ministÉrio da ciÊncia e tecnologia e inovaÇÃo - mcti ... da silva goncalves.pdf · each tree,...
TRANSCRIPT

MINISTÉRIO DA CIÊNCIA E TECNOLOGIA E INOVAÇÃO - MCTI INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM ENTOMOLOGIA
FLUTUAÇÃO POPULACIONAL DA MOSCA-NEGRA-DOS-CITROS,
Aleurocanthus woglumi (HEMIPTERA: ALEYRODIDAE) E DE SEUS INIMIGOS
NATURAIS EM UM PLANTIO DE CITROS, MANAUS, AMAZONAS, BRASIL
Maiara da Silva Gonçalves
Manaus, Amazonas
Junho, 2013

Maiara da Silva Gonçalves
FLUTUAÇÃO POPULACIONAL DA MOSCA-NEGRA-DOS-CITROS,
Aleurocanthus woglumi (HEMIPTERA: ALEYRODIDAE) E DE SEUS INIMIGOS
NATURAIS EM UM PLANTIO DE CITROS, MANAUS, AMAZONAS, BRASIL
Orientador(a): Dra. Beatriz Ronchi-Teles
Co-orientador(a): Dra. Márcia Reis Pena
Dissertação apresentada ao
Instituto Nacional de Pesquisas da
Amazônia como parte dos
requisitos para obtenção do título
de Mestre em Ciências Biológicas
(Entomologia).
Manaus, Amazonas
Junho, 2013

ii

iii
BANCA EXAMINADORA
Dr. Neliton Marques da Silva
Universidade Federal do Amazonas
Membro titular
Dr. Geraldo José Nascimento de Vasconcelos
Universidade Estadual do Amazonas
Membro titular
Dra. Rachel da Silva Corrêa
Escola Superior Batista do Amazonas
Membro titular
Dr. José Wellington de Morais
Instituto Nacional de Pesquisas da Amazônia
Membro suplente
Dra. Vivian de Siqueira Dutra
Universidade Federal do Amazonas
Membro suplente

iv
Dedico
Aos meus pais Nelma Shirley e David Benedito, que me deram os bens mais
preciosos: a vida e a liberdade de escolher meu caminho.

v
Ofereço
Ao meu querido Álvaro César por ser companheiro de todas as horas, pelo
carinho, dedicação, incentivos diários e pela fé inabalável em mim.

vi
AGRADECIMENTOS
Ao Instituto Nacional de Pesquisas da Amazônia - INPA pela oportunidade de
realizar este projeto e aprimorar minha formação.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico - CNPq, pela
bolsa de estudos concedida para realização do Mestrado.
Ao Sr. Osiris Silva, proprietário do sítio Brejo do Matão por permitir a realização
deste trabalho em um dos plantios de Laranja Pera.
À Dra. Beatriz Ronchi Teles, pela orientação e confiança em mim depositada.
À Dra. Márcia Reis Pena, por todas as excelentes sugestões, pelo incentivo
constante, por sempre acreditar no meu potencial, pela amizade ao longo desses
anos: muitíssimo obrigada por tudo!
À Malu Christine Barbosa Feitosa pelo enorme auxílio com as lâminas de
parasitóides e identificação, companhia, conselhos e sugestões.
Ao Dib Mady Diniz Gomes pela companhia em laboratório e auxílio imprescindível
nas coletas e triagens.
Ao Bruno Garcia Oliveira, que em pouco tempo ajudou tanto com boas idéias e
bibliografias indispensáveis e bom humor diário.
Ao amigo Pedro Aurélio Costa Lima Pequeno pela imensurável ajuda nas análises
estatísticas, gráficos e boas sugestões.
Ao Willians Lourenço Porto pelas incontáveis ajudas com questões estatísticas,
companhia e desabafos em „mineirês‟ no laboratório, além do sagrado café e
bolachas de toda tarde.
Aos colegas de curso pelas companhias durante as disciplinas e bons momentos
de descontração nos corredores do prédio de Entomologia.
Aos professores do curso de Entomologia por passarem valiosos ensinamentos
que só a vasta experiência pode transmitir.
Aos queridos amigos de longa data Ives Montefusco, Aline Castro, Luciana
Frazão, Elis Perrone, Kim Machado e Ed Barros pelas visitas, ligações, palavras
constantes de incentivo e torcida diária para que este trabalho fosse bem
sucedido, muito obrigada mesmo!

vii
Às amigas da FAK, Márcia Marques, Natália Wagner, Mayana Nobre e Marianna
Sarmanho por ouvirem minhas dificuldades e torcerem sempre por mim.
A todos que de alguma forma auxiliaram na realização deste trabalho.

viii
RESUMO
A mosca-negra-dos-citros, Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae) é uma importante praga asiática que tem causado danos à citricultura brasileira desde que foi introduzida no país, em 2001. Objetivou-se neste estudo verificar a flutuação populacional da mosca-negra-dos-citros, A. woglumi, de seus possíveis inimigos naturais e a influência de fatores bióticos e abióticos na flutuação no município de Manaus/AM. As coletas foram realizadas quinzenalmente em um pomar de Laranja Pêra (Citrus sinensis L.), no sítio Brejo
do Matão (0251‟12.6‟‟ S; 06002‟40.6‟‟ O) durante o período de dezembro de 2011 a novembro de 2012. Para o estudo de flutuação populacional de mosca-negra e seus inimigos naturais associados, foram selecionadas ao acaso dez plantas, em caminhamento zigue zague. Em cada planta foi colocada uma armadilha adesiva amarela. Nestas plantas foi realizado o estudo da flutuação populacional dos imaturos (ovos e ninfas) de mosca-negra e sua distribuição por quadrante (norte, sul, leste e oeste). Foi escolhido aleatoriamente um ramo por quadrante em cada planta, e de cada ramo foram retiradas seis folhas - da base, meio e ápice do ramo - totalizando 24 folhas por planta. O maior pico populacional de mosca-negra-dos-citros ocorreu no período chuvoso. Não houve diferença significativa na quantidade de adultos, ovos e ninfas de A. woglumi entre período chuvoso e seco. Não houve diferença significativa na quantidade de ovos e ninfas de A. woglumi entre os quadrantes. Foram coletados os inimigos naturais de A. woglumi com representantes das ordens Neuroptera: Chrysopidae e Coleoptera: Coccinelidae. Estes predando ninfas de mosca-negra. Foi verificada a ação parasitária de insetos da ordem Hymenoptera pertencentes às famílias Aphelinidae e Encyrtidae. Dentre as populações de parasitóides encontradas, Cales sp. e Arrenophagus sp. foram as que estiveram mais relacionadas com a flutuação populacional da mosca-negra. Os resultados deste trabalho refletem a viabilidade de se estabelecer um programa de manejo integrado de A. woglumi, sendo necessários estudos complementares sobre a flutuação populacional da mosca-negra e biologia de seus parasitóides para que o programa de MIP seja realizado de forma efetiva. Palavras chave: Citricultura, parasitóides, Aphelinidae, Encyrtidae, Chrysopidae

ix
ABSTRACT
The citrus blackfly, Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae) is an important Asiatic pest which has been damaging the brazilian citriculture since it was introduced in the country, in 2001. This study had the aim to verify the populational fluctuation of citrus blackfly, A. woglumi, and of its possible natural enemies and the influence of biotic and abiotic factors on the fluctuation in Manaus/AM. The evaluations occurred fortnightly in a Sweet Orange (Citrus
sinensis L.) crop, at Sítio Brejo do Matão (0251‟12.6‟‟ S; 06002‟40.6‟‟ O), during the period of December 2011 to November 2012. For the population fluctuation study of citrus blackfly and its associated and efective natural enemies, ten plants from the crop were selected at random. In each plant was placed a yellow sticky trap. On these same plants was conducted the population fluctuation study of citrus blackfly immatures (eggs and nymphs) and their distribution per quadrants (north, south, east and west). It was chosen at random a branch per quadrant in each tree, and from each branch were removed six leaves – from the base, middle and apex of the branch – totalizing 24 leaves per tree. The highest citrus blackfly peak occurred during the period of higher rainfall. There were no difference on the quantity of adults, eggs and nymphs of A. woglumi during rainy and dry period. There were no difference on the quantity of eggs and nymphs of A. woglumi among the quadrants. There were collected the natural enemies of A. woglumi from Neuroptera order, Chrysopidae family and Coleoptera: Coccinelidae. These preying on blackfly nymphs. The parasitic action of insects from Hymenoptera order belonging to Aphelinidae and Encyrtidae families was verified. Among the populations of parasitoids found, the genus Cales sp. and Arrenophagus sp. were the most correlated to the population fluctuation of citrus blackfly. The results of this work reflect the viability of establishing an integrated managing program of A. woglumi, being necessary complementary studies about the population fluctuation of A. woglumi and the biology of its natural enemies in a way the MIP program can be done effectively. Key words: Citriculture, parasitoids, Aphelinidae, Encyrtidae, Chrysopidae

10
SUMÁRIO
FOLHA DE ROSTO.................................................................................................ii
AGRADECIMENTOS..............................................................................................vi
RESUMO...............................................................................................................viii
ABSTRACT.............................................................................................................ix
LISTA DE FIGURAS..............................................................................................11
1.0 – INTRODUÇÃO..............................................................................................15
2.0 - REVISÃO DE LITERATURA.........................................................................17
2.1 – Citricultura.....................................................................................................17
2.2 – Mosca-negra-dos-citros, Aleurocanthus woglumi.........................................18
2.2.1 – Taxonomia e aspectos gerais....................................................................18
2.2.2 - Distribuição Geográfica e Histórico............................................................19
2.2.3 – Biologia......................................................................................................19
2.2.4 - Danos.........................................................................................................22
2.2.5 - Plantas hospedeiras ..................................................................................22
2.2.6 – Controle.....................................................................................................23
2.2.6.1 – Controle biológico...................................................................................23
2.2.6.1.1 – Predadores..........................................................................................23
2.2.6.1.2 – Parasitóides.........................................................................................24
2.2.6.1.3 - Fungos entomopatogênicos.................................................................26
2.2.6.2 – Controle químico.....................................................................................27
2.2.6.3 – Controle alternativo com extratos botânicos...........................................28
2.2.7 - Flutuação populacional...............................................................................28
3.0 - OBJETIVOS...................................................................................................30
3.1 - Objetivo Geral................................................................................................30
3.2 - Objetivos específicos......................................................................................30
4.0 - MATERIAL E MÉTODOS..............................................................................31
4.1 - Local de estudo..............................................................................................31
4.2 – Amostragens da mosca-negra-dos-citros, A. woglumi e de seus inimigos
naturais associados...............................................................................................32
4.2.1 - Amostragem com armadilhas adesivas amarelas......................................32

11
4.2.2 - Coleta de Imaturos de A. woglumi nas folhas.............................................33
4.2.3 - Coleta e identificação de inimigos naturais.................................................33
4.2.3.1 - Predadores .............................................................................................33
4.2.3.2 – Parasitóides............................................................................................33
4.3 Registro das variáveis abióticas......................................................................35
4.4 - Análise dos dados.........................................................................................35
4.4.1. Testes de normalidade e homocedasticidade.............................................35
4.4.2. Influência das variáveis abióticas na flutuação populacional da mosca-negra
e inimigos naturais.................................................................................................35
4.4.3. População da mosca-negra em período chuvoso e período seco...............36
4.4.4. Distribuição de imaturos da mosca-negra por quadrantes e período..........36
4.4.5. Relação entre mosca-negra e novos brotamentos foliares..........................36
4.4.6. Relação entre predadores, parasitóides e ninfas de mosca-negra..............36
4.4.7. Distribuição de parasitóides por quadrantes e período................................36
5.0 – RESULTADOS E DISCUSSÃO....................................................................37
5.1 Flutuação populacional de adultos e imaturos de A. woglumi e sua distribuição
espacial nos quadrantes leste, oeste, norte e sul da planta..................................37
5.1.1 - Flutuação populacional de adultos e imaturos de A. woglumi, coletados em
armadilhas adesivas amarelas e em folhas, respectivamente, e sua correlação
com os fatores abióticos........................................................................................37
5.1.2. – Distribuição espacial de ovos e ninfas (I, II, III e IV) de A. woglumi
coletados em folhas durante período chuvoso e seco...........................................40
5.1.2.1 – Relação entre adultos e imaturos de A. woglumi e fatores bióticos
(ocorrência de brotações)......................................................................................42
5.2 - Inimigos naturais associados, da mosca-negra-dos-citros, Aleurocanthus
woglumi em um pomar de Laranja Pêra (Citrus sinensis L.)
...............................................................................................................................43
5.2.1 Registro dos agentes de controle biológico..................................................43
5.2.1.1 – Fungos entomopatogênicos....................................................................43
5.2.1.2 – Predadores..............................................................................................44
5.2.1.3 – Parasitóides............................................................................................46

12
5.2.2 – Flutuação populacional de inimigos naturais da mosca-negra-dos-citros, A.
wolumi em um pomar de Laranja Pêra (Citrus sinensis L.)...................................50
5.2.2.1 Predadores................................................................................................50
5.2.2.2 Parasitóides...............................................................................................52
5.2.2.2.1 Relação entre flutuação populacional de A. woglumi e parasitóides....52
5.2.2.2.2 Distribuição espacial dos parasitóides da mosca-negra-dos-citros,
A.wolumi nos diferentes quadrantes da planta (norte, sul, leste e oeste) em um
pomar de Laranja Pêra (Citrus sinensis L.). ..........................................................57
6.0 – CONCLUSÕES.............................................................................................58
REFERÊNCIAS BIBLIOGRÁFICAS......................................................................59
LISTA DE FIGURAS
Figura 1 – Imagem de satélite do talhão onde foram realizadas as coletas.........31
Figura 2 – Croqui da área de estudo, no sítio Brejo do Matão, BR174 – km 15,
caminhamento em zigue-zague e plantas marcadas.............................................32
Figura 3 – Flutuação populacional de adultos de A. woglumi capturados em
armadilhas adesivas amarelas e sua correlação com o índice pluviométrico (mm)
(teste de regressão simples, p = 0,67), temperatura média (25,5±0,1°C)
(regressão simples, p = 0,51) e umidade relativa média (67,6±0,8%) (regressão
simples, p = 0,37) coletados no período de dezembro de 2011 a novembro de
2012 em um pomar de Laranja Pêra (Citrus sinensis L.).......................................38
Figura 4 – Distribuição temporal de adultos, ovos e ninfas de todos os ínstares de
Aleurocanthus woglumi Ashby (Aleyrodidae) durante período chuvoso (dezembro
de 2011 a maio de 2012) e período seco (junho a novembro de 2012), utilizando
teste T para adultos (p = 0,45) e ovos (p = 0,22) e teste Wilkoxon para ninfas (p =
0,81).......................................................................................................................39
Figura 5 – Distribuição espacial de ovos por quadrantes no período chuvoso
(ANOVA, p = 0,23) e no período seco (ANOVA, p = 0,15)....................................41
Figura 6 – Distribuição espacial de ninfas de A. woglumi em período chuvoso
(Kruskal-Wallis, p = 0,45) e período seco (Kruskal-Wallis, p = 0,3).......................41

