fortaleza 2010 - petagronomia.ufc.br lineker.pdf · palavras chaves: vigna unguiculata. interação...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO CEARÁ
CENTRO DE CIÊNCIAS AGRÁRIAS
CURSO DE AGRONOMIA
LINEKER DE SOUSA LOPES
EFEITO AMBIENTAL SOBRE A RESERVA PROTÉICA E CAPACIDADE DE
HIDRATAÇÃO EM DIFERENTES VARIEDADES DE FEIJÃO CAUPI
FORTALEZA
2010

LINEKER DE SOUSA LOPES
EFEITO AMBIENTAL SOBRE A RESERVA PROTÉICA E CAPACIDADE DE
HIDRATAÇÃO EM DIFERENTES VARIEDADES DE FEIJÃO CAUPI
Monografia submetida à coordenação do curso
de Agronomia, da Universidade Federal do
Ceará, como requisito parcial necessário à
obtenção do titulo de Engenheiro Agrônomo.
Orientadora: Profa. Dra. Maria Izabel Gallão.
Co-orientadora: Profa. Dra. Cândida Hermínia
Campos de Magalhães Bertini
FORTALEZA
2010

L854e Lopes, Lineker de Sousa
Efeito ambiental sobre a reserva protéica e capacidade de hidratação em
sementes de feijão caupi / Lineker de Sousa Lopes. -- Fortaleza, 2010.
59 f. ; il. color. enc.
Orientadora: Profa. Dra. Maria Izabel Gallão
Co-orientadora: Profa. Dra. Cândida Hermínia Campos de Magalhães
Bertini
Monografia (Graduação) - Universidade Federal do Ceará, Centro de
Ciências Agrárias, Fortaleza, 2010.
1. Vigna unguiculata. 2. Fisiologia vegetal. I. Gallão, Maria Izabel,
(Orient.). II. Bertine, Cândida Hermínia Campos de Magalhães. III.
Universidade Federal do Ceará – Graduação em Agronomia. IV. Título.
CDD 631

LINEKER DE SOUSA LOPES
EFEITO AMBIENTAL SOBRE A RESERVA PROTÉICA E CAPACIDADE DE
HIDRATAÇÃO EM DIFERENTES VARIEDADES DE FEIJÃO CAUPI
Monografia submetida à coordenação do curso de Agronomia, da Universidade
Federal do Ceará, como requisito parcial necessário à obtenção do titulo de Engenheiro
Agrônomo.
A citação de qualquer parte dessa monografia é permitida, desde que seja feita em
conformidade com as normas da ética cientifica.
LINEKER DE SOUSA LOPES
Monografia aprovada em 24 de Novembro de 2010.

Aos meus Pais,
Francisco José Lopes e
Maria Gêrda Araújo de Sousa Lopes
DEDICO

AGRADECIMENTOS
A Deus, por ter me dado força e discernimento durante toda a minha vida, sempre me
conduzindo pelos caminhos corretos, mas nem sempre fáceis.
Aos meus Pais, que a cada dia lutam para nos dar o pão de cada dia sempre com
dificuldades, mas nunca desistindo e nunca me deixando desistir.
À SESu, especialmente ao Programa de Educação Tutorial (PET), pelo auxílio
financeiro e concessão da bolsa.
Projeto do CNPq Universal (5311156954827847), pelo auxílio financeiro.
A professora Izabel Gallão, pela convivência, ensinamentos e sem duvida pela amizade.
A professora Cândida Bertini, pelos ensinamentos na área da genética e melhoramento
de plantas, sempre com dedicação e boa vontade na hora de ensinar.
Ao professor Ervino Bleicher, pelo seu empenho em nos orientar no Programa de
ensino tutorial (PET) introduzindo-nos no mundo da pesquisa, ensino e extensão por
meio de uma visão critica da realidade.
Ao Dr. Edy Brito, pela disponibilidade em ajudar.
Aos colegas do curso de agronomia, dos quais compartilhamos alegrias e dificuldade na
busca de uma formação acadêmica de qualidade, especialmente meus colegas Everton
Pinheiro, Gabriel Nuto e José Moacir.
A grande família PET-Agronomia da qual tenho imenso orgulho de participar e que sem
dúvida nunca deixarei de fazer parte.
A grande família LAS da qual tem imenso orgulho de participar.
Aos amigos que me ajudaram na realização desta monografia, sem os quais teria sido
mais difícil essa caminhada: Wener e Stellamaris. A todos que contribuíram de uma
forma ou de outra, meu eterno obrigado!

"Toda a nossa ciência, comparada
com a realidade, é primitiva e infantil - e, no
entanto, é a coisa mais preciosa que temos. "
(Albert Einstein)

RESUMO
EFEITO AMBIENTAL SOBRE A RESERVA PROTÉICA E CAPACIDADE DE
HIDRATAÇÃO EM DIFERENTES VARIEDADES DE FEIJÃO CAUPI
As reservas de nutrientes armazenados pelas plantas nas sementes são provisões
para o desenvolvimento do embrião e estabelecimento da nova planta perpetuando
assim a espécie. As proteínas são nutrientes essenciais ao desenvolvimento de embrião e
a alimentação humana. O trabalho foi realizado com o objetivo de avaliar o valor
protéico de variedades de ciclo precoce, o efeito ambiental sobre o acumulo de reservas
protéicas, a atuação do conteúdo protéico como componente matricial responsável pelo
movimento de água no início da embebição das sementes e determinar a curva de
embebição dos lotes de sementes de vinte e duas variedades oriundos de Quixadá e
Cascavel. Vinte e duas variedades de feijão caupi escolhidos por serem precoce e
estarem disponíveis no banco de germoplasma de caupi do Centro de Ciências Agrárias
(CCA) na Universidade Federal do Ceará. Sementes de cada variedade de caupi para os
dois ambientes foram submetidas a extração de proteínas, quantificada por
espectrofotometria. As sementes colocadas para germinar em condições de germinação
foram controladas por câmaras de germinação tipo BOD, com temperatura a 25°C e
fotoperíodo de 12h de luz e 12h de escuro. As variedades CNCx 251-11E, Marataóa
apresentaram maior valor protéico, em Quixadá. As condições ambientais de Quixadá
mostraram-se favoráveis ao acúmulo de reservas protéicas nas sementes de feijão
caupi.A variedade Pitiúba mostrou-se estável à ação do ambiente sobre o acúmulo de
reserva protéica. Recomenda-se o cruzamento das variedades CNCx 251-11E ou
Marataóa com 7919-Dixie hee, Paulistinha, Purple knuckie hull-55, CNCx 251-76E,
BRS-Rouxinol, MNC 03-731C-21, Milagroso ou Setentão e o uso da variedade Pitiúba,
para conferir estabilidade ambiental, para formação de novas variedades no programa de
melhoramento para aumento do valor nutricional do feijão caupi O modelo trifásico de
Bewley e Black (1978) não se adéqua as curvas de embebição de todas as variedades de
feijão caupi. As condições ambientais de cultivo influenciam adequação ao modelo. O
conteúdo protéico não age como principal componente mátrico em sementes de feijão
caupi.
Palavras chaves: Vigna unguiculata. Interação genótipo x ambiente. Curva de
embebição.

ABSTRACT
ENVIRONMENTAL EFFECT ON RESERVE PROTEIN AND HYDRATION
CAPACITY IN DIFFERENT VARIETIES OF COWPEA BEANS
The nutrient reserves stored in the seed plants are provisions for the development
of embryo and establishment of new plant, like this perpetuating the species. Proteins
are essential nutrients to the developing embryo and human nutrition. The study was
conducted to evaluate the protein value of early maturity genotypes, the environmental
effect on the accumulation of protein reserves, the performance of protein content as a
matric component responsible for the movement of water in the beginning of seed
imbibition and determine the imbibition curves of seed lots of twenty-two cowpea
varieties from the Quixadá and Cascavel. Twenty-two cowpea varieties selected by
early maturity and by being available in the cowpea germplasm collection at the Center
for Agricultural Sciences (CCA) at the Federal University of Ceará. Composite samples
of each varieties of cowpea of the two environments were crushed and 40 mg of them
were put to react with 1 mL of NaOH 0.1 M. The protein was quantified by
spectrophotometry. The seeds were put to germinate in conditions controlled by a
germination chamber, BOD, with temperature 25 °C and a photoperiod of 12h light and
12h dark. The varieties CNCx 251-11E and Marataóa have higher protein, in Quixadá.
The environmental conditions of Quixadá were favorable the accumulation of protein
reserves in the cowpea seeds. The variety Pitiúba was stable to the action of the
environment on the accumulation of protein reserve. It is recommended that the
crossing of varieties CNCx 251-11E or Marataóa with 7919-Dixie hee, Paulistinha,
Purple hull knuckie-55, CNCx 251-76E, BRS-Rouxinol, MNC 03-731C-21, Milagorso
or Setentão and use of variety Pitiúba, to provide environmental stability, for formation
of new varieties in a breeding program for enhancing the nutritional value of
cowpea.The triphasic model of Bewley and Black (1978) does not fit the curves of
absorption of all varieties of cowpea. The environmental conditions influence the model
adequacy. The protein content is not acting as principal component matric in cowpea
seeds.
Keywords: Vigna unguiculata. Interaction genotype x environment. Imbibition curve.

SUMÁRIO
1. INTRODUÇÃO GERAL .............................................................................................. 12
2. CAPITULO I - REFERENCIAL TEÓRICO ............................................................... 14
2.1. Origem do feijão caupi .................................................................................................. 14
2.2. Importância socioeconômica do feijão caupi ................................................................ 15
2.3. Melhoramento do feijão caupi no Brasil ....................................................................... 15
2.4. Proteínas do feijão caupi ............................................................................................... 16
2.5. Avaliação de proteínas .................................................................................................. 17
2.6. Acúmulo de reservas nutritivas em sementes ............................................................... 19
2.7. Modelo de embebição de Bewley e Black (1978) ........................................................... 20
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 22
3. CAPITULO II - INFLUÊNCIA AMBIENTAL NO ACÚMULO DE PROTEÍNAS
EM SEMENTES DE DIFERENTES VARIEDADES DE FEIJÃO CAUPI ....................... 26
RESUMO .............................................................................................................................. 26
ABSTRACT .......................................................................................................................... 27
3.1. Introdução ..................................................................................................................... 28
3.2. Material e métodos ........................................................................................................ 29
3.2.1. Material vegetal e delineamento experimental ............................................................... 29
3.2.2. Condições edafoclimáticas ............................................................................................ 30
3.2.3. Preparo das amostras ..................................................................................................... 31
3.2.4. Determinação química................................................................................................... 31
3.2.5. Analise estatística.......................................................................................................... 31
3.3. Resultados e discussão ................................................................................................... 32
3.4. Conclusão ....................................................................................................................... 37
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 38
4. CAPITULO 3 - CAPACIDADE DE HIDRATAÇÃO DE SEMENTES EM
VARIEDADES FEIJÃO CAUPI .......................................................................................... 40
RESUMO .............................................................................................................................. 40
ABSTRACT .......................................................................................................................... 41
4.1. Introdução ..................................................................................................................... 42
4.2. Material e métodos ........................................................................................................ 43
4.2.1. Material vegetal e delineamento experimental ............................................................... 43
4.2.2. Preparo da amostra e curva de embebição ...................................................................... 44
4.2.3. Determinação química................................................................................................... 45

4.2.4. Análise estatística.......................................................................................................... 45
4.3. Resultados e discussão ................................................................................................... 46
4.4. Conclusão ....................................................................................................................... 53
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 54
ANEXOS ............................................................................................................................... 56

12
1. INTRODUÇÃO GERAL
O Feijão caupi é uma excelente fonte de proteínas (23-25% em média),
apresentando todos os aminoácidos essenciais, carboidratos (62%, em média), vitaminas
e minerais, além de possuir grande quantidade de fibras dietéticas, baixa quantidade de
gordura (teor de óleo de 2%, em média) e não conter colesterol. O feijão caupi
representa alimento básico para as populações de baixa renda, e geração de empregos e
renda no Nordeste brasileiro.
As frações que compõem a fibra alimentar exercem efeitos fisiológicos
importantes no organismo. As fibras solúveis têm a propriedade de reduzir a absorção
de glicose e do colesterol plasmático, enquanto que as fibras insolúveis aceleram o
tempo de trânsito intestinal, auxiliando na prevenção de doenças do trato
gastrointestinal (OLSON et al., 1987; MOORE et al., 1998; VANDERHOOF, 1998). O
feijão é um dos poucos alimentos que possui conteúdo balanceado de fibras solúvel e
insolúvel, por isso pode e deve compor os cardápios (HUGHES et al., 1993).
Adicionalmente, o feijão constitui a base alimentar da maioria dos brasileiros, sobretudo
nas classes sociais de baixa renda, devido ao menor custo quando comparado, por
exemplo, às fontes de proteínas de origem animal.
A característica de precocidade do ciclo em feijão caupi é importante por permitir
o melhor aproveitamento das poucas chuvas das regiões semi-áridas pelo produtor no
Nordeste brasileiro para cultivo de feijão caupi. Essa característica reduz a exposição às
intempéries, reduzindo os riscos de perdas na safra se comparadas com variedades de
ciclo longo.
O ambiente tem ação modificadora das características ou balanço de reservas das
sementes. Durante o processo de maturação das sementes, existem duas fases que
exigem condições climáticas completamente diferentes. Na primeira fase, as sementes
acumulam rapidamente matéria seca e a presença de umidade em quantidade adequada é
indispensável. A falta de chuva nesta fase, de forma geral, tornará menos eficiente à
deposição de substâncias nutritivas no interior das sementes, tornando-as menos
vigorosas e com menor potencial de armazenamento. Na segunda fase as sementes se
desidratam rapidamente e se ocorrer muita chuva, a desidratação será lenta. Isto fará
com que o teor de umidade das sementes seja mantido em nível elevado, conduzindo as

13
sementes a uma rápida deterioração. Estas sementes terão o seu potencial de
armazenamento reduzido (MARCOS FILHO, 2005).
Após um período de armazenamento o potencial fisiológico das sementes é
reduzido, mas este potencial pode ser recuperado através do condicionamento
fisiológico, uma técnica de pré-umedecimento que usa a água como re-ativador
metabólico preparatório para o futuro processo germinativo. O pré-umedecimento deve
ser realizado com uma quantidade de água e tempo que permita a semente chegar até no
máximo à segunda fase do modelo trifásico de Bewley e Black, antes da secagem das
sementes condicionadas fisiologicamente (MARCOS FILHO, 2005).
A identificação das causas determinantes dos diversos efeitos imediatos do
condicionamento fisiológico é ainda relativamente obscuro. A maioria das pesquisas
sugere ou conclui que a hidratação controlada promove a atividade de mecanismos de
reparo das membranas, de ácidos nucléicos, de outras macromoléculas e componentes
da estrutura celular, sem, no entanto, identificá-los completamente, mas ressaltando a
redução da liberação de exsudados durante a embebição e a menor ocorrência de
microrganismos associados às sementes. (MARCOS FILHO, 2005)
O presente trabalho foi constituído de duas pesquisas com o objetivo de fornecer
informações do efeito ambiental sobre formação de reservas protéicas, da capacidade
das proteínas de alterarem processo absortivo de água durante a germinação, e sobre
curvas de embebição de diferentes variedades de feijão caupi produzidos em diferentes
ambientes e seu ajuste ao modelo trifásico de Bewley e Black (1978). Vinte e duas
variedades precoces de feijão caupi plantados em Quixadá e Cascavel em regime de
sequeiro foram avaliados na busca das informações acima citadas.
Estas informações servirão de base para programas de melhoramento de feijão
caupi com objetivo de melhorar a qualidade nutricional dos grãos e para o
condicionamento fisiológico de sementes armazenadas em bancos de germoplasma.

