efeito de compostos presentes na dieta e de drogas de ... · a relevância do transporte celular de...
TRANSCRIPT

Efeito de compostos presentes na dieta e de
drogas de abuso na captação de glicose numa
linha celular de trofoblastos humanos (células
BeWo)
João Ricardo Diniz de Araújo
Orientado por: Prof. Doutora Maria de Fátima Moreira Martel
Trabalho de Investigação
Porto, 2007


iii


i
AGRADECIMENTOS
Agradeço a preciosa orientação da Prof. Doutora Maria de Fátima Moreira Martel.
Agradeço também o apoio técnico fundamental do Dr. Pedro Gonçalves.
Trabalho financiado pela FCT (Programa Ciência, Tecnologia e Inovação do
Quadro Comunitário de Apoio e POCTI/SAU-FCF/59382/2004).

ii
ÍNDICE
Lista de Abreviaturas ......................................................................................... iii
Resumo e palavras-chave em português e inglês ............................................. iv
Introdução .......................................................................................................... 1
Objectivos .......................................................................................................... 9
Material e Métodos........................................................................................... 10
Resultados ....................................................................................................... 16
Discussão......................................................................................................... 35
Conclusão ........................................................................................................ 49
Bibliografia ....................................................................................................... 51
Anexos ............................................................................................................. 62
Índice de anexos .............................................................................................. 63

iii
LISTA DE ABREVIATURAS
Amax : acumulação de 3H-DG no equilíbrio
ATP: trifosfato de adenosina
BeWo: linha celular de coriocarcinoma humano
DG: 2-desoxiglicose
DG-6-P: 2-desoxiglicose-6-fosfato
DMSO: dimetilsulfóxido
EGCG: epigalocatequina-3-galato
EPM: erro padrão da média
GLUT: transportador de glicose independente do sódio
GF-HBS: solução amortecedora salina com HEPES sem glicose
HMIT: transportador de mio-inositol acoplado ao ião H+
3H-DG: 2-desoxi-D-glicose marcada com o isótopo radioactivo (trício)
kin: taxa constante do transporte para dentro da célula
kout: taxa constante do transporte para fora da célula
MDMA: 3,4-metilenodioximetanfetamina
MTT: brometo de 3-(4,5-dimetil-2-tiazolil)-2,5-difenil-2H-tetrazólio
SGLT: transportador activo de glicose dependente do sódio
SERT: transportador de serotonina
THC: ∆9-tetrahidrocanabinol

iv
RESUMO EM PORTUGUÊS
Os mecanismos que regulam o transporte placentar de glicose são ainda
pouco conhecidos. Por isso o objectivo deste trabalho, foi caracterizar a captação
de glicose da circulação materna e estudar a modulação desse processo por
diversas substâncias (polifenóis, metilxantinas, etanol e o seu metabolito
acetaldeído), presentes em bebidas alcoólicas e não-alcoólicas de consumo
habitual (como o vinho, chá e café) e por drogas de abuso. Para tal, determinou-
se o efeito agudo (26 minutos) e crónico (48 horas) desses compostos na
captação de desoxiglicose marcada radioactivamente com trício (3H-DG), numa
linha celular de trofoblastos humanos (BeWo).
Verificamos que a captação de 3H-DG (1 �M) ocorre principalmente por um
mecanismo facilitado (GLUT), e é modulada de modo diferente pelos compostos
das bebidas e pelas drogas de abuso. Agudamente, o acetaldeído (30 e 100 mM),
o resveratrol, o xanto-humol, a epigalocatequina-3-galato (100 �M), e a quercetina
e crisina (ambos a 10 e 100 �M) reduziram o transporte apical de 3H-DG,
enquanto que a rutina, a catequina (ambos a 10 e 100 �M), a epicatequina (100
�M) e o etanol (10 mM) aumentaram-no. A catequina e a epicatequina em
conjunto (ambas a 100 �M) reduziram a captação de 3H-DG, enquanto que a
epicatequina e o xanto-humol (ambos a 100 �M) contrabalançaram os efeitos que
cada um tinha isoladamente. Todos estes compostos parecem interagir com o
GLUT para exercerem os seus efeitos; contudo o acetaldeído parece exercer o
seu efeito por outro mecanismo. A quercetina e o xanto-humol são inibidores não
competitivos da captação de 3H-DG, ao passo que a epigalocatequina-3-galato e
o acetaldeído reduziram ambos o valor de Km e Vmax.

v
Cronicamente, a rutina e a miricetina isoladamente (0,1 e 1 �M) e em
conjunto (1 �M) aumentaram a captação de 3H-DG, enquanto que a teoflina (0,1 e
1 �M) e as drogas de abuso 3,4-metilenodioximetanfetamina, anfetamina (ambas
a 0,25 e 2,5 �M) e o ∆9-tetrahidrocanabinol (1 nM) reduziram-na. Para exercerem
os seus efeitos crónicos, estes compostos parecem actuar ao nível do GLUT.
Os resultados obtidos neste trabalho, vêem aprofundar o conhecimento da
bioactividade e a interferência na saúde humana, de alguns polifenóis,
metilxantinas e drogas de abuso.
Palavras-chave: glicose, captação apical, linha celular BeWo, polifenóis,
metilxantinas, etanol, acetaldeído, anfetamina, ∆9-tetrahidrocanabinol, 3,4-
metilenodioximetanfetamina.
RESUMO EM INGLÊS
Very little is known concerning the mechanisms responsible for the
regulation of the placental transfer of glucose. So, the aim of this work was to
characterize the placental uptake of glucose from the maternal circulation, and to
study the modulation of that process by several compounds (polyphenols,
methylxanthines, ethanol and its metabolite acetaldehyde), present in alcoholic
and nonalcoholic drinks (such as wine, tea and coffee) and by drugs of abuse. For
this, the acute (26 min) and chronic (48 h) effect of these compounds upon the

vi
uptake of 3H-2-deoxy-D-glucose (3H-DG) by a human trophoblast cell line (BeWo)
was determined.
We verified that the uptake of 3H-DG occurs mainly by a facilitated
mechanism (GLUT), that is differentially modulated by compounds present in
drinks and by drugs of abuse. Acutely, acetaldehyde (30-100 mM), resveratrol,
xanthohumol, epigallocatechin-3-gallate (100 µM), chrysin and quercetine (both at
10-100 µM), decreased 3H-DG uptake, whereas rutin, catechin (both at 10-100
µM), epicatechin (100 µM) and ethanol (10 mM) increased it. Together, catechin
and epicatechin (both at 100 µM) decreased the uptake of 3H-DG, whereas
epicatechin and xanthohumol (both at 100 µM) counterbalanced each one’s
isolated effect. All these compounds seem to interact with GLUT in order to exert
their effects, except acetaldehyde that seems to exert its effect by a different
mechanism. Quercetin and xanthohumol are non-competitive inhibitors of 3H-DG
uptake, whereas both epigallocatechin-3-gallate and acetaldehyde decreased both
the Km and Vmax values.
Chronically, rutin and miricetin increased the uptake of 3H-DG both isolated
(both at 0,1-1 �M) and in combination (both at 1 �M), whereas theophylline (0,1-1
µM) and the drugs amphetamine, 3,4-methylenedioxymethamphetamine (0,1-2,5
�M) and ∆9-tetrahydrocannabinol (1 nM) decreased it. In order to exert their
chronic effects, these compounds seem to interact with GLUT.
The results of these work, will generate a deeper knowledge about the
bioactivity and health interference of polyphenols, methylxantines and drugs of
abuse.

vii
Palavras-chave em inglês: glucose, apical uptake, BeWo cell line, polyphenols,
methylxantines, ethanol, acetaldehyde, ∆9-tetrahydrocannabinol, amphetamine,
3,4-methylenedioxymethamphetamine.


1
INTRODUÇÃO
As membranas plasmáticas são estruturas laminares constituídas por
moléculas proteicas, lipídicas e glicídicas (1), que funcionam como uma barreira
semi-permeável entre o citoplasma e o fluido extracelular (2).
O transporte de substâncias (moléculas ou iões) para dentro ou para fora
da célula, pode ser classificado de dois modos: activo ou passivo e mediado ou
não por transportadores (2). O transporte activo requer o consumo de ATP
(trifosfato de adenosina) ou outro intermediário metabólico, sendo as moléculas
transportadas contra o seu gradiente de concentração. Pelo contrário, o
transporte passivo não requer energia sendo as moléculas transportadas a favor
do seu gradiente de concentração (2,3).
O transporte mediado por transportadores ocorre na presença de uma
proteína membranar integral, que permite o movimento de moléculas com
determinadas características. (2) Nestas, incluem-se moléculas hidrofílicas,
polares e de alto peso molecular (geralmente superior a 200 Da) como glícidos,
aminoácidos e neurotransmissores, que são incapazes de atravessar livremente a
membrana plasmática por difusão simples (3). Existem dois tipos de transporte
mediados por proteínas: o transporte facilitado, que não requer energia pois
transporta moléculas a favor do seu gradiente de concentração e o transporte
activo (2). Em geral, as proteínas transportadoras exibem especificidade para um
determinado soluto, saturação (à medida que a concentração de soluto aumenta,
a sua taxa de transporte aumenta mas só até certo ponto) e velocidade máxima
de transporte (quando todos os transportadores estão ocupados, a presença de

2
mais substância não aumenta a taxa de transporte pois os transportadores estão
saturados) (2).
A relevância do transporte celular de glicose decorre do facto de esta ser
uma fonte de energia fundamental e um importante substrato metabólico nos
mamíferos.
A glicose é obtida directamente da dieta e através da síntese de novo a
partir de outros substratos gliconeogénicos existentes em órgãos como o fígado.
A glicose é transportada da corrente sanguínea para as células, por dois
tipos de transportadores estruturalmente distintos, cujos membros têm sido
identificados ao longo dos últimos anos. Um deles é um transportador activo
dependente do ião sódio. Pelo menos três e até seis transportadores de glicose
dependentes do sódio, (SGLT1-SGLT6, genes da família SLC5A) já foram
identificados. Através destes transportadores, o gradiente electroquímico do ião
sódio gerado pela bomba de sódio e potássio, é utilizado para transportar a
glicose contra o seu gradiente de concentração (4). O outro transportador, é um
mecanismo de transporte facilitado de glicose que engloba uma família com treze
membros (GLUT1-GLUT12 e HMIT, genes da família SLC2), subdivididos em três
subclasses (4). Estes são independentes do sódio e utilizam o gradiente de
concentração da glicose (ou de outros açúcares), para realizar a sua função de
transporte (4, 5). Os primeiros transportadores a serem clonados foram o GLUT1,
existente sobretudo nos eritrócitos, cérebro e barreira placentária, e o SGLT1
predominante na membrana apical dos enterócitos e nas células renais do tubo
contornado proximal (4). Todos estes transportadores de glicose exibem
diferentes especificidades para o substrato e diferentes propriedades cinéticas e

3
padrões de expressão tecidular, o que indica que a captação celular de glicose é
um processo complexo (4).
O papel que os transportadores de glicose placentários desempenham
ainda antes da implantação do embrião, é essencial para garantir que o
metabolismo da glicose seja adequado (6). Mais tarde, o transporte facilitado
glicose da mãe para o feto será a principal fonte de energia para o crescimento e
metabolismo dos tecidos feto-placentários (7, 8), uma vez que a gliconeogénese
fetal não ser suficiente para assegurar as necessidades de glicose fetais (9).
A placenta humana é um órgão temporário especializado, de origem fetal
(placa coriónica e vilosidades coriónicas) e materna (decídua basalis),
constituindo a principal ligação entre a mãe e o feto (9). A corrente sanguínea
materna e fetal estão separadas pela estrutura da placenta, que é formada por
três camadas tecidulares: o sinciciotrofoblasto, estrutura multinucleada que
resulta da fusão dos citotrofoblastos com morfologia polarizada, cuja membrana
apical está em contacto directo com o sangue materno e cuja membrana basal
está voltada para a circulação fetal; uma fina camada de tecido conjuntivo e o
endotélio fetal (10) (ver figura 1).
Figura 1 - Estrutura da placenta humana e distribuição das isoformas dos transportadoresGLUT. Legenda: syncytiotrophoblast - sinciciotrofoblasto, cytotrophoblast - citotrofoblasto,microvillous membrane - membrana apical, basal membrane - membrana basal, stromal cells -células estromais, endothelial cell - célula endotelial, fetal capillary - capilares fetais eintervillous space – espaço intervilosidades. � - GLUT 1, � - GLUT 3 (11).

4
A placenta é fundamental para o desenvolvimento fetal normal, pois
desempenha numerosas funções: é o local de transferência de nutrientes (glicose,
aminoácidos, ácidos gordos e nucleosídeos) e oxigénio da mãe para o feto, e é o
local de eliminação e remoção de produtos do metabolismo da circulação fetal (9,
12). Estas funções primordiais são reguladas por uma rede complexa de
mecanismos de transporte, que se distribuem de uma forma polarizada
essencialmente no sinciciotrofoblasto. Além destas, são também funções da
placenta: sintetizar e segregar para as circulações materna e fetal hormonas,
enzimas, factores de crescimento, citoquinas e outras moléculas responsáveis
pelo desenvolvimento fetal normal (12).
A placenta não é contudo uma barreira estanque, já que moléculas nocivas
circulantes podem atingir a circulação fetal. De facto, os mecanismos de
transporte de nutrientes da mãe para o feto não têm especificidade restrita aos
seus substratos fisiológicos, pelo que qualquer xenobiótico estruturalmente
semelhante ao substrato fisiológico, pode ser transportado para a circulação fetal,
ou mesmo sem ser transportado interferir com o transporte de nutrientes (12).
O transporte de glicose ao nível da placenta é mediado por transportadores
estereoespecíficos (selectivos para a D-glicose relativamente à L-glicose),
independentes do sódio, sensíveis à inibição pela citocalasina B e floretina, e que
apresentam características cinéticas semelhantes. Esse transporte ocorre
essencialmente na zona vásculo-sincicial, (7) preferencialmente na direcção
materno-fetal e em menor extensão na direcção oposta (9).
O principal transportador envolvido no movimento transplacentar de glicose
é o GLUT1, o qual está presente essencialmente no sinciciotrofoblasto e em

5
menor grau nas células vasculares fetais (11). A sua distribuição entre a
membrana apical e basal é assimétrica, com um grau de expressão e actividade 4
a 5 vezes superior no lado apical em virtude da sua maior área de superfície (13).
Por isso, em condições normais, os transportadores de glicose presentes na
membrana basal são o passo limitante do transporte, permitindo através dos seus
níveis de expressão controlar o fornecimento de glicose para o feto (11). O
aumento da expressão do GLUT1 na membrana basal, que ocorre durante o
segundo e terceiro trimestres de gestação, está pois de acordo com o
crescimento fetal e necessidade de glicose máximos característicos deste
período (14). Em sinciciotrofoblastos de placentas de termo, a sobre-expressão
de transportadores GLUT1 ao nível da membrana basal, pode ser responsável
por situações clínicas de diabetes e macrossomia fetal (13). Contudo, os
mecanismos desencadeantes dessa sobre-expressão não estão ainda totalmente
esclarecidos (11).
Em virtude de condições experimentais variadas e de diferenças na
interpretação de resultados, (13, 15) existe alguma contradição no que diz
respeito aos restantes transportadores de glicose expressos na placenta e
respectiva localização celular (11). Os transportadores GLUT3 são expressos
somente no endotélio vascular e funcionam como scavengers de glicose (11).
Contudo, durante a fase inicial da gestação, é possível que citotrofoblastos
mitoticamente activos expressem o GLUT3, ao invés daqueles que já não se
dividem ou que já se tenham diferenciado em sinciciotrofoblastos característicos
de uma idade gestacional mais avançada. Seguindo o mesmo padrão de
expressão e localização do GLUT 3, estão também descritas outras isoformas da
família GLUT: o GLUT4 (transportador sensível à insulina) e o GLUT12 (13).

6
Com o objectivo de obter um conhecimento mais profundo da intensa
actividade transportadora que ocorre na placenta, é importante comparar dados
obtidos de sistemas in vitro, como linhas celulares imortalizadas e culturas
primárias (ambas de sinciciotrofoblastos), com sistema in vivo de transporte em
tecidos (9). Estes estudos comparativos são absolutamente necessários (9), daí
que este seja um dos motivos científicos para a realização deste trabalho.
As células BeWo são uma linha celular de coriocarcinoma humano, que
exibe muitas das características morfológicas e bioquímicas, descritas em
trofoblastos humanos (9). Estas células ao se diferenciarem, formam um tecido
semelhante aos sinciciotrofoblastos, incluindo os aspectos relacionados com a
expressão e actividade de transportadores (5). Como tal, as células BeWo exibem
uma distribuição assimétrica dos transportadores de glicose, expressando
principalmente o GLUT1 (14). Esta linha celular pode ser cultivada em
monocamada polarizada aderente ao suporte, (14) sendo considerada um modelo
de trofoblasto válido e conveniente para estudar actividades celulares e processos
de transporte transplacentar de nutrientes e fármacos (9).
A relação existente entre os mais de 8000 polifenóis presentes na dieta e a
prevenção de doenças no homem, tem sido um campo de investigação intenso
durante os últimos anos (16). De acordo com as suas estruturas, as principais
classes de polifenóis incluem os flavonóides, os mais abundantes na nossa dieta,
e os menos comuns estilbenos (como o resveratrol) e linhanos.
Uma das mais importantes fontes alimentares de polifenóis são as bebidas
de consumo habitual como sumos de fruta, vinho, café, bebidas à base de cacau
e cerveja (17). Algumas destas bebidas, especialmente o vinho tinto e o chá

7
verde, evidenciaram em estudos bioquímicos e epidemiológicos um efeito
protector de várias doenças degenerativas como cancro, doenças
cardiovasculares, doenças neurodegenerativas e diabetes (18, 19, 20, 21, 22, 23).
Devido à riqueza destes alimentos em polifenóis, estes últimos têm sido alvo de
numerosíssimos estudos os quais lhes têm atribuído imensas propriedades
biológicas protectoras (24, 25, 26). Entre essas propriedades destacam-se a
actividade anti-oxidante, anti-carcinogénica, anti-inflamatória, anti-estrogénica,
anti-proliferativa, anti-alérgica, anti-trombótica e moduladora de actividade
enzimática (27, 24, 25, 26).
Contudo, interessa referir que existem também estudos epidemiológicos,
que não atribuem efeito protector ou que sugerem mesmo um efeito prejudicial
dos polifenóis na saúde humana (27).
Através da dieta, e igualmente por via de bebidas de consumo habitual
como o café, chá, bebidas de cola, e através de medicamentos, o público em
geral consome uma grande variedade de produtos contendo cafeína (28). Cerca
de 75% das mulheres grávidas norte-americanas, consomem quantidades
moderadas ou baixas de cafeína (29). Contudo, as estimativas do consumo diário
são muito variáveis, oscilando em média entre os 150 a 300 mg ou menos (29,
30). Apesar de ainda permanecerem dúvidas acerca do consumo materno de
cafeína representar ou não um risco para o crescimento intrauterino do feto, (28)
desde 1980 que a Food and Drug Administration recomenda que as mulheres
grávidas evitem o consumo de cafeína (29).

8
O uso de drogas ilícitas e lícitas durante a gravidez é um grave problema
de saúde pública, devido aos potenciais efeitos adversos para a saúde materna e
fetal (31). Nas mulheres em idade fértil, sobretudo em adolescentes, a prevalência
do consumo de drogas de abuso tem vindo a crescer. (32, 33). No final dos anos
90, 5 a 6% das mulheres grávidas norte-americanas (mais de duzentas mil
mulheres), afirmou ter consumido drogas ilícitas durante o período gestacional
(32, 32). Destas, 28% abstiveram-se de consumir durante o primeiro trimestre, e
93% apenas durante o último trimestre, após terem conhecimento da sua gravidez
(34). A marijuana (dois terços) seguida da cocaína (um décimo) eram as drogas
ilícitas mais consumidas (34).
Adicionalmente, metade das mulheres grávidas norte-americanas (mais de
2 milhões de mulheres) consumiram tabaco e álcool durante a gravidez, sendo
por isso as substâncias de abuso alvo de maior consumo (34, 35). O consumo
materno abusivo de álcool provoca uma série de complicações graves que
constituem a síndrome alcoólica fetal (30), enquanto que o tabaco e seus
constituintes (sobretudo a nicotina) podem provocar: aborto, parto prematuro,
atraso de crescimento intra-uterino e baixo peso à nascença (36). No que diz
respeito às consequências adversas do consumo materno de drogas ilícitas,
existem ainda inconsistências (32). Alguns estudos demonstram uma associação
entre uso de drogas e efeitos adversos na saúde materna (aborto e parto
prematuro) e fetal (atraso de crescimento intra-uterino, malformações congénitas
e alterações comportamentais à nasçenca) (34, 37), enquanto outros estudos não
mostram associação (32).
É de referir ainda que na população consumidora de drogas, existe uma
tendência crítica para o consumo de múltiplas substâncias. Assim, o consumo de

9
tabaco e álcool está normalmente associado ao consumo de uma série de
estimulantes e alucinogéneos que incluem a cocaína, marijuana, anfetaminas e
seus derivados como o ecstasy (3,4-metilenodioximetamfetamina - MDMA) (38).
OBJECTIVOS
Os mecanismos que regulam o transporte placentar de glicose a partir da
membrana apical do sinciciotrofoblasto e o seu efluxo na membrana basal do
sinciciotrofoblasto, são ainda escassamente conhecidos tanto em sistemas in vivo
como in vitro. Por isso, o principal objectivo deste trabalho de investigação foi
caracterizar o transporte de glicose na placenta, através do estudo da captação
apical de desoxiglicose (DG) marcada radioactivamente com trício (3H-DG), na
linha celular de coriocarcinoma humano BeWo.
Dado que a ingestão de glicose na dieta raramente ocorre de forma isolada
mas sim no contexto do alimento, foi objectivo deste trabalho determinar o efeito
de alguns polifenóis, metilxantinas, do etanol e do composto decorrente do seu
metabolismo hepático – o acetaldeído, no transporte de 3H-DG em células BeWo.
Para tal, estudou-se o efeito agudo e crónico de diferentes concentrações dos
seguintes polifenóis: catequina, crisina, epicatequina, epigalocatequina-3-galato
(EGCG), miricetina, quercetina, resveratrol, rutina e xanto-humol, das
metilxantinas cafeína e teofilina, bem como do etanol e acetaldeído.

