efeito do sorgo sobre o desempenho zootÉcnico ... · para conseguir um rápido desenvolvimento...
TRANSCRIPT

UNIVERSIDADE ESTADUAL PAULISTA “JULIO DE MESQUITA FILHO” FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS
CÂMPUS DE JABOTICABAL
EFEITO DO SORGO SOBRE O DESEMPENHO ZOOTÉCNICO, CARACTERÍSTICAS DA CARCAÇA E O DESENVOLVIMENTO
DA MUCOSA INTESTINAL DE FRANGOS
Daniel Mendes Borges Campos
Zootecnista
JABOTICABAL - SÃO PAULO - BRASIL
2006

UNIVERSIDADE ESTADUAL PAULISTA “JULIO DE MESQUITA FILHO” FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS
CÂMPUS DE JABOTICABAL
EFEITO DO SORGO SOBRE O DESEMPENHO ZOOTÉCNICO, CARACTERÍSTICAS DA CARCAÇA E O DESENVOLVIMENTO
DA MUCOSA INTESTINAL DE FRANGOS
Daniel Mendes Borges Campos
Orientador: Prof. Dr. Marcos Macari
Co-orientador: Renato Luis Furlan
Dissertação apresentada à Faculdade de Ciências Agrárias e Veterinárias – Unesp, Câmpus de Jaboticabal, como parte das exigências para a obtenção do título de Mestre em Zootecnia (Produção Animal).
Maio de 2006
Jaboticabal – SP

ii
DADOS CURRICULARES DO AUTOR
DANIEL MENDES BORGES CAMPOS – nascido em Jaboticabal – SP, no dia 10
de julho de 1979. Em março de 1998 ingressou no curso de Zootecnia da Faculdade de
Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal (FCAV–
UNESP), concluindo em novembro de 2002. No período de 20 janeiro de 2003 até 21 de
janeiro de 2004, trabalhou como “Trainee” de produção na empresa Agroceres Nutrição
Animal, no município de Patos de Minas – MG. Em março de 2004 iniciou o curso de
Mestrado em Zootecnia, na Faculdade de Ciências Agrárias e Veterinárias,
Universidade Estadual Paulista, Jaboticabal (FCAV–UNESP).

iii
Aos meus queridos Pais,
Ivomar Borges Campos e Sandra Maria Mendes Borges Campos
Aos meus irmãos,
Eduardo Mendes Borges Campos e Emiliano Mendes Borges Campos
A minha babá,
Vera Lúcia Rodrigues da Silva,
A minha noiva,
Thaisa Cristina Devós Ganga
Obrigado pelo apoio, incentivo, amor e exemplo de vida
Dedico

iv
AGRADECIMENTOS
A Deus pela vida e seus encantos.
Aos Professores Marcos Macari e Renato Luis Furlan pela orientação e exemplo
como profissionais.
Aos amigos da Graduação e Pós-Graduação, em especial, Aiani Maria Vaz,
Bruno Serpa Vieira, Carla Heloisa Faria Domingues, Daniel Emygdio Faria Filho, Karoll
Alfonso Torres, Paulo Sergio Rosa, Tiago Urbano pela colaboração na condução das
etapas de campo e discussão do presente trabalho. Um conselho “MENOS CHARLA
MAS TRABAJO” (Comandante Alfonso).
Aos funcionários Clara, Damaris, Euclides, Shirley e em especial o Sr. Orandi
(Departamento de Morfologia e Fisiologia Animal), Robson, Isildo, Sr. João, e Vicente
(Setor de Avicultura), Sandra e Oswaldo (Fábrica de Ração) e a Tânia (Departamento
de Tecnologia) pelo auxílio e amizade.
Aos professores Isabel Boleli e Vera Maria Barbosa de Moraes (Exame Geral de
Qualificação) pelas preciosas contribuições desta dissertação.
À Fundação de Amparo à Pesquisa do Estado de São Paulo, FAPESP, pela
concessão de bolsa e estudo durante 2004 e 2005 (Processo 04/05058-8).
Aos meus amigos e colegas que não estão citados neste texto, mas contribuíram
de alguma forma para eu chegasse a este momento.

v
SUMÁRIO
Página
CAPÍTULO 1 - CONSIDERAÇÕES GERAIS........................................................................... 1 CAPÍTULO 2 - EFEITO DA SUBSTITUIÇÃO DE MILHO POR SORGO SOBRE O DESEMPENHO, AS CARACTERÍSTICAS DA CARCAÇA E O CUSTO DE ARRAÇOAMENTO EM FRANGOS DE CORTE...................................................................... 6
Resumo.................................................................................................................. 6
Introdução.............................................................................................................. 7 Material e Métodos................................................................................................ 8 Resultados............................................................................................................. 11 Discussão.............................................................................................................. 13 Conclusão.............................................................................................................. 15
CAPÍTULO 3 - CARACTERÍSTICAS MORFOLÓGICAS DA MUCOSA INTESTINAL DE FRANGOS DE CORTE ALIMENTADOS COM SORGO BAIXO TANINO............................... 16
Resumo.................................................................................................................. 16 Introdução.............................................................................................................. 17 Material e Métodos................................................................................................ 18 Resultados............................................................................................................. 22 Discussão.............................................................................................................. 32 Conclusão.............................................................................................................. 35
CAPÍTULO 4 - USO DA REVISÃO SISTEMÁTICA DA LITERATURA E MODELOS MATEMÁTICOS PARA PREDIZER O DESEMPENHO DE FRANGOS DE CORTE ALIMENTADOS COM DIETAS À BASE DE MILHO E SORGO.............................................. 36
Resumo.................................................................................................................. 36 Introdução.............................................................................................................. 37 Material e Métodos................................................................................................ 38 Resultados e Discussão........................................................................................ 40 Conclusão.............................................................................................................. 43
REFERÊNCIAS........................................................................................................................ 43

vi
EFEITO DO SORGO SOBRE O DESEMPENHO ZOOTÉCNICO, CARACTERÍSTICAS
DA CARCAÇA E O DESENVOLVIMENTO DA MUCOSA INTESTINAL DE FRANGOS
RESUMO - A variação nos preços dos principais ingredientes da alimentação
avícola (milho e soja), fazem da utilização de ingredientes alternativos a resposta viável
para a obtenção de resultados econômicos satisfatórios. O sorgo (Sorghum bicolor (L.)
Moench) é tido como uns dos principais ingredientes energéticos substitutos ao milho,
todavia, existe certa resistência na utilização de sorgo em rações pré-iniciais devido à
possibilidade deste cereal afetar o desempenho dos pintainhos. Neste sentido, foram
realizadas três pesquisas para analisar o desempenho, as características da carcaça e o
desenvolvimento da mucosa intestinal de frangos alimentados com sorgo baixo tanino
em substituição ao milho. Concluiu-se que: (1) o sorgo baixo teor em tanino pode
substituir completamente o milho em dietas para frangos de corte a partir do primeiro dia
de idade sem comprometimento do desempenho e o rendimento de carcaça e cortes
comerciais; (2) A altura dos vilos e o número de células caliciformes do intestino não são
afetados pelos níveis de substituição de milho por sorgo baixo tanino, no entanto, a
profundidade de cripta do duodeno aumenta com a inclusão de sorgo nas rações; (3) a
idade apresenta efeito significativo sobre o número de células caliciformes onde ocorre
aumento na quantidade de células produtoras de mucina no período de 1 a 21 dias de
idade. A altura dos vilos no duodeno atinge seu desenvolvimento máximo aos 21 dias de
idade.
Palavras-chave: altura de vilo, células caliciformes, dieta pré-inicial, modelos
matemáticos, profundidade de cripta

vii
SORGHUM EFFECT ON PERFORMANCE, CARCASS CHARACTERISTICS AND
MUCOSAL DEVELOPMENT OF BROILERS
SUMMARY – Prices variation of main grains of poultry production (corn and soybean),
make the utilizations of alternative feeds an important way to obtain good economics
results. Sorghum (Sorghum bicolor (L.) Moench) is the most common alternative for
corn, therefore, there is a resistance to use sorghum in pre-starter diets due the
possibility of this cereal affects the chicks performance. In these sense, three studies,
were carried out to assess the performance, carcass characteristics and mucosal
development in broilers fed with sorghum in place of corn. In conclusion: (1) sorghum
with low tannin content can substitute completely corn in broilers diets since the first day
of life without impair performance and carcass yield; (2) villus height and goblet cells
numbers of small gut are not affected by levels of substitutions of corn by sorghum with
low tannin content, however, crypt depth of duodenum increases with sorghum inclusion;
(3) goblet cells are significative affected by age, with a the higher amount of these cells
from 1 to 21 days of age. Villus heights on duodenum reach the maximum development
at 21 days of age.
Keywords: crypt depth, goblet cells, mathematical models, villus height, pre-starter diet

1
CAPÍTULO 1 – CONSIDERAÇÕES GERAIS
As grandes oscilações de preços dos ingredientes das rações conduzem a
avicultura de corte à busca por substitutos dos tradicionais ingredientes utilizados para o
arraçoamento, buscando assim minimizar custos de produção. No entanto, esta
substituição pode ser limitada pela qualidade de determinados ingredientes alternativos,
devido à presença de fatores antinutricionais ou a sua composição bromatológica.
A nutrição da primeira semana de vida do pintainho de corte tem sido o foco de
diversas pesquisas, pois representa cerca de 17% do período de vida da ave, além de
ocorrer a maior taxa de crescimento do trato gastrintestinal (TGI) (GÔNZALEZ &
SALDANHA, 2001; UNI & FERKET, 2004).
Os períodos pré e pós-eclosão são críticos para o desenvolvimento e
sobrevivência de frangos de corte. Durante esse período, os pintainhos sofrem uma
transição metabólica e fisiológica em função da troca da alimentação dos nutrientes do
saco vitelínico para um alimento exógeno, uma fonte de nutrientes baseada
principalmente em lipídeos para uma dieta sólida, baseada em carboidratos e proteínas.
Imediatamente após a eclosão, os pintainhos utilizam suas limitadas reservas corporais
para conseguir um rápido desenvolvimento físico e funcional do trato gastrintestinal, a
fim de desenvolver a capacidade de digerir alimentos e assimilar nutrientes (UNI &
FERKET, 2004). Esta imaturidade do TGI na fase pré-inicial reduz a capacidade de
utilização dos nutrientes, o que parece acarretar decréscimo dos valores de energia
metabolizável (EM) do alimento (LONGO et al., 2005). O intestino delgado é o principal
órgão responsável pela digestão e absorção de nutrientes, assim quanto antes os
pintainhos alcançarem sua capacidade funcional, mais cedo poderão utilizar os
nutrientes da dieta,, crescer eficientemente e demonstrar seu potencial genético, resistir
a infecções e a doenças metabólicas (UNI & FERKET, 2004).
O peso do intestino em comparação com o peso do embrião aumenta de 1% do
décimo sétimo dia de incubação para 3,5% no dia da eclosão. Logo após a eclosão, o
proventrículo, moela e intestino delgado crescem mais rapidamente do que outros

2
órgãos e tecidos. Nos pintainhos, maior tamanho relativo destes órgãos digestivos
ocorre entre 3 e 7 dias de idade (UNI & FERKET, 2004).
Na eclosão o TGI dos pintainhos está anatomicamente completo. No entanto, a
superfície de absorção e taxa de proliferação dos enterócitos aumenta após a eclosão,
sendo esse rápido desenvolvimento diferente ao longo do intestino delgado. No
duodeno, o crescimento dos vilos está praticamente completo aos 7 dias de vida,
enquanto que no jejuno e íleo o desenvolvimento continua além do 14° dia de vida (UNI,
1999).
O desenvolvimento da mucosa intestinal envolve o aumento da altura e
quantidade dos vilos, o que corresponde a um aumento em número de suas células
epiteliais, enterócitos, células caliciformes e enteroendócrinas. Esse processo decorre
primariamente de dois eventos citológicos associados: renovação e extrusão celular. A
renovação consiste na proliferação e diferenciação resultante das divisões mitóticas
sofridas por células totepotentes (“stem cells”) localizadas na cripta e ao longo dos vilos,
e a extrusão é a perda de células, que ocorre normalmente no ápice dos vilos. O
equilíbrio entre esses dois processos determina o “turnover” (síntese-migração-extrusão)
constante na mucosa, o qual garante a manutenção do tamanho dos vilos e, portanto, a
manutenção da capacidade digestiva e de absorção intestinal. Quando a mucosa
intestinal responde a algum agente com um desequilíbrio no “turnover” a favor de um
desses processos, ocorre modificação na altura dos vilos. A manutenção da mucosa
intestinal, em condições fisiológicas normais, tem custo energético elevado para o
frango, segundo MCBRIDE & KELLY (1990) a manutenção do epitélio intestinal e
estruturas de suporte anexas corresponde a cerca de 20% da energia bruta consumida
pelo animal, além da redução da quantidade de substrato digerido e absorvido, há ainda
o custo da restauração do epitélio. A energia conservada pelo reduzido “turnover” de
células no epitélio intestinal poderá ser utilizada para o desenvolvimento de massa
muscular. Assim, o rendimento econômico do lote será seriamente comprometido
quando o “turnover” intestinal for alterado (MAIORKA et al., 2002).
As aves neste período pré-inicial possuem limitações na atividade enzimática e
digestiva. Aparentemente são necessários três a quatro dias entre o nascimento e a

3
capacidade máxima enzimática para a digestão de carboidratos. A amilase pancreática,
maltase e sacarase atingem a atividade específica máxima dentro deste período
(MARCHAIN & KULKA, 1997). NOY & SKLAN (1995) trabalhando com pintainhos de
corte com quatro dias de idade, demonstraram que a digestão do amido aumentou
pouco do 4° para o 21° dia de vida das aves, passando de 82 para 89%,
respectivamente. NITSAN et al. (1991) observaram que o valor máximo de amilase no
pâncreas ocorre no 8° dia de vida, e no intestino delgado o valor máximo foi encontrado
no 17°dia. A digestão de gordura também melhora com o aumento da idade,
(SAKOMURA et al. 2004) concluíram que a digestibilidade da gordura aumentou com a
idade e que tal variação foi associada com a atividade enzimática da lipase. NITSAN et
al. (1991) encontraram valores máximos de lipase no pâncreas no 8° dia e no intestino
delgado no 4° dia de vida das aves. Em peruzinhos a digestibilidade de sebo bovino e
uma mistura de óleos vegetais passou de 66,4 e 83,7% na 2ª semana de vida para 90,8
e 96,5% na 8ª semana (SELL et al., 1986). NITSAN et al. (1991) confirmaram a
presença de quimiotripsina e de tripsina no pâncreas de pintainhos na eclosão, contudo,
a atuação de ambas as enzimas foram baixas até o 5° ou 6° dia de idade quando
tiveram um aumento significativo, sendo o ápice da quimiotripsina no 10° dia de vida. No
geral, a digestão protéica passa de 78 para 90% do 4° para o 21° dia de vida (NOY &
SKLAN, 1995).
Assim sendo, devido a características anatômicas e fisiológicas do TGI pertinente
a esta primeira semana de vida, tais como: as necessidades nutricionais existentes e
suas limitações em digerir e absorver determinados nutrientes, o rápido crescimento do
animal, sua dificuldade de sobrevivência em ambientes frios e a alta correlação entre o
peso vivo aos sete dias de vida com o peso de abate, fazem com que os ingredientes
utilizados para a formulação de sua dieta sejam diferenciados.
O valor nutricional dos alimentos depende basicamente de seu conteúdo em
nutrientes, e da sua disponibilidade biológica. Depende ainda, da presença e dos níveis
de substâncias tóxicas e/ou antinutricionais, as quais podem alterar essa composição ou
tornar indisponíveis esses nutrientes (PEZZATO, 1995). O sorgo (Sorghum bicolor (L.)
Moench) tem sido avaliado como substituto do milho nas rações para aves e suínos,

