dissertação biodiversidade besouros longicórneos
TRANSCRIPT

UNIVERSIDADE FEDERAL DE OURO PRETO
INSTITUTO DE CIÊNCIAS EXATAS E BIOLÓGICAS
DEPARTAMENTO DE BIODIVERSIDADE, EVOLUÇÃO E MEIO AMBIENTE
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA DE BIOMAS TROPICAIS
Biodiversidade de Besouros Longicórneos (Coleoptera: Cerambycidae) da
Estação Ecológica do Tripuí, Ouro Preto, MG.
FELIPE DONATELI GATTI
OURO PRETO - MG
2015

UNIVERSIDADE FEDERAL DE OURO PRETO
INSTITUTO DE CIÊNCIAS EXATAS E BIOLÓGICAS
DEPARTAMENTO DE BIODIVERSIDADE, EVOLUÇÃO E MEIO AMBIENTE
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA DE BIOMAS TROPICAIS
Biodiversidade de Besouros Longicórneos (Coleoptera: Cerambycidae) da
Estação Ecológica do Tripuí, Ouro Preto, MG.
FELIPE DONATELI GATTI
Dissertação apresentada ao Programa de Pós Graduação em Ecologia de Biomas Tropicais do Departamento de Biodiversidade, Evolução e Meio ambiente do Instituto de Ciências Exatas e Biológicas da Universidade Federal de Ouro Preto como requisito parcial para obtenção do título de Mestre em Ecologia.
ORIENTADOR: PROF. Dr. MARCO ANTONIO A. CARNEIRO
OURO PRETO - MG
2015



Agradecimentos
Meus sinceros agradecimentos:
Aos meus pais, Tadeu e Edina, e a minha irmã Mariana, pelo apoio
incondicional e por sempre serem um porto seguro para mim. Sem vocês eu não teria
chegado nem na metade do caminho que já trilhei até agora.
A todos os meus familiares que de alguma forma contribuíram para minha
formação. Em especial a minha tia Maria que sempre me apoiou e demostrou confiança
em todas as minhas escolhas.
Ao professor Marcão, pela orientação e por me ensinar a tratar a ciência de
forma metódica e ética.
Aos meus amigos Leonardo Rodrigues, Sebastião Zarantonelli, Ricardo Brandão
e Bruno Gomes, não só por toda a ajuda para realizar este trabalho, mas por terem sido
como irmãos para mim ao longo de todos os anos que passei em Ouro Preto.
A Sarah pela paciência, companheirismo e por sempre estar ao meu lado me
acolhendo em todos os momentos.
A todos os meus amigos do Laboratório de Entomologia Ecológica, em especial
ao Luiz Figueiredo, Taís Araujo e Henrique Glater, que foram imprescindíveis para a
realização deste trabalho.
A todos os meus amigos do Biomas, em especial a Ana Clara Magalhães, Iuri
Martins, Fernando Pinho e Filipe Moura, por me ajudarem de inúmeras formas, além de
dividirem comigo seus conhecimentos e experiências e terem sido muito importantes
para a realização deste trabalho.
A todos os professores que de alguma forma contribuíram para minha formação,
seja em aula ou em uma conversa descontraída na cantina e corredores.
Ao Rubens, secretário do Biomas, por se mostrar à disposição e ter me ajudado
sempre que precisei.
A Universidade Federal de Ouro Preto, por me fornecer uma bolsa de estudos.
Ao setor de transporte da UFOP, em especial aos motoristas Seu Hélio e Sapo.
Aos membros da banca por suas valiosas sugestões.
Este trabalho foi realizado com o apoio financeiro da Universidade Federal de
Ouro Preto e com recursos do autor e do professor orientador.

“If the doors of perception were cleansed everything would appear to man as it is: infinite.”
William Blake

Sumário
Apresentação ................................................................................................. 4
CAPÍTULO I: Riqueza, abundância e sazonalidade de besouros
longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí, Ouro Preto, MG. ........................ 8
Resumo ....................................................................................................... 9
Introdução ................................................................................................. 10
Metodologia ............................................................................................. 13
Resultados ................................................................................................ 16
Discussão ................................................................................................. 44
CAPÍTULO II: A influência da heterogeneidade estrutural do habitat na
comunidade de besouros longicórneos (Coleoptera: Cerambycidae) em um
fragmento de Mata Atlântica no Brasil. ...................................................... 48
Resumo ..................................................................................................... 49
Introdução ................................................................................................. 50
Metodologia ............................................................................................. 52
Resultados ................................................................................................ 58
Discussão ................................................................................................. 67
Conclusão ................................................................................................... 71
Referências .............................................................................................. 72

4
Apresentação
A degradação e a fragmentação do habitat natural em Ouro Preto começou com
o início do extrativismo do ouro na região. Para atender a grande demanda de pessoas
que aqui chegavam nesse período, foi necessário realizar a abertura de estradas, a
implantação de lavouras e pastos, a retirada de madeira, o reflorestamento de espécies
de interesse econômico e a consequente urbanização (Paniago, 1983; Paula et al., 2002).
Entretanto, o cenário começou a piorar com o declínio da exploração do ouro no final
do século XIX e início do século XX. A partir daí, as monoculturas de café e chá, e a
criação de gado, sempre pautadas no desmatamento predatório, passaram a se expandir
rapidamente pela região (Golfari, 1975; Paniago, 1983).
Dentro desse contexto, surgiram as formações vegetais secundárias, compostas
por indivíduos resultantes de todos esses processos de ocupação antrópica (Paula et al.,
2002). Há muito tempo Gómez-Pompa e Wiechers (1979) haviam alertado que, em um
futuro próximo, as florestas primárias estariam restritas as unidades de conservação.
Esse futuro já é uma realidade para grande parte da Mata Atlântica.
Na Estação Ecológica do Tripuí (EET), um fragmento de Mata Atlântica situado
em Ouro Preto onde o presente trabalho foi realizado, é possível observar reflexos de
todas essas atividades antrópicas, fato que faz da EET um mosaico de vegetação com
diferentes graus de impacto e estágios sucessionais (Werneck et al., 2000, 2001),
tornando-a um excelente laboratório de pesquisa para quem procura entender como a
heterogeneidade estrutural das diferentes formações vegetais pode afetar a
biodiversidade.
Entretanto, entender como a mudança estrutural do habitat afeta a biodiversidade
demanda tempo e recursos, e muitas vezes nós não temos isso a nosso dispor. Assim, as
abordagens práticas das questões que envolvem os estudos ecológico dos fragmentos

5
ambientais requerem a seleção de espécies, ou grupos de espécies, que possam de forma
rápida, barata e eficiente representar a biodiversidade e os processos ecológicos que
envolvem o ecossistema (Fahrig, 2003, 2001; Feinsinger, 2001). Dentro desse contexto,
os insetos surgem como um excelente grupo de bioindicadores para essa finalidade. Eles
estão intimamente associados a cobertura vegetal da área e suas comunidades são
sensíveis a praticamente qualquer intensidade de perturbação. Assim, os insetos se
tornaram os melhores indicadores da qualidade ambiental de uma região, e
consequentemente, de outros grupos taxonômicos, sendo portanto bons bioindicadores
de todo ecossistema (Brown, 1997a, 1997b; New, 1995).
Os besouros longicórnios pertencem a família Cerambycidae, que encontra-se
dentro da superfamília Chrysomeloidea, inserida na ordem Coleoptera. Eles se
distinguem dos outros insetos principalmente por apresentarem tarsos pseudotetrâmeros
e pela grande maioria de suas espécies possuírem longas antenas, que estão inseridas
entre a fronte e olhos (Costa Lima, 1955; Vives, 2000). As antenas são importantes
órgãos sensoriais que permitem aos besouros encontrar sua planta hospedeira e um
parceiro para acasalar através de compostos voláteis transportados pelo ar (Vives,
2000). Seus olhos são compostos e conforme o hábito da espécie, diurno, crespuscular
ou noturno, podem ser fina, intermediária ou grosseiramente facetados (J. R. M.
Mermudes em comunicação pessoal, 2015).
Esses besouros são organismos conspícuos em florestas, onde ocupam desde o
solo até a copa das árvores (Monné et al. 2009). Podem ser polífagos ou monófagos, e
apresentam, em vários níveis, uma alta especificidade com sua planta hospedeira. Isso
foi extremamente importante para moldar a evolução do grupo (Linsley, 1959).
Podemos ver reflexos desse processo evolutivo no comportamento desses besouros,
como por exemplo, as espécies pertencentes as subfamílias mais primitivas, como

6
Parandrinae, Prioninae e Lepeturinae, são em sua maioria polífagas, podendo ter como
planta hospedeira tanto gimnospermas como angiospermas. Enquanto que as espécies
pertencentes a subfamília Lamiinae e as mais especializadas de Cerambycinae parecem
estarem restritas a Angiospermae (Linsley, 1959).
O processo de acasalamento dos besouros longicórneos ocorre na planta
hospedeira que foi selecionada para a postura dos ovos pelos progenitores (Vives,
2000). Os ovos são postos em galhos vivos, mortos ou recém cortados, conforme o
comportamento da espécie (Costa Lima, 1955). As suas larvas são xilófagas e
broqueiam a madeira, entretanto, existem algumas espécies em que as larvas podem se
alimentar externamente de raízes, fazendo um túnel no solo (Morillo, 2007) e também
existem larvas espermófogas, que se desenvolvem em sementes (Costa Lima, 1955).
Esses besouros desempenham importantes papéis no ecossistema florestal.
Muitos organismos dependem do seu comportamento, de serrar galhos e árvores e
brocar a madeira, para encontrar um habitat adequado para se desenvolverem. Esse fato
faz os besouros longicórnos serem considerados “engenheiros de ecossistemas” (Buse et
al., 2008; Calderón-Cortés et al., 2011). Grande parte desses besouros dependem da
madeira morta durante parte do seu ciclo de vida, e por isso, são classificados como
insetos saproxílicos (Speight, 1989; Stokland et al., 2012). Essas espécies participam
ativamente do processo de decomposição da madeira morta e da ciclagem de nutrientes
nos ecossistemas florestais (Cobb et al., 2010).
A diversidade dos besouros longicórneos tem sido alterada por distúrbios
ambientais em várias partes do mundo (Meng et al., 2013). Assim, alguns pesquisadores
têm usado esses besouros como indicadores da biodiversidade e do estado de
conservação dos habitats, compostos por mosaicos vegetacionais, que sofreram as mais

7
diversas atividades antrópicas (Holland, 2007; Jansson et al., 2009; Sugiura et al.,
2009a).
Tudo isso mostra como os besouros longicórneos estão diretamente vinculados
as formações vegetais e são fortemente influenciados pelo estado de conservação da
floresta (Martins, 1999a; Raje et al., 2012). Nesse contexto, a composição da
comunidade dos besouros longicórneos vai responder de forma diferente à
heterogeneidade estrutural do habitat em áreas de uma floresta, as quais sofreram
diferentes tipos de impactos ambientais no passado, e se encontram em diferentes
estágios sucessionais.
Desta forma, este estudo fez uma descrição inicial da composição de espécies de
besouros longicórneos presentes na EET. No primeiro capítulo foi descrita a riqueza,
abundância e a sazonalidade da comunidade de besouros longicórneos na EET. No
segundo capítulo foi avaliado o efeito da heterogeneidade estrutural do habitat de três
áreas da EET, que se encontram em diferentes estágios sucessionais, sobre essa
comunidade.

