lljiiipime ;aqç,.,. - usp · a sra. mirian o. marques que além de ter colaborado em uma das fases...
TRANSCRIPT


llJIIipIMe IJ'-I'J .4 ;aqç,.,.

AGRADECIMENTOS
Ourante a elaboração desta tese houve sempre a colabo
ração de inúmeras pessoas e Instituições a quem expresso o meu
mais profundo agradecimento. Entre elas gostaria de salientar:
-- O Prof. Or. Francisco J.S. Lara, orientador desta
tese, que não poupou esforços em criar tôdas as condições ne
cessárias para o desenvolvimento dêste trabalho, assim como, pe
lo continuo estimulo a mim proporcionado.
-- O Sr. Antônio G. de Bianchi com quem dividi o en
tusiasmo e as frustações da maior parte dêste trabalho.
-- O Sr. Ângelo G. Gambarini com quem realizei os es
tágios iniciais dêste trabalho.
-- A srta. Yolanda Tavares e Sra. ~alkyria H. Lara, a~
bas do Instituto Adolfo Lutz, pela colaboração em campos de sua
especialidade.
O Or. Gerhard Malnic, Ora. Rebeca C. de Angelis e
Sra. Ilza C. M. Terra, todos do Instituto de Ciências Biomédicas
da Universidade de são Paulo e Or. Herval P. Ribeiro do Hospital
Central da Santa Casa de Misericôrdia de são Paulo, pela preste
za com que me emprestaram equipamentos de sua responsabilidade.
-- Os meus colegas do Instituto de Química da Univer
sidade de são Paulo: Or. Fernando Galembeck, Jane Balsamo, Júlia
M. Alves, Manuel T. Pueyo, Marila Cordeiro, Nelson Marques, Or.
Rogério Meneghini, Roberto V. Santelli, que colaboraram em algu
mas das fases experimentais ou contribuíram com valiosas .críti
cas e sugestões.
-- A Ora. Eline s. Prado em cujo curso de pés-gradua
çao em química de proteínas aprendi várias técnicas que se torn~
ram úteis durante a realização dêste trabalho.

-- A Ora. Carrninda da C. Landim que através de seu cur
so de Morfologia de Insetos muito contribuiu para uma boa inter
pretação de muitos resultados por mim obtidos.
o Or. Gilberto B. Oomont do Instituto de Química da
Universidade Federal do Rio de Janeiro que me introduziu nas téc
nicas de cromatografia de aminoácidos.
A sra. Mirian O. Marques que além de ter colaborado
em uma das fases experimentais, forneceu numerosas informações'
nao publicadas de seu trabalho.
-- A srta. Hiromi Tamaki e Sr. José Coelho sem cuja
assistência técnica teria sido muito difícil a realização desta
tese.
-- A Fundação de Amparo à Pesquisa do Estado de são PaE
lo que através de uma bôlsa de Iniciação Científica e duas bôlsas
de Doutoramento me possibilitou ingressar na vida cientIfica.
-- A srta. Yvone de Abreu responsável pela eficiente da
tilografia desta tese.
List
INT~
MeTO

através de seu cur
para uma boa inter- íNDICE
1. Considerações iniciais " .•••. " • . • • • . • . . • . • • . . . • • • • • . 2
2. Contrôle hormonal do desenvolvimento dos insetos.... 6
3. Origem das proteínas do inseto adulto ...•••••..•.•. 9
4. Sêdas e casulos ••.••••.•.•...•.•••.••..•••'......... 14
5. Finalidades dêste trabalho •....•....•.••••....•••.• 16
tituto de Química da
introduziu nas téc-
m de ter colaborado
umerosas informações-
sé Coelho sem cuja
1 a realização desta
Lista de Abreviaturas emoregadas
INTRODt!ÇÃO •••.••••• ~ •••••••••••.•••.••.••••.•..•••.••.•••••
pág.
1
2
ia Estado de são PaE
tIfica e duas bÔ1sas
"ida cientIfica.
~l pela eficiente da
r~TODOS •...•.•••..••.•••••.••..••.••.•....•••.•........••. 20
1. Cultura de Rhynchosciata ••••••••...••.•••••••.••.••.. 20
2. Determinação da idade fisiológica larval .•.....•.'.. 20
3. Coleta de hemolinfa .•.••...•••••.•••.•.••...•....•. 23
4. Determinação do volume ocupado pelos hemocitos na
hemolinfa •••••••••••••••••.•••.••••••.••.•....•...• 23
5. Métodos de estudo da coagulação e escurecimento
da hemolinfa •• • • • • • . . • . • • • . • • • • . • • • . . • • . • . • . . • . • . . 24
6. Determinação da osmolaridade, densidade, volume
e pêso-sêco da hemolinfa . • • • • • . • • • . • . • • . • . • . . . . . . . 24
7. Determinação de p02' pC02 , pH e HC03- na hemo-
linfa ••••••••.•••••....•••..•.•••••••..•••.•••.•.•• 25
8. Determinações químicas na hemolinfa ••••..•.......•• 25
9. Cromatografia de troca iônica dos compostos ni~~i-
drina-po~itivos ácidos~solúveis na hemolinfa •....•• 29
10. Preparação dos pigmentos da hemolinfa •.•••••••.•••• 30
11. Identificação dos carotenóides •.••••••.••.•...•.••• 31
12. Preparação do casulo I, II e III e determinação da
massa produzida por indivíduo •••.••••.•••••..•••••• 32
13. Dete~inação da solubilidade do casulo .•••..••••••. 32
14. Determinações químicas do casulo •••••...•••.••••..• 33
15. Cromatografia em papel de carboidratos do casulo .•• 35
16. Cromatografia de troca iônica de aminoácidos da
fração insolúvel do casulo ••••..••••••••..••.•.•••• 36

pág.
REAGEh'TES •..•••.•.•.•.•...•.•..•....•.•.•....•..•.••.•..• 38
A. Química da hemolinfa •••••.•..•..••.••..•••••...•••.. 85
A. Química da hemolinfa de larvas de R. "americana ..•.• 39
B.
REFERt
RESUMO
ABSTRA
85
39
58
37
37
1. Apresentação geral dos resultados 58
2. Solubilidade, conteúdo de cinzas e metais 59
3 . Carboidratos •..................•........•........ 6].
4. Compost9s nitrogenados .....•..................•.. 65
5. Produção e composição do casulo em função do tp.mpo 68
1. Resultados gerais ....•......•••.....•.......•.... 39
2. Cromatografia dos compostos ninhidrina-positivos
solúveis em ácido ....••. , ....•....•......•...... ' 40
3. Apresentação dos resultados das análises químicas. 48
4. Pigmentos .....•....•......•..•.•................. 50
1. Extensão da análise realizada .••.........•...•... 85
2. Componentes orgânicos da hemolinfa ••.•...•....... 87
3. Composição molar da hemolinfa e seus componentes
inorgânicos. • . . • . . • • • . . • . . . . . . . . . . . . . . • • . . • . . . • . • . 93
4. Coagulação e escurecimento .•.....•.....••.•...... 99
5, Pigmentos ...••.•..............•••.....•......•... 100
1. Massa corpórea e volume de hemolinfa •.•........•. 72
2. Determinações químicas na hemolinfa .••.•.•....•.• 76
3. pêso-sêco e glicogênio no corpo gorduroso •....... 81
4. pêso-sêco e metais nos túbulos de Malpighi •...... 82
B. Química do casulo coletivo de R. americana
C. Aspectos químicos e fisiológicos durante o 49 estádio
de R. americana ..... , •. . . . . • •. .• •. . •. ..•. . •.. ... .. • 72
17. Determinação de pêso-sêco e metais nos tubos de
Malpighi .••.....•......•.•.•.•••.••••..•...•......
18. Determinação de glicogênio e pêso-sêco do corpo
gorduroso ...••.........••.....••..•................
RESULTADOS ..•.••••••.•.•..........•.••••..........•......
DISCUSSÃO

pág.
~ubos de
pág.
· .~o corpo
• a••••••••••
· .
· .
37
37
38
39
B. Produção do casulo coletivo por ~arvas de R. ame-
ricana
1. Quími ca do casulo ...•••.••.••..•......•.••..•••
2. Origem dos componentes do casulo ....••.........
3. Mecanismos e regulação da produção do casulo ...
RESUMO •••••••••••••••• •••••••••••••• '. =• • • • • • • • • • • • •• • • ••
102
102
104
111
114~icana ....• 39
· . 39 ABSTRACT 116
l-positivos
• . •• • • • • . • • . 40
;es químicas. 48
••••• " • .. .. 50
~ .......... 58
·........... 58ais ........ 59·........... 61
· ............. 65:rão do tAmpo 68
) 49 estádio
·................. 72
, ............. 72
·.............. 76)So ........ 81.ghi •...... 82
• . . . . . . • . • . 85
• . • . • . . • • • . 85
· . • . • . . • . . • 85
• • •• • • • • • • • 87
nponentes
· • • • • • . • . • • 93
· • • • • • • • • •. 99
.•••••.••• ' 100
REFERtNéIAS BIBLIOGRÂFICAS •••••••••••••••••••••••••••••• 118

ABREVIATURAS
TCA - Âcido tricloroacético
19P - Primeiro período do 49 estádio de R. americana (ver Tabe
la I). Da mesma forma 29P, 39P, 49P, 59P e 69P, corres
pondem aos demais períodos do 49 estádio.
p.e. - POnto de ebulição
O.V. - Luz ultravioleta
RNA - Âcido ribonucléico
DNA - Âcido desoxirribonucléico
Cf. - Confrontar com
PAS - Periodic Acid Schiff, teste para carboidratos
A - Comprimento de onda
p.ex.- Por exemplo
p/p - péso por pêso
p/V - pêso por volume
V/V - Volume por volume
SDS - Dodecil sulfato de sódio
.Ác. - Âcido
EC - Numeração das enzimas conforme recomendações feitas pela
Enzyme Commission da International Union of Biochemistry
em 1964.

INTRODUÇÃO
1. CONSIDERAÇÕES INICIAIS
A classe Insecta constitui o grupo com o maior número
conhecido de espécies no reino animal; cêrca de 900.000 para um
total descrito de 1.200.000. Sua distribuição abrange tôda a
superfície do planêta, com exceção das profu~dezas do oceano e
está representada mesmo entre os poucos animais que habitam per
manentemente a região do Polo Sul. Essa enorme distribuição ec~
lógica e a variedade de formas que assume, a torna excelente pa
ra o estudo das relações organismo-meio e certamente seria mui~o
interessante conhecer as razões morfológicas, fisiológicas e bio
químicas de seu sucesso.
A importáncia econômica dos insetos, pode ser avalia
da quando se considera o montante de prejuízos causados por êles
anualmente; sõmente nos Estados Unidos, como consequência de
seus ataques a colheitas, produtos armazenados, domésticos e sim~
lares, além das doenças e desconfôrto causados ho homem e nos a
nimais: 15 biliões de dólares! (ROSS, 1965). Ainda dentro do
prisma econômico, sua importância se faz sentir também na indús
'tria da sêda natural, do mel, da cêra, etc.
O estudo dos insetos não tem sido estimulado apenas
pelo seu número e sucesso biológico ou pelas possibilidades dês
se conhecimento contribuir para o aperfeiçpamento dos métodos de
contrôle de pragas. Os insetos tem-se mostrado excelentes mode
los para estudos de problemas genéticos e citogenéticos (SINNOT,
DUNN & DOBZHANSKY, 1958), bioquímicos (GILMOUR, 1961), fisioló
gicos (WIGGLESWORTH, 1948, 1965), etc.
Todos os insetos, com exceção dos Ametabola, sofrem
uma metamorfose. Esta pode ser definida, segundo WIGGLESWORTH
(1965), como a mudança mais ou menos marcada de form~, que ocor
re quando o inseto torna-se adulto; independente do fato dessa
modificação requerer duas mudas e um estágio pupal como nos Holo

-3-
metabola, ou uma única muda final, como nos Hemimetabola.
Existem várias teorias para explicar a origem da me
tamorfose completa em insetos. Aquela de maior aceitação en
tre os especialistas é a de POYARKOFF-HINTON (Cf. NOVÃK, 1966).Segundo esta teoria a adaptação da larva e adulto a nichos ecol2
gicos distintos os tornou muito diferentes morfolõgicamente. A
pupa poderia ser interpretada corno um meio de intercalar uma se
gunda muda entre a larva de último estádio e o adulto, permitin
do o estabelecimento das ligações musculares imaginais em apóde
mas cuticulares do adulto, que são completamente diferentes da
queles da larva, em duas etapas. A primeira muda permitiria o
descolamento e histólise da musculatura larval e a segunda seria
necessária para a ligação dos músculos imaginais.
Segundo WIGGLESWORTH (1965, 1970) a metamorfose, do
ponto de vista ontogenético, poderia ser considerada como um ti
po particular de polimorfismo. Os insetos Holometabola possui
riam três sistemas genéticos distintos, enquanto que os Hernime
tabola p~ssuiriam dois sistemas; correspondentes no primeiro ca
so à larva, pupa e adulto e no segundo caso à ninfa e adulto. Du
rante a metamorfose ocorreria a inibição de um sistema genético
e a indução de outro.
Ó fato da profunda reorganização de tecidos e órgãos
que caracteriza a metamorfose de insetos estar sob contrõle hor
monal (ver item ~ abaixo), tem levado a se considerar êsse fe
nõmeno um excelente modélo para o estudo da reguYação horrnonal
da expressão genética e seu papel na morfogênese. Isso tem ins
pirado um número crescente de trabalhos em bioquímica do desen
volvimento de insetos,que foram revistos recentemente por alguns
autores (AGRELL, 1964; GILBERT, 1967b; WYATT, 1968).
A metamorfose nos insetos Holometabola inicia-se por
ocasião da muda pupal, embora naqueles insetos que produzem um
pupar~o ou tecem um casulo para a pupa, a metamorfose inicia-se
mais precisamente com a produção dessas estruturas (WIGGLESWORTH
1965),
A respiração dos insetos durante a metamorfose segue
uma curva em forma de U, indicando uma respiração mais intensa
no iní
inicia
gênese
tológi
os doi
indica
mente
níveis
portan
adulto
nor de
maior
as est
metamo
ocorre
biossí
larva,
protei
tuição
cie te
devido
balhar
manho.
forese
difere
visao
CHI, 1
* HINT
que es
mo "ph
to por
design
confor
(1946)
dentro

-4-
s Hemimetabola.
plicar a origem da me
3 maior aceitação en
ON (Cf. NOVÃK, 1966).
adulto a nichos ecol§
morfolõgicamente. A
de intercalar uma se
e o adulto, permitin-
!s imaginais em apóde
~ente diferentes da
:a muda permitiria o
:val e a segunda seria
~nais.
I) a metamorfose, do
'nsiderada corno um ti
Holometabola possui
uantoque os Hemime
entes no primeiro ca
à ninfa e adulto. D~
e um sistema genético
o de tecidos e órgãos
tar sob contrõle hor
3 considerar êsse fe
a reguTação hormonal
~nese. Isso tem ins
)ioquímica do desen
~entemente por alguns
r, 1968).
~tabola inicia-se por
ietos que produzem um
~tamorfose inicia-se
~uturas (WIGGLESWORTH
a me~amorfose segue
ipiração mais intensa
no início e no fim do processo. Êste fenõmeno foi interpretado
inicialmente corno refletindo sucessivamente a histólise e histo
gênese dos tecidos (Cf. AGRELL, 1964). Entretanto, estudos his
tológicos mostraram que essa interpretação não era correta, pois
os dois fenômenos ocorriam simultâneamente. Estudos posteriores
indicaram que a curva em U da taxa de respiração era principal
mente uma consequência do contrôle das vias respiratórias pelos
níveis de ADP, ATP e fosfato inorgânico celular, refletindo,
portanto, as demandas energéticas do inseto. Como a pupa ou o*adulto faratopossuem pequena atividade muscular, a maior ou m~
nor demanda energética na metamorfose tem sido relacionada com a
maior ou menor atividade biossintética de proteínas(~NATT,1968).
Os resultados comentados acima indicam, portanto, que
as estruturas imaginais começam a ser formadas desde o início da
metamorfose e que subjacentemente à formação dessas estruturas
ocorrem processos que demandam energia e que devem incluir a
biossíntese de proteínas.
As estruturas imaginais diferem bastante daquelas da
larva, sugerindo que deve haver uma diferença notável entre as
proteínas imaginais e as larvais. A extensão em que a consti
tuição protéica do adulto difere daquela qa larva da mesma espé
cie tem sido estudada principalmente nas proteínas da hemolinfa,
devido às facilidades de sua coleta e pela dif~culdade_de se tr~
halhar com órgãos de larvas de insetos, devido ao seu pequeno t~
manho. Êsses estudos foram realizados principalmente com eletr~
forese de gel de poliacrilamida e mostraram que existe algumas
diferenças de padrão protéico entre a larva e o adulto (ver re
visão em ~NATT, 1968. No caso de R. americana, consultar BIAN
CHI, 1972).
* HINTON (1946) propôs o têrmo "pharate" para designar o estádio
que está fechado dentro da cutícula do estádio anterior. O têr
mo "pharate" é derivado do grego "pharata" que significa "cober
to por um véu". Como não existe nenhum têrmo em português para
designar "pharate" será usado aqui o têrmo "farata" ou "farato",
conforme o caso, que utiliza a mesma raiz grega usada por HINTON
(1946). Dessa forma adulto farato corresponde ao adulto fechado
dentro da cutícula pupal.

~~
-5-
Eletroferogramas em gel de poliacrilamida de homogen~
tos de tecido, que devem ser mais significativos do que a hemo
linfa na representação da constituição protéica do inseto, mos
traram a existência de algumas diferenças entre larvas e adultos.
Entretanto, devido ao número relativw~ente pequeno de bandas el~
troforetican~nte resolvidas (cêrca de 20), em relação às cente
nas de proteínas qüe devem estar presentes nas células, êsses r~
sultados são pouco ccnclusivos na avaliação do grau de diferença
existente entre a constituição protéica larva1 e imaginaI.
Uma outra maneira de atacar o problema é isolar pro
teínas homólogas de diversos estágios do desenvolvimento do ins~
to e compará-las quimicamente. Poucos estudos foram realizados
nesse sentido para se ter uma idéia clara a respeito (ver revi
são em WYATT, 1968). A questão da origem das proteínas do adul
to, isto é, se elas originam-se integralmente por síntese "de
novo" a partir de aminoácidos livres ou se parte das proteínas
larvais são conservadas intactas no adulto, está intimamente re
lacionada ao problema da extensão em que a~ proteínas do inseto
adulto diferem daquela~ larvais. O problema encontra-se ainda
relativamente pouco estudado e os resultados obtidos até o pres~n
te, err~ora conflitantes, sugerem que as proteínas imaginais po
deriam originar-se em :9arte por síntese "de novo" e em parte a
partir de proteínas lar:vais que se conservariam intactas no adul
to (ver ítem 3, a seguir).
Abaixo será mostrado que a produção do casulo em B.*americana representa um modelo interessante para o estudo da o-
rigem das proteínas do adulto durante a metamorfose e quais obj~
tivos dentro desse estudo se pretende atingir especificamente com
esta tese. Antes disso, porém, será conveniente um comentário
breve a respeito dos conhecimentos atuais sóbre o contróle horm~
nal do desenvolvimento dos insetos, origem das proteínas do inse
to adulto e sôbre a química ~e casulos e sedas naturais.
* Rhynchosciara americana Wiedernann, 1821 (Diptera Nematocera
Sciaridae), foi redescrita por NONATO & PAVAN (1951) como ange
~ e essa denominação foi usada em numerosas publicações até
ser demonstr~a sua sinonímia com americana por BREUER (1969).

-6-
2. CONTROLE HORMONAL DO DESE~TVOLVlMENTO DOS INSETOS
O estudo da regulação hormonal do desenvolvimento pós
embrionário dos insetos foi iniciado há mais de 50 anos. Os es
forços conjugados de numerosos pesquisadores, permitiram estabele
cer na década de 50 ma modêlo, hoje clássico, dessa regulação.
Posteriormente êste modêlo foi ampliado para todos os insetos e
enriquecido de detalhes porém sem nenhuma alteração substancial.
As pesquisas básicas que levaram ã ?roposição dêsse modêlo foram
revistas minuciosamente por vários autores (o. ex.,GILBERT, 1964;
SCHNEIDERMAN & GILBERT, 1964; HIGGLESI·mRTH, 1964 ; NOVÃK, 1966; HER
MAN, 1968; lvIGGLESWORTH, 1970) e não serão apreciadas aqui. O mo
dêlo clássico da regulação hormonal dos insetos pode ser resumido
como segue abaixo.
As células neurossecretoras localizadas no "pars inter
cerebralis" secretam o "hormônio cerebral", que se acumula nas
"corpora cardiaca", de onde se difunde para a hemolinfa, estimu
lando as glândulas protoráxicas a secretar a ecdisona (hormônio
da muda). Sob influência dêste hormônio o processo de muda se
inicia. O nível de hormônio juvenil, que é produzido nas "cor
pora allata", determina o tipo de muda. A presença de um título
elevado de hormônio juvenil e ecdisona resulta em muda larva-lar
va; um decréscimo no tItulo de hormônio juvenil leva à muda lar
va-pupa am Holometabola e as mudas para adulto ocorrem quando
ecdisona está presente, mas o hormônio juvenil ausente.
Depois de estabelecido o modêlo descrito acima, grande
esfôrço foi realizado na identificação química dos três hormônios
envolvidos. Graças aos trabalhos de vários pesquisadores, entre
os quais se destacou KARLSON, a ecdisona foi cristalizada, iden
tificada como sendo um esteróide e essa identificação foi confir
mada por síntese. Estas descobertas foram amplamente revistas por
KARLSON & SEKERIS (1966). Mais tarde foi identificado nas prepa
rações de ecdisona um componente presente em menores quantidades,
com atividade similar à ecdisona, ao qual se denominou inicialmen
te de B-ecdisona e que, após a determinação de sua estrutura, ve
rificou-se tratar-se da 20-hidroxi-ecdisona. Esta substância foi
demonstrada ser idêntica ao composto denominado ecdisterona e ao
hormônio da muda de crustáceos, a crustecdisona(WIGGLESWORTH,1970).

-7-
: INSETOS
lizadas no "pars inte!
que se acumula nas
a hemolinfa, estimu
a ecdisona (hormônio
processo de muda se
é produzido nas "cor
presença de um título
lta em muda larva-lar
enil leva à muda lar-
.0 desenvolvimento pós
s de 50 anos. Os es
s, permitiram estabel~
o, dessa regulação.
a todos os insetos e
Iteração substancial.
ão dêsse modêlo foram
(D. eX.,GILBERT, 1964;
1964;NOVÂK,1966; HER
apreciadas aqui. O mo
etos pode ser resumido
descrito acima, grande
ica dos três hormônios
pesquisadores, entre
i cristalizada, iden
ntificação foi confir
amplamente revistas por
dentificado nas prepa-
TI menores quantidades,
= denominou inicialme~
de sua estrutura, ve
Esta substância foi
nado ecdisterona e ao
sona (I"lIGGLESWORTH,1970).
do hormônio juvenil de extratos de
derivado terpenóide foi realizada
sua confirmação por síntese por
A identificação
Hyalophora cecropia como um
por ROLLER et alii (1967) e
DAHM, TROST & ROLLER (1967).
Além dos três hormônios mencionados acima são conheci
dos numerosos outros, mas como não estão diretamente envolvidos
no contrôle do desenvolvimento e metamorfose, não serão discuti
dos aqui. Os conhecimentos existentes sôbre êsses hormônios fo
ram revistos. por: GILMOUR (1965), NOVÂK (1966), \\'IGGLES\·lORTH,
(1970), LAW & REGNIER (1971).
A natureza química do hormônio cerebrál ainda está m~
definida, embora resultados recentes indicam tratar-se de um peE
tídeo ou proteína de baixo pêso-molecular e que talvez seja for-
mado por várias espécies de proteínas ou polipeptídeos (Cf.
WIGGLESWORTH, 1970).
O modo de ação dos hormônios morfogenéticos dos ins~
tos tem sido muito estudado. Entre êles o melhor estudado é a
quêle da ecdisona e nêsse sentido merecem uma ênfase especial os
estudos realizados em glândulas salivares de Diptera. Estas
glândulas, possuem cromossomos politênicos que apresentam um pa
drão de "puffs" (expansões globulares dos cromossomos politêni
cos) que são típicos do estágio de desenvolvimento do inseto,a~
sim como, do tecido em que se encontram (Cf. BREUER & PAVAN,1955;
BEERMAN,1956). Êsses " puffs" cromossômicos parecem representar
gens em atividade (BEERMAN, 1963) e alguns " puffs" podem ser i~
,duzidos especificamente por injeção de ecdisona em larvas de
Chironomus (CLEVER & KARLSON, 1960). Essa indução de "puffs"
cromossômicos pela ecdisona parece ocorrer mesmo nas condições
normais de desenvolvimento larval. Isso foi demonstrado pelo f~
to de larvas de Drosoohila divididas em dois compartimentos por
uma ligadura apresentar um padrão de "puffs" da época de forma
ção do pupário, sõmente nos cromossomos de células das glândulas
salivares presentes no mesmo comnartimento que contém as glându
las protoráxicas. Por outro lado implantes de glândulas saliva
res de pré-pupas de Drosophila em larvas de último estádio reve!
tem a um padrão de "puffs" correspondente à época do início da
formação do pupârio (BECKER, 1962). Mais recentemente AMABIS &
quandolto ocorrem
nil ausente.

:"8-
CABRAL (1970) mostraram, em experimentos similares aos de BECKER
(1962), que os "puffs" da glândula salivar de R. americana tam
bém devem estar sob contrô1es hormonal.
KARLSON e colaboradores conseguiram, em uma série de
trabalhos, mostrar que ecdisona injetada em larvas de Calliphora
erythrocephala dirige-se ao núcleo das células epidérmicas, onde
in~uz a síntese preferencial de RNAs mensageiros para a síntese
de dopa descarboxilase (EC 4.1.26), enzima chave para o início'
do processo de esclerotização na formação do pupário e, baseando
se nêsses resultados e nos discutidos anteriormente, propuse
ram um modêlo para o mecanismo de ação da ecdisona (ver revisão
em KARLSON & SEKERIS,1966). ~ste modêlo consiste em assumir um
mecanismo de regulação das células de :nsetos similar ao propos
to por JACOB & MONOD para bactérias e o papel da ecdisona seria
combinar-se com repressores específicos, liberando secções espe
cíficas do DNA para transcrição. Êsse modêlo apresenta várias
dificuldades, entre elas a não indução de síntese de dopa descaE
boxilase (EC 4.1.26) na pupa de Calliphora, quando há elevação
do título de eC01sona, o que torna mais provável admitir que a
ação da ecdisona sôbre o material genético seja mais complexa que
a proposta por KARLSON & SEKERIS (1966), denendendo do tecido e
do seu estágio de desenvolvimento (ver discussão mais ampla em
WIGGLESWORTH, 1970).
o hormônio juvenil parece agir sôbre o metabolismo de
ácidos nucléicos, embora não exista ainda informação precisa sô
bre o seu mecanismo (sLÁMA, 1971).
Os estudos de endocrinologia de insetos descritos aci
ma, além de terem permitido o estabelecimento de um excelente mo
dêlo para estudo da regulação hormonal da expressão genética,que
é o desenvolvimento dos insetos, estão fadados a criarem as ba
ses de uma revolucionária tecnologia de combate aos insetos dani
nhos. Esta tecnologia está assentada no fato do hormônio juve
nil impedir o desenvolvimento da pupa em adulto. Assim, pupas
de insetos submetidas ao contato, ou mesmo vapores do hormônio
juvenil, acabam morrendo por não serem canazes de se desenvolve
rem até adultos ou, caso se desenvolvam, originam adultos esté
reís (sLÂMA, 1971).
.-'I
;
no SE
logo!
vos,
nham
homeIl
1971;
juver
ticic
os ar
ção cagem
arnbiE
do fé
:va cc
mentE
sito
nomir
975 (
3. CF
blemé
nismc
do en
feché
que é
a his
paI,
aadas
nais
ção creprE
tese
dos r
de cc
·::"l.ntc

-9-
; similares aos de BECKER
'ar de R. americana tam-
,guiram, em uma série de
em larvas de Cal1iphora
'é1ulas epidérmicas, onde
sageiros para a síntese
ma chave para o início
o do pupário e, baseand~
nteriormente, propuse
a ecdisona (ver revisão
consiste em assumir um
setos similar ao propos
papel da ecdisona seria
liberando secções espe
~dêlo apresenta várias
e síntese de dopa descaE
~, quando há elevação
?rovável admitir que a
:0 seja mais complexa que
deDendendo do tecido e
Intensas pesquisas estão sendo realizadas atualmente
no sentido de se conseguir uma síntese química econômica de aná
logos do hormônio juvenil, quimicamente estáveis e altamente at~
vos, e assim criar uma nova geração de inseticidas que não te
nham efeitos tóxicos sóbre as plantas e animais, incluindo o
homem, mas que sejam altamente eficazes contra os insetos (sLÃMA
1971~ CRUICKSHANK. e PALMERE, 1971)0 Os análogos do hormônio
juvenil possuem, entretanto, uma desvantagem em relação aos ins~
ticidas porque, enquanto êstes matam os insétos em poucas horas,
os análogos do hormónio juvenil simplesmente impedem a reprodu
ção da oraga. Em comoensação, o~ análogos do hormônio juvenil
agem em oeaueníssimas quantidades, o que permitirá uma ooluição
ambiental bem men9r do que ocorre com os inseticidas atuais,além
do fato de alguns análogos do hormônio juvenil terem ação selet!
va contra determinados insetos daninhos (SLÂMA, 1971). Recente
mente uma firma industrial foi constituída com o exclusivo prop~
sito de desenvolver essa nova geração de inseticidas que ela de
nominou "Entocon" CCf. boletins técnicos da Zoecon Corporation
975 California Avenue, PaIo Alto, Califórnia 94304).
Lscussão mais ampla em3. ORIGEM DAS FROTErNAS DO INSETO ADULTO
pazes de se desenvolve
originam adultos esté-
- sôbre o metabolismo de
i informação precisa sô-
A origerndas proteínas do inseto adulto é um dos pr2
blemas bioquímicos mais fundamentais da metamorfose dêstes orga
nismos. A maneira mais simples de considerar o problema (levan
do em conta o fato de que a pupa e o adulto faratosão sistemas
fechados que trocam apenas gases com o meio ambiente), é admitir
que as proteínas larvais são hiurolizadas paulatinamente durante
a histólise dos tecidos larvais, desde o início do estágio pu
paI, e que concomitantemente as proteínas do adulto são sinteti
3adas "de novo" a partir de aminoácidos livres nos tecidos imag.:!:.
nais em desenvolvimento. Visto assim, esquemàticamente a solu
ção do problema' se resumiria' na demonstração de como é feita a
repressão paulatina dos' sistema gen~ticos responsáveis pela siQ
tese das proteínas larvais e a indução daquêles sistemas envolvi
dos na síntese das proteínas imaginais, assim como, do mecanismo
de contrôle da hidrôlise das proteínas larvais. Existem, entre
::"tnto, algumas evidências a indicar que o processo poderia ser
juve-
pupas
hormônio
le insetos descritos ac!
lento de um excelente m2
l expressão genética,que
ldados a criarem as ba
dani
10 val?ores do
:ombate aos insetos
fato do hormônio
adulto. Assim,

-10-
mais complexo, envolvendo a utilização de proteínas larvais na
formação das proteínas do adulto sem hidró1ise prévia.
As considerações acima mencionadas passaram a serem
feitas com base experimental ao se verificar o papel das proteí
nas da hemo1infa na vitelogênese. ~ste estudo iniciou-se quando
TELFER (1954) mostrou que uma proteína de hemo linfa é tomada pe
los ovócitos em desenvolvimento e usada diretamente na constitui
çao do vitela em Hya10phora (Lepidoptera). Resultados similares
foram obtidos por numerosos autores em difo.rentes ordens de inse
tos, mostrando que o fenômeno é geral em insetos (Co1eoptera: De
LOOF e WILDE, 1970. Dictyoptera: ADIYODI, 1967; NIELSEN e
MILLS, 1968. Orthoptera: HILL, 1962. Diptera: STRANGWAYS
DIXON, 1962; ORR, 1954a, b). O contrô1e hormona1 dêsse processo
foi estudado principalmente em Orthoptera, onde as células neu
rossecretoras do protocérebro estão implicadas no estímulo à sí~
tese de proteínas da hemo1infa pelo corpo gorduroso, enquanto
que a "corpora allata" estimula a tomada pinocitótica dessas prQ
teínas pelos ovócitos em desenvolvimento (TELFER, 1965). Estudos
realizados em outras ordens de insetos mostraram, entretanto,
que pode haver variações no mecanismo proposto em Orthoptera,
havendo uma alteração na importância relativa das células neuros
secretoras e "corpora a11ata" na vite10gênese, conforme o inseto
estudado (WIGGLESWORTH, 1970).
As proteínas da hem01infa, além de serem utilizadas na
constituição do vitelo, são tomadas por um processo pinocitótico
para vârios outros órgãos, no inicio da pupação (LOUGHTON e t~ST
1965; LOCKE e COLLINS, 1966, 1967; TOBE e LOUGHTON, 1969;
CHIPPENDALE e KILBY, 1969) onde forneceriam aminoácidos para a
síntese "de novo" de proteínas imaginais (LOCKE e COLLINS,
1966, 1967) e/ou transportariam nutrientes essenciais que lhes
estão conjugados corno: carboidratos, 1ipídeos. ou cromóforos
(LOUGHTON e WEST, 1965). Alguns resultados sugerem que a tomada
pelos tecidos, pelo menos no caso do corpo gorduroso em Lepidop
tera, seria estimulada pela ecdisona (COLLINS, 1969).
Os trabalhos referidos acima sôbre metabolismo de pro
teínas durante a metamorfose dão uma idéia apenas qualitativa
dos fenômenos envolvidos. Um estudo quantitativo dêsse metabo
lismo foi realizado por BOYD e MITCHELL (1966). ~stes autores
injetaI
de~
era ba~
da hemc
ria a li
tamorfc
em PhOI
injetaI
dultos
dação é
injeçãc
Phormié
3/4 da
A partj
fa pass
na forn
ticada
mi tir é
tras pI
resulté
pelos t
tes peI
tilizac
alaniné
maram é
apenas
esteja
minoácl
forma ~
teínas
e ress3
1ismo (
dos tré
nicas r
cífica~
I!letamoJtudos l

-11-
~ proteínas larvais na
Use prévia.
~r o papel das proteí
:udo iniciou-se quando
lemolinfa é tomada pe
:etamente na constitu~
Resultados similares
!rentes ordens de ins~
lsetos (Coleoptera: De
1967; NIELSEN e
Diptera: STRANGWAYS
'rmonal desse processo
onde as células neu-
das no estímulo à sí~
gorduroso, enquanto
nocitótica dessas prQ
ELFER, 1965). Estudos
straram, entretanto,
oposto em Orthoptera,
va das células neuros
se, conforme o inseto
:las passaram a serem
injetaram proteínas da hemolinfa previamente marcadas em larvas
de Drosophila melanogaster e verificaram que o. seu "turnover"
era bastante elevado, o que os levou a propor que as proteínas
da hemolinfa seriam intermediários importantes pelos quais pass~
ria a maior parte do "turnover" protéico total do inseto em me
tamorfose. Uma situação completamente diferente foi encontrada
em Phormia regina por CHEN e LEVENBOOK (1966). Êstes autores
injetaram proteínas marcadas da hemolinfa em larvas, pupas e a
dultos de Phormia regina e verificaram que a velocidade de degr~
dação das proteínas era sempre baixa e demonstraram também que a
injeção de proteínas da hemolinfa marcadas em larvas maduras de
Phormia regina pouco antes de puparem leváva à recuperação de
3/4 da radioatividade total em proteínas nos tecidos do adulto.
A partir dêsse resultado propuseram que as proteínas da hemolin
fa passavam para os tecidos e eram em seguida usadas intactas
na formação das estruturas do adulto. Essa proposição foi cri
ticada por WYATT (1968) que argumentou que não é necessário ad
mitir a transformação direta de proteínas da hemolinfa (ou ou
tras proteínas larvais) em proteínas do adulto para explicar os
resultados obtidos, bastando admitir que as proteínas tomadas
pelos tecidos seriam ali degradadas e os aminoácidos resultan
tes permaneceriam dentro dos tecidos, onde seriam ràoidamente u
tilizados na ress!ntese protéica.
~ serem utilizadas na
?rocesso pinocitótico
~ção (LOUGHTON e WEST
e LOUGHTON, 1969;
un aminoácidos para a
(LOCKE e COLLINS,
essenciais que lhes
:deos. ou cromóforos
sugerem que a tomada
rorduroso em Lepidop-
rS,1969).
! metabolismo de pro
a apenas qualitativa
tativo desse metabo
.966). Êstes autores
l~.
DINAMARCA e LEVENBOOR (1966) medindo a incorporação de
alanina e lisina durante a metamorfose de Phormia regina, esti
maram a síntese protéica "de novo" durante a sua metamorfose em
apenas 2% do total das proteínas do adulto. Embora esse cálculo
esteja baseado na hipótese simplificativa de que o "pool" de á
minoácidos é homogêneo em todo o inseto,os autores sugeriram, de
forma similar a CHEN e LEVENBOOK (1966), que uma parte das pro
teínas larvais seria conservada intacta no adulto sem degradação
e ressíntese.
Devido às dificuldades envolvidas no estudo do metabo
lismo de proteínas durante a metamorfose, como pode-se avaliar
dos trabalhos comentados acima, seria interessante o uso de téc
nicas mais refinadas, envolvendo o isolamento de proteínas esp~
cíficas de tecidos individuais e o seu acompanhamento durante a
~etamorfose. Dentro dêsse prisma são muito interessantes os es
tudos relacionados com a califorina.