10
Determinou-se também o efeito agudo e crónico de algumas drogas como
a anfetamina, cocaína, MDMA, e ∆9-tetrahidrocanabinol (THC) e de drogas lícitas
como a nicotina, na captação de 3H-DG em células BeWo.
MATERIAIS E MÉTODOS
Materiais
Desoxi-D-glicose, 2-[1,2-3H] (hidrocloreto; actividade específica 40-50
Ci/mmol) (American Radiolabeled Chemicals, Saint Louis, MO, USA); cafeína
(BDH Laboratory Chemicals Ltd., Poole, England); 2-desoxi-D-glicose (Fluka
Chemie GmbH, Buchs, USA); (+)-catequina hidratada, citocalasina B obtida de
Helminthosporum dematiodium, colagénio tipo 1, crisina, epicatequina, EGCG ((-
)epigalocatequina-3-galato), Ham’s F12 K (Nutrient mixture F12-Ham Kaighn’s
modification), HEPES (ácido N-2-hidroxietilpiperazina-N’-2-etanesulfónico),
miricetina, MTT (brometo de 3-(4,5-dimetil-2-tiazolil)-2,5-difenil-2H-tetrazólio); (-)-
nicotina (sal de tartarato hidrogenado), quercetina di-hidratada, resveratrol, rutina,
solução de antibiótico/antimicótico (100 unidades ml-1 penicilina; 100 �g ml-1
estreptomicina e 0.25 �g ml-1 anfotericina B), solução de tripsina-EDTA, soro
bovino fetal, teofilina (Sigma, St. Louis, MO, USA). DMSO (dimetilsulfóxido), Triton
X-100, Tris (hidrocloreto de tris-(hidroximetil)-aminometano) (Merck, Darmstadt,
Germany); etanol absoluto (Panreac Química SA, Barcelona, Espanha);
acetaldeído (May & Baker, Dagenham, UK); (±)-anfetamina, (±)-MDMA (ecstasy;
3,4-metilenodioximetamfetamin), THC ((-)-∆9-tetrahidrocanabinol) (Cerilliant

11
Corporation, Round Rock, TX, USA); cocaína (hidrocloreto) (Uquipa, Lisboa,
Portugal); xanto-humol (gentilmente cedido pelo Eng. Machado Cruz, iBesa-
Instituto de Bebidas e Saúde, S. Mamede Infesta, Portugal).
Cultura de Células BeWo
A linha celular BeWo foi obtida da American Type Culture Collection (ATCC
CCL-98, Rockville, MD, EUA) e foi utilizada entre as passagens 34 e 65. As
células foram mantidas numa atmosfera humidificada de 5% CO2–95% ar e foram
cultivadas em meio Ham’s F12K suplementado com 2,5 g/l bicarbonato de sódio,
10% de soro bovino fetal inactivado pelo calor e solução de 1%
antibiótico/antimicótico. O meio de cultura era mudado a cada 2-3 dias e as
células eram sub-cultivadas (divisão da cultura) a cada 7 dias na razão de 1:2.
Para realizar uma sub-cultura, as células eram removidas das placas por acção
enzimática (0,25 % tripsina-EDTA, 5 minutos, 37ºC) e eram cultivadas em placas
de Petri (21 cm2; Ø 60 mm; TPP®, Trasadingen, Switzerland). Para as
experiências de transporte, as células BeWo eram semeadas em placas de 24
poços (2 cm2; Ø 16 mm; TPP®) previamente tratadas com colagénio, para uma
maior aderência das células ao suporte. Após 3-5 dias de cultura (até atingirem
90-100% de confluência), as células eram usadas nas experiências. Nesta altura
cada cm2 continha cerca de 60 �g de proteína celular.
Estudos de transporte
A captação de glicose nas células BeWo foi medida usando o análogo da
glicose não metabolizável (3H-DG) como marcador.

12
As experiências de transporte foram realizadas em meio tampão GF-HBS
(solução amortecedora salina sem glicose) com a seguinte composição (em mM):
140 NaCl, 5 KCl, 1 CaCl2, 2,5 MgSO4, 20 HEPES e pH 7,4. Em cada experiência
as células BeWo em monocamadas nos poços, eram tratadas do seguinte modo:
inicialmente o meio de cultura era aspirado e as células eram lavadas com 0.3 ou
0,5 ml de meio GF-HBS a 37ºC. Depois as células eram pré-incubadas durante 20
minutos com 0,3 ml de meio GF-HBS a 37ºC. A captação de 3H-DG pelas células,
iniciava-se através da incubação com 0,3 mL de meio GF-HBS a 37ºC contendo 1
�M de 3H-DG (excepto nas experiências de determinação da cinética de captação
de 3H-DG). Após 6 minutos, procedia-se à paragem da incubação por meio de
lavagem das células com 0,3 ou 0,5 mL de meio GF-HBS gelado (4ºC). As células
eram depois solubilizadas com 0,3 mL de Triton X-100 a 0,1% (em tris-HCL 5 mM,
pH 7,4). As células eram deixadas durante a noite à temperatura de 4ºC. A
radioactividade captada pelas células era então medida por cintilometria líquida.
Tratamento agudo e crónico das células com os compostos a testar
Para testar o efeito agudo dos compostos na captação de 3H-DG em
células BeWo, estes estavam presentes durante o período de pré-incubação (20
minutos) e incubação com 3H-DG 1 �M (6 minutos).
Quanto ao estudo do efeito crónico dos compostos na captação de 3H-DG,
este foi realizado em células BeWo cultivadas à 3 dias (com 90-95% de
confluência), em que o meio de cultura continha os compostos a testar. Os
compostos eram diluídos 1000 vezes no meio de cultura de forma a minimizar o
efeito do solvente, com excepção da citocalasina B que foi diluída 100 vezes. O
meio era renovado após as primeiras 24 horas, e no final das 48 horas realizou-se

13
as experiências de transporte. As experiências de transporte eram idênticas às
descritas em “Estudos de transporte”, mas sem período de pré-incubação, sendo
as células unicamente incubadas com a 3H-DG durante 6 minutos na ausência do
composto a testar.
As concentrações dos compostos a testar quer a nível agudo quer em
efeito crónico, foram escolhidas com base em trabalhos anteriormente realizados
pelo nosso grupo (39, 40).
Os controlos para os compostos testados foram feitos na presença do
respectivo solvente. Para os compostos cujos solventes são DMSO
(dimetilsulfóxido), etanol, metanol ou água, a concentração final destes solventes
foi de 1% no meio GF-HBS de pré-incubação e incubação para os tratamentos
agudos, ou de 0,1% no meio de cultura para os tratamentos crónicos (com
excepção do solvente da citocalasina B que foi 1%).
Determinação da viabilidade celular
O efeito dos diferentes compostos testados na viabilidade das células
Bewo foi determinado pelo teste do MTT (brometo de 3-(4,5-dimetil-2-tiazolil)-2,5-
difenil-2H-tetrazólio) (41).
Para determinar se os compostos que tiveram efeito agudo (26 minutos) na
captação de 3H-DG afectaram a viabilidade celular, as células Bewo foram
incubadas durante 3 horas a 37ºC em 500 �L de meio de cultura contendo 50 �L
de solução MTT (5 mg/ml). Nos últimos 26 minutos deste período, os compostos a
serem testados foram adicionados a cada poço. Para determinar se os compostos
que tiveram efeito crónico (48 horas) na captação de 3H-DG afectaram a

14
viabilidade celular, as células Bewo foram cronicamente tratadas com os
compostos. Após 45 horas de tratamento, 50 �L de solução MTT (5 mg/ml) foram
adicionados a cada poço e as células foram posteriormente incubadas durante 3
horas a 37ºC.
A solução de MTT foi removida após o período de incubação de 3 horas e
as células foram solubilizadas através da adição de 200 �l de DMSO, seguida de
agitação da placa durante 10 minutos à temperatura ambiente. A densidade
óptica para as soluções de cada poço foi determinada a 550 nm e a 650 nm no
leitor de placas. A densidade óptica a 650 nm, que corresponde a uma absorção
inespecífica de luz, foi subtraída da densidade óptica a 550 nm, para calcular o
valor específico da densidade óptica para os cristais de formazan derivados da
redução do MTT. Esse valor será proporcional ao número de células viáveis com
mitocôndrias activas.
Doseamento de proteínas
A quantidade de proteínas existentes nas células em monocamada foi
determinada segundo o protocolo descrito por Bradford (42), utilizando como
padrão a albumina de soro bovino.
Cálculos e tratamento estatístico
Para analisar a dependência da captação de 3H-DG em função do tempo,
os parâmetros da equação (1) foram ajustados aos dados experimentais por
análise de regressão não linear utilizando um programa informático (Muzyka,
Tarkany, Yelizanof, Sergienko and Boichuk (2005) (43).

15
(1) A(t)=k in/kout (1-e-kout.t)
A(t) representa a acumulação de 3H-DG no instante t, kin e kout representam as
taxas constantes do transporte para dentro e para fora da célula,
respectivamente, e t representa o tempo de incubação. Amax corresponde por
definição, à acumulação de 3H-DG no equilíbrio (t��). kin é expresso em pmol
mg proteina-1 min-1 (sendo depois convertido em �L mg proteina-1 min-1) e kout em
min-1.
Para o cálculo do IC50, os parâmetros da equação de Hill para inibição em
múltiplos sítios foi ajustada aos dados experimentais por análise de regressão não
linear, utilizando um programa informático (Muzyka, Tarkany, Yelizanof, Sergienko
and Boichuk (2005) (43).
Para analisar a curva de saturação, os parâmetros da equação de
Michaelis-Menten foram ajustados aos dados experimentais por análise de
regressão não linear, utilizando um programa informático (Muzyka, Tarkany,
Yelizanof, Sergienko and Boichuk (2005) (43).
As médias aritméticas são apresentadas com o erro padrão da média
(EPM) e as médias geométricas com intervalos de confiança a 95%. O significado
estatístico da diferença entre grupos, foi avaliado pelo teste t de Student. As
diferenças são consideradas significativas sempre que P<0.05.
O valor de n indica o número de réplicas de pelo menos duas actividades
experimentais realizadas em dias diferentes.

16
RESULTADOS
O análogo não metabolizável da glicose (3H-DG) foi utilizado nestes
estudos de transporte, pois é captado pelos GLUT sendo fosforilado pela
hexocínase a 2-desoxiglicose-6-fosfato (DG-6-P), que não será utilizada
posteriormente nas vias de metabolismo glicolítico. Além disso, o facto da DG-6-P
ter carga eléctrica, torna-a incapaz de ser transportada para fora da célula por
difusão passiva. Por isso a acumulação de DG-6-P radioactiva nas células, é um
indicador fiável da actividade transportadora da glicose (44).
1. Caracterização da captação de 3H-DG em células BeWo
Na primeira fase do trabalho experimental, caracterizou-se a captação
apical de 3H-DG em células BeWo, no que diz respeito à dependência do tempo e
à sua especificidade.
1.1 Dependência do tempo
Inicialmente determinou-se a variação da captação de 3H-DG em função do
tempo (time-course). Para tal, as células BeWo foram incubadas com 3H-DG (1
µM) na presença ou ausência de citocalasina B (um potente inibidor do transporte
facilitado de glicose), durante diferentes períodos de tempo (figura 2).

17
A análise dos resultados, revelou que na ausência de citocalasina B o kin foi
de 24,08 ± 4.13 �l mg proteina-1 min-1, o kout de 0,045 ± 0.010 min-1 e a Amax de
535,70 ± 46,84 pmol mg proteina-1. Ou seja, uma quantidade de células
correspondente a 1 mg de proteína celular, removeu a 3H-DG presente em 24,1 �l
de meio de incubação por minuto, e simultaneamente 4,5% da 3H-DG intracelular
acumulada saiu das células por minuto. Na presença de 50 µM de citocalasina B,
ocorreram diferenças significativas na captação de 3H-DG em função do tempo. A
citocalasina B reduziu o kin para 8,93 ± 1,40 �l mg proteina-1 min-1 e a Amax para
339,00 ± 35,67 pmol mg proteina-1 (P<0,05). Contudo, não alterou
significativamente o kout (0.026 ± 0.006 min-1).
De referir ainda, que também se realizaram experiências com 3H-DG a 100
µM. Quer na ausência (kin de 26,60 ± 3,10 �l mg proteina-1 min-1, kout de 0,046 ±
0,007 min-1 e Amax de 566,30 ± 32,90 pmol mg proteina-1) quer na presença de
citocalasina B (kin de 10,10 ± 1,20 �l mg proteina-1 min-1 (P<0,05); kout de 0,019 ±
0,004 min-1 e Amax de 531,00 ± 57,57 pmol mg proteina-1) os resultados obtidos
foram semelhantes aos anteriores. Contudo, o efeito inibitório da citocalasina B foi
0 10 20 30 40 50 60 70 80 90 1000
100
200
300
400
500
600 ControloCito B 50 µM
T ime-course 1 µM
Tempo de incubação (min)C
apta
ção
de
3 H-D
G(p
mo
l/m
gp
rote
ína)
Figura 2 - Variação da captação apical de 3H-DG em função dotempo de incubação (time-course). As células foram pré-incubadasdurante 20 minutos e posteriormente incubadas a 37ºC com 3H-DG 1µM, na ausência (controlo, n=6) ou presença de citocalasina B 50 µM(Cito B 50 µM, n=5-6). Os resultados são apresentados sob a formade média ± EPM.

18
menor na concentração de 100 µM 3H-DG do que na de 1 µM, pois o valor de kin
obtido na presença de citocalasina B foi superior (figura 3 – anexo 1).
Em virtude destes resultados as experiências subsequentes foram
realizadas com 3H-DG a 1 µM, à excepção das experiências de cinética de
captação.
De acordo com as figuras 2 e 3, a captação de 3H-DG nas células BeWo
ocorreu de uma forma dependente do tempo aumentando linearmente até aos 6
minutos de incubação, à qual se seguiu uma fase de estabilização. Com base
nesta informação, as experiências subsequentes foram realizadas utilizando
períodos de incubação de 6 minutos.
1.2 Especificidade da captação de 3H-DG
Com o objectivo de investigar a especificidade do mecanismo envolvido na
captação de 3H-DG, determinou-se o efeito de diferentes concentrações de
citocalasina B (figura 4).
Figura 4. Efeito de concentrações crescentes de citocalasina B (cito B 1,10, 50 e 100 µM; n=6-9), na captação de 3H-DG. As células BeWo forampré-incubadas durante 20 minutos e incubadas posteriormente com 3H-DG1 µM durante 6 minutos a 37ºC, na presença de citocalasina B ou dorespectivo solvente (controlo, n=14, correspondendo a 100%). Osresultados são apresentados sob a forma de média ± EPM.
-7 -6 -5 -4 -3
0102030405060708090
100110
Log [ cito B ]
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)

19
A presença deste composto exerceu um efeito inibitório dependente da
concentração, quando comparado com o controlo (ausência de citocalasina B,
correspondendo a 100% de captação). Na concentração de 10 µM a citocalasina
B reduziu significativamente a captação de 3H-DG em 22% (78,3 ± 1,7% do
controlo), enquanto que nas concentrações de 50 e 100 µM a redução foi ainda
mais significativa, cerca de 60% (40,3 ± 3,3% e 41,8 ± 1,3% do controlo,
respectivamente), obtendo-se um efeito inibitório máximo.
Apesar da inibição da captação não ter sido mais significativa, o valor do
IC50 (concentração necessária para provocar 50 % de inibição da captação
facilitada de 3H-DG) calculado para a citocalasina B foi de 17,04 µM (8,92-32,6
µM, n=6-9).
2. Modulação nutricional da captação de 3H-DG
Na segunda fase do trabalho, investigou-se a modulação da captação de
3H-DG nas células BeWo, por diferentes concentrações de compostos bioactivos
presentes em bebidas: polifenóis, metilxantinas, etanol e o seu metabolito
acetaldeído. Para tal, realizaram-se tratamentos agudos (26 minutos) e
tratamentos crónicos (48 horas de exposição).
2.1 Efeito agudo
A exposição aguda das células BeWo a diferentes concentrações de etanol
e acetaldeído (0,1-100 mM), resultou num efeito diferente para cada um deles
(figura 5).

20
O etanol aumentou significativamente a captação de 3H-DG apenas na
concentração de 10 mM (111,5 ± 3,3% do controlo). Quanto ao acetaldeído,
diminuiu significativamente a captação de 3H-DG nas concentrações mais
elevadas, 30 mM (71,2 ± 6,7% do controlo) e 100 mM (35,3 ± 4,6% do controlo).
Testou-se também o efeito agudo de vários polifenóis, todos nas
concentrações de 1, 10 e 100 µM. O resveratrol, a EGCG e o xanto-humol
diminuíram significativamente a captação celular de 3H-DG na concentração de
100 µM (84,5 ± 4,7%, 61,1 ± 2,0% e 74,0 ± 8,4% do controlo, respectivamente)
(figuras 6, 7 e 8).
Figura 5 - Efeito da exposição aguda das células BeWo a diferentes concentrações(0.1, 3, 10, 30 e 100 mM) de etanol (ETOH, n=5-6) e acetaldeído (ACA, n=3-4), nacaptação de 3H-DG. As células BeWo foram pré-incubadas durante 20 minutos eposteriormente incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, na presença docomposto ou do respectivo solvente (controlo, n=8, correspondendo a 100%). Osresultados são apresentados sob a forma de média ± EPM. *Significativamentediferente do controlo (P < 0,05).
0.1 3 10 30 100 0.1 3 10 30 1000
20
40
60
80
100
120 Controlo
ETOH
ACA
*
*
*
Concentração (mM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
21
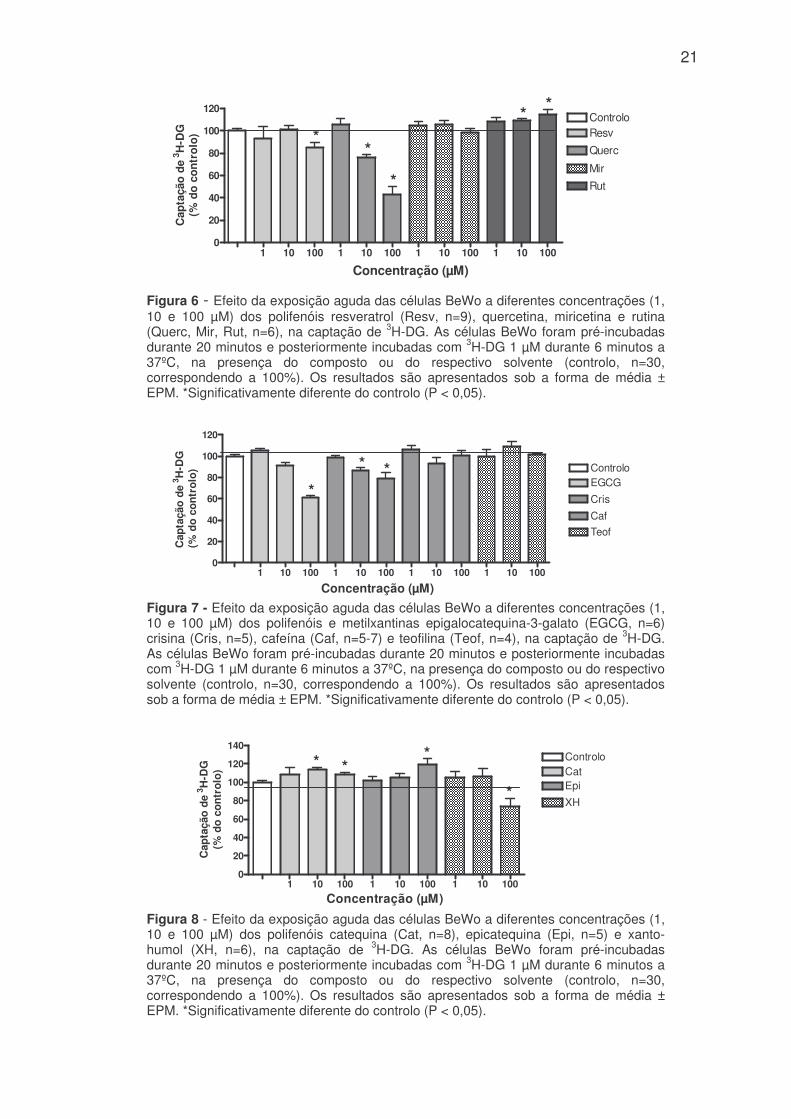
Figura 6 - Efeito da exposição aguda das células BeWo a diferentes concentrações (1,10 e 100 µM) dos polifenóis resveratrol (Resv, n=9), quercetina, miricetina e rutina(Querc, Mir, Rut, n=6), na captação de 3H-DG. As células BeWo foram pré-incubadasdurante 20 minutos e posteriormente incubadas com 3H-DG 1 µM durante 6 minutos a37ºC, na presença do composto ou do respectivo solvente (controlo, n=30,correspondendo a 100%). Os resultados são apresentados sob a forma de média ±EPM. *Significativamente diferente do controlo (P < 0,05).
1 10 100 1 10 100 1 10 100 1 10 1000
20
40
60
80
100
120ControloResv
Querc
Mir
Rut
*
*
*
**
Concentração (µM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 7 - Efeito da exposição aguda das células BeWo a diferentes concentrações (1,10 e 100 µM) dos polifenóis e metilxantinas epigalocatequina-3-galato (EGCG, n=6)crisina (Cris, n=5), cafeína (Caf, n=5-7) e teofilina (Teof, n=4), na captação de 3H-DG.As células BeWo foram pré-incubadas durante 20 minutos e posteriormente incubadascom 3H-DG 1 µM durante 6 minutos a 37ºC, na presença do composto ou do respectivosolvente (controlo, n=30, correspondendo a 100%). Os resultados são apresentadossob a forma de média ± EPM. *Significativamente diferente do controlo (P < 0,05).
1 10 100 1 10 100 1 10 100 1 10 1000
20
40
60
80
100
120
ControloEGCG
Cris
Caf
Teof
*
* *
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Concentração (µM)
Figura 8 - Efeito da exposição aguda das células BeWo a diferentes concentrações (1,10 e 100 µM) dos polifenóis catequina (Cat, n=8), epicatequina (Epi, n=5) e xanto-humol (XH, n=6), na captação de 3H-DG. As células BeWo foram pré-incubadasdurante 20 minutos e posteriormente incubadas com 3H-DG 1 µM durante 6 minutos a37ºC, na presença do composto ou do respectivo solvente (controlo, n=30,correspondendo a 100%). Os resultados são apresentados sob a forma de média ±EPM. *Significativamente diferente do controlo (P < 0,05).
1 10 100 1 10 100 1 10 1000
20
40
60
80
100
120
140ControloCatEpi
XH
* **
*
Concentração (µM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)

