metabolismo do carbono em plantas de palma de óleo ... filemetabolismo do carbono em plantas de...
TRANSCRIPT

1
Metabolismo do carbono em plantas de palma de óleo submetidas à deficiência hídrica.
Jéssica Taynara da Silva Martins(1)
; Liliane Correa Machado(2)
; Thays
Corrêa Costa(3)
; Mayra Taniely Ribeiro Abade(4); Ana Ecídia de Araújo Brito(5); Cândido Ferreira de Oliveira Neto(6).
(1)Estudante de graduação em Agronomia; Universidade Federal Rural da Amazônia (UFRA); Belém, PA;
Estudante de graduação em Agronomia;
(UFRA);[email protected];(3)
Estudante de graduação em Agronomia;
(UFRA);[email protected]; (4)
Estudante de mestrado em Agronomia; Universidade Estadual do
Oeste do Paraná (UNIOESTE); [email protected];(5)
Estudante de Doutorado em Agronomia
(UFRA); [email protected]; (6)
Professor/Orientador da UFRA/Campus Belém; [email protected].
RESUMO: A palma de óleo apresenta uma boa capacidade de adaptação contribuindo à sua
dispersão respondendo bem as modificações do meio ambiente. O objetivo deste trabalho foi avaliar os aspectos fisiológicos e bioquímicos no metabolismo do carbono da palma de óleo sob condições de plena irrigação e deficiência hídrica. O experimento foi conduzido em casa de vegetação na Universidade Federal Rural da Amazônia, campus Capitão Poço, realizado em delineamento inteiramente casualizado com os tratamentos dispostos em fatorial 2 x 2 [duas condições hídricas (controle e deficiência hídrica) x dois genótipos de dendê (2501 e 2528)], com cinco repetições por tratamento. O conteúdo relativo de água reduziu em ambos os genótipos submetidos à deficiência hídrica, possivelmente pelo fechamento estomático. O Amido nas raízes foi reduzido e nas folhas quase não sofreu alteração quando se submeteu os genótipos em deficiência hídrica. Entretanto, os resultados mostraram que o teor de carboidrato foi menor no tratamento com estresse hídrico, indicando que em condições severas de secas, as plantas submetidas ao estresse têm seu teor de carboidratos reduzido, possivelmente relacionado à menor assimilação de CO2. Em relação à Sacarose observa-se um maior teor nas folhas para os 2 genótipos, do que nas raízes, este resultado mostra redução na taxa de translocação de carboidratos das folhas para as raízes e diminuição do crescimento e das atividades metabólicas das raízes, que por sua vez, passam a demandar por menos carboidratos. A deficiência hídrica durante o período do experimento provocou alterações no metabolismo do carbono da palma de óleo. Termos de indexação: Elaeis guineensis, fisiologia, bioquímica.
INTRODUÇÃO
O dendezeiro, Elaeis guineensis Jacq. também é conhecido como palma de óleo, esta espécie apresenta uma boa capacidade de adaptação que contribuiu para a dispersão desta cultura em outras partes do mundo, passando a integrar a flora local tanto através da formação dos dendezais subespontâneos quanto na forma de exploração comercial. É cultivado principalmente na Ásia, na África e nas Américas Central e do Sul (Ramalho Filho et al., 2010). É uma oleaginosa de grande importância em diversos aspectos, tendo uma das maiores produtividades de óleo conhecida no mundo, com um rendimento de 4 a 6 toneladas de óleo/há (Muller et al., 2006).