-12-
Califorina é uma proteína isolada de Calliphora ery
~rocephala, sintetisada pelo corpo gorduroso larval, armazenada
principalmente na hemolinfa e que corresponde a 60, 40 e 10% das
proteínas extraíveis de insetos com 4 dias após a eclosão, 2
dias após a formação do pupário e 3 dias após a emergência do
adulto, respectivamente, indicando que deve ser uma proteína de
armazenamento usada na formação dos tecidos imaginais (MUNN,
FEIN5TEIN e GREVILLE, 1967; MUNN, PRICE e GREVILLE, 1969; MUNN e
GREVILLE, 1969).
A presença de califorina foi pesquisada em outrcos in
setos, utilizando-se técnicas imunológicas e os resultados indi
cam a ocorrência de reações de identidade completa ou parcial em
4 gêneros de outrOG Diptera Cyclorrhapha, de identidade parcial
em 4 gêneros de Brachycera, apenas uma reação de identidade par
cial em 3 gêneros de Nematocera e nenhuma reação em insetos de
ordem diferente de Diptera, embora ocorressem part~culas seme
lhantes (ao microscópio eletrônico) à califorina em várias espé
cies de insetos, especialmente Lepidoptera (MUNN e GREVILLE,
1969). ~sses resultados sugerem a ocorrência generalizada de
proteínas do tipo da califorina, que seriam armazenadas princi
palmente na hemolinfa e mais tarde seriam tornadas pelos tecidos
no início da pupação (como discutido acima) e, após hidrólise,
forneceriam os precursores para a síntese das proteínas do adul
to. Essas proteínas poderiam então ser vistas como uma grande
reserva de aminoácidos, que ocorreriam na forma de proteínas e
nao livres, para evitar aumentar a osmolaridade do animal.
Embora os estudos com a califorina indiquem claramente
a existência em vários insetos de proteínas larvais com função
de simples reServa aminoácidos, nao excluem a possibilidade da
ocorrência de proteínas larvais que permaneçam intactas no adul
to, como comentado anteriormente.
A utilização de proteínas larvais específicas, sem hi
drólise prévia, é demonstrada nos dois trabalhos que serão comen
tados a seguir.
LAUFER e NAKA5E (1965) na base de estudos imunoquími
cos e radioautográficos propuseram que a maioria das proteínas
da l
ali
sul o
pele
se e
sec:;; c.onM
liv.
fa
·0 Dua:.;
j A
na
ass·
sej
ces
de
fut
te,
Cf.
é p
va
195
de
vid
9llção
gin
qua
mui
ima

era (MUNN
::>rrência
iso~ada de Calliphora ery
juroso larval, armazenada
~ponde a 60, 40 e 10% das
dias após a eclosão, 2
2S após a emergência do
deve ser uma proteína de
tecidos imaginais (Mm~N,
~ e GREVILLE, 1969; MUNN e
lesquisada em outrcos in
:as e os resultados ind!
:de completa ou parcial em
de identidade parcial
reação de identidade par
uma reação em insetos de
rressem partLculas seme
aliforina em várias espé-
e GREVILLE,
generalizada de
eriam armazenadas princi
3.1TI tornadas pelos tecidos
na) e, após hidrõlise,
,e das proteínas do adul
ristas como uma grande
i forma de proteínas e
iridade do animal.
:ina indiquem claramente
dnas lar.rais com função
:luem a possibilidade da
meçam intactas no adul-
.is específicas, sem hi
rabalbos que serão comen
de estudos imunoquími
maioria das proteínas
i....
r-13-
da secreçao da glândula salivar de Chironomus tentans nao sao
ali sintetizadas, mas são sequestradas da hemolinfa. Êsses re
sultados foram confirmaóos e a evidência a seu favor extendida
pelo estudo das proteínas da hemolinfa e glândula por eletrofor~
se de disco em gel de po~iacrilarnida, por estudos imunológicos da
secreção salivar e por experimentos de incorooração de aminoáci~
dos marcauos realizados por DOYLE e LAUFER (1969).
Os resultados evidenciam que o processo de secreçao s~
livar'em Chironomus envolve o sequestro de proteínas da hemolin
fa de forma similar àquela que ocorre durante a vitelogênese'~
Duas diferenças importantes devem ser, entretanto, assinaladas.
A primeira delas é que o sequestro de proteínas da hemolinfa
na formação da secreção salivar tem que ser específico, afim àe
assegurar que as propriedades químicas e físicas dessa secreção
sejam as mesmas, enquanto que na vitelogênese isso não é tão ne
cessário, porque o papel das proteínas sequestradas parece ser o
de compor uma reserva de aminoácidos para o desenvclviment? do
futuro embrião (geralmente uma proteína é tomada específicamen~"
te, enquanto que as outras são tomadas com menor especificidade,
Cf. TELFER, 1965). A outra diferença notável é que a secreção
é produzida e utilizada na confecção do casulo em que vive a laE
va do Chironomus enquanto, êle ainda estâ se alimentando (S~GUY,
1951; IMMS, 1957), não há necessidade, portanto, dêle lançar mão
de suas reservas, como ocorre na vitelogênese que se realiza na
vida pupal ou no imago recém emergido.
Essa mesma diferença entre a formação da secreção em
Chironomus e a vitelogênese é também um obstáculo na considera
ção dessa formação como ID~ modêlo da produção das proteínas ima
ginais porque, diferentemente destas~ a secreção é formada en
quanto as larvas estão se alimentando e em um ambiente hormonal
muito distinto daquele em que se inicia a formação das proteínas
imaginais •

::\;
-14-
4. StDAS E CASULOS
As sédas, definidas como um material fibroso secreta
do por certas espécies de Insecta e Arachnida (SEIFTER e GALLOP,
1966), apresentam vários tipos estruturais conhecidos. Segundo
RUDALL (1962) é possível distinguir pelo menos 3 tipos princi
pais de sédas. Existem sêdas que são constituídas principalme~
te por fibroína (comum em Leoidoptera), colágeno (ocorrem em
Hymenoptera) ou quitina (observado em Dictyoptera). Os estudos
dêsse último tipo ioram realizados apenas com difração de
raios-X, mas revelam a existência de um-componente protéico as
sociado à qui tina que deve-se asemelhar à proteína cuticular
(RUDALL, 1962).
Um quarto e um quinto tipo de sêda foram reconheci
dos mais tarde em Neuroptera cujos componentes principais sao:
um composto do tipo cuticulina e uma póliglicina, respectivamente
(RUDALL e KENCHINGTON, 1971).
As sêdas melhor estudadas sao aquelas constituídas
principalmente de e-fibroína e produzidas por Lepidoptera. Aná
lises de aminoácidos realizadas em fibroínas de 74 Arthrcpoda,
principalmente Lepidoptera, mostraram que o conteúdo molar de
aminoácidos pequenos (alanina, glicina e serina) varia entre
46,2 e 94,6% do total (LUCAS, SHAlv e SMITH, 1958, 1960) o que e~
plica uma série de características típicas de difração de raios
X que apresentam, e sua conformação e com cadeias polipeptídicas
antiparalelas.
Além da conformação e, outras conformações foram reco
nhecidas nas fibroínas, usando difração de raios-X: conformação
a, observada em Hymenoptera e "cross-e" t::ffi alguns Hymenoptera
e em Arachnocampa luminosa (Diptera). O primeiro tipo de con
formação referido permitiu a demonstração de outro caso de tran
sição de conformação a para e por estiramento, além da queratina
e o segundo forneceu o primeiro caso conhecido de transformação
"cross-e" para - e-paralela (RUDALL e KENCHINGTON, 1971).
Análises de aminoácidos realizadas nas sêdas com cons
tituição diferente das e-fibroínas já referidas, permitiram uma
melhor compreensão das sêdas tipo colágeno, poliglicina,fibroína,
"cross·
zam pOl
de hid,
de 66%
teúdo (
em reli
caractc
do toti
portê'.n
da con
Os tral
ceram ,
das, a:gas de
lhos d,
foram
gueada
aderne:
da tra:
Por fi:
proteí:
hidrox.
KENCHIl
afirma.
conven
vidas:
trar t
geno e
tina, :
dados,
determ
em 196
bém sa

-15-
Além da sêdn, os casulos de insetos podem conter t~
bém sais, geralmente originaõos dos túbulos de Malpighi. Êsses
Os resultados acima referidos levaram RUDALL (1962)a
afirmar que as proteínas da sêda representam modêlos biológicos,
convenientes para o estudo dos mecanismos evolucionários envol
vidos nas transições de conformação de proteínas, além de ilus
trar tipos intermediários de proteínas estruturais entre o colá
geno e o grupo das proteínas fibrosas afins formado pela quera
tina, miosina, epidermina e fibrinogênio.
Os estudos sôbre as proteínas da sêda tiveram e têm i~
portante papel no conhecimento da confo~ação de proteínas, além
da contribuição ao conhecimento dos produtos naturais de insetos.
Os trabalhos de MEYER e MARK (1928) em fibroína da séda estabel~
ceram o conceito de cadeias polipept!dicas completamente extend!
das, além de fortalecer a visão das proteínas como cadeias lon
gas de a-aminoácidos unidos por ligações peptldicas. Os traba
lhos de MA.R8H,PATJL:NG e COREY (1955a, b) com o mesmo material
foram importantes, no estabelecimento do conceito da fôlha :ore
gueada em estrutura protéica. A séda rara de Chrysopa permitiu
a demcnstração da existência da confcrma.ção protéica "Cross 13" e
da transição "Cross 13" -->B-paralp.la (PARKER e RUDAL:S, 1957) •
Por fim a sêda de Nematus ribesii, permitiu a demonstração de
proteínas com conformação de colágeno serr. apresentar, entretanto,
hidroxiprolina na molécula (LUCAS e PUDALL, 1968; RUDALL e
KENCHINGTON, 1971).
"cross-B" e a-fibroína. As sidas do tipo colágeno se caracteri
zam por um alto conteúdo de glicina, prolina e pela ocorrência
de hidroxilisina; aquelas de tipo poliglicina ãpresentarn cêrca
de 66% dos aminoácidos corno glicina; a-fibroínas mostram um con
teúdo de glicina baixo e um alto conteúdo de aminoácidos ácidos
em relação às B-fibroínas e por .fim, as sêdas do tipo "cross-B li
caracterizam-se por ~~ alto sonteúdo de serina que chega a 43%
do total (LUCAS e RUDALL, 1968).
Os componentes não protéicos da sêda forfu~ pouco est~
dados, como pode-se ver pelo fato das hexosaminas só terem sido
determinadas na sêda de Bo~byx mori, que é a sêda mais estudada,
em 1965 (SMITH e CLARK, 1965).
conformação
Hymenoptera
tipo de con-
erial fibroso secreta
da (SEIFTER e GALLOP,
conhecidos. Segundo
nos 3 tipos princi
tituídas principalme~
colágeno (ocorrem em
optera). Os estudos
nas com difração de
omponente protéico as-
protelna cuticular
~da foram reconheci
1tes principais são:
lcina, respectivamente
I alguns
imeiro
outro caso de tran
o, além da queratina
do de transformação
NGTON, 1971).
lformações foram reco
raios-x:
s nas sêdas com con~
das, permitiram uma
poliglicina,fibroína,
aquelas constituídas
)r Lepidoptera. Aná
las de 74 Arthrcpoda,
o conteúdo molar de
serina) varia entre
1958, 1960) o que e~
ledifração de raios
:adeias polipeptídicas

-16-
sais podem ser de oxalato de cálcio(OHNISHI et alii, 1968) ou
carbonato de cálcio (CLARK, 1958; WIGGLESWORTH, 1965).
5. FINALIDADES D~STE TRAB~~HO
o estudo da origem das proteínas imaginais seria muito
facilitada se fôsse conhecida alguma estrutura do adulto formada
por um pequeno número de proteínas, porém presentes em massa su
ficiente, para poderem ser isoladas, caracterizadas e ter a sua
origem traçada com segurança ao longo da metamorfose. No desco
nhecimento de uma estrutura imaginaI com as características des
critas, seria talvez proveitosa a utilização de um modêlo.
Um modêlo adequado parece ser a produção do casulo por
R. americana. Êsse casulo possui uma massa considerável, deve
possuir um número pequeno de proteínas e é feito depois que a
larva parou de comer. Como a glândula salivar da larva nao pos
sui um reservatôrio para acúmulo de secreção, com a qual é feita
pelo menos em parte o casulo, essa tem de ser sintetizada a par
tir das reservas do animal. Portanto, esta síntese dá-se de for
ma análoga àquela que ocorre durante a metamorfose, quando o ani
mal sintetiza tôdas as proteínas imaginais a partir de reservas
(não importa de que origem), porque nesta ocasião o inseto é um
sistema fechado. A produção das proteínas da secreção, ainca de
forma análoga àquela na metamorfose, deve ser específica a fim
de manter inalteradas as propriedades físicas, químicas e bioló
gicas do casulo.
Um outro aspecto importante do modêlo em questão é a
produção do casulo ter início no fim do estágio larval, no come
ço da metamorfose, em ambiente hormonal similar àquele em que
tem início a produção das proteínas imaginais (ver ítem l,acima).
~ste aspecto é muito importante porque, considerando-se a meta
- morfose um tipo particular de polimorfismo (ver ítem 1), onde os
sistemas ge!'léticos da larva, pUJ:)a e adulto seriam ativados um a
pós o outro pelo ambiente hormonal interno do animal, se houves
se uma diferença muito grande entre a cronologia da produção do
casulo e do início da formação das proteínas imaginais,o que nao
acontece neste caso, isso poderia corresponder também a um ambi-
entl
ces!
sin<
e s\
bilj
vime
do n
de l:
"' da 1=
.. glâr.
ment
sao't
larv~(,
do d
tar
vime
inse
nho
de Ó
gãos
vel
suem
seta
a pr
um m
imag
a vi
estu
modê
Entr
cos
duçã
tenç
fora

-17-
~SHI et alii, 1968) ou
SlVORTH,1965).
~s imaginais seria muito
rutura do adulto formada
n presentes em ~assa su
icterizadas e ter a sua
metamorfose. No desco
\ as características de~
lção de um modêlo.
l produção do casulo por
ssa considerável, deve
é feito àepois que a
livar da larva não pos
ção, com a qual é feita
ser sintetizada a par
ta síntese dá-se de for
tamorfose, qUwldo o an!
~ a partir de reservas
ocasião o inseto é um
; da secreção, ainca de
ser específica a fim
.cas, químicas e hioló-
lodêlo em questão é a
;tágio larval, no come
milar àquele em que
ais (ver ítem l,acima).
nsiderando-se a met~
(ver ítem 1), onde os
seriam ativados um a
do animal, se houves-
ologia da produção do
as imaginais,o que não
~der também a um ambi-
L
ente hormonal muito distinto e, provàvelmente, implicaria em pr~
cessos radicalmente 4iferentes.
A produção do casulo em R. americana é feita de forma
sincrônica e coordenada por larvas oriundas de uma mesma postura
e sujeitas às mesmas condições ambientais, o que garante a poss!
bilidade de se estudar larvas em diferentes estágios do desenvo!
vimento, apresentando um mínimo de variações individuais. Além
do ma~s, a correlação de resultados é facilitada pela existência
de um método de datar fisiolõgicamente as larvas, após o início
da produção do casulo, baseado nos "puffs" . cromossômicos das
glândulas salivares (GUARACIABA e TOLEDO, 1967) que foi ligeira
mente reformulado nesta tese (ver Métodos). Como êsses "puffs"
são regulados por hormônios, assim como todo o desenvolvimento
larval,conforme foi comentado no ítem 2 acima, o referido méto
do de datação é extremamente adequado. E' importante acrescen
tar ainda o fato de que todos os estudos bioquímicos do desenv01
vimento de R. americana são facilitados, em relação aos demais
insetos Holometabola, com exceção de Lepidoptera, devido ao tama
nho de sua larva, que permite com relativa facilidade o estudo
de órgãos isolados por dissecção. A dificuldade de se estudar §rgãos isolados em larvas da maioria dos Holometabola é a respons~
vel pela incidência maior dêsses estudos em Lepidoptera, que po~
suem larvas maiores que as encontradas nas demais ordens de in
setos (Cf. r~ATT, 1968).
R~ vista de tôdas as características comentadas acima,
a produção do casulo por R. americana representa, provàvelmente,
~~ modêlo mais prorrQssor para o estudo da formação das proteínas
imaginais do que a formação da secreção salivar em Chironomus e
a vitelogênese referidos no ítem 3, acima.
A intenção básica dêste trabalho é contribuir para o
estudo da origem das proteínas do inseto adulto, utilizando como
modêlo a produção do casulo coletivo por larvas de R. americana.
Entretanto, devido à carência quase abas luta de dados fisiológ!
cos e bioquímicas fundamentais em R. americana durante essa pro
dução, êste trabalho orientou-se mais especificamente para a ob
tenção dêstes dados. Com êsse objetivo, dois tópicos principais
foram selecionados para estudo:

"';18-
a) a composição química da hemolinfa ao longo do de
senvolvimento larval;
b) composição química do casulo coletivo e suas alte
rações durante a sua proGução.
~nfase foi dada ao estudo da hemolinfa por várias ra-
zoes:
a) a hemolinfa corresponde a porção considerável do
animal (37% do seu pêso-Úffiido, 26% de seu peso
sêco) e é fácil de se obter;
b) a hemolinfa deve refletir as principais ~lterações
bioquímicas dos tecidos, porque os insetos pos
suem um sistema de circulação aberto, em que o único fluído corpóreo é a hemo linfa e sõ~ente atr~
vés da qual muitos órgãos podem trocar compostos
entre si;
c) a participação da hemolinfa em muitos eventos bi2
químicos no final da vida larval já foi documenta
da em vários insetos (WYATT, 1961; "YATT, 1968;
ver tarooém ítem 3), o que abre a possibilidade d~
la participar na produção do casulo de R. ameri
cana.
o estudo químico da hemolinfa foi feito com mais deta
lhe em larvas no fim da vida larval, pouco antes de iniciarem a
produção do casulo (idade de 45-48 dias, quando mantidas a 20oC)
pozque nessa ocasião o volume de hemolinfa é maior.
o conhecimento da composição do casulo orienta a pes
quisa sôbre a origem de seus componentes e eventualmente sôbre o
mecanismo e regulação de sua síntese. Além disso, devido ao fa
to do c~8ulo ser constituído oelo menos em parte por séda, puacomposição química apresenta um elevado interêsse, conforme foi
chamado a atenção no ítem 4 acima. ~ste interêsse ainda é mais
acentuado porque êsse tipo de produto natural foi muito po~co e~
tudado em Diptera, se fôr excluído o trabalho de GROSSBACH(1969), realizado com a secreção salivar de duas espécies de
Ca~ptochironomus. Nêste caso a secreção é responsável pela pro-
dução
rios I
a lar'
lo ve:
que o
mal (:

-19-
,linfa ao longo do de-
I coletivo e suas alte~.
,linfa por várias ra-
,rção considerável do
lo, 26% de seu pêso-
principais aIterações
:que os insetos pos
io aberto, em que o ú
lolinfa e sõ;nente atr~
,dem trocar compostos
em muitos eventos bi2
rval já foi docurnent~
1961; HYATT, 1968;
re a possibilidade ã~
casulo de R. ameri-
feito co~ mais deta
antes de iniciarem a
ando mantidas a 20oC)
é maior.
~sulo orienta a pcs
~ve~tualmente sôbre o
disso, devido ao fa
parte por sêda, pua
~rêsse, conforme foi
:erêsse ainda é mais
.1 foi muito pouco e~
abalho de GROSSBACHduas espécies de
"esponsável pela pro-
~,t
L
dução de um fio, semelhante à sêda, que prende partículas de v~
rios materiais, fcrmando um tubo no interior do qual permanece
a larva dêsse animal, não se constituindo, portanto, em um cas~
lo verdadeiro, em contraste com o que ocorre em R.americana,poE
que o tubo não é constituído inteiramente de secreções do ani
mal (S~GUY, 1951; IMMS, 1957).

·1
~TODOS
1. Cultura de Rhynchosciara
Rhynchosciara americana (Diptera Nematocera Sciaridae)
foi cultivada nas condições previamente descritas por LARA, TAMA
KI e PAVAN (1965). Só foram utilizadas larvas de um mesmo sexoem tôdas as determinações para simplificar a interpretação dos
resultados. O sexo utilizado foi sempre o feminino devido ao
seu tamanho maior.
2. Determinação da idade fisiológica larval
O presente trabalho procura entre outras coisas o cor
reiacionamento de eventos fisiológicos com alterações químicas
em alguns tecidos. Torna-se imperioso então a utilização de mé
todos adequados para a comparaçao de resultados obtidos em lar
vas oriundas·de grupos de postura diferentes. Embora a compara
çao por idade cronológica seja a mais fácil de ser realizada, não
é a mais adequada, mesmo em animais submetidos a idénticas condi
çôes de cultura. Pequenas diferenças na velocidade de desenvol
vimento em grupos da mesma idade cronológica são frequentemente
observadas. De maneira geral essas pequenas diferenças não sao
de importância, entretanto, em alguns casos, nos quais se inclu
em muitos daquêles descritos nesse trabalho, essas diferenças EOdem levar a resultados de interpretação difícil.
Um método de datação fisiológica para R. americana foi
desenvolvido por GUARACIABA e TOLED0 (1967) para o 49 estádio,
o qual corresponde a 70% da ,vida larval e onde ocorrem os princ!
pais e mais vislveis eventos larvais. Abaixo será apresentado o
ciclo vital de R. americana afim de facilitar as discussões pos
teriores e uma tabela das subdivisões do 49 estádio segundo GUA
RACIABA e TOLEDO (1967) com modificações baseadas em observações
feitas durante a realização desta tese. A notação aqui apresen
tada será empregada em tôda a extensão dêste trabalho.
ecll
vai:
mudo
pec1
dia:
49 I
ra (
cio
aprl
gia
~
-. vem
aquj
esq\
sere
da (
cutj
da j
últj
mo n
nestvime
de 4
uma
mude
res
da i
dem
LED(

-21-
,a Nematocera Sciaridae)
iescritas por LARA, T~
Larvas de um mesmo sexo
Ir a interpretação dos
o feminino devido ao
~
re outras coisas o cor
om alterações químicas
tão a utilização de mé
ltados obtidos em lar
teso Embora a compara
il de ser realizada, não
tidos a idênticas condi
~elocidade de desenvol
ica são frequentemente
)as diferenças não são
)s, nos quais se inclu
lho, essas diferenças EOLfícil.
l para R. americana foi
1) para o 49 estádio,
onde ocorrem os princ!
lixo será apresentado o
.tar as discussões pos
9 estádio segundo GUA
,aseadas em observações
. notação aqui apresen
te trabalho.
t
As fêmeas de R. americana poem de 1500 a 2000 ovos que
eclodem cêrca de 13 dias após a postura, a 200 C. As mudas lar
vais ocorrem aos 7, 13 e 21 dias após a eclosão, enquanto que a
muda pupal e a emergência do adulto se dão aos 64 e 70 dias res-.
pectivamente. A postura das fêmeas fecundadas inicia-se aos 75
dias. A Tabela I descreve os principais eventos fisiológicos do
49 estádio e a nomenclatura usada para a sua datação. De manei
ra geral, os eventos fisiológicos descritos (mudança de côr, in!
cio da produção de casulo, etc.) oscilam em tôrno da cronologia
apresentada nessa tabela de cêrca de 1 a 2 dias. Essa cronolo
gia corresponde a animais se desenvolvendo a 200 C.
Algumas observações quanto à terminologia empregada d~
vem ser feitas,em relação à tabela I. O têrmo "apélise·" usado
aqui corresponde ao descolamento das células epidérmicas do exo
esqueleto antigo, corno propôsto por JENKIN e HlNTON (1966). Muda
será entendida aqui corno o processo pelo qual o inseto se despe
da cutícula antiga.
Pupa farata corresponde à pupa fechada no interior da
cutícula do último estádio larval, corno foi comentado no rodapé
da introdução (p. 3). Apupa farata é formada, portanto, após a
última apólise larval e é chamada também, muitas vêzes, pelo têr·
mo menos preciso de pré-pupa.
A expressão larva madura que será usada frequentemente
nesta tese, refere-se, à larva que atingiu o seu máximo desenvol
vimento, isto é, uma larva que está no fim do 29 período (cêrca
de 45-48 dias de idade).
GUARAClABA e TOLEDO (1967) reportaram a existência de
uma muda entre o 59 período e a muda pupal. Entretanto, nenhuma
muda verdadeira parece ocorrer nesse intervalo de tempo; os autQ
res devem estar chamando de muda à última apólise larval referi
da acima .
O~tras informações sôbre a biologia de R.americana po
dem ser encontradas em DREYFUS et alii (1951) e GUARACIABA e TO
LEDO (1967) .

Subdivisão do 49 estádio de R.
Idade Cronológica
aproximada (dias)
21
32
49
54
57
58
59
61-64
-22-
TABELA I
Eventos Fisiológicosou
Aparência da larva
3~ muda larva1, lar
vas claras
Larvas começam a se
tornarem avermelha
das
Larvas iniciam a te
cer o casulo
Início da formação
do "puff" B-2 da
glândula salivar. Ca
sulo consistente
Máximo desenvolvimen
to do "puff" B-2 lar
vas quietas, em po
sição curva
Máximo desenvolvimen
to do "puff" C-3.
Larvas curvas, mas
já envolvidas por c~.
lulas individuais
do casulo
Última apólise lar
vaI. Túbulos de
Malpighi livre~ de
caC0 3
Muda pupal
americana
Correspondente ao
início do período
19
29
39
49
59
69
Pré-Pupal
(pupa farata)
3. Coleta de hemolinfa
As larvas era
cada para ficarem imóve
picadas na porção termi
fa era recolhida em tub
centrifugação a 3.200g
tos tissulares, a hemol
ções químicas.
Para a deterrn
hemolinfa era recolhida
noS tubos de centrífuga
(EC 1.10.3.1) endógena
tais, o estilete usado
vidraria usada era prev
Hcl e âgua bidestilada
4. Determinação do volu
Hemolinfa col
minação com fragmentos
rada para capilares de
Microcaps), conservand
dos. Por.êsse lado, os
cool e em seguida, cent
ças de centrífuga com t
correspondentes a 1500g
da da altura do capilar
ta ocupada pelos hemoci
permitia calcular o vol
hemocitos corresponden
permitiram calcular fàc
hemocitos na hemolinfa.

-23-
3. Coleta de hemolinfa
americana
Correspondente ao
início do período
19
29
39
49
59
69
Pré-Pupal
(pupa farata)
l
As larvas eram lavadas com água, dispostas sóbre gêlo pi
cado para ficarem imóveis, sê"cas em papel de filtro e em seguida
picadas na porção terminal do abdome com um estilete. A hemolin
fa era recolhida em tubos de centrífuga mantidos em gêlo. Após
centrifugação a 3.200g a 40 C, para sedimentar hemocitos e fragmen
tos tissulares, a hemolinfa sobrenadante era usada nas determina
ções quimi cas .
Para a determinação de compostos ninhidrina-positivos, a
hemolinfa era recolhida sôbre cristais de feniltiouréia colocados
nos tubos de centrífuga a fim de inibir a atividade da tirosinase
(EC 1.10.3.1) endógena (LEVENBOOK, 1966). Nas determinações de m~
tais, o estilete usado para picar a larva era de vidro e tóda a
vidraria usada era previamente lavada extensivamente com H2S04 ,
HCl e água bidestilada e deionizada.
4. Determinação do volume ocupado pelos hemocitos na hemolinfa
Hemolinfa coletada com todo o cuidado para evitar conta
minação com fragmentos tissulares e sem ser centrifugada foi aspi
rada para capilares de 25 lJl ("disposable micropipettes", DRUMOND
Microcaps), conservando-se cêrca de 4 mm de altura não preenchi
dos. Por êsse lado, os capilares foram selados em lâmpada a ál
cool e em seguida, centrifugados envolvidos por algodão, em cabe
ças de centrífuga com tubos basculantes. Foram usad?s rotações
correspondentes a 1500g ou 3200g com resultados idênticos. A medi
da da altura do capilar ocupado pela hemolinfa e a altura da calo
ta ocupada pelos hemocitos, realizada sob uma lupa esteroscópica,
permitia calcular o volume de hemolinfa no capilar e o volume de
hemocitos correspondente a essa hemolinfa. Êsses dois resultados
permitiram calcular fàcilmente o volume porcentual ocupado pelos
hemocitos na hemolinfa.

I,·
".,.
I',
-24-
5. Métodos de estudo da coagulação e escurecimento da hemolinfa
Cinco capilares de 15 111 ("disposablp. micropipettes" ,
DRUMMOND Microcaps) foram preenchidos com hemolinfa total e cin
co outros preenchidos com hernoliL~a cent=ifugada de larvas de 29
P, 45 dias de idade. Após 1, 2, 5, 10 e 15 horas, os capilares
eram observados quanto à cór, presença de sedimentos e fluidez.
Paralelamente eram f~itas observações similares em urro tubo de
ensaio com cêrca àe e,5 ml de hemolinfa total.
6. Determinação da osmolaridade, densidade, volume e pêso-sêco
da hemolinfa
As determinações de osmolaridade foram tôdas realiza
das utilizando-se o método crioscépico em um Fiske Osmometer,ge~
tilrnente cedido pelo Dr. Gerhard Malnic.
A densidade foi seropre determinada pesanão-se "dispo
sable micropipettes" ,(Drurnrnond Microcaps) de 15 111 com e sem he
molinfa. Os desvios observados entre as determinações foram in
feriores a 1%.
Os volumes de hemolinfa foram todos determinados pelo
método de exanguinação (RICFJUmSON, BURDETTE e EAGLESON, 1931).0
método de diluição de corante (PRICE, 1961) foi também, tentado,
produzindo-se nos casos favoráveis (larvas jovens), resultados
muito semelhantes aos de exanguinacão corno pode ser visto em re
sultados (Tabela XVII). Êsse método foi entretanto abandonado
porque em larvas maduras os resultados variavam muito dependendo
do intervalo de tempo entre a injeção e a coleta de hemolinfa.Es
sa variabilidade deve ser consequência de excreção do corante
pelos tUbulos de Malpighi, corno demonstrado em Schistocerca gre
garia por LEE (1961). Corno nenhuma excreção de corante foi ob
servada em larvas jovens isso parece sugerir que a eliminação de
corante varia com a idade. e que portanto, seria necessário de
terminar cuidadosamente os tempos máximos a serem usados entre i~
jeção de corante e coleta de hemolinfa para cada idade. Em vis
ta disso preferiu-se utilizar sõmente o método de exanguinação.
A massa de sólidos da hemolinfa foi obtida aquecendo-
se amostras de hemolinfa centrifugada a 800 C at~ atingir pêso
çonstante.
7. Determinação de p
Amostras· d,
vas, limpas corno des
de óleo de parafina.
para a determinação .
Analyser Model 113" I
Massachusetts", cedi.
determinaçoes foram :
370C~
As concent,
das determinações de
do-se que as constan1
plasma sanguíneo hum,
e 6,10 respectivamen1
moles por litro :e pC
1958).
pH =
O pH de hen
dido a 250 C, em condi
de vidro de um "Metrc
lo Dr. Gerhard Malnic
B. Determinações guín
Aminoácidos
damente no item 9.
a-amino-N t
hemolinfa foram adiei
da a 45009 por 10 min
se 0,5ml de HCI04 IN
se 10 minu~os a 11500
ml de HCI04 0,5N. Os
venientemente e usado
cido solúvel pelo rnét
feita com DL-leucina.

:urecimento da hemolinfa
:posablp. micropipettes" ,
Im hemolinfa total e cin
,=ifugada de larvas de 29
15 horas, os capilares
e sedimentos e fluidez.
milares em UIT. tubo detotal.
-25-
7. Determinação de P02' pC02 , pH e HC0 3- na hemolinfa
Amostras de hemolinfa foram colhidas picando-se as laE
vas, limpas como descrito anter~ormente, sob uma espêssa camada
de óleo de parafina. Essas amostras foram em seguida utilizadas
para a determinação anaeróbica de p02' pC02 e pH em um "pH/Gas
Analyser Model 113" da "Instrumentation Laboratory Inc., Boston,
Massachusetts", cedido pelo Dr. Herval P. Ribeiro. Tódas essas
determinações foram realizadas com o aparelho termoestatisado a
370C~
As concentrações de HC03 foram calculadas.a partir
das determinações de pH e pc02 usando a equação abaixo, assumin
do-se que as constantes ~ e pK são idênticas na hemolinfa e
plasma sanguíneo humano. Neste os valores de ~ e ~ são 0,0310
e 6,10 respectivamente, a 370 C, quando HC03- é medido em mili
moles por litro :e pC02 em milímetros de mercúrio (DAVENPORT,
1958).
ie, volume e pêso-sêco
1e foram tódas realiza
1 um Fiske Osmometer,ge~
,ada pesandO-se "dispo
de 15 ~l com e sem he
determinações foram in-pH = pK + log
[HC03-]
a (pC02 )
~dos determinados pelo
rTE e EAGLESON, 1931).0
1) foi também, tentado,
jovens), resultados
) pode ser visto em re
!ntretanto abandonado
"iavam muito dependendo
coleta de hemolinfa.Es
excreção do corante
o em Schistocerca gre
ão de corante foi ob
rir que a eliminação de
seria necessário de
a serem usados entre ig
a cada idade. Em vis
todo de exanguinação.
:oi obtida aquecendo-
)C at~ atingir pêsoII,
II
~
° pH de hemolinfa coletada como descrito acima foi me
dido a 25°C, em condições anaeróbicas, usando um microeletrodo
de vidro de um "Metroiun Herisau pH Meter", gentilmente cedido p~
lo Dr. Gerhard Malnic.
8. Determinações guímicas na hemolinfa
Aminoácidos e peptideos livres serão discutidos separ~
damente no item 9.
a-amino-N total-ácido solúvel. Amostras de 80 vl de
hemolinfa foram adicionadas a 0,72 ml de NaCl O,lM e centrifuga
da a 4500g por 10 minutos, 'para a remoção de hemocitos. Juntouse 0,5ml de HCI0 4 lN a 0,5ml do sobrenadante acima. Centrifugou-
se 10 minutos a 1l500g e lavou-se duas vêzes o sedimento com 0,5
ml de HCI0 4 0,5N. Os sobrenadantes reunidos foram diluídos con
venientemente e usados para a determinação de a-amino N total-á
cido solúvel pelo método de ROSEN (1957). A padronização foi
feita com DL-leucina.

"I-,-r-
-26-
Carboidrato e trealose. Alíquotas de 0,15 ml de hemo
linfa centrifugada eram reunidas com 1 ml de etanol 60% v/v, dei
xados a 75 0 C por 10 minutos e após atingir a temperatura ambien
te eram centrifugadas. Após recolher o sobrenadante, o sedimen
to era novamente aquecido oom etanol 60% v/v e .centrifugado como
acima. Essa operação era repeti~a mais uma vez. Os sobrenadan
tes reunidos eram diluídos convenientemente com etanol 60% v/v e
alíquotas dêsses eram usadas, depois de evaporadas em. estufa a
600 C para a determinação de carboidratos com reagente de antro
na segundo MOKRASCH (1954). A massa de carboidratos era calcul~
da como glicose.
O extrato alcoólico preparado acima foi usado também
para o isolamento químico e determinação quantitativa de trealo
se de acôrdo com a metodologia de \VYATT e KALF (1957).
Cloreto. Tôdas as determinações foram feitas com hemQ
linfa centrifugada total usando a técnica de SHALES e SHALES
(1941) •
Cinzas. Essas foram determinadas por incineração de ~
mostras de 1 rol de hemolinfa centrifugada em cápsulas de plati~
na, em mufla a 560oC, até produzir cinzas brancas.
Citrato. Foi determinado em amostras de hemolinfa ce~
trifugada e desproteinizacas de acôrdo com ETTINGER, GOLDBAUM e
SMITH Jr. (1952).
Fôsforo total, inorgânico, ácido solúvel, lipídico e
protéico. O método empregado para a ceterminação de fósforo to
tal, inorgânico, lipídico e- ácido solúvel foi aquêle descrito
por CHEN, TORIBARA e WARNER (1956). As amostras para a determi
n~ção de fósforo protéico foram preparadas por duas técnicas di
férentes: uma envolvendo precipitação das proteínas por TCA e
outra.por sulfato de amônia. A primeira preparação consistia em
tomar alíquotas de 0,2 ml de hemolinfa centrifugadae adicionar
0,8 ml de TCA 10% P/V. Após a centrifugação e desprêzo do sobr~
nadante, o sedimento era lavado com 1 ml de TCA 5% P/V e o sobre
nadante novar.~nte desprezado. O sedimento ~ra então suspenso em
1,5 ml de etanol-éter (3:l, .v/v), aquecido à ebulição, centrifu-
o sobrenadante dI
vêzes. O sedimento era
depois incinerado e usal
técnica referida acima.
A determinaçã.
tação das proteínas com
alíquotas de O, 2 ml de J
0,6 ml de sulfato de am'
230 C). Após a centrifu
mento foi lavado com 1,'
pós nova centrifugação
foi suspenso em 3,8 ml
ebulição, centrifugado
passo foi repetido uma
ção, foi usado para a d
O conteúdo do
raçao das proteínas por
ma, é o mesmo.
Glicogênio. A
da eram desproteinizada
obtido por centrifugaçâ
todos os sobrenadantes
tanol absoluto e incube
Em seguida, o material
do. O sedimento era er
80%, evaporado em estui
carboidratos pela reaç~
massa de glicogênio eré
a fração assim isolada
na-positivas diferentes
e CLARINGBOLD, 1956).
Lipídeos. ts1
mostras centrifugadas (
e SLOANE-STANLEY (1957:
vaporação do solvente (
tura ambiente, até se (

..i
-27-
IS de 0,15 rol de hemo
e etanol 60% v/v, de.!.
a temperatura ambien
renadante, o sedimen
v e .centrifugado como
vez. Os sobrenadan
com etanol 60% v/v e
poradas em. estufa a
~ reagente de antro
boidratos era calCUla
Lma foi usado também
~ntitativa de trealo
CALF (1957).
:oram feitas com hemo
gado e o sobrenadante desprezado. O último passo era repetido 3- "
vêzes. O sedimento era em seguida lavado com 1,5 ml de acetona e
depois incinerado e usado para a determinação de fósforo pela
técnica referida acima.
A determinação de fósforo protéico, envolvendo precipi
tação das proteínas com sulfato de amônio foi realizada tomando
alíquotas de 0,2 ml de hemolinfa centrifugada e lhes adicionando
0,6 ml de sulfato de amônio saturado (à temperatura ambiente,
230C). Após a centrifugação e abandono do sobrenadante, o sedi
mento foi lavado com 1,0 ml de sulfato de amônio 75% saturado. A
pós nova centrifugação e desprêzo do sobrenadante, o sedimento
foi suspenso em 3,8 ml de etanol-éter (3:1, v/v), aquecido até a
ebulição, centrifugado e o sobrenadante, descartado. O último
passo foi repetido uma vez e o sobrenadante final, após incinera
ção, foi usado para a determinação de fósforo.
ras de hemolinfa ceg
TTINGER, GOLDBAUM e
por incineração de ~
I cápsulas de plati..,.
"ancas.
olúvel, lipídico e
nação de fósforo to
i aquéle descrito
~ras para a determi
)r duas técnicas di
)teínas por TCA e
)aração consistia em
.fugadae adicionar
e desprezo do sobr~
~CA 5% P/V e o sobr~
·a então suspenso em
ebulição, centrifu-
SP.ALES e SHALES O conteúdo do fósforo protéico determinado após a prep~
ração das proteínas por qualquer das duas técnicas descritas aci
ma, ê o mesmo.
Glicogênio. Amostras de 0,4 ml de hemolinfa centrifuga
da eram desproteinizadas com 0,1 ml de HCI04 2,5N. O sedimento
obtido por centrifugação era lavado com 0,5 ml de HCl04 0,5N e
todos os sobrenadantes reunidos. A éstes juntava-se 2,5 ml de e
tanol absoluto e incubava-se a preparação a 370 C por 15 minutos.
Em seguida, o material era centrifugado e o sobrenadante desprez~
do. O sedimento era então lavado duas vézes com 0,5 ml de etanol
80%, evaporado em estufa a 600C e usado para as determinações de
carboidratos pela reação de antrona segundo MOKRASCH (1954). A
massa de glicogenio era calculada como giicose. E l possível que
a fração assim isolada e determinada contenha substâncias antro
na-positivas diferentes do verdadeiro glicogenio (BALMAIN,BIGGERS
e CLARINGBOLD, 1956).
Lipídeos. Êstes foram isolados quantitativamente de a
mostras centrifugadas de hemolinfa pela técnica de FOLCH, LEE
e SLOANE-STANLEY (1957) e determinados gravimetricamente, após e
vaporação do solvente da extração por pressão reduzida, à temper~
tura ambiente, até se obter peso-constante.

I''i!i
11111
-28-
Metais. A matéria orgânica de amostras de hemolinfa cen
trifugada com as precauções já descritas, era destruída e'o mate
rial remanescente, após diluições convenientes em água bidestila
da e deionizada era usado para as determinações de metais por es
pectroscopia de absorção atõmica. Para essa finalidade foi empr~
gado um "Atomic Spectrometer" da "Perkin-Elmer" operàdo pela srta
Yolanda Tav~res.
As determinações de sódio e potássio, entretanto, .foram
realizadas sem destruição prévia da matéria orgânica, por espec
troscopia de chamas, em um "Flame Photometer Evans" da "Electro
selenium Ltd.", gentilmente cedido pelo Dr. Gerhard Malnic.
Três técnicas distintas foram utilizadas para a destr~
ição da matéria orgânica. A primeira delas consistia em incine
rar a hemolinfa em cápsula de platina ou porcelana (os resultados
foram idênticos) em uma mufla a 5600 C até a formação de cinzas
brancas. Outra técnica consistia em destruir a matéria orgânica
da mesma forma que a descrita por CHEN, TORIBARA e WARNER (1956)
antes de determinar fõsforo total. Por fim outra técnica utiliza
da consistia em aplicar a hemolinfa em quadrados de papel \Vhatman
n91 de cêrca de 4cm de lado e colocar o sistema em um frasco de"combustão de Schoniger para a sua destruição por radiação infra-
vermelha.
Como os resultados obtidos com as três técnicas de des
truição foram idênticos passou-se a utilizar a técnica que fôsse
mais conveniente na ocasião.
Proteínas. Estas foram determinadas utilizando-se o
procedimento de LOvlRY et ali i (1951). Afim de se eliminar inter
ferências de compostos não protêicos, a absorbância de cada amos
tra foi corrigida pela absorbância do sobrenadante de uma duplic~
ta desproteinizada com HCl0 4, em concentração final de 0,5Ni am
bas absorbâncias medidas após a reação colorimétrica. Os títulos
de proteína foram calculados tendo albumina cristalina de sôro
bovino como padrão.
9. cromatografia de .
vos ácido-solúvei:
A preparaç;
descri to antes, para
tos ninhidrina-posit:
to não hidrolizadas,
A, cromatogJ
Analyser, Model 120C'
Angelis, utilizando c
necido junto com o eç
pequena modificação c
A identific
ta baseando-se em se.
sorbância 440/570 m~
tamento na hidrõlise.
permitia a sua identi
lizados com uma amost
ros adquiridos de "Ma
gramas pub2icados (SP
TALLEY, 1962; HAMILTO
to dos compostos na h
cima) e cromatogramas
les de amostras não h
~os não foi possIvel
sos, a identificação
Os resultad
çao da área sob os pi
tes de cor para cada
gramas padrões. No c
mistura padrão, usou
fatôres fornecidos pe
cada modêlo de equipa
caso de.peptídeos uso
te média é obtida tom
cor para os aminoácid
na, prolina e cisteIn

-29-
~ostras de hemolinfa ceg
era destruída e'o mat~
entes em água bidestila
nações de metais por es
'ssa finalidade foi empr~
Elrner" operádo pela srta
ássio, entretanto, foram
ia orgânica, por espec
ter Evans" da "Electro
r. Gerhard Malnic.
tilizadas para a destr~
~s consistia em incine
Jorcelana (os resultados
a formação de cinzas
ruir a matéria orgânica
)RIBARA e WARNER (1956)
Lm outra técnica utiliz~
tdrados de papel ~Vhatman
;istema em um frasco de
;ão por radiação infra-
lS três técnicas de des
:ar a técnica que fôsse
9. Cromatografia de troca iônica dos compostos ninhidrina-positi
vos ácido-solúveis da hernolinfa
A preparação das amostras de hemolinfa, coletadas corno
descrito antes, para a identificação e quantificação dos compos
tos ninhidrina-positivos ácido solúveis, tanto hidrolizadas quan
to não hidrolizadas, foi feita segundo LEVENBOOK e OINAMARCA(1966)
A, cromatografia fo~ realizada em um "Beckman Arninoacid
Analyser, Model 120C", gentilmente cedido pela Ora. Rebeca C. de
Angelis, utilizando o procedi~ento para fluídos fisiológicos for
necido junto com o equipamento da Beckman e que consiste em uma
pequena modificação do-método de SPACKMAN, STEIN e MOORE(195B).
A identificação de cada pico nos cromatogramas foi fei
ta baseando-se em seu tempo relativo de eluição, sua razão de ab
sorbância 440/570 m~ (ZACHARIUS e TALLEY, 1962) e em seu compor
tamento na hidrólise. O tempo relativo de eluição dos compostos
permitia a sua identificação por comparação com cromatogramas re~
lizados com uma amostra de 17 aminoácidos, cromatogràficamente p~
ros adquiridos. de "Mann' Research Laboratories~', ou/e com cromato
gramas pub2icados (SPACKMAN, STEIN e MOORE, 1958; ZACHARIUS e
TALLEY, 1962; HAMILTON, 1963). Para a observação do comportamen
to dos compostos na hidrólise, amostras eram hidrolizadas (ver a
cima) e cromatogramas obtidos com essas eram comparados com aquê-.
les de amostras não hidrolizadas. Na identificação de vários pi~os não foi possível utilizar todos êsses critérios. Nesses ca
sos, a identificação foi considerada apenas tentativa.
.adas utilizando-se o
m de se eliminar inter
sorbância de cada amos
enadante de uma duplic~
ção final de 0,5N; am
orirnétrica. Os títulos
a cristalina de sôro
i!!.,\l.
Os resultados quantitativos foram obtidos por integra
çao da área sob os picos dos cromatogramas e utilizando constan
tes de côr para cada aminoácido, conforme determinado nos cromat2
gramas padrões. No caso de compostos que não estavam presentes na
mistura padrão, usou-se a constante de côr média, corrigida por
fatôres fornecidos pela "Beckrnan Instruments Inc." adequados para
cada modêlo de equipamento e correspondentes a cada composto. No
caso depeptídeos usou-se a constante de côr média. Essa consta~
te média é obtida tornando-se a média de tôdas as constantes de
côr para os aminoácidos comuns exceto: histidina, lisina, argini
na, prolina e cisteína.

i 11
I1:·::lii I,
IIlIn
-30-
As concentrações de serina, treonina, tirosina e proli
na nas amostras hidrolizadas foram elevadas de 15, 5, 6 e 5% re~
pectivamente para compensar as perdas durante a hidrólise ácida.
As.correções foram calculadas para o tempo de hidrólise de 18
horas utilizado aqui, a partir dos resultados de HIRS, STEIN e
MOORE (1954).
10. Preparação dos pigmentos da hemolinfa
Um volume de hemolinfa centrifugado era reunido a 19
volumes de ~lorofór~io-metanol'2:1 (v/v, ambos redestilados) be~
agitados e filtrados em papel de filtro quantitativo, pré-lavado
com clorofórmio;metanol. Ao filtrado adicionou-se 0,2 volumes
de água deionizada e após agitação o material foi centrifu~ado a
11500g por 10 minutos. A fase superior aquosa continha um pig
mento de côr limão, enquanto que a fase lipídica apresentava uma
côr alaranjada. A fase aquosa após ser evaporada ã pressão red~
zida e ao abrigo da luz, teve o seu resíduo dissolvido em meta
nol 90% (v/v) e lavado por agitação com igual volume de éter de
petróleo (p.e. 30-S00 C) duas vezes, DeP9is disso o pigmento cor
de limão teve o seu solvente novamente evaporado e foi conserva
do no vácuo e ao abrigo da luz até a sua utilização. Essa ~rep~
ração é chamada daqui para diante de pigmento limão.
A fase lipídica foi evaporada à pressão reduzida e ao
abrigo da luz e em seguida, seu resíduo foi dissolvido em éter
de petróleo (p.e. 30-S00 C). Essa di~solução foi imediata e com
pleta o que indicava, juntamente com a côr alaranjada, que se
tratava de carotenóides. O material dissolvido em éter de petr~
leo, após duas lavagens por agitação com igual volume de água
deionizada, foi recolhido, evaporado e conservado no vacuo, ao
abrigo da luz até a sua utilização. Essa preparação é denomina
da daqui para diante de carotenóides da hemolinfa.
A saponificação dos carotenóides da hemolinfa foi rea
lizada essencialmente como recomendado por KARRER e JUCKER(1950).
A amostra foi dissolvida em 2 mlde ROH 12% em metanol,transferi
do para tubo de ensaio, ao qual,juntou-se alguns cristais de pi
rogalol e deixou-se 2S minutos a 7S oC,após fechar o tubo com al
dogão. Esfriou-se o tubo em gêlo e adicionou-se 6ml de água deio
nizada e 4 ml de éter d
gitar bem, a fase do ét
agitações com água e us
abrigo da luz,. após eva
11.'I'de'ntific'ação dos c
A identificaç
tada utilizando~se: par
sorção em hidroxiapatit
Partição. A s
caroten5ides epifásicos
treéter de petróleo (p
e JUCRER,19S0). Com ess
ter de petróleo (p.e. 3
metanol 90% v/v, agitad
Cromatografia
foi feita como se segue
metro foi preenchida in
da de 2 em de altura.
acetona e §ter de petró
adicionou-se uma suspen
leo, suficiente para pr
de altura. Seguiu-se a
ter de petróleo, para p
As amostras d
dfsselvidas em éter de
coluna. A separação do
droxiapatita era feita
quanto que a sua eluiçã
solventes aplicados na
-SOoC), acetona 7% em é
Teste qualita
zado como descrito por
de potássio usado no te$CHNEIDER (19631.

