modificaÇÃo fenotÍpica do melanoma murino …livros01.livrosgratis.com.br/cp049076.pdf · rica,...
TRANSCRIPT

ELIZABETH CRISTINA PÉREZ HURTADO
MODIFICAÇÃO FENOTÍPICA DO MELANOMA MURINO INDUZIDA PELA
INTERAÇÃO COM CÉLULAS B-1 – MECANISMOS DE TRANSDUÇÃO DE
SINAL
São Paulo 2008
Tese apresentada à Universidade Federal de São Paulo – Escola Paulista de Medicina para obtenção do Título de Doutor em Ciências da Saúde. Orientador: Prof. Dr. José Daniel Lopes Co-orientador: Joel Jr. Machado

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
Pérez, Elizabeth Cristina Modificação fenotípica do melanoma murino induzida pela interação com células B-1 – Mecanismos de Transdução de Sinal. / Elizabeth Cristina Pérez. – São Paulo, 2008. xxi, 109f. Tese (Doutorado) – Universidade Federal de São Paulo. Escola Paulista de Medicina. Programa de Pós-Graduação em Microbiologia e Imunologia Título em Inglês: B-1 lymphocytes increase metastatic behavior of melanoma cells through the extracellular signal-regulated kinase pathway. 1. Células B16. 2. Linfócitos B-1. 3. ERK. 4. Metástases.

iii
Universidade Federal de São Paulo
Escola Paulista de Medicina
Departamento de Microbiologia, Imunologia e Parasitologia
Chefe do Departamento: Prof. Dr. José Daniel Lopes
Coordenador do Curso de Pós-graduação: Prof. Dr. Renato Arruda Mortara

iv
ELIZABETH CRISTINA PÉREZ HURTADO
MODIFICAÇÃO FENOTÍPICA DO MELANOMA MURINO INDUZIDA PELA
INTERAÇÃO COM CÉLULAS B-1 – MECANISMOS DE TRANSDUÇÃO DE
SINAL
Presidente da banca: Prof. Dr. José Daniel Lopes
BANCA EXAMINADORA Profa. Dra. Maria Mitzi Brentani
Dra. Nathalie Cella
Profa. Dra. Célia Regina Whitaker Carneiro
Prof. Dr. Hugo Pequeno Monteiro
Aprovada em: 23/01/2008

v
Este trabalho foi desenvolvido na disciplina de
Imunologia do Departamento de Microbiologia,
Imunologia e Parasitologia da Universidade
Federal de São Paulo, com auxílios financeiros
concedidos pela Fundação de Amparo a
Pesquisa do Estado de São Paulo (FAPESP
processo 03/05176-8)

vi
Dedicatória
Este trabalho é dedicado :
As memórias da minha mãe Carmen e da minha avó Rackel, Aos meus adorados homens: meu esposo Harold e meus filhos Juan David
e Luis Felipe
A meu pai Guillermo e meus irmãos Carolina e Guillermo

vii
Agradecimentos
Ao Prof. Dr. José Daniel Lopes, pelo seu carinhoso acolhimento, dedicação,
amizade, apoio, ensinamentos e auxílio na redação desta tese.
Ao Prof. Dr. Joel Machado pelo espírito crítico no desenvolvimento deste
trabalho, auxílio na redação desta tese e pela valiosa contribuição na realização
do artigo científico.
Ao Prof. Dr. Mario Mariano pela amizade e valiosas contribuições.
Às Profs. Dras. Célia Regina Whitaker Carneiro, Ieda Maria Longo Maugéri,
Miriam Galvonas Jasiulionis e Zulma Fernandes Peixinho, pela amizade, carinho e
valiosas contribuições.
À Prof. Dra. Marimélia Porcionatto (INFAR) pelo carinho, amizade e
grandes ensinamentos.
À Dra. Valderes Bastos Valero e sua equipe: Amarildo, Luis, Renato do
Laboratório de Experimentação animal do INFAR por seu grande carinho, amizade
e disposição no fornecimento de animais.
À Prof. Dra. Elaine Guadalupe Rodrigues, pela amizade e colaboração no
fornecimento de camundongos,
À Prof. Dra. Edna Freymüller (CEME) e sua equipe: André, Isabel, Márcia e
Patrícia, pelo carinho, amizade, disposição e colaboração nas análises por
microscopia eletrônica.

viii
Ao Prof. Dr. João Bosco Pesqueiro e sua equipe: Felipe, Raphael, Giuliana,
Jõao e demais estudantes, pela amizade, disposição e valiosa colaboração na
realização dos ensaios e análises por RT-PCR em tempo real,
Ao Prof. Dr. Sergio Schenkman e sua equipe pela amizade e colaboração
no empréstimo do sistema de foto-documentação utilizado nas análises por RT-
PCR realizadas durante este trabalho.
Aos funcionários da Disciplina de Imunologia: Aparecido Mendes, Creusa
Marina, Creusa Rosa, Eraldina do Nascimento, Geová dos Santos, Gisélia Lopes,
Ivone Mozat, Naide, Neta e Zélia Pereira, pela amizade, colaboração e convívio
alegre durante estes anos.
À Mércia, secretária da pós-graduação do departamento, pela amizade e
auxílio constante.
Aos professores, funcionários e colegas da disciplina de Biologia Celular
(oitavo andar) pela colaboração e boa disposição.
As amigas do “Lab 2”, Érika Kioshima (Londri), Fabiana Aliperti (Fabico),
Juliana Maricato (Juju), Lívia Pugliese e Patrícia Xander (Patlicia), pela
cumplicidade, carinho, apoio, clima alegre e grande ajuda na correção desta tese.
A todos os amigos do “Lab1 e Lab2”, Adriana, Aline (pulga), Bruno, Camila,
Creusa, Nadia, Marcela, Márcia, Patrícia, Renata, Toshie pela amizade, carinho e
convívio alegre e estimulante durante a realização deste trabalho.
À Fernanda Staquicini, colega que defendeu doutorado 3 anos atrás,
obrigada pela amizade e acolhida carinhosa ao chegar na Imuno assim como por
me ensinar o mundo das B-1.

ix
Aos amigos da Imuno, Inferno (Ronni Brito), Gugu (André), Carla Squaiella,
Juliana Mussalem, Tatiana, Adriana, Alice, Fernanda Molognoni, Karina, Tatiana
Rica, Ana Flavia Popi, Felipe, Fernanda, Helena, Maiko, Mauro, Ricardo, Carol,
Janaina, pelo carinho, amizade e simpatia.
Às amigas e amigos com quem tive oportunidade de trabalhar e que há
algum tempo já trilham seu próprio caminho: Ana Cristina Campos, Ana Flavia
Vigna (Oxigenada), Beatriz Helena Pizarro, Daniela Grosso, Fernanda Staquicini,
Hellen Fuzii, Ludmila Chinen, Luciana, Luciano Feitosa, Luiz Cláudio Godoy,
Priscila Penteado, Renata Ananias, Thalita Abrão; obrigada pelo carinho, amizade
e feliz convivência.
À FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo) por
financiar este trabalho.
À minha “Virgencita” e aos meus Anjos protetores pela companhia,
iluminação e proteção de sempre.
E por suposto a minha família, meu esposo Harold e meus dos filhotes Juan
David e Luis Felipe por seu grande apoio, alegria, amor e carinho durantes estes
anos de correria, muitíssimo obrigada.

x
Sumário
Dedicatória..............................................................................................................VI
Agradecimentos......................................................................................................VII
Lista de Gráficos....................................................................................................XIII
Lista de Tabelas....................................................................................................XIV
Lista de Figuras......................................................................................................XV
Lista de Abreviaturas e Símbolos........................................................................XVII
RESUMO...............................................................................................................XX
1. INTRODUÇÃO......................................................................................................1
2. OBJETIVOS........................................................................................................16
2. MATERIAIS E MÉTODOS..................................................................................18
2.1 MATERIAIS.......................................................................................................19
2.1.1 Soluções Rotineiras.......................................................................................19
2.1.1.1 Soluções para SDS-PAGE.........................................................................19
2.1.1.2 Soluções para Coloração por Comassie Blue............................................20
2.1.1.3 Soluções para Immunoblotting...................................................................20
2.1.1.4 Soluções para ELISA.................................................................................20
2.1.1.5 Soluções para cultura de células................................................................21
2.1.2 Animais..........................................................................................................21
2.2 MÉTODOS........................................................................................................22
2.2.1 Cultura de células e tratamentos...................................................................22
2.2.2 Análises de moléculas de superfície por citometria de fluxo.........................24

xi
2.2.3 Ensaio de viabilidade celular por Trypan blue...............................................24
2.2.4 Ensaio de inibição da adesão........................................................................25
2.2.5 Ensaio de metástases experimental..............................................................25
2.2.6 Preparação de extratos celulares..................................................................26
2.2.7 Dosagem protéica pelo micrométodo de Bradford........................................26
2.2.8 Eletroforese em gel de poliacrilamida com dodecilsulfato de sódio (SDS-
PAGE).....................................................................................................................26
2.2.9 Transferência de proteínas de nitrocelulose (Immunoblot ou Western
blot).........................................................................................................................28
2.2.10 Microscopia eletrônica de transmissão........................................................29
2.2.11 Ensaios de proliferação celular....................................................................30
2.2.11.1 Incorporação de timidina tritiada...............................................................30
2.2.11.2 MTT (tetrazolium salt3-(-4,4-dimethylthiazol-2-yl_-2,5-diphenyltetrazolium
bromide...................................................................................................................30
2.2.12 Dosagem de citocinas no sobrenadante de cultura…………………............31
2.2.13 Obtenção de RNA total................................................................................32
2.2.14 RT-PCR (Reverse Transcriptase – Polymerase Chain Reaction)...............33
2.2.14.1 Análise qualitativa por RT-PCR................................................................34
2.2.14.2 Análise quantitativa por RT-PCR em tempo real......................................36
2.2.15 Análise Estatística.......................................................................................37
3 RESULTADOS....................................................................................................39
3.1 Caracterização do sistema de co-cultivo..........................................................40
3.2 Células B-1 e B16 fisicamente interagem durante o co-cultivo........................41

xii
3.3 Fosforilação de ERK está aumentada nas células B16 após interação com
células B-1..............................................................................................................43
3.4 Aumento no potencial metastático de células B16 é dependente dos níveis de
ERK fosforilado.......................................................................................................44
3.5 Células B-1 não induzem aumento na proliferação de células B16.................46
3.6 Células B-1 induzem a expressão de genes relacionados com metástases nas
células B16.............................................................................................................46
3.7 Níveis basais de fosforilação de ERK nas células B-1 são necessários para
induzir aumento no potencial metastático das células B16....................................47
3.8 Níveis basais de fosforilação de ERK nas células B16 são inalterados pelo
estímulo com IL-10.................................................................................................50
4. DISCUSSÃO....................................................................................................,,.67
5. CONCLUSÕES...................................................................................................75
6. REFERÊNCIAS BIBLIOGRÁFICAS...................................................................92
ABSTRACT
ANEXOS

xiii
Lista de Gráficos
Gráfico 1. Via canônica de Ras-Raf-MEK-ERK.....................................................15

xiv
Lista de Tabelas
Tabela 1. Classificação clínica de melanoma........................................................11
Tabela 2. Composição de gel para SDS-PAGE.....................................................27
Tabela 3. Seqüências oligo-nucleotídicas dos primers sense (S) e anti-sense (AS),
temperatura de anelamento (°C) e o tamanho do produto do PCR em pares de
bases (BP), dos transcritos estudados por RT-PCR qualitativo.............................35
Tabela 4. Seqüências oligo-nucleotídicas dos primers sense (S) e anti-sense (AS)
e tamanho do produto do PCR em pares de bases (bp), dos transcritos estudados
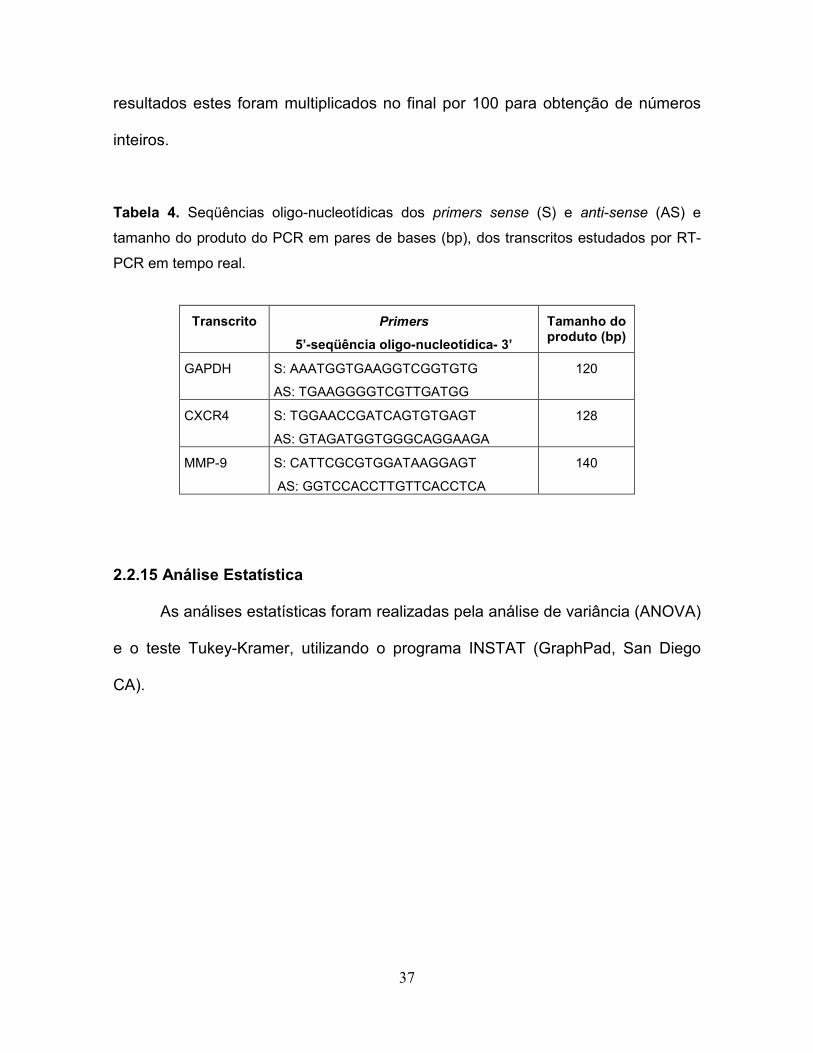
por RT-PCR em tempo real....................................................................................37

xv
Lista de Figuras
Figura 1. Células B-1 são eliminadas eficientemente das co-culturas após
aspiração do sobrenadante....................................................................................51
Figura 2. Células B-1 aumentam o potencial metastático das células B16...........52
Figura 3. Células B-1 induzem aumento no potencial metastático de células B16
independentemente da adesão celular...................................................................53
Figura 4. Fatores solúveis das células B-1 não contribuem para o aumento da
capacidade metastática das células B16................................................................54
Figura 5. Células B16 e B-1 interagem fisicamente durante o co-cultivo..............55
Figura 6. Células B-1 induzem aumento nos níveis de fosforilação de ERK nas
células B16.............................................................................................................56
Figura 7. Níveis basais de fosforilação de ERK em células B16 e B-1 são inibidos
pelo tratamento com PD98059...............................................................................57
Figura 8. O aumento no potencial metastático das células B16 induzido pelo
contato com células B-1 é dependente dos níveis de ERK fosforilado..................58
Figura 9. Células B-1 não induzem aumento na proliferação das células B16......59
Figura 10. Qualidade do RNA total após extração.................................................60
Figura 11. Células B-1 induzem a expressão de CXCR4 e MMP-9 nas células
B16..........................................................................................................................61
Figura 12. Produção de IL-10 por células B-1.......................................................62
Figura 13. Níveis basais de fosforilação de ERK nas células B-1 são necessários
para induzir aumento no potencial metastático das células B16............................63

xvi
Figura 14. Níveis de fosforilação de Stat3 nas células B16 antes e após co-cultivo
com células B-1......................................................................................................64
Figura 15. Níveis de ERK fosforilado e comportamento metastático das células
B16 não são aumentados por IL-10 recombinante.................................................65
Figura 16. Células B-1 de animais knock-out para IL-10 na são capazes de induzir
aumento no potencial metastático das células B16, mesmo na presença de IL-10
recombinante..........................................................................................................66

xvii
Lista de Abreviaturas e Símbolos
% Porcentagem
µg Micrograma
µL Microlitro
µM Micromolar
BCR B cell receptor
bp Pares de bases
BSA Soro albumina bovina
cDNA DNA complementar
CXCR4 Chemokine (C-X-C motif) receptor
DEPC Dietil pirocarbonato
DMSO Dimetilsulfóxido
DO Densidade óptica
EDTA Ácido etilenodiamino tetracético
ELISA Enzyme linked Immunosorbent Assay
ERK Extracellular signal-regulated kinase
FACS Fluorescence Activated Cell Sorting
FITC Isotiocianato de fluoresceína
g Gramas
GM-CSF Granulocyte-macrophage colony-stimulating factor
GPDH Gliceraldeído-3-fosfato desidrogenase
H Hora

xviii
IgD Imunoglobulina D
IgD Imunoglobulina D
IgM Imunoglobulina M
IL Interleucina
JAKs Janus-activated kinase
JNK June N-terminal kinase
kDA Kilo Dalton
LPS Lipopolissacarídeo
MAPK Mitogen activated protein kinase
MCP-1/CCL2 Monocyte chemotactic protein -1/ chemokine (C-C motif)
M-CSF Monocyte-colony-stimulating factor
MDSC Myeloid-derived suppressor cells
mL Mililitro
mM Milimolar
MMPs Matrix Metalloproteinases
MMP-9 Matrix Metalloproteinase-9
NK Natural killer
NO Óxido nítrico
PBS Tampão salino fosfato
PKC Protein kinase C
PMSF Fluoreto de fenilmetanosulfonato
q.s.p quantidade suficiente para
RGP Radial growth phase
ROI Reativos intermediários de oxigênio

xix
rpm Rotação por minuto
RTKs Receptores tirosina kinase
RTKs Receptores tirosina kinase
RT-PCR Reverse transcriptase polymerase chain reaction
SDF-1/CXCL12 Stromal cell-derived factor 1
SDS-PAGE Sodium dodecyl sulphte polyacrylamide gel
STAT Signal Transducers and Activators of Transcription
Stat3 Signal Transducer and Activator of Transcription 3
TAMs Tumour-associated macrophages
TNF Tumour necrosis factor
V Volts
v/v Volume a volume
VEGF-A Vascular endothelial growth factor - A
VGP Vertical growth phase
W Wats
W Wats
X Vezes

xx
Resumo
Evidências indicam que tumores necessitam constante influxo de células mielo-
monocíticas para sustentar seu comportamento maligno. Esse fato é devido a
fatores derivados do tumor, os quais recrutam e induzem diferenciação funcional
de células mielo-monocíticas, das quais a maioria são macrófagos. Embora os
precursores clássicos de macrófagos sejam linhagens mielóides, linhagens
linfóides como células B-1, subtipo de linfócitos B encontrados
predominantemente nas cavidades pleural e peritoneal são também capazes de
migrar para focos inflamatórios e se diferenciar em fagócitos mononucleares
apresentando fenótipo semelhante ao dos macrófagos. No presente trabalho foi
avaliada a interação entre células B16 de melanoma murino e células B-1 durante
o co-cultivo e se esta interação influencia ativação de vias de transdução de sinal
envolvidas com progressão e metástases. Utilizando sistema de co-cultura in vitro,
foi mostrado que células B16 e células B-1 interagem fisicamente após 48 horas
de co-cultura. Além disso, esta interação resulta em aumento da expressão de
genes associados com metástases como MMP-9 e CXCR4 nas células B16 que
favorecem o aumento na capacidade metastática destas células, como revelado
por ensaios experimentais de metástases in vivo. Este trabalho também revela
evidências de que células B16 apresentam marcado aumento na fosforilação da
quinase regulada por sinais extracelulares (ERK) após contato com células B-1. A
inibição da fosforilação de ERK com inibidor farmacológico da quinase ascendente
de ERK, MEK1/2, suprime fortemente a expressão de MMP-9 e CXCR4 e inibe o
aumento da capacidade metastática das células B16 induzido por contato com

xxi
células B-1. Adicionalmente, níveis constitutivos de ERK fosforilado nas células B-
1 são necessários para que estas células sejam capazes de induzir aumento no
potencial metastático das células B16. Nossos resultados em conjunto mostram
que células B-1 podem contribuir na aquisição de um fenótipo mais agressivo das
células tumorais.


1. INTRODUÇÃO

1. INTRODUÇÃO
Uma das principais características no desenvolvimento do câncer é a
formação de metástases na qual células tumorais provenientes do tumor primário
invadem e colonizam tecidos ou órgãos distantes do sítio de origem (Hanahan &
Weinberg, 2000; Balkwill & Mantovani, 2001; Langley & Fidler, 2007; Aguirre-
Ghiso, 2007).
Embora grandes avanços tenham sido realizados na prevenção,
diagnóstico, técnicas cirúrgicas e terapias adjuvantes, a maior parte das mortes
em câncer são resultado da formação de metástases resistentes a terapias
convencionais. Por esta razão, o entendimento dos mecanismos celulares e
moleculares envolvidos na formação de metástases é importante para o
desenvolvimento de novos alvos terapêuticos
A formação de metástases é um processo complexo e seletivo que consiste
em vários passos seqüenciais e relacionados, nos quais células tumorais precisam
adquirir a capacidade de invadir tecidos adjacentes para formar tumores
secundários ou metástases (Poste & Fidler, 1980; Fidler, 2002a). Neste processo,
células com capacidade de metastatizar separam-se do tumor primário, entram na
circulação ao invadirem a parede dos vasos sanguíneos ou linfáticos, disseminam-
se e, posteriormente, aderem a órgãos determinados. Para que todos estes
eventos ocorram, o aumento na expressão de moléculas de adesão celular como
integrinas, proteases como metaloproteinases (MMPs) e moléculas quimio-

2
atraentes como citocinas e quimiocinas são essenciais para invasão e
metástatização das células tumorais (Sierra, 2005; Deryugina & Quigley, 2006).
Observações clínicas em pacientes com câncer e estudos experimentais
em camundongos têm demonstrado que certos tipos de tumores metastatizam
para órgãos específicos independentemente da anatomia vascular, do tamanho do
tumor ou do número de células tumorais recebidas em cada órgão (Sierra, 2005;
Fokas et al., 2007). Para explicar essa especificidade, a teoria soil and seed,
proposta por Paget em 1889, sustenta que órgãos diferentes (soil) provêm
condições de crescimento ótimas para cânceres específicos (seed; revisado por
Fidler, 2002b).
Assim como sugerido por Paget, outros autores (Balkwill & Mantovani,
2001; Coussens & Werb, 2002; Croci et al., 2007) têm demonstrado que durante a
progressão tumoral e metástase ocorre comunicação ativa (cross-talk) entre
células tumorais e seu estroma. Esta comunicação pode ser mediada tanto pelo
contato direto entre as células, como pela produção de citocinas, quimiocinas,
fatores de crescimento e metaloproteinases, que favorecem o avanço das células
tumorais para sítios distantes do tumor primário (Lehmann et al., 1987; Christofori
& Semb, 1999; McGary et al., 2002; Li et al., 2002). A interação entre células
estromais e tumorais sugere que o tumor não é apenas uma massa composta de
células malignas, e sim, um tecido com ampla diversidade celular que incluem
células residentes como adipócitos, fibroblastos, populações de células
hematopoiéticas migratórias como monócitos, macrófagos, neutrófilos, mastócitos,
eosinófilos, linfócitos T e B (Coussens et al., 1999; Coussens et al., 2000; Kuper et
al., 2000; Balkwill & Mantovani, 2001; Di Carlo et al., 2001; Coussens & Werb,

3
2002; Joyce, 2005; Inoue et al., 2006). Essa diversidade celular produz assim um
único micro-ambiente o qual é capaz de modificar as propriedades neoplásicas
das células tumorais (Coussens & Werb, 2002; Croci et al., 2007).
Macrófagos associados ao tumor (TAMs, tumor-associated
macrophages)
Macrófagos associados ao tumor (TAMs) representam o maior componente
do infiltrado inflamatório da maioria dos tumores (Mantovani et al., 1992, Balkwill &
Mantovani, 2001). No entanto, a alta freqüência destas células no infiltrado de
muitos cânceres humanos está associada a mau prognóstico (Mantovani et al.,
2006). TAMs são derivados de monócitos circulantes os quais são recrutados ao
sítio do tumor por fatores quimiotáticos como a proteína quimiotática para
monócitos - 1 [MCP-1/CCL2; monocyte chemotactic protein-1/chemokine (C-C
motif) ligand 2], o fator estimulador de colônias de monócitos (M-CSF; monocyte-
colony-stimulating factor) e o fator de crescimento do endotélio vascular – A
(VEGF-A; vascular endothelial growth factor – A; Mantovani et al., 1992; Barleon
et al., 1996).
Macrófagos são células versáteis que representam a primeira linha de
defesa da resposta imune inata e não requerem adaptação ou imunização para
funcionar. Estas células têm papel importante no recrutamento e ativação de
linfócitos por meio da apresentação de antígenos (Sica et al., 2002). Entretanto,
devido a sua funcionalidade heterogênea, podem também realizar funções
supressoras (Rutherford et al., 1993). Em câncer, os macrófagos são importantes

4
células efetoras na imunidade contra tumores e, dependendo de seu fenótipo,
podem tanto prevenir como promover a progressão tumoral.
Fenotipicamente dois tipos de macrófagos têm sido descritos: macrófagos
ativados pela via clássica e macrófagos ativados pela via alternativa. Os
macrófagos ativados pela via clássica têm fenótipo IL-12high, IL-23high, IL-10low e
são ativados por IFN-γ e lipopolissacarídeo (LPS; Mantovani et al., 2004a). Em
contraste, os macrófagos ativados pela via alternativa têm fenótipo IL-12low, IL-
23low, IL-10high e são ativados por IL-4 e IL-13 (Gordon, 2003). Pelo fato de
possuírem perfil de citocinas IL-12 e IL-10 similares aos das células T CD4+ Th1 e
Th2, os macrófagos ativados classicamente e os ativados alternativamente têm
sido chamados de M1 e M2, respectivamente (Mills et al., 2000; Mantovani et al.,
2002; Mantovani et al., 2005).
Quanto à função destas células no câncer, macrófagos M1 são células
efetoras potentes que eliminam células tumorais e promovem a rejeição do tumor
(Mantovani et al., 1992, Guiducci et al., 2005). Entretanto, os macrófagos M2
favorecem crescimento e disseminação tumoral provavelmente por favorecer um
ambiente imunossupressor pela produção de IL-10 (Ibe et al., 2001), assim como
por promover proliferação celular, angiogênese, remodelação tissular e reparo
(Sica et al., 2002; Mantovani et al., 2002; Gordon, 2003; Mantovani et al., 2004b;
Condeelis & Pollard, 2006; Porta et al., 2007).

5
Células supressoras derivadas da linhagem mielóide (MDSC: myeloid-
derived suppressor cells)
Além dos TAMs, recentes estudos mostraram o acumulo de células
mielóides imaturas na medula óssea, linfonodos e baço tanto em pacientes com
câncer como em camundongos com tumores implantados (Young et al., 1987;
Garrity et al., 1997; Almand et al., 2001; Zea et al., 2005). Estas células,
denominadas atualmente como células supressoras derivadas da linhagem
mielóide (MDSC; myeloid-derived suppressor cells), expressam marcadores de
células mielóides Mac-1 (CD11b) e Gr-1+ (Ly6G) e são induzidas por fatores de
crescimento como o fator de crescimento do endotélio vascular (VEGF), o fator
estimulador de colônias granulocítica e monocítica (GM-CSF) e por citocinas pro -
inflamatórias como IL-1β (Gabrilovich et al., 1996; Young et al., 1997; Bronte et al.,
1999; Song et al., 2005; Bunt et al., 2006). MDSC interferem com a imunidade
tumoral e promovem crescimento tumoral por inibir a citotoxidade das células
tumorais mediada por células NK (Susuki et al., 2005), e por bloquear a ativação
de linfócitos T CD4+ e CD8+ reativos ao tumor (Kusmartsev et al., 2000; Bronte et
al., 2001; Melani et al., 2003; Sinhá et al., 2005). Além disso, recentes estudos
mostraram que MDSC também interferem na imunidade anti-tumoral tanto por
induzir o acúmulo de células T reguladoras imunossupressoras (Gabrilovich et al.,
2001; Huang et al., 2006) como por interagir com macrófagos, os quais aumentam
a produção de IL-10 e decrescem os níveis de IL-12, favorecendo assim um
ambiente imunossupressor propício para a progressão tumoral (Sinhá et al.,
2007).

6
Células B-1
Embora esteja bem estabelecido que linhagens mielóides são os
precursores clássicos de macrófagos, recentes evidências sugerem que a
linhagem linfóide pode também gerar células fagocíticas com características de
macrófagos (macrófago-like), como as células B-1 (Almeida et al., 2001).
As células B-1, subtipo de células B, foram inicialmente identificadas como
células B CD5+ que participam de processos autoimunes em camundongos
(Hayakawa et al., 1983), com características similares às das células responsáveis
pela Leucemia Linfocítica Crônica em humanos (Boumsell et al., 1978). Estas
células diferem das células B-2 convencionais por sua localização anatômica,
expressão de marcadores de superfície e funcionalidade (Hayakawa et al., 1985,
Herzenberg et al., 1986; Martin et al., 2001).
Estas células representam somente 1-5% das células B totais em
camundongos, mas são a principal população de células B nas cavidades
peritoneal e pleural, podendo ser também encontradas no baço, linfonodos e
intestinos (Kantor & Herzenberg, 1993). Células B-1 diferem das células B-2
convencionais por expressar altos níveis de IgM (IgMhi), baixos níveis de IgD
(IgDlo) e B220 (B220lo) e por expressar CD43 (sialoforina, marcador de leucócitos,
exceto nas células B). A maioria das células B-1 das cavidades peritoneal e
pleural expressam CD11b ou Mac-1, marcador de macrófagos/granulócitos que é
parte do receptor do complemento CR3. Entretanto, a maioria das células B-1 do
baço não expressam este marcador (Kantor & Herzenberg, 1993). Além disso, as
células B-1 presentes na cavidade peritoneal podem ser subdividas em 3

7
populações pela expressão de CD5 (marcador de linfócitos T) e CD11b. As células
B-1a expressam CD5 e CD11b; células B-1b não expressam CD5 mas expressam
CD11b; e, mais recentemente descritas, as células B-1c expressam CD5 mas não
expressam CD11b (Herzenberg et al., 1986; Kantor et al., 1992; Hastings et al.,
2006).
As diferenças funcionais entre as células B-1 e B-2 são numerosas e têm
sido revisadas extensamente (Kantor & Herzenberg, 1993; Martin et al., 2001;
Berland and Wortis, 2002; Hardy, 2006). O repertório de anticorpos produzidos
pelas células B-1 é menos diverso, comparado ao das células B-2 convencionais,
refletindo assim na capacidade de resposta ao antígeno destas duas populações
celulares (Hardy et al., 1989; Baumgarth et al., 2005). Células B-2 respondem
eficientemente a antígenos protéicos e participam da resposta imune adaptativa
por sofrer hipermutação somática dos genes das imunoglobulinas e maturação da
afinidade do receptor. Em contraste, células B-1 respondem rapidamente a
antígenos timo-independentes, razão pela qual têm sido reconhecidas como
veículos da imunidade inata (Hardy, 1992; Herzenberg, 2000; Baumgarth et al.,
2005). Além disso, recentes observações mostraram diferenças funcionais mesmo
entre os subtipos de células B-1 (Alugupalli et al., 2004; Haas et al., 2005;
Alugupalli & Gerstein, 2005; Hsu et al., 2006). Células B-1a secretam de forma
espontânea IgM, provendo a primeira barreira contra bactérias encapsuladas
como Streptococcus pneumoniae. Em contraste, a produção de anticorpos por
células B-1b é induzida e está envolvida na remoção final de patógenos e na
proteção a longo prazo (Haas et al., 2005).

8
Por outro lado, diferentemente das células B-2, que têm período de vida
curto e são continuamente geradas por progenitores na medula óssea, as células
B-1 são mantidas por proliferação homeostática ou auto-renovação, como
mostram os experimentos de transferência adotiva de células B-1 em animais
imunodeficientes (Hayakawa et al., 1985; Forster & Rajewisky, 1987). Além disso,
células B-1 independem da sinalização via receptor de células B (BCR, B cell
receptor) para proliferar (Berland & Wortis, 2002; Casola et al., 2004). Karras e
colaboradores (1997) mostraram que esta propriedade de crescimento das células
B-1 é devida à ativação constitutiva da proteína Stat3 (Signal transducer and
activator of transcription 3), que substitui o sinal de proliferação mediado pela
ligação BCR-antígeno. Adicionalmente, células B-1 também expressam níveis
constitutivos da quinase regulada por sinais extracelulares (ERK: extracellular
signal-regulated kinase 1/2). Entretanto, o papel desta quinase nestas células,
ainda é desconhecido, uma vez que o bloqueio da ativação de ERK não afeta a
capacidade de auto-renovação das células B-1 (Wong et al., 2002).
Células B-1 também produzem grandes quantidades de interleucina 10 (IL-
10; O’Garra et al., 1992; Gieni et al., 1997), uma citocina do tipo Th2,
antiinflamatória, que participa da supressão de respostas pro-inflamatórias (Moore
et al., 1993). As células B-1 utilizam esta citocina como fator autócrino de
crescimento (O’Garra et al., 1992) e, por esta razão, a eliminação de células B-1
induzida pelo tratamento com anticorpos anti-IL-10 é acompanhada por drástica
diminuição dos títulos de imunoglobulinas séricas (Ishida et al., 1992).
A participação das células B-1 tem sido observada em várias patologias,
tanto em humanos como em camundongos (Aramaki et al., 1998; Mohan et al.,

9
1998; Hayakawa et al., 2000; Antsiferova et al., 2005; Hardy, 2006; Duan & Morel,
2006). A expansão de células B-1 autorreativas foi associada ao desenvolvimento
de doenças auto-imunes (Murakami et al., 1992; Murakami et al., 1995; Mohan et
al., 1998) e leucemias de células B (Chevallier et al., 1998). Além disso, Minoprio e
colaboradores (1993) demonstraram que na infecção chagásica experimental
ocorre aumento significativo de células B-1 esplênicas na fase aguda da doença e
ainda, camundongos Xid, geneticamente modificados e desprovidos de células B-
1, resolvem com maior eficiência a infecção por esse protozoário. Resultados
semelhantes foram obtidos também em infecções causadas por Paracoccidioides
brasiliensis e por filarias linfáticas (Paciorkowoski et al., 2000; Marcelino França et
al., 2006). Por outro lado, estudos realizados em nosso laboratório demonstraram
que estas células são capazes de migrar para focos inflamatórios inespecíficos
induzidos por corpo estranho, diferenciando-se em fagócitos mononucleares com
características similares a macrófagos (Almeida et al., 2001; Bogsan et al., 2005).
Além disso, outros trabalhos do nosso grupo, alguns deles em andamento,
mostraram a participação das células B-1 na evolução da infecção causada por P.
brasiliensis (Godoy et al., 2003), no lupus eritematoso sistêmico murino (E Brito et
al., 2007), na cicatrização (Oliveira et al., in preparation), na formação de células
gigantes (Bogsan et al., 2005), e de granulomas (Vigna et al., 2006; Russo, in
preparation).
Além do papel imuno-modulador das células B-1 nas patologias descritas
acima, resultados ainda não publicados pelo nosso grupo (Staquicini, 2004)
mostraram que o contato prévio de células B-1 com células de melanoma murino
da linhagem B16 foi capaz de induzir aumento no potencial metastático dessas

10
células tumorais sugerindo assim, a participação das células B-1 também na
regulação de mecanismos que controlam metástases.
Melanoma
Melanoma é um tipo comum e letal de câncer de pele que acomete
principalmente populações relativamente jovens (Parker et al., 1997). O melanoma
tem sido catalogado como a terceira forma mais comum de câncer de pele; porém,
devido à sua alta taxa de recorrência e capacidade metastática, é a forma que
maior número de mortes produz. Além disso, o melanoma é altamente resistente
aos métodos convencionais de tratamento como quimioterapia e radioterapia
(Sober et al., 1991; Li et al., 2002; Tarhini & Agarwala, 2006).
O melanoma origina-se de células pigmentadas denominadas melanócitos,
células derivadas de melanoblastos neuro-ectodermais que migraram durante a
embriogênese para a epiderme, derme e outros sítios, sendo encontradas na
camada basal da pele e em outras superfícies epiteliais como olhos e ouvido
interno (Herlyn et al., 2000; Smith et al., 2002).
De acordo com sua localização anatômica e padrões de crescimento, o
melanoma pode ser dividido em diferentes subtipos: melanoma superficial,
nodular, lentigo acral, lentigo maligno e melanoma não-cutâneo (Tabela 1).
Quanto à progressão, cinco etapas têm sido propostas baseadas no curso clínico
e nas características histopatológicas: 1) nevo congênito, com melanócitos
estruturalmente normais os quais têm curto período de vida e geralmente
carregam anormalidades citogenéticas; 2) nevo displásico, caracterizado por

11
células com atipia estrutural e, em alguns casos, é precursor da neoplasia; 3)
etapa de crescimento radial (RGP), fase inicial do melanoma primário,
caracterizado pelo crescimento horizontal da epiderme podendo ou não alcançar
crescimento vertical (VGP), e pode ser curado por excisão cirúrgica; 4) VGP, fase
na qual as células invadem a derme e têm potencial para metastatizar; 5)
melanoma metastático, com a formação de metástases em sítios distantes do
local de origem do tumor primário (Clark et al., 1984; Meier et al., 1998; Rusciano,
2000; Chudnovsky et al., 2005).
Tabela 1. Classificação clínica de melanoma.
Subtipo Freqüência Sitio comum Características gerais
Melanoma expansivo
superficial (MES)
70% Tronco nos homens
Pernas nas mulheres
RGP, evolução de 1–5 anos
Melanoma nodular
(MN)
10–25% Tronco nos homens
Pernas nas mulheres
RGP, evolução de 6–18 meses.
Sexo masculino, na proporção
2:1
Melanoma
lentiginoso acral
(MLA)
5% Regiões palmoplantares,
extremidades digitais
(unhas)
Não relacionado à exposição
solar.
Não tem predileção por sexo.
Mais freqüente em não brancos
(30 a 70%).
Melanoma lentigo
maligno (MLM)
<1% Cabeça e pescoço de
idosos
Associada com exposição
crônica ao sol
RGP, evolução de 3–15 anos
Melanoma não
cutâneo
5% Ocular, mucosa Não relacionado à exposição
solar.
Prognóstico e tratamento
deferem dos subtipos cutâneos
Chudnovsky et al., 2005

12
Para compreender os mecanismos biológicos envolvidos no processo
metastático em melanomas, estudos experimentais vêm sendo realizados com
células de melanoma murino B16 (Johnson et al., 1987). Estas células foram
geradas espontaneamente em camundongos C57BL/6 e isoladas por Isaiah Fidler
(1973). A partir da linhagem parental B16, Fidler (1978) obteve variantes com
diferentes potenciais de malignidade. Para tanto, células B16 foram injetadas
endovenosamente em camundongos singênicos e, após três semanas, as
primeiras colônias formadas no pulmão foram dissecadas e adaptadas à cultura
celular. A primeira variante selecionada in vivo, B16F1, foi re-injetada em novos
animais e, após três semanas, as colônias formadas no pulmão foram novamente
dissecadas para posterior cultivo. Assim, dez linhagens foram selecionadas in vivo
com crescente potencial metastático, uma vez que células B16F10 apresentaram
capacidade superior à linhagem parental para metastatização. Em conjunto este
estudo demonstrou claramente que células B16 são heterogêneas e que poucas
células de potencial altamente maligno pré-existem na linhagem parental.
Recentes avanços para determinação dos mecanismos envolvidos na
transformação e malignização de células tumorais como o melanoma mostraram
aumento nos níveis de ativação de várias vias de sinalização intracelular
envolvidas com invasão e progressão metastática, dentre as quais se destaca a
via das MAPKs (Mitogen-activated protein kinases; Buettner et al., 2002;
Chudnovsky et al., 2005; Meier et al., 2005, Lopez-Bergami et al., 2007; Meier et
al., 2007). O aumento na ativação destas vias induz expressão de genes que
codificam integrinas, fatores de crescimento, metaloproteinases, quimiocinas,
citocinas, oncogenes, entre outras moléculas que favorecem o avanço e

13
instalação das células tumorais em sítios distantes do tumor primário (Lewis et al.,
2005; Axelsen et al., 2007). Sendo assim, o estudo do papel das vias de
transdução de sinal envolvidas com invasão e metástases torna-se indispensável
para esclarecer os mecanismos celulares e moleculares pelos quais células
tumorais como o melanoma adquirem o fenótipo metastático.
Quinase regulada por sinais extracelulares (ERK: Extracellular-signal
Regulated Kinase)
A via das proteínas quinases ativadas por mitógenos (MAPKs: mitogen-
activated protein kinase) é uma cascata de sinalização intracelular essencial para
numerosos eventos celulares. Esta via é ativada diferencialmente por fatores de
crescimento, hormônios, citocinas, stress celular (irradiação, choque térmico,
desigualdade osmótica, dano ao DNA) e por produtos bacterianos como o
lipopolissacarídeo (LPS). A ativação das MAPKs em resposta a estes estímulos
está relacionada ao controle de diversos eventos biológicos como resposta
inflamatória, metabolismo, diferenciação, ciclo celular, proliferação e apoptose. As
três principais vias das MAPKs incluem a quinase regulada por sinais
extracelulares (ERK1/2) também referida como via Ras-Raf-MEK-ERK; a quinase
Jun N-terminal (JNK: June N-terminal kinase) e p38 (Cobb & Goldsmith, 1995;
Weston & Davies, 2002; Zebisch et al., 2007).
As MAPK são quinases serina/treonina as quais são ativadas pela
fosforilação de seus resíduos serina e/ou treonina localizados em regiões
específicas (Crews et al., 1992). A via canônica de Ras-Raf-MEK-ERK é
desencadeada pela interação de ligantes extracelulares com os receptores tirosina

14
quinase (RTKs) na superfície celular. Esta ligação induz o recrutamento
intracelular de Ras GTP, envolvido na ativação de Raf-1. Raf-1 ativado fosforila
MEK que fosforila ERK, o qual dimeriza e transloca para o núcleo celular, onde
fosforila e estimula diversos fatores de transcrição que regulam a expressão de
genes associados ao ciclo celular, proliferação, sobrevivência, diferenciação,
adesão, remodelamento de matriz extracelular, entre outros eventos (Viala &
Pouyssegur, 2004; Zebisch et al., 2007; Gráfico 1).
Nos últimos anos o papel da via Ras/Raf/MEK/ERK na progressão do
melanoma tem recebido grande atenção devido à identificação de mutações no
gene B-Raf em 70% dos casos de melanomas ocasionados por exposição
constante ao sol (Davies et al., 2002; Maldonado et al., 2003). Esta mutação
confere super-ativação da quinase ERK a qual tem sido associada com expansão
metastática de melanomas (Klafter & Arbiser, 2000; Cohen et al., 2002;
Govindarajan et al., 2003; Smalley, 2003; Lopez-Bergami et al., 2007). Além das
mutações no gene B-Raf, mutações no gene N-Ras, verificadas em quase 20% de
melanomas humanos, podem também conferir ativação constitutiva da via das
quinases Raf/MEK/ERK em melanoma (Mansour et al., 1994; Davies et al., 2002;
Gorden et al., 2003).
Devido ao fato desta via estar envolvida com várias funções celulares,
numerosos estudos experimentais em modelos murinos e humanos mostraram
correlação direta entre a ativação da via das MAPKs com angiogênese,
tumorigênese (D´Angelo et al., 1995; Ilan et al., 1998; Cohen et al., 2002) e
metástases (Krueger et al., 2001; Sahai et al., 2001; Choo et al., 2005), razão pela

15
qual esta via vem sendo estudada como alvo terapêutico (Chudnovsky et al.,
2005; Alvarado & Giles, 2007; Lopez-Bergami et al., 2007).
Assim, o estudo da participação da via ERK no aumento do potencial
metastático das células B16 induzido pelo contato com células B-1, poderia
contribuir para explicar os mecanismos envolvidos neste fenômeno.
Gráfico 1. Via canônica de Ras-Raf-MEK-ERK .
RTKRTK

2. OBJETIVOS

2. OBJETIVOS
1.1 OBJETIVOS
Considerando que células B-1 são capazes de induzir aumento no potencial
metastático de células de melanoma murino da linhagem B16, este estudo tem
como objetivo principal determinar os mecanismos celulares envolvidos nesse
fenômeno.
Objetivo específico
1. Investigar a participação de ERK no aumento do potencial metastático de
células de melanoma B16 induzido pelo contato com células B-1.

3. MATERIAIS E MÉTODOS

3. MATERIAIS E MÉTODOS
2.1. MATERIAIS
2.1.1 Soluções Rotineiras
Todas as soluções aqui descritas foram preparadas com água bi-destilada
ou com padrão Mili-Q e com reagentes qualidade P.A.
• Solução estoque de tampão fosfato-salina 20X (PBS): NaCl 2,7378 M,
KCl 0,053 M, Na2HPO4 0,1301 M e K2HPO4 0,0229 M. Para uso, esta solução foi
diluída 20X (PBS 1X).
• Solução estoque de tampão tris-salina 10X (TBS): Tris base 24,2 g; NaCl
80g; pH 7.6 e 1000 mL de água q.s.p. Para uso, esta solução foi diluída 10X.
2.1.1.1 Soluções para SDS-PAGE
• Tampão de amostra para SDS-PAGE: Tris-HCl 62 mM, pH 8,8, contendo
0,2% (p/v) de SDS, β-mercaptoetanol 50 mM, 0,005% (p/v) de azul de bromofenol
e 10% (v/v) de glicerol.
• Solução estoque de acrilamida/bis para gel de proteína: 30% de
acrilamida, 0,8% de bis-acrilamida e água bi-destilada 100 mL q.s.p.
• Tampão para gel de proteína (separação) desnaturante: Tris-HCl 1,5 M,
pH 8,8, contendo 0,4% de SDS (p/v).
• Tampão para gel de proteína (empilhamento) desnaturante: Tris-HCl 0,5
M, pH 6,8, contendo 0,4% de SDS (p/v).
• Tampão de corrida para SDS-PAGE: Tris 25 mM, glicina 190 mM, pH 8,3,
e 0,1% de SDS (p/v).

20
2.1.1.2 Soluções para coloração por Comassie blue
• Corante de azul de Coomassie: 0,2% de Coomassie R250 (p/V) em 50%
de metanol e 10% de ácido acético (v/v).
• Descorante forte para Coomassie: 50% de metanol, 10% de ácido acético
e 40% de água destilada (v/v).
2.1.1.3 Soluções para Immunoblotting
• Tampão de transferência: Tris-HCl 25 mM, glicina 192 mM e 20% de
metanol (v/v).
• Corante de Ponceau: 0,5% (p/v) de Ponceau S em 5% de ácido acético
(v/v).
• Tampão de bloqueio: TBS 1X, 0,1% de Tween-20 com 5% de leite
desnatado em pó (p/v).
• Tampão de lavagem: 0,1% de Tween-20 em TBS 1X.
• Tampão striping: 1,87 g de glicina pH 2.5, 100 mL de SDS 10% e 1000
mL de água q.s.p.
2.1.1.4 Soluções para ELISA (Enzyme Linked Immuno Sorbent Assay)
• Tampão de lavagem (PBS-T): 0,05% de Tween-20 em PBS 1X.
• PBS-BSA (soro albumina bovina): 3% de BSA em PBS 1X.
• Peróxido de Hidrogênio: 3% de Peróxido de Hidrogênio em PBS 1X.
• Tampão para OPD: Na2PO4 0,4 M; Ácido Cítrico 0,4 M; pH 5,8, contendo
5.05% (V/V) de H2O2 (Perhidrol, Merk).

