o objectivo deste documento é que o usem como revisão de ...º... · duas moléculas de piruvato...
TRANSCRIPT

O objectivo deste documento é que o usem como revisão de conteúdos e através dele
percebam quais os pontos mais importantes da matéria. Recomendo os casos clínicos do
Devlin porque vos ajudam a consolidar conhecimentos mas não são necessariamente para
decorar.
Bom estudo! Boa sorte!
Joana Franco
2010/2011

1 – Metabolismo Glicídico
A – Glicólise
Introdução
A glicólise é a sequência de reacções que metabolizam uma molécula de glicose em
duas moléculas de piruvato com produção concomitante de duas moléculas de ATP.
O piruvato pode depois ser fermentado a lactado (fermentação láctica) ou etanol
(fermentação alcoólica)
A glicose é o único combustível utilizado pelo cérebro em situações normais e o único
utilizado pelos glóbulos vermelhos de todo.
Em situações aeróbias, o piruvato é metabolizado a dióxido de carbono e água através
do ciclo de Krebs e a cadeia transportadora de electrões.
Na ausência de oxigénio, o piruvato é convertido em lactato.
o No músculo, a fermentação láctica é utilizada quando a necessidade de
energia suplanta a capacidade de transporte de oxigénio (por exemplo,
durante o exercício físico intenso). O músculo funciona em anaerobiose até ao
ponto em que a acumulação de lactato faz descer o pH até ao ponto em que a
própria via de produção de lactato é inibida.
Fig.1
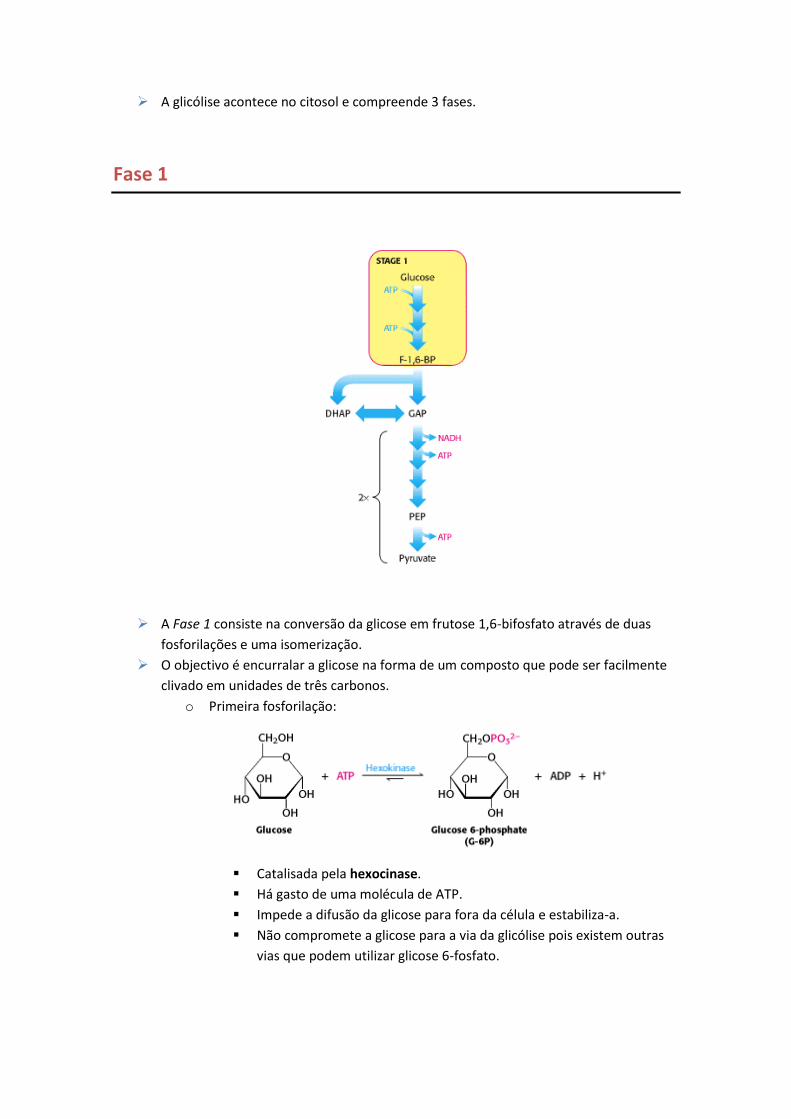
A glicólise acontece no citosol e compreende 3 fases.
Fase 1
A Fase 1 consiste na conversão da glicose em frutose 1,6-bifosfato através de duas
fosforilações e uma isomerização.
O objectivo é encurralar a glicose na forma de um composto que pode ser facilmente
clivado em unidades de três carbonos.
o Primeira fosforilação:
Catalisada pela hexocinase.
Há gasto de uma molécula de ATP.
Impede a difusão da glicose para fora da célula e estabiliza-a.
