instituto nacional de pesquisas da amazÔnia - inpa...
TRANSCRIPT

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DE ÁGUA DOCE E
PESCA INTERIOR
Mapeamento cromossômico comparativo de ciclídeos amazônicos utilizando sequências repetitivas de DNA
Carlos Henrique Schneider
Manaus, Amazonas Fevereiro, 2013

ii
CARLOS HENRIQUE SCHNEIDER
Mapeamento cromossômico comparativo de ciclídeos amazônicos utilizando sequências repetitivas de DNA
Orientador: Eliana Feldberg, Dra.
Co-orientador: Cesar Martins, Dr. Unesp/Botucatu-SP
Tese apresentada ao Instituto
Nacional de Pesquisas da
Amazônia como parte dos
requisitos para obtenção do título
de Doutor em Biologia de Água
Doce e Pesca Interior.
Manaus, Amazonas Fevereiro, 2013

iii
FICHA CATALOGRÁFICA
S358 Schneider, Carlos Henrique
Mapeamento cromossômico comparativo de ciclídeos amazônicos utilizando sequências repetitivas de DNA / Carlos Henrique Schneider. --- Manaus : [s.n.], 2013.
xviii, 121 f. : il. color.
Tese (doutorado) --- INPA, Manaus, 2013.
Orientadora : Eliana Feldberg
Coorientador : Cesar Martins
Área de Concentração : Biologia de Água Doce e Pesca Interior
1. Evolução genômica – Ciclídeos – Amazônia. I. Título.
CDD 19. ed. 597.580415
Sinopse:
Análise da organização genômica comparativa de cinco espécies de
cinco tribos de ciclídeos neotropicais foram realizadas com uma
abordagem citogenética clássica (coloração convencional, detecção da
heterocromatina e regiões organizadoras de nucléolo) e molecular (DNAr
18 e 5S, sequência telomérica, retrotransposons Rex1, 3 e 6, sequências
microssatélites e fração repetitiva Cot-1 DNA). As análises evidenciaram
ampla diversidade genômica e cariotípica existente em ciclídeos
neotropicais e sugerem que grande parte desta ocorre em função de
sequências repetitivas.
Palavras-chave: FISH, retrotransposons, composição genômica,
evolução cariotípica

iv
Dedico esta Tese aos meus pais João Carlos e Edite, às minhas irmãs Marielle e Silvia e especialmente a minha esposa Maria Claudia Gross.

v
“O primeiro pecado da humanidade foi
a fé; a primeira virtude foi a dúvida.”
Carl Sagan
“Uma vida não questionada não
merece ser vivida.”
Platão

vi
A realização deste projeto foi possível devido:
Ao Programa de Pós-graduação em Biologia de Água Doce e Pesca
Interior, do INPA.
Ao laboratório de Genética Animal do INPA, Coordenação de Pesquisas
em Biodiversidade, onde a maior parte deste trabalho foi desenvolvido, com
financiamento proporcionado pelos Projetos de Pesquisas Institucionais (PPIs),
Fundação de Amparo à Pesquisa do Amazonas/Conselho Nacional de
Desenvolvimento Científico e Técnológico (PRONEX FAPEAM/CNPq
003/2009) e convênios entre estas agências e o Ministério de Ciências e
Tecnologia (INCT ADAPTA, FAPEAM/CNPq 573976/2008-2).
Aos laboratórios Genômica Integrativa da Universidade Estadual
Paulista “Júlio de Mesquita Filho” – Botucatu/SP, Genética Animal da
Universidade Estadual de Ponta Grossa – Ponta Grossa/PR onde análises da
FISH foram realizadas, e ao Laboratório de Tecnologias de DNA da
Universidade Federal do Amazonas – Manaus/AM, onde as clonagens dos
DNAs repetitivos foram desenvolvidas.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq) pela concessão da bolsa de estudo e pela taxa de bancada (processo
140816/2009-7) durante a realização deste trabalho.

vii
Agradecimentos Agradeço a todas as pessoas, colegas e amigos que, direta ou
indiretamente, possibilitaram a realização deste trabalho.
À Dra Eliana Feldberg, minha orientadora, que possibilitou a minha vinda
à Manaus, sempre deixou seu laboratório de portas abertas e topou encarar
este trabalho. Muito obrigado pelos inúmeros ensinamentos, apoio, conselhos,
atenção, críticas, sugestões, incentivo e amizade. Obrigado também por estar
sempre pronta a sanar dúvidas, ensinar, ler e reler rapidamente tudo que é
necessário para o bom andamento do trabalho.
Ao Dr Cesar Martins, meu co-orientador, que mesmo distante me
ensinou muito. Obrigado pela amizade, discussões, críticas, sugestões
agilidade nas correções, e também pelos momentos de descontração e muitas
risadas comendo costelas de tambaqui.
Ao Dr Jorge Ivan Rebelo Porto, por sua amizade pelos estímulos,
atenção, por sanar minhas dúvidas e por plantar muitas outras dúvidas na
minha cabeça que possibilitaram uma outra visão do meu trabalho.
Aos amigos do Laboratório de Genética Animal do INPA: Leandra, Leila,
Eduardo “Belugão”, Érica, Heidi, Carlos Faresin, Maelin, Lucas, Denise, Patrick,
Marajó, Masseo, Vanessa Susan, Cebola e tantos os outros que passaram por
lá. Valeu pelo bate-papo (científico ou não), pelas críticas e sugestões, bem
como pelo convívio diário e festas. Em especial aos amigos Leandra, Edu
(Belugão) e Leilinha, sem os vocês o Laboratório seria mais triste.
Aos amigos do Laboratório de Citogenômica da UFAM: Claudia, Natália,
Rafael, Renan, Ana Carolina “Fotula”, Thais, Francy, Marilia, Camila, Karol e a
agregada Jennifer “Janafa”, por me acolherem no reta final da tese. A família
LACA é demais.
À Carminha e Elany, super secretárias do BADPI. Sem elas os alunos
iriam sofrer muito.
Ao Robertão, Mara, Marcelo, Vivi, Leonardo, Miguel e demais colegas do
Laboratório de Citogenética Animal da Universidade Estadual de Ponta Grossa,
por me iniciarem na vida científica e pela imensa ajuda na reta final deste
trabalho.

viii
Ao MSc Edson Junior do Carmo, “o cara”, pela grande ajuda em todas
as clonagens. Edson, você é muito gente fina, sempre achando tempo para
ajudar todo mundo que te procura. Valeu mesmo.
A Leandra, minha parceira de bancada, amiga e nossa família aqui em
Manaus. Valeu pela paciência (desde a graduação), pelas inúmeras risadas e
pelos momentos de tensão divididos (que não foram poucos). Sempre esteve
presente em todas as conquistas e fracassos científicos desde a UEPG, rindo
ou chorando junto e sempre largando uma que mata todo mundo de rir. Você
fará falta em Manaus. Valeu Lê, uma bolsa de PIBIC divida em quatro deu
resultado.
Ao Lineu Gross e Marilda Delfrate Gross, grandes amigos que mesmo
longe estão sempre perto. É bom fazer parte da vida de vocês.
À Dra Maria Claudia Gross, pelo exemplo profissional, pela orientação,
pelo apoio, pelas críticas e sugestões, pela inúmeras revisões em todos os
capítulos e na versão final da tese, bem como pela acolhida na UFAM. E à
minha esposa Claudinha, obrigado pelo companheirismo e amizade nas horas
alegres e tristes, por aguentar a choradeira e os inúmeros estresses quando eu
achava que nada daria certo. Sempre com seu amor e carinho me incentivou a
seguir adiante e mostrou o ponto positivo de tudo. Claudinha: “Diante da
vastidão do tempo e da imensidão do universo, é um imenso prazer para mim
viver uma época e compartilhar a vida com você”. É maravilhoso ter você ao
meu lado. Amo você.
As minhas irmãs, Marielle e Silvia, pelo incentivo, apoio e momentos de
descontração nos finais de ano no Paraná. É ótimo ser irmão e amigo de
vocês.
Aos meus pais João Carlos Schneider (in memorian) e Edite Maria
Schneider, que sempre serão meus exemplos de vida. Obrigado pelo esforço e
ajuda que me fizeram chegar até aqui. Pai, cada dia sem você é triste, cada
volta ao Paraná, sem ter você para conversar e dar risadas é horrível. Obrigado
por você ter feito parte da minha vida. Mãe, obrigado por tudo, broncas e
palavras de apoio. Mãe, Mari e Silvinha passei os melhores dias da minha vida
com vocês e, depois que vim para Manaus, valorizo muito mais cada momento
que passamos juntos. Sem o apoio e incentivo de vocês, eu não teria crescido
pessoal e profissionalmente. Por isso esta conquista é nossa! Amo vocês!

ix
Resumo Os ciclídeos, além de serem explorados na pesca, no comércio ornamental e
na aquicultura, são considerados modelos para estudos de genética e
evolução. Distribuem-se nas Américas Central e do Sul, África, Madagascar e
sul da Índia. Os ciclídeos neotropicais, todos pertencem à subfamília Cichlinae,
que é subdividida em sete tribos: Cichlini, Retroculini, Astronotini,
Chaetobranchini, Geophagini, Cichlasomatini, e Heroini. Abordagens
citogenéticas clássicas têm sido realizadas no grupo neotropical, contudo
faltam informações sobre a mapeamento físico cromossômico, bem como
análise comparativa de sequências repetitivas, sendo que estas abordagens
são fundamentais para a identificação de rearranjos cromossômicos e melhor
interpretação da organização genômica deste grupo de peixes. O presente
estudo investigou de maneira comparativa, a organização genômica de
exemplares de tribos basais (Cichlini e Astronotini), intermediária (Geophagini)
e derivadas (Heroini) com uma abordagem citogenética clássica e molecular,
utilizando DNAr 18 e 5S, sequências teloméricas, retrotransposons Rex1, 3 e 6,
sequências microssatélites e fração repetitiva Cot-1 DNA, para mapeamento
cromossômico. Análises das sequências nucleotídicas foram realizadas.
Variações nos diversos marcadores citogenéticos foram evidenciadas, porém
nenhuma tendência com relação a aumento, diminuição ou manutenção do
número diploide basal de 2n=48 foi verificada nestas tribos. Algumas espécies
possuem dupliçações gênicas do DNAr 18S, bem como, sítios simples ou
múltiplos. Na maioria dos táxons analisados o DNAr 5S foi localizado na região
intersticial de um par de cromossomos homólogos, porém variações neste
padrão foram verificadas. Sítios teloméricos intersticiais (ITS) estão presentes
em cromossomos de algumas espécies, os quais foram relacionados com
eventos de rearranjos cromossômicos. O mapeamento físico dos
retroelementos evidenciou abundantes marcações dispersas nos cromossomos
da maioria das espécies. Algumas espécies apresentaram marcações
conspícuas em porções terminais e centroméricas, que estão marcações estão
associadas à regiões heterocromáticas, e às porções cromossômicas
eucromáticas. O elemento Rex1 apresentou maior quantidade de marcações
em espécies derivadas. As sequências nucleotídicas de Rex1 e 3 foram mais
conservadas em espécies basais que em derivadas, entretanto este padrão

x
não foi verificado em Rex6. Ainda, a presença de diferentes linhagens foram
observadas em Rex1, bem como possíveis eventos de transferência horizontal
marcaram a evolução de Rex1 e 3 no genoma de ciclídeos neotropicais. O
mapeamento do Cot-1 DNA e de sequências microssatélites evidenciou sinais
em porções centroméricas e teloméricas em algumas espécies e dispersos em
outras. Foi possível verificar ainda para as análises de Cot-1 DNA, que
espécies mais próximas filogeneticamente apresentam maior similaridade na
organização da fração repetitiva nos cromossomos. Os dados demonstram
ampla diversidade genômica e cariotípica existente em ciclídeos neotropicais e
sugerem que grande parte desta ocorre em função de sequências repetitivas,
as quais parecem estar influenciando a evolução destes peixes.

xi
Abstract In addition to their importance as food sources and for ornamental purposes
and aquiculture, cichlids are considered models in genetic and evolutionary
studies. These fish are distributed throughout Central and South America,
Africa, Madagascar, and southern India. All of the neotropical cichlids belonging
to the subfamily Cichlinae, which is in turn subdivided into seven tribes: Cichlini,
Retroculini, Astronotini, Chaetobranchini, Geophagini, Cichlasomatini, and
Heroini. Classical cytogenetic investigations have focused on the neotropical
group, but more information is still needed concerning their physical
chromosomal mapping as well as comparative analyses of their repetitive
sequences – fundamental approaches for the identification of chromosomal
rearrangements and for more precise interpretations of the genomic
organization of this group of fish. In that sense, the present study undertook a
comparative study of the genomic organization of specimens of the basal
(Cichlini and Astronotini), intermediate (Geophagini), and derived (Heroini)
tribes of Cichlinae using both classical and molecular cytogenetic techniques,
utilizing 18 and 5S DNAr, telomeric sequences, retrotransposons Rex1, 3 and
6, microsatellite sequences, and repetitive Cot-1 DNA fractions as probes for
chromosomal mapping. Analyses of nucleotide sequences were also
performed. Variations were seen in diverse cytogenetic markers, although no
tendency was confirmed in these tribes in terms of increasing, decreasing, or
maintaining the basal diploid number 2n=48. Some species demonstrated
genetic duplications of 18S DNAr, as well as simple or multiple sites. 5S DNAr
was located in the interstitial region of a homologous chromosome pair in most
of the taxa analyzed, although variations on this pattern were seen. Interstitial
telomeric sites (ITS) were observed on the chromosomes of some species and
were related to chromosomal rearrangement events. In terms of retroelements,
physical mapping indicated abundant markers dispersed along the
chromosomes of most species. Some species demonstrated conspicuous
markings in their terminal and centromeric portions that were associated with
heterochromatic as well as euchromatic chromosomal regions. The Rex1
element demonstrated greater numbers of markings in derived species. The
nucleotide sequences of Rex1 and 3 were more conserved in basal species
than in derived species, although this same pattern was not observed for Rex6.

xii
Evidence for different lineages was observed in Rex1, as well as possible
horizontal transfer events marking the evolution of Rex1 and 3 in the cichlid
genome. The mapping of the Cot-1 DNA repetitive fraction and microsatellite
sequences indicated centromeric and telomeric sites in some species but
dispersed sites in others. It was also possible to determine through Cot-1 DNA
analyses that the phylogenetically more closely related species demonstrated
greater similarity in the organizations of the repetitive fractions of their
chromosomes. Our data indicated ample genomic and karyotypic diversity
among Neotropical cichlids, suggesting that most of this diversity occurred due
to repetitive sequences that influenced the evolution of these fish.

xiii
SUMÁRIO
1 Introdução ..................................................................................................... 19
1.1 Objetivos ................................................................................................. 26
1.1.1 Objetivo Geral ................................................................................... 26
1.1.2 Objetivos específicos ........................................................................ 26
2 Material e Métodos........................................................................................ 27
2.1 Material ................................................................................................... 27
2.2 Métodos .................................................................................................. 29
2.2.1 Indução de mitoses........................................................................... 29
2.2.2 Obtenção de cromossomos mitóticos ............................................... 29
2.2.3 Detecção da heterocromatina constitutiva (Banda C)....................... 30
2.2.4 Detecção das regiões organizadoras de nucléolo – RONs............... 30
2.2.5 Extração de DNA .............................................................................. 31
2.2.6 Isolamento de sequências repetitivas por PCR (Polymerase Chain
Reaction) ................................................................................................... 31
2.2.7 Isolamento de sequências repetitivas através da técnica de Cot-1
DNA ........................................................................................................... 32
2.2.8 Clonagem ......................................................................................... 33
2.2.9 Sequenciamento dos fragmentos de DNA repetitivo ........................ 33
2.2.10 Hibridação in situ por fluorescência - FISH..................................... 34
2.2.11 Análise das Sequências.................................................................. 35
3. Resultados e Discussão............................................................................... 36
3.1 Artigo 1 – Evolução cromossômica de ciclídeos Neotropicais: o papel das
sequências de DNA repetitivo na organização e estrutura do cariótipo. ....... 37
3.1.1 Introdução......................................................................................... 39
3.1.2 Material e Métodos ........................................................................... 40
3.1.3 Resultados........................................................................................ 43
3.1.4 Discussão ......................................................................................... 52

xiv
3.2 Artigo 2 - Dinâmica evolutiva de elementos retrotransponíveis Rex1, Rex3
e Rex6 no genoma de ciclídeos Neotropicais. .............................................. 57
3.2.1 Introdução......................................................................................... 58
3.2.2 Material e Métodos ........................................................................... 60
3.2.3 Resultados........................................................................................ 62
3.2.4 Discussão ......................................................................................... 73
3.3 Artigo 3 - Mapeamento cromossômico e sequenciamento nucleotídico de
DNAs repetitivos de ciclídeos Neotropicais................................................... 78
3.3.1 Introdução......................................................................................... 79
3.3.2 Material e Métodos ........................................................................... 81
3.3.3 Resultados........................................................................................ 83
3.3.4 Discussão ......................................................................................... 89
3.4 Artigo 4 – Distribuição de sequências microssatélites em cromossomos
de ciclídeos amazônicas. .............................................................................. 95
3.4.1 Introdução......................................................................................... 96
3.4.2 Material e Métodos ........................................................................... 97
3.4.3 Resultados........................................................................................ 98
3.4.4 Discussão ....................................................................................... 101
4 Conclusões Gerais ...................................................................................... 104
5 Referências bibliográficas ........................................................................... 105

xv
Lista de figuras
Introdução
Figura 1 Árvore de máxima parcimônia, com base dados genéticos e
morfológicos proposta por Smith et al. 2008. ................................................... 21
Figura 2 Imagem de satélite da região da bacia amazônica central,
evidenciando os pontos de coleta; 1=Barcelos; 2=rio Branco; 3=Anavilhanas;
4=rio Manacapuru; 5=lago Catalão; 6=Careiro; 7=lago de Balbina.................. 28
Capítulo 1
Figura 1 Cariótipos das espécies pertencentes as tribos de Cichlinae (Cichlini,
Astronotini, Geophagini, Cichlasomatini e Heroini) em banda-C. Os
cromossomos portadores da RON são mostrados na caixa. ........................... 45
Figura 2 Cariótipos das espécies pertencentes as tribos de Cichlinae (Heroini)
em banda-C. Os cromossomos portadores da RON são mostrados na caixa. 46
Figura 3 Cariótipos das espécies pertencentes as tribos da subfamília
Cichlinae, mostrando os sítios de DNAr 18S (vermelho) e 5S (verde). Os
cromossomos foram contra corados com DAPI. .............................................. 48
Figura 4 Cariótipos das espécies pertencentes as tribos da subfamília
Cichlinae, mostrando os sítios de DNAr 18S (vermelho) e 5S (verde). Os
cromossomos foram contra corados com DAPI. .............................................. 49
Figura 5 Cariótipos das espécie pertencentes as tribos de Cichlinae,
mostrando sinais de hibridização com sonda telomérica (vermelho). Os
cromossomos foram contra corados com DAPI. .............................................. 50
Figura 6 Cariótipos das espécie pertencentes as tribos de Cichlinae,
mostrando sinais de hibridização com sonda telomérica (vermelho). Os
cromossomos foram contra corados com DAPI. .............................................. 51
Capítulo 2
Figura 1 Alinhamento das sequências de aminoácidos dos diferentes
retroelementos entre ciclideos: A) Região correspondente a transcriptase

xvi
reversa de Rex1 das espécies analisadas em comparação com O. niloticus
(GenBank: AJ288473); B) Região correspondente a transcriptase reversa de
Rex3 em comparação com O. niloticus (GenBank: AJ400370) e C) Região da
endonuclease de Rex6 das espécies analisadas em comparação com O.
niloticus (GenBank: AJ293545). ....................................................................... 64
Figura 2 Árvore de máxima verossimilhança dos 49 clones de Rex1, revelando
duas linhages monofiléticas deste retroelemento. Valores de bootstrap maiores
que 60 são mostrado sobre os ramos. ............................................................. 65
Figura 3 Árvore de máxima verossimilhança dos 50 clones do retroelemento
Rex3. Valores de bootstrap maiores que 60 são mostrado sobre os ramos. ... 67
Figura 4 Árvore de máxima verossimilhança dos 59 clones do retroelemento
Rex6. Valores de bootstrap maiores que 60 são mostrado sobre os ramos. ... 69
Figura 5 Mapeamento cromossômico do retrotransposon Rex1 (sinais em
vermelho). Os cromossomos foram contra corados com DAPI........................ 71
Figura 6 Mapeamento cromossômico do retrotransposon Rex3 (sinais em
vermelho). Os cromossomos foram contra corados com DAPI........................ 72
Figura 7 Mapeamento cromossômico do retrotransposon Rex6 (sinais em
vermelho). Os cromossomos foram contra corados com DAPI........................ 73
Capítulo 3
Figura 1 Hibridização da fração Cot-1 DNA em quatro espécies de Cichlinae;
(A, E, I, M) cromossomos contra corados com DAPI das espécies C.
monoculus, A. ocellatus, G. proximus e S. discus, respectivamente; (B, F, J, N)
Sonda Cot-1 DNA isolada do genoma de C. monoculus hibridizada em seus
próprios cromossomos e contra os cromossomos das demais espécies, em (F)
cabeça de seta evidencia par sem sinais de hibridização, e alguns sinais
restringem-se a regiões teloméricas (seta); (C, G, K, O) Sonda Cot-1 DNA
isolada do genoma de S. discus hibridizada contra os cromossomos das
espécies analisadas e em seus próprios cromossomos, em (O) seta evidencia
cromossomos sem sinais de hibridização; (D, H, L, P) Double FISH da fração
Cot-1 DNA. ....................................................................................................... 88
Capítulo 4

xvii
Figura 1 Cromossomos metafásicos de C. monoculus, P. scalare e S. discus
hibridizados com sequências microssatélites (a – m). Setas evidenciam
ausência de sinais de hibridizações. Os cromossomos foram contra corados
com DAPI. ...................................................................................................... 101

xviii
Lista de tabelas Capítulo 1
Tabela 1 Espécies de Cichlinae analisadas..................................................... 41
Capítulo 3
Tabela 1 Similaridade entre sequências da fração Cot-1 DNA de C. monoculus
e sequências conhecidas em banco de dados públicos................................... 84
Tabela 2 Similaridade entre sequências da fração Cot-1 DNA de S. discus e
sequências conhecidas em banco de dados públicos...................................... 85
Capítulo 4
Tabela 1 Sequências dos microssatélites hibridizados nos cromossomos dos
ciclideos. (+) presença de sinal de hibridização; (-) ausência de sinal de
hibridização. ..................................................................................................... 99

19
1 Introdução
Biologia e citogenética de ciclídeos Sul Americanos
A família Cichlidae pertence à ordem Perciformes e é uma das mais
ricas em espécies de peixes de água doce do mundo, com aproximadamente
3.000 espécies. Possui ampla distribuição geográfica, sendo encontrada nas
Américas Central e do Sul, África, Madagascar e sul da Índia. Sua maior
diversidade ocorre nos lagos do leste africano, seguida da região Neotropical,
onde a região amazônica é a principal responsável por esta riqueza de
espécies (Kullander 2003).
Na Amazônia, os ciclídeos destacam-se tanto como fonte protéica,
quanto para fins ornamentais e pesca esportiva. No que diz respeito à
alimentação, os peixes são a base alimentar em regiões ribeirinhas e nos
grandes centros amazônicos, sendo umas das espécies de ciclídeos mais
consumidas o acará-açu (Astronotus spp.), o papa-terra (Geophagus spp.) e o
tucunaré (Cichla spp.), onde dados pesqueiros chegam a registrar um
desembarque de mais de 5.000 toneladas por ano da última espécie (Cabral
Júnior e Almeida 2006).
Além disso, os tucunarés também vêm atraindo aficionados pela pesca
esportiva de todo o mundo devido ao seu comportamento agressivo. Este tipo
de pesca tem aumentado no decorrer dos anos, principalmente na região que
abrange o médio rio Negro e seus afluentes (Freitas e Rivas 2006). Nesta
mesma região ocorre a exploração de peixes ornamentais. Segundo Anjos et
al. (2007) foram exportados mais de 700.000 exemplares de ciclídeos no ano
de 2005, com destaque para o acará-bandeira (Pterophyllum spp.), com mais
de 6.000 exemplares exportados legalmente em 2007 e o acará-disco
(Symphysodon spp.), com mais de 42.000 exportações na mesma época
(IBAMA 2008). O destino da maioria destes peixes é a Ásia e a Europa.
Os ciclídeos são considerados um grupo monofilético (Kullander 1998;
Farias et al. 1999, Smith et al. 2008), que se destacam pelo grande sucesso
evolutivo e alta taxa de especiação e especialização (Lowe-McConnell 1969;
Thompson 1979). Farias et al. (2000), combinando caracteres morfológicos e
genéticos (sequências de DNA de genes mitocondriais e nucleares) sugeriram
que as subfamílias de ciclídeos africanos Heterochromidinae e

20
Pseudocrenilabrinae seria grupo irmão dos ciclídeos neotropicais. De acordo
com Kullander (1998), os ciclídeos neotropicais pertencem à família Cichlidae,
e esta é subdividida nas seguintes subfamílias: Retroculinae, Cichlinae,
Astronotinae, Geophaginae e Cichlasomatinae. Entretanto, um gênero africano
(Heterochromis) era agrupado com os ciclídeos neotropicais. Na última revisão
filogenética realizada neste grupo, baseada em diferentes marcadores
genéticos e morfológicos, todos os ciclídeos neotropicais foram agrupados na
subfamília Cichlinae subdividida em sete tribos: Cichlini, Retroculini, Astronotini,
Chaetobranchini, Geophagini, Cichlasomatini, e Heroini (Smith et al. 2008)
(Figura 1).