8
CAPÍTULO I
Riqueza, abundância e sazonalidade de besouros
longicórneos (Coleoptera: Cerambycidae) coletados
com armadilha luminosa na Estação Ecológica do
Tripuí, Ouro Preto, MG.

9
RESUMO. Riqueza, abundância e sazonalidade de besouros longicórneos
(Coleoptera: Cerambycidae) coletados com armadilha luminosa na Estação
Ecológica do Tripuí, Ouro Preto, MG. Este estudo fez uma descrição inicial da
composição de espécies de besouros longicórneos presentes na Estação Ecológica do
Tipuí. Foram coletados 581 indivíduos distribuídos em 145 espécies e 3 subfamílias. A
subfamília mais rica e abundante foi Lamiinae, seguida por Cerambycinae e Prioninae.
Entre as espécies amostradas, 46,2% foram consideradas singletons, 13,8% doubletons
e 90,3% raras. As curvas de acumulação de espécies não estabilizaram, o que já era
esperado, tendo em vista que foram amostradas mais espécies e indivíduos de besouros
longicórneos nos últimos meses de nossa coleta. As estimativas de riqueza apresentaram
valores muito mais elevados do que a riqueza observada. Isso foi reflexo da alta
proporção de espécies raras presentes na amostra. Este trabalho mostrou que inventários
faunísticos de grupos hiperdiversos, realizados com apenas um ano de coleta, pode
subestimar a riqueza de espécies de uma região. Assim, séries temporais maiores,
associadas a mais métodos de coletas, é essencial para uma amostragem mais precisa.
PALAVRAS-CHAVE: entomologia florestal, espécies raras, estimadores de riqueza.

10
INTRODUÇÃO
Os besouros longicórneos pertencem à família Cerambycidae, incluída na ordem
Coleoptera. Essa é uma das maiores famílias de besouros, e consequentemente de
insetos (Monné et al., 2010, 2009), e possui oito subfamílias: Aseminae, Cerambycinae,
Lamiinae, Lepturinae, Oxypeltinae, Parandrinae, Prioninae e Spondylidinae (Monné et
al., 2010; Napp, 1994). A família Cerambycidae apresenta uma alta diversidade, com
pelo menos 4.000 gêneros e 35.000 espécies em todo o mundo (Costa, 2000). Nas
Américas compreende 1.600 gêneros com 9.000 espécies (Monné e Bezark, 2009) e o
Brasil conta com aproximadamente 1.000 gêneros com 4.000 espécies (Costa, 2000;
Monné e Bezark, 2009).
Esses besouros podem ser xilófagos, fitófagos ou saprófagos, e suas espécies
podem variar de monófagas a polífagas. Os adultos se alimentam de madeira viva ou
morta, folhas, flores, pólen, frutos, seiva, sementes e raízes (Costa Lima, 1955; Lieutier
et al., 2004; Linsley, 1959; Martins, 1997; Morillo, 2007). São normalmente de vida
curta, direcionados especialmente para cópula e nidificação, geralmente morrendo após
executar essas etapas. Esses besouros são capazes de nidificar tanto em árvores sadias e
pouco estressadas quanto em árvores com deficiências nutricionais e minerais (Lieutier
et al., 2004). Algumas espécies, como por exemplo, os serra-paus matam o caule para
torná-lo adequado para postura de ovos. Para isso, eles cortam um anel em volta da
planta, e quando o caule é anelado, todos os tecidos acima do cinturão morrem e as
gemas laterais começam a se desenvolver abaixo do cinturão. Isso afeta muito a
arquitetura das árvores, reduzindo o crescimento em altura, causando má formação e
aumentando a taxa de mortalidade (Lieutier et al., 2004). Suas larvas são, na grande
maioria, xilófagas e broqueiam os troncos e galhos. Alimentam-se, principalmente da
casca, floema, seiva e cerne das plantas, podendo causar prejuízos e eventualmente

11
matar sua planta hospedeira (Martins, 1997). Como vimos, muitas espécies de besouros
longicórneos dependem da madeira morta ou decadente em parte do seu ciclo de vida,
fato que faz essas espécies serem consideradas insetos saproxílicos (Speight, 1989;
Stokland et al., 2012).
Os besouros longicórneos atendem muito bem aos sete critérios propostos por
Pearson (1994) para serem bons bioindicadores da qualidade do habitat. Eles possuem a
taxonomia estável (Martins, 2011, 2010, 2008, 2007, 2006, 2005, 2003a, 2003b, 2002,
1999b, 1998, 1997); a sua história natural é bem conhecida (Costa Lima, 1955; Lieutier
et al., 2004; Linsley, 1959); são facilmente amostrados por diferentes técnicas
entomológicas, como armadilhas luminosas (Wolda, 1978a, 1978b), armadilhas de
Malaise (Makino et al., 2007; Meng et al., 2013), guarda-chuva entomológico e rede
entomológica (Monné et al., 2012, 2010, 2009); são amplamente distribuídos em todo o
mundo, porém, possuem espécies especialistas de habitat (Lieutier et al., 2004; Wagner
et al., 2008); são sensíveis a mudanças ambientais e apresentam rápidas respostas a
estas mudanças (Baur et al., 2005, 2002; Collinge et al., 2001; Hjältén et al., 2012;
Maeto et al., 2002; Saint-Germain et al., 2004; Sugiura et al., 2009b; Taki et al., 2010;
Vodka et al., 2009); seus padrões de biodiversidade refletem a diversidade de outros
táxons (Buse et al., 2008; Calderón-Cortés et al., 2011); e possuem espécies com
potencial importância econômica em grandes culturas como eucalipto (Zanúncio et al.,
2009), milho (Pires et al., 2011) e tamarindo (Orozco-Santos et al., 2011). Além disso,
existem espécies pirófagas, que preferem se alimentar de árvores mortas por fogo e são
indicadoras de áreas que sofreram queimada recente (Cobb et al., 2010), e outras que só
nidificam em árvores perenes já estabelecidas (Grove, 2002a) podendo assim serem
indicadoras de florestas primárias.

12
No Brasil existem muitos trabalhos taxonômicos envolvendo os besouros
longicórneos, mas poucos estudos com ênfase ecológica. Entretanto, esses trabalhos
taxonômicos ainda relatam, com frequência, novas espécies e novos registros de
ocorrência desses besouros (Barreto et al., 2013; Galileo e Martins, 2010; Galileo et al.,
2014; Martins et al., 2011; Martins et al., 2009a, 2009b; Martins et al., 2006; Monné et
al., 2012, 2010, 2009; Morillo, 2007; Rodrigues et al., 2010; Souza e Silva, 2012).
Assim, embora bem estudados em relação a outros grupos, a descrição frequente dessas
novas espécies para a família, mostra que ainda há muito a se conhecer sobre a
diversidade dos besouros longicórneos.
Neste capítulo foram descritas a riqueza, a abundância e a sazonalidade de
besouros longicórneos amostrados na Estação Ecológica do Tripuí, ao longo de doze
meses consecutivos de coleta.

