mts del huso mts de interfase - iib - instituto de ... · en las células el estado de...
TRANSCRIPT


axonema
MTs de interfase
MTs del huso
Microtúbulos
vesícula
Cuando una célula entra en mitosis
los microtúbulos citoplasmáticos se
desensamblan y se reensamblan
para formar el huso mitótico.

El dímero de tubulina une dos moléculas de GTP. Los
heterodímeros de tubulina se ensamblan para formar los
microtúbulos. La molécula de GTP unida a la subunidad de
tubulina beta puede hidrolizarse afectando la dinámica de los
microtúbulos. La orientación constante de los monómeros en
los polímeros determina su polaridad estructural. Las
velocidades de crecimiento diferentes de los extremos (+) y
(-) son consecuencia de esta polaridad.
Estructura de las subunidades de tubulina y los microtúbulos
la orientación de los ganchos de
tubulina en dirección horaria
indican que el extremo (+) apunta
hacia el observador.
extremo (-) hacia el frente
Análisis al microscopio
electrónico de la polaridad de
microtúbulos seccionados
transversalmente
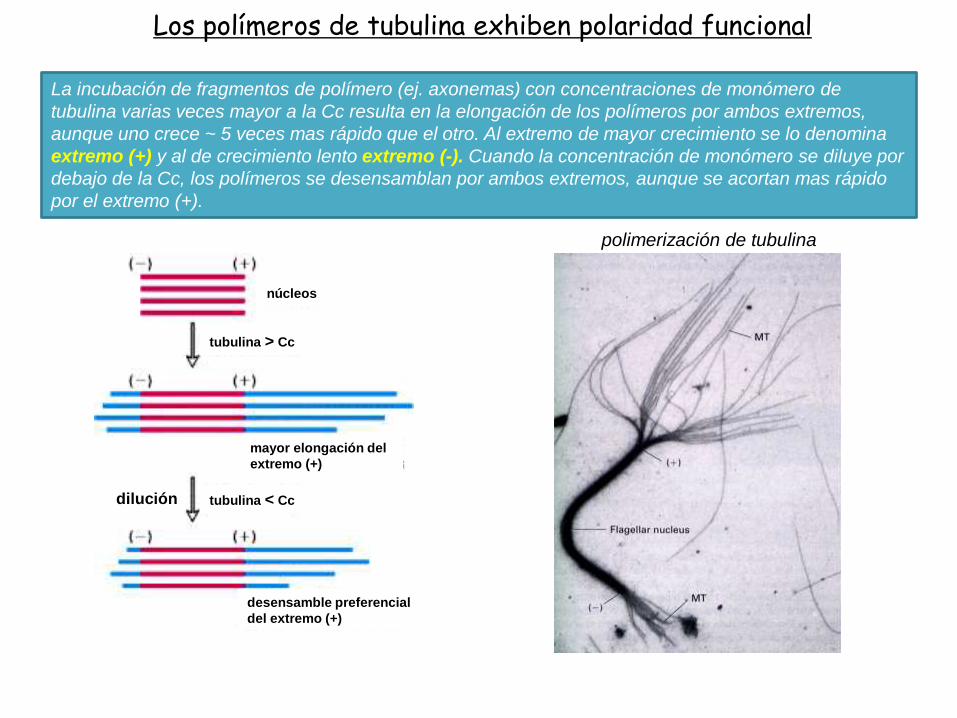
La incubación de fragmentos de polímero (ej. axonemas) con concentraciones de monómero de
tubulina varias veces mayor a la Cc resulta en la elongación de los polímeros por ambos extremos,
aunque uno crece ~ 5 veces mas rápido que el otro. Al extremo de mayor crecimiento se lo denomina
extremo (+) y al de crecimiento lento extremo (-). Cuando la concentración de monómero se diluye por
debajo de la Cc, los polímeros se desensamblan por ambos extremos, aunque se acortan mas rápido
por el extremo (+).
Los polímeros de tubulina exhiben polaridad funcional
polimerización de tubulina
tubulina > Cc
tubulina < Cc
mayor elongación del
extremo (+)
desensamble preferencial
del extremo (+)
dilución
núcleos

tiempo 0 tiempo 2 min tiempo 5 min tiempo 11 min
10 mm
A diferencia del treadmilling, la
inestabilidad dinámica se caracteriza
por la alternancia entre fases de
crecimiento lento y de acortamiento
rápido en los extremos (+). La transición
de la fase de crecimiento a la de
acortamiento se denomina catástrofe y
la transición de acortamiento a
crecimiento rescate.
Los microtúbulos además exhiben un comportamiento dinámico conocido como inestabilidad dinámica
Lodish et al MCB2004
La inestabilidad dinámica de los microtúbulos puede visualizarse en células vivas.

La inestabilidad dinámica es una consecuencia de la presencia de tubulina-GDP/GTP en los extremos (+) de los MTs
La inestabilidad dinámica
de los microtúbulos
depende de la presencia
de tubulina-GDP o tubulina-
GTP en los extremos (+).
En una población de
microtúbulos, existe una
mezcla de ambos estados.
Cuando la velocidad de
adición de monómeros
supera a la de la hidrólisis
del GTP se acumula
tubulina-GTP ("GTP cap"),
condición que favorece la
elongación. Cuando el cap
de GTP se pierde, la
velocidad de disociación de
monómeros incrementa
dramáticamente, causando
el acortamiento rápido del
MT.
ex
trem
o
ine
sta
ble
ex
trem
o
es
tab
le
Alberts et al., MBC2002

