“junÇÕes comunicantes e migraÇÃo celular em ... (2).pdf · células derivadas da zona...
TRANSCRIPT

“JUNÇÕES COMUNICANTES E MIGRAÇÃO CELULAR EM EXPLANTES DE ZONA SUBVENTRICULAR PÓS-NATAL DE
ROEDORES”
MÔNICA MARINS
TESE DE DOUTORADO SUBMETIDA À UNIVERSIDADE
FEDERAL DO RIO DE JANEIRO
Universidade Federal do Rio de Janeiro Centro de Ciências da Saúde Programa de Pós-Graduação em Ciências Morfológicas 2006

i
Mônica Marins
JUNÇÕES COMUNICANTES E MIGRAÇÃO CELULAR EM EXPLANTES DE ZONA SUBVENTRICULAR DE ROEDORES
Tese de Doutorado, apresentada ao Programa de Pós-Graduação em Ciências Morfológicas (Neurobiologia do Desenvolvimento), Departamento de Anatomia, no Instituto de Ciências Biomédicas, Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Doutora em Ciências morfológicas (Neurobiologia do desenvolvimento).
Esta tese foi desenvolvida, no Laboratório de Neuroanatomia Celular, sob a orientação do Prof. João Ricardo Lacerda de Menezes e co-orientação da Profa. Maira Monteiro Fróes e contou com ao apoio financeiro das seguintes entidades: CAPES, CNPq, CNPq/PRONEX , FAPERJ, FUJB.
Rio de Janeiro 2006

ii
assinaturas da banca

iii
Marins, Mônica
Junções comunicantes e migração celular em explantes de zona
subventricular pós-natal de roedores / Mônica Marins. – Rio de Janeiro:
UFRJ / Instituto de Ciências Biomédicas, 2006.
xi, 127 f. : il. ; 31 cm Orientadores: João Ricardo Lacerda de Menezes e Maira Monteiro Fróes
Ferrão Tese (doutorado) -- UFRJ, Instituto de Ciências Biomédicas,
Programa de Pós-graduação em Ciências Morfológicas, 2006. Referências bibliográficas: f. 93-117
1. Telencéfalo - citologia. 2. Telencéfalo – crescimento & desenvolvimento. 3. Neurônios - citologia. 4. Neurônios – fisiologia. 5. Comunicação celular. 6. Junções gap. 7. Movimento celular - fisiologia. 8. Cultura de células. 9. Carbenoxolona – análogos & derivados. 10. Microscopia de vídeo. 11. Imunohistoquímica. 12. Animais recém -

iv
Em memória de
Odilon Ribeiro da Silva (Pai)
e
Graciela de Oliveira Silva (Vó)

v
Agradecimentos
Agradeço primeiramente a Deus, pois sem a permissão Dele nada acontece.
Especialmente à minha mãe, Helenice Marins da Silva, o esteio de minha vida. À
minha filha Ana Clara Marins, que deu um sentido para minha vida e me ensinou a
ter mais garra ainda.
Aos meus dois orientadores, João Ricardo Lacerda de Menezes e Maira Monteiro Fróes, pela "paciência de jó" com minhas urgências (nem sempre tão
urgentes) e principalmente por terem sido muito mais que orientadores, me
presenteando com seu humor impagável, amizade, carinho e exemplo.
Ao Fábio da Silva de Azevedo Fortes, pelo auxílio imprescindível com os
experimentos de injeção intracelular.
Ao Professor Antônio Carlos Campos de Carvalho por ter cedido os
animais nocaute para conexina 43 e à Sylvia Suadicani responsável pelos
experimentos com estes animais.
Ao Professor Marcelo Pelajo Machado por ter disponibilizado o Microscópio
Confocal, do Departamento de Patologia da Fundação Oswaldo Cruz, e ao Pedro Paulo de Abreu Manso, pelo excelente trabalho de operação do Confocal.
Ao apoio técnico impecável e eficiência de Carla Moreira Furtado, Elizabeth C. P. Moraes, Antônia Lima Carvalho, Adiel B. do Nascimento e Alexandra F. dos Santos. Não sei o que seria de mim sem vocês.
Aos membros do Departamento de Anatomia, especialmente o prof. Vivaldo Moura Neto e o pessoal de seu laboratório.
Às amigas Rosenilde C. Holanda e Mair Machado de Oliveira pelas dicas
valiosíssimas e amizade inestimável.
A TODOS os membros passados e presentes dos laboratórios de Neuroplasticidade e Neuroanatomia Celular, não esquecendo das convidadas
especiais Rose e Mair, que fazem do dia-a-dia profissional uma convivência em
família, com muito bom humor, companheirismo e paciência, resultando em nossos
almoços festivos de todos os dias. Foi uma grande honra ter convivido com todos
vocês.
Por fim aos ratinhos que perderam generosa e literalmente as cabeças.

vi
Resumo: MARINS, Mônica. Biblioteca e memória: o efeito do bloqueio juncional com carbenoxolone em explantes de zona subventricular pós-natal de roedores. Rio de Janeiro, 2001. Dissertação (Mestrado em Ciências Morfológicas)—Instituto de Ciências Biomédicas, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2001.
Células derivadas da zona subventricular telencefálica pós-natal (do inglês, SVZ, subventricular zone) participam de uma massiva migração tangencial para o bulbo olfatório em um processo conhecido como fluxo migratório rostral (do inglês, RMS, rostral migratory stream). Os mecanismos subjacentes a esta migração ainda são pouco esclarecidos. Em estudos prévios demonstramos que células da SVZ/RMS apresentam acoplamento celular mediado por junções comunicantes. No presente trabalho investigamos os pares acoplados, revelados por injeção intracelular de corantes, em células do halo migratório de explantes de SVZ. A transferência de corantes mostrou que ambos neuroblastos e astroglia participam de acoplamento homo- e heterocelular. Em ensaios in vitro de cultura de explantes de SVZ testamos a ação de agentes desacoplantes sobre a migração celular. Em resumo, explantes de SVZ de ratos de 4 dias pós-natais foram cultivados sobre poli-L-lisina por 3 a 5 dias (3-5DIV), na presença e ausência de Carbenoxolone (CBX) e tiveram seu halo migratório medido. O halo migratório, a partir destes explantes, foi significativamente reduzido na presença do CBX, sem qualquer sinal de efeito inespecífico ou tóxico. Realizamos ensaios de videomicroscopia em explantes cultivados sobre poli-L-lisina ou matrigel por 2-3 DIV. Estes explantes eram observados por 3-5 horas nas condições pré-tratamento, CBX (ou halotano) e pelo período de recuperação, após a retirada da droga. A migração foi bloqueada fortemente em um curso temporal consistente com a cinética do bloqueio de acoplamento por estes fármacos. Além disto, este efeito foi plenamente reversível. Nossos resultados sugerem que, tanto o acoplamento homo- como heterocelular estão presentes na SVZ/RMS e que este acoplamento é importante para a inicialização e manutenção da migração na SVZ pós-natal.

vii
Abstract: MARINS, Mônica. Biblioteca e memória: o efeito do bloqueio juncional com carbenoxolone em explantes de zona subventricular pós-natal de roedores. Rio de Janeiro, 2001. Dissertação (Mestrado em Ciências Morfológicas)—Instituto de Ciências Biomédicas, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2001.
Cells derived from the anterior subventricular zone (SVZ) participate in a massive tangential migration to the olfactory bulb known as the rostral migratory stream (RMS). The mechanisms underlying this migration are still poorly understood. We have previously demonstrated that cells throughout the SVZ/RMS are coupled through functional gap junctions. In the present work the coupled cell partners were revealed by intracellular dye injection assays throughout the migratory halo in SVZ explants. Intracellular injections have shown that both neuroblast and astroglial cells participate in homo- and heterocellular coupling. We have then tested the effects of uncoupling agents on cell migration in SVZ explants. Briefly, SVZ explants from 3-4 postnatal rats were cultured for 3 days (3DIV) onto poly-L-lisine, in presence or absence of the uncoupler carbenoxolone (CBX), and the migratory halo was measured. Significant reduction was recorded in the presence of CBX, with no signs of unspecific or cytotoxic actions. Cell migration was also analysed using time-lapse video-microscopy in explants plated onto poly-L-lysine or matrigel for 2-3 DIV. These explants were observed for 3-5 hours under each condition: pre-treatment, CBX (or halothane) and recovery conditions, after drug washout. Pharmacological uncoupling reversibly blocked cell migration, with inhibitory kinetics’ consistent with the distinct modes of action of the agents employed. Our results indicate that both homocellular and heterocellular coupling are present in the postnatal SVZ and that junctional coupling is important for initialization and maintenance of cell migration in this neurogenic zone.

viii
Lista de Abreviaturas
βIII-Tubulina = Classe III β-tubulina
BMP = proteína morfogênica dos ossos (bone morphogenetic protein)
CBX = carbenoxolone
Cx = conexina
DIV = dias in vitro ou dias em cultura
gap junction = junção comunicante
GFAP = proteína glial fibrilar acídica (glial fibrillary acidic protein)
GZA = Ácido glicirrízico
LY = Lucifer Yellow (sem tradução em português)
P0 = Dia do nascimento
PBS = solução de tampão fosfato (phosphate buffered saline)
P120ctn = p120 catenina (Discussão p.88)
RD = Rodamina dextran
RMS = corrente migratória rostral (rostral migratory zone)
RNA = ácido ribonucléico
SVZ = zona subventricular (subventricular zone)
SVZb = zona subventricular contida no bulbo olfatório
SVZc = zona subventricular contida no córtex cerebral
SNC = sistema nervoso central
VZ = zona ventricular (ventricular zone)

ix
Sumário
I – Introdução......................................................................................................... 01
1 – Formação do telencéfalo ................................................................................ 01
1.1 – Camadas germinativas ..................................................................... 03
1.1.1 – A zona ventricular (VZ) ....................................................... 03
1.1.2 – A zona subventricular (SVZ) ............................................... 04
1.1.2.1 – A zona subventricular anterior (SVZa) .................. 07
1.1.2.2 – A migração na SVZa ............................................. 10
1.2 – Comunicação juncional ..................................................................... 12
1.2.1 – A comunicação juncional no Sistema Nervoso Central em
desenvolvimento................................................................... 15
1.2.1.1 – As junções comunicantes nas camadas
germinativas.......................................................................... 15
1.2.1.1.1 – As junções comunicantes na SVZ............ 17
1.2.2 – Junções comunicantes e migração celular.......................... 20
2 – O principal bloqueador juncional utilizado....................................................... 23
II – Objetivos......................................................................................................... 26
III – Materiais e Métodos....................................................................................... 27
1 – Preparação de explantes...................................................................... 27
2 – Cultura de explantes de zona subventricular em matrigel.................... 27
3 – Cultura de explantes de camundongos nocaute de Cx43.................... 29
4 – Bloqueio da comunicação juncional..................................................... 30
5 – Injeção de corante................................................................................ 31
6 – Imuno-histoquímica............................................................................... 32

x
7 – Teste de especificidade do bloqueador juncional................................. 32
8 – Análise e quantificação da migração.................................................... 33
9 – Vídeo microscopia de tempo intervalado.............................................. 34
10 – Análise da vídeo microscopia de tempo intervalado.......................... 35
IV – Resultados..................................................................................................... 38
1 – Migração celular a partir de explantes de SVZb e SVZc...................... 38
2 – Caracterização do acoplamento no explante....................................... 40
3 – A Cx43 está presente em diferentes tipos celulares no explante......... 48
4 – Apesar da expressão elevada, a Cx43 não afeta significativamente a
migração celular nos explantes............................................................. 53
5 – Bloqueio farmacológico crônico por CBX nos explantes...................... 53
5a – A especificidade do CBX.......................................................... 60
5b – O efeito do CBX é reversível.................................................... 62
6 – O bloqueio farmacológico inibe a migração diretamente...................... 65
V – Discussão........................................................................................................ 80
1 – A cultura de explantes parece reproduzir a fisologia da SVZ/RMS
in vivo................................................................................................... 81
2 – Acoplamento celular nos explantes de SVZ......................................... 83
3 - Fatores envolvidos na migração celular da SVZ/RMS.......................... 85
4 – O acoplamento e a migração celular.................................................... 86
5 – O possível papel da conexina 43 com a migração celular................... 89
6 - A função do acoplamento juncional na SVZ/RMS................................ 91
VI – Conclusões ................................................................................................... 98
VII – Referências .................................................................................................. 99
VIII – Anexos......................................................................................................... 126

xi
Anexo I – Filmes de vídeo microscopia de tempo intervalado.................... 126
Anexo II – Menezes et. al., 2002................................................................. 127

1
I - Introdução:
Durante o desenvolvimento do Sistema Nervoso Central (SNC), existe intensa
movimentação celular de precursores, ou células imaturas, em direção aos seus
destinos finais no tecido maduro. Este fenômeno é vital para o estabelecimento da
complexa circuitaria neural. As moléculas e mecanismos celulares, envolvidos com a
correta migração e posicionamento destas células, ainda são amplamente
desconhecidos, apesar dos estudos intensivos sobre a migração celular de algumas
regiões. Nesta tese estudamos a influência do acoplamento celular, mediado por
junções comunicantes, sobre a extensa migração tangencial, que ocorre em uma
das camadas germinativas do cérebro pós-natal. Este fenômeno possui
características importantes para o entendimento da plasticidade e regeneração do
SNC adulto.
1 – Formação do Telencéfalo:
A formação do telencéfalo envolve uma série de eventos seqüenciais:
proliferação, migração, diferenciação e morte celular. A proliferação em geral ocorre
em camadas especializadas, denominadas germinativas. Uma vez gerados, os
neurônios não retornam ao ciclo mitótico, e migram destas camadas germinativas
para seu destino final. Essa migração pode ser radial e/ou tangencial à superfície
pial. A migração radial caracteriza-se por apresentar neurônios jovens “escalando”
células gliais especializadas, comuns no período embrionário, conhecidas como
células da glia radial (Rakic, 1972, 1990; Edmondson e Hatten, 1987; Alvarez-Buylla
e Nottebohm, 1988). Alternativamente foi proposto que alguns neurônios herdam a

2
fibra radial do precursor, com morfologia radial, e migram por translocação nuclear
(Miyata et al., 2001). Já na migração tangencial, os neurônios imaturos se movem
paralelamente à superfície pial e perpendicular às fibras da glia radial (Walsh e
Cepko, 1990; Luskin, 1993; Lois e Alvarez-Buylla, 1994; O’Rourke et al., 1995).
Neste tipo de migração as pistas de direcionamento utilizadas pelas células ainda
não são conhecidas, embora existam alguns possíveis candidatos (Hu et al., 1996;
Hu e Rutishauser, 1996; Mendez-Otero e Cavalcante, 1996; Zhu et al., 1999;
Conover et al., 2000; Menezes et al., 2002; Ming e Song, 2005).
Finalmente, à medida que chegam a seu destino, seja no bulbo olfatório (BO),
no córtex cerebral, ou no subpallium, os neurônios jovens começam a diferenciar-se
expressando marcadores fenotípicos e morfologia complexa, conseqüentemente
conferindo ao tecido a organização histotípica característica, como no caso das
camadas corticais.
Normalmente o número de células geradas no sistema nervoso central (SNC)
é maior que o número final de células na maturidade. Isso ocorre devido ao processo
de morte celular programada (apoptose), ajudando a definir a forma final do cérebro
e possibilitando a correção dos erros que ocorrem ao longo do desenvolvimento.
Acreditava-se que a apoptose ocorria principalmente no destino final das células, no
entanto sabe-se que está presente inclusive nas zonas germinativas (Blaschke et
al.,1996, 1998; Haydar et al., 1999), indicando uma concomitância destes vários
processos durante o desenvolvimento (Blaschke et al., 1996).
Neste trabalho nosso interesse estará centralizado na migração celular.
Estudaremos a influência de interações intercelulares mediadas por acoplamento
juncional neste fenômeno. Para tanto, utilizamos um modelo in vitro de uma das
camadas germinativas presentes no período pós-natal, a zona subventricular.

3
1.1 – Camadas germinativas:
De modo geral, as camadas germinativas possuem as seguintes
características: alto índice de proliferação, grande densidade celular,
homogeneidade populacional e ausência de prolongamentos axonais. No período
embrionário, as células proliferativas que darão origem ao telencéfalo são
encontradas principalmente em duas regiões (chamadas de zonas ou camadas
germinativas): a zona ventricular (do inglês, Ventricular Zone - VZ) e a subventricular
(do inglês, Subventricular Zone - SVZ; Boulder Committee, 1970; McConnell, 1988;
Jacobson, 1991; Bayer e Altman, 1994; Takahashi et al., 1995). Em ambas são
gerados neurônios e células gliais (Temple e Quian, 1996; Kriegstein and Noctor,
2004). Algumas regiões corticais apresentam proliferação celular persistente no
período pós-natal como por exemplo: VZ (Tramontin et al., 2003), SVZ (Allen, 1912),
hipocampo (Altman e Das, 1965) camada granular externa do cerebelo (External
Granular Layer - EGL - Miale e Sidman, 1961), dentre outras (Gould et al., 1999;
Horner et al., 2000). Mas no adulto apenas a SVZ telencefálica e a camada
subgranular do giro denteado persistem como zonas neurogênicas ativas (Altman e
Das, 1965; Altman,1969; Kaplan e Bell, 1983; Emsley et al., 2005).
1.1.1 – A Zona Ventricular (VZ):
A VZ margeia os ventrículos, apresentando-se como uma camada
pseudoestratificada de células alinhadas radialmente, cujos núcleos posicionam-se
de acordo com a fase do ciclo celular (migração nuclear intercinética – MNI; Sauer,
1935, 1937; Sauer e Walker, 1959; Angevine e Sidman, 1961; Berry et al., 1964;

4
Caviness e Sidman, 1973; Shoukimas e Hinds, 1978; Nowakowski e Rakic, 1981;
Rakic, 1988; Bayer e Altman, 1991; Takahashi et al., 1992). As células da VZ
atingem o pico proliferativo durante a embriogênese, continuam proliferando
residualmente na primeira semana pós-natal quando a glia radial começa a se
transformar em astrócitos da SVZ e em células ependimárias que circundam os
ventrículos (Tramontin et al., 2003; Spassky et al., 2005; Menezes et al., 2005). Com
exceção da microglia, de origem extraneural (Cuadros e Navascués, 1998), a VZ é a
fonte de todos os tipos celulares no SNC, gerando astrócitos, oligodendrócitos,
células ependimárias e neurônios. Além disso, existem evidencias de que a glia
radial, originária da VZ subcortical, produz as células-tronco da SVZ do adulto, que
mantém a linhagem neurogênica no cérebro adulto (Tramontin et al., 2003, Merkle et
al., 2004). O destino das células de glia radial cortical parece ser restrito a formação
de linhagens gliais ou a simples persistência residual no córtex pos-natal (Alves et
al., 2002; Menezes et al. 2005).
1.1.2 – A Zona Subventricular (SVZ):
A SVZ localiza-se suprajacente à VZ (Fig. 1). Seu aparecimento é
concomitante à neurogênese cortical (Altman, 1969). No rato, distingue-se da VZ a
partir do décimo sexto dia embrionário (E16 – Pencea e Luskin, 2003) e no final da
gestação e em idades pós-natais seus limites já estão claramente distintos
(Takahashi et al., 1992, 1995). De modo peculiar, a SVZ mantém atividade
proliferativa, estendendo-se à idade adulta (Allen, 1912; Smart, 1961; Altman, 1969;
Sturrock e Smart, 1980, Tropepe et al., 1997). A SVZ caracteriza-se pela presença
de figuras de mitose abventriculares e não apresenta migração nuclear intercinética

5
(Sturrock e Smart, 1980; Takahashi et al. 1992; 1995). Devido ao aumento tardio da
proliferação na SVZ acreditava-se que esta gerava principalmente células de
linhagem glial (Privat e Leblond, 1972; Takahashi et al., 1992). Contudo, atualmente
sabe-se que a SVZ embrionária é um importante sítio de origem de neurônios para
as camadas supragranulares do córtex (Tarabykin et al., 2001; Kriegstein e Noctor,
2004; Nieto et al., 2004). Além disto, em idades pós-natais sua porção anterior é
fonte de interneurônios que colonizam o bulbo olfatório (Luskin, 1993). O caráter
multipotente desta camada pode ser evidenciado inclusive em modelos in vitro (Lois
e Alvarez-Buylla, 1993; Luskin et al., 1997). Luskin (1993) propôs um modelo
compartimentado, onde a porção mais anterior da SVZ (SVZa) do recém-nato seria
especializada na produção de precursores neuronais para o bulbo olfatório,
enquanto a mais caudal apresentaria altos índices de morte celular (Morshead e van
der Kooy, 1992) ou geraria células de linhagem glial (Luskin e McDermott, 1994).
Este modelo atualmente desacreditado foi parcialmente corroborado pelo trabalho
de Lim e colaboradores (2000), que mostra que a expressão de BMPs e Noguina na
SVZ perinatal faz de sua porção anterior um compartimento neurogênico; e por
Marshall e colaboradores (2003), que sugere a existência de um compartimento glial
na porção dorso-lateral da SVZ perinatal. Com o uso de antigos vetores retrovirais
de baixa titulação foi demonstrado que os tipos celulares predominantes que surgem
da porção posterior da SVZ in vivo e in vitro são astrócitos e oligodendrócitos (Privat,
1975; Vaysse e Goldman, 1990; Reynolds e Weiss, 1992; Levison e Goldman, 1993;
Levison et al., 1993; Luskin e McDermott, 1994). No entanto, com o uso de um vetor
de alta titulação, foi possível demonstrar a presença de precursores
neuronais,destinados ao bulbo, provenientes de porções caudais da SVZ neonatal
(Suzuki e Goldman, 2003). Este resultado demonstrou que a aparente

6
Figura 1. Desenho esquemático de algumas camadas celulares do período embrionário. No telencéfalo, o lúmen do tubo neural, chamado de ventrículo (V) está rodeado por células proliferativas da zona ventricular (VZ). Em volta da VZ começa a se formar uma outra camada de células proliferativas, a zona subventricular (SVZ). Por último, em posições mais periféricas temos uma camada celular genericamente chamada de manto, que contém células em diferenciação ou diferenciadas. Interposta entre as camadas germinativas e a zona do manto, temos a zona intermediária, não apontada no esquema, por onde percorrem as celulas em migração e prologamentos axonais da circuitaria neural em desenvolvimento.

7
compartimentação da SVZ neonatal seria apenas fruto de uma proporção mais
elevada de precursores gliais na porção posterior nestas idades. Assim, o animal
imaturo aproxima-se do adulto, no qual os precursores neuronais, encontrados em
“ilhas” de neuroblastos, se espalham inclusive para porções mais caudais da SVZ
(Doetsch et al., 1997).
1.1.2.1 – A Zona Subventricular anterior (SVZa):
A porção rostral da SVZ (SVZa) circunda os ventrículos laterais até o bulbo
olfatório (Fig. 2; Hinds, 1968a,b; Altman, 1969; Kishi, 1987; Frazier-Cierpial e
Brunjes, 1989; Kishi et al., 1990). Virtualmente todas as células geradas na SVZa do
cérebro pós-natal precoce se diferenciarão em neurônios, e em particular se
tornarão interneurônios do bulbo olfatório (células granulares e periglomerulares;
Luskin, 1993; Betarbet et al., 1996; Zigova et al., 1996). Para isto, neurônios
imaturos e neuroblastos migram tangencialmente e de forma independente de
células gliais em direção ao bulbo olfatório pelo interior da SVZ/RMS (Fig. 3). Este
fenômeno migratório é conhecido como a “corrente migratória rostral” (RMS, rostral
migratory stream; Altman, 1969; Luskin, 1993). Apesar do pico de geração dessas
células ocorrer nas duas primeiras semanas pós-natais, a neurogênese e a RMS
continuam até a idade adulta (Hinds, 1968a,b; Lois e Alvarez-Buylla, 1994; Tropepe
et al., 1997). Durante o pico de neurogênese, a SVZ pode ser facilmente
reconhecida por apresentar grande densidade e uniformidade celular e por ser
altamente proliferativa (Altman, 1969). Apesar da aparente homogeneidade na
densidade celular, células em fase S apresentam-se mais concentradas na periferia
da RMS, indicando uma possível segregação entre células proliferativas e

8
Figura 2. Vista dorsal do cérebro do rato adulto. O ventrículo (V) está ladeado por células ependimárias. No cérebro adulto, ainda persiste uma pequena zona subventricular (SVZ, pontilhado e reta maior). A zona subventricular anterior (SVZa, com limites aproximados indicados pela reta menor) é quase contínua com a corrente migratória rostral, que invade o bulbo olfatório (OB). Os hemisférios cerebrais (CH) também estão indicados. Por esta via rostral navegam neurônios imaturos e precursores para popular o bulbo olfatório durante toda vida do animal. (Modificada da Figura 1 de Smart, 1961).

9
migratórias (Menezes et al., 1998). Além disso, o acoplamento celular mediado por
junções comunicantes distribui-se de modo semelhante ao das células em fase S
(Menezes et al., 2002); a glia radial em transformação se concentra em volta da
SVZ, fornecendo uma espécie de revestimento (Alves et al., 2002), também
semelhante ao animal adulto, onde os neuroblastos migram em cadeias revestidas
por túneis astrocitários (Lois et al., 1996; Peretto et al., 1997). Em conjunto, estas
evidências sugerem a existência de discretos compartimentos celulares no interior
da SVZ.
As células da SVZ/RMS neonatal expressam propriedades especiais que as
distinguem de outros progenitores do SNC. Por exemplo, sua proliferação e
migração podem ser concomitantes e acontecem acompanhadas de expressão de
fenótipo neuronal desde o ponto de origem destas células até seu destino final (o
bulbo olfatório; Menezes et al., 1995). Em contraste, sabe-se que em qualquer outro
local do SNC, os neurônios migratórios imaturos são pós-mitóticos (Menezes e
Luskin, 1994). Na SVZ/RMS do adulto (e provavelmente no jovem) encontramos
cinco tipos celulares: células indiferenciadas, neuroblastos, neurônios imaturos,
astrócitos e microglia. No jovem, somam-se a estes as células de glia radial
Figura 3. Vista para-sagital do cérebro do rato neonato (entre o nascimento e o terceiro dia de vida –P0-P3). O ventrículo (V) em preto ladeado pela zona subventricular anterior (SVZa). Acima da SVZ está o corpo caloso (CC), ambos contidos no córtex (CTX) do animal. A corrente migratória rostral (RMS) coincide com a SVZa e é constituída pelo fluxo migratório de suas células em direção ao bulbo olfatório (OB). Modificada da Figura 1 de Menezes et al., 1995.