-31-
nina, tirosina e pro11
s de 15, 5, 6 e 5% re~
nte a hidrólise ácida.
de hidrólise de 18
dos de HIRS, STEIN e
- - ( 0 5 0) . dnizada e 4 ml de eter de petroleo p.e. 30 - O C. Dep9~s e a-
gitar bem, a fase do éter de petróleo foi recuperada, lavada por
agitações com água e usada em seguida ou conservada no vácuo, ao
abrigo da luz, após evaporação do solvente.
se
petrQ
águaAs amostras de carotenóides saponificadas ou nao eram
digselvida~ em éter de petróleo (p.e. 300 -50oC) e aplicadas na
coluna. A separação dos carotenóides ao longo da coluna de hi
droxiapatitaera feita com éter de petróleo (p.e. 30o-50oC), en
quanto que a sua eluição para fora da coluna era realizada com
solventes aplicados na seguinte ordem: éter de petróleo (p.e.30o0__
-50 C), acetona 7% em eter de petroleo e acetona.
Partição. A separação dos carotenóides da hemolinfa em
carotenóides epifásicos e hipofásicos foi feita por partição en
treéter de petróleo (p.e.300 -50oC) e metanol 90% v/v (Cf.KARRER
e JUCKER,1950). Corrl essa finalidade, a amostra dissolvida em éter de petróleo (p.e. 30o -500 C) foi juntada a igual volume de
metanol 90% v/v, agitada e deixada a repousar.
ll.'I'dentificação dos carotenó:ides
A identificação dos 9arotenóides da hemolinfa foi ten
tada utilizando~se: partição em duas fases, cromatografia de ad
sorção em hidroxiapatita e um teste qualitativo para astaxantina.
Teste qualitativo para astaxantina. O teste foi real!
zado como descrito por GOODWIN e SRISUKH (1949). O t-butóxido
de potássio usado no teste foi preparàdo segUndo JOHNSON e$CHNEIDER (1963).
. Cromatografia. A montagem da coluna de hidroxiapatita
foi feita como se segue. Uma coluna de vidro de 1,2 em de diâ
metro foi preenchida inferiormente com algodão formando uma cam~
da de 2 ~ de altura. ~ste algodão foi previamente lavado com
acetona e ~ter de petróleo. Após fechar a extremidade inferior,
adicionou-se uma suspensão de hidroxiapatita em éter de petró
leo, suficiente para produzir uma coluna de sedimento de 8,0 cm
de altura. Seguiu-se a adição de urnà suspensão de Na2S04 em é·ter de petróleo, para produzir um sedimento de 1,0 em de altura.
com-
e ao
éter
ao
reparação é denominaolinfa.
lressão reduzida
dissolvido em
:0 foi imediata e
alaranjada, que
vida em éter de
ual volume de
ervado no vácuo,
da hemolinfa foi rea
KARRER e JUCKER(1950).
em metanol,transfer!
19uns cristais de pi
fechar o tubo com al
u-se 6ml de água deio
ado era reunido a 19
nbos redestilados) bem
antitativo, pré-lavado
Lonou-se 0,2 volumes
lal foi centrifuqado a
losa continha um pig
)idica apresentava uma
,parada à pressão red~
) dissolvido e~ meta
lal volume de éter de
l disso o pigmento côr
>orado e foi conserva
:ilização. Essa prep~
ltO limão.

PI'li I
I,ll' i,
~ i'
Iil
, ;:,iillll,,~:i[i:H,:::;
-32-
12. Preparação do casulo I, 11 e 111 e determinação da massa pro
duzida por indivíduo
Diferentes grupos de larvas maduras limpas em número co
nhecido foram colocadas em recipientes forrados com um papel de
filtro mantido sempre úmido. Os grupos após iniciar a produção
de casulo tinham as suas larvas removidas em tempos adequados, o
pêso-sêco do casulo produzido até a ocasião era determinado e ~~~
partir do número de larvas presentes era calculada a produção
por larva. Para facilitar o estudo, o período de produção do ca
sulo foi dividido em 3 fases. A primeira fase corresponde à pro
dução do casulo do início até o começo do 49 período do 49 está
dio; a segunda, do começo do 49 período até 59 período e a tercei
ra fase do 59 período até a muda pupal. O casulo produzido do iní
cio até o fim da primeira fase foi denominado de casulo I; do iní
cio até o fim da segunda fase, de casulo 11; e do início até o
fim da terceira fase, de casulo 111. Evidentemente, pela nomen
clatura usada o casulo 111 corresponde ao casulo terminado, en
quanto que os demais correspondem ao casulo em fases anteriores
de produção;
13. Determinação da solubilidade do casulo
Amostras sêcas e trituradas de casulo pesando cêrca de
100 mg foram colocadas em tubos de centrífuga previamente levados
a pêso-constante. Essas amostras eram lavadas 3 vêzes com 5 ml
de água deionizada cada vez. A recuperação do material insolúvel
foi sempre realizada por centrifugação a 11500g. O material re
manescente após as lavagens foi pesado depois de ter sidosêco a- - o -peso-constante em estufa a 100 e. A diferença de peso observada
em relação às amostras antes de serem lavadas, foi considerada co
mo a massa de material hidrossolúvel.
Amostras de cêrca de 100 mg de casulo, lavadas com água
Como descrito acima, foram colocadas em tubos de centrífuga pre
viamente levados a pêso-constante. Adicionou-se em cada tubo 5ml
de ácido acético 10% (v/v), agitando com bastão até a efervescên
cia observada desaparecer. Após centrifugação a 11500 g, o sedi
mento foi lavado duas vêzes com 1 ml de ácido acético 10% e uma
vez com 1 ml de água. O sedimento remanescente das lavagens foi
sêco a pêso consté
inicial foi assumi
do acético solúvel
nado fração insolú
14. Determinações'
Aminoáci.
ítem 16.
lI-amino-l
50 mg da fração in~
hidrolizadas em lOC
diluídas do hidroli
tico glacial e o cc
nica de ROSEN (1957
"carboidra
Amostras sêcas de c
ram hidrolizadas em
2 ml de Hel 3N, a 1
nada a hidrólise, a
e filtradas em lã &ra a determinação d,
tros e para a purif:
As hexosaI
uma modificação do r
Dowex-50, como descl
drão galactosamina 1
Os açúcarE
hidrolizados pela rE
DUBOIS et alii (195l
na presença e ausênc
tra foi a diferença
tomado para eliminal
cífica de côr resu11
utilizada como padri

-33-
jeterrninacão da massa pro-
iduras limpas em número c~
:orrados com um papel de
após iniciar a produção
IS em tempos adequados, o
:ião era determinado e ~~a
-a calculada a produção
'errado de produção do ca
'a fase corresponde à pro
a 49 período do 49 está
até 59 período e a terce~
O casulo produzido do ini
inado de casulo I; do ini
lI; e do início até o
identemente, pela nomen
ia casulo terminado, en
;ulo em fases anteriores
,o
casulo pesando cêrca de
fuga previamente levados
vadas 3 vêzes com 5 ml
ão do material insolúvel
11500g. O material re
?ois de ter sido sêco a
rença de pêso observada
idas, foi considerada co
:asulo, lavadas com água
mos de centrífuga prê
'nou-se em cada tubo 5ml
astão até a efervescên
ação a 11500 g, o sedi
ido acético 10% e uma
cente das lavagens foi'
sêco a pêso constante e pesado. A diferença do último pêso e o
inicial foi assumido como massa de material hidroinsolúvel e áci
do acético solúvel. O sedimento remanescente no final foi denomi
nado fração insolúvel do casulo.
14. Determinações químicas do casulo
Aminoácidos da fração insolúvel serão discutidos no
ítem 16.
a-amino-N total insolúvel. Amostras sêcas de cêrca de
50 mg da fração insolúvel, preparadas como descrito acima foram
hidrolizadas em 100ml de NaOH 5N a lOOoe por 8 horas. Alíquotas
diluídas do hidrolizado eram acertadas para pH 6,0 com.ácido acé
tico glacial e o conteúdo de a-amino N foi determinado pela téc
nica de ROSEN (1957), usando como padrão DL-leucina.
Carboidratos insolúveis e purificação de melanoidinas.
Amostras sêcas de cêrca4e 10 mg da fra~ão insolúvel do casulo f2
ram hidrolizadas em balões volumétricos de lO ml, com cêrca de
2 ml de Hel 3N, a loooe, durante vários períodos de temoo. Termi
nada a hidrólise, as amostras eram acertadas para lO ml com água
e filtradas em lã de vidro. ~sses filtrados foram utilizados pa
ra a determinação de hexosaminas, ácidos urônicQs, açúcares neu
tros e para a purificação e isoiamento de melanoidinas.
As hexosaminas foram determinadas nos filtrados usando
uma modificação do méto~o de ELSON-MORGN~, após purificação em
Dowex-SO, como descrito por BOAS (1953). Foi utilizado como pa
drão gaIactosamina HeI purificada em Dowex-SO como as amostras.
Os açúcares neutros foram determinad~s nos filtrados dos
hidrolizados pela reação de fenol-H 2S04 , segundo a técnica de
DUBOIS et alii (19S6). Tôdas as determinações foram realizadas
na presença e ausencia de fenol. O valor aceito para cada amos
tra foi a diferença entre as duas determinações. ,~sse cuidado foi
tomado para eliminar as interferências devido à ~odução inespe
cífica de côr resultante do aquecimento com H2S04 . Glicose foi
utilizada como padrão nessas determinações. Deve-se notar que o

1
J·.ill"
:111.
Iilüll:
IIIIJO
-34-
método de fenol-H 2S04 é empregável para açúcares neutros e ácidos
uron1COS. Entretanto, corno no presente material o conteúdo de ácidos urônicos é insignificante em relação ao conteúdo de carboi
dratos total medido com fenol-H 2S04 (ver Resultados), a presente
determinação pode ser considerada uma determinação de açúcares
neutros.
Os ácidos urônicos foram determinados nos filtrados dos
hidrolizados após reação com carbazol, segundo GALAMBOS (1967)
utilizando ácido glicurônico corno padrão.
o isolamento de melanoidinas dos filtrados dos hidroli
zados foi conseguido corno se segue. Amostras dos filtrados fo
ram aplicados em uma coluna de Dowex-50 preparada. como descri
to por BOAS (1953) e eluídas com 10 ml de água, seguido de 5 ml
de HCl 2N. Nessas condições os carboidratos ácidos e neutros, a
minoácidos e hexosaminas são liberados da coluna, enquanto que o
pigmento ainda fica retido. ~sse foi então eluído com NaOH 2N.
Após neutralização o eluato foi submetido à reaçao de ELSON-MOR
GAN na modificação proposta por BOAS (1953) e seu espectro de ab
sorção realizado. A absorbância em cada comprimento de onda foi
corrigida pela absorbância do pigmento purificado e na mesma di
luição queaquêle medido após a reaçao de ELSON-MORGAN.
Carboidratos solúveis. Foram determinados segundo DUBOIS
et alii (1956) usando o mesmo cuidado para eliminar interferênci
as descrito acima.
Carbonato. Foi identificado no casulo como descrito em
seguida. Cêrca de 50 mg de casulo foi colocado em um sistema fe
chado, 2 ml de H2S04
O,lN foram adicionados e o frasco aquecido
em banho-maria. O gás liberado foi recolhido em solução saturada
de Ca(OH)2' Afim de evitar qualquer dúvida quanto à identidade do
precipitado branco formado na solução de Ca(OH)2' cêrca de 50 mg
de casulo foi previamente tratada com H20 2 100 volumes e em se
guida o procedimento acima foi repetido.
Cinzas. Foram determinadas após o aquecimento em cadi
nho de porcelana a aoooc, em mufla, até produzir cinzas brancas.
Foram usadas amostras de casulo sêco (determinação de cinzas to-
de casulo apó 9(determinação dI
Lipídeos. Fo:
li:
Metais. Foral
linfa, com exceção de K'
tais, isto é, por espec'
truição da matéria orgã:
Nitrogênio-to
terminado segundo o mét
ALBANESE e ORTO (1963).
Proteínas sol
por LOWRY et ali i (19~1
vina corno padrão.
15. Cromatografia em pa
!Unostras de c
ácido (ver Item 3) for
horás a 100oC. Os hidr
plicados diretamente no
O sistema de
te em cuba pré-saturada
senvolver o cromatogram
solventes: um dêles con
v/v/v) com um tempo cor
piridina-água (3:1:1, v
papel para cromatografi
casO foi usado papel Wh
coso
A revelação cligeira modificação de
por pAR'!'RIDGE e WESTALI

r-35-
~úcares neutros e ácidos
iterial o conteúdo de á
) ao conteúdo de carboi
~esultados), a presente
~rminação de açúcares
Lnados nos filtrados dos
~gundo GALAMBOS (1967)
; filtrados dos hidroli
.tras dos filtrados fo
:eparada. como descri
água, seguido de 5 ml
~s ácidos e neutros, a
coluna, enquanto que o
.0 eluído com NaOH 2N.
à reação de ELSON-MOR-
e seu esPectro de ab
omprimento de onda foi
ificado e na mesma di
ELSON-MORGAN.
=rminados segundo DUBOIS
eliminar interferênci
lsulo como descrito em
lado em um sistema fe
; e o frasco aquecido
do em solução saturada
quanto à identidade do
. (OH) 2' cêrca de 50 mg
100 volumes e em se-
aquecimento em cadi
juzir cinzas brancas.
ninação de cinzas to-
tais) e de casulo apó~ lavagens em água e ácido como descrito no
ítem 13 (determinação de cinzas insolúveis).
Lipídeos. Foram determinados como descrito para hemo
linfa.
Metais. Foram determinados como descri~o para hemo
linfa, com exceção de K+ que foi determinado como os outros me
tais, isto é, por espectroscopia de absorção atômica, após des
truição da matéria orgânica.
Nitrogênio-total da fração insolúvel do casulo foi de
terminado segundo o método de Kjeldahl, como recomendado por
ALBANESE e ORTO (1963).
Proteínas solúveis. Foram determinadas como descrito
por LOWRY et alii (19~1), usando albumina cristalina de sôro bo
vina como padrão.
15. Cromatografia em papel de carboidratos do casulo
~ostras de cêrca de 7 mg de casulo lavado em água e
ácido (ver ítem 3) foram hidrolizados em 1 ml de HCl 3N por 2
horás a 1000C. Os hidrolizados .filtrados em lã de vidro foram ~
plicados diretamente no papel de cromatografia.
O sistema de cromatografia usado foi sempre o descenden
te em cuba pré-saturada com o ~olvente a ser empregado para de
senvolver o cromatograma. Foram usados dois tipos de si~temas
solventes: um.dêles consistia embutanol-etanol-ág~a (10:1:2,
v/v/v) com um tempo corrido de 92-94 horas e o outro em butanol
piridina-âgua (3:1:1, v/v/v) com uma corrida de 27-29 horas. O
papel para cromatografia usado foi Whatman n9 1 e em apenas um
caso.foi usado papel Whatman 3MM que produziu resultados idênti
cos.
A revelação dos cromatogramas foi realizada usando uma
ligeira modificação do sistema de nitrato de prata introduzido
por PARTRIDGE e WESTALL (1948). Foi·usado também um sistema de

rl!~II::
111 IIli li
-36:"
revelação específico para hexosaminas, baseando-se em uma modifi
cação do método de ELSON-MORGAN desenvolvido por PARTRIDGE e
WESTALL (1948).
A identificação dos carboidratos foi realizada corren
do amostras de carboidratos conhecidos no mesmo cromatograma e
comparando-se as migrações. A comparação da migração de um com
posto em cromatogramas diferentes foi feita pelo seu RG, defini
do pela razão entre sua migração e a da glicose no mesmo cromat~
grama.
16. Cromatografia na troca iônica de aminoácidos da fracão inso
lúvel do casulo
Amostras de 6-9 mg da fração insolúvel do casulo secas
e finamente pulverizadas, foram pesadas em ampolas. Adicionou
se 2 ml de HCI 6N em cada ampola. Estas foram seladas à vácuo
(110 mrn Hg) e 3 delas aquecidas a 10SoC por 22 horas, enquanto
que 3 outras por 70 horas. Então 0;( conteúdo das ampo·las e as
lavagens dessas foram filtrados em lã de vidro. Os filtrados fo
ram recolhidos, liofilizados e depois dissolvidos em 4,0 ml de
tampão citrato de sódio pH 2,20 (0,2N em sódio). Alíquotas de
0,25 ml dêsses foram aplicados em um "Beckman Aminoacid Analyser
Model 120C", gentilmente cedido pela Dra. Rebeca C. de Angelis, e
as cromatografias foram realizadas usando o procedimento de 2
horas para hidrolizados protéicos, descritos em boletins técni
cos fornecidos juntamente com o equipamento.
A identificação e quantificação de cada pico presente
nos cromatogramas foi feita como descrito no ítem 9 acima. Os
títulos dos aminoácidos instáveis em ácido foram extrapolados p~
ra tempo zero de hidrólise, como descrito por HIRS, STEIN e
MOORE (1954), a partir dos resultados obtidos nos dois tempos de
hidrólise empregados.
17. Determinação de
As larvas E
Malpighi removidos,
mitir o escorrimento
eram colocados em pef
tante e sêcos até pÉ
lninação de seu pêSC-f
man n9 1 e usados paI
no ítem 8, com exceçc
pectroscopia de absel
gânica.
18. Determinação de 9
As larvas e
durosos centrais eram
dos de maneira a perrn
em tubos mantidos no
as determinações de 9
sêcos até pêso-consta
seu pêso-sêco.
O glicogêni
tir de hidrolizado aI
descrita por BELL e Y
gênio foi quantificad
(1954), usando glicos
sim isolada e deterrni
diferentes do verdade
BOLD, 1956).

-37-
e
baseando-se em uma modif!
,lvido por PARTRIDGE e
tos foi realizada corren
no mesmo cromatograma e
io da migração de um com
~ita pelo seu RG, defini-
glicose no mesmo cromat2
noácidos da fracão inso-
nsolúvel do casulo secas
em ampolas. Adicionou-
foram seladas à vácuo
)or 22 horas, enquanto
lteúdo das ampolas e as
vidro. Os filtrados f2
solvidos em 4,0 ml de
sódio). Alíquotas de
kman Aminoacid Analyser
Rebeca C. de Angelis, e
J o procedimento de 2
:os em boletins técni:0.
• de cada pico presente
no ítem 9 acima. Os
foram extrapolados p~
por HIRS, STEIN
jos nos dois tempos de
17. Determinação de peso-sêco e metais rios tubos de Malpighi
As larvas eram dissecadas em NaCl 0,1M, os túbulos de
Malpighi removidos, lavados em NaCl e deixados de maneira a per
mitir o escorrimento da solução de lavagem. Depois disso eles
eram colocados em pesa-filtros previamente levados a peso-cons-- - - otante e secos ate peso-constante, em estufa a 100 C, para deteE
lrinação de seu pêso-seco ou aplicado em quadrados de papel What
man n9 1 e usados para a determinação de metais, como descrito
no item 8, com excp.ção do K, que também foi determinado por es
pectroscopia de abserção atômica, após destruiçao da matéria or
gâr.ica.
18. Determinação de glicogenio e peso-sêco do corpo gorduroso
As larvas eram dissecadas em NaCl O,lM, os corpos gor
durosos centrais eram removidos, lavados- em NaCl 0,1M e deixa
dos de maneira a permitir o escorrimento da solução de lavagem,
em tubos mantidos no gêlo. Depois disso eles eram usados para
as determinações de glicogênio ou colocados em Pesa-filtros e
secos até pêso-constante em estufa a lOOoC, para determinação de
seu peso-sêco.
o glicogênio do corpo gorduroso foi preparado a par
tir de hidrolizado alcalino do tecido de forma similar àquela
descrita por BELL e YOUNG (1934). Após a sua preparação o glic~
gênio foi quantificado por reação com antrona, segundo MOKRASCH
(1954), usando glicose como padrão. E' possível que a fração ~
sim isolada e determinada contenha subsêâncias antrona-positivas
diferentes do verdadeiro glicogênio (BALMAIN, BIGGERS e CLARING
BOLD, 1956).

,li
: :111
li!!
ili;;t:1IIIIijHI
REAGENTES
Os aminoácidos usados eram produtos cromatogràficamen
te puros da Mann Research Labor~tories.
A hidroxiapatita empregada era produto especial para
cromatografia em coluna (Bio Gel HTP) fornecida pelo Bio Rad La
boratories, Richmond, California.
tter de petróleo foi obtido por destilação fracionada
(fração de p.e. 30o-S0o C) do produto técnico.
Etanol também foi obtido do produto técnico por desti
lação simples.
Clorofórmio, metanol e butanol eram produtos analíti
cos da British Drug Houses e eram redestilados antes de usar.
As resinas de troca iônica Dowex empregados foram obti
das da Sigma Chemical Company.
Todos os demais reagentes empregados foram produtos a
nalíticos obtidos principalmente da British Drug Houses, E.Merck
A. G. e J. T. Baker.
Qulmica da bemolinfa
1. Resultados Gerais
A hemolinfa d
quido escurc, avermelha
, de substâncias sólidas.
com o ar é de 7,27, aoutros Diptera Neli
o abaixamentc
-o, 4020C, que
~sse abaixe
setos, com exceção de
1953J. TOKUMARU, FERI
laridade correspondentE
R. americana, utilizan<
tamanho de" gotas de 1
das por camadas de ar c
çóes conhecidas por 24
de valor duvidoso porq"
drollticas como amil
etc. (FLORKIN e JEUNIA
tenha havido intensa h
tão longo e à tempe
osmolaridade de cêrca
com sucesso para a inc
ricana (LARA e TOLED
1969; TERRA et a1ii, rn
A densidade
1,043 que é um valor e
A hemolinfa
em circulação que pode

RESULTADOS
utos cromatogràficamen_
)roduto especial para
lecida pelo Bio Rad La-
destilação fracionada.co.
luto técnico por desti-
ram produtos analíti
ados antes de usar.
empregados foram obti
idos foram produtos a
1 Drug Houses, E.Merck
A. Química da hemolinfa de larvas de R. americana
1. Resultados Gerais
A hemolinfa de larvas maduras de R. americana é um lí
quido escure, avermelhado, formado por 89% em massa de água e 11%
de substâncias sólidas. Seu pH medido sem entrar em contacto
com o ar é de 7,27, a 250 C ou 370 C e é similar ao determinadopara outros Diptera Nematocera (Cf. BUCK, 1953).
O abaixamento do ponto de congelação observados em m~
dia é de -0,4020 C, que corresponde a uma osmolaridade de 216 mi
liosmoles. ~sse abaixamento é pequeno em relação aos demais in
setos, com exceção de larvas aquáticas de Diptera (Cf. BUCK,
1953~. TOKUMARU, FERRI eWORSMANN (1968) encontraram uma osmo
laridade correspondente a 320 miliosmoles para a hemolinfa de
R. americana, utilizando método baseado na alteração ou não do
tamanho de· gotas de hemolinfa fechadas em capilares e separa
das por camadas de ar de gotas de soluções de KCl de concentra
Qões conhecidas por 24 horas. ~sses resultados são, entretanto ,
de valor duvidoso porque a hemolinfa possui numerosas enzimas h!
drolíticas como amilases, esterases, proteases, fosfatases,
etc. (FLORKIN e JEUNIAUX, 1964) e é difícil se assegurar que não
tenha havido intensa hidrólise de compostos durante um período
tão longo e à temperatura ambiente Além do mais, meios com
osmolaridade de cêrca de 216 miliosmoles foram feitos e usados
com sucesso para a incubação de glândulas salivares de R. ame
ricana (LARA e TOLEDO, citado em ARMELIN, MENEGHINI e LARA,
1969; TERRA et alii, manuscrito em preparação).
A densidade média da hemolinfa de larvas maduras é de
1,043 que é um valor elevado entre insetos (Cf. BUCK, 1953).
A hemolinfa da maioria dos insetos apresenta hemocitos
em circulação que podem ocupar volumes de até 15% do volume to~

:~
.~ I
:--;;
-40-
tal da hemolinfa, em certas ocasiões como na muda (BUCK, 1953).
Em R. americana, o volume ocupado pelos hemocitos é de cerca
de 0,3% do volume total da hemolinfa, o que dispensa a necessi
dade de se corrigir os tItulos dos compostos determinados no
plasma (hemolinfa centrifugada) para hemolinfa total. O valor
de 0,3% obtido concorda com a maioria dos determinados para he
molinfa de insetos (Cf. BUCK, 1953).
A hemolinfa de larvas de R. americana deixada em re
pouso por períodos de até 15 horas à temperatura ambiente, na
auséncia ou presença de hemocitos, nao coagula e escurece muito
pouco. Por outro lado, amostras de hemolinfa centrifugadas e
diluídas 10 vézes com água deionizada, mesmo mantidas no gêlo,
escurecem e floculam intensamente após 2 horas, resultando em
espêsso sedimento cinzento escuro. Entretanto, se uma centri
fugação fór' feita logo após a diluição, resulta em sedimento ama
relado, correspondente a apenas 3% das proteInas da hemolinfa.
2. Crbmatografia dos compostos ninhidrina-positivos solúveis
em ácido
A figura 1 mostra um cromatograma típico de uma amos
tra de compostos ninhidrina-positivos ácidos solúveis, não hidr~
lizada, de hemolinfa de larvas de R. americana do 29 período
(45-48 dias de idade). Para facilitar a visualização, alguns
picos foram ampliados, ficando fora da escala que é 1:4.
Aidentificaçao dos picos apresentados a seguir foi
feita utilizando os critérios referidos em Métodos: 7, ácido
aspártico; 8, treonina; 9,. serina; 12, prolinai 13, ácido glu
tâmico; 14, glicinai 15, alanina; 18, va1inai 19, cistatio
nina; 20, metionina; 21, isoleucinai 22, leucina; 25, tiro
sina; 26, fenilalanina; 29, ornitina; 30, amônia; 32, lisi
na; 34, histidinai 35, triptofano; 36, arginina (Ver figura
1) •
O pico n9 10 (figura 1) poderia ser identificado como
asparagina e/ou glutamina porque o método utilizado não permite
a resolução dos dois componentes. Entretanto, como TUNDISI et
alii (1964), encontraram em um estudo qualitativo da hemolinfa de
R. americana os dois compostos, utilizando cromatografia em pa
pel, o pico foi atribuldo a ambos os compostos.
O.
tídeos num
A identifi
relativos
nhidrina-p
ridos em M
O
mente base
foserina;
tioninas
amino-n-bu
y-amino-bu
p,
zada como
dos result
vio médio
sos entre
observada::
setas (LE\
cser inferj
fre destn
fa, na pn
Os tItulO!
mente SUpE
timadcs, (
te glutam:
(MOORE e :

la (EDeR, 1953).
:os é de cérca
msa a necessi
leterminados no
:otal. O valor
.nados para he-
deixada em re
'a ambiente, na
escurece muito
:entrifugadas e
ltidas no gélo,
resultando em
:e uma centri
I sedimento ama
la hemolinfa .
tivos solúveis
:0 de uma amos
'eis, não hidr~
. do 29 periodo
zação, alguns
é 1:4.
,s a seguir foi
dos: 7, ácido
13, ácido glu-
19, cistatio-
na: 25, tiro
.i 32, 1isi
na (Ver figura
.entificado como
ado não permite
como TUNDISI et
da hemo1infa de
.tografia em pa-
Os picos n9 5, 11, 23, 31 e 33 foram atribuídos a pep
tídeos numerados respectivamente 1, 2, 3, 4e 5 (ver figura 1).
A identificação désses picos baseia-se no fato de seus tempos
relativos de eluição não corresponderem a nenhum composto ni
nhidrina-positivo encontrável nos cromatogramas publicados refe
ridos em Métodos e desaparecem por hidrólise.
Os picos que se seguem foram identificados tentativa
mente baseando-se nos seus tempos relativos de eluição: 1, fo~
foserina: 2, f06foetanolamina: 3, taurina; 4, uréia i 6, m~
tioninas sulfóxidos: 16, ácido a-amino-adípicoi 17, ácido a_
amino-n-butíricoi 24, glicosaminai 27, B-alanina; 28, ácido
y-arndno-butírico (figura 1).
A quantificação dos picos dos cromatogramas foi reali
zada corno descrito em Métodos e a Tabela II apresenta a média
dos resultados obtidos em 3 determinações independentes. O des
vio médio observados nas determinações ficou na maioria dos ca
sos entre 5 e 15%. Variações dessa ordem e até. maiores foram
observadas em determinações semelh~rites realizadas em outros in
setos (LEVENBOOR e DINAMARCA, 1966).
O valor apresentado para o titulo de triptofano deve
ser inferior ao que ocorre "in vivo", porque ésse aminoácido s2
fre destruição parcial durante as desproteinizações da hemolin
fa, na preparação da amostra para análise (MOORE e STEIN,l954).
Os títulos encontrados para ácido glutâmico devem estar ligeir~
mente superestimados e aquéles da glutamina + asparagina, sube~
timados, devido a hidrólise parcial das amidas, especialmen
te glutamina, durante a preparação e análise das ~IDstras
(MOORE e STEIN, 1954).
••I'I.
1-


TABELAII
Compostos ninhidrina-positivos, solúveis em ácidos, da hemo1infa
de larvas de R. americana, 29 período, de 45-48 dias de idade 1
1. Para maiores detalhes, consultar o texto.
2. Determinado na fração ácido solúvel da hemolinfa como descritoem Métodos.
3. Traços são denominados os compostos que nas diluições empregadas (ver Métodos) não podem ser convenientemente estimados.Correspondem a concentrações inferiores a 0,05 mM. O sinal(+++) indica que o composto foi encontrado 3 vêzes em 3 análises e (++) indica que foi encontrado 2 vêzes em 3 análises.
ConcentraçuomM
"
'.'I;.
+++
++
++
++
+++
+++
+++
++
94,0
0,20
1,18
9,01
0,95
1,38
12,78
26,25
1,31
102,96
Q-amino-N tota12
Tra-ços3
Composto
Taurina
Uréia
Metionina<3 su1fóxidos
ÂC., Q-amino adípico
Âc. Q-amino-n-butírico
Glicosamina
a-alanina
Âc. y-amino-butírico
Amônia
Fosfoetanolamina
total geral
Peptídeo-l
PeptIdeo-2
Peptídeo-3
Peptídeo-4
PeDtIdeo-5
total de peptIdeos
0,07
0,35
13,15
5,30
8,05
7,65
0,96
2,11
3,92
0,41
0,61
1,78
2,06 .
0,27
2,42
4,67
2,76
1,40
0,89
0,63
3,22
62,62
concentraçãomM
Composto
Fosfoserina
Âc. aspático
Treonina
Serina
Prolina
Âc.glutâmico
Glicina
Alanina
Valina
Metionina
Isoleucina
Leucina
Tirosina
Feni1alanina
Ornitina
Lisina
Histidina
Triptofano
Arginina
Cistationina
Glutamina+asparagina
Total de aminoácidos
'M I <tilI~r--N-.-I~,
~ ~ " ~ OI ttS"r-t-IJ, ••.•• I::4JO'I <ti O--i ='-.-1IS:::UU..Q~, -.-I -ri =' I 4Ju01o.QJOI <ti '.-I ~ s::: 'u~'O -.-ILtl
<ti <ti 'I:; M- o. O N <ti, Ul s::: N I "
<tI-.-I ,~<tI
- E:" s:::I + <ti <ti 0-.-1I I I:: 'C 'C) <ti IS -.-I -ri -ri,s:: U U 4J'-.-10 =,,<tI Ul
E:'OQJ -ri• <ti -ri ~ ,.c:'4JUOlX>
=' '<ti Ul N '.~ -.-I ~
lO" ··M: \o ... n::s~ '~~S::"~O N-riLtl1 ~.. s:::1 <ti •• <ti OJ •• s::: <ti ~ QJ
<ti -ri I:: <ti 'C, s::: s::: -.-I I '.-l1 -ri r.l s::: a:>. 4J~ ~ O O-
'QJ<tI-ri'QJ1 Ul 4J r-- O-: ' QJ N~,LtlE: ': 0'1 ~ ··Ml '<tIMj " " O s:::) <ti <ti N -ri ": s::: I:: s:: <til ..-1..-1 " <ti s:::I s::: U <tI~..-I
) 0..-1 s::: <ti Ul •) QJ~-ri~-.-I Ot ~ 01S:::..-I~4J) 4J O s::: X) '..-IQJ •• QJI '~4J~N4JCO~<tI M
• 4J' OI " " Ul \C> "OO-.-IN~~
-UUU QJ1..-1..-1 •• O >:4JE: '<tlQJI ~.ltlO'l S:::'O Ul"'ltl4J~-.-I'.-I QJ) o. =' Ul 4J .c:I Ul ~ •• O o.~
I<tI01<tl~QJltlI s:: -.-I 0-4J100-ri4J QJ''O'O~ ''C1..-1..-1 ltl ,~
UU>LtlMu:ol<tl 1l'Cl N QJI ••• ~
• 'CO " ltl Or--M~<tI..-I..-I
I ~ S::S::: <ti••• ··..-1.0 I:;: tJl " O I:; E:: O tO U <tl ltl ltl''C s::: -.-I Ul lol'..-1..-1 ~ O '<ti'X~~ UOQ.,"0 O 4J -.-I M~ l-I =' ~:~o....QO'l""
=' I <tl <tiUl 'S::: • s:: s:::
N 1 ~-.-I-ri
tJl~ON4JS:::
<ti s::: -.-I-ris::: •• ..-1 " s:: 01-.-IN eM lollols::: tO O<tlO O I O-.-IQJISQJ' ,4J'O '00>\C>Q)'.-lO .... NMe4J'04J
o. -.-I 0.," "'Q)OQJOO
\C> 0." ro 0., u s::

:;:1
",]I
A concentração dos ~~noácidos é elevada, como é ca
racterístico de insetos, especialmente os Endopterygota (FLORKIN
e JEUNIAUX, 1964). Os aminoácidos predominantes na hemolinfa
são: treonina; prolina, ácido glutámico e serina, que juntos
perfazem mais do que 50% da concentração total de aminoácidos
livres da hernolinfa. A ocorrência de ácido glutârnico e prolina
entre os aminoácidos mais concentrados é típico de inse
tos Endopterygota (FLORKIN e JEUNIAUX, 1964). E '. interessante
assinalar a ausência, até mesmo de traços,d~ cisteína ou cistina
e citrulina. Cisteína, entretanto, tambérn não foi detectado
em vários outros Diptera como: Gastrophilus intestinalis (LEVEN
BOOK, 1950a), Phormia regina (LEVENBOOK, 1966). E' notável tam
bém a concentração relativamente elevada da ornitina e a ocor
rência de cistationina na hemolinfa, embora esta já tenha sido
identificada em outros insetos (WYATT, 1961).
Os peptídeos ocorrem em pequeno número, embora a sua
concentração total seja considerável. Dois dêles são ácidos e
3 dêles, representando cêrca de 87% do total dos peptídeos, sao
neutros ou básicos, a julgar pelos seus tempos relativos de e
luição. A concentração do peptídeo 3, que corresponde a 70%
do total dos peptídeos, só é superada entre os aminoácidos pela
treonina.
Vários insetos apresentam concentrações muito baixas
de amônia na hemolinfa circulante, mas depois que essa é retira
da da larva a sua concentração cresce ràpidamente a partir de
precursores desconhecidos (WIGGLESWORTH, 1965;LEVENBOOK, 1950a).
Como nenhum estudo foi feito nesse sentido em R. americana, a e
levada concentração de amônia atribuída à hemolinfa nesta tese
deve ser considerada com certa reserva. Outro motivo para essa
reserva é o fato da concentração de amônia determinada na hemo
linfa, 26,3 ~~, exceder de cinco vêzes aquela concentração consi
derada tóxica, que é 5 mM (CRAIG, 1960).
A razoável concordáncia observada entre a soma dos
compostos ninhidrina-positivos e a determinação direta de a-ami
no N, indica que nenhuma fração quantitativamente importante da
queles compostos deixou de ser analisada.
Aminoáci
americana
Aminoácid
Histidina
Âcido aspár:
Âcido glutâJ
Lisina
Prolina
Glicina
Leucina
Ornitina
Serina
l.
2.
A
cido solúvel
cida é de 95
ácida aument,
Assumindo qUI
peptídeos da
peptídeos na
aminoácl dos 1
ponde a uma !
identificado!
número médio
rando as cone
estimar o n1Ír1
ficados. Ass
deve ser um caminoácidos s
so, como já f
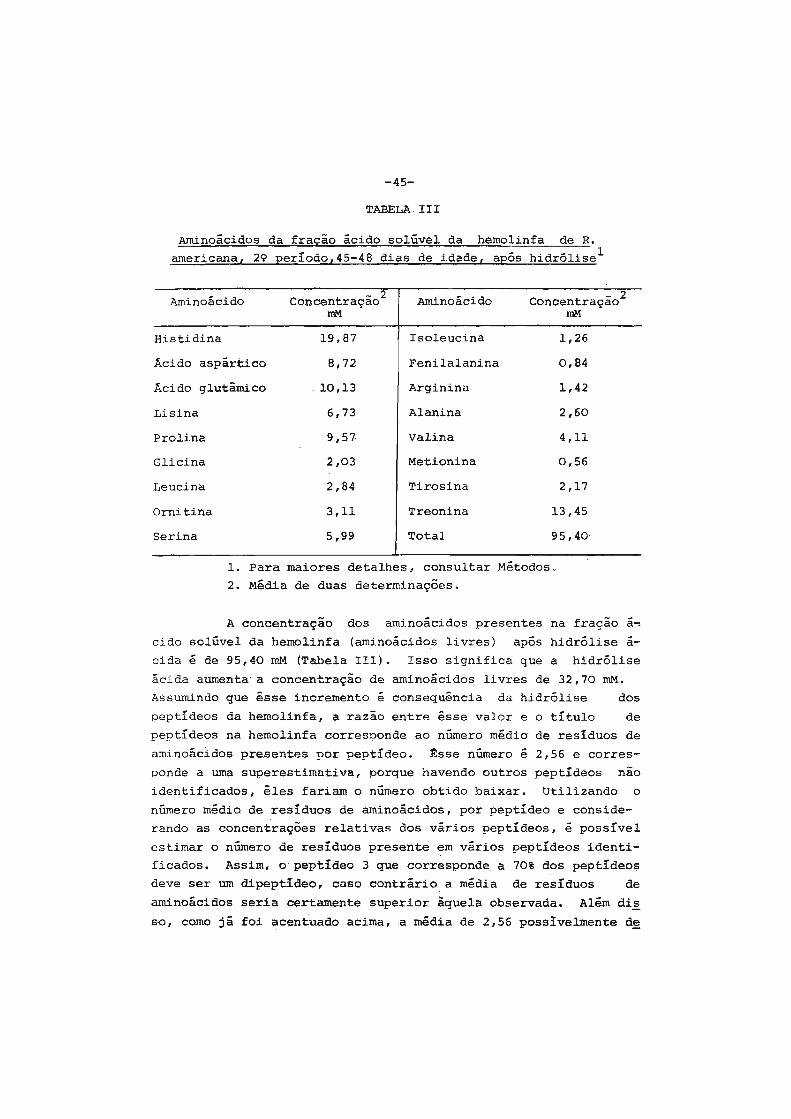
rada, como é ca
~rygota (FLORKIN
na hemolinfa
la, que juntos
de aminoácidos·
ltâmico e prolina
[pico de inse-
I. interessante
:eína ou cistina
foi detectado
~tinalis (LEVEN
E' notável tam
nitina e a ocor
~ já tenha sido
~, embora a sua
les são ácidos e
peptídeos, são
relativos de e
~rresponde a 70%
~inoácidos pela
s muito baixas
3 essa é retira
3nte a partir de
ilENBOOK, 1950a).
americana, a e-
nfa nesta tese
~tivo para essa
cminada na hemo-
ncentração consi
re a soma dos
direta de a-ami
e importante da-
TABELA !II
Aminoácidos da fração ácido solúvel da hemolinfa de R.
americana, 29 período,45-4B dias de idade, após hidrõlise l
Aminoácido - 2 Aminoácido - 2Concentraçao ConcentraçaomM mM
Histidina 19,87 . Isoleucina 1,26
Âcido aspárt:ico 8,72 Fenilalanina 0,84
Âcido glutâmico 10,13 Arginina 1,42
Lisina 6,73 Alanina 2,60
Prolina 9,57. Valina 4,11
Glicina 2,03 Metionina 0,56
Leucina 2,84 I Tirosina 2,17
Ornitina 3,11
ITreonina 13,45
Serina 5,99 Total 95,40
1. Para maiores detalhes, consultar Métodos.
2. Média de duas determinações.
A concentração dos aminoácidos presentes na fração á~
cido solúvel da hemolinfa (aminoácidOS livres) após hidrólise á
cida é de 95,40 mM (Tabela 111). Isso significa que ahidrólise
ácida aumenta a concentração de aminoácidos livres de 32,70 mM.
Assumindo que êsse incremento. é consequência da hidrólise dos
peptídeos da hemolinfa, a razão entre êsse valor e o título de
peptídeos na hemoiinfa corresponde ao número médio de resíduos de
aminoácidos presentes por peptídeo. tsse número é 2,56 e corres
ponde a uma superestimativa, porque havendo outros peptídeos não
identificados, êles fariam o núrr.ero obtido baixar. Utilizando o
número médio de resíduos de aminoácidos, por peptídeo e conside
rando as concentrações relativas dos vários peptídeos, é possível
estimar o número de resíduos presente em vários peptídeos identi
ficados. Assim, opeptídeo 3 que corresponde ã 70% dos peptídeos
deve ser um dipeptídeo, caso contrário a média de resíduos de
aminoácidos seria certamente superior àquela observada~ Além di~
so, como já foi acentuado acima, a média de 2,56 possivelmente d~
J
I••Ii
~-.,

