michelle karen brasil serafim definiÇÃo de um meio de cultivo para o … · 2014-02-19 ·...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DO CEARÁ
FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS- PPGCV
MICHELLE KAREN BRASIL SERAFIM
DEFINIÇÃO DE UM MEIO DE CULTIVO PARA O DESENVOLVIMENTO IN
VITRO DE FOLÍCULOS PRÉ-ANTRAIS CANINOS ISOLADOS
Fortaleza
2013

2
MICHELLE KAREN BRASIL SERAFIM
DEFINIÇÃO DE UM MEIO DE CULTIVO PARA O DESENVOLVIMENTO IN
VITRO DE FOLÍCULOS PRÉ-ANTRAIS CANINOS ISOLADOS
Tese apresentada ao Programa de Pós-Graduação em
Ciências Veterinárias da Faculdade de Veterinária da
Universidade Estadual do Ceará, como requisito parcial
para obtenção do título de Doutor em Ciências
Veterinárias.
Área de Concentração: Reprodução e Sanidade Animal
Linha de pesquisa: Reprodução e Sanidade de
carnívoros, onívoros, herbívoros e aves.
Orientador: Prof. Dr. José Ricardo de Figueiredo.
Fortaleza
2013

3
Dados Internacionais de Catalogação na Publicação
Universidade Estadual do Ceará
Biblioteca Central Prof. Antônio Martins Filho
Bibliotecário Responsável – Francisco Welton Silva Rios – CRB-3/919
S482a Serafim, Michelle Karen Brasil
Definição de um meio de cultivo para o desenvolvimento in vitro de
folículos pré-antrais caninos isolados / Michelle Karen Brasil Serafim . –
2013.
CD‘ROM. 140 f. : il. (algumas color.) ; 4 ½ cm.
―CD-ROM contendo o arquivo no formato PDF do trabalho
acadêmico, acondicionado em caixa de DVD Slim (19 x 14 cm x 7 mm)‖.
Tese (doutorado) – Universidade Estadual do Ceará, Faculdade de
Veterinária, Programa de Pós-Graduação em Ciências Veterinárias,
Curso de Doutorado em Ciências Veterinárias, Fortaleza, 2013.
Área de Concentração: Reprodução e Sanidade Animal.
Orientação: Prof. Dr. José Ricardo de Figueiredo.
Co-orientação: Prof.ª Dr.ª Lúcia Daniel Machado da Silva.
1. Cadela. 2. Folículo ovariando. 3. Cultivo in vitro. 4. Oócito. I.
Título.
CDD: 572.8

4

5
Dedico esta tese aos meus pais por todo o amor incondicional,
respeito, dedicação e por sempre se fazerem presentes em minha vida.
E dedico também à minha família, materna e paterna,
por sempre me darem a certeza de que nunca estou sozinha.

6
AGRADECIMENTOS
Á Força maior que guia, ilumina, protege, ensina e alimenta meu espírito diariamente.
Seja por intermédio de meu anjo da guarda e/ou guia espiritual, ou via meus pais e
familiares. Ao Nosso Senhor Deus, meu mais sincero e humilde obrigado.
Agradeço inicialmente com todo respeito e humildade, a todos as cadelas que foram
utilizadas em meus experimentos, principalmente as oriundas do Centro de Controle de
Zoonozes de Fortaleza (CCZ). Em que todas as vezes após anestesiadas, vinham até
mim, e se deitavam aos meus pés, por me verem como proteção e segurança. Como
sempre sussurrei após cada coleta, a cada uma das doadoras: "Muito obrigada,
Princesinha!".
A todos os funcionário do CCZ, por todo apoio e ajuda, meu muito obrigado
Agradeço À Universidade Estadual do Ceará, ao Programa de Pós-Graduação em
Ciências Veterinárias e a todos os seus funcionários Adriana Albuquerque e Antônio
Cesar Camelo (que sempre me supriu de café e sorrisos), coordenadores, ex-
coordenadores e professores, em especial ao Prof. Dácio Teixeira e Davide Rondina.
Aos órgãos de fomento CAPES e CNPq, pelos auxílios em forma de bolsa de estudos,
tanto no Brasil, quanto no exterior (Estados Unidos).
Ao meu orientador, por todos os conselhos, ensinamentos, e por 4 anos de intensas
tentativas de fazer de mim uma profissional melhor. Agradeço imensamente cada
"carão", cada reunião de "10 min", cada palavra de incentivo ou crítica, pois me fizeram
crescer. Que Deus o proteja, Mestre.
A todos os meus colegas da equipe LAMOFOPA. Em especial por todo apoio e carinho,
agradeço a Laritza Lima, Débora Magalhães, Allana Pessoa, Hudson Correia, Jamily
Bruno, Valdevane Araújo, Franciele Lunardi (minha Guria), Débora Sales ("chapa" de
pensionato e caronas repletas de "aventuras") e a Profa. Ana Paula Ribeiro Rodrigues. E
aos ex-LAMOFOPA, Victor Galindo, minha Paula Correia, Aglailson Pinheiro, Valesca
Luz e Ana Beatriz Duarte. A pessoa que dentro do LAMOFOPA compartilha comigo
todo o amor pelo universo "NERD/GEEK", e se demonstrou um amigo verdadeiro,
obrigado ao Fco
Léo Aguiar.
Aos meus amigos Priscila Ramos (paraense que amo), Ana Kelen Lima (minha
"Camelete" preferida), Cláudio Ferro (Abraço da Ursa que te adora), Ticiana Franco
(―Branca de Neve‖), Henna Roberta (Henninha), Antônio Mota (Toim), Rosivaldo
Júnior, Maia Júnior, Liduína Modesto (minha tia), Lívia e Lígia Costa, e Gerlane

7
Modesto (A Nega) por todos os momentos compartilhados em Fortaleza e amizade,
agradeço.
Em principal agradeço aos meus pais Damião Serafim (um coração puro, lutador,
persistente e emotivo), e Mª José Brasil (que agrega serenidade, resistência, equilíbrio e
imparcialidade de uma forma harmônica e inspiradora). Vocês são meu exemplo de Yin
Yang, a razão do meu viver, e meu sempre eterno e verdadeiro amor. Aos meus irmãos,
Diego Brasil (meu Otário) e Karol Brasil (minha Filha), sem vocês isso não seria
possível, com e por vocês tudo tem mais sentido, vocês são minha estrutura, respiração,
meus presentes de Deus. Aos meus dois grandes bebês e sobrinhos, que "Nenem" ama,
Diego Filho e Mariana Brasil. Agradeço às minhas cadelas Saphira e Lulu (in
memorian), e aos demais componentes da "matilha Brasil Serafim".
Aos meus familiares, por respeitarem minhas ausências constantes (mesmo a
contragosto), e serem todo meu apoio e companheirismo. Agradeço em especial aos
primos/irmãos Laura Brasil, Kharen Brasil, Kharina Brasil, Leonides Júnior, Karize
Brasil, Jaqueline e Jéssica Brasil e Débora Arnaut (um dos et.al. omitidos em meus
artigos), meus tios Rui Figueira, Aiene Brasil, Laura Brasil, Fco Serafim, Edvis
Serafim, e meus avós Maria Santíssima e José Serafim.
Ao meu namorado David Figueira, obrigada por toda paciência, compreensão,
companheirismo, risadas, e por todo o amor, "You are my home, my all, my love".
Thank to all people from Smithsonian Biology Conservation Institute, for the company,
parties and studies, you all did my days in USA one of the best moments of my life. In
special, thank to Dr. Nucharin Songsasen, for the personal and scientific teachings,
patience and all great moments. Thank you ka, you are an amazing and young person,
when I grow up, I want to be like you. Thank to Chommanart Thongkittidilok (my little
sister Giftfy Timon) you are the best part of USA to me, thank you for every moment ka.
And thank to Jennifer Nagashima and Mayaco Fujihara, work with you was a pleasure
and a gift.
Obrigado a todos os componentes da banca examinadora, por terem aceitado o convite
de forma tão imediata.
E finalizo agradecendo in memorian ao meu Anjo de Luz (Daniel F. de Cavalho), e a
todos os outros que do outro plano me protegem, guiam e iluminam me dando a plena
certeza da presença de Deus ao meu lado.

8
RESUMO
A presente tese teve como objetivo avaliar o efeito de diferentes concentrações de 1)
insulina, 2) GH, associados ou não ao FSH sobre o cultivo de folículos pré-antrais
caninos e sua influência na esteriodogênese, e ainda o efeito do 3) EGF na presença ou
ausência de FSH, e a influência do meio de base (MEM ou αMEM) em diferentes
sistema de cultivo (2D ou 3D). Para tanto, folículos pré-antrais e antrais avançados
foram isolados por microdissecção e cultivados por 18 dias em gotas de 100 μL de α-
MEM ou MEM suplementado. Para as condições experimentais, nas fases I, II, e III,
foram avaliados respectivamente os efeitos da insulina (5, 10 ng/ml ou 10g/mL), GH
(10 ou 50ng/ml) e EGF (50 ou 100 ng/ml) na ausência ou presença de concentrações
crescentes de FSH (100, 500 e 1000 ng/mL), adicionados sequencialmente a partir dos
dias 0, 6 e 12 de cultivo, respectivamente. Na Fase 3, foi ainda avaliado os meio de base
MEM ou αMEM, cultivados no sistema 2D, 3D em matriz de alginato ou 3D em matriz
de alginato associado à fibrina. Durante o cultivo foram avaliados os seguintes
parâmetros: crescimento folicular, formação de antro, extrusão e crescimento oocitário.
Após o cultivo, foi analisada a viabilidade folicular (marcadores calceína e etídio
homodímero), bem como a integridade da cromatina com o Hoechst 33342. Na Fase I,a
suplementação com insulina a 10g/mL associada ao FSH apresentou média do
diâmetro folicular, taxa de formação de antro e sobrevivência significativamente
superiores aos demais tratamentos (P <0,05). Referente à Fase II, o crescimento
folicular no grupo com GH a 50ng/ml associado ao FSH apresentou maior diâmetro
folicular (P˂ 0,05), bem como taxas de formação de antro significativamente superior
ao Controle. Os folículos oriundos do Controle e do GH 50ng/ml associado ao FSH
apresentaram um aumento significativo da produção de estradiol, do dia 6 ao dia 18 de
cultivo. Na Fase III, a taxa de crescimento folicular, bem como o percentual de oócitos
com cromatina intacta ao fim do cultivo foram superiores (P˂ 0,05) no tratamentos
MEM acrescido de EGF 100ng/ml associado ao FSH, quando comparado aos demais
tratamentos. Os folículos cultivados em meio base αMEM demonstraram uma taxa de
crescimento significativamente superior (P< 0.05) aos demais tratamentos. Vale
salientar que não foi observada influência do sistema de cultivo (2D ou 3D) sobre o
desenvolvimento folicular. Concluindo, este estudo mostrou que a insulina, GH, e EGF
de forma dose dependente beneficiam a manutenção da morfologia folicular e sua
associação ao FSH estimulou o desenvolvimento folicular in vitro.
Palavras chaves: cadela, folículo ovariando, cultivo in vitro, oócito

9
ABSTRACT
The objective of this study was to evaluate the effect of different concentrations of 1)
Insulin, 2) GH, associated or not with FSH, on culture of domestic dog preantral
follicles and the influence on the steroidogenesis, also the effect of 3) EGF in presence
or absence of FSH, and the influence of basic media (MEM or αMEM) in different
culture systems (2D or 3D). Secondary follicles and early antral follicles were isolated
by microdissection, and then, cultured for 18 days in MEM or αMEM supplemented.
The experimental conditions, at phase 1, 2, and 3, were evaluated the effect of insulin
(5, 10 ng/ml, 10 g/ml), GH (10 or 50 ng/ml) and EGF (50 or 100 ng/ml), respectively,
in the presence or absence of sequentially added FSH (100, 500 and 1000 ng/ml) at days
0, 6 and 12 of culture, respectively. At phase 3, was rated yet, the influence of basic
media MEM or αMEM, cultured in 2D, 3D system with alginate matrix or 3D with
matrix of alginate and fibrin. During the culture, these parameter were assessed:
follicular growth, antrum formation rate, oocyte extrusion and growth. At the end of
culture, viability parameters was evaluated using fluorescence cell viability assay
(calcein-AM and ethidium homodimer-1), also the oocyte chromatin integrity was
evaluated by Hoechst 33342 staining. At phase 1, supplementation of insulin at 10 μg
with FSH significantly increased rate of follicular growth, antrum formation, and
percentage of viable follicles, compared to other treatments (P <0.05). In phase 2, GH at
50 ng/ml supplemented with FSH resulted in the highest average of follicular diameter
and antrum formation rate than the control group (P<0.05). Follicles from both the
control and the GH50+FSH treatment groups actively and increasingly secreted
estradiol from days 6 to 18 of culture (P<0.05). At phase 3, group of MEM plus EGF
100ng/ml in the presence of FSH, exhibited a higher percentage (P< 0.05) of oocytes
with intact chromatin and rates of follicular growth when compare to the other groups
evaluated. The use of α-MEM was more beneficial on the maintenance of chromatin
integrity as well as follicle growth compared with MEM. Neither 2D nor 3D culture
systems were affecting on follicular growth. In conclusion, our study demonstrates that
insulin, GH and EGF have beneficial effect on the maintenance of follicular
morphology in a dose-dependent manner and, in association with FSH, stimulates in
vitro follicular development. However, the three dimensional culture systems did not
promote dog‘s in vitro folliculogenesis.
Key words: Bitch, ovarian follicle, culture, oocyte

10
LISTA DE FIGURAS
Revisão Bibliográfica
Figura 01: Desenho esquemático da localização anatômica do ovário canino, e
estruturas adjacentes. .............................................................................................22
Figura 02: Esquematização da oogênese na espécie canina.........................................23
Figura 03: Folículo pré-antral canino, com oócito de Grau 1, com grande quantidade de
gotícula de lipídio em seu interior...................................................................................24
Figura 04:Folículo pré-antral canino incluso em gota de alginato (Fonte:N. Songsasen)
(A) e alginato acrescido de fibrina (B). As setas vermelhas indicam a ação da fibrina ao
degradar o alginato ao longo do cultivo..........................................................................31
Capítulo 2
Fig. 1. Canine ovarian preantral follicle diameter measured on Day 0 of culture (a) and a
follicle without an antrum (b). O, oocyte; C.C, cumulus cells; G.C, granulosa
cells......................................................................................................................74
Fig. 2. (a) A non-viable canine ovarian follicle, (b) labelled with ethidium homodimer-1,
after 18 days of in vitro culture in absence of FSH and insulin. (c) Viable canine ovarian
follicle showing the antrum cavity and (d ) the same follicle labelled with calcein-AM
after 18 days of in vitro culture in the presence of FSH and 10 mgmL-1
insulin..............75
Fig. 3. Follicle diameter after in vitro culture in the absence (CtrlMEM) or presence of 5
ngmL-1
,10 ngmL-1
or 10 mgmL-1
insulin (Ins5ng, Ins10ng, Ins10mg, respectively) for 18
days in the absence of FSH (Experiment 1). The number of follicles cultured is given at
the bottom of the histograms.....................................................................................76
Fig. 4. Follicle diameter after in vitro culture for 18 days in medium containing 10
mgmL-1
,insulin alone (Ins10mg), FSH alone (CtrlFSH) or FSH-containing medium
supplemented with 5 ngmL-1
,10 ngmL-1
or 10 mgmL-1
insulin (Ins5ngF, Ins10ngF,
Ins10mgF, respectively; Experiment 2). The number of follicles cultured is given at the
bottom of the histograms...........................................................................................77

11
Fig. 5. Antrum formation of canine preantral follicles cultured in the absence (CtrlMEM)
or presence of 5 ngmL-1
, 10ngmL-1
or 10 mgmL-1
insulin (Ins5ng, Ins10ng, Ins10mg,
respectively) for 18 days in the absence of FSH (Experiment 1). The number of follicles
cultured is given at the bottom of the histograms.........................................................77
Fig. 6. Antrum formation of canine preantral follicles culture in medium containing 10
mgmL-1
insulin alone (Ins10mg), FSH alone (CtrlFSH) or FSH-containing medium
supplemented with 5 ngmL-1
,10 ngmL-1
or 10 mgmL 1 insulin (Ins5ngF, Ins10ngF,
Ins10mgF, respectively; Experiment 2). The number of follicles cultured is given at the
bottom of the histograms...........................................................................................78
Capítulo 3
Figure 1: Percentage of morphologically normal canine follicles after in vitro culture in
the absence (αMEM+, αMEM+FSH) or presence of different GH concentrations (50
ng/mL or 10 ng/mL) with (GH10+FSH and GH50+FSH) or without FSH (GH10 and
GH50) for 18 days..................................................................................................94
Figure 2: Canine follicular diameter (mean ± S.D.) after in vitro culture in the absence
(αMEM+, αMEM+FSH) or presence of different GH concentrations (50 ng/mL or 10
ng/mL) with (GH10+FSH and GH50+FSH) or without FSH (GH10 and GH50) for 18
days.....................................................................................................................95
Figure 3: Antrum formation rates (mean ± S.D.) after in vitro culture in the absence
(αMEM+, αMEM+FSH) or presence of different GH concentrations (50 ng/mL or 10
ng/mL) with (GH10+FSH and GH50+FSH) or without FSH (GH10 and GH50) for 18
days.......................................................................................................................96
Figure 4: Domestic dogs follicle after 18 days of culture in media αMEM added of GH
(50 ng/mL) in presence of FSH (a), and the same follicle stained with calcein-AM and
classified as viable (b)...............................................................................................97

12
Capítulo 4
Figure 01: Domestic dogs oocytes stained with Hoechst 33342, and classified chromatin
configutation as Intact (a), Fragmented (b) and Unclassifiable (c)...............................115
Figure 02: Domestic dog preantral follicles in different in vitro culture systems for 18
days.............................................................................................................................................115

13
LISTA DE TABELAS
Capítulo 2
Table 1. Follicular viability after 18 days culture of canine preantral follicles in the
presence or absence of different concentrations of insulin (Experiment 1).....................75
Table 2. Follicular viability after 18 days culture of canine preantral follicles in the
presence or absence of different concentrations of insulin with or without sequential
addition of 100, 500 and 1000 ngmL21 FSH on Days 0, 6 and 12 of culture (Experiment
2)...........................................................................................................................76
Capítulo 3
Table 1: Follicular viability after 18 days of culture of canine preantral follicles in the
presence or absence of different concentrations of growth hormone (GH), with or
without the addition of FSH (100, 500 and 1000 ng/mL).............................................97
Table 2: Evaluation of estradiol concentration (mean ± S.D.) in canine preantral follicle
culture medium between days 6 (D6) and 18 (D18) of culture for control medium
(αMEM) or medium supplemented with high concentrations of GH (50 ng/ml) associated
with FSH (GH50+FSH)..................................................................................................97
Capítulo 4
Table 01:, Antrum formation, follicular growth and oocyte diameter of domestic dog
preantral follicles cultured for 18 days in the presence or absence of FSH and EGF (50 or
100 ng/ml)....................................................................................................................116
Table 02: Antrum formation, follicular growth and oocyte diameter of domestic dog
preantral follicles cultured for 18 days of culture in media MEM or αMEM, and in
different culture systems: under mineral oil (2D), enclosed in a alginate-only matrix
(ALG) or in a fibrin- alginate hydrogel (FA-IPN).........................................................116
TABLE 03: Chromatin integrity of domestic dog oocytes obtained from ovarian follicles
after 18 days of culture in media in the presence or absence of FSH and EGF (50 or 100
ng/ml)..............................................................................................................................116

14
TABLE 04: Chromatin integrity of domestic dog oocytes obtained from ovarian follicles
after 18 days of culture in media MEM or αMEM, and in different culture systems:
under mineral oil (2D), enclosed in an alginate-only matrix (ALG) or in a fibrin- alginate
hydrogel (FA-IPN)....................................................................................................117

15
LISTA DE ABREVIATURAS E SIGLAS
ALG - Alginate (Alginato)
ANOVA - Analysis of variance (Análise de variância)
ART - Assisted reproduction techniques (Técnicas de reprodução assistida)
BSA - bovine serum albumin (Albumina sérica bovina)
cAMP - AMP cíclico
CAPES - Coordenação de Aperfeiçoamento do Pessoal de Nível Superior
CCO - Complexo cumulus oócitos
CCZ - Centro de Controle de Zoonoses
CE - Ceará
CG - Células da granulosa
CGP - Células germinativas primordiais
CIV - Cultivo in vitro
CNPq - Conselho Nacional de Desenvolvimento Científico e Tecnológico
CO2 - Dióxido de carbono
COCs - Cumulus-oocyte complexes (Complexos cúmulus oócito)
E2- Estradiol
EGF - Epidermal growth factor (Fator de crescimento epidermal)
FAVET - Faculdade de Veterinária
FA-IPN: fibrin- alginate hydrogel ( Hidrogel de alginato e fibrina)
FINEP - Financiadora de Estudos e Projetos
FIV - Fecundação in vitro
FOPA - preantral ovarian follicle (Folículo ovariano pré-antral)
FSH - Follicle stimulating hormone (Hormônio folículo estimulante)
FSHr - Recombinant follicle stimulating hormone (Hormônio folículo estimulante
FUNCAP - Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico
g - Gramas
GC - Granulosa cells (células da granulosa)
GDF-9 - Growth diferentiation factor-9 (Fator de crescimento e diferenciação-9)
GH - Growth hormone (Hormônio do crescimento)
GnRH - Gonadotropin-releasing hormone (Hormônio liberdor de gonadrotofina)
GT - granulosa and thecal cells
GV - Germinal vesicle (Vesícula germinativa)

16
GVBD - Germinal vesicle break down (Quebra da vesícula germinativa)
h - Hours (horas)
HC - Classic histology (Histologia clássica)
HSD: honestly significant difference test (Teste Honestly de diferença significativa)
IGF-I, II - Insulin-like growth factor-I, -II (Fator de crescimento semelhante à insulina -
I, II)
ITS - Insulin, tranferrin and selenium (Insulina, transferrina e selênio)
IVM - In vitro maturation (Maturação in vitro)
KL - Kit ligand
l - Litros
LAMOFOPA - Laboratório de Manipulação de Oócitos e Folículos Pré-Antrais
LH - Luteinizing hormone (Hormônio luteinizante)
MEM - Minimal essential medium (Meio essencial mínimo)
MEM+ - Supplemented minimal essential medium (Meio essencial mínimo
suplementado)
mg - Miligramas
MII - Metaphase II (Metáfase II)
min. - Minutos
mL, ml - Mililitros
mm - Milímetro
mM - Milimolar
MOIFOPA - Manipulação de Oócitos Inclusos em Folículos Ovarianos Pré-Antrais
n - número de amostras
ng - Nanograma
P<0.05 - Probabilidade de erro menor do que 5%
P≤0.1 - Probabilidade de erro menor ou igual a 1%
PBS - Phosphate buffered saline (tampão fosfato-salino)
pH - potencial hidrogeniônico
PPGCV - Programa de Pós-Graduação em Ciências Veterinárias
s - Segundos
SAS - Statistical analysis system
SCBI - Smithsonian Conservation Biology Institute
SEM - Standart error of the mean (Erro padrão médio)
TC - Theca cells (células da teca)

17
TCM-199 - Tissue Culture Medium - 199 (Meio de cultivo de tecido - 199)
TGF-α - Transforming growth factor alfa (Fator de crescimento transformante alfa)
TGF-β - Transforming growth factor beta (Fator de crescimento transformante beta)
UECE - Universidade Estadual do Ceará
USA - United States of America (Estados Unidos da América)
VEGF - Vascular endothelial growth factor (Fator de crescimento do endotélio vascular)
Vol. - Volume
ZP - Zona pellucida (Zona pelúcida)
% - Percentage (Porcentagem)
< - Menor
= - Igual
> - Maior
± - Mais ou menos
≤ - Menor ou igual
≥ - Maior ou igual
°C - Graus Celsius
2D - Two dimensional system of culture (Sistema bidimensional de cultivo)
3D - Three dimensional system of culture (Sistema tridimensional de cultivo)
α-MEM - Alpha minimal essential medium (Meio essencial mínimo alfa)
α-MEM+ - Supplemented alpha minimal essential medium (Meio essencial mínimo alfa
Δ - Delta
μg - Microgramas
μL, μl - Microlitros
μm - Micrômetros
μM - Micromolar

18
SUMÁRIO
1.INTRODUÇÃO.............................................................................................................19
2. REVISÃO BIBLIOGRÁFICA.....................................................................................20
2.2. OVÁRIO CANINO........................................................................................20
2.3. OOGÊNESE..................................................................................................22
2.4. FOLICULOGÊNESE.....................................................................................24
2.4. BIOTÉCNICA DE MOIFOPA......................................................................27
2.5. SISTEMA DE CULTIVO BIDIMENSIONAL.............................................29
2.6. SISTEMA DE CULTIVO TRIDIMENSIONAL...........................................30
2.7. IMPORTÂNCIA DA COMPOSIÇÃO DO MEIO DE CULTIVO...............32
a) Insulina..................................................................................................34
b) Hormônio do Crescimento (GH)...........................................................36
c) Fator de Crescimento Epidermal (EGF)................................................37
2.8. TÉCNICAS PARA AVALIAÇÃO DE FOLÍCILOS PRÉ-ANTRAIS PÓS
CULTIVO IN VITRO........................................................................................................38
a) Microscopia de fluorescência................................................................39
b) Dosagem de estradiol............................................................................41
3.JUSTIFICATIVA .........................................................................................................43
4. HIPÓTESE CIENTÍFICA ...........................................................................................44
5. OBJETIVO ..................................................................................................................44
5.1. Objetivo Geral ...........................................................................................................44
5.2. Objetivos Específicos ................................................................................................44
6. CAPÍTULO 1 ...............................................................................................................45
7. CAPÍTULO 2 ...............................................................................................................71
8. CAPÍTULO 3 ...............................................................................................................81
9. CAPÍTULO 4 .............................................................................................................102
10. CONCLUSÕES .......................................................................................................124
11. PERSPECTIVAS .....................................................................................................124
12. REFERÊNCIAS BIBLIOGRÁFICAS......................................................................126

19
1. INTRODUÇÃO
A utilização de biotécnicas como a maturação in vitro (MIV), fertilização in
vitro (FIV), criopreservação de gametas, dentre outros, são importantes ferramentas por
auxiliar no conhecimento acerca da biologia reprodutiva do cão doméstico Bem como,
permitir que o cão venha a servir de modelo experimental no auxílio da conservação de
espécies de canídeos selvagens em extinção, considerando-se as similaridades
reprodutivas entre estas espécies (COMIZZOLE, et al., 2000; SONGSASEN, et al.,
2007). Porém, o maior obstáculo para a aplicabilidade e desenvolvimento de biotécnicas
na espécie canina reside nas peculiaridades existentes na fisiologia reprodutiva da
cadela, com consequente baixas taxas de MIV de oócitos caninos, comparado a outras
espécies de mamíferos domésticos (FARSTAD, 2000; EVENCE, et al., 2011).
Entretanto, essa baixa taxa de ooócitos maturados in vitro pode vir a ser otimizada a
partir da utilização de oócitos oriundos de folículos pré-antrais (FOPA) existentes em
um grande número nos ovários canino (DURRANT, et al.,1998; FUJIHARA et al.,
2012).
Nesse contexto, a biotécnica de Manipulação de Oócitos Inclusos em Folículos
Ovarianos Pré-antrais (MOIFOPA), também denominada de Ovário Artificial,
(FIGUEIREDO et al., 2009), surge como uma ferramenta atrativa para que pequenos
oócitos inclusos em FOPA, recuperados dos ovários, possam sobreviver, crescer, ser
maturados e posteriormente fertilizados in vitro. A biotécnica de MOIFOPA
compreende as etapas de isolamento e cultivo in vitro, bem como a criopreservação dos
oócitos inclusos em FOPA. A MOIFOPA visa minimizar o impacto da perda folicular
originada pelo processo de atresia que ocorre largamente nos ovários. Em associação
com outras tecnologias reprodutivas, como a fertilização in vitro (FIV), o cultivo in
vitro e transferência de embriões (CIV e TE), a MOIFOPA permitiria ainda, não
somente otimizar, como também conservar o material genético de animais valiosos e de
espécies em vias de extinção (FIGUEIREDO et al., 2008; FIGUEIREDO et al., 2011;
SONGSASEN et al.,2011).
A literatura já relata a obtenção de embriões a partir do cultivo de FOPA em
algumas espécies domésticas como suínos (WU et al., 2007), bubalinos (GUPTA et al.,
2008) e caprinos (SARAIVA et al.,2010), bovinos (THOMAS et al. 2007,
MCLAUGHLIN & TELFER 2010), ovinos (ARUNAKUMARI et al. 2010) e felinos
(JEWGENOW & PARIS 2006). Entretanto, somente em murinos foi observado o

20
nascimento de crias saudáveis após maturação e fertilização de oócitos oriundos de
FOPA cultivados in vitro (O´BRIEN et al., 2003).
No que diz respeito aos caninos domésticos, maiores estudos são necessários
para incrementar o desenvolvimento in vitro dos FOPA, visto que os resultados
referentes a essa espécie ainda se limitam ao crescimento folicular e observação de
formação da cavidade antral (DURRANT et al., 1998; BOLAMBA et al., 1998;
SONGSASEN et al, 2011). Nesse sentido, torna-se necessário a elaboração de um meio
de cultivo que possa preencher os requisitos imprescindíveis ao estabelecimento de um
sistema eficiente de cultivo in vitro de FOPA caninos na tentativa se obter oócitos
competentes, ao ponto de tornarem-se aptos à produção in vitro de embriões (PIVE).
Nesse contexto, vários sistemas (bi ou tridimensional) e meios de cultivo in vitro
têm sido propostos para as diferentes espécies visando fornecer o suporte necessário
para o desenvolvimento folicular in vitro (EPPIG & O´BRIEN, 1996; O´BRIEN et al.,
2003; DEMEESTERE et al., 2005, TELFER, 2008). Dentre as substâncias utilizadas na
suplementação dos meios de cultivo que apresentaram efeito positivo no
desenvolvimento in vitro de FOPA pode-se destacar a atuação do Hormônio Folículo
Estimulante (FSH) (em caprinos, SARAIVA et al., 2008; bovinos, ROSSETO et al.,
2013; e caninos, SERAFIM et al., 2010), insulina (caprinos, CHAVES et al., 2012;
bovinos, ITOH et al., 2002; e ovinos ARUNAKUMARI et al., 2010), o Fator de
Crescimento Epidermal (EGF) (caprinos, SILVA et al., 2013; bubalinos, GUPTA et al.,
2002; e ovinos ANDRADE, et al., 2005) e o Hormônio do Crescimento (GH) (caprinos,
MAGALHÃES et al., 2011; camundongos KOBAYASHI et. al. 2000; e ovinos,
KHALID et. al., 2000).
Para uma melhor compreensão do presente trabalho será abordado, a seguir, na
revisão de literatura aspectos relacionados ao ovário canino, oogênese, foliculogênese,
biotécnica de MOIFOPA, sistemas de cultivo, importância da composição do meio no
desenvolvimento folicular, bem como, a atuação do GH, insulina e EGF, no cultivo in
vitro de FOPA.

21
2. REVISÃO BIBLIOGRÁFICA
2.1.Ovário canino
Na espécie canina, os ovários são estruturas de formato ovaladoo, se encontram
envoltos pela Bursa ovarica e suspensos pelo ligamento mesovário, estando localizados
à altura das 3ª e 4ª vértebras lombares, em situação caudal aos rins (RODRIGUES,
2003; DAVIDSON & BAKER, 2009) (Figura 01). Considerado o órgão reprodutivo
primário da fêmea, o ovário recebe essa denominação por realizar duas funções
primordiais: (1) formar a célula germinativa feminina e (2) produzir os hormônios
sexuais e diferentes peptídeos. Este papel duplo é complementar, interdependente e
necessário para o sucesso da reprodução (HAFEZ 2004). Além disso, o ovário é
formado por duas regiões: a medular e a cortical, que podem ser diferenciadas em torno
de 40 dias de desenvolvimento fetal, na espécie canina (KARLSON et al., 1982,
MONTEIRO et al., 2008).
A região medular é responsável pela nutrição e sustentação, está localizada mais
internamente, e contêm nervos, vasos sanguíneos, linfáticos, assim como tecido
conjuntivo de conformação firme (KARLSON et al., 1982). Na região cortical,
evidencia-se uma porção externa com uma camada de tecido conjuntivo denso, chamada
túnica albugínea ovariana, localizada entre a porção externa do córtex e o epitélio
germinativo. Ainda no córtex, podem ser encontradas diferentes estruturas tais como
corpo lúteo, corpo hemorrágico e corpo albicans além das células germinativas também
denominadas oócitos, que por sua vez, estão inclusos nos folículos ovarianos em
diferentes fases de desenvolvimento (dependendo da fase reprodutiva do indivíduo)
(BEARDEN & FUQUAY, 1997).