10
(Stensaas e Gilson, 1972; Kishi et al., 1990; Luskin, 1993; Doetsch et al., 1997;
Menezes et al, 2000).
1.1.2.2 - A migração na SVZa:
A maioria das células migratórias é bipolar apresentando um prolongamento
“líder”, com um grande cone de crescimento na ponta, outro curto, chamado de
“seguidor” (Kishi, 1987, Luskin, 1993). No adulto estas células migram como uma
rede de cadeias tangencialmente orientadas (Doetsch e Alvarez-Buylla, 1996; Lois et
al., 1996) e que convergem na RMS. Essa organização em cadeias foi relacionada à
expressão da N-CAM polisialilada (PSA-N-CAM; Rousselot et al., 1995), do
gangliosídeo 9-O-acetil GD3 (Mendez-Otero e Cavalcante, 1996; Miyakoshi et al.,
2001) e da p120catenina através da regulação das RhoGTPases (Chauvet et al.,
2003). O fato das cadeias migratórias se restringirem ao interior da RMS tem sido
relacionado à presença de pistas direcionadoras como fatores difusíveis, moléculas
de matriz celular, interações intercelulares e de células com substrato (Menezes et
al., 2002). Mais recentemente, outros fatores também foram descritos funcionando
como pistas de direcionamento para migração tangencial na SVZ, tais como: um
fator atrator liberado pelo bulbo olfatório (Liu e Rao, 2003), semaforinas-neuropilinas,
e neuregulinas (Kriegstein e Noctor, 2004). Acredita-se que estes fatores possam se
combinar para criar um corredor permissivo para direcionar neurônios durante a
migração tangencial (Wichterle et al., 2003).
Os neuroblastos, após terem alcançado o bulbo olfatório, passam a migrar
radialmente, como células individualizadas, para camadas mais externas (Kishi,
1987; Luskin, 1993), possivelmente sobre tenascina-R, que parece funcionar como

11
molécula atratora ou permissiva, além de fornecer o sinal para que os neuroblastos
abandonem as cadeias migratórias (Saghatelyan et al., 2004). Além disso, o gene
homeobox Arx parece estar ligado à entrada das células da RMS no bulbo olfatório
(Yoshihara et al., 2005). Também no adulto as cadeias migratórias avançam através
de tubos definidos por processos astrocitários que proliferam lentamente. Apesar da
existência de células de glia radial no animal pós-natal jovem, a migração na
SVZ/RMS parece não depender da glia radial ou de direcionamento axonal
(Rutishauser et al., 1985; Kishi, 1987; Rutishauser e Landmesser,1991; Luskin,
1993, Menezes et al., 1995; Wichterle et al., 1997; Law et al., 1999; Menezes et al.,
2000). No neonato, as células de glia radial são facilmente diferenciadas dos
neuroblastos, os quais não apresentam morfologia radial neste estágio, e se
posicionam ortogonais às células de glia (Kishi et al. 1990; Menezes et al., 2000).
Em um trabalho anterior, nosso grupo demonstrou a presença de extenso
acoplamento juncional por corante na SVZ e RMS pós-natal (Menezes et al., 2000,
vide adiante), de forma coincidente com o período de maior atividade proliferativa e
migratória desta região. Demonstrações anteriores da importância do acoplamento
celular em outras camadas germinativas, [como: na regulação da proliferação na VZ
embrionária (Bittman et al., 1997; Bittman et al., 1999; Weissman et al., 2004); a
correlação negativa com a iniciação da migração celular (Bittman et al., 1997;
Rozental et al., 1995) e mesmo a expressão da conexina 43 em células quiescentes
da SVZ pós-natal (Miragall et al., 1997)] nos levaram a investigar a influencia do
acoplamento celular na proliferação e migração das células presentes na SVZ pós-
natal (Marins, 2001). No presente trabalho focalizamos na contra-intuitiva correlação
positiva do acoplamento juncional com a migração celular.

12
1.2 – Comunicação Juncional:
A comunicação via gap junctions, ou comunicação juncional, é realizada pelo
alinhamento de conjuntos de canais protéicos transmembranares, chamados de
placas juncionais, presentes entre células apostas. Cada placa juncional pode conter
até milhares destes canais protéicos. Estes canais são formados pelo alinhamento
de dois conjuntos de proteínas transmembrana, chamadas conexinas (Cx), cada
grupo de seis compondo um hemicanal, ou conexon (Fig. 4B e C).
A
B
C
D
3,5nm 20nm
Canal formadopor poros emcada membrana
Espaço extracelularnormal
6 conexinas = 1 conexon
61 2
345
61 2
345
fechadoaberto
Meio Extracelular
Meio Citoplasmático
Citoplasma
Transmembrana
Extracelular
Cana
l
Conexon
CanalConexon homomérico heteromérico homomérico heteromérico
homotípico homotípico heterotípico heterotípico
Citoplasmapré-sináptico
Citoplasmapós-sináptico
M1 M2 M3 M4
Figura 4. Modelos tridimensionais dos canais juncionais baseados em estudos de difração de Raios X. A) As células apostas contribuem com um hemicanal para completar o canal juncional. Cada hemicanal é formado por seis conexinas. B) Modelo de um único conexon, com representação da mudança de conformação das conexinas para abrir ou fechar o canal. O sombreado indica a porção transmembrana das conexinas. C) Modelo topológico da conexina com os cilindros representando os domínios transmembrana (M1-M4). As alças entre o primeiro e o segundo (E1) e entre o terceiro e o quarto (E2) são extracelulares. D)Desenho esquemático de possíveis combinações de hemicanais para formar um canal juncional completo. Os hemicanais podem ser homoméricos (formados por seis conexinas idênticas) ou heteroméricos (compostos de mais de uma isoforma de conexina). Os canais formados pelo alinhamento dos hemicanais podem ser homotípicos (os dois conexons são idênticos) ou heterotípicos (com dois conexons diferentes). Modificada da Figura 1 de Willecke et al., 2002.

13
Conexons compostos por conexinas do mesmo tipo formam canais
homotípicos, enquanto que aqueles compostos por conexinas de tipos diferentes
formarão canais heterotípicos, com propriedades diferentes daquelas esperadas
para canais homotípicos (Willecke et al., 2002; Goodenough e Paul, 2003). A
variedade de canais aumenta quando dentro de cada conexon encontramos
diferentes tipos de conexinas (Fig. 4D – Willecke et al., 2002). Canais intercelulares
funcionais são formados quando células adjacentes expressam hemicanais
complementares que, quando alinhados, permitem o intercâmbio bidirecional de íons
e pequenas moléculas entre as células, determinando o acoplamento metabólico e
elétrico. Inicialmente pensava-se que somente moléculas de até 1 kDa
atravessariam os canais juncionais (Loewenstein, 1981; Kumar e Gilula,1996),
atualmente sabe-se que dependendo da conformação, é possível que moléculas
com mais de 1,5, e com até 4 kDa, passem pelo poro juncional (Neijssen et al.,
2005; Valiunas et al., 2005).
Mais importante, o acoplamento pode ainda ser classificado como
homocelular, portanto envolvendo células do mesmo tipo, ou heterocelular, quando
estabelecido entre células de tipos diferentes (Rouach et al., 2002). Muitas vezes o
acoplamento heterocelular envolve conexinas ou conexons de natureza diversa,
canais heterotípicos, que combinam características de seletividade e permeabilidade
dos diferentes conexons. Desta forma, é possível prever que acoplamentos
heterocelulares podem apresentar características mais complexas do que as
geralmente presentes nos acoplamentos homocelulares.
O agrupamento dos canais juncionais na membrana depende de contato
intercelular e resulta na formação da placa juncional (normalmente composta de
centenas de canais), região onde o espaço extracelular fica bastante reduzido

14
embora as membranas não cheguem a se tocar (Fig. 4A – Lampe et al., 1991; Kistler
et al., 1994). A placa juncional não é uma estrutura estática e sim dinâmica, isto é,
pode ser desagregada e reagrupada em seguida numa outra região da membrana
celular em resposta à estímulos fisiológicos (Evans e Martin, 2002; Thomas et al.,
2005). Além disso, cada conexon possui meia-vida curta (1-5h, dependendo do tipo
de conexina) e é continuamente renovado na membrana, isolado ou agrupado nas
placas juncionais (Evans e Martin, 2002; Salameh e Dhein, 2005; Thomas et al.,
2005). Estudos recentes mostraram que os hemicanais isolados também podem
funcionar como via de intercâmbio entre a célula e o meio extracelular (Goodenough
e Paul, 2003; Ye et al., 2003; Contreras et al., 2004).
Atualmente vinte tipos de conexinas já foram identificados em camundongos e
vinte e um em humanos (Söhl e Willecke, 2003). Presentes em diversos tecidos,
estas proteínas são nomeadas de acordo com o peso molecular deduzido de seu
cDNA. Pertencem a uma única família gênica e exibem antigenicidades específicas,
permitindo-nos acompanhar seu perfil de expressão e distribuição in situ. No sistema
nervoso de mamíferos foram encontradas as seguintes conexinas: Cx26, Cx29,
Cx30, Cx31, Cx32, Cx36, Cx37, Cx43, Cx45, Cx47 e Cx57 (Nagy et al., 2004).
Algumas conexinas apresentam certo grau de predominância celular, p. ex.: os
astrócitos apresentam canais juncionais formados principalmente pela Cx43
(também descrita em neurônios imaturos e alguns neurônios já diferenciados), Cx30,
Cx45 e talvez a Cx26; Cx36 é expressa exclusivamente em neurônios, embora
alguns tipos de neurônios também expressem as conexinas Cx32, Cx37, Cx45,
Cx47 e Cx57; a Cx26, predomina entre precursores, células leptomeningeais,
ependimócitos, astrócitos e neurônios maduros; e, finalmente os oligodendrócitos
acoplam entre si via Cx32, Cx29 e Cx47 (Bittman et al., 2002 e V. revisão em Spray

15
e Dermitzel, 1996; Naus e Bani-Yaghoub, 1998; Rouach et al., 2002; Söhl et al.,
2004; Montoro e Yuste, 2004, Söhl et al, 2005; Sutor e Hagerty, 2005).
1.2.1 – A comunicação juncional no Sistema Nervoso Central em
desenvolvimento:
As junções comunicantes são comuns desde o desenvolvimento inicial do
tubo neural (Ruangvoravat e Lo, 1991; Yancey et al., 1992) e parecem cumprir
diversos papéis desde o controle de proliferação e da diferenciação, como também
na indução de padrões histotípicos regionais (Loewenstein, 1979; Fraser et al., 1987;
Warner et al., 1992; Mellitzer et al., 1999). O acoplamento celular está presente
desde a fase proliferativa (da fase S até G2, as células desacoplam na fase M e
tornam a se acoplar em G1 - Bittman et al., 1997) até o estabelecimento das redes
neurais (período no qual acredita-se que o acoplamento seja responsável pela
sincronização neuronal, necessária para o estabelecimento adequado das primeiras
sinapses - Peinado et al., 1993; Personius e Balice-Gordon, 2000; Sutor e Hagerty,
2005). Algumas conexinas, presentes no cérebro embrionário, continuam sendo
expressas nas primeiras semanas pós-natal, p. ex., Cx26, Cx43 e Cx45 (Montoro e
Yuste, 2004).
1.2.1.1 – As junções comunicantes nas camadas germinativas:
Resultados prévios demonstraram (direta e indiretamente) o acoplamento
celular na VZ telencefálica (LoTurco e Kriegstein, 1991; Bittman et al., 1997;
Nadarajah et al., 1997; Bahrey e Moody, 2003; Weissman et al., 2004). As conexinas

16
descritas na VZ são as Cx26, Cx43 e Cx45 (Dermietzel, 1996; Nadarajah et al.,
1997; Montoro e Yuste, 2004). Estas junções também foram identificadas na SVZ,
por microscopia eletrônica (Miragall et al., 1992; Yamamoto et al., 1990), e por
imuno-histoquímica (Miragall et al., 1997; Peretto et al., 2005). As proteínas
específicas destas junções, as conexinas (Cxs. 43 e 26), foram detectadas nesta
região tanto no período embrionário quanto em idades pós-natais (Miragall et al.,
1997; Nadarajah et al., 1997; Peretto et al., 2005). Os ensaios que finalmente
confirmariam a funcionalidade destas junções foram apresentados por Menezes e
cols. (2000), que demonstraram não somente que a comunicação juncional na
SVZ/RMS envolve vastas populações celulares, como também se encontra
compartimentada, predominando em regiões associadas a nichos de células
proliferativas em fase S (vide adiante).
Alguns autores acreditam que o acoplamento por junção comunicante está
relacionado com a entrada e/ou permanência da célula no ciclo mitótico, e o
desacoplamento com a saída do ciclo (LoTurco e Kriegstein, 1991; Bittman et al.,
1997, 2002; Duval et al., 2002; Bahrey e Moody, 2003; Scemes et al., 2003;
Weissman et al., 2004). Em concordância, Rozental e Cols. (1995) relaciona o
desacoplamento com o início da diferenciação celular, em neuroblastos hipocampais
imortalizados. Por outro lado, Miragall e cols. (1997) sugerem que na SVZ/RMS o
acoplamento é inversamente proporcional à proliferação, semelhante ao que ocorre
em certas linhagens de células tumorais (Loewenstein et al., 1979; Zhu et al., 1991;
Lee et al., 1992; Naus et al., 1992; Tabernero et al., 2001; Sánchez-Alvarez et al.,
2001; Trosko, 2003). Outras possíveis funções das conexinas durante o
desenvolvimento são a participação na formação de barreiras e regionalização do
tubo neural (Naus e Bani-Yaghoub, 1998; Mellitzer et al., 1999).

17
Em algumas situações a comunicação juncional tem sido implicada
positivamente na migração celular (Bannerman et al., 2000; Lin et al., 2002; Fushiki
et al., 2003; Scemes et al., 2003), como na migração de células cardíacas
simpáticas da crista neural (Huang et al., 1998). Este trabalho propõe que o
acoplamento favorece a migração por comunicar o sinal migratório para todas as
células da via, coordenando sua resposta às pistas migratórias. Além disso, estudos
recentes com neuroesferas relacionam o acoplamento, mediado por Cx43, com a
manutenção da proliferação e da migração celular (Duval et al., 2002; Scemes et al.,
2003).
1.2.1.1.1 – As junções comunicantes na SVZ:
Miragall e colaboradores (1997) encontraram imunomarcação de Cx43
abundante na RMS, levando-os a supor que tanto neuroblastos quanto precursores
gliais a expressariam. Contudo os níveis de marcação de Cx43 variaram durante o
desenvolvimento pós-natal, i. e., sendo baixos nos primeiros estágios pós-natais e
aumentando em estágios tardios, incluindo o adulto, num gradiente inverso ao da
atividade proliferativa da SVZ/RMS. Além disto, Miragall descreve a expressão da
Cx43 apenas em células pós-mitóticas da SVZ/RMS, e conseqüentemente uma
correlação inversa entre acoplamento celular e proliferação.
Menezes e cols. (2000), além de demonstrarem a funcionalidade dos contatos
juncionais na SVZ/RMS, identificaram dois padrões de acoplamento celular: um
extenso, distribuído ao longo da periferia da SVZ/RMS(especialmente na sua borda
dorsal); e outro restrito, presente homogeneamente pela espessura da SVZ/RMS
(Fig. 5). Este padrão é reminiscente da distribuição de células em fase S observada

18
anteriormente (Menezes et al., 1998). A correlação espacial entre a distribuição das
células proliferativas e do acoplamento celular enfraquece a proposta de Miragall e
colaboradores (1997), formulada sobre evidências puramente estruturais, e sugere
uma correlação positiva entre o acoplamento juncional e a proliferação celular na
SVZ jovem. Coerentemente, o acoplamento celular apresentou um gradiente
decrescente no sentido caudo-rostral, similar ao gradiente de proliferação celular
encontrado na SVZ/RMS (Menezes et al., 1998). A extensa distribuição de
acoplamento juncional na SVZ/RMS, também sugere que esta comunicação
intercelular estaria relacionada com outros processos celulares desta camada,
incluindo a migração e a diferenciação celulares bem como a compartimentação da
SVZ/RMS.
Durante o Mestrado, desenvolvemos o modelo de cultura de micro-explantes
para estudar os efeitos do acoplamento juncional sobre a proliferação e migração
dos progenitores da SVZ. O fato das células da SVZ apresentarem comportamento
migratório variável em seu trajeto até o bulbo (V. item 1.1.2.2 da introdução), nos
levou a separar os explantes conforme sua origem: SVZb (aqueles provenientes do
bulbo olfatório) e SVZc (porção contida abaixo do manto do córtex cerebral). Neste
trabalho nosso primeiro objetivo foi estabelecer o modelo de cultura de explantes e
estudar o possível efeito do bloqueio do acoplamento juncional sobre a migração
celular (Fig. 6A e B). O próximo passo foi identificar a presença do acoplamento
funcional através da técnica de “scrape loading” (do inglês, carregamento por corte),
na qual utilizamos a mistura de dois corantes: um permeante à junções
comunicantes (Lucifer Yellow, verde) e outro não permeante (Rodamina Dextran,
vermelha). Desse modo, as células duplamente marcadas foram preenchidas no
momento do corte e aquelas somente verdes receberam o corante via acoplamento
19
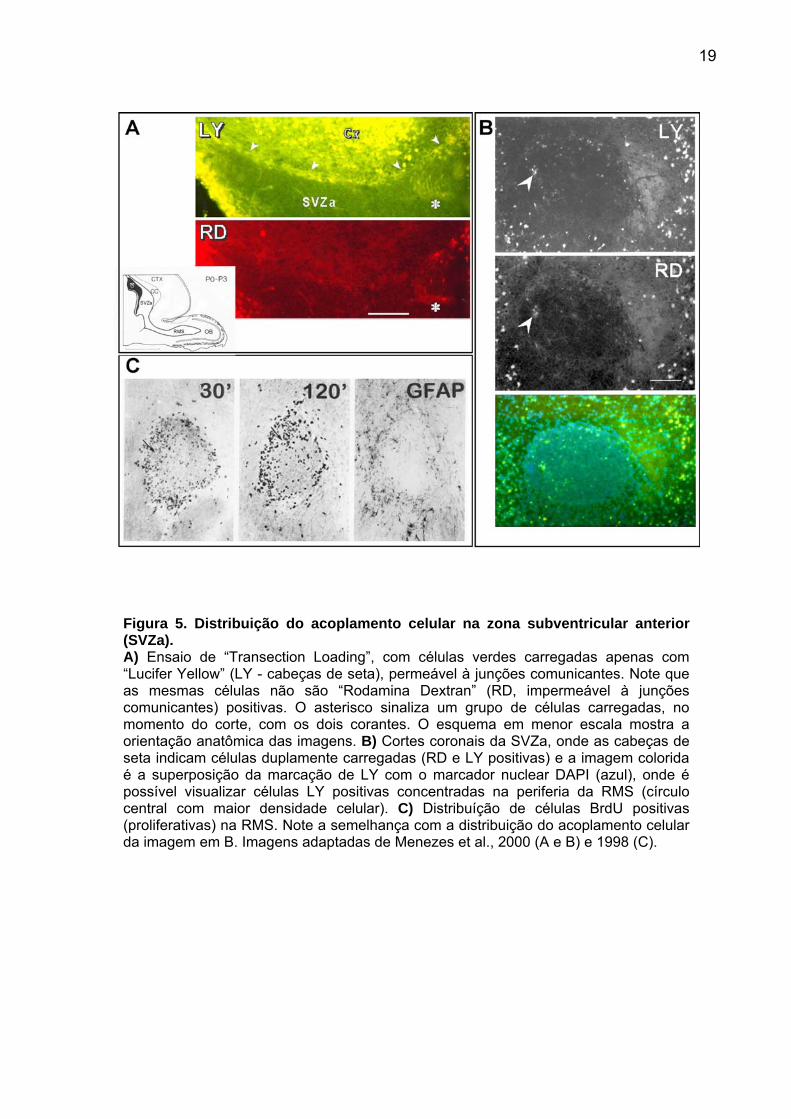
Figura 5. Distribuição do acoplamento celular na zona subventricular anterior (SVZa). A) Ensaio de “Transection Loading”, com células verdes carregadas apenas com “Lucifer Yellow” (LY - cabeças de seta), permeável à junções comunicantes. Note que as mesmas células não são “Rodamina Dextran” (RD, impermeável à junções comunicantes) positivas. O asterisco sinaliza um grupo de células carregadas, no momento do corte, com os dois corantes. O esquema em menor escala mostra a orientação anatômica das imagens. B) Cortes coronais da SVZa, onde as cabeças de seta indicam células duplamente carregadas (RD e LY positivas) e a imagem colorida é a superposição da marcação de LY com o marcador nuclear DAPI (azul), onde é possível visualizar células LY positivas concentradas na periferia da RMS (círculo central com maior densidade celular). C) Distribuíção de células BrdU positivas (proliferativas) na RMS. Note a semelhança com a distribuição do acoplamento celular da imagem em B. Imagens adaptadas de Menezes et al., 2000 (A e B) e 1998 (C).

20
juncional (Fig. 6C-F). Em resumo, neste trabalho anterior mostramos que a
presença do acoplamento celular mediado por junções comunicantes é importante
para a manutenção da proliferação (Fig. 6G e H) e da migração nos explantes (Fig.
6A e B) e parece ser vital para a inicialização da migração em toda a SVZ. No
entanto, apenas nos explantes de SVZc o bloqueio do acoplamento juncional foi
revertido, para níveis migratórios comparáveis aos controles. Este dado confirmou
nossa suspeita da existência de diferenças regionais. Apesar dos resultados
apontarem para o envolvimento de junções comunicantes com os fenômenos
estudados, algumas questões ainda precisavam ser melhor esclarecidas: quais
seriam as conexinas presentes nos explantes, quais tipos celulares estariam
envolvidos no acoplamento, como ficaria a migração diante do uso de outros
bloqueadores juncionais, o efeito do bloqueio juncional sobre a migração é imediato
ou indireto, por exemplo, via proliferação celular?
1.2.2 – Junções comunicantes e migração celular:
Apesar do deslocamento celular ser aparentemente incompatível com o
estabelecimento de contatos juncionais estáveis, diversos estudos têm sugerido, que
os níveis de comunicação juncional afetam ou são afetados pela atividade migratória
celular (Huang et al., 1998; Menezes et al., 2002).
Estudos utilizando células e tecidos neoplásicos têm sido particularmente
úteis como modelos de migração celular. Inicialmente, estes estudos tinham em
comum o estabelecimento de uma correlação negativa entre o grau de acoplamento
juncional homocelular e a invasividade/proliferação tumorais (Elias e Friend, 1976;
Navolotski et al., 1997; Hamada et al., 1998; Naus e Bani-Yaghoub, 1998;

21
Figura 6. Efeito do acoplamento celular sobre a migração e proliferação nos explantes de SVZ. A) Explante de SVZ contida no córtex (SVZc) em situação controle com 3 dias in vitro (DIV). B) Explante da mesma região em presença de Carbenoxolone (CBX) bloqueador de acoplamento juncional. C e D) Ensaio de “Scrape Loading”, com células verdes carregadas apenas com “Lucifer Yellow” (LY - cabeças de seta), permeável à junções comunicantes. Note que, em presença de CBX (D) o acoplamento foi bastante reduzido. E e F) Imagens dos mesmos campos de C e D (respectivamente) onde as células LY positivas não são “Rodamina Dextran” (RD, impermeável à junções comunicantes) positivas. G) Explante de SVZ contida no bulbo olfatório (SVZb), com 3DIV em situação controle, imunomarcado para BrdU. H) Explante de SVZb, com 3DIV em presença de CBX, com poucas células BrdU positivas. Barra de calibração 10µm.Modificado de Marins, 2001.

22
McDonough et al., 1999; Li e Herlyn, 2000). Em gliomas, os níveis de conexina 43 e
conseqüentemente o grau de acoplamento intercelular encontram-se reduzidos (Zhu
et al, 1991; McDonough et al., 1999), enquanto a superexpressão de conexina 43
induz à reversão do fenótipo tumoral (Naus et al., 1992). Ao comparar o grau de
severidade metastásica entre diferentes linhagens tumorais humanas, Navolotski e
cols. (1997) demonstraram que o desenvolvimento tumoral (migração e proliferação)
estava realcionado à redução dos níveis de comunicação celular.
Estudos recentes, desvincularam as junções comunicantes da possível
função de supressores tumorais (Lin et al., 2002; Zhang et al., 2003; Pollman et al.,
2005). Pollman e cols. (2005) demonstraram que o acoplamento heterocelular (via
Cx43) entre células de tumor de mama e células endoteliais aumenta a motilidade
das primeiras e conseqüentemente a invasibilidade do tumor. Correlações positivas
semelhantes foram extraídas de estudos realizados com tumores de próstata
(Pollman et al., 2005; Miekus et al., 2005) e em modelos de invasividade do córtex
por glioma (Lin et al., 2002).
Em outros sistemas, tem-se evidenciado uma correlação negativa entre o
acoplamento e a migração celular, como relatado durante a cicatrização epitelial (Qiu
et al., 2003) ou no desenvolvimento de resposta inflamatória (Zahler et al., 2003). A
dicotomia persiste, no entanto, quanto ao papel da comunicação juncional sobre a
migração celular em sistemas não neurais. Células não-neurais, como células
epiteliais (Pawar et al., 1995; Lampe et al., 1998; Chanson et al., 2005), fibroblastos
(Nagy et al., 1996; Ehrlich e Diez, 2003) e células musculares lisas (Blackburn et al,
1997; Déglise et al., 2005) apresentam também uma correlação positiva entre
acoplamento juncional e migração celular. Pawar e cols. (1995) demonstraram a
existência de um mecanismo de seleção das células epiteliais renais, que

23
desenvolviam a atividade migratória, mas não a proliferativa, em resposta à necrose
tubular. Na interpretação dos autores, este movimento estava na dependência de
um gradiente de sinalizadores intercelulares, que partiam do ponto da necrose
através dos canais juncionais (Pawar et al., 1995). Este estudo ilustra, portanto, a
possibilidade de independência entre os processos proliferativo e migratório.
Estudos recentes sugerem que a comunicação juncional permanece ausente
durante a migração (Bittman et al., 1997; Bittman e Lo Turco, 1999) só retornando
transitoriamente durante a formação da circuitaria cortical (Yuste et al., 1992;
Peinado et al., 1993; Yuste et al., 1995). Desta forma, o acoplamento parece
precisar ser desligado durante a migração neuronal. No entanto, existe uma
evidência de que, durante o desenvolvimento, possa ocorrer uma implicação direta e
positiva de junções comunicantes sobre o fenômeno migratório. Huang e cols.
(1998) demonstraram esta relação utilizando um modelo de migração de precursores
de neurônios simpáticos cardíacos derivados da crista neural. De forma interessante,
tal como a migração presente na SVZ/RMS, estes precursores neurais também
migram em grupo, “em masse”, com células intimamente apostas umas as outras. É
possível que esta seja uma caracteristica comum àqueles processos migratórios que
apresentem uma participação positiva de junções comunicantes.
2 – O principal bloqueador juncional utilizado:
O carbenoxolone (CBX; ácido 3β-hidroxi-11-oxooleano-12-eno–30-óico 3-
hemissuccinato) foi utilizado como principal bloqueador juncional neste trabalho. O
CBX é um derivado sintético hemisuccinato do ácido β-glicirretínico (β-GA), princípio
ativo da raiz do alcaçuz (Fig. 7). Apesar do β-GA ser muito utilizado para o bloqueio

24
de junções comunicantes (Davidson e Baumgarten, 1988), o CBX foi considerado
mais adequado, para o tratamento crônico de nossas culturas de explantes, por ser
hidrofílico e menos tóxico. Além disso, o CBX é inespecífico quanto ao isotipo das
conexinas, isto é, bloqueia o acoplamento mediado por quaisquer conexinas (Spray
et al., 2002, Salameh e Dhein, 2005). O primeiro trabalho relacionando o CBX com o
bloqueio de junções comunicantes foi publicado em 1986, por Davidson e cols.
Atualmente, tem sido muito utilizado no bloqueio juncional de regiões do sistema
nervoso em modelos in vivo (Pacha et al., 2003; Masaki et al., 2004) ex-vivo
(Menezes et al., 2000) e in vitro (Alvarez-Maubecin et al., 2000; Marins, 2001; Pais et
al., 2003; Ye et al., 2003; Bittman et al., 2004; Kano et al., 2005). Dentre os
mecanismos imaginados para a ação dos bloqueadores juncionais, inclui-se a
possibilidade de estarem agindo por modificação das estruturas terciária e
quaternária do oligômero juncional, através da adsorção e intercalação molecular na
região de camada lipídica anular do conexon. Guan e colaboradores (1996)
relacionaram β-GA com desfosforilação da Cx43 e desorganização das placas
juncionais. O CBX também foi capaz de desagregar as placas juncionais,
possivelmente por causar mudanças conformacionais ao se ligar ao conexon,
alterando a permeabilidade dos canais e com isso, dificultando sua organização em
placas juncionais (Goldberg et al., 1996). Além disto existem evidências de outros
efeitos intra-celulares, tais como: bloqueio da enzima 11-β-dehidrogenase (Monder
et al., 1989). Embora o CBX seja um potente bloqueador juncional, sua ação não
inibe completamente o acoplamento celular, apenas o reduz em torno de 70-90%
(Van Haarst et al., 1996; Menezes et al., 2000; Marins, 2001; Pais et al., 2003).

