luciana dos santos gomes barbosa - ppgn.ufrj.br · À denise, por morrer e “vê deus” comigo...
TRANSCRIPT

Universidade Federal do Rio de Janeiro
Centro de Ciências da Saúde
Instituto de Nutrição Josué de Castro
Programa de Pós-Graduação em Nutrição
Luciana dos Santos Gomes Barbosa
Microdomínios de membrana e perfil lipídico de células de
câncer de mama humano MCF-7 e MDA-MB-231 sob a ação
do resveratrol
Rio de Janeiro
Março/2017

Microdomínios de membrana e perfil lipídico de células de câncer de
mama humano MCF-7 e MDA-MB-231 sob a ação do resveratrol
Luciana dos Santos Gomes Barbosa
Orientadoras: Prof. ª Dr. ª Eliane Fialho de Oliveira
Prof. ª Dr. ª Georgia Correa Atella
Rio de Janeiro
Março/2017
Tese apresentada ao Programa de Pós-Graduação em Nutrição
(PPGN), do Instituto de Nutrição Josué de Castro da
Universidade Federal do Rio de Janeiro, como parte dos
requisitos necessários à obtenção do título de doutor em
Ciências Nutricionais.


MICRODOMÍNIOS DE MEMBRANA E PERFIL LIPÍDICO DE CÉLULAS DE
CÂNCER DE MAMA HUMANO MCF-7 E MDA-MB-231 SOB A AÇÃO DO
RESVERATROL
Luciana dos Santos Gomes Barbosa
TESE SUBMETIDA AO CORPO DOCENTE DO PROGRAMA DE PÓS-GRADUAÇÃO EM
NUTRIÇÃO DO INSTITUTO DE NUTRIÇÃO JOSUÉ DE CASTRO DA UNIVERSIDADE FEDERAL DO RIO DE JANEIRO COMO PARTE DOS REQUISITOS NECESSÁRIOS
PARA A OBTENÇÃO DO GRAU DE DOUTOR EM CIÊNCIAS NUTRICIONAIS.
Banca Examinadora:
___________________________________________________
Professora Doutora Eliane Fialho de Oliveira
Professora Associada do Instituto de Nutrição Josué de Castro/UFRJ
Orientadora
___________________________________________________
Professora Doutora Georgia Correa Atella
Professora associada do Instituto de Bioquímica Médica/UFRJ
Coorientadora
___________________________________________________
Professora Doutora Evelize Folly das Chagas
Professora Adjunta do Departamento de Biologia Celular e Molecular/UFF
Revisora - titular externo
___________________________________________________
Professor Doutor Julio Beltrame Daleprane
Professor Adjunto do Instituto de Nutrição/UERJ
Titular externo
___________________________________________________
Professor Doutor Robson de Queiroz Monteiro
Professor Associado do Instituto de Bioquímica Médica/UFRJ
Titular externo

___________________________________________________
Professora Doutora Eliane Lopes Rosado
Professora associada do Instituto de Nutrição Josué de Castro/UFRJ
Titular interno
___________________________________________________
Professora Doutora Kelse Tibau de Albuquerque
Professora Adjunto do Instituto de Nutrição Josué de Castro/UFRJ
Titular interno
___________________________________________________
Professora Doutora Fátima Lucia de Carvalho Sardinha
Professora Associada do Instituto de Nutrição Josué de Castro/UFRJ
Suplente interno
___________________________________________________
Professora Doutora Danielly Cristiny Ferraz da Costa
Professora Adjunta do Instituto de Nutrição/UERJ
Suplente externo

RIO DE JANEIRO
MARÇO/2017
Este trabalho foi realizado no Laboratório de Alimentos Funcionais e Sementes
(LabAFs), do Instituto de Nutrição Josué de Castro, no Centro Nacional de Bioimagem
(CENABIO) e no laboratório de Bioquímica de Lipídeos e Lipoproteínas, do Instituto
de Bioquímica Médica (IBqM), todos localizados no Centro de Ciências da Saúde, da
Universidade Federal do Rio de Janeiro, sob orientação da Professora Eliane Fialho de
Oliveira e da Professora Georgia Correa Atella. O projeto contou com a colaboração da
Professora Claudia dos Santos Mermelstein, pertencente ao Instituto de Ciências
Biomédicas e do Professor Marcos Henrique Ferreira Sorgine, do IBqM e o auxílio
financeiro da Fundação do Câncer, do Conselho Nacional de Desenvolvimento
Científico e Tecnológico (CNPq) e da Fundação Carlos Chagas Filho de Amparo à
Pesquisa do Estado do Rio de Janeiro (FAPERJ).

Dedico esta tese a todos os amigos que me acompanharam nessa caminhada,
e a todos aqueles que estão na busca por respostas em auxilio aos que sofrem física e
moralmente.

AGRADECIMENTOS
Com muita satisfação eu registro nessas poucas linhas a minha gratidão.
À minha orientadora Lili. É muito difícil descrever o que você representa para mim,
pois faltam as palavras e sobram as lágrimas...mas, são de alegria e gratidão por você ter
me acolhido como sua aluna, lá no final de 2008. Desde então, foram muitos os desafios
que você me proporcionou e saiba que eu tentei aproveitar todos da melhor forma
possível, mesmo com minhas limitações (rsrsrs). Precisei sair da minha zona de
conforto muitas vezes, e você sempre foi a “bússola” que direcionava e sempre
mostrava com seus exemplos, suas atitudes como agir nas adversidades dos
experimentos (nada me aborrece!) e nas relações com o próximo. Muito obrigada por
tudo. Estou terminando o doutorado, mas, você será sempre minha querida “chefa”!
À minha coorientadora “prof. Ge”, como eu frequentemente a procurava pelos
corredores do bloco H. A frase que os seus alunos mais ouviram de mim foi “você viu a
prof. Ge hoje?”. Agradeço por toda a sua generosidade cedendo momentos preciosos
para me ensinar a montar figuras e me acompanhar em experimentos. Nosso começo foi
meio tumultuado, já que eu estava tendo problemas com a adorável centrifuga Beckman,
mas, nada como uma procura por uma Beckman funcionando para quebrar o gelo.
Obrigada por manter as portas do seu laboratório sempre abertas para mim e minhas
células. Meu carinho por você é muito grande!
À Nuccia querida, que me ensinou o passo a passo do mundo do isolamento dos
microdomínios de membrana, lá no comecinho dessa história. Mesmo na reta final para
a sua defesa de tese teve paciência para me explicar e passar os protocolos de todas as
técnicas e soluções. Eu não poderia ter tido melhor professora, todo o agradecimento
será pouco.
À minha revisora Evelize Folly, por aceitar essa tarefa e se colocar à disposição para
ajudar desde o nosso primeiro contato na disciplina SAT II. Agradeço muito por cada
contribuição naquela época, elas ajudaram muito no preparo deste documento.

Ao professor Julio Beltrame, por aceitar prontamente participar da banca. Só os alunos
em reta final de defesa entenderão o valor dessa atitude (rsrsrs), muito obrigada!
Ao professor Robson Monteiro, por aceitar participar da banca, mas, especialmente, por
sempre nos apoiar. Perdi a conta de quantas vezes fui ao seu laboratório em busca de
ajuda, e em todas recebi “sim” do senhor ou dos seus alunos. Gratidão!
À professora Eliane Rosado, por participar da banca, e por suas palavras seja ao longo
da graduação, seja no meu Seminário de Atualização Científica, aquele dia foi um
divisor de águas e a senhora contribuiu muito para o dia de hoje.
À professora Kelse Tibau, por aceitar participar da banca e contribuir com o meu
trabalho.
À professora Fátima Sardinha, por ter aceitado participar da banca e por todos os
ensinamentos desde o mestrado, eles me auxiliaram muito na confecção desse trabalho.
À professora Danielly Ferraz, por aceitar participar da banca com muitos pontos de
exclamação. Dany querida, obrigada por sempre responder minhas dúvidas via
whatsapp. Muito sucesso para você!
Ao professor Mário, que como a Lili diz, é meu avô científico. Obrigada por sempre me
receber muito bem quando eu o abordo nos corredores ou na JIC para tirar dúvidas
sobre meu projeto, ou simplesmente para saber o que o senhor acha desse ou daquele
resultado. Sempre lembrarei da gentileza em me esprestar as chaves do seu laboratório
para que eu pudesse revelar blotting em “horários alternativos”.
À professora Claudia Mermelstein, por aceitar colaborar conosco e por disponibilizar
seu espaço, anticorpos e equipamentos. Aos seus queridos alunos John e Ivone, por toda
ajuda com os ensaios de imunofluorescência, aprendi muito com vocês.
Ao professor Marcos Sorgine, por também colaborar conosco, analisar meus resultados
e se interessar pelo meu trabalho, mesmo estando em outro continente! À sua aluna
Vivian, que dividiu por muitas vezes sua bancada comigo e me auxilio no PCR no

período em o professor estava fora. E aos outros alunos do seu laboratório que
esclaresceram muitas dúvidas minhas.
Aos alunos do laboratório do professor Robson. À Araci que me ajudou muitas vezes a
desvendar o PCR e me apoio emocionalmente quando as coisas não estavam muito bem
(rsrsrs). Ao Fausto querido, sempre muito solícito, fazia o intercâmbio entre mim e o
prof. Robson quando eu não conseguia encontrá-lo, sempre com um sorriso no rosto e
palavras de incentivo, mesmo eu falando para ele que as duas células estavam
contaminadas. Um exemplo de bom ânimo! À Vanessa, eu que pensei que não
perturbaria mais você, depois de um pouco mais de dois anos de proteômica...eis que
você surge nessa reta final doando células e me ajudando a preparar meio, muita
gratidão!
Aos alunos do laboratório do professor Mário, em especial ao Willy (que pertubo desde
os meus tempos de mestrado), ao Filipe, ao Carlos e à Ana, valeu por tudo!
Aos alunos do laboratório da prof. Ge, George, Isabela e Cinara e ao Iron, por me
tratarem como se eu fosse de casa.
À Michele, por tudo. Acho que não tenho como enumerar aqui todas as vezes que pude
contar com você. Gostaria de retribuir a atenção e o cuidado, será que consigo algum
dia? Como não nos conhecemos antes menina??? Gratidão por tudo.
À Karol, por ser a menina do 32Pi! Obrigada por me ensinar a técnica e pela
preocupação com a minha segurança. Sua tranquilidade foi essencial durantes os
experimentos. Você mora no meu coração!
Ao professor Masuda e todos do seu laboratório, Lilian, Helô e professora Isabela, que
quando me viam...sabiam que era para pedir (rsrsrs) e mesmo assim mantinham as
portas abertas. Muito obrigada!
À Mileane, por analisar minhas amostras no GC/MS com tanto carinho, e estar sempre
disposta a responder meus emails e mensagens por whatsapp.

À Maria Clara, técnica do Cenabio, por muitas vezes salvar a vida preparando o
precioso meio de cultura nos momentos mais críticos.
À Julia, por ter muita paciência comigo. Eu sei que no começo e no meio foi difícil, não
é?! Mas agora está tudo mara! Me perdoe pelos momentos que lhe causei aflição e
angustia. Foram momentos de aprendizado para mim. Obrigada por querer que eu faça
parte da sua vida e não desistir. Estamos juntas nessa!
À Patsy, por ser minha revisora de inglês! Gratidão pela sua ajuda, mesmo em pleno
domingo...talvez mais de um...rsrsrs. Sua moleca que está virando borboleta, desejo um
caminho colorido e brilhante para você!
À Iris, pelo incentivo, por sempre me lembrar a Chris, por ficar tão feliz com um pedaço
de brownie, bolo de coco ou pão! Minha parceira de loucuras no aeroporto, desejo que
continuemos assim, afinal, nosso fã club Chris Angel tem que continuar!
À Denise, por morrer e “vê Deus” comigo por muitas e muitas vezes! Obrigada pela
amizade, pelo abraço sincero e pelo carinho de sempre, querida professora!
Ao Christian e ao Luan, meus amigos de outras vidas, pela sintonia desde os primeiros
instantes que nos conhecemos e o incentivo de sempre. O tempo foi curto, mas o
suficiente para eu aprender muito com vocês. Queridos, que nossos caminhos
continuem se cruzando!
Às professoras Maria Lúcia, Ana Luisa e Mariana e ao Marcelooooo pelas palavras de
incentivo e amizade ao longo desses anos.
À todas as amigas do LabAFs Ju, Kim, Marina, Mariana, Samara, Cláudia, Geannetti,
Carol Duque, Carol França, Bia, Paula e Paola pelo calor humano em nosso laboratório
acolhedor, e pelo apoio, principalmente, nos momentos pré journal, sei que vocês me
entendem!
À Chris, minha queridona, por participar e me ajudar mesmo dizendo “Lu eu não sei
nada sobre o seu trabalho”, ainda bem que eu não me importava. Foi com o seu apoio

que consegui alcançar o objetivo de terminar a tese dentro do prazo (foi quase lá). Horas
no trânsito dentro do 913 ou 616 serviram para pesquisarmos casas na Ilha, e não é que
deu certo? Você sabe o quanto foi doído ver você indo embora, mas você nasceu para
ser professora, seja aqui, ou em Curitiba, onde você continua sendo presente na minha
vida, até mesmo nesse finalzinho! Gratidão à Deus por você existir!
À Fabi, por me ensinar, com prazer, os segredos (alguns...) do Western Blotting
(alguns...), e dessa vez sem me sabotar. Ligações durante a noite para saber como
estavam os experimentos e para me incentivar quando o ânimo estava fraco e eu não via
luz no fim do túnel, tudo isso você fez, então, como não te agradecer?! Espero retribuir
ficando um pouquinho com o meu sobrinho para você descansar!
À Kenita, pela praticidade e por me trazer à terra algumas vezes, ainda que com
palavras bem diretas. Obrigada por segurar as pontas com nossas crianças, quando eu
precisei me ausentar. Gratidão pelo carinho e pela amizade.
À Leidi, minha aluna de IC, por ser como você é! Precisei aprender e refletir muito para
poder te orientar de forma produtiva, espero não ter traumatizado você (rsrsrs). Minha
menina da extração de lipídeos, agora nutricionista, sua ajuda foi essencial. Desejo o
melhor para você!
À Paulete, pela amizade de todas as horas, risos e choros. A gente não se larga e que
seja sempre assim. Gratidão por me apoiar, alegrar e torcer por mim ao longo desses
anos, você me inspira!
À Rê, minha lindona, pela amizade e a companhia de sempre, nesse ou em outro país
(amo a internet por permitir isso)! Gratidão por compartilhar comigo todos os
momentos, na alegria e na tristeza (isso é quase um casamento rsrsrs). Nossa amizade é
infinita.
À Thaís, laranjeira do meu quintal, pelo companheirismo nesses anos. Sempre lembrarei
do dia em que você me ofereceu ajuda, mesmo sem nunca ter tido experiência em
laboratório. Sua amizade vale ouro!

Aos amigos do Bezerra pelas preces, em especial, à Luzia e ao Daniel, por
compreenderem a minha ausência e me incentivarem à distância, sempre com muito
carinho.
Aos meus pais Tiana e Celso e irmão Júnior, pelo amor incondicional, pelo carinho e
cuidado quando eu passava a maior parte do dia (e noite) no computador. Ao meu pai,
por me ajudar a descontrair fazendo pão comigo. À minha mãe, por sempre me
empurrar quando eu tinha vontade de parar. Suas palavras são um alento todos os dias e
seu sorriso é meu sol!
Ao marido Julio, por ter topado ir morar comigo na Ilha, só para eu ficar mais perto do
Fundão, que no último ano foi minha primeira casa. Agradeço pelos momentos de
compreensão (em torno de 90% rsrsrs) e pela participação em tudo relacionado ao nosso
lar. Sem a nossa parceria tudo isso teria sido muito mais difícil.
Aos meus amores Sansão e Dalila por sempre me arrancarem sorrisos, mesmos nos
momentos de cansaço e estresse.
Aos meus familiares, que mesmo sem compreender o que significa esse universo,
sempre me deram apoio e carinho.
À Deus, Jesus, Dr. Bezerra e todos os amigos que me acompanharam, me inspirando,
fortalecendo, iluminando o caminho para que tudo ocorresse dessa forma. Gratidão!

RESUMO
O câncer é a segunda maior causa de morte no mundo. Para o biênio 2016-2017, a
estimativa para o Brasil indica a ocorrência de, aproximadamente, 600 mil novos casos,
e o câncer de mama feminino representa 28,1%. O resveratrol (Resv) é um fitoquímico
que pode atuar nos três estágios da carcinogênese apresentando alvos moleculares
diversos. Frequentemente seu efeito é relacionado a sua captação por microdomínios de
membrana. Essas regiões são compostas, por colesterol, esfingolipídeos e proteínas,
como as flotilinas (flot), que são associadas ao mau prognóstico para o câncer, quando
estão superexpressas. No presente estudo, foi investigado o efeito do Resv sobre os
microdomínios de membrana e o perfil lipídico das linhagens de câncer de mama
humano MCF-7 e MDA-MB-231. Verificou-se a citotoxicidade de Resv por redução de
MTT (methylthiazolyldiphenyl-tetrazolium bromide), após 24 h de tratamento. O efeito
do Resv sobre a flot-1 foi analisado por dot-blotting após tratamento por 24 h com
Resv. A expressão de flot-1 e flot-2 foi verificada por Western Blotting, e a regulação da
expressão gênica foi analisada por reação em cadeia da polimerase com transcriptase
reversa quantitativa (RT-qPCR). Microscopia de imunofluorescência foi empregada
para a pesquisa de flot-2 na célula MDA-MB-231. O estudo do perfil lipídico ocorreu
por cromatografia em camada fina de alta afinidade (HPTLC) e cromatografia em
camada fina (TLC). Os ácidos graxos (FA) foram identificados por cromatografia
gasosa com espectometria de massa (GC/MS). A biossíntese de fosfolipídeos (PL) foi
analisada utilizando-se o fosfato inorgânico radioativo e TLC. O metabolismo de
lipídeos foi pesquisado por Western Blotting, verificando-se as enzimas acil-
CoA:diacilglicerol aciltransferase 2, ácido graxo sintase, acetil CoA carboxilase
fosforilada (ρACCβ), proteína quinase ativada por adenosina monofosfato alfa
constitutiva (AMPKα) e fosforilada (ρAMPKα). Os resultados demonstram que para a
MCF-7 houve efeito citotóxico do Resv a partir de 50 µM e para a MDA-MB-231 a
partir de 100 µM. As concentrações de 80 e 200 µM foram utilizadas nos ensaios
posteriores, para MCF-7 e MDA-MB-231, respectivamente. Os dados obtidos por
Western Blotting não indicaram diferenças expressivas para flot-1 e flot-2, no entanto,
na microscopia de imunofluorescência nota-se o aumento de 13,4 vezes da área
correspondente a flot-2. O RT-qPCR indicou aumento acentuado da expressão gênica de
flot-1 e flot-2 na MDA-MB-231. Houve redução (p < 0,05) de éster de colesterol na
MCF-7 e aumento de FA (2,9 vezes). Na MDA-MB-231 FA aumentou 5,3 vezes. Os
FA saturados predominantes em ambas as células foram o ácido palmítico e o ácido
esteárico e os insaturados foram o ácido oleico e o ácido elaídico. Na MCF-7 houve
aumento em 65% de fosfatidiletanolamina. Esfingomielina e lisofosfatidilcolina (LPC)
demonstraram o maior percentual de redução, 57% e 40%, respectivamente. A MDA-
MB-231 exibiu redução de fosfatidilcolina (63%) e LPC (52%). O Resv reduziu ρACCβ
(3,3 vezes; p < 0,05) e AMPKα/ρAMPKα (1,5 vezes; p < 0,04) na MCF-7 e, também na
MDA-MB-231 em 111,8 (p < 0,03) e 1,2 (p < 0,005) vezes, para ρACCβ e
AMPKα/ρAMPKα, respectivamente. O conjunto de resultados obtido demonstra que o
Resv aumenta a expressão de flot e de FA nas células de câncer de mama MCF-7 e
MDA-MB-231, e o efeito citotóxico do fitoquímico se mantém. Isso indica que, embora
o composto esteja favorecendo eventos relacionados ao avanço tumoral, outros alvos
moleculares do Resv podem estar ativos, contribuindo para sua ação anticâncer. Com
isso, são necessários novos estudos que analisem por quais mecanismos o Resv atua
nessas condições.
Palavras-chave: Resveratrol, flotilina, detergent-resistant membranes, câncer de
mama, metabolismo lipídico

ABSTRACT
Cancer is the second largest cause of death worldwide. For the biennium 2016-2017 the
estimate for Brazil indicates the occurrence of approximately 600 thousand new cases,
with female breast cancer representing 28.1%. Resveratrol (Resv) is a phytochemical
which can act at the three stages of carcinogenesis through several molecular targets.
Frequently its effect is related to its uptake by membrane microdomains. These regions
are composed of cholesterol, sphingolipids and proteins, such as flotillins (flot), which
are associated with cancer’s poor prognosis when overexpressed. Thus, the aim of this
study was to investigate the effect of Resv on the membrane microdomains and the lipid
profile of human breast cancer cell lines MCF-7 and MDA-MB-231. The cytotoxicity
was checked by MTT (methylthiazolyldiphenyl-tetrazolium bromide) assay, after 24
hours of treatment. Resv effect on flot-1 was analyzed by dot-blotting after 24 hours of
treatment. Flot-1 and flot-2 expression was measured by Western Blotting analysis, and
the regulation of the gene expression was analyzed by quantitative reverse transcriptase
polymerase chain reaction (RT-qPCR). Immunofluorescence microscopy was used to
screen for flot-2 in MDA-MB-231 cell. Study of the lipid profile was performed by high
performance thin-layer chromatography (HPTLC) and thin-layer chromatography
(TLC). Fatty acids (FA) identification occurred by gas chromatography with mass
spectrometry (GC/MS). Phospholipid (PL) biosynthesis was performed using
radioactive inorganic phosphate and TLC. Lipid metabolism were analyzed by Western
Blotting, checking diacylglycerol O-acyltransferase 2 (DGAT2), fatty acid synthase
(FAS), acetyl-CoA carboxylase β phosphorylated (ρACCβ), 5' adenosine
monophosphate-activated protein kinase (AMPKα) and phosphorylated (ρAMPKα)
enzymes. Results demonstrate that for MCF-7 there was cytotoxic effect of Resv from
50 μM and for MDA-MB-231 from 100 μM. Concentrations of 80 and 200 μM were
used in subsequent assays for MCF-7 and MDA-MB-231, respectively. The data
obtained by Western Blotting did not indicate significant differences for flot-1 and flot-
2, however, in immunofluorescence microscopy it was observed a 13.4-fold increase in
the area corresponding to flot-2. RT-qPCR indicated a sharp increase in the gene
expression of flot-1 and flot-2 in MDA-MB-231. Significant reduction (p < 0.05) of
cholesterol ester and an increase of FA (2.9-fold) was observed im MCF-7. In MDA-
MB-231 FA increased 5.3-fold. The predominant saturated FA in both cells were
palmitic acid and stearic acid and unsaturated FA were oleic acid and elaidic acid.
MCF-7 cells showed an increase of 65% of phosphatidylethanolamine. Sphingomyelin
and lysophosphatidylcholine (LPC) showed the highest reduction percentage, 57% and
40%, respectively. MDA-MB-231 showed reduction of phosphatidylcholine (63%) and
LPC (52%). Resv reduced ρACCβ (3.3-fold, p < 0.05) and AMPKα / ρAMPKα (1.5-
fold, p <0.04) in MCF-7 and in MDA-MB- 231 at 111.8 (p < 0.03) and 1.2 (p < 0.005)
times, for ρACCβ and AMPKα / ρAMPKα, respectively. The results obtained allow us
to verify that Resv increases the expression of flot and FA in MCF-7 and MDA-MB-
231, and that the cytotoxic effect of the phytochemical remains. This indicates that
although the compound is favoring events related to tumor progression, other molecular
targets of Resv might be active, contributing to its anticancer action. With this, new
studies are needed to analyze by what mechanisms Resv acts under these conditions.
Keywords: Resveratrol, flotillin, detergent-resistant membranes, breast cancer, lipid
metabolism

Lista de figuras
Descrição Página
Figura 1: Representação dos estágios da carcinogênese, incluindo iniciação,
promoção e progressão. 24
Figura 2: Os dez tipos de cânceres mais incidentes estimados no Brasil, para
2016, por sexo, exceto pele não melanoma. 25
Figura 3: Representação espacial das taxas brutas de incidência por 100 mil
mulheres da neoplasia de mama, para o ano de 2016. 26
Figura 4: Microscopia representativa das células de câncer de mama MCF-7 e
MDA-MB-231 28
Figura 5: Estrutura química do trans-resveratrol. 30
Figura 6: Resveratrol e sua ação sobre a AMPK e SIRT1. 32
Figura 7: Esquema representativo de microdomínios de membrana e seus
componentes presentes na membrana plasmática. 36
Figura 8: Efeito do resveratrol sobre a viabilidade celular das linhagens MCF-
7 e MDA-MB-231. 60
Figura 9: Isolamento de microdomínios de membrana e análise do perfil do
gradiente de sacarose, após tratamento com resveratrol. 62
Figura 10: Resveratrol altera a distribuição dos microdomínios lipídicos de
membrana nas frações do gradiente de sacarose da célula MCF-7. 63
Figura 11: Resveratrol altera a distribuição dos microdomínios lipídicos de
membrana nas frações do gradiente de sacarose da célula MDA-MB-231. 64
Figura 12: Efeito do resveratrol sobre os níveis de flot-1 e flot-2 em células
MCF-7 e MDA-MB-231. 65
Figura 13: Imunofluorescência para flot-2 em células MDA-MB-231 após
tratamento com resveratrol. 67
Figura 14: Efeito do resveratrol na expressão de flot-1 e flot 2 nas linhagens
celulares MCF-7 e MDA-MB-231. 69
Figura 15: Efeito do resveratrol sobre o perfil de lipídeos neutros em células
MCF-7 e MDA-MB-231, por HPTL. 71
Figura 16: Resveratrol altera o perfil de lipídeos neutros em células MCF-7 e
MDA-MB-231. 73

Figura 17: Efeito do resveratrol sobre lipídeos totais, colesterol e
triacilglicerol em células MCF-7 e MDA-MB-231. 75
Figura 18: Efeito do resveratrol sobre os fosfolipídeos totais de células MCF-
7 e MDA-MB-231. 81
Figura 19: Efeito do tempo de tratamento com resveratrol sobre a biossíntese
de fosfolipídeos em células MCF-7 e MDA-MB-231. 82
Figura 20: Tempo de evolução do efeito do resveratrol na biossíntese de
fosfolipídeos em linhagens celulares MCF-7. 84
Figura 21: Tempo de evolução do efeito do resveratrol na biossíntese de
fosfolipídeos em linhagens celulares MDA. 85
Figura 22: Efeito do resveratrol sobre as enzimas participantes do
mestabolismo lipídico DGAT2, FAS, ρACCβ e AMPK em células MCF-7 e
MDA-MB-231. 87
Figura 23: Esquema representando o efeito do resveratrol sobre as flotilinas e
as enzimas do metabolismo lipídico e as possíveis vias de sinalização inibidas,
por ação do fitoquímico. 103

Lista de tabelas
Descrição Página
Tabela 1: Resveratrol altera perfil de ácidos graxos saturados em células
MCF-7 e MDA-MB-231. 77
Tabela 2: Resveratrol altera perfil de ácidos graxos insaturados em células
MCF-7 e MDA-MB-231. 79

Lista de abreviaturas e siglas
32Pi Fosfato inorgânico radioativo (Radioactive inorganic phosphate)
Akt Proteína serina/treonina quinase, também conhecida como proteína
quinase B (protein kinase B)
AMPKα Proteína quinase ativada por adenosina monofosfato alfa (5' adenosine
monophosphate-activated protein kinase)
BSTFA:TMCS Bistrimetilsililtrifluoroacetamida:trimetilclorosilano
CHO Colesterol livre (Cholesterol)
CHOE Éster de colesterol (Cholesterol ester)
CPM Contagens por minuto
DAPI Marcador fluorescente de ácidos nucléicos (4’,6-diamidino-2-
fenilindol-dihidrocloreto)
DG Diacilglicerol
DGAT acil-CoA:diacilglicerol aciltransferase
DMEM do inglês Dulbecco´s modified eagle médium
DMSO Dimetil sulfóxido
EDTA Ácido etilenodiaminotetracético
EGTA Ácido etilenoglicoltetracético
ER+ Receptor de estrogênio positivo (Estrogen receptors)
FA Ácido graxo (Fatty acids)
FADD Proteína de domínio de morte associada ao Fas (Fas-associated protein
with death domain)
FAS Ácido graxo sintase (Fatty acid synthase)
Fas/CD95 Receptor de superfície celular da superfamília dos receptores fator de
necrose tumoral
Flot-1 Flotilina-1
Flot-2 Flotilina-2
GAPDH Gliceraldeído-3-fosfato desidrogenase (Glyceraldehyde 3-phosphate
dehydrogenase)
GC/MS Cromatografia Gasosa com Espectrometria de Massa (Gas
chromatography with mass spectrometry)