Não compromete a glicose para a via da glicólise pois existem outras
vias que podem utilizar glicose 6-fosfato.
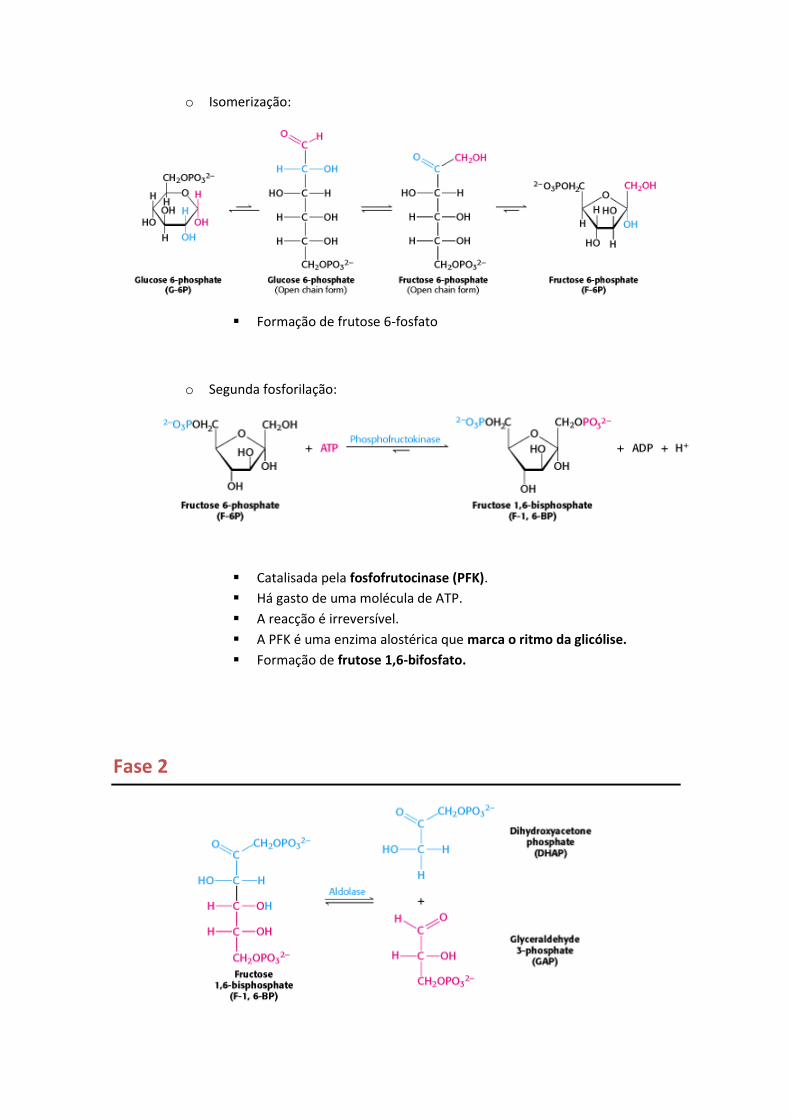
o Isomerização:
Formação de frutose 6-fosfato
o Segunda fosforilação:
Catalisada pela fosfofrutocinase (PFK).
Há gasto de uma molécula de ATP.
A reacção é irreversível.
A PFK é uma enzima alostérica que marca o ritmo da glicólise.
Formação de frutose 1,6-bifosfato.
Fase 2
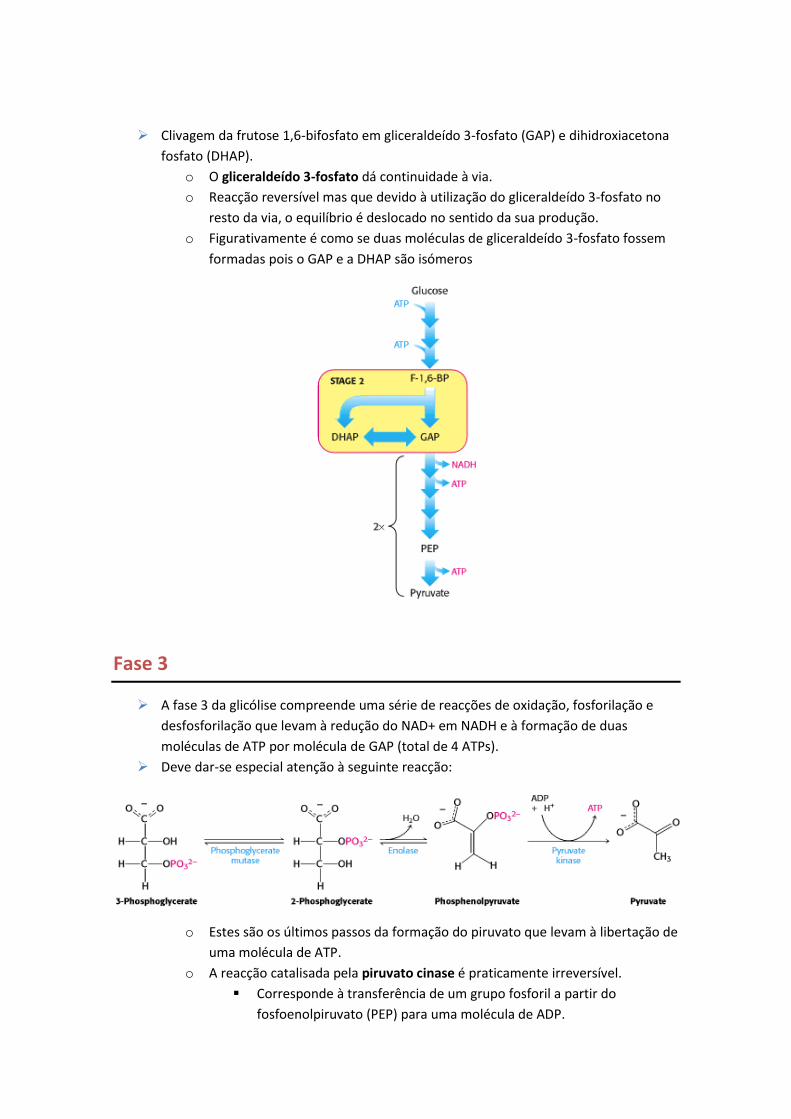
Clivagem da frutose 1,6-bifosfato em gliceraldeído 3-fosfato (GAP) e dihidroxiacetona
fosfato (DHAP).
o O gliceraldeído 3-fosfato dá continuidade à via.
o Reacção reversível mas que devido à utilização do gliceraldeído 3-fosfato no
resto da via, o equilíbrio é deslocado no sentido da sua produção.
o Figurativamente é como se duas moléculas de gliceraldeído 3-fosfato fossem
formadas pois o GAP e a DHAP são isómeros
Fase 3
A fase 3 da glicólise compreende uma série de reacções de oxidação, fosforilação e
desfosforilação que levam à redução do NAD+ em NADH e à formação de duas
moléculas de ATP por molécula de GAP (total de 4 ATPs).
Deve dar-se especial atenção à seguinte reacção:
o Estes são os últimos passos da formação do piruvato que levam à libertação de
uma molécula de ATP.
o A reacção catalisada pela piruvato cinase é praticamente irreversível.
Corresponde à transferência de um grupo fosforil a partir do
fosfoenolpiruvato (PEP) para uma molécula de ADP.
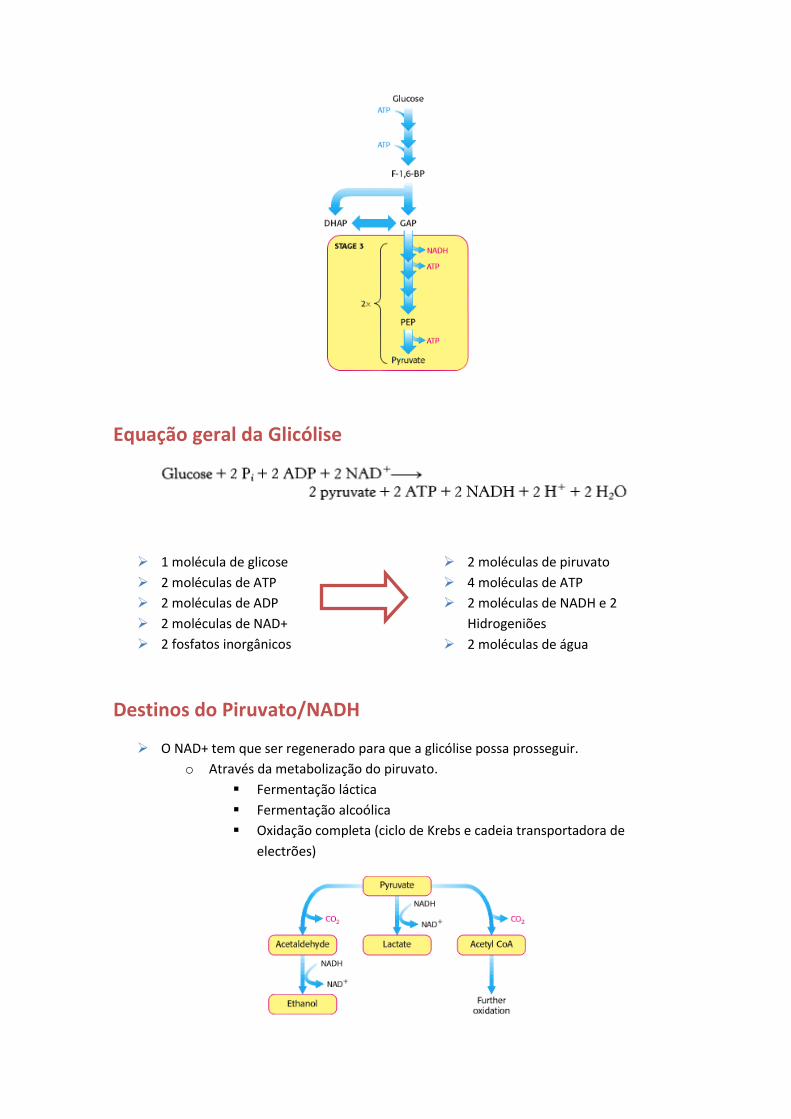
Equação geral da Glicólise
1 molécula de glicose
2 moléculas de ATP
2 moléculas de ADP
2 moléculas de NAD+
2 fosfatos inorgânicos
2 moléculas de piruvato
4 moléculas de ATP
2 moléculas de NADH e 2
Hidrogeniões
2 moléculas de água
Destinos do Piruvato/NADH
O NAD+ tem que ser regenerado para que a glicólise possa prosseguir.
o Através da metabolização do piruvato.
Fermentação láctica
Fermentação alcoólica
Oxidação completa (ciclo de Krebs e cadeia transportadora de
electrões)