25
Figura 7. Bloquedores do acoplamento celular (A e B) e análogo sem efeito (C).Modelos moleculares tridimensionais e fórmulas estruturais planas de bloqueadores juncionais. A) Princípio ativo extraído da raiz do alcaçus, o ácido glicirretínico (aGA) é um dos bloqueadores juncionais mais utilizados, embora seja lipossolúvel. B) Análogo sintético do αGA, o Carbenoxolone (CBX), preferido em trabalhos que envolvam culturas teciduais e celulares, é hidrossolúvel e possui características anfipáticas. C)Análogo sintético do αGA, mas sem qualquer efeito sobre junções comunicantes, o ácido glicirrízico (GZA) é comumente usado como controle dos previsíveis efeitos colaterais. Funciona como controle da especificidade do efeito dos bloqueadores sobre as junções comunicantes. As fórmulas estruturais planas foram obtidas no site www.chemfinder.com e os modelos tridimensionais foram gerados com o software ChemOffice2004 da CambridgeSoft Corporation.

26
II - Objetivos:
Neste trabalho, focalizaremos nossa atenção na determinação do substrato
celular envolvido com a inibição da migração celular a partir de explantes de SVZ
após o bloqueio do acoplamento mediado por junções comunicantes. Para tanto,
utilizaremos as seguintes estratégias:
1. Determinação do padrão de acoplamento individual das células do
explante inclusive com a identificação dos pares celulares acoplados;
2. Análise da expressão da conexina 43, a mais abundante da SVZ, nos
explantes;
3. Análise da especificidade e toxicidade do carbenoxolone com o
emprego de outros bloqueadores juncionais, análogo inativo e ensaios
de curso temporal da ação do fármaco;
4. Ensaios de vídeo microscopia de tempo intervalado para observar se o
acoplamento interfere direta ou indiretamente com a migração.
5. Comparar o comportamento migratório e a importância do acoplamento
juncional entre SVZb e SVZc.

27
III - Materiais e Métodos:
1 – Preparação de explantes:
Utilizamos ratos P3 (o primeiro dia pós-natal é P0) ou P4, para gerar os
explantes de SVZ. Os animais foram decapitados, os cérebros retirados e fatiados
(V. Fig. 8) utilizando um McIlwain Tissue Chopper, à espessura de 400 µm. As fatias
foram transferidas para placas de Petri com Gey’s (solução salina balanceada -
GIBCO, USA) gelado para a dissecção da SVZ (Fig. 8). Os explantes obtidos foram
plaqueados, individualmente sobre lamínulas de vidro previamente distribuídas em
placas de cultura de 24 poços, tratadas com poli-L-lisina (Sigma, USA; 10 µg/mL), e
recobertas com meio de cultura contendo 60% de D-MEM (GIBCO), 30% de solução
Hank's (GIBCO), 10% de soro fetal bovino (Fazenda Pigue, Brasil), 1% de solução
de penicilina (10.000 U/L) + Streptomicina (10mg/mL – GIBCO), 1% de Fungizon
(Bristol Myers Squibb, Brasil) e 1% de glicose. Os explantes foram cultivados a 37o C
por 3-5 dias em incubadora com injeção de 5% de CO2, o meio foi trocado no dia
seguinte de cada experimento e em dias alternados até o quinto dia em cultura,
quando foram fixados com HistoChoice (AMRESCO, USA). Separamos explantes
provenientes de duas porções distintas da SVZ, de modo que a SVZ contida no
bulbo olfatório foi chamada de bulbar (SVZb) e a subjacente ao córtex, de cortical
(SVZc), conforme o esquema da Figura 8 (2a).
2 - Cultura de explantes de zona subventricular em Matrigel:
Para uma parte dos experimentos de videomicroscopia utilizamos como

28
Figura 8. Metodologia de preparação de explantes P4 de SVZb e SVZc. Os cérebros dissecados foram cortados coronalmente à 400 µm (1); as fatias eram transferidas para uma placa de Petri contendo Gey's a 4 0C, colocada sobre fundo preto para melhor visualização da SVZ, (que, por diferença de densidade celular, ficava mais escura que o tecido adjacente - 2); separamos a SVZ em duas porções: uma contida no bulbo olfatório (SVZb) e outra contida no córtex (SVZc - 2a); a SVZ era dissecada e cortada em explantes (3-4); os explantes provenientes das duas regiões da SVZ, eram plaqueados separada-e individualmente sobre lamínulas de vidro, previamente colocadas em placas de cultura com 24 poços, tratadas com Poly-L-Lisina e recobertas com meio de cultura (5-6).

29
substrato o Matrigel (Collaborative Biomedical Products, USA) devido a maior
velocidade e homogeneidade das células que migram para fora do explante neste
substrato tridimensional (Wichterle et al., 1997). O Matrigel, é um gel tridimensional
composto de colágeno tipo IV, laminina e proteoglicanos de matriz extracelular
(Kleinman et al, 1982). As idades dos animais utilizados, os procedimentos de
dissecção e plaquemento são idênticos aos da cultura de explantes sem matrigel, à
exceção do tratamento prévio das lamínulas com poli-L-lisina. No entanto,
atendendo às especificações do Matrigel, foi utilizado meio Neurobasal (GIBCO)
sem soro, acrescido de suplemento B-27 (GIBCO), L-Glutamina e antibióticos. Após
o plaqueamento, era aplicada uma fina camada do Matrigel, diluído em meio
neurobasal em uma concentração 1:3, sobre os explantes. Em seguida, os explantes
foram colocados na estufa por cerca de 20 minutos, para permitir a solidificação do
Matrigel. Após este período, foram adicionados 200 µl de meio Neurobasal em cada
poço. Estas culturas foram mantidas na estufa por períodos de 2 a 3 dias, quando
alguns explantes foram selecionados para um dos ensaios de vídeo microscopia de
tempo intervalado, enquanto o restante dos explantes foi fixado com HistoChoice.
3 – Cultura de explantes de camundongos nocaute de Cx43:
Explantes de SVZ, foram obtidos a partir dos cérebros de camundongos
mutantes (com deleção completa da Cx43), gentilmente cedidos pelo Prof. Antônio
Carlos Campos de Carvalho. No final do período embrionário (E19), os animais
foram retirados do útero materno por meio de cesariana, as cabeças foram
decapitadas e imediatamente transferidas para recipientes identificados e
previamente preenchidos com Gey’s gelado. Após dissecção, os cérebros foram

30
cortados em McIlwain Tissue Chopper, a uma espessura de 300 µm. Neste caso os
cortes foram mais finos que aqueles feitos para os explantes de rato porque o
cérebro do camundongo era bem menor e precisávamos de mais fatias contendo
SVZ para poder dissecar os explantes. Em seguida as fatias foram submetidas aos
mesmos procedimentos dos explantes de ratos e plaqueadas nas mesmas
condições com a devida identificação de procedência. No momento da cesariana, foi
colhido material dos animais para verificação do genótipo de cada um deles. O
Laboratório do Prof. Antônio Carlos forneceu o resultado da identidade genotípica
dos animais obtida por análise de RT-PCR. Os explantes foram cultivados nas
mesmas condições dos explantes de rato, sendo que somente em situação controle
até o quinto dia in vitro, quando foram fixados com HistoChoice.
4 - Bloqueio da Comunicação Juncional:
Utilizamos o bloqueador carbenoxolone (CBX; Sigma – Fig. 8), com
características anfipáticas, a uma concentração de 100µM (Davidson e Baumgarten,
1988, Van Haarst et al., 1996) sobre a metade dos explantes cultivados nas
lamínulas, a partir das primeiras 14-18 horas de cultura. Todos os explantes,
controles e tratados, foram submetidos a trocas de meio a cada 48h até sua fixação.
O CBX foi mantido até o término da cultura. A remoção ou adição de CBX para as
situações de reversão ou CBX no terceiro dia (V. adiante) foi feita no terceiro dia in
vitro (3DIV) e os explantes submetidos às novas condições foram mantidos até o
quinto e último dia em cultura.

31
5 – Injeção de corante:
Lamínulas contendo explantes de SVZb e SVZc, com 3 dias de cultivo, foram
transferidas para placas de cultura de plástico e algumas células, dos halos
migratórios, foram injetadas com “Lucifer Yellow” CH (5 % em 150 mM LiCl sem
ajuste de pH), com o auxílio de microeletrodos de vidro (resistência entre 40-70 MΩ)
por aplicação de pequenos pulsos de corrente hiperpolarizante (0.1 nA, 100 ms),
usando um amplificador WPI (modelo 7060). A transferência de corante foi
observada no microscópio invertido Axiovert 100 (Carl Zeiss, Oberkochen,
Germany), equipado com arco de iluminação de xenônio, as fotografias foram
tiradas, usando filmes Kodak Gold 400, cerca de 2 minutos após cada injeção (Fróes
et al. 1999) e posteriormente por sistema de fotografia digital acoplado ao mesmo
microscópio. As imagens de microscopia confocal foram obtidas com a utilização do
microscópio confocal Zeiss LSM 510 Meta (Carl Zeiss; Depto de Patologia,
Fundação Oswaldo Cruz, Fiocruz). Os tipos celulares injetados e os acoplados foram
identificados por observação de sua morfologia, sendo que os corpos celulares com
núcleos pequenos e citoplasma escasso, foram considerados como neurônios ou
neuroblastos; e as células com núcleos grandes e citoplasma abundante, como
células gliais de natureza astrocitária. Estes critérios são baseados em nossos
resultados prévios de ensaios de imuno-histoquímica, com a expressão de GFAP e
βIII tubulina, freqüentemente associados à morfologia acima descrita. A cada
injeção, as informações foram registradas e posteriormente os dados foram
quantificados para produção de gráficos que revelaram a incidência do acoplamento
e sua extensão. Os dados foram submetidos à análise estatística com teste Qui
quadrado, apresentando significância para valores de α < 0,05.

32
6 – Imuno-histoquímica:
Para as reações de imuno-histoquímica, os explantes foram fixados com
HistoChoice (AMRESCO, USA) por 3-8 dias. Depois de várias lavagens com solução
salina e tampão fosfato (PBS, 10mM e pH 7.4), incubamos os anticorpos primários
diluídos em PBS com 0,03% de Triton-X 100 (Reagen, USA) contendo 10% de soro
de cabra (solução de bloqueio). Para detecção da presença neuronal utilizamos o
anticorpo monoclonal contra proteínas do citoesqueleto, classe III β-tubulina (Sigma;
1:200), para identificação glial usamos anticorpo policlonal contra GFAP (proteína
glial fibrilar acídica – Dako, USA; 1:200), a presença da glia radial foi detectada com
anticorpo monoclonal contra Nestina (marcador do filamento intermediário das
células tronco neurais – Pharmingen, USA; 1:100), a conexina Cx43 foi identificada
com o anticorpo policlonal contra Cx43 (Zymed, USA; 1:200). Após 24h em solução
de anticorpo primário a 4o C o tecido foi lavado três vezes com PBS e incubado por
2h, à temperatura ambiente, em solução de bloqueio com anticorpos secundários.
Utilizamos os seguintes anticorpos secundários: anticorpo de cabra contra IgG de
coelho conjugado à FITC (Accurate, USA; 1:100); contra IgG de camundongo
conjugado à Cy3 (Jackson ImmunoResearch, USA; 1:50 e 1:200); ao Alexa Fluor
488 (Molecular Probes, USA; 1:200) e ao Alexa Fluor 633 (Molecular Probes; 1:200).
7 - Teste de especificidade do bloqueador juncional:
Cultivamos alguns explantes, até o 3DIV, em presença do ácido glicirrízico
[GZA; sal de amônio do ácido 3-O-(2-O-β-D-glicopiranuronosil-α-D-
glicopiranuronosil)-18β-glicirretínico – 100µM; Sigma - Fig. 8], análogo sem efeito do

33
CBX. O ácido glirrízico não é hidrossolúvel como o CBX, por isso, foi diluído em uma
mistura de Dimetil sulfóxido (DMSO) e etanol (3:2, vol:vol - Duval et al., 2002). Uma
parte dos explantes (SVZb=4; SVZc=6) foi cultivada apenas na presença do DMSO
e do etanol (situação controle) e outra parte igual foi exposta ao ácido glicirrízico. No
terceiro dia de cultivo os explantes foram fixados com HistoChoice.
8 - Análise e quantificação da migração:
A comparação e a quantificação da migração entre os explantes tratados e
nao-tratados se deram por uma razão entre a área de explantes fotografados nas
primeiras 14-18 horas (1o dia), antes da formação de um halo migratório significativo,
e a área de distribuição das células destes mesmos explantes fotografados no 3o e
5o (e último) dias em cultura. Estes valores são, portanto, adimensionais e referidos
como medidas fracionárias 3/1 e 5/1, conforme indicadas nos gráficos (Figs. 17 à
21). Desta forma, a razão 3/1 é o índice migratório do primeiro até o terceiro dia, do
mesmo modo que a 5/1 mostra o índice migratório do período total do experimento,
ou seja, do primeiro ao quinto dia in vitro. Nos casos de adição de CBX no 3DIV e
reversão, os explantes foram fotografados antes da execução destes procedimentos.
Note que são os mesmos explantes submetidos às diferentes condições
experimentais. Os negativos foram projetados no papel através da utilização do
Aristophoto (Eleitz Weizlar, no 10545), seus contornos externos foram desenhados e
os desenhos, com as respectivas barras de escala, foram digitalizados (HP Precision
Scan) e editados (Adobe Photostyler). Durante a última etapa de experimentos as
imagens dos explantes foram adquiridas, por sistema de vídeo captura, em um
microscópio invertido Nikon (TE200). Quantificamos a área total de explantes

34
somada ao halo migratório utilizando o software Scion (NIH). Para cada condição
foram analisados de 4 a 12 explantes, de 3 ou mais experimentos independentes,
com exceção dos explantes de camundongo nocaute Cx43, que foi único. Os
resultados foram submetidos aos testes Mann-Witney, à exceção dos seguintes
experimentos: nocaute Cx43, submetido ao teste Kruskal-Wallis; GZA e seu veículo,
que ao serem comparados com a situação controle e CBX foram submetidos ao
teste ANOVA; e dos de reversão e CBX no terceiro dia in vitro, que foram
submetidos ao teste T de Student pareado. Todos os testes apresentaram
significância para valores de p<0,05. Os gráficos foram gerados com as barras de
desvio padrão para todas as situações experimentais.
9 - Vídeo Microscopia de Tempo Intervalado:
Nosso ensaio teve início com a seleção prévia de explantes com 2DIV,
cultivados sobre Matrigel, que foram transferidos para placas de cultura de 4 ou 24
poços e cobertos com 400µL de meio de cultura. Em seguida, os explantes de SVZ
foram individualmente monitorados, em um sistema de vídeo microscopia, que
reproduz as condições da estufa de cultura, composto de microscópio invertido
Axiovert 135 (Carl Zeiss), equipado com: CCD Axio Cam, CTI Controler 3.700
(controlador de injeção de CO2, mantida a 5%), TRZ 3.700 (controlador de
temperatura da placa aquecida que forma a base do microscópio, constante a 370C).
Além disso, o sistema de vídeo microscopia está ligado a um computador, e é
controlado pelo programa AxioVision (versão 3.1), onde escolhemos os intervalos de
tempo nos quais as imagens dos explantes deveriam ser capturadas e a duração de
cada seção de captura. As imagens foram obtidas a cada dois minutos, por 3h em

35
situação controle (na presença de meio de cultura controle). Após este período, o
meio foi substituído pela mesma quantidade de meio contendo CBX. Depois de 3h
de observação, o bloqueador foi substituído por meio controle. Três horas depois, os
explantes foram fixados com HistoChoice por 3-8 dias. O mesmo procedimento foi
feito para explantes de 3DIV cultivados com Halotano (borbulhado no meio de
cultura vol:vol), GZA (100µM) e seu veículo (DMSO e etanol - 3:2, vol:vol), utilizado
como controle.
10 – Análise da vídeo microscopia de tempo intervalado:
Devido a problemas com o foco nos ensaios de vídeo microscopia, resultante
da tridimensionalidade das cadeias, optamos por fazer dois tipos diferentes de
análise das imagens:
1 – Na primeira etapa, acompanhamos algumas células individualmente para estimar
sua movimentação e velocidade média de deslocamento. Escolhemos quinze células
aleatórias, localizadas nas cadeias migratórias que apresentaram maior
movimentação. As células foram marcadas pela superposição de pequenos círculos
coloridos sobre a porção mais volumosa de seu corpo a cada imagem. Na passagem
de uma imagem para a próxima, círculos de mesma cor foram posicionados nos
corpos das mesmas células, localizadas por sua posição relativa ao plano de fundo.
As células deixavam de ser acompanhadas quando houvesse dúvida sobre sua
localização ou dificuldade na identificação de seu contorno em relação às demais. A
área onde as células estavam contidas foi delimitada por um retângulo, alinhado com
as bordas esquerda e inferior da imagem, portanto regiões fixas. Os retângulos e os
círculos foram agrupados e duplicados a cada imagem e os círculos receberam um

36
número da imagem de onde foram extraídas. Em seguida, grupos de 10 retângulos
(intervalos de 18 minutos), foram superpostos, na mesma ordem em que as imagens
foram produzidas, e alinhados pelas bordas esquerda e inferior (pontos de referência
fixos). Ao final do procedimento, o deslocamento das células se tornava visível, bem
como sua trajetória, marcada pela ordem numérica de cada círculo. Na etapa
seguinte, traçamos uma linha, passando pelo centro de cada círculo, ligando a
primeira à última da trajetória de cada célula. A reta foi partida no centro de cada
círculo e os segmento de reta foram medidos e convertidos em micrômetros. As
distâncias percorridas em cada hora de filmagem, para as três situações
experimentais, foram correlacionadas com os intervalos de tempo em planilha do
programa Excel (versão 5.1.2600). Calculamos a velocidade média através da razão
da distância total percorrida pelo intervalo de tempo total de movimentação para
cada hora de filmagem. As velocidades médias foram comparadas internamente
dentro de cada condição experimental e entre as condições experimentais utilizando
o teste T de Student não pareado.
2 – O segundo método de análise foi desenvolvido para estudar a direção migratória
das células, classificada em centrífuga (para fora do explante) ou centrípeta (para
dentro do explante). Três investigadores independentes contaram o movimento
aparente das células através da colocação de uma barra, com aproximadamente
5µm, sobre as cadeias migratórias a serem analisadas. Novamente escolhemos as
cadeias onde havia maior movimentação e nitidez de foco, ao longo das três horas
de filmagem de cada etapa do experimento. As “células” que ultrapassassem a barra
para um dos lados eram contabilizadas de acordo com a direção de seu movimento.
Os números fornecidos por cada investigador foram submetidos a teste de
correlação, com valor de p entre -1 e 1 (sendo a maior correlação possível igual a 1)

37
e comparados através do teste não paramétrico Kruskal-Wallis, com significância
para valores de p<0,05.

38
IV - Resultados:
1 – Migração celular a partir de explantes de SVZb e SVZc:
Anteriormente estabelecemos o modelo de cultura de explantes de SVZ para
estudar a migração de neuroblastos derivados desta região (Marins, 2001). Nosso
modelo apresenta as seguintes características:
a) Os explantes cultivados sobre lamínulas (3 DIV e 5 DIV) recobertas com poli-L-
lisina ou Matrigel, apresentaram características semelhantes às anteriormente
descritas na literatura (Hockberger et al., 1987; Hu et al., 1996; Wichterle et al.
1997 e 1999, Mason et al., 2001);
b) Os explantes de SVZb apresentaram halos migratórios maiores que os explantes
de SVZc com idades equivalentes. Além disso, apenas os explantes de SVZc
conseguiram atingir a reversão completa dos efeitos do agente desacoplante;
c) Explantes cultivados sobre poli-L-lisina e Matrigel apresentaram características
semelhantes: as cadeias migratórias se formaram normalmente e a morfologia
dos tipos celulares presentes nos explantes mostrou-se bastante semelhante
(Fig.9). No entanto, observamos algumas diferenças: no matrigel as cadeias
migratórias eram mais numerosas, as células pareciam migrar de modo mais
independente de glia subjacente que na poli-L-lisina e a velocidade migratória
aumentada;
d) As reações de imunohistoquímica contra GFAP, Nestina, proteína Classe-IIIβ-
tubulina (βIII) e BrdU, em explantes controle, revelaram a presença de células
gliais, neuronais e de precursores neurais, respectivamente.
Com este modelo, demonstramos anteriormente que a migração e

39
Figura 9. Características dos explantes de SVZ cultivados sobre diferentes substratos. Explantes de SVZc com três dias in vitro (3DIV) cultivados sobre poli-L-lisina (A) e com matrigel (B). Note que as cadeias migratórias se formaram normalmente em ambas situações. No entanto, o substrato tridimensional conferido pelo matrigel aumenta o número de cadeias e parece aumentar também a velocidade migratória. Explantes representativos de 600 (A) e de 6 (B). Barra de escala 100µm.

40
proliferação celular em explantes de SVZ parecem ser influenciados pelo
acoplamento juncional. Nesta tese pretendemos caracterizar a influência do
acoplamento celular sobre a migração.
2 – Caracterização do acoplamento nos explantes:
Anteriormente, detectamos a presença de acoplamento funcional mediado por
junções comunicantes nos explantes, mas não havíamos identificado os tipos
celulares envolvidos e nem a extensão do acoplamento.
Realizamos injeções intracelulares do corante “Lucifer Yellow”, permeante a
junções comunicantes, em células identificadas por sua morfologia e localizadas nas
extremidades do halo migratório. A densidade celular, nos explantes, decresce em
direção ao halo migratório, facilitando a visualização e a identificação morfológica
das células injetadas. Encontramos acoplamento homo- e heterocelular nos
explantes de SVZc e SVZb (Fig. 10), como já havia sido descrito anteriormente para
modelos in vitro (Nedergaard, 1994; Fróes et al., 1999; Duval et al., 2002). Cada
injeção resultou em três possibilidades: a) ausência de acoplamento, com nenhuma
outra célula preenchida pelo corante além daquela injetada; b) acoplamento
homocelular, entre células com a mesma morfologia; e c) acoplamento heterocelular,
envolvendo células com morfologia claramente diferente (Fig. 10). Em alguns casos,
a célula injetada estava acoplada com outras das imediações (Fig. 10A), em outros
havia pelo menos uma célula mais afastada preenchida pelo corante,
possivelmemente via prolongamentos (Fig. 10B-E). Ao detectarmos acoplamento,
contabilizamos o número de células preenchidas pelo corante e as dividimos usando
critérios morfológicos (V. Mat. e Met.) entre células com morfologia glial (Astroglia)

41
Figura 10. Exemplos dos pares acoplados em explantes de SVZ com 3DIV (A-E). Fotomicrografias de contraste de fase e fluorescência de injeção iontoforética de Lucifer Yellow (LY- verde; insets em A e B) em explantes de SVZb (A-E). A. A morfologia das células acopladas sugere a presença de acoplamento homocelular (apenas entre precursores neurais). B. A célula acoplada à distância apresenta morfologia semelhante à da glia radial, sugerindo a presença de acoplamento heterocelular entre precursores neuronais e célula glial (seta). A micropipeta de injeção pode ser observada em todos as figuras; as cabeças de seta indicam a extremidade da micropipeta. C-E. Neste exemplo, a célula injetada apresenta morfologia glial, e está acoplada com outra mais afastada e com morfologia semelhante à de um precursor neural. Imagens representativas de 14 injeções realizadas em 3 explantes. Barra de calibração, 50µm (A e B) e 20µm (C - E).

42
e não glial (Neurônios; estes podendo representar também neuroblastos e raros e
hipotéticos precursores indiferenciados com forma indefinida). Deste modo
obtivemos um gráfico com as seguintes variáveis: para cada neurônio injetado
identificamos o acoplamento homocelular como NN, o heterocelular como NA
(neurônio acoplado com astrócito) e ausência de acoplamento como N0 (neurônio
mais zero); o mesmo critério foi adotado para os astrócitos injetados com AA para
acoplamento homocelular, AN para heterocelular e A0 para ausência de
acoplamento. Utilizamos o teste Qui quadrado para comparar a distribuição de
incidência de pares acoplados em explantes de SVZb e SVZc e não encontramos
diferença entre as duas regiões. Registramos, no entanto, valores de incidência de
pareamentos NN em explantes de SVZb que praticamente dobram aqueles obtidos
em explantes SVZc. Apesar de não terem forçado a significância estatística no
conjunto de pareamentos entre as duas sub-regiões, acreditamos que estes reflitam
de fato diferentes propriedades de comunicação juncional entre
neurônios/neuroblastos destas duas categorias de explantes e que estas, por sua
vez possam indicar graus maturacionais distintos nestas duas populações celulares.
A análise destes dados trouxe-nos às seguintes conclusões: a) a incidência
do acoplamento heterocelular não parece diferir significativamente entre as duas
regiões da SVZ; b) injeções em neurônios levaram a incidências de acoplamento
heterocelular maiores, quando comparadas com injeções em células astrogliais; uma
interpretação para esta aparente assimetria é de que os altos níveis de acoplamento
interastrocitário determinariam vias de menor resistência para o escoamento do
corante através do sincício glial (Fróes et al., 1999); c) Os pareamentos
homocelulares diferem em explantes de SVZb e SVZc, de forma a privilegiar, nestes
últimos, o acoplamento astroglial sobre o neuronal (Fig. 11).

43
Figura 11. Incidência dos tipos de acoplamento nos explantes da SVZb (A) e SVZc (B). Os diferentes tipos de acoplamento estão indicados na abcissa. O primeiro elemento de cada par representa o tipo celular injetado (A – astroglia; N – neurônio). Pareamentos homocelulares classificam-se como neuronais (NN – n = 6 em A e 17 em B) e astrogliais (AA – n = 8 em A e 11 em B). Ausência de acoplamento está indicada por A0 e N0. Os pareamentos heterocelulares estão indicados como NA ou AN, conforme derivados de injeções em neurônios ou em células astrogliais, respectivamente. A e B) Independente da região de origem do explante, oacoplamento homocelular apresentou maior incidência que os demais, enquanto que o heterocelular mostrou-se equivalente à ausência de acoplamento. Em todos os casos o pareamento heterocelular apresentou maior incidência quando o neurônio foi injetado. Explantes derivados de SVZb (n = 3 - A) apresentaram maior incidência de acoplamento homocelular entre astrócitos que entre neurônios, o contrário acontece em explantes derivados de SVZc (n = 4 - B). Essa diferença pode refletir dos diferentes graus de maturação celular entre as duas regiões.

44
Em um dos experimentos de injeções intracelulares realizamos imuno-
histoquímica contra GFAP (proteína acídica fibrilar glial) para identificar pelo menos
um dos tipos celulares envolvidos com o acoplamento. Econtramos exemplos de
acoplamento heterocelular e homocelular tanto na SVZb quanto na SVZc (Fig. 12).
Os resultados de imuno-histoquímica confirmaram a relação da morfologia com o
fenótipo celular, aumentando a confiabilidade de nossas análises.
Além da quantificação dos tipos de acoplamento, também nos preocupamos
em saber se a sua extensão (o número de células acopladas para cada injeção)
revelaria diferenças entre a SVZb e a SVZc. As duas regiões apresentaram-se muito
semelhantes no que se refere à extensão do acoplamento, por isso agrupamos
todos os resultados e analisamos os dados conforme a célula injetada (Fig. 13). O
acoplamento homocelular ocorreu mais entre astrócitos que entre neurônios e foi
mais extenso que o heterocelular (Fig. 13A), resultado compatível com o alto índice
de acoplamento homocelular característico destas células (Rouach et al., 2002;
Theis et al, 2005; Sutor e Hagerty, 2005). Mais ainda, o tratamento estatístico
revelou que a extensão do acoplamento homocelular astroglial é significativamente
diferente do homocelular neuronal (p=0,0138) e dos pareamentos heterocelulares
(p≤0,0001). A extensão do acoplamento heterocelular foi maior quando um neurônio
era injetado que quando a célula injetada era um astrócito (p=0,0001), proporção
inversa à do acoplamento homocelular (Fig.13B). Por fim, o acoplamento
homocelular neuronal é significativamente diferente do heterocelular definido por
injeção em neurônios (p=0,0133) e em astrócitos (p<0,0001).