21
Figura 1 Árvore de máxima parcimônia, com base em dados genéticos e
morfológicos proposta por Smith et al. 2008.
O número diploide mais frequente para espécies da subfamília Cichlinae
é 2n=48 cromossomos, número considerado ancestral para esta subfamília e
também para a ordem Perciformes (Brum 1995; Feldberg et al. 2003). Porém,
de acordo com Feldberg et al. (2003), três tendências de evolução
cromossômica são evidenciadas nos ciclídeos: a primeira é a manutenção do

22
número diploide ancestral, a segunda é a diminuição do número cromossômico
chegando até 2n=32 e a terceira é o aumento do número diploide atingindo
2n=60 cromossomos.
Estas três tendências foram evidenciadas na tribo Cichlasomatini por
Santos (2006), creditadas à ocorrência de rearranjos Robertsonianos (fissão e
fusão cêntrica) e não Robertsonianos (inversão pericêntrica) durante a
evolução cariotípica dos ciclídeos amazônicos. Um exemplo de diminuição do
número diploide é visualizado no gênero Pterophyllum, onde P. scalare e P.
leopoldi apresentam número diploide igual a 48 cromossomos e a espécie P.
altum, 2n=46 cromossomos, diminuição esta que ocorreu devido à fusão de
quatro cromossomos. Além disso, inversões pericêntricas também são
evidentes no gênero e teriam modificado o número de braços cromossômicos.
Para o gênero Cichla, destaca-se o trabalho realizado por Alves-Brinn et
al. (2004), onde, analisando espécimes provenientes da Usina Hidroelétrica de
Balbina (bacia do rio Uatumã), evidenciaram uma possível hibridização natural
entre C. monoculus e C. temensis. Os indivíduos híbridos (Cichla sp.)
apresentaram características morfológicas e cariotípicas intermediárias. Todos
os indivíduos (C. monoculus, C. temensis e Cichla sp.) apresentavam cariótipo
com 2n=48 cromossomos acrocêntricos, porém com relação à região
organizadora de nucléolo (RON) os exemplares de C. monoculus e cinco
indivíduos híbridos apresentaram marcação no segundo par cromossômico,
enquanto para C. temensis e três híbridos esta foi evidenciada no terceiro par.
Como C. monoculus e C. temensis são encontrados em simpatria em muitos
locais da bacia amazônica, os autores sugerem que o ancestral de Cichla sp.
seria o resultado do cruzamento destas espécies.
Com objetivo de conhecer o cariótipo de ciclídeos neotropicais Felberg e
Bertollo (1985) analisaram Geophagus brasiliensis, G. surinamensis e
Astronotus ocellatus. Com relação à G. brasiliensis e G. surinamensis os
autores encontraram 2n=48 cromossomos para ambos, porém evidenciaram
uma diferença na morfologia cromossômica, onde a primeira espécie apresenta
apenas um par de metacêntrico enquanto a segunda dois pares. No que se
refere à Astronotus ocellatus, todos os espécimens apresentaram 2n=48
cromossomos, porém com seis pares de cromossomos meta-
submetacêntricos. Estes dados evidenciam novamente que, embora o número

23
cromossômico da subfamília seja conservado, algumas espécies tiveram
rearranjos estruturais durante a sua evolução cromossômica propiciando assim
a alteração da fórmula cariotípica.
Os acarás-discos do gênero Symphysodon foram analisados
citogeneticamente e revelaram dados interessantes. As três espécies do
gênero (S. aequifasciatus, S. discus e S. haraldi) apresentaram número
diploide igual a 2n=60 cromossomos, com a maioria dos cromossomos meta-
submetacêntricos (Mesquita et al. 2008; Gross et al. 2009a). Além desta
característica derivada, S. aequifasciatus apresentou também uma cadeia
cromossômica meiótica em anel contendo 20 elementos e mais 20 bivalentes
em células diplotênicas, provavelmente originada de translocações múltiplas e
seriadas, enquanto S. discus apresentou apenas 30 bivalentes, o que sugere
que S. aequifasciatus diferenciou-se cromossomicamente num período mais
recente que S. discus (Gross et al. 2009a).
DNAs repetitivos
Genes de cópia única ou de poucas cópias correspondem a uma
pequena parte do genoma dos vertebrados (2-10%), quando comparado às
regiões repetitivas (Horvath et al. 2001). Eucariotos têm dois tipos de DNA
altamente repetitivos: sequências de DNA repetitivos dispersas e sequências
de DNA repetidas em tandem. As sequências de DNA repetidas em tandem
incluem os DNAs satélite, enquanto as sequências repetitivas dispersas
englobam os elementos transponíveis (Phillips e Reed 1996; Martins 2007).
DNAs satélites podem variar de 1.000 a 100.000 cópias por genoma,
comumente com repetições entre 100 a 300 pares de bases. Estas sequências
são organizadas em grandes grupos, principalmente na região telomérica e
centromérica dos cromossomos e são os principais componentes das
heterocromatinas (Martins 2007). Esta classe de DNA repetitivo é uma
ferramenta útil no mapeamento físico do genoma, contribuindo para o
desenvolvimento de marcadores genéticos de fundamental importância para o
conhecimento da estrutura cromossômica e da evolução em muitas espécies
de peixes (Azevedo et al. 2005).
Os elementos transponíveis (transposons e retrotransposons) são
encontrados dispersos pelo genoma, com um número de cópias que pode

24
variar de 10 a milhares, dependendo do elemento transponível e da espécie
que se encontra (Tafalla et al. 2006). Com base na estrutura e em seu
mecanismo de transposição estes elementos são divididos em duas classes: a
primeira engloba os três tipos de transposons que têm um RNA como
intermediário (retrotransposon), sendo eles: elementos longos intercalados
(long interspersed elements-LINEs), elementos curtos intercalados (short
intersperser elements-SINEs) e os que possuem longas repetições terminais
(long terminal repeats-LTRs); a segunda inclui todos os que são transpostos
diretamente de DNA para DNA, ou seja, sem outra molécula intermediária (Van
Sluys et al. 2001; Tafalla et al. 2006; Martins 2007). Nos genomas dos peixes
são encontradas as duas classes de elementos transponíveis (Volff et al. 2003;
Tafalla et al. 2006).
Elementos transponíveis podem estar associados a regiões
heterocromáticas, como demonstrado por Capriglione et al. (2002), em
Chionodraco hamatus, família Channichthyidae da região Antártica. Os autores
estudaram o transposon Tc1 e evidenciaram que ele está localizado em
regiões heterocromáticas, particularmente no cromossomo sexual (Y)
sugerindo que este elemento transponível esteve envolvido na diferenciação
deste cromossomo.
Analisando retrotransposons (Rex1, Rex3, Rex6) em peixes da
Antártica, Ozouf-Costaz et al. (2004) sugeriram uma possível correlação entre
rearranjos cromossômicos e atividade dos retrotransposons (Rex3), tendo em
vista que a espécie que possui o cariótipo derivado tem um padrão de
hibridização mais compartimentalizada, com acúmulo destes elementos nas
regiões pericentroméricas dos cromossomos, enquanto as espécies basais
apresentam distribuição homogênea.
Mapeamento citogenético
Apesar da citogenética estar proporcionando dados interessantes a
respeito dos ciclídeos amazônicos, estudos envolvendo a localização de
sequências específicas de DNA em cromossomos são ainda incipientes nos
peixes desta região. A citogenética molecular é utilizada para identificação de
sequências de DNA específicas no cromossomo, consistindo basicamente no

25
pareamento de determinado segmento de DNA ou RNA com uma sequência de
nucleotídeos complementar situado na célula alvo (Guerra 2004).
Na citogenética de peixes a hibridização fluorescente in situ (FISH)
revela-se importante, pois tem permitido novas interpretações da diversidade
cariotípica, em particular sobre a origem de cromossomos sexuais e de
supranumerários, bem como sobre a organização genômica de segmentos
cromossômicos (Galetti Jr e Martins 2004).
As sequências repetitivas de DNA tem sido amplamente exploradas
como sondas para FISH uma vez que este tipo de DNA gera sinais de fácil
visualização nos cromossomos. Tais abordagens vem sendo aplicadas a
estudos que visam o entendimento da carioevolução e organização genômica
em peixes, uma vez que permite a localização de sequências específicas de
DNA no cariótipo de uma espécie e a comparação destas com grupos
relacionados. Tendo em vista que os ciclídeos amazônicos são altamente
explorados, estudos que visem o conhecimento do genoma destes peixes
fazem-se necessários. Dessa forma, o mapeamento cromossômico de
sequências repetitivas de DNA em ciclídeos pode propiciar uma melhor visão
da organização do genoma e, portanto, gerar subsídios para programas de
melhoramento genético assistido na piscicultura, além de ser um marcador
interessante para inferências sobre rearranjos cromossômicos que ocorreram
na evolução do grupo. Para tanto foram escolhidas espécies de ciclídeos
neotropicais de interesse econômico, englobando as diferentes tribos, desde
basais até derivadas, com ampla distribuição e elevada abundância na bacia
amazônica.

26
1.1 Objetivos
1.1.1 Objetivo Geral
Identificar e mapear sequências de DNA repetitivo em ciclídeos
amazônicos, englobando espécies consideradas basais e derivadas,
contribuindo para o entendimento da organização genômica dos ciclídeos
neotropicais e sua evolução cariotípica.
1.1.2 Objetivos específicos
Isolar e caracterizar sequências repetitivas de DNA presentes no genoma
das espécies de diferentes tribos de Cichlinae.
Comparar o mapeamento cromossômico destas espécies, utilizando as
sequências repetitivas isoladas.
Inferir quais foram os principais rearranjos que ocorreram na carioevolução
dos ciclídeos neotropicais.
Analisar comparativamente as sequências nucleotídicas entre as espécies
modelo.

27
2 Material e Métodos
2.1 Material
Foram estudados citogeneticamente 12 exemplares de Cichla
monoculus (Cichlini), 10 de Astronotus ocellatus (Astronotini), 12 Geophagus
proximus (Geophagini), 6 de Acaronia nassa e 1 de Bujurquina peregrinabunda
(Cichlasomatini), 6 de Hoplarchus psittacus, 4 de Hypselecara coryphaenoides,
5 de Hypselecara temporalis, 3 de Caquetaia spectabilis, 6 de Uaru
amphiacanthoides, 4 de Pterophyllum leopoldi, 12 de Pterophyllum scalare e 12
de Symphysodon discus (Heroini). Os indivíduos foram coletados em diferentes
localidades da bacia amazônica (Figura 2), com redes de pesca (malhas 70, 80
e 90), e seguindo as normatizações do IBAMA de acordo com a autorização
SISBIO 10609-1/2007.
Os peixes foram mantidos vivos em aquário aerado durante o transporte
até o laboratório de campo ou para o Laboratório de Genética Animal do
Instituto Nacional de Pesquisa da Amazônia - Manaus, onde foram realizadas
as análises.
Os exemplares foram anestesiados em água gelada. Para a obtenção de
preparações cromossômicas foram extraídos os órgãos hematopoiéticos e para
extração do DNA amostras de tecido muscular e fígado de todos os indivíduos,
sendo os animais numerados, registrados, fixados em formol 10% durante 24h
e acondicionados posteriormente em recipientes contendo álcool 70%.
Exemplares testemunhos foram depositados na Coleção Ictiológica do INPA
sob números: INPA 26451, INPA 26452, INPA 26453, INPA 26459, INPA
26462, INPA 26466

28
Figura 2 Imagem de satélite da região da bacia amazônica central, evidenciando os pontos de coleta; 1=Barcelos; 2=rio Branco;
3=Anavilhanas; 4=rio Manacapuru; 5=lago Catalão; 6=Careiro; 7=lago de Balbina.

29
2.2 Métodos
2.2.1 Indução de mitoses
Para se obter um maior número de células em metáfase, foi utilizada a
técnica para indução de mitoses, descrita por Oliveira et al. (1988), na qual
uma solução de fermento biológico foi aplicada nos espécimes.
A solução de fermento biológico foi preparada com 0,3 g de fermento
biológico comercial, 0,3 g de açúcar e 10 mL de água destilada, sendo esta
mantida em estufa (37 °C) por um período de 15-20 minutos, e posteriormente
injetada na região dorso-lateral do peixe, na proporção de 1mL para cada 100 g
de peso do animal. Os peixes submetidos ao tratamento com solução de
fermento biológico foram colocados em aquários bem aerados por 24 horas.
2.2.2 Obtenção de cromossomos mitóticos
As preparações citogenéticas foram obtidas seguindo-se a metodologia
descrita por Bertollo et al. (1978), com algumas modificações, que consiste em
injetar intraperitonealmente colchicina 0,0125% na proporção de 1 mL para
cada 100 g de peso do animal. Os peixes permaneceram em aquário bem
aerado por 40 minutos. Em seguida foram sacrificados por imersão em água
gelada e a porção anterior do rim foi retirada, sendo transferida para uma
solução hipotônica de KCL 0,075 M (6-8 mL). O tecido foi divulcionado com o
auxílio de uma seringa de vidro e o sobrenadante (suspensão celular) foi
transferido com o auxílio de uma pipeta Pasteur para um tubo de centrífuga. A
suspensão celular obtida foi incubada em estufa a 37 °C por 30 minutos.
Transcorrido este tempo a suspensão foi pré-fixada com 6 gotas de
metanol:ácido acético (3:1) e ressuspendida. Após 5 minutos, foi adicionado
fixador até encher o tubo. A suspensão foi centrifugada por 10 minutos a 900
rpm. O sobrenadante foi desprezado e 6 mL de fixador foi adicionado, sendo o
material ressuspendido novamente e centrifugado por 10 minutos a 900 rpm. O
sobrenadante foi desprezado e 6 mL de fixador foi adicionado, e essa lavagem
foi repetida por mais duas vezes. Após a última centrifugação e eliminação do
sobrenadante, 1,5 mL de fixador foram adicionados e o material foi
ressuspendido com cuidado. Essa suspensão celular foi então estocada em
micro tubo e mantida em freezer para posterior utilização.

30
Para a preparação das lâminas as mesmas foram lavadas, secas ao ar e
posteriormente imersas em água destilada a 55 ºC, em banho-maria. Após
cinco minutos, as lâminas foram retiradas da água de forma a manter uma
película de água sobre a sua superfície, na qual foi gotejada a suspensão
celular em diferentes regiões. As lâminas secaram diretamente ao ar.
2.2.3 Detecção da heterocromatina constitutiva (Banda C)
Para a detecção da heterocromatina constitutiva foi utilizada a técnica
descrita por Sumner (1972), com algumas modificações. As lâminas com a
suspensão celular foram tratadas em solução de HCl 0,2N a 45 °C, por 2
minutos e lavadas rapidamente em água destilada à temperatura ambiente.
Após secas as lâminas foram incubadas por 1 minuto e 20 segundos em
solução de hidróxido de bário a 5%, a 42 °C, recém preparada e filtrada. A
ação do hidróxido de bário foi interrompida imergindo-se rapidamente a lâmina
em solução de HCl 0,2N, lavada em água destilada e deixada secar ao ar.
Após secas, as lâminas foram incubadas em solução 2xSSC (cloreto de sódio
0,3M e citrato trisódico 0,03M, pH 6,8) em banho-maria a 60 °C, por 15
minutos. Em seguida, as lâminas foram lavadas várias vezes em água
destilada e secas ao ar, sendo posteriormente coradas com solução de Giemsa
(diluída a 5% em tampão fosfato 0,06M e pH 6,8) durante 10 minutos, lavadas
em água destilada e secas.
2.2.4 Detecção das regiões organizadoras de nucléolo – RONs
Para a detecção das regiões organizadoras de nucléolo (RONs) foi
utilizada a técnica descrita por Howell e Black (1980), com modificações. Sobre
as lâminas com a suspensão foram adicionadas 2 gotas de uma solução
coloidal de gelatina (2 g de gelatina comercial sem sabor, dissolvida em 100
mL de água destilada, acrescida de 1 mL de acido fórmico) e, sobre cada gota
de gelatina, duas gotas de solução aquosa de nitrato de Prata (AgNO3) a 50%,
agitando-se levemente a lâmina, que foi coberta com lamínula. A lâmina foi
colocada em câmara úmida, em banho-maria a 60 °C, até adquirir uma
coloração marrom dourada, sendo posteriormente lavada em água destilada.

31
2.2.5 Extração de DNA
Para extração do DNA foram descartadas as vísceras e as escamas,
utilizando-se somente o tecido muscular. O protocolo de extração de DNA
utilizado foi o descrito por Sambrook et al. (1989), com algumas modificações.
O tampão de lise utilizado foi (Tris-HCl pH 8,0 em 10 mM, NaCl 0,3 M, EDTA
10 mM), posteriormente foram acrescentados: 15 μL de proteinase K (10
mg/mL) e 6 μL de RNAse A (10 mg/mL). As amostras foram incubadas para
que o tecido fosse totalmente digerido.
A seguir foram feitas lavagens sucessivas com fenol clorofórmio e
clorofórmio hidratado (500μL de cada um destes reagentes). Após a lise, o
DNA foi separado das proteínas por precipitação salina juntamente com
centrifugação a 14000 rpm. O sobrenadante foi então precipitado com 600 μL
de isopropanol 100% gelado, também com o auxílio de centrifugação. Ao final,
o DNA foi hidratado com aproximadamente 100 μL de água milli-Q,
dependendo do tamanho do pellet (DNA precipitado) formado.
Para possibilitar a análise da quantidade e integridade do material, o
DNA extraído foi quantificado por comparação com marcador de concentração
conhecida, em eletroforese padrão (com tampão Tris-Borato-EDTA 0,5X e
corrida a 70 V por 40 minutos) em gel de agarose 0,8% e corado com GelRed
Acid Gel Stain Biotium (1:500). A visualização e análise do DNA no gel foram
feitas no fotodocumentador Easy Doc 100 (BioAgency), o qual possui acoplado
um transluminador de luz ultravioleta (260 nM). Adicionalmente, quantificações
em espectrofotômetro NanoVue Plus (GE Healthcare) foram realizadas.
2.2.6 Isolamento de sequências repetitivas por PCR (Polymerase Chain
Reaction)
Elementos transponíveis Rex1, Rex3 e Rex6, bem como as sequências
teloméricas e os DNAs ribossomais 18S e 5S foram amplificados por PCR. Os
primers utilizados, as condições de PCR e os perfis encontram-se detalhados
nas seções Material e Métodos de cada capítulo. Depois de isolados, os
elementos transponíveis foram clonados e sequenciados. Os produtos de PCR
foram verificados e quantificados por comparação com o marcador Low Mass
DNA Ladder (Invitrogen) em eletroforese em gel de agarose 1,0% corado com

32
GelRed Acid Gel Stain (Biotium 1:500). A visualização e análise do DNA no gel
foram feitas no fotodocumentador Easy Doc 100 (BioAgency), o qual possui
acoplado um transluminador de luz ultravioleta (260 nM). Adicionalmente,
quantificações em espectrofotômetro NanoVue Plus (GE Healthcare) foram
realizadas.
Após isolados e caracterizados por sequenciamento, os elementos
foram utilizados como sondas para hibridização dos cromossomos metafásicos.
2.2.7 Isolamento de sequências repetitivas através da técnica de Cot-1 DNA
O isolamento de sequências repetitivas baseado na cinética de
reassociação do DNA foi conduzido segundo Zwick et al. (1997) e adaptado por
Ferreira e Martins (2008). O DNA genômico foi diluído a 100-500 ng/µl em NaCl
0,3 M. Em seguida, em tubos de 1,5 ml foi colocado 50 µl de DNA e
autoclavado por 3 minutos a 1.4 atm/120 oC. Foi aplicado 3 µl do DNA
autoclavado em gel de agarose 1% para checar o tamanho dos fragmentos
obtidos. Após, 3 alíquotas (tubos 0, 3 e 5) com 50 µl do DNA autoclavado foram
desnaturadas em banho a 95 oC por 10 minutos. Os tubos foram colocados em
gelo por 10 segundos. O tubo 0 foi tratado imediatamente com enzima S1
nuclease. Os tubos 1 e 5 foram colocados em banho a 65 oC para renaturação.
Após 3 minutos, foi retirado o tubo 1 e tratado com S1 nuclease e após 5
minutos, foi retirado o tubo 5 e tratado com a mesma enzima. Para o
tratamento com S1 nuclease, foi utilizado 1U da enzima para cada 1 µg de
DNA (5,5 µl tampão 10X para o volume final de 50 µl). As amostras foram
incubadas a 37 oC por 8 minutos e posteriormente congeladas imediatamente
em nitrogênio líquido. Foi adicionado volume igual de fenol/clorofórmio (1:1) e
centrífugado por 5 minutos a 13.000 rpm. Em seguida, as amostras foram
transferidas para um novo tubo e o DNA foi precipitado com 2,5 volumes de
etanol absoluto gelado. O material foi deixado no freezer a 80 oC negativos por
30 minutos e posteriormente centrifugado por 15 minutos a 15.000 rpm a 4 oC,
o álcool foi descartado e o pellet foi seco e estufa. O material foi ressuspendido
em 50 µl de água ultra-pura estéril autoclavada e o tamanho do fragmento foi
checado por comparação com marcador de Low Mass DNA 1 Kb, em

33
eletroforese padrão (com tampão Tris-Borato-EDTA 0,5X e corrida a 70 V por
40 minutos) em gel de agarose 0,8%, corado com GelRed Acid Gel Stain
(Biotium 1:500). A visualização e análise do DNA no gel foram feitas no
fotodocumentador Easy Doc 100 (BioAgency), o qual possui acoplado um
transluminador de luz ultravioleta (260 nM).
2.2.8 Clonagem
Os elementos transponíveis obtidos por PCR e as sequências repetitivas
obtidas por Cot-1 foram inseridos no vetor plasmidial pGem-T easy (Promega)
e pMOS (GE HealthCare) respectivamente. As construções recombinantes
obtidas foram utilizadas para transformar bactérias competentes de Escherichia
coli linhagem DH5α. Para isso foram pipetadas 50 µL de células competentes
em um tubo eppendorf; e adicionados 10 µL do produto obtido na ligação às
células competentes, misturando tudo suavemente. As células foram incubadas
no gelo por 30 minutos, e em seguida foram submetidas a um choque térmico
por 40 segundos no banho-maria a 42 ºC e colocadas no gelo por 2 minutos.
Foi adicionado 80 µL de meio cultura LB (Luria-Bertani) em cada tubo e os
mesmos foram colocados para agitar a 150 rpm por uma hora a 37 ºC.
Enquanto isso, 30 µL de X-Gal (50 mg/mL) e 50 µL IPTG (0,1 M) foram
espalhados em cada placa de meio de cultura sólido contendo ampicilina (100
mg/mL), para a seleção dos clones recombinantes. As bactérias foram
plaqueadas e incubadas a 37 ºC durante uma noite (aproximadamente 16
horas).
2.2.9 Sequenciamento dos fragmentos de DNA repetitivo
Para confirmar e identificar os produtos isolados os fragmentos foram
sequenciados pelo método de Sanger et al. (1977), com terminadores
marcados com fluorescência.
Para a realização do sequenciamento foram utilizados entre 150 e 300
ng de DNA, 5 pmol de cada primer em reações separadas; 4 μL do premix (kit)
e água mili-Q para completar o volume de 10 μL. Essas reações foram
processadas em um termociclador em 30 ciclos sendo: desnaturação a 95 °C

34
por 20 segundos, anelamento a 50 °C por 15 segundos e extensão a 72 °C por
1 minuto e 30 segundos. Os primers utilizados no sequenciamento foram
primers universais que se anelam nos plasmídios pMOS e pGEM-T utilizados
como vetores de clonagem. Depois de sequenciado, os fragmentos foram
submetidos a um tratamento de precipitação para a eliminação do produto não
incorporado durante a reação de sequenciamento e logo após foi realizada a
leitura automática do fragmento no sequenciador automático ABI 3130 XL
(Applied Biosystem) nas condições de injeção e corrida recomendadas pelo
fabricante.
2.2.10 Hibridização fluorescente in situ (FISH)
Foi realizada a técnica de hidridização fluorescente in situ (FISH)
descrita por Pinkel et al. (1986) com alguns ajustes.
Tratamento das lâminas:
As lâminas foram lavadas em tampão PBS 1x durante 5 minutos em
temperatura ambiente, sendo posteriormente desidratadas em série alcoólica
70, 85 e 100%, 5 minutos cada e secas ao ar. As lâminas foram incubadas em
90 μl de RNAse (0,4% RNAse/2xSSC) a 37 °C por 1h em câmara úmida.
Transcorrido este tempo, as lâminas foram lavadas três vezes em 2xSSC por 5
minutos cada e em PBS 1x pelo mesmo tempo. As lâminas foram fixadas em
formaldeídeo 1% em PBS 1x/50mM MgCl2 durante 10 minutos à temperatura
ambiente e em seguida lavadas em PBS 1x por 5 minutos, sendo então
desidratadas em série alcoólica gelada (70,85, 100 %) por 5 minutos cada.
Simultaneamente a desidratação da lâmina em série alcoólica, a solução de
hibridização contendo formamida 100% (concentração final 50%), sulfato de
dextrano 50%, 20xSSC (concentração final de 2xSSC), sondas marcadas e
água milli-Q foi desnaturada a 99 ºC por um período de 10 minutos, sendo
passada imediatamente ao gelo.
O DNA cromossômico foi desnaturado com formamida 70% em 2xSSC a 70
ºC por 5 minutos e desidratado em etanol gelado 70%, 85% e 100% por 5
minutos cada, secando ao ar. A solução de hibridização foi colocada sobre a
lâmina com os cromossomos desnaturados e incubada em câmara úmida a 37
oC por aproximadamente 18 horas.

35
Após o tempo de hibridização, as lâminas foram lavadas em formamida
15% a 42 oC durante 10 minutos e posteriormente lavadas em solução Tween
0,5%, por 5 minutos. Para detecção do sinal as lâminas foram incubadas em
tampão NFDM por 15 minutos e lavar 2 vezes com Tween 0,5% temperatura
ambiente por 5 minutos cada. As lâminas foram incubadas com o anticorpo
antidigoxigenina ou com estreptavidina, por 60 minutos em câmara úmida a 37 oC. Após este tempo as lâminas foram lavadas 3 vezes com Tween 0,5%
temperatura ambiente por 5 minutos cada, desidratadas em série alcoólica
gelada 70, 85 e 100% durante 5 minutos cada e após secas montadas com
antifade e DAPI, sendo cobertas com lamínulas.
2.2.11 Análise das Sequências
As amostras sequenciadas foram salvas (formato abd) e transferidas do
sequenciador para outro computador. Todas as sequências foram alinhadas
utilizando-se o programa de alinhamento múltiplo Clustal W (Thompson et al.
1994), que está incluso no BioEdit 7 (Hall 1999). Para identificação de
possíveis homologias, as sequências nucleotídicas obtidas foram submetidas a
buscas online BLAST (“Basic Local Alignment Search Tool”) (Altschul et al.
1990) através do National Center for Biotechnology Information (NCBI) (USA),
“website” (http://www.ncbi.nlm.nih.gov/blast/) e na base de dados Repbase do
Genetic Information Research Institute (http://www.girinst.org/censor/) utilizando
o programa on-line Censor (Kohany et al. 2006). As sequências geradas foram
depositadas no banco de dados do GenBank e os números de acessos
encontram-se em cada capítulo no tópico Resultados e Discussão.