21
• Bloqueio da Revelação: Ácido Sulfúrico (H2SO4) 4 N em água destilada.
2.1.1.5 Soluções para cultura de células
• Meio RPMI: RPMI 1640 (Sigma, St Louis, MO) acrescido de ácido N-2-
hiroxietilpiperazina-N-2-etanosulfônico 10 mM (HEPES, da Sigma), bicarbonato de
sódio 24 mM (Sigma) e 40 µg/mL de garamicina (Schering Plough, N.J, USA).
Após preparação o meio foi esterilizado por filtração positiva em filtro de 0,22 µM.
• Meio completo (R-10): meio RPMI suplementado com 10% de soro bovino
fetal (SBF, da Cultilab, Campinas, SP).
• Meio para congelamento de células: 45% de RPMI, 45% de soro fetal
bovino e 10% de DMSO.
• PBS-EDTA (ácido etileno diaminoacético): 2,0 mM de EDTA em PBS 1X.
Após preparada esta solução foi esterilizada em autoclave a 120°C por 15
minutos.
2.1.2 Animais
Camundongos C57BL/6 wild-type (wt) fêmeos de 2 a 3 meses de idade
foram cedidos pelo Laboratório de experimentação animal do INFAR da
Universidade Federal de São Paulo, UNIFESP. Camundongos C57BL/6 knock-out
para IL-10 (KO) fêmeos de 2 a 3 meses de idade foram cedidos gentilmente pelo
Dr. João Santana da Silva (Departamento de Farmacologia, Escola de Medicina,
USP, Riberão Preto, SP). Os animais foram mantidos livres de germes, em micro-
isoladores com água e ração autoclavadas. Todos os procedimentos realizados

22
com os animais foram executados de acordo com normas estabelecidas pelo
Comitê de Ética em Experimentação Animal da Universidade Federal de São
Paulo.
2.2 MÉTODOS
2.2.1 Cultura de células e tratamentos
Células de melanoma B16 (B16): Células de melanoma de baixo potencial
metastático, obtidas em nosso laboratório a partir da linhagem de melanoma
murino B16F10 (Staquicini et al., 2003), foram cultivadas como monocamadas em
meio completo (RPMI-1640 suplementado com 10% de soro fetal bovino, SBF) a
37oC em atmosfera úmida de 5% de CO2. O descolamento das células para
posterior sub-cultivo foi realizado por curta exposição à PBS-EDTA 2 mM ou
Tripsina (Cultilab). Nos ensaios de Western blott ou Immunoblot, as células B16
utilizadas para extratos foram previamente plaqueadas em garrafas de cultura de
75 cm2 ou placas de Petri 100 X 90 mm. Após de alcançar confluência de 80%, o
meio foi aspirado e as células aderidas foram lavadas 3 vezes com PBS e re-
cultivadas com RPMI sem soro por 24 horas. Em seguida, estas células foram ou
não estimuladas com 10% de SBF ou tratadas com inibidor de MEK1/2 (PD98059,
Cell Signalling, Beverly, MA, USA) ou com IL-10 recombinante (Pharmigen, San
Diego, CA), durante diferentes períodos de tempo (entre 15 e 120 minutos).
Células B-1: Linfócitos B-1 foram obtidos a partir de cultura de células
aderentes de lavado peritoneal de camundongos C57BL/6 como descrito
previamente (Almeida et al., 2001). Assim, células peritoneais de camundongos

23
wild-type (B-1wt) e knock-out para IL-10 (B-1KO), foram coletadas por lavagens
sucessivas com RPMI e incubadas em garrafas de cultura por 40 minutos a 37oC.
Em seguida, o sobrenadante foi desprezado e as células aderidas foram re-
cultivadas até 7 dias, em na presença de meio completo. Após este período a
maior porcentagem de células presentes no sobrenadante destas culturas (células
flutuantes) corresponde às células B-1 (Almeida et al., 2001). Para ensaios de
Western blot, monocultura de células B-1 com seis dias de incubação, presentes
no sobrenadante da cultura acima descrita, foram centrifugadas, lavadas 3 vezes
com PBS e re-cultivadas com RPMI sem soro por 24 horas. Em seguida, estas
células foram estimuladas ou não com SBF ou tratadas com inibidor farmacológico
de MEK1/2 (PD98059) de 1 até 24 horas.
Co-culturas de células B16 e B-1: Co-culturas de células B16 e B-1 foram
realizadas em placas de 6 orifícios ou em garrafas para ensaio de metástases in
vivo ou para Western blot, respectivamente. Brevemente, 1X105 células B16 foram
plaqueadas em meio completo e incubadas por 24 horas para adesão. Após este
período, o meio foi aspirado e 1X106 células de B-1 com 5 dias de cultura foram
adicionadas juntamente com seu meio condicionado (mB1) e incubadas por
diferentes períodos na ausência ou presença de PD98059. Logo após os
diferentes períodos de incubação, as células B-1 foram removidas por aspiração e
as células B16 aderidas foram lavadas 3 vezes com PBS para remover qualquer
resíduo de células B-1. Em seguida, células B16 foram coletadas e submetidas a
análises por citometria de fluxo (FACS), utilizando-se os marcadores B220 e Mac-
1 de células B-1 objetivando confirmar a pureza das células B16 após co-cultivo.
Assim células B16 livres de células B-1 após co-cultivo (B16+B-1) foram usadas

24
nos ensaios experimentais in vivo e na preparação de extratos celulares para
análises de Western Blot.
2.2.2 Análises de moléculas de superfície por citometria de fluxo
Monoculturas de células B16 ou B-1 ou células B16 após co-cultivo com B-1
foram coletadas e lavadas 2 vezes com PBS e ajustadas à concentração 1X106
células/amostra. Estas células foram marcadas duplamente com anticorpos anti-
B220 conjugado com ficoeritrina (PE) e anti CD11b (Mac-1) conjugado a
fluoresceína (FITC), ambos da Pharmingen (San Diego, CA, USA), na diluição de
1:100 em PBS acrescido de 1% de BSA e incubados no escuro por 1 hora, no
gelo. Após este período as células foram lavadas e ressuspendidas em 500 µl de
PBS e, em seguida, analisadas por citometria de fluxo (FACSCalibur da Becton
Dickinson, Mountain View, CA) usando o programa CellQuest.
2.2.3 Ensaio de viabilidade celular por Trypan blue
Solução de Trypan blue foi preparada em PBS e filtrada antes do uso.
Células B16 ou B-1 de monoculturas ou co-culturas após diferentes tratamentos
descritos anteriormente, foram coletadas e ressuspendidas em PBS. Dez
microlitros de cada suspensão celular foram diluídos com 10 µL do reagente
Trypan blue e, em seguida, 10 µl desta diluição foram contados em câmara de
Neubauer para estabelecer o porcentual de células coradas (mortas) em cada
amostra. Depois de verificada a viabilidade celular, as suspensões celulares com
mais de 80% de células viáveis foram utilizadas para ensaios experimentais in vivo

25
e para a preparação dos extratos celulares utilizados nas análises de Western
Blot.
2.2.4 Ensaio de inibição da adesão
Para criar a condição de cultura não-aderente, monoculturas de células B16
ou B-1 ou co-culturas de ambas as células foram plaqueadas em placas de Petri
cobertas previamente com uma camada fina de agarose 1% e incubadas a 37°C
com 5 % de CO2 por 72 horas. Após este período, células em suspensão foram
coletadas e ressuspendidas em PBS. Agregados celulares foram mecanicamente
separados por agitação vigorosa e as células individuais foram submetidas a
análise de viabilidade por Trypan blue e em seguida a ensaio experimental de
metástase in vivo.
2.2.5 Ensaio de metástase experimental
Células B-1 ou B16 de monoculturas ou células B16 após co-cultivadas com
células B-1 foram coletadas e ressuspendidas em PBS. Logo, 1X105 células
diluídas em 100 µL de PBS foram injetadas na veia da cauda de camundongos
C57BL/6 wild-type. Para cada grupo celular analisado foram feitas injeções em
pelo menos três camundongos. Após 14 dias de inoculação os animais foram
sacrificados e seus pulmões removidos cirurgicamente para estabelecer
macroscopicamente o número de nódulos metastáticos instalados.

26
2.2.6 Preparação de extratos celulares
Para extração de proteínas totais, células foram lavadas 2 vezes com PBS
a 4°C e, em seguida, lisadas com 100 µL de tampão de lise (contendo, 20 mM de
Tris-HCl pH7.4, 100 mM de NaCl, 1 mM de Na3VO4, 40 mM de NaF, o,5% de NP-
40, 1 mM de PMSF, 10 µg/mL de leupeptina e 10 µg/mL de aprotinina), por 10
minutos a 4°C. Após este período, os lisados celulares foram centrifugados por 15
minutos a 4°C para remoção de resíduos não protéicos. Em seguida, os
sobrenadantes foram coletados e a concentração protéica determinada pelo
método de Bradford (1976).
2.2.7 Dosagem protéica pelo micrométodo de Bradford
As determinações das concentrações protéicas foram realizadas segundo
método proposto por Bradford (1976), que utiliza Coomassie brilliant blue (CBB)
G-250 (Sigma) como reativo e albumina bovina (BSA) 1,0 mg/mL como padrão. A
leitura da curva padrão foi realizada em comprimento de onda de 570 nm em leitor
EIA Reader Titertek, Multiskan, MCC340 (Labsystem, Santa Fé, USA).
2.2.8 Eletroforese em gel de poliacrilamida com dodecilsulfato de sódio
(SDS-PAGE)
Os procedimentos para SDS-PAGE foram executados em equipamentos
Mini-Protean II (Bio-Rad Laboratories, Hercules, CA) conforme descrito
anteriormente (Laemmli, 1970). Para as proteínas em estudo, o gel de separação
foi feito na concentração de 10% de acrilamida e o gel de empilhamento na
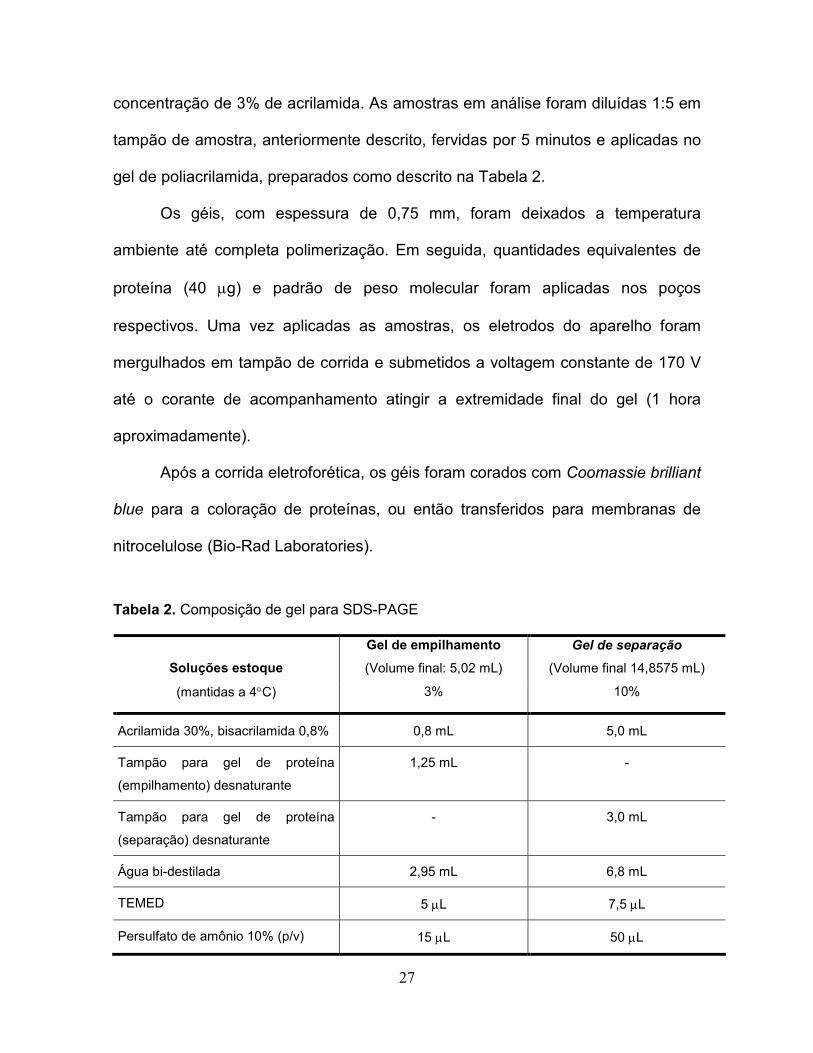
27
concentração de 3% de acrilamida. As amostras em análise foram diluídas 1:5 em
tampão de amostra, anteriormente descrito, fervidas por 5 minutos e aplicadas no
gel de poliacrilamida, preparados como descrito na Tabela 2.
Os géis, com espessura de 0,75 mm, foram deixados a temperatura
ambiente até completa polimerização. Em seguida, quantidades equivalentes de
proteína (40 µg) e padrão de peso molecular foram aplicadas nos poços
respectivos. Uma vez aplicadas as amostras, os eletrodos do aparelho foram
mergulhados em tampão de corrida e submetidos a voltagem constante de 170 V
até o corante de acompanhamento atingir a extremidade final do gel (1 hora
aproximadamente).
Após a corrida eletroforética, os géis foram corados com Coomassie brilliant
blue para a coloração de proteínas, ou então transferidos para membranas de
nitrocelulose (Bio-Rad Laboratories).
Tabela 2. Composição de gel para SDS-PAGE
Gel de empilhamento
(Volume final: 5,02 mL)
Gel de separação
(Volume final 14,8575 mL)
Soluções estoque
(mantidas a 4°C) 3% 10%
Acrilamida 30%, bisacrilamida 0,8% 0,8 mL 5,0 mL
Tampão para gel de proteína
(empilhamento) desnaturante
1,25 mL -
Tampão para gel de proteína
(separação) desnaturante
- 3,0 mL
Água bi-destilada 2,95 mL 6,8 mL
TEMED 5 µL 7,5 µL
Persulfato de amônio 10% (p/v) 15 µL 50 µL

28
2.2.9 Transferência de proteínas para membranas de nitrocelulose
(Immunoblot ou Western blot)
A transferência eletroforética dos géis de poliacrilamida para membranas de
nitrocelulose foi efetuada conforme descrito por Towbin e colaboradores (1979),
usando equipamento Trans-Blot System (Bio-Rad Laboratories). Após a
eletroforese os géis foram postos sobre membranas de nitrocelulose (Bio-Rad
Laboratories) ou PVDF (Amersham Pharmacia Biotech, Buckinghamshire, UK),
recobertos com papel de filtro e comprimidos com esponjas. Todos os materiais
foram embebidos em tampão de transferência e em seguida, encaixados em
placas acrílicas perfuradas e mergulhados na câmara de eletroforese contendo o
mesmo tampão. A transferência foi realizada na voltagem constante de 100 V por
1 hora e meia. Em seguida, as membranas foram coradas com Ponceau S para a
visualização das bandas protéicas e depois bloqueadas com tampão de bloqueio
por 1 hora a temperatura ambiente. Após lavagem, as membranas foram
incubadas overnight a 4°C com anticorpo primário específico para ERK fosforilado
[p-ERK, (E-4): sc-7383), ERK total [(K-23): sc-94], Stat3 total [(C-20): sc-482]
todos da Santa Cruz Biotechnology, INC (Santa Cruz, CA, USA) ou Stat3
fosforilado (p-Stat3, da BioVision, CA, USA), diluídos em tampão de bloqueio
segundo instruções do fabricante. Após este período, as membranas foram
lavadas e incubadas por 1 hora a temperatura ambiente, com o respectivo
anticorpo secundário (Ig-G) conjugado também a peroxidase. Em seguida, bandas
reativas foram visualizadas por quimiluminescência (ECL-Amersham-Pharmacia)
em filmes Kodak e/ou Amersham (Hyperfilm ECL). Após a detecção, as

29
membranas foram lavadas e incubadas com tampão de stripping para serem re-
incubadas com outros anticorpos.
2.2.10 Microscopia eletrônica de transmissão
Células B-1 ou células B16 co-cultivadas ou não com células B-1 foram
incubadas por 24, 48 e 72 horas sobre lamínulas de plástico contidas em placas
de doze (12) orifícios (Costar). Após estes períodos de incubação, as células
foram fixadas em 50% de glutaraldeído, 4% de formaldeído, 0,2 M de cacodilato
de sódio em pH 7.2 por 2 horas. Em seguida, foram lavadas por 4 vezes (1 hora
cada lavagem) com tampão cacodilato 0,1 M e logo fixadas novamente com
tetróxido de ósmio 2% em tampão cacodilato 0,1 M. As amostras foram então
lavadas 3 vezes e desidratadas em concentrações crescentes de etanol e óxido
de propileno. Em seguida, as amostras foram incluídas em resina Araldite® e
mantidas a vácuo por 4 horas e logo a 60°C por 40 horas para polimerização. Uma
vez polimerizadas, as amostras incluídas em Araldite® foram cortadas em seções
de 90 a 95 µm de espessura e coradas com 2% de uranil-acetato aquoso durante
8 minutos. Estas preparações foram subsequentemente coradas com citrato de
chumbo diluído em água destilada por 4 minutos. Amostras foram analisadas e
fotografadas em microscópio eletrônico (JEOL JEM 1200 EX II).

30
2.2.11 Ensaios de proliferação celular
2.2.11.1 Incorporação de timidina tritiada
Células B16 (2X103 células por poço em um volume final de 200 µl) foram
cultivadas com meio completo em placas de 96 poços (uma placa para cada
tempo de incubação: 24, 48, 72 e 120 horas) e incubadas por 6 horas em estufa a
37oC e 5% de CO2. Em seguida, células aderidas foram lavadas com PBS e
ressuspendidas em RPMI sem soro e incubadas overnight nestas condições. Após
este período, o sobrenadante foi retirado e substituído por meio completo ou meio
condicionado de cultura de células B-1 na presença (B16+B-1) ou ausência
(B16+mB-1) de 2X104 células B-1. Células submetidas a estas condições foram
incubadas durante 24, 48 e 72 horas a 37oC e 5% de CO2.. Dezoito horas antes do
término de cada incubação, 1 µCi de timidina tritiada (Amersham) foi adicionado
às culturas. Posteriormente, cada amostra foi adsorvida em papel de filtro e cada
filtro foi colocado em tubos com 2 ml de líquido de cintilação para a leitura em
aparelho Beckman LS 5801. O experimento foi realizado em triplicata para cada
grupo celular estudado.
2.2.11.2 MTT [tetrazolium salt3-(4,5-dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromide]
Células B16 (2,5X103 células por poço em volume final de 100 µl) foram
cultivadas com meio completo em placas de 96 poços (uma placa para cada
tempo de incubação: 0, 24, 48, 72 horas) e incubadas 12 horas em estufa a 37oC
e 5% de CO2. Em seguida, células aderidas foram lavadas com PBS

31
ressuspendidas com RPMI sem soro e incubadas overnight nestas condições.
Após este período, o sobrenadante foi retirado e substituído por meio completo ou
meio condicionado de cultura de células B-1 na presença (B16+B-1) ou ausência
(B16+mB-1) de 2X104 células B-1. Células submetidas a estas condições foram
então incubadas durante 24, 48 e 72 horas a 37oC e 5% de CO2. Posteriormente,
2,5 horas antes do término de cada incubação, foram adicionados 10 µl de MTT (5
mg/mL) às culturas e deixados por 1,5 horas a 37oC e 5% de CO2. Em seguida, o
sobrenadante de cada poço foi aspirado e foram adicionados 100 µl de
isopropanol (Merk) e mantidas a temperatura ambiente por 15 minutos. Após este
período, as placas foram analisadas em comprimento de onda de 570 nm em leitor
de ELISA, EIA Reader Titertek, Multiskan, MCC340 (Labsystem). O experimento
foi realizado em triplicata para cada grupo celular estudado.
2.2.12 Dosagem de citocinas no sobrenadante de cultura
A produção de IL-10 no sobrenadante de cultura de células B-1 tanto de
animais wt como KO foi quantificada por ELISA (Enzymed linked
immunoabsorvent assay) seguindo as instruções do fabricante (Duoset mouse IL-
10, R&D System, Minneapolis, MN, USA). Brevemente, células B-1 foram
cultivadas como descrito acima e os sobrenadantes foram coletados após 7 dias
de cultura e mantidos a 70°C até o momento da dosagem. Para a determinação
da citocina, placas de ELISA de 96 poços foram sensibilizadas overnight a
temperatura ambiente com anticorpos monoclonais anti-IL-10. Após este período,
as placas foram lavadas por 4 vezes com PBS-T e logo incubadas com 200 µL de

32
PBS-BSA a temperatura ambiente durante 2 horas, para prevenir ligações
inespecíficas. Placas foram lavadas 4 vezes com OS-T e incubadas 2 horas com
100 µL por orifício de cada sobrenadante (amostras em triplicatas) ou com
standard de IL-10 diluído serialmente em PBS-BSA. Em seguida, placas foram
lavadas 4 vezes com PBS-T e incubadas 2 horas com 100 µL/poço de anti-IL-10 a
temperatura ambiente. As placas foram então novamente lavadas e a revelação
foi realizada com 50 µL/poço de ο-fenilenodiamina (OPD, 1 mg/mL, Sigma)
dissolvida em tampão citrato-fosfato 100 mM pH5,8, contendo 0,05% (v/v) de H2O2
(água oxigenada). A reação foi interrompida após o aparecimento de cor com 50
µL/poço de H2SO4 4 N (50 µL/poço). Os resultados foram avaliados por leitura
espectrofotométrica em comprimento de onda 492 nm em leitor de ELISA
(Labsystems). A sensibilidade do ensaio foi de 20 pg/mL. Resultados são
apresentados como medianas ±SD.
2.2.13 Obtenção de RNA total
O RNA total foi extraído de células B-1 e de células B16 tanto de
monoculturas como co-culturas utilizando Trizol (Invitrogen, Carlsbad, CA) de
acordo com as instruções do fabricante. Brevemente, 1,5 mL de Trizol foi
adicionado às células e após homogeneização por 5 minutos, o lisado celular foi
transferido para tubos de propilpropileno de 2 mL, e adicionados 300 µL de
clorofórmio a cada tubo. O conteúdo foi agitado vigorosamente e centrifugado a
14.000 rpm por 15 minutos a 4°C. A fase aquosa (superior) foi transferida para
microtubos de 1,5 mL, contendo 750 µL de isopropanol e em seguida

33
homogeneizado. Após precipitação por 24 horas a -20°C, centrifugou-se a 14.000
rpm por 15 minutos a 4°C. O RNA precipitado foi lavado 2 vezes com etanol 70%
em água DEPC 0,001% (dietil pirocarbonato), desidratado por evaporação a
temperatura ambiente e diluído de 30 a 50 µL de água DEPC.
A quantificação do RNA foi feita por espectrofotometria a 260 nm. O
resultado da densidade óptica (DO) foi multiplicado pelo fator 40, o que fornece a
concentração em µg/mL, uma vez que uma unidade de DO a 260 nm equivale a
40 µg/mL.
A qualidade do RNA foi avaliada de duas maneiras: em primeiro lugar, com
relação à contaminação por proteína, a qual é dada pela razão entre as leituras a
260 e 280 nm, que precisa estar entre 1,6 e 1,8. Em segundo lugar, foi avaliada a
integridade do RNA utilizando gel de agarose. Nesta análise a amostra é
considerada íntegra e livre de contaminação quando no gel estão presentes duas
bandas, 28S e 18S, sendo que a banda de 28S é mais intensa que a de 18S. A
ocorrência de outra banda, de tamanho superior a de 28S, denota contaminação
por DNA genômico e, neste caso, a amostra deve ser submetida à re-extração por
Trizol.
2.2.14 RT-PCR (Reverse Transcriptase – Polymerase Chain Reaction)
A sínteses de cDNA a partir de amostras de RNA total (transcrição reversa)
para estudos de expressão gênica por RT-PCR, foi realizada com o kit SuperScript
III (Invitrogen- Life Technology) de acordo com as instruções do fabricante. Assim,
a transcrição reversa foi realizada a partir de 3 µg de RNA total, tratado

34
previamente com DNase 1 (Invitrogen- Life Technology), para eliminar qualquer
tipo de contaminação com DNA genômico, conforme verificado pela não
amplificação do gene de controle interno GAPDH.
Após preparados os cDNAs foram submetidos a RT-PCR qualitativo e a
RT–PCR quantitativo em tempo real.
2.2.14.1 Análise qualitativa por RT-PCR
Cada amplificação foi realizada em volume total de 25 µl contendo 100 ng
de cada primer (sense e anti-sense), 1,5 mM de solução de 25 mM de Mg2+, 10 ul
de MasterMix (2,5x) (Eppendorf) e 2 µl de cDNA diluído 1:2 em água bi-destilada
estéril. Como controle negativo de cada reação de PCR foi utilizada água bi-
destilada ao invés de cDNA .
As condições de amplificação comuns para cada par de primers foram:
desnaturação 94°C por 30 segundos, anelamento por 30 segundos (temperatura
específica para cada par de primers), extensão 72°C por 30 segundos, número de
ciclos 35. A temperatura de anelamento de cada par de primers foi escolhida após
o teste de gradiente de temperatura utilizando o termo-ciclador Mastercycler
Gradient (Eppendorf). Na Tabela 3 estão descritas as seqüências de
oligonucleotídeos utilizadas para cada transcrito e, também a temperatura de
anelamento, e o tamanho dos produtos do PCR em número de pares de bases
(bp).

35
Tabela 3. Seqüências oligo-nucleotídicas dos primers sense (S) e anti-sense (AS),
temperatura de anelamento (°C) e o tamanho do produto do PCR em pares de bases
(BP), dos transcritos estudados por RT-PCR qualitativo.
Transcrito Primers
5’-seqüência oligo-nucleotídica- 3’
Temperatura de anelamento (°C)
Tamanho do produto (bp)
GAPDH S: ACCACAGTCCATGCCATCAC AS: TCCACCACCCTGTTGCTGTA
62 500
CXCR4 S: AGTTTTCACTCCAGCTAACCCTTAT AS: TGGAATGTTCAGTTTCTTCCTCTAC
58 321
MMP-9 S: TTCTCTGGACGTCAAATGTGG AS: CAAAGAAGGAGCCCTAGTTCAAGG
60 413
Para a análise da amplificação, aos produtos de PCR foram adicionados 5
µl de loading buffer, constituído por solução de glicerol a 50% (Sigma Chemical
Company) acrescida de 1mg/ml de vermelho de cresol (Sigma Chemical
Company) em 0,5x TBE (Gibco BRL, Gaithersburg, MD, USA). Cada amostra
diluída 1:6 com este tampão foi aplicada em volume de 10 µl ao gel de agarose
1,5 % (Gibco BRL) preparado em 0,5x TBE com 0,75 µg/ml de brometo de etídio
(Sigma Chemical Company) e submetida a eletroforese com 100 V por 50
minutos. Adicionalmente, também foi submetido a eletroforese o marcador de
massa molecular (Low mass, Invitrogen). As bandas foram visualizadas sob luz
ultravioleta. Uma vez verificada a positividade no controle e a ausência de
contaminação (controle negativo), a análise do mRNA foi realizada através do
sistema de documentação de imagem do gel de eletroforese “Kodak Digital
Science – Electrophoresis Documentation and Analysis System 120” (Eastman
Kodak Co,, Rochester, NY, USA).

36
2.2.14.2 Análise quantitativa por RT-PCR em tempo real
A reação de PCR em tempo real foi realizada com o reagente Sybr Green
(Applied Biosystems). A partir da reação de transcrição reversa, utilizou-se 1 mL
de cDNA, 7,5 mL de master mix Sybr Green, 0,5 mL de cada oligonucleotídeo (18
µM): sense e anti-sense (Tabela 4), e água Milli Q autoclavada q.s.p 15 mL. Em
seguida, as reações para PCR em tempo real foram realizadas utilizando ABI
7000 Real Time PCR System (Applied Biosystems), nas seguintes condições: 10
minutos a 50°C para ativação da enzima, desnaturação por 5 minutos a 95°C, e 40
ciclos de 95°C por 30 segundos e 60°C por 1 minuto. Após avaliar a qualidade da
reação com base nas curvas de dissociação, os resultados foram analisados
utilizando o programa ABI 7000 SDS software (Applied Biosystems). Para a
quantificação relativa, foi realizado o seguinte cálculo: inicialmente determinou-se
o cycle threshold (CT), dado pelo número do ciclo em que o sinal de fluorescência
atingiu a linha limiar (threshold line), ou seja, a linha em que a emissão de
fluorescência está acima do ruído de fundo (background). O CT encontrou-se
invariavelmente na região correspondente à fase exponencial da amplificação, o
que torna mais acurada a estimativa de quantificação dos transcritos na amostra
original. Os valores de CT dos genes de interesse foram normalizados em relação
ao CT do gene constitutivo (GAPDH), resultando o ∆CT, representado pelo
CTgene - CTconstitutivo. Por fim, calculou-se o 2-∆CT, sendo este o valor a ser
trabalhado como representante da expressão relativa para cada gene
(Vandesompele et al., 2002; Bustin et al., 2005). Para melhor interpretação dos
37
resultados estes foram multiplicados no final por 100 para obtenção de números
inteiros.
Tabela 4. Seqüências oligo-nucleotídicas dos primers sense (S) e anti-sense (AS) e
tamanho do produto do PCR em pares de bases (bp), dos transcritos estudados por RT-
PCR em tempo real.
Transcrito Primers
5’-seqüência oligo-nucleotídica- 3’
Tamanho do produto (bp)
GAPDH S: AAATGGTGAAGGTCGGTGTG
AS: TGAAGGGGTCGTTGATGG
120
CXCR4 S: TGGAACCGATCAGTGTGAGT
AS: GTAGATGGTGGGCAGGAAGA
128
MMP-9 S: CATTCGCGTGGATAAGGAGT
AS: GGTCCACCTTGTTCACCTCA
140
2.2.15 Análise Estatística
As análises estatísticas foram realizadas pela análise de variância (ANOVA)
e o teste Tukey-Kramer, utilizando o programa INSTAT (GraphPad, San Diego
CA).

4. RESULTADOS

4. RESULTADOS
3.1 Caracterização do sistema de co-cultivo
Estudos anteriores realizados no nosso laboratório mostraram que o co-
cultivo de células B-1 com células de melanoma murino da linhagem B16, foi
capaz de induzir aumento no potencial metastático destas células (Staquicini,
2004). Com o intuito de determinar quais mecanismos moleculares controlam este
efeito, experimentos iniciais foram realizados para caracterizar o sistema de co-
cultivo e avaliar o tipo de interação entre estas células. Para isso, células B-1
foram isoladas de lavados peritoneais de camundongos e co-cultivadas por 24, 48
e 72 horas com uma sublinhagem de células B16 caracterizada pelo seu baixo
potencial metastático (Staquicini et al., 2003). Após estes períodos, células B-1
foram eliminadas por aspiração e em seguida, células B16 livres de células B-1
foram utilizadas em ensaios experimentais de metástase in vivo. O método de
eliminação das células B-1 após os diferentes períodos de co-cultivo foi eficiente,
como evidenciado por citometria de fluxo (Fig. 1). Esta análise mostrou que
populações celulares duplamente marcadas para os marcadores fenotípicos B220
e Mac-1 de células B-1, foram detectados em células B-1, porém não em
monoculturas de células B16 ou em células B-16 livres de células B-1 após o co-
cultivo (B16+B1). Após 14 dias da inoculação, o número de nódulos metastáticos
presentes nos pulmões dos animais foi determinado macroscopicamente.
Conforme demonstrado na Figura 2, a inoculação de células B16 submetidas ao
co-cultivo com células B-1 por 48 ou 72 horas, aumentou significativamente o

40
número de nódulos metastáticos quando comparado com animais injetados com
células B16 ou B-1 isoladas de monoculturas.
Uma vez que no sistema de co-cultivo as células B16 são aderentes e
células B-1 não são, foi determinado se o aumento do potencial metastático das
células B16, induzido pela presença das células B-1, dependia da adesão das
células B16 ao plástico. Para testar isto, os dois tipos celulares foram co-
cultivados sobre substrato não adesivo (agarose) por 72 horas e após este
período um pool de células B16+B1 foi submetido a ensaios experimentais de
metástases in vivo. Conforme mostrado na Figura 3, animais injetados com células
B16 co-cultivadas com B-1 em suspensão (sB16+B1) produziram
significativamente mais colônias metastáticas no pulmão comparado aos grupos
controles injetados com células B16 (sB16) ou B-1 (sB1) coletadas de
monoculturas mantidas nas mesmas condições (Fig. 3A). Adicionalmente, ensaios
de viabilidade celular por Trypan blue mostraram a manutenção da viabilidade das
células B16 e B-1 mantidas em suspensão por 72 horas (Fig. 3B). Em conjunto,
estes resultados demonstram que células B-1 são capazes de aumentar o
potencial metastático das células B16 e que a adesão das células B16 não é
necessária para este efeito.
3.2 Células B-1 e B16 fisicamente interagem durante o co-cultivo.
Em seguida, foi determinado se o aumento no potencial metastático das
células B16 era dependente do contato físico entre as duas células ou se fatores
solúveis produzidos pelas células B-1 poderiam estar envolvidos neste feito. Para

41
esta análise, células B16 foram cultivadas por 72 horas em meio condicionado
proveniente tanto de monoculturas de células B-1 como de células B16 co-
cultivadas com células B-1 por 72 horas. Como apresentado na Figura 4, não
foram observadas diferenças no número de nódulos metastáticos nos pulmões de
animais injetados com células B16 cultivadas em seu próprio meio comparado
com aquelas cultivadas em meio condicionado tanto de monoculturas de células
B-1 (B16+mB1) como do co-cultivo de células B16 com B-1 [B16+m(B16+B1)]. Por
outro lado, quando células B16 submetidas ao co-cultivo com células B-1 (B16+B-
1) foram injetadas, o número de nódulos metastáticos presentes nos pulmões
desses animais foi significativamente maior comparado com os grupos acima
descritos.
Desta forma, os resultados acima descritos mostram que fatores solúveis
produzidos pelas células B-1 são insuficientes para aumentar a capacidade
metastática das células B16, sugerindo que o contato físico entre as duas células
ocorre durante o co-cultivo. De fato, análises por microscopia eletrônica de
transmissão revelaram a existência de interação física entre as células B16 e B-1.
Na Figura 5A, podemos observar uma célula B-1 caracterizada por sua atípica
morfologia nuclear com reentrâncias delimitando dois lóbulos distintos, descrita
por Abrahão e colaboradores em 2004. Na Figura 5B, não observamos interação
física entre células B-1 e células B16 co-cultivadas por 24h. Entretanto, quando
estas células foram co-cultivadas por 48 e 72h, vários pontos de interação entre as
membranas plasmáticas destas células foram detectados (Fig. 5C e D). Juntos
estes resultados demonstram que estes dois tipos celulares interagem fisicamente

42
quando co-cultivados por tempos superiores a 48h, observação esta que
correlaciona com o tempo de co-cultivo necessário para aumentar o potencial
metastático das células B-16 (Fig. 2).
3.3 Fosforilação de ERK está aumentada nas células B16 após
interação com células B-1
Considerando que células B16 e B-1 interagem fisicamente, foi
hipotetizado que as células B-1 poderiam influenciar no aumento do potencial
metastático das células B16 através da interferência em vias de sinalização celular
que regulam invasão e metástase.
Em melanomas há relatos mostrando que a ativação da quinase regulada
por sinais extracelulares ERK 1/2, da família das MAPKs (proteínas quinases
ativadas por mitógenos), está envolvida de forma positiva na regulação da
proliferação celular, assim como no aumento do potencial metastático destes
tumores (Davies et al, 2002; Zebish et al., 2007; Lopez –Bergami et al., 2007). Por
esta razão, a participação de ERK foi avaliada no aumento do potencial
metastático das células B16 após o co-cultivo com células B-1.
Inicialmente, foi determinado o perfil de fosforilação basal de ERK nas
monoculturas tanto de células B16 como de células B-1. Para isso foi utilizado um
anticorpo que reconhece especificamente as duas formas (p42/p44 kDa) de ERK
fosforilada/ativada. Como pode ser observado na Figura 6A, monoculturas de
células B16 ou de células B-1 cultivadas overnight na ausência de soro bovino
fetal (SBF) apresentaram níveis basais de fosforilação de ERK. Enquanto a adição

43
de SBF induziu um leve aumento na fosforilação de ERK nas células B16, a
presença de SBF não alterou o perfil basal de fosforilação desta quinase nas
células B-1, mostrando que ambos os tipos celulares apresentam ERK
constitutivamente fosforilado.
Para avaliar se células B-1 poderiam aumentar os níveis basais de
fosforilação de ERK nas células B16 durante o co-cultivo, células B16 foram
mantidas por 24 horas na ausência de soro e então estimuladas ou não com SBF
por 1 hora (1h) na presença ou ausência de células B-1. Após este período,
extratos celulares de monoculturas de células B16 ou de células B16 livres de
células B-1 após o co-cultivo foram produzidos e submetidos a Immunobloting
para investigar os níveis de ERK fosforilado. Conforme mostrado na Figura 6B, um
aumento considerável nos níveis de fosforilação de ERK foi observado nas células
B16 após o co-cultivo com células B-1 (B16+B1), independente da presença de
soro, comparado com células B16 de monoculturas.
3.4 Aumento no potencial metastático de células B16 é dependente
dos níveis de ERK fosforilado
Para avaliar se o aumento na ativação de ERK nas células B16 após o co-
cultivo com células B-1 é um evento importante no aumento do potencial
metastático das células B16, foi utilizado o inibidor farmacológico de MEK1/2
(PD98059) que é a quinase up-stream a ERK na via de sinalização.
Como observado na Figura 7, os níveis basais de ERK fosforilado foram
completamente inibidos na presença de 40 µM de PD98059. Além disso, as

44
análises por Trypan blue não indicaram perda significativa da viabilidade destas
células na presença do inibidor (dados não mostrados).
Em seguida, foi avaliado se o tratamento das células B16 com PD98059
antes ou durante o co-cultivo poderia prevenir o aumento da fosforilação de ERK
induzido pelas células B-1. Para isto, células B16 foram pré-tratadas ou não com
40 µM de PD98059 por 24 horas e co-cultivadas ou não com células B-1 por 72
horas. Adicionalmente, ambos os tipos celulares foram co-cultivados por 72 horas
na presença do inibidor. Como mostra a Figura 8A, independentemente se as
células B16 foram pré-tratadas com PD98059 antes do co-cultivo [(B16+PD)+B1]
ou co-cultivadas com células B-1 na presença do inibidor [(B16+B1)+PD], os
níveis de ERK fosforilado permaneceram similares aos encontrados nas
monoculturas de células B16 não tratadas (B16). Em contraste, os níveis de ERK
fosforilado aumentarão nas células B16 co-cultivadas com B-1 na ausência do
inibidor (B16+B1). Interessantemente, prevenindo o aumento na fosforilação de
ERK induzido pelas células B-1, o número de nódulos metastáticos produzidos
pelas células B16 não apresentou diferença estatística daqueles produzidos por
células B16 cultivadas sozinhas na ausência do inibidor, enquanto que células B16
provenientes de co-culturas não tratadas desenvolveram mais nódulos
metastáticos (Fig. 8B). Notavelmente, apesar da quase completa inibição da
fosforilação de ERK nas células B16 cultivadas sozinhas na presença de
PD98059, esta inibição não refletiu quantitativamente em redução de nódulos
pulmonares, comparado com células B16 cultivadas sozinhas na ausência do
inibidor (Fig. 8B). Juntos estes resultados mostram que existe uma clara

45
associação entre aumento da fosforilação de ERK induzido pelas células B-1 e
aumento no potencial metastático das células B16.
3.5 Células B-1 não induzem aumento na proliferação de células B16
Uma vez que a quinase ERK além de regular invasão e metástase também
está envolvida no controle da proliferação celular (Raman et al., 2007), foi
investigado se o co-cultivo com as células B-1 poderia induzir aumento na
proliferação das células B16. Entretanto, ensaios de MTT e timidina tritiada
realizados por 24, 48, e 72 horas não mostraram aumento na proliferação de
células B16 após co-cultivo com células B-1 (B16+B1) ou quando as células B16
foram cultivadas em meio condicionado proveniente de monoculturas de células B-
1 (B16+mB1) (Fig. 9).
3.6 Células B-1 induzem a expressão de genes relacionados com
metástases nas células B16.
Para explorar em maior detalhe a influência biológica das células B-1 no
aumento do potencial metastático das células B16, a expressão de genes
relacionados com metástase como CXCR-4 [Chemokine (C-X-C motif) receptor] e
MMP-9 (Matrix metalloproteinase-9) foi avaliada nestas células antes e após o co-
cultivo com células B-1. Para isto, RNA total de células B16 co-cultivadas ou não
com células B-1 na ausência ou presença de PD98059 foi extraído. Uma vez
verificada nas amostras de RNA, após tratamento com DNase, a integridade do
RNA (Fig. 10A) e a não amplificação de GAPDH, gene de controle interno, (Fig.

46
10B) foi preparado cDNA a partir destas amostras para análises de expressão
gênica por RT-PCR qualitativo e RT-PCR em tempo real.
Conforme demonstrado na Figura 11A e B, células B16 co-cultivadas com
células B-1 (B16+B1) apresentaram considerável aumento na expressão de MMP-
9 e CXCR4 comparado com monoculturas de células B16 (B16).
Interessantemente, o aumento na expressão destes genes foi inibido nas células
B16 provenientes de co-culturas com células B-1 tratadas com PD98059
[(B16+B1)+PD]. Em conjunto estes resultados sugerem que o aumento na
fosforilação de ERK induzido pelas células B-1 regula a expressão de genes
relacionados a metástase nas células B16, os quais por sua vez podem contribuir
para aumentar o potencial metastático destas células.
3.7 Níveis basais de fosforilação de ERK nas células B-1 são
necessários para induzir aumento no potencial metastático das células
B16.
Resultados descritos na Figura 6A mostraram que células B-1 ex vivo
apresentam níveis constitutivos de ERK fosforilado. Considerando esta evidência,
foi avaliada se a ativação constitutiva desta quinase nas células B-1 poderia estar
relacionada no comprometimento destas células para induzir aumento no potencial
metastático das células B16.
Trabalhos na literatura mostraram que células B-1 produzem e utilizam IL-
10 como fator de crescimento autócrino (O`Garra et al., 1992). Além disso,
existem vários relatos mostrando que esta citocina foi capaz de promover ativação

47
de ERK, enquanto outros relacionaram a eficiente transcrição de IL-10 com a
atividade de ERK (Zhou et al., 2001; Lee et al., 2002; Niemand et al., 2003; Mathur
et al., 2004). Baseado nestas premissas, foi determinado o perfil de fosforilação de
ERK em células B-1 provenientes de camundongos knock-out para IL-10 (B-1
KO). Como pode ser observado na Figura 12, níveis de IL-10 foram detectados no
sobrenadante de culturas de células B-1 de animais wilde-type (B1 wt), enquanto
que no sobrenandante de culturas de células B-1 KO (B1KO) essa citocina não
apresentou níveis detectáveis. Interessantemente, os níveis de fosforilação de
ERK foram notavelmente menores em células B-1 KO, comparado com os níveis
observados em células B-1 wt (Fig. 13A), mostrando assim diferença nos níveis
constitutivos de ERK fosforilado nestas duas células.
Por outro lado, levando em conta relatos da literatura que mostram a
contribuição de ERK para a fosforilação do resíduo serina 727 do fator de
transcrição Stat3 (Signal transducer and activator of transcription 3; Kuroki &
O`Flaherty, 1999; Kakisis et al., 2005) foi avaliado se os níveis baixos de ERK
fosforilado encontrados nas células B-1 KO poderiam influenciar os níveis de
fosforilação de Stat3 nestas células. Entretanto, conforme mostrado na Figura 13B
os níveis de Stat3 fosforilado (resíduo serina727) encontrados nas células B-1 KO
foram equivalentes aos níveis das células B-1 wt.
Em seguida, foi investigado o efeito das células B-1 KO nas células B16
após co-cultivo de ambas as células. Quando os níveis de fosforilação de ERK nas
células B16 submetidas ao co-cultivo com células B-1 KO (B16+B1KO) foram
avaliados estes permaneceram basais, enquanto que nas células B16 de co-
culturas com células B-1 wt (B16+B1wt) os níveis de fosforilação de ERK foram

48
aumentados (Fig. 13C). Por outro lado, o incremento nos níveis de fosforilação de
ERK foi abolido quando as células B-1 wt foram pré-tratadas com PD98059 antes
do co-cultivo com células B16 (Fig. 13C).
Corroborando com os resultados acima, ensaios experimentais de
metástase (Fig. 13D) demonstraram que injeção de células B16 co-cultivadas com
células B-1 wt (B16+B1wt), mas não com células B-1 KO (B16+B1KO) ou células
B-1 wt pré-tratadas com PD98059 antes do co-cultivo [B16+(B1+PD)], induziram
aumento no número de nódulos metastáticos pulmonares. Estes resultados
sustentam a hipótese que níveis constitutivos de ERK fosforilado nas células B-1
participam no comprometimento destas células para induzir aumento no potencial
metastáticos das células B16.
Adicionalmente, o status de ativação de Stat3 nas células B16 co-cultivadas
ou não com células B-1 wt ou B-1 KO foi analisado também por Immunoblot.
Conforme mostrado na Figura 14, níveis basais equivalentes de fosforilação de
Stat3 foram observados nas células B16 cultivadas tanto na ausência quanto na
presença de soro. Entretanto, quando células B16 foram co-cultivadas com células
B-1 wt ou KO, aumento nos níveis de fosforilação de Stat3 foi observado nas
células B16. Juntos estes resultados mostram que, a pesar do co-cultivo com
células B-1 wt e B-1 KO aumentarem os níveis de fosforilação de Stat3 nas células
B16, este aumento não correlaciona com o aumento do potencial metastático das
células B16, uma vez que o co-cultivo destas células com células B-1 KO não
potencializou a capacidade metastática das células B16 (Fig. 13D).