Regulação da Glicólise
Nas vias metabólicas, em geral, todas as reacções essencialmente irreversíveis são
potenciais locais de controlo.
Na glicólise os locais de controlo correspondem às reacções catalisadas pela:
o Hexocinase
o Fosfofrutocinase
o Piruvato Cinase
A actividade destas enzimas é controlada pela ligação reversível de efectores
alostéricos ou por modificação covalente.
A quantidade destas enzimas varia segundo a regulação da sua transcrição
(hormonas).
o Fosfofrutocinase (PFK)
É o elemento de controlo mais importante.
A reacção que catalisa corresponde ao passo que compromete a
glicose para a via da glicólise pois esta reacção é exclusiva desta via.
No fígado, o ATP inibe-a alostericamente (diminui a sua afinidade para
a frutose 6-fosfato.
O AMP reverte o efeito inibitório do ATP.
O citrato (intermediário do ciclo de Krebs) também inibe a PFK
aumentando o efeito inibitório do ATP.
Um nível elevado de citrato significa que os precursores
biossintéticos são abundantes e que a glicose não deve ser
degradada na glicólise.
A glicólise é estimulada quando a quantidade de energia diminui.
o Hexocinase
É inibida pelo seu produto, glicose 6-fosfato (sinaliza que a célula já
não precisa de mais glicose para energia ou como precursor
biossintético).
A inibição da fosfofrutocinase leva à inibição da hexocinase por
acumulação de substratos a montante.
No fígado:
Existe uma isoforma da hexocinase, a glicocinase (ou
hexocinase IV).

A hexocinase IV só fosforila a glicose quando esta é abundante
pois tem 50 vezes menos afinidade para a glicose que a
hexocinase.
O papel da hexocinase IV no fígado é fornecer glicose para
produção de glicogénio (molécula de armazenamento de
glicose) e de ácidos gordos
A pouca afinidade da hexocinase IV para a glicose faz com que
esta esteja disponível principalmente para o cérebro e
músculos em períodos de escassez de glicose e que ela possa
ser armazenada em períodos de abundância.
o Piruvato cinase
Catalisa o terceiro passo irreversível da glicólise.
Controla a velocidade de saída de piruvato.
A frutose 1,6-bifosfato (produto do passo irreversível anterior) activa a
piruvato cinase.
O ATP inibe-a alostericamente de forma a abrandar a glicólise.
A alanina (um aminoácido que pode ser sintetizado em um passo para
formar piruvato – ver metabolismo proteico) pode inibir
alostericamente a piruvato cinase para sinalizar que os precursores
biossintéticos são abundantes.
Inibida por fosforilação reversível
Cascata de sinalização desencadeada pela glucagina e mediada
pelo AMPc leva à fosforilação da piruvato cinase diminuindo a
sua actividade.

B - Gliconeogénese
É a síntese de glicose a partir de precursores não-glicídicos.
Converte piruvato em glicose.
Os precursores principais são:
o Lactato
o Aminoácidos (proveniente da proteólise)
o Glicerol (proveniente da hidrólise de triacilglicerois)
Acontece principalmente no fígado e alguma quantidade no rim.
Ajuda a manter os níveis de glicose no sangue a um nível que permita que o cérebro e
os músculos consigam ter glicose suficiente para as suas necessidades metabólicas.
A gliconeogénese não é o reverso da glicólise devido à existência de passos muito
exergónicos (passos irreversíveis catalisados pela hexocinase, fosfofrutocinase e
piruvato cinase) que são contornados por outras enzimas.

Regulação da Gliconeogénese/Glicólise
A glicólise e a gliconeogénese são controladas de forma coordenada para que dentro
da célula uma via esteja relativamente inactiva enquanto a outra está muito activa.
O ritmo da glicólise é determinado pela concentração de glicose enquanto o da
gliconeogénese é determinado pela concentração de lactato e outros precursores da
glicose.
Locais de controlo:
o Interconversão de frutose 6-fosfato em frutose 1,6-bifosfato (último passo da
Fase 1 da glicólise)
Fosfofrutocinase na glicólise
Frutose 1,6-bifosfatase na gliconeogénese.
Inibida pelo AMP
Activada pelo ATP e citrato
Um nível elevado de AMP indica que existe pouca carga energética e
sinaliza a necessidade de formação de ATP e consequentemente de
activação da glicólise e inibição da gliconeogénese.
Um nível elevado de ATP ou citrato indica que existe uma carga
energética alta e que os precursores biossintéticos estão elevados e
portanto a glicólise deve ser inibida e a gliconeogénese deve ser
activada.
o Interconversão do fosfoenolpiruvato em piruvato (último passo da glicólise,
primeiro da gliconeogénese a partir do piruvato)
Piruvato cinase na glicólise
Piruvato carboxilase na gliconeogénese
Activada pelo acetil-CoA
Inibida pelo ADP
A gliconeogénese é assim favorecida quando os precursores
metabólicos estão abundantes.
Regulação hormonal:
o Através da alteração do ritmo da expressão genética (transcrição ou
degradação do mRNA) das enzimas intervenientes nas vias.
o Insulina:
A concentração aumenta no período pós-prandial.
Estimula a expressão da fosfofrutocinase e piruvato cinase.
o Glucagina:
A concentração aumenta em períodos de jejum.
Inibe a expressão da fosfofrutocinase e piruvato cinase.
Estimula a produção de enzimas gliconeogénicas, a fosfoenolpiruvato
carboxilase e a frutose 1,6-bifosfatase

C – Ciclo de Krebs (ciclo do ácido cítrico)
Generalidades
O processamento aeróbio da glicose começa com a oxidação completa dos seus
derivados a dióxido de carbono – ciclo de Krebs.
É a via final comum à oxidação de todas as moléculas combustíveis (aminoácidos,
ácidos gordos e glícidos) sendo que a maior parte delas entra no ciclo sob a forma de
acetil-CoA.
Acontece na mitocôndria.
A função do ciclo de Krebs é conseguir electrões de alta energia a partir de compostos
de carbono e usá-los na formação de NADH e FADH2. O NADH e FADH2 são depois
utilizados na cadeia transportadora de electrões para a formação de ATP.
Formação de Acetil-CoA a partir do piruvato
Na matriz mitocondrial.
Descarboxilação oxidativa.
o Catalisada pelo complexo piruvato desidrogenase.
o Reacção irreversível.
o É a ligação entre a glicólise e o ciclo de Krebs.
A coenzima A é um transportador de muitos grupos acil para o ciclo.
Ciclo de Krebs propriamente dito
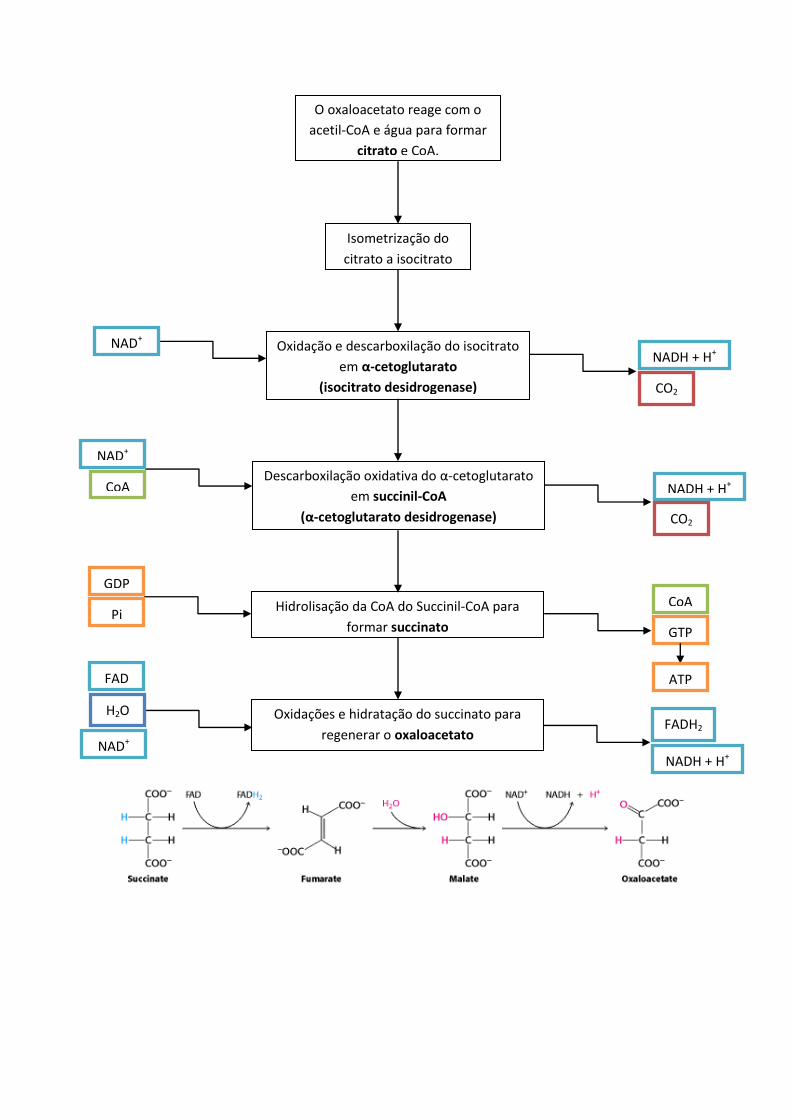
O oxaloacetato reage com o
acetil-CoA e água para formar
citrato e CoA.
Isometrização do
citrato a isocitrato
Oxidação e descarboxilação do isocitrato
em α-cetoglutarato
(isocitrato desidrogenase)
Descarboxilação oxidativa do α-cetoglutarato
em succinil-CoA
(α-cetoglutarato desidrogenase)
Hidrolisação da CoA do Succinil-CoA para
formar succinato
Oxidações e hidratação do succinato para
regenerar o oxaloacetato
NAD+
NADH + H+
CO2
NAD+
CoA
NADH + H+
CO2
GDP
Pi
GTP
CoA
ATP
NAD+
H2O
FAD
NADH + H+
FADH2