4
principalmente nas regiões semi-áridas e tropicais, onde sua cultura apresenta melhor
rendimento por unidade de área (ROSTAGNO et al., 2000). No entanto, de acordo com
a classificação apresentada por FARIA FILHO et al. (2005), que estudaram o perfil dos
aminoácidos dos alimentos utilizados na avicultura em relação ao padrão de
aminoácidos da proteína ideal, concluíram que a proteína do sorgo apresenta o pior
perfil de aminoácidos dos cereais utilizados na avicultura. Segundo esta classificação, o
sorgo só apresenta um perfil melhor de aminoácido que o glúten de milho. As
simulações de rações elaboradas com a respectiva classificação criada pelos autores,
demonstraram que a substituição do milho por sorgo baixo tanino afetou o balanço de
aminoácidos da ração. Dispõe ainda, de nível muito baixo de pigmentos e nível inferior
de extrato etéreo (SCHEUERMANN, 2003).
O sorgo contém vários compostos fenólicos que podem afetar a cor, a aparência
e qualidade nutricional. Esses compostos podem ser classificados em três grupos
básicos: ácidos fenólicos, flavonóides e taninos. Os ácidos fenólicos são encontrados
em todos os tipos de sorgo enquanto os flavonóides não o são. O fenol, conhecido como
tanino, encontra-se concentrado na testa da semente, que é um tecido altamente
pigmentado localizado logo abaixo do pericarpo. Os taninos são classificados em
hidrolisáveis e condensados. Os taninos condensados ou proantocianidinas são
polímeros de flavonóides, cujos monômeros (flavan-3-ol) são unidos carbono-carbono.
Os taninos hidrolisáveis são ésteres de ácido gálico e de ácido hexahidroxidifênico e
glicose, além de outros polióis (AGOSTNI-COSTA, 2003). O grau de toxidade do tanino
depende do seu tipo, se hidrolisável ou condensado, das suas proporções na dieta, dos
produtos finais da hidrólise no intestino e da espécie animal (QUINTERO PINTO, 2000).
As proantocianidinas estão ligadas à coagulação e precipitação de enzimas e alguns
minerais, formação de complexos com os carboidratos da dieta, inibição da atividade de
algumas enzimas digestivas, como a tripsina e α-amilase. Quando ingeridos em certas
quantidades, reduzem a taxa de crescimento, uma vez que diminui o aproveitamento
energético e protéico da dieta levando a maior excreção de nitrogênio nas fezes como
resultado da interação tanino-proteínas, formadas por múltiplas pontes de hidrogênio.
Os taninos condensados também causam necroses e distorções nos vilos da mucosa

5
intestinal e aumento no número de células caliciformes (MITJAVILA et al., 1977;
TREVINO, 1992; CHANG et al., 1994; ORTIZ et al., 1994; OLIVEIRA, 1998). Os ácidos
fenólicos não têm efeito adverso na qualidade nutricional, porém podem causar cor
indesejável aos alimentos, quando processados sob condições alcalinas. Os
flavonóides, a exemplo dos ácidos fenólicos, também não causam problemas na
digestibilidade e palatabilidade do sorgo. Constituem um amplo grupo de compostos
fenólicos encontrados nas plantas, sendo que alguns deles estão entre os principais
pigmentos presentes em vegetais (BUTULO, 2003).
Alguns trabalhos relataram os efeitos negativos nas dietas que contêm níveis
elevados de tanino sobre o ganho de peso, consumo de ração e conversão alimentar
dos frangos de corte (CHANG & FULLER, 1964; ROSTAGNO et al., 1973; DOUGLAS &
SULLIVAN, 1991; ORTIZ et al., 1994;). No entanto, em trabalhos no quais foram
utilizados sorgo com baixo teor em tanino, não foram observados efeitos negativos nos
índices de desempenho. Os autores SILVA (2003) e GARCIA et al. (2005) trabalhando
com níveis de 0, 25, 50, 75 100% de substituição de milho por sorgo baixo teor em
tanino, verificaram que os frangos apresentaram médias similares para consumo de
ração, ganho de peso e conversão alimentar nas fases inicial e crescimento. DOUGLAS
& SULLIVAN (1991) ao substituírem milho por sorgo baixo teor em tanino na fase inicial
de frangos de corte não observaram efeito significativo para ganho de peso e conversão
alimentar. Contudo, mesmo quando o sorgo possui níveis baixos de tanino existe uma
resistência de se utilizar este cereal em dietas na fase pré-inicial de frangos de corte,
devido à possibilidade de afetar o desenvolvimento da mucosa intestinal e a atividade
enzimática do animal e assim afetar o desempenho zootécnico na primeira semana de
vida.
Assim, o objetivo do trabalho foi estudar o efeito da substituição do milho por
sorgo baixo tanino sobre o desempenho, as características da carcaça, o
desenvolvimento da mucosa intestinal e elaborar modelos de superfície de resposta
para a predição do desempenho de frangos de corte alimentados com sorgo e/ou milho
como ingredientes energéticos em rações de frangos de corte.

6
CAPÍTULO 2 – EFEITO DA SUBSTITUIÇÃO DE MILHO POR SORGO SOBRE O
DESEMPENHO, AS CARACTERÍSTICAS DA CARCAÇA E O CUSTO DE
ARRAÇOAMENTO EM FRANGOS DE CORTE.
RESUMO – O experimento foi conduzido com o objetivo de avaliar a influência do sorgo
baixo teor em tanino sobre o desempenho e o rendimento de carcaça e cortes
comerciais nos frangos de corte de 1 – 42 dias de idade. Foram utilizados 750
pintainhos de um 1 dia de idade Cobb-500®, em um delineamento inteiramente
casualizado. A dieta controle foi à base de milho e farelo de soja e outras quatro dietas
com substituições de 25, 50, 75 e 100% do milho por sorgo baixo teor em tanino, com 6
repetições por tratamento totalizando 30 unidades experimentais. As rações
experimentais foram isonutrientes dentro de cada fase de criação: inicial (1-21 dias de
idade) e crescimento (22-42 dias de vida). O consumo de ração, o ganho de peso e a
conversão alimentar não foram afetados (p>0,05) pelos níveis de inclusão de sorgo nos
períodos 1 a 7, 1 a 14, 1 a 21 e 1 a 42 dias de vida das aves. A viabilidade criatória, o
índice de eficiência produtiva e o rendimento de carcaça e cortes comerciais aos 42 dias
de idade também não foram influenciados (p>0,05) pelos níveis crescentes de sorgo.
Assim, o sorgo baixo teor em tanino pode substituir completamente o milho em dietas
para frangos de corte a partir do primeiro dia de idade sem comprometimento do
desempenho e o rendimento de carcaça e cortes comerciais.
Palavras-Chave: sorgo, frangos de corte, desempenho, características de carcaça.

7
Introdução
A nutrição do pintainho de corte em sua primeira semana de vida tem sido o foco
de diversas pesquisas, devido a forte correlação entre o peso dos animais no sétimo dia
de vida com o peso de abate. Esta fase representa cerca de 17% do período de vida da
ave, além de ocorrer a maior taxa de crescimento relativo do trato gastrintestinal (UNI &
FERKET, 2004; GONZÁLEZ & SALDANHA, 2001). Esse período é considerado uma
transição entre a vida embrionária (dependente dos nutrientes do saco vitelínico) para a
vida adulta, baseada em uma dieta sólida constituída basicamente por proteínas e
carboidratos. Imediatamente após a eclosão, os pintainhos utilizam suas limitadas
reservas corporais para conseguir um rápido desenvolvimento físico e funcional do trato
gastrintestinal (TGI), a fim de desenvolver a capacidade de digerir alimentos e assimilar
nutrientes (UNI & FERKET, 2004). As aves neste período possuem limitações na
atividade enzimática e digestiva. Diversos autores demonstraram melhora significativa
na digestibilidade de carboidratos, gorduras e proteínas com o aumento da idade (SELL
et al., 1986; NITSAN et al., 1991; NOY & SKLAN, 1995; MARCHAIN & KULKA, 1997).
A necessidade de minimizar custos de alimentação das aves mesmo que
pequena resulta em grande impacto na avicultura devido ao seu volume de produção,
por isso, o sorgo (Sorghum bicolor (L.) Moench) tem sido avaliado como possível
substituto do milho nas rações para aves e suínos para tal objetivo. De acordo com
DOWLING et al. (2002) o sorgo se torna viável economicamente quando seu preço
equivale a 95% do preço do milho. O sorgo contém vários compostos fenólicos que
podem afetar a cor, a aparência e qualidade nutricional da dieta. Esses compostos
podem ser classificados em três grupos básicos: ácidos fenólicos, flavonóides e taninos.
No entanto, somente os taninos podem afetar a qualidade nutricional da dieta. Os
taninos estão ligados à coagulação e precipitação de enzimas e alguns minerais. Eles
podem formar complexos com os carboidratos da dieta e inibir a atividade de algumas
enzimas digestivas, como a tripsina e α-amilase. Quando ingeridos em certas
quantidades, causam redução no ganho de peso e uma pior conversão alimentar de

8
frangos de corte, uma vez que diminui o aproveitamento energético e protéico da dieta
levando a maior excreção nitrogênio nas fezes como resultado da interação tanino-
proteínas, formada por múltiplas pontes de hidrogênio (CHANG & FULLER, 1964;
ROSTAGNO et al., 1973; DOUGLAS & SULLIVAN, 1991; TREVINO, 1992; CHANG et
al., 1994; ORTIZ et al., 1994; OLIVEIRA, 1998).
No entanto em trabalhos nos quais foram utilizados sorgos com baixos teores em
tanino, não foram observados efeitos negativos nos índices de desempenho (DOUGLAS
& SULLIVAN, 1991; POUR-REZA & EDRISS, 1997; DINIZ et al., 2002; SILVA, 2003 e
GARCIA et al., 2005). Contudo, mesmo quando o sorgo possui níveis baixos de tanino,
existe resistência de se utilizar este cereal em dietas na fase pré-inicial devido à
possibilidade de afetar a digestibilidade dos nutrientes através das interações do tanino
com a ração ou com as limitadas enzimas digestivas, além de atuar como possível fator
de perturbação da mucosa intestinal principalmente na fase pré-inicial ou acarretar lotes
não homogêneos.
Assim, o presente trabalho teve como objetivo avaliar o custo do arraçoamento
de dietas com sorgo baixo teor em tanino, o desempenho zootécnico e o rendimento de
carcaça e cortes comerciais de frangos de corte alimentados com sorgo baixo teor em
tanino em substituição ao milho.
Material e Métodos
O experimento foi conduzido no setor de Avicultura do Departamento de
Zootecnia da Faculdade de Ciências Agrárias e Veterinárias, UNESP, Jaboticabal-SP.
Foram utilizadas duas câmaras climáticas com 16 unidades experimentais de 2,5 x
1,6m.
Foram utilizados 750 frangos machos Cobb-500®, distribuídos em um
delineamento com blocos casualizados. O objetivo dos blocos foi controlar diferenças de
temperatura, umidade e gases nas diferentes câmaras. Na chegada dos pintainhos foi

9
realizada pesagem de uma amostra do lote para a determinação do peso médio e foram
montadas unidades experimentais com animais pesando em média 42,48 ± 0,56g. Os
tratamentos consistiram na administração ad-libitum de uma dieta controle a base de
milho e farelo de soja e outras quatro dietas com substituições de 25, 50, 75 e 100% do
milho por sorgo baixo teor em tanino, com 6 repetições por tratamento totalizando 30
unidades experimentais. As rações experimentais foram isonutrientes dentro de cada
fase de criação: inicial (1-21 dias de idade) e crescimento (22-42 dias de vida). Foram
formuladas seguindo a composição nutricional dos ingredientes e os níveis nutricionais
proposto por ROSTAGNO et al. (2000) (Tabela1).
O sorgo utilizado no presente experimento teve 4,52 mg de catequina por grama
de amostra. O conteúdo total de tanino foi analisado pelo método da Vanilina – HCl, este
método baseia-se na reação da leucoantocianidina (catequina) e proantocianidina
(tanino condensado) com vanilina na presença de HCl, é um método específico para a
determinação de tanino condensado e alguns flavonóides.
Os animais foram vacinados contra doença de Marek e Bouba Aviária no
incubatório. No 8° dia de idade as aves foram vacinadas (via ocular) contra as doenças
de Newcastle e Gumboro e no 14° dia contra a doença de Gumboro na água de bebida.

10
Tabela 1 – Composição centesimal das rações experimentais do período inicial (1 - 21 dias de idade) e crescimento (22 - 42 dias de idade).
* Suplementadas a fim de alcançar os níveis nutricionais de ROSTAGNO et al. (2000). 1 Níveis de garantia por quilograma do produto: selênio 54,6mg; ácido nicotínico 6930mg; cobre 25000mg; pantotenato de cálcio 1900mg; biotina 32mg; manganês 15252mg; DL-metionina (mínimo) 340g; iodo 260mg; coccidiostático 25000mg; antioxidante 100mg; colina 120mg; promotor de crescimento e eficiência alimentar 10000mg; vitamina A 1400000U.I.; vitamina B1 356mg; vitamina B12 2000mcg; vitamina B2 1920mg; vitamina B6 693mg; vitamina D3 600000U.I.; vitamina E 5000mg; vitamina K 196mg; zinco 18250mg. 2 Níveis de garantia por quilograma do produto: selênio 54,6mg; ácido nicotínico 6930mg; cobre 25000mg; pantotenato de cálcio 1900mg; biotina 32mg; manganês 15252mg; DL-metionina (mínimo) 270g; iodo 260mg; coccidiostático 22000mg; antioxidante 100mg; colina 120mg; promotor de crescimento e eficiência alimentar 10000mg; vitamina A 1400000U.I.; vitamina B1 356mg; vitamina B12 2000mcg; vitamina B2 1920mg; vitamina B6 693mg; vitamina D3 600000U.I.; vitamina E 5000mg; vitamina K 196,5mg; zinco 18250mg.
As aves e as sobras de ração nos comedouros foram pesadas, permitindo
determinar o consumo de ração (CR), o ganho de peso (GP) e a conversão alimentar
(CA) no 7°, 14°, 21° e 42° dia de vida das aves.
A viabilidade criatória (VC) foi calculada pela seguinte formula: VC = 100 – {(NAM
/ NAI) x 100} onde, NAM é o número de aves mortas por unidade experimental e NAI é o
número de aves inicial por unidade experimental ao final de 42 dias de vida.
O índice de eficiência produtiva (IEP) foi calculado pela seguinte formula: IEP =
(GMD x VC) / (CA x10) onde, GMD é o ganho médio diário (g).
INICIAL CRESCIMENTO Ingredientes % 0% 25% 50% 75% 100% 0% 25% 50% 75% 100%
Milho 54,89 41,17 27,45 13,72 - 60,33 45,25 30,17 15,08 - Sorgo baixo tanino - 13,72 27,45 41,17 54,89 - 15,08 30,17 45,25 60,33 Farelo de Soja, 45 36,31 36,22 36,12 36,03 35,94 30,61 30,51 30,40 30,30 30,20 Óleo de soja 3,72 4,02 4,32 4,63 4,93 4,24 4,57 4,90 5,24 5,57 Fosfato bicálcico 1,82 1,82 1,81 1,80 1,80 1,62 1,61 1,61 1,60 1,59 Calcário Calcítico 0,99 0,99 0,99 0,99 0,99 0,94 0,94 0,94 0,94 0,94 Sal Comum 0,46 0,46 0,46 0,46 0,47 0,39 0,39 0,39 0,40 0,40 DL-metionina* 0,06 0,07 0,08 0,08 0,09 0,08 0,09 0,09 0,10 0,11 L-lisina* 0,15 0,16 0,17 0,17 0,18 0,20 0,21 0,22 0,23 0,24 Suplemento mineral e vitamínico 0,501 0,50 0,50 0,50 0,50 0,502 0,50 0,50 0,50 0,50
Caulim 1,10 0,88 0,65 0,43 0,21 1,10 0,85 0,60 0,36 0,12 Total 100 100 100 100 100 100 100 100 100 100 Energia e Nutrientes Composição Calculada Energia Metabolizável(Kcal) 3000 3000 3000 3000 3000 3100 3100 3100 3100 3100 Proteína bruta (%) 21,40 21,40 21,40 21,40 21,40 19,31 19,31 19,31 19,31 19,31 Cálcio (%) 0,96 0,96 0,96 0,96 0,96 0,87 0,87 0,87 0,87 0,87 Fósforo Disponível (%) 0,45 0,45 0,45 0,45 0,45 0,41 0,41 0,41 0,41 0,41 Sódio (%) 0,22 0,22 0,22 0,22 0,22 0,19 0,19 0,19 0,19 0,19 Lisina (%) 1,26 1,26 1,26 1,26 1,26 1,16 1,16 1,16 1,16 1,16 Metionina (%) 0,56 0,56 0,56 0,56 0,56 0,51 0,51 0,51 0,51 0,51 Aminoácidos sulfurados (%) 0,90 0,90 0,90 0,90 0,90 0,82 0,82 0,82 0,82 0,82