49
3.8 Níveis basais de fosforilação de ERK nas células B16 são
inalterados pelo estímulo com IL-10
Considerando que a IL-10 pode promover aumento nos níveis de
fosforilação de ERK via ativação do receptor de IL-10 (Zhou et al., 2001; Lee et al.,
2002), foi descartada a possível participação da IL-10, produzida pelas células B-1
wt, no aumento dos níveis de fosforilação de ERK nas células B16 submetidas ao
co-cultivo. Para checar isto, células B16 foram tratadas com diferentes
concentrações de IL-10 recombinante (IL-10r) por diferentes tempos. Como
apresentado na Figura 15A, os tratamentos com IL-10r não foram capazes de
aumentar os níveis basais de fosforilação de ERK nas células B16. Além disso, a
inabilidade da IL-10r em incrementar os níveis de fosfo-ERK nas células B-16,
correlacionou com a incapacidade desta citocina ou do meio condicionado de
culturas de células B-1 wt de aumentar a capacidade metastática das células B16
(Fig. 15B). Comparativamente, células B16 submetidas ao co-cultivo com células
B-1 wt produziram maior número de nódulos pulmonares.
Adicionalmente, foi verificado se o tratamento de células B-1 KO na
presença de IL-10r poderia mimetizar os efeitos das células B-1 wt no aumento do
potencial metastático das células B16 durante o co-cultivo. Para avaliar esta
hipótese, células B16 (B16) foram co-cultivadas ou não com células B-1 wt
(B16+B1wt) ou com células B-1 KO na ausência (B16+B1KO) ou presença de 10
ng/ml de IL-10r [(B16+B1KO)+IL-10r] por 72 horas. Adicionalmente, células B-1
KO foram pré-tratadas com 10 ng/ml de IL-10r [B16+(B1KO+IL-10r)] ou com meio
condicionado de culturas de células B-1 wt [B16+(B1KO+mB1wt)] antes do co-

50
cultivo. Após estes tratamentos, células B16 foram coletadas e submetidas a
ensaios de metástase experimental in vivo. Conforme observado na Figura 16A,
células B16 co-cultivadas com células B-1 KO submetidas aos tratamentos com
IL-10r ou com meio condicionado de células B-1 wt, produziram número de
nódulos metastáticos similares àqueles produzidos por células B16 de
monoculturas, enquanto células B16 co-cultivadas com células B-1 wt foram
significativamente mais metastáticas.
Uma vez que o tratamento de células B-1 KO com IL-10r não mimetizou o
efeito das células B-1 wt no aumento do potencial metastático das células B16, foi
verificado se existiam diferenças na expressão do receptor de IL-10 entre essas
células. Para isso, utilizando RT-PCR qualitativo foi determinada a expressão de
mRNA do receptor de IL-10 nas células B-1 wt e KO. Conforme observado na
Figura 16B a expressão de mRNA do receptor de IL-10 (IL-10R) nas células B-1
KO é quase inexistente comparado com os níveis de expressão das células B-1
wt.
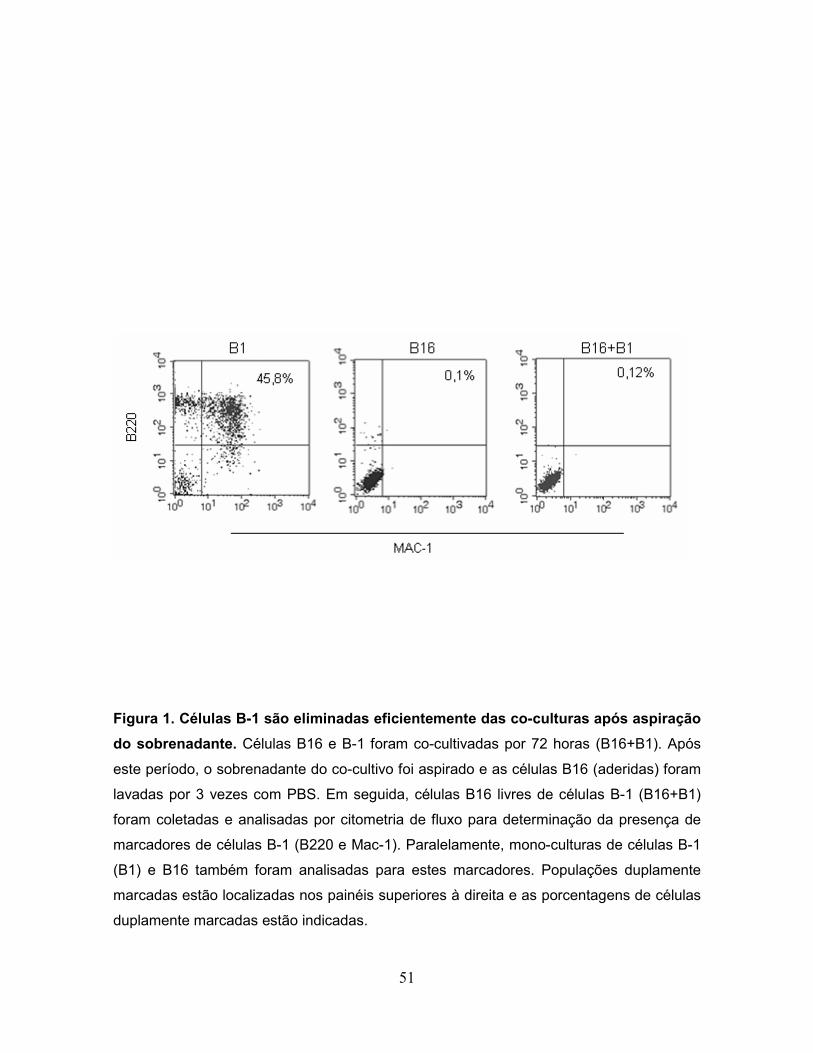
51
Figura 1. Células B-1 são eliminadas eficientemente das co-culturas após aspiração
do sobrenadante. Células B16 e B-1 foram co-cultivadas por 72 horas (B16+B1). Após
este período, o sobrenadante do co-cultivo foi aspirado e as células B16 (aderidas) foram
lavadas por 3 vezes com PBS. Em seguida, células B16 livres de células B-1 (B16+B1)
foram coletadas e analisadas por citometria de fluxo para determinação da presença de
marcadores de células B-1 (B220 e Mac-1). Paralelamente, mono-culturas de células B-1
(B1) e B16 também foram analisadas para estes marcadores. Populações duplamente
marcadas estão localizadas nos painéis superiores à direita e as porcentagens de células
duplamente marcadas estão indicadas.
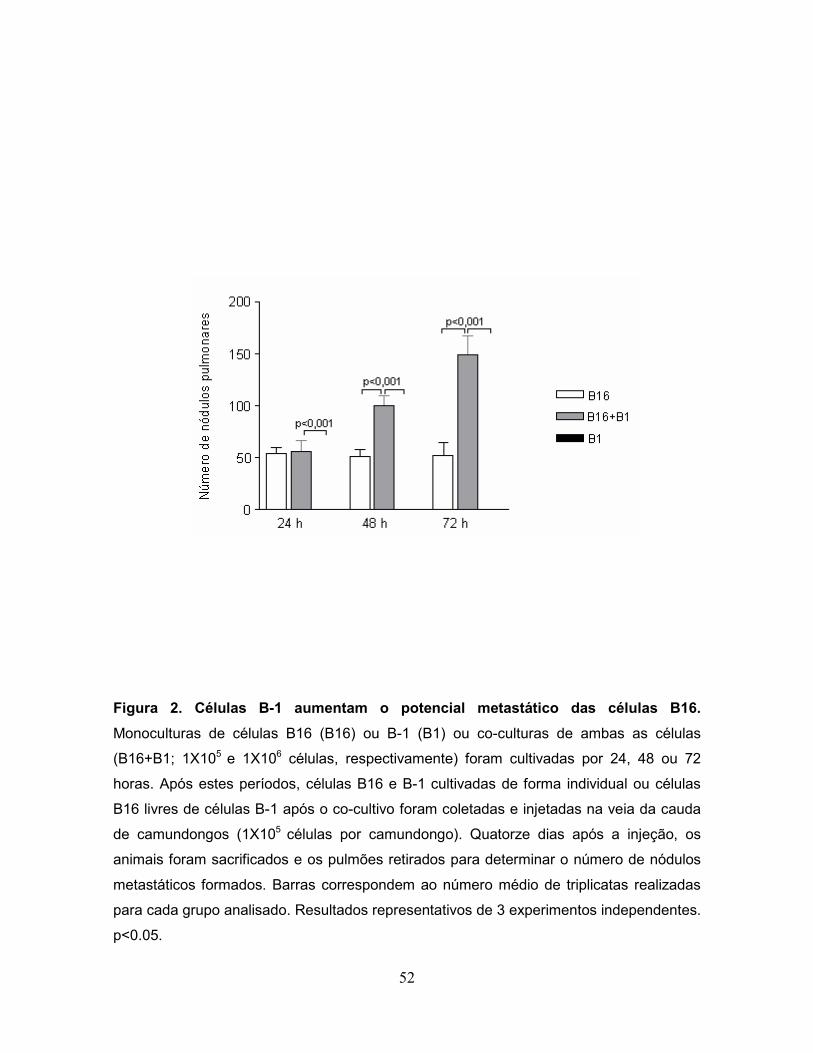
52
Figura 2. Células B-1 aumentam o potencial metastático das células B16.
Monoculturas de células B16 (B16) ou B-1 (B1) ou co-culturas de ambas as células
(B16+B1; 1X105 e 1X106 células, respectivamente) foram cultivadas por 24, 48 ou 72
horas. Após estes períodos, células B16 e B-1 cultivadas de forma individual ou células
B16 livres de células B-1 após o co-cultivo foram coletadas e injetadas na veia da cauda
de camundongos (1X105 células por camundongo). Quatorze dias após a injeção, os
animais foram sacrificados e os pulmões retirados para determinar o número de nódulos
metastáticos formados. Barras correspondem ao número médio de triplicatas realizadas
para cada grupo analisado. Resultados representativos de 3 experimentos independentes.
p<0.05.
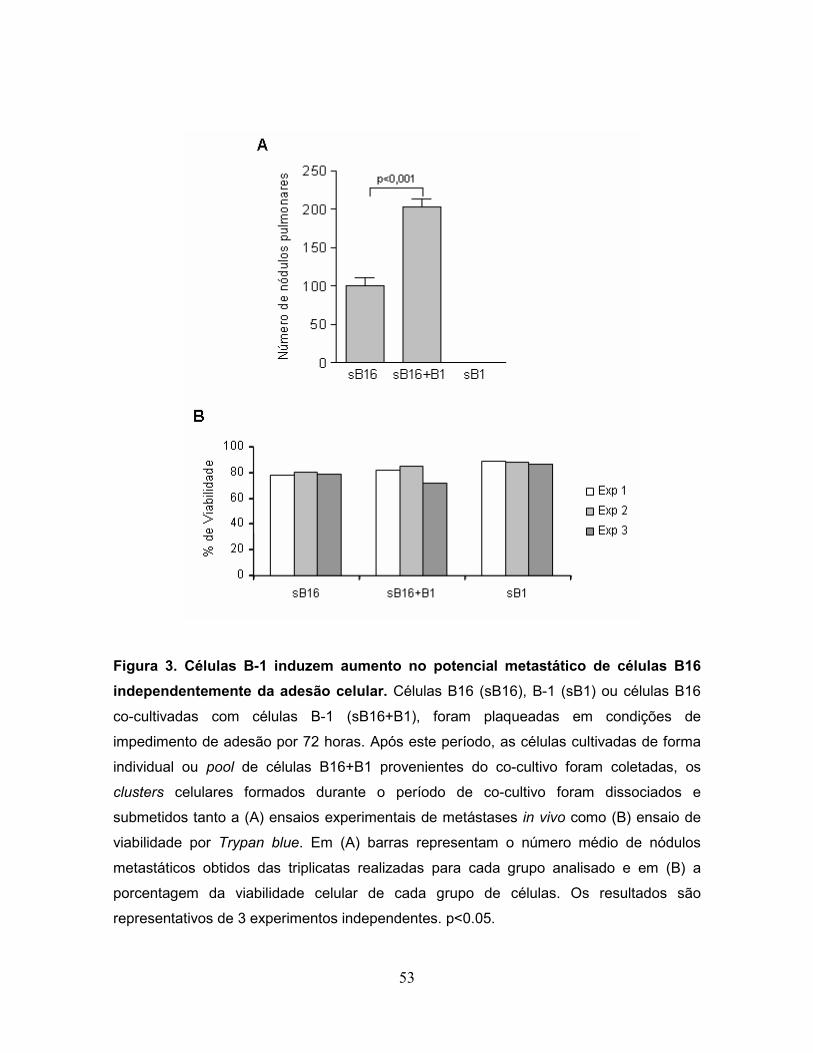
53
Figura 3. Células B-1 induzem aumento no potencial metastático de células B16
independentemente da adesão celular. Células B16 (sB16), B-1 (sB1) ou células B16
co-cultivadas com células B-1 (sB16+B1), foram plaqueadas em condições de
impedimento de adesão por 72 horas. Após este período, as células cultivadas de forma
individual ou pool de células B16+B1 provenientes do co-cultivo foram coletadas, os
clusters celulares formados durante o período de co-cultivo foram dissociados e
submetidos tanto a (A) ensaios experimentais de metástases in vivo como (B) ensaio de
viabilidade por Trypan blue. Em (A) barras representam o número médio de nódulos
metastáticos obtidos das triplicatas realizadas para cada grupo analisado e em (B) a
porcentagem da viabilidade celular de cada grupo de células. Os resultados são
representativos de 3 experimentos independentes. p<0.05.

54
Figura 4. Fatores solúveis das células B-1 não contribuem para o aumento da
capacidade metastática das células B16. Células B16 foram cultivadas por 72 horas em
seu próprio meio (B16) ou co-cultivadas com células B-1 (B16+B1) ou cultivadas em meio
condicionado tanto de monoculturas de 5 dias de células B-1 (B16+mB1) como de co-
cultura de células B16 com B-1 [B16+m(B16+B1)]. Após este período, células B16 de
monoculturas ou do co-cultivo com células B-1 foram coletadas e submetidas a ensaios
experimentais de metástases in vivo. Barras correspondem ao número médio de
triplicatas realizadas para cada grupo analisado. Resultados são representativos de 3
experimentos independentes. p<0.05.
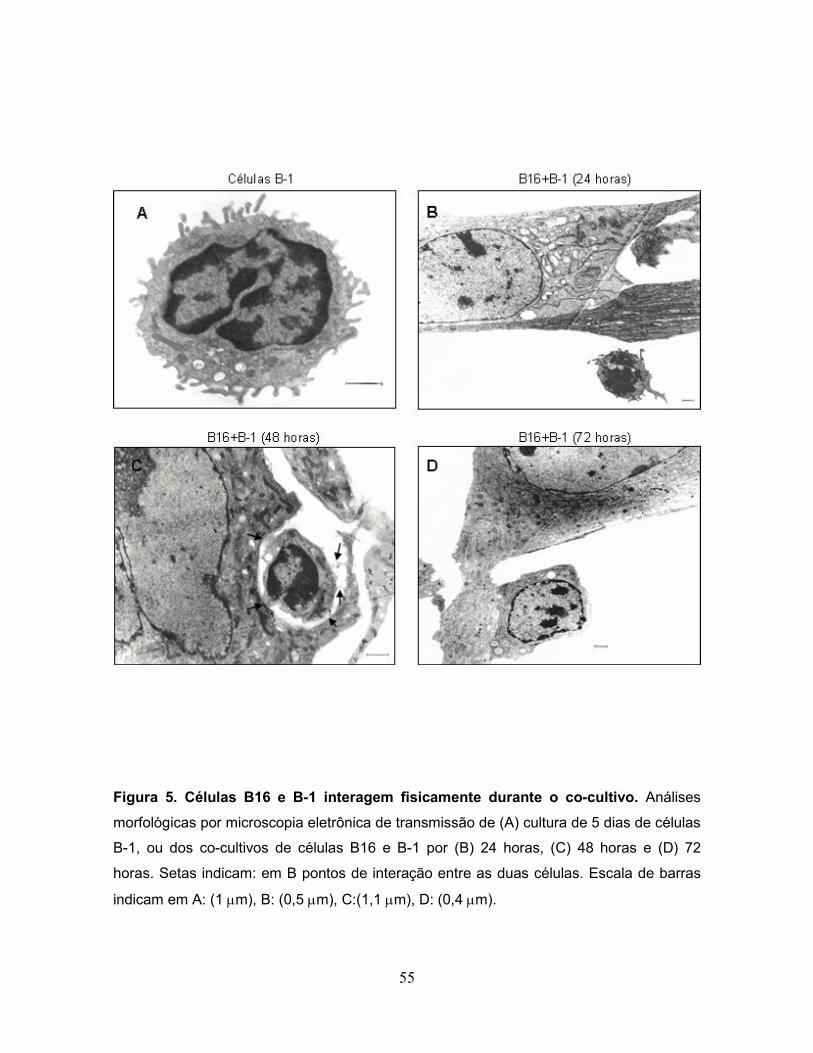
55
Figura 5. Células B16 e B-1 interagem fisicamente durante o co-cultivo. Análises
morfológicas por microscopia eletrônica de transmissão de (A) cultura de 5 dias de células
B-1, ou dos co-cultivos de células B16 e B-1 por (B) 24 horas, (C) 48 horas e (D) 72
horas. Setas indicam: em B pontos de interação entre as duas células. Escala de barras
indicam em A: (1 µm), B: (0,5 µm), C:(1,1 µm), D: (0,4 µm).

56
Figura 6. Células B-1 induzem aumento nos níveis de fosforilação de ERK nas
células B16. (A) Monoculturas de células B16 e B-1 foram mantidas por 24 horas em
meio sem soro e estimuladas ou não com 10% de soro bovino fetal (SBF) por 1 hora (1 h),
conforme indicado. (B) Células B16 mantidas por 24 h sem soro foram estimuladas ou
não com 10% SFB por 1 h e em cada uma destas condições as células B16 foram co-
cultivadas ou não com células B-1 (B16+B-1), como indicado. Após este período, extratos
totais de monoculturas de células B16 e de células B16 livres de células B-1 após o co-
cultivo foram produzidos e submetidos a Immunobloting para detecção dos níveis de ERK
fosforilado (p-ERK). Níveis de ERK total foram analisados como controle interno dos
níveis protéicos aplicados no gel.

57
Figura 7. Níveis basais de fosforilação de ERK em células B16 e B-1 são inibidos
pelo tratamento com PD98059. (A) Monoculturas de células B16 e B-1 foram cultivadas
em meio completo e tratadas ou não com 20 ou 40 µM de PD98059 por 24 horas. Após
este período extratos totais foram preparados e analisados por Western Blot para a
detecção dos níveis de ERK fosforilado. Níveis de ERK total foram analisados como
controle interno dos níveis protéicos aplicados no gel.

58
Figura 8. O aumento no potencial metastático das células B16 induzido pelo contato
com células B-1 é dependente dos níveis de ERK fosforilado. Monoculturas de células
B16 foram mantidas em meio completo na ausência (B16) ou presença de 40 µM de
PD98059 (B16+PD) por 72 horas (72 h). Para co-culturas, células B16 e B-1 foram co-
cultivadas na ausência (B16+B-1) ou presença de 40 µM de PD98059 por 72 h [(B16+B1)
+PD]. Adicionalmente, monoculturas de células B16 foram pré-tratadas 24 h com 40 µM
de PD98059 e em seguida, co-cultivadas com células B-1 por 72 h na ausência do inibidor
[(B16+PD) +B1]. Após este período, células B16 das monoculturas ou células B16 livres
de células B-1 após o co-cultivo foram coletadas e submetidas tanto a (A) análises por
Western blot para detecção dos níveis de ERK fosforilado, como (B) ensaios
experimentais de metástases in vivo. Barras correspondem ao número médio de nódulos
obtidos das triplicatas realizadas para cada grupo analisado. Estes resultados são
representativos de 3 experimentos independentes. p<0.05.

59
Figura 9. Células B-1 não induzem aumento na proliferação das células B16. Células
B16 cultivadas em meio completo na ausência (B16) ou presença de células B-1
(B16+B1) ou do meio condicionado de cultura de células B-1 de 5 dias (B16+mB1) foram
incubadas durante 24, 48 e 72 horas. Após este período, células B16 (aderidas) foram
submetidas a ensaios de proliferação: (A) MTT e (B) incorporação de timidina tritiada.
Barras correspondem ao valor médio das triplicatas. Resultados representativos de 4
experimentos independentes. DO=Densidade Ótica, cpm=contas por minuto.
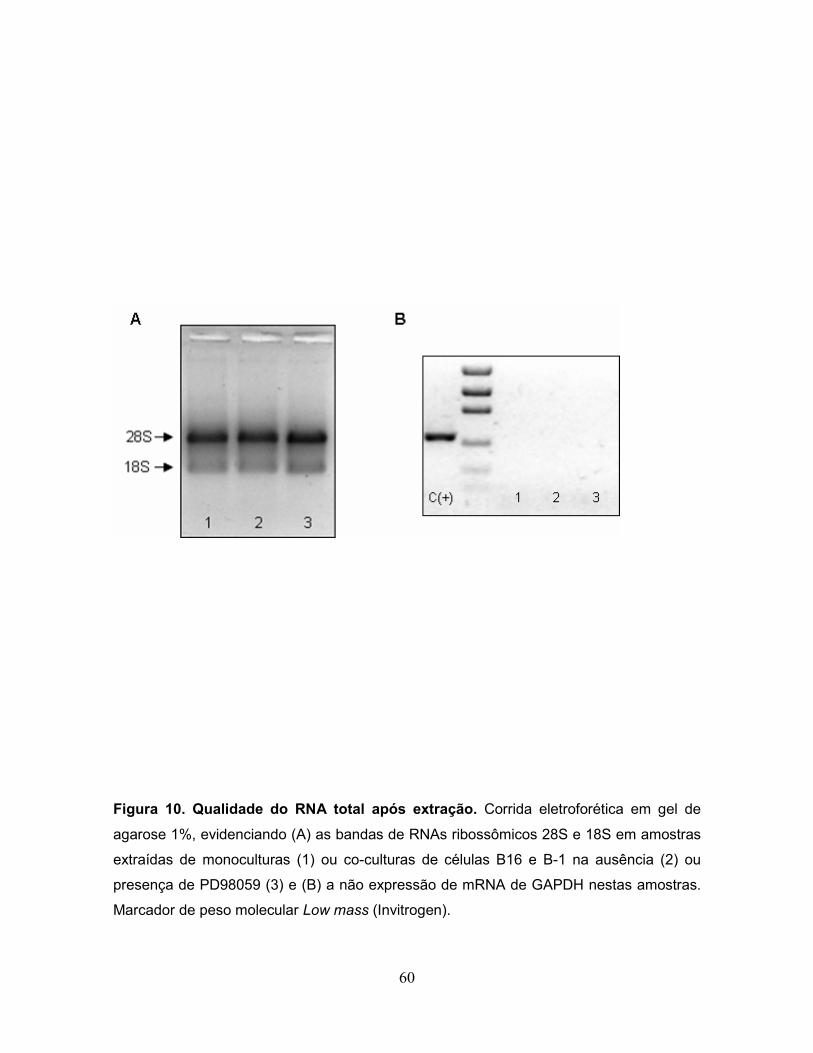
60
Figura 10. Qualidade do RNA total após extração. Corrida eletroforética em gel de
agarose 1%, evidenciando (A) as bandas de RNAs ribossômicos 28S e 18S em amostras
extraídas de monoculturas (1) ou co-culturas de células B16 e B-1 na ausência (2) ou
presença de PD98059 (3) e (B) a não expressão de mRNA de GAPDH nestas amostras.
Marcador de peso molecular Low mass (Invitrogen).
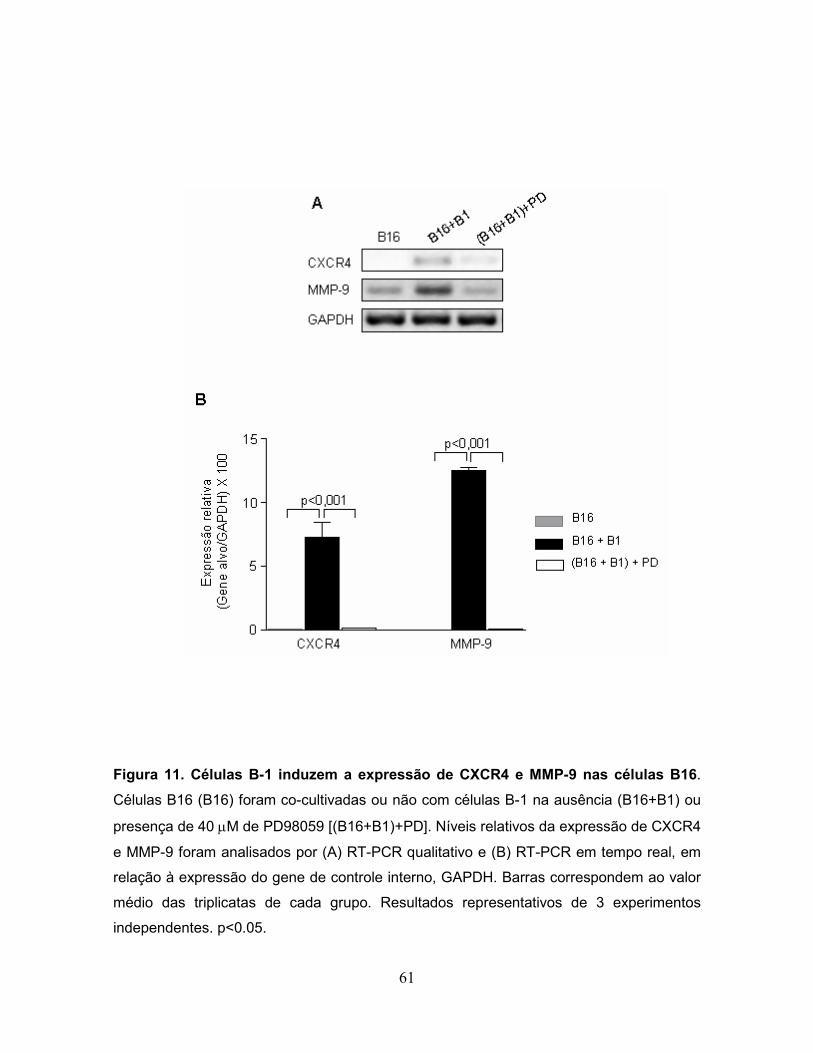
61
Figura 11. Células B-1 induzem a expressão de CXCR4 e MMP-9 nas células B16.
Células B16 (B16) foram co-cultivadas ou não com células B-1 na ausência (B16+B1) ou
presença de 40 µM de PD98059 [(B16+B1)+PD]. Níveis relativos da expressão de CXCR4
e MMP-9 foram analisados por (A) RT-PCR qualitativo e (B) RT-PCR em tempo real, em
relação à expressão do gene de controle interno, GAPDH. Barras correspondem ao valor
médio das triplicatas de cada grupo. Resultados representativos de 3 experimentos
independentes. p<0.05.
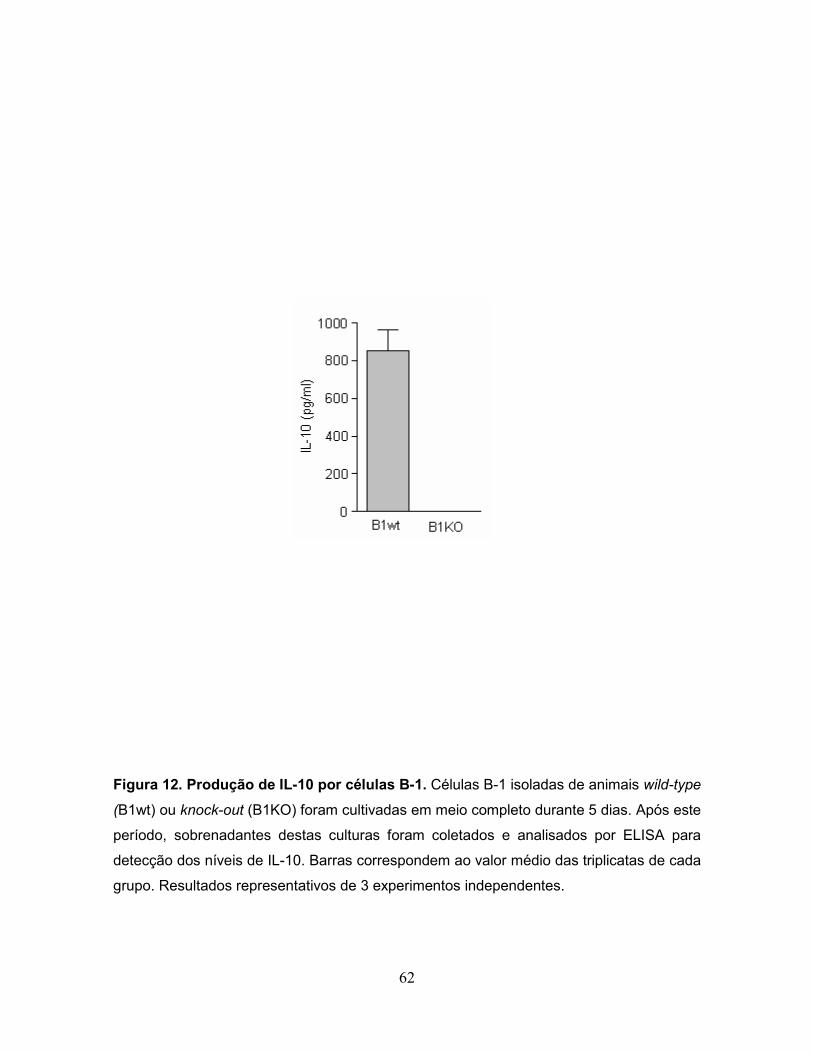
62
Figura 12. Produção de IL-10 por células B-1. Células B-1 isoladas de animais wild-type
(B1wt) ou knock-out (B1KO) foram cultivadas em meio completo durante 5 dias. Após este
período, sobrenadantes destas culturas foram coletados e analisados por ELISA para
detecção dos níveis de IL-10. Barras correspondem ao valor médio das triplicatas de cada
grupo. Resultados representativos de 3 experimentos independentes.

63
Figura 13. Níveis basais de fosforilação de ERK nas células B-1 são necessários
para induzir aumento no potencial metastático das células B16. Extratos totais de (A
e B) monoculturas de 5 dias de células B-1 tanto de animais wild-type (B1wt) como knock-
out para IL-10 (B1KO) ou (C) células B16 de monoculturas (B16) ou co-cultivadas por 72
horas com células B-1wt (B16+B1wt), B-1KO (B16+B1KO) ou com B-1wt pré-tratadas
com 40 µM de PD98059 [B16+(B1wt+PD)], foram submetidas à análises por Western blot
para detecção dos níveis de ERK fosforilado (A e C) ou fosforilação do resíduo serina 727
da Stat3 (B). Níveis de ERK total e Stat3 total foram usados para confirmar quantidades
iguais dos extratos celulares. (D) Células B16 foram tratadas como descrito em C e
posteriormente submetidas a ensaio de metástases experimental in vivo. Barras
correspondem ao número médio de nódulos obtidos das triplicatas realizadas para cada
grupo analisado. Resultados representativos de 3 experimentos independentes. p<0.05.

64
Figura 14. Níveis de fosforilação de Stat3 nas células B16 antes e após co-cultivo
com células B-1. Extratos totais de células B16 mantidas 24 horas na ausência de soro
(SBF) e em seguida estimuladas com SBF ou co-cultivadas com células B-1 de animais
tanto wild-type (B16+B1wt) como knock-out para IL-10 (B16+B1KO) foram submetidos a
Western blot para verificar a fosforilação da serina 727 da Stat3. Níveis de Stat3 total
foram usados para confirmar quantidades iguais dos extratos celulares. Resultados
representativos de 3 experimentos independentes.
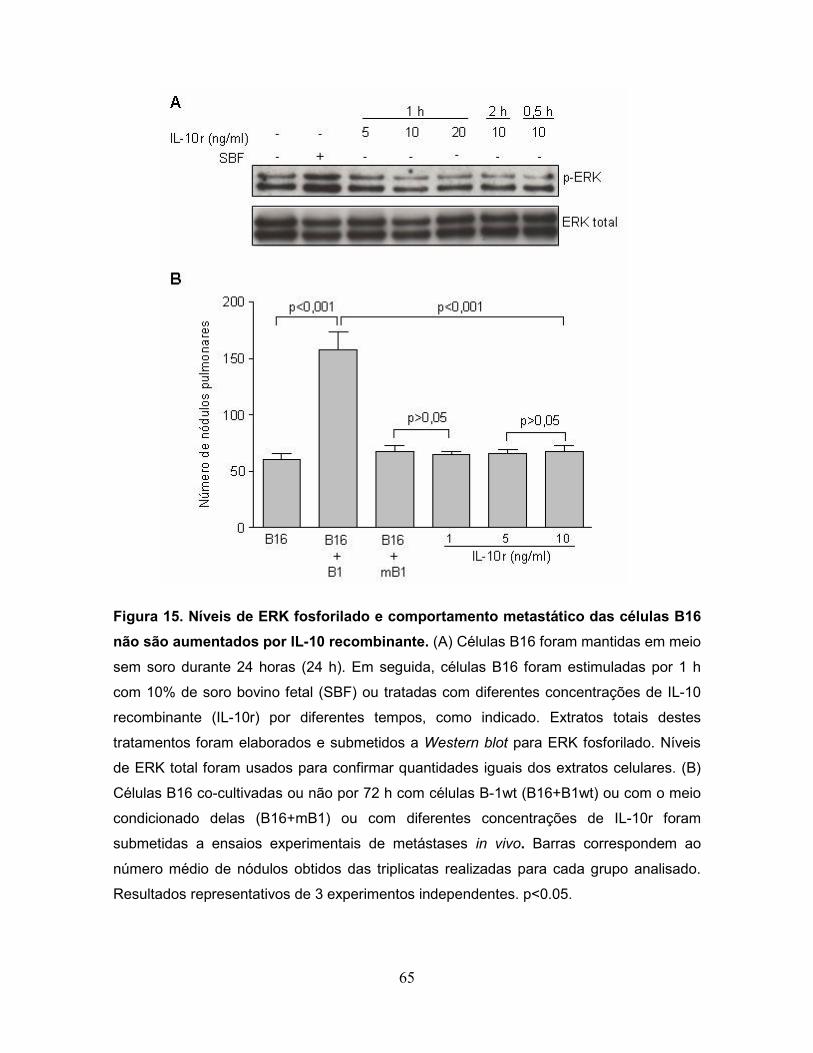
65
Figura 15. Níveis de ERK fosforilado e comportamento metastático das células B16
não são aumentados por IL-10 recombinante. (A) Células B16 foram mantidas em meio
sem soro durante 24 horas (24 h). Em seguida, células B16 foram estimuladas por 1 h
com 10% de soro bovino fetal (SBF) ou tratadas com diferentes concentrações de IL-10
recombinante (IL-10r) por diferentes tempos, como indicado. Extratos totais destes
tratamentos foram elaborados e submetidos a Western blot para ERK fosforilado. Níveis
de ERK total foram usados para confirmar quantidades iguais dos extratos celulares. (B)
Células B16 co-cultivadas ou não por 72 h com células B-1wt (B16+B1wt) ou com o meio
condicionado delas (B16+mB1) ou com diferentes concentrações de IL-10r foram
submetidas a ensaios experimentais de metástases in vivo. Barras correspondem ao
número médio de nódulos obtidos das triplicatas realizadas para cada grupo analisado.
Resultados representativos de 3 experimentos independentes. p<0.05.
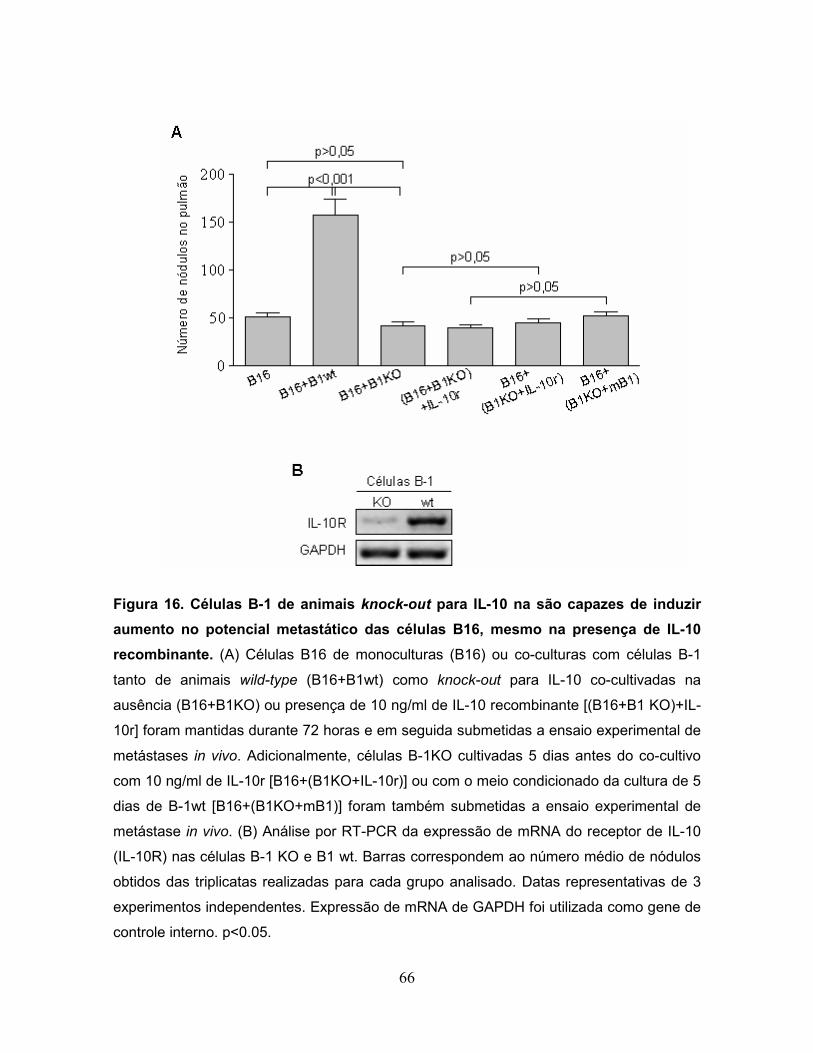
66
Figura 16. Células B-1 de animais knock-out para IL-10 na são capazes de induzir
aumento no potencial metastático das células B16, mesmo na presença de IL-10
recombinante. (A) Células B16 de monoculturas (B16) ou co-culturas com células B-1
tanto de animais wild-type (B16+B1wt) como knock-out para IL-10 co-cultivadas na
ausência (B16+B1KO) ou presença de 10 ng/ml de IL-10 recombinante [(B16+B1 KO)+IL-
10r] foram mantidas durante 72 horas e em seguida submetidas a ensaio experimental de
metástases in vivo. Adicionalmente, células B-1KO cultivadas 5 dias antes do co-cultivo
com 10 ng/ml de IL-10r [B16+(B1KO+IL-10r)] ou com o meio condicionado da cultura de 5
dias de B-1wt [B16+(B1KO+mB1)] foram também submetidas a ensaio experimental de
metástase in vivo. (B) Análise por RT-PCR da expressão de mRNA do receptor de IL-10
(IL-10R) nas células B-1 KO e B1 wt. Barras correspondem ao número médio de nódulos
obtidos das triplicatas realizadas para cada grupo analisado. Datas representativas de 3
experimentos independentes. Expressão de mRNA de GAPDH foi utilizada como gene de
controle interno. p<0.05.

5. DISCUSSÃO

5. DISCUSSÃO
Relatos da literatura têm demonstrado que a expansão metastática de
células de melanoma é um processo altamente complexo dependente de
interações entre as células tumorais e seu estroma (Langley & Fidler, 2007;
Aguirre-Ghiso, 2007). Embora os macrófagos associados ao tumor (TAMs) sejam
a principal população de células inflamatórias presentes no micro-ambiente
tumoral, outros tipos celulares podem também favorecer a formação de
metástases ao interagir de forma direta com as células tumorais e ou indireta pela
produção de fatores solúveis (Balkwill & Mantovani, 2001; Inoue et al., 2006; Croci
et al., 2007; Mantovani et al., 2007; Hofmeister et al., 2008).
Evidências apresentadas no presente estudo mostram que interações in
vitro entre células de melanoma murino B16 e células B-1, subtipo de linfócitos B
presentes nas cavidades pleural e peritoneal, resultam em aumento do potencial
metastático das células de melanoma (Fig.2). Os resultados apresentados
mostraram inequivocamente que interações físicas entre células B16 e B-1, e não
fatores solúveis produzidos pelas células B-1, representam a principal condição
para aumentar a capacidade metastática das células B16 (Fig. 4 e 5),
corroborando dados prévios de nosso grupo, ainda não publicados. Análises de
microscopia eletrônica (Fig. 5), realizadas com intuito de investigar com mais
detalhe o tipo de interação entre essas células durante o co-cultivo, demonstraram
estreita comunicação física entre elas. Esta interação, entretanto, não sugeriu

69
fusão celular, já que os contornos das membranas plasmáticas de cada célula
apresentaram-se claramente delimitados.
Um dos resultados inéditos deste trabalho mostrou que o aumento do
potencial metastático das células B16 induzido pelo contato com células B1 está
associado a aumento na ativação de ERK. Análises de Western blot evidenciaram
que o contato de células B-1 induz aumento nos níveis de fosforilação de ERK nas
células B16 (Fig. 6B). Interessantemente, quando o aumento da fosforilação de
ERK foi bloqueada pelo inibidor farmacológico de MEK (PD98059), quinase up-
stream de ERK, o aumento da capacidade metastática das células B16 foi inibido
(Fig. 8B). Em contraste, monoculturas de células B16 apresentam níveis basais
constitutivos de ERK (Fig. 6A), e a completa inibição desta quinase nestas células
não inibiu sua capacidade natural de formar nódulos metastáticos no pulmão (Fig.
8B). Esta observação sugere que o fenótipo intrínseco da linhagem celular B16 de
baixo potencial metastático pode ser sustentado por outra via de sinalização que
independe de background mínimo de ERK fosforilado. Por outro lado, o fato de
que células B-1 não serem capazes de elevar os níveis de ERK fosforilado nem o
potencial metastático das células B16 tratadas previamente com PD98059 sugere
que, os efeitos desencadeados pelas células B-1 dependem de níveis basais de
ERK fosforilado nas células B16.
Vários relatos da literatura sustentam que os substratos de ERK incluem
alvos presentes no citoplasma como também fatores de transcrição tais como as
famílias ELK/SAP, Ets/PEA3, AP-1 e Stat (Hill & Treisman, 1996; Harper &
LoGrasso, 2001; Viala & Pouyssegur, 2004). Algumas destas famílias, como AP-1
e Ets são importantes na regulação da expressão de várias metaloproteinases

70
(MMPs) como MMP-9 e quimiocinas como CXCR4, as quais estão associadas
com progressão metastática em diversos tipos de cânceres entre eles o melanoma
(John & Tuszynski, 2001; Hofmann et al., 2003; Kulbe et al., 2004; Tower et al.,
2002; Kim et al., 2006; Maroni et al., 2007; Ruffini et al., 2007).
Análises de expressão gênica para CXCR4 e MMP-9 por RT-PCR
qualitativo e em tempo real realizadas neste trabalho mostraram aumento
substancial na expressão destes transcritos nas células B16 após contato com
células B-1 (Fig. 11). Interessantemente, estes resultados também mostraram que
o aumento na expressão de mRNA de CXCR4 e MMP-9 é inibido pela adição do
inibidor farmacológico PD98059. Assim como os achados de outros grupos, este
resultado sustenta a participação de ERK na regulação da expressão destes
transcritos (Gum et al., 1997; Simon et al., 1998; Lakka et al., 2000; Maroni et al.,
2007) e ainda, sugere que o aumento na expressão de mRNA de MMP-9 e
CXCR4 nas células B16 induzida pelo co-cultivo com células B-1 pode contribuir
para potencializar a capacidade metastática das células B16 ao favorecer o
extravasamento e migração destas células aos pulmões.
Outro dado importante e inédito de nossa pesquisa foi a necessidade de
níveis constitutivos de ERK fosforilado nas células B-1 para o comprometimento
destas células na indução do aumento no potencial metastático das células B16.
Resultados obtidos mostram que a inibição da fosforilação de ERK nas células B-1
pelo tratamento com PD98059, prejudica consideravelmente a capacidade destas
células de induzir aumento tanto nos níveis de ERK fosforilado como na
capacidade metastática das células B16. Esta observação foi também sustentada
por estudos com células B-1 provenientes de camundongos knock-out para IL-10

71
(KO), as quais apresentaram níveis basais de ERK fosforilado menores dos níveis
encontrados nas células B-1 de animais wild-type (wt) (Fig. 13A). Resultados
obtidos também com células B-1 KO mostraram que estas células não têm
capacidade para aumentar os níveis de ERK fosforilado nem o potencial
metastático das células B16 quando comparadas com células B-1 wt.
Considerando dados da literatura que evidenciam aumento nos níveis de
ERK fosforilado via ativação do receptor de IL-10 (Yi et al., 2002; Mathur et al.,
2004; Liu et al., 2006; Chanteux et al., 2007), é possível especular que os níveis
baixos de ERK fosforilado observados nas células B-1 KO sejam devidos a níveis
também baixos de expressão de mRNA do receptor de IL-10 nestas células (Fig.
16). Embora o nível de fosforilação de ERK nas células B-1 wt após estímulo com
IL-10 recombinante (IL-10r) não tenha sido analisado, os resultados acima
descritos sugerem que o estímulo das células B-1 com IL-10, poderia contribuir
com os níveis basais de fosforilação de ERK observados nestas células. Em
contraste, células B-16 não apresentaram aumento nos níveis de fosforilação de
ERK nem aumento do seu potencial metastático após estímulo com IL-10r ou com
o sobrenadante de cultura de células B1wt (Fig. 15). Estes resultados, em
conjunto, sugerem que IL-10 pode ser um fator importante para manutenção de
níveis constitutivos de ERK nas células B-1 wt, os quais podem favorecer a
expressão de prováveis moléculas de membrana, que ao interagir no co-cultivo
com receptores presentes nas células B16 promovem o crescimento metastático
destas células.
Fosforilação constitutiva de ERK nas células B-1 foi previamente
demonstrada por Wong e colaboradores (2002). Embora a atividade de ERK não

72
seja necessária para a sobrevida destas células em cultura, ainda são
desconhecidos quais aspectos da fisiologia das células B-1 são regulados pela
quinase ERK (Wong et al., 2002). Com base nos resultados aqui apresentados
ainda não foi possível estabelecer porque as células B-1 precisam de altos níveis
de ERK fosforilado para potencializar a capacidade metastática das células B16.
Entretanto, relatos da literatura têm demonstrado que diferenças na duração,
magnitude e distribuição sub-celular da atividade de ERK determinam programas
celulares específicos (Balmanno & Cook, 1999; Sasagawa et al., 2005; Ebisuya et
al., 2005). Assim, considerando os resultados acima descritos que sustentam a
necessidade do contato físico entre células B16 e B-1, é possível especular que
níveis basais de ERK nas células B-1 são requeridos para regular a expressão de
moléculas de superfície nestas células, as quais ao interagirem com células B16
durante o co-cultivo, formam uma rede interativa de sinalização célula-célula.
Por outro lado, além da ativação constitutiva de ERK, células B-1 também
apresentam ativação constitutiva de Stat3 (Signal transducer and activator of
transcription 3), o qual tem papel importante na capacidade de auto-renovação
destas células (Karras et al., 1997). As proteínas STATs são fatores de transcrição
ativados por fosforilação de uma tirosina (Y705) ou uma serina (S727) em
resposta a um ligante extracelular, que pode ser citocina ou fatores de
crescimento (Darnell, et al., 1997; Bromberg, et al., 1999; Ihle, 2001; Chen, et al.,
2004). Considerando relatos da literatura, nos quais foi mostrado que ERK pode
contribuir com a fosforilação do resíduo serina da Stat3 (Kuroki & O`Flaherty,
1999; Kakisis et al., 2005), investigou-se nas células B-1 KO os níveis de
fosforilação de Stat3, visto que estas células apresentam níveis de ERK fosforilado

73
menores quando comparados com os níveis observados nas B-1 wt (Fig. 13A). O
fato que os níveis de Stat3 nas células B-1 KO foram semelhantes aos observados
nas células B-1 wt (Figura 13B) sugere que a ativação do resíduo serina (727) da
Stat3 nestas células independe da atividade de ERK. Por outro lado, considerando
que aumento na ativação de Stat3 foi associado também com progressão
metastática em vários tipos de cânceres, entre eles o melanoma (Gabrilovich et
al., 1996; Mora et al., 2002; Kortylewisky et al., 2005; Lassmann et al., 2007), o
status de ativação deste fator de transcrição foi avaliado nas células B16 antes e
após contato com células B-1. Conforme mostrado na Figura 14, células B16
apresentam níveis basais de Stat3 (resíduo serina 727) e estes níveis são
aumentados após interação com células B-1, sejam KO ou wt. Entretanto, células
B-1 KO não têm capacidade de aumentar nem os níveis de fosforilação de ERK
nem a capacidade metastática das células B16 (Fig 13 C e D, respectivamente).
Embora outros experimentos sejam necessários para excluir a participação de
Stat3 neste modelo de estudo, estes resultados sugerem fortemente que a via da
Stat3 não tem participação no aumento do potencial metastático induzido pelo
contato com células B-1.
Adicionalmente, outras vias de transdução de sinal que atuam sobre
mensageiros intermediários como PKC e Fak (Kahana, et al., 2002; Oka & Kikawa,
2005), cuja up-regulation tem sido associada com invasão e metástases de células
de melanoma, também foram analisadas. Entretanto, resultados obtidos com estas
quinases não levaram a resultados conclusivos da sua participação neste modelo
de estudo.

74
Em resumo os resultados apresentados neste trabalho fornecem as
primeiras evidências do envolvimento da via ERK no aumento da capacidade
metastática das células B16 induzido pelo contato com células B-1. Sugerem que
a up-regulation desta via é um dos mecanismos pelos quais linfócitos B-1 podem
influenciar diretamente no potencial metastático de células tumorais. Além disso,
este trabalho reforça a idéia que as células B-1 podem estar presentes no micro-
ambiente tumoral realizando funções similares aos TAMs ao favorecer a formação
de metástases. Para confirmar esta última hipótese, atualmente nosso grupo
investiga a presença das células B-1 em isolados de tecidos tumorais humanos
com o intuito de determinar se o efeito das células B-1 sobre o comportamento
metastático das células de melanoma apresentadas neste estudo ocorre também
in vivo no contexto dinâmico do micro-ambiente tumoral.

6. CONCLUSÕES

6. CONCLUSÕES
1 – Células B-1 interagem fisicamente com células de melanoma murino
B16 após 48 horas de co-cultivo e esta interação é necessária para o aumento do
potencial metastático das células B16;
2 – A participação da citocina IL-10 ou outro fator solúvel produzido pelas
células B-1 como responsáveis pela indução do aumento tanto dos níveis de
fosforilação de ERK como da capacidade metastática das células B16 foi excluída;
3 – Células B-1 induzem aumento na capacidade metastática das células
B16 ao induzir up-regulation dos níveis basais de ERK fosforilado nestas células;
4 – Níveis basais de ERK fosforilado tanto nas células B16 como nas B-1
são necessários para potencializar o crescimento metastático das células B16
induzido pelo contato com células B-1;
5 – Células B-1 induzem aumento na expressão de genes associados a
progressão metastática, como MMP-9 e CXCR-4 nas células B16;

77
6 – Altos níveis de foforilação de ERK nas células B-1 são necessários para
o comprometimento destas células para induzir aumento no potencial metastático
das células B16;
7 – A fosforilação do resíduo serina da Stat3 nas células B-1 independe dos
níveis de ativação de ERK, nestas células.