13
Figura 7 – Relação entre quantidade de ovos (p = 0,6), ninfas (p = 0,31) e adultos
de A. woglumi (p = 0,26) e presença (S) ou ausência (N) de novas brotações
utilizando teste T de duas amostras......................................................................42
Figura 8 – Fungos entomopatogênicos Aschersonia aleyrodis e outro não
identificado sobre ovos ninfas de A. woglumi em um pomar de Laranja Pêra
(Citrus sinensis L.)..................................................................................................44
Figura 9 – A) Larva de Scymnus predando ninfas de III e IV ínstar de A. woglumi;
B) Vista dorsal do adulto de Scymnus; C) Vista ventral do adulto............................44
Figura 10 – A) Ceareochrysa sp. 1; B) Ceareochrysa sp. 2; C) Leucochrysa.
.............................................................................................................................. 45
Figura 11 – Larvas de crisopídeos com restos de ninfas de A. woglumi. A) Larva
de crisopídeo ainda esbranquiçada, com poucos restos de ninfas de A. woglumi
na carapaça; B) Larva de crisopídeo com carapaça mais enegrecida; C) Larva de
crisopídeo em campo com carapaça completamente cheia de restos de ninfas de
A. woglumi...............................................................................................................46
Figura 12 – Parasitóides da família Aphelinidae emergidos de ninfas de III e IV
ínstar de A. woglumi. A) Aphelinus cf.; B) Fêmea de Cales cf.; D) Macho de Cales
cf.............................................................................................................................49
Figura 13 - Parasitóides da família Encyrtidae emergidos de ninfas de III e IV
ínstar de A. woglumi. A) Fêmea de Arrenophagus cf.; B) Fêmea de Encyrtidae
cf.............................................................................................................................49
Figura 14 – Análise de Regressão Simples entre crisopídeos adultos e ninfas de
A. woglumi (r2 = 0,01, p = 0,68)..............................................................................50
Figura 15 – Correlação entre Precipitação acumulada no período de dezembro de
2011 a novembro de 2012 (teste de Regressão Simples, r2 = 0,13, p = 0,23),
Temperatura média (teste de Regressão Simples, r2 = 0,06, p = 0,41), Umidade
Relativa (teste de Regressão Simples, r2 = 0,01, p = 0,66) e o número total de
crisopídeos adultos coletados................................................................................51
Figura 16 – Flutuação populacional de parasitóides encontrados em armadilhas
adesivas e ninfas de todos os estádios de A. woglumi..........................................53
Figura 17. Flutuação populacional dos parasitóides emergidos de ninfas III e
ninfas IV de A. woglumi e ninfas de todos os estádios de A. woglumi..................53

14
Figura 18. Análise de regressão simples da relação entre ninfas de A. woglumi e
parasitóides Aphelinus sp. encontrados em armadilhas adesivas (r2 = 0,07, p =
0,37) e emergidos (r2 = 0,004, p = 0,83)................................................................54
Figura 19 – Análise de regressão simples da correlação entre parasitóides
Arrenophagus sp. encontrados em armadilhas adesivas e ninfas de A. woglumi (r2
= 0,06, p = 0,003) e Arrenophagus sp emergidos e ninfas de A. woglumi (r2 = 0,09,
p = 0,34).................................................................................................................54
Figura 20 – Análise de regressão simples da relação entre Cales sp. encontrados
em armadilhas adesivas e ninfas de A. woglumi (r2 = 0,78, p = 0,0001) e Cales sp
emergidos e ninfas de A. woglumi.........................................................................55
Figura 21 – Distribuição espacial por quadrantes dos Parasitóides emergidos de
ninfas de A. woglumi durante o período chuvoso (ANOVA, p = 0,94) e durante
período seco (ANOVA, p = 0,7) em um pomar de Laranja Pêra (Citrus sinensis L.)
no período de dezembro de 2011 a novembro de 2012........................................57

15
1.0 - Introdução
O Brasil é o maior produtor e exportador do suco de laranja e detém 37%
da produção mundial. Em 2012 foram produzidas 19.901.149 toneladas de laranja
(Agrianual, 2013). Em cinco décadas o país se tornou referência mundial na
produção de citros, com mudas e viveiros certificados, cultivares de laranja bem
estruturados e industrialização do suco para abastecer tanto o mercado
consumidor interno quanto os mercados consumidores europeu, asiático e norte-
americano, gerando lucros de 1 a 2 bilhões de dólares por ano ao Brasil (Neves et
al. 2010).
A mosca-negra-dos-citros, Aleurocanthus woglumi Ashby (Hemiptera:
Aleyrodidae) causa grandes prejuízos à citricultura em muitos países. Essa praga,
de origem asiática, foi detectada no Brasil em 2001, no estado do Pará, atingindo
o estado do Amazonas em 2004 e São Paulo, o maior polo citrícola do mundo em
2008 (Dietz e Zetek, 1920, Lemos et al. 2006, Jordão e Silva 2006, Pena et al.
2008, Ronchi-Teles et al. 2009, ADAB 2011, IDAF 2010, Correia et al. 2011).
Atualmente encontra-se amplamente disseminada no Brasil, ocorrendo em quase
todas as regiões, com exceção dos estados da Região Sul.
É uma praga de hábito alimentar polífago tendo os citros como hospedeiros
primários, ou seja, são os preferidos (Oliveira et al. 1999). No entanto, se estiver
em elevada densidade populacional, os adultos de A. woglumi podem dispersar
para outras plantas hospedeiras que estiverem próximas (Nguyen e Hamon,
2003), como por exemplo: Abacate (Persea americana Mill.), Bananeira (Musa
spp.), Café (Coffea arabica L.), Caju (Anacardium occidentale L.), Carambola
(Averrhoa carambola L.), Gengibre (Zingiber officinale Roscoe), Goiaba (Psidium
guajava L.), Graviola (Annona muricata L.), Mamão (Carica papaya L.), Manga
(Mangifera indica L.), Maracujá (Passiflora edulis Sims.), Pêra (Pyrus spp.), Rosa
(Rosa spp.), Sapoti (Manilkara zapota L.) e Uva (Vitis vinifera L.), plantas onde foi
observada a presença de A. woglumi (MAPA, 2011).
A alimentação das ninfas e adultos prejudica a planta pela sucção da seiva
nas folhas, causando danos celulares ao removerem nutrientes e pelos danos
indiretos pela produção de “honeydew” que são excretas açucaradas (Dowell et
al. 1981) que proporcionam o crescimento do fungo fumagina (Capnodium sp.),

16
fazendo com que as folhas não consigam fotossintetizar e os frutos se tornem
enegrecidos (Nguyen et al. 2003).
De acordo com Martinez (1982), o controle químico da mosca-negra-dos-
citros é oneroso e pouco eficiente. O controle biológico tem sido mais eficiente
que o controle químico, em diversas partes do mundo, e é realizado utilizando
predadores, parasitóides e fungos entomopatogênicos.
Os principais inimigos naturais da mosca-negra são os coleópteros Azya
trinitatis Marshall, Azya sp., Pentilia castanea Mulsant, Curinus sp., Delphastus
sp., Diomus sp. e Stethorus sp., os crisopídeos Chrysopa sp. e Nodita sp., os
fungos Aegerita webberi Fawcett e Aschersonia aleyrodis Webber (Martínez,
1979) e os parasitóides Amitus hesperidum Silvestri e Encarsia opulenta Silvestri
(Nguyen et al., 1983).
Os estudos de flutuação populacional de insetos praga servem como
parâmetros importantes para programas de Manejo Integrado de Pragas (MIP)
(Gazzoni et al. 1988), pois permitem conhecer melhor como as populações se
comportam ao longo do tempo e quais fatores externos bióticos e abióticos têm
correlação mais estreita, influenciando positiva ou negativamente a flutuação
populacional (Fernandes et al. 2003).
As informações obtidas com estudos de flutuação populacional de insetos
pragas permitem a previsão de picos populacionais (Cividanes, 2000) e a
definição de métodos de controle (Silveira Neto et al. 1976, Rodrigues et al.
2010).
Como o MIP é caracterizado como um manejo que altere o mínimo
possível o ambiente, é necessário fazer uma amostragem rápida e eficiente das
pragas e seus inimigos naturais (Fernandes et al. 2003). Estas informações são
muito importantes para que os produtores possam planejar melhor a necessidade
de controle e evitar desequilíbrios ecológicos no pomar.
2.0 - Revisão de literatura
2.1 - Citricultura
Atualmente o Brasil é o terceiro maior produtor de frutas do mundo, ficando
atrás da China (122 milhões de toneladas) e da Índia (84 milhões). Dos 39

17
milhões de toneladas de frutas produzidas no Brasil, 48% é de laranja, com 19,1
milhões de toneladas, estabelecendo-se como o país que mais produz laranja e
suco de laranja no mundo (FAO, 2012, Agrianual, 2013).
Além de ser o maior produtor de suco de laranja, detendo 37% da
produção mundial, o Brasil é também o maior exportador de suco congelado e
não congelado. Em 2012 foram produzidas 19.901.149 toneladas de laranja
(Agrianual, 2013). O Estado de São Paulo é o que mais tem municípios
produtores de laranja e melhor área plantada, sendo responsável por 76,3% da
produção brasileira (IBGE, 2012).
A maior parte do suco congelado é exportado para a Bélgica, Japão e
países baixos e o suco não congelado é exportado principalmente para a Bélgica
também, países baixos, Estados Unidos e Suíça além de outros 45 países. A
exportação de fruta in natura é pequena e, somada à fruta comercializada
internamente, representa 30% da produção (Agrianual, 2013).
No Amazonas, a citricultura é uma atividade que aos poucos vem se
incorporando ao processo de integração agroindustrial, por se adaptar bem às
condições edafoclimáticas da região, ser portadora de alta densidade-renda e
possuir mercado tanto para o produto in natura, quanto para seus subprodutos,
como suco concentrado, no resto do Brasil (Benedito, 1995, Gasparotto et al.
1998). A área destinada a produção de citros no Amazonas é de cerca de 2.951
hectares, com 343.642 frutos produzidos, entre as culturas de laranja, limão e
tangerina (IDAM, 2010).
A produção de laranja no Estado despertou o interesse dos produtores em
virtude do bom preço que a fruta tem alcançado no mercado (Silva e Souza,
2002). A área de produção está concentrada nos municípios de Rio Preto da Eva,
Manaus, Iranduba, Itacoatiara, Beruri, Presidente Figueiredo, Manacapuru e
Manaquiri, em ordem decrescente de produção (IDAM, 2010).
A citricultura nacional tem se mostrado bastante vulnerável aos diversos
problemas fitossanitários que têm surgido ao longo da sua história e que são
responsáveis por grandes perdas para a cultura (Santos, 2009).
Entre estes problemas fitossanitários, estão os causados por vírus e
bactérias, transmitidos por insetos e ácaros vetores e também por insetos e

18
ácaros pragas. Estas limitações têm sido uma constante na história da citricultura
nos principais centros produtores mundiais, o que ocasiona variações nos
sistemas de manejo como um todo (Müller et al. 2005).
A rápida propagação de doenças e pragas eleva o custo de produção
(Agrianual, 2003). As pragas são responsáveis pelos altos custos de implantação
e manutenção dos pomares cítricos, além de causar prejuízos decorrentes da
perda de colheitas e mercados (Maia, 2008).
Dentre as pragas dos citros que mais causam prejuízos no Amazonas
estão: o ácaro da leprose (Brevipalpus phoenicis Geijskes), que transmite o vírus
da leprose, pulgão preto (Toxoptera citricida Kirkaldy), vetor do vírus da tristeza
dos citros, cigarrinhas dos gêneros Acrogonia sp., Oncometopia sp., Homalodisca
sp., vetores da bactéria Xylella fastidiosa Wells, que causa a doença Clorose
Variegada de Citros (CVC) e a mosca-negra-dos-citros, que ao sugar nutrientes
da planta e deixar uma excreta açucarada, permite a formação de fumagina,
impedindo que ocorra respiração e fotossíntese, reduzindo a frutificação em até
80% (Bentes et al. 2007).
2.2 – Mosca-negra-dos-citros, Aleurocanthus woglumi
2.2.1 – Taxonomia e aspectos gerais
A mosca-negra-dos-citros, Aleurocanthus woglumi Ashby foi descrita de
forma técnica pela primeira vez por Quaintance e Baker em um manuscrito,
utilizado por Ashby posteriormente para descrever detalhadamente, mesmo de
forma não técnica, os estágios de vida do inseto (Dietz e Zetek, 1920). É um
inseto da ordem Hemiptera, subordem Stenorryncha e pertencente à família
Aleyrodidae. Possui as sinonímias Aleurocanthus punjabensis Corbett e
Aleurocanthus husain Corbett (Raga e Costa, 2008). Os adultos têm o corpo
vermelho alaranjado e as asas azuladas, mas na incidência da luz parecem
enegrecidas. As fêmeas medem cerca de 1,3 mm e os machos 0,9 mm de
comprimento (Silva et al. 2011). Os ovos são amarelados, as ninfas de primeiro
ínstar são bem clara, as de segundo ínstar tem coloração marrom e as ninfas de
terceiro e quarto ínstar são de fato enegrecidas (Dietz e Zetek, 1920).

19
2.2.2 - Distribuição Geográfica e Histórico
A mosca-negra-dos-citros está disseminada por quatro continentes:
Américas, Ásia, África e Oceania (Heu e Nagamine, 2001, Cosave, 1999).
Na América do Norte está presente nos Estados Unidos, Bermudas e México. Na
América Central e Caribe foi encontrada na Bahamas, Barbados, Costa Rica,
Cuba, República Dominicana, El Salvador, Guatemala, Haiti, Ilhas Caimã,
Jamaica, Nicarágua, Panamá, Porto Rico, Antilhas Holandesas e Trinidad. Na
América do Sul está na Colômbia, Venezuela, Equador, Peru, Guiana Francesa,
Suriname e Brasil (Hart et al. 1978, EPPO, 2008, Nguyen e Hamon, 1983 e Heu e
Nagamine, 2001, Maia et al. 2005, MAPA, 2011).
Foi constatada pela primeira vez no Brasil, em 2001, no estado do Pará
(Maia et al. 2005), em seguida, detectada em 2003 no Maranhão (Lemos et al.
2006); em 2004 foi registrada a primeira ocorrência no Amazonas, nos municípios
de Rio Preto da Eva, Itacoatiara e Iranduba (Ronchi-Teles et al. 2009); em
Tocantins (MAPA, 2008) e no Amapá em 2006 (Jordão e Silva, 2006), Goiás e
São Paulo em 2008 (Pena et al. 2008), Roraima em 2009 (Raga et al. 2011),
Paraíba em 2009 (Lopes et al. 2010), na Bahia em 2010 (ADAB, 2010), Ceará em
2010 (ADAGRI, 2010) e Pernambuco em 2010 (ADAGRO, 2010). Era
considerada praga quarentenária presente de alerta máximo (A2) (MAPA, 2008),
no entanto, como está disseminada em mais da metade dos estados brasileiros,
já não pode mais ser considerada praga exótica limitada a apenas algumas
regiões.
2.2.3 - Biologia
A mosca-negra-dos-citros realiza a oviposição na superfície abaxial das
folhas, geralmente em forma de espiral, mas pode não haver nenhum formato
caso a fêmea adulta esteja sob condições de estresse e o número de ovos em
uma espiral pode variar tanto quanto a forma e a média é de 32 ovos por espiral,
podendo haver até 50 espirais numa mesma folha (Dietz e Zetek, 1920).
Entre 11 a 20 dias, as ninfas de primeiro ínstar eclodem. Têm formato
ovalado e alongado, são esbranquiçadas e possuem pernas e antenas curtas. É a
única fase além da adulta em que há mobilidade. No final desta fase, que tem