stathmin
stathmin-P
proteínas que se unen a las
subunidades no ensambladas *
tubulina actina
thymosin
profilin
*las proteínas promueven la depolimerizacion (↓)o
polimerizacion (↑) de los respectivos polímeros
proteínas que causan la fragmentación
de los polímeros
katanin
microtúbulos microfilamentos
gelsolin
proteínas que se asocian a las paredes
de los polímeros y los estabilizan
MAP2, Tau
XMAP215
tropomyosin
microtúbulos microfilamentos
XMAP215-P
KIF2
cofilin
microtúbulos microfilamentos
proteínas que se unen a los
extremos de los polímeros
g-TuRC (-), nucleación
+ TIPs (CLIP170, Op18)
CapZ (+), capping
Arp2/3 (-), nucleación
microtúbulos microfilamentos
proteínas que se asocian a las paredes
de los polímeros y los desestabilizan
En las células el estado de polimerización de la actina y tubulina es regulado por diversas proteínas asociadas
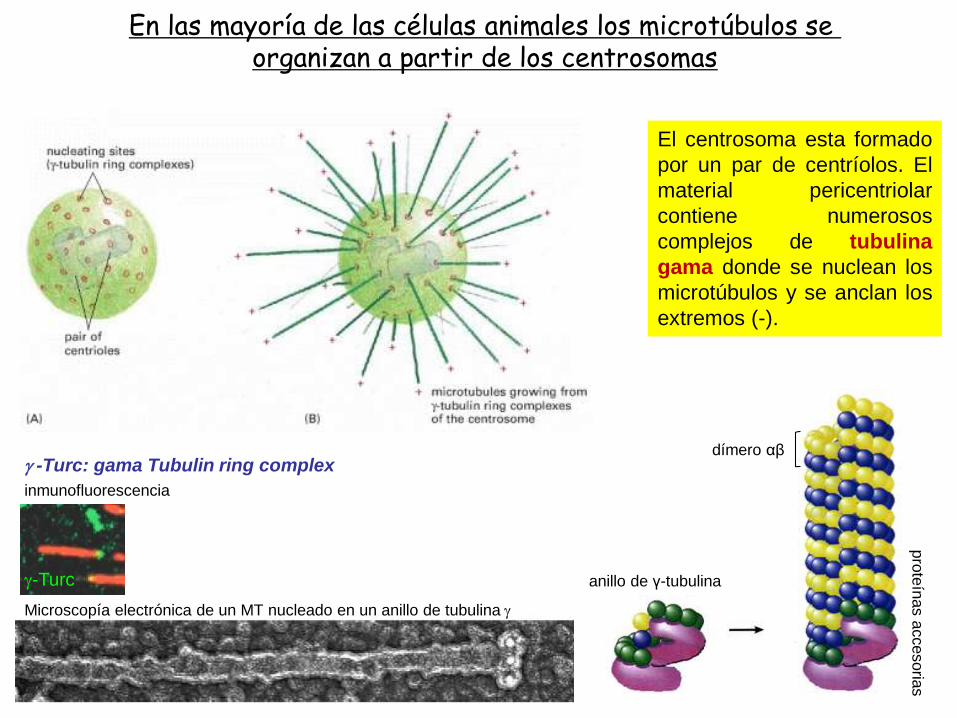
En las mayoría de las células animales los microtúbulos se organizan a partir de los centrosomas
pro
teín
as
accesoria
s
El centrosoma esta formado
por un par de centríolos. El
material pericentriolar
contiene numerosos
complejos de tubulina
gama donde se nuclean los
microtúbulos y se anclan los
extremos (-).
g -Turc: gama Tubulin ring complex
Microscopía electrónica de un MT nucleado en un anillo de tubulina g
inmunofluorescencia
g-Turc
dímero αβ
anillo de γ-tubulina
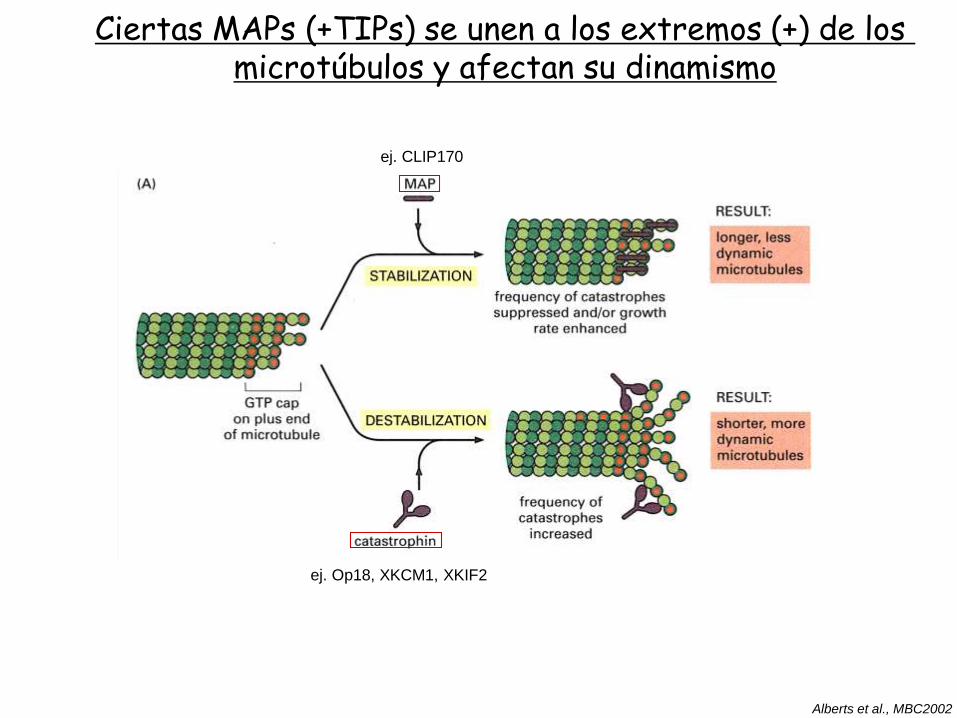
Ciertas MAPs (+TIPs) se unen a los extremos (+) de los microtúbulos y afectan su dinamismo
ej. Op18, XKCM1, XKIF2
Alberts et al., MBC2002
ej. CLIP170