36
3. Resultados e Discussão
Os resultados obtidos com a análise cromossômica comparativa de
ciclídeos amazônicos e a discussão destes dados são apresentados na forma
de quatro artigos científicos, mencionado a seguir:
3.1 Artigo 1 – Evolução cromossômica de ciclídeos Neotropicais: o papel das
sequências de DNA repetitivo na organização e estrutura do cariótipo.
Publicado no periódico Reviews in Fish Biology and Fisheries, doi:
10.1007/s11160-012-9285-3.
3.2 Artigo 2 – Dinâmica evolutiva de elementos retrotransponíveis Rex1, Rex3
e Rex6 no genoma de ciclídeos Neotropicais. Enviado para publicação no
periódico BMC Evolutionary Biology, em 10/09/2012 (MS ID:
9875706468235423).
3.3 Artigo 3 – Mapeamento cromossômico e sequenciamento nucleotídico de
DNAs repetitivos de ciclídeos Neotropicais. Enviado para publicação no
periódico Genetica, em 14/11/2012 (MS ID: GENE-D-12-02244).
3.4 Artigo 4 – Distribuição de sequências microssatélites em cromossomos de
ciclídeos amazônicos. Enviado para publicação no periódico Hydrobiologia, em
27/12/2012 (MS ID: HYDR-D-12-08297)

37
3.1 Artigo 1 – Evolução cromossômica de ciclídeos Neotropicais: o papel das sequências de DNA repetitivo na organização e estrutura do cariótipo

38
RESEA RCH PA PER
Chromosomal evolution of neotropical cichlids: the roleof repetitive DNA sequences in the organizationand structure of karyotype
Car los Henr ique Schneider • Mar ia Claudia Gross •
Mar ia Leandra Terencio • Rober to Ferreira Ar toni •
Marcelo Ricardo Vicar i • Cesar Mar tins • Eliana Feldberg
Received: 15 June 2012 / Accepted: 20 August 2012Ó Springer Science+Business Media B.V. 2012
Abstract Cichlids are important in the aquacultureand ornamental fish trade and are considered modelsfor evolutionary biology. However, most studies ofcichlids have investigated African species, and theSouth American cichlids remain poorly characterized.Studies in neotropical regions have focused almostexclusively on classical cytogenetic approaches with-out investigating physical chromosomal mapping ofspecific sequences. The aim of the present study is toinvestigate the genomic organization of speciesbelonging to different tribes of the subfamilyCichlinae (Cichla monoculus, Astronotus ocellatus,
Geophagus proximus, Acaronia nassa, Bujurquinaperegrinabunda, Hoplarchus psittacus, Hypselecaracoryphaenoides, Hypselecara temporalis, Caquetaiaspectabilis, Uaru amphiacanthoides, Pterophyllumleopoldi , Pterophyl lum scalare, and Symphysodondiscus) and reexamine the karyotypic evolutionarypatterns proposed for this group. Variations in somecytogenetic markers were observed, although notrends were found in terms of the increase, decrease,or maintenance of the basal diploid chromosomenumber 2n = 48 in the tribes. Several species wereobserved to have 18S rDNA genetic duplications, aswell as multiple rDNA loci. In most of the taxaanalyzed, the 5S rDNA was located in the interstitialregion of a pair of homologous chromosomes,although variations from this pattern were observed.Interstitial telomere sites were also observed andappear to be involved in chromosomal rearrangementevents and the accumulation of repeat-rich satelli teDNA sequences. Our data demonstrated the karyo-typic diversity that exists among neotropical cichlids,suggesting that most of this diversity is due to therepetitive sequences present in heterochromaticregions and that repeat sequences have greatly influ-enced the karyotypic evolution of these fishes.
Keywor ds Karyotype evolutionInterstitial telomeric sitesFluorescent in situ hybridizationRibosomal DNA
C. H. Schneider (& ) M. L. Terencio E. FeldbergLaboratorio de Genetica Animal, Instituto Nacional dePesquisas da Amazonia (INPA), Av. Andre Araujo, 2936,Petropolis, Manaus, Amazonas 69011-970, Brazile-mail: [email protected]
M. C. GrossLaboratorio de Citogenomica, Departamento de Biologia,Instituto de Ciencias Biologicas, Universidade Federal doAmazonas, Manaus, AM, Brazil
R. F. Artoni M. R. VicariLaboratorio de Citogenetica e Evolucao, Departamento deBiologia Estrutural, Molecular e Genetica, UniversidadeEstadual de Ponta Grossa, Ponta Grossa, PR, Brazil
C. MartinsLaboratorio de Genomica Integrativa, Departamento deMorfologia, Instituto de Biociencias, UniversidadeEstadual Paulista Julio de Mesquita Filho (UNESP),Botucatu, SP, Brazil
123
Rev Fish Biol FisheriesDOI 10.1007/s11160-012-9285-3
Author's personal copy

39
3.1.1 Introdução
Ciclideos pertencem a uma das mais ricas famílias em número de
espécies, de peixes de água doce mundo e uma das principais famílias de
vertebrados, com mais de 3.000 espécies distribuídas nas Américas Central e
do Sul, na África, Madagascar e sul da Índia (Kullander 1998; Kocher 2004).
Muitas de suas espécies são importantes recursos para pesca e aquicultura e
bem como são amplamente explorados no comércio de peixes ornamentais.
A família Cichlidae é composta por quatro subfamílias: Etroplinae,
Ptychochrominae, Pseudocrenilabrinae e Cichlinae. Esta última subfamília é
restrita à região Neotropical e é composta por sete tribos: Cichlini, Retroculini,
Astronotini, Chaetobranchini, Geophagini, Cichlasomatini, e Heroini (Sparks e
Smith 2004; Smith et al. 2008).
Ciclídeos têm sido descritos como um excelente grupo para estudos
evolutivos devido a diversidade de nichos ecológicos ocupados, estratégias de
vida, adaptações morfológicas e comportamentais (Lowe-McConnell 1991;
López-Fernández et al. 2005). Muitos estudos genéticos estão sendo
realizados para investigar a história evolutiva dos ciclídeos e muitas espécies já
tiveram seu genoma sequenciado (Cichlid Genome Consortium Program em
http://cichlid.umd.edu/cichlidlabs/kocherlab/bouillabase.html). Estas
informações genômicas abrem oportunidades para integração entre sequências
nucleotídicas e dados cromossômicos (Cabral-de-Mello et al. 2012). A maioria
das informações disponíveis acerca da organização genômica em Cichlidae é
restrita a poucas espécies do Velho Mundo, entretanto diferentes classes de
DNAs repetitivos foram utilizadas para a compreensão da estrutura cariotípica
destes peixes (Oliveira et al. 1999; Martins et al. 2004; Ferreira e Martins 2008;
Poletto et al. 2010a). Por outro lado, muitos ciclídeos neotropicais apresentam
alguma informação citogenética, embora a maioria dos estudos enfoque
apenas descrições clássicas da macroestrutura cariotípica (Thompson 1979;
Feldberg e Bertollo 1985), faltando informações sobre a localização física
cromossômica de sequências de interesse. A maioria dos estudos de
citogenética molecular em ciclídeos Neotropicais tem mapeado genes de DNAr
45S (Gross et al. 2009b; Gross et al. 2010; Poletto et al. 2010b) e elementos
transponíveis para poucas espécies (Mazzuchelli e Martins 2009; Teixeira et al.
2009). Tendo em vista que muitas espécies de ciclídeos terão seu genoma

40
totalmente sequenciado nos próximos anos, o mapeamento físico
cromossômico de sequências de DNA pode contribuir para a identificação de
rearranjos cromossômicos, embasar estudos evolutivos e permitir melhor
interpretações do panorama genômico das espécies.
Assim, o objetivo do presente estudo foi analisar a organização
genômica dos DNAr 5S e 18S e sequências teloméricas em cromossomos
mitóticos de 13 espécies de ciclídeos Amazônicos, pertencentes a cinco tribos
da subfamília Cichlinae. Os resultados obtidos permitiram rediscutir os padrões
de evolução cariotípica propostos para o grupo, com uma abordagem
citogenética clássica e molecular, enfatizando o papel das sequências
repetitivas de DNA.
3.1.2 Material e Métodos
Foram analisados exemplares pertencentes a cinco tribos da subfamília
Cichlinae coletados na Amazônia Central (Tabela 1). Todos os espécimes
foram anestesiados por imersão em água gelada até a morte. Exemplares
testemunhos foram depositados na Coleção de Peixes do Instituto Nacional de
Pesquisas da Amazônia (INPA), Manaus, Amazonas, Brasil (números dos
exemplares INPA 26451, INPA 26452, INPA 26453, INPA 26459, INPA 26462
e INPA 26466).
3.1.2.1 Preparação cromossômica e localização da heterocromatina constitutiva e região organizadora de nucléolo (RON)
Cromossomos mitóticos foram obtidos a partir de células renais
utilizando o protocolo de “air drying” (Bertollo et al. 1978). A heterocromatina foi
analisada pela técnica de banda C (Sumner 1972), e RONs foram detectadas
pela impregnação pelo nitrato de prata (Howell e Black 1980).
41
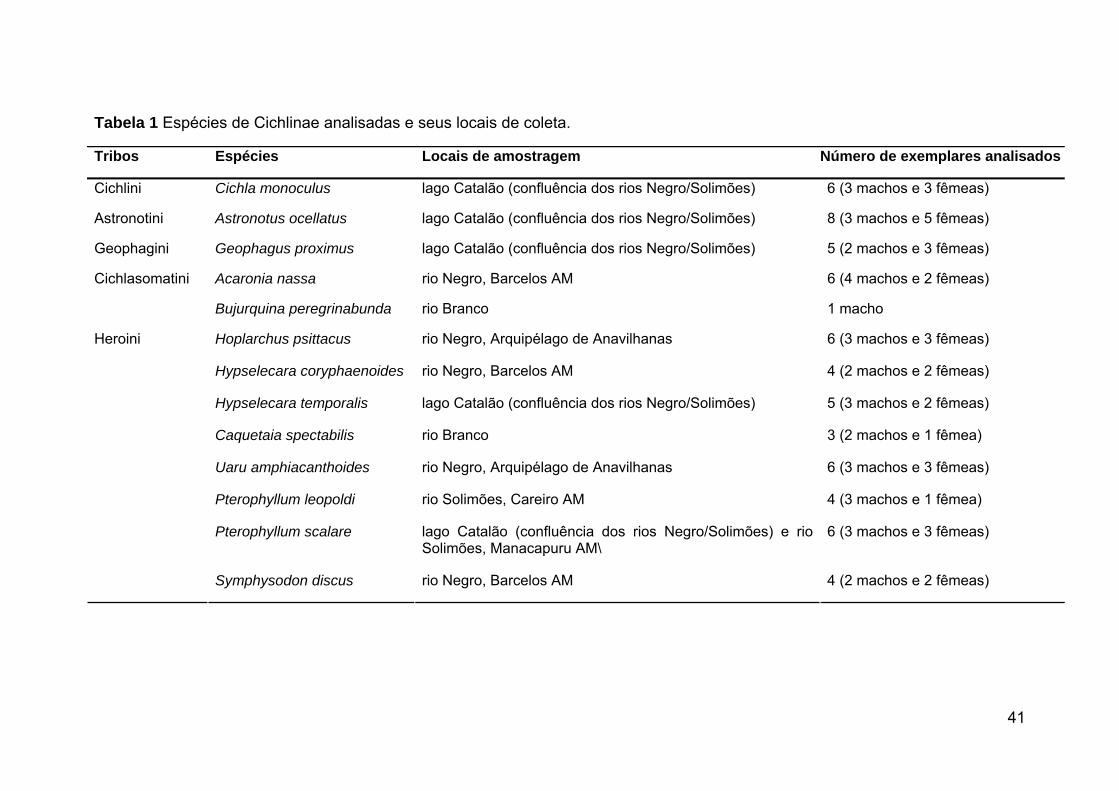
Tabela 1 Espécies de Cichlinae analisadas e seus locais de coleta.
Tribos Espécies Locais de amostragem Número de exemplares analisados
Cichlini Cichla monoculus lago Catalão (confluência dos rios Negro/Solimões) 6 (3 machos e 3 fêmeas)
Astronotini Astronotus ocellatus lago Catalão (confluência dos rios Negro/Solimões) 8 (3 machos e 5 fêmeas)
Geophagini Geophagus proximus lago Catalão (confluência dos rios Negro/Solimões) 5 (2 machos e 3 fêmeas)
Acaronia nassa rio Negro, Barcelos AM 6 (4 machos e 2 fêmeas) Cichlasomatini
Bujurquina peregrinabunda rio Branco 1 macho
Hoplarchus psittacus rio Negro, Arquipélago de Anavilhanas 6 (3 machos e 3 fêmeas)
Hypselecara coryphaenoides rio Negro, Barcelos AM 4 (2 machos e 2 fêmeas)
Hypselecara temporalis lago Catalão (confluência dos rios Negro/Solimões) 5 (3 machos e 2 fêmeas)
Caquetaia spectabilis rio Branco 3 (2 machos e 1 fêmea)
Uaru amphiacanthoides rio Negro, Arquipélago de Anavilhanas 6 (3 machos e 3 fêmeas)
Pterophyllum leopoldi rio Solimões, Careiro AM 4 (3 machos e 1 fêmea)
Pterophyllum scalare lago Catalão (confluência dos rios Negro/Solimões) e rio Solimões, Manacapuru AM\
6 (3 machos e 3 fêmeas)
Heroini
Symphysodon discus rio Negro, Barcelos AM 4 (2 machos e 2 fêmeas)

42
3.1.2.2 Extração de DNA e reação em cadeia da polimerase (PCR)
O DNA total foi extraído do tecido muscular utilizando o protocolo de
fenol-clorofórmio descrito por Sambrook et al. (1989) e quantificado em gel de
agarose por comparação com marcador lambda padrão. As amplificações dos
genes de RNAr 18S e 5S foram realizadas por reação em cadeia da polimerase
(PCR) utilizando os oligonucleotídeos iniciadores (primers) 18Sf (5’-CCG CTT
TGG TGA CTC TTG AT-3’) e 18Sr (5’-CCG AGGACC TCA CTA AAC CA-3’)
(Gross et al. 2010), 5Sa (5’-TAC GCC CGA TCT CGT CCG ATC-3’) e 5Sb (5’-
CAGGCT GGT ATG GCC GTA AGC-3’) (Gross et al. 2010; Martins e Galetti
1999, respectivamente). As reações de PCR foram realizadas em um volume
final de 25 µL contendo DNA genômico (200 ng), tampão 10x com 1,5 mM de
MgCl2, Taq DNA polimerase (5 U/µL), dNTPs (1 mM), par de primers (5 mM), e
água Milli-Q. O perfil de reação para o DNAr 18S foi 1 min 95 °C; 35 ciclos de 1
min a 94 °C, 1 min a 56 °C e 1 min 30 s a 72 °C; seguidos por 5 min a 72 °C.
As condições para amplificação do DNAr 5S foram 1 min 95 °C; seguidos por
30 ciclos de 1 min a 94 °C, 1 min a 59 °C, e 1 min 30 s a 72 °C; e uma
extensão final de 5 min a 72 °C.
PCR da sequência telomérica (TTAGGG)n foi realizada em um volume
final de 25 µL contendo tampão 10x com 1,5 mM de MgCl2, dNTPs (1 mM), 0,2
µL primer (TTAGGG)5, 0.2 µL primer (CCCTAA)5 (Ijdo et al. 1991), e 2 U of Taq
DNA polymerase. A primeira parte da amplificação foi realizada com baixa
estringência (4 min a 94 °C; seguidos por 12 ciclos de 1 min a 94 °C, 45 s a 52
°C e 1 min 30 s a 72 °C), seguidos por 35 ciclos de alta estringência (1 min a
94 °C, 1 min 30 s a 60 °C e 1 min 30 s a 72 °C). Hibridizações de sequências
teloméricas em posições cromossômicas intersticiais foram consideradas ITS
(sítios teloméricos intersticiais) quando sinais estavam presentes em todas as
metáfases analisadas.
3.1.2.3 Hibridização fluorescente in situ (FISH)
Os produtos de PCR do DNAr 18S e das sequências teloméricas, foram
marcados com digoxigenina-11-dUTP (Dig Nick Translation mix; Roche)
enquanto o DNAr 5S foi marcado com biotina-14-dATP (Biotin Nick Translation
mix; Roche). Ambos por reações de nick translation de acordo com instruções

43
dos fabricantes. Os anticorpos Avidina-FITC (Sigma-Aldrich), anti-avidina
biotinilada (Sigma-Aldrich), e anti-digoxigenina rodamina (Roche) foram usados
para detectar e amplificar o sinal da sonda. Hibridizações homólogas e
heterólogas foram realizadas de acordo com o protocolo descrito por Pinkel et
al. (1986) em elevadas condições de estringência 77% (2,5 ng/µL de DNAr
18S, DNA 5S ou sondas teloméricas, 50% formamida, 10% sulfato de dextrano,
e 2x SSC a 37 °C por 18 h). Os cromossomos foram contra corados com DAPI
(2 mg/ml) em meio de montagem VectaShield (Vector).
3.1.2.4 Microscopia/Processamento das imagens
Os cromossomos foram analisados em microscópio de epifluorescência
Olympus BX51 e as imagens foram capturadas com câmera digital acoplada
(Olympus DP71) utilizando o software Image-Pro MC 6.3. As metáfases foram
processadas no programa Adobe Photoshop CS3 e os cromossomos foram
medidos no programa Image J. Os cromossomos foram classificados de acordo
com nomenclatura proposta por Levan et al. (1964).
3.1.3 Resultados
Nenhum cromossomo sexual diferenciado foi observado em qualquer um
dos exemplares analisados, embora variações interespecíficas em número
diploide, padrão de heterocromatina constitutiva, número e localização de sítios
ribossomais, bem com na localização das sequências foram observadas.
3.1.3.1 Número diploide, heterocromatina e RONs
O menor número diploide observado foi de 42 cromossomos (16
meta/submetacêntricos [m-sm] e 26 subtelo/acrocêntricos [st-a]) em
Hypselecara coryphaenoides. A única espécie que apresentou número diploide
de 46 cromossomos (16 [m-sm] + 30 [st-a]) foi Uaru amphiacanthoides.
Astronotus ocellatus, Cichla monoculus, Geophagus proximus, Hoplarchus
psittacus, Hypselecara temporalis, Pterophyllum leopoldi, e Pterophyllum
scalare apresentaram número diploide com 48 cromossomos, embora existam

44
diferenças na fórmula cariotípica. Cichla monoculus possui apenas
cromossomos st-a, enquanto A. ocellatus, H. psittacus, H. temporalis, P.
leopoldi, e P. scalare possuem cariótipo com 16 cromossomos m-sm + 32 st-a.
A fórmula cariotípica de Geophagus proximus é de 12 cromossomos m-sm + 36
st-a. Número diploide de 50 cromossomos foi observado em Acaronia nassa (4
m-sm + 46 st-a), Bujurquina peregrinabunda (20 m-sm + 30 st-a), e Caquetaia
spectabilis (12 m-sm+ 38 st-a). Symphysodon discus foi a espécie que
apresentou o maior número diploide de 2n=60 (50 m-sm+ 10 st-a) (Figuras 1 e
2).
Todas as espécies analisadas apresentaram heterocromatina localizada
predominantemente na região centromérica de todos os cromossomos.
Algumas bandas-C em porções intersticiais foram observadas em C.
monoculus e H. temporalis, bem como em regiões terminais em A. nassa, H.
temporalis e C. spectabilis, e em regiões pericentroméricas em P. leopoldi, A.
ocellatus, H. coryphaenoides, C. spectabilis e U. amphiacanthoides. Blocos
conspícuos de heterocromatina foram mais evidentes em cromossomos de
algumas espécies tais como em A. ocellatus, P. scalare, U. amphiacanthoides,
S. discus e B. peregrinabunda (Figuras 1 e 2).
Com exceção de Uaru amphiacanthoides, que possui múltiplas
marcações de RONs (região terminal do braço curto dos pares 7, 8, 16 e
intersticial do braço longo do par 14), as demais espécies possuem apenas um
par cromossômico marcado pela AgNO3. O primeiro par cromossômico foi o
portador da RON em B. peregrinabunda, P. scalare, A. ocellatus, G. proximus,
e H. psittacus, enquanto que em C. monoculus e H. temporalis foi o segundo
par cromossômico que carreou a RON. Ainda, em H. coryphaenoides, P.
leopoldi, C. spectabilis e S. discus as RONs foram localizadas nos pares 5, 9,
11 e 17, respectivamente. A. ocellatus, G. proximus, H. psittacus e A. nassa
apresentaram duplicação da RON em um homólogo do par. (Figura 1 e 2).

45
Figura 1 Cariótipos das espécies pertencentes às tribos de Cichlinae (Cichlini
Astronotini, Geophagini, Cichlasomatini e Heroini) em banda-C. Os
cromossomos portadores da RON são mostrados na caixa.

46
Figura 2 Cariótipos das espécies pertencentes às tribos de Cichlinae (Heroini)
em banda-C. Os cromossomos portadores da RON são mostrados na caixa.
3.1.3.2 Mapeamento dos DNAr 18S e 5S e sequências teloméricas
Os resultados das hibridizações fluorescentes in situ (FISH) utilizando a
sonda de DNAr 18S foram coincidentes com as RONs, entretanto em Uaru
amphiacanthoides, apenas dois pares de cromossomos (7 e 8) exibiram sinais
positivos de hibridização com esta sonda em contraste com os quatro pares
impregnados pelo nitrato de prata. P. leopoldi apresentou resultados

47
contrastantes com as demais espécies devido ao grande número de sítios de
DNAr 18S. Além do braço curto do par 9, o qual foi o portador da RON, outros
vinte pares de cromossomos apresentaram sinais de hibridização, tanto em
regiões teloméricas, quanto em regiões intersticiais (Figura 3 e 4).
Quanto aos sítios de DNAr 5S, apenas Caquetaia spectabilis possui
múltiplos sítios e eles foram localizados em regiões intersticiais (homólogos do
par 8) e terminais (homólogos do par 9). Ainda, sítios de DNAr 5S foram
observados nas regiões terminais dos cromossomos de Bujurquina
peregrinabunda (par 12), Hoplarchus psittacus (par 11), and Symphysodon
discus (par 18). As demais espécies exibiram sítios em porções terminais e
pericentroméricas, embora com variações nos pares que portaram estes sítios
(Figura 3 e 4).
Os telômeros de todos os cromossomos das 13 espécies de Cichlinae
apresentaram sinais de hibridização com a sonda telomérica, sendo que sítios
teloméricos intersticiais (ITS) foram observados em espécies com números
diploides de 42, 46, 48 e 50 cromossomos. Variações no número de ITS foram
observadas entre as espécies analisadas. Os ITS foram visualizados como
sinais pontuais em alguns cromossomos, ou em blocos em outros, podendo
marcar totalmente os braços curtos de alguns cromossomos porém, este
padrão de hibridização foi visualizado em todas as metáfases analisadas.
Alguns destes ITS foram localizados adjacentes aos sítios de DNAr 18S, como
em Astronotus ocellatus, Acaronia nassa, Bujurquina peregrinabunda e
Hypselecara temporalis. O menor número de ITS foi observado em
Hypselecara coryphaenoides (pares 4 e 19) e Uaru amphiacanthoides (pares
14 e 15), seguidos por Pterophyllum scalare (pares 13, 15 e 16), Astronotus
ocellatus (pares 1, 2, 3, 7 e 17), Bujurquina peregrinabunda (pares 1, 3, 4, 6, 8
e 9), Hypselecara temporalis (pares 1, 2, 3, 7, 8 e 12), e Acaronia nassa (pares
2, 3, 4, 5, 6, 7, 8, 9, 22, 24 e 25). Todos os ITS foram coincidentes com blocos
heterocromáticos. Cichla monoculus, Geophagus proximus, Hoplarchus
psittacus, Symphysodon discus, Pterophyllum leopoldi, Caquetaia spectabilis
não mostraram evidência de ITS (Figuras 5 e 6).

48
Figura 3 Cariótipos das espécies pertencentes às tribos da subfamília
Cichlinae, mostrando os sítios de DNAr 18S (vermelho) e 5S (verde). Os
cromossomos foram contra corados com DAPI.

49
Figura 4 Cariótipos das espécies pertencentes às tribos da subfamília
Cichlinae, mostrando os sítios de DNAr 18S (vermelho) e 5S (verde). Os
cromossomos foram contra corados com DAPI.

50
Figura 5 Cariótipos das espécie pertencentes às tribos de Cichlinae,
mostrando sinais de hibridização com sonda telomérica (vermelho). Os
cromossomos foram contra corados com DAPI.

51
Figura 6 Cariótipos das espécie pertencentes às tribos de Cichlinae,
mostrando sinais de hibridização com sonda telomérica (vermelho). Os
cromossomos foram contra corados com DAPI.

52
3.1.4 Discussão Com base em dados sobre macroestrutura cariotípica, durante muito
tempo acreditou-se que os ciclídeos neotropicais eram um grupo conservado
do ponto de vista citogenético (Kornfield 1984; Feldberg e Bertollo 1985).
Contudo, com o avanço do conhecimento acerca do genoma de diferentes
espécies de Cichlinae, ficou evidente que o mesmo é composto por diferentes
classes de DNAs repetitivos e que estes tiveram um papel importante na
evolução cariotípica destes peixes, podendo inclusive ter favorecido rearranjos
cromossômicos (Gross et al. 2009b; Teixeira et al. 2009; Mazzuchelli e Martins
2009; Gross et al. 2010; Valente et al. 2011).
A presença de rearranjos cromossômicos robertsonianos e não
robertsonianos em Cichlinae proporcionou que três caminhos evolutivos
fossem sugeridos para este grupo: (i) manutenção do 2n=48 cromossomos
considerado basal, (ii) diminuição do 2n ou (iii) aumento do número diploide
(Feldberg et al. 2003). Porém, estes caminhos evolutivos parecem ser
adotados independentemente dentro das tribos, com exceção das tribos basais
(Cichlini, Astronotini e Chaetobanchini) onde todas as espécies analisadas
apresentam 2n=48 cromossomos. Nas demais tribos, os números diploides
variam de 38 a 60, não havendo uma tendência em aumento, redução ou
manutenção do número diploide, tais como visto em Cichlasomatini, variando
de 44 a 50 cromossomos e em Heroini, de 42 a 60 cromossomos (presente
estudo; para revisão de número diploide ver Feldberg et al. 2003; Poletto et al.
2010b). Além disso, se considerado o padrão de distribuição da
heterocromatina constitutiva, o número e localização de sítios ribossomais e a
presença de sequências teloméricas intersticiais, novamente não é possível
verificar tendências dentro das tribos.
Em uma revisão citogenética efetuada para os ciclídeos neotropicais foi
proposto que as espécies que possuem um menor número diploide têm mais
cromossomos meta-submetacêntricos, enquanto que as espécies com número
diploide igual a 48 cromossomos possuem poucos cromossomos de dois
braços. Rearranjos do tipo inversão cromossômica poderiam promover a
existência de diferentes fórmulas cariotípicas (Feldberg et al. 2003). Porém,
nossos dados não corroboram totalmente estas predições, pois tanto nas
espécies com 2n=48 (A. ocellatus, H. psittacus, H. temporalis, P. leopoldi, P.