45

46
Figura 12. Identificação fenotípica de um dos componentes do par acoplado. Imagens (microscopia confocal) de injeção iontoforética de “Lucifer Yellow” (LY; verde) feita em explantes de SVZc (A-D) e SVZb (E-G) com imunomarcação contra GFAP (vermelho). A) A seta indica a célula injetada, a coloração verde das outras células revela a presença do acoplamento. B) O grupo de células acopladas não apresentou marcação para GFAP, isto sugere a possibilidade desse acoplamento ser homocelular (entre células com fenótipo semelhante). C) Superposição da injeção de LY com a imunomarcação contra GFAP. O retângulo indica a região selecionada para a reconstrução tridimensional. D) Reconstrução tridimensional do retângulo contido em C. Os pequenos retângulos em volta da foto maior representam o eixo Z do bloco de fatias cortado ao nível das linhas brancas, com separação dos reagentes (acima e à direita) e com a superposição das duas cores (à esquerda e abaixo). Note que as células acopladas aparecem escuras nas imagens contendo apenas GFAP. E) A seta indica a célula injetada e o sombreado verde do fundo da imagem reflete a presença de autofluorescência. F) Neste exemplo, a célula injetada é imunopositiva para GFAP, mas seu par não. G) A reconstrução tridimensional nos permite evidenciar a morfologia de célula glial, confirmada pela imunomarcação para GFAP, ao mesmo tempo em que a ausência de morfologia glial estava associada com a ausência de GFAP neste caso particular. Barra de calibração 10µm.

47
Figura 13. Extensão do acoplamento celular na SVZ. Os gráficos representam a freqüência com que “clusters” (grupo de células carregadas com o permeante juncional a partir de uma única injeção) com diferentes tamanhos (número de células acopladas) aparecem no total de injeções*∗. As medidas não discriminam entre as injeções realizadas em explantes de SVZb e SVZc. A) Acoplamento homocelular NN, à esquerda (total de 17 injeções), e AA, à direita (total de 11 injeções). B) Acoplamento heterocelular NA, à esquerda (total 17 injeções), e AN, à direita (total de 11 injeções). A extensão do acoplamento homocelular astroglial prevaleceu sobre os demais, e é significativamente diferente do acoplamento heterocelular (p≤0,0001) e do acoplamento homocelular neuronal (p=0,0138). Acoplamentos homo e heterocelular definidos por injeções em neurônios praticamente se equivalem, mas são significativamente diferentes (p=0,0133) ainda que menos freqüentes relativamente ao acoplamento astroglial. O acoplamento homocelular neuronal também é significativamente diferente do acoplamento heterocelular definido por injeção astroglial (p<0,0001).
∗ A primeira barra em cada gráfico representa acoplamento nulo (por ausência de espalhamento transcelular do fluorocromo em níveis detectáveis) somado aos casos em que não aconteceu acoplamento homocelular (A) ou heterocelular (B).

48
3 – A Cx43 está presente em diferentes tipos celulares no explante:
O fato da Cx43 ser a principal conexina dos astrócitos somado à grande incidência
de astrócitos em nossos explantes e à possibilidade de neurônios imaturos
expressarem esta conexina, nos levou a elegê-la para um ensaio de
imunocitoquímica. Utilizamos também um marcador para fenótipo celular, a nestina,
proteína presente nas células progenitoras, glia radial e em alguns astrócitos
diferenciados, quando em cultura. Os explantes apresentaram grande densidade de
células contendo os dois marcadores, contudo a Cx43 também está presente nas
células negativas para nestina (Fig. 14). O padrão de distribuição celular destas duas
marcações é compatível com os compartimentos subcelulares esperados para as
duas proteínas (Fig. 15). No exemplo mostrado na Figura 15, duas regiões do halo
migratório de um dos explantes (Fig. 15A) estão representadas. No primeiro
exemplo, encontramos uma célula, em mitose, e prolongamentos nestina-positivos,
repletos de Cx43, enquanto que outras células apostas, nestina-negativas, também
eram Cx43-positivas (Fig.15B). Na outra região selecionada, encontramos uma
célula fortemente marcada para nestina e igualmente positiva para Cx43, no interior
de uma cadeia migratória contendo células negativas para nestina e marcadas
apenas para Cx43 (Fig. 15C-F). Este resultado nos levou a crer que a Cx43 pode ser
a principal envolvida no acoplamento heterocelular e principalmente com a migração
em nossos explantes.
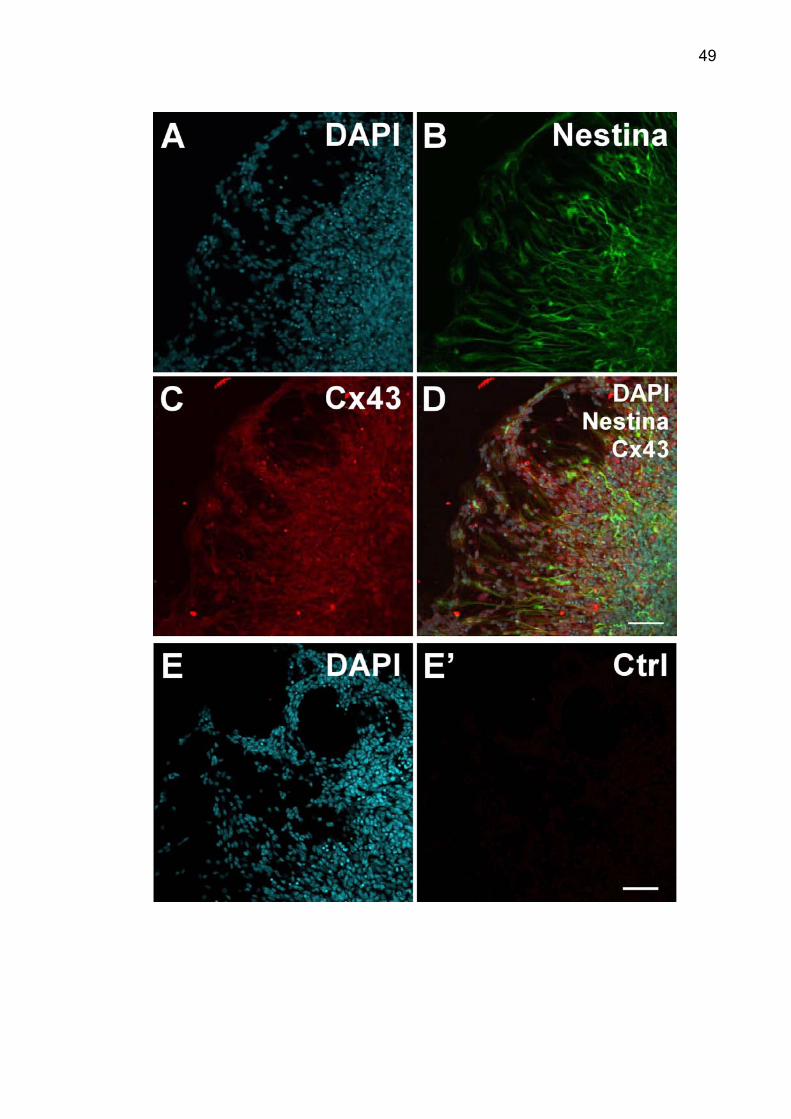
49

50
Figura 14. Imagens de microscopia confocal de explante de SVZb no 3DIV (A-E’). Fotomicrografias de fluorescência de imunomarcação contra Nestina (verde) e conexina 43 (Cx43 - vermelho). A. A contracoloração com DAPI revela a forma e a densidade celular do halo migratório do explante. B. A marcação com Nestina revela a presença abundante de glia radial e astrócitos em nossos explantes. C. Grande parte, senão todas as células apresentaram-se marcadas para Cx43. D. Tripla exposição para Nestina, Cx43 e DAPI. E. Contracoloração do explante utilizado como controle do experimento de imuno-histoquímica. E’. Imagem da fluorescência do controle (ausência dos anticorpos primários). Barra de calibração 50µm.

51

52
Figura 15. Imagens de microscopia confocal de explante de SVZb no 3DIV (A-F). Fotomicrografias de fluorescência de imuno-marcação contra Nestina (verde) e conexina 43 (Cx43 - vermelho). A. Imagem de tripla exposição (vermelho, verde e azul, contracoloração com DAPI) do halo migratório do explante. Os retângulos indicam as regiões selecionadas para as imagens de maior aumento. B. Célula mitótica, com características de glia radial, imuno-positiva para Nestina e Cx43. O retângulo indica a região selecionada para separação da marcação de cada um dos reagentes. B’. Imuno-marcação de Nestina. B’’. Imuno-marcação para Cx43. Note que as células negativas para Nestina também apresentam marcação para Cx43. B’’’. Contracoloração com DAPI. C. Região contendo célula fortemente marcada para Nestina (seta). D. Imuno-marcação para Nestina. O sombreado verde que envolve alguns corpos celulares é marcação inespecífica. As células marcadas apresentam prolongamentos fortemente fluorescentes. E. A marcação da Cx43 revela um pontilhado abundante que reveste praticamente todas as células do explante. F. Dupla exposição de Nestina e Cx43. Note que embora todas as células sejam Cx43 positivas, nem todas apresentam reatividade para Nestina (cabeças de seta). Barra de calibração 50µm (A) e 10µm (B-F).

53
4 – Apesar da expressão elevada, a Cx43 não afeta significativamente a
migração celular nos explantes:
Realizamos dois experimentos de cultura de explantes de SVZ de
camundongos mutantes que não possuíam a Cx43. Devido aos altos níveis de
expressão desta conexina nos mais diversos sistemas do organismo, atingindo
principalmente o coração, os animais nascem com inúmeras deficiências e morrem
em poucos minutos. Para contornar o problema, utilizamos animais embrionários
pouco antes do término da gestação (décimo nono dia embrionário – E19). O
cérebro diminuto e a SVZ precoce impediram a separação das duas regiões
estudadas neste trabalho, fornecendo apenas explantes de SVZ.
O primeiro experimento foi perdido quando passaria pela terceira rodada de
medição dos halos migratórios, ainda sem acusar qualquer discrepância entre
homozigotos sem Cx43 (n=3), heterozigotos ou animais selvagens. O segundo
experimento foi feito em uma ninhada com característica mendeliana (um
homozigoto sem Cx43, dois heterozigotos e um selvagem). Os halos migratórios dos
explantes foram fotografados e medidos nas mesmas condições que os demais
experimentos e a análise estatística não revelou qualquer diferença entre os
explantes dos diferentes animais (Fig. 16).
5 – Bloqueio farmacológico crônico por CBX nos explantes:
No presente trabalho estendemos nossos estudos anteriores sobre o bloqueio
do acoplamento celular com CBX, adicionando mais experimentos e refinando as
análises estatísticas do efeito sobre a migração. Sendo assim, a droga foi adicionada
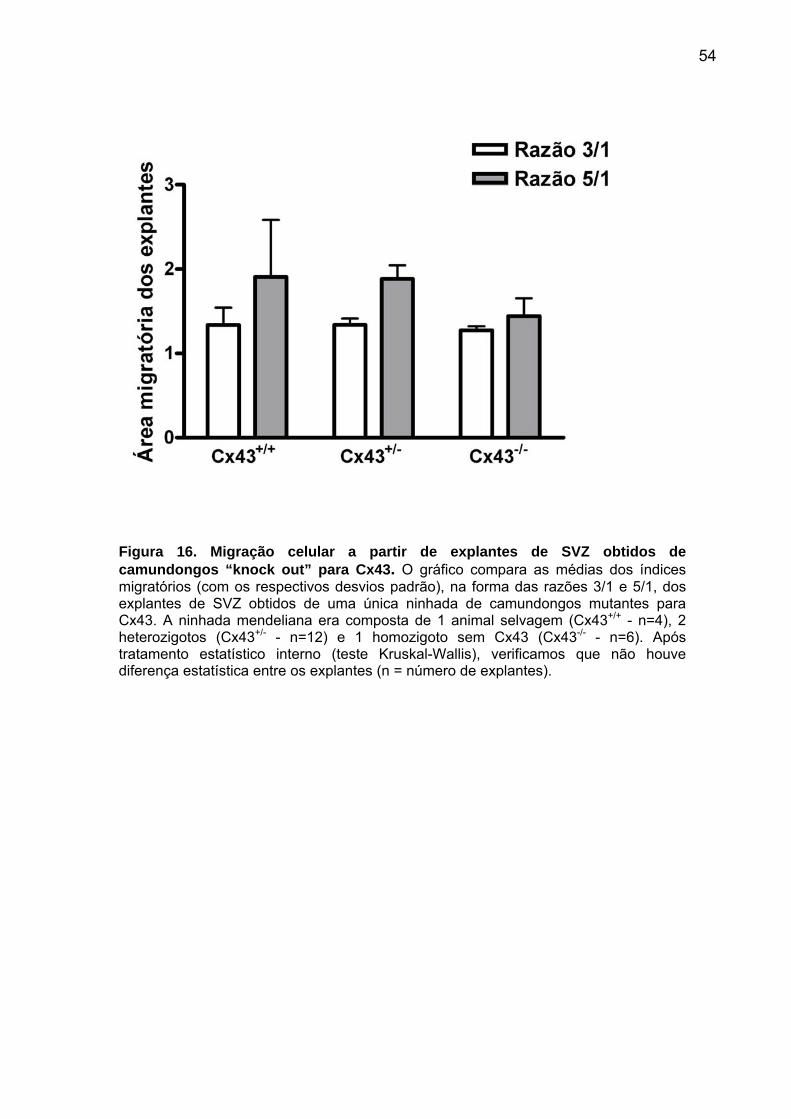
54
Figura 16. Migração celular a partir de explantes de SVZ obtidos de camundongos “knock out” para Cx43. O gráfico compara as médias dos índices migratórios (com os respectivos desvios padrão), na forma das razões 3/1 e 5/1, dos explantes de SVZ obtidos de uma única ninhada de camundongos mutantes para Cx43. A ninhada mendeliana era composta de 1 animal selvagem (Cx43+/+ - n=4), 2 heterozigotos (Cx43+/- - n=12) e 1 homozigoto sem Cx43 (Cx43-/- - n=6). Após tratamento estatístico interno (teste Kruskal-Wallis), verificamos que não houve diferença estatística entre os explantes (n = número de explantes).

55
cerca de 14-18 horas após o plaqueamento e incubação dos explantes, diluída em
meio de cultura normal para uma concentração final de 100µM, e mantida por até
5DIV. A superfície migratória dos explantes foi mensurada em dois ou três
momentos diferentes do seu cultivo in vitro, que incluem obrigatoriamente o final do
primeiro dia, do terceiro e/ou quinto dias após o plaqueamento inicial. A redução da
área de superfície migratória, nas duas regiões estudadas, foi bastante significativa,
como podemos observar nos histogramas da Figura 17. Mais ainda, os explantes de
zona subventricular mostraram-se equivalentemente susceptíveis às ações do
desacoplante (Fig. 17A e B). Este resultado mostrou-se consistente visto que a
variação interna dos explantes dentro de cada experimento foi consideravelmente
baixa, como demonstram os histogramas apresentados nas Figuras 18 e 19 (A e B).
O bloqueio do acoplamento reduziu a média dos halos migratórios em todos os
experimentos e durante todo o tempo da cultura. Um ou outro explante escapa do
tamanho médio esperado para mais ou para menos, mas a maioria permaneceu
bem próxima.
Finalmente, avaliamos as possíveis conseqüências de uma administração
mais tardia de CBX sobre a migração celular em explantes de SVZb e SVZc. O
agente desacoplante foi adicionado ao meio de cultura no 3o DIV e mantido até o 5o
dia, quando a área de superfície migratória foi novamente mensurada e comparada
àquelas registradas no 1o e 3o DIV. A administração de CBX no 3o DIV não afetou o
índice migratório dos explantes, a razão 5/1, que fornece o índice migratório de todo
o período em cultura, indica um aumento significativo do halo migratório em relação
aos três primeiros dias em cultura (Fig. 20A e B). De fato, quando comparamos as
razões 5/1 da administração de CBX no 3o DIV com a razão 5/1 da situação controle,

56
Fig. 17. Migração celular a partir de explantes de SVZ de 3 e 5DIV e os efeitos do agente desacoplante CBX. No quadro superior estão representados o perfil de explantes da SVZb e SVZc em situações controle e em presença de CBX, após 5 dias in vitro (DIV). Os valores das áreas apresentados abaixo foram obtidos a partir destes perfis. Nos gráficos abaixo, os valores são expressos como razões das áreas medidas ao final do 1o, 3o e 5oDIV, tomando como normalizador o 1oDIV. Representam portanto, valores adimensionais e são referidos como quocientes 3/1 e 5/1, conforme indicado nos gráficos. A e B) Medidas comparativas de explantes de SVZb (A) e SVZc (B), nas condições controle e CBX, expressas como razões 3/1 e 5/1. As razões representam médias e respectivos desvios padrão, calculados a partir de 4 a 12 explantes diferentes, de um total de 3 experimentos independentes. Os asteriscos indicam significância estatística na condição CBX 100µM frente aos respectivos controles (teste Mann-Witney; H0 para valores de p<0,05). Desta forma temos: p=0,0004 (SVZb) e p<0,0001 (SVZc) para razões 3/1; p=0,0059 (SVZb) e p=0,0124 (SVZc) para razões 5/1. Barra de escala 100µm.

57
Figura 18. Comparação da variação dos índices de crescimento dos explantes de SVZb, entre experimentos independentes. Símbolos diferentes (quadrado, triângulo e círculo pretos) identificam os diferentes experimentos e as cores (quadrados verde e vermelho) indicam as condições experimentais (controle ou CBX), a superposição das cores resulta na coloração preta. A) Até o 3oDIV, a maioria dos explantes controle (símbolos verdes) variou entre 2 e 4, com apenas três explantes acima deste patamar. Já os explantes CBX (símbolos vermelhos) apresentaram índice de crescimento ainda mais uniforme, com a maioria abaixo de 2. B) Os explantes controle até 5DIV apresentaram índices de crescimento bastante variáveis, com a maioria concentrada entre 3 e 9. Aqueles que ficaram em presença de CBX até o 5oDIV apresentaram índice de crescimento variável, com a maioria entre 1 e 3. As barras coloridas horizontais representam as médias das respectivas razões dentro de cada experimento. Note que apesar de alguma sobreposição entre as razões migratórias dos explantes tratados e controle, tanto as médias das razões 3/1 quanto as 5/1 dos explantes com CBX ficaram sempre abaixo dos controles equivalentes.

58
Figura 19. Comparação da variação dos índices de crescimento dos explantes de SVZc, entre experimentos independentes. Os símbolos identificam os diferentes experimentos e as cores (quadrados coloridos) indicam as condições experimentais (a superposição das cores resulta na coloração preta). A) Os explantes controle até o 3oDIV apresentaram crescimento menor que os de SVZb (V. Fig. 19), com a maioria se concentrada entre 1 e 3, e com apenas um acima de 4. Os explantes CBX variaram pouco entre si, da mesma forma que os de SVZb. Com índices bastante semelhantes, a maioria se concentrou abaixo de 2. B) Explantes controle até 5DIV apresentaram crescimento e variabilidade menores que os de SVZb, com a maioria se fixando entre 2 e 6. Enquanto que aqueles que ficaram em presença de CBX até o 5oDIV variaram da mesma forma que os de SVZb. Com índices bastante semelhantes, a maioria também se concentrou entre 1 e 3. Novamente as médias dos explantes com CBX (barras vermelhas) ficaram sempre abaixo das médias controle (barras verdes) dentro de cada experimento.

59
Figura 20. O bloqueio do acoplamento celular a partir do 3oDIV não tem efeito sobre a migração celular a partir de explantes de SVZ. A-B) Medidas comparativas (com os respectivos desvios padrão) de explantes da SVZb (A) e SVZc (B) em situações controle, do 1o ao 3o dia em cultura e em presença de CBX, do 3o ao 5o dia in vitro (DIV). C) Comparação da razão 5/1 obtida para os explantes de SVZb e SVZc nas condições controle e CBX 100µM ministrado no 3oDIV. Os asteriscos indicam significância estatística da razão 5/1 frente à razão 3/1 correspondentes. A e B) Teste T de Students pareado (H0 para valores de p<0,05), com p=0,0022 (A) e p<0,0001 (B). C) Teste Mann-Witney (H0 para valores de p<0,05)revelou ausência de diferença significativa entre as condições experimentais.

60
confirmamos a ausência de alterações significativas no crescimento migratório dos
explantes de SVZ (Fig. 20C).
Em conjunto, estes resultados fornecem um forte indício de que o
acoplamento pode ser importante para o fenômeno de inicialização da migração em
nossos explantes.
5a – A especificidade do CBX:
Para testar especificidade dos efeitos do CBX, utilizamos seu análogo sem
efeito, o ácido glicirrízico (GZA – V. Mat. e Met. Item 6), para tratamento crônico até
o 3oDIV. Em razão do GZA não ser hidrossolúvel como o CBX, utilizamos seu
veículo, uma mistura de DMSO e etanol, adicionado ao meio de cultura para
situação controle. A presença do GZA ou de seu veículo não afetou
significativamente os explantes de SVZ, quando comparados às situações controle e
CBX (Fig. 21A e B). Apenas na condição CBX os explantes migraram
significativamente menos que em presença de GZA ou de seu veículo. Além disso,
ensaios com vídeo-microscopia de tempo intervalado (V. adiante), utilizando
halotano, outro conhecido bloqueador juncional, também confirmaram o efeito do
CBX sobre o acoplamento mediado por junções comunicantes, além de permitirem a
monitoração da baixa toxicidade do CBX e do GZA (não mostrado) sobre explantes
da SVZ. Dessa forma, este conjunto de resultados sugere que o tratamento com
CBX afetou o acoplamento juncional e não alterou a viabilidade dos explantes.

61
Figura 21. O análogo inativo do agente desacoplante CBX, o GZA, não interfere com a migração celular a partir de explantes de SVZ. A-B) Os explantes cultivados com GZA (100µm) ou seu veículo (DMSO/Etanol) apresentaram índices migratórios muito semelhantes aos observados na situação controle. Note que as razões 3/1 CBX são as mesmas daquelas apresentadas nos histogramas da figura 17. Cada condição experimental apresenta os respectivos desvios padrão. Os asteriscos indicam significância estatística na condição CBX frente às demais condições de cultura (teste ANOVA; H0 para valores de p<0,05).

62
5b - O efeito do CBX é reversível:
Embora resultados prévios com “Acridine Orange”, marcador de viabilidade
celular, não revelarem qualquer diferença, nos níveis de morte celular, entre
explantes controle e em presença de CBX (Marins, 2001), Mantivemos e
completamos os experimentos para os ensaios de reversão. Tais ensaios são muito
importantes quando o experimento envolve a utilização de fármacos, pois quando a
reversão é possível, vemos diminuir a chance de um efeito devido a toxicidade da
droga. Para avaliar as conseqüências da remoção do CBX sobre a migração celular
nos explantes de SVZ,o tratamento com a droga foi mantido até o final do 3o DIV,
quando o meio com CBX foi substituído por meio normal. Todos os explantes de
SVZ apresentaram reversão significativa dos efeitos do bloqueio juncional (Fig. 22A
e B). O halo migratório apresentou recuperação considerável desde a retirada da
droga até o último dia de cultivo (Fig. 22A e B). No entanto, a reversão da área
migratória para níveis equivalentes aos níveis controle pôde ser constatada em
razões 5/1 somente nos explantes de SVZc (Fig. 22C). Embora os explantes de
SVZb tenham apresentado reversão do bloqueio do acoplamento, a velocidade ou
capacidade migratória das células foi aparentemente menor que nos explantes de
SVZc, impossibilitando o retorno dos halos migratórios a níveis próximos dos
controles (Fig. 22C). Sendo assim, a razão média de área migratória 5/1 registrada
na condição CBX reversão para SVZb equivale àquela determinada na condição
CBX mantido por 5 dias, conforme podemos verificar pela comparação com a figura
17A.

63

64
Figura 22. Remoção do agente desacoplante CBX no 3o dia in vitro (CBX reversão) reverte seus efeitos sobre a migração celular nos explantes de SVZ. Fotomicrografias: A) Fotomicrografia de contraste de fase de um explante controle de 3DIV. B, B’) Imagem de contraste de fase de um mesmo explante tratado com CBX até 3DIV (B) e dois dias após a retirada do CBX (B’). Gráficos: A-B) Medidas comparativas de explantes de SVZb (A) e SVZc (B), nas condições CBX 100µM (do 1° ao 3° DIV), e a variação total da área do explante (do 1° ao 5° DIV) expressas como razões 3/1 e 5/1, respectivamente. C) Comparação da razão 5/1 obtida para os explantes de SVZb e SVZc nas condições controle e reversão. As razões representam médias e respectivos desvios padrão, calculados a partir de 4 a 12 explantes diferentes, de um total de 3 experimentos independentes. Embora os explantes de SVZb não retomem os índices migratórios esperados para um explante controle, como os explantes de SVZc, a migração celular é reestabelecida, contudo lentamente. Os asteriscos indicam significância estatística entre a razão 5/1 em relação à razão 3/1 (teste T de Student pareado; Ho para valores de p<0,05), sendo p<0,0001 (A) e p= 0,0004 (B). Em C, utilizamos o teste T de Student não pareado (Ho para valores de p<0,05), com p=0,0054. Barras de escala de 100µm.

65
6 – O bloqueio farmacológico inibe a migração diretamente:
Para responder se o acoplamento estaria interferindo diretamente na
migração, ou indiretamente via proliferação celular realizamos ensaios de vídeo
microscopia de tempo intervalado, que possibilita acompanhar o efeito do
bloqueador juncional, sobre o comportamento migratório das células dos explantes,
em tempo real. Se as células pararem de migrar em poucos minutos poderemos
concluir que o acoplamento interfere diretamente na migração, caso o efeito do
bloqueador só possa ser observado após algumas horas, teremos grandes chances
do acoplamento agir sobre outros processos celulares, como a proliferação, que
eventualmente poderim intervir sobre a a migração ou sobre o número de células
que iniciam o processo de migração. Imagens de explantes representativos de SVZb
e SVZc, cultivados sobre poli-L-lisina, foram capturadas por aproximadamente nove
horas (9h), sendo 3h em situação controle, 3h em presença do CBX e 3h após a
remoção do agente desacoplante (V. Mat. e Mét. Item 9). A adição de CBX,
interrompeu a migração celular sem provocar, no entanto, grandes alterações
morfológicas. Após a lavagem do CBX, a migração foi restabelecida em alguns
minutos (Fig. 23).
De forma semelhante, explantes tratados com halotano (vol/vol) também
apresentaram um importante arresto da migração celular. Diferentemente do CBX, a
instalação deste efeito foi quase que instantânea, refletindo as diferentes ações
destes agentes sobre as junções comunicantes (Spray et al., 2002). Nos
experimentos com halotano a migração retornou a um ritmo aparentemente normal
após uma hora de monitoramento, de forma espontânea e progressiva. Acreditamos
que esta recuperação seja devido à grande volatilidade deste agente desacoplante.

