efeitos do abrasamento elétrico da amígdala basolateral em … · 2019. 6. 3. · rugby clube. i...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
DEPARTAMENTO DE NEUROCIÊNCIAS E CIÊNCIAS DO COMPORTAMENTO
Efeitos do abrasamento elétrico da amígdala
basolateral em padrões oscilatórios durante o sono
LEONARDO RAKAUSKAS ZACHARIAS
RIBEIRÃO PRETO – SP
2018

LEONARDO RAKAUSKAS ZACHARIAS
Efeitos do abrasamento elétrico da amígdala basolateral em padrões
oscilatórios durante o sono
Dissertação apresentada à Faculdade de
Medicina de Ribeirão Preto da Universidade
de São Paulo, como parte das exigências
para a obtenção do título de Mestre em
Ciências
Área de concentração: Neurociências
Orientador: Prof. Dr. João Pereira Leite
Co-orientador: Prof. Dr. Cleiton Lopes Aguiar
RIBEIRÃO PRETO – SP
2018

Autorizo a reprodução e divulgação total ou parcial deste trabalho, por qualquer meio convencional ou eletrônico, para fins de estudo e pesquisa, desde que citada a fonte.
Zacharias, Leonardo Rakauskas Efeitos do abrasamento elétrico da amígdala basolateral em padrões
oscilatórios durante o sono– Ribeirão Preto, 2018
Dissertação de Mestrado, apresentada à Faculdade de Medicina de Ribeirão Preto/USP. Área de concentração: Neurologia, Eletrofisiologia e Comportamento. Orientador: Leite, João Pereira. Co-orientador: Lopes-Aguiar, Cleiton. 1. Epilepsia 2. Aprendizado. 3. Sono. 4. Eletrofisiologia. 5. Hipocampo. 6. Córtex pré-frontal. 7. Potenciais locais de campo.

NOME: ZACHARIAS, Leonardo Rakauskas
TÍTULO: Efeitos do abrasamento elétrico da amígdala basolateral em padrões
oscilatórios durante o sono
Dissertação apresentada à Faculdade de
Medicina de Ribeirão Preto da Universidade
de São Paulo, como parte das exigências
para a obtenção do título de Mestre em
Neurociências
Aprovado em:
Banca examinadora
Prof. Dr.___________________________ Instituição:___________________
Julgamento:________________________ Assinatura:__________________
Prof. Dr.___________________________ Instituição:___________________
Julgamento:________________________ Assinatura:__________________
Prof. Dr.___________________________ Instituição:___________________
Julgamento:________________________ Assinatura:__________________


Agradecimentos
Agradeço primeiramente ao Prof. Dr. João Pereira Leite, por me aceitar como
seu aluno de mestrado e possibilitar a realização deste trabalho. Agradeço ao Prof.
Dr. Cleiton Lopes Aguiar pelo convite a ingressar no mestrado e participar
ativamente na realização deste trabalho, sempre disposto a ensinar e orientar. Este
trabalho não seria possível sem a orientação destes dois professores.
Agradeço também aos membros do Laboratório de Investigação em Epilepsia,
sempre abertos à discussão e dispostos em contribuir com os trabalhos
desenvolvidos no laboratório. Nesse sentido agradeço especialmente ao Lucas
Barone Peres e Rafael Naime Ruggiero, que juntamente com o Cleiton participaram
ativamente no desenvolvimento técnico e teórico deste trabalho, desde a coleta de
dados à discussão e interpretação dos resultados. Gostaria de agradecer também
aos eletrofisiologistas Matheus, Danilo, Lézio e Fernando, que contribuíram neste
trabalho ensinando e discutindo as melhores técnicas para a realização dos
experimentos. Agradeço à Fernanda e Isabella por toda a ajuda nos experimentos
comportamentais, e também à Renata e Dani, pelos ensinamentos e proatividade
nas análises histológicas. Agradeço ao Renato Meirelles e Marco Antonio, sempre
dispostos a resolver qualquer tipo de problema em nossos laboratórios e biotérios.
Agradeço também aos colegas do Departamento de Neurologia e Neurociências,
principalmente durante as pausas para o cafezinho.
Dos amigos de Ribeirão Preto gostaria de agradecer aos membros do
Laboratório de Biomagnetismo, por além de iniciar minha carreira científica ainda
manter as portas abertas para discussões e sugestões, mesmo em diferentes áreas
de pesquisa. Agradeço especialmente ao Fernando Torrieri, Renan Matsuda, André
Salles e Victor Souza por também auxiliar no planejamento técnico e sugestões
principalmente em questões experimentais essenciais no desenvolvimento deste
trabalho. Também agradeço aos amigos da República da Gruta e do Ribeirão Preto
Rugby Clube.
I would like to thank Dr. Marie Carlén for accepting me as a Research
Assistant at the Carlén Lab during my master. A special thanks to Nicolas Guyón,
Sofie Ährlund, and Hoseok Kim, who dedicated part of their time to including me at

their projects, providing essential knowledge to my scientific development. I want to
thank all the other friends at the DMC Lab for the collaboration for my scientific and
personal formation.
Agradeço também à Coordenação de Aperfeiçoamento de Pessoal de Nível
Superior (CAPES), à Swedish Foundation for International Cooperation in Research
and Higher Education (STINT), ao Conselho Nacional de Desenvolvimento Científico
e Tecnológico (CNPq) e à Fundação de Amparo à Pesquisa do Estado de São Paulo
(FAPESP) pelo financiamento necessário ao desenvolvimento deste projeto.
Por ultimo, porém não menos importante, gostaria de agradecer à minha
família. Agradeço aos meus pais, Fernando e Shirley, por todo o apoio e
compreensão desde meu ingresso no mestrado. Também agradeço aos meus
irmãos Ligia, Lucas e Luiz, que sempre me apoiaram em todas as questões
necessárias e também compreenderam os momentos de ausência durante esse
período. Obrigado a cada um de vocês, pois sem o seu apoio não seria possível a
realização deste trabalho.


RESUMO
ZACHARIAS, L.R. Efeitos do abrasamento elétrico da amígdala
basolateral em padrões oscilatórios durante o sono. Dissertação (Mestrado) −
Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão
Preto, 2018
Na epilepsia do lobo temporal (ELT), alterações morfofuncionais em estruturas
límbicas são classicamente acompanhadas de déficits cognitivos. Estudos anteriores
revelaram que disfunções eletrofisiológicas em circuitos hipocampo-corticais são
observadas durante o sono NREM (non rapid eye movement), onde eventos
patológicos como fast ripples e IEDs (interictal epileptiform discharges) substituem
gradativamente eventos fisiológicos, como as sharp-wave ripples (SWR). Tal
substituição pode estar por trás dos prejuízos cognitivos observados tanto nos
modelos animais como em pacientes, já que as SWRs são fundamentais para a
transferência de informação do hipocampo ao córtex durante a consolidação de
memórias. De maneira complementar, o sono REM também parece exercer um
papel fundamental em processos mnemônicos, facilitando eventos de plasticidade
sináptica e coordenando regiões encefálicas distantes por meio de acoplamento
entre diferentes frequências oscilatórias, tais como teta e gama. Entretanto, as
alterações no sono REM durante os processos de epileptogênese ainda foram
pouco exploradas. Neste trabalho testamos a hipótese de que disfunções na
coordenação rítmica durante o sono REM estariam associadas a prejuízos de
memória que se manifestam na epileptogênese. Para isso, submetemos ratos Wistar
adultos machos a protocolo de abrasamento rápido da amígdala basolateral,
possibilitando a avaliação de alterações eletrofisiológicas gradativas durante o
processo de epileptogênese. Foram realizados implantes crônicos de eletrodos para
registro do potencial local de campo (LFP, Local Field Potential) nas regiões de CA1
e do córtex pré-frontal medial (mPFC, medial prefrontal cortex), além de eletrodos
bipolares para estímulo na amígdala basolateral. Os protocolos de abrasamento
foram realizados durante um período de 3 dias, aplicando-se diariamente 10 trens de
estímulos a 50 Hz com duração de 10 segundos. Para avaliarmos os prejuízos
cognitivos, os animais foram submetidos a testes de reconhecimento de objetos
antes do início a após o término dos protocolos de abrasamento. O sono

subsequente às sessões de reconhecimento de objetos e aos protocolos de
estimulações foram registrados diariamente. Além do prejuízo no reconhecimento de
objetos e alterações eletrofisiológicas durante o sono NREM, como a substituição
gradativa de ripples por IEDs, os animais submetidos ao abrasamento elétrico
apresentaram um aumento na comodulação fase-amplitude entre oscilações teta e
gama durante o sono REM após as estimulações, exibindo também uma correlação
negativa entre a comodulação e a duração das pós-descargas induzidas pelos
estímulos elétricos do abrasamento durante o sono subsequente a aplicação dos
protocolos. Nossos achados ampliam a compreensão vigente sobre como alterações
de oscilações cerebrais durante o sono, especialmente da fase REM, poderiam estar
subjacentes a prejuízos de memória que ocorrem na ELT.
Palavras-chave: epilepsia; epileptogênese; abrasamento elétrico; sono;
memória; hipocampo; amígdala; córtex pré-frontal; eletrofisiologia; potenciais de
campo locais;

ABSTRACT
ZACHARIAS, L.R. Effects of basolateral amygdala kindling on oscillatory
patterns during sleep. Dissertation (Master Degree) − Faculdade de Medicina de
Ribeirao Preto, Universidade de São Paulo, Ribeirão Preto, 2018
Morphofunctional changes in limbic structures are classically followed by
cognitive deficits in Temporal Lobe Epilepsy (TLE) patients. Previous studies
revealed that electrophysiological dysfunctions in hippocampal-cortical circuits are
observed during NREM (non-rapid eye movement) sleep, where pathological events
such as fast ripples and IEDs (interictal epileptiform discharges) gradually replace
physiological events, such as Sharpwave Ripples (SWR). This replacement seems to
describe the cognitive impairments observed in animal models and TLE patients
since SWRs are fundamental for information transfer from the hippocampus to cortex
during memory consolidation. Complementary, REM sleep also plays a significant
role in mnemonic processes, facilitating synaptic plasticity events and coordinating
distant brain regions by coupling different frequencies, such as theta and gamma.
However, alterations in REM sleep during the epileptogenesis processes are poorly
investigated. In this study, we tested the hypothesis that dysfunctions on rhythmic
coordination during REM sleep would be associated with memory deficits showed
during epileptogenesis. For this, we submitted adult Wistar rats to a rapid kindling
protocol on basolateral amygdala (BLA), allowing the evaluation of progressive
electrophysiological changes during the epileptogenic process. Chronic electrodes
were implanted for the local field potentials (LFP) recording in the CA1 and medial
prefrontal cortex (mPFC), as well as bipolar electrodes for BLA stimulation. The
kindling protocols were performed during three days, applying ten trains of 50 Hz
stimulations with ten seconds duration. Object recognition tasks were performed
before and after the kindling protocol to evaluate cognitive impairment. Sleep
recordings were performed daily after the object recognition or kindling application.
Along with object recognition impairment and electrophysiological changes during
NREM sleep, such as progressive SWR substitution by IEDs, kindled rats presented
an increase in phase-amplitude comodulation between theta and gamma oscillations
during REM sleep after stimulation sessions, which also correlates negatively with
after-discharges (AD) duration induced by the kindling stimulation. Our findings

expand the comprehension about how changes in brain oscillations during REM
sleep underlies observed memory deficits in TLE.
Keywords: epilepsy; epileptogenesis; animal models of epilepsy; electric
kindling; sleep; memory; hippocampus; amygdala; prefrontal cortex;
electrophysiology; local field potentials;

LISTA DE ABREVIATURAS E SIGLAS
AD - After Discharges
AP - ântero-posterior
BLA - Basolateral Amygdala
CE - Córtex Entorrinal
CEUA - Comissão de Ética em Uso de Animais
CTRL - Controle (grupo experimental)
DP - Desvio Padrão
DV - Dorso-ventral
ELT - Epilepsia do Lobo Temporal
EMG – Eletromiograma
FRs - Fast Ripples
GD - Giro Denteado
HFO - High Frequency Oscillation
HG - High Gamma
ID - Índice de Discriminação
IEDs - Interictal Epileptiform Discharges
KD - Kindling (grupo experimental)
LFP - Local Field Potential
LG - Low Gamma
MI - Modulation Index
ML - Médio-lateral
mPFC - medial Prefrontal Cortex
NREM - non-rapid eye movement

PV+ - Parvalbumina Positivos
REM - rapid eye movement
RMS - Root Mean Square
SC - Colateral de Schaffer
Septo Medial – SM
SL - Septo Lateral
SM-DB -Septo Medial/Banda Diagonal de Broca
SWR - Sharpwave Ripples

SUMÁRIO
1 INTRODUÇÃO---------------------------------------------------------------------------------------------------------------- 1
2 OBJETIVOS-------------------------------------------------------------------------------------------------------------------11
2.1 OBJETIVO GERAL --------------------------------------------------------------------------------------------------------11
2.2 OBJETIVOS ESPECÍFICOS ------------------------------------------------------------------------------------------------11
3 MATERIAIS E MÉTODOS -------------------------------------------------------------------------------------------------12
3.1 ANIMAIS -----------------------------------------------------------------------------------------------------------------12
3.2 DESENHO EXPERIMENTAL ----------------------------------------------------------------------------------------------13
3.3 GRUPOS EXPERIMENTAIS -----------------------------------------------------------------------------------------------14
3.4 CIRURGIA ESTEREOTÁXICA ----------------------------------------------------------------------------------------------14
3.5 RECONHECIMENTO DE OBJETOS ---------------------------------------------------------------------------------------16
3.6 PROTOCOLO DE ABRASAMENTO RÁPIDO -------------------------------------------------------------------------------18
3.7 REGISTRO DE SONO -----------------------------------------------------------------------------------------------------21
3.8 MÉTODOS PARA ANÁLISES ELETROFISIOLÓGICAS ----------------------------------------------------------------------21
3.8.1 Determinação dos estágios de sono ----------------------------------------------------------------------22
3.8.2 Métodos para detecção de eventos eletrofisiológicos durante o sono NREM: Ondas Delta,
Spindles, Ripples e IEDs -------------------------------------------------------------------------------------------------------------25
3.8.2.1 Detecção de Ripples e IEDs --------------------------------------------------------------------------------------- 26
3.8.2.2 Detecção de Ondas Delta ----------------------------------------------------------------------------------------- 29
3.8.2.3 Detecção de Spindles ---------------------------------------------------------------------------------------------- 32
3.8.3 Análise da incidência de eventos eletrofisiológicos durante o sono NREM ---------------------32
3.8.4 Análise do acoplamento de eventos hipocampais com eventos corticais durante o sono
NREM 32
3.9 SONO REM: ANÁLISES DE DENSIDADES DE POTÊNCIA E COMODULAÇÃO ENTRE OSCILAÇÕES TETA E GAMA ------33
3.9.1 Análise da densidade de potência durante sono REM -----------------------------------------------33
3.9.2 Análise da comodulação entre fase em teta e amplitude em gama -----------------------------34
3.10 ANÁLISES ESTATÍSTICAS ----------------------------------------------------------------------------------------------35
3.10.1 Teste de Mann-Whitney ------------------------------------------------------------------------------------35
3.10.2 Análise de tendência de Mann-Kendall -----------------------------------------------------------------35
3.10.3 Análise por reamostragem - Bootstrap -----------------------------------------------------------------35
3.10.4 Correlação entre os efeitos do abrasamento e alterações em sono NREM e REM----------36
3.11 HISTOLOGIA ---------------------------------------------------------------------------------------------------------36
4 RESULTADOS ----------------------------------------------------------------------------------------------------------------37
4.1 EFEITOS COMPORTAMENTAIS DO ABRASAMENTO ELÉTRICO RÁPIDO DA BLA ---------------------------------------37
4.2 RECONHECIMENTO DE OBJETOS ----------------------------------------------------------------------------------------38

4.3 EFEITOS DO ABRASAMENTO ELÉTRICO SOBRE O SONO ----------------------------------------------------------------40
4.3.1 Alterações na Arquitetura do Sono durante a aplicação do abrasamento ---------------------40
4.3.2 Efeitos do abrasamento sobre o sono NREM: substituição de Ripples por IEDs ---------------41
4.3.3 Acoplamento hipocampo-cortical: probabilidade de Ondas Delta e Spindles após eventos
hipocampais 42
4.4 SONO REM: ALTERAÇÕES NAS OSCILAÇÕES LOCAIS DE CA1 ---------------------------------------------------------45
4.4.1 Potência de oscilações teta e gama -----------------------------------------------------------------------45
4.4.1.1 Dinâmica oscilatória: comodulação de fase-amplitude entre teta e gama --------------------------- 47
4.5 CORRELAÇÃO ENTRE OS EFEITOS DO ABRASAMENTO, SONO E MEMÓRIA --------------------------------------------49
4.6 HISTOLOGIA -------------------------------------------------------------------------------------------------------------51
5 DISCUSSÃO ------------------------------------------------------------------------------------------------------------------52
5.1 VANTAGENS DO MODELO DE ABRASAMENTO RÁPIDO DA AMÍGDALA BASOLATERAL --------------------------------53
5.2 ALTERAÇÕES COGNITIVAS INDUZIDAS PELO ABRASAMENTO ELÉTRICO -----------------------------------------------54
5.3 EFEITOS DO ABRASAMENTO SOBRE EVENTOS ELETROFISIOLÓGICOS DURANTE SONO NREM ----------------------55
5.4 ALTERAÇÃO DE OSCILAÇÕES TETA E GAMA DURANTE O SONO REM -------------------------------------------------57
5.4.1 Alterações na comodulação de fase-amplitude de oscilações teta e gama --------------------59
6 LIMITAÇÕES E POSSIBILIDADES FUTURAS --------------------------------------------------------------------------60
7 CONCLUSÕES ---------------------------------------------------------------------------------------------------------------62
8 REFERÊNCIAS ---------------------------------------------------------------------------------------------------------------63