7. REFERÊNCIAS BIBLIOGRÁFICAS

7. REFERÊNCIAS BIBLIOGRÁFICAS
- Abrahão TB, Freymüller E, Mortara RA, Lopes JD, Mariano M.Morphological
characterization of mouse B-1 cells.Immunobiology. 2003;208(4):401-11. - Aguirre-Ghiso JA. Models, mechanisms and clinical evidence for cancer dormancy.
Nat Rev Cancer. 2007 Nov;7(11):834-46. - Almand B, Clark JI, Nikitina E, van Beynen J, English NR, Knight SC, Carbone DP,
Gabrilovich DI. Increased production of immature myeloid cells in cancer patients: a mechanism of immunosuppression in cancer. J Immunol. 2001 Jan 1;166(1):678-89.
- Almeida SR, Aroeira LS, Frymuller E, Dias MA, Bogsan CS, Lopes JD, Mariano M.
Mouse B-1 cell-derived mononuclear phagocyte, a novel cellular component of acute non-specific inflammatory exudate. Int Immunol. 2001 Sep;13(9):1193-201.
- Alugupalli KR, Leong JM, Woodland RT, Muramatsu M, Honjo T, Gerstein RM. B1b
lymphocytes confer T cell-independent long-lasting immunity. Immunity. 2004 Sep;21(3):379-90.
- Alugupalli KR, Gerstein RM. Divide and conquer: division of labor by B-1 B cells.
Immunity. 2005 Jul;23(1):1-2. - Alvarado Y, Giles FJ. Ras as a therapeutic target in hematologic malignancies.
Expert Opin Emerg Drugs. 2007 May;12(2):271-84. - Antsiferova YS, Sotnikova NY, Posiseeva LV, Shor AL. Changes in the T-helper
cytokine profile and in lymphocyte activation at the systemic and local levels in women with endometriosis. Fertil Steril. 2005 Dec;84(6):1705-11.
- Aramaki M, Nagasawa T, Koseki T, Ishikawa I. Presence of activated B-1 cells in
chronic inflamed gingival tissue. J Clin Immunol. 1998 Nov;18(6):421-9. - Axelsen JB, Lotem J, Sachs L, Domany E. Genes overexpressed in different
human solid cancers exhibit different tissue-specific expression profiles. Proc Natl Acad Sci U S A. 2007 Aug 7;104(32):13122-7.
- Balkwill F, Mantovani A. Inflammation and cancer: back to Virchow? Lancet. 2001
Feb 17;357(9255):539-45. - Balmanno K, Cook SJ.Sustained MAP kinase activation is required for the
expression of cyclin D1, p21Cip1 and a subset of AP-1 proteins in CCL39 cells. Oncogene. 1999 May 20;18(20):3085-97.
- Barleon B, Sozzani S, Zhou D, Weich HA, Mantovani A, Marmé D. Migration of
human monocytes in response to vascular endothelial growth factor (VEGF) is mediated via the VEGF receptor flt-1. Blood. 1996 Apr 15;87(8):3336-43.

80
- Baumgarth N, Tung JW, Herzenberg LA. Inherent specificities in natural antibodies: a key to immune defense against pathogen invasion. Springer Semin Immunopathol. 2005 Mar;26(4):347-62.
- Berland R, Wortis HH. Origins and functions of B-1 cells with notes on the role of
CD5. Annu Rev Immunol. 2002;20:253-300. - Bogsan CS, Novaes e Brito RR, Palos Mda C, Mortara RA, Almeida SR, Lopes JD,
Mariano M. B-1 cells are pivotal for in vivo inflammatory giant cell formation. Int J Exp Pathol. 2005 Aug;86(4):257-65.
- Boumsell L, Bernard A, Lepage V, Degos L, Lemerle J, Dausset J. Some chronic
lymphocytic leukemia cells bearing surface immunoglobulins share determinants with T cells. Eur J Immunol. 1978 Dec;8(12):900-4.
- Bradford MM.A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976 May 7;72:248-54.
- Bromberg JF, Wrzeszczynska MH, Devgan G, Zhao Y, Pestell RG, Albanese C,
Darnell JE Jr. Stat3 as an oncogene. Cell. 1999 Aug 6;98(3):295-303. - Bronte V, Chappell DB, Apolloni E, Cabrelle A, Wang M, Hwu P, Restifo NP.
Unopposed production of granulocyte-macrophage colony-stimulating factor by tumors inhibits CD8+ T cell responses by dysregulating antigen-presenting cell maturation. J Immunol. 1999 May 15;162(10):5728-37.
- Bronte V, Serafini P, Apolloni E, Zanovello P. Tumor-induced immune dysfunctions
caused by myeloid suppressor cells. J Immunother (1997). 2001 Nov-Dec;24(6):431-46. - Buettner R, Mora LB, Jove R. Activated STAT signaling in human tumors provides
novel molecular targets for therapeutic intervention. Clin Cancer Res. 2002 Apr;8(4):945-54.
- Bunt SK, Sinha P, Clements VK, Leips J, Ostrand-Rosenberg S. Inflammation
induces myeloid-derived suppressor cells that facilitate tumor progression. J Immunol. 2006 Jan 1;176(1):284-90.
- Bustin SA, Benes V, Nolan T, Pfaffl MW. Quantitative real-time RT-PCR--a
perspective. J Mol Endocrinol. 2005 Jun;34(3):597-601. - Casola S, Otipoby KL, Alimzhanov M, Humme S, Uyttersprot N, Kutok JL, Carroll
MC, Rajewsky K. B cell receptor signal strength determines B cell fate. Nat Immunol. 2004 Mar;5(3):317-27.
- Chanteux H, Guisset AC, Pilette C, Sibille Y. LPS induces IL-10 production by
human alveolar macrophages via MAPKinases- and Sp1-dependent mechanisms. Respir Res. 2007 Oct 4;8(1):71.

81
- Chen W, Daines MO, Khurana Hershey GK.Turning off signal transducer and activator of transcription (STAT): the negative regulation of STAT signaling.J Allergy Clin Immunol. 2004 Sep;114(3):476-89; quiz 490.
- Chevallier N, Berthelemy M, Le Rhun D, Laine V, Levy D, Schwartz-Cornil I. Bovine
leukemia virus-induced lymphocytosis and increased cell survival mainly involve the CD11b+ B-lymphocyte subset in sheep. J Virol. 1998 May;72(5):4413-20.
- Choo MK, Sakurai H, Koizumi K, Saiki I. Stimulation of cultured colon 26 cells with
TNF-alpha promotes lung metastasis through the extracellular signal-regulated kinase pathway. Cancer Lett. 2005 Dec 8;230(1):47-56.
- Christofori G, Semb H. The role of the cell-adhesion molecule E-cadherin as a
tumour-suppressor gene. Trends Biochem Sci. 1999 Feb;24(2):73-6. - Chudnovsky Y, Khavari PA, Adams AE. Melanoma genetics and the development
of rational therapeutics. J Clin Invest. 2005 Apr;115(4):813-24. - Clark WH Jr, Elder DE, Guerry D 4th, Epstein MN, Greene MH, Van Horn M. A
study of tumor progression: the precursor lesions of superficial spreading and nodular melanoma. Hum Pathol. 1984 Dec;15(12):1147-65.
- Cobb MH, Goldsmith EJ. How MAP kinases are regulated. J Biol Chem. 1995 Jun
23;270(25):14843-6. - Cohen C, Zavala-Pompa A, Sequeira JH, Shoji M, Sexton DG, Cotsonis G,
Cerimele F, Govindarajan B, Macaron N, Arbiser JL. Mitogen-actived protein kinase activation is an early event in melanoma progression. Clin Cancer Res. 2002 Dec;8(12):3728-33.
- Condeelis J, Pollard JW. Macrophages: obligate partners for tumor cell migration,
invasion, and metastasis. Cell. 2006 Jan 27;124(2):263-6. - Coussens LM, Raymond WW, Bergers G, Laig-Webster M, Behrendtsen O, Werb
Z, Caughey GH, Hanahan D. Inflammatory mast cells up-regulate angiogenesis during squamous epithelial carcinogenesis. Genes Dev. 1999 Jun 1;13(11):1382-97.
- Coussens LM, Tinkle CL, Hanahan D, Werb Z. MMP-9 supplied by bone marrow-
derived cells contributes to skin carcinogenesis. Cell. 2000 Oct 27;103(3):481-90. - Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002 Dec 19-
26;420(6917):860-7. - Cowley S, Paterson H, Kemp P, Marshall CJ. Activation of MAP kinase kinase is
necessary and sufficient for PC12 differentiation and for transformation of NIH 3T3 cells. Cell. 1994 Jun 17;77(6):841-52.
- Crews CM, Alessandrini A, Erikson RL. The primary structure of MEK, a protein
kinase that phosphorylates the ERK gene product. Science. 1992 Oct 16;258(5081):478-80.

82
- Croci DO, Zacarias Fluck MF, Rico MJ, Matar P, Rabinovich GA, Scharovsky OG. Dynamic cross-talk between tumor and immune cells in orchestrating the immunosuppressive network at the tumor microenvironment. Cancer Immunol Immunother. 2007 Nov;56(11):1687-700.
- D'Angelo G, Struman I, Martial J, Weiner RI. Activation of mitogen-activated protein
kinases by vascular endothelial growth factor and basic fibroblast growth factor in capillary endothelial cells is inhibited by the antiangiogenic factor 16-kDa N-terminal fragment of prolactin. Proc Natl Acad Sci U S A. 1995 Jul 3;92(14):6374-8.
- Darnell JE Jr. STATs and gene regulation. Science. 1997 Sep 12;277(5332):1630-
5. - Davies H, Bignell GR, Cox C, Stephens P, Edkins S, Clegg S, Teague J, Woffendin
H, Garnett MJ, Bottomley W, Davis N, Dicks E, Ewing R, Floyd Y, Gray K, Hall S, Hawes R, Hughes J, Kosmidou V, Menzies A, Mould C, Parker A, Stevens C, Watt S, Hooper S, Wilson R, Jayatilake H, Gusterson BA, Cooper C, Shipley J, Hargrave D, Pritchard-Jones K, Maitland N, Chenevix-Trench G, Riggins GJ, Bigner DD, Palmieri G, Cossu A, Flanagan A, Nicholson A, Ho JW, Leung SY, Yuen ST, Weber BL, Seigler HF, Darrow TL, Paterson H, Marais R, Marshall CJ, Wooster R, Stratton MR, Futreal PA. Mutations of the BRAF gene in human cancer. Nature. 2002 Jun 27;417(6892):949-54.
- Deryugina EI, Quigley JP. Matrix metalloproteinases and tumor metastasis. Cancer
Metastasis Rev. 2006 Mar;25(1):9-34. - Di Carlo E, Forni G, Lollini P, Colombo MP, Modesti A, Musiani P. The intriguing
role of polymorphonuclear neutrophils in antitumor reactions. Blood. 2001 Jan 15;97(2):339-45.
- Duan B, Morel L. Role of B-1a cells in autoimmunity. Autoimmun Rev. 2006
Jul;5(6):403-8. - Ebisuya M, Kondoh K, Nishida E. The duration, magnitude and
compartmentalization of ERK MAP kinase activity: mechanisms for providing signaling specificity. J Cell Sci. 2005 Jul 15;118(Pt 14):2997-3002.
- E Brito RR, De Lorenzo BH, Xander P, Godoy LC, Lopes JD, da Silva NP, Sampaio
SC, Mariano M. Role of distinct immune components in the radiation-induced abrogation of systemic lupus erythematosus development in mice. Lupus. 2007;16(12):947-54.
- Fidler IJ. Selection of successive tumour lines for metastasis. Nat New Biol. 1973
Apr 4;242(118):148-9. - Fidler IJ.Tumor heterogeneity and the biology of cancer invasion and
metastasis.Cancer Res. 1978 Sep;38(9):2651-60. - Fidler IJ. Critical determinants of metastasis. Semin Cancer Biol. 2002a
Apr;12(2):89-96. - Fidler IJ. The organ microenvironment and cancer metastasis. Differentiation.
2002b Dec;70(9-10):498-505.

83
- Fidler IJ. The pathogenesis of cancer metastasis: the 'seed and soil' hypothesis
revisited. Nat Rev Cancer. 2003 Jun;3(6):453-8. - Fokas E, Engenhart-Cabillic R, Daniilidis K, Rose F, An HX. Metastasis: the seed
and soil theory gains identity. Cancer Metastasis Rev. 2007 Dec;26(3-4):705-715. - Forster I, Rajewsky K. Expansion and functional activity of Ly-1+ B cells upon
transfer of peritoneal cells into allotype-congenic, newborn mice. Eur J Immunol. 1987 Apr;17(4):521-8.
- Gabrilovich DI, Chen HL, Girgis KR, Cunningham HT, Meny GM, Nadaf S,
Kavanaugh D, Carbone DP. Production of vascular endothelial growth factor by human tumors inhibits the functional maturation of dendritic cells. Nat Med. 1996 Oct;2(10):1096-103.
- Gabrilovich DI, Velders MP, Sotomayor EM, Kast WM. Mechanism of immune
dysfunction in cancer mediated by immature Gr-1+ myeloid cells. J Immunol. 2001 May 1;166(9):5398-406.
- Garrity T, Pandit R, Wright MA, Benefield J, Keni S, Young MR. Increased
presence of CD34+ cells in the peripheral blood of head and neck cancer patients and their differentiation into dendritic cells. Int J Cancer. 1997 Nov 27;73(5):663-9.
- Gieni RS, Umetsu DT, DeKruyff RH. Ly1- (CD5-) B cells produce interleukin (IL)-
10. Cell Immunol. 1997 Feb 1;175(2):164-70. - Godoy LC, Mariano M, Lopes JD. Immunity and hypersensitivity to gp43 antigen in
susceptible and resistant mice infected with Paracoccidioides brasiliensis. Med Mycol. 2003 Oct;41(5):427-36.
- Gorden A, Osman I, Gai W, He D, Huang W, Davidson A, Houghton AN, Busam K,
Polsky D. Analysis of BRAF and N-RAS mutations in metastatic melanoma tissues. Cancer Res. 2003 Jul 15;63(14):3955-7.
- Gordon S. Alternative activation of macrophages. Nat Rev Immunol. 2003
Jan;3(1):23-35. - Govindarajan B, Bai X, Cohen C, Zhong H, Kilroy S, Louis G, Moses M, Arbiser JL.
Malignant Transformation of Melanocytes to Melanoma by Constitutive Activation of Mitogen-activated Protein Kinase Kinase (MAPKK) Signaling. J Biol Chem. 2003 Mar 14;278(11):9790-5.
- Guiducci C, Vicari AP, Sangaletti S, Trinchieri G, Colombo MP. Redirecting in vivo
elicited tumor infiltrating macrophages and dendritic cells towards tumor rejection. Cancer Res. 2005 Apr 15;65(8):3437-46.
- Gum R, Wang H, Lengyel E, Juarez J, Boyd D.Regulation of 92 kDa type IV collagenase expression by the jun aminoterminal kinase- and the extracellular signal-regulated kinase-dependent signaling cascades.Oncogene. 1997 Mar 27;14(12):1481-93.

84
- Haas KM, Poe JC, Steeber DA, Tedder TF. B-1a and B-1b cells exhibit distinct developmental requirements and have unique functional roles in innate and adaptive immunity to S. pneumoniae. Immunity. 2005 Jul;23(1):7-18.
- Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000 Jan 7;100(1):57-70. - Hardy RR, Carmack CE, Shinton SA, Riblet RJ, Hayakawa K. A single VH gene is
utilized predominantly in anti-BrMRBC hybridomas derived from purified Ly-1 B cells. Definition of the VH11 family. J Immunol. 1989 May 15;142(10):3643-51.
- Hardy RR. Variable gene usage, physiology and development of Ly-1+ (CD5+) B
cells. Curr Opin Immunol. 1992 Apr;4(2):181-5. - Hardy RR. B-1 B cells: development, selection, natural autoantibody and leukemia.
Curr Opin Immunol. 2006 Oct;18(5):547-55. - Harper SJ, LoGrasso P. Signalling for survival and death in neurones: the role of
stress-activated kinases, JNK and p38. Cell Signal. 2001 May;13(5):299-310. - Hastings WD, Gurdak SM, Tumang JR, Rothstein TL. CD5+/Mac-1- peritoneal B
cells: a novel B cell subset that exhibits characteristics of B-1 cells. Immunol Lett. 2006 May 15;105(1):90-6.
- Hayakawa K, Hardy RR, Parks DR, Herzenberg LA. The "Ly-1 B" cell
subpopulation in normal immunodefective, and autoimmune mice. J Exp Med. 1983 Jan 1;157(1):202-18.
- Hayakawa K, Hardy RR, Herzenberg LA, Herzenberg LA. Progenitors for Ly-1 B
cells are distinct from progenitors for other B cells. J Exp Med. 1985 Jun 1;161(6):1554-68.
- Hayakawa K, Shinton SA, Asano M, Hardy RR. B-1 cell definition. Curr Top
Microbiol Immunol. 2000;252:15-22. - Herlyn M, Berking C, Li G, Satyamoorthy K. Lessons from melanocyte development
for understanding the biological events in naevus and melanoma formation. Melanoma Res. 2000 Aug;10(4):303-12.
- Herzenberg LA, Stall AM, Lalor PA, Sidman C, Moore WA, Parks DR, Herzenberg
LA. The Ly-1 B cell lineage. Immunol Rev. 1986 Oct;93:81-102. - Herzenberg LA. B-1 cells: the lineage question revisited. Immunol Rev. 2000
Jun;175:9-22. - Hill CS, Treisman R. Transcriptional regulation by extracellular signals:
mechanisms and specificity. Cell. 1995 Jan 27;80(2):199-211. - Hofmann UB, Eggert AA, Blass K, Brocker EB, Becker JC. Expression of matrix
metalloproteinases in the microenvironment of spontaneous and experimental melanoma metastases reflects the requirements for tumor formation. Cancer Res. 2003 Dec 1;63(23):8221-5.

85
- Hofmeister V, Schrama D, Becker JC. Anti-cancer therapies targeting the tumor
stroma.Cancer Immunol Immunother. 2008 Jan;57(1):1-17. - Houghton AN, Gold JS, Blachere NE. Immunity against cancer: lessons learned
from melanoma. Curr Opin Immunol. 2001 Apr;13(2):134-40. - Hsu MC, Toellner KM, Vinuesa CG, Maclennan IC. B cell clones that sustain long-
term plasmablast growth in T-independent extrafollicular antibody responses. Proc Natl Acad Sci U S A. 2006 Apr 11;103(15):5905-10.
- Huang B, Pan PY, Li Q, Sato AI, Levy DE, Bromberg J, Divino CM, Chen SH. Gr-
1+CD115+ immature myeloid suppressor cells mediate the development of tumor-induced T regulatory cells and T-cell anergy in tumor-bearing host. Cancer Res. 2006 Jan 15;66(2):1123-31.
- Ibe S, Qin Z, Schuler T, Preiss S, Blankenstein T. Tumor rejection by disturbing
tumor stroma cell interactions. J Exp Med. 2001 Dec 3;194(11):1549-59. - Ihle JN. The Stat family in cytokine signaling. Curr Opin Cell Biol. 2001
Apr;13(2):211-7. - Ilan N, Mahooti S, Madri JA. Distinct signal transduction pathways are utilized
during the tube formation and survival phases of in vitro angiogenesis. J Cell Sci. 1998 Dec 18;111 ( Pt 24):3621-31.
- Inoue S, Leitner WW, Golding B, Scott D. Inhibitory effects of B cells on antitumor
immunity. Cancer Res. 2006 Aug 1;66(15):7741-7. - Ishida H, Hastings R, Kearney J, Howard M. Continuous anti-interleukin 10
antibody administration depletes mice of Ly-1 B cells but not conventional B cells. J Exp Med. 1992 May 1;175(5):1213-20.
- John A, Tuszynski G. The role of matrix metalloproteinases in tumor angiogenesis
and tumor metastasis. Pathol Oncol Res. 2001;7(1):14-23. - Johnson CW, Barth RF, Adams D, Holman B, Price JE, Sautins I. Phenotypic
diversity of murine B16 melanoma detected by anti-B16 monoclonal antibodies. Cancer Res. 1987 Feb 15;47(4):1111-7.
- Joyce JA. Therapeutic targeting of the tumor microenvironment. Cancer Cell. 2005
Jun;7(6):513-20. - Kahana O, Micksche M, Witz IP, Yron I. The focal adhesion kinase (P125FAK) is
constitutively active in human malignant melanoma. Oncogene. 2002 Jun 6;21(25):3969-77.
- Kakisis JD, Pradhan S, Cordova A, Liapis CD, Sumpio BE. The role of STAT-3 in
the mediation of smooth muscle cell response to cyclic strain. Int J Biochem Cell Biol. 2005 Jul;37(7):1396-406.

86
- Kantor AB, Stall AM, Adams S, Herzenberg LA, Herzenberg LA. Differential development of progenitor activity for three B-cell lineages. Proc Natl Acad Sci U S A. 1992 Apr 15;89(8):3320-4.
- Kantor AB, Herzenberg LA. Origin of murine B cell lineages. Annu Rev Immunol.
1993;11:501-38. - Karras JG, Wang Z, Huo L, Howard RG, Frank DA, Rothstein TL. Signal transducer
and activator of transcription-3 (STAT3) is constitutively activated in normal, self-renewing B-1 cells but only inducibly expressed in conventional B lymphocytes. J Exp Med. 1997 Mar 17;185(6):1035-42.
- Kim J, Mori T, Chen SL, Amersi FF, Martinez SR, Kuo C, Turner RR, Ye X, Bilchik
AJ, Morton DL, Hoon DS. Chemokine receptor CXCR4 expression in patients with melanoma and colorectal cancer liver metastases and the association with disease outcome. Ann Surg. 2006 Jul;244(1):113-20.
- Klafter R, Arbiser JL. Regulation of angiogenesis and tumorigenesis by signal
transduction cascades: lessons from benign and malignant endothelial tumors. J Investig Dermatol Symp Proc. 2000 Dec;5(1):79-82.
- Klemke RL, Cai S, Giannini AL, Gallagher PJ, de Lanerolle P, Cheresh DA.
Regulation of cell motility by mitogen-activated protein kinase. J Cell Biol. 1997 Apr 21;137(2):481-92.
- Kortylewski M, Kujawski M, Wang T, Wei S, Zhang S, Pilon-Thomas S, Niu G, Kay
H, Mulé J, Kerr WG, Jove R, Pardoll D, Yu H. Inhibiting Stat3 signaling in the hematopoietic system elicits multicomponent antitumor immunity. Nat Med. 2005 Dec;11(12):1314-21.
- Krueger JS, Keshamouni VG, Atanaskova N, Reddy KB. Temporal and quantitative
regulation of mitogen-activated protein kinase (MAPK) modulates cell motility and invasion. Oncogene. 2001 Jul 12;20(31):4209-18.
- Kulbe H, Levinson NR, Balkwill F, Wilson JL The chemokine network in cancer--
much more than directing cell movement. Int J Dev Biol. 2004;48(5-6):489-96. - Kuper H, Adami HO, Trichopoulos D. Infections as a major preventable cause of
human cancer. J Intern Med. 2000 Sep;248(3):171-83. - Kuroki M, O'Flaherty JT. Extracellular signal-regulated protein kinase (ERK)-
dependent and ERK-independent pathways target STAT3 on serine-727 in human neutrophils stimulated by chemotactic factors and cytokines. Biochem J. 1999 Aug 1;341 ( Pt 3):691-6.
- Kusmartsev SA, Li Y, Chen SH. Gr-1+ myeloid cells derived from tumor-bearing
mice inhibit primary T cell activation induced through CD3/CD28 costimulation. J Immunol. 2000 Jul 15;165(2):779-85.
- Laemmli UK.Cleavage of structural proteins during the assembly of the head of
bacteriophage T4.Nature. 1970 Aug 15;227(5259):680-5.

87
- Langley RR, Fidler IJ. Tumor cell-organ microenvironment interactions in the
pathogenesis of cancer metastasis.Endocr Rev. 2007 May;28(3):297-321. - Lakka SS, Jasti SL, Kyritsis AP, Yung WK, Ali-Osman F, Nicolson GL, Rao JS.
Regulation of MMP-9 (type IV collagenase) production and invasiveness in gliomas by the extracellular signal-regulated kinase and jun amino-terminal kinase signaling cascades. Clin Exp Metastasis. 2000;18(3):245-52.
- Lassmann S, Schuster I, Walch A, Göbel H, Jütting U, Makowiec F, Hopt U,
Werner M. STAT3 mRNA and protein expression in colorectal cancer: effects on STAT3-inducible targets linked to cell survival and proliferation. J Clin Pathol. 2007 Feb;60(2):173-9.
- Lee CH, Kakinuma T, Wang J, Zhang H, Palmer DC, Restifo NP, Hwang ST.
Sensitization of B16 tumor cells with a CXCR4 antagonist increases the efficacy of immunotherapy for established lung metastases. Mol Cancer Ther. 2006 Oct;5(10):2592-9.
- Lee TS, Chau LY.Heme oxygenase-1 mediates the anti-inflammatory effect of
interleukin-10 in mice.Nat Med. 2002 Mar;8(3):240-6. - Lehmann JM, Holzmann B, Breitbart EW, Schmiegelow P, Riethmüller G, Johnson
JP. Discrimination between benign and malignant cells of melanocytic lineage by two novel antigens, a glycoprotein with a molecular weight of 113,000 and a protein with a molecular weight of 76,000. Cancer Res. 1987 Feb 1;47(3):841-5.
- Lehmann JM, Riethmuller G, Johnson JP. MUC18, a marker of tumor progression
in human melanoma, shows sequence similarity to the neural cell adhesion molecules of the immunoglobulin superfamily. Proc Natl Acad Sci U S A. 1989 Dec;86(24):9891-5.
- Lewis TB, Robison JE, Bastien R, Milash B, Boucher K, Samlowski WE, Leachman
SA, Dirk Noyes R, Wittwer CT, Perreard L, Bernard PS. Molecular classification of melanoma using real-time quantitative reverse transcriptase-polymerase chain reaction. Cancer. 2005 Oct 15;104(8):1678-86.
- Li G, Satyamoorthy K, Herlyn M. Dynamics of cell interactions and communications
during melanoma development. Crit Rev Oral Biol Med. 2002;13(1):62-70. - Liu YW, Chen CC, Tseng HP, Chang WC. Lipopolysaccharide-induced
transcriptional activation of interleukin-10 is mediated by MAPK- and NF-kappaB-induced CCAAT/enhancer-binding protein delta in mouse macrophages. Cell Signal. 2006 Sep;18(9):1492-500.
- Locati M, Riboldi E, Otero K, Martinez FO, Riva F, Perrier P, Baviera S, Signorelli
P, Bonecchi R, Allavena P, Sozzani S, Mantovani A. Regulation of the chemokine system at the level of chemokine receptor expression and signaling activity. Immunobiology. 2001 Dec;204(5):536-42.
- Lopez-Bergami P, Huang C, Goydos JS, Yip D, Bar-Eli M, Herlyn M, Smalley KS,
Mahale A, Eroshkin A, Aaronson S, Ronai Z. Rewired ERK-JNK signaling pathways in melanoma. Cancer Cell. 2007 May;11(5):447-60.

88
- Luster AD. Chemokines--chemotactic cytokines that mediate inflammation. N Engl
J Med. 1998 Feb 12;338(7):436-45. - MacDougall JR, Matrisian LM. Contributions of tumor and stromal matrix
metalloproteinases to tumor progression, invasion and metastasis. Cancer Metastasis Rev. 1995 Dec;14(4):351-62.
- Maldonado JL, Fridlyand J, Patel H, Jain AN, Busam K, Kageshita T, Ono T,
Albertson DG, Pinkel D, Bastian BC. Determinants of BRAF mutations in primary melanomas. J Natl Cancer Inst. 2003 Dec 17;95(24):1878-90.
- Mansour SJ, Matten WT, Hermann AS, Candia JM, Rong S, Fukasawa K, Vande
Woude GF, Ahn NG. Transformation of mammalian cells by constitutively active MAP kinase kinase.Science. 1994 Aug 12;265(5174):966-70.
- Mantovani A, Bottazzi B, Colotta F, Sozzani S, Ruco L. The origin and function of
tumor-associated macrophages. Immunol Today. 1992 Jul;13(7):265-70. - Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization:
tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002 Nov;23(11):549-55.
- Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine
system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004a Dec;25(12):677-86.
- Mantovani A, Sozzani S, Locati M, Schioppa T, Saccani A, Allavena P, Sica A.
Infiltration of tumours by macrophages and dendritic cells: tumour-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Novartis Found Symp. 2004b;256:137-45; discussion 146-8, 259-69.
- Mantovani A, Sica A, Locati M. Macrophage polarization comes of age. Immunity.
2005 Oct;23(4):344-6. - Mantovani A, Schioppa T, Porta C, Allavena P, Sica A. Role of tumor-associated
macrophages in tumor progression and invasion. Cancer Metastasis Rev. 2006 Sep;25(3):315-22.
- Mantovani A, Marchesi F, Porta C, Sica A, Allavena P. Inflammation and cancer:
Breast cancer as a prototype. Breast. 2007 Aug 29; [Epub ahead of print]. - Marcelino França K, Vericimo MA, Retamal C, Kipnis TL, Arnholdt AC. B1 cells
contribution to susceptibility in experimental paracoccidioidomycosis: immunoglobulin isotypes and repertoire determination. Med Mycol. 2006 Dec;44(8):755-66.
- Maroni P, Bendinelli P, Matteucci E, Desiderio MA. HGF induces CXCR4 and
CXCL12-mediated tumor invasion through Ets1 and NF-kappaB. Carcinogenesis. 2007 Feb;28(2):267-79.

89
- Martin F, Oliver AM, Kearney JF. Marginal zone and B1 B cells unite in the early response against T-independent blood-borne particulate antigens. Immunity. 2001 May;14(5):617-29.
- Mathur RK, Awasthi A, Wadhone P, Ramanamurthy B, Saha B. Reciprocal CD40
signals through p38MAPK and ERK-1/2 induce counteracting immune responses. Nat Med. 2004 May;10(5):540-4. Erratum in: Nat Med. 2004 Jul;7(10):755.
- McGary EC, Lev DC, Bar-Eli M. Cellular adhesion pathways and metastatic
potential of human melanoma. Cancer Biol Ther. 2002 Sep-Oct;1(5):459-65. - Meier F, Satyamoorthy K, Nesbit M, Hsu MY, Schittek B, Garbe C, Herlyn M.
Molecular events in melanoma development and progression. Front Biosci. 1998 Sep 15;3:D1005-10.
- Meier F, Schittek B, Busch S, Garbe C, Smalley K, Satyamoorthy K, Li G, Herlyn
M. The RAS/RAF/MEK/ERK and PI3K/AKT signaling pathways present molecular targets for the effective treatment of advanced melanoma. Front Biosci. 2005 Sep 1;10:2986-3001.
- Meier F, Busch S, Lasithiotakis K, Kulms D, Garbe C, Maczey E, Herlyn M, Schittek
B. Combined targeting of MAPK and AKT signalling pathways is a promising strategy for melanoma treatment. Br J Dermatol. 2007 Jun;156(6):1204-13.
- Melani C, Chiodoni C, Forni G, Colombo MP. Myeloid cell expansion elicited by the
progression of spontaneous mammary carcinomas in c-erbB-2 transgenic BALB/c mice suppresses immune reactivity. Blood. 2003 Sep 15;102(6):2138-45.
- Mills CD, Kincaid K, Alt JM, Heilman MJ, Hill AM. M-1/M-2 macrophages and the
Th1/Th2 paradigm. J Immunol. 2000 Jun 15;164(12):6166-73. - Minoprio P, el Cheikh MC, Murphy E, Hontebeyrie-Joskowicz M, Coffman R,
Coutinho A, O'Garra A. Xid-associated resistance to experimental Chagas' disease is IFN-gamma dependent. J Immunol. 1993 Oct 15;151(8):4200-8.
- Mohan C, Morel L, Yang P, Wakeland EK. Accumulation of splenic B1a cells with
potent antigen-presenting capability in NZM2410 lupus-prone mice. Arthritis Rheum. 1998 Sep;41(9):1652-62.
- Moore KW, O'Garra A, de Waal Malefyt R, Vieira P, Mosmann TR. Interleukin-10.
Annu Rev Immunol. 1993;11:165-90. - Mora LB, Buettner R, Seigne J, Diaz J, Ahmad N, Garcia R, Bowman T, Falcone R,
Fairclough R, Cantor A, Muro-Cacho C, Livingston S, Karras J, Pow-Sang J, Jove R. Constitutive activation of Stat3 in human prostate tumors and cell lines: direct inhibition of Stat3 signaling induces apoptosis of prostate cancer cells. Cancer Res. 2002 Nov 15;62(22):6659-66.
- Murakami M, Tsubata T, Okamoto M, Shimizu A, Kumagai S, Imura H, Honjo T.
Antigen-induced apoptotic death of Ly-1 B cells responsible for autoimmune disease in transgenic mice. Nature. 1992 May 7;357(6373):77-80.

90
- Murakami M, Honjo T. Involvement of B-1 cells in mucosal immunity and
autoimmunity. Immunol Today. 1995 Nov;16(11):534-9. - Murakami T, Maki W, Cardones AR, Fang H, Tun Kyi A, Nestle FO, Hwang ST.
Expression of CXC chemokine receptor-4 enhances the pulmonary metastatic potential of murine B16 melanoma cells. Cancer Res. 2002 Dec 15;62(24):7328-34.
- Nagase H, Woessner JF Jr. Matrix metalloproteinases. J Biol Chem. 1999 Jul
30;274(31):21491-4. - Niemand C, Nimmesgern A, Haan S, Fischer P, Schaper F, Rossaint R, Heinrich
PC, Müller-Newen G. Activation of STAT3 by IL-6 and IL-10 in primary human macrophages is differentially modulated by suppressor of cytokine signaling 3. J Immunol. 2003 Mar 15;170(6):3263-72.
- O'Garra A, Chang R, Go N, Hastings R, Haughton G, Howard M. Ly-1 B (B-1) cells
are the main source of B cell-derived interleukin 10. Eur J Immunol. 1992 Mar;22(3):711-7. - Oka M, Kikkawa U. Protein kinase C in melanoma. Cancer Metastasis Rev. 2005
Jun;24(2):287-300. - Paciorkowski N, Porte P, Shultz LD, Rajan TV. B1 B lymphocytes play a critical role
in host protection against lymphatic filarial parasites. J Exp Med. 2000 Feb 21;191(4):731-6.
- Paget S: The distribution of secondary growths in cancer of the breast. Lancet
1889, 1:571-573. - Parker SL, Tong T, Bolden S, Wingo PA. Cancer statistics, 1997. CA Cancer J Clin.
1997 Jan-Feb;47(1):5-27. - Pollard JW. Tumour-educated macrophages promote tumour progression and
metastasis. Nat Rev Cancer. 2004 Jan;4(1):71-8. - Porta C, Subhra Kumar B, Larghi P, Rubino L, Mancino A, Sica A. Tumor
promotion by tumor-associated macrophages. Adv Exp Med Biol. 2007;604:67-86. - Poste G, Fidler IJ. The pathogenesis of cancer metastasis. Nature. 1980 Jan
10;283(5743):139-46. - Raman M, Chen W, Cobb MH. Differential regulation and properties of MAPKs.
Oncogene. 2007 May 14;26(22):3100-12. - Ruffini PA, Morandi P, Cabioglu N, Altundag K, Cristofanilli M. Manipulating the
chemokine-chemokine receptor network to treat cancer. Cancer. 2007 Jun 15;109(12):2392-404.
- Rusciano D. Differentiation and metastasis in melanoma. Crit Rev Oncog.
2000;11(2):147-63.

91
- Rutherford MS, Witsell A, Schook LB. Mechanisms generating functionally heterogeneous macrophages: chaos revisited. J Leukoc Biol. 1993 May;53(5):602-18.
- Sahai E, Olson MF, Marshall CJ. Cross-talk between Ras and Rho signalling
pathways in transformation favours proliferation and increased motility. EMBO J. 2001 Feb 15;20(4):755-66.
- Sasagawa S, Ozaki Y, Fujita K, Kuroda S. Prediction and validation of the distinct
dynamics of transient and sustained ERK activation. Nat Cell Biol. 2005 Apr;7(4):365-73. .- Sica A, Saccani A, Mantovani A. Tumor-associated macrophages: a molecular
perspective. Int Immunopharmacol. 2002 Jul;2(8):1045-54. - Sierra A. Metastases and their microenvironments: linking pathogenesis and
therapy. Drug Resist Updat. 2005 Aug;8(4):247-57. - Simon C, Goepfert H, Boyd D. Inhibition of the p38 mitogen-activated protein
kinase by SB 203580 blocks PMA-induced Mr 92,000 type IV collagenase secretion and in vitro invasion. Cancer Res. 1998 Mar 15;58(6):1135-9.
- Sinha P, Clements VK, Ostrand-Rosenberg S. Reduction of myeloid-derived
suppressor cells and induction of M1 macrophages facilitate the rejection of established metastatic disease. J Immunol. 2005 Jan 15;174(2):636-45.
- Sinha P, Clements VK, Bunt SK, Albelda SM, Ostrand-Rosenberg S. Cross-talk
between myeloid-derived suppressor cells and macrophages subverts tumor immunity toward a type 2 response. J Immunol. 2007 Jul 15;179(2):977-83.
- Smalley KS. A pivotal role for ERK in the oncogenic behaviour of malignant
melanoma? Int J Cancer. 2003 May 1;104(5):527-32. - Smith SH, Goldschmidt MH, McManus PM. A comparative review of melanocytic
neoplasms. Vet Pathol. 2002 Nov;39(6):651-78. - Sober AJ, Lew RA, Koh HK, Barnhill RL. Epidemiology of cutaneous melanoma. An
update. Dermatol Clin. 1991 Oct;9(4):617-29. - Song X, Krelin Y, Dvorkin T, Bjorkdahl O, Segal S, Dinarello CA, Voronov E, Apte
RN. CD11b+/Gr-1+ immature myeloid cells mediate suppression of T cells in mice bearing tumors of IL-1beta-secreting cells. J Immunol. 2005 Dec 15;175(12):8200-8.
- Staquicini FI, Moreira CR, Nascimento FD, Tersariol IL, Nader HB, Dietrich CP,
Lopes JD. Enzyme and integrin expression by high and low metastatic melanoma cell lines. Melanoma Res. 2003 Feb;13(1):11-8.
- Staquicini FI. Participação de Linfócitos B-1 no Desenvolvimento do Melanoma
Murino Experimental [tese]. São Paulo: Universidade Federal de São Paulo – Escola
Paulista de Medicina; 2004.

92
- Sumimoto H, Imabayashi F, Iwata T, Kawakami Y. The BRAF-MAPK signaling pathway is essential for cancer-immune evasion in human melanoma cells. J Exp Med. 2006 Jul 10;203(7):1651-6.
- Suzuki E, Kapoor V, Jassar AS, Kaiser LR, Albelda SM. Gemcitabine selectively
eliminates splenic Gr-1+/CD11b+ myeloid suppressor cells in tumor-bearing animals and enhances antitumor immune activity. Clin Cancer Res. 2005 Sep 15;11(18):6713-21.
- Tarhini AA, Agarwala SS. Cutaneous melanoma: available therapy for metastatic
disease. Dermatol Ther. 2006 Jan-Feb;19(1):19-25. - Tower GB, Coon CC, Benbow U, Vincenti MP, Brinckerhoff CE. Erk 1/2
differentially regulates the expression from the 1G/2G single nucleotide polymorphism in the MMP-1 promoter in melanoma cells. Biochim Biophys Acta. 2002 Apr 24;1586(3):265-74.
- Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A. 1979 Sep;76(9):4350-4.
- Treisman R. Regulation of transcription by MAP kinase cascades. Curr Opin Cell
Biol. 1996 Apr;8(2):205-15. - Tsung K, Dolan JP, Tsung YL, Norton JA. Macrophages as effector cells in
interleukin 12-induced T cell-dependent tumor rejection. Cancer Res. 2002 Sep 1;62(17):5069-75.
- Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A,
Speleman F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002 Jun 18;3(7):RESEARCH0034.
- Viala E, Pouysségur J.Regulation of tumor cell motility by ERK mitogen-activated
protein kinases.Ann N Y Acad Sci. 2004 Dec;1030:208-18. - Vigna AF, Almeida SR, Xander P, Freymüller E, Mariano M, Lopes JD. Granuloma
formation in vitro requires B-1 cells and is modulated by Paracoccidioides brasiliensis gp43 antigen. Microbes Infect. 2006 Mar;8(3):589-97.
- Wen Z, Zhong Z, Darnell JE. Maximal activation of transcription by statl and stat3
requires both tyrosine and serine phosphorylation. Cell, Vol 82, 251-260, 28 July 1995. - Weston CR, Davis RJ. The JNK signal transduction pathway. Curr Opin Genet Dev.
2002 Feb;12(1):14-21. - Wong SC, Chew WK, Tan JE, Melendez AJ, Francis F, Lam KP. Peritoneal CD5+
B-1 cells have signaling properties similar to tolerant B cells. J Biol Chem. 2002 Aug 23;277(34):30707-15.
- Yi AK, Yoon JG, Yeo SJ, Hong SC, English BK, Krieg AM. Role of mitogen-
activated protein kinases in CpG DNA-mediated IL-10 and IL-12 production: central role of

93
extracellular signal-regulated kinase in the negative feedback loop of the CpG DNA-mediated Th1 response. J Immunol. 2002 May 1;168(9):4711-20.
- Yin Z, Bahtiyar G, Zhang N, Liu L, Zhu P, Robert ME, McNiff J, Madaio MP, Craft
J. IL-10 regulates murine lupus. J Immunol. 2002 Aug 15;169(4):2148-55. - Young MR, Newby M, Wepsic HT. Hematopoiesis and suppressor bone marrow
cells in mice bearing large metastatic Lewis lung carcinoma tumors. Cancer Res. 1987 Jan 1;47(1):100-5.
- Young MR, Wright MA, Lozano Y, Prechel MM, Benefield J, Leonetti JP, Collins
SL, Petruzzelli GJ. Increased recurrence and metastasis in patients whose primary head and neck squamous cell carcinomas secreted granulocyte-macrophage colony-stimulating factor and contained CD34+ natural suppressor cells. Int J Cancer. 1997 Feb 20;74(1):69-74.
- Zea AH, Rodriguez PC, Atkins MB, Hernandez C, Signoretti S, Zabaleta J,
McDermott D, Quiceno D, Youmans A, O'Neill A, Mier J, Ochoa AC. Arginase-producing myeloid suppressor cells in renal cell carcinoma patients: a mechanism of tumor evasion. Cancer Res. 2005 Apr 15;65(8):3044-8.
- Zebisch A, Czernilofsky AP, Keri G, Smigelskaite J, Sill H, Troppmair J. Signaling
through RAS-RAF-MEK-ERK: from basics to bedside. Curr Med Chem. 2007;14(5):601-23.
- Zhou JH, Broussard SR, Strle K, Freund GG, Johnson RW, Dantzer R, Kelley KW.
IL-10 inhibits apoptosis of promyeloid cells by activating insulin receptor substrate-2 and phosphatidylinositol 3'-kinase. J Immunol. 2001 Oct 15;167(8):4436-42.
- Zou W, Machelon V, Coulomb-L'Hermin A, Borvak J, Nome F, Isaeva T, Wei S,
Krzysiek R, Durand-Gasselin I, Gordon A, Pustilnik T, Curiel DT, Galanaud P, Capron F, Emilie D, Curiel TJ.Stromal-derived factor-1 in human tumors recruits and alters the function of plasmacytoid precursor dendritic cells. Nat Med. 2001 Dec;7(12):1339-46.

ABSTRACT

Summary
Increasing evidence indicates that tumors require a constant influx of
myelomonocytic cells to support their malignant behavior. This is caused by tumor-
derived factors, which recruit and induce functional differentiation of
myelomonocytic cells, most of which are macrophages. Although myeloid lineages
are the classical precursors of macrophages, B lymphoid lineages such as B-1
cells, a subset of B lymphocytes found predominantly in pleural and peritoneal
cavities, are also able to migrate to inflammatory sites and differentiate into
mononuclear phagocytes exhibiting macrophage-like phenotype. Here we
examined the interplay of B-1 cells and tumor cells and checked whether this
interaction provides signals to influence melanoma cells metastases. Using in vitro
coculture experiments we showed that B16, a murine melanoma cell line, and B-1
cells physically interact. Moreover, interaction of B16 with B-1 cells leads to
upregulation of metastasis-related genes expression (MMP-9 and CXCR-4),
increasing its metastatic potential, as revealed by experimental metastases assays
in vivo. We also provide evidences that B16 cells exhibit markedly upregulated
phosphorylation of the extracellular signalregulated kinase (ERK) when co-cultured
with B-1 cells. Inhibition of ERK
phosphorylation induced by B-1 cells with an inhibitor of MEK1/2 strongly
suppressed the induction of MMP-9 and CXCR-4 mRNA expression and impaired
the increased metastatic behavior of B16. In addition, constitutive levels of ERK1/2
phosphorylation in B-1 cells are necessary for their commitment to affect the
metastatic potential of B16 cells. Our findings show for the first time, that B-1

80
lymphocytes can contribute to tumor cell properties required for invasiveness
during metastatic spread.