11
Aos 42 dias foram sacrificadas duas aves por unidade experimental para
avaliação do rendimento de carcaça inteira, peito, coxa+sobrecoxa, pés, asas, dorso,
cabeça mais pescoço e gordura abdominal, todos os valores percentuais foram
expressos em relação ao peso vivo.
O modelo matemático utilizado para VC, IEP, RC e cortes comercias foi: yij = µ + ti
+ bj + eij. Onde yij = é o valor observado na parcela que recebeu tratamento i no bloco j, µ
é o efeito da média, ti é o efeito do tratamento i, bj é o efeito do bloco j e eij é o erro
experimental na parcela que recebeu o tratamento i no bloco j.
Os dados foram verificados quanto à presença de “outliers” e testou-se as
pressuposições de normalidade dos erros studentizados e de homogeneidade das
variâncias. Após constada a não violação dessas pressuposições, os dados foram
submetidos à análise de variância através do procedimento GLM do programa SAS®
(SAS Institute, 2002) e em caso de diferença significativa (5%) as médias foram
comparadas pelo teste de Tukey (5%).
Resultados
Os valores médios obtidos para o consumo de ração, o ganho de peso e a
conversão alimentar nos períodos de 1 a 7, 1 a 14, 1 a 21 e 1 a 42 dias de idade
encontram-se na Tabela 2.
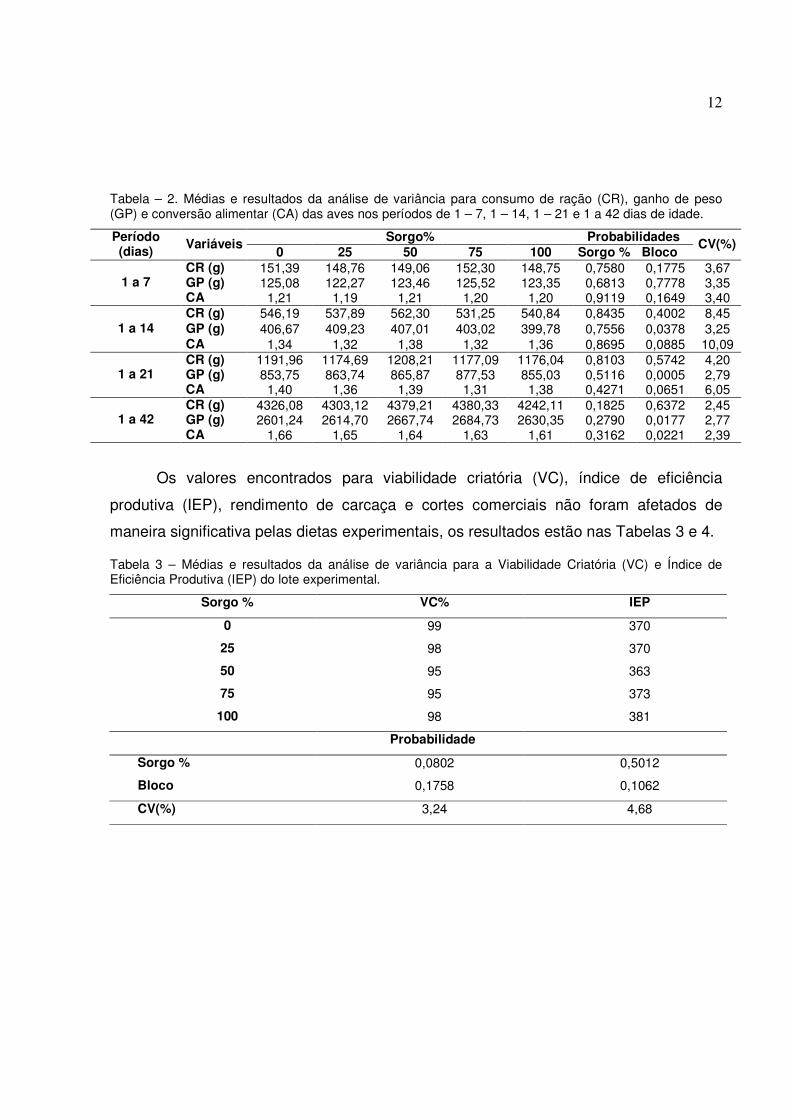
Os níveis de inclusão de sorgo baixo teor em tanino não afetaram de maneira
significativa os valores médios para consumo de ração, ganho de peso e conversão
alimentar nos períodos de 1 a 7, 1 a 14, 1 a 21 e 1 a 42 dias de idade.
12
Tabela – 2. Médias e resultados da análise de variância para consumo de ração (CR), ganho de peso (GP) e conversão alimentar (CA) das aves nos períodos de 1 – 7, 1 – 14, 1 – 21 e 1 a 42 dias de idade.
Sorgo% Probabilidades Período (dias) Variáveis
0 25 50 75 100 Sorgo % Bloco CV(%)
CR (g) 151,39 148,76 149,06 152,30 148,75 0,7580 0,1775 3,67 GP (g) 125,08 122,27 123,46 125,52 123,35 0,6813 0,7778 3,35 1 a 7 CA 1,21 1,19 1,21 1,20 1,20 0,9119 0,1649 3,40 CR (g) 546,19 537,89 562,30 531,25 540,84 0,8435 0,4002 8,45 GP (g) 406,67 409,23 407,01 403,02 399,78 0,7556 0,0378 3,25 1 a 14 CA 1,34 1,32 1,38 1,32 1,36 0,8695 0,0885 10,09 CR (g) 1191,96 1174,69 1208,21 1177,09 1176,04 0,8103 0,5742 4,20 GP (g) 853,75 863,74 865,87 877,53 855,03 0,5116 0,0005 2,79 1 a 21 CA 1,40 1,36 1,39 1,31 1,38 0,4271 0,0651 6,05 CR (g) 4326,08 4303,12 4379,21 4380,33 4242,11 0,1825 0,6372 2,45 GP (g) 2601,24 2614,70 2667,74 2684,73 2630,35 0,2790 0,0177 2,77 1 a 42 CA 1,66 1,65 1,64 1,63 1,61 0,3162 0,0221 2,39
Os valores encontrados para viabilidade criatória (VC), índice de eficiência
produtiva (IEP), rendimento de carcaça e cortes comerciais não foram afetados de
maneira significativa pelas dietas experimentais, os resultados estão nas Tabelas 3 e 4.
Tabela 3 – Médias e resultados da análise de variância para a Viabilidade Criatória (VC) e Índice de Eficiência Produtiva (IEP) do lote experimental.
Sorgo % VC% IEP
0 99 370
25 98 370
50 95 363
75 95 373
100 98 381
Probabilidade
Sorgo % 0,0802 0,5012
Bloco 0,1758 0,1062
CV(%) 3,24 4,68
13
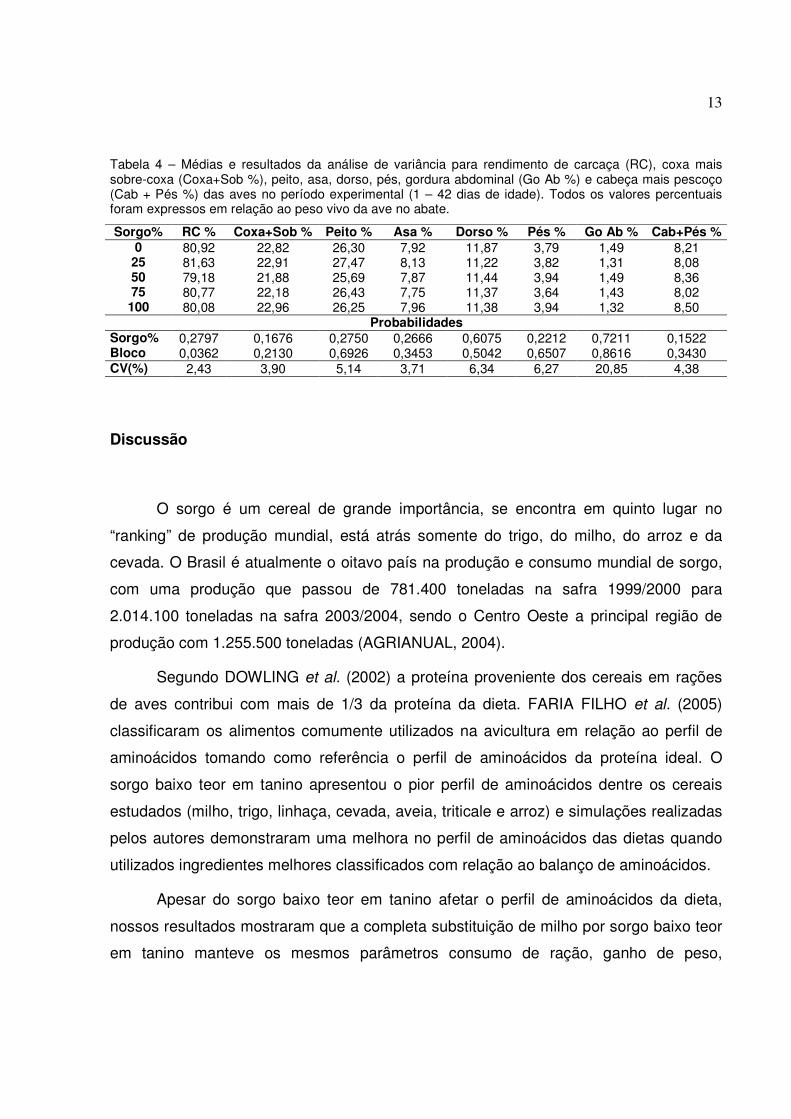
Tabela 4 – Médias e resultados da análise de variância para rendimento de carcaça (RC), coxa mais sobre-coxa (Coxa+Sob %), peito, asa, dorso, pés, gordura abdominal (Go Ab %) e cabeça mais pescoço (Cab + Pés %) das aves no período experimental (1 – 42 dias de idade). Todos os valores percentuais foram expressos em relação ao peso vivo da ave no abate.
Sorgo% RC % Coxa+Sob % Peito % Asa % Dorso % Pés % Go Ab % Cab+Pés % 0 80,92 22,82 26,30 7,92 11,87 3,79 1,49 8,21
25 81,63 22,91 27,47 8,13 11,22 3,82 1,31 8,08 50 79,18 21,88 25,69 7,87 11,44 3,94 1,49 8,36 75 80,77 22,18 26,43 7,75 11,37 3,64 1,43 8,02 100 80,08 22,96 26,25 7,96 11,38 3,94 1,32 8,50
Probabilidades Sorgo% 0,2797 0,1676 0,2750 0,2666 0,6075 0,2212 0,7211 0,1522 Bloco 0,0362 0,2130 0,6926 0,3453 0,5042 0,6507 0,8616 0,3430 CV(%) 2,43 3,90 5,14 3,71 6,34 6,27 20,85 4,38
Discussão
O sorgo é um cereal de grande importância, se encontra em quinto lugar no
“ranking” de produção mundial, está atrás somente do trigo, do milho, do arroz e da
cevada. O Brasil é atualmente o oitavo país na produção e consumo mundial de sorgo,
com uma produção que passou de 781.400 toneladas na safra 1999/2000 para
2.014.100 toneladas na safra 2003/2004, sendo o Centro Oeste a principal região de
produção com 1.255.500 toneladas (AGRIANUAL, 2004).
Segundo DOWLING et al. (2002) a proteína proveniente dos cereais em rações
de aves contribui com mais de 1/3 da proteína da dieta. FARIA FILHO et al. (2005)
classificaram os alimentos comumente utilizados na avicultura em relação ao perfil de
aminoácidos tomando como referência o perfil de aminoácidos da proteína ideal. O
sorgo baixo teor em tanino apresentou o pior perfil de aminoácidos dentre os cereais
estudados (milho, trigo, linhaça, cevada, aveia, triticale e arroz) e simulações realizadas
pelos autores demonstraram uma melhora no perfil de aminoácidos das dietas quando
utilizados ingredientes melhores classificados com relação ao balanço de aminoácidos.
Apesar do sorgo baixo teor em tanino afetar o perfil de aminoácidos da dieta,
nossos resultados mostraram que a completa substituição de milho por sorgo baixo teor
em tanino manteve os mesmos parâmetros consumo de ração, ganho de peso,

14
conversão alimentar, rendimento de carcaça e cortes comerciais dentro dos valores
apresentados por aves alimentadas com ração à base de milho e soja.
Estes achados positivos corroboram com outros autores que não observaram
diferença significativa no desempenho de aves alimentadas com sorgo baixo teor em
tanino. DOUGLAS & SULLIVAN (1991), ao substituírem milho por sorgo baixo teor em
tanino na fase inicial (1-21 dias) não observaram diferença estatística para ganho de
peso e conversão alimentar. POUR-REZA & EDRISS (1997) relataram que a
substituição de milho por sorgo baixo teor em tanino não afetou o desempenho das
aves, contudo o autor trabalhou a partir do 7° até o 49° dia de vida, não fornecendo
sorgo na primeira semana. DINIZ et al. (2002) trabalharam com a fase pré-inicial (tempo
que a parcela experimental levou para consumir 300 g/ave) até os 49 dias de idade, com
uma substituição de 50 e 100% do milho por sorgo baixo teor em tanino observaram que
os níveis de inclusão não afetaram a conversão alimentar aos 49 dias de idade. SILVA
(2003) não verificou diferença significativa no consumo de ração e conversão alimentar
aos 21 e 42 dias de idade quando o milho foi substituído por 25, 50, 75 e 100% de sorgo
baixo teor em tanino, resultados semelhantes foram encontrados por GARCIA et al.
(2005) que não encontraram diferença significativa ao trabalhar com sorgo baixo teor em
tanino nos níveis 0, 25, 50, 75 e 100% em substituição ao milho para as variáveis
consumo de ração, ganho de peso e conversão alimentar aos 21, 35 e 42 dias de idade.
Com relação ao rendimento de carcaça e cortes comerciais resultados
semelhantes foram encontrados por GARCIA et al. (2005) que não encontraram
diferença estatística para os valores de carcaça inteira, perna, asas, peito e dorso.
DINIZ et al. (2002) trabalharam com lote misto. Os machos não foram afetados pela
substituição do milho por sorgo para as porcentagens de peito, asa e dorso. No entanto,
os rendimentos de carcaça inteira e coxa + sobre-coxa foram maiores para os
tratamentos contendo 50 e 100% de sorgo baixo tanino em substituição ao milho. Nas
fêmeas as médias observadas não diferiram para carcaça inteira, peito e dorso, porém a
porcentagem de asa apresentou uma média superior para a dieta à base de sorgo e
soja.