2
O dendezeiro responde bem as modificações do meio ambiente, mostrando variações no desempenho dos genótipos quando submetidos a diferentes condições ambientais (Rafii et al., 2002). A disponibilidade hídrica é um dos principais mecanismos que regulam o desenvolvimento e sobrevivência de plantas (Tobe et al., 2005). A água é um recurso que possibilita o crescimento, pois estimula o alongamento celular, síntese e hidratação de proteínas, e a entrada de nutrientes solúveis do solo. Segundo Pimentel (2004) os efeitos da seca sobre o desenvolvimento dos vegetais estão relacionados a intensidade, referente à duração do estresse, fenologia e genética da planta. Na palma de óleo, a deficiência hídrica provoca aborto de inflorescência e redução do menor número de cachos. Entretanto, não está claro se essas respostas são um resultado direto da redução da oferta de carboidratos da fotossíntese durante a seca, ou uma consequência de alterações no balanço de fitormônios (Corley & Tinker, 2003). Outro parâmetro importante é a diminuição da condutância estomática, uma das primeiras respostas da planta ao déficit a fim de limitar a perda de água por transpiração (Suresh et al., 2010).Segundo Suresh et al. (2010), a deficiência hídrica não altera de forma irreversível as trocas gasosas em mudas de dendê, pois estas exibem recuperação total após a reidratação. Nesse contexto, o objetivo deste trabalho foi avaliar os aspectos fisiológicos e bioquímicos no metabolismo do carbono da palma de óleo (Elaeis guineensis Jacq) sob condições de plena irrigação e deficiência hídrica.
MATERIAL E MÉTODOS As plantas jovens de Elaeis guineensis Jacq, com 10 meses de idade provenientes da Embrapa Amazônia Oriental que apresentaram aspectos e tamanhos semelhantes foram selecionadas e colocadas em vasos de 20 L preenchidos com substrato constituído de solo e esterco bovino curtido (na proporção de 3:1 v/v). O experimento foi conduzido em casa de vegetação na Universidade Federal Rural da Amazônia, campus Capitão Poço (Latitude 01º 44’ 47’’ e Longitude 47º 03’34’’), Brasil. As plantas foram submetidas a dois regimes hídricos: irrigado (controle) e deficiência hídrica (suspensão total da irrigação no início do experimento), em um período de 30 dias. Durante o período experimental as plantas chamadas controle foram irrigadas diariamente para repor a água perdida. O experimento foi realizado em delineamento inteiramente casualizado com os tratamentos dispostos em fatorial 2 x 2 [duas condições hídricas (controle e deficiência hídrica) x dois genótipos de dendê (2501 e 2528)], com cinco repetições por tratamento. Os dados foram analisados por meio de ANOVA utilizando SISVAR 5.3 e os gráficos por Excel (Microsoft Office 2010). As médias foram comparadas pelo teste de Tukey ao nível de 5% de significância. O conteúdo relativo de água (C.R.A) foi determinado conforme Slavick (1979). A determinação do amido foi descrita por Dubois et al. (1956) através da extração etanólica. O método descrito por Dubois et al.(1956) também foi utilizado para determinar os carboidratos solúveis totais. A sacarose foi determinada segundo o método de Van Handel (1968).
RESULTADOS E DISCUSSÃO Conteúdo relativo de água O teor do conteúdo relativo de água (C.R.A) (Figura1) sob controle da disponibilidade hídrica apresentou médias de 85,5 no genótipo 2501 e 81,55 no 2528. Quando submetidas à deficiência hídrica o C.R.A mostrou redução no seu comportamento, com médias de 63,57 para o genótipo 2501 e 62,55 para o 2528, essa redução é causada pelo possível fechamento dos estômatos, sendo tomado como estratégia para a manutenção da planta no período de suspensão hídrica. Assim, a manutenção do status hídrico das plantas, por meio do fechamento estomático, ocorre em resposta à sinalização desencadeada pelo ácido abscísico (Hong Bo et al., 2008). Fioreze et al. (2011) estudando plantas de soja encontraram resultados semelhantes aos do presente estudo afirmando que as plantas utilizam outra estratégia para a manutenção do turgor celular, a qual pode estar associada ao ajustamento osmótico, por meio do acúmulo de compostos osmoticamente ativos e não nocivos nas células.