ANEXOS

80
----- Mensagem encaminhada de [email protected] ----- Data: Tue, 15 Jan 2008 19:19:14 -0500 (EST) De: [email protected] Reponder para: [email protected] Assunto: Cancer Science (CAS-OA-0602-2007.R1): Acceptance Para: [email protected] Date: 15-Jan-2008 RE: Ms No.: CAS-OA-0602-2007.R1 Title: B-1 lymphocytes increase metastatic behavior of melanoma cells through the extracellular signal-regulated kinase pathway. Author(s): Pérez, Elizabeth; Machado Jr, Joel; Aliperti, Fabiana; Freym�ler, Edna; Mariano, Mario; Lopes, Jos� Dear Prof. Joel Machado Jr: We are pleased to inform you that your revised paper has been accepted for publication in Cancer Science and will be published in Cancer Science. The comments of the Associate Editor and reviewer(s) who reviewed your manuscript are included at the foot of this letter. Thank you for your fine contribution. On behalf of the Editors of Cancer Science, we look forward to your continued contributions to the Journal. We would like to take this opportunity to explain our recently launched functions to assist authors with tracking their manuscripts through the production process. To minimise publication time of your manuscript it is important that all electronic artwork is supplied to the editorial office in the correct format and resolution. I recommend that you consult the Illustration guidelines at http://www.blackwellpublishing.com/bauthor/digill.asp if you need advice on any aspect of preparing your artwork. Copyright - If you havenuft already sent your completed Copyright Assignment Form for your manuscript at the time of submission, please sign and return to this office within two weeks. Accepted articles canuft go into production for an issue until a completed form has been received. Author Services - Production status tracking - You can now track your article via the publisherufs Author Services. Once your paper is with the Production Editor, you will receive an e-mail with a unique code that automatically adds your article to the system when you register. With Author Services you can check the status of your article online and choose to receive automated e-mails at key stages of production. Therefore, please ensure that we have your complete e-mail address. There may be a short delay whilst the article is sent to the Production Editor and logged into the production tracking system. Additional services will be added to the website soon. Proofing your manuscript – Cancer Science is included in a new electronic service, uge-proofinguh. You will receive an e-mail from the typesetter when your article is ready for proofing. Youufll receive instructions about how to download your paper and how to return your corrections. Your e-mail address is needed for this vital step, too. OnlineEarly – Cancer Science operates a system called OnlineEarly, whereby articles are published online ahead of assignment to an issue and publication in print. If your article is eligible for this service (Review Articles and Original Articles only), you can track the progress of your article and learn when it is published online by registering for Author Services. Please note that in order to publish your article as quickly as possible, and if your article is received very close to the copy deadline for an issue, it may be

81
incorporated directly into that issue without first appearing OnlineEarly. The Blackwell Synergy Website for Cancer Science is http://www.blackwell-synergy.com/loi/cas. Reprints and Offprints - You will receive instructions for ordering offprints when you are notified that your proofs are ready for review. Sincerely yours, Dr. Setsuo Hirohashi, Editor Cancer Science --- Cancer Science Editorial Office TEL: 81-3-3830-1266 FAX: 81-3-5689-7278 [email protected] http://www.blackwellpublishing.com/cas Associate Editor's decision: Acceptance Comments to Author: Associate Editor: 1 Comments to the Author: The authors have properly revised the manuscript according to the reviewers' comments. I recommend to accept this manuscript for publication in Cancer Science. ----- Finalizar mensagem encaminhada -----


ELIZABETH CRISTINA PÉREZ HURTADO
MODIFICAÇÃO FENOTÍPICA DO MELANOMA MURINO INDUZIDA PELA
INTERAÇÃO COM CÉLULAS B-1 – MECANISMOS DE TRANSDUÇÃO DE
SINAL
São Paulo 2008
Tese apresentada à Universidade Federal de São Paulo – Escola Paulista de Medicina para obtenção do Título de Doutor em Ciências da Saúde. Orientador: Prof. Dr. José Daniel Lopes Co-orientador: Joel Jr. Machado

ii
Pérez, Elizabeth Cristina Modificação fenotípica do melanoma murino induzida pela interação com células B-1 – Mecanismos de Transdução de Sinal. / Elizabeth Cristina Pérez. – São Paulo, 2008. xxi, 109f. Tese (Doutorado) – Universidade Federal de São Paulo. Escola Paulista de Medicina. Programa de Pós-Graduação em Microbiologia e Imunologia Título em Inglês: B-1 lymphocytes increase metastatic behavior of melanoma cells through the extracellular signal-regulated kinase pathway. 1. Células B16. 2. Linfócitos B-1. 3. ERK. 4. Metástases.

iii
Universidade Federal de São Paulo
Escola Paulista de Medicina
Departamento de Microbiologia, Imunologia e Parasitologia
Chefe do Departamento: Prof. Dr. José Daniel Lopes
Coordenador do Curso de Pós-graduação: Prof. Dr. Renato Arruda Mortara

iv
ELIZABETH CRISTINA PÉREZ HURTADO
MODIFICAÇÃO FENOTÍPICA DO MELANOMA MURINO INDUZIDA PELA
INTERAÇÃO COM CÉLULAS B-1 – MECANISMOS DE TRANSDUÇÃO DE
SINAL
Presidente da banca: Prof. Dr. José Daniel Lopes
BANCA EXAMINADORA Profa. Dra. Maria Mitzi Brentani
Dra. Nathalie Cella
Profa. Dra. Célia Regina Whitaker Carneiro
Prof. Dr. Hugo Pequeno Monteiro
Aprovada em: 23/01/2008

v
Este trabalho foi desenvolvido na disciplina de
Imunologia do Departamento de Microbiologia,
Imunologia e Parasitologia da Universidade
Federal de São Paulo, com auxílios financeiros
concedidos pela Fundação de Amparo a
Pesquisa do Estado de São Paulo (FAPESP
processo 03/05176-8)

vi
Dedicatória
Este trabalho é dedicado :
As memórias da minha mãe Carmen e da minha avó Rackel, Aos meus adorados homens: meu esposo Harold e meus filhos Juan David
e Luis Felipe
A meu pai Guillermo e meus irmãos Carolina e Guillermo

vii
Agradecimentos
Ao Prof. Dr. José Daniel Lopes, pelo seu carinhoso acolhimento, dedicação,
amizade, apoio, ensinamentos e auxílio na redação desta tese.
Ao Prof. Dr. Joel Machado pelo espírito crítico no desenvolvimento deste
trabalho, auxílio na redação desta tese e pela valiosa contribuição na realização
do artigo científico.
Ao Prof. Dr. Mario Mariano pela amizade e valiosas contribuições.
Às Profs. Dras. Célia Regina Whitaker Carneiro, Ieda Maria Longo Maugéri,
Miriam Galvonas Jasiulionis e Zulma Fernandes Peixinho, pela amizade, carinho e
valiosas contribuições.
À Prof. Dra. Marimélia Porcionatto (INFAR) pelo carinho, amizade e
grandes ensinamentos.
À Dra. Valderes Bastos Valero e sua equipe: Amarildo, Luis, Renato do
Laboratório de Experimentação animal do INFAR por seu grande carinho, amizade
e disposição no fornecimento de animais.
À Prof. Dra. Elaine Guadalupe Rodrigues, pela amizade e colaboração no
fornecimento de camundongos,
À Prof. Dra. Edna Freymüller (CEME) e sua equipe: André, Isabel, Márcia e
Patrícia, pelo carinho, amizade, disposição e colaboração nas análises por
microscopia eletrônica.

viii
Ao Prof. Dr. João Bosco Pesqueiro e sua equipe: Felipe, Raphael, Giuliana,
Jõao e demais estudantes, pela amizade, disposição e valiosa colaboração na
realização dos ensaios e análises por RT-PCR em tempo real,
Ao Prof. Dr. Sergio Schenkman e sua equipe pela amizade e colaboração
no empréstimo do sistema de foto-documentação utilizado nas análises por RT-
PCR realizadas durante este trabalho.
Aos funcionários da Disciplina de Imunologia: Aparecido Mendes, Creusa
Marina, Creusa Rosa, Eraldina do Nascimento, Geová dos Santos, Gisélia Lopes,
Ivone Mozat, Naide, Neta e Zélia Pereira, pela amizade, colaboração e convívio
alegre durante estes anos.
À Mércia, secretária da pós-graduação do departamento, pela amizade e
auxílio constante.
Aos professores, funcionários e colegas da disciplina de Biologia Celular
(oitavo andar) pela colaboração e boa disposição.
As amigas do “Lab 2”, Érika Kioshima (Londri), Fabiana Aliperti (Fabico),
Juliana Maricato (Juju), Lívia Pugliese e Patrícia Xander (Patlicia), pela
cumplicidade, carinho, apoio, clima alegre e grande ajuda na correção desta tese.
A todos os amigos do “Lab1 e Lab2”, Adriana, Aline (pulga), Bruno, Camila,
Creusa, Nadia, Marcela, Márcia, Patrícia, Renata, Toshie pela amizade, carinho e
convívio alegre e estimulante durante a realização deste trabalho.
À Fernanda Staquicini, colega que defendeu doutorado 3 anos atrás,
obrigada pela amizade e acolhida carinhosa ao chegar na Imuno assim como por
me ensinar o mundo das B-1.

ix
Aos amigos da Imuno, Inferno (Ronni Brito), Gugu (André), Carla Squaiella,
Juliana Mussalem, Tatiana, Adriana, Alice, Fernanda Molognoni, Karina, Tatiana
Rica, Ana Flavia Popi, Felipe, Fernanda, Helena, Maiko, Mauro, Ricardo, Carol,
Janaina, pelo carinho, amizade e simpatia.
Às amigas e amigos com quem tive oportunidade de trabalhar e que há
algum tempo já trilham seu próprio caminho: Ana Cristina Campos, Ana Flavia
Vigna (Oxigenada), Beatriz Helena Pizarro, Daniela Grosso, Fernanda Staquicini,
Hellen Fuzii, Ludmila Chinen, Luciana, Luciano Feitosa, Luiz Cláudio Godoy,
Priscila Penteado, Renata Ananias, Thalita Abrão; obrigada pelo carinho, amizade
e feliz convivência.
À FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo) por
financiar este trabalho.
À minha “Virgencita” e aos meus Anjos protetores pela companhia,
iluminação e proteção de sempre.
E por suposto a minha família, meu esposo Harold e meus dos filhotes Juan
David e Luis Felipe por seu grande apoio, alegria, amor e carinho durantes estes
anos de correria, muitíssimo obrigada.

x
Sumário
Dedicatória..............................................................................................................VI
Agradecimentos......................................................................................................VII
Lista de Gráficos....................................................................................................XIII
Lista de Tabelas....................................................................................................XIV
Lista de Figuras......................................................................................................XV
Lista de Abreviaturas e Símbolos........................................................................XVII
RESUMO...............................................................................................................XX
1. INTRODUÇÃO......................................................................................................1
2. OBJETIVOS........................................................................................................16
2. MATERIAIS E MÉTODOS..................................................................................18
2.1 MATERIAIS.......................................................................................................19
2.1.1 Soluções Rotineiras.......................................................................................19
2.1.1.1 Soluções para SDS-PAGE.........................................................................19
2.1.1.2 Soluções para Coloração por Comassie Blue............................................20
2.1.1.3 Soluções para Immunoblotting...................................................................20
2.1.1.4 Soluções para ELISA.................................................................................20
2.1.1.5 Soluções para cultura de células................................................................21
2.1.2 Animais..........................................................................................................21
2.2 MÉTODOS........................................................................................................22
2.2.1 Cultura de células e tratamentos...................................................................22
2.2.2 Análises de moléculas de superfície por citometria de fluxo.........................24

xi
2.2.3 Ensaio de viabilidade celular por Trypan blue...............................................24
2.2.4 Ensaio de inibição da adesão........................................................................25
2.2.5 Ensaio de metástases experimental..............................................................25
2.2.6 Preparação de extratos celulares..................................................................26
2.2.7 Dosagem protéica pelo micrométodo de Bradford........................................26
2.2.8 Eletroforese em gel de poliacrilamida com dodecilsulfato de sódio (SDS-
PAGE).....................................................................................................................26
2.2.9 Transferência de proteínas de nitrocelulose (Immunoblot ou Western
blot).........................................................................................................................28
2.2.10 Microscopia eletrônica de transmissão........................................................29
2.2.11 Ensaios de proliferação celular....................................................................30
2.2.11.1 Incorporação de timidina tritiada...............................................................30
2.2.11.2 MTT (tetrazolium salt3-(-4,4-dimethylthiazol-2-yl_-2,5-diphenyltetrazolium
bromide...................................................................................................................30
2.2.12 Dosagem de citocinas no sobrenadante de cultura…………………............31
2.2.13 Obtenção de RNA total................................................................................32
2.2.14 RT-PCR (Reverse Transcriptase – Polymerase Chain Reaction)...............33
2.2.14.1 Análise qualitativa por RT-PCR................................................................34
2.2.14.2 Análise quantitativa por RT-PCR em tempo real......................................36
2.2.15 Análise Estatística.......................................................................................37
3 RESULTADOS....................................................................................................39
3.1 Caracterização do sistema de co-cultivo..........................................................40
3.2 Células B-1 e B16 fisicamente interagem durante o co-cultivo........................41

xii
3.3 Fosforilação de ERK está aumentada nas células B16 após interação com
células B-1..............................................................................................................43
3.4 Aumento no potencial metastático de células B16 é dependente dos níveis de
ERK fosforilado.......................................................................................................44
3.5 Células B-1 não induzem aumento na proliferação de células B16.................46
3.6 Células B-1 induzem a expressão de genes relacionados com metástases nas
células B16.............................................................................................................46
3.7 Níveis basais de fosforilação de ERK nas células B-1 são necessários para
induzir aumento no potencial metastático das células B16....................................47
3.8 Níveis basais de fosforilação de ERK nas células B16 são inalterados pelo
estímulo com IL-10.................................................................................................50
4. DISCUSSÃO....................................................................................................,,.67
5. CONCLUSÕES...................................................................................................75
6. REFERÊNCIAS BIBLIOGRÁFICAS...................................................................92
ABSTRACT
ANEXOS

xiii
Lista de Gráficos
Gráfico 1. Via canônica de Ras-Raf-MEK-ERK.....................................................15

xiv
Lista de Tabelas
Tabela 1. Classificação clínica de melanoma........................................................11
Tabela 2. Composição de gel para SDS-PAGE.....................................................27
Tabela 3. Seqüências oligo-nucleotídicas dos primers sense (S) e anti-sense (AS),
temperatura de anelamento (°C) e o tamanho do produto do PCR em pares de
bases (BP), dos transcritos estudados por RT-PCR qualitativo.............................35
Tabela 4. Seqüências oligo-nucleotídicas dos primers sense (S) e anti-sense (AS)
e tamanho do produto do PCR em pares de bases (bp), dos transcritos estudados
por RT-PCR em tempo real....................................................................................37

xv
Lista de Figuras
Figura 1. Células B-1 são eliminadas eficientemente das co-culturas após
aspiração do sobrenadante....................................................................................51
Figura 2. Células B-1 aumentam o potencial metastático das células B16...........52
Figura 3. Células B-1 induzem aumento no potencial metastático de células B16
independentemente da adesão celular...................................................................53
Figura 4. Fatores solúveis das células B-1 não contribuem para o aumento da
capacidade metastática das células B16................................................................54
Figura 5. Células B16 e B-1 interagem fisicamente durante o co-cultivo..............55
Figura 6. Células B-1 induzem aumento nos níveis de fosforilação de ERK nas
células B16.............................................................................................................56
Figura 7. Níveis basais de fosforilação de ERK em células B16 e B-1 são inibidos
pelo tratamento com PD98059...............................................................................57
Figura 8. O aumento no potencial metastático das células B16 induzido pelo
contato com células B-1 é dependente dos níveis de ERK fosforilado..................58
Figura 9. Células B-1 não induzem aumento na proliferação das células B16......59
Figura 10. Qualidade do RNA total após extração.................................................60
Figura 11. Células B-1 induzem a expressão de CXCR4 e MMP-9 nas células
B16..........................................................................................................................61
Figura 12. Produção de IL-10 por células B-1.......................................................62
Figura 13. Níveis basais de fosforilação de ERK nas células B-1 são necessários
para induzir aumento no potencial metastático das células B16............................63

xvi
Figura 14. Níveis de fosforilação de Stat3 nas células B16 antes e após co-cultivo
com células B-1......................................................................................................64
Figura 15. Níveis de ERK fosforilado e comportamento metastático das células
B16 não são aumentados por IL-10 recombinante.................................................65
Figura 16. Células B-1 de animais knock-out para IL-10 na são capazes de induzir
aumento no potencial metastático das células B16, mesmo na presença de IL-10
recombinante..........................................................................................................66

xvii
Lista de Abreviaturas e Símbolos
% Porcentagem
µg Micrograma
µL Microlitro
µM Micromolar
BCR B cell receptor
bp Pares de bases
BSA Soro albumina bovina
cDNA DNA complementar
CXCR4 Chemokine (C-X-C motif) receptor
DEPC Dietil pirocarbonato
DMSO Dimetilsulfóxido
DO Densidade óptica
EDTA Ácido etilenodiamino tetracético
ELISA Enzyme linked Immunosorbent Assay
ERK Extracellular signal-regulated kinase
FACS Fluorescence Activated Cell Sorting
FITC Isotiocianato de fluoresceína
g Gramas
GM-CSF Granulocyte-macrophage colony-stimulating factor
GPDH Gliceraldeído-3-fosfato desidrogenase
H Hora

xviii
IgD Imunoglobulina D
IgD Imunoglobulina D
IgM Imunoglobulina M
IL Interleucina
JAKs Janus-activated kinase
JNK June N-terminal kinase
kDA Kilo Dalton
LPS Lipopolissacarídeo
MAPK Mitogen activated protein kinase
MCP-1/CCL2 Monocyte chemotactic protein -1/ chemokine (C-C motif)
M-CSF Monocyte-colony-stimulating factor
MDSC Myeloid-derived suppressor cells
mL Mililitro
mM Milimolar
MMPs Matrix Metalloproteinases
MMP-9 Matrix Metalloproteinase-9
NK Natural killer
NO Óxido nítrico
PBS Tampão salino fosfato
PKC Protein kinase C
PMSF Fluoreto de fenilmetanosulfonato
q.s.p quantidade suficiente para
RGP Radial growth phase
ROI Reativos intermediários de oxigênio

xix
rpm Rotação por minuto
RTKs Receptores tirosina kinase
RTKs Receptores tirosina kinase
RT-PCR Reverse transcriptase polymerase chain reaction
SDF-1/CXCL12 Stromal cell-derived factor 1
SDS-PAGE Sodium dodecyl sulphte polyacrylamide gel
STAT Signal Transducers and Activators of Transcription
Stat3 Signal Transducer and Activator of Transcription 3
TAMs Tumour-associated macrophages
TNF Tumour necrosis factor
V Volts
v/v Volume a volume
VEGF-A Vascular endothelial growth factor - A
VGP Vertical growth phase
W Wats
W Wats
X Vezes

xx
Resumo
Evidências indicam que tumores necessitam constante influxo de células mielo-
monocíticas para sustentar seu comportamento maligno. Esse fato é devido a
fatores derivados do tumor, os quais recrutam e induzem diferenciação funcional
de células mielo-monocíticas, das quais a maioria são macrófagos. Embora os
precursores clássicos de macrófagos sejam linhagens mielóides, linhagens
linfóides como células B-1, subtipo de linfócitos B encontrados
predominantemente nas cavidades pleural e peritoneal são também capazes de
migrar para focos inflamatórios e se diferenciar em fagócitos mononucleares
apresentando fenótipo semelhante ao dos macrófagos. No presente trabalho foi
avaliada a interação entre células B16 de melanoma murino e células B-1 durante
o co-cultivo e se esta interação influencia ativação de vias de transdução de sinal
envolvidas com progressão e metástases. Utilizando sistema de co-cultura in vitro,
foi mostrado que células B16 e células B-1 interagem fisicamente após 48 horas
de co-cultura. Além disso, esta interação resulta em aumento da expressão de
genes associados com metástases como MMP-9 e CXCR4 nas células B16 que
favorecem o aumento na capacidade metastática destas células, como revelado
por ensaios experimentais de metástases in vivo. Este trabalho também revela
evidências de que células B16 apresentam marcado aumento na fosforilação da
quinase regulada por sinais extracelulares (ERK) após contato com células B-1. A
inibição da fosforilação de ERK com inibidor farmacológico da quinase ascendente
de ERK, MEK1/2, suprime fortemente a expressão de MMP-9 e CXCR4 e inibe o
aumento da capacidade metastática das células B16 induzido por contato com

xxi
células B-1. Adicionalmente, níveis constitutivos de ERK fosforilado nas células B-
1 são necessários para que estas células sejam capazes de induzir aumento no
potencial metastático das células B16. Nossos resultados em conjunto mostram
que células B-1 podem contribuir na aquisição de um fenótipo mais agressivo das
células tumorais.

1. INTRODUÇÃO

1. INTRODUÇÃO
Uma das principais características no desenvolvimento do câncer é a
formação de metástases na qual células tumorais provenientes do tumor primário
invadem e colonizam tecidos ou órgãos distantes do sítio de origem (Hanahan &
Weinberg, 2000; Balkwill & Mantovani, 2001; Langley & Fidler, 2007; Aguirre-
Ghiso, 2007).
Embora grandes avanços tenham sido realizados na prevenção,
diagnóstico, técnicas cirúrgicas e terapias adjuvantes, a maior parte das mortes
em câncer são resultado da formação de metástases resistentes a terapias
convencionais. Por esta razão, o entendimento dos mecanismos celulares e
moleculares envolvidos na formação de metástases é importante para o
desenvolvimento de novos alvos terapêuticos
A formação de metástases é um processo complexo e seletivo que consiste
em vários passos seqüenciais e relacionados, nos quais células tumorais precisam
adquirir a capacidade de invadir tecidos adjacentes para formar tumores
secundários ou metástases (Poste & Fidler, 1980; Fidler, 2002a). Neste processo,
células com capacidade de metastatizar separam-se do tumor primário, entram na
circulação ao invadirem a parede dos vasos sanguíneos ou linfáticos, disseminam-
se e, posteriormente, aderem a órgãos determinados. Para que todos estes
eventos ocorram, o aumento na expressão de moléculas de adesão celular como
integrinas, proteases como metaloproteinases (MMPs) e moléculas quimio-

2
atraentes como citocinas e quimiocinas são essenciais para invasão e
metástatização das células tumorais (Sierra, 2005; Deryugina & Quigley, 2006).
Observações clínicas em pacientes com câncer e estudos experimentais
em camundongos têm demonstrado que certos tipos de tumores metastatizam
para órgãos específicos independentemente da anatomia vascular, do tamanho do
tumor ou do número de células tumorais recebidas em cada órgão (Sierra, 2005;
Fokas et al., 2007). Para explicar essa especificidade, a teoria soil and seed,
proposta por Paget em 1889, sustenta que órgãos diferentes (soil) provêm
condições de crescimento ótimas para cânceres específicos (seed; revisado por
Fidler, 2002b).
Assim como sugerido por Paget, outros autores (Balkwill & Mantovani,
2001; Coussens & Werb, 2002; Croci et al., 2007) têm demonstrado que durante a
progressão tumoral e metástase ocorre comunicação ativa (cross-talk) entre
células tumorais e seu estroma. Esta comunicação pode ser mediada tanto pelo
contato direto entre as células, como pela produção de citocinas, quimiocinas,
fatores de crescimento e metaloproteinases, que favorecem o avanço das células
tumorais para sítios distantes do tumor primário (Lehmann et al., 1987; Christofori
& Semb, 1999; McGary et al., 2002; Li et al., 2002). A interação entre células
estromais e tumorais sugere que o tumor não é apenas uma massa composta de
células malignas, e sim, um tecido com ampla diversidade celular que incluem
células residentes como adipócitos, fibroblastos, populações de células
hematopoiéticas migratórias como monócitos, macrófagos, neutrófilos, mastócitos,
eosinófilos, linfócitos T e B (Coussens et al., 1999; Coussens et al., 2000; Kuper et
al., 2000; Balkwill & Mantovani, 2001; Di Carlo et al., 2001; Coussens & Werb,

3
2002; Joyce, 2005; Inoue et al., 2006). Essa diversidade celular produz assim um
único micro-ambiente o qual é capaz de modificar as propriedades neoplásicas
das células tumorais (Coussens & Werb, 2002; Croci et al., 2007).
Macrófagos associados ao tumor (TAMs, tumor-associated
macrophages)
Macrófagos associados ao tumor (TAMs) representam o maior componente
do infiltrado inflamatório da maioria dos tumores (Mantovani et al., 1992, Balkwill &
Mantovani, 2001). No entanto, a alta freqüência destas células no infiltrado de
muitos cânceres humanos está associada a mau prognóstico (Mantovani et al.,
2006). TAMs são derivados de monócitos circulantes os quais são recrutados ao
sítio do tumor por fatores quimiotáticos como a proteína quimiotática para
monócitos - 1 [MCP-1/CCL2; monocyte chemotactic protein-1/chemokine (C-C
motif) ligand 2], o fator estimulador de colônias de monócitos (M-CSF; monocyte-
colony-stimulating factor) e o fator de crescimento do endotélio vascular – A
(VEGF-A; vascular endothelial growth factor – A; Mantovani et al., 1992; Barleon
et al., 1996).
Macrófagos são células versáteis que representam a primeira linha de
defesa da resposta imune inata e não requerem adaptação ou imunização para
funcionar. Estas células têm papel importante no recrutamento e ativação de
linfócitos por meio da apresentação de antígenos (Sica et al., 2002). Entretanto,
devido a sua funcionalidade heterogênea, podem também realizar funções
supressoras (Rutherford et al., 1993). Em câncer, os macrófagos são importantes

4
células efetoras na imunidade contra tumores e, dependendo de seu fenótipo,
podem tanto prevenir como promover a progressão tumoral.
Fenotipicamente dois tipos de macrófagos têm sido descritos: macrófagos
ativados pela via clássica e macrófagos ativados pela via alternativa. Os
macrófagos ativados pela via clássica têm fenótipo IL-12high, IL-23high, IL-10low e
são ativados por IFN-γ e lipopolissacarídeo (LPS; Mantovani et al., 2004a). Em
contraste, os macrófagos ativados pela via alternativa têm fenótipo IL-12low, IL-
23low, IL-10high e são ativados por IL-4 e IL-13 (Gordon, 2003). Pelo fato de
possuírem perfil de citocinas IL-12 e IL-10 similares aos das células T CD4+ Th1 e
Th2, os macrófagos ativados classicamente e os ativados alternativamente têm
sido chamados de M1 e M2, respectivamente (Mills et al., 2000; Mantovani et al.,
2002; Mantovani et al., 2005).
Quanto à função destas células no câncer, macrófagos M1 são células
efetoras potentes que eliminam células tumorais e promovem a rejeição do tumor
(Mantovani et al., 1992, Guiducci et al., 2005). Entretanto, os macrófagos M2
favorecem crescimento e disseminação tumoral provavelmente por favorecer um
ambiente imunossupressor pela produção de IL-10 (Ibe et al., 2001), assim como
por promover proliferação celular, angiogênese, remodelação tissular e reparo
(Sica et al., 2002; Mantovani et al., 2002; Gordon, 2003; Mantovani et al., 2004b;
Condeelis & Pollard, 2006; Porta et al., 2007).

5
Células supressoras derivadas da linhagem mielóide (MDSC: myeloid-
derived suppressor cells)
Além dos TAMs, recentes estudos mostraram o acumulo de células
mielóides imaturas na medula óssea, linfonodos e baço tanto em pacientes com
câncer como em camundongos com tumores implantados (Young et al., 1987;
Garrity et al., 1997; Almand et al., 2001; Zea et al., 2005). Estas células,
denominadas atualmente como células supressoras derivadas da linhagem
mielóide (MDSC; myeloid-derived suppressor cells), expressam marcadores de
células mielóides Mac-1 (CD11b) e Gr-1+ (Ly6G) e são induzidas por fatores de
crescimento como o fator de crescimento do endotélio vascular (VEGF), o fator
estimulador de colônias granulocítica e monocítica (GM-CSF) e por citocinas pro -
inflamatórias como IL-1β (Gabrilovich et al., 1996; Young et al., 1997; Bronte et al.,
1999; Song et al., 2005; Bunt et al., 2006). MDSC interferem com a imunidade
tumoral e promovem crescimento tumoral por inibir a citotoxidade das células
tumorais mediada por células NK (Susuki et al., 2005), e por bloquear a ativação
de linfócitos T CD4+ e CD8+ reativos ao tumor (Kusmartsev et al., 2000; Bronte et
al., 2001; Melani et al., 2003; Sinhá et al., 2005). Além disso, recentes estudos
mostraram que MDSC também interferem na imunidade anti-tumoral tanto por
induzir o acúmulo de células T reguladoras imunossupressoras (Gabrilovich et al.,
2001; Huang et al., 2006) como por interagir com macrófagos, os quais aumentam
a produção de IL-10 e decrescem os níveis de IL-12, favorecendo assim um
ambiente imunossupressor propício para a progressão tumoral (Sinhá et al.,
2007).

6
Células B-1
Embora esteja bem estabelecido que linhagens mielóides são os
precursores clássicos de macrófagos, recentes evidências sugerem que a
linhagem linfóide pode também gerar células fagocíticas com características de
macrófagos (macrófago-like), como as células B-1 (Almeida et al., 2001).
As células B-1, subtipo de células B, foram inicialmente identificadas como
células B CD5+ que participam de processos autoimunes em camundongos
(Hayakawa et al., 1983), com características similares às das células responsáveis
pela Leucemia Linfocítica Crônica em humanos (Boumsell et al., 1978). Estas
células diferem das células B-2 convencionais por sua localização anatômica,
expressão de marcadores de superfície e funcionalidade (Hayakawa et al., 1985,
Herzenberg et al., 1986; Martin et al., 2001).
Estas células representam somente 1-5% das células B totais em
camundongos, mas são a principal população de células B nas cavidades
peritoneal e pleural, podendo ser também encontradas no baço, linfonodos e
intestinos (Kantor & Herzenberg, 1993). Células B-1 diferem das células B-2
convencionais por expressar altos níveis de IgM (IgMhi), baixos níveis de IgD
(IgDlo) e B220 (B220lo) e por expressar CD43 (sialoforina, marcador de leucócitos,
exceto nas células B). A maioria das células B-1 das cavidades peritoneal e
pleural expressam CD11b ou Mac-1, marcador de macrófagos/granulócitos que é
parte do receptor do complemento CR3. Entretanto, a maioria das células B-1 do
baço não expressam este marcador (Kantor & Herzenberg, 1993). Além disso, as
células B-1 presentes na cavidade peritoneal podem ser subdividas em 3

7
populações pela expressão de CD5 (marcador de linfócitos T) e CD11b. As células
B-1a expressam CD5 e CD11b; células B-1b não expressam CD5 mas expressam
CD11b; e, mais recentemente descritas, as células B-1c expressam CD5 mas não
expressam CD11b (Herzenberg et al., 1986; Kantor et al., 1992; Hastings et al.,
2006).
As diferenças funcionais entre as células B-1 e B-2 são numerosas e têm
sido revisadas extensamente (Kantor & Herzenberg, 1993; Martin et al., 2001;
Berland and Wortis, 2002; Hardy, 2006). O repertório de anticorpos produzidos
pelas células B-1 é menos diverso, comparado ao das células B-2 convencionais,
refletindo assim na capacidade de resposta ao antígeno destas duas populações
celulares (Hardy et al., 1989; Baumgarth et al., 2005). Células B-2 respondem
eficientemente a antígenos protéicos e participam da resposta imune adaptativa
por sofrer hipermutação somática dos genes das imunoglobulinas e maturação da
afinidade do receptor. Em contraste, células B-1 respondem rapidamente a
antígenos timo-independentes, razão pela qual têm sido reconhecidas como
veículos da imunidade inata (Hardy, 1992; Herzenberg, 2000; Baumgarth et al.,
2005). Além disso, recentes observações mostraram diferenças funcionais mesmo
entre os subtipos de células B-1 (Alugupalli et al., 2004; Haas et al., 2005;
Alugupalli & Gerstein, 2005; Hsu et al., 2006). Células B-1a secretam de forma
espontânea IgM, provendo a primeira barreira contra bactérias encapsuladas
como Streptococcus pneumoniae. Em contraste, a produção de anticorpos por
células B-1b é induzida e está envolvida na remoção final de patógenos e na
proteção a longo prazo (Haas et al., 2005).

8
Por outro lado, diferentemente das células B-2, que têm período de vida
curto e são continuamente geradas por progenitores na medula óssea, as células
B-1 são mantidas por proliferação homeostática ou auto-renovação, como
mostram os experimentos de transferência adotiva de células B-1 em animais
imunodeficientes (Hayakawa et al., 1985; Forster & Rajewisky, 1987). Além disso,
células B-1 independem da sinalização via receptor de células B (BCR, B cell
receptor) para proliferar (Berland & Wortis, 2002; Casola et al., 2004). Karras e
colaboradores (1997) mostraram que esta propriedade de crescimento das células
B-1 é devida à ativação constitutiva da proteína Stat3 (Signal transducer and
activator of transcription 3), que substitui o sinal de proliferação mediado pela
ligação BCR-antígeno. Adicionalmente, células B-1 também expressam níveis
constitutivos da quinase regulada por sinais extracelulares (ERK: extracellular
signal-regulated kinase 1/2). Entretanto, o papel desta quinase nestas células,
ainda é desconhecido, uma vez que o bloqueio da ativação de ERK não afeta a
capacidade de auto-renovação das células B-1 (Wong et al., 2002).
Células B-1 também produzem grandes quantidades de interleucina 10 (IL-
10; O’Garra et al., 1992; Gieni et al., 1997), uma citocina do tipo Th2,
antiinflamatória, que participa da supressão de respostas pro-inflamatórias (Moore
et al., 1993). As células B-1 utilizam esta citocina como fator autócrino de
crescimento (O’Garra et al., 1992) e, por esta razão, a eliminação de células B-1
induzida pelo tratamento com anticorpos anti-IL-10 é acompanhada por drástica
diminuição dos títulos de imunoglobulinas séricas (Ishida et al., 1992).
A participação das células B-1 tem sido observada em várias patologias,
tanto em humanos como em camundongos (Aramaki et al., 1998; Mohan et al.,

9
1998; Hayakawa et al., 2000; Antsiferova et al., 2005; Hardy, 2006; Duan & Morel,
2006). A expansão de células B-1 autorreativas foi associada ao desenvolvimento
de doenças auto-imunes (Murakami et al., 1992; Murakami et al., 1995; Mohan et
al., 1998) e leucemias de células B (Chevallier et al., 1998). Além disso, Minoprio e
colaboradores (1993) demonstraram que na infecção chagásica experimental
ocorre aumento significativo de células B-1 esplênicas na fase aguda da doença e
ainda, camundongos Xid, geneticamente modificados e desprovidos de células B-
1, resolvem com maior eficiência a infecção por esse protozoário. Resultados
semelhantes foram obtidos também em infecções causadas por Paracoccidioides
brasiliensis e por filarias linfáticas (Paciorkowoski et al., 2000; Marcelino França et
al., 2006). Por outro lado, estudos realizados em nosso laboratório demonstraram
que estas células são capazes de migrar para focos inflamatórios inespecíficos
induzidos por corpo estranho, diferenciando-se em fagócitos mononucleares com
características similares a macrófagos (Almeida et al., 2001; Bogsan et al., 2005).
Além disso, outros trabalhos do nosso grupo, alguns deles em andamento,
mostraram a participação das células B-1 na evolução da infecção causada por P.
brasiliensis (Godoy et al., 2003), no lupus eritematoso sistêmico murino (E Brito et
al., 2007), na cicatrização (Oliveira et al., in preparation), na formação de células
gigantes (Bogsan et al., 2005), e de granulomas (Vigna et al., 2006; Russo, in
preparation).
Além do papel imuno-modulador das células B-1 nas patologias descritas
acima, resultados ainda não publicados pelo nosso grupo (Staquicini, 2004)
mostraram que o contato prévio de células B-1 com células de melanoma murino
da linhagem B16 foi capaz de induzir aumento no potencial metastático dessas

10
células tumorais sugerindo assim, a participação das células B-1 também na
regulação de mecanismos que controlam metástases.
Melanoma
Melanoma é um tipo comum e letal de câncer de pele que acomete
principalmente populações relativamente jovens (Parker et al., 1997). O melanoma
tem sido catalogado como a terceira forma mais comum de câncer de pele; porém,
devido à sua alta taxa de recorrência e capacidade metastática, é a forma que
maior número de mortes produz. Além disso, o melanoma é altamente resistente
aos métodos convencionais de tratamento como quimioterapia e radioterapia
(Sober et al., 1991; Li et al., 2002; Tarhini & Agarwala, 2006).
O melanoma origina-se de células pigmentadas denominadas melanócitos,
células derivadas de melanoblastos neuro-ectodermais que migraram durante a
embriogênese para a epiderme, derme e outros sítios, sendo encontradas na
camada basal da pele e em outras superfícies epiteliais como olhos e ouvido
interno (Herlyn et al., 2000; Smith et al., 2002).
De acordo com sua localização anatômica e padrões de crescimento, o
melanoma pode ser dividido em diferentes subtipos: melanoma superficial,
nodular, lentigo acral, lentigo maligno e melanoma não-cutâneo (Tabela 1).
Quanto à progressão, cinco etapas têm sido propostas baseadas no curso clínico
e nas características histopatológicas: 1) nevo congênito, com melanócitos
estruturalmente normais os quais têm curto período de vida e geralmente
carregam anormalidades citogenéticas; 2) nevo displásico, caracterizado por

11
células com atipia estrutural e, em alguns casos, é precursor da neoplasia; 3)
etapa de crescimento radial (RGP), fase inicial do melanoma primário,
caracterizado pelo crescimento horizontal da epiderme podendo ou não alcançar
crescimento vertical (VGP), e pode ser curado por excisão cirúrgica; 4) VGP, fase
na qual as células invadem a derme e têm potencial para metastatizar; 5)
melanoma metastático, com a formação de metástases em sítios distantes do
local de origem do tumor primário (Clark et al., 1984; Meier et al., 1998; Rusciano,
2000; Chudnovsky et al., 2005).
Tabela 1. Classificação clínica de melanoma.
Subtipo Freqüência Sitio comum Características gerais
Melanoma expansivo
superficial (MES)
70% Tronco nos homens
Pernas nas mulheres
RGP, evolução de 1–5 anos
Melanoma nodular
(MN)
10–25% Tronco nos homens
Pernas nas mulheres
RGP, evolução de 6–18 meses.
Sexo masculino, na proporção
2:1
Melanoma
lentiginoso acral
(MLA)
5% Regiões palmoplantares,
extremidades digitais
(unhas)
Não relacionado à exposição
solar.
Não tem predileção por sexo.
Mais freqüente em não brancos
(30 a 70%).
Melanoma lentigo
maligno (MLM)
<1% Cabeça e pescoço de
idosos
Associada com exposição
crônica ao sol
RGP, evolução de 3–15 anos
Melanoma não
cutâneo
5% Ocular, mucosa Não relacionado à exposição
solar.
Prognóstico e tratamento
deferem dos subtipos cutâneos
Chudnovsky et al., 2005

12
Para compreender os mecanismos biológicos envolvidos no processo
metastático em melanomas, estudos experimentais vêm sendo realizados com
células de melanoma murino B16 (Johnson et al., 1987). Estas células foram
geradas espontaneamente em camundongos C57BL/6 e isoladas por Isaiah Fidler
(1973). A partir da linhagem parental B16, Fidler (1978) obteve variantes com
diferentes potenciais de malignidade. Para tanto, células B16 foram injetadas
endovenosamente em camundongos singênicos e, após três semanas, as
primeiras colônias formadas no pulmão foram dissecadas e adaptadas à cultura
celular. A primeira variante selecionada in vivo, B16F1, foi re-injetada em novos
animais e, após três semanas, as colônias formadas no pulmão foram novamente
dissecadas para posterior cultivo. Assim, dez linhagens foram selecionadas in vivo
com crescente potencial metastático, uma vez que células B16F10 apresentaram
capacidade superior à linhagem parental para metastatização. Em conjunto este
estudo demonstrou claramente que células B16 são heterogêneas e que poucas
células de potencial altamente maligno pré-existem na linhagem parental.
Recentes avanços para determinação dos mecanismos envolvidos na
transformação e malignização de células tumorais como o melanoma mostraram
aumento nos níveis de ativação de várias vias de sinalização intracelular
envolvidas com invasão e progressão metastática, dentre as quais se destaca a
via das MAPKs (Mitogen-activated protein kinases; Buettner et al., 2002;
Chudnovsky et al., 2005; Meier et al., 2005, Lopez-Bergami et al., 2007; Meier et
al., 2007). O aumento na ativação destas vias induz expressão de genes que
codificam integrinas, fatores de crescimento, metaloproteinases, quimiocinas,
citocinas, oncogenes, entre outras moléculas que favorecem o avanço e

13
instalação das células tumorais em sítios distantes do tumor primário (Lewis et al.,
2005; Axelsen et al., 2007). Sendo assim, o estudo do papel das vias de
transdução de sinal envolvidas com invasão e metástases torna-se indispensável
para esclarecer os mecanismos celulares e moleculares pelos quais células
tumorais como o melanoma adquirem o fenótipo metastático.
Quinase regulada por sinais extracelulares (ERK: Extracellular-signal
Regulated Kinase)
A via das proteínas quinases ativadas por mitógenos (MAPKs: mitogen-
activated protein kinase) é uma cascata de sinalização intracelular essencial para
numerosos eventos celulares. Esta via é ativada diferencialmente por fatores de
crescimento, hormônios, citocinas, stress celular (irradiação, choque térmico,
desigualdade osmótica, dano ao DNA) e por produtos bacterianos como o
lipopolissacarídeo (LPS). A ativação das MAPKs em resposta a estes estímulos
está relacionada ao controle de diversos eventos biológicos como resposta
inflamatória, metabolismo, diferenciação, ciclo celular, proliferação e apoptose. As
três principais vias das MAPKs incluem a quinase regulada por sinais
extracelulares (ERK1/2) também referida como via Ras-Raf-MEK-ERK; a quinase
Jun N-terminal (JNK: June N-terminal kinase) e p38 (Cobb & Goldsmith, 1995;
Weston & Davies, 2002; Zebisch et al., 2007).
As MAPK são quinases serina/treonina as quais são ativadas pela
fosforilação de seus resíduos serina e/ou treonina localizados em regiões
específicas (Crews et al., 1992). A via canônica de Ras-Raf-MEK-ERK é
desencadeada pela interação de ligantes extracelulares com os receptores tirosina

14
quinase (RTKs) na superfície celular. Esta ligação induz o recrutamento
intracelular de Ras GTP, envolvido na ativação de Raf-1. Raf-1 ativado fosforila
MEK que fosforila ERK, o qual dimeriza e transloca para o núcleo celular, onde
fosforila e estimula diversos fatores de transcrição que regulam a expressão de
genes associados ao ciclo celular, proliferação, sobrevivência, diferenciação,
adesão, remodelamento de matriz extracelular, entre outros eventos (Viala &
Pouyssegur, 2004; Zebisch et al., 2007; Gráfico 1).
Nos últimos anos o papel da via Ras/Raf/MEK/ERK na progressão do
melanoma tem recebido grande atenção devido à identificação de mutações no
gene B-Raf em 70% dos casos de melanomas ocasionados por exposição
constante ao sol (Davies et al., 2002; Maldonado et al., 2003). Esta mutação
confere super-ativação da quinase ERK a qual tem sido associada com expansão
metastática de melanomas (Klafter & Arbiser, 2000; Cohen et al., 2002;
Govindarajan et al., 2003; Smalley, 2003; Lopez-Bergami et al., 2007). Além das
mutações no gene B-Raf, mutações no gene N-Ras, verificadas em quase 20% de
melanomas humanos, podem também conferir ativação constitutiva da via das
quinases Raf/MEK/ERK em melanoma (Mansour et al., 1994; Davies et al., 2002;
Gorden et al., 2003).
Devido ao fato desta via estar envolvida com várias funções celulares,
numerosos estudos experimentais em modelos murinos e humanos mostraram
correlação direta entre a ativação da via das MAPKs com angiogênese,
tumorigênese (D´Angelo et al., 1995; Ilan et al., 1998; Cohen et al., 2002) e
metástases (Krueger et al., 2001; Sahai et al., 2001; Choo et al., 2005), razão pela

15
qual esta via vem sendo estudada como alvo terapêutico (Chudnovsky et al.,
2005; Alvarado & Giles, 2007; Lopez-Bergami et al., 2007).
Assim, o estudo da participação da via ERK no aumento do potencial
metastático das células B16 induzido pelo contato com células B-1, poderia
contribuir para explicar os mecanismos envolvidos neste fenômeno.
Gráfico 1. Via canônica de Ras-Raf-MEK-ERK .
RTKRTK

2. OBJETIVOS

2. OBJETIVOS
1.1 OBJETIVOS
Considerando que células B-1 são capazes de induzir aumento no potencial
metastático de células de melanoma murino da linhagem B16, este estudo tem
como objetivo principal determinar os mecanismos celulares envolvidos nesse
fenômeno.
Objetivo específico
1. Investigar a participação de ERK no aumento do potencial metastático de
células de melanoma B16 induzido pelo contato com células B-1.

3. MATERIAIS E MÉTODOS

3. MATERIAIS E MÉTODOS
2.1. MATERIAIS
2.1.1 Soluções Rotineiras
Todas as soluções aqui descritas foram preparadas com água bi-destilada
ou com padrão Mili-Q e com reagentes qualidade P.A.
• Solução estoque de tampão fosfato-salina 20X (PBS): NaCl 2,7378 M,
KCl 0,053 M, Na2HPO4 0,1301 M e K2HPO4 0,0229 M. Para uso, esta solução foi
diluída 20X (PBS 1X).
• Solução estoque de tampão tris-salina 10X (TBS): Tris base 24,2 g; NaCl
80g; pH 7.6 e 1000 mL de água q.s.p. Para uso, esta solução foi diluída 10X.
2.1.1.1 Soluções para SDS-PAGE
• Tampão de amostra para SDS-PAGE: Tris-HCl 62 mM, pH 8,8, contendo
0,2% (p/v) de SDS, β-mercaptoetanol 50 mM, 0,005% (p/v) de azul de bromofenol
e 10% (v/v) de glicerol.
• Solução estoque de acrilamida/bis para gel de proteína: 30% de
acrilamida, 0,8% de bis-acrilamida e água bi-destilada 100 mL q.s.p.
• Tampão para gel de proteína (separação) desnaturante: Tris-HCl 1,5 M,
pH 8,8, contendo 0,4% de SDS (p/v).
• Tampão para gel de proteína (empilhamento) desnaturante: Tris-HCl 0,5
M, pH 6,8, contendo 0,4% de SDS (p/v).
• Tampão de corrida para SDS-PAGE: Tris 25 mM, glicina 190 mM, pH 8,3,
e 0,1% de SDS (p/v).

20
2.1.1.2 Soluções para coloração por Comassie blue
• Corante de azul de Coomassie: 0,2% de Coomassie R250 (p/V) em 50%
de metanol e 10% de ácido acético (v/v).
• Descorante forte para Coomassie: 50% de metanol, 10% de ácido acético
e 40% de água destilada (v/v).
2.1.1.3 Soluções para Immunoblotting
• Tampão de transferência: Tris-HCl 25 mM, glicina 192 mM e 20% de
metanol (v/v).
• Corante de Ponceau: 0,5% (p/v) de Ponceau S em 5% de ácido acético
(v/v).
• Tampão de bloqueio: TBS 1X, 0,1% de Tween-20 com 5% de leite
desnatado em pó (p/v).
• Tampão de lavagem: 0,1% de Tween-20 em TBS 1X.
• Tampão striping: 1,87 g de glicina pH 2.5, 100 mL de SDS 10% e 1000
mL de água q.s.p.
2.1.1.4 Soluções para ELISA (Enzyme Linked Immuno Sorbent Assay)
• Tampão de lavagem (PBS-T): 0,05% de Tween-20 em PBS 1X.
• PBS-BSA (soro albumina bovina): 3% de BSA em PBS 1X.
• Peróxido de Hidrogênio: 3% de Peróxido de Hidrogênio em PBS 1X.
• Tampão para OPD: Na2PO4 0,4 M; Ácido Cítrico 0,4 M; pH 5,8, contendo
5.05% (V/V) de H2O2 (Perhidrol, Merk).

21
• Bloqueio da Revelação: Ácido Sulfúrico (H2SO4) 4 N em água destilada.
2.1.1.5 Soluções para cultura de células
• Meio RPMI: RPMI 1640 (Sigma, St Louis, MO) acrescido de ácido N-2-
hiroxietilpiperazina-N-2-etanosulfônico 10 mM (HEPES, da Sigma), bicarbonato de
sódio 24 mM (Sigma) e 40 µg/mL de garamicina (Schering Plough, N.J, USA).
Após preparação o meio foi esterilizado por filtração positiva em filtro de 0,22 µM.
• Meio completo (R-10): meio RPMI suplementado com 10% de soro bovino
fetal (SBF, da Cultilab, Campinas, SP).
• Meio para congelamento de células: 45% de RPMI, 45% de soro fetal
bovino e 10% de DMSO.
• PBS-EDTA (ácido etileno diaminoacético): 2,0 mM de EDTA em PBS 1X.
Após preparada esta solução foi esterilizada em autoclave a 120°C por 15
minutos.
2.1.2 Animais
Camundongos C57BL/6 wild-type (wt) fêmeos de 2 a 3 meses de idade
foram cedidos pelo Laboratório de experimentação animal do INFAR da
Universidade Federal de São Paulo, UNIFESP. Camundongos C57BL/6 knock-out
para IL-10 (KO) fêmeos de 2 a 3 meses de idade foram cedidos gentilmente pelo
Dr. João Santana da Silva (Departamento de Farmacologia, Escola de Medicina,
USP, Riberão Preto, SP). Os animais foram mantidos livres de germes, em micro-
isoladores com água e ração autoclavadas. Todos os procedimentos realizados

22
com os animais foram executados de acordo com normas estabelecidas pelo
Comitê de Ética em Experimentação Animal da Universidade Federal de São
Paulo.
2.2 MÉTODOS
2.2.1 Cultura de células e tratamentos
Células de melanoma B16 (B16): Células de melanoma de baixo potencial
metastático, obtidas em nosso laboratório a partir da linhagem de melanoma
murino B16F10 (Staquicini et al., 2003), foram cultivadas como monocamadas em
meio completo (RPMI-1640 suplementado com 10% de soro fetal bovino, SBF) a
37oC em atmosfera úmida de 5% de CO2. O descolamento das células para
posterior sub-cultivo foi realizado por curta exposição à PBS-EDTA 2 mM ou
Tripsina (Cultilab). Nos ensaios de Western blott ou Immunoblot, as células B16
utilizadas para extratos foram previamente plaqueadas em garrafas de cultura de
75 cm2 ou placas de Petri 100 X 90 mm. Após de alcançar confluência de 80%, o
meio foi aspirado e as células aderidas foram lavadas 3 vezes com PBS e re-
cultivadas com RPMI sem soro por 24 horas. Em seguida, estas células foram ou
não estimuladas com 10% de SBF ou tratadas com inibidor de MEK1/2 (PD98059,
Cell Signalling, Beverly, MA, USA) ou com IL-10 recombinante (Pharmigen, San
Diego, CA), durante diferentes períodos de tempo (entre 15 e 120 minutos).
Células B-1: Linfócitos B-1 foram obtidos a partir de cultura de células
aderentes de lavado peritoneal de camundongos C57BL/6 como descrito
previamente (Almeida et al., 2001). Assim, células peritoneais de camundongos

23
wild-type (B-1wt) e knock-out para IL-10 (B-1KO), foram coletadas por lavagens
sucessivas com RPMI e incubadas em garrafas de cultura por 40 minutos a 37oC.
Em seguida, o sobrenadante foi desprezado e as células aderidas foram re-
cultivadas até 7 dias, em na presença de meio completo. Após este período a
maior porcentagem de células presentes no sobrenadante destas culturas (células
flutuantes) corresponde às células B-1 (Almeida et al., 2001). Para ensaios de
Western blot, monocultura de células B-1 com seis dias de incubação, presentes
no sobrenadante da cultura acima descrita, foram centrifugadas, lavadas 3 vezes
com PBS e re-cultivadas com RPMI sem soro por 24 horas. Em seguida, estas
células foram estimuladas ou não com SBF ou tratadas com inibidor farmacológico
de MEK1/2 (PD98059) de 1 até 24 horas.
Co-culturas de células B16 e B-1: Co-culturas de células B16 e B-1 foram
realizadas em placas de 6 orifícios ou em garrafas para ensaio de metástases in
vivo ou para Western blot, respectivamente. Brevemente, 1X105 células B16 foram
plaqueadas em meio completo e incubadas por 24 horas para adesão. Após este
período, o meio foi aspirado e 1X106 células de B-1 com 5 dias de cultura foram
adicionadas juntamente com seu meio condicionado (mB1) e incubadas por
diferentes períodos na ausência ou presença de PD98059. Logo após os
diferentes períodos de incubação, as células B-1 foram removidas por aspiração e
as células B16 aderidas foram lavadas 3 vezes com PBS para remover qualquer
resíduo de células B-1. Em seguida, células B16 foram coletadas e submetidas a
análises por citometria de fluxo (FACS), utilizando-se os marcadores B220 e Mac-
1 de células B-1 objetivando confirmar a pureza das células B16 após co-cultivo.
Assim células B16 livres de células B-1 após co-cultivo (B16+B-1) foram usadas

24
nos ensaios experimentais in vivo e na preparação de extratos celulares para
análises de Western Blot.
2.2.2 Análises de moléculas de superfície por citometria de fluxo
Monoculturas de células B16 ou B-1 ou células B16 após co-cultivo com B-1
foram coletadas e lavadas 2 vezes com PBS e ajustadas à concentração 1X106
células/amostra. Estas células foram marcadas duplamente com anticorpos anti-
B220 conjugado com ficoeritrina (PE) e anti CD11b (Mac-1) conjugado a
fluoresceína (FITC), ambos da Pharmingen (San Diego, CA, USA), na diluição de
1:100 em PBS acrescido de 1% de BSA e incubados no escuro por 1 hora, no
gelo. Após este período as células foram lavadas e ressuspendidas em 500 µl de
PBS e, em seguida, analisadas por citometria de fluxo (FACSCalibur da Becton
Dickinson, Mountain View, CA) usando o programa CellQuest.
2.2.3 Ensaio de viabilidade celular por Trypan blue
Solução de Trypan blue foi preparada em PBS e filtrada antes do uso.
Células B16 ou B-1 de monoculturas ou co-culturas após diferentes tratamentos
descritos anteriormente, foram coletadas e ressuspendidas em PBS. Dez
microlitros de cada suspensão celular foram diluídos com 10 µL do reagente
Trypan blue e, em seguida, 10 µl desta diluição foram contados em câmara de
Neubauer para estabelecer o porcentual de células coradas (mortas) em cada
amostra. Depois de verificada a viabilidade celular, as suspensões celulares com
mais de 80% de células viáveis foram utilizadas para ensaios experimentais in vivo

25
e para a preparação dos extratos celulares utilizados nas análises de Western
Blot.
2.2.4 Ensaio de inibição da adesão
Para criar a condição de cultura não-aderente, monoculturas de células B16
ou B-1 ou co-culturas de ambas as células foram plaqueadas em placas de Petri
cobertas previamente com uma camada fina de agarose 1% e incubadas a 37°C
com 5 % de CO2 por 72 horas. Após este período, células em suspensão foram
coletadas e ressuspendidas em PBS. Agregados celulares foram mecanicamente
separados por agitação vigorosa e as células individuais foram submetidas a
análise de viabilidade por Trypan blue e em seguida a ensaio experimental de
metástase in vivo.
2.2.5 Ensaio de metástase experimental
Células B-1 ou B16 de monoculturas ou células B16 após co-cultivadas com
células B-1 foram coletadas e ressuspendidas em PBS. Logo, 1X105 células
diluídas em 100 µL de PBS foram injetadas na veia da cauda de camundongos
C57BL/6 wild-type. Para cada grupo celular analisado foram feitas injeções em
pelo menos três camundongos. Após 14 dias de inoculação os animais foram
sacrificados e seus pulmões removidos cirurgicamente para estabelecer
macroscopicamente o número de nódulos metastáticos instalados.