22
Figura 01: Desenho esquemático da localização anatômica do ovário canino, e
estruturas adjacentes. Fonte: O'Meara Pet Informed.
2.2. Oogênese
A oogênese tem início em mámíferos do sexo feminino durante o
desenvolvimento embrionário com a migração das células germinativas primordiais
(CGP) oriundas do saco vitelínico para a crista gonadal. Imediatamente após a
diferenciação das gônadas, em que no feto canino acontece por volta do 42º dia pós-
coito, ocorre a transformação das CGP em oogônias mitoticamente ativas e, então, em
oócitos primários. As oogônias são células grandes e arredondadas que migram e
colonizam a crista, perdendo a motilidade e iniciando a gametogênese por meio de
divisões mitóticas. Na cadela, quando a mitose é cessada pouco depois do nascimento,
inicia-se a meiose (BRISTOL-GOULD & WOODRUFF; 2006; SONGSASEN, et al.,
2007).
No período correspondente às duas semanas após o nascimento, as oogônias
começam a se dividir, ingressam na meiose I no estádio de pré-leptóteno e evoluem para
oócitos primários. Em seguida, uma camada de células somáticas planas conhecidas
como células da pré-granulosa, originárias do epitélio celômico, circundam os oócitos

23
primários formando os folículos primordiais e dando início à foliculogênese (SUH et al,
2002; RODRIGUES, 2003).
Figura 02: Esquematização da oogênese na espécie canina.
A partir desse evento, os oócitos primários passam para a segunda etapa da
meiose compreendida pelos sucessivos estádios da prófase I, constituindo assim os
folículos primordiais que são observados nos caninos com cerca de três semanas após o
nascimento. Alguns meses após o nascimento, todos os folículos primordiais com o
número definitivo de oócitos já estarão formados e entram em um período de
quiescência (BUCCIONE et al., 1990; ENGLAND e HEWITT, 1999) (Figura 02).
Na maioria das espécies mamíferas, com a chegada da puberdade, ocorre a
liberação do pico pré-ovulatório de FSH e do hormônio luteinizante (LH), que promove

24
a progressão da divisão meiótica, formação do oócitos secundários e outra parada na
fase de metáfase II. Dessa forma, a meiose é retomada novamente após a fecundação do
oócito pelo espermatozóide, originando o oócito haplóide fecundado e finalizando a
oogênese (FIGUEIREDO et al., 2008).
Entretanto, na espécie canina,, os oócitos de coloração enegrecida devido a
grande quantidade de gotículas de lipídios (Figura 03), apresentam como particularidade
o fato de serem ovulados imaturos, em estádio de vesícula germinativa, enquanto as
outras espécies ovulam oócitos em metáfase II (MII). Dessa forma, é necessário um
período de dois a três dias de permanência no oviduto para completar a maturação
meiótica do oócito (LUVONI et al, 2005; HATOYA, et al. 2009; BUKOWSKA et al.,
2012).
Figura 03: Folículo pré-antral canino, com oócito de Grau 1, com grande quantidade de
gotícula de lipídio em seu interior.
2.3. Foliculogênese
A foliculogênese pode ser definida como o processo de ativação, crescimento, e
maturação folicular, iniciando com a formação do folículo primordial e culmina com o
estádio de folículo maduro ou pré-ovulatório (FIGUEIREDO et al., 2002;
SONGSASEN et al, 2009). Esse processo é contínuo e envolve o recrutamento diário de
folículos do pool de reserva, os quais crescem e regridem em sua maioria, estando desse
modo o conteúdo ovariano em constante modificação. Na cadela, a foliculogênese
Oócito

25
inicia-se com a formação dos folículos primordiais após o nascimento, em um intervalo
situado entre 2 e 12 semanas (ANDERSEN & SIMPSON, 1973).
A população folicular presente no ovário mamífero é bastante heterogênea e
localiza-se no córtex ovariano. De acordo com o grau de evolução, os folículos podem
ser divididos em folículos pré-antrais ou não cavitários que representam cerca de 90%
da população folicular e compreendem os folículos primordiais, primários e
secundários; folículos antrais ou cavitários, englobando os folículos terciários e os
folículos de De Graaf. Estes últimos são também denominados de folículos maduros ou
pré-ovulatórios (FIGUEIREDO et al.; 2002; LUVONE et al., 2005)
O crescimento folicular se inicia com a ativação dos folículos primordiais, em
que os folículos do pool de reserva ou folículos quiescentes iniciam um processo
irreversível de desenvolvimento e passam a fazer parte do pool de folículos em
crescimento. O primeiro sinal de ativação dos folículos primordiais é a retomada da
proliferação das células da granulosa, ou seja, o folículo primordial transforma-se em
folículo primário (VAN DER HURK et al., 1997; ERICKSON & SHIMASAKI,2003).
Na espécie canina os folículos primários medem cerca de 59.0 ± 13.8 µm de diâmetro, e
abrigam pequenos oócitos de 1ª ordem, com diâmetro de 31.0 ± 3.1µm, com uma única
camada de células do cumulus (BARBER et al.,2001; SONGSASEN et al., 2009).
O início da ativação folicular pode ocorrer de forma espontânea (FORTUNE et
al., 2003) ou via estímulo de substâncias produzidas pelo oócito e células da granulosa,
que atuam no desenvolvimento folicular de forma autócrina e parácrina (ALBERTINI
& BARRETT, 2003). Dentre os inúmeros fatores que atuam na ativação folicular, pode-
se destacar o Kit ligand (PARROTT & SKINNER (1999), insulina (ENGELMAN, et al
2006), o fator de crescimento derivado de plaqueta (PDGF) (NAGAO et al., 2006), o
fator de crescimento do hepatócito (HGF) (TAHER et al., 2002), o fator de crescimento
queratinócito (KGF) (CHANG et al., 2005; BAO et al., 2005), e o fator de crescimento
do endotélio vascular (VEGF) (ALON et al., 1995), dente outros.
Em caninos, Songsasen et al. (2009) detectaram o Fator de Crescimento
Fibroblástico 2 (FGF-2) em oócitos e células da granulosa de folículos primordiais a
secundários, em todas as fases reprodutivas, exceto no anestro. Já em cadelas adultas, os
autores identificaram o Fator de Crescimento Fibroblástico 7 (FGF-7) nas células da
granulosa de folículos primários e secundários, também ocorrendo nas camadas das
células da teca de folículos antrais durante o proestro e estro, comprovando dessa forma

26
que, na espécie canina, o FGF-2 está implicado na ativação do folículo no início do
desenvolvimento folicular, enquanto que a atividade do FGF-7 parece estar relacionada
com a foliculogênese mais tardia.
Em continuação à foliculogênese, através de divisões mitóticas celulares o
folículo primário evolui para estágio de folículo secundário, que é constituído por
diversas camadas adicionais de células da granulosa de formato cúbico. Na cadela, o
diâmetro de um folículo secundário varia entre 152 a 304µm (211,4±1,5µm)
(DURRANT et al., 1998; DOLEZEL et al., 2003). Com o crescimento folicular, uma
cavidade repleta de líquido é formada dentro da camada de células da granulosa e a teca
torna-se mais estratificada. Esses folículos são chamados de folículos terciários ou
folículos antrais (FIGUEREDO et al. 2002; ABIR,et al., 2006).
No folículo antral, observam-se as células da granulosa do cumulus oophorus
que envolvem o oócito e as células que constituem as camadas internas do folículo,
denominadas de células murais. As células do cumulus fornecem nutrientes aos oócitos
durante seu crescimento, participam na formação da zona pelúcida e sintetizam a matriz
extracelular composta por proteínas e pelo polissacarídeo ácido hialurônico, de
importância no transporte e na atração dos espermatozóides no oviduto (BEDFORD &
KIM, 1993; TOSHIMORI, 2000).
Uma vez iniciado o crescimento, os folículos podem chegar à ovulação, evento
esse que se destina ao um número mínimo de folículos, ou podem sofrer morte celular
por atresia podendo esta ocorrer, em qualquer fase do crescimento folicular (LIMA,
2006; MONTEIRO et al., 2008). De acordo com Fugihara et al. (2012), cerca de 99,9%
dos folículos não atinge a ovulação, pois são eliminados por atresia, fazendo com que o
desenvolvimento de um folículo pré-ovulatório a partir de um folículo primordial seja
um evento biológico extremamente raro.
A atresia pode ocorrer pelas vias degenerativa e/ou apoptótica. A primeira pode
ser causada por isquemia, que resulta em algumas alterações na permeabilidade da
membrana celular e conseqüente degeneração. Já a segunda, ocorre quando o ambiente
parácrino ou endócrino não é apropriado para suportar o crescimento folicular e/ou
diferenciação das células da granulosa (SAUMANDE, 1991; FIGUEIREDO et al.,
1995; SILVA et al., 2002).

27
2.4. A biotécnica de MOIFOPA
Durante muito tempo, as pesquisas na área de biotecnologia da reprodução se
restringiram a estudos com animais de produção e laboratoriais, e somente a partir da
década de 90, com o interesse da comunidade científica pela preservação da
biodiversidade é que a espécie canina mereceu atenção. Esse aspecto deveu-se ao fato
da fêmea canina constituir-se em um modelo experimental para canídeos selvagens,
dentre eles o cachorro vinagre (Spheothos venaticus) e a raposinha-do-campo
(Lycalopex vetulus), que são espécies ameaçadas da fauna brasileira (COMIZZOLI et
al., 2000; EVECEN et al., 2011; The IUCN Red List of Threatened Species, 2013). O
modelo canino também visou auxililar a cinofilia, aperfeiçoando biotecnologias que
melhorassem o potencial reprodutivo no cão doméstico e permitissem a preservação de
germoplasma das cadelas domésticas portadoras de características zootécnicas
superiores ou de raças raras (DURRANT et al., 1998; STRÖM, 1997, RODRIGUES &
RODRIGUES, 2002; SONGSASEN et al., 2011) (VER CAPITÚLO 2 DA TESE).
Dentre as biotécnicas estudadas, a MOIFOPA surgiu como uma importante
alternativa, por permitir o resgate dos FOPA antes que estes se tornem atrésicos e
cultiva-los in vitro até a maturação, preservando-os do processo da atresia (GUPTA et
al., 2008; FIGUEIREDO et al., 2011). Além disso, consiste em uma das principais
ferramentas utilizadas atualmente para a elucidação da foliculogênese inicial. A
MOIFOPA pertence ao grupo das biotécnicas fundamentais ligadas à reprodução, que se
encontra em franco desenvolvimento, e compreende três fases: o resgate de FOPA, a
conservação folicular através do resfriamento e/ou criopreservação, e o cultivo in vitro
até o estádio de maturação folicular (FIGUEIREDO et al., 1999; GUPTA & NANDI.,
2012).
O cultivo in vitro dos FOPA, visa avaliar o efeito de diferentes substâncias, em
diversas concentrações e em várias fases do desenvolvimento folicular, com intuito de
mimetizar in vitro o que ocorre no ovário com uns poucos folículos que escapam à
atresia e ovulam (FORTUNE, 2003; WOODRUFF, 2007).
Nesse contexto, diversos sistema de cultivo vem sendo estudados com intuito de
promover o adequado desenvolvimento folicular in vitro. Um sistema de cultivo ideal
deve preservar a viabilidade folicular, conservar a aparência morfológica pré-existente

28
in vivo, assim como assegurar o crescimento e a maturação folicular (FIGUEIREDO et
al., 2008; PICTON et al, 2008).
Dessa forma, existem basicamente duas formas de cultivo folicular: 1) incluso
em tecido ovariano (fragmentos de tecido ovariano ou ovário inteiro) também
denominado cultivo in situ ou 2) utilizando folículos isolados (ARAÚJO in press).
Outra modalidade de cultivo é a associação entre esses dois métodos de cultivo (cultivo
em dois passos), em que inicialmente se realiza o cultivo in situ dos folículos
primordiais, e ,em seguida, isolam-se os folículos secundário obtidos para então serem
cultivados isoladamente até a obtenção dos folículos antrais (KREEGER et al., 2006;
TELFER et al., 2008; JIN et al., 2010).
No que se refere a espécie canina, em um estudo recente, Fujihara et al. (2012),
realizaram com sucesso o cultivo de fragmentos de tecido ovariano, sob um substrato de
gel de agarose, com obtenção de significativa viabilidade folicular por um período de
até 15 dias de cultivo.
Porém, quando se deseja avaliar o metabolismo folicular, esteroidogênese,
desenvolvimento do oócito ou mesmo a influência hormonal sobre o cultivo de
folículos, o método de cultivo de folículos isolados tem se demonstrado mais
apropriado. Neste tipo de cultivo é possível visualizar detalhadamente a influência que
determinado fator irá exercer sobre o desenvolvimento do folicular, a difusão do meio
ocorre de forma mais eficaz, e observa-se um maior sucesso na formação de antro com
posterior desenvolvimento (PEDERSEN, 1970; ABIR et al., 1999, GUPTA & NANDI.,
2012).
Basicamente, o cultivo in vitro de folículos isolados pode ser realizado no
modelo bidimensional (2D), ou ainda em modelo tridimensional (3D), ambos os
modelos apresentando variações dentro do mesmo sistema de cultivo (DEMEESTERE
et al., 2005).

29
2.4. Sistema de cultivo bidimensional
O sistema de cultivo folicular bidimensional é caracterizado pelo cultivo dos
folículos pré-antrais isolados diretamente sobre uma superfície, podendo esta ser a
própria superfície plástica da placa, uma membrana hidrofóbica que inibe a aderência e
migração celular, ou sobre uma matriz extracelular (por exemplo: colágeno, ágar, etc).
Alternativamente, este cultivo pode ser realizado em placas de micro-poços contendo o
meio de cultivo, ou em microgotas de meio de cultivo sob óleo mineral, com trocas de
meio frequente que previnem dentre outros fatores a aderência (NAYUDU &
OSBORN,1992; BOLAND et al., 1993; EPPIG & O'BRIEN, 1996; EPPIG et al., 1998,
PICTON, et al., 2008).
Em um estudo realizado com folículos pré-antrais de humanos, associando-se o
cultivo in situ seguido do cultivo isolado na presença de activina, Telfer et al. (2008),
realizaram um cultivo bidimensional de folículos isolados em placas de micro-poços
com fundo em formato de "V", para garantir uma forma igualitária o contato com os
nutrientes e trocas gasosas nas diferentes extremidades foliculares. Os autores relatam
um satisfatório e acelerado desenvolvimento folicular, com o surgimento de cavidade
antral. Este sistema foi também utilizado na espécie bovina permitindo a manutenção da
estrutura do folículo e promoveu o crescimento e diferenciação das células foliculares
(GUTIERREZ et al., 2000; WALTERS et al., 2007; TELFER & MCLAUGHLIN 2012)
Alguns autores relatam que o sistema bidimensional apresenta como
desvantagens o fato de não permitir a preservação da estrutura espacial folicular, muitas
vezes promovendo o aderência das células da granulosa ao fundo da placa, tornando o
folículo propício a ruptura das junções gap, e consequente perda da conexão entre as
células somáticas e o oócito, interação essa importante para o desenvolvimento oocitário
(WEST et al., 2007; DESAI et al, 2010; JIN et al, 2010). Entretanto, esse sistema vem
sendo empregado com sucesso no cultivo de FOPA em diversas espécies, fornecendo
oócitos viáveis e aptos a serem submetidos a FIV e PIVE, incluindo a obtenção de
embriões a partir desses oócitos, em bovinos (THOMAS et al. 2007; MCLAUGHLIN
& TELFER, 2010), camundongo (EPPIG & O‘BRIEN, 1996), ovino
(ARANAKUMARI et al., 2010), caprinos (SARAIVA et al., 2010; MAGALHÃES et
al., 2011), suínos (WU et al., 2007) e bubalinos (GUPTA et al., 2008). Alguns autores
ao realizarem comparações referentes ao sistema de cultivo 2D e 3D para o cultivo de

30
FOPA, relataram que em suas condições de cultivo, ambos os sistemas de cultivo
suportaram de forma similar e satisfatória o desenvolvimento folicular (ZHOU &
ZHANG, 2005; ARUNAKUMARI, et al., 2010).
Na espécie canina, o cultivo bidimensional foi realizado com o cultivo em placas
de 24 poços contendo de 5 a 10 ml de meio de cultivo por poço (BOLAMBA et al.,
1998; BOLAMBA et al, 2002,) ou em placas contendo microgotas de 100µl de meio,
cobertas com óleo mineral (SERAFIM et al., 2010) que permitiu o acompanhamento do
crescimento folicular até a formação de antro.
2.5. Sistema de cultivo tridimensional
Um sistma de cultivo ideal, deve assegurar, dentre outros fatores, a manutenção
da morfologia pré-existente do folículo, tendo em vista que o desenvolvimento normal
do folículo e a maturação oocitária são altamente dependente da estrutura folicular
(BERKHOLTZ, et al., 2006; WOODRUFF, et al., 2007).
Nesse contexto, o sistema de cultivo tridimensional (3D), vem sendo
amplamente estudado com objetivo de recriar um ambiente in vitro que permitia a
manutenção da estrutura tridimensional do folículo, preservando a interação célula-
célula/célula-estroma, bem como evitando a aderência das células da granulosa a
superfície da placa e consequentemente beneficiando o desenvolvimento folicular, (JIN
et al., 2010, DESAI et al, 2012; TELFER & MCLAUGHLIN 2012).
Para tanto, estudos foram realizados para solucionar questões envolvendo a
avaliação das propriedades físicas e respostas celulares do tipo de biomaterial a ser
utilizado no encapsulamento folicular.visando mimetizanr com maior fidedignidade a
matriz extracelular do ovário (EISELT , et al, 2000; DESAI et al, 2010).
O uso da matriz de colágeno para o cultivo de folículos pré-antrais de bubalinos
(SHARMA et al, 2009), bovinos (ALM et al., 2006) e humanos (ABIR et al., 1999)
bem como do hidrogel hialurônico para o cultivo de FOPA em murinos (DESAI et al.,
2012) tem sido relatado com sucesso na literatura. Porém, o uso da matriz de alginato
vem sendo o sistema de encapsulamento mais difundido e utilizado devido seu

31
promissor e encorajador sucesso no desenvolvimento folicular em diversas espécies
como bubalinos (GUPTA et al., 2008); camundongos (KREEGER et al, 2006),
humanos (XU et al., 2010) e primatas não humanos (XU et al., 2006; XU et al., 2009,
TING et al., 2011), até mesmo com a obtenção de oócitos meioticamente competentes
com essa espécie (XU et al, 2011), bem como a obtenção de crias viáveis em
camundongos (XU et al., 2006)
O alginato é um polissacarídeo linear copolímero do ácido β-D-manurônico e do
ácido α-L-gulurônico, que são produzidos pelas algas marrons. A redução da rigidez da
matriz de alginato pela redução de sua concentração, permitiu a diferenciação de tipo de
células foliculares e da teca, formação de cavidade antral, maturação oocitária e
produção hormonal (XU et al., 2006, WEST et al 2007; PICTON et al., 2008) (Figura
04, A).
Figura 04: Folículo pré-antral canino incluso em gota de alginato (Fonte: N.
Songsasen) (A) e alginato acrescido de fibrina (B). As setas vermelhas indicam a ação
da fibrina ao degradar o alginato ao longo do cultivo.
Na espécie canina, Songsasen et al. (2011) realizaram o cultivo de FOPA
caninos comparando o encapsulamento em duas diferentes concentrações de alginato, a
0,5 ou 1,5%. Os autores observaram que os folículos caninos cultivados em alginato
mantiveram a integridade estrutural, apresentaram crescimento ao longo do cultivo, bem
como foram hormonalmente ativos. Entretanto, o baixo percentual de alginato (0,5%)
A B

32
foi o mais adequado para ser utilizado na espécie canina, por possibilitar um
crescimento mais significativo quando comparado a 1,5% de alginato.Entretanto, uma
recente questionamento surgiu referente ao fato do hidrogel de alginato não ser
degradável e tornar-se mais rígido nas regiões adjacentes da gota de alginato, à medida
que o folículo aumentaria de tamanho. Por essa razão, foi proposto com sucesso a
associação do alginato e fibrina, objetivando criar um ambiente dinâmico que forneça
um maior suporte para o crescimento folicular (WEST et a., al, 2007, SHIKANOV et
al., 2009).
A fibrina é um derivado natural, e consiste do fibrinogênio (crioprecipitado do
plasma sanguíneo) polimerizado através da ação da trombina na presença do cálcio (JIN
et al., 2010; XU et al., 2013). Durante o cultivo, o folículo encapsulado ativa proteases
que degradam a fibrina presente na gota de hidrogel, o que resulta na redução gradativa
da rigidez do gel, e uma maior acomodação do folículo dentro da gota, e maior suporte
mecânico a medida que o folículo aumenta de tamanho (SHIKANV et al., 2011) (Figura
03, B). Esse sistema foi utilizado com sucesso em camundongos (KREEGER et al.,
2006; JIN et al., 2010; SHIKANV et al, 2009) e quando comparado ao cultivo com
alginato sozinho, resultou em uma taxa significativa e satisfatória de folículos viáveis, e
até mesmo a produçção de embriões de duas células.
Em um estudo recente XU et al. (2013), compararam a matriz composta somente
de alginato, com o alginato acrescido de fibrina para o cultivo de folículos primários de
primatas não-humanos. Os autores observaram que a presença de fibrina no alginato
possibilitou um aumento das taxas de crescimento folicular, produção de estradiol,
hormônio anti-mulleriano e do VEGF, fato este não observado para o cultivo de
folículos secundários nas mesmas condições. De acordo com os autores, devido ao fato
do folículo secundário apresentar uma expansão de volume mais rápida quando
comparada ao primário nessa espécie. Possivelmente, a fibrina não acompanhou a
redução sua rigidez de forma adequada para assegurar o rápido crescimento folicular.
2.6. Importância da composição do meio de cultivo
No desenvolvimento de um sistema de cultivo eficiente é essencial que ocorra
um controle irrestrito de todas as variáveis que possam afetar direta ou indiretamente a
sobrevivência folicular, tais como, temperatura, CO2, tensão de oxigênio, controle de

33
contaminações fúngicas e bacterianas, e fatores referentes ao meio de cultivo como a
presença de nutrientes e suplementos (SILVA et al.; 2004,PICTON, et al., 2008).
Dessa forma, a composição do meio é uma importante ferramenta para a
obtenção do sucesso do cultivo de FOPA in vitro, visto que, é no meio de cultivo que os
folículos encontrarão subsídios necessários para dar suporte ao seu crescimento
(TELFER et al., 2000). Assim, muitos autores têm investigado o efeito de vários meios
de cultivo e seus componentes, no cultivo in vitro de FOPA, tanto de animais de
laboratórios, como animais domésticos (DANFORTH et al., 2003; MATOS et al.,
2007).
Nesse sentido, diversas substâncias podem compor o meio de cultivo e
influenciar no crescimento e no desenvolvimento folicular (HOVATTA, et al., 1997;
TELFER et al., 2000). Segundo Guérin (1998), na composição do meio deve conter
ainda suplementos como albumina sérica bovina (BSA) ou soro fetal bovino (SFB); e
dentre outros, hormônios como GH, fatores de crescimento como o EGF, ,ou
gonadotrofinas como o FSH, para garantir uma melhor similaridade com o ambiente in
vivo, e melhor suporte ao desenvolvimento folicular
Em relação à espécie canina de forma geral, as pesquisas conduzidas utilizam
meios de cultivo semelhantes aos empregados para as outras espécies (GUÉRIN, 1998).
Serafim et al.(2010) realizaram o cultivo de FOPA caninos, por 18 dias, em meio
αMEM, suplementado com ITS, ácido ascórbico e BSA. Os autores puderam observar o
adequado crescimento folicular, com formação de antro e viabilidade de todos os
folículos ao final do cultivo.
Bolamba et al. (1998) realizaram o cultivo de FOPA avançados em meio base
Meio Essencial Mínimo Dulbelco‘s (DMEM) suplementado, e concluíram que é
possível obter-se retomada de meiose em oócitos recuperados de FOPA nesse estádio.
Também foi observado que o cultivo de FOPA de cadelas em meio SOF resultou em
uma baixa percentagem de oócitos caninos maturos, e tanto o SFB, quanto o BSA não
foram efetivos quando adicionados ao meio de cultivo (BOLAMBA et al., 2002). No
entanto, sabe-se que a suplementação do meio com BSA ou SFB é benéfica por ser uma
importante fonte de proteína, suprindo as necessidades de aminoácidos essenciais e
fornecendo energia oriunda de carboidratos. Além disso, o acréscimo de BSA aos meios

34
pode facilitar a aderência de fatores de crescimento e evitar decréscimo na concentração
de oxigênio do meio (HEWITT & ENGLAND, 1999).
Em um estudo recendo, Fugihara et al.,(2012), realizando o cultivo in situ de
folículos pré-antrais caninos, realizaram uma comparação entre dois diferentes meios de
base (MEM e αMEM) para fragmentos de ovário cultivados sob uma superfície de gel
de agarose. Os autores observaram que o meio de base αMEM, fornecia um melhor
suporte para o desenvolvimento in situ dos folículos caninos, sendo o percentual de
viabilidade no meio αMEM, significativamente superior ao MEM.
Vale resaltar que ambos meios αMEM e MEM são meios ricos, porém, que
apresentam formulações diferenciada em determinados fatores, como a presença de
aminoácidos não essenciais. O αMEM,a apresenta maiores concentrações de
aminoácidos não essenciais como L-alanina, L-asparaginea, L-ácido aspartico, L-
cisteina, L-ácido glutâmico, glicina, L-prolina and L-serina, que são fontes de
energéticas de grande importância para os estágios tardios do desenvolvimento folicular
e oocitário (LANE & GARDNER 2007; FUJIHARA et al., 2012).
Além disso, a adição de compostos antioxidantes e substratos energéticos ao
meio de cultivo também é aconselhável para manutenção da sobrevivência celular.
Nesse sentido, suplementos como a combinação de insulina, transferrina e selênio (ITS),
podem ser utilizados para o fim acima descrito (AUGUSTIN et al., 2003, PICTON et
al., 2008, TELFER & MCLAUGHLIN 2012).
A seguir será abordado o papel de algumas substâncias importantes que para o
desenvolvimento folicular in vitro.
a) Insulina
A insulina é uma pequena proteína de baixo peso molecular (5808 kDa)
produzida nas ilhotas de Langerhans, células do pâncreas endócrino. Inicialmente é
produzida como um pré-pró hormônio no retículo endoplasmático, que é clivado no
próprio retículo para formar a pró-insulina. A pró-insulina é homóloga aos fatores de
crescimento semelhante à insulina do tipo I e II (IGF-I e IGF-II). Em seguida, a pró-
insulina é convertida no complexo de Golgi em insulina, composta por duas cadeias de

35
aminoácidos, conectadas por pontes dissulfeto (YARAK et al.,2005; SILVA et
al.,2009).
Quando se liga aos receptores localizados na membrana plasmática da célula
alvo, aumenta a captação de glicose, que é rapidamente fosforilada, tornando-se
substrato para todas as funções metabólicas celulares. Acredita-se que o transporte
aumentado da glicose resulte da translocação de múltiplas vesículas intracelulares para
as membranas celulares. Essas vesículas transportam em suas próprias membranas
múltiplas moléculas de proteínas transportadoras de glicose, que se ligam á membrana
celular e facilitam a captação de glicose para o interior da célula (CARVALHEIRAS et
al., 2002; CHAVES et al., 2012).
A presença da insulina também torna a membrana celular mais permeável a íons
de potássio e íons e fosfato, assim como a muitos aminoácidos, resultando no transporte
aumentado dessas substâncias para o interior da célula, característica essa que
compartilha com o GH, além de aumentar a tradução do RNA-mensageiro com a
consequente síntese de novas proteínas (VAN DEN HURK & ZHAO, 2005).
A insulina é normalmente adicionada ao meio de cultivo in vitro em
concentrações supra fisiológicas (10 µg/ml) associada ao selênio, que atua como
estimulador da síntese de glutationa (GSH), a qual age como sistema de defesa na célula
contra os radicais livres. A transferrina também foi identificada como um dos maiores
constituintes protéicos nas células da granulosa e no fluido folicular e ampolar, tendo
ação de quelar radicais hidroxila, facilitando o transporte de ferro e outros metais para
dentro da célula além de atuar como fator de crescimento. Tal associação de insulina,
transferrina e selênio recebem a denominação de ITS (FRESHNEY, 1994; LIM &
HANSEL, 2000).
Estudos in vitro têm demonstrado que a suplementação do meio de cultivo com
ITS favorece o crescimento bem como a sobrevivência folicular com uma baixa taxa de
degeneração em bovinos (KATSKA et al., 2000, MACHADO et al., 2006), humanos
(LOUHIO et al.,2000), felinos (JEWGENOW & GORITZ, 1995) e murinos (LIU et al.,
2002). Entretanto, Eppig et al. (1998) demonstraram que, na ausência de soro, a insulina
não apresentou nenhum efeito sobre o desenvolvimento das células da granulosa de
camundongos e sobre a competência do oócito. O mesmo autor observou efeitos

36
deletérios da combinação de insulina e FSH no meio de cultivo. Em contrapartida,
outros autores mostraram que a associação de gonadotrofinas e insulina foi considerada
ótima em cultivo de FOPA intactos, de modo a favorecer a proliferação e diferenciação
folicular, gerando oócitos maduros (LIU et al., 2002).
b) Hormônio do crescimento (GH)
O GH também denominado de hormônio somatotrófico, é uma molécula
pequena de proteína que contém 191 aminoácidos em cadeia única. Este hormônio
induz a proliferação de quase todos os tecidos do corpo que tem capacidade de crescer,
promove o aumento de tamanho das células e do número de mitoses, bem como a
diferenciação específica de certos tipos celulares. Além do efeito geral sobre a
estimulação do crescimento, o GH exerce numerosos efeitos metabólicos específicos,
potencializando quase todos os aspectos da captação de aminoácidos e da síntese de
proteínas pelas células, reduzindo ao mesmo tempo a degradação das proteínas (SILVA
et al., 2009, MAGALHÂES, et al, 2011).
Foi constatado que o GH induz o fígado e outros tecidos, em grau bem menor, a
sintetizar várias proteínas pequenas, denominadas somatomedinas, ou fator de
crescimento semelhante à insulina (IGF) (FORTUNE, 2003).
O GH pode influenciar a função ovariana através do aumento do transporte para
o ovário de IGF-I produzido no fígado ou em outros locais, podendo também agir
diretamente sobre as células da granulosa que expressam receptores de GH ou ainda
promover um aumento da esteroidogênese. Adicionalmente, o GH pode afetar
indiretamente a ação do IGF ovariano, alterando níveis intrafoliculares de uma ou mais
proteínas de ligação de IGF (RECHLER, 1993).
Estudos têm sugerido que esse hormônio desempenha um importante papel no
crescimento folicular e função luteal. Em ovários de coelhos perfundidos, o GH
promoveu o estímulo do crescimento folicular, a maturação de oócitos, e produção
ovariana de estradiol (YOSHIMURA et al., 2003). Além disso, o receptor do GH já foi
localizado nas células foliculares de murinos (LOBIE et al., 1990), bovinos (LUCY et
al., 1993; IZADYAR et al., 1997), ovinos (ECKERY et al., 1997), suínos (QUESNEL,
1999) e humanos (SHARARA & NIEMAN, 1994). Na espécie caprina, o GH foi capaz
de promover o crescimento, maturação de oócitos oriundos de FOPA e o
desenvolvimento de um embrião oriundo de um oócito FOPA crescido in vitro

37
(MAGALHÃES ,et al., 2011). Já na espécie equina, seu emprego resultou na indução da
expansão do cumulus, sem ter influência, no entanto, sobre a produção embrionária
nessa espécie, bem como em suínos (MARCHAL et al., 2003).
Dessa forma, maiores estudos são necessários para que se possa compreender a
função do GH na foliculogênese, e sua ação no meio de cultivo de FOPA caninos.
c) Fator de crescimento Epidermal (EGF)
O EGF foi um dos primeiros fatores a serem identificado em mamíferos como
fator promotor de crescimentos e tem suas propriedades extensivamente caracterizadas.
Este fator faz parte da grande família de proteínas pertencentes ao fator de crescimentos
transformado alpha (TGF-α), que inclue a amfirregulina (AREG), betarregulina (BTC),
epirregulina e neuregulina (CARPENTER et al., 1993; CONTI et al, 2006; SILVA et
al., 2013).
O EGF é considerado um fator de crescimento protéico cuja atividade biológica
é mediada por receptores de membrana EGF-R (ErbB1) do tipo tirosina-quinase,
pertencentes à superfamília ErbB (CONTI et al., 2006; CELESTINO et al.,2009,
SILVA et al., 2013). Estudos in vitro demonstraram que o EGF tem ação na regulação
de diversos processos no ovário, como a ativação folicular e estímulo de mitoses nas
células foliculares, proliferação e diferenciação das células da granulosa em bovinos
(SAHA et al., 2000; WANG et al., 2007), e estímulo do crescimento folicular em
humanos (ROY et. al.,1998). Além disso, exerce uma importante função na
foliculogênese e esteriodogênese nas células da granulosa e apresenta um efeito
estimulante na maturação oocitária e expansão das células do cumulus in vitro por
ambas as vias, autócrina e parácrina, em suínos, bovinos, roedores e humanos (RIEGER
et al., 1998; ABEYDEERA et al, 1998, JEZOVA et al., 2001; SONG et al, 2011)
Na espécie canina, o EGF já foi utilizado no meio de maturação de oócitos
crescidos in vivo, resultando em um efeito positivo na progressão da meiose (CUI et al.,
2006) bem como benefício na expansão das células do cumulus (BOLAMBA et al.,
2006) sem no entanto apresentar um efeito estimulatório na produção de EGFr na
maturação in vitro (SONG et. al, 2011).