66
Figura 23. Imagens de vídeo-microscopia de tempo intervalado de explante de SVZc. Na seqüência controle, três células escolhidas tiveram sua movimentação acompanhada durante 2h, em intervalos de 20 min. Uma hora após a adição de CBX, outras três células foram acompanhadas por uma hora e dez minutos, com intervalos de 10 min. Após a retirada do CBX, acompanhamos as mesmas três células da sequência anterior até 40 min., quando a célula mais externa se confundiu com as que estavam à sua frente. Uma hora depois da retirada do CBX outra célula (verde) foi adicionada às demais. Vinte minutos depois, mais uma célula foi adicionada e acompanhada até o final da análise (após 2h com intervalos de 20 min). Note que a movimentação celular presente na situação controle cessou em presença do bloqueador juncional e foi recuperada após o retorno à condição controle. Experimento representativo de 4. Barra de escala 50µm.

67
(Estes resultados estão apresentados de forma qualitativa nos filmes do material
suplementar em anexo).
O mesmo procedimento de captura de imagens foi utilizado para explantes de
SVZc cultivados sobre Matrigel, que fornece um substrato tridimensional para o
explante. Esses explantes pareceram ter sua velocidade migratória aumentada em
relação àqueles cultivados sobre poli-L-lisina (substrato bidimensional). Além disso,
as cadeias migratórias foram muito mais numerosas sobre o matrigel e praticamente
sem glia subjacente, apesar do aspecto geral do explante não ter sofrido qualquer
alteração. A migração foi afetada pelo CBX da mesma forma que nos ensaios
anteriores, no entanto, o aumento aparente da velocidade migratória tornou mais
evidente a parada drástica da migração quando em presença do agente
desacoplante.
Problemas de foco na vídeo microscopia (resultantes da tridimensionalidade
das cadeias), fizeram com que a investigação do movimento individual dos
neuroblastos foi feita em um número reduzido de células, o que pode causar um erro
devido ao viés causado pelo tamanho da amostra. Por isso, analisamos também o
movimento e a direção total das células em secções das cadeias dos explantes.
Acompanhamos a movimentação individual de algumas células (n=15) a cada hora
de cada uma das três situações estudadas na vídeo microscopia do explante sobre
Matrigel (Fig. 24). As células escolhidas estavam nas cadeias que apresentaram
maior mobilidade e precisavam estar visíveis na primeira imagem (V. Mat. e Met.
item 10). A medição do deslocamento de cada célula permitiu o cálculo da
velocidade média e a representação gráfica da progressão do movimento ao longo
do tempo (Figs. 25 à 27). Em situação controle, as células apresentaram uma
grande variação em sua movimentação, com algumas lentas e outras rápidas

68

69
Figura 24. O CBX afeta diretamente a movimentação celular nos explantes de SVZ. Imagens de ensaio de vídeo microscopia de tempo intervalado com explante de SVZc cultivado por 2 DIV com Matrigel. As fotomicrografias maiores revelam o aspecto inicial do explante em cada etapa do experimento. Os pequenos retângulos vermelhos indicam uma das regiões escolhidas para acompanhamento individual da movimentação celular e representam a origem das fotos menores. As fotos menores compõem os primeiros dezoito minutos da primeira hora de cada situação experimental, cada círculo colorido indica a célula cuja movimentação foi acompanhada. No período pré-carbenoxolone (Pré-CBX), algumas células apresentaram movimentação enquanto outras permaneceram praticamente paradas. Na presença do CBX, a movimentação da maioria das células ficou bastante reduzida. Já na situação Pós-CBX, as células tornam a se movimentar de modo semelhante ao observado na situação controle. Algumas células não puderam ser acompanhadas por todo o período do experimento por que com sua movimentação e mudança de plano focal estas desaparecem a medida que se confundem com outras, impedindo sua identificação. As barras de escala são de 50 (foto maior) e 20 µm (foto menor).

70

71
Figura 25. Trajetórias e deslocamento das células no halo migratório de um explante de SVZc em matrigel, antes da adição de CBX. Análises de movimentação celular individual a partir de ensaio de vídeo microscopia de tempo intervalado. Os retângulos à esquerda contêm as trajetórias das células analisadas em cada hora (indicada pelo número acima e à esquerda) de filmagem durante o período pré-CBX. A posição das células a cada 2 minutos está representada por círculos de cores diferentes para cada célula individual analisada. O maior círculo representa a posição inicial de cada célula. As células escolhidas para análise, durante a primeira hora, não foram as mesmas da segunda ou terceira hora. Os gráficos da coluna da direita contém o deslocamento das células dos respectivos retângulos. As cores das linhas nos gráficos correspondem às cores dos círculos. As médias das velocidades médias de cada grupo celular estão indicadas em µm/min dentro da área dos gráficos. Enquanto algumas células se deslocam mais rapidamente, outras o fazem de modo um pouco mais lento, acarretando uma grande variação no comportamento migratório.

72

73
Figura 26. Trajetórias e deslocamento das células de explante de SVZc em matrigel, na presença de CBX. Análises de movimentação celular individual a partir de ensaio de vídeo microscopia de tempo intervalado. As células escolhidas para análise, durante a primeira hora, foram as mesmas da segunda e terceira hora. O número de células foi diminuindo a medida que tornou-se impossível identificá-las com segurança. As cores das linhas nos gráficos correspondem às cores das bolinhas. As médias das velocidades médias de cada grupo celular estão indicadas em µm/min dentro da área dos gráficos. A presença do CBX reduziu significativamente a movimentação celular e homogeneizou o comportamento migratório, reduzindo drásticamente a variação. O rápido curso temporal deste efeito é consistente com a ação direta das junções comunicantes na migração celular.

74
presentes em cada uma das três horas de filmagem (Fig. 25). Do momento em que é
colocado até surgir seu efeito, o CBX leva aproximadamente 20 minutos para
bloquear a migração. Este retardo na iniciação do bloqueio fica claro ao
compararmos o gráfico da primeira hora com os gráficos das outras duas horas de
filmagem (Fig. 26). Além de bloquear a migração celular, o CBX reduziu a variação
do comportamento migratório das células, homogeneizando-o (V. segunda e terceira
hora da Fig. 26). Quando o CBX é retirado e o meio controle restituído, as células
começam a se movimentar quase que imediatamente e não pudemos observar
nenhum tipo de atraso na inicialização da movimentação. Contudo, a variação do
comportamento migratório permaneceu reduzida, com as células se movimentando
homogeneamente mais rápido (Fig. 27).
As culturas crônicas dos explantes de SVZ revelaram a presença de migração
centrífuga, nos levando a crer que esta seria a direção migratória adotada pelas
células. No entanto, os ensaios de vídeo-microscopia possibilitaram acompanhar os
movimentos migratórios de células individuais, de modo que as direções migratórias
não indicaram nenhum tipo de preferência. Mesmo assim restava-nos analisar a
direcionalidade do movimento no explante com Matrigel. Devido ao grande número
de células em movimento, o aspecto tridimensional das cadeias (dificultando a
individualização das células) e o longo tempo intervalo (2 minutos), utilizamos um
critério subjetivo de análise de movimento aparente. Para diminuir o erro, três
investigadores independentes contaram células das mesmas regiões com alta
correlação entre eles: 0,87 e 0,75 em situação controle; 0,97 e 0,96 em presença de
CBX; e 0,91 e 0,77 após a retirada do CBX (onde -1≤ p ≤1). Além disso, o teste
estatístico Kruskal-Wallis não apontou qualquer diferença significativa entre eles. O
movimento para fora do explante foi chamado de migração centrífuga e o inverso,
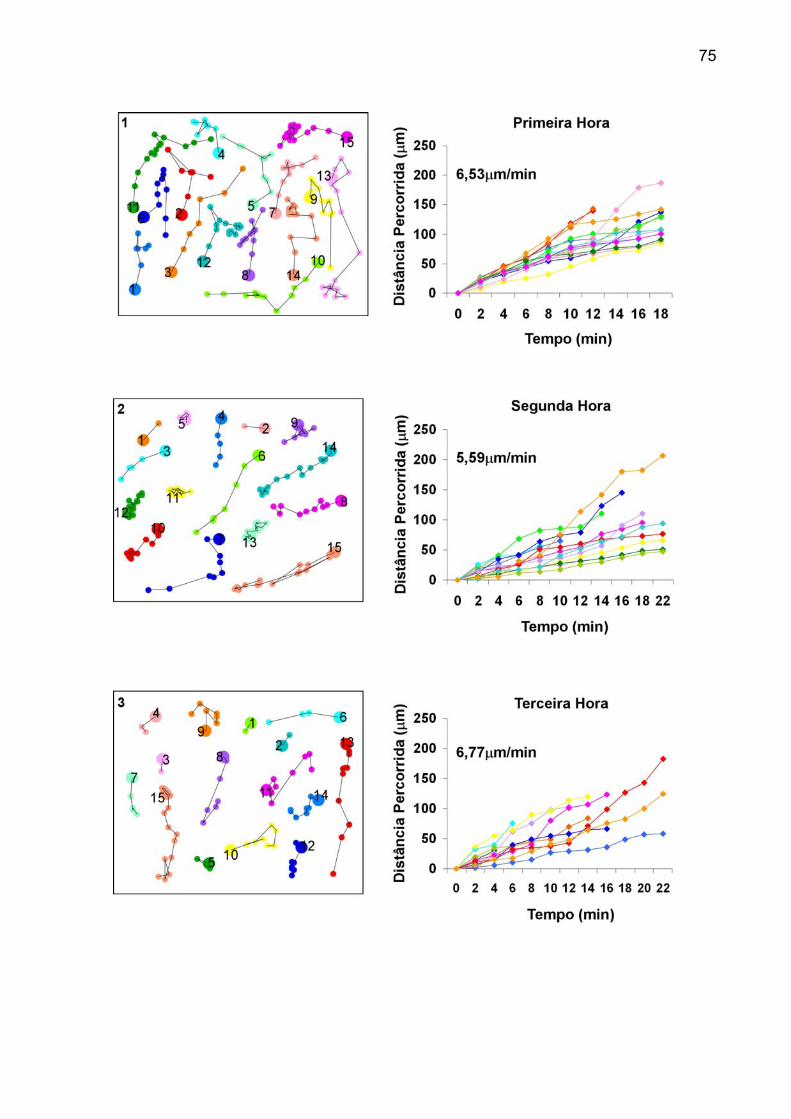
75

76
Figura 27. Trajetórias e deslocamento das células de um explante de SVZc em matrigel, após a retirada do CBX. Análises de movimentação celular individual a partir de ensaio de vídeo microscopia de tempo intervalado. As células escolhidas para análise, durante a primeira hora, não foram as mesmas da segunda ou terceira hora. Logo após a retirada do CBX as células retornaram a sua atividade migratória, de forma generalizada, mas não aos níveis pré-CBX, com a velocidade média ligeiramente inferior ao controle. No conjunto as células apresentaram reduzida variação no comportamento migratório visto que a grande maioria passou a se movimentar rapidamente.

77
migração centrípeta. Apesar de haver uma tendência para a migração centrífuga em
todas as condições experimentais, esta direção é significativamente preferencial
apenas após a retirada do CBX (Fig. 28). Este resultado explica a retomada da
migração dos explantes de SVZc, após a reversão, para índices semelhantes ao
esperado para explantes controles (V. Fig. 21). Em conjunto, estes resultados
sugerem uma relação direta do acoplamento celular com a migração, nos explantes
de SVZ.
Demonstramos uma relação direta do acoplamento com a migração celular na
SVZ, aliada à baixa toxicidade dos agentes farmacológicos. Temos, no entanto,
indícios de que agentes desacoplantes como o CBX, testado em nossos ensaios,
interfira em outros eventos celulares. Nos ensaios de vídeo microscopia, utilizando
explantes cultivados sobre poli-L-lisina, acompanhamos modificações morfológicas
sofridas por células gliais subjacentes à extremidade das cadeias migratórias. Em
situação controle, a dinâmica de alterações morfológicas nestas células restringiu-se
a raras expansões e retrações protoplasmáticas, típicas de astrócitos mantidos em
cultura (Fig. 29). No entanto, com a adição de CBX, freqüentemente observa-se
transições morfológicas dramáticas de células gliais em extremidade de cadeias,
durante as quais, estas assumiam aspecto fusiforme a fibroso. Por fim, a retirada do
desacoplante, restaurava o aspecto protoplasmático, testemunhando a
reversibilidade destas alterações.

78
Figura 28. O bloqueio do acoplamento juncional diminui a atividade migratória em geral e sem apresentar preferência direcional neste efeito. Contagens de movimento celular aparente, feitas em ensaio de vídeo microscopia de tempo intervalado com explante de SVZc, cultivado por 2 DIV sobre Matrigel. O movimento celular aparente foi avaliado por três investigadores independentes em cinco cadeias migratórias para cada situação experimental. Foi contabilizada a movimentação total que ultrapassasse uma barra, de aproximadamente 5 µm de comprimento e espessura negligivel, colocada transversalmente sobre a cadeia a ser avaliada. A correlação entre os investigadores foi de 0,87 e 0,75; 0,97 e 0,96; e 0,97 e 0,77 nas condicoes pré-CBX, CBX e pós-CBX, respectivamente. Além disso, a comparação estatística entre eles (teste Kruskal-Wallis) não apontou qualquer diferença significativa Parece haver uma tendência para a migração centrífuga em detrimento da centrípeta, mas só a condição pós-CBX apresentou diferença significativa entre as duas direções. Note que, novamente a presença do CBX diminui fortemente a movimentação celular no explante. O asterísco indica significância estatística entre as direções centrífuga e centrípeta (p=0,0199; teste T de Student nãopareado; H0 para valores de p<0,05), que ocorreu apenas na condição Pós-CBX.

79
Figura 29. Transformação de célula glial nos explantes de SVZc sobre poli-L-lisina tratados com CBX. Seqüência de cinco imagens de um ensaio de vídeo microscopia de tempo intervalado focalizando um grupo de cadeias no halo miratório de um explante de SVZc, com 3 DIV, cultivado sobre poli-L-lisina. Durante as três primeiras horas de filmagem, antes da adição do CBX, a morfologia de uma célula glial (cabeça de seta) localizada na extremidade de uma das cadeias migratórias apresenta uma grande mobilidade de seus prolongamentos, mas mantém uma morfologia protoplasmática típica de astrócitos em cultura. A adição de CBX modifica a morfologia da célula para um aspecto fusiforme alongado, concomitante com um colapso da cadeia a que se relacionava. Após a retirada do CBX, a morfologia da célula glial retoma um aspecto protoplasmático. Barra de escala de 50µm.

80
V – Discussão:
Nesta tese, apresentamos as primeiras evidências diretas de envolvimento
positivo de junções comunicantes na migração celular de neuroblastos no SNC.
Utilizando uma preparação in vitro, confirmamos o envolvimento das junções
comunicantes na migração das células oriundas da porção anterior da SVZ pós-
natal. Numa primeira etapa, a mediação de junções comunicantes neste fenômeno
foi demonstrada, em culturas de longa duração, pela importante redução da
migração celular em presença do agente desacoplante carbenoxolone (CBX). A
especifidade do CBX foi confirmada pela utilização de seu análogo sem efeito sobre
junções comunicantes, ácido glicirrízico (GZA). Além disto, o efeito deste agente
desacoplante foi reversível e nenhuma alteração de viabilidade foi detectada
(Marins, 2001), afastando a possibilidade de ação inespecífica. Numa segunda
etapa, em ensaios de vídeo-microscopia com o tratamento agudo com CBX e outro
agente desacoplante, o halotano, demonstramos que o acoplamento juncional afeta
a migração celular diretamente. Nestes ensaios, foi possível observar a suspensão
da migração em uma cinética correspondente à cinética de ação farmacológica dos
agentes empregados sobre as junções comunicantes. As parcerias celulares
estabelecidas nos sincícios de comunicação juncional definidos na SVZ foram
reveladas em explantes derivados de várias porções da SVZ pós-natal mediante a
combinação de técnicas de injeção iontoforética de corantes com imuno-
histoquímica para marcadores fenotípicos. Além disto demonstramos, com a técnica
de imuno-histoquímica, que a Cx43, a conexina mais abundante durante o
desenvolvimento do SNC, distribui-se por todos os tipos celulares presentes nos
explantes. Apesar de sua expressão abundante, outras conexinas podem estar
envolvidas na formação das redes de acoplamento juncional na SVZ, já que nossos

81
ensaios com camundongos nocaute Cx43 não revelaram alterações significativas do
comportamento migratório em explantes da SVZ. Em conjunto com a descrição
prévia da presença de conexinas e de acoplamento celular na SVZ/RMS (Menezes
et al., 2000; Marins, 2001, Liu et al., 2006) estes resultados sugerem fortemente que
estas junções cumprem um importante papel na fisiologia desta região neurogênica.
1 – A cultura de explantes parece reproduzir a fisologia da SVZ/RMS in vivo:
Nosso modelo experimental é amplamente descrito e utilizado por vários
grupos (Hu et al., 1996; Wichterle et al., 1997; Zhu et al., 1999; Lim et al., 1999,
Mason et al., 2001; Bolteus e Bordey, 2004; Sailer et al., 2005). A descrição de uma
grande migração centrífuga a partir do explante inicial é uma característica comum a
todos estes grupos. Este modelo já foi utilizado para estudar a dinâmica desta
migração (Wichterle et al., 1997; Mason et al., 2001, Bolteus e Bordey, 2004, Bolteus
et al., 2005, Koizumi et al., 2006) bem como os fatores moleculares que influenciam
no fenômeno (Hu et al., 1996; Zhu et al., 1999; Miyakoshi et al., 2001; Bolteus e
Bordey, 2004, Bolteus et al., 2005, Koizumi et al., 2006). Apesar desta característica
em comum, pequenas variações experimentais produzem explantes com diferenças
de extensão do halo de migração, tipos celulares e forma da cadeia migratória.
Como exemplo, nos explantes cultivados sobre matrigel (fórmula comercial de
substrato adesivo tridimensional para cultura de células composta de uma mistura de
colágeno, vitronectina e outros fatores), um ambiente tridimensional é fornecido e
supostamente favorece a formação de extensas cadeias celulares a partir do
explante, com pouca ou nenhuma presença de células gliais (Wichterle et al., 1997).
Estas características também foram observadas em nossos explantes cultivados

82
com matrigel. Além disso, a velocidade migratória aumentou, o que é uma resposta
celular característica de células de origem neural para este substrato, como
recentemente descrito na literatura (Wichterle et al., 1997; Kleinman e Martin, 2005).
Nos ensaios utilizando a poli-L-lisina como substrato, observamos a formação inicial
de um halo migratório homogêneo, com cadeias migratórias individualizadas apenas
em sua porção mais externa e proporção significativa de células gliais. Acreditamos
que este padrão é conseqüência do plaqueamento sobre lamínulas pré-tratadas com
poli-L-lisina, que não fornece um ambiente tridimensional. Este padrão é mais
parecido com o obtido por Mason e cols. (2001) com explantes cultivados sobre poli-
D-lisina.
Em nossos explantes conseguimos repetir várias das particularidades
publicadas e principalmente reproduzimos as muitas características da SVZ durante
a primeira semana pós-natal, sendo elas: 1. extensa migração celular; 2. formação
de cadeias migratórias compostas de células semelhantes entre si (neuroblastos
βIII+); 3. presença de células de glia radial; 4. acoplamento celular mediado por
junções comunicantes; 5. proliferação celular mantida (pelo tempo de duração da
cultura; Marins, 2001). Em estudo prévio os tipos celulares majoritariamente
encontrados nos explantes foram neuroblastos (células βIII+, alguns nestina*) e
células gliais (GFAP+, fortemente nestina+), ambos proliferativos (BrdU+) como já
descrito na literatura (Menezes et al., 1995; Doetsch et al., 1997; Luskin et al., 1997;
Lim et al., 1999).
O uso de agentes farmacológicos como principal método de bloqueio do
acoplamento celular, leva ao questionamento sobre a especificidade da droga. A
ausência de inespecificidade do CBX tem sido defendida por sua ação direta sobre
os conexons (Goldberg et al., 1996). Verificamos a especificidade do CBX através de

83
ensaios crônicos e agudos com o ácido glicirrízico (GZA), análogo sem efeito sobre
o acoplamento. Tanto o CBX como o GZA são conhecidos por interagirem com
receptores para glicocorticóides e mineralocorticóides (Bani-Yaghoub et al., 1999),
mas esses receptores não medeiam a redução do acoplamento causada pelo CBX
(Davidson et al., 1986). Além disso, o tratamento com CBX não afetou a viabilidade
dos explantes porque: 1. as células continuaram migrando e proliferando, mesmo
que em menor proporção; 2. o tratamento com CBX foi reversível; 3. o acoplamento
celular também estava presente, sendo que de forma reduzida; 4. Em estudos
prévios não encontramos sinais de degeneração ou morte apoptótica nas células
dos explantes tratados ou não (Marins, 2001).
Em conjunto estes dados sugerem que o modelo de explante, apesar de
perder a complexa organização tridimensional encontrada in situ reproduz muitas
características encontradas in vivo, e sugerem fortemente que as junções
comunicantes devem cumprir um importante papel na regulação da migração
tangencial desta região.
2 - Acoplamento celular nos explantes de SVZ:
Resultados anteriores de nosso grupo mostraram a presença do acoplamento
funcional na SVZ in situ e in vitro (apenas no halo migratório dos explantes foi
analisado) (Menezes et al., 2000; Marins, 2001). Em ambos os casos o extenso
acoplamento parecia envolver células astrogliais e precursores neuronais. A técnica
de injeção intracelular de fluorocromos (tb. apenas realizada no halo migratório),
utilizada neste trabalho, veio esclarecer os possíveis pareamentos celulares
presentes em nossos explantes de SVZ. Utilizando critérios morfológicos aliados a

84
critérios de imunorreatividade célula-específica, encontramos acoplamento homo- e
heterocelulares, calculamos a incidência relativa destes pareamentos e verificamos
que: o acoplamento homocelular foi mais freqüente que o heterocelular; a incidência
do acoplamento heterocelular foi eqüivalente à ausência de acoplamento; e o
acoplamento homocelular entre neurônios foi maior na SVZb que na SVZc,
ocorrendo o contrário com o acoplamento homocelular astroglial.
A presença de acoplamento homo- e heterocelular nos explantes sugere que
estes possam também estar presentes na SVZ/RMS in situ, por exemplo,
neuroblastos-neuroblastos e/ou neuroblastos-glia radial ou mesmo envolvendo
outros precursores. No entanto, apesar da clara diferença morfológica entre as
células classificadas como astroglia e neurônios/neuroblastos, e da ratificação
parcial por imuno-histoquímica, nossos critérios de identificação celular podem
introduzir um grau de incerteza em nossas análises, quanto à determinação precisa
do fenótipo. Mesmo assim, resultados in situ de nosso laboratório (não publicados) e
de eletrofisiologia em fatias agudas (Liu et al., 2006) revelam a presença do
acoplamento homo- e heterocelular na SVZ.
Não é surpresa detectar a presença de acoplamento homo- e heterocelular
nos explantes ou in situ. O acoplamento celular é uma propriedade marcante de
células neuronais imaturas e de células gliais, durante o desenvolvimento do SNC
(Fróes et al., 1999; Fróes e Menezes, 2002; Bittman et al., 2002; Montoro e Yuste,
2004; Sutor e Hagerty, 2005). Está presente em diversos estágios de diferenciação
celular e maturação histoarquitetônica, pois é detectável desde precursores neurais
(gliais) e neuroblastos, em sítios neurogênicos (Bittman et al., 1997; Bittman e
LoTurco, 1999), até neurônios jovens em fase de sinaptogênese e estabelecimento
da circuitaria do córtex telencefálico (Peinado et al., 1993; Yuste et al., 1995; Bittman

85
et al., 2002; Rouach et al., 2002; Kosaka e Kosaka, 2003). Neurônios pós-
migratórios do córtex cerebral, a princípio em redes homocelulares, participando da
construção da circuitaria sináptica cortical, estabeleceriam domínios, aparentemente
colunares, de acoplamento juncional, com correspondência territorial com os
domínios de co-ativação definidos por flutuações espontâneas de cálcio (Yuste e
Katz, 1991; Yuste et al., 1992; Peinado et al., 1993; Yuste et al., 1995; Katz e Shatz,
1996; Crowley e Katz, 2000; Okada et al., 2003), que contribuiriam para a
estabilização de vias sinápticas envolvendo neurônios de projeção e interneurônios
(Roerig e Katz, 1997, Katz e Crowley, 2002, Fukuda et al., 2006).
No nosso caso, parcerias de acoplamento envolvendo elementos gliais entre
si, neuroblastos ou ambos foram detectadas nos explantes de SVZ, provavelmente
reproduzindo o que ocorre in situ. Apesar de não termos encontrado evidências da
contribuição relativa das diferentes parcerias para o comportamento migratório na
SVZ, apresentamos indícios de que, em seu conjunto, o sincício pancelular na SVZ
convive com a formação do halo migratório e sugerimos que este seja um importante
modulador positivo da migração em cadeia, típica da SVZ (V. adiante).
3 – Fatores envolvidos na migração celular da SVZ/RMS:
A migração na SVZ/RMS tem sido extensivamente estudada in vivo e in vitro
(Doetsch et al., 1997; Fricker et al., 1999; Chazal et al., 2000; Wichterle et al., 2001;
Hoehn et al., 2002; Kakita et al., 2003; Suzuki e Goldman, 2003; Sanai et al., 2004;
Wang et al., 2006; Lim et al., 2006; Gashghaei et al., 2006). A Zona Subventricular é
conhecida por sua migração em cadeias, independente de glia radial e com as
células migrando umas sobre as outras com destino às camadas granular e

86
periglomerular do bulbo olfatório. Vários fatores têm sido implicados na migração
celular desta região: como a netrina, slit/Robo, EGF, FGF, TGFβ, BDNF, PSA-
NCAM, efrinas e seus receptores, gangliosídeo 9-O-acetil GD3, tenascina,
integrinas, BMP/noggina, PTEN, eritropoietina, ativador de proteína G (AGS3),
metaloproteinases (MMP2 eMMP9), GDNF e até mesmo o fluxo de líquido cérebro-
espinhal (Miyakoshi et al., 2001; Menezes et al., 2002; Marshall et al., 2003; Li et al.,
2003; Tsai et al., 2006; Taymans et al., 2006; Sawamoto et al., 2006; Wang et al.,
2006; Paratcha et al., 2006). Esta miríade de moléculas, muitas das quais quando
manipuladas inibem a atividade migratória completamente, sugere uma complexa
regulação desta migração, seja em níveis de interação com o meio extracelular,
interações inter-celulares, de regulação da maquinaria motora celular, do
direcionamento ou da manutenção do estado de diferenciação, entre outros.
Acreditamos que as conexinas interferem principalmente na interação intercelular
que permite a migração “en masse”, típica da migração encontrada na SVZ/RMS,
células de crista neural cardíacas (Huang et al., 1998), e em alguns outros tipos
celulares em processo de invasão metastásica (Friedl et al., 1995; Navolotski et al.,
1997; Lin et al., 2002).
4 – O acoplamento e a migração celular:
De um modo geral o CBX bloqueou a migração celular nos explantes de
SVZb e SVZc sobre lamínulas cobertas com poli-L-lisina ou com matrigel. Este
resultado sugere que o acoplamento celular é um importante fator para a migração
destes neuroblastos. É importante frisar que a ausência de migração pode indicar
não apenas um efeito na migração per se, mas na sua inicialização também.