20
duração de 7 a 16 dias, as ninfas perdem as pernas e fixam o aparelho bucal na
folha para o início da alimentação (Raga e Costa, 2008).
No segundo ínstar, o inseto tem um formato mais ovalado e convexo,
medindo 0,40 mm x 0,20 mm e de coloração marrom escura com espinhos
dorsais mais numerosos e proeminentes (Raga e Costa, 2008). Essa fase dura de
5 a 30 dias em indivíduos de laboratório, enquanto que no campo, o tempo
máximo observado foi de 21 dias (Dietz e Zetek, 1920).
Os indivíduos que estão entrando no terceiro ínstar levam de meia hora a
duas para se tornarem completamente escurecidos, de cor preto brilhante. Os
espinhos são mais numerosos e resistentes que no segundo, com convexidade
mais pronunciada e de contorno mais ovalado. Neste ínstar os sexos podem ser
definidos, sendo os machos pelo menos 1/3 menores do que as fêmeas, que são
sempre mais largas. A duração da fase é de 6 a 20 dias (Dietz e Zetek, 1920).
No quarto ínstar, também chamado de pupa, os insetos demoram de uma
hora à uma hora e meia para terem cor preta mais brilhante e com uma cera
branca ao redor do corpo. Os machos geralmente secretam muito mais cera que
as fêmeas. A duração deste período é irregular, variando de 16 a 80 dias em
laboratório. Em campo, o maior tempo registrado foi de 48 dias (Dietz e Zetek,
1920).
Após emergirem da pupa, e deixarem uma abertura em forma de “T”, os
adultos geralmente descansam ao lado dela até que várias partes do corpo já
estejam mais rígidas. Medem cerca de 0,99 a 1,24 mm, sendo as fêmeas maiores
que os machos (Raga e Costa, 2008). A cabeça, tórax e abdome são da cor
“vermelho tijolo”, sendo o vértice da cabeça amarelo pálido. As pernas e antenas
são esbranquiçadas e os olhos são vermelhos ou marrom avermelhados. As asas
são escuras, de cor azul ardósia com manchas incolores, parecendo uma faixa
branca (Dietz e Zetek, 1920; Dowell et al. 1981).
Após o enrijecimento do corpo, os adultos deixam sua posição inicial e
começam a buscar alimento para depois acasalar. No campo, o acasalamento
pode ocorrer a qualquer hora do dia, sendo mais freqüente no fim da tarde, devido
ao aumento no número de adultos que se juntam ao longo do dia. Segundo as
observações de Dietz e Zetek (1920) os adultos são fototrópicos positivos, pois

21
sempre se juntavam no lado da gaiola onde a luz era mais forte e a duração da
vida foi de, em média, uma semana, podendo chegar a até 12 dias.
O ciclo de ovo a adulto é de 45 a 133 dias, dependendo da temperatura
(Nguyen e Hamon, 1993). No Pará, em estudo realizado em laboratório, o ciclo da
mosca-negra-dos-citros foi de 59 a 88 dias, com média de 74 dias, sendo cerca
de 4,9 gerações ao ano. Foi observada uma baixa taxa de sobrevivência de ovos
e ninfas, relacionada com presença de predadores, parasitóides e altas
precipitações (Cunha, 2003).
No Maranhão, Lopes (2009) observou que não houve diferença estatística
na duração do ciclo biológico de A. woglumi nos hospedeiros laranjeira e
mangueira com 66,71 e 69 dias, respectivamente, valores inferiores aos
encontrados por Pena et al. (2009) e Cunha (2003) que observaram durante três
meses a biologia da mosca-negra em condições brasileiras.
Martinez (1982), em trabalho realizado na Venezuela em condições de
campo, verificou uma taxa de 16,8% de viabilidade dos ovos enquanto que em
laboratório observou maior taxa de viabilidade dos ovos (44,7%). A redução da
taxa observada em campo foi atribuída à possível influência de variações
climáticas e inimigos naturais.
De acordo com Dowell et al. (1981), chuva, vento e baixa umidade são
fatores ambientais importantes para a mosca-negra-dos-citros, assim como para
qualquer inseto. O desenvolvimento favorecido pela faixa de temperatura de 20 a
34C e umidade relativa de 70 a 80% (EPPO, 2008), o que facilita sua dispersão
por quase todos os Estados do país.
Observações feitas por Lopes (2010) indicam que no estudo do ciclo
biológico da mosca-negra, tanto a duração como a viabilidade das fases de
desenvolvimento foram muito afetadas pelo controle natural, pois nas fases de
ovo até ninfa de segundo ínstar observou-se grande mortalidade das mesmas
devido à presença de agentes externos, como os ácaros predadores Amblyseius
acalyphus Denmark e Muma, 1973 e Amblyseius aerialis Muma, 1955 (Acari:
Phytoseiidae).

22
2.2.4 - Danos
Tanto os adultos como as ninfas causam injúrias às plantas por se
alimentarem no floema, sugando a seiva. Eliminam uma secreção açucarada que
causa o aparecimento de fungos saprófitos, formando a fumagina (Oliveira et al.
2001) que diminui a quantidade de luz incidente sobre os órgãos clorofilados da
planta, reduzindo a capacidade de fotossíntese. O fato de se posicionarem na
parte abaxial da folha também interfere com as trocas gasosas feitas através dos
estômatos.
Do ponto de vista econômico, as fumaginas podem prejudicar a qualidade
dos produtos agrícolas, alterando a aparência dos mesmos (Bergamin Filho et al.
1995). A produção de frutos é reduzida em mais de 90% quando as infestações
excedem 5 a 7 ninfas por cm2 em todas as folhas. São necessárias de 50 a 100
ninfas por folhas para que haja redução dos níveis de nitrogênio para abaixo de
2,2% (Reitz, 1972).
2.2.5 - Plantas hospedeiras
A mosca-negra é uma praga de hábito alimentar polífago, sendo as plantas
cítricas as hospedeiras favoritas. A habilidade de suportar o desenvolvimento
completo da mosca-negra é a definição de “planta hospedeira” (Dowell et al.
1981).
Como os adultos de mosca-negra são atraídos por objetos que refletem luz
na faixa de 500-600 nm (nanômetros) e que é a faixa de todas as plantas verdes,
teoricamente os adultos poderiam infestar qualquer planta. No entanto, assim que
as fêmeas pousam na folha, são capazes de distinguir entre uma planta boa ou
fraca hospedeira e não hospedeira (Dowell, 1979).
A ocorrência do inseto foi relatada em cerca de trezentas espécies de
plantas com destaque para: manga, uva, citros, caju, abacate, goiaba, maçã, figo,
banana, mamão, pêra, romã, marmelo, café, rosas, entre outras (Nguyen et al.
2003). Em Belém foram registradas como plantas hospedeiras: laranja, tangerina,
pomelo, limão-tahiti, limão-galego, laranja “da terra”, manga e jambo (Cunha,
2003). Na região urbana de Manaus, tem sido encontrada infestando folhas de
citros, mangueiras e café em menor quantidade (Ronchi-Teles et al. 2009) e na

23
região rural dos municípios de Rio Preto da Eva, Iranduba e Itacoatiara há
infestações em pomares de citros (Pena, 2009).
Em testes com e sem chance de escolha de hospedeiros, A. woglumi
apresentou preferência por ovipositar nas espécies cítricas, mantendo um padrão
de não-preferência em cajueiro e goiabeira (Lopes, 2009).
2.2.6 – Controle
2.2.6.1 – Controle biológico
2.2.6.1.1 - Predadores
A mosca-negra possui muitos inimigos naturais relatados em outros países,
dentre os quais insetos predadores das famílias Chrysopidae (Ordem Neuroptera,
apenas imaturos), Coccinelidae (Ordem Coleoptera, tanto as larvas como
adultos), Dolichopodidae (Ordem Diptera, apenas imaturos) e Araneidae (Ordem
Araneae) (Cherry e Dowell, 1979).
As espécies de predadores registradas na Ásia foram Acletoxenus indica
Malloch, Acletoxenus sp., Chrysopa sp., Cryptoblabes gnidiella Millière,
Cryptognatha sp., Scymnus smithianus Silvestri, Scymnus sp., Cybocephalus sp.
(Clausen, 1934), em El Salvador foram utilizados os predadores Delphastus sp. e
Chrysopa sp. (Quezada, 1974), nos Estados Unidos, Delphastus pusillus
LeConte, Delphastus pallidus LeConte, Cycloneda sanguinea Linnaeus e
Chilocorus stigma Say (Cherry e Dowell, 1979), na Venezuela foram encontrados
Azya trinitatis Marshall, Curinus sp., Delphastus sp., Diomus sp., Pentilia castanea
Mulsant, Stethorus sp, Chrysopa sp., Nodita sp e Mantispilla sp. (Martínez et al.,
1979), e no Brasil foram relatados os predadores Amblyseius aerialis Muma, D.
pusillus, Ceraeochrysa cubana Hagen (Silva, 2005, Lemos, 2010).
Cherry e Dowell (1979) estudaram a abundância sazonal e preferência
alimentar dos predadores da mosca-negra no sudeste da Florida e registraram a
ocorrência dos predadores D. pusillus e D. pallidus, que se alimentavam dos ovos
e ninfas (entre 50 e 67%), e mais de 80% dos adultos presentes e também C.
sanguinea e C. stigma que foram atraídos por armadilhas amarelas e tiveram
ocorrência em maior quantidade no quadrante sul da planta (Dowell e Cherry,
1981).

24
Martínez et al. (1979) registraram predadores da mosca-negra das ordens
Coleoptera, Neuroptera e Mantodea. Da ordem Coleoptera foi encontrado um
inseto da família Dermestidae e as seguintes espécies da família Coccinelidae: A.
trinitatis, Curinus sp., Delphastus sp., Diomus sp., P. castanea, Stethorus sp.,
todos predadores de ovos e ninfas de primeiro ínstar. Da ordem Neuroptera,
foram encontrados os gêneros Chrysopa sp. e Nodita sp., da família Chrysopidae,
predando ninfas de todos os ínstares. Os insetos da ordem Mantodea são da
família Mantispidae, gênero Mantispilla sp. e predam ninfas de mosca-negra.
Lemos (2010) verificou a presença de dois predadores, A. aerialis (Acari:
Phytoseiidae) e D. pusillus (Coleoptera: Coccinellidae) associados à mosca-
negra.
Dentre os predadores da ordem Neuroptera, o bicho lixeiro da espécie C.
cubana é freqüentemente encontrado se alimentando das ninfas de mosca-negra.
Em Capitão Poço, no Pará, foi observado que sua carcaça feita de exúvias dos
insetos dos quais se alimenta, geralmente de cor clara, passou a ser escura por
conta das exúvias de mosca-negra das quais estava se alimentando (Silva, 2005).
Gonçalves et al. (2007) observaram a presença dos crisopídeos C. cubana,
Ceraeochrysa cincta, Ceraeochrysa claveri, Ceraeochrysa sanchezi e
Leucochrysa sp. em um pomar de citros em Manaus infestado pela mosca-negra.
Os predadores geralmente podem se alimentar de um grande número de
ninfas e adultos, mas sozinhos não são capazes de fazer a população decrescer
porque a fecundidade natural destas é alta o suficiente para compensar a perda
de indivíduos (Dowell et al. 1981).
2.2.6.1.2 - Parasitóides
Os parasitóides mais utilizados e conhecidos mundialmente são Encarsia
opulenta Silvestri (Hymenoptera: Aphelinidae) e Amitus hesperidum Silvestri
(Hymenoptera: Platygastridae). E. opulenta possui uma taxa de reprodução mais
lenta que A. hesperidum, mas tem melhor habilidade de procura (Nguyen et al.
2003).
Além destas duas espécies de parasitóides, mais registros foram feitos em
vários locais, os parasitóides registrados na Ásia foram Encarsia merceti Silvestri,

25
Eretmocerus serius Silvestri, Prospaltella divergens Silvestri, Prospaltella smithi
Silvestri, Prospaltella sp. (Clausen, 1934). Nos EUA, além de E. opulenta e A.
hesperidum, há também Prospaltella opulenta Silvestri (Cherry e Pastor, 1980,
Nguyen, 2003). Na Venezuela, Cales sp., Prospaltella sp. e Eretmocerus sp
(Martínez et al.,1979), na Nicarágua E. opulenta (Sweezey e Vasquez, 1991), no
México, Encarsia clypealis Silvestri, Encarsia smithi Silvestri e A. hesperidum
(Sagar, 1999) e no Brasil Cales noacki Howard e Encarsia sp. (Rossato, 2007,
Lemos, 2010).
Em campo os parasitóides adultos se alimentam do “honeydew”, a excreta
doce de pulgões e de outros insetos fitófagos como a própria mosca-negra, a qual
é rica em carboidratos e proteinas (Stary, 1988) e também pode agir como um
cairomônio de localização para que os parasitóides encontrem o hospedeiro
(Hagvar e Hofsvang, 1991).
Uma maneira simples de verificar se está ocorrendo o parasitismo é
através da observação das exúvias (pupas) da mosca-negra com auxílio de lupa
de aumento de 10 a 20 vezes. Se a abertura for em formato de “T” o inseto
emergido foi a mosca-negra, caso seja uma abertura circular, a emergência é de
parasitóide, no caso de haver dois círculos na mesma exúvia, significa
emergência de A. hesperidum (Silva et al. 2011).
Cherry e Pastor (1980) verificaram durante um programa de erradicação da
mosca-negra na Florida, que duas espécies de parasitóides introduzidas, A.
hesperidum e P. opulenta, se estabeleceram bem e foram responsáveis pela
redução da população de A. woglumi na Flórida, havendo alto índice de
parasitismo (80%) no primeiro ano, apenas 30% no segundo e 25% no terceiro
ano de pesquisa. Dowell e Cherry (1981) também observaram a ocorrência do
parasitóide P. opulenta na Florida.
No Texas, Summy et al. (1985) observaram que com a presença de E.
opulenta nos plantios onde havia grande densidade de mosca-negra, ocorreu um
controle biológico muito efetivo, onde hospedeiro e parasitóide coexistiam em
uma interação bastante estável de baixa densidade populacional.
Sweezey e Vasquez (1991) verificaram que E. opulenta foi responsável
pelo declínio da população de A. woglumi na Nicarágua, apesar de o tamanho da