22
A quercetina (à excepção da concentração mais baixa) e a crisina
reduziram a captação de 3H-DG de uma forma dependente da concentração. Este
efeito foi significativo para as concentrações mais elevadas, sendo a redução de
aproximadamente 24 e 57 % para a quercetina 10 e 100 µM e de 13 e 21% para a
crisina 10 e 100 µM (figuras 6 e 7). Em suma, os compostos com efeito inibitório
agudo mais significativo, foram o acetaldeído 100 mM (65% de inibição), a EGCG
100 µM (40% de inibição) e a quercetina 100 µM (57% de inibição).
Por outro lado, a rutina aumentou a captação de 3H-DG nas concentrações
de 10 e 100 µM (109,2 ± 1,6% e 114,4 ± 5,0% do controlo, respectivamente) e a
epicatequina aumentou apenas na concentração de 100 µM (119,4 ± 6,4% do
controlo) (figuras 6 e 8). A catequina, nas concentrações de 10 e 100 µM também
aumentou significativamente a captação apical de 3H-DG (113,5 ± 3,0% e 108,3 ±
2,5% do controlo para as concentrações de 10 e 100 µM respectivamente) (figura
8). Em suma, os compostos que aumentaram mais significativamente a captação
de 3H-DG foram a rutina (14,4%) e epicatequina (19,5%) ambas nas
concentrações mais elevadas (100 µM).
A miricetina e as metilxantinas (cafeína e teofilina) nas concentrações
testadas, não modificaram significativamente a captação de 3H-DG nas células
BeWo (figuras 6 e 7).
2.2 Efeito crónico
Todos os compostos testados nas experiências de efeito agudo foram
também testados cronicamente, sendo que a maioria deles não alterou
significativamente a captação de 3H-DG nas células BeWo (figuras 9 e 10, e
figuras 11 e 12 – anexo 1). No entanto nas concentrações testadas, a rutina e a
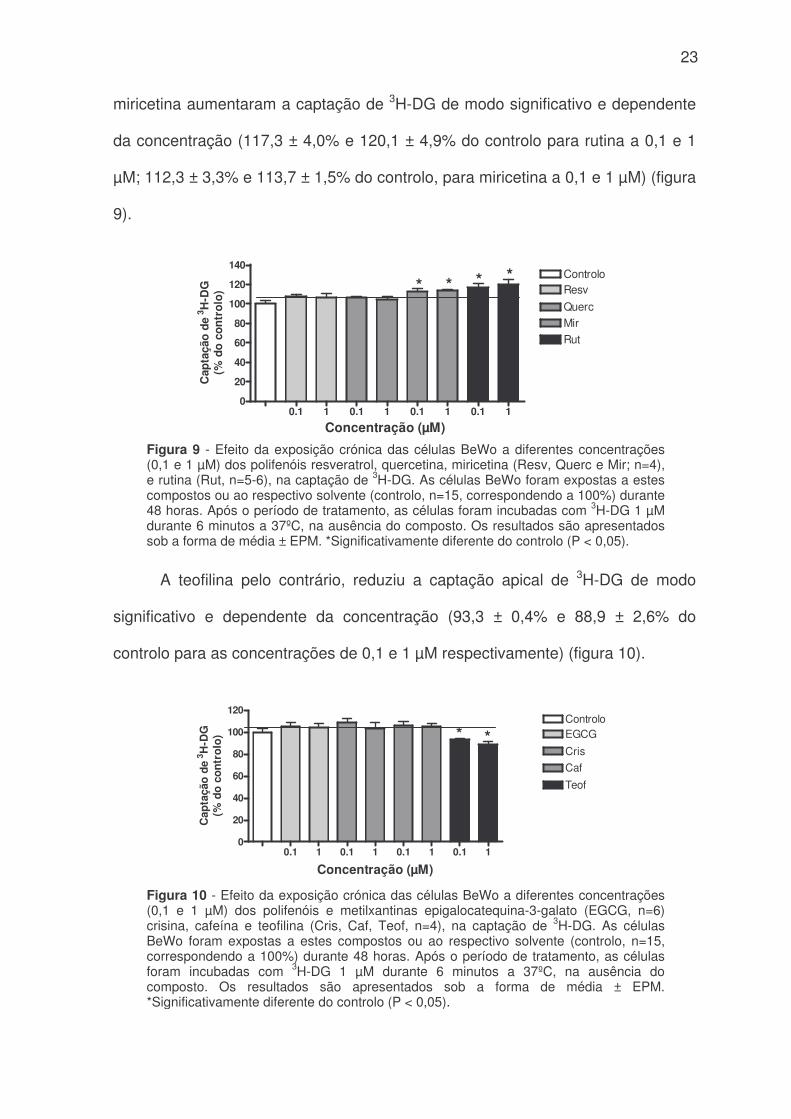
23
miricetina aumentaram a captação de 3H-DG de modo significativo e dependente
da concentração (117,3 ± 4,0% e 120,1 ± 4,9% do controlo para rutina a 0,1 e 1
µM; 112,3 ± 3,3% e 113,7 ± 1,5% do controlo, para miricetina a 0,1 e 1 µM) (figura
9).
A teofilina pelo contrário, reduziu a captação apical de 3H-DG de modo
significativo e dependente da concentração (93,3 ± 0,4% e 88,9 ± 2,6% do
controlo para as concentrações de 0,1 e 1 µM respectivamente) (figura 10).
Figura 9 - Efeito da exposição crónica das células BeWo a diferentes concentrações(0,1 e 1 µM) dos polifenóis resveratrol, quercetina, miricetina (Resv, Querc e Mir; n=4),e rutina (Rut, n=5-6), na captação de 3H-DG. As células BeWo foram expostas a estescompostos ou ao respectivo solvente (controlo, n=15, correspondendo a 100%) durante48 horas. Após o período de tratamento, as células foram incubadas com 3H-DG 1 µMdurante 6 minutos a 37ºC, na ausência do composto. Os resultados são apresentadossob a forma de média ± EPM. *Significativamente diferente do controlo (P < 0,05).
0.1 1 0.1 1 0.1 1 0.1 10
20
40
60
80
100
120
140ControloResv
QuercMirRut
* * * *
Concentração (µM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 10 - Efeito da exposição crónica das células BeWo a diferentes concentrações(0,1 e 1 µM) dos polifenóis e metilxantinas epigalocatequina-3-galato (EGCG, n=6)crisina, cafeína e teofilina (Cris, Caf, Teof, n=4), na captação de 3H-DG. As célulasBeWo foram expostas a estes compostos ou ao respectivo solvente (controlo, n=15,correspondendo a 100%) durante 48 horas. Após o período de tratamento, as célulasforam incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, na ausência docomposto. Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P < 0,05).
0.1 1 0.1 1 0.1 1 0.1 10
20
40
60
80
100
120ControloEGCG
Cris
Caf
Teof
* *
Concentração (µM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)

24
Os restantes compostos (resveratrol, quercetina, EGCG, crisina, cafeína,
catequina, epicatequina, xanto-humol, etanol e acetaldeído) não modificaram a
captação de 3H-DG nas células BeWo (figuras 9 e 10, e figuras 11 e 12 – anexo
1).
Apesar disso, o resveratrol (na concentrações de 0,1 e 1 µM) evidenciou
uma tendência para aumentar a captação de 3H-DG enquanto o xanto-humol na
concentração de 1 µM mostrou uma tendência para reduzir (figuras 9 e figura 11 -
anexo 1).
3. Modulação da captação de 3H-DG por drogas de abuso
Da mesma forma que se investigou a modulação nutricional da captação
de 3H-DG em células BeWo, investigou-se também o efeito agudo e crónico de
diferentes concentrações de drogas de abuso.
3.1 Efeito Agudo
Nenhuma das drogas de abuso testadas agudamente (nicotina, cocaína,
MDMA, THC e anfetamina) alterou significativamente a captação de 3H-DG
(figuras 13 e 14 – anexo 1).
3.2 Efeito Crónico
O MDMA e a anfetamina inibiram o transporte apical de 3H-DG de forma
estatisticamente significativa nas duas concentrações testadas, 0,25 e 2,5 µM
(88,7 ± 2,2% e 86,7 ± 2,5% do controlo para o MDMA 0,25 e 2,5 µM
respectivamente; 91,8 ± 1,7% do controlo e 92 ± 2,3% do controlo para a
anfetamina 0,25 e 2,5 µM respectivamente). Quanto ao THC, apenas a

25
concentração mais baixa provocou uma diminuição significativa da captação (para
93,4 ± 1,8% do controlo) (Figura 15).
De realçar que estes efeitos crónicos inibitórios foram quantitativamente
semelhantes, situando-se em torno dos 10%.
A nicotina e a cocaína tiveram um comportamento diferente, não
provocando alterações significativas na captação de 3H-DG (Figura 16 – anexo 1).
4. Efeito de compostos presentes na dieta e de drogas de abuso na
viabilidade celular
Após a realização dos estudos de transporte, determinou-se o efeito agudo
e crónico dos compostos testados na viabilidade celular. Este procedimento, teve
o objectivo de analisar se os efeitos inibitórios ou estimuladores dos compostos
na captação de 3H-DG, estavam relacionados ou não com possíveis efeitos
citotóxicos ou proliferativos nas células BeWo.
*
0.25 1 0.25 1 0.001 0.01 0.10
20
40
60
80
100
120ControloMDMA
ANF
THC
* * * *
Concentração (µM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 15 - Efeito da exposição crónica das células BeWo a diferentes concentraçõesde 3,4-metilenodioximetanfetamina (MDMA; 0,25 e 1 µM; n=6), anfetamina (ANF; 0,25e 1 µM; n=6) e ∆
9-tetrahidrocanabinol (THC; 0,001, 0,01 e 0,1 µM; n=6-9) na captaçãode 3H-DG. As células BeWo foram expostas a estes compostos ou ao respectivosolvente (controlo, n=12, correspondendo a 100%) durante 48 horas. Após o período detratamento, as células foram incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, naausência do composto. Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P < 0,05).
26
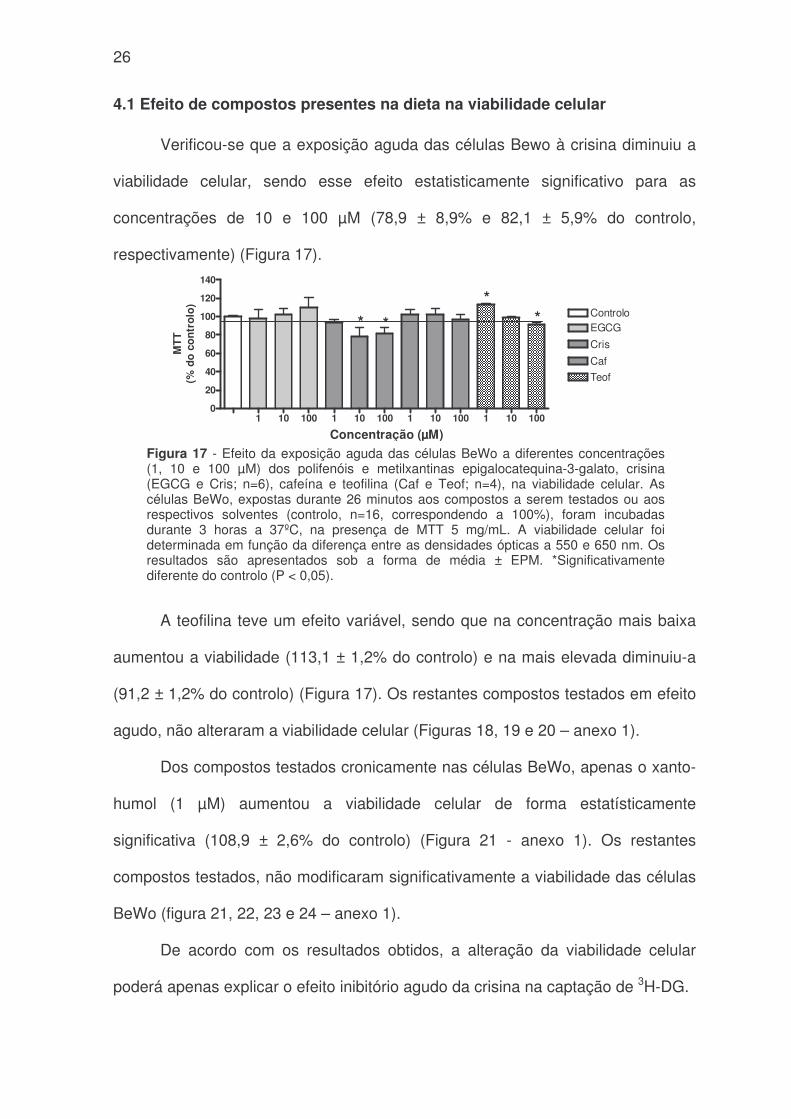
4.1 Efeito de compostos presentes na dieta na viabilidade celular
Verificou-se que a exposição aguda das células Bewo à crisina diminuiu a
viabilidade celular, sendo esse efeito estatisticamente significativo para as
concentrações de 10 e 100 µM (78,9 ± 8,9% e 82,1 ± 5,9% do controlo,
respectivamente) (Figura 17).
A teofilina teve um efeito variável, sendo que na concentração mais baixa
aumentou a viabilidade (113,1 ± 1,2% do controlo) e na mais elevada diminuiu-a
(91,2 ± 1,2% do controlo) (Figura 17). Os restantes compostos testados em efeito
agudo, não alteraram a viabilidade celular (Figuras 18, 19 e 20 – anexo 1).
Dos compostos testados cronicamente nas células BeWo, apenas o xanto-
humol (1 µM) aumentou a viabilidade celular de forma estatísticamente
significativa (108,9 ± 2,6% do controlo) (Figura 21 - anexo 1). Os restantes
compostos testados, não modificaram significativamente a viabilidade das células
BeWo (figura 21, 22, 23 e 24 – anexo 1).
De acordo com os resultados obtidos, a alteração da viabilidade celular
poderá apenas explicar o efeito inibitório agudo da crisina na captação de 3H-DG.
1 10 100 1 10 100 1 10 100 1 10 1000
20
40
60
80
100
120
140
ControloEGCG
Cris
Caf
Teof
* *
**
Concentração (µM)
MT
T(%
do
con
tro
lo)
Figura 17 - Efeito da exposição aguda das células BeWo a diferentes concentrações(1, 10 e 100 µM) dos polifenóis e metilxantinas epigalocatequina-3-galato, crisina(EGCG e Cris; n=6), cafeína e teofilina (Caf e Teof; n=4), na viabilidade celular. Ascélulas BeWo, expostas durante 26 minutos aos compostos a serem testados ou aosrespectivos solventes (controlo, n=16, correspondendo a 100%), foram incubadasdurante 3 horas a 37ºC, na presença de MTT 5 mg/mL. A viabilidade celular foideterminada em função da diferença entre as densidades ópticas a 550 e 650 nm. Osresultados são apresentados sob a forma de média ± EPM. *Significativamentediferente do controlo (P < 0,05).

27
4.2 Efeito de drogas de abuso na viabilidade celular
A exposição aguda das células BeWo à cocaína, diminuiu a viabilidade
celular de forma significativa e dependente da concentração (91,9 ± 2,2%, 84,2 ±
4,8%, e 80,8 ± 3,8% do controlo, nas concentrações de 0,1, 1 e 10 µM
respectivamente) (figura 25 - anexo 1).
A nicotina e o MDMA nas concentrações mais baixas (1 e 0,1 µM
respectivamente), reduziram a viabilidade das células BeWo em cerca de 12%
(88,6 ± 4,6 % e 88,8 ± 2,3% do controlo) comparativamente ao controlo. As
restantes concentrações não alteraram significativamente a viabilidade celular
(figura 25 e 26 - anexo 1). Quanto à anfetamina, apenas na concentração de 1 µM
reduziu ligeiramente, mas com significado estatístico, a viablilidade celular (94.4 ±
1.7% do controlo) (figura 26 - anexo 1). O THC comparativamente ao controlo,
não modificou a viabilidade das células BeWo (figura 26 - anexo 1).
Das drogas de abuso testadas cronicamente, nenhuma delas alterou
significativamente a viabilidade celular (figuras 27 e 28 – anexo 1), logo o efeito
inibitório crónico do MDMA, anfetamina e THC na captação de 3H-DG, não se
deve a efeitos citóxicos nas células BeWo.
5. Caracterização do efeito dos compostos testados
Na última fase do trabalho, caracterizou-se o efeito agudo e crónico dos
compostos testados sobre a captação de 3H-DG em células BeWo.
Para tal, realizaram-se tratamentos agudos ou crónicos na presença
simultânea de cada composto, cuja concentração tinha inibido ou estimulado