38
2.7. Técnicas para avaliação de folículos pré-antrais pós-cultivo in vitro
Diversas técnicas podem vir a ser utilizadas para avaliação estrutural e de
viabilidade dos folículos pré-antrais após o cultivo in vitro. Através delas é possível
avaliar as possíveis alterações ocorridas ao longo do cultivo, bem como, monitorar a
qualidade e funcionalidade folicular, para possibilitar melhorias nos sistemas de cultivo.
Dentre as técnicas existentes, iremos abordar a microscopia de fluorescência e dosagem
de hormônios esteroides.
a) Microscopia de fluorescência
A microscopia de fluorescência é uma ferramenta analítica muito poderosa que
combina as propriedades de aumento de microscopia de luz com a visualização da
fluorescência. É realizada em conjunção com o microscópio de luz básico pela adição
de uma fonte potente de luz, filtros especializados, e um marcador específico de
fluorescência na amostra, que permite avaliar dentre outros fatores, a viabilidade celular
(LIM & DANUSER, 2009; BLACKBURN et al, 2011).
O uso da microscopia de fluorescência para a análise dos folículos pré-antrais
após o cultivo in vitro, vem sendo empregado com sucesso em diversas espécies,
principalmente para avaliação da viabilidade celular após cultivo (BRUNO et al., 2009;
SARAIVA et al., 2011; SILVA et al., 2013). Para essa finalidade, pode-se fazer uso dos
marcadores Hoescht, calceína AM e etídio homodímero-1.
O marcador Hoescht penetra em células vivas e tem sido largamente empregado
em citologia com a finalidade de marcar cromossomos, avaliar a integridade da
membrana e configuração da cromatina de oócitos oriundos de folículos pré-antrais
(SAHA et al., 2000; MATOS et al., 2007). Em felinos, Jewgenow (1996) observou uma
mudança na aparência morfológica dos folículos isolados após cinco dias de cultivo em
M199 suplementado com soro fetal bovino. Além disso, o autor relata que foi observado
a substituição da cromatina normal (estrutura com aparência reticular) por uma
cromatina degenerada (estrutura com aparência circular ou com distribuição compacta).
Para a espécie canina, antes do marcador Hoescht, a literatura relata a
necessidade prévia de fixação do oócito em paraformaldeído, para uma melhor
visualização do ooplasma (KIM et al., 2005; LOPES et al., 2007; SONG et al., 2011).
Visto que, devido à grande quantidade de gotículas de lipídios existente no interior do
oócito dos caninos, também observado em outras espécies como porcinos (STURMEY

39
et al., 2006) e bubalinos (SHARMA et al., 2009), a visualização da cromatina pode ser
dificultada na microscopia de fluorescência, necessitando dessa forma ser fixado
(HEWITT et al., 1998)
A calceína AM e o etídio homodímero, conferem uma marcação dupla,
identificando simultaneamente células viáveis e não viáveis, respectivamente. A
Calceína-AM, atravessa passivamente a membrana celular, sendo clivada por esterases
nas células vivas, e como produto dessa clivagem o componente fluorescente de
coloração verde é então visualizada (VAN DEN HURK et al., 1998; LOPES et al.,
2009). Já o etídio homodímero, é evidenciado somente quando ocorre a perda da
integridade da membrana, cessa a atividade da esterase, e o etídio reage com os ácidos
nucléicos, produzindo a fluorescência de coloração vermelha (SANTO et al., 2006;
MATOS et al., 2007).
b) Dosagem de estradiol
O desenvolvimento folicular e ovulação são dependentes da proliferação e
diferenciação ocorridas nas células da granulosa e da teca, as quais iniciam a
esteriodogênese mediante estímulos gonadotróficos e por citocinas. O estradiol ou 17β-
estradiol é sintetizado por enzimas esteroidogênicas como o citocromo P450 nas células
da teca e pelo citocromo P450aromatase nas células da granulosa. Tais enzimas
convertem de colesterol a pregnenolona e a síntese de estradiol a partir de precursores
de andrógenos, respectivamente. Na fase final do desenvolvimento folicular, essa
produção é mais acentuada, sendo esse evento necessário para manutenção da
sobrevivência folicular (SCARAMUZZI et al., 1993; WANDJI et al., 1996; TAMURA
et al., 2007). De acordo com Telfer et al., (2000), no cultivo in vitro de folículos
ovarianos é possível realizar o monitoramento do meio de cultivo e dos fatores
secretados pelo folículo e oócito, possibilitando dessa forma, fazer uso dessa análise
como uma importante técnica de avaliação do desenvolvimento folicular. Estudos
prévios tem verificado a produção de estradiol em folículos pré-antrais em outras
espécies como em caprinos (SILVA et al., 2013), ovinos: (CECCONI et al., 1999)
humanos (TELFER et al., 2008) e bovinos (SPICER et al., 1996).
Em um estudo recente, Songsasen et al., (2011); avaliou a produção de estradiol
em folículos pré-antrais caninos cultivados no sistema tridimensional com diferentes

40
concentrações de alginato, e concentrações crescentes de FSH. Os autores observaram
que o sistema de cultivo possibilitou a manutenção da capacidade esteriodogênica
folicular, em ambas as concentrações de alginato avaliadas. Dessa forma, a habilidade
de produzir estradiol in vitro, indica que o folículo se encontra em condições favoráveis
para a multiplicação e diferenciação das células da teca e granulosa, sendo portanto, um
bom indicador para o desenvolvimento folicular in vitro (WANDJI et al., 1996;
DRIANCOURT, 2001).

41
3. JUSTIFICATIVA
Experimentos realizados com gametas de cães (Canis familiaris) são
considerados importantes, tendo em vista que o cão, sendo usado como modelo
experimental, pode responder a uma série de questões da fisiologia reprodutiva de
canídeos selvagens, considerando-se as similaridades entre as espécies. Dessa forma, a
definição de um meio de cultivo e sistema adequado de cultivo in vitro possibilitaria a
expansão de técnicas de reprodução assistida, facilitando a PIVE de embriões, e a
conservação de gametas (COMIZZOLI,et al, 2000; ABDEL-GHANI, et al., 2012).
Essa por sua vez, poderá constituir-se em uma etapa para salvaguardar a diversidade
genética em populações mantidas em cativeiro ou sob ameaça de extinção. Não obstante
o caráter futuro, a perspectiva de aplicação dessas técnicas em caninos é bastante real
(RODRIGUES & RODRIGUES, 2003). Porém um fator limitante para o
desenvolvimento e eficiência dessas biotécnicas é a ausência de um número satisfatório
de oócitos a serem fertilizados (TELFER, 2001). Este problema pode vir a ser
solucionado por meio do uso de uma grande fonte de oócitos inclusos em FOPA,
obtidos com a técnica de MOIFOPA (FIGUEIREDO et al., 2011).
Além disso, o uso da MOIFOPA pode vir a auxiliar na solução de inúmeras
questões que ainda existem em relação à foliculogênese na espécie canina, assim como
a ação dos fatores envolvidos no crescimento, diferenciação e seleção folicular. Tendo
em vista que o folículo ovariano pré-antral é o precursor da população de folículos
antrais que chegarão ao estádio ovulatório, faz-se necessário o seu estudo para elucidar
os mecanismos responsáveis pelo desenvolvimento dessa grande fração da população
ovariana.
Neste sentido, para que ocorra o sucesso do cultivo, é necessário que o sistema
de cultivo utilizado (2D ou 3D), bem como a compisição do meio de cultivo
proporcionem as condições favoráveis para subsidiar o desenvolvimento folicular.
Nesse sentido, estudos têm sido desenvolvidos visando verificar a influência da adição
de diversas substâncias que beneficiem o desenvolvimento folicular, merecendo
destaque a insulina (KATSKA et al., 2000, MACHADO et al., 2006) GH
(HARTSHORE, 1997; TELFER et al., 2000) e o EGF (BOLAMBA et al., 2006,
CELESTINO et al.,2009). Na espécie canina, a literatura relata o uso do cultivo
bidimensional em poços, com meios suplementados com FSH, hCG e estradiol no

42
cultivo oócitos de FOPA avançados e antrais precoces, porém, com um baixo percentual
de oócitos que progrediram de MI à MII. Além disso, não foi avaliado o envolvimento
desses hormônios sobre o desenvolvimento folicular (BOLAMBA et al., 1998). Por
outro lado, Serafim et al. (2010) realizaram o cultivo bidimensional de FOPA em
microgotas de 100µl de meio submersas em óleo mineral, e determinaram que a adição
de FSH de forma sequencial é benéfico para o desenvolvimento folicular, já que foi
evidenciada formação de antro e sobrevivência ao fim do cultivo. Songsasen. et
al.(2011), analisou a influência do FSH no cultivo de FOPA caninos encapsulados com
matriz de alginato, e observou que o FSH é de fato essencial para o desenvolvimento
folicular, e que a concentração de 0,5% de alginato era a mais adequada para suportar o
desenvolvimento folicular.
Entretanto, um sistema de cultivo ideal bem como o meio a ser utilizado que
venha a promover incremento no desenvolvimento folicular de FOPA caninos ainda não
foi determinado. Dessa forma, faz-se necessária a realização de estudos mais
aprofundados, visando identificar o melhor sistema a ser empregado no cultivo (2D ou
3D), quais substâncias devem ser adicionadas ao meio, assim como suas concentrações
e momento ideal de adição. Assim, será possível identificar os componentes ideais para
o crescimento e desenvolvimento adequado dos FOPA caninos, e a partir disso, será
possível a obtenção de oócitos viáveis, os quais serão destinados à MIV. Esta opção se
faz relevante, frente ao baixo índice de oócitos maturados in vitro na espécie canina.

43
4. HIPÓTESE CIENTÍFICA
A adição de insulina, GH e EGF, beneficia a sobrevivência, o crescimento e a
maturação in vitro de oócitos inclusos em de FOPA caninos
O meio de base αMEM sustenta de forma mais satisfatória o crescimento
folicular em cães domésticos do que o MEM.
O tipo de sistema de cultivo (2D ou 3D) influencia o desenvolvimento in vitro
de FOPA.

44
5. OBJETIVOS
5.1.Objetivo geral:
Avaliar o efeito de diferentes concentrações de insulina, EGF e GH, na presença
ou ausência de FSH sobre o cultivo in vitro de FOPA caninos isolados.
5.2.Objetivos específicos:
Analisar o efeito de diferentes concentrações de insulina (5, 10 ng/mL e 10
µg/ml), de GH (10 e 50 ng/ml) e de EGF (50 ou 100 ng/ml),sobre o crescimento,
formação de antro e viabilidade dos FOPA caninos isolados e cultivados por 18
dias.
Investigar a interação do FSH com as substâncias insulina, GH, e EGF, no
cultivo in vitro de FOPA caninos.
Avaliar a influência do meio de cultivo de base (αMEM ou MEM) no
desenvolvimento folicular de cães
Verificar o comportamento dos FOPA nos sistemas de cultivo bidimensional e
tridimensional, e como esses sistemas agem sobre o desenvolvimento folicular

45
6. CAPÍTULO 01
AVANÇOS NO CRESCIMENTO, MATURAÇÃO E FECUNDAÇÃO IN VITRO
DE OÓCITOS CANINOS.
Periódico :Ciência Animal/in press

46
RESUMO- A produção in vitro de embriões em cães pode auxiliar na compreensão de
inúmeros fenômenos fisiológicos como o crescimento, a maturação e a fecundação de
oócitos e ainda se apresenta como alternativa para contornar obstáculos reprodutivos
inerentes à espécie. Além disso, os conhecimentos adquiridos com esta técnica podem
ser utilizados para a preservação de cães selvagens ameaçados de extinção. Entretanto
diferenças na fisiologia reprodutiva canina quando comparada às demais espécies
domésticas, aliadas a uma baixa eficiência na maturação in vitro, resulta em uma
dificuldade de maiores avanços na produção in vitro de embriões caninos. Dessa forma,
este artigo tem como objetivo, discorrer sobre os avanços obtidos até o momento no
crescimento e maturação in vitro de oócitos de cães domésticos.
Palavras chaves: Canino, maturação in vitro, oócito

47
AVANÇOS NO CRESCIMENTO, MATURAÇÃO E FECUNDAÇÃO IN VITRO
DE OÓCITOS CANINOS.
(Advances on in vitro growth, maturation and fecundation of canine oocyte )
Michelle Karen Brasil Serafim1*; Gerlane Modesto da Silva1; Ana Kelen Felipe Lima 1;
Ticiana Franco Pereira da Silva1, Deborah de Melo Magalhães Padilha1, Lúcia Daniel
Machado da Silva2, José Ricardo de Figueiredo1
1Faculdade de Medicina Veterinária, Laboratório de Manipulação de Oócitos e
Folículos Pre-antrais (LAMOFOPA), PPGCV, Universidade Estadual do Ceará,
Fortaleza, CE, Brasil
2 Faculdade de Medicina Veterinária, Laboratório de Reprodução de Carnívoros (LRC),
PPGCV, Universidade Estadual do Ceará, Fortaleza, CE, Brasil
RESUMO- A produção in vitro de embriões em cães pode auxiliar na compreensão de
inúmeros fenômenos fisiológicos como o crescimento, a maturação e a fecundação de
oócitos e ainda se apresenta como alternativa para contornar obstáculos reprodutivos
inerentes à espécie. Além disso, os conhecimentos adquiridos com esta técnica podem
ser utilizados para a preservação de cães selvagens ameaçados de extinção. Entretanto
diferenças na fisiologia reprodutiva canina quando comparada às demais espécies
domésticas, aliadas a uma baixa eficiência na maturação in vitro, resulta em uma
dificuldade de maiores avanços na produção in vitro de embriões caninos. Dessa forma,
este artigo tem como objetivo, discorrer sobre os avanços obtidos até o momento no
crescimento e maturação in vitro de oócitos de cães domésticos.
Palavras chaves: Canino, maturação in vitro, oócito
ABSTRACT- The in vitro embryo production in dogs can help the understanding of
several physiological processes such as growth, maturation and fertilization of oocytes,
and also represents an alternative to overcome reproductive obstacles in this species.

48
Moreover, the knowledge obtained from this technique can be used for the preservation
of endangered wild dogs. However, the existence of differences between dogs and other
domestic species regarding the reproductive physiology as well as the low efficiency in
the in vitro maturation in this species difficult major advances in the in vitro embryo
production in dogs. Thus, this review aims to address the progress obtained so far in the
in vitro growth and maturation of oocytes from domestic dogs.
Key-worlds: Canine, in vitro maturation, oocyte
1. INTRODUÇÃO
A produção in vitro de embriões, que envolve as etapas de maturação (MIV),
fecundação (FIV) e cultivo de embriões (CIV), é de extrema importância em caninos
domésticos uma vez que possibilita sua aplicação na multiplicação de espécies de
canídeos selvagens vulneráveis ou ameaçados de extinção, como o Lobo Guará
(Chrysocyon brachyurus), a Raposinha do Campo (Pseudalopex sp.) e o Cachorro
Vinagre (Speothos venaticus). Adicionalmente, estas técnicas também poderão
contribuir para a compreensão da própria fisiologia reprodutiva canina, bem como
contornar obstáculos reprodutivos em animais de alto valor zootécnico (SILVA et al.,
2004a; SONGSASEN et al., 2007, HATOYA et al., 2009).
Entretanto, o maior obstáculo para a aplicabilidade e desenvolvimento dessas
biotécnicas consiste nas diferenças da fisiologia reprodutiva da cadela e na baixa
eficiência da MIV em oócitos caninos, comparadas às outras espécies de mamíferos
(OTOI et al., 2007). Diante disso, inúmeros estudos relacionados à foliculogênese e
oogênese têm sido conduzidos com a finalidade de determinar quais fatores afetam a
viabilidade, crescimento e a maturação dos oócitos caninos. Para tanto, a presente
revisão tem como objetivo, discorrer sobre as estratégias utilizadas até o presente
momento para aumentar a produção in vitro de embriões caninos, utilizando-se oócitos
oriundos tanto de folículos pré-antrais como de folículos antrais.

49
2. A biotécnica de Manipulação de Oócitos Inclusos em Folículos Pré-antrais
(MOIFOPA) em cadelas
De acordo com Figueiredo et al. (2007), os mamíferos domésticos nascem com
uma reserva folicular de milhares a milhões de folículos primordias quiescentes. Porém,
esse número é progressivamente reduzido ao longo da vida reprodutiva da fêmea, por
um processo fisiológico denominado de atresia.
Uma vez que 99,9% de toda a população folicular presente no ovário mamífero é
perdida por este fenômeno, a biotécnica de Manipulação de Oócitos Inclusos em
Folículos Ovarianos Pré-antrais (MOIFOPA), recentemente designado de ovário
artificial, surge como uma importante ferramenta para evitar essa grande perda folicular
que ocorre naturalmente ―in vivo”, pois se propõe a resgatar um grande número de
folículos pré-antrais (FOPA), cultivá-los ―in vitro‖ até a completa maturação oocitária
para utilização posterior na PIV de embriões. Além disso, devido à possibilidade de
proporcionar o fornecimento de uma população homogênea de oócitos oriundos de um
mesmo animal (FIGUEIREDO et al., 2009), essa biotécnica contribuirá também para a
padronização de técnicas, como a clonagem, a FIV e a transgenia, uma vez que será
utilizada uma população de oócitos em um mesmo estádio de desenvolvimento e de
mesma origem (FIGUEIREDO et al., 2008).
As etapas da MOIFOPA consistem na conservação visando à estocagem de
FOPA por um curto (resfriamento) ou longo período (criopreservação); no isolamento
ou resgate de FOPA a partir de ovários e no cultivo, que tem como finalidade promover
o crescimento, a MIV e FIV dos oócitos previamente inclusos em FOPA
(FIGUEIREDO et al., 2008). Através dessa biotécnica já foram obtidos embriões em
algumas espécies como bubalinos (GUPTA, et al, 2008), suínos (WU et al., 2001) e
caprinos (SARAVIVA et al., in press) bem como crias vivas, conforme relatado em
camundongos (O‘BRIEN et al., 2003). Nas sessões a seguir serão abordados os
principais avanços referentes ao resfriamento, isolamento e cultivo in vitro de FOPA
caninos.
2.1.RESFRIAMENTO
De acordo com Silva et al. (2004b), o tempo decorrido entre a coleta do ovário e
o início do cultivo in vitro é uma questão fundamental. Quando este tempo é muito
longo, especialmente quando o animal doador está longe do laboratório de reprodução,

50
como é o caso dos animais selvagens, a preservação da viabilidade folicular é
diminuída, pois as células após a coleta encontram-se sob uma situação de isquemia.
Nesse sentido, fatores como meio de transporte, temperatura e tempo de preservação,
devem ser meticulosamente avaliados, pois podem afetar diretamente as células
ovarianas e resultar na ocorrência de autólise celular (NAKAO & NAKATSUJI, 1992;
TAS et al., 2006).
Poucos estudos avaliam os efeitos do meio e temperatura de transporte para
folículos pré-antrais caninos. Geralmente, o foco maior está nos efeitos dessas variáveis
sobre a maturação in vitro dos oócitos oriundos de folículos antrais, e os achados
geralmente são controversos (TAS et al., 2006; LOPES et al., 2009).
Durrant et al. (1998) e Bolamba et al. (1998) realizaram o cultivo de folículos
pré-antrais caninos avançados, para obtenção de oócitos a serem destinados à MIV.
Essa equipe realizou o transporte dos ovários em solução salina a 4°C acrescida de
antibióticos. Em seguida, os ovários foram armazenados por 24h a 4°C em solução
salina contendo antibióticos e antifúngicos.
Em um estudo recente, Lima et al. (2010) comprovaram que o transporte de
ovários caninos realizados em meio água de coco em pó (ACP) em baixas temperaturas,
é adequado para manutenção da morfologia dos folículos por até 24h.
Lopes et al. (2009) concluíram que a temperatura, o tempo de estocagem e meio
no qual os ovários são acondicionados influencia a preservação da morfologia e
viabilidade de folículos pré-antrais caninos. Os resultados obtidos nesse estudo
mostraram que os ovários podem ser estocados com sucesso em hipotermia, ou seja, a
4°C por no máximo 12 horas, em solução de NaCl 0,9% ou meio essencial mínimo
(MEM). Serafim et al. (2010) fizeram uso com sucesso desse protocolo para o
transporte de folículos pré-antrais caninos destinados ao cultivo de folículos isolados
por um longo período, confirmando sua eficiência.
De acordo com Kirklin & Barratt-Boyes (1993), a hipotermia tem demonstrado
ser um meio excepcional de proteção de qualquer órgão, com a queda exponencial da
taxa metabólica em cerca de 50%. Isso pode ser explicado devido à redução do
metabolismo seguido de uma diminuição da exigência de oxigênio e nutrientes assim

51
como da produção de metabólitos, que é criticamente importante para células em
isquemia.
2.2. ISOLAMENTO FOLICULAR
Outro ponto importante para o sucesso da MOIFOPA é o isolamento folicular,
que consiste na dissociação ou separação dos FOPA dos demais componentes do
estroma ovariano (fibroblastos, fibras de colágeno e elásticas, fibronectina, etc). Para
isso, podem ser utilizados procedimentos mecânicos associados ou não aos enzimáticos
(FIGUEIREDO et al., 2008).
O método enzimático de isolamento consiste na incubação de tecido ovariano
com pronase, colagenase ou desoxiribonuclease (DNAse), para dissociar e desagregar a
matriz de tecido seguido pela liberação dos folículos do estroma com auxílio de agulhas
(EPPIG & O‘BRIEN, 1996). Essa técnica permite isolar um grande número de
pequenos FOPA a partir de ovários mamíferos, e tem demonstrado ser dependente do
tempo de exposição e das concentrações enzimáticas utilizadas, tendo em vista que
podem provocar danos, como a degradação da membrana basal e a remoção das
camadas de células da teca, uma vez que essas estruturas apresentam os mesmos
constituintes que as enzimas degradam (NAYUDU et al, 2001; PARK et al., 2005).
Na tentativa de desenvolver um protocolo enzimático de recuperação de FOPA
caninos fazendo uso de DNAse (75 UI/ml) e colagenase (1000 UI/ml), Durrant et al.
(1998) submeteram tecidos ovarianos de cadelas a três diferentes protocolos. No
primeiro protocolo, os ovários foram submetidos ao processo enzimático e recuperação
dos FOPA dentro de 3horas após remoção dos ovários; no segundo protocolo, os
ovários inicialmente foram armazenados a 4°C por 18 à 24h, e só então submetidos à
digestão enzimática e isolamento folicular; já no terceiro protocolo, o tratamento
enzimático dos ovários foi realizado no dia da coleta, e em seguida, armazenado a 4°C
por 18h, para finalmente ser realizado o isolamento dos FOPAs. Apesar de terem sido
obtidos folículos em diversos estádios de desenvolvimento, as taxas de degeneração
foram elevadas em todos os protocolos avaliados.
A fim de contornar os problemas inerentes à utilização do processo enzimático,
métodos mecânicos têm sido utilizados. A técnica mecânica de isolamento de FOPA
permite recuperar uma maior quantidade de folículos intactos, sendo relativamente
barata simples e rápida (HARTSHORNE, 1997). Essa técnica já foi utilizado com

52
sucesso em ovários felinos (LIMA et al., 2003), bovinos (FIGUEIREDO et al.,1993;
HULSHOF et al.,1994), caprinos (LUCCI et al., 1999, SILVA et al., 2010) e ovinos
(AMORIM et al., 1998). Os FOPAs podem ser isolados mecanicamente através de
aparelhos como tissue chopper ou utilizando agulhas para o procedimento de
microdissecção. (FIGUEIREDO et al., 2008).
A microdissecção consiste em dissecar os FOPA de pequenos fragmentos do
córtex ovariano com auxílio de agulhas de fino calibre ou fórceps (GUTIERREZ et al.,
2000; PICTON, et al., 2003). Segundo KURVILA (2007), apesar da microdissecção
não ser um método adequado para se obter um grande número de folículos, esse tipo de
isolamento permite garantir a qualidade folicular, visto que mantém a integridade da
estrutura folicular, e da membrana basal, bem como as interações entre as células da
teca e da granulosa. Portanto, é um importante método para determinar as condições de
cultivo ideal para FOPA (RALPH et al., 1995).
A técnica de microdissecção já foi utilizada com sucesso em caprinos (SILVA et
al., 2010), camundongos (SPEARS et al., 1994; LIU et al., 2002), bubalinos
(SHARMA et al., 2009), ovinos (TAMILMANI et al., 2005; ARUNAKUMARI et al.,
2010), bovinos (GUTIERREZ et al., 2000), suínos (MAO et al., 2002), humanos (ABIR
et al., 1999) e caninos (LOPES et al., 2009). Serafim et al. (2010) isolaram
manualmente por meio de microdissecção, com auxílio de agulhas 26G acopladas a
seringas de 1 ml, FOPA secundários com diâmetro superior a 200 µm, e posteriormente
cultivaram por 18 dias. A viabilidade folicular foi avaliada ao término do cultivo,
através de marcadores fluorescentes para detecção de células vivas ou mortas, sendo
que 100% dos folículos encontravam-se viáveis e com estrutura morfológica.
2.3. CULTIVO
O sucesso do desenvolvimento folicular in vitro depende do sistema de cultivo
empregado, das condições de cultivo e o meio utilizado (HARTSHORNE, 1997).
Basicamente existem dois sistemas de cultivo de FOPA, o cultivo in situ e o
isolado. No primeiro deles, os folículos são cultivados inclusos no fragmento ovariano
ou no ovário inteiro; tendo essa técnica, a vantagem de manter a estrutura folicular e ter
uma maior praticidade. Enquanto que no segundo sistema, o cultivo isolado, os folículos

53
são cultivados isoladamente sobre a placa ou um substrato, caracterizando um sistema
bidimensional, ou inserido no substrato, caracterizando o sistema tridimencional. O
cultivo de FOPA isolados permite uma maior perfusão do meio bem como a
possibilidade de cultivar individualmente folículos em diferentes categorias (primordial,
primário, secundário) (HARTSHORNE, 1997; FIGUEIREDO et al., 2008).
Bolamba et al., (1998) e Bolamba et al.,(2002) avaliaram a maturação nuclear de
oócitos caninos derivados de folículos cultivados in vitro em placas de plástico de 24
poços revestidas com 0,6% de ágar estéril, para evitar a perda de células da granulosa.
Os resultados mostraram a progressão oocitária para os estádios de metáfase I e II,
porém os percentuais de maturação foram muito baixos e o desenvolvimento folicular
não foi avaliado.
De acordo com Isobel & Terada (2001), um dos sistemas de mais comumente
utilizado para diferentes espécies, seja para o cultivo de folículos ou de oócitos, é o
sistema de gotas de meio cobertas com óleo mineral. Neste sistema, o principal fator
biofísico é representado pela proporção de volume de meio em relação ao número de
estruturas por gota. Tal sistema também tem sido utilizado com sucesso para o cultivo
de FOPA de caprinos (SILVA et al., 2010) e ovinos (ARUNAKUMARI et al., 2010).
Segundo Silva et al. (2004b), no desenvolvimento de um sistema de cultivo
eficiente também é essencial que ocorra um controle irrestrito de todas variáveis que
possam afetar direta ou indiretamente o desenvolvimento folicular, tais como,
temperatura, CO2, tensão de oxigênio, controle de contaminações fúngicas e
bacterianas, e fatores referentes ao meio de cultivo, como a presença de nutrientes e
suplementos.
Além disso, a composição do meio é uma importante ferramenta para a obtenção
do sucesso do cultivo de FOPA in vitro, visto que, é no meio de cultivo que os folículos
encontrarão os subsídios necessários para dar suporte ao seu crescimento (TELFER et
al., 2000). Assim, muitos autores têm investigado o efeito de vários componentes no
meio de cultivo in vitro de FOPA, tanto de animais de laboratórios, como animais
domésticos (LIU et al., 2002; DANFORTH et al., 2003).

54
2.4. IMPORTÂNCIA DO MEIO NO CULTIVO IN VITRO DE
FOLÍCULOS PRÉ-ANTRAIS CANINOS
Existem poucos relatos sobre o cultivo in vitro de FOPA caninos. Dessa forma,
ainda não foi desenvolvido um protocolo padrão ou um sistema de cultivo definido a ser
utilizado nessa espécie (SERAFIM et al., 2010). Os meios de cultivo utilizados em cães
são adaptações dos empregados para as outras espécies, tendo geralmente como meio de
base o TCM199, MEM, Meio Essencial Mínimo Dulbelco‘s (DMEM), fluido sintético
ovidutário (SOF) dentre outros (GUÉRIN, 1998). Bolamba et al. (1998) realizaram o
cultivo de FOPA caninos avançados em DMEM suplementado, e concluíram que é
possível obter retomada de meiose em oócitos recuperados de FOPA avançados. Ainda
em caninos, Bolamba et al. (2002), observaram que o meio SOF pode suportar a
maturação nuclear de oócitos oriundo de FOPA avançados cultivados por 72h sem a
presença de suplementação protéica. Os autores afirmam que a presença de Soro fetal
bovino (SFB) ou Albumina Sérica Bovina (BSA), não melhoraram as taxas de
maturação. Apesar disso, estudos têm demonstrado que a suplementação do meio de
cultivo com BSA ou SFB é benéfica por ser uma importante fonte de proteína, suprindo
as necessidades de aminoácidos essenciais, fornecendo energia oriunda de carboidratos,
funcionando também como uma fonte de íons dióxido de carbono para manutenção de
um pH ideal, além de favorecer a captação de fatores de crescimento (HEWITT &
ENGLAND, 1999).
Além da adição dessas fontes proteicas, é aconselhável a suplementação com
compostos antioxidantes como a combinação de insulina, transferrina e selênio (ITS) e
o ácido ascórbico (AUGUSTIN et al., 2003; LUVONI et al., 2004). Os antioxidantes
têm papel fundamental na redução dos danos causados pela ação de espécies reativas de
oxigênio e complexos metal-oxigênio ao DNA, proteínas, carboidratos, lipídios e
membranas celulares (SIES et al., 1992).
A adição de outras substâncias como fatores de crescimento e/ou hormônios
incluindo gonadotrofinas como o FSH (HARTSHORNE, 1997), também tem sido
realizado. Serafim et al. (2010) realizaram o cultivo in vitro de folículos pré-antrais
isolados caninos em meio MEM suplementado (α-MEM), na ausência ou presença de
diferentes concentrações e formas de adição de FSH. Os autores comprovaram que o
FSH adicionado de forma seqüencial (100, 500 e 1000 ng/ml, nos dias 0, 6 e 12 de

55
cultivo respectivamente) beneficiou diretamente o cultivo, promoveu a manutenção da
sobrevivência folicular, além de aumentar as taxas de crescimento folicular e formação
de antro.
Entretanto, maiores estudos são necessários para se determinar quais os
substâncias devem ser adicionadas ao meio de cultivo dos FOPA caninos, de modo a
manter a viabilidade, crescimento e maturação de oócitos inclusos em FOPA para
posterior PIV de embriões. Os principais avanços relativos à PIV de embriões caninos
serão abordados a seguir
2. PRODUÇÃO in vitro (PIV) EM CADELAS
As diferenças na fisiologia canina quando comparadas aos demais animais
domésticos como os bovinos, dificultam a extrapolação, para espécie canina, de
protocolos desenvolvidos para ruminantes. O primeiro entrave na PIV de embriões
caninos é a MIV e por esta razão, vários estudos têm sido realizados para testar
diferentes condições de cultivo, no entanto, os resultados da maturação nuclear ainda
permanecem baixos (SANTOS et al., 2006; RODRIGUES et al., 2009).
A produção in vitro de embriões envolve as etapas de: maturação, fecundação e
cultivo in vitro que serão abordadas a seguir.
2.1.MATURAÇÃO IN VITRO
A MIV é um processo complexo, cujo objetivo é mimetizar as alterações
dinâmicas que ocorrem no folículo ovariano durante o período pré-ovulatório. A
ativação do oócito ocorre com o desenvolvimento do folículo, no interior do ovário com
um preciso controle físico e hormonal. No cultivo in vitro, o oócito é submetido a uma
série de injúrias que podem interferir neste processo, prejudicando assim, o seu
desenvolvimento e competência meiótica, bem como a sua utilização para a FIV no
futuro (FIGUEIREDO et al.,2008; RODRIGUES et al., 2009).
Protocolos para MIV na espécie canina, com o objetivo de promover a
competência dos complexos cumulus oophorus (CCOs) para a fecundação e clivagem,
ainda não foram completamente desenvolvidos. As principais razões da falha no cultivo

56
residem em identificar os suplementos necessários e suas concentrações ideais para o
desenvolvimento oocitário (FARSTAD, 2000; RODRIGUES et al., 2003). Diante disso,
alguns estudos têm avaliado tanto as etapas iniciais, levando em consideração desde a
fase do ciclo estral em que a fêmea se encontra, até a coleta e transporte dos ovários,
bem como o sistema de cultivo utilizado, avaliando principalmente o efeito da adição ou
não de várias substâncias aos meios de cultivo (LUVONI et al., 2004).
Para Rodrigues e Rodrigues (2003), a MIV não é influenciada pelo estádio do
ciclo reprodutivo da fêmea doadora de oócitos, não apresentando diferença significativa
entre fases em que se encontrem as fêmeas, seja folicular, luteal ou mesmo em anestro.
Entretanto, Lopes et al. (in press) confirmaram a existência de diferenças marcantes na
morfologia ultra-estrutural de oócitos caninos de grau I, oriundos de folículos antrais
coletados de cadelas em diferentes fase do ciclo estral (anestro e diestro) como
diferenças na localização nuclear que são indicativas de estádios finais do
desenvolvimeto oócitário, e na quantidade e distribuição de organelas no citoplasma.
Já Evecen et al. (2010), avaliando o efeito do estádio do ciclo estral e as
temperaturas de transporte dos ovários sobre a MIV de oócitos caninos, obtiveram uma
maior taxa de retomada da meiose (metáfase I e II, 62,9%) a partir de oócitos de ovários
colhidos em anestro e transportados à 4°C, enquanto que uma menor taxa de MIV foi
obtida com o transporte dos ovários a 37 °C, em todas as fases do ciclo estral.
Sabe-se que a eficiência da MIV é afetada diretamente durante o transporte e
estocagem dos ovários, influenciada principalmente pela temperatura do meio em que
esses ovários são acondicionados (NAKAO & NAKATSUJI, 1992). Estudos anteriores
têm mostrado resultados controversos acerca da temperatura ideal de transporte. Na
espécie canina, Bolamba et al. (2002) e Tas et al. (2006), verificaram que o transporte
realizado por 2, 4 ou 24h a 4°C tem efeito positivo ou não afeta as taxas de MIV canina.
Já Lee et al. (2006), observaram uma redução significativa na viabilidade folicular após
o transporte a 4°C por 5h.
Outros estudos têm relatado que a manutenção da viabilidade de oócitos de cães
domésticos é maior sob temperaturas de transporte mais próximas da temperatura
corporal, independentemente do meio ou método de transporte (HANNA et al., 2008).
Isso ocorre, possivelmente, como consequência do maior teor de lipídios presentes no
oócito canino, que resulta em uma maior intolerância à temperatura. Tal fato é