13
METODOLOGIA
Área de estudo. As coletas foram realizadas na Estação Ecológica do Tripuí
(EET), um fragmento de Mata Atlântica Estacional Semidecídua Montana, com uma
área de 337 ha, localizado na cidade de Ouro Preto, zona metalúrgica do estado de
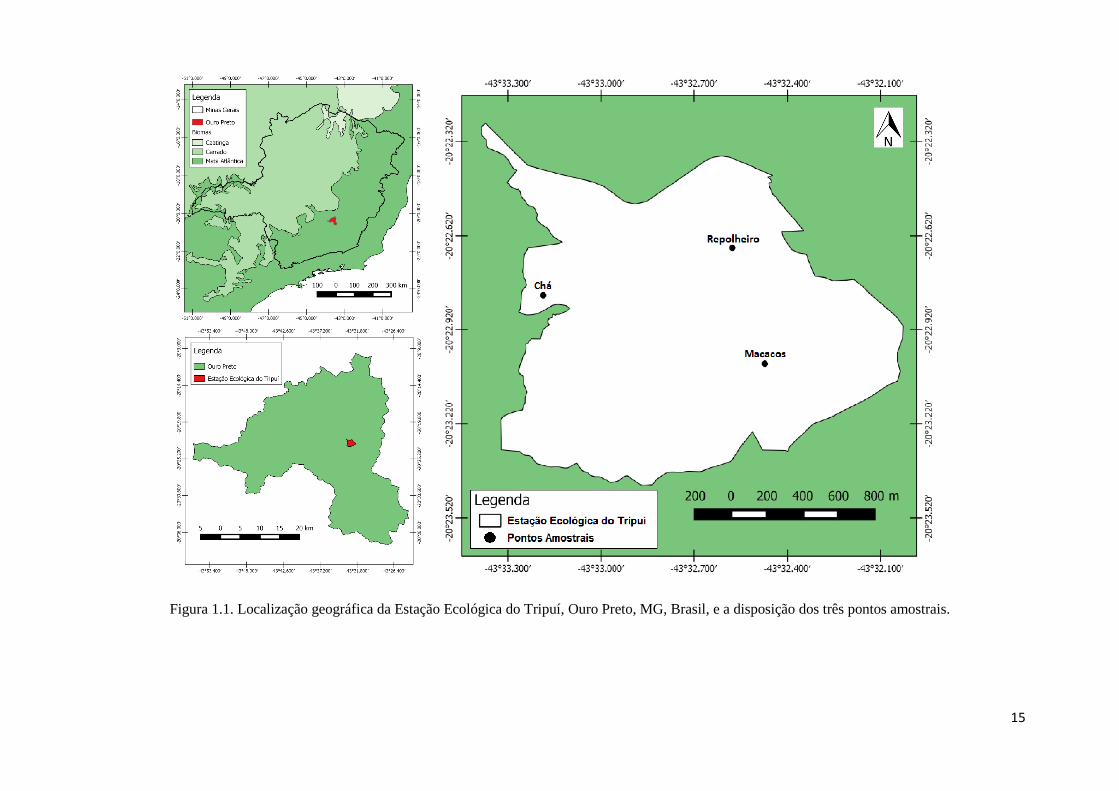
Minas Gerais, Brasil (20°23’45” S e 43°34’33” W) (Fig. 1.1). Encontra-se na sub-bacia
do córrego Tripuí, e possui um gradiente altitudinal variando entre 1280 e 1450 m.
Segundo a classificação de Köppen, seu clima é Cwb, com verão temperado e o inverno
seco, índice pluviométrico anual próximo de 1.600 mm, e temperatura média anual de
18ºC. Seus solos são litólicos, de natureza álica com textura areno-siltosa (Alvares et
al., 2013; Werneck et al., 2001, 2000).
Na EET foram escolhidas três áreas, já estudadas previamente por Werneck et
al. (2000), que sofreram diferentes impactos no passado e agora se encontram em
diferentes estágio sucessionais. Uma dessas áreas é conhecida como Repolheiro, que
corresponde ao trecho de floresta mais preservado da EET, onde não há relatos de
perturbação antrópica ou fogo nos últimos 45 anos; outra área é conhecida como
Macacos, que corresponde a um trecho intermediário de floresta na EET onde não há
relato de retirada de madeira ou fogo nos últimos 45 anos, mas que nos últimos anos
vem sofrendo perturbação sistêmica por equinos de moradores da EET e de fazendas
vizinhas; e uma área conhecida como Chá, onde havia uma plantação de chá preto
(Camellia sinensis (L.) Kuntze) que foi abandonada à aproximadamente 55 anos e
corresponde ao trecho da floresta em estágio sucessional mais jovem da EET (Werneck
et al., 2000).
Coleta de Besouros Longicórneos. Os besouros foram coletados através de
armadilhas luminosas nas três áreas de estudo durante doze meses consecutivos, de
fevereiro de 2014 a janeiro de 2015. Foram realizadas duas campanhas de coleta por

14
mês, sendo que cada campanha correspondeu a três noites de coleta consecutivas,
totalizando assim seis noites de coleta por mês. A armadilha luminosa utilizada para a
coleta foi a modelo Luiz de Queiroz, com fonte luminosa fluorescente ultravioleta de 15
W e 100 V, fabricada pela empresa Biocontrole®. O funcionamento das armadilhas é
crepuscular noturno–vespertino, entre 18–06 hs. As armadilhas foram posicionadas a
1,80 m de altura do solo para permitir uma maior dispersão de luz (De Paula e Ferreira,
2000, 1998). Os insetos foram sacrificados com acetato de etila ainda no campo. No
laboratório, os espécimes coletados foram montados e identificados até o menor nível
taxonômico possível. Todos os espécimes coletados estão depositados na coleção
entomológica do Laboratório de Entomologia Ecológica do Departamento de
Biodiversidade, Evolução e Meio Ambiente da Universidade Federal de Ouro Preto. As
fotos dos besouros foram tiradas com uma lente Canon® Ef 100 mm Macro acoplada a
uma câmera Canon® T1i.
Análises estatísticas. Para avaliar o esforço amostral da coleta foram feitas três
curvas de acumulação de espécies: a curva do coletor, que adiciona as espécies na
medida que elas vão aparecendo na amostra; método exato, que encontra a expectativa
média da riqueza de espécies em cada amostra; e a curva de rarefação, que encontra a
média da acumulação de indivíduos (Oksanen et al., 2015). Para estimar a riqueza de
besouros longicórneos foram usados dois estimadores não-paramétricos baseados na
abundância das espécies, o Chao1 (Chao, 1987, 1984) e o ACE (Chao e Lee, 1992;
Chao e Yang, 1993). Todas essas análises foram realizadas através dos pacotes ‘vegan’
(Oksanen et al., 2015) e ‘BiodiversityR’ (Kindt e Coe, 2005) desenvolvido para o
programa estatístico R (R Development Core Team 2012).
15
Figura 1.1. Localização geográfica da Estação Ecológica do Tripuí, Ouro Preto, MG, Brasil, e a disposição dos três pontos amostrais.