28
significativamente a captação de 3H-DG, e de citocalasina B na concentração de
50 µM (que tinha provocado um efeito inibitório máximo na captação de 3H-DG).
O objectivo foi testar se o efeito inibitório ou estimulador dos compostos, era
mantido ou alterado na presença de citocalasina B.
Realizaram-se também tratamentos agudos e crónicos na presença
simultânea de dois compostos (de consumo conjunto habitual por serem
abundantes nas mesmas bebidas), cujas concentrações tinham provocado efeitos
significativos na captação de 3H-DG. O objectivo consistiu em verificar se esses
compostos, mantinham ou alteravam (de um modo somativo por exemplo) a
captação de 3H-DG, quando eram testados em conjunto.
Por fim, determinou-se o efeito agudo inibitório de alguns compostos
presentes na dieta, sobre os parâmetros cinéticos da captação de 3H-DG em
células BeWo. Especificamente, determinou-se os efeitos desses compostos nos
valores de Vmax (velocidade da captação de 3H-DG para concentrações de DG
ditas saturantes) e Km (concentração de substrato que origina uma velocidade de
captação correspondente a metade da Vmax). Este último parâmetro é considerado
uma medida de afinidade do substrato para o transportador (45).
5.1 Caracterização do efeito agudo
Isoladamente, e tal como tínhamos previsto, a citocalasina B inibiu
consideravelmente a captação de 3H-DG (figura 29, e figuras 30 e 31 – anexo 1).
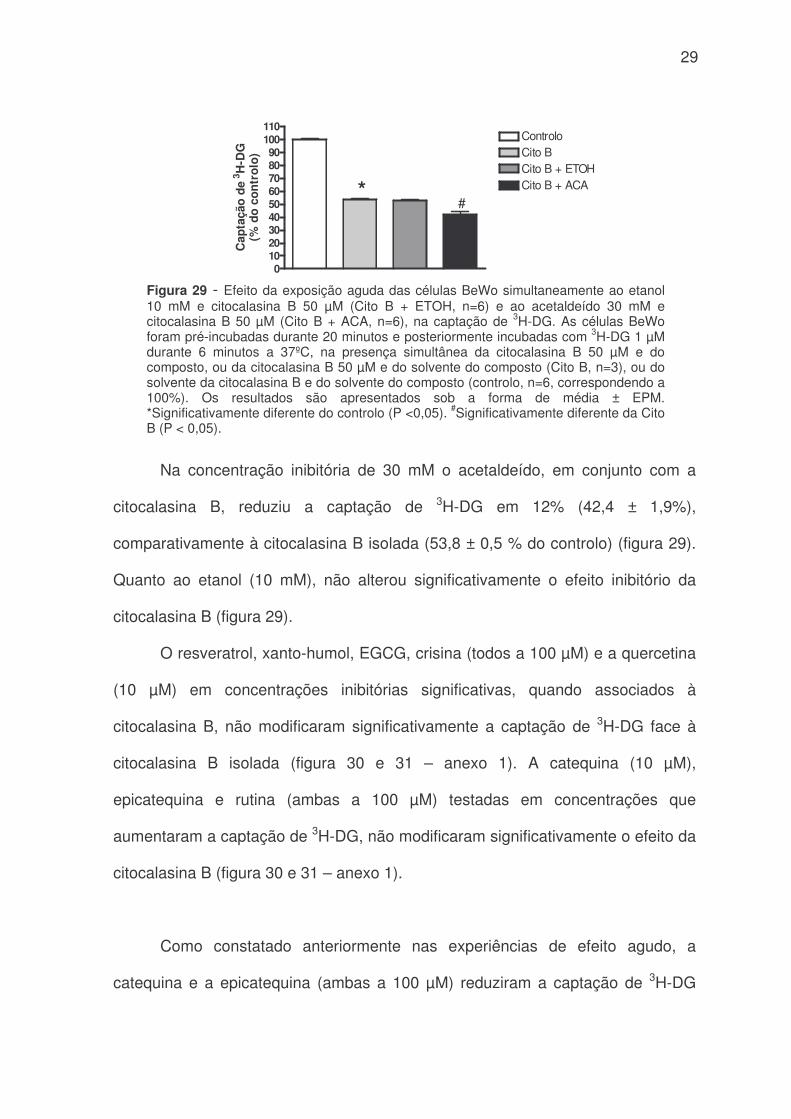
29
Na concentração inibitória de 30 mM o acetaldeído, em conjunto com a
citocalasina B, reduziu a captação de 3H-DG em 12% (42,4 ± 1,9%),
comparativamente à citocalasina B isolada (53,8 ± 0,5 % do controlo) (figura 29).
Quanto ao etanol (10 mM), não alterou significativamente o efeito inibitório da
citocalasina B (figura 29).
O resveratrol, xanto-humol, EGCG, crisina (todos a 100 µM) e a quercetina
(10 µM) em concentrações inibitórias significativas, quando associados à
citocalasina B, não modificaram significativamente a captação de 3H-DG face à
citocalasina B isolada (figura 30 e 31 – anexo 1). A catequina (10 µM),
epicatequina e rutina (ambas a 100 µM) testadas em concentrações que
aumentaram a captação de 3H-DG, não modificaram significativamente o efeito da
citocalasina B (figura 30 e 31 – anexo 1).
Como constatado anteriormente nas experiências de efeito agudo, a
catequina e a epicatequina (ambas a 100 µM) reduziram a captação de 3H-DG
0102030405060708090
100110
Cito BCito B + ETOHCito B + ACA
Controlo
*#
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 29 - Efeito da exposição aguda das células BeWo simultaneamente ao etanol10 mM e citocalasina B 50 µM (Cito B + ETOH, n=6) e ao acetaldeído 30 mM ecitocalasina B 50 µM (Cito B + ACA, n=6), na captação de 3H-DG. As células BeWoforam pré-incubadas durante 20 minutos e posteriormente incubadas com 3H-DG 1 µMdurante 6 minutos a 37ºC, na presença simultânea da citocalasina B 50 µM e docomposto, ou da citocalasina B 50 µM e do solvente do composto (Cito B, n=3), ou dosolvente da citocalasina B e do solvente do composto (controlo, n=6, correspondendo a100%). Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P <0,05). #Significativamente diferente da CitoB (P < 0,05).
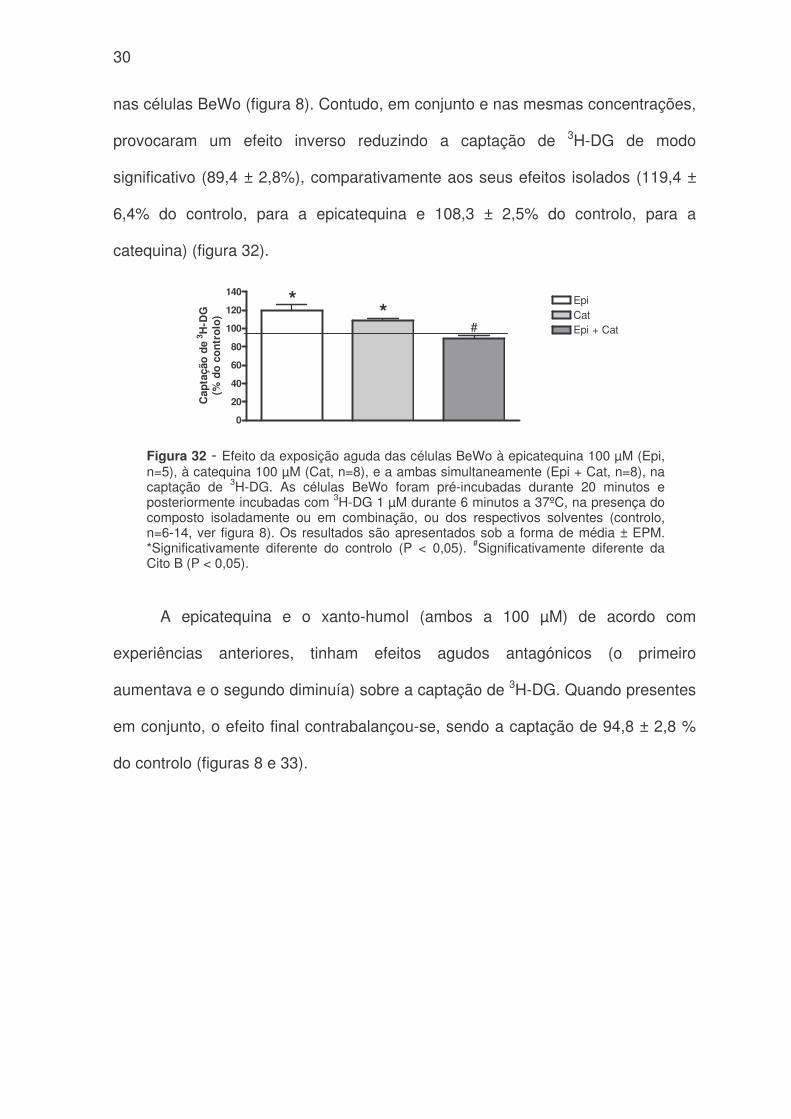
30
nas células BeWo (figura 8). Contudo, em conjunto e nas mesmas concentrações,
provocaram um efeito inverso reduzindo a captação de 3H-DG de modo
significativo (89,4 ± 2,8%), comparativamente aos seus efeitos isolados (119,4 ±
6,4% do controlo, para a epicatequina e 108,3 ± 2,5% do controlo, para a
catequina) (figura 32).
A epicatequina e o xanto-humol (ambos a 100 µM) de acordo com
experiências anteriores, tinham efeitos agudos antagónicos (o primeiro
aumentava e o segundo diminuía) sobre a captação de 3H-DG. Quando presentes
em conjunto, o efeito final contrabalançou-se, sendo a captação de 94,8 ± 2,8 %
do controlo (figuras 8 e 33).
Figura 32 - Efeito da exposição aguda das células BeWo à epicatequina 100 µM (Epi,n=5), à catequina 100 µM (Cat, n=8), e a ambas simultaneamente (Epi + Cat, n=8), nacaptação de 3H-DG. As células BeWo foram pré-incubadas durante 20 minutos eposteriormente incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, na presença docomposto isoladamente ou em combinação, ou dos respectivos solventes (controlo,n=6-14, ver figura 8). Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P < 0,05). #Significativamente diferente daCito B (P < 0,05).
0
20
40
60
80
100
120
140EpiCatEpi + Cat
**
#
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)

31
5.2 Caracterização do efeito crónico
A miricetina e rutina que isoladamente e na concentração de 1 µM,
aumentaram a captação de 3H-DG, não modificaram significativamente o efeito da
citocalasina B. (figura 34 – anexo 1). O mesmo aconteceu com a teofilina na
concentração inibitória de 1 µM (figura 34 – anexo 1). Por último, a anfetamina,
THC e MDMA em concentrações que inibiram a captação de 3H-DG em efeito
crónico, quando associadas à citocalasina B deixaram de evidenciar esse efeito
(figura 35 – anexo 1).
De referir ainda que, em efeito crónico a citocalasina B apenas inibiu
moderadamente a captação de 3H-DG (aproximadamente em 15%) por
comparação com o controlo (figuras 34 e 35 – anexo 1).
Por fim, estudou-se o efeito crónico decorrente da combinação da
miricetina e rutina (ambas a 1 µM) na captação de 3H-DG. Verificou-se que esta
combinação aumentou a captação (130,0 ± 5,4 %, n=9) comparativamente ao
Figura 33 - Efeito da exposição aguda das células BeWo à epicatequina 100 µM (Epi,n=5), ao xantohumol 100 µM (XH, n=6), e a ambos simultaneamente (Epi + XH, n=6),na captação de 3H-DG. As células BeWo foram pré-incubadas durante 20 minutos eposteriormente incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, na presença docomposto isoladamente ou em combinação, ou dos respectivos solventes (controlo,n=6-14, ver figura 8). Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P <0,05). #Significativamente diferente da CitoB (P <0,05).
0
20
40
60
80
100
120
140EPIXHEPI + XH
*
*#
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
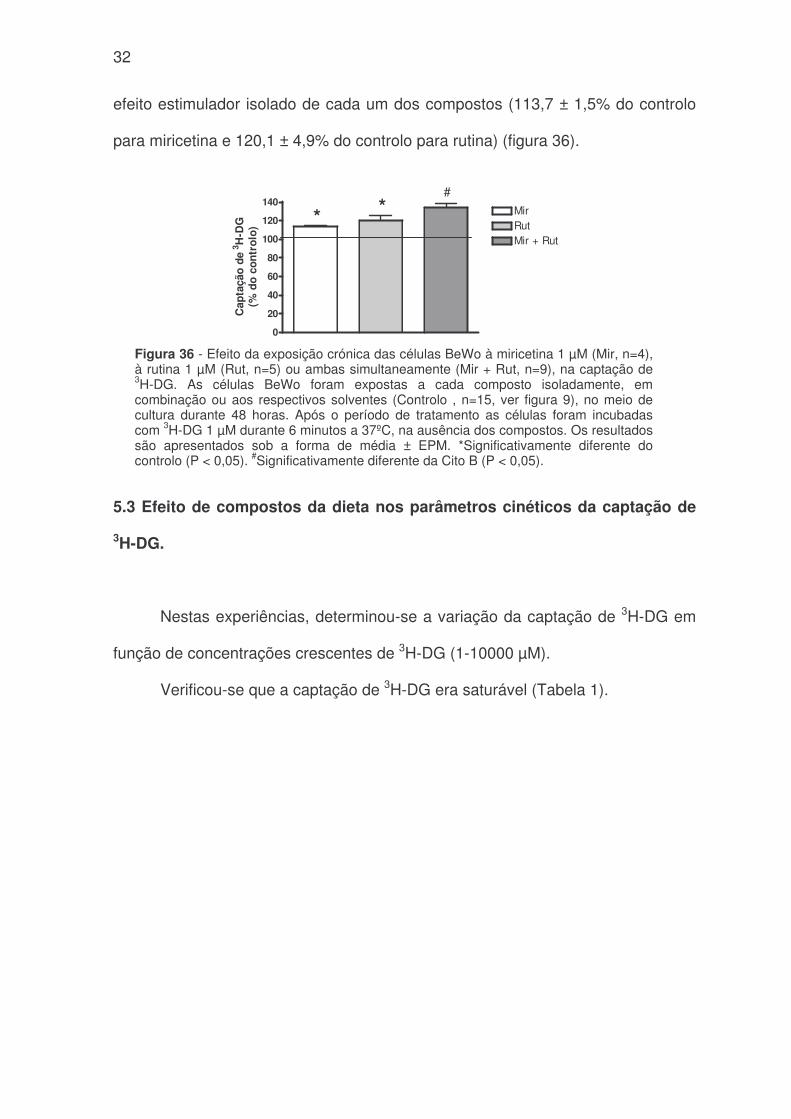
32
efeito estimulador isolado de cada um dos compostos (113,7 ± 1,5% do controlo
para miricetina e 120,1 ± 4,9% do controlo para rutina) (figura 36).
5.3 Efeito de compostos da dieta nos parâmetros cinéticos da captação de
3H-DG.
Nestas experiências, determinou-se a variação da captação de 3H-DG em
função de concentrações crescentes de 3H-DG (1-10000 µM).
Verificou-se que a captação de 3H-DG era saturável (Tabela 1).
0
20
40
60
80
100
120
140MirRutMir + Rut
* *#
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 36 - Efeito da exposição crónica das células BeWo à miricetina 1 µM (Mir, n=4),à rutina 1 µM (Rut, n=5) ou ambas simultaneamente (Mir + Rut, n=9), na captação de3H-DG. As células BeWo foram expostas a cada composto isoladamente, emcombinação ou aos respectivos solventes (Controlo , n=15, ver figura 9), no meio decultura durante 48 horas. Após o período de tratamento as células foram incubadascom 3H-DG 1 µM durante 6 minutos a 37ºC, na ausência dos compostos. Os resultadossão apresentados sob a forma de média ± EPM. *Significativamente diferente docontrolo (P < 0,05). #Significativamente diferente da Cito B (P < 0,05).
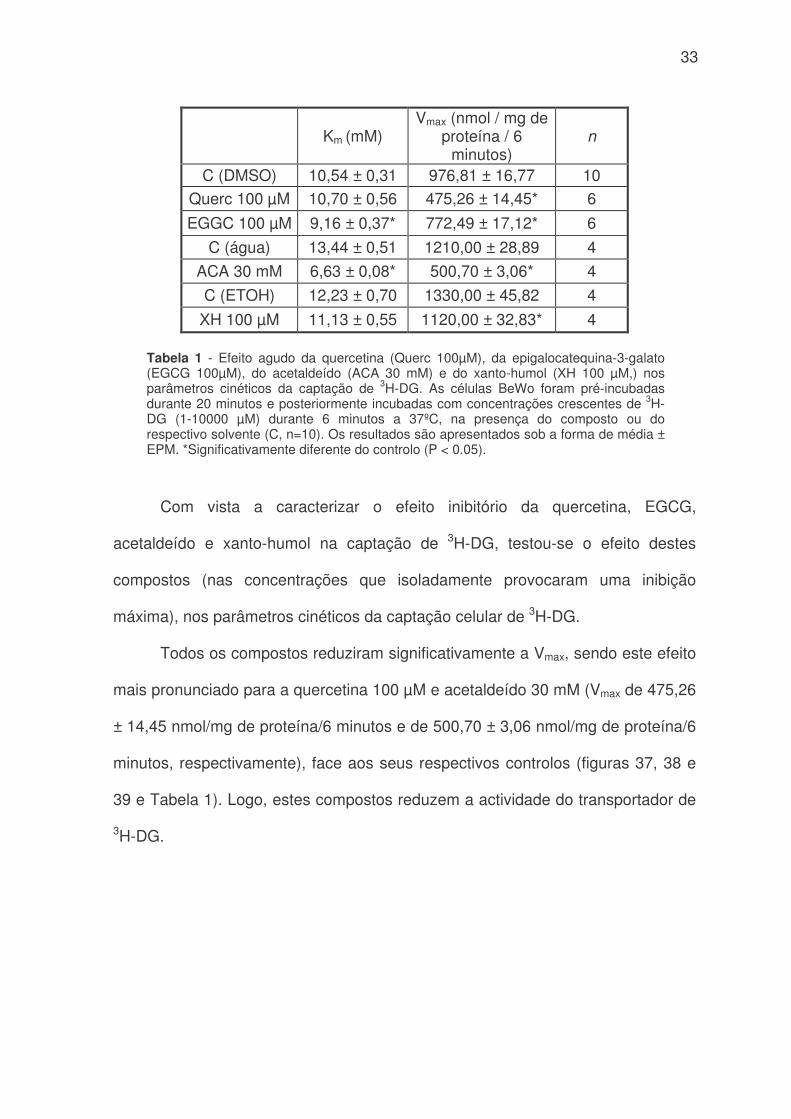
33
Com vista a caracterizar o efeito inibitório da quercetina, EGCG,
acetaldeído e xanto-humol na captação de 3H-DG, testou-se o efeito destes
compostos (nas concentrações que isoladamente provocaram uma inibição
máxima), nos parâmetros cinéticos da captação celular de 3H-DG.
Todos os compostos reduziram significativamente a Vmax, sendo este efeito
mais pronunciado para a quercetina 100 µM e acetaldeído 30 mM (Vmax de 475,26
± 14,45 nmol/mg de proteína/6 minutos e de 500,70 ± 3,06 nmol/mg de proteína/6
minutos, respectivamente), face aos seus respectivos controlos (figuras 37, 38 e
39 e Tabela 1). Logo, estes compostos reduzem a actividade do transportador de
3H-DG.
Km (mM)Vmax (nmol / mg de
proteína / 6minutos)
n
C (DMSO) 10,54 ± 0,31 976,81 ± 16,77 10
Querc 100 µM 10,70 ± 0,56 475,26 ± 14,45* 6
EGGC 100 µM 9,16 ± 0,37* 772,49 ± 17,12* 6
C (água) 13,44 ± 0,51 1210,00 ± 28,89 4
ACA 30 mM 6,63 ± 0,08* 500,70 ± 3,06* 4
C (ETOH) 12,23 ± 0,70 1330,00 ± 45,82 4
XH 100 µM 11,13 ± 0,55 1120,00 ± 32,83* 4
Tabela 1 - Efeito agudo da quercetina (Querc 100µM), da epigalocatequina-3-galato(EGCG 100µM), do acetaldeído (ACA 30 mM) e do xanto-humol (XH 100 µM,) nosparâmetros cinéticos da captação de 3H-DG. As células BeWo foram pré-incubadasdurante 20 minutos e posteriormente incubadas com concentrações crescentes de 3H-DG (1-10000 µM) durante 6 minutos a 37ºC, na presença do composto ou dorespectivo solvente (C, n=10). Os resultados são apresentados sob a forma de média ±EPM. *Significativamente diferente do controlo (P < 0.05).
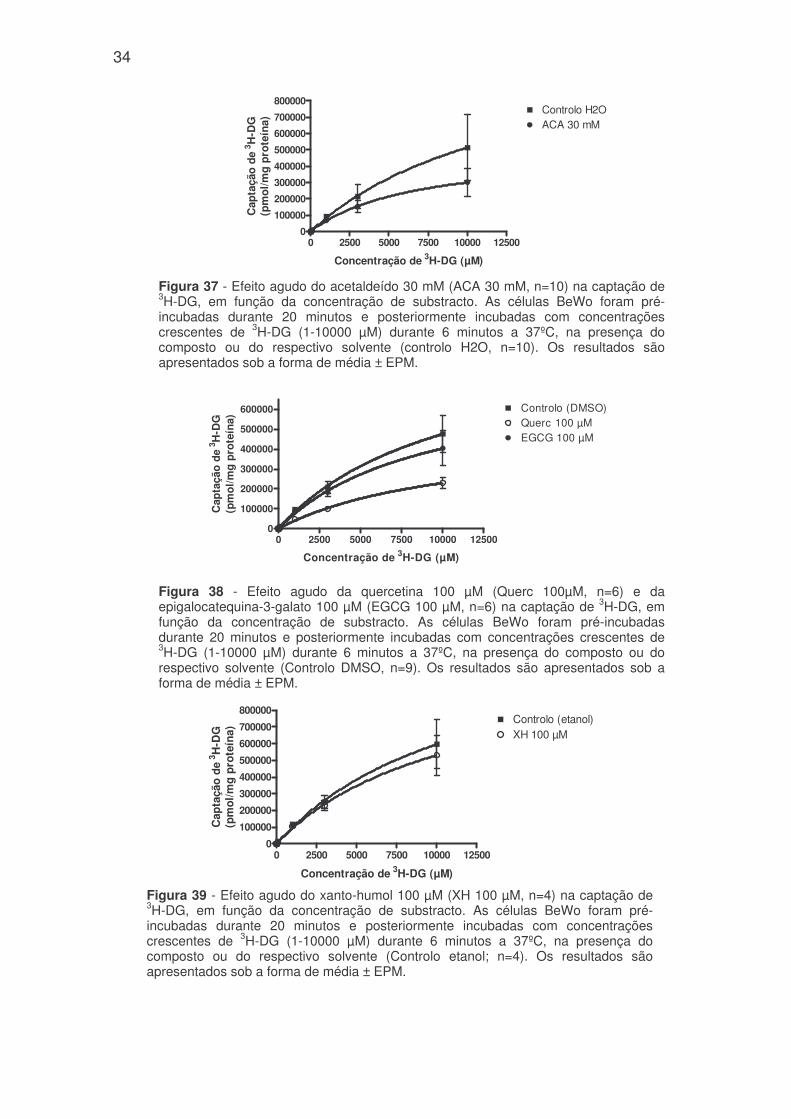
34
Figura 37 - Efeito agudo do acetaldeído 30 mM (ACA 30 mM, n=10) na captação de3H-DG, em função da concentração de substracto. As células BeWo foram pré-incubadas durante 20 minutos e posteriormente incubadas com concentraçõescrescentes de 3H-DG (1-10000 µM) durante 6 minutos a 37ºC, na presença docomposto ou do respectivo solvente (controlo H2O, n=10). Os resultados sãoapresentados sob a forma de média ± EPM.
0 2500 5000 7500 10000 125000
100000
200000
300000
400000
500000
600000
700000
800000Controlo H2OACA 30 mM
Concentração de 3H-DG (µM)C
apta
ção
de
3 H-D
G(p
mo
l/m
gp
rote
ína)
Figura 39 - Efeito agudo do xanto-humol 100 µM (XH 100 µM, n=4) na captação de3H-DG, em função da concentração de substracto. As células BeWo foram pré-incubadas durante 20 minutos e posteriormente incubadas com concentraçõescrescentes de 3H-DG (1-10000 µM) durante 6 minutos a 37ºC, na presença docomposto ou do respectivo solvente (Controlo etanol; n=4). Os resultados sãoapresentados sob a forma de média ± EPM.
0 2500 5000 7500 10000 125000
100000
200000
300000
400000
500000
600000
700000
800000Controlo (etanol)XH 100 µM
Concentração de 3H-DG (µM)
Cap
taçã
od
e3 H
-DG
(pm
ol/
mg
pro
teín
a)
0 2500 5000 7500 10000 125000
100000
200000
300000
400000
500000
600000 Controlo (DMSO)Querc 100 µMEGCG 100 µM
Concentração de 3H-DG (µM)
Cap
taçã
od
e3 H
-DG
(pm
ol/
mg
pro
teín
a)
Figura 38 - Efeito agudo da quercetina 100 µM (Querc 100µM, n=6) e daepigalocatequina-3-galato 100 µM (EGCG 100 µM, n=6) na captação de 3H-DG, emfunção da concentração de substracto. As células BeWo foram pré-incubadasdurante 20 minutos e posteriormente incubadas com concentrações crescentes de3H-DG (1-10000 µM) durante 6 minutos a 37ºC, na presença do composto ou dorespectivo solvente (Controlo DMSO, n=9). Os resultados são apresentados sob aforma de média ± EPM.