87
A correlação entre migração e acoplamento juncional não é intuitiva, já que o
deslocamento celular é aparentemente incompatível com o estabelecimento de
contatos juncionais estáveis. Mesmo assim diversos estudos têm sugerido, ao longo
das duas últimas décadas, que os níveis de comunicação juncional afetam ou são
afetados pela atividade migratória celular (Huang et al., 1998; Menezes et al., 2002).
De fato, estudos utilizando células e tecidos neoplásicos têm sido
particularmente úteis como modelos de migração celular. Inicialmente, estes estudos
tinham em comum o estabelecimento de uma correlação negativa entre o grau de
acoplamento juncional homocelular e a invasividade/proliferação tumorais (Elias e
Friend, 1976; Navolotski et al., 1997; Hamada et al., 1998; Naus e Bani-Yaghoub,
1998; McDonough et al., 1999; Li e Herlyn, 2000). Em gliomas, os níveis de conexina
43 e conseqüentemente o grau de acoplamento intercelular encontram-se reduzidos
(Zhu et al, 1991; McDonough et al., 1999), enquanto a superexpressão de conexina
43 induz à reversão do fenótipo tumoral (Naus et al., 1992). Ao comparar o grau de
severidade metastásica entre diferentes linhagens tumorais humanas, Navolotski e
cols. (1997) demonstraram que a redução significativa dos níveis de comunicação
juncional era necessária à ativação de programas de desenvolvimento tumoral.
Estes incluíam não somente a motilidade celular, de caráter migratório, como
também o posterior crescimento (proliferação) das espécies colonizantes.
Estudos recentes, no entanto, colocaram em xeque a hipótese das junções
comunicantes como supressores tumorais (Lin et al., 2002; Zhang et al., 2003;
Pollman et al., 2005). Pollman e cols. (2005) demonstraram que o acoplamento
heterocelular (via Cx43) entre células de tumor de mama e células endoteliais
aumenta a motilidade das primeiras e conseqüentemente a invasibilidade do tumor.
Correlações positivas semelhantes foram extraídas de estudos realizados com

88
tumores de próstata (Pollman et al., 2005; Miekus et al., 2005) e em modelos de
invasividade do córtex por glioma (Lin et al., 2002).
Em outros sistemas, tem-se evidenciado uma correlação negativa entre o
acoplamento e a migração celular, como relatado durante a cicatrização epitelial (Qiu
et al., 2003) ou no desenvolvimento de resposta inflamatória (Zahler et al., 2003). A
dicotomia persiste, no entanto, quanto ao papel da comunicação juncional sobre a
migração celular em sistemas não neurais. Células não-neurais, como células
epiteliais (Pawar et al., 1995; Lampe et al., 1998; Chanson et al., 2005), fibroblastos
(Nagy et al., 1996; Ehrlich e Diez, 2003) e células musculares lisas (Blackburn et al,
1997; Déglise et al., 2005) apresentam também uma correlação positiva entre
acoplamento juncional e migração celular. Pawar e cols. (1995) demonstraram a
existência de um mecanismo de seleção das células epiteliais renais, que
desenvolviam a atividade migratória, mas não a proliferativa, em resposta à necrose
tubular. Na interpretação dos autores, este movimento estava na dependência de
um gradiente de sinalizadores intercelulares, que partiam do ponto da necrose
através dos canais juncionais (Pawar et al., 1995). Este estudo ilustra, portanto, a
possibilidade de independência entre os processos proliferativo e migratório.
Evidências recentes sugerem que a comunicação juncional homo- e/ou
heterocelular, na histogênese do SNC, precede a atividade migratória (Bittman et al.,
1997; Nadarajah et al., 1997). O acoplamento permanece ausente durante a
migração (Bittman et al., 1997; Bittman e Lo Turco, 1999) só retornando
transitoriamente durante a formação da circuitaria cortical (Yuste et al., 1992;
Peinado et al., 1993; Yuste et al., 1995). Desta forma, o acoplamento parece
precisar ser desligado durante a migração neuronal. No entanto, como mencionado
anteriormente Huang e cols. (1998) demonstraram uma relacao positiva entre as

89
conexinas e migracao celular utilizando um modelo de migração de precursores de
neurônios simpáticos cardíacos derivados da crista neural.
5 –O possível papel da conexina 43 com a migração celular:
Nesta tese verificamos a presença abundante da Cx43 tanto em glia quanto
em neurônios/neuroblastos, no halo e nas cadeias migratórias. Estudamos
principalmente a Cx43 por representar o isotipo mais abundante em mamíferos,
estar presente durante o desenvolvimento do SNC, e por termos à época
disponibilidade de animal nocaute Cx43. A Cx43 está presente em quase todos os
tecidos: é a principal conexina do tecido cardíaco e dos astrócitos cerebrais (Beyer
et al., 1987; Giaume et al., 1991; Dermietzel et al., 1991). Diversos trabalhos a
relacionam com eventos do desenvolvimento, como: formação de cartilagens (Zhang
et al., 2002), proliferação celular (Sánchez-Alvarez et al., 2001; Tabernero et al.,
2001; Solan et al., 2003), migração celular (Huang et al., 1998; Li et al., 2002;
Scemes et al., 2003; Fushiki et al., 2003; Oliveira et al., 2005) e diferenciação
(Venance et al., 1995; Duval et al., 2002). Além disso, ainda está relacionada com
outras funções independentes do acoplamento celular, como: a participação de
hemicanais na neuroproteção (Contreras et al., 2004; Lin et al., 2003).
A Cx43 aparece em nichos proliferativos e na placa cortical em formação
durante o desenvolvimento do telencéfalo. Apesar de ser majoritária em astrócitos,
também tem sido relatada em neurônios e neuroblastos, com algumas controvérsias
na literatura (Belliveau e Naus, 1995; Nadarajah et al., 1996; Venance et al., 2000;
Duval et al., 2002; Fonseca et al., 2002).

90
Esta conexina tem sido implicada na formação do sincício astroglial e no
estabelecimento de fluxos transmembrana através do funcionamento na forma de
hemicanais em astrócitos submetidos a estresse experimental (isquemia,
envenenamento metabólico – Contreras et al., 2004). Hemicanais astrocitários
formados por Cx43 têm sido implicados na secreção de neurotransmissores, como
glutamato e GABA e neuromoduladores, como ATP e derivados, NAD+, entre outros
(Goodenough e Paul, 2003). Nesta tese, não há evidência de formação de
hemicanais, ao menos nas condições experimentais testadas, uma vez que não foi
observado o extravasamento de corante para o meio extracelular durante as
injeções iontoforéticas de LY, nem captação inespecífica de corante nos ensaios de
scrape-loading. Apesar disto, não podemos excluir completamente a formação de
hemicanais em nossas preparações, que abrindo-se em número reduzido ou muito
transitoriamente, tenham se tornado indetectáveis por nossos métodos funcionais.
A ampla distribuição da Cx43 em todos os tipos celulares presentes no
explante nos levou a supor que esta poderia constituir um substrato molecular
importante para o estabelecimento de pareamentos homo- e heterocelulares,
reconhecidos em nossos ensaios funcionais. Além disso, por ter sido anteriormente
relacionada à migração celular em outros modelos (Huang et al., 1998; Duval et al.,
2002; Lin et al., 2002; Scemes et al., 2003; Fushiki et al., 2003), poderia ser o isotipo
mais diretamente relacionado com a migração celular em nossos explantes de SVZ.
Contudo, esta relação não foi confirmada em ensaios que realizamos com os
animais transgênicos (Cx43-/- e -/+). É possível que a ausência de Cx43 seja
compensada por outras conexinas (Fushiki et al., 2003; Theis et al., 2005).
Infelizmente não pudemos confirmar a presença de outras conexinas em nosso
modelo até o momento. Acreditamos que outras conexinas estejam presentes em

91
nossos explantes, conforme tem sido constatado em células astrogliais e neuronais
(Dermietzel et al., 2000; Montoro e Yuste, 2004; Sutor e Hagerty, 2005). Portanto,
dadas às limitações inerentes a modelos nocaute, nos quais outras transcrições
podem ser afetadas em cascatas gênicas, e à possibilidade de expressão
compensatória de outros isotipos de conexinas, não eliminamos a possibilidade da
Cx43 ter um papel fisiologicamente importante na migração celular em explantes de
SVZ. O papel da Cx43 poderá ser mais bem estudado através de ensaios de
ablação genética direcionada em células de animais normais, como as técnicas de
anti-senso (Qiu et al., 2003) e uso de RNAi (Zhang et al., 2006).
6 - A função do acoplamento juncional na SVZ/RMS:
A SVZ/RMS é particular no sentido de que proliferação, migração e
diferenciação acontecem simultaneamente, revelando-se, portanto, como processos
intimamente intrincados ao longo do desenvolvimento (Menezes et al., 1995).
Coerentemente, nossos dados apontam para o envolvimento da comunicação
juncional não apenas na migração, mas também na proliferação e na diferenciação
(Marins, 2001 e resultados não mostrados) das células oriundas da SVZ/RMS. Por
exemplo, quanto à diferenciação, observamos que além do bloqueio da migração
celular nos explantes de SVZ (em especial nos explantes de SVZb), o tratamento
crônico com CBX produziu um maior número de células com morfologia mais
complexa (semelhante a astrócitos maduros e/ou neurônios) nas extremidades do
halo migratório, em comparação com os explantes não tratados (resultados não
mostrados). Um outro possível papel para o acoplamento celular na SVZ/RMS é a
formação de compartimentos e bordas teciduais, como sugerido para outras regiões

92
(Naus e Bani-Yaghoub, 1998; Melitzer et al., 1999; Yoshida e Colman, 2000; Sutor e
Hagerty, 2005).
Encontramos uma correlação positiva entre a comunicação juncional e a
migração celular. Isto sugere que o intercâmbio metabólico e elétrico, mediado por
junções homo- e/ou heterocelulares do tipo gap, seria importante na migração de
precursores neurais. Nosso principal resultado neste trabalho foi a demonstração do
envolvimento direto do acoplamento celular com a migração das células de SVZ.
Esta conclusão é fruto de três observações: 1. A rápida instalação da parada
migratória, após a adição dos agentes desacoplantes CBX e halotano, em um curso
temporal consistente com a cinética de ação destas drogas sobre as junções
comunicantes (Spray et al., 2002); 2. Alta incidência de acoplamento homocelular
entre neuroblastos evidenciado por meio de injeção intracelular de corante (Figs. 10,
11, 12 e 13); 3. Abundante expressão de Cx43 entre neuroblastos (Figs. 14 e 15).
Além disto, este fenômeno de acoplamento homocelular dirigindo a migração em
massa de coortes de células é muito semelhante ao descrito para as células da
crista neural cardíaca, no qual o envolvimento de Cx43 foi claramente demonstrado
(Huang et al., 1998).
Os resultados da adição de CBX apenas no 3DIV, nos levam a concluir que a
importância do acoplamento celular para a migração pode repousar em parte no
processo de sinalização para iniciá-la. Apesar do CBX ser capaz de bloquear a
migração estabelecida das células da SVZ (Figs. 20, 23, 24 e 26), o tratamento
iniciado depois de 3DIV, não apresenta diferença significativa entre os índices
migratórios destes em relação aos índices totais dos controles no 5DIV. Isto sugere
que, pelo menos dentro da janela de tempo analisada, uma vez tendo as células

93
iniciado o seu processo migratório anteriormente, a influência do acoplamento
celular não se faz mais imprescindível.
Alguns aspectos distintos da migração entre os explantes de SVZc e SVZb
podem indicar diferentes atuações do acoplamento celular sobre a fisiologia local.
Sabemos que apesar de muitas semelhanças, a SVZ bulbar é povoada por
diferentes precursores da região cortical (Hack et al., 2005). Além disto, podemos
predizer que alguns neuroblastos na SVZb, estão próximos ou iniciando o processo
de mudança de direção migratória de tangencial para radial. Apesar destas
importantes diferenças na composição celular e dos explantes de SVZb terem
desenvolvido os maiores halos migratórios, ambos os explantes apresentaram um
nível similar de sensibilidade migratória ao bloqueio do acoplamento juncional. Os
explantes de SVZb e SVZc são, portanto, pouco distinguíveis quanto à sensibilidade
aos efeitos anti-migratórios do CBX, com redução da área de migração celular para
cerca de 50% do controle. Diferem, no entanto, quanto à reversão dentro do período
analisado. Apesar dos efeitos do CBX terem sido revertidos (com a retomada da
migração) em todos os explantes, apenas explantes de SVZc recuperaram índices
migratórios semelhantes aos índices totais dos controles. Isto sugere que os efeitos
do desacoplamento nas células oriundas da SVZb, não seriam apenas sobre a
migração mas em outras fases do desenvolvimento celular, como por exemplo, na
indução precoce da diferenciação destas células.
A presença, de forma majoritária, do acoplamento homocelular neuronal e
astroglial indica que pelo menos dois mecanismos diferentes de sinalização celular
podem potencialmente interferir com a migração em nossos explantes.
O acoplamento homocelular entre neuroblastos parece ser um substrato para
o efeito rápido da inibição da migração pelos agentes desacoplantes, conforme

94
sugerido em nossos ensaios de videomicroscopia. Como referido anteriormente,
essa interação celular pode influenciar a migração por um mecanismo semelhante
ao observado no efeito da Cx43 sobre a migração das células de crista neural
(Huang et al., 1998, Lin et al., 2002). Neste modelo de nocaute da Cx43 a migração
das células da crista neural é impedida, causando neste animal malformação
conotruncal cardíaca e obstrução do fluxo sanguíneo. Este grupo demonstrou que,
apesar da ausência da Cx43 inibir a migração das células de crista neural cardíaca,
aparentemente o acoplamento celular não é essencial para este fenômeno ocorrer
(Xu et al., 2001). A influência da Cx43 na migração parece ser mediada por sua
interação com a p120catenina (p120ctn) através da formação de complexos
moleculares de membrana com a N-caderina (Xu et al., 2001) Curiosamente, a
p120ctn é abundantemente expressa por células da SVZ/RMS (Chauvet et al.,
2003), sugerindo que esta via pode ser importante também em nosso modelo. A
p120ctn é uma proteína de ancoragem, da família de proteínas armadillo, que forma
complexos com a N-caderina. Além disso, a p120ctn está envolvida na regulação da
atividade das proteínas RhoGTPases, relacionadas com a migração, bem como com
outros fenômenos envolvendo a dinâmica do citoesqueleto: crescimento axonal,
dendrítico e plasticidade sináptica (Luo, 2000; Dickson, 2001). Além de poder atuar
sinergisticamente com proteínas do citoesqueleto através destes complexos
protéicos em células migratórias, possíveis interações das conexinas, principalmente
da Cx43, com outras moléculas do maquinário celular, principalmente descritas para
Cx43, no exercício de funções extra-juncionais poderiam influenciar o citoesqueleto
(Giepmans, 2004) e conseqüentemente a migração.
Durante os ensaios de vídeo-microscopia observamos a alteração morfológica
de células gliais em resposta ao tratamento agudo com CBX. Este era um fenômeno

95
que consistentemente observávamos associado ao colapso das cadeias migratórias
nos explantes cultivados sobre poli-L-lisina. Como estas células em geral se
apresentavam abaixo do plano de foco usado nos experimentos dirigidos para a
observação das cadeias não analisamos este fenômeno de forma sistemática.
Apesar disto, esta observação fortuita poderia apontar para um possível
envolvimento envolvimento da comunicação juncional na manutenção de uma
morfofisiologia da população astroglial importante para a regulação da migração
celular da SVZ/RMS. Se o acoplamento homocelular glial tem alguma influência
sobre a atividade migratória, esta, no entanto, terá de ser necessariamente indireta,
modulando a interação neurônio-glia presente na SVZ/RMS. De fato é sugerido que
o acoplamento homocelular glial poderia atuar na manutenção da diferenciação de
células gliais (Duval et al., 2002; Leung et al., 2002). Estas por sua vez podem,
direta ou indiretamente, liberar fatores tróficos para a manutenção da atividade
proliferativa e migratória da SVZ. Coerentemente com esta possibilidade existem
evidências demonstrando a importância da população astroglial da SVZ/RMS sobre
aspectos da migração celular desta camada (Mason et al., 2001; Bolteus e Bordey,
2004). A outra função deste tipo de acoplamento pode ser a formação de barreiras e
compartimentos celulares (Naus e Bani-Yaghoub, 1998; Melitzer et al., 1999;
Yoshida e Colman, 2000, Sutor e Hagerty, 2005) por mecanismos ainda
desconhecidos.
Por fim, o acoplamento heterocelular pode auxiliar na migração das células da
RMS para o interior do bulbo olfatório, através de um mecanismo semelhante ao
descrito para invasividade de células tumorais (Miekus et al., 2005; Pollman et al.,
2005).

96
A migração em cadeia, que parece ocorrer sem o auxílio da glia radial e as
por longas distâncias percorridas, dos precursores neurais da SVZ, tem intrigado
diversos grupos de pesquisa. Alguns grupos acreditam na possibilidade de fatores
secretados pelo bulbo olfatório (aliados às diversas moléculas descritas
anteriormente) serem os prováveis responsáveis pela atração das células da SVZ
para seu destino final. Este tema permanece controverso na literatura, visto que,
dentre os trabalhos que descrevem os efeitos da retirada do bulbo olfatório sobre as
células da SVZ, alguns afirmam não haver qualquer efeito (Hu e Rutishauser, 1996;
Kirschenbaum et al., 1999), enquanto outros mostram a existência de efeito atrator
(Murase e Horvitz, 2002 e 2004; Liu e Rao, 2003). Dentre os possíveis candidatos a
fatores direcionadores provenientes do bulbo olfatório, estão alguns
neurotransmissores expressos nesta região, principalmente a dopamina (Baker et
al., 2001; Coronas et al., 2004; Popolo et al., 2004; VanKampen et al., 2004; Baker
et al., 2004; Saino-Saito et al., 2004) e o GABA (Stewart et al., 2002; Belluzzi et al.,
2003; Nguyen et al., 2003; Bolteus e Bordey, 2004; De Marchis et al., 2004; Liu et
al., 2005; Represa e Ben-Ari, 2005; Luján et al., 2005).
Trabalhos recentes encontraram acoplamento mediado por junções
comunicantes entre interneurônios corticais GABA+ e entre outros tipos neuronais
(Rouach et al., 2002; Fukuda e Kosaka, 2003). Além disto, o acoplamento celular
tem sido relacionado com o estabelecimento de sincronização neuronal antes da
formação da circuitaria cortical e a do bulbo olfatório (Rouach et al., 2002; Owens e
Kriegstein, 2002; Zhang e Restrepo, 2003; Kosaka e Kosaka, 2003; Söhl et al., 2005;
Theis et al., 2005). Diante destes achados, acreditamos ser possível que o
acoplamento participe na propagação da sinalização dos neurotransmissores,
presentes no bulbo olfatório, para as células migratórias da SVZ/RMS. A maior

97
proximidade in situ das células dos explantes de SVZb, com aquelas que são a fonte
destes neurotransmissores do bulbo olfatório, sugere uma maior influência destes
sobre o comprometimento fenotípico dos neuroblastos desta região da SVZ. Se esta
suposição for verdadeira, poderia explicar em parte os diferentes efeitos do bloqueio
do acoplamento juncional sobre a migração celular nos explantes oriundos de SVZc
e b, encontrados neste trabalho.
Em conclusão, é possível que o acoplamento celular, tanto homo- como
heterocelular, seja um dos mecanismos responsáveis pela manutenção da rota
migratória de longa distância em direção ao bulbo olfatório.

98
VI - Conclusões:
Neste trabalho concluímos que:
1. Os acoplamentos homo- e heterocelular coexistem nos explantes e poderiam
influenciar as propriedades proliferativas e migratórias, características da
neurogênese da SVZ;
2. Apesar dos altos índices de expressão em nossos explantes, a conexina 43 não
parece ser a principal mediadora do acoplamento celular na SVZ e pode ter sua
ausência compensada por outras conexinas;
3. O CBX e o halotano não afetaram a viabilidade dos explantes, seus efeitos são
reversíveis e o CBX parece atuar especificamente sobre o acoplamento celular;
4. A migração foi bloqueada em ambos os explantes, SVZc e SVZb. O bloqueio
parece privilegiar a inicialização da migração em nosso modelo experimental;
5. Os ensaios de vídeo-microscopia de tempo intervalado confirmam que o
bloqueio da comunicação juncional interfere diretamente na migração celular;
6. As regiões SVZb e SVZc apresentam comportamentos distintos pós-bloqueio,
revelando uma possível heterogeneidade regional.

99
VII - Referências:
ALLEN, E. (1912). The cessation of mitosis in the central nervous system of the albino rat. J. Comp. Neurol. 22: 547-568;
ALTMAN, J. & DAS, G. D. (1965). Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 124: 319-335;
ALTMAN, J. (1969). Autoradiographic and histological studies of postnatal neurogenesis IV. cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. J. Comp. Neurol. 137: 433-457;
ALVAREZ-BUYLLA, A. & NOTTEBOHM, F. (1988). Migration of young neurons in adult avian brain. Nature 335:353-354;
ALVAREZ-MAUBECIN, V., GARCÍA-HERNÁNDEZ, F., WILLIAMS, J. T. & BOCKSTAELE, E. J. V. (2000). Functional coupling between neurons and glia. J. Neurosci. 20: 4091-4098;
ALVES, J. A. J., BARONE, P., ENGELENDER, S., FRÓES, M. M. & MENEZES, J. R. L. (2002). Initial stages of radial glia astrocytic transformation in the early postnatal anterior subventricular zone. J. Neurol. 52: 251-265;
ANGEVINE, J. B. & SIDMAN, R. l. (1961). Autoradiographic study of cell migration during histogenesis of the cerebral cortex in the mouse. Nature 192: 766-768;
BAHREY, H. L. & MOODY, W. J. (2003). Voltage-gated currents, dye and electrical coupling in the embryonic mouse neocortex. Cereb. Cortex 13: 239-251;
BAKER, H.; LIU, N.; CHUN, H. S.; SAINO, S.; BERLIN, R.; VOLPE, B. & SON, J. H. (2001). Phenotypic differentiation during migration of dopaminergic progenitor cells to the olfactory bulb. J. Neurosci. 21: 8505-8513;
BAKER, S. A.; BAKER ,K. A. & HAGG, T. (2004). Dopaminergic nigrostriatal projections regulate neural precursor proliferation in the adult mouse subventricular zone. Eur. J. Neurosci. 20: 575-579;

100
BANI-YAGHOUB, M., BECHBERGER, J. F., UNDERHILL, T. M. & NAUS, C. C. G. (1999). The effects of gap junction blockage on neuronal differentiation of human NTera2/Clone D1 cells. Exp. Neurol. 156: 16-32;
BANNERMAN, P., NICHOLS, W., PUHALLA, S., OLIVER, T., BERMAN, M. & PLEASURE, D. (2000). Early migratory rat neural crest cells express functional gap junctions: evidence that neural crest cell survival requires gap junction function. J. Neurosci. Res. 61: 605-615;
BAYER, S. A. & ALTMAN, J. (1991). Neocortical development. Raven Press: New York;
BAYER, S. A. & ALTMAN, J. (1994). Neocortex development. Raven Press: New York;
BELLIVEAU, D. J. & NAUS, C. C. (1995). Cellular localization of gap junction mRNAs in developing rat brain. Dev. Neurosci. 17: 81-96;
BELLUZZI, O.; BENEDUSI, M.; ACKMAN, J. & LOTURCO, J. J. (2003). Electrophysiological differentiation of new neurons in the olfactory bulb. J. Neurosci. 23: 10411-10418;
BERRY, M., ROGERS, A. W. & EAYRES, J. T. (1964). Pattern of cell migration during cortical histogenesis. Nature 203: 591-593;
BETARBET, R., ZIGOVA, T., BAKAY, R. A. E. & LUSKIN, M. B. (1996). Dopaminergic and GABAergic interneurons of the olfactory bulb are derived from the neonatal subventricular zone. Int. J. Dev. Neurosci. 14: 921-930;
BEYER, E. C., PAUL, D. L. & GOODENOUGH, D. A. (1987). Connexin43: a protein from rat heart homologous to a gap junction protein from liver. J. Cell Biol. 105: 2621-2629;
BITTMAN, K., OWENS, D. F., KRIEGSTEIN, A. R. & LOTURCO, J. J. (1997). Cell coupling and uncoupling in the ventricular zone of the developing cortex. J. Neurosci. 17 (18): 7037-7044;
BITTMAN, K. & LOTURCO, J. J. (1999). Differential regulation of connexin 26 and 43 in murine neocortical precursors. Cereb.Cortex 9: 188-195;

101
BITTMAN, K., BECKER, D. L., CICIRATA, F. & PARNAVELAS, J. G. (2002). Connexin expression in homotypic and heterotypic cell coupling in the developing cerebral cortex. J. Comp. Neurol. 443: 201-212;
BITTMAN, K. S., PANZER, J. A. & BALICE-GORDON, R. J. (2004). Patterns of cell-cell coupling in embryonic spinal cord studied via ballistic delivery of gap-junction-permeable dyes. J. Comp. Neurol. 477: 273-285;
BLACKBURN, J. P., CONNAT, J. L., SEVERS, N. J. & GREEN, C. R. (1997). Connexin43 gap junction levels during development of the thoracic aorta are temporally correlated with elastic laminae deposition and increased blood pressure. Cell Biol. Int. 21: 87-97;
BLASCHKE, A. J., STALEY, K. & CHUN, J. (1996). Widespread programmed cell death in proliferative and postmitotic regions of the fetal cerebral cortex. Development 122: 107-127;
BLASCHKE, A. J.; WEINER, J. A. & CHUN, J. (1998). Programmed cell death is a universal feature of embryonic and postnatal neuroproliferative regions throughout the central nervous system. J. Comp. Neurol. 396: 39-50;
BOLTEUS, A. J. & BORDEY, A. (2004). GABA release and uptake regulate neuronal precursor migration in the postnatal subventricular zone. J. Neurosci. 24: 7623-7631;
BOLTEUS, A. J., GARGANTA, C. & BORDEY, A. (2005). Assays for measuring extracellular GABA levels and cell migration rate in acute slices. Brain Res. Brain Res. Protoc. 14: 126-134;
BOULDER COMMITTEE (1970). Embryonic vertebrate central nervous system: revised terminology. Anat. Rec. 166, 257;
CAVINESS, V. S. & SIDMAN, R. L. (1973). Time of origin of corresponding cell classes in the cerebral cortex of normal and reeler mutant mice: an autoradiographic analysis. J. Comp. Neurol. 148: 141-152;
CHANSON, M., DEROUETTE, J. P., ROTH, I., FOGLIA, B., SCERRI, I., DUDEZ, T. & KWAK, B. (2005). Gap junctional communication in tissue inflammation and repair. Biochim. Biophys. Acta. 1711: 197-207;

102
CHAUVET, N., PRIETO, M., FABRE, C., NOREN, N. K. & PRIVAT, A. (2003). Distribution of p120 catenin during rat brain development: potential role in regulation of cadherin-mediated adhesion and actin cytoskeleton organization. Mol. Cell. Neurosci. 22: 467-486;
CHAZAL, G., DURBEC, JANKOVSKI, A., ROUGON, G. & CREMER, H. (2000). Consequences of neural cell adhesion molecule deficiency on cell migration in the rostral migratory stream of the mouse. J. Neurosci. 20: 1446-1457;
CONOVER, J. C., DOETSCH, F., GARCIA-VERDUGO, J. -M., GALE, N. W., YANCOPOULOS, G. D. & ALVAREZ-BUYLLA, A. (2000). Disruption of Eph/ephrin signaling affects migration and proliferation in the adult subventricular zone. Nature Neurosci. 3: 1091-1097;
CONTRERAS, J. E., SANCHEZ, H. A., VELIZ, L. P., BUKAUSKAS, F. F., BENNETT, M. V. & SAEZ, J. C. (2004). Role of connexin-based gap junction channels and hemichannels in ischemia-induced cell death in nervous tissue. Brain Res. Brain Res. Rev. 47: 290-303;
CORONAS, V.; BANTUBUNGI, K.; FOMBONNE, J.; KRANTIC, S.; SCHIFFMANN, S. N. & ROGER, M. (2004). Dopamine D3 receptor stimulation promotes the proliferation of cells derived from the post-natal subventricular zone. J. Neurochem. 91: 1292-1301;
CROWLEY, J. C. & KATZ, L. C. (2000). Early development of ocular dominance columns. Science 290: 1321-1324;
CUADROS, M. A. & NAVASCUÉS, J. (1998). The origin and differentiation of microglial cells during development. Prog. Neurobiol. 56: 173-189;
DAVIDSON, J. S., BAUMGARTEN, I. M. & HARLEY, E. H. (1986). Reversible inhibition of intercellular junctional communication by glycyrrhetinic acid. Biochem. Biophys. Res. Commun. 134: 29-36;
DAVIDSON, J. S. & BAUMGARTEN, I. M. (1988). Glycyrrhetinic acid derivatives: a novel class of inhibitors of gap-junctional intercellular communication. Structure-activity relationships. J. Neuropharmacol. 246: 1104-1107;
DE MARCHIS, S.; TEMONEY, S.; ERDELYI, F.; BOVETTI, S.; BOVOLIN, P.; SZABO, G. & PUCHE, A. C. (2004). GABAergic phenotypic differentiation of a