Regulação
Complexo piruvato desidrogenase
o A formação de acetil-CoA a partir de piruvato compromete os átomos de
carbono provenientes da glicose para o ciclo de Krebs ou para a formação de
ácidos gordos.
o Altas concentrações dos produtos do complexo inibem a reacção:
Acetil-CoA
NADH
o A fosforilação do complexo inactiva-o. Uma fosfatase (retira o fosfato) reactiva
o complexo.
o Piruvato e ADP activam o complexo inactivando a fosfatase.
O piruvato e o ADP são moléculas sinalizadoras de que a carga
energética da célula está baixa.
Isocitrato desidrogenase
o Estimulada por:
ADP
o Inibida por:
ATP
NADH
α-cetoglutarato desidrogenase
o Inibida por:
Succinil-CoA
NADH
ATP
O ritmo do ciclo de Krebs é diminuído quando a célula tem uma alta concentração de
ATP.
A isocitrato desidrogenase e a α-cetoglutarato desidrogenase são os locais primários
de controlo do ciclo de Krebs.

D – Cadeia transportadora de electrões e fosforilação oxidativa
Introdução
Cadeia transportadora de electrões é um sistema de transportadores de electrões
localizados na membrana interna da mitocôndria
A cadeia transportadora de electrões é responsável pela oxidação de NADH e FADH2
em presença de O2 (aceitador final de electrões) com aproveitamento da energia livre
para a formação de ATP (fosforilação oxidativa).
A oxidação completa de 1 mol NADH resulta na produção de aproximadamente 2,5
moles de ATP.
A oxidação completa de 1 mol FADH2 resulta na produção de aproximadamente 1,5
moles de ATP.
A cadeia transportadora de electrões não é mais do que um sistema de reacções de
oxidação-redução ligadas.
Complexos da cadeia transportadora de electrões
A energia libertada durante a remoção dos electrões do NADH e FADH2 e durante a
sua transferência entre os complexos da cadeia transportadora (reacções exergónicas)
é usada para bombear protões através da membrana interna da mitocôndria da matriz
para o espaço intermembranar.
A cadeia é constituída por 4 complexos enzimáticos que catalisam reacções parciais.
o Complexo I:
NADH-ubiquinona óxido-redutase
Catalisa a transferência de electrões do NADH para a ubiquinona (UQ).
Bombeia 4 protões por cada 2 electrões transferidos para a
ubiquinona.
o Complexo II:
Succinato-ubiquinona óxido-redutase (succinato desidrogenase)
Catalisa a transferência de electrões do succinato para a ubiquinona
(UQ)
o Complexo III:
Ubiquinol-citocromo c redutase
Catalisa a transferência de electrões do ubiquinol (forma reduzida da
ubiquinona após os passos anteriores) para o citocromo c.
Espaço intermembranar
Matriz

Bombeia 4 protões por cada 2 electrões transferidos do ubiquinol para
ao citocromo c.
O citocromo c vai funcionar como um transportador de electrões.
o Complexo IV:
Citocromo c oxidase
Catalisa a transferência de electrões do citocromo c para o O2
formando água havendo acoplamento do bombeamento de protões
(4 electrões/4 protões).
Inibido por cianeto (CN-) que impede a transferência de electrões
ligando-se ao complexo.
Fosforilação oxidativa
A ATP sintetase (“complexo V”) usa a energia do gradiente electroquímico (diferença
de concentração de protões nos dois lados da membrana interna da mitocôndria)
produzido pelos protões resultantes da transferência de electrões para a produção de
energia sob a forma de ATP.
A ATP sintetase dirige a síntese de ATP pelo movimento de protões a favor do
gradiente electroquímico.
Espaço intermembranar
Matriz

O movimento de protões através da ATP sintetase provoca uma alteração
conformacional com alteração do local de ligação do ATP, libertando-o e permitindo a
ligação de um ADP e um Pi à subunidade catalítica.
A velocidade de síntese de ATP pela ATP sintetase é determinada pela velocidade de
utilização de ATP pela célula que por sua vez regula a velocidade de funcionamento da
cadeia transportadora de electrões.
Quando as necessidades energéticas da célula são baixas, o ATP acumula-se e o
gradiente de protões deixa de ser usado para a síntese de ATP.
o A magnitude do gradiente de protões aumenta até que a energia necessária
para bombear protões através da membrana, a partir da matriz, contra o
gradiente eléctrico existente, fique igual à energia libertada durante a
transferência dos electrões de NADH para O2. Neste ponto é atingido o
equilíbrio e o transporte de electrões pára.
O ADP e o NAD+ estimulam a ATP sintetase.
À medida que o ATP é sintetizado, a magnitude do gradiente de protões diminui uma
vez que os protões se movem através da ATP sintetase.
Shuttle malato-aspartato
No coração e no fígado, os electrões do NADH citosólico são levados para a
mitocôndria através deste shuttle.
1- Os electrões são transferidos do NADH para o oxaloacetato formando malato
(redução do oxaloacetato).
2- O malato atravessa a membrana mitocondrial e é reoxidado pelo NAD+ para
formar NADH e oxaloacetato (malato desidrogenase).
3- Para poder voltar ao citosol, o oxaloacetato tem que ser transaminado (ver
metabolismo proteico) a aspartato.
4- O aspartado volta ao citosol onde é desaminado (ver metabolismo proteico) para
regenerar o oxaloacetato.