1
1 INTRODUÇÃO
A epilepsia é considerada uma desordem neurológica, afetando
aproximadamente 50 milhões de pessoas em todo o mundo, e é caracterizada pela
incidência de duas ou mais crises convulsivas espontâneas. As principais causas da
epilepsia estão relacionadas à alteração de estruturas cerebrais tanto durante sua
formação (anormalidades genéticas, danos cerebrais durante o desenvolvimento por
hipóxia ou crises febris) como também após o amadurecimento do encéfalo, como
traumatismos cranioencefálicos graves, acidentes vascular-encefálicos, infecções ou
tumores (WHO, 2018), levando a alterações como redução do volume de estruturas
subcorticais e atrofia da substância cinzenta em regiões como o neocórtex e o
tálamo, assim como também à esclerose hipocampal (WHELAN et al., 2018).
Apesar das crises epilépticas aumentarem os riscos de lesões corporais e
mortalidade, as comorbidades psiquiátricas observadas na epilepsia afetam
diretamente qualidade de vida dos pacientes, levando a distúrbios psicológicos como
ansiedade e depressão (SUURMEIJER; REUVEKAMP; ALDENKAMP, 2002;
KANDRATAVICIUS et al., 2012; WHO, 2018) mas também a impactos
socioeconômicos relacionados principalmente a prejuízos cognitivos (HERMANN et
al., 2008; MELBOURNE CHAMBERS et al., 2014).
Estes prejuízos cognitivos são observados frequentemente na Epilepsia do
Lobo Temporal (ELT), o tipo mais comum de epilepsia focal famacorresistente em
adultos. Contudo, ainda não está claro como tais alterações cognitivas estariam
relacionadas a padrões de atividade patológicos que ocorrem durante períodos
interictais (BRAGIN; PENTTONEN; BUZSÁKI, 1997; KANDRATAVICIUS et al., 2014;
GELINAS et al., 2016).
Prejuízos cognitivos estão principalmente relacionados a alterações
morfofuncionais, incluindo a perda de neurônios excitatórios e inibitórios em regiões
como o hipocampo e o córtex entorrinal (CE) (HOLMES, 2015; JANZ et al., 2017).
Juntamente com a perda neuronal, também ocorre a reorganização sináptica dessas
redes, com o crescimento aberrante de fibras musgosas na região de CA3 e a
ocorrência de neurogênese no giro denteado (GD) (HOLMES, 2015) do hipocampo.
Essas alterações hipocampais levam a um aumento da excitabilidade principalmente
por vias glutamatérgicas (BARKER-HALISKI; WHITE, 2015), e que através de um

2
circuito de retroalimentação podem potencializar os danos por excitotoxicidade e as
reorganizações sinápticas, exemplificados na Figura 1 (HOLMES, 2015):
Figura 1: Alterações fisiológicas em diferentes intervalos após as crises epilépticas e
possíveis mecanismos envolvidos no déficit cognitivo – adaptado de (HOLMES, 2015)
Essa reorganização estrutural leva à alteração dos padrões de atividades
eletrofisiológicas no hipocampo e também em suas projeções, como a geração de
oscilações patológicas de alta frequência e a redução na precisão de disparos
neuronais (FOFFANI et al., 2007; STALEY, 2007).
Para compreender melhor os efeitos de tais mudanças sobre as alterações
em estruturas do Lobo Temporal, podemos utilizar modelos animais que não apenas
reproduzam as características da ELT, mas que também nos permitam acessar os
efeitos durante o período latente, onde ocorrem os processos de epileptogênese
(KANDRATAVICIUS et al., 2014). Neste sentido, é possível modelar a
epileptogênese através de estimulações elétricas repetitivas sobre estruturas do
Lobo Temporal, gerando o acúmulo de disparos epilépticos pós-estímulo e,
consequentemente, levando a crises espontâneas e alterações histopatológicas
semelhantes aos observados em pacientes (GODDARD, 1967; RACINE, 1972a;
KANDRATAVICIUS et al., 2014; MORALES et al., 2014). Esta técnica, conhecida
como abrasamento elétrico, é uma potente ferramenta para estudar mecanismos de
epileptogênese, reproduzindo os aspectos da alteração da plasticidade sináptica
decorrentes da atividade anormal em focos epiléticos e as alterações graduais que
3
podem levar às crises generalizadas e comorbidades da ELT (GELINAS et al.,
2016), além de apresentar prejuízos cognitivos semelhantes aos observados em
outros modelos animais e em pacientes de ELT.
Estes prejuízos cognitivos observados emergem em conjunto com eventos
patológicos de alta frequência hipocampais, como as Descargas Interictais
Epileptiformes (Interictal Epileptiform Discharges, IEDs) e as Fast Ripples (FRs, 200
– 600 Hz), detectadas principalmente em zonas epileptogênicas e, apesar de
comumente associadas à perda neuronal, também foram reportadas mesmo em
modelos de ELT não-lesionais, como a infusão de toxina tetânica intrahipocampal
(JIRUSKA et al., 2010; LI et al., 2018) ou a aplicação de protocolos de abrasamento
sobre a amígdala basolateral (SMITH et al., 2005; MORALES et al., 2014).
Durante o processo de epileptogênese em modelos por infusão de toxina
tetânica intrahipocampal (JIRUSKA et al., 2010) e por abrasamento elétrico do
hipocampo (GELINAS et al., 2016), a emergência de IEDs e FRs decorre de um
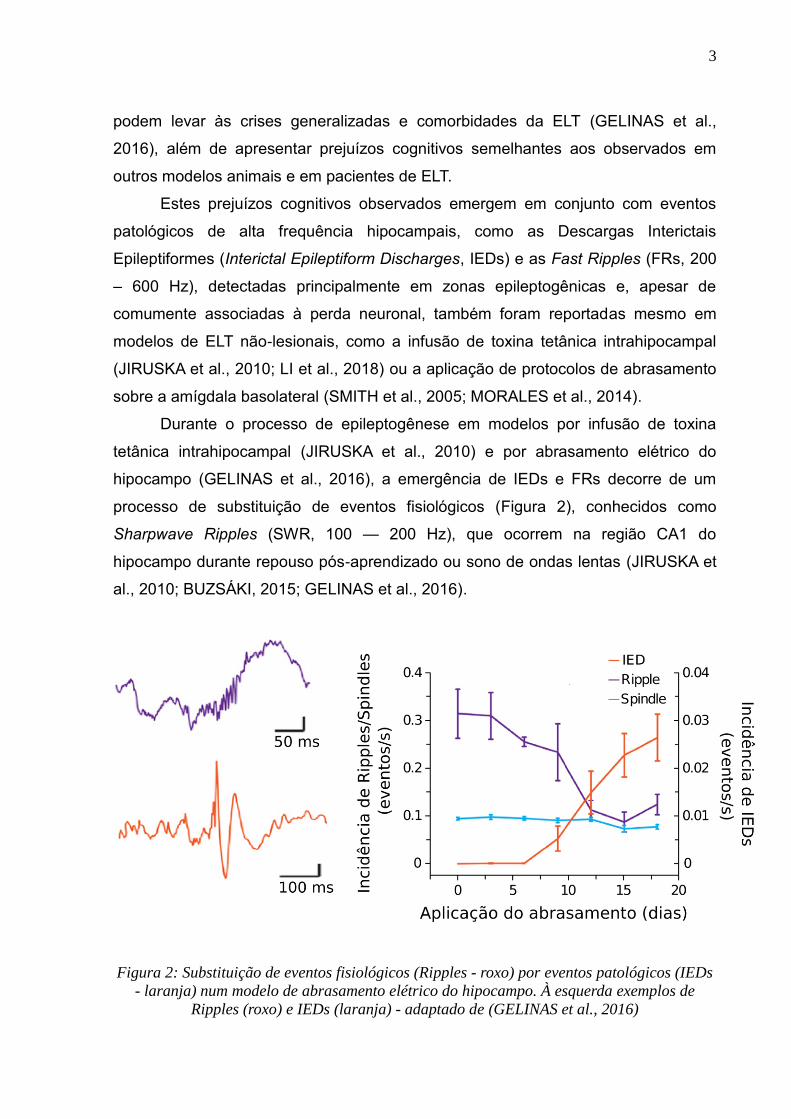
processo de substituição de eventos fisiológicos (Figura 2), conhecidos como
Sharpwave Ripples (SWR, 100 — 200 Hz), que ocorrem na região CA1 do
hipocampo durante repouso pós-aprendizado ou sono de ondas lentas (JIRUSKA et
al., 2010; BUZSÁKI, 2015; GELINAS et al., 2016).
Figura 2: Substituição de eventos fisiológicos (Ripples - roxo) por eventos patológicos (IEDs
- laranja) num modelo de abrasamento elétrico do hipocampo. À esquerda exemplos de
Ripples (roxo) e IEDs (laranja) - adaptado de (GELINAS et al., 2016)

4
As SWR são estruturas eletrofisiológicas de campo altamente precisas,
compostas por (1) uma massiva salva de disparos em CA3 e (2) oscilações de alta
frequência síncronas (Ripples, 100 — 200 Hz), decorrentes de atividade altamente
organizada de interneurônios parvalbumina positivos (PV+) presentes em CA1
(BUZSÁKI, 2015). As SWR que ocorrem no sono de ondas lentas ou repouso pós-
aprendizado eventualmente aparecem associadas ao replay de sequências de
disparos exibidas pelas place-cells de CA1 durante o comportamento exploratório
(ROUX et al., 2017). A reativação de representações espaciais, associada ao alto
acoplamento temporal entre eventos de SWR e spindles (12 a 18 Hz) tálamo-
corticais, parece fundamentar processos que dependam da transferência de
informação hipocampo-cortical, como a consolidação de memória espacial
(BUZSÁKI, 2015; TAXIDIS et al., 2015; GELINAS et al., 2016). Sendo assim, a
substituição gradativa de SWR por FRs e IEDs durante os processos de
epileptogênese parece estar subjacente aos mecanismos responsáveis pela
manifestação de prejuízos de memória em modelos animais de ELT, pois afeta
diretamente processos de transferência de informação hipocampo-cortical
(GELINAS et al., 2016).
Porém essas alterações estão relacionadas apenas à atividade
eletrofisiológica registrada durante os períodos de repouso ou sono NREM (non-
rapid eye movement) após o aprendizado. De acordo com as teorias de
consolidação sistêmica da memória, outros estágios do sono, como o sono REM,
também apresentam aspectos fundamentais na consolidação da memória durante o
sono pós aprendizado (DIEKELMANN; BORN, 2010). Nesse sentido, o sono REM
atuaria de maneira complementar ao sono NREM na regulação das atividades
neuronais, como exemplificado na Figura 3:

5
Figura 3 - Diferentes estágios da consolidação sistêmica de memórias. Primeiramente as
informações adquiridas durante os processos de aprendizado (encoding) são armazenadas
tanto no córtex (roxo) quando nas regiões hipocampais (amarelo). Durante o sono NREM, as
projeções hipocampo-corticais fortalecem as sinapses desenvolvidas durante o aprendizado
através da facilitação sináptica, enquanto que durante o sono REM estes processos
aconteceriam de maneira independentes em cada uma das regiões – adaptado de
(DIEKELMANN; BORN, 2010).
Marcado principalmente pela presença de oscilações teta e gama, o sono
REM promove a diminuição das taxas de disparos durante as transições de
episódios de sono NREM (Figura 4), reorganizando a excitabilidade sináptica das
regiões hipocampais (GROSMARK et al., 2012). Essa reorganização depende
também da potência das oscilações teta presentes durante este estágio (MIYAWAKI;
DIBA, 2016), com evidências causais de que a diminuição da potência dessas
oscilações durante o sono REM pode levar a prejuízos cognitivos (BOYCE et al.,
2016).

6
Figura 4 - Efeitos do sono REM sobre a taxa de disparos neuronais entre dois episódios de
sono NREM – adaptado de (GROSMARK et al., 2012)
O acoplamento entre oscilações teta e gama é sugerido como um possível
mecanismo de consolidação de memórias, ocorrendo durante os processos de
aprendizado e também durante o sono REM (LISMAN; IDIART, 1995; LISMAN;
BUZSAKI, 2008; TORT et al., 2009; LISMAN; JENSEN, 2013), onde uma sequência
de oscilações gama sincroniza a atividade de diferentes grupos neuronais durante
um período das oscilações teta (Figura 5), reacessando padrões de ativação
observados durante o aprendizado.

7
Figura 5 - Possível mecanismo de consolidação da memória através das oscilações teta e
gama. Diferentes padrões de ativação seriam armazenados em cada oscilação gama (A-G),
acopladas a fase positiva de teta – adaptado de (LISMAN; BUZSAKI, 2008).
Assumindo este papel complementar entre diferentes estágios do sono na
consolidação da memória, podemos sugerir que os prejuízos cognitivos devido a
alterações eletrofisiológicas durante o período da epileptogênese não estejam
restritos apenas à substituição de SWR por eventos patológicos durante o sono
NREM, mas também a alterações nas oscilações teta e gama durante o sono REM,
uma vez que a ELT pode levar a mudanças morfológicas e funcionais em estruturas
responsáveis pela geração e/ou manutenção destas oscilações (para uma revisão,
ver SHUMAN; AMENDOLARA; GOLSHANI, 2017)
Dentro dessas estruturas podemos destacar circuitos osciladores em teta
intrahipocampais, como as regiões CA1, CA3 e o GD, e também circuitos
extrahipocampais, como o CE e o septo medial (BUZSÁKI, 2002; PATEL et al., 2012;
COLGIN, 2016; SHUMAN; AMENDOLARA; GOLSHANI, 2017). As oscilações teta
emergem de uma complexa interação entre estes diferentes circuitos osciladores,
envolvendo projeções GABAérgicas, glutamatérgicas e colinérgicas dentro de um
circuito septo-hipocampo-entorrinal retroalimentável (Figura 6).

8
Figura 6 – Modelo do circuito septo-hipocampal-entorrinal envolvido na geração e
manutenção de teta, exemplificando projeções GABAérgicas (azul), colinérgicas (verde) e
glutamatérgicas (vermelho) entre as regiões do Septo Medial/Banda diagonal de Broca (SM-
DB), o Septo Lateral (SL), as regiões de CA1 e CA3, conectadas pela via Colateral de
Schaffer (SC), o giro denteado (GD) e diferentes camadas do Córtex Entorrinal (CE) –
adaptado de (KITCHIGINA et al., 2013).
Neste sistema, o septo lateral aparece como principal coordenador das
oscilações teta, e a manipulação das atividades septais ou a perturbação das
projeções septo-hipocampais podem reduzir a amplitude das oscilações teta
registradas no hipocampo durante períodos interictais (COLOM et al., 2006;
BUTUZOVA; KITCHIGINA, 2008; KITCHIGINA et al., 2013; BOYCE et al., 2016;
SHUMAN; AMENDOLARA; GOLSHANI, 2017).
Em modelos animais de ELT é observado alterações na dinâmica das
atividades eletrofisiológicas do septo medial, assim como também sua fisiologia.
Enquanto o abrasamento elétrico da Via Perfurante altera os rítmos de ativação
neuronal no septo medial e leva a uma sincronização destes disparos com IEDs
(KITCHIGINA; BUTUZOVA, 2009), a injeção intraperitonial de pilocarpina pode levar
a perda de neurônios GABAérgicos nessa região, e consequentemente a uma
diminuição da potência de oscilações teta registradas no hipocampo (COLOM et al.,
2006), como destacado na Figura 7.

9
Figura 7 - Alterações nos circuitos entre septo medial (SM), septo lateral (SL) e hipocampo
(H). Em preto, neurônios GABAérgicos. Em cinza e branco, neurônios colinérgicos e
glutamatérgicos, respectivamente. Neurônios piramidais estão representados por triângulos.
A perda neuronal de neurônios GABAérgicos e piramidais está representada por neurônios
tracejados, enquanto que as fibras musgosas anormais estão representadas pelos traçados
finos – adaptado de (COLOM et al., 2006)
Alterações intrahipocampais também são capazes de modificar as oscilações
teta (SHUMAN; AMENDOLARA; GOLSHANI, 2017). Em modelos por injeção
intrahipocampal de ácido caínico é observado uma diminuição da coerência de
oscilações teta entre as regiões do CE e do giro denteado no hipocampo, porém
com perdas neuronais verificadas apenas na região hipocampal (FRORIEP et al.,
2012), e principalmente nas projeções hipocampais de CA1 ao CE dorsal (JANZ et
al., 2017). Neste mesmo trabalho também foi verificado que as projeções entorrinais
ao giro denteado através da via perfurante apresentam alterações morfológicas
semelhantes às observadas em estudos de potenciações sinápticas de longa
duração, como o aumento do número e tamanho de terminais sinápticos, o que
poderia induzir a geração de atividades epileptiformes hipocampais e também
contribuir para a desincronização das oscilações teta nestas regiões (JANZ et al.,
2017). Neste caso, as alterações morfológicas e funcionais dessas estruturas levam
a uma desincronização das oscilações teta, sendo refletida como uma diminuição de
sua potência (BUZSÁKI, 2002; SHUMAN; AMENDOLARA; GOLSHANI, 2017).

10
A interação entre as regiões CA1 e o CE também parecem fundamentais
durante o processo de aprendizado de memórias espaciais. O CE, além de atuar
como uma das principais interfaces entre o hipocampo e outras regiões corticais,
também oferece suporte à codificação das place cells hipocampais através das grid
cells, integrando informações espaciais provenientes do córtex. A transferência e
consolidação de informações durante o aprendizado entre o CE e o hipocampo
ocorre principalmente com a sincronização de disparos neuronais com oscilações
gama, induzindo a facilitação sináptica nas projeções CE-CA1 através das via
temporoamônica durante o aprendizado (para uma revisão ver NAKAZONO et al.,
2018 - Figura 8). Portanto, as alterações na comunicação entre o CE e CA1
poderiam afetar o aprendizado e a consolidação de memórias não somente devido à
desincronização de oscilações teta, mas também à alteração na sincronização dos
disparos neuronais com oscilações gama.
Figura 8 - Aumento da sincronia de disparos neuronais com oscilações gama durante o
processo de aprendizado, induzindo a facilitação sináptica na via entorrinal-CA1 – adaptado
de (NAKAZONO et al., 2018)

11
Portanto, considerando os possíveis mecanismos de consolidação da
memória no sono REM através da interação entre oscilações teta e gama e a
alteração nos mecanismos responsáveis por sua geração e manutenção, nossa
hipótese é de que o processo de epileptogênese altera também a relação entre
essas duas oscilações, acompanhando os prejuízos cognitivos observados.
Para testar esta hipótese, registramos o Potencial de Campo Local (Local
Field Potential - LFP) nas regiões de CA1 do hipocampo e do córtex pré-frontal
medial (medial prefrontal cortex – mPFC) durante o sono subsequente a aplicação
de protocolos de abrasamento elétrico rápido na amígdala basolateral (basolateral
amygdala - BLA), verificando possíveis alterações na atividade eletrofisiológica
destas áreas durante o sono NREM e sono REM, e se tais alterações são suficientes
para induzir déficits cognitivos nos animais durante testes de reconhecimento de
objetos.
2 OBJETIVOS
2.1 Objetivo geral
Investigar os efeitos do abrasamento elétrico sobre padrões oscilatórios e
coordenação entre hipocampo e córtex pré-frontal durante diferentes estágios do
sono e sua possível relação com a consolidação de memórias de reconhecimento de
objetos.
2.2 Objetivos específicos
1. Verificar os efeitos de um modelo de abrasamento elétrico rápido da BLA sobre
os processos de consolidação de memória espacial em testes de
reconhecimento de objetos;
2. Caracterizar a incidência de eventos eletrofisiológicos e patológicos em sono
NREM durante o processo de epileptogênese, identificando e calculando a
incidência de IEDs e ripples na região CA1 do hipocampo e de Ondas Delta e
Spindles no mPFC;

12
3. Caracterizar as alterações nas oscilações teta e gama na região CA1 do
hipocampo em sono REM durante o processo de epileptogênese, verificando a
densidade de potência relativa destas frequências e também o acoplamento
entre fase e amplitude, indicados pelo Índice de Comodulação (Modulation Index
– MI) (TORT et al., 2010).
3 MATERIAIS E MÉTODOS
3.1 Animais
Foram utilizados ratos Wistar com pesos iniciais entre 200 e 350 gramas (7 a
8 semanas de idade) fornecidos pelo Biotério Central do campus da Universidade de
São Paulo em Ribeirão Preto – SP. Os animais foram habituados ao ambiente do
biotério experimental e manipulados durante um período de 7 dias, mantidos em
grupos de 2 a 3 animais em caixas de polipropileno (40x33x18 cm) forradas com
raspas de madeira, com livre acesso a água e alimentação. O biotério experimental
possui alternação de ciclos de claro/escuro com duração de 12 horas, com início de
ciclos claros às 7h00 e temperaturas controladas de 23º a 25º C.
Os procedimentos experimentais utilizados neste trabalho foram aprovados
pela Comissão de Ética em Uso de Animais (CEUA) da FMRP sob o protocolo
016/2016.