26
sua população ter diminuído após períodos de chuvas intensas e aplicação de
inseticidas.
Também no Texas, após um período de severo congelamento que antingiu
e enfraqueceu os plantios, um trabalho de restabelecimento do controle biológico
de mosca-negra teve que ser feito, utilizando os parasitóides A. hesperidum e E.
opulenta., os quais conseguiram diminuir a população de mosca-negra após um
ano (Meagher e French, 2004).
No México, o controle biológico da mosca-negra é feito com três espécies
de parasitóides: E. clypealis, E. smithi e A. hesperidum. São liberadas em plantios
que tenham muitas ninfas de mosca-negra e em dias nos quais o vento não
ultrapasse a velocidade de 10 km/h e nem esteja chovendo (Sagar, 1999).
Em Belém, no estado do Pará, verificou-se um bom controle de A. woglumi
pelo parasitismo de microhimenópteros ainda não identificados, com relatos de
parasitismo da ordem de 90% (Silva 2005, Maia, 2008). Rossato (2007) verificou
parasitismo de mosca – negra por C. noacki (Hymenoptera: Aphelinidae) e
Encarsia sp. (Hymenoptera: Aphelinidae), considerando C. noacki um parasitóide
eficiente. Em municípios da Região Norte do Maranhão Lemos (2010) também
verificou a presença dos parasitoides, C. noacki e Encarsia sp.
2.2.6.1.3 - Fungos entomopatogênicos
Além de predadores e parasitóides, fungos entomopatogênicos também
podem ser utilizados no controle da mosca-negra. Martínez et al. (1979)
observaram que os fungos Aegerita webberi Fawcett e Aschersonia aleyrodis
Webber cresciam sobre as ninfas de mosca-negra e Pena et al. (2009)
comprovaram a eficiência no controle de mosca-negra utilizando o fungo
entomopatogênico Aschersonia aleyrodis na concentração de 2,3x107, onde ovos
e ninfas de primeiro e segundo ínstar foram mais suscetíveis.
Em outros países mais fungos entomopatogênicos foram registrados
crescendo sobre ninfas de mosca-negra. No Panamá foram Aschersonia sp. e A.
webberi (Dietz e Zetek, 1920), na Florida Paecilomyces farinosus Brown & Smith
(Fawcett, 1944), na Colômbia, Aschersonia sp. (Agudelo e Falcon, 1977), em El
Salvador e Costa Rica A. aleyrodis (Quezada, 1974, Elizondo e Quezada, 1990),

27
em Cuba Verticillium lecanii Zimmermann (Oppenheim, 2001), e no Brasil são os
fungos Aschersonia aleyrodis, Fusarium sp. e Aegerita webberi (Raga e Costa,
2008, Pena, 2009).
2.2.6.2 – Controle químico
De acordo com Martinez (1982), o controle químico da mosca-negra-dos-
citros é oneroso e pouco eficiente. O controle biológico tem sido mais eficiente
que o controle químico, em diversas partes do mundo, e é realizado utilizando
predadores, parasitóides e fungos entomopatogênicos.
O controle químico da mosca-negra é pouco utilizado mundialmente, e
quando é feito serve mais para o controle da fumagina. Como os produtos
utilizados para controlar a fumagina são a base de cobre, fungos
entomopatogênicos são prejudicados e, portanto, a população da mosca-negra
volta a crescer (Maia, 2008).
No Texas após um surto populacional de mosca-negra durante uma geada,
o programa de controle biológico utilizando os parasitóides E. opulenta e A.
hesperidum inicialmente não foi bem sucedido, portanto French e Meagher (1992)
testaram quatro inseticidas e obtiveram 90% de mortalidade das ninfas aplicando
Orthene 75S® e Talstar 10WP®.
O controle químico de mosca-negra-dos-citros no Brasil é feito utilizando os
inseticidas Kohinor 200 SC®, Provado 200 SC®, Timon 200 SC®, que possuem
imidacloprido na fórmula e Ampligo que tem os grupos químicos antranilamida e
piretróide na composição (Agrofit, 2003).
2.2.6.3 – Controle alternativo com extratos botânicos
Apesar de existirem muitos produtos químicos voltados para o controle de
pragas agrícolas, os métodos de controle alternativo têm ganhado espaço devido
principalmente à crescente preocupação com os efeitos negativos de agrotóxicos
tanto para o ambiente como para o corpo humano e pelos altos custos na
utilização de produtos químicos em plantios. Alguns trabalhos realizados com
mosca branca (B. tabaci) e mosca-negra (A. woglumi) mostram que o método de
controle alternativo pode ser eficaz.

28
Katole et al. (1996) verificaram que a pulverização de monocrotofos
0,025% mais óleo de nim 1% (emulsificado) num estágio ninfal inicial da mosca-
negra, seguido de uma pulverização de 1,25% de óleo de nim após 15 dias, foi o
tratamento mais efetivo.
Costa (2010) verificou que o extrato aquoso de Melia azedarach na
concentração de 8% possibilita um controle intermediário (47,3% de mortalidade)
sobre ovos de mosca-negra e Gonçalves (2010) observou que os extratos de
Melia azedarach em etanol a 0,56% exerceram tanto atividade ovicida (79% de
mortalidade) quanto ninficida (82,2% de mortalidade) sobre a mosca-negra e o
extrato de folhas de Azadirachta indica em etanol 0,56% exerceu atividade
ninficida (75,8% de mortalidade) sobre a praga.
Santos et al. (2010) verificaram a atividade ninficida de Carapa guianensis
(Meliaceae) em ninfas de segundo ínstar de mosca-negra, com mortalidade de
44,2% utilizando o extrato aquoso na concentração de 8%.
Silva Neto et al. (2010) também observaram atividade ninficida em ninfas
de segundo ínstar com o extrato aquoso de amêndoas de A. indica nas
concentrações 4 e 8%.
Castro et al. (2010) relataram a atividade ovicida de extratos aquosos de
folhas de Piper aduncum (Piperaceae) na concentração de 4%, causando 54,8%
de mortalidade nos ovos de mosca-negra.
2.2.7 - Flutuação populacional
Bellotti e Farias (1979) afirmam que picos populacionais de moscas-
brancas estão associados com a estação chuvosa, época em que as plantas
geralmente estão mais vigorosas, pois os níveis de população podem depender
mais das condições fisiológicas da planta que do clima.
Gomez et al. (2005) observaram que as populações de Bemisia sp. foram
maiores nas épocas chuvosas, resultado similar ao de Araújo (2005), onde a
população de Aleurothrixus aepim (Goeldi) e Bemisia tuberculata (Bondar) foi
maior nos meses de precipitação mediana e umidade relativa ainda elevada.
Em um estudo realizado no Maranhão, Vieira (2008) constatou que nos
meses de janeiro a março, correspondentes a estação chuvosa, a infestação de

29
mosca-negra foi ascendente, com o número máximo de 800 indivíduos por
armadilha. No entanto, em abril, mês de maior índice pluviométrico, houve uma
queda brusca, com cerca de 100 indivíduos por armadilha, e os picos
populacionais na estação seca tiveram uma periodicidade bimestral.
As informações obtidas com estudos de flutuação populacional de insetos
pragas permitem a previsão de picos populacionais (Cividanes, 2000) e a
definição de métodos de controle (Silveira Neto et al., 1976, Rodrigues et al.,
2010).
De acordo com Srinivasan et al. (2012), a população de moscas brancas
da espécie B. tabaci pode variar de um ano para outro, entre um pomar e outro, e
ter picos populacionais tanto na primavera quanto no outono, e que isto depende
de fatores climáticos e da nutrição das plantas.
Andrade e Ottati (2008) observaram no município de São José do Ribamar,
Maranhão, um pico populacional de mosca-negra em janeiro de 2008, na estação
chuvosa do estado, quando foram coletados 29% do total de 10.460 adultos em
um período de seis meses.
Santos et al. (2008) em um estudo da flutuação populacional de A. woglumi
em Capitão Poço, no Maranhão, relataram picos populacionais de mosca-negra
nos meses de outubro e novembro de 2007 (final da estação seca), sendo
observada ainda elevada população em dezembro de 2007, no início da estação
chuvosa.
EMBRATER/CIAT (1982) verificaram que as altas populações de moscas
brancas, estão associadas com a estação chuvosa porque é a época em que as
plantas estão mais fortes e que os possíveis elementos envolvidos na flutuação
populacional podem ser uma combinação de fatores ecológicos, como a fisiologia
das plantas, predadores e parasitóides.
Miklasiewics e Walker (1990) observaram que os parasitóides C. noacki e
Eretmocerus sp. foram responsáveis pela redução substancial na população da
mosca-branca Aleurothrixus floccosus. Também relatam que houve substituição
de população de parasitóides, inicialmente Eretmocerus sp. estava em maior
quantidade mas a população de C. noacki passou a ser maior.
Cherry et al. (1980) em um estudo de três anos na Flórida, constatou que

30
os parasitóides A. hesperidum e E. opulenta se estabeleceram em um plantio de
citros e mantiveram os índices populacionais de mosca-negra baixos. Também
verificaram que inicialmente a população de A. hesperidum era mais abundante e
foi ultrapassada por E. opulenta, mas ambas as espécies continuaram coexistindo
devido a disponibilidade de alimento. Luna-Salas et al. (2004) relataram dois picos populacionais do parasitóide
E. perplexa logo após os picos populacionais de mosca-negra, nos períodos de
março a junho e abril a maio, época da primavera na região de Tamaulipas, no
México.
3.0 - Objetivos
3.1 - Objetivo Geral
Verificar a flutuação populacional da mosca-negra-dos-citros, A. woglumi e
de seus possíveis inimigos naturais em um cultivo de citros no município de
Manaus, AM, identificando estes quando presentes.
3.3 - Objetivos específicos
1 - Verificar a flutuação populacional de A. woglumi e de seus possíveis inimigos
naturais em um pomar de citros.
2 - Diagnosticar a época de maior e menor incidência A. woglumi e de seus
possíveis inimigos naturais em um pomar de citros.
3 - Identificar as espécies de parasitóides e predadores associados e atuantes no
controle natural de A. woglumi em um pomar de citros.
4 - Relacionar os fatores abióticos (temperatura, umidade e precipitação) e
bióticos (brotação de ramos novos) na população de A. woglumi e de seus
inimigos naturais em um pomar de citros.
4.0 - Material e Métodos
4.1 - Local de estudo
O trabalho foi desenvolvido em um pomar de citros localizado na Rodovia
BR 174 (0251‟12.6‟‟ S; 06002‟40.6‟‟ O; Altitude: 101 m), km 15, Sítio Brejo do
Matão, área rural da região Metropolitana de Manaus, Amazonas, Brasil (Clima
tropical, de monção – Classificação de Kopen).

31
Figura 1. Imagem de satélite do talhão onde foram realizadas as coletas (Fonte: Google Earth).
As coletas foram realizadas em um plantio de laranja Pêra Rio (Citrus
sinensis L.) enxertados sobre Limão Cravo (Citrus limonia O.) com idade média
de seis anos. Esta área consiste de 1,2 hectares, onde as plantas tiveram os
tratamentos fitossanitários suspensos por um período de pelo menos 6 meses
anteriores ao início do projeto e continuaram suspensos durante a realização
deste .
4.2 - Amostragens da mosca-negra-dos-citros, A. woglumi e de seus
inimigos naturais associados.
As coletas foram realizadas quinzenalmente de dezembro de 2011 a
novembro de 2012, sendo amostradas dez plantas em caminhamento zigue
zague, para contagem de adultos de mosca-negra-dos citros capturados em
armadilhas adesivas amarelas e por meio da coleta de ovos e ninfas nas folhas.
Os inimigos naturais foram amostrados nas mesmas folhas utilizadas para
contagem dos imaturos de mosca-negra. As plantas amostradas foram marcadas
com faixas coloridas para controle das coletas.

32
A contagem dos indivíduos, triagem e identificação do material coletado foi
feita no Laboratório de Entomologia Agrícola do INPA – Campus II com auxílio de
um contador de volume, uma lupa, microscópio e chaves de identificação.
4.2.1.- Amostragem com armadilhas adesivas amarelas
Para a coleta dos adultos de mosca-negra foram utilizadas dez armadilhas
adesivas amarelas, nas dimensões de 24,5 x 10 cm, as quais foram colocadas
nas dez plantas escolhidas, sendo uma armadilha para cada planta, trocadas a
cada quinze dias.
4.2.2 - Coleta de Imaturos de A. woglumi nas folhas
Em cada planta, selecionada anteriormente, foram escolhidos ao acaso
quatro galhos, um de cada quadrante (norte, sul, leste e oeste), dos quais foram
retiradas seis folhas (da base, meio e ápice) por ramo, ao acaso, para contagem
de ovos e ninfas. Para a análise destes imaturos foi delimitado em cada folha uma
área de 8 cm² na região central desta. A contagem dos imaturos foi feita utilizando
microscópio estereoscópico e contador de volume.
Figura 2. Croqui da área de estudo, no sítio Brejo do Matão, BR174 – km 15, caminhamento em
zigue-zague e plantas marcadas.

33
4.2.3 - Coleta e identificação de inimigos naturais
4.2.3.1 - Predadores
Para confirmar a ocorrência de predação, as larvas dos potenciais
predadores, encontradas junto das folhas coletadas no item 4.2.2, foram
acondicionadas em placas de Petri cobertas com filme plástico contendo folhas
que foram coletadas apenas para alimentação dos predadores, sem ovos e
adultos de mosca-negra, apenas ninfas. Estas foram acondicionadas em BOD
(temperatura média de 25,5±0,1°C e 67,6±0,8% de UR média) por duas semanas,
até a emergência dos adultos, que foram montados e etiquetados.
Os predadores adultos encontrados em campo foram coletados utilizando-
se frasco morteiro, levados ao Laboratório de Entomologia Agrícola, onde foram
conservados em álcool 70% e etiquetados.
4.2.3.2 – Parasitóides
Para verificar a ocorrência de parasitismo, as folhas utilizadas na contagem
de imaturos de mosca-negra no item 4.2.2 foram previamente avaliadas ao
microscópio estereoscópio para retirada de todos os insetos existentes que não
fossem mosca-negra-dos-citros e assim assegurar a certeza das emergências
dos parasitóides da mosca-negra.
Em seguida as folhas foram acondicionadas em placas de Petri cobertas
com filme plástico e diariamente, durante quinze dias, a emergência dos
parasitóides foi avaliada. Estas foram acondicionadas em BOD (temperatura
média de 25,5±0,1°C e 67,6±0,8% de UR média).
A identificação dos parasitóides foi realizada por meio da preparação de
lâminas em Bálsamo do Canadá, e as chaves de identificação utilizadas foram “A
review of the genera of Neotropical Encyrtidae (Hymenoptera: Chalcidoidea)” de
John S. Noyes, 1980 e “Hymenoptera of the World: An identification guide to
families” de Henri Goulet e John T. Huber, 1993. A identificação dos predadores
da ordem Neuroptera foi feita utilizando-se a chave “The Green Lacewing of
Brazilian Agro-ecosystems” de Sérgio de Freitas e Norman D. Penny, 2001. A
identificação dos predadores da ordem Coleoptera foi feita utilizando-se a chave

34
contida no livro “O estudo dos insetos” de Charles A. Triplehorn e Norman F.
Johnson, 2011.
A metodologia utilizada no preparo de lâminas foi a elaborada pelo grupo
de estudos do especialista em parasitóides Dr. Serguei Triapitsyn, da
Universidade da California. Foram preparadas lâminas permanentes, utilizando-se
hidróxido de potássio (KOH) para a diafanização dos tecidos moles como
gorduras e musculatura e em seguida mergulhando os parasitóides em água
destilada para diluir o hidróxido de potássio, a fim de diminuir a atividade deste
sobre o inseto e depois em ácido acético Glacial para neutralizar o hidróxido de
potássio. Após este processo houve o mergulho em diferentes concentrações de
álcool (70, 80, 90 e 100%), para desidratar os insetos gradualmente. Depois de
desidratar em álcool, os insetos foram banhados em eugenol (produto alternativo
ao xilol, que é uma substância cancerígena) por no mínimo 10 minutos, para
amolecer as articulações e facilitar a montagem da lâmina. Por fim, cada inseto foi
posicionado em uma lâmina e uma gota de bálsamo-do-Canadá foi aplicada para
fixar e conservar o inseto na lâmina. Os insetos foram montados dorso-
ventralmente, com as asas e antenas esticadas, depois as lâminas foram
mantidas em estufa por cerca de três dias para secarem.
Os parasitóides emergidos foram comparados com os parasitóides adultos
observados nas armadilhas, para verificar se eram os mesmos.
4.3 Registro das variáveis abióticas
Foram utilizados os dados de temperatura, umidade e pluviosidade
registrados pelo Instituto Nacional de Meteorologia (INMET), Estação Manaus e
do Instituto Nacional de Pesquisas da Amazônia, Clima e Ambiente (CLIAMB),
dados referentes ao período de dezembro de 2011 a Julho de 2012 (CLIAMB) e
agosto de 2012 a novembro de 2012 (INMET).
4.4 - Análise dos dados
Os dados quinzenais para cada mês foram agrupados para análise,
registrando a quantidade mensal de ovos, ninfas, adultos de mosca-negra e

35
inimigos naturais (parasitóides e predadores) encontrados. O programa utilizado
nas análises foi o software livre R, versão 2.15.
4.4.1. Testes de normalidade e homocedasticidade
Os dados foram submetidos aos testes estatísticos de normalidade (teste
Shapiro-Wilk) e homocedasticidade (teste Bartlett). Quando necessário, os dados
foram transformados em log de base 10. Quando mesmo após as transformações
os dados não apresentavam homocedasticidade, foram utilizados os testes não
paramétricos equivalentes, Wilkoxon e Kruskal-Wallis.
4.4.2. Influência das variáveis abióticas na flutuação populacional da mosca-
negra e inimigos naturais
Foram utilizadas análises de Regressão Simples para verificar a influência
das variáveis abióticas como a pluviosidade, temperatura média e umidade
relativa média na flutuação populacional de adultos da mosca-negra e na
população de inimigos naturais. Os coeficientes de correlação significativos
obtidos foram comparados para determinar qual fator abiótico teve maior relação
com a flutuação populacional da mosca-negra e de crisopídeos.
4.4.3. População da mosca-negra em período chuvoso e período seco
Para verificar diferença na quantidade de adultos e ovos de mosca-negra
por período, foram feitos Testes T pareados. Para as ninfas, foi utilizado o teste
Wilkoxon.
4.4.4. Distribuição de imaturos da mosca-negra por quadrantes e período
Para verificar diferença na quantidade de ovos entre quadrantes, os dados
foram submetidos à ANOVA. Na análise para verificar a diferença na quantidade
de ninfas entre quadrantes foi utilizado o teste Kruskal-Wallis.