26
2.2.6 Preparação de extratos celulares
Para extração de proteínas totais, células foram lavadas 2 vezes com PBS
a 4°C e, em seguida, lisadas com 100 µL de tampão de lise (contendo, 20 mM de
Tris-HCl pH7.4, 100 mM de NaCl, 1 mM de Na3VO4, 40 mM de NaF, o,5% de NP-
40, 1 mM de PMSF, 10 µg/mL de leupeptina e 10 µg/mL de aprotinina), por 10
minutos a 4°C. Após este período, os lisados celulares foram centrifugados por 15
minutos a 4°C para remoção de resíduos não protéicos. Em seguida, os
sobrenadantes foram coletados e a concentração protéica determinada pelo
método de Bradford (1976).
2.2.7 Dosagem protéica pelo micrométodo de Bradford
As determinações das concentrações protéicas foram realizadas segundo
método proposto por Bradford (1976), que utiliza Coomassie brilliant blue (CBB)
G-250 (Sigma) como reativo e albumina bovina (BSA) 1,0 mg/mL como padrão. A
leitura da curva padrão foi realizada em comprimento de onda de 570 nm em leitor
EIA Reader Titertek, Multiskan, MCC340 (Labsystem, Santa Fé, USA).
2.2.8 Eletroforese em gel de poliacrilamida com dodecilsulfato de sódio
(SDS-PAGE)
Os procedimentos para SDS-PAGE foram executados em equipamentos
Mini-Protean II (Bio-Rad Laboratories, Hercules, CA) conforme descrito
anteriormente (Laemmli, 1970). Para as proteínas em estudo, o gel de separação
foi feito na concentração de 10% de acrilamida e o gel de empilhamento na
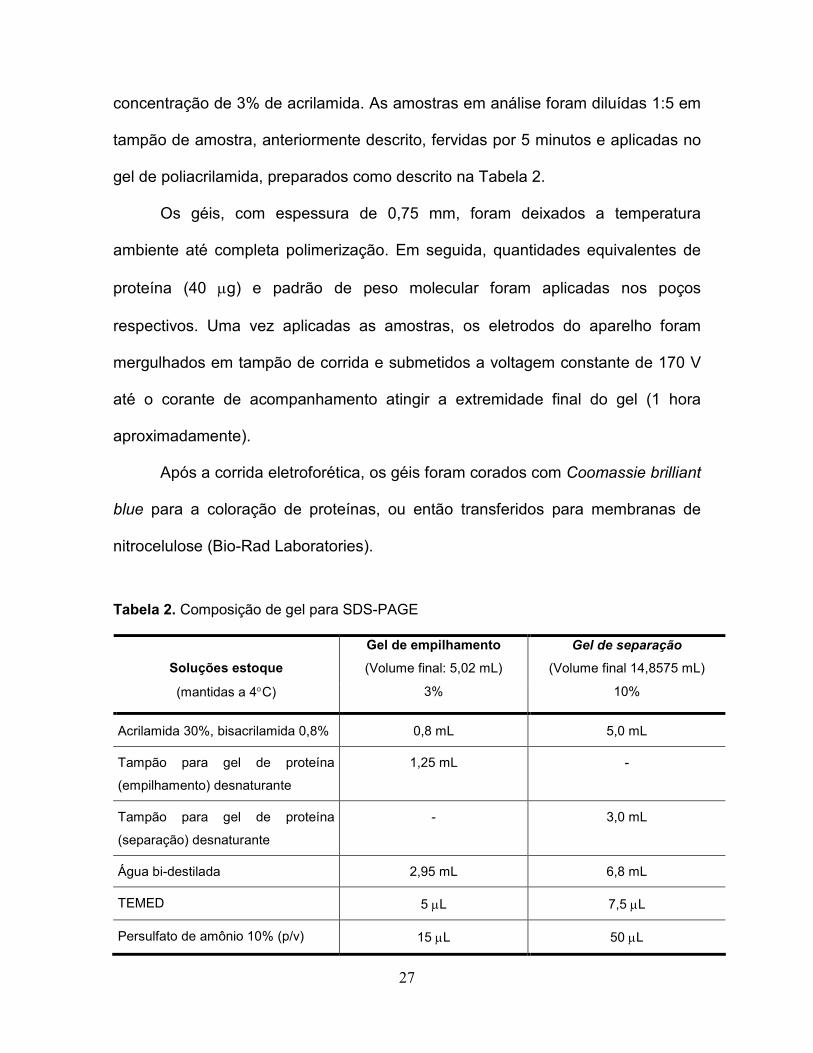
27
concentração de 3% de acrilamida. As amostras em análise foram diluídas 1:5 em
tampão de amostra, anteriormente descrito, fervidas por 5 minutos e aplicadas no
gel de poliacrilamida, preparados como descrito na Tabela 2.
Os géis, com espessura de 0,75 mm, foram deixados a temperatura
ambiente até completa polimerização. Em seguida, quantidades equivalentes de
proteína (40 µg) e padrão de peso molecular foram aplicadas nos poços
respectivos. Uma vez aplicadas as amostras, os eletrodos do aparelho foram
mergulhados em tampão de corrida e submetidos a voltagem constante de 170 V
até o corante de acompanhamento atingir a extremidade final do gel (1 hora
aproximadamente).
Após a corrida eletroforética, os géis foram corados com Coomassie brilliant
blue para a coloração de proteínas, ou então transferidos para membranas de
nitrocelulose (Bio-Rad Laboratories).
Tabela 2. Composição de gel para SDS-PAGE
Gel de empilhamento
(Volume final: 5,02 mL)
Gel de separação
(Volume final 14,8575 mL)
Soluções estoque
(mantidas a 4°C) 3% 10%
Acrilamida 30%, bisacrilamida 0,8% 0,8 mL 5,0 mL
Tampão para gel de proteína
(empilhamento) desnaturante
1,25 mL -
Tampão para gel de proteína
(separação) desnaturante
- 3,0 mL
Água bi-destilada 2,95 mL 6,8 mL
TEMED 5 µL 7,5 µL
Persulfato de amônio 10% (p/v) 15 µL 50 µL

28
2.2.9 Transferência de proteínas para membranas de nitrocelulose
(Immunoblot ou Western blot)
A transferência eletroforética dos géis de poliacrilamida para membranas de
nitrocelulose foi efetuada conforme descrito por Towbin e colaboradores (1979),
usando equipamento Trans-Blot System (Bio-Rad Laboratories). Após a
eletroforese os géis foram postos sobre membranas de nitrocelulose (Bio-Rad
Laboratories) ou PVDF (Amersham Pharmacia Biotech, Buckinghamshire, UK),
recobertos com papel de filtro e comprimidos com esponjas. Todos os materiais
foram embebidos em tampão de transferência e em seguida, encaixados em
placas acrílicas perfuradas e mergulhados na câmara de eletroforese contendo o
mesmo tampão. A transferência foi realizada na voltagem constante de 100 V por
1 hora e meia. Em seguida, as membranas foram coradas com Ponceau S para a
visualização das bandas protéicas e depois bloqueadas com tampão de bloqueio
por 1 hora a temperatura ambiente. Após lavagem, as membranas foram
incubadas overnight a 4°C com anticorpo primário específico para ERK fosforilado
[p-ERK, (E-4): sc-7383), ERK total [(K-23): sc-94], Stat3 total [(C-20): sc-482]
todos da Santa Cruz Biotechnology, INC (Santa Cruz, CA, USA) ou Stat3
fosforilado (p-Stat3, da BioVision, CA, USA), diluídos em tampão de bloqueio
segundo instruções do fabricante. Após este período, as membranas foram
lavadas e incubadas por 1 hora a temperatura ambiente, com o respectivo
anticorpo secundário (Ig-G) conjugado também a peroxidase. Em seguida, bandas
reativas foram visualizadas por quimiluminescência (ECL-Amersham-Pharmacia)
em filmes Kodak e/ou Amersham (Hyperfilm ECL). Após a detecção, as

29
membranas foram lavadas e incubadas com tampão de stripping para serem re-
incubadas com outros anticorpos.
2.2.10 Microscopia eletrônica de transmissão
Células B-1 ou células B16 co-cultivadas ou não com células B-1 foram
incubadas por 24, 48 e 72 horas sobre lamínulas de plástico contidas em placas
de doze (12) orifícios (Costar). Após estes períodos de incubação, as células
foram fixadas em 50% de glutaraldeído, 4% de formaldeído, 0,2 M de cacodilato
de sódio em pH 7.2 por 2 horas. Em seguida, foram lavadas por 4 vezes (1 hora
cada lavagem) com tampão cacodilato 0,1 M e logo fixadas novamente com
tetróxido de ósmio 2% em tampão cacodilato 0,1 M. As amostras foram então
lavadas 3 vezes e desidratadas em concentrações crescentes de etanol e óxido
de propileno. Em seguida, as amostras foram incluídas em resina Araldite® e
mantidas a vácuo por 4 horas e logo a 60°C por 40 horas para polimerização. Uma
vez polimerizadas, as amostras incluídas em Araldite® foram cortadas em seções
de 90 a 95 µm de espessura e coradas com 2% de uranil-acetato aquoso durante
8 minutos. Estas preparações foram subsequentemente coradas com citrato de
chumbo diluído em água destilada por 4 minutos. Amostras foram analisadas e
fotografadas em microscópio eletrônico (JEOL JEM 1200 EX II).

30
2.2.11 Ensaios de proliferação celular
2.2.11.1 Incorporação de timidina tritiada
Células B16 (2X103 células por poço em um volume final de 200 µl) foram
cultivadas com meio completo em placas de 96 poços (uma placa para cada
tempo de incubação: 24, 48, 72 e 120 horas) e incubadas por 6 horas em estufa a
37oC e 5% de CO2. Em seguida, células aderidas foram lavadas com PBS e
ressuspendidas em RPMI sem soro e incubadas overnight nestas condições. Após
este período, o sobrenadante foi retirado e substituído por meio completo ou meio
condicionado de cultura de células B-1 na presença (B16+B-1) ou ausência
(B16+mB-1) de 2X104 células B-1. Células submetidas a estas condições foram
incubadas durante 24, 48 e 72 horas a 37oC e 5% de CO2.. Dezoito horas antes do
término de cada incubação, 1 µCi de timidina tritiada (Amersham) foi adicionado
às culturas. Posteriormente, cada amostra foi adsorvida em papel de filtro e cada
filtro foi colocado em tubos com 2 ml de líquido de cintilação para a leitura em
aparelho Beckman LS 5801. O experimento foi realizado em triplicata para cada
grupo celular estudado.
2.2.11.2 MTT [tetrazolium salt3-(4,5-dimethylthiazol-2-yl)-2,5-
diphenyltetrazolium bromide]
Células B16 (2,5X103 células por poço em volume final de 100 µl) foram
cultivadas com meio completo em placas de 96 poços (uma placa para cada
tempo de incubação: 0, 24, 48, 72 horas) e incubadas 12 horas em estufa a 37oC
e 5% de CO2. Em seguida, células aderidas foram lavadas com PBS

31
ressuspendidas com RPMI sem soro e incubadas overnight nestas condições.
Após este período, o sobrenadante foi retirado e substituído por meio completo ou
meio condicionado de cultura de células B-1 na presença (B16+B-1) ou ausência
(B16+mB-1) de 2X104 células B-1. Células submetidas a estas condições foram
então incubadas durante 24, 48 e 72 horas a 37oC e 5% de CO2. Posteriormente,
2,5 horas antes do término de cada incubação, foram adicionados 10 µl de MTT (5
mg/mL) às culturas e deixados por 1,5 horas a 37oC e 5% de CO2. Em seguida, o
sobrenadante de cada poço foi aspirado e foram adicionados 100 µl de
isopropanol (Merk) e mantidas a temperatura ambiente por 15 minutos. Após este
período, as placas foram analisadas em comprimento de onda de 570 nm em leitor
de ELISA, EIA Reader Titertek, Multiskan, MCC340 (Labsystem). O experimento
foi realizado em triplicata para cada grupo celular estudado.
2.2.12 Dosagem de citocinas no sobrenadante de cultura
A produção de IL-10 no sobrenadante de cultura de células B-1 tanto de
animais wt como KO foi quantificada por ELISA (Enzymed linked
immunoabsorvent assay) seguindo as instruções do fabricante (Duoset mouse IL-
10, R&D System, Minneapolis, MN, USA). Brevemente, células B-1 foram
cultivadas como descrito acima e os sobrenadantes foram coletados após 7 dias
de cultura e mantidos a 70°C até o momento da dosagem. Para a determinação
da citocina, placas de ELISA de 96 poços foram sensibilizadas overnight a
temperatura ambiente com anticorpos monoclonais anti-IL-10. Após este período,
as placas foram lavadas por 4 vezes com PBS-T e logo incubadas com 200 µL de

32
PBS-BSA a temperatura ambiente durante 2 horas, para prevenir ligações
inespecíficas. Placas foram lavadas 4 vezes com OS-T e incubadas 2 horas com
100 µL por orifício de cada sobrenadante (amostras em triplicatas) ou com
standard de IL-10 diluído serialmente em PBS-BSA. Em seguida, placas foram
lavadas 4 vezes com PBS-T e incubadas 2 horas com 100 µL/poço de anti-IL-10 a
temperatura ambiente. As placas foram então novamente lavadas e a revelação
foi realizada com 50 µL/poço de ο-fenilenodiamina (OPD, 1 mg/mL, Sigma)
dissolvida em tampão citrato-fosfato 100 mM pH5,8, contendo 0,05% (v/v) de H2O2
(água oxigenada). A reação foi interrompida após o aparecimento de cor com 50
µL/poço de H2SO4 4 N (50 µL/poço). Os resultados foram avaliados por leitura
espectrofotométrica em comprimento de onda 492 nm em leitor de ELISA
(Labsystems). A sensibilidade do ensaio foi de 20 pg/mL. Resultados são
apresentados como medianas ±SD.
2.2.13 Obtenção de RNA total
O RNA total foi extraído de células B-1 e de células B16 tanto de
monoculturas como co-culturas utilizando Trizol (Invitrogen, Carlsbad, CA) de
acordo com as instruções do fabricante. Brevemente, 1,5 mL de Trizol foi
adicionado às células e após homogeneização por 5 minutos, o lisado celular foi
transferido para tubos de propilpropileno de 2 mL, e adicionados 300 µL de
clorofórmio a cada tubo. O conteúdo foi agitado vigorosamente e centrifugado a
14.000 rpm por 15 minutos a 4°C. A fase aquosa (superior) foi transferida para
microtubos de 1,5 mL, contendo 750 µL de isopropanol e em seguida

33
homogeneizado. Após precipitação por 24 horas a -20°C, centrifugou-se a 14.000
rpm por 15 minutos a 4°C. O RNA precipitado foi lavado 2 vezes com etanol 70%
em água DEPC 0,001% (dietil pirocarbonato), desidratado por evaporação a
temperatura ambiente e diluído de 30 a 50 µL de água DEPC.
A quantificação do RNA foi feita por espectrofotometria a 260 nm. O
resultado da densidade óptica (DO) foi multiplicado pelo fator 40, o que fornece a
concentração em µg/mL, uma vez que uma unidade de DO a 260 nm equivale a
40 µg/mL.
A qualidade do RNA foi avaliada de duas maneiras: em primeiro lugar, com
relação à contaminação por proteína, a qual é dada pela razão entre as leituras a
260 e 280 nm, que precisa estar entre 1,6 e 1,8. Em segundo lugar, foi avaliada a
integridade do RNA utilizando gel de agarose. Nesta análise a amostra é
considerada íntegra e livre de contaminação quando no gel estão presentes duas
bandas, 28S e 18S, sendo que a banda de 28S é mais intensa que a de 18S. A
ocorrência de outra banda, de tamanho superior a de 28S, denota contaminação
por DNA genômico e, neste caso, a amostra deve ser submetida à re-extração por
Trizol.
2.2.14 RT-PCR (Reverse Transcriptase – Polymerase Chain Reaction)
A sínteses de cDNA a partir de amostras de RNA total (transcrição reversa)
para estudos de expressão gênica por RT-PCR, foi realizada com o kit SuperScript
III (Invitrogen- Life Technology) de acordo com as instruções do fabricante. Assim,
a transcrição reversa foi realizada a partir de 3 µg de RNA total, tratado

34
previamente com DNase 1 (Invitrogen- Life Technology), para eliminar qualquer
tipo de contaminação com DNA genômico, conforme verificado pela não
amplificação do gene de controle interno GAPDH.
Após preparados os cDNAs foram submetidos a RT-PCR qualitativo e a
RT–PCR quantitativo em tempo real.
2.2.14.1 Análise qualitativa por RT-PCR
Cada amplificação foi realizada em volume total de 25 µl contendo 100 ng
de cada primer (sense e anti-sense), 1,5 mM de solução de 25 mM de Mg2+, 10 ul
de MasterMix (2,5x) (Eppendorf) e 2 µl de cDNA diluído 1:2 em água bi-destilada
estéril. Como controle negativo de cada reação de PCR foi utilizada água bi-
destilada ao invés de cDNA .
As condições de amplificação comuns para cada par de primers foram:
desnaturação 94°C por 30 segundos, anelamento por 30 segundos (temperatura
específica para cada par de primers), extensão 72°C por 30 segundos, número de
ciclos 35. A temperatura de anelamento de cada par de primers foi escolhida após
o teste de gradiente de temperatura utilizando o termo-ciclador Mastercycler
Gradient (Eppendorf). Na Tabela 3 estão descritas as seqüências de
oligonucleotídeos utilizadas para cada transcrito e, também a temperatura de
anelamento, e o tamanho dos produtos do PCR em número de pares de bases
(bp).

35
Tabela 3. Seqüências oligo-nucleotídicas dos primers sense (S) e anti-sense (AS),
temperatura de anelamento (°C) e o tamanho do produto do PCR em pares de bases
(BP), dos transcritos estudados por RT-PCR qualitativo.
Transcrito Primers
5’-seqüência oligo-nucleotídica- 3’
Temperatura de anelamento (°C)
Tamanho do produto (bp)
GAPDH S: ACCACAGTCCATGCCATCAC AS: TCCACCACCCTGTTGCTGTA
62 500
CXCR4 S: AGTTTTCACTCCAGCTAACCCTTAT AS: TGGAATGTTCAGTTTCTTCCTCTAC
58 321
MMP-9 S: TTCTCTGGACGTCAAATGTGG AS: CAAAGAAGGAGCCCTAGTTCAAGG
60 413
Para a análise da amplificação, aos produtos de PCR foram adicionados 5
µl de loading buffer, constituído por solução de glicerol a 50% (Sigma Chemical
Company) acrescida de 1mg/ml de vermelho de cresol (Sigma Chemical
Company) em 0,5x TBE (Gibco BRL, Gaithersburg, MD, USA). Cada amostra
diluída 1:6 com este tampão foi aplicada em volume de 10 µl ao gel de agarose
1,5 % (Gibco BRL) preparado em 0,5x TBE com 0,75 µg/ml de brometo de etídio
(Sigma Chemical Company) e submetida a eletroforese com 100 V por 50
minutos. Adicionalmente, também foi submetido a eletroforese o marcador de
massa molecular (Low mass, Invitrogen). As bandas foram visualizadas sob luz
ultravioleta. Uma vez verificada a positividade no controle e a ausência de
contaminação (controle negativo), a análise do mRNA foi realizada através do
sistema de documentação de imagem do gel de eletroforese “Kodak Digital
Science – Electrophoresis Documentation and Analysis System 120” (Eastman
Kodak Co,, Rochester, NY, USA).

36
2.2.14.2 Análise quantitativa por RT-PCR em tempo real
A reação de PCR em tempo real foi realizada com o reagente Sybr Green
(Applied Biosystems). A partir da reação de transcrição reversa, utilizou-se 1 mL
de cDNA, 7,5 mL de master mix Sybr Green, 0,5 mL de cada oligonucleotídeo (18
µM): sense e anti-sense (Tabela 4), e água Milli Q autoclavada q.s.p 15 mL. Em
seguida, as reações para PCR em tempo real foram realizadas utilizando ABI
7000 Real Time PCR System (Applied Biosystems), nas seguintes condições: 10
minutos a 50°C para ativação da enzima, desnaturação por 5 minutos a 95°C, e 40
ciclos de 95°C por 30 segundos e 60°C por 1 minuto. Após avaliar a qualidade da
reação com base nas curvas de dissociação, os resultados foram analisados
utilizando o programa ABI 7000 SDS software (Applied Biosystems). Para a
quantificação relativa, foi realizado o seguinte cálculo: inicialmente determinou-se
o cycle threshold (CT), dado pelo número do ciclo em que o sinal de fluorescência
atingiu a linha limiar (threshold line), ou seja, a linha em que a emissão de
fluorescência está acima do ruído de fundo (background). O CT encontrou-se
invariavelmente na região correspondente à fase exponencial da amplificação, o
que torna mais acurada a estimativa de quantificação dos transcritos na amostra
original. Os valores de CT dos genes de interesse foram normalizados em relação
ao CT do gene constitutivo (GAPDH), resultando o ∆CT, representado pelo
CTgene - CTconstitutivo. Por fim, calculou-se o 2-∆CT, sendo este o valor a ser
trabalhado como representante da expressão relativa para cada gene
(Vandesompele et al., 2002; Bustin et al., 2005). Para melhor interpretação dos
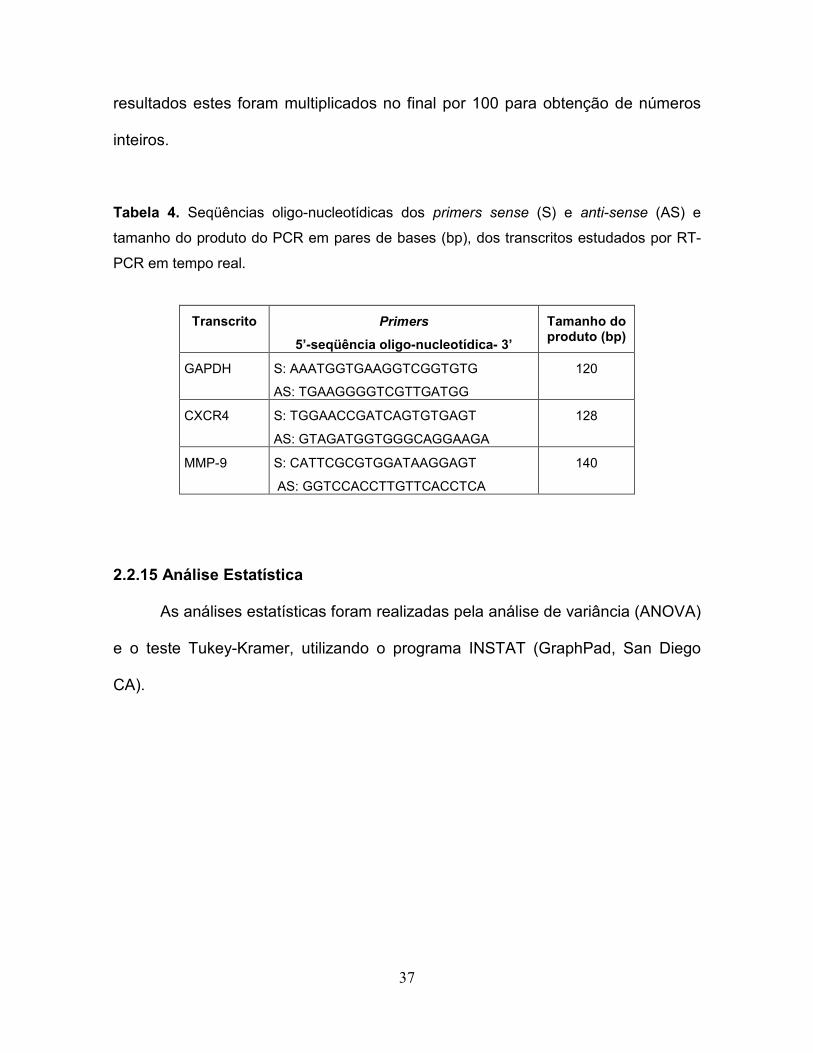
37
resultados estes foram multiplicados no final por 100 para obtenção de números
inteiros.
Tabela 4. Seqüências oligo-nucleotídicas dos primers sense (S) e anti-sense (AS) e
tamanho do produto do PCR em pares de bases (bp), dos transcritos estudados por RT-
PCR em tempo real.
Transcrito Primers
5’-seqüência oligo-nucleotídica- 3’
Tamanho do produto (bp)
GAPDH S: AAATGGTGAAGGTCGGTGTG
AS: TGAAGGGGTCGTTGATGG
120
CXCR4 S: TGGAACCGATCAGTGTGAGT
AS: GTAGATGGTGGGCAGGAAGA
128
MMP-9 S: CATTCGCGTGGATAAGGAGT
AS: GGTCCACCTTGTTCACCTCA
140
2.2.15 Análise Estatística
As análises estatísticas foram realizadas pela análise de variância (ANOVA)
e o teste Tukey-Kramer, utilizando o programa INSTAT (GraphPad, San Diego
CA).

4. RESULTADOS

4. RESULTADOS
3.1 Caracterização do sistema de co-cultivo
Estudos anteriores realizados no nosso laboratório mostraram que o co-
cultivo de células B-1 com células de melanoma murino da linhagem B16, foi
capaz de induzir aumento no potencial metastático destas células (Staquicini,
2004). Com o intuito de determinar quais mecanismos moleculares controlam este
efeito, experimentos iniciais foram realizados para caracterizar o sistema de co-
cultivo e avaliar o tipo de interação entre estas células. Para isso, células B-1
foram isoladas de lavados peritoneais de camundongos e co-cultivadas por 24, 48
e 72 horas com uma sublinhagem de células B16 caracterizada pelo seu baixo
potencial metastático (Staquicini et al., 2003). Após estes períodos, células B-1
foram eliminadas por aspiração e em seguida, células B16 livres de células B-1
foram utilizadas em ensaios experimentais de metástase in vivo. O método de
eliminação das células B-1 após os diferentes períodos de co-cultivo foi eficiente,
como evidenciado por citometria de fluxo (Fig. 1). Esta análise mostrou que
populações celulares duplamente marcadas para os marcadores fenotípicos B220
e Mac-1 de células B-1, foram detectados em células B-1, porém não em
monoculturas de células B16 ou em células B-16 livres de células B-1 após o co-
cultivo (B16+B1). Após 14 dias da inoculação, o número de nódulos metastáticos
presentes nos pulmões dos animais foi determinado macroscopicamente.
Conforme demonstrado na Figura 2, a inoculação de células B16 submetidas ao
co-cultivo com células B-1 por 48 ou 72 horas, aumentou significativamente o

40
número de nódulos metastáticos quando comparado com animais injetados com
células B16 ou B-1 isoladas de monoculturas.
Uma vez que no sistema de co-cultivo as células B16 são aderentes e
células B-1 não são, foi determinado se o aumento do potencial metastático das
células B16, induzido pela presença das células B-1, dependia da adesão das
células B16 ao plástico. Para testar isto, os dois tipos celulares foram co-
cultivados sobre substrato não adesivo (agarose) por 72 horas e após este
período um pool de células B16+B1 foi submetido a ensaios experimentais de
metástases in vivo. Conforme mostrado na Figura 3, animais injetados com células
B16 co-cultivadas com B-1 em suspensão (sB16+B1) produziram
significativamente mais colônias metastáticas no pulmão comparado aos grupos
controles injetados com células B16 (sB16) ou B-1 (sB1) coletadas de
monoculturas mantidas nas mesmas condições (Fig. 3A). Adicionalmente, ensaios
de viabilidade celular por Trypan blue mostraram a manutenção da viabilidade das
células B16 e B-1 mantidas em suspensão por 72 horas (Fig. 3B). Em conjunto,
estes resultados demonstram que células B-1 são capazes de aumentar o
potencial metastático das células B16 e que a adesão das células B16 não é
necessária para este efeito.
3.2 Células B-1 e B16 fisicamente interagem durante o co-cultivo.
Em seguida, foi determinado se o aumento no potencial metastático das
células B16 era dependente do contato físico entre as duas células ou se fatores
solúveis produzidos pelas células B-1 poderiam estar envolvidos neste feito. Para

41
esta análise, células B16 foram cultivadas por 72 horas em meio condicionado
proveniente tanto de monoculturas de células B-1 como de células B16 co-
cultivadas com células B-1 por 72 horas. Como apresentado na Figura 4, não
foram observadas diferenças no número de nódulos metastáticos nos pulmões de
animais injetados com células B16 cultivadas em seu próprio meio comparado
com aquelas cultivadas em meio condicionado tanto de monoculturas de células
B-1 (B16+mB1) como do co-cultivo de células B16 com B-1 [B16+m(B16+B1)]. Por
outro lado, quando células B16 submetidas ao co-cultivo com células B-1 (B16+B-
1) foram injetadas, o número de nódulos metastáticos presentes nos pulmões
desses animais foi significativamente maior comparado com os grupos acima
descritos.
Desta forma, os resultados acima descritos mostram que fatores solúveis
produzidos pelas células B-1 são insuficientes para aumentar a capacidade
metastática das células B16, sugerindo que o contato físico entre as duas células
ocorre durante o co-cultivo. De fato, análises por microscopia eletrônica de
transmissão revelaram a existência de interação física entre as células B16 e B-1.
Na Figura 5A, podemos observar uma célula B-1 caracterizada por sua atípica
morfologia nuclear com reentrâncias delimitando dois lóbulos distintos, descrita
por Abrahão e colaboradores em 2004. Na Figura 5B, não observamos interação
física entre células B-1 e células B16 co-cultivadas por 24h. Entretanto, quando
estas células foram co-cultivadas por 48 e 72h, vários pontos de interação entre as
membranas plasmáticas destas células foram detectados (Fig. 5C e D). Juntos
estes resultados demonstram que estes dois tipos celulares interagem fisicamente

42
quando co-cultivados por tempos superiores a 48h, observação esta que
correlaciona com o tempo de co-cultivo necessário para aumentar o potencial
metastático das células B-16 (Fig. 2).
3.3 Fosforilação de ERK está aumentada nas células B16 após
interação com células B-1
Considerando que células B16 e B-1 interagem fisicamente, foi
hipotetizado que as células B-1 poderiam influenciar no aumento do potencial
metastático das células B16 através da interferência em vias de sinalização celular
que regulam invasão e metástase.
Em melanomas há relatos mostrando que a ativação da quinase regulada
por sinais extracelulares ERK 1/2, da família das MAPKs (proteínas quinases
ativadas por mitógenos), está envolvida de forma positiva na regulação da
proliferação celular, assim como no aumento do potencial metastático destes
tumores (Davies et al, 2002; Zebish et al., 2007; Lopez –Bergami et al., 2007). Por
esta razão, a participação de ERK foi avaliada no aumento do potencial
metastático das células B16 após o co-cultivo com células B-1.
Inicialmente, foi determinado o perfil de fosforilação basal de ERK nas
monoculturas tanto de células B16 como de células B-1. Para isso foi utilizado um
anticorpo que reconhece especificamente as duas formas (p42/p44 kDa) de ERK
fosforilada/ativada. Como pode ser observado na Figura 6A, monoculturas de
células B16 ou de células B-1 cultivadas overnight na ausência de soro bovino
fetal (SBF) apresentaram níveis basais de fosforilação de ERK. Enquanto a adição

43
de SBF induziu um leve aumento na fosforilação de ERK nas células B16, a
presença de SBF não alterou o perfil basal de fosforilação desta quinase nas
células B-1, mostrando que ambos os tipos celulares apresentam ERK
constitutivamente fosforilado.
Para avaliar se células B-1 poderiam aumentar os níveis basais de
fosforilação de ERK nas células B16 durante o co-cultivo, células B16 foram
mantidas por 24 horas na ausência de soro e então estimuladas ou não com SBF
por 1 hora (1h) na presença ou ausência de células B-1. Após este período,
extratos celulares de monoculturas de células B16 ou de células B16 livres de
células B-1 após o co-cultivo foram produzidos e submetidos a Immunobloting
para investigar os níveis de ERK fosforilado. Conforme mostrado na Figura 6B, um
aumento considerável nos níveis de fosforilação de ERK foi observado nas células
B16 após o co-cultivo com células B-1 (B16+B1), independente da presença de
soro, comparado com células B16 de monoculturas.
3.4 Aumento no potencial metastático de células B16 é dependente
dos níveis de ERK fosforilado
Para avaliar se o aumento na ativação de ERK nas células B16 após o co-
cultivo com células B-1 é um evento importante no aumento do potencial
metastático das células B16, foi utilizado o inibidor farmacológico de MEK1/2
(PD98059) que é a quinase up-stream a ERK na via de sinalização.
Como observado na Figura 7, os níveis basais de ERK fosforilado foram
completamente inibidos na presença de 40 µM de PD98059. Além disso, as

44
análises por Trypan blue não indicaram perda significativa da viabilidade destas
células na presença do inibidor (dados não mostrados).
Em seguida, foi avaliado se o tratamento das células B16 com PD98059
antes ou durante o co-cultivo poderia prevenir o aumento da fosforilação de ERK
induzido pelas células B-1. Para isto, células B16 foram pré-tratadas ou não com
40 µM de PD98059 por 24 horas e co-cultivadas ou não com células B-1 por 72
horas. Adicionalmente, ambos os tipos celulares foram co-cultivados por 72 horas
na presença do inibidor. Como mostra a Figura 8A, independentemente se as
células B16 foram pré-tratadas com PD98059 antes do co-cultivo [(B16+PD)+B1]
ou co-cultivadas com células B-1 na presença do inibidor [(B16+B1)+PD], os
níveis de ERK fosforilado permaneceram similares aos encontrados nas
monoculturas de células B16 não tratadas (B16). Em contraste, os níveis de ERK
fosforilado aumentarão nas células B16 co-cultivadas com B-1 na ausência do
inibidor (B16+B1). Interessantemente, prevenindo o aumento na fosforilação de
ERK induzido pelas células B-1, o número de nódulos metastáticos produzidos
pelas células B16 não apresentou diferença estatística daqueles produzidos por
células B16 cultivadas sozinhas na ausência do inibidor, enquanto que células B16
provenientes de co-culturas não tratadas desenvolveram mais nódulos
metastáticos (Fig. 8B). Notavelmente, apesar da quase completa inibição da
fosforilação de ERK nas células B16 cultivadas sozinhas na presença de
PD98059, esta inibição não refletiu quantitativamente em redução de nódulos
pulmonares, comparado com células B16 cultivadas sozinhas na ausência do
inibidor (Fig. 8B). Juntos estes resultados mostram que existe uma clara

45
associação entre aumento da fosforilação de ERK induzido pelas células B-1 e
aumento no potencial metastático das células B16.
3.5 Células B-1 não induzem aumento na proliferação de células B16
Uma vez que a quinase ERK além de regular invasão e metástase também
está envolvida no controle da proliferação celular (Raman et al., 2007), foi
investigado se o co-cultivo com as células B-1 poderia induzir aumento na
proliferação das células B16. Entretanto, ensaios de MTT e timidina tritiada
realizados por 24, 48, e 72 horas não mostraram aumento na proliferação de
células B16 após co-cultivo com células B-1 (B16+B1) ou quando as células B16
foram cultivadas em meio condicionado proveniente de monoculturas de células B-
1 (B16+mB1) (Fig. 9).
3.6 Células B-1 induzem a expressão de genes relacionados com
metástases nas células B16.
Para explorar em maior detalhe a influência biológica das células B-1 no
aumento do potencial metastático das células B16, a expressão de genes
relacionados com metástase como CXCR-4 [Chemokine (C-X-C motif) receptor] e
MMP-9 (Matrix metalloproteinase-9) foi avaliada nestas células antes e após o co-
cultivo com células B-1. Para isto, RNA total de células B16 co-cultivadas ou não
com células B-1 na ausência ou presença de PD98059 foi extraído. Uma vez
verificada nas amostras de RNA, após tratamento com DNase, a integridade do
RNA (Fig. 10A) e a não amplificação de GAPDH, gene de controle interno, (Fig.

46
10B) foi preparado cDNA a partir destas amostras para análises de expressão
gênica por RT-PCR qualitativo e RT-PCR em tempo real.
Conforme demonstrado na Figura 11A e B, células B16 co-cultivadas com
células B-1 (B16+B1) apresentaram considerável aumento na expressão de MMP-
9 e CXCR4 comparado com monoculturas de células B16 (B16).
Interessantemente, o aumento na expressão destes genes foi inibido nas células
B16 provenientes de co-culturas com células B-1 tratadas com PD98059
[(B16+B1)+PD]. Em conjunto estes resultados sugerem que o aumento na
fosforilação de ERK induzido pelas células B-1 regula a expressão de genes
relacionados a metástase nas células B16, os quais por sua vez podem contribuir
para aumentar o potencial metastático destas células.
3.7 Níveis basais de fosforilação de ERK nas células B-1 são
necessários para induzir aumento no potencial metastático das células
B16.
Resultados descritos na Figura 6A mostraram que células B-1 ex vivo
apresentam níveis constitutivos de ERK fosforilado. Considerando esta evidência,
foi avaliada se a ativação constitutiva desta quinase nas células B-1 poderia estar
relacionada no comprometimento destas células para induzir aumento no potencial
metastático das células B16.
Trabalhos na literatura mostraram que células B-1 produzem e utilizam IL-
10 como fator de crescimento autócrino (O`Garra et al., 1992). Além disso,
existem vários relatos mostrando que esta citocina foi capaz de promover ativação

47
de ERK, enquanto outros relacionaram a eficiente transcrição de IL-10 com a
atividade de ERK (Zhou et al., 2001; Lee et al., 2002; Niemand et al., 2003; Mathur
et al., 2004). Baseado nestas premissas, foi determinado o perfil de fosforilação de
ERK em células B-1 provenientes de camundongos knock-out para IL-10 (B-1
KO). Como pode ser observado na Figura 12, níveis de IL-10 foram detectados no
sobrenadante de culturas de células B-1 de animais wilde-type (B1 wt), enquanto
que no sobrenandante de culturas de células B-1 KO (B1KO) essa citocina não
apresentou níveis detectáveis. Interessantemente, os níveis de fosforilação de
ERK foram notavelmente menores em células B-1 KO, comparado com os níveis
observados em células B-1 wt (Fig. 13A), mostrando assim diferença nos níveis
constitutivos de ERK fosforilado nestas duas células.
Por outro lado, levando em conta relatos da literatura que mostram a
contribuição de ERK para a fosforilação do resíduo serina 727 do fator de
transcrição Stat3 (Signal transducer and activator of transcription 3; Kuroki &
O`Flaherty, 1999; Kakisis et al., 2005) foi avaliado se os níveis baixos de ERK
fosforilado encontrados nas células B-1 KO poderiam influenciar os níveis de
fosforilação de Stat3 nestas células. Entretanto, conforme mostrado na Figura 13B
os níveis de Stat3 fosforilado (resíduo serina727) encontrados nas células B-1 KO
foram equivalentes aos níveis das células B-1 wt.
Em seguida, foi investigado o efeito das células B-1 KO nas células B16
após co-cultivo de ambas as células. Quando os níveis de fosforilação de ERK nas
células B16 submetidas ao co-cultivo com células B-1 KO (B16+B1KO) foram
avaliados estes permaneceram basais, enquanto que nas células B16 de co-
culturas com células B-1 wt (B16+B1wt) os níveis de fosforilação de ERK foram

48
aumentados (Fig. 13C). Por outro lado, o incremento nos níveis de fosforilação de
ERK foi abolido quando as células B-1 wt foram pré-tratadas com PD98059 antes
do co-cultivo com células B16 (Fig. 13C).
Corroborando com os resultados acima, ensaios experimentais de
metástase (Fig. 13D) demonstraram que injeção de células B16 co-cultivadas com
células B-1 wt (B16+B1wt), mas não com células B-1 KO (B16+B1KO) ou células
B-1 wt pré-tratadas com PD98059 antes do co-cultivo [B16+(B1+PD)], induziram
aumento no número de nódulos metastáticos pulmonares. Estes resultados
sustentam a hipótese que níveis constitutivos de ERK fosforilado nas células B-1
participam no comprometimento destas células para induzir aumento no potencial
metastáticos das células B16.
Adicionalmente, o status de ativação de Stat3 nas células B16 co-cultivadas
ou não com células B-1 wt ou B-1 KO foi analisado também por Immunoblot.
Conforme mostrado na Figura 14, níveis basais equivalentes de fosforilação de
Stat3 foram observados nas células B16 cultivadas tanto na ausência quanto na
presença de soro. Entretanto, quando células B16 foram co-cultivadas com células
B-1 wt ou KO, aumento nos níveis de fosforilação de Stat3 foi observado nas
células B16. Juntos estes resultados mostram que, a pesar do co-cultivo com
células B-1 wt e B-1 KO aumentarem os níveis de fosforilação de Stat3 nas células
B16, este aumento não correlaciona com o aumento do potencial metastático das
células B16, uma vez que o co-cultivo destas células com células B-1 KO não
potencializou a capacidade metastática das células B16 (Fig. 13D).

49
3.8 Níveis basais de fosforilação de ERK nas células B16 são
inalterados pelo estímulo com IL-10
Considerando que a IL-10 pode promover aumento nos níveis de
fosforilação de ERK via ativação do receptor de IL-10 (Zhou et al., 2001; Lee et al.,
2002), foi descartada a possível participação da IL-10, produzida pelas células B-1
wt, no aumento dos níveis de fosforilação de ERK nas células B16 submetidas ao
co-cultivo. Para checar isto, células B16 foram tratadas com diferentes
concentrações de IL-10 recombinante (IL-10r) por diferentes tempos. Como
apresentado na Figura 15A, os tratamentos com IL-10r não foram capazes de
aumentar os níveis basais de fosforilação de ERK nas células B16. Além disso, a
inabilidade da IL-10r em incrementar os níveis de fosfo-ERK nas células B-16,
correlacionou com a incapacidade desta citocina ou do meio condicionado de
culturas de células B-1 wt de aumentar a capacidade metastática das células B16
(Fig. 15B). Comparativamente, células B16 submetidas ao co-cultivo com células
B-1 wt produziram maior número de nódulos pulmonares.
Adicionalmente, foi verificado se o tratamento de células B-1 KO na
presença de IL-10r poderia mimetizar os efeitos das células B-1 wt no aumento do
potencial metastático das células B16 durante o co-cultivo. Para avaliar esta
hipótese, células B16 (B16) foram co-cultivadas ou não com células B-1 wt
(B16+B1wt) ou com células B-1 KO na ausência (B16+B1KO) ou presença de 10
ng/ml de IL-10r [(B16+B1KO)+IL-10r] por 72 horas. Adicionalmente, células B-1
KO foram pré-tratadas com 10 ng/ml de IL-10r [B16+(B1KO+IL-10r)] ou com meio
condicionado de culturas de células B-1 wt [B16+(B1KO+mB1wt)] antes do co-

50
cultivo. Após estes tratamentos, células B16 foram coletadas e submetidas a
ensaios de metástase experimental in vivo. Conforme observado na Figura 16A,
células B16 co-cultivadas com células B-1 KO submetidas aos tratamentos com
IL-10r ou com meio condicionado de células B-1 wt, produziram número de
nódulos metastáticos similares àqueles produzidos por células B16 de
monoculturas, enquanto células B16 co-cultivadas com células B-1 wt foram
significativamente mais metastáticas.
Uma vez que o tratamento de células B-1 KO com IL-10r não mimetizou o
efeito das células B-1 wt no aumento do potencial metastático das células B16, foi
verificado se existiam diferenças na expressão do receptor de IL-10 entre essas
células. Para isso, utilizando RT-PCR qualitativo foi determinada a expressão de
mRNA do receptor de IL-10 nas células B-1 wt e KO. Conforme observado na
Figura 16B a expressão de mRNA do receptor de IL-10 (IL-10R) nas células B-1
KO é quase inexistente comparado com os níveis de expressão das células B-1
wt.
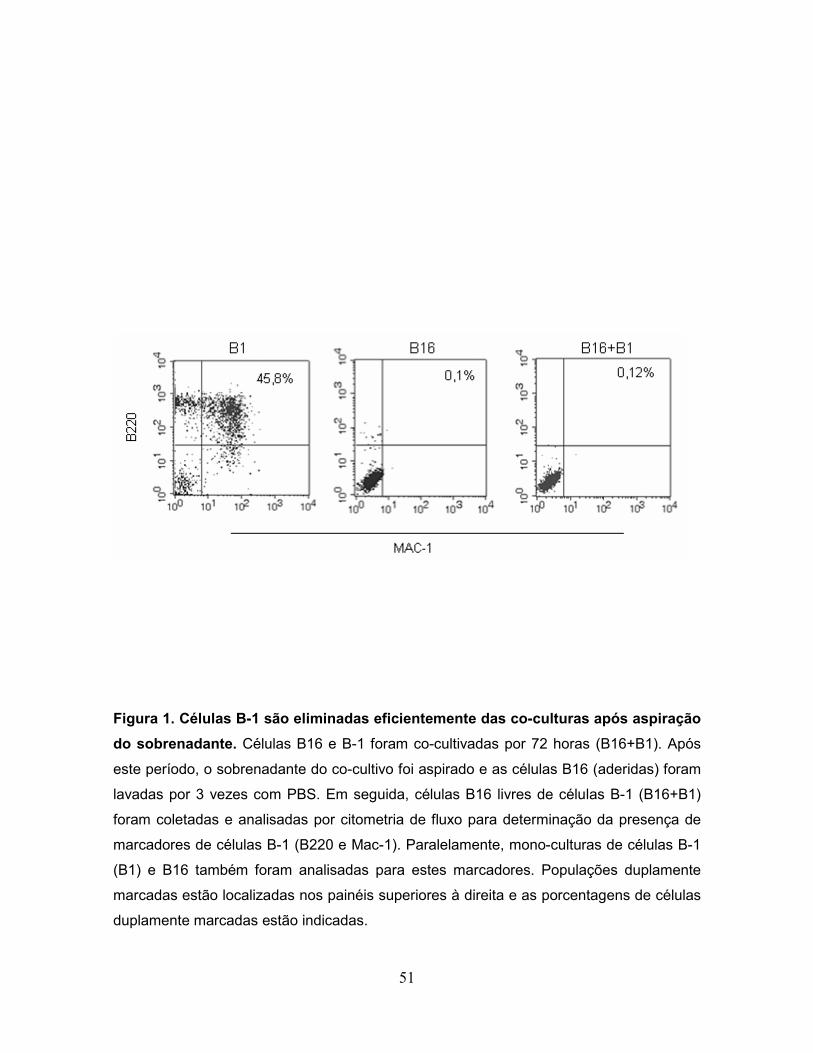
51
Figura 1. Células B-1 são eliminadas eficientemente das co-culturas após aspiração
do sobrenadante. Células B16 e B-1 foram co-cultivadas por 72 horas (B16+B1). Após
este período, o sobrenadante do co-cultivo foi aspirado e as células B16 (aderidas) foram
lavadas por 3 vezes com PBS. Em seguida, células B16 livres de células B-1 (B16+B1)
foram coletadas e analisadas por citometria de fluxo para determinação da presença de
marcadores de células B-1 (B220 e Mac-1). Paralelamente, mono-culturas de células B-1
(B1) e B16 também foram analisadas para estes marcadores. Populações duplamente
marcadas estão localizadas nos painéis superiores à direita e as porcentagens de células
duplamente marcadas estão indicadas.
52
Figura 2. Células B-1 aumentam o potencial metastático das células B16.
Monoculturas de células B16 (B16) ou B-1 (B1) ou co-culturas de ambas as células
(B16+B1; 1X105 e 1X106 células, respectivamente) foram cultivadas por 24, 48 ou 72
horas. Após estes períodos, células B16 e B-1 cultivadas de forma individual ou células
B16 livres de células B-1 após o co-cultivo foram coletadas e injetadas na veia da cauda
de camundongos (1X105 células por camundongo). Quatorze dias após a injeção, os
animais foram sacrificados e os pulmões retirados para determinar o número de nódulos
metastáticos formados. Barras correspondem ao número médio de triplicatas realizadas
para cada grupo analisado. Resultados representativos de 3 experimentos independentes.
p<0.05.