57
observado com mesma intensidade nos oócitos de suínos, que também apresentam
lipídios no citoplasma, e sensibilidade às baixas temperaturas durante o transporte (KIM
et al., 2006). Já a composição dos meios de transporte é variável de acordo com o autor,
sendo os meios mais comumente utilizados a solução salina ou o PBS acrescidos de
antibióticos na temperatura de 35 a 39°C (HAY et al., 1997; OTOI et al., 2000).
O oócito canino necessita ter determinadas características morfológicas
específicas para ser selecionado à MIV, devendo possuir um diâmetro superior a 120
µm, pigmentos enegrecidos, citoplasma homogêneo e ser completamente cercado por
duas ou mais camadas de células do cumulus, apresentando dessa forma competência
para os estádios finais da maturação nuclear (SAINT-DIZIER et al., 2001; KIM et al.,
2006).
No tocante ao tempo necessário para o cultivo in vitro, experimentos realizados
com oócitos caninos submetidos à MIV revelaram similaridade com os eventos
manifestados in vivo, confirmando, portanto, a necessidade de cultivo dos gametas
femininos por um período situado ao redor de 48-72 horas (FULTON et al., 1998).
O meio de maturação utilizado para a maioria dos estudos é o TCM199,
modificado ou não, sob condições de temperatura e umidade variáveis (HEWITT &
ENGLAND, 1999). Rota & Cabianca (2004) demonstraram que o TCM199 apresentou
resultados superiores ao SOF, em relação ao número de oócitos que retomaram a meiose
e desenvolveram-se até os estádios de metáfase I (MI), anáfase I (AI) e metáfase II
(MII). Entretanto, Saikhun et al. (2008) observaram que não havia diferença
significativa entre os meios de cultivo complexos (TCM199, DMEM/F12) e simples
(SOF, HAM-F10) com relação à retomada da meiose ou maturação nuclear de oócitos
de cadela.
A suplementação do meio de cultivo também é um importante fator a ser
avaliado, e pode ser realizado com a adição de algumas substâncias que visem favorecer
a progressão da maturação até o estádio de MII. (NICKSON et al., 1993; RODRIGUES
& RODRIGUES, 2003). Inúmeros suplementos já foram testados no meio de MIV
canina, dentre eles o fator de crescimento epidermal (EGF) (CUI et al, 2006, SONG et.
al., in press), estrógeno associado ao EGF (HATOY et al., 2009), fator de crescimento
semelhante à insulina (IGF-I) (OLIVEIRA et al., 2009), SFB ou BSA (HEWITT &
ENGLAND, 1997; BOLAMBA et al., 2002), SOF(HEWITT & ENGLAND, 1999),

58
gonadotrofina coriônica humana (hCG) (DE LOS REYES et al., 2005), estradiol e
progesterona (KIM et al, 2005), somatotrofina humana (hST) (RODRIGUES et al.,
2003), água de coco em pó (ACP-318®) (SILVA et al., 2009), polivinil-pirrolidona
(PVA) (SANTOS et al., 2006), associações de ITS (ROTA & CABIANCA., 2004),
hormônio do crescimento (GH) (CHIGIONI et al., 2008), dentre muitos outros.
Entretanto, os resultados ainda apresentam-se limitados, não fornecendo uma taxa
satisfatória de oócitos que possam ser destinados à FIV e à PIV.
A grande maioria dos estudos com MIV de oócitos caninos oriundo de folículos
antrais, não reportam taxas de maturação nuclear (MII) superiores a 15-20% (RIBEIRO
et al, 2010). Dessa forma, novos estudos devem ser realizados no intuito de melhorar as
taxas de MIV nesta espécie (SERAFIM et al., 2010). O cultivo in vitro de oócitos
inclusos em folículos pré-antrais se apresenta, então, como alternativa para ampliar os
estudos em caninos possibilitando futuramente o fornecimento de um grande número de
oócitos maturos a serem destinados para esta finalidade.
2.2. FECUNDAÇÃO IN VITRO
A FIV é uma das importantes etapas da PIV de embriões. O teste crucial que
avalia o sucesso da MIV é a habilidade do oócito de dar continuidade ao
desenvolvimento embrionário após a penetração espermática e a fecundação. Como
conseqüência dos resultados limitados da maturação oocitária canina, o uso da FIV
nessa espécie ainda não se apresenta eficiente (RODRIGUES & RODRIGUES, 2006).
Os primeiros relatos com FIV em cães foram descritos por Mahi &
Yanagimachi em 1978, onde foram observadas taxas de 20 a 30% de fecundação,
julgados pela presença de espermatozóide dentro dos oócitos. Entretanto, a fecundação
é caracterizada através da observação de penetração e fusão do espermatozóide ao
oócito, extrusão do 2º corpúsculo polar ou formação de pró-núcleo masculino e
feminino no ooplasma (POPE et al., 1993). Logo em seguida, Renton et al. (1991) e
Yamada et al. (1993) obtiveram o sucesso comprovado da FIV canina com o
desenvolvimento embrionário até o estádio de blastocisto. Já Songasasen et al. (2002)
inseminaram oócitos cultivados in vitro, e encontraram 34% desses oócitos penetrados
por espermatozóides. Fecundações normais, julgadas pela presença de dois pró-núcleos

59
e uma única cauda de espermatozóide dentro do citoplasma somente foram observadas
em 4% dos oócitos.
Os protocolos de FIV canina são geralmente adaptados daqueles utilizados
rotineiramente na espécie bovina, fazendo uso do sêmen fresco ou congelado. Do ponto
de vista do esperma, o sucesso da clivagem requer o recrutamento e seleção da
população espermática, bem como uma concentração adequada para fecundar os oócitos
in vitro. Variações temporais na co-incubação de espermatozóide-oócito mostraram
penetração e incorporação do espermatozóide ao ooplasma, e formação de pró-núcleos,
iniciados algumas horas após a fecundação. Uma exposição em torno de 24 h ao
espermatozóide foi necessária para se obter 10-25% de taxa de penetração oócitária
(RODRIGUES et al., 2003). O co-cultivo espermatozóide-oócito geralmente ocorre em
um período de 12 a 24h, após o qual os presumíveis zigotos formados são avaliados e
aqueles de boa qualidade são transferidos para gotas de meio de CIV (SAIKHUN et al.,
2008).
O co-cultivo da FIV canina geralmente é realizado em meio de capacitação
canino (CCM) desenvolvido por Mahi e Yanagimachi (1978), ou o Fert-TALP
adaptado, suplementado com heparina, sendo este muito semelhante ao CCM. Tais
meios são suficientes para capacitar de forma satisfatória o espermatozóide canino,
suprimindo a necessidade de uma capacitação prévia, especialmente quando se trata do
espermatozóide canino criopreservado (KAWAKAMI et al., 1993; DE LOS REYES et
al. , 2006; SAIKHUN et al., 2008).
2.3.CULTIVO IN VITRO DE EMBRIÕES
A PIV de embriões não tem apresentado sucesso e aplicabilidade na espécie
canina. As taxa de desenvolvimento embrionário são ainda muito baixas tendo sido
obtidos embriões no estádio de 2-12 células (SONGASASEN et al., 2002); oito células
(YAMADA et al., 1992; RODRIGUES & RODRIGUES, 2007), 8-16 células
(SAIKHUN et al., 2008) ou blastocisto (OTOI et al., 2000). Neste último estudo, dos
217 oócitos que foram inseminados, apenas um blastocisto foi produzido. Hatoya et al.
(2006) afirmam que embriões co-cultivados com fibroblastos embrionários caninos
(CEF), desenvolveram-se até estádio de 16 células. Dos 147 embriões co-cultivados

60
com fibroblastos embrionários de camundongo, um desenvolveu-se até o estádio de
mórula.
Até o presente momento, o mais surpreendente sucesso do CIV canino foi
obtido por England et al. (2001). Os autores obtiveram oócitos de cadelas em estro, os
maturaram por 24-72h e em seguida realizaram a FIV. Foram transferidos 88 embriões
de uma ou duas células para as receptoras, e foi relatada uma única gestação por ultra-
sonografia no dia 20 após a transferência. No entanto, a gestação foi perdida dois dias
mais tarde por reabsorção.
De acordo com Rodrigues et al. (2006), poucos estudos enfatizam a
influência das condições de cultivo de embriões caninos. Um aspecto fundamental diz
respeito à influência da temperatura de cultivo para o desenvolvimento embrionário. Os
embriões caninos são comumente produzidos na temperatura padronizada dos
programas de reprodução in vitro (37 a 39 °C), já tendo sido relatado anteriormente que
as temperaturas elevadas têm um efeito deletério sobre o crescimento do oócito, a
síntese de proteínas ou a formação de transcrições necessárias para posterior
desenvolvimento embrionário.
No entanto, ainda não se sabe como os suplementos adicionados ao meio
afetam o desenvolvimento in vitro do embrião canino. Rodrigues et al. (2007)
observaram um bloqueio da clivagem dos embriões em torno do estádio 6-8 células, que
pode ter sido resultado tanto da maturação incompleta dos oócitos, como também
devido às condições insuficientes do meio de cultivo em sustentar o desenvolvimento
embrionário. Saikhun et al. (2008), realizaram um comparativo entre o CIV de embriões
em meio TCM199 e meio SOF, comprovando que o meio SOF fornece um maior
suporte para o desenvolvimento embrionário canino. Entretanto, mais estudos ainda são
necessários para identificar as condições ótimas para o CIV embrionário na espécie
canina.
3. CONSIDERAÇÕES FINAIS
Muitos estudos ainda precisam ser realizados na tentativa de se esclarecer os
complexos processos envolvidos no crescimento e maturação dos oócitos caninos,
visando melhorar a produção in vitro de embriões e, posteriormente, a obtenção de crias
até o presente não relatado na literatura.

61
Neste contexto, dentre as biotécnicas temos a MOIFOPA, que surge como uma
importante ferramenta para se obter uma população mais homogênea de oócitos, com
objetivo de otimizar os resultados da MIV e FIV, e garantir um número suficiente de
oócitos a serem destinados a transferência de embriões. A realização desses estudos
resultará em novas abordagens sobre a reprodução assistida canina, que poderão ser
aplicadas não somente aos animais de companhia, como aos cães de alto valor
comercial na cinofilia, e também aos canídeos selvagens ameaçados de extinção.
4. REFERÊNCIAS
ABIR R.; ROIZMAN P.; FISCH B.; NITKE S.; OKON E.; ORVIETO R.; BEN R.Z.
Pilot study of isolated early human follicles cultured in collagen gels for 24 h. Hum
Reprod, v.14,n. 5, p.1299 –301, 1999.
AMORIM, C.A.; RODRIGUES, A.P.R.; LUCCI,C.M.; FIGUEIREDO, J.R.;
GONÇALVES, P.B.D. Mechanical method for isolation of preantral follicles from adult
ovine ovaries, Arq. Fac. Vet., v. 26, n.1, p. 215, 1998.
ARUNAKUMARI G.; SHANMUGASUNDARAM N.; RAO V.H. Development of
morulae from the oocytes of cultured sheep preantral follicles; Theriogenology v.74,
n.5, p.884–894, 2010.
AUGUSTIN, R.; POCAR, P.; WRENZYCKI, C.; NIEMANN, H.; FICHER, B.;
Mitogenic and anti-apoptotic activity of insulin on bovine embryos produced in vitro.
Reproduction, v.126, p.91-99, 2003.
BOLAMBA D.; BORDEN-RUSS K.D.; DURRANT B.S.; In vitro maturation of
domestic dog oocytes cultures in advanced preantral and early antral follicles.
Theriogenology. v.49, p.933-942, 1998.
BOLAMBA, D.; BORDEN-RUSS, K.D.; DURRANT, B.S. In vitro maturation of bitch
oocytes from advanced preantral follicle in synthetic oviduct fluid medium: serum is not
essencial. Theriogenology, v.58, p.1689-1703, 2002.

62
CHIGIONI, S.; SECCHI, C.; BORROMEO, V.; MODINA S.; BERETTA, M. S.;
LUVONI G. C. Effects of growth hormone on oocyte in vitro maturation and its
localization in the canine cumulus-oocyte complexes. Vet Res Commun. v. 32, p.131–
134. 2008.
CUI, X.S.; JIN Y.X.; SHEN, X.H.; LEE , J. Y.; LEE H.S.; YIN X.J.; KONG I.K.; KIM,
N.H. Epidermal growth factor enhances meiotic resumption of canine oocytes in the
presence of BSA. Theriogenology. v.66, p.267–274, 2006.
DANFORTH, D.R.; ARBOGAST, L.K.; GHOSH, S.; DICKERMAN, A.; ROFAGHA,
R.; FRIEDMAN, C.I. Vascular Endothelial Growth Factor Stimulates Preantral Follicle
Growthin the Rat Ovary, Biology of Reproduction, v.68, p.1736–1741, 2003.
DE LOS REYES M.; CARRION R.; BARROS C. In vitro fertilization of in vitro
matured canine oocytes using frozen–thawed dog semen. Theriogenology. v.66,
p.1682–1684, 2006.
DE LOS REYES M.; LANGEA J. D.E.; MIRANDA P.; PALOMINOS J.; BARROS C.
Effect of human chorionic gonadotrophin supplementation during different culture
periods on in vitro maturation of canine oocytes. Theriogenology. v.64, p.1–11, 2005.
DURRANT B.S.; PRATT N.C.: RUSS K.D.; BOLAMBA D. Isolation and
characterization of canine advanced, preantral and early antral follicles.
Theriogenology. v.49, p.917-932, 1998.
ENGLAND, G.C.W.; VERSTEGEN, J.P.; HEWITT, D.A. Pregnancy following in vitro
fertilization of canine oocytes. Veterinary Record. v.148,p.20-22, 2001.
EPPIG J. J., O'BRIEN M. J. Development In Vitro of Mouse Oocytes from Primordial
Follicles. Biology of reproduction. v.54, p.197-207, 1996.
EVECEN M.; CIRIT U.; DEMIRA K., OZDAS O.B.; TAS, M.; BIRLER S.,
PABUCCUO S. Effects of estrous cycle stage and transport temperature of ovaries on in
vitro maturation of canine oocytes. Animal Reproduction Science. v.117, p.160–165,
2010.
FARSTAD, W. Current state in biotechnology in canine and feline reproduction.
Animal Reproduction Science. v.61, p.375-387, 2000.

63
FIGUEIREDO J R.; Essential role of follicle stimulating hormone in the maintenance of
caprine preantral follicle viability in vitro. Zygote;v.15, p.173– 82, 2007.
FIGUEIREDO, J.R.; HULSHOF, S.C.J.; VAN DEN HURK, R.; ECTORS, F. J.;
FONTES, I.L.L.; NUSGENS, B.; BEVERS, M.M.; BECKERS, J.F. Development of a
new mechanical and enzymatic method for the isolation of intact preantral follicles from
fetal, calf and adult bovine ovaries. Theriogenology. v.40, p.789-799, 1993.
FIGUEIREDO, J.R.; RODRIGUES, A. P. R.; LOPES, C. A. P. Importância do ovário
artificial na reprodução medicamente assistida e bem-estar animal, Revista CFMV, Ano
XV, n.45, p.72-75, 2009.
FIGUEIREDO, J.R.; RODRIGUES, A.P.R.; AMORIM, C.A.; SILVA, J.R.V.
Manipulacão de Oocitos Inclusos em Foliculos Ovarianos Pre-antrais – MOIFOPA. In:
GONCALVES, P.B.D.; FIGUEIREDO, J.R.; FREITAS, V.J.F. (Eds). Biotécnicas
Aplicadas à Reprodução Animal. São Paulo, Roca, 2008, p.303-327.
FULTON, R.; DURRANT B.; KESKINTEPE L.; BRANDON C.; FAYRER-HOSKEN
R. Assisted reproductive techniques for the treatment of canine infertility. Society for
Theriogenology Newsletter. v.21, p.2-7, 1998.
GUÉRIN C. Fecondation in vitro chez la chienne.Ou en est-on? Pratique Médicale et
Chirurgicale des.Animaux de Compagnie. v.33, p.151-161, 1998.
GUPTA P.S.P.; NANDI,S.; RAVINDRANATHA, B.M. ; SARMA, P.V. In vitro
culture of buffalo (Bubalus bubalis) preantral follicles.Theriogenology. v.57, p.1839-
1854, 2008.
GUTIERREZ, C.G., RALPH, J.H., TELFER, E.E., WILMUT, I.; WEBB, R. Growth
and antrum formation of bovine preantral follicles in long-term culture in vitro, Biol
Reprod. v.62, p.1322–1328, 2000.
HANNA, C.; LONG C.; HINRICHS K.; WESTHUSIN M.; KRAEMER D. Assessment
of canine oocyte viability after transportation and storage under different conditions.
Anim Reprod Sci, v.105, p.451–456, 2008.
HARTSHORNE, G. In vitro culture of ovarian follicles. Rev Reprod, v.2, p.94-104,
1997.

64
HATOYA, S.; SUGIYAMA, Y.; NISHIDA, H.; OKUNO, T.; TORII, R.; SUGIURA,
K.; KIDA, K.; KAWATE, N.; TAMADA, H.; INABA, T. Canine oocyte maturation in
culture: Significance of estrogen and EGF receptor gene expression in cumulus cells,
Theriogenology. v.71, p.560–567, 2009.
HATOYA, S.; SUGIYAMA, Y.; TORII, R.; WIJEWARDANA, V.; KUMAGAI, D.;
SUGIURA, K.; KIDA, K.; KAWATE, N.; TAMADA, H.; SAWADA, T.; INABA, T.
Effect of co-culturing with embryonic fibroblasts on IVM, IVF and IVC of canine
oocytes. Theriogenology. v.66, p.1083–1090, 2006.
HAY, M.A.; KING, W.A.; GARTLEY, C.J.; LEIBO, S.P.; GOODROWE, K.L. Canine
spermatozoa cryopreservation and evaluation of gamete interaction. Theriogenology.
v.48, p.1329–42, 1997.
HEWITT, D.A.; ENGLAND, G.C.W. The canine oocyte penetration assay; its use as an
indicator of dog spermatozoal performance in vitro. Animal Reproduction Science. v.50,
p.123–139, 1997.
HEWITT, D.A.; ENGLAND, G.C.W. Influence of gonadotrophin supplementation on
the in vitro maturation of bitch oocytes. Veterinary Record. London. v.144, p.237-239,
1999.
HULSHOF, S.C.J.; FIGUEIREDO, J.R.; BECKRES, J.B.; BEVERS, M.M.; VAN DER
DONK, J.A.; VAN DEN HURK, R. Effects of fetal bovine serum, FSH and 17-β-
estradiol on the culture of bovine preantral follicles. Theriogenology. v.44, p.217– 226,
1995.
ISOBEL N.; TERADA T. Effect of the factor inhibiting germinal vesicle breakdown on
the disruption of gap junctions and cumulus expansion of pig cumulus–oocyte
complexes cultured in vitro. Reproduction. v.121, p.249–257, 2001.
KAWAKAMI, E.; VANDEVOOT, C.A.; MAHI, C.A.; OVERSTREET, J.W. Induction
of acrosome reaction of canine sperm by homologous zona pellucida. Biol. Reprod.
v.48, p.841–845, 1993.
KIM, M.K.; FIBRIANTO, Y. H.; OH, H.J.; JANG, G.; KIM, H.J.; LEE, K.S.; KANG,
S.K.; LEE, B.C.; HWANG, W.S. Effects of estradiol-17b and progesterone

65
supplementation on in vitro nuclear maturation of canine oocytes. Theriogenology, v.63,
p.1342–1353, 2005.
KIM, H.J.; CHOI, S.H.; SON, D.S.; CHO, S.R..; CHOE, C.Y.; KIM, Y.K.; HAN, M.H.;
RYU, I.S.; KIM, I.C.; KIM, I.H.; IM, K.S.; NAGAI, T. Effect of exposure duration of
ovaries and oocytes at ambient temperature on parthenogenetic development of porcine
follicular oocytes. J. Reprod. Dev. v.52, p.633–638, 2006.
KIRKLIN, J.W.; BARRATT-BOYES, B.G. Cardiac Surgery; Churchill Livingstone,
New York, USA, 1993, p. 91.
KURVILA, A.; Cloning and sequencing of FSH receptor gene from buffalo preantral
follicles, 2007. M.V.Sc. thesis submitted to division of physiology and climatology,
Deemed University, IVRI, Izatnagar.
LEE, H.S.; YIN, X.J.; KONG, I.K. Sensitivity of canine oocytes to low temperature.
Theriogenology. v.66, p.1468–1470, 2006.
LIMA, G.L.; COSTA, L.L.M.;. CAVALCANTI, D.M.L.P.; RODRIGUES, C.M.F;
FREIRE, F.A.M.; FONTENELE-NETO, J.D.; SILVA, A.R. Short-term storage of
canine preantral ovarian follicles using a powdered coconut water (ACP1)-based
medium. Theriogenology. v.74, p.146–152, 2010.
LIMA, A. K. F.; BEZERRA, M. B. ; OLIVEIRA, L. C. ; FIGUEIREDO, J. R. ; SILVA,
L. D. M. Isolamento e caracterização de folículos ovarianos pré-antrais em gatas
domésticas (Felis catus). Revista Brasileira de Reprodução Animal. v.27, p.396-397,
2003.
LIU, H.; HE, Z.; ROSENWAKS, Z. In vitro culture and in vitro maturation of mouse
preantral follicles with recombinant gonadotropins. Fertility and sterility. v.77, p.373-
83, 2002.
LOPES, C.A.P.; SANTOS, R.R.; CELESTINO, J.J.H.; MELO, M.A.P.; CHAVES, R.
N.; CAMPELLO, C.C.; SILVA, J.R.V.; BAO, S.N.; JEWGENOW, K., FIGUEIREDO,
J.R. Short-term preservation of canine preantral follicles: Effects of temperature,
medium and time. Animal Reproduction Science. v.115, p.201–214, 2009.

66
LOPES, G.; ROCHA A.; SOUSA M. Comparative ultrastructural analysis of diestrous
and anestrous canine Grade 1 cumulus-oocyte complexes. Anim. Reprod. Sci., in press.
2010.
LUCCI, C.M., AMORIM, C.A., RODRIGUES, A.P.R.; FIGUEIREDOA J.R.; BÁO
S.N.;. SILVA J.R.V; GONÇALVES, P.B.D. Study of preantral follicle population in
situ and after mechanical isolation from caprine ovaries at different reproductive stages.
Anim. Reprod. Sci. v.56, p.223-236, 1999.
LUVONI, G. C., CHIGIONI, S., ALLIEVI, E., MACIS, D. Factors involved in vivo
and in vitro maturation of canine oocytes. Theriogenology. v. 63, p.41–59, 2004.
MAHI, C.A.; YANAGIMACHO, R. Capacitacion, acrosome reaction, and egg
penetration by canine spermatozoa in a simple defined medium. Gamete Res. v1, p.101-
109, 1978.
MAO, J.; WU, G.; SMITH, M.F.; MCCAULEY, T.C.; CANTLEY, T.C.; PRATHER,
R.S.; DIDION, B.A.; DAY, B.N. Effects of culture medium, serum type, and various
concentrations of follicle-stimulating hormone on porcine preantral follicular
development and antrum formation in vitro. Biol Reprod. v.67, p.1197–203, 2002.
NAKAO, H.; NAKATSUJI, N. Effect of storage conditions of bovine ovaries and
oocytes on the success rate of in vitro fertilization and culture. J. Reprod. Dev. v.38,
p.11–13, 1992.
NAYUDU, P. L.; FEHRENBACHA, A.; KIESELA, P.; VITTA. U.A.;
PANCHARATNAA, K. S. Progress toward understanding follicle development in
vitro, Arch Med Res. v.32, n.6, p.587-594, 2001.
O‘BRIEN, M. J.; PENDOLA, J. K.; EPPIG, J. J. A revised protocol for in vitro
development of nouse oocytes from primordial follicles dramatically improves their
developmental competence. Biology of Reproduction. v.68, p.1682–1686, 2003.
OLIVEIRA, K. S.; MACHADO, M. A.; TONIOLLO, G. H. Avaliação do fator de
crescimento IGF-I na maturação in vitro de oócitos caninos no meio tcm 199, Ciência
Animal Brasileira. v.10, p.560-567, 2009.

67
OTOI, T.; SHIN, T.; KRAEMER, D.C.; WESTHUSIN, M.E. Role of cumulus cells on
in vitro maturation of canine oocytes. Reprod Domest Anim. V.42, p.184–9, 2007.
OTOI, T.; MURAKAMI, M.; FUJII, M.; TANAKA, M.; OOKA, A.; UNE, S.;
SUZUKI T. Development of canine matured and fertilized in vitro. Veterinary Record.
v.146, p.52-53, 2000.
PARK, K.S.; LEE, T.H.; PARK, Y.K.; SONG, H.B.; CHUN, S.S. Effects of isolating
methods (mechanical or enzymatical) on structure of pre-antral follicles in mouse. J
Assisted Reprod Genet. v.22, p.9–10, 2005.
PICTON, H.M.; DANFOUR, M.; HARRIS, S.E.; CHAMBERS, E.; HUNTRISS, J. In
vitro growth and maturation of oocyte. Reproduction Supplement, v.61, p.445–462
2003.
POPE, C.E.; KELLER, G.L.; DRESSER, B.L. In vitro fertilization in domestic and non-
domestic cats including sequences of early nuclear events, developments in vitro,
cryopreservation and successful intra- and interspecies embryo transfer. J. Reprod.
Fertil. Suppl. v.47, p.189-201, 1993.
RALPH, JI-I.; WILMUT, I.; TELFER, E.E. In vitro growth of bovine preantral follicles
and the influence of FSH on follicular and oocyte diameters. J Reprod Fert. Abstract
series, v.15, p.6, 1995.
RENTON, J.P.; BOYD, J.S.; ECKERSALL, P.D.; PERRY, B. Ovulation and early
embryogenic development in the bitch (Canis familiaris). Journal of Reproduction and
Fertility. v.93, p.221-231, 1991.
RIBEIRO, A.P.C.; PIRES, E.A.; APPARÍCIO, M.; ALVES, A.E.; CARARETO, R.;
VICENTE, W.R.R. Maturação in vitro de oócitos caninos: aspectos fisiológicos e sua
relação com a evolução da técnica. Rev. Bras. Reprod. Anim. v.34, p.50-57, 2010.
RODRIGUES, B. A; SANTOS, L.C.; RODRIGUES, J.L. Effect of maturation medium
on in vitro cleavage of canine oocytes fertilized with fresh and cooled homologous
semen. Zygote. v.15, p.43–53, 2007.
RODRIGUES, B.A.; SILVA, A.E.F.; RODRIGUEZ P.; CAVALCANTE L.F.;
RODRIGUES J.L. Cumulus cell features and nuclear chromatin configuration of in

68
vitro matured canine COCs and the influence of in vivo serum progesterone
concentrations of ovary donors. Zygote. p.1-13, 2009.
RODRIGUES, B.A.; RODRIGUES, J.L. Meiotic Response of In vitro matured Canine
Oocytes under different proteins and heterologous hormone supplementation. Reprod
Dom. Anim. v.38, p.58–62, 2003.
RODRIGUES, B.A.; RODRIGUES, J.L. Responses of canine oocytes to in vitro
maturation and in vitro fertilization outcome. Theriogenology. v.66, p.1667–1672, 2006.
ROTA, A.; CABIANCA, G. In vitro maturation rates of canine oocytes from anoestrous
bitches in simple media. Reprod. Nutr. Dev. v.44, p.105–109, 2004.
SAIKHUN, J.; SRIUSSADAPORN, S.; THONGTIP, N.; PINYOPUMMIN, A.;
KITIYANANT Y. Nuclear maturation and development of IVM/IVF canine embryos in
synthetic oviductal fluid or in co-culture with buffalo rat liver cells. Theriogenology.
v.69, p.1104–1110, 2008.
SAINT-DIZIER, M.; RENARD J.P.; CHASTANT-MAILLARD, S. In vitro maturation
of bitch oocytes: effect of sperm penetration. Journal of Reproduction and Fertilility.
Supplement. v.57, p.147-150, 2001.
SANTOS, L. C.; RODRIGUES, B. A.; RODRIGUES, J. L. In vitro nuclear maturation
of bitch oocytes in the presence of polyvinyl-pyrrolidone. Anim. Reprod. v.3, p.70-75,
2006.
SARAIVA, M.V.A.; ROSSETTO, R.; BRITO, I.; CELESTINO, J.J.H; SILVA, C.;
FAUSTINO, L.; ALMEIDA, A.; BRUNO, J.; MAGALHÃES, D.M.; MATOS, M.H.;
CAMPELLO, C.C.; FIGUEIREDO J.R. Dynamic medium produces caprine embryo
from preantral follicles grown in vitro; Reproductive Sciences, in press.
SERAFIM, M.K.B.; ALMEIDA, A.P.; DUARTE, A.B.G.; SILVA, G.M.; ARAUJO,
V.R.; CHAVES, R.N.; SILVA, L.D.M; FIGUEIREDO, J.R.; CAMPELLO, C.C.;
LOPES, C.A.P. Canine preantral follicles culture at different concentrations of follicle-
stimulating hormone (FSH). Theriogenology. v.74, p.749–755, 2010.

69
SHARMA, G.T.; DUBEY, P.K.; MEUR, S.K. Effect of different mechanical isolation
techniques on developmental competence and survival of buffalo ovarian preantral
follicles. Livestock Science .v.123, p.300-305, 2009.
SIES, H.; STAHL, W.; SUNDQUIST, A.R. Antioxidant functions of vitamins. Ann NY
Acad Sci. v.669, p.7-201992.
SILVA, A.R.; MORATO, R.G.; SILVA, L.D.M. The potential for gamete recovery
from non-domestic canids and felids. Anim Reprod. v.81, p.159–75, 2004a.
SILVA, A.E.F.; CAVALCANTE, L.F.; RODRIGUES B.A.; RODRIGUES J.L. The
influence of powdered coconut water (ACP-318®) in vitro maturation of canine
oocytes. Reprod Dom Anim. 2009.
SILVA, C.M.G.; MATOS, M.H.T.; RODRIGUES, G.Q.; FAUSTINO, L.R.; PINTO,
L.C.; CHAVES, R.N; ARAÚJO, V.R.; CAMPELLO, C.C.; FIGUEIREDO J.R. In vitro
survival and development of goat preantral follicles in two different oxygen tensions;
Animal Reproduction Science. v.117, p.83–89, 2010.
SILVA, J.R.V.; VAN DEN HURK; MATOS, M.H.T.; SANTOS, R.R.; PESSOA, C.;
MORAES, M.O.; FIGUEIREDO, J.R. Influences of FSH and EGF on primordial
follicles during in vitro culture of caprine ovarian cortical tissue. Theriogenology. v.61,
p.1691-1704, 2004b.
SONG, H.J.; KANG, E.J.; MAENG , G.H.; OCK , S.A.; LEE, S.L.; YOO, J.G; JEON,
B.G.; RHO, G.J. Influence of epidermal growth factor supplementation during in vitro
maturation on nuclear status and gene expression of canine oocytes. Research in
Veterinary Science. in press.
SONGSASEN, N.; WILDT, D.E. Oocyte biology and challenges in developing in vitro
maturation systems in the domestic dog. Animal Reproduction Science. v.98, p.2–22,
2007.
SONGSASEN, N.; YU, I.; LEIBO, S.P. Nuclear maturation of canine oocytes cultured
in protein-free media. Mol. Reprod. Dev. v.62, p.407–415, 2002.
SPEARS, N.; BOLAND, N.I.; MURRAY, A.A.; GOSDEN, R.G. Mouse oocytes
derived from in vitro growth primary ovarian follicles are fertile. Hum. Reprod., v.9,
p.527–532, 1994.

70
TAMILMANI, G.; RAO, B.S.; VAGDEVI, R.; AMARNATH, D.; NAIK, B.R.,
MUTHARAO, M.; RAO, V.H. Nuclear maturation of ovine oocytes from sheep
preantral follicles cultured in vitro. Small Rum. v.60, p.295–305, 2005.
TAS, M.; EVECEN, M.; OZDAS, O. B.; CIRIT, U.; DEMIR, K.; BIRLER, S.;
PABUCCUOGLU, S. Effect of transport and storage temperature of ovaries on in vitro
maturation of bitch oocytes. Anim. Reprod. Sci.v.96, p.30–34, 2006.
TELFER, E.E.; BINNIE, J.P.; MCCAFFERY, F.H.; CAMPBELL, B.K. In vitro
development of oocytes from porcine and bovine primary follicles. Mol. Cell Endo. v.
163, p.117–123 2000.
WU, J.; EMERY, B.R.; CARRELL, D.T. In vitro growth, maturation, fertilization, and
embryonic development of oocytes from porcine preantral follicles. Biology of
Reproduction. v.64, p.375–381, 2001.
YAMADA, S.; SHIMAZU, Y.; KAWAO, Y.; NAKAZAWA, M.; NAITO, K.;
TOYODA, Y. In vitro maturation and fertilization of pre-ovulatory canine oocytes. J
Reprod Fertil. v.47, p.227-229, 1993.