14
2. CAPITULO I - REFERENCIAL TEÓRICO
2.1. Origem do feijão caupi
O feijão caupi é uma planta Dicotiledonea, que pertence à ordem Fabales, família
Fabaceae, subfamília Famboideae, Tribo Phaseoleae, subtribo Phaseolina, gênero
Vigna, subgênero Vigna, espécie Vigna unguiculata (L.) Walp., subespécie unguiculata
(VERDCOURT, 1970; MARÉCHAL et. al., 1978; PADULOSI; NG, 1997).
O oeste da África, mais precisamente a Nigéria, é citado por Steele e Mehra
(1980) e Ng e Maréchal (1985) como sendo o centro primário de diversidade da espécie
Vigna unguiculata (L.) Walp. e Padulosi e Ng (1997) relatam que, provavelmente, a
região de Transvaal, na República da África do Sul, seja a região de especiação da
espécie.
O feijão caupi tem vários nomes vulgares dependendo da região. No Nordeste,
por exemplo, é conhecido como feijão de corda, feijão de macáçar ou macaça. O nome
feijão de corda advém do ramo produzido por algumas de suas variedades, sendo estas
classificadas ramadoras. Já o nome feijão de macáçar ou macaça, não vem de uma
estrutura morfológica do vegetal e sim de uma cidade indonésia, Macaçar, hoje
conhecida como Ujung Pandang, que funcionava como entreposto comercial português,
nossos patrícios de outrora (BARRACLOUGH, 1995).
Em 1568, já havia indicação da presença de muitos tipos de feijão no Brasil fato
que se confirmou em 1578, quando foi relatado que uma grande variedade de feijões e
favas era cultivada no Estado da Bahia, sendo os grãos e as vagens usados na
alimentação humana do mesmo modo como o era em Portugal e na Espanha. Embora
nenhuma citação indicasse as cultivares de feijão utilizado, é bastante provável que o
feijão-caupi estivesse entre elas. Nessa época, o comércio entre o Brasil e o Oeste da
África, de Guiné e Angola, era intenso, tornando-se ainda mais importante a partir de
1549, com a fundação da Bahia como capital administrativa (GANDAVO, 2001). Freire
Filho (1988) apresenta várias evidências de que o feijão caupi foi introduzido na
America Latina, no século 16, pelos colonizadores espanhóis e portugueses,
primeiramente nas colônias espanholas e em seguida no Brasil, provavelmente no
Estado da Bahia.

15
2.2. Importância socioeconômica do feijão caupi
Apresenta alta rusticidade e adaptabilidade às condições de estiagem
prolongadas e capacidade de se desenvolver em solo de baixa fertilidade, podendo fixar
nitrogênio atmosférico, por meio da simbiose com bactérias do gênero Rhizobium.
(OLIVEIRA; CARVALHO, 1988). É uma cultura autógama com taxa média de
cruzamento natural, igual ou inferior a 1%, variando com a variedade, condições
ambientais e mais particularmente, com a população de insetos, especialmente as
abelhas (BLACKHURST; MILLER, 1980).
A área ocupada com feijão-caupi, no mundo, está em torno de 12,5 milhões de ha,
com 8 milhões (64% da área mundial) na parte oeste e central da África. Entre todos os
países, os principais produtores mundiais são Nigéria, Niger e Brasil (QUIN, 1997). No
Nordeste, os maiores produtores são os Estados do Ceará, Piauí, Bahia e Maranhão e em
2009, a safra do estado do Ceará representou 15% do total de 866.258 t (IBGE, 2010).
Totalizando o feijão caupi é responsável pela geração de 1.451.578 empregos/ano no
Brasil, com o valor de produção estimado em US$ 249.142.582,00/ano (EMBRAPA,
2010).
Em condições experimentais Bezerra (1997) relatou produtividades de grãos secos
acima de 3 t/ha, tendo a expectativa de que seu potencial genético permita ultrapassar
produtividade de 6 t/ha. Por tanto, o uso de variedades melhoradas e condições de
manejo apropriadas, a cultura de feijão caupi pode proporcionar altos rendimentos.
2.3. Melhoramento do feijão caupi no Brasil
Se comparado a outras culturas, o feijão caupi tem seu potencial genético ainda
pouco explorado. Tendo em vista o valor deste potencial genético, faz-se necessária a
quantificação da variabilidade genética, no pré-melhoramento, para posterior utilização
em programas de melhoramento.
Os objetivos do melhoramento de feijão caupi tem sido, em curto prazo, aumentar
a produtividade e melhorar a qualidade visual, culinária e nutricional dos grãos;
aumentar a adaptabilidade, a estabilidade e a tolerância a estresses hídricos; desenvolver

16
variedades com arquitetura para cultivo mecanizado; desenvolver variedades com
arquitetura moderna mais adequada à agricultura familiar; incorporar resistência
múltipla a doenças; aumentar resistência a insetos-praga, e em médio prazo,
desenvolver variedades com grãos de cor verde persistente à secagem para enlatamento
e congelamento; desenvolver variedades com características para processamento
industrial, para fabricação de farinha e sopa pré-cozida (FREIRE FILHO et al., 2005).
Smartt (1990) afirma que, em relação ao uso, o feijão caupi é muito semelhante
ao feijoeiro comum, porém é mais bem adaptado às condições climáticas dos trópicos
semi-árido, úmido e subúmido. Portanto, graças a essa capacidade de se desenvolver
bem em locais onde o feijão comum não se desenvolve satisfatoriamente e cumprir a
mesma função alimentar, fornecendo os dez aminoácidos essenciais ao ser humano e
excelente valor calórico, é que o feijão caupi deve ser considerado uma cultura
complementar, não competidora do feijão comum.
Segundo Freire Filho et al. (2000), em razão do aumento da importância dos
ciclos mais precoces, o feijão caupi passou a ser classificado segundo o ciclo como:
Superprecoce, aquele que alcança a maturidade com até 60 dias após semeadura,
Precoce, aquele que alcança a maturidade entre 61 e 70 dias após semeadura, Médio-
precoce, aquele que alcança a maturidade entre 71 e 80 dias após semeadura, Médio-
tardio, aquele que alcança a maturidade entre 81 e 90 dias após semeadura, Tardio,
aquele que alcança a maturidade a partir de 91 dias após semeadura.
Outra importante classificação do feijão caupi é quanto a seu porte. Apesar da
classificação de Rachie e Rawal (1976) e Freire filho et al. (1981) com sete e seis
classificações, constata-se que ocorrem quatro principais tipos de porte, os quais seriam
Ereto, ramos principal e secundários curtos; Semi-ereto, ramos principal e secundários
curto a médio; Semiprostrado, ramos principal e secundários médio; e Prostrado, ramos
principal e secundários longos.
2.4. Proteínas do feijão caupi
As proteínas das sementes das leguminosas são geralmente deficientes em
aminoácidos sulfurados (cisteína e metionina), mas, ao contrário dos cereais, seu

17
conteúdo de lisina é satisfatório. As globulinas são as principais proteínas de reserva das
sementes de leguminosas, perfazendo até 70% ou mais do conteúdo total de nitrogênio
da semente. No feijão caupi, a composição de aminoácidos das sementes revela a
presença de todos os aminoácidos essenciais (treonina, valina, isoleucina, leucina,
lisina, fenilalanina, metionina, triptofano, arginina), denominados assim por não serem
sintetizados no organismo de animais (GRAGEIRO et al., 2005).
A simples determinação química é insuficiente para caracterizar o valor nutritivo
de um alimento, pois este se relaciona diretamente com a concentração e ao balanço de
nutrientes, à biodisponibilidade e à presença de componentes tóxicos e/ou
antinutricionais (SGARBIERI, 1987). O feijão caupi possui algumas proteínas que
agem como fator antinutricionais como os inibidores de proteases e as lectinas.
Os inibidores de proteases são proteínas largamente encontradas em vegetais e
outros organismos, que inibem com alta especificidade a ação de enzimas proteolíticas,
como tripsina e quimiotripsina, do trato digestivo de insetos e animais (NORTON,
1991).
As lectinas podem representar até 10% das proteínas armazenadas nos
cotilédones e são glicoproteínas que possuem a capacidade de reconhecer com alta
especificidade e de se ligar de forma reversível a resíduos de carboidratos, que exercem
a função de defesa contra ataques de fitófagos e animais herbívoros, e estão envolvidas
na interação simbiótica entre planta e as bactérias fixadoras de nitrogênio do gênero
Rhizobium (HIRSCH, 1999). Os efeitos tóxicos e antinutricionais das lectinas resultam
da sua interação com glicoconjugados da superfície celular das células epiteliais do
VARo digestório, promovendo desarranjos estruturais e morfológicos da borda em
escova, estimulação da secreção de nitrogênio endógeno e redução da atividade
enzimática intestinal (OLIVEIRA et al., 1988; GRANT, 1991; CAVADA et al., 2001).
2.5. Avaliação de proteínas
Muitos métodos espectrofotométricos, ao longo dos anos, têm sido propostos para
determinação de proteínas totais como o método de Lowry, o método de Smith, o
método de Bradford, mas também existem metodologias não espectrofotométricas

18
muito usadas para este mesmo fim como a de Kjeldahl. Não há uma metodologia
considerada de uso universal, todas têm um determinado nível de sensibilidade e
algumas características que podem ser vantajosas ou não.
O método de Lowry, ou método do folin-fenol, tem por principio a produção de
um composto com absorção máxima em um comprimento de onda de 720nm, a partir da
redução de uma mistura contendo molibdato, tungstato e ácido fosfórico (reagente de
Folin-Ciocalteau) quando reage com proteínas, na presença do catalisador cobre (II). A
principal vantagem do método de lowry é sua alta sensibilidade e as desvantagens desse
método são as muitas substâncias interferentes (Barbital, Tampões, Cloreto do césio,
CiVARo, Cysteine, Diethanolamine, Dithiothreitol, EDTA, EGTA, HEPES,
Mercaptoethanol, Nonidet p-40, Fenol, Pyrrolidone do polyvinyl, Deoxycholate do
sódio, Salicylate de sódio, Thimerosol, Tricine, TRIS, Triton x-100), apresentar longo
tempo de análise, possuir absortividade específica altamente variável para diferentes
proteínas e seguir a lei de Beer-Lambert apenas numa pequena faixa de concentração de
proteínas (ZAIA, et al., 1998).
O método de Smith se baseia na reação de cobre (II) com proteínas, em meio
alcalino, produzindo cobre (I) e formando um complexo com o ácido bicinchoninico,
que absorve luz fortemente no comprimento de onda de 560nm. Apesar de ser fácil o
preparo dos reagentes e possui a mesma sensibilidade do método de Lowry, tem como
desvantagens a dependência da temperatura de reação e a variação da absorbância com
o tempo (ZAIA, et al., 1998).
O método de Bradford, 1976, se baseia na interação entre o corante BG-250 e
macromoléculas de proteínas que contém aminoácidos de cadeias laterais básicas ou
aromáticas. No pH da reação, a interação entre proteínas de alto peso molecular e o
corante BG-250 provoca o deslocamento do equilíbrio do corante para a forma aniônica,
que absorve fortemente em 595nm. O método de Bradford é mais rápido e sensível que
o de Lowry e tem sido utilizado para a determinação de proteínas totais em tecidos de
plantas (ZAIA, et al., 1998).
O método de Kjeldahl é baseado na decomposição da matéria orgânica através da
digestão a 400 °C com ácido sulfúrico concentrado, em presença de sulfato de cobre
como catalisador que acelera a oxidação da matéria orgânica. O nitrogênio presente na
solução ácida resultante é determinado por destilação por arraste de vapor, seguida de

19
titulação com ácido diluído. O método de Kjeldahl permite a determinação indireta de
proteínas em várias amostras biológicas, assim como o nitrogênio em plantas para a
avaliação do estado nutricional (GALVANI; GAERTNER, 2006).
2.6. Acúmulo de reservas nutritivas em sementes
Segundo o modelo proposto por Dure III (1975), o acúmulo de reservas
nutricionais ou matéria seca nas sementes se dá logo após a histodiferenciação, divisão
e expansão celular pós-fecundação da oosfera e se estende até a maturidade fisiológica
quando há o corte de conexão vascular iniciando sua desidratação (Figura 1).
O modelo de Dure III dividido em quatro fases. As fases I e II compreendem os
processos de divisão, histodiferenciação e expansão celular. Na fase II, a habilidade das
células de aumentar de volume pode ser determinante do tamanho final atingido pelas
sementes, indicando o importante papel regulador do desenvolvimento desempenhado
pela relação água/sementes (EGLI, 1998). A fase III é caracterizada pelo intenso
acúmulo de matéria seca que ocorre através da digestão de tecidos, principalmente
nucela e tecidos adjacentes, e da transferência de reservas da planta para a semente. A
fase IV representa o período pós-maturidade fisiológica em que a semente perde a
conexão vascular com planta mãe e sofre rápida desidratação (MARCOS FILHO,
2005).
Apesar do intenso acúmulo de matéria seca durante a fase III, a semente não uma
simples “depósito” passivo das reservas oriundas da planta mãe. Pode regular seu
próprio crescimento pela capacidade de sintetizar moléculas complexas, em quantidades
relativamente precisas. No entanto, isto não a semente independente do fornecimento de
substâncias da planta mãe (EGLI, 1998).