15
Estes resultados de desempenho e características de carcaça se devem
provavelmente à suplementação de aminoácidos sintéticos 100% digestíveis como a
DL-metionina e L-lisina. No presente estudo, as diferenças na suplementação destes
aminoácidos chegaram a 50% para DL-metionina e 20% de L-lisina para a fase inicial e
37,5% de DL-metionina e 20% de L-lisina para a fase de crescimento entre as dietas 0 e
100% de sorgo baixo teor em tanino. A dieta 100% de sorgo forneceu as seguintes
quantidades de aminoácidos digestíveis 1,06% de lisina, 0,51% de metionina e 0,75%
de metionina+cistina, e a dieta controle forneceu 1,05% de lisina, 0,49% de metionina e
0,82% de metionina+cistina.
A viabilidade criatória, índice que avalia o número de aves alojadas e o número
de aves retiradas para o abate, esta relacionada à genética da ave, ao manejo utilizado,
às condições sanitárias e ambientais e também as condições nutricionais fornecidas ao
lote. No presente experimento a viabilidade criatória não sofreu efeito significativo para
os níveis de inclusão de sorgo baixo teor em tanino, indicando que este cereal não
afetou o desenvolvimento das aves nas condições em que foram criadas. GARCIA et al.
(2005) não observaram diferença significativa para mortalidade quando trabalhou com
sorgo baixo teor em tanino.
O índice de eficiência produtiva, variável que pondera o ganho de peso diário,
conversão alimentar e a viabilidade criatória também não apresentou diferença
significativa, indicando os bons resultados de desempenho encontrados nos tratamentos
experimentais.
Conclusão
O sorgo baixo teor em tanino pode substituir completamente o milho em dietas
para frangos de corte a partir do primeiro dia de idade sem comprometimento do
desempenho e o rendimento de carcaça e cortes comerciais.

16
CAPÍTULO 3 – CARACTERÍSTICAS MORFOLÓGICAS DA MUCOSA INTESTINAL
DE FRANGOS DE CORTE ALIMENTADOS COM SORGO BAIXO TANINO.
RESUMO – O experimento foi conduzido com o objetivo de avaliar a influência do
sorgo baixo teor em tanino sobre a altura dos vilos, profundidade das criptas e número
de células caliciformes no epitélio intestinal de frangos de corte de 1 – 42 dias de idade.
Foram utilizados 750 pintainhos de um dia de idade Cobb-500®, em um delineamento
em blocos casualizados. A dieta controle foi a base de milho e farelo de soja e outras
quatro dietas com substituições de 25, 50, 75 e 100% do milho por sorgo baixo teor em
tanino, com 6 repetições por tratamento totalizando 30 unidades experimentais. As
rações experimentais foram isonutrientes dentro de cada fase de criação: inicial (1-21
dias de idade) e crescimento (22-42 dias de vida). Ajustou-se o modelo de superfície de
resposta para as variáveis analisadas, considerando os efeitos lineares e quadráticos da
porcentagem do sorgo e idade bem como a interação entre eles. Os parâmetros foram
estimados pelo método dos quadrados mínimos. A altura dos vilos nos diferentes
segmentos intestinais não foi afetada pelos níveis crescentes de sorgo. A altura dos
vilos no duodeno apresentou um crescimento contínuo durante a fase de inicial de
criação. A profundidade das criptas do duodeno foram maiores para os tratamentos com
maiores níveis de sorgo na fase final de criação. O número de células caliciformes não
foi influenciado pelas dietas experimentais. O fator idade apresentou efeito significativo
sobre o número de células caliciformes onde ocorreu aumento na quantidade de células
produtoras de mucina no período de 1 a 21 dias de idade.
Palavras-Chave: sorgo, frangos de corte, células caliciformes, vilos e criptas intestinais.

17
Introdução
Oscilações nos preços dos alimentos comumente utilizados nas rações de
animais de produção levam os nutricionistas a utilizarem ingredientes alternativos para
que possam alcançar resultados de desempenho e econômicos satisfatórios, contudo
existem limitações que vão desde fatores antinutricionais à composição nutricional do
alimento.
O estudo do desenvolvimento da mucosa intestinal de frangos de corte tem sido o
foco de diversas pesquisas devido a sua crucial importância no processo de digestão e
absorção dos alimentos. Após o nascimento, os pintainhos dependentes dos nutrientes
do saco vitelínico passam a utilizar os nutrientes provenientes da ração, passando de
uma fonte de nutrientes baseada em gorduras para uma fonte baseada principalmente
em carboidratos e proteínas. Assim, os pintainhos utilizam suas limitadas reservas
corporais (saco vitelínico) para conseguir um rápido desenvolvimento físico e funcional
do trato gastrintestinal, a fim de desenvolver a capacidade de digerir alimentos e
assimilar nutrientes (UNI & FERKET, 2004). De acordo com MAIORKA et al. (2002) o
desenvolvimento da mucosa intestinal envolve o crescimento dos vilos, o que
corresponde a um aumento em número de células epiteliais (enterócitos, células
caliciformes e enteroendócrinas).
As células caliciformes, responsáveis pela produção de uma camada de muco
composta principalmente por glicoproteínas que são conhecidas como mucinas, que
contêm uma longa porção polissacarídica tornando-as hidrofílicas e viscosas, as
funções das mucinas são: de proteger a membrana em escova contra agressões
químicas e efeitos abrasivos da digesta, atuar como barreira contra microrganismos,
pois integrada ao muco encontra-se a flora intestinal natural e imunoglobulinas e
influenciar o transporte entre o conteúdo luminal e a membrana em escova (BOLELI, et
al., 2002; UNI et al., 2003). A hiperplasia e/ou hipertrofia das células caliciformes no
intestino delgado é visto como resposta a algum tipo de agressão visando manter a
integridade da mucosa. Diversos trabalhos (MITJAVILA et al., 1977; ORTIZ et al., 1994;

18
OLIVEIRA et al., 2000 e UNI et al., 2003) relataram este efeito por diferentes razões, tais
como os fatores antinutricionais dos alimentos ou o atraso no fornecimento de alimento
após eclosão.
O sorgo (Sorghum bicolor (L.) Moench) é tido como um dos principais substitutos
ao milho em rações de aves e suínos. Este cereal contém vários compostos fenólicos
que podem afetar a cor, a aparência e qualidade nutricional. Esses compostos podem
ser classificados em três grupos básicos: ácidos fenólicos e flavonóides (inócuos aos
animais) e o composto fenólico antinutricional conhecido como tanino. As
proantocianidinas ou tanino estão ligados à coagulação e precipitação de enzimas e
alguns minerais, formação de complexos com os carboidratos da dieta, inibição da
atividade de algumas enzimas digestivas, como a tripsina e α-amilase. Quando
ingeridos em certas quantidades, reduzem a taxa de crescimento do animal, uma vez
que diminui o aproveitamento energético e protéico da dieta levando a maior excreção
de nitrogênio nas fezes como resultado da interação tanino-proteínas, formadas por
múltiplas pontes de hidrogênio. Os taninos condensados também causam necroses e
distorções nos vilos da mucosa intestinal e aumento no número de células caliciformes
(MITJAVILA et al., 1977; TREVINO, 1992; CHANG et al., 1994; ORTIZ et al., 1994;
OLIVEIRA, 1998). De acordo com JIMENEZ et al. (1994), os taninos condensados não
são absorvidos pela mucosa intestinal de frangos, mas os demais compostos fenólicos
são absorvidos e distribuídos em vários tecidos do organismo.
Os cultivares de sorgo que são vendidos atualmente no Brasil possuem pequena
quantidade de tanino condensado nos grãos. Assim, o objetivo deste trabalho foi avaliar
o desenvolvimento da mucosa intestinal de frangos de corte alimentados com níveis
crescentes de sorgo baixo teor em tanino em substituição ao milho.
Material e Métodos
O experimento foi conduzido no setor de Avicultura do Departamento de
Zootecnia da Faculdade de Ciências Agrárias e Veterinárias, UNESP, Jaboticabal-SP.

19
Foram utilizadas duas câmaras climáticas com 16 unidades experimentais com 2,5 x
1,6m.
Foram utilizados 750 frangos machos Cobb-500®, distribuídos em um
delineamento com blocos casualizados. O objetivo dos blocos foi controlar diferenças de
temperatura, umidade e gases nas diferentes câmaras. Na chegada dos pintainhos foi
realizada uma pesagem de uma amostra do lote para a determinação do peso médio e
foram montadas unidades experimentais com animais pesando em média 42,48 ± 0,56g.
Os tratamentos consistiram na administração ad-libitum de uma dieta controle a base de
milho e farelo de soja e outras quatro dietas com substituições de 25, 50, 75 e 100% do
milho por sorgo baixo teor em tanino, com 6 repetições por tratamento totalizando 30
unidades experimentais. As rações experimentais foram isonutrientes dentro de cada
fase de criação: inicial (1-21 dias de idade) e crescimento (22-42 dias de vida). Foram
formuladas seguindo a composição nutricional dos ingredientes e os níveis nutricionais
de acordo com ROSTAGNO et al. (2000) (Tabela1).
O sorgo utilizado no presente experimento teve 4,52 mg de catequina por grama
de amostra. O conteúdo total de tanino foi analisado pelo método da Vanilina – HCl, este
método baseia-se na reação da leucoantocianidina (catequina) e proantocianidina
(tanino condensado) com vanilina na presença de HCl, é um método específico para a
determinação de tanino condensado e alguns flavonóides.
Os animais foram vacinados contra doença de Marek e Bouba Aviária no
incubatório. No 8° dia de idade as aves foram vacinadas (via ocular) contra as doenças
de Newcastle e Gumboro e no 14° dia contra a doença de Gumboro na água de bebida.

20
Tabela 1 – Composição centesimal das rações experimentais do período inicial (1 - 21 dias de idade) e crescimento (22 - 42 dias de idade).
* Suplementadas a fim de alcançar os níveis nutricionais de ROSTAGNO et al. (2000). 1 Níveis de garantia por quilograma do produto: selênio 54,6mg; ácido nicotínico 6930mg; cobre 25000mg; pantotenato de cálcio 1900mg; biotina 32mg; manganês 15252mg; DL-metionina (mínimo) 340g; iodo 260mg; coccidiostático 25000mg; antioxidante 100mg; colina 120mg; promotor de crescimento e eficiência alimentar 10000mg; vitamina A 1400000U.I.; vitamina B1 356mg; vitamina B12 2000mcg; vitamina B2 1920mg; vitamina B6 693mg; vitamina D3 600000U.I.; vitamina E 5000mg; vitamina K 196mg; zinco 18250mg. 2 Níveis de garantia por quilograma do produto: selênio 54,6mg; ácido nicotínico 6930mg; cobre 25000mg; pantotenato de cálcio
1900mg; biotina 32mg; manganês 15252mg; DL-metionina (mínimo) 270g; iodo 260mg; coccidiostático 22000mg; antioxidante 100mg; colina 120mg; promotor de crescimento e eficiência alimentar 10000mg; vitamina A 1400000U.I.; vitamina B1 356mg; vitamina B12 2000mcg; vitamina B2 1920mg; vitamina B6 693mg; vitamina D3 600000U.I.; vitamina E 5000mg; vitamina K 196,5mg; zinco 18250mg.
No 2°, 8°, 14°, 21° e 41° dia de vida, uma ave por repetição foi sacrificada por
deslocamento cervical e coletadas duas partes de cada região do intestino delgado
(duodeno: a partir do piloro até a porção distal da alça duodenal; jejuno: a partir da
porção distal da alça duodenal até o divertículo de Meckel e íleo: entre o divertículo de
Meckel e a abertura dos cecos). Após a coleta, um dos segmentos foi fixado em Bouin
por 24 horas para a determinação de mucina neutra, e o outro em Paraformaldeído por
48 horas para a determinação de mucina ácida, altura de vilo e profundidade de cripta.
Após a fixação ambos os segmentos foram desidratados, em uma série de
concentração crescente de etanol (50, 60, 70, 80, 90 e 100%), diafanizados (xilol I e xilol
II), infiltrados e incluídos em parafina e finalmente cortados.
INICIAL CRESCIMENTO Ingredientes % 0% 25% 50% 75% 100% 0% 25% 50% 75% 100%
Milho 54,89 41,17 27,45 13,72 - 60,33 45,25 30,17 15,08 - Sorgo baixo tanino - 13,72 27,45 41,17 54,89 - 15,08 30,17 45,25 60,33 Farelo de Soja, 45 36,31 36,22 36,12 36,03 35,94 30,61 30,51 30,40 30,30 30,20 Óleo de soja 3,72 4,02 4,32 4,63 4,93 4,24 4,57 4,90 5,24 5,57 Fosfato bicálcico 1,82 1,82 1,81 1,80 1,80 1,62 1,61 1,61 1,60 1,59 Calcário Calcítico 0,99 0,99 0,99 0,99 0,99 0,94 0,94 0,94 0,94 0,94 Sal Comum 0,46 0,46 0,46 0,46 0,47 0,39 0,39 0,39 0,40 0,40 DL-metionina* 0,06 0,07 0,08 0,08 0,09 0,08 0,09 0,09 0,10 0,11 L-lisina* 0,15 0,16 0,17 0,17 0,18 0,20 0,21 0,22 0,23 0,24 Suplemento mineral e vitamínico 0,50
1 0,50 0,50 0,50 0,50 0,502 0,50 0,50 0,50 0,50
Caulim 1,10 0,88 0,65 0,43 0,21 1,10 0,85 0,60 0,36 0,12 Total 100 100 100 100 100 100 100 100 100 100
Energia e Nutrientes Composição Calculada Energia Metabolizável(Kcal) 3000 3000 3000 3000 3000 3100 3100 3100 3100 3100 Proteína bruta (%) 21,40 21,40 21,40 21,40 21,40 19,31 19,31 19,31 19,31 19,31 Cálcio (%) 0,96 0,96 0,96 0,96 0,96 0,87 0,87 0,87 0,87 0,87 Fósforo Disponível (%) 0,45 0,45 0,45 0,45 0,45 0,41 0,41 0,41 0,41 0,41 Sódio (%) 0,22 0,22 0,22 0,22 0,22 0,19 0,19 0,19 0,19 0,19 Lisina (%) 1,26 1,26 1,26 1,26 1,26 1,16 1,16 1,16 1,16 1,16 Metionina (%) 0,56 0,56 0,56 0,56 0,56 0,51 0,51 0,51 0,51 0,51 Aminoácidos sulfurados (%) 0,90 0,90 0,90 0,90 0,90 0,82 0,82 0,82 0,82 0,82