71
7. CAPÍTULO 2
Altas concentrações de insulina promovem o crescimento in vitro e a viabilidade de
folículos preantrais caninos
High insulin concentrations promote the in vitro growth and viability of canine
preantral follicles
Periódico : Reproduction, Fertility and Development 25(6) 927-934
http://dx.doi.org/10.1071/RD12074

72
RESUMO: Este trabalho tem como objetivo avaliar o efeito da adição de
diferentes concentrações de insulina, associados ou não ao FSH no desenvolvimento dos
folículos pré-antrais caninos no cultivo in vitro. Para tanto, folículos secundários (>200
μm) foram isolados por microdissecção e posteriormente cultivados por 18 dias em
meio α-MEM+
(CtrlMEM) suplementado, e avaliadas várias concentrações de insulina
(5 (Ins5ng) or 10 ng/ml (Ins10ng), ou a 10µg/ml (Ins 10µg)) na ausência (Experimento
1) ou presença (Experimento 2) de FSH acrescido de forma sequencial (Ins5ngF,
Ins10ngF and 10µgF).Os parâmetros avaliados foram manutenção da estrutura
morfológica ao longo do cultivo, viabilidade e diâmetro folicular, e taxa de formação de
antro. Pode-se observar que na manutenção da morfologia folicular ao fim do cultivo,
no Experimento 1, o tratamento CtrlMEM apresentou maior percentual de folículos
intactos (86,79%), quando comparado ao grupo de alta concentração de insulina
(Ins10g: 58,18%) (P < 0.05). Já no Experimento 2, não houve diferença significativa
entre os tratamentos avaliados. Quanto à viabilidade, no Experimento 1, o grupo
Ins10µg, apresentou percentual de folículos viáveis significativamente superior
(91,23%) aos tratamentos sem insulina (CtrlMEM¸39,39%), Ins5ng (75,68%) e Ins10ng
(68,63%). Porém, no Experimento 2, o tratamento CtrlFSH apresentou 100% dos
folículos viáveis, enquanto o meio contendo somente insulina (Ins10µg) apresentou
91,43% de folículos viáveis ao fim do cultivo (P < 0.05). No diâmetro folicular, o grupo
Ins10g apresentou-se significativamente superior (334,18 ± 74,12) quando comparado
aos demais tratamentos, CtrlMEM (217,83 ± 39,50); Ins5ng (238,64 ± 35,41) e Ins10ng
(244,76 ± 45,74) (P<0.05). Já no Experimento 2, a alta concentração de insulina
associada ao FSH (Ins10gF) apresentou média do diâmetro folicular
significativamente superior (411.05 ± 87.24), as demais concentrações de insulina
testadas (Ins5ngF: 347.38 ± 79.43; Ins10ngF: 336.10 ± 69.78). Para a formação de antro
no Experimento 1, não foi observado diferença entre os tratamento testados.. Entretanto,
no Experimento 2, o grupo com maior concentração de insulina (Ins10µgF: 59,26%)
apresentou taxa de formação de antro significativamente superior em relação aos demais
tratamentos (CtrlFSH: 15.87%; Ins5ngF:34,43% e Ins10ngF: 30,16%; P<0.05). Dessa
forma, diante do exposto, pode-se concluir que a associação de uma alta concentração
de insulina (10µg/ml) ao FSH seqüencial, é benéfica para a sobrevivência e
crescimento in vitro de folículos pré-antrais caninos.

73

74

75
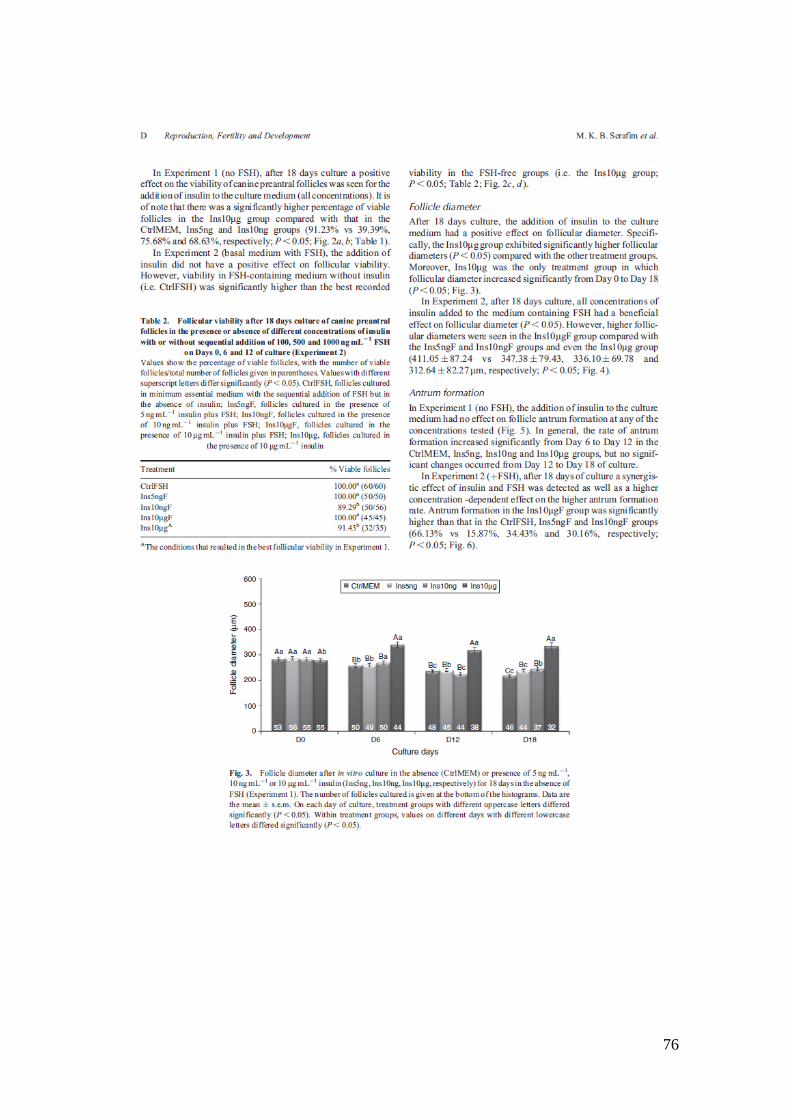
76

77
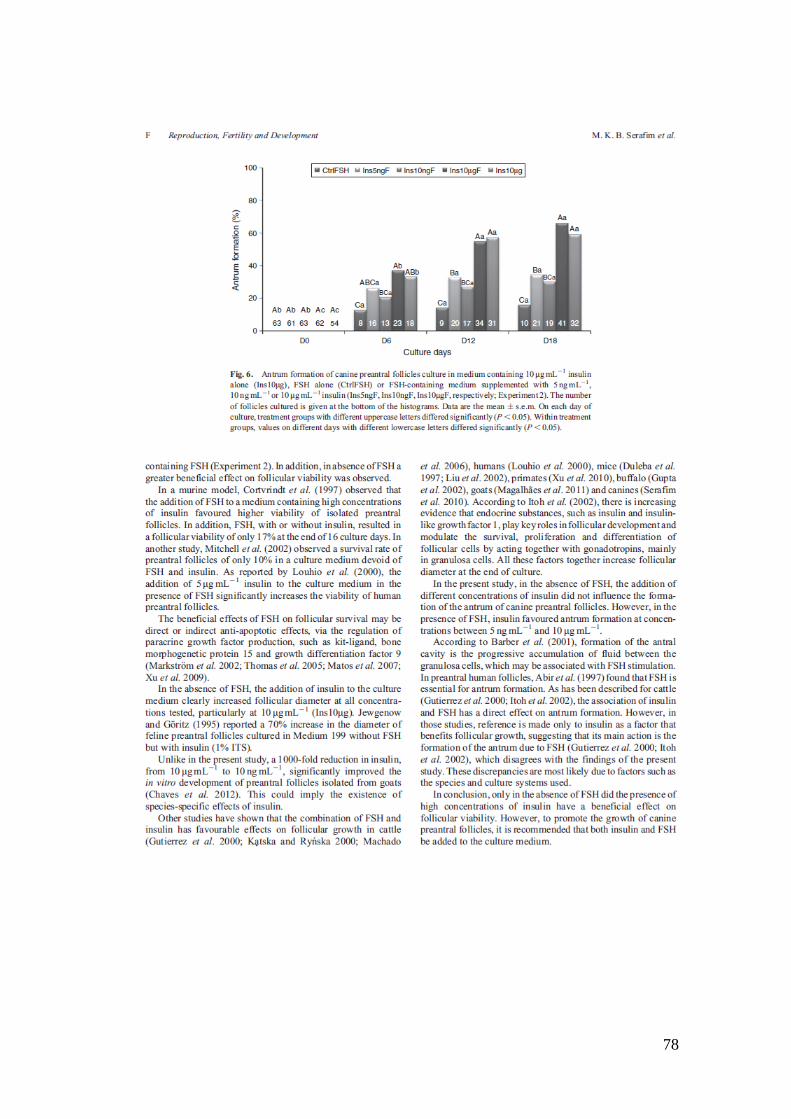
78

79

80

81
8. CAPÍTULO 3
Folículos pré-antrais caninos cultivado em meio suplementado com diferentes
concentrações do hormônio do crescimento (GH)
Canine preantral follicle culture in medium containing different concentrations of
growth hormone (GH)
Periódico: Biopreservation and Biobanking, submetido

82
RESUMO
O objetivo do presente estudo foi avaliar o efeito do hormônio do crescimento
sobre o desenvolvimento in vitro de folículos pré-antrais caninos na presença ou
ausência de FSH. Para tanto, folículos secundários isolados por microdissecção foram
cultivados em seis tratamentos: meio controle (αMEM+) constituído por αMEM
suplementado com BSA; grutamina; hipoxantina; insulina; tranferrina; selênio e ácido
ascórbico; αMEM+ acrescido de GH nas concentrações de 10 (GH10) ou 50 ng/mL
(GH50); e essas concentrações adicionadas de FSH (GH10+FSH e GH50+FSH). Foram
avaliados parâmetros como percentual de folículos intactos; formação de antro;
diâmetro e viabilidade folicular com marcadores de fluorescência, bem como análise da
produção de estradiol. Quanto à manutenção da morfologia normal dos folículos ao
longo do cultivo, o grupo GH50 foi o único tratamento que manteve o percentual de
folículos intactos do dia zero ao dia 18 de cultivo (P˂ 0,05). No tocante a viabilidade
folicular, com exceção do Controle, em todos os tratamentos os folículos se
apresentaram viáveis (P˂ 0,05). No que se refere ao crescimento folicular, o tratamento
GH50+FSH apresentou o maior diametro (P˂ 0,05) a partir do dia 12, mantendo essa
condição até o fim do cultivo. Além disso, o referido tratamento foi único que resultou
em taxas de formação de antro significativamente superior ao Controle. Os folículos
oriundos do Controle e do GH50+FSH foram esteroigenicamente ativos observando-se
aumento significativo da produção de estradiol do dia 6 ao dia 18 de cultivo, em ambos
os tratamentos analisados (P< 0.05). Concluindo, este estudo mostrou que o GH de
forma dose dependente beneficiou a manutenção da morfologia folicular e sua
associação ao FSH estimulou o crescimento folicular in vitro.
Palavras chaves: Folículo ovariano, GH, FSH, cão doméstico

83
Canine preantral follicle culture in medium containing different concentrations of
growth hormone (GH)
Michelle Karen Brasil Serafim A, D, Ana Beatriz Graça Duarte A; Gerlane Modesto da
Silva A, Carlos Eduardo Azevedo Souza C,Arlindo de A. Araripe Moura A, Lúcia
Daniel Machado da SilvaB, Cláudio Cabral Campielo A, José Ricardo de Figueiredo A
A Faculty of Veterinary Medicine, Laboratory of Manipulation of Oocytes and Preantral
Follicles (LAMOFOPA), Veterinary Science Post Graduation Program (VSPGP), State
University of Ceara,Av. Paranjana, 1700, Campus do Itaperi, Fortaleza, CE 60.740-000,
Brazil.
B Faculty of Veterinary Medicine, Laboratory of Carnivore Reproduction (LRC), State
University of Ceara, Av. Paranjana, 1700, Campus do Itaperi, Fortaleza, CE 60.740-000,
Brazil.
C Laboratory of Animal Physiology; Department of Animal Science, Federal University of
Ceará, Av. Mister Hull S/N, Campus Pici, Fortaleza – CE – Brazil, CE 60021-970
D Corresponding author. Email: [email protected]

84
ABSTRACT
The purpose of this study was to evaluate the effect of different concentrations
of growth hormone (GH) on the in vitro development of domestic dog (Canis lupus
familiaris) preantral follicles in the presence or absence of follicle stimulating hormone
(FSH). Secondary preantral follicles, isolated by microdissection, were cultured in
αMEM+ (control), a medium composed of αMEM with bovine serum albumin (BSA),
glutamine, hypoxantine, insulin, transferrin, selenium and ascorbic acid. The other
treatments included αMEM+ with GH added at concentrations of 10 ng/mL or 50
ng/mL, with (GH10+FSH, GH50+FSH) or without FSH (GH10, GH50). Follicle
development was evaluated on the basis of the percentage of intact follicles, antrum
formation, follicular diameter, follicular viability using fluorescent markers and
estradiol production. GH50 was the only treatment that maintained the same percentage
of morphologically normal follicles from day 0 to day 18 of culture (P<0.05). For all
treatments except the control treatment, follicles were viable throughout the 18 days of
culture (P<0.05). GH50 supplemented with FSH (GH50+FSH) resulted in the highest
average follicular diameter (P<0.05) from days 12 to 18; in other words, GH50+FSH
was the only treatment to yield higher antral formation rates than the control treatment.
Follicles from both the control and the GH50+FSH treatment groups actively and
increasingly secreted estradiol from days 6 to 18 of culture (P<0.05). In conclusion, our
study demonstrates that GH benefits the maintenance of follicular morphology in a
dose-dependent manner and, in association with FSH, stimulates in vitro follicular
growth.
Keywords: Ovarian follicles, GH, FSH, domestic dog,

85
INTRODUCTION
In vitro culture of preantral follicles is an important tool for understanding
folliculogenesis and oogenesis in domestic mammals, and the knowledge acquired
could also be applied in the efforts to conserve endangered canine species (1). Although
in vitro embryo production has been described in different species such as pigs (2),
water buffaloes (3), sheep (4) and goats (5), the birth of live offspring generated from
oocytes recovered from the in vitro growth of follicles has only been achieved in mice
(6). In domestic dogs (Canis lupus familiaris), the best results of preantral follicles
cultured in vitro are still limited to follicular growth and antrum formation (7, 8, 9)
Culture of preantral follicles in canines has been accomplished in medium
supplemented with hormones, in particular with FSH, in both 2D and 3D systems (7;
10; 11). However, other hormones and growth factors have been used as supplements in
follicle culture media in different species, including the Somatotrophic Hormone,
otherwise known as Growth Hormone (GH) (12, 13).
Studies that have stimulated in vitro growth of both preantral and antral follicles in
several species (12, 14), including rabbits (15), mares and sows (16), and goats (17),
have consistently shown that GH plays an important role in the regulation of ovarian
function. Kobayashi et. al. (2000) (18) and Apa et. al. (1994) (19) reported a significant
increase in theca and granulosa cell proliferation after in vitro culture of mice preantral
follicles in the presence of GH. In this same species, joint stimulation with GH and FSH
resulted in increased follicle diameter after four days of in vitro culture (20; 21). In
sheep, both FSH and GH act as regulators of insulin-like growth factor I (IGF-I) and
stimulate estrogen production in a dose-dependent manner (22). A recent study

86
described the first goat embryo generated from in vitro cultured oocytes grown in
medium containing GH and FSH (5).
Currently, only modest advances have been achieved in the culture of domestic dog
preantral follicles. Despite promising results showing increased follicular diameter of
follicles cultured in medium supplemented with GH in several species, no information
is available on the influence of GH on follicle growth in the canine species. Therefore,
the aim of this research was to investigate the effects of medium supplementation with
different concentrations of GH on the in vitro growth of domestic dog preantral follicles
in the presence or absence of FSH.
1. MATERIAL AND METHODS
2.1. Collection and transport of ovaries
Ovaries (n= 20) from bitches (undefined crossbreds) in various stages of the estrous
cycle were collected immediately after euthanasia in the Center for Animal Disease
Control in Fortaleza, CE, Brazil (approval by Animal Care and Use Committee; numb:
09172558-5/49, UECE/FORTALEZA/BRAZIL). After collection, ovaries were washed
once in 70% alcohol for 10 s and twice in HEPES-buffered Minimum Essential Medium
(MEM-HEPES) supplemented with penicillin (100 µg/mL) and streptomycin (100
µg/mL); ovaries were then transported to the laboratory within 1 h at 4 ºC (23).
2.2. Isolation of domestic dog advanced preantral follicles
After removing the ovarian bursa and other adjacent tissues and ligaments, the
ovarian cortex was sliced (approximately 1 mm thick). Then, secondary preantral
follicles <200 µm in diameter were manually isolated by microdissection using 27.5 G

87
needles attached to 1-mL syringes under a stereomicroscope dissecting microscope
(SMZ 645 Nikon, Tokyo, Japan). After isolation, follicles were immersed in drops of
culture medium and washed three times before being transferred to culture plates. Only
follicles considered morphologically normal (i.e., with a centrally located spherical and
dark oocyte, surrounded by two or three compact layers of granulosa cells and with no
apparent damage to the basal membrane) were used.
2.3. In vitro culture
Follicles were cultured individually for 18 days in 100 μL drops of medium under
mineral oil, using 60-mm diameter Petri dishes (Corning Inc., Lowell, MA, USA,
catalog no. 430166). The basic culture medium consisted of MEM supplemented with
bovine serum albumin (3 mg/mL), glutamine (2 mM), hypoxanthine (2 mM), ITS
supplement (Sigma-Aldrich, USA; 10 µg/mL), insulin (10 µg/mL), transferrin (5,5
µg/ml), selenium (5 ng/mL), and ascorbic acid (50 µg/mL); this medium was defined as
α-MEM+ (Control). The domestic dog preantral follicles were allocated into six
treatments: control medium alone (α-MEM+) or control medium supplemented with
recombinant bovine FSH (rFSH) (MEM+FSH), with GH (growth hormone from bovine
pituitary gland) at 10 ng/mL (GH10) or 50 ng/mL (GH50) alone, or with GH associated
with rFSH (GH10+FSH and GH50+FSH, respectively). The rFSH (rFSH, Nanocore,
Campinas, São Paulo, Brazil) was added sequentially at appropriate growing
concentrations (100 ng/mL until Day 6, 500 ng/mL until Day 12, and 1000 ng/mL until
Day 18 of culture) (9). Follicles were cultured at 39 ºC with 5% CO2, and 60 µL of the
medium was changed every 2 days, except for days 6 and 12, when the entire medium
was changed.

88
At days 0, 6, 12 and 18 of culture, follicular diameter, morphology, antrum
formation and oocyte extrusion rate were evaluated. Follicles showing morphological
signs of degeneration, such as whitening of oocytes and of the surrounding granulosa
cells, misshapen oocytes, rupture of the basement membrane and/or oocyte extrusion,
were classified as morphologically abnormal. Follicles were measured every 6 days
using an ocular micrometer attached to a stereomicroscope (SMZ 645 Nikon, Tokyo,
Japan), and the average of two measurements (vertical + horizontal l/2) was recorded. In
addition, the growth rate was calculated as the variation in diameter throughout the
culture period. To evaluate the antrum formation rate, the emergence of a translucent
cavity filled with follicular fluid was noted.
2.4. Analysis of follicular viability
At the end of the culture period, a two-color fluorescence cell viability assay based
on the simultaneous determination of live and dead cells by calcein-AM and ethidium
homodimer-1 was used. The first probe detected intracellular esterase activity of viable
cells, whereas the latter probe labeled nucleic acids of non-viable cells with plasma
membrane disruption. The test was performed by incubating cells in a PBS medium
supplemented with 4 µM calcein-AM and 2 µM ethidium homodimer-1(Molecular
Probes, Invitrogen, Karlsruhe, Germany) at 37°C for 15 min. Afterwards, follicles were
washed three times in HEPES-buffered TCM199, mounted between a slide and a
coverslip, and evaluated (100x) with the aid of an epifluorescence microscope
(Olympus). The emitted fluorescent signals of calcein-AM and ethidium homodimer
were collected at 488 and 568 nm, respectively. Oocytes and granulosa cells were

89
considered viable if the cytoplasm was stained positively with calcein-AM (green) and
if chromatin was not labeled with ethidium homodimer (red).
2.5. Evaluation of estradiol production
To evaluate estradiol production on days 6 and 18 of culture, the medium used to
culture each domestic dog follicle was aspirated and stored at -80°C until being shipped
to the laboratory for hormone analysis. Medium samples were thawed at room
temperature and used for the determination of estradiol concentration using the
ARCHITECT®
platform (Abbott Diagnostics, Abbott Park, IL, USA, sensitivity of 2.5
pg/mL) according to the manufacturer‘s instructions. This platform uses CMIA
(chemiluminescence microparticle immunoassay) and has a detection limit between 10
and 1000 pg/mL. Medium samples used for follicle culture were diluted with standard
medium between 100 and 400 times to bring concentrations below the detection limit of
the kit. A sample of standard medium was used as a negative control. Intra and inter-
assay CVs were below 10%.
2.6. Statistical analysis
Data were initially evaluated for homoscedasticity and normal distribution of the
residues by Bartlett‘s and Shapiro-Wilk tests, respectively. Upon confirming both of
these requirements for an analysis of variance, the effects of treatment, time and a
treatment by time interaction term were analyzed using the PROC MIXED command of
SAS (2002) with a repeated statement to account for autocorrelation between sequential
measurements. The model used was represented by the equation
Yijk=µ+Ri+Fj+Tk+(RT)ik+eijk, where Yijk is the observation of the jth follicle in the ith

90
treatment at the kth time of culture, µ is the overall mean, Ri is the ith treatment, Fj is the
random effect of the jth follicle within the ith treatment, Tk is the kth time of culture,
(RT)ik is the treatment by time interaction term and eijk is the random residual effect.
Comparisons between treatments or times were further analyzed by the LSD test. A
probability of P<0.05 indicated a significant difference, and results were expressed as
the mean ± S.D.
2. RESULTS
3.1 Follicular morphology
A total of 240 domestic dog preantral follicles were selected for culture and
randomly allocated to one of six treatments. Figure 1 shows the percentage of
morphologically normal follicles with an intact basal membrane throughout the 18 days
of culture. Over the course of culture, we found a decrease in the percentage of normal
follicles from day 0 to day 18 for all treatments except GH50. However, after 18 days of
culture, there were no differences among treatments (Figure 1; P<0.05).
3.2 Follicular viability
After 18 days of culture, the viability of all follicles was examined. With the
exception of the control group (αMEM+: 80%), all follicles in all treatment groups were
viable (Table 1) (P< 0.05) (Figure 4).
3.3 Follicular growth after in vitro culture
As culture progressed, all treatments displayed increases in follicular diameter
(P˂0.05) from day 0 to 18 (Figure 2). At day 12 of culture, the follicles in the treatment

91
with GH and FSH (GH50+FSH) had a higher follicular diameter than the follicles in the
other treatments groups (P˂0.05).
3.4 Antrum Formation
All treatments resulted in the formation of an antral cavity starting at day 6 of
culture (Figure 3). Between days 6 and 12 of culture, we found an increase in the antral
formation rates for all treatments except GH50 (P< 0.05). There were no further
significant changes between days 12 and 18.
At day 6 of culture, groups GH50 (76.92%) and GH50+FSH (77.5%) had similar
antral formation rates, and these rates were significantly higher than those of the other
treatments. It is important to note that at day 12 of culture, group GH50+FSH reached a
rate of antrum formation of 100%, but the rate of this treatment did not differ from that
of the GH10+FSH treatment (92.5%). However, at the end of the 18 days of culture,
only the GH50+FSH treatment had rates that were significantly different from those of
the αMEM+ (90%), αMEM+FSH (90%), GH10 (89.47%) and GH50 (87.18%)
treatments (P< 0.05).
3.5 Estradiol assays
Table 2 shows the estradiol concentrations in the recovered culture media of
domestic dog preantral follicles cultured for 18 days in the control and GH50+FSH
groups. There was a significant increase in estradiol secretion between days 6 and 18 for
both treatments (P<0.05), but there were no differences between treatments.
3. DISCUSSION
For the first time, the present study shows the effect of GH with or without
sequential addition of FSH on the in vitro development of domestic dog preantral

92
follicles. Only culture with GH alone at a concentration of 50 ng/mL (GH50) was
sufficient to maintain the percentage of morphologically normal follicles throughout the
experiment. This treatment also yielded 100% viable follicles by day 18 of culture.
Therefore, our results suggest that GH in the above mentioned concentration (GH50)
favors in vitro culture survival. Our findings are supported by those of other studies
suggesting that GH might promote follicular survival by preventing apoptosis either via
promotion of IGF-I gene expression or stimulation of anti-apoptotic PI-3K/Akt
signaling (24, 25).
Our results also demonstrated that GH (50 ng/mL) with FSH (GH50+FSH)
promoted follicular growth in a dose-dependent manner, with both hormones possibly
acting synergistically. To the best of our knowledge, this treatment (GH50+FSH) also
resulted in the largest follicular diameter yet reported (almost 500 µm) and an antrum
formation rate of 100% in domestic dog preantral follicles after long-term culture.
Several studies showed that GH, acting alone or in concert with FSH, affects
ovarian function, promoting cellular growth by stimulating granulosa cell proliferation
and differentiation (26, 27). Similar results have been reported in mice, where
significant increases in the diameter of preantral follicles after 4 days of culture in the
presence of GH (1 mg/mL) and FSH were observed (20, 21). In sheep, both FSH and
GH (200 ng/mL) regulate ovarian function in a dose-dependent manner, stimulating the
proliferation of granulosa cells and IGF-I secretion by these cells after 96 h of culture
(22).
Others studies published by our research group on domestic dog follicles
cultured in the presence of FSH and insulin (10 µg/mL) described an antrum formation
rate of 60% after 18 days of culture (11). In the present report, supplementation of the
culture medium with 50 ng/mL of GH likely enhanced this effect, resulting in the

93
formation of antra in all follicles. This result might be related to the ability of GH to
stimulate the production of fluid during follicular development (27, 28).
Other studies have reported that the addition of GH to culture medium stimulates
proliferation and differentiation of granulosa cells, which results in increased estrogen
secretion (21,26,29). Our study, however, found that the estradiol concentration in the
medium was not significantly affected by supplementation with GH or FSH. This
finding clearly shows that the increases in follicular diameter seen in the GH50+FSH
group were not attributable to greater estradiol secretion. Similar results were reported
by Songsasen et. al. (2011) (10) for enclosed domestic dog follicles cultured under
different alginate concentrations. The authors observed substantial increases in follicular
size after 240 hours of culture but no change in estradiol secretion throughout
incubation.
In conclusion, this study shows that supplementation of culture medium with
GH clearly produces dose-dependent beneficial effects on follicular morphology, while
the addition of FSH further enhances in vitro follicular growth.
Acknowledgements
This research was supported by grants from the National Council for Scientific and
Technological Development (CNPq; grant no. 142065/2010–2) and the Brazilian
Innovation Agency (FINEP). MKB received financial support from CNPq. The authors
express their appreciation to the Center for Zoonosis Control (CCZ) of Fortaleza
Municipality (Brazil).

94
Fig. 1: Percentage of morphologically normal canine follicles after in vitro culture in the
absence (αMEM+, αMEM+FSH) or presence of different GH concentrations (50 ng/mL
or 10 ng/mL) with (GH10+FSH and GH50+FSH) or without FSH (GH10 and GH50)
for 18 days.
Capital letters: difference among treatments on each day of culture (P< 0.05).
Lower case letters: difference among days of culture within each treatment (P <0.05).

95
Fig. 2: Canine follicular diameter (mean ± S.D.) after in vitro culture in the absence
(αMEM+, αMEM+FSH) or presence of different GH concentrations (50 ng/mL or 10
ng/mL) with (GH10+FSH and GH50+FSH) or without FSH (GH10 and GH50) for 18
days.
Capital letters: difference among treatments on each day of culture (P< 0.05).
Lower case letters: difference among days of culture within each treatment (P <0.05).

96
Fig. 3: Antrum formation rates (mean ± S.D.) after in vitro culture in the absence
(αMEM+, αMEM+FSH) or presence of different GH concentrations (50 ng/mL or 10
ng/mL) with (GH10+FSH and GH50+FSH) or without FSH (GH10 and GH50) for 18
days.
Capital letters: difference among treatments on each day of culture (P< 0.05).
Lower case letters: difference among days of culture within each treatment (P <0.05).
Fig. 4: Domestic dogs follicle after 18 days of culture in media αMEM added of GH (50
ng/mL) in presence of FSH (a), and the same follicle stained with calcein-AM and
classified as viable (b).
a b

97
TABLE 1: Follicular viability after 18 days of culture of canine preantral follicles in the
presence or absence of different concentrations of growth hormone (GH), with or
without the addition of FSH (100, 500 and 1000 ng/mL).
TREATMENT FOLLICULAR VIABILITY
αMEM+ 80% (24/30) B
αMEM+FSH 100% (20/20) A
GH10 100%(25/25) A
GH50 100% (20/20) A
GH10+FSH 100% (24/24) A
GH50+FSH 100% (28/28) A
Values with different superscript letters differ (P <0.05)
TABLE 2: Evaluation of estradiol concentration (mean ± S.D.) in canine preantral
follicle culture medium between days 6 (D6) and 18 (D18) of culture for control
medium (αMEM) or medium supplemented with high concentrations of GH (50 ng/ml)
associated with FSH (GH50+FSH).
TREATMENT D6 (pg/ml) D18 (pg/ml)
αMEM 7.29 ± 2.92a 41.61 ± 16.87
b
GH50+FSH 4.66 ± 3.43a 66.21 ± 38.91
b
a, b- Indicates a significant difference between the days of culture within each
treatment (P <0.05).
REFERENCES
(1) Figueiredo, J.R., Rodrigues, A.P.R., Silva, J.R.V., et al. Cryopreservation and in
vitro culture of caprinepreantral follicles Reprod. Fertil. Develop. , 2011; 23: 40–47.
(2) Wu, J., Tian, Q. Role of follicle stimulation hormone and epidermal growth factor in
the development of porcine preantral follicle in vitro. Zygote 2007; 15: 233–240.
(3) Gupta, P.S., Ramesh, H.S., Manjunatha, B.M., et al. Production of buffalo embryos
using oocytes from in vitro grown follicles Zygote 2008; 16: 57–63.

98
(4) Arunakumari, G., Shanmugasundaram, N., Rao, V.H. Development of morulae from
the oocytes of cultured sheep preantral follicles Theriogenology 2010; 74: 884–894.
(5) Magalhães, D.M., Duarte, A.B.G., Araújo, V.R., et al. In vitro production of a
caprine embryo from a preantral follicle cultured in media supplemented with growth
hormone Theriogenology 2011; 75: 182–188.
(6) O‘brien, M.J., Pendola, J.K., Eppig, J.J. A revised protocol for in vitro development
of mouse oocytes from primordial follicles dramatically improves their developmental
competence. Biol. Reprod. 200368, 1682– 1686.
(7) Bolamba, D, Borden-Russ, K.D., Durrant, B.S. In vitro maturation of domestic dog
oocytes cultured in advanced preantral and early antral follicles Theriogenology 1998;
49: 933–942.
(8) Durrant, B.S., Pratt, N.C., Russ. K.D., et al. Isolation and characterization of canine
advanced preantral and early antral follicles Theriogenology 1998; 49: 917–932.
(9) Serafim, M.K., Araújo, V.R., Silva, G.M., Duarte. A.B.G. Canine preantral follicles
cultured with various concentrations of folliclestimulating hormone (FSH)
Theriogenology 2010; 74: 749–755.
(10) Songsasen, N., Woodruff, T.K., Wild D.E. In vitro growth and steroidogenesis of
dog follicles are influenced by the physical and hormonal microenvironment.
Reproduction 2011; 142: 113–122.
(11) Serafim, M.K.B., Silva, G.M., Duarte, A.B.G. et al. High insulin concentrations
promote the in vitro growth and viability of canine preantral follicles. Reprod. Fertil.
Develop. in press.

99
(12) Shimizu, T., Murayama, C., Sudo, N., et al. Involvement of insulin and growth
hormone (GH) during follicular development in the bovine ovary. Anim. Reprod. Sci.
2008;106: 143-52.
(13) Sirotkin, A.V., Makarevich, A.V. GH regulates activity and apoptosis in cultured
bovine granulosa celle through the activation of cAMP/protein kinase a system. J.
Endocrinol. 1999; 163: 317-327.
(14) Zhao, J., Taverne, M.A., Van Deweijden, G.C., et al. Immunohistochemical
localization of growth hormone (GH), GH receptor (GHR), insulin-like growth factor I
(IGF-I) and type I IGF-I receptor, and gene expression of GH and GHR in rat pre-antral
follicles. Zygote 2002; 10: 85–94.
(15) Yoshimura, Y., Iwashita, M., Karube, M.,et al. Growth hormonestimulates
follicular development by stimulating ovarian production of insulin-like growth factor-I.
Endocrinology 1994; 135: 887–894.
(16) Marchal, R., Caillaud, M., Martoriati, A., et al. Effect of Growth Hormone (GH) on
In vitro Nuclear and Cytoplasmic Oocyte Maturation, Cumulus Expansion, Hyaluronan
Synthases, and Connexins 32 and 43 Expression, and GH Receptor Messenger RNA
Expression in Equine and Porcine. Species. Biol. Reprod. 2003; 69:1013–1022.
(17) Martins, F.S., Celestinom J.J.H., Saraiva, M.V.A.,et al. Interaction between growth
differentiation factor 9, insulin-like growth factor I and growth hormone on the in vitro
development and survival of goat preantral follicles. Braz. J. Med. Biol. Res. 2010; 43:
728-736.

100
(18) Kobayashi, J., Mizunuma, H., Kikuchi, N., et al. Morphological assessment of the
effect of growth hormone on preantral follicles from 11-day-old mice in an in vitro
culture system. Biochem. Biophys. Res. Commun. 2000; 268: 36–41.
(19) Apa, R., Lanzone, A., Micelia, F., et al., Growth hormone induces in vitro
maturation, Mol. Cell. Endocrinol. 1994; 106: 207-212.
(20) Liu, X., Andoh, K., Yokota, H., et al. Effects of growth hormone, activin, and
follistatin on the development of preantral follicle from immature female mice.
Endocrinology 1998; 139: 2342-2347.
(21) Kikuchi, N.; Andoh, K., Abe, Y., Inhibitory action of leptin on early follicular
growth differs in immature and adult female mice. Biol. Reprod. 2001; 65: 66-71.
(22) Khalid, M., Haresign, W., Luck, M.R., Secretion of IGF-I by ovine granulosa cells:
effects of growth hormone and follicle stimulating hormone. Anim. Reprod. Sci. 2000;
58: 261-72.
(23) Lopes, C.A.P., Santos, R.R., Celestino, J.J. H., et al. Short-term preservation of
canine preantral follicles: effects of temperature, medium and time. Anim. Reprod. Sci.
2009; 115: 201–214.
(24) Eisenhauer, K.M., Chun, S.Y., Billig, H., et al. Growth hormone suppression of
apoptosis in preovulatory rat follicles and partial neutralization by insulin-like growth
factor binding protein. Biol. Reprod. 1995; 53: 13-20.
(25) Costoya, J.A., Finidori, J., Moutoussamy, S. et al. Activation of growth hormone
receptor delivers an antiapoptotic signal: evidence for a role of akt in this pathway.
Endocrinology 1999; 140: 5937- 5943.