GPI Glicofosfatidilinositol (Glycophosphatidylinositol)
HEPES 4-(2-Hydroxyethyl)piperazine-1-ethanesulfonic acid
HER Receptor do fator de crescimento epidermal humano (Human
epidermal growth factor receptors)
HPTLC Cromatografia em camada fina de alta afinidade (High performance
thin-layer chromatography)
IC50 Concentração inibitória de 50%
LC-MS Cromatografia líquida com espectrometria de massa (Liquid
chromatography - mass spectrometry)
LPC Lisofosfatidilcolina (Lysophosphatidylcholine)
MAPK Proteína quinase ativada por mitógeno (Mitogen activated protein
kinase)
MG Monoacilglicerol (Monoacylglycerol)
MS Espectrometria de massa (mass spectrometry)
mTOR Proteína alvo da rapamicina em mamíferos (mammalian target of
rapamycin)
MTT do inglês Methylthiazolyldiphenyl-tetrazolium bromide
MβCD Metil-β-ciclodextrina
ND Não determinado
NF-κB Factor nuclear kappa B (nuclear factor- κB)
PA Ácido fosfatídico (Phosphatidic acid)
PBS Tampão fosfato de sódio pH 7,4 (Phosphate buffered saline)
PC Fosfatidilcolina (Phosphatidylcholine)
PE Fosfatidiletanolamina (Phosphatidylethanolamine)
PI Fosfatidilinositol (Phosphatidylinositol)
PI3K Fosfatidilinositol-3-quinase (Phosphatidylinositol-3-kinase)
PL Fosfolipídeos (Phospholipids)
PR+ Receptor de progesterona positivo (Progesterone receptors)
PVDF Difluoreto de polivinilideno (polyvinylidene difluoride)
Resv Trans-resveratrol
RMD Resistência a múltiplas drogas
rpm Rotações por minuto
RT-qPCR Reação em cadeia da polimerase com transcriptase reversa quantitativa

(Quantitative reverse transcriptase polymerase chain reaction)
SD Desvio padrão (Standard deviation)
SEM Erro padrão das médias (Standard error mean)
SM Esfingomielina (Sphingomyelin)
TBS Tampão Tris + NaCl pH 7,6 (Tris buffered saline)
TG Triacilglicerol (Triacylglycerol)
TLC Cromatografia em camada fina (Thin-layer chromatography)
TNBC Câncer de mama triplo negativo (Triple negative breast cancer)
TNE Tampão Tris + NaCl + EGTA pH 7,5
u.m.a Unidade de massa atômica
WHO Organização Mundial de Saúde (World Health Organization)
ρACCβ Acetil-CoA carboxilase β fosforilada (Acetyl-CoA carboxylase β
phosphorylated)
ρAMPKα Proteína quinase ativada por adenosina monofosfato α fosforilada (5'
adenosine monophosphate-activated protein kinase phosphorylated)

Sumário
1. INTRODUÇÃO 22
1.1. Câncer de mama: breve relato sobre o cenário mundial e nacional 23
1.2. As células de câncer de mama MCF-7 e MDA-MB-231 e suas
peculiaridades moleculares 27
1.3. Resveratrol: um composto com potencial anticâncer e suas moléculas alvo 29
1.4. Microdomínios de membrana e sua função no câncer 34
1.5. Metabolismo lipídico no câncer 39
2. OBJETIVOS 43
2.1. Objetivo geral 44
2.2. Objetivos específicos 44
3. MATERIAL & MÉTODOS 45
3.1. Reagentes 46
3.2. Culturas de células 46
3.3. Ensaio de viabilidade celular 47
3.4. Isolamento dos microdomínios de membrana 47
3.5. Análise da densidade do gradiente de sacarose 48
3.6. Análise dos efeitos do resveratrol sobre os microdomínios de membrana por
dot-blotting 48
3.7. Investigação do efeito do resveratrol sobre expressão das proteínas flotilina-
1 e flotilina-2 por Western Blotting 50
3.7.1. Preparo do extrato proteico 50
3.7.2. Eletroforese unidimensional em gel de poliacrilamida em condições
desnaturantes 50
3.7.3. Western Blotting 51
3.8. Ensaio de imunofluorescência para flotilina-2 em células MDA-MB-231
tratadas com resveratrol 52
3.9. Análise do efeito do resveratrol sobre a expressão de flotilina-1 e flotilina-2
por reação em cadeia da polimerase com transcriptase reversa quantitativa (RT-
qPCR) 52
3.10. Estudo da ação do resveratrol sobre perfil lipídico das células de câncer de 54

mama MCF-7 e MDA-MB-231
3.10.1. Extração de lipídeos 54
3.10.2. Análise de lipídeos neutros e ácidos graxos 54
3.10.2.1. Estudo do perfil de lipídeos neutros por cromatografia em camada fina
de alta afinidade (HPTLC) e cromatografia em camada fina (TLC) 54
3.10.2.2. Identificação e quantificação de ácidos graxos por cromatografia gasosa
com espectometria de massa (GC/MS) 55
3.10.2.3. Dosagem de lipídeos totais, colesterol e triacilglicerol 56
3.10.3. Análise de composição de fosfolipídeos 57
3.10.3.1. Ensaio com fosfato inorgânico radioativo (32Pi) 57
3.10.3.2. Cromatografia em camada fina (TLC) 57
3.11. Investigação da ação do resveratrol sobre enzimas do metabolismo lipídico
por Western Blotting 58
3.12. Análise estatística 58
4. RESULTADOS 59
4.1. Citotoxicidade do resveratrol em células de câncer de mama MCF-7 e
MDA-MB-231 60
4.2. Efeito do resveratrol sobre microdomínios de membrana e expressão de
flotilina-1 e flotilina-2 em células de câncer de mama MCF-7 e MDA-MB-231 61
4.3. Efeito do resveratrol sobre perfil de lipídeos neutros, ácidos graxos e
fosfolipídeos de células de câncer de mama MCF-7 e MDA-MB-231 70
5. DISCUSSÃO 89
5.1. Citotoxicidade do resveratrol em células de câncer de mama MCF-7 e
MDA-MB-231 90
5.2. Efeito do resveratrol sobre microdominíos de membrana em células de
câncer de mama MCF-7 e MDA-MB-231 91
5.3. Efeito do resveratrol sobre a expressão de flotilina-1 e flotilina-2 em células
de câncer de mama MCF-7 e MDA-MB-231 93
5.4. Efeito do resveratrol sobre perfil de lipídeos neutros, ácidos graxos e
fosfolipídeos de células de câncer de mama MCF-7 e MDA-MB-231 95
6. CONCLUSÃO 101
7. REFERÊNCIAS 104
8. ANEXOS 114

22
1. INTRODUÇÃO

23
1.1. Câncer de mama: breve relato sobre o cenário mundial e nacional
Câncer é uma palavra de origem grega, karkinos, que significa caranguejo,
utilizada pela primeira vez por Hipócrates (viveu entre 460 e 377 antes de Cristo). Nos
dias atuais, esse vocábulo abrange um conjunto de mais de 100 doenças caracterizada
pela proliferação descontrolada de células anormais e o mau funcionamento de
mecanismos reguladores. A gênese tumoral inicia-se com o acúmulo progressivo de
mutações no genoma (danos ao DNA) e alterações na expressão ou função de genes
essenciais para a manutenção da homeostasia celular (FREITAS et al., 2010; INCA,
2016).
A carcinogênese ocorre em três estágios principais: iniciação, promoção e
progressão (Figura 1). A iniciação caracteriza-se pela interação de agentes
cancerígenos com o DNA resultando em mutação irreversível. A promoção é uma etapa
reversível, a qual pode ocorrer muitos anos após a iniciação, em que acontece a
proliferação de células tumorais iniciadas. Durante a progressão, as células
gradualmente se transformam, até o estabelecimento do câncer e, em estágios mais
avançados, a metástase (DAYEM et al., 2016).

24
Figura 1: Representação dos estágios da carcinogênese, incluindo iniciação,
promoção e progressão. Fonte: Adaptado de Dayem et al. (2016).
Considerado um problema de saúde pública, o câncer é a segunda maior causa
de morte no mundo, sendo responsável por 8,8 milhões de óbitos em 2015, isto é, 1 a
cada 6 mortes é devido a essa doença. A previsão é de que o número de novos casos
aumente em 70% até o ano de 2037. Vale ressaltar que mais de 60% do total de novos
casos no mundo ocorrem na África, Ásia, América Central e América do Sul, e essas
regiões são responsáveis por 70% das mortes por câncer no âmbito mundial (WHO,
2017a).

25
Em populações em desenvolvimento, o impacto do câncer corresponderá a 80%
dos mais de 20 milhões de novos casos estimados para 2025. Para o biênio 2016-2017, a
estimativa para o Brasil indica a ocorrência de, aproximadamente, 600 mil novos casos,
incluindo o câncer de pele não melanoma, o mais incidente e com menor índice de
mortalidade (cerca de 180 mil). Excetuando-se esse tumor, os três tipos mais frequentes
(Figura 2) serão os de próstata (28,6%) e pulmão (8,1%) entre os homens e, os de
mama (28,1%) e intestino (8,6%) entre as mulheres (INCA, 2016).
Figura 2: Os dez tipos de cânceres mais incidentes estimados no Brasil, para 2016,
por sexo, exceto pele não melanoma. Fonte: INCA, 2016.
O câncer de mama tem como característica a maior incidência e mortalidade na
população feminina independente das condições socioeconômicas, isto é, ocorre tanto
em países em desenvolvimento quanto naqueles desenvolvidos. Para o ano de 2016
foram esperados 57.960 novos caso de câncer de mama em todo o Brasil. Sua maior
frequência é observada em mulheres das regiões Sul, Sudeste, Centro-Oeste e Nordeste
(INCA, 2016).

26
Figura 3: Representação espacial das taxas brutas de incidência por 100 mil
mulheres da neoplasia de mama, para o ano de 2016. Fonte: INCA, 2016.
As perspectivas para o cenário nacional são alarmantes (Figura 3), o Estado do
Rio de Janeiro (RJ) apresenta o segundo maior número de novos casos (8.020), sendo
antecedido por São Paulo (15.570). Somente na capital do RJ, estimou-se 112,75 novos
casos de neoplasia maligna da mama feminina por 100 mil mulheres. Apesar dos
números expressivos, o câncer de mama quando detectado em fases iniciais, na maioria
dos casos, apresenta chances aumentadas de tratamento e cura (INCA, 2016).
Recentemente, a Organização Mundial da Saúde publicou o guia para o
diagnóstico precoce do câncer, enfatizando o avanço na capacidade de detecção da
doença nos estágios iniciais como importante estratégia para o seu controle em todos os
contextos. Tais melhorias podem ter como repercussão vidas salvas e redução do custo
individual, social e econômico que os cuidados necessários no câncer demandam.
(WHO, 2017b).

27
1.2. As células de câncer de mama MCF-7 e MDA-MB-231 e suas peculiaridades
moleculares
São diversos os subtipos moleculares dos tumores de mama, os principais são
luminal A, luminal B, superexpressão de HER2, basaloide, mama-normal símile e
claudin-low. Essa heterogeneidade confere aos tumores diferentes prognósticos e
respostas às terapias. No presente estudo, foram utilizadas duas linhagens distintas, a
MCF-7, pertencente ao subtipo luminal A, e a MDA-MB-231, integrante do subtipo
claudin-low (PRAT et al., 2010; CIRQUEIRA et al., 2011).
A linhagem celular MCF-7 (Figura 4, superior) apresenta como características
o fato de serem positivas para o receptor de estrogênio (ER+) e/ou receptor de
progesterona (PR+/-) e negativas para o receptor tipo 2 do fator de crescimento
epidermal humano (HER2-). Adicionalmente, exibe expressão reduzida de Ki67, um
antígeno marcador de proliferação celular. Devido a isso, é frequentemente responsiva a
quimioterapia com tamoxifeno e inibidores de aromatases, além de intervenções alvo-
específicas, por terapias hormonais (antiestrogênicos). Representando,
aproximadamente, 60% dos casos de carcinoma mama, tumores desse subtipo possuem
melhor prognóstico quando comparado aos demais (CIRQUEIRA, 2011; HOLLIDAY
& SPEIRS, 2011).
Por outro lado, a linhagem MDA-MB-231 (Figura 4, inferior) encontra-se no
subtipo claudin-low e anteriormente era classificada como basal. Essa modificação
ocorreu em função da identificação da baixa expressão de proteínas claudina 3, 4 e 7,
ocludinas, E-caderina e Ki67, em conjunto com aumento de marcadores relacionados a
transição epitelial-mesenquimal, o que pode favorecer o potencial invasivo observado
nesse tipo celular. Além de apresentar características associadas às células-tronco, com
fenótipo CD44+/CD24-, seu imunofenótipo triplo negativo (TNBC, do inglês triple
negative breast cancer) ER-, PR- e HER2-, é relacionado ao mau prognóstico. Até o
momento, não foi definida intervenção terapêutica específica e as existentes obtêm
resposta intermediária (CIRQUEIRA, 2011; HOLLIDAY & SPEIRS, 2011).

28
Figura 4: Microscopia representativa das células de câncer de mama MCF-7
(superior) e MDA-MB-231 (inferior). Fonte: ATCC, 2017a; ATCC, 2017b.

29
As diferenças apresentadas entre as duas células refletem não somente em sua
resposta aos quimioterápicos, mas também àquelas induzidas por compostos bioativos.
MDA-MB-231 é mais sensível a genisteina (isoflavona encontrada na soja, com
estrutura química similar ao estrogênio) do que a MCF-7, por exemplo. Embora a
MDA-MB-231 seja TNBC, a MCF-7 apresenta deficiência na expressão de caspase 3,
importante na regulação da morte celular por apoptose e determinante para a ação da
isoflavona (YANG et al., 2007).
O resveratrol (Resv) é outro fitoquímico que apresenta efeito célula-dependente,
em relação à viabilidade, proliferação, apoptose e ciclo celular. Resv em concentrações
reduzidas (50 µM), na MCF-7, diminui a proliferação por bloqueio do ciclo celular,
enquanto que na MDA-MB-231 isso ocorre em decorrência da morte celular.
Adicionalmente, o Resv (50 a 150 µM) induz a MCF-7 a apoptose, mas não a MDA-
MB-231. O fato de as células responderem distintamente pode ser consequência de, na
MCF-7, o Resv promover o aumento de p21, p27 e p53, entretanto, na MDA-MB-231, a
expressão desses genes supressores de tumor não é detectada. Isso indica que o Resv
pode regular diferencialmente o processo de morte nas duas linhagens (POZO-
GUISASO et al., 2002). Ademais, p53 desempenha importante função no
desencadeamento de apoptose (mediada por caspases) induzida por Resv (COSTA et
al., 2012).
1.3. Resveratrol: um composto com potencial anticâncer e suas moléculas alvo
Raros são os casos de câncer causados por fatores hereditários, em sua grande
maioria eles são ocasionados por estilo de vida pouco saudáveis e fatores ambientais
prejudiciais, sendo assim evitáveis. Entre os fatores ambientais, a dieta inadequada
contribui cerca de 30%. Diante desse cenário nutricional e alimentar, em 2003, a OMS
lançou a Iniciativa de Promoção de Frutas e Vegetais, na qual, estimulava o consumo
diário de, no mínimo, 400 g de frutas e hortaliças com o objetivo de reduzir o risco de
doenças crônicas não transmissíveis (DCNT), entre elas o câncer (redução de 5 a 12%).
Em conjunto, foi iniciado o Programa 5 ao Dia, inclusive no Brasil, que além de
incentivar a ingestão 5 porções de frutas e/ou hortaliças diariamente, mostrava a
importância da variedade de cores desses alimentos (WHO, 2003). Ainda nesse
contexto, recentemente, foi divulgado o novo Guia Alimentar para a População
Brasileira, no qual alimentação é abordada em aspectos que vão além da ingestão de

30
nutrientes. Nesse guia constam os dez passos para uma alimentação adequada e
saudável, baseados, principalmente, no consumo de alimentos in natura ou
minimamente processados, em sua ampla diversidade, com a finalidade de promover a
saúde e prevenir as DCNT (BRASIL, 2014).
A diversidade de cores encontradas nos alimentos ocorre em função da presença
de compostos que podem ser nutrientes ou não nutrientes. Entre esses, está o Resv,
presente principalmente nas uvas roxas, vinho tinto, oleaginosas, dentre outras plantas,
sendo encontrado em mais de 70 espécies de plantas (AGGARWAL et al., 2004). Esse
fitoquímico é um metabólito secundário e produzido a partir de injúrias sofridas pela
planta, para atuar em seu mecanismo de defesa contra patógenos (fitoalexina) ou até
mesmo em consequência de alterações climáticas e características do solo. Classificado
como um polifenol, da categoria dos estilbenos, o resveratrol possui dois anéis
aromáticos e três grupos hidroxilas, nas formas cis e trans (Figura 5). Mais comumente
encontrado, o isômero trans é a configuração que apresenta atividade biológica (LIU,
2004; SIGNORELLI & GHIDONI, 2005).
Figura 5: Estrutura química do trans-resveratrol. Fonte: NCBI, 2017.
Um dos grandes desafios em estudos in vivo com Resv é seu rápido metabolismo
e baixa biodisponibilidade. Após a administração oral do composto ocorre a detecção no
plasma, seguido por redução da concentração e a identificação de um segundo pico,
referente a recirculação do resveratrol, e em poucas horas é verificada sua presença nos

31
tecidos. Estudo in vivo, com ratos, indica que 75% do Resv ingerido é absorvido
(SIGNORELLI & GHIDONI, 2005).
No entanto, em humanos a concentração detectada no plasma é menor. A
administração de 5 ou 50 mg de trans-resveratrol, duas vezes ao dia por 4 e 12 semanas,
em mulheres que apresentavam risco aumentado para o câncer de mama, resultou na
elevação significativa da concentração sérica de Resv e seus metabólitos após as
semanas de tratamento. O isômero trans foi predominante, enquanto que a forma cis
representou apenas 5% ou menos do total detectado. O efeito do Resv foi de acordo com
seu aumento, que se mostrou diretamente relacionado à redução da metilação do gene
supressor tumoral RASSF-1α e da expressão de prostaglandina E2, sugerindo sua
atuação como agente quimiopreventivo (ZHU et al., 2012).
Após consumo de doses diárias de 0,5 ou 1,0 g de Resv (por oito dias), por
pacientes com câncer de cólon retal, antes da ressecção cirúrgica, foram identificados no
tecido intestinal os metabólitos resveratrol-3-O-glucuronídeo, resveratrol-4′-O-
glucuronídeo, resveratrol-3-O-sulfato, resveratrol-4′-O-sulfato, resveratrol sulfato
glucuronídeo e resveratrol disulfato. A concentração de Resv no tecido tumoral após a
ingestão de 0,5 ou 1,0 g foi de 8,3 e 94,1 nmol/g, respectivamente. Por outro lado, no
tecido não cancerígeno, localizado nas adjacências ao lado direito, o teor foi de 18,6 e
674 nmol/g, respectivamente. No plasma, o Resv foi encontrado no limite de detecção,
sendo a maior concentração média de 22,3 nmol/mL, em pacientes que consumiram 1,0
g de Resv. Mesmo nessas condições, o Resv reduziu em 5% a proliferação dessas
células de câncer de cólon retal (PATEL et al., 2010).
Considerado um composto anticâncer, o fitoquímico pode inibir as fases de
iniciação, promoção e progressão da carcinogênese, como demonstrado por Jang et al.
(1997) em um estudo pioneiro. Os mecanismos de ação do Resv incluem sua
capacidade antioxidante, indução de apoptose, modulação do factor nuclear kappa B
(NF-κB) e da enzima ciclooxigenase (COX) e inibição da isoenzima citocromo P450
(CYPIA1). O desencadeamento de apoptose está associado a ativação do supressor
tumoral p53 e caspases, além do estímulo ao Fas/CD95 (COSTA et al., 2012; HWANG
& CHOI, 2015).
São diversos os alvos moleculares em células de câncer descritos para o Resv,
assim como são várias as moléculas influenciadas por esses alvos, a partir da ação do
Resv, como exemplificado na Figura 6. Sirtuina 1 (SIRT1) e proteína quinase ativada
por adenosina monofosfato (AMPK) são duas proteínas moduladas por Resv.

32
Importantes na regulação da p53, na biogênese tumoral e na manutenção da homeostase
de lipídeos e glicose, SIRT1 e AMPK atuam em conjunto quando ativadas por Resv,
sugerindo que possam participar da mesma via de sinalização (KULKARNI & CANTÓ,
2015).
Figura 6: Resveratrol e sua ação sobre a AMPK e SIRT1. Fonte: Adaptado de
Kulkarni & Cantó (2015).
O efeito do Resv sobre a AMPK ocorre independentemente da SIRT1, porém, o
contrário não se verifica. No entanto, parece que esse comportamento é dose e célula
dependente. Por outro lado, SIRT1 pode modular a AMPK desacetilando a quinase do
fígado B1 (LKB1, do inglês liver kinase B1), essencial para a ativação da AMPK. Tanto
SIRT1 quanto AMPK são efetores centrais da ação do Resv e suas atividades estão
intimamente ligadas (KULKARNI & CANTÓ, 2015).
Outras moléculas podem ser influenciadas direta ou indiretamente pelo Resv e
seus alvos, como ocorre com a COX, que ao ser inibida pelo Resv não catalisa a reação
de conversão do ácido araquidônico a prostaglandinas, reduzindo assim processos
inflamatórios e o estímulo ao crescimento tumoral. O mesmo efeito é encontrado para as
lipoxigenases, indicando a participação do fitoquímico na regulação do metabolismo de
eicosanoides (JANG et al., 1997).

33
O NF-κB pode ter a concentração reduzida ou a atividade transcricional alterada
por ação do Resv. Sugere-se que o Resv ao diminuir estímulos pró-inflamatórios, assim
como o fator de necrose tumoral alfa (TNF-α) ou peróxido de hidrogênio, prejudique a
translocação nuclear do NF-κB, impedindo que ele exerça sua função. Além disso, a
SIRT1 ao ser ativada por Resv, interage com uma das subunidades (RelA/p65) do NF-
κB, desatilando-a, e assim inibindo seu potencial de transativação (YEUNG et al.,
2004).
A enzima fosfatidilinositol-3-quinase (PI3K) pode apresentar sinalização
diretamente inibida por ação do Resv em seu sítio catalítico de ligação de adenosina
trifosfato. Essa inibição acarreta em não ativação da proteína quinase serina/treonina
(Akt). PI3K é um componente essencial para a transdução de sinal de diversas proteínas
quinases da via PI3K/Akt/mTOR (proteína alvo da rapamicina em mamíferos), que
regula o metabolismo, a proliferação, a migração, a sobrevivência, a angiogênese e a
resistência a quimioterapia. Em razão de sua importância para diversos eventos
celulares, modificações na ativação da via de sinalização mediada por PI3K são o
principal foco no desenvolvimento de quimioterápicos. O gene PI3KCA,
frequentemente, apresenta-se com mutação em cânceres de mama, em maior proporção
nos tumores do tipo ER+ e HER- (FRÖJDÖ et al., 2007; TURNER et al., 2016; ZHAO
et al., 2017).
Com participação crítica em processos de proliferação, sobrevivência e
diferenciação celular, a proteína quinase ativada por mitógeno (MAPK) tem a atividade
influenciada pelo Resv, incluindo as quinases reguladas por sinal extracelular 1 e 2
(ERK1 e 2) e as quinase c-Jun n-terminal (JNK). No entanto, a concentração de Resv
determina qual será o efeito sobre as MAPK. Doses menores que 50 µM podem
aumentar a sinalização de ERK, por outro lado, quando acima de 50 µM, o Resv inibe
MAPK (BOWERS et al., 2000; KULKARNI & CANTÓ, 2015).
O fenótipo de resistência a múltiplas drogas (RMD), encontrado em diversos
tipos de cânceres, é um dos grandes obstáculos durante o tratamento da doença. Em
células de osteosarcomas, NF-κB e p38 MAPK estão envolvidos na RMD, modulando a
resistência regulada pela glicoproteína-P (P-gp) contra o quimioterápico adriamicina
(doxorrubicina). No entanto, 25, 50 e 100 µM de Resv podem reverter a RMD ao
fármaco, por meio da inibição da MAPK, consequentemente, não ocorre a ativação de
NF-κB, e assim, a P-gp não é estimulada (ZHANG et al,. 2016). A participação do Resv
nas vias mencionadas acima, com efeito direto ou indireto, demonstra que a ação do

34
composto é complexa, não podendo ser analisada de um único aspecto, visto que, as
moléculas reguladas pelo fitoquímico atuam em cascata e formam uma rede de
sinalização que contribuem para seu efeito anticâncer.
1.4. Microdomínios de membrana e sua função no câncer
Microdomínios de membrana são pequenos (10 - 200 nm), heterogêneos,
altamente dinâmicos, domínios enriquecidos de esfingolipídeos (SL) e esteróis que
compartimentalizam processos celulares. Pequenas plataformas podem algumas vezes
ser estabilizadas para formar grandes plataformas por meio de interações proteína-
proteína e proteína-lipídeo. Essa foi a definição consensual obtida para os lipid rafts
durante o Keystone Symposium on Lipid Rafts and Cell Function, realizado em 2006, no
Colorado (PIKE, 2006).
Nesse mesmo encontro foi decidido substituir o termo lipid raft por membrane
raft, pois o primeiro passaria o conceito de que apenas as interações entre lipídeos
determinariam a formação das plataformas lipídicas, enquanto a segunda, surgiu do
entendimento de que proteínas e lipídeos contribuem para a origem desses
microdomínios de membrana. Com a justificativa de que proteínas ou complexos
podem ser isolados por meio de técnicas que utilizam detergentes e ainda assim fazerem
parte das plataformas, o uso do termo detergent-resistant, para membranas resistentes a
detergentes (detergent-resistant membranes), foi desencorajado (PIKE, 2006).
Desde muito antes da realização do simpósio as funções dos microdomínios de
membrana são discutidas, sendo atribuídas a transdução de sinais, o tráfego de
moléculas, a organização do citoesqueleto e a entrada de patógenos (BROWN &
LONDON, 1998; MUNRO, 2003). Questionamentos sobre sua existência são realizados
até os dias atuais (LEVENTAL & VEATCH, 2016). Em parte, isso se deve a
dificuldade em se visualizar os microdomínios em células vivas e ao processo de
isolamento utilizando-se o detergente não iônico Triton X-100 e baixas temperaturas,
favorecendo o rearranjo da bicamada lipídica (MUNRO, 2003). García-Marcos et al.
(2006) ao analisarem dois métodos diferentes para caracterizar microdomínios de
membrana, sendo um baseado na solubilização seletiva com Triton X-100 a 4 °C e o
outro no processo de sonicação, encontraram similar composição de lipídeos nas
amostras obtidas por ambos os métodos.

35
Entretanto, identificou diferenças no conteúdo proteico, sugerindo que o
detergente pode afetar as interações entre proteínas e microdomínios de membrana, mas
pode ser utilizado. Porém, vale ressaltar que o processo inicial de obtenção das amostras
não foi igual, enquanto em um protocolo as células foram ressuspendidas diretamente
em HEPES, no outro, elas foram homogeneizadas em tampão contendo inibidores de
diferentes classes de proteases, o que pode ter sido um fator determinante para os
resultados encontrados. Ademais, o emprego do Triton X-100, seguido por
ultracentrifugação do gradiente de sacarose é o protocolo usualmente adotado para o
estudo dos microdomínios de membrana (GAJATE & MOLLINEDO, 2017) em
conjunto com técnicas de imagem, em especial, a microscopia de fluorescência de alta
resolução (OWEN et al., 2012).
Microdomínios de membrana são compostos principalmente por colesterol
(CHO) e SL, apresentam diferentes proteínas em sua estrutura (Figura 7), as quais são
utilizadas como marcadores dessas regiões. Entre elas, podem ser salientadas as
proteínas ancoradas por glicosilfosfatidilinositol (GPI), as caveolinas (cav) e as
flotilinas (flot). As caveolinas são encontradas em duas principais isoformas, cav-1 e
cav-2, que conferem à membrana plasmática invaginações, com função de ancoramento,
importante no processo de endocitose e transporte de moléculas. As flotilinas, também
denominadas reggies, presentes como flot-1/reggie 2 e flot-2/reggie 1, são pertencentes
a família das proteínas estomatinas, proibitinas, flotilina e Hflk/C (SPFH). Conferem
aos microdomínios característica planar, sendo ancoradas na bicamada lipídica por meio
de palmitoilização (flot-1 e flot-2) e miristoilização (flot-2), e apesar de não
atravessarem a membrana, se associam com outras proteínas residentes nos
microdomínios de membrana (STAUBACH & HANISCH, 2011; ZHAO et al., 2011).
Entre as proteínas mencionadas nessa tese, as flotilinas serão destaque.