21
Para a determinação das mucinas neutras (PAS+), cortes semi-seriados com 6
µm de espessura, em um total de cinco cortes por ave, foram desparafinizadas (Xilol I e
II), hidratadas (álcool absoluto I e II), coradas com ácido periódico 5% (cinco minutos),
um rápido banho de água destilada, seguido de um banho de água destilada à 50ºC,
incubação com reativo de Schiff por quinze minutos (acompanhando a coloração do
tecido), um banho de água sulfurosa (10mL de metabissulfito de sódio, mais 10 mL
ácido clorídrico 1N com 200 mL de água destilada), um banho com água corrente
seguido de um banho de água destilada. Após esta bateria de coloração as lâminas
foram desidratadas, diafanizadas e finalmente montadas.
Para determinação de mucinas ácidas (AB+), altura de vilo e profundidade de
cripta, cortes semi-seriados com 5 µm de espessura, em um total de cinco cortes por
ave, foram desparafinizadas, re-hidratadas e incubadas em uma solução de 3% ácido
acético por três minutos, seguidas de uma incubação de dez minutos em uma solução
de Alcian Blue (AB 1% em 3% de ácido acético com pH 2,5). Sendo lavadas com água
destilada e um rápido banho de eosina. Após sofrerem novo banho com água destilada,
as lâminas foram desidratadas, diafanizadas e montadas.
O número de células caliciformes foi contado ao longo de um segmento de
epitélio do vilo equivalente a 200 µm, em um total de oito vilos por ave (nº células
caliciformes / 200 µm de epitélio do vilo), através de microscopia de luz.
As imagens das lâminas AB+ foram digitalizadas e mensuradas a altura dos vilos
(µm) e a profundidade das criptas (µm) para cada segmento do intestino delgado
através do programa Image J®, onde cada repetição teve um mínimo de 30 medidas de
altura de vilo e profundidade de cripta.
Ajustou-se o modelo de superfície de resposta para as variáveis analisadas,
considerando os efeitos lineares e quadráticos de porcentagem de substituição de milho
por sorgo baixo tanino (sorgo%) e idade bem como a interação entre eles. Os
parâmetros foram estimados pelo método dos quadrados mínimos. Os dados foram
verificados quanto à presença de “outliers” e testou-se as pressuposições de
normalidade dos erros studentizados e de homogeneidade de variância. As análises
foram realizadas pelo general linear models do programa Statistica® 6.0.
22
Resultados
As estimativas dos parâmetros do modelo de superfície de resposta para altura
do vilo e profundidade da cripta do duodeno encontram-se na Tabela 2.
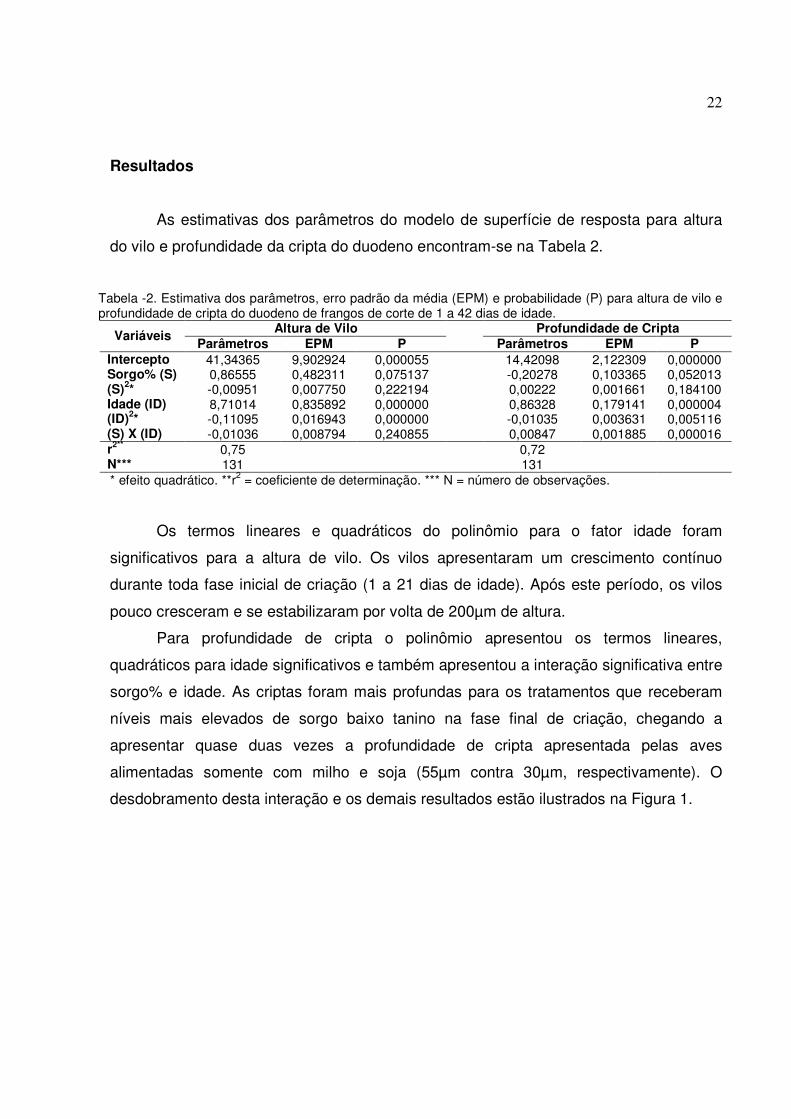
Tabela -2. Estimativa dos parâmetros, erro padrão da média (EPM) e probabilidade (P) para altura de vilo e profundidade de cripta do duodeno de frangos de corte de 1 a 42 dias de idade.
Altura de Vilo Profundidade de Cripta Variáveis Parâmetros EPM P Parâmetros EPM P Intercepto 41,34365 9,902924 0,000055 14,42098 2,122309 0,000000 Sorgo% (S) 0,86555 0,482311 0,075137 -0,20278 0,103365 0,052013 (S)2* -0,00951 0,007750 0,222194 0,00222 0,001661 0,184100 Idade (ID) 8,71014 0,835892 0,000000 0,86328 0,179141 0,000004 (ID)2* -0,11095 0,016943 0,000000 -0,01035 0,003631 0,005116 (S) X (ID) -0,01036 0,008794 0,240855 0,00847 0,001885 0,000016 r2** 0,75 0,72 N*** 131 131 * efeito quadrático. **r2 = coeficiente de determinação. *** N = número de observações.
Os termos lineares e quadráticos do polinômio para o fator idade foram
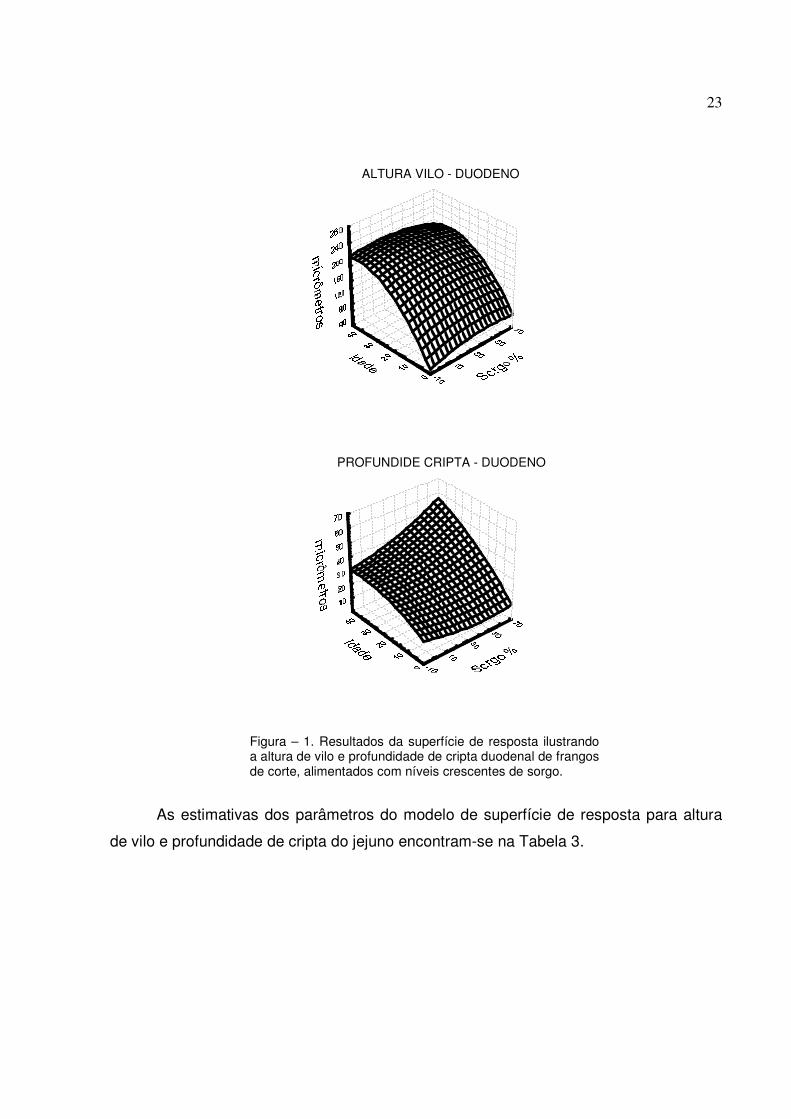
significativos para a altura de vilo. Os vilos apresentaram um crescimento contínuo
durante toda fase inicial de criação (1 a 21 dias de idade). Após este período, os vilos
pouco cresceram e se estabilizaram por volta de 200µm de altura.
Para profundidade de cripta o polinômio apresentou os termos lineares,
quadráticos para idade significativos e também apresentou a interação significativa entre
sorgo% e idade. As criptas foram mais profundas para os tratamentos que receberam
níveis mais elevados de sorgo baixo tanino na fase final de criação, chegando a
apresentar quase duas vezes a profundidade de cripta apresentada pelas aves
alimentadas somente com milho e soja (55µm contra 30µm, respectivamente). O
desdobramento desta interação e os demais resultados estão ilustrados na Figura 1.
23
ALTURA VILO - DUODENO
PROFUNDIDE CRIPTA - DUODENO
Figura – 1. Resultados da superfície de resposta ilustrando a altura de vilo e profundidade de cripta duodenal de frangos de corte, alimentados com níveis crescentes de sorgo.
As estimativas dos parâmetros do modelo de superfície de resposta para altura
de vilo e profundidade de cripta do jejuno encontram-se na Tabela 3.
24
Tabela -3. Estimativa dos parâmetros, erro padrão da média (EPM) e probabilidade (P) para altura de vilo e profundidade de cripta do jejuno de frangos de corte de 1 a 42 dias de idade.
Altura de Vilo Profundidade de Cripta Variáveis Parâmetros EPM P Parâmetros EPM P Intercepto 35,36205 6,952254 0,000001 5,620623 1,743696 0,001619 Sorgo% (S) -0,18261 0,362608 0,615426 -0044692 0,090946 0,624002 (S)2* 0,00517 0,006428 0,422967 0,001440 0,001612 0,373584 Idade (ID) 7,91334 0,634315 0,000000 1,729393 0,159093 0,000000 (ID)2* -0,12117 0,013466 0,000000 -0,022561 0,003377 0,000000 (S) X (ID) -0,00924 0,008220 0,263063 -0,001450 0,002062 0,483170 r2** 0,70 0,73 N*** 130 130 * efeito quadrático. **r2 = coeficiente de determinação. *** N = número de observações.
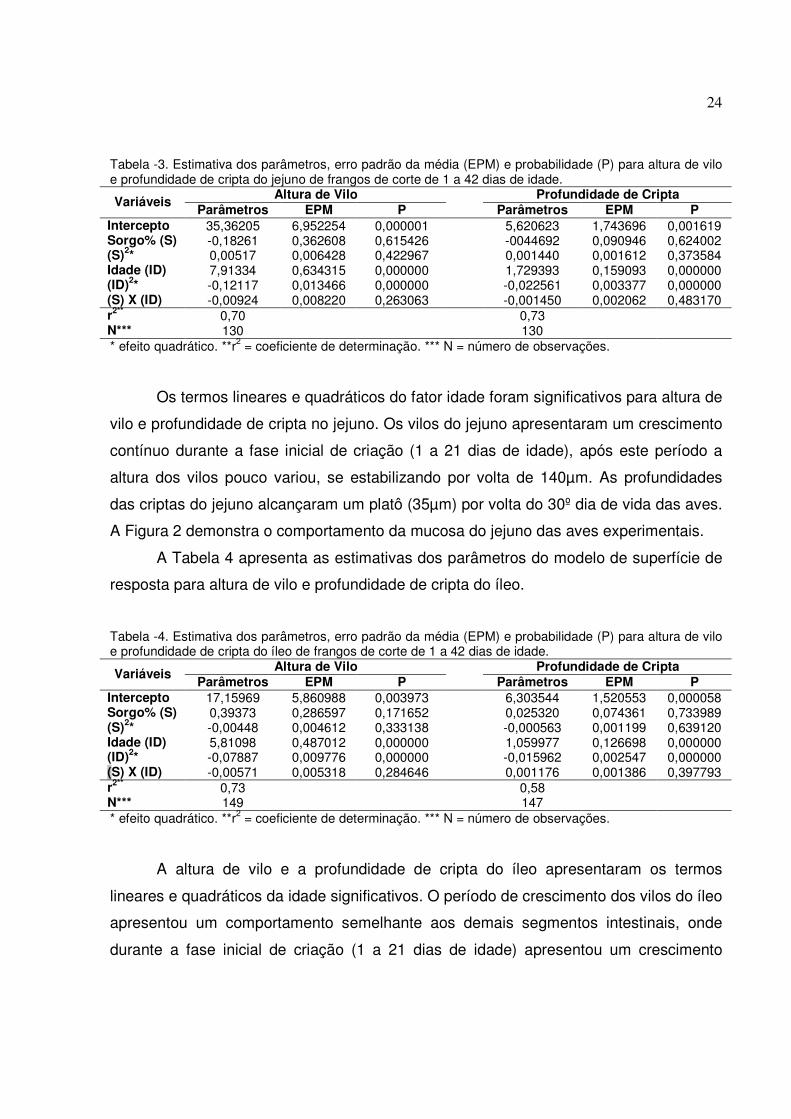
Os termos lineares e quadráticos do fator idade foram significativos para altura de
vilo e profundidade de cripta no jejuno. Os vilos do jejuno apresentaram um crescimento
contínuo durante a fase inicial de criação (1 a 21 dias de idade), após este período a
altura dos vilos pouco variou, se estabilizando por volta de 140µm. As profundidades
das criptas do jejuno alcançaram um platô (35µm) por volta do 30º dia de vida das aves.
A Figura 2 demonstra o comportamento da mucosa do jejuno das aves experimentais.
A Tabela 4 apresenta as estimativas dos parâmetros do modelo de superfície de
resposta para altura de vilo e profundidade de cripta do íleo.
Tabela -4. Estimativa dos parâmetros, erro padrão da média (EPM) e probabilidade (P) para altura de vilo e profundidade de cripta do íleo de frangos de corte de 1 a 42 dias de idade.
Altura de Vilo Profundidade de Cripta Variáveis Parâmetros EPM P Parâmetros EPM P Intercepto 17,15969 5,860988 0,003973 6,303544 1,520553 0,000058 Sorgo% (S) 0,39373 0,286597 0,171652 0,025320 0,074361 0,733989 (S)2* -0,00448 0,004612 0,333138 -0,000563 0,001199 0,639120 Idade (ID) 5,81098 0,487012 0,000000 1,059977 0,126698 0,000000 (ID)2* -0,07887 0,009776 0,000000 -0,015962 0,002547 0,000000 (S) X (ID) -0,00571 0,005318 0,284646 0,001176 0,001386 0,397793 r2** 0,73 0,58 N*** 149 147 * efeito quadrático. **r2 = coeficiente de determinação. *** N = número de observações.
A altura de vilo e a profundidade de cripta do íleo apresentaram os termos
lineares e quadráticos da idade significativos. O período de crescimento dos vilos do íleo
apresentou um comportamento semelhante aos demais segmentos intestinais, onde
durante a fase inicial de criação (1 a 21 dias de idade) apresentou um crescimento
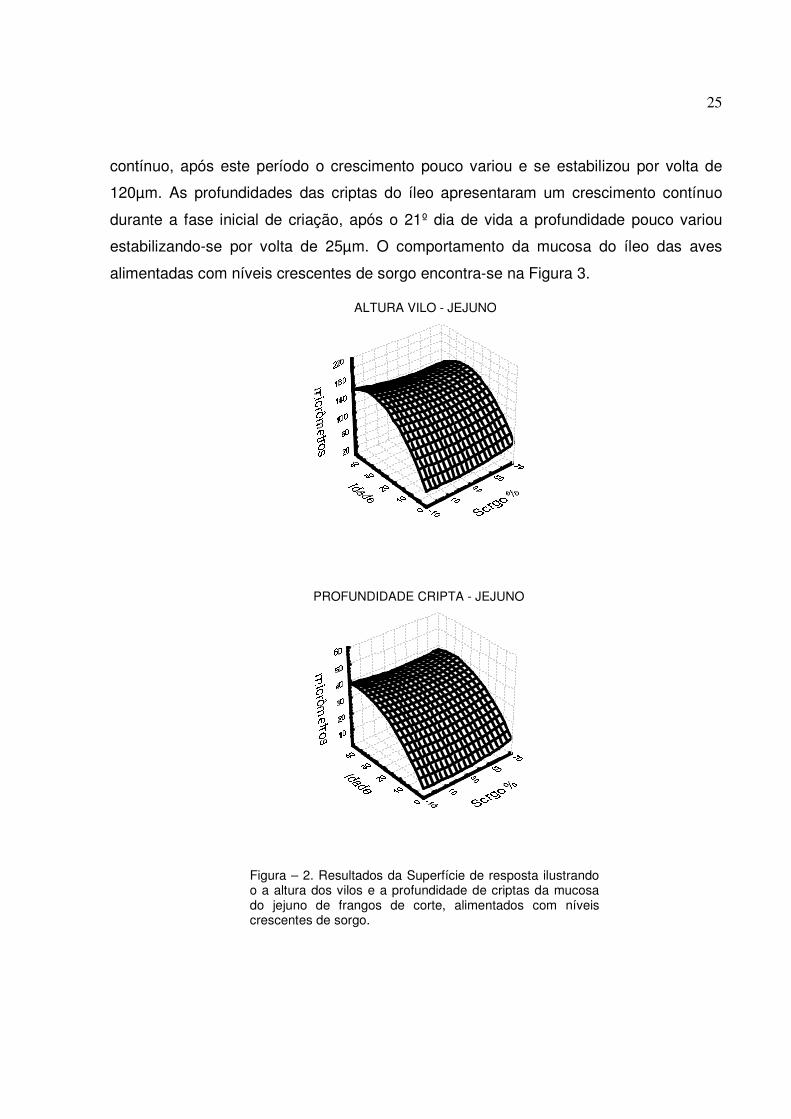
25
contínuo, após este período o crescimento pouco variou e se estabilizou por volta de
120µm. As profundidades das criptas do íleo apresentaram um crescimento contínuo
durante a fase inicial de criação, após o 21º dia de vida a profundidade pouco variou
estabilizando-se por volta de 25µm. O comportamento da mucosa do íleo das aves
alimentadas com níveis crescentes de sorgo encontra-se na Figura 3.
ALTURA VILO - JEJUNO
PROFUNDIDADE CRIPTA - JEJUNO
Figura – 2. Resultados da Superfície de resposta ilustrando o a altura dos vilos e a profundidade de criptas da mucosa do jejuno de frangos de corte, alimentados com níveis crescentes de sorgo.

26
ALTURA VILO - ÍLEO
PROFUNDIDADE CRIPTA - ÍLEO
Figura – 3. Resultados da superfície de resposta ilustrando a altura dos vilos e a profundidade das criptas da mucosa do íleo de frangos de corte, alimentados com níveis crescentes de sorgo.
A Tabela 5 apresenta as estimativas dos parâmetros do modelo de superfície de
resposta para o número de células caliciformes que produzem mucina neutra (PAS+) e
mucina ácida (AB+) por 200 µm de epitélio duodenal.
Os termos lineares e quadráticos do fator idade apresentaram efeito significativo
para o número de células que produzem mucina neutra e ácida. Houve um aumento no
número de ambas as células durante a fase inicial de criação, seguido de um

27
decréscimo no número de células depois deste período. Estes resultados estão
ilustrados na Figura 4.
Tabela -5. Estimativa dos parâmetros, erro padrão da média (EPM) e probabilidade (P) para o número de células caliciformes que produzem mucina neutra (PAS+) e mucina ácida (AB+) por 200 µm de epitélio duodenal de frangos de corte de 1 a 42 dias de idade.
Células PAS+ Células AB+ Variáveis Parâmetros EPM P Parâmetros EPM P
Intercepto 11,47492 0,722849 0,000000 7,176561 0,487889 0,000000 Sorgo% (S) 0,01393 0,033175 0,675304 -0,002573 0,026122 0,921695 (S)2* -0,00006 0,000537 0,911209 0,000116 0,000437 0,791825 Idade (ID) 0,31977 0,058861 0,000000 0,489407 0,043771 0,000000 (ID)2* -0,00701 0,001162 0,000000 -0,010046 0,000915 0,000000 (S) X (ID) 0,00019 0,000644 0,771982 -0,000245 0,000522 0,639748 r2** 0,25 0,53 N*** 122 109 * efeito quadrático. **r2 = coeficiente de determinação. *** N = número de observações.
As estimativas dos parâmetros do modelo de superfície de resposta para o
número de células caliciformes que produzem mucina neutra (PAS+) e mucina ácida
(AB+) por 200 µm de epitélio de jejuno encontram-se na Tabela 6.
Tabela - 6. Estimativa dos parâmetros, erro padrão da média (EPM) e probabilidade (P) para o número de células caliciformes que produzem mucina neutra (PAS+) e mucina ácida (AB+) por 200 µm de epitélio de jejuno de frangos de corte de 1 a 42 dias de idade.
Células PAS+ Células AB+ Variáveis Parâmetros EPM P Parâmetros EPM P Intercepto 12,63138 0,682035 0,000000 7,733326 0,465564 0,000000 Sorgo% (S) 0,04697 0,033679 0,165674 -0,022422 0,026189 0,393881 (S)2* -0,00123 0,000552 0,027201 0,000468 0,000449 0,300314 Idade (ID) 0,33428 0,059070 0,000000 0,556361 0,043988 0,000000 (ID)2* -0,00793 0,001194 0,000000 -0,011684 0,000930 0,000000 (S) X (ID) 0,00070 0,000676 0,304764 0,000124 0,000526 0,814125 r2** 0,31 0,64 N*** 127 109 * efeito quadrático. **r2 = coeficiente de determinação. *** N = número de observações.
Os termos quadráticos para o fator sorgo% e idade e o termo linear do fator idade
apresentaram efeitos significativos para o número de células caliciformes produtoras de
mucina neutra (PAS+). Para o número de células produtoras de mucina ácida (AB+)
somente o fator idade apresentou os termos do polinômio linear e quadrático
significativo.

28
Os números de células caliciformes que produzem mucina neutra e ácida
aumentaram durante os primeiros 21 dias de vida, após este período, o número de
células diminui acentuadamente.
As médias de células produtoras de mucina neutra (PAS+) no jejuno foram
maiores para os tratamentos que apresentaram quantidades similares de sorgo e milho,
para as dietas contento somente milho soja ou sorgo soja as médias foram menores. As
ilustrações destes resultados estão na Figura 5.
Número de Células PAS+ DUODENO
Número de Células AB+ DUODENO
Figura – 4. Gráfico de superfície de resposta ilustrando o número de células caliciformes produtoras de mucina por 200µm de epitélio duodenal das aves alimentadas com níveis crescentes de sorgo.

29
Na Tabela 7 encontram-se as estimativas dos parâmetros do modelo de superfície
de resposta para o número de células caliciformes que produzem mucina neutra (PAS+)
e mucina ácida (AB+) por 200 µm de epitélio de íleo.
Tabela -7. Estimativa dos parâmetros, erro padrão da média (EPM) e probabilidade (P) para o número de células caliciformes que produzem mucina neutra (PAS+) e mucina ácida (AB+) por 200µm de epitélio de íleo de frangos de corte de 1 a 42 dias de idade.
Células PAS+ Células AB+ Variáveis Parâmetros EPM P Parâmetros EPM P Intercepto 11,71879 0,738472 0,000000 7,575651 0,524105 0,000000 Sorgo% (S) -0,00397 0,038797 0,918587 0,017454 0,027177 0,521955 (S)2* 0,00037 0,000643 0,565068 0,000115 0,000472 0,807726 Idade (ID) 0,52183 0,061442 0,000000 0,583985 0,046061 0,000000 (ID)2* -0,01038 0,001248 0,000000 -0,011568 0,000963 0,000000 (S) X (ID) -0,00089 0,000764 0,247061 -0,000747 0,000553 0,178845 r2** 0,38 0,59 N*** 133 124 * efeito quadrático. **r2 = coeficiente de determinação. *** N = número de observações.
Os termos lineares e quadráticos do polinômio foram significativos para fator
idade para o número de células caliciformes que produzem mucina neutra e ácida. A
quantidade de células caliciformes do íleo apresentou um comportamento semelhante
ao número de células do duodeno, onde as médias aumentaram nos primeiros 21 dias
de idade e sofreram uma queda após este período. O número de células caliciformes
durante o período experimental no íleo está ilustrado na Figura 6.

30
Número de Células PAS+ JEJUNO
Número de Células AB+ JEJUNO
Figura – 5. Gráfico de superfície de resposta ilustrando o número de células caliciformes produtoras de mucina por 200µm de epitélio de jejuno das aves alimentadas com níveis crescentes de sorgo.

31
Número de Células PAS+ ÍLEO
Número de Células AB+ ÍLEO
Figura – 6. Gráfico de superfície de resposta ilustrando o número de células caliciformes produtoras de mucina por 200µm de epitélio de íleo das aves alimentadas com níveis crescentes de sorgo.

32
Discussão
Após o nascimento, as aves passam por uma abrupta mudança na fonte de
suprimentos, os nutrientes do saco vitelínico são substituídos por nutrientes de uma
dieta exógena rica em carboidratos e proteínas. A ingestão desta nova fonte é seguida
por um rápido crescimento do trato gastrintestinal e órgãos anexos para assimilar o
alimento que está sendo ingerido (UNI et al., 1998). A massa intestinal aumenta mais
rápido do que os demais órgãos da ave, alcançando um pico por volta do sexto dia de
vida (UNI et al., 1999). A interação entre o crescimento intestinal, funções digestivas e a
dieta são críticas durante este período. Diversas pesquisas sobre nutrição durante a
primeira semana tem sido intensificadas e difundidas devido à alta correlação entre o
peso dos pintainhos no sétimo dia de vida e seu respectivo peso ao abate (ROCHA et
al., 2003). O sorgo atualmente comercializado em território nacional não possui altas
concentrações do composto fenólico antinutricional conhecido como tanino, apesar
desta evidência existe uma resistência de se utilizar este cereal em altas concentrações
em dietas pré-iniciais. A manutenção da mucosa intestinal, em condições fisiológicas
normais, tem custo energético elevado para o frango. Quando ocorrem lesões, além da
redução da quantidade de substrato digerido e absorvido, há ainda o custo da
restauração desse epitélio. A energia conservada pelo reduzido “turnover” de células no
epitélio intestinal poderá ser utilizada para o desenvolvimento da massa muscular
(MAIORKA et al., 2002).
No presente estudo os níveis crescentes de sorgo baixo tanino não afetou a
altura dos vilos do duodeno, jejuno e íleo em nenhuma idade avaliada. No entanto, a
profundidade de cripta do duodeno foi afetada significativamente pela interação entre os
níveis de inclusão de sorgo e a idade, onde as criptas foram mais profundas nos
tratamentos com os maiores níveis de inclusão de sorgo na fase final de criação.
Devemos ressaltar que a cripta é responsável por 55% da capacidade de proliferação
celular do intestino (BOLELI et al., 2002) e uma cripta maior nos indica um maior
potencial de proliferação celular. COOK & BIRD (1973) avaliando parâmetros
morfométricos e a taxa de migração das células de criptas de aves “germ-free” e aves

33
criadas em um ambiente convencional, encontraram que as aves convencionais
apresentaram criptas 54% maiores e maiores taxas de migração celular, e que o
aumento na taxa de extrusão de algum modo é um estímulo à capacidade de
proliferação e crescimento das criptas destas aves. Portanto, no duodeno onde
encontramos criptas com tamanhos diferentes e vilos com alturas semelhantes inferimos
que ocorreu uma maior proliferação e extrusão nos tratamentos com maiores criptas, ou
seja, ocorreu um maior “turnover” e um maior gasto energético para manutenção deste
epitélio.
O duodeno e o jejuno são as porções do intestino delgado que apresentam
maiores taxas de proliferação e migração celular, maiores vilos e criptas e respondem a
agressões e a estímulos da mucosa de maneira mais acentuada (UNI et al., 1998;
OLIVEIRA et al., 2000; GEYRA et al., 2001). Dessa forma, qualquer tipo de perturbação
no intestino delgado leva a alterações morfológicas nestas duas regiões, assim a
alteração das criptas do duodeno na fase final de criação não é um resultado ilógico.
Porém, alguns autores relatam a capacidade de adaptação de aves mais velhas a
fatores estressantes como lectinas e tanino (D’MELLO &THOMAS, 1978; CHUBB, 1982;
OLIVEIRA et al., 2000). Portanto, a alteração morfométrica das criptas do duodeno na
fase final de criação pode ser uma resposta adaptativa da mucosa intestinal a um efeito
acumulativo da presença de proantocianidinas do sorgo.
A altura dos vilos e a profundidade das criptas aumentaram conforme a idade. No
entanto, o crescimento dos vilos no duodeno teve um comportamento diferente do
encontrado na literatura. De acordo com UNI (1999), o crescimentos dos vilos do
duodeno está praticamente completo aos sete dias de vida. Neste experimento, a altura
dos vilos para todos os tratamentos apresentou um crescimento contínuo durante toda
fase inicial (de 1 até 21 dias de idade) das aves.
A camada de mucina no intestino delgado possui uma importante função
protetora para as células da mucosa e de transporte entre o lúmen e a membrana em
escova, tendo extensivas implicações nas funções intestinais. A relevância das distintas
mucinas produzidas ainda não é bem entendida, no entanto, tem sido sugerido que a
mucina ácida funciona como uma barreira bacteriana dificultando a locomoção e a

34
mucina neutra parece ser menos degradável pelas glicosidades bacterianas e proteases
endógenas. O alimento fornecido, a taxa de migração das células da mucosa intestinal e
a população microbiana podem afetar o tipo de mucina e a quantidade produzida (UNI et
al., 2003).
As análises em superfície de resposta dos diferentes segmentos intestinais
durante o período experimental, mostraram que os números de células caliciformes que
produzem mucina neutra e ácida não foram afetados pelos níveis crescentes de sorgo
baixo tanino. Estes resultados evidenciaram que os maiores níveis de inclusão de sorgo
não fornece quantidade suficiente de tanino condensado para aumentar o número de
células caliciformes, mesmo na fase pré-inicial onde o animal se encontra em processo
de maturação do trato gastrintestinal. Diversos autores trabalhando com alimentos que
possuem concentrações elevadas de fatores antinutricionais, tais como: lectinas e tanino
e diferentes modelos biológicos (ratos e frangos), apresentaram um aumento no número
de células caliciformes no epitélio do trato gastrintestinal como resposta às agressões
causadas por tais substâncias (OLIVEIRA et al., 2000; ORTIZ et al., 1994; MITJAVILA et
al., 1977).
O fator idade demonstrou ter grande importância no número de células
caliciformes que produzem mucina neutra e ácida. Durante o período de maior
crescimento do intestino delgado (as primeiras três semanas) as aves apresentaram um
número crescente de células caliciformes. Quando a velocidade de crescimento do
epitélio diminuiu, o número de células caliciformes também decresceu. UNI et al. (2003)
demonstraram que após a eclosão até o 8° dia a proporção de células caliciformes
aumentou com a idade, porém em uma constante proporção aos enterócitos. No
presente trabalho, dentro de um mesmo tamanho de epitélio intestinal (200µm) o
número de células caliciformes variou com a idade. Talvez a necessidade da produção
de mucina não seja a mesma com o avançar da idade devido a mudanças na taxa de
proliferação celular, capacidade digestiva, mudança na flora intestinal ou a adaptação à
dieta utilizada.

35
Conclusão
A altura dos vilos e o número de células caliciformes do intestino não são
afetados pelos níveis de substituição de milho por sorgo baixo tanino, no entanto, a
profundidade de cripta do duodeno aumenta com a inclusão de sorgo nas rações.
A idade apresenta efeito significativo sobre o número de células caliciformes onde
ocorre aumento na quantidade de células produtoras de mucina no período de 1 a 21
dias de idade. A altura dos vilos no duodeno atinge seu desenvolvimento máximo aos 21
dias de idade.

36
CAPÍTULO 4 – USO DA REVISÃO SISTEMÁTICA DA LITERATURA E MODELOS
MATEMÁTICOS PARA PREDIZER O DESEMPENHO DE FRANGOS DE CORTE
ALIMENTADOS COM DIETAS À BASE DE MILHO E SORGO.
RESUMO – Objetivo desta pesquisa foi criar modelos de superfície de resposta, com
base nos dados selecionados pela revisão sistemática da literatura, para as variáveis
independentes, níveis de substituição do milho por sorgo e idade, para predição do
desempenho zootécnico. Localizaram-se nove trabalhos através do método de revisão
sistemática da literatura, no período de 1995 a janeiro de 2006. Três trabalhos da
literatura mais um trabalho não publicado do nosso grupo de pesquisa se adequaram
aos critérios de inclusão estabelecidos: 1) efeito da substituição do milho por sorgo
baixo teor em tanino; 2) envolveram frangos de corte de linhagem comercial; 3)
apresentaram número de repetições, número de aves por repetição, níveis nutricionais
empregados (proteína bruta e energia metabolizável) e apresentaram resultados de
desempenho em tabela; 4) experimentos em baterias não foram aceitos; 5) utilizaram
somente milho, soja e sorgo como ingredientes e as dietas com sorgo foram fornecidas
já na primeira semana de vida. O banco de dados para consumo de ração e ganho de
peso foi composto por um total de 10900 aves. Ajustou-se um modelo de superfície de
resposta com duas variáveis independentes (sorgo% e idade). Os parâmetros foram
estimados pelo método dos quadrados mínimos. Somente o termo quadrático da idade
apresentou efeito significativo para consumo de ração (r2 = 0,98) e ganho de peso (r2 =
0,97). Os demais termos do polinômio não apresentaram efeito significativo. Estes
resultados demonstram que o nível de substituição de milho por sorgo não prejudica o
consumo de ração e o ganho de peso nas fases pré-inicial, inicial, crescimento ou
terminação.
Palavras-chave: análise de regressão, idade, sorgo baixo tanino, revisão sistemática da
literatura, superfície de resposta.

37
Introdução
A grande flutuação no cambio dos principais ingredientes (milho e soja) da
alimentação avícola, fazem da utilização de ingredientes alternativos a resposta viável
para a obtenção de resultados econômicos satisfatórios. O sorgo (Sorghum bicolor (L.)
Moench) é tido como uns dos principais ingredientes energéticos substitutos ao milho,
todavia, existe certa resistência na utilização de sorgo em rações pré-iniciais devido à
possibilidade deste cereal afetar o desempenho dos pintainhos neste período.
A determinação precisa da relação benefício/custo na criação de frangos de corte
está na dependência do desenvolvimento de modelos matemáticos capazes de predizer
o crescimento e o consumo de ração dos frangos em função dos fatores que influenciam
essas variáveis (OVIEDO-RONDÓN & WALDROUP, 2002). Os modelos matemáticos
são ferramentas que fornecem subsídios para tomadas de decisões que envolvem
diversas variáveis, mostrando qual caminho seguir para obtenção do melhor retorno
econômico da atividade desenvolvida. Dentre os possíveis modelos, os de superfície de
resposta são de grande aplicabilidade prática, pois permitem modelar mais de um fator
simultaneamente através de polinômios de primeira, segunda ou mais ordens, além de
testarem a interação entre os fatores envolvidos na pesquisa (FREUND & LITTELL,
2000). EITS et al. (2005) ressaltaram que os modelos devem ser práticos (incluir
variáveis conhecidas pelo nutricionista), flexíveis (envolver variáveis como sexo, níveis
nutricionais, ingredientes, etc.) e precisos (derivados de vários conjuntos de dados).
Neste contexto, a utilização de modelos que demonstrem a variação do
desempenho das aves quando são submetidas às dietas compostas por ingredientes
alternativos é de grande valia para embasar os nutricionistas em suas decisões.
Para obtenção de dados consistentes, utilizou-se uma revisão sistemática da
literatura (WANG & BUSHMAN, 1999), uma técnica simples capaz de identificar,
selecionar e avaliar as pesquisas a serem utilizadas na elaboração do banco de dados a
ser analisado. Portanto deve-se definir previamente a estratégia de busca, as bases de
dados utilizadas na pesquisa, período da revisão e os critérios de seleção de estudos. A

38
revisão deve mostrar os trabalhos selecionados e também os trabalhos excluídos e a
razão para tal procedimento. O objetivo de se realizar uma revisão sistemática da
literatura é propiciar uma revisão reprodutível e não viesada (pois critérios de inclusão
devem ser explícitos e coerentes impossibilitando uma seleção tendenciosa dos dados).
Assim, o objetivo desta pesquisa foi criar modelos de superfície de resposta, com
base nos dados selecionados pela revisão sistemática da literatura, para as variáveis
independentes, níveis de substituição do milho por sorgo e idade, para predição do
desempenho zootécnico.
Material e métodos
Localização dos estudos
A revisão sistemática da literatura foi realizada por buscas eletrônicas com as
palavras chaves sorgo e frangos de corte nas bases de dados “Biological Abstracts”,
“Cab Abstracs”, “Medline”, “Scielo” e “Athena” no período de 1995 a janeiro de 2006.
Foram inseridos dados de trabalhos não publicados de nosso grupo de pesquisa
(Universidade Estadual Paulista, Faculdade de Ciências Agrárias e Veterinárias,
Departamento de Morfologia e Fisiologia Animal).
Critérios de inclusão
Os trabalhos selecionados apresentaram: 1) efeito da substituição do milho por
sorgo baixo teor em tanino; 2) envolveram frangos de corte de linhagem comercial; 3)
apresentaram número de repetições, número de aves por repetição, níveis nutricionais
empregados (proteína bruta e energia metabolizável) e apresentaram resultados de
desempenho em tabela; 4) os dados de aves criadas em baterias não foram aceitos; 5)
utilizou-se somente milho, soja e sorgo como ingredientes e as dietas com sorgo foram
fornecidas já na primeira semana de vida.

39
Coleta de dados
Nos trabalhos inclusos foram coletadas as médias para consumo de ração e
ganho de peso. Esses dados foram armazenados no Microsoft Excel® com os
respectivos valores de substituição de milho por sorgo e idade (dias) e número de
repetições e de aves por repetição.
Análise estatística
Os modelos de superfície resposta (FREUND & LITTELL, 2000) utilizados foram:
Yi = �0 (Xi0) + �1 (Xi1) + �2 (Xi2) + �3 (Xi3) + �4 (Xi4) + �5 (Xi1 Xi3) + �i
Yi = ganho de peso (g) ou consumo de ração (g);
Xi0 = 1 (intercepto);
Xi1 = idade média (dias);
Xi2 = Xi12;
Xi3 = inclusão de sorgo (%);
Xi4 = Xi32;
�0, �1, �2, �3, �4 e �5, = parâmetros que foram estimados pelo método dos
quadrados mínimos ponderados, considerando como fator de ponderação o número
total de aves com que cada média foi obtida (número de repetições*número de aves por
repetição) expresso como porcentagem do total de aves do conjunto de dados. Assim,
as médias obtidas com o maior número de aves tiveram maior peso na estimativa dos
parâmetros.
�i =
erro aleatório associado a observação i.

40
Resultados e Discussão
Revisão sistemática da literatura
A revisão sistemática da literatura localizou nove trabalhos, sendo que seis não
se enquadraram dentro dos critérios de inclusão: - utilizaram cultivares de sorgo com
altos teores em tanino (JACOB et al., 1996 a; NYACHOTI et al., 1996; NYACHOTI et al.,
1998); utilizaram outros cereais e/ou leguminosas além do sorgo, soja e milho (JACOB
et al., 1996 b; LÓPEZ et al., 2001 a); as aves foram criadas em baterias (CUEVAS et al.,
2002).
Os trabalhos que se enquadram nos critérios de seleção foram: LÓPEZ et al.
(2001 b), SILVA (2003), GARCIA et al. (2005) e um trabalho de nossa equipe
(CAMPOS, dados não publicados). Esses trabalhos foram realizados com frangos
sexados (machos e/ou fêmea), com as linhagens comerciais Arbor Acres, Coob e Ross,
com os níveis de proteína bruta variando de 19 a 23% e os níveis de energia
metabolizável variando de 2900 a 3200 Kcal. O banco de dados formado com os
trabalhos selecionados para as variáveis, consumo de ração e ganho de peso,
apresentaram o total 10900 aves.
Superfície de resposta
As estimativas dos parâmetros do modelo de superfície de resposta para
consumo de ração e conversão alimentar estão na Tabela 1.
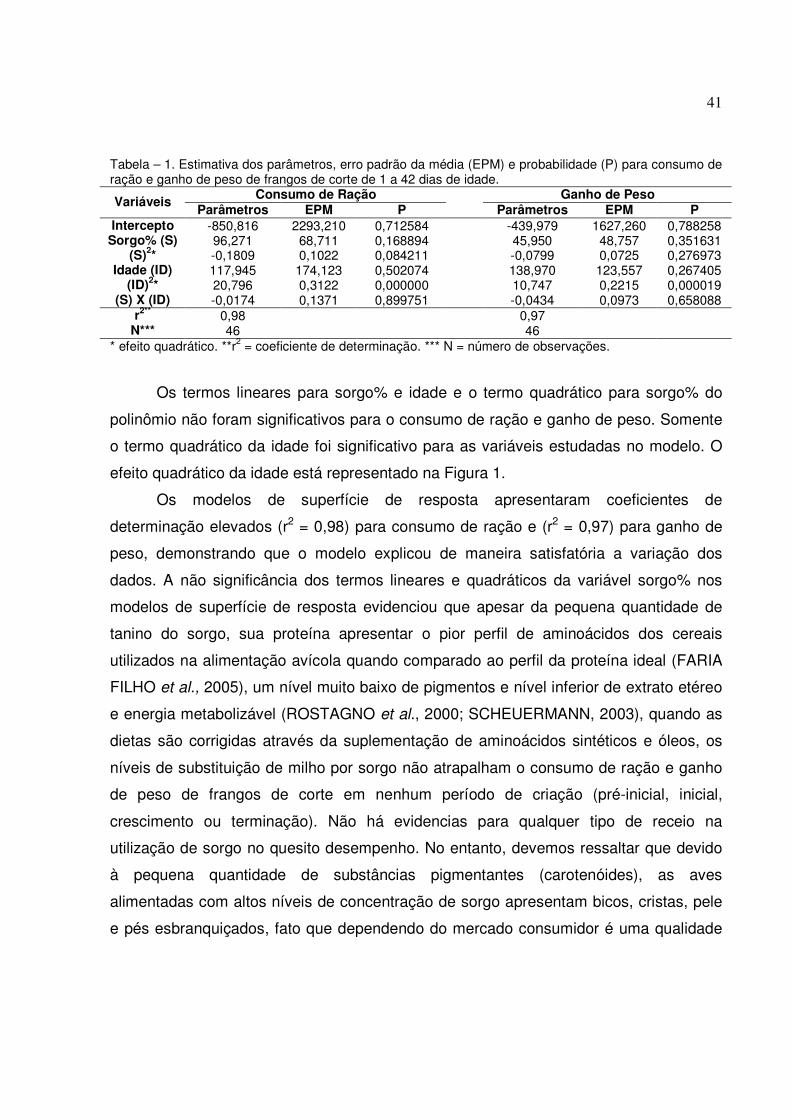
41
Tabela – 1. Estimativa dos parâmetros, erro padrão da média (EPM) e probabilidade (P) para consumo de ração e ganho de peso de frangos de corte de 1 a 42 dias de idade.
Consumo de Ração Ganho de Peso Variáveis Parâmetros EPM P Parâmetros EPM P Intercepto -850,816 2293,210 0,712584 -439,979 1627,260 0,788258
Sorgo% (S) 96,271 68,711 0,168894 45,950 48,757 0,351631 (S)2* -0,1809 0,1022 0,084211 -0,0799 0,0725 0,276973
Idade (ID) 117,945 174,123 0,502074 138,970 123,557 0,267405 (ID)2* 20,796 0,3122 0,000000 10,747 0,2215 0,000019
(S) X (ID) -0,0174 0,1371 0,899751 -0,0434 0,0973 0,658088 r2** 0,98 0,97
N*** 46 46 * efeito quadrático. **r2 = coeficiente de determinação. *** N = número de observações.
Os termos lineares para sorgo% e idade e o termo quadrático para sorgo% do
polinômio não foram significativos para o consumo de ração e ganho de peso. Somente
o termo quadrático da idade foi significativo para as variáveis estudadas no modelo. O
efeito quadrático da idade está representado na Figura 1.
Os modelos de superfície de resposta apresentaram coeficientes de
determinação elevados (r2 = 0,98) para consumo de ração e (r2 = 0,97) para ganho de
peso, demonstrando que o modelo explicou de maneira satisfatória a variação dos
dados. A não significância dos termos lineares e quadráticos da variável sorgo% nos
modelos de superfície de resposta evidenciou que apesar da pequena quantidade de
tanino do sorgo, sua proteína apresentar o pior perfil de aminoácidos dos cereais
utilizados na alimentação avícola quando comparado ao perfil da proteína ideal (FARIA
FILHO et al., 2005), um nível muito baixo de pigmentos e nível inferior de extrato etéreo
e energia metabolizável (ROSTAGNO et al., 2000; SCHEUERMANN, 2003), quando as
dietas são corrigidas através da suplementação de aminoácidos sintéticos e óleos, os
níveis de substituição de milho por sorgo não atrapalham o consumo de ração e ganho
de peso de frangos de corte em nenhum período de criação (pré-inicial, inicial,
crescimento ou terminação). Não há evidencias para qualquer tipo de receio na
utilização de sorgo no quesito desempenho. No entanto, devemos ressaltar que devido
à pequena quantidade de substâncias pigmentantes (carotenóides), as aves
alimentadas com altos níveis de concentração de sorgo apresentam bicos, cristas, pele
e pés esbranquiçados, fato que dependendo do mercado consumidor é uma qualidade
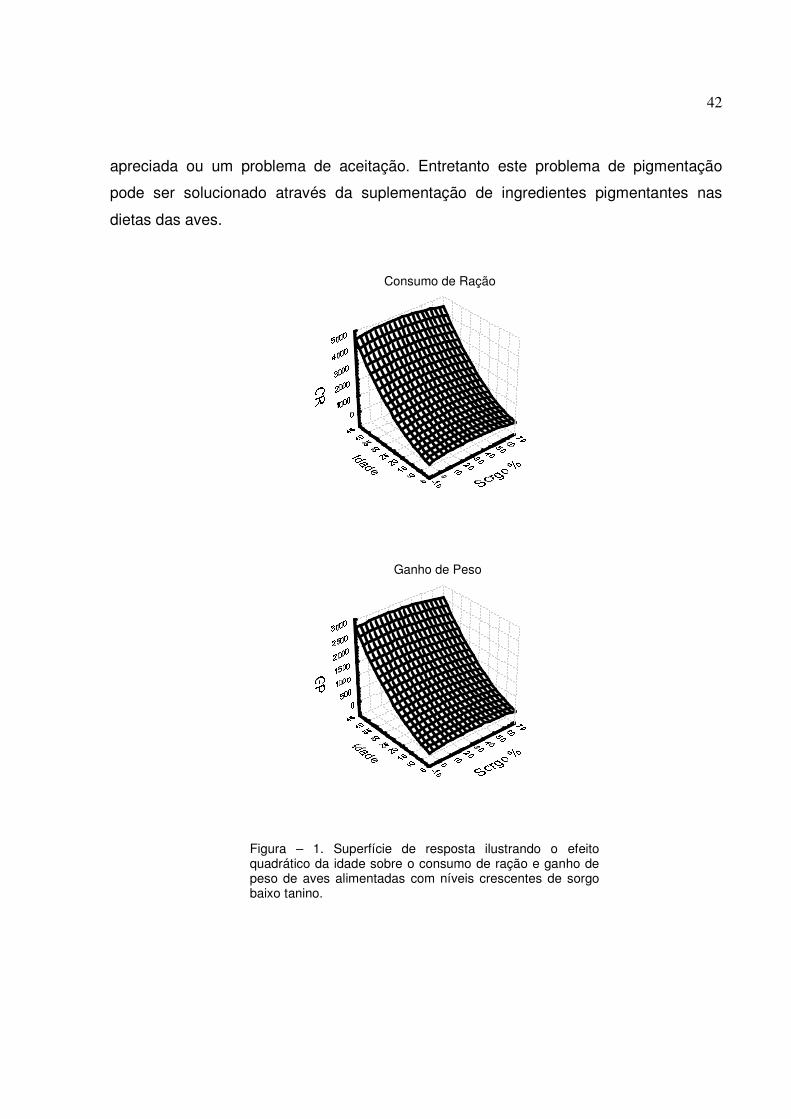
42
apreciada ou um problema de aceitação. Entretanto este problema de pigmentação
pode ser solucionado através da suplementação de ingredientes pigmentantes nas
dietas das aves.
Consumo de Ração
Ganho de Peso
Figura – 1. Superfície de resposta ilustrando o efeito quadrático da idade sobre o consumo de ração e ganho de peso de aves alimentadas com níveis crescentes de sorgo baixo tanino.