;:11
-46-
ve estar superestimada, o que reforça ainda mais a conclusão
acima. Assumindo que o peptídeo 3 é um dipeptídeo, os peptídeos
2, 4 e 5 devem ter no máximo 10 resíduos, enquanto que o peptí
deo 1, por ocorrer em baixa concentração, não permite nenhuma es
timativa segura.
A Tabela IV,mostra a participação porcentual dos vá
rios aminoácidos no incremento arninoacídico total, após a hidró
lise ácida da porção ácido solúvel da hemolinfa. Se fôr admiti
do, corno foi feito acima, que o referido incremento é resultan
te da hidrólise dos peptídeos da hemolinfa, a Tabela IV é uma
boa estimativa da composição porcentual molar dos aminoácidos pep
tídicos.
A presença de ornitina, na Tabela IV deve ser conse
quência de variações amostrais, porque a ornitina é um aminoáci
do tipicamente livre. Variações amostrais devem ~er responsá
veis também pela presença na Tabela de vários aminoácidos cuia
participação no incremento é pequena corno: tirosina, metionina,
etc. A participação do ácido aspártico e glutâmico no incremen
to rea~ é menor do ~ue o indicado na Tabela, porque parte dêles
se originaram da hidrólise das aroidas correspondentes. Apesar
de tôdas as restrições acima, a Tabela IV permite concluir que
os peptídeos são compostos principalmente de histidina e ácido
aspártico, e em menor grau de ácido glutámico, lisina e outros.
A ocorrência generalizada de histidina explica porque os peptí
deos presentes em concentrações mais significativas sao neutros
ou básicos, a julgar pelos seus tempos relativos de eluição.
Participaç
Aminoácido
Histidina
Acido aspártico
Acido glutâmico
Lisina
Prolina
Glicina
Leucina
Ornitina
Serina
* Dados calcula,
O pl
peptídeo e,
hidrólise dev,
tais liberado:
iguais deverá
l:e 55% dos am:
que se os doil
nos doi s aminc
tais. Uma inl
sultados são :
consti tuido p<
histidina e 01
ção do peptidE
j a formado pOJ
co.
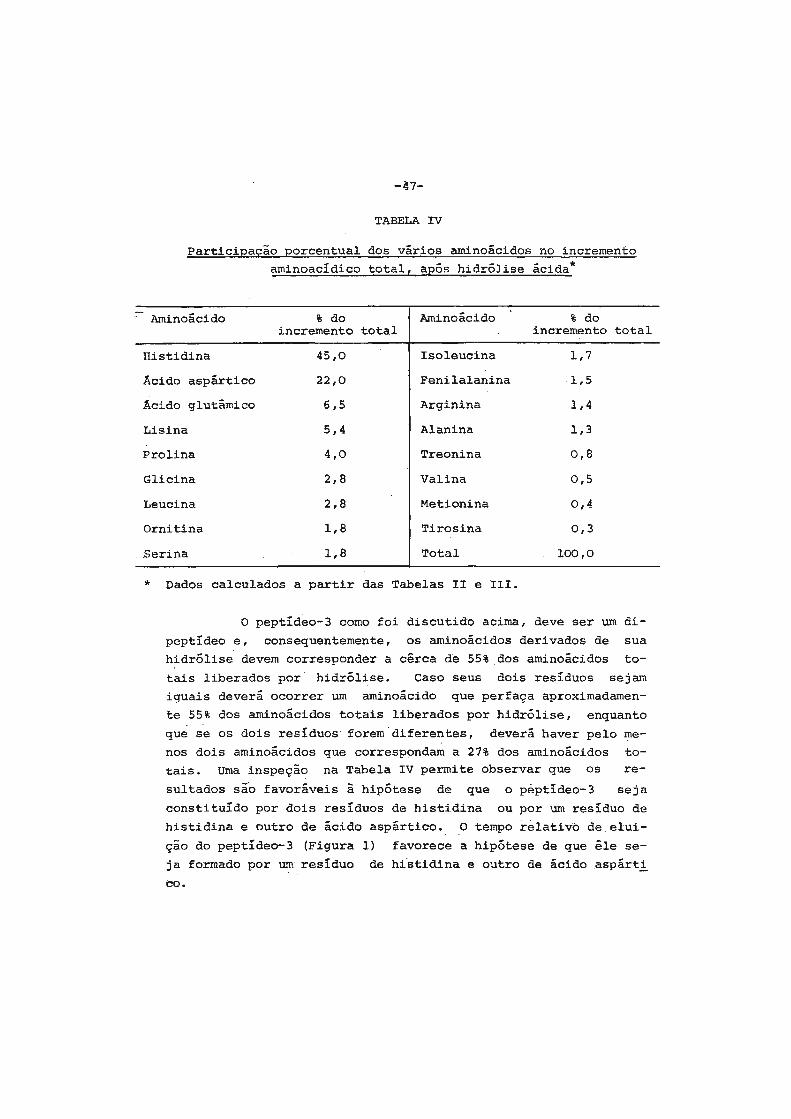
a conclusão
's oeptídeos
le o peptí
, nenhwna es
ltual dos va
'ós a hidró
fôr admiti-
é resultan
IV é wna
.noácidos peE
! ser conse
un aminoáci
responsá
:idcs cuia
rnetionina,
10 incremen
larte dêles
~s. Aoesar
lCluir que
la e ácido
l e outros.
os peotí
lao neutros
!luição.
TABELA IV
ParticiEação porcentual dos vários aminoácidos no incremento
aminoacldico total, após hidrólise ácida*
Aminoácido % do Aminoácido % doincremento total incremento total
Histidina 45,0 Isoleucina 1,7
Acido aspártico 22,0 I Fenilalanina 1,5
Acido glutâmico 6,5 Arginina 1,4
Lisina 5,4 Alanina 1,3
Prolina 4,0 Treonina 0,8
Glicina 2,8 Valina 0,5
Leucina 2,8 Metionina 0,4
Ornitina 1,8 Tirosina 0,3
Serina 1,8 Total 100,0
* Dados calculados a partir das Tabelas II e III.
o peptídeo-3 como foi discutido acima, deve ser um di
peptídeo e, consequentemente, os aminoácidos derivados de sua
hidrólise devem corresponder a cerca de 55% dos aminoácidos to
tais liberados por hidrólise. Caso seus dois resíduos sejam
iguais deverá ocorrer um aminoácido que perfaça aproximadamen
te 55% dos aminoácidos totais liberados por hidrólise, enquanto
que se os dois resíduos· forem diferentes, deverá haver pelo me
nos dois aminoácidos que correspondam a 27% dos aminoácidos to
tais. Uma inspeção na Tabela IV permite observar que os re
sultados são favoráveis à hipótese de que o peptldeo~3 seja
constituído por dois reslduos de histidina ou por um resíduo de
histidina e outro de ácido aspártico. O tempo relativo de elui
ção do peptídeo-3 (Figura 1) favorece a hipótese de que ele se
ja formado por um resíduo de h1"stidina e outro de ácidoaspárti
co.

TABELA V
-48--
Composiçãoguímica da hemolinfa da larva de R.
americana, 29 período, 45-48 dias de idade*
o (~ é elevac
gânicos ácidc
ta (WYATT, l~
do solúvel OJ
O resultado
perestimado
da proteína c
sentar o mesn
ácidos, indic
taminado com
dições salim
te em concent
conforme foi
citoplasmátic
Tre
O t
tretanto, tít
e em outros i
uma caracteri
HOLLIS, 1961)
como ocorre
e JEUNIAUX,
concentração.
As
larvas de ins
de grandeza d
A c
de R. americ
larvas dos de
o que já pode
densidade e
outros inseto
110
12,9
0,49
0,81
1,60
77,5
67,6
9,7
55,4
10,2
2,8
65,5
42,1
47,6
6,9 g/lOO ml
201
1401,4
520
391
460
mg/lOO mlComposto
Mg
Ca
Cu
Zn
Fe
Na
K
P-inorgânico
P-ácido-solúvel
P-lipídico
P-protéico
P-total
HC03
- (pH 7,270)
Cl
Proteína
Carboidrato total
Trealose
Glicogênio
Lipídeo total
Ácido cítrico**Cinzas
3. Apresentação dos resultados das análises químicas
As concentrações de Ca, Na, K e Cl na hemolinfa de
larvas maduras de R. americana (Tabela V) são similares às de
terminadas para grande número de insetos, enquanto que a concen
tração de Mg é muito elevada, sõmente comparável com aquelasen
contradas em alguns Lepidoptera e Coleoptera (BUCK, 1953; PROS
SER e BROWN, 1961). Concentrações semelhantes às apresentadas
na Tabela V para Cu e Zn foram determinadas em outro Diptera :
Gastrophilus -intestinalis (LEVENBOOK, 1950a).
'*
**
Os resultados referentes a ~mpostos ninhidrinapositivos encontram-se nas Tabelas 11 e 111. Para maiores detalhes consultar o texto.
Apenas uma determinação.
As
terminações)
lado (ver Mét

(micas
la hemolinfa de
ni lares às de
) que a concen
:om aquelas ·en
C, 1953: PROS-
apresentadas
lutro Diptera :
le R.
~
o conteúdo de fósforo total na hemolinfa de R. ameri
~ é elevado e a maior parte dêste está ligado a compostos or
gânicos ácido-solúveis, o que é característico da classe Insec
ta (WYATT, 1961). Só foram identificados na fração fósforo áci
do solúvel orgânico: fosfoetanolamina e fosfoserina (Tabela 11).•
O resultado apresentado para fósforo protéico poderia estar su
perestimado se ocorressem ácidos nucléicos na hemo1infa. O fato
da proteína obtida por precipitação com sulfato de amônio apre
sentar o mesmo conteúdo em fósforo que aquela precipitada por
ácidos, indica que o fósforo considerado protéico não está con
taminado com fósforo de ácidos nucléicos. Isso porque nas con
dições salinas empregadas DNA não se precipitaria e RNA presen
te em concentrações de pelo menos até 100 pg/ml, também não,
conforme foi observado em exp~riência contrôle utilizando RNA
citoplasmático de glândula salivar de R. americana.
O título de citrato encontrado é bastante elevado, en
tretanto, títulos elevados como êsse ocorrem em outros Diptera
e em outros insetos, o que levou a se considerar ésse fenômeno
uma característica de larvas da Classe Insecta (LEVENBOOK e
HOLLIS, 1961).
Trealose é o carboidrato predominante da hemo1infa,
como ocorre em grande número de insetos (~~ATT, 1961: FLORKIN,
e JEUNIAUX, 1964), enquanto que glicogênio ocorre em pequena
concentração.
00 ml
As determinações para
larvas de insetos (BUCK, 1953:
de grandeza do valor encontrado
lipídeos totais na hemo1infa de
WYATT, 1961) são da mesma ordem
em R.americana.
["inaPa
A concentração de proteínas encontrada na hemo1infa
de R. americana é superior à média determinada na hemo1infa de
larvas dos demais insetos (Cf. BUCK, 1953: ENGLE e WOODS, 1960),
o que já poderia ser antecipado por sua osmo1aridade baixa, mas
densidade e conteúdo de sólidos mais elevados que a média dos
outros insetos.
As determinações de pC02 a 37°C deram em média (3 de
terminações) 15,7 mmHg. A partir dessa determinação foi calcu
lado (ver Métodos) o valor para bicarbonato apresentado na Ta-


A
HEMOLINI'A TOTAL
SOBRENAOAt,lTE ~CIO ...
SOBRENAOANTE A(;IOO
SULI'OSSALICILI CO
\\\\ ,.,~ \
\
\ \ ,-.,,-....' " .....--, .,,~, 0••-', " ....................
......... _-..... .. ............-.._--- --_.~.::::...-_-- -.._-
0,200
0.800
~ \
~, 0,/300
,)--~
0....00
!m média (3 de
; um período de
!ffi equilíbrio.
.ei turas não se
l deve ser cau
.tes na hemolin
lfa é retirada,
medidas corre
liretamente no
ferir um pouco
ácido carbôni
.ssumidos serem
pode não ser
em que foi de
animal normal
'alores obtidos
fornecidos por
'ivo" seria 46%
o pH da hemo
a temperatura
admitido nêsse
depende apenas
a temperatura,
'in vivo'" seria
!ssa forma, em
.ferir do valor
loa estimativa
"
11li
I'
+004580a202804-e0 &20
A (m)! I
Figura 2 - Espectro de absorção da hemolinfa de larvas madurasde R. americana. O espectro da hemolinfa total foi feito apósdiluição de 50 vêzes em água deionizada e aquêles dos. sobrenadantes de desproteinização com HCl0 4 0,5N ou ácido sulfossalicílico 3% (P/V) foram realizados apos neutralização com NaOH
e numa diluição final de 50 vêzes.
.1 de larvas de
o: um em 270
o a 270-280 mil
e triptofano
s aminoácidos,
eia de um pico

~IH
-52-
nessa mesma posição em amostras desproteinizadas com HCI04 O,SN
(figura 2). O pico a 310 m~ deve estar ligado ã existência de
proteínas, pois êle quase desaparece após a desproteinização. O
Dica a 460 m~, parece corresponder a carotenóides ligados a pro
teínas, conforme será mostrado abaixo. E' interessante observar
ainda o fato de que o sobrenadante da desproteinização com áci
do sulfossalicílico apresenta absorção nos comprimentos de onda
mais curtos do espectro visível, contràriamente ao sobrenadante
de HCI04 (figura 2). A referida absorção confere ao sobrena
dante ácido sulfossalicílico uma cor limão, enquanto que o so
brenadante HCI0 4 é incolor.
o sobrenadante de amostras desproteinizadas com TCA
10% (p/v) ou com ácido tungístico (Van SLYKE e HA~VKINS, 1928),
apresenta um espectro de absorção semelhante àquele obtido após
precipitação das proteínas com HCI04 , O,SN, enquanto que aque
las amostras desproteinizadas com 100% de saturação em (NH4)2S04
ou com um volume de clorofórmio apresentam sobrenadantes cujos
espectros são idênticos àqueles obtidos após precipitação com
ácido sulfossalicílico 3% (P/V). ~sses resultados indicam que
a maior parte dos pigmentos deve estar ligada a proteínas, pois
são precipitadas por todos precipitantes de proteínas usados.
Deve-se assinalar, entretanto, o fato de um pigmento de cor li~
mao nao ser precipitado por ácido sulfossalicílico 3% (ver fi
gura 2), sulfato de amônio saturado e clorofórmio.
A figura 3 mostra o espectro de absorção do .pigmento
de cor limão após a sua preparação e purificação como descrito
em Métodos. O seu resíduo obtido após a evaporação de seu sol
vente por pressão reduzida ao abrigo da luz é solúvel em: água
deionizada, NaCl O,lM, etanol 60% (V/V), ROH 5% (P/V) em meta
nol e NaOH O,lN. Oresiduo nao é solúvel em éter de petróleo
(p.e. 30o-S0oC) e acetona.
O resíduo do pigmento cor de limão solubilizado em a
gua é precipitável em HCI04 O,SN, mas mantém-se solúvel em eta
nol 60% após aquecimento a 7SoC por lO minutos ou após adição de
um volume de clorofórmio.
L O'
O'toN

-53-
oIIUo-com HCI0
4 O,SNlU1-1xistência de lUo.'teini zação. 0- (I)° ~::t.1-1o.ligados a pro-
~Ul
'0sante observar0-< o.
lUzação áci- If)com10
mentos de onda:>
.0 sobrenadanteo ~
sobrena- iõaodi'otanto que o so- m
O .-IOI O.,. CU!
lUO401'0lizadas com TCA(1)0
o 8401A\'JI<INS, 1928) , '(1),...8::E:.,.le obtido após(I) e0(1)nto que aque- IlU
o 8010 .... 401o em (NH4 ) 2S04 ... .-I ....
!-lnadantes cujos !-lO.0 U!cipitação 0(1)como 'Otf) CIls indicam que~ '00
Eoternas, 00pois4010cternas usados. o (1)0
i EII\l0">0-nto de cor li.,.
.... lU0.0::0 3% (ver fi- ....
o 0'1-1OI 'O ....tf) l-l
0::1IlU o-o- lllI:> do 'pigmento 1-1(1)
o odescrito ,... U! I.comoIf) .Cl
lU JIIio de seu s01-CIllúve1 água 'O
f
era:o11) o(P/V) em meta-l') l-l401
~r de petróleo [)Glo. 11o U! 'II')~o o o o ,.,o o o o
.1izado em á- ~ ct III 6!.IIN N
~~úve1 em eta-
<t
I'IpÓS adição de

I
1111. -54-
'111
IHll
o pigmento côr de limão após solubilização em água,
adição de um volume de HCl04 4N produz um sedimento que após la
vagens com HCl0 4 0,5N dá reação positiva em testes colorimétri
cos para proteína (LOWRY et alii, 1951) e para carboidrato (DU
BOIS et alii, 1956).
As preparações do pigmento de cor limão apresentam uma
intensa fluorescência amarelo-esverdeado quando expostos a U.V.
de comprimento de onda longo (315-380 mp), enquanto que com U.V.
de comprimento de onda curto (200-280 mp), nenhuma fluorescên
cia é observada.
A fluorescência das preparações é mantida mesmo em
H2S0 4 0,5N, NaOH O,lN ou em meios aquosos com ditionito de só
dio em quantidades supersaturantes.
s ao predomina
quena concent
Amo
ou não, separ
de hidroxiapa
tróleo. A cc
o isolamento
pondentes às
gundo migraçã
petróleo elui
enquanto que
IV•.. ~sses re
te com inforrn
posição das b
após desenvo
e 6 são muito
mas, embora
nenhuma tenta
ções 111 e IV
NQ da bandal
1
2
.3
4
5
6
L Numeraçãopetróleo
A b
A banddas.
O pigmento que permanece no sobrenadante após as des
proteinizações da hemolinfa com ácido sulfossalicílico, 3% ou
sulfato de amônto 100% saturado, depois de purificado por preci
pitação com HCl04 0,5N, lavagens com o mesmo ácido e dissolução
em etanol 60% produz um espectro de absorção idêntico àquele
do pigmento côr de limão, discutido acima. Além disso, êsse pi~
mento apresenta o mesmo tipo de fluorescência que o pigmento li
ma0. Isso permite concluir que o pigmento da hemolinfa precipi
tável com sõmente alguns precipitantes de proteína deve ser o
pigmento cor de limão.
As propriedades observadas do pigmento de côr limão su
gerem que êsse seja uma cromoglicoproteína. Os resultados mais
indicativos dessa hipótese são: a sua solubilidade em etanol 60%
e sulfato de amônio saturado, além da reação positiva observada
para proteínas e carboidratos. Os resultados indicam ainda que
a molécula cromógena do pigmento deve ser muito estável e que
grupos ionizáveis não devem estar comprometidos com a estrutura
responsável pela fluorescência, porque essa não é al~erada na
presença de um redutor como ditionito de sódio e nem em meios
ácidos ou básicos.
Os resultados obtidos com a partição dos carotenóides
da hemolinfa entre éter de petróleo (p.e. 30o-50oC)e metano1
90% corno descrito em Métodos, indicam' que êsses carotenóides
.11'1n

ao em agua,
que após la
olorimétri
idrato (DU-
resentam uma
stos a U.V.
que com U.V.
fluorescên-
ida mesmo em
rüto de só-
-55-
são predominantemente epifásicos, embora ocorra também uma p~
quena concentração de pigmentos amarelados hipofásicos.
Amostras de carotenóides da hemolinfa, saponificadas
ou nao, separam-se em 6 bandas coloridas ao longo de uma coluna
de hidroxiapatita (ver Métodos), durante eluição com éter de pe
tróleo. A continuação da eluição com éter de petróleo permite
o isolamento de duas frações distintas (Fração I e 11) corres
pondentes às bandas 1 e 2 respectivamente (bandas numeradas se
gundo migração decrescente). A adição d~ acetona 7% em éter de
petróleo elui as bandas 3 e 4 em uma única fração (Fração III),
enquanto que adição de acetona elui as bandas 5 e 6 na fração
IV•.. Êsses resultados estão sumarizados na Tabela VI, juntamerr
te com informações sôbre as côres correspondentes a cada banda.
1. Numeração segundo ordem decrescente de migração com éter depetróleo (p.e. 300 -500 C).
A banda 2 é bem mais intensa que tôdas as demais reuni
das. A banda 1 é de intensidade intermediária, as bandas 3, .~e 6 são muito fracas e a banda 4 é mais intensa do que as últi
mas, embora menos que a banda 1. Em vista dess~observações
nenhuma tentativa foi feita de se isolar os pigmentos das fra
ções III e IV.
:l.pÓS as des
lico, 3% ou
::> por preci
~ dissolução
Lco àquele
;0, êsse pi.9:
)igmento li
lfa precipi
~ve ser o
:or limão su
Ltados mais
n etanol 60%
'l observada
:l.ffi ainda que
;tável e que
a estrutura
al~erada na
am em meios
=arotenóides
::lC)e metanol
:l.rotenóides
fraç,ões
N9 da bandal Côr da banda
1 amarela
2 vermelha
3 amarela
4 alaranjada
5 amarela
6 alaranjada
TABELA VI
Eluente
éter de petróleo
éter de petróleo
acetona 7% em éter
acetona 7% em éter
acetona
acetona
Fração recolhida
I
Ir
IIr
IIr
IV
IV
u
j

li -56-
I~
xantofilas,
tuintes à h
hipofásicos
tanol 90%.
;I~
UI
A
I e lI, ap
e ao abrigo
vamente. A
carotenóide
suposição di
posição é fi
ficar adsor'
Nenhuma idel
ça ao pigmel
lhança de SE
aplicado à
traídos da
cia visava
astaxantina
ção II não t
xantina e ne
sultado foi5/0 550
J..lm)))470430390
---- l"'RAÇÃO I__ FRAçÃon li.
I \I \
I \I \
I \I ,_," '-' ,
" ",--'--~/" \I I
I \I I/ ,
I ,I I
I I// I
/ ,I \
/ \/ \.. " \.. \
~.. \
/ \...... _--*' "\
350
0,020
A
0,100
O,OGO
0,060
•
11. Figura 4 - Espectro de absorção daê frações I e II dos carote-!~ noides da hemolinfa, após preparaçao e cromatografia em hi.dro-
xiapatita, como descrito em Métodos. O espectro da fração Ifoi realizado em hexano e o da fração II em piridina.
n

'I
;10 660
A ( mJJ)
dos carote.a em hj,drola fração Iridina.
A fração IV e possIvelmente também a fração 111 contém
xantofilas, devido a intensa adsorção de seus pigmentos' consti
tuintes à hidroxiapatita, que devem corresponder aos pigmentos
hipofásicos observados na partição entre éter de petróleo e me
tanol 90%.
A figura 4 mostra os espectros de absorção 'das frações
I e lI, após evaporação do éter de petróleo a pressão reduzida
e ao abrigo da luz e redissolução em pexano e piridina, respecti
vamente. A confrontação dos espectros obtidos com aquêles dos
carotenóides mais conhecidos (KARRER e JUCKER,l950)perrnite a
suposição de que o pigmento da fração I seja a-caroteno. Essa su
posição é fortalecida pelo fato daquele pigmento pràticarnente não
ficar adsorvido na coluna, como seria de se esperar do fl-caroteno ..
Nenhuma identidade póde, entretanto, ser atribuída com seguran
ça ao pigmento da fração 11. E' interessante assinalar aseme
lhança de seu espectro com aquêle do astaceno em piridina.
Um teste qualitativo para astaxantina (ver Métodos) foi
aplicado à fração 11 (fração predominante) de carotenóides ex
traídos da hemolinfa e. cromatografados no mesmo dia. A experiên
cia visava testar a afirmação de TUNDISI et alii (1964) de que a
astaxantina é o pigmento predominante na hemolinfa, embora a fr~
ção 11 não tivesse o comportamento na coluna esperado para asta
xantina e nem o seu espectro (Cf. KUHN e SORENSEN, 1938). O re
sultado foi negativo.
t

-58-
B. Química do casulo coletivo de R. americana
1. Apresentação geral dos resultados
A composição geral do casulo 111 calculada a partir
das análises quantitativas apresentadas nos ítens seguintes, émostrada na Tabela VII. Os valores apresentados sao médias de
3 determinações.
TABELA VII
composição química do casulo coletivo de R. americanal
A ansulo, exceçãcpresentes em q
notável do cas
e carboidratosxo).
2. Solubili
Apena(Tabela VIII).
do pêso sêco à
terial lúdrossConstituinte
Material hidrossolúvel nao identificado
Carboidrato solúve12
Proteína solúve12
3CaC03Galactosamina insolúve1 4
Âcidos ur6nicos insolúveis4
Açúcares neutros insolúveis 4
Proteína insolúveIS
~, Cinzas insolúveis 6
Lipídeos
Total 7
% docasulo em pêso
4,3
2,4
4,4
43,S
1,9
0,2
8,2
33,6
4,3
<1,0
102,8
A maa lavagem com
0,8%,em pêso s
é proteína. 11
deve correspon
Após45,4% do casul
casulo (ve~ Mé
guintes solven
peratura ambie50% (P/V), HCl
Solubi
1. A composição se refere ao casulo 111, isto é, ao casulo acabado. Maiores detalhes no texto.
2. Proteínas ou carboidratos determinados na, lavagem com água adicionados àqueles medidos na lavagem com ácido acético.
3. Assumindo corresponder ao material hidroinsolúvel e ácido acético solúvel. A massa de proteínas e carboidratos solúvelnessas condições foi subtraída.
4. Extraoolado para tempo zero de hidrólise ácida da fração docasulo remanescente após as lavagens em água e ácido (fraçãoinsolúvel do casulo).
5. Calculado a partir jos aminoácidos de hidrolizado ácido dafração insolúvel, extrapolado para tempo zero de hidrólise.
6. Determinadas na fração insolúvel do casulo.7. Excluído lipídeos.
Massa hidross
Massa hidroin
Âcido acético
Massa insolúvl
Cinzas totais
Cinzas insolú'* Para maiores
de :3 determil** Determinadas

a partir
lintes, é
lédias de
-59-
A análise abrangeu a totalidade dos componentes do ca
sulo, exceção feita naturalmente a compostos que possam estar
presentes em quantidades insignificantes. A característica mais
notável do casulo é o seu alto contéudo em CaC03 e em proteínas
e carboidratos insolúveis em todos solventes utilizados (ver abai
xo).
2. Solubilidade, conteúdo dei cinzas e metais
~1
Apenas uma pequena parcela do cas~lo é hidrossoluve1
(Tabela VIII). Dessa fração 2,5% correspondem a proteínas e 1,6%
do pêso sêco do casulo a carboidratos, remanescendo 4,3% de ma
terial hidrossolúvel não identificado.
!m peso A massa de material solúvel em ácido acético 10% após
a lavagem com água é de 46,2% do pêso-sêco do casulo. Cêrca de
0,8%,em pêso sêco do casulo, dessa fração é carboidrato e 1,9~
é proteína. A massa resultante de 43,5% do pêso sêco do casulo
deve corresponder a caC0 3 (ver abaixo).
Após as lavagens com água e ácido acético 10% sobram
45,4% do casulo. Essa fração foi denominada fração insolúvel do
casulo (ver Métodos). ~sse material permanece insolúvel nos ~e··
guintes solventes empregados, tanto a quente (37oC) quanto ~ tem
peratura ambiente (22 0 C): SDS 10% (p/V), uréia 6M, ácido formico
50% (P/V), HCl 2N, KOH 1M e NaHC03 1M.
TABELA VIII
Solubilidade e conteúdo de cinzas do casulo 111 de
*. Rhynchosciara americana
% emEéso-sêco do casulo
,
sao mediãs
8,4
45,4
32,5
4 L 3
46,2
resultados
Determinação
Massa hidrossolúvel
Massa hidroinsolúvel,
Âcido acético solúvel
Massa insolúvel
Cinzas totais**Cinzas insolúveis
* Para maiores detalhes consultar Metodos. Osde 3 determinações.
** Determinadas na fração insolúvel do casulo.
J
;cido daldrólise.
3
fração doio (fração
::om água a;tico.
:! ácido a:>s solúvel
:lsulo acaba

-60-
Os resultados mostram que existe massa considerá
vel de cinzas no casulo e que a maior parte dessas é solúvel em
ácido acético 10%, enquanto que a fração insolúvel é essencial
mente orgânica. As análises de metais do casulo (Tabela IX)
indicam que cálcio e o metal predominante, correspondendo
a 19% do pêso-sêco do casulo~ consequentemente, as cinzas das
lavagens em ácido acético devem ser correspondentes a compostos
de cálcio.
TABELA IX
*Metais no casulo 111 de R. americana
Metal % do casulo (P/P)
Ca 19,0
Fe 0,108
Zn 0,024
Mg 0,602
Cu 0,002
K 0,001
*
3.~
A
ronzeamento ,
5) dá uma idi
amarronzado.
menta é form;
O
hidrólise e
guintes propJ
seu comportai
U. V. de compJ
cência fraca,
curto {200- 21
com reagente
e spectro de c
As propriedac
"reação de rem uma reaçãc
menta. Desse
rante a hidré
uma destruiçã
pois de 6 hOI
zero de hidré
amostra de ca
da hidrÓlise.
das a aquecirr
HANNAN e GREl\
A f
carboidratos
rios tempos d
aoos 4 horas
admitirmos g
locidade desd
vel, a melhor
la extrapolaç
de hidrólise.
neutros prese
'"
Para maiores detalhes consultar Métodos.Os resultados são médias de 3 determinaçoes.
Um teste qualitativo para carbonatos realizado como
descrito em Métodos, indicou a presença de quantidades conside
ráveis de carb~natos no casulo. ~sse resultado juntamente com
os apresentados acima sugerem que porção hidroinsolúvel e ácido
acético solúvel do casulo é carbonato de cálcio. Os cálculos a
presentados a seguir mostram que a hipótese referida é bastante
verossimel. Calculando todo o Ca presente no casulo como CaC03resulta em 47,5%,que é similar à massa determinada como material
hidroinsolúvel e ácido acético solúvel, após subtrair a massa
de proteínas e carboidratos solúveis. Por outro lado, calculan
do as cinzas correspondentes a todo Ca presente no casulo resul
ta em 26,6%, que é bem próximo dos 28,8% presente nas lavagens
do casulo, como seria de se esperar se pràticamente todo Ca es
tivesse na forma de CaC0 3 •
dina. E' pc

considerá
é solúvel em
é essencial-
=> (Tabela IX)
)rrespondendo
cinzas das
a compostos
Lizado como
'ies conside
'ltamente com
ível e ácido
cálculos a
:l. é bastante
) como CaC0 3)mo material
rair a massa
), calculan
:l.sulo resul-
lavagens
:odo Ca es-
-61-
3. Carboidratos
A hidrólise ácida do casulo é acompanhada por um amar
ronzeamento da solução. A curva de absorbância a 400 m~ (Figura
5) dá uma idéia da progressão de formação de um pigmento amarelo
amarronzado. E' interessante observar que a maior parte do pig~
mento é formado nas primeiras 6 horas de hidrólise.
o pigmento amarelado (melanoidina) produzido durante a
hidrólise e purificado como descrito em Métodos apresenta as se
guintes propriedades: caráter altamente básico, a julgar pelo
seu comportamento em Dowex-50, fluorescência azulada intensa em
U.V. de comprimento de onda longo (315-380 m~) e uma fluores
cência fraca, mas também azulada em U.V. de comprimento de onda
curto (200-280 m~). Além disso o pigmento dá reação positiva
com reagente de ELSON-MORGAN para hexosaminas, apresentando um
espectro de absorção nessas condições com dois picos (Figura 6)
As propriedades dêsse pigmento são similares aos produtos da
"reação de Maillard" (Cf. ELLIS, 1959). Essa reação consiste
em uma reação entre proteínas e carboidratos durante o aqueci
mento. Dessa form~, o incremento em melanoidinas observado d~
rante a hidrólise do casulo (Figura 5) deve ser acompanhado de
uma destruição désses, mais intensa no início e decrescendo de
pois de 6 horas. E' interessante observar que mesmo no tempo
zero de hidrólise já existe uma pequena quantidade de melanoi
dina. E' possível que essa tenha sido formada ao se secar a
amostra de casulo afim de se determinar o seu pêso-sêco antes
da hidrólise. Reação de Maillard em amostras úmidas submeti
das a aquecimento é fenómeno conhecido há já algum tempo (LEA,
HANNAN e GREAVES, 1950; ELLIS, 1959).
A figura 5 mostra o resultado das determinações de
carboidratos neutros na fração insolúvel do casulo III após vá
rios tempos de hidrólise. A destruição dos açúcares neutros
aoós 4 horas de hidrólise .segue uma cinética de l~ ordem. Se
admitirmos que a destruição dos carboidratos segue a mesma ve
locidade desde o início da hidrólise, o que é bastante prov~
vel, a melhor determinação para açúcares neutros seria dada pe
la extrapolação gráfica da curva de cinética para tempo zero
de hidrólise. Dessa forma a porcentagem em massa de açúcares
neutros presentes na fração insolúvel seria de cêrca de 18%. A
1

-62-
OIt>o.o
J>CIlUlo
0,200:llCil1>,ZoI>
I>0,100
~
oo
3-c:
0,050
0,030
0,020
o
12
o
AÇUCARES NEUTROS
I
4 e 10
TE M PO OE HID ROLIS~ (HORAS)
o
ÁCIDOS URÔNICOS
ti
2
Â
,,,,,,,,,,,\ ,,,
\\ ,,,,
~
20,0
10,0O.J::lcn~(,.)
O 5,0o.JllJ>::l.JOCIlZ
~ 0,7
~S
0,5:;!llJ
:lillJ(!)
~z...Lo)
c::~ 0,2
Oc~
(,.)o
~
a:lL
~ 1,0o
Figura 5 - Recuoeração de açúcares neutros e ácidos urônicosapôs vários tempos de hidrólise_da fra~ão insolúvel do casulo 111 de R. americana e produçao de melanoidinas nos mes-
mos tempos. Ver texto.

A
Figura 6 - Espectro de absorção do pigmento amarronzado (melanoidina) formado durante a hidróliseácida da fração insolúvel do casulo 111, após purificação em Dowex-50 e reação de Elson-Morgan.
Ver detalhes em Métodos.
I0'1'wI
690
A (m}J1
670650630610590570550530510490470450
::l ~u ~o o _o ~oo Q, ~ o o ã o NUI O ti N C>I (Jl OO, O O O O OO ::lri W OOt>!3 llJ 1-'.
\' VIONVS!:tOSev(1) UI O"UI ~ O
I I UI
0,025~
0,050

••
-64-
5,0
--~----.,• ---..,.~. a ~
210L---'L.----I.---6L..---!.---,l.O--.J'2:---:'~4---:1:6--~'8:---;:20TEhlPO DE HIDRÓLISE (HORAS)
Figura 7 - Recuperação da galactosamina após var10S temposde hidrólise da fração insolúvel do casulo 111 de R. ameri
cana .
validade deltar de acôrcção das mel,
carboidrato!
ofica a aceit
se, como a
galactosamir.
casulo (ver
cos e galaet
insolúvel dc
Os
dois sistema
sistema) per
1actosamina
sulo. Uma
togramas e CI
mo nenhum di
g1icose, gali
ti19li cosamil
lactosamina I
confirmada eI
MORGAN.
4. Compos1
A1.É
rer nenhum cc
nos hidrolizc
fato" da soma
cados excedel
ção de dois :I
do de Kj eldar.
uma recuperaç
o método do t
recuperações
método de KjE

-J
18 20
( HORAS)
-65-
validade dessa extrapolação é apoiada ainda pelo fato dela es
tar de acôrdo com a previsão feita acima, baseando-se na for.m2
ção das melanoidinas, de que a maior parte da destruição dos
carboidratos deveria ocorrer antes de 6 horas de hidrólise.
o mesmo tipo de argumentação apresentado acima justi
fica a aceitação das extrapolações, para tempo zero de hidróli
se, como a melhor determinação de ácidos urônicos (Figura 5) e
galactosamina (Figura 7), que é a única hexosamina presente no
casulo (ver abaixo). Os valores extrapolados para ácidos urôni
cos e galactosamina são, respectivamente: 0,45% .. e 4,1% da fração
insolúvel do casulo.
Os resultados de cromatografias em papel feitas em
dois sistemas solventes diferentes (2 cromatogramas em cada
sistema) permitiram a identificação de glicose, galactose e ga
lactosamina nos hidrolizados ácidos da fração insolúvel do ca
sulo. Uma mancha pouco intensa, mas constante de todos croma
togramas e com elevada migração, não ·._pôde ser identificada co
mo nenhum dos carboidratos: ribose, xilose, manose, frutose,
glioose, galactose, galactosamina, glicosamina, sorbose, N-ace
tilglicosaminae N-acetilgalactosamina. A identificação de ga
lactosamina como única hexosamina presente.nos ·hidrolizados foi
confirmada em 2 cromatogramas revelados com reagente de ELSON
MORGAN.
I
ios tempose R. ameri-
4. Compostos nitroqenados
Além de aminoácidos e galactosamina parece não ocor- ,li
rer nenhum composto ni~ogenado em quantidades significantes
nos hidrolizados da fração insolúvel do casulo (Tabela X). O ,
fato da soma do nitrogênio dos compostos nitrogenados identifi
cados exceder o conteúdo de N total deve ser devido a conjug~
ção de dois fatôres. O primeiro dêles é o fato de que o méto
do de Kjeldàhl clássico empregado, de tubo aberto, não permitir
uma recuperação de N rigorosamente quantitativa, como corre com
o método do tubo selado e nem tão precisa (JACOB,196S). Aliás,
recuperações relativamente baixas de nitrogênio determinado pelo
método de Kjeldahl foram também observados éIn cutIculas de inse-

••
-66-
tos (HACKMAN e GOLDBERG, 1971) e em vários outros produtos naturais
(STITCHER, citado em HACKMAN e GOLDBERG, 1971). Em vista disso, os
valores para N-total mais exatos seriam mais elevados do que aquê
les obtidos. O segundo fator a ser considerado é uma possível su
perestimativa do nitrogênio de arninoácidcscomo consequência de va
riações amostrais.
TABELA X
Compostos nitrogenados da fração insolúvel do casulo 111
de R. americana
Determinação % em N da fração insolúvel
N de aminoácidos 1 10,0
N de galactosamina 2 0,3
Soma dos anteriores 10,3
N-total (Kjeldahl) 3 8,7
1. Calculado a partir da análise de aminoácidos, apóscorreção para tempo zero de hidrólise. Ver Métodos.
2. Calculado a partir do valor extrapolado para zeroda Figura 7.
3. Média de 3 determinações realizadas como descritoem Métodos.
Acido glutâmico, ácido aspártico, serina e treoni
na são os aminoácidos mais abundantes da fração insolúvel do ca
sulo, juntos correspondendo a 42% do total de aminoácidos (Tabela
XI). A composição de aminoácidos da fração insolúvel do casu
lo assemelha-se mais àquela do pupário de L. cuprina do que a
qualquer outra apresentada na Tabela XI.
Composicão
do casulo
Cal
Aminoácido ~
. Ac. glutàmico
Ac.aspártico
Serina
Treonina
Glicina
Leucina
Alanina
Valina
Isoleucina
1/2 Cistina
Lisina
Tirosina
Fenilalanina
Prolina
Arginina
Histidina
Metionina
a-alanina
Hidroxilisi~a
L As determinedas por pere
2. Calculado a
3. Segundo GROS
4. Calculado a
5. Segundo LUCJl

Composicão porcentual molar dos aminoácidos da fração insolúvel
do casulo IIr de R. americana e de outros produtos de insetos
TABELA XI
~67-
% em moles
secreção Fibroína4Salivar 3 P.cynthia
C.tentans
PupárioL: cU2~
Casulo 111R. amertca
naAminoácido
dutos naturais
-ista disso, os
s do que aquê-
possível su
uência de va-
ulo 111
solúvel
, aoósétodos.
zero
crito
a e treoni
Lúvel do ca
:::idos (Tabela
.,el do casu-
do que a
- Ác. glutâmico 11,6 10,9 12,79 0,8 9,8
Ác.aspártico 10,9 9,1, 6,18 3,5 5,8
Serina 10,2 8,3 9,39 - - 5,1 11,0
Treonin.a 9,3 4,7 2,96 0,6 3,5
Glicina 8,1 13,1 4,51 31,4 27,5
Leucina 7,3 4,1 1,88 cr,2 2,2
Alanina 6,4 8,0 10,54 47,8 13,8
Valina 6,0 ~,6 2,84 0,6 3,8
Isoleucina 5,5 2,3 -1,96 0,3 2,0
1/2 Cistina 4-,5 1,5 12,88 ...
Lisina 4,2 2,9 11,09 0,3 1,7
Tirosina 4,0 3,2 - 5,6 2,5
Fenilalanina 3,4 3,1 - 0,1 0,4
Prolina 3,1 7,8 2,49 0,3 9,7li
Arginina 2,5 3,8 13,02 1,9 4,0 ill
Histidina 2,0 3,9 --- 1,5 0,5
Metionína 1,0 0,3 7,42
e-alanina - 6,4 --ªidroxilisi~a - - - - 1,8
1. As determinações foram realizadas em hidro1izados ácidos e corrigidas por perdas devido à hidró1ise como descrito em Métodos.
2. Calculado a partir de GILBY e McKELLAR (1970).
3. Segundo GROSSBACH (1969) •
4. Calculado a partir de-LUCAS, SHAW e SMITH (1960) •
5. Segundo LUCAS e RUDALL (1968).