36
4.4.5. Relação entre mosca-negra e novos brotamentos foliares
Foi testada a relação de imaturos (ovos e ninfas) e adultos de mosca-negra
com a presença ou ausência de novos brotamentos utilizando Teste T de duas
amostras.
4.4.6. Relação entre predadores, parasitóides e ninfas de mosca-negra
Para testar a influência da população de predadores e parasitóides na
população de ninfas da mosca-negra, foram realizados testes de Regressão
Simples, com correlação entre cada grupo de inimigos naturais e número total de
ninfas de mosca-negra.
4.4.7. Distribuição de parasitóides por quadrantes e período
Os dados de parasitóides emergidos por quadrante durante período seco e
período chuvoso foram submetidos à ANOVA para verificar diferenças.
5.0 – Resultados e Discussão
5.1 Flutuação populacional de adultos e imaturos de A. woglumi e sua
distribuição espacial nos quadrantes leste, oeste, norte e sul da planta.
5.1.1 - Flutuação populacional de adultos e imaturos de A. woglumi,
coletados em armadilhas adesivas amarelas e em folhas, respectivamente, e
sua correlação com os fatores abióticos.
Durante o período de coletas não foi observada influência direta dos fatores
abióticos na flutuação populacional dos adultos de mosca-negra.
Na observação da temperatura e flutuação de adultos de A. woglumi não
foi encontrada relação significativa (p = 0,51), pois a temperatura variou pouco
durante os meses de coleta e não exerceu influencia na flutuação populacional da
mosca-negra (Figura 1).
Quanto à umidade relativa média, esta também não exerceu influência (p =
0,37) e também não variou, decrescendo de 80% em média para 70% apenas
durante o período seco. Apesar de o índice pluviométrico não influenciar
diretamente na quantidade de adultos de mosca-negra (p = 0,67), é possível
observar que o pico populacional ocorreu em janeiro de 2012, durante o período

37
de chuvas e decaiu bruscamente nos meses de maior quantidade de precipitação
acumulada (fevereiro a maio) (Figura 3). A população de adultos de mosca-negra
se manteve baixa durante todo o período chuvoso (dezembro de 2011 a maio de
2012), voltando a crescer apenas durante o período seco (a partir de junho de
2012) (Figura 3).
Figura 3. Flutuação populacional de adultos de A. woglumi capturados em armadilhas adesivas
amarelas e sua correlação com o índice pluviométrico (mm) (teste de regressão simples, p = 0,67),
temperatura média (25,5±0,1°C) (regressão simples, p = 0,51) e umidade relativa média
(67,6±0,8%) (regressão simples, p = 0,37) coletados no período de dezembro de 2011 a novembro
de 2012 em um pomar de Laranja Pêra (Citrus sinensis L.) na Região Metropolitana de Manaus,
Amazonas.
O pico populacional de adultos de mosca-negra capturados em armadilhas
adesivas amarelas ocorreu no período chuvoso, mas as populações de adultos,
ovos e ninfas coletadas não diferiram estatisticamente entre período chuvoso e
seco (p = 0,81, p = 0,22, p = 0,81, respectivamente) (Figura 4).

38
Figura 4. Distribuição temporal de adultos, ovos e ninfas de todos os ínstares de Aleurocanthus
woglumi Ashby (Aleyrodidae) durante período chuvoso (dezembro de 2011 a maio de 2012) e
período seco (junho a novembro de 2012), utilizando teste T para adultos (p = 0,45) e ovos (p =
0,22) e teste Wilkoxon para ninfas (p = 0,81).
Viera (2008) no Maranhão, ao observar a flutuação populacional e
dependência espacial de A. woglumi em Citrus latifolia e constatar que o nível
populacional de mosca-negra foi maior durante o período chuvoso (março, maio e
julho), sendo o pico populacional também logo no início das chuvas e decaindo
bruscamente no mês com maior índice pluviométrico, em abril.

39
Patel e Patel (2006) em Gujarat, India, concluíram que a população de
mosca-negra se manteve ativa durante o ano inteiro, mas ocorreram três picos
populacionais de ninfas, nos meses de abril (verão), agosto e novembro (estação
chuvosa), diminuindo drasticamente em fevereiro (inverno), corroborando com os
resultados deste trabalho, onde também houve pico populacional de mosca-negra
na estação chuvosa.
Maia (2008), em Capitão Poço – Pará, verificou que a densidade
populacional de mosca-negra foi maior durante os meses de dezembro, janeiro,
fevereiro e outubro (estação chuvosa), com média de 90% de infestação e menor
durante os meses de abril (final da estação chuvosa), maio, junho e julho (estação
seca), com média de 25% de infestação.
No entanto, Gonzaga Filho et al. (2011) obtiveram resultados diferentes
dos deste estudo ao verificar a sazonalidade de A. woglumi em um cultivo de
citros em Manaus. O período com maior incidência de adultos de mosca-negra foi
de março a julho de 2010, final da estação chuvosa e início da estação seca, com
média de 193 indivíduos e média de 818 mm de precipitação. O período de menor
incidência de A. woglumi foi de agosto de 2010 a fevereiro de 2011, abrangendo o
período seco e início do período chuvoso, com precipitação média de 1165,7 mm
e média de 34,9 indivíduos de mosca-negra.
Medeiros (2009) ao observar uma grande influência da precipitação sobre a
densidade populacional da mosca-negra no Maranhão, também obteve resultados
diferentes. A densidade de ovos e ninfas foi alta durante o período seco (de
julho/2006 a início de janeiro/2007) e decaiu durante todo o período chuvoso
(fevereiro/2007 a junho/2007).
5.1.2. – Distribuição espacial de ovos e ninfas (I, II, III e IV) de A. woglumi
coletados em folhas durante período chuvoso e seco.
Não houve diferença significativa na distribuição espacial de ovos entre os
diferentes quadrantes (norte, sul, leste e oeste) tanto durante o período chuvoso
como durante o período seco (Figura 5).

40
Figura 5. Distribuição espacial de ovos por quadrantes no período chuvoso (ANOVA, p = 0,23) e
no período seco (ANOVA, p = 0,15).
Também não houve diferença estatística na distribuição de ninfas (I, II, III e
IV ínstares) entre os quadrantes (leste, oeste, norte e sul) tanto no período
chuvoso quanto no período seco (Figura 6).
Figura 6. Distribuição espacial de ninfas de A. woglumi em período chuvoso (Kruskal-Wallis, p =
0,45) e período seco (Kruskal-Wallis, p = 0,3)
Medeiros (2009) também não observou diferença na distribuição espacial
de ovos e ninfas de A. woglumi durante o período chuvoso, mas constatou que no
período seco houve maior concentração dos ovos e ninfas nos quadrantes leste e
oeste.

41
5.1.2.1 – Relação entre adultos e imaturos de A. woglumi e fatores bióticos
(ocorrência de brotações)
Foram testadas as relações entre a presença de brotações novas e
quantidade de ovos, ninfas e adultos de A. woglumi.
Os meses em que ocorreram brotações foliares e floração foram fevereiro,
abril, junho e novembro. Aparentemente, o crescimento vegetativo foi bimestral no
período chuvoso, ocorrendo apenas uma vez no período seco. Não houve
diferença na quantidade de ovos, ninfas e adultos entre o período com novas
brotações e sem novas brotações (p = 0,6, p = 0,31 e p = 0,26, respectivamente)
(Figura 7).
Figura 7. Relação entre quantidade de ovos (p = 0,6), ninfas (p = 0,31) e adultos de A. woglumi (p
= 0,26) e presença (S) ou ausência (N) de novas brotações utilizando teste T de duas amostras.
Silva (2010) observou em Capitão Poço – Pará, que em abril e maio/2009,
período imediatamente posterior aos meses de maior precipitação acumulada,
houve um alto índice de plantas com presença de mosca-negra em ambos os

42
plantios, o que foi relacionado à um maior crescimento vegetativo das plantas de
citros.
Ainda que os efeitos de maior incidência de chuvas sejam diferentes entre
as culturas, geralmente quanto maior a disponibilidade hídrica, maior a
produtividade em plantios (CONAB, 2010), havendo maior oferta de alimento para
insetos fitófagos.
A chuva pode influenciar direta ou indiretamente na população de insetos
fitófagos, sua ação pode ser direta e mecânica, ao “lavar” os insetos das folhas e
assim diminuir a quantidade, mas também pode beneficiá-los ao facilitar a
emissão de novas brotações foliares, que são a preferência de insetos fitófagos
como a mosca-negra para ovopositar.
A falta de relação entre ovos, ninfas e adultos de mosca-negra neste
trabalho provavelmente se dá pelo curto período de avaliação, que interfere nos
resultados estatísticos.
5.2 - Inimigos naturais da mosca-negra-dos-citros, Aleurocanthus woglumi
em um pomar de Laranja Pêra (Citrus sinensis L.)
5.2.1 Registro dos agentes de controle biológico
5.2.1.1 – Fungos entomopatogênicos
Foi registrada a ocorrência de duas espécies de fungos entomopatogênicos
crescendo sobre ovos e ninfas de A. woglumi, principalmente durante a época
chuvosa e com umidade mais alta. Um dos fungos é Aschersonia aleyrodis e o
outro não foi possível identificar (Figura 8).

43
Figura 8. Fungos entomopatogênicos Aschersonia aleyrodis e outro não identificado sobre ovos e
ninfas de A. woglumi em um pomar de Laranja Pêra (Citrus sinensis L.).
5.2.1.2 – Predadores
Os predadores registrados neste trabalho foram seis indivíduos da família
Coccinelidae (Coleoptera), gênero Scymnus (Figura 9) e quarenta insetos da
família Chrysopidae (Neuroptera), gêneros Ceareochrysa (9 indivíduos) (Figura
10.A e 10.B) e Leucochrysa (31 indivíduos) (Figura 10.C).
Figura 9. A) Larva de Scymnus predando ninfas de III e IV ínstar de A. woglumi; B) Vista dorsal do
adulto de Scymnus; C) Vista ventral do adulto.
Aschersonia aleyrodis
Não identificado
identificado
44
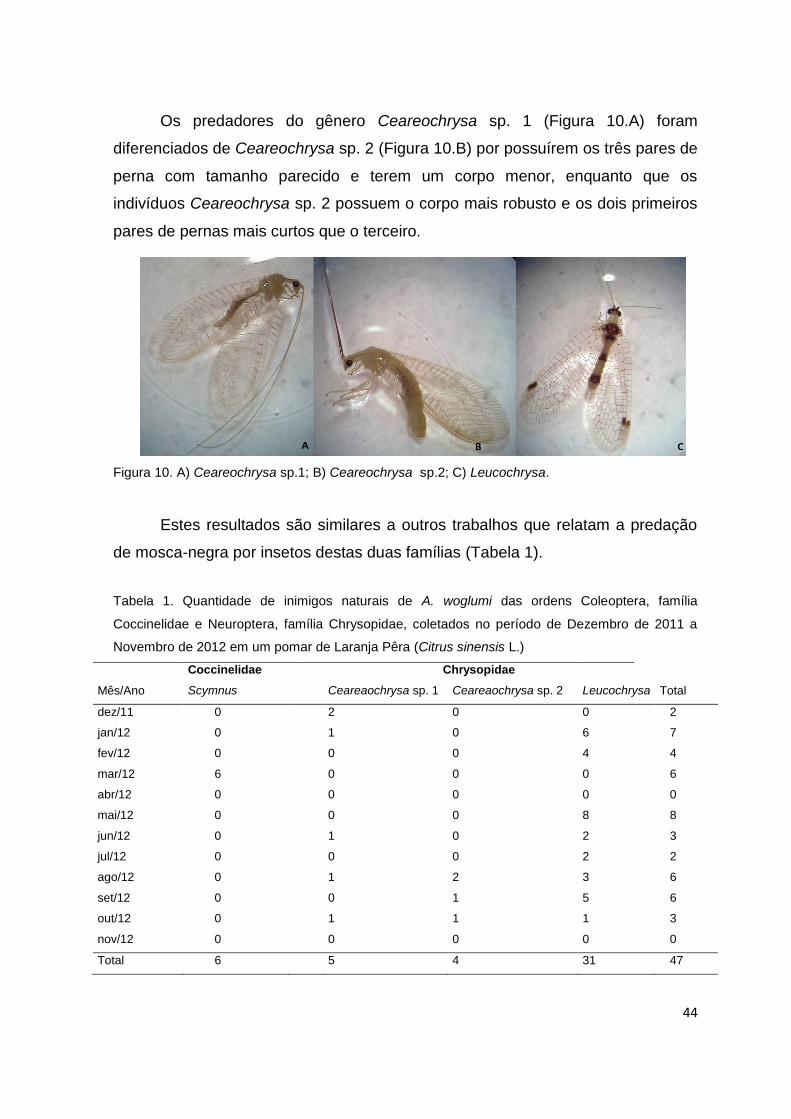
Os predadores do gênero Ceareochrysa sp. 1 (Figura 10.A) foram
diferenciados de Ceareochrysa sp. 2 (Figura 10.B) por possuírem os três pares de
perna com tamanho parecido e terem um corpo menor, enquanto que os
indivíduos Ceareochrysa sp. 2 possuem o corpo mais robusto e os dois primeiros
pares de pernas mais curtos que o terceiro.
Figura 10. A) Ceareochrysa sp.1; B) Ceareochrysa sp.2; C) Leucochrysa.
Estes resultados são similares a outros trabalhos que relatam a predação
de mosca-negra por insetos destas duas famílias (Tabela 1).
Tabela 1. Quantidade de inimigos naturais de A. woglumi das ordens Coleoptera, família
Coccinelidae e Neuroptera, família Chrysopidae, coletados no período de Dezembro de 2011 a
Novembro de 2012 em um pomar de Laranja Pêra (Citrus sinensis L.)
Coccinelidae Chrysopidae
Mês/Ano Scymnus
Ceareaochrysa sp. 1 Ceareaochrysa sp. 2 Leucochrysa Total
dez/11 0
2 0 0 2
jan/12 0
1 0 6 7
fev/12 0
0 0 4 4
mar/12 6
0 0 0 6
abr/12 0
0 0 0 0
mai/12 0
0 0 8 8
jun/12 0
1 0 2 3
jul/12 0
0 0 2 2
ago/12 0
1 2 3 6
set/12 0
0 1 5 6
out/12 0
1 1 1 3
nov/12 0
0 0 0 0
Total 6
5 4 31 47