E- Via das fosfopentoses
Introdução
Funciona paralelamente à glicólise e ao ciclo de Krebs.
Produz poder redutor na forma de NADPH e pentoses/hexoses intermediárias.
O NADPH serve como dador de hidrogénio e electrões para a biossíntese.
A via é altamente influenciada pelas necessidades de NADPH.
Ocorre no citosol.
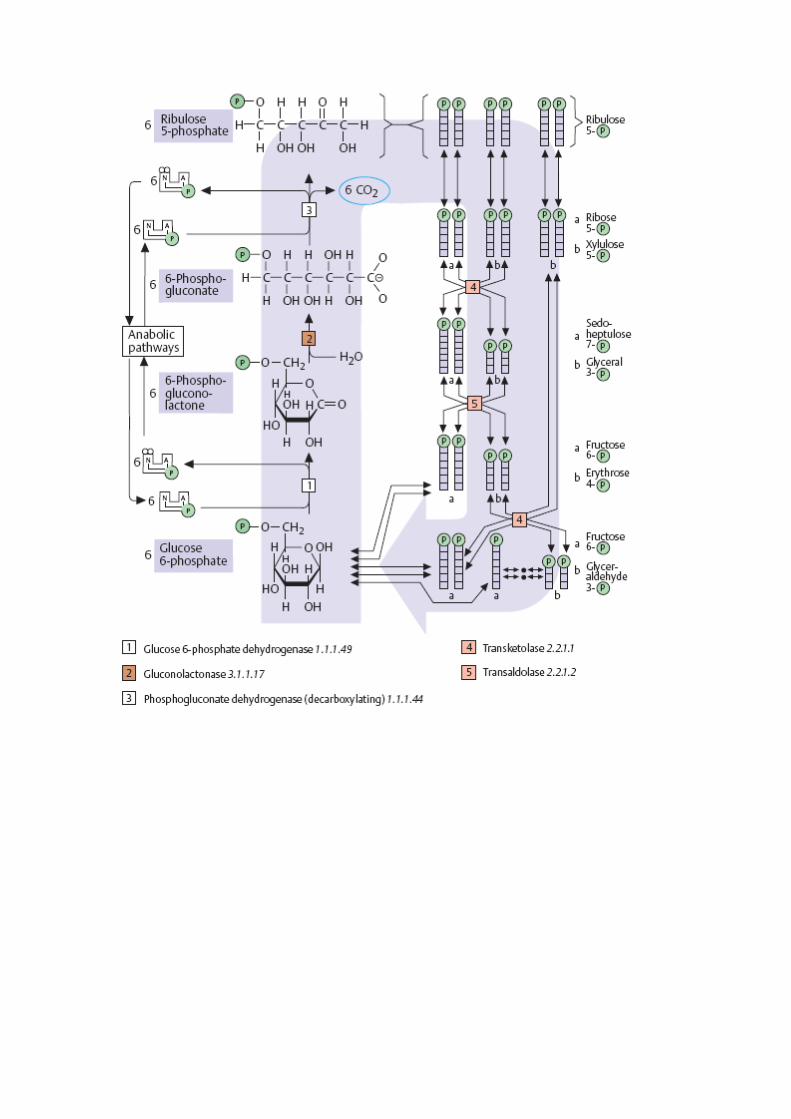
Tem duas fases: oxidativa e não oxidativa
Fase oxidativa
É uma forma de cortar a cadeia de carbonos de uma molécula glicídica removendo um
carbono de cada vez.
o Oxidação e descarboxilação da glicose 6-fosfato para formar ribose 5-fosfato e
NADPH.
o Reacção, em parte, catalisada pela glicose 6-fosfato desidrogenase (G6P).
A via pode parar neste ponto:
o NADPH pode ser usado para reacções biossintéticas redutoras.
o Ribose 5-fosfato pode ser usada como precursor para a síntese de nucleótidos.
Fase não-oxidativa
Quando o NADPH é mais necessário para a célula do que a ribose 5-fosfato, esta é
rearranjada para formar intermediários da glicólise e consequentemente ser
reaproveitada ajuste das necessidades da célula.
Para que as interconversões aconteçam, forma-se primeiramente xilulose 5-fosfato a
partir da isomerização da ribose 5-fosfato. Estas duas moléculas são então utilizadas
nas interconversões.
Esta fase corresponde a 3 reacções de transcetolisação e transaldolisação que levam à
formação de alguns intermediários da glicólise:
o Frutose 6-fosfato glicose 6-fosfato
o Gliceraldeído 3-fosfato
Vias de destino do NADPH:
o Biossíntese de ácidos gordos
o Biossíntese de colestrol
o Biossíntese de nucleótidos
o Biossíntese de neurotransmissores
o Redução do glutatião (molécula
causadora de stress oxidativo) a
glutationa
o Monooxigenases do citocromo
P450 (enzima hepática)

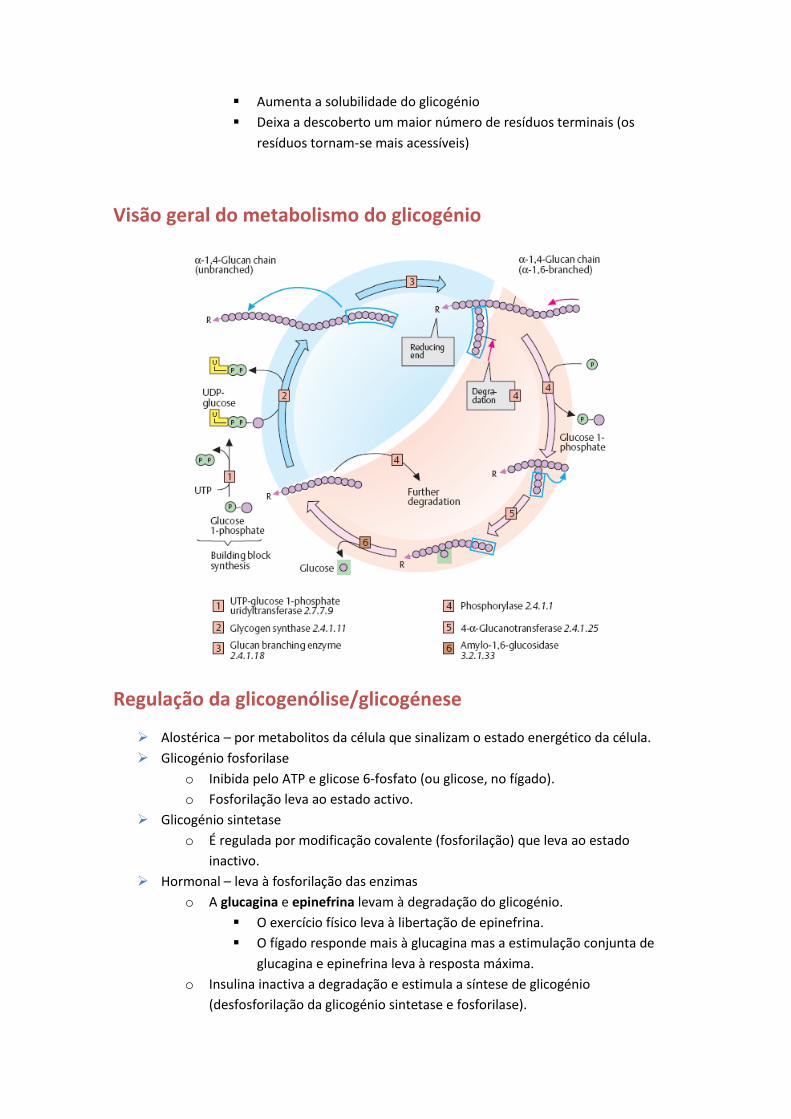
F – Metabolismo do glicogénio
Introdução
O glicogénio é uma forma de armazenamento de glicose facilmente mobilizável.
Os resíduos de glicose estão unidos por ligações α-1,4 glicosídicas e a cada 10 resíduos
é criada uma ramificação através de ligações α-1,6 glicosídicas.
Responsável por manter estável a quantidade de glicose no sangue entre as refeições.
O glicogénio encontra-se no fígado e no músculo (concentração maior no fígado).
o Fígado regula a glicemia em relação às necessidades de todo o organismo.
o Músculo utiliza o glicogénio para consumo energético próprio.
Glicogenólise
Consiste na degradação do glicogénio a glicose.
A glicogénio fosforilase é a enzima chave da glicogenólise.
o Cliva as ligações entre os resíduos glicosil (glicose 1-fosfato) que compõem o
glicogénio entre C-1 de um resíduo glicosil e C-4 do seguinte (tendo em conta
que os resíduos estão unidos por ligações α-1,4) através da adição de um
fosfato inorgânico (Pi).
o A glicose 1-fosfato não se difunde para fora da célula.
o A glicose 1-fosfato é facilmente convertida em glicose 6-fosfato
(fosfoglucomutase).
A razão pela qual a glicogénio fosforilase utiliza o fosfato inorgânico para clivar os
resíduos de glicose em vez de os hidrolisar é que desta forma já não há necessidade de
gastar ATP para fosforilar a glicose (o Pi fica na molécula – glicose 1-fosfato).
A glicogénio fosforilase só consegue efectuar este processo até encontrar um
obstáculo – a ramificação.
o As ligações α-1,6 não são clivadas pela glicogénio fosforilase.
o Termina a clivagem a uma distância de 4 resíduos da ramificação.