35
Quanto ao Km, verificou-se que somente o acetaldeído e a EGCG reduziram
significativamente esse valor sendo o efeito do primeiro mais pronunciado (Km de
6,63 ± 0,08 mM e Km de 9,16 ± 0,37 mM, respectivamente), comparativamente
aos controlos. Logo, estes compostos aumentam a afinidade do transportador
para a 3H-DG (figuras 37, 38 e 39 e Tabela 1).
DISCUSSÃO
A placenta humana expressa vários mecanismos de transporte,
nomeadamente transportadores de nutrientes, cujo funcionamento e expressão
são conhecidos noutros tecidos, mas que nela são ainda alvo de alguma
especulação (13). A placenta tem uma função fulcral no fornecimento de
nutrientes ao feto em desenvolvimento. Nutrientes como a glicose, aminoácidos,
vitaminas e minerais, atravessam a placenta através de mecanismos de
transporte presentes essencialmente na membrana apical e basolateral do
sinciciotrofoblasto (13). O transporte transplacentar de glicose, fundamental para
o crescimento e desenvolvimento fetal, ocorre principalmente através de um
mecanismo de transporte facilitado, GLUT1, que permite o movimento de glicose
a favor do seu gradiente de concentração. As restantes isoformas do GLUT
implicadas neste processo (GLUT3, 4 e 12) são ainda pouco conhecidas, em
virtude da sua expressão proteica e localização celular não serem consensuais
(11).

36
A linha celular BeWo, é um modelo celular de sinciciotrofoblasto humano
que exibe muitas das características morfológicas e bioquímicas dos trofoblastos,
durante o último trimestre de gravidez (15). Além disso, forma rapidamente uma
monocamada confluente com morfologia polarizada e aderente ao suporte (15), o
que a torna particularmente atractiva para o estudo do transporte transplacentar
de nutrientes (9).
Tal como no sinciciotrofoblasto humano, o principal transportador de
glicose expresso nas células BeWo é o GLUT 1. Apesar de estarem descritos
outros transportadores, existe alguma controvérsia quanto à sua expressão na
membrana (11, 15, 14).
Na maioria dos estudos realizados em culturas celulares, onde se
pretendeu estudar o transporte placentar de glicose, a escolha do substrato recai
sobre a DG (11, 15, 44). Este substrato ao ser captado pelas células, é fosforilado
pela hexocínase a DG-6-P não sendo posteriormente metabolizado. Isto permite a
manutenção do gradiente de DG, aumentando o tempo necessário para que se
atinja um equilíbrio entre os dois lados da membrana (11, 15).
Pelo facto do transporte materno-fetal de glicose ser ainda pouco
conhecido, e pela sua modulação por compostos da dieta e drogas de abuso não
estar descrita, os objectivos deste trabalho foram: caracterizar a captação apical
de 3H-DG em células BeWo, e determinar especificamente o efeito agudo e
crónico de alguns compostos presentes em bebidas e de algumas drogas de
abuso, nesse processo.
Na primeira fase do trabalho caracterizou-se a captação apical de 3H-DG,
no que diz respeito à dependência do tempo, saturabilidade e especificidade.

37
A análise dos resultados, mostrou que a captação de 3H-DG nas células
BeWo é dependente do tempo e saturável.
A determinação dos parâmetros cinéticos de captação de 3H-DG, revelou
valores de Km e Vmax da ordem dos 13,5 mM e dos 1200 nmol/mg prot,
respectivamente. O valor de Km obtido, foi intermédio face aos descritos por Shah
et al (1999) (15) (Km de 1,5 mM) e por Carruthers et al (1990) (46) (Km de 20 mM)
ambos para o GLUT1 (43).
Além disso a citocalasina B, um conhecido inibidor do transporte facilitado
de glicose (11, 15), provocou um efeito inibitório máximo na captação de 3H-DG
de cerca de 60%. Strobel et al (2005) e Martin et al (2003) referem que
citocalasina B se liga com alta afinidade a um domínio transmembranar interno do
GLUT e não aos mesmos locais de ligação da glicose (domínio interno conhecido
por região QLS) (47, 48).
Então, de acordo com os nossos resultados, cerca 60% do transporte de
glicose nas células BeWo é facilitado. Contudo, outros autores referem valores
um pouco diferentes: Shah et al (1999) afirma que esse transporte pode ser da
ordem dos 85% (15) e Pratibshary et al (2002) (14) afirma que é de 50%. De
qualquer forma os 60% obtidos, são suficientes para investigar o transporte
apical. Nas células BeWo, apesar de terem sido identificados, além do GLUT1,
outros transportadores de glicose, nomeadamente o GLUT3 (transportador de
baixo Km) e GLUT5 (transportador de frutose), e da citocalasina B não ter
especificidade para se ligar somente ao GLUT1, é pouco provável que estes
tenham um papel relevante na captação de 3H-DG (11, 15, 14).
A restante percentagem do transporte (40%) que não foi inibido pela
citocalasina B, pode corresponder a adsorção não-específica de 3H-DG às células

38
ou ao material plástico da placa, ou corresponder a um mecanismo de difusão
passiva de 3H-DG para o interior das células (o que não parece provável devido à
natureza polar desta molécula), ou pode eventualmente corresponder à captação
por um transportador adicional (49) dependente do sódio (SGLT). Contudo,
actualmente não está descrito outro tipo de transportadores a nível placentário,
além dos GLUT (5).
Em suma, podemos concluir que o principal mecanismo funcionalmente
relevante para o transporte de glicose nas células BeWo é o GLUT1, coincidindo
com o mecanismo identificado em trofoblastos de placentas de termo (5, 8, 11,
15,14).
Os polifenóis constituem um grupo complexo e extenso de compostos,
presentes em bebidas como sumos de fruta, vinho, chá e cerveja (17, 50).
Na população norte-americana, a ingestão média diária de polifenóis foi
avaliada como sendo 1 g por dia e as concentrações plasmáticas máximas como
sendo não superiores a 1 �M, após a ingestão de 10-100 mg de polifenóis.
Contudo a produção de metabolitos nos tecidos e flora intestinal e a
suplementação dietética, podem aumentar essa concentração (16).
Numerosos estudos têm atribuído a estes compostos efeitos benéficos
para a saúde humana (ver introdução) (24, 25, 26, 27, 50). Alguns desses efeitos
estão relacionados com a capacidade de regularem os mecanismos de transporte
de glicose (51).
A nível placentar, desconhece-se a influência dos polifenóis no transporte
de glicose. Contudo, está descrito que estes compostos têm capacidade para
atravessar a barreira placentária (52). Por isso, um dos objectivos deste trabalho

39
foi investigar a modulação nutricional, por diferentes compostos bioactivos
presentes em bebidas alcoólicas e não-alcoólicas, do transporte de 3H-DG em
células BeWo. Neste contexto, testou-se o efeito de polifenóis, mas também de
metilxantinas como a cafeína e teofilina e do etanol e do seu metabolito
acetaldeído.
A análise dos resultados obtidos permite concluir que agudamente o
resveratrol, xanto-humol, EGCG (todos na concentração de 100 �M), e a
quercetina e crisina (ambas nas concentrações de 10 e 100 �M) diminuíram
significativamente a captação de 3H-DG, sendo o efeito inibitório mais
pronunciado para o xanto-humol (26% de inibição), EGCG (40% inibição) e
quercetina (57 % de inibição). Estes resultados sugerem que a exposição aguda a
quantidades elevadas destes polifenóis, só atingidas por suplementação, poderá
reduzir os níveis de glicose fetais causando efeitos adversos.
Nenhum destes compostos alterou o efeito inibitório da citocalasina B, por
isso conclui-se que actuam no mesmo transportador (GLUT) para exercerem os
seus efeitos inibitórios.
Em comparação com trabalhos publicados noutros tipos celulares, os
resultados obtidos revelaram-se de forma geral concordantes. Em linhas celulares
mielocíticas e linfocíticas (células U937 e Jurkat respectivamente) a quercetina e
miricetina foram os polifenóis que nas mais baixas concentrações (9-12 �M),
reduziram para metade a captação de 3H-DG mediada pelos GLUT (53).
Do mesmo modo Johnston et al (2004) verificaram que a miricetina e a
quercetina, ambas na concentração de 100 �M, reduziram em 82% e 75%
respectivamente, o transporte de glicose mediado pelos GLUT, em células Caco-2

40
(50). Em adipócitos e ovócitos de Xenopus Laevis, a quercetina teve um efeito
semelhante ao nível do GLUT4 e 2 (54, 47). Segundo Park et al (1999) a
capacidade inibitória da quercetina pode estar relacionada com a presença de
grupos de hidroxiflavonas na sua estrutura (53).
Relativamente ao resveratrol, os resultados estão de acordo com os
obtidos por Park (2000) que demonstrou que nas linhas celulares U937 e HL-60, o
resveratrol inibiu competitivamente a captação de 3H-DG pelos GLUT1 e 3 (55).
A crisina, como diminuiu a viabilidade celular de modo estatísticamente
significativo, nas concentrações de 10 e 100 �M, é provável que reduza a
captação de 3H-DG devido a citotoxicidade celular. Os efeitos agudos dos
restantes polifenóis, não são explicados por alteração da viablilidade celular.
A análise dos efeitos inibitórios dos polifenóis sobre os parâmetros
cinéticos da captação de 3H-DG, revelou que a quercetina e o xanto-humol
reduziram a Vmax mas não alteraram o valor de Km, enquanto que a EGCG reduziu
ambos. Logo, concluiu-se que a quercetina e o xanto-humol são inibidores não
competitivos, e que provavelmente se ligam a um sítio alostérico do GLUT
induzindo uma alteração conformacional no sítio activo (onde se liga a 3H-DG),
que diminui a actividade do transportador para altas concentrações de substrato.
No fundo, o efeito assemelha-se à redução do número de transportadores na
superfície celular.
Os estudos realizados neste campo são controversos, pois alguns
evidenciam que a quercetina comporta-se como um inibidor competitivo do
transporte de 3H-DG, enquanto outros evidenciam o contrário (53, 56, 47, 54, 48).
Quanto ao xanto-humol, uma chalcona prenilada que ocorre na planta do
lúpulo e na cerveja, estudos realizados em células BeWo, evidenciaram que este

41
composto não tem qualquer efeito agudo na captação de ácido fólico ou de
vitamina B1 (40). Paralelamente aos nossos resultados, este aspecto não deixa de
ser interessante, visto que na mesma cultura celular o xanto-humol teve efeitos
distintos sobre a captação placentar de diferentes nutrientes.
Relativamente à EGCG, esta mostra um comportamento atípico, contudo
pode-se pressupor que o seu efeito resulta da ligação a um sítio alostérico,
induzindo uma alteração conformacional no sítio activo do transportador, cujo
resultado é o aumento da afinidade para o substrato e simultaneamente
diminuição da actividade do transportador. Vários estudos demonstraram, noutros
tipos celulares (50, 56, 57), que no grupo das catequinas apenas a EGCG teve
um potente efeito inibidor do transporte de glicose mediado pelos GLUT (56, 50).
Em células de epitélio intestinal, concentrações de 0,1 e 1 mM de EGCG
reduziram o transporte em cerca 35-37%, sendo o resíduo galato importante para
esse efeito (50, 57).
No que diz respeito ao seu efeito nos parâmetros cinéticos da captação de
glicose, os estudos são ainda contraditórios. (56, 57). O efeito inibitório da EGCG
pode dever-se à ligação a um domínio externo do GLUT1, estando de acordo com
os nossos resultados, ou a um bloqueio da ligação da glicose ao transportador
(56, 57).
Ainda relativamente aos resultados do efeito agudo de polifenóis, a rutina a
catequina (ambas a 10 e 100 �M) e a epicatequina (100 �M) aumentaram de
modo significativo a captação de 3H-DG. Estes compostos não alteraram o efeito
da citocalasina B, por isso ambos interagem com o GLUT de modo a exercem os
seus efeitos. De acordo com Park et al (1999) e Johnston et al (2004) a catequina
e epicatequina (ambas a 100 �M), demonstraram um efeito estimulador sobre a

42
captação facilitada de glicose em várias linhas celulares (53, 50). Quanto à rutina
(em concentrações inferiores a 100 �M), estudos evidenciaram que não teve
efeito sobre o transporte facilitado de glicose, em células Caco-2, U937 e Jurkat
(50, 53). Estes estudos contrariam os nossos resultados provavelmente porque o
efeito da rutina varia conforme a cultura celular e, como iremos ver mais à frente,
com o tempo de exposição (50, 53).
As catequinas são muito abundantes no chá (sobretudo no verde) sendo a
EGCG o principal componente (cerca de 60% das catequinas), e a catequina e
epicatequina componentes existentes em quantidades mais modestas (inferiores
a 20%) (56).
Relativamente à associação de polifenóis, verificou-se que a catequina e
epicatequina (ambas a 100 �M) contrariamente ao seu efeito isolado, reduziram a
captação de 3H-DG quando associadas, e a epicatequina e o xanto-humol (ambos
a 100 �M) em conjunto contrabalançaram os efeitos antagónicos que tinham
isoladamente. Estes resultados evidenciam que nas bebidas, em que há uma
associação destes compostos, os efeitos observados podem ser muito distintos
dos efeitos de cada um compostos isoladamente.
Cronicamente, a análise dos resultados revelou que a rutina (tal como
agudamente) e a miricetina, aumentaram de um modo dependente da
concentração a captação de 3H-DG. Como nenhum dos compostos alterou a
viabilidade celular, estes resultados não podem ser explicados por efeitos
proliferativos sobre as células BeWo. A miricetina e a rutina não alteraram o efeito
crónico da citocalasina B, por isso estes polifenóis interagem com o GLUT para
exercerem os seus efeitos.

43
Tal como visto anteriormente, existem estudos que indicam que a
miricetina tem um efeito inibitório na captação facilitada de glicose, em algumas
linhas celulares (53, 54, 47). Apesar de contrariarem os nossos resultados, é
importante salientar que esses estudos avaliam apenas o efeito agudo da
miricetina e como tal, um maior tempo de exposição das células ao composto
pode ter influência no seu efeito sobre a captação de glicose.
A mesma explicação aplica-se à rutina, pois como visto anteriormente, não
teve efeito agudo sobre o transporte facilitado de glicose, em algumas linhas
celulares. (50, 53)
As flavonas quercetina, um dos principais polifenóis presente na dieta, a
rutina (forma glicosilada da primeira) e a miricetina (26) existem principalmente no
vinho tinto, chá e cerveja (17, 40). Quando associadas cronicamente, verificou-se
que a miricetina e a rutina (ambas a 1 �M) evidenciaram um efeito estimulador
somativo (de 30%) na captação de 3H-DG, face ao seus efeitos isolados.
Os riscos associados ao consumo materno de cafeína (presente em
bebidas como o chá ou café) nomeadamente o aborto espontâneo e atraso de
crescimento intra-uterino, são ainda controversos (28, 29).
Os resultados obtidos com as metilxantinas, revelaram que apenas a
teofilina alterou a captação de 3H-DG. A teoflina (0,1 e 1 �M) quando testada em
efeito crónico, reduziu em 10% o transporte apical de 3H-DG de um modo
dependente da concentração. Este resultado não está relacionado com efeitos
citotóxicos nas células BeWo, visto que a teofilina não alterou a viabilidade
celular. Pelo facto deste composto não modificar o efeito inibitório da citocalasina
B, conclui-se que interage com o GLUT para exercer o seu efeito inibitório.

44
Já foi demonstrado em alguns estudos (58, 59), que as metilxantinas
cafeína e teofilina têm capacidade para inibir o transporte facilitado de glicose.
Contudo, os mecanismos responsáveis por esse efeito não são ainda totalmente
conhecidos. De acordo com Steinfelder et al (1990), a teofilina e a cafeína
inibiram a captação facilitada de glicose em adipócitos, através de dois modos:
por interação com o mesmo local de ligação da citocalasina B no transportador, e
por inibição dos receptores A1 da adenosina (58).
Ho et al (2001) refere um mecanismo de actuação diferente, afirmando que
a cafeína e a teofilina (em concentrações inferiores ou iguais a 10 mM) inibem a
captação de glicose pelo GLUT1 em eritrócitos, reduzindo a Vmax do transportador
(59). Apesar dos estudos mencionados revelarem um efeito inibitório de ambas as
metilxantinas sobre o transporte de glicose, Keating et al (2006) demonstraram
que apenas a teofilina, nas concentrações de 10 e 100 �M, reduziu a captação
apical de ácido fólico em células BeWo (40). E no presente trabalho, também se
verificou que somente a teofilina teve efeito inibitório sobre o transporte de
glicose.
Como tal, os nossos resultados sugerem que a exposição crónica à
teofilina, poderá ter um efeito adverso no feto, devido à redução dos níveis fetais
de glicose.
O etanol é reconhecidamente um potente teratogénio nos seres humanos,
apesar de ser a droga mais consumida em todo mundo (39, 40). O seu consumo
abusivo por grávidas, pode estar associado a um conjunto de manifestações
clínicas fetais conhecidas como Síndrome alcoólica fetal (32). Os efeitos
teratogénicos do etanol estão relacionados com vários factores, entre os quais a

45
formação de acetaldeído no feto, na placenta e na própria mãe (60, 61). O
acetaldeído é formado principalmente a partir da oxidação hepática do etanol,
pela acção da desidrogénase do aldeído e em menor grau pelo sistema
enzimático do citocromo P-450 (através da enzima CYP2E1 ou MEOS -
“microssomal ethanol oxidizing system”) (62). Este metabolito pode atingir
concentrações fetais superiores às da circulação materna (60, 61).
Os resultados obtidos, evidenciaram que o etanol e o acetaldeído apenas
exerceram um efeito agudo, na captação de 3H-DG em células BeWo. O etanol
teve um efeito estimulador de 10%, na concentração de 10 mM. O acetaldeído
pelo contrário, reduziu a captação de 3H-DG de modo dependente da
concentração, provocando reduções de 30% e 60% nas concentrações mais
elevadas (30 e 100 mM, respectivamente). Verificou-se também que os efeitos de
ambos os compostos, não se devem a alterações na viabilidade celular. O etanol
não alterou o efeito da citocalasina B, por isso interage com o GLUT para exercer
o seu efeito estimulador na captação de 3H-DG.
Actualmente existe alguma evidência científica, de que a teratogenicidade
do etanol está associada a alterações do transporte de nutrientes na placenta,
nomeadamente a uma redução do transporte de aminoácidos, ácido fólico e
tiamina (40, 60, 63). Nesse contexto, não deixa de ser interessante verificar que
não teve efeito, ou demonstrou uma ligeira tendência para aumentar a captação
de 3H-DG.
Adicionalmente, devido aos efeitos dos polifenóis e do etanol serem
distintos, não é correcto extrapolar os efeitos isolados do etanol para os efeitos de
bebidas alcoólicas.

