, R.J. Brasil. Tese de
Mestrado. Universidade Federal do Rio de Janeiro – UFRJ, Rio de Janeiro. 163 p.
MONTÚ, M. & GLOEDEN, I.M. 1986. Atlas dos Cladocera e Copepoda (Crustacea) do
estuário da Lagoa dos Patos (Rio Grande, Brasil). Nerítica, Pontal do Sul, PR, vol. 1, no.
2, p. 1-134.
MONTÚ, A.M. 1980. Zooplâncton do estuário da Lagoa dos Patos I. Estrutura e variações
temporais e especiais da comunidade. Atlântica 4:53-72. MOREIRA, G.S. 1973. On the diurnal vertical migration of hydromedusae off Santos, Brazil.
Publs Seto mar. biol. Lab. 20: 537-566.
MUXAGATA, E. 1999. Avaliação da biomassa e distribuição zooplanctônica na plataforma
continental sudeste brasileira durante o inverno de 1995. Dissertação de mestrado.
Fundação Universidade do Rio Grande. 168 p.
NUNES, R. D. 2007. Variação espaço-temporal da comunidade zooplanctônica na enseada da
Armação do Itapocoroy, litoral norte de Santa Catarina (Brasil). Trabalho de Conclusão
de Curso (Graduação em Oceanografia) – Centro de Ciências Tecnológicas da Terra e do
Mar, Universidade do Vale do Itajaí, Itajaí. 30 p.
Oliveira, M. R. 1999. Zooplâncton da plataforma continental sudeste do Brasil, entre
Mongaguá-SP (24º 05'S) e o Cabo de Santa Marta Grande-SC (28º 40'S), com especial

68
referência aos copépodos Eucalanidae. Unpublished Ph.D. Dissertation, Universidade de
São Paulo, São Paulo. 147p. OMORI, M. & IKEDA, T. 1984. Methods in marine zooplancton ecology, New Tork: Publ.
John Wiley & Sons, 332p.
ONBÉ, T. 1999. Ctenopoda and Onychopoda (Cladocera). In: D. Boltovskoy, (ed.). South
Atlantic zooplankton. Backhuys Publishers Leiden, The Netherlands, pp. 797-813.
PALÁCIO, F.J. (1982): Revision zoogeografica marina del sur del Brasil. Bol. Inst Ocean
USP. 31(1):69-92.
PARSONS, T.R.; TAKAHASHI, M.; HARGRAVE, B. 1984. Biological oceanographic
processes. Pergamon Press. Oxford. 330 p.
PIELOU, E.C. 1984. The interpretation of ecological data. A primer on classification and
ordination, New York: Publ. John Wiley & Sons, 263p.
PIELOU, E.C. The measure of diversity in different types of biological collections. J. Theor.
Biol., 13:133-144, 1966.
PIOLA AR, CAMPOS EJD, MÖLLER Jr OO, CHARO M AND MARTINEZ C. 2000.
Subtropical shelf front off eastern south America. J. Geoph. Res. 105(C3): 6565-6578.
PIOLA AR, MATANO RP, PALMA E, MÖLLER Jr OO AND CAMPOS EJD. 2005. The
influence of the Plata river discharge on the western South Atlantic Shelf. Geoph. Res.
Letters. 32: 1-4.
POSTA, P.D. 2006. Influência de Fatores Ambientais sobre a Distribuição Espacial e
Temporal de Cladóceros Marinhos (Crustacea, Branchiopoda) em um Estuário de
Ubatuba, SP. Dissertação de Mestrado. Universidade de Taubaté, São Paulo. 68 p.
RAMÍREZ, F.C. 1981. Cladocera. In: Atlas del Zooplancton del Atlántico Sudoccidental y
métodos de trabajo con el zooplancton marino, Boltovskoy, D.(ed.).Pub. Esp. INIDEP.
Mar del Plata, Argentina. 533-542.
RAYMONT, J.E.G. 1983. Plankton and productivity in the oceans - Volume 2 - Zooplankton.
Pergamon Press. Oxford.824 p.
RESGALLA Jr. C AND MONTÚ M. 1993. Cladóceros marinhos da plataforma continetal do
Rio Grande do Sul – Brasil. Nauplius. 1: 63-79.
RESGALLA Jr. C, DE LA ROCHA C AND MONTÚ M. 2001. The influence of Ekman
transport on zooplankton biomass variability off southern Brazil. J. Plankton Res. 23(1):
1191-1216.