36
Figura 7: Esquema representativo de microdomínios de membrana e seus
componentes presentes na membrana plasmática. Fonte: Adaptado de El-Sayed &
Harashima (2013).
Microdomínios de membrana são considerados plataformas de sinalização para
diversos eventos celulares, incluindo a apoptose, que é crucial na carcinogênese. Nesse
processo, participam moléculas que se encontram co-localizadas nos microdomínios de
membrana, como exemplo o receptor Fas, que ao ser ativado ou apenas redistribuído ao
longo da plataforma, inicia a cascata de ativação para a morte celular programada,
dependente de caspases. As quinases, frequentemente são mobilizadas nessas regiões
para promover ou inibir esse processo, como é o caso da Akt, da JNK e da proteína
quinase C (PKC). Já foi demonstrado que a desorganização dos microdomínios de
membrana, utilizando metil-β-ciclodextrina (MβCD) resulta em prejuízo à fosforilação
da Akt, com consequente redução de sua atividade, e aumento de apoptose em células
de macrófagos de rato, de adenocarcinoma epitelial de pulmão e de leucemia (GEORGE
& WU, 2012).
Para que microdomínios de membrana atuem como plataforma de sinalização é
necessário que haja a integridade dos mesmos, em relação ao conteúdo de lipídeos e
proteínas, principalmente, receptores de membrana, proteínas quinases e canais de
cálcio. Concentrações específicas de cada um desses componentes, incluindo as

37
diferenças de lipídeos, determinam a função dos microdomínios. A desorganização dos
microdomínios de membrana altera a homeostase celular, modificando assim suas vias
de morte e sobrevivência (GEORGE & WU, 2012). Células TNBC MDA-MB-231
tratadas com MβCD 0,5 mM por 48 h, mostraram redução de 52% da adesão, de 48%
da invasão, e de 52% da formação de capilar endotelial em cultura, enquanto que as
células na fase G2M do ciclo celular (pode culminar em apoptose) aumentaram de 5%
(controle) para 25%. Após 72 h de tratamento, 58% das células estavam em apoptose.
Ao suplementar as células com CHO os resultados obtidos foram revertidos, indicando
que a integridade dos microdomínios de membrana é determinante para a sobrevivência
e metástase dessa linhagem TNBC (BADANA et al., 2016).
Alguns quimioterápicos e compostos bioativos também podem suprimir o
crescimento e induzir a apoptose de células de câncer por meio a interferência no
conteúdo dos microdomínios de membrana, assim como ocorre com o resveratrol
(GEORGE & WU, 2012). Como descrito na seção anterior, o Resv pode induzir a morte
celular influenciando diversas vias. Uma delas ocorre por meio dos microdomínios de
membrana, a partir do seu estímulo para a redistribuição do receptor Fas associado a
recrutamento da proteína de domínio de morte associada ao Fas (FADD),
desencadeando a apoptose em células de carcinoma de cólon, após 48 h de tratamento,
utilizando concentrações de Resv que variaram entre 10 a 100 µM (DELMAS et al.,
2003).
Colin et al. (2011) a partir do questionamento de qual mecanismo o Resv “entra”
na célula e como inicia seus efeitos, verificaram que em linhagens de câncer de cólon é
captado rapidamente, por endocitose. Entretanto, a internalização do composto depende
da integridade dos microdomínios de membrana, como observado após utilizar
diferentes inibidores de endocitose (monensina, óxido de fenilarsina, clorpromazina,
monodansilcadaverina e amilorida) ou os desorganizadores de microdomínios de
membrana MβCD, filipina e nistatina, constatando a redução de Resv intracelular para a
MβCD e esse efeito foi dose dependente. Isso resultou na diminuição da ativação de
AMPK, principalmente, após as células serem expostas à monensina, e a MβCD
impediu a redistribuição do receptor de Resv, integrina β3, ao longo dos microdomínios
de membrana. A análise das frações oriundas do isolamento dos microdomínios, por
meio do gradiente de sacarose, demonstrou o acúmulo do fitoquímico principalmente
nas frações de baixa densidade 3 e 4, co-localizado com as proteínas flotilina e cav-2.

38
Embora os resultados associando microdomínios de membrana e a sinalização
para a morte de células tumorais cause entusiasmo, nos últimos anos tem sido crescente
o número de estudos atribuindo à superexpressão de flotilinas, em especial a flot-2,
condições indesejáveis na carcinogênese. Flotilinas são proteínas altamente conservadas
e usualmente empregadas como marcadores de microdomínios de membrana ou de
ancoramento (STAUBACH & HANISCH, 2011). Entretanto, elas apresentam
atividades que vão além, possuem unidades dinâmicas com suas próprias funções
regulatórias. Formando homo ou hetero-oligômeros, as reggies atuam como um
ancoramento estável para a organização de complexo multiproteico, no entanto, a
estabilidade de flot-1 está fortemente ligada à presença de flot-2 (LANGHORST et al.
2005).
Não obstante ao desconhecimento das funções das flotilinas, elas têm sido
associadas a transdução de sinais, endocitose, tráfego de vesículas e reorganização do
citoesqueleto de actina, além da interação com ampla variedade de proteínas
(LANGHORST et al. 2005). Tais interações foram identificadas em maior número para
flot-1 do que para a flot-2 (BANNING et al., 2011). Asp et al. (2014) demonstraram em
células de mama que flot-1 e flot-2 encontram-se co-localizadas com HER, e a depleção
das flotilinas acarreta em redução da expressão de HER, bem como a redução da
migração celular e da formação de dímeros por esse receptor. Nessas células, flot-2
parece regular positivamente a ativação de Akt e de ERK 1 e 2.
Por outro lado, em uma linhagem de carcinoma esofágico de células escamosas,
a flot-1 superexpressa foi atribuída a progressão e ao mau prognóstico da doença, por
aumentar o potencial de agressividade das células in vitro e contribuir para a progressão
tumoral in vivo. Além disso, o aumento de flot-1 foi diretamente relacionado a maior
atividade de NF-κB (SONG et al., 2012). Semelhantes associações entre a expressão
elevada de flot-2 e mau prognóstico para o câncer são relatados (WEN et al., 2015; LIU
et al., 2015a; LIU et al., 2015b).
Nas buscas bibliográficas realizadas ao longo do doutorado, e para a confecção
desta tese, não foram encontrados estudos que relacionassem o aumento da expressão de
flotilinas, induzida por Resv, e a carcinogênese. No entanto, Touzet & Philips (2010),
verificaram que, sob condições de estresse, o Resv protege os microdomínios de
membrana presentes no retículo sarco/endoplasmático dos danos causados por espécies
reativas de oxigênio. Entre os danos evitados observou-se a inibição da expressão de

39
cav-3 e flot-1, e esse efeito do fitoquímico foi atribuído a sua atividade antioxidante.
Porém, se tratavam de células não tumorais.
1.5. Metabolismo lipídico no câncer
Atualmente, de acordo com a International Lipid Classification and
Nomenclature Committee, os lipídeos são classificados em oito categorias: ácidos
graxos (FA), glicerolipídeos (GL), glicerofosfolipídeos (GPL), SL, esteróis, prenol,
sacarolipídeos e policetídeos. Os lipídeos com maior relevância para o desenvolvimento
do câncer e para a quimioterapia são os FA, GPL, SL e esteróis (HUANG & FRETER,
2015).
FA são compostos por uma cadeia de hidrocarboneto com um grupo carboxila
terminal e produzidos pela ação da enzima ácido graxo sintase (FAS) a partir dos
precursores acetil-CoA e malonil-CoA, ou pela degradação de GL por lipases, ou por
fosfolipases ao quebrarem GPL. Os GPL são os principais componentes das membranas
biológicas, que podem ser encontrados em diversos tipos quando apresentam um grupo
adicional, tal como, colina, etanolamina, serina, inositol e glicerol (HUANG &
FRETER, 2015).
A biossíntese ou degradação desses lipídeos é regulada por diferentes vias de
sinalização. Adicionalmente, o metabolismo de GPL fornece para a célula uma
variedade de moléculas bioativas, incluindo inositol trifosfato, diacilglicerol (DG),
ácido araquidônico, ácido fosfatídico (PA) e ácido lisofosfatídico (LPC). As SM podem
ser hidrolisadas por esfingomielinases produzindo ceramidas e fosfocolinas. A
biossíntese de CHO envolve mais de trinta reações enzimáticas diferentes (HUANG &
FRETER, 2015).
Metabolismo de lipídeos abrange captação, transporte, síntese e degradação das
moléculas e constitui um processo complexo. Tanto a biossíntese quanto a degradação
são regulados por diferentes vias de sinalização, que podem variar de acordo com o
tecido onde ocorrem, incluindo as condições fisiológicas no momento. Esses fatores
determinam a ativação ou inibição das vias, pois, fornecem informações sobre as
necessidades da célula e possíveis alterações em seu microambiente (HUANG &
FRETER, 2015).
Como mencionado na seção anterior, microdomínios de membrana são,
particularmente, ricos em CHO e SL. Nessas regiões, as cadeias laterais de fosfolipídeos

40
(PL) são enriquecidas com FA saturados, ao contrário das regiões não raft. Essa
composição confere às plataformas lipídicas característica mais ordenada e menos fluida
em relação a membrana circundante, além de determinar qual será a sua função
(CALDER & YAQOOB, 2007; GEORGE & WU, 2012). Sabe-se que durante a
carcinogênese o metabolismo lipídico sofre reprogramação, para que seja gerada
energia suficiente para a expansão e disseminação das células tumorais, pois,
apresentam elevada taxa de proliferação (BELORIBI-DJEFAFLIA et al., 2016).
Modificações no fenótipo metabólico são diretamente direcionadas por ativação
de oncogenes e redução de supressores tumorais, e por condições do microambiente,
tais como pouca oxigenação e escassez de nutrientes. O primeiro evento adaptativo
observado no metabolismo tumoral é a captação exacerbada de glicose e consequente
glicólise, causando o aumento da produção de lactato. O aumento da expressão de
enzimas lipogênicas como a acetil-CoA carboxilase (ACC), a FAS e a ATP citrato liase
(ACLY), representa a quase totalidade das alterações fenotípicas na maioria dos
tumores (BELORIBI-DJEFAFLIA et al., 2016). O processo de lipogênese também pode
ser promovido por mTOR, aumentando a síntese de novo e reduzindo a lipólise
(CHAKRABARTI et al., 2010).
A expressão aumentada de FAS é um preditor de mau prognóstico, relacionada
ao surgimento de lesões pré-cancerosas e a metástase de tumores de mama e próstata.
Informações sobre a abundância, composição de FA e a distribuição espacial das
diferentes classes de lipídeos em tumores contribuem para a sua distinção em relação a
células benignas, bem como para a identificação do estadiamento da doença. No câncer
de mama é bem caracterizado o aumento acentuado de fosfatidiletanolamina (PE) e
fosfatidilcolina (PC). Esse último, quando enriquecido em FA saturados é
correlacionado ao alto grau tumoral e menor sobrevida global (BELORIBI-DJEFAFLIA
et al., 2016).
Hopperton et al. (2014) demonstraram que células de câncer de mama MCF-7 e
MDA-MB-231 incorporaram de 2 a 3 vezes mais acetato radioativo do que a linhagem
não tumoral MCF-10A, indicando taxa muito mais elevada de síntese de FA endógenos.
Os autores associam esse resultado à superexpressão de FAS. Além disso, FA
sintetizados endogenamente são excretados pelas células tumorais, sugerindo que eles
são produzidos em excesso. Ainda nesse contexto, Azordegan et al. (2013), analisaram
três frações de tecidos de câncer de mama (normal, marginal e tumoral). Os autores
observaram que os fosfolipídeos PC, PE, fosfatidilinositol (PI), lisofosfatidilcolina

41
(LPC), SM e fosfatidilserina (PS) da fração tumoral apresentaram maior percentual de
incorporação de ácido araquidônico (PC, PE e PI), ácido oleico (PI e PS) e ácido
palmítico (LPC, PC e SM). Na fração normal, somente PS exibiu maior incorporação de
ácido palmítico, seguida pela fração marginal e tumoral.
Alterações lipídicas identificadas em células de câncer podem, claramente,
distinguí-las das não neoplásicas, tanto para FA quanto para PL. Vale enfatizar que para
os PL, além de determiná-los, é importante caracterizar os FA que o constituem
(CADENAS et al., 2012; AZORDEGAN et al., 2013; WAKI et al., 2014). Além disso,
esses mesmos lipídeos podem ser utilizados como biomarcadores específicos de
diferentes tipos de tumores e potencial metastático (BANDU et al., 2016; MISTRY &
FRENCH, 2016; KIM et al., 2016). Por outro lado, eles também podem ser utilizados
em estratégias para o tratamento do câncer, como propõe Arlia-Ciommo et al. (2016).
Esses pesquisadores, após um extenso estudo da literatura sobre morte celular
programada provocada por mudanças em lipídeos de membrana, a qual denominam
liponecrose, em células de leveduras (modelo de organismo com alta taxa proliferativa)
e câncer, sugeriram que o aumento do índice de dessaturação (relação entre FA
monoinsaturados e FA saturados) dos FA na membrana plasmática pode promover a
liponecrose, seletiva em células tumorais, por dois mecanismos. O primeiro baseia-se na
modificação da estrutura da membrana do retículo endoplasmático, gerando estresse
excedente ao limiar citotóxico da célula. E o segundo, fundamenta-se na inibição da
enzima estearoil-CoA dessaturase, provocando a redução da atividade da ACC
(ARLIA-CIOMMO et al., 2016).
Fármacos empregados na quimioterapia podem apresentar como alvo os
lipídeos. Entre eles pode-se destacar o tamoxifeno, um anti-estrogênico, utilizado em
terapias endócrinas de primeira linha para câncer de mama ER+, atuando na inibição do
metabolismo de PL, levando as células à morte por apoptose (MORAD & CABOT,
2015). Compostos bioativos como o Resv também podem atuar na modulação do
metabolismo lipídico. Embora sejam escassos os estudos que demonstrem a relação
entre RESV e alterações lipídicas em células de câncer de mama (SCARLATTI et al.,
2003), já se tem descrito que o fitoquímico pode ser incorporado a membrana
plasmática, principalmente em microdomínios de membrana, causando o enrijecimento
transitório da membrana (DELMAS & LI, 2011). No entanto, em outro momento, ao
promover a síntese de ceramidas, o Resv favorece o aumento da fluidez e a apoptose
(SCARLATTI et al., 2003; DELMAS & LI, 2011).

42
Em células de leucemia mieloide, o Resv promove a translocação da enzima
esfingomiosina quinase-1 (SphK1) para o citoplasma e a diminuição de sua atividade, a
redução de esfingosina-1-fosfato (S1P) e o aumento de ceramida. Além disso, induziu a
apoptose e inibiu a proliferação. Assim como a ceramida, a S1P é derivada do
metabolismo de SL, intimamente relacionado com a proliferação celular e apoptose,
sendo os efeitos da S1P antagônicos, pois promove a progressão tumoral (TIAN & YU,
2015). Outro mecanismo pelo qual o Resv poderia regular a síntese ou degradação de
lipídeos seria via ativação de AMPK (KULKARNI & CANTÓ, 2015), como descrito na
seção anterior.

43
2. OBJETIVOS

44
2.1. Objetivo geral
Investigar o efeito do composto bioativo resveratrol sobre os microdomínios de
membrana e o perfil lipídico das linhagens de câncer de mama humano MCF-7 e MDA-
MB-231.
2.2. Objetivos específicos
• Isolar os microdomínios de membrana;
• Caracterizar a distribuição da proteína flotilina-1 nos microdomínios de membrana;
No extrato celular total:
• Analisar a expressão de flotilina-1 e flotilina-2;
• Verificar a composição de lipídeos neutros e de ácidos graxos;
• Avaliar a biossíntese de fosfolipídeos;
• Examinar a expressão de enzimas envolvidas no metabolismo lipídico.

45
3. MATERIAL & MÉTODOS

46
3.1. Reagentes
Os anticorpos flotilina-1 (#25506), acil-CoA:diacilglicerol aciltransferase 2
(#32399), ácido graxo sintase (#20140), acetil CoA carboxilase fosforilada (#30446),
proteína quinase ativada por adenosina monofosfato alfa constitutiva (#398861) e
gliceraldeído-3-fosfato desidrogenase (#0411) foram adquiridos da Santa Cruz
Biotechnology Inc. (Dallas, TE, USA). A proteína quinase ativada por adenosina
monofosfato alfa fosforilada foi obtida da Cell Signaling Technology (#2535), o DAPI
da Molecular Probes (#3571, Carlsbad, CA, USA) e a Flotilina-2 da BD Transduction
(#610384). O fosfato inorgânico radioativo foi obtido da Comissão Nacional de Energia
Nuclear (CNEN, São Paulo, SP, Brazil) e os lipídeos utilizados como padrão foram
adquiridos da Sigma-Aldrich Inc. (St. Louis, MO, USA) e da Avanti Polar Lipids Inc.
(Alabaster, AL, USA).
Os primers foram adquiridos da Invitrogen (Carlsbad, CA, USA), com as
seguintes sequencias:
Proteína Forward Reverse
Flotilina-1 CTCCACCCCACCTCAACTTATTTA TCCAGCCCATCCCTCAGTCT
Flotilina-2 CCCCAGATTGCTGCCAAA TCCA-CTGAGGACCACAATCTCA
Actina GTCCGCGATATCAAGGAAAA GTGTTGGCGTACAGGTCCTT
3.2. Culturas de células
As linhagens de células epiteliais de câncer de mama humano MCF-7 e MDA-
MB-231 foram obtidas da ATCC (American Type Culture Collection). MCF-7 e MDA,-
MB-231 foram mantidas em meio DMEM (Dulbecco´s Modified Eagle Medium),
suplementado com 10% de soro fetal bovino e 100 unidades/mL de penicilina e 100
μg/mL de sulfato de estreptomicina. As duas linhagens celulares foram mantidas a 37
°C em uma atmosfera umidificada de 5% de CO2 (modificado de Casanova et al., 2012).

47
3.3. Ensaio de viabilidade celular
Para analisar a citotoxicidade do resveratrol, utilizou-se o método de redução do
MTT (methylthiazolyldiphenyl-tetrazolium bromide) (MOSMANN, 1983). As linhagens
celulares MCF-7 e MDA-MB-231 foram cultivadas em placas de 24 poços (2 x 105) e
tratadas por 24 h com 50, 80, 100, 150, 200, 250 e 300 µM de Resv. O Resv foi
solubilizado em DMSO (dimetilsulfóxido) e para todos os tratamentos com o
fitoquímico padronizou-se a concentração do solvente em 0,1%. As amostras controle
foram tratadas com DMSO 0,1%. Após o tratamento, o meio de cultura foi retirado e as
células foram lavadas três vezes com PBS (fosfato de sódio 10 mM e NaCl 150 mM;
pH 7,4).
Em seguida, adicionou-se a solução de MTT em PBS (0,5 mg/mL), incubou-se
por 3 h em estufa a 37 °C, em atmosfera umidificada de 5% de CO2. Posteriormente, a
solução de MTT foi retirada cuidadosamente e adicionou-se isopropanol 100%,
homogeneizando-se, logo em seguida, para a solubilização dos cristais de formazan (de
coloração roxa), produzidos a partir da redução do MTT por ação de desidrogenases
mitocondriais (MOSMANN, 1983).
Após centrifugação por 5 min a 5000 rpm, o sobrenadante foi transferido para
microplaca de 96 poços e submetido a análise colorimétrica, em espectrofotômetro a
570 nm e 650 nm. Para a determinação da viabilidade celular considerou-se a produção
de cristais de formazan (quanto maior a intensidade da coloração roxa, maior a
proporção de células viáveis), em relação ao controle (equivalente a 100%). Foram
realizados três experimentos independentes, em triplicata, e os resultados expressos em
percentual.
3.4. Isolamento dos microdomínios de membrana
A partir do ensaio de citotoxicidade foram definidas as concentrações de Resv
em 80 e 200 µM para o tratamento da MCF-7 e da MDA-MB-231, respectivamente. As
amostras controle foram tratadas com DMSO 0,1%. Após 24 h de tratamento, as células
(1 x 108) foram lavadas três vezes com PBS. Em seguida, ao pellet adicionou-se 1 mL
do tampão TNE (Tris 100 mM, NaCl 150 mM e EGTA 0,2 mM; pH 7,5) e 2 µL de
coquetel de inibidores de proteases 10 vezes diluído (benzenosulfonil fluoreto

48
hidrocloreto, aprotinina, bestatina hidrocloreto, E-64, leupeptina e pepstatina A; Sigma-
Aldrich Inc, St. Louis, MO, USA, #P8340).
As amostras foram, então, sonicadas a 10% da amplitude total, em 3 ciclos de 10
s cada, com intervalos de 3 s entre cada ciclo, usando-se o sonicador Branson Sonifier
250 (Analog Ultrasonic Homogenizer, Branson Ultrasonics Corp., Danbury, CT, USA).
Posteriormente, foram incubadas com Triton X-100 2% de em tampão TNE pH 7,5, por
20 minutos a 4 °C (BROWN & ROSE, 1992).
Após o tempo de incubação, adicionou-se às amostras 2 mL de solução de
sacarose 80%, resultando na concentração final de 40%. Transferiu-se esta solução para
o tubo de centrífuga específico para o rotor SW41 Ti (Beckman Coulter Inc, Brea, CA,
USA). Em seguida, foram adicionados, lentamente, 4 mL de solução de sacarose 30% e
posteriormente 4 mL de solução de sacarose 5%, formando três fases distintas. Todas as
soluções de sacarore foram preparadas em tampão TNE pH 7,5 (1:1 p/v). O gradiente
foi centrifugado a 38000 rpm, por 20 horas a 4 °C (rotor SW41 Ti, Beckman Coulter
Inc, Brea, CA, USA). Após esse período, foram coletadas 12 frações de 1 mL de cada
tubo, iniciando-se no topo até o fundo do tubo.
3.5. Análise da densidade do gradiente de sacarose
Imediatamente após a coleta das doze frações do gradiente de sacarose, 10 µL de
cada fração foi analisado em refratômetro modelo 10406 (American Optical Company,
Buffalo, NY, USA). Os índices de refração obtidos foram convertidos em percentual de
sacarose utilizando-se a tabela de conversão (GRIFFITH, 2010) para a determinação do
percentual de sacarose das frações.
3.6. Análise dos efeitos do resveratrol sobre os microdomínios de membrana por
dot-blotting
Com o intuito de verificar possíveis alterações nos microdomínios de membrana,
ocasionadas pelo tratamento com Resv, foi investigada uma das principais proteínas
marcadoras de microdomínios de membrana, a flot-1, empregando-se a técnica dot-
blotting. Para isso, as frações obtidas do gradiente de sacarose foram submetidas ao
processo de diálise para a retirada de sacarose, em tampão fosfato 25 mmol e EDTA 1

49
mmol pH 7,5, com duas trocas a cada 4 h e um período overnight. Foram utilizadas
membranas de diálise de 6 a 8 kDa.
O dot-blotting foi realizado utilizando-se o equipamento Dot-blot Minifold I, de
acrílico, com 96 poços (GE Healthcare Life Sciences Corp., Buckinghamshire, GBR,
UK). O sistema foi montado colocando-se no interior do aparelho papel filtro e
membrana de nitrocelulose Hybond-ECL, com 0,45 μm de espessura (GE Healthcare
Life Sciences Corp., Buckinghamshire, GBR, UK), previamente umedecidos em
Tampão TBS (Tris-HCl 10 mM, NaCl 150 mM; pH 7,6). Em seguida, o equipamento
foi conectado a bomba de sucção e foi aplicado em seus poços o volume de 500 μL de
cada fração do gradiente. Após 10 minutos de sucção, as proteínas aderidas a membrana
de nitrocelulose foram bloqueadas em tampão TBS-Tween-Leite pH 7,6 (Tris-HCl 10
mM, NaCl 150 mM, Tween-20 0,1% (v/v); pH 7,6 e leite em pó desnatado 5% (p/v)),
por 2 h em temperatura ambiente.
Logo após, a membrana foi incubada com o anticorpo contra flot-1 diluído
(1:800) em tampão TBS-Tween-Leite, por 18 h a 4 °C. Posteriormente, a membrana foi
lavada cinco vezes, a cada 5 min, com tampão TBS-Tween pH 7,6, seguida por
incubação com anticorpo secundário conjugado a peroxidase diluído (1:1000) em TBS-
Tween-Leite, por 2 h em temperatura ambiente. Terminado esse período, a membrana
foi lavada cinco vezes, a cada 5 min, com tampão TBS-Tween pH 7,6 e três vezes, a
cada 5 min, com tampão TBS pH 7,6. O complexo antígeno-anticorpo formado foi
revelado por meio do método de intensificação de quimioluminescência pelo luminol
(Millipore, Darmstadt, HE, GER). O filme Kodak X-OMAT (Eastman Kodak Comp.,
Rochester, NY, USA) foi exposto por 30 s, revelado e fixado adequadamente em
câmara escura. A análise densitométrica foi realizada após digitalização, com auxílio do
programa ImageJ versão 1.5 (National Institute of Health, USA) e convertida em
percentual.

50
3.7. Investigação do efeito do resveratrol sobre expressão das proteínas flotilina 1 e
flotilina-2 por Western Blotting
3.7.1. Preparo do extrato proteico
As linhagens MCF-7 e MDA-MB-231 foram tratadas com 80 e 200 µM de Resv,
respectivamente, por 24 h. Utilizou-se DMSO 0,1% para tratar o controle. Em seguida,
retirou-se o meio e as células foram lavadas três vezes com PBS. Posteriormente,
adicionou-se nitrogênio líquido e, após sua evaporação, o tampão de lise composto por
Tris-HCl 5 mM pH 7,4, EDTA 10 mM, ortovanadato 1 mM, fluoreto de sódio 5 mM,
óxido de fenilarsina 1 mM, ácido ocadaico 1 µM, PMSF 1 mM e coquetes de inibidores
de proteases 100 vezes diluído (benzenosulfonil fluoreto hidrocloreto, aprotinina,
bestatina hidrocloreto, E-64, leupeptina e pepstatina A; Sigma-Aldrich Inc, St. Louis,
MO, USA, #P8340). Em seguida, as amostras foram sonicadas em banho de ultrassom
por 10 min e centrifugadas a 8000 rpm, por 5 min a 4 °C. O sobrenadante foi
armazenado a -80 °C para as análises posteriores. A determinação da concentração de
proteínas do extrato proteico ocorreu de acordo com o método descrito por Lowry et al.
(1951), utilizando-se albumina bovina sérica 0,1 g% como padrão.
3.7.2. Eletroforese unidimensional em gel de poliacrilamida em condições
desnaturantes
Oitenta microgramas de proteínas de cada amostra foram submetidos à
eletroforese em gel de poliacrilamida a 10%, na presença de dodecil sulfato de sódio
(SDS) (LAEMMLI, 1970). O gel de corrida em condições desnaturantes foi constituído
por Tris-HCl 375 mM, SDS 0,1 g%, acrilamida 9,7 g%, bisacrilamida 0,3 g%,
persulfato de amônio 0,033 g%, tetrametiletilenodiamina (TEMED) 0,055 % (v/v) pH
8,8. O gel de empacotamento foi composto por Tris-HCl 83 mM, SDS 0,07 g%,
acrilamida 3,2 g%, bisacrilamida 0,09 g%, persulfato de amônio 0,073 g%, TEMED
0,123 % (v/v) pH 6,8. O tampão de corrida apresentou a seguinte composição Tris-HCl
24,7 mM, glicina 190 mM, SDS 0,1 g% pH 8,3.
Antes da realização da eletroforese, adicionou-se às proteínas tampão de amostra
contendo Tris-HCl 62,5 mM, glicerol 10% (v/v), azul de bromofenol 0,001 g%, SDS 2
g%, β- mercaptoetanol 10%. Os padrões de massa molecular utilizados foram: β-

51
galactosidase (120 kDa), albumina bovina sérica (85 kDa), ovoalbumina (50 kDa),
anidrase carbônica (35 kDa), β-lactoglobulina (25 kDa) e lisozima (20 kDa) (Thermo
Fisher Scientific Inc, Muskego, WI, USA, #26612). A corrida ocorreu sob voltagem
constante e corrente de 15 mA.
3.7.3. Western Blotting
Após eletroforese, as proteínas foram eletrotransferidas do gel para uma
membrana de fluoreto de polivinilideno (PVDF), anteriormente ativada em metanol
100%, por 10 min. A eletrotransferência foi realizada em tampão de corrida adicionado
de metanol 20% (v/v) pH 8,3, em 210 mA, pelo período de 90 minutos a 4 ºC
(TOWBIN, et al., 1979), utilizando-se o sistema de transferência da Bio-Rad (Bio-Rad
Laboratories, Hercules, CA, USA). Em seguida, a membrana foi corada com o reagente
Ponceau para confirmar a transferência das proteínas. TBS pH 7,6 foi utilizado para
lavar a membrana, até a retirada completa do Ponceau.
Posteriormente, a membrana contendo as proteínas aderidas foi bloqueada em
tampão TBS-Tween-Leite pH 7,6, overnight a 4 °C. Logo após, foi incubada com o
anticorpo contra flot-1, flot-2 ou GAPDH (controle interno de carregamento de
proteínas) diluídos (1:1000) em tampão TBS-Tween-Leite, por 2 h em temperatura
ambiente. Em seguida, foi lavada cinco vezes, a cada 5 min, com tampão TBS-Tween
pH 7,6, seguida por incubação com anticorpo secundário conjugado a peroxidase
diluído (1:1000) em TBS-Tween-Leite, por 2 h em temperatura ambiente.
Terminado esse período, a membrana foi lavada cinco vezes, a cada 5 min, com
tampão TBS-Tween pH 7,6 e, três vezes, a cada 5 min, com tampão TBS pH 7,6. O
complexo antígeno-anticorpo formado foi revelado por meio do método de
intensificação de quimioluminescência pelo luminol (Millipore, Darmstadt, HE, GER),
com auxílio do equipamento C-Digit Blot Scanner (Li-Cor Inc., Lincoln, NE, USA). A
análise densitométrica foi realizada após digitalização, com auxílio do programa ImageJ
versão 1.5 (National Institute of Health, USA). Para o descarregamento, as membranas
foram lavadas duas vezes, por 10 min, com tampão TBS pH 7,6 e, incubadas com a
solução de descarregamento composta por glicina 200 mM, SDS 0,1% e Tween 20 1%
pH2,2. Em seguida, foram lavadas duas vezes, por 10 min, com água deionizada, três
vezes, por 10 min com TBS pH 7,6 e três vezes, por 10 min com TBS-Tween.