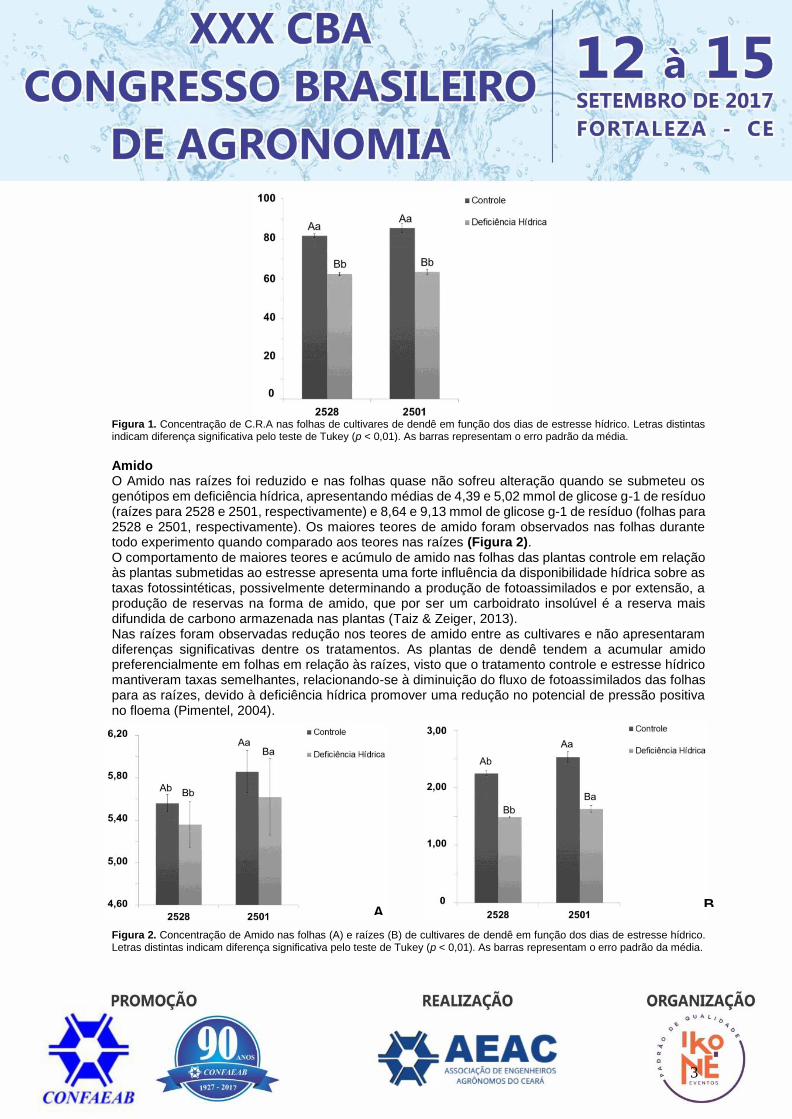
3
Figura 1. Concentração de C.R.A nas folhas de cultivares de dendê em função dos dias de estresse hídrico. Letras distintas indicam diferença significativa pelo teste de Tukey (p < 0,01). As barras representam o erro padrão da média.
Amido O Amido nas raízes foi reduzido e nas folhas quase não sofreu alteração quando se submeteu os genótipos em deficiência hídrica, apresentando médias de 4,39 e 5,02 mmol de glicose g-1 de resíduo (raízes para 2528 e 2501, respectivamente) e 8,64 e 9,13 mmol de glicose g-1 de resíduo (folhas para 2528 e 2501, respectivamente). Os maiores teores de amido foram observados nas folhas durante todo experimento quando comparado aos teores nas raízes (Figura 2). O comportamento de maiores teores e acúmulo de amido nas folhas das plantas controle em relação às plantas submetidas ao estresse apresenta uma forte influência da disponibilidade hídrica sobre as taxas fotossintéticas, possivelmente determinando a produção de fotoassimilados e por extensão, a produção de reservas na forma de amido, que por ser um carboidrato insolúvel é a reserva mais difundida de carbono armazenada nas plantas (Taiz & Zeiger, 2013). Nas raízes foram observadas redução nos teores de amido entre as cultivares e não apresentaram diferenças significativas dentre os tratamentos. As plantas de dendê tendem a acumular amido preferencialmente em folhas em relação às raízes, visto que o tratamento controle e estresse hídrico mantiveram taxas semelhantes, relacionando-se à diminuição do fluxo de fotoassimilados das folhas para as raízes, devido à deficiência hídrica promover uma redução no potencial de pressão positiva no floema (Pimentel, 2004). Figura 2. Concentração de Amido nas folhas (A) e raízes (B) de cultivares de dendê em função dos dias de estresse hídrico. Letras distintas indicam diferença significativa pelo teste de Tukey (p < 0,01). As barras representam o erro padrão da média.