13
3.2 Desenho experimental
A Figura 9 ilustra o desenho experimental realizado neste trabalho.
Primeiramente os animais foram submetidos a cirurgia para implante de
eletrodos para registro eletrofisiológico crônico do LFP nas regiões do mPFC pré-
límbico e CA1 e estimulações elétricas na BLA. Após a cirurgia, os animais
passaram por um período de recuperação de 7 dias, sendo mantidos em caixas
individuais com grades elevadas para preservar o implante.
O experimento foi conduzido durante um período total de 6 dias após a
recuperação cirúrgica, incluindo 3 dias de habituação, onde os animais foram
habituados às caixas de registro de sono e a arenas de reconhecimento de objetos.
No primeiro dia de experimento foi iniciado o protocolo de reconhecimento de
objetos através das sessões de encoding, com registros de sono subsequente
estendidos até às 19h00. No segundo dia foram realizadas as sessões de decoding,
seguido pelo início da aplicação dos protocolos de abrasamento rápido, com
duração média de 3 horas por dia, durante 3 dias, baseados em (MORALES et al.,
2014). As sessões de sono foram registradas após as estimulações, novamente até
às 19h00. Nos grupos controle apenas o registro de sono foi realizado durante o
período relativo à aplicação do abrasamento.
Figura 9 - Desenho experimental, indicando os testes de reconhecimento de objetos, registro
de sono e aplicação dos protocolos de abrasamento elétrico (kindling).

14
No quinto e sexto dia foram realizadas novas sessões para o reconhecimento
de objeto, com registro de sono subsequente ao encoding. Ao final do experimento,
os animais foram sacrificados e perfundidos em soluções de PBS e PFA 4%, com
extração dos cérebros para posterior verificação do posicionamento dos eletrodos.
Todos os procedimentos experimentais serão descritos em detalhes nas
próximas seções.
3.3 Grupos experimentais
Os animais foram divididos em 2 grupos principais: grupo KD (Kindling, n = 8)
e grupo CTRL (Controle, n = 6), onde apenas os animais do grupo KD foram
submetidos a protocolos de abrasamento elétrico rápido.
Cada grupo foi subdividido em 2 novos grupos (‘a’ e ‘b’) nos animais dos
grupos KD-a (n = 5) e CTRL-a (n = 3) foram implantados eletrodos de registro
monopolares nas regiões do mPFC (1 eletrodo) e CA1 (3 eletrodos), e eletrodos
bipolares para estimulação na BLA. Nos grupos KD-b (n = 3) e CTRL-b (n = 3) foram
implantados eletrodos de registro monopolares somente em CA1 (1 eletrodo), e
eletrodos bipolares para estimulação na BLA. Em todos os animais foram
implantados eletrodos para medir a atividade muscular (eletromiograma - EMG) sob
a musculatura da nuca.
Nos primeiros subgrupos (KD-a e CTRL-a) os experimentos foram conduzidos
individualmente. Já nos grupos KD-b e CTRL-b, o menor número de canais de
registros permitiu a realização de experimentos simultâneos, utilizando diferentes
caixas de registro de sono.
Os registros de sono e a aplicação do protocolo de abrasamento foram
realizadas na mesma caixa, sem acesso a água e comida durante as sessões de
estímulos.
3.4 Cirurgia estereotáxica
Os eletrodos implantados foram construídos manualmente utilizando fios de
tungstênio encapados com teflon (d = 45 µm; AM-Systems, EUA) soldados em
conectores torneados (Figura 10). Quatro microparafusos foram implantados sobre

15
ossos frontais e parietais para fixação do implante, e um microparafuso fixado sobre
o osso occipital para atuar como referência e terra para o registro eletrofisiológico.
Figura 10- Exemplo de eletrodos utilizados nas cirurgias. a) eletrodos de registro monopolar
para implante em mPFC. b) Eletrodos bipolares de estímulo para implante na BLA. c)
Eletrodos para registro de eletromiograma sob a musculatura da nuca.
Os animais foram anestesiados com injeções de ketamina (100mg/Kg, i.p. em
NaCl 0,15M) e xylazina (10mg/Kg, i.p. em NaCl 0,15M), com verificação do grau de
anestesia por pinçamento da cauda durante toda a cirurgia. Uma vez anestesiados,
animais foram submetidos à tricotomia do escalpo, fixação do crânio em
estereotáxico (KOPF; KOPF Instruments; EUA) e assepsia do escalpo com iodo e
água oxigenada, seguida de injeção subcutânea de anestésico local (lidocaína 2%
em NaCl 0,15M) para incisão do escalpo e então exposição e limpeza da superfície
do crânio.
Tomando-se o bregma como referência ântero-posterior (AP) e médio-lateral
(ML), foram realizadas craniotomias ipsilaterais nas coordenadas respectivas às
regiões de implante indicadas na Tabela 1, baseadas em (PAXINOS; WATSON,
2006; MORALES et al., 2014) e adaptadas de acordo com procedimentos já
estabelecidos em nosso laboratório.

16
Tabela 1 – Coordenadas esteteotáxicas relativas ao Bregma para implante de eletrodos de registro e estimulação
mPFC CA1 BLA
AP (mm) +3,0 -5,4 -2,5
ML (mm) -0,5 -4,6 -5,1
DV (mm) -3,5 -2,5 -7,2
3.5 Reconhecimento de Objetos
O reconhecimento de objetos foi baseado nos procedimentos descritos em
(MAINGRET et al., 2016), e tem como objetivo verificar a consolidação de memórias
espaciais dependentes do hipocampo (WARBURTON; BROWN, 2015) antes e após
a aplicação de protocolos de abrasamento.
Os experimentos foram realizados numa caixa de madeira retangular
(40x35x80 cm) ventilada e iluminada, com uma base de borracha (arena) removível
para limpeza. O protocolo de reconhecimento de objetos foi dividido em duas
sessões: encoding e decoding, separadas por um período de 24 horas. Durante o
encoding os animais são colocados na arena, explorando dois objetos idênticos
posicionados em cantos adjacentes durante 20 minutos (Figura 11-a). No decoding,
um dos objetos é posicionado no canto oposto, e os animais podem explorá-los
durante um período de 5 minutos (Figura 11-b). Em todos os grupos o teste foi
realizado no período da manhã. Todas as sessões foram registradas em vídeo
através de câmeras posicionadas dentro da arena, na parte superior.
Figura 11 - a) Representação do animal explorando os objetos durante a sessão de encoding.
b) Representação do animal explorando o objeto deslocado, durante a sessão de decoding.

17
Uma das paredes da caixa conta com uma janela de vidro escurecido,
servindo como um marcador espacial para os animais. Para que a exploração seja
voltada principalmente aos objetos utilizados, os animais foram habituados à arena
durante 3 dias precedendo a primeira sessão de encoding, permitindo uma
exploração de 20 minutos por dia sem objetos.
Para avaliarmos a eficácia nessa consolidação da memória avaliamos o
tempo de exploração despendido pelo animal sobre cada objeto na arena,
calculando o Índice de Discriminação (ID), dado por:
𝐼𝐷 =𝑡𝑂𝑏𝑗2
(𝑡𝑂𝑏𝑗1 + 𝑡𝑂𝑏𝑗2)
Onde tObj1 e tObj2 corresponde ao tempo total de exploração do Objeto 1 e
do Objeto 2, respectivamente. Foram considerados como tempos de exploração
situações onde o animal manteve a cabeça orientada ao objeto, numa distância de
pelo menos 2 cm. Situações como subir ou sentar-se sobre os objetos não foram
contabilizadas como exploração.
Os períodos de exploração dos objetos foram analisados utilizando o sofware
Open Source BORIS (FRIARD; GAMBA, 2016), que permite o registro de eventos
comportamentais durante análise de vídeos. Dessa maneira, o ID pode ser calculado
para cada minuto do experimento.
Caso a consolidação da memória espacial não esteja afetada, é esperado que
o tempo de exploração do Objeto 2 seja maior em relação ao Objeto 1, resultando
num ID maior que 0,5. Numa situação aonde a consolidação possa ter sido
comprometida, os tempos de exploração destes objetos tendem a ser os mesmos,
levando a um ID igual a 0,5.
O teste de reconhecimento de objetos foi realizado precedendo a aplicação
do abrasamento e no dia seguinte ao término das sessões de estimulação, dividindo
as análises em 4 grupos: Kindling pré-estimulação (KD-PRE), Kindling pós-
estimulação (KD-POS), Controle pré-estimulação (CTRL-PRE) e Controle pós-
estimulação (CTRL-PRE). Os grupos PRÉ e PÓS são compostos pelos mesmos
animais, e diferentes objetos foram utilizados nas sessões PRÉ e PÓS.

18
As comparações de ID entre os grupos foram realizadas utilizando testes não-
paramétricos de Mann-Whitney, com n = 8 animais no Grupo KD e n = 6 no Grupo
CTRL.
3.6 Protocolo de abrasamento rápido
Para verificarmos os efeitos de oscilações patológicas sobre a consolidação
da memória durante o sono utilizamos um modelo de abrasamento rápido sobre a
BLA, descrito por (MORALES et al., 2014). O protocolo adotado para o abrasamento
rápido consiste em 3 dias de estimulação. Os trens de pulsos (ondas quadradas
bifásicas, largura: 1 ms e frequência: 50 Hz) tem duração total de 10 segundos, e
são aplicados 10 vezes a cada dia, com intervalo inter-trem de 20 minutos, ilustrado
na Figura 12:
Figura 12 - Protocolo de abrasamento rápido, adaptado de (MORALES et al., 2014)
Os limiares de intensidade de estimulação são definidos durante o primeiro
dia. Iniciando em 100 µA, um trem de pulso é aplicado a cada 10 minutos,
aumentando-se a corrente num passo de 25 µA até que ocorram pós-descargas com
duração mínima de 5 segundos. Identificadas as pós-descargas, a intensidade de
estimulação é então reduzida em 20%, sendo utilizada durante os próximos 3 dias.
Para garantir que não haja o acúmulo de carga sobre os eletrodos de
estimulação durante a aplicação do protocolo de abrasamento, os estímulos

19
elétricos foram gerados por uma unidade isoladora bifásica SIU-BI acoplada a um
gerador S88X (Grass Instruments, EUA).
A progressão dos efeitos convulsivos durante a aplicação dos estímulos
abrasivos foi avaliada com base na Escala de Racine (RACINE, 1972b; MORALES
et al., 2014), analisando comportamentos característicos após a aplicação de cada
trem de pulsos. As respostas foram classificas posteriormente através da análise dos
vídeos registrados, sendo classificadas de acordo com a Tabela 2:
Tabela 2 – Comportamentos característicos em diferentes estágios de Racine
Estágio de Racine Comportamento observado
R1 Movimentos orofaciais
R2 Movimentos de cabeça
R3 Clonia de patas dianteiras
R4 Levantamento
R5 Levantamento e queda, perda de equilíbrio
Os animais que apresentaram R5 pelo menos 3 vezes foram considerados
totalmente abrasados. No entanto, todos os animais foram utilizados para análises
comportamentais e eletrofisiológicas.
Juntamente com os efeitos comportamentais relacionados à aplicação dos
trens de estímulos, calculamos também a duração das Pós Descargas (AD, de After
Discharges) geradas após cada estimulação (EBERT; RUNDFELDT; LÖSCHER,
1995; BRAGIN; PENTTONEN; BUZSÁKI, 1997). Para isso calculamos o envelope
do LFP (não filtrado) nos períodos pós-estimulação e identificamos automaticamente
valores do envelope superiores a duas vezes o RMS do LFP no intervalo de 5
minutos após o início da AD (Figura 13). A duração da AD é calculada como o
intervalo entre o início da AD principal (primeira identificada) e o final das AD
secundárias (último bloco identificado, exemplo na Figura 14). Junto com o cálculo
da duração das AD, também calculamos o número total de descargas ictais (Spikes),
identificando os picos com amplitudes acima do limiar.

20
Figura 13 - Exemplo da detecção das AD (vermelho) após as estimulações. O período de
estimulação é evidenciado pelo artefato de estímulo presente no registro (preto, -10 a 0
segundos). O envelope (azul) e o limiar para detecção (linha vermelha pontilhada) são
calculados relativos ao período de 5 minutos após a estimulação. Em cinza, os Spikes ictais
detectados.
Figura 14 – Exemplo de detecção das AD e Spikes com ocorrência de ADs secundárias.

21
3.7 Registro de sono
Para avaliarmos os efeitos dos protocolos de abrasamento sobre os
mecanismos eletrofisiológicos relacionados à consolidação da memória é essencial
registrarmos o sono subsequente ao aprendizado e também às estimulações
decorrentes do abrasamento. Para isso, o LFP coletado pelos eletrodos de registro
foi amplificado 1000 vezes e filtrado analogicamente nas bandas de 0,1 a 1000 Hz, e
digitalizado com frequência de 5 kHz (grupos KD-a e CTRL-a) ou 4 kHz (grupos KD-
b e CTRL-b).
Nos grupos KD-a e CTRL-a o registro foi realizado utilizando um sistema
Open Ephys (Open Science Brasil, BR - SIEGLE et al., 2017), juntamente com um
headstage Intan (Intan Technologies, EUA), que permite a amplificação e
digitalização dos dados diretamente sobre a cabeça do animal. Nestes grupos foram
registrados os LFPs do mPFC (1 canal) e de CA1 (3 canais), além do registro do
EMG e dos acelerômetros incluídos no headstage.
Os animais dos grupos KD-b e CTRL-b foram registrados. utilizando um
amplificador e filtro analógico A-M Systems 3600 (A-M Systems, EUA) e um
digitalizador PowerLab Daq 16/30 (ADInstruments, NZ). Apesar do primeiro sistema
apresentar facilidades, como a digitalização diretamente sobre cabeça do animal e
filtros e amplificadores integrados no mesmo sistema, este segundo setup
possibilitou o registro de 6 animais simultaneamente.
O sono dos animais foi registrado durante o ciclo claro, entre 8h00 e 19h00,
visando obter o maior tempo de registro possível após os procedimentos de
reconhecimento de objeto e da aplicação dos protocolos de abrasamento. Os
registros foram iniciados logo após as sessões de encoding do reconhecimento de
objetos (8h30) e também após a aplicação dos protocolos de abrasamento (12h00).
3.8 Métodos para análises eletrofisiológicas
Nesta sessão descreveremos os métodos desenvolvidos neste trabalho para
a separação de estágios de sono, análises oscilatórias e detecção de eventos
eletrofisiológicos durante diferentes estágios de sono. Todas as rotinas foram
desenvolvidas em Matlab 2015a (MathWorks, EUA).

22
3.8.1 Determinação dos estágios de sono
Para avaliarmos as possíveis alterações eletrofisiológicas induzidas pelo
modelo de abrasamento estagiamos o sono-vigília em três principais categorias:
Acordado, Sono NREM e Sono REM. Os estágios REM e Acordado possuem
oscilações hipocampais características nas faixas de teta (5 – 12 Hz), enquanto que
estágios NREM possuem oscilações características em frequências mais lentas,
como delta (0.5 – 4 Hz). Essa diferença entre as frequências características permite
a diferenciação dos estágios através da análise das densidades de potência das
oscilações em diferentes frequências.
Desenvolvemos uma rotina onde o sono é estagiado de forma semi-
automatizada baseado na atividade oscilatória do LFP em CA1 e na atividade
locomotora extraída pelo EMG, acelerômetros ou análise de vídeos.
O primeiro passo consiste na separação do registro em épocas de 10
segundos. Para cada época são estimadas as densidades de potências totais do
LFP através do Método de Welch (com janelas de 1,25 segundos e nfft igual a 212 -
WELCH, 1967), em CA1 e então separada nas seguintes faixas de frequência: 0,5 –
4,5 Hz; 0,5 – 9 Hz; 0,5 – 20 Hz e 0,5 – 55 Hz. A partir dessas densidades de
potência, calculamos as seguintes razões:
𝑅𝑎𝑡𝑖𝑜1 =0.5– 4.5𝐻𝑧
0.5– 9𝐻𝑧
𝑅𝑎𝑡𝑖𝑜2 =0.5– 20𝐻𝑧
0.5– 55𝐻𝑧
Calculando as razões Ratio 1 e Ratio 2 para cada época de 10 segundos,
obtivemos um Scatterplot (Figura 15) onde podemos manualmente selecionar
Clusters de estados Ativados (REM/Acordado, sem informação do EMG) e
Desativados (NREM, sem informação do EMG):

23
Figura 15 - Exemplo de clusterização manual de estados Ativados e Desativados num registro
de aproximadamente 8 horas. O Cluster azul foi classificado como Desativado, enquanto que
o vermelho classificado como Ativado.
Em seguida adicionamos as informações sobre a locomoção do animal,
criando novos gráficos de entre as Razões 1 e 2 pelo RMS da atividade locomotora
(Figura 16):
Figura 16 - Scatterplots tridimensionais (a) e bidimensionais (b e c) entre Ratio 1, Ratio 2 e o
EMG. Nesta etapa é possível excluir épocas selecionadas como estados Ativado ou
Desativado, mas que possuem altos valores de EMG, indicando movimentação do animal.
0.8 0.85 0.9 0.95 1
Ratio 2
0
0.2
0.4
0.6
0.8
1R
atio 1
Cluster manual
épocas de 10s
0.8 0.85 0.9 0.95 1
Ratio 2
0
0.2
0.4
0.6
0.8
1
Ratio 1
Cluster manual
épocas de 10s
Cluster Desativado
Cluster Ativado