52
3.8. Ensaio de imunofluorescência para flotilina-2 em células MDA-MB-231
tratadas com resveratrol
As células MDA-MB-231 (5 x 105) foram cultivadas em lamínulas com poli-L-
lisina 0,1% e tratadas com Resv 200 µM e DMSO 0,1% (controle) por 24 h. Em
seguida, foram lavadas 3 vezes com PBS, fixadas com metanol 4% em PBS por 15 min
em temperatura ambiente (ASP et al., 2014) e, novamente lavadas três vezes com PBS.
Posteriormente, descartou-se o PBS e as amostras foram lavadas três vezes por 10 min,
com PBS-Triton X-100 0,5% e incubadas, por 1 h a 37 °C, com anticorpo contra flot-2
(1:70) diluído em PBS-Triton X-100 0,5%.
Logo após, foram novamente lavadas com PBS-Triton X-100 0,5%, três vezes
por 10 min, seguida de incubação, por 1 h a 37 °C, com anticorpo secundário (1:100)
conjugado ao fluoróforo Alexa flúor 488. Após nova lavagem com PBS-Triton X-100
0,5%, três vezes por 10 min, as células foram incubadas, sob agitação, com a solução de
NaCl 0,9% por 10 min. A seguir, foram mantidas com DAPI (1:2000) em NaCl 0,9%,
em agitação por três min para corar os núcleos celulares. Subsequentemente, as imagens
foram adquiridas utilizando-se microscópio confocal modelo Axion Vert 100 (Zeiss
Corporate, Oberkochen, BW, GER), com câmera modelo Olympus DP71 (Olympus
Corporation, Shinjuku, TYO, JP). Para a amostra controle foram fotografados cinco
campos diferentes e nove para as tratadas. A média do valor das áreas encontradas foi
utilizada para o cálculo da área correspondente a flot-2.
3.9. Análise do efeito do resveratrol sobre a expressão de flotilina-1 e flotilina-2 por
reação em cadeia da polimerase com transcriptase reversa quantitativa (RT-
qPCR)
Para o ensaio de RT-qPCR, após tratamento por 24 h com 80 e 200 µM de Resv,
as células MCF-7 e MDA-MB-231, respectivamente foram lavadas três vezes com PBS.
Em seguida, adicionou-se 1 mL do reagente Trizol (Invitrogen, Life Tecnologies,
Carlsbad, CA, USA, #15596-026) e as amostras foram mantidas overnight, a -20 °C em
tubos livres de RNase. Posteriormente, a extração de RNA foi realizada seguindo as
instruções do fabricante. Inicialmente, foram acrescentados 200 µL de clorofórmio,
seguida por agitação vigorosa, por 3 min em temperatura ambiente.

53
Após centrifugação a 12000 g, por 15 min a 4 °C, a fase aquosa foi transferida
para novo tubo, onde foram adicionados 500 µL de isopropanol, que permaneceram
incubados por 10 min em temperatura ambiente. Logo após, houve nova centrifugação a
12000 g, por 15 min a 4 °C e o sobrenadante foi removido cuidadosamente, para não
ocorrer prejuízo ao precipitado. Em seguida, o pellet foi lavado com etanol 70% a 4 °C,
centrifugado a 7500 g, por 5 min a 4 °C, o sobrenadante descartado e o restante
evaporado em banho seco a 56 °C por 5 min.
O precipitado foi solubilizado em 20 µL de água autoclavada e a concentração
de RNA foi determinado no NanoDrop. Para a síntese de cDNA utilizou-se 2 µg do
total de RNA de cada amostra, previamente submetidas ao tratamento com DNase, na
qual foram incubadas por 30 min a 37 °C com 1 µL de DNase e 1 µL tampão de reação
10x (volume final da reação foi de 10 µL, ajustado com água autoclavada) e, por 10 min
a 65 °C com 1µL de EDTA 25 mM. As amostras de RNA foram armazenadas a -80 °C.
Posteriormente, para a reação de síntese de cDNA, nas amostras de RNA (10
µL) foram adicionados 2 µL de tampão RT 10x, 0,8 µL de dNTP 25x (100 mM), 2 µL
de RT Random Primers 10x, 1 µL de MultiScribeTMReverse Transcriptase (Applied
Biosystems, Waltham, MA, USA, #4368813) e o volume ajustado para 10 µL com água
autoclavada (volume final da reação foi de 20 µL). A reação foi conduzida por 10 min a
25 °C, seguida por 120 min a 37 °C e 5 s a 85 °C.
Para condução da RT-qPCR, o cDNA das amostras (1:50) e os primers de flot-1,
flot-2 e actina (1:10, cada) foram diluídos em água autoclavada. A actina foi empregada
como controle interno. A reação para cada proteína foi constituída de uma mistura com
0,5 µL de água autoclavada, 1 µL de primer forward e reverse (10 µM), 7,5 µL de
PowerUpTM SYBR® Green Master Mix (Applied Biosystems, Waltham, MA, USA,
#A25742) e 5 µL de cDNA da amostra.
Cada gene foi quantificado em triplicata utilizando-se o StepOne Real-Time PCR
Systems (Applied Biosystems, Waltham, MA, USA), tendo como parâmetros a pré-
incubação por 10 min a 95 °C, seguido por 40 ciclos de 15 s a 95 °C, 60 s a 60 °C, com
curva de fusão de 15 s a 95 °C, 60 s a 60 °C e 15 s a 95 °C. Os valores encontrados para
cada gene foram normalizados considerando-se a expressão de actina. Foram realizados
três experimentos independentes.

54
3.10. Estudo da ação do resveratrol sobre perfil lipídico das células de câncer de
mama MCF-7 e MDA-MB-231
3.10.1. Extração de lipídeos
A extração de lipídeos das céulas MCF-7 e MDA-MB-231 (1 x 107) tratadas
com Resv 80 e 200 µM, respectivamente, ou DMSO 0,1%, por 24 h, foi realizada
conforme descrito por Bligh & Dyer (1959). Após o período de incubação, as células
foram lavadas três vezes com PBS, em seguida, adicionou-se clorofórmio:metanol:água
(1:2:0,8, v/v). Após agitação a cada 5 min, por 1 hora, as amostras foram centrifugadas
a 3000 rpm, por 20 min a 4 °C e, o sobrenadante foi separado do precipitado, sendo
reservado em novo tubo.
O precipitado foi submetido a segunda extração com a mesma mistura de
solventes, tempo de agitação e condições de centrifugação. Posteriormente, os
sobrenadantes foram unificados e a eles adicionados água deionizada:clorofórmio (1:1).
Após agitação por 30 s, o material foi centrifugado a 3000 rpm, por 30 minutos a 4 °C, e
a fase orgânica foi transferida para tubos resistentes a solventes orgânicos, previamente
pesados 5 vezes. O solvente foi evaporado por arraste de nitrogênio gasoso e os tubos
foram pesados novamente. A quantidade de lipídeos foi determinada por gravimetria, na
qual considerou-se a diferença entre as pesagens realizadas. Em seguida, os lipídeos
foram ressuspendidos em clorofórmio:metanol (1:1 v/v) para as análises.
3.10.2. Análise de lipídeos neutros e ácidos graxos
3.10.2.1. Estudo do perfil de lipídeos neutros por cromatografia em camada fina de
alta afinidade (HPTLC) e cromatografia em camada fina (TLC)
Os ensaios de HPTLC foram conduzidos com 380 µg de lipídeos, enquanto que
para a TLC aplicou-se a quantidade total de lipídeos obtidos em 1 x 107 de células. Em
ambas as cromatografias utilizou-se placa de sílica (Merck, Frankfurt, DS, GER). O
sistema de solvente foi composto por n-hexano:éter dietílico:ácido acético (30:20:0,5
para a HPTLC e 60:40:1 v/v para a TLC) (VOGEL et al., 1962). Os lipídeos foram
idetificados com auxílio dos padrões de monoacilglicerol (MG), diacilglicerol (DG),

55
triacilglicerol (TG), colesterol livre (CHO), éster de colesterol (CHOE) e ácido graxo
livre (FA) (Avanti Polar Lipids Inc., Alabaster, AL, USA).
Após o arraste de lipídeos com o sistema de solventes e evaporação, as placas
foram borrifadas com reagente de Charring (sulfato de cobre 10% e ácido fosfórico 8%)
(RUIZ & OCHOA, 1997). Aguardou-se a secagem do reagente para só então colocar as
placas de sílicas entre placas de vidro, que foram colocadas em estufa a 200 °C para a
revelação dos lipídeos, por tempo variável entre 15 e 20 min. Após digitalização das
placas, a análise densitométrica foi realizada com auxílio do programa ImageJ versão
1.5 (National Institute of Health, USA), e os resultados expressos em µg/µL após
normalização com os padrões utilizados. Para a identificação dos lipídeos por HPTLC,
foram realizados dois experimentos independentes.
3.10.2.2. Identificação e quantificação de ácidos graxos por cromatografia gasosa
com espectometria de massa (GC/MS)
Após a extração, os lipídeos foram transesterificados (CHRISTIE, 1989). Para
isso, foram ressuspendidos em 1 mL de tolueno, em seguida adicionou-se 2 mL de
ácido sulfúrico 1% (em metanol) e agitou-se de forma intermitente por 1 min. As
amostras foram mantidas overnight a 50 °C. Posteriormente, foi adicionado 1 mL de
NaCl 5% e agitou-se vigorosamente. Para a extração dos ésteres adicionou-se hexano,
em seguida, as amostras foram centrifugadas a 3000 rpm, por 20 min. Esse processo foi
realizado duas vezes. Logo após, o solvente foi seco por arraste de nitrogênio. As
amostras foram ressuspendidas na solução com o padrão interno C19 (0,5 µg/µL).
Injetou-se 1 µL de cada amostra ao cromatógrafo acoplado ao MS Shimadzu,
modelo GP2010 Plus, com a coluna HP Ultra 2 (5% fenil - metilpolisiloxano), com 25
m x 0,20 mm x 0,33 μm de dimensões. O injetor foi mantido a 250 °C, com o primeiro
minuto no modo splitless. A temperatura do forno da coluna permaneceu a 40 °C por 1
minuto e elevada a 160 °C, com taxa de aquecimento de 30 °C/min e de 233-300 °C,
com taxa de aquecimento de 1 °C/min, mantida por 10 min. Hélio foi utilizado como
gás de arraste com velocidade linear de 36,0 cm/s.
Para a detecção por espectrometria de massas utilizou-se detector contendo uma
fonte de ionização por elétrons (EI-70 eV) e um analisador de massas quadrupolo,
operado em varreduras de 40 a 440 u.m.a. A interface e a fonte de íons foram mantidas
a 240°C. A identificação dos constituintes da mistura foi feita por comparação dos seus

56
espectros de massas com aqueles da biblioteca NIST05, contida no banco de dados
digital do equipamento, assim como com seus tempos de retenção com padrão Supelco
37 Component FAME Mix (Sigma-Aldrich Inc, St. Louis, MO, USA). A quantificação
foi realizada por meio de curva de calibração utilizando-se o padrão interno metil
nonadecanoato (Sigma-Aldrich Inc, St. Louis, MO, USA). Para a detecção de possíveis
alterações causadas pelo Resv, foram realizados dois experimentos independentes e os
resultados expressos em µg/µL. Os cromatogramas de cada amostra encontram-se
representados no Anexo 2 e Anexo 3.
3.10.2.3. Dosagem de lipídeos totais, colesterol e triacilglicerol
Para esse ensaio foi utilizado o extrato celular total, sem a realização da extração
de lipídeos. Dessa forma, as células MCF-7 e MDA-MB-231 (1 x 107) foram tratadas
com Resv 80 e 200 µM, respectivamente, ou DMSO 0,1%, mantidas em PBS com
coquetel de inibidores de proteases 100 vezes concentrado 0,1% (Sigma-Aldrich Inc, St.
Louis, MO, USA, #P8340) e sonicadas a 10% da amplitude total, em 3 ciclos de 10 s
cada, com intervalos de 3 s entre cada ciclo, usando-se o sonicador Branson Sonifier
250 (Analog Ultrasonic Homogenizer, Branson Ultrasonics Corp., Danbury, CT, USA).
Para cada amostra determinou-se a concentração de proteínas (LOWRY et al., 1951),
que foi utilizada para a normalização dos teores de lipídeos totais, CHO e triacilglicerol
encontrados.
A determinação de lipídeos totais baseou-se no método descrito por Knight et al.
(1972). As amostras (5 µL) foram colocadas em tubos de ensaio e mantidas em banho-
seco a 100 °C por 10 min, após esse período foram resfriadas e adicionou-se 100 µL de
ácido sulfúrico e acondicionou-se novamente em banho-seco a 100 °C por 10 min. Em
seguida, os tubos foram novamente resfriados e acrescentou-se 1 mL do reagente de
vanilina (1,98 g de vanilina e 668 mL de ácido orto-fosfórico 85% para 1L de reagente).
Logo após, os tubos foram agitados e permaneceram ao abrigo da luz por 30 min.
Posteriormente, realizou-se análise colorimétrica, utilizando-se espectrofotômetro com
leitura a 546 nm.
Para a dosagem de CHO, utilizou-se o método descrito por Zlatkis et al. (1953).
Em 5 µL de amostra adicionou-se 0,75 mL de ácido acético e 0,5 mL de cloreto de ferro
2,5% (0,05 g de cloreto de ferro diluído em 2 mL de ácido fosfórico, com volume
ajustado com ácido sulfúrico para 25 mL). Após agitados, os tubos com as amostras

57
foram mantidos por 30 min ao abrigo da luz. Em seguida, realizou-se a leitura em
espectrofotômetro a 560 nm. Tanto para a dosagem de lipídeos totais quanto para a de
determinação de CHO de utilizou-se como padrão o CHO (0,5 mg/mL) em clorofórmio,
que foi empregado nos cálculos de determinação dos teores desses lipídeos.
A análise do conteúdo de triacilglicerol foi realizada por meio do kit comercial
Triglicérides 120 (Doles, Goiânia, BRA), seguindo as instruções do fabricante. Em 10
µL de amostra adicionou-se 200 µL de tampão e 10 µL de água, que foram incubados
por 10 min a 37 °C. A leitura foi realizada em espectrofotômetro a 510 nm.
Para a três quantificações foram realizados três experimentos independentes, em
triplicata. E os resultados expressos em µg de lipídeo/µg de proteína de cada amostra.
3.10.3. Análise de composição de fosfolipídeos
3.10.3.1. Ensaio com fosfato inorgânico radioativo (32Pi)
Para a análise dos fosfolipídeos marcados com 32Pi, as linhagens MCF-7 e
MDA-MB-231 (2 x 105) foram cultivadas em placas de 24 poços e tratadas com Resv
(DMSO 0,1% foi o controle) em meio DMEM sem soro fetal bovino e 20 µL de 32Pi
(1,5 mCi/mL) e o volume de incubação foi de 500 µL. O início da exposição ao Resv
ocorreu em conjunto com a incubação com 32Pi e os intervalos de tempo de 0, 2, 8 e 24
h foram verificados. As células foram submetidas à extração de lipídeos conforme
descrição anterior. Em seguida, 10 µL foi utilizada para determinação dos fosfolipídeos
associados ao 32Pi por cintilação líquida. Os resultados foram expressos em contagens
por minuto (CPM). (XIMENES et al., 2015)
3.10.3.2. Cromatografia em camada fina (TLC)
A análise de fosfolipídeos por TLC ocorreu submetendo-se as amostras,
aplicadas na placa de sílica, ao conjunto de solventes constituído por
clorofórmio:acetona:metanol:ácido acético:água destilada (40:15:13:12:8 v/v) (revisado
por FUCHS et al., 2011). Posteriormente, os lipídeos foram visualizados após exposição
com filme Kodak X-OMAT (Eastman Kodak Comp., Rochester, NY, USA) por,
aproximadamente, 2 semanas a -70 °C, com auxílio de um TranScreeen phospho
(Eastman Kodak Comp., Rochester, NY, USA).

58
Os spots foram marcados na placa de sílica e raspados para a análise por
cintilação líquida. Os resultados foram expressos em contagens por minuto (CPM).
Foram utilizados os seguintes padrões para a identificação dos fosfolipídeos por
comparação: lisofosfatidilcolina (LPC), fosfatidiletanolamina (PE), fosfatidilcolina
(PC), fosfatidilinositol (PI), ácido fosfatídico (PA), fosfatidilserina (PS),
fosfatidilglicerol (PG) e esfingomielina (SM). A imagem representativa da TLC
encontra-se no Anexo 4.
3.11. Investigação da ação do resveratrol sobre enzimas do metabolismo lipídico
por Western Blotting
As possíveis alterações da via lipídica foram avaliadas por Western Blotting
(TOWBIN, et al., 1979). O preparo do extrato proteico, a realização da eletroforese
unidimensional em gel de poliacrilamida em condições desnaturantes e o Western
Blotting ocorreram conforme descrito nos itens 3.7.1., 3.7.2. e 3.7.3., respectivamente.
Entretanto, o tempo de bloqueio com TBS-Tween-Leite e de incubação com os
anticorpos primários e secundários foi de 2 h em temperatura ambiente.
As enzimas analisadas foram acil-CoA:diacilglicerol aciltransferase 2 (DGAT2),
ácido graxo sintase (FAS), acetil CoA carboxilase fosforilada (ρACCβ), proteína
quinase ativada por adenosina monofosfato alfa constitutiva (AMPKα) e fosforilada
(ρAMPKα). Foram realizados três experimentos independentes e os resultados
expressos em unidades arbitrárias normalizadas pelo GAPDH.
3.12. Análise estatística
O erro padrão foi calculado por meio da média dos experimentos. O nível de
significância das diferenças entre as médias foi avaliado pelo método de análise
estatística Teste-t não pareado (p < 0,05) e por análise de variância (ANOVA) seguido
dos testes de comparações múltiplas Tukey, Sidak e Dunnett (p < 0,05). Para os testes
estatísticos utilizou-se o programa GraphPad Prism, versão 5.0 e versão 6.0 (GraphPad
Software, Inc, La Jolla, CA, USA)

59
4. RESULTADOS

60
4.1. Citotoxicidade do resveratrol em células de câncer de mama MCF-7 e MDA-
MB-231
A análise do impacto do Resv sobre a viabilidade das células MCF-7 e MDA-
MB-231 foi avaliado utilizando-se o ensaio de redução do MTT, após 24 h de exposição
a concentrações de 50, 80, 100, 150, 200, 250 e 300 µM do fitoquímico. Nas células
MCF-7, observou-se o efeito citotóxico a partir de 50 µM (Figura 8A), com diminuição
de 38%, nos demais pontos da curva o decréscimo da viabilidade foi de 45 (80 µM) a
58% (300 µM) (p < 0,05). Por outro lado, na MDA-MB-231 a citotoxicidade ocorreu,
inicialmente, com 100 µM do composto (Figura 8B) (p < 0,05), e os percentuais de
queda na viabilidade foram menores, variando entre 30 (50 µM), 33 (80 µM) e 52%
(300 µM).
Contr
ole
50,0
80,0
100,0
150,0
200,0
250,0
300,0
0
20
40
60
80
100
120
* *
R e s v e r a tr o l (M )
Via
bil
ida
de
ce
lula
r (
%)
* * * * * * * * * * * *
* * *
* * *
A
Contr
ole
50,0
80,0
100,0
150,0
200,0
250,0
300,0
0
20
40
60
80
100
120
R e s v e r a tr o l (M )
Via
bil
ida
de
ce
lula
r (
%)
*
*
*
*
*
B
Figura 8: Efeito do resveratrol sobre a viabilidade celular das linhagens MCF-7 e MDA-MB-231.
Células (A) MCF-7 e (B) MDA-MB-231 (2x105) foram incubadas na presença de diferentes
concentrações (0 a 300 µM) de resveratrol, ou DMSO 0,1% (controle), por 24 h a 37 °C. Em seguida,
foram lavadas e mantidas com MTT (0,5 mg/mL) por 3 h a 37 °C, para a formação dos cristais de
formazan e a densidade ótica associada foi analisada a 570 e 650 nm. Os resultados foram expressos em
média ± SD, n = 3. */**/*** Significativamente diferentes (p < 0,05) dos valores correspondentes ao
controle, utilizando-se One-way ANOVA, seguido por teste de comparações múltiplas Dunnett.

61
4.2. Efeito do resveratrol sobre microdominíos de membrana e expressão de
flotilina-1 e flotilina-2 em células de câncer de mama MCF-7 e MDA-MB-231
Com o intuito de isolar os microdomínios lipídicos de membrana utilizou-se o
Triton X-100 a 4°C e as células MCF-7 e MDA-MB-231, após 24 h de tratamento com
Resv, foram submetidas ao gradiente sacarose com ultracentrifugação. Ao final das 22 h
(20 h de centrifugação + 2 h até parar de girar) foi possível visualizar material
esbranquiçado flutuante em todas as condições de tratamento (Figura 9A). Quando as
amostras foram aliquotadas em doze frações com volume de 1 mL cada, observou-se
que este material se concentrou nas frações 4 e 5, conforme confirmado posteriormente
(Figuras 10 e 11). Em seguida, o índice de refração foi determinado e convertido em
percentual de sacarose. Nota-se que houve a formação gradual do gradiente para as
células MCF-7 controle (Figura 9B) e tratada (Figura 9C) e, MDA-MB-231 controle
(Figura 9D) e tratada (Figura 9E). O percentual de sacarose mínimo foi de 5% e o
máximo de 50%, confirmando a formação do gradiente.
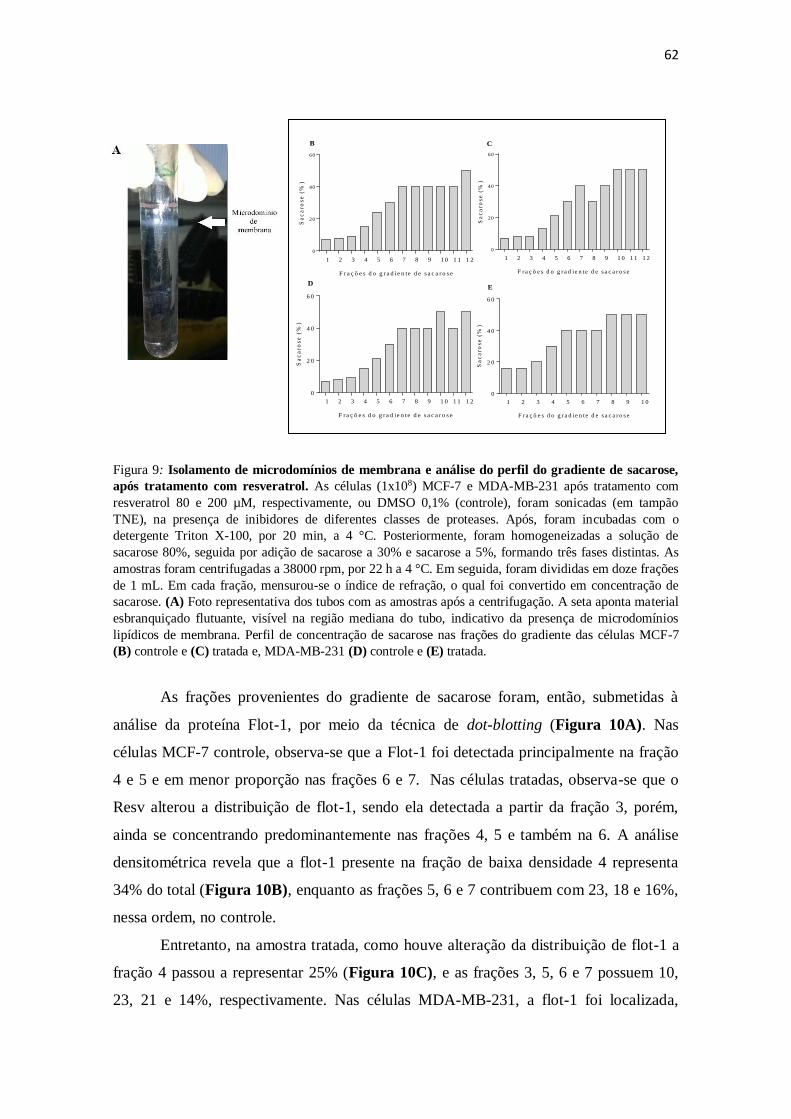
62
Figura 9: Isolamento de microdomínios de membrana e análise do perfil do gradiente de sacarose,
após tratamento com resveratrol. As células (1x108) MCF-7 e MDA-MB-231 após tratamento com
resveratrol 80 e 200 µM, respectivamente, ou DMSO 0,1% (controle), foram sonicadas (em tampão
TNE), na presença de inibidores de diferentes classes de proteases. Após, foram incubadas com o
detergente Triton X-100, por 20 min, a 4 °C. Posteriormente, foram homogeneizadas a solução de
sacarose 80%, seguida por adição de sacarose a 30% e sacarose a 5%, formando três fases distintas. As
amostras foram centrifugadas a 38000 rpm, por 22 h a 4 °C. Em seguida, foram divididas em doze frações
de 1 mL. Em cada fração, mensurou-se o índice de refração, o qual foi convertido em concentração de
sacarose. (A) Foto representativa dos tubos com as amostras após a centrifugação. A seta aponta material
esbranquiçado flutuante, visível na região mediana do tubo, indicativo da presença de microdomínios
lipídicos de membrana. Perfil de concentração de sacarose nas frações do gradiente das células MCF-7
(B) controle e (C) tratada e, MDA-MB-231 (D) controle e (E) tratada.
As frações provenientes do gradiente de sacarose foram, então, submetidas à
análise da proteína Flot-1, por meio da técnica de dot-blotting (Figura 10A). Nas
células MCF-7 controle, observa-se que a Flot-1 foi detectada principalmente na fração
4 e 5 e em menor proporção nas frações 6 e 7. Nas células tratadas, observa-se que o
Resv alterou a distribuição de flot-1, sendo ela detectada a partir da fração 3, porém,
ainda se concentrando predominantemente nas frações 4, 5 e também na 6. A análise
densitométrica revela que a flot-1 presente na fração de baixa densidade 4 representa
34% do total (Figura 10B), enquanto as frações 5, 6 e 7 contribuem com 23, 18 e 16%,
nessa ordem, no controle.
Entretanto, na amostra tratada, como houve alteração da distribuição de flot-1 a
fração 4 passou a representar 25% (Figura 10C), e as frações 3, 5, 6 e 7 possuem 10,
23, 21 e 14%, respectivamente. Nas células MDA-MB-231, a flot-1 foi localizada,
F ra ç õ e s d o g ra d ie n te d e s a c a r o s e
Sa
ca
ros
e (
%)
1 2 3 4 5 6 7 8 9 1 0
0
2 0
4 0
6 0
E
F ra ç õ e s d o g ra d ie n te d e s a c a r o s e
Sa
ca
ros
e (
%)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
0
20
40
60
C
F ra ç õ e s d o g ra d ie n te d e s a c a r o s e
Sa
ca
ros
e (
%)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
0
2 0
4 0
6 0
D
F ra ç õ e s d o g ra d ie n te d e s a c a r o s e
Sa
ca
ros
e (
%)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
0
20
40
60
B

63
principalmente, nas frações 4 e 5, assim como ocorreu com a MCF-7 (Figura 11A),
representando 29 e 31%, respectivamente, nas amostras não tratadas (Figura 11B). As
frações 6 e 7 contribuíram com 21 e 18%. Após o tratamento, houve redução de flot-1
nas frações 4 (28%) e 7 (8%) (Figura 11C), com aumento de flot-1 nas frações 5 (38%)
e 6 (25%).
F ra ç õ e s d o g ra d ie n te d e s a c a r o s e
De
ns
ito
me
tria
(%
)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
0
5
10
15
20
25
30
35
B
F ra ç õ e s d o g ra d ie n te d e s a c a r o s e
De
ns
ito
me
tria
(%
)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
0
5
10
15
20
25
30
35
C
Figura 10: Resveratrol altera a distribuição dos microdomínios lipídicos de membrana nas frações
do gradiente de sacarose da célula MCF-7. O volume de 500 µL de cada fração foi aplicado na
membrana de nitrocelulose. Em seguida, a membrana foi bloqueada com TBS-Tween-Leite, por 2 h em
temperatura ambiente. Posteriormente, foi incubada com anticorpo contra flot-1 diluído (1:800) em TBS-
Tween-Leite, por 18 h a 4 C. Após, a membrana foi lavada e incubada com anticorpo secundário
conjugado a peroxidase (1:1000), por 2 h em temperatura ambiente. A detecção de flot-1 realizou-se por
quimioluminescência. (A) Digitalização da foto obtida após a revelação. Densitometria, em percentual, da
imagem digitalizada para as amostras (B) controle (DMSO 0,1%) e (C) tratada (Resv 200 µM).