53
Figura 3. Células B-1 induzem aumento no potencial metastático de células B16
independentemente da adesão celular. Células B16 (sB16), B-1 (sB1) ou células B16
co-cultivadas com células B-1 (sB16+B1), foram plaqueadas em condições de
impedimento de adesão por 72 horas. Após este período, as células cultivadas de forma
individual ou pool de células B16+B1 provenientes do co-cultivo foram coletadas, os
clusters celulares formados durante o período de co-cultivo foram dissociados e
submetidos tanto a (A) ensaios experimentais de metástases in vivo como (B) ensaio de
viabilidade por Trypan blue. Em (A) barras representam o número médio de nódulos
metastáticos obtidos das triplicatas realizadas para cada grupo analisado e em (B) a
porcentagem da viabilidade celular de cada grupo de células. Os resultados são
representativos de 3 experimentos independentes. p<0.05.
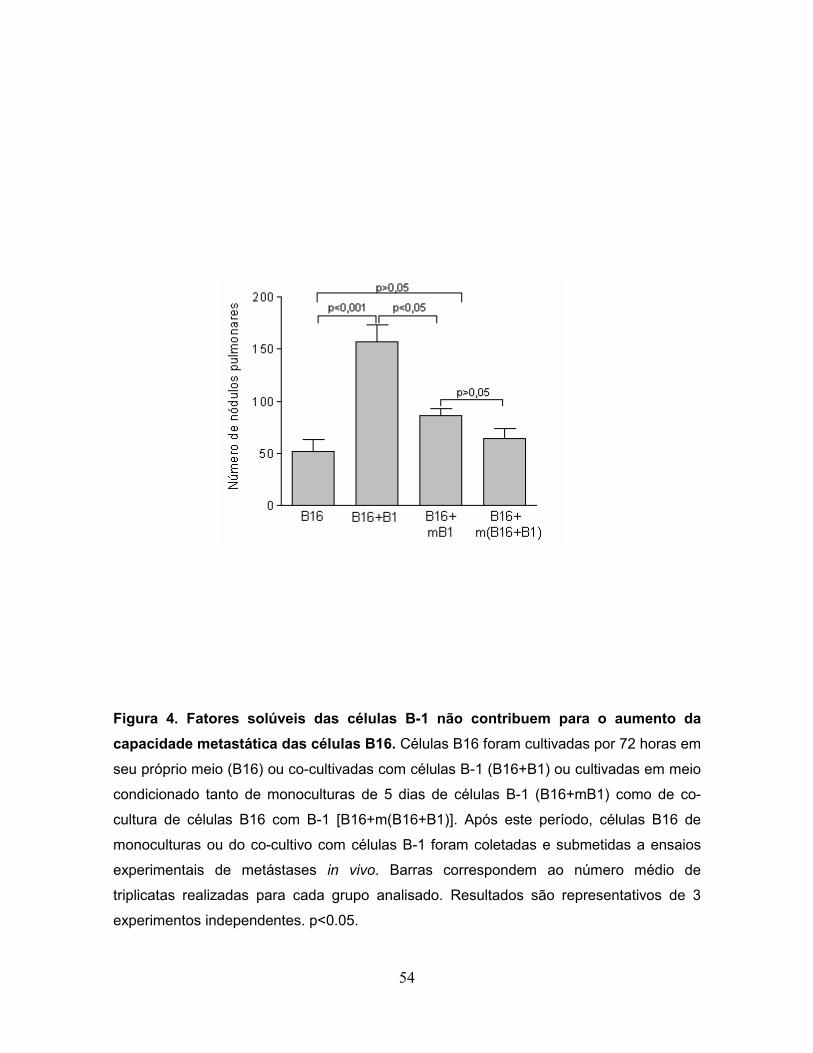
54
Figura 4. Fatores solúveis das células B-1 não contribuem para o aumento da
capacidade metastática das células B16. Células B16 foram cultivadas por 72 horas em
seu próprio meio (B16) ou co-cultivadas com células B-1 (B16+B1) ou cultivadas em meio
condicionado tanto de monoculturas de 5 dias de células B-1 (B16+mB1) como de co-
cultura de células B16 com B-1 [B16+m(B16+B1)]. Após este período, células B16 de
monoculturas ou do co-cultivo com células B-1 foram coletadas e submetidas a ensaios
experimentais de metástases in vivo. Barras correspondem ao número médio de
triplicatas realizadas para cada grupo analisado. Resultados são representativos de 3
experimentos independentes. p<0.05.
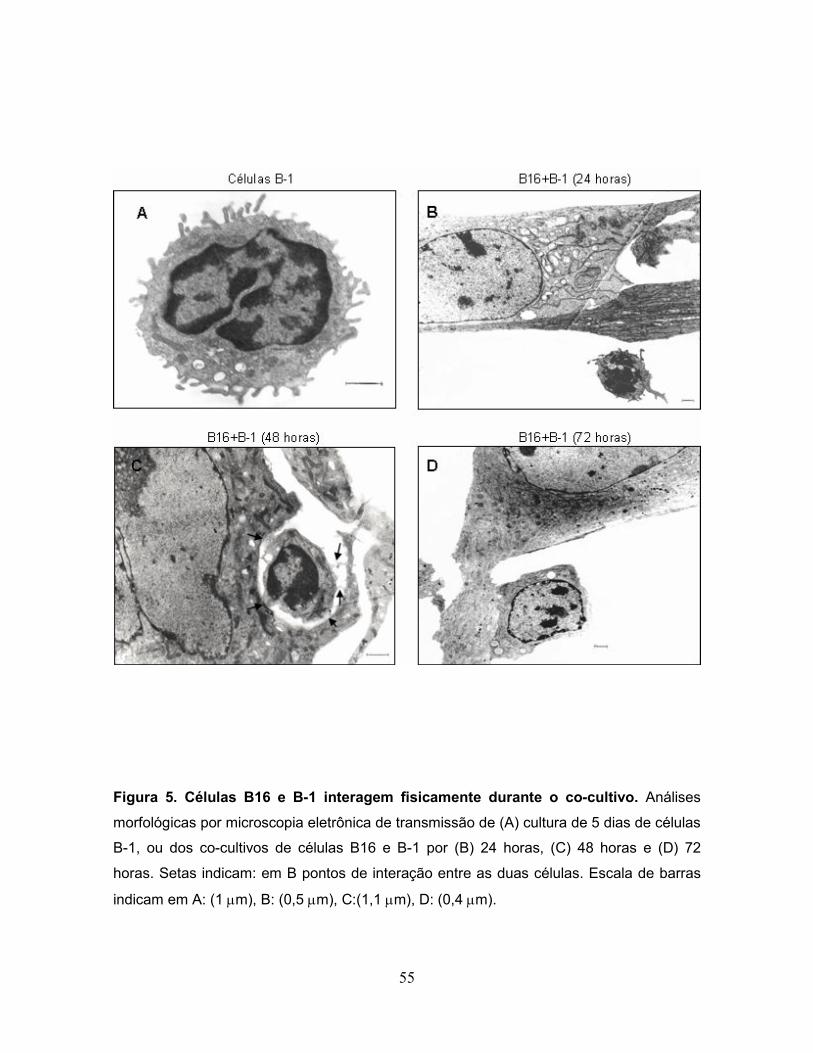
55
Figura 5. Células B16 e B-1 interagem fisicamente durante o co-cultivo. Análises
morfológicas por microscopia eletrônica de transmissão de (A) cultura de 5 dias de células
B-1, ou dos co-cultivos de células B16 e B-1 por (B) 24 horas, (C) 48 horas e (D) 72
horas. Setas indicam: em B pontos de interação entre as duas células. Escala de barras
indicam em A: (1 µm), B: (0,5 µm), C:(1,1 µm), D: (0,4 µm).
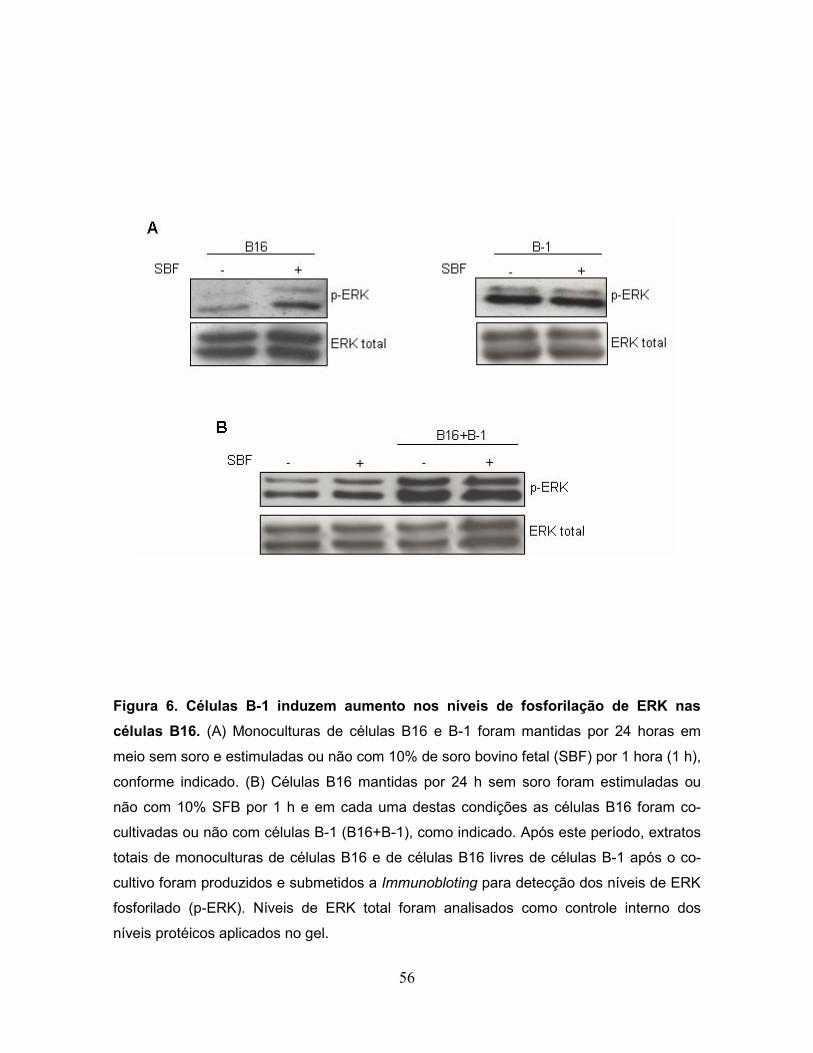
56
Figura 6. Células B-1 induzem aumento nos níveis de fosforilação de ERK nas
células B16. (A) Monoculturas de células B16 e B-1 foram mantidas por 24 horas em
meio sem soro e estimuladas ou não com 10% de soro bovino fetal (SBF) por 1 hora (1 h),
conforme indicado. (B) Células B16 mantidas por 24 h sem soro foram estimuladas ou
não com 10% SFB por 1 h e em cada uma destas condições as células B16 foram co-
cultivadas ou não com células B-1 (B16+B-1), como indicado. Após este período, extratos
totais de monoculturas de células B16 e de células B16 livres de células B-1 após o co-
cultivo foram produzidos e submetidos a Immunobloting para detecção dos níveis de ERK
fosforilado (p-ERK). Níveis de ERK total foram analisados como controle interno dos
níveis protéicos aplicados no gel.

57
Figura 7. Níveis basais de fosforilação de ERK em células B16 e B-1 são inibidos
pelo tratamento com PD98059. (A) Monoculturas de células B16 e B-1 foram cultivadas
em meio completo e tratadas ou não com 20 ou 40 µM de PD98059 por 24 horas. Após
este período extratos totais foram preparados e analisados por Western Blot para a
detecção dos níveis de ERK fosforilado. Níveis de ERK total foram analisados como
controle interno dos níveis protéicos aplicados no gel.
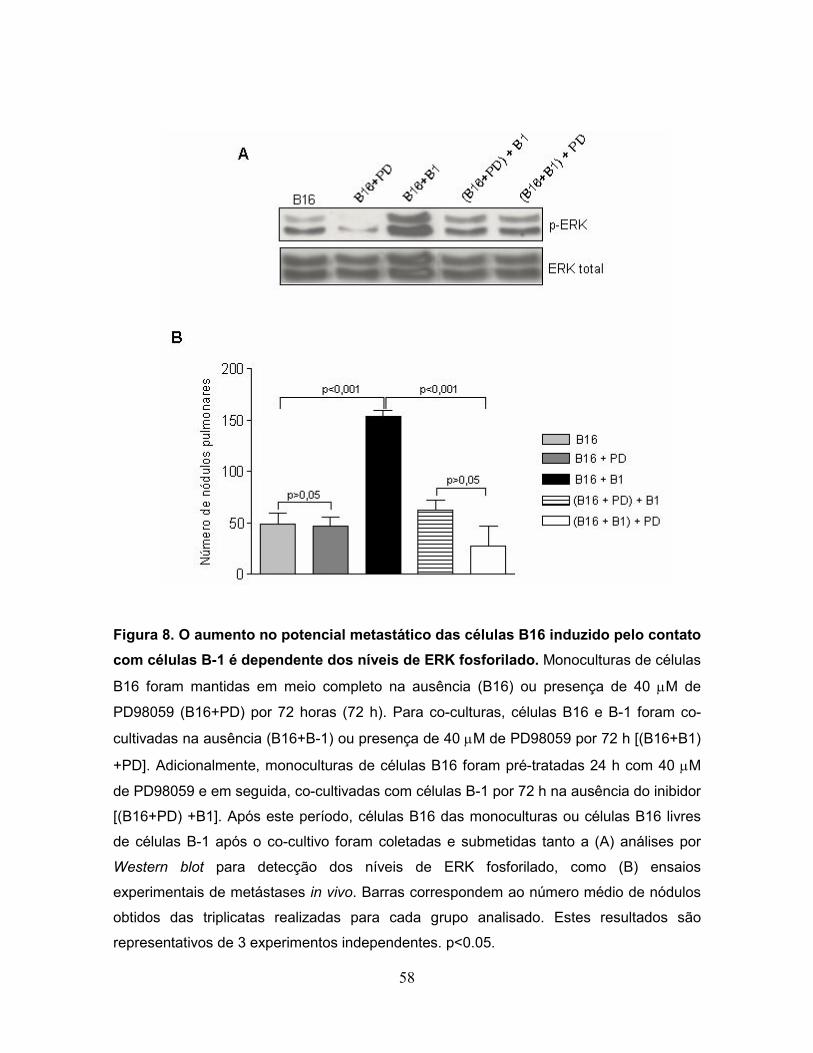
58
Figura 8. O aumento no potencial metastático das células B16 induzido pelo contato
com células B-1 é dependente dos níveis de ERK fosforilado. Monoculturas de células
B16 foram mantidas em meio completo na ausência (B16) ou presença de 40 µM de
PD98059 (B16+PD) por 72 horas (72 h). Para co-culturas, células B16 e B-1 foram co-
cultivadas na ausência (B16+B-1) ou presença de 40 µM de PD98059 por 72 h [(B16+B1)
+PD]. Adicionalmente, monoculturas de células B16 foram pré-tratadas 24 h com 40 µM
de PD98059 e em seguida, co-cultivadas com células B-1 por 72 h na ausência do inibidor
[(B16+PD) +B1]. Após este período, células B16 das monoculturas ou células B16 livres
de células B-1 após o co-cultivo foram coletadas e submetidas tanto a (A) análises por
Western blot para detecção dos níveis de ERK fosforilado, como (B) ensaios
experimentais de metástases in vivo. Barras correspondem ao número médio de nódulos
obtidos das triplicatas realizadas para cada grupo analisado. Estes resultados são
representativos de 3 experimentos independentes. p<0.05.
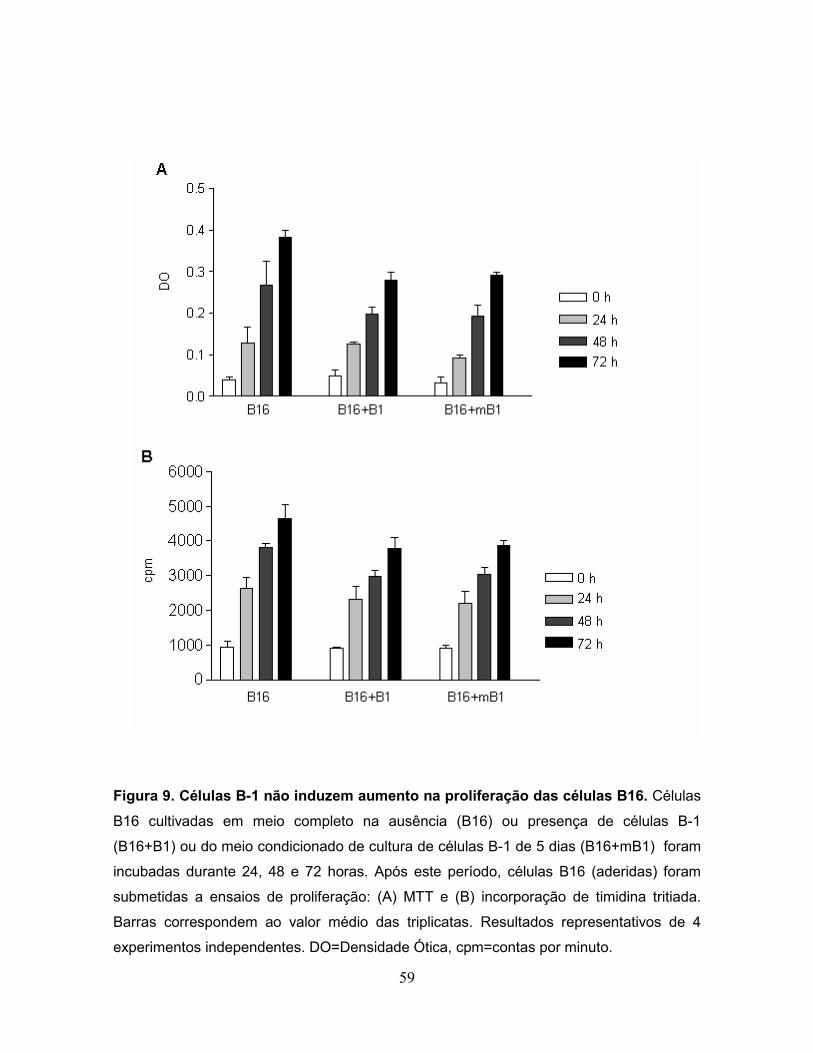
59
Figura 9. Células B-1 não induzem aumento na proliferação das células B16. Células
B16 cultivadas em meio completo na ausência (B16) ou presença de células B-1
(B16+B1) ou do meio condicionado de cultura de células B-1 de 5 dias (B16+mB1) foram
incubadas durante 24, 48 e 72 horas. Após este período, células B16 (aderidas) foram
submetidas a ensaios de proliferação: (A) MTT e (B) incorporação de timidina tritiada.
Barras correspondem ao valor médio das triplicatas. Resultados representativos de 4
experimentos independentes. DO=Densidade Ótica, cpm=contas por minuto.
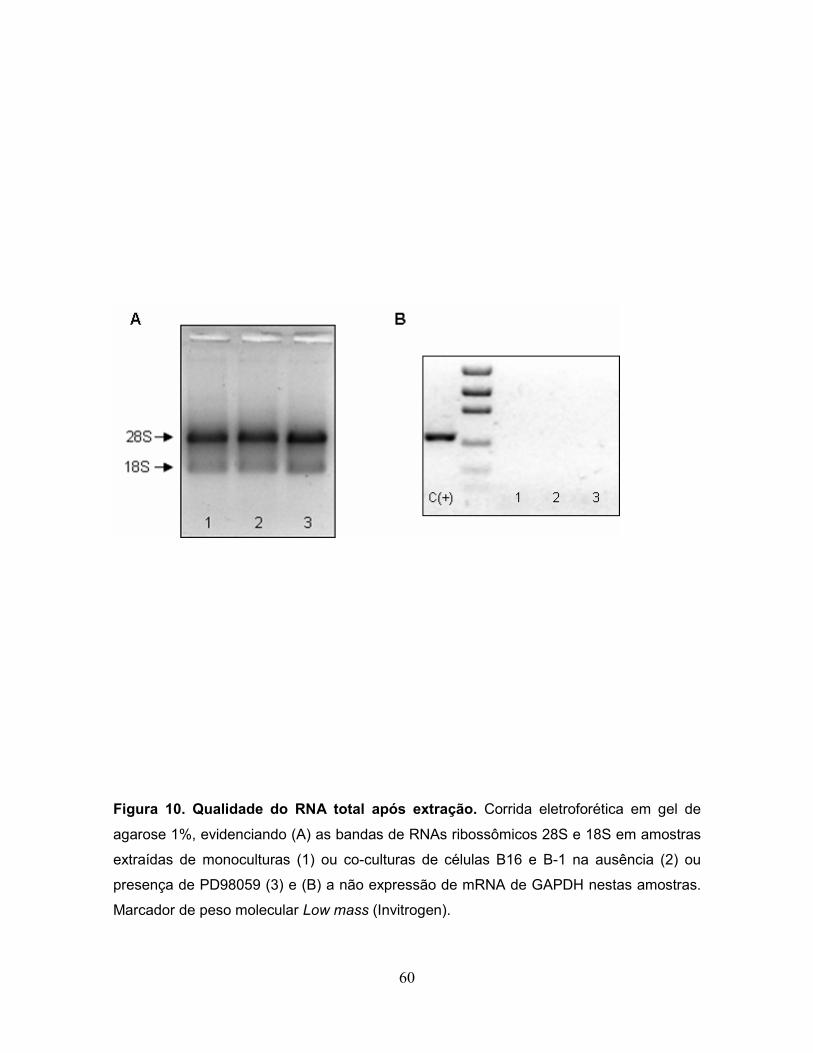
60
Figura 10. Qualidade do RNA total após extração. Corrida eletroforética em gel de
agarose 1%, evidenciando (A) as bandas de RNAs ribossômicos 28S e 18S em amostras
extraídas de monoculturas (1) ou co-culturas de células B16 e B-1 na ausência (2) ou
presença de PD98059 (3) e (B) a não expressão de mRNA de GAPDH nestas amostras.
Marcador de peso molecular Low mass (Invitrogen).

61
Figura 11. Células B-1 induzem a expressão de CXCR4 e MMP-9 nas células B16.
Células B16 (B16) foram co-cultivadas ou não com células B-1 na ausência (B16+B1) ou
presença de 40 µM de PD98059 [(B16+B1)+PD]. Níveis relativos da expressão de CXCR4
e MMP-9 foram analisados por (A) RT-PCR qualitativo e (B) RT-PCR em tempo real, em
relação à expressão do gene de controle interno, GAPDH. Barras correspondem ao valor
médio das triplicatas de cada grupo. Resultados representativos de 3 experimentos
independentes. p<0.05.

62
Figura 12. Produção de IL-10 por células B-1. Células B-1 isoladas de animais wild-type
(B1wt) ou knock-out (B1KO) foram cultivadas em meio completo durante 5 dias. Após este
período, sobrenadantes destas culturas foram coletados e analisados por ELISA para
detecção dos níveis de IL-10. Barras correspondem ao valor médio das triplicatas de cada
grupo. Resultados representativos de 3 experimentos independentes.
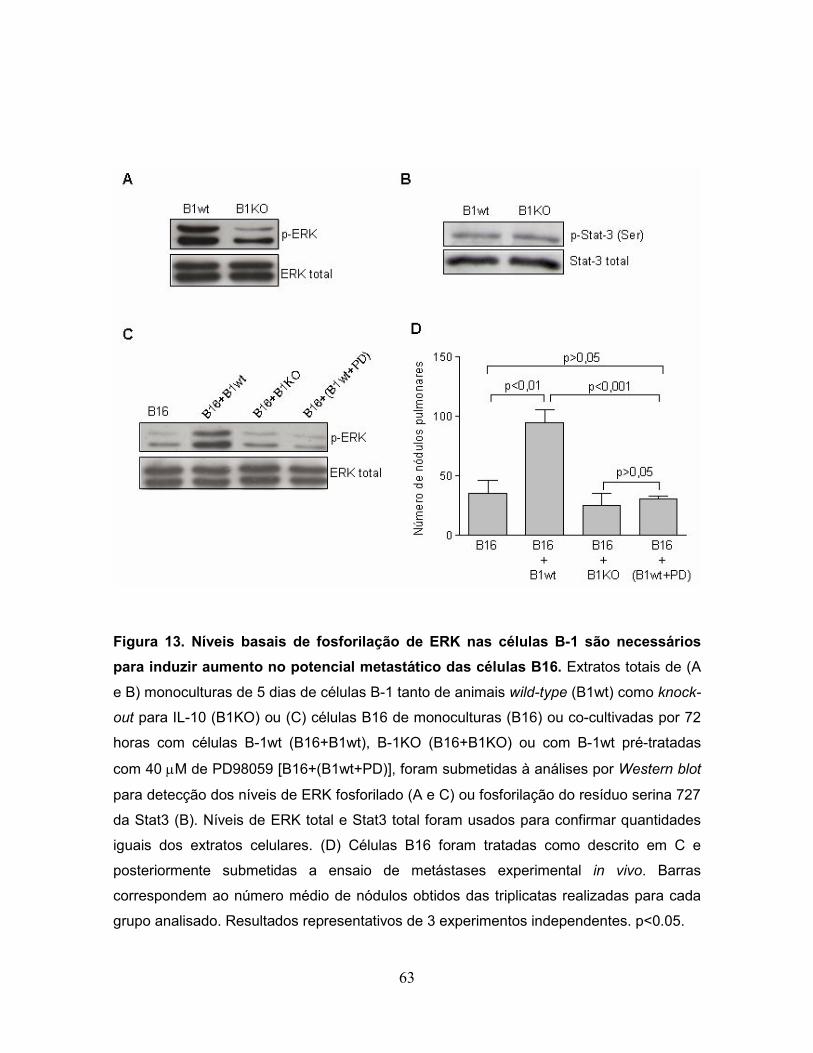
63
Figura 13. Níveis basais de fosforilação de ERK nas células B-1 são necessários
para induzir aumento no potencial metastático das células B16. Extratos totais de (A
e B) monoculturas de 5 dias de células B-1 tanto de animais wild-type (B1wt) como knock-
out para IL-10 (B1KO) ou (C) células B16 de monoculturas (B16) ou co-cultivadas por 72
horas com células B-1wt (B16+B1wt), B-1KO (B16+B1KO) ou com B-1wt pré-tratadas
com 40 µM de PD98059 [B16+(B1wt+PD)], foram submetidas à análises por Western blot
para detecção dos níveis de ERK fosforilado (A e C) ou fosforilação do resíduo serina 727
da Stat3 (B). Níveis de ERK total e Stat3 total foram usados para confirmar quantidades
iguais dos extratos celulares. (D) Células B16 foram tratadas como descrito em C e
posteriormente submetidas a ensaio de metástases experimental in vivo. Barras
correspondem ao número médio de nódulos obtidos das triplicatas realizadas para cada
grupo analisado. Resultados representativos de 3 experimentos independentes. p<0.05.

64
Figura 14. Níveis de fosforilação de Stat3 nas células B16 antes e após co-cultivo
com células B-1. Extratos totais de células B16 mantidas 24 horas na ausência de soro
(SBF) e em seguida estimuladas com SBF ou co-cultivadas com células B-1 de animais
tanto wild-type (B16+B1wt) como knock-out para IL-10 (B16+B1KO) foram submetidos a
Western blot para verificar a fosforilação da serina 727 da Stat3. Níveis de Stat3 total
foram usados para confirmar quantidades iguais dos extratos celulares. Resultados
representativos de 3 experimentos independentes.
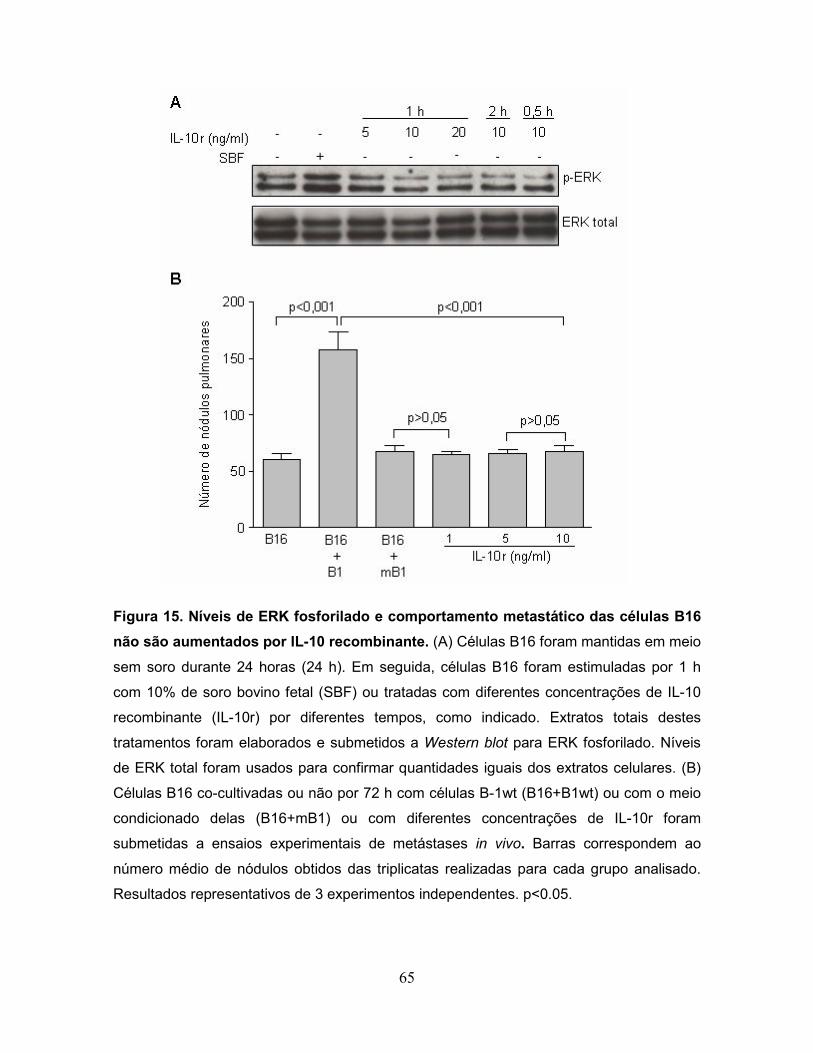
65
Figura 15. Níveis de ERK fosforilado e comportamento metastático das células B16
não são aumentados por IL-10 recombinante. (A) Células B16 foram mantidas em meio
sem soro durante 24 horas (24 h). Em seguida, células B16 foram estimuladas por 1 h
com 10% de soro bovino fetal (SBF) ou tratadas com diferentes concentrações de IL-10
recombinante (IL-10r) por diferentes tempos, como indicado. Extratos totais destes
tratamentos foram elaborados e submetidos a Western blot para ERK fosforilado. Níveis
de ERK total foram usados para confirmar quantidades iguais dos extratos celulares. (B)
Células B16 co-cultivadas ou não por 72 h com células B-1wt (B16+B1wt) ou com o meio
condicionado delas (B16+mB1) ou com diferentes concentrações de IL-10r foram
submetidas a ensaios experimentais de metástases in vivo. Barras correspondem ao
número médio de nódulos obtidos das triplicatas realizadas para cada grupo analisado.
Resultados representativos de 3 experimentos independentes. p<0.05.
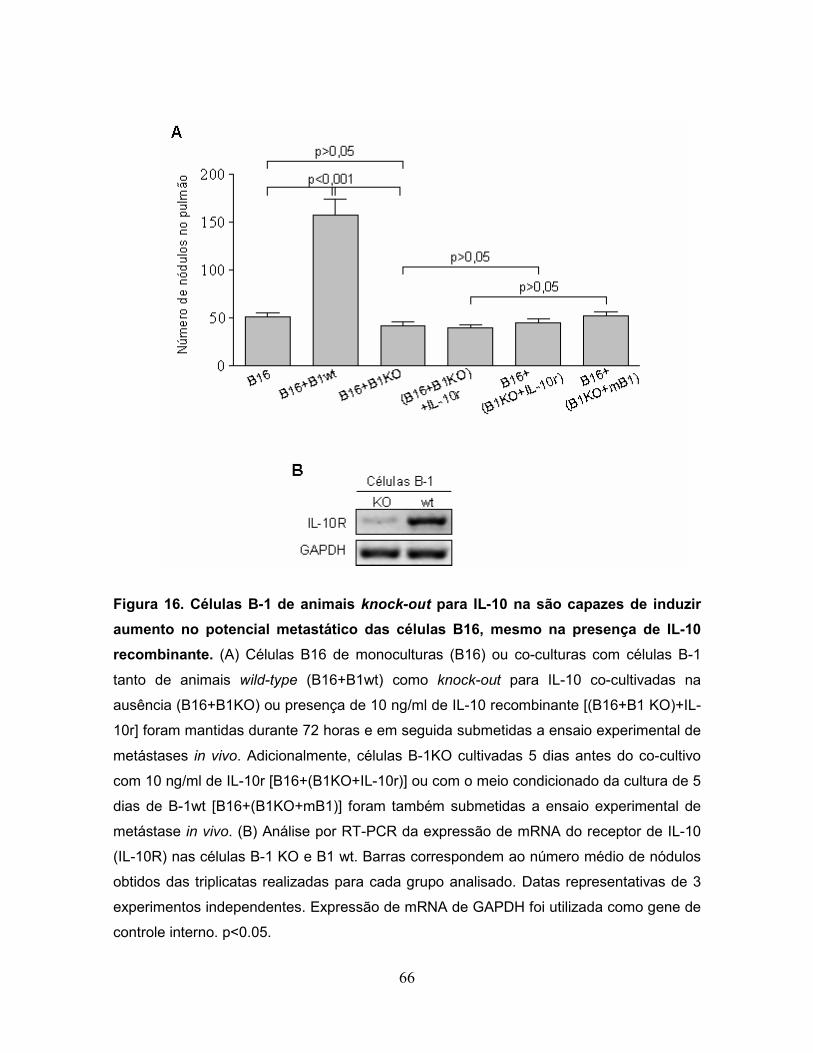
66
Figura 16. Células B-1 de animais knock-out para IL-10 na são capazes de induzir
aumento no potencial metastático das células B16, mesmo na presença de IL-10
recombinante. (A) Células B16 de monoculturas (B16) ou co-culturas com células B-1
tanto de animais wild-type (B16+B1wt) como knock-out para IL-10 co-cultivadas na
ausência (B16+B1KO) ou presença de 10 ng/ml de IL-10 recombinante [(B16+B1 KO)+IL-
10r] foram mantidas durante 72 horas e em seguida submetidas a ensaio experimental de
metástases in vivo. Adicionalmente, células B-1KO cultivadas 5 dias antes do co-cultivo
com 10 ng/ml de IL-10r [B16+(B1KO+IL-10r)] ou com o meio condicionado da cultura de 5
dias de B-1wt [B16+(B1KO+mB1)] foram também submetidas a ensaio experimental de
metástase in vivo. (B) Análise por RT-PCR da expressão de mRNA do receptor de IL-10
(IL-10R) nas células B-1 KO e B1 wt. Barras correspondem ao número médio de nódulos
obtidos das triplicatas realizadas para cada grupo analisado. Datas representativas de 3
experimentos independentes. Expressão de mRNA de GAPDH foi utilizada como gene de
controle interno. p<0.05.

5. DISCUSSÃO

5. DISCUSSÃO
Relatos da literatura têm demonstrado que a expansão metastática de
células de melanoma é um processo altamente complexo dependente de
interações entre as células tumorais e seu estroma (Langley & Fidler, 2007;
Aguirre-Ghiso, 2007). Embora os macrófagos associados ao tumor (TAMs) sejam
a principal população de células inflamatórias presentes no micro-ambiente
tumoral, outros tipos celulares podem também favorecer a formação de
metástases ao interagir de forma direta com as células tumorais e ou indireta pela
produção de fatores solúveis (Balkwill & Mantovani, 2001; Inoue et al., 2006; Croci
et al., 2007; Mantovani et al., 2007; Hofmeister et al., 2008).
Evidências apresentadas no presente estudo mostram que interações in
vitro entre células de melanoma murino B16 e células B-1, subtipo de linfócitos B
presentes nas cavidades pleural e peritoneal, resultam em aumento do potencial
metastático das células de melanoma (Fig.2). Os resultados apresentados
mostraram inequivocamente que interações físicas entre células B16 e B-1, e não
fatores solúveis produzidos pelas células B-1, representam a principal condição
para aumentar a capacidade metastática das células B16 (Fig. 4 e 5),
corroborando dados prévios de nosso grupo, ainda não publicados. Análises de
microscopia eletrônica (Fig. 5), realizadas com intuito de investigar com mais
detalhe o tipo de interação entre essas células durante o co-cultivo, demonstraram
estreita comunicação física entre elas. Esta interação, entretanto, não sugeriu

69
fusão celular, já que os contornos das membranas plasmáticas de cada célula
apresentaram-se claramente delimitados.
Um dos resultados inéditos deste trabalho mostrou que o aumento do
potencial metastático das células B16 induzido pelo contato com células B1 está
associado a aumento na ativação de ERK. Análises de Western blot evidenciaram
que o contato de células B-1 induz aumento nos níveis de fosforilação de ERK nas
células B16 (Fig. 6B). Interessantemente, quando o aumento da fosforilação de
ERK foi bloqueada pelo inibidor farmacológico de MEK (PD98059), quinase up-
stream de ERK, o aumento da capacidade metastática das células B16 foi inibido
(Fig. 8B). Em contraste, monoculturas de células B16 apresentam níveis basais
constitutivos de ERK (Fig. 6A), e a completa inibição desta quinase nestas células
não inibiu sua capacidade natural de formar nódulos metastáticos no pulmão (Fig.
8B). Esta observação sugere que o fenótipo intrínseco da linhagem celular B16 de
baixo potencial metastático pode ser sustentado por outra via de sinalização que
independe de background mínimo de ERK fosforilado. Por outro lado, o fato de
que células B-1 não serem capazes de elevar os níveis de ERK fosforilado nem o
potencial metastático das células B16 tratadas previamente com PD98059 sugere
que, os efeitos desencadeados pelas células B-1 dependem de níveis basais de
ERK fosforilado nas células B16.
Vários relatos da literatura sustentam que os substratos de ERK incluem
alvos presentes no citoplasma como também fatores de transcrição tais como as
famílias ELK/SAP, Ets/PEA3, AP-1 e Stat (Hill & Treisman, 1996; Harper &
LoGrasso, 2001; Viala & Pouyssegur, 2004). Algumas destas famílias, como AP-1
e Ets são importantes na regulação da expressão de várias metaloproteinases

70
(MMPs) como MMP-9 e quimiocinas como CXCR4, as quais estão associadas
com progressão metastática em diversos tipos de cânceres entre eles o melanoma
(John & Tuszynski, 2001; Hofmann et al., 2003; Kulbe et al., 2004; Tower et al.,
2002; Kim et al., 2006; Maroni et al., 2007; Ruffini et al., 2007).
Análises de expressão gênica para CXCR4 e MMP-9 por RT-PCR
qualitativo e em tempo real realizadas neste trabalho mostraram aumento
substancial na expressão destes transcritos nas células B16 após contato com
células B-1 (Fig. 11). Interessantemente, estes resultados também mostraram que
o aumento na expressão de mRNA de CXCR4 e MMP-9 é inibido pela adição do
inibidor farmacológico PD98059. Assim como os achados de outros grupos, este
resultado sustenta a participação de ERK na regulação da expressão destes
transcritos (Gum et al., 1997; Simon et al., 1998; Lakka et al., 2000; Maroni et al.,
2007) e ainda, sugere que o aumento na expressão de mRNA de MMP-9 e
CXCR4 nas células B16 induzida pelo co-cultivo com células B-1 pode contribuir
para potencializar a capacidade metastática das células B16 ao favorecer o
extravasamento e migração destas células aos pulmões.
Outro dado importante e inédito de nossa pesquisa foi a necessidade de
níveis constitutivos de ERK fosforilado nas células B-1 para o comprometimento
destas células na indução do aumento no potencial metastático das células B16.
Resultados obtidos mostram que a inibição da fosforilação de ERK nas células B-1
pelo tratamento com PD98059, prejudica consideravelmente a capacidade destas
células de induzir aumento tanto nos níveis de ERK fosforilado como na
capacidade metastática das células B16. Esta observação foi também sustentada
por estudos com células B-1 provenientes de camundongos knock-out para IL-10

71
(KO), as quais apresentaram níveis basais de ERK fosforilado menores dos níveis
encontrados nas células B-1 de animais wild-type (wt) (Fig. 13A). Resultados
obtidos também com células B-1 KO mostraram que estas células não têm
capacidade para aumentar os níveis de ERK fosforilado nem o potencial
metastático das células B16 quando comparadas com células B-1 wt.
Considerando dados da literatura que evidenciam aumento nos níveis de
ERK fosforilado via ativação do receptor de IL-10 (Yi et al., 2002; Mathur et al.,
2004; Liu et al., 2006; Chanteux et al., 2007), é possível especular que os níveis
baixos de ERK fosforilado observados nas células B-1 KO sejam devidos a níveis
também baixos de expressão de mRNA do receptor de IL-10 nestas células (Fig.
16). Embora o nível de fosforilação de ERK nas células B-1 wt após estímulo com
IL-10 recombinante (IL-10r) não tenha sido analisado, os resultados acima
descritos sugerem que o estímulo das células B-1 com IL-10, poderia contribuir
com os níveis basais de fosforilação de ERK observados nestas células. Em
contraste, células B-16 não apresentaram aumento nos níveis de fosforilação de
ERK nem aumento do seu potencial metastático após estímulo com IL-10r ou com
o sobrenadante de cultura de células B1wt (Fig. 15). Estes resultados, em
conjunto, sugerem que IL-10 pode ser um fator importante para manutenção de
níveis constitutivos de ERK nas células B-1 wt, os quais podem favorecer a
expressão de prováveis moléculas de membrana, que ao interagir no co-cultivo
com receptores presentes nas células B16 promovem o crescimento metastático
destas células.
Fosforilação constitutiva de ERK nas células B-1 foi previamente
demonstrada por Wong e colaboradores (2002). Embora a atividade de ERK não

72
seja necessária para a sobrevida destas células em cultura, ainda são
desconhecidos quais aspectos da fisiologia das células B-1 são regulados pela
quinase ERK (Wong et al., 2002). Com base nos resultados aqui apresentados
ainda não foi possível estabelecer porque as células B-1 precisam de altos níveis
de ERK fosforilado para potencializar a capacidade metastática das células B16.
Entretanto, relatos da literatura têm demonstrado que diferenças na duração,
magnitude e distribuição sub-celular da atividade de ERK determinam programas
celulares específicos (Balmanno & Cook, 1999; Sasagawa et al., 2005; Ebisuya et
al., 2005). Assim, considerando os resultados acima descritos que sustentam a
necessidade do contato físico entre células B16 e B-1, é possível especular que
níveis basais de ERK nas células B-1 são requeridos para regular a expressão de
moléculas de superfície nestas células, as quais ao interagirem com células B16
durante o co-cultivo, formam uma rede interativa de sinalização célula-célula.
Por outro lado, além da ativação constitutiva de ERK, células B-1 também
apresentam ativação constitutiva de Stat3 (Signal transducer and activator of
transcription 3), o qual tem papel importante na capacidade de auto-renovação
destas células (Karras et al., 1997). As proteínas STATs são fatores de transcrição
ativados por fosforilação de uma tirosina (Y705) ou uma serina (S727) em
resposta a um ligante extracelular, que pode ser citocina ou fatores de
crescimento (Darnell, et al., 1997; Bromberg, et al., 1999; Ihle, 2001; Chen, et al.,
2004). Considerando relatos da literatura, nos quais foi mostrado que ERK pode
contribuir com a fosforilação do resíduo serina da Stat3 (Kuroki & O`Flaherty,
1999; Kakisis et al., 2005), investigou-se nas células B-1 KO os níveis de
fosforilação de Stat3, visto que estas células apresentam níveis de ERK fosforilado

73
menores quando comparados com os níveis observados nas B-1 wt (Fig. 13A). O
fato que os níveis de Stat3 nas células B-1 KO foram semelhantes aos observados
nas células B-1 wt (Figura 13B) sugere que a ativação do resíduo serina (727) da
Stat3 nestas células independe da atividade de ERK. Por outro lado, considerando
que aumento na ativação de Stat3 foi associado também com progressão
metastática em vários tipos de cânceres, entre eles o melanoma (Gabrilovich et
al., 1996; Mora et al., 2002; Kortylewisky et al., 2005; Lassmann et al., 2007), o
status de ativação deste fator de transcrição foi avaliado nas células B16 antes e
após contato com células B-1. Conforme mostrado na Figura 14, células B16
apresentam níveis basais de Stat3 (resíduo serina 727) e estes níveis são
aumentados após interação com células B-1, sejam KO ou wt. Entretanto, células
B-1 KO não têm capacidade de aumentar nem os níveis de fosforilação de ERK
nem a capacidade metastática das células B16 (Fig 13 C e D, respectivamente).
Embora outros experimentos sejam necessários para excluir a participação de
Stat3 neste modelo de estudo, estes resultados sugerem fortemente que a via da
Stat3 não tem participação no aumento do potencial metastático induzido pelo
contato com células B-1.
Adicionalmente, outras vias de transdução de sinal que atuam sobre
mensageiros intermediários como PKC e Fak (Kahana, et al., 2002; Oka & Kikawa,
2005), cuja up-regulation tem sido associada com invasão e metástases de células
de melanoma, também foram analisadas. Entretanto, resultados obtidos com estas
quinases não levaram a resultados conclusivos da sua participação neste modelo
de estudo.

74
Em resumo os resultados apresentados neste trabalho fornecem as
primeiras evidências do envolvimento da via ERK no aumento da capacidade
metastática das células B16 induzido pelo contato com células B-1. Sugerem que
a up-regulation desta via é um dos mecanismos pelos quais linfócitos B-1 podem
influenciar diretamente no potencial metastático de células tumorais. Além disso,
este trabalho reforça a idéia que as células B-1 podem estar presentes no micro-
ambiente tumoral realizando funções similares aos TAMs ao favorecer a formação
de metástases. Para confirmar esta última hipótese, atualmente nosso grupo
investiga a presença das células B-1 em isolados de tecidos tumorais humanos
com o intuito de determinar se o efeito das células B-1 sobre o comportamento
metastático das células de melanoma apresentadas neste estudo ocorre também
in vivo no contexto dinâmico do micro-ambiente tumoral.

6. CONCLUSÕES

6. CONCLUSÕES
1 – Células B-1 interagem fisicamente com células de melanoma murino
B16 após 48 horas de co-cultivo e esta interação é necessária para o aumento do
potencial metastático das células B16;
2 – A participação da citocina IL-10 ou outro fator solúvel produzido pelas
células B-1 como responsáveis pela indução do aumento tanto dos níveis de
fosforilação de ERK como da capacidade metastática das células B16 foi excluída;
3 – Células B-1 induzem aumento na capacidade metastática das células
B16 ao induzir up-regulation dos níveis basais de ERK fosforilado nestas células;
4 – Níveis basais de ERK fosforilado tanto nas células B16 como nas B-1
são necessários para potencializar o crescimento metastático das células B16
induzido pelo contato com células B-1;
5 – Células B-1 induzem aumento na expressão de genes associados a
progressão metastática, como MMP-9 e CXCR-4 nas células B16;

77
6 – Altos níveis de foforilação de ERK nas células B-1 são necessários para
o comprometimento destas células para induzir aumento no potencial metastático
das células B16;
7 – A fosforilação do resíduo serina da Stat3 nas células B-1 independe dos
níveis de ativação de ERK, nestas células.