24
Nesta etapa podemos excluir manualmente épocas que foram incluídas nos
Clusters Ativados ou Desativados, mas que indicam movimento, resultando na
Figura 17:
Figura 17 - Scatterplot tridimensional e bidimensional de Ratio1 e Ratio2 contra o EMG (a).
Somente os blocos que apresentam Atonia foram mantidos (b e c), resultando na figura d).
Ao final, podemos verificar a densidade espectral média dos Clusters
resultantes (Figura 18):
Figura 18 – Exemplo do espectro de potência para um animal entre 0 e 100 Hz para os
estados NREM (laranja) e REM (azul) determinados pela Clusterização (à esquerda), e a
potência normalizada das principais frequências que diferenciam os estados (à direita).
100
102
Frequencia (Hz)
10 5
10 4
10 3
10 2
10 1
Log10 (P
otê
ncia
)
REM
NREM
Delta Theta L. Gamma H. Gamma Gamma
Bandas de frequência
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Potê
ncia
norm
aliz
ada (%
of to
tal)
REM
NREM

25
A maior densidade de potência nas frequências mais baixas (de 0 a 5 Hz)
caracterizam oscilações do Sono de NREM (em laranja), enquanto que o pico em
torno de 5 a 12 Hz (teta) caracteriza o sono REM (azul), o que nos indica que a
separação dos estágios de sono foi bem-sucedida.
3.8.2 Métodos para detecção de eventos eletrofisiológicos durante o sono
NREM: Ondas Delta, Spindles, Ripples e IEDs
As detecções de eventos eletrofisiológicos foram baseadas na análise da
amplitude de sinais filtrados em bandas de frequências específicas, verificando
intervalos temporais com amplitudes e intervalos dentro de limiares sugeridos pela
literatura (GELINAS et al., 2016; MAINGRET et al., 2016), apresentados na Tabela 3.
Para refinar a detecção destes eventos optamos por adotar limiares iguais ou
ligeiramente abaixo dos recomendados pela literatura e utilizar variáveis auxiliares
que tragam informações que não estejam relacionadas somente às bandas de
frequência utilizadas, mas também sobre dinâmica da amplitude dos eventos
detectados, como o número de cruzamentos em zero, o desvio padrão da amplitude
sobre o tempo e a integral da amplitude sobre a curva. Neste segundo passo
utilizamos estas variáveis auxiliares para criar Clusters a partir dos eventos
detectados, visualizando e selecionamento manualmente os que mais se adequarem
ao evento estudado, ignorando possíveis falsos positivos. A seguir exemplificaremos
o processo de detecção para Ripples, IEDs, Ondas-Delta e Spindles.
Tabela 3 – Bandas de frequência utilizadas na detecção de eventos eletrofisiológicos
Evento Ripples IEDs Ondas-Delta Spindles
Frequência (Hz) 100 – 250 50 – 90** 0.5 – 4 9 – 17
Intervalo de duração (ms)
25 – 200 20 – 100 150 – 500 400 – 3000
Limiar adotado *Média + 3*DP Média + 3*DP Média + 2*DP ***Média + 1.5*DP
*Adotado Média + 2*DP para alguns animais; ** Intervalo sugerido: 60 – 90 Hz; ***Limiar sugerido: Média + 3*DP;

26
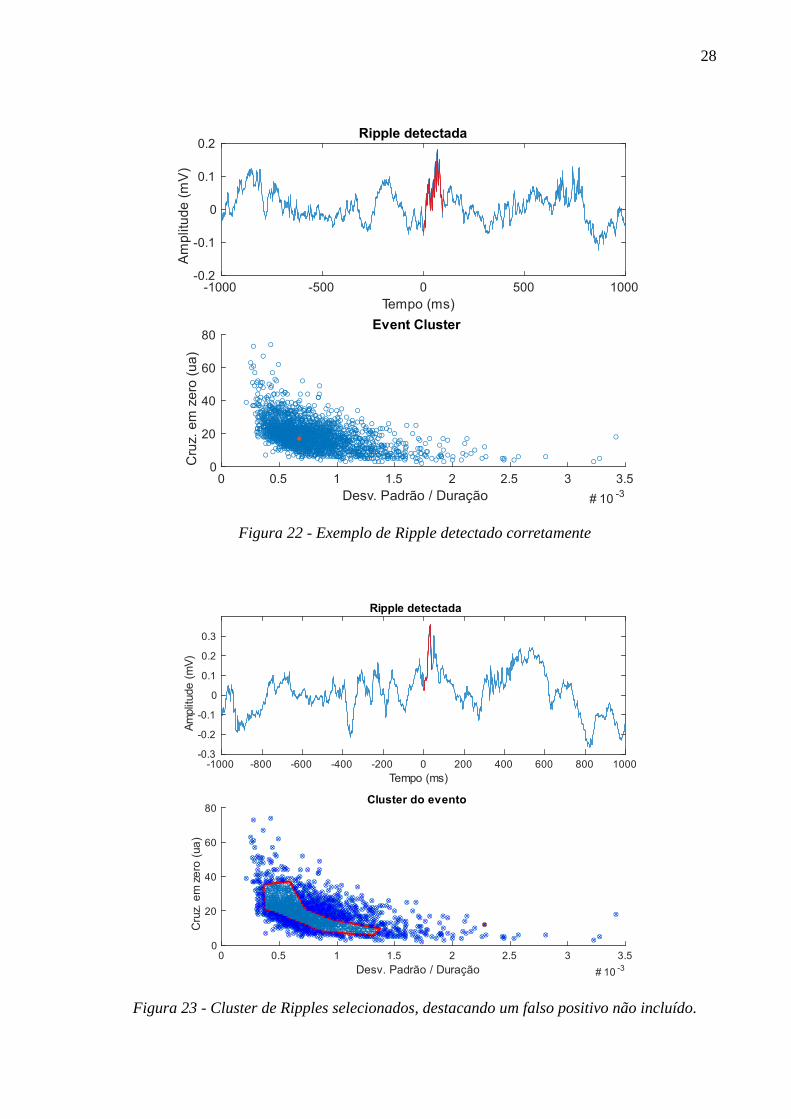
3.8.2.1 Detecção de Ripples e IEDs
A clusterização de eventos de alta frequência detectados em CA1 foi
realizada contabilizando o número de cruzamentos em zero (eixo y) e o desvio
padrão da amplitude sobre o tempo total do evento detectado (eixo x), fornecendo
informações sobre a frequência de oscilação e sobre a variação da amplitude,
respectivamente. As Figura 19 a Figura 23 ilustram o processo de seleção de
eventos de IEDs e Ripples, exemplificando também falsos positivos detectados. A
Figura 23 exemplifica um Cluster de eventos Ripples selecionado manualmente,
com um evento falso positivo excluído da seleção.
Figura 19 - Exemplo de IED detectada corretamente.
500 0 500
Tempo (ms)
0.4
0.2
0
0.2
0.4
0.6
Am
plit
ude (m
V)
IED Detectada
0 1 2 3 4 5 6
Desv. Padrão / Duração (ua) 10 3
0
10
20
30
40
Cru
zam
ento
s e
m z
ero
(ua)
Cluster do evento

27
Figura 20 - Exemplo de evento falso positivo durante a detecção de IEDs
Figura 21 - Exemplo de falso Ripple detectado
500 0 500
Tempo (ms)
0.4
0.2
0Am
plit
ude (m
V)
IED detectada
0 1 2 3 4 5 6
Desv. Padrão / Duração (ua) 10 3
0
10
20
30
40
Cru
z. e
m z
ero
(ua)
Cluster do evento
1000 800 600 400 200 0 200 400 600 800 1000
Tempo (ms)
0.3
0.2
0.1
0
0.1
0.2
0.3
Am
plit
ude (m
V)
Ripple detectada
0 0.5 1 1.5 2 2.5 3 3.5
Desv. Padrão / Duração 10 3
0
20
40
60
80
Cru
z. e
m z
ero
(ua)
Cluster do evento
28
Figura 22 - Exemplo de Ripple detectado corretamente
Figura 23 - Cluster de Ripples selecionados, destacando um falso positivo não incluído.
1000 500 0 500 1000
Tempo (ms)
0.2
0.1
0
0.1
0.2
Am
plitu
de (m
V)
Ripple detectada
0 0.5 1 1.5 2 2.5 3 3.5
Desv. Padrão / Duração 10 3
0
20
40
60
80
Cru
z. em
zero
(ua)
Event Cluster
1000 800 600 400 200 0 200 400 600 800 1000
Tempo (ms)
0.3
0.2
0.1
0
0.1
0.2
0.3
Am
plit
ude (m
V)
Ripple detectada
0 0.5 1 1.5 2 2.5 3 3.5
Desv. Padrão / Duração 10 3
0
20
40
60
80
Cru
z. e
m z
ero
(ua)
Cluster do evento

29
Apesar de utilizarmos diferentes faixas de frequência para a detecção de cada
evento, as IEDs podem surgir como falsos positivos em Ripples. Sendo assim,
Ripples detectadas durante o mesmo intervalo que IEDs foram descartadas, como
sugerido em (GELINAS et al., 2016).
3.8.2.2 Detecção de Ondas Delta
Além da detecção de amplitudes do sinal filtrado a partir dos limiares
sugeridos, a detecção de ondas delta ainda conta com outro processo anterior a
clusterização. Semelhante a (MAINGRET et al., 2016), identificamos o ponto máximo
dentro dos intervalos que se destacam acima dos limiares do sinal filtrado. A partir
deste ponto máximo, calculamos o diferencial num intervalo de -250 a + 250 ms,
sendo 0 o ponto máximo detectado previamente. Calculando o máximo e o mínimo
do diferencial nos intervalos de -250 – 0 ms e 0 – 250 ms, respectivamente, obtemos
o ponto inicial e final da onda delta, como exemplificado na Figura 24:
Figura 24- Exemplo de cálculo do início e fim de onda Delta a partir da derivada. Em azul
temos o envelope do sinal filtrado e retificado. Em cinza, o sinal filtrado. As linhas verticais
indicam o início e fim da onda delta detectada.
250 0 250
Tempo (ms)
0.04
0.02
0
0.02
0.04
0.06
0.08
Am
plit
udes
Delta envelope
Derivada
LFP Filtrado
Início de Delta
Fim de Delta

30
Como a detecção dos limiares de sinais filtrados é realizada através do
envelope do sinal retificado, podemos obter falsos negativos como ondas delta de
curvatura negativa, como na Figura 25:
Figura 25 - Exemplo de falso positivo na detecção de onda delta: curvatura negativa
Para evitar este tipo de erro, introduzimos em nossa Clusterização o cálculo
da integral numérica do sinal filtrado do evento detectado, como exemplificado na
Figura 26 e Figura 27:
250 0 250
Tempo (ms)
0.1
0.08
0.06
0.04
0.02
0
0.02
0.04
0.06
0.08
0.1
Am
plit
udes
Delta envelope
Derivada
LFP Filtrado
Início de Delta
Fim de Delta

31
Figura 26 - Exemplo do processo de clusterização de ondas delta, identificando falsos
positivos detectados pela curvatura negativa através do valor da integral numérica,
calculada somente no intervalo destacado em vermelho.
Figura 27 - Exemplo de onda delta corretamente classificada
1000 500 0 500 1000Tempo (ms)
0.1
0.05
0
0.05
0.1A
mplitu
de (m
V)
Onda delta detectada
0 1 2 3 4 5 6 7
Desv. Padrão / Duração 10 4
0.2
0.1
0
0.1
Inte
gra
l
Cluster do evento
1000 500 0 500 1000
Tempo (ms)
0.1
0.05
0
0.05
0.1
0.15
Am
plitu
de (m
V)
Onda delta detectada
0 1 2 3 4 5 6 7
Desv. Padrão / Duração 10 4
0.2
0.1
0
0.1
Inte
gra
l
Cluster do evento

32
3.8.2.3 Detecção de Spindles
A clusterização dos eventos Spindles através dos valores de Cruzamentos em
zero, desvio padrão ou integral numérica não apresentou resultados eficientes em
termos de clusterização, principalmente devido à grande variação de intervalos em
que os eventos podem ocorrer, indo de 400 ms a 3 segundos. Com isso, apenas a
detecção de eventos acima do limiar foi utilizada. No entanto, a ocorrência de IEDs e
ondas Delta também podem contaminar o sinal filtrado na faixa de spindles (9 – 17
Hz), podendo levar a detecção de falsos positivos. Sendo assim, todos os eventos
spindles que foram detectados juntamente com IEDs ou ondas delta foram
descartados.
3.8.3 Análise da incidência de eventos eletrofisiológicos durante o sono
NREM
A incidência de IEDs, Ripples, Ondas Delta e Spindles durante o sono NREM
foi avaliada através da análise de tendência de Mann-Kendall (HAMED;
RAMACHANDRA RAO, 1998; exemplo de aplicação em GELINAS et al., 2016),
verificando se a Tendência da incidência ao longo dos dias de experimento é
significante (sessão 3.10.2).
3.8.4 Análise do acoplamento de eventos hipocampais com eventos corticais
durante o sono NREM
Nesta etapa verificamos se o acoplamento entre eventos eletrofisiológicos de
CA1 e do mPFC podem ser alterados em animais submetidos ao abrasamento. Para
isso, verificamos a probabilidade de ocorrência de Ondas Delta ou Spindles
detectados no mPFC após eventos Ripples ou IEDs.
Para cada evento hipocampal (Ripple ou IED) procuramos a ocorrência mais
próxima de um evento cortical (Delta ou Spindle), calculando o intervalo temporal
entre os eventos. Os intervalos calculados foram separados entre grupos KD e
CTRL, e reamostrados aleatoriamente através da técnica de bootstrap com 5000
iterações (sessão 3.10.3).

33
3.9 Sono REM: análises de densidades de potência e comodulação entre oscilações teta e gama
Foram analisadas as densidades de potência do LFP em CA1 e calculados os
MI (TORT et al., 2010) entre as fases de teta e amplitudes de gama para cada dia de
experimento, dividindo os episódios de sono REM em épocas contínuos de 30
segundos.
Os animais dos grupos KD-b e CTRL-b não foram incluídos nas análises de
sono REM, pois o tempo total de registro deste segundo sub-grupo foi relativamente
menor comparado ao grupo ‘a’ devido a questões técnicas relacionadas às
estimulações e registros simultâneos, resultando em poucos episódios de sono REM
registrados de maneira adequada.
3.9.1 Análise da densidade de potência durante sono REM
Analisamos as densidades de potência média através da estimação dos
espectros pelo Método de Welch (WELCH, 1967), permitindo a utilização de uma
janela de 2 segundos com sobreposição das estimativas em 50%, e resolução em
frequência (via Transformada de Fourier Discreta) de 213 pontos, na faixa de 0 a 500
Hz.
Foram calculadas as densidades de potência para cada época de 30
segundos durante todos os dias, e verificadas as potências relativas em 4 diferentes
bandas de frequência: teta (5 – 12 Hz), gama baixo (low-gamma, LG: 30 – 55 Hz),
gama alto (high-gamma, HG: 55 – 120 Hz) e oscilações de alta frequência (high
frequency oscillation, HFO: 100 – 120 Hz).
Separando as épocas em dois grupos, KD e CTRL, as amostras foram
normalizadas em relação ao primeiro dia de experimento e comparadas entre os
grupos através de reamostragem estatística por Bootstrap com 5000 iterações
(sessão 3.10.3), verificando diferenças na distribuição das potências entre os
animais controle e nos animais submetidos ao abrasamento elétrico.

34
3.9.2 Análise da comodulação entre fase em teta e amplitude em gama
A amplitude de frequências gama moduladas pela fase de teta dependem das
estruturas hipocampais avaliadas, com variações características no eixo dorso-
ventral (SCHEFFER-TEIXEIRA; TORT, 2017). Para determinarmos os canais mais
adequados para os cálculos do MI geramos comodulogramas bidimensionais
envolvendo diferentes frequências para a análise de fase e diferentes frequências
para análise de amplitude, como mostra a Figura 28:
Figura 28 - Exemplo de diferentes acoplamentos de fase-amplitude durante o sono REM num
animal do grupo KD. As linhas representam os comodulogramas gerados entre diferentes
canais de registro no hipocampo (1, 2 e 3, respectivamente) para o cálculo da frequência de
amplitude, e um canal fixo (3) para a frequência de fase. As colunas representam os dias de
experimento. Os eletrodos 1 e 2 apresentam comodulações entre teta e HFOs, enquanto que o
eletrodo 3 apresenta comodulações em gama. Em cada gráfico o eixo ‘x’ apresenta as
frequências para o cálculo da fase, e o eixo ‘y’ as frequências para o cálculo da amplitude.
De acordo com os comodulogramas gerados, determinamos que as bandas
em gama com alterações mais significantes durante a aplicação do abrasamento
foram em LG (30 – 55 Hz), utilizada portanto para o cálculo de MI.