A B
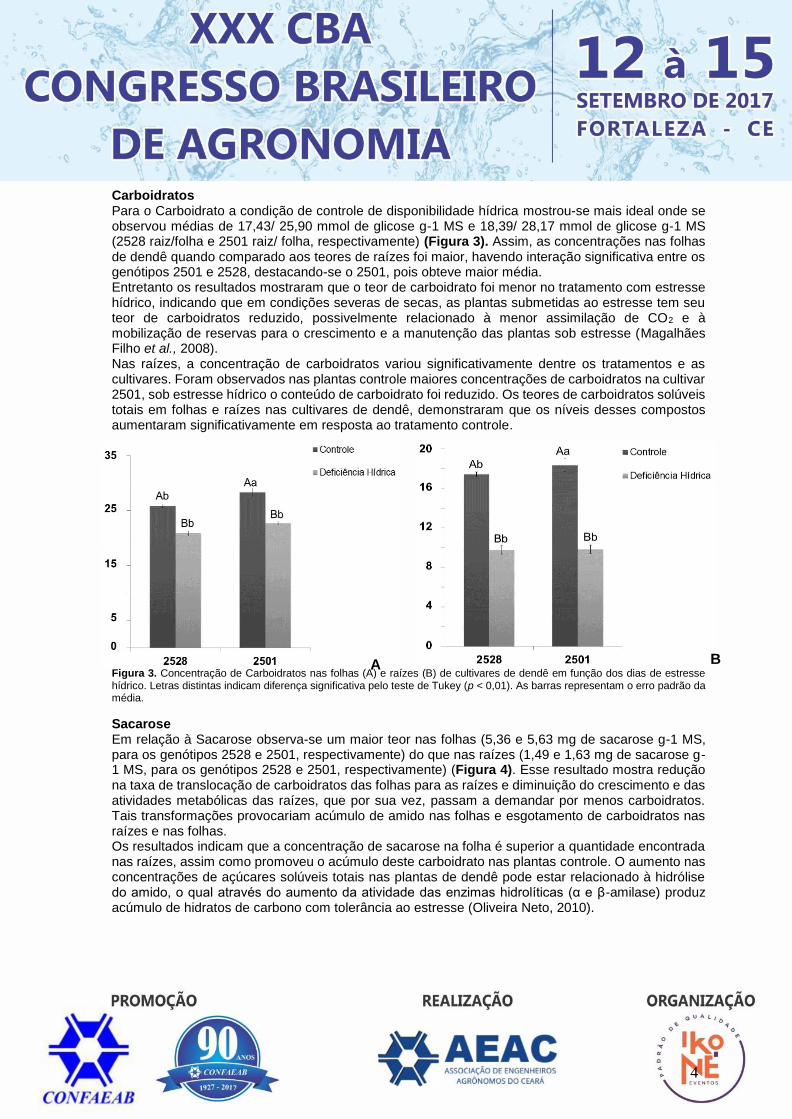
4
Carboidratos Para o Carboidrato a condição de controle de disponibilidade hídrica mostrou-se mais ideal onde se observou médias de 17,43/ 25,90 mmol de glicose g-1 MS e 18,39/ 28,17 mmol de glicose g-1 MS (2528 raiz/folha e 2501 raiz/ folha, respectivamente) (Figura 3). Assim, as concentrações nas folhas de dendê quando comparado aos teores de raízes foi maior, havendo interação significativa entre os genótipos 2501 e 2528, destacando-se o 2501, pois obteve maior média. Entretanto os resultados mostraram que o teor de carboidrato foi menor no tratamento com estresse hídrico, indicando que em condições severas de secas, as plantas submetidas ao estresse tem seu teor de carboidratos reduzido, possivelmente relacionado à menor assimilação de CO2 e à mobilização de reservas para o crescimento e a manutenção das plantas sob estresse (Magalhães Filho et al., 2008). Nas raízes, a concentração de carboidratos variou significativamente dentre os tratamentos e as cultivares. Foram observados nas plantas controle maiores concentrações de carboidratos na cultivar 2501, sob estresse hídrico o conteúdo de carboidrato foi reduzido. Os teores de carboidratos solúveis totais em folhas e raízes nas cultivares de dendê, demonstraram que os níveis desses compostos aumentaram significativamente em resposta ao tratamento controle.