-6-8-
5. Produção e composição do casulo em funçã~ do tempo
° pêso-sêco do casulo em distintas fases de sua pro
dução é mostrado na TABELA XII. Conforme já foi apresentado emMétodos, o casulo III correspondente ao casulo já terminado, en
quanto que o casulo 11 e casulo I correspondem ao casulo produ
zido até o início do 59 período e início do 49 período, respec
tivaroente.
TABELA XII
- - *Aumento do peso-seco do casulo ao longo do desenvolvimento
Deterr
Material
Ca
a-aroino-N
* Os valores determinados são médiasde 3 determinações. Detalhes sãoencontrados em Métodos.
A alteração mais evidente que ocorre no casulo durante
o desenvolvimento é um aumento percentual da fração hidroinso
lúvel-ácido acético solúvel, que foi admitida corresponder ao
caC03
(Tabela XIII). Ocorre também um grande aumento no conteú
do de Ca, como seria de se esperar na base do que já foi assumido
acima e dos resultados apresentados no ítem B.2 anterior. Isso
parece indicar que a deposição de CaC03 no casulo ocorre prefe
rencialmente após o 49 período do 49 estádio.
••
pêso-sêco do casulo
mq!produzido/larva
0,43
1,53
3,47
Casulo
I
Ir
IIl
Fração im
1. Tôdas metrês det
2. Assumidcacéticolúvel ne
3. Determi~
drólisetados safoi apli
4. MaterialáCido co
E
drossolúvel
pode estar
fim do peri
de remoção
sarnento de
Dessa forma
derivado do
o animal el
que, conseq
111. Confo
componente
sa forma as

tempo
:s de sua pro
.presentado em
rminado, en
'asulo produ
Iodo, respec-
*::>lvimento
:ulo durante
LO hidroinso
londer ao
~o no conteú
foi assumido
'ior. Isso
,rre prefe-
TABELA XIII
composição química do casulo em três- 1
etapas do desenvolvimento
Porcentagem do pêso-sêco do casulo
Determinação Casulo I Casulo 11 Casulo 111
Material hidrossolúvel 3,0 3,1 8,4
CaC032 11,0 40,4 43,5
Ca 7,5 19,5 19,0
a-amino-N insolúve1 3 5,18 2,49 2,46
Fração insolúvel 4 84,2· 54,5 45,4
1. Tôdas medidas, salvo indicação em contrário, são médias detrês determinações. Procure Métodos para detalhes.
2. Assumido corresponder ao material hidroinsolúvel e ácidoacético solúvel. A massa de proteínas e carboidratos solúvel nessas condições já foi subtraída.
3. Determinado por reação de ninhidrina após 8 horas de hidrólise alcalina da fração insolúvel do casulo. Os resultados são médias de duas determinações e nenhuma correcãofoi aplicada devido a destruição de aminoácidos sensívêis.
4. Material do casulo remanescente após lavagens com água eácido conforme descrito em Métodos.
E' interessante notar também um aumento na fração hi
drossolúvel do casulo 111 quando comparado com o 11 ou I. Isso
pode estar relacionado com o fato do casulo 111 ser obtido no
fim do período pré-pupal (pupa~farata), quando as larvas são
de remoção difícil do casulo, sendo impossível evitar extrava
samento de flu~exuvial de umas ou de hemolinfa de outras.
Dessa forma, o incremento em material hidrossolúvel poderia ser
derivado dos fluIdos acima referidos, assim como das fezes que
o animal elimina normalmente durante todo o feitio do casulo e
que, consequentemente, ocorrem em maior quantidade no casulo
111. Conforme já foi apresentado anteriormente, proteína é o
componente majoritário da fração insolúvel do casulo 111. Des
sa forma as determinações de a-amino N insolúvel, permitem uma
I
,.'•

-70-
comparação preliminar da composição da fracão insolúvel nas
três fases estudadas. Os resultados da Tabela XIII permitem
calcular que a porcentagem de a-amino N na fração insolúvel dos
casulos I, 11 e 111 é: 6,16, 4,57 e 5,40 respectivamente. Dessa
forma pode-se admitir tentativamente que a com~osição da fração
insolúvel do casulo varia um pouéo com o desenvolvimento, sendo
a fração do casulo I mais rica em proteínas e aquela do casulo
11 a mais pobre porcentualmente.
Tentar obter valores para o conteúdo de proteínas in
solúveis a partir dêsses dados de a-amino N é arriscado. Entre
tanto, partindo dos valores de a-amino N para o casulo 111, ass~
mindo um pêso-molecular médio para o resíduo de aminoácido de
120 e considerando que a recuperação em um hidrolizado alcalino
de aibumina realizado e calculado corno essas amostras de casulo
é de 74%, obtemos um valor de cêrca de 28,6% para as proteínas
insolúveis. Êsse valor concorda de forma razoável com aquêle
obtido de forma mais rigorosa apresentado na Tabela VII.
A Tabela XIV mostra a massa produzida dos principais
componentes do casulo nas três fases estudadas, enquanto que a
Tabela XV_ mostra a velocidade de produção désses componentes,
nas mesmas fases.
TABELA XIV
Produção dos componentes do casulo ao lonqo do*desenvolvimento
Intervalos estudados
Composto 39P--149P 49P-159P 59P-lpupa Total
Proteína insolúvel 268 350 552 1170
Carboidrato insolúvel 82 106 170 358
CaC0 3 47 572 891 1510
* Os dados foram calculados das Tabelas XII e XIII, assumindoque a fração insolúvel dos casulos I eII tem a mesma composição que aquela do casulo 111. Todos os resultados estãoexpressos em ~g produzido/larva.
Oiz
dia anterior ide velocidade
tervalo do 59
deração baseie
na~se uma pupc
corno uma pupa
a produzir caE
los: 39P -:., 4<;
dias respecti"
dura 3 dias.
Tabela XV cor!
superestimadas
Velocidade Ir
Composto
Proteína inso
Carboidrato i
cac03
* Dados calculvem.·.ser procem lIg produz
** ~sses valore
Os res
ponentes do casu
mente entre o 59
de que enquanto
duas vêzes maior
a produção de Ca'
entre os process,
teínas e carbOid
dução de CaC0 3 , !
maneira idênticapu~e.. farata.

olúvel nas
11 permitem
solúvel dos
fite. Dessa
éla fração
nto, sendo
do casulo
oteínas in
do. Entre
o 111, ass~
noácido de
do alcalino
s de casulo
as proteínas
com aquêle
II .
principais
anto que a
:lmponentes,
° intervalo de produção de casulo do 59 periodo até o
dia anterior à.muda pupal foi considerado, para fins de cálculo
de velocidade de produção do casulo, como correspondente ao in
tervalo do 59 periodo até a última apólise larval. Essa consi
deração baseia-se no fato de que após essa apólise o inseto tor
na~se uma pupa farata, funcionando do ponto de vista fisiológico
como uma pupa (HINTON, 1946), . sendo en tão incapaz de continuar
a produzir casulo. ° número de dias correspondente aos interva
los: 39P --I 49P, 49P --I 59P e 59P -4 pupa farata são: 5, 3 e 2
dias respectivamente. Às vêzes o inter~alo 59P ~ pupa farata
dura 3 dias. Consequentemente as velocidades apresentadas na
Tabela XV correspondentes a êsse intervalo podem estar um pouco
superestimadas.
TABELA XV
Velocidade média de produção dos diferentes constituintes do
casulo em três etapas de sua formação*
Intervalos estudados
39P=-J 49P 49P=-J 59P 59P -4 pupa farata**Composto
Proteina insolúvel 54 117 276
Carboidrato insolúvel 16 35 85
CaC03 9,4 191 446
I
:lo*
**
Dados calculados a partir da Tabela XIV. Maiores informações devem ser procuradas no texto. Todos os resultados estão expressõsem ~g produzidos/individuo/dia.
~sses valores podem estar um pouco superestimados. Ver texto.
:Idos--
-Ipupa Total
552 1170
l70 358
391 1510
3.ssumindo3. compo-
estão
Os resultados indicam que a produção de todos os com
ponentes do casulo é mais intensa em tõrno do 59 periodo, especial
mente entre o 59P e pupa farata. E' interessante assinalar o fato
de que enquanto a produção de proteinas e carboidratos insolúveis é
duas vêzes maior no intervalo 49P ~ 59P em relação ao 39P ~ 49P,
a produção de caC03 é 20 vêzes maior. Isso sugere uma independência
entre os processos de produção de CaC0 3 e aquêles de produçao de PEo
teinas e carboidratos insolúveis. Entretanto, a velocidade de pro
dução de caC0 3 , proteínas e carboidratos insolúveis é aumentada de
maneira idêntica ao se passar do intervalo 49P ~ 59P para 59P -1pu~a farata.
!.

-12-
C. Aspectos químicos e fisiológicos durante o 4~ estádio de
R. americana
1. Massa cor~órea e volume de hemolinfa
A larva de R. americana aumenta ràpidamente de pêso
até alguns dias depois da 3~ muda, isto é, até cêrca de 26 dias
de idade (Figura 8). Em seguida ainda cresce logaritmicamente
até ficar madura, isto é, atingir o máximo desenvolvimento lar
vaI. A~partir de então o inseto pára de comer e começa a prod~
zir o casulo e a perder massa até a emergência do adulto (70
dias de idade). Uma inspecção na figura 8 mostra que os desvios
das médias de massas medidas em larvas com menos do que 48 dias
(com uma exceção) são pequenos. Êsse fato torna possível a uti
lização da presente curva para a estimativa da idade fisiológi
ca de larvas, a partir da medida de sua massa. Essa estimativa
é muito útil, especialmente para insetos recolhidos na natureza,
porque antes dos 48 dias de idade existe apenas um marcador da
idade fisiológica larval: a mudança de côr da larva, que ocor
re em tôrno dos 32 dias de idade.
TOKU~~U, FERRI e WORSM1U~ (1968) obtiveram resulta
dos muito discrepanteS dos aqui apresentados, chegando a pe
sar indivi~u0s ainda na fase larval com 80 dias de idade, em
contraste também com as observações de outros autores (Cf. GUA~
CIABA e TOLEDO, 1969). Essas diferenças encontradas por TOKUMA
RU, FERRI e WORSMANN, podem ser explicados por uma má termoesta
tização do ambiente de criação das larvas.
A determinação dos volumes de hemolinfa presente nas
larvas e pupas de ~_americana foi sempre realizado pelo método
de exanguinação (Tabela XVI). Isto justifica-se pelo fato (ver
comentários em Métodos) de que os resultados obtidos por esta
técnica e pela técnica de diluição de corante serem semelhantes
em larvas com idades inferiores a 35 dias (Tabela XVII). Figura 8 - COs traços vecadas, obtidnimo de 5 deidade em diaperíodos do
assim CI

:;> estádio de I
~, que ocor-
presente nas
pelo método
1,0
0.7~
Jli
o.'f I39/4i1fp: IPv~110,4- II~ I 22
0.3
10 2:0 JO +O 60 60 70
OiAS APOS A fCLOSÃO
+0,0
30,0
60,0
20,0
70,0
10,0
-.c:n 7,0E...,
6,0<If)
~of)
<I
E",O
2,0
Figura 8 - Crescimento e~ massa de fêmeas de R. americana a 200 C.Os traços verticais representam o desvio padrão das médias indicadas, obtidas em 3 grupos independentes de insetos, após um mínimo de 5 determinações em cada. Embaixo são indicados,além daidade em dias após a eclosão. os intervalos correspondentes aosperíodos do 49 estádio; 19, 29, 39, 49, 59, 69, prê-pupal (PP),- assim como do período pupal (ver detalhes em Métodos).
(ver.0 fato
por esta
semelhantes
'Ir) •
~ram resulta
~gando a pe
~ idade, em
!S (Cf. GUAR?;
; por TOKUMA
lá termoesta-
nte de peso
ca de 26 dias
aritmicamente
lvimento lar
omeça a prod~
::> adulto (70
Lle os desvios
::> que 48 dias
:;sível a uti-
:1e fisiológi
:;a estimativa
na natureza,
n marcador da

-74-
TABELA XVI
Volume, proteína, a-amino-N ácido solúvel e densidade na hemo*linfa de R. americana durante o desenvolvimento larval
Est! Período Idade Volume de Massa de ~-aminb N Densidade da
dio (dias) hemo1infa proteína p moles hemo1infa Idade\l11 larva ;\lg/larva larva g/ml
IIr 20 3,28 18 0,32 1,013
**19P 22 6,0 32 0,53
19P 27 9,5 54 0,91 1,023 *** De
-19P 30 13,6
29P 32 14,8 96
** A ra29P 34 16,8 151 2,02
(0,401 :!:** tante29P 35 17,2 201 tádio, isto é,29P 38 16,7 267 fato permite a
29P 41 19,2 542 1,53 1,026 vas nao madura
IV29P 44 20,6 1,89 A ra
29P 45 20,6 887 2,00 1,032 vas maduras é
e 59 ela é de29P 48 24,6 1700 2,31 1,043 de-se concluir
39P 49 17,9 1790 2,29 1,037 damente do que
49P 54 14,3 1890 1,30 1,058 nidades. Isso
ra pupa deve h59P 57 9,4 1190 0,93 1,057 tecidos. Ess
64 4,0 488 1,025 vantada abaixopupa
de água para o
* A metodologia empregada está descrita em Métodos.
** Volumes estimados usando a razão volume de hemolinfa (vermassa da larva
texto) •

TABELA XVII
:idade na hemo*I larval
Densidade dahemolinfa
g/ml
1,013
Volumes de hemolinfa em larvas de R. americana*usando duas técnicas difere~tes
Volume (~l/larva)
Idade (dias) Exanguinação Diluição de corante
27 9,5 9,3
32 14,8 17,7
1,023* Detalhes em Métodos.
1,026
1,032
1,043
1,037
1,058
1,057
1,025
~ (ver'a
A razão volume de hemolinfa e massa de larva é cons
tante (0,401 : 0,009 ~l/mg) durante considerável parte do 49 es
tádio, isto é, de larvas com 22 dias até 42 dias de idade. ~sse
fato permite a estimativa rápida do volume de hemolinfa de lar
vas não maduras, conhecendo sua massa.
A razão volume de hemolinfa e massa corpórea de lar
vas maduras é de 0,31 - 0,39 ~l/mg, durante os períodos 39, 49
e 59 ela é de 0,24 - 0,32 ~l/mg e na pupa 0,11 - 0,15 ~l/mg. P~
de-se concluir então que o volume de hemolinfa diminui mais ràp!
damente do que o pêso-sêco do animal em pelo menos duas oportu
nidades. Isso indica que de larvas- maduras para 39P e de 59P p~
ra pupa deve haver transferência de água da hemolinfa para os
tecidos. Essa interpretação está de acôrdo com a hipótese le
vantada abaixo da hemolinfa servir como importante reservatório
de água para o animal.
11
IÚ

TABELA XVIII
pêso-sêco e conteúdo de água das larvas de R. americana imedia
tamente antes do início e após o término do casulo
2. Determinações químicas na hemolinfa
Os resultados das determinações de proteína, carboi
dratos e a-amino-N ácido solúvel e densidade da hemolinfa en
contram-se na Tabela XVI e Tabela XIX. ~sses mesmos· resulta
dos estão também representados na figura 9.
Os resultados apresentados na Tabela XVIII permitem
calcular que a hemolinfa é respónsável por 37% do pêso-fresco
e 26% do pêso-séco de larvas maduras, o que lhe garante urna
imoortância, pelo menos quantitativa, na vida do inseto. Além
disso, êsses resultados levam à conclusão de que a hemolinfa
é importante reservatório de água para o animal, pois contém
39% da água das larvas maduras e contribui com 53% da água
perdida pela larva durante a produção do casulo, isto e, entre
o fim do 29 período e o período pré-pupal.
OnO}II'O"'1
onO)l\lowl/6n
ono} 1\1 o NII S3' o
000 ..0'<co-
'o Olli ....~~
-::r-::::>~
' ...D:...
-..--'"TOIot
o 01... li: '"<Il: oQ CI
Õz ~
UI(D :E 0-I>: <t o.. I a:<) 't Q.
1.,II
I.. OI
'"
3,7
6,5
22,4
pré-pupal, 63
10,7
58,7
22,9
Período e idade (dias)
29P, 48
Média de duas determinações. Resultados expressos em . mg/larva.
- Calculado a partir da diferença entre pêsofresco e pêso-sêco da larva ou hemolinfa.
1<Determinação
pêso-sêco da larva
*
**Âgua, total
**Âgua, hemolinfa
**

o o o oOnOllllOWI 16(( "1l13.l01:ld o o o o. UI ~ co •
I I
:eína, I o o o ocarboi- 000)11101111 6rl 0.1"1:1010911\1:> ... N -ot
lemolinfa en-;mos· resulta- • o O
OnO!1II0WI/U'Ol'll'f N-ONIH1f,. ~ q.OI
o+lX(l)+l
~(l):>
P=;,
(l)'O
co'l-Is::..;.-Io(:;(l)
.c::coc[J)
o+lco~
'O..;o.o~cot.J
(l)
j
coccou''';~(l)(:;co
cos::
*1(l)+lo~p..
.-I<ll:>
'::l.-IoUl
o'O..-iU
1/01ZIos::''';
~Il:l
oC'l
oUI
O
'"
O
'"
o
""
C1
filO'o -<I
Ao lD<I o
..Icn C)
<I ...Õ
'.
"" ,-,.,",
'-.,,I
II,
\ Il, ., \
r( '.\'" .......<J.-...._~
.............. --..... .-., .. -"<1" ",
",' ..,,,,,
",8,,,
,:<fl/
./"/
/"
1'"i
\"\,\
\\
OIot
~
?:,
OIOI
00
~.~Q'"
'o OIã<1LI'" o'Q.'"~
~::>~
' ...a:...-"r-
~.
o... z<c o <IQ
ÕZ Z- lU
'" :lO t-a: ... o<I I a:u 't n.
1.,II
I..
(dias)
6,5
2,4
3,7
t:ados ex-
lupal, 63
XVIII permitem
lo pêso-fresco
garante uma
inseto. Além
e a hemolinfa
pois contém
53% da água
isto é, entre
unericana imedia
) do casulo
re pêsolinfa.

-78-
TABELA XIX
Conteúdo de carboidratos na hemolinfa durante o
desenvolvimento de R. americana
Idade jJg/indivíduoPerIodo (dias) Carboidrato Trealose Glicogênio
19P 30 13.,1 li,4 0,0
29P 32 32,3 26,2 0,2
29P 48 49,4 34,4 0,3
39P 49 35,8 24,4 0,3
49P 54 46,5 28,2 0,2
59P 57 47,5 29,4 0,3
Pupa 64 34,6 27,6
Durante o 39 estádio e 49 estádio até a idade de 48
dias ocorre um acúmulo generalizado de compostos na hemolin
fa, resultando em um aumento de sua densidade. Após o início da
produção do casulo ocorre ainda um ligeiro aumento na massa de
proteínas, seguido de uma queda grande até o fim do período est~
dado (pupa). Os compostos ninhidrina-positivos ácido solúveis (a
amino-N ácido solúvel} diminuem em massa desde o início da produ
ção do casulo até a fase de pupa. Carboidrato total cai no iní
cio da produção do casulo (39P), em seguida retorna ao valor ini
cial e se mantém estável até a transformação em pup~ quando cai
novamente. Trealose é o carboidrato predominante na hemolinfa de
R. americana durante todo o desenvolvimento larval, enquanto
que glicogênio ocorre em massa insignificante.
o início da elevação do conteúdo de proteínas na hem~
linfa (cêrca de 30 dias de idade), coincide com a acentuação da
côr avermelhada das larvas o que marca o inIcio do 29 período do
49 estádio.
Composição de ar
de R. amE
Componente
Arginina
Âcido aspárticl
Treonina
Serina
Prolina
Âcido glutâmic·
Glicina
Alanina
Valina
Metionina
Isoleucina
Leucina
Tirosina
Fenilalanina
Glutamina + Ai!:
Amônia
Lisina
Histidina
Triptofano
Fosfoetanolamj
Cistationina
Fosfoserina
Ornitina
Peptídeos
Total de amin<
Total geral
* Os valores
** Os numeros I
em dias apó
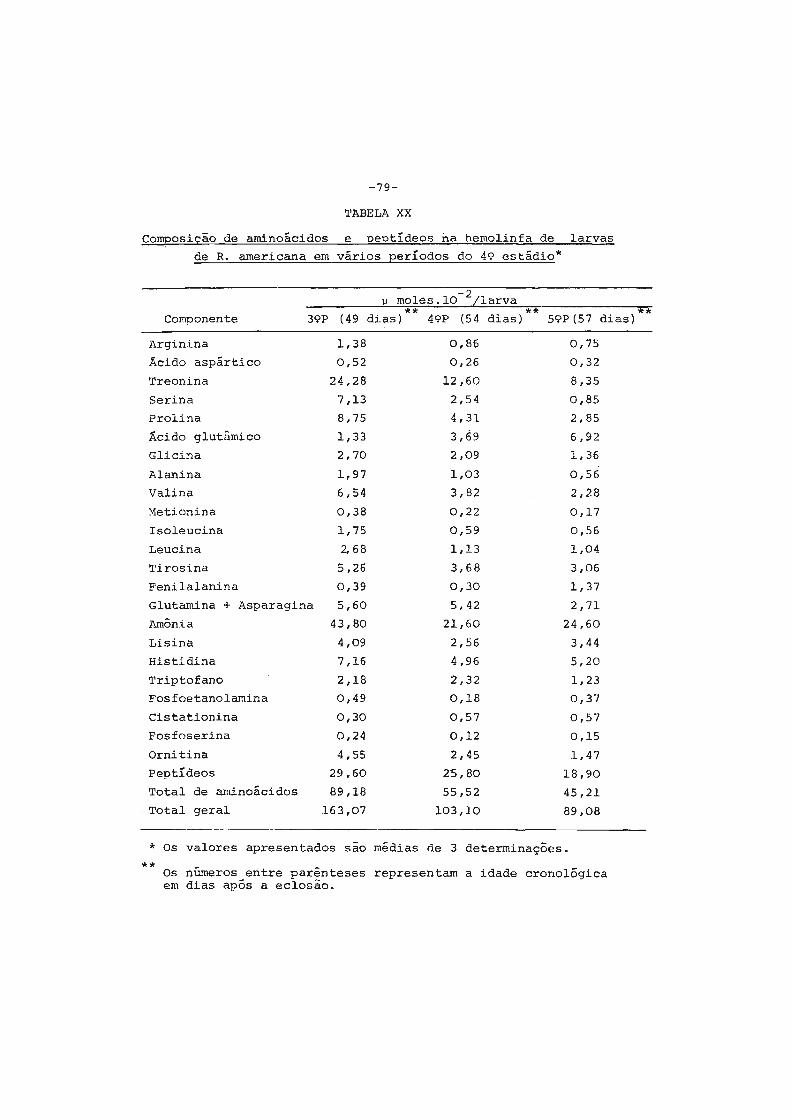
! o
3"ênio
3
ldade de 48
na hemolin
início da
massa de
1:!ríodo est~
solúvois(a
j da produ
:li no iní-
valor ini
uando cai
:amo1infa de
enquanto
nas na hemo
:antuação da
período do
-79-
TABELA XX
Composição de aminoácidos e peptídeos na hemo1infa de larvas
de R. americana em vários períodos do 49 estádio*
-2~ moles.lO jlarva
** ** **Componente 39P (49 dias) 49P (54 dias) 59P(57 dias)
Arginina 1,38 0,86 0,75
Âcido aspártico 0,52 0,26 0,32
Treonina 24,28 12,60 8,35
Serina 7,13 2,54 0,85
Prolina 8,75 4,31 2,85
Âcido glutâmico 1,33 3,69 6,92
Glicina 2,70 2,09 1,36
Alanina 1,97 1,03 0,56
Valina 6,54 3,82 2,28
Metionina 0,38 0,22 0,17
Isoleucina 1,75 0,59 0,56
Leucina 2,68 1,13 1,04
Tirosina 5,26 3,68 3,06
Fenilalanina 0,39 0,30 1,37
Glutamina + Asparagina 5,60 5,42 2,71
Amônia 43,80 21,60 24,60
Lisina 4,09 2,56 3,44
Histidina 7,16 4,96 5,20
Triptofano 2,18 2,32 1,23
Fosfoetanolarnina 0,49 0,18 0,37
Cistationina 0,30 0,57 0,57
Fosfoserina 0,24 0,12 0,15
Ornitina 4,55 2,45 1,47
Peptídeos 29,60 25,80 18,90
Total de aminoácidos 89,18 55,52 45,21
. Total geral 163,07 103,10 89,08
* Os valores apresentados são médias de 3 determinações.
** ~ - -Os numeros entre parenteses representam a idade cronologicaem dias apôs a eclosão.
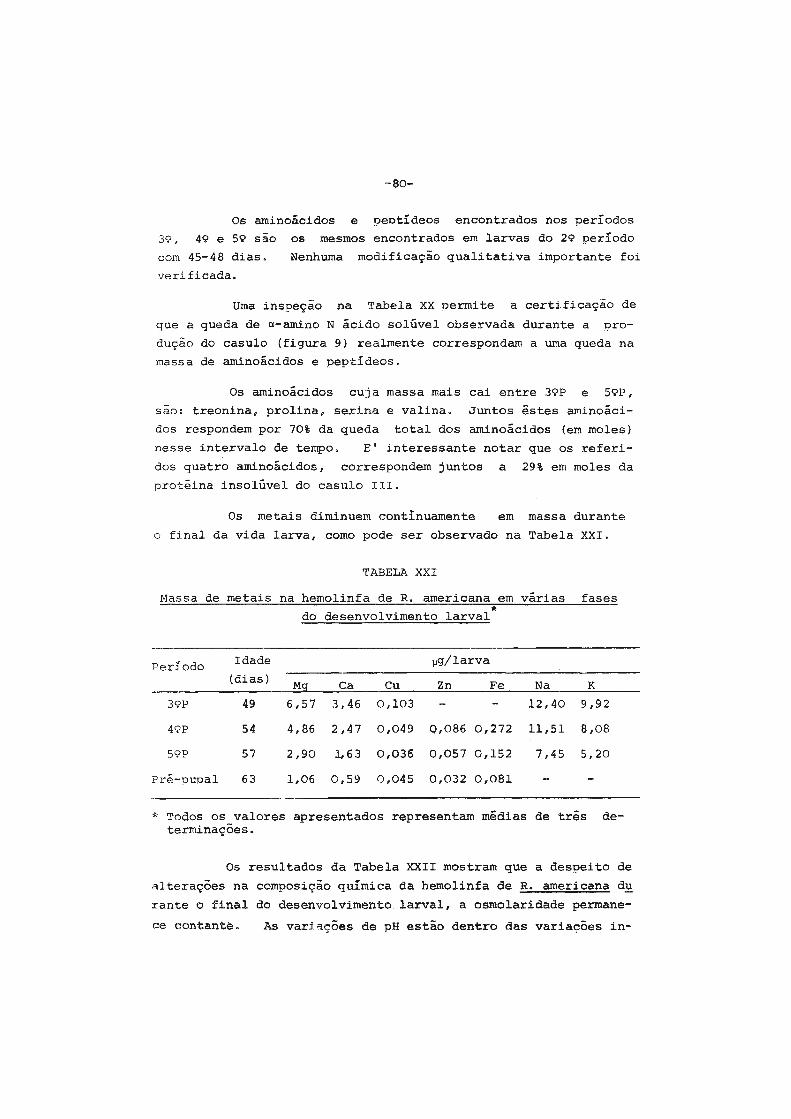
1
-80-
Os aminoácidos e peptídeos encontrados nos períodos
39, 49 e 59 sao os mesmos encontrados em larvas do 29 período
com 45-48 dias. Nenhuma modificação qualitativa importante foi
verificada.
Urna inspeção na Tabela xx permite a certificação de
que a queda de a-amino N ácido solúvel observada durante a pro
dução do casulo (figura 9) realmente correspondam a uma queda na
massa de aminoácidos e peptídeos.
dividuais, c
ce realmente
Os
rante o dese
Osmolaridade
Os aminoácidos cuja massa mais cai entre 39P e 59P,
sao: treonina, prolina, serina e valina. Juntos êstes aminoáci
dos respondem por 70% da queda total dos aminoácidos (em moles)
nesse intervalo de tempo. E' interessante notar que os referi
dos quatro aminoácidos, correspondem juntos a 29% em moles da
protéina insolúvel do casulo 111.
Período
39P
49P
59P
I,(d.
TABELA XXI
o final da vida larva, como pode ser observado na Tabela XXI.
Os metais diminuem continuamente em massa durante"I<
Todos os v,naçoes comminação re,
Massa de metais na hemolinfa de R. americana em várias fases*do desenvolvimento larval 3. pêso-sÉ
Os resultados da Tabela XXII mostram que a despeito de
alterações na composição química da hernolinfa de R. americana d~
rante o final do desenvolvimento larval, a osmolaridade permane-
Período Idade )Jg/larva
(dias) Mg Ca Cu Zn Fe Na K
39P 49 6,57 3,46 0,103 12,40 9,92
49P 54 4,86 2,47 0,049 0,086 0,272 11,51 8,08
59P 57 2,90 1, 63 0,036 0,057 0,152 7,45 5,20
pré-pupal 63 1,06 0,59 0,045 0,032 0,081
* Todos os valores apresentados representam médias de três de-terminações.
ce contante. As variações de pH estão dentro das variações in-
o c
porções princ
férica dispos
segmentarment
pequena em re
di fici I em la
roso central
todo o desenv
quando então
(Tabela XXIII
bola (WIGGLES
o c
vel, crescenà
responde a 18
guida até pup

nos períodos
lo 29 período
.mportante foi
~tificação de
'ante a pro
uma queda na
-81-
dividuais, com a possível exceção daquela do 49 P, cujo pH pare
ce realmente ser ligeiramente mais baixo que dos demais períodos.
Os valores par.a cloreto e amônia sofrem alterações du
rante o desenvolvimento.
TABELA XXII
Osmolaridade, pH, cloreto e amônia na hemolinfa de R. americana*em diferentes idades
Concentração (mM)Cloreto NH3
39P e 59P,
es aminoáci-
s (em moles)
e os referi
em moles da
Periodo
39P
49P
59P
Idade(dias)
49
54
57
18,3
25,0
23,0
24,5
15,1
26,2
Osmolaridade(m. osmoles)
217
216
209
pH
7,31
7,18
7,28ssa durante
Jela XXI.
Las fases
K
40 9,92
51 8,08
45 5,20
três de-
despeito de
mericana d,!!
de permane
riações in-
* - -Todos os valores da Tabela representam medias de tres determi-nações com exceção de pH, que corresponde a apenas uma determinação realizada a 250 C.
3. pêso-sêco e glicogênio no corpo gorduroso
O corpo gorduroso de R. americana é formado por duas
porções principais: uma central bastante volumosa e outra peri
férica disposta em pequenos aglomevados laterais organizados
segmentarmente. Essa porção periférica apresenta massa muito
pequena em relação ao corpo gorduroso central e é de dissecção
dificil em larvas jovens. .Devido a isso, sômente o corpo gordu
roso central foi pesquisado. ~sse aumenta em pêso-sêco duran
todo o desenvolvimento da larva até essa atingir a maturidade,
quando então passa a corresponder a 38% do pêso-sêco da larva
(Tabela XXIII). Valores dessa magnitude são comuns em Holomet~.
bola (WIGGLESWORTH, 1965).
O conteúdo de glicogênio do corpo. gorduroso é apreciá
vel, crescendo até atingir um máximo no 49 período, quando cor
responde a 18% do pêso-sêco do corpo gorduroso, declinando em s~
guida até pupa (Tabela XXIII).
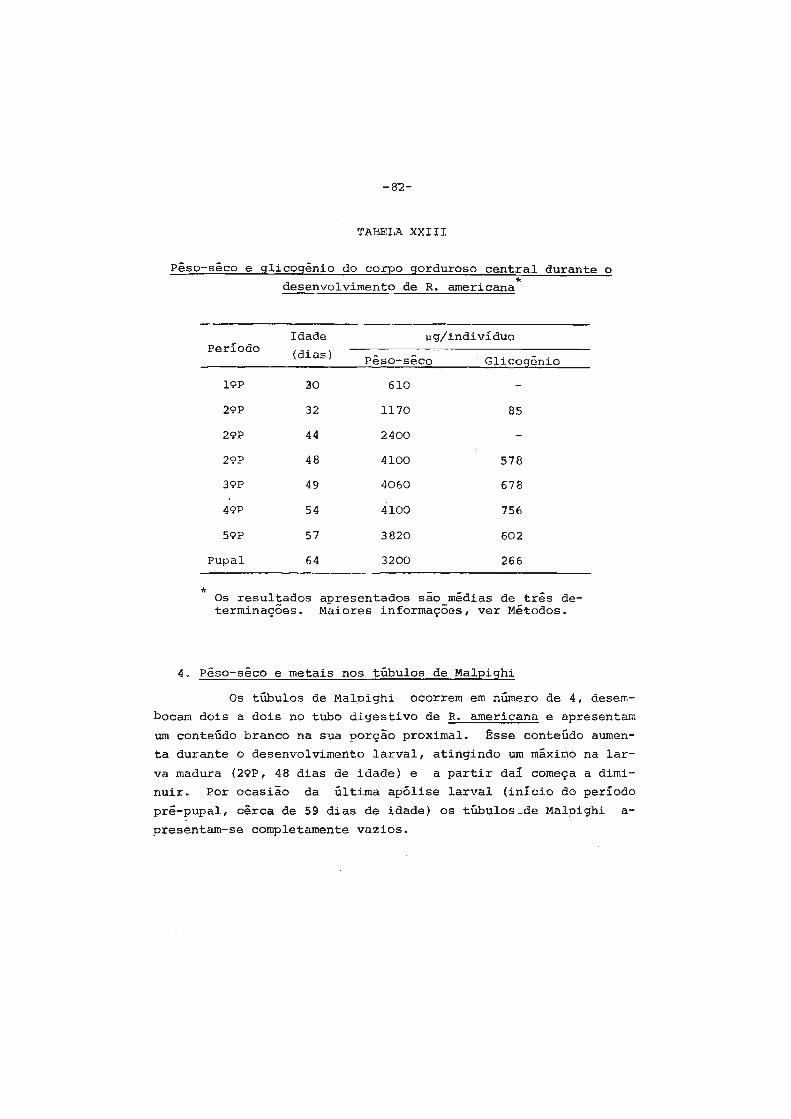
-82-
TABELA XXIII
pêso-sêco e glicogênio do corpo gorduroso central durante o*desenvolvimento de R. americana
Pe.
Pré·
pêso-sêc
Glicogênio
\lg/indivíduo
pêso-sêco
Idade
(dias)Período
19P
29P
30
32
610
1170 85
* CóSI
29P
29P
39P
49P
44
48
49
54
2400
4100
4060
4100
578
678
756
A Te
pighi repleto!
seu total eSVi
rença entre OI
branco: 1,30 r
o/<Os resultados apresentados são médias de três de-terminações. Maiores informações, ver Métodos.
59P
Pupal
57
64
3820
3200
602
266o c(
fervescente ell
bonatos. A o(
Malpighi já f(
clusive Diptel
4. pêso-sêco e metais nos túbulos de Malpighi
Os túbulos de Malpighi ocorrem em número de 4, desem
bocam dois a dois no tubo digestivo de R. americana e apresentam
um conteúdo branco na sua porção proximal. ~sse conteúdo aumen
ta durante o desenvolvimento larval, atingindo um máximo na lar
va madura (29P, 48 dias de idade) e a partir daI começa a dimi
nuir. Por ocasião da última apólise larval (início do período
pré-pupal, cêrca de 59 dias de idade) os túbulos~de Malpighi a
presentam-se completamente vazios.
A me;
bastante granê
dessa determir.
··sente nos túb\;
cia dêsse vale
referido (Tab
realmente CaCO
A ma
presente nos t
conteúdo de Ca
sulo (Tabela X
lo 111 e nos t
'cerm1nada expe.
ver os túbulos
seu conteúdo.

lrante o
de-
4, desem
apresentam
údo aumen
mo na lar
ça a dimi
do período
lpighi a-
-il3
TABELA XXIV
pêso-sêco dos tUbulos de Malpighi antes e após a formação
do casulo
Período Idade mg/indivíduo*(dias) pêso-sêco caC0
32QP 48 1,39 1,35
Pré-pupal í3 0,09
* Calculado a partir do conteúdo de Ca apresentado na Tabela XXV.
A Tabela XXIV mostra o pêso-sêco dos tUbulos de Mal
pighi repletos com o conteúdo branco na larva madura, e após o
seu total esvaziamento, depois da última apólise larval. A dif~
rença entre os dois valores permite estimar a massa do conteúdo
branco: 1,30 mg/indivíduo.
o conteúdo branco dos tUbulos de Malpighi torna-se e
fervescente em ácido acético 10%, o que sugere a presença de ca~
bonatos. A ocorrência de carbonato de cálcio em tUbulos de
Malpighi já foi documentada em larvas de numerosos insetos, in
clusive Diptera (CLARK, 1958).
A massa de cálcio presente nos túbulos de Malpighi é
bastante grande, como pode ser vista na Tabela XXV. A partir
dessa determinação de Ca pode-se estimar a massa de CaC03, pre-
··sente nos tUbulos de Malpighi: 1,35 mg/indivíduo. A concordân
cia dêsse valor com a massa de conteúdo dos túbulos de Malpighi
referido (Tabela XXIV), reforça a hipótese de que o conteúdo érealmente CaC0 3 .
A massa de Ca presente no casulo III é similar àquela
presente nos túbulos de Malpighi e muito superior à queda no
conteúdo de Ca verificada na hemolinfa durante a produção do ca
sulo (Tabela XXV). A semelhança entre o conteúdo de Ca no cas~
lo III e nos tllbulos de Malpighi deve ser ainda maior que a de
term~nada experimentalmente, devido à impossibilidade de se rem2
ver os túbulos de Malpighi sem perda de uma pequena fração de
seu conteúdo.