45
Martínez (1979), encontrou na Venezuela, entre outros predadores, os
crisopídeos Chrysopa sp. e Nodita sp. (Neuroptera: Chrysopidae) e o coleóptero
Delphastus sp. (Coleoptera: Coccinellidae). No Pará foram identificados os
predadores Ceraeochrysa caligata Banks (Neuroptera: Chrysopidae) e Delphastus
pusillus (Coleoptera: Coccinellidae) (Maia, 2004).
Uma observação importante sobre as larvas de crisopídeos é que
geralmente possuem a carapaça de coloração branca (Figura 11.A), devido a
predação de outros insetos como algumas cochonilhas, e neste estudo foi
verificada uma coloração enegrecida da carapaça (Figura 11.B) em laboratório
assim como também em campo (Figura 11.C), confirmando a predação de ninfas
de mosca-negra.
Figura 11. Larvas de crisopídeos com restos de ninfas de A. woglumi. A) Larva de crisopídeo
ainda esbranquiçada, com poucos restos de ninfas de A. woglumi na carapaça; B) Larva de
crisopídeo com carapaça mais enegrecida; C) Larva de crisopídeo em campo com carapaça
completamente cheia de restos de ninfas de A. woglumi.
5.2.1.3 – Parasitóides
Neste trabalho foram identificadas duas famílias de parasitóides:
Encyrtidae com 689 indivíduos em armadilhas adesivas (Tabela 2) e 25
emergidos (Tabela 3) e Aphelinidae com 3680 indivíduos em armadilhas adesivas
(Tabela 2) e 195 emergidos (Tabela 3), que são as duas entre as famílias mais
importantes da superfamília Chalcidoidea para o controle biológico. Apresentam
espécies endoparasitóides ou ectoparasitóides, e algumas são hiperparasitas de

46
insetos pertencentes às famílias Aleyrodidae, Aphididae, Psyllidae e superfamília
Coccoidea, todos da ordem Hemiptera (Goulet e Huber, 1993).
Neste estudo observou-se que a população dos parasitóides da família
Aphelinidae ocorreu em todos os meses de coleta e em maior quantidade que os
da família Encyrtidae.
Tabela 2. Parasitóides associados a mosca-negra-dos-citros coletados em armadilhas adesivas
amarelas no período de dezembro de 2011 a novembro de 2012 em um pomar de Laranja Pêra
(Citrus sinensis L.).
Encyrtidae Aphelinidae
Mês/Ano Encyrtidae sp.1 Arrenophagus sp. ♀ Cales sp. ♀ Cales sp. ♂ Aphelinus sp. ♀ Total
dez/11 107 81 190 207 121 706
jan/12 29 56 139 29 183 436
fev/12 23 33 45 23 135 259
mar/12 2 23 17 2 95 139
abr/12 1 6 9 1 131 148
mai/12 0 7 22 0 239 268
jun/12 5 17 35 5 407 469
jul/12 15 37 51 15 639 757
ago/12 4 21 50 4 327 406
set/12 33 62 40 33 121 289
out/12 13 67 109 13 86 288
nov/12 7 40 92 7 58 204
Total 239 450 799 339 2542 4369

47
Tabela 3. Parasitóides emergidos de ninfas de A. woglumi no período de dezembro de 2011 a
novembro de 2012 em um pomar de Laranja Pêra (Citrus sinensis L.) na região metropolitana de
Manaus, Amazonas.
Encyrtidae Aphelinidae
Mês/Ano Encyrtidae sp.1 Arrenophagus sp. ♀ Cales sp. ♀ Cales sp. ♂ Aphelinus sp. ♀ Total
dez/11 0 4 0 0 1 5
jan/12 0 4 26 0 9 39
fev/12 0 11 23 3 99 136
mar/12 1 0 10 5 7 23
abr/12 0 1 0 0 1 2
mai/12 0 0 0 0 0 0
jun/12 0 0 0 0 0 0
jul/12 0 1 0 0 1 2
ago/12 0 0 0 0 1 1
set/12 0 0 0 0 2 2
out/12 0 2 2 0 3 7
nov/12 0 1 0 0 2 3
Total 1 24 61 8 126 220
Os parasitóides encontrados da família Aphelinidae, provavelmente,
pertencem aos gêneros Aphelinus cf. apenas com fêmeas (Figura 12.A e B),
sendo 2542 indivíduos em armadilhas adesivas (Tabela 2) e 126 emergidos
(Tabela 3), Cales cf., com fêmeas (Figura 12.C) e machos (Figura 12.D)
totalizando 1138 indivíduos em armadilhas adesivas (Tabela 2) e 69 emergidos
(Tabela 3). Foram obtidos dois gêneros da família Encyrtidae, sendo que um
provavelmente pertence ao gênero Arrenophagus cf. (Figura 12.E e F), com 450
indivíduos em armadilhas adesivas (Tabela 2) e 24 emergidos (Tabela 3) e o
outro ainda não foi identificado (Figura 12.G e H).
Em Capitão Poço, no estado do Pará, Rossato (2007) também encontrou
parasitóides da família Aphelinidae, a espécie Cales noacki e espécimes do
gênero Encarsia sp. A espécie C. noacki é um endoparasitóide de insetos da
família Aleyrodidae que pode parasitar ninfas de segundo, terceiro e quarto
ínstares e teve maior abundância que Encarsia sp. nos municípios de Belém,
Capitão Poço e Irituia, no Pará.

48
No Maranhão, Machado et al. (2012) assim como Rossato (2007), também
verificaram a presença de C. noacki e Encarsia sp. em São Luís com ocorrências
de 46,42% e 66,66%, respectivamente.
De acordo com Maia et al. (2005), C. noacki é mais eficiente que Encarsia
sp. em condições de maiores infestações de mosca-negra, mas o contrário ocorre
quando a densidade populacional de A. woglumi é menor, sugerindo substituição
de espécies de parasitóides em campo, conforme a flutuação populacional de A.
woglumi.
Figura 12. Parasitóides da família Aphelinidae emergidos de ninfas de III e IV ínstar de A. woglumi.
A) Aphelinus cf.; B) Fêmea de Cales cf.; D) Macho de Cales cf.
Figura 13. Parasitóides da família Encyrtidae emergidos de ninfas de III e IV ínstar de A. woglumi.
A) Fêmea de Arrenophagus cf.; B) Fêmea de Encyrtidae cf.

49
5.2.2 – Flutuação populacional de inimigos naturais da mosca-negra-dos-
citros, A. wolumi em um pomar de Laranja Pêra (Citrus sinensis L.) na região
metropolitana de Manaus, Amazonas
5.2.2.1 Predadores
Os insetos predadores da família Chrysopidae apesar de serem
constantemente encontrados com restos de ninfas de A. woglumi em suas
carapaças e a predação ter sido confirmada em laboratório, exerceram baixa
influência na flutuação populacional destas (r2 = 0,01, p = 0,68) (Figura 14),
provavelmente por haver outras opções de alimentos como cochonilhas de
carapaça e a cochonilha Praelongaorthezia praelonga, encontradas com
frequência no local de coleta.
Figura 14. Análise de Regressão Simples entre crisopídeos adultos e ninfas de A. woglumi (r2
=
0,01, p = 0,68).

50
As variáveis abióticas precipitação (r2 = 0,13, p = 0,23), umidade relativa (r2
= 0,01, p = 0,66) e a temperatura média (r2 = 0,06, p = 0,41) não exerceram
influência na flutuação populacional de crisopídeos (Figura 15).
Figura 15. Relação entre Precipitação acumulada no período de dezembro de 2011 a novembro
de 2012 (teste de Regressão Simples, r2
= 0,13, p = 0,23 ), Temperatura média (teste de
Regressão Simples, r2 = 0,06, p = 0,41), Umidade Relativa (teste de Regressão Simples, r
2 = 0,01,
p = 0,66) e o número total de crisopídeos adultos coletados.
Estes insetos são considerados importantes agentes de controle biológico
devido a sua voracidade e adaptabilidade ecológica, podendo predar diferentes
pragas (Bortoli et al. 2009)
Outro fator que pode está relacionado à baixa predação é a temperatura,
que de acordo com os estudos de Maia et al. (2000) e Boregas et al. (2003) se for
em média de 30°C, afeta negativamente a capacidade predatória por agilizar o

51
desenvolvimento dos crisopídeos, diminuindo a duração das fases de vida e por
conseguinte, a quantidade de presas consumida é reduzida.
De acordo com Silva (2004), a população dos adultos de C. externa é
influenciada negativamente pela temperatura e precipitação, observação feita
também por Cardoso et al. (2003) que registraram picos populacionais de
crisopídeos em dezembro, fevereiro e março, meses correspondentes à época
mais quente no Paraná.
5.2.2.2 Parasitóides
5.2.2.2.1 Relação entre flutuação populacional de A. woglumi e parasitóides
Neste estudo foi verificado que apenas a atuação dos parasitóides não é o
único fator responsável pela queda na população de A. woglumi após o pico
populacional. Nos testes de regressão simples entre a quantidade de adultos de
mosca-negra e parasitóides encontrados nas armadilhas adesivas, o valor de r2 foi
0,01, ou seja, aparentemente os parasitóides são responsáveis apenas por 1% da
diminuição dos adultos de A. woglumi, no entanto, o valor de p = 0,23 mostra que
há dúvidas quanto a este resultado. Na regressão simples entre ninfas de A.
woglumi e parasitóides nas armadilhas, o valor de r2 foi 0,06, significando que
estes parasitóides são responsáveis por 6% do decréscimo na população de
ninfas, com p = 0,02, assegurando que o resultado é confiável.
Utilizando testes de Regressão Simples foi possível observar que o
parasitóides mais dependentes da flutuação populacional das ninfas de A.
woglumi foram Arrenophagus cf. e Cales cf. (Figura 16).
Apesar de a população de Aphelinus sp. ter sido maior e ocorrido durante
todo o período avaliado, a flutuação populacional de Cales cf. e Arrenophagus cf.
acompanhou de forma mais congruente a das ninfas de mosca-negra (Figura 17).

52
Figura 16. Flutuação populacional de parasitóides encontrados em armadilhas adesivas e ninfas
de todos os estádios de A. woglumi.
Figura 17. Flutuação populacional dos parasitóides emergidos de ninfas III e ninfas IV de A.
woglumi e ninfas de todos os estádios de A. woglumi.
Como é possível observar nos gráficos de regressão linear entre Aphelinus
cf. e ninfas de A. woglumi (Figura 18), a relação é negativa para os parasitóides
em armadilha (r2 = 0,07, p = 0,37) e praticamente nula para os emergidos (r2 =
0,004, p = 0,83), ou seja, aparentemente pouco interferem na flutuação
populacional das ninfas.

53
Este alto índice populacional de Aphelinus cf. pode ser explicado em
função desse parasitóide ter à disposição outras fontes de alimento além da
mosca-negra ou então por ovopositar em ínstares mais novos, inviabilizando a
ovoposição de outras espécies (exceto se estas puderem aproveitar um
hospedeiro já utilizado), como foi proposto por Bográn et al. (2002).
Figura 18. Análise de regressão simples da relação entre ninfas de A. woglumi e parasitóides
Aphelinus cf. encontrados em armadilhas adesivas (r2 = 0,07, p = 0,37) e emergidos (r
2 = 0,004, p
= 0,83).
Os parasitóides do gênero Arrenophagus cf. encontrados em armadilha
tem uma relação positiva e mais direta com as ninfas (r2 = 0,6, p = 0,003) e os
emergidos (r2 = 0,09, p = 0,34) (Figura 19).
Figura 19. Análise de regressão simples da correlação entre parasitóides Arrenophagus cf.
encontrados em armadilhas adesivas e ninfas de A. woglumi (r2 = 0,06, p = 0,003) e Arrenophagus
cf. emergidos e ninfas de A. woglumi (r2 = 0,09, p = 0,34).

54
Os parasitóides do gênero Cales cf. encontrados em armadilhas foram os
que tiveram maior relação com as ninfas de A. woglumi (r2 = 0,78, p = 0,0001) e
os emergidos (r2 = 0,06, p = 0,41) estavam relacionados às ninfas em 6%, um
pouco abaixo da correlação de Arrenophagus cf. emergidos (Figura 20).
Figura 20. Análise de regressão simples da relação entre Cales cf. encontrados em armadilhas
adesivas e ninfas de A. woglumi (r2
= 0,78, p = 0,0001) e Cales cf. emergidos e ninfas de A.
woglumi.
A sincronia existente entre A. woglumi e parasitóides também foi observada
por Rao et al. (1996), com os parasitóides Encarsia spp. e Eretmocerus indicus,
além de haver preferência alimentar pelo segundo ínstar ninfal apesar de haver
parasitismo dos ínstares posteriores, resultado diferente deste estudo, onde foi
observado parasitismo apenas em ninfas de terceiro e quarto ínstar.
Thompson et al. (1987) observaram sucessão de populações de
parasitóides da mosca-negra na Florida, em um estudo de quatro anos, sendo
que inicialmente a dominância era de A. hesperidum, depois foi de Encarsia smithi
e no último ano o parasitóide dominante foi E. opulenta.
A sucessão nas populações de parasitóides comumente se deve às
relações de competição entre espécies, que ocorrem frequentemente em
ambiente natural (Haccou e Alphen, 2008).
É possível observar nos gráficos que a população de Aphelinus cf. se
sobressai em relação às outras. Os parasitóides deste gênero podem estar se

55
desenvolvendo em outros hospedeiros além da mosca-negra ou então as fêmeas
podem ter o comportamento de superparasitóides, deixando ovos em ninfas
previamente parasitadas, ou por elas ou por outras espécies. O superparasitismo
é uma forma comum de competição entre parasitóides, esse comportamento já foi
registrado para várias espécies de himenópteros e dipteros. Em um hospedeiro
superparasitado, geralmente os insetos que se desenvolvem são da fêmea
superparasita (Godfray, 1994).
Os parasitóides dos gêneros Cales cf. e Arrenophagus cf. que
acompanham a flutuação da mosca-negra sem suprimir a população da praga,
podem estar se comportando de acordo com o modelo de crescimento
populacional dependente da densidade. Essa dependência ocorre quando há
agregação de parasitóides em locais onde exista uma densidade maior de
hospedeiros ou quando há aumento na procura dos parasitóides por áreas de alta
densidade de hospedeiros. Os dois gêneros de parasitóides provavelmente
competem entre si e quanto mais tempo as fêmeas permanecerem em um
mesmo local com outras competidoras, menor será a recompensa. No entanto, a
espécie que conseguir permanecer até que a população da outra deixe o local,
terá mais vantagens e poderá ser utilizada de forma mais efetiva em um programa
de controle biológico (Heimpel e Casas, 2008).
As relações ecológicas entre os parasitóides em si são complexas por
dependerem de estudos mais aprofundados sobre a biologia e comportamento de
cada espécie para poder compreender melhor sua interação com a mosca-negra-
dos-citros. No entanto, a diversidade de parasitóides encontrada neste trabalho é
um resultado promissor para futuros programas de manejo integrado de pragas,
sendo necessário decidir, após estudos de biologia, comportamento e parasitismo
qual ou quais parasitóides são mais apropriados para criação massal e o uso no
MIP.
Sendo o presente estudo realizado em pomar comercial, o não uso de
agrotóxicos ou, em última alternativa, ser necessário a aplicação de agrotóxico
para controle das pragas associadas aos citros, é recomendável priorizar o uso de
agrotóxicos seletivos e de baixo impacto sobre os inimigos naturais.