101
(26) Kaiser, G.G., Sinowatz, F., Palma, G.A. et al. Effects of growth hormone on
female reproductive organs. Anat. Histol. Embryol 2001; 30: 265–271.
(27) Bachelot, A.; Monget, P., Imbert-Bolloré, P., et al. Growth hormone is required for
ovarian follicular growth. Endocrinology 2002; 143: 4104- 4112.
(27) Apa, R., Caruso, A., Andreani, C.L., Grown hormone stimulates androsterone
synthesis by rat theca-interstitial cell. Mol. Cell. Endocrinol. 1996; 118:95-101.
(28) Mendoza, C., Ruiz-Requena, E., Ortege, E., et al. Follicular fluid markers of
oocyte development potential. Hum.Reprod. 2002; 17: 1017–22.
(29) Semiz, O., Evirgen, O. The effect of growth hormone on ovarian follicular
response and oocyte nuclear maturation in young and aged mice. Acta Histochem. 2009;
111: 104—111.

102
9. CAPÍTULO 4
Influência da suplmentação com o fator de crescimento epidermal e do microambiente
na sobrevivência e crescimento de folículos ovarianos caninos.
Influence of epidermal growth factor supplementation and in vitro microenvironment on
the survival and growth of dog ovarian follicles.
Periódico: Theriogenology

103
Resumo
O objetivo deste estudo foi investigar o efeito do fator de crescimento epidermal (EGF),
hormônio folículo estimulante (FSH), tipo de meio (MEM ou αMEM), e condições de
cultivo (2D vs 3D) no cultivo in vitro de folículos ovariando pré-antrais e antrais iniciais
de cadelas domésticas. Para tanto, no Experimento I, folículos ovarianos foram obtidos
e incubados por 18 dias em um dos segunites meios:1)MEM sozinho (Controle), 2)
MEM + FSH. 3) MEM + 50 ng/ml EGF, 4) MEM + 100 ng/ml EGF, 5) MEM + FSH +
50 ng/ml EGF e 6) MEM + FSH + 100 ng/ml EGF O FSH foi adicionado
sequencialmente nas concentrações de 100, 500 e 1,000 ng/mL, nos dias 0, 6, e 12 de
cultivo,respectivamente No Experimento II, os folículos foram cultivados por 18 dias
em meio 1)α-MEM ou MEM suplementado.Dessa forma, os folículos foram cultivados
em três diferentes sistemas de cultivo: sistema bidimensional (2D), tridimensional com
matriz hidrogel de alginato (ALG) or matriz de alginato-fibrina. Foram avaliadas as
taxas de formação de antro, modificações no diâmetro folicular e oocitário, e
integridade da cromatina oocitária após o cultivo.No Experimento I, o grupo
MEM+EGF100+FSH, exibiu um maior percentual (P< 0.05) de oócitos com cromatina
intacta, bem como taxas de crescimento folicular. Para o Experimento II, o uso do α-
MEM foi mais benéfico na manutenção da integridade da cromatina e crescimento
folicular do que o MEM. Taxas de expanção da cavidade antral e diâmentro oocitário
nos Experimento I e II forma similares (P< 0.05) entre os tratamentos, independente do
período de cultivo em ambos os parâmentros avaliados. Nenhum efeito do sistema de
cultivo (2D ou 3D) no cresimento folicular foi identificado. Em conclusão, o
crescimento, desenvolvimento e integridade da cromatina de folículos ovarianos do cão
doméstico in vitro apresenta um aumento de uma forma dose-dependente quando
associado ao FSH e EGF a 50ng/ml, e com o uso do αMEM como meio de base.
Entretanto, o sistema de cultivo tridimensional não conferiu nenhum benefício adicional
na foliculogênese in vitro do cão.

104
Influence of epidermal growth factor supplementation and in vitro microenvironment on
the survival and growth of dog ovarian follicles.
SERAFIM, M.K.B.; NAGASHIMA, J.; FIGUEIREDO, J. R., SONGSASEN, N.
Abstract
The purpose of this research was to investigate the effects of epidermal growth factor
(EGF), follicle stimulating hormone(FSH), medium type (MEM or αMEM), and culture
condition (2D vs 3D) on in vitro growth of dog advanced preantral and early antral
follicles. In Experiment I, ovarian follicles were obtained and incubated for 18 d in one
of the following medium: 1)MEM alone (Control), 2) MEM + FSH, 3) MEM + 50
ng/ml EGF, 4) MEM + 100 ng/ml EGF, 5) MEM + FSH + 50 ng/ml EGF and 6) MEM
+ FSH + 100 ng/ml EGF. The FSH was added sequentially at concentration of 100, 500
and 1,000 ng/mL, on days 0, 6, and 12 of culture, respectively. In Experiment II,
follicles were cultured for 18 d in either Eagle‘s α-MEM or 2) MEM, supplemented..
Then, follicles were cultured in three different systems: two dimensional (2D), three
dimensional with alginate hydrogel matrix (ALG) or in a fibrin–alginate matrix (FA-
IPN). Rates of antrum formation, change in follicular and oocyte diameter, and oocyte
chromatin integrity after culture were evaluated. In Experiment I, the group
MEM+EGF100+FSH, exhibited a higher percentage (P< 0.05) of oocytes with intact
chromatin and rates of follicular growth. For Experiment II, the use of α-MEM was
more beneficial on the maintenance of chromatin integrity as well as follicle growth
compared with MEM. Rates of antrum expansion and oocyte diameter in Experiments I
and II were similar (P< 0.05) among treatments, independent of culture period in both

105
parameters evaluated. No effect of culture system (2D or 3D) on follicular growth was
detected. In conclusion, the growth, development, and chromatin integrity of domestic
dog ovarian follicles in vitro dose-dependently improved in the presence of FSH
associated to EGF a 50ng/ml,, as well as with αMEM as the basic media. However, the
three dimensional culture systems did not confer any additional benefit to in vitro
folliculogenesis in the dog.
1. Introduction
Studies regarding culture of domestic dog follicles have been conducted in an attempt to
improve our ability to grow follicles and oocytes in vitro, by identifying an optimum
culture condition supportive of follicular development (BOLAMBA et. al., 1998,
DURRANT et. al., 1998, COMIZZOLI, et al, 2007). Previous studies have shown that
biochemical (hormone, growth factors and culture media) and physical
microenvironment (culture condition) significantly affect follicular development
(HARTSHORNE, et al, 1997; JEWGENOW, 1998; TELFER, 2001, DING et .al, 2012).
Specifically, FSH has been shown to be essential for in vitro growth of ovarian follicles
in many mammalian species, including the domestic dog (SERAFIM et al., 2010;
SONGSASEN et al., 2011). Our team has observed that the presence of follicle
stimulating hormone (FSH), added sequentially, maintained the survival of isolated
canine preantral follicles and promoted an increased rate of follicular growth and
antrum formation when compared to a fixed concentration of 100 ng/mL throughout the
entire culture period (SERAFIM et al, 2010).
EGF is on member of a large family of transforming growth factor-α (TGF-α) proteins
(CONTI et al., 2006), also is a known mitogenic polypeptide that has been shown to
stimulate proliferation of granulosa cells, recovered from primary and secondary
follicles in the bovine (WANDJI et al., 1996; ROY AND KOLE, 1998) and pigs
(MORBECK et al., 1993) models. Moreover, in culture medium containing FSH, EGF
promotes antrum formation, follicular growth and oocyte viability in goats (SILVA et

106
al., 2013), pigs (MAO et al, 2004; WU & TIAN, 2007) and buffalo (GUPTA et al.,
2008) isolated preantral follicles. With regard to domestic dogs, to our knowledge, the
influence of EGF has only been evaluated during in vitro maturation (IVM), and has
been shown to improve expansion of cumulus cells and rates of meiotic resumption
(CUI et al., 2006; BOLAMBA et al., 2006)
Concerning the culture of isolated domestic dog follicles, previous studies have reported
the use of Modified Eagle's medium (MEM) (BOLAMBA et al, 1998) or α-Minimum
Essential Medium (α-MEM) (SONGSASEN et al, 2011; SERAFIM et al., 2012) as a
basic media. Alpha-MEM has been used satisfactorily to culture goats (MAGALHÃES
et al., 2011), ewes (ARUNAKUMARI et al., 2010) and mouse (NONOWAKI et al.,
2010) preantral follicles. However, for preantral follicles of buffalo (GUPTA et al.,
2008) and cows (ANDRADE et al., 2012), MEM better supports follicular growth and
activation, respectively. These basic media differ largely in their composition, for
example, α-MEM contains higher concentrations of non-essential amino acids, which
have been shown to serve as energy sources, precursors for protein synthesis, as well as
regulate intracellular pH and osmotic pressure, than MEM (LANE & GARDNER 2007;
FUJIHARA et al., 2012). Recently, Fujihara et al., (2012), demonstrated that α-MEM
better sustained viability of primordial dog follicles enclosed in an ovarian tissue after
15 days of culture. However, no comparisons have been made between MEM and
αMEM for the culture of more advanced follicular stages in this species. As other
requirements for in vitro folliculogenesis have been shown to be stage-specific
(KREEGER et al., 2005; ROSSETO et al., 2012), the need for non-essential amino
acids for dog preantral follicle culture should also be evaluated.
Other relevant factor which can affect follicular development is the culture system used.
The culture of domestic dog preantral follicles has been undertaken using a two
dimensional system (2D), wherein follicles were cultured in a direct contact with plastic
superficies of multi-well plates (BOLAMBA et al., 1998) or in microdrop culture
(SERAFIM et al., 2010). These studies observed follicular growth and antrum
formation. However, some studies describe that the 2D approach may not be suitable for
in vitro culture of isolated follicles, because the granulosa cells could migrate and cause
disrupt communication with the adjacent oocyte that, in turn, compromises gamete
development (WEST et al. 2007, WOODRUFF et al., 2007). Accordingly, three-
dimensional (3D) systems, including alginate hydrogel have been developed showing

107
encouraging results in the mouse (XU et al., 2006), buffalo (GUPTA et al., 2008),
human (XU et al., 2009), and non-human primate (XU et al., 2011) follicle culture. Live
offspring have been produced from embryos derived from oocytes recovered from
follicles cultured in alginate hydrogel (XU et al., 2006; GUPTA et al., 2008). Although
offspring have not been produced in other species, including domestic dog
(SONGSASEN et al., 2011) and monkeys (XU et al., 2010; XU et al., 2011) follicles
cultured in alginate hydrogels increase in size, form antrum and produce steroids
Nevertheless, a more recent study suggests that the combination of fibrin and alginate in
a matrix provides a more dynamic microenvironment for follicle growth (JIN et al.,
2010, XU et al., 2011). In this system, fibrin is degraded by proteases secreted from
growing follicles, thereby softening the hydrogel and allowing further expansion by the
follicle while retaining the ability to mechanically support it with the presence of the
alginate (SHIKANOV et al., 2011; XU et al., 2013).
Therefore, the aim of this research was to investigate the effects of EGF and its
interaction with FSH, basic media, and culture conditions on the growth and
development of domestic dog advanced preantral and early antral follicles
2. Material and Methods
2.1. Recovery of ovarian follicles
Thirty-three ovaries were recovered from bitches (age of 6 months to 8 years old) in
various stages of the estrous cycle were collected immediately after undergoing routine
ovariohysterectomy at local veterinary clinics. The ovaries were transported to the
laboratory at 4ºC within 4–6 hours of surgery in L-15 medium (Leibovitz; Sigma,
Chemical Co., St Louis, MO, USA) with 10 mg/ml penicillin G sodium and
streptomycin sulfate.
In the laboratory, the ovarian bursa and other adjacent tissues were removed and the
ovarian cortex was sliced (approximately 1 mm thick). Then, advanced preantral (˃200
to 300 µm; n = 402) and early antral follicles (˃300µm to 420µm, n= 98) (DURRANT,
et al.,1998, SONGSASEN et al, 2007, SONGSASEN et al, 2009) were manually
isolated by microdissection, using 25 - 27 G needles attached to 1-mL syringes under a

108
dissecting microscope (SMZ 645 Nikon, Tokyo, Japan). After isolation, follicles were
washed in culture medium before being transferred to a randomly assigned culture
system. Only follicles considered morphologically normal, i.e., with a centrally located
spherical and dark oocyte, surrounded by two or three compact layers of granulosa cells,
and with no apparent damage to the basal membrane were selected for culture
(BOLAMBA et al., 1998).
2.3. In vitro culture
All follicles were individually cultured 39 ºC with 5% CO2 for 18 days in 100 μL drops
of culture medium (see Experimental design) under mineral oil, using 60-mm diameter
Petri dishes (Corning Inc., Lowell, MA, USA, catalog no. 430166). Half of culture
medium was exchanged every 2 days with except of Days 6 and 12 where the medium
was exchanged completely (100 µl).
2.4. Follicle Encapsulation
For the alginate culture system, each follicle was encapsulated in a bead of alginate
solution as previously described (SONGSASEN et al., 2011) with minor modifications.
Briefly, isolated follicles were individually transferred into a 2–3 µl droplet of 0.5%
(w/v) alginate. The droplets were then pipetted in a solution containing 50 mM CaCl2
and 140 mM NaCl, allowed to cross-link for 2 min, then rinsed for 10 min in culture
medium. The alginate matrix contained 0.5% (w/v) sterile sodium alginate (FMC
BioPolymers, Philadelphia, PA, USA)–in phosphate-buffered saline (PBS -Invitrogen,
Carlsbad, CA)
For follicle encapsulation in FA-IPN, fibrinogen (25 mg/ml; Tisseel [Fibrin Sealant],
Baxter Healthcare, Inc., Deerfield, IL, USA) mixed with alginate (0.25% w/v) was
dropped into a cross-linking solution containing 50 mg/ml thrombin (Tisseel [Fibrin
Sealant]), after that, rinsed for 10 min in culture medium, then each encapsulated
follicle was transferred into to culture drop (XU et al., 2013).
2.5. Assessment of follicle morphology, antrum formation and growth
Follicular diameter, morphology, and antrum expansion were evaluated on Day 0, 6, 12
and 18 of in vitro culture. Follicles showing morphological signs of degeneration, such

109
as clearing of the oocyte‘s cytoplasm and the surrounding granulosa cells, misshapen
oocytes, rupture of the basement membrane and/or oocyte extrusion were classified as
abnormal morphology (DURRANT et al., 1998). To evaluate growth, follicles were
measured every 6 days under an inverted microscope (Leitz DM-IL, Research
Instrument Limited, Falmouth, Cornwall, UK) equipped with a heated stage and a
calibrated optical micrometer, Each follicle was sized from the outer layer of the
somatic cells, and the measurements included the widest diameter and the perpendicular
width to the initial assessment. The mean of these two metrics then was calculated and
reported as ‗follicle diameter‘. Growth rate was calculated as the change in diameter
during the culture period, compared with Day 0 size. Antrum formation (for preantral)
or expansion (for early antral) was noted when the emergence or enlargement of a
translucent cavity filled with follicular fluid was observed, respectively.
2.6. Evaluation of cultured oocytes
Oocyte diameter including the zona pellucida was assessed Day 0 and 18, under an
inverted microscope (Leitz DM-IL, Research Instrument Limited, Falmouth, Cornwall,
UK) equipped with a heated stage and a calibrated optical micrometer. At the end of
culture, all healthy follicles were mechanically opened with 26 G needles under a
stereomicroscope and enclosed oocytes recovered. Cumulus cells associated with
oocytes were removed by gentle pipetting, and the gamete were then fixed in 4%
paraformaldehyde overnight, washed and stored in PBS solution until further analysis.
Oocytes were labeled with Hoechst 33342 (1.9µM Hoechst + 90% glycerol/PBS) to
assess chromatin integrity. Chromatin integrity was classified as: Intact (with intact
chromatin structure), Fragmented (oocytes with dispersed DNA fragments or chromatin
aggregates) or Unclassifiable (no DNA staining was visible) (LOPES et al., 2007).
2.7. Experimental design
In the Experiment I; MEM was used as the basic culture medium,supplemented with
bovine serum albumin (BSA, 3 mg/mL), glutamine (2 mM), hypoxanthine (2 mM), ITS
(10 µg/ml insulin, 5,5 g/mL transferrin and 5 ng/mL selenium), and ascorbic acid (50
µg/mL), named as MEM (Control). Canine preantral and early antral follicles were
allocated into six treatments: control medium alone (MEM) or control medium
sequentially supplemented with FSH (MEM+FSH); EGF alone at 50 ng/mL

110
(MEM+EGF50) or 100 ng/mL (MEM+EGF100) (GUPTA et al., 2002; ZHOU &
ZHANG, 2005; SILVA,et al., 2013,) or EGF with sequential FSH (MEM+EGF50+FSH
and MEM+EGF100+FSH). FSH (Folltropin-V, Bioniche Animal Health, Belleville,
ON, Canada) was added sequentially at previously determined concentrations (100
ng/mL until Day 6, 500 ng/mL until Day 12, and 1000 ng/mL until Day 18 of culture)
(SERAFIM et.al., 2010).
In the Experiment II, two different basic media were compared 1) Eagle‘s α-MEM
(Irvine Scientific, Santa Ana, CA, USA) and 2) MEM. The MEM plus 100 ng/ml EGF
and sequential FSH (MEM+EGF100+FSH) from the Experiment I, was used in the
Experiment II as a Control group. The same concentration of EGF (100ng/ml) and FSH
were used in all treatments groups in Experiment II. The follicles were cultured in three
different systems, 1) two dimensional system (2D) (SERAFIM et. al, 2012) with the
follicles in a drops of medium in direct contact with a culture dish, 2) in a three
dimensional system with alginate hydrogel matrix (ALG) (SONGSASEN et al., 2012)
or 3) in a three dimensional fibrin–alginate matrix (FA-IPN) (XU et al., 2013).
All follicles in both Experiments (I and II), were cultured at 39 ºC with 5% CO2, and 60
µL of the medium was changed every 2 days with except of days 6 and 12 where the
medium was exchanged completely (100 µl).
3. Statistical analysis
Antrum formation and chromatin integrity data were analyzed using a Chi square test
with the results expressed as percentages. Follicle and oocyte/zona pellucida diameter
results were first evaluated by Shapiro-Wilk and Levene‘s test to confirm normality and
the homogeneity of variances, respectively, using Sigma Stat software (SigmaStat;
SPSS Inc, Chicago, IL, USA). Follicular diameters met both criteria and were
subsequently analyzed via Analysis of Variance (ANOVA), followed by the Tukey's
honestly significant difference test. A probability of P < 0.05 indicated a significant
difference and results were expressed as mean ± S.E.M.
4. Results

111
4.1 Study 1: Influence of EGF
4.1.1. Follicular growth
In Experiment I, the rate of follicular growth at the end of culture (Day 18), was
significantly higher in MEM with 100 ng/ml EGF and FSH than other treatments
groups, except MEM with 50 ng/ml EGF (P˂0.05) (Table 01).
4.1.2. Antrum expansion
Regarding the percentage of antrum expansion in Experiment I, the addition of EGF to
the culture medium had no effect at any concentrations tested (Table 1). In general, the
rate of antrum expansion increased significantly from Day 6 to Day 18 in all the
treatments, but no significant changes occurred among treatments independent of
culture period (P<0.05).
4.1.3. Oocyte diameter
Tables 1 display oocyte diameter including the zona pellucida, in the Experiments I at
days 0 and 18 of culture. Although a significant increase in oocyte diameter was seen
between the onset of culture and day 18, there were no significant differences among
treatments evaluated in either Experiment I and II. (overall mean 114 µm).
4.1.4. Chromatin configuration
Representative oocytes with chromatin Intact, Fragmented and Unclassifiable are shown
in Figure 01 a, b and c, respectively. In the Experiment I, the addition of EGF at 100
ng/ml plus FSH (MEM+EGF100+FSH), exhibited a high percentage (P< 0.05) of
oocytes with intact chromatin when compared to the other treatments (Table 03)
4.2. Study 2: Influence of culture media and condition
4.2.1. Follicular growth

112
In Experiment II, the follicles cultured in αMEM showd a significantly higher
percentage of growth from day 0 to day 18 of culture compared to those cultured in
MEM (Table 02). Furthermore, no effect of culture systems (2D or 3D) on follicular
growth was detected (Figure 02).
4.2.2. Antrum expansion
Between days 6 and 18 of culture, we found an increase in the antrum formation rates
for all treatments evaluated (P< 0.05). However, in general, there is no effect of basic
media and system of culture on rate of antrum formation, since there are no differences
among the treatments when compared each other in the same day of culture (P< 0.05)
(Table 2).
4.2.3. Oocyte diameter
The oocyte diameter significant increased from the day 0 at the end of culture in all
treatments evaluated (Table 2). However, after 18 days of culture, all the conditions of
culture submitted (two or three dimensions) and both basic media (MEM and αMEM)
when compare all treatments within the same day of culture, there is no difference
(P<0.05) among the treatments evaluated.
4.2.4. Chromatin configuration
For Experiment II, the use of α-MEM as basic media, was more beneficial in
maintaining chromatin integrity than with MEM (P<0.05), independent of culture
system used (see Table 04). Furthermore, domestic dog preantral follicles were not
affected by culture system used, since the 2D system, 3D in alginate hydrogel and fibrin
and alginate matrix did not differ in the percentages of oocytes with intact chromatin at
the end of 18 days of culture.
5. Discussion

113
To our knowledge, this is the first study to evaluate the effect of different concentrations
of EGF, and its interaction with FSH, on domestic dog ovarian follicles. Further, is the
first comparison of three types of culture systems with two different basic media on dog
in vitro folliculogenesis. The concentrations of EGF tested in our study (50 and 100
ng/ml) were chosen based on studies in other species, including the goat (ZHOU &
ZHANG, 2005; SILVA et al., 2013), buffalo (GUPTA et al., 2002), and sheep
(ANDRADE et al., 2005).
In the current study, our data demonstrated the combination high concentrations of EGF
(100 ng/ml) and sequentially supplemented FSH (EGF100+FSH), was beneficial to dog
follicular growth. Several studies have shown that EGF plays an important role in
folliculogenesis, acting as a potent mitotic factor for follicular cells, and promoting
follicular growth in goats (CELESTIBO et al., 2009, SILVA et al., 2013), cows
(GUTIERREZ et al., 2000), sheep (HEMAMALINI et al., 2003); rats (DEMEESTERE
et al., 2005) and buffalos (GUPTA et al., 2008). In addition, the presence of FSH added
sequentially used in this study mimics the pulsatile and increasing patterns of FSH
secretion that occurs in vivo in the bitch. (KOOISTRA et al., 1999; ENGLAND et al.,
2009), and is possibly accompanied by an increase in FSH receptors that will support
the higher concentrations during the final phases of follicular growth (EVENCEN et al.,
2011, SARAIVA et al., 2011).
Results also show that the combination of EGF and FSH (100 ng/ml) added to the
culture medium promoted rates of follicular growth, as well as oocyte meiotic
competence, in swine (MAO et al, 2004), buballine (GUPTA et al., 2002) and goats
(SILVA et al., 2002). In the domestic dog, the presence of EGF at 100 ng/ml along with
FSH (1 µg/ml) in IVM media promotes an increase in the meiotic resumption rates of
oocytes (CUI et al., 2006; SONG et al, 2011). Previous studies have suggested that both
EGF and FSH can effectively activate the mitogen-activated protein kinase (MAPK)
cascade in granulosa cells to promote cell proliferation (MAIZELS et al., 1998).
Furthermore, studies have demonstrated that the expression of the EGF gene in
follicular cells is under FSH control, through stimulating the expression of EGF genes,
and therefore the two play synergistic roles in the promotion of follicular development
(ROY & HARRIS 1994; WU et al., 2007).

114
In both Experiments (I and II) there was no effect of EGF, FSH, basic media (MEM or
αMEM) or culture system (2D or 3D) on expansion of the antral cavity and oocyte
diameter at the end of culture. It is possible that differences among treatment groups
may have been obscured by the efficiency of other supplements added to basic media,
based on previous work by this laboratory for their role in stimulating follicle
development (SARAIVA et al., 2010, SILVA et al., 2013). For example, the addition of
ascorbic acid to goat preantral follicles improved rates of antrum formation (SILVA et
al., 2011), and has also been used in the culture media of domestic dog preantral
follicles (SERAFIM et al., 2010). The probable role of ascorbic acid is on the
preservation of basement membrane integrity, providing support for follicular
development and consequent antrum expansion. Further, the presence of insulin (10
µg/ml) in the basic media has been reported to be an important factor for the stimulation
of antrum formation in this species (SERAFIM et al., 2012).
Recently, research with the alginate hydrogel 3D culture system has provided
encouraging results in the culture of follicles from the buffalo (GUPTA et al., 2008),
non-human primate (XU et al. 2009a, 2011), human (XU et al., 2009b), and the
domestic dogs (SONGSASEN et al, 2011). Even more recently, the use of fibrin-
alginate interpenetrating network demonstrated the importance of providing a dynamic
cell-responsive mechanical microenvironment for cultured mouse follicles through
degradation of fibrin by follicle-secreted proteases (SHIKANOV et al., 2011; XU et al,
2013). However, in our present study, there was no effect of culture system (2D, ALG
or FA-IPN) on follicular growth, antrum formation or chromatin integrity. Likely, our
culture conditions (basic media and supplements), provided a propitious environment to
support the follicular development, independent of the system selected for culture.
Regarding the two basic culture media evaluated, our results demonstrated that αMEM
better supported the in vitro growth of domestic dog follicles compared with MEM.
This was likely due to an increased need for more non-essential amino acids for
folliculogenesis in this species, as observed by Fujihara, et al, (2012), in a recent study
culturing domestic dog ovarian tissue.
In conclusion, the growth, development and chromatin integrity of domestic dog
ovarian follicles grown in vitro was improved in the presence of FSH associated to EGF
in a dose-dependent way, and with αMEM as the basic media. However, the three

115
dimensional culture systems did not confer any additional benefit to in vitro
folliculogenesis in the dog.
Figure 01: Domestic dogs oocytes stained with Hoechst 33342, and classified chromatin
configutation as Intact (a), Fragmented (b) and Unclassifiable (c).
Figure 02: Domestic dog preantral follicles in different in vitro culture systems for 18 days.

116
TABLE 01:, Antrum formation, follicular growth and oocyte diameter of domestic dog preantral follicles cultured for
18 days in the presence or absence of FSH and EGF (50 or 100 ng/ml).
Treatments Antrum Formation (%)
Follicular
Growth (%)
Oocyte plus ZP diamater (µm)
D6 D18 D0-D18 D0 D18
MEM 13,95Ab
58.14 Aa
26.35±2.60 CB
60.54±3,60 Ab
118.18±9.07 Aa
MEM+ FSH 29,41 Ab
47.05 Aa
23.88±2.73 C 70.25±2,64
Ab 113.37±5.2,6
Aa
MEM+EGF50 27,45 Ab
54.90 Aa
31.70±3.89 AB
61.70±2,84 Ab
121. 48±3,02 Aa
MEM+EGF100 19.23 Ab
44.23 Aa
20.93±2.62 C 67.45±2,79
Ab 113,13±2,28
Aa
MEM+EGF50+FSH 30.35 Ab
53.57 Aa
22.71±2.24 C 65,21±2,79
Ab 113,04±2,81
Aa
MEM+EGF100+FSH 39.62 Ab
41.50 Aa
35.44±4.01 A 70.22±2,93
Ab 104,43±6,66
Aa
(A,B,C): Significant difference among treatments in the same column (P<0.05)
(a,b): Significant difference among treatments in the same days of culture at the same parameter of evaluation (P<0.05)
TABLE 02: Antrum formation, follicular growth and oocyte diameter of domestic dog preantral follicles cultured for 18
days of culture in media MEM or αMEM, and in different culture systems: under mineral oil (2D), enclosed in a
alginate-only matrix (ALG) or in a fibrin- alginate hydrogel (FA-IPN).
Treatments
Antrum Formation (%) Follicular
Growth (%)
Oocyte/ZP diamater (µm)
D6 D18 D0-D18 D0 D18
MEM 2D
(CONTROL)
36,21 Ab
64,91Aa
33.19±3.39
B 66.81±2.75
Ab 97.72±2.07
Aa
MEM ALG 26,32 Ab
70,00Aa
33.72±3.39 B 68.37±2.54
Ab 99.28±4.33
Aa
MEM FA-IPN 18,18 Ab
75,00 Aa
41.36± 5.46 B 75.38±3.09
Ab 105.00±2.49
Aa
αMEM 2D 28,57 Ab
52,27 Aa
50.63±5.10 A 68.73±1.94
Ab 111.53±21,87
Aa
αMEM ALG 35,29 Ab
60,00 Aa
50.43±3.04 A 72.64±2.61
Ab 103.23±2,09
Aa
αMEM FA-IPN 34,38 Ab
76,67 Aa
59.45±3,2 A 69.64±66.81
Ab 99.28±4.33
Aa
(A,B,C): Significant difference among treatments in the same column (P<0.05)
(a,b): Significant difference among treatments in the same days of culture at the same parameter of evaluation (P<0.05)
TABLE 03: Chromatin integrity of domestic dog oocytes obtained from ovarian follicles
after 18 days of culture in media in the presence or absence of FSH and EGF (50 or 100
ng/ml).
Treatment Intact Fragmented Unclassifiable TOTAL
MEM 6 (15.78%) B 29 (76,31%)
A 3 (7,89%)
B 38
MEM+ FSH 6 (17.14%) B 24 (68,57%)
B 5 (15,28%)
A 35
MEM+EGF50 6 (16.21%) B 29 (78,37%)
A 2 (5,4%)
B 37
MEM+EGF100 9 (19.56%) B 32 (69,56%)
B 5 (10,86%)
A 46
MEM+EGF50+FSH 3 (7.89%) C 32 (84,21%)
A 3 (7,89%)
B 38
MEM+EGF100+FSH 16 (44,44%) A 17 (47.22%)
C 3 (8,33%)
B 36
(A,B,C): Significant difference among treatments in the same column (P<0.05)

117
TABLE 04: Chromatin integrity of domestic dog oocytes obtained from ovarian follicles after
18 days of culture in media MEM or αMEM, and in different culture systems: under mineral oil
(2D), enclosed in an alginate-only matrix (ALG) or in a fibrin- alginate hydrogel (FA-IPN).
Treatment Intact Fragmented Unclassifiable TOTAL
MEM 2D
(CONTROL)
7 (29.16%)B 15 (62.50%)
B 10(41.66%)
A
32
MEM ALG 6 (28.57%)B 18 (85.71%)
A 6(28.57%)
AB 30
MEM FA-IPN 5 (21.73%)B 13 (56.52%)
AB 12(52.17%)
A 30
αMEM 2D 17 (62.96%)A 11 (40.74%)
A 5(18.51%)
AB 33
αMEM ALG 18 (81.81%)A 6 (27.27%)
B 8(36.36%)
A 32
αMEM FA-IPN 17 (77.27%)A 4 (18.18%)
C 9(40.90%)
A 30
(A,B,C): Significant difference among treatments in the same column (P<0.05)
Referências
ANDRADE, E.R., SENEDA, M.M., ALFIERI, A.A., OLIVEIRA, J.A.,
BRACARENSE, A.P.F.R.L., FIGUEIREDO, J.R., TONIOLLI, R. Interactions of
indole acetic acid with EGF and FSH in the culture of ovine preantral follicles.
Theriogenology 64, 1104– 1113, 2005.
ANDRADE, E.R.; VAN DER HURK, R.; LISBOA, L.A.; HERTEL, M.F.; MELO-
STERZA, F.A.; MORENO, K.; BRACARENSE, A.P.F.R.L.; LANDIM-
ALVARENGA, F.C.; SENEDA, M.M.; ALFIERI, A.A. Effects of ascorbic acid on in
vitro culture of bovine preantral follicles. Zygote, p.1-10, 2012.
ARUNAKUMARI, G., SHANMUGASUNDARAM, N., RAO, V.H. Development of
morulae from the oocytes of cultured sheep preantral follicles Theriogenology 2010; 74:
884–894.
BOLAMBA, D.; BORDEN-RUSS, K.D.; DURRANT, B.S. In vitro maturation of bitch
oocytes from advanced preantral follicle in synthetic oviduct fluid medium: serum is not
essencial. Theriogenology, v.58, p.1689-1703, 2002.
BOLAMBA, D.; RUSS, K.D.; HARPER, S.A.; SANDLER, J.L.; DURRANT, B.S.
Effects of epidermal growth factor and hormones on granulosa expansion and nuclear
maturation of dog oocytes in vitro. Theriogenology, v. 65, p. 1037-1047, 2006.
BOLAMBA,D.; BORDEN-RUSS, K. D.; DURRANT, B. S. In vitro maturation of
domestic dog oocytes cultures in advanced preantral and early antral follicles.
Theriogenology, v. 49, n.5, p. 933-942, 1998.