43
Conclusão
Conclui-se que os níveis de substituição de milho por sorgo não prejudica o
consumo de ração e o ganho de peso nas fases pré-inicial, inicial, crescimento ou
terminação.
Referências
AGOSTINI-COSTA, T. S. Determinação de tanino em pedúnculo de caju: método da
vanilina versus método do butanol ácido. Química Nova, v. 5, p. 763-765, 2003.
AGRIANUAL. anuário da agricultura brasileira. São Paulo: FNP Consultoria e
Agroinfornativos, 2004.
BOLELI, I. C. MAIORKA, A., MACARI, M. Estrutura Funcional do Trato Digestório. In:
MACARI, M.; FURLAN, R.L.; GONZALES, E. (ed.). Fisiologia Aviária: Aplicada a
frangos de corte. 2 ed. Jaboticabal: FUNEP, 2002. p. 75-95.
BUTULO, J., E. Qualidade de Ingredientes de Alimentação Animal. CBNA, 2003.
CHANG, J.J., BAILEY, J. W., COLLINS, J. L. Dietary tannnis from cowpeas and tea
transiently alter apparent calcium absorption and utilization of protein in rats. Journal of
Nutrition, v.124, p.283-88, 1994.
CHANG, S. I., FULLER, H. L. Effect of tannin contents of grain sorghum for growing
chicks. Poultry Science, v. 43, p. 30-36, 1964.
CHUBB, L. G. Anti-nutritive factors in animal feedstuffs. In: HARESING, W. Studies in
agricultural food sciences. Butterworks, p. 21-23, 1982.

44
COOK, R. H., BIRD, F. Duodenal villus area and epithelial cellular migration in
conventional and germ-free chicks. Poultry Science, v. 52, p. 2276-2280, 1973.
CUEVAS, A. C., GUTIÉRREZ, A. C., GONZÁLEZ, E. A., BARRERA, E. M. Valor
nutrimental de cuatro pastas de soya procesadas en diferentes estados de la república
mexicana. Veterinaria México. v. 33, n. 3, p. 209-217, 2002.
D’MELLO, J. P. F., THOMAS, D. The nutritive value of the leucena leaf meal from
Malawi studies with young chicks. Tropical Agricultural Trinidad, v. 55, p. 45-50, 1978.
DINIZ, F; V., FERNANDES, E. A., MUNDIM, S. A. P., ALVES, M. B. R., AQUINO, C. A.
M., CHIARELI, D. Desempenho de frangos de corte submetidos a dietas formuladas a
base de milho e sorgo. Revista Brasileira de Ciência Avícola, s.4, p.60, 2002.
DOUGLAS, J. H., SULLIVAN, T. W. Influence of infrared (micronization) treatment on the
nutritional value of corn and low and high-tannin sorghum. Poultry Science, v.70.
p.1534-1539, 1991.
DOWLING, L. F., ARNDT, C., HAMAKER, B. R. Economic Viability of high digestibility
sorghum as feed for market broilers. Agronomy Journal, v. 94, p. 1050-1058, 2002.
EITS, R.M.; KWAKKEL, R.P.; VERSTEGEN, M.W.A. et al. Dietary balanced protein in
broiler chickens. 1. A flexible and practical tool to predict dose-response curves. British
Poultry Science, v. 46, p. 300-309, 2005.
FARIA FILHO, D. E., TORRES, K. A. A., CAMPOS, D. M. B., VIEIRA, B. S., URBANO,
T., ROSA, P. S., FERRAUDO, A. S. Ingredient Classification According to the Digestible

45
Amino Acid Profile: An Exploratory Analysis. Brazilian Journal of Poultry Science, v.7,
n.3, p. 185-193, 2005.
FREUND, R.J.; LITTELL, R.C. SAS® System for Regression. SAS Institute, 2000.
Fourth edition, SAS Institute Inc, Cary, NC. 235p.
GARCIA, R. G., MENDES A. A., COSTA, C., PAZ, I. C. L. A., TAKAHASHI, S. E.,
PELÍCIA, K. P., KOMIYAMA, C. M., QUINTEIRO, R. R. Desempenho e qualidade de
carne de corte alimentados com diferentes níveis de sorgo em substituição ao milho.
Arquivo Brasileiro Medicina Veterinária e Zootecnia, v.57, n.5, p.634-643, 2005.
GEYRA, A., UNI, Z., SKLAN, D. The effect of fasting at different ages on growth and
tissue dynamics in the small intestine of the young chick. British Journal of Nutrition, v.
86, p. 53-61, 2001.
GÔNZALEZ, E., SALDANHA, E. S. P. B. Os primeiros dias de vida do frango e a
produtividade futura: In: CONGRESSO BRASILEIRO DE ZOOTECNIA – ZOOTEC, 11.,
2001, Goiânia. Anais...Goiânia: UCG, 2001. p. 310-327.
JACOB, J. P., MITARU, B. M., MBUGUA, P. N., BLAIR, R. The effect of substituting
Kenyan Serena Sorghum for maize in broiler starter diets with different dietary crude
protein and methionine levels. Animal Feed technology, v. 61, p. 27-39, 1996 a.
JACOB, J. P., MITARU, B. M., MBUGUA, P. N., BLAIR, R. The feeding value of Kenyan
sorghum, sunflower seed cake and sesame seed cake for boilers and layers. Animal
Feed Science Technology. v. 61, p. 41-56, 1996 b.
JIMENEZ, R. L. M., ROGLER, J. C., HOUSLEY, T. L., BUTLER, L. G., ELKIN, R. G.
Absorption and distribution of 14C-labeled condensed tannins and related sorghum
phenolics in chickens. Journal Agriculture Food Chemistry, v. 42, p. 963-967, 1994.

46
LONGO, F. A., MENTEN, J. F. M., PEDROSO, A. A., FIGUEIREDO, A. N., RACANICCI,
A. M. C., GAIOTTO, J. B., SORBARA, J. O. B. Carboidratos na Dieta Pré-Inicial de
Frangos de Corte. Revista Brasileira de Zootecnia, v. 34, n.1, p.123-133, 2005.
LÓPEZ, R. M., HUGUENIN, T. C., GONZÁLEZ, E. A. Lesiones macroscópicas
gastrointestinales y renales em pollos de engorda, criados com niveles altos de proteína
de soya em la dieta. Veterinaria México, v. 32, n. 3, p. 175-182, 2001 a.
LÓPEZ, R. M., MÉNDEZ, T. J., GONZÁLEZ, E. A., AMEZCUA, C. M., Necessidades de
treonina em pollos sometidos a dos calandários de vacunación. Veterinaria México. V.
32, n. 3, p. 189-194, 2001 b.
MAIORKA, A., BOLELI, I. C., MACARI, M. Desenvolvimento e reparo da mucosa
intestinal. In: MACARI, M.; FURLAN, R.L.; GONZALES, E. (ed.). Fisiologia Aviária:
Aplicada a frangos de corte. 2 ed. Jaboticabal: FUNEP, 2002. p. 113-123.
MARCHAIM, U., KULKA R. G., The non-parallel increase of amylase, chymotripsinogen
and procarboxypeptidase in the developing chick pancreas. Biochimica and
Biophysica Acta. 146:553-559, 1967.
MCBRIDE, B. W., KELLY, J. M., Energy cost of absorption and metabolism in the
ruminant gastrointestinal tract and liver: A review. Journal Animal Science, 68: 2997-
3010, 1990.
MITJAVILA, S., LACOMBE, C., CARRERA, G., DERACHE, R. Tannic acid and oxidized
tannic acid on the functional state of rat intestinal epithelium. Journal of Nutrition. v.
107, 2113-2121. 1977.

47
NITSAN, Z. I., BEN-AVRAHAN, G., ZOREF, Z., et al. Growth and development of the
digestive organs and some enzymes in broiler chicks after hatching. British Poultry
Science, v. 32, p. 515-523, 1991.
NOY, Y. SKALN, D. Digestion and absorption in the young chick, Poultry Science, v.
47, p.366-373, 1995.
NYACHOTI, C. M., ATKINSON, J. L., LEESON, S. Evaluation of Magadi soda-treated
sorghum grain for young broilers. Animal Feed Science Technology. v. 70, p. 295-304,
1998.
NYACHOTI, C. M., ATKINSON, J. L., LEESON, S. Response of broiler chicks fed a
high-tannin sorghum diet. Journal Applied Poultry Science. v. 5, p. 239-245, 1996.
OLIVEIRA, P. B. Influência de fatores antinutricionais da leucena (Leucaena
leucocephala e Leucaena eunningan) e do feijão guandu (Cajanus cajan) sobre o
epitélio intestinal e o desempenho de frangos de corte. Dissertação (Mestrado). –
UEM (Universidade Estadual de Maringá), Maringá, 1998.
OLIVEIRA, P. B., MURAKAMI, A. E., GARCIA, E. R. M., MACARI, M., SCAPINELLO, C.
Influência de Fatores Antinutricionais da Leucena (Leucaena leucocephala e Leucaena
eunningan) e do Feijão Guandu (Cajanus cajan) Sobre o Epitélio Intestinal e o
Desempenho de Frangos de Corte. Revista Brasileira de Zootecnia, 29(6): 1759-1769,
2000.
ORTIZ, L. T., ALZUETA, C., TREVINO, J., CASTANO, M. Effects of faba bean on the
growth and histological structure of the intestinal tract and liver of chicks and rats. British
Poultry Science, v. 35, p. 743-754, 1994.

48
OVIEDO-RONDÓN, E.O.; WALDROUP, P.W. Model to estimate amino acid
requirements for broiler chickens: a review. International Journal of Poultry Research,
v. 1, p. 106-113, 2002.
PEZZATO, L. E. Alimentos convencionais e não convencionais disponíveis para
indústria da nutrição de peixes no Brasil. In: SIMPÓSIO INTERNACIONAL SOBRE
NUTRIÇÃO DE PEIXES E CRUSTÁCEOS, 1995, Campos do Jordão.
Proceedings...Campos do Jordão, Brasil: CNBA, 1995, p. 33-52.
POUR-REZA, J.; EDRISS, M. A. Effects of dietary sorghum of different tannin
concentrations and tallow supplementation on the performance of broiler chicks. British
Poultry Science, v. 38, p.512-17, 1997.
QUINTERO PINTO, L. G. Tanino em rações para peixes tropicais. p55. Dissertação
(mestrado em Aqüicultura) – Centro de Aqüicultura, UNESP (Universidade Estadual
Paulista), Jaboticabal, 2000.
ROCHA, P. T., STRINGHINI, J. H., ANDRADE, M. A., LEANDRO, N. S. M., ANDRADE,
M. L., CAFÉ, M. B. Desempenho de frangos de corte alimentados com rações pré-
iniciais contendo diferentes níveis de proteína e energia metabolizável. Revista
Brasileira de Zootecnia, v. 32, n. 1, p. 162-170, 2003.
ROSTAGNO, H. S., FEATHERSTON, W. R., ROGLER, J. C. Studies on the nutritional
value of sorghum grains with varying tannin contents for chicks: Growth studies. Poultry
Science, 52:765-772, 1973.
ROSTAGNO, H. S., SILVA, D.J., COSTA, P.M. Composição de alimentos e
exigências nutricionais de aves e suínos. Universidade Federal de Viçosa, Viçosa.
M.G., p. 11-13, 2000.

49
SAKOMURA, N. K., BIANCHI, M. D., PIZAURO Jr., J. M., CAFÉ, M. B., FREITAS, E. R.
Efeito da idade dos frangos de corte sobre a atividade enzimática e digestibilidade dos
nutrientes do farelo de soja e da soja integral. Revista Brasileira de Zootecnia, v. 33,
n. 4, p. 924-935, 2004.
SAS Institute, 2002. SAS® User's Guide: Statistics, SAS Institute Inc, Cary, NC.
SCHEUERMANN, G. N. Utilização do sorgo em rações para frangos de corte. UBA -
Informa (Informativo Técnico – União brasileira de Avicultura), p. 95-96, 2003.
SELL, J. L., KRODGAHL, A., HANYU, N. Influence of age in utilizations supplemental
fats by young turkeys. Poultry Science, 65 (3), p. 546-554, 1986.
SILVA, J. D. T. Uso de sorgo com baixo teor em tanino na alimentação de frangos
de corte. 2003. Trabalho de conclusão de curso (Graduação em Zootecnia) - Faculdade
de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal, 2003.
UNI, Z. Functional development of the small intestine in domestic birds: Cellular and
molecular aspects. Poultry and Biology Reviews. p. 167-179, 1999.
UNI, Z., FERKET, R. P. methods for early nutrition and their potential. Poultry Science,
v. 60, p. 101-111, 2004.
UNI, Z., GANOT, S., SKLAN, D. Posthatch development of mucosal function in the small
intestine. Poultry Science, v. 77, p. 75-77, 1998.
UNI, Z., NOY, Y., SKLAN, D. Posthatch development of small intestinal function in the
poult. Poultry Science, v. 78, p. 215-222, 1999.

50
UNI, Z., SMIRNOV, A., SKALAN, D. Pre and Posthatch of Development of Goblet Cells
in the Broiler Small Intestine: Effect of Delayed Access to Feed. Poultry Science, v.82,
p. 320-327, 2003.
WANG, M.C.; BUSHSMAN, B.J. Integrating results through meta-analytic review
using SAS software. 1 ed. Cary: SAS Institute Inc.; 1999.