53
scalare), quanto nas que possuem 2n=42 (H. coryphaenoides) e 46 (U.
amphiacanthoides), são visualizados oito pares de cromossomos
meta/submetacêntricos, com exceção de Geophagus proximus, que apresenta
2n=48 cromossomos, sendo seis pares meta-submetacêntricos. Contudo, a
descrição de diferentes fórmulas cariotípicas para uma mesma espécie é
comumente encontrada em ciclídeos, como Pterophyllum scalare, que tem
descritas as fórmulas: como possuindo 4 m-sm + 44 st-a (Thompson 1979), 12
m-sm + 36 st-a (Nascimento et al. 2006), 6 m-sm + 42 st-a (Poletto et al.
2010b) ou 16 m-sm +32 st-a (presente estudo). A maioria desta variação não é
devido a rearranjos cromossômicos que ocasionaram diferença intraespecífica,
mas provavelmente à qualidade da preparação cromossômica analisada, ao
padrão de compactação do DNA e erros na medida dos braços cromossômicos
que geram divergência na interpretação do cariótipo.
Contudo, inversões cromossômicas podem ter ocorrido em alguns
ciclídeos que mantiveram o número diploide basal (tais como A. ocellatus e H.
temporalis) uma vez que sítios teloméricos intersticiais (ITS) foram encontrados
em pares cromossômicos destas espécies. ITS também foram encontrados em
H. coryphaenoides (2n=42 cromossomos), sugerindo a presença de rearranjos
do tipo fusão neste grupo. Entretanto em Uaru amphiacanthoides que
apresenta 2n= 46 cromossomos, nenhum ITS foi observado. A ausência de ITS
em cromossomos que foram originados por eventos de fusões, pode indicar
que estes ITS foram perdidos devido a processos de erosão molecular de
sequências de DNA repetitivo, bem como pequenos sítios teloméricos podem
não ser detectados pela FISH (Mandrioli et al. 1999). Ainda, essas fusões são
devido a mecanismos de encurtamento do telômero, à inativação do telômero,
a regiões descompactadas de DNA ou a presença de sequências similares às
teloméricas no DNA repetitivo centromérico tais como transposons (Shampay
et al. 1995; Slijepcevic 1998).
Braços curtos inteiros foram marcados em FISH utilizando sondas
teloméricas, para diversas espécies analisadas. Provavelmente, outros DNAs
repetitivos podem estar associados às sequências teloméricas nestas
espécies. Este tipo de associação já foi relatado anteriormente em Drosophila,
onde foi caracterizada uma família de DNA satélite associada ao telômero com
repetições idênticas às encontradas na região centromérica (Bachmann et al.

54
1990). Este mesmo padrão foi identificado em algumas plantas (Navajas-Pérez
et al. 2009), refletindo a complexidade dos padrões evolutivos relacionados
com sequências de DNA repetitivo. Considerando esta associação entre
sequências teloméricas e DNA satélite, é possível que em algum momento
ambas foram carreadas para outras porções do genoma por meio de trocas
desiguais durante processos de crossing over, transposições ou duplicações
dessas sequências e que atualmente estão localizadas em regiões
heterocromáticas. Estas regiões heterocromáticas podem estar protegendo
DNAs repetitivos de pressões seletivas que atuam em regiões eucromáticas,
possibilitando uma evolução diferencial destas porções genômicas (Grewal e
Jia 2007).
A heterocromatina na maioria dos ciclídeos analisados apresenta-se
localizada na região centromérica ou pericentromérica da maioria dos
cromossomos, sendo o padrão de distribuição diferencial da mesma muitas
vezes utilizado como um marcador espécie-específico ou populacional
(Feldberg et al. 2003; Vicari et al. 2006; Benzaquem et al. 2008; Perazzo et al.
2011). Estudos recentes mostram que o centrômero é uma porção
cromossômica dinâmica que têm capacidade de reposicionar-se devido a
respostas epigenômicas, podendo assim modificar a fórmula cariotípica sem
reposicionamento de genes (Rocchi et al. 2012). Esta pode ser a explicação
para a existência de heteromorfismo cromossômico em alguns pares de
homólogos, comum em ciclídeos neotropicais, o que gera a descrição de
citótipos diferenciados para algumas espécies, como relatado para
Symphysodon (Mesquita et al. 2008), bem como pode ocasionar divergência na
fórmula cariotípica de indivíduos de uma mesma espécie, proveniente de
diferentes localidades. Atualmente, a heterocromatina é reconhecida por
desempenhar atividades importantes, tais como auxiliar na segregação
cromossômica, organização nuclear e regulação da expressão gênica
associada a respostas às mudanças no ambiente, também podendo afetar o
processo de recombinação gênica (Grewal e Jia 2007; Varriale et al. 2008;
Bühler 2009). Assim, a associação entre regiões heterocromáticas e sítios
ribossomais, como encontrada nos ciclídeos neotropicais bem como em
diversos outros grupos de peixes, pode estar relacionada com a variabilidade
na localização e número das RONs ativas.

55
Vários trabalhos que inferiram evolução cariotípica em ciclídeos levaram
em conta apenas dados da citogenética clássica, o que pode ter levado a
incongruências em alguns resultados, mascarando a real organização
genômica. Isso pode ser evidenciado em Satanoperca jurupari, onde quatro
sítios de NORs foram descritos por meio da impregnação com nitrato de prata
(Mendonça et al. 1999), porém apenas um par de homólogos apresentou sítios
de DNAr 18S (Poletto et al. 2010b). Ainda, estudos anteriores sugerem a
presença de região organizadora de nucléolo múltipla em Caquetaia spectabilis
(Salgado et al. 1995), porém dentre as espécies analisadas no presente
trabalho, tanto com impregnação por nitrato de prata quanto por localização
dos sítios ribossomais da família 45S, apenas Uaru amphiacanthoides
apresentou sítios múltiplos de RONs. Contudo, semelhante ao que foi
visualizado em Satanoperca jurupari, em U. amphiacanthoides oito sítios foram
evidenciados pela Ag-RON e apenas quatro sítios de DNAr 18S foram
detectados. Estes resultados divergentes entre os sítios Ag-RON positivos e os
de DNAr 18S, podem estar relacionados à afinidade que algumas regiões
heterocromáticas têm pelo nitrato de prata, mascarando assim o real número
de RONs (Sumner 2003). Em peixes, frequentemente ocorre associação entre
RONs e regiões heterocromáticas. Em S. discus, estudos anteriores indicaram
a presença de variações intra-específicas quanto à localização dos genes de
RNAr 18S. Estes puderam ser identificados entre dois a cinco sítios, sempre
envolvidos com regiões terminais dos braços curtos dos pares cromossômicos
6, 17 e 23 (Gross et al. 2010). Estas observações foram corroboradas no
presente estudo.
Vinte e um sítios de DNAr 18S foram observados em Pterophyllum
leopoldi. Este grande número de sítios de DNAr é incomum em peixes, embora
já tenha sido descrito em outros vertebrados (Cazaux et al. 2011). Artefatos de
técnica que implicariam em erros durante o processo podem ser descartados,
uma vez que as hibridizações foram feitas em condições de alta extringência e
com sondas homólogas. Estes múltiplos sítios ribossomais provavelmente são
derivados de segmentos de DNAr 18S que foram duplicados e dispersos no
genoma de Pterophyllum leopoldi, como evidenciado para o DNAr 5S em
outras espécies de peixes (Martins et al. 2006; Silva et al. 2011). Contudo,
podemos inferir que o sítio de DNAr 18S no par cromossômico nove apresenta

56
atividade transcricional, pois foram os únicos que impregnaram com o nitrato
de prata. Assim, os sítios adicionais de DNAr 18S podem estar relacionados a
processos naturais de nascimento e morte de sequências repetitivas no
genoma (Rooney e Ward 2005).
Diferenças na localização do DNAr 5S foram observadas entre as tribos
de ciclídeos analisadas. Na maioria das espécies, os sítios de DNAr 5S foram
localizados em posição intersticial de um par de cromossomos, entretanto C.
spectabilis apresentou quatro sítios de DNAr 5S, dois na região intersticial e
dois na porção telomérica. Os DNAr 18S e 5S foram localizados em pares
cromossômicos distintos, e isto é um padrão comum para peixes (Martins e
Wasko 2004). Martins (2007) atribui este padrão a diferentes processos
transcricionais nos cístrons ribossomais, onde os genes de DNAr 45S (o qual
engloba entre outros o DNAr 18S) são transcritos pela RNA polimerase I,
enquanto o DNAr 5S é transcrito pela RNA polimerase III.
Dessa forma, nossos dados indicam que os ciclídeos neotropicais
possuem grande variação cariotípica, como evidenciado pela distinta
organização do cariótipo. Provavelmente, uma grande parte desta variação é
devido as diferentes classes de sequências repetitivas, que apresentam
diferentes quantidades e distribuição nos cromossomos. Estudos estão sendo
realizados para elucidar a composição da heterocromatina, para refinar ainda
mais a análise da organização genômica dos ciclídeos neotropicais, e mostrar
o real panorama evolutivo deste grupo.

57
3.2 Artigo 2 - Dinâmica evolutiva de elementos retrotransponíveis Rex1, Rex3 e Rex6 no genoma de ciclídeos Neotropicais

58
3.2.1 Introdução
Elementos transponíveis (TEs) têm potencial para produzir grandes
alterações no genoma de seus hospedeiros (Kidwell e Lisch 2000; 2001),
agindo como uma importante ferramenta evolutiva, gerando uma gama de
novas regiões regulatórias e sequências codificantes (Volff 2006; Bourque
2008). TEs tem o potencial de gerar biodiversidade, transições evolutivas
através de mutações, e domesticação molecular (Blass et al. 2012).
Eventualmente, TEs podem ser transferidos entre espécies isoladas
reprodutivamente, através de transferência horizontal, independente da
clássica herança de pais para prole, e também desempenha papel importante
na diversidade genômica entre espécies (Schaack et al. 2010).
Com base na estrutura e em seu mecanismo de transposição, os
elementos móveis são divididos em duas classes: classe I engloba os
elementos transponíveis que têm um RNA como intermediário
(retrotransposons) copiado em um cDNA por uma transcriptase reversa e
integrado em uma nova região genômica; classe II inclui todos os que são
transpostos diretamente de DNA para DNA, sem outra molécula intermediária
(Van Sluys et al. 2001; Tafalla et al. 2006; Martins 2007; Levin e Moran 2011).
Os retrotransposons podem ser agrupados de acordo com a presença ou
ausência de longas repetições terminais (LTR long terminal repeats). As LTRs
são necessárias para a transcrição e integração do cDNA depois da transcrição
reversa, sendo que são encontrados, na sequência dos LTRs domínios para
proteinase, integrase, transcriptase reversa e RNAse. Já, os retrotransposons
não-LTRs utilizam promotores internos para a sua transposição e codificam
uma transcriptase reversa e uma RNAse, sendo também conhecidos como
LINES (long interspersed elements) ou retrotransposons TP (target-primed).
Além disso, dentro da categoria dos não-LTRs, existem também os SINES
(short interspersed nucleotide elements), que não codificam a maquinaria
enzimática necessária para a transposição, mas obtém esta provavelmente por
meio dos elementos LINES (Böhne et al. 2008).
Diversos elementos transponíveis são encontrados nos genomas tanto
em níveis quantitativos como qualitativos e o mesmo tipo de TE pode ter
diferente sucesso em invadir diferentes espécies (Böhne et al. 2008). Acredita-

59
se que TEs possam estar associados à biodiversidade e especiação de peixes
teleósteos. A diversidade presente em teleósteos é também refletida na
diversidade do tamanho e estrutura de seu genoma, o qual foi
consideravelmente influenciado por retroelementos (Volff et al. 2003; Volff
2005). Todos os tipos de retroelementos já descritos foram encontrados nos
genomas de peixes (Okada et al. 1997, Poulter et al. 1999; Volff et al. 1999;
Aparicio et al. 2002) e entre os non-LTR retrotransposons destacam-se Rex1,
Rex3 e Rex6, os quais estiveram ativos durante a evolução deste grupo de
vertebrados (Volff et al. 1999; 2000; 2001a).
Os peixes da família Cichlidae (Perciformes) apresentam grande
importância na pesca de subsistência e esportiva, na aquicultura e aquariofilia
e se destacam como excelente modelo evolutivo devido a sua radiação
adaptativa, diversidade ecológica e comportamental (Kocher 2004; López-
Fernández et al. 2005). Dentre os ciclídeos, apenas a subfamília Cichlinae é
encontrada na região Neotropical. Análises, combinando caracteres
morfológicos e dados de sequências nucleotídicas, agrupam a subfamília
Cichlinae em um clado monofilético dividido em sete tribos: Astronotini,
Chaetobranchini, Cichlasomatini, Cichlini, Geophagini, Heroini, e Retroculini.
Chaetobranchini + Geophagini é grupo irmão de Heroini + Cichlasomatini.
Astronotini é uma tribo monogenérica e grupo irmão destas quatro tribos.
Finalmente, o clado que engloba Cichlini + Retroculini é o grupo basal, irmão
das demais tribos (Smith et al. 2008).
Diferentes classes de TEs foram mapeadas no genoma dos ciclídeos
neotropicais, tais como transposon Tc1 que foi mapeado em Cichla kelberi
evidenciando blocos conspícuos em regiões centroméricas e marcações
dispersas nos cromossomos (Teixeira et al. 2009). Três novas classes de
retroelementos foram descritas: RCk que apresenta marcações dispersas nos
cromossomos de C. kelberi; AoRex3 e AoLINE que são agrupados na
heterocromatina centromérica de todos os cromossomos de Astronotus
ocellatus (Mazzuchelli e Martins 2009). O mapeamento de diferentes classes
do retroelemento Rex também tem sido realizado neste grupo visando
compreender a organização genômica e os resultados têm mostrado um
padrão de distribuição compartimentalizado na heterocromatina
pericentromérica entretanto sinais dispersos e agrupados em regiões

60
eucromáticas também foram observados (Gross et al. 2009b; Mazzuchelli e
Martins 2009; Teixeira et al. 2009; Valente et al. 2011). Contudo, as
informações restringem-se ao mapeamento físico cromossômico, não sendo
conhecida a diversidade genética destes retroelementos. Portanto, análises
comparativas englobando espécies consideradas basais e derivadas, são de
particular interesse para o entendimento da dinâmica dos retroelementos na
evolução genômica dos ciclídeos do novo mundo. Assim, a organização
genômica dos non-LTR retrotransposons Rex1, Rex3 e Rex6 em cinco
espécies de ciclídeos amazônicos foi avaliada por meio do mapeamento físico
cromossômico e sequenciamento destes elementos móveis, o que permitiu o
entendimento de como estes retroelementos se comportam no genoma deste
grupo de peixes.
3.2.2 Material e Métodos
Foram analisados exemplares pertencentes a quatro tribos da subfamília
Cichlinae coletados na Amazônia Central. As espécies analisadas foram:
Cichla monoculus (3 machos e 3 fêmeas) tribo Cichlini, Astronotus ocellatus (3
machos e 5 fêmeas) Astronotini, Geophagus proximus (2 machos e 3 fêmeas)
Geophagini, Pterophyllum scalare (3 machos e 3 fêmeas) e Symphysodon
discus (2 machos e 2 fêmeas) Heroini. Os exemplares foram amostrados com
autorização de coleta ICMBio SISBIO 10609-1/2007. Todos os espécimes
foram anestesiados por imersão em água gelada até a morte. Os cromossomos
mitóticos foram obtidos de células renais utilizando o protocolo de “air drying”
(Bertollo et al. 1978).
3.2.2.1 Extração de DNA e reação em cadeia da polimerase (PCR)
O DNA total foi extraído do tecido muscular utilizando o protocolo de
fenol-clorofórmio descrito por Sambrook et al. (1989) e quantificado em
espectrofotômetro NanoVue Plus (GE Healthcare). A reação em cadeia da
polimerase (PCR) para amplificação dos retroelementos, foi realizada utilizando
os primers RTX1-F1 (5’-TTC TCC AGT GCC TTC AAC ACC-3’) e RTX1-R1 (5’-
TCC CTC AGC AGA AAG AGT CTG CTC-3’) para amplificação do segmento

61
que corresponde ao gene da transcriptase reversa do Rex1 (Volff et al. 2000);
os primers RTX3-F3 (5’-CGG TGA YAA AGG GCA GCC CTG-3’) e RTX3-R3
(5’-TGG CAG ACN GGG GTG GTG GT-3’) para amplificar a região que codifica
o gene da transcriptase reversa do retrotransposon Rex3 (Volff et al. 1999); e
os primers Rex6-Medf1 (5’-TAA AGC ATA CAT GGA GCG CCA C-3’) e Rex6-
Medr2 (5’-GGT CCT CTA CCA GAG GCC TGG G-3’) para amplificar a parte C-
terminal da endonuclease do retrotransposon Rex6 (Volff et al. 2001a).
As reações de PCR foram realizadas em um volume final de 25 μL
contendo DNA genômico (200 ng), tampão 10x com 1,5 mM de MgCl2, Taq
DNA polimerase (5 U/μL), dNTPs (1 mM), par de primers (5 mM), e água Milli-
Q. Os perfis das reações para o Rex1, Rex3, e Rex6 incluem os seguintes
passos: 95 °C por 5 min; 35 ciclos de 95 °C por 1 min, 55 °C por 40 s, e 72 °C
por 2 min; seguidos por uma extensão final de 72 °C por 5 min. Os produtos de
PCR foram checados em gel de agarose 1% quantificados em
espectrofotômetro NanoVue Plus (GE Healthcare) e utilizados para clonagem e
como sondas para a FISH.
3.2.2.2 Clonagem, sequenciamento e análise das sequências
Os produtos de amplificação dos retrotransposons Rex1, Rex3 e Rex6
foram inseridos em plasmídeo pGEM-T easy (Promega). Os produtos de
ligação foram inseridos em células competentes da linhagem DH5α de
Escherichia coli. Clones que possuíam o inserto foram sequenciados em
sequenciador automático de DNA ABI 3130 XL (Applied Biosystem). Cada
clone foi identificado por buscas na base de dados do NCBI
(http://www.ncbi.nlm.nih.gov) com a ferramenta BLASTn e na base do Repbase
(Jurka et al. 2005) no Genetic Information Research Institute (Giri)
(http://www.girinst.org/repbase) utilizando o software CENSOR (Kohany et al.
2006). As sequências obtidas foram depositadas no GenBank (NCBI) sob os
seguintes números de acessos: Rex1, GenBank: JX576302-JX576350; Rex3,
GenBank: JX576351-JX576400; e Rex6, GenBank: JX576401-JX576459. As
sequências nucleotídicas foram alinhadas utilizando o programa ClustalW
(Thompson et al. 1994) incluso no programa BioEdit 7 (Hall 1999). As árvores
filogenéticas dos retroelementos foram obtidas por busca heurística de máxima

62
verossimilhança. A robustez da topologia encontrada foi testada com 1000
replicatas de bootstrap. Estas análises foram realizadas utilizando o programa
MEGA5 (Tamura et al. 2011). Para análise de máxima verossimilhança foram
inseridas sequências de retroelementos Rex1, 3 e 6 de outros ciclídeos,
disponíveis no GenBank. As sequências dos aminoácidos foram deduzidas a
partir das sequências nucleotídicas utilizando o programa BioEdit 7 (Hall 1999).
3.2.2.3 Hibridização fluorescente in situ (FISH)
Os produtos de PCR dos retroelementos (Rex1, 3 e 6) foram marcados
com digoxigenina-11-dUTP (Dig Nick Translation mix; Roche) por reações de
nick translation de acordo com instruções do fabricante e o anticorpo anti-
digoxigenina rodamina (Roche) foi usado para detectar o sinal da sonda.
Hibridizações homólogas e heterólogas foram realizadas de acordo com o
protocolo descrito por Pinkel et al. (1986) em condições de estringência de 77%
(2,5 ng/µL de DNA, 50% formamida, 10% sulfato de dextrano, e 2x SSC a 37
°C por 18 h). Os cromossomos foram contra corados com DAPI (2 mg/ml) em
meio de montagem VectaShield (Vector).
3.2.2.4 Microscopia/Processamento das imagens
Os cromossomos foram analisados em microscópio de epifluorescência
Olympus BX51 e as imagens foram capturadas com câmera digital acoplada
(Olympus DP71) utilizando o software Image-Pro MC 6.3. As metáfases foram
processadas no programa Adobe Photoshop CS3 e os cromossomos foram
medidos no programa Image J. Os cromossomos foram classificados de acordo
com nomenclatura proposta por Levan et al. (1964).
3.2.3 Resultados
3.2.3.1 Diversidade molecular dos TEs analisados
Um total de 49 clones de Rex1 foram sequenciados sendo 11 de C.
monoculus, 8 de A. ocellatus, 10 de G. proximus, 12 de P. scalare e 8 de S.
discus. Sequências de 573 a 575 pb (pares de bases) foram obtidas.

63
Substituições nucleotídicas foram mais frequentes em espécies derivadas (P.
scalare e S. discus) e em menor número em espécies basais (C. monoculus e
A. ocellatus). Nenhuma substituição foi observada em C. monoculus, enquanto
que nas demais espécies as substituições nucleotídicas foram: uma transição
em A. ocellatus, duas transições em G. proximus, duas transições e três
transversões em P. scalare e 18 transições, sete transversões e 4 indels
(insersões/deleções) em S. discus. Quando comparado com sequências de
Rex1 de outros ciclídeos disponíveis no Genbank observou-se 91% de
identidade genética com Hemichromis bimaculatus (GenBank: AJ288479) e
94% com Oreochromis niloticus (GenBank: AJ288473). Foi possível identificar
blocos conservados correspondentes à sequência da transcriptase reversa,
sendo a sequência de S. discus similar à de O. niloticus, assim como as
demais são semelhantes entre si (Figura 1a). A árvore filogenética dos clones
do elemento Rex1 indicou separação entre Symphysodon discus e as demais
espécies, evidenciada por dois ramos na árvore suportados por elevados
valores de bootstrap. Os clones de cada espécie foram agrupados em ramos
separados. O ramo que engloba os clones das espécies Cichla monoculus,
Astronotus ocellatus, Geophagus proximus e Pterophyllum scalare teve um
clone de P. scalare basal a todos os demais e os clados que englobam as
tribos Heroini, Cichlini, Astronotini e Geophagini apresentaram-se como grupos
irmãos (com exceção de S. discus) (Figura 2), não refletindo a relação
filogenética das espécies analisadas.

64
Figura 1 Alinhamento das sequências de aminoácidos dos diferentes
retroelementos entre ciclídeos: A) Região correspondente à transcriptase
reversa de Rex1 das espécies analisadas em comparação com O. niloticus
(GenBank: AJ288473); B) Região correspondente à transcriptase reversa de
Rex3 em comparação com O. niloticus (GenBank: AJ400370) e C) Região da
endonuclease de Rex6 das espécies analisadas em comparação com O.
niloticus (GenBank: AJ293545).

65
Figura 2 Árvore de máxima verossimilhança dos 49 clones de Rex1, revelando duas linhages monofiléticas deste retroelemento.
Valores de bootstrap maiores que 60 são mostrado sobre os ramos.

66
Sequências entre 418 e 475 pb foram obtidas entre os 50 clones de
Rex3 analisados. Ao total foram 6 clones de cada espécie (C. monoculus, A.
ocellatus, G. proximus e P. scalare) e 26 clones de S. discus. O retroelemento
Rex3 apresentou-se muito váriavel em todas as espécies analisadas, com
várias substituições nucleotídicas, sendo: 113 em C. monoculus (79 transições,
25 transversões e 9 indels), 159 em A. ocellatus (83 transições, 19
transversões e 57 indels), 215 em G. proximus (89 transições, 34 transversões
e 92 indels), 242 em P. scalare (130 transições, 58 transversões e 54 indels) e
245 em S. discus (133 transições, 58 transversões e 54 indels). Quando
comparado com sequências de Rex3 de outros ciclídeos disponíveis no
Genbank observou-se 87% de identidade genética com Oreochromis niloticus
(GenBank: AJ400370), 91% com Cichla kelberi (GenBank: FJ687588) e 92%
com Geophagus surinamensis (GenBank: HM535302). Mesmo com ampla
variação nas sequências dos clones, foi possivel identificar blocos conservados
correspondentes à sequência da transcriptase reversa de Rex3 (Figura 1b). A
árvore filogenética dos clones do elemento Rex3 não agrupou os clones por
espécie, com exceção de um clado que engloba clones de S. discus. As
demais tribos não foram separadas em clados exclusivos (Figura 3).

67
Figura 3 Árvore de máxima verossimilhança dos 50 clones do retroelemento Rex3. Valores de bootstrap maiores que 60 são
mostrado sobre os ramos.

68
Fragmentos entre 459 e 505 pb foram gerados para os 59 clones
sequenciados do retroelemento Rex6. Ao total foram sequenciados oito clones
de C. monoculus, sete clones de A. ocellatus, sete de G. proximus, cinco de P.
scalare e 32 de S. discus. C. monoculus e S. discus apresentaram elevada taxa
de substituição nucleotídica: 213 (80 transições, 63 transversões e 70 indels) e
214 (103 transições, 88 transversões e 23 indels) respectivamente. As
substituições nucleotídicas evidenciadas nas demais espécies foram: 62 (41
transições, 17 transversões e 4 indels) em A. ocellatus, 66 (47 transições, 19
transversões) em G. proximus e 34 (26 transições, 7 transversões e 1 indels)
em P. scalare. Quando comparado com sequências de Rex6 de outros
ciclídeos disponíveis no Genbank observou-se 81% de identidade genética
com Oreochromis niloticus (GenBank: AJ293545), 83% com Melanochromis
auratus (GenBank: HM535303) e 88% com Crenicichla sp. (GenBank:
HM535301). As sequências apresentaram região de homologia com a
endonuclease do Rex6 (Figura 1c). Os clones de cada espécie foram
agrupados em ramos distintos na árvore filogenética dos clones do elemento
Rex6, sendo que A. ocellatus aparece como grupo irmão de alguns clones de
C. monoculus. E todos os C. monoculus e A. ocellatus são grupo irmão das
demais espécies derivadas. Geophagus proximus aparece como grupo irmão
de Crenicichla sp., sendo este clado basal ao que engloba P. scalare e S.
discus (Figura 4). Estes dados apresentam similaridade com a árvore
filogenética proposta para as espécies.

69
Figura 4 Árvore de máxima verossimilhança dos 59 clones do retroelemento Rex6. Valores de bootstrap maiores que 60 são
mostrado sobre os ramos.