Una serie de proteínas denominadas +TIPs se unen al extremo (+) de los microtúbulos y regulan su función
EB1, CLIP170 y otras, son ejemplos de proteínas que interaccionan con el extremo (+) de los
microtúbulos en crecimiento.
Cumplen diversas funciones, promueven la estabilidad del extremo (+), promueven el anclaje
de microtúbulos con membranas, filamentos de actina, interaccionan con moléculas motoras,
moléculas de señalización, etc.
La interacción de estos factores con el extremo (+) permite que la célula emplee la energía
que libera la polimerización de microtúbulos para generar fuerzas que pueden usarse para
posicionar el huso, cromosomas u organelas.
Galjart 2005 Nat Rev Mol Cell Biol

stathmin secuestra
tubulina no ensamblada,
favoreciendo la frecuencia
de catastrofes de los
extremos (+)
Otras MAPs se unen a los monómeros y promuevenla depolimerización de los MTs
Alberts et al., MBC2002

MAP2 y tau son proteínas de
entrecruzamiento que regulan
el espaciamiento entre MTs.
MAP2 Tau
MAP2 y Tau se unen a la pared de los MTs y contribuyen a suestabilización y espaciamiento en fascículos
Microscopía electrónica de células transfectadas con MAP2 o Tau.
Alberts et al., MBC2002

En neuronas MAP2 se localiza principalmente en dendritas y Tau en axones
Neurona de hipocampo. La inmunofluorescencia
revela la localización de MAP2 en dendritas
(anaranjado) y de la tau en axones (verde).
Microscopía electrónica de un axón (A) y de una
dendrita (B). Observe el diferente espaciamiento
de los microtúbulos.
A B
MAP2tau

Drogas que alteran la dinámica de microtúbulos se usan comoagentes quimioterápicos para el tratamiento de tumores
Las drogas que afectan la polimerización detubulina son agentes citotóxicos. Las drogas queestabilizan los microtúbulos (taxol y epotilonas)se unen al sitio taxano en la subunidad β detubulina estabilizando los contactoslongitudinales y laterales. Laulimalida ypelorusida A estabilizan los microtúbulosuniendo dos subunidades β protofilamentosadyacentes. Los agentes desestabilizantes seunen al sitio de colchicina ubicado entre lassubunidades α y β de tubulina en el mismodímero, o al sitio de vinca, que se forma entresubunidades α y β de dos dímeros distintosalineados longitudinalmente. Los complejostubulina-colchicina pueden incorporarse durantela elongación alterando los extremos delmicrotúbulo. Los compuestos dirigidos al sitiomaitansina se unen a tubulina β en el extremomás del microtúbulo inhibiendo la elongación.
Akhmanova (2015) Nat Rev Mol Cell Biol

Los microtúbulos y las moléculas motoras son esenciales para eltransporte intracelular
Galjart 2005 Nat Rev Mol Cell Biol
Los microtúbulos de fibroblastos
y de conos de crecimiento
axonal son dinámicos y alternan
fases de elongación y
acortamiento. Durante la
mitosis, el treadmilling de
microtúbulos del cinetocoro (B)
facilita la segregación de
cromosomas.
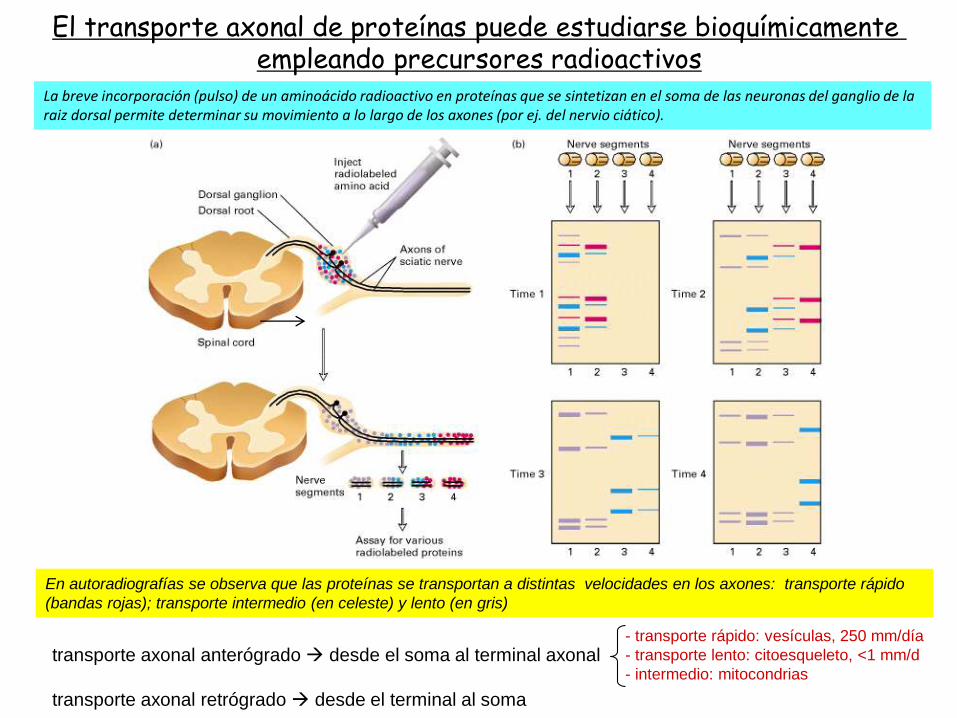
El transporte axonal de proteínas puede estudiarse bioquímicamenteempleando precursores radioactivos
transporte axonal anterógrado desde el soma al terminal axonal
transporte axonal retrógrado desde el terminal al soma
- transporte rápido: vesículas, 250 mm/día
- transporte lento: citoesqueleto, <1 mm/d
- intermedio: mitocondrias
En autoradiografías se observa que las proteínas se transportan a distintas velocidades en los axones: transporte rápido
(bandas rojas); transporte intermedio (en celeste) y lento (en gris)
La breve incorporación (pulso) de un aminoácido radioactivo en proteínas que se sintetizan en el soma de las neuronas del ganglio de la raiz dorsal permite determinar su movimiento a lo largo de los axones (por ej. del nervio ciático).