16
RESULTADOS
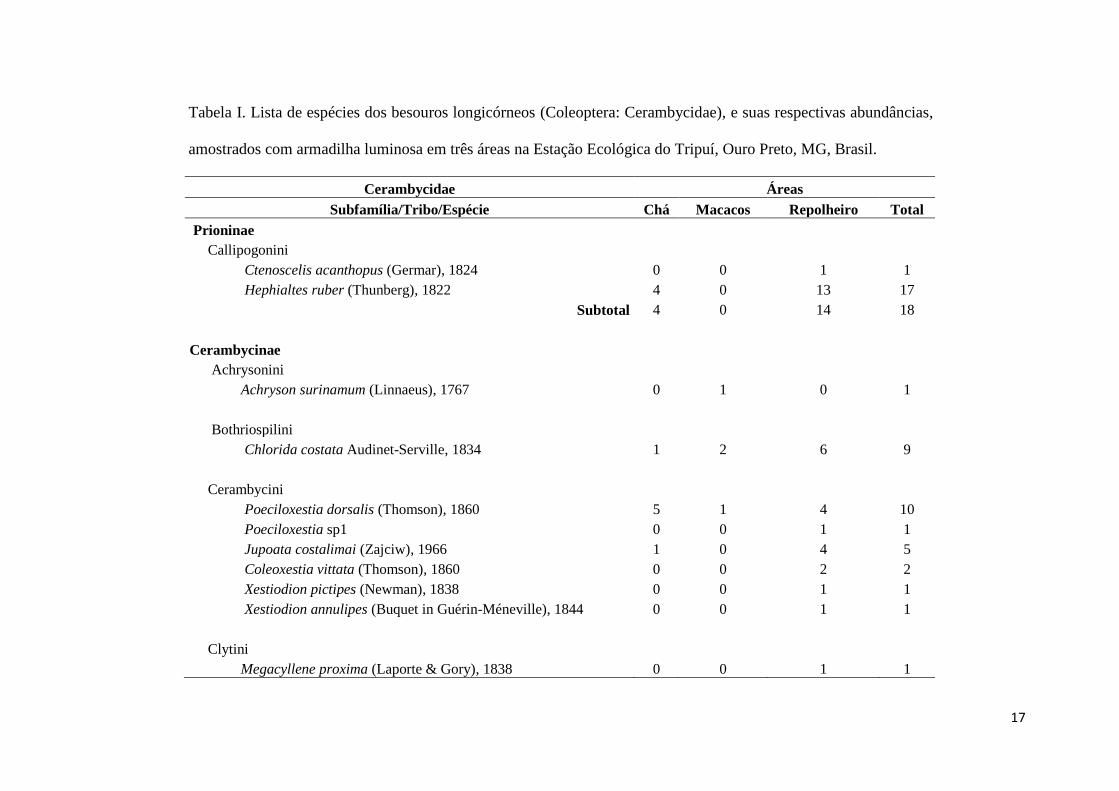
Foram coletados 581 indivíduos de besouros longicórneos, distribuídos em 145
espécies e 3 subfamílias (Tab. I e Fig. 1.2 a 1.14). A subfamília mais rica e abundante
foi Lamiinae (91 espécies e 347 indivíduos), seguida por Cerambycinae (52 espécies e
216 indivíduos) e Prioninae (2 espécies e 18 indivíduos). A comunidade foi
caracterizada por apresentar um grande número de espécies raras. Sessenta e sete
(46,2%) espécies foram consideradas “singletons” (representadas por apenas um
indivíduo na amostra) e 20 (13,8%) espécies “doubletons” (representadas por apenas
dois indivíduos na amostra). Setenta e uma (48,95%) espécies ocorreram em uma única
amostra, sendo assim consideradas “unicatas” e 22 (15,15%) espécies ocorreram em
duas amostras, sendo assim consideradas “duplicatas”. Cento e trinta e uma espécies, ou
90,3% da amostra, apresentaram abundância entre 1 e 10 indivíduos, sendo assim,
consideradas raras. As espécies mais abundantes foram Sp87 com 42 (7,2%) indivíduos,
Trypanidius dimidiatus Thomson 1861 com 28 (4,8%) indivíduos e Sp85 com 22
(3,8%) indivíduos.
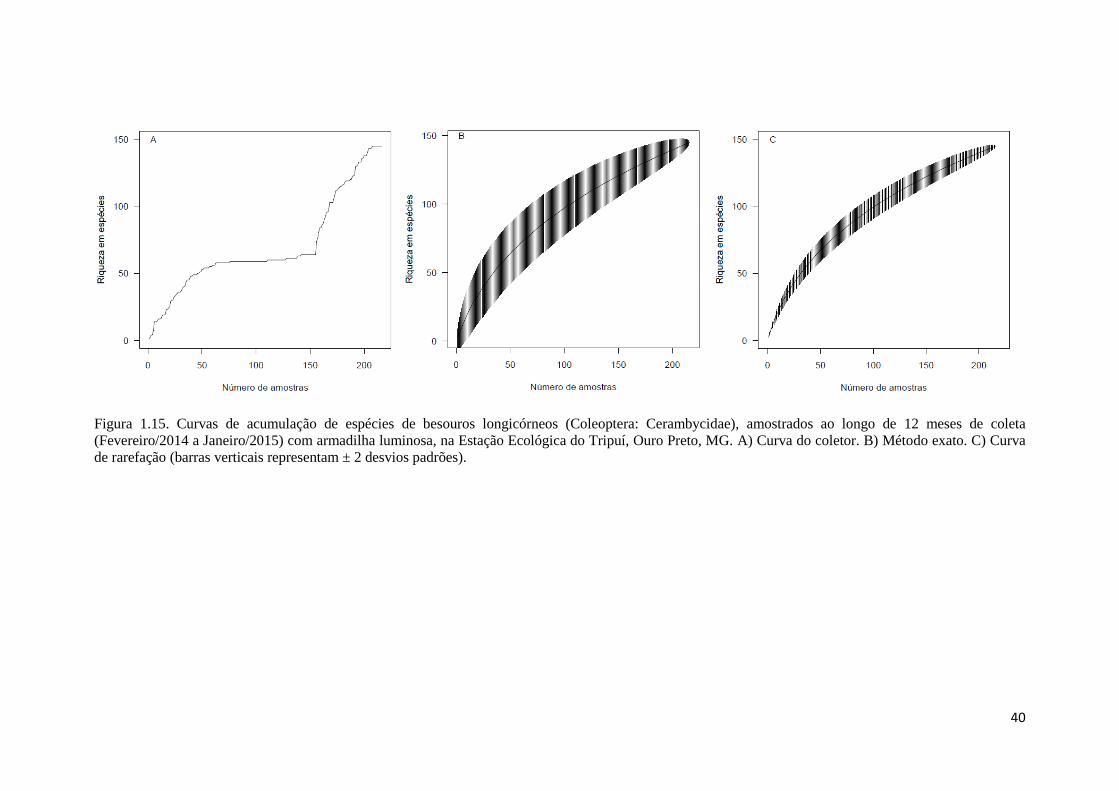
Independentemente do método de ajuste utilizado, as curvas de acumulação de
espécies não estabilizaram (Fig. 1.15). Esse resultado já era esperado, tendo em vista
que os últimos meses de coleta (outubro de 2014 a janeiro de 2015) compreenderam o
ápice da estação chuvosa, e nesse período foram coletadas mais espécies (81 ou 55,85%
do total da amostra) e indivíduos (428 ou 73,7% do total da amostra) de besouros
longicórneos (Fig. 1.16 e 1.17). As estimativas da riqueza foram mais elevadas (SChao1=
250 ± 3,87; SACE= 236 ± 1,72) que o número de espécies observadas (Sobs = 145 ± 1,59)
e as suas curvas não se estabilizaram (Fig. 1.18). O que também era esperado, já que as
estimativas realizadas pelo Chao1 e o ACE dependem da proporção de espécies raras na
amostra.
17
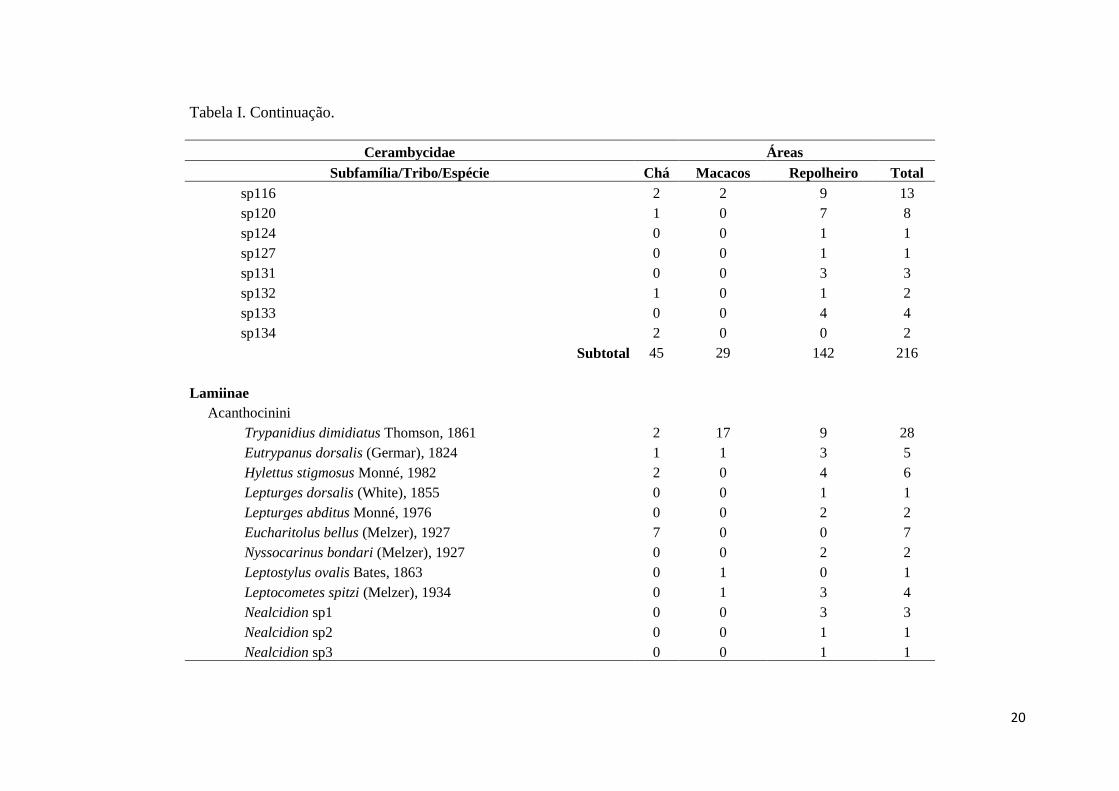
Tabela I. Lista de espécies dos besouros longicórneos (Coleoptera: Cerambycidae), e suas respectivas abundâncias,
amostrados com armadilha luminosa em três áreas na Estação Ecológica do Tripuí, Ouro Preto, MG, Brasil.
Cerambycidae Áreas
Subfamília/Tribo/Espécie Chá Macacos Repolheiro Total
Prioninae
Callipogonini
Ctenoscelis acanthopus (Germar), 1824 0 0 1 1
Hephialtes ruber (Thunberg), 1822 4 0 13 17
Subtotal 4 0 14 18