118
CELESTINO, J.J.H.; BRUNO, J.B.; LIMA-VERDE, I.B.; MATOS, M.H.T.;
SARAIVA, M.V.A.; CHAVES, R.N.; MARTINS, F.S.; LIMA, L.F.; NAME, K.P.O.;
CAMPELLO, C.C.; SILVA, J.R.V.; BÁO, S.N.; FIGUEIREDO, J.R. Recombinant
epidermal growth factor maintains follicular ultrastructure and promotes the transition
to primary follicles in caprine ovarian tissue cultured in vitro. Reproductive Science, v.
16, p. 239-246, 2009.
COMIZZOLI, P.; MERMILLOD, P.; MAUGET R.; Reproductive biotechnologies for
endangered mammalian species. Reprod. Nutr. Dev.; v.40, p.493–504, 2000.
CONTI, M.; HSIEH, M.; PARK, J.Y.; SU, Y.Q. Role of the epidermal growth factor
network in ovarian follicles. Molecular Endocrinology, v. 20, p. 715-723, 2006.
CUI, X.S.; JIN Y.X.; SHEN, X.H.; LEE , J. Y.; LEE H.S.; YIN X.J.; KONG I.K.; KIM,
N.H.; Epidermal growth factor enhances meiotic resumption of canine oocytes in the
presence of BSA. Theriogenology v.66, p.267–274, 2006.
DEMEESTERE, I.; CENTNER, J.; GERVY, Y.; DELBAERE, A. Impact of various
endocrine and paracrine factors on in vitro culture of preantral follicles in rodents.
Reproduction, v. 130, p. 147-156, 2005.
DING T., LUO A., YANG S., LAI Z., WANG Y., SHEN W., JIANG J., LU Y., MA,
D., WANG, S. Effects of basal media and supplements on diethylstilbestrol-treated
immature mouse primary granulosa cell growth and regulation of steroidogenesis in
vitro. Reprod Dom Anim v.47, p.355–364, 2012.
DURRANT B.S.; PRATT N.C.: RUSS K.D.; BOLAMBA D.; Isolation and
characterization of canine advanced, preantral and early antral follicles.
Theriogenology; v.49, p.917-932, 1998.
EGF REFERÊNCIAS
ENGLAND G.C.W., RUSSO, M., Freeman S.L. Follicular dynamics, ovulation and
conception rates in bitches. Reproduction in Domestic Animals v.44, p.53–58, 2009.
EVECEN M., CIRITB U., DEMIRA KHAMZAOGLUA., A. I., BAKIRERA G.,
PABUCCUOGLUA S., BIRLERA S.; Adding hormones sequentially could be an
effective approach for IVM of dog oocytes, Theriogenology, v.75, p.1647-51, 2011.
FUJIHARA M., COMIZZOLI P., WILDT, D.E. SONGSASEN N.; Cat and Dog
Primordial Follicles Enclosed in Ovarian Cortex Sustain Viability after In vitro Culture
on Agarose Gel in a Protein-Free Medium. Reprod Dom Anim, v.47, p.102–108, 2012.
GUPTA P.S.P.; NANDI,S.; RAVINDRANATHA, B.M. ; SARMA, P.V. In vitro
culture of buffalo (Bubalus bubalis) preantral follicles.Theriogenology v.57, p. 1839-
1854., 2008.

119
GUPTA P.S.P.; NANDI; S.; RVINDRANATHA B.M.; SARMA P.V.; In vitro culture
of buffalo (Bubalus bubalis) pre antral follicles. Theriogenology, 57, 1839-1854, 2002.
GUTIERREZ, C.G.; RALPH, J.H.; TELFER, E.E.; WILMUT, I.; WEBB, R. Growth
and antrum formation of bovine preantral follicles in long-term culture in vitro. Biology
of Reproduction, v. 62, p. 1322-1328, 2000.
HARTSHORNE G.;. In vitro culture of ovarian follicles. Rev Reprod, v.2, p.94-104.
1997
HEMAMALINI, N.C.; RAO, B.S.; TAMILMANI, G.; AMARNATH, D.; VAGDEVI,
R.; NAIDU, K.S.; REDDY, K.K.; RAO, V.H. Influence of transforming growth factor-
α, insulin-like growth factor-II, epidermal growth factor or follicle stimulating hormone
on in vitro development of preantral follicles in sheep. Small Rumin. Res., v. 50, p. 11-
22, 2003.
JEWGENOW K. Impact of peptide growth factors on the culture of small preantral
follicles of domestic cats. Theriogenology, v.45, p.889-895, 1996.
JEWGENOW, K.; PARIS, M.C.J.; Preservation of female germ cells from ovaries of
cat species. Theriogenology. v.66. p. 93–100, 2006.
JIN, S.Y.; LEI, L.; SHIKANOV, A.; SHEA, L.D.; WOODRUFF, T.K. A novel two-
step strategy for in vitro culture of early-stage ovarian follicles in the mouse. Fertility
and Sterility. v. 93, n. 8, p. 2633-9, 2010.
KOOISTRA KS, OKKENS AC, BEVERS MM, Concurrent pulsatile secretion of
luteinizing hormone and follicle-stimulating hormone during different phases of the
estrous cycle and anestrus in beagle bitches. Biol Reprod; v.60, p.65-71, 1999.
KREEGER PK, DECK JW, WOODRUFF TK, SHEA LD. The in vitro regulation of
ovarian follicle development using alginate-extracellular matrix gels. Biomaterials;v.27,
p.714–723, 2006.
LANE, M., GARDNER, D.K., Embryo culture medium: which is the best? Best Pract
Res Clin Obstet Gynaecol v.21,p. 83–100, 2007.
LOPES G., SOUSA M., LUVONI C.G., ROCHA A. Recovery rate, morphological
quality and nuclear maturity of canine cumulus-oocyte complexes collected from
anestrous or diestrous bitches of different ages, Theriogenology v.68, p.821–825, 2007.
MAGALHÃES D.M., DUARTE A.B.G., ARAÚJO V.R., BRITO I.R., SOARES T.G.,
LIMA I.M.T., LOPES C.A.P., CAMPELLO C.C., RODRIGUES A.P.R.,
FIGUEIREDO J.R.;In vitro production of a caprine embryo from a preantral follicle
cultured in media supplemented with growth hormone, Theriogenology, v.75, p.182–
188, 2011.

120
MAIZELS, E.T., COTTOM. J,, JONES, J.C., HUNZICKER-DUN0N, M. Follicle
stimulating hormone (FSH) activates the p38 mitogen-activated protein kinase pathway,
inducing small heat shock protein phosphorylation and cell rounding in immature rat
ovarian granulosa cells. Endocrinology. v.139, p.3353-6, 1998.
MAO, J.; SMITH, M.F.; RUCKER, E.B.; WU, G.M.; McCAULEY, T.C.; CANTLEY,
T.C.; PRATHER, R.S.; DIDIONS, B.A.; DAY, B.N. Effect of epidermal growth factor
and insulin-like growth factor I on porcine preantral follicular greowth, antrum
formation, and stimulation of granulosal cell proliferation and supperssion of apoptosis
in vitro. Journal of Animal Science, v. 82, p. 1967-1975, 2004.
MAO, J.; SMITH, M.F.; RUCKER, E.B.; WU, G.M.; McCAULEY, T.C.; CANTLEY,
T.C.; PRATHER, R.S.; DIDIONS, B.A.; DAY, B.N. Effect of epidermal growth factor
and insulin-like growth factor I on porcine preantral follicular greowth, antrum
formation, and stimulation of granulosal cell proliferation and supperssion of apoptosis
in vitro. Journal of Animal Science, v. 82, p. 1967-1975, 2004.
MORBECK, D.E.; FLOWERS, W.L.; BRITT, J.H. Response of porcine granulosa cells
isolated from primary and secondary follicles to FSH, 8-bromo-cAMP and epidermal
growth factor in vitro. Journal of Reproduction and Fertility, v. 99, p. 577–584, 1993.
NONOWAKI, S.; TAKAHASHI, K.; HORIUCHI, T .Culture Media Affect Follicle
Survival and Oocyte Maturation in Preantral Mouse Follicle Cultures. Journal of
Mammalian Ova Research, v. 27, p. 35-41, 2010.
ROSSETO,R.; SARAIVA, M.V.A.; SANTOS,R.R.; SILVA,C.M.G.; FAUSTINO,L.R.;
CHAVES,R.N.; BRITO,I.R.; RODRIGUES,G.Q.; LIMA,I.M.T.; DONATO, M.A.M.;
PEIXOTO, C.A.; FIGUEIREDO, J.R.Effect of medium composition on the in vitro
culture of bovine pre-antral follicles: morphology and viability do not guarantee
functionality.Zygote, v.21, p.125-128, 2012.
ROY, S.K. & HARRIS, S.G. Antisense epidermal growth factor oligodeoxynucleotides
inhibit follicle-stimulating hormone-induced in vitro DNA and progesterone, synthesis
in hamster preantral follicles. Mol Endocrinol v.8, p.1175–81,1994.
ROY, S.K.; KOLE, A.R. Ovarian transforming growth factor-beta (TGF-beta)
receptors: in vitro effects of follicle stimulating hormone, epidermal growth factor and
TGF-beta on receptor expression in human preantral follicles. Molecular Human
Reproduction, v. 3, p. 207-214, 1998.
SARAIVA, M.V.A., ROSSETTO, R., BRITO, I.R., CELESTINO, J.J.H., SILVA,
C.M.G., FAUSTINO, L.R., ALMEIDA, A.P., BRUNO, J.B., MAGALHÃES, D.M.,
MATOS, M.H.T., CAMPELLO, C.C., FIGUEIREDO, J.R. Dynamic medium produces
caprine embryo from preantral follicles grown in vitro. Reproductive Science v.12,
p.1135–1143, 2010.

121
SARAIVA, M.V.A.; CELESTINO, J.J.H.; ARAÚJO, V.R.; CHAVES, R.N.;
ALMEIDA, A.P.; LIMA-VERDE, I.B.; DUARTE, A.B.G.; SILVA, G.M.; MARTINS,
F.S.; BRUNO, J.B.; MATOS, M.H.T.; CAMPELLO, C.C.; SILVA, J.R.V.;
FIGUEIREDO, J.R. Expression of follicle-stimulating hormone receptor (FSH-R) in
goat ovarian follicles and the impact of sequential culture medium on in vitro
development of caprine preantral follicles. Zygote, v. 19, p. 205-214, 2011.
SERAFIM, M. K.; ARAUJO, V. R.; SILVA, G. M.; DUARTE, A. B.; ALMEIDA, A.
P.; CHAVES, R. N.; CAMPELLO, C. C.; LOPES, C. A.; FIGUEIREDO, J. R.; SILVA,
L. D. Canine preantral follicles cultured with various concentrations of follicle-
stimulating hormone (FSH). Theriogenology, v. 74, p. 749-55, 2010.
SERAFIM, M. K.; SILVA, G. M.; DUARTE, A. B.; ARAÚJO, V. R.; SILVA, T. F.;
LIMA, A. K.; CHAVES, R. N.; CAMPELLO, C. C.; SILVA, L. D.; FIGUEIREDO, J.
R. High insulin concentrations promote the in vitro growth and viability of canine
preantral follicles. Reproduction, Fertility and Development, 2012.
SHIKANOV A., SMITH R. M., XU M., WOODRUFF T. K., SHEA, L.D. Hydrogel
network design using multifunctional macromers to coordinate tissue maturation in
ovarian follicle culture. Biomaterials, v.32, p 2524-2531, 2011.
SHIKANOV, A., XU, M., WOODRUFF, T.K., SHEA, L.D. Interpenetrating
fibrinalginate matrices for in vitro ovarian follicle development. Biomaterials v.30,
p.5476–5485, 2009.
SILVA, C. M. G.; CASTRO, S. V.; FAUSTINO, L. R.; RODRIGUES, G. Q.; BRITO,
I. R.; ROSSETTO, R.; SARAIVA, M. V.; CAMPELLO, C. C.; LOBO, C. H.; SOUZA,
C. E.; MOURA, A. A.; DONATO, M. A.; PEIXOTO, C. A.; FIGUEIREDO, J. R. The
effects of epidermal growth factor (EGF) on the in vitro development of isolated goat
secondary follicles and the relative mRNA expression of EGF, EGF-R, FSH-R and
P450 aromatase in cultured follicles. Research in Veterinary Science, v. 94, p. 453-461,
2013.
SILVA, G.M.; ARAÚJO, V.R.; DUARTE, A.B.G.; CHAVES, R.N.; SILVA, C.M.G.;
LOBO, C.H.; ALMEIDA, A.P.; MATOS, M.H.T.; TAVARES, L.M.T.; CAMPELO,
C.C.; FIGUEIREDO, J.R. Ascorbic acid improves the survival and in vitro growth of
isolated caprine preantral follicles. Animal Reproduction, v. 8, p. 14-24, 2011.
SILVA, J.R.V; FERREIRA, M.A.L.; COSTA, S.H.F.; SANTOS, R.R.; CARVALHO,
F.C.A; RODRIGUES, A.P.R.; LUCCI, C.M.; BÁO, S.N.; FIGUEIREDO, J.R.

122
Degeneration rate of preantral follicles in the ovaries of goats. Small Ruminant
Research, v. 43, p. 203-209, 2002.
SONG, H.J.; KANG, E.J.; MAENG, G.H.; OCK, S.A.; LEE, S.L.; YOO, J.G.; JEON,
B.G.; RHO, G.J. Influence of epidermal growth factor supplementation during in vitro
maturation on nuclear status and gene expression of canine oocytes. Research in
Veterinary Science, v. 91, p. 439-445, 2011.
SONGSASEN, N., FICKES, A., PUKAZHENTHI B.S., WILDT, D.E. Follicular
Morphology, Oocyte Diameter and Localisation of Fibroblast Growth Factors in the
Domestic Dog Ovary. Reprod Dom Anim v.44, p. 65–70, 2009.
SONGSASEN, N., WILDT, D.E.; Oocyte biology and challenges in developing in vitro
maturation systems in the domestic dog; Animal Reproduction Science, v.98, p.2–22.
2007.
SONGSASEN, N., WOODRUFF, T.K., WILD D.E. In vitro growth and steroidogenesis
of dog follicles are influenced by the physical and hormonal microenvironment.
Reproduction v142, p.113–122, 2011.
TELFER, E.E. In vitro development of pig preantral follicles. Reprod. Suppl. 58, 81–
90, 2001.
WANDJI, S.A., SRSEN, V., VOSS, A.K., EPPIG, J.J., FORTUNE, J.E..; Initiation in
vitro of growth of bovine primordial follicles. Biol Reprod 55:942–948. 1996.
WEST, E. R.; XU, M.; WOODRUFF, T. K.; SHEA, L. D. Physical properties of
alginate hydrogels and their effects on in vitro follicle development. Biomaterials, v. 28,
p. 4439-4448, 2007.
WOODRUFF T. K., SHEA L. D. The Role of the The Role of the Extracellular Matrix
in Ovarian Follicle Development, Reproductive Sciences, v. 14, p. 6, 2007.
WU, J., XU, B., WANG, W.; Effects of luteinizing hormone and follicle stimulating
hormone on the developmental competence of porcine preantral follicle oocytes grown
in vitro. Journal of Assisted Reproduction and Genetics v. 24. p.419–424, 2007.
WU, J.; TIAN, Q. Role of follicle stimulating hormone and epidermal growth factor in
the development of porcine preantral follicle in vitro. Zygote, v. 15, p. 233-240, 2007.
XU, J., BERNUCI, M.P., LAWSON, M.S., YEOMAN, R.R., FISHER, T.E.,
ZELINSKI, M.B., STOUFFER, R.L. Survival, growth, and maturation of secondary
follicles from prepubertal, young, and older adult rhesus monkeys during encapsulated
three-dimensional culture: effects of gonadotropins and insulin. Reproduction v.140, p.
685–697, 2010.
XU, J., LAWSON, M.S., YEOMAN, R.R., MOLSKNESS T.A., TING, A.Y.,
STOUFFER, R.L., ZELINSK, M.B. Fibrin promotes development and function of

123
macaque primary follicles during encapsulated three-dimensional culture. Human
Reproduction, v.0, p.1–14, 2013.
XU, J., LAWSON, M.S., YEOMAN, R.R., MOLSKNESS T.A., TING, A.Y.,
STOUFFER, R.L., ZELINSK, M.B. Fibrin promotes development and function of
macaque primary follicles during encapsulated three-dimensional culture. Human
Reproduction, v.0, p.1–14, 2013.
XU, J., LAWSON, M.S., YEOMAN, R.R., PAU, K.Y., BARRETT, S.L., ZELINSKI,
M.B., STOUFFER, R.L. Secondary follicle growth and oocyte maturation during
encapsulated three-dimensional culture in rhesus monkeys: effects of gonadotrophins,
oxygen and fetuin. Hum. Reprod. v.26, p.1061–1072, 2011.
XU, M., BARRETT, S.L.,WEST-FARRELL, E., KONDAPALLI, L.A.,
KIESEWETTER, S.E., SHEA, L.D., WOODRUFF, T.K. In vitro grown human ovarian
follicles from cancer patients support oocyte growth. Human Reproduction v.24,
p.2531–2540, 2009a.
XU, M., WEST-FARRELL, E.R., STOUFFER, R.L., SHEA, L.D., WOODRUFF ,T.K.,
ZELINSKI, M.B. Encapsulated three-dimensional culture supports development of non-
human primate secondary follicles. Biology of Reproduction v.81, p.587–594, 2009b.
XU, M.; WEST, E.; SHEA, L.D.; WOODRUFF, T.K. Identification of a stage-specific
permissive in vitro culture environment for follicle growth and oocyte development.
Biology of Reproduction, v. 75, p. 916-923, 2006.
XU, M.; WEST-FARRELL, E.R.; STOUFFER, R.L.; SHEA, L.D.; WOODRUFF, T.K.;
ZELINSKI, M.B. Encapsulated three-dimensional culture supports development of
nonhuman primate secondary follicles. Biology of Reproduction, v. 81, p. 587-594,
2009.
ZHOU, H.M.; ZHANG, Y. Impact of growth factors on in vitro development of caprine
oocytes at pre-antral stage. Reproduction in Domestic Animals, v. 40, p. 161-165, 2005.

124
9.CONCLUSÕES
Somente na ausência de FSH, a presença de altas concentrações de insulina teve
efeito benéfico sobre a viabilidade folicular. Entretanto, para a promoção do
crescimento de folículos pré-antrais caninos, recomenda-se a associação da
insulina e do FSH.
O GH de forma dose dependente beneficiou a manutenção da morfologia
folicular e sua associação ao FSH garantiu um incremento no crescimento
folicular in vitro.
O desenvolvimento folicular e a manutenção da integridade da cromatina nos
folículos ovarianos caninos domésticos foram melhores na presença do FSH
associado ao EGF e utilizando-se o α-MEM como meio de base.
O tipo de sistema de cultivo (2D vs 3D) não influenciou o desenvolvimento in
vitro de FOPA caninos isolados.

125
10. PERSPECTIVAS
Diante dos resultados obtidos através dos estudos da presente tese, foi possível
observar que a adição de substâncias como a insulina, GH e EGF, associados ao FSH
possibilitou um incremento no desenvolvimento folicular in vitro de folículos de
caninos domésticos. Porém, maiores estudos ainda são necessários na tentativa de se
definir um meio cultivo para folículos pré-antrais caninos que venha a proporcionar
maiores taxa de oócitos aptos a serem maturados. Aliado a isso, torna-se de suma
importância o desenvolvimento de meios específicos para a maturação in vitro de
oócitos, já que os meios disponíveis são ainda bastante ineficientes.

126
11. REFERÊNCIAS
ABDEL-GHANI, M.A., SHIMIZU T., ASANO T., SUZUKI H. In vitro maturation of
canine oocytes co-cultured with bovine and canine granulosa cell monolayers.
Theriogenology v.77, p.347–355, 2012.
ABEYDEERA, L.R., WANG, W.H., CANTLEY, T.C., RIEKE, A., PRATHER, R.S.,
DAY, B.N., Presence of epidermal growth factor during in vitro maturation of pig
oocytes and embryo culture can modulate blastocyst development after in vitro
fertilization. Mol. Reprod. Dev. v.51, p.395–401, 1998.
ABIR, R. NITKE, A. BEN-HAROUSH, A., FISCH, B. In vitro maturation of
primordial ovarian follicles: Clinical significance, progress in mammals, and methods
for growth evaluation. Histology and Histopatology. V. 21, p. 887-898, 2006.
ABIR, R.; ROIZMAN, P.; FISCH, B.; NITKE, S.; OKON, E.; ORVIETO, R.; BEN
RAFAEL, Z. Pilot study of isolated early human follicles cultured in collagen gels for
24 hours. Human Reproduction, v. 14, p. 1299-1301, 1999.
ALBERTINI, D. F.; BARRETT, S. L. Oocyte-somatic cell communication.
Reproduction Supplement, v. 61, n. 3, p. 49–54, 2003.
ALM, H.; KATSKA-KSIAZKIEWICZ, L.; RYŃSKA, B.; TUCHSCHERER, A.
Survival and meiotic competence of bovine oocytes originating from earlyantral ovarian
follicles. Theriogenology, v. 65, p. 1422–1434, 2006.
ALON, T.; HEMO, I.; ITIN, A.; PE‘ER, J.; STONE, J.; KESHET, E. Vascular
endothelial growth factor acts as a survival factor for newly formed retinal vessels and
has implications for retinopathy of prematurity. Nature Medicine, v.1, p.1024-8, 1995.
ANDERSEN, A. C.; SIMPSON, M.E. The ovary and Reproductive Cycle of the dog
(Beagle). Los Altos: Geron- X, Inc., 1973
ANDRADE, E.R., SENEDA, M.M., ALFIERI, A.A., OLIVEIRA, J.A.,
BRACARENSE, A.P.F.R.L., FIGUEIREDO, J.R., TONIOLLI, R. Interactions of
indole acetic acid with EGF and FSH in the culture of ovine preantral follicles.
Theriogenology 64, 1104– 1113, 2005.
ARAÚJO V.R., GASTAL M.O., FIGUEIREDO J.R., GASTAL, E.L. In vitro culture of
bovine preantral follicles: A review. Molecular Reproduction and Development, in
press.

127
ARUNAKUMARI, G., SHANMUGASUNDARAM, N., RAO, V.H., Development of
morula from the oocytes of cultured sheep preantral follicles. Theriogenology v.74, p.
884-894; 2010.
AUGUSTIN, R.; POCAR, P.; WRENZYCKI, C.; NIEMANN, H.; FICHER, B.;
Mitogenic and anti-apoptotic activity of insulin on bovine embryos produced in vitro.
Reproduction, v.126, v.91-99, 2003.
BAO, S.; WANG, Y.; SWEENEY, P.; CHAUDHURI, A.; DOSEFF, A. I.; MARSH, C.
B.; KNOELL, D. L. Keratinocyte growth factor induces Akt kinase activity and inhibits
Fas-mediated apoptosis in A549 lung epithelial cells. American Journal of Physiology -
Lung Cellular and Molecular, v. 288, n. 1, p. 36-42, 2005.
BARBER, M.R. LEE S.M., STEFFENS W.L., ARD M, FAYRER-HOSKEN R.A.,
Immunolocalization of zona pellucida antigens in the ovarian follicle of dogs, cats,
horses and elephants. Theriogenology. Los Altos. v. 55, n. 8, p. 1705-1917, 2001.
BEARDEN, H.J. & FUQUAY, J.W.; Applied Animal Reproduction. 4ª ed. Prentice
Hall. 351p. 1997.
BEDFORD, J.M.; KIM, H.H. Cumulus oophorus as sperm sequestering device in vivo;
Journal of Exp. Zoologica. v. 265, p. 321-328, 1993.
BERKHOLTZ, C. B., LONNIE D. S., WOODRUFF, T. K. Extracellular Matrix
Functions in Follicle Maturation. Semin Reprod Med., v 24, p 262–269, 2006.
BLACKBURN, J. S., LIU, S., LANGENAU, D. M. Quantifying the Frequency of
Tumor-propagating Cells Using Limiting Dilution Cell Transplantation in Syngeneic
Zebrafish; J. Vis. Exp., v: 53, p.2790, 2011.
BOLAMBA, D.; BORDEN-RUSS, K.D.; DURRANT, B.S. In vitro maturation of bitch
oocytes from advanced preantral follicle in synthetic oviduct fluid medium: serum is not
essencial. Theriogenology, v.58, p.1689-1703, 2002.
BOLAMBA, D.; RUSS, K.D.; HARPER, S.A.; SANDLER, J.L.; DURRANT, B.S.
Effects of epidermal growth factor and hormones on granulosa expansion and nuclear
maturation of dog oocytes in vitro. Theriogenology, v. 65, p. 1037-1047, 2006.
BOLAMBA,D.; BORDEN-RUSS, K. D.; DURRANT, B. S. In vitro maturation of
domestic dog oocytes cultures in advanced preantral and early antral follicles.
Theriogenology, v. 49, n.5, p. 933-942, 1998.
BOLAND, N.I., HUMPHERSON, P.G., LEESE, H.J., GOSDEN, R.G. Pattern of
lactate production and steroidogenesis during growth and maturation of mouse ovarian
follicles in vitro. Biology of Reproduction v.48, p.798–806, 1993.

128
BRISTOL-GOULD, S., WOODRUFF, T.K. Folliculogenesis in the domestic cat (Felis
catus). Theriogenology, v.66, p.5-13, 2006.
BRUNO, J. B.; CELESTINO, J. J. H.; LIMA-VERDE, I. B.; LIMA, L. F.; MATOS, M.
H. T.; ARAÚJO, V. R.; SARAIVA, M. V. A.; MARTINS, F. S.; NAME, K. P. O.;
CAMPELLO, C. C.; BÁO, S. N.; SILVA, J. R. V.; FIGUEIREDO, J. R. Expression of
vascular endothelial growth factor (VEGF) receptor in goat ovaries and improvement of
in vitro caprine preantral follicle survival and growth with VEGF. Reproduction
Fertility and Development, v. 21, p. 679-687, 2009.
BUCCIONE, R.; SCHROEDER, A.C.; EPPIG, J.J. Interation between somatic cells and
germ cells throughout mammalian oogenesis. Biology of Reproduction, v. 43, p. 543-
547, 1990.
BUKOWSKA, D., KEMPISTY, B., PIOTROWSKA, H., ZAWIERUCHA, P.,
BRUSSOW, K.P., JAŚKOWSKI, J.M., NOWICKI, M. The in vitro culture supplements
and selected aspects of canine oocytes maturation. Polish Journal of Veterinary Sciences
v15, v. 199, p. 205, 2012.
CARPENTER, C.L.; AUGER, K.R.; CHANUDHURI, M.; YOAKIM, M.;
SCHAFFHAUSEN, B.; SHOELSON, S.; CANTLEY, L.C. Phosphoinositide 3-kinase
is activated by phosphopeptides that bind to the SH2 domains of the 85-kDa subunit.
The Journal of Biological Chemistry, v. 268, p. 9478-9483, 1993.
CARVALHEIRAS, J.B.C., ZECCHIN, H.G. SAAD M.J.A. Vias de sinalização da
insulina. Arquivo Brasileiro de Endocrinologia e Metabolismo. v.46, p. 419-425, 2002.
CECCONI, S.; BARBONI, B.; COCCIA, M.; MATTIOLI, M. In vitro development of
sheep preantral follicles. Biology of Reproduction, v. 60, p. 594-601, 1999.
CELESTINO, J. J. H.; BRUNO, J. B.; LIMA-VERDE, I. B.; MATOS, M. H. T.;
SARAIVA, M. V. A.; CHAVES, R. N.; MARTINS, F. S.; ALMEIDA, P.; CUNHA, R.
M. S.; LIMA, L. F.; NAME, K. P. O.; CAMPELLO, C. C.; SILVA, J. R. V.; BAO, S.
N.; FIGUEIREDO, J. R. Steady-state level of kit ligand mrna in goat ovaries and the
role of kit ligand in preantral follicle survival and growth in vitro. Molecular
Reproduction & Development, v. 77, p. 231-240, 2010.
CELESTINO, J.J.H.; BRUNO, J.B.; LIMA-VERDE, I.B.; MATOS, M.H.T.;
SARAIVA, M.V.A.; CHAVES, R.N.; MARTINS, F.S.; LIMA, L.F.; NAME, K.P.O.;
CAMPELLO, C.C.; SILVA, J.R.V.; BÁO, S.N.; FIGUEIREDO, J.R. Recombinant
epidermal growth factor maintains follicular ultrastructure and promotes the transition

129
to primary follicles in caprine ovarian tissue cultured in vitro. Reproductive Science, v.
16, p. 239-246, 2009.
CHANG, Y.; WANG, J.; LU, X.; THEWKE, D. P.; MASON, R. J. KGF induces
lipogenic genes through a PI3K and JNK/SREBP-1 pathway in H292 cells. The Journal
of Lipid Research, v. 46, n. 12, p. 2624-2635, 2005.
CHAVES, R. N.; DUARTE, A. B.; RODRIGUES, G. Q.; CELESTINO, J. J.; SILVA,
G. M.; LOPES, C. A.; ALMEIDA, A. P.; DONATO, M. A.; PEIXOTO, C. A.;
MOURA, A. A.; LOBO, C. H.; LOCATELLI, Y.; MERMILLOD, P.; CAMPELLO, C.
C.; FIGUEIREDO, J. R. The effects of insulin and follicle-simulating hormone (FSH)
during in vitro development of ovarian goat preantral follicles and the relative mRNA
expression for insulin and FSH receptors and cytochrome P450 aromatase in cultured
follicles. Biology of Reproduction, v. 87,2012.
CHAVES, R.N.; DUARTE, A.B.G.; RODRIGUES, G.Q.; CELESTINO, J.J.H.;
SILVA, M.G.; LOPES, C.A.P.; ALMEIDA, A.P.; DONATO, M.A.M.; MOURA,
A.A.A.; LOBO, C.; LOCATELLI, Y.; MERMILLOD, P.; CAMPELLO, C.C.;
FIGUEIREDO, J.R. The Effects of Insulin and FSH During In vitro Development of
Ovarian Goat Preantral Follicles and the Relative mRNA Expression for Insulin and
FSH Receptors and Cytochrome P450 Aromatase in Cultured Follicles. Biology of
Reproduction, v. 87, p. 1-11, 2012.
COMIZZOLI, P.; MERMILLOD, P.; MAUGET R.; Reproductive biotechnologies for
endangered mammalian species. Reprod. Nutr. Dev.; v.40, p.493–504, 2000.
CONTI, M.; HSIEH, M.; PARK, J.Y.; SU, Y.Q. Role of the epidermal growth factor
network in ovarian follicles. Molecular Endocrinology, v. 20, p. 715-723, 2006.
CUI, X.S.; JIN Y.X.; SHEN, X.H.; LEE , J. Y.; LEE H.S.; YIN X.J.; KONG I.K.; KIM,
N.H.; Epidermal growth factor enhances meiotic resumption of canine oocytes in the
presence of BSA. Theriogenology v.66, p.267–274, 2006.
DANFORTH, D. R.; ARBOGAST, L. K.; GHOSH, S.; DICKERMAN, A.;
ROFAGHA, R.; FRIEDMAN, C. I. Vascular endothelial growth factor stimulates
preantral follicle growth in the rat ovary. Biology of Reproduction, v. 68, p. 1736-1741,
2003.
DAVIDSON P.; BAKER T. W.; Reproductive Ultrasound of the Bitch and Queen;
Topics in Companion Animal Medicine, v.24, Number 2, 2009.

130
DEMEESTERE, I.; CENTNER, J.; GERVY, Y.; DELBAERE, A. Impact of various
endocrine and paracrine factors on in vitro culture of preantral follicles in rodents.
Reproduction, v. 130, p. 147-156, 2005.
DESAI N., ABDELHAFEZ F., CALABRO, A. FALCONE T. Three dimensional
culture of fresh and vitrified mouse pre-antral follicles in a hyaluronan-based hydrogel:
a preliminary investigation of a novel biomaterial for in vitro follicle maturation.
Reproductive Biology and Endocrinology, v10, p.29, 2012.
DESAI N., ALEX A., HAFEZ F.A., CALABRO A., GOLDFARB J., FLEISCHMAN
A., FALCONE, A. T. Three-dimensional in vitro follicle growth: overview of culture
models, biomaterials, design parameters and future directions. Reproductive Biology
and Endocrinology, v.8, p.119,2010.
DESAI, N., ABDELHAFEZ, F., CALABRO, A., FALCONE, T. Three dimensional
culture of fresh and vitrified mouse pre-antral follicles in a hyaluronan-based hydrogel:
a preliminary investigation of a novel biomaterial for in vitro follicle maturation.
Reprod Biol Endocrinol v.10, p.1–12, 2012.
DESAI, N., ALEX, A., ABDELHAFEZ, F., CALABRO, A., GOLDFARB, J.,
FLEISCHMAN, A., FALCONE, T. Three-dimensional in vitro follicle growth:
overview of culture models, biomaterials, design parameters and future directions.
Reproductive Biology and Endocrinology 8 119, 2010.
DOLEZEL, R ; KYLIANKOVA R., KUMMER V., MASKOVA J.; STARA P.,
VITASEK R.; Follicular population and oestrogen receptor alpha in canine ovaries
during the oestrus cycle. Reproduction in Domestic Animals. Berlin, v. 38, p. 347, 2003
DRIANCOURT, M.A. Regulation of ovarian follicular dynamics in farm animals:
Implications for manipulation of reproduction. Theriogenology, v. 55, p. 1211-1239,
2001.
DURRANT B. S; PRATT, N. C., RUSS; K. D.; BOLAMBA D.; Isolation and
characterization of canine advanced, preantral and early antral follicles, Theriogenology
v.49, p.917-932, 1998.
ECKERY, D.C.; MOELLER, C.L.; NETT, T.M.; SAWYER, H.R. Localization and
quantification of binding sites for follicle-stimulating hormone, luteinizing hormone,
growth hormone, and insulin-like growth factor I in sheep ovarian follicles. Biology of
Reproduction; v.57, p.507–513, 1997.