As ligações α-1,6 exigem a acção de outras duas enzimas:
o Transferase – transfere um bloco de 3 resíduos de uma ramificação (que está a
ser degradada) para outra mais exterior. Resta apenas um único resíduo nesta
ramificação que está unido à molécula de glicogénio por uma ligação α-1,6.
o α-1,6-glucosidase ou enzima desramificadora do glicogénio – hidrolisa a
ligação α-1,6 libertando glicose.
Esta molécula de glicose é fosforilada pela hexocinase formando
glicose 6-fosfato.
No fígado:
o O fígado contem a enzima hidrolítica glicose 6-fosfatase que cliva o grupo fosforil
da glicose 6-fosfato para formar glicose livre e o fosfato inorgânico.
Esta enzima é a mesma que conclui a gliconeogénese.
Não existe na maioria dos outros tecidos o que faz com que a glicose 6-
fosfato fique retida para a produção de ATP
A actividade da glicogénio fosforilase utiliza o fosfato piridoxal (PLP), um derivado da
vitamina B6.
Glicogénese
Consiste na síntese do glicogénio a partir de glicose.
Utiliza uridina difosfato glicose (UDP-glicose) em vez da glicose 1-fosfato.
o A UDP-glicose é uma forma activa de glicose tal como o acetil-CoA é uma
forma activa de acetato.
o A UDP-glicose é sintetizada a partir da glicose 1-fosfato e uridina trifosfato
(UTP) numa reacção catalisada pela UDP-glicose pirofosforilase.
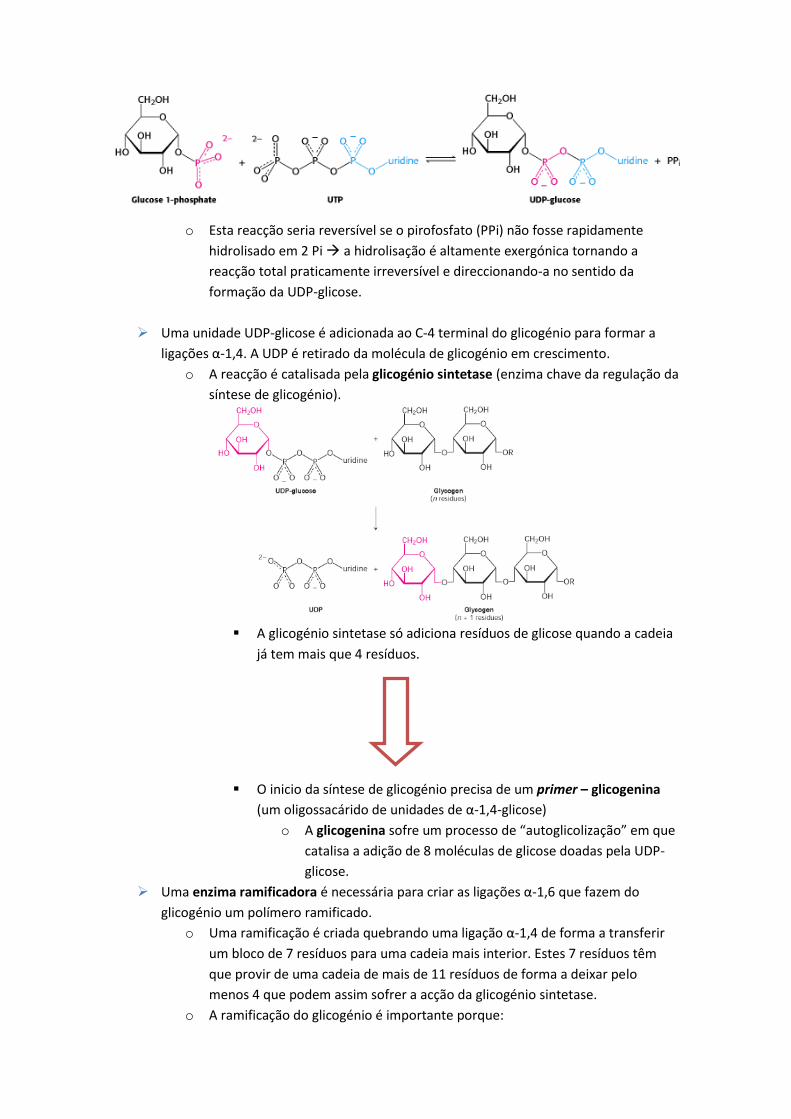
o Esta reacção seria reversível se o pirofosfato (PPi) não fosse rapidamente
hidrolisado em 2 Pi a hidrolisação é altamente exergónica tornando a
reacção total praticamente irreversível e direccionando-a no sentido da
formação da UDP-glicose.
Uma unidade UDP-glicose é adicionada ao C-4 terminal do glicogénio para formar a
ligações α-1,4. A UDP é retirado da molécula de glicogénio em crescimento.
o A reacção é catalisada pela glicogénio sintetase (enzima chave da regulação da
síntese de glicogénio).
A glicogénio sintetase só adiciona resíduos de glicose quando a cadeia
já tem mais que 4 resíduos.
O inicio da síntese de glicogénio precisa de um primer – glicogenina
(um oligossacárido de unidades de α-1,4-glicose)
o A glicogenina sofre um processo de “autoglicolização” em que
catalisa a adição de 8 moléculas de glicose doadas pela UDP-
glicose.
Uma enzima ramificadora é necessária para criar as ligações α-1,6 que fazem do
glicogénio um polímero ramificado.
o Uma ramificação é criada quebrando uma ligação α-1,4 de forma a transferir
um bloco de 7 resíduos para uma cadeia mais interior. Estes 7 resíduos têm
que provir de uma cadeia de mais de 11 resíduos de forma a deixar pelo
menos 4 que podem assim sofrer a acção da glicogénio sintetase.
o A ramificação do glicogénio é importante porque:
Aumenta a solubilidade do glicogénio
Deixa a descoberto um maior número de resíduos terminais (os
resíduos tornam-se mais acessíveis)
Visão geral do metabolismo do glicogénio
Regulação da glicogenólise/glicogénese
Alostérica – por metabolitos da célula que sinalizam o estado energético da célula.
Glicogénio fosforilase
o Inibida pelo ATP e glicose 6-fosfato (ou glicose, no fígado).
o Fosforilação leva ao estado activo.
Glicogénio sintetase
o É regulada por modificação covalente (fosforilação) que leva ao estado
inactivo.
Hormonal – leva à fosforilação das enzimas
o A glucagina e epinefrina levam à degradação do glicogénio.
O exercício físico leva à libertação de epinefrina.
O fígado responde mais à glucagina mas a estimulação conjunta de
glucagina e epinefrina leva à resposta máxima.
o Insulina inactiva a degradação e estimula a síntese de glicogénio
(desfosforilação da glicogénio sintetase e fosforilase).