7. REFERÊNCIAS BIBLIOGRÁFICAS

7. REFERÊNCIAS BIBLIOGRÁFICAS
- Abrahão TB, Freymüller E, Mortara RA, Lopes JD, Mariano M.Morphological
characterization of mouse B-1 cells.Immunobiology. 2003;208(4):401-11. - Aguirre-Ghiso JA. Models, mechanisms and clinical evidence for cancer dormancy.
Nat Rev Cancer. 2007 Nov;7(11):834-46. - Almand B, Clark JI, Nikitina E, van Beynen J, English NR, Knight SC, Carbone DP,
Gabrilovich DI. Increased production of immature myeloid cells in cancer patients: a mechanism of immunosuppression in cancer. J Immunol. 2001 Jan 1;166(1):678-89.
- Almeida SR, Aroeira LS, Frymuller E, Dias MA, Bogsan CS, Lopes JD, Mariano M.
Mouse B-1 cell-derived mononuclear phagocyte, a novel cellular component of acute non-specific inflammatory exudate. Int Immunol. 2001 Sep;13(9):1193-201.
- Alugupalli KR, Leong JM, Woodland RT, Muramatsu M, Honjo T, Gerstein RM. B1b
lymphocytes confer T cell-independent long-lasting immunity. Immunity. 2004 Sep;21(3):379-90.
- Alugupalli KR, Gerstein RM. Divide and conquer: division of labor by B-1 B cells.
Immunity. 2005 Jul;23(1):1-2. - Alvarado Y, Giles FJ. Ras as a therapeutic target in hematologic malignancies.
Expert Opin Emerg Drugs. 2007 May;12(2):271-84. - Antsiferova YS, Sotnikova NY, Posiseeva LV, Shor AL. Changes in the T-helper
cytokine profile and in lymphocyte activation at the systemic and local levels in women with endometriosis. Fertil Steril. 2005 Dec;84(6):1705-11.
- Aramaki M, Nagasawa T, Koseki T, Ishikawa I. Presence of activated B-1 cells in
chronic inflamed gingival tissue. J Clin Immunol. 1998 Nov;18(6):421-9. - Axelsen JB, Lotem J, Sachs L, Domany E. Genes overexpressed in different
human solid cancers exhibit different tissue-specific expression profiles. Proc Natl Acad Sci U S A. 2007 Aug 7;104(32):13122-7.
- Balkwill F, Mantovani A. Inflammation and cancer: back to Virchow? Lancet. 2001
Feb 17;357(9255):539-45. - Balmanno K, Cook SJ.Sustained MAP kinase activation is required for the
expression of cyclin D1, p21Cip1 and a subset of AP-1 proteins in CCL39 cells. Oncogene. 1999 May 20;18(20):3085-97.
- Barleon B, Sozzani S, Zhou D, Weich HA, Mantovani A, Marmé D. Migration of
human monocytes in response to vascular endothelial growth factor (VEGF) is mediated via the VEGF receptor flt-1. Blood. 1996 Apr 15;87(8):3336-43.

80
- Baumgarth N, Tung JW, Herzenberg LA. Inherent specificities in natural antibodies: a key to immune defense against pathogen invasion. Springer Semin Immunopathol. 2005 Mar;26(4):347-62.
- Berland R, Wortis HH. Origins and functions of B-1 cells with notes on the role of
CD5. Annu Rev Immunol. 2002;20:253-300. - Bogsan CS, Novaes e Brito RR, Palos Mda C, Mortara RA, Almeida SR, Lopes JD,
Mariano M. B-1 cells are pivotal for in vivo inflammatory giant cell formation. Int J Exp Pathol. 2005 Aug;86(4):257-65.
- Boumsell L, Bernard A, Lepage V, Degos L, Lemerle J, Dausset J. Some chronic
lymphocytic leukemia cells bearing surface immunoglobulins share determinants with T cells. Eur J Immunol. 1978 Dec;8(12):900-4.
- Bradford MM.A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976 May 7;72:248-54.
- Bromberg JF, Wrzeszczynska MH, Devgan G, Zhao Y, Pestell RG, Albanese C,
Darnell JE Jr. Stat3 as an oncogene. Cell. 1999 Aug 6;98(3):295-303. - Bronte V, Chappell DB, Apolloni E, Cabrelle A, Wang M, Hwu P, Restifo NP.
Unopposed production of granulocyte-macrophage colony-stimulating factor by tumors inhibits CD8+ T cell responses by dysregulating antigen-presenting cell maturation. J Immunol. 1999 May 15;162(10):5728-37.
- Bronte V, Serafini P, Apolloni E, Zanovello P. Tumor-induced immune dysfunctions
caused by myeloid suppressor cells. J Immunother (1997). 2001 Nov-Dec;24(6):431-46. - Buettner R, Mora LB, Jove R. Activated STAT signaling in human tumors provides
novel molecular targets for therapeutic intervention. Clin Cancer Res. 2002 Apr;8(4):945-54.
- Bunt SK, Sinha P, Clements VK, Leips J, Ostrand-Rosenberg S. Inflammation
induces myeloid-derived suppressor cells that facilitate tumor progression. J Immunol. 2006 Jan 1;176(1):284-90.
- Bustin SA, Benes V, Nolan T, Pfaffl MW. Quantitative real-time RT-PCR--a
perspective. J Mol Endocrinol. 2005 Jun;34(3):597-601. - Casola S, Otipoby KL, Alimzhanov M, Humme S, Uyttersprot N, Kutok JL, Carroll
MC, Rajewsky K. B cell receptor signal strength determines B cell fate. Nat Immunol. 2004 Mar;5(3):317-27.
- Chanteux H, Guisset AC, Pilette C, Sibille Y. LPS induces IL-10 production by
human alveolar macrophages via MAPKinases- and Sp1-dependent mechanisms. Respir Res. 2007 Oct 4;8(1):71.

81
- Chen W, Daines MO, Khurana Hershey GK.Turning off signal transducer and activator of transcription (STAT): the negative regulation of STAT signaling.J Allergy Clin Immunol. 2004 Sep;114(3):476-89; quiz 490.
- Chevallier N, Berthelemy M, Le Rhun D, Laine V, Levy D, Schwartz-Cornil I. Bovine
leukemia virus-induced lymphocytosis and increased cell survival mainly involve the CD11b+ B-lymphocyte subset in sheep. J Virol. 1998 May;72(5):4413-20.
- Choo MK, Sakurai H, Koizumi K, Saiki I. Stimulation of cultured colon 26 cells with
TNF-alpha promotes lung metastasis through the extracellular signal-regulated kinase pathway. Cancer Lett. 2005 Dec 8;230(1):47-56.
- Christofori G, Semb H. The role of the cell-adhesion molecule E-cadherin as a
tumour-suppressor gene. Trends Biochem Sci. 1999 Feb;24(2):73-6. - Chudnovsky Y, Khavari PA, Adams AE. Melanoma genetics and the development
of rational therapeutics. J Clin Invest. 2005 Apr;115(4):813-24. - Clark WH Jr, Elder DE, Guerry D 4th, Epstein MN, Greene MH, Van Horn M. A
study of tumor progression: the precursor lesions of superficial spreading and nodular melanoma. Hum Pathol. 1984 Dec;15(12):1147-65.
- Cobb MH, Goldsmith EJ. How MAP kinases are regulated. J Biol Chem. 1995 Jun
23;270(25):14843-6. - Cohen C, Zavala-Pompa A, Sequeira JH, Shoji M, Sexton DG, Cotsonis G,
Cerimele F, Govindarajan B, Macaron N, Arbiser JL. Mitogen-actived protein kinase activation is an early event in melanoma progression. Clin Cancer Res. 2002 Dec;8(12):3728-33.
- Condeelis J, Pollard JW. Macrophages: obligate partners for tumor cell migration,
invasion, and metastasis. Cell. 2006 Jan 27;124(2):263-6. - Coussens LM, Raymond WW, Bergers G, Laig-Webster M, Behrendtsen O, Werb
Z, Caughey GH, Hanahan D. Inflammatory mast cells up-regulate angiogenesis during squamous epithelial carcinogenesis. Genes Dev. 1999 Jun 1;13(11):1382-97.
- Coussens LM, Tinkle CL, Hanahan D, Werb Z. MMP-9 supplied by bone marrow-
derived cells contributes to skin carcinogenesis. Cell. 2000 Oct 27;103(3):481-90. - Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002 Dec 19-
26;420(6917):860-7. - Cowley S, Paterson H, Kemp P, Marshall CJ. Activation of MAP kinase kinase is
necessary and sufficient for PC12 differentiation and for transformation of NIH 3T3 cells. Cell. 1994 Jun 17;77(6):841-52.
- Crews CM, Alessandrini A, Erikson RL. The primary structure of MEK, a protein
kinase that phosphorylates the ERK gene product. Science. 1992 Oct 16;258(5081):478-80.

82
- Croci DO, Zacarias Fluck MF, Rico MJ, Matar P, Rabinovich GA, Scharovsky OG. Dynamic cross-talk between tumor and immune cells in orchestrating the immunosuppressive network at the tumor microenvironment. Cancer Immunol Immunother. 2007 Nov;56(11):1687-700.
- D'Angelo G, Struman I, Martial J, Weiner RI. Activation of mitogen-activated protein
kinases by vascular endothelial growth factor and basic fibroblast growth factor in capillary endothelial cells is inhibited by the antiangiogenic factor 16-kDa N-terminal fragment of prolactin. Proc Natl Acad Sci U S A. 1995 Jul 3;92(14):6374-8.
- Darnell JE Jr. STATs and gene regulation. Science. 1997 Sep 12;277(5332):1630-
5. - Davies H, Bignell GR, Cox C, Stephens P, Edkins S, Clegg S, Teague J, Woffendin
H, Garnett MJ, Bottomley W, Davis N, Dicks E, Ewing R, Floyd Y, Gray K, Hall S, Hawes R, Hughes J, Kosmidou V, Menzies A, Mould C, Parker A, Stevens C, Watt S, Hooper S, Wilson R, Jayatilake H, Gusterson BA, Cooper C, Shipley J, Hargrave D, Pritchard-Jones K, Maitland N, Chenevix-Trench G, Riggins GJ, Bigner DD, Palmieri G, Cossu A, Flanagan A, Nicholson A, Ho JW, Leung SY, Yuen ST, Weber BL, Seigler HF, Darrow TL, Paterson H, Marais R, Marshall CJ, Wooster R, Stratton MR, Futreal PA. Mutations of the BRAF gene in human cancer. Nature. 2002 Jun 27;417(6892):949-54.
- Deryugina EI, Quigley JP. Matrix metalloproteinases and tumor metastasis. Cancer
Metastasis Rev. 2006 Mar;25(1):9-34. - Di Carlo E, Forni G, Lollini P, Colombo MP, Modesti A, Musiani P. The intriguing
role of polymorphonuclear neutrophils in antitumor reactions. Blood. 2001 Jan 15;97(2):339-45.
- Duan B, Morel L. Role of B-1a cells in autoimmunity. Autoimmun Rev. 2006
Jul;5(6):403-8. - Ebisuya M, Kondoh K, Nishida E. The duration, magnitude and
compartmentalization of ERK MAP kinase activity: mechanisms for providing signaling specificity. J Cell Sci. 2005 Jul 15;118(Pt 14):2997-3002.
- E Brito RR, De Lorenzo BH, Xander P, Godoy LC, Lopes JD, da Silva NP, Sampaio
SC, Mariano M. Role of distinct immune components in the radiation-induced abrogation of systemic lupus erythematosus development in mice. Lupus. 2007;16(12):947-54.
- Fidler IJ. Selection of successive tumour lines for metastasis. Nat New Biol. 1973
Apr 4;242(118):148-9. - Fidler IJ.Tumor heterogeneity and the biology of cancer invasion and
metastasis.Cancer Res. 1978 Sep;38(9):2651-60. - Fidler IJ. Critical determinants of metastasis. Semin Cancer Biol. 2002a
Apr;12(2):89-96. - Fidler IJ. The organ microenvironment and cancer metastasis. Differentiation.
2002b Dec;70(9-10):498-505.

83
- Fidler IJ. The pathogenesis of cancer metastasis: the 'seed and soil' hypothesis
revisited. Nat Rev Cancer. 2003 Jun;3(6):453-8. - Fokas E, Engenhart-Cabillic R, Daniilidis K, Rose F, An HX. Metastasis: the seed
and soil theory gains identity. Cancer Metastasis Rev. 2007 Dec;26(3-4):705-715. - Forster I, Rajewsky K. Expansion and functional activity of Ly-1+ B cells upon
transfer of peritoneal cells into allotype-congenic, newborn mice. Eur J Immunol. 1987 Apr;17(4):521-8.
- Gabrilovich DI, Chen HL, Girgis KR, Cunningham HT, Meny GM, Nadaf S,
Kavanaugh D, Carbone DP. Production of vascular endothelial growth factor by human tumors inhibits the functional maturation of dendritic cells. Nat Med. 1996 Oct;2(10):1096-103.
- Gabrilovich DI, Velders MP, Sotomayor EM, Kast WM. Mechanism of immune
dysfunction in cancer mediated by immature Gr-1+ myeloid cells. J Immunol. 2001 May 1;166(9):5398-406.
- Garrity T, Pandit R, Wright MA, Benefield J, Keni S, Young MR. Increased
presence of CD34+ cells in the peripheral blood of head and neck cancer patients and their differentiation into dendritic cells. Int J Cancer. 1997 Nov 27;73(5):663-9.
- Gieni RS, Umetsu DT, DeKruyff RH. Ly1- (CD5-) B cells produce interleukin (IL)-
10. Cell Immunol. 1997 Feb 1;175(2):164-70. - Godoy LC, Mariano M, Lopes JD. Immunity and hypersensitivity to gp43 antigen in
susceptible and resistant mice infected with Paracoccidioides brasiliensis. Med Mycol. 2003 Oct;41(5):427-36.
- Gorden A, Osman I, Gai W, He D, Huang W, Davidson A, Houghton AN, Busam K,
Polsky D. Analysis of BRAF and N-RAS mutations in metastatic melanoma tissues. Cancer Res. 2003 Jul 15;63(14):3955-7.
- Gordon S. Alternative activation of macrophages. Nat Rev Immunol. 2003
Jan;3(1):23-35. - Govindarajan B, Bai X, Cohen C, Zhong H, Kilroy S, Louis G, Moses M, Arbiser JL.
Malignant Transformation of Melanocytes to Melanoma by Constitutive Activation of Mitogen-activated Protein Kinase Kinase (MAPKK) Signaling. J Biol Chem. 2003 Mar 14;278(11):9790-5.
- Guiducci C, Vicari AP, Sangaletti S, Trinchieri G, Colombo MP. Redirecting in vivo
elicited tumor infiltrating macrophages and dendritic cells towards tumor rejection. Cancer Res. 2005 Apr 15;65(8):3437-46.
- Gum R, Wang H, Lengyel E, Juarez J, Boyd D.Regulation of 92 kDa type IV collagenase expression by the jun aminoterminal kinase- and the extracellular signal-regulated kinase-dependent signaling cascades.Oncogene. 1997 Mar 27;14(12):1481-93.

84
- Haas KM, Poe JC, Steeber DA, Tedder TF. B-1a and B-1b cells exhibit distinct developmental requirements and have unique functional roles in innate and adaptive immunity to S. pneumoniae. Immunity. 2005 Jul;23(1):7-18.
- Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000 Jan 7;100(1):57-70. - Hardy RR, Carmack CE, Shinton SA, Riblet RJ, Hayakawa K. A single VH gene is
utilized predominantly in anti-BrMRBC hybridomas derived from purified Ly-1 B cells. Definition of the VH11 family. J Immunol. 1989 May 15;142(10):3643-51.
- Hardy RR. Variable gene usage, physiology and development of Ly-1+ (CD5+) B
cells. Curr Opin Immunol. 1992 Apr;4(2):181-5. - Hardy RR. B-1 B cells: development, selection, natural autoantibody and leukemia.
Curr Opin Immunol. 2006 Oct;18(5):547-55. - Harper SJ, LoGrasso P. Signalling for survival and death in neurones: the role of
stress-activated kinases, JNK and p38. Cell Signal. 2001 May;13(5):299-310. - Hastings WD, Gurdak SM, Tumang JR, Rothstein TL. CD5+/Mac-1- peritoneal B
cells: a novel B cell subset that exhibits characteristics of B-1 cells. Immunol Lett. 2006 May 15;105(1):90-6.
- Hayakawa K, Hardy RR, Parks DR, Herzenberg LA. The "Ly-1 B" cell
subpopulation in normal immunodefective, and autoimmune mice. J Exp Med. 1983 Jan 1;157(1):202-18.
- Hayakawa K, Hardy RR, Herzenberg LA, Herzenberg LA. Progenitors for Ly-1 B
cells are distinct from progenitors for other B cells. J Exp Med. 1985 Jun 1;161(6):1554-68.
- Hayakawa K, Shinton SA, Asano M, Hardy RR. B-1 cell definition. Curr Top
Microbiol Immunol. 2000;252:15-22. - Herlyn M, Berking C, Li G, Satyamoorthy K. Lessons from melanocyte development
for understanding the biological events in naevus and melanoma formation. Melanoma Res. 2000 Aug;10(4):303-12.
- Herzenberg LA, Stall AM, Lalor PA, Sidman C, Moore WA, Parks DR, Herzenberg
LA. The Ly-1 B cell lineage. Immunol Rev. 1986 Oct;93:81-102. - Herzenberg LA. B-1 cells: the lineage question revisited. Immunol Rev. 2000
Jun;175:9-22. - Hill CS, Treisman R. Transcriptional regulation by extracellular signals:
mechanisms and specificity. Cell. 1995 Jan 27;80(2):199-211. - Hofmann UB, Eggert AA, Blass K, Brocker EB, Becker JC. Expression of matrix
metalloproteinases in the microenvironment of spontaneous and experimental melanoma metastases reflects the requirements for tumor formation. Cancer Res. 2003 Dec 1;63(23):8221-5.

85
- Hofmeister V, Schrama D, Becker JC. Anti-cancer therapies targeting the tumor
stroma.Cancer Immunol Immunother. 2008 Jan;57(1):1-17. - Houghton AN, Gold JS, Blachere NE. Immunity against cancer: lessons learned
from melanoma. Curr Opin Immunol. 2001 Apr;13(2):134-40. - Hsu MC, Toellner KM, Vinuesa CG, Maclennan IC. B cell clones that sustain long-
term plasmablast growth in T-independent extrafollicular antibody responses. Proc Natl Acad Sci U S A. 2006 Apr 11;103(15):5905-10.
- Huang B, Pan PY, Li Q, Sato AI, Levy DE, Bromberg J, Divino CM, Chen SH. Gr-
1+CD115+ immature myeloid suppressor cells mediate the development of tumor-induced T regulatory cells and T-cell anergy in tumor-bearing host. Cancer Res. 2006 Jan 15;66(2):1123-31.
- Ibe S, Qin Z, Schuler T, Preiss S, Blankenstein T. Tumor rejection by disturbing
tumor stroma cell interactions. J Exp Med. 2001 Dec 3;194(11):1549-59. - Ihle JN. The Stat family in cytokine signaling. Curr Opin Cell Biol. 2001
Apr;13(2):211-7. - Ilan N, Mahooti S, Madri JA. Distinct signal transduction pathways are utilized
during the tube formation and survival phases of in vitro angiogenesis. J Cell Sci. 1998 Dec 18;111 ( Pt 24):3621-31.
- Inoue S, Leitner WW, Golding B, Scott D. Inhibitory effects of B cells on antitumor
immunity. Cancer Res. 2006 Aug 1;66(15):7741-7. - Ishida H, Hastings R, Kearney J, Howard M. Continuous anti-interleukin 10
antibody administration depletes mice of Ly-1 B cells but not conventional B cells. J Exp Med. 1992 May 1;175(5):1213-20.
- John A, Tuszynski G. The role of matrix metalloproteinases in tumor angiogenesis
and tumor metastasis. Pathol Oncol Res. 2001;7(1):14-23. - Johnson CW, Barth RF, Adams D, Holman B, Price JE, Sautins I. Phenotypic
diversity of murine B16 melanoma detected by anti-B16 monoclonal antibodies. Cancer Res. 1987 Feb 15;47(4):1111-7.
- Joyce JA. Therapeutic targeting of the tumor microenvironment. Cancer Cell. 2005
Jun;7(6):513-20. - Kahana O, Micksche M, Witz IP, Yron I. The focal adhesion kinase (P125FAK) is
constitutively active in human malignant melanoma. Oncogene. 2002 Jun 6;21(25):3969-77.
- Kakisis JD, Pradhan S, Cordova A, Liapis CD, Sumpio BE. The role of STAT-3 in
the mediation of smooth muscle cell response to cyclic strain. Int J Biochem Cell Biol. 2005 Jul;37(7):1396-406.

86
- Kantor AB, Stall AM, Adams S, Herzenberg LA, Herzenberg LA. Differential development of progenitor activity for three B-cell lineages. Proc Natl Acad Sci U S A. 1992 Apr 15;89(8):3320-4.
- Kantor AB, Herzenberg LA. Origin of murine B cell lineages. Annu Rev Immunol.
1993;11:501-38. - Karras JG, Wang Z, Huo L, Howard RG, Frank DA, Rothstein TL. Signal transducer
and activator of transcription-3 (STAT3) is constitutively activated in normal, self-renewing B-1 cells but only inducibly expressed in conventional B lymphocytes. J Exp Med. 1997 Mar 17;185(6):1035-42.
- Kim J, Mori T, Chen SL, Amersi FF, Martinez SR, Kuo C, Turner RR, Ye X, Bilchik
AJ, Morton DL, Hoon DS. Chemokine receptor CXCR4 expression in patients with melanoma and colorectal cancer liver metastases and the association with disease outcome. Ann Surg. 2006 Jul;244(1):113-20.
- Klafter R, Arbiser JL. Regulation of angiogenesis and tumorigenesis by signal
transduction cascades: lessons from benign and malignant endothelial tumors. J Investig Dermatol Symp Proc. 2000 Dec;5(1):79-82.
- Klemke RL, Cai S, Giannini AL, Gallagher PJ, de Lanerolle P, Cheresh DA.
Regulation of cell motility by mitogen-activated protein kinase. J Cell Biol. 1997 Apr 21;137(2):481-92.
- Kortylewski M, Kujawski M, Wang T, Wei S, Zhang S, Pilon-Thomas S, Niu G, Kay
H, Mulé J, Kerr WG, Jove R, Pardoll D, Yu H. Inhibiting Stat3 signaling in the hematopoietic system elicits multicomponent antitumor immunity. Nat Med. 2005 Dec;11(12):1314-21.
- Krueger JS, Keshamouni VG, Atanaskova N, Reddy KB. Temporal and quantitative
regulation of mitogen-activated protein kinase (MAPK) modulates cell motility and invasion. Oncogene. 2001 Jul 12;20(31):4209-18.
- Kulbe H, Levinson NR, Balkwill F, Wilson JL The chemokine network in cancer--
much more than directing cell movement. Int J Dev Biol. 2004;48(5-6):489-96. - Kuper H, Adami HO, Trichopoulos D. Infections as a major preventable cause of
human cancer. J Intern Med. 2000 Sep;248(3):171-83. - Kuroki M, O'Flaherty JT. Extracellular signal-regulated protein kinase (ERK)-
dependent and ERK-independent pathways target STAT3 on serine-727 in human neutrophils stimulated by chemotactic factors and cytokines. Biochem J. 1999 Aug 1;341 ( Pt 3):691-6.
- Kusmartsev SA, Li Y, Chen SH. Gr-1+ myeloid cells derived from tumor-bearing
mice inhibit primary T cell activation induced through CD3/CD28 costimulation. J Immunol. 2000 Jul 15;165(2):779-85.
- Laemmli UK.Cleavage of structural proteins during the assembly of the head of
bacteriophage T4.Nature. 1970 Aug 15;227(5259):680-5.

87
- Langley RR, Fidler IJ. Tumor cell-organ microenvironment interactions in the
pathogenesis of cancer metastasis.Endocr Rev. 2007 May;28(3):297-321. - Lakka SS, Jasti SL, Kyritsis AP, Yung WK, Ali-Osman F, Nicolson GL, Rao JS.
Regulation of MMP-9 (type IV collagenase) production and invasiveness in gliomas by the extracellular signal-regulated kinase and jun amino-terminal kinase signaling cascades. Clin Exp Metastasis. 2000;18(3):245-52.
- Lassmann S, Schuster I, Walch A, Göbel H, Jütting U, Makowiec F, Hopt U,
Werner M. STAT3 mRNA and protein expression in colorectal cancer: effects on STAT3-inducible targets linked to cell survival and proliferation. J Clin Pathol. 2007 Feb;60(2):173-9.
- Lee CH, Kakinuma T, Wang J, Zhang H, Palmer DC, Restifo NP, Hwang ST.
Sensitization of B16 tumor cells with a CXCR4 antagonist increases the efficacy of immunotherapy for established lung metastases. Mol Cancer Ther. 2006 Oct;5(10):2592-9.
- Lee TS, Chau LY.Heme oxygenase-1 mediates the anti-inflammatory effect of
interleukin-10 in mice.Nat Med. 2002 Mar;8(3):240-6. - Lehmann JM, Holzmann B, Breitbart EW, Schmiegelow P, Riethmüller G, Johnson
JP. Discrimination between benign and malignant cells of melanocytic lineage by two novel antigens, a glycoprotein with a molecular weight of 113,000 and a protein with a molecular weight of 76,000. Cancer Res. 1987 Feb 1;47(3):841-5.
- Lehmann JM, Riethmuller G, Johnson JP. MUC18, a marker of tumor progression
in human melanoma, shows sequence similarity to the neural cell adhesion molecules of the immunoglobulin superfamily. Proc Natl Acad Sci U S A. 1989 Dec;86(24):9891-5.
- Lewis TB, Robison JE, Bastien R, Milash B, Boucher K, Samlowski WE, Leachman
SA, Dirk Noyes R, Wittwer CT, Perreard L, Bernard PS. Molecular classification of melanoma using real-time quantitative reverse transcriptase-polymerase chain reaction. Cancer. 2005 Oct 15;104(8):1678-86.
- Li G, Satyamoorthy K, Herlyn M. Dynamics of cell interactions and communications
during melanoma development. Crit Rev Oral Biol Med. 2002;13(1):62-70. - Liu YW, Chen CC, Tseng HP, Chang WC. Lipopolysaccharide-induced
transcriptional activation of interleukin-10 is mediated by MAPK- and NF-kappaB-induced CCAAT/enhancer-binding protein delta in mouse macrophages. Cell Signal. 2006 Sep;18(9):1492-500.
- Locati M, Riboldi E, Otero K, Martinez FO, Riva F, Perrier P, Baviera S, Signorelli
P, Bonecchi R, Allavena P, Sozzani S, Mantovani A. Regulation of the chemokine system at the level of chemokine receptor expression and signaling activity. Immunobiology. 2001 Dec;204(5):536-42.
- Lopez-Bergami P, Huang C, Goydos JS, Yip D, Bar-Eli M, Herlyn M, Smalley KS,
Mahale A, Eroshkin A, Aaronson S, Ronai Z. Rewired ERK-JNK signaling pathways in melanoma. Cancer Cell. 2007 May;11(5):447-60.

88
- Luster AD. Chemokines--chemotactic cytokines that mediate inflammation. N Engl
J Med. 1998 Feb 12;338(7):436-45. - MacDougall JR, Matrisian LM. Contributions of tumor and stromal matrix
metalloproteinases to tumor progression, invasion and metastasis. Cancer Metastasis Rev. 1995 Dec;14(4):351-62.
- Maldonado JL, Fridlyand J, Patel H, Jain AN, Busam K, Kageshita T, Ono T,
Albertson DG, Pinkel D, Bastian BC. Determinants of BRAF mutations in primary melanomas. J Natl Cancer Inst. 2003 Dec 17;95(24):1878-90.
- Mansour SJ, Matten WT, Hermann AS, Candia JM, Rong S, Fukasawa K, Vande
Woude GF, Ahn NG. Transformation of mammalian cells by constitutively active MAP kinase kinase.Science. 1994 Aug 12;265(5174):966-70.
- Mantovani A, Bottazzi B, Colotta F, Sozzani S, Ruco L. The origin and function of
tumor-associated macrophages. Immunol Today. 1992 Jul;13(7):265-70. - Mantovani A, Sozzani S, Locati M, Allavena P, Sica A. Macrophage polarization:
tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002 Nov;23(11):549-55.
- Mantovani A, Sica A, Sozzani S, Allavena P, Vecchi A, Locati M. The chemokine
system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004a Dec;25(12):677-86.
- Mantovani A, Sozzani S, Locati M, Schioppa T, Saccani A, Allavena P, Sica A.
Infiltration of tumours by macrophages and dendritic cells: tumour-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Novartis Found Symp. 2004b;256:137-45; discussion 146-8, 259-69.
- Mantovani A, Sica A, Locati M. Macrophage polarization comes of age. Immunity.
2005 Oct;23(4):344-6. - Mantovani A, Schioppa T, Porta C, Allavena P, Sica A. Role of tumor-associated
macrophages in tumor progression and invasion. Cancer Metastasis Rev. 2006 Sep;25(3):315-22.
- Mantovani A, Marchesi F, Porta C, Sica A, Allavena P. Inflammation and cancer:
Breast cancer as a prototype. Breast. 2007 Aug 29; [Epub ahead of print]. - Marcelino França K, Vericimo MA, Retamal C, Kipnis TL, Arnholdt AC. B1 cells
contribution to susceptibility in experimental paracoccidioidomycosis: immunoglobulin isotypes and repertoire determination. Med Mycol. 2006 Dec;44(8):755-66.
- Maroni P, Bendinelli P, Matteucci E, Desiderio MA. HGF induces CXCR4 and
CXCL12-mediated tumor invasion through Ets1 and NF-kappaB. Carcinogenesis. 2007 Feb;28(2):267-79.

89
- Martin F, Oliver AM, Kearney JF. Marginal zone and B1 B cells unite in the early response against T-independent blood-borne particulate antigens. Immunity. 2001 May;14(5):617-29.
- Mathur RK, Awasthi A, Wadhone P, Ramanamurthy B, Saha B. Reciprocal CD40
signals through p38MAPK and ERK-1/2 induce counteracting immune responses. Nat Med. 2004 May;10(5):540-4. Erratum in: Nat Med. 2004 Jul;7(10):755.
- McGary EC, Lev DC, Bar-Eli M. Cellular adhesion pathways and metastatic
potential of human melanoma. Cancer Biol Ther. 2002 Sep-Oct;1(5):459-65. - Meier F, Satyamoorthy K, Nesbit M, Hsu MY, Schittek B, Garbe C, Herlyn M.
Molecular events in melanoma development and progression. Front Biosci. 1998 Sep 15;3:D1005-10.
- Meier F, Schittek B, Busch S, Garbe C, Smalley K, Satyamoorthy K, Li G, Herlyn
M. The RAS/RAF/MEK/ERK and PI3K/AKT signaling pathways present molecular targets for the effective treatment of advanced melanoma. Front Biosci. 2005 Sep 1;10:2986-3001.
- Meier F, Busch S, Lasithiotakis K, Kulms D, Garbe C, Maczey E, Herlyn M, Schittek
B. Combined targeting of MAPK and AKT signalling pathways is a promising strategy for melanoma treatment. Br J Dermatol. 2007 Jun;156(6):1204-13.
- Melani C, Chiodoni C, Forni G, Colombo MP. Myeloid cell expansion elicited by the
progression of spontaneous mammary carcinomas in c-erbB-2 transgenic BALB/c mice suppresses immune reactivity. Blood. 2003 Sep 15;102(6):2138-45.
- Mills CD, Kincaid K, Alt JM, Heilman MJ, Hill AM. M-1/M-2 macrophages and the
Th1/Th2 paradigm. J Immunol. 2000 Jun 15;164(12):6166-73. - Minoprio P, el Cheikh MC, Murphy E, Hontebeyrie-Joskowicz M, Coffman R,
Coutinho A, O'Garra A. Xid-associated resistance to experimental Chagas' disease is IFN-gamma dependent. J Immunol. 1993 Oct 15;151(8):4200-8.
- Mohan C, Morel L, Yang P, Wakeland EK. Accumulation of splenic B1a cells with
potent antigen-presenting capability in NZM2410 lupus-prone mice. Arthritis Rheum. 1998 Sep;41(9):1652-62.
- Moore KW, O'Garra A, de Waal Malefyt R, Vieira P, Mosmann TR. Interleukin-10.
Annu Rev Immunol. 1993;11:165-90. - Mora LB, Buettner R, Seigne J, Diaz J, Ahmad N, Garcia R, Bowman T, Falcone R,
Fairclough R, Cantor A, Muro-Cacho C, Livingston S, Karras J, Pow-Sang J, Jove R. Constitutive activation of Stat3 in human prostate tumors and cell lines: direct inhibition of Stat3 signaling induces apoptosis of prostate cancer cells. Cancer Res. 2002 Nov 15;62(22):6659-66.
- Murakami M, Tsubata T, Okamoto M, Shimizu A, Kumagai S, Imura H, Honjo T.
Antigen-induced apoptotic death of Ly-1 B cells responsible for autoimmune disease in transgenic mice. Nature. 1992 May 7;357(6373):77-80.

90
- Murakami M, Honjo T. Involvement of B-1 cells in mucosal immunity and
autoimmunity. Immunol Today. 1995 Nov;16(11):534-9. - Murakami T, Maki W, Cardones AR, Fang H, Tun Kyi A, Nestle FO, Hwang ST.
Expression of CXC chemokine receptor-4 enhances the pulmonary metastatic potential of murine B16 melanoma cells. Cancer Res. 2002 Dec 15;62(24):7328-34.
- Nagase H, Woessner JF Jr. Matrix metalloproteinases. J Biol Chem. 1999 Jul
30;274(31):21491-4. - Niemand C, Nimmesgern A, Haan S, Fischer P, Schaper F, Rossaint R, Heinrich
PC, Müller-Newen G. Activation of STAT3 by IL-6 and IL-10 in primary human macrophages is differentially modulated by suppressor of cytokine signaling 3. J Immunol. 2003 Mar 15;170(6):3263-72.
- O'Garra A, Chang R, Go N, Hastings R, Haughton G, Howard M. Ly-1 B (B-1) cells
are the main source of B cell-derived interleukin 10. Eur J Immunol. 1992 Mar;22(3):711-7. - Oka M, Kikkawa U. Protein kinase C in melanoma. Cancer Metastasis Rev. 2005
Jun;24(2):287-300. - Paciorkowski N, Porte P, Shultz LD, Rajan TV. B1 B lymphocytes play a critical role
in host protection against lymphatic filarial parasites. J Exp Med. 2000 Feb 21;191(4):731-6.
- Paget S: The distribution of secondary growths in cancer of the breast. Lancet
1889, 1:571-573. - Parker SL, Tong T, Bolden S, Wingo PA. Cancer statistics, 1997. CA Cancer J Clin.
1997 Jan-Feb;47(1):5-27. - Pollard JW. Tumour-educated macrophages promote tumour progression and
metastasis. Nat Rev Cancer. 2004 Jan;4(1):71-8. - Porta C, Subhra Kumar B, Larghi P, Rubino L, Mancino A, Sica A. Tumor
promotion by tumor-associated macrophages. Adv Exp Med Biol. 2007;604:67-86. - Poste G, Fidler IJ. The pathogenesis of cancer metastasis. Nature. 1980 Jan
10;283(5743):139-46. - Raman M, Chen W, Cobb MH. Differential regulation and properties of MAPKs.
Oncogene. 2007 May 14;26(22):3100-12. - Ruffini PA, Morandi P, Cabioglu N, Altundag K, Cristofanilli M. Manipulating the
chemokine-chemokine receptor network to treat cancer. Cancer. 2007 Jun 15;109(12):2392-404.
- Rusciano D. Differentiation and metastasis in melanoma. Crit Rev Oncog.
2000;11(2):147-63.

91
- Rutherford MS, Witsell A, Schook LB. Mechanisms generating functionally heterogeneous macrophages: chaos revisited. J Leukoc Biol. 1993 May;53(5):602-18.
- Sahai E, Olson MF, Marshall CJ. Cross-talk between Ras and Rho signalling
pathways in transformation favours proliferation and increased motility. EMBO J. 2001 Feb 15;20(4):755-66.
- Sasagawa S, Ozaki Y, Fujita K, Kuroda S. Prediction and validation of the distinct
dynamics of transient and sustained ERK activation. Nat Cell Biol. 2005 Apr;7(4):365-73. .- Sica A, Saccani A, Mantovani A. Tumor-associated macrophages: a molecular
perspective. Int Immunopharmacol. 2002 Jul;2(8):1045-54. - Sierra A. Metastases and their microenvironments: linking pathogenesis and
therapy. Drug Resist Updat. 2005 Aug;8(4):247-57. - Simon C, Goepfert H, Boyd D. Inhibition of the p38 mitogen-activated protein
kinase by SB 203580 blocks PMA-induced Mr 92,000 type IV collagenase secretion and in vitro invasion. Cancer Res. 1998 Mar 15;58(6):1135-9.
- Sinha P, Clements VK, Ostrand-Rosenberg S. Reduction of myeloid-derived
suppressor cells and induction of M1 macrophages facilitate the rejection of established metastatic disease. J Immunol. 2005 Jan 15;174(2):636-45.
- Sinha P, Clements VK, Bunt SK, Albelda SM, Ostrand-Rosenberg S. Cross-talk
between myeloid-derived suppressor cells and macrophages subverts tumor immunity toward a type 2 response. J Immunol. 2007 Jul 15;179(2):977-83.
- Smalley KS. A pivotal role for ERK in the oncogenic behaviour of malignant
melanoma? Int J Cancer. 2003 May 1;104(5):527-32. - Smith SH, Goldschmidt MH, McManus PM. A comparative review of melanocytic
neoplasms. Vet Pathol. 2002 Nov;39(6):651-78. - Sober AJ, Lew RA, Koh HK, Barnhill RL. Epidemiology of cutaneous melanoma. An
update. Dermatol Clin. 1991 Oct;9(4):617-29. - Song X, Krelin Y, Dvorkin T, Bjorkdahl O, Segal S, Dinarello CA, Voronov E, Apte
RN. CD11b+/Gr-1+ immature myeloid cells mediate suppression of T cells in mice bearing tumors of IL-1beta-secreting cells. J Immunol. 2005 Dec 15;175(12):8200-8.
- Staquicini FI, Moreira CR, Nascimento FD, Tersariol IL, Nader HB, Dietrich CP,
Lopes JD. Enzyme and integrin expression by high and low metastatic melanoma cell lines. Melanoma Res. 2003 Feb;13(1):11-8.
- Staquicini FI. Participação de Linfócitos B-1 no Desenvolvimento do Melanoma
Murino Experimental [tese]. São Paulo: Universidade Federal de São Paulo – Escola
Paulista de Medicina; 2004.

92
- Sumimoto H, Imabayashi F, Iwata T, Kawakami Y. The BRAF-MAPK signaling pathway is essential for cancer-immune evasion in human melanoma cells. J Exp Med. 2006 Jul 10;203(7):1651-6.
- Suzuki E, Kapoor V, Jassar AS, Kaiser LR, Albelda SM. Gemcitabine selectively
eliminates splenic Gr-1+/CD11b+ myeloid suppressor cells in tumor-bearing animals and enhances antitumor immune activity. Clin Cancer Res. 2005 Sep 15;11(18):6713-21.
- Tarhini AA, Agarwala SS. Cutaneous melanoma: available therapy for metastatic
disease. Dermatol Ther. 2006 Jan-Feb;19(1):19-25. - Tower GB, Coon CC, Benbow U, Vincenti MP, Brinckerhoff CE. Erk 1/2
differentially regulates the expression from the 1G/2G single nucleotide polymorphism in the MMP-1 promoter in melanoma cells. Biochim Biophys Acta. 2002 Apr 24;1586(3):265-74.
- Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A. 1979 Sep;76(9):4350-4.
- Treisman R. Regulation of transcription by MAP kinase cascades. Curr Opin Cell
Biol. 1996 Apr;8(2):205-15. - Tsung K, Dolan JP, Tsung YL, Norton JA. Macrophages as effector cells in
interleukin 12-induced T cell-dependent tumor rejection. Cancer Res. 2002 Sep 1;62(17):5069-75.
- Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De Paepe A,
Speleman F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002 Jun 18;3(7):RESEARCH0034.
- Viala E, Pouysségur J.Regulation of tumor cell motility by ERK mitogen-activated
protein kinases.Ann N Y Acad Sci. 2004 Dec;1030:208-18. - Vigna AF, Almeida SR, Xander P, Freymüller E, Mariano M, Lopes JD. Granuloma
formation in vitro requires B-1 cells and is modulated by Paracoccidioides brasiliensis gp43 antigen. Microbes Infect. 2006 Mar;8(3):589-97.
- Wen Z, Zhong Z, Darnell JE. Maximal activation of transcription by statl and stat3
requires both tyrosine and serine phosphorylation. Cell, Vol 82, 251-260, 28 July 1995. - Weston CR, Davis RJ. The JNK signal transduction pathway. Curr Opin Genet Dev.
2002 Feb;12(1):14-21. - Wong SC, Chew WK, Tan JE, Melendez AJ, Francis F, Lam KP. Peritoneal CD5+
B-1 cells have signaling properties similar to tolerant B cells. J Biol Chem. 2002 Aug 23;277(34):30707-15.
- Yi AK, Yoon JG, Yeo SJ, Hong SC, English BK, Krieg AM. Role of mitogen-
activated protein kinases in CpG DNA-mediated IL-10 and IL-12 production: central role of

93
extracellular signal-regulated kinase in the negative feedback loop of the CpG DNA-mediated Th1 response. J Immunol. 2002 May 1;168(9):4711-20.
- Yin Z, Bahtiyar G, Zhang N, Liu L, Zhu P, Robert ME, McNiff J, Madaio MP, Craft
J. IL-10 regulates murine lupus. J Immunol. 2002 Aug 15;169(4):2148-55. - Young MR, Newby M, Wepsic HT. Hematopoiesis and suppressor bone marrow
cells in mice bearing large metastatic Lewis lung carcinoma tumors. Cancer Res. 1987 Jan 1;47(1):100-5.
- Young MR, Wright MA, Lozano Y, Prechel MM, Benefield J, Leonetti JP, Collins
SL, Petruzzelli GJ. Increased recurrence and metastasis in patients whose primary head and neck squamous cell carcinomas secreted granulocyte-macrophage colony-stimulating factor and contained CD34+ natural suppressor cells. Int J Cancer. 1997 Feb 20;74(1):69-74.
- Zea AH, Rodriguez PC, Atkins MB, Hernandez C, Signoretti S, Zabaleta J,
McDermott D, Quiceno D, Youmans A, O'Neill A, Mier J, Ochoa AC. Arginase-producing myeloid suppressor cells in renal cell carcinoma patients: a mechanism of tumor evasion. Cancer Res. 2005 Apr 15;65(8):3044-8.
- Zebisch A, Czernilofsky AP, Keri G, Smigelskaite J, Sill H, Troppmair J. Signaling
through RAS-RAF-MEK-ERK: from basics to bedside. Curr Med Chem. 2007;14(5):601-23.
- Zhou JH, Broussard SR, Strle K, Freund GG, Johnson RW, Dantzer R, Kelley KW.
IL-10 inhibits apoptosis of promyeloid cells by activating insulin receptor substrate-2 and phosphatidylinositol 3'-kinase. J Immunol. 2001 Oct 15;167(8):4436-42.
- Zou W, Machelon V, Coulomb-L'Hermin A, Borvak J, Nome F, Isaeva T, Wei S,
Krzysiek R, Durand-Gasselin I, Gordon A, Pustilnik T, Curiel DT, Galanaud P, Capron F, Emilie D, Curiel TJ.Stromal-derived factor-1 in human tumors recruits and alters the function of plasmacytoid precursor dendritic cells. Nat Med. 2001 Dec;7(12):1339-46.

ABSTRACT

Summary
Increasing evidence indicates that tumors require a constant influx of
myelomonocytic cells to support their malignant behavior. This is caused by tumor-
derived factors, which recruit and induce functional differentiation of
myelomonocytic cells, most of which are macrophages. Although myeloid lineages
are the classical precursors of macrophages, B lymphoid lineages such as B-1
cells, a subset of B lymphocytes found predominantly in pleural and peritoneal
cavities, are also able to migrate to inflammatory sites and differentiate into
mononuclear phagocytes exhibiting macrophage-like phenotype. Here we
examined the interplay of B-1 cells and tumor cells and checked whether this
interaction provides signals to influence melanoma cells metastases. Using in vitro
coculture experiments we showed that B16, a murine melanoma cell line, and B-1
cells physically interact. Moreover, interaction of B16 with B-1 cells leads to
upregulation of metastasis-related genes expression (MMP-9 and CXCR-4),
increasing its metastatic potential, as revealed by experimental metastases assays
in vivo. We also provide evidences that B16 cells exhibit markedly upregulated
phosphorylation of the extracellular signalregulated kinase (ERK) when co-cultured
with B-1 cells. Inhibition of ERK
phosphorylation induced by B-1 cells with an inhibitor of MEK1/2 strongly
suppressed the induction of MMP-9 and CXCR-4 mRNA expression and impaired
the increased metastatic behavior of B16. In addition, constitutive levels of ERK1/2
phosphorylation in B-1 cells are necessary for their commitment to affect the
metastatic potential of B16 cells. Our findings show for the first time, that B-1

80
lymphocytes can contribute to tumor cell properties required for invasiveness
during metastatic spread.

ANEXOS

80
----- Mensagem encaminhada de [email protected] ----- Data: Tue, 15 Jan 2008 19:19:14 -0500 (EST) De: [email protected] Reponder para: [email protected] Assunto: Cancer Science (CAS-OA-0602-2007.R1): Acceptance Para: [email protected] Date: 15-Jan-2008 RE: Ms No.: CAS-OA-0602-2007.R1 Title: B-1 lymphocytes increase metastatic behavior of melanoma cells through the extracellular signal-regulated kinase pathway. Author(s): Pérez, Elizabeth; Machado Jr, Joel; Aliperti, Fabiana; Freym�ler, Edna; Mariano, Mario; Lopes, Jos� Dear Prof. Joel Machado Jr: We are pleased to inform you that your revised paper has been accepted for publication in Cancer Science and will be published in Cancer Science. The comments of the Associate Editor and reviewer(s) who reviewed your manuscript are included at the foot of this letter. Thank you for your fine contribution. On behalf of the Editors of Cancer Science, we look forward to your continued contributions to the Journal. We would like to take this opportunity to explain our recently launched functions to assist authors with tracking their manuscripts through the production process. To minimise publication time of your manuscript it is important that all electronic artwork is supplied to the editorial office in the correct format and resolution. I recommend that you consult the Illustration guidelines at http://www.blackwellpublishing.com/bauthor/digill.asp if you need advice on any aspect of preparing your artwork. Copyright - If you havenuft already sent your completed Copyright Assignment Form for your manuscript at the time of submission, please sign and return to this office within two weeks. Accepted articles canuft go into production for an issue until a completed form has been received. Author Services - Production status tracking - You can now track your article via the publisherufs Author Services. Once your paper is with the Production Editor, you will receive an e-mail with a unique code that automatically adds your article to the system when you register. With Author Services you can check the status of your article online and choose to receive automated e-mails at key stages of production. Therefore, please ensure that we have your complete e-mail address. There may be a short delay whilst the article is sent to the Production Editor and logged into the production tracking system. Additional services will be added to the website soon. Proofing your manuscript – Cancer Science is included in a new electronic service, uge-proofinguh. You will receive an e-mail from the typesetter when your article is ready for proofing. Youufll receive instructions about how to download your paper and how to return your corrections. Your e-mail address is needed for this vital step, too. OnlineEarly – Cancer Science operates a system called OnlineEarly, whereby articles are published online ahead of assignment to an issue and publication in print. If your article is eligible for this service (Review Articles and Original Articles only), you can track the progress of your article and learn when it is published online by registering for Author Services. Please note that in order to publish your article as quickly as possible, and if your article is received very close to the copy deadline for an issue, it may be

81
incorporated directly into that issue without first appearing OnlineEarly. The Blackwell Synergy Website for Cancer Science is http://www.blackwell-synergy.com/loi/cas. Reprints and Offprints - You will receive instructions for ordering offprints when you are notified that your proofs are ready for review. Sincerely yours, Dr. Setsuo Hirohashi, Editor Cancer Science --- Cancer Science Editorial Office TEL: 81-3-3830-1266 FAX: 81-3-5689-7278 [email protected] http://www.blackwellpublishing.com/cas Associate Editor's decision: Acceptance Comments to Author: Associate Editor: 1 Comments to the Author: The authors have properly revised the manuscript according to the reviewers' comments. I recommend to accept this manuscript for publication in Cancer Science. ----- Finalizar mensagem encaminhada -----


Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo