efeitos da tricostatina a sobre a acetilação de histonas ... 958.pdf · efeitos da tricostatina a...
TRANSCRIPT

1
C.S.C.S.C.S.C.S.C.S. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, L.Z. L.Z. L.Z. L.Z. L.Z. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, N.Z. N.Z. N.Z. N.Z. N.Z. S S S S Sarararararaivaivaivaivaiva,a,a,a,a, F F F F F.M..M..M..M..M. M M M M Monononononttttteireireireireiro & J.M.o & J.M.o & J.M.o & J.M.o & J.M. G G G G Garararararcia.cia.cia.cia.cia. 2011. 2011. 2011. 2011. 2011. Efeitos da tricostatina A sobre a acetilação dehistonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
Acta Scientiae Veter inariae, 2011. 39(2) : 958.Acta Scientiae Veter inariae, 2011. 39(2) : 958.Acta Scientiae Veter inariae, 2011. 39(2) : 958.Acta Scientiae Veter inariae, 2011. 39(2) : 958.Acta Scientiae Veter inariae, 2011. 39(2) : 958.
ORIGINAL ARTICLEPub. 958Pub. 958Pub. 958Pub. 958Pub. 958
ISSN 1679-9216 (Online)
Received: September 2010 www.ufrgs.br/actavet Accepted: January 2011
*Trabalho realizado com apoio financeiro da FAPESP. Laboratório de Reprodução Animal, Departamento de Medicina Veterinária Preventivae Reprodução Animal, FCAV, UNESP, Jaboticabal, Brasil. CORRESPONDÊNCIA: C.S. Oliveira [[email protected] - FAX: + 55 (16)3209-2633]. Departamento de Medicina Veterinária Preventiva e Reprodução Animal, FCAV, UNESP Jaboticabal, Brasil. Via de Acesso Prof.Paulo Donato Castellane s/n., CEP 14884-900 Jaboticabal, SP, Brasil.
Efeitos da tricostatina A sobre a acetilação de histonas, proliferação celular eEfeitos da tricostatina A sobre a acetilação de histonas, proliferação celular eEfeitos da tricostatina A sobre a acetilação de histonas, proliferação celular eEfeitos da tricostatina A sobre a acetilação de histonas, proliferação celular eEfeitos da tricostatina A sobre a acetilação de histonas, proliferação celular ediferenciação de células tronco embrionárias murinas*diferenciação de células tronco embrionárias murinas*diferenciação de células tronco embrionárias murinas*diferenciação de células tronco embrionárias murinas*diferenciação de células tronco embrionárias murinas*
Effects of Trichostatin A on Histone Acetylation, Cell Proliferation and Differentiation of MurineEmbryonic Stem Cells
CCCCClarlarlarlarlara Sa Sa Sa Sa Slade Olade Olade Olade Olade Olllll iviviviviveireireireireira ,a ,a ,a ,a , L L L L Letícia Zetícia Zetícia Zetícia Zetícia Zoooooccccccccccolarolarolarolarolaro Oo Oo Oo Oo Olivl ivl ivl ivl iveireireireireira,a ,a ,a ,a , N N N N Naiaraiaraiaraiaraiara Za Za Za Za Zoooooccccccccccal Sal Sal Sal Sal Sarararararaivaivaivaivaiva ,a ,a ,a ,a , F F F F Fabio Mabio Mabio Mabio Mabio Morororororaaaaattttto Mo Mo Mo Mo Monononononttttteireireireireiro &o &o &o &o &Joaquim Mansano GarciaJoaquim Mansano GarciaJoaquim Mansano GarciaJoaquim Mansano GarciaJoaquim Mansano Garcia
ABSTRACTBackground: Embryonic stem cells are cells derived from early-stage embryos that are characterized by pluripotency and self-renewal capacity. The in vitro cultured murine embryonic stem cells can indefinitely propagate in an undifferentiated state inthe presence of leukemia inhibitory factor (LIF). However, when stimulated, these cells can differentiate into cell lines derivedfrom all three embryonic germ layers. The trichostatin A (TSA) is an epigenetic modifier agent and several studies have usedthe TSA to stimulate cellular differentiation. However, most of these studies only assessed one TSA concentration. Therefore,this study aimed to evaluate the effects of different TSA concentrations on histone hyperacetylation during in vitro celldifferentiation of murine pluripotent embryonic stem cells, cultured with or without LIF, in the quest of to standardize theirapplication on early cultures of embryonic stem cells.Materials, Methods & Results: Undifferentiated murine embryonic stem cells were plated in the presence of different TSAconcentrations (0 nM, 15 nm, 50 nM and 100 nM) in the presence or absence of LIF. Thus, the treatments were evaluated inundifferentiated embryonic stem cells cultured in the presence of LIF (Control group: 0 nM LIF+; Group 15 nM LIF+; Group50 nM LIF+ and Group 100 nM LIF+), and in embryonic stem cells cultured in the absence of LIF (Control group: 0 nM LIF-
; Group 15 nM LIF-; Group 50 nM LIF- and Group 100 nM LIF-). Treatment with TSA was performed for 24 h. After that themedium was replaced with fresh medium without TSA. Samples were collected at 0, 12, 24, 36 and 48 h after the beginning ofthe experiment. Three replicates were performed in each experimental group. The relative amount of Histone H3 lysine 9acetylation was analyzed in all groups, as well as the cell proliferation in the embryonic stem cells cultured in the presence ofLIF. In the control group (0 nM), the absence of LIF resulted in higher levels (P <0.05) of H3lys9ac compared to thecultures supplemented with LIF. In the embryonic stem cells cultured in the presence of LIF, the 50 nM and 100 nM treatmentsresulted in higher levels (P <0.05) of H3lys9ac when compared with 0 nM and 15 nM treatments. Evaluating the Hoechstarea in the 0 nM group, it was observed that the number of cells increased (P <0.05) according to the time of culture. Treatmentwith 15 nM also reflected a similar distribution, but the Hoechst area in 15 nM group was lower (P <0.05) at 24 and 48h whencompared to the observed in the control group. In the 100 nM treatment, was observed that the area of Hoechst was lower (P<0.05) to that obtained in the control group at 12, 24 and 48h. In addition, it was observed that treatment with TSA inducesgreater cellular differentiation when compared to control groups in stem cells cultured in the presence of LIF as well as in theabsence of LIF.Discussion: In the present study it was observed that TSA treatment increased the levels of histone acetylation in murineembryonic stem cells at a 50 nM concentration, making it possible to reduce the concentration recommended in the literature(100 nM). In addtion, it was concluded that the lower TSA concentrations utilized (15 nm and 50 nM) was less harmful tocellular proliferation than the 100 nM TSA concentration.
Keywords: embryonic stem cells, histone acetylation, murine, trichostatin A.
Descritores: células tronco embrionárias, acetilação de histonas, murinos, tricostatina A.

2
C.S .C .S .C .S .C .S .C .S . O O O O Ol ivl ivl ivl ivl ive i re i re i re i re i ra ,a ,a ,a ,a , L .Z . L .Z . L .Z . L .Z . L .Z . O O O O Ol ivl ivl ivl ivl ive i re i re i re i re i ra ,a ,a ,a ,a , N .Z . N .Z . N .Z . N .Z . N .Z . S S S S Sa ra ra ra ra ra i va i va i va i va i va ,a ,a ,a ,a , F F F F F. M .. M .. M .. M .. M . M M M M Mo no no no no nttttt e i re i re i re i re i ro & J .M.o & J .M.o & J .M.o & J .M.o & J .M. G G G G Ga ra ra ra ra rc ia .c ia .c ia .c ia .c ia . 2011 2011 2011 2011 2011..... Efeitos da tricostatina A sobre aacetilação de histonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
INTRODUÇÃO
Células tronco embrionárias (CTE) são célu-las oriundas de embriões em estádios iniciais de de-senvolvimento, caracterizadas pela pluripotência ecapacidade de auto-renovação [18].
Quando cultivadas in vitro, as CTE murinaspodem propagar-se indefinidamente no estadoindiferenciado, na presença do fator inibidor deleucemia (LIF) e/ou de uma monocamada defibroblastos embrionários murinos [17,19]. Quandoestimuladas, essas células são capazes de diferenci-ar-se em linhagens celulares oriundas das três cama-das germinativas [4].
O processo de reprogramação celular queocorre durante a diferenciação de CTE é controladoem grande parte por eventos epigenéticos, incluindoa acetilação de histonas [16]. A tricostatina A (TSA)é uma droga modificadora epigenética que promovehiperacetilação das histonas por inibir a enzimahistona desacetilase [21]. Nas CTE, o uso da TSAinduz modificações rápidas de ordem morfológicano perfil de expressão gênica, semelhantes às ocorri-das na diferenciação inicial das CTE induzida pelaretirada do LIF [11].
Existem na literatura diversos trabalhos queutilizam a TSA para estimular a diferenciação celular[1,5-7,10,11]. Entretanto, a maioria destes avaliou osefeitos de apenas uma concentração de TSA[1,5,7,11]. Salienta-se ainda que os efeitos da TSApodem variar conforme a concentração da droga nomeio de cultivo [14].
Assim sendo, o presente estudo teve comoobjetivo avaliar os efeitos de diferentes concentra-ções da droga modificadora epigenética TSA nahiperacetilação das histonas durante a diferenciaçãocelular in vitro de CTE murinas pluripotentes, culti-vadas com ou sem LIF, buscando padronizar suaaplicação em cultivos iniciais de CTE.
MATERIAS E MÉTODOS
Delineamento experimentalNo presente experimento, CTE indife-
renciadas foram plaqueadas na presença de diferen-tes concentrações de TSA (0 nM, 15 nM, 50 nM e100 nM), na presença ou ausência de LIF. Desta for-ma, os tratamentos foram avaliados em CTEindiferenciadas, na presença de LIF [grupos experi-mentais: 0 nM LIF+ (Controle CTE indiferenciadas);15 nM LIF+; 50 nM LIF+; 100 nM LIF+] e em CTEno início da diferenciação celular, na ausência de LIF[grupos experimentais: 0 nM LIF- (Controle CTE emdiferenciação); 15 nM LIF-; 50 nM LIF-; 100 nMLIF-]. O tratamento com TSA foi realizado por 24 h,e após o meio foi substituído por meio fresco sem adroga. Os grupos experimentais se encontra detalha-dos na Tabela 1.
Amostras foram coletadas 0, 12, 24, 36 e 48h após o início do experimento. Três replicatas fo-ram realizadas em cada grupo experimental. Foramanalisadas a quantificação relativa de histona H3acetilada na lisina 9 (H3lys9ac) em todos os grupos,e a proliferação celular nos cultivos na presença deLIF.
GRUPOS LIF Meio de cultivo 0-24h Meio de cultivo 24-48h
0nM LIF+(Controle indiferenciado) Presente DMEN CTE DMEM CTE
0nM LIF-(Controle em diferenciação) Ausente DMEM CTE DMEM CTE
15nM TSA LIF+ Presente DMEM TSA 15nM DMEM CTE
15nM TSA LIF- Ausente DMEM TSA 15nM DMEM CTE
50nM TSA LIF+ Presente DMEM TSA 50nM DMEM CTE
50nM TSA LIF- Ausente DMEM TSA 50nM DMEM CTE
100nM TSA LIF+ Presente DMEM TSA 100nM DMEM CTE
100nM TSA LIF- Ausente DMEM TSA 100nM DMEM CTETSA: Tricostatina A; LIF: fator inibidor de leucemia.
Tabela 1. Descrição dos sistemas de cultivo de células tronco embrionárias (CTE) utilizados para avali-ação dos efeitos da TSA sobre a acetilação de histonas, proliferação e diferenciação celular.

3
C.S.C.S.C.S.C.S.C.S. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, L.Z. L.Z. L.Z. L.Z. L.Z. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, N.Z. N.Z. N.Z. N.Z. N.Z. S S S S Sarararararaivaivaivaivaiva,a,a,a,a, F F F F F.M..M..M..M..M. M M M M Monononononttttteireireireireiro & J.M.o & J.M.o & J.M.o & J.M.o & J.M. G G G G Garararararcia.cia.cia.cia.cia. 2011. 2011. 2011. 2011. 2011. Efeitos da tricostatina A sobre a acetilação dehistonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
Cultivo das CTE pluripotentesOs reagentes e meios de cultura não especifi-
cados foram adquiridos da Sigma Chemical1.A linhagem de CTE H106, utilizada no pre-
sente estudo, foi obtida e caracterizada pelo Prof.Joaquim Mansano Garcia (UNESP, Jaboticabal, SãoPaulo). As células foram mantidas em monocamadasde fibroblastos primários murinos previamenteinativados com motomicina-C, em placas gela-tinizadas de poliestireno2, em meio DMEM3
suplementado com 15% de Knockout SerumReplacement4, 1% aminoácidos não essenciais, 10mM â-mercaptoetanol, 1mM piruvato sódico, 2 mML-glutamina e 83.4 ug/mL amicacina5. De acordocom o grupo experimental, o fator inibidor deleucemia (LIF) murino recombinante (1000 IU/mL)6
foi adicionado. As placas foram incubadas a 37oCem atmosfera de 5% CO2, e as células foram repicadasa cada 48 h.
Ao início dos experimentos, os cultivos fo-ram repicados para placas de 96 poços gelatinizadasna ausência de fibroblastos primários murinos.
Tratamento dos cultivos com tricostatina A (TSA)As CTE foram plaqueadas no d 0, e o meio
de cultivo foi empregado de acordo com os gruposexperimentais. A TSA foi diluída previamente emetanol e a concentração no meio de cultivo ajustadapara 15 nM, 50 nM e 100 nM de TSA, de forma quea concentração final de etanol não ultrapassasse0.01%. O meio foi substituído 24 h após o início dotratamento, por meio fresco sem TSA.
Avaliação dos níveis de acetilação de histonas eproliferação celular
Em todos os grupos, as amostras foram fixa-das 0, 12, 24, 36 e 48 h após a suplementação deTSA. Para tanto, foi adicionada solução deparaformaldeído a 4% em PBS por 20 min a 37º C, eas placas foram armazenadas a 4ºC por até uma se-mana.
No momento da reação, as amostras foramlavadas novamente com PBS-L, e incubadas em câ-mara úmida por 5 min, em solução de 0,5% TritonX-100. Posteriormente, a solução de triton X-100 foiretirada e os cultivos foram incubados em soluçãode 5% BSA por uma hora. Após retirar a solução deBSA, o anticorpo primário monoclonal mouse anti-
acLys9H3 (1:200) foi adicionado, o qual permane-ceu por uma noite reagindo com as células. Os culti-vos foram lavados três vezes com PBS-L, e oanticorpo secundário anti-camundongo conjugado aFITC (1:200)7 foi adicionado. As placas foram incu-badas protegidas da luz, a temperatura ambiente, por2 h. Posteriormente, o anticorpo secundário foi reti-rado e o corante HOECHST 33342 foi adicionadopor 10 min, para a coloração do núcleo das células.Os cultivos foram lavados três vezes com PBS-L.
Os cultivos foram avaliados sob fluorescênciaem microscópio óptico invertido Olympus IX-708,sob os filtros WV (340-380nm) para HOECHST33342, e WIB para FITC (478-495nm). Em cada pla-ca, foram reservados poços para a realização do con-trole negativo, nos quais apenas o anticorpo secun-dário era adicionado. Utilizou-se o sistema de capta-ção de imagens Kontron Elektronik Imaging Systeme o Software KS-300 3.09.
Após a captura das imagens, estas foram pro-cessadas no software ImageJ1.41º (NIH, USA). Asimagens foram transformadas em 8bit e avaliadasseguindo os mesmos parâmetros de threshold. As fra-ções de área positivas para H3lys9ac (FITC) foramdivididas pelas frações de área do HOECHST, ob-tendo-se a razão FITC/HOECHST. A intensidade defluorescência obtida para H3lys9ac (FITC) foi divi-dida pelo número de células do campo, obtendo-se aintensidade de FITC/ N total de células.
A proliferação celular foi estimada pela ana-lise da área preenchida pelos núcleos das células (co-rados com HOECHST) em cada imagem.
Análise EstatísticaOs dados obtidos foram analisados utilizan-
do o programa Statistical Analysis System (SASInstitute, Cary, NC, USA). Os resultados referentes àrazão da intensidade do FITC sobre o número decélulas nos quatro tratamentos (0 nM, 15 nM, 50 nMe 100 nM), com ou sem LIF, foram analisados peloteste de Mann Whitney e Wilcoxon Rank. Os resul-tados referentes à área total de HOECHST foram sub-metidos à análise de variância (ANOVA) One-way eao teste de comparação de médias de Tukey, consi-derando mesma concentração de TSA, e em outraanálise considerando mesmo tempo de exposição àdroga TSA. Para todas as análises, foi estabelecidonível de significância de 5%.
4
C.S .C .S .C .S .C .S .C .S . O O O O Ol ivl ivl ivl ivl ive i re i re i re i re i ra ,a ,a ,a ,a , L .Z . L .Z . L .Z . L .Z . L .Z . O O O O Ol ivl ivl ivl ivl ive i re i re i re i re i ra ,a ,a ,a ,a , N .Z . N .Z . N .Z . N .Z . N .Z . S S S S Sa ra ra ra ra ra i va i va i va i va i va ,a ,a ,a ,a , F F F F F. M .. M .. M .. M .. M . M M M M Mo no no no no nttttt e i re i re i re i re i ro & J .M.o & J .M.o & J .M.o & J .M.o & J .M. G G G G Ga ra ra ra ra rc ia .c ia .c ia .c ia .c ia . 2011 2011 2011 2011 2011..... Efeitos da tricostatina A sobre aacetilação de histonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
RESULTADOS
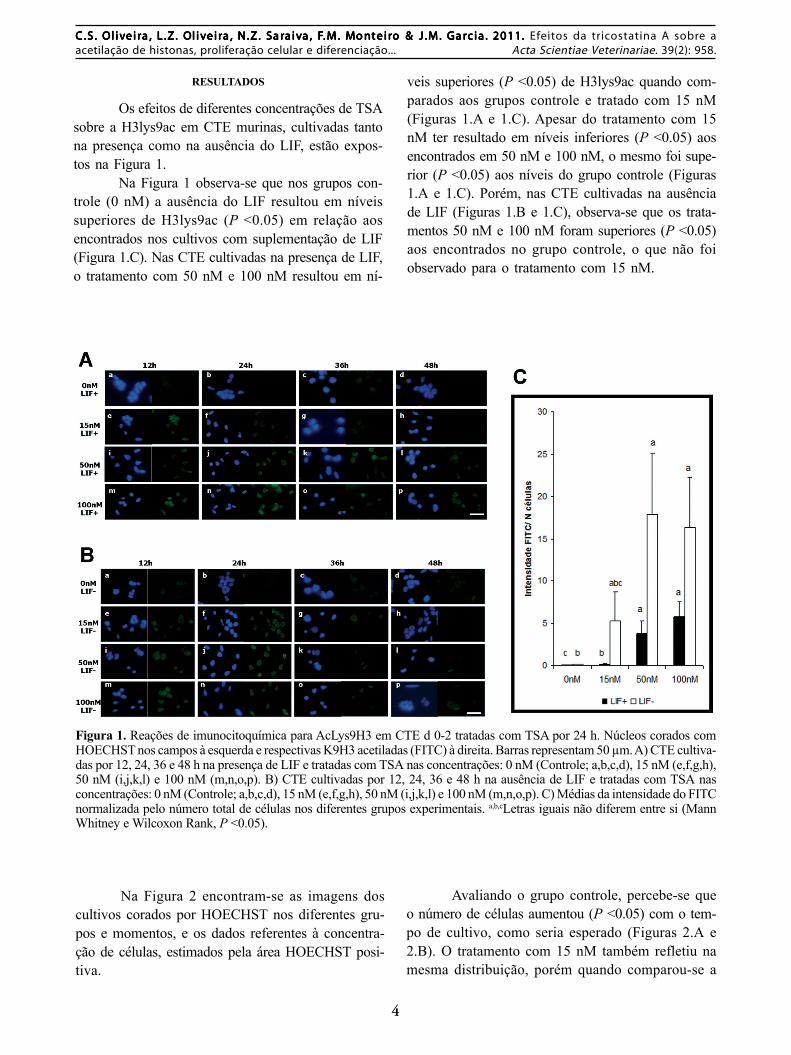
Os efeitos de diferentes concentrações de TSAsobre a H3lys9ac em CTE murinas, cultivadas tantona presença como na ausência do LIF, estão expos-tos na Figura 1.
Na Figura 1 observa-se que nos grupos con-trole (0 nM) a ausência do LIF resultou em níveissuperiores de H3lys9ac (P <0.05) em relação aosencontrados nos cultivos com suplementação de LIF(Figura 1.C). Nas CTE cultivadas na presença de LIF,o tratamento com 50 nM e 100 nM resultou em ní-
veis superiores (P <0.05) de H3lys9ac quando com-parados aos grupos controle e tratado com 15 nM(Figuras 1.A e 1.C). Apesar do tratamento com 15nM ter resultado em níveis inferiores (P <0.05) aosencontrados em 50 nM e 100 nM, o mesmo foi supe-rior (P <0.05) aos níveis do grupo controle (Figuras1.A e 1.C). Porém, nas CTE cultivadas na ausênciade LIF (Figuras 1.B e 1.C), observa-se que os trata-mentos 50 nM e 100 nM foram superiores (P <0.05)aos encontrados no grupo controle, o que não foiobservado para o tratamento com 15 nM.
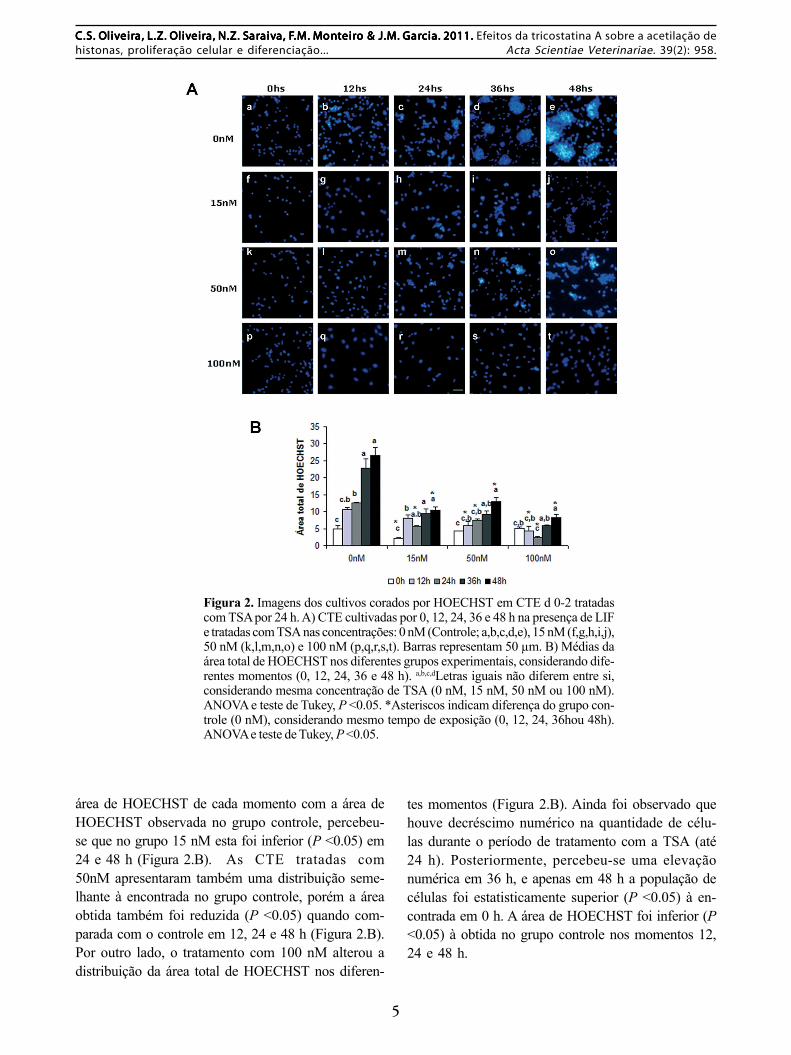
Na Figura 2 encontram-se as imagens doscultivos corados por HOECHST nos diferentes gru-pos e momentos, e os dados referentes à concentra-ção de células, estimados pela área HOECHST posi-tiva.
Figura 1. Reações de imunocitoquímica para AcLys9H3 em CTE d 0-2 tratadas com TSA por 24 h. Núcleos corados comHOECHST nos campos à esquerda e respectivas K9H3 acetiladas (FITC) à direita. Barras representam 50 µm. A) CTE cultiva-das por 12, 24, 36 e 48 h na presença de LIF e tratadas com TSA nas concentrações: 0 nM (Controle; a,b,c,d), 15 nM (e,f,g,h),50 nM (i,j,k,l) e 100 nM (m,n,o,p). B) CTE cultivadas por 12, 24, 36 e 48 h na ausência de LIF e tratadas com TSA nasconcentrações: 0 nM (Controle; a,b,c,d), 15 nM (e,f,g,h), 50 nM (i,j,k,l) e 100 nM (m,n,o,p). C) Médias da intensidade do FITCnormalizada pelo número total de células nos diferentes grupos experimentais. a,b,cLetras iguais não diferem entre si (MannWhitney e Wilcoxon Rank, P <0.05).
Avaliando o grupo controle, percebe-se queo número de células aumentou (P <0.05) com o tem-po de cultivo, como seria esperado (Figuras 2.A e2.B). O tratamento com 15 nM também refletiu namesma distribuição, porém quando comparou-se a
5
C.S.C.S.C.S.C.S.C.S. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, L.Z. L.Z. L.Z. L.Z. L.Z. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, N.Z. N.Z. N.Z. N.Z. N.Z. S S S S Sarararararaivaivaivaivaiva,a,a,a,a, F F F F F.M..M..M..M..M. M M M M Monononononttttteireireireireiro & J.M.o & J.M.o & J.M.o & J.M.o & J.M. G G G G Garararararcia.cia.cia.cia.cia. 2011. 2011. 2011. 2011. 2011. Efeitos da tricostatina A sobre a acetilação dehistonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
área de HOECHST de cada momento com a área deHOECHST observada no grupo controle, percebeu-se que no grupo 15 nM esta foi inferior (P <0.05) em24 e 48 h (Figura 2.B). As CTE tratadas com50nM apresentaram também uma distribuição seme-lhante à encontrada no grupo controle, porém a áreaobtida também foi reduzida (P <0.05) quando com-parada com o controle em 12, 24 e 48 h (Figura 2.B).Por outro lado, o tratamento com 100 nM alterou adistribuição da área total de HOECHST nos diferen-
Figura 2. Imagens dos cultivos corados por HOECHST em CTE d 0-2 tratadascom TSA por 24 h. A) CTE cultivadas por 0, 12, 24, 36 e 48 h na presença de LIFe tratadas com TSA nas concentrações: 0 nM (Controle; a,b,c,d,e), 15 nM (f,g,h,i,j),50 nM (k,l,m,n,o) e 100 nM (p,q,r,s,t). Barras representam 50 µm. B) Médias daárea total de HOECHST nos diferentes grupos experimentais, considerando dife-rentes momentos (0, 12, 24, 36 e 48 h). a,b,c,dLetras iguais não diferem entre si,considerando mesma concentração de TSA (0 nM, 15 nM, 50 nM ou 100 nM).ANOVA e teste de Tukey, P <0.05. *Asteriscos indicam diferença do grupo con-trole (0 nM), considerando mesmo tempo de exposição (0, 12, 24, 36hou 48h).ANOVA e teste de Tukey, P <0.05.
tes momentos (Figura 2.B). Ainda foi observado quehouve decréscimo numérico na quantidade de célu-las durante o período de tratamento com a TSA (até24 h). Posteriormente, percebeu-se uma elevaçãonumérica em 36 h, e apenas em 48 h a população decélulas foi estatisticamente superior (P <0.05) à en-contrada em 0 h. A área de HOECHST foi inferior (P<0.05) à obtida no grupo controle nos momentos 12,24 e 48 h.
6
C.S .C .S .C .S .C .S .C .S . O O O O Ol ivl ivl ivl ivl ive i re i re i re i re i ra ,a ,a ,a ,a , L .Z . L .Z . L .Z . L .Z . L .Z . O O O O Ol ivl ivl ivl ivl ive i re i re i re i re i ra ,a ,a ,a ,a , N .Z . N .Z . N .Z . N .Z . N .Z . S S S S Sa ra ra ra ra ra i va i va i va i va i va ,a ,a ,a ,a , F F F F F. M .. M .. M .. M .. M . M M M M Mo no no no no nttttt e i re i re i re i re i ro & J .M.o & J .M.o & J .M.o & J .M.o & J .M. G G G G Ga ra ra ra ra rc ia .c ia .c ia .c ia .c ia . 2011 2011 2011 2011 2011..... Efeitos da tricostatina A sobre aacetilação de histonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
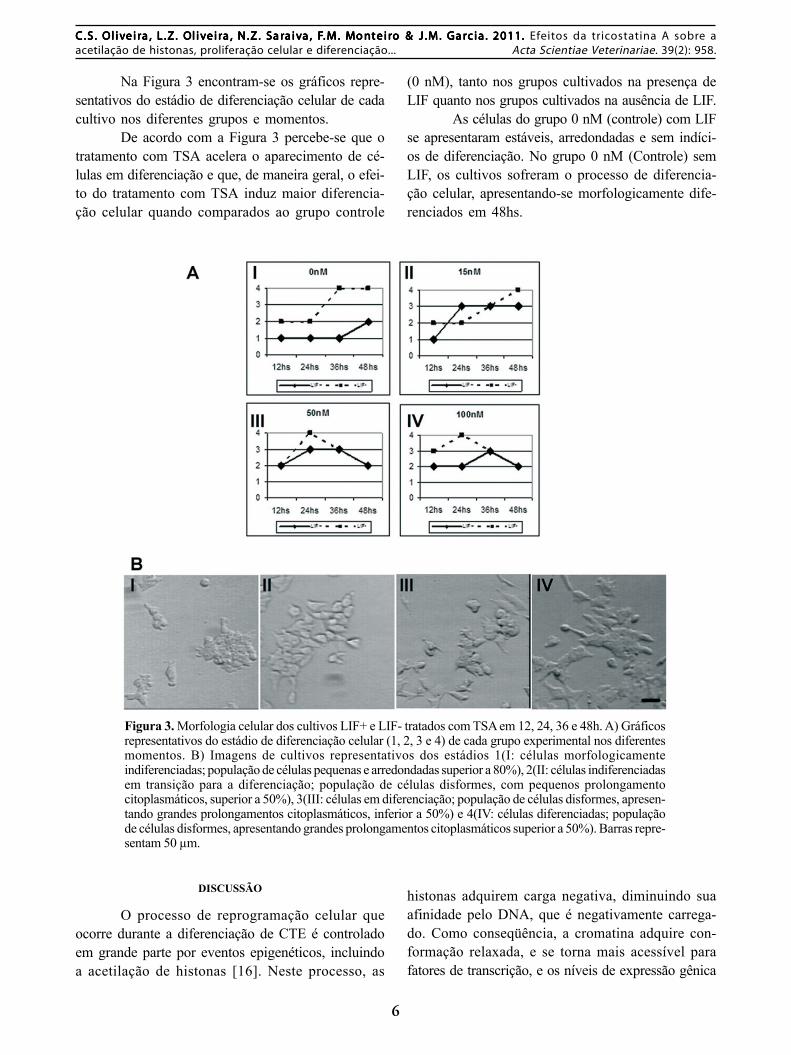
Na Figura 3 encontram-se os gráficos repre-sentativos do estádio de diferenciação celular de cadacultivo nos diferentes grupos e momentos.
De acordo com a Figura 3 percebe-se que otratamento com TSA acelera o aparecimento de cé-lulas em diferenciação e que, de maneira geral, o efei-to do tratamento com TSA induz maior diferencia-ção celular quando comparados ao grupo controle
(0 nM), tanto nos grupos cultivados na presença deLIF quanto nos grupos cultivados na ausência de LIF.
As células do grupo 0 nM (controle) com LIFse apresentaram estáveis, arredondadas e sem indíci-os de diferenciação. No grupo 0 nM (Controle) semLIF, os cultivos sofreram o processo de diferencia-ção celular, apresentando-se morfologicamente dife-renciados em 48hs.
Figura 3. Morfologia celular dos cultivos LIF+ e LIF- tratados com TSA em 12, 24, 36 e 48h. A) Gráficosrepresentativos do estádio de diferenciação celular (1, 2, 3 e 4) de cada grupo experimental nos diferentesmomentos. B) Imagens de cultivos representativos dos estádios 1(I: células morfologicamenteindiferenciadas; população de células pequenas e arredondadas superior a 80%), 2(II: células indiferenciadasem transição para a diferenciação; população de células disformes, com pequenos prolongamentocitoplasmáticos, superior a 50%), 3(III: células em diferenciação; população de células disformes, apresen-tando grandes prolongamentos citoplasmáticos, inferior a 50%) e 4(IV: células diferenciadas; populaçãode células disformes, apresentando grandes prolongamentos citoplasmáticos superior a 50%). Barras repre-sentam 50 µm.
DISCUSSÃO
O processo de reprogramação celular queocorre durante a diferenciação de CTE é controladoem grande parte por eventos epigenéticos, incluindoa acetilação de histonas [16]. Neste processo, as
histonas adquirem carga negativa, diminuindo suaafinidade pelo DNA, que é negativamente carrega-do. Como conseqüência, a cromatina adquire con-formação relaxada, e se torna mais acessível parafatores de transcrição, e os níveis de expressão gênica

7
C.S.C.S.C.S.C.S.C.S. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, L.Z. L.Z. L.Z. L.Z. L.Z. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, N.Z. N.Z. N.Z. N.Z. N.Z. S S S S Sarararararaivaivaivaivaiva,a,a,a,a, F F F F F.M..M..M..M..M. M M M M Monononononttttteireireireireiro & J.M.o & J.M.o & J.M.o & J.M.o & J.M. G G G G Garararararcia.cia.cia.cia.cia. 2011. 2011. 2011. 2011. 2011. Efeitos da tricostatina A sobre a acetilação dehistonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
elevam-se [revisado em 13] e o fenótipo das célulasé alterado, uma vez que as mesmas adquirem com-prometimento com linhagens específicas.
A tricostatina A (TSA) é uma substância utili-zada em estudos envolvendo alterações epigenéticas,pois promove inibição das enzimas histona desa-cetilases, que retiram os grupamentos acetil da cau-da das histonas [21]. As propriedades da TSA comoagente promotor de elevação na acetilação dashistonas já foram demonstradas em CTE em diver-sos estudos [3,10,11]. Dentre suas vantagens sobreas demais drogas inibidoras da enzima histonadesacetilase, pode-se destacar sua reduzida toxicidadee a necessidade de baixas concentrações para induçãodo efeito desejado [8].
No presente estudo, buscou-se determinaruma concentração ideal de TSA para aplicação emestudos com CTE, que resultasse em aumentossatisfatórios nos níveis de acetilação de histonas semcausar grandes prejuízos na proliferação celular. Pri-meiramente, foram coletadas amostras a cada 12 hpara avaliar os níveis de acetilação da lisina 9 dehistonas H3 (H3Lys9ac) nos diferentes grupos expe-rimentais. Como o objetivo dos estudos empregan-do a TSA é promover hiperacetilação, e é sabido quea substância apresenta efeito residual, avaliaram-seas médias de H3Lys9ac considerando os quatro pe-ríodos estudados (12, 24, 36 e 48 h). Sendo assim, ascoletas foram estendidas 24 h além do período detratamento, pois os níveis de H3Lys9ac se mantêmelevados após a retirada da droga.
Os resultados observados na Figura 1 estãode acordo com outros autores [11], que observaramonda de acetilação global das histonas H3 e H4 dosdias 1 a 5 após a retirada do LIF em CTE pluripotentes.Portanto, com o início da diferenciação celular as CTEsofrem uma onda de acetilação correspondente à ati-vação de diversos genes tecido-específico [12]. Nogrupo cultivado na presença de LIF, como o LIF ageinibindo a diferenciação celular, os níveis de acetilaçãopermanecem baixos, correspondentes aos de CTEpluripotentes.
Uma vez que na ausência de LIF as CTE ini-ciam a diferenciação celular e naturalmente elevamseus níveis de acetilação, nesta condição a concen-tração de 15 nM não resultou em um aumento emH3lys9ac. Já no caso de CTE indiferenciadas, queapresentam níveis baixos de acetilação [12], o resul-tado do tratamento com 15 nM foi satisfatório. Ape-
sar de não serem estatisticamente relevantes, os ní-veis de H3lys9ac observados nos grupos sem LIFforam numericamente superiores aos observados nosgrupos cultivados na presença de LIF. Esta observa-ção está de acordo com outros relatos [3], onde seobservou, após tratamento com TSA, aumentos trêsvezes maiores nos níveis de acetilação de CTEpluripotentes comparados aos de fibroblastos (célu-las diferenciadas).
Portanto, considerando os efeitos da TSAsobre os níveis de acetilação de histonas, para CTEcultivadas na presença ou não de LIF, a concentra-ção 50 nM de TSA resultou em aumentos nos níveisde H3lys9ac semelhantes aos obtidos para a concen-tração 100nM. Para a concentração 15 nM, não hou-ve aumento (LIF-) ou o aumento foi inferior (LIF+)ao observado nas concentrações 50nM e 100nM.Desta forma, considerando os níveis de acetilação, aconcentração 50nM foi mais indicada na presençaou não de LIF.
Após avaliação da acetilação das histonas,observou-se como os diferentes tratamentos propos-tos afetam a proliferação celular. Para tanto, conside-rou-se os grupos tratados na presença de LIF, poisestudos em CTE mostraram que células indi-ferenciadas são mais sensíveis aos efeitos da TSAque as células diferenciadas [3].
Em relação aos resultados expostos na Figu-ra 2 do presente estudo, observa-se que houve acen-tuada morte celular nos cultivos tratados com 100nM de TSA. Os efeitos anti-proliferativos da TSA jáforam demonstrados em diversos estudos [2,10,15],e estão relacionados com a ativação de genes pró-apoptoticos [20] e com a exposição da fita de DNA,que se torna mais aberta com a acetilação das histonas,a endonucleases [9].
Neste sentido, observou-se neste experimen-to que concentrações de TSA menores que 100nM,como a concentração 50nM, devem ser usadas parao tratamento de CTE a fim de se obter altos níveis deacetilação sem comprometer a sobrevivência dasmesmas.
Por último, avaliou-se ainda os efeitos da TSAsobre a morfologia celular. A perda do estadoindiferenciado ocorre após a exposição a um sinalde diferenciação que resulta em uma perdairreversível do fenótipo das células tronco embrio-nárias, caracterizadas pela perda do agrupamento dacolônia de crescimento, por alteração na auto-reno-

8
C.S .C .S .C .S .C .S .C .S . O O O O Ol ivl ivl ivl ivl ive i re i re i re i re i ra ,a ,a ,a ,a , L .Z . L .Z . L .Z . L .Z . L .Z . O O O O Ol ivl ivl ivl ivl ive i re i re i re i re i ra ,a ,a ,a ,a , N .Z . N .Z . N .Z . N .Z . N .Z . S S S S Sa ra ra ra ra ra i va i va i va i va i va ,a ,a ,a ,a , F F F F F. M .. M .. M .. M .. M . M M M M Mo no no no no nttttt e i re i re i re i re i ro & J .M.o & J .M.o & J .M.o & J .M.o & J .M. G G G G Ga ra ra ra ra rc ia .c ia .c ia .c ia .c ia . 2011 2011 2011 2011 2011..... Efeitos da tricostatina A sobre aacetilação de histonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
vação, e por um padrão de expressão gênica que nãomais se assemelha ao de CTE indiferenciadas [11,13].Assim, a perda da morfologia indiferenciada e mu-danças dramáticas na expressão de alguns genesmarcadores caracterizam o fenótipo associado comum estágio bem inicial de diferenciação de CTEmurinas em resposta a retirada do LIF e também aotratamento com TSA [11].
Alterações morfológicas também foram ob-servadas na Figura 3 do presente experimento. Nosgrupos tratados com TSA na presença de LIF, emgeral, foram observadas alterações morfológicas quese reverteram quando a droga foi retirada, e as célu-las retornaram a um fenótipo mais semelhante aoindiferenciado. Esta observação está de acordo comoutros autores [11], que também observaram regres-são do estádio de diferenciação após retirada da TSAem CTE cultivadas na presença do LIF. Nos grupostratados com TSA na ausência de LIF, observou-seque o processo de diferenciação foi acelerado emrelação ao grupo controle.
A retomada do fenótipo indiferenciado, ob-servado depois da retirada da TSA, nos grupos culti-vados com LIF demonstra que este fator dificulta adiferenciação celular. A propriedade do LIF de difi-cultar e/ou retardar a diferenciação parece ser eficazmesmo que a TSA induza e/ou estimule o aumentoda expressão gênica [11]. Neste sentido, o presenteestudo demonstrou que, quando na presença de LIF,mesmo que a TSA promova alteração de fenótipocelular, ela não promove comprometimento com ti-pos celulares diferenciados.
A onda de acetilação global provocada pelaTSA se assemelha à observada nos estádios iniciaisde diferenciação celular. Porém, observou-se que oscultivos tratados com TSA na presença de LIF retoma-
vam seu fenótipo original, indiferenciado, após a re-tirada da TSA. Portanto, não pode ser afirmado que aonda de acetilação provocada pela TSA promovediferenciação celular, mas talvez que ela possa esti-mular e antecipar o processo.
CONCLUSÃO
Diante do exposto, os principais achados desteestudo são: i) a TSA promove elevação dos níveis deacetilação de histonas em CTE já na concentração 50nM, tornando possível a redução da concentração pre-conizada na literatura (100 nM), e ii) que estas concen-trações inferiores (15 nM e 50 nM) não provocam preju-ízos na proliferação celular como observado para 100nM.
Embora a elucidação dos processos que re-gem a diferenciação de CTE e o consequente domí-nio destes eventos seja objetivo de numerosos estu-dos, permanecemos ainda distante do entendimentopleno do processo de diferenciação celular. Assim,outros estudos avaliando diferenciação celular sãonecessários para melhor entendimento dos efeitos dahiperacetilação de histonas sobre o comprometimentocom linhagens diferenciadas em CTE.