20
Figura 1 – Modelo de Dure III (1975) para acúmulo de matéria seca.
2.7. Modelo de embebição de Bewley e Black (1978)
O processo de embebição das sementes é físico, relacionado basicamente a
propriedades coloidais dos seus constituintes e às diferenças de potencial hídrico (Ѱw)
entre a semente e o meio externo. O potencial hídrico é composto pelo potencial de
pressão (Ѱp), potencial osmótico (Ѱ ), potencial gravitacional (Ѱg) e o potencial
matricial (Ѱm). O potencial osmótico (Ѱ ) representa à capacidade dos solutos, quando
dissolvidos, de reduzir a energia livre do sistema e o potencial matricial (Ѱm)
representa a capacidade de sólidos ou substâncias insolúveis, de reduzirem o Ѱw. O
componente matricial é particularmente importante em estágios iniciais de absorção de
água pelas sementes secas, enquanto ainda há pouca atuação do componente osmótico
por não haver plena embebição da semente (PIMENTA, 2004).
O processo de embebição de sementes foi padronizado por Bewley e Black
(1978), segundo o modelo trifásico de curva polinomial de terceiro grau onde f(x) = ax³
± bx² ± cx ± d, e a > 0. A primeira fase é caracterizada por uma rápida embebição,
graças à diferença acentuada entre os potenciais hídricos. A segunda fase é o momento
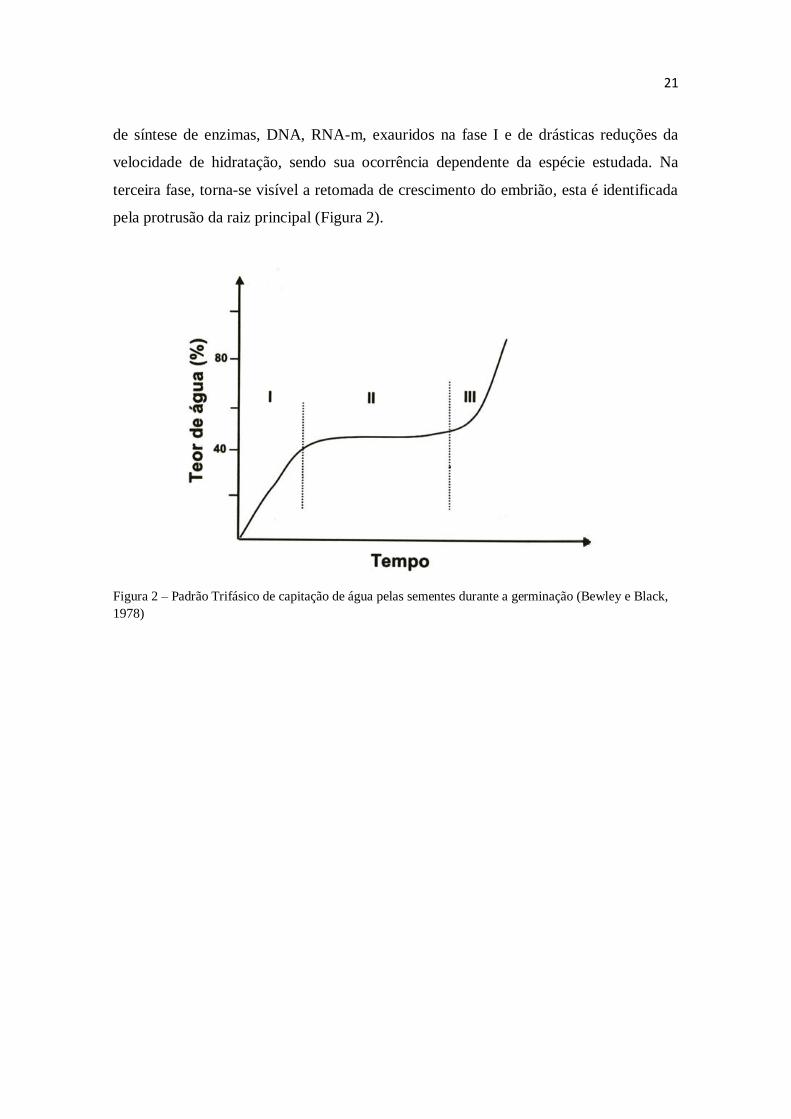
21
de síntese de enzimas, DNA, RNA-m, exauridos na fase I e de drásticas reduções da
velocidade de hidratação, sendo sua ocorrência dependente da espécie estudada. Na
terceira fase, torna-se visível a retomada de crescimento do embrião, esta é identificada
pela protrusão da raiz principal (Figura 2).
Figura 2 – Padrão Trifásico de capitação de água pelas sementes durante a germinação (Bewley e Black,
1978)

22
REFERÊNCIAS BIBLIOGRÁFICAS
BARRACLOUGH, G. Altas da história do mundo da Folha de São Paulo/Times. 4. ed.
rev. São Paulo: Folha da Manhã. 1995. p. 154-157.
BEWLEY, J.D.; BLACK, M. Physiology and Biochemistry of seed in relation to
Germination. Berlin: Springer Verlag, v.1, 306p. 1978.
BEZERRA, A. A. de C. Variabilidade e diversidade genética em caupi (Vigna
unguiculata (L.) Walp.) precoce, de crescimento determinado e porte ereto e semi-
ereto. 1997. 105 f. Dissertação (Mestrado em Melhoramento Genético Vegetal)
Universidade federal Rural de Pernambuco, Recife.
BLACKHURST, H. T.; MILLER JUNIOR, J. C. Cowpea. In: FEHR, W. R.; HADLEY,
H. H. (Ed.). Hybridization of crop plants. Mandson, Wisconsin: American Society of
Agronomy, 1980. p. 327-337.
BRADFORD, M.M. A rapid and sensitive method for the quantifications of microgram
quantities of protein utilizing the principle of protein-dye binding. Analitycal
Biochemistry, v. 72, p.248-254, 1976.
CAVADA, B. S.; BARBOSA, T.; ARRUDA, S.; GRANEIRO, T. B.; BARRAL-
NETO, M. Revisting proteus: Do minor changes in lectin structure matter in biological
activity? Lessons from and potencial biotechnological uses of the Diocleinae subtribe
lectins. Current Protein and Peptide Science, Hilversum, holanda, v. 2, n. 2, p. 123-
135, 2001.
EGLI, D. B. Seed biology and the yield of grain crops. New York, CABI
International. 178 p., 1998.
EMBRAPA. Sistemas de Produção. Disponível em:
http://sistemasdeproducao.cnptia.embrapa.br. Acesso: 18 agost. 2010.
FREIRE FILHO, F. R. Origem, evolução e domesticação do caupi (Vigna unguiculata
(L.) Walp.) In: ARAÚJO, J. P. P. de; WATT, E. E. (Org.) O caupi no Brasil. Goiânia:
Embrapa-CNPAF; Ibadan: IITA, 1988. p. 25-46.

23
FREIRE FILHO, F. R., LIMA, J. A. de A., RIBEIRO, V. Q. Feijão-caupi avanços
tecnológicos. In: FREIRE FILHO, F. R., RIBEIRO, V. Q., BARRETO, P. D.,
SANTOS, A. A. Melhoramento genético. 1. ed. Brasília; Embrapa Informação
Tecnológica, 2005. Cap. 1, p. 29-92.
FREIRE FILHO, F. R.; CARDOSO, M. J.; ARAÚJO, A. G. de; SANTOS, A. A. dos;
SILVA, P. H. S. da. Características botânicas e agronômicas de variedades de feijão
macáçar ( Vigna unguiculata (L.) walp.). Teresina: Embrapa-UEPAE de Teresina,
1981. 40 p. (Boletim de Pesquisa, 4).
FREIRE FILHO, F. R.; RIBEIRO, V. Q.; SANTOS, A. A. Variedades de caupi para
região Meio-norte do Brasil. In: CARDOSO, M. J. (Org.). A cultura do feijão caupi
no Meio-norte do Brasil. Teresina: Embrapa-CPAMN, 2000. P. 67-88. Embrapa-
CPAMN, (Circular Técnica, 28).
GALVANI, F.; GAERTNER, E. Adequação da metodologia Kjeldahl para
determinação de nitrogênio total e proteína bruta. Embrapa, Corumbá, Maio, 2006,
(Circular Técnica 63).
GANDAVO, P. de M. Dos mantimentos de terra. In: GANDAVO, P. de M. VARado da
terra do Brasil. Disponível em : < http://bndigital.bn.br/>. Acesso em: 17. Agost. 2010.
GRAGEIRO, T. B. et al. Composição bioquímica da semente. In: FREIRE FILHO, F.
R.; LIMA, J. A. A.; RIBEIRO, V. Q. Feijão caupi: avanços tecnológicos. Brasília:
Embrapa, 2005. p. 339-365.
GRANT, G. L. In: D’MELLO, J. P. F.; DUFUS, C. M. (Ed.). Toxic substances in crop
plants. Great Britain: The Royal Society of Chemistry, 1991. P. 49-67.
HIRSCH, A. M. Role of lectins (and rhizobial exopoly-saccharides) in legume
nodulation. Current Opinion in Plant Biology, London, v. 2, n. 4, p. 320-326, 1999.
HUGHES, P.E. et al. Coefficient of parentage and cluster analysis of north American
dry bean variedades. Crop Science, v.33, n.1, p.190-193, 1993.
IBGE. Levantamento Sistemático da produção Agrícola: pesquisa mensal de previsão e
acompanhamento das safras agrícolas no ano civil.Levant. Sistem. Prod. Agríc. Rio de
Janeiro v.23 n.09 p.1-80 set.2010.

24
MARCOS FILHO, J. Fisiologia de Sementes de Plantas Cultivadas. 12ed. Editora
Fealq. Piracicaba, 2005. 495 p.
MARÉCHAL, R.; MASCHERPA, J. M.; STAINIER, F. Étude taxonomique d’um
groupe complexe d’espècies de genes Phaseolus et vigna (Papilionaceae) sur la base de
donneés morplhologiques et polliniques, traitées par I’analyse informatique. Boissiera,
Genebra, n. 28, p. 1-273, 1978.
MOORE, M.A. et al. Soluble and insoluble fiber influences on cancer development.
Critical reviews in Oncology/Hematology, v.27, n.3, p.229-242, 1998.
NG, N. Q.; MARÉCHAL, R. Cowpea taxonomy, origin germ plasm. In: SINCH, S. R.;
RACHIE, K. O. (Ed.). Cowpea research, production and utilization. Cheichecter:
Johm Wiley, 1985. p. 11-21.
NORTON, G. Proteinase inhibitors. In: D’MELLO, J. P. F.; DUFUS, C. M. (Ed.).
Toxic substances in crop plants. Great Britain: The Royal Society of Chemistry, 1991.
OLIVEIRA, I. P.; CARVALHO, A. M. A de. A cultura do caupi nas condições de clima
e solo dos trópicos úmidos e semi-áridos do Brasil. In: ARAÚJO, J. P. de; WATT, E. A
(Org.) O caupi no Brasil. Brasília: IITA/EMBRAPA, 1988. p.65-95.
OLIVEIRA, J. T. A.; PUSZTAI, A.; GRANT, G. Changes in organs and tissues induced
by feeding of purified kidney bean (Phaseolus vulgaris) lectins. Nutrition Research,
Tarrytown, v. 8, p. 943-947, 1988.
OLSON, A. et al. Chemistry and analysis of soluble dietary fiber. Food Technology,
v.4, n.2, p.71-82, 1987.
PADULOSI, S.; NG, N. Q. Origin taxonomy, and morphology of Vigna unguiculata
(L.) Walp. In: SINGH, B. B.; MOHAN, R.; DASHIELL, K. E.; JACKAI, L. E. N.
(Ed.). Advances in Cowpea Research. Tsukuba: IITA: JIRCAS, 1997. p. 1-12.
QUIN, F. M. Introduction. In: SING, B. B.; MOHAN RAJ, D. R.; DASHIEL, K. E.;
JACKAI, L. E. N. (Ed.) Advances in cowpea research. Ibadan: IITA-JIRCAS, 1997.
p. ix-xv.

25
RACHIE, K. O.; RAWAL, K. M. Integrated approaches to improving cowpeas
vigna unguiculata (L.) Walp. Ibadan: IITA, 1976. 36 p. (IITA. Techincal Bulletin, 5).
SGARBIERI, V. C. Composição e valor nutritivo dos alimentos. In: SGARBIERI, V. C.
Alimentação e nutrição: fator de saúde e desenvolvimento. São Paulo: Almed, 1987. p.
213-242.
SMARTT, J. Grain legumes: evolution and genetic resources. Cambridge, great
Britain: Cambrige University Press, 1990. 333 p.
STEELE, W. M.; MEHRA, K. L. Structure, evolution and adaptation to farming system
and inveronment in Vigna. In: SUMMERFIELD, D. R.; BUNTING, A. H. (Ed.)
Advances in legume science. England: Royal Botanic Gardens, 1980.
VANDERHOOF, J.A. Immunonutrition: the role of carbohydrates. Nutrition, v.14,
n.7/8, p.595-598, 1998.
VERDCOURT, B. Studies in the leguminosae – Papilonoidea for the flora of tropical
East Africa. IV. Kew Bulletin, Londres, v. 24, p. 597-569, 1970.
ZAIA, D. A. M.; ZAIA, C. T. B. V.; LICHTIG, J. Determinação de proteínas totais via
espectrofotometria: vantagens e desvantagens dos métodos existentes. Química Nova,
21(6), 1998.

26
3. CAPITULO II - INFLUÊNCIA AMBIENTAL NO ACÚMULO DE
PROTEÍNAS EM SEMENTES DE DIFERENTES VARIEDADES DE
FEIJÃO CAUPI
RESUMO
O feijão caupi é muito cultivado e consumido no Nordeste brasileiro, sendo assim
acessível à grande parte da população, representando a principal fonte de proteína
vegetal. Os riscos de perda total ou redução de produção e da qualidade dos grãos de
feijão caupi podem ser reduzidos por técnicas de cultivo que permitem o convívio com
a seca e com as chuvas irregulares, uma delas é o uso de variedades adaptadas e
precoces. O trabalho foi realizado com o objetivo de avaliar o valor protéico e o efeito
ambiental sobre o acúmulo de reservas protéicas de variedades de feijão caupi de ciclo
precoce. As variedades de feijão caupi foram plantadas em 2010, nas Fazendas Lavoura
seca e Boa água localizadas em Quixadá e Cascavel, respectivamente. Foram avaliados
22 variedades, os quais foram escolhidas por serem precoces e estarem disponíveis no
banco de germoplasma de caupi do Centro de Ciências Agrárias (CCA) na Universidade
Federal do Ceará. Amostras composta de cada variedade de caupi para os dois
ambientes foram maceradas e de 40 mg de cada amostra foram extraídas proteínas com
1 mL de NaOH a 0,1M, sendo em seguida quantificada por espectrofotometria. Os
dados foram estatisticamente estudados através da análise de variância e a comparação
de médias pelo teste de Scott-Knott. Na quantificação da divergência genética entre as
variedades, foram utilizadas as técnicas de aglomeração com base na distância
generalizada de Mahalanobis (D2). As variedades CNCx 251-11E, Marataóa
apresentaram maior valor protéico, em Quixadá. As condições ambientais de Quixadá
mostraram-se favoráveis ao acúmulo de reservas protéicas nas sementes de feijão
caupi.A variedade Pitiúba mostrou-se estável à ação do ambiente sobre o acúmulo de
reserva protéica. Recomenda-se o cruzamento das variedades CNCx 251-11E ou
Marataóa com 7919-Dixie hee, Paulistinha, Purple knuckie hull-55, CNCx 251-76E,
BRS-Rouxinol, MNC 03-731C-21, Milagroso ou Setentão e o uso da variedade Pitiúba,
para conferir estabilidade ambiental, para formação de novas variedades no programa de
melhoramento para aumento do valor nutricional do feijão caupi.
Palavras chaves – Vigna unguiculata. Valor protéico. Sementes.

27
ABSTRACT
The cowpea is widely grown and consumed in the Brazilian Northeast. It's
accessible to the majority of the population and representing the main source of
vegetable protein. The risks of total loss or reduction of production and quality of the
beans can be reduced by cultivation techniques that allow interaction with drought and
irregular rainfall, one of them is the use of early cowpea varieties. The study was
conducted to evaluate the protein value of genotypes early-maturing and environmental
effect on the accumulation of protein reserves. The cowpea was planted in 2010, in
farms located in Quixadá and Cascavel. The 22 varieties were chosen to be early
matured and be available in the cowpea germplasm collection at the Center for
Agricultural Sciences (CCA) at the Federal University of Ceará. Composite samples of
each varieties of cowpea of the two environments were macerated and from 40 mg of
sample were extracted proteins with 1 mL of 0.1 M NaOH, and then quantified by
spectrophotometry. The data were statistically analyzed by analysis of variance and
comparisons of means by Scott-Knott test. In the quantification of genetic divergence,
we have used clustering based on the Mahalanobis distance (D²). Varieties CNCx 251-
11E and Marataóa have higher protein, in Quixadá. The environmental conditions of
Quixadá were favorable the accumulation of protein reserves in the cowpea seeds. The
variety Pitiúba was stable to the action of the environment on the accumulation of
protein reserve. It is recommended that the crossing of varieties CNCx 251-11E or
Marataóa with 7919-Dixie hee, Paulistinha, Purple hull knuckie-55, CNCx 251-76E,
BRS-Rouxinol, MNC 03-731C-21, Milagorso or Setentão and use of variety Pitiúba, to
provide environmental stability, for formation of new varieties in a breeding program
for enhancing the nutritional value of cowpea.
Keywords - Vigna unguiculata. Protein value. Seeds.