64
F ra ç õ e s d o g ra d ie n te d e s a c a r o s e
De
ns
ito
me
tria
(%
)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
0
5
10
15
20
25
30
35
40
B
F ra ç õ e s d o g ra d ie n te d e s a c a r o s e
De
ns
ito
me
tria
(%
)
1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
0
5
10
15
20
25
30
35
40
C
Figura 11: Resveratrol altera a distribuição dos microdomínios lipídicos de membrana nas frações
do gradiente de sacarose da célula MDA-MB-231. O volume de 500 µL de cada fração foi aplicado na
membrana de nitrocelulose. Em seguida, a membrana foi bloqueada com TBS-Tween-Leite, por 2 h em
temperatura ambiente. Posteriormente, foi incubada com a flot-1 diluída 1:800 em TBS-Tween-Leite, por
18 h a 4 C. Após, a membrana foi lavada e incubada com anticorpo secundário conjugado a peroxidase
(1:1000), por 2 h em temperatura ambiente. A detecção de flot-1 realizou-se por quimioluminescência.
(A) Digitalização da foto obtida após a revelação. Densitometria, em percentual, da imagem digitalizada
para as amostras (B) controle (DMSO 0,1%) e (C) tratada (Resv 200 µM).
Como a análise de flot-1 revelou resultados interessantes, indicando que o Resv
altera a distribuição da proteína nas regiões de microdomínios lipídicos de membrana,
também foi verificado seu efeito sobre a flot-2, pela técnica de Western blot. Tanto flot-
1 quanto flot-2 foram detectadas nas células MCF-7 e MDA-MB-231 controle e tratadas
com Resv (Figura 12A). No entanto, após o tratamento com Resv, os níveis de flot-1 e
flot-2 mantiveram-se similares aos do controle, sem alterações expressivas observadas
por esta técnica. Na linhagem MCF-7 (Figura 12B) obteve-se para a flot-1 e flot-2 a
redução de 1,06 e 1,07 vezes, respectivamente, e o aumento de 0,85 e 0,93 vezes para
flot-1 e flot-2, respectivamente, na MDA-MB-231 (Figura 12C).

65
Figura 12: Efeito do resveratrol sobre os níveis de flot-1 e flot-2 em células MCF-7 e MDA-MB-231.
As células (1x108) MCF-7 e MDA-MB-231 após 24 h de tratamento com 80 e 200 µM de resveratrol,
respectivamente, ou DMSO 0,1% (controle), foram congeladas em nitrogênio líquido. Em seguida,
adicionou-se tampão de lise, composto por inibidores de proteases de diferentes classes. Posteriormente,
as amostras foram sonicadas, centrifugadas a 8.000 rpm, por 5 min a 4 °C. O volume do sobrenadante
(extrato proteico) equivalente a 80 µg de proteínas foi submetido a eletroforese em gel de poliacrilamida
10%, em condições desnaturantes. Após, as proteínas do gel foram transferidas para a membrana PVDF, a
qual foi bloqueada com TBS-Tween-Leite, overnight a 4 °C, posteriormente, foi incubada com flot-1 ou
flot-2 diluída (1:1000) em TBS-Tween-Leite, por 2 h em temperatura ambiente. O GAPDH (1:1000) foi
utilizado como controle de carregamento interno de proteínas. Após, a membrana foi lavada e incubada
com anticorpo secundário conjugado a peroxidase (1:1000), por 2 h em temperatura ambiente. A detecção
de flot-1 e flot-2 realizou-se por quimioluminescência. (A) Imagens obtidas e a densitometria da MCF-7
(B) e MDA-MB-231 (C), expressa em unidades arbitrárias.
Com os resultados obtidos por dot-blotting e Western blotting observou-se que o
Resv alterou a distribuição de flot-1 nas regiões de microdomínio de membrana
(Figuras. 10 e 11), mas não modificou os níveis de flot-1 e flot-2 (Figura 12). Nesse
contexto, para complementar os resultados obtidos, realizou-se o ensaio de
imunofluorescência para a pesquisa de flot-2 nas células MDA-MB-231 cultivadas em
lamínulas e tratadas com Resv 200 µM por 24 h. Posteriormente, foram fixadas com
metanol 4% e tratadas com anticorpo contra flot-2, em seguida, com anticorpo
secundário conjugado ao fluoróforo e marcadas com DAPI. Os resultados obtidos em
microscópio confocal revelaram que flot-2 apresenta-se em quantidade expressivamente
aumentada nas células tratadas com Resv (Figura 13D, E e F), quando comparadas ao
0.0
0.5
1.0
1.5
MCF-7
Flot-1 Flot-2
Controle Resv Controle Resv
Flo
t-1 e
Flo
t-2/G
AP
DH
(unid
ades
arb
itrá
rias
)
0.0
0.5
1.0
1.5
Flot-1 Flot-2
MDA-MB-231
Controle Resv Controle ResvF
lot-
1 e
Flo
t-2/G
AP
DH
(unid
ades
arb
itrá
rias
)

66
controle (Figura 13A, B e C). Após expostas ao Resv houve elevação de cerca de 13,4
vezes da área correspondente a flot-2.

67
Resveratrol
Controle

68
0
2
4
6
8
1 0
1 2
Áre
a c
orre
sp
on
de
nte
à f
lot-
2 (
%)
C o n t r o l
R e s v
G
Figura 13: Imunoflurescência para flot-2 em células MDA-MB-231 após tratamento com
resveratrol. As células (5 x 105) foram tratadas com resveratrol 200 µM ou DMSO 0,1% (controle), por
24 h. Em seguida, foram lavadas com PBS-Triton X-100 0,5% e incubadas com flot-2 (diluição1:70), por
1 h a 37 °C. Após, lavou-se e incubou-se as amostras com o anticorpo secundário (diluição 1:100)
conjugado ao fluoróforo, por 1 h a 37 °C. Posteriormente, as células foram lavadas e incubadas com
DAPI (diluição 1:2000), por 3 min. As imagens obtidas em microscópio confocal (aumento de 40 vezes)
mostram a marcação para flot-2 (verde) na amostra controle (A) e tratada (D). O núcleo nas amostras
controle (B) e tratada (E) foi corado com DAPI (azul). (C) e (F) são a sobreposição das imagens das
amostras controle (A e B) e tratadas (D e E). (G) representação em percentual da área correspondente a
flot-2. Os resultados foram expressos em média ± S.D, n = 5 para a amostra controle e n = 9 para a
amostra tratada.
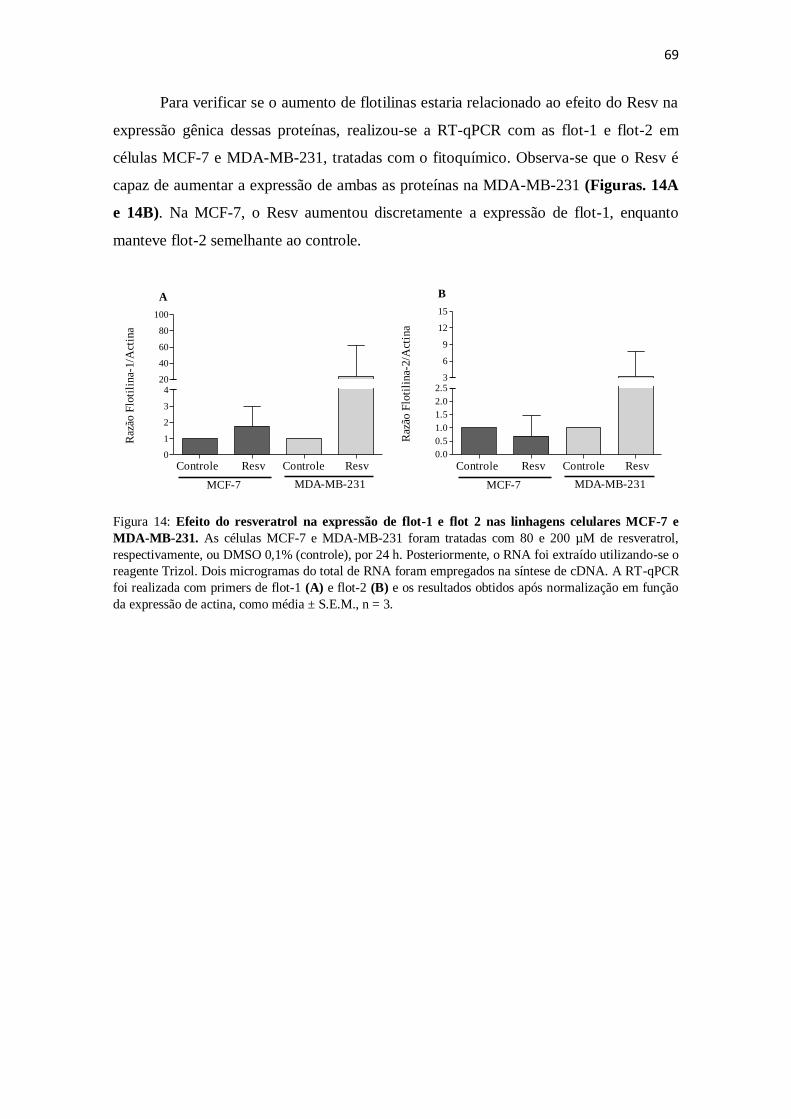
69
Para verificar se o aumento de flotilinas estaria relacionado ao efeito do Resv na
expressão gênica dessas proteínas, realizou-se a RT-qPCR com as flot-1 e flot-2 em
células MCF-7 e MDA-MB-231, tratadas com o fitoquímico. Observa-se que o Resv é
capaz de aumentar a expressão de ambas as proteínas na MDA-MB-231 (Figuras. 14A
e 14B). Na MCF-7, o Resv aumentou discretamente a expressão de flot-1, enquanto
manteve flot-2 semelhante ao controle.
0
1
2
3
420
40
60
80
100
MCF-7 MDA-MB-231
Controle ResvControleResv
A
Raz
ão F
loti
lina-
1/A
ctin
a
0.0
0.5
1.0
1.5
2.0
2.53
6
9
12
15
MCF-7 MDA-MB-231
Controle ResvControleResv
B
Raz
ão F
loti
lina-
2/A
ctin
a
Figura 14: Efeito do resveratrol na expressão de flot-1 e flot 2 nas linhagens celulares MCF-7 e
MDA-MB-231. As células MCF-7 e MDA-MB-231 foram tratadas com 80 e 200 µM de resveratrol,
respectivamente, ou DMSO 0,1% (controle), por 24 h. Posteriormente, o RNA foi extraído utilizando-se o
reagente Trizol. Dois microgramas do total de RNA foram empregados na síntese de cDNA. A RT-qPCR
foi realizada com primers de flot-1 (A) e flot-2 (B) e os resultados obtidos após normalização em função
da expressão de actina, como média ± S.E.M., n = 3.

70
4.3. Efeito do resveratrol sobre perfil de lipídeos neutros, ácidos graxos e
fosfolipídeos de células de câncer de mama MCF-7 e MDA-MB-231
Dada a importância que o fenótipo lipídico de membrana em células de câncer e
as alterações no perfil de lipídeos apresentam para a determinação de seu início,
progressão e grau de malignidade (HE et al., 2015), além da escassez de informações
sobre o tema associando a ação do Resv, foram analisados os efeitos desse fitoquímico.
Para isso, as linhagens MCF-7 e MDA-MB-231 foram expostas ao Resv por 24 h, e
diferentes classes de lipídeos foram investigadas. Inicialmente, extratos lipídicos das
duas linhagens celulares foram submetidos à HPTLC (Figura 15A), para a análise de
lipídeos neutros, com quantidades padronizadas em 380 µg de lipídeos. Identificou-se
éster de colesterol (CHOE), triacilglicerol (TG), ácido graxo (FA) colesterol (CHO) e
monoacilglicerol (MG). Nesses experimentos não foram considerados os PL presentes
na origem da HPTLC.
As células MCF-7 (Figura 15B), após exposição ao resveratrol, apresentaram
redução de CHOE de 8,1 para 7,1 µg/µL (p > 0,05), com ligeira diminuição de TG (4,2
para 2,9 µg/µL) e FA (2,3 para 1,9 µg/µL), por outro lado, CHO (5,8 µg/µL - controle;
5,9 µg/µL - Resv) e MG (5,6 µg/µL - controle; 5,9 µg/µL - Resv) mantiveram
conteúdos similares ao controle. Na linhagem MDA-MB-231 (Figura 15C) a redução
mais expressiva observada foi para o TG, de 5,2 para 3,7 µg/µL. Tanto CHOE (9,3
µg/µL - controle; 9,2 µg/µL - Resv), quanto FA (2,2 µg/µL -controle; 2,0 µg/µL -
Resv), CHO (6,4 µg/µL - controle; 5,9 µg/µL - Resv) e MG (5,7 µg/µL - controle; 6,2
µg/µL - Resv) apresentaram concentrações próximas as do controle.

71
0.0
2.5
5.0
7.5
10.0
12.5
15.0
*
CHOE TG FA CHO MG CHOE TG FA CHO MG
Controle Resv
MCF-7
B
Co
nte
údo
de
lipíd
eos
(
g/
L)
0.0
2.5
5.0
7.5
10.0
12.5
15.0
CHOE TG FA CHO MG CHOE TG FA CHO MG
Controle Resv
MDA-MB-231
C
Conte
údo d
e li
píd
eos
( g/
L)
Figura 15: Efeito do resveratrol sobre o perfil de lipídeos neutros em células MCF-7 e MDA-MB-
231, por HPTLC. As células (1x107) após tratamento com resveratrol 80 e 200 µM, respectivamente, ou
DMSO 0,1% (controle), por 24 h, foram submetidas à extração de lipídeos. Posteriormente, 380 µg de
lipídeos totais (determinados por gravimetria (Anexo 1)) foram submetidos a HPTLC, em conjunto de
solventes composto por n-hexano:éter dietílico:ácido acético (30:20:0,5 v/v). Em seguida a evaporação
dos solventes, a HPTLC foi revelada com o reagente de Charring e aquecimento a 200 °C. (A) Imagem
representativa de dois experimentos independentes para as células (B) MCF-7 e (C) MDA-MB-231. O
conteúdo de CHOE (éster de colesterol), TG (triacilglicerol), FA (ácido graxo), CHO (colesterol livre) e
MG (monoacilglicerol) foram expressos em µg/µL, após normalização com os respectivos padrões. A
barra representa média ± S.E.M. * Significativamente diferente (p < 0,05) dos valores correspondentes ao
controle, utilizando-se Two-way ANOVA, seguido por teste de comparações múltiplas Sidak.

72
Após analisar o perfil de lipídeos neutros padronizando a quantidade aplicada à
HPTLC em 380 µg, foi verificado se ao utilizar a quantidade total de lipídeos extraídos
em 1 x 107 de células seria possível observar alterações não detectadas anteriormente.
Dessa forma, pode-se considerar a variabilidade do teor de lipídeos da própria célula
não tratada e àquela em consequência da exposição ao Resv. Com isso, o conteúdo total
de lipídeos obtidos em 1 x 107 de células, tanto para a MCF-7 (1020 µg - controle; 1700
µg - Resv) quanto para a MDA-MB-231 (1120 µg - controle; 900 µg - Resv), foi
aplicado à TLC para a análise de lipídeos neutros, como demonstrado na Figura 16A.
Foram encontrados CHOE, TG, FA e CHO.
Diferentemente do observado nos resultados obtidos por HPTLC, as células
MCF-7 (Figura 16B) apresentaram alterações sensíveis nas concentrações de FA (2,45
µg/µL - controle; 7,05 µg/µL - Resv) e CHO (11,33 µg/µL - controle; 14,93 µg/µL -
Resv), as quais foram elevadas em 188 e 32%, respectivamente, após tratamento com
Resv. CHOE e TG mantiveram teores semelhantes aos do controle. Na linhagem MDA-
MB-231 (Figura 16C), notou-se aumento acentuado de FA em 435% (1,49 µg/µL -
controle; 7,97 µg/µL - Resv). TG (18,3 µg/µL - controle; 15,34 µg/µL - Resv) e CHO
(19,93 µg/µL - controle; 17,45 µg/µL - Resv) demonstraram redução sutil de 16 e 12%,
respectivamente, enquanto o CHOE (21,59 µg/µL - controle; 20,59 µg/µL - Resv)
manteve-se similar ao controle.
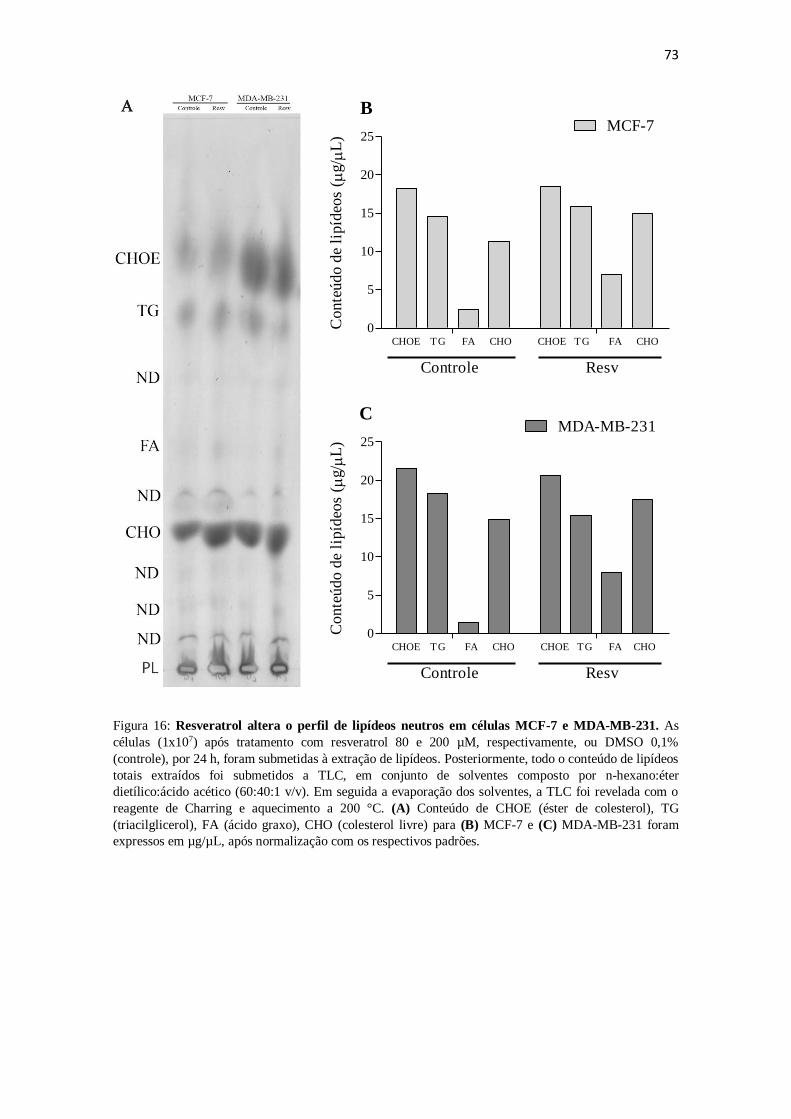
73
Figura 16: Resveratrol altera o perfil de lipídeos neutros em células MCF-7 e MDA-MB-231. As
células (1x107) após tratamento com resveratrol 80 e 200 µM, respectivamente, ou DMSO 0,1%
(controle), por 24 h, foram submetidas à extração de lipídeos. Posteriormente, todo o conteúdo de lipídeos
totais extraídos foi submetidos a TLC, em conjunto de solventes composto por n-hexano:éter
dietílico:ácido acético (60:40:1 v/v). Em seguida a evaporação dos solventes, a TLC foi revelada com o
reagente de Charring e aquecimento a 200 °C. (A) Conteúdo de CHOE (éster de colesterol), TG
(triacilglicerol), FA (ácido graxo), CHO (colesterol livre) para (B) MCF-7 e (C) MDA-MB-231 foram
expressos em µg/µL, após normalização com os respectivos padrões.
0
5
10
15
20
25
CHOE TG FA CHO CHOE TG FA CHO
Controle Resv
MCF-7B
Conte
údo d
e li
píd
eos
( g/
L)
0
5
10
15
20
25
CHOE TG FA CHO CHOE TG FA CHO
Controle Resv
MDA-MB-231C
Conte
údo d
e li
píd
eos
( g/
L)

74
Outras técnicas foram utilizadas para a determinação do conteúdo de lipídeos
nas células MCF-7 e MDA-MB-231. Empregou-se ácido acético e cloreto de ferro
(ZLATKIS et al., 1953) para análise de CHO (Figura 17B), o kit comercial (Doles®,
Ltda, Gôiania, GO, Brasil) para dosagem de TG (Figura 17C), além da quantificação
de lipídeos totais (Figura 17A) por meio do método sulfo-fosfo-vanilina (KNIGHT et
al., 1972). Todos os resultados foram expressos em relação ao teor de proteínas
encontrado em cada extrato celular utilizado.
Na linhagem MCF-7, pode-se observar que após o tratamento com Resv houve
discreto aumento de lipídeos totais (0,130 para 0,159 µg/µg de proteína), redução de TG
(0,009 para 0,001 µg/µg de proteína) e CHO manteve-se similar ao controle (0,013 e
0,012 µg/µg de proteína). Nas células MDA-MB-231 o Resv ocasionou o aumento do
conteúdo de lipídeos totais de 0,147 para 0,308 µg/µg de proteína (p < 0,004) e a
redução de CHO de 0,072 para 0,036 (p < 0,05), entretanto, para o TG há a tendência ao
aumento da concentração, porém a diferença não foi significativa.

75
0.0
0.1
0.2
0.3
0.4
MCF-7 MDA-MB-231
Controle
Resv**
Lip
ídeo
s to
tais
( g/
g d
e p
rote
ína)
0.00
0.01
0.020.03
0.05
0.07
0.09
MCF-7 MDA-MB-231
*
Controle
Resv
B
CH
O
( g/
g d
e pro
teín
a)
0.00
0.01
0.02
0.03
MCF-7 MDA-MB-231
Controle
Resv
C
TG
( g/
g d
e p
rote
ína)
Figura 17: Efeito do resveratrol sobre lipídeos totais, colesterol e triacilglicerol em células MCF-7 e
MDA-MB-231. Após tratamento com resveratrol, as células (1 x 107) foram lavadas e sonicadas em PBS
com inibidores de proteases. Em seguida, foram submetidas as análises das concentrações de (A) lipídeos
totais utilizando-se o método Sulfo-fosfo-vanilina, de (B) colesterol total com ácido acético e cloreto de
ferro e, (C) TG (triacilglicerol) por meio de kit comercial (Doles®). Os resultados obtidos foram
expressos em µg de lipídeo/ µg de proteína da amostra. A barra representa média ± S.E.M., n = 2 - 4. *
Significativamente diferente (p < 0,004; p < 0,05) dos valores correspondentes ao controle, utilizando-se
Teste-t não pareado e teste de comparações múltiplas Tukey.

76
Para verificar o efeito do Resv sobre o perfil de FA das células de câncer de
mama MCF-7 e MDA-MB-231, após a extração de lipídeos, as amostras foram
submetidas à análise por cromatografia gasosa com espectrometria de massas (Anexo 2
e Anexo 3). O total de treze FA saturados foram detectados (Tabela 1), desses, as
células MCF-7 apresentaram doze, em concentrações que variaram de 0,003 a 0,745
µg/µL e de 0,009 a 0,946 µg/µL antes e após o tratamento com Resv, respectivamente.
Observou-se, em maior teor, o ácido palmítico (C16:0), seguido pelo ácido esteárico
(C18:0), ácido láurico (C12:0) e ácido mirístico (C14:0), com 0,745, 0,583, 0,212 e
0,158 µg/µL, respectivamente. O tratamento com Resv reduziu em 52% o ácido
tridecanoico (C13:0), enquanto que os demais FA saturados aumentaram seu conteúdo,
especialmente o C18:0 em 45% e o C12:0 em 43%. Os FA ácido heneicosanoico
(C21:0) e ácido tricosanoico (C23:0) foram encontrados apenas nas células não tratadas,
nas concentrações de 0,004 e 0,003 µg/µL, respectivamente, enquanto que o ácido
hexacosanoico (C26:0) não foi detectado em ambas condições (amostras controle e
tradada com Resv).
A linhagem MDA-MB-231, apresentou concentrações de FA saturados que
variam de 0,010 a 0,997 e 0,013 a 1,125 µg/µL, para as células controle e expostas ao
Resv, respectivamente. O C18:0 foi encontrado em maior teor, acompanhado pelo
C16:0, com concentrações de 0,997 e 0,814 µg/µL, respectivamente. Entretanto, esses
dois FA evidenciaram os menores percentuais de aumento de seu conteúdo após
tratamento com Resv, com 13% para C18:0 e 14% para C16:0. Por outro lado, o ácido
lignocérico (C24:0), o C26:0 e o ácido docosanoico (C22:0), exibiram aumentos
acentuados de seus teores em 87, 53 e 50%, nessa ordem. Ao contrário do observado
com as células MCF-7, na MDA-MB-231 não foram detectados os FA C13:0, C21:0 e
C23:0 nas células tratadas e controle.