;;'84-
TABELA XXV
Determinação de metais nos túbulos de Malpighi em larvas maduras
de R. americana e comparação com o conteúdo de metais do casulo
111 e com a queda dêsses verificada na hemolinfa durante a pro-
dução do casulo
Jlg!individuo
Metais Hemolinfa 1 Malpighi 2 Casulo 111 3
Zn 1,30 0,83
Cu 0,058 0,17 0,056
Ca 2,61 540 660
K 36 0,031
Mg 5,51 76 0,056
Fe 5,0 3,75
1. Queda durante a produção do casulo. Calculado a partir das Tabelas V, XVI e XXI.
2. Determinado em animais do 29 período, 48 dias de idade. Maiores informações, ver Métodos.
3. Calculado a partir das Tabelas IX e XII.
A. Quími ca de
1. Extens~
A '1
já" apresentac
a extensão cc
maduras de B..de 88% do pês
tre os inse
mori e Hyal
cerca de 96
1"961) .
A f
na deve corre
dos orgânicos
proteínas. E:
proteínas con:
vadas em inse"
ser responsávl
te como g li co<
LHO (1969) qUE
cas separadas
1arvas de R. e

larvas maduras
ais do casulo
durante a pro-
:lsulo III 3
0,83
0,056
i60
0,031
0,056
3,75
llado a par-
ias de ida-
DISCUSSÃO
A. Química da hemolinfa
1. Extensão da análise realizada
A Tabela XXVI apresenta uma compilação de resultados
já apresentados, porém agora na forma adequada para se estimar
a extensão coberta pela análise realizada da hemolinfa de larvas
maduras de R. americana. Como se vê, a análise abrangeu cêrca
de 88% do pêso-sêco da hemolinfa, o que coloca R. americana en
tre os insetos de hemolinfa melhor conhecida, próximo a Bombyx
mori e Hyalophora cecropia cujas hemolinfas são conhecidas· em
cêrca de 96 e 92% de seu pêso-sêco, respectivamente (WYATT,
1"961) .
A fração não identificada da hemolinfa de R. america
~ deve corresponder, entre possíveis outros compostos, a áci
dos orgânicos (ver abaixo, ítem 3) ea carboidratos ligados às
proteínas. Esta última suposição está baseada no fato de que·
proteínas conjugadas a carboidratos ocorrem em concentrações el~
vadas em insetos (SIAKOTOS, 1960a). Tais carboidratos parecem
ser responsáveis por parte dos componentes determinados usualme~
te como glicogênio (WYATT, 1961) e pela observação de TOLEDO FI
LHO (1969) que encontrou reação PAS positiva nas 9 bandas protéi
cas separadas poreletroforese em gel de agar da hemolinfa de
larvas de R. americana.

composição porcentual em massa do resíduo sêco da hemolinfa de
larvas maduras de R.americana*
Os valores apresentados foram calculados a partir das tabelas 11 e V. A concentração dos compostos fosforiladosorgânicos não identificados foi calculada a partir de dedeterminação de P-orgânico ácido· solúvel (P-ácido solúvel-P-inorgânico) da Tabela V, assumindo que êste represen-~té:15% em pêso dos referidos compostos, seguido da subtração do título de fosfoetanolamina e fosfoserina. Ver discussão no texto.
-86-
TABELA XXVI
Composto
Proteína
Aminoácidos
Lipídeos
Peptídeos
.Citrato
Compostos fosforilados orgânicos
não identificados
Metais
Carboidratos
Cloreto
Ortofosfato
Fosfoetanolamina
Glicogênio
. Total
pêso-sêco
*
mg/ml % pêso-sêco
69;00 61,5
8,22 7,3
5,20 4,6
4,23 3,8
3,91 3,5
2,77 2,5
2,71 2,4
2,01 1,8
0,48 0,4
0,30 0,3
0,18 0,2
0,01 0,0
99,02 88,3
112,30 100,0
2. componentes
TUNDI
aminoácidos liv
matografia em 1=
Seus resultados
gura 1, embora
tídeos por êles
figura. ° fate
nina, fenilalan
nina alêm de 11
te se deve à me
liti co em relaç
° coné bastante elev
les merecem uma
Cista
em concentraçõe
não se detecta
11). Isso pode
deriva-se da me
moserina devido
o caso de Boinby
teina é um amin
demonstrado no
1948).
Argin
dos, podendo se
por ornitina, o
ornitina -->
tina completo e:
por INOKUCHI, Ri
dão uma evidênc.
lo menos em Bom
ser explicada, ,
na exógena, pore
rosos de vários
deria ser forma,

olinfa de
êso-sêco
61,5
7,3
4,6
3,8
3,5
2,5
2,4
1,8
0,4
0,3
0,2
0,0
88,3
00,0
s tari1adosde . de-
solúpresensubtrar dis-
-87-
2. Componentes orgânicos da hemolinfa
TUNDISI et alii (1964) em uma análise qualitativa dos
aminoácidos livres da hemolinfa de R. americana utilizando cro
matografia em papel, identificaram 15 aminoácidos e 2 peptídeos.
Seus resultados estão de acõrdo com aquêles apresentados na fi
gura 1, embora nenhuma correlação possa ser feita entre os pep
tideos por êles identificados e· aquêles apresentados na referida
figura. O fato dos referidos autores não terem encontrado metiQ
nina, fenilalanina, triptofano, arginína, fosfoserina, cistatio
nina além de 11 outros compostos ninhidrina-positivos, certamen
te se deve à menor resolução e sensibilidade de seu sistema ana
lítico em relação ao utilizado nesta tese.
O conteúdo de aminoácidos na hemolinfa de R. americana
é bastante elevado, como foi indicado em Resultados e vários dê
les merecem uma discussão particular.
Cistationina acumula-se na hemolinfa de R. americana
em concentrações superiores até que ametionina, enquanto que
não se detecta nem mesmo traços de cisteína ou cistina (Tabela
lI). Isso poderia ser explicado admitindo-se que a cistationina
deriva-se da metionina e que ela não é clivada em cisteina e ho
moserina devido a ausência da enzima apropriada, como parece ser
o caso de Bombyx mori (GILMOUR, 1965). Isso indicaria que cis
teina é um aminoácido essencial para R. americana como já foi
demonstrado no Diptera Nematocera~· (GOLDBERG e De MEILLON,
1948) •
Arginina é ~equerida na dieta de todos insetos estuda
dos, podendo ser substituída parcialmente por citrulina, mas não
por ornitina, o que levou a se propor a não ocorrência do passo
ornitina ----> citrulina e consequentemente de um ciclo da orni
tina completo em insetos (GILMOUR, 1961). Os resultados obtidos
por INOKUCHI, HORIE e ITO (1969) usando marcadores radioativos
dão uma evidência direta de que o referido passo não ocorre, pe
lo menos em Bombyx mori. A presença de ornitina e uréia poderia
ser explicada, entretanto, pela ação da arginase sõbre a argini
na exégena, porque arginase (EC 3.5.3.1) ocorre nos corpos gord~
rosos de vários insetos pesquisados (GILMOUR, 1965). A uréia po
deria ser formada ainda a partir de ác~do a1antéico sob ação da
J••

-88-
alantoicase (EC 3.5.3.4), como ocorre com o grilo Acheta domes
~ (GILMOUR, 1965). Nenhuma informação existe sôbre a possi
bilidade da ornitina se derivar do ácido glutâmico ou prolina,
como acorre em vários animais (Cf. GLIMOUR, 1965).
A partir do exp.:'sto acima, pode-se admitir que o acú
mulo de ornitina verificado na hemolinfa de R. americana e a au
sência de traços de citrulina seja consequência da formação de
ornitina a p~rtir da arginina exógena e sua impossibilidade de
poder dar origem à arginina, devido a inexistência do passo ornl
tina ---> citrulina. A uréia presente em traços de hemolinfa
poderia ter surgido pela arginase (EC 3.5.3.1) ou, menos provà
velmente, pela alantoicase (EC 3.5.3.4), como descrito acima.
As concentrações de tirosina.na hemolinfa de R. ameri
~ sao bastante elevadas, chegando a 3,26 mM no 59 período,
que corresponde aproximadamente ao limite de solubilidade da ti
rosina em água a 250 C (0,05% p/P). Entretanto, concentrações
ainda mais elevadas foram encontradas na hemolinfa de larvas de
outros Diptera como Calliphora auqur (7,41 mM, segundo HACKMAN,
1956) e na larva inteira de Phormia regina, onde a concentra
ção da tirosina livre é tão elevada, cêrca de lO vêzes o limite
de solubilidade da tirosina em água, que LEVENBOOK e DINAMARCA
(1966) propuseram que "in vivo" a tirosina deve ocorrer no esta
do sólido. Essas concentrações elevadas de tirosina devem estar
relacionadas com o papel dêsse aminoácido como precursor cos fe
nóis que intervém na formação do pupário e/ou de nova cutícula
(GILMOUR, 1965). Como Ca11iphora e Phormia produzem um pupário,
enquanto que R. americ~~a não o faz, seria mesmo de se esperar
que as primeiras possuíssem um conteúdo maior de tirosina em re
lação a essa última.
Taurina é encontrada frequentemente na hemolinfa ce iQ
setos e deve-se originar a partir da descarboxilação do derivado
oxidado da cisteína, o ácido cisteico, e não possui nenhuma fun
ção conhecida em insetos (GILMOUR, 1965). Recentemente vários
eXperimentos tem sugerido a possibilidade de taurina atuar como
um neurotransmissor semelhante ao ácido y-amino butírico em ma
míferos (Cf. DAVISON e KACZI~REK, 1971). E' possível que o mes
mo ocorra em insetos, ewbora ainda não exista nenh~~ ,estudo a
,respeito.
o ãtação vegetal
tionina (GILM
o ádo ácido glut
da biossíntes,
missoras do s.
e S- alanina t,
tização de cu'
Fos:
devem estar 1
deos. E I intl
tídeo predoro.:
~ (BIEBER et
1963).
A f\
das concentraç
(CHEN~ 1966),
culares como t
ram' entretant
e, segundo CE
participação e
morregulação.
dos livres da
na biossíntese
aminoácidos n
fato dos inset
gânicos (FLORK
Adiante, ítem
que os aminoá
pam na síntese
A oc
setos é fenô
existem, entre
presentes. En
te ácidos,
SIMMONS, 1962)

) Acheta domes
sôbre a possi
) ou prolina I
.tir que o acú
,ricana e a au
da formação de
ssibilidade de
do passo orn.!.
de hemolinfa
menos provã-
rito acima.
a de R. ameri
) 59 período,
Llidade da ti-
concentrações
1 de larvas de
fundo HACKMAN,
! a concentra
'êzes o limite
'K e DINAMARCA
'orrer no est~
a devem estar
ursor dos fe~
nova cutícula
m um pupário,
de se esperar
rosina em re-
nolinfa de i12
) do derivado
nenhuma fun
!mente vários
I atuar como
:írico em ma
,1 que o mes-
estudo a
-89-
o ácido a-amino-n-butírico deve originar-se da alimen
tação vegetal, embora possa surgir a partir da degradação da me
tionina (GILMOUR, 1965).
o ácido y-amino-n-butírico, resulta da descarboxilação
do ácido glutâmico, enquanto que nenhum dado existe a respeito
da biossíntese'da B-alanina; ambos devem ser substâncias trans
missoras do sistema nervoso central em insetos (GILMOUR,1965)
e e-alanina t~ sido implicada também nos processos de esclero
tização de cutículas (HACKMAN e GOLDBERG, 1971).
Fosfoetanolamina e possivelmente fosfoserina também
devem estar relacionados com a síntese e degradação de fosfatí
deos. E' interessante notar que fosfatidiletanolamina é o fosf~
tídeo predominante em Diptera, como por exemplo: Phormia regi
~ (BIEBER et alii, 1961) e Musca domestica (CRONE e BRIDGES,
1963) .
A função dos aminoácidos livres presentes em tão elev~
das concentrações na hemolinfa ainda não está bem esclarecida
(CHEN~ 1966), exceptuando-se aquela de alguns aminoácidos parti
culares como tirosina, como já foi comentado. Algumas funções fQ
ram, entretanto, atribuídas aos aminoácidos livres da hemolinfa
e, segundo CHEN (1966), são: precursores de síntese protéica,
participação em mecanismos de detoxificação, tamponamento e os
morregulação. O caso mais conhecido da utilização dos aminoáci
dos livres da hemolinfa como precursores de síntese protéica é
na biossíntese da sêda por Bombyx (WYATT, 1961). O papel dos
aminoácidos na osmorregulação parece estar relacionado com o
fato dos insetos possuírem uma baixa concentração de íons inor
gânicos (FLORKIN e JEUNIAUX,1964; ver também ítem A3, abaixo).
Adiante, ítem B2, será apresentada alguma evidência indireta de
que os aminoácidos livres da hemolinfa de R. americana partici
pam na síntese do seu casulo.
A ocorréncia de peptídeos na hemolinfa de larvas de i12setos é fenômeno generalizado (CHEN, 1966). Entre os Diptera
existem, entretanto, diferenças quanto aos tipos de peptídeos
presentes. Enquanto que em Drosophila os peptídeos são geralme12
te ácidos, de cadeia re1ativamente longa (Cf. MITCHELL e
SIMMONS, 1962), aquêles presentes em Phormia reqina são de cará-

-90-
(1950b) as proteínas da hemo
62% do poder tamoonante to-
ter básico ou neutro e predominantemente compostos de dois ami
noácidos (Cf. LEVENBOOK, 1966).
Os peptídeos oresentes na hemolinfa de R. americana
são do tipo observado em Phormia regina, isto é, predominante
mente básicos ou neutros e compostos de pequeno número de resí
duos de aminoácidos.
O significado bioquímico dos peptídeos da hemolinfa
não é ,conhecido com segurança. LEVENBOOK (1966) a oartir da
observação do desaparecimento dêsses peptídeos em Phormia regina
por ocasião da pupação sugeriu a possibilidade dêstes funciona
rem como uma reserva de aminoácidos, que seriam utilizados nos
processos de histogênese na pupa. E' possível, entretanto,
que êsses peptídeos tenham também funções mais específicas, como
por exemplo o peptídeo sexual de Drosophila melanogaster,que é
produzido nas glândulas ;acessórias do macho e transferido para
as fêmeas na cópula, aumentando a ovoposição nessas (CHEN e"BUHLER, 1970).
As proteínas da hemolinfa sao sintetizadas principal
mente pelo corpo gorduroso (SHIGEMATSU, 1958; 1960; PRICE e
BOSMAN, 1966), embora outros órgãos como o intestino aparente
mente sejam também responsáveis pela síntese de pelo menos parte
dessas proteínas (ROTH e PORTER, ·1964; CHIPPENDALE, 1970).
Várias funções tem sido atribuídas às proteínas da he
molinfa. Segundo SIAKOTOS (1960a, b) as proteínas da hemolinfa
transportam carboidratos e lipídeos dos sítios que os armazenam
para outros em que serão utilizados. Pelo menos em alguns inse
tos, as proteínas da hemolinfa funcionam corno fornecedoras de
aminoácidos para a manutenção da osmolaridade da hemolinfa
(WYATT, 1961).
De acôrdo com LEVENBOOK
linfa sao responsáveis por cêrca de
tal da hemolinfa de Gastrophilus.
A presença de numerosas enzimas entre as bandas prote!
cas separadas por eletroforese de amostras de hemolinfa foi de
monstrada primeiramente por LAUFER {1960a, b), embora atividades
enzimáticas já tivessem sido detectadas na hemolinfa por numero-
sos outros in
1964; CHEN, 196
um complexo muI
ce ser tomado i
para a degradaç
tanto, uma das
atraído a atenç
como precursora
durante a metam
contribuírem.na
to, conforme já
Em B.:..proteínas da he
que será discut
dos de BIANCHI
proteínas da he:
se, como ocorre
demonstração po
cas da hemolinf.
gel de agar dão
dicar uma parti
rido por SIAKOTI
Os li]ciados a proteíl
te na forma de I
1967a), embora (
dos principalmel
tados desta tesE
predominante na
de fosfolipídeo!
Os fo!
parte do P-lipic
aproximadamente
de fosfolipídeo~
mente multipliccV) por 25. ~s~
de hemolinfa.

de dois ami-
R. americana
redominante
ero de resí-
a hemolinfa
a partir da
ormia regina
es funciona
ilizados nos
entretanto,
íficas, como
aster ,que éferido para
(CHEN e
s principal
60; PRICE e
no aparente
menos parte
1970).
eínas da he
da hemolinfa
os armazenam
alguns inse
necedoras de
a hemolinfa
nas da hemo
oonante to-
andas prote!.
nfa foi de
:i atividades
por -numero-
sos outros investigadores anteriores (Cf. FLORKIN e JEUNIAUX,
1964; CHEN, 1966). Mais recentemente foi detectado e purificado
um complexo multienzimático da hemolinfa de maripôsas, que pare
ce ser tomado intacto por tecidos em histólise, contribuíndo ali
para a degradação do DNA (RIECHERS, MEYERS e BERRY, 1969). Entr~
tanto, uma das funções das proteínas da hemolinfa que mais tem
atraído a atenção dos pesquisad~res recentemente é a de servirem
como precursoras diretas ou indiretas das proteínas tissulares
durante a metamorfose, para a constituição do adulto, além de
contribuírem na formação do vitelo dos ovócitos em desenvolvimen
to, conforme já foi comentado na Introdução.
Em R. americana a função melhor estabelecida para as
proteínas da hemolinfa é sua participação na produção do casulo,
que será discutida em detalhe, no ítem 2B, abaixo. Os resulta
dos de BIANCHI (1972) sugerem ainda a possibilidade de uma das
proteínas da hemolinfa de R. americana participar na vitelogêne
se, como ocorre com grande número de insetos. Por outro lado a
demonstração por TOLEDO FILHO (1969) de que as 9 bandas protéicas da hemolinfa de R. americana separáveis por eletroforese em
gel de agar dão reações positivas para carboidratos poderiam in
dicar uma participação no transporte de carboidratos, como suge
rido por SIAKOTOS (l960a, b) em Periplaneta americana.
Os lipídeos encontram-se na hemolinfa de insetos asso
ciados a proteínas, através de ligações específicas principalmen
te na forma de diglicerídeos (CHINO e GILBERT, 1965; GILBERT,
1967a), embora ocorra exceções onde os lipídeos são transporta
dos principalmente como triglicerídeos (MARTIN, 1969). Os resu!
tados desta tese não permitem concluir qual é a forma lipídica
predominante na hemolinfa de R. americana,embora uma estimativa
de fosfolipídeos e lipídeos neutros possa ser feita.
Os fosfatídeos são de regra responsáveis pela. maior
parte do P-lipídicoem Diptera (GILBY, 1965) e como êles contém
aproximadamente 4% em pêso de fósforo é possível estimar a massa
de fosfolipídeos presentes na hemolinfa de R. americana, simple~
mente multiplicando o conteúdo de P-lipídico determinado (Tabela
v) por 25. tsse cálculo resulta em 2,55 mg de fosfolipídeos!ml
de hemolinfa.

Os compostos fosforilados presentes na hemolinfa de
larvas maduras de R. americdna são fosfolipIdeos (discutidos
acima), ortofosfato, fosfOProteInas e componentes da fração or
gânica ácido solúvel. Nesta fração sõmente dois compostos fos
forilados foram identificados e quantificados nesta tese; fosf~
serina e fosfoetanolamina (Tabela 11): Os compostos organ~cos
fosforilados ácidos solúveis não identificados podem ser, entr~
tanto, estimados se for assumido que êles possuem um conteúdo
médio de P similar ao observado em compostos dessa fração em o~
tros insetos, isto é, 15% em pêso (WYATT, 1961). O resultado
dêsse cálculo é apresentado na Tabela XXVI.
-92-
A concentração de lipIdeos neutros por sua vez,
ser calculada a partir do conteúdo lipídico total (Tabela
da estimativa de fosfolipídeos feita acima, resultando em
mg de lipídeos neutros/ml de hemolinfa.
pode
v) e
2,05
3. Composiç~
A
tribuição pc
tal da hemo]
dos para a h
f.0InPosição IIi
Aminoácidos
Mg
Na
Os compostos fosforilados orgãn~éos.e ácido solúveis
sao muito importantes para o tamponamento da hemolinfa de inse
tos (LEVENBOOK, 1950b) e para o seu balanço ãnion-cátion
(WYATT, LOUGHHEED e ~VYA'rl', 1956; LEVENBOOK, 1950a). A importãg
cia dêsses compostos no balanço ânion-cátion da hemolinfa de ~.
americana pode ser apreciada na. Tabela XXVIII uma vez que fosf~
to ácido solúvel é um dos principais ãnions da hemolinfa e éconstituído principalmente de compostosorgân~cos como pode ser
demonstrado a partir da Tabela V.
A concentração de citrato na hemolinfa de larvas de
R. americana é bastante elevada e êste ânion corresponde a cêr
ca de 47% de todos ânions detectados na hemolinfa dêste inseto
(Tabela XXVIII) •. Concentrações elevadas de citrato na hemolinfa
foram observadas em vários outros insetos e trabalhos realiza
dos em Prodenia eridaniaindicam que o citrato é de origem eg
dógena e que seria acumulado juntamente com outros ácidos orgâ~
nicos devido a uma desproporção entre a biossIntese e a oxida
ção e/ou utilização dêsses ácidos {LEVENBOOK e HOLLIS, 1961 ;
LEVENBOOK, 1961).
1\.ci do cI tri
K
Cloreto
Compos tos fI
nao;
Peptideos
HC03
Trealose
Carboidratol
Ca
Ortofosfato
Fos foetanoli
Total
Osmolaridad!
1. Os valorel11 e V, cc
2. Calculado
3. Calculadotrações dEbela V.

sua vez, pode
L (Tabela v) e
:ando em 2 ,05
hemolinfa de
)s (discutidos
da fração or
:ompostos fos
:a tese~ fosf9
:05 orgânicos
tem ser, entr~
I um conteúdo
fração em 0O!!
O resultado
cido solúveis
infa de inse
ânion-cátion
A importâ!!
molinfa de R.
vez que fosf~
9molinfa e écomo pode ser
de larvas de
3ponde a cêr
dêste inseto
) na hemolinfa
Lhos realiza
de origem e!!
ácidos orgâ;"
;e e a oxida-
lLLIS, 1961 ~
3. Composição molar da hemolinfa e seus componentes inorgânicos
A composição molar da hemolinfa permite estimar a con
tribuiçãoporcentual de cada componente para a osmolaridade to
tal da hemolinfa. A Tabela XXVII mostra os resultados calcula
dos para a hemolinfa de R.americana.
TABELA XXVII
C9mposição molar da hemolinfa de larvas maduras de R. americanal
Composto mM % do total
Aminoácidos 62,62 26,2
Mg 45,30 19,0
Na 33,70 14,1
Acido cítrico 18,60 7,8
K 17,30 7,2
Cloreto 13,40 5,6
Compostos fosforilados orgânicos
não identificados2 13,32 5,5
Peptídeos 12,78 5,4
HC03- 6,90 2,9
Trealose 4,10 1,7
Carboidratos não identificados 3 3,39 1,4
Ca 3,23 1,4
Ortofosfato 3,13 1,3
Fosfoetanolamina 1,31 0,5
Total 239,08 100,0
Osmolaridade (mi liosmoles) 216
1. Os valores apresentados foram calculados a partir das tabelas11 e V, com as exceções indicadas. Ver discussão no texto.
2. Calculado a partir da Tabela XXVI~
3. Calculado como glicose, a partir da diferença entre as concentrações de carboidrato total e trealose, apresentadas na Ta=bela V.
f,ll

-94-
Amônia nao se encontra representada na Tabela XXVII,
devido à possibilidade de que o seu elevado título seja um arte
fato (ver Tabela 11 e comentários a ela referente em Resultados).
Similarmente ali não se encontram os valores para Fe, Cu e Zn,
por ocorrerem em concentrações muito baixas (ver Tabela V). As
protefnas, embora presentes em elevada concentração, quando ex
pressas em massa por volume, são osmõticamente de efeito desprezi
vel, em relação aos componentes representados na Tabela XXVII e
por isto também nao foram ali apresentadas.
A soma das concentrações molares dos componentes da he
molinfa é maior do que a sua osmolaridade total. Essa diferença
deve ser real, porque possíveis superestimativas dos compos
tos não determinados diretamente, como os compostos fosforilados
orgânicos não identificados e os carboidratos não identificados,
nao seriam suficientes para corresponder à diferença observada.
Além disso, se amônia 0correr realmente na concentração determi
nada, ou o seu eventual precurser, a diferença entre a concen
tração molar total e a osmolaridade da hemolinfa seria ainda
maior. A explicação mais simples para essa diferença é admitir
que alguns componentes não sejam osmõticamente ativos por esta
rem complexados a outros. Essa possibilidade voltará a ser ex
plorada abaixo ao se discutir o balanço ânion-cátion da hemolinfa.
A contribuição osmótica dos íons inorgânicos e amino
ácidos da hemolinfa pode ser analisada~ com maior facilidade
através da figura 10. Nesta ~lgura A, B, e D representam os
principais padrões iônicos inorgânicos nos insetos pterygota,
segundo SUTCLIFFE, conforme citado por FLORKIN e JEUNIAUX (1964).
Segundo êsse mesmo autQr, A representa o tipo considerado mais
primitivo, caracterizado por concentração elevada de íons inor
gânicos e baixa concentração de arninoácidoS1 D é considerado o
tipo mais evoluído, apresentando elevada concentração de amino
ácido~ e baixa concentração de íons inorgânicos e, finalmente,B
é um tipo intermediário que é usual nas ordens: Megaloptera,
Neuroptera, Mecoptera, Trichoptera e Diptera. R. americana re
presentada por C, possui um padrãq bem de acôrdo com aquêle u
sual para Diptera (B) isto e, quase 50% da pressão osmótica é
devido a cátions inorgânicos, cloreto'é um ânion de importância
relativamente pequena e o papel osmótico dos aminoácidos é ele
vado.
A
Atol 11'10 Ácroos !
Nn
k
P04
H-
ei
EPHE 111EROPTEI
OOONATAPl.EOOPTERAOICTVOPTERAHETEROPTERA
Figura lO - Ef,dos como porce:de insetos pteduas seções veconcentração ID<tions são ilusà direita. Osvel em igual prespondem a CO!
tados de acôrd,C f,

da hemolinfa.
abela XXVII,
eja um arte
Resultados) .
, Cu e Zn,
ela V) • As
quando ex
eito desprezi
ela XXVII e
nentes da he
5sa diferença
dos compos
fosforilados
lentificados,
~a observada.
~ção determi
:re a concen
~ seria ainda
lça é admitir
TOS por esta-
~á a ser ex-
A
,AMINO ACID05
k
04-
ti.
çe
B
,AMINO ACIDOS
Mg
e.",
k
N~
P04
cf
c
,AMINO ACIDOS
l1li9
Ca
K
N~
cf
o
I
AMINO ACIDOS
NaCá
De POA
e!Na
111'JII:
::~lI"
r,:
Figura lO - Efeitos osmóticos de co~ponentes inorgânicos ilustr~dos como porcentagens da concentraçao molar total da hemolinfade insetos pterygota. Cada bloco na figura é visualizado comoduas seções verticais, cada uma destas correspondendo a 50% daconcentração molar total. As contribuições porcentuais dos cátions são ilustrados na seção à esquerda, enquanto que os ânionsà direita. Os aminoácidos são representados, na medida do possível em igual proporção em ambas seções. As áreas em branco cor=respondem a componentes não indicados. A, B e D estão representados de acôrdo com SUTCLIFFE segundo FLORKIN e JEUNIAUX(1964).
C foi calculado a partir da Tabela XXVII.
)s e amino
facilidade
~presentam os
Pterygota,
IIAUX (1964).
derado mais
.e íons inor
'onsiderado o
ão de amino
finalmente,B
Megaloptera,
mericana re
om aquêle u
o osmótica é
importância
cidos é ele-
EPHEMEROPTERA
OOONATA
PLECOPTERA
DICTYOPTERAHETEROPTERA
MEGALOPTERA
HEUROPTIERA
IIIIECOPTERA
TIIUCHOPTERADIPTER A
R. AWI ERICANALEPIDOPTERA
HYMENOPTERA
:11'

a maior parte dos cá
devem ser neutrali-
-96-
Algumas diferenças notáveis existem, entretanto, entre
R. americana e o padrão de Diptera apresentado. são elas: um ní
vel muito elevado de Mg e K,e um nível relativamente baixo de
Na. Segundo BON~ (1944), insetos carnívoros tenderiam a ter um
conteúdo elevado de Na, enquanto que os insetos fitófagos, um
conteúdo elevado de K e Mg em sua hemolinfa de tal forma que a
razão" molar Na/K nos insetos fi tófagos é menor ou próximo de 1,
e nos carnívoros bem acima de 1 (cêrca de 5 a 15 de regra). Essa
explicação -dietética para os diferentes padrões catiônicos "exis
tentes encontra, entretanto, várias exceções, conforme ressaltado
pelo próprio BO~. Apesar disso, essa hipótese é aplicável a nu
merosos insetos e é possível que o alto conteúdo de K e Mg na he
molinfa de R. americana, em relação ao padrão de Diptera apresen
tado (figura 10,B), seja devido aos seus hábitos fitôfagos, en
quanto que a maioria dos insetos utilizados para a elaboração do
padrão de Diptera apresentado são carnívoros (Cf. FLORKIN e JEU
NIAUX, 1964).
A razão molar Na/Kna hemolinfa de_ larvas maduras de
R. americana é de 1,95, que é relativamente próximo da unidade,
como seria de se esperar para um inseto fitôfago.
A elevada concentração de K presente na hemolinfa de
muitos insetos parece incompatível com uma função neuromuscular
normal, especialmente porque, segundo CARRINGTON e TENNEY (1959),
todo K encontra-se na forma livre e fisiolõgicamente ativo na
hemolinfa. Experimentos electrofisiológicos indicaram, entretan
to, que a bainha em tôrno das fibras nervosas dos insetos a
tua como eficiente barreira de difusão, fazendo com que o fluído
imediatamente em contacto com as fibras nervosas possua uma comp~
sição iónica diferente daquela da hemolinfa (Cf. NARAHASHI, 1963).
Mais tarde foi demonstrado que essa diferen~e composição era man
tida pela bainha não por ser esta impermeável aos íons, mas por
ser capaz de manter um equilíbrio dinámico adequado entre os íons
da hemolinfa e do fluído circundante das fibras nervosas (Cf.
NARAHASHI, 1963). Entretanto, mesmo a existência dessa bainha
nao explica a insensibilidade de certos insetos a variações iônicas extremas (GILMOUR, 1965).
A figura 10 permite observar que
tions presentes na hemolinfa de R. americana
zados por' ân:gânicos é mu:
tanto, me:Lho:
Balanço ânior
Composto
Mg++
Na+
Ca++
Fe++
Total
1. Os valoresXXVII e ar
2. Calculadosecundária
3. Estimado ada hemolinda hemolinta. Nos cáTER (1960)
As
deradas apena
taria na form
razões que nã
da alta conce
mostra clar
dos orgânicos
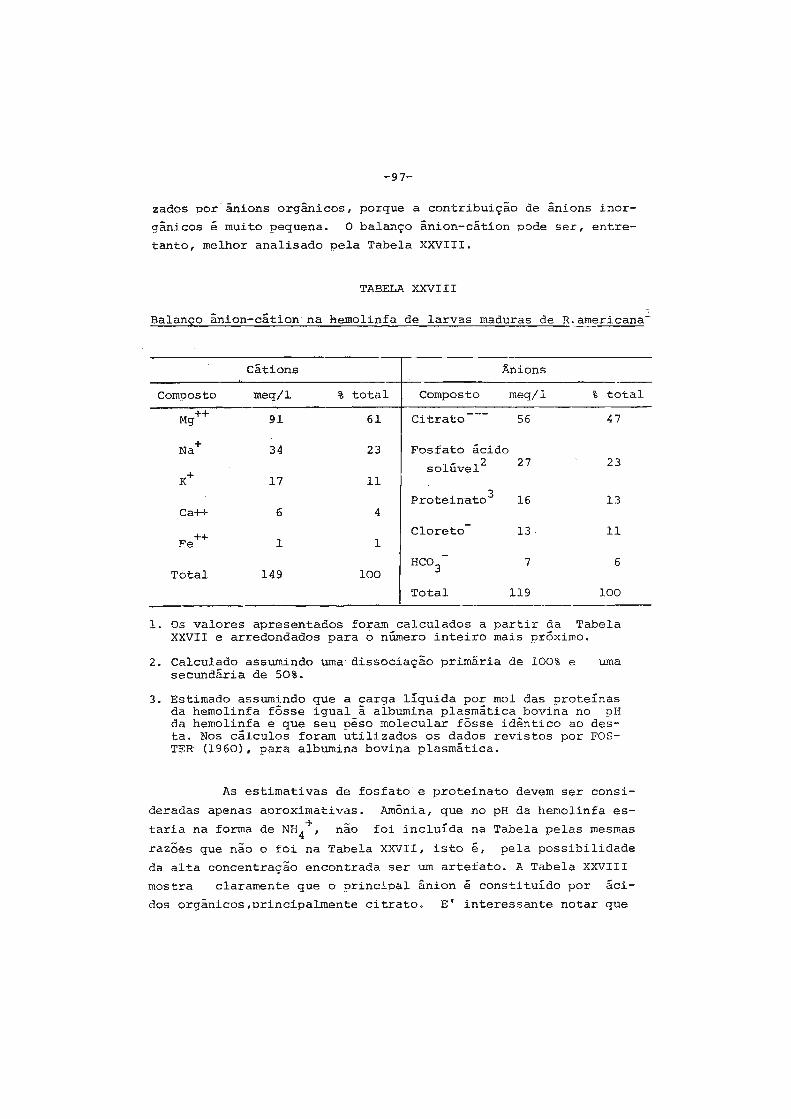
TABELA XXVIII
Balanço ânion-cátionna hemolinfa de larvas maduras de R.americana-
zados por"ânions orgânicos, porque a contribuição de ânions inor
gânicos é muito pequena. O balanço ânion-cátion pode ser, entre
tanto, melhor analisado pela Tabela XXVIII.
23
13
47
% totalComposto meq/l
Ânions
proteinat03 16
Citrato 56
Fosfato ácido
solúve12 27
4
23
11
61
% total
6
34
17
91
meq/l
Cátions
Na+
Mg++
Ca++
K+
Composto
3 fitófagos, um
:al forma que a
)U próximo de 1,
ntretanto, entre
são elas: um níarnente baixo de
~deriam a ter um
ieregra) . Essa
ltiônicos ,-_exis
:orme ressaltado
aplicável a nu
"e K e Mg na he
'iptera apresem
itófagos, en-
elaboração do
FLORl<IN e JEU-
\Tas maduras de
imo da unidade,
++Fe
Total
1
149
1
100
Cloreto-
HC03
-
13
7
11
6
Total 119 100
1 hemolinfa de
) neuromuscular1. Os valores apresentados foram calculados a partir da Tabela
XXVII e arredondados para o número inteiro mais próximo.
3. Estimado assumindo que a carga líquida por mal das proteínasda hemolinfa fôsse igual â albumina plasmática bovina no pHda hemolinfa e que seu pêso molecular fôsse idêntico ao desta. Nos cálculos foram utilizados os dados revistos por FOSTER (1960), para albumina bovina plasmática.
As estimativas de fosfato e proteinato devem ser consi
deradas apenas aproximativas. Amônia, que no pH da hemolinfa es
taria na forma de NH4+, não foi incluída na Tabela pelas mesmas
razõés que não o foi na Tabela XXVII, isto é, pela possibilidade
da alta concentração encontrada ser um artefato. A Tabela XXVIII
mostra claramente que o principal ânion é constituído por áci
dos orgânicos,principalmente citrato. E' interessante notar que
TENNEY (1959),
lmente ativo na
lram, entretan
los insetos a
l que o fluído
,s s ua uma comp~
RAHASHI, 1963).
sição era man-
íons, mas por
entre os íonsnervosas (Cf.
a dessa bainha
ariaçóes iôni-
2. Calculado assumindo uma dissociação primária de 100% esecundária de 50%.
uma
~1111
parte dos cã
ser neutrali-

-98-
hâ um excesso de câtions em relação ao de ânions,que poderia ser
exolicado pela ocorrência de cátions ligados a proteínas·ou ami
noácidos e pela ocorrência de compostos ácidos não determinados.
Os aminoácidos poderiam contribuir muito pouco no valor do pH da
hemolinfa :essa contribuição seria para os cátions, não para os
ânions. A demonstração de que 15-20% do cálcio e magnésio da
hemolinfa estão ligados a proteínas em Telea polyphemus (CARRINQ
TON e TENNEY, 1959) favorece à idéia de que o desbalanço iônico
observado é devido principalmente a uma fração dos cátions que
nao estaria livre.
Se fôr admitido que a diferença observada entre cátions
e ânions da Tabela XXVIII corresponde a cátions que não estão l~
vres, seria possível explicar também a discrepância entre a con
centração molar total estimada da hemolinfa e sua determinação
direta, apresentada na Tabela XXVII.
A osmolaridade da hemolinfa permanece constante duran
te a produção do casulo, embora haja uma queda generalizada em
massa por indivIduo dos comoonentes da hemolinfa (Tabela XXII) .
Essa constância parece ser consequência da pequena variação, que
é ainda mutuamente compensada, da concentração molar dos compo
nentes que juntos perfazem cérca de 80% da osmolaridade total da
hemolinfa (Tabela XXIX). Deve-se notar, entretanto, que os me
tais sofrem uma redução, nao balanceada pelo aumento de outros cgn
ponentes da hemolinfa, entre o 29P e 39P sem haver uma queda cor
respondente na osmolaridade da hemolinfa. ~ste resultado estâ
de acôrdo com a hipótese levantada acima de que parte dos cátions
não está na forma livre, mas ligada a outras moléculas e seria
explicado se o decréscimo no conteúdo de metais ocorrer devido à
perda dos cátions ligados.
1
Osmolaridade e
mente mais ati-
Período Idade
(dias) -Ml
29 P 48 1<
39 p 49
49 p 54
59 p 57
* Valores reprodl
4. Coagulaç;
A heI
pontâneamente.
reações observi
cri to por G::lliGC
teroptera, al~
adultos, embori
outro padrão, E
TAGNON, 1962).
A hen
cipalmente peJ
forma aparenten
mamíferos atrc
TAGNON, 1962).
O esc
çao com água de
de B.:.. americanc
1971), mas o f

poderia ser
.nas . ou ami
,terminados.
or do pH da
não para os
nésio da
~ (CARRINQ
nço iônico
tions que
ntre cátions
não estão l!ntre a con
determinação
TABELA XXIX
Osmolaridade e concentracão rnilirnolar dos comoonentes osmôtica~
mente maiS ativos da hemolinfa de larvas de R. americana duran
te o desenvolvimento*
Período Idade m!'1 miliosmoles
(dias) Metais Aminoácidos Peptídeos Cloreto Osmolaridade
29 P 48 100,0 62,6 12,8 13,4 216
39 P 49 64,3 56,1 16,5 18,3 217
49 P 54 68,3 38,8 18,0 25,0 216
59 P 57 66,2 48,1 20,1 23,0 209
* Valores reproduzidos ou calculados das Tabelas lI, V, XX, XXI e XXII.
II
llltl
i1~1Ij~ t
maduras
YOUNG,explic~
4. Coagulação e escurecimentu
/li
dilui-
A hemolinfa de larvas de R. americana nao coagula es
pontâneamente. Êsse comportamento é semelhant~ ao padrão IV de
reações observadas na hemolinfa após a sua retirada do inseto de~
crito por G~GO~RE (1951). Êsse padrão é ccrr.~~ em ~emipt€ra He
teroptera, alguns Coleoptera, alguns Hymenoptera e em Diptera
adultos, embora as poucas larvas de Diptera estudadas apresentam
outro padrão, envolvendo coagulação da hemolinfa (Cf. GRÉGOlRE e
TAGNON, 1962).
A hemostasia no padrão IV de insetos é realizada prin
cipalmente pela formação de hemocitos aglutinados na ferida, de
forma aparentemente análoga àquela verificada na hemostasia de
mamíferos através da ag~utinação de plaquetas (GRÉGOIRE e
TAGNON, 1962).
o escurecimento e floculação da hemolinfa após
ção com água destilada, similar ao observado em larvas
de ~ americana, foi reportado em Choristoneura (SCKMIDT e
1971), mas o fenômeno permanece obscuro. Uma hipótese
ante duran
ralizada em
abela XXII) .
riação, que
dos compo
3.e total da
iue os me
3.e outros can
3. queda. cor
:ado está
dos cátions
e seria
~r devido à

-100-
tiva, pelo menos para o escurecimento, é admitir a ocorrência em
condições normais de um inibidor da tirosinase (EC 1.10.3.1) que
deixaria de ser efe~Lvo após a diluição, de forma análoga ao ini
bidor de trealase (EC 3.2.1.28) da hemolinfa da Phormia regina
por FRIEDMAN (1961).
5. Pigmentos
A hemolinfa de larvas de R. americana maduras possuem
pelo menos 7 pigmentos ligados a proteínas. Os mais importan
tes quantitativamente são: uma cromoglicooroteína de côr limão,
cujo pigmento fluorescente de natureza desconhecida está forte
mente ligado à proteína e dois carotenóides prêsos por ligações
fracas a proteinas. Um dêsses carotenóides parece ser o S-car~
teno e o outro, mais abundante, possui um espectro similar, mas
não idêntico,ao do astaceno. TUNDISI et alii (1964) propuseram
baseando-se em resultados cromatográficos, e espectrográficos
que o principal pigmento da hemolinfa de larvas maduras seria
astaxantina e outro pigmento também importante seria um similar
ao e-caroteno. A presença dêsse último foi confir~ado no pre
sente trabalho, enquanto que a ocorrência de astaxantina não foi
observada. Urna explicação possível para a discrepância poderia
ser uma má interpretação dos resultados por TUNDISI et alii
(1964), corno consequência de ter feito os espectros em dissulf~
to de carbono, onde astaxantina e astaceno apresentam espectros
similares de uma única banda e não em oiridina, onde astaceno
apresenta um único oico de absorcão, enquanto que astaxantinali •
apresenta três (KUHN e SORENSEN, 1938). Urna outra possibilidade
seria da astaxantina ter sido oxidada a astaceno nas condições
usadas de extração e cromatografia nesta tese, o mesmo nao ocor
rendo com os citados autores. Esta possibilidade, entretanto, émuito remota porque a oxidação da astaxantina oelo ar é favoreci
da sàmente em meio alcalinq e, com exceção das experiências em
que as amostras foram previamente saponificadas os meios usados
eram sempre neutros.
BASILE et alii (1970) identificaram três pigmentos na
hemolinfa de larvas selvagens de R. americana, usando eletrofo
rese em agarose, cujas côres eram: limão, tijolo e violeta.
Isolou também um mutante recessivo, ligado ao sexo que apresen-
1tava apenas
côr de limão
cor de limão
tijolo e vio
nas associad
no. A refer
alii (1970)
cretas e com
Os
diretamente,
1954; CROMAR
~ provàvelm
tenóide simi
ele maneira a
é derivada d
ce to- caroten,
derivados do
1970). A
veis para
J:)rimeira int,
tação tenha ,
pigmentoS ca:
mente espeeí
mutação tenh,
possível ê ai
capacidade d,
to. Por fim
eia de uma m'
roteno, admi
seja derivad
seu favor o
toda a situa,
portamento d,
Os
nhecida em il
devido a sua
valor duvido:
lO ão amare10-,

ocorrência em
1.10.3.1) que
náloga ao in~
ormia regina
tava apenas o pigmento cór de limão da hemolinfa. O pigmento
côr de limão observado deve corresponder à cromoglicoproteína
côr de limão descrita nesta tese, enquanto que o pigmento côr de
tijolo e violeta devem corresoonder, respectivamente, a prote!
nas associadas ao l3-caroteno e ao carotenóide similar a astace
no. A referidn associação deve ser específica, porque BAST.LE et
alii (1970) encontraram os pigmentos distribuídos em bandas dis
cretas e com migração ~eproduzível na eletroforese.
~~lfl' .
~ll~~I
W'MII1(,1,
pigmentos na
do eletrofo
e violeta.
uras possuem
ais importan
de côr limão,
a está forte
por ligações
ser o l3-car!i
similar, mas
propuseram
~ctrográficos
naduras seria
La um similar
ltina não foi
leia poderia
SI et alii
; em dissulf~
:am espectros
Ide astaceno
astaxantina
>ossibilidade
las condições
:mo nao ocor
:ntretanto, é
Ir é favorec~
-iências em
leios usados
~ill:
",W-~I'"
,
Os carotenóid~s de insetos são derivados do alimento
diretamente, ou após ligeiras alterações na molécula (GOODWIN,
1954; CROMARTIE, 1959). O l3-caroteno de hemolinfa de R. america
~ provàvelmente deve provir da dieta, enquanto que o caro
tenóide similar- ao astaceno poderia ser derivado do e-caroteno,
ne maneira análoga à astaxantina que, segundo GOODWIN (1949),
é derivada do l3-caroteno em Schistocerca gregaria ou como os
ceto-carotenóides de Leptinotarsa decemlineata que são todos
derivados do B-caroteno recebido na dieta (LEUENBERGER e THOMMEN,
1970). A partir dêsses fatos, três interpretações são possí
veis para a mutação limão de R. americana, referida acima. A
primeira interpretação pode ser imaginada admitindo-se que a mu
tação tenha afetado a capacidade da larva de absorver os dois
pigmentos carotenóides. Entretanto, corno essa absorçao é geral
mente específica (GOOpWIN, 1954) é pouco provável que uma única
mutação tenha causado essa situação. A segunda interpretação
possível é admitir que se trate de urna dupla mutação afetando a
capacidade da larva de absorver os dois carotenóides do alimen
to. Por fim, a terceira interpretação é considerá-la consequên
cia de uma mutação afetando a capacidade da larva absorv~r l3-ca~
roteno, admitindo ainda, que o carotenóide similar ao astaceno
seja derivado do B-caroteno. Esta última interpretação tem a
seu favor o fato de admitir apenas uma mutação para explicar
toda a situação encontrada, o que está mais de acôrdo com o com
portamento de fator mendeliano simples dessa mutação.
Os carotenóides não tem nenhuma função específica co
nhecida em insetos, embora seja admitido um paoel na reprodução,
devido a sua presença nos ovos (GOODl1IN, 1954). Êsse papel é de
valor duvidoso em R. americana, porque no mutante limão, os ovos
são amarelo-limão (BASILE, 1970), denunciando a ausência de caro
pre-~do no
que apresen-