28
3.1. Introdução
As proteínas são acumuladas no interior das sementes com o objetivo de fornecer
aminoácidos para desenvolvimento do embrião, no entanto pode ser aproveitado pelo
homem e animais como fonte de proteínas. Assim, do ponto de vista nutricional, a
identificação e a quantificação das proteínas contidas nas sementes de diferentes
variedades de feijão caupi permitem selecionar aquelas de alto valor nutricional.
O feijão caupi, além de aumentar o conteúdo protéico na alimentação, contribui
para melhorar a qualidade da dieta, quando constituída predominantemente de
leguminosas e cereais, porque além de ser uma excelente fonte de proteínas (23 a 25%
em média) apresenta todos os aminoácidos essenciais, em especial, lisina, uma vez que
os cereais são deficientes neste aminoácido (IQBAL; KHALIL; SHAH, 2003).
A qualidade nutricional das proteínas das leguminosas é influenciada pelo gênero,
espécie, variedade botânica, concentração de fatores antinutricionais, tempo de
estocagem, tratamento térmico e, em geral, é inferior àquela da proteína de origem
animal (BRESSANI, 1993; CRUZ et al., 2003).
O acúmulo de matéria seca pode ser influenciado por fitormônios, pelo ambiente
(oferta de água, fertilidade do solo, temperatura, luminosidade) e até pela posição da
semente na planta (MARCOS FILHO, 2005).
Dentre todos os fitormônios, as auxinas e as giberelinas atuam no acúmulo de
matéria seca. As auxinas são responsáveis por atividades de assimilação de compostos
provenientes da planta-mãe enquanto as giberelinas direcionam a síntese de reservas na
semente (MARCOS FILHO, 2005).
Têm sido observadas reduções no teor de proteínas, em estações agrícolas com
alta precipitação pluvial ou irrigação mal controlada, nas culturas de trigo, cevada e
aveia, enquanto o oposto, ou seja, elevação no proteína total em plantas expostas a
períodos de baixa disponibilidade hídrica ocorre em soja, milho, feijão e, também, em
trigo. (BEWLEY; BLACK, 1994; COPELAND; MCDONALD, 1995).
A fertilidade do solo pode contribuir para alteração da composição protéica das
sementes. Os íons de potássio presente em maiores quantidade são importantes para o
metabolismo normal de proteínas e para a atividade enzimática, assim como o
nitrogênio que pode ser fornecido pelo solo ou pela simbiose rizóbio - leguminosa. O

29
íon sulfato (SO4-2
) absorvido pelas raízes fornece o enxofre necessário à formação de
aminoácidos sulfurados (cisteína e metionina), mas em leguminosas não há grande
presença de aminoácidos sulfurados, indicando assim pouca expressividade do íon
sulfato (SO4-2
) na alteração da quantidade de proteína total em feijão caupi. O íon Mg2+
atua como elemento-ponte conferindo estabilidade a conformação de proteínas e
enzimas, contribuindo para o acréscimo de N-protéico (FURLANI, 2004).
A temperatura e luminosidade atuam sobre a fotossíntese e consequente sucesso
no acúmulo de matéria seca. Marcos Filho (2005) afirma que um número relativamente
pequeno de pesquisas sobre estresse por temperatura na formação da semente, sugere
que temperaturas superiores a 30 °C na fase de transferência de matéria seca pode trazer
sérios prejuízos à produção e qualidade das sementes devido à redução fotossintética em
espécies de leguminosas. A temperatura ótima para a germinação de sementes da maioria
das espécies está entre 15 e 30°C, a máxima varia de 30 a 40°C, podendo a mínima se
aproximar do ponto de congelamento (COPELAND; MCDONALD, 1995).
Considerando-se que o feijão caupi é uma tradicional fonte de proteína para a
população nordestina, objetivou-se com esse trabalho avaliar o valor protéico de
diferentes variedades de ciclo precoce em dois ambientes do Ceará e o efeito ambiental
sobre o acúmulo de proteínas.
3.2. Material e métodos
3.2.1. Material vegetal e delineamento experimental
Para a execução do trabalho, vinte e duas variedades foram cultivadas no primeiro
semestre do ano de 2010, nos meses de abril, maio e junho, em Quixadá-CE e em
Cascavel-CE sob condições de sequeiro. O plantio seguiu o delineamento experimental
de blocos casualizados (DBC), com quatro repetições e com vinte e duas variedades
constituindo os Tratamentos. As parcelas foram constituídas de 24 plantas espaçadas a
0,8 x 0,6 m e uma área útil constituída por 12 plantas.
As vinte e duas variedades foram Sempre verde (CE-25), Pitiúba (CE-31),
Milagroso (CE-46), Rouxinho 1 (CE-73), Rouxinho 2 (CE-77), 7919-Dixie hee (CE-
104), Purple knuckie hull-55 (CE-113), Setentão (CE-596), CNCx 251-11E (CE-790),

30
CNCx 251-60E (CE-796), CNCx 251-76E (CE-798), Pingo de ouro (CE-930),
Marataóa (CE-933), BRS-Rouxinol (CE-937), Paulistinha (CE-939), BRS-Punjante
(CE-940), CB-3 (ainda sem registro no Banco de germoplasma de feijão caupi, do
CCA/UFC), AU 94-418-07-01 (CE-176), UCR-95-701 (CE-493), MNC 03-731C-21
(CE-390), MNC 03-720-11 (CE-178), MNC 01-627D-65-1 (CE-497), escolhidas pelo
critério de precocidade, já que variedades precoces são mais adaptados ao ambiente
semi-árido, resultando assim maior rendimento ao agricultor sertanejo.
Em Quixadá, o plantio foi realizado na Fazenda Lavoura Seca, pertencente à
Universidade Federal do Ceará e em Cascavel, o plantio foi realizado na Fazenda Boa
Água, pertencente ao Eng° Agrônomo José Mesquita L. Almeida, numa área de 1200
m2.
3.2.2. Condições edafoclimáticas
As condições edafoclimáticas do sertão central e tabuleiro costeiro do Ceará
foram representados por Quixadá e Cascavel, respectivamente. O ambiente de Cascavel
é típico tropical com inverno seco e verão subúmido, enquanto o ambiente de Quixadá é
típico tropical semi-árido com inverno seco e verão com chuvas irregulares pela ação
inconstante das massas de ar. Durante o ano de cultivo observou-se que, no estado do
Ceará, as chuvas se normalizaram tardiamente, sendo o mês de abril o que mais choveu
e o que se iniciou uma boa distribuição das chuvas no período, o que representou uma
boa disponibilidade de água no inicio do desenvolvimento da cultura (Tabela 1).
Os solos dos locais de plantio foram quimicamente corrigidos com base na análise
de solos e segundo a recomendação de adubação para cultivo de feijão caupi para o
estado do Ceará (AQUINO et al, 1993).
Tabela 1 – Pluviosidade mensal nos ambientes Quixadá e Cascavel no primeiro
semestre de 2010
LOCAL PLUVIOSIDADE (mm)
Janeiro Fevereiro Março Abril Maio Junho
QUIXADÁ 47,4 0,0 57,8 288,4 5 36,2
CASCAVEL 80 33 76 179 75 60
Fonte: FUNCEME 2010.

31
3.2.3. Preparo das amostras
A partir das sementes colhidas e beneficiadas foram coletadas amostras 1000
sementes em suas repetições de cada parcela e homogeneizadas para compor uma
amostra composta representativa do ambiente.
Subamostras das amostras compostas das sementes dos vinte e duas variedades
destinadas as análises de proteínas foram obtidas por trituração em liquidificador
industrial, sendo o material triturado peneirado com uma peneira de 18 mesh, para a
uniformização da amostra e para facilitar o contato soluto com o solvente, reagente
extrator. Foram realizadas três repetições de cada amostra para quantificação protéica,
segundo o delineamento inteiramente casualizado (DIC).
3.2.4. Determinação química
Foram pesadas 40 mg de cada amostra e colocadas para reagir com 1 mL de
NaOH a 0,1 M, por 20 minutos, sendo o sobrenadante removido com centrifugação a
5000 rpm por 4 minutos, esta operação foi repetida com cada amostra até exauri-la de
proteínas. O sobrenadante foi reservado para quantificação protéica, segundo o método
proposto por Bradford (1976). Neste método utiliza-se uma solução albumina (BSA)
como padrão e um espectrofotômetro regulado para mensurar a absorbância da luz com
o comprimento de onda de 595 nm pela solução de 100 µL da amostra/mL de solução
de Bradford. A quantificação das proteínas existentes se dá a partir da absorbância
obtida, processada segundo a equação de regressão da solução padrão de albumina
(BSA) com absorbância registrada.
3.2.5. Analise estatística
A análise de variância foi realizada segundo ANOVA para o delineamento
inteiramente casualizado (DIC) com esquema fatorial simples (2x22), onde se tem
constituindo o primeiro fator os dois ambiente e o segundo as 22 variedades. Após a
verificação do nível de significância pelo teste F da ANOVA, realizou-se a comparação
de médias pelo teste Scott e Knott (1974).

32
Com o intuito de identificar similaridades e diferenças entre as variedades quanto
o valor de proteína total nos diferentes ambientes avaliados, realizou-se a quantificação
da divergência genética, utilizando-se a técnica de aglomeração com base na distância
generalizada de Mahalanobis (D2), proposta por Mahalanobis (1936). Após a obtenção
da matriz de dissimilaridade genética por meio das distâncias de Mahalanobis (D²)
obtidas para os diferentes pares de variedades, realizou-se o agrupamento das
variedades pelo método de agrupamento de ligação média entre grupo (UPGMA). Foi
utilizado o programa estatístico GENES (CRUZ, 2006).
3.3. Resultados e discussão
A análise de variância de proteína total mostra que houve diferença significativa
entre as variedades de feijão caupi e entre os ambientes, além de uma interação entre
variedade e ambiente significativa e que não houve diferença significativa entre a
variedade Pitiúba (VAR 2) plantada em Quixadá e a plantada em Cascavel (Tabela 2).
Tabela 2 – Análise de variância de proteína total para esquema fatorial 2x22
FV GL SQ QM F P
VARIEDADES 21 24634,95748 1173,09321 2,08367
AMBIENTES 1 63055,66089 63055,66089 112,00091
VARxAMB 21 11822,84076 562,99242 6,05011
VAR/AMB 42 36457,79823 868,04282 9,32828 P=0,0
VAR/AMB 1 21 17591,40861 837,68612 9,00206 P=0,0
VAR/AMB 2 21 18866,38962 898,39951 9,6545 P=0,0
AMB/VAR 22 74878,50165 3403,56826 36,57589 P=0,0
AMB/VAR 1 1 1701,04683 1701,04683 18,28002 P=0,00005
AMB/VAR 2 1 1,82854 1,82854 0,01965 P=100,0
AMB/VAR 3 1 4702,42513 4702,42513 50,53384 P=0,0
AMB/VAR 4 1 3035,81678 3035,81678 32,62391 P=0,0
AMB/VAR 5 1 3656,73225 3656,73225 39,29647 P=0,0
AMB/VAR 6 1 5572,4278 5572,4278 59,88318 P=0,0
AMB/VAR 7 1 6976,42216 6976,42216 74,97097 P=0,0
AMB/VAR 8 1 2996,62768 2996,62768 32,20277 P=0,0
AMB/VAR 9 1 5776,79956 5776,79956 62,07943 P=0,0
AMB/VAR 10 1 1767,6275 1767,6275 18,99552 P=0,00003