70
3.2.3.2 Mapeamento físico cromossômico dos TEs
O retroelemento Rex1 apresentou-se disperso nos cromossomos de
Cichla monoculus, Astronotus ocellatus, Geophagus proximus, Pterophyllum
scalare e Symphysodon discus. Ainda, Astronotus ocellatus apresentou os
pares 11, 17 e 23 com ampla distribuição deste retroelemento, assim como os
pares 1, 5, 7, 8, 15, 19 e 23 de Geophagus proximus e a maioria dos pares de
Symphysodon discus. Contudo, todas as espécies apresentaram marcações
mais conspícuas em regiões terminais e/ou centroméricas, havendo maior
quantidade de marcações em espécies derivadas, tais como Pterophyllum
scalare (Figura 5 – sinais em vermelho).
Múltiplos e intensos sinais de hibridização foram obtidos para o
retroelemento Rex3 nas cinco espécies analisadas. Estas marcações
conspícuas localizaram-se em regiões terminais da maioria dos cromossomos
das cinco espécies, bem como em regiões centroméricas, pericentroméricas e
intersticiais. Contudo, em ambos os homólogos do par 21 de Cichla monoculus
e pares 4, 5, 8, 9 e 12 de Symphysodon discus não foram evidenciados sinais
de hibridização (Figura 6 – sinais em vermelho). O Rex6 foi visualizado, de maneira geral, em marcações conspícuas na
maioria dos cromossomos de todas as espécies, em posições terminais,
intersticiais, centroméricas e/ou pericentroméricas. Em Geophagus proximus
ambos os homólogos dos pares 2, 21, 23 e 24 não apresentaram sinais de
hibridização, bem como os pares 2, 8, 13, 20 e 25 de Symphysodon discus
(Figura 7 – sinais em vermelho). Marcações de Rex1, 3 e 6 também foram
detectadas em regiões eucromáticas.

71
Figura 5 Mapeamento cromossômico do retrotransposon Rex1 (sinais em
vermelho). Os cromossomos foram contra corados com DAPI.

72
Figura 6 Mapeamento cromossômico do retrotransposon Rex3 (sinais em
vermelho). Os cromossomos foram contra corados com DAPI.

73
Figura 7 Mapeamento cromossômico do retrotransposon Rex6 (sinais em
vermelho). Os cromossomos foram contra corados com DAPI.
3.2.4 Discussão
3.2.4.1 Distribuição cromossômica dos TEs
Estudos recentes têm mostrado que ciclídeos possuem cariótipo
dinâmico, com variação na organização e estrutura do genoma destes peixes,
sendo que diversas classes de DNA repetitivos podem influenciar e propiciar
este dinamismo (Gross et al. 2009b; Poletto et al. 2010b; Valente et al. 2011;
Schneider et al. 2012a). Dentre estes elementos repetitivos destacam-se os

74
TEs, que podem conduzir a evolução de genomas por diferentes processos,
incluindo inativação gênica, combinando diferentes éxons e por conversão
gênica (para revisão ver Kazazian 2004). Ainda, a recombinação entre cópias
não homólogas de TEs pode conduzir a formação de deleções, duplicações,
inversões e translocações, bem como o próprio processo de transposição pode
ocasionar vários rearranjos (Volff 2005). Deste modo, TEs parecem ter papel
importante na diversificação cariotípica dos ciclídeos.
Elementos transponíveis acumulam-se principalmente em regiões
heterocromáticas do genoma em diversos grupos, incluindo peixes (Fischer et
al. 2004; Mazzuchelli e Martins 2009; Gross et al. 2009b; Valente et al. 2011).
Entretanto, em algumas espécies estes elementos transponíveis são
encontrados em regiões eucromáticas (Teixeira et al. 2009; Terencio et al.
2012a). As espécies de ciclídeos analisadas apresentam acúmulo de Rex1,
Rex3 e Rex6 em maior quantidade em regiões heterocromáticas, mas algumas
marcações também foram detectadas em porções eucromáticas. Estes
resultados indicam ao menos duas hipóteses: i) que a localização
cromossômica destes TEs é dirigida por outros mecanismos evolutivos
podendo evoluir de forma diferencial de outras sequências repetitivas
presentes na heterocromatina; ii) que estes TEs possuem alguma função
estrutural/regulatória e seu acúmulo em regiões eucromáticas pode trazer
vantagens para o genoma do hospedeiro.
O movimento e o acúmulo de TEs exercem grande influência na
trajetória evolutiva do hospedeiro e parece evidente que TEs são ferramentas
regulatórias do genoma, com papel importante no direcionamento da
expressão gênica (Brookfield 2005; Slotkin e Martienssen 2007), porém a
relação entre TEs e função regulatória nem sempre é direta e facilmente
visualizada. Contudo, correlação entre rearranjos cariotípicos e atividade de
retrotransposons foi evidenciada em Perciformes marinhos, onde distribuição
compartimentalizada com acúmulo em regiões pericentroméricas são
observadas em espécies derivadas, tais como Notothenia coriiceps (Ozouf-
Costaz et al. 2004).
Rearranjos cromossômicos são evidentes na evolução dos Cichlinae e
sítios teloméricos intersticiais (ITS) são reconhecidos em Astronotus ocellatus
(Schneider et al. 2012a). Estes ITS provavelmente estão associados a

75
diferentes classes de DNA repetitivo e possivelmente estes TEs podem ter
atuado nos rearranjos cromossômicos de A. ocellatus, uma vez que os
retroelementos Rex1, 3 e 6 estão nas mesmas porções dos cromossomos
onde os ITS são visualizados (Schneider et al. 2012a). O acúmulo de TEs em
regiões específicas do genoma de ciclídeos foi evidenciada e associada a
possíveis eventos de alterações cromossômicas (Teixeira et al. 2009;
Mazzucheli e Martins 2009; Gross et al. 2009b; Ferreira et al. 2010; Valente et
al. 2011).
Virtualmente os TEs podem inserir-se em qualquer porção do genoma
do hospedeiro, entretanto algumas insersões em porções preferenciais já foram
relatadas, tais como em genes ribossomais (Xiong e Eickbush 1993; Silva et al.
2011). Associação entre Rex1, Rex3 e Rex6 e sítios ribossomais 18S e 5S foi
evidenciada em todas as espécies analisadas no presente estudo. Esta
associação entre TE e DNAr em ciclídeos pode ser a causa dos múltiplos sítios
de DNAr 18S encontrados em P. scalare (Schneider et al. 2012a), bem como
pode estar relacionado à variabilidade de sítios de DNAr 18S em Symphysodon
aequifasciatus (Gross et al. 2010). Análise da distribuição do gene de RNAr
18S no genoma de Oreochromis niloticus tem demonstrado que algumas
cópias de 18S são sempre ladeadas por TEs, sugerindo que estes elementos
podem ter atuado na dispersão de cópias de 18S, gerando assim variabilidade
e um grande número de sítios (Nakajima et al. 2012).
3.2.4.2 Diversidade dos TEs
Apesar de pouco se saber sobre a evolução e o efeito genômico de
retroelementos em peixes, estes parecem ser responsáveis pelo dinamismo
cariotípico recentemente revelado para os ciclídeos (Gross et al. 2009b; Poletto
et al. 2010b; Valente et al. 2011; Schneider et al. 2012a). Este dinamismo
também pode ser notado com base nas sequências dos clones dos ciclídeos
neotropicais, onde Rex1 apresentou duas linhagens evolutivas. A presença de
diferentes linhagens de Rex1 já foi observada anteriormente em Oreochromis
niloticus, onde foi verificado a presença de quatro linhagens deste
retroelemento (Volff et al. 2000). Os clones analisados no presente estudo
correspondem à linhagem 4, a qual é amplamente detectada no genoma dos

76
peixes da superordem Acanthopterygii (Volff et al. 2000). O surgimento de
linhagens pode estar associado a resultados de perdas frequentes ou rápida
divergência destes retroelementos (Rouzic e Capy 2006). O elevado nível de
similaridade entre as sequências de Rex de algumas espécies de peixes
teleósteos sugere ainda atividade relativamente recente deste retroelemento
(Volff et al. 2001a).
A presença de stop codons nas sequências de aminoácidos indica que
estas correspondem a elementos inativos, porém foi possível observar blocos
conservados que correspondem ao fragmento da transcriptase reversa para
clones de Rex1 e 3 e da endonuclease em Rex6. As sequências nucleotídicas
de Rex1 e 3 foram mais conservadas em espécies basais (C. monoculus) que
em derivadas (S. discus). Possivelmente este fato está associado a forças
seletivas que tendem a estabilizar o genoma, em muitos casos eliminado
famílias de TEs do genoma de algumas espécies (Böhne et al. 2008), pois os
mecanismos de repressão em cada hospedeiro podem ser diferentes (Capy et
al. 1994). A evolução genômica de Symphysodon é complexa e engloba
eventos de hibridização entre as espécies (Amado et al. 2011), o que pode ter
causado instabilidade nos TEs, levando a eventos de transposição que
resultaram em vários rearranjos cromossômicos (Gross et al. 2009b). Por outro
lado, Rex6 apresentou elevadas taxas de substituições em C. monoculus
(basal) similares a de S. discus (derivado). Este acúmulo de mutações pode
estar associado à senescência do TE, que normalmente é caracterizada pelo
acúmulo de mutações seguida da provável perda estocástica do elemento pelo
genoma (Pinsker et al. 2001).
Estudos demostraram que as sequências de TEs evoluem de forma
independente no genoma de seus hospedeiros (Lerat et al. 2000; Lerat et al.
2002) e uma das principais razões é a possibilidade de transferência horizontal,
que ocasiona incongruência entre as filogenias do hospedeiro e do TE (Silva e
Kidwel 2000). Com relação às árvores filogenéticas dos Rex1 e Rex3, estas
não refletem a árvore filogenética das espécies (Smith et al. 2008) e
provavelmente eventos de transferência horizontal podem ter ocorrido nestes
retroelementos durante a história evolutiva das espécies analisadas.
Entretanto, polimorfismo ancestral e diferentes taxas de evolução de TEs
entre hospedeiros, incluindo perda de cópias de TEs, não podem ser

77
descartados. Por outro lado, para Rex6 as espécies formaram clados
monofiléticos e a árvore filogenética do elemento apresenta congruência com a
proposta para os ciclídeos neotropicais (Smith et al. 2008).
Assim, nossos dados mostram que os ciclideos são um grupo
particularmente interessante para estudos de evolução dos TEs, os quais
atuaram efetivamente na estruturação do genoma e do cariótipo. A distribuição
de Rex no genoma dos ciclídeos sugere que estes retroelementos estão sob
ação de mecanismos evolutivos que tendem a acumula-los em regiões
cromossômicas específicas (principalmente em heterocromatinas). Além disso,
nossos dados indicam que a presença de transferência horizontal, surgimento
ou eliminação de linhagens de TEs podem ter tido papel importante na história
evolutiva de Rex no genoma dos ciclídeos.

78
3.3 Artigo 3 - Mapeamento cromossômico e sequenciamento nucleotídico de DNAs repetitivos de ciclídeos Neotropicais

79
3.3.1 Introdução
Genes de cópia única ou de poucas cópias correspondem a uma
pequena parte do genoma dos vertebrados (2-10%), quando comparado às
regiões repetitivas (Horvath et al. 2001). Estes DNAs repetitivos podem
apresentar-se organizados em tandem ou dispersos pelo genoma
(Charlesworth et al. 1994), sendo que DNAs satélites, minisatélites e
microssatélites normalmente apresentam o primeiro padrão de organização. Os
DNAs satélites são organizados em repetições com centenas ou milhares de
cópias, localizados normalmente em porções cromossômicas
heterocromáticas, preferencialmente próximos aos centrômeros ou em regiões
subteloméricas ou intersticiais (Adega et al. 2009; Kunh et al. 2008). A
importância biológica dos DNAs satélites é intrigante e em muitos casos sem
explicação, pois alguns satélites são conservados evolutivamente por um longo
período de tempo. A presença de DNA satélites em sequências expressas,
bem como as interações com proteínas específicas dos centrômeros, sugere
um papel biológico para alguns destes satélites, embora isto não esteja
claramente elucidado (revisado por Ugarkovic 2005).
Com relação às sequências repetitivas que se encontram dispersas no
genoma destacam-se os elementos transponíveis (TEs), que podem ser
classificados em transposons ou retrotransposons. Estes TEs diferem de outras
sequências repetitivas pela sua capacidade de se mover no genoma do
hospedeiro, gerando uma gama de polimorfismos como consequência da
inserção, bem como pela variação do número de cópias destas sequências
dentro e entre espécies (Biémont e Vieira 2006; Biémont 2010). Devido a
dinâmica dos TEs, acredita-se que eles têm impulsionado a evolução do
genoma de diversas maneiras, incluindo inativação gênica, combinando
diferentes éxons, por conversão e duplicação gênica, além de transposições. A
recombinação entre cópias não homólogas de TEs pode ainda conduzir à
formação de deleções, duplicações, inversões e translocações no genoma do
hospedeiro (Volff et al. 2003; Kazazian 2004; Rebollo et al. 2012).
Estudos envolvendo sequências repetitivas de DNA em peixes ciclídeos
têm indicado a participação ativa destas na carioevolução das espécies, porém
a maioria das informações refere-se ao mapeamento físico de genes

80
ribossomais e alguns poucos estudos focam nos elementos transponíveis e
sequências teloméricas (Gross et al. 2009b; Gross et al. 2010; Polleto et al.
2010b; Valente et al. 2011; Schneider et al. 2012a). Assim, informações sobre
a composição genômica destas regiões repetitivas de DNA são escassas e
para ciclídeos focam principalmente nas espécies africanas (Ferreira e Martins
2008).
Entre as diferentes metodologias de isolamento de sequências
repetitivas a que utiliza cinética de reassociação do DNA (Cot-1 DNA) sido tem
sido empregada com sucesso em peixes (Ferreira e Martins 2008; Vicari et al.
2010). Para os ciclídeos africanos, a utilização desta técnica, aliada ao
mapeamento físico das sequências obtidas permitiu identificar principalmente
DNAs satélites, microssatélites e TEs na fração repetitiva do genoma, os quais
apresentaram-se distribuídos predominantemente em regiões centroméricas e
teloméricas, bem como ao longo do maior par cromossômico de diversas
espécies de ciclídeos africanos da subfamília Pseudocrenilabrinae (Ferreira e
Martins 2008; Ferreira et al. 2010; Fantinatti et al. 2011). Porém, o
conhecimento da porção repetitiva do genoma dos ciclídeos neotropicais ainda
representa uma lacuna que precisa ser elucidada para auxiliar na compreensão
acerca da evolução genômica deste grupo de peixes.
Assim, o objetivo deste trabalho foi avaliar a composição genômica da
fração repetitiva de duas espécies pertencentes à sub-família Cichlinae, que
agrupa os ciclídeos neotropicais: uma espécie que é considerada basal e
apresenta heterocromatina restrita a blocos centroméricos (Cichla monoculus),
enquanto a outra é considerada uma espécie derivada e que apresenta blocos
heterocromáticos conspícuos (Symphysodon discus) (para relações
filogenéticas ver Smith et al. 2008; para dados cariotípicos ver Schneider et al.
2012a). A fração repetitiva foi acessada pela obtenção de Cot-1 DNA (DNA
enriquecido com sequências alta e moderadamente repetitivas), que foi
utilizada para avaliar a distribuição das mesmas em cromossomos metafásicos
de espécies de diferentes tribos de Cichlinae: C. monoculus (Cichlini),
Astronotus ocellatus (Astronotini), Geophagus proximus (Geophagini) e S.
discus (Heroini), o que permitiu o entendimento da organização destas na
carioevolução destes ciclideos. Além disso, o conteúdo genômico da fração

81
Cot-1 DNA foi determinado através de clonagem e sequenciamento
nucleotídico.
3.3.2 Material e Métodos
Foram analisados exemplares pertencentes a quatro tribos da subfamília
Cichlinae coletados na Amazônia Central. As espécies analisadas foram Cichla
monoculus (3 machos e 3 fêmeas), Astronotus ocellatus (3 machos e 5
fêmeas), Geophagus proximus (2 machos e 3 fêmeas), Pterophyllum scalare (3
machos e 3 fêmeas) e Symphysodon discus (2 machos e 2 fêmeas). Os
exemplares foram amostrados com autorização de coleta ICMBio SISBIO
10609-1/2007. Todos os espécimes foram anestesiados por imersão em água
gelada até a morte. Os cromossomos mitóticos foram obtidos de células renais
utilizando o protocolo de “air drying” (Bertollo et al. 1978).
3.3.2.1 Isolamento do DNA repetitivo por meio da cinética de reassociação Cot-1 DNA
Amostras de DNA enriquecido com sequências repetitivas de Cichla
monoculus e Symphysodon discus, foram obtidas por meio da cinética de
renaturação Cot-1 DNA (DNA enriquecido com sequências alta e
moderadamente repetitivas), de acordo com o protocolo descrito por Zwick et
al. (1997), e adaptado por Ferreira e Martins (2008). O DNA total foi extraído à
partir do tecido muscular utilizando o protocolo de fenol-clorofórmio de
Sambrook et al. (1989) e quantificado em espectrofotômetro NanoVue Plus (GE
Healthcare). As amostras de DNA (50 μl de DNA 100-500 ng/μl diluídos em
NaCl 0.3 M) foram autoclavadas (121 ºC) por 5 minutos para obter fragmentos
entre 100 a 2.000 pares de bases. Estas alíquotas de DNA foram desnaturados
por 10 minutos à 95 ºC, colocadas em gelo por 10 segundos e posteriormente
colocadas à 65 ºC por 3 minutos para renaturar. As amostras foram incubadas
por 8 minutos à 37 ºC com 1 U da enzima S1 nuclease, para digerir o DNA fita
simples. A fração repetitiva do DNA foi recuperada por congelamento em
nitrogênio líquido e em seguida re-extraída, utilizando o método e fenol-

82
clorofórmio (Sambrook et al. 1989). Os fragmentos de DNA resultantes, foram
utilizados como sondas para FISH e posteriormente clonados e sequenciados.
3.3.2.2 Clonagem e sequenciamento do DNA repetitivo
Os fragmentos da fração repetitiva Cot-1 DNA foram inseridos em
plasmídeo pMOS Blue Blunt Ended Cloning Kit (GE Healthcare) e os produtos
de ligação foram transformados em células competentes de Escherichia coli
linhagem DH5α. Os clones positivos foram sequenciados em sequenciador
automático de DNA ABI 3130 XL (Applied Biosystem). Cada clone foi
identificado por buscas na base de dados do NCBI
(http://www.ncbi.nlm.nih.gov) com a ferramenta BLASTn e na base do Repbase
(Jurka et al. 2005) no Genetic Information Research Institute (Giri)
(http://www.girinst.org/repbase/) utilizando o software CENSOR (Kohany et al.
2006). As sequências obtidas foram depositadas no banco de dados do NCBI
sob os números: GenBank JX993730 até JX993748. As sequências
nucleotídicas foram alinhadas utilizando o programa ClustalW (Thompson et al.
1994) incluso no programa BioEdit 7 (Hall 1999).
3.3.2.3 Hibridização fluorescente in situ (FISH)
Os fragmentos da fração repetitiva Cot-1 DNA isolados do genoma de C.
monoculus e S. discus foram marcados com Digoxigenina-11-dUTP e Biotina-
16-dUTP (Dig-Nick Translation mix e Biotin-Nick Translation mix; Roche)
respectivamente, por reações de nick translation, conforme instruções do
fabricante. O anticorpo anti-digoxigenina rodamina (Roche) e a estreptavidina
(Life Technologies) foram utilizados para detectar o sinal da FISH (Pinkel et al.
1986). Hibridizações homólogas e heterólogas foram realizadas utilizando
estas sondas nas metáfases de Cichla monoculus, Astronotus ocellatus,
Geophagus proximus e Symphysodon discus. As hibridizações foram feitas
com 77% de estringência (2,5 ng/µL de DNA, 50% formamida, 10% sulfato de
dextrano, e 2x SSC a 37 °C por 18 h). Os cromossomos foram contra corados
com DAPI (2 mg/ml) em meio de montagem VectaShield (Vector).

83
3.3.2.4 Microscopia/Processamento das imagens
Os cromossomos foram analisados em microscópio de epifluorescência
Olympus BX51 e as imagens foram capturadas com câmera digital acoplada
(Olympus DP71) utilizando o software Image-Pro MC 6.3.
3.3.3 Resultados
3.3.3.1 Composição da fração repetitiva do genoma de S. discus e C.
monoculus
Foram sequenciados 77 clones de Cot-1 DNA de C. monoculus e estes
mostraram similaridade com diversas sequências depositadas no GenBank.
Diversos tipos de famílias de DNAs repetitivos foram identificados na fração
repetitiva do genoma de C. monoculus, dentre eles: 30 classes de non-LTR
retrotransposons, 21 de transposons, 6 de microssatélites, 5 de DNAs satélites.
Ainda, alguns fragmentos (15) apresentaram similaridade com genes ou
pseudogenes (Tabela 1).
Entre os 105 clones de Cot-1 DNA de S. discus sequenciados, foram
identificados 38 classes de non-LTR retrotransposons, 23 de transposons, 22
de microssatélites, 11 fragmentos de genes ou pseudogenes, 8 classes de
DNAs satélites e 3 de LTR retrotransposon (Tabela 2). Ainda, alguns clones de
C. monoculus e S. discus apresentaram similaridade a dois diferentes tipos de
sequência, dependendo do banco de dados em qual foi realizada a busca
(Tabela 1 e 2).

84
Tabela 1 Similaridade entre sequências da fração Cot-1 DNA de C. monoculus e sequências conhecidas em banco de dados
públicos
Clone Homologia Similaridade IdentidadeCm1 DNA satélite SATB de Tilapiinae e Haplochrominae (RepBase/GIRI*) 85% Cm2 retrotransposon não-LTR LINE de Oreochromis niloticus (GenBank AF016498.1) 97% Cm3 retrotransposon não-LTR MAUI de Takifugu rubripes (RepBase/GIRI*) 72% Cm4 retrotransposon não-LTR Rex6 (RepBase/GIRI*) 79% Cm5 Transposon Tc1 de Labeo rohita (Genbank AY083617.1) 83% Cm6 retrotransposon não-LTR MAUI de Takifugu rubripes (RepBase/GIRI*) 70% Cm7 retrotransposon não-LTR LINE de Astronotus ocellatus (GenBank FJ164043.1) 84% Cm8 retrotransposon não-LTR EXPANDER (RepBase/GIRI*) 82% Cm9 retrotransposon não-LTR LINE de Oreochromis niloticus (GenBank AF074486.1) 85%
Cm10 Transposon transposon da superfamília P (RepBase/GIRI*) 78% Cm11 retrotransposon não-LTR Rck de Cichla kelberi (GenBank FJ687586.1) 94% Cm12 Transposon Tc1 Mariner de pufferfish (RepBase/GIRI*) 90% Cm13 retrotransposon não-LTR LTR 1 de Xiphophorus (GenBank S77502.1) 88% Cm14 Microssatélite Cyprinus carpio (GenBank JN764809.1) 77% Cm15 retrotransposon não-LTR MAUI de Haplochromis (GenBank AB601486.1) 92% Cm16 retrotransposon não-LTR EXPANDER2 de Takifugu rubripes (RepBase/GIRI*) 81% Cm17 retrotransposon não-LTR Rex3 de Cichla kelberi (GenBank FJ687588.1) 92% Cm18 Microssatélite Parambassis siamensis (GenBank AB720623.1) 100% Cm19 Transposon Tc1 from Takifugu rubripes (RepBase/GIRI*) 84% Cm19 Gene gene Hox de Astatotilapia (GenBank EF594313.1) 90% Cm20 Transposon Tc1 de Rana pipiens (RepBase/GIRI*) 76% Cm20 sequência desconhecida sequência desconhecida de Oreochromis niloticus (GenBank AF533976.1) 83% Cm21 Microssatélite Eleutheronema tetradactylum (GenBank AB697180.1) 96% Cm22 Transposon Tc1 Pogo de Takifugu rubripes (RepBase/GIRI*) 80% Cm22 Gene gene da opsina de Oreochromis niloticus (GenBank JF262086.1) 83% Cm23 Microssatélite Astronotus ocellatus (GenBank GQ302874.1) 90% Cm24 Transposon Tc1 Mariner de Xenopus tropicalis (RepBase/GIRI*) 94% Cm24 Pseudogene Histone H3 pseudogene de Salmo salar (GenBank HM159473.1) 96% Cm25 Satélite DNA satélite de Astronotus ocellatus (GenBank FJ164040.1) 77% Cm26 Satélite DNA satélite de Astronotus ocellatus (GenBank FJ164039.1) 77% Cm27 Satélite DNA satélite de Astronotus ocellatus (GenBank FJ164035.1) 85% Cm28 retrotransposon não-LTR EXPANDER (RepBase/GIRI*) 93% Cm29 Transposon Mariner1 (RepBase/GIRI*) 88% Cm29 Pseudogene Gene da proteína ribosomal de Cyprinus carpio (GenBank AY115478.1) 89% Cm30 Transposon Tc1 DR3 (RepBase/GIRI*) 88% Cm31 retrotransposon não-LTR EXPANDER1 DR de Danio rerio (RepBase/GIRI*) 93% Cm32 retrotransposon não-LTR locus para inserção SINE Tilapia rendalli (GenBank AB100537.1) 85% Cm33 Transposon Helitron2 de Danio rerio (RepBase/GIRI*) 86%