46
O acetaldeído (30 mM) reduziu em 12% o efeito da citocalasina B na
captação de 3H-DG. Logo parece exercer o seu efeito inibitório, ou pelo menos
parte dele, por outro mecanismo que não envolve o GLUT.
O efeito do acetaldeído nos parâmetros cinéticos da captação de 3H-DG,
resultou numa redução dos valores de Km e Vmax. Em virtude destes resultados,
pressupõe-se que o efeito do acetaldeído pode também resultar da ligação a um
sítio alostérico do GLUT, com a consequente alteração conformacional do sítio
activo do transportador. O resultado é um aumento da afinidade para o substrato
e simultaneamente uma diminuição da actividade do transportador. Pelo facto de
atravessar livremente a barreira placentária (61), o acetaldeído também pode
exercer o seu efeito inibitório, influenciando a actividade de enzimas do
metabolismo intracelular da 3H-DG, como a hexocínase. O acetaldeído pode
diminuir a actividade da hexocínase, conduzindo a uma redução da captação da
3H-DG.
Através destes resultados, podemos concluir que um dos possíveis
mecanismos envolvidos nos efeitos nocivos do acetaldeído, será a redução da
captação apical de glicose no sinciciotrofoblasto. Até porque, em concentrações
semelhantes às testadas (20 mM) o acetaldeído também reduziu a actividade
transportadora de aminoácidos (64).
Como nota, interessa referir que os efeitos inibitórios dos polifenóis e do
acetaldeído na captação de 3H-DG, foram estudados utilizando concentrações de
3H-DG (1 �M) claramente inferiores aos valores de Km obtidos.

47
Nos Estados Unidos, estima-se que anualmente mais de duzentas mil
mulheres sejam expostas, pelo menos uma vez, a drogas ilícitas durante o
período gestacional (34).
Nenhuma das drogas de abuso testadas agudamente, alterou a captação
de 3H-DG nas células BeWo. No entanto cronicamente o MDMA, a anfetamina
(0,25 e 2,5 �M) e o THC (apenas na concentração mais baixa, 1 nM) reduziram
aproximadamente em 10% a captação de 3H-DG. Estes resultados não estão
relacionados com efeitos citotóxicos, pois a viabilidade celular não foi alterada.
Adicionalmente, nas concentrações inibitórias estas drogas de abuso não
alteraram o efeito da citocalasina B, o que pressupõe que interagam com o GLUT
para exercer esses efeitos. Estes resultados poderão explicar o efeito crónico do
MDMA, anfetamina e THC. Contudo, não temos nenhuma explicação para o facto
do THC apenas inibir a captação de 3H-DG na concentração mais baixa. No seu
conjunto, estes resultados sugerem que a exposição crónica ao MDMA,
anfetamina e THC, poderão ter um efeito adverso no feto devido à redução dos
níveis fetais de glicose.
Estão já descritos, efeitos adversos de drogas de abuso noutros
transportadores placentários, nomeadamente de nutrientes e de catecolaminas
(13, 65, 66). Segundo Ganapathy et al (1999), os transportadores das
monoaminas serotonina, noradrenalina e dopamina são os principais alvos de
drogas ilícitas na placenta (62). A cocaína é um potente inibidor competitivo do
transportador de serotonina (SERT), pois os valores plasmáticos de consumidores
habituais (1-2.5 �M) são muito superiores à concentração que inibe 50% da
captação de serotonina (0,1 �M). Apesar de ser um inibidor competitivo, a cocaína

48
e seus análogos não são substratos do SERT. Igualmente, mas de forma menos
potente, a cocaína também inibe o transportador de noradrenalina (13).
A anfetamina é um potente inibidor do transportador de noradrenalina, e os
valores plasmáticos em doses terapêuticas e em consumidores habituais (0,3-0,8
�M e 1-2 �M, respectivamente), são muito superiores à concentração responsável
por inibir 50% da captação (0,1 �M) (13). Em relação ao SERT, a anfetamina é
também um inibidor, mas menos potente (13). Como é um substrato alternativo de
ambos os transportadores, a anfetamina pode acumular-se no sinciciotrofoblasto,
alterando funções intracelulares entre as quais a mitocondrial (13, 65). Quanto ao
THC, inibe a função do SERT de forma não competitiva (13).
A inibição do transporte materno-fetal de monoaminas, resulta num
aumento dos seus níveis no fluxo sanguíneo placentário, causando
vasoconstrição, redução do fluxo sanguíneo, e em última instância diminuição do
aporte de nutrientes ao feto (13).
Além dos transportadores de monoaminas, estão descritos outros possíveis
alvos de drogas de abuso (13, 65). A anfetamina, metanfetamina, cocaína e
nicotina inibem competitivamente a função do transportador de catiões orgânicos
do tipo 3 (OCT3) (13). Também Rytting et al (2004) demonstraram que a
anfetamina e seus derivados inibiram o transporte de carnitina, por via do
transportador de catiões orgânicos (OCTN2), em células BeWo (67).
O transporte de aminoácidos, principalmente o dependente do ião sódio
(como o dos aminoácidos glutamina e ácido �-aminoisobutírico) é inibido de forma
não-competitiva pela cocaína, nicotina e canabinóides (13, 66). Esse efeito deve-
se ao facto de dissiparem o gradiente electroquímico do ião sódio, sendo as
concentrações inibitórias muito variáveis (13).

49
Por fim, e com base nos resultados obtidos neste trabalho, podemos
especular sobre o impacto dos compostos testados, na proliferação tumoral. Uma
vez que a glicose é um substrato energético importante para o controlo do
crescimento e proliferação celular, a inibição e/ou activação do seu transporte
para as células neoplásicas será um alvo terapêutico a considerar.
CONCLUSÃO
Com este trabalho, concluímos que o principal mecanismo de captação
apical de 3H-DG nas células BeWo é o transporte facilitado (GLUT). A captação
de 3H-DG é modulada de modo diferente, por compostos bioactivos presentes na
dieta (polifenóis e metilxantinas) e por drogas de abuso.
Agudamente o resveratrol, xanto-humol, EGCG (100 �M), quercetina e
crisina (ambos a 10 e 100 �M) reduziram o transporte apical de 3H-DG, enquanto
que a rutina, catequina (ambos a 10 e 100 �M), epicatequina (100 �M) e etanol
(10 mM) aumentaram-no. A catequina e epicatequina em conjunto (ambas a 100
�M) reduziram a captação de 3H-DG, enquanto que a epicatequina e o xanto-
humol (ambos a 100 �M) contrabalançaram os efeitos antagónicos que tinham
isoladamente. Todos estes compostos testados em efeito agudo, parecem
interagir com o GLUT para exercerem os seus efeitos.
A quercetina e o xanto-humol reduziram a actividade do transportador sem
alterar a afinidade para o substrato, ao passo que a EGCG simultaneamente

50
aumentou a afinidade para o substrato e diminuiu a actividade do transportador. O
acetaldeído teve um efeito inibitório nas concentrações mais elevadas (30 e 100
mM), que é explicado em parte pela ligação a um sítio alostérico do GLUT
aumentando a afinidade para o substrato e diminuindo a actividade do
transportador.
Cronicamente, a rutina e a miricetina isoladamente (0,1 e 1 �M) e em
conjunto (1 �M) aumentaram a captação de 3H-DG, enquanto que a teoflina (0,1 e
1 �M) e as drogas de abuso MDMA, anfetamina (ambas a 0,25 e 2,5 �M) e THC
(1 nM) reduziram essa mesma captação. Para exercerem os seus diferentes
efeitos estes compostos parecem interagir com o GLUT.
Pelo facto dos efeitos agudos e crónicos destes compostos serem distintos,
não é correcto extrapolar efeitos crónicos a partir dos agudos e vice-versa.
Os resultados obtidos neste trabalho, vêm aprofundar o conhecimento da
bioactividade e a influência na saúde materno-fetal de alguns polifenóis,
metilxantinas e drogas de abuso.

51
REFERÊNCIAS BIBLIOGRÁFICAS
1. Stryer L. (1996); Biochemistry. 4th Ed. New York. W. H: Freeman and
Company.
2. Saladin K. (2001); Anatomy and Physiology: the unit of form and function.
2nd Ed. New York. MacGraw-Hill.
3. Guyton AC, Hall JE. (2000); Textbook of Medical physiology. 10th Ed.
Philadelphia. WB Saunders.
4. Wood I, Trayhurn P. Glucose transporters (GLUT and SGLT): expanded
families of sugar trnasport proteins. Br J Nutr. 2007; 89:3-9.
5. Baumann M, Zamudio S, Illsley N. Hypoxic upregulation of glucose
transporters in BeWo choriocarcinoma cell is mediates by hipoxia-inducible factor-
1 (HIF-1). Am J Cell Physiol. 2007; (article in press).
6. Von Wolf M, Ursel S, Hahn U, Steldinger R, Strowitzki T. Glucose
transporter proteins (GLUT) in human endometrium: expression, regulation, and
function throughout the menstrual cycle. J Clin Endocrinol Metab. 2003;
88(8):3885-3892.
7. Scneider H, Reiber W, Sager R, Malek A. Asymetrical transport of glucose
across the in vitro perfused human placenta. Placenta. 2002; 24:27-33.

52
8. Leonce J, Brockton N, Robinson S, Venkatesan S, Bannister P, Raman V,
Murphy K, Parker K, Pavitt, D Teoh T, Regan L, Burchell A, Steer P, Johnston D.
Glucose production the human placenta. Placenta. 2006; Trophoblast Research
(vol. 20), 27(suppl. A).
9. Fuchus N, Ellinger I. Endocytic and transcytotic process in villous
syncyciotrophoblast: role in nutrient transport to the human fetus. Traffic. 2004;
5:725-738.
10 Sastry, BVR. Techniques to study human placental transport. Adv Drug
Deliv Rev. 1999; 38:17-39.
11. Illsley N. Glucose transporters in the human placenta. Placenta. 2000; 21:
14-22.
12. Ganapathy V, Prasad PD, Ganapathy ME, Leibach FH. Drugs of abuse and
placental transport. Adv Drug Deliv Ver. 1999; 38:99-110.
13. Hormonal regulation of glucose and system A amino acid transport in first
trimester placental villous fragments. Am J Physiol Regul Integr Comp Physiol.
2005; 288:R656-662.
14. Vardhana P, Illsley N. Transepithelial glucose transport and metabolism in
BeWo choriocarcinoma cells. Placenta. 2002; 23:653-660.

53
15. Shah S, Zhao H, Low S, MacArdle H, Hundal H. Characterization of glucose
transport and glucose transporters in the human choriocarcinoma cell line, BeWo.
Placenta. 1999; 20:651-659.
16. Ito H, Gonthier MP, Manach C, Morand C, Mennen L, Rémésy C, Scalbert
A. Polyphenol levels in human urine after intake of six different polyphenol-rich
beverages. Br J Nutr. 2005; 94:500-509.
17. Scalbert A, Williamson G. Dietary intake and bioavalibility of polyphenols. J
Nutr. 2000; 13:2073S-2085S.
18. Scalbert A, Manach C, Morand C, Rémésy C, Jimanez L. Dietary
polyphenols and the prevention of diseases. Crit Rev Food Sci Nutr. 2005; 45:287-
386.
19. Soleas GJ, Diamandis EP, Goldberg DM. Wine as a biological fliud: history,
production, and role in disease prevention. J Clin Lab Anal 1997; 11(5):287-213.
20. Mukhtar H, Ahmad N. Tea polyphenols: prevention of cancer and optimizing
health. Am J Clin Nutr. 2000; 71 (suppl. 6):1698S-702S.
21. Trevisanato SI, Kim YI. Tea and health. Nutr Rev. 2000; 58(1):1-10.

54
22. Van de Wiel A, Van Golde PHM, Hart HC. Blessings of the grape. Eur J Int
Med. 2001; 12:484-9.
23. Yang CS, Malakial P, Meng X. Inhibition of carcinogenesis by tea. Annu
Rev Pharmacol. 2002; 42:25-54.
24. Bravo L. Polyphenols: chemistry, dietary sources, metabolism and
nutricional significance. Nutr Rev 1998;56(11):317-33.
25. Middleton E Jr, Kandaswami C, Theoharides TC. The effects of plant
flavonoids on mamalian cells: implications for inflammation, heart disease, and
cancer. Pharmacol Rev 2000; 52(4):673-751.
26. Nijveldt RJ, Van Nood E, Van Hoorn DE, Boelens PG, Van Norren K, Van
Leeuwen PA. Flavonoids: a review of probable mechanisms of action and
potencial applications. Am J Clin Nutr. 2001; 74 (4):418-25.
27. Ross J, Kasum C. Dietary flavonoids: bioavailiabiity, metabolic effects, and
safety. Annu Rev Nutr. 2002; 22:29-34.
28. Hinds TS, West WL, Knight EM, Harland BF. The effect of caffeine on
pregnancy outcomes variables. 1996; Nutr Rev. 54(4):203-7.

55
29. Bracken M, Triche E, Belanger K, Hellenbrand K, Leaderer B. Association
of maternal caffeine consumption with decrements in fetal growth. Am J Epidemiol.
2003;157:456-466.
30. Vik T, Bakketeig S, Trygg K, Ling-Larsen K, Jacobsen G. High caffeine
consumption in the third trimester of pregnancy: gender-specific effects on fetal
growth. Pedr Perinat Epidemiol. 2003; 17:324-331.
31. Thadani P, Strauss J, Dey S, Andersen V, Audus K, Coats K, Cross J,
Erlebacher A, Ganapathy V, Linzer D, Miller R, Novak D, Rapaka S, Sadowsky Y,
Salafia C, Soares M, Unadkt J. National institute on drug abuse conference report
on placental proteins, drug transport, and fetal development. Am J Obstet
Gynecol. 2004; 191:1858-62.
32. Howell E, Heiser N, Harrington M. A review of recent findings on substance
abuse treatment for pregnant women. J Subs Ab Treat.1999; 16(3):195-219.
33. Cornelius M, Goldschmidt L, Day N, Larkby C. Alcohol, tobacco and
marijuana use among pregnant teenagers: 6-year follow-up of offspring growth
effects. Neurotoxicol Teratol. 2002; 24:703-710.
34. Ebrahim S, Gfroerer J. Pregnancy-related substance abuse in the United
States during 1996-1998. Obstet Gynecol. 2003; 101:374-9.

56
35. Bennet AD. Perinatal substance abuse and the drug-exposed. Adv Nurse
Pract. 1999; 7(5):32-36.
36. Rash V. Cigarrete, alcohol, and caffeine consuption:risk factors for
spontaneous abortion. Acta Obstet Gynecol Scand. 2003; 82:182-188.
37. Fergusson D, Horwood L, Northstone, K, ALSPAC study team. Maternal
use of cannabis and pregnancy outcome. Br J Obstet Gynaecol. 2002; 109:21-27.
38. Ho E, Karimi-Tabesh L, Koren G. Characteristics of pregnant women who
use excstasy (3,4-methylenedioximethamphetamine). Neurotoxicol. Teratol. 2001;
23:561-567.
39. Keating E, Lemos C, Azevedo I, Martel F. Characteristics of thiamine
uptake by the BeWo human trophoblast cell line. J Biochem Mol Biol. 2006;
34(9):383-393.
40. Keating E, Lemos C, Gonçalves P, Martel F. Acute and chronic effect of
some dietary bioactive compounds on folic acid uptake and on the expression of
folic acid transporters by the human trphoblast cell line BeWo. J Nutr Biochem.
2006 (article in press).
41. Mosman T. Rapid colorimetric assay for celular growth and survival:
application to proliferation and cytotoxicity assays. J Immunol Methods 1983;
65:55-63

57
42. Bradford MM. A rapid method for the quantitation of microgram quantities of
protein utilizing the principle of protein dye-binding. Anal Biochem. 1976; 72:248-
54.
43. Muzyka A, Tarkany O, Yelizanov V, Sergienko U, Boichuk A. 2005-
GraphPad Prism (version 4.03) for Windows, San Diego, CA.
44. Shubert P, Gordon M, Landon M, Gabbe S, Kniss D. Ketoacids attenuate
glucose uptake in human trophoblasts isolated form first trimester chorionic villi.
Am J Obstet Gynecol 1996; 175:56-62.
45. Hipólito-Reis C, Alçada M, Azevedo I. (2002) Práticas de Bioquímica para
as ciências da saúde. Lisboa. Lidel - edições técnica lda.
46. Strobel P, Allard C, Perz-acle T, Calderon R, Aldunate R, Leighton F.
Miricetin, quercetin and catechin gallate inhibit glucose uptake in isolated rat
adipocytes. Biochem J. 2005; 386:471-478.
47. Carruthers A. Facilitated diffusion of glucose. Physiol Rev. 1990; 70:1135-
1176.
48. Martin HJ, Kornmann F, Fuhrmann G. The inhibitory effects of flavonoids
and antiestrogens on the Glut1 glucose transporter in human erythrocytes. Chem-
Biol Int. 2003; 146:225-235.

58
49. Keating E, Lemos C, Azevedo I, Martel F. Comparision of folic acid uptake
characteristics by human placental choriocarcinoma cells at acidic and
physiological pH. Can J Physiol Pharmacol. 2006; 84(2):245-255.
50. Johnston K, Sharp P, Clifford M, Morgan L. Dietary polyphenols decrease
glucose uptake by human intestinal Caco-2 cells. FEBS letters. 2005; 579:1653-
1657.
51. Purintrapiban J, Suttajit M, Forsberg N. Differential activation of glucose
transport in cultured muscle cells byb poliphenolic compounds from Canna Indica
L. root. Biol Pharm Bull. 2006; 29(10):19956-1998.
52. Schroder-van der Elst JP, van der Heide D, Rochos H, Morreale de
Escobar G, Kohrle J. Synthetic flavonoids cross the placenta in the rat and are
found in fetal brain. Am J Physiol. 1998; 274(2 pt 1):E253-E256.
53. Park J. Flavonoids are potencial inhibitors of glucose uptake in U937 cells.
Biochem Biophys Res Commun. 1999; 260:568-574.
54. Kwon O, Eck P, Chen S, Corpe C, Lee JH, Kruhulak M, Levine M.
Inhibition of the intestinal glucose transporter GLUT 2 by flavonoids. FASEB J.
2007; 21:366-377.

59
55. Park J. Inhibition of glucose and dehydroascorbic acid uptakes by
resveratrol in human transformed myelocytic cells. J Nat Prod. 2001; 64:381-384.
56. Naftalin R, Azfal I, Cunningham P, Halai M, Ross C, Salleh N, Miligan S.
Interactions of androgens, green tea catechins and the antiadrgen flutamide with
the external glucose-binding site of the human erythrocyte glucose transporter
GLUT1. Br J Pharmacol. 2003; 140:487-499
57. Kobayashi Y, Suzuki M, Satsu H, Arai S, Hara Y, Suzuki K, Miyamoto S,
Shimizu M. Green tea polyphenols inhibit the sodium-dependent glucose
transporter of intestinal epithelial cells by a competitive mechanism. J Agic Food
Chem. 2000; 48:5618-5623.
58. Steinfelder HJ, Petho-Schramm. Methylxantines inhibit glucose transport in
rat adipocytes by two independent mechanisms. Biochem Pharmacol. 1990; 40
(5):1154-7.
59. Ho YY, Yang H, Klepper J, Fischbarg J, Wang D, Vivo D. Glucose
transporter type 1 deficiency syndrome (Glut1ds): methylxantines potentiate glut 1
haploinsuficiency in vitro. Pediatr Res. 2001; 50(2):254-260.
60. Chaudhuri J. Alcohol and the developing fetus - a review. Med Sci Monit.
2000; 6(5):1031-1041.

60
61. Hard M, Iqbal U, Brien J, Koren G. Binding of acetaldehyde to human and
guinea pig placentae in vitro. Placenta. 2003. 24:149-154.
62. Timbrell J. (2000); Principles of Biochemical Toxicology. 3rd Ed.
Philadelphia. Taylor and Francis Inc.
63. Jones C, Srinivas S, Devoe L, Ganapathy V, Prasad P. Inhibition of system
A amino acid transport activity by ethanol in BeWo choriocarcinoma cells. Am J
Obstet Gynecol 2002; 187:209-16.
64. Asai M, Narita A, Kashiwamata S. Effects of acetaldehyde and/or ethanol
on neutral aminoacid transport systems in microvillous brush border menbrane
vesicles prepared form human placenta. Experientia. 1985; 41(12):1566-1568.
65. Rytting E, Audus K. Novel organic cation transporter 2-Mediated Carnitine
uptake in placental choriocarcinoma (BeWo) cells. J Pharmacol Exp Ther.
2004;312(1):192-197.
66. Pastrakujulic A, Derewlany L. Knie B, Koren G. Effects of cocaine and
nicotine on amino acid transport across the human placental cotyledon perfused in
vitro. J Pharmacol Exp Ther. 2000; 294(1):141-146.