33
AMB/VAR 11 1 3148,64573 3148,64573 33,8364 P=0,0
AMB/VAR 12 1 3984,74578 3984,74578 42,82142 P=0,0
AMB/VAR 13 1 5300,87621 5300,87621 56,96499 P=0,0
AMB/VAR 14 1 4252,19336 4252,19336 45,6955 P=0,0
AMB/VAR 15 1 2165,23513 2165,23513 23,26834 P=0,0
AMB/VAR 16 1 12082,81857 12082,81857 129,84602 P=0,0
AMB/VAR 17 1 1292,42903 1292,42903 13,88888 P=0,00034
AMB/VAR 18 1 639,93982 639,93982 6,87701 P=0,01029
AMB/VAR 19 1 1195,32313 1195,32313 12,84534 P=0,00055
AMB/VAR 20 1 541,62161 541,62161 5,82045 P=0,01791
AMB/VAR 21 1 2427,64515 2427,64515 26,08829 P=0,0
AMB/VAR 22 1 1659,27388 1659,27388 17,83111 P=0,00006
RESÍDUO 88 8188,83794 93,05498
TOTAL 131 107702,2971
MÉDIA 139,680
CV(%) 6,9
Em Quixadá, as variedades CNCx 251-11E, Marataóa se mostram com maior
reserva de proteínas na semente enquanto as variedades Sempre verde, Pitiúba,
Milagroso, Setentão, CNCx 251-11E, CNCx 251-60E, CNCx 251-76E, Marataóa, BRS-
Rouxinol, Paulistinha, AU 94-418-07-01, UCR-95-701, MNC 03-731C-21, MNC 03-
720-11 mostram maior capacidade de armazenar proteínas de reserva em Cascavel
(Tabela 3).
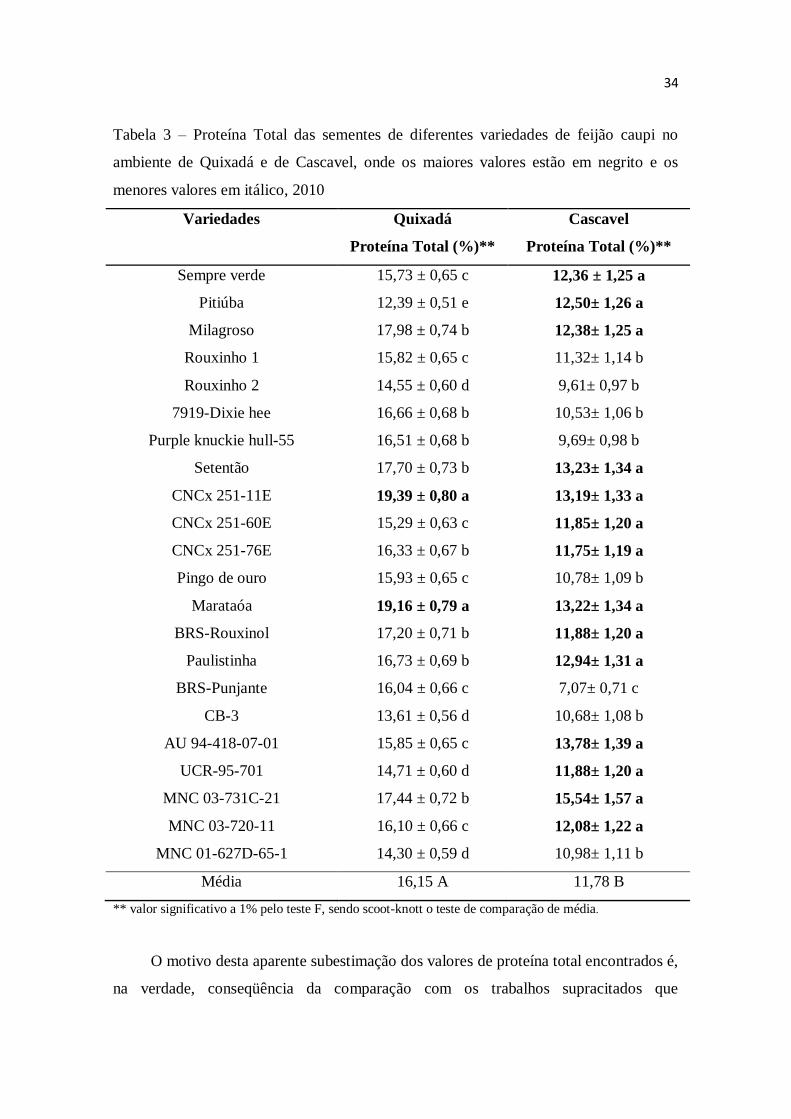
Na Tabela 3, observam-se os valores para de proteína total de cada variedade nos
dois ambientes de cultivo. De modo geral, pode-se observar que o valor de proteína
total máximo foi de 19,39% e o mínimo foi de 7%, obtidos com o variedade CNCx 251-
11E em Quixadá e com BRS-Punjante em Cascavel, respectivamente. Mesmo o valor
máximo de proteína total obtido no presente trabalho é inferior a média protéica em
feijão caupi, que é entre 23 e 25% segundo a EMBRAPA Meio Norte (2003), assim
como aos teores protéicos, variando de 19,5% a 26,1%, encontrados por Maia et al.
(2000) em variedades Epace-10, Epace-11, Pitiúba, TVu-1888, IPA-2006 e Olho-de-
ovelha, no Nordeste.
34
Tabela 3 – Proteína Total das sementes de diferentes variedades de feijão caupi no
ambiente de Quixadá e de Cascavel, onde os maiores valores estão em negrito e os
menores valores em itálico, 2010
Variedades Quixadá
Proteína Total (%)**
Cascavel
Proteína Total (%)**
Sempre verde 15,73 ± 0,65 c 12,36 ± 1,25 a
Pitiúba 12,39 ± 0,51 e 12,50± 1,26 a
Milagroso 17,98 ± 0,74 b 12,38± 1,25 a
Rouxinho 1 15,82 ± 0,65 c 11,32± 1,14 b
Rouxinho 2 14,55 ± 0,60 d 9,61± 0,97 b
7919-Dixie hee 16,66 ± 0,68 b 10,53± 1,06 b
Purple knuckie hull-55 16,51 ± 0,68 b 9,69± 0,98 b
Setentão 17,70 ± 0,73 b 13,23± 1,34 a
CNCx 251-11E 19,39 ± 0,80 a 13,19± 1,33 a
CNCx 251-60E 15,29 ± 0,63 c 11,85± 1,20 a
CNCx 251-76E 16,33 ± 0,67 b 11,75± 1,19 a
Pingo de ouro 15,93 ± 0,65 c 10,78± 1,09 b
Marataóa 19,16 ± 0,79 a 13,22± 1,34 a
BRS-Rouxinol 17,20 ± 0,71 b 11,88± 1,20 a
Paulistinha 16,73 ± 0,69 b 12,94± 1,31 a
BRS-Punjante 16,04 ± 0,66 c 7,07± 0,71 c
CB-3 13,61 ± 0,56 d 10,68± 1,08 b
AU 94-418-07-01 15,85 ± 0,65 c 13,78± 1,39 a
UCR-95-701 14,71 ± 0,60 d 11,88± 1,20 a
MNC 03-731C-21 17,44 ± 0,72 b 15,54± 1,57 a
MNC 03-720-11 16,10 ± 0,66 c 12,08± 1,22 a
MNC 01-627D-65-1 14,30 ± 0,59 d 10,98± 1,11 b
Média 16,15 A 11,78 B
** valor significativo a 1% pelo teste F, sendo scoot-knott o teste de comparação de média.
O motivo desta aparente subestimação dos valores de proteína total encontrados é,
na verdade, conseqüência da comparação com os trabalhos supracitados que

35
superestimação os valores protéicos devido à metodologia empregada na quantificação
protéica. A metodologia usada no presente trabalho é de quantificação indireta através
de espectrofotometria conforme proposto por Bradford (1976), enquanto a metodologia
dos trabalhos supracitados é conforme o método Kjeldhal.
Os valores superiores de proteína em feijão caupi em muitos trabalhos é,
claramente, conseqüência da metodologia Kjeldhal que superestima o valor de proteína
total (GUIMARÃES; MARQUEZ, 2002; MORRISON, 1992). O método Kjeldhal foi
desenvolvido para determinação de nitrogênio total, sendo realizada uma conversão de
nitrogênio total (NT) em proteína total (PT) que se dá por meio da equação teórica PT =
NT x 6,25. O fator de conversão 6,25 baseia-se na afirmação de que em 100g de
proteínas há 16g de nitrogênio, o que é apenas uma estimativa já que os aminoácidos
têm um átomo de nitrogênio em sua estrutura básica, mas podem ter mais em seu
radical, e que o tipo e a quantidade de aminoácidos que compõem uma proteína podem
variar muito. Além disso, outro fator que torna superestimador o método é a ocorrência
de reações de digestão com ácido sulfúrico que disponibilizam todo nitrogênio dos
tecidos, sejam oriundos de proteína, ácido nucléico ou outros compostos nitrogenados.
O ambiente Quixadá apresentou-se favorável ao armazenamento de proteínas de
reserva nas sementes de feijão caupi, exceto sobre a variedade Pitiúba que não
respondeu ao ambiente com significativo acréscimo de proteína total como as demais
variedades (Tabela 2 e 3).
Os principais fatores ambientais que podem ter contribuído para que as sementes
produzidas em Quixadá tenham maior valor protéico foram a disponibilidade de água e
de rizóbios no solo.
A diferença pluviométrica entre os ambientes estudados foi de apenas 15,6 mm
(Tabela 1). No entanto, com a irregularidade das chuvas nos meses de cultivo, o
feijoeiro foi submetido a uma limitação hídrica em Quixadá durante o período de
floração e enchimento dos grãos, que corresponde ao mês de maio (Tabela 1).
Conforme Bewley e Black (1985) há uma tendência a elevação no valor de proteína
total em sementes de plantas de feijão expostas a períodos de baixa disponibilidade
hídrica, assim como observado no ambiente de Quixadá.
As áreas de cultivo tinham diferenças quanto ao uso. Em Quixadá a área era
continuamente cultivada com várias culturas, inclusive a de feijão caupi. Em Cascavel a

36
área estava em pousio há pelo menos dez anos. Por isso, a ação dos rizóbios pode ter
sido mais pronunciada em Quixadá, onde a carga de rizóbios tende a ser maior pelos
vários ciclos de cultivo de feijão caupi no campo, contribuindo para o incremento
protéico nas sementes.
A ação do incremento de nitrogênio no solo sobre a elevação de proteína pode ser
exemplificado pelo trabalho de Calarota e Carvalho (1984) que concluíram, em girassol,
que o fornecimento de doses mais altas de N, até um limite que parece estar próximo de
8g por planta (1g na semeadura e 7g em cobertura), provoca a formação de sementes
com conteúdos crescentes de proteína e decrescentes de óleo.
O agrupamento das variedades com base na dissimilaridade pelo método
UPGMA, em conjunto com um teste de comparação de médias permite identificar os
pares de genótipos mais divergentes quanto ao valor protéico e com valores acima da
média, ou seja, identificar combinações híbridas entre genótipos permita a obtenção de
populações segregantes com teores de proteína acima da média.
Observando as médias de proteína total em Quixadá e em Cascavel percebe-se
que a expressão fenotípica em Quixadá aproxima-se mais do potencial possível e
indeterminado para acúmulo de proteína em sementes de feijão caupi. Com base nisto,
para fins de melhoramento iremos analisar o máximo potencial expresso, tomando
assim, como referência os valores de proteína total obtidos para cada variedade em
Quixadá.
Com base na matriz de dissimilaridade, os pares de variedades mais dissimilares
para a característica de protéica total nas sementes nas condições de Quixadá foram
CNCx 251-11E e Pitiúba (111,1) (Anexos 1). O dendrograma formado com o
agrupamento de ligação média entre grupo (UPGMA) mostra a formação de dois grupos
no maior nível de fusão (30,98), onde o grupo 1 é formado por todas as variedades
exceto as variedades CNCx 251-11E e Marataóa que compõem o segundo grupo
(Anexo 2).
Se analisar junto a Tabela 3 e o Anexo 2, observa-se que as variedades do grupo 2
foram as que mais acumularam reservas protéicas e que no grupo 1 há dois subgrupos
formados pelas variedades 7919-Dixie hee, Paulistinha, Purple knuckie hull-55, CNCx
251-76E (1° subgrupo) e BRS-Rouxinol, MNC 03-731C-21, Milagroso, Setentão (2°
subgrupo) que têm proteína total acima da média. Além disso, a variedade Pitiúba que

37
acumulou proteína a baixo da média está agrupado no grupo 1 e apresenta estabilidade
para a característica proteína total.
Com base na dissimilaridade e reserva de proteína total pode-se recomendar o
cruzamento das variedades CNCx 251-11E ou Marataóa com 7919-Dixie hee,
Paulistinha, Purple knuckie hull-55, CNCx 251-76E, BRS-Rouxinol, MNC 03-731C-
21, Milagroso ou Setentão em programa de melhoramento para aumento do valor
nutricional do feijão caupi e o uso da variedade Pitiúba para conferir estabilidade as
novas variedades em formação no programa de melhoramento.
3.4. Conclusão
As variedades CNCx 251-11E, Marataóa apresentaram maior valor protéico em
Quixadá.
As condições edafoclimáticas encontradas no ambiente de Quixadá mostraram-se
favoráveis ao acúmulo de reservas protéicas nas sementes de feijão caupi.
A variedade Pitiúba mostrou-se estável à ação do ambiente sobre o acúmulo de
reserva protéica, mas esta variedade compõe o grupo inferior quanto à capacidade de
armazenar proteína em Quixadá.
Recomenda-se o cruzamento das variedades CNCx 251-11E ou Marataóa com
7919-Dixie hee, Paulistinha, Purple knuckie hull-55, CNCx 251-76E, BRS-Rouxinol,
MNC 03-731C-21, Milagroso ou Setentão e o uso da variedade Pitiúba para conferir
estabilidade ambiental para formação de novas variedades no programa de
melhoramento para aumento do valor nutricional do feijão caupi.

38
REFERÊNCIAS BIBLIOGRÁFICAS
AQUINO, A. B.; AQUINO, B. F.; HERNANDEZ, F. F. F.; HOLANDA, F. J. M.;
FREIRE, J. M.; CRISÓSTOMO, L. A.; COSTA, R. I.; UCHÔA, S. C. P.; FENANDES,
V. L. B.Recomendações de adubação e calagem para o estado do Ceará.Editora UFC.
Fortaleza, 1993. 248p.
BEWLEY, J. D.; BLACK, M. Seeds – Physiology of development and germination.
New York, Plenum Press. 2Ed. 367p., 1994.
BRADFORD, M.M. A rapid and sensitive method for the quantifications of microgram
quantities of protein utilizing the principle of protein-dye binding. Analitycal
Biochemistry, v. 72, p.248-254, 1976.
BRESSANI, R. Grain quality of common beans. Food Reviews International,
Philadelphia, v. 9, n. 2, p. 237-297, 1993.
CALAROTA, N. E.;CARVALHO, N. M. Efeitos da adubação nitrogenada em
cobertura sobre os conteúdos de óleo e de proteína e a qualidade fisiológica de sementes
de girassol (Helianthus annuus). Revista Brasileira de Sementes, vol. 06, nº 3, p. 41-
50, 1984.
COPELAND, L. O.; MCDONALD, M. B. Principles of seed science and technology.
New York, Chapman & Hall. 3ª ed. 409p., 1995.
CRUZ, C.D. Programa Genes: Biometria. Editora UFV. Viçosa (MG). 382p. 2006.
CRUZ, G. A. D. R. et al. Protein quality and in vivo digestibility of different varieties of
bean (Phaseolus vulgaris L.). Brazilian Journal of Food Technology, Campinas, v. 6,
n. 2, p. 157-162, 2003.
DURE III, L. S. Seed formation. Annual Review of Plant Physiology, v. 26, p. 259-
278, 1975.
EMBRAPA MEIO-NORTE. Cultivo de feijão caupi. Jul/2003. Disponível em:
<http://www.cpamn.embrapa.br/pesquisa/graos/FeijaoCaupi/referencias.htm>. Acesso
em: 8 set. 2010.

39
FURLANI, A. M. C. Nutricão mineral. In: KERBAUY, G.B., Fisiologia Vegetal,
Guanabara-Koogan, 2004. Cap. 2, p. 40-93.
GUIMARÃES, C. P.; MARQUEZ, U. M. L. Composição química de tabletes de caldo
de carne: nitrogênio protéico, não-protéico e fenilalanina. Ciência e Tecnologia de
Alimentos vol. 22 no. 3 Campinas, 2002.
IQBAL, A.; KHALIL, I. A.; SHAH, H. Nutritional yield and amino acid profile of rice
protein as influenced by nitrogen fertilizer. Sarhad Journal of Agriculture, v. 19, n. 1,
p. 127-134, 2003.
MAHALANOBIS, P. C. On the generalized distance in statistics. Proceedings of the
National Institute of Sciences of India, New Delhi, v.2, p.49-55, 1936.
MAIA, F. M. M.; OLIVEIRA, J. T. A.; MATOS, M. R. T.; MOREIRA, R. A.;
VASCONCELOS, I. M. Proximate composition, amino acid content and
haemagglutinating and trypsin-inhibiting activies of some Brazilian Vigna unguiculata
(L.) Walp. Variedades. Journal of the Science of Food and Agriculture, Lodon, v. 80,
n. 4, p. 453-458, 2000.
MARCOS FILHO, J. Fisiologia de Sementes de Plantas Cultivadas. 12ed. Editora
Fealq. Piracicaba, 2005. 495 p.
MORRISON, W.R. Analysis of cereal starches. In: LINSKENS, H.F.; JACKSON, J.F.
(eds.) Seed analysis. Berlin: Springer-Verlag, 1992. p.199-215.
SCOTT, A.J.; KNOTT, M.A. A cluster analysis method for grouping means in the
analysis of variance. Biometrics, v.30, n.3, p.507-512, 1974.