Cerambycinae
Achrysonini
Achryson surinamum (Linnaeus), 1767 0 1 0 1
Bothriospilini
Chlorida costata Audinet-Serville, 1834 1 2 6 9
Cerambycini
Poeciloxestia dorsalis (Thomson), 1860 5 1 4 10
Poeciloxestia sp1 0 0 1 1
Jupoata costalimai (Zajciw), 1966 1 0 4 5
Coleoxestia vittata (Thomson), 1860 0 0 2 2
Xestiodion pictipes (Newman), 1838 0 0 1 1
Xestiodion annulipes (Buquet in Guérin-Méneville), 1844 0 0 1 1
Clytini
Megacyllene proxima (Laporte & Gory), 1838 0 0 1 1
18
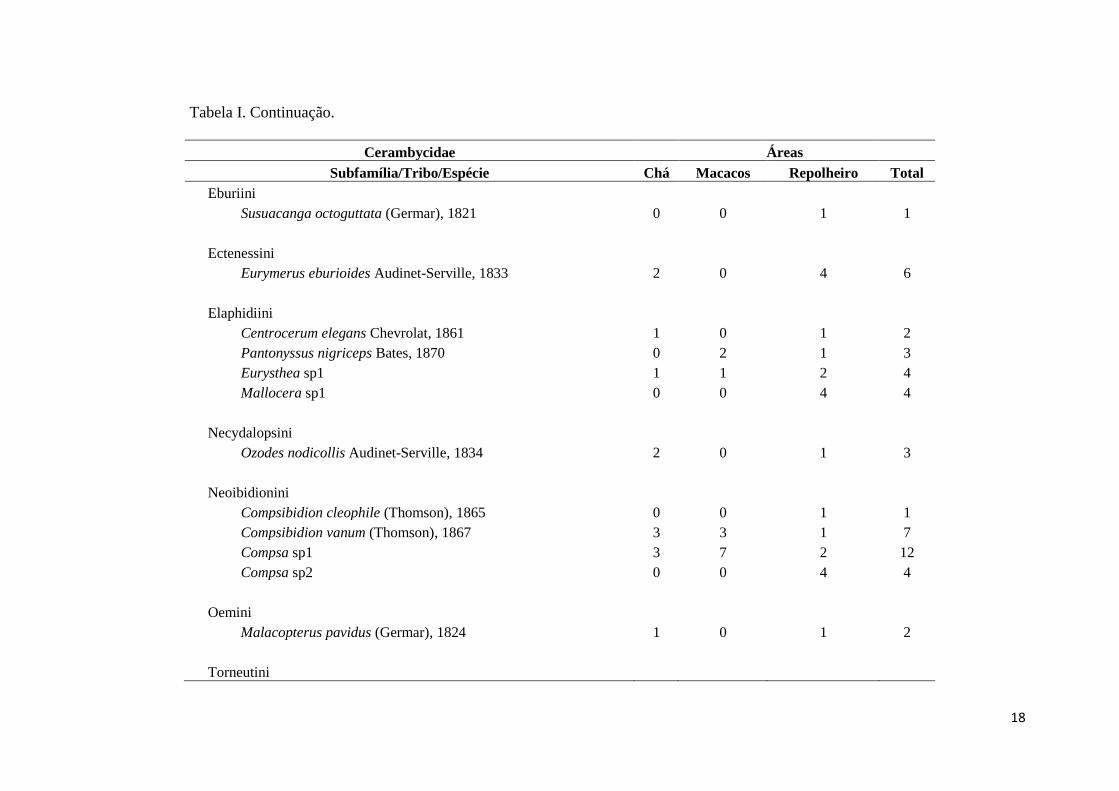
Tabela I. Continuação.
Cerambycidae Áreas
Subfamília/Tribo/Espécie Chá Macacos Repolheiro Total
Eburiini
Susuacanga octoguttata (Germar), 1821 0 0 1 1
Ectenessini
Eurymerus eburioides Audinet-Serville, 1833 2 0 4 6
Elaphidiini
Centrocerum elegans Chevrolat, 1861 1 0 1 2
Pantonyssus nigriceps Bates, 1870 0 2 1 3
Eurysthea sp1 1 1 2 4
Mallocera sp1 0 0 4 4
Necydalopsini
Ozodes nodicollis Audinet-Serville, 1834 2 0 1 3
Neoibidionini
Compsibidion cleophile (Thomson), 1865 0 0 1 1
Compsibidion vanum (Thomson), 1867 3 3 1 7
Compsa sp1 3 7 2 12
Compsa sp2 0 0 4 4
Oemini
Malacopterus pavidus (Germar), 1824 1 0 1 2
Torneutini
19
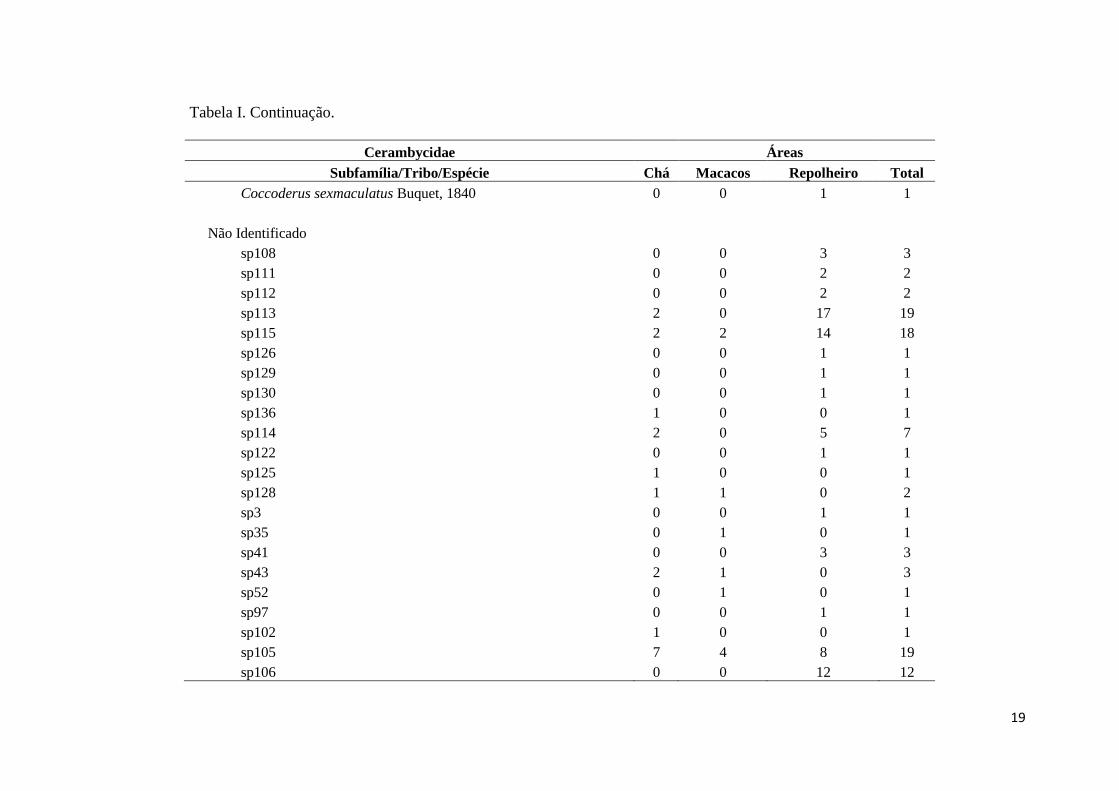
Tabela I. Continuação.
Cerambycidae Áreas
Subfamília/Tribo/Espécie Chá Macacos Repolheiro Total
Coccoderus sexmaculatus Buquet, 1840 0 0 1 1
Não Identificado
sp108 0 0 3 3
sp111 0 0 2 2
sp112 0 0 2 2
sp113 2 0 17 19
sp115 2 2 14 18
sp126 0 0 1 1
sp129 0 0 1 1
sp130 0 0 1 1
sp136 1 0 0 1
sp114 2 0 5 7
sp122 0 0 1 1
sp125 1 0 0 1
sp128 1 1 0 2
sp3 0 0 1 1
sp35 0 1 0 1
sp41 0 0 3 3
sp43 2 1 0 3
sp52 0 1 0 1
sp97 0 0 1 1
sp102 1 0 0 1
sp105 7 4 8 19
sp106 0 0 12 12
20
Tabela I. Continuação.
Cerambycidae Áreas
Subfamília/Tribo/Espécie Chá Macacos Repolheiro Total
sp116 2 2 9 13
sp120 1 0 7 8
sp124 0 0 1 1
sp127 0 0 1 1
sp131 0 0 3 3
sp132 1 0 1 2
sp133 0 0 4 4
sp134 2 0 0 2
Subtotal 45 29 142 216
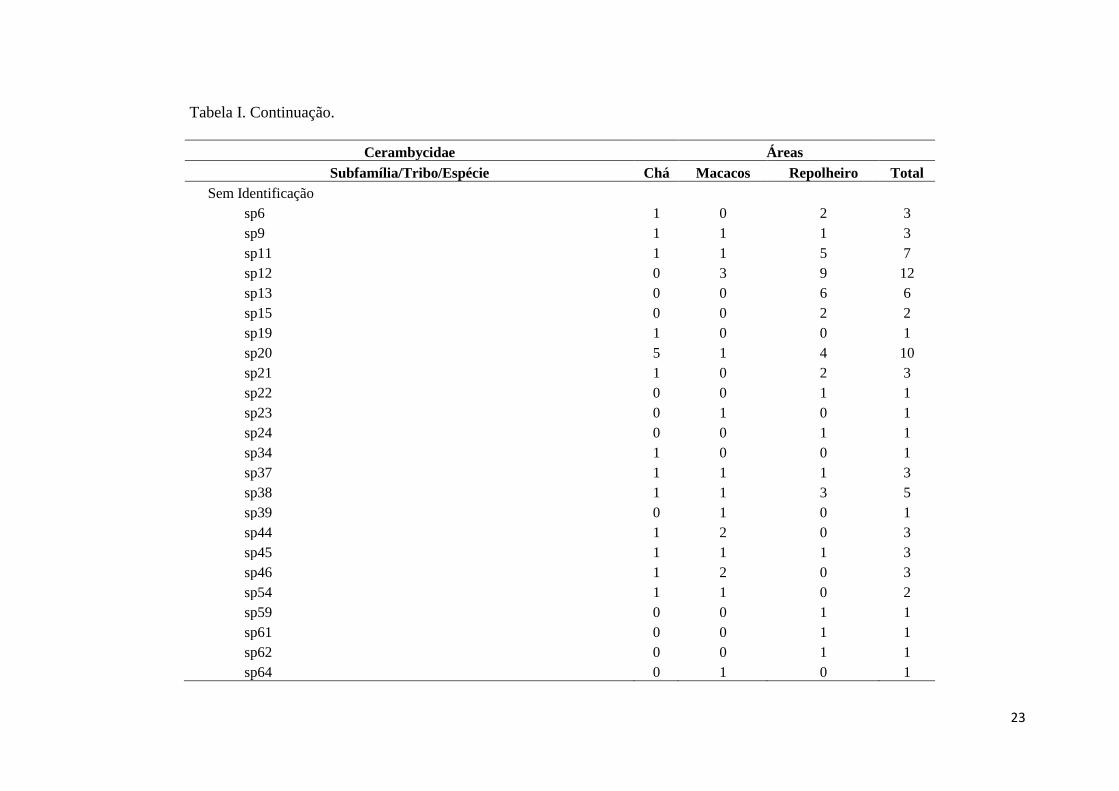
Lamiinae
Acanthocinini
Trypanidius dimidiatus Thomson, 1861 2 17 9 28
Eutrypanus dorsalis (Germar), 1824 1 1 3 5
Hylettus stigmosus Monné, 1982 2 0 4 6
Lepturges dorsalis (White), 1855 0 0 1 1
Lepturges abditus Monné, 1976 0 0 2 2
Eucharitolus bellus (Melzer), 1927 7 0 0 7
Nyssocarinus bondari (Melzer), 1927 0 0 2 2
Leptostylus ovalis Bates, 1863 0 1 0 1
Leptocometes spitzi (Melzer), 1934 0 1 3 4
Nealcidion sp1 0 0 3 3
Nealcidion sp2 0 0 1 1
Nealcidion sp3 0 0 1 1