Figura 3. Concentração de Carboidratos nas folhas (A) e raízes (B) de cultivares de dendê em função dos dias de estresse hídrico. Letras distintas indicam diferença significativa pelo teste de Tukey (p < 0,01). As barras representam o erro padrão da média.
Sacarose Em relação à Sacarose observa-se um maior teor nas folhas (5,36 e 5,63 mg de sacarose g-1 MS, para os genótipos 2528 e 2501, respectivamente) do que nas raízes (1,49 e 1,63 mg de sacarose g-1 MS, para os genótipos 2528 e 2501, respectivamente) (Figura 4). Esse resultado mostra redução na taxa de translocação de carboidratos das folhas para as raízes e diminuição do crescimento e das atividades metabólicas das raízes, que por sua vez, passam a demandar por menos carboidratos. Tais transformações provocariam acúmulo de amido nas folhas e esgotamento de carboidratos nas raízes e nas folhas. Os resultados indicam que a concentração de sacarose na folha é superior a quantidade encontrada nas raízes, assim como promoveu o acúmulo deste carboidrato nas plantas controle. O aumento nas concentrações de açúcares solúveis totais nas plantas de dendê pode estar relacionado à hidrólise do amido, o qual através do aumento da atividade das enzimas hidrolíticas (α e β-amilase) produz acúmulo de hidratos de carbono com tolerância ao estresse (Oliveira Neto, 2010).
A B
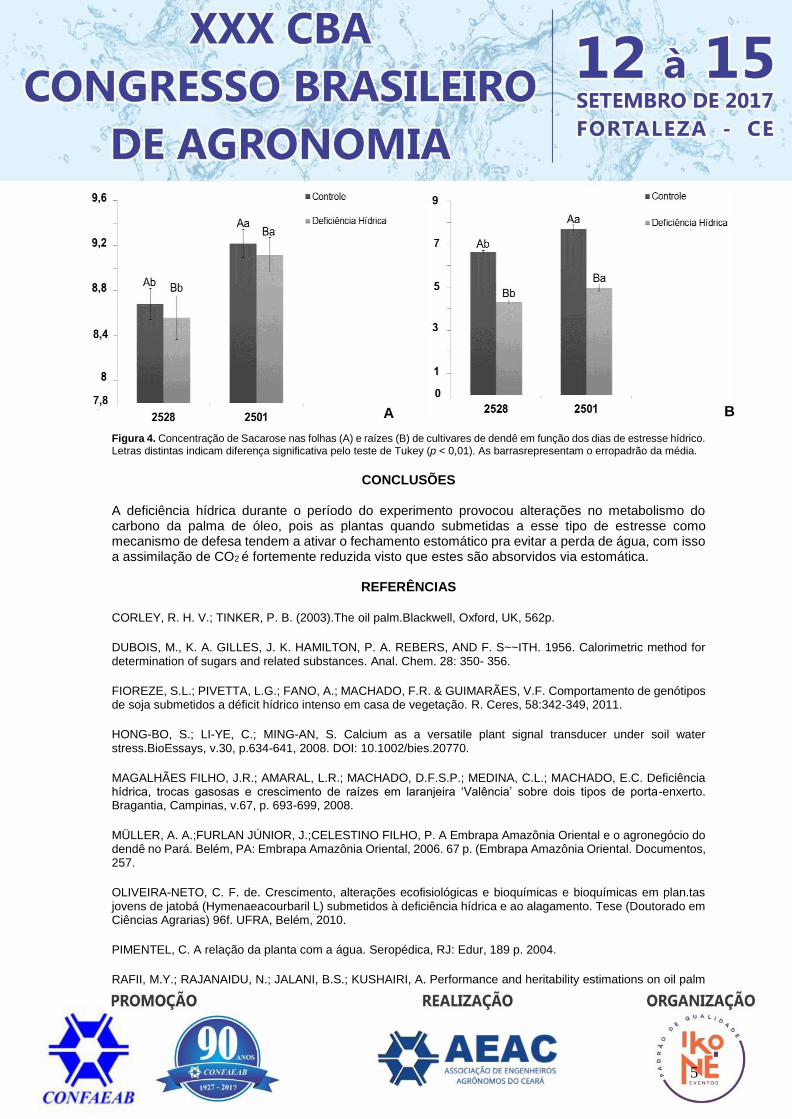
5
Figura 4. Concentração de Sacarose nas folhas (A) e raízes (B) de cultivares de dendê em função dos dias de estresse hídrico. Letras distintas indicam diferença significativa pelo teste de Tukey (p < 0,01). As barrasrepresentam o erropadrão da média.