102-
tenóides (os ovos de fêmeas selvagens sao alaranjados).
A origem e função do pigmento fluorescente de côr li
mão é completamente obscura,principalmente porque a sua natureza
química é desconhecida. E' interessante notar.que a sua presen
ça foi detectada na secreção salivar de larvas de R. americana
(BIANCHI, 1972) e é possível que se encontre também nos ovos,
a julgar pela coloração dêstes no mutante limão.
B. Produção do casulo coletivo por larvas de R. americana
1. Química do casulo
A fiação do casulo inicia-se com a formação de fila
mentos bem definidos junto aos quais a larva deposita um mate
rial semelhante a fezes. Mais tarde o espaço entre fios sao pr~
enchidos por um material amorfo, membranoso, visível mesmo de
pois da retirada de Cac03
por ácido acético. Êste material mem
branoso, juntamente com o filamentar fazem parte, portanto, do
que tem sido chamado fração insolúvel do casulo. Esta fração c~
responde à sêda de R. americana (ver ítem 2 abaixo), e a Tabela
XI representa a sua composição aminoacídica. BIANCHI (1972)
fêz a determinação da composição aminoacídica da secreção sali
var da larva madura de R. americana. Esta secreção é responsá
vel pela origem dos filamentos que são formados logo no início
da produção do casulo, referidos acima. Como os seus resultados
sao similares àqueles obtidos com o casulo terminado isso sugere
que a parte fibrosa e membranosa da fração insolúvel do casulo
tem a mesma composição ou que a parte fibrosa predomina de tal
forma sôbre a amorfa que mascara a eventual composição peculiar
deSta última.
Os tipos principais de sêda conhecidos sao constituí
dos principalmente de fibroína, colágeno, qui tina, cuticulina ou
poliglicina (RUDALL e KENCHINGTON, 1971). A composição da sêda
de R. americana, representada pela composição da fração insolúvel
do casulo, é principalmente protéica, não podendo portanto, ser
do tipo cuticulina ~ ou quitina. A Tabela XI mostra que a sêda
de R. american
menos do que 6
ser também do
a semelhança d
res, a julgar
do pupário de
rio de Dipte
tícula externa
que a sêda de
to de sêda, em
sêdas qui tinos
adas com prote
de R. american
hexosaminas au
te seja galact
levado em cons
carboidratos n
A se
acíclica mui to
bora os dois g
te. Isso talv
mus não produz
quáticos bem d
gundo LUCAS, S
te em que o in
que a diversid
A fu
pupaçao é a de
variações de
(IMMS, 1957).
do casulo em R
baseando-se no
as larvas dêst
procedéncia, p
tes úmidos, os
em locais sêco

,} .
~ de côr li
;ua natureza
sua presen
americana
nos ovos,
~
de fila-
um mate
ios são pr~
mesmo de
terial mem
tanto, do
3. fração cQ:
= a Tabela
(l972)
~çao sali
responsá
lO início
resultados
.sso sugere
lo casulo
la de tal
peculiar
constituí
iculina ou
o da sêda
insolúvel
tanto, ser
que a sêda
-103-
de R. americana não pode ser do tipo poliglicina, pois tem muito
menos do que 66% dos aminoácidos como glicina, bem como não pode
ser também do tipo fibroína ou colágeno. E' notável, entretanto,
a semelhança de sua composição com aquela de proteínas cuticula
res, a julgar pela sua semelhança com a composição aminoacídica
do pupário de Lucilia cuprina (Tabela XI). Como se sabe o pupá
rio de Diptera Cyclorrhapha é derivado da epicutícula e endocu
ticula externa da larva madura (WHITEN, 1957). Parece, portanto,
que a sêda de R. americana representa um tipo ainda não descri
to de sêda, embora guarde semelhanças com a porção protéica das
sêdas quitinosas, as quais segundo RUDALL .(l962) estariam assoc!
adas com proteínas similares àquelas da cutícula. O fato da sêda
de R. americana apresentar um conteúdo relativamente elevado de
hexosaminas aumenta estas semelhanças, embora a hexosamina prese~
te seja galactosamina e não N-acetilglicosamina e deva ainda ser
levado em consideração o seu conteúd~ relativamente elevado de
carboidratos neutros.
A sêda de Camptochironomus possui uma composição aminQ
acidica muito diferente daquela de R. americana (Tabela XI), em
bora os dois gêneros estejam relativamente próximos taxonómicame~
te. Isso talvez esteja relacionado com o fato ãe Camptochirono
~ não produzir um casulo sedoso verdadeiro e possuir hábitos .a
quáticos bem distintos, portanto, daquêles de R. americana. Se
gundo LUCAS, SHAW e SMITH (1960) é possível que o tipo de ambien
te em que o inseto pupe e suas características peculiares expli
que a diversidade das sêdas.
A função dos casulos de insetos formados por ocasião da
pupaçao é a de oferecer proteção à pupa contra choques mecânicos,
variações de temperatura, dessecamento e ataques de predadores
(IMMS, 1957). DREYFUS et alii (1951) puseram em dúvida a função
do casulo em R. americana como um protetor contra o dessecamento,
baseando-se no ambiente úmido em que são normalmente encontradas
as larvas dêste inseto. Entretanto, a sua objeção parece não ter
procedência, porque embora as larvas sejam encontradas. em ambien
tes úmidos, os casulos muitas vêzes têm sido encontrados no campo
em locais sêcos.
~~.
fdR,Jmlllil':
IQlq
!:llll1~,1111
I~·j
',i:

-104-
2. Orígem dos componentes do casulo
A produção do casulo exige um enorme esfôrço da larva
de "R. americana, pois durante a sua produção essa perde 62% de
seu conteúdo aquoso e 39% de seu pêso-sêco. Sômente a constitu!
çao do casulo em si consome cêrca de 29% de seu pêso-sêco. Re
sultados surpreendentes como êsse foram observados também em ou
tros insetos, como Bombyx mori, em que cêrca da metade das pro
teínas da larva madura é usada para fazer o casulo (WIGGLES
WORTR, 1965).
Urna consequência do expôsto acima é que a observação
das alterações químicas na hemolinfa e em vários órgãos da lar
va deve permitir a identificação dos precursores dos componentes
do casulo. Essa identificação seria dificultada se a maior par
te das secreções formadoras do casulo fóssem sintetizadas e ar
mazenadas durante o período de alimentação da larva. Entretan
to, corno será mostrado abaixo, e já foi brevemente referido na
Introdução, êsse não é o caso emR. americana.
Os componentes hidrossolúveis do casulo devem provir,
pelo menos em parte, de contaminações pelo fluído exuvial e/ou
hemolinfa das larvas durante a coleta dos casulos, além das fe
zes eliminadas pelo animal durante o feitio do casulo, conforme
já comentado em Resultados. Devido a isto, êsses componentes não
mais serão considerados aqui. Os principais componentes do caSE
lo c~ja origem deve então ser traçada são: cac03 e as proteínas
e carboidratos insolúveis~
A hipótese formulada em Resultados de que o composto
efervescente, contendo cálcio, observado nos túbulos de Malpighi
deve ser CaC0 3 , encontrou confirmação nos estudos histoquímicos
de MARQUES (informação pessoal). Esta mostrou a ocorrência de
cristais de carbonato de cálcio no lúrnen da porção proximal dos
túbulos de Malpighi das la~vas de R. americana. A ocorrência
dêsses cristais e com essa mesma localização já havia sido de
monstrada em outros insetos, inclusive em Diptera (CLARK, 1958).
A partir dessa demonstração e urna inspeção nas Tabelas XXIV e
XXV, torna-se possível admitir com segurança que o CaC03 do
casulo provém dos túbulos de Malpighi. ° acúmulo de "carbonato
de cálcio nos túbulos de Malpighi durante o desenvolvimento lar
vaI e a sua posterior eliminação encontra paralelo em vários in-
setos e tem:
cálcio, (CLARJ
As
vir tainbém d<
postos de cá:
com base no E
do casulo qUi
terial hidro!
VIII "e IX e
provável a cc
que já foi er
insetos (CLAI
AJ
americana, pl
resulta da 01
produzida pOJ
similar a fe,
pelo menos er
ghi, enquantc
ção do casul,
disto é o fat
a fezes e e c
pri.ncipalmen1
a composição
da secreção
fiação" (fim cproteínas con
nas insolúvej
sa média de 1=
em vários pel
com os incren
nos mesmos PE
39P --I 49P,
276 Ilg/dia/lé
massas de sec
livar; nos meE
pectivamente
respondentes

fôrço da larva
3. perde 62% de
te a constitui
~so-sêco. Re
também em ou
;ade das pro
;ulo (WIGGLES-
! a observação
irgãos da lar
IS componentes
! a maior par
!tizadas e ar
L. Entretan
:e refe'rido na
.evem provir,
:uvial e/ou
além das fe
.10, conforme
'omponentes não
entes do cas~
as proteínas
ue O composto
s de Malpighi
istoquímicos
orrência. de
proximal dos
ocorrência
ia sido de
CLARK, 1958).
elas XXIV e
o Cac03 do
e . carbonato
lvimento lar
em vários in-
-105-
setos e tem sido interpretado como um mecanismo de excreção de
cálcio. (CLARK, 1958).
As cinzas insolúveis, determinadas no casulo devem pro
vir também dos túbulos de Malpighi. E' possível que sejam com
postos de cálcio diferentes de caC03 . Essa hipôtese é levantada
com base no excesso de caC0 3 calculado a partir do conteúdo de Ca
do casulo quando comparado com o caC03 calculado a partir do ma
terial hidroinsolúvel e ácido acético solúvel do casulo (Tabela
VIII'e IX e texto correspondente). ° composto de cálcio mais
provável a corresponder às cinzas insolúveis é fosfato de cálcio,
que já foi encontrado nos tUbulos de Malpighi de várias larvas de
insetos (CLARK, 1958).
A fração insolúvel do casulo corresponde à sêda de ~.
americana, produzida pelas glândulas salivares. Essa conclusão
resulta da observação direta de que o casulo é feito por urna sêda
produzida por essas glândulas salivares (labiais) e um material
similar a fezes. Êste material similar a fezes deve corresponder
pelo menos em parte, ao CaC03 , que provém dos tÚbulos de Malpi
ghi, enquanto que a séda deve corresponder ao restante dacomposi
ção do casulo, isto é, à fração insolúv~l. Uma evidência a favor
disto é o fato do casulo I, que apresenta pouco material similar
a fezes e é quase que sômente feito por fios de sêda, ser formado
pr~ncipalmente pela f~ação insolúvel (Tabela XIII). Além do mais,
a composição da sêda, obtida a partir de determinações' químicas
da secreção salivar colhida pouco antes do início do processo de
fiação (fim do 29 período do 49 estádio) mostra que elà é rica em
proteínas com composição aminoacídica similar àquela das protéi
nas insolúveis do casulo 111 (BIANCHI, 1972). Além disso, a mas
sa média de proteínas secretadas pela glândula salivar, por dia,
em vários perlodos da produção do casulo, concorda bastante bem
com os incrementos médios diários da proteína do casulo, medidos
nos mesmos períodos. ~sses, incrementos para os intervalos de
39P ~ 49P, 49P ~ 59P e 59P ~ Pupa farata são: 54, 117 e.
276 ~g/dia/larva, respectivamente (Tabela XV), enquanto que as
massas de secreção protéica, produzidas por dia, pela glândula s~
liva~ nos mesmos intervalos são: 42, 102 e 90 ~g/dia/larva, res
pectivamente (BIANCHI, 1972). A discrepância nos resultados cor
respondentes ao -último intervalo seguramente é inferior à apre-
III1I1
111'1I~U
U!II~'I
1~11_11,
i1lij
l;!ltI~I'"
1111
~~,
II,,~iil!~
,ri

-106-
sentada, porque o resultado obtido no casulo pode estar superesti
mado (Tabela XV) enquanto que aauêle da glândula salivar deve es
tar subestimado (BIANCHI, 1972).
o conteúdo de carboidratos da secreçao e, entretanto,
be~ inferior ao determinado no casulo III (BIANCHI, 1972). Essa
discrepância parece ser consequência de uma mudança na composi
ção da fração insolúvel durante o desenvolvimento, no sentido de
aumentar o seu conteúdo de carboidratos (Tabela XIII e texto cOE
respondente). Urna outra hipótese para ~xplicar a diferença no
conteúdo de carboidratos encontrada entre o casulo e secreção s~
livar seria admitir que a larva de R. americana produzisse sêda
também pelo túbulo de Malpighi. Êsse fenômeno ocorre em vários
Coleoptera e Neuroptera (WIGGLESWORTH, 1965) e está relacionado
com modificações do túbulo de Malpighi e no reto, corno as descri
tas por MAZZI e BACCETTI (1956) para Donus crinitus (Coleoptera)~
Corno essas modificações nao ocorrem em R. americana (MARQUES
informação pessoal), o túbulo de Malpighi em R. americana não d~
ve ter nenhuma atividade secretora de compostos para o casulo,
além do CaC0 3 e, portanto, tôda a sêda deve ser produzida nas
glândulas salivares.
Segundo BIANCHI (1972) a glândula salivar no início do
processo de fiação possui no máximo 136 ug de proteínas de secr~
ção acumulada por larva. Corno o casulo produzido por larva con
tém cêrca de 1170 ug de proteínas na fração insolúvel (Tabela XIV)
pode-se concluir que o animal acumula apenas cerca de 10% de tô
da sêda que vai utilizar para fazer o casulo antes de parar de
comer e iniciar a fiar. A larva de B. mori comporta-se de manei
ra radicalmente diferente, a maior parte d~ secreção de sua sêda
(cêrca de 70%) é sintetizada e armazenada antes do animal parar
de comer (FUKUDA et alii, 1959). Êsse comportamento diferencial
deve estar ligado , pelo menos em parte, às diferenças anatômi
cas entre as glândulas labiais dos dois insetos. Enquanto que
as glândulas labiais (sericígenas) de B. mori apresentam na sua
porção mediana um reservatório considerável para a secreção (LU
CAS, SHAW e SMITH, 1958), as glândulas labiais (salivares) de ~.
americana apresentam apenas um lúrnen relativamente estreito e de
diâmetro pouco variável ao longo da glândula (DREYFUS et alii,
1951).
o fa,
após o animal
car a origem (
na hemolinfa e
valo mais adeq'
tes do 49P os
cia de reserva
gir alterações
mação do anima:
A Tal
precursores na
correspondem a
túbulos de Mal!
são ·formados q\
discussão acim,
seus precursorE
16% do pêso-sêe
mente a cutícu:
vo. Entretant<
apresentam cêr<
196 4), e as tJ
tina (WIGGLESWC
não estudada q\
abaixo de 16~
como· representé
seus precursore
A Té
fícil admitir
vem, pelo menOE
cimo de todos E
é de 199 ug/laI
dos 350 ug/lan
Se cj
~, corno foi Se
teínas insolúvE
adequadamente,

I~I
ar superesti
\lar deve es-
:ntretanto,
n2l. Essa
la comoosi
sentido de
! texto co!.
.ferença no
:ecreção s~
lzisse sêda
, em vários
'elacionado
as descri
oleoptera):
(MARQUES
~ não d~
casulo,
zida nas
início do
3 de secr~
larva co!}.
rabela XIV)
lO% de tô-
)arar de
de manei
sua sêda
\al parar
ferencial
anatômi
,anto que
am na sua
eção (LU
es) de ~.
eito e de
et alii,
-107-
o fato de 90% da sêda de R. americana ser sintetizada
após o animal parar de comer, torna razoável procurar identifi
car a origem de seus componentes através de análises químicas
na hemolinfa e em vários órgãos durante a sua produção. O inte!.
valo mais adequado para êsse estudo é entre o 49P e o 59P. An
tes do 49P os resultados tendem a ser prejudicados pela existê!}.
cia de reserva de secreção na glândula e após o 59P devem sur
gir alterações químicas relacionadas com a apólise e transfor
mação do animal em uma pupa farata .
A Tabela XXX mostra o balanço de proteínas ou seus
precursores na hemolinfa e em órgãos de R. americana que juntos
correspondem a 71% do pêso-sêco total da larva madura. Corno os
tUbulos de Malpighi correspondem a 13% do pêso-sêco da larva e
são 'formados quase que exclusivamente de CaC03 (Tabela XXIV e
discussão acima), pode-se dizer que o balanço de proteínas ou
seus precursores é conhecido em 84% do pêso-sêco da larva. Os
16% do pêso-sêco da larva não estudados correspondem principal
mente a cutículas, traquéias, parede muscular e tubo digesti
vo. Entretanto, como as cutículas moles de larvas de Diptera
apresentam cêrca de 50-60% em quitina (RUDALL, 1963, HACKMAN,
1964), e as t~aquéias e o revestimento do intestino contém qui
tina (WIGGLESWORTH, 1965), a porcentagem do pêso-sêco larval
não estudada que pode fornecer precursores de proteínas desce
abaixo de 16%. Dessa forma a Tabela XXX pode ser considerada
como representando um balanço bem completo das proteínas e
seus precursores entre os períodos indicados.
A Tabela XXX e aargurnentação acima tornam muito di
fícil admitir que as proteínas insolúveis do casulo não deri
vem, pelo menos em parte, das proteínas da hemolinfa. O decré~
cimo de todos precursores, excluído as proteínas da hemolinfa
é de 199 ~g/larva, enquanto que nêsse período são sintetiza
dos 350 ~g/larva de proteínas insolúveis do casulo.
Se cisteína for aminoácido essencial para R. america
~, como foi sugerido no ítem A2 acima, sua presença nas pro
teínas insolúveis do casulo (Tabela XI) só poderia ser explicada
adequadamente, admitindo-se que essas proteínas s~o feitas, pelo
l~i
1111"1IIHIi,
I~I'
11;t:
I'~II,111;
WIH
Iflll'lili):
11'11..,11
ltl~'""bil
~"i,

-108-
menos parcialmente, a partir das proteínas da hemolinfa. Isso é
necessário porque a hemolinfa não possui cisteína livre (Tabela
11) e nem ligada a peptídeos (Tabela 111).
TABELA XXX
Balanço dos possíveis contribuintes de precursores das proteínas
insolúveis do casulo e incremento dessas, no intervalo do 49P ao
S9P do 49 estádio deR. americanal
BI
secreção sal
em diferente
de incorpora
salivar em à
teínas da
do casulo,
linfa, enqua
glândula.
glângula sal
pos a sua hi
1. Discussão no texto.
2. Segundo BIANCHI (1972).
3. Calculado a partir da hipótese de que a concentração de aminoácidos livres da glândula salivar é a mesma da hemolinfa eque a densidade dessa glândula é 1,00.
4. Calculado a partir da Tabela xx.5. Decrescimo do pêso-sêco do corpo gorduroso, após subtração
do decréscimo do conteudo-de glicogênio no mesmo período.Calculado a partir da Tabela XXIII.
6. Calculado a partir da Tabela XVI.
7. Retirado da Tabela XIV.
Possível contribuinteOu
Proteínas do casulo
Proteína da glândula salivar2
Aminoácidos da glândula salivar3
Aminoácidos da hemolinfa4
peptídeos da hemolinfa4
Corpo gordurosoS
Proteínas da hemolinfa6
Proteínas insolúveis do casul07
llg/larva
decréscimo incremento
o
38
12
23
126
700
350
Se
feita apart
um reflexo d
Em Resultado
ja queda em .
do total da
quêles que p
fôrem consid
dos tornarn-s·
que mais dim
na hemolinfa
leucina (Tab·
que ocorrem '
cente: treon.
se vê, os re
síntese de p:
vres da hemo
nas da hemo1
A·
das larvas d
ria das la
de uma adapt
utilizadas e
A
glândula sal
carboidratos
demais carbo

-109-
Linfa. Isso élivre (Tabela
das proteínas
valo do 49P ao
rva
incremento
350
:ação de amin~lemolinfa e
subtração, período.
BIANCHI (1972) propôs a partir de eletroferogramas da
secreç.ão salivar e da glândula salivar "in vivo" e "in vitro",
em diferentes períodos larvais, assim corno a partir de cinéticas
de incorporação de aminoácidos marcados em proteínas da glândula
salivar em diferentes circunstâncias, que cêrca de 60% das pro
teínas da secreção salivar (que corresponde à fração insolúvel
do casulo, ver acima) provém diretamente das proteínas da hem~
linfa, enquanto que os outros 40% são sintetizados na própria
glândula. Esta síntese utilizaria os aminoácidos livres da
glângula salivar e/ou hemolinfa ou as proteínas da hemolinfa a
pós a sua hidrólise.
Se parte da síntese de proteínas da secreção salivar éfeita a partir dos .aminoácidos da hemolinfa, seria de se esperar
um reflexo dessa síntese na composição aminoacídica da hemolinfa.
Em Resultados foi ressaltado o fato de queaquêles aminoácidos cE
ja queda em moles, ao passar do 39P ao 59P correspondem a 70%
do total da queda dos aminoácidos da hemolinfa são justamente a
quêles que perfazem 29% da proteína insolúvel do casulo 111. Se
fôrem considerados apenas os aminoácidos essenciais, os resulta
dos tornam-se ainda mais reveladores. Os aminoácidos essenciais
que mais diminuem em moles, por ordem decrescente de grandeza,
na hemolinfa entre o 39P e 59P são: treonina, valina, isoleucina,
leucina (Tabela XX); enquanto que aquêles aminoácidos essenciais
que ocorrem em maior quantidade no casulo são, por ordem decres
cente: treonina, leucina, valina e isoleucina (Tabela XI). Corno
se vê, os resultados são indicativos de que pelo menos parte da
síntese de proteínas do casulo é feita a partir de aminoácidos li
vres da hemolinfa, enquanto que a outra parte deriva das proteí
nas da hernolinfa.
A ocorrência de uma concentração protéica na hemolinfa
das larvas de R. americana maior do que a apresentada pela maio
ria das larvas de insetos (Tabela V), talvez seja consequência
de uma adaptação dêsse animal, pelo fato dessas proteínas serem
utilizadas em parte na produção do casulo.
A síntese dos carboidratos insolúveis do casulo pela
~lli
nu:~iI1
:'II[III!
111111,:
~III
IK~I'I!lll
'1glândula salivax' tem que
carboidratos presentes na
demais carboidratos antes
utilizar, corno precursores imediatos.
glândula salivar ou na hemolinfa. Os
de poderem ser utilizados na síntese

-110-
tem que ser primeiro liberados na hemolinfa. Os carboidratos
que podem ser liberados na hemolinfa após hidrólise incluem a
qui tina cuticular e o glicogênio presente nos vários tecidos co
rno corpo gorduroso, masculatura e células do tubo digestivo. A
quitina cuticular, entretanto, só é hidrolizada sob a açao do
fluído exuvial e seus produtos transferidos para a hemolinfa,
nas vésperas da muda pupal (BADE e WYATT, 1961). enquanto que o
glicogênio que é mais fàci1mente mobilizado para a hemolinfa na
forma de trealose, pertence ao corpo gorduroso (KILBY, 1963).
·.TABELA XXXI
ga~ta, mantend
linfa (Cf. HOR
hormônio da "c
tração de trea
gorduroso (STE:
do glucagon e:
1965; WIENS e
A ut
facilitada pel
enzima que cli·
1963) •
Balanço dos posslveis contribuintes de precursores dos carboi
dratos insolúveis do casulo e incremento dêsses no intervalo do
49P ao S9P do 49 estádio de R.americanal 3. Mecanism
1. Ver discussão.2. Segundo BIANCHI (1912)3. Calculado a partir da Tabela XIX4. Calculado a partir da Tabela XXIII5. Retirado da Tabela XIV
\Jg/larva
Glicogênio do
.. ·decrés·cimo
A fi,
lar àquela des
liberação da
da hemolinfa s
tração da musc
te demonstrado
na glàndula sa
secreção por c
secreção da s,
res àquelas da
é, a secreçao
vários solvent,
altamente inso
10,7
106
incremento
18
154.. 4
corpo· ·gorduroso
insolúveis do casuloS
Possível.contribuinteou
Cqrboidratos do casulo
Carboidratos
~lândula salivar 2
Hemolinfa3
A Tabela XXXI mostra o balanço dos possíveis contribu
intes de precursores dos carboidratos insolúveis do casulo.
Conforme pode-se concluir da Tabela XXXI e da a~gurnentação aci
ma, oglicogênio do corpo gorduroso é o grande fornecedor de
precursores de carboidratos.
O corpo gorduroso é capaz de sintetisar t.realose e
liberá-la para a hemolinfa na mesma velocidade com que essa é
Estu,
noptera mostra:
te organizada,
raleIas (RUDAL:
ção no fio de :
dêsses filamen·
drodinâmi.cas
tivas e produz.
levanta ainda,
sêda por Bomby:
bre a estrutur,

::arboidratos
~ incluem a
tecidos co
Lgestivo. A
a ação do
l hemolinfa,
Iuanto que o
!molinfa na
~, 1963).
los carboi
ntervalo do
ncremento
10,7
106
; contribu
lo casulo.
ltação aci
lecedor de
t.realose e
ue essa é
-111-
gaeta, mantendo constante a concentração de carboidratos ha hem~
linfa (Cf. HORIE,l960). Essa conversão está sob contrôle de um
hormônio da "corpora cardíaca" que é capaz de aumentar a concen
tração de trealose na hemolinfa às custas do glicogênio do corpo
gorduroso (STEELE, 1961) e cujos efeitos são similares àqueles
do glucagon em fígado de mamíferos (STEELE, citado em KILBY,
1965; WIENS e GILBERT, 1967).
A utilização da trealose da hemolinfa pelos tecidos é
facilitada pela ocorrência generalizada da trealase (EC3.2.l.28)
enzima que cliva a trealose em duas moléculas de glicose (KILBY,
1963) •
3. Mecanismo e regUlação da produção do casulo
A fiação da sêda em R. americana ocorre de forma simi
lar àquela descrita para Bombyx(LUCAS, SHAWe SMITH, 1958). A
liberação da secreção da sêda é feita por pressão hidrostática
da hemolinfa sôbre a glândula salivar como consequência de con
tração da musculatura corporal da larva. Isso pode ser fàcilmeg
te demonstrado pela constatação da inexistência de musculatura
na glândula salivar (MARQUES; informação pessoal), e obtenção de
secreção por compressão do corpo da larva (BIANCHI, 1972). A
secreção da sêda de R. americana apresenta propriedades simila
res àquelas da sêda de Bombyx (LUCAS, SHAW e SMITH, 1958), isto
é, a secreção é expelida cómo um fluído esverdeado solúvel em
vários solventes, mas que após ser submetido a tensões torna-se
altamente insolúvel.
Estudos realizados com glândulas sericígenas de Hyme
noptera mostraram que a secreção da sêda encontra-se ali altameg
te organizada, na forma de filamentos dispostos em fileiras pa
ralelas (RUDALL, 1965). Segundo êsse mesmo autor, a transforma
ção no fio de sêda insolúvel seria consequência do deslizamento
dêsses filamentos, uns sôbre os outros, sob ação de fôrças hi
drodinâmicas e tensões permitindo· o contato de extremidades re~
tivas e produzindo novas ligações entre os filamentos. O autor
levanta ainda a possibilidade de ocorrer o mesmo na produçãó da
sêda por Bombyx mori, embora não exista estudos nêsse animal sô
bre a estrutura da sêda dentro das glândulas sericígenas e o cog
Ilil
IIII
~:;
~ll~
;!:~
:iII
!lUII
mil"h,
I"!II.;~
~Il~
111"

-112-
ceito mais aceito para a transformação da secreção da sêda em fio
insolúvel nêsse animal seja baseado na alteração da conformação
das moléculas protéicas, de uma forma dobrada, para outra completamente distendida.
Observações preliminares realizadas, durante a elabora
ção desta tese, com microscópio de fase na glândula salivar de
R. americana, indicaram a presença ali de uma certa organização
do material, embora não no nível observado em Hymenoptera (Cf.
RUDALL, 1965). Essas observações sugerem que a transformação
da secreção da sêda em um fio insolúvel em R.americana pode ocor
rer segundo o mesmo mecanismo descrito por RUDALL (J.965).
A regulação hormonal do processo de fiação é pouco co
nhecida, embora haja evidências em Lepidoptera de que ecdistero
na (20-hidroxi-ecdisona) seja um fator importante para a promo
ção e manutenção da síntese de fibroína na divisão posterior da
glândula sericlgena no período larval (SHIGEMATSU e MORIYAMA,
1970) e de que o início da fiação dependa de um baixo título de
hormônio juvenil (WIGGLESWORTH, 1970).
A liberação de cac03 pelos tÚbulos de Malpighi de ~
ricana poderia ocorrer pela conjugação de dois fat?rea: pressão
hidrostática da hemolinfa sôbre os. túbulos como consequência de
contraçôes corporais da larva e ação da musculatura presente nos
dutos comuns de cada par de túbulos de Malpighi •. tste tipo de
musculatura é comum entre os Diptera (PATTON, 1953) e,R. america
na não é exceção (MARQUES, informação pessoal). No início da
fiação, os dutos comuns dos túbulos de Malpighi estariam fechados,
impedindo que a pressão hidrostática da hemolinÍa, consequente de
contrações corporais da larva, ocasionasse a liberação de CaC03 .
Posteriormente, os dutos se abririam, permitindo a saída de cris
tais de Cac03 , sob ação da pressão hidrostáti~a da hemolinfa. t~
te mecanismo é perfeitamente compatível com os resultados apreseQ
tados na Tabela XV, que mostram uma liberação de caC03 muito pe
quena no início da fiação, seguido de um grande aumento alguns di
as depois. A Tabela mostra também que a produção da sêda é inte~
sa durante tôda a fiação, o que indica a ocorrência de elevada
pressão hidrostática durante todo êste período, devido ao fato
de que essa pressão é necessária para a liberação da secreção da
sêda (ver acima).
A:
par de túbulc
estado atual
colha entre
é i.nteressan1de Malpighi (
nas "corpora
xemplo, em Lc
WM
túbulos de Mê
intestino con
p.:los túbulos
lar também pc
de loialpighi à
A o
guardar, entre
túbulos de Ma

Il'~,'
da sêda em fio
da conformação
i outra comple-
mte a elabora
lula salivar de
·ta organização
lenoptera (Cf.
transformação
~ pode oco!:J.965) •
ão ê pouco co
que ecdistero
para a promo
posterior da
e MORIYAMA,
üxo tí tulo de
)ighi de R. ame
~es: pressão
;equência de
presente nos
:ste tipo de
e iR. america-
I início da
,riam fechados,
'onsequente de
ão de Caco3 .
aída de cris
hemolinfa. ts
tados apresen
D3 muito pe
nto alguns di
sêda é inten
:!e elevada
:!o ao fato
secreção da
I
-113-
A regulação da musculatura dos dutos comuns de cada
par de tUbulos de Malpighi poderia ser nervosa ou hormonal. °estado atual dos conhecimentos em R.americana não permite a es
colha entre nenhum dos dois processos de regUlação. Contudo,
é j,nteressante notar que a regulação da musculatura dos tUbulos
de Malpighi de alguns insetos é feita por um hormônio produzido
nas "corpora allata", similar à adrenalina, corno ocorre, por e
xemplo, em Locusta migratoria (CAMERON, 1953)~
WATERHOUSE (1950) propôs que os grânulos presentes nos
tUbulos de Malpighi de Lucilia cuprina seriam descarregados no
intestino como consequência de um aumento na tomada de fluídos
pelos tUbulos de Malpighi no estágio pupal. Um mecanismo simi
lar também poderia explicar a liberação de caC0 3 pelos tUbulos
de Malpighi de R. americana.
A opçao entre os dois mecanismos propostos deverá a-
guardar,entretanto, a expansão dos estudos de fisiologia dos
tUbulos de Malpighi em R. americana.
illil:
\li'l
:I::~
11''''1

~S~O
Os resultados obtidos nesta tese podem ser distribuídos
em três grandes grupos: composição química da hemolinfa e do casu
lo coletivo e determinação química de alguns componentes princi
pais do corpo gorduroso e tUbulos de Malpighi ao longo do desen
volvimento.
Os principais resultados referentes à química da hemo
linfa de larvas maduras são:
1) A hemolinfa possui uma densidade 1,043, pH = 7,27,
osmolaridade 216 miliosmoles e corresponde a 37% do pêso-úmido
do animal e 26% de seu pêso-sêco. A hemolinfa não se coagula e
possui um volume de células correspondente a 0,3% de seu volume
total.
2) A análise química realizada deu conta de 88% do pe
so-sêco total da hemolinfa e revelou que entre os componentes pre
sentes mais importantes em massa estão as proteínas, seguidas dos
aminoácidos livres, enquanto que os osmõticamente mais ativos sao* +os aminoácidos livres seguido de Mg e Na. Entre os aminoáci-
dos é notável a presença de ornitina e cistationina em concentra
ções relativamente elevadas e a ausência, mesmo em traços, de cis
teína e/ou cistina e citrulina.
3) Os peptídeos ocorrem em concentrações eleyadas, mas
em pequeno número, e são compostos de 2 a 3 resíduos de aminoáci
dos em média; o mais abundante dos quais deve ser U~ dipeptídeo
de histidina e ácido aspártico.
4) Citrato é o ânion mais importante da hemolinfa, se
guido dos fosfatos orgânicos, enquanto que trealose é-o principal
carboidrato presente.
5) Existem pelo menos 6 carotenóides ligados de forma
não covalente a proteínas da hemol~nfa e uma cromoglicoproteína,
de côr amarelo-limão, fluorescente, de natureza desconhecida. Os
1
carotenóides mai
e um similar ao
A compo
da muda pupal é
34% de proteínas
dos solventes ut
HCl 2N, KOH 1M e
4% de cinzas nao
parcialmente ide
enquanto que 4%
O corre
casulo, tUbulos
volvimento, poss
1) A fr
dratos) correspc
da larva, enquan
Malpighi.
2) As p
nos em parte, da
boidratos devem
Os res~
veis significac
~nfase é dada na
orgânicos e inor
de secreção da s

iIn'
distribuídos
nfa e do cas!:,!
entes princi
ngo do desen-
mica da hemo-
3, pH = 7,27,
do pêso-úmido
se coagula e
seu volume
:le 88% do pê
nponentes pr~
seguidas dos
Ls ativos são
)s aminoáci
~m concentra
~aços, de cis
Leyadas, mas
de aminoáci
lm dipeptídeo
!molinfa, se
,- O principal
dos de forma
icoproteína,
nhecida. Os
l
~1l5-
carotenóides mais importantes quantitativamente sao: a-caroteno
e um similar ao astaceno.
A composição em massa do casulo coletivo na véspera
da muda pupal é a seguinte, em números inteiros: 44% de CaC03
;
34% de protelnas e 10% de carboidratos~ambos insolúveis em to
dos solventes utilizados (SDS 10%, uréia 6M, ácido fórmico 50%,
HCl 2N, KOH 1M e Na HC03 1M); 10% de material hidrossolúvel e
4% de cinzas não identificadas. O material hidrossolúvel foi
parcialmente identificado como: proteína (4%), carboidratos (2%) ,
enquanto que 4% ainda permanece desconhecido.
O correlacionamento da análise química da hemolinfa,
casulo, túbulos de Malpighi e corpo gorduroso ao longo do desen
volvimento, possibilitou ainda as seguintes conclusões:
1) A fração insolúvel do casulo (protelnas e carboi
dratos) corresponde à sêda secretada pelas glândulas salivares
da larva, enquanto que o caC03 presente provém dos túbulos de
Malpighi.
2) As proteínas do casulo devem originar-se, pelo me
nos em parte, das proteínas da hemolinfa, enquanto que seus car
boidratos devem provir do glicogênio do corpo gorduroso.
Os resultados são considerados em têrmos de seus possi
veis significados metabólicos e eventualmente fisiológicos.
~nfase é dada na discussão do papel metabólico dos componentes
orgânicos e inorgânicos da hemolinfa, assim corno dos mecanismos
de secreção da sêda e Cac03
e sua possível regulação hormonal.
II~'
1",1
b
ir

ABSTRACT
Theresults of this thesis are concerned to three main
lines of research: the chemical composition of the hemolymph, of
the cornrnunal cocoon, and the chernical deterrnination of some
components in the rat body and Malpighian tubules along larval
development.
The chief results on the hemolymph chemistry of mature
larvae are:
1) The hemolymph has a density = 1.043, pH = 7.27,
osmolarity = 2l~ rniliosmols and corresponds to 37% of the larva
(Wet-Weight) or to 26% ofthe larva (dry weight). The hemolymph
does not clott and the volume of the hemocytes is 0.3% of blood
total volume.
2) The chemical analysis dealt with 88% (dry-weight)
of the hemolymph and sh0wed that proteins are the most important
component in mass, while free arnino acids, Mg++ and Na+ are· the
most osmotically active substances. Arnong the amino acids
is rernarkable the titres of ornithine.and cystathionine, and the
complete absenceof cysteine and/or cystine and citrulline.
3) There are few peptides, but present in high titres.
They are built of two to' three arnino acids residues,and the most
concentrated of them must be a dipeptide of histidine and as
partic acid.
4) Citrate and organic phosphates are the most im
portant anions in the hemolymph. Trehalose is the chief car
bohydrate present.
5) Thereare at least 6 non-covalentely protein-bound
carotenoids and one lemon-yellow, fluorescent chromoglycopro
tein of unknown nature.
The chemical composition of the communal cocoon of
R. americana expressed in percentage of its total dry weight
(nurnbers r01
proteins an(
used (10% SI
NaHC03 ) ~ 10\
nown nature.
as: protein
nown.
T~
analy'sis of
body during
1)
carbohydrate
livary glanc
2)
from the h
from the fat
Th
tabolic and
in the discu
ganic compon
the silk sec
monal regula

!d to three main
le hemolymph, of
lation of some
.es along larva1
listry of mature
'43, pH = 7.27,
7% of the larva
The hemolymph
s 0.3% of blood
8% (dry-weight)
most important+.
and Na are· the
amino acids
ionine, and the
itrulline.
in high titres.
~s , and the most
Une and as-
the most im
the chief car-
protein-bound
:hromoglycopro-
Inal cocoon of
.1 dry weight
J
(numbers rounded to the nearest unity) are: 44% of caC03
; 34% of
proteins and 10% of carbohydrates both insoluble in alI solvents
used (10% 50S, 6M urea, 50% forroic acid, 2N HCl, 1M KOH and 1M
NaHC03 ); 10% of water soluble material and 4% of ashes of unk
nown nature. The water soluble material was identified in part
as: protein· (4%} and carbohydrates (2%), while 4% remained unk
nown.
The interrelationship among the results of the chemical
analysis of the hemolymph, cocoon, Malpighian tubules and fat
body during larval developmentwas used to draw the conclusions:
1) The insoluble fraction of the cocoon (proteins and
carbohydrates) corresponds to the silk produced by the larval sa
livary glands, while caC03 comes from Malpighian tubules.
2) The cocoon proteins must come, at least .in part,
from the hemolymph proteins, while its carbohydrates must come
from the fat body glycogen.
The resu~ts are discusseuin terms of its possible me
tabolic and eventually physiological meanings. Emphasis is given
in the discussion ofthe metabolic role of the inorganic and or
ganic components of the hemolymph, as yet in the mecanisms of
the silk secretion and caC03 deposition and their possLble hor
monal regulation.
I1:~I:
li
IHiIIHII
:~
I"
I