69
RESGALLA Jr., C.; COUTINHO DE SOUZA, V.G.; RORIG, L.R. & SCHETTINI, C.A.F.
2008. Spatial and temporal variation of the zooplâcton community in the area of
influence of the Itajaí-açú river, SC (Brazil). Braz. J. Oceanogr. 56(3):211-224.
RESGALLA Jr, C. 2008. Pteropoda, Cladocera, and Chaetognatha associations as
hydrological indicators in the southern Brazilian Shelf. Lat. Am. J. Aquat. Res., (36)2: 0-
0.
RESGALLA Jr. (no prelo). The holoplankton of the Santa Catarina coast, southern Brazil.
An. Acad. Bras. Cienc.
ROCHA, C.E.F. 1985. The occurrence of Pleopis schmackeri (poppe) in the southern atlantic
and other marine cladocerans on the brazilian coast. Crustaceana, 49(2):202-204.
ROCHA, C.E.F. 1977. Distribuição dos Cladócero e Ostracoda (Crustácea) planctônicos
marinhos ao largo de Santos, Brasil. Tese de Mestrado IBUSP. 100pp.
RODRIGUES, R.F. 1977. Evolução da Massa d’água durante a Ressurgência de Cabo Frio.
Publicações. Inst. Pesq. Mar.,(115): 1-31.
SANT’ANNA, E. M. E. & BJÖNBERG, T. K. S. 2006. Seasonal dynamics of
mesozooplankton in Brazilian coastal waters. Hydrobiologia 563:253-268.
SARTORI, L.P. & LOPES, R.M. 2000. Seasonal variability of pelagic copepod assemblages
on the inner continental shelf off Paraná, Brazil. Nauplius 8:79-88.
SCHETTINI CAF, RESGALLA Jr. C, PEREIRA FILHO J, SILVA MAC, TRUCCOLO EC
AND RÖRIG LR. 2005. Variabilidade temporal das características oceanográficas e
ecológicas da região de influência fluvial do rio Itajaí-Açu. Braz. J. Aquat. Sci. Technol.
9(2): 93-102.
SILVA, A.C.da, ARAÚJO, M. & BOURLÈS, B. Variação sazonal da estrutura de massas de
água da Plataforma Continental do Amazonas e área oceânica adjacente. Revista
Brasileira de Geofísica, v. 23 n. 2, p.145-157, 2005.
SILVA, L.S.; MIRANDA, L.B. de & CASTRO FILHO, B.M. 2004. Estudo numérico da
circulação e da estrutura termohalina na região adjacente à Ilha de São Sebastião (SP).
Revista Brasileira de Geofísica, 22(3): 197-221.
STERZA, J. M. & LOUREIRO FERNANDES , L. F. Zooplankton community of the Vitória
bay estuarine system (Southeastern Brazil). Characterization during a three-year study.
Brazilian Journal of oceanography, v. 2/3, n. 54, p. 95-105, 2006.

70
VALENTIN, J.L., MONTEIRO-RIBAS, W.M., MUREB, M.A., PESSOTTI, E. 1976. A
origem das massas de água na ressurgência de Cabo Frio (Brasil) vista através do estudo
das comunidades de copépodos. Publ. Inst. Pesq. Marinha, 97: 1-35.
VALENTIN JL. 1988. A dinâmica do plâncton da ressurgência de Cabo Frio – RJ. In:
BRANDINI FP (Ed.) Mem. III Encontro Brasileiro de Plâncton. Caiobá. p. 25-35.
VALENTIN JL, GAETA AS, SAPCH HL, MONTÚ M AND ODEBRECHT C. 1994.
Diagnóstico ambiental oceânico e costeiro das regiões sul e sudeste do Brasil. Volume 4.
Oceanografia Biológica: Plâncton. Brasília, Petrobras, 321p.
VAN DER SPOEL, S. & BOLTOVSKOY, D. 1981. Pteropoda, p. 493-533. In: D.
Boltovskoy (Ed.). Atlas del zooplancton del Atlántico sudoccidental y métodos de trabajo
com el zooplancton mariño. Mar del Plata, Publicación Especial del Intituto Nacional de
Investigatión y Desarrollo Pesquero, 936p.
VEADO, L. D. V. 2002. Alteração da comunidade zooplanctônica do Saco dos Limões após
impacto das obras da Via Expressa Sul - Baía Sul da ilha de Santa Catarina. Trabalho de
Conclusão de Curso – Centro de Ciências Tecnológicas da Terra e do Mar, Universidade
do Vale do Itajaí, Itajaí. 65 p.
VEADO, L.D.V. & RESGALLA Jr. C. 2005. Alteração da comunidade zooplanctônica do
saco dos Limões após impacto das obras da via expressa sul - Baía Sul da Ilha de Santa
Catarina. Braz. J. Aquatic Sci. Technol. 9(2): 65 - 73.
VEGA-PÉREZ, L.A. & BOWMAN, T.E. 1992. Description of the Pelagic Copepod,
Ctenocalanus heronae Vega-Pérez and Bowman, from off São Paulo, Brazil (Calanoida,
Clausocalanidae). Proc. Biol. Soc. Washington, 105:97-101.
VEGA-PÉREZ, L.A. 1993. Estudo do zooplâncton da região de Ubatuba, Estado de São
Paulo. Pub. esp. Inst. oceanogr. 10: 65-84.
VEGA-PÉREZ, L.A. & HERNÁNDEZ, S. 1997. Composição e Distribuição da Família
Paracalanidae (Copepoda, Calanoida) ao Largo de São Sebastião, Estado de São Paulo –
Brasil, com Ênfase em três espécies de Paracalanus. Rev. Bras. Oceanogr., 45: 61-75.
WIAFE, G. & FRID, C. L. J. 1996. Short-term temporal variation in coastal zooplankton
communities: the relative importance of physical and biological mechanisms. Journal of
Plankton Research, 8(8): l485-1501.
WISHNER, K. F., M. M. GOWING, AND C. GELFMAN. 1998. Mesozooplankton biomass
in the upper 1000 m in the Arabian Sea: Overall seasonal and geographic patterns, and
relationship to oxygen gradients. Deep-Sea Res. II 45: 2405–2432.

71
YAMASHITA, C. 1977. Efeitos combinados de temperatura e salinidade na sobrevivência do
copépode Euterpina acutifrons (Dana, 1847), da região de Santos e do canal de São
Sebastião. Boletim de Fisiologia Animal 1: 91-120.
YAMASHITA, C. & MOREIRA, G.S. 1981. Variação anual em tamanho de Euterpina
acutifrons (Dana) (Copepoda, Harpacticoida) do Canal de São Sebastião e da região de
Santos, Brasil. Boletim de Fisiologia Animal 5: 29-43.