CONCLUSÕES
A deficiência hídrica durante o período do experimento provocou alterações no metabolismo do carbono da palma de óleo, pois as plantas quando submetidas a esse tipo de estresse como mecanismo de defesa tendem a ativar o fechamento estomático pra evitar a perda de água, com isso a assimilação de CO2 é fortemente reduzida visto que estes são absorvidos via estomática.
REFERÊNCIAS
CORLEY, R. H. V.; TINKER, P. B. (2003).The oil palm.Blackwell, Oxford, UK, 562p.
DUBOIS, M., K. A. GILLES, J. K. HAMILTON, P. A. REBERS, AND F. S~~ITH. 1956. Calorimetric method for determination of sugars and related substances. Anal. Chem. 28: 350- 356.
FIOREZE, S.L.; PIVETTA, L.G.; FANO, A.; MACHADO, F.R. & GUIMARÃES, V.F. Comportamento de genótipos de soja submetidos a déficit hídrico intenso em casa de vegetação. R. Ceres, 58:342-349, 2011.
HONG-BO, S.; LI-YE, C.; MING-AN, S. Calcium as a versatile plant signal transducer under soil water stress.BioEssays, v.30, p.634-641, 2008. DOI: 10.1002/bies.20770.
MAGALHÃES FILHO, J.R.; AMARAL, L.R.; MACHADO, D.F.S.P.; MEDINA, C.L.; MACHADO, E.C. Deficiência hídrica, trocas gasosas e crescimento de raízes em laranjeira ‘Valência’ sobre dois tipos de porta-enxerto. Bragantia, Campinas, v.67, p. 693-699, 2008.
MÜLLER, A. A.;FURLAN JÚNIOR, J.;CELESTINO FILHO, P. A Embrapa Amazônia Oriental e o agronegócio do dendê no Pará. Belém, PA: Embrapa Amazônia Oriental, 2006. 67 p. (Embrapa Amazônia Oriental. Documentos, 257.
OLIVEIRA-NETO, C. F. de. Crescimento, alterações ecofisiológicas e bioquímicas e bioquímicas em plan.tas jovens de jatobá (Hymenaeacourbaril L) submetidos à deficiência hídrica e ao alagamento. Tese (Doutorado em Ciências Agrarias) 96f. UFRA, Belém, 2010.
PIMENTEL, C. A relação da planta com a água. Seropédica, RJ: Edur, 189 p. 2004.
RAFII, M.Y.; RAJANAIDU, N.; JALANI, B.S.; KUSHAIRI, A. Performance and heritability estimations on oil palm
A B

6
progenies tested in different environments. JournalofOil Palm Research, v.14, p.15-24, 2002.
RAMALHO FILHO. A.; MOTTA, P. E. F. da; NAIME, U.J.e; BACA, J. F. M. Avaliação da Aptidão das Terras. In: Zoneamento Agroecológico do Dendezeiro para as Áreas Desmatadas da Amazônia Legal. Rio de Janeiro: Embrapa - CNPS, 2010. 44p.
SLAVICK, B. Methods of studyng plant water relations. New York: Springer Verlag, 1979. 449 p.
SURESH, K; NAGAMANI, C; RAMACHANDRUCU, K.; MATHUR, R. K. (2010) Gas Exchange characteristics, leaf water potential and chlorophyll a fluorescence in oil palm (ElaeisguineensisJacq.) seedlings under water stress and recovery. Photosynthetica, 48: 430-436.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 3. ed.Porto Alegre: Artmed, 2004. 719 p.
TOBE, K.; ZHANG, L.; OMASA, K. 2005. Annals of Botany, 95: 649-659.