Epinefrina:
Verde – enzima activa Rosa – enzima inactiva
Nota: a fosforilase cinase também pode ser parcialmente activada pelo cálcio iónico e o efeito
é conjunto com o da epinefrina importante no músculo
Síntese Degradação

2 – Metabolismo lipídico
Introdução
Papel metabólico dos ácidos gordos:
o São precursores biossintéticos dos fosfolípidos e glicolípidos.
o Modificação de proteínas.
o São moléculas combustíveis.
o Os seus derivados podem ser hormonas ou mensageiros intracelulares.
A degradação e síntese de ácidos gordos são processos praticamente contrários um ao
outro.
A maior parte dos lípidos ingeridos são triacilglicerois.
A – Lípidos na dieta
O local de maior acumulação de lípidos (triacilglicerois) é o citoplasma das células do
tecido adiposo. Estas células são especializadas para a síntese e armazenamento de
triacilglicerois.
Os triacilglicerois provenientes da dieta são degradados a ácidos gordos e um
monoacilglicerol para que sejam absorvidos pelo epitélio intestinal e posteriormente
passarem para o sistema linfático e depois para a corrente sanguínea.
o No lúmen intestinal, são incorporados em micelos formados com o ácido dos
sais biliares e que propiciam a digestão dos triacilglicerois pelas enzimas
pancreáticas.
o Nas células do epitélio intestinal, os triacilglicerois são ressintetisados e
transferidos para os quilomicra - uma lipoproteína transportadora de
triacilglicerois.
A parte proteica das lipoproteínas é chamada apolipoproteína.
Os quilomicra ligam-se a lipoproteínas lipases primeiramente no
tecido adiposo e no músculo – no músculo podem ser oxidados para
formação de energia.

B – Utilização de ácidos gordos como combustível
Introdução
3 fases:
o Triacilglicerois são degradados a ácidos gordos e glicerol que são libertados do
tecido adiposo para a corrente sanguínea e transportados para os tecidos com
necessidade energética.
o Nos tecidos, os ácidos gordos são activados e transportados para a
mitocôndria para serem degradados.
o Os ácidos gordos são degradados em acetil-CoA que pode ser utilizado no ciclo
de Krebs.
Fase 1
Lipólise:
o Corresponde à hidrólise dos triacilglicerois por lipases formando ácidos gordos
e glicerol.
o As lipases são activadas pela epinefrina e glucagina e inibidas pela insulina.
Os ácidos gordos não são solúveis no plasma e por isso ligam-se à albumina (proteína
transportadora presente no plasma) para serem transportados para os tecidos.
O glicerol é absorvido pelo fígado, fosforilado e oxidado para formar dihidroxiacetona
fosfato que é isomerada a gliceraldeído 3-fosfato (intermediário da glicólise e
gliconeogénese). Pode ser convertido a glicose ou piruvato.
Fase 2
A oxidação dos ácidos gordos acontece na mitocôndria.
Antes de entrarem na matriz mitocondrial, os ácidos gordos são activados através da
ligação à Co-enzima A formando acil-CoA isto acontece na membrana externa da
mitocôndria e a reacção é catalisada pela acil-CoA sintetase.
Há gasto de uma molécula de ATP. No entanto a reacção é bastante
favorável pois o equivalente a duas moléculas de ATP é hidrolisado com
Ácido gordo Acil-CoA
libertação de um pirofosfato (PPi) que é rapidamente hidrolisado tornando
a reacção irreversível.
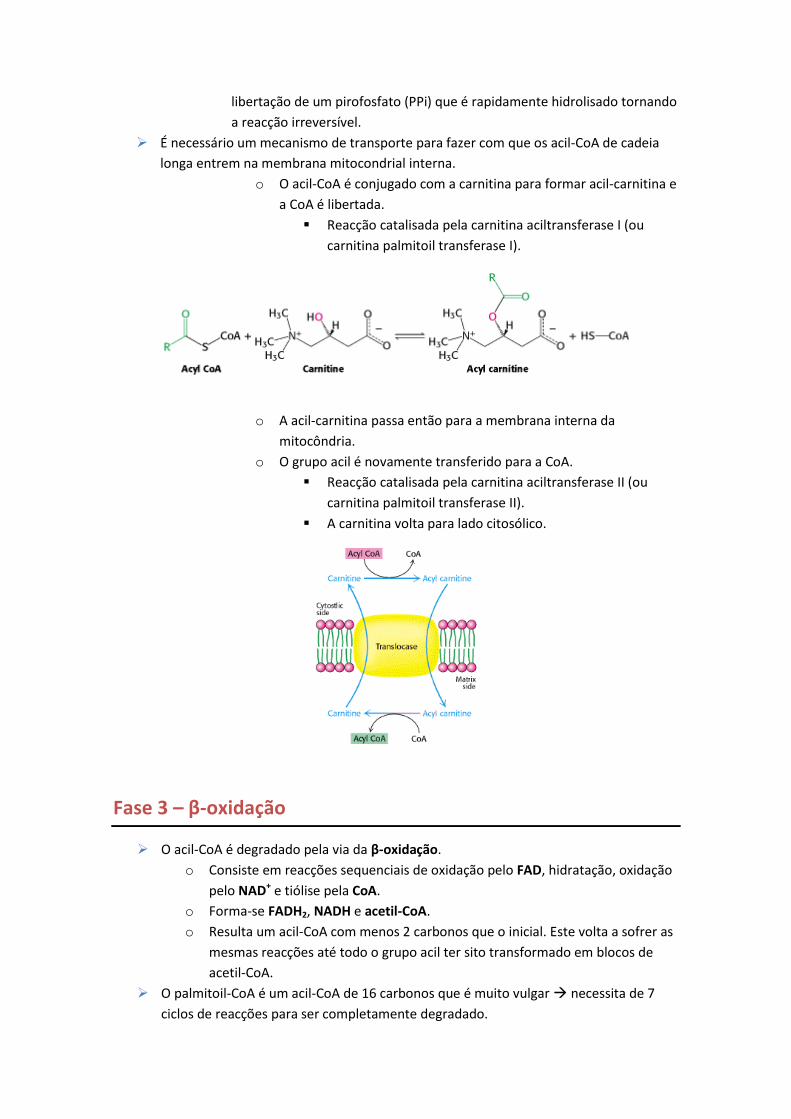
É necessário um mecanismo de transporte para fazer com que os acil-CoA de cadeia
longa entrem na membrana mitocondrial interna.
o O acil-CoA é conjugado com a carnitina para formar acil-carnitina e
a CoA é libertada.
Reacção catalisada pela carnitina aciltransferase I (ou
carnitina palmitoil transferase I).
o A acil-carnitina passa então para a membrana interna da
mitocôndria.
o O grupo acil é novamente transferido para a CoA.
Reacção catalisada pela carnitina aciltransferase II (ou
carnitina palmitoil transferase II).
A carnitina volta para lado citosólico.
Fase 3 – β-oxidação
O acil-CoA é degradado pela via da β-oxidação.
o Consiste em reacções sequenciais de oxidação pelo FAD, hidratação, oxidação
pelo NAD+ e tiólise pela CoA.
o Forma-se FADH2, NADH e acetil-CoA.
o Resulta um acil-CoA com menos 2 carbonos que o inicial. Este volta a sofrer as
mesmas reacções até todo o grupo acil ter sito transformado em blocos de
acetil-CoA.
O palmitoil-CoA é um acil-CoA de 16 carbonos que é muito vulgar necessita de 7
ciclos de reacções para ser completamente degradado.