40
4. CAPITULO 3 - CAPACIDADE DE HIDRATAÇÃO DE SEMENTES EM
VARIEDADES FEIJÃO CAUPI
RESUMO
As sementes possuem a capacidade de embebição governada pelas diferenças
entre o potencial hídrico dos tecidos da semente e do substrato fornecedor de água. Com
base em estudos com curva de embebição, um gráfico que relaciona tempo e peso
hidratado da semente, e mudanças fisiológicas das sementes em germinação os
pesquisadores Bewley e Black propuseram um modelo trifásico que reflete o
comportamento comum durante o processo de hidratação. Objetivou-se com o trabalho
determinar as curvas de embebição dos lotes de sementes das 22 variedades oriundas de
Quixadá e de Cascavel, e avaliar a atuação do conteúdo protéico como componente
matricial responsável pelo movimento de água no início da embebição. Os feijões caupi
foram plantados em 2010, nas Fazendas Lavoura seca e Boa água localizadas em
Quixadá e em Cascavel, respectivamente. As 22 variedades foram escolhidas por serem
precoce e estarem disponíveis no banco de germoplasma de caupi do Centro de Ciências
Agrárias (CCA) na Universidade Federal do Ceará. As sementes foram colocadas para
germinar em rolos papel germitest 28x38 cm com três repetições de cada variedade e
com sementes oriundas de apenas um ambiente. As condições de germinação foram
controladas por câmaras de germinação tipo BOD, com temperatura a 25 °C e
fotoperíodo de 12h de luz e 12h de escuro. As avaliações, para formação das curvas de
embebição, se deram no início da embebição (0h) e às 2h, 4h, 6h, 12h, 18h, 24h, 30h,
36h, 42h, 50h, 58h, 66h, 74h, 82h após o início da embebição. Amostras compostas de
cada variedade de caupi para os dois ambientes foram maceradas e de 40 mg de amostra
foram extraídas proteínas com 1 mL de NaOH a 0,1 M, sendo em seguida quantificadas
por espectrofotometria. O modelo trifásico de Bewley e Black (1978) não se adéqua as
curvas de embebição de todas as variedades de feijão caupi. As condições ambientais de
cultivo influenciam adequação ao modelo. O conteúdo protéico não age como principal
componente mátrico em sementes de feijão caupi.
Palavras chaves - Curva de embebição. Priming. Proteína.

41
ABSTRACT
Seeds have the ability to absorb water or imbibition governed by differences in
tissue water potential of the seed and substrate water supplier different. Based on
studies with imbibition curves and physiological changes of the germinating seeds
researchers Bewley and Black (1978) proposed a triphasic model that reflects the
common behavior during the hydration process. The objective of the work to determine
the imbibition curve of seed lots of twenty-two varieties from the Quixadá and
Cascavel, and evaluate the performance of the protein content as a matrical component
responsible for the movement of water in the beginning of imbibition. The cowpeas
were planted in 2010, in farms located in Quixadá and Cascavel. The varieties were
chosen to be early matured and be available in the cowpea germplasm collection at the
Center for Agricultural Sciences (CCA) at the Federal University of Ceará. The seeds
germinated in paper rolls germitest 28x38cm with three replications of each varieties of
one environmental. The germination conditions were controlled by a germination
chamber, BOD, temperature 25 °C and a photoperiod of 12h light and 12h dark. The
assessments, training for imbibition curves, occurred at the beginning of imbibition (0h)
and at 2h, 4h, 6h, 12h, 18h, 24h, 30h, 36h, 42h, 50h, 58h, 66h, 74h, 82h after beginning
of imbibitions. Composite samples of each varieties of cowpea of the two environments
were macerated and from 40 mg of sample were extracted proteins with 1mL of 0.1 M
NaOH, and then quantified by spectrophotometry. The triphasic model of Bewley and
Black (1978) does not fit the curves of absorption of all varieties of cowpea. The
environmental conditions influence the model adequacy. The protein content is not
acting as principal component matric in cowpea seeds.
Keywords - Imbibition curve. Priming. Protein.

42
4.1. Introdução
A captação de quantidade considerável de água é imprescindível para o reinicio
de atividades metabólicas da semente após a maturidade. A deficiência hídrica
geralmente é considerada o fator limitante da germinação de sementes não-dormentes,
afetando a percentagem, a velocidade e a uniformidade de germinação. A embebição é
um tipo especial de difusão provocada pela atração entre moléculas de água e a
superfície matricial, governada pelas diferenças entre o potencial hídrico dos tecidos da
semente e do substrato fornecedor de água (MARCOS FILHO, 2005).
A água é fundamental para o metabolismo celular durante a germinação por
permitir a atividade enzimática, a solubilização e transporte de reservas e agir como
reagente em si. Além disso, a água torna tenros os tecidos facilitando a protrusão
radicular (MARCOS FILHO, 2005).
O componente matricial é o principal responsável pelo movimento de água no
início da embebição, mas com o aumento do nível de água livre o componente osmótico
torna-se também muito participativo (CARDOSO, 2004).
Em nível celular as proteínas atuam como retentora de água e componente
mátrico, já que possuem grupos iônicos e polares, como amina (NH2), hidroxila (OH),
carboxila (COOH) e outros que permitem ligações de hidrogênio com a água e sua
conformação. Assim, modificações na atividade metabólica da semente, em resposta a
variações do teor de água, parecem estar ligadas a alteração de propriedades da água
(VILLELA; MARCOS FILHO, 1998).
Marcos Filho (2005) cita sementes de ervilha, feijão comum, milho, soja, alface,
rabanete e amendoim como espécies com relativa pausa caracterizando a segunda fase
do modelo trifásico de Bewley e Black (1978), mas também cita espécies que não
exibem esta segunda fase como trigo, mamona, cevada, arroz e aveia.
A expressão seed enhancement é utilizada no meio industrial para nomear um
conjunto de técnicas ou processos destinados a realçar a qualidade ou beneficiar o
desempenho de lotes de sementes e/ou plântulas produzidas (TAYLOR, 1998). Não
havendo, ainda, uma proposta concreta para a tradução da expressão seed enhancement
Marcos Filho (2005), sugere o uso da expressão condicionamento de sementes para
nominar este conjunto de técnicas. O condicionamento fisiológico (priming) é apenas

43
uma técnica do condicionamento de sementes, que visa elevar o potencial fisiológico da
semente através de um pré-umedecimento. Kidd e West (1919 apud Hegarty, 1978),
relataram que as sementes pré-umedecidas germinavam mais rapidamente se fossem
embebidas em quantidade limitada de água. Heydecker e Coolbear (1977) comentando
a técnica relataram reversão dos benefícios do condicionamento, durante a secagem,
indicando a necessidade de fazer logo a semeadura. Entretanto, Bewley e Black (1982)
sugeriram que os efeitos da aplicação do condicionamento fisiológico não desaparecem
com a secagem, se respeitado o grau de umidade seguro para o armazenamento.
Um dos sintomas mais claros do declínio no potencial fisiológico é a redução na
velocidade de germinação, identificada pelo aumento do tempo entre a semeadura e o
início da germinação e também entre a primeira e a última germinação de uma
população de sementes. O potencial fisiológico pode ser recuperado com absorção de
água pela semente durante a fase I e II, sem permitir a protrusão ou inicio da fase III. A
hidratação ativa a digestão das reservas e sua translocação e assimilação, levando as
sementes do lote a um estado metabólico relativamente uniforme (BEWLEY; BLACK,
1982).
A curva de embebição pode ser usada para referenciar a aplicação do
condicionamento fisiológico, indicando a quantidade de água e tempo de pré-
umedecimento a ser aplicado.
Objetivou-se com o trabalho determinar a curva de embebição dos lotes de
sementes de vinte e duas variedades oriundos de Quixadá e Cascavel, e avaliar a
atuação do conteúdo protéico como componente matricial responsável pelo movimento
de água no início da embebição.
4.2. Material e métodos
4.2.1. Material vegetal e delineamento experimental
Para a execução do trabalho, vinte e duas variedades foram cultivadas no primeiro
semestre do ano de 2010, nos meses de abril, maio e junho, em Quixadá-CE e em
Cascavel-CE sob condições de sequeiro. O plantio seguiu o delineamento experimental
de blocos inteiramente casualizados (DBC), com quatro repetições e com vinte e duas

44
variedades constituindo os Tratamentos. As parcelas foram constituídas de 24 plantas
espaçadas a 0,8 x 0,6 m e uma área útil constituída por 12 plantas.
As vinte e duas variedades foram Sempre verde (CE-25), Pitiúba (CE-31),
Milagroso (CE-46), Rouxinho 1 (CE-73), Rouxinho 2 (CE-77), 7919-Dixie hee (CE-
104), Purple knuckie hull-55 (CE-113), Setentão (CE-596), CNCx 251-11E (CE-790),
CNCx 251-60E (CE-796), CNCx 251-76E (CE-798), Pingo de ouro (CE-930),
Marataóa (CE-933), BRS-Rouxinol (CE-937), Paulistinha (CE-939), BRS-Punjante
(CE-940), CB-3 (ainda sem registro no Banco de germoplasma de caupi, do
CCA/UFC), AU 94-418-07-01 (CE-176), UCR-95-701 (CE-493), MNC 03-731C-21
(CE-390), MNC 03-720-11 (CE-178), MNC 01-627D-65-1 (CE-497), escolhidas pelo
critério de precocidade, já que variedades precoces são mais adaptados ambiente semi-
árido, resultando assim maior rendimento ao agricultor sertanejo.
Em Quixadá, o plantio foi realizado na Fazenda Lavoura Seca, pertencente à
Universidade Federal do Ceará e em Cascavel, o plantio foi realizado na Fazenda Boa
Água, pertencente ao Eng° Agrônomo José Mesquita L. Almeida, numa área de 1200
m2.
4.2.2. Preparo da amostra e curva de embebição
A partir das sementes colhidas e beneficiadas foram coletadas amostras de 1000
sementes em suas repetições de cada parcela e homogeneizadas para compor uma
amostra composta de cada variedade representativa do ambiente.
Sementes retiradas das 22 amostras compostas foram dispostas segundo o
delineamento inteiramente casualizado (DIC). As sementes foram colocadas para
germinar em rolos de papel germitest 28x38cm com três repetições de cada variedade e
com sementes oriundas de apenas um ambiente.
As sementes em germinação estavam sob condições controladas por câmaras de
germinação tipo BOD, com temperatura a 25 °C e fotoperíodo de 12 h de luz e 12 h de
escuro simulando as condições fotoperiodicas encontradas próximo a linha do Equador
durante todo ano.
As avaliações se deram no início da embebição (0 h) e às 2 h, 4 h, 6 h, 12 h, 18
h, 24 h, 30 h, 36 h, 42 h, 50 h, 58 h, 66 h, 74 h, 82 h após o início da embebição. No

45
momento de cada avaliação de peso úmido de cada semente, dois rolos eram retirados
um com sementes do ambiente Quixadá e outro com sementes de Cascavel.
4.2.3. Determinação química
Foram pesadas 40mg de cada amostra e colocadas para reagir com 1 mL de NaOH
a 0,1M, por 20 minutos, sendo o sobrenadante removido com centrifugação a 5000 rpm
por 4 minutos, esta operação foi repetida com cada amostra até exauri-la de proteínas. O
sobrenadante foi reservado para quantificação protéica, segundo o método proposto por
Bradford (1976). Neste método utiliza-se uma solução albumina (BSA) como padrão e
um espectrofotômetro regulado para mensurar a absorbância da luz com o comprimento
de onda de 595 nm pela solução de 100 µL da amostra/mL de solução de Bradford. A
quantificação das proteínas existentes se dá a partir da absorbância obtida, processada
segundo a equação de regressão da solução padrão de albumina (BSA) com absorbância
registrada.
4.2.4. Análise estatística
A análise de variância realizada seguiu o modelo de delineamento inteiramente
ao acaso (DIC) e as variâncias foram testadas quanto a sua homogeneidade pelo Teste
de Cochran para avaliação de valores discrepantes de peso. Logo após foi realizado teste
de regressão para cada variedade nos dois ambientes para obtenção das curvas de
embebição.
O teste de correlação pelo coeficiente de Pearson, seguido pelo teste de Mantel
(MANTEL, 1967) foi realizado para verificar a correlação entre valor protéico total e
peso de semente seca após 2h, 4h e 6h de embebição e da distância espacial entre as
unidades experimentais (mensurada através da distância euclidiana) com suas
respectivas flutuações na densidade (baseada na correlação de Pearson), como medida
da heterogeneidade espacial no tratamento com ambientes.
O teste de Mantel (Z) é dado por:

46
onde, Xij e Yij são os elementos das matrizes X e Y a serem comparadas. A significância
estatística da correlação matricial foi obtida com 5000 permutações aleatórias.
Os programas estatísticos utilizados foram o ASSISTAT e o GENES (SILVA,
1996; SILVA; AZEVEDO, 2002; SILVA; AZEVEDO 2006; SILVA; CRUZ, 2006;
AZEVEDO, 2009).
4.3. Resultados e discussão
A análise de variância mostrou-se significativa e o teste de Cochran foi positivo
para a hipótese de homogeneidade das variâncias, indicando não haver valores
discrepantes de peso.
Com base nas curvas de embebição representadas nas Figuras 3-24, o modelo
trifásico proposto por Bewley e Black (1978) foi observado na curva de embebição dos
seguintes variedades: Sempre verde cultivado em Cascavel, Pitiúba cultivado em
Quixadá, Milagroso cultivado em Cascavel, Rouxinho 1 cultivado em Quixadá, 7919-
Dixie hee cultivado em Quixadá, Setentão cultivado em Cascavel, CNCx 251-11E
cultivado em Cascavel, CNCx 251-60E cultivado em Quixadá e Cascavel, CNCx 251-
76E cultivado em Quixadá e Cascavel, Marataóa cultivado em Quixadá e Cascavel,
BRS-Rouxinol cultivado em Quixadá, Paulistinha cultivado em Cascavel, BRS-
Punjante cultivado em Quixadá e Cascavel, CB-3 cultivado em Cascavel, AU 94-418-
07-01 cultivado em Quixadá e Cascavel, UCR-95-701 cultivado em Quixadá e
Cascavel, MNC 03-731C-21 cultivado em Quixadá, MNC 03-720-11 cultivado em
Quixadá, MNC 01-627D-65-1 cultivado em Quixadá e Cascavel, os demais variedades
avaliados não apresentaram claramente a segunda fase do modelo trifásico através da
redução relativa na absorção de água, mas isto não implica que não houve síntese de
enzimas, de DNA e de RNA-m em algum momento na transição da fase I para fase III.
Em feijão comum se relata a adequação plena ao modelo trifásico. No entanto, a
falta aparente da fase II na curva de embebição é relatada em outras culturas, como
trigo, mamona, cevada, arroz e aveia (MARCOS FILHO, 2005). Por meio do presente
trabalho podendo-se afirmar a falta aparente da fase II, ocorre em algumas variedades
de Vigna unguiculata (L.) Walp e que as condições ambientais durante a formação da
semente pode influenciar a presença ou não da fase II (Figuras 3-24).

47
As curvas de embebição representadas nas Figuras 3-24 podem servir de base para
uma hidratação temporalmente controlada em regime de embebição lenta (em papel
úmido) para o futuro condicionamento fisiológico dos lotes de sementes estudados e
armazenados no banco de germoplasma da Universidade Federal do Ceará.
Pandey (1988) enfatizou a eficiência da embebição lenta, entre folhas de papel-
toalha umedecido a 25 °C durante 48 horas para feijão francês (Phaseolus vulgaris L.),
para recuperação do potencial fisiológico de sementes armazenadas por até três anos.
Balbinot e Lopes (2006), concluíram que a secagem das sementes de cenoura após o
condicionamento osmótico não afetou a sua germinação e o condicionamento das
sementes em água destilada, por até 48 horas em sistema aerado seguido de secagem,
contribuiu para aumentar o vigor das sementes de cenoura.
A correlação foi moderada entre a proteína total, e o peso da semente seca (0,60, a
1% de significância), e peso da semente com duas horas de embebição (0,43, a 1% de
significância), e peso da semente com quatro horas de embebição (0,38, a 5% de
significância), e peso da semente com seis horas de embebição (0,36, a 5% de
significância). As correlações indicam que a proteína total, apesar de sua contribuição
como componente matricial por suas propriedades mátricas em nível celular, não
representa o componente matricial que determina a hidratação inicial das sementes de
feijão caupi.
Observa-se uma redução na correlação entre proteína total e o peso da semente
nas primeiras horas de embebição. Isto pode ser resultado da redução da
responsabilidade do componente matricial pelo movimento de água no início da
embebição, já que com o aumento do nível de água livre o componente osmótico torna-
se também muito participativo conforme Marcos filho (2005) e Cardoso (2004).