85
Tabela 2 Similaridade entre sequências da fração Cot-1 DNA de S. discus e sequências conhecidas em banco de dados públicos
Clone Homologia Similaridade IdentidadeSd1 Microssatélite Symphysodon discus (GenBank EU109271.1) 100% Sd2 Transposon Mariner de Takifugu rupripes (RepBase/GIRI*) 83% Sd3 Retrovírus ERV3 de Echinops telfairi (RepBase/GIRI*) 85% Sd3 receptor de lipoproteína receptor de lipoproteína deletedo em tumor de D. rerio (GenBank XM001920004.3) 96% Sd4 retrotransposon não-LTR MAUI de Takifugu rupripes (RepBase/GIRI*) 76% Sd4 Gene GnRH1 de Oreochromis niloticus (GenBank AF467291.1) 85% Sd5 Microssatélite Cyprinus carpio (GenBank JN779151.1) 86% Sd6 Microssatélite Symphodus melops (GenBank GQ922115.1) 95% Sd7 Transposon Tc1 de Xenopus tropicalis (RepBase/GIRI*) 92% Sd8 Microssatélite Cyprinus carpio (GenBank JN777259.1) 100% Sd9 Satélite Hinfl 10 de Astronotus ocellatus (GenBank FJ164040.1) 88%
Sd10 Satélite Hinfl 8 de Astronotus ocellatus (GenBank FJ164039.1) 88% Sd11 Microssatélite Cyprinus carpio (GenBank JN782703.1) 100% Sd12 retrotransposon não-LTR Coprina Cc1 (RepBase/GIRI*) 73% Sd13 retrotransposon não-LTR MAUI de Haplochromis chilotes (GenBank AB601474.1) 100% Sd14 Transposon RNAm da transposase Tcb1 de Salmo salar (GenBank BT059762.1) 77% Sd15 Transposon Tc1 Pogo de Takifugu rubripens (RepBase/GIRI*) 86% Sd16 Gene Proteina fosfatase de Oreochromis niloticus (GenBank XM003451427.1) 91% Sd17 retrotransposon não-LTR Rex1 de Cichlasoma labridens (GenBank AJ288469.1) 97% Sd18 retrotransposon não-LTR Rex3 de Tetraodon nigroviridis (GenBank AJ621035.1) 86% Sd19 retrotransposon não-LTR EXPANDER de Haplochromis chilotes (GenBank AB601473.1) 88% Sd20 retrotransposon não-LTR AoRex3 de Astronotus ocellatus (GenBank FJ164041.1) 86% Sd21 retrotransposon não-LTR Rex1 de Haplochomis chilotes (GenBank AB6073.1) 86% Sd22 Transposon Helitron8 de Drosophila rhopaloa (RepBase/GIRI*) 75% Sd23 retrotransposon não-LTR CiLINE de Oreochromis niloticus (GenBank AF016499.1) 93% Sd24 retrotransposon não-LTR LINE de Haplotaxodon microlepis (GenBank AB017056.1) 92% Sd25 retrotransposon não-LTR Rex1 de lampréia marinha (RepBase/GIRI*) 76% Sd25 Gene gene vasa de Oreochromis niloticus (GenBank AB649033.1) 84% Sd26 Gene 18S de Dictyosoma burger (GenBank AB375573.1) 96% Sd27 Microssatélite Pristipomoides filamentosus (GenBank GQ998938.1) 91% Sd28 retrotransposon não-LTR Copia8 de pêssego (RepBase/GIRI*) 87% Sd28 Gene Histone desacetilase 1-B de Salmo salar 92% Sd29 Microssatélite Pangasius (GenBank AF378285.1) 85% Sd30 Satélite ONSAT de tilapia (RepBase/GIRI*) 70% Sd31 retrotransposon não-LTR retrotransposon Penelope de ciclídeos do lago Malawi (RepBase/GIRI*) 80% Sd32 retrotransposon não-LTR Poseidon de Tetraodon nigroviridis (GenBank AJ621377.1) 74% Sd33 Pseudogene LSU rRNA (RepBase/GIRI*) 94% Sd33 Gene Gene ribossomal 28S de Centropristis striata (GenBank EF120947.1) 98% Sd34 Satélite SATA de Sarotherodon galilaeus (GenBank SGU02717) 74%

86
Tabela 2 Continuação
Clone Homologia Similaridade IdentidadeSd35 retrotransposon não-LTR SINE2 de ciclídeos do lago Malawi (RepBase/GIRI*) 90% Sd36 Transposon Mariner3 de musgo (RepBase/GIRI*) 93% Sd37 Gene gene da Opsina de Oreochromis niloticus (GenBank JF262087.1) 85% Sd38 Microssatélite Trachinotus ovatus (GenBank JF964929.1) 97% Sd39 Gene Anoctamina 1 de Oreochromis niloticus (GenBank XM003455720.1) 100% Sd40 retrotransposon não-LTR EXPANDER2 de Takifugu rubripes (RepBase/GIRI*) 82% Sd41 retrotransposon não-LTR Rex1 de Haplochromis chilotes (GenBank AB601473.1) 83% Sd42 Transposon Tc1 de Labeo rohita (GenBank AY083617.1) 86% Sd43 Transposon Helitron N1 de Danio rerio (RepBase/GIRI*) 89% Sd44 Satellite SATB de Oreochromis niloticus (GenBank S57288.1) 88% Sd45 Gene proteina zinc finger de Oreochromis niloticus (GenBank XM003448688.1) 86% Sd46 Transposon Tc1-4Xt (RepBase/GIRI*) 88% Sd47 Microssatélite Gobiocypris rarus (GenBank DQ490152.1) 76% Sd48 retrotransposon não-LTR Rex3 de Xiphophorus maculatus (GenBank AY298859.1) 92% Sd49 retrotransposon LTR Copia 21 de pêssego (RepBase/GIRI*) 77% Sd50 Transposon Mariner 1 de Danio rerio (RepBase/GIRI*) 94% Sd51 retrotransposon não-LTR SINE de Hypophthalmichthys molitrix (GenBank FJ171630.1) 97%

87
3.3.3.2 Mapeamento físico cromossômico da fração repetitiva Cot-1 DNA
Cichla monoculus (Cichlini), Astronotus ocellatus (Astronotini) e
Geophagus proximus (Geophagini) apresentaram número diploide igual a 48
cromossomos (Figura 1A, 1E, 1L, respectivamente) e S. discus (Heroini)
apresentou 60 cromossomos (Figura 1M). A hibridização da sonda Cot-1 DNA
de Cichla monoculus nos cromossomos da própria espécie evidenciou que a
fração repetitiva do genoma encontra-se localizada na região centromérica e
terminal de todos os cromossomos (Figura 1B). A hibridização heteróloga desta
sonda em A. ocellatus (Figura 1F), G. proximus (Figura 1J) e S. discus (Figura
1N) evidenciou sinais centroméricos, teloméricos e dispersos na maioria dos
cromossomos. Contudo em A. ocellatus um par cromossômico não apresentou
nenhuma região de homologia e em outro as mesmas restringiram-se a sítios
teloméricos.
Hibridização com sonda enriquecida com fração repetitiva do DNA de S.
discus (Cot-1 DNA) na própria espécie evidenciou que a fração repetitiva do
genoma encontra-se localizada em grandes blocos na região centromérica de
todos os cromossomos (Figura 1O), com exceção dos dois menores
cromossomos do complemento que não apresentaram nenhuma região de
homologia. A hibridização cruzada da sonda Cot-1 DNA de S. discus nos
cromossomos de C. monoculus (Figura 1C), A. ocellatus (Figura 1G), G.
proximus (Figura 1K) revelou sítios dispersos, com algumas marcações
conspícuas na região centromérica e telomérica de alguns pares de
cromossomos.
A sobreposição de ambas as sondas nos cromossomos de todas as
espécies ressaltou que regiões teloméricas e centroméricas apresentam
grande quantidade de regiões repetitivas de DNA, havendo homologia entre as
espécies (Fig 1D, 1H, 1L, 1P). Contudo, estas regiões repetitivas não são
exclusivas destas regiões, sendo observadas dispersas ao longo dos braços
cromossômicos, especialmente nas hibridações heterólogas.

88
Figura 1 Hibridização da fração Cot-1 DNA em quatro espécies de Cichlinae;
(A, E, I, M) cromossomos contra corados com DAPI das espécies C.
monoculus, A. ocellatus, G. proximus e S. discus, respectivamente; (B, F, J, N)
Sonda Cot-1 DNA isolada do genoma de C. monoculus hibridizada em seus
próprios cromossomos e contra os cromossomos das demais espécies, em (F)
cabeça de seta evidencia par sem sinais de hibridização, e alguns sinais
restringem-se a regiões teloméricas (seta); (C, G, K, O) Sonda Cot-1 DNA
isolada do genoma de S. discus hibridizada contra os cromossomos das
espécies analisadas e em seus próprios cromossomos, em (O) seta evidencia
cromossomos sem sinais de hibridização; (D, H, L, P) Double FISH da fração
Cot-1 DNA.

89
3.3.4 Discussão
3.3.4.1 Organização do DNA repetitivo nos cromossomos dos ciclídeos
Diversas classes de DNAs compõem a fração repetitiva do genoma de
C. monoculus e S. discus, tais como DNAs satélites, microssatélites,
retrotransposons, transposons, genes e pseudogenes. Assim, como na maioria
dos organismos, se compararmos o mapeamento físico cromossômico destes
elementos repetitivos que foram isolados de C. monoculus e S. discus com o
padrão heterocromático descrito para as mesmas, a maior parte destes está
alocado em regiões de heterocromatina (Schneider et al. 2012a).
As regiões heterocromáticas tendem a ser mais conspícuas e
proeminentes em espécies derivadas quando comparadas com espécies
basais de ciclídeos neotropicais, estando predominantemente alocadas em
regiões centroméricas e pericentroméricas (Schneider et al. 2012a). Contudo,
algumas bandas heterocromáticas são visualizadas em C. monoculus, que é
um táxon basal e grupo irmão do ramo que engloba A. ocellatus. O clado que
abriga G. proximus tem posição intermediária e S. discus é agrupado em um
clado derivado (Smith et al. 2008).
O mapeamento físico cromossômico comparativo do Cot-1 DNA
utilizando sondas homólogas em C. monoculus e S. discus revelou que a
espécie basal apresenta a maior parte dos seus elementos repetitivos alocados
em pequenos blocos teloméricos e centroméricos, enquanto que a espécie
derivada apresenta grandes blocos centroméricos. Ou seja, as regiões
repetitivas isoladas da própria espécie não estão restritas às regiões
heterocromáticas em C. monoculus.
Ainda, a hibridização da fração repetitiva do DNA de C. monoculus nos
cromossomos das outras espécies mostrou o mesmo padrão encontrado na
hibridização homóloga, sugerindo que esta parte do genoma deve apresentar-
se conservada ao longo da evolução mantendo possíveis papéis funcionais. Já
a hibridização da fração repetitiva do DNA de S. discus nos cromossomos das
outras espécies mostrou que a compartimentalização destas sequências na
região centromérica ou pericentromérica é uma característica derivada, uma
vez que nas demais espécies estas sequências repetitivas apresentam-se
predominantemente dispersas nos cromossomos, ficando evidente que

90
espécies mais próximas filogeneticamente apresentam maior similaridade na
organização da fração repetitiva do genoma e que as sequências repetitivas
não são exclusivas de regiões heterocromáticas.
Processos de formação de heterocromatina são influenciados por
diversos fatores, entre eles a presença e quantidade de elementos repetitivos
de DNA, como clusters de DNA satélite e elementos transponíveis (Weiler e
Wakimoto 1995; Birchler et al. 2000; Grewal e Jia 2007). Contudo as
sequências repetitivas de DNA alocadas nesta região, provavelmente
desempenham papel relevante na biologia celular, estrutura genômica,
regulação da expressão gênica e evolução adaptativa (Ganley e Kobayashi
2007; Varriale et al. 2008; Bühler 2009; Schneider et al. 2012a). Porém, as
sequências repetitivas não estão restritas às regiões heterocromáticas e já
foram localizadas anteriormente em porções eucromáticas do genoma de
ciclídeos neotropicais (Teixeira et al. 2009; Schneider et al. 2012b), o que não
significa que estas estão sendo constantemente expressas, uma vez que
outras formas de inativação podem ocorrer, tais como RNA de interferência
(RNAi) onde pequenos RNAs agem silenciando a expressão pós-transcricional
(Fire et al. 1998; Djupedal et al. 2005; Jinek e Doudna 2009; Moazed 2009).
3.3.4.2 Diversidade dos DNAs repetitivos em ciclídeos
Independente das sequências repetitivas estarem alocadas em regiões
heterocromáticas ou eucromáticas, estas possivelmente estão envolvidas na
evolução dos grupos, tais como os DNAs satélites SATA e SATB, que foram
isolados do genoma de Oreochromis niloticus, espécie esta que pertence à
subfamília Pseudocrenilabrinae, clado irmão dos ciclídeos Neotropicais (Franck
e Wright 1993; Franck et al. 1994; Smith et al. 2008). O SATB não é tão
abundante no genoma de O. niloticus quanto o SATA, mas havia sido
encontrado apenas em espécies africanas proximamente relacionadas,
estando ausente no genoma de ciclídeos neotropicais (Franck e Wright 1993;
Franck et al. 1994). Entretanto, um clone de C. monoculus apresentou 85% de
identidade genética com este DNA satélite, mostrando assim uma conservação
de SATB com espécies basais dos ciclídeos neotropicais.

91
Dentre os ciclídeos neotropicais também foi possível verificar
similaridades entre DNAs satélites de espécies distantes filogeneticamente, tais
como clones isolados de S. discus (espécie derivada) que possuem
similaridade ao DNA satélite AoSAT isolado de A. ocellatus (Mazzuchelli e
Martins 2009), uma espécie que ocupa uma posição basal na filogenia dos
ciclídeos (Smith et al. 2008). Ainda, estes AoSAT apresentam similaridades
com famílias de satélites centroméricos de outras espécies de peixes,
evidenciando assim uma interessante conservação desta porção genômica. A
conservação de alguns DNAs satélites, em diferentes grupos ao longo do
tempo evolutivo, corrobora a hipótese que esta classe de DNA possa ter papel
regulatório em eucariotos. Entretanto, para compreender a complexa evolução
dos DNAs satélites é necessário a elucidação das bases moleculares que
afetam as mudanças nestas classes de DNA (Ugarkovic e Plohl 2002;
Ugarkovic 2005).
Além dos DNAs satélites, também foram encontrados microssatélites na
porção repetitiva do genoma das espécies estudadas. Os microssatélites
identificados apresentaram similaridade com sequências isoladas de diversas
espécies, entre elas A. ocellatus e S. discus (Amado et al. 2008; Sousa et al.
2009). Devido ao elevado grau de polimorfismo, sequências microssatélites são
comumente utilizadas em estudos populacionais, entretanto nos últimos anos,
o emprego destas sequências tem possibilitado inferências acerca da evolução
cariotípica (Schmidt e Heslop-Harrison 1996; Fillon et al. 1998; Jiang et al.
2011). Estudos futuros que visem mapear microssatélites nos cromossomos de
ciclídeos neotropicais, poderão auxiliar a compreender o papel destas
sequências na evolução cromossômica deste grupo de peixes.
Além dos DNAs repetidos em tandem, os que normalmente se
apresentam dispersos no genoma também têm sido mapeados no genoma dos
ciclídeos neotropicais. Entretanto, a maior parte dos estudos é focado em
retrotransposons não-LTR da família Rex (Mazzuchelli e Martins 2009; Gross et
al. 2009b; Teixeira et al. 2009; Valente et al. 2011; Schneider et al. 2012b). No
presente estudo diversas classes de TEs foram identificadas no genoma de C.
monoculus e S. discus e para ambas espécies as sequências mais abundantes
correspondem a retrotransposons não-LTR. Provavelmente, isto se deve ao
método de transposição destes elementos, pois transposons são excisados de

92
um sítio genômico e integrado a outro, por um processo de “corta e cola”,
enquanto retrotransposons são transcritos em um RNA intermediário copiado
em um cDNA por uma transcriptase reversa e integrado em uma nova região
genômica, dessa forma duplicando o elemento (Burt e Trivers 2006). Ainda, a
maior abundância de retrotransposons não-LTR pode estar associada ao fato
de que o mecanismo de retrotransposição é mais simples, se comparado aos
complexos processos dos retrotransposons LTR (para revisão veja Kazazian
2004).
Diferentes classes de retrotransposons não-LTR foram identificadas no
genoma de C. monoculus e de S. discus, tais como: MAUI, Expander, LINEs,
SINEs, Penelope, Poseidon e Rex. O retroelemento Rex vem sendo
amplamente estudado em peixes, onde diferentes linhagens foram descritas
(Volff et al. 2000). Em ciclídeos neotropicais estes TEs estão associados à
mobilização de sequências não autônomas, e eventos de transferência
horizontal parecem ter ocorrido durante a evolução destes peixes (Schneider et
al. 2012b). Por outro lado, esta é a primeira descrição do retroelemento MAUI
em ciclídeos neotropicais. Este TE foi descrito no genoma de Takifugu rubripes
e pertence ao grupo dos CR1 LINEs (long interspersed elements) (Poulter et al.
1999).
Uma classe específica do elemento LINE denominada CiLINE2 foi
isolada do genoma de Oreochromis niloticus e mapeamentos comparativos
entre os gêneros Oreochromis, Tilapia, e Sarotherodon mostraram padrão
diferencial de localização cromossômica (Oliveira et al. 1999). No presente
estudo foram identificadas sequências isoladas de S. discus que apresentaram
93% de similaridade com CiLINE2 de O. niloticus, mostrando uma manutenção
de sequências repetitivas mesmo em espécies distantes filogeneticamente.
Ainda, este mesmo padrão foi observado para as demais classes de DNAs
repetitivos isoladas, onde o genoma de S.discus apresentou maior quantidade
e diversidade de sequências repetitivas.
Os retrotransposons Penelope e Poseidon foram detectados na fração
repetitiva do genoma de S. discus, porém parecem estar ausentes em C.
monoculus. Contudo, esses retroelementos podem estar presentes no genoma
de C. monoculus e não terem sido amostrados, ou então terem sido eliminados
do genoma desta espécie. Ambos retroelementos, Penelope e Poseidon, são

93
formas ancestrais de retrotransposons presentes em diferentes espécies de
peixes. Penelope foi primeiramente identificado no genoma de Drosophila virilis
na qual apresenta papel importante na evolução cromossômica, onde os sítios
de inserções estão significativamente associados a rearranjos cromossômicos.
Entretanto, este retroelemento está ausente em espécies próximas como
Drosophila melanogaster (Evgen’ev et al. 2000; Volff et al. 2001b; Evgen’ev e
Arkhipova 2004).
As classes de transposons isolados do genoma de S. discus e C.
monoculus foram em sua maioria variantes de Tc1/mariner. Esta superfamília
de transposons é amplamente encontrada no genoma de diversas espécies de
peixes (Radice et al. 1994). A distribuição deste elemento em diferentes
espécies parece ter sido marcada por eventos de transferência horizontal,
devido a incongruências entre relações filogenéticas das espécies e do
transposon. Ainda, a maioria destes elementos é defeituosa em seu gene de
transposase devido a deleções, inserções ou substituições de bases levando a
terminação da matriz de leitura. Isto causa inativação do transposon, então
este elemento será um componente permanente do genoma do hospedeiro,
como um fóssil genético, acumulando diversas mutações (Ivics et al. 1996;
Leaver 2001; Pocwierz-Kotus et al. 2007). Provavelmente esta deve ser a
explicação para a grande diversidade do transposon Tc1, que foi isolados da
fração repetitiva de S. discus e C. monoculus. Ainda, em Cichla kelberi Tc1 foi
localizado em porções centroméricas e análises de sequências mostraram que
no genoma desta espécie estão presentes Tc1 e MITEs (miniature inverted
repeat transposable elements), os quais são um variantes deste transposon
(Teixeira et al. 2009).
Alguns clones de S. discus e de C. monoculus apresentaram
similaridade a dois tipos de sequência, tais como o clone Cm19 que mostrou
84% de identidade com Tc1 de Takigugu rubripes, quando feita a busca no
banco de dados Repbase (Jurka et al. 2005) e 90% de similaridade com parte
do gene Hox de Astatotilapia, quando feita a busca com sequências
depositadas no GenBank. O gene Hox codifica fatores de transcrição que em
metazoários, irão especificar segmentos do corpo ao longo do eixo antero-
posterior. Este gene ocorre em clusters, os quais foram gerados por séries de
duplicações em tandem em um gene ancestral (Stellwag 1999; Ravi et al.

94
2009). Estudos indicam que muitas espécies de peixes apresentam cópias não
funcionais deste gene, com presença de stop códons em alguns clusters
(Hoegg et al. 2007). Por outro lado, a participação de TEs na evolução do gene
Hox já foi descrita em vertebrados (Di-Poï et al. 2009), mostrando assim
complexa relação entre TEs e o genoma do hospedeiro.
Ainda, o clone de S. discus Sd25 apresentou similaridade tanto com
Rex1 em busca realizada no Repbase, quanto com segmento do gene vasa em
busca no GenBank. Este gene desempenha um papel essencial no
desenvolvimento de linhagens de células germinativas (Raz 2000) e
acreditava-se, que em Oreochromis niloticus, este gene era cópia única com
diferentes isoformas geradas por splicing alternativos (Kobayashi et al. 2002).
Entretanto, estudos recentes têm mostrado que o gene vasa é composto ao
menos por três cópias: uma original que foi duplicada gerando duas cópias
adicionais. Ainda, foi encontrado um fragmento de um Expander, um
retrotranspon não-LTR junto a uma destas cópias, o qual pode ter mediado a
duplicação de um locus do gene vasa em O. niloticus (Fujimura et al. 2011).
Assim, é possivel que o retrotransposon não-LTR Rex1 possa ter atuado na
evolução do gene vasa em S. discus, tal qual Expander atuou em O. niloticus,
porém análises adicionais são necessárias.
A utilização Cot-1 DNA foi eficiente para estudar a fração repetitiva do
genoma de C. monoculus e S. discus e evidenciou uma ampla gama de
sequências repetitivas inéditas para ciclídeos neotropicais, bem como, foi
possível verificar que espécies mais próximas filogeneticamente, apresentam
maior similaridade na organização da fração repetitiva nos cromossomos.
Ainda, foi possível verificar a interessante similaridade entre sequências
gênicas e elementos transponíveis, cuja interpretação é complexa. Entretanto,
podemos inferir que estes elementos repetitivos tiveram papel fundamental na
evolução genômica deste grupo de peixes e estudos futuros que venham a
sequenciar o genoma total de ciclídeos neotropicais poderão gerar subsídios
para avançar no entendimento da composição, organização e evolução
genômica deste grupo de peixes.

95
3.4 Artigo 4 – Distribuição de sequências microssatélites em cromossomos de ciclídeos amazônicos
\

96
3.4.1 Introdução
Cichlidae é uma das famílias mais ricas em espécies de peixes de todo
mundo, com aproximadamente 3.000 espécies distribuídas na América Central
e do Sul, África, Madagascar e sul da Índia (Kullander 1998; Kocher 2004). A
evolução deste grupo é marcada por repetidos eventos de radiação adaptativa
e especiações simpátricas (Schliewen et al. 1994; Seehausen 2006) e estes
peixes são reconhecidos como excelente modelo para estudos evolutivos
devido a diversidade de nichos explorados, bem como adaptações
morfológicas e comportamentais (Lowe-McConnell 1991). Na região
Neotropical cerca de 400 espécies já foram descritas e estas se distribuem por
uma gama de hábitats, sendo que a maior diversidade encontra-se na região
Amazônica, com mais de 300 espécies descritas (Kullander 2003).
Em ciclídeos neotropicais a diversidade de formas e adaptações
morfológicas não é reflexo de variações no número diploide, uma vez que a
grande maioria possui 48 cromossomos (Feldberg et al. 2003). Contudo, as
espécies deste grupo apresentam um genoma dinâmico, com variação na sua
estrutura, composição e organização cariotípica evidenciadas pelo
sequenciamento de DNA e mapeamento físico cromossômico de diferentes
sequências de DNA repetitivo, tais como sequências teloméricas,
retrotransposons Rex1, Rex3, Rex6, AoRex3, AoLINE e RCk, transposons Tc1
e sequências gênicas de RNAs ribossomais 18 e 5S, bem como U1 snRNAs
(Vicari et al. 2006; Gross et al. 2009b; Mazzuchelli e Martins, 2009; Teixeira et
al. 2009; Gross et al. 2010; Poletto et al. 2010b; Cabral-de-Mello et al. 2011;
Valente et al. 2011; Schneider et al. 2012a; Schneider et al. 2012b). Além
disso, DNAs repetitivos, tais como elementos transponíveis, estão co-
localizados ou associados com DNAs ribossomais neste grupo de peixes e
parecem estar atuando em processos de duplicação e dispersão das
repetições de DNAr (Gross et al. 2010; Schneider et al. 2012a; Schneider et al.
2012b; Nakajima et al. 2012).
As sequências de DNA repetitivo apresentam um elevado grau de
polimorfismo devido a variação no número de unidades de repetição,
consequência da sua particular dinâmica evolutiva. Contudo, dentre estes
elementos os microssatélites (ou Short Tandem Repeat) são os mais

97
polimórficos, sendo estes formados por pequenas sequências com 1 a 6
nucleotídeos, repetidas em tandem e encontradas ao longo do DNA (Tautz e
Renz 1984). Estes marcadores moleculares têm sido muito utilizados em
genética de populações, na identificação de limites taxonômicos e casos de
hibridização, bem como na análise forense devido a uma suposição implícita de
que estes evoluem de forma neutra (Goldstein e Pollock 1997; Filcek et al.
2005; Racey et al. 2007; McCusker et al. 2008). No entanto, estudos recentes
têm identificado que alguns microssatélites são funcionais e podem afetar a
regulação de genes (Kashi e King 2006; Gemayel et al. 2010), assim como
estarem envolvidos com rearranjos cromossômicos (Kamali et al. 2011).
A utilização do mapeamento físico de sequências microssatélites em
cromossomos tem sido pouco explorada, sendo esta abordagem
principalmente aplicada para o entendimento da evolução de diferentes
sistemas cromossômicos de determinação sexual (Li et al. 2010; Pokorná et al.
2011; Terencio et al. 2012b). Levando-se em conta que microssatélites
representam o componente mais dinâmico do genoma, o entendimento da sua
organização nos cromossomos mostra-se promissora, no sentido de avançar
no conhecimento do papel dos elementos repetitivos de DNA nos mecanismos
de evolução cromossômica. Dessa forma, sequências microssatélites foram
mapeadas em cromossomos de três espécies de ciclídeos neotropicais (Cichla
monoculus, Pterophyllum scalare e Symphysodon discus) que ocupam
diferentes posições filogenéticas visando uma melhor compreensão da
organização e evolução cromossômica neste grupo de peixes.
3.4.2 Material e Métodos
Foram analisados exemplares de Cichla monoculus (4 machos e 4
fêmeas), Pterophyllum scalare (3 machos e 3 fêmeas) e Symphysodon discus
(2 machos e 3 fêmeas) coletados no rio Negro. As coletas foram realizadas
com autorização do ICMBio SISBIO 10609-1/2007. Todos os indivíduos foram
anestesiados em água gelada até a morte. Os cromossomos mitóticos foram
obtidos pela técnica de “air drying” (Bertollo et al. 1978).