Análisis por videomicroscopía revela el transporte bi-direccionaly saltatorio de las vesículas
El transporte axonal ha sido visualizado en extractos de axon gigante de calamar incubados en presencia de ATP. En el
panel de la izquierda se muestra una serie de imágenes adquiridas a distintos tiempos por una cámara de video acoplada a
un microscopio óptico. Las flechas negra y amarilla señalan vesículas moviéndose en dirección opuesta sobre los MTs. (B):
Una región similar a (A) fue procesada para microscopía electrónica. Se observan 2 vesículas unidas a un MT
BA
videos disponibles

En los melanóforos, los microtúbulos son requeridos parala redistribución de vesículas con pigmentos
El movimiento de los gránulos de pigmento sobre los MTs es regulado por señales extracelulares.
movimiento centrípeto
dependiente de dineínas
movimiento centrífugo
dependiente de kinesinas
Hong & Laverty, Proc Assoc Biol Lab 2011
melanóforos
killifish
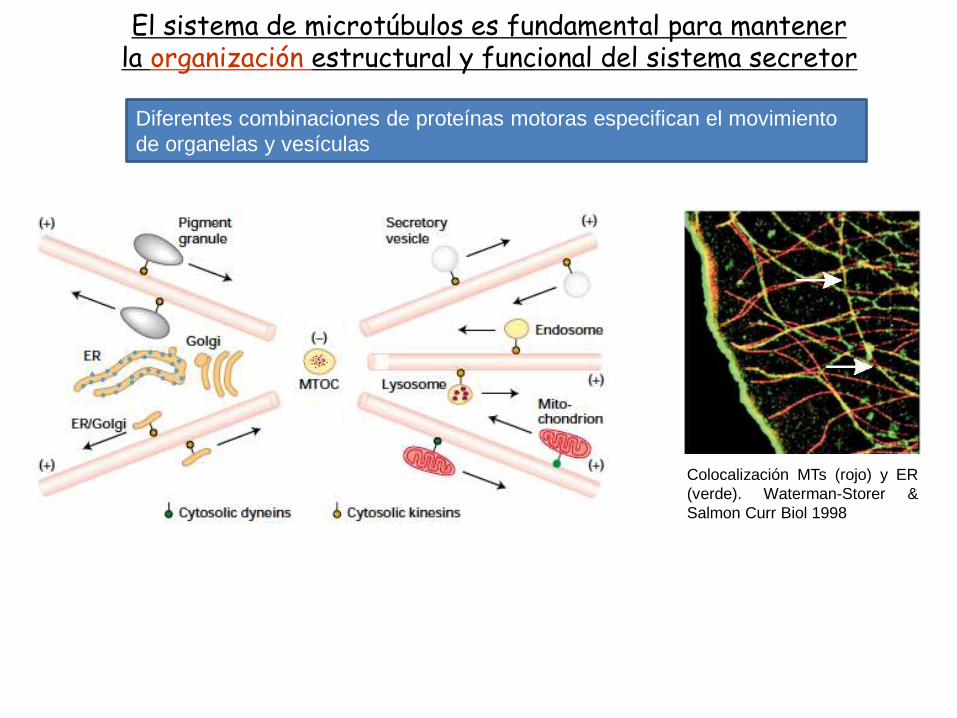
El sistema de microtúbulos es fundamental para mantenerla organización estructural y funcional del sistema secretor
Diferentes combinaciones de proteínas motoras especifican el movimiento
de organelas y vesículas
Colocalización MTs (rojo) y ER
(verde). Waterman-Storer &
Salmon Curr Biol 1998

MT
ER
Golgi
ER
MTs
Golgi
En células tratadas con colchicina el ER colapsa alrededor del núcleo y el aparato de Golgi se
fragmenta. Cuando se remueve la colchicina, los microtúbulos se reensamblan y las proteína
motoras transportan las organelas a sus posiciones originales.
núcleo
MTOC
Los microtúbulos contribuyen al posicionamiento de organelas

Las dineínas y kinesinas son moléculas motoras que emplean energíade la hidrólisis de ATP para translocarse sobre microtúbulos
Lodish et al MCB 2000
* Movement of motor protein toward the (+) end or (−) end of microtubules.† Also known as kinesin related proteins (KRPs).‡ Outer-arm dyneins have three heavy chains, and inner-arm dyneins have two heavy chains.
Class Members Cargo Direction of Movement *
Cytosolic kinesins Kinesin, Unc-104 Cytosolic vesicles (+)
Spindle kinesins† Ncd/KAR3, BimC/Eg5, CENP-E
Spindle and astral MTs, centrosomes, kinetochores
(+) or (−)
Cytosolic dyneins Cytoplasmic dynein Cytosolic vesicles, kinetochores during mitosis and meiosis
(−)
Axonemal dyneins Outer-arm dyneins, inner-arm dyneins‡
A tubule of doublet microtubules in cilia and flagella
(−)