103
subpopulation of subventricular derived migrating progenitors. Eur. J. Neurosci. 20: 1307-1317;
DEGLISE, S., MARTIN, D., PROBST, H., SAUCY, F., HAYOZ, D., WAEBER, G.,
NICOD, P., RIS, H. B., CORPATAUX, J. M. & HAEFLIGER, J. A. (2005). Increased connexin43 expression in human saphenous veins in culture is associated with intimal hyperplasia. J. Vasc. Surg. 41: 1043-1052;
DERMIETZEL, R., HERTBERG, E. L., KESSLER, J. A. & SPRAY, D. C. (1991). Gap junctions between cultured astrocytes: immunocytochemical, molecular, and electrophysiological analysis. J. Neurosci. 11:1421-1432;
DERMIETZEL, R. (1996). Molecular diversity and plasticity of gap junctions in the nervous system. In: Gap junctions in the nervous system (Spray, D., Dermietzel, R., eds.), pp 13-38. New York: Chapman and Hall;
DERMIETZEL, R., GAO, Y., SCEMES, E., VIEIRA, D., URBAN, M., KREMER, M., BENNETT, M. V. & SPRAY, D. C. (2000). Connexin43 null mice reveal that astrocytes express multiple connexins. Brain Res. Brain Res. Rev. 32: 45-56;
DICKSON, B. J. (2001). Rho GTPases in growth cone guidance. Curr. Opin. Neurobiol. 11: 103-110;
DOETSCH, F. & ALVAREZ-BUYLLA, A. (1996). Network of tangential pathways for neuronal migration in adult mammalian brain. Proc. Natl. Acad. Sci. U. S. A. 93: 14895-14900;
DOETSCH, F., GARCIA-VERDUGO, J. M. & ALVAREZ-BUYLLA, A. (1997). Cellular composition and three-dimensional organisation of the subventricular germinal zone in the adult mammalian brain. J. Neurosci. 17: 5046-5061;
DUVAL, N., GOMÈZ, D., CALAORA, V., CALABRESE, A., MEDA, P. & BRUZZONE, R. (2002). Cell coupling and Cx43 expression in embryonic mouse neural progenitor cells. J. Cell Sci. 115: 3241-3251
EDMONDSON, J. C. & HATTEN, M. E. (1987). Glial-guided neuronal migration in vitro: A high resolution timelapse video microscopic study. J. Neurosci. 7: 1928-1934:
EHRLICH, H. P. & DIEZ, T. (2003). Role for gap junctional intercellular communications in wound repair. Wound Repair Regen. 11: 481-489;

104
ELIAS, P. M. & FRIEND, D.S. (1976). Vitamin-A-induced mucous metaplasia. An in vitro system for modulating tight and gap junction differentiation. J Cell Biol. 68:173-188;
EMSLEY, J. G., MITCHELL, B. D., KEMPERMANN, G. & MACKLIS,J. D. (2005). Adult neurogenesis and repair of the adult CNS with neural progenitors, precursors, and stem cells. Prog. Neurobiol. 75: 321-341;
EVANS, W. H. & MARTIN, P. E. M. (2002). Gap junctions: structure and function (Review). Mol. Membr. Biol. 19: 121-136;
FONSECA, C.G., GREEN, C. R. & NICHOLSON, L. F. (2002). Upregulation in astrocytic connexin 43 gap junction levels may exacerbate generalized seizures in mesial temporal lobe epilepsy. Brain Res. 929: 105-116;
FRASER, S. E., GREEN, C. R., BODE, H. R. & GILULA, N. B. (1987). Selective disruption of gap junctional communication interferes with a patterning process in Hydra. Science 237: 49-55;
FRAZIER-CIERPIAL, L. & BRUNJES, P. C. (1989). Early postnatal cellular proliferation and survival in the olfactory bulb and rostral migratory stream of normal and unilaterally odor-deprived rats. J. Comp. Neurol. 289: 481-492;
FRICKER, R. A., CARPENTER, M. K., WINKLER, C., GRECO, C., GATES, M. A. & BJORKLUND A. (1999). Site-specific migration and neuronal differentiation of human neural progenitor cells after transplantation in the adult rat brain. J. Neurosci. 19: 5990-6005;
FRIEDL, P., NOBLE, P. B., WALTON, P. A., LAIRD, D. W., CHAUVIN, P. J., TABAH, R. J., BLACK, M. & ZANKER, K. S. (1995). Migration of coordinated cell clusters in mesenchymal and epithelial cancer explants in vitro. Cancer Res. 55:4557-4560;
FRÓES, M. M., CORREIA, A. H. P., GARCIA-ABREU, J., SPRAY, D. C., CAMPOS DE CARVALHO, A. C. & MOURA-NETO, V. (1999). Gap-junctional coupling between neurons and astrocytes in primary central nervous system cultures. Proc. Natl. Acad. Sci. U. S. A. 96: 7541-7546;
FRÓES, M. M. & MENEZES, J. R. L. (2002). Coupled heterocellular arrays in the brain. Neurochem. Int. 41: 367-375;

105
FUKUDA, T. & KOSAKA, T. (2003). Ultrastructural study of gap junctions between dendrites of parvalbumin-containing GABAergic neurons in various neocortical areas of the adult rat. Neuroscience 120: 5-20;
FUKUDA, T., KOSAKA, T., SINGER, W. & GALUSKE, R. A. (2006). Gap junctions among dendrites of cortical GABAergic neurons establish a dense and widespread intercolumnar network. J. Neurosci. 26: 3434-3443;
FUSHIKI, S., PEREZ VELAZQUEZ, J. L., ZHANG, L., BECHBERGER, J. F., CARLEN, P. L. & NAUS, C. C. (2003). Changes in neuronal migration in neocortex of connexin43 null mutant mice. J. Neuropathol. Exp. Neurol. 62: 304-314;
GASHGHAEI, H. T., WEBER, J., PEVNY, L., SCHMID, R., SCHWAB, M. H., LLOYD, K. C., EISENSTAT, D. D., LAI, C. & ANTON, E. S. (2006). The role of neuregulin-ErbB4 interactions on the proliferation and organization of cells in the subventricular zone. Proc. Natl. Acad. Sci. U. S. A. 103: 1930-1935;
GIAUME, C., FROMAGET, C., AOUMARI, A., CORDIER, J., GLOWINSKI, J. & GROS, D. (1991). Gap junctions in cultured astrocytes: single-channel currents and characterization of channel-forming protein. Neuron 6: 133-143;
GIEPMANS, B. N. (2004). Gap junctions and connexin-interacting proteins. Cardiovasc. Res. 62: 233-245;
GOLDBERG, G. S., MORENO, A. P., BECHBERGER, J. F., HEARN, S. S., SHIVERS, R. R., MACPHEE, D. J., ZHANG, Y. –C. & NAUS, C. C. G. (1996). Evidence that disruption of connexon particle arrangements in gap junction plaques is associated with inhibition of gap junctional communication by a glycyrrhetinic acid derivative. Exp. Cell Res. 222: 48-53;
GOODENOUGH, D. A. & PAUL, D. L. (2003). Beyond the gap: functions of unpaired connexon channels. Nature Rev. Mol. Cell Biol. 4: 285-294;
GOULD, E., REEVES, A. J., GRAZIANO, M. S. A. & GROSS, C. G. (1999). Neurogenesis in the neocortex of adult primates. Science 286: 548-552;
GUAN, X., WILSON, S., SCHLENDER, K. K. & RUCH, R. J. (1996). Gap-junction disassembly and connexin 43 dephosphorylation induced by 18 beta-glycyrrhetinic acid. Mol. Carcinog. 16: 157-164;

106
HACK, M. A.; SAGHATELYAN, A.; DE CHEVIGNY, A.; PFEIFER, A.; ASHERY-PADAN, R.; LLEDO, P. M. & GOTZ, M. (2005). Neuronal fate determinants of adult olfactory bulb neurogenesis. Nature Neurosci. 8: 865-872;
HAMADA, J., NAGAYASU, H., KAWANO, T., MIZUTANI, T., NAKATA, D., HOSOKAWA, M. & TAKEICHI, N. (1998). Characterization of the progressive sublines derived from a weakly malignant cloned cell line, ER-1, co-inoculated subcutaneously with a foreign body. Clin. Exp. Metastasis. 16: 291-8.
HAYDAR, T. F., KUAN, C-Y., FLAVEL, R.A. & RAKIC, P. (1999). The role of cell death in regulating the size and shape of the mammalian forebrain. Cereb. Cortex 9: 621-626;
HINDS, J. W. (1968a). Autoradiograpphic study of histogenesis in the mouse olfactory bulb. I. Time of origin of neurons and glia. J. Comp. Neurol. 134: 287-304;
HINDS, J. W. (1968b). Autoradiograpphic study of histogenesis in the mouse olfactory bulb. II. Cell proliferation and migration. J. Comp. Neurol. 134: 305-322;
HOCKBERGER, P. E., TSENG, H.-Y. & CONNOR, J. A. (1987). Immunocytochemical and electrophysiological differentiation of rat cerebellar granule cells in explant cultures. J. Neurosci. 7: 1370-1383;
HOEHN, M., KUSTERMANN, E., BLUNK, J., WIEDERMANN, D., TRAPP, T., WECKER, S., FOCKING, M., ARNOLD, H., HESCHELER, J., FLEISCHMANN, B. K., SCHWINDT, W. & BUHRLE, C. (2002). Monitoring of implanted stem cell migration in vivo: a highly resolved in vivo magnetic resonance imaging investigation of experimental stroke in rat. Proc. Natl. Acad. Sci. U. S. A. 99: 16267-16272;
HORNER, P. J., POWER, A. E., KEMPERMANN, G., KUHN, H. G., PALMER, T. D., WINKLER, J., THAL, L. J. & GAGE, F. H. (2000). Proliferation and differentiation of progenitor cells throughout the intact adult rat spinal cord. J. Neurosci. 20: 2218-2228;
HU, H., TOMASIEWICZ, H., MAGNUSON, T. & RUTISHAUSER, U. (1996). The role of polysialic acid in migration of olfactory bulb interneuron precursors in the subventricular zone. Neuron 16: 735-743;

107
HU, H. Y. & RUTISHAUSER, U. (1996). A septum-derived chemorepulsive factor for migrating olfactory interneurons precursors. Neuron 16: 933-940;
HUANG, G. Y., COOPER, E. S., WALDO, M. L., GILULA, N. B. & LO, C. W. (1998). Gap junction-mediated cell-cell communication modulates mouse neural crest migration. J. Cell Biol. 143: 1725-1734;
JACOBSON, M. (1991). Developmental Neurobiology. Plenum, New York;
KAKITA, A., ZERLIN, M., TAKAHASHI, H. & GOLDMAN, J. E. (2003). Some glial progenitors in the neonatal subventricular zone migrate through the corpus callosum to the contralateral cerebral hemisphere. J. Comp. Neurol. 458: 381-388;
KANO, T., INABA, Y. & AVOLI, M. (2005). Periodic oscillatory activity in parahippocampal slices maintained in vitro. Neuroscience 130; 1041-1053;
KAPLAN, M. S. & BELL, D. H. (1983). Neuronal proliferation in the 9-month-old rodent-radioautographic study of granule cells in the hippocampus. Exp. Brain Res. 52: 1-5;
KATZ, L. C. & SHATZ, C. J. (1996). Synaptic activity and the construction of cortical circuits. Science 274: 1133-1138;
KATZ, L. C. & CROWLEY, J. C. (2002). Development of cortical circuits: lessons from ocular dominance columns. Nat.Rev.Neurosci. 3: 34-42;
KIRSCHENBAUM, B.; DOETSCH, F.; LOIS, C. & ALVAREZ-BUYLLA, A. (1999). Adult subventricular zone neuronal precursors continue to proliferate and migrate in the absence of the olfactory bulb. J. Neurosci. 19: 2171-2180;
KISHI, K. (1987). Golgi studies on the development of granule cells of the rat olfactory bulb with reference to migration in the subependymal layer. J. Comp. Neurol. 258: 112-124;
KISHI, K., PENG, J. Y., KAKUTA, S., MURAKAMI, K., KURODA, M., YUKOTA, S., HAYAKAWA, S., KUGE, T. & ASAYAMA, T. (1990). Migration of bipolar subependymal cells, precursors of the granule cells of the rat olfactory bulb, with reference to the arrangement of the radial glial fibers. Arch. Histol. Cytol. 53:219-226;

108
KISTLER, J., GOLDIE, K., DONALDSON, P. & ENGEL, A. (1994). Reconstitution of native-type noncrystalline lens fiber gap junctions from isolated hemichannels. J. Cell Biol. 126: 1047-1058;
KLEINMAN, H. K., MCGARVEY, M. L., LIOTTA, L. A., ROBEY, P. G., TRYGGVASON, K. & MARTIN, G. R. (1982). Isolation and characterization of type IV procollagen, laminin, and heparan sulfate proteoglycan from the EHS sarcoma. Biochemistry 21: 6188-6193;
KLEINMAN, H. K. & MARTIN, G. R. (2005). Matrigel: basement membrane matrix with biological activity. Semin. Cancer Biol. 15: 378-386;
KOIZUMI, H., HIGGINBOTHAM, H., POON, T., TANAKA, T., BRINKMAN, B. C. & GLEESON, J. G. (2006). Doublecortin maintains bipolar shape and nuclear translocation during migration in the adult forebrain. Nature Neurosci. 9: 779-786;
KOSAKA, T. & KOSAKA, K. (2003). Neuronal gap junctions in the rat main olfactory bulb, with special reference to intraglomerular gap junctions. Neurosci. Res. 45:189-209;
KRIEGSTEIN, A. R. & NOCTOR, S. C. (2004). Patterns of neuronal migration in the embryonic cortex. Trends Neurosci. 27: 392-399;
KUMAR, N. M. & GILULA, N. B. (1996). The gap junction communication channel. Cell 84: 381-388;
LAMPE, P. D., KISTLER, J., HEFTI, A., BOND, J., MÜLLER, S., JOHNSON, R. G. & ENGEL, A. (1991). In vitro assembly of gap junctions. J. Struct. Biol. 107: 281-290;
LAMPE, P. D., NGUYEN, B. P., GIL, S., USUI, M., OLERUD, J., TAKADA, Y. & CARTER, W. G. (1998). Cellular interaction of integrin alpha3beta1 with laminin 5 promotes gap junctional communication. J. Cell Biol. 143: 1735-1747;
LAW, A. K. T., PENCEA, V., BUCK, C. R. & LUSKIN, M. B. (1999). Neurogenesis and neuronal migration in the neonatal rat forebrain anterior subventricular zone do not require GFAP-positive astrocytes. Dev. Biol. 216: 622-634;

109
LEE S., TOMASETTO, C., PAUL D., KEYOMARSI, K. & SAGER, R. (1992). Transcriptional downregulation of gap-junction proteins blocks junctional communication in human mammary tumor cell lines. J. Cell Biol. 118: 1213-1221;
LEUNG, D. S.; UNSICKER, K. & REUSS, B. (2002). Expression and developmental regulation of gap junction connexins cx26, cx32, cx43 and cx45 in the rat midbrain-floor. Int. J. Dev. Neurosci. 20: 63-75;
LEVISON, S. W., CHUANG, C., ABRAMSON, B. J. & GOLDMAN, J. E. (1993). The migrational patterns and developmental fates of glial precursors in the rat subventricular zone are temporally regulated. Development 119: 611-622;
LEVISON, S. W. & GOLDMAN, J. E. (1993). Both oligodendrocytes and astrocytes develop from progenitors in the subventricular zone of postnatal rat forebrain. Neuron 10: 201-212;
LI, G. & HERLYN, M. (2000). Dynamics of intercellular communication during melanoma development. Mol. Med. Today. 6: 163-169;
LI, H., BRODSKY, S., KUMARI, S., VALIUNAS, V., BRINK, P., KAIDE, J., NASJLETTI, A. & GOLIGORSKY, M.S. (2002). Paradoxical overexpression and translocation of connexin43 in homocysteine-treated endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 282: H2124-2133;
LI, L., LIU, F. & ROSS, A. H. (2003). PTEN regulation of neural development and CNS stem cells. J. Cell Biochem. 88: 24-28;
LIM, D. A. & ALVAREZ-BUYLLA, A. (1999). Interaction between astrocytes and adult subventricular zone precursors stimulates neurogenesis. Proc. Natl. Acad. Sci. U. S. A. 96:7526-7531;
LIM, D. A., TRAMONTIN, A. D., TREVEJO, J. M., HERRERA, D. G., GARCIA-VERDUGO, J. M. & ALVAREZ-BUYLLA, A. (2000). Noggin antagonizes BMP signaling to create a niche for adult neurogenesis. Neuron 28: 713-726;
LIM, D. A., SUAREZ-FARINAS, M., NAEF, F., HACKER, C. R., MENN, B., TAKEBAYASHI, H., MAGNASCO, M., PATIL, N. & ALVAREZ-BUYLLA, A. (2006). In vivo transcriptional profile analysis reveals RNA splicing and chromatin remodeling as prominent processes for adult neurogenesis. Mol. Cell. Neurosci. 31: 131-148;

110
LIN, J. H. -C., TAKANO, T., COTRINA, M. L., ARCUINO, G., KANG, J., LIU, S., GAO, Q., JIANG, L., LI, F., LICHTENBERG-FRATE, H., HAUBRICH, S., WILLECKE, K., GOLDMAN, S. A. & NEDERGAARD, M. (2002). Connexin 43 enhances the adhesivity and mediates the invasion of malignant glioma cells. J. Neurosci. 22: 4302-4311;
LIN, J. H. -C.; YANG, J.; LIU, S.; TAKANO, T.; WANG, X.; GAO, Q.; WILLECKE, K. & NEDERGAARD, M. (2003). Connexin mediates gap junction-independent resistance to cellular injury. J. Neurosci. 23: 430-441;
LIU, G. & RAO, Y. (2003). Neuronal migration from the forebrain to the olfactory bulb requires a new attractant persistent in the olfactory bulb. J. Neurosci. 23: 6651-6659;
LIU, X.; WANG, Q.; HAYDAR, T. F. & BORDEY, A. (2005). Nonsynaptic GABA signaling in postnatal subventricular zone controls proliferation of GFAP-expressing progenitors. Nat. Neurosci. 8: 1179-1187;
LIU, X., BOLTEUS, A. J., BALKIN, D. M., HENSCHEL, O. & BORDEY, A. (2006). GFAP-expressing cells in the postnatal subventricular zone display a unique glial phenotype intermediate between radial glia and astrocytes. Glia 54: 394-410;
LO TURCO, J. J. & KRIEGSTEIN, A. R. (1991). Clusters of coupled neuroblasts in neocortex. Science 252: 563-566;
LOEWENSTEIN, W. R. (1979). Junctional intercellular communication and the control of growth. Biochem. Biophys. Acta. 560: 1-65;
LOEWENSTEIN, W. R. (1981). Junctional intercellular communication: the cell-to-cell membrane channel. Physiol Rev. 61: 829-913;
LOIS, C. & ALVAREZ-BUYLLA, A. (1993). Proliferating subventricular zone cells in the adult mammalian forebrain can differentiate into neurons and glia. Proc. Natl. Acad. Sci. U. S. A. 90: 2074-2077;
LOIS, C. & ALVAREZ-BUYLLA, A. (1994). Long distance neuronal migration in the adult mammalian brain. Science 264: 1145-1148;

111
LOIS, C., GARCIA-VERDUGO, J. M. & ALVAREZ-BUYLLA, A. (1996). Chain migration of neuronal precursors. Science 271: 978-981;
LUJAN, R.; SHIGEMOTO, R. & LOPEZ-BENDITO, G. (2005). Glutamate and GABA receptor signalling in the developing brain. Neuroscience 130: 567-580;
LUO, L. (2000). Rho GTPases in neuronal morphogenesis. Nat. Rev. Neurosci. 1: 173-180;
LUSKIN, M. B. (1993) Restricted proliferation and migration of postnatally generated neurons derived from the forebrain subventricular zone. Neuron 11: 173-189;
LUSKIN, M. B. & MCDERMOTT, K. (1994). Divergent cell lineages for oligodendrocytes and astrocytes originating in the neonatal forebrain subventricular zone. Glia 11: 211-226;
LUSKIN, M. B., ZIGOVA, T., SOTERES, B. J. & STEWART, R. R. (1997). Neuronal progenitor cells derived from the anterior subventricular zone of the neonatal rat forebrain continue to proliferate in vitro and express a neuronal phenotype. Mol. Cell. Neurosci. 8: 351-366;
MARINS, M. (2001). O efeito do bloqueio juncional com carbenoxolone em explantes de zona subventricular pós-natal de roedores. Dissertação de Mestrado – Universidade Federal do Rio de Janeiro, UFRJ;
MARSHALL, C. A. G., SUZUKI, S. O. & GOLDMAN, J. E. (2003). Gliogenic and neurogenic progenitors of the subventricular zone: who are they, where did they come from, and where are they going? Glia 43: 52-61;
MASAKI, E., KAWAMURA, M. & KATO, F. (2004). Attenuation of gap-junction-mediated signaling facilitated anesthetic effect of sevoflurane in the central nervous system of rats. Anesth. Analg. 98: 647-652, table;
MASON, H. A., ITO, S. & CORFAS, G. (2001). Extracellular signals that regulate the tangential migration of olfactory bulb neuronal precursors: inducers, inhibitors, and repellents. J. Neurosci. 21: 7654-7663;
MCCONNELL, S. K. (1988).Development and decision making in the mammalian cerebral neocortex. Brain Res. Rev. 13 (1): 1-23;

112
MCDONOUGH, W. S., JOHANSSON, A., JOFFEE, H., GIESE, A. & BERENS, M. E. (1999). Gap junction intercellular communication in gliomas is inversely related to cell motility. Int. J. Dev. Neurosci. 17: 601-11.
MELLITZER, G., XU, Q. & WILKINSON, D. G. (1999). Eph receptors and ephrins restrict cell intermingling and communication. Nature 400: 77-81;
MENDES-OTERO, R. & CAVALCANTE, L. A. (1996). Expression of 9-o-acetylated gangliosides is correlated with tangential cell migration in the rat brain. Neurosci. Letters 204: 97-100;
MENEZES, J. R. L. & LUSKIN, M. B. (1994). Expression of neuron-specific tubulin defines a novel population of neurons in the proliferative layers of the developing telencephalon. J. Neurosci. 14: 5399-5416;
MENEZES, J. R. L., SMITH, C. M., NELSON, K. C. & LUSKIN, M. B. (1995). The division of neuronal progenitor cells during migration in the neonatal mammalian forebrain. Mol. Cell. Neurosci. 6: 496-508;
MENEZES, J. R. L., DIAS, F., GARSON, A. V. B. & LENT, R. (1998). Restricted distribution of S-phase cells in the anterior subventricular zone of the postnatal mouse forebrain. Anat. Embryol. 198: 205-211;
MENEZES, J. R. L., FRÓES, M. M., MOURA NETO, V. & LENT, R. (2000). Gap junction-mediated coupling in the postnatal anterior subventricular zone. Dev. Neurosci. 22: 34-43;
MENEZES, J. R. L., MARINS, M., ALVES, J. A. J., FRÓES, M. M. & HEDIN-PEREIRA, C. (2002). Cell migration in the postnatal subventricular zone. Braz. J. Med. Biol. Res. 35:1411-1421;
MENEZES, J. R. L., EMSLEY, J. G. & MACKLIS, J. D. (2005). Persistence of radial glia in the cortical subventricular zone of the adult mouse. Society for Neuroscience Annual Meeting prog. (Wednesday, Nov. 16) 831-11;
MERKLE, F. T., TRAMONTIN, A. D., GARCÍA-VERDUGO, J. M. & ALVAREZ-BUYLLA, A. (2004). Radial glia give rise to adult neural stem cells in the subventricular zone. Proc. Nat. Acad. Sci. 101: 17528-17532;

113
MIALE, I. L. & SIDMAN, R. L. (1961). An autoradiographic analysis of histogenesis in the mouse cerebellum. Exp. Neurol. 4: 277-296;
MIEKUS, K., CZERNIK, M., SROKA, J., CZYZ, J. & MADEJA, Z. (2005). Contact stimulation of prostate cancer cell migration: the role of gap junctional coupling and migration stimulated by heterotypic cell-to-cell contacts in determination of the metastatic phenotype of Dunning rat prostate cancer cells. Biol. Cell 97: 893-903;
MING, G. & SONG, H. (2005). Adult neurogenesis in the mammalian central nervous system. Annu. Rev. Neurosci. 28:223-250;
MIRAGALL, F., HWANG, T. K., TRAUB, O., HERTZBERG, E. L. & DERMIETZEL, R. (1992). Expression of connexins in the developing olfactory system of the mouse. J. Comp. Neurol. 325: 359-378;
MIRAGALL, F., ALBIEZ, P., BARTELS, H., DeVRIES, U. & DERMIETZEL, R. (1997). Expression of the gap junction protein connexin43 in the subependymal layer and the rostral migratory stream of the mouse: evidence for an inverse correlation between intensity of connexin43 expression and cell proliferation activity. Cell Tissue Res. 287: 243-253;
MIYAKOSHI, L. M., MENDEZ-OTERO, R. & HEDIN-PEREIRA, C. (2001). The 9-O-acetyl GD3 gangliosides are expressed by migrating chains of subventricular zone neurons in vitro. Braz. J. Med. Biol. Res. 34: 669-673;
MIYATA, T., KAWAGUCHI, A. OKANO, H. & OGAWA, M. (2001). Assymetric inheritance of radial glial fibers by cortical neurons. Neuron 31: 727-741;
MONDER, C., STEWART, P. M., LAKSHMI, V., VALENTINO, R., BURT, D. & EDWARDS, C. R. (1989). Licorice inhibits corticosteroid 11 beta-dehydrogenase of rat kidney and liver: in vivo and in vitro studies. Endocrinology 125: 1046-1053;
MONTORO, R. J. & YUSTE, R. (2004). Gap junctions in developing neocortex: a review. Brain Res. Rev. 47: 216-226;
MORSHEAD, C. M. & VAN DER KOOY, D. (1992). Postmitotic death is the death of constitutively proliferating cells in the subependymal layer of the adult mouse brain. J. Neurosci. 12: 249-256;