35
A comparação dos MI entre os grupos KD-a e CTRL-a foram realizadas
utilizando técnicas de bootstrap para intervalos de confiança, calculados para cada
dia de experimento.
3.10 Análises estatísticas
3.10.1 Teste de Mann-Whitney
Para a comparação entre os ID e a proporção dos estágios de sono entre os
grupos KD e CTRL foram realizados testes não-paramétricos de Mann-Whitney, com
valores de alfa iguais a 0,05.
3.10.2 Análise de tendência de Mann-Kendall
O teste de Mann-Kendall aparece como uma opção aos testes de Correlação
de Pearson/Spearman, que exigem a linearidade entre as variáveis observadas. Os
resultados do teste podem ser interpretados através dos valores de tau, sendo -1
indicando Tendência Negativa perfeita, e +1 indicando Tendência Positiva perfeita
(HAMED; RAMACHANDRA RAO, 1998). A significância do teste é verificada através
do teste Z, analisando se a Tendência da incidência ao longo dos dias de
experimento é significante. O teste Z é similar ao teste-t de Student, porém
considera os valores de variância da amostra normalizada durante a comparação.
Distribuições das Tendências que estejam centradas em zero são indicadas como
não significativas (p > 0,05).
3.10.3 Análise por reamostragem - Bootstrap
A estratégia de Bootstrap consiste na reamostragem aleatória repetitiva com
reposição de um conjunto de dados experimentais aplicados a uma função
desejada, como o cálculo da média de uma distribuição ou a comparação entre a
diferença entre médias de dois grupos. O caráter aleatório das reamostragens leva à
redução de vieses estatísticos que acompanham a comparação entre diversos
indivíduos num mesmo grupo (EFRON, 1992).
A partir da aplicação do Bootstrap nossas variáveis passam a ser descritas
como distribuições, não apenas como um conjunto de dados observados. Neste
trabalho, aplicamos o Boostrap para a caracterização dos valores médios de um

36
grupo e também para a diferença média entre dois grupos. No primeiro caso, foram
realizadas 5000 reamostragens do conjunto de dados experimentais, e a cada
iteração foi calculada a média dos valores selecionados aleatoriamente, obtendo
assim uma distribuição das médias em cada reamostragem aleatória. Para a
comparação entre dois grupos foram calculadas as diferenças entre as médias de
cada iteração, obtendo assim uma distribuição das diferenças entre as médias. Para
avaliar essa diferença foram calculados os intervalos de confiança com alfa de 0.05.
As distribuições onde o intervalo de confiança contém o valor zero não foram
consideradas estatisticamente diferentes. As técnicas de Bootstrap foram aplicadas
nas análises de probabilidade de ocorrência entre eventos durante sono NREM e
também para a análise do MI entre oscilações teta-gama e análise de potência de
oscilações teta, LG, HG e HFO durante sono REM.
3.10.4 Correlação entre os efeitos do abrasamento e alterações em sono NREM
e REM
Para avaliarmos quais os efeitos da aplicação do abrasamento elétrico e
verificar a dependência das alterações nos padrões oscilatórios e os déficits
cognitivos, calculamos as correlações de Pearson entre o ID, a duração das AD, o
número de Spikes, a incidência de IEDs durante sono NREM, a variação das
potências de Teta e o MI durante o sono REM.
3.11 Histologia
Ao final dos experimentos os animais foram anestesiados com Uretana (1,5
mg/kg, solução a 50%). Em seguida foram aplicadas correntes de 1 mA durante 1
segundo para indução da lesão eletrolítica nos tecidos para a verificação do
posicionamento dos eletrodos, sendo perfundidos em soluções de tampão fosfato-
salina (PBS – 350 ml)0 e paraformaldeído (PFA 4% - 250 ml) logo em seguida. Após
a perfusão os encéfalos foram extraídos e imersos em solução de PFA 4% durante
12 horas. Durante os 3 dias seguintes foram realizados os procedimentos para
crioproteção através da desidratação dos encéfalos, sendo transferidos em soluções
de 10%, 20% e 30% de sacarose a cada 24 horas, respectivamente. Ao final, foram
37
congelados por imersão em solução de Isopentano e armazenados em freezer a -
80º C.
Para verificação do posicionamento das lesões eletrolíticas foram realizados
cortes transversais em criostato com 30 µm de espessura em torno das
coordenadas respectivas ao mPFC, BLA e CA1. Em seguida, as lâminas foram
coradas utilizando Cresil-violeta para identificação das lesões.
4 RESULTADOS
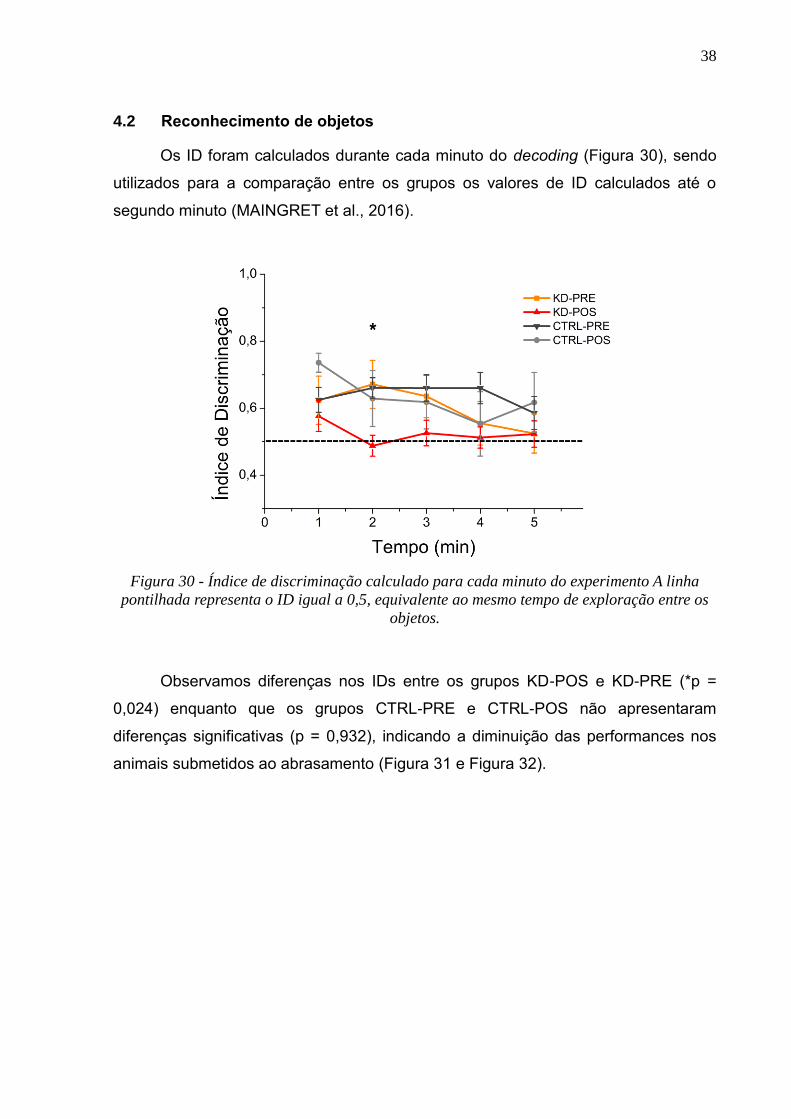
4.1 Efeitos comportamentais do abrasamento elétrico rápido da BLA
Durante o protocolo de indução de pós-descargas as respostas
comportamentais após cada trem de estímulos foram analisadas por vídeo e
classificadas de acordo com a Escala de Racine (Tabela 2, RACINE, 1972b). A
Figura 29 traz os valores máximos na escala alcançados pelos animais dentro de
cada dia de aplicação, com no mínimo 3 repetições para cada comportamento.
Apenas os animais K1, K7 e B7 não alcançaram o estágio 5.
Figura 29 – Estágios de Racine máximo alcançado por cada animal em cada dia de
estimulação, aplicados nos dias 1, 2 e 3. O estágio de Racine reportados para cada animal é
relativo ao estágio máximo alcançado em cada dia, com o mínimo de 3 repetições.
38
4.2 Reconhecimento de objetos
Os ID foram calculados durante cada minuto do decoding (Figura 30), sendo
utilizados para a comparação entre os grupos os valores de ID calculados até o
segundo minuto (MAINGRET et al., 2016).
Figura 30 - Índice de discriminação calculado para cada minuto do experimento A linha
pontilhada representa o ID igual a 0,5, equivalente ao mesmo tempo de exploração entre os
objetos.
Observamos diferenças nos IDs entre os grupos KD-POS e KD-PRE (*p =
0,024) enquanto que os grupos CTRL-PRE e CTRL-POS não apresentaram
diferenças significativas (p = 0,932), indicando a diminuição das performances nos
animais submetidos ao abrasamento (Figura 31 e Figura 32).

39
Figura 31 - Boxplot indicando a distribuição dos ID para cada grupo, com diferença entre os
grupos KD-PRE e KD-POS (*p = 0,024; Teste de Mann-Whitney). Os grupos CTRL-PRE e
CTRL-POS não apresentaram diferenças (p = 0,936; Teste de Mann-Whitney)
Figura 32 - Médias e erro padrão dos ID calculados até 2 minutos para todos os grupos, com
diferença entre os grupos KD-PRE e KD-POS (*p = 0,024; Teste de Mann-Whitney). Os
grupos CTRL-PRE e CTRL-POS não apresentaram diferenças (p = 0,932; Teste de Mann-
Whitney)

40
4.3 Efeitos do abrasamento elétrico sobre o sono
4.3.1 Alterações na Arquitetura do Sono durante a aplicação do abrasamento
Após a classificação dos estágios do sono (Figura 33, exemplo da densidade
média de potência espectral por grupos durante o primeiro dia de experimento)
verificamos que a proporção entre os estados Acordado, NREM e REM não foi
alterada entre os grupos KD e CTRL (Figura 34 - Teste de Mann-Whitney – p = 0,714
(Acordado), p = 0,714 (NREM) e p = 0,341 (REM)):
Figura 33 – Densidade espectral dos estágios de sono nos grupos KD e CTRL durante o
primeiro dia
Figura 34 - Proporção média entre os estágios de sono para cada grupo. A proporção entre
os grupos não apresentou diferenças significativas (Teste de Mann-Whitney – p = 0,714
(Acordado), p = 0,714 (NREM) e p = 0,341 (REM)).
1 10 100
Frequencia (Hz)
10 6
10 5
10 4
10 3
10 2
Log10 (Potê
ncia
)
Densidade Espectral nos estados Grupo KD
1 10 100
Frequencia (Hz)
10 6
10 5
10 4
10 3
10 2
Log10 (Potê
ncia
)
Densidade Espectral nos estados Grupo CTRL
REM
NREM
Acordado
0 5
Dias
20
30
40
50
60
70
Pro
porç
ão d
o e
stá
gio
(%
)
Acordado
0 5
Dias
20
30
40
50
60
70NREM
0 5
Dias
0
10
20
30
40
50REM
Grupo KD
Grupo CTRL

41
4.3.2 Efeitos do abrasamento sobre o sono NREM: substituição de Ripples
por IEDs
Nesta etapa calculamos a incidência de eventos (eventos/segundo) durante o
sono NREM (Figura 35 e Figura 36). As incidências de IEDs entre os dias 1 e 4
apresentaram uma Tendência Positiva significativa (tau = 0,83; p < 0,001 – teste Z),
enquanto que ripples e spindles apresentaram uma Tendência Negativa significativa
(tau = -0,70; p < 0,01 e tau = -0,73; p < 0,001 – teste Z, respectivamente). Os
eventos de onda delta no grupo KD e os eventos de onda delta, spindle e ripple no
grupo Controle não apresentaram Tendências significativas (tau = 0,2; p = 0,4 para
Ondas Delta (KD), tau = 0,06; p = 0,8 para Ripples (CTRL), tau = 0,04; p = 0,17
para Spindles (CTRL) e tau = 0,02; p = 0,5 para Ondas Delta (CTRL)).
Figura 35 - Incidência de eventos durante o sono NREM em animais submetidos ao
abrasamento
0 1 2 3 4 5 6
Dias
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Incid
ência
de O
ndas D
elta
e S
pin
dle
s (evento
s/s
)
0
0.05
0.1
0.15
0.2
0.25
0.3
Incid
ência
de IE
Ds e
Rip
ple
s (e
vento
s/s
)
Incidência de eventos Grupo KD
Ondas Delta
Spindles
IEDs
Ripples

42
Figura 36 - Incidência de eventos Delta, Spindle e Ripples em animais controle.
4.3.3 Acoplamento hipocampo-cortical: probabilidade de Ondas Delta e
Spindles após eventos hipocampais
A Figura 37 traz as distribuições de Probabilidade para cada evento
analisado, detalhadas na Tabela 5:
0 1 2 3 4 5 6
Dia
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Incid
ência
de O
ndas D
elta
e S
pin
dle
s (evento
s/s
)
0
0.05
0.1
0.15
0.2
0.25
0.3
Incid
ência
de R
ipple
s (evento
s/s
)
Incidência de eventos Grupo CTRL
Ondas delta
Spindles
Ripples

43
Figura 37 - Probabilidade de ocorrência de ondas delta após IEDs e Ripples. b)
Probabilidade de ocorrência de Spindles após IEDs e Ripples. c) Probabilidade de
ocorrência de Spindles após ondas delta.

44
Tabela 5 – Distribuição do intervalo de ocorrência de eventos corticais em relação a eventos hipocampais.
Eventos
Densidade de probabilidade
máxima
Intervalo de ocorrência
Número total de eventos
Média ± desvio padrão (s)
Intervalo de confiança (s)
IED – delta 0,026 0,11 ± 0,018 [-0,07; 0,14] 8343
Ripple (KD) – delta 0,007 0,37 ± 0,08 [-0,20; 0,55] 8411
Ripple (CT) – delta 0,011 0,33 ± 0,05 [0,25; 0,42] 12457
IED – Spindle 0,009 3,0 ± 0,6 [1,80; 4,32] 8343
Ripple (KD) – Spindle 0,001 1,5 ± 0,5 [0,40; 2,52] 13421
Ripple (CT) – Spindle 0,011 2,6 ± 0,5 [1,68; 3,70] 12457
Delta (KD – Spindle 0,024 0,7 ± 0,2 [0,31; 1,08] 83813
Delta (CT) – Spindle 0,044 0,9 ± 0,1 [0,70; 1,08] 73619
A Tabela 6 compara as distribuições das probabilidades de eventos calculadas
através da análise do Intervalo de Confiança, sendo a hipótese H0 de que as
distribuições de probabilidade não possuem diferença (h = 1).
Tabela 6 – Comparações entre as distribuições dos intervalos de ocorrências.
Probabilidade de ocorrência de onda delta
Média ± Desvio Padrão da Diferença
(s)
Intervalo de confiança da
diferença
Teste de hipótese: igualdade
entre os grupos N
IEDs vs Ripples (KD) -0,27 ± 0,09 [-0,43; -0,10] h = 1 8343
IEDs vs Ripples (CT) -0,22 ± 0,06 [-0,35; -0,12] h = 1 8343
Ripples (CT) vs Ripples (KD)
-0,04 ± 0,10 [-0,24; 0,16] h = 0 8411
Probabilidade de ocorrência de Spindles
IEDs vs Ripples (KD) 1,61 ± 0,94 [-0,19; 3,42] h = 0 8343
IEDs vs Ripples (CT) 0,43 ± 0,90 [-1,38; 2,14] h = 0 8343
Ripples (KD) vs Ripples (CT)
-1,17 ± 0,77 [-2,72; 0,33] h = 0 12457
Delta (KD) vs Delta (CT) -0,20 ± 0,24 [-0,66; 0,26] h = 0 73619
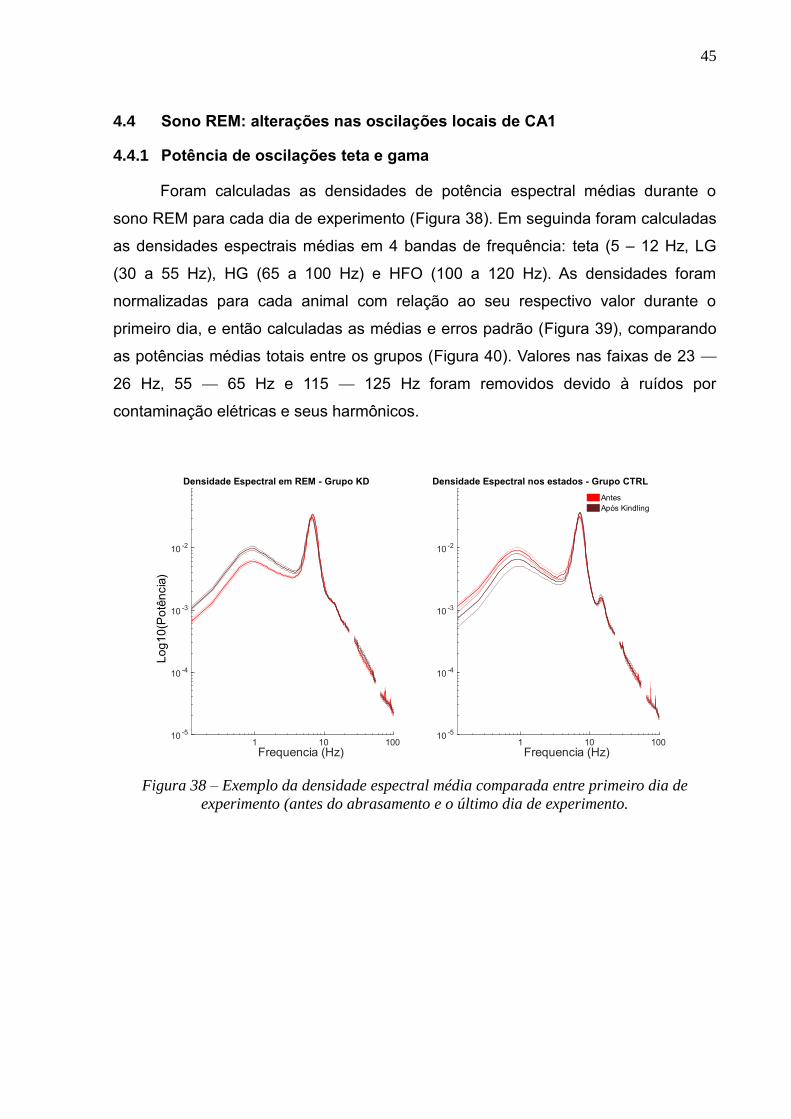
45
4.4 Sono REM: alterações nas oscilações locais de CA1
4.4.1 Potência de oscilações teta e gama
Foram calculadas as densidades de potência espectral médias durante o
sono REM para cada dia de experimento (Figura 38). Em seguinda foram calculadas
as densidades espectrais médias em 4 bandas de frequência: teta (5 – 12 Hz, LG
(30 a 55 Hz), HG (65 a 100 Hz) e HFO (100 a 120 Hz). As densidades foram
normalizadas para cada animal com relação ao seu respectivo valor durante o
primeiro dia, e então calculadas as médias e erros padrão (Figura 39), comparando
as potências médias totais entre os grupos (Figura 40). Valores nas faixas de 23 —
26 Hz, 55 — 65 Hz e 115 — 125 Hz foram removidos devido à ruídos por
contaminação elétricas e seus harmônicos.
Figura 38 – Exemplo da densidade espectral média comparada entre primeiro dia de
experimento (antes do abrasamento e o último dia de experimento.
1 10 100
Frequencia (Hz)
10 5
10 4
10 3
10 2
Densidade Espectral em REM Grupo KD
1 10 100
Frequencia (Hz)
10 5
10 4
10 3
10 2
Densidade Espectral nos estados Grupo CTRL
Antes
Após Kindling
Log10(P
otê
ncia
)