48
Figura 3 – Curva de embebição de feijão caupi cv. Sempre verde cultivado em Quixadá e Cascavel, 2010.
( ) simboliza protrusão radicular.
Figura 4 – Curva de embebição de feijão caupi cv. Pitiúba cultivado em Quixadá e Cascavel, 2010. ( )
simboliza protrusão radicular.
Figura 5 – Curva de embebição de feijão caupi cv. Milagroso cultivado em Quixadá e Cascavel, 2010. ( )
simboliza protrusão radicular.
Figura 6 – Curva de embebição de feijão caupi cv. Rouxinho 1 cultivado em Quixadá e Cascavel, 2010.
( ) simboliza protrusão radicular.

49
Figura 7 – Curva de embebição de feijão caupi cv. Rouxinho 2 cultivado em Quixadá e Cascavel, 2010.
( ) simboliza protrusão radicular.
Figura 8 – Curva de embebição de feijão caupi cv. 7919-Dixie hee cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
Figura 9 – Curva de embebição de feijão caupi cv. Purple knuckie hull-55 cultivado em Quixadá e
Cascavel, 2010. ( ) simboliza protrusão radicular.
Figura 10 – Curva de embebição de feijão caupi cv. Setentão cultivado em Quixadá e Cascavel, 2010. ( )
simboliza protrusão radicular.
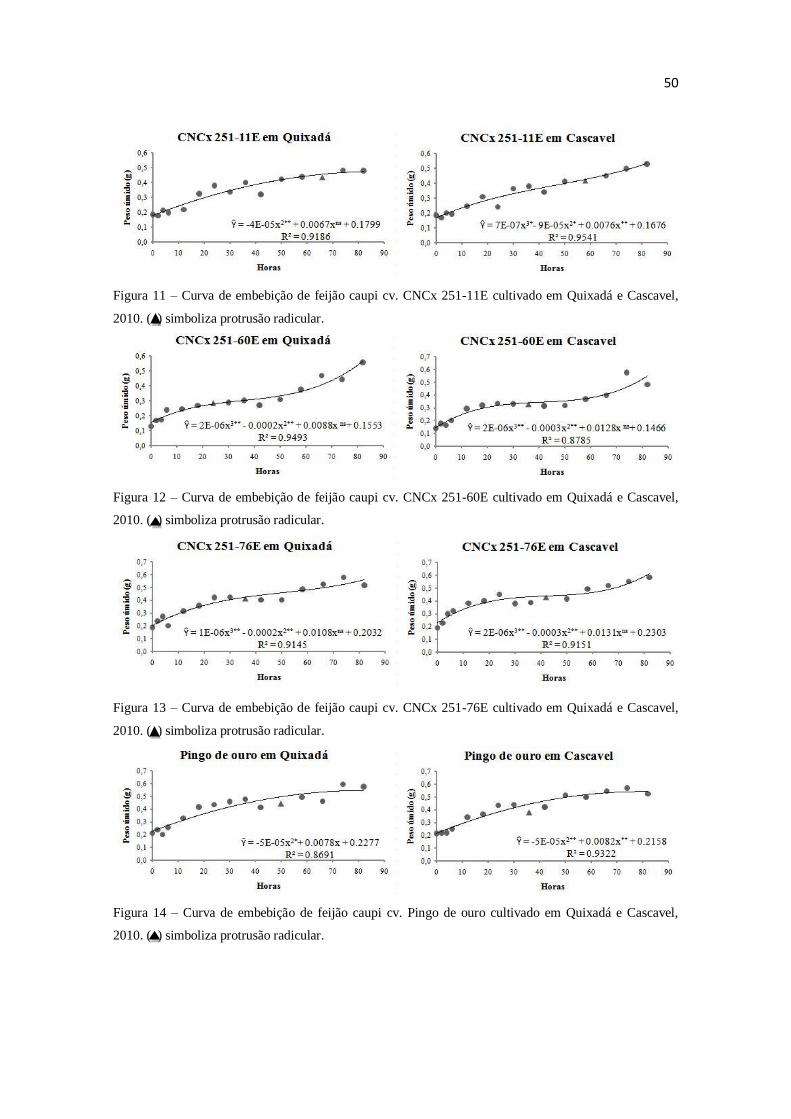
50
Figura 11 – Curva de embebição de feijão caupi cv. CNCx 251-11E cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
Figura 12 – Curva de embebição de feijão caupi cv. CNCx 251-60E cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
Figura 13 – Curva de embebição de feijão caupi cv. CNCx 251-76E cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
Figura 14 – Curva de embebição de feijão caupi cv. Pingo de ouro cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
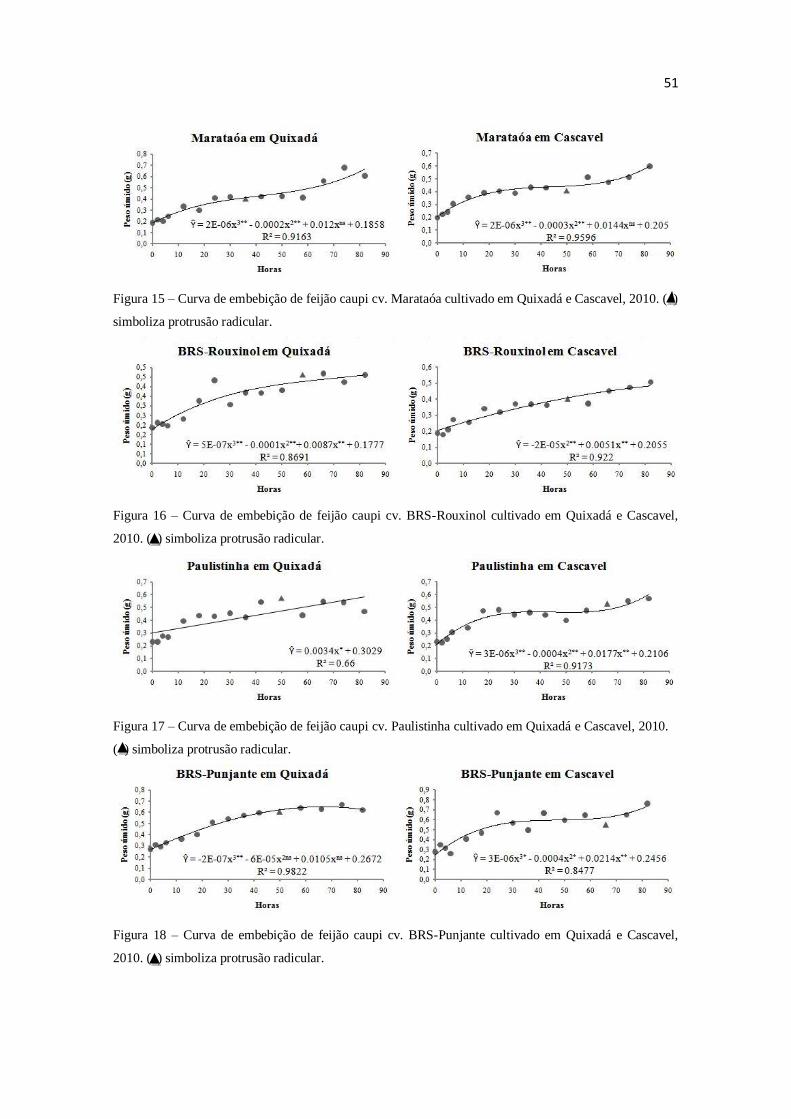
51
Figura 15 – Curva de embebição de feijão caupi cv. Marataóa cultivado em Quixadá e Cascavel, 2010. ( )
simboliza protrusão radicular.
Figura 16 – Curva de embebição de feijão caupi cv. BRS-Rouxinol cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
Figura 17 – Curva de embebição de feijão caupi cv. Paulistinha cultivado em Quixadá e Cascavel, 2010.
( ) simboliza protrusão radicular.
Figura 18 – Curva de embebição de feijão caupi cv. BRS-Punjante cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
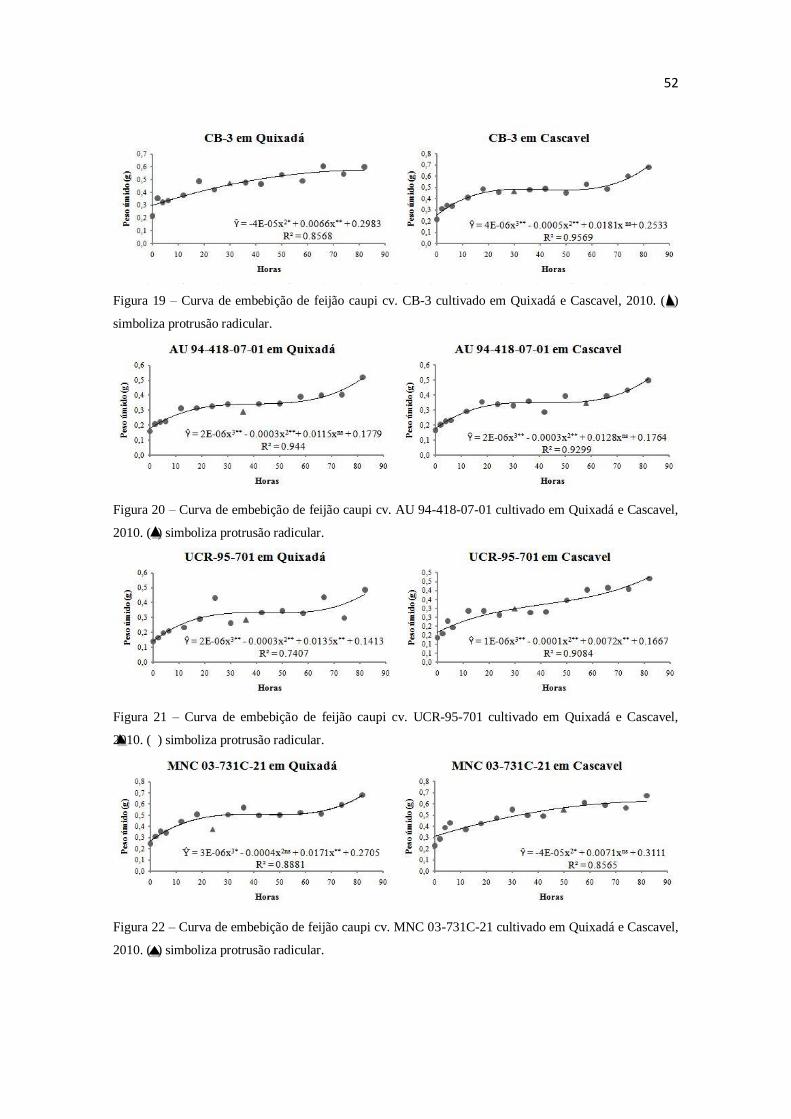
52
Figura 19 – Curva de embebição de feijão caupi cv. CB-3 cultivado em Quixadá e Cascavel, 2010. ( )
simboliza protrusão radicular.
Figura 20 – Curva de embebição de feijão caupi cv. AU 94-418-07-01 cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
Figura 21 – Curva de embebição de feijão caupi cv. UCR-95-701 cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
Figura 22 – Curva de embebição de feijão caupi cv. MNC 03-731C-21 cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
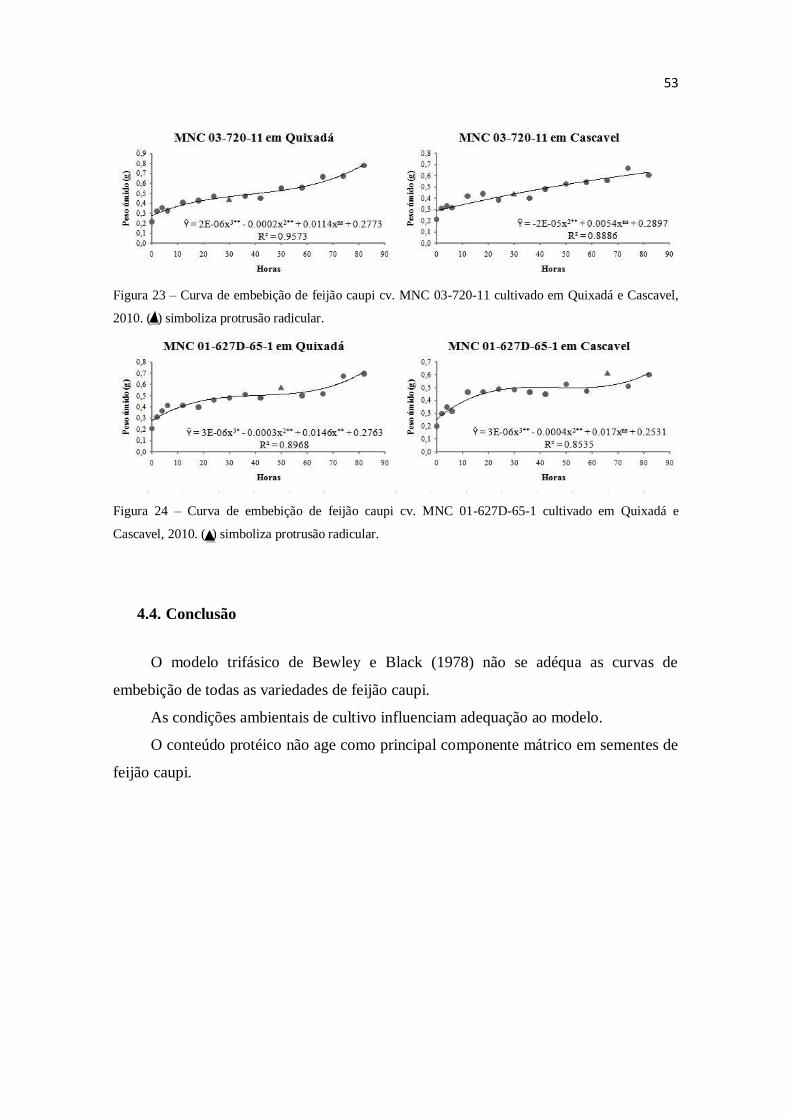
53
Figura 23 – Curva de embebição de feijão caupi cv. MNC 03-720-11 cultivado em Quixadá e Cascavel,
2010. ( ) simboliza protrusão radicular.
Figura 24 – Curva de embebição de feijão caupi cv. MNC 01-627D-65-1 cultivado em Quixadá e
Cascavel, 2010. ( ) simboliza protrusão radicular.
4.4. Conclusão
O modelo trifásico de Bewley e Black (1978) não se adéqua as curvas de
embebição de todas as variedades de feijão caupi.
As condições ambientais de cultivo influenciam adequação ao modelo.
O conteúdo protéico não age como principal componente mátrico em sementes de
feijão caupi.