Las dineínas forman complejos multimoleculares que transportan diversoscargos hacia los extremos (-) de los microtúbulos
Las dineínas citosólicas son
responsables del transporte axonal
retrógrado, transporte de vesículas
endocíticas hacia lisosomas,
posicionamiento perinuclear del Golgi,
elongación y posicionamiento del huso
mitótico, movimiento de cromosomas,
movimiento radial centrípeto de
pigmentos en melanóforos, etc. La
velocidad de transporte varía entre 1-2
mm/seg. Las numerosas subunidades que
componen al complejo dineína-dinactina
explicaría la amplia diversidad de cargos
transportados por las dineínas.
microscopía electrónica."freeze-etch"
Las dineínas son proteínas multiméricas compuestas de
2-3 cadenas pesadas y varias subunidades intermedias y
livianas. Las subunidades pesadas son ATPasas que
interaccionan con el microtúbulo. Las cadenas livianas
interaccionan con un complejo multiproteico denominado
dinactina que se asocia al cargo.
cabezas
globulares
de las cadenas
pesadas
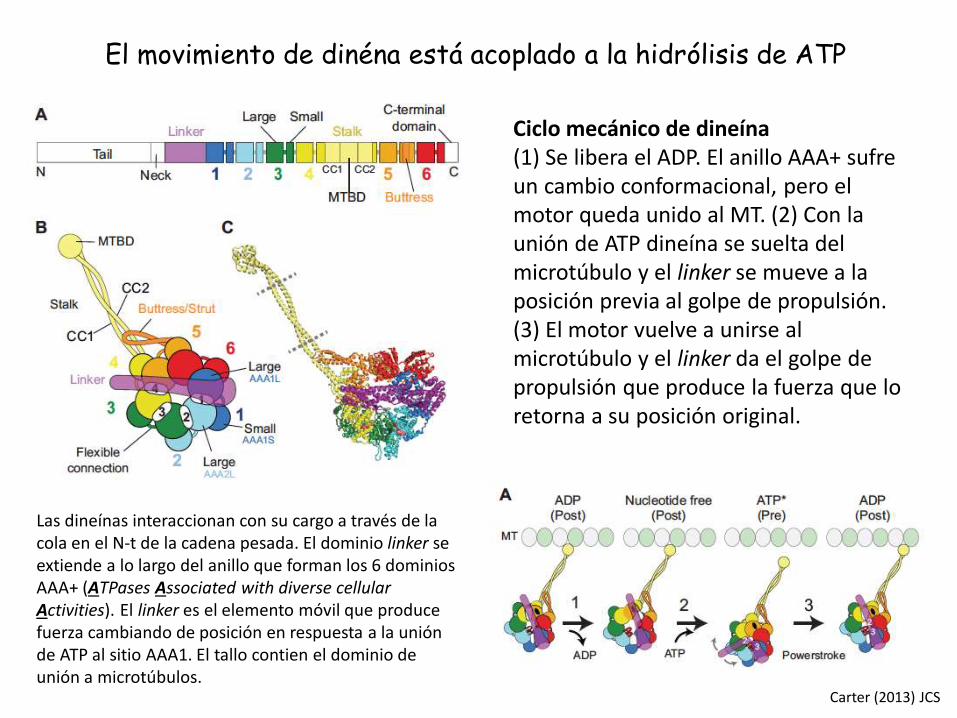
Las dineínas interaccionan con su cargo a través de la cola en el N-t de la cadena pesada. El dominio linker se extiende a lo largo del anillo que forman los 6 dominiosAAA+ (ATPases Associated with diverse cellular Activities). El linker es el elemento móvil que produce fuerza cambiando de posición en respuesta a la uniónde ATP al sitio AAA1. El tallo contien el dominio de unión a microtúbulos.
Ciclo mecánico de dineína(1) Se libera el ADP. El anillo AAA+ sufreun cambio conformacional, pero el motor queda unido al MT. (2) Con la unión de ATP dineína se suelta del microtúbulo y el linker se mueve a la posición previa al golpe de propulsión. (3) El motor vuelve a unirse al microtúbulo y el linker da el golpe de propulsión que produce la fuerza que lo retorna a su posición original.
El movimiento de dinéna está acoplado a la hidrólisis de ATP
Carter (2013) JCS
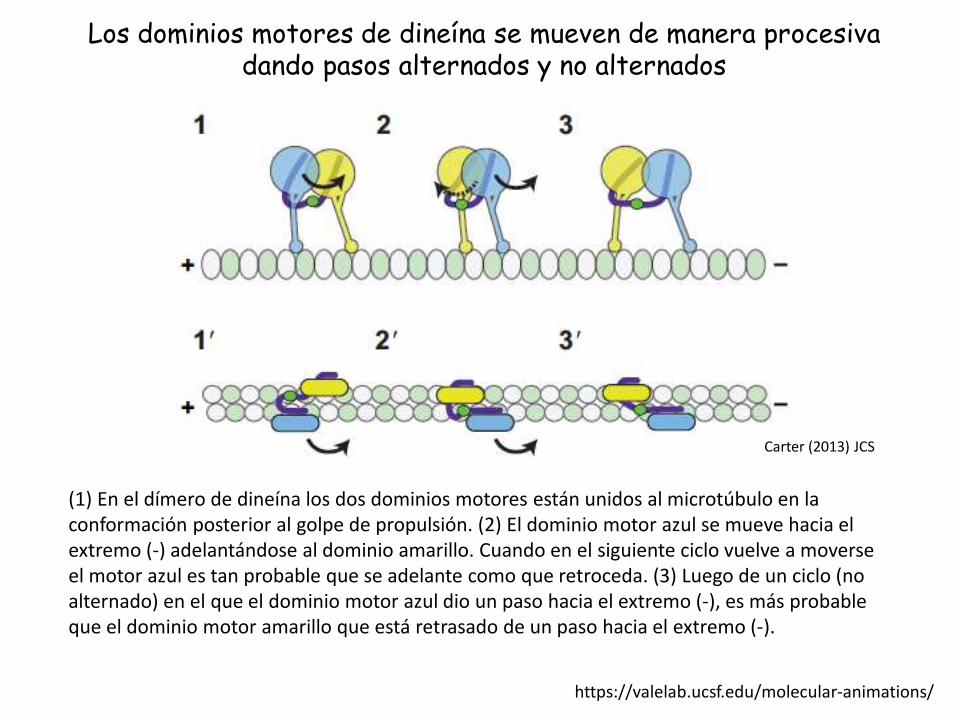
Los dominios motores de dineína se mueven de manera procesivadando pasos alternados y no alternados
(1) En el dímero de dineína los dos dominios motores están unidos al microtúbulo en la conformación posterior al golpe de propulsión. (2) El dominio motor azul se mueve hacia el extremo (-) adelantándose al dominio amarillo. Cuando en el siguiente ciclo vuelve a moverseel motor azul es tan probable que se adelante como que retroceda. (3) Luego de un ciclo (no alternado) en el que el dominio motor azul dio un paso hacia el extremo (-), es más probable que el dominio motor amarillo que está retrasado de un paso hacia el extremo (-).
Carter (2013) JCS
https://valelab.ucsf.edu/molecular-animations/
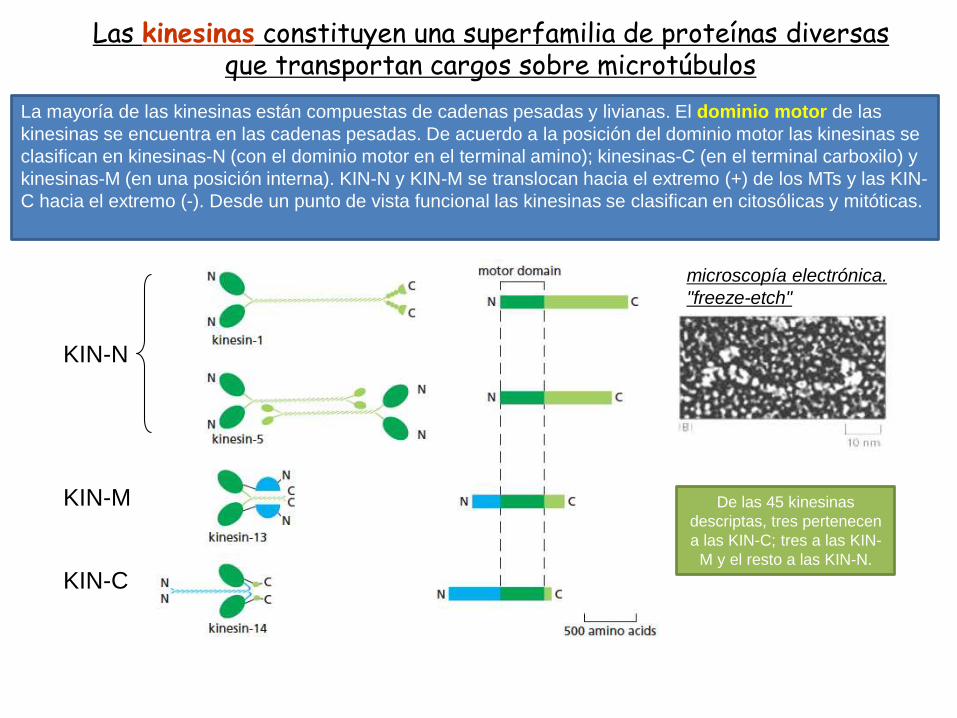
Las kinesinas constituyen una superfamilia de proteínas diversasque transportan cargos sobre microtúbulos
La mayoría de las kinesinas están compuestas de cadenas pesadas y livianas. El dominio motor de las
kinesinas se encuentra en las cadenas pesadas. De acuerdo a la posición del dominio motor las kinesinas se
clasifican en kinesinas-N (con el dominio motor en el terminal amino); kinesinas-C (en el terminal carboxilo) y
kinesinas-M (en una posición interna). KIN-N y KIN-M se translocan hacia el extremo (+) de los MTs y las KIN-
C hacia el extremo (-). Desde un punto de vista funcional las kinesinas se clasifican en citosólicas y mitóticas.
microscopía electrónica.
"freeze-etch"
KIN-N
KIN-M
KIN-C
De las 45 kinesinas
descriptas, tres pertenecen
a las KIN-C; tres a las KIN-
M y el resto a las KIN-N.