21
Tabela I. Continuação.
Cerambycidae Áreas
Subfamília/Tribo/Espécie Chá Macacos Repolheiro Total
Acanthoderini
Macropophora accentifer (Olivier), 1795 0 0 2 2
Oreodera charisoma Lane, 1955 8 5 7 20
Oreodera sp1 0 1 0 1
Eupromerella propinqua (Melzer), 1931 0 5 0 5
Psapharochrus jaspideus (Germar), 1824 2 0 1 3
Psapharochrus sp1 0 1 0 1
Aerenicini
Phaula lichenigera (Perty), 1832 0 0 1 1
Agapanthiini
Pachypeza pennicornis (Germar), 1824 1 1 1 3
Compsosomatini
Compsosoma perpulchrum (Vigors), 1825 0 0 2 2
Compsosoma phaleratum Thomson, 1857 6 6 4 16
Desmiphorini
Desmiphora sp1 1 0 1 2
Desmiphora sp2 0 0 1 1
Desmiphora sp3 0 1 0 1
Estola sp1 2 0 1 3
Estola sp2 1 0 1 2

22
Tabela I. Continuação.
Cerambycidae Áreas
Subfamília/Tribo/Espécie Chá Macacos Repolheiro Total
Hemilophini
Gagarinia mniszechii (Chabrillac), 1857 4 0 0 4
Phoebe phoebe (Lepeletier & Audinet-Serville in Latreille), 1825 0 0 1 1
Phoebe cava (Germar), 1824 0 0 1 1
Lycaneptia sp1 1 0 0 1
Onciderini
Oncideres dejeanii Thomson, 1868 0 1 2 3
Oncideres sp1 1 0 0 1
Oncideres sp2 2 2 0 4
Psyllotoxus griseocinctus Thomson, 1868 0 0 2 2
Trestonia capreola (Germar), 1824 4 1 0 5
Trachysomus fragifer (Kirby), 1818 0 0 1 1
Pseudobeta doris (Thomson), 1868 1 0 0 1
Chitron mniszechi (Buquet), 1859 0 1 0 1
Glypthaga sp1 1 0 0 1
Polyrhaphidini
Polyrhaphis grandini Buquet, 1853 1 1 1 3
Pteropliini
Rhaphiptera affinis Thomson, 1868 1 1 3 5
Rhaphiptera nodifera Audinet-Serville, 1835 2 0 0 2
Rhaphiptera pallens Gounelle, 1908 1 0 5 6
23
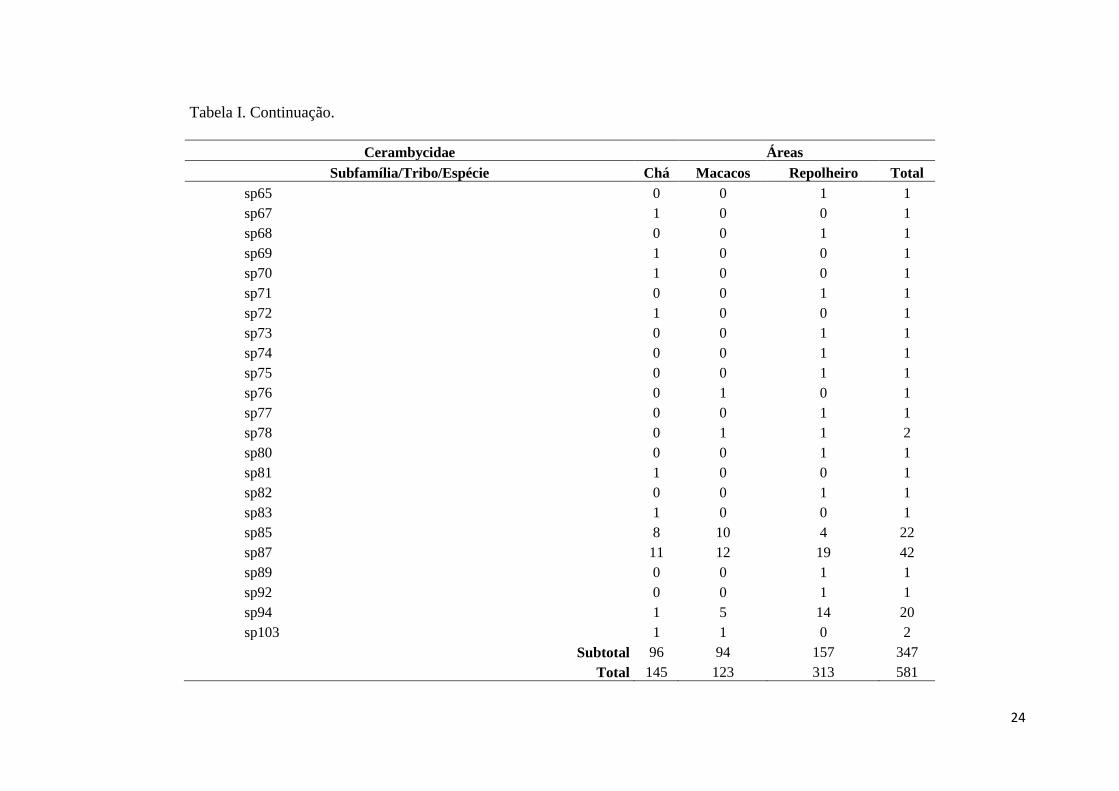
Tabela I. Continuação.
Cerambycidae Áreas
Subfamília/Tribo/Espécie Chá Macacos Repolheiro Total
Sem Identificação
sp6 1 0 2 3
sp9 1 1 1 3
sp11 1 1 5 7
sp12 0 3 9 12
sp13 0 0 6 6
sp15 0 0 2 2
sp19 1 0 0 1
sp20 5 1 4 10
sp21 1 0 2 3
sp22 0 0 1 1
sp23 0 1 0 1
sp24 0 0 1 1
sp34 1 0 0 1
sp37 1 1 1 3
sp38 1 1 3 5
sp39 0 1 0 1
sp44 1 2 0 3
sp45 1 1 1 3
sp46 1 2 0 3
sp54 1 1 0 2
sp59 0 0 1 1
sp61 0 0 1 1
sp62 0 0 1 1
sp64 0 1 0 1
24
Tabela I. Continuação.
Cerambycidae Áreas
Subfamília/Tribo/Espécie Chá Macacos Repolheiro Total
sp65 0 0 1 1
sp67 1 0 0 1
sp68 0 0 1 1
sp69 1 0 0 1
sp70 1 0 0 1
sp71 0 0 1 1
sp72 1 0 0 1
sp73 0 0 1 1
sp74 0 0 1 1
sp75 0 0 1 1
sp76 0 1 0 1
sp77 0 0 1 1
sp78 0 1 1 2
sp80 0 0 1 1
sp81 1 0 0 1
sp82 0 0 1 1
sp83 1 0 0 1
sp85 8 10 4 22
sp87 11 12 19 42
sp89 0 0 1 1
sp92 0 0 1 1
sp94 1 5 14 20
sp103 1 1 0 2
Subtotal 96 94 157 347
Total 145 123 313 581

25
Figura 1.2.

26
Figura 1.3.

27
Figura 1.4.

28
Figura 1.5.

29
Figura 1.6.

30
Figura 1.7.

31
Figura 1.8.

32
Figura 1.9.

33
Figura 1.10.

34
Figura 1.11.

35
Figura 1.12.

36
Figura 1.13.

37
Figura 1.14.
Legendas das figuras 1.2 a 1.14:
Figura 1.2. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Prioninae: a) Ctenoscelis
acanthopus; b) Hephialtes ruber. Subfamília Cerambycinae: c) Achryson surinamum; d)
Chlorida costata; e) Poeciloxestia dorsalis; f) Poeciloxestia sp1; g) Jupoata costalimai; h)
Coleoxestia vittata; i) Xestiodion pictipes; j) Xestiodion annulipes; k) Megacyllene
proxima; l) Susuacanga octoguttata.
Figura 1.3. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Cerambycinae: a) Eurymerus
eburioides; b) Centrocerum elegans; c) Centrocerum elegans; d) Pantonyssus nigriceps; e)
Eurysthea sp1; f) Mallocera sp1; g) Ozodes nodicollis; h) Compsibidion cleophile; i)
Compsibidion vanum; j) Compsa sp1; k) Compsa sp2; l) Malacopterus pavidus.
Figura 1.4. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Cerambycinae: a) Coccoderus
sexmaculatus; b) sp108; c) sp111; d) sp112; e) sp113; f) sp115; g) sp126; h) sp129; i)
sp130; j) sp136; k) sp114; l) sp122.
Figura 1.5. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Cerambycinae: a) sp125; b) sp128; c)
sp3; d) sp35; e) sp41; f) sp43; g) sp52; h) sp97; i) sp102; j) sp105; k) sp106; l) sp116.

38
Figura 1.6. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Cerambycinae: a) sp120; b) sp124; c)
sp127; d) sp131; e) sp132; f) sp133; g) sp134. Subfamília Lamiinae: h) Trypanidius
dimidiatus; i) Eutrypanus dorsalis; j) Hylettus stigmosus; k) Lepturges dorsalis; l)
Lepturges abditus.
Figura 1.7. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Lamiinae: a) Eucharitolus bellus; b)
Nyssocarinus bondari; c) Leptostylus ovalis; d) Leptocometes spitzi; e) Nealcidion sp1; f)
Nealcidion sp2; g) Nealcidion sp3; h) Macropophora accentifer; i) Oreodera charisoma; j)
Oreodera sp1; k) Eupromerella propinqua; l) Psapharochrus jaspideus.
Figura 1.8. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Lamiinae: a) Psapharochrus sp1; b)
Phaula lichenigera; c) Pachypeza pennicornis; d) Compsosoma perpulchrum; e)
Compsosoma phaleratum; f) Desmiphora sp1; g) Desmiphora sp2; h) Desmiphora sp3; i)
Estola sp1; j) Estola sp2; k) Gagarinia mniszechii; l) Phoebe phoebe.
Figura 1.9. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Lamiinae: a) Phoebe cava; b)
Lycaneptia sp1; c) Oncideres dejeanii; d) Oncideres sp1; e) Oncideres sp2; f) Psyllotoxus
griseocinctus fêmea; g) Psyllotoxus griseocinctus macho; h) Trestonia capreola; i)
Trachysomus fragifer; j) Pseudobeta doris; k) Chitron mniszechi; l) Glypthaga sp1.
Figura 1.10. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Lamiinae: a) Polyrhaphis grandini;
b) Rhaphiptera affinis; c) Rhaphiptera nodifera; d) Rhaphiptera pallens; e) sp6; f) sp9; g)
sp11; h) sp12; i) sp13; j) sp15; k) sp19; l) sp20.
Figura 1.11. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Lamiinae: a) sp21; b) sp22; c) sp23;
d) sp24; e) sp34; f) sp37; g) sp38; h) sp39; i) sp44; j) sp45; k) sp46; l) sp54.
Figura 1.12. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Lamiinae: a) sp59; b) sp61; c) sp62;
d) sp64; e) sp65; f) sp67; g) sp68; h) sp69; i) sp70; j) sp71; k) sp72; l) sp73.