SOURCES AND MANUFACTURERS1Sigma Chemical, St. Louis, MO, USA.2NUNC, Thermo Fisher Scientific, Rochester, NY, USA. 3Gibco, Life Technologies, Carlsbad, CA, USA.4Invitrogen-Life Technologies, Carlsbad, CA, USA.5Instituto Biochimico, Rio de Janeiro, RJ, Brasil.6Chemicon-Millipore, Billerica, MA, USA.7SCBT, Santa Cruz, CA, USA.8Olympus, Tokyo, Japão.9Carl Zeiss, Jena, Germany.
Declaration of interest. The authors report no conflicts ofinterest. The authors alone are responsible for the contentand writing of the paper.
REFERÊNCIAS
1 Baqir S. & Smith L.C. 2006. Inhibitors of histone deacetylases and DNA methyltransferases alter imprinted gene regulationin embryonic stem cells. Cloning Stem Cells. 8(3): 200-213.
2 Chuikin I.A., Lianguzova M.S. & Pospelov V.A. 2006. Anti-proliferative effect of histone deacetylase inhibitors on murineembryonic stem cells. Tsitologiia. 48(8): 674-683.
3 Dai B. & Rasmussen T.P. 2007. Global epiproteomic signatures distinguish embryonic stem cells from differentiated cells.Stem Cells. 25: 2567-2574.
4 Gadue P., Huber T.L., Nostro M.C., Kattman S. & Keller G.M. 2005. Germ layer induction from embryonic stem cells.Experimental Hematology. 33: 955-964.
5. Gregory R.I., O’neil L.P., Randall T.E., Fournier C., Khosla S., Turner B.M. & Feil R. 2002. Inhibition of histone

9
C.S.C.S.C.S.C.S.C.S. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, L.Z. L.Z. L.Z. L.Z. L.Z. O O O O Olivlivlivlivliveireireireireira,a,a,a,a, N.Z. N.Z. N.Z. N.Z. N.Z. S S S S Sarararararaivaivaivaivaiva,a,a,a,a, F F F F F.M..M..M..M..M. M M M M Monononononttttteireireireireiro & J.M.o & J.M.o & J.M.o & J.M.o & J.M. G G G G Garararararcia.cia.cia.cia.cia. 2011. 2011. 2011. 2011. 2011. Efeitos da tricostatina A sobre a acetilação dehistonas, proliferação celular e diferenciação... Acta Scientiae Veterinariae. 39(2): 958.
deacetylases alters allelic chromatin conformation at the imprinted U2áf1-rs1 locus in mouse embryonic stem cells.Journal of Biological Chemistry. 277(14): 11728-11734.
6. Hattori N., Nishino K., Ko Y.G., Hattori N., Ohgane J., Tanaka S. & Shiota K. 2004. Epigenetic control of mouse Oct-4gene expression in embryonic stem cells and trophoblast stem cells. Journal of Biological Chemistry. 279: 17063-17069.
7. Hosseinkhani M., Hasegawa K. & Ono K. 2007. Trichostatin A induces myocardial differentiation of monkey ES cells.Biochemical and Biophysical Research Communications. 356: 386-391.
8. Johnstone R.W. 2002. Histone-deacetylase inhibitors: novel drugs for the treatment of cancer. Nature Reviews DrugDiscovery. 1: 287-299.
9. Koyama Y., Adachi M., Sekiya M., Takekawa M. & Imai K. 2000. Histone deacetylase inhibitors suppress IL-2-mediatedgene expression prior to induction of apoptosis. Blood. 96: 1490-1495.
10. Lee J.H., Hart S.R.L. & Skalnik G. 2004. Histone deacetylase activity is required for embryonic stem cell differentiation.Genesis. 38: 32-38.
11. McCool K.W., Xu X., Singer D.B., Murdoch F.E. & Fritsch M.K. 2007. The Role of Histone Acetylation in RegulatingEarly Gene Expression Patterns during Early Embryonic Stem Cell Differentiation. Journal of Biological Chemistry.282(9): 6696-6706.
12. Meshorer E. & Misteli T. 2006. Chromatin in pluripotent embryonic stem cells and differentiation. Na-ture Reviews Molecular Cell Biology. 7: 540-546.
13. Monteiro F.M., Oliveira C.S., Oliveira L.Z., Saraiva N.Z., Mercadante M.E.Z., Lopes F.L., Arnold D.R. & Garcia J.M.2011. Chromatin modifying agents in the in vitro production of bovine embryos. Veterinary Medicine International(Recent Advances in Reproductive Technologies): 1-9.
14. Oliveira C.S., Saraiva N.Z., De Souza M.M., Tetzner T.A.D., De Lima M.R. & Garcia J.M. 2010. Effects of histonehyperacetylation on the preimplantation development of male and female bovine embryos. Reproduction, Fertility andDevelopment. 22(6): 1041-1048.
15. Saunders N., Dicker A., Popa C., Jones S. & Dahler A. 1999. Histone deacetylase inhibitors as potential anti-skin canceragents. Cancer Research. 59: 399-404.
16. Saraiva N.Z., Oliveira C.S. & Garcia J.M. 2010. Histone acetylation and its role in embryonic stem cell differentiation.World J Stem Cells. 2(6): 121-126.
17. Smith A. 1998. Cell therapy: in search of pluripotency. Current Biology. 8: 802-804.18. Solter D. 2006. From teratocarcinomas to embryonic stem cells and beyond: a history of embryonic stem cell research.
Nature Review Genetics. 7: 319-327.19. Williams R.L., Hilton D.J., Pease S., Willson T.A., Stewart C.I., Gearing D.P., Wagner E.F., Metcalf D., Nicola N.A. &
Gough N.M. 1988. Myeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cells.Nature. 36: 684-687.
20. Yakovlev A., Khafizova M., Abdullaev Z., Loukinov D. & Kondratyev A. 2010. Epigenetic regulation of caspase-3 geneexpression in rat brain development. Gene. 450(1-2): 103-108.
21. Yoshida M., Kijima M., Akita M. & Beppu T. 1990. Potent and specific inhibition of mammalian histone deacetylase bothin vivo and in vitro by trichostatin A. Journal of Biological Chemistry. 265: 17174-17179.
Pub. 958Pub. 958Pub. 958Pub. 958Pub. 958www.ufrgs.br/actavet