56
5.2.2.2.2 Distribuição espacial dos parasitóides da mosca-negra-dos-citros,
A.wolumi nos diferentes quadrantes da planta (norte, sul, leste e oeste) em
um pomar de Laranja Pêra (Citrus sinensis L.) na região metropolitana de
Manaus, Amazonas
Não houve diferença significativa na distribuição de parasitóides entre os
diferentes quadrantes tanto no período chuvoso (p = 0,94, f = 0,12) quanto no
período seco (p = 0,7, f = 0,93) (Figura 21), sendo que no período chuvoso
houveram alguns registros pontuais de maior quantidade de parasitóides
emergidos, provavelmente acompanhando a quantidade de ninfas de A. woglumi
disponíveis, pois conforme a população de hospedeiros aumenta, cresce também
a de parasitóides (Heimpel e Casas, 2008).
Figura 21. Distribuição espacial por quadrantes dos Parasitóides emergidos de ninfas de A.
woglumi durante o período chuvoso (ANOVA, p = 0,94) e durante período seco (ANOVA, p = 0,7)
em um pomar de Laranja Pêra (Citrus sinensis L.) na Região Metropolitana de Manaus no período
de dezembro de 2011 a novembro de 2012.

57
6.0 – Conclusões
O pico populacional de A. woglumi ocorreu no mês de menor índice
pluviométrico do período chuvoso mas não houve diferença flutuação
populacional entre período chuvoso e período seco.
Os fatores abióticos temperatura média, umidade relativa média e
pluviosidade não influenciaram diretamente a população de A. woglumi.
Os crisopídeos encontrados não exerceram influência na população de A.
woglumi.
Os parasitóides encontrados apresentaram relação com A. woglumi, no
entanto, devido à complexidade das relações ecológicas entre estes insetos, mais
estudos biológicos são necessários para determinar quais parasitóides são mais
adequados em programas de controle biológico.

58
Referências Bibliográficas
ADAB. 2010 .Agência estadual de defesa agropecuária da Bahia (http://www.adab.ba.gov.br).
Acesso em 11/10/2011.
ADAGRI. 2010. Agência de defesa agropecuária do estado do Ceará
(http://www.adagri.ce.gov.br). Acesso em 27/11/2011.
ADAGRO. 2010. Agência de defesa e fiscalização agropecuária de Pernambuco
(http://www.adagro.pe.gov.br). Acesso em 27/11/2011.
Agudelo, F.; Falcon, L.A. 1977. Some naturally occurring insect pathogens in Colombia. Turrialba.
27: 423 – 424.
Agrianual. 2013. Anuário da Agricultura Brasileira. São Paulo, FNP Consultoria &
Agroinformativos. 247-275.
Agrianual. 2003. Anuário da Agricultura Brasileira. São Paulo, FNP Consultoria &
Agroinformativos. 295-332.
Agrofit. 2003. Sistema de Agrotóxicos Fitossanitários.
(http://extranet.agricultura.gov.br/agrofit_cons/principal_agrofit_cons). Acesso em: 04/05/2013.
Andrade, A.F.; Ottati, A.L.T. 2008. Distribuição espacial intraplanta e flutuação populacional da
mosca-negra-dos-citros, Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae), em pomar de
limão. Resumo do XXII Congresso Brasileiro de Entomologia – Uberlândia, Minas Gerais.
Araújo, R.O.C. 2005. Ocorrência e flutuação populacional de Aleurothrixus aepim (Goeldi, 1886),
Bemisia tuberculata (Bondar) (Hemiptera: Aleyrodidae) e Iatrophobia brasiliensis (Rübsaamen,
1907) (Diptera: Cecidomyiidae) em genótipos de mandioca em Teresina, Piauí. Dissertação de
Mestrado, Universidade Federal do Piauí, Teresina, Piauí. 62p.
Belloti, A. C.; Farias, A. R. N. Pragas importantes na cultura da mandioca (Manihot esculenta
Crantz) e meios de controle. 1979. Trabalho apresentado no III Curso Intensivo Nacional de
Mandioca. Cruz das Almas, Bahia, 05 a 16/05/1979.
Benedito, L. 1995. Dinâmica do complexo agroindustrial. Ed. BASA, Belém, Pará, 53p.

59
Bentes, J.L.S.; Silva, N.M.; Ronchi-Teles, B.; Verás, S.; Jansen, T.C. 2007. Citros: Pragas e
doenças. Ed. EDUA, Manaus, Amazonas, 60p.
Bergamin Filho, A.; Lopes, D.B.; Amorim, L.; Godoy, C.V.; Berger, R.D. 1995. Avaliação de danos
causados por doenças de plantas. Revisão Anual de Patologia de Plantas. 3: 133-184.
Bográn, C.E.; Heinz, K.M.; Ciomperlik, M.A. 2002. Interspecific competition among insect
parasitoids: field experiments with whiteflies as hosts in cotton. Ecology. 83: 653 – 668.
Boregas, K.G.B.; Carvalho, C.F.; Souza, B. 2003. Aspectos biológicos de Chrysoperla externa
(Hagen, 1861) (Neuroptera: Chrysopidae) em casa de vegetação.Ciência e agrotecnologia. 27: 7-
16.
Bortoli, S.A.; Murata, A.T.; Brito, C.H.; Narciso, R.S. 2009. Aspectos biológicos de Ceraeochrysa
cincta (Neuroptera, Chrysopidae), em condições de laboratório. Revista de Biologia e Ciências da
Terra. 9: 101-106.
Cardoso, J.T.; Lázzari, S.N.M.; Freitas, S.; Iede, E.T. 2003. Ocorrência e fluturação populacional
de Chrysopidae (Neuroptera) em áreas de plantio de Pinus taeda (L.) (Pinacea) no sul do Paraná.
Revista Brasileira de Entomologia. 47: 473-475.
Castro, R.S.; Pena, M.R.; Silva, N.M.; Vendramim, J.D.; Costa, I.B. 2010. Atividade ovicida de
extratos aquosos de folhas de Piper aduncum L. sobre a mosca-negra-dos-citros, Aleurocanthus
woglumi Ashby (Aleyrodidae) em condições de laboratório. Resumo do XXIII Congresso Brasileiro
de Entomologia. Natal – Rio Grande do Norte.
Cherry, R.; Dowell, R.V. 1979. Predators of citrus blackfly (Hemiptera: Aleyrodidae). Entomophaga.
24: 385-391.
Cherry, R.; Pastor Jr., S. 1980. Variations in population levels of Citrus Blackfly, Aleurocanthus
woglumi (Hom.: Aleyrodidae) and parasites during an eradication program in Florida.
Entomophaga. 25: 365-368.
Cividanes, F.J. 2000. Uso de graus-dia em entomologia: Com particular referência ao controle de
percevejos pragas da soja. Ed. FUNEP, Jaboticabal, São Paulo, 31p.
Clausen, C.P. 1934. The natural enemies of Aleyrodidae in Tropical Asia. Philippine Journal of
Science, 253- 265.

60
CONAB - Companhia Nacional de Abastecimento. 2010. Boletim agroclimático para avaliação de
impactos nas culturas: trimestre 2010: janeiro, fevereiro e março. Conab, Brasília, Distrito Federal,
50p.
Correia, R.G.; Lima, A.C.S.; Farias, P.R.S.; Maciel, F.C.S.; Silva, M.W.; Silva, A.G. 2011. Primeiro
registro da ocorrência de mosca-negra-dos-citros, Aleurocanthus woglumi Ashby, 1915
(Hemiptera: Aleyrodidae) em Roraima (www.agroambiente.ufrr.br). Acesso em 28/01/2011.
Cosave: Comitê de sanidade vegetal do Cone Sul (http://www.cosave.org.pv) 1999. Acesso em
26/11/2011.
Costa, I.B. 2010. Atividade ovicida e ninficida de extratos aquosos de folhas de Melia azedarach L.
(Meliaceae) sobre a mosca-negra-dos-citros, Aleurocanthus woglumi Ashby (Hemiptera:
Aleyrodidae) em condições de laboratório. Trabalho de conclusão de curso. Manaus: Universidade
Federal do Amazonas. 37p.
Cunha, M.L.A. 2003. Distribuição geográfica, aspectos biológicos e controle químico da mosca-
negra dos citros, Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae), nas condições
ambientais do estado do Pará. Dissertação de mestrado, Universidade Federal Rural da
Amazônia, Belém, Pará. 57p.
Dietz, H.F.; Zetek, J. 1920. The blackfly of citrus and other subtropical plants. USDA Bulletin. 885:
1-55.
Dowell, R.V. 1979. Host selection by the citrus blackfly Aleurocanthus woglumi (Homoptera:
Aleyrodidae). Entomologia Experimentalis et Appliccata. 25: 289-296.
Dowell, R.V.; Cherry, R.H. 1981. Survey traps for parasitoids, and coccinellid predators of the citrus
blackfly, Aleurocanthus woglumi. Amsterdam. Nederlandse Entomologische Vereniging. 29: 356-
362.
Dowell, R.V.; Cherry, R.H.; Fitzpatrick, G.E.; Reinert, J.A.; Knapp, J.L. 1981. Biology, plant-insect
relations and control of the Citrus Blackfly. Technical Bulletin 818, Agricultural Experiment Station,
University of Florida. Gainesville, 49p.
Elizondo, J.M.; Quezada, J.R. 1990. Identificación y evaluación de los enemigos naturales de la
mosca prieta de los cítricos Aleurocanthus woglumi Ashby (Homoptera: Aleyrodidae) em cuatro
zonas citrícolas de Costa Rica. Turrialba. 40: 190 – 197p.

61
EMBRATER/CIAT. 1982. Descrição das pragas que atacam a mandioca (Manihot esculenta
Crantz) e características de seus prejuízos. EMBRATER/DIRET, 04, Brasília, Distrito Federal,
1982. 47p.
EPPO n 103. 2008.
(http://www.eppo.org/QUARANTINE/insects/Aleurocanthus_woglumi/ALECWO_ds.pd). Acesso em
20/11/2011.
FAO – Food and Agriculture Organization, 2012 (http://faostat.fao.org). Acesso em 20/01/ 2013.
Fawcett, H.S. 1944. Fungus and bacterial diseases of insects as factors in biological control.
Botanical Review. 10: 327 – 348.
Fernandes, M.G.; Busoli, A.C.; Barbosa, J.C. 2003. Distribuição espacial de Alabama argillace
(Hubner) (Lepdoptera: Noctuidae) em Algodoeiro. Neotropical Entomology. 32: 107 – 115.
French, V.J.; Meagher, R.L.J. 1992. Citrus Blackfly: Chemical control on nursery citrus. Subtropical
Plant Science. 45: 7-10.
Gasparotto, L.; Junqueira, N.T.V.; Pereira, J.C.R. 1998. Doenças de citros no Estado do
Amazonas. Ed. EMBRAPA-CPAA, Circular Técnica N 6, Manaus, Amazonas, 20p.
Gazzoni, D.; Oliveira, E.B.; Corso, I.C.; Ferreira, B.S.C.; Vilas Boas, G.L.; Moscardi, F.; Panizzi,
A.R. 1988. Manejo de pragas da soja. EMBRAPA/CNPSo, Londrina, Paraná. 44p.
Godfray, H.C.G. 1994. Parasitoids: behavioral and evolutionary ecology. Princeton University
Press, New Jersey. 461p.
Gomez; S.A.; Duarte, M.M.; Rohden, V. da S. 2005. Flutuação Populacional de Bemisia sp.
(Homoptera: Aleyrodidae) em mandiocais de Mato Grosso do Sul. Anais do XI Congresso
Brasileiro de Mandioca. (www.cpao.embrapa.br/11cbm/_html/trabalhos/.../pasta43.PDF). Acesso
em 30/01/2012.
Gonçalves, M.S.; Pena, M.R.; Silva, N.M. 2007. Entomofauna de inimigos naturais da mosca-
negra-dos-citros, Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae), na região de Manaus,
Amazonas. Resumo do XXII Congresso Brasileiro de Entomologia, Uberlândia, Minas Gerais.

62
Gonçalves, M.S. 2010. Atividade ovicida e ninficida de diferentes extratos de meliáceas sobre a
mosca-negra-dos-citros, Aleurocanthus woglumi Ashby (Aleyrodidae) em condições de laboratório.
Trabalho de conclusão de curso. Manaus: Universidade Federal do Amazonas. 34p.
Gonzaga Filho, S.; Lima, S.C.; Pena, M.R.; Vasconcelos, G.J.N.; Silva, N.M. 2011. Flutuação
populacional da mosca-negra-dos-citros Aleurocanthus woglumi (Hemiptera: Aleyrodidae) em
pomar de citros em Manaus – AM. Resumo do XXIII SICONBIOL - Simpósio de Controle Biológico.
São Paulo.
Goulet, H.; Huber, J. T. 1993. Hymenoptera of the World. Ottawa, Canada Communication Group.
678p.
Haccou, P.; Alphen, J.J.M. 2008. Competition and asymmetric wars of attrition in insect parasitoids.
In: Wajnberg, E., Bernstein, C., Alphen, J.J.M. Behavioural ecology of insect parasitoids: from
theoretical approaches to field application. Oxford: Blackwell Publishing Ltd. 193-211p.
Hagvar, E.B.; Hofsvang, T. 1991. Aphid parasitoids (Hymenoptera, Aphidiidae): biology, host
selection and use in biological control. Biocontrol News and Information.12: 13-41.
Hart, W.G.; Selhime, A.G.; Harlan, D.P.; Ingle, S.J.; Sanches, R.M.; Rhode, R.H.; Garcia, C.A.;
Caballero, J.; Garcia, R.L. 1978. The introduction and establishment of parasites of citrus blackfly,
Aleurocanthus woglumi Ashby, in Florida. Entomophaga. 23: 361-366.
Heu, R.A.; Nagamine, W.T. 2001. Citrus Blackfly, Aleurocanthus woglumi Ashby (Homoptera,
Aleyrodidae). New Pest Advisory, Division of Plant Industry, 99: 1-3.
Heimpel, G.E.; Casas, J. 2008. Parasitoid foraging and oviposition behaviour in the field. In:
Wajnberg, E.; Berstein, C; Alphen, J. (Ed.) Behavioral ecology of insect parasitoids – from
theoretical approaches to field applications.445p.
IBGE. 2012. Instituto Brasileiro de Geografia e Estatística (http://www.ibge.gov.br). Acesso em
18/10/2012.
IDAF. 2010. Instituto de defesa agropecuária e florestal do Espírito Santo
(http://www.idaf.es.gov.br). Acesso em 11.10.2011.
IDAM. Instituto de desenvolvimento agropecuário e florestal sustentável do Estado do Amazonas.
2010. Relatório de acompanhamento trismestral – Produção vegetal, 3p.