114
MURASE, S. & HORWITZ, A. F. (2002). Deleted in colorectal carcinoma and differentially expressed integrins mediate the directional migration of neural precursors in the rostral migratory stream. J. Neurosci. 22: 3568-3579;
MURASE, S. & HORWITZ, A. F. (2004). Directions in cell migration along the rostral migratory stream: the pathway for migration in the brain. Curr. Top. Dev. Biol. 61: 135-152;
NADARAJAH, B.; THOMAIDOU, D.; EVANS, W. H. & PARNAVELAS, J. G. (1996). Gap junctions in the adult cerebral cortex: regional differences in their distribution and cellular expression of connexins. J. Comp. Neurol. 376: 326-342;
NADARAJAH, B., JONES, A., EVANS, W. & PARNAVELAS, J. (1997). Differential expression of connexins during neocortical development and neuronal circuit formation. J. Neurosci. 17: 3096-3111;
NAGY, J. I., HOSSAIN, M. Z., LYNN, B. D., CURPEN, G. E., YANG, S. & TURLEY, E. A. (1996). Increased connexin-43 and gap junctional communication correlate with altered phenotypic characteristics of cells overexpressing the receptor for hyaluronic acid-mediated motility. Cell Growth Differ. 7: 745-751;
NAGY, J. I., DUDEK, F. E. & RASH, J. E. (2004). Update on connexins and gap junctions in neurons and glia in the mammalian nervous system. Brain Res. Brain Res. Rev. 47: 191-215;
NAUS, C. C. G., ELISEVICH, K., ZHU, D., BELLIVEAU, D. J. & DEL MAESTRO, R. F. (1992). In vivo growth of C6 glioma cells transfected with Cx43 cDNA. Cancer Res. 52: 4208-4213;
NAUS, C. C. G. & BANI-YAGHOUB, M. (1998). Gap junctional communication in the developing central nervous system. Cell Biol. Int. 22: 751-763;
NAVOLOTSKI, A., RUMJNZEV, A., LU, H., PROFT, D., BARTHOLMES, P. & ZANKER, K. S. (1997). Migration and gap junctional intercellular communication determine the metastatic phenotype of human tumor cell lines. Cancer Lett. 118:181-187;
NEDERGAARD, M. (1994). Direct signaling from astrocytes to neurons in cultures of mammalian brain cells. Science 263: 1768-1771;

115
NEIJSSEN, J., HERBERTS, C., DRIJFHOUT, J. W., REITS, E., JANSSEN, L. & NEEFJES, J. (2005). Cross-presentation by intercellular peptide transfer through gap junctions. Nature 434: 83-88;
NGUYEN, L.; MALGRANGE, B.; BREUSKIN, I.; BETTENDORFF, L.; MOONEN, G.; BELACHEW, S. & RIGO, J. M. (2003). Autocrine/paracrine activation of the GABA(A) receptor inhibits the proliferation of neurogenic polysialylated neural cell adhesion molecule-positive (PSA-NCAM+) precursor cells from postnatal striatum. J. Neurosci. 23: 3278-3294;
NIETO, M., MONUKI, E. S., TANG, H., IMITOLA, J.,; HAUBST, N., KHOURY, S. J., CUNNINGHAM, J., GOTZ, M. & WALSH, C. A. (2004). Expression of Cux-1 and Cux-2 in the subventricular zone and upper layers II-IV of the cerebral cortex. J. Comp. Neurol. 479: 168-180;
NOWAKOWSKI, R. S. & RAKIC, P. (1981). The site of origin and route and rate of migration of neurons to the hippocampal region of the rhesus monkey. J. Comp. Neurol. 196: 129-154;
O’ROURKE, N. A., SULLIVAN, D. P., KAZNOWSKI, C. E., JACOBS, A. A. & MCONNELL, S. K. (1995). Tangential migration of neurons in the developing cerebral cortex. Development 121: 2165-2176;
OKADA, Y., MIYAMOTO, T. & TODA, K. (2003). Dopamine modulates a voltage-gated calcium channel in rat olfactory receptor neurons. Brain Res. 968: 248-255;
OLIVEIRA, R., CHRISTOV, C., GUILLAMO, J. S., DE BOUARD, S., PALFI, S., VENANCE, L., TARDY, M. & PESCHANSKI, M. (2005). Contribution of gap junctional communication between tumor cells and astroglia to the invasion of the brain parenchyma by human glioblastomas. BMC Cell Biol. 6: 7;
OWENS, D. F. & KRIEGSTEIN, A. R. (2002). Developmental neurotransmitters? Neuron 36: 989-991;
PACHA, J., VAGNEROVA, R. & BRYNDOVA, J. (2003). Carbenoxolone accelerates maturation of rat intestine. Pediatr. Res. 53; 808-813;
PAIS, I., HORMUZDI, S. G., MONYER, H., TRAUB, R. D., WOOD, I. C., BUHL, E. H., WHITTINGTON, M. A. & LEBEAU, F. E. N. (2003). Sharp wave-like activity

116
in the hippocampus in vitro in mice lacking the gap junction protein connexin 36. J. Neurophysiol. 89: 2046-2054;
PARATCHA, G., IBANEZ, C. F. & LEDDA, F. (2006). GDNF is a chemoattractant factor for neuronal precursor cells in the rostral migratory stream. Mol. Cell. Neurosci. 31:505-14;
PAWAR, S., KARTHA, S. & TOBACK, F. G. (1995). Differential gene expression in migrating renal epithelial cells after wounding. J. Cell Physiol. 165: 556-565;
PEINADO, A., YUSTE, R. & KATZ, L. C. (1993). Gap junctional communication and the development of local circuits in neocortex. Cereb. Cortex 3: 488-498;
PENCEA, V. & LUSKIN, M. B. (2003).Prenatal development of the rodent rostral migratory sream. J. Comp. Neurol. 463: 402-418;
PERETTO, P., MERIGHI, A., FASOLO, A. & BONFANTI, L. (1997). Glial tubes in the rostral migratory stream of the adult rat. Brain Res. Bull. 42: 9-21;
PERETTO, P., GIACHINO, C., AIMAR, P., FASOLO, A. & BONFANTI, L. (2005). Chain formation and glial tube assembly in the shift from neonatal to adult subventricular zone of the rodent forebrain. J. Comp. Neurol. 487: 407-427;
PERSONIUS, K. E. & BALICE-GORDON, R. J. (2000). Activity-dependent editing of neuromuscular synaptic connections. Brain Res. Bull. 53: 513-522;
POLLMANN, M. A., SHAO, Q., LAIRD, D. W. & SANDIG, M. (2005). Connexin 43 mediated gap junctional communication enhances breast tumor cell diapedesis in culture. Breast Cancer Res. 7: R522-534;
POPOLO, M.; MCCARTHY, D. M. & BHIDE, P. G. (2004). Influence of dopamine on precursor cell proliferation and differentiation in the embryonic mouse telencephalon. Dev. Neurosci. 26: 229-244;
PRIVAT, A. & LEBLOND, C. P. (1972). The subependymal layer and neighbouring region in the brain of the young rat. J. Comp. Neurol. 146: 277-302;
PRIVAT, A. (1975). Postnatal gliogenesis in the mammalian brain. Int. Rev. Cytol. 40: 281-323;

117
QIU, C., COUTINHO, P., FRANK, S., FRANKE, S., LAW, L. Y., MARTIN, P., GREEN, C. R. & BECKER, D. L. (2003). Targeting connexin43 expression accelerates the rate of wound repair. Curr. Biol. 13: 1697-1703;
RAKIC, P. (1972). Mode of cell migration to the superficial layers of fetal monkey neocortex. J. Comp. Neurol. 145: 61-84;
RAKIC, P. (1988). Specification of cerebral cortical areas. Science 241: 170-176;
RAKIC, P. (1990). Principles of neural cell migration. Experientia 46 (9): 882-891;
REPRESA, A. & BEN ARI, Y. (2005). Trophic actions of GABA on neuronal development. Trends Neurosci. 28:278-283;
REYNOLDS, B. A. & WEISS, S. (1992). Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science 255: 1707-1710;
ROERIG, B. & KATZ, L. C. (1997). Modulation of intrinsic circuits by serotonin 5-HT3 receptors in developing ferret visual cortex. J. Neurosci. 17: 8324-8338;
ROUACH, N., AVIGNONE, E., MÊME, W., KOULAKOFF, A., VENANCE, L., BLOMSTRAND, F. & GIAUME, C. (2002). Gap junctions and connexin expression in the normal and pathological central nervous system. Biol. Cell 94: 457-475;
ROUSSELOT, P., LOIS, C. & ALVAREZ-BUYLLA, A. (1995). Embryonic (PSA) N-CAM reveals chains of migrating neuroblasts between the lateral ventricle and the olfactory bulb of adult mice. J. Comp. Neurol. 351: 51-61;
ROZENTAL, R., MEHLER, M. F., MORALES, M., ANDRADE-ROZENTAL, A. F., KESSLER, J. A. & SPRAY, D. C. (1995). Differentiation of hippocampal progenitor cells in vitro: temporal expression of intercellular coupling and voltage- and ligand-gated responses. Dev. Biol. 167: 350-362;
RUANGVORAVAT, C. P. & LO, C. W. (1991). Restrictions in gap junctional communication in the Drosophila larval epidermis. Dev. Dyn. 193: 70-82;

118
RUTISHAUSER, U., WATANABE, M., SILVER, J., TROY, F. A. &VIMR, E. R. (1985). Specific alteration of NCAM-mediated cell adhesion by an endoneuraminidase. J. Cell Biol. 101: 1842-1849;
RUTISHAUSER, U. & LANDMESSER, L. (1991). Polysialic acid on the surface of axons regulates patterns of normal and activity-dependent innervation. Trends Neurosci. 14: 528-532;
SAGHATELYAN, A., CHEVIGNY, A., SCHACHNER, M. & LLEDO, P. –M. (2004). Tenascin-R mediates activity-dependent recruitment of neuroblastos in the adult mouse forebrain. Nature Neurosci. 7: 347-356;
SAILER, M. H., HAZEL, T. G., PANCHISION, D. M., HOEPPNER, D. J., SCHWAB, M. E. & MCKAY, R. D. (2005). BMP2 and FGF2 cooperate to induce neural-crest-like fates from fetal and adult CNS stem cells. J. Cell Sci. 118: 5849-5860;
SAINO-SAITO, S.; SASAKI, H.; VOLPE, B. T.; KOBAYASHI, K.; BERLIN, R. & BAKER, H. (2004). Differentiation of the dopaminergic phenotype in the olfactory system of neonatal and adult mice. J. Comp. Neurol. 479: 389-398;
SALAMEH, A. & DHEIN, S. (2005). Pharmacology of gap junctions. New pharmacological targets for treatment of arrhythmia, seizure and cancer? Biochim. Biophys. Acta 1719: 36-58;
SANAI, N., TRAMONTIN, A. D., QUIÑONES-HINOJOSA, A., BARBARO, N. M., GUPTA, N., KUNWAR, S., LAWTON, M. T., McDERMOTT, M. W., PARSA, A. T., MANUEL-GARCIA VERDUGO, J., BERGER, M. S. & ALVAREZ-BUYLLA, A. (2004). Unique astrocyte ribbon in adult human brain contains neural stem cells but lacks chain migration. Nature 427: 740-744;
SÁNCHEZ-ALVAREZ, R, TABERNERO, A., SÁNCHEZ-ABARCA, L. I., ORFAO, A., GIAUME, C. & MEDINA, J. M. (2001). Proliferation of C6 glioma cells is blunted by the increase in gap junction communication caused by tolbutamide. FEBS Lett. 509: 202-206;
SAUER, F. C. (1935). Mitosis in the neural tube. J. Comp. Neurol. 62: 377-405;
SAUER, F. C. (1937). Some factors in the morphogenesis of vertebrate embryonic epithelia. J. Morphol. 61: 563-579;

119
SAUER, M. E. & WALKER, B. E. (1959). Radioautographic study of interkinetic nuclear migration in the neural tube. Proc. Soc. Exp. Biol. Med. 101: 557-560;
SAWAMOTO, K., WICHTERLE, H., GONZALEZ-PEREZ, O., CHOLFIN, J. A., YAMADA, M., SPASSKY, N., MURCIA, N. S., GARCIA-VERDUGO, J. M., MARIN, O., RUBENSTEIN, J. L., TESSIER-LAVIGNE, M., OKANO, H. & ALVAREZ-BUYLLA, A. (2006). New neurons follow the flow of cerebrospinal fluid in the adult brain. Science 311:629-32;
SCEMES, E., DUVAL, N & MEDA, P. (2003). Reduced expression of P2Y1 receptors in connexin43-null mice alters calcium signaling and migration of neural progenitor cells. J. Neurosci. 23: 11444-11452;
SHOUKIMAS, G. M. & HINDS, J. W. (1978). The development of the cerebral cortex in the embryonic mouse: an electron microscopic serial section analysis. J. Comp. Neurol. 179: 795-830;
SMART, I. (1961). The subependimal layer of the mouse brain and its cell production as shown by radioautography after thymidine-H3 injection. J. Comp. Neurol. 116: 325-348;
SÖHL, G. & WILLECKE, K. (2003). An update on connexin genes and their nomenclature in mouse and man. Cell Commun. Adhes. 10: 173-180;
SÖHL, G., ODERMATT, B., MAXEINER, S., DEGEN, J. & WILLECKE, K. (2004). New insights into the expression and function of neural connexins with transgenic mouse mutants. Brain Res. Rev. 47: 245-259;
SÖHL, G., MAXEINER, S. & WILLECKE, K. (2005). Expression and functions of neural gap junctions. Nat. Rev. Neurosci. 6: 191-200;
SOLAN, J. L., FRY, M. D., TENBROEK, E. M. & LAMPE, P. D. (2003). Connexin43 phosphorylation at S368 is acute during S and G2/M and in response to protein kinase C activation. J. Cell Sci. 116: 2203-2211;
SPASSKY, N.; MERKLE, F. T.; FLAMES, N.; TRAMONTIN, A. D.; GARCIA-VERDUGO, J. M. & ALVAREZ-BUYLLA, A. (2005). Adult ependymal cells are postmitotic and are derived from radial glial cells during embryogenesis. J. Neurosci. 25: 10-18;

120
SPRAY, D. C. & DERMIETZEL, R. (1996). X-linked dominant Charcot-Marie-Tooth disease and other potential gap-junction diseases of the nervous system. Trends Neurosci. 18: 256-262;
SPRAY, D. C., ROZENTAL, R. & SRINIVAS, M. (2002). Prospects for rational development of pharmacological gap junction channel blockers. Curr. Drug Targets. 3: 455-464;
STENSAAS, L. J. & GILSON, B. C. (1972). Ependymal and subependymal cells of the caudato-pallial junction in the lateral ventricle of the neonatal rabbit. Z. Zelforsh 132: 297-322;
STEWART, R. R.; Hoge, G. J.; ZIGOVA, T. & LUSKIN, M. B. (2002). Neural progenitor cells of the neonatal rat anterior subventricular zone express functional GABA(A) receptors. J. Neurobiol. 50: 305-322;
STURROCK, R. R. & SMART, I. H. M. (1980). A morphological study of the mouse subependimal layer from embryonic life to old age. J. Anat. 130: 391-415;
SUTOR, B. & HAGERTY, T. (2005). Involvement of gap junctions in the development of the neocortex. Biochim. Biophys. Acta 1719: 59-68;
SUZUKI, S. O. & GOLDMAN, J. E. (2003). Multiple cell populations in the early postnatal subventricular zone take distinct migratory pathways: a dynamic study of glial and neuronal progenitor migration. J. Neurosci. 23: 4240-4250;
TABERNERO, A., JIMÉNEZ, C., VELASCO, A., GIAUME, C. & MEDINA, J. M. (2001). The enhancement of glucose uptake caused by the collapse of gap junction communication is due to an increase in astrocyte proliferation. J. Neurochem. 78: 890-898;
TAKAHASHI, T., NOWAKOWSKI, R. S. & CAVINESS, V. S. (1992). BudR as an S-phase marker for quantitative studies of cytokinetic behaviour in the murine cerebral ventricular zone. J. Neurocytol. 21: 185-197;
TAKAHASHI, T., NOWAKOWSKI, R. S., CAVINESS, V. S. (1995). Early ontogeny of secondary proliferative population of the embryonic murine cerebral wall. J. Neurocytol. 21: 185-197;

121
TARABYKIN, V., STOYKOVA, A., USMAN, N. & GRUSS, P. (2001). Cortical upper layer neurons derive from the subventricular zone as indicated by Svet1 gene expression. Development 128:1983-1993;
TAYMANS, J. M., KIA, H. K. & LANGLOIS, X. (2006). Activator of G protein signaling type 3 mRNA is widely distributed in the rat brain and is particularly abundant in the subventricular zone-olfactory bulb system of neural precursor cell proliferation, migration and differentiation. Neurosci. Lett. 391:116-21;
TEMPLE, S. & QUIAN, X. (1996). Vertebrate neural progenitor cells: subtypes and regulation. Curr. Opin. Neurobiol. 6: 11-17;
THEIS, M., SOHL, G., EIBERGER, J. & WILLECKE, K. (2005). Emerging complexities in identity and function of glial connexins. Trends Neurosci. 28: 188-195;
THOMAS, T., JORDAN, K., SIMEK, J., SHAO, Q., JEDESZKO, C., WALTON, P. & LAIRD, D. W. (2005). Mechanisms of Cx43 and Cx26 transport to the plasma membrane and gap junction regeneration. J. Cell Sci. 118: 4451-4462;
TRAMONTIN, A. D., GARCÍA-VERDUGO, J. M., LIM, D. A. & ALVAREZ-BUYLLA, A. (2003). Postnatal development of radial glia and the ventricular zone (VZ): a continuum of the neural stem cell compartment. Cereb. Cortex 13: 580-587;
TROPEPE, V., CRAIG, C. G., MORSHEAD, C. M. & VAN DER KOOY, D. (1997). Transforming growth factor- null and senescent mice show decreased neural progenitor cell proliferation in the forebrain subependyma. J. Neurosci. 17: 7850-7859;
TROSKO, J. E. (2003). The role of stem cells and gap junctional intercellular communication in carcinogenesis. J. Biochem. Mol. Biol. 36: 43-48;
TSAI, P. T., OHAB, J. J., KERTESZ, N., GROSZER, M., MATTER, C., GAO, J., LIU, X., WU, H. & CARMICHAEL, S. T. (2006). A critical role of erythropoietin receptor in neurogenesis and post-stroke recovery. J. Neurosci. 26:1269-1274;
VALIUNAS, V., POLOSINA, Y. Y., MILLER, H., POTAPOVA, I. A., VALIUNIENE, L., DORONIN, S., MATHIAS, R. T., ROBINSON, R. B., ROSEN, M. R., COHEN, I. S. & BRINK, P. R. (2005). Connexin-specific cell-to-cell transfer of short interfering RNA by gap junctions. J. Physiol. 568: 459-468;

122
VAN HAARST, A. D., WELBERG, L. A., SUTANTO, W., OITZL, M. S. & DE KLOET, E. R. (1996). 11 beta-Hydroxysteroid dehydrogenase activity in the hippocampus: implications for in vivo corticosterone receptor binding and cell nuclear retention. J. Neuroendocrinol. 8: 595-600;
VAN KAMPEN, J. M.; HAGG, T. & ROBERTSON, H. A. (2004). Induction of neurogenesis in the adult rat subventricular zone and neostriatum following dopamine D receptor stimulation. Eur. J. Neurosci. 19: 2377-2387;
VAYSSE, P. J. –J., & GOLDMAN, J. E. (1990). A clonal analysis of glial lineages in neonatal forebrain development in vitro. Neuron 4: 833-845;
VENANCE, L., CORDIER, J., MONGE, M., ZALC, B., GLOWINSKI, J. & GIAUME, C. (1995). Homotypic and heterotypic coupling mediated by gap junctions during glial cell differentiation in vitro. Eur. J. Neurosci. 7: 451-461;
VENANCE, L.; ROZOV, A.; BLATOW, M.; BURNASHEV, N.; FELDMEYER, D. & MONYER, H. (2000). Connexin expression in electrically coupled postnatal rat brain neurons. Proc. Natl. Acad. Sci. U. S. A 97: 10260-10265;
WALSH, C. & CEPKO, C. L. (1990). Cell lineage and cell migration in the developing cerebral cortex. Experientia 46 (9): 940-947;
WANG, L., ZHANG, Z. G., ZHANG, R. L., GREGG, S. R., HOZESKA-SOLGOT, A., LETOURNEAU, Y., WANG, Y. & CHOPP, M. (2006). Matrix metalloproteinase 2 (MMP2) and MMP9 secreted by erythropoietin-activated endothelial cells promote neural progenitor cell migration. J. Neurosci. 26: 5996-6003;
WARNER, A. (1992). Gap junctions in development--a perspective. Semin. Cell Biol. 3: 81-91;
WEISSMAN, T. A., RIQUELME, P. A., IVIC, L., FLINT, A. C. & KRIEGSTEIN, A. R. (2004). Calcium waves propagate through radial glial cells and modulate proliferation in the developing neocortex. Neuron 43: 647-661;
WICHTERLE, H., GARCÍA-VERDUGO, J. M. & ALVAREZ-BUYLLA, A. (1997). Direct evidence for homotypic, glia-independent neuronal migration. Neuron 18: 779-791;

123
WICHTERLE, H.; GARCIA-VERDUGO, J. M.; HERRERA, D. G. & ALVAREZ-BUYLLA, A. (1999). Young neurons from medial ganglionic eminence disperse in adult and embryonic brain. Nature Neurosci. 2: 461-466;
WICHTERLE, H., TURNBULL, D. H., NERY, S., FISHELL, G. & ALVAREZ-BUYLLA, A. (2001). In utero fate mapping reveals distinct migratory pathways and fates of neurons born in the mammalian basal forebrain. Development 128: 3759-3771;
WICHTERLE, H., ALVAREZ-DOLADO, M., ERSKINE, L. & ALVAREZ-BUYLLA, A. (2003). Permissive corridor and diffusible gradients direct medial ganglionic eminence cell migration to the neocortex. Proc. Natl. Acad. Sci. USA 100: 727-732;
WILLECKE, K., EIBERGER, J., DEGEN, J., ECKARDT, D., ROMUALDI, A., GÜLDENAGEL, M., DEUTSCH, U. & SÖHL, G. (2002). Structural and functional diversity of connexin genes in the mouse and human genome. Biol. Chem. 383:725-737;
XU, X.; LI, W. E.; HUANG, G. Y.; MEYER, R.; CHEN, T.; LUO, Y.; THOMAS, M. P.; RADICE, G. L. & LO, C. W. (2001). Modulation of mouse neural crest cell motility by N-cadherin and connexin 43 gap junctions. J. Cell Biol. 154: 217-230;
YAMAMOTO, T., OCHALSKY, A., HERTZBERG, E. L., & NAGY, J. I. (1990). On the organisation of astrocytic gap junctions in rat brain as suggested by LM. and EM immunohistochemistry of connexin 43 expression. J. Comp. Neurol. 302: 853-883;
YANCEY, S. B., BISWAL, S. & REVEL, J. P. (1992). Spatial and temporal patterns of distribution of the gap junction protein connexin43 during mouse gastrulation and organogenesis. Development (Camb.) 114: 203-212;
YE, Z. -C., WYETH, M. S., BALTAN-TEKKOK, S. & RANSOM, B. R. (2003). Functional hemichannels in astrocytes: a novel mechanism of glutamate release. J. Neurosci. 23: 3588-3596;
YOSHIDA, M. & COLMAN, D. R. (2000). Glial-defined rhombomere boundaries in developing Xenopus hindbrain. J. Comp. Neurol. 424: 47-57;

124
YOSHIHARA, S., OMICHI, K., YANAZAWA, M., KITAMURA, K. & YOSHIHARA, Y. (2005). Arx homeobox gene is essential for development of mouse olfactory system. Development 132: 751-762;
YUSTE, R., & KATZ, L. C. (1991). Control of postsynaptic Ca2+ influx in developing neocortex by excitatory and inhibitory neurotransmitters. Neuron 6: 333-344;
YUSTE, R., PEINADO, A. & KATZ, L. C. (1992). Neuronal domains in developing neocortex. Science 257: 665-669;
YUSTE, R., NELSON, D. A., RUBIN, W. W. & KATZ, L. C. (1995). Neuronal domains in developing neocortex: mechanisms of coactivation. Neuron 14: 7-17;
ZAHLER, S., HOFFMANN, A., GLOE, T. & POHL, U. (2003). Gap-junctional coupling between neutrophils and endothelial cells: a novel modulator of transendothelial migration. J. Leukoc. Biol. 73: 118-126;
ZHANG, C. & RESTREPO, D. (2003). Heterogeneous expression of connexin 36 in the olfactory epithelium and glomerular layer of the olfactory bulb. J. Comp. Neurol. 459: 426-439;
ZHANG, W., GREEN, C. & STOTT, N. S. (2002). Bone morphogenetic protein-2 modulation of chondrogenic differentiation in vitro involves gap junction-mediated intercellular communication. J. Cell Physiol. 193: 233-243;
ZHANG, X., CHEN, X., WU, D., LIU, W., WANG, J., FENG, Z., CAI, G., FU, B., HONG, Q. & DU, J. (2006). Downregulation of connexin 43 expression by high glucose induces senescence in glomerular mesangial cells. J. Am. Soc. Nephrol. 17: 1532-1542;
ZHANG, Z. Q., HU, Y., WANG, B. J., LIN, Z. X., NAUS, C. C. & NICHOLSON, B. J. (2003). Effective asymmetry in gap junctional intercellular communication between populations of human normal lung fibroblasts and lung carcinoma cells. Carcinogenesis 25: 473-482;
ZHU, D., CAVENEY, S., KIDDER, G. & NAUS, C. (1991). Transfection of C6 glioma cells with connexin 43 cDNA: analysis of expression, intercellular coupling, and cell proliferation. Proc. Natl. Acad. Sci. USA 88: 1883-1887;

125
ZHU, Y., LI, H., ZHOU, L., WU, J. Y. & RAO, Y. (1999). Cellular and molecular guidance of GABAergic neuronal migration from an extracortical origin to the neocortex. Neuron 23: 473-485;
ZIGOVA, T., BETARBET, R., SOTERES, B. J., BROCK, S., BAKAY, R. A. E. & LUSKIN, M. B. (1996). A comparison of the pattern of migration and destination of homotopically transplanted telencephalic ventricular zone cells. Dev. Biol. 173: 459-474.

126
VIII - Anexos:
Anexo I – Filmes de videomicroscopia de tempo intervalado:
Filme 1 – Situação controle do experimento com Halotano em explante cultivado sobre poli-
L-lisina (representativo de 3 experimentos); duração de 2h com imagens
registradas em intervalos de 1 minuto; observe a movimentação das células
localizadas na principal cadeia do explante;
Filme 2 – Explante do Filme 1 após tratamento com halotano; Note que embora ainda exista
bastante movimentação celular; duração de 3h com imagens registradas em
intervalos de 2 minutos; as células presentes na cadeia em evidência
movimentam-se em direção ao explante e, assim que o bloqueador evapora
(após 1h), elas tornam a se mover em direção à placa de cultura;
Filme 3 – Situação controle do experimento com CBX em explante cultivado sobre matrigel
(representativo de 3 experimentos); duração de 3h com imagens registradas em
intervalos de 2 minutos; as células movimentam-se intensamente tanto para fora
quanto para dentro do explante;
Filme 4 – Explante do Filme 3 após tratamento com CBX; duração de 3h com imagens
registradas em intervalos de 2 minutos; a presença do bloqueador juncional
reduziu consideravelmente a movimentação celular presente no explante; (Note
que devido ao tempo de troca do meio os primeiros 8 minutos após a troca não
foram registrados);
Filme 5 – Explante do Filme 3 após a retirada do CBX; duração de 3h com imagens
registradas em intervalos de 2 minutos; a retirada do bloqueador juncional
restabeleceu a movimentação observada na situação controle. (V nota do Filme
4).

127
Anexo II – Artigo de revisão:
MENEZES, J. R. L., MARINS, M., ALVES, J. A. J., FRÓES, M. M. & HEDIN-PEREIRA, C.
(2002). Cell migration in the postnatal subventricular zone. Braz. J. Med. Biol. Res.
35:1411-1421;