61
ANEXOS

62

63
ÍNDICE DE ANEXOS
Anexo 1 - Figuras nº 3, 11, 12, 13, 14, 16, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27,
28, 30, 31, 34 e 35. ......................................................................................... a1

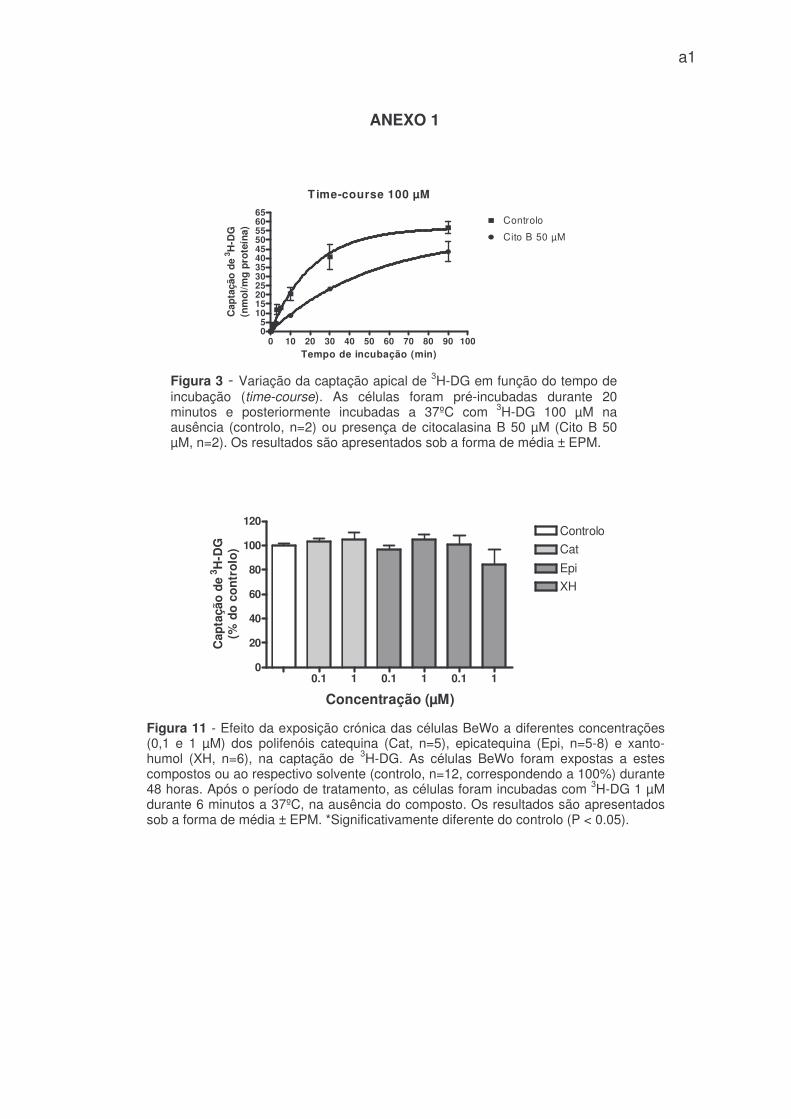
a1
ANEXO 1
Time-course 100 µM
0 10 20 30 40 50 60 70 80 90 10005
101520253035404550556065
Controlo
Cito B 50 µM
Tempo de incubação (min)
Cap
taçã
od
e3 H
-DG
(nm
ol/
mg
pro
teín
a)
Figura 3 - Variação da captação apical de 3H-DG em função do tempo deincubação (time-course). As células foram pré-incubadas durante 20minutos e posteriormente incubadas a 37ºC com 3H-DG 100 µM naausência (controlo, n=2) ou presença de citocalasina B 50 µM (Cito B 50µM, n=2). Os resultados são apresentados sob a forma de média ± EPM.
Figura 11 - Efeito da exposição crónica das células BeWo a diferentes concentrações(0,1 e 1 µM) dos polifenóis catequina (Cat, n=5), epicatequina (Epi, n=5-8) e xanto-humol (XH, n=6), na captação de 3H-DG. As células BeWo foram expostas a estescompostos ou ao respectivo solvente (controlo, n=12, correspondendo a 100%) durante48 horas. Após o período de tratamento, as células foram incubadas com 3H-DG 1 µMdurante 6 minutos a 37ºC, na ausência do composto. Os resultados são apresentadossob a forma de média ± EPM. *Significativamente diferente do controlo (P < 0.05).
0.1 1 0.1 1 0.1 10
20
40
60
80
100
120Controlo
Cat
Epi
XH
Concentração (µM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
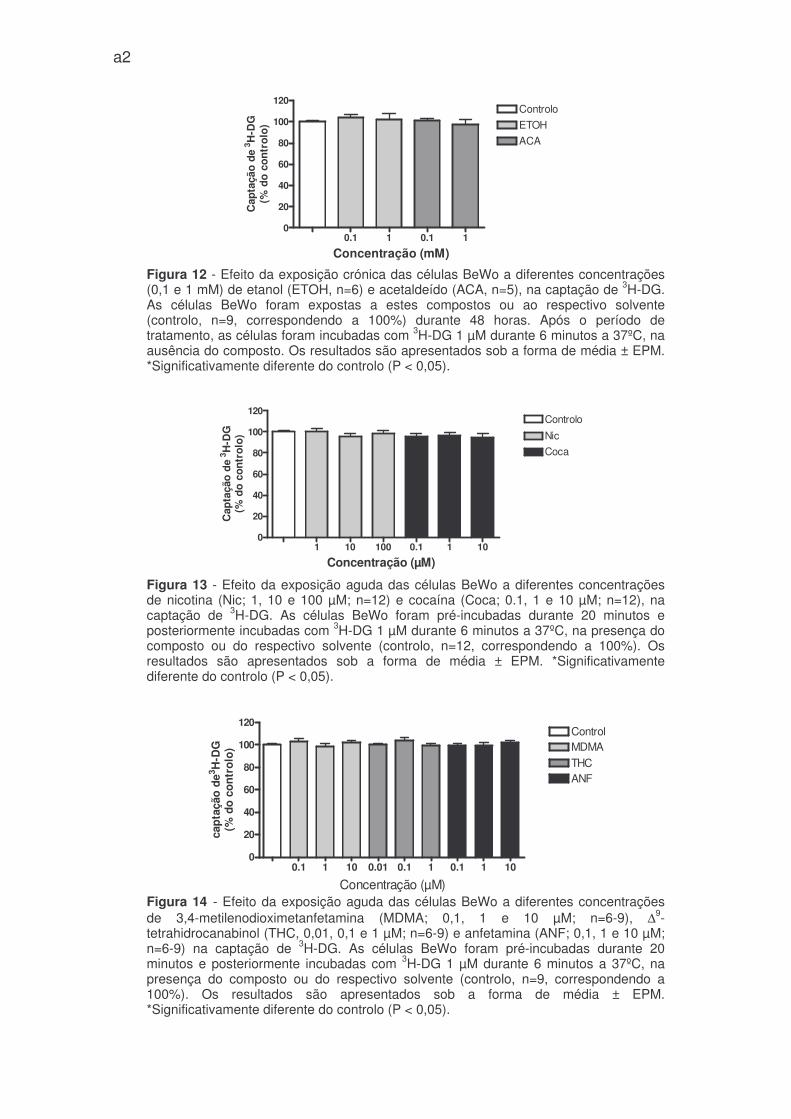
a2
Figura 12 - Efeito da exposição crónica das células BeWo a diferentes concentrações(0,1 e 1 mM) de etanol (ETOH, n=6) e acetaldeído (ACA, n=5), na captação de 3H-DG.As células BeWo foram expostas a estes compostos ou ao respectivo solvente(controlo, n=9, correspondendo a 100%) durante 48 horas. Após o período detratamento, as células foram incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, naausência do composto. Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P < 0,05).
0.1 1 0.1 10
20
40
60
80
100
120Controlo
ETOH
ACA
Concentração (mM)C
apta
ção
de
3 H-D
G(%
do
con
tro
lo)
Figura 13 - Efeito da exposição aguda das células BeWo a diferentes concentraçõesde nicotina (Nic; 1, 10 e 100 µM; n=12) e cocaína (Coca; 0.1, 1 e 10 µM; n=12), nacaptação de 3H-DG. As células BeWo foram pré-incubadas durante 20 minutos eposteriormente incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, na presença docomposto ou do respectivo solvente (controlo, n=12, correspondendo a 100%). Osresultados são apresentados sob a forma de média ± EPM. *Significativamentediferente do controlo (P < 0,05).
1 10 100 0.1 1 100
20
40
60
80
100
120Controlo
Nic
Coca
Concentração (µM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 14 - Efeito da exposição aguda das células BeWo a diferentes concentraçõesde 3,4-metilenodioximetanfetamina (MDMA; 0,1, 1 e 10 µM; n=6-9), ∆
9-tetrahidrocanabinol (THC, 0,01, 0,1 e 1 µM; n=6-9) e anfetamina (ANF; 0,1, 1 e 10 µM;n=6-9) na captação de 3H-DG. As células BeWo foram pré-incubadas durante 20minutos e posteriormente incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, napresença do composto ou do respectivo solvente (controlo, n=9, correspondendo a100%). Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P < 0,05).
0.1 1 10 0.01 0.1 1 0.1 1 100
20
40
60
80
100
120ControlMDMATHCANF
Concentração (µM)
cap
taçã
od
e3 H-D
G(%
do
con
tro
lo)
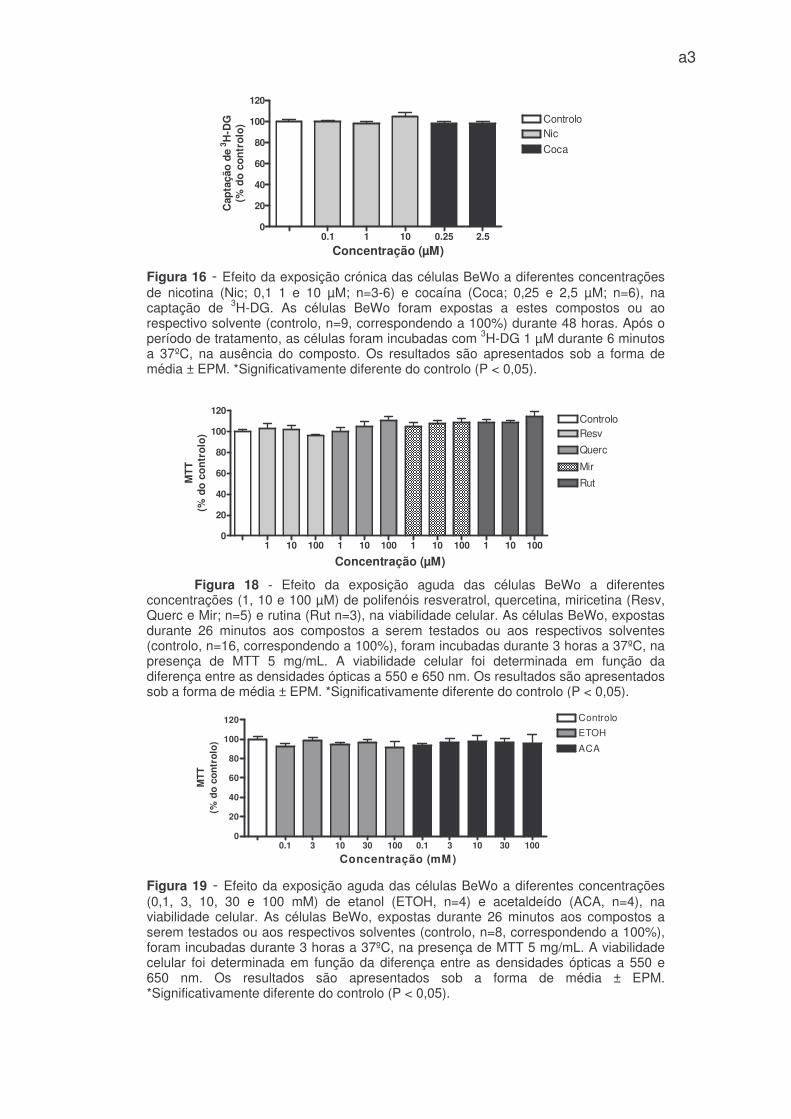
a3
0.1 1 10 0.25 2.50
20
40
60
80
100
120
ControloNic
Coca
Concentração (µM)
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 16 - Efeito da exposição crónica das células BeWo a diferentes concentraçõesde nicotina (Nic; 0,1 1 e 10 µM; n=3-6) e cocaína (Coca; 0,25 e 2,5 µM; n=6), nacaptação de 3H-DG. As células BeWo foram expostas a estes compostos ou aorespectivo solvente (controlo, n=9, correspondendo a 100%) durante 48 horas. Após operíodo de tratamento, as células foram incubadas com 3H-DG 1 µM durante 6 minutosa 37ºC, na ausência do composto. Os resultados são apresentados sob a forma demédia ± EPM. *Significativamente diferente do controlo (P < 0,05).
Figura 18 - Efeito da exposição aguda das células BeWo a diferentesconcentrações (1, 10 e 100 µM) de polifenóis resveratrol, quercetina, miricetina (Resv,Querc e Mir; n=5) e rutina (Rut n=3), na viabilidade celular. As células BeWo, expostasdurante 26 minutos aos compostos a serem testados ou aos respectivos solventes(controlo, n=16, correspondendo a 100%), foram incubadas durante 3 horas a 37ºC, napresença de MTT 5 mg/mL. A viabilidade celular foi determinada em função dadiferença entre as densidades ópticas a 550 e 650 nm. Os resultados são apresentadossob a forma de média ± EPM. *Significativamente diferente do controlo (P < 0,05).
1 10 100 1 10 100 1 10 100 1 10 1000
20
40
60
80
100
120ControloResv
Querc
Mir
Rut
Concentração (µM)
MT
T(%
do
con
tro
lo)
Figura 19 - Efeito da exposição aguda das células BeWo a diferentes concentrações(0,1, 3, 10, 30 e 100 mM) de etanol (ETOH, n=4) e acetaldeído (ACA, n=4), naviabilidade celular. As células BeWo, expostas durante 26 minutos aos compostos aserem testados ou aos respectivos solventes (controlo, n=8, correspondendo a 100%),foram incubadas durante 3 horas a 37ºC, na presença de MTT 5 mg/mL. A viabilidadecelular foi determinada em função da diferença entre as densidades ópticas a 550 e650 nm. Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P < 0,05).
0.1 3 10 30 100 0.1 3 10 30 1000
20
40
60
80
100
120 Controlo
ETOH
ACA
Concentração (mM)
MT
T(%
do
con
tro
lo)
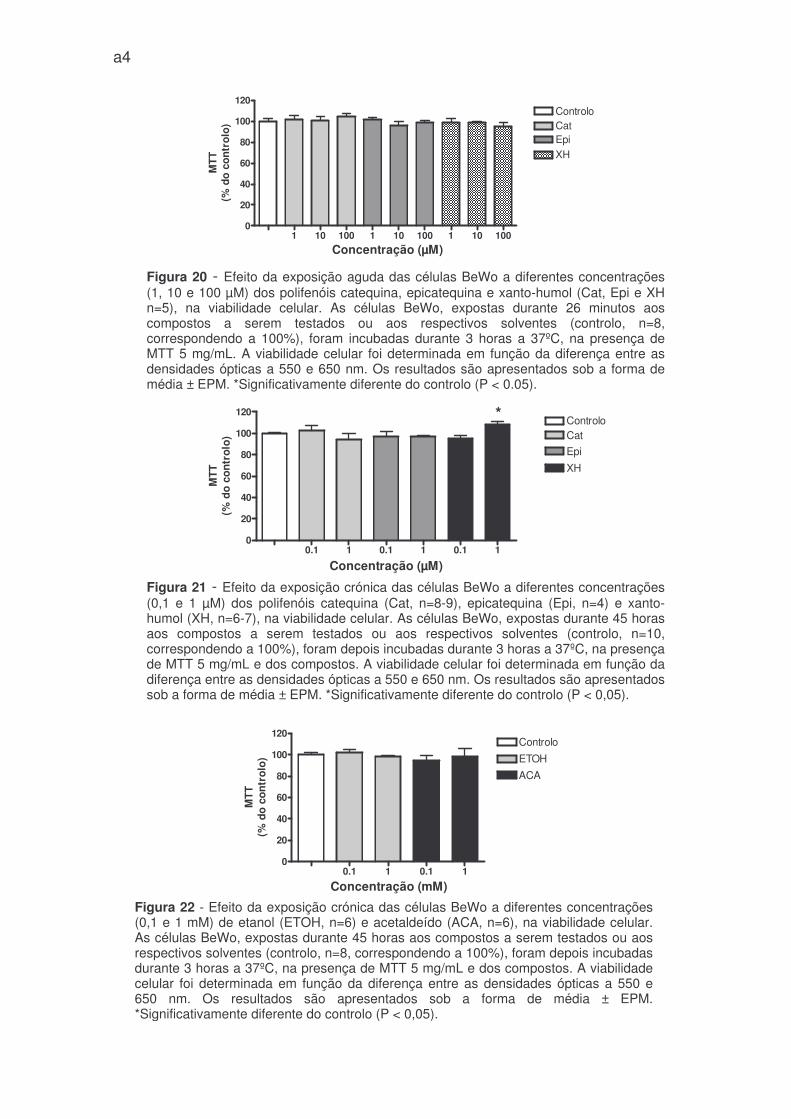
a4
Figura 20 - Efeito da exposição aguda das células BeWo a diferentes concentrações(1, 10 e 100 µM) dos polifenóis catequina, epicatequina e xanto-humol (Cat, Epi e XHn=5), na viabilidade celular. As células BeWo, expostas durante 26 minutos aoscompostos a serem testados ou aos respectivos solventes (controlo, n=8,correspondendo a 100%), foram incubadas durante 3 horas a 37ºC, na presença deMTT 5 mg/mL. A viabilidade celular foi determinada em função da diferença entre asdensidades ópticas a 550 e 650 nm. Os resultados são apresentados sob a forma demédia ± EPM. *Significativamente diferente do controlo (P < 0.05).
1 10 100 1 10 100 1 10 1000
20
40
60
80
100
120ControloCatEpiXH
Concentração (µM)
MT
T(%
do
con
tro
lo)
0.1 1 0.1 10
20
40
60
80
100
120Controlo
ETOH
ACA
Concentração (mM)
MT
T(%
do
con
tro
lo)
Figura 22 - Efeito da exposição crónica das células BeWo a diferentes concentrações(0,1 e 1 mM) de etanol (ETOH, n=6) e acetaldeído (ACA, n=6), na viabilidade celular.As células BeWo, expostas durante 45 horas aos compostos a serem testados ou aosrespectivos solventes (controlo, n=8, correspondendo a 100%), foram depois incubadasdurante 3 horas a 37ºC, na presença de MTT 5 mg/mL e dos compostos. A viabilidadecelular foi determinada em função da diferença entre as densidades ópticas a 550 e650 nm. Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P < 0,05).
Figura 21 - Efeito da exposição crónica das células BeWo a diferentes concentrações(0,1 e 1 µM) dos polifenóis catequina (Cat, n=8-9), epicatequina (Epi, n=4) e xanto-humol (XH, n=6-7), na viabilidade celular. As células BeWo, expostas durante 45 horasaos compostos a serem testados ou aos respectivos solventes (controlo, n=10,correspondendo a 100%), foram depois incubadas durante 3 horas a 37ºC, na presençade MTT 5 mg/mL e dos compostos. A viabilidade celular foi determinada em função dadiferença entre as densidades ópticas a 550 e 650 nm. Os resultados são apresentadossob a forma de média ± EPM. *Significativamente diferente do controlo (P < 0,05).
0.1 1 0.1 1 0.1 10
20
40
60
80
100
120ControloCat
Epi
XH
*
Concentração (µM)
MT
T(%
do
con
tro
lo)
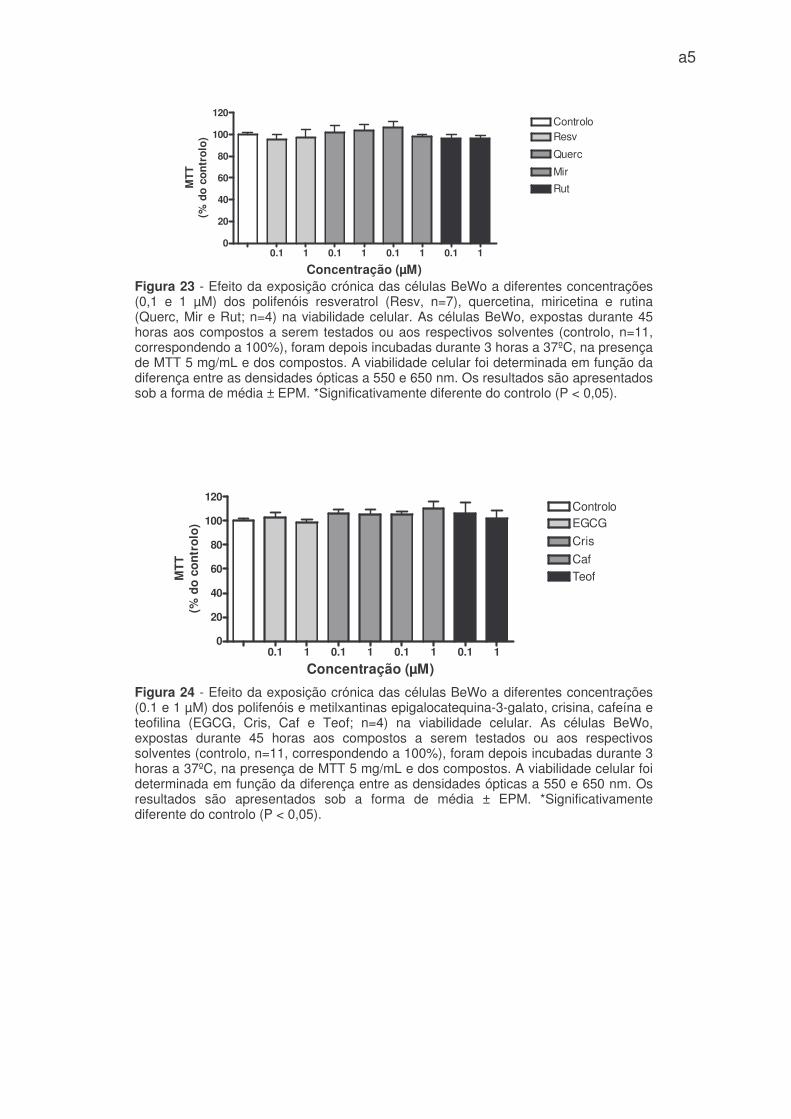
a5
Figura 23 - Efeito da exposição crónica das células BeWo a diferentes concentrações(0,1 e 1 µM) dos polifenóis resveratrol (Resv, n=7), quercetina, miricetina e rutina(Querc, Mir e Rut; n=4) na viabilidade celular. As células BeWo, expostas durante 45horas aos compostos a serem testados ou aos respectivos solventes (controlo, n=11,correspondendo a 100%), foram depois incubadas durante 3 horas a 37ºC, na presençade MTT 5 mg/mL e dos compostos. A viabilidade celular foi determinada em função dadiferença entre as densidades ópticas a 550 e 650 nm. Os resultados são apresentadossob a forma de média ± EPM. *Significativamente diferente do controlo (P < 0,05).