63
Jordão A.L.; Silva R.A. 2006. Guia de pragas agrícolas para o manejo integrado no estado do
Amapá. Ribeirão Preto. Editora Holos. 182p.
Katole, S. R.; Mahajan, R. K.; Tayde, G. S. Gawande, R. B. 1996. Neem oil: an environmentally
safer biocide in the management of citrus blackfly (Aleurocanthus woglumi Ashby). PKV Research
Journal. 20: 28-30p.
Lemos R.N.S.; Silva, G.S.; Araújo, J.R.G.; Chagas, E.F.; Moreira, A.A.; Soares, A.T.M. 2006.
Ocorrência de Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae) no Maranhão. Neotropical
Entomology. 35: 558-559p.
Lopes, G.S. 2010. Preferência de oviposição e ciclo de vida da mosca negra dos citros,
Aleurocanthus woglumi Ashby, 1915 (Hemiptera: Aleyrodidae) em diferentes plantas hospedeiras.
Dissertação de Mestrado: Universidade Estadual do Maranhão, São Luís, Maranhão. 56p.
Lopes, E.B.; Brito, C.H.; Batista, J.L.; Silva, A.B. 2010. Ocorrência da mosca-negra-dos-citros na
Paraíba. Tecnologia e Ciência Agropecuária. 4: 19-22p.
Luna-Salas, J.F.; Myartseva, S.N.; Ruiz-Cancino, E.; Varela-Fuentes, S.E.; Coronado-Blanco, J.M.
2004. Population dynamics of the citrus blackfly parasitoids on Valencia oranges in Central
Tamaulipas, Mexico. The 2004 ESA Annual Meeting and Exhibition,
(http://esa.confex.com/esa/2004/techprogram/paper_15882.htm). Acesso em: 15/02/2013.
Machado, K.K.G.; Lemos, R.N.S.; Pereira, C.F.M.; Medeiros, F.R.; Araujo, J.R.G.; Costa, V.A.
2012. Ocorrência de parasitóides da mosca-negra-dos-citros Aleurocanthus woglumi Ashby
(Hemiptera: Aleyrodidae) na região norte do Estado do Maranhão. Resumo do XXIV Congresso
Brasileiro de Entomologia. Curitiba, Paraná.
Maia, W.J.M.S.; Carvalho, C.F.; Souza, B. 2000. Exigências térmicas de Chrysoperla externa
(Hagen, 1861) (Neuroptera: Chrysopidae) alimentada com Schizaphis graminum (Rondani, 1852)
(Homoptera: Aphididae) em condições de laboratório. Ciência e Agro-tecnologia. 24: 74-80p.
Maia, W.J.M.S.; Souza, J.C.; Marques, L.C.; Silva, L.M.S.; Benaduce, R.V.; Gentil R.M. 2005.
Infestação em citros por Aleurocanthus woglumi (Ashby) e perspectivas de controle biológico
aplicado no Pará. Anais do 9º Simpósio de Controle Biológico. Recife, Pernambuco. 183p.
Maia, P.S.P. 2008. Caracterização da distribuição espacial da mosca-negra-dos-citros
(Aleurocanthus woglumi Ashby, 1915) em pomar georreferenciado para determinar um plano de

64
amostragem seqüencial. Dissertação de mestrado, Universidade Rural da Amazônia, Belém, Pará.
77p.
MAPA. 2008. Ministério da Agricultura, Pecuária e Abastecimento (http://www.agricultura.gov.br/).
Acesso em 28/01/2012.
MAPA. 2011. Ministério da Agricultura, Pecuária e Abastecimento. Lista de pragas quarentenárias
presentes - (A2). (http://extranet.agricultura.gov.br/sislegis-
consulta/servlet/VisualizarAnexo?id=14560). Acesso em 28/01/2012.
Martínez, N.B.; Terán, J.; Geraud, F. 1979. Enemigos naturales de la mosca prieta de los cítricos
Aleurocanthus woglumi Ashby (Homoptera: Aleyrodidae) em Venezuela. Agronomia Tropical. 29:
453-458.
Martínez, N.B. 1982. Biologia de la mosca prieta de los cítricos Aleurocanthus woglumi Ashby
(Homoptera: Aleyrodidae) em el campo. Agronomia Tropical. 31: 211-218.
Meagher, R.L.; French, J.V. 2004. Augmentation of parasitoids for biological control of Citrus
Blackfly in Southern Texas. Florida Entomologist. 87: 186–193.
Medeiros, F.R.; Lemos, R.N.S.; Ottati, A.L.T.; Araújo, J.R.G.; Machado, K.K.G.; Rodrigues, A.A.C.
2009. Dinâmica populacional da mosca-negra-dos-citros Aleurocanthus woglumi Ashby
(Hemiptera: Aleyrodidae) em Citrus spp. no município de São Luís – MA. Revista Brasileira de
Fruticultura, Jaboticabal. 31: 1016-1021.
Miklasiewicz, T.J.; Walker, G.P. 1990. Environmental Entomology. 19: 1485-1490.
Müller, G.W.; Targon, M.L.P.N.; Carvalho, S.A.; Souza, A.A.; Rodrigues, J.C.V. 2005 In: Mattos,
S.J.D.; Negri, J.R.; Pio, R.M.; Pompeu Junior, J. Citros. Campinas: Instituto Agronômico e Fundag,
p. 147 - 196.
Neves, M.F.; Trombin, V.G.; Milan, P.; Lopes, F.F.; Cressoni, F.; Kalaki, R. 2010. O retrato da
citricultura brasileira. Markestrat, FEA/USP. (http://www.citrusbr.com.br/exportadores-
citricos/saiba-mais/o-retrato-da-citricultura-brasileira-189513-1.asp). Acesso em 21/01/2013.
Nguyen, R.; Hamon, A.B. 2003. Citrus Blackfly, Aleurocanthus woglumi Ashby (Homoptera:
Aleyrodidae). CIR 360, University of Florida.

65
Nguyen, R.; Brasil, Jr.; Poacher, C. 1983. Population density of the citrus blackfly, Aleurocanthus
woglumi Ashby (Homoptera: Aleyrodidae), and its parasites in urban Florida in 1979-81.
Environmental Entomology. 12: 878-884.
Oliveira, M.R.V.; Silva, C.C.A. da; Navia, D. 1999. Praga quarentenária A1: A mosca-negra dos
citros, Aleurocanthus woglumi Ashby (Hemiptera: Aleyrodidae). EMBRAPA – Comunicado Técnico
40. 7p.
Oliveira, R.P. de; Scivittaro, W.B.; Borges, R. de Sá; Nakasu, B.H. 2001. Mudas de citros. Pelotas:
Embrapa Clima Temperado. 32p.
Oppenheim, S. 2001. Alternative agriculture in Cuba. American Entomologist. 47: 216 – 227.
Patel, P.S.; Patel, G.M. 2006. Population Dynamics of important pests of citrus, Citrus aurantifolia,
Swingle. Indian Journal of Entomology. 68: 255-259.
Pena, M.R.; Vendramim, J.D.; Lourenção, A.L.; Silva, N.M.; Yamamoto, P. T.; Gonçalves, M.S.
2008. Ocorrência da mosca-negra-dos-citros, Aleurocanthus woglumi Ashby (Hemiptera:
Aleyrodidae) no estado de São Paulo. Revista de Agricultura. 83: 61-65.
Pena, M.R.; Silva, N.M.; Bentes, J.L.S.; Alves, S.B.; Bezerra, E.J.S.; Vendramim, J.D.; Lourenção,
A.L.; Humber, R.A. 2009. Inibição do desenvolvimento de Aleurocanthus woglumi Ashby
(Hemiptera: Aleyrodidae) por Aschersonia CF. Aleyrodis Webber (Deuteromycotina:
Hyphomycetes). Arquivos do Instituto Biológico. São Paulo. 76: 619-625.
Quezada, J.R. 1974. Biological control of Aleurocanthus woglumi (Homoptera: Aleyrodidae) in El
Salvador. Entomophaga. 19: 243 – 254.
Raga, A.; Costa, V.A. 2008. Mosca-negra dos Citros. Instituto Biológico – APTA Documento
Técnico 001. (www.biologico.sp.gov.br). Acesso em 27/01/2012.
Raga, A.; Marsaro Junior, A.L.; Racca Filho, F.; Costa, V.A. 2011 Novos Registros de Espécies de
Aleyrodidae (Hemiptera) no Estado de Roraima, Brasil. Arquivos do Instituto Biológico. 78: 439-
441.
Rao, N.G.V.; Lokhande, R.K.; Punit, M. 1996. Parasitoid of citrus aleyrodids: their behaviour and
seasonal occurrence. Advances in Plant Sciences. 9: 147-149.

66
Reitz, H.J.; Leonard, C.D.; Stewart, I.; Koo, R.C.J.; Anderson, C.A.; Reese, R.L.; Calvert, D.V.;
Smith, P.F. 1972. Recommended fertilizers and nutritional sprays for citrus. Cooperative Extension
Bulletin. 26.
Rodrigues, T.R.; Fernandes, M.G.; Santos, H.R. 2010. Distribuição espacial de Aphis gossypii
(Glover) (Hemiptera, Aphididae) e Bemisia tabaci (Gennadius) biótipo B (Hemiptera, Aleyrodidae)
em algodoeiro Bt e não-Bt. Revista Brasileira de Entomologia. 54: 136-143.
Ronchi-Teles, B.; Pena, M.R.; Silva, N.M. 2009. Observações sobre a ocorrência de mosca-negra-
dos-citros, Aleurocanthus woglumi Ashby, 1915 (Hemiptera: Sternorrhyncha: Aleyrodidae) no
estado do Amazonas. Acta Amazonica. 39: 241 – 244.
Rossato, V. 2007. Ocorrência de parasitóides de Aleurocanthus woglumi Ashby, 1903 (Hemiptera:
Aleyrodidae) e seu parasitismo por Cales noacki Howard, 1907 (Hymenoptera: Aphelinidae) nos
municípios de Belém, Capitão Poço e Irituia no estado do Pará. Dissertação de Mestrado,
Universidade Federal Rural da Amazônia, Belém, Pará. 40 pp.
SAGAR. 1999. Ficha Técnica. Control Biológico de la Mosca Prieta de los Cítricos. Centro
Nacional de Referencia de Control Biológico, Tecomán, Colima, México. 4p.
Santos, R.S.; Maia, W.J.M.S.; Alves, G.A.R.; Gentil, R.M.; Silva, C.S.; Magalhães, T.L. 2008.
Flutuação Populacional de Aleurocanthus woglumi (Hemiptera: Aleyrodidae) em plantio
convencional e orgânico em Capitão Poço, Pará. Resumo do XXII Congresso Brasileiro de
Entomologia. Uberlândia, Minas Gerais.
Santos, M.S. 2009. Resistência de genótipos de citros a lagarta-minadora-dos-citros Phyllocnistis
citrella Stainton, 1856 (Lepidoptera: Gracillariidae). Tese de Doutorado em Entomologia. Ed.
ESALQ, Piracicaba., São Paulo, 133p.
Santos, A.B.; Pena, M.R.; Vendramim, J.D.; Castro, R.S.; Silva Neto, M.R. 2010. Atividade
ninficida de extratos aquosos de folhas de andiroba, Carapa guianensis (Meliaceae), sobre mosca-
negra-dos-citros, Aleurocanthus woglumi (Aleyrodidae) em laboratório. Resumo do XXIII
Congresso Brasileiro de Entomologia, Natal, Rio Grande do Norte.
Silva, S.E.L.; Souza, A.G.C. 2002. Produção de Mudas de Laranja. Circular Técnica, 14. Embrapa
Amazônia Ocidental, Manaus, Amazonas.

67
Silva, A de B. 2005. Mosca-negra dos citros, (Aleurocanthus woglumi Ashby) uma praga potencial
para a citricultura brasileira in: Poltronieri, L. S.; Trindade, D. R.; Santos, I. P. dos. Pragas e
doenças de cultivos amazônicos. Embrapa Amazônia Oriental, Belém, Pará, 483p.
Silva, A.G., 2010. Dinâmica populacional de mosca-negra-doscitros (Aleurocanthus woglumi
Ashby, 1915) em pomares de citros em sistema agroflorestal e monocultura. Dissertação de
Mestrado, Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista,
Jaboticabal. 79p.
Silva Neto, M.R.; Pena, M.R.; Vendramim, J.D.; Castro, R.S.; Santos, A.B. 2010. Bioatividade de
extratos aquosos de amêndoas de nim, Azadirachta indica (Meliacea) sobre a mosca-negra-dos-
citros, Aleurocanthus woglumi (Aleyrodidae) em condições de laboratório. Resumo do XXIII
Congresso Brasileiro de Entomologia. Natal – Rio Grande do Norte.
Silva, A.G.; Farias, P.R.S.; Boiça Júnior, A.L.; Souza, B.H.S. 2011. Mosca-negra-dos-citros:
Características gerais, bioecologia e métodos de controle dessa importante praga quarentenária
da citricultura brasileira. Jaboticabal, São Paulo. Entomobrasilis. 4: 85-91.
Silveira Neto, S.; Nakano, O.; Bardini, D.; Villa Nova, N.A. 1976. Manual de ecologia dos insetos.
Editora Agronômica Ceres LTDA, São Paulo, 419p.
Srinivasan, R.; Riley, D.; Diffie, S.; Sparks, A.; Adkins, S. 2012. Whitefly population dynamics and
evaluation of whitefly-transmitted Tomato yellow leaf curl virus (TYLCV)-resistant tomato
genotypes as whitefly and TYLCV reservoirs. Journal of Economic Entomology. 105: 1447 – 1456.
Starý, P. 1988. Aphidiidae. In: Aphids. Their Biology, Natural Enemies and Control. World Crop
Pests (eds A.K. Minks and P. Harrewijn), Volume 2B, 171-184p. Elsevier, Amsterdam, p. 364.
Summy, K.R.; Gilstrap, F.E.; Hart, W.G. 1985. Aleurocanthus woglumi [Hom.: Aleyrodidae] and
Encarsia opulent [Hym.: Encyrtidae] density-dependent relationship between adult parasite
aggregation and mortality of the host. Entomophaga. 30: 107-112.
Swezey, S.L.; Vasquez, E.C. 1991. Biological control of Citrus blackfly (Homoptera: Aleyrodidae) in
Nicaragua. Environmental Entomology. 20: 1691-1698.
Thompson, C.R.; Cornell, J.A.; Sailer, R.I. 1987. Interactions of parasites and a hyperparasite in
Biological control of Citrus Blackfly, Aleurocanthus woglumi (Homoptera: Aleyrodidae), in Florida.
Environmental Entomology. 16: 140 – 144.

68
Vieira, Daniela Lavra. 2008. Flutuação populacional e dependência espacial de Aleurocanthus
woglumi Ashby, 1915 (Hemiptera: Aleyrodidae) em Citrus latifolia. Dissertação de Mestrado,
Universidade Estadual do Maranhão, São Luís, Maranhão. 41p.