46
Figura 39 - Potência normalizada para teta, LG, HG e HFO durante o sono REM.
Figura 40 - Histogramas das diferenças entre densidades de potência entre o Grupo KD e
Grupo CTRL obtidas por meio de Bootstrap. Todas as diferenças rejeitam a hipótese de que
as distribuições entre os grupos sejam iguais, exceto para Teta e HG. Porém, apenas Teta
possui uma alteração razoável, com uma diminuição média de 7,32%
A variação da densidade de potência com relação ao primeiro dia foi
significativa nas frequências de Teta e HG. Em teta temos uma diminuição de 7,32 ±
0.40% no grupo KD em relação ao grupo CTRL, enquanto que as outras frequências
apresentam variações que, apesar de significantes, são relativamente baixas, sendo
a maior observada em HG (-0,10 ± 0,01 %).
As comparações realizadas com o Bootstrap das diferenças entre os Grupo
KD e Grupo CTRL podem ser conferidas na Tabela 7:
0 1 2 3 4 5
Dias
20
10
0
10
20
Potê
ncia
norm
aliz
ada(%
)
Teta (5 12 Hz)
0 1 2 3 4 5
Dias
5
2.5
0
2.5
5LG (3 55 Hz)
0 1 2 3 4 5
Dias
1
0.5
0
0.5
1HG (65 1 Hz)
0 1 2 3 4 5
Dias
1
0.5
0
0.5
1HFO (1 12 Hz)
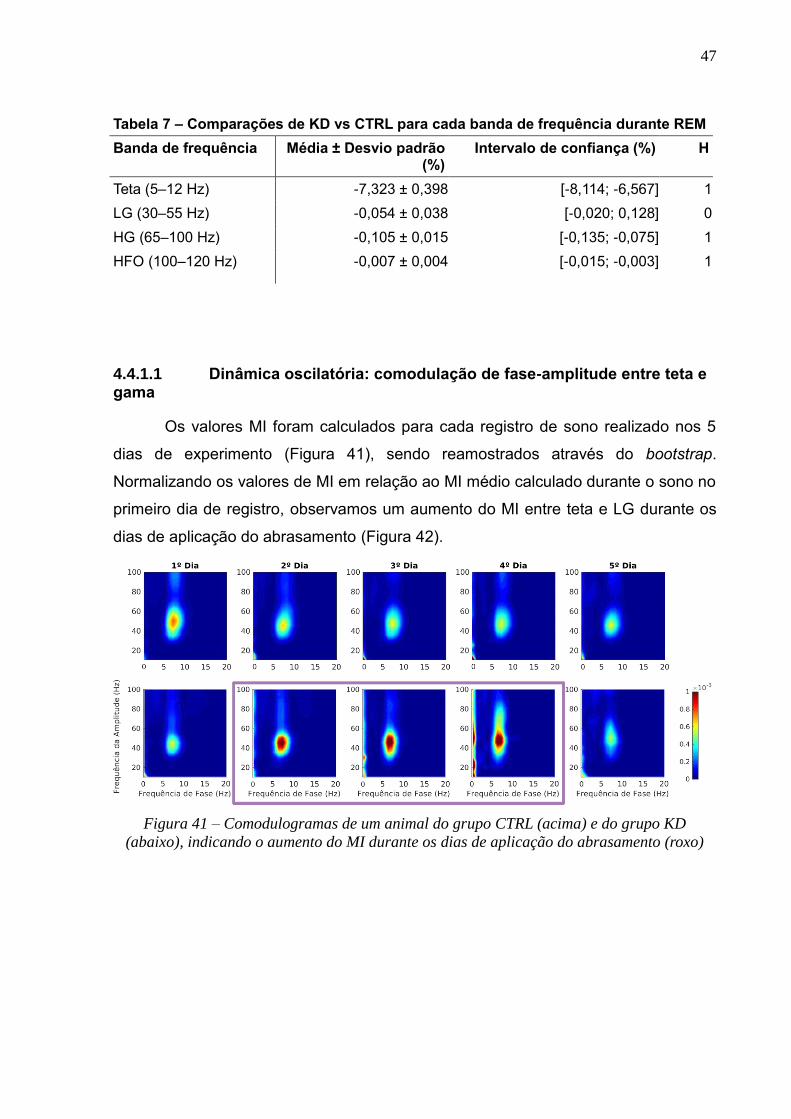
47
Tabela 7 – Comparações de KD vs CTRL para cada banda de frequência durante REM
Banda de frequência Média ± Desvio padrão (%)
Intervalo de confiança (%) H
Teta (5–12 Hz) -7,323 ± 0,398 [-8,114; -6,567] 1
LG (30–55 Hz) -0,054 ± 0,038 [-0,020; 0,128] 0
HG (65–100 Hz) -0,105 ± 0,015 [-0,135; -0,075] 1
HFO (100–120 Hz) -0,007 ± 0,004 [-0,015; -0,003] 1
4.4.1.1 Dinâmica oscilatória: comodulação de fase-amplitude entre teta e gama
Os valores MI foram calculados para cada registro de sono realizado nos 5
dias de experimento (Figura 41), sendo reamostrados através do bootstrap.
Normalizando os valores de MI em relação ao MI médio calculado durante o sono no
primeiro dia de registro, observamos um aumento do MI entre teta e LG durante os
dias de aplicação do abrasamento (Figura 42).
Figura 41 – Comodulogramas de um animal do grupo CTRL (acima) e do grupo KD
(abaixo), indicando o aumento do MI durante os dias de aplicação do abrasamento (roxo)

48
Figura 42 - Média e desvio padrão das distribuições de MI calculados para cada dia de
experimento nos grupos CTRL (Preto) e KD (vermelho), destacando o aumento do MI
durante a aplicação do abrasamento (dias 2, 3 e 4).
Para verificarmos a diferença entre os grupos para cada dia, calculamos a
distribuição da diferença entre as médias de cada grupo pelo bootstrap, indicada
pelos histogramas da Figura 43:
Figura 43 - Histogramas com a distribuição da diferença entre as médias do grupo KD e do
grupo CTRL repetidas 5000 vezes.
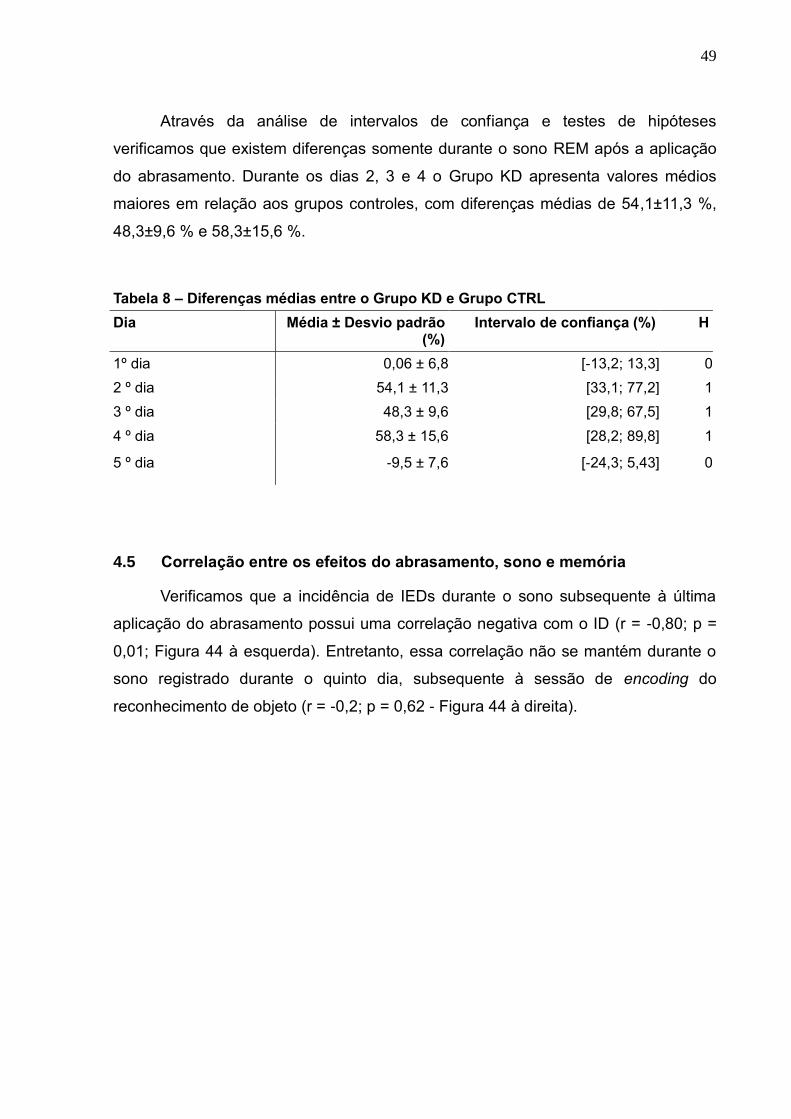
49
Através da análise de intervalos de confiança e testes de hipóteses
verificamos que existem diferenças somente durante o sono REM após a aplicação
do abrasamento. Durante os dias 2, 3 e 4 o Grupo KD apresenta valores médios
maiores em relação aos grupos controles, com diferenças médias de 54,1±11,3 %,
48,3±9,6 % e 58,3±15,6 %.
Tabela 8 – Diferenças médias entre o Grupo KD e Grupo CTRL
Dia Média ± Desvio padrão (%)
Intervalo de confiança (%) H
1º dia 0,06 ± 6,8 [-13,2; 13,3] 0
2 º dia 54,1 ± 11,3 [33,1; 77,2] 1
3 º dia 48,3 ± 9,6 [29,8; 67,5] 1
4 º dia 58,3 ± 15,6 [28,2; 89,8] 1
5 º dia -9,5 ± 7,6 [-24,3; 5,43] 0
4.5 Correlação entre os efeitos do abrasamento, sono e memória
Verificamos que a incidência de IEDs durante o sono subsequente à última
aplicação do abrasamento possui uma correlação negativa com o ID (r = -0,80; p =
0,01; Figura 44 à esquerda). Entretanto, essa correlação não se mantém durante o
sono registrado durante o quinto dia, subsequente à sessão de encoding do
reconhecimento de objeto (r = -0,2; p = 0,62 - Figura 44 à direita).
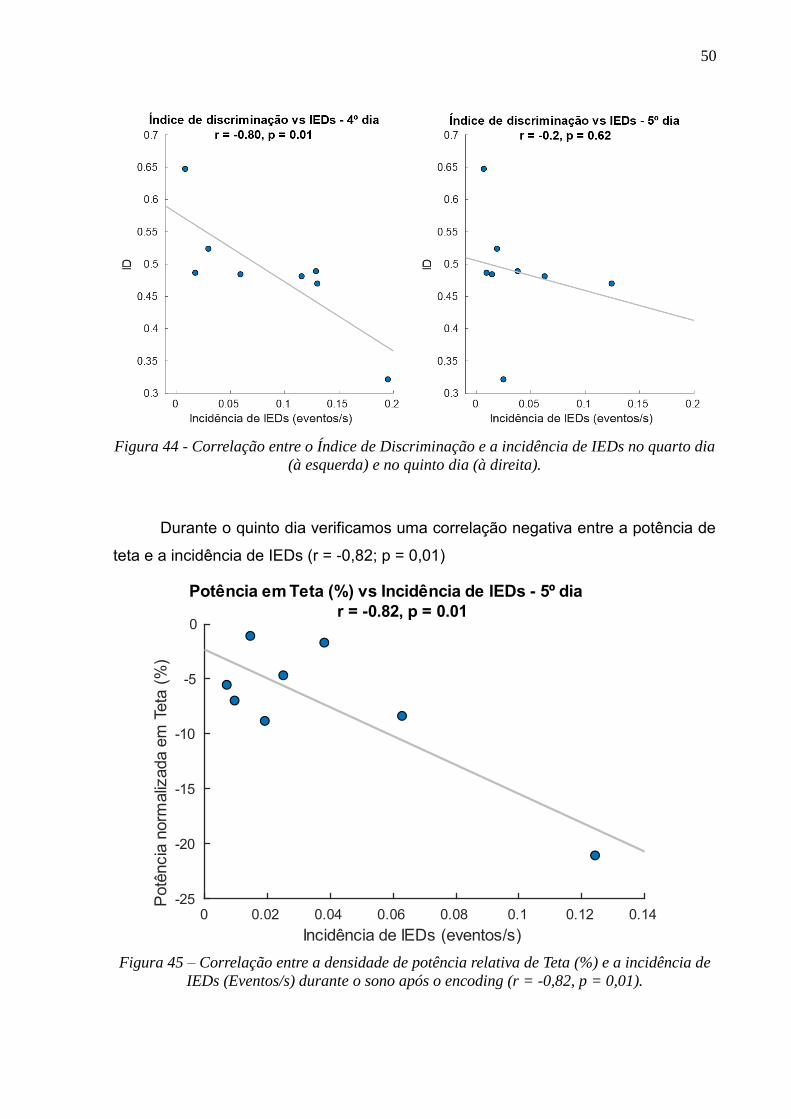
50
Figura 44 - Correlação entre o Índice de Discriminação e a incidência de IEDs no quarto dia
(à esquerda) e no quinto dia (à direita).
Durante o quinto dia verificamos uma correlação negativa entre a potência de
teta e a incidência de IEDs (r = -0,82; p = 0,01)
Figura 45 – Correlação entre a densidade de potência relativa de Teta (%) e a incidência de
IEDs (Eventos/s) durante o sono após o encoding (r = -0,82, p = 0,01).
0 0.02 0.04 0.06 0.08 0.1 0.12 0.14
Incidência de IEDs (eventos/s)
25
20
15
10
5
0
Potê
ncia
norm
aliz
ada e
m T
eta
(%
)
Potência em Teta (%) vs Incidência de IEDs 5 dia
r .82, p . 1

51
Observamos também uma correlação negativa entre os valores de MI médios
por animal a cada dia e a duração das pós-descargas médias para cada dia:
Figura 46 – Correlação negativa entre os valores médios de MI (%) e os valores médios de
AD(s) (r = -0,55; p = 0,03).
4.6 Histologia
As lesões eletrolíticas realizadas no mPFC geraram lesões menores quando
comparadas às lesões na BLA e CA1. Como implantamos um fio único no mPFC, a
referência para as estimulações foram o parafuso terra posicionado sobre o osso
occipital, gerando uma menor quantidade de corrente em torno do eletrodo. Em
CA1, no entanto, as lesões foram extensas, uma vez que foram aplicados correntes
sobre cada um dos três eletrodos.
0 20 40 60 80 100 120 140
Duração das AD (s)
0
100
200
300
400
Índic
e d
e C
om
odula
ção M
I (%
)
ndice de Comodulação (%) vs Duração das ADs (s)
r .55, p . 3

52
Figura 47 – Lesões identificadas a partir de cortes coronais do atlas estereotáxico (adaptado
de (PAXINOS; WATSON, 2006) nas regiões do mPFC, BLA e CA1. As lesões em CA1 foram
representadas por marcadores maiores, correspondendo à extensão da lesão.
5 DISCUSSÃO
Uma das mais complexas comorbidades presentes na ELT são os prejuízos
cognitivos verificados tanto em pacientes como em modelos animais. As alterações
morfofuncionais das estruturas límbicas envolvidas tanto na ELT como na
consolidação de memórias durante o sono pode descrever estes efeitos, porém os
mecanismos que levam a estas alterações não foram completamente elucidados
(HOLMES, 2015).
Neste trabalho buscamos caracterizar como oscilações típicas durante o sono
são alteradas na ELT e quais as possíveis implicações destas alterações nos
processos de consolidação de memória. Utilizando um modelo de abrasamento
rápido da BLA, verificamos que além do prejuízo no reconhecimento de objetos e
alterações eletrofisiológicas durante o sono NREM, como a substituição gradativa de
ripples por IEDs, os animais submetidos ao abrasamento elétrico apresentaram um
aumento na comodulação fase-amplitude entre oscilações teta e gama durante o
sono REM após as estimulações, exibindo também correlação negativa entre a

53
comodulação e a duração das pós-descargas induzidas pelos estímulos do
abrasamento durante o sono subsequente a aplicação dos protocolos. Estes
resultados podem levar a identificação de novos marcadores eletrofisiológicos que
possam esclarecer como o estabelecimento de circuitos epilépticos recrutam
mecanismos fisiológicos relacionados à consolidação de memória, como discutido
nos próximos itens.
5.1 Vantagens do modelo de abrasamento rápido da amígdala basolateral
O estudo dos processos epileptogênicos através de modelos de abrasamento
possibilita um controle preciso da progressão da epilepsia e também dos focos
epilépticos (GODDARD, 1967; RACINE, 1972b; KANDRATAVICIUS et al., 2014;
MORALES et al., 2014), se tornando um dos principais modelos utilizados neste tipo
de estudo. Essa popularidade também levou ao desenvolvimento de diversos
métodos de aplicação, incluindo modelos de abrasamento elétrico, químicos e até
por indução de traumas (BUTUZOVA; KITCHIGINA, 2008; MORALES et al., 2014;
ASTASHEVA; ASTASHEV; KITCHIGINA, 2015). Porém grande parte destes
protocolos são realizados diariamente durante períodos de 14 até 30 dias de
estimulações, tornando-se um processo despendioso.
A redução do período de estimulações é interessante para estudos
envolvendo registros eletrofisiológicos, principalmente de alta densidade, uma vez
que reações do tecido cerebral aos eletrodos implantados podem levar a perda da
estabilidade do registro ao longo do tempo (WINSLOW; TRESCO, 2010). Diversos
protocolos de abrasamento rápido propostos dependem de estimulações acima do
limiar de evocação de ADs e da necessidade de aplicação sobre as mesmas
estruturas a serem estudadas, o que pode levar a alterações como astrogliose e
perda neuronal não somente devido à epileptogênese, mas também ao próprio
protocolo de estimulação (MORALES et al., 2014). A aplicação do protocolo de
abrasamento rápido da BLA sugerido por (MORALES et al., 2014) possibilita a
estimulação de correntes abaixo do limiar para evocação de ADs, levando a
alterações comportamentais e também na morfologia e funcionalidade de células
hipocampais semelhante a um modelo de abrasamento lento. Apesar de demonstrar
as semelhanças entre estes protocolos, a caracterização realizada por (MORALES

54
et al., 2014) não avalia os prejuízos na consolidação de memória e as alterações
eletrofisiológicas envolvidas nesse déficit.
Além de reproduzimos os efeitos comportamentais da aplicação do protocolo
de abrasamento rápido sugerido por (MORALES et al., 2014), onde 5 animais
atingiram o estágio 5 de Racine (enquanto que 2 permaneceram no estágio 4 e
apenas 1 no estágio 3 - Figura 29), também caracterizamos os prejuízos cognitivos e
disfunções eletrofisiológicas resultantes deste modelo, evidenciando tanto sua
reprodutibilidade como também ampliando sua aplicabilidade, introduzindo as
possibilidades de sua utilização em estudos sobre as alterações cognitivos e
fisiológicas durante os processos de epileptogênese.
5.2 Alterações cognitivas induzidas pelo abrasamento elétrico
Ratos tendem naturalmente a explorar mudanças que observam ao seu redor,
desde novos objetos inseridos num ambiente familiar como também objetos já
conhecidos, porém em novas posições ou durante novos contextos ambientais
(MUMBY, 2002). A consolidação do aprendizado relativo a cada um desses modelos
de exploração pode ser atribuído a diferentes estruturas cerebrais, como a
dependência do Córtex Perirrinal durante a exploração de novos objetos e a
dependência do Hipocampo na exploração de objetos conhecidos, porém
deslocados ou em novos contextos ambientais (WARBURTON; BROWN, 2015).
O teste de reconhecimento de objetos utilizado neste experimento permite
verificar a eficácia na consolidação de informações dependentes do hipocampo, uma
vez que envolvem todo o contexto ambiental/espacial em torno dos objetos. Nesse
sentido, os animais submetidos aos protocolos de abrasamento elétrico
apresentaram prejuízos na consolidação dessa memória espacial, indicando assim
possíveis alterações nos mecanismos de consolidação de memória dependentes do
hipocampo. Prejuízos em testes semelhantes também foram observados em
modelos animais de ELT por administração de pilocarpina (JIANG et al., 2018),
enquanto que em modelos de abrasamento elétrico do hipocampo os animais
apresentaram um menor desempenho em labirintos circulares (CheeseBoard Maze,
GELINAS et al., 2016), sugerindo que os efeitos observados neste trabalho
corroboram com a literatura a respeito dos prejuízos cognitivos em modelos de ELT.