No caso dos ácidos gordos de cadeia impar:
o São oxidados da mesma forma mas no final resultam uma molécula de acetil-
CoA (em vez de duas) e uma molécula de propionil-CoA (composto de 3
carbonos).
o O propionil-CoA entra no ciclo de Krebs depois de ter sido convertido a
succinil-CoA (reacção dependente da vitamina B12 ou cobalamina).
C – Síntese de corpos cetónicos
O acetil-CoA produzido na β-oxidação só entra no ciclo de Krebs se a degradação de
ácidos gordos e glícidos estiver equilibrada porque a sua entrada depende da
disponibilidade de oxaloacetato (normalmente formado a partir do piruvato).
Podemos encarar os corpos cetónicos como uma forma transportável de unidades de
acetil-CoA.
Corpos cetónicos:
o Compostos formados a partir do acetil-CoA no fígado, passam para a corrente
sanguínea e são transportados para os tecidos periféricos.
Acetoacetato
3-hidroxibutirato
Acetona
o O músculo cardíaco e o córtex renal usam acetoacetato preferencialmente à
glicose.
o O cérebro pode adaptar-se à sua utilização em períodos de jejum prolongado
ou diabetes.
o Acetoacetato e 3-hidroxibutirato:
Condensação de duas moléculas de acetil-CoA para formar
acetoacetil-CoA.
Acetoacetil-CoA condensa-se com mais uma molécula de acetil-CoA
com formação de HMG-CoA e CoA.
O HMG-CoA é então clivado para formar acetoacetato e 3-
hidroxibutirato
o Acetona:
Proveniente da descarboxilação espontânea do acetoacetato.
Provoca o “hálito cetónico”.
HMG-CoA

Quando a quantidade de acetoacetato em circulação está elevada, a lipólise é inibida
no tecido adiposo.
D – Síntese de ácidos gordos
Introdução
Acontece no citosol.
O ácido gordo é aumentado a partir da adição sucessiva de unidades de carbonos –
acetil-CoA e malonil-ACP.
O poder redutor para a síntese é dado pelo NADPH.
Nos organismos mais complexos, a síntese de ácidos gordos é levada a cabo por um
sistema enzimático chamado ácidos gordos sintetase.
Formação do malonil-CoA
Carboxilação do acetil-CoA a malonil-CoA
o Reacção irreversível.
o Catalisada pela acetil-CoA carboxilase (tem como grupo prostético a biotina).
É a enzima reguladora do metabolismo dos ácidos gordos.
o É necessária a hidrólise de uma molécula de ATP.
o O CO2 activado (HCO3-) é transferido para o acetil-CoA.
Ciclo de enlongação na síntese de ácidos gordos
Os intermediários estão ligados a uma proteína transportadora de grupos acil – ACP.
o Podemos pensar na ACP como uma CoA gigante.
o Forma-se acetil-ACP e malonil-ACP.
A enlongação de ácidos gordos é catalisada pela ácidos gordos sintetase e consiste na
adição sucessiva de unidades acetil-ACP e malonil-ACP com reduções e
descarboxilações concomitantes.
O ciclo de enlongação termina no composto C16 – o palmitato.
Posterior enlogação é conseguida através de outros sistemas enzimáticos acessórios.

Equações gerais da síntese de ácidos gordos
Síntese do malonil-CoA:
Síntese do palmitato:
Total:
Transporte de acetil-CoA e fontes de NADPH para a síntese de
ácidos gordos
A síntese do palmitato requer 8 moléculas de acetil-CoA.
o O acetil-CoA é formado a partir do piruvato na mitocôndria por isso tem que
ser transportado para o citosol para ser utilizado na síntese de ácidos gordos.
Esta barreira é ultrapassada através de um bypass pelo citrato que transporta grupos
acil através da membrana interna da mitocôndria.
o O citrato é formado na matriz mitocondrial através da condensação do
oxacloacetato com o acetil-CoA.
o O citrato pode atravessar a membrana interna da mitocôndria e no citosol é
clivado pela ATP-citrato liase para voltar a formar acetil-CoA e oxaloacetato.
o Há gasto de uma molécula de ATP.
O oxaloacetato formado no citosol tem que voltar à mitocôndria e a membrana
interna é impermeável necessidade de um sistema de bypass de transporte:
o Primeiro, o oxaloacetato é reduzido a malato pelo NADH .
Reacção catalisada pela malato desidrogenase.

o Segundo, o malato sofre uma descarboxilação oxidadativa pelo NAD+.
Reacção catalisada pela enzima málica.
Há formação de NADPH
o Por último, o piruvato formado volta espontaneamente para a mitocôndria
onde é carboxilado a oxaloacetato.
O sistema de transporte gera NADPH (que pode ser utilizado na síntese de ácidos
gordos) por cada molécula de acetil-CoA transferida para o citosol.
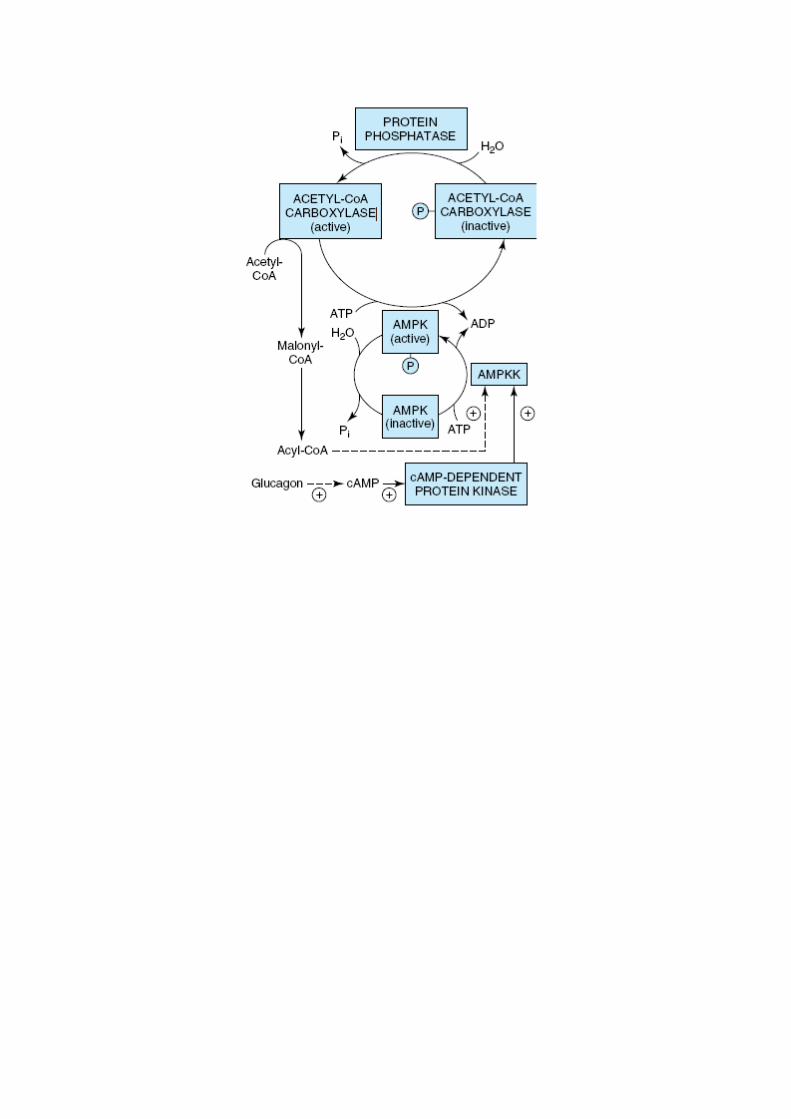
Regulação do metabolismo dos ácidos gordos
A acetil-CoA carboxilase (catalisa a síntese de malonil-CoA a partir de acetil-CoA –
passo inicial da síntese de ácidos gordos) é a enzima mais importante na regulação do
metabolismo dos ácidos gordos.
o É controlada por 3 sinais globais:
Glucagina
Epinefrina
Insulina
o A insulina estimula a síntese de ácidos gordos enquanto a glucagina e
epinefrina têm o efeito inverso.
o Pode também ser controlada pelos níveis de metabolitos:
Citrato – sinaliza que os precursores biossintéticos estão abundantes e
por isso activa a acetil-CoA carboxilase para dar inicio à síntese de
ácidos gordos.
Palmitoil-CoA e AMP – levam à inibição da acetil-CoA carboxilase.
o Fosforilação – inactiva a acetil-CoA carboxilase.