REFERENCIAS BIBLIOGRÁFICAS'
ADIYODI, K.G. - The nature of haernolymph proteins in relation
to the ovarian cycle in the viviparous cockroach, Nauphoeta
cinerea. J. Insect Physiol., 13: 1189-1195, 1967.
AGRELL, I. - Physio1ogical and biochemical changes during insect
development. In: ROCKSTEIN, M., ed. ~e physiology of Insec
~, New York, Academic Press, 1964, v.l, p. 91-148.
ALBANESE, A.A. & ORTO, I •. A. - Proteins and amino acids. In: AL
BANESE, A.A., ed. Newer roethods of nutri ti anal biochernistry,
New York, Academic Press, 1963, v.1, p. 1-112.
AMABIS, J.M. & CABRAL, D. - RNA and DNA puffs in polytene chromo
somes of Rhynchosciara: inhibition by extirpation of prothorax.
Science, 169: 692-694, 1970.
ARMELIN, H.A.i MENEGHINI, R. & LARA, F.J.S. - Extraction and cha-
racterization of newly synthesized RNA from who1e ce11s and
cel1u1ar fractions of Rhynchosciara angelae salivary glands.
Biochim. biophys. Acta, 190: 358-367, 1969.
BADE, M.L. & vNATT, G.R. - Metabolic conversions during pupation
of the cecropia si1kworm. I. Deposition and utilization of
nutrient reserves. Biochem. J., 83: 470-478, 1961.
BALMAIN, J.H.i BIGGERS, J.D. & CHARINGBOLD, P.J. - Micromethod
for the estimation of glycogen in the genital organs of the
mouse. Aust. J. biol. Sci., ~: 139-146, 1956.
BASILE, R.i TOLEDO FILHO, S.A.i CUNHA, A.B. dai MORGANTE, J.S. &
MARQUES, J. - Análises da mutação que afeta a bigmentacão de lar
vas de Rhynchosciara angelae Nonato & Pavan, 1951 (Diptera
Sciaridae). Rev. bras. Biol., 30: 471-475, 1970.
* De acôrdo com as normas preconizadas pela ABNT (Associação Bra
sileira de Normas Técnicas). Abreviaturas dos títulos de peri§
dicas de acôrdo com World Medical Periodical, 3~ ed, London,
1961.
TBECKER, H.J.
phi la melan.
Puffbildungli
der Ringdru:
BEERMAN, W. - 1
of chromosOl
232, 1956.
BEERMAN, H. - \
ce11ular di:
BELL, D.J. & Yl
g1ycogen.
BIANCHI, A.G. (
ra american;
presentada é
mica da USP.
BIEBER, L. L. i
BURGH, R. W. - 1
~ (Meigen)
BOAS, N. F. - ME
J. biol. ChE
Bom, G.J. - LE
ques des in!
1944.
BOYD, J. B. & M:
in Drosop}
BREUER, M.E.
tera, Sciari
lo, 17: 167·
BREUER, M.E. &
angelae at (
7: 317-386,

-119-
relation
Nauphoeta
ing insect
of Insec-
In: AL
1emistry,
~ne chromo-
E prothorax.
m and cha
!lls and
glands.
"BECKER, H.J. - Die Puffs der Speiche1drusenchromosomen von Droso-. ,,---
phila me1anogaster. 11. Mitteilung. Die Auslosung der"Puffbildung, ihre Spezifitat und ihre Beziehung zur Funktion
"der Ringdruse. Chromosoma, ll: 341-384, 1962.
BEERMAN, W. - Nuclear differentiation and functiona1 morpho10gy
of chromosomes. Co1d Spr. Harb. Symp. quanto Biol., 21: 217
232, 1956.
BEERMAN, \~. - Cyto10gica1 aspects of information transfer in
ce11u1ar differentiation. Amer. Zool., 1: 23-32, 1963.
BELL, D.J. & YOUNG, F.G. - Observations on the chemistry of 1iver
glycogen. Biochem.·J., 28:882-889, 1934.
BIANCHI, A.G. de - Bioqu{rniep da secreção salivar de Rhynchoscia
ra americana, são Paulo, 1972 [Tese de doutoramento a ser a
presentada ao Departamento de Bioquímica do Instituto de Quí
mica da Usp].
BIEBER, L.L.; HODGSON, E.; CHELDELIN, V.H.; BROOKES, V.J. & NEW
BURGH, R.W. - Phosph01ipids patterns in the b10wfly Phormia reqi
na (Meigen). J. biol. Chem., 236: 2590-2595, 1961.
BOAS, N.F. - Method for the determination of hexosarnines in tissues.
J. biol. Chem., ~l 553-563, 1953.
i~jj
III
1III
:00
r pupation
ltion
·omethod
of
of
the
BONt, G.J. -Le rapport sodium/potassium dans le liquide coelomi
ques des insectes. Ann. Soe. roy. Zool. Belg., 75: 123 -132,
1944.
BOYD, J.B. & MITCHELL, H.R. - Turnover of the hemolymph proteins
in Drosophi1a. Arch. Biochem., 117: 310-3~9, 1966.
11111
!~
'E, J .S. &
ão de lar
Diptera
iação Bra
s de peri§
London,
J
"BREUER, M.E. - Revision of the genus Rhynchosciara Rubsaarnen (DiE
tera, Sciaridae) in the neotropical region. Arch. Zo01.S.Pau
lo, 17: 167-198, 1969.
BREUER, M.E. & PAVAN, C. - Behaviour of polytene chromosomes of B.anqe1ae at different stages of larva1 development. Chromosoma,
7: 317-386, 1955.
111i'1!tll:1
i~

12'0-
BUCK, J.B. - Physica1 prooerties and chemica1 composition of in
sect blood. In: ROEDER, K.D., ed. Insect physiology. New
York, John Wiley, 1953, p. 147-190.
CLEVER, U. &
Speicheld:
Exp. Cel1
CAMERON, M.L. - Secretion of an orthodiphenol in the corpus car
diacurn of the insect. Nature, 172: 349-350, 1953.
CARRINGTON, C.B. & TENNEY, S.M. - Chernical constituents of haemo
1yrnph and tissue in Te1ea oolyphemus Cram. with particular r~
ference to the question of ion binding. J. Insect Physiol.,
3: 402-413, 1959.
CHEN, P.S. - Amino acid and protein metabo1ism in insect develop
ment. Advanc. Insect Physiol., 1: 53-131, 1966.
"CHEN, P.S. & BUHLER, R. - Paragonial substance (sex peptide) and
other free ninhydrin-positive components in male and female
adults of Drosophi1a melanogaster. J. Insect Physiol., 16:
615-627, 1970.
COLLINS, J. V,
Ca1podes t
12: 341-3!
CRAIG, R. - ~
tomol., 5:
CROMARTIE, R,
76, 1959.
CRONE, H. D. I
Musca domE
CRUICKSHANK,
juvenile t
CHEN JUNIOR, P.S.; TORIBARA, T.Y. & WARNER, H. - Microdeterrnina
tion of phosphorus. Analyt. Chem., 28: 1756-1758, 1956.
CHINO, H. & GILBERT, L.I. - Lipid release and transport in in
sects. Biochim. biophys. Acta, 2l!:94-110; 1965.
CHIPPENDALE, G.M. - Metamorphic changes in haemo1ymph and midgut
oroteins of the southwestern cornborer, Diatraea grandiose11a.
J. InsectPhysiol., li: 1909-1920, 1970.
CHEN, P.S. & LEVENBOOK, L. - Studies on the haemolyrnph proteins
of the blowf1y Phorrnia regina. 11. Synthesis and breakdown
as reveàled by isotopic 1abel1ing. J. Insect Physiol., 12:
1611-1627, 1966.
DAHM, K. H.; 'J
Synthesis
89: 5292-~
DAVENPORT, H.
go, Univex
DE LOOF, A. 8
proteins a
sa decem1i
DAVISQN, A.N rrnitter ?
DINAMARCA, M.
corooratia
sis of the
110-119, 1
pro
of
CHIPPENDALE, G.M. & KILBY, B.A. - Re1ationship between the
teins of the haemolymph and fat body during deve10pment
Pieris brassicae. J. Insect Physio1., 15: 905-926, 1969.
CLARK, E.l~. - A review of 1iterature on ca1ciurn and magnesiurn in
insects. Ann. entornaI. Soe. Amer., 51: 142-154, 1958.
DOYLE, D. & L
tion in th
61-78, 196

1"1
'sition of in
l2.gy. New
e corpus car
3.
ents of haem~
particular r~
ct Physio1.,
-121-
"CLEVER, U. & KARLSON, P. - Induction von Puff-Veranderung in den
Speicheldrüsenchromosomen von Chironomus tentans durch Ecdyson.
Exp. Cell Res., 20: 623-626, 1960.
COLLINS, J.V. - The hormonal control of fat body develoornent in
Calrodes ethlius (Lepidoptera, Hesneriidae). J. Insect Physiol.,
12: 341-352, 1969.
CRAIG, R. - The physiology of excretion in insects. Ann. Rev. En
tomol., ~: 53-68, 1960.
I'q
,,";fi
\'
I!I
~sect developCROMARTIE, R.I.T •. - Insect pigments. Ann. Rev. Entorno1., 4:
76, 1959.
59-
peptide) and
~ and fema1e
CRONE, H.D. & BRIDGES, R.G. ~ The phosph01ipids of the housef1y,
Musca domestica. Biochem." J., 89: 11-12~ 1963.
{si01. , 16: CRUICKSHANK, P.A. & PALMERE, R.M. - Terpenoid arnines as
juvenile horrnones~ Nature, 233= 488-489, 1971.
insect
nph proteins
1 breakdown
'si01. , 12:
n
DAHM, K.H.; TROST, B.M. & ROLLER, H. - The juveni1e horrnone. V.
Synthesis of the racemic juvenile horrnone. J. Am. chem. Soe.,
~: 5292-5294, 1967.
DAVISQN, A.N r & KACZMAREK, L.K. - Taurine - a possib1e neurotrans
mitter? Nature, lli:l07-10a, 1971.
DAVENPORT, H.N. - TheABC of acid-basic chemistry. 4th ed., Chica
go, Universtty Press, 1958, 118p.:rodeterrnina
I, 1956.
lort in in-
,h and midgut
grandiosella.
DE LOOF, A. & DE WILDE, J.
proteins and vitellogenesis
sa decemlineata. J. Insect
The relation between haemolyrnph
in the colorado beetle, Lentinotar
Physiol., 16: 157-169, 1970.
11111
•
en the pro
oprnent of
6, 1969.
magnesi um in
1958.
I
DINAMARCA, M.L. & LEVENBOOK, L. - Oxidation, utilization and in
cornoration into protein,of alanine and lysine durinq rnetamornh~
sis of the blowf1y Phorrnia regina (Meigen). Arch. Biochern. ,1!I:110-119, 1966.
DOYLE, D. & LAUFER, H. - Sources of larva1 salivary g1and secre
tion in the Dipteran Chironornus tentans. J. Ce11 Biol., 40:
61-78, 1969.
111/u::"

-122-
DREYFUS, A.; NONATO, E.; BREUER, M.E. & PAVAN, C. - Cromosomas P2litênicos em vários órgãos de "Rhynchosciara angelae" Nonato &
Pavan (Diptera). Rev. bras. Biol., 11: 439-450, 1951.
DUBOIS, M.; GILLES, R.A.; FJlliILTON, J.K.; REBERS, P.A. & SMITH,F.
Colorimetric method for determination of sugars and related
substances. Analyt. Chem., ~: 350- 356, 1956.
ELLIS, G.P. - The Maillard reaetion. Advanc. Carbohyd. Chem., 14:
63-134, 1959.
GAMBINO, S.R.
345, 1961.
GILBERT, L.I.
aspects. Ir:
York, Acaden
GILBERT, L.I.
ltisect Pnysi
ENGLE JUNIOR, R.L. & WOODS, K.R. - Comparative biochemistry
embriology. In: PUTNAM, F.W., ed. The plasma proteins.
Yórk, Academic Press, 1960, v.2, p. 183-265.
and
New
GILBERT, L.I.
In: FLORKIN,
Amsterdam,
ETTINGER, R.H.; GOLDBAUM, L.R. & SMITH JUNIOR, L.H. - A simplified
photometric method for the determination of citric acid in
biological fluids. J. bio1. Chem., 199: 531-536, 1952.
FLORKIN, M. & JEUNIAUX, C. - Hemolymph: composition. In: ROCKSTEIN,
M.,ed. The physiology of Insecta, NeN York, Academic Press,
1964, v. 3, p. 109-152.
FOLCH, J.; LEES, M. & SLOANE-STANLEY, G.H. - A simple method for
the isolation and purification of total lipids from animal
tissues. J. biol. Chem., 226: 497-509,1957.
FOSTER, J.E. - Plasma albumin. In: PUTNAM, F.I'l. ,ed., The olasma
proteins. New York, Academic Press, 1960, v.l, p.179-239.
FRIEDMAN, S. - Inhibition of treha1ase activity in the hemo1ymph
of Phormia regina Meig. Arch. Biochem., 93: 550-554, 1961.
FUKUDA, T.; SUDO, M.; MATUDA, M.; HAYASHI, T.; KUROSE, T.; HORIU
CHI, Y. & FLORKIN, M. - Formation of the silkproteins during the
growth of the silkworm larva (Bombyx mori). In: LEVENBOOK,L.
ed. Biochemistry of insects. London, Pergamon Press, 1959.
GALAMBOS, J.T. - The reaction of carbazole with carbohydrates. I.
Effect of borate and sulfamate on the carbazole colar of
sugars. Analyt. Biochem., 19: 119-132, 1967.
GILBY, A.R. - I
Entomo1., 1C
GILBY, A. R. & lJ
ria of a blc
GILMOUR, D. - 1Press, 1961,
GILMOUR, D. - '1
1965, 215 p.
GOLBERG, L. & [
des aegypti
Biochem. J.,
GOom·lIN, T. W.
tribution in
gratory locu
the desert 1
45: 472-479,
GOODWIN, T. Iv. New York, Ch
GOODWIN, T.W. &
carotenoids
migratoria Ir,
Forsk.) . E

-1:23-
:omosomas P2
~" Nonato &
151.
& SMITH,F.
1 re1ated
, Chem., 14:
GAMBINO, S.R. - Determination of b100d pC0 2 . C1in. Chem., 7:336
345, 1961.
GILBERT, L.I. - Physiology of growth and development: endocrine
aspects. In: ROCKSTEIN, M.,ed. The physio1ogy Insecta. New
York, Academic Press, 1964. v.l, p. 194-225.'
GILBERT, L.I. - Lipid metabo1ism and function in insects.Advanc.
Ihsect Physíol., !: 69-211, 1967a.
\ simplified
.stry
?ins.
acid
and
New
in
GILBERT, L.I. - Biochemical corre1ationsin insect metarnorphosis.
In: FLORKIN, M. & STOTZ, E.H., eds. Comprehensive biochemistry.
Amsterdam, Elsevier, 1967. v.28, p. 199~252.
GILBY, A.R. - Lipids and their metabolism in insects. Ann. Rev.
Entomol.,12: 141-160, 1965.
L952.
1: ROCKSTEIN,
müc Press,
nethod for
n animal
~he plasma
.79-239.
hemo1ymph
;4, 1961.
T.; HORIU
luring the
WENBOOK,L.
;5, 1959.
'drates. I.
GILBY, A.R. & McKELLAR, S.N. - The composition of the emp~y pupa
ria of a b1owfly. J. Insect Physiol., 16: 1517-1529, 1970.
GILMOUR, D. - The biochemistry of insects. New York, Academic
Press, 1961, 343 p.
GILMOUR, D. - The metabolism of insects. London, Oliver and Boy.d,
1965, 215 p.
GOLBERG, L. & DE MEILLON, B. - The nutrition of the larva of Aê
des aegypti Linnaeus. 4. Protein and amino-acid requirements.
Biochem. J., 43: 379-387, 1948 .
GOODWIN, T.W. - The biochemistry of locust. 2. Carotenoid àis
tribution in solitary and gregarions phases of the African mi
gratory locust (Locusta migratoria migratorioides R & F) and
the desert locust (Schistocerca gregaria Forsk). Biochem. J.,
45: 472-479, 1949.
GOOmHN, T. \'l. - The comparative biochemistry of the carotenoids.
New York, Chemical· .Publishing, 1954, 356 p.
.or of GOOmvIN, T.W. & SRISUKH, S. - The biochemistry of locust. I. The
carotenoids of the integument of two locust species (Locusta
migratoria migratorioides R. and F and Schistocerca gregaria
Forsk.) . Biochem. J., 45: 263- 268, 1949.

-124-
G~GOlRE,' C. - Blood eoagu1ation in arthropods. 11. Phase oon~
trast mieroseopie observations on hemo1ymph eoagu1ation in
sixty-one speeies of inseets. B1ood,~: 1173-1198, 1951.
~RtGOlRE, C. & TAGNON, H.J. - B100d eoagulation. In: FLORKIN,M.
& MASON, H.S., eds. comparative bioehemistry. New York, Aea
demie Press, 1962. v.4, p. 435-482.
"GROSSBACH, U. - Chromosomen-Aktivitat und bioehemisehe Ze11-"differenzierung in den Speieheldrusen von Camptoehironomus.
Chromosoma, 28: 136-187, 1969.
GUARACIABA, H.L. & TOLEDO, L.F.A. - Age determination of "Rhyn
ehoseiara angelae 11 1arvae. Rev. bras. Biolo, 27: 321-332,
1967.
HORIE, Y;
Bombyx mo.
IMMS, A.D.
"O. W. Ri e:886 !?
INOKUCHI, T.
Bombvx mo
JACOB, S.
Meth. bio
JENKIN, P.M.
eles. Nat
HACKMAN, R.H. - Changes in the free
blowf1y larvae at metamorphosis.
405, 1956.
amino aeids of the blood of
Aust. J. bio1. Sei., 9:400-
JOr."NSON, i'l. S
aeetie ae
HACKMAN, R.H. & GOLDBERG, M. - Studies on the hardening and dark
ening of inseet eutieles. J. Insect Physio1., 17:335-347,1971.
HACK~~, R.H. - Chemistry of the insect eutie1e. In: ROCKSTEIN,
M., ed. The physio1ogy of Insecta. New York, Aeademic Press,
1964. v.3, p. 471-506.
HAMILTON, P.B. - Ion exehange ehromatography of aminoacids,
sing1e eo1umn, high resolving, ful1y automatic proeedure.
Analyt. Chem., 35: 2055- 2064, 1963.
a
KARLSON, P.
ne, and i
473-502,
KARRER, P. &
384 p.
KILBY, B.A.
Insect Ph
KILBY, B.A.
In: Goom,
HERMAN, W.S. - Control of hormone production in insects. In:ETKIN,
W. & GILBERT, L.I., eds. Metarnorphosis: a orob1ern in deve10prnent
bio1ogy. New York, App1eton Century-Crofts, 1968, p. 107-141.
HILL, L. - Neurosecretory eontrol of hemo1ymph protein coneentra
tion during ovarian deve10pment in the desert locust. J. Inseet
Physio1., 8: 609-621, 1962.
HINTON, H.E. - Coneea1ed phases in the rnetamorphosis of inseets.
Nature, 157: 552-553, 1946.
HIRS, M.W.; STEIN, W.H. & MOORE, S. - The amino acid cornposition
of ribonuc1ease. J. bio1. Chem., 211: 941-950, 1954.
York, Acc
KUHN, R. 81 ~
Chem. Bel
LARA, F.J.S,
anqe1ae.
LAUFER, H•.
proteins
torno1. ,
200.

-l:25-
by
1957,
'. Phase oon~
'ulation in
.98, 1951.
n: FLORKIN,M.
w York, Aea-
isehe Zell
oehironomus.
)n Df "Rhvn
7: 321-332,
:he blood of
Sei., 2.: 400-
I: ROCKSTEIN,
.demie Press,
ing and dark
:335-347,1971.
HORIE, Y; - Blood trehalose and fat body glyeogen in the silkworm,
Bombyx mori. Nature, 188: '583-584, 1960 .
IMMS, A.D. - A general textbook of entom2lY. 9th ed. Rev.
U. W. Riehards and R. G. Davies. New York, ~lethuen,
886 ~.
INOKUCHI, T.; HORIE, Y. & ITO, T. - Urea eye1e in the silkworm,
Bo~hvx mori: Bioehem. biophys. Res. Commun., 35: 783-787,1969.
JACOB, S. - The determination Df nitrogen in biolcgiea1 materiaIs.
Meth. bioehem. Anal., 13: 241-263, lS65.
JENKIN, P.M. & HINTON, H.E. - Apolysis in arthropod moulting ey
eles. Nature~ 211: 871, 1966. .,JOENSON, ~·].S. & SCffi-lEIDER, H.P. - B-earbethoxy-y,y-dipheny1viny,!
aeetie aeid. Org. synth. Rev. ed., ,1: 132-136, 1963.
KARLSON, P. & SEKERIS, C.E. - Eedyso~e, an inseet steroid hormo
ne, and its mode of aetion. Reeent. Progr. Hormone Res., 22:
473-502, 1966.
KARRER, P. & JUCKER, E. - Carotenoids. :Jew York, Elsevier,1950,
384 p.
"II!!
Illi
li!
oaeids,
oeedure.a
KILBY, B.A. - The bioehemistry of the inseet fat body.
Inseet Physiol., 1: 111-174, 1963.
Advane.
::ts. In:ETKIN,
in development
, p. 107-141.
Ln concentra
1St. J. Insect
of insects.
comoosition
54.
KILBY, B.A. - Intermediary metacolism and the inseet fat body.
In: GOODNIN, T.N.,ed. Aspects of Insect biochemistrv. New
York, Acadernic Press, 1965, P. 39-48.
» "KUHN, R. & SORENSEN, N.A. Uber Astaxanthin und Ovoverdin.
Ch~~. Ber., 11: 1879-1888, 1938.
LARA, F.J.S.; TAMAKI, H. &.PAVAN, C. - Laboratory eu1ture of R.
anqelae. Amer. Natur., ~: 189-191, 1965.
LAUFER, H. - Studies Df ehanges in enzynatie activities Df blood
proteins in the developing si1k moth. Proe. Int. Congr. En
torno1., 11th , Vienna, 1960. Proc. Pavia, 1960a. v.3, p. 194
200.

-1:26-
LAUFER, H. - Blood proteins in insect development. Ann. N.Y.Acad.
Sei., 89: 490-515, 1960b.
LAUFER, H. & NAKASE, Y. - Developmental studies of the Dipteran
salivary gland. 11. DNase activity in Chircno~us thummi. J.
Cell Biol., 25: 97-102, 1965.
LAW, J.H. & REGNIER, F.E. - Pheromones. Ann. Rev. Bioc~ern., 40:
533-:-548, 1971.
LEA, C.H.; HANNAN, R.S. & GREAVES, R.I.N.· - 'l'he reaction bet-
ween proteins and reducing sugars in the "dry"state. Biochem.
J., i2: 626-629, 1950.
LEE, R.M. - The variation of blood volume with age in the desert
locust. (Schistocerca qregaria Forsk.). J. Insect Physiol.,
.§.: 36-51, 1961.
LEUENBERGER, F. & TOMMEN, H. - Ke:to-carotenoids in the Colcrv.do
potato beetle, Leotinotarsa decemlineata. J. Insect Physicl.,
lf: 1855-1858, 1970.
LEVENBOOK, L. - The composition of horse bot fly (Gastroohilus in
testinalis) larva blood. Biochem. J., 47: 336-346, 1950a.
LEVENBOOK, L. - The physiology of carbon dioxide transoort in in
sect blood. 111. The buffer capacity of Gastroohilus bIood.
J. exp. Biol., l2: 184-191, 1950b.
LEVENBOOK, L. - Organic acids in insects. 11. Tricarboxilic acid
cycle and related enzymes in the larva of the southern army
worm, Prodenia eridania. pxch. Biochem., 92: 114-121, 1961.
LEVENBOOK, L. - Hemolymph aminoacids and peptides during lanral
grovlth of '.he blowfly, Pbormia regina. Comp. Biochem. Physiol.,
18: 341-351, 1966.
LE\~NBOOK, L. & DINAMARCA, M.L. - Free amino acids and related
compounds during metamorphosis of the blowfly Phormia regina.
J. Insect Physiol., 12: 1343-1362, 1966.
LEVENBOOK, L. & HOLLIS JUNIOR, V.W. - Organic acid in insects. I.
Citric acid. J. Insect physiol., .§.: 52-61, 1961.
LOCKE, M. & COl
body of an :
LOCKE, M. & COl
dies' in the
467-469, 191
LOUGHTON, B.G.
of haemolymI
11: 919-932,
Lm1RY, o. H.; R(
tein meaSUrE
ill: 265-27!
LUCAS, F. & RUI
silks. In:
chemistry.
LUCAS, F.; SHA\
Protein Cher
LUCAS, F.; SHAI'
fibroins.
and i ts sigr
and taxononl)
I-IARSH, R.E.; CC
si1k fibroir
ne). Acta C
MARSH, R.E.; CC
structure oí
1955b.
MARTIN, J.S.
of lipids b)
rus. J. Ins
MAZ ZI, V. & BAC
de11a seta r
Curcu1ionidc

-127-
n. N.Y.Acad.
Dipter.'ln
thummi. J.
c~em., 40:
on bet-
Biochem.
the desert
Physiol. ,
Colorudo
t Physic1.,
roohilus in
1950a.
port in in
~ blood.
oxilic acid
ern army
21, 1961.
og larval
m. Physiol.,
:l related
ia regina.
insects. I.
J
LOCKE, M. & COLLlNS, J.V. - Sequestration of protein by the fat
body of an insect. Nature, 210: 552-553, 1966.
LOCKE, M. & COLLlNS, J.V. - Protein uptake in multivesicular bo
dies'in the molt-intermolt cycle of an insect. Science, 155:
467-469, 1967.
LOUGHTON, B.G. & ~mST, A.S. - The development and distribution
of haemolymph proteins in Lepidoptera. J. Insect Physiol.,
11: 919-932, 1965.
LO~~Y, O.H.; ROSEBROUGH, N.S.; FARR, A.L. & RANDAL, R.J. - Pro
tein measurement with the folin phenol reagent. J. biol.Chern.,
193: 265-275, 1951.
LUCAS, F. & RUDALL, K. - Extracellular fibrcus proteins: the
silks. In: FLORKlN, M. & STOTZ, E.H., eds. Comprehensive bio
chernistry. Amsterdarn, Elsevier, 1968. v. 26B, p. 475-558.
LUCAS, F.; SHAW, J.T.B. & SMITH, S.G. - The silk fibroins. Advanc.
Protein Chern., 11: 107-242, 1958.
LUCAS, F.; SHAW, J.T.B. & SMlTH, S.G. - Cornparative studies of
fibroins. I. The aminoacid cornposition of various fibroins
and its significance in re1ation to their crystal structure
and taxonon,y. J. mo1ec. llio1., ~: 339-349, 1960.
V~RSH, R.E.; COREY, R.B. & PAULING, L. - The structure of Tussah
si1k fibroin (with a note on the structure of ~-po1y-L-a1ani
ne). Acta Cryst., 8: 710-715, 1955a.
~~RSH, R.E.; COREY, R.B. & PAULING, L. - An investigation of the
structure of si1k fibroin. Biochim. biophys. Acta, 16:1-34,
1955b.
MARTIN, J.S. - Studies on assimi1ation, mobi1ization and transoort
of lipids by the fat body and haemo1ymph of Pyrrhocoris apte
rus. J. lnsect Physio1., 15: 2319-2344, 1969.
MAZZI, V. & BACCETTI, B. - I tubi Ma1pighiani e la secrezione
de11a seta ne11e 1arve di Donus crinitus Boheman (Coleoptera,
Curculionidae, Hyperini) Redia, 41: 315-341, 1956.

-128-
OI
MEYER, K.H. & MARK, H. - Uber den Aufbau des Seiden-Fibroins.
Chem. Ber., i!: 1932-1936, 1928.
MITCHELL, H.K. & SIMMONS, J.R. - Amino acids and derivatives in
Drosophi1a. In: HOLDEN. J.T., ed. Amino acid poo1s. Amster
dam, E1sevier, 1962, P. 136-146.
MOKRASCH, L.C. - Ana1ysis of hexose ohosphates and sugar mixtures
with the anthrone reagent. J. bio1. Chem., 208: 55-59,1954.
MOORE, S. & STEIN, H.H. - Procedures for the chromatographic de
termination of amino acids on four per cent cross-linked su1
fonated po1ystyrene resins. J. bio1. Chem., 211: 893-906,
1954.
MUNN, E.A.; FEINSTEIN, A. & GREVILLE, G.D. - A major protein
consti tuent of pupae of the b1mlfly Cal1iphora ervthrocepha1a
(Diptera). Biochem. J., 102: SP-6P, 1967.
MUNN, E.A. & GREVILLE, G.D. - The soluble oroteins of developing
Cal1iphora erythrocepha1a, particular1y cR11iphorin, and sim~
lar proteins in other insect. J. Insect Physiol., 15,: 1935
1950, 1969.
~1UNN, E.A. ;PRICE, G.M. & GREVILLE, G.D. - The synthesis "in vi
tro" of the protein calliphorin by fat body from the larva of
the blovTfly, Calliphora erythrocephala. J. Insect Physio1., 15:
16Ól-160S, 1969.
NARAHASHI, T. - The properties of insect axons. Advanc. Insect
Physio1., 1: 175-256, 1963.
NIELSEN, D.J. & MILLS, R.R. - Changes in electrophoretic proper-"ties of haenolymph and terminal oocyte proteins d~ring vite11~
genesis in the american cockroach. J. Insect Physiol., 14:
163-170, 1968.
"NONATO, E. & PAVAN, C. ~ A new species of "R-l-)ynchosciara" Rubsaa
men, 1894 (Diptera, Mycetophi1idae). Rev. bras. Biol., 11:
435-437, 1951.
NOVÂK, V.J.A. - Insect hQrmones. London, Methuen, 1966. 478p.
OHNISHI, -E
from COI
783-784
ORR, C. W.M.
on egg (
Insect I
ORR, C.W.M.
ori the c
b10wfly,
119, 19€
PARKER, K.rgreen I"
PARTRIDGE,
tography
to the q
white an
PATTON, R.L
New York
PRICE, G.M.
house fI
PRIq:, G.M.
teins is
throceph
PROSSER, C.ndgy. 2
RICHARDSON,
1arva1.
RIECHERS, I
of "mu1t
J. Insec
"RÓi..LER, H.;
ture of

-129-
-Fibroins.
rivatives in
~. Arnster-
,ugar mixtures
55-59,1954.
;ographic de
;-linked sul
893-906,
OHNISHI, -E.; TAKAHASHI, S.Y.; SONOBE, H. & HAYASHI, T. - Crys~al
from cocoon of Ma1acosoma neustria testacea. Science,~:
783-784~ 1968.
ORR, C.W.M. - The influence of nutritional and hormonal factors
on egg deve10pment in the b10wfly Phormia regina (meig.). J.
Insect Physiol., 12: 53-64, 1964a.
ORR, C.W.M. - The inf1uence of nutritiona1 and hormona1 factor
on the chemistry of the fat body, b100d and ovaries of the
b1owfly, Phormia regina, Meig. J. Insect Physiol., 10: 103
119, 1964b.
'r orotein
ythroceohala
f developing
in, and simi
PARKER, K.D. & RUDALL, K.M. - The silk of the egg-stalk of the
green 1ace-wing fly. Nature, 111: 905-906, 1957.
PARTRIDGE, S.M. & ~mSTALL, R.G. - Filter-paper partition chroma
tography of sugars. I. General description and apolication~
to the qua1itative ana1ysis of sugars in apple juice, egg
white and foeta1 blood of sheep. Biochem. J., 42:238-248,1948.
, 15: 1935- PATTON, R.L. - Excretion. In: ROEDER, K.D., ed. Insect ohysiology.
New York, John Wi1ey, 1953, p. 387-403.
:osis "in vi-
the larva of
Physiol. , 15:
mc. Insect
PRICE, G.M. - Some aspects of amino acid metabo1ism in the adu1t
house f1y, Musca domestica. Biochem. J., 80: 420-428,1961.
PRIC~, G.M. & BOSMAN, T. - The e1ectrophoretic separation of pro
teins isolated from the larva of the blowfly, Ca11iohora ery~
throcephala. J. Insect Physiol., 12: 741-745, 1966.
!tic proper
~ring vi te11Q
.io1. , 14:
PROSSER, C.L. & BRO\m JUNIOR, F.A. - Comoarative animal ohysiolo
91. 2nd ed., Philadelphia, Saunders, 1961, 688p.
RICHAROSON, C.H.: BURDETTE, R.C. &.EAGLESON, C.W. - Blood volume
larval. Ann. entomol. Soe. Amer., li: 503-506, 1931.
"~" Rubsa2-io1. , 11:
RIECHERS, L.A.; MEYERS, F.H. & BERRY, S.J. - DNase as a component
of "multi-enzyme complexes" in the blood of saturniid moths.
J. Insect Physiol., 15: 743-753, 1969 •
..Rd~LER, H.; DAHM, K.H.; SvffiELEY, C.C. & TROST, B.M. - The struc-
ture of the juvenile hormone. Angew. Chem., i:179-1BO,1967.I____1 _
66. 47Bp.

1-130-
ROSEN, H. - A modified ninhydrin eo1orimetrie ana1ysis for amino
aeids. Areh. Bioehem., 67: 10-15,1957.
SHIGEMATSU, )
the silkwc
ROSS, H.H. - A textbook of entomo1ogy. New York, John Wi1ey,1965,
539 p.
ROTH, T.F. & PORTER, R.K. - Yo1k protein uptake in ooey±e of the
mosquito Aedes aegypti L. J. Cell Biol., 20: 313-332, 1964.
RUDALL, K.M. - Silk al1d other eoeoon proteins.
MASON, H.S., eds; Comparative bioehemistry.
mie Press, 1962. v. 4, p. 397-433.
In: FLORKIN, M. &
New York, Aeade-
SHIGEMATSU, l
worm, ~
170, 1960
SHIGEMATSU, l
synthesis
si1kworm,
1970.
RUDALL, K.M. - The ehitinprotein eomplexes of inseet eutieles.
Advane. inseet Physiol., 1: 257-313, 1963.
SIAKOTOS, A. reoekroaeh
1013, 196C
RUDALL, K.M. - Ske1eta1 strueture in inseets. In: GOODHIN, T.!'l.,
ed. Aspeets of inseet bioehemistry. London, Academic Press,
1965. _p. 83~92.
SIAKOTOS, A. r
coekroach.
cesses.
RUDAI.L, K. M. & KENCHINGTON, \-7. - Arthropod
of fibrous proteins in animal tissues.
73~96, 1971.
silks: the prob1em
Ann. Rev. entorno1.,16:SINNOT, E. W. l
ties. 5 t l'
SCHNEIDERMAN, H.A. & GILBERT, L.I. - Control of growth and deve
loprnent in inseets. Scienee, 143: 325-333, 1964.
SCHMIDT, F.H. & YOUNG, C.L. - Larva1 eoloration in Choristoneura
~. (Lepidoptera; Tortrieidae). Bi1e pigment in haemo1yrnph
J. Inseet Physiol., 17: 843-855, 1971.
SEIFTER, S. & GALLOP, P.M. - The structure proteins. In: NEU
RATH, H., ed. The proteins. New York, Aeadernic Press, 1966.
v. 4, p. 155-458.
stGUY, E. - Ordre des dipteres. In: GRASSt, P.P., ed. Traité de
zoologie: anatomie, systématique, bio1ogie. Paris, Masson,
1951, v. lO, p. 448-744.
12.: 293-3C
ovaries,
STEELE, J.E.
eardiaeum
SMITH-JUNIOR,
Nature, 20;
SPACKMAN, D.F
apparatus
Chem., .2Q:
sLÂMA, K.
40: 1079-J
~
STRANGIvAYS-DJ
nes and re
(Meig. )for
biol.
SHALES, O.S. & SHLAES, S.S. - A simple and aecurate method
the determination of ehloride in biological fluids. J.
Chern., 140: 879-884, 1941.
I
1,

-Bl-
for amino SHIGEMATSU, H. - Synthesis of blood proteins by the fat body in
the silkworm, Bombyx mori. Nature,~: 880-882, 1958.
SHIGEMATSU, H. - Protein metabolism in the fat body of the silk
worm, Bombyx mori L. BulI. Seric. Exp. Stn. Japan, 16: 141
170, 1960.
Wiley,1965,
yt:é of the
32, 1964.
,ORKIN, M. &
rk, Acade-
SHIGEMATSU, H. & MORIYAMA, H; - Effect of ecdysterone on
synthesis in the posterior division of the si1k gland
silkworm, Bombyx mori. J. Insect Physioi., 16: 2015
1970.
fibroin
of the
- 2022,
cutic1es.SIAKOTOS, A.N.
cockroach I.
1013, 1960a.
The conjugated plasma proteins of the American
The normal state. J. gen. Physiol., 43: 999 -
DT-nN, T. vl. ,
mic Press,
problem
."ltomol. , 16:
~istoneura
haemo1ymph
and deve-
Traité de
[asson,
In: NEU-
55, 1966.
hod for
J. bio1.
i
r
~
SIAKOTOS, A.N. - The conjugated plasma proteins of the American
cockroach. lI. Changes during the mou1ting and clotting pro
cesses. J. gen. Physiol ., 43: 1015-1030, 1960b.
SINNOT, E.W.; DUN~, L.C. & DOBZHANSKY, T. - PrincipIes of gene
tics. 5 th ed. New York, McGraw Hil1, 1958, 581p.
sLÂMA, K. - Insect juveni1e hormone ana1ogues. Ann. Rev.Biochem.,
40: 1079-1102, 1971.
SMITH-JUNIOR, J.G. & CLARK, R.D. - Hexosamine in Bombyx mori si1k.
Nature, ~: 860-861, 1965.
SPACKMAN, D.H.; STEIN, W.H. & MOORE, S. - Automatic recording
apparatus for use in the chromatography of amino acids. Ana1yt.
Chem., 30: 1190-1206, 1958.
STEELE, J.E. ~ Ocurrence of a hyperglycemic factor in the corpus
cardiacum of an insect. Nature,~: 680-681, 1961.
'A'STRANGWAYS-DIXON, J. - The relationship between nutrition, hormo
nes and reproduction in the blowfly Ca11iphora erythrocephala
(Meig. ) lI!. The corpusal1atum in -relation to nutri tion, the
avaries, innervation and the corpus cardiacum. J. exp.Biol.,
,22: 293-306, 1962.

-];-32-
TELFER, W.H. - Immuno1ogica1 studies of insect metamorphosis. II.
The role of a sex-1imited fema1e protein in egg formation by
the cecropia si1kworm. J. gen. Physiol., 37: 539-558, 1954.
TELFER, W.H. - The mechanism.:. and control of yo1k formation. Ann·.
Rev. Entomo1., 10: 161-184, 1965.
TOBE, S.S. & LOUGHTON, B.G. - An autoradiographic study of haemo
lymph protein uptake by the tissues of the fifth instar 1ocust.
J. Insect Physiol., 15: 1331-1346, 1969.
TOKUMARU, M.; FERRI, M.O.G. & l'10RSMANN,T.U. - Pressão osmótica
da hemolinfa da "Rhynchosciara angelae" Nonato & Pavan, 1951
(Oiptera, Mycetophilidael. Rev. bras. Biol., 28: 435 - 445,
1968.
TOLEOO FILHO, S.A. - Oifferentiation of sibling species of "R..hvn
chcsciara" by hemolyrnph electrophoresis (Oiptera, Sciaridae).
Rev. bras. Biol., 29: 289-294, 1969.
TUNOIsr, J.; KOKROM, M.I.; DTAZ, G.; M/,GALOI, J.B. & LP~RA, F.J.S.
Composição da hemo1infr. de "Rhynchosciara angelae" Cienc. e
Cu1t., 16: 185, 1964.
V&~SLYKE, 0.0. & HA~~INS, J.A. - A gasometric method for deter~i
nation of raduc~ng sugars, and its application to analysis of'
b100d dnd urine. J. bio1. Chem., 79: 739-767, i928.
~ATERHOUSE, O.F. - Stuãies of the physiology and toxico1ogy of
blowf1ies. XIV. The composition, formation and fate of the
granules in the Ma1pighian tubu1es of Lucilia cuprina 1arvae.
Aust. J. Sei. Res. B, l: 75-112, 1950.
iVEINER, B.A. - A..microe1ectrode to measure dissolved oxygen in in
sect 1arvae. J. Insect Physiol., 11: 817-830, 1965.
~mITTEN, J.M.- The supposed pre-pupa in cyclorrhaphous Oiptera.
Quart. J. micro Sei., 98: 241-250,1957.
WIENS, A.W. & GILRERT, L.I. - Regu1ation of carbohydrate mobili
zation and utilization in Leucophaea maderae. J. Insect Fhy
sio1., 13: 779-794, 1967.
j,
IIi
J
WIGGLESWOR'J
the stuc
1948.
WIGGLESWOR'I
producti
1964.
WIGGLESWOR'I
London,
WIGGLESWORT
1970, 15
í'1YATT, G. R.
EntomoI.
í'1YATT, G.R.
W. & GIL
lopmenta
p. 143-1
WYATT,G.R.
II. Treh
833-847,
WYP.TT, G. R.
insect h,
si1kworm
39: 853-
ZACHARIUS,
ocuring I
chromato.

-133-
lorphosis. 11.
'ormation by
'-558, 1954.
,rmatiori. Ann·.
udy of haemo
instar locust.
WIGGLESWORTH, V.B. - Croonian lecture: the insect as a medium for
the study of physiology. Proc. roy. Soe. B, 135: 430 - 446,
1948.
WIGGLESWORTH, V.B. - The hormona1 regulation of growth and re
production in insects. Advanc. Insect Physiol., l: 244-332,
1964.
WIGGLESWORTH, V.B. - The principIes of insect physiology. 6th ed.
London, Methuen, 1965, 741 p.
WIGGLESWORTH, V.B. - Insect hormones. Edinburgh, 01ivar &
1970, 159 p.ao osmótica
Pavan, 1951
435 - 445,~VYATT, G.R. - The biochemistry of insect hemolymph.
Entomol., ~: 75 - 102, 1961.
Ann.
Boyd,
Rev.
ies of "Rhvn
Sciaridae) .
LP~RA, F.J.S.
'I Cienc. e
1 for determi
analysis of .
l28.
.cology of
:e.te of the
lrina larvae.
oxygen in in'1:,-'o
,us Diptera.
ate mobili
Insect Fhy-
'1II
f
j
íVYATT, G.R. - Biochemistry of insect metamorphosis. In: ETKIN,
W. & GILBERT, L.I., eds. Metamorphosis: a prob1em in deve
1opmenta1 bio1ogy. New York, Appleton Century-Crofts,1968.
p. 143-184.
WYATT,G.R. & KALF, G.F. - The chemistry of insect hemo1ymph.
11. Trehalose and other carbohydrates. J.gen.Physio1., 40:
833-847, 1957.
WYATT, G.R.; LOUGHHEED, T.C. & vNATT, S.S. - The chemistry of
insect hemolymph. Organic components of the hemolymph of the
si1kworm, Bornbyx mori, and two other species. J.gen. Physio1.,
39: 853-868, 1956.
ZACHARIUS, R.M. & TALLEY, E.A. - E1ution pattern of natura11y
ocuring ninhydrin positive compounds during ion exchange
chromatography. Analyt. Chem., 34: 1551-1556, 1962.