131
EISELT, P., YEH, J., LATVALA, R,K., SHEA, L.D., MOONEY, D.J.: Porous carriers
for biomedical applications based on alginate hydrogels. Biomaterials , v.21, p.1921-
1927, 2000.
ENGELMAN, J. A.; LUO, J.; CANTLEY, L. C. The evolution of phosphatidylinositol
3-kinases as regulators of growth and metabolism. Nature Reviews Genetics, v. 7, n. 8,
p. 606-619, 2006.
ENGLAND, G.C.W.; HEWITT, D.A. Follicular growth and ovulation in dogs. In:
EVSSAR Annual Symposium. Lyon, p. 51. 1999.
EPPIG, J. J., O'BRIEN M. J.; Development In Vitro of Mouse Oocytes from Primordial
Follicles. Biol. Reprod. v.54, p.197-207, 1996.
EPPIG, J.J.; O‘BRIEN, M.J.; PENDOLA, F.L.; WATANABE, S.; Factors affecting the
developmental competence of mouse oocytes grown in vitro: follicle-stimulating
hormone and insulin. Biology of Reproduction v.59, p.1445–1453, 1998.
ERICKSON, G.F.; SHIMASAKI, S. The spatiotemporal expression pattern of the bone
morphogenic protein family in rat ovary cell types during the estrous cycle.
Reproduction Biology and Endocrinology, v.1, p. 1-20, 2003.
EVECEN M., CIRITB U., DEMIRA KHAMZAOGLUA., A. I., BAKIRERA G.,
PABUCCUOGLUA S., BIRLERA S.; Adding hormones sequentially could be an
effective approach for IVM of dog oocytes, Theriogenology, v.75, p.1647-51, 2011.
FARSTAD, W.; Current state in biotechnology in canine and feline reproduction.
Animal Reproduction Science. v.61, p. 375-387, 2000.
FIGUEIREDO J.R, AMORIM CA, LUCCI CM, GONÇALVES PBD. Isolation and in
vitro culture of ruminant preantral follicles. Arq. Fac. Vet. UFRGS, v. 27, p. 11-31,
1999.
FIGUEIREDO, J.R., RODRIGUES, A.P.R., SILVA, J.R.V., SANTOS, R.R.
Cryopreservation and in vitro culture of caprinepreantral follicles. Reprod. Fertil.
Develop.v. 23,p. 40–47, 2011.
FIGUEIREDO, J.R.; FREITAS, V.J.F. Biotécnicas aplicadas à reprodução animal. São
Paulo: Livraria Varela, p. 227-256, 2002.
FIGUEIREDO, J.R.; HULSHOF, S.C.J.; THIRY, M.; VAN DEN HURK, R.; BEVERS,
M.M.; NUSGENS, B.; BECKERS, J.F. Extracellular matrix proteins and basement
membrane: their identification in bovine ovaries and significance for the attachment of
cultured preantral follicles. Theriogenology, v. 43, p. 845-858, 1995.

132
FIGUEIREDO, J.R.; RODRIGUES, A. P. R.; LOPES, C. A. P.; Importância do ovário
artificial na reprodução medicamente assistida e bem-estar animal, Revista CFMV, Ano
XV, n.45, p.72-75, 2009.
FIGUEIREDO, J.R.; RODRIGUES, A.P.R.; AMORIM, C.A.; SILVA, J.R.V.
Manipulação de Oócitos Inclusos em Folículos Ovarianos Pré-antrais – MOIFOPA. In:
GONÇALVES, P.B.D.; FIGUEIREDO, J.R.; FREITAS, V.J.F. (Eds). Biotécnicas
Aplicadas à Reprodução Animal. Editora Roca, São Paulo, p.303-327, 2008.
FORTUNE, J. E. The early stages of follicular development: activation of primordial
follicles and growth of preantral follicles. Animal Reproduction Science,. v. 78, p. 135–
163, 2003.
FRESHNEY, I.R.; Culture of Animal Cells: A Manual of Basic Technique. 3rd ed. New
York: Wiley-Liss, 93, 1994.
FUJIHARA M., COMIZZOLI P., WILDT, D.E. SONGSASEN N.; Cat and Dog
Primordial Follicles Enclosed in Ovarian Cortex Sustain Viability after In vitro Culture
on Agarose Gel in a Protein-Free Medium. Reprod Dom Anim, v.47, p.102–108, 2012.
GUÉRIN C. Fecondation in vitro chez la chienne.Ou en est-on? Pratique Médicale et
Chirurgicale des.Animaux de Compagnie; v.33, p.151-161, 1998.
GUPTA P.S.P.; NANDI,S.; RAVINDRANATHA, B.M. ; SARMA, P.V. In vitro
culture of buffalo (Bubalus bubalis) preantral follicles.Theriogenology v.57, p. 1839-
1854., 2008.
GUPTA P.S.P.; NANDI; S.; RVINDRANATHA B.M.; SARMA P.V.; In vitro culture
of buffalo (Bubalus bubalis) pre antral follicles. Theriogenology, 57, 1839-1854, 2002.
GUPTA, P.S.P., NANDI S.; Isolation and Culture of Preantral Follicles for Retrieving
Oocytes for the Embryo Production: Present Status in Domestic Animals, Reproduction
in Domestic Animals, v. 47, p. 513–519, 2012
HAFEZ, E.S.E. Reprodução Animal, 7ª ed., São Paulo, Manole, 513p. 2004.
HARTSHORE, G.M.; In vitro culture of ovarian follicles; Reviews of Reproduction,
v.2, p.94–104; 1997.
HATOYA S., SUGIYAMA Y., NISHIDA H. , OKUNO T. , TORII R. , SUGIURA K.,
KIDA K., KAWATE N., TAMADA H. , INABA T. Canine oocyte maturation in
culture: Significance of estrogen and EGF receptor gene expression in cumulus cells,
Theriogenology v.71, p.560–567, 2009.
HATOYA S.; SUGIYAMA Y.; TORII R.; WIJEWARDANA V.; KUMAGAI D.;
SUGIURA K.; KIDA K.; KAWATE N.; TAMADA H.; SAWADA T.; INABA T..

133
Effect of co-culturing with embryonic fibroblasts on IVM, IVF and IVC of canine
oocytes. Theriogenology. v.66, v.1083–1090, 2006.
HEWITT, D.A., WATSON, P.F., ENGLAND, G.C. Nuclear staining and culture
requirements for in vitro maturation of domestic bitch oocytes. Theriogenology.
v.15;p.1083-101, 1998.
HEWITT, D.A.; ENGLAND, G.C.W.; Influence of gonadotrophin supplementation on
the in vitro maturation of bitch oocytes. Veterinary Record. London. v.144, p.237-239,
1999.
HOVATTA, O., SILYE, R., ABIR, R.; Extracellular matrix improves survival of both
stored and fresh human primordial and primary ovarian follicles in long-term culture.
Hum. Reprod., v.12, p.1032–1036, 1997.
HOVATTA, O.; SILYE, R.; ABIR, R.; KRAUSZ, T.; WINSTON, R. M. L.
Extracellular matrix improves survival of both stored and fresh human primordial and
primary ovarian follicles in long-term culture. Human Reproduction, v. 12, p. 1032-
1036, 1997.
ITOH, T., KACCHI, M., ABE, H., SENDAI, Y., HOSHI, H.; Growth, antrum
formation, and estradiol production of bovine preantral follicles cultured in a serum-free
medium. Biology of Reproduction v.67, p.1099-1105, 2002.
IZADYAR, F.; VAN TOL, H.T.A.; COLENBRANDER, B.; BEVERS, M.M.
Stimulatory effect of growth hormone on in vitro maturation of bovine oocytes is
exerted through cumulus cells and not mediated by IGF-I. Molecular Reproduction and
Development, v.47, p.175–180, 1997.
JEWGENOW K. Impact of peptide growth factors on the culture of small preantral
follicles of domestic cats. Theriogenology, v.45, p.889-895, 1996.
JEWGENOW, K.; GÖRITZ, F. The recovery of preantral follicles from ovaries of
domestic cats and their characterization before and after culture. Animal Reproduction
Science. v. 39, p. 285-297, 1995.
JEWGENOW, K.; PARIS, M.C.J.; Preservation of female germ cells from ovaries of
cat species. Theriogenology. v.66. p. 93–100, 2006.
JEZOVA, M., SCSUKOVA, S., NAGYOVA, E., VRANOVA, J., PROCHAZKA, R.,
KOLENA, J. Effect of intraovarian factors on porcine follicular cells: cumulus
expansion, granulosa and cumulus cell progesterone production. Anim. Reprod. Sci.
v.65, p.115–126, 2001.

134
JIN, S.Y.; LEI, L.; SHIKANOV, A.; SHEA, L.D.; WOODRUFF, T.K. A novel two-
step strategy for in vitro culture of early-stage ovarian follicles in the mouse. Fertility
and Sterility. v. 93, n. 8, p. 2633-9, 2010.
KARLSON, P., GEROK, W., GROSS, W., HOXTER, G. Patobioquímica, 1ª ed.,
Editora Guanabara Koogan S.A., Rio de Janeiro, 321p., 1982.
KATSKA, L.; ALM, H.; RYNSKA, B. Nuclear configuration of bovine oocytes from
fresh and in vitrocultured preantral and early antral ovarian follicles. Theriogenology, v.
54, n. 2, p. 247-260, 2000.
KHALID, M., HARESIGN, W., LUCK, M.R. Secretion of IGF-I by ovine granulosa
cells: effects of growth hormone and follicle stimulating hormone. Anim. Reprod. Sci.;
v.58, p.261-72, 2000.
KIM, M. K.,. FIBRIANTO Y. H, OH H. J., JANG G., KIM H. J., LEE K. S., KANG S.
K., LEE B. C., HWANG. W. S.; Effects of estradiol-17b and progesterone
supplementation on in vitro nuclear maturation of canine oocytes, Theriogenology
v.63,p. 1342–1353, 2005.
KOBAYASHI, J., MIZUNUMA, H., KIKUCHI, N., LIU, X., ANDOH, K., ABE, Y.,
YOKOTA, H., YAMADA, K., IBUKI, Y., HAGIWARA, H. Morphological assessment
of the effect of growth hormone on preantral follicles from 11-day-old mice in an in
vitro culture system. Biochem. Biophys. Res. Commun. v.268,p.36–41, 2000.
KREEGER PK, DECK JW, WOODRUFF TK, SHEA LD. The in vitro regulation of
ovarian follicle development using alginate-extracellular matrix gels. Biomaterials;
v.27, p.714–723, 2006.
LANE, M., GARDNER, D.K., Embryo culture medium: which is the best? Best Pract
Res Clin Obstet Gynaecol v.21,p. 83–100, 2007.
LIM, J., DANUSER, G. Live Cell Imaging of F-actin Dynamics via Fluorescent
Speckle Microscopy (FSM); J. Vis. Exp. v:30), p.1325, 2009.
LIM, J.M.; HANSEL, W. Exogeneus substances affecting development of in
vitroderived bovine embryous before and after embryonic genome activation.
Theriogenology, v.53, p.1081-1091, 2000.
LIMA, A.K.F.; Determinação da população folicular, criopreservação e cultivo de
oócitos inclusos em folículos ovarianos pré-antrais de gata doméstica, Fortaleza, 75
p.Tese (Doutorado em Ciências Veterinárias) Universidade Estadual do Ceará,
Faculdade de Veterinária. 2006.

135
LIU, H.C.; HE, Z.; ROSENWAKS, Z. In vitro culture and in vitro maturation of mouse
preantral follicles with recombinant gonadotropins; Fertility and Sterility, v.77, p.373–
383, 2002.
LOBIE, P.E.; BREIPOHL, W.; ARAGON, J.G.; WATERS, M.J. Cellular localization
of the growth hormone receptor/binding protein in the male and female reproductive
systems. Endocrinology; v.126, p.2214–2221, 1990.
LOPES G., SOUSA M., LUVONI C.G., ROCHA A. Recovery rate, morphological
quality and nuclear maturity of canine cumulus-oocyte complexes collected from
anestrous or diestrous bitches of different ages, Theriogenology v.68, p.821–825, 2007.
LOPES, C.A.P.; SANTOS, R.R.; CELESTINO, J.J.H.; MELO, M.A.; CHAVES, R.N.;
CAMPELLO, C.C.; SILVA, J.R.; BÁO, S.N.; JEWGENOW, K.; FIGUEIREDO, J.R.
Short-term preservation of canine preantral follicles: Effects of temperature, medium
and time. Animal Reproduction Science, v. 115, p. 201-214, 2009.
LOUHIO, H.; HOVATTA, O.; SJÖBERG, J.; TUURI, T. The effects of insulin, and
insulin-like growth factors I and II on human ovarian follicles in long-term culture. Mol
Human Reproduction; v.6, p.694–8, 2000.
LUCY, M.C.; COLLIER, R.J.; KITCHELL, M.L.; DIBNER, J.J.; HAUSER, S.D.;
KRIVI, G.G. Immunohistochemical and nucleic acid analysis of somatotropin receptor
populations in the bovine ovary. Biology of Reproduction, v.48, p.1219– 1227, 1993.
LUVONI G. C., CHIGIONI S., ALLIEVI E., MACIS D.Factors involved in vivo and in
vitro maturation of canine oocytes, Theriogenology,v. 63, p.41–59, 2005.
MACHADO, G.M.; AMORIM, C.A.; BÁO, S.N.; LUCCI, C.M. Cultivo in vitro de
folículos pré-antrais isolados a partir de ovários de vacas zebuínas (Bos taurus indicus),
Vet. Not., Uberlândia, v. 12, p. 11-16, 2006.
MAGALHÃES D.M., DUARTE A.B.G., ARAÚJO V.R., BRITO I.R., SOARES T.G.,
LIMA I.M.T., LOPES C.A.P., CAMPELLO C.C., RODRIGUES A.P.R.,
FIGUEIREDO J.R.;In vitro production of a caprine embryo from a preantral follicle
cultured in media supplemented with growth hormone, Theriogenology, v.75, p.182–
188, 2011.
MARCHAL, R.; CAILLAUD, M.; MARTORIATI, A.; GÉRARD, N.; MERMILLOD,
P.; GOUDET, G. Effect of Growth Hormone (GH) on In vitro Nuclear and Cytoplasmic
Oocyte Maturation, Cumulus Expansion, Hyaluronan Synthases, and Connexins 32 and
43 Expression, and GH Receptor Messenger RNA Expression in Equine and Porcine
Species, BIOLOGY OF REPRODUCTION 69, 1013–1022, 2003.

136
MARTINS, F. S.; Papel do GDF-9, IGF-I e GH sobre o desenvolvimento in vitro de
folículos pré-antrais caprinos. Fortaleza, 2009 75 p.Tese (Doutorado em Ciências
Veterinárias) Universidade Estadual do Ceará, Faculdade de Veterinária.
MATOS, M.H.T.; LIMA-VERDE, I.B.; LUQUE, M.C.A.; MAIA JR, J.E.; SILVA,
J.R.; CELESTINO, J.J.H.; MARTINS, F.S.; BÁO, S.N.; LUCCI, C.M.; FIGUEIREDO,
J.R. Essential role of follicle stimulating hormone in the maintenance of caprine
preantral follicle viability in vitro. Zygote, v. 15, p. 173-182, 2007.
MCLAUGHLIN, M.; TELFER, E.E. Oocyte development in bovine primordial follicles
is promoted by activin and FSH within a two-step serum-free culture system.
Reproduction, v.139, p.971–978, 2010.
MONTEIRO C. M. R.; PERRI, S. H. V.; CARVALHAL R. ; CARVALHO R. G.;
Estudo morfológico comparativo dos ovários de vacas e novilhas da raça elore (Bos
taurus indicus), ARS VETERINARIA,Jaboticabal,SP ,v.24, 122-126, 2008.
NAGAO, S.; MURAO, K.; IMACHI, H.; CAO, W.M.; YU, X.; LI, J.; MATSUMOTO,
K.; NISHIUCHI, T.; AHMED, R. A.; WONG, N. C.; UEDA, K.; ISHIDA, T. Platelet
derived growth factor regulates ABCA1 expression in vascular smooth muscle cells.
FEBS Letters, v. 580, n. 18, p. 4371-4376, 2006.
NAYUDU, P.L., OSBORN, S.M. Factors influencing the rate of preantral and antral
growth of mouse ovarian follicles in vitro. Journal of Reproduction and Fertility, v. 95
p.349–362, 1992.
O‘BRIEN, M.J., PENDOLA, J.K., EPPIG, J.J. A revised protocol for in vitro
development of mouse oocytes from primordial follicles dramatically improves their
developmental competence. Biol. Reprod. v.68, p.1682– 1686, 2003.
PARROTT, J.A.; SKINNER, M.K. Kit-ligand/stem cell factor induces primordial
follicle development and initiates folliculogenesis. Endocrinology, v. 140, p. 4262-
4271, 1999.
PEDERSEN T. Determination of follicle growth rate in the ovary of the immature
mouse. Journal of Reproduction and Fertility, v. 21, p.81–93, 1970.
PICTON, H.M.; HARRIS, S.E.; MURUVI, W.; CHAMBERS, E.L. The in vitro growth
and maturation of follicles. Reproduction, v. 136, p. 703-715, 2008.
QUESNEL, H. Localization of binding sites for IGF-I, insulin and GH in the sow ovary.
Journal of Endocrinol; v.163, p.363–372, 1999.
RECHLER, M.M. Insulin-like growth factor binding proteins. Vitam Horm. v.47, p.1–
91, 1993.

137
RIEGER, D., LUCIANO, A.M., MODINA, S., POCAR, P., LAURIA, A., GANDOLFI,
F. The effects of epidermal growth factor and insulin-like growth factor I on the
metabolic activity, nuclear maturation and subsequent development of cattle oocytes in
vitro. J. Reprod. Fertil. v.112, p.123–130, 1998.
RODRIGUES B.A.; RODRIGUES J.L.; Meiotic Response of In vitro matured Canine
Oocytes under different proteins and heterologous hormone supplementation, Reprod
Dom Anim. v.38, p.58–62, 2003.
RODRIGUES, B.A.; RODRIGUES, J.L. Reproductive endocrinology in the bitch,
Clínica Veterinária, VII, v.40, p.50-58, 2002.
RODRIGUES, BERENICE DE ÁVILA; Maturação e fecundação "in vitro" de ovócitos
de caninos domésticos (canis familiaris) Tese de doutorado em Ciências Veterinárias na
área de Reprodução Animal PORTO ALEGRE 2003.
ROSSETTO R., SANTOS, R.R., SILVA, G.M., DUARTE, A.B.G., SILVA, C.M.G.,
CAMPELLO, C.C., FIGUEIREDO, J.R.. Comparative study on the in vitro
development of caprine and bovine preantral follicles. Small Ruminant Res, v.113,
p.167-170, 2013.
ROY, S.K.; KOLE, A.R. Ovarian transforming growth factor-beta (TGF-beta)
receptors: in vitro effects of follicle stimulating hormone, epidermal growth factor and
TGF-beta on receptor expression in human preantral follicles. Molecular Human
Reproduction, v. 3, p. 207-214, 1998.
SAHA S. SHIMIZU M, GESHI M, IZAIKE Y. In vitro culture of bovine preantral
follicles. Anim. Reprod. Sci. v.63, p.27-39, 2000.
SANTOS, R.R.; THARASANIT, T.; FIGUEIREDO, J.R.; VAN HAEFTEN, T.; VAN
DEN HURK, R. Preservation of caprine preantral follicle viability after
cryopreservation in sucrose and ethylene glycol. Cell and Tissue Research, v. 25, p.
523–531, 2006.
SARAIVA, M.V.A., . CELESTINO J.J.H., CHAVES R.N., MARTINS F.S., BRUNO,
J.B., LIMA VERDE, I.B., MATOS M.H.T., SILVA G.M., PORFIRIO E.P., BÁO, S.N.,
CAMPELLO C.C., SILVA, J.R.V., FIGUEIREDO, J.R. Influence of different
concentrations of LH and FSH on in vitro caprine primordial ovarian follicle
development. Small Ruminant Research v.78, p.87–95, 2008.
SARAIVA, M.V.A., ROSSETTO, R., BRITO, I.R., CELESTINO, J.J.H., SILVA,
C.M.G., FAUSTINO, L.R., ALMEIDA, A.P., BRUNO, J.B., MAGALHÃES, D.M.,
MATOS, M.H.T., CAMPELLO, C.C., FIGUEIREDO, J.R. Dynamic medium produces

138
caprine embryo from preantral follicles grown in vitro. Reproductive Science v.12,
p.1135–1143, 2010.
SARAIVA, M.V.A.; CELESTINO, J.J.H.; ARAÚJO, V.R.; CHAVES, R.N.;
ALMEIDA, A.P.; LIMA-VERDE, I.B.; DUARTE, A.B.G.; SILVA, G.M.; MARTINS,
F.S.; BRUNO, J.B.; MATOS, M.H.T.; CAMPELLO, C.C.; SILVA, J.R.V.;
FIGUEIREDO, J.R. Expression of follicle-stimulating hormone receptor (FSH-R) in
goat ovarian follicles and the impact of sequential culture medium on in vitro
development of caprine preantral follicles. Zygote, v. 19, p. 205-214, 2011.
SAUMANDE, J. La folliculogenèse chez les ruminants. Rec. Vét., v. 167, p. 205-218,
1991.
SCARAMUZZI, R.J.; ADAMS, N.R.; BAIRD, D.T.; CAMPBELL, B.K.; DOWNING,
J.A.; FINDLAY, J.K.; HENDERSON, K.M.; MARTIN, G.B.; MCNATTY, K.P.;
MCNEILLY, A.S.; TSONIS, C.G. A model for follicle selection and the determination
of ovulation rate in the ewe. Reproduction, Fertility and Development, v. 5, p. 459-478,
1993.
SERAFIM, M. K.; ARAUJO, V. R.; SILVA, G. M.; DUARTE, A. B.; ALMEIDA, A.
P.; CHAVES, R. N.; CAMPELLO, C. C.; LOPES, C. A.; FIGUEIREDO, J. R.; SILVA,
L. D. Canine preantral follicles cultured with various concentrations of follicle-
stimulating hormone (FSH). Theriogenology, v. 74, p. 749-55, 2010.
SERAFIM, M.K.; ARAUJO, V.R.; SILVA, G.M.; DUARTE, A.B.; ALMEIDA, A.P.;
CHAVES, R.N.; CAMPELLO, C.C.; LOPES, C.A.; FIGUEIREDO, J.R.; SILVA, L.D.
Canine preantral follicles cultured with various concentrations of follicle-stimulating
hormone (FSH). Theriogenology, v. 74, n. 5, p. 749-755, 2010.
SERAFIM, M.K.B., SILVA, G.M., DUARTE, A.B.G., ARAÚJO, V.R., SILVA, T.F.P.,
LIMA, A.K.F., CHAVES, R.N., CAMPELLO, C.C., SILVA, L.D.M., FIGUEIREDO,
J.R. High insulin concentrations promote the in vitro growth and viability of canine
preantral follicles. Reprod Fert Develop, v.25, p 927-934, 2012.
SHARARA, F.I., NIEMAN, L.K. Identification and cellular localization of growth
hormone receptor gene expression in the human ovary. J Clin Endocrinol Metab;
79:670–672, 1994.
SHARMA, G.T., DUBEY, P.K.; MEUR, S.K. Effect of different mechanical isolation
techniques on developmental competence and survival of buffalo ovarian preantral
follicles. Livestock Science, v. 123, p. 300-305, 2009.

139
SHIKANOV, A., SMITH R. M., XU M., WOODRUFF T. K., SHEA, L.D. Hydrogel
network design using multifunctional macromers to coordinate tissue maturation in
ovarian follicle culture. Biomaterials, v.32, p 2524-2531, 2011.
SHIKANOV, A., XU, M., WOODRUFF, T.K., SHEA, L.D. Interpenetrating
fibrinalginate matrices for in vitro ovarian follicle development. Biomaterials v.30,
p.5476–5485, 2009.
SILVA, C. M. G.; CASTRO, S. V.; FAUSTINO, L. R.; RODRIGUES, G. Q.; BRITO,
I. R.; ROSSETTO, R.; SARAIVA, M. V.; CAMPELLO, C. C.; LOBO, C. H.; SOUZA,
C. E.; MOURA, A. A.; DONATO, M. A.; PEIXOTO, C. A.; FIGUEIREDO, J. R. The
effects of epidermal growth factor (EGF) on the in vitro development of isolated goat
secondary follicles and the relative mRNA expression of EGF, EGF-R, FSH-R and
P450 aromatase in cultured follicles. Research in Veterinary Science, v. 94, p. 453-461,
2013.
SILVA, J. R. V.; VAN DEN HURK, R.; VAN TOL, H. T. A.; ROELEN, B. A. J.;
FIGUEIREDO, J. R. V. Expression of growth differentiation factor 9 (GDF-9), bone
morphogenetic protein 15 (BMP-15) and BMP receptors in the ovaries of goats.
Molecular Reproduction and Development, v. 70, p. 11-19, 2004.
SILVA, J.R.V., FIGUEIREDO J.R., HURK R. VAN DEN. Involvement of growth
hormone (GH) and insulin-like growth factor (IGF) system in ovarian folliculogenesis.
Theriogenology, v.71, p 1193–1208, 2009.
SILVA, J.R.V; FERREIRA, M.A.L.; COSTA, S.H.F.; SANTOS, R.R.; CARVALHO,
F.C.A; RODRIGUES, A.P.R.; LUCCI, C.M.; BÁO, S.N.; FIGUEIREDO, J.R.
Degeneration rate of preantral follicles in the ovaries of goats. Small Ruminant
Research, v. 43, p. 203-209, 2002.
SONG, H.J.; KANG, E.J.; MAENG, G.H.; OCK, S.A.; LEE, S.L.; YOO, J.G.; JEON,
B.G.; RHO, G.J. Influence of epidermal growth factor supplementation during in vitro
maturation on nuclear status and gene expression of canine oocytes. Research in
Veterinary Science, v. 91, p. 439-445, 2011.
SONGSASEN N., WOODRUFF, T. K., WILDT, D. E. In vitro growth and
steroidogenesis of dog follicles are influenced by the physical and hormonal
microenvironment, Reproduction, v. 142, p.113–122, 2011.
SONGSASEN, N., FICKES, A., PUKAZHENTHI B.S., WILDT, D.E. Follicular
Morphology, Oocyte Diameter and Localisation of Fibroblast Growth Factors in the
Domestic Dog Ovary. Reprod Dom Anim v.44, p. 65–70, 2009.

140
SONGSASEN, N., WILDT, D.E.; Oocyte biology and challenges in developing in vitro
maturation systems in the domestic dog; Animal Reproduction Science, v.98, p.2–22.
2007.
SONGSASEN, N., WOODRUFF, T.K., WILD D.E. In vitro growth and steroidogenesis
of dog follicles are influenced by the physical and hormonal microenvironment.
Reproduction v142, p.113–122, 2011.
SPICER, L.J.; STEWART, R.E. Interaction among basic fibroblast growth factor;
epidermal growth factor; insulin and insulin-like growth factor-I (IGF-I) on cell
numbers and steroidogenesis of bovine thecal cells: role of IGF-I receptors. Biology of
Reproduction, v. 54, n. 1, p. 255-263, 1996.
STRÖM B., ROTA A., LINDE-FORSBERG C., In vitro characteristics of canine
spermatozoa subjected to two methods of cryopreservation. Theriogenology, v.48,
p.247-256, 1997.
STURMEY, R. G. TOOLE, P. J. O‘; LEESE H. J.; Fluorescence resonance energy
transfer analysis of mitochondrial:lipid association in the porcine oocyte, Society for
Reproduction and Fertility, v:1470, p.1626; 2006.
SUH, C.S., SONNTAG, B., ERICKSON, G.F. The ovarian life cycle: a contemporany
view. Reviews in Endocrine and Metabolic Disorders, v. 3, p. 5-12, 2002.
TAHER, T. E.; DERKSEN, P. W.; DE BOER, O. J.; SPAARGAREN, M.; TEELING,
P.; VAN DER WAL, A. C.; PALS, S. T. Hepatocyte growth factor triggers signaling
cascades mediating vascular smooth muscle cell migration. Biochemical and
Biophysical Research Communications, v. 298, n. 1, p. 80-86, 2002.
TAMURA, K.; MATSUSHITA, M.; ENDO, A.; KUTSUKAKE, M.; KOGO, H. Effect
of insulin-like growth factor-binding protein 7 on steroidogenesis in granulosa cells
derived from equine chorionic gonadotropin-primed immature rat ovaries. Biology of
Reproduction, v. 77, n. 3, p. 485-491, 2007.
TELFER, E.E. In vitro development of pig preantral follicles. Reprod. Suppl., v.58,
p.81–90, 2001.
TELFER, E.E., MCLAUGHLIN, M., DING, C., THONG, K.J. A two-step serum free
culture system supports development of human oocytes from primordial follicles in the
presence of activin. Human Reproduction v.23, p.1151–1158, 2008.
TELFER, E.E.; BINNIE, J.P., MCCAFFERY, F.H.; CAMPBELL, B.K.; In vitro
development of oocytes from porcine and bovine primary follicles; Molecular and
Cellular Endocrinology; v.163; p. 117–123, 2000.

141
TELFER, E.E.; MCLAUGHLIN M., Strategies to support human oocyte development
in vitro, Int. J. Dev. Biol. v.56, p. 901-907, 2012.
THOMAS, F.H.; CAMPBELL, B.K.; AMSTRONG, D.G.; TELFER, E.E. Effects of
IGF-I bioavailability on bovine preantral follicular development in vitro. Reproduction,
v.133, p.1121–1128, 2007.
TING, A.Y., YEOMAN, R.R. LAWSON MS, ZELINSKI MB (2011) In vitro
development of secondary follicles from cryopreserved rhesus macaque ovarian tissue
after slow-rate freeze or vitrification. Hum Reprod 26: 2461-2472.
TOSHIMORI, K. Sperm plasma membrane modifications associated with fertilization
in mammals Journal of Reproduction and Development. Coolingwood. v. 46, p. 65-78,
2000.
VAN DEN HURK, R.; BEVERS, M.M.; BECKER, J.F. In vivo and in vitro
development of preantral follicles. Theriogenology, v. 47, p. 73-82, 1997.
VAN DEN HURK, R.; SPEK, E. R.; HAGE, W. J.; FAIR, T., RALPH, J. H.;
SCHOTANUS, K. Ultrastructure and viability of isolated bovine preantral follicles.
Human Reproduction, v. 4, n. 6, p. 833-841, 1998.
VAN DEN HURK, R.; ZHAO, J. Formation of mammalian oocyte and their growth,
differentiation and maturation within ovarian follicles. Theriogenology. v. 63, p. 1717-
1751, 2005.
WALTERS, K.A., CAMPBELL, B.K., ARMSTRONG, D.G., TELFER, E.E. The
effects of IGF-I on bovine follicle development and IGFBP-2 expression are dose and
stage dependent. Reproduction v.131, p. 515-523, 2007.
WANDJI, S.A., SRSEN, V., VOSS, A.K., EPPIG, J.J., FORTUNE, J.E..; Initiation in
vitro of growth of bovine primordial follicles. Biol Reprod 55:942–948. 1996.
WANG, Y.; LI, J.; WANG, C.Y.; KWOK, A.H.Y.; LEUNG, F.C. Epidermal growth
factor (EGF) receptor ligands in the chicken ovary. I. Evidence for heparin-binding
EGF-like growth factor (HB-EGF) as a potential oocyte-derived signal to control
granulose cell proliferation and HB-EGF and kit ligand expression. Endocrinology, v.
148, p. 3426-3440, 2007.
WEST, E.R., XU, M., WOODRUFF, T.K., SHEA, L.D.; Physical properties of alginate
hydrogels and their effects on in vitro follicle development. Biomaterials;v.28, p.4439–
4448, 2007.
WOODRUFF, T. K., SHEA, L. D. The Role of the Extracellular Matrix in Ovarian
Follicle Development, Reproductive Sciences v.14, p.6, 2007.

142
WU, J., XU, B., WANG, W.; Effects of luteinizing hormone and follicle stimulating
hormone on the developmental competence of porcine preantral follicle oocytes grown
in vitro. Journal of Assisted Reproduction and Genetics v. 24. p.419–424, 2007.
XU, J., BERNUCI, M.P., LAWSON, M.S., YEOMAN, R.R., FISHER, T.E.,
ZELINSKI, M.B., STOUFFER, R.L. Survival, growth, and maturation of secondary
follicles from prepubertal, young, and older adult rhesus monkeys during encapsulated
three-dimensional culture: effects of gonadotropins and insulin. Reproduction v.140, p.
685–697, 2010.
XU, J., LAWSON, M.S., YEOMAN, R.R., MOLSKNESS T.A., TING, A.Y.,
STOUFFER, R.L., ZELINSK, M.B. Fibrin promotes development and function of
macaque primary follicles during encapsulated three-dimensional culture. Human
Reproduction, v.0, p.1–14, 2013.
XU, J., LAWSON, M.S., YEOMAN, R.R., PAU, K.Y., BARRETT, S.L., ZELINSKI,
M.B., STOUFFER, R.L. Secondary follicle growth and oocyte maturation during
encapsulated three-dimensional culture in rhesus monkeys: effects of gonadotrophins,
oxygen and fetuin. Hum. Reprod. v.26, p.1061–1072, 2011.
XU, M.; WEST, E.; SHEA, L.D.; WOODRUFF, T.K. Identification of a stage-specific
permissive in vitro culture environment for follicle growth and oocyte development.
Biology of Reproduction, v. 75, p. 916-923, 2006.
XU, M.; WEST-FARRELL, E.R.; STOUFFER, R.L.; SHEA, L.D.; WOODRUFF, T.K.;
ZELINSKI, M.B. Encapsulated three-dimensional culture supports development of
nonhuman primate secondary follicles. Biology of Reproduction, v. 81, p. 587-594,
2009.
YARAK S., BAGATIN E., HASSUN K.M., PARADA M.O.A.B. & TALARICO
FILHO S. Hiperandrogenismo e pele: síndrome do ovário policístico e resistência
periférica à insulina. Anais Brasileiros de Dermatologia. v.80. p.395-410, 2005.
YOSHIMURA, Y. Insulin-like growth factors and their binding proteins: potential
relevance to reproductive physiology. Reproductive Medicine and Biology, Boston, v.2,
p.1-24, 2003.
ZHOU, H.M.; ZHANG, Y. Impact of growth factors on in vitro development of caprine
oocytes at pre-antral stage. Reproduction in Domestic Animals, v. 40, p. 161-165, 2005.