Estructura de las kinesinas convencionales
Lodish et al MCB2004
La kinesina I o convencional consiste en dos cadenas pesadas, cada una de las cuales se asocia a una
cadena liviana. El dominio globular (cabeza) de las cadenas pesadas une ATP e interaccionan con la tubulina
en el microtúbulo. Mediante una region flexible (cuello) las cabezas se conectan a un dominio ¨coiled coil¨ o
tallo. El cuello flexible acopla los ciclos de hidrólisis de ATP con cambios conformacionales que se trasladan al
tallo. Las cadenas livianas se asocian al extremo del tallo y contribuyen a las interacciones con el cargo.
Las kinesinas I son responsables del transporte axonal anterógrado, el transporte de vesículas secretoras hacia la membrana
plasmática y el movimiento radial centrífugo de gránulos con pigmentos en melanóforos.

El transporte mediado por kinesinas puede ser visualizado en ensayos in vitro
MT pegado al cubreobjeto
las vesículas siempre
se mueven hacia
el extremo (+)
kinesinas pegadas
al cubreobjeto
El movimiento direccional mediado por kinesinas ha sido estudiado fijando microtúbulos (a)
o kinesinas (b) a cubreobjetos. El movimiento de esferas cubiertas con kinesinas (a) o
microtúbulos (b) requiere de la presencia de ATP en el buffer de incubación.
https://valelab.ucsf.edu/motility/

Las vesículas pueden translocarse en direcciones opuestas sobre el mismo microtúbulo
Hancock 2014 Nat Rev Mol Cell Biol

Paradoja de la co-dependencia entre motores antagonistas
Experimentos de pérdida de función revelan la co-dependencia entre motoresantagonistas. El transporte bidireccional de vesículas (PrP, lipid droplets), mitocondrias y gránulos requiere de la actividad de kinesinas y dineínas. La inhibición de un motor también inhibe la actividad del motor que se mueve en dirección opuesta.
Quimografos del transporte anterógrado y retrógrado de vesículas de PrP celular fusionada a YFP en neuronas: (a) knock out para la cadena liviana de kinesina 1 (KLC1) y knock down para KLC2, o (b) knock down para la subunidad p150 de la proteína adaptadora de dineína dinactina