54
REFERÊNCIAS BIBLIOGRÁFICAS
BALBINOT, E.; LOPES, H. M. Efeitos do condicionamento fisiológico e da secagem
na germinação e no vigor de sementes de cenoura. Revista Brasileira de
sementes, vol.28, n° 1, Pelotas, Apr. 2006.
BEWLEY, J. D.; BLACK, M. Physiology and biochemistry of seeds in relation to
germination. New York, Springer-Verlag. V. 2, 375p., 1982.
BEWLEY, J.D.; BLACK, M. Physiology and Biochemistry of seed in relation to
Germination. Berlin: Springer Verlag, v.1, 306p. 1978.
BRADFORD, M.M. A rapid and sensitive method for the quantifications of microgram
quantities of protein utilizing the principle of protein-dye binding. Analitycal
Biochemistry, v. 72, p.248-254, 1976.
CARDOSO, V. J. M. Germinação. In: KERBAUY, G.B., Fisiologia Vegetal,
Guanabara-Koogan, 2004. Cap. 17, p. 386-407.
CRUZ, C.D. Programa Genes: Biometria. Editora UFV. Viçosa (MG). 382p. 2006.
HEGARTHY, T. W. The physiology of seed hydration and dehydration, and relation
between water stress and the control of germination. Plant, Cell and Enviroment, v. 1,
n. 1, p. 101-109, 1978.
HEYDECKER, W.; COOLBEAR, P. Seed treatments for improved performance –
survery and attempted prognosis. Seed Science and Technology, v. 5, n. 2, p. 353-425,
1977.
MANTEL, N. The detection of disease clustering and a generalized regression
approach. Cancer, 1967. Res. 27: 209 – 220.
MARCOS FILHO, J. Fisiologia de Sementes de Plantas Cultivadas. 12ed. Editora
Fealq. Piracicaba, 2005. 495 p.
PANDEY, D. K. Priming induced repair in French bean seeds. Seed Science and
Technology, v. 16, n. 2, p. 527-532, 1988.

55
PIMENTA, J. A. Relações hidricas. In: KERBAUY, G.B., Fisiologia Vegetal,
Guanabara-Koogan, 2004. Cap. 1, p. 1-39.
SILVA, F. de A. S. The ASSISTAT Software: statistical assistance. In: International
conference on computers in agriculture, 6, Cancun, 1996. Anais... Cancun: American
Society of Agricultural Engineers, 1996. p.294-298.
SILVA, F. de A. S.; AZEVEDO, C. A. V. de. A New Version of The Assistat-
Statistical Assistance Software. In: WORLD CONGRESS ON COMPUTERS IN
AGRICULTURE, 4, Orlando-FL-USA: Anais... Orlando: American Society of
Agricultural and Biological Engineers, 2006. p.393-396.
SILVA, F. de A. S.; AZEVEDO, C. A. V. de. Versão do programa computacional
Assistat para o sistema operacional Windows. Revista Brasileira de Produtos
Agroindustriais, Campina Grande, v.4,n.1, p71-78,2002.
SILVA, F. de A. S.; AZEVEDO, C. A. V. de. Principal Components Analysis in the
Software Assistat-Statistical Attendance. In: World congress on computers in
agriculture, 7, Reno-NV-USA: American Society of Agricultural and Biological
Engineers, 2009.
TAYLOR, A. G.; ALLEN, P. S.; BENNETT, M. A.; BRADFORD, K. J.; BURRIS, J.
S.; MISRA, M. K. Seed enhancements. Seed Science Research, v. 8, n. 2, p. 245-256,
1998.
VILLELA, F. A.; MARCOS FILHO, J. Estados energéticos e tipos de água na semente.
Revista Brasileira de Sementes, vol. 20, n° 2, p.79-83 – 1998.

56
ANEXOS
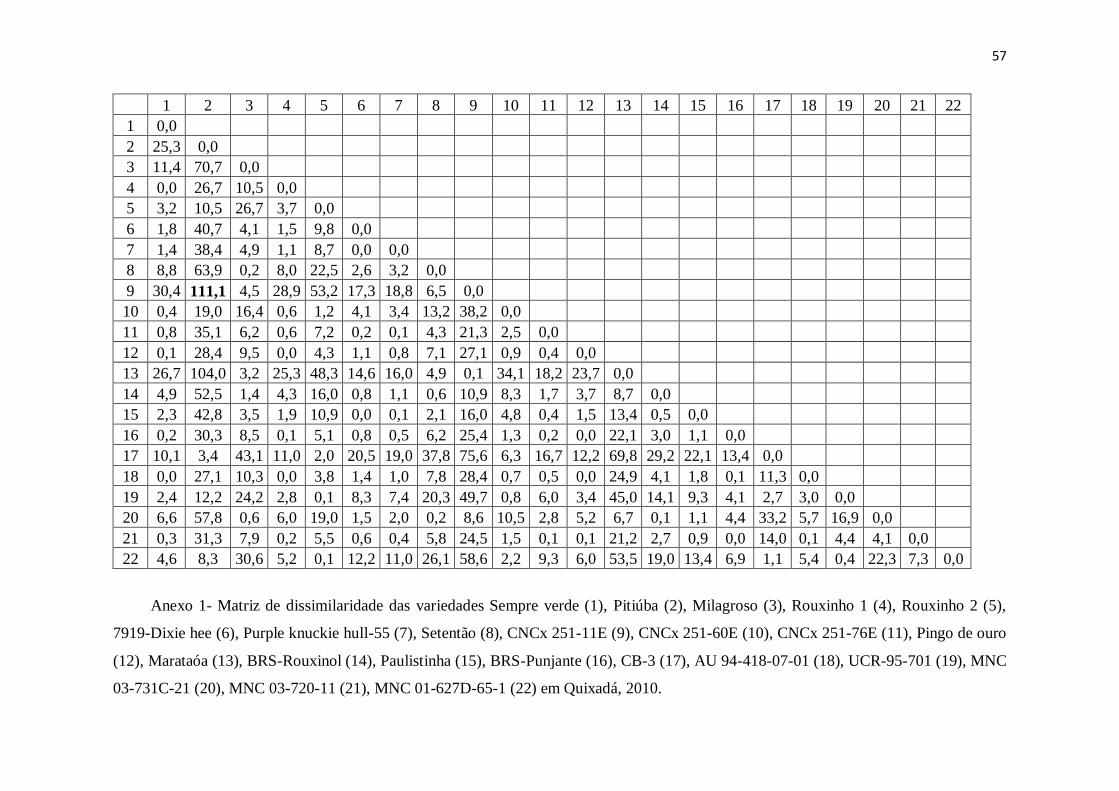
57
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
1 0,0
2 25,3 0,0
3 11,4 70,7 0,0
4 0,0 26,7 10,5 0,0
5 3,2 10,5 26,7 3,7 0,0
6 1,8 40,7 4,1 1,5 9,8 0,0
7 1,4 38,4 4,9 1,1 8,7 0,0 0,0
8 8,8 63,9 0,2 8,0 22,5 2,6 3,2 0,0
9 30,4 111,1 4,5 28,9 53,2 17,3 18,8 6,5 0,0
10 0,4 19,0 16,4 0,6 1,2 4,1 3,4 13,2 38,2 0,0
11 0,8 35,1 6,2 0,6 7,2 0,2 0,1 4,3 21,3 2,5 0,0
12 0,1 28,4 9,5 0,0 4,3 1,1 0,8 7,1 27,1 0,9 0,4 0,0
13 26,7 104,0 3,2 25,3 48,3 14,6 16,0 4,9 0,1 34,1 18,2 23,7 0,0
14 4,9 52,5 1,4 4,3 16,0 0,8 1,1 0,6 10,9 8,3 1,7 3,7 8,7 0,0
15 2,3 42,8 3,5 1,9 10,9 0,0 0,1 2,1 16,0 4,8 0,4 1,5 13,4 0,5 0,0
16 0,2 30,3 8,5 0,1 5,1 0,8 0,5 6,2 25,4 1,3 0,2 0,0 22,1 3,0 1,1 0,0
17 10,1 3,4 43,1 11,0 2,0 20,5 19,0 37,8 75,6 6,3 16,7 12,2 69,8 29,2 22,1 13,4 0,0
18 0,0 27,1 10,3 0,0 3,8 1,4 1,0 7,8 28,4 0,7 0,5 0,0 24,9 4,1 1,8 0,1 11,3 0,0
19 2,4 12,2 24,2 2,8 0,1 8,3 7,4 20,3 49,7 0,8 6,0 3,4 45,0 14,1 9,3 4,1 2,7 3,0 0,0
20 6,6 57,8 0,6 6,0 19,0 1,5 2,0 0,2 8,6 10,5 2,8 5,2 6,7 0,1 1,1 4,4 33,2 5,7 16,9 0,0
21 0,3 31,3 7,9 0,2 5,5 0,6 0,4 5,8 24,5 1,5 0,1 0,1 21,2 2,7 0,9 0,0 14,0 0,1 4,4 4,1 0,0
22 4,6 8,3 30,6 5,2 0,1 12,2 11,0 26,1 58,6 2,2 9,3 6,0 53,5 19,0 13,4 6,9 1,1 5,4 0,4 22,3 7,3 0,0
Anexo 1- Matriz de dissimilaridade das variedades Sempre verde (1), Pitiúba (2), Milagroso (3), Rouxinho 1 (4), Rouxinho 2 (5),
7919-Dixie hee (6), Purple knuckie hull-55 (7), Setentão (8), CNCx 251-11E (9), CNCx 251-60E (10), CNCx 251-76E (11), Pingo de ouro
(12), Marataóa (13), BRS-Rouxinol (14), Paulistinha (15), BRS-Punjante (16), CB-3 (17), AU 94-418-07-01 (18), UCR-95-701 (19), MNC
03-731C-21 (20), MNC 03-720-11 (21), MNC 01-627D-65-1 (22) em Quixadá, 2010.

58
Anexo 2 - Dendrograma segundo método de agrupamento de ligação média entre grupo (UPGMA) das variedades de Feijão caupi em
Quixadá, 2010.
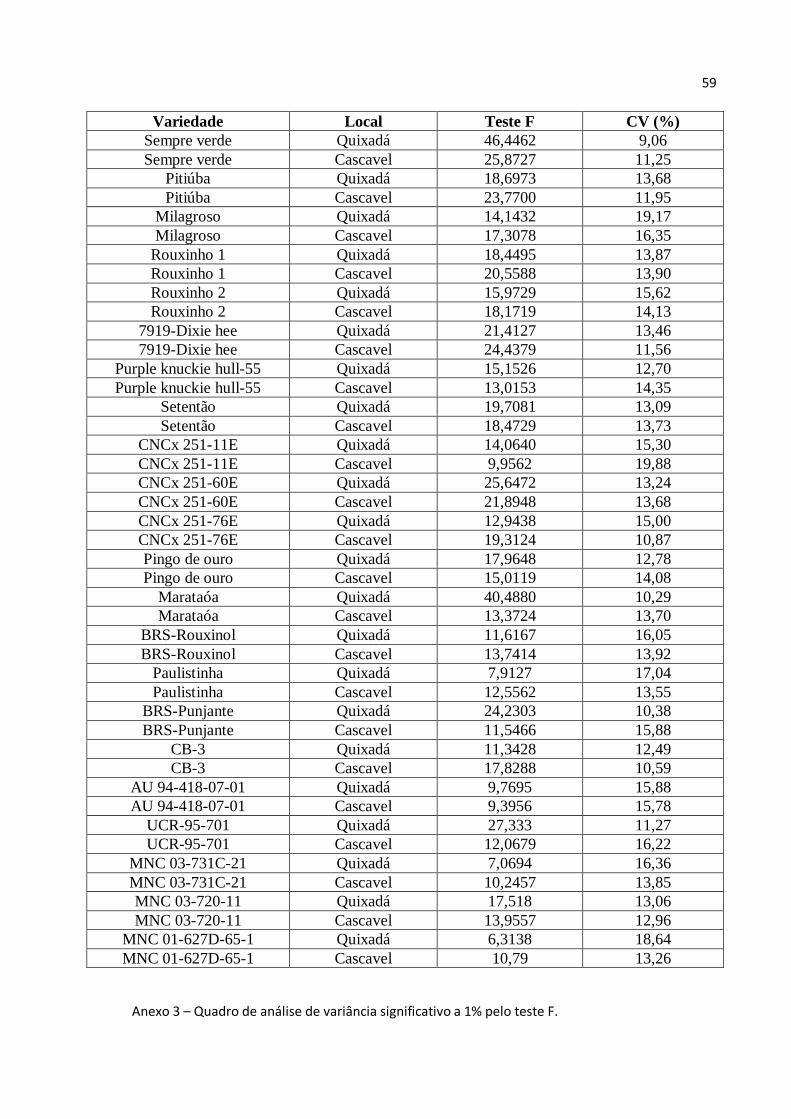
59
Variedade Local Teste F CV (%)
Sempre verde Quixadá 46,4462 9,06
Sempre verde Cascavel 25,8727 11,25
Pitiúba Quixadá 18,6973 13,68
Pitiúba Cascavel 23,7700 11,95
Milagroso Quixadá 14,1432 19,17
Milagroso Cascavel 17,3078 16,35
Rouxinho 1 Quixadá 18,4495 13,87
Rouxinho 1 Cascavel 20,5588 13,90
Rouxinho 2 Quixadá 15,9729 15,62
Rouxinho 2 Cascavel 18,1719 14,13
7919-Dixie hee Quixadá 21,4127 13,46
7919-Dixie hee Cascavel 24,4379 11,56
Purple knuckie hull-55 Quixadá 15,1526 12,70
Purple knuckie hull-55 Cascavel 13,0153 14,35
Setentão Quixadá 19,7081 13,09
Setentão Cascavel 18,4729 13,73
CNCx 251-11E Quixadá 14,0640 15,30
CNCx 251-11E Cascavel 9,9562 19,88
CNCx 251-60E Quixadá 25,6472 13,24
CNCx 251-60E Cascavel 21,8948 13,68
CNCx 251-76E Quixadá 12,9438 15,00
CNCx 251-76E Cascavel 19,3124 10,87
Pingo de ouro Quixadá 17,9648 12,78
Pingo de ouro Cascavel 15,0119 14,08
Marataóa Quixadá 40,4880 10,29
Marataóa Cascavel 13,3724 13,70
BRS-Rouxinol Quixadá 11,6167 16,05
BRS-Rouxinol Cascavel 13,7414 13,92
Paulistinha Quixadá 7,9127 17,04
Paulistinha Cascavel 12,5562 13,55
BRS-Punjante Quixadá 24,2303 10,38
BRS-Punjante Cascavel 11,5466 15,88
CB-3 Quixadá 11,3428 12,49
CB-3 Cascavel 17,8288 10,59
AU 94-418-07-01 Quixadá 9,7695 15,88
AU 94-418-07-01 Cascavel 9,3956 15,78
UCR-95-701 Quixadá 27,333 11,27
UCR-95-701 Cascavel 12,0679 16,22
MNC 03-731C-21 Quixadá 7,0694 16,36
MNC 03-731C-21 Cascavel 10,2457 13,85
MNC 03-720-11 Quixadá 17,518 13,06
MNC 03-720-11 Cascavel 13,9557 12,96
MNC 01-627D-65-1 Quixadá 6,3138 18,64
MNC 01-627D-65-1 Cascavel 10,79 13,26
Anexo 3 – Quadro de análise de variância significativo a 1% pelo teste F.