98
3.4.2.1 Hibridização fluorescente in situ (FISH)
Oito microssatélites (Amado et al. 2008; Passos et al. 2010) foram
mapeados por hibridização fluorescente in situ (FISH) em metáfases mitóticas
de Cichla monoculus, Pterophyllum scalare e Symphysodon discus (Tabela 1).
As sequências microssatélites foram marcadas com Digoxigenina-11-dUTP e
Biotina-16-dUTP (Dig-Nick Translation mix e Biotin-Nick Translation mix;
Roche), por reações de nick translation, conforme instruções do fabricante. O
anticorpo anti-digoxigenina rodamina (Roche) e a estreptavidina (Life
Technologies) foram utilizados para detectar o sinal da FISH, realizada
seguindo o protocolo de Pinkel et al. 1986. As hibridizações foram feitas com
77% de estringência (2,5 ng/µL de DNA, 50% formamida, 10% sulfato de
dextrano, e 2x SSC a 37 °C por 18 h). Os cromossomos foram contra corados
com DAPI (2 mg/ml) em meio de montagem VectaShield (Vector).
3.4.3 Resultados
Quatro microssatélites apresentaram marcações positivas no genoma de
Cichla monoculus, sendo quatro dinucleotídicos e um pentanucleotídico
(Tabela 1). Hibridizações com o microssatélite pentanucleotídico (GAATA)8
mostraram sinais dispersos em todos os cromossomos e marcações
conspícuas foram verificadas em alguns pares. Contudo, um par não
apresentou nenhuma homologia com a sonda (Figura 1a). O microssatélite
(CA)16 apresentou-se distribuído em todos os cromossomos de C. monoculus,
com exceção de um par. A maioria dos cromossomos apresentou o
microssatélite distribuído de forma dispersa, sendo que em alguns
cromossomos as marcações foram conspícuas (Figura 1b). Padrão disperso foi
verificado quando foi hibridizado o microssatélite (GT)13, entretanto sinais fortes
em porções teloméricas, intersticiais e/ou centroméricas foram visualizadas nos
cromossomos (Figura 1c). Por outro lado, hibridizações utilizando o
microssatélite (AC)7 evidenciaram apenas dois pares marcados, um com
marcações na região telomérica do braço curto e na região intersticial do braço
longo e o outro com ambas regiões teloméricas marcadas (Figura 1d).

99
Tabela 1 Sequências dos microssatélites hibridizados nos cromossomos dos
ciclídeos. (+) presença de sinal de hibridização; (-) ausência de sinal de
hibridização.
Motivo de repetição C. monoculus P. scalare S. discus (CA)16 + + + (AC)7 + - - (GT)13 + + - (GA)12 - + -
(GAATA)8 + + + (GAGAA)12 - + -
(GT)9CA(GT)7CG(GT)19 - - + (CT)14GT(GT)5(CG)2(CT)9 - - +
Com relação a Pterophyllum scalare, cinco microssatélites foram
mapeados no genoma desta espécie: três dinucletídicos e dois
pentanucleotídicos (Tabela 1). Os microssatélites (GAATA)8, (CA)16 e (GT)13,
que se apresentaram dispersos em C. monoculus, apresentaram sinais
compartimentalizados em P. scalare. O microssatélite (CA)16 foi mapeado em
blocos centroméricos de cinco pares cromossômicos (Figura 1f), enquanto os
demais foram localizados em porções teloméricas, centroméricas e intersticiais
na maioria dos cromossomos (Figura 1e e 1g). Ainda, os microssatélites (GA)12
e o (GAGAA)12 apresentaram padrões semelhantes com marcações
conspícuas nos centrômeros, entretanto com (GAGAA)12 um par cromossômico
não apresentou marcação (Figura 1h e 2i).
Quatro microssatélites foram mapeados em Symphysodon discus. Os
microssatélites (GAATA)8 e (CA)16 apresentaram padrões similares a C.
monoculus, com sinais dispersos e com algumas marcações conspícuas na
região centromérica (Figura 1j e 1k). Apenas um par cromossômico apresentou
marcações com os microssatélites (GT)9CA(GT)7CG(GT)19 e
(CT)14GT(GT)5(CG)2(CT)9, entretanto o primeiro foi localizado na região
telomérica do braço longo e o segundo em regiões centroméricas e teloméricas
(Figura 1l e 1m).

100

101
Figura 1 Cromossomos metafásicos de C. monoculus, P. scalare e S. discus
hibridizados com sequências microssatélites (a – m). Setas evidenciam
ausência de sinais de hibridizações. Os cromossomos foram contra corados
com DAPI.
3.4.4 Discussão
A fração repetitiva do genoma normalmente escapa da pressão seletiva
que atua sobre os segmentos não-repetitivos e a maior parte das sequências
microssatélites evolui de forma neutra, não afetando o fenótipo dos indivíduos
(Schlötterer 2000). No entanto, estudos recentes têm identificado
microssatélites funcionais que afetam a forma física de um indivíduo (Kashi e
King 2006; Gemayel et al. 2010). Estes microssatélites putativamente
funcionais estão localizados principalmente em regiões gênicas ou perto delas
e há alteração do número de vezes que o motivo é repetido, estando esta
repetição relacionada à capacidade de modificar a expressão do gene ou
ocasionar uma mudança na sequência de proteínas (Wren et al. 2000; Li et al.
2004; Vinces et al. 2009; Gemayel et al. 2010). Além deste aspecto funcional,
os variantes de DNAs repetitivos incluindo microssatélites podem ser agentes
eficientes de evolução adaptativa (King e Kashi 2009).
Para todas as classes de DNA repetitivo, parece haver uma tendência
geral para o aumento no comprimento da matriz no tempo evolutivo. As
sequências altamente repetitivas tendem ainda a acumular-se nas regiões de
baixa recombinação, como centrômeros e telômeros, enquanto que as
repetições que ocorrem na eucromatina são muito mais susceptíveis a
crossing-over (Pathak e Ali 2012).
De maneira geral, a hibridização cromossômica dos microssatélites
evidenciou padrões contrastantes de abundância e localização destas
sequências nos cromossomos de Cichla monoculus, Pterophyllum scalare e
Symphysodon discus, ficando evidente que estas sequências repetitivas se
acumularam diferencialmente no genoma destas espécies.
Apesar das três espécies apresentarem ampla distribuição dos
microssatélites (GAATA)8 e (CA)16 no seu genoma, maior

102
compartimentalização destes marcadores foi evidenciada em P. scalare, que é
uma espécie derivada na filogenia de Cichlinae (Smith et al. 2008). Esta
compartimentalização de sequências repetitivas em espécies derivadas
também foi evidenciada em elementos transponíveis para este grupo de peixes
(Gross et al. 2009b; Valente et al. 2011; Schneider et al. 2012b). A associação
entre microssatélites e abundância de elementos retrotransponíveis já foi
proposta como um dos mecanismos que pode gerar aumento nos
microssatélites (Nadir et al. 1996), entretanto, em muitos organismos, não há
uma relação entre alta densidade de elementos transponíveis e elevada taxa
de microssatélites.
As sequências microssatélites mapeadas no presente trabalho,
apresentaram localização similar ao padrão descrito para retroelementos nas
mesmas espécies, com algumas marcações dispersas ao longo dos
cromossomos e outras compartimentalizadas em regiões terminais e
centroméricas (Schneider et al. 2012b), o que indica que estas sequências
podem estar envolvidas na formação estrutural do centrômero e telômero. Além
disso, os microssatélites presentes nas regiões centroméricas e subteloméricas
são diferentes entre as espécies de ciclídeos, o que ressalta a importância
destas sequências na evolução deste grupo.
O centrômero é uma estrutura essencial, que tem vários papéis
funcionais para a segregação dos cromossomos replicados para as células
filhas. Esses papéis incluem marcação genética/epigenética e montagem do
complexo proteico do cinetócoro durante o ciclo celular, pontos de controle da
mitose, coesão e liberação das cromátides irmãs, migração cromossômica para
os pólos celulares e citocinese (Santaguida e Musacchio 2009; Allshire e
Karpen 2008; Perpelescu e Fukagawa 2011). Centrômeros são compostos por
longos trechos de repetições em tandem de DNA satélites e microssatélites,
fundamentais para ligação com complexos proteicos (Verdaasdonk e Bloom
2011; Kalitsis e Choo 2012). A região centromérica dos cromossomos de P.
scalare é rica em sequências microssatélites e a composição desta região
cromossômica é variável, uma vez que todos os centrômeros desta espécie
apresentaram sinais de hibridização com o microssatélite (GA)12, mas quando
foi utilizada a sonda (GAGAA)12 um par não apresentou sinal de hibridização.
Ainda, em Cichla monoculus e Symphysodon discus a região centromérica não

103
é rica em (GA)12 e (GAGAA)12. Isso ocorre porque as sequências de DNA
centromérico normalmente possuem elevada taxa de evolução e é comum a
variação entre espécies e até mesmo entre cromossomos da mesma espécie
(Plohl et al. 2008).
Outra região cromossômica que apresenta alta taxa de evolução é a
subterminal dos cromossomos. Normalmente esta região é formada por
diferentes classes de DNA repetitivo, as quais podem auxiliar na estabilização
da porção final dos cromossomos devido a possibilidade de amplificação
destas sequências, mesmo na ausência de telomerase (Jain et al. 2010; Torres
et al. 2011). Em S. discus, são evidenciados na região subterminal de dois
pares de cromossomos os microssatélites (GT)9CA(GT)7CG(GT)19 e
(CT)14GT(GT)5(CG)2(CT)9, os quais não são verificados nos cromossomos de
C. monoculus e P. scalare. O mesmo ocorre com o microssatélite (AC)7 que é
evidenciado apenas em C. monoculus. Já os microssatélites (GAATA)8 e (GT)13
aparecem em regiões subterminais de Cichla monoculus e P. scalare, mas não
são encontrados nesta região em Symphysodon discus. Ainda, em P. scalare,
uma marcação conspícua do microssatélite (GT)13 foi evidenciada na região
terminal no maior par cromossômico, local onde fica localizado o sítio do gene
ribossomal 18S (Schneider et al. 2012a), mostrando assim sintenia entre esta
duas classes de DNA repetitivo.
Além das diferentes sequências que compõem as regiões centroméricas
e teloméricas nos ciclídeos neotropicais, ressalta-se que as sequências
repetitivas não são exclusivas de regiões heterocromáticas (Schneider et al.
2012c). Muitas marcações em regiões eucromáticas foram verificadas com
vários microssatélites, sugerindo que estas podem ter desempenhado papel na
evolução deste grupo de peixes, bem como pode ser possível que estas
sequências tenham alguma função no genoma dos ciclídeos neotropicais.
Nossos dados de mapeamentos comparativos geraram novas informações
acerca da estrutura e organização da porção repetitiva do genoma dos
ciclídeos, contribuindo ainda para o entendimento da composição genômica
destes peixes.

104
4 Conclusões Gerais
A evolução cromossômica dos ciclídeos neotropicais é marcada por
eventos de rearranjos cromossômicos, entretanto nenhuma tendência quanto
aumento, manutenção ou diminuição do número diploide foi verificada, pois
estes eventos ocorrem independentemente nas diferentes tribos de Cichlinae.
A distribuição dos retroelementos Rex1, 3 e 6 no genoma dos ciclídeos
sugere que estes retroelementos estão sob mecanismos evolutivos que tendem
à acumula-los em regiões cromossômicas específicas, principalmente em
heterocromatinas.
Duas linhagens evolutivas de Rex1 foram evidenciadas no genoma dos
exemplares analisados.
As sequências nucleotídicas de Rex1 e 3 foram mais conservadas em
espécies basais como C. monoculus, que em derivadas como S. discus.
Elevadas taxas de substituições nucleotídicas foram verificadas no
retroelemento Rex6 nas espécies analisadas.
A evolução genômica dos ciclídeos neotropicais foi marcada por eventos
de transferência horizontal, surgimento ou eliminação de linhagens de
elementos transponíveis.
Espécies mais próximas filogeneticamente apresentam maior
similaridade na organização da fração repetitiva nos cromossomos.
A fração repetitiva do genoma de S. discus (espécie derivada) apresenta
maior diversidade de sequências quando comparada à de uma espécie basal
(C. monoculus).

105
5 Referências bibliográficas
Adega, F.; Guedes-Pinto, H.; Chaves, R. 2009. Satellite DNA in the Karyotype
Evolution of Domestic Animals – Clinical Considerations. Cytogenetic and
Genome Research, 126:12-20.
Allshire, R.C; Karpen, G.H. 2008. Epigenetic regulation of centromeric
chromatin: old dogs, new tricks? Nature Reviews Genetics, 9: 923-937.
Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. 1990. Basic local
alignment search tool. Journal of Molecular Biology, 215: 403–410.
Alves-Brinn, M.N.; Porto, J.I.R.; Feldberg, E. 2004. Karyological evidence for
interspecific hybridization between Cichla monoculus and C. temensis
(Perciformes, Cichlidae) in the Amazon. Hereditas, 141: 252-257.
Amado, M.V.; Farias, I.P.; Hrbek, T. 2011. A molecular perspective on
systematics, taxonomy and classification amazonian discus fishes of the
genus Symphysodon. International Journal of Evolutionary Biology, 1-16.
Amado, M.V.; Hrbek, T.; Gravena, W.; Fantin, C.; Assunção, E.M.; Astolfi-Filho,
S.; Farias, I.P. 2008. Isolation and characterization of microsatellite markers
for the ornamental discus fish Symphysodon discus and cross-species
amplification in other Heroini cichlid species. Molecular Ecology Resources,
8:1451–1453.
Anjos, H.D.B.; Siqueira, J.A.; Amorim, R.M.S. 2007. Comércio de peixes
ornamentais do Estado do Amazonas, Brasil. Boletim Sociedade Brasileira
de Ictiologia, 4-5.
Aparicio, S.; Chapman, J.; Stupka, E.; Putnam, N.; Chia, J.M.; Dehal, P. et al.
2002. Whole genome shotgun assembly and analysis of the genome of Fugu
rubripes. Science, 297: 1301–1310.
Azevedo, M.F.C.; Oliveira, C.; Martins, C.; Wasko, A.P.; Foresti, F. 2005.
Isolation and characterization of a satellite DNA family in Achirus lineatus
(Teleostei: Pleuronectiformes: Achiridae). Genetica, 125: 205–210.
Bachmann, L.; Raab. M.; Sperlich, D. 1990. Evolution of a telomere associated
satellite DNA sequence in the genome of Drosophila tristis and related
species. Genetica, 83:9-16.

106
Benzaquem, D.C.; Feldberg, E.; Porto, J.I.R.; Gross, M.C.; Zuanon, J.A.S.
2008. Cytotaxonomy and karyoevolution of the genus Crenicichla
(Perciformes, Cichlidae). Genetics and Molecular Biology, 31:250-255.
Bertollo, L.A.C.; Takahashi, C.S.; Moreira-Filho. O. 1978. Cytotaxonomic
considerations on Hoplias lacerdae (Pisces, Erythrinidae). Brazilian Journal
of Genetics, 1:103–120.
Biémont, C. 2010. A brief history of the status of transposable elements: from
junk DNA to major players in evolution. Genetics, 186:1085–1093.
Biémont, C.; Vieira, C. 2006. Junk DNA as an evolutionary force. Nature, 443:
521-524.
Birchler, J.A.; Bhadra, M.P.; Bhadra, U. 2000. Making noise about silence:
repression of repeated genes in animals. Current Opinion in Genetics &
Development, 10, 211–216.
Blass, E.; Bell, M.; Boissinot, S. 2012. Accumulation and rapid decay of non-
LTR retrotransposons in the genome of the three-spine stickleback. Genome
Biology and Evolution, 4:687–702.
Böhne, A.; Brunet, F.; Galiana-Arnoux, D.; Schultheis, C.; Volff, J.N. 2008.
Transposable elements as drivers of genomic and biological diversity in
vertebrates. Chromosome Research, 16:203–215.
Bourque, G.; Leong, B.; Vega, V.B.; Chen, X.; Lee, Y.L.; Srinivasan, K.G.;
Chew, J.L.; Ruan, Y.; Wei, C.H.; Ng, H.H.; Liu. E.T. 2008. Evolution of the
mammalian transcription factor binding repertoire via transposable elements.
Genome Research, 18:1752–1762.
Brookfield, J. 2005. The ecology of the genome-mobile DNA elements and their
hosts. Nature Reviews Genetics, 5:128–136.
Brum, M.J.I. 1995. Correlações entre a filogenia e a citogenética dos peixes
Teleósteos. Séries Monografias, 2: 5-42. Sociedade Brasileira de Genética,
Ribeirão Preto, SP, Brasil.
Bühler, M. 2009. RNA turnover and chromatin-dependent gene silencing.
Chromosoma, 118:141–151.
Burt, A.; Trivers, R. 2006. Genes in conflict: the biology of selfish genetic
elements. Cambridge: Harvard University Press, pp 228-300.
Cabral Júnior, W.; Almeida, O.T. 2006. Avaliação do mercado da indústria
pesqueira na Amazônia. In: Almeida, O.T. (Ed). A indústria pesqueira na

107
Amazônia. Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis/ProVárzea. p. 17-40.
Cabral-de-Mello, D.C.; Valente, G.T.; Nakajima, R.T.; Martins, C. 2012.
Genomic organization and comparative chromosome mapping of the U1
snRNA gene in cichlid fish, with an emphasis in Oreochromis niloticus.
Chromosome Research, 20: 279-292.
Capriglione, T.; Odierna, G.; Caputo, V.; Canapa, A.; Olmo, E. 2002.
Characterization of a Tc1-like transposon in the Antarctic ice-fish,
Chionodraco hamatus. Gene, 295:193-198.
Capy, P.; Login, T.; Bigot, Y.; Brunet, F.; Daboussi, M.J.; Periquet, G.; David, J.;
Hartl, D. 1994. Horizontal transmission versus ancient origin: mariner in the
witness box. Genetica, 93:161-170.
Cazaux, B.; Catalan, J.; Veyrunes, F.; Douzery, E.J.P.; Britton-Davidian, J.
2011. Are ribosomal DNA clusters rearrangement hotspots? A case study in
the genus Mus (Rodentia, Muridae). BMC Evolutionary Biology, 11:124-137.
Charlesworth, B.; Sniegowski, P.; Stephan, W. 1994. The evolutionary
dynamics of repetitive DNA in eukaryotes. Nature, 371:215-220.
Di-Poï, N.; Montoya-Burgos, J.I.; Duboule, D. 2009. A typical relaxation of
structural constraints in Hox gene clusters of the green anole lizard. Genome
Research, 19:602-610.
Djupedal, I.; Portoso, M.; Spahr, H.; Bonilla, C.; Gustafsson, C.M.; Allshire,
R.C.; Ekwall, K. 2005. RNA Pol II subunit Rpb7 promotes centromeric
transcription and RNAi directed chromatin silencing. Genes & Development,
19:2301–2306.
Evgen’ev, M.; Zelentsova, H.; Poluectova, H.; Lyozin, G.; Velikodvorskaya, V.;
Pyatkov, K.; Zhivotovsky, L.; Kidwell, M. 2000. Mobile elements and
chromosomal evolution in the virilis group of Drosophila. Proceedings of the
National Academy of Sciences of United States of America, 97:11337–
11342.
Evgen’ev, M.B.; Arkhipova, I.R. 2004. Penelope-like elements – a new class of
retroelements: distribution, function and possible evolutionary significance.
Cytogenetic and Genome Research, 110:510–521.
Fantinatti, B.E.A.; Mazzuchelli, J.; Valente, G.T.; Cabral-de-Mello, D.C.; Martins,
C. 2011. Genomic content and new insights on the origin of the B

108
chromosome of the cichlid fish Astatotilapia latifasciata. Genetica, 139:1273–
1282.
Farias, I.P.; Ortí, G.; Meyer, A. 2000. Total evidence: molecules, morphology,
and the phylogenetics of cichlid fishes. Journal of Experimental Zoology, 288:
76–92.
Farias, I.P.; Ortí, G.; Sampaio, I.; Schneider H.; Meyer, A. 1999. Mitochondrial
DNA phylogeny of the family Cichlidae: monophyly and fast molecular
evolution of the Neotropical assemblage. Journal of Molecular Evolution, 48:
703–711.
Felberg, E.; Bertollo, L.A.C. 1985. Karyotypes of 10 species of Neotropical
cichlids (Pisces, Perciformes). Caryologia, 38 (3-4): 257-268.
Feldberg, E.; Porto, J.I.R.; Bertollo, L.A.C. 2003. Chromosomal changes and
adaptation of cichlid fishes during evolution. In: Val, A.L., Kapoor, B.G. (Eds).
Fish adaptation. Science Publishers, Enfield, USA. p. 285-308.
Ferreira, I.A.; Martins, C. 2008. Physical chromosome mapping of repetitive
DNA sequences in Nile tilapia Oreochromis niloticus: Evidences for a
differential distribution of repetitive elements in the sex chromosomes.
Micron, 39:411–418.
Ferreira, I.A.; Poletto, A.B.; Kocher, T.D.; Mota-Velasco, J.C.; Penman, D.J.;
Martins, C. 2010. Chromosome Evolution in African Cichlid Fish:
Contributions from the Physical Mapping of Repeated DNAs. Cytogenetic
and Genome Research, 129:314–322.
Filcek, K.B.; Gilmore, S.A.; Scribner, K.T.; Jones, M.L. 2005. Discriminating
lamprey species using multilocus microsatellite genotypes. North American
Journal of Fisheries Management, 25:502–509.
Fillon, V.; Morisson, M.; Zoorob, R.; Auffray, C.; Douaire, M.; Gellin, J.; Vignal,
A. 1998. Identification of 16 chicken microchromosomes by molecular
markers using two-colour fluorescence in situ hybridization (FISH).
Chromosome Research, 6:307-313.
Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. 1998.
Potent and specific genetic interference by double-stranded RNAin
Caenorhabditis elegans. Nature, 391:806-810.
Fischer, C.; Bouneau, L.; Coutanceau, J.P.; Weissenbach, J.; Volff, J.N.; Ozouf-
Costaz, C. 2004. Global heterochromatic colocalization of transposable

109
elements with minisatellites in the compact genome of the pufferfish
Tetraodon nigroviridis. Gene 336 175–183.
Franck, J.P.C.; Kornfield, I.; Wright, J.M. 1994. The utility of SATA satellite DNA
sequences for inferring phylogenetic relationships among the three major
genera of tilapiine cichlid fishes. Molecular Phylogenetics and Evolution,
3:10–16.
Franck, J.P.C.; Wright, J.M. 1993. Conservation of a satellite DNA sequence
(SATB) in the tilapiine and haplochromine genome (Pisces: Cichlidae).
Genome, 36:187–194.
Freitas, C.E.C.; Rivas, A.A.F. 2006. A pesca e os recursos pesqueiros na
Amazônia Ocidental. Ciência e Cultura, 58 (3): 30-32.
Fujimura, K.; Conte, M.A.; Kocher, T.D. 2011. Circular DNA Intermediate in the
Duplication of Nile Tilapia vasa Genes. PLoS One, 6: 29477.
Galetti Jr, P.M.; Martins, C. 2004. Contribuição da hibridação in situ para o
conhecimento dos cromossomos dos peixes. In Guerra. M. (Ed) FISH:
Conceitos e Aplicações na Citogenética. Editora da Sociedade Brasileira de
Genética, p. 61-88.
Ganley, A.R.D.; Kobayashi, T. 2007. Highly efficient concerted evolution in the
ribosomal DNA repeats: Total rDNA repeat variation revealed by whole-
genome shotgun sequence data. Genome Research, 2007 17: 184-191.
Gemayel, R.; Vinces, M.D.; Legendre, M.; Verstrepen, K.J. 2010. Variable
tandem repeats accelerate evolution of coding and regulatory sequences.
Annual Reviews Genetics, 44:445-477.
Goldstein, D.B.; Pollock, D.D. 1997. Lauching microsatellities. Jounal of
Heredity, 88:335-342.
Grewal, S.I.S.; Jia, S. 2007. Heterochromatin revised. Nature Reviews
Genetics, 8:35–46.
Gross, M.C.; Feldberg, E.; Cella, D.M.; Schneider, M.C.; Schneider, C.H.; Porto,
J.I.R.; Martins. C. 2009a. An intriguing evidence of translocations in discus
fish (Symphysodon, Cichlidae) and report of the biggest meiotic
chromosomal chain in vertebrates. Heredity, 102:435–441.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Porto, J.I.R.; Martins, C.; Feldberg,
E. 2009b. Comparative Cytogenetic Analysis of the Genus Symphysodon
(Discus Fishes, Cichlidae): Chromosomal Characteristics of

110
Retrotransposons and Minor Ribosomal DNA. Cytogenetic and Genome
Research, 127:43-53.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Porto, J.I.R.; Martins, C.; Feldberg
E. 2010. Variability of 18S rDNA locus among Symphysodon fishes:
chromosomal rearrangements. Journal of Fish Biology, 76:1117–1127.
Guerra, M. 2004. Hibridização in situ: Princípios básicos. In: Guerra, M. (Ed)
FISH: Conceitos e Aplicações na Citogenética. Editora da Sociedade
Brasileira de Genética, pp. 1-32.
Hall, T.A. 1999. BioEdit: a user-friendly biological sequence alignment editor
and analysis program for Windows 95/96/NT. Nucleic Acids Symposium
Series 1999, 41:95–98.
Hoegg, S.; Boore, J.L.; Kuehl, J.V.; Meyer, A. 2007. Comparative phylogenomic
analyses of teleost fish Hox gene clusters: lessons from the cichlid fish
Astatotilapia burtoni. BMC Genomics, 8:317.
Horvath, J.E.; Bailey, J.A.; Locke, D.P.; Eichler, E.E. 2001. Lessons from the
human genome: transitions between euchromatin and heterochromatin.
Human Molecular Genetics, 10: 2215-2223.
Howell, W.M.; Black, D.A. 1980. Controlled silver-staining of nucleolus
organizer regions with a protective colloidal developer: a 1-step method.
Experientia, 36:1014–1015.
Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. 1991. Improved telomere
detection using a telomere repeat probe (TTAGGG)n generated by PCR.
Nucleic Acids Research, 19:4780
Ivics, Z.; Izsák, Z.; Minter, A.; Hackett, P.B. 1996. Identification of functional
domains and evolution of Tc1-like transposable elements. Proceedings of the
National Academy of Sciences of United States of America, 93:5008-5013.
Jain, D.; Hebden, A.K.; Nakamura, T.M.; Miller, K.M.; Cooper, J.P. 2010. HAATI
survivors replace canonical telomeres with blocks of generic
heterochromatin. Nature, 467: 223–227.
Jiang, B.; Lou, Q.; Wu, Z.; Zhang, W.; Wang, D.; Mbira, K.G.; Weng, Y.; Chen,
J. 2011. Retrotransposon-and microsatellite sequence-associated genomic
changes in early generations of a newly synthesized alletetraploid Cucumis x
hytivus Chen & Kirkbride. Plant Molecular Biology, 77:225-233.