La dispersión de melanóforos correlaciona con altos niveles de AMPc, activación de kinesina y miosina V paraeventos de transporte coordinado que resulta en la distribución uniforme de gránulos de pigmento en el citoplasma. La agregación de melanóforos correlacionacon disminución en los niveles de AMPc y activación de dineínas que resulta en la agregación en el centro de la célula.
El AMPc regula diferencialmente la actividad de motoresasociados a la superficie de gránulos de pigmento
Modelo para la activación del transporte dirigido a los extremo (-) de microtúbulos
Kashina & Rodinov (2005) TiCB
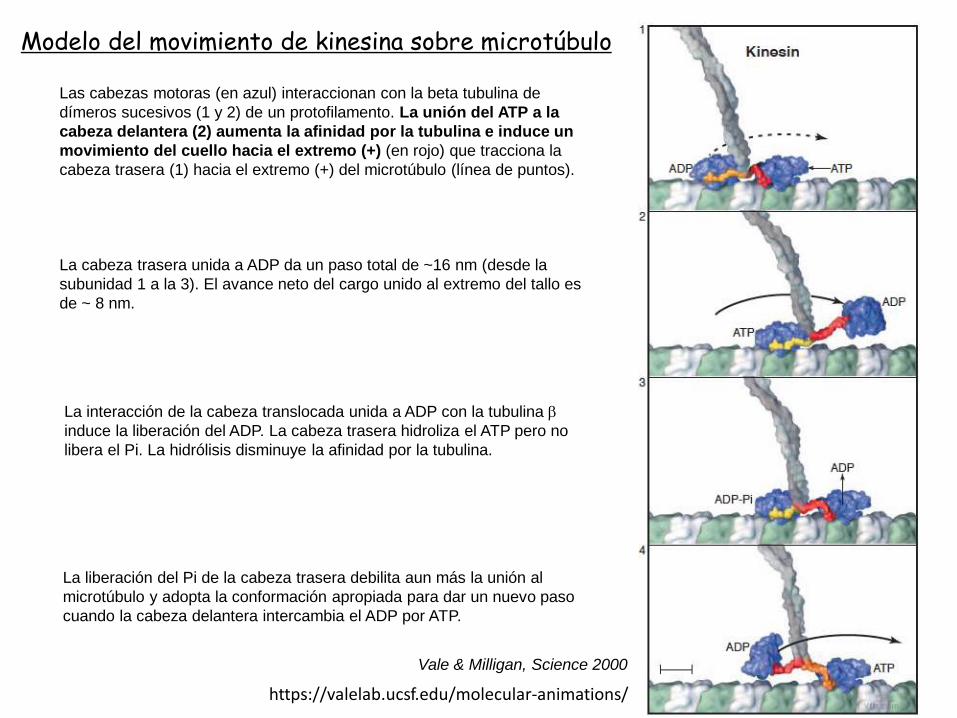
Las cabezas motoras (en azul) interaccionan con la beta tubulina de
dímeros sucesivos (1 y 2) de un protofilamento. La unión del ATP a la
cabeza delantera (2) aumenta la afinidad por la tubulina e induce un
movimiento del cuello hacia el extremo (+) (en rojo) que tracciona la
cabeza trasera (1) hacia el extremo (+) del microtúbulo (línea de puntos).
La interacción de la cabeza translocada unida a ADP con la tubulina b
induce la liberación del ADP. La cabeza trasera hidroliza el ATP pero no
libera el Pi. La hidrólisis disminuye la afinidad por la tubulina.
La liberación del Pi de la cabeza trasera debilita aun más la unión al
microtúbulo y adopta la conformación apropiada para dar un nuevo paso
cuando la cabeza delantera intercambia el ADP por ATP.
La cabeza trasera unida a ADP da un paso total de ~16 nm (desde la
subunidad 1 a la 3). El avance neto del cargo unido al extremo del tallo es
de ~ 8 nm.
Modelo del movimiento de kinesina sobre microtúbulo
Vale & Milligan, Science 2000
https://valelab.ucsf.edu/molecular-animations/

Comparación de los ciclos mecano-químicos de kinesina y miosina
Vale & Milligan, Science 2000
Miosina ejecuta un paso y se disocia del filamento. En contraste, kinesina es un motor
altamente procesivo y ejecuta varios cientos de pasos antes de disociarse del microtúbulo.

kinesin 1kinesin 14 (Ncd)
Kinesina 1 se mueve hacia el extemo (+) de microtúbulosporque el linker en el cuello apunta hacia adelante en el motor más retrasado (naranja) y hacia atrás en el que vaadelante (rojo). Cuando el linker del dominio motor másadelantado encastra (amarillo), empuja al otro dominiomotor hacia adelante. En cambio, la unión de ATP haceque el elemento mecánico de Ncd rote hacia el extremo(-) (amarillo). Luego el motor se libera y vuelve a la posición previa al golpe de propulsión y el ciclocomienza de nuevo.
El elemento mecánico de kinesina 14 sufre un golpe de propulsiónhacia el extremo (-)

El huso mitótico separa las cromátids durante la anafase
El huso mitótico es un arreglo bipolar de microtúbulos. Los extremos (+) de losmicrotúbulos se proyectan hacia afuera de los polos del huso, mientras que losextremos (-) quedan anclados en los polos. En el ecuador del huso mitótico sesuperponen los extremos (+) de los microtúbulos interpolares. Los microtúbulos delcinetocoro conectan los polos del huso con los cinetocoros de las cromátideshermanas. Los microtúbulos astrales conectan los polos con la actina cortical.

La función del huso mitótico depende de proteínas motorasasociadas a microtúbulos
Kinesina 5 interacciona con dos filamentos antiparalelos y su movimiento hacia los extremos (+)resulta en la separación de los polos.La kinesina 14 tiene un solo dominio motor que se mueve hacia el extremo (-). Al interaccionar conmicrotúbulos interpolares antiparalelos tiende a acercar los polos.Las kinesisnas 4 y 10 se asocian a los brazos de los cromosomas alejándolos de los polos.Las dineínas conectan el extremo (+) de microtúbulos astrales con la actina cortical. El movimientode dineína hacia el extremo (-) tira de los polos hacia la actina cortical.

- Son polímeros no polares extremadamente estables
- Los monómeros son proteínas fibrosas ricas en alfa hélice
- Los monómeros no unen nucleótidos
- El ensamblaje no requiere de hidrólisis de nucleótidos
- No participan en motilidad celular ni se conocen motores asociados a IFs
- Proveen resistencia al estrés mecánico

Los filamentos intermedios se forman a partir de una familia de subunidades
Lodish et al MCB 2016

Todas las subunidades de los filamentos intermedios comparten un dominiorígido rico en alfa hélice, y difieren en las regiones amino y carboxilo
El dominio central conservado posee repeticiones de 7 aminoácidos hidrofóbicos (héptadas)
involucradas en la formación de estructuras coiled-coil (super hélices) entre monómeros.

Los filamentos intermedios son polímeros no polares que se ensamblan por interacciones laterales de dímeros y tetrámeros
monómero
2 monómeros se asocian
por sus a-hélice y forman
un dímero coiled coil
2 dímeros se asocian
lateralmente en orientación
antiparalela. Tetrámero
2 tetrámeros se asocian
lateralmente y forman
octámeros. 4 octámeros
se asocian lateralmente
y forman el filamento.