55
5.3 Efeitos do abrasamento sobre eventos eletrofisiológicos durante sono NREM
As alterações nas oscilações eletrofisiológicas durante o sono NREM
sugerem que a substituição de ripples hipocampais por oscilações patológicas, como
as IEDs, estejam relacionados a déficits na consolidação da memória espacial,
alterando principalmente a comunicação hipocampo-cortical (GELINAS et al., 2016).
Relações causais já foram estabelecidas entre a eficiência da consolidação
de memória e o acoplamento entre ripples com ondas delta corticais e spindles
tálamo-corticais. Em MAINGRET et al., 2016, o acoplamento entre essas oscilações
foi aumentado artificialmente através de estimulações elétricas sobre regiões
corticais, gerando ondas deltas e spindles sincronizados com a detecção de ripples
em CA1. Este acoplamento hipocampo-cortical artificial melhorou o desempenho dos
animais em tarefas de reconhecimento de objetos, enquanto que a estimulação
elétrica não-sincronizada com ripples não modificou o desempenho. Como
observamos neste estudo, as IEDs levam a um acoplamento aberrante entre o
hipocampo e o córtex pré-frontal, com intervalos entre IEDs e ondas delta até 0.27 ±
0.09 segundos menores quando comparados a ripples (Figura 37, Tabela 5). Essa
diminuição no intervalo de ocorrência de ondas delta após eventos hipocampais se
assemelha à estimulação elétrica realizada em (MAINGRET et al., 2016) (Figura 48),
porém com a ausência de informações relevantes contidas em ripples e
possivelmente com a introdução de informações irrelevantes ou distorcidas
provenientes das IEDs.

56
Figura 48 – Probabilidade de ocorrência de uma onda delta no mPFC após a detecção de
ripples em CA1. Em verde, a probabilidade de ocorrência durante o acoplamento artificial
por estimulação elétrica, e em roxo a probabilidade de ocorrência com estimulação elétrica
não pareada aos ripples. A diminuição no intervalo entre os eventos hipocampais e corticais
se assemelha ao observado durante IEDs e ondas delta – adaptado de (MAINGRET et al.,
2016).
De fato, a geração de IEDs ocorre de maneira semelhante à geração de
ripples em CA3, porém com a diminuição da atividade das basket cells
parvalbuminérgicas, levando a um aumento da atividade de neurônios excitatórios
(GULYÁS; FREUND, 2015). Essa diminuição na regulação da atividade neuronal
durante a geração dos ripples pode explicar os déficits cognitivos observados na
ELT, sendo as IEDs uma representação distorcida dos padrões de disparos
neuronais em consolidação durante o sono NREM.
As análises de probabilidade de eventos mostram os efeitos do abrasamento
principalmente sobre o acoplamento hipocampo-cortical através das ondas delta,
uma vez que a probabilidade de ocorrência de spindles após IEDs ou Ripples (grupo
KD e CTRL), e a probabilidade de spindles após ondas delta não possuem
diferenças significativas entre os grupos (Tabela 5). Este resultado sugere que o
acoplamento entre eventos hipocampais (IEDs ou Ripples) e as ondas delta possa
se iniciar através de estruturas hipocampais, indicado principalmente pelo curto
intervalo de 0,11 ± 0,02 segundos entre IEDs e ondas delta.
Neste trabalho também observamos que a incidência de IEDs durante o sono
subsequente ao último dia de abrasamento está correlacionada ao Índice de

57
Discriminação de objetos (r = -0,8; p = 0,01; Figura 44). Esta correlação, entretanto,
pode indicar o efeito das aplicações do abrasamento sobre a performance nas
tarefas comportamentais, e não necessariamente o efeito das IEDs durante a
consolidação da memória, uma vez que a incidência durante o sono subsequente ao
aprendizado no quinto dia não apresentou correlação com o desempenho dos
animais durante o teste.
5.4 Alteração de oscilações teta e gama durante o sono REM
A geração de oscilações teta no hipocampo emerge da interação entre
estruturas intrahipocampais e também extrahipocampais, envolvendo diferentes
circuitos osciladores teta ao longo do eixo septo-temporal no hipocampo como
também a interação do hipocampo com estruturas corticais, como o CE, e
subcorticais, como o septo medial (BUZSÁKI, 2002; PATEL et al., 2012; SHUMAN;
AMENDOLARA; GOLSHANI, 2017). Alterações nestes circuitos podem levar tanto à
diminuição das potências como também da sincronia em teta, e as correlações
destas alterações eletrofisiológicas com déficits em tarefas envolvendo memórias
dependentes do hipocampo sugerem que estas alterações podem levar à
perturbação da transferência e integração de informações através destes circuitos
(SHUMAN; AMENDOLARA; GOLSHANI, 2017).
Já a manipulação destes circuitos osciladores em teta pode levar a condições
semelhantes a modelos de ELT e até mesmo reverter os efeitos cognitivos
observados. A administração repetitiva de antagonistas de receptores GABAa ou
agonistas de receptores inespecíficos de glutamato no septo-medial, por exemplo,
são capazes de induzir atividades epileptiformes no hipocampo (BUTUZOVA;
KITCHIGINA, 2008; ASTASHEVA; ASTASHEV; KITCHIGINA, 2015), enquanto que a
inibição optogenética de neurônios GABAérgicos do septo-medial durante o sono
REM leva à diminuição da potência de teta, acompanhado por prejuízos na
consolidação da memória (BOYCE et al., 2016). Já a estimulação elétrica da mesma
região em frequências teta é capaz de promover uma melhora na performance de
animais tratados com pilocarpina em tarefas de memória dependentes do
hipocampo (LEE et al., 2017).
É interessante notar que a diminuição da potência em teta observada em
nossos experimentos possui uma correlação negativa com a incidência de IEDs

58
durante o sono pós-aprendizado (r = -0,82; p = 0,01; Figura 45). Uma das
interpretações da correlação observada envolveria os possíveis efeitos de
abrasamento elétrico sobre o septo-medial. Resultados semelhantes ao nosso
trabalho foram verificados em animais submetidos ao abrasamento elétrico da via
perfurante, induzindo o surgimento de IEDs e também uma tendência à diminuição
de teta hipocampal. Entretanto, durante os eventos de IEDs registrados em CA1
também foram detectados disparos neuronais no septo-medial sincronizados com os
eventos patológicos (KITCHIGINA et al., 2013). O septo-medial, por sua vez, pode
influenciar diretamente o processo de epileptogênese. A administração repetitiva de
antagonistas de receptores GABAa, como a infusão intraseptal de bicuculina, é
capaz de induzir atividades epileptiformes no hipocampo e comportamentos de
crises epilépticas (BUTUZOVA; KITCHIGINA, 2008), enquanto que o mesmo
procedimento utilizando o agonista de receptores glutamatérgicos L-glutamato leva
aos mesmos efeitos (ASTASHEVA; ASTASHEV; KITCHIGINA, 2015). Em ambos
experimentos também é observado a diminuição das oscilações teta registradas no
hipocampo durante períodos interictais. A indução destes fenômenos epilépticos
ocorre principalmente através das projeções septo-hipocampais, levando a
alterações no equiíbrio excitatório/inibitório das estruturas hipocampais (BUTUZOVA;
KITCHIGINA, 2008). Com base nestas evidências, podemos sugerir que a
correlação entre IEDs e a diminuição das potências em teta observadas em nosso
experimento pode estar relacionada a alterações nos circuitos septo-hipocampal-
entorrinal observadas nestes modelos, levando tanto ao surgimento de IEDs como
também a diminuição da potência em teta e prejuízos cognitivos.
Uma hipótese alternativa seria de que a diminuição das oscilações teta é
resultado da desincronização das atividades eletrofisiológicas entre diferentes
estruturas hipocampais, devido principalmente a alterações nas projeções
hipocampo-entorinais. Em animais tratados com injeções intraperiotoniais de ácido
caínico, o prejuízo na consolidação de memórias episódicas (relacionadas à
representação espaço-temporal) ocorre junto à diminuição da coerência de
oscilações teta entre as regiões de CA1 e do giro denteado (INOSTROZA et al.,
2013). Analisando a influência das projeções entre o hipocampo e o CE, LAURENT
et al., 2015 sugere que essa desincronização ocorre principalmente devido à
reorganização das vias temporoamônica e perfurante após a esclerose hipocampal
característica da ELT. Da mesma forma que alterações morfológicas podem induzir a

59
desincronização das oscilações teta entre as regiões hipocampais, as IEDs
poderiam surgir principalmente devido à diminuição da inibição em CA3 e à
modificação dos circuitos geradores de ripples (MA et al., 2006; GULYÁS; FREUND,
2015), associando a correlação entre IEDs e a potência de teta a possíveis
mecanismos comuns de modifições morfológicas das regiões hipocampais.
5.4.1 Alterações na comodulação de fase-amplitude de oscilações teta e gama
No presente estudo, não observamos diferenças em relação a densidade de
potência em gama durante os dias de experimento. Entretanto, o acoplamento entre
as oscilações teta e LG (indicados pelo MI) mostrou um aumento durante o sono
REM subsequente ao abrasamento (Figura 42). O aumento na comodulação de
fase-amplitude nestas frequências é comumente associada aos processos de
aprendizado e consolidação de memória (TORT et al., 2009; COLGIN, 2015), e o
fato de não apresentar correlação com a incidência de IEDs ou com as densidades
de potência em teta pode sugerir que esse acoplamento seja reflexo de outros
mecanismos afetados durante a epileptogênese, como a reativação de circuitos
relacionados aos padrões de disparos epileptiformes pelo hipocampo.
A correlação negativa entre o MI e a duração das ADs (Figura 46, r = -0,55; p
= 0,03) sugere que possa haver certa intersecção entre mecanismos de formação de
memória e de epileptogênese. A duração das ADs se torna maior à medida em que
as crises deixam de ser parciais (locais) e passam a influenciar outras regiões
(crises generalizadas) (MORALES et al., 2014). Durante este processo, as redes
hipocampais gradativamente se tornam hiperexcitáveis, podendo levar a crises
recorrentes, sendo considerado um aprendizado do circuito à redução da inibição
local (GODDARD, 1967; MUSTO; SAMII; HAYES, 2009). Neste sentido, a correlação
encontrada mostra que durante os períodos de menor duração das ADs ocorrem
eventos eletrofisiológicos semelhantes aos observados durante a consolidação de
informações durante o sono REM. Podemos sugerir que, neste período, os circuitos
modulados pela estímulação elétrica estariam sendo reativados durante o sono,
passando assim por um processo de plasticidade sináptica, indicando diretamente o
processo de epileptogênese na alteração de excitabilidade hipocampal. Já os
períodos com maiores durações de ADs podem estar relacionados às crises
generalizadas, indicando um circuito epiléptico já consolidado.

60
Resultados semelhantes foram observados em pacientes epilépticos durante
o sono NREM. Utilizando uma combinação de macro e microeletrodos para registro
da atividade eletrofisiológica tanto no foco epiléptico como em regiões temporais
adjacentes, BOWER et al., 2015 mostraram que o padrão de ativação de grupos
neuronais que disparam minutos antes da ocorrência de crises epilépticas são
repetidos durante o sono NREM. Além disso, esses grupos não necessariamente
estão localizados na região do foco epiléptico, demonstrando a ativação de uma
rede neuronal relacionada às descargas epilépticas distribuída espacialmente. Além
da ativação destes grupos neuronais, também foi observado que as alterações na
forma de onda das IEDs podem ocorrer tanto após episódios de crises epilépticas
como também após o sono NREM (BOWER et al., 2017). Essa alteração das
características de onda das IEDs estariam relacionadas à modificação e
incorporação de novos circuitos geradores de descargas interictais, tornando-se um
indicador dos processos de epileptogênese, e sua alteração durante o sono NREM
indica que essas mudanças podem ocorrer principalmente através de circuitos
relacionados à consolidação da memória (BOWER et al., 2017). Dessa forma,
podemos sugerir que o aumento da comodulação de fase-amplitude em teta-gama
observado em nossos experimentos também emerge como um marcador
eletrofisiológico dos processos de epileptogênese através dos mecanismos de
consolidação de memória empregados durante o sono REM, complementando as
evidências apresentadas por (BOWER et al., 2017).
6 LIMITAÇÕES E POSSIBILIDADES FUTURAS
Os resultados obtidos neste trabalho permitem uma abordagem observacional
sobre as alterações de oscilações em diferentes estágios de sono durante a
aplicação de um protocolo de abrasamento elétrico, não possibilitando o
estabelecimento de relações causais entre os eventos observados e correlatos
comportamentais, como prejuízos cognitivos ou indução de crises epilépticas.
Através da utilização deste modelo podemos apenas inferir que os efeitos
comportamentais e eletrofisiológicos estão relacionados ao processo de
epileptogênese, e que apesar de descritos em (MORALES et al., 2014) não

61
descrevemos alterações características de tecidos epilépticos, como a excitabilidade
neuronal ou astrogliose reativa em tecidos hipocampais, ou ainda crises
espontâneas (para uma revisão ver KANDRATAVICIUS et al., 2014).
As sugestões acerca da origem da correlação entre mecanismos envolvidos
na diminuição das potências de teta em CA1 e a incidência de IEDs são baseados
principalmente em evidências da literatura, analisando os efeitos de diferentes
modelos de ELT sobre circuitos oscilatórios hipocampais/extrahipocampais
responsáveis pela coordenação das oscilações teta. Nossos resultados são relativos
apenas às oscilações locais em CA1, enquanto que para determinar quais
mecanismos estão engajados tanto na diminuição de teta como na incidência de
IEDs seriam necessários registros dos vários componentes dos circuitos septo-
hipocampo-entorrinal, assim como uma análise histológica para verificar alterações
celulares em diferentes instâncias deste circuito, como a diminuição de projeções
GABAérgicas do septo-medial ao hipocampo (COLOM et al., 2006) e também
alterações no giro denteado (FRORIEP et al., 2012) e nas projeções entre o CE e
Hipocampo (INOSTROZA et al., 2013; LAURENT et al., 2015; JANZ et al., 2017).
Outra limitação nas considerações sobre a diminuição das potências de teta e
as alterações morfológicas envolvidas se dá ao fato de que grande parte das
caracterizações presentes na literatura são conduzidas durante tarefas
comportamentais ou comportamentos exploratórios (INOSTROZA et al., 2013;
LAURENT et al., 2015; LOPEZ-PIGOZZI et al., 2016), e não necessariamente
durante períodos de sono REM, o que pode refletir como uma caracterização dos
prejuízos durante o aprendizado, e não necessariamente na consolidação das
memórias durante o sono. O melhor desempenho de animais epilépticos em tarefas
dependentes do hipocampo através da estimulação elétrica do septo-medial em
frequências de teta durante as tarefas comportamentais (LEE et al., 2017)
confirmaria essa hipótese. Para verificar a influência dessa diminuição da potência
durante o sono REM na ELT podemos propor intervenções semelhantes a (LEE et
al., 2017), porém estimulando os circuitos osciladores em teta durante o sono REM
pós-aprendizado.
O aumento da comodulação de fase-amplitude entre oscilações teta-gama
durante o sono REM aparece como um dos principais achados de nosso trabalho.
Para verificarmos de fato quais tipos de processos essa comodulação pode estar
influenciando podemos propor experimentos induzindo perturbações nesse

62
acoplamento. Caso esse acoplamento esteja relacionado ao processo de
epileptogênese, podemos dificultar a propagação dos efeitos num modelo de
abrasamento elétrico, levando a um aumento do número de estímulos necessários
para levar ao abrasamento completo.
7 CONCLUSÕES
Neste trabalho caracterizamos os efeitos comportamentais e eletrofisiológicos
de um modelo de abrasamento rápido da BLA, reproduzindo os déficits cognitivos
em um teste de reconhecimento de objetos e alterações nas oscilações durante o
sono NREM observados em modelos tradicionais de abrasamento elétrico, como a
substituição de ripples por IEDs e alterações no acoplamento hipocampo-cortical
através da diminuição do intervalo entre ondas delta corticais geradas após IEDs.
Nossos resultados mostram que a incidência de IEDs durante a consolidação
de memória está correlacionada à diminuição da densidade de potência de
oscilações teta durante o sono REM. Uma hipótese plausível é que os possíveis
mecanismos que podem levar a estas alterações eletrofisiológicas em conjunto
envolvem tanto a alteração da atividade do septo-medial, capaz de induzir atividades
epilépticas e a redução de oscilações teta no hipocampo (COLOM et al., 2006;
BUTUZOVA; KITCHIGINA, 2008; GARCÍA-HERNÁNDEZ et al., 2010; KITCHIGINA
et al., 2013; ASTASHEVA; ASTASHEV; KITCHIGINA, 2015), como também a
alteração morfológica de circuitos osciladores de teta intrahipocampais, levando à
desincronização de teta e também à geração de IEDs (MA et al., 2006; INOSTROZA
et al., 2013; LAURENT et al., 2015). Apesar de sugerirmos estes possíveis
mecanismos nas alterações observadas durante o sono REM, nossos dados
refletem eventos registrados apenas em CA1, não possibilitando uma delimitação
refinada das circuitarias envolvidas nestes processos.
Além das alterações relacionadas aos processos de consolidação de
memória, o sono REM também fornece informações sobre o processo de
epileptogênese através do acoplamento de fase-amplitude entre oscilações teta e
gama. O aumento do MI durante o sono após a aplicação dos protocolos de
abrasamento sugere que a comodulação de fase-amplitude em teta-gama possa ser
um marcador eletrofisiológico dos processos de epileptogênese. A correlação dos

63
valores médios de MI e os valores médios de AD durante os dias de aplicação do
abrasamento pode indicar a dinâmica da epileptogênese sobre os mecanismos de
consolidação de memória empregados durante o sono REM, complementando as
evidências verificadas durante o sono NREM (BOWER et al., 2015, 2017).
Em conclusão, nossos achados ampliam a compreensão vigente sobre como
alterações de oscilações cerebrais durante o sono, especialmente da fase REM,
poderiam estar subjacentes a prejuízos de memória que ocorrem na ELT .
8 REFERÊNCIAS
ASTASHEVA, E.; ASTASHEV, M.; KITCHIGINA, V. Changes in the behavior and oscillatory
activity in cortical and subcortical brain structures induced by repeated l-glutamate injections to the
medial septal area in guinea pigs. Epilepsy Research, v. 109, p. 134–145, jan. 2015. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0920121114003015>.
BARKER-HALISKI, M.; WHITE, H. S. Glutamatergic Mechanisms Associated with Seizures
and Epilepsy. Cold Spring Harbor Perspectives in Medicine, v. 5, n. 8, p. a022863, ago. 2015.
Disponível em: <http://perspectivesinmedicine.cshlp.org/lookup/doi/10.1101/cshperspect.a022863>.
BOWER, M. R. et al. Evidence for Consolidation of Neuronal Assemblies after Seizures in
Humans. Journal of Neuroscience, v. 35, n. 3, p. 999–1010, 21 jan. 2015. Disponível em:
<http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.3019-14.2015>.
BOWER, M. R. et al. Reactivation of seizure-related changes to interictal spike shape and
synchrony during postseizure sleep in patients. Epilepsia, v. 58, n. 1, p. 94–104, jan. 2017. Disponível
em: <http://doi.wiley.com/10.1111/epi.13614>.
BOYCE, R. et al. Causal evidence for the role of REM sleep theta rhythm in contextual
memory consolidation. Science, v. 352, n. 6287, p. 812–816, 13 maio 2016. Disponível em:
<http://www.sciencemag.org/cgi/doi/10.1126/science.aad5252>.
BRAGIN, A.; PENTTONEN, M.; BUZSÁKI, G. Termination of Epileptic Afterdischarge in the
Hippocampus. The Journal of Neuroscience, v. 17, n. 7, p. 2567–2579, 1 abr. 1997. Disponível em:
<http://www.jneurosci.org/lookup/doi/10.1523/JNEUROSCI.17-07-02567.1997>.
BUTUZOVA, M. V.; KITCHIGINA, V. F. Repeated blockade of GABAA receptors in the medial
septal region induces epileptiform activity in the hippocampus. Neuroscience Letters, v. 434, n. 1, p.
133–138, mar. 2008. Disponível em: <http://linkinghub.elsevier.com/retrieve/pii/S0304394008001183>.
BUZSÁKI, G. Theta Oscillations in the Hippocampus. Neuron, v. 33, n. 3, p. 325–340, jan.
2002. Disponível em: <http://linkinghub.elsevier.com/retrieve/pii/S089662730200586X>.