0.1 1 0.1 1 0.1 1 0.1 10
20
40
60
80
100
120ControloResv
Querc
Mir
Rut
Concentração (µM)
MT
T(%
do
con
tro
lo)
0.1 1 0.1 1 0.1 1 0.1 10
20
40
60
80
100
120ControloEGCG
Cris
CafTeof
Concentração (µM)
MT
T(%
do
con
tro
lo)
Figura 24 - Efeito da exposição crónica das células BeWo a diferentes concentrações(0.1 e 1 µM) dos polifenóis e metilxantinas epigalocatequina-3-galato, crisina, cafeína eteofilina (EGCG, Cris, Caf e Teof; n=4) na viabilidade celular. As células BeWo,expostas durante 45 horas aos compostos a serem testados ou aos respectivossolventes (controlo, n=11, correspondendo a 100%), foram depois incubadas durante 3horas a 37ºC, na presença de MTT 5 mg/mL e dos compostos. A viabilidade celular foideterminada em função da diferença entre as densidades ópticas a 550 e 650 nm. Osresultados são apresentados sob a forma de média ± EPM. *Significativamentediferente do controlo (P < 0,05).
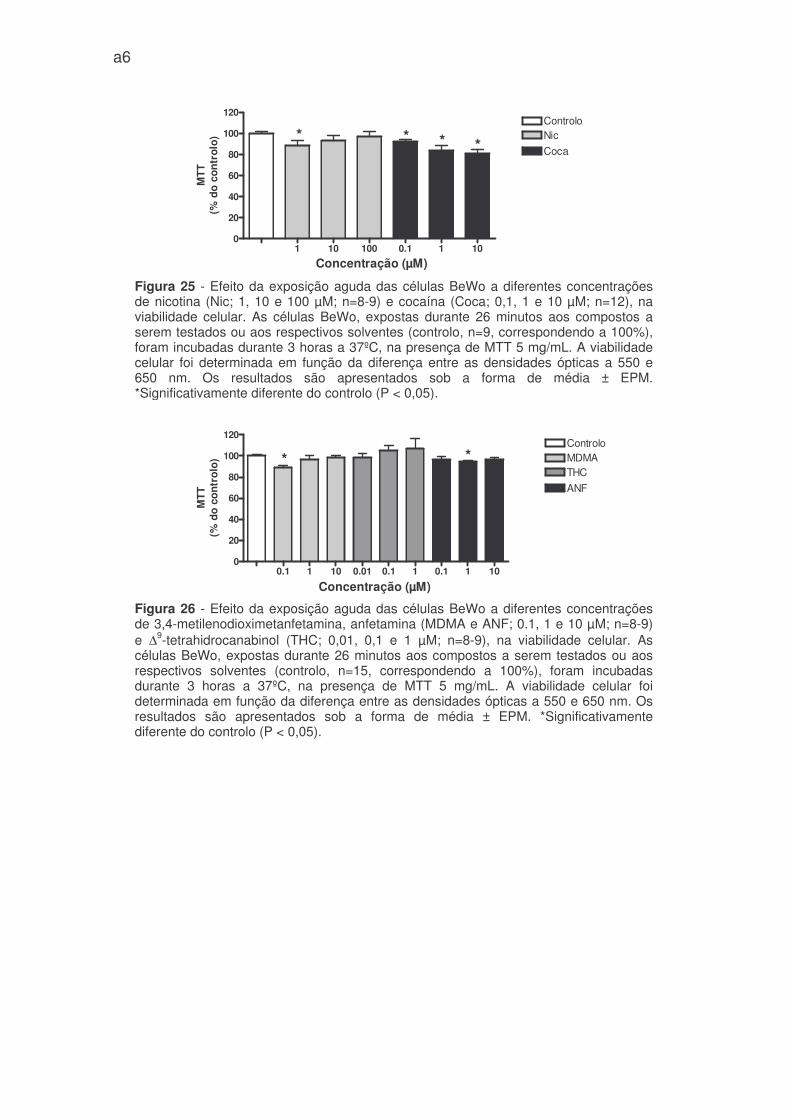
a6
Figura 26 - Efeito da exposição aguda das células BeWo a diferentes concentraçõesde 3,4-metilenodioximetanfetamina, anfetamina (MDMA e ANF; 0.1, 1 e 10 µM; n=8-9)e ∆
9-tetrahidrocanabinol (THC; 0,01, 0,1 e 1 µM; n=8-9), na viabilidade celular. Ascélulas BeWo, expostas durante 26 minutos aos compostos a serem testados ou aosrespectivos solventes (controlo, n=15, correspondendo a 100%), foram incubadasdurante 3 horas a 37ºC, na presença de MTT 5 mg/mL. A viabilidade celular foideterminada em função da diferença entre as densidades ópticas a 550 e 650 nm. Osresultados são apresentados sob a forma de média ± EPM. *Significativamentediferente do controlo (P < 0,05).
0.1 1 10 0.01 0.1 1 0.1 1 100
20
40
60
80
100
120ControloMDMATHC
ANF
* *
Concentração (µM)
MT
T(%
do
con
tro
lo)
1 10 100 0.1 1 100
20
40
60
80
100
120ControloNic
Coca* * * *
Concentração (µM)
MT
T(%
do
con
tro
lo)
Figura 25 - Efeito da exposição aguda das células BeWo a diferentes concentraçõesde nicotina (Nic; 1, 10 e 100 µM; n=8-9) e cocaína (Coca; 0,1, 1 e 10 µM; n=12), naviabilidade celular. As células BeWo, expostas durante 26 minutos aos compostos aserem testados ou aos respectivos solventes (controlo, n=9, correspondendo a 100%),foram incubadas durante 3 horas a 37ºC, na presença de MTT 5 mg/mL. A viabilidadecelular foi determinada em função da diferença entre as densidades ópticas a 550 e650 nm. Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P < 0,05).
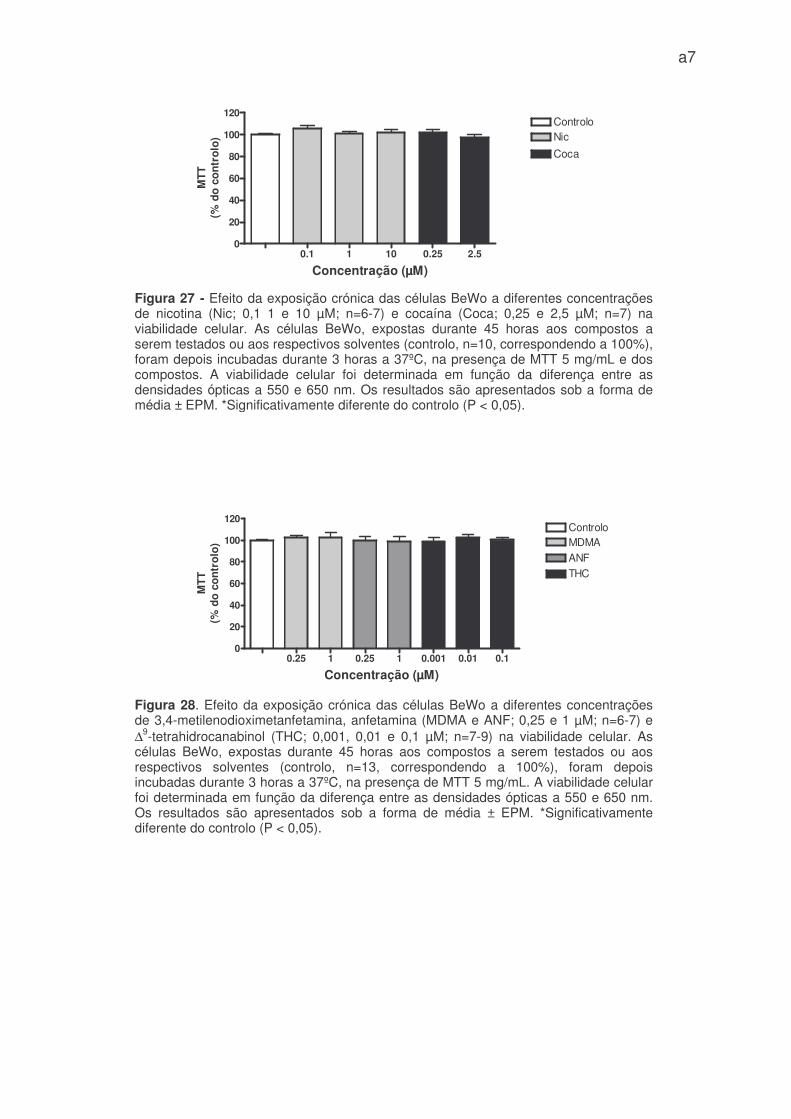
a7
Figura 27 - Efeito da exposição crónica das células BeWo a diferentes concentraçõesde nicotina (Nic; 0,1 1 e 10 µM; n=6-7) e cocaína (Coca; 0,25 e 2,5 µM; n=7) naviabilidade celular. As células BeWo, expostas durante 45 horas aos compostos aserem testados ou aos respectivos solventes (controlo, n=10, correspondendo a 100%),foram depois incubadas durante 3 horas a 37ºC, na presença de MTT 5 mg/mL e doscompostos. A viabilidade celular foi determinada em função da diferença entre asdensidades ópticas a 550 e 650 nm. Os resultados são apresentados sob a forma demédia ± EPM. *Significativamente diferente do controlo (P < 0,05).
0.1 1 10 0.25 2.50
20
40
60
80
100
120ControloNic
Coca
Concentração (µM)
MT
T(%
do
con
tro
lo)
Figura 28. Efeito da exposição crónica das células BeWo a diferentes concentraçõesde 3,4-metilenodioximetanfetamina, anfetamina (MDMA e ANF; 0,25 e 1 µM; n=6-7) e∆
9-tetrahidrocanabinol (THC; 0,001, 0,01 e 0,1 µM; n=7-9) na viabilidade celular. Ascélulas BeWo, expostas durante 45 horas aos compostos a serem testados ou aosrespectivos solventes (controlo, n=13, correspondendo a 100%), foram depoisincubadas durante 3 horas a 37ºC, na presença de MTT 5 mg/mL. A viabilidade celularfoi determinada em função da diferença entre as densidades ópticas a 550 e 650 nm.Os resultados são apresentados sob a forma de média ± EPM. *Significativamentediferente do controlo (P < 0,05).
0.25 1 0.25 1 0.001 0.01 0.10
20
40
60
80
100
120ControloMDMAANFTHC
Concentração (µM)
MT
T(%
do
con
tro
lo)
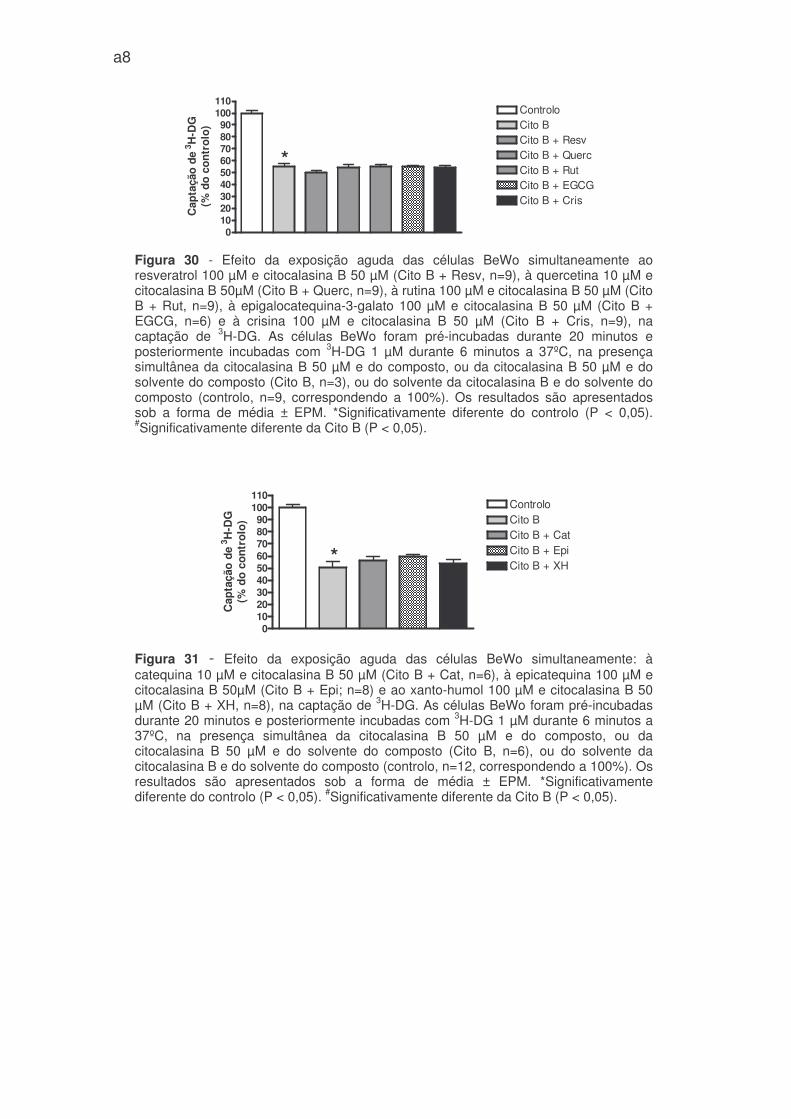
a8
Figura 30 - Efeito da exposição aguda das células BeWo simultaneamente aoresveratrol 100 µM e citocalasina B 50 µM (Cito B + Resv, n=9), à quercetina 10 µM ecitocalasina B 50µM (Cito B + Querc, n=9), à rutina 100 µM e citocalasina B 50 µM (CitoB + Rut, n=9), à epigalocatequina-3-galato 100 µM e citocalasina B 50 µM (Cito B +EGCG, n=6) e à crisina 100 µM e citocalasina B 50 µM (Cito B + Cris, n=9), nacaptação de 3H-DG. As células BeWo foram pré-incubadas durante 20 minutos eposteriormente incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, na presençasimultânea da citocalasina B 50 µM e do composto, ou da citocalasina B 50 µM e dosolvente do composto (Cito B, n=3), ou do solvente da citocalasina B e do solvente docomposto (controlo, n=9, correspondendo a 100%). Os resultados são apresentadossob a forma de média ± EPM. *Significativamente diferente do controlo (P < 0,05).#Significativamente diferente da Cito B (P < 0,05).
0102030405060708090
100110
ControloCito BCito B + ResvCito B + QuercCito B + RutCito B + EGCGCito B + Cris
*
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
0102030405060708090
100110
ControloCito BCito B + CatCito B + EpiCito B + XH
*
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 31 - Efeito da exposição aguda das células BeWo simultaneamente: àcatequina 10 µM e citocalasina B 50 µM (Cito B + Cat, n=6), à epicatequina 100 µM ecitocalasina B 50µM (Cito B + Epi; n=8) e ao xanto-humol 100 µM e citocalasina B 50µM (Cito B + XH, n=8), na captação de 3H-DG. As células BeWo foram pré-incubadasdurante 20 minutos e posteriormente incubadas com 3H-DG 1 µM durante 6 minutos a37ºC, na presença simultânea da citocalasina B 50 µM e do composto, ou dacitocalasina B 50 µM e do solvente do composto (Cito B, n=6), ou do solvente dacitocalasina B e do solvente do composto (controlo, n=12, correspondendo a 100%). Osresultados são apresentados sob a forma de média ± EPM. *Significativamentediferente do controlo (P < 0,05). #Significativamente diferente da Cito B (P < 0,05).
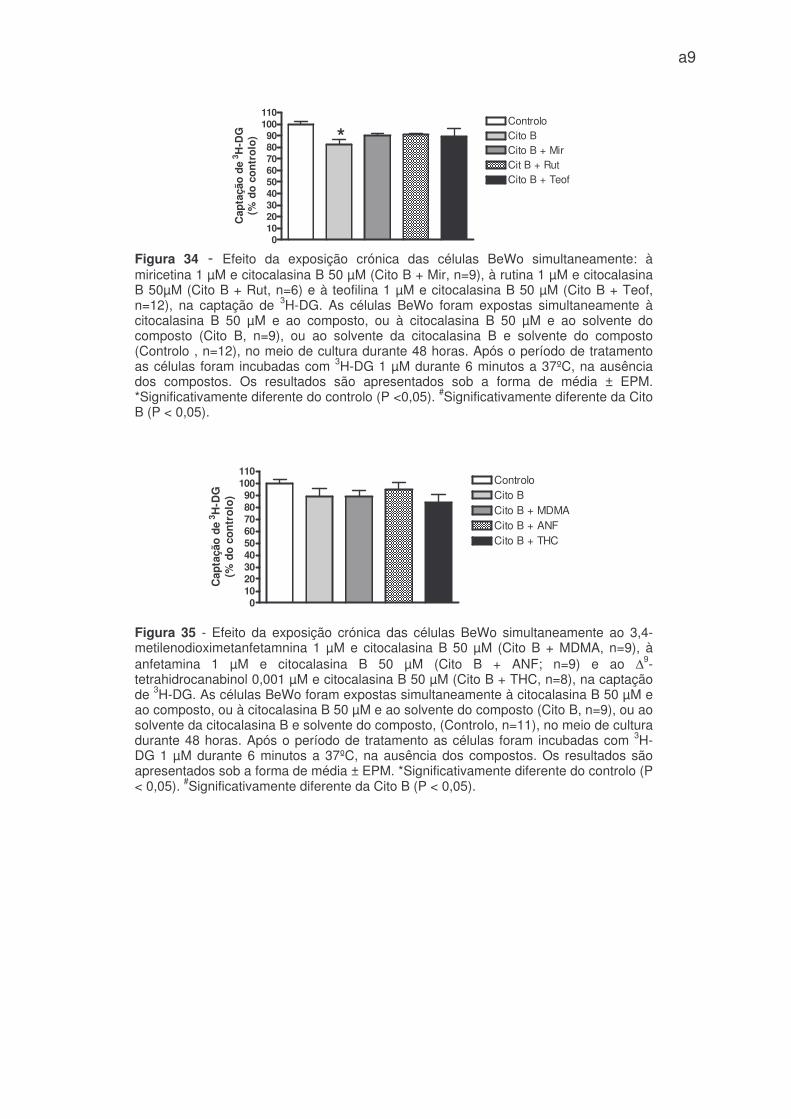
a9
Figura 34 - Efeito da exposição crónica das células BeWo simultaneamente: àmiricetina 1 µM e citocalasina B 50 µM (Cito B + Mir, n=9), à rutina 1 µM e citocalasinaB 50µM (Cito B + Rut, n=6) e à teofilina 1 µM e citocalasina B 50 µM (Cito B + Teof,n=12), na captação de 3H-DG. As células BeWo foram expostas simultaneamente àcitocalasina B 50 µM e ao composto, ou à citocalasina B 50 µM e ao solvente docomposto (Cito B, n=9), ou ao solvente da citocalasina B e solvente do composto(Controlo , n=12), no meio de cultura durante 48 horas. Após o período de tratamentoas células foram incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, na ausênciados compostos. Os resultados são apresentados sob a forma de média ± EPM.*Significativamente diferente do controlo (P <0,05). #Significativamente diferente da CitoB (P < 0,05).
0102030405060708090
100110
ControloCito BCito B + MirCit B + RutCito B + Teof
*
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
0102030405060708090
100110
ControloCito BCito B + MDMACito B + ANFCito B + THC
Cap
taçã
od
e3 H
-DG
(%d
oco
ntr
olo
)
Figura 35 - Efeito da exposição crónica das células BeWo simultaneamente ao 3,4-metilenodioximetanfetamnina 1 µM e citocalasina B 50 µM (Cito B + MDMA, n=9), àanfetamina 1 µM e citocalasina B 50 µM (Cito B + ANF; n=9) e ao ∆
9-tetrahidrocanabinol 0,001 µM e citocalasina B 50 µM (Cito B + THC, n=8), na captaçãode 3H-DG. As células BeWo foram expostas simultaneamente à citocalasina B 50 µM eao composto, ou à citocalasina B 50 µM e ao solvente do composto (Cito B, n=9), ou aosolvente da citocalasina B e solvente do composto, (Controlo, n=11), no meio de culturadurante 48 horas. Após o período de tratamento as células foram incubadas com 3H-DG 1 µM durante 6 minutos a 37ºC, na ausência dos compostos. Os resultados sãoapresentados sob a forma de média ± EPM. *Significativamente diferente do controlo (P< 0,05). #Significativamente diferente da Cito B (P < 0,05).

a10