Las células epiteliales poseen filamentos de la proteína queratina

Los filamentos de queratinas se anclan en desmosomas y hemidesmosomasestabilizando la estructura del epitelio

Los desmosomas son áreas especializadas de contacto célula-célula
Los desmosomas se asocian en su lado citosólico a proteínas adaptadoras de la familia de las
plaquinas que anclan los filamentos intermedios de queratina. Proteínas transmembrana
relacionadas con las cadherinas median el contacto entre células adyacentes

Los hemidesmosomas son sitios de anclaje de la célula a la matriz
Un tipo particular de integrina media el anclaje de la célula a laminina en la lámina basal y a
filamentos intermedios de queratina a tavés de proteínas adaptadoras

Sección normal de pielSección de piel de ratón mutante
en un gen de queratina
Mutaciones en la queratina desestabilizan la estructura de la epidermis
epidermis
Esquema de epitelio
mutante. La flecha indica
la ruptura de las células
debido a la fragilidad de
los filamentos de
queratina
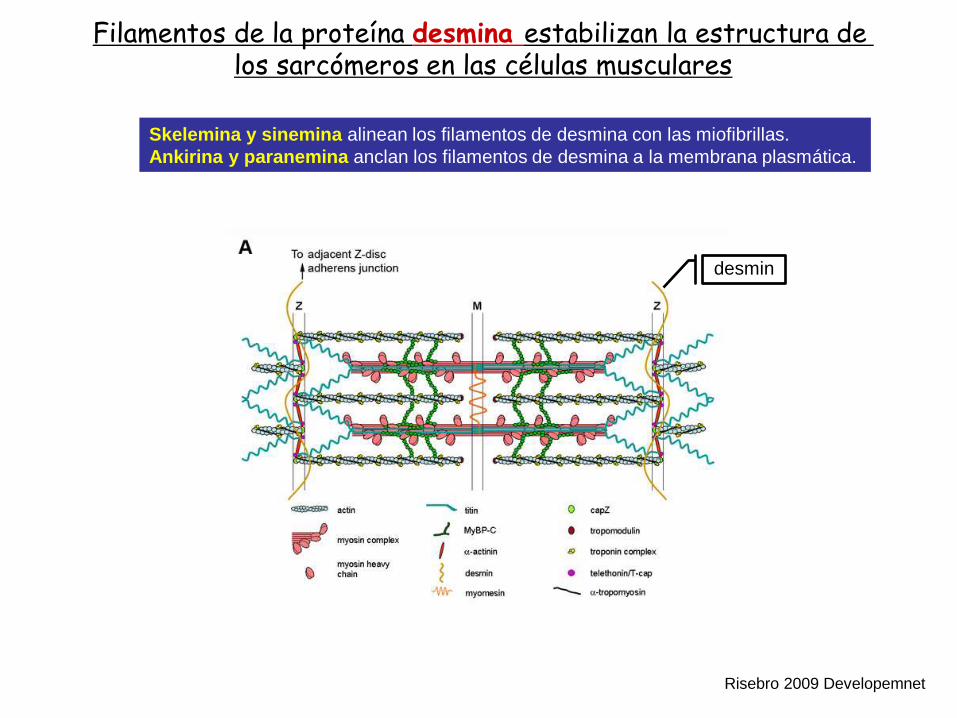
Filamentos de la proteína desmina estabilizan la estructura de los sarcómeros en las células musculares
Skelemina y sinemina alinean los filamentos de desmina con las miofibrillas.
Ankirina y paranemina anclan los filamentos de desmina a la membrana plasmática.
Risebro 2009 Developemnet
desmin

Los neurofilamentos (NFs) son esenciales para la estructura del axon
Tres tipos de proteínas de neurofilamentos (NF-L, NF-M y NF-H) se ensamblan para formar
heteropolímeros. Los NFs axonales estan áltamente interconectados por puentes de NF-H
Alteraciones en proteínas de neurofilamentos en neuronas motoras estan asociadas
a la enfermedad neurodegenerativa amyotrophic lateral sclerosis (ALS).
N-t C-tα-helical rod domain

Los filamentos de laminas estabilizan la envoltura nuclear
Hay tres tipos de laminas: A, B y C. Las laminas son fosforiladas por Cdks-M al comienzo
de la mitosis provocando el desensamble de los filamentos y el desarmado de la red.

Los filamentos intermedios se asocian con microtúbulos mediante proteínas puente: plectina
microscopía electrónica de barrido

Materila de Estudio
Libros
Alberts, Molecular Biology of the Cell
Lodish, Molecular Cell Biology
Artículos
Thomas D. Pollard, et al. Actin, a Central Player in Cell Shape and Movement. Science 326, 1208
(2009)
Thomas D. Pollard, et al. Cellular Motility Driven by Assembly and Disassembly of Actin Filaments.
Cell 112, 453–465 (2003)
Ronald D. Vale, et al. The Way Things Move: Looking Under the Hood of Molecular Motor Proteins.
Science 288, 88 (2000)
Andre P. Carter. Crystal clear insights into how the dynein motor moves. Journal of Cell Science 126,
705–713 (2013)
Hancock. Bidirectional cargo transport: moving beyond tug of war Bidirectional cargo transport: moving
beyond tug of war. Nature Reviews Molecular Cell Biology (2014)
iBioseminars
JULIE THERIOT: CELL MOTILITY AND THE CYTOSKELETON
Protein Polymers, Crawling Cells and Comet Tails: http://www.ibiology.org/ibioseminars/cell-biology/julie-
theriot-part-1.html
THOMAS POLLARD: CELL MOTILITY AND CYTOKINESIS
I. Mechanism of cell motility 1: http://www.ibiology.org/ibioseminars/thomas-pollard-part-1.html
II. Mechanism of cell motility 2: http://www.ibiology.org/ibioseminars/thomas-pollard-part-2.html
RON VALE: MOLECULAR MOTOR PROTEINS
Molecular Motor Proteins: http://www.ibiology.org/ibioseminars/cell-biology/ron-vale-part-1.html