64
BUZSÁKI, G. Hippocampal sharp wave-ripple: A cognitive biomarker for episodic memory and
planning. Hippocampus, v. 25, n. 10, p. 1073–1188, out. 2015. Disponível em:
<http://doi.wiley.com/10.1002/hipo.22488>.
COLGIN, L. L. Theta–gamma coupling in the entorhinal–hippocampal system. Current
Opinion in Neurobiology, v. 31, p. 45–50, abr. 2015. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0959438814001639>.
COLGIN, L. L. Rhythms of the hippocampal network. Nature Reviews Neuroscience, v. 17,
n. 4, p. 239–249, 10 abr. 2016. Disponível em: <http://www.nature.com/articles/nrn.2016.21>.
COLOM, L. V. et al. Septo-Hippocampal Networks in Chronically Epileptic Rats: Potential
Antiepileptic Effects of Theta Rhythm Generation. Journal of Neurophysiology, v. 95, n. 6, p. 3645–
3653, jun. 2006. Disponível em: <http://www.physiology.org/doi/10.1152/jn.00040.2006>.
DIEKELMANN, S.; BORN, J. The memory function of sleep. Nature Reviews Neuroscience,
v. 11, n. 2, p. 114–126, 4 fev. 2010. Disponível em: <http://www.nature.com/articles/nrn2762>.
EBERT, U.; RUNDFELDT, C.; LÖSCHER, W. Development and Pharmacological Suppression
of Secondary Afterdischarges in the Hippocampus of Amygdala-kindled Rats. European Journal of
Neuroscience, v. 7, n. 4, p. 732–741, abr. 1995. Disponível em: <http://doi.wiley.com/10.1111/j.1460-
9568.1995.tb00677.x>.
EFRON, B. Bootstrap Methods: Another Look at the Jackknife. In: KOTZ, S.; JOHNSON, N. L.
(Ed.). Breakthroughs in Statistics: Methodology and Distribution. New York, NY: Springer New
York, 1992. p. 569–593.
FOFFANI, G. et al. Reduced Spike-Timing Reliability Correlates with the Emergence of Fast
Ripples in the Rat Epileptic Hippocampus. Neuron, v. 55, n. 6, p. 930–941, set. 2007. Disponível em:
<http://linkinghub.elsevier.com/retrieve/pii/S0896627307005879>.
FRIARD, O.; GAMBA, M. BORIS: a free, versatile open-source event-logging software for
video/audio coding and live observations. Methods in Ecology and Evolution, v. 7, n. 11, p. 1325–
1330, 2016.
FRORIEP, U. P. et al. Altered theta coupling between medial entorhinal cortex and dentate
gyrus in temporal lobe epilepsy. Epilepsia, v. 53, n. 11, p. 1937–1947, nov. 2012. Disponível em:
<http://doi.wiley.com/10.1111/j.1528-1167.2012.03662.x>.
GARCÍA-HERNÁNDEZ, A. et al. Septo-hippocampal networks in chronic epilepsy.
Experimental Neurology, v. 222, n. 1, p. 86–92, mar. 2010. Disponível em:
<http://linkinghub.elsevier.com/retrieve/pii/S0014488609005032>.
GELINAS, J. N. et al. Interictal epileptiform discharges induce hippocampal-cortical coupling in
temporal lobe epilepsy. Nature Medicine, v. 22, n. 6, p. 641–648, 2016.
GODDARD, G. V. Development of epileptic seizures through brain stimulation at low intensity.
Nature, v. 214, n. 5092, p. 1020–1, 3 jun. 1967. Disponível em:

65
<http://www.ncbi.nlm.nih.gov/pubmed/6055396>.
GROSMARK, A. D. et al. REM Sleep Reorganizes Hippocampal Excitability. Neuron, v. 75, n.
6, p. 1001–1007, set. 2012. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0896627312007520>.
GULYÁS, A. I.; FREUND, T. T. Generation of physiological and pathological high frequency
oscillations: the role of perisomatic inhibition in sharp-wave ripple and interictal spike generation.
Current Opinion in Neurobiology, v. 31, p. 26–32, abr. 2015. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0959438814001573>.
HAMED, K. H.; RAMACHANDRA RAO, A. A modified Mann-Kendall trend test for
autocorrelated data. Journal of Hydrology, v. 204, n. 1–4, p. 182–196, jan. 1998. Disponível em:
<http://linkinghub.elsevier.com/retrieve/pii/S002216949700125X>.
HERMANN, B. P. et al. Growing up with epilepsy: A two-year investigation of cognitive
development in children with new onset epilepsy. Epilepsia, v. 49, n. 11, p. 1847–1858, nov. 2008.
Disponível em: <http://doi.wiley.com/10.1111/j.1528-1167.2008.01735.x>.
HOLMES, G. L. Cognitive impairment in epilepsy: the role of network abnormalities. Epileptic
disorders : international epilepsy journal with videotape, v. 17, n. 2, p. 101–16, jun. 2015.
Disponível em: <http://www.ncbi.nlm.nih.gov/pubmed/25905906>.
INOSTROZA, M. et al. Specific Impairment of “What-Where-When” Episodic-Like Memory in
Experimental Models of Temporal Lobe Epilepsy. Journal of Neuroscience, v. 33, n. 45, p. 17749–
17762, 6 nov. 2013. Disponível em: <http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.0957-
13.2013>.
JANZ, P. et al. Synaptic Remodeling of Entorhinal Input Contributes to an Aberrant
Hippocampal Network in Temporal Lobe Epilepsy. Cerebral Cortex, v. 27, n. 3, p. 2348–2364, 1 mar.
2017. Disponível em: <https://academic.oup.com/cercor/article/27/3/2348/3056317>.
JIANG, Y. et al. Abnormal hippocampal functional network and related memory impairment in
pilocarpine-treated rats. Epilepsia, v. 59, n. 9, p. 1785–1795, set. 2018. Disponível em:
<http://doi.wiley.com/10.1111/epi.14523>.
JIRUSKA, P. et al. Epileptic high-frequency network activity in a model of non-lesional
temporal lobe epilepsy. Brain, v. 133, n. 5, p. 1380–1390, maio 2010. Disponível em:
<https://academic.oup.com/brain/article-lookup/doi/10.1093/brain/awq070>.
KANDRATAVICIUS, L. et al. Psychiatric Comorbidities in Temporal Lobe Epilepsy: Possible
Relationships between Psychotic Disorders and Involvement of Limbic Circuits. Revista Brasileira de
Psiquiatria, v. 34, n. 4, p. 454–466, dez. 2012. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S1516444612000396>.
KANDRATAVICIUS, L. et al. Animal models of epilepsy: use and limitations. Neuropsychiatric
disease and treatment, v. 10, p. 1693–705, 2014. Disponível em:

66
<http://www.ncbi.nlm.nih.gov/pubmed/25228809>.
KITCHIGINA, V. et al. Disturbances of septohippocampal theta oscillations in the epileptic
brain: Reasons and consequences. Experimental Neurology, v. 247, p. 314–327, set. 2013.
Disponível em: <https://linkinghub.elsevier.com/retrieve/pii/S0014488613000423>.
KITCHIGINA, V. F.; BUTUZOVA, M. V. Theta activity of septal neurons during different
epileptic phases: The same frequency but different significance? Experimental Neurology, v. 216, n.
2, p. 449–458, abr. 2009. Disponível em:
<http://linkinghub.elsevier.com/retrieve/pii/S0014488609000065>.
LAURENT, F. et al. Proximodistal Structure of Theta Coordination in the Dorsal Hippocampus
of Epileptic Rats. Journal of Neuroscience, v. 35, n. 11, p. 4760–4775, 18 mar. 2015. Disponível em:
<http://www.jneurosci.org/cgi/doi/10.1523/JNEUROSCI.4297-14.2015>.
LEE, D. J. et al. Stimulation of the medial septum improves performance in spatial learning
following pilocarpine-induced status epilepticus. Epilepsy Research, v. 130, p. 53–63, fev. 2017.
Disponível em: <https://linkinghub.elsevier.com/retrieve/pii/S0920121117300116>.
LI, L. et al. Extrahippocampal high-frequency oscillations during epileptogenesis. Epilepsia, v.
59, n. 4, p. e51–e55, abr. 2018. Disponível em: <http://doi.wiley.com/10.1111/epi.14041>.
LISMAN, J.; BUZSAKI, G. A Neural Coding Scheme Formed by the Combined Function of
Gamma and Theta Oscillations. Schizophrenia Bulletin, v. 34, n. 5, p. 974–980, 21 jul. 2008.
Disponível em: <https://academic.oup.com/schizophreniabulletin/article-
lookup/doi/10.1093/schbul/sbn060>.
LISMAN, J. E.; IDIART, M. A. Storage of 7 +/- 2 short-term memories in oscillatory subcycles.
Science (New York, N.Y.), v. 267, n. 5203, p. 1512–5, 10 mar. 1995. Disponível em:
<http://www.ncbi.nlm.nih.gov/pubmed/7878473>.
LISMAN, J. E.; JENSEN, O. The Theta-Gamma Neural Code. Neuron, v. 77, n. 6, p. 1002–
1016, mar. 2013. Disponível em: <https://linkinghub.elsevier.com/retrieve/pii/S0896627313002316>.
LOPEZ-PIGOZZI, D. et al. Altered Oscillatory Dynamics of CA1 Parvalbumin Basket Cells
during Theta-Gamma Rhythmopathies of Temporal Lobe Epilepsy. eNeuro, v. 3, n. 6, 22 nov. 2016.
Disponível em: <http://eneuro.org/cgi/doi/10.1523/ENEURO.0284-16.2016>.
MA, D. L. et al. Reorganization of CA3 area of the mouse hippocampus after pilocarpine
induced temporal lobe epilepsy with special reference to the CA3-septum pathway. Journal of
Neuroscience Research, v. 83, n. 2, p. 318–331, 1 fev. 2006. Disponível em:
<http://doi.wiley.com/10.1002/jnr.20731>.
MAINGRET, N. et al. Hippocampo-cortical coupling mediates memory consolidation during
sleep. Nature Neuroscience, v. 19, n. 7, p. 959–964, 16 jul. 2016. Disponível em:
<http://www.nature.com/articles/nn.4304>.
MELBOURNE CHAMBERS, R. et al. Cognition, academic achievement, and epilepsy in

67
school-age children: A case–control study in a developing country. Epilepsy & Behavior, v. 33, p. 39–
44, abr. 2014. Disponível em: <https://linkinghub.elsevier.com/retrieve/pii/S1525505014000389>.
MIYAWAKI, H.; DIBA, K. Regulation of Hippocampal Firing by Network Oscillations during
Sleep. Current Biology, v. 26, n. 7, p. 893–902, abr. 2016. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0960982216300768>.
MORALES, J. C. et al. A new rapid kindling variant for induction of cortical epileptogenesis in
freely moving rats. Frontiers in Cellular Neuroscience, v. 8, 23 jul. 2014. Disponível em:
<http://journal.frontiersin.org/article/10.3389/fncel.2014.00200/abstract>.
MUMBY, D. G. Hippocampal Damage and Exploratory Preferences in Rats: Memory for
Objects, Places, and Contexts. Learning & Memory, v. 9, n. 2, p. 49–57, 1 mar. 2002. Disponível em:
<http://www.learnmem.org/cgi/doi/10.1101/lm.41302>.
MUSTO, A. E.; SAMII, M. S.; HAYES, J. F. Different phases of afterdischarge during rapid
kindling procedure in mice. Epilepsy Research, v. 85, n. 2–3, p. 199–205, ago. 2009. Disponível em:
<http://linkinghub.elsevier.com/retrieve/pii/S0920121109000722>.
NAKAZONO, T. et al. Gamma oscillations in the entorhinal-hippocampal circuit underlying
memory and dementia. Neuroscience Research, v. 129, p. 40–46, abr. 2018. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0168010217306570>.
PATEL, J. et al. Traveling Theta Waves along the Entire Septotemporal Axis of the
Hippocampus. Neuron, v. 75, n. 3, p. 410–417, ago. 2012. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0896627312006666>.
PAXINOS, G.; WATSON, C. The Rat Brain in Stereotaxic Coordinates Sixth Edition by.
Academic press, v. 170, p. 547612, 2006.
RACINE, R. J. Modification of seizure activity by electrical stimulation: I. after-discharge
threshold. Electroencephalography and Clinical Neurophysiology, v. 32, n. 3, p. 269–279, mar.
1972a. Disponível em: <http://linkinghub.elsevier.com/retrieve/pii/0013469472901769>.
RACINE, R. J. Modification of seizure activity by electrical stimulation: II. Motor seizure.
Electroencephalography and Clinical Neurophysiology, v. 32, n. 3, p. 281–294, mar. 1972b.
Disponível em: <http://linkinghub.elsevier.com/retrieve/pii/0013469472901770>.
ROUX, L. et al. Sharp wave ripples during learning stabilize the hippocampal spatial map.
Nature Neuroscience, v. 20, n. 6, p. 845–853, 10 abr. 2017. Disponível em:
<http://www.nature.com/doifinder/10.1038/nn.4543>.
SCHEFFER-TEIXEIRA, R.; TORT, A. B. L. Unveiling Fast Field Oscillations through
Comodulation. Eneuro, v. 4, n. 4, p. ENEURO.0079-17.2017, 2017. Disponível em:
<http://eneuro.sfn.org/lookup/doi/10.1523/ENEURO.0079-17.2017>.
SHUMAN, T.; AMENDOLARA, B.; GOLSHANI, P. Theta Rhythmopathy as a Cause of
Cognitive Disability in TLE. Epilepsy Currents, v. 17, n. 2, p. 107–111, mar. 2017. Disponível em:

68
<http://epilepsycurrents.org/doi/10.5698/1535-7511.17.2.107>.
SIEGLE, J. H. et al. Open Ephys: an open-source, plugin-based platform for multichannel
electrophysiology. Journal of Neural Engineering, v. 14, n. 4, p. 045003, 1 ago. 2017. Disponível
em: <http://stacks.iop.org/1741-
2552/14/i=4/a=045003?key=crossref.72f4c42751bd1cca996f430a5a7a3758>.
SMITH, P. D. et al. Seizures, not hippocampal neuronal death, provoke neurogenesis in a
mouse rapid electrical amygdala kindling model of seizures. Neuroscience, v. 136, n. 2, p. 405–415,
jan. 2005. Disponível em: <http://linkinghub.elsevier.com/retrieve/pii/S0306452205008067>.
STALEY, K. J. Neurons Skip a Beat during Fast Ripples. Neuron, v. 55, n. 6, p. 828–830, set.
2007. Disponível em: <http://linkinghub.elsevier.com/retrieve/pii/S0896627307006733>.
SUURMEIJER, T. P. B. M.; REUVEKAMP, M. F.; ALDENKAMP, B. P. Social Functioning,
Psychological Functioning, and Quality of Life in Epilepsy. Epilepsia, v. 42, n. 9, p. 1160–1168, 12 jan.
2002. Disponível em: <http://doi.wiley.com/10.1046/j.1528-1157.2001.37000.x>.
TAXIDIS, J. et al. Local Field Potentials Encode Place Cell Ensemble Activation during
Hippocampal Sharp Wave Ripples. Neuron, v. 87, n. 3, p. 590–604, ago. 2015. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0896627315006364>.
TORT, A. B. L. et al. Theta-gamma coupling increases during the learning of item-context
associations. Proceedings of the National Academy of Sciences, v. 106, n. 49, p. 20942–20947, 8
dez. 2009. Disponível em: <http://www.pnas.org/cgi/doi/10.1073/pnas.0911331106>.
TORT, A. B. L. et al. Measuring Phase-Amplitude Coupling Between Neuronal Oscillations of
Different Frequencies. Journal of Neurophysiology, v. 104, n. 2, p. 1195–1210, ago. 2010.
Disponível em: <http://www.physiology.org/doi/10.1152/jn.00106.2010>.
WARBURTON, E. C.; BROWN, M. W. Neural circuitry for rat recognition memory.
Behavioural Brain Research, v. 285, p. 131–139, maio 2015. Disponível em:
<https://linkinghub.elsevier.com/retrieve/pii/S0166432814006512>.
WELCH, P. The use of fast Fourier transform for the estimation of power spectra: A method
based on time averaging over short, modified periodograms. IEEE Transactions on Audio and
Electroacoustics, v. 15, n. 2, p. 70–73, jun. 1967. Disponível em:
<http://ieeexplore.ieee.org/document/1161901/>.
WHELAN, C. D. et al. Structural brain abnormalities in the common epilepsies assessed in a
worldwide ENIGMA study. Brain, v. 141, n. 2, p. 391–408, 1 fev. 2018. Disponível em:
<https://academic.oup.com/brain/article/141/2/391/4818311>.
WHO. Epilepsy - World Health Organization. Disponível em: <http://www.who.int/news-
room/fact-sheets/detail/epilepsy>.
WINSLOW, B. D.; TRESCO, P. A. Quantitative analysis of the tissue response to chronically
implanted microwire electrodes in rat cortex. Biomaterials, v. 31, n. 7, p. 1558–1567, mar. 2010.

69
Disponível em: <http://linkinghub.elsevier.com/retrieve/pii/S0142961209012770>.