77
Tabela 1: Resveratrol altera perfil de ácidos graxos saturados em células MCF-7 e MDA-MB-231.
MCF-7 MDA-MB-231
Controle Resv Controle Resv
ID FA Lipídeo T.R. (µg/µL) (µg/µL)
C12:0 Ácido láurico 9,963 0,212 a 0,304
a 0,015 a 0,013
a
C13:0 Ácido tridecanoico 11,986 0,023 a 0,011
a ND ND
C14:0 Ácido mirístico 15,442 0,158±0,011 0,184±0,096 0,081±0 0,106±0,014
C15:0 Ácido pentadecanoico 19,425 0,035±0,002 0,036±0,020 0,013±0,009 0,018±0,012
C16:0 Ácido palmítico 24,417 0,745±0,100 0,946±0,538 0,814±0,235 0,926±0,184
C17:0 Ácido margárico 30,250 0,036±0,016 0,044±0,030 0,049±0,036 0,064±0,045
C18:0 Ácido esteárico 36,800 0,583±0,285 0,846±0,611 0,997±0,587 1,125±0,572
C20:0 Ácido eicosanoico 51,017 0,012±0,001 0,014±0,003 0,015±0 0,019±0
C21:0 Ácido heneicosanoico 56,921 0,004 a ND ND ND
C22:0 Ácido docosanoico 65,542 0,011±0,007 0,013±0,003 0,010±0,007 0,015±0,003
C23:0 Ácido tricosanoico 71,133 0,003 a ND ND ND
C24:0 Ácido lignocérico 79,517 0,009±0,006 0,010±0,001 0,023±0,002 0,043±0,010
C26:0 Ácido hexacosanoico 82,342 ND ND 0,015±0,002 0,023±0
Conteúdo total de lipídeos saturados 1,831 2,408 2,033 2,352
ID FA: Identificação do ácido graxo; Resv: resveratrol; T,R,: Tempo de retenção; ND: Não determinado. a Representa o valor obtido em apenas um experimento independente.
As células MCF-7 e MDA-MB-231 (1x107) após tratamento com resveratrol 80 e 200 µM,
respectivamente, ou DMSO 0,1% (controle), por 24 h, foram submetidas à extração de lipídeos.
Posteriormente, todo o conteúdo de lipídeos totais extraídos foi submetido à GC/MS. Resultados
expressos em média ± S.E.M., n = 2.
Foram identificados quinze FA insaturados (Tabela 2). A linhagem MCF-7,
exibiu como FA insaturado principal o ácido oleico (C18:1n9c), seguido pelo ácido
elaídico (C18:1n9t) e ácido palmitoleico (C16:1n7) que apresentaram concentrações de
0,542, 0,419 e 0,223 µg/µL, respectivamente. Após o tratamento com resveratrol, esses
três lipídeos continuaram predominantes, com elevação de seus teores de 53% para o
C18:1n9c e de 13% para o C16:1n7, enquanto o C18:1n9t apresentou redução de 20%.
Os demais FA foram encontrados em menores teores que variaram de 0,005 a 0,073
µg/µL. Apesar disso, esses FA exibiram aumento acentuado de seu conteúdo, na
presença do Resv, com destaque para o EPA (C20:5n3) com acréscimo de 73% e o
DHA (C22:6n3) com 65% a mais.

78
As células MDA-MB-231 apresentaram também como FA insaturados
principais o C18:1n9c, seguido pelo C18:1n9t e o ácido araquidônico (C20:4n6) com
teores de 0,460, 0,162 e 0,123 µg/µL, respectivamente. O tratamento com resveratrol
favoreceu alterações consideráveis no conteúdo de FA com aumentos expressivos de
141% para o ácido cis-8,11,14-eicosadienóico (C20:3n6), de 91% para o C20:4n6, de
90% para o DHA (C22:6n3) e 65% para o ácido cis-10-heptadecenóico (C17:1n7).
Observou-se redução do ácido 7-hexadecanóico (C16:1n9) e do ácido cis-11-
eicosenóico (C20:1n9), ambas em 15%. O FA C20:1 foi detectável somente após o
tratamento, na concentração de 0,002 µg/µL.
Embora tenha ocorrido o aumento de FA saturados para as duas células de
câncer de mama, e eles representem o maior conteúdo dos ácidos FA totais encontrados,
vale ressaltar que na MDA-MB-231 o Resv proporcionou o aumento superior de FA
insaturados (53%), quando comparados ao FA saturados (16%). Com isso, as
concentrações de FA insaturados das células MCF-7 e MDA-MB-231 ficaram similares,
nos valores de 1,881 e 1,882 µg/µL, respectivamente. Entretanto, o Resv ocasionou a
elevação em 32% do teor de FA saturados na MCF-7, ultrapassando, dessa forma, o
encontrado para a MDA-MB-231 (2,408 e 2,352 µg/µL, respectivamente).

79
Tabela 2: Resveratrol altera perfil de ácidos graxos insaturados em células MCF-7 e MDA-MB-231.
MCF-7 MDA-MB-231
Controle Resv Controle Resv
ID FA Lipídeo T.R. (µg/µL) (µg/µL)
C16:1n9 Ácido 7-hexadecanoico 23,321 0,027±0 0,040±0,025 0,112a 0,095
a
C16:1n7 Ácido palmitoleico 23,486 0,223±0,133 0,252±0,061 0,093±0,005 0,131±0,030
C17:1n7 Ácido cis-10-heptadecenoico
28,908 0,027±0,02 0,026±0,01 0,023±0,005 0,038±0,013
C18:2n6 Ácido linoleico 34,492 0,036±0,012 0,038±0,002 0,038±0,012 0,056±0,001
C18:1n9c Ácido oleico 34,958 0,542±0,076 0,831±0,478 0,460±0,014 0,718±0,206
C18:1n9t Ácido elaídico 35,433 0,419±0,168 0,334±0,002 0,162±0,005 0,234±0,036
C20:4n6 Ácido araquidônico 45,675 0,073±0,014 0,102±0,064 0,123±0,071 0,235±0,159
C20:5n3 Ácido eicosapentaenoico (EPA)
46,117 0,011±0,001 0,019±0,016 0,040±0,020 0,060±0,036
C20:3n6 Ácido cis-8,11,14-eicosatrienoico
47,067 0,016±0,001 0,025±0,020 0,022±0,008 0,053±0,031
C20:2n6 Ácido cis-11,14-
eicosadienoico 48,508 0,005
a 0,007±0,004 0,007a 0,012±0,003
C20:1n9 Ácido cis-11-eicosenoico
48,975 0,038±0,013 0,042±0,030 0,013a 0,011±0,007
C20:1 49,165 0,036±0,025 0,027±0,012 ND 0,002a
C22:6n3
Ácido cis-4,7,10,13,16,19-docosahexaenoico (DHA)
59,433 0,052±0,010 0,086±0,057 0,084±0,049 0,160±0,108
C22:1n9 Ácido erúcico 63,500 0,014±0,010 0,014±0,004 0,010±0,004 0,008±0,004
C24:1n9 Ácido nervônico 77,792 0,034±0,019 0,038±0,013 0,012±0,006 0,019±0,002
Conteúdo total de lipídeos insaturados 1,553 1,881 1,199 1,832
ID FA: Identificação do ácido graxo; Resv: resveratrol; T,R,: Tempo de retenção; ND: Não determinado. a Representa o valor obtido em apenas um experimento independente.
As células MCF-7 e MDA-MB-231 (1x107) após tratamento com resveratrol 80 e 200 µM,
respectivamente, ou DMSO 0,1% (controle), por 24 h, foram submetidas à extração de lipídeos.
Posteriormente, todo o conteúdo de lipídeos totais extraídos foi submetido à GC/MS. Resultados
expressos em média ± SD, n = 2.

80
O efeito do Resv no conteúdo de PL foi analisado utilizando-se PL marcado com
fosfato inorgânico radioativo (32Pi), que ao serem incorporados permitem a
identificação desses lipídeos presentes em menor concentração nas células. Após a
incubação com Resv e 32Pi, por 0, 2, 8, e 24 h, foi possível observar a incorporação de
32Pi e a biossíntese de PL ao longo do tempo (Figura 18). Nas células MCF-7, a síntese
de PL totais foi ligeiramente aumentada com 2 h de tratamento com Resv (77 CPM),
quando comparada ao controle (51 CPM). Nos tempos de 8 e 24 h, os valores
encontrados foram similares aos do controle, sendo de 1067 CPM (8 h) e 4153 CPM (24
h) para a célula não tratada e, 1035 CPM (8 h) e 3637 CPM (24 h) após o tratamento
com Resv. Por outro lado, o Resv alterou o perfil de biossíntese de PL nas células
MDA-MB-231. Com 8 h de tratamento com o composto bioativo observa-se a redução
(518 CPM) de PL em relação ao controle (1155 CPM), a qual é acentuada com 24 h
(5181 CPM para a célula não tratada e 1992 CPM para a tratada).

81
0 2 8 24 0 2 8 240
50100150200250300500
2000
3500
5000
6500
Controle Resv
MCF-7
Tempo (horas)
32P
i-fo
sfoli
píd
eos
(CP
M)
0 2 8 24 0 2 8 240
50100150200250300500
2000
3500
5000
6500
Controle Resv
MDA-MB-231
Tempo (horas)
32P
i-fo
sfoli
píd
eos
(CP
M)
Figura 18: Efeito do resveratrol sobre os fosfolipídeos totais de células MCF-7 e MDA-MB-231. As
linhagens celulares MCF-7 e MDA-MB-231 (2 x 105) foram incubadas por 0, 2, 8, 24 h com resveratrol
80 e 200 µM, respectivamente, ou DMSO 0,1% (controle) e acrescidas de 32Pi. Posteriormente, foram
submetidas à extração de lipídeos, seguida por análise por cintilação líquida. Os resultados foram
expressos como média ± SD, n = 2 -3, de contagens por minuto (CPM) de 32Pi-fosfolipídeo sintetizado
nos tempos indicados.
Para identificar quais PL apresentam sua biossíntese alterada pelo Resv, após
incubação das células MCF-7 e MDA-MB-231 com o fitoquímico e com 32Pi, os
lipídeos foram extraídos e submetidos à TLC (Anexo 4). Os PL sintetizados foram PA,
PE, PC, PI, SM e LPC para a MCF-7 e PE, PC, PI, SM e LPC para a MDA-MB-231.
Nas células MCF-7 os PL sintetizados em maior concentração foram PE, PC e PI
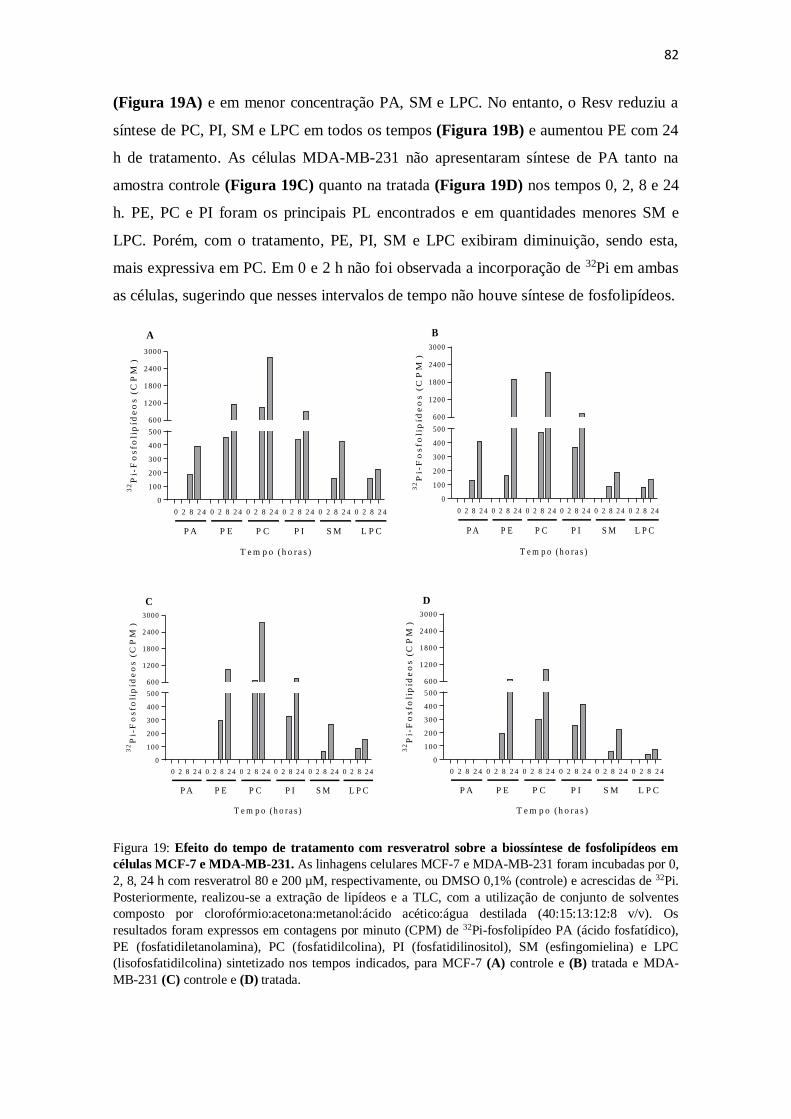
82
(Figura 19A) e em menor concentração PA, SM e LPC. No entanto, o Resv reduziu a
síntese de PC, PI, SM e LPC em todos os tempos (Figura 19B) e aumentou PE com 24
h de tratamento. As células MDA-MB-231 não apresentaram síntese de PA tanto na
amostra controle (Figura 19C) quanto na tratada (Figura 19D) nos tempos 0, 2, 8 e 24
h. PE, PC e PI foram os principais PL encontrados e em quantidades menores SM e
LPC. Porém, com o tratamento, PE, PI, SM e LPC exibiram diminuição, sendo esta,
mais expressiva em PC. Em 0 e 2 h não foi observada a incorporação de 32Pi em ambas
as células, sugerindo que nesses intervalos de tempo não houve síntese de fosfolipídeos.
T e m p o (h o ra s )
32P
i-F
os
fo
lip
íde
os
(C
PM
)
0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4
0
100
200
300
400
500
600
1200
1800
2400
3000
P A P E P C P I S M L P C
A
T e m p o (h o ra s )
32P
i-F
os
fo
lip
íde
os
(C
PM
)
0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4
0
100
200
300
400
500
600
1200
1800
2400
3000
P A P E P C P I S M L P C
B
T e m p o (h o ra s )
32P
i-F
os
fo
lip
íde
os
(C
PM
)
0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4
0
100
200
300
400
500
600
1200
1800
2400
3000
P A P E P C P I S M L P C
C
T e m p o (h o ra s )
32P
i-F
os
fo
lip
íde
os
(C
PM
)
0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4 0 2 8 2 4
0
100
200
300
400
500
600
1200
1800
2400
3000
P A P E P C P I S M L P C
D
Figura 19: Efeito do tempo de tratamento com resveratrol sobre a biossíntese de fosfolipídeos em
células MCF-7 e MDA-MB-231. As linhagens celulares MCF-7 e MDA-MB-231 foram incubadas por 0,
2, 8, 24 h com resveratrol 80 e 200 µM, respectivamente, ou DMSO 0,1% (controle) e acrescidas de 32Pi.
Posteriormente, realizou-se a extração de lipídeos e a TLC, com a utilização de conjunto de solventes
composto por clorofórmio:acetona:metanol:ácido acético:água destilada (40:15:13:12:8 v/v). Os
resultados foram expressos em contagens por minuto (CPM) de 32Pi-fosfolipídeo PA (ácido fosfatídico),
PE (fosfatidiletanolamina), PC (fosfatidilcolina), PI (fosfatidilinositol), SM (esfingomielina) e LPC
(lisofosfatidilcolina) sintetizado nos tempos indicados, para MCF-7 (A) controle e (B) tratada e MDA-
MB-231 (C) controle e (D) tratada.

83
Ao analisar a biossíntese de cada PL isoladamente (Figuras 20 e 21), pode-se
verificar a evolução da biossíntese ao longo do tempo detalhadamente. A síntese de PA
manteve-se similar entre células MCF-7 controle e tratadas, (Figura 20) com ligeira
redução em 8 h de tratamento (183 CPM - controle; 130 CPM - Resv) e aumento de
17% em 24 h (390 CPM - controle; 407 CPM - Resv). PE exibiu aumento 65% em 24 h
de tratamento (1156 CPM - controle; 1905 CPM - Resv). PC e PI apresentaram
biossíntese menor em 23% (2796 CPM - controle; 2141 CPM - Resv) e 20% (892 CPM
- controle; 710 CPM - Resv), respectivamente, em relação ao controle. SM e LPC além
de apresentarem-se em menores quantidades de síntese quando comparados aos PL
encontrados, também demonstraram o maior percentual de redução, 57% (424 CPM -
controle; 184 CPM - Resv) e 40% (223 CPM - controle; 133 CPM - Resv),
respectivamente, em 24 h, na presença de Resv.
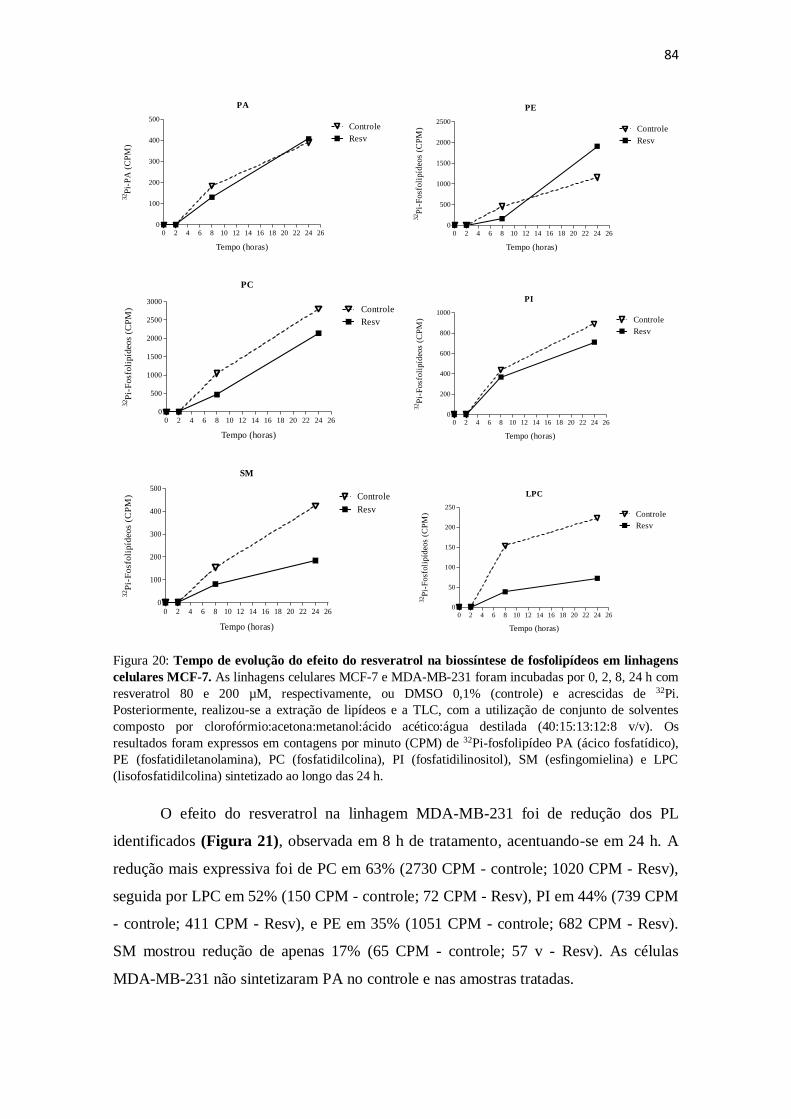
84
PA
0 2 4 6 8 10 12 14 16 18 20 22 24 260
100
200
300
400
500Controle
Resv
Tempo (horas)
32P
i-P
A (
CP
M)
PE
0 2 4 6 8 10 12 14 16 18 20 22 24 260
500
1000
1500
2000
2500
Resv
Controle
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
PC
0 2 4 6 8 10 12 14 16 18 20 22 24 260
500
1000
1500
2000
2500
3000
Resv
Controle
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
PI
0 2 4 6 8 10 12 14 16 18 20 22 24 260
200
400
600
800
1000
Resv
Controle
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
SM
0 2 4 6 8 10 12 14 16 18 20 22 24 260
100
200
300
400
500
Resv
Controle
Tempo (horas)
32P
i-F
osfo
lipíd
eos
(CP
M)
LPC
0 2 4 6 8 10 12 14 16 18 20 22 24 260
50
100
150
200
250Controle
Resv
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
Figura 20: Tempo de evolução do efeito do resveratrol na biossíntese de fosfolipídeos em linhagens
celulares MCF-7. As linhagens celulares MCF-7 e MDA-MB-231 foram incubadas por 0, 2, 8, 24 h com
resveratrol 80 e 200 µM, respectivamente, ou DMSO 0,1% (controle) e acrescidas de 32Pi.
Posteriormente, realizou-se a extração de lipídeos e a TLC, com a utilização de conjunto de solventes
composto por clorofórmio:acetona:metanol:ácido acético:água destilada (40:15:13:12:8 v/v). Os
resultados foram expressos em contagens por minuto (CPM) de 32Pi-fosfolipídeo PA (ácico fosfatídico),
PE (fosfatidiletanolamina), PC (fosfatidilcolina), PI (fosfatidilinositol), SM (esfingomielina) e LPC
(lisofosfatidilcolina) sintetizado ao longo das 24 h.
O efeito do resveratrol na linhagem MDA-MB-231 foi de redução dos PL
identificados (Figura 21), observada em 8 h de tratamento, acentuando-se em 24 h. A
redução mais expressiva foi de PC em 63% (2730 CPM - controle; 1020 CPM - Resv),
seguida por LPC em 52% (150 CPM - controle; 72 CPM - Resv), PI em 44% (739 CPM
- controle; 411 CPM - Resv), e PE em 35% (1051 CPM - controle; 682 CPM - Resv).
SM mostrou redução de apenas 17% (65 CPM - controle; 57 v - Resv). As células
MDA-MB-231 não sintetizaram PA no controle e nas amostras tratadas.

85
PE
0 2 4 6 8 10 12 14 16 18 20 22 24 260
250
500
750
1000
1250
1500Controle
Resv
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
PC
0 2 4 6 8 10 12 14 16 18 20 22 24 260
500
1000
1500
2000
2500
3000Controle
Resv
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
PI
0 2 4 6 8 10 12 14 16 18 20 22 24 260
200
400
600
800
Resv
Controle
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
SM
0 2 4 6 8 10 12 14 16 18 20 22 24 260
50
100
150
200
250
300Controle
Resv
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
LPC
0 2 4 6 8 10 12 14 16 18 20 22 24 260
50
100
150
200Controle
Resv
Tempo (horas)
32P
i-F
osf
oli
píd
eos
(CP
M)
Figura 21: Tempo de evolução do efeito do resveratrol na biossíntese de fosfolipídeos em linhagens
celulares MDA-MB-231. As linhagens celulares MCF-7 e MDA-MB-231 foram incubadas por 0, 2, 8,
24 h com resveratrol 80 e 200 µM, respectivamente, ou DMSO 0,1% (controle) e acrescidas de 32Pi.
Posteriormente, realizou-se a extração de lipídeos e a TLC, com a utilização de conjunto de solventes
composto por clorofórmio:acetona:metanol:ácido acético:água destilada (40:15:13:12:8 v/v). Os
resultados foram expressos em contagens por minuto (CPM) de 32Pi-fosfolipídeo PE
(fosfatidiletanolamina), PC (fosfatidilcolina), PI (fosfatidilinositol), SM (esfingomielina) e LPC
(lisofosfatidilcolina) sintetizado ao longo das 24 h, após normalização com os respectivos padrões.

86
Para verificar se o resveratrol modifica o perfil lipídico das células MCF-7 e
MDA-MB-231 por modulação da via de metabolismo lipídico, foram investigadas as
enzimas acil-CoA:diacilglicerol aciltransferase 2 (DGAT2), ácido graxo sintase (FAS),
acetil-CoA carboxilase β fosforilada (ρACCβ) e proteína quinase ativada por adenosina
monofosfato α fosforilada/constitutiva (AMPKα/ρAMPKα), após 24 h de tratamento
com Resv. Na linhagem MCF-7 o resveratrol reduziu discretamente a DGAT2 (Figura
22 A) e a FAS (Figura 22 B), enquanto que para a ρACCβ e AMPKα/ρAMPKα
(Figuras 22 D e E) a diminuição apresentada após o tratamento com o Resv foi de 3,3
vezes e 1,5 vezes, respectivamente (p < 0,05 e p < 0,04). Em células MDA-MB-231 o
efeito do Resv sobre a DGAT2 foi de ligeiro aumento (Figura 22 A), no entanto, a
ρACCβ e AMPKα/ρAMPKα (Figuras 22 D e E) foram reduzidas 111,8 vezes e 1,2
vezes, respectivamente (p < 0,03 e p < 0,005).

87
0.0
0.5
1.0
1.5
MCF-7 MDA-MB-231
Controle Resv Controle Resv
B
DG
AT
2/G
AP
DH
(un
idad
es a
rbit
rári
as)
0.0
0.5
1.0
1.5
MCF-7 MDA-MB-231
Controle Resv Controle Resv
C
FA
S/G
AP
DH
(un
idad
es a
rbit
rári
as)
0.0
0.5
1.0
1.5
MCF-7 MDA-MB-231
*
*
Controle Resv Controle Resv
D
A
CC
/GA
PD
H
(arb
itra
ry u
nit
s)
0.0
0.5
1.0
1.5
**
MCF-7 MDA-MB-231
Controle Resv Controle Resv
E
A
MP
K
/AM
PK
1
(arb
itra
ry u
nit
s)
Figura 22: Efeito do resveratrol sobre as enzimas participantes do mestabolismo lipídico DGAT2,
FAS, ρACCβ e AMPK em células MCF-7 e MDA-MB-231. As células MCF-7 e MDA-MB-231 após

88
24 h de tratamento com 80 e 200 µM de resveratrol, respectivamente, ou DMSO 0,1% (controle), foram
congeladas em nitrogênio líquido. Em seguida, adicionou-se tampão de lise, composto por inibidores de
proteases de diferentes classes. Posteriormente, as amostras foram sonicadas, centrifugadas a 8.000 rpm,
por 5 min a 4 °C. O volume do sobrenadante (extrato proteico) equivalente a 80 µg de proteínas foi
submetido a eletroforese em gel de poliacrilamida 10%, em condições desnaturantes. Após, as proteínas
do gel foram transferidas para a membrana PVDF, a qual foi bloqueada com TBS-Tween-Leite, por 2 h,
posteriormente, foi incubada com as enzimas da via lipídica diluídas 1:1000 em TBS-Tween-Leite, por 2
h. O GAPDH foi utilizado como controle de carregamento. Após, a membrana foi lavada e incubada com
anticorpo secundário conjugado a peroxidase (1:1000), por 2 h. A detecção das enzimas realizou-se por
quimioluminescência. (A) Digitalização da imagem obtida e a análise densitométrica das enzimas (B)
DGAT2 - acil-CoA:diacilglicerol aciltransferase 2, (C) FAS - Ácido graxo sintase, (D) ρACCβ - Acetil-
CoA carboxylase β fosforilada (E) ρAMPKα/AMPKα - Proteína quinase ativada por adenosina
monofosfato α fosforilada/constitutiva. Os resultados foram expressos em unidades arbitrárias
normalizadas pelo GAPDH. A barra representa média ± S.E.M, n = 3. * Significativamente diferente (p <
0,005; p < 0,03; p < 0,04; p < 0,05) dos valores correspondentes ao controle, utilizando-se Teste-t não
pareado.

89
5. DISCUSSÃO

90
5.1. Citotoxicidade do resveratrol em células de câncer de mama MCF-7 e MDA-
MB-231
Ao analisar a citotoxicidade do Resv, por MTT (Figura 8), observamos que as
duas linhagens responderam diferente ao tratamento. Na MCF-7 (Figura 8A) o
composto reduziu a viabilidade a partir de 50 µM e na MDA-MB-231 (Figura 8B) o
efeito só se iniciou com 100 µM. Em estudo previamente realizado pelo grupo, foi
verificado que o Resv apresenta efeitos citotóxicos a MCF-7 e MDA-MB-231, em 24 h
de tratamento, com valores de IC50 de 120 e 370 µM, respectivamente (CASANOVA et
al., 2012). Tais dados corroboram os resultados apresentados, demonstrando que a
MCF-7 apresenta maior sensibilidade ao fitoquímico. Enquanto que a MDA-MB-231
devido ao seu imunofenótipo TNBC e ao seu subtipo claudin-low, os quais conferem a
essa linhagem maior potencial de agressividade (CIRQUEIRA, 2011; HOLLIDAY &
SPEIRS, 2011), necessita de maior concentração de Resv para a redução da sua
viabilidade.
Um dos benefícios do Resv é o reduzido ou ausente efeito citotóxico em células
não tumorais. Casanova et al. (2012) verificaram que em células BHK-21 (fibroblasto
de rim de hamster), por exemplo, não houve redução da viabilidade no tratamento com
até 200 µM, e na VERO (células epiteliais de rim de macaco) a mesma concentração
reduziu em 23%, aproximadamente. Por outro lado, em células de câncer de bexiga
humana com fenótipo RMD, foi demonstrado que o Resv reduz a viabilidade de 6,9 a
76,1% em 24 h de tratamento com concentrações variando de 10 a 350 µM, com IC50 de
265 (WANG et al., 2017). Em tumor de pulmão de não pequenas células o Resv
também apresenta efeito citotóxico, reduzindo a viabilidade e alterando a morfologia
dessas células na concentração de 50 µM (LI et al., 2016b).
Considerando-se a diferença de resposta ao Resv e o percentual de redução de
viabilidade exibido pelas duas células e as informações sobre sua citotoxicidade em
linhagens não cancerosas, as concentrações de 80 e 200 µM foram selecionadas para
serem utilizadas em todos os ensaios posteriores, para MCF-7 e MDA-MB-231,
respectivamente.