39
Figura 1.13. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Lamiinae: a) sp74; b) sp75; c) sp76;
d) sp77; e) sp78; f) sp80; g) sp81; h) sp82; i) sp83; j) sp85; k) sp87; l) sp89.
Figura 1.14. Besouros longicórneos (Coleoptera: Cerambycidae) coletados com armadilha
luminosa na Estação Ecológica do Tripuí. Subfamília Lamiinae: a) sp92; b) sp94; c) sp103.
40
Figura 1.15. Curvas de acumulação de espécies de besouros longicórneos (Coleoptera: Cerambycidae), amostrados ao longo de 12 meses de coleta
(Fevereiro/2014 a Janeiro/2015) com armadilha luminosa, na Estação Ecológica do Tripuí, Ouro Preto, MG. A) Curva do coletor. B) Método exato. C) Curva
de rarefação (barras verticais representam ± 2 desvios padrões).
41
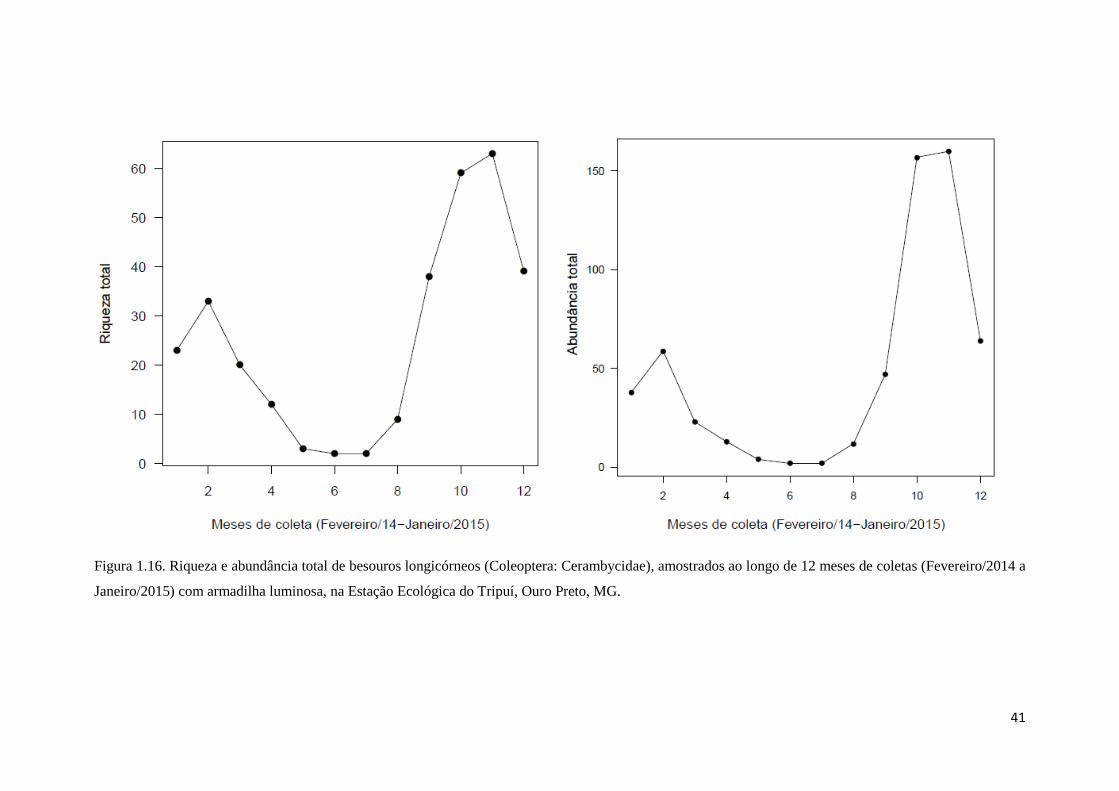
Figura 1.16. Riqueza e abundância total de besouros longicórneos (Coleoptera: Cerambycidae), amostrados ao longo de 12 meses de coletas (Fevereiro/2014 a
Janeiro/2015) com armadilha luminosa, na Estação Ecológica do Tripuí, Ouro Preto, MG.
42
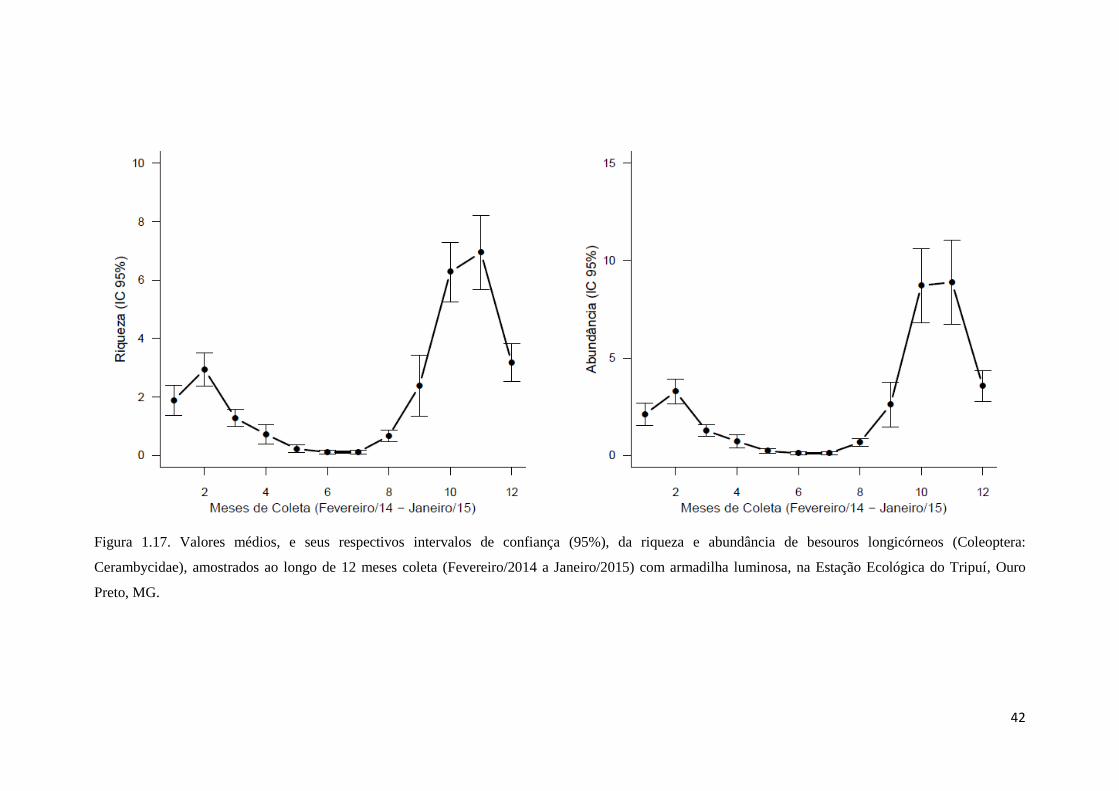
Figura 1.17. Valores médios, e seus respectivos intervalos de confiança (95%), da riqueza e abundância de besouros longicórneos (Coleoptera:
Cerambycidae), amostrados ao longo de 12 meses coleta (Fevereiro/2014 a Janeiro/2015) com armadilha luminosa, na Estação Ecológica do Tripuí, Ouro
Preto, MG.

43
Figura 1.18. Curvas dos estimadores da riqueza de besouros longicórneos (Coleoptera: Cerambycidae), amostrados
ao longo de 12 meses de coleta (Fevereiro/2014 a Janeiro/2015) com armadilha luminosa, na Estação Ecológica do
Tripuí, Ouro Preto, MG (barras verticais representam ± 1 desvio padrão).

44
DISCUSSÃO
Apesar das diferenças entre os desenhos amostrais, tempo de duração e métodos
de coleta dificultarem comparações com os dados dos levantamentos faunísticos
realizados até o momento, a EET apresentou uma considerável diversidade
(Abundância: 581, Riqueza: 145) de besouros longicórneos quando comparada com a
encontrada por Rodrigues et al. (2010) na Ilha Grande, RJ (Riqueza: 50) e por Souza e
Silva (2012) no Parque Natural Municipal de Porto Velho, RO (Abundância: 171,
Riqueza: 61). Entretanto, com exceção do trabalho realizado por Morillo (2007) na
Mata do Paraíso, Viçosa, MG (Abundância: 307, Riqueza: 105), as regiões onde os
levantamentos faunísticos vêm sendo realizados a mais anos por especialistas do grupo,
como por exemplo, o Parque Nacional do Itatiaia, RJ (Monné et al., 2012, 2010, 2009) e
o Estado do Maranhão (Martins et al., 2011, 2009a, 2009b; Monné, 2005; Zajciw,
1967), apresentam uma riqueza muito mais elevada, 648 e 367 espécies
respectivamente, do que a encontrada na EET. Assim como nesses estudos citados
anteriormente (Monné et al., 2012, 2010, 2009; Morillo, 2007; Rodrigues et al., 2010;
Souza e Silva, 2012), neste trabalho, as subfamílias Lamiinae e Cerambycinae também
apresentaram maior riqueza e abundância de besouros longicórneos.
Como esperado, os meses de novembro e dezembro de 2014, que correspondem
à primavera no hemisfério sul e compreendem o ápice da estação chuvosa, possuindo
naturalmente temperatura e pluviosidade muito elevadas, foram os meses onde mais
espécies e indivíduos de besouros foram amostrados (Fig. 1.16 e 1.17). Esse
sincronismo da diversidade de besouros longicórneos com a estação chuvosa
provavelmente está relacionada com o período de maior disponibilidade de recursos. É
nesse período que, na grande maioria das plantas, surgem novas folhas e flores, e
também aumentam a quantidade de galhos mortos na floresta (Bullock e Solís-