111
Jinek, M.; Doudna, J.A. 2009. A three-dimensional view of the molecular
machinery of RNA interference. Nature, 457:405-412.
Jurka, J.; Kapitonov, V.V.; Pavlicek, A.; Klonowski, P.; Kohany, O.;
Walichiewicz, J. 2005. Repbase Update, a database of eukaryotic repetitive
elements. Cytogenetic and Genome Research, 110:462–467.
Kalitsis, P.; Choo, K.H.A. 2012. The evolutionary life cycle of the resilient
centromere. Chromosoma, 121:327–340.
Kamali, M.; Sharakhova, M.V.; Baricheva, E.; Karagodin, E.; Tu, Z.; Sharakhov,
I.V. 2011. An Integrated Chromosome Map of Microsatellite Markers and
Inversion Breakpoints for an Asian Malaria Mosquito, Anopheles stephensi.
Journal of Heredity, 102:719–726.
Kashi Y,; King D. 2006. Simple sequence repeats as advantageous mutators in
evolution. Trends in Genetics, 22:253-259.
Kazazian Jr, H.H. 2004. Mobile elements: drivers of genome evolution. Science,
303:1626–1632.
Kidwell, M.G.; Lisch, D.R. 2000. Transposable elements and host genome
evolution. Tree, 15: 95-98.
Kidwell, M.G.; Lisch, D.R. 2001. Perspective: transposable elements, parasitic
DNA, and genome evolution. Evolution, 55:1-24.
King, D.G.; Kashi, Y. 2009. Heretical DNA sequences? Science, 326:229-230.
Kobayashi, T.; Kajiura-Kobayashi, H.; Nagahama, Y. 2002. Two isoforms of
vasa homologs in a teleost fish: their differential expression during germ cell
differentiation. Mechanisms of Development, 111:167–171.
Kocher, T.D. 2004. Adaptive evolution and explosive speciation: the cichlid fish
model. Nature Reviews Genetics, 5:288-298.
Kohany, O.; Gentles, A.J.; Hankus, L.; Jurka, J. 2006. Annotation, submission
and screening of repetitive elements in Repbase: Repbase Submitter and
Censor. BMC Bioinformatics, 7:474.
Kornfield, I.L. 1984. Descriptive Genetics of Cichlid fishes. In: Turner, B.J. (Ed).
Evolutionary Genetics of Fishes. Plenum Press, New York. p. 591-616.
Kuhn, G.C.S.; Sene, F.M.; Moreira-Filho, O.; Schwarzacher, T.; Heslop-
Harrison, J.S. 2008. Sequence analysis, chromosomal distribution and long-
range organization show that rapid turnover of new and old pBuM satellite

112
DNA repeats leads to different patterns of variation in seven species of the
Drosophila buzzatii cluster. Chromosome Research, 16:307-324.
Kullander, S.O. 1998. A phylogeny and classification of the South America
Cichlidae (Teleostei: Perciformes). In: Malabarba, L.R.; Reis, R.E.; Vari, R.P.;
Lucena, Z.M.S.; Lucena, C.A.S. (Eds). Phylogeny and classification of
neotropical fishes. Editora da Pontifícia Universidade Católica, Porto Alegre,
Brasil. p. 461-498.
Kullander, S.O. 2003. Family Cichlidae. In: Reis, R.R.; Kullander, S.O.; Ferraris.
C.J. (Eds). Check list of the freshwater of South and Central America. Editora
da Universidade Católica, Porto Alegre, Brasil. p. 605-654.
Leaver, M.J. 2001. A family of Tc1-like transposons from the genomes of fishes
and frogs: evidence for horizontal transmission. Gene, 271:203-214.
Lerat, E.; Biémont, C.; Capy, P. 2000. Codon usage and the origin of P
elements. Molecular Biology and Evolution, 17: 467–468.
Lerat, E.; Capy, P.; Biémont, C. 2002. Codon usage by transposable elements
and their host genes in five species. Journal of Molecular Evolution, 54: 625–
637.
Levan, A.; Fredga, K.; Sandberg, A.A. 1964. Nomenclature for centromeric
position on chromosomes. Hereditas, 52:201–220.
Levin, H.L.; Moran, J.V. 2011. Dynamic interactions between transposable
elements and their hosts. Nature Reviews Genetics, 12 615-627.
Li, J.; Phillips, R.B.; Harwood, A.S.; Koop, B.F.; Davidson, W.S. 2010.
Identification of the Sex Chromosomes of�Brown Trout (Salmo trutta) and
Their Comparison with the Corresponding Chromosomes in Atlantic Salmon
(Salmo salar) and Rainbow Trout (Oncorhynchus mykiss). Cytogenetic and
Genome Research, 133:25–33.
Li, Y.C.; Korol, A.B.; Fahima, T.; Nevo, E. 2004. Microsatellites within genes:
structure, function, and evolution. Molecular Biology and Evolution, 21:991-
1007.
López-Fernández, H.; Honeycutt, R.L.; Winemiller, K.O. 2005. Molecular
phylogeny and evidence for an adaptive radiation of geophagine cichlids from
South America (Perciformes: Labroidei). Molecular and Phylogenetics
Evolution, 34:227–244.

113
Lowe-McConnell, R.H. 1969. The cichlid fishes of Guyana, South America, with
notes on their ecology and breeding behavior. Zoological Journal of the
Linnean Society, 48:255-302.
Lowe-McConnell, R.H. 1991. Ecology of cichlids in South American and African
waters, excluding the African Great Lakes. In: Keenleyside MHA (Ed). Cichlid
Fishes – Behavior, Ecology and Evolution. Chapman and Hall, London, p.
60–85.
Mandrioli, M.; Cuoghi, B.C.; Marini, M.; Manicardi, G.C. 1999. Localization of
the (TTAGGG)n telomeric repeat in the chromosomes of the pufferfish
Tetraodon fluviatilis (Hamilton Buchanan) (Osteichthyes). Caryologia,
52:155-157.
Martins, C. 2007. Chromosomes and repetitive DNA: a contribution to the
knowledge of the fish genome. In: Pisano, E.; Ozouf-Costaz, C.; Foresti, F.;
Kappor, B.G. (Eds) Fish cytogenetics. Science Publisher, Enfield, p. 421–
453.
Martins, C.; Ferreira, I.A.; Oliveira, C.; Foresti, F.; Galetti Jr, P.M. 2006. A
tandemly repetitive centromeric DNA sequence of the fish Hoplias
malabaricus (Characiformes: Erythrinidae) is derived from 5S rDNA.
Genetica, 127:133-141.
Martins, C.; Galetti, P.M. 1999. Chromosomal localization of 5S rDNA genes in
Leporinus fish (Anostomidae, Characiformes). Chromosome Research,
7:363-367.
Martins, C.; Oliveira, C.; Wasko, A.P.; Wright, J.M. 2004. Physical mapping of
the Nile tilapia (Oreochromis niloticus) genome by fluorescent in situ
hybridization of repetitive DNAs to metaphase chromosomes a review.
Aquaculture, 231:37–49.
Martins, C.; Wasko, A.P. 2004. Organization and evolution of 5S Ribosomal
DNA in the fish genome. In: Williams, C.R. (Ed). Focus on Genome
Research. Nova Science Publishers, Inc, Hauppauge, p. 335-363.
Mazzuchelli, J.; Martins, C. 2009. Genomic organization of repetitive DNAs in
the cichlid fish Astronotus ocellatus. Genetica, 136:461–469.
McCusker, M.R.; Paterson, I.G.; Bentzen, P. 2008. Microsatellite markers
discriminate three species of North Atlantic wolffishes (Anarhichas spp.).
Journal of Fish Biology, 72:375–385.

114
Mendonça, M.N.C.; Porto, J.I.R.; Feldberg, E. 1999. Ocorrência de três citótipos
em Satanoperca aff. jurupari (Perciformes, Cichlidae) no Catalão, Manaus,
AM. Genetics and Molecular Biology, 22: 69.
Mesquita, D.R.; Porto, J.I.R.; Feldberg, E. 2008. Chromosomal variability in the
wild ornamental variability fish Symphysodon spp. (Perciformes, Cichlidae)
from amazon. Neotropical Ichthyology, 6:181–190.
Moazed, D. 2009. Small RNAs in transcriptional gene silencing and genome
defence. Nature, 457:413-420.
Nadir, E.; Margalit, H.T.; Gallily, T.; Ben-Sasson, S.A. 1996. Microsatellite
spreading in the human genome: Evolutionary mechanisms and structural
implications. Proceedings of the Natural Academy of Science, 93:6470-6475.
Nakajima, R.T.; Cabral-de-Mello, D.C.; Valente, G.T.; Venere, P.C.; Martins, C.
2012. Evolutionary dynamics of rRNA gene clusters in cichlid fish. BMC
Evolutionary Biology, 12:198.
Nascimento, A.L.; Souza, A.C.P.; Feldberg, E.; Carvalho, J.R.; Barros, R.M.S.;
Pieczarka, J.C.; Nagamachi, C.Y. 2006. Cytogenetic analysis on
Pterophyllum scalare (Perciformes, Cichlidae) from Jari River, Pará state.
Caryologiai, 59:138-143.
Navajas-Pérez, R.; Schwarzacher, T.; Ruiz, R.M.; Garrido-Ramos, M.A. 2009.
Characterization of RUSI, a telomere-associated satellite DNA, in the genus
Rumex (Polygonaceae). Cytogenetic and Genome Research, 124:81–89.
Okada, N.; Hamada, M.; Ogiwara, I.; Ohshima, K. 1997. SINEs and LINEs
share common 39 sequences: a review. Gene, 205:229–243.
Oliveira, C.; Almeida-Toledo, L.F.; Foresti, F.; Toledo-Filho, S.A. 1988.
Supernumerary Chromosomes, Robertsonian rearrangement and multiple
NORs in Corydora aeneas (Pisces, Siluriformes, Callichthyidae). Caryologia,
41: 227-236.
Oliveira, C.; Chew, J.S.K.; Porto-Foresti, F.; Dobson, M.J.; Wright, J.M. 1999. A
LINE2 repetitive DNA sequence from the cichlid fish, Oreochromis niloticus:
sequence analysis and chromosomal distribution. Chromosoma, 108:457–
468.
Ozouf-Costaz, C.; Brandt, J.; Körting, C.; Pisano, E.; Bonillo, C.; Coutanceau,
J.P.; Volff, J.N. 2004. Genome dynamics and chromosomal localization of the

115
non-LTR retrotransposons Rex1 and Rex3 in Antarctic fish. Antarctic Science
16: 51-57.
Passos, K.B.; Leão, A.S.A.; Oliveira, D.P.; Farias, I.P.; Hrbek, T. 2010.
Polymorphic microsatellite markers for the overexploited Amazonian fish,
Semaprochilodus insignis (Jardine�and Schomburgk 1841). Conservation
Genetics Resources, 2:231–234.
Pathak, D.; Ali, S. 2012. Repetitive DNA: A Tool to Explore Animal Genomes/
Transcriptomes. In: Germana, M,; Petrera, F. (Eds). Functional Genomics.
InTech, Published, p.155-180.
Perazzo, G.; Noleto, R.B.; Vicari, M.R.; Machado, P.C.; Gava, A.; Cestari, M.M.
2011. Chromosomal studies in Crenicichla lepidota and Australoheros
facetus (Cichlidae, Perciformes) from extreme Southern Brazil. Reviews in
Fish Biology and Fisheries, 21:509–515.
Perpelescu, M.; Fukagawa, T. 2011. The ABCs of CENPs. Chromosoma,
120:425–446.
Phillips, R.B.; Reed, K.M. 1996. Application of fluorescence in situ hybridization
(FISH) techniques to fish genetics: a review. Aquaculture, 140: 197-216.
Pinkel, D.; Straume, T.; Gray, J.W. 1986. Cytogenetic analysis using
quantitative, high-sensitivity, fluorescence hybridization. Proceedings of the
Natural Academy of Science, 83: 2934-2938.
Pinsker, W.; Haring, E.; Hagemann, S.; Miller, W.J. 2001. The evolutionary life
history of P transposons: From horizontal invaders to domesticated
neogenes. Chromosoma, 110:148-158.
Plohl, M.; Mesltrovin, N.; Bruvo, B.; Ugarkovic, D. 2008. Similarity of structural
features and evolution of satellite DNAs from Palorus subdepressus
(Coleoptera) and related species. Journal of Molecular Evolution, 46, 234-
239.
Pocwierz-Kotus, A.; Burzynski, A.; Wenne, R. 2007. Family of Tc1-like elements
from fish genomes and horizontal transfer. Gene, 390:243–251.
Pokorná, M.; Kratochvíl, L.; Kejnovský, E. 2011. Microsatellite distribution on
sex chromosomes at different stages of heteromorphism and
heterochromatinization in two lizard species (Squamata: Eublepharidae:
Coleonyx elegans and Lacertidae: Eremias velox). BMC Genetics, 12:90.

116
Poletto, A.B.; Ferreira, I.A.; Martins, C. 2010a. The B chromosomes of the
African cichlid fish Haplochromis obliquidens harbour 18S rRNA gene copies.
BMC Genetics, 11:1.
Poletto, A.B.; Ferreira, I.A.; Cabral-de-Mello, D.C.; Nakajima, R.T.; Mazzuchelli,
J.; Ribeiro, H.B.; Venere, P.C.; Nirchio, M.; Kocher, T.D.; Martins, C. 2010b.
Chromosome differentiation patterns during cichlid fish evolution. BMC
Genetics, 11:50-62.
Poulter, R,; Butler, M.; Ormandy, J. 1999. A LINE element from the pufferfish
(fugu) Fugu rubripes which shows similarity to the CR1 family of non-LTR
retrotransposons. Gene, 1999, 227:169–179.
Racey, P.A.; Barratt, E.M.; Burland, T.M.; Deaville,� R.; Gotelli, D.; Jones, G.;
Piertney S.B 2007. Microsatellite DNA polymorphism confirms reproductive
isolation and reveals differences in population genetic structure of cryptic
pipistrelle bat species. Biological Journal of the Linnean Society, vol. 90:539–
550.
Radice, A.D.; Bugaj, B.; Fitch, D.A.H.; Emmons, S.W. 1994. Widespread
occurrence of the Tcl transposon family: Tcl.like transposons from teleost
fish. Molecular and General Genetics, 244:606-612.
Ravi, V.; Lam, K.; Tay, B.H.; Tay, A.; Brenner, S.; Venkatesh, B. 2009. Elephant
shark (Callorhinchus milii) provides insights into the evolution of Hox gene
clusters in gnathostomes. Proceedings of the National Academy of Sciences,
106:16327–16332.
Raz, E. 2000. The function and regulation of vasa-like genes in germ-cell
development. Genome Biology, 1:017.1–1017.6.
Rebollo, R.; Romanish, M.T.; Mager, D.L. 2012. Transposable Elements: An
Abundant and Natural Source of Regulatory Sequences for Host Genes.
Annual Reviews of Genetics, 46:21–42.
Rocchi, M.; Archidiacono, N.; Schempp, W.; Capozzi, O.; Stanyon, R. 2012.
Centromere repositiong in mammals. Heredity, 108:59-67.
Rooney, A.P.; Ward, T.J. 2005. Evolution of a large ribosomal RNA multigene
family in filamentous fungi: Birth and death of a concerted evolution
paradigm. Proceedings of the National Academy of Sciences, 102:5084–
5089.

117
Rouzic, A.; Capy, P. 2006. Theoretical approaches to the dynamics of
transposable elements in genomes, populations, and species. In: Lankenau,
D.H.; Volff, J.N. (Eds). Transposons and dynamic genome. Springer Verlag
Berlin Heidelberg, p. 1-19.
Salgado, S.M., Feldberg, E., Porto, J.I.R. 1995. Estudos citogenéticos em cinco
espécies da família Cichlidae (Perciformes-Labroidei), da bacia amazônica
central. Brazilian Journal of Genetics, 18: 463.
Sambrook, J.; Fritsch, E.F.; Maniatis, T. 1989. Molecular Cloning: a Laboratory
Manual. Cold Springs Harbor Laboratory Press, Cold Springs Harbor, NY.
Sanger, F.; Nicklen, S.; Coulson, A.R. 1977. DNA Sequencing with chain
terminating inhibitors. Proceedings of National Academy of Science 74: 5463-
5467.
Santaguida, S.; Musacchio, A. 2009. The life and miracles of kineto- chores.
EMBO Journal, 28:2511–2531.
Santos, A.C. 2006. Caracterização cromossômica de espécies de acarás da
subfamília Cichlasomatinae (Perciformes: Cichlidae) da Amazônia Central.
Dissertação de mestrado, Instituto Nacional de Pesquisas da Amazônia/
Universidade Federal do Amazonas, Manaus, Brasil. 72pp.
Schaack, S.; Choi, E.; Lynch, M.; Pritham, E.J. 2010. DNA transposons and the
role of recombination in mutation accumulation in Daphnia pulex. Genome
Biology, 11:R46.
Schliewen, U.K.; Tautz, D.; Paabo, S. 1994. Sympatric �speciation suggested
by monophyly of crater lake cichlids. �Nature, 368:629–632.
Schlötterer, C. 2000. Evolutionary dynamics of microsatellite DNA.
Chromosoma, 109:365–371.
Schmidt, T.; Heslop-Harrison, J.S. 1996. The physical and genomic
organization of microsatellites in sugar beet. Proceedings of the National
Academy of Sciences of United States of America, 93:8761-8765.
Schneider, C.H.; Gross, M.C.; Terencio, M.L.; Artoni, R.F.; Vicari, M.R.; Martins,
C.; Feldberg, E. 2012a. Chromosomal evolution of Neotropical cichlids: the
role of repetitive DNA sequences in the organization and structure of
karyotype. Reviews in Fish Biology and Fisheries, doi:10.1007/s11160-012-
9285-3.

118
Schneider, C.H.; Gross, M.C.; Terencio, M.L.; Carmo, E.J.; Martins, C.;
Feldberg, E. 2012b. Evolutionary dynamics of retrotransposable elements
Rex1, Rex3 and Rex6 in Neotropical cichlids genome. BMC Evolutionary
Biology, submitted.
Schneider, C.H.; Gross, M.C.; Terencio, M.L.; Carmo, E.J.; Martins, C.;
Feldberg, E. 2012c. Chromosome mapping and nucleotide sequencing of
repetitive DNAs of Neotropical cichlids. Genetica, submitted.
Seehausen, O. 2006. African cichlid fish: a model system in adaptive radiation
research. Proceedings of the Natural Academy of Science, 273:1987–1998.
Shampay, J.; Schmitt, M.; Bassham, S. 1995. A novel minisatellite at a cloned
hamster telomere. Chromosoma, 104:29-38.
Silva, J.C.; Kidwel, M.G. 2000. Horizontal transfer and selection in the evolution
of P elements. Molecular Biology and Evolution, 17:1542-1557.
Silva, M.; Matoso, D.A.; Vicari, M.R.; Almeida, M.C.; Margarido, V.P.; Artoni,
R.F. 2011. Physical Mapping of 5S rDNA in Two Species of Knifefishes:
Gymnotus pantanal and Gymnotus paraguensis (Gymnotiformes).
Cytogenetic and Genome Research, 134:303-307.
Slijepcevic, P. 1998. Telomeres and mechanisms of Robertsonian fusion.
Chromosoma, 107:136-140.
Slotkin, R.K.; Martienssen, R. 2007. Transposable elements and the epigenetic
regulation of the genome. Nature Reviews Genetics, 8:272-85.
Smith, W.L.; Chakrabarty, P.; Sparks, J.S. 2008. Phylogeny, taxonomy, and
evolution of Neotropical cichlids (Teleostei: Cichlidae: Cichlinae). Cladistics
24:625–641.
Sousa, C.F.S.; Santos, C.H.A.; Sousa, C.B.; Paula-Silva, M.N.; Souza, P.;
Farias, I.P.; Ferreira-Nozawa, M.S.; Almeida-Val, V.M.F. 2009. Development
and characterization of microsatellite markers in Astronotus crassipinis
(Heckel, 1840). Conservation Genetics Resources, 1:277–280.
Sparks, J.S.; Smith, W.L. 2004. Phylogeny and biogeography of cichlid fishes
(Teleostei: Perciformes: Cichlidae). Cladistics 20:501-517.
Stellwag, E.J. 1999. Hox gene duplication in fish. Cell & Developmental Biology, 10:531-540.
Sumner, A.T. 1972. A simple technique for demonstrating centromeric
heterocromatin. Experimentall Cell Research, 75:304–306.

119
Sumner, A.T. 2003. Chromosomes organization and function. Blackwell
Publishing, Malden.
Tafalla, C.; Estepa, A.; Coll, J.M. 2006. Fish transposons and their potential use
in aquaculture. Journal of Biotechnology, 123: 397–412.
Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. 2011.
MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum
Likelihood, Evolutionary Distance, and Maximum Parsimony Methods.
Molecular and Biology Evolution, 28:2731–2739.
Tautz, D.; Renz, M. 1984. Simple Sequences Are Ubiquitous Repetitive
Components Of Eukaryotic Genomes. Nucleic Acids Research, 25, 4127-
4138.
Teixeira, W.G.; Ferreira, I.A.; Cabral-de-Mello, D.C.; Mazzuchelli, J.; Valente,
G.T.; Pinhal, D.; Poletto, A.B.; Venere, P.C.; Martins, C. 2009. Organization
of repeated DNA elements in the genome of the cichlid fish Cichla kelberi
and its contributions to the knowledge of fish genomes. Cytogenetic and
Genome Research, 125:224–234.
Terencio, M.L.; Schneider, C.H.; Gross, M.C.; Nogaroto, V.; Almeida, M.C.;
Artoni, R.F.; Vicari, M.R.; Feldberg, E. 2012a. Influence of repetitive
sequences in the evolution of the W chromosome in fish of the family
Prochilodontidae. Genetica, in press.
Terencio, M.L.; Schneider, C.H.; Gross, M.C.; Vicari, M.R.; Farias, I.P.; Passos,
K.B.; Feldberg, E. 2012b. Evolutionary dynamics of repetitive DNA and sex
chromosomes: Semaprochilodus (Characiformes, Prochilodontidae) as a
model. Sexual Development, submitted.
Thompson, J.D.; Higging, D.G.; Gibson, T.J. 1994. ClustalW: improving the
sensitivity of progressive multiple sequence alignment through sequence
weighting, position specific gap penalties and matrix choice. Nucleic Acids
Research, 22:4673–4680.
Thompson, K.W. 1979. Cytotaxonomy of 41 species of Neotropical Cichlidae.
Copeia, 4:679-691
Torres, G.A.; Gong, Z.; Iovene, M.; Hirsch, C.D.; Buell, C.R.; Bryan,G.J.; Novák,
P.; Macas,J.; Jiang, J. 2011. Organization and Evolution of Subtelomeric
Satellite Repeats in the Potato Genome. Genetics, 1:85-92.

120
Ugarkovic, D. 2005. Functional elements residing within satellite DNAs. EMBO
reports, 11:1035-1039.
Ugarkovic, D.; Plohl, M. 2002. Variation in satellite DNA profiles-causes and
effects. The EMBO Journal, 21:5955-5959.
Valente, G.T.; Mazzuchelli, J.; Ferreira, I.A.; Poletto, A.B.; Fantinatti, B.E.A.;
Martins, C. 2011. Cytogenetic mapping of the retroelements Rex1, Rex3 and
Rex6 among cichlid cish: new insights on the chromosomal distribution of
transposable elements. Cytogenetic and Genome Research, 133:34-42.
Van Sluys, M.A.; Scortecci, K.C.; Costa, A.P.P. 2001. O genoma instável,
sequências genéticas móveis. In: Matioli, S.R. (Ed) Biologia Molecular e
Evolução. Holos Editora, Ribeirão Preto, São Paulo. p. 70-81.
Varriale, A.; Torelli, G.; Bernardi, G. 2008. Compositional properties and
thermal adaptation of 18S rRNA in vertebrates. RNA, 14:1492–1500.
Verdaasdonk, J.S.; Bloom, K. 2011. Centromeres: unique chromatin structures
that drive chromosome segregation. Nature Reviews Molecular Cell Biology,
12:320-332.
Vicari, M.R.; Artoni, R.F.; Moreira-Filho, O.; Bertollo, L.A.C. 2006. Basic and
molecular cytogenetics in freshwater Cichlidae (Osteichthyes, Perciformes).
Karyotypic conservationism and divergence. Caryologia, 59:260-266.
Vicari, M.R.; Nogaroto, V.; Noleto, R.B.; Cestari, M.M.; Cioffi, M.B.; Almeida,
M.C.; Moreira-Filho, O.; Bertollo, L.A.C.; Artoni, R.F. 2010. Satellite DNA and
chromosomes in Neotropical fishes: methods, applications and perspectives.
Journal of Fish Biology, 76:1094–1116.
Vinces, M.; Legendre, M.; Caldara, M.; Hagihara, M.; Verstrepen, K. 2009.
Unstable tandem repeats in promoters confer transcriptional evolve ability.
Science, 324:1213-1216.
Volff, J.F.; Bouneau, L.; Ozouf-Costaz, C.; Fischer, C. 2003. Diversity of
retrotransposable elements in compact pufferfish genomes. Trends in
Genetics, 19:674-678.
Volff, J.N. 2005. Genome evolution and biodiversity in teleost fish. Heredity, 94,
280–294.
Volff, J.N. 2006. Turning junk into gold: domestication of transposable elements
and the creation of new genes in eukaryotes. Bioessays, 28:913-922.

121
Volff, J.N.; Körting, C.; Froschauer, A.; Sweeney, K.; Schartl, M. 2001a. Non-
LTR retrotransposons encoding a restriction enzyme-like endonuclease in
vertebrates. Journal of Molecular Evolution, 52: 351–360.
Volff, J.N., Hornung, U., Schartl, M. 2001b. Fish retroposons related to the
Penelope element of Drosophila virilis define a new group of
retrotransposable elements. Molecular Genetics and Genomics 265: 711–
720.
Volff, J.N.; Korting, C.; Schartl, M. 2000. Multiple lineages of the non-LTR
retrotransposon Rex1 with varying success in invading fish genomes.
Molecular Biology and Evolution, 17:1673–1684.
Volff, J.N.; Korting, C.; Sweeney, K.; Schartl, M. 1999. The non-LTR
retrotransposon Rex3 from the fish Xiphophorus is widespread among
teleosts. Molecular Biology and Evolution, 16: 1427–1438.
Weiler, K.S.; Wakimoto, B.T. 1995. Heterochromatin and gene expression in
Drosophila. Annual Reviews of Genetics, 29:577–605.
Wren, J.; Forgacs, E.; Fondon, J.; Pertsemlidis, A.; Cheng, S.; Gallardo, T.;
Williams, R.; Shohet, R.; Minna, J.; Garner, H. 2000. Repeat polymorphisms
within gene regions: phenotypic and evolutionary implications. American
Journal of Human Genetics, 67:345-356.
Xiong, Y.; Eickbush, T.H. 1993. Dong, a non-long terminal repeat (non-LTR)
retrotransposable element from Bombyx mori. Nucleic Acids Research,
21:1318.
Zwick, M.S.; Hanson, R.E.; McKnight, T.D.; Nurul-Islam-Faridi, M.; Stelly, D.M.
1997. A rapid procedure for the isolation of Cot-1 DNA from plants. Genome,
40:138-142.