91
5.2. Efeito do resveratrol sobre microdominíos de membrana em células de câncer
de mama MCF-7 e MDA-MB-231
Microdomínios de membrana, também conhecidos como lipid rafts ou
membranas resistentes a detergentes (Detergent-resistant membranes), são plataformas
lipídicas flutuantes capazes de ativar vias de sinalização pró- e anti-apoptóticas.
Alterações em sua composição, principalmente as relacionadas ao CHO, podem
favorecer a ativação ou inativação de suas proteínas associadas. Dentre essas estão
receptores de morte, proteínas quinases e canais de cálcio, tais constituintes tornam
esses domínios dinâmicos e heterogêneos, capazes de participar de amplo número de
sinalizações que são traduzidas da membrana extracelular para as organelas presentes
no citoplasma ou no núcleo (GEORGE & WU, 2012).
A análise dos microdomínios de membrana pode ocorrer de diferentes formas,
tradicionalmente, realiza-se seu isolamento utilizando-se Triton X-100 a 4 °C, uma vez
que são insolúveis nesse detergente. Embora seja afetado pela capacidade do Triton X-
100 em promover a fusão dos microdomínios de membrana heterogêneos, esse método
é o mais comumente utilizado (GEORGE & WU, 2012; GAJATE & MOLLINEDO,
2017). Técnicas de microscopia têm sido empregadas para a observação de
modificações e de interações físicas de proteínas dos microdomínios de membrana
(GEORGE & WU, 2012), tornando-se opção como método complementar para as
análises realizadas a partir do isolamento com Triton X-100, por gradiente de sacarose.
No presente trabalho nós empregamos as duas técnicas, isolamos os
microdomínios de membrana por gradiente de sacarose, utilizando Triton X-100 a 4°C
(Figura 9) e, em outro momento, analisamos nas células íntegras por microscopia de
imunofluorescência (Figura 9). Com isolamento por gradiente de sacarose foi possível
observar material flutuante (Figura 9A), que se concentrou entre as frações de baixa
densidade 4 e 5. Esse resultado está de acordo com o encontrado na literatura, no qual
caracteriza os microdomínios de membrana nessa faixa densidade (GAJATE &
MOLLINEDO, 2017).
Em nosso estudo, analisamos a presença de flotilina-1 (flot-1) nas frações do
gradiente de sacarose, por dot-blotting para verificar se o tratamento com Resv alteraria
as regiões de microdomínos de membrana. Observamos que o fitoquímico modifica a
distribuição de flot-1, deslocando a proteína para a direita do gradiente, conforme pode

92
ser observado em ambas as células MCF-7 (Figuras 10A e B) e MDA-MB-231
(Figuras 11A e B).
Verificar a integridade dos microdomínios de membrana, fornece informações
de alterações que podem direcionar para a ativação de vias de sinalização para a
sobrevivência ou para morte, uma vez que essas plataformas participam da homeostase
celular. Compostos com efeitos anticâncer podem inibir o crescimento e induzir a
apoptose em tumores malignos, por meio de mudanças na distribuição e composição
dos microdomínios de membrana. Entre esses compostos pode-se citar a edelfosina
(alquil lisofofolipídeos sintético) e o Resv (GAJATE et al., 2009; GEORGE & WU,
2012).
Ambos induzem a morte celular programada por meio da ativação de Fas
presente nos microdomínios de membrana, no entanto, a edelfosina provoca o rearranjo
da plataforma lipídica redistribuindo seus componentes proteicos, em especial, os
componentes do complexo DISC (death-inducing signaling complex) e apoptossomo,
enquanto o mecanismo de ação proposto para o Resv indica que também ocorre a
ativação simultânea de receptores do ligante indutor de apoptose relacionado ao TNF
(TRAIL) (GAJATE et al., 2009; GEORGE & WU, 2012).
O efeito do Resv sobre os microdomínios de membrana inicia-se com sua
captação por endocitose, conforme descrito por Colin et al. (2011). Esses pesquisadores,
ao incubarem células de câncer de cólon e leucemia com diferentes inibidores de
endocitose e do desorganizador de microdomínios de membrana, a MβCD, verificaram
que houve a redução da captação do fitoquímico. Porém, quando os microdomínios de
membrana foram mantidos íntegros e, então, a endocitose ocorreu por esta plataforma
lipídica, houve a ativação de vias de sinalização dependentes de quinase e o
desencadeamento de apoptose dependente de caspases. Eles ainda observaram que 30
µM de Resv por 10 min, foi o suficiente para alterar a distribuição de flotilina entre as
frações do gradiente de sacarose 4, 5 e 6, porém, os autores não informaram qual
isoforma da proteína identificaram.

93
5.3. Efeito do resveratrol sobre a expressão de flotilina-1 e flotilina-2 em células de
câncer de mama MCF-7 e MDA-MB-231
Para investigar se o Resv, além de alterar a distribuição de flotilina, também
poderia influenciar na expressão da mesma, nós tratamos as células com o composto
bioativo, preparamos o extrato proteico, conforme descrito no item 4.7.1 da seção
material e métodos. Em seguida, realizamos a eletroforese unidimensional em condições
desnaturantes e a técnica de western blotting para a pesquisa de flot-1 e flot-2.
Observamos conteúdos similares das duas proteínas em ambas as células, quando
comparadas ao controle, com sutil tendência à redução na MCF-7 (Figuras 12A e B) e
ao aumento na MDA-MB-231 (Figuras 12A e C).
Apesar de verificarmos que o Resv modificou a distribuição de flot-1 nos
microdomínios de membrana, não constatamos o aumento do conteúdo dessa proteína e
de sua isoforma flot-2 nas duas linhagens de câncer de mama. Então, com o intuito de
complementar os resultados obtidos, submetemos a MDA-MB-231 ao ensaio de
microscopia de imunofluorescência, para pesquisa de flot-2. Para isso, as células foram
cultivadas em lamínulas e tratadas com Resv 200 µM por 24 h. Os resultados obtidos
por microscopia confocal (Figuras 13D, E e F) demonstram o efeito acentuado do
fitoquímico sobre a flot-2, aumentando a área correspondente à proteína em 13,4 vezes.
Interessados em esclarecer se o Resv atuaria sobre a expressão gênica de
flotilinas, submetemos as células MCF-7 e MDA-MB-231 à análise por RT-qPCR para
flot-1 e flot-2. Os resultados encontrados confirmaram as informações obtidas por
microscopia de imunofluorescência para flot-2 na MDA-MB-231, com aumento
acentuado de sua expressão (Figura 14B). Curiosamente, a expressão de flot-1 é muito
mais elevada do que a de flot-2 nessa célula (Figura 14A). Nas células MCF-7, o Resv
demonstrou discreta tendência em elevar a expressão de flot-1 (Figura 14A), enquanto
que manteve a flot-2 similar ao controle (Figura 14B). Vale ressaltar que a estabilidade
de flot-1 é dependente da presença de flot-2 (LANGHORST et al., 2005). A diferença
que observamos entre os resultados obtidos pela técnica de western blotting,
microscopia de imunofluorescência e RT-qPCR, poderia ser justificada pelo fato das
técnicas possuírem graus de sensibilidade distintos. Apesar disso, microscopia de
imunofluorescência e RT-qPCR corroboram o aumento acentuado de flot-1 e flot-2 nas
células MDA-MB-231, induzido por Resv.

94
Flotilinas conferem aos microdomínios de membrana característica planar, e
podem até mesmo substituir a caveolina em células deficientes em cav-1, como é o caso
da MCF-7 (STAUBACH & HANISCH, 2011). Entretanto, não são apenas marcadores
de microdomínios de membrana ou de ancoramento (STAUBACH & HANISCH,
2011), mas possuem unidades dinâmicas com suas próprias funções regulatórias
(LANGHORST et al. 2005). Atualmente, elevadas concentrações de flot em células de
diferentes tipos de cânceres têm sido associadas a metástase e mau prognóstico. Esse
aumento de expressão tornou-se um novo fator de risco e marcador de progressão e
agressividade tumoral. Liu et al. (2015b) sugerem que a flot-2 pode ser utilizada como
um oncogene no desenvolvimento de câncer cervical devido a sua super expressão
nessas células, enquanto que Li et al. (2016c) classificam a flot-1 como um preditor de
metástase em câncer do colo do útero em estágios iniciais.
Embora em nossos resultados tenha ocorrido o aumento de expressão de flot-1 e
flot-2 na MDA-MB-231, foi observada redução da viabilidade dessas células (Figura
8B). Somente a elevação de flotilinas não contribui para o crescimento e progressão
tumoral, no caso da flot-1, por exemplo, é necessário que ocorra simultaneamente a
ativação de NF-κB (SONG et al., 2012; LIU et al., 2015a). No entanto, um dos efeitos
anticâncer do Resv é suprimir a ativação de NF-κB, resultado obtido em células com
fenótipo de resistência a múltiplas drogas (ZHANG et al., 2016).
Vale salientar que os estudos in vitro e in vivo que associam o aumento de
expressão das flot-1 e flot-2 ao aumento do grau de malignidade com impacto negativo
no prognóstico do paciente, o fazem de forma isolada (ASP et al., 2014; WANG et al.,
2015; LIU et al., 2015a; LIU et al., 2015b; LI et al., 2016c), não associando esses
efeitos à exposição de fitoquímicos ou a quimioterápicos, isto é, não se trata do aumento
induzido por um composto, mas sim, aquele proveniente de fatores intrínsecos à própria
célula de câncer. Lembrando que flotilinas contribuem para a homeostase celular e a
formação do tumor, um processo de desregulação/desequilíbrio de diferentes
mecanismos, pode ser um estímulo independente para a superexpressão dessas
proteínas.
Resv é extensivamente relacionado a vias de sinalização que contribuem para a
inibição da iniciação, proliferação e metástase, induzindo as células de câncer de mama
à morte ou a parada do ciclo celular (CASANOVA et al., 2012; DAYEM et al., 2016;
DANDAWATE et al., 2016) e por isso, possui diferentes alvos moleculares, entre eles
podem ser destacados AMPK, NF-κB, PI3K/Akt, ER e MAPK (YEUNG et al., 2004;

95
FRÖJDÖ et al., 2007; KULKARNI & CANTÓ, 2015; TURNER et al., 2016; ZHAO et
al., 2017), que podem estar co-localizados em microdomínios de membrana e interagir
com as flotilinas, assim como ocorre com a proteína de domínio de morte associada ao
Fas (FADD) e a flot-2 (WEI et al., 2013), Dessa forma, é necessário estudar a relação
entre esses integrantes dos microdomínios de membrana e o aumento da expressão de
flot-1 e flot-2 induzida pelo Resv. Vale ressaltar que não foram encontrados estudos que
relacionassem a superexpressão de flotilinas na presença de Resv com a progressão da
carcinogênese.
5.4. Efeito do resveratrol sobre perfil de lipídeos neutros, ácidos graxos e
fosfolipídeos de células de câncer de mama MCF-7 e MDA-MB-231
Para investigar as possíveis alterações do Resv sobre o perfil de lipídeos neutros,
as amostras tratadas com Resv por 24 h foram submetidas à extração de lipídeos e em
seguida realizou-se a HPTLC (Figura 15). Foram identificados CHO, TG, FA e MG.
Desses lipídeos, a MCF-7 (Figura 15A) apresentou redução discreta de CHOE, TG e
FA, enquanto que CHO e MG mantiveram-se similares ao controle. A MDA-MB-231
(Figura 15B) exibiu concentrações de CHOE, FA, CHO e MG próximas as do controle,
no entanto, houve redução expressiva de TG.
As alterações pouco expressivas do conteúdo de lipídeos, não coerente as
informações obtidas por gravimetria (Anexo 1), nos fizeram refletir sobre a
padronização da quantidade de lipídeos aplicados para a HPTLC, se o efeito não estaria
minimizado. Com isso, realizamos a TLC (Figura 16) utilizando o conteúdo total de
lipídeos em 1x107 de células. Os resultados obtidos indicaram modificações mais
acentuadas das concentrações de lipídeos após tratamento com Resv, para as duas
células.
A MCF-7 (Figura 16A) aumentou em 188 e 32% os teores de FA e CHO,
respectivamente, e a MDA-MB-231 (Figura 16B) elevou em 435% a concentração de
FA, por outro lado houve redução de TG e CHO em 16 e 12%, respectivamente. A
utilização do material lipídico total, obtido após a extração de lipídeos, demonstrou que
dessa forma é possível observar modificações não verificadas com a padronização de
peso do extrato lipídico e comprovou que o Resv pode alterar a composição de lipídeos
neutros dessas células.

96
A composição dos microdomínios de membrana, contendo principalmente CHO
e SL, além de proteínas ancoradoras é drasticamente alterada no câncer, impactando na
atividade de quinases, no tráfego de proteínas, na adesão e apoptose, tornando-se pré-
requisito para a sua rápida proliferação. Essa modificação pode ocorrer em decorrência
da reprogramação metabólica, comum na carcinogênese (BAENKE et al., 2013;
GUÉGUINOU et al., 2015).
A reprogramação metabólica inclui maior captação de glicose e biossíntese de
FA. A produção de FA e CHO é restrita a um conjunto de tecidos, como hepatócito,
adipócito e tecido mamário em lactante. Entretanto, a reativação da síntese de lipídeos é
frequentemente observada em tumores e sua exata função para a manutenção e
desenvolvimento da doença não está completamente esclarecida. Mas, sabe-se que
contribui também para fornecer energia em períodos de depleção de nutrientes e
participam da transformação celular (BAENKE et al., 2013).
Ao analisarmos as concentrações de lipídeos totais, CHO e TG (Figura 17) do
extrato celular, sem a prévia extração de lipídeos, também observamos modificações em
decorrência do tratamento com o Resv, em especial para a linhagem MDA-MB-231.
Essa célula apresentou significativo aumento de lipídeos totais (Figura 17A) e redução
de CHO (Figura 17B), por outro lado, esses, se mantiveram similares ao controle na
MCF-7. Embora se observe diferenças em relação a concentração de TG em ambas as
células, os valores encontrados variaram acentuadamente entre os experimentos
independentes e os teores foram muito reduzidos, não sendo possível verificar
diferenças significativas entre as amostras.
Alterações no conteúdo de lipídeos podem ocasionar modificações na
organização dos microdomínios de membrana e, consequentemente, em suas funções, as
quais podem induzir ou inibir vias de sinalização pró- e anti-apoptóticas. Isso ocorre por
influenciarem na biologia do câncer, uma vez que a presença ou abundância de alguns
lipídeos na membrana celular, tais como DHA e EPA, modificam a sinalização do HER
e dos canais iônicos KCa, ClCa e Ca2+ localizados nos microdomínios de membrana, os
quais possuem importantes funções na migração, invasão e metástase de células de
câncer, incluindo o de mama (GUÉGUINOU et al., 2015).
Para analisar o efeito do Resv sobre os FA, após tratamento com Resv, por 24 h,
as células foram submetidas à extração de lipídeos e à cromatografia gasosa com
espectrometria de massas. Foram encontradas diferenças entre os treze FA saturados
(Tabela 1) e quinze FA insaturados (Tabela 2) identificados. Tanto a MCF-7, quanto a

97
MDA-MB-231 exibiram aumento dos FA ao serem expostas ao Resv, exceto o C13:0
(reduziu 52%) para a MDA-MB-231.
Em ambas as linhagens, os FA saturados predominantes foram C16:0 e C18:0.
Entre os FA insaturados o C18:1n9c e o C18:1n9t foram principais. Os FA da série n-3
EPA e DHA mostraram os maiores percentuais de aumento na MDA-MB-231, 73 e
65%, respectivamente. Tais alterações na composição de lipídeos podem ser explicadas
pelo fato de tecidos cancerosos serem capazes de gerar FA, e também PL por lipogênese
de novo, que em sua maioria são lipídeos específicos, necessários para a rápida
proliferação da célula. Lipídeos também podem participar de mudanças pós-
translacionais de proteínas, atuando no tráfego e localização sub celular de proteínas.
Modificação protéica com cadeias acil saturadas podem promover sua associação com
os microdomínios de membrana, enquanto que FA insaturados excluem proteínas dessas
plataformas (BAENKE et al., 2013).
Utilizamos 32Pi para investigar o efeito do Resv nos PL nos períodos de 0, 2, 8 e
24 h. Ao longo do tempo é possível verificar a incorporação de 32Pi na amostra controle
(Figura 18), indicando que há ocorrência de biossíntese nas células. Após tratamento
com Resv, a MCF-7 manteve o conteúdo de PL totais semelhante ao controle, por outro
lado, a MDA-MB-231 demonstrou redução de 55% em 8 h, acentuando-se com 24 h
(62%) de exposição ao Resv. Com o intuito de investigar as principais classes de PL,
realizamos a TLC das amostras marcadas com 32Pi e verificamos a evolução do efeito
do Resv na biossíntese (Figura 20 e Figura 21). A MCF-7 apresentou aumento
expressivo de PE e redução acentuada de SM e LPC, enquanto a MDA-MB-231 não
exibiu PA e diminuiu todos os PL encontrados, pronunciada em PC, LPC, PI e PE.
Além de sua utilização para a sobrevivência das células, os lipídeos são
importantes moléculas de sinalização. Dentre esses, estão os fosfoinositídeos, segundo
mensageiros que retransmitem informações do HER para a maquinaria celular,
recrutando proteínas efetoras para compartimentos específicos da membrana celular. O
principal representante dessa classe é o fosfatidilinositol 3,4,5-trifosfato, produzido pela
PI3K, em resposta ao sinal do fator de crescimento mediante ativação de Akt. Vale
relembrar que tanto HER, quanto PI3K/Akt, são alvos de ação do Resv (BAENKE et
al., 2013; KULKARNI & CANTÓ, 2015).
Atualmente, diversos estudos têm surgido com a proposta de elucidar as
diferenças existentes entre células tumorais e não tumorais, com técnicas que permitem
a detecção e caracterização de mudanças sutis em componentes lipídicos, como MS/MS

98
e LC-MS e, assim, empregá-los como biomarcadores no diagnóstico precoce de
variados tipos de cânceres (DÓRIA et al., 2013; NIEVA et al., 2012; HE et al., 2015).
A linhagem de câncer de mama humano MDA-MB-231, por sua característica TNBC, é
alvo de pesquisas nesse sentindo. Comparada com outros dois tipos celulares, um não
maligno e outro ER+ com baixo potencial metastático, mostrou tendência semelhante de
perfil de PL, com maiores percentuais de PC, PE, PS, PI e SM, nessa ordem, e a LPC
foi o menos abundante PL encontrado.
Algumas diferenças significativas em relação à célula não tumoral foram
percebidas, como o menor percentual de PE e a exclusividade em apresentar PA
(DÓRIA et al., 2013). Esse dado difere dos resultados que encontramos, no qual, não
houve detecção de PA nessas células, além disso, observamos que o Resv aumentou o
conteúdo de PE na MCF-7, mas reduziu na MDA-MB-231 (Figura 19), entretanto não
é possível verificar comparativamente as alterações, pois, Dória et al. (2013) utilizou
método de análise diferente.
Porém, seria interessante avaliar se o Resv alterou o PL para valores de
abundância próximos ou não aos de células não tumorais. Ainda de acordo com Dória et
al. (2013), as diferenças expressivas entre as três células que ele analisou ocorreu no
perfil de composição de FA de cada PL, por exemplo, o potencial metastático foi
associado a PC (O-16:0/18:1), PC (O-16:0/20:1) e PI (22:5/18:0), mas elevados na
MDA-MB-231. Por outro lado, a capacidade migratória foi atribuída a redução de PC
(18:0/18:1), SM (18:1/24:0), SM (18:1/22:0) e SM (18:1/24:1).
He et al., (2015) encontraram conteúdos aumentados de PC e reduzidos de SM e
PI em seis células de câncer de mama (incluindo a MCF-7 e a MDA-MB-231), em
relação com o controle, uma linhagem não tumoral. Os autores enfatizam que o
aumento significativo de lipídeos monoinsaturados pode ser associado com o grau de
malignidade de células de câncer de mama, assim como ocorre com FA (NIEVA et al.,
2012; HE et al., 2015).
Embora seja essencial caracterizar as diferenças existentes entre células tumorais
e não tumorais, pesquisa que auxiliem a elucidar o efeito de compostos bioativos, como
o Resv, com potencial de ser utilizado como adjuvante na quimioterapia (CASANOVA
et al, 2012) sobre a composição de lipídeos, são necessárias. Grande parte dos trabalhos
publicados nesse contexto verificam a ação de lipídeos adicionados à cultura de células
(GRAMMATIKOS et al., 1994; CHAMRAS et al., 2002; HOFMANOVÁ, et al., 2012)
e não somente aqueles sintetizados por elas mesma, a partir de um estímulo não

99
lipídico. Com isso, não é possível atribuir as consequências apresentadas somente às
modificações intrínsecas, uma vez que os FA acrescentados são incorporados à
membrana celular.
Utilizando-se a técnica na qual se forma uma monocamada celular, Olas &
Holmsen (2012), analisaram a interação do Resv com glicerofosfolipídeos de
membrana. O composto bioativo a 2,5 e 50 µg/mL parece localizar-se entre as
moléculas da monocamada de fosfolipídeos, penetrando mais facilmente entre serinas e
etanolaminas. É possível que uma das hidroxilas do Resv seja atraída pelo grupo
carboxila da serina, explicando sua maior afinidade por PS do que por PE e PC. No
entanto, tal condição é influenciada pelo comprimento do grupo acila, pelo grau de
instauração e pela natureza da cabeça hidrofóbica do PL.
Em hepatócitos de ratos o efeito do Resv é dose dependente sobre a redução da
síntese de FA em concentrações de 10, 25, 50 e 100 µM, por 1 h, não modificando a
síntese de CHO. Nesses tecidos, 25 µM de Resv por 30 min, diminuem diacilglicerol
(DG) e TG, além de PE, PI e PC. Dentre as enzimas que participam do metabolismo de
lipídeos, a atividade da ACC foi significativamente afetada pelo fitoquímico, o mesmo
ocorreu para os FA C14:0, C16:0, C18:0 e C18:1 (GNONI & PAGLIALONGA, 2009).
Frequentemente, o Resv está relacionado à inibição da lipogênese em células não
tumorais, como em adipócitos. Seus alvos moleculares, nesse caso, podem ser a ACC, a
Akt e a AMPK (LASA et al., 2012; LI et al., 2016a). No entanto, nossos resultados
demosntram que o Resv não inibiu a síntese de lipídeos em células de câncer de mama
MCF-7 e MDA-MB-231.
A partir dos resultados de perfil lipídico obtidos, analisamos por western blotting
a expressão de cinco enzimas envolvidas no metabolismo de lipídeos, a DGAT2, FAS,
ρACCβ, ρAMPKα/AMPKα (Figura 22A). Nossos resultados demonstraram que o Resv
reduziu a fosforilação de ACC (Figura 22D) em ambas as células, sendo mais
acentuada na MDA-MB-231, além disso, o fitoquímico regulou negativamente a
AMPKα (Figura 22E). A expressão de DGAT2 e FAS não foi alterada
significativamente (Figuras 22B e C).
Pandey et al. (2011) investigaram a FAS, sob o efeito do Resv, nas linhagens
MCF-7 e MDA-MB-231 e encontraram resultados divergentes dos nossos. Verificaram
que ao tratarem as células com 50 e 100 µM de Resv, houve redução significativa de
FAS e, consequentemente, do conteúdo total de lipídeos, adicionalmente, a diminuição

100
de FAS ocasionou a indução dos pró-oncogenes DAPK2 e BNIP3, elevando
expressivamente o percentual de células em apoptose.
Nosso estudo revelou que o tratamento com Resv reduziu, significativamente, a
ρACCβ e a ρAMPKα/AMPKα, componentes chave no metabolismo de lipídeos que,
dessa forma, foram ativados e inativados, respectivamente, o que propiciou a síntese de
FA, como foi observado. Embora o aumento na biossíntese de FA seja uma condição
indesejável em células tumorais, com íntima relação com sua sobrevivência,
proliferação, metástase e malignidade, outras questões devem ser levadas em
consideração.
As vias de sinalização para tais condições, em sua maioria, apresentam
integrantes modulados por Resv. O FA C20:4n6, que em nosso estudo apresentou
elevação de sua concentração na MDA-MB-231 na presença de Resv, é um indutor de
migração e invasão nessas mesmas células. Porém, essa ação ocorre por meio da via
dependente de PI3K/Akt, duas moléculas inibidas por ação do Resv (VILLEGAS-
COMONFORT et al., 2014). Dessa forma, é necessário investigar outras moléculas
envolvidas no metabolismo de lipídeos e moduladas por Resv, para esclarecer qual o
efeito da alteração de perfil lipídico induzida pelo fitoquímico, nas células MCF-7 e
MDA-MB-231.

101
6. CONCLUSÕES

102
De acordo com os resultados obtidos concluiu-se que:
• A viabilidade das células de câncer de mama MCF-7 e MDA-MB-231 foi
reduzida após o tratamento com Resv, porém a MDA-MB-231 necessita de maior
concentração do composto para o efeito citotóxico ser observado.
• O Resv altera a distribuição de flot-1 e flot-2 nos microdomínios de membrana
de células de câncer de mama MCF-7 e MDA-MB-231;
• Além disso, o fitoquímico aumenta a quantidade de flot-2 na célula MDA-MB-
231. Esse fato pode ser em decorrência do efeito do composto na regulação gênica da
proteína, sendo o mesmo efeito observado para a flot-1. Porém, não houve
modificação expressiva de flot-1 e flot-2 na linhagem MCF-7;
• O perfil de lipídeos neutros é modificado pelo Resv, principalmente, devido ao
aumento de ácidos graxos livres; sendo esse efeito melhor observado por TLC,
aplicando-se o conteúdo total de lipídeos presentes em 1 x107 de células;
• As concentrações de ácidos graxos aumentaram em ambas as células, sobretudo
os insaturados, nas células MDA-MB-231. Os saturados principais foram C18:0 e
C16:0 e os insaturados predominantes foram C18:1n9c e C18:1n9t;
• O Resv elevou expressivamente as concentrações de DHA e EPA;
• O perfil de fosfolipídeos é influenciado pelo Resv. Apesar de não haver
modificações acentuadas na biossíntese de fosfolipídeos totais na MCF-7, o Resv
reduziu o seu conteúdo na MDA-MB-231 em 8 e 24 h de tratamento;
• Além disso, nas células MCF-7 o Resv aumentou PE e reduziu expressivamente
PC, LPC, PI e PE na MDA-MB-231;
• O fitoquímico regulou o metabolismo de lipídeos, e um dos mecanismos
observados que pode ter contribuído para esse efeito foi a modulação das enzimas
ACC e AMPK, principalmente, na linhagem MDA-MB-231.
O conjunto de resultados obtido nos permite verificar que o Resv foi citotóxico
para as células de câncer de mama MCF-7 e MDA-MB-231. O aumento da
expressão de flotilinas, do conteúdo de ácidos graxos e a modificação da composição
lipídica observados, não repercutiram em aumento da viabilidade celular, nas
condições estudas. Embora o composto esteja favorecendo eventos relacionados ao
avanço tumoral, outras vias de sinalização podem estar sendo reguladas pelo Resv,
contribuindo para sua ação anticâncer, como representado na Figura 23. Com isso, é
necessário elucidar por quais mecanismos o Resv atua nessas condições.

103
Figura 23: Esquema representando o efeito do resveratrol sobre as flotilinas e as enzimas do
metabolismo lipídico e as possíveis vias de sinalização inibidas, por ação do fitoquímico.

104
7. REFERÊNCIAS

105
Aggarwal, BB; Bhardwaj, A; Aggarwal, RS; Seeram, NP; Shishodia, S; Takada, Y.
Role of resveratrol in prevention and therapy of cancer: preclinical and clinical studies.
Anticancer Reserch, 24, 2783-2840, 2004.
Arlia-Ciommo, A; Svistkova, V; Mohtashami, S; Titorenko, VI. A novel approach to
the discovery of anti-tumor pharmaceuticals: searching for activators of liponecrosis.
Oncotarget, 7, 5204-5225, 2016. doi: 10.18632/oncotarget.6440.
Asp, N; Pust, S; Sandvig, K. Flotillin depletion affects ErbB protein levels in different
human breast cancer cells. Biochimica et Biophysica Acta, 1843, 1987-1996. 2014. doi:
10.1016/j.bbamcr.2014.04.013.
ATCC - American Type Culture Collection, 2017a. Disponível em:
<https://www.atcc.org/Products/All/HTB-22.aspx#characteristics>. Acesso em 08 de
fevereiro de 2017.
ATCC - American Type Culture Collection, 2017b. Disponível em:
<https://www.atcc.org/Products/All/HTB-26.aspx#characteristics>. Acesso em 08 de
fevereiro de 2017.
Azordegan, N; Fraser, V. Khuong, L; Hillyer, LM; Ma, DWL; Fischer, G;
Moghadasian, MH. Carcinogenesis alters fatty acid profile in breast tissue. Molecular
and Cellular Biochemistry, 374, 223-232, 2013. doi 10.1007/s11010-012-1523-4.
Badana, A; Chintala, M; Varikuti, G; Pudi, N; Kumari, S; Kappala, VR; Malla, RR.
Lipid raft integrity is required for survival of triple negative breast cancer cells. Journal
of Breast Cancer, 19, 372-384, 2016. doi.org/10.4048/jbc.2016.19.4.372.
Baenke, F; Peck, B; Miess, H; Schulze, A. Hooked on fat: the role of lipid synthesis in
cancer metabolism and tumour development. Disease Models & Mechanisms 6, 1353-
1363, 2013. doi:10.1242/dmm.011338.
Bandu, R; Mok, HJ; Kim, KP. Phospholipids as cancer biomarkers: mass spectrometry-
based analysis. Mass Spectrometry Reviews, 2016. doi: 10.1002/mas.21510.
Banning, A; Tomasovic, A; Tikkanen, R. Functional aspects of membrane association
of reggie/flotillin proteins. Current Protein and Peptide Science, 12, 725-735, 2011.
Beloribi-Djefaflia, S; Vasseur, S; Guillaumond, F. Lipid metabolic reprogramming in
cancer cells. Oncogenesis, 2016. doi:10.1038/oncsis.2015.49.
Bligh, EG & Dyer, WJ. A rapid method of total lipid extraction and purification.
Canadian Journal of Biochemistry and Physiology, 37, 911-917, 1959. doi:
10.1139/o59-099.
Bowers, JL; Tyulmenkov, VV; Jernigan, SC; Klinge, CM. Resveratrol acts as a mixed
agonist/antagonist for estrogen receptors alpha and beta. Endocrinology, 141, 3657-
3667, 2000. doi: 10.1210/endo.141.10.7721.

106
Brasil. Ministério da Saúde. Secretaria de Atenção à Saúde. Departamento de Atenção
Básica. Guia alimentar para a população brasileira, 2014. Disponivel em:
<http://bvsms.saude.gov.br/bvs/publicacoes/guia_alimentar_populacao_brasileira_2ed.p
df>. Acesso em: 18 de fevereiro de 2017.
Brown, DA & London, E. Functions of lipid rafts in biological membranes. Annual
Review of Cell and Developmental Biology, 14, 111-136, 1998. doi:
10.1146/annurev.cellbio.14.1.111.
Brown, DA & Rose, JK. Sorting of GPI-anchored proteins to glycolipid-enriched
membrane subdomains during transport to the apical cell surface. Cell, 68, 533-544,
1992.
Cadenas, C; Vosbeck, S; Hein, EM; Hellwig, B; Langer, A; Hayen, H; Franckenstein,
D; Büttner, B; Hammad, S; Marchan, R; Hermes, M; Selinski, S; Rahnenführer, J;
Peksel, B; Török, Z; Vígh, L; Hengstler, JG. Glycerophospholipid profile in oncogene-
induced senescence. Biochimica et Biophysica Acta, 1821, 1256-1268, 2012.
Calder, PC & Yaqoob, P. Lipid rafts—composition, characterization, and controversies.
Journal of Nutrition, 137, 548-553, 2007.
Casanova, F; Quarti, J; Costa, DC; Ramos, CA; Silva, JL; Fialho, E. Resveratrol
chemosensitizes breast cancer cells to melphalan by cell cycle arrest. Journal of Cellular
Biochemistry, 113, 2586-2596, 2012. doi: 10.1002/jcb.24134.
Chakrabarti, P; English, T; Shi, J; Smas, CM; Kandror, KV. Mammalian target of
rapamycin complex 1 suppresses lipolysis, stimulates lipogenesis, and promotes fat
storage. Diabetes, 59, 775-781, 2010. doi: 10.2337/db09-1602.
Chamras, H; Ardashian, A; Heber, D; Glaspy, JA. Fatty acid modulation of MCF-7
human breast cancer cell proliferation, apoptosis and differentiation. Journal of
Nutritional Biochemistry, 13, 711- 716, 2002.
Christie, WW. Gas Chromatography and Lipids. A practical guide. Editora The Oily
Press, 1989.
Cirqueira, MB; Moreira, MAR; Soares, LR; Freitas-Júnior, R. Molecular subtypes of
breast câncer. FEMINA, 39, 499-503, 2011.
Colin, D; Limagne, E; Jeanningros, S; Jacquel, A; Lizard, G; Athias, A; Gambert, P;
Hichami, A; Latruffe, N. Endocytosis of resveratrol via lipid rafts and activation of
downstream signaling pathways in cancer cells. Cancer Prevention Research, 4, 1095-
1106, 2011. doi: 10.1158/1940-6207.CAPR-10-0274.
Costa, DCF; Casanova, FA; Quarti, J; Malheiros, MS, Sanches, S; Santos, OS; Fialho,
E; Silva, JL. Transient transfection of a wild-type p53 gene triggers resveratrol-induced
apoptosis in cancer cells. PloS One, 2012. doi: 10.1371/journal.pone.0048746.

107
Dandawate, PR; Subramaniam, D; Jensen, RA; Anant, S. Targeting cancer stem cells
and signaling pathways by phytochemicals: novel approach for breast cancer therapy.
Seminars in Cancer Biology, 2016. doi: 10.1016/j.semcancer.2016.09.001.
Dayem, AA; Choi, HY; Yang, GM; Kim, K; Saha, SK; Cho, SG. The anti-cancer effect
of polyphenols against breast cancer and cancer stem cells: molecular mechanisms.
Nutrients, 2016. doi:10.3390/nu8090581.
Delmas, D & Li, HY. Role of membrane dynamics processes and exogenous molecules
in cellular resveratrol uptake: consequences in bioavailability and activities. Molecular
Nutrition & Food Research, 55, 1142-1153, 2011. doi: 10.1002/mnfr.201100065.
Delmas, D; Rébé, C; Lacour, S; Filomenkos, R; Athias, A; Gambert, P; Cherkaoui-
Malki, M; Jannin, B; Dubrez-Daloz, L; Latruffe, N; Solary, E. Resveratrol-induced
apoptosis is associated with Fas redistribution in the rafts and the formation of a death-
inducing signaling complex in colon cancer cells. The Journal of Biological Chemistry,
278, 41482-41490, 2003. doi: 10.1074/jbc.M304896200.
Dória, ML; Cotrim, CZ; Simões, C; Macedo, B; Domingues, P; Domingues, MR;
Helguero, LA. Lipidomic analysis of phospholipids from human mammary epithelial
and breast cancer cell lines. Journal of Cellular Physiology. 228, 457-468, 2013. doi:
10.1002/jc24152.
El-Sayed, A & Harashima, H. Endocytosis of gene delivery vectors: from clathrin-
dependent to lipid raft-mediated endocytosis. Molecular Therapy, 21, 1118-1130, 2013.
doi: 10.1038/mt.2013.54.
Freitas, MS; Costa, DCF; Casanova, FA. Principais mecanismos de oncogênese. Em:
Bases moleculares em clínica médica (Alves, PCC; Da Poian, AT; Abrahão, A). Editora
Atheneu, São Paulo, 1, 555-571, 2010.
Fröjdö, S; Cozzone, D; Vidal, H; Pirola, L. Resveratrol is a class IA phosphoinositide 3
kinase inhibitor. Biochemical Journal, 406, 511-518, 2007. doi: 10.1042/BJ20070236.
Fuchs, B; Süss, R; Teuber, K; Eibisch, M; Schiller, J. Lipid analysis by thin-layer
chromatography--a review of the current state. Journal of Chromatography A, 1218,
2754-2774, 2011. doi: 10.1016/j.chroma.2010.11.066.
Gajate, C & Mollinedo, F. Isolation of lipid rafts through discontinuous sucrose gradient
centrifugation and Fas/CD95 death receptor localization in raft fractions. Methods and
Protocols, Methods in Molecular Biology, 1557, 125-138, 2017. doi: 10.1007/978-1-
4939-6780-3_13.
Gajate, C; Gonzalez-Camacho, F; Mollideno, F. Lipid raft connection between extrinsic
and intrinsic apoptotic pathways. Biochemical and Biophysical Research
Communications, 380, 780-784, 2009. doi:10.1016/j.bbrc.2009.01.147.

108
García-Marcos, M; Pochet, S; Tandel, S; Fontanils, U; Astigarraga, E; Fernández-
González, JA; Kumps, A; Marino, A; Dehaye, JP. Characterization and comparison of
raft-like membranes isolated by two different methods from rat submandibular gland
cells. Biochimica et Biophysica Acta, 1758, 796-806, 2006. doi:
10.1016/j.bbamem.2006.05.008.
George, K & Wu, S. Lipid raft: a floating island of death or survival. Toxicology and
Applied Pharmacology, 259, 311-319, 2012. doi:10.1016/j.taa2012.01.007.
Gnoni, GV & Paglialonga, G. Resveratrol inhibits fatty acid and triacylglycerol
synthesis in rat hepatocytes. European Journal of Clinical Investigation, 39, 211-218,
2009. doi: 10.1111/j.1365-2362.2008.02077.x.
Grammatikos, SI; Subbaiah, PV; Victor, TA; Miller, WM. n-3 and n-6 fatty acid
processing and growth effects in neoplastic and non-cancerous human mammary
epithelial cell lines. British Journal of Cancer, 70, 219-227, 1994.
Griffith, OM. Practical techniques for centrifugal separations. Application Guide, 2010.
Disponível em:
<https://thermofisher.co.nz/Uploads/file/Scientific/Applications/Equipment-
Furniture/Practical-Techniques-for-Centrifugal-Separations.pdf>. Acesso em: 03 de
fevereiro de 2017.
Guéguinou, M; Gambade, A; Félix, R; Chantôme, A; Fourbon, Y; Bougnoux, Weber,
G; Potier-Cartereau, M; Vandier, C. Lipid rafts, KCa/ClCa/Ca2+ channel complexes and
EGFR signaling: novel targets to reduce tumor development by lipids? Biochimica et
Biophysica Acta, 1848, 2603-2620, 2015. doi: 10.1016/j.bbamem.2014.10.036.
He, M; Guo, S; Li, Z. In situ characterizing membrane lipid phenotype of breast cancer
cells using mass spectrometry profiling, 2015. Scientific Reports, doi:
10.1038/srep11298.
Hofmanová, J; Ciganek, M; Slavík, J; Kozubík, A; Stixová, L; Vaculová, A; Dušek, L;
Machala, M. Lipid alterations in human colon epithelial cells induced to differentiation
and/or apoptosis by butyrate and polyunsaturated fatty acids. Journal of Nutritional
Biochemistry, 23, 539-548, 2012. doi:10.1016/j.jnutbio.2011.02.010.
Holliday, DL & Speirs, V. Choosing the right cell line for breast cancer research. Breast
Cancer Research, 13, 2011. doi: 10.1186/bcr2889.
Hopperton, KE; Duncan, RE; Bazinet, RP; Archer, MC. Fatty acid synthase plays a role
in cancer metabolism beyond providing fatty acids for phospholipid synthesis or
sustaining elevations in glycolytic activity. Experimental Cell Research, 320, 302-310,
2014. doi: 10.1016/j.yexcr.2013.10.016.
Huang, C & Freter, C. Lipid metabolism, apoptosis and cancer therapy. International
Journal of Molecular Sciences, 16, 924-949, 2015. doi:10.3390/ijms16010924.

109
Hwang, KA & Choi, KC. Anticarcinogenic effects of dietary phytoestrogens and their
chemopreventive mechanisms. Nutrition and Cancer, 67, 796-803, 2015. doi:
10.1080/01635581.2015.1040516.
INCA - Instituto Nacional de Câncer/Ministério da Saúde. Estimativa 2016: incidência
de câncer no Brasil, 2015. Disponível em: <2017
http://www.inca.gov.br/estimativa/2016/estimativa-2016-v11.pdf>. Acesso em: 08 de
junho de 2016.
Jang, M; Cai, L; Udeani, GO; Slowing, KV; Thomas, CF; Beecher, CW; Fong, HH;
Farnsworth, NR; Kinghorn, AD; Mehta, RG; Moon, RC; Pezzuto, JM. Cancer
chemopreventive activity of resveratrol, a natural product derived from grapes. Science,
10, 218-220, 1997. doi: 10.1126/science.275.5297.218.
Kim, HY; Lee, KM; Kim, SH; Kwon, YJ; Chun, YJ; Choi, HK. Comparative metabolic
and lipidomic profiling of human breast cancer cells with different metastatic potentials.
Oncotarget, 7, 67111-67128, 2016. doi: 10.18632/oncotarget.11560.
Knight, JA; Anderson, S; Rawie, JM. Chemical basis of the sulfo-phospho-vanillin
reaction for estimating total serum lipids. Clinical Chemistry, 18, 199-202, 1972.
Kulkarni, SS; Cantó, C. The molecular targets of resveratrol. Biochimica et Biophysica
Acta, 1852, 1114-1123, 2015. doi: 10.1016/j.bbadis.2014.10.005.
Laemmli, UK. Cleavage of Structural Proteins during the Assembly of the Head of
Bacteriophage T4. Nature, 227, 680-685, 1970. doi:10.1038/227680a0.
Langhorst, MF; Reuter, A; Stuermer, CAO. Scaffolding microdomains and beyond: the
function of reggie/flotillin proteins. Cellular and Molecular Life Sciences, 62, 2228-
2240, 2005. doi: 10.1007/s00018-005-5166-4.
Lasa, A; Schweiger, M; Kotzbeck, P; Churruca, I; Simón, E; Zechner, R; Portillo, MP.
Resveratrol regulates lipolysis via adipose triglyceride lipase. Journal of Nutritional
Biochemistry, 23, 379-384, 2012. doi: 10.1016/j.jnutbio.2010.12.014.
Levental, I; Veatch, SL. The continuing mystery of lipid rafts. Journal of Molecular
Biology, 428, 4749-4764, 2016. doi: 10.1016/j.jmb.2016.08.022.
Li, S; Bouzar, C; Cottet-Rousselle, C; Zagotta, I; Lamarche, F; Wabitsch, M; Tokarska-
Schlattner, M; Fischer-Posovszky, P; Schlattner, U; Rousseau, D. Resveratrol inhibits
lipogenesis of 3T3-L1 and SGBS cells by inhibition of insulin signaling and
mitochondrial mass increase. Biochimica et Biophysica Acta, 1857, 643-652, 2016a.
doi: 10.1016/j.bbabio.2016.03.009.
Li, X; Wang, D; Zhao, QC; Shi, T; Chen, J. Resveratrol inhibited non-small cell lung
cancer through inhibiting STAT-3 signaling. American Journal of the Medical Sciences,
352, 524-530, 2016b. doi: 10.1016/j.amjms.2016.08.027.

110
Li, Z; Yang, Y; Gao, Y; Wu, X; Yang, X; Zhu, Y; Yang, H; Wu, L; Yang, C; Song, L.
Elevated expression of flotillin-1 is associated with lymph node metastasis and poor
prognosis in early-stage cervical cancer. American Journal of Cancer Research, 6, 38-
50, 2016c.
Liu, J; Huang, W; Ren, C; Wen, Q; Liu, W; Yang, X; Wang, L; Zhu, B; Zeng, L; Feng,
X; Zhang, C; Chen, H; Jia, W; Zhang, L; Xia, X; Chen, Y. Flotillin-2 promotes
metastasis of nasopharyngeal carcinoma by activating NF-κB and PI3K/Akt3 signaling
pathways. Scientific Reports, 2015a. doi: 10.1038/srep11614.
Liu, RH. Potential synergy of phytochemicals in cancer prevention: mechanism of
action. Journal of Nutrition, 134, 3479S-3485S, 2004.
Liu, Y; Lin, L; Huang, Z; Ji, B; Mei, S; Lin, Y; Shen, Z. High expression of flotillin-2 is
associated with poor clinical survival in cervical carcinoma. International Journal of
Clinical and Experimental Pathology, 8, 622-628, 2015b.
Lowry, OH; Rosebrough, NJ; Farr, AL; Randall, RJ. Protein measurement with the
Folin phenol reagent. Journal of Biological Chemistry, 193, 265-275, 1951.
Mistry, DAH & French, PW. Circulating phospholipids as biomarkers of breast cancer:
a review. Breast Cancer: Basic and Clinical Research, 10, 191-196, 2016.
doi:10.4137/BCBCR.S40693.
Morad, SAF & Cabot, MC. Tamoxifen regulation of sphingolipid metabolism—
therapeutic implications. Biochimica et Biophysica Acta, 1851, 1134-1145, 2015.
doi:10.1016/j.bbalip.2015.05.001.
Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to
proliferation and cytotoxity assays. Journal of Immunological Methods, 65, 55-63,
1983.
Munro, S. Lipid Rafts: Elusive or Illusive? Cell, 115, 377-388, 2003.
NCBI - National Center for Biotechnology Information. PubChem Substance Database.
SID= 445154. 2017. Disponível em:
<https://pubchem.ncbi.nlm.nih.gov/compound/resveratrol#section=Top>. Acesso em:
08 dde fevereiro de 2017.
Nieva, C; Marro, M; Santana-Codina, N; Rao, S; Petrov, D; Sierra. The lipid phenotype
of breast cancer cells characterized by raman microspectroscopy: towards a
stratification of malignancy. PLoS One, 2012. doi: 10.1371/journal.pone.0046456.
Olas, B & Holmsen, H. Interaction of resveratrol with membrane glycerophospholipids
in model system in vitro. Food and Chemical Toxicology, 50, 4028-4034, 2012.
doi.org/10.1016/j.fct.2012.07.066.

111
Owen, DM; Magenau, A; Williamson, D; Gaus, K. The lipid raft hypothesis revisited -
New insights on raft composition and function from super-resolution fluorescence
microscopy. Bioessays, 34, 739-747, 2012. doi: 10.1002/bies.201200044.
Pandey, PR; Okuda, H; Watabe, M; Pai SK; Liu, W; Kobayashi, A; Xing, F; Fukuda, K;
Hirota, S; Sugai, T; Wakabayashi, G; Koeda, K; Kashiwaba, M; Suzuki, K; Chiba, T;
Endo, M; Fujioka, T; Tanji, S; Mo, YY; Cao, D; Wilber, AC; Watabe, K. Resveratrol
suppresses growth of cancer stem-like cells by inhibiting fatty acid synthase. Breast
Cancer Research and Treatment, 130, 387-398, 2011. doi: 10.1007/s10549-010-1300-6.
Patel, KR; Brown, VA; Jones, DJ; Britton, RG; Hemingway, D; Miller, AS; West, KP;
Booth, TD; Perloff, M; Crowell, JA; Brenner, DE; Steward, WP; Gescher, AJ; Brown,
K. Clinical pharmacology of resveratrol and its metabolites in colorectal cancer patients.
Cancer Research, 70, 7392-7399, 2010. doi: 10.1158/0008-5472.CAN-10-2027.
Pike, LJ. Rafts defined: a report on the Keystone symposium on lipid rafts and cell
function. Journal of Lipid Research, 47, 1597-1598, 2006. doi: 10.1194/jlr.E600002-
JLR200.
Pozo-Guisado, E; Alvarez-Barrientos, A; Mulero-Navarro; Santiago-Josefat, B;
Fernandez-Salguero, PM. The antiproliferative activity of resveratrol results in
apoptosis in MCF-7 but not in MDA-MB-231 human breast câncer cells: cell-specific
alteration of the cell cycle. Biochemical Pharmacology, 64, 1375-1386, 2002.
Prat, A; Parker, JS; Karginova, O; Fan, C; Livasy, C; Herschkowitz, JI, He, X; Perou,
CM. Phenotypic and molecular characterization of the claudin-low intrinsic subtype of
breast cancer. Breast Cancer Research, 12, 2010. doi: 10.1186/bcr2635.
Ruiz, JI & Ochoa, B. Quantification in the subnanomolar range of phospholipids and
neutral lipids by monodimensional thin-layer chromatography and image analysis.
Journal of Lipid Research, 38, 1482-1489, 1997.
Scarlatti, F; Sala, G; Somenzi, G; Signorelli, P; Sacchi, N; Ghidoni, R. Resveratrol
induces growth inhibition and apoptosis in metastatic breast cancer cells via de novo
ceramide signaling. FASEB Journal, 17, 2339-2341, 2003. doi: : 10.1096/fj.03-0292fje.
Signorelli, P & Ghidoni, R. Resveratrol as an anticancer nutrient: molecular basis, open
questions and promises. Journal of Nutritional Biochemistry, 16, 449-466, 2005. doi:
10.1016/j.jnutbio.2005.01.017.
Song, L; Gong, H; Lin, C; Wang, C; Liu, L; Wu, J; Li, M; Li, J. Flotillin-1 promotes
tumor necrosis factor-α receptor signaling and activation of NF-κB in esophageal
squamous cell carcinoma cells. Gastroenterology, 143, 995-1005, 2012. doi:
10.1053/j.gastro.2012.06.033.
Staubach, S & Hanisch, FG. Lipid rafts: signaling and sorting platforms of cells and
their roles in cancer. Expert Review of Proteomics, 8, 263-277, 2011. doi:
10.1586/epr.11.2.

112
Tian, H & Yu, Z. Resveratrol induces apoptosis of leukemia cell line K562 by
modulation of sphingosine kinase-1 pathway. International Journal of Clinical and
Experimental Pathology, 8, 2755-2762, 2015.
Touzet, O; Philips, A. Resveratrol protects against protease inhibitor-induced reactive
oxygen species production, reticulum stress and lipid raft perturbation. AIDS, 24, 1437-
1447, 2010. doi:10.1097/QAD.0b013e32833a6114.
Towbin, H; Staehelin, T; Gordon, J. Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets: procedure and some applications.
Proceedings of the National Academy of Sciences, 76, 4350-4354, 1979.
Turner, NC; Neven, P; Loibl, S; Andre, F. Advances in the treatment of advanced
oestrogen-receptor-positive breast cancer. Lancet, 2016. doi: 10.1016/S0140-
6736(16)32419-9.
Villegas-Comonfort, S; Castillo-Sanchez, R; Serna-Marquez, N; Cortes-Reynosa, P;
Salazar EP. Arachidonic acid promotes migration and invasion through a PI3K/Akt-
dependent pathway in MDA-MB-231 breast cancer cells. Prostaglandins, Leukotrienes
and Essential Fatty Acids, 90, 169-177, 2014. doi: 10.1016/j.plefa.2014.01.007.
Vogel, WC; Zieve, L; Carleton, RO. Measurement of serum lecithin, lysolecithin, and
sphingomyelin by a simplified chromatographic technique. Journal of Laboratory and
Clinical Medicine, 59, 335-344, 1962.
Waki, M; Ide, Y; Ishizaki, I; Nagata, Y; Masaki, N; Sugiyama, E; Kurabe, N;
Nicolaescu, D; Yamazaki, F; Hayasaka, T; Ikegami, K; Kondo, T; Shibata, K; Hiraide,
T; Taki, Y; Ogura, H; Shiiya, N; Sanada, N; Setou, M. Single-cell time-of-flight
secondary ion mass spectrometry reveals that human breast cancer stem cells have
significantly lower content of palmitoleic acid compared to their counterpart non-stem
cancer cells. Biochimie, 107, 73-77, 2014. doi: 10.1016/j.biochi.2014.10.003.
Wang, S; Meng, Q; Xie, Q; Zhang, M. Effect and mechanism of resveratrol on drug
resistance in human bladder cancer cells. Molecular Medicine Reports, 15, 1179-1187,
2017. doi: 10.3892/mmr.2017.6111.
Wang, YL; Yao, WJ; Guo, L; Xi, HF; Li, SY; Wang, ZM. Expression of flotillin-2 in
human non-small cell lung cancer and its correlation with tumor progression and patient
survival. International Journal of Clinical and Experimental Pathology, 8, 601-607,
2015.
Wei, S; Moon, HG; Zheng, Y; Liang, X; As, CH; Jin, Y. 2 Modulates Fas signaling
mediated apoptosis after hyperoxia in lung epithelial cells. PLoS ONE, 2013. doi:
10.1371/journal.pone.0077519.
Wen, Q; Alnemah, MM; Luo, J; Wang, W; Chu, S; Chen, L; Li, J; Xu, L; Li, M; Zhou,
J; Fan, S. Flot-2 is an independent prognostic marker in oral squamous cell carcinoma.
International Journal of Clinical and Experimental Pathology, 8, 8236-8243, 2015.

113
WHO - World Health Organization. Cancer. Fact sheet updated february 2017, 2017a.
Disponível em: < http://www.who.int/mediacentre/factsheets/fs297/en/>. Acesso em: 06
de fevereiro de 2017.
WHO - World Health Organization. Fruit and vegetable promotion initiative - report of
the meeting, 2003. Disponível em:
<http://www.who.int/dietphysicalactivity/publications/f&v_promotion_initiative_report.
pdf>. Acesso em: 06 de fevereiro de 2017.
WHO - World Health Organization. Guide to cancer early diagnosis, 2017b. Disponível
em: <http://apps.who.int/iris/bitstream/10665/254500/1/9789241511940-
eng.pdf?ua=1>. Acesso em: 06 de fevereiro de 2017.
Ximenes, AA; Silva-Cardoso, L; De Cicco, NNT; Pereira, MG; Lourenço, DC; Fampa,
P; Folly, E; Cunha-e-Silva, NL; Silva-Neto, MAC; Atella, GC. Lipophorin drives lipid
incorporation and metabolism in insect trypanosomatids. Protist, 166, 297-309, 2015.
doi: 10.1016/j.protis.2015.04.003.
Yang, S; Zhou, Q; Yang, X. Caspase-3 status is a determinant of the differential
responses to genistein between MDA-MB-231 and MCF-7 breast cancer cells.
Biochimica et Biophysica Acta, 1773, 903-911, 2007. doi:
10.1016/j.bbamcr.2007.03.021.
Yeung, F; Hoberq, JE; Ramsey, CS; Keller, MD; Jones, DR; Frye, RA; Mayo, MW.
Modulation of NF-kappaB-dependent transcription and cell survival by the SIRT1
deacetylase. EMBO Journal, 23, 2369-2380, 2004. doi: 10.1038/sj.emboj.7600244.
Zhang, R; Lu, M; Zhang, Z; Tian, X; Wang, S; LV, D. Resveratrol reverses P-
glycoprotein-mediated multidrug resistance of U2OS/ADR cells by suppressing the
activation of the NF-κB and p38 MAPK signaling pathway. Oncology Letters, 12,
4147-4154, 2016. doi: 10.3892/ol.2016.5136.
Zhao, F; Zhang, J; Liu, YS; Li,L; He, YL. Research advances on flotillins. Virology
Journal, 2011. doi: 10.1186/1743-422X-8-479.
Zhao, W; Qiu, Y; Kong, D. Class I phosphatidylinositol 3-kinase inhibitors for cancer
therapy. Acta Pharmaceutica Sinica B, 7, 27-37, 2017. doi: 10.1016/j.apsb.2016.07.006.
Zhu, W; Qin, W; Zhang, K; Rottinghaus, GE; Chen, YC; Kliethermes, B; Sauter, ER.
Trans-resveratrol alters mammary promoter hypermethylation in women at increased
risk for breast cancer. Nutrition and Cancer, 64, 393-400, 2012.
doi:10.1080/01635581.2012.654926.
Zlatkis, A; Zak, B; Boyle, AJ. A new method for the direct determination of serum
cholesterol. Journal of Laboratory and Clinical Medicine, 68, 486-492, 1953.

114
8. ANEXOS

115
Anexo 1
Teor de lipídeos totais mensurado por gravimetria.
0
1000
2000
3000
4000
M C F - 7
M D A
C o n tr o l R e s v C o n tr o l R e s v
Lip
íde
os
to
tais
(
g)

116
Anexo 2
MCF-7 - Cromatografia ácidos graxos Controle (superior) e resveratrol (inferior)
117
Anexo 3
MDA-MB-231 - Cromatografia ácidos graxos Controle (superior) e resveratrol (inferior)
118
Anexo 4
Identificação do perfil de fosfolipídeos de células de câncer de mama MCF-7 e MDA-
MB-231, após tratamento com resveratrol por 0, 2, 8 e 24 horas, por TLC e marcação
com 32Pi.