45
Magallanes, 1990; Martínez-Yrízar, 1995). Assim, a estação chuvosa é marcada pela
emergência dos adultos a procura de comida, plantas hospedeiras, troncos mortos e do
sexo oposto para realizar o acasalamento e a nidificação (Makino et al., 2007;
Rodrigues et al., 2010; Toledo et al. 2002).
Levando em consideração que as curvas de acumulação de espécies não
estabilizaram (Fig. 1.15), devido ao acréscimo de espécies nos meses finais da coleta, e
que os resultados dos estimadores de riquezas mostraram que pode haver mais de cem
espécies além das quais foram coletadas (Fig. 1.18), a lista de espécies apresentada
neste trabalho ainda é incompleta, e mais espécies de besouros longicórneos devem ser
amostradas na EET com a continuação dos estudos.
Esses resultados corroboram com a sugestão de outros autores de que,
inventários faunísticos mais precisos necessitam de séries temporais maiores, em
projetos de longa duração com combinações de diferentes métodos de coleta (Longino
et al., 2002; Wolda, 1978a, 1978b, 1988). Considerando que os besouros longicórneos
pertencem a um grupo hiperdiverso, com cerca de 35.000 espécies (Costa, 2000),
possuindo táxons que habitam os mais variados habitats florestais (Monné et al., 2009) e
podem apresentar grandes diferenças no período de desenvolvimento larval, variando de
1 a 3 anos (Beeson e Bhatia, 1939; Lieutier et al., 2004), é esperado que a composição
da comunidade desses besouros varie não só sazonalmente como também anualmente.
Assim, sempre seriam incrementadas novas espécies ano após ano de coleta, deixando o
padrão da curva do coletor em forma de “S” (ver Figura 1.15a) (Glater et al., em
preparação). Esses dados demonstram que inventários de grupos hiperdiversos,
realizados ao longo de um ano de coleta, subestimam a riqueza e a composição de
espécies, diminuindo a eficácia de ações práticas que visam a conservação e a
manutenção da biodiversidade.

46
A ideia de que uma boa amostragem deve causar uma estabilização nas curvas
de acumulação, entendida como o ponto em que a inclusão de unidades amostrais não
culminará na adição significativa de novas espécies, é um ponto de debate para os
trabalhos realizados com grupos de organismos hiperdiversos que habitam florestas
tropicais (Schilling e Batista, 2008). Pois essas florestas aportam a maior biodiversidade
do mundo, o que torna a completa estabilização das curvas algo irrealista (Longino et
al., 2002).
Os altos valores encontrados para as estimativas de riqueza (Fig. 1.18) são
reflexos da alta proporção de espécies raras presente na amostra. É importante deixar
claro que o fato dessas espécies serem raras na amostra não quer dizer, necessariamente,
que elas sejam raras na natureza. Elas podem ser espécies especialistas com baixo nível
populacional, generalistas mal amostradas ou espécies turistas (Longino et al., 2002;
Novotný e Basset, 2000; Schoener, 1987). O fato é que espécies raras são componentes
importantes das comunidades de insetos herbívoros nas florestas tropicais, e nunca
devem ser excluídas ou marginalizadas nos estudos dessas comunidades (Novotný e
Basset, 2000).
Como se pode observar, o valor de riqueza estimado pelo ACE é menor que o do
Chao1 (Fig. 1.18). Como o valor dado por Chao1 é uma estimativa mínima da riqueza
(Chao 1984), o ACE parece estar subestimando a riqueza dos besouros longicórneos da
EET. O fato é que diferentes estimadores vão estimar diferentes riquezas de espécies, e
sendo assim, é essencial e mais seguro usar mais de um estimador para fazer inferências
sobre a riqueza de uma área (O’hara, 2005). Os estimadores são uma ferramenta muito
útil para a realização de trabalhos ecológicos, mas é importante deixar claro que eles
também possuem seus vieses matemáticos e metodológicos e é preciso ser prudente ao
usá-los. Nenhum deles infere com certeza quantas espécies podem existir em uma área,

47
e o aconselhado é tratar essas estimativas como estimativas mínimas da riqueza (ver
Longino et al., 2002; Ohara, 2005).
Este trabalho mostrou que as florestas das montanhas mineiras aportam uma
grande diversidade de besouros longicórneos, e que a continuidade do estudo, com a
implementação de outros métodos de coletas, é essencial para a obtenção de novos
registros de espécies para a região. Além disso, anos consecutivos de estudos
acarretariam em informações mais claras que permitiriam entender melhor as flutuações
temporais da comunidade desses besouros.

48
CAPÍTULO II
A influência da heterogeneidade estrutural do
habitat na comunidade de besouros longicórneos
(Coleoptera: Cerambycidae) em um fragmento de
Mata Atlântica.

49
RESUMO. A influência da heterogeneidade estrutural do habitat na comunidade
de besouros longicórneos (Coleoptera: Cerambycidae) em um fragmento de Mata
Atlântica. Entender a influência da mudança massiva dos habitats nas comunidades
biológicas, principalmente nas espécies que participam de processos ecossistêmicos, se
tornou extremamente importante para o desenvolvimento de estratégias que visam a
proteção da biodiversidade nos remanescentes florestais. Os besouros longicórneos
desempenham importantes funções nos processos ecossistêmicos da floresta e sua
diversidade é afetada por muitos distúrbios ambientais em todo o mundo. Este trabalho
teve como objetivo mostrar a influência da heterogeneidade estrutural do habitat, de três
áreas de uma floresta que sofreram diferentes impactos no passado e agora se encontram
em diferentes estágios sucessionais, sobre a comunidade de besouros longicórneos. A
área em estágio sucessional mais avançado apresentou, principalmente, árvores mais
grossas e maior disponibilidade de madeira morta, importantes componentes florestais
que ajudam a manter a diversidade estrutural do habitat. Nessa área também foi
observada a maior riqueza e abundância de besouros longicórneos. Esses resultados
mostram que esses besouros estão intimamente associados à heterogeneidade estrutural
das florestas e podem ser bons indicadores para a avaliação da diversidade e da
qualidade do habitat.
PALAVRAS-CHAVE: biodiversidade, biologia da conservação, entomologia florestal,
glmmADMB, serviços ecossistêmicos.

50
INTRODUÇÃO
A fragmentação, degradação e a perda de habitat observadas em todas as regiões
do globo é um reflexo negativo das atividades antrópicas. No Brasil a Mata Atlântica foi
reduzida a “ilhas” esparsas de vegetação ao longo do tempo, sendo que a maioria delas
se encontra bastante perturbadas pela ação antrópica (Fundação SOS Mata Atlântica e
INPE, 2011; Oliveira-Filho e Machado, 1993; Teixeira et al., 2009). Como reflexo
dessa perturbação, grande parte desses fragmentos de Mata Atlântica se tornaram
mosaicos vegetacionais com diferentes graus de impacto e estágios sucessionais
(Teixeira et al., 2009; Werneck et al., 2001, 2000).
Com a crescente conscientização da importância da preservação e conservação
dos remanescentes florestais, entender a influência da mudança massiva dos habitats nas
comunidades biológicas, especialmente nas espécies que participam ativamente dos
processos ecossistêmicos, se tornou extremamente importante para o desenvolvimento
de estratégias que visam à proteção da biodiversidade desses remanescentes florestais.
Os besouros longicórneos (Coleoptera: Cerambycidae) desempenham papéis
importantes no ecossistema florestal. Eles participam ativamente do processo de
coevolução entre a flora e a comunidade de insetos, exercendo papéis importantes como
o de polinizadores (Makino et al., 2007; Meng et al., 2013). Grande parte desses
besouros são insetos saproxílicos (Speight, 1989; Stokland et al., 2012) e estão
diretamente associados a disponibilidade e condições da madeira presente no habitat
(Grove, 2002a; Hanks, 1999; Hjältén et al., 2012; Lachat et al., 2006; Siitonen, 2001;
Stokland et al., 2012). Esses besouros utilizam árvores e troncos mortos como recursos
e são importantes componentes ecológicos da comunidade de insetos que contribuem
para a degradação de madeira morta, contribuindo assim para a ciclagem de nutrientes
(Buse et al., 2008; Cobb et al., 2010).

51
Os besouros longicórneos possuem o comportamento de serrar galhos e árvores
para torná-los adequados a nidificação, afetando assim a arquitetura da planta
hospedeira (Duval e Whitford, 2008; Feller, 2002) e a dinâmica de clareiras na floresta.
Suas larvas são, na grande maioria, xilófagas e broqueiam a madeira fazendo galerias
nela. Esses hábitos comportamentais alteram fisicamente o ambiente e proporcionam
novos habitats para outros organismos, assim esses besouros são considerados
“engenheiros de ecossistemas” (Buse et al., 2008; Calderón-Cortés et al., 2011).
Estudos mostram que as interações mediadas por besouros longicórneos engenheiros
são fatores que aumentam a riqueza e a estrutura da comunidade de artrópodes
brocadores (Calderón-Cortés et al., 2011).
Estudos mostram que a diversidade dos besouros longicórneos é afetada por
muitos distúrbios diferentes, como por exemplo: a fragmentação e destruição do habitat
(Baur et al., 2005, 2002; Collinge et al., 2001), o fogo (Muona e Rutanen, 1994; Saint-
Germain et al., 2004), diferentes práticas de manejos florestais (Grove, 2002b; Hjältén
et al., 2012; Lachat et al., 2006; Maeto et al., 2002; Meng et al., 2013; Taki et al., 2010;
Vodka et al., 2009) e plantas invasoras (Sugiura et al., 2009b). Grande parte desses
trabalhos mostram que florestas primárias não perturbadas apresentam maior
diversidade de besouros longicórneos quando comparadas à florestas secundárias e
plantações artificiais (Meng et al., 2013).
Assim, este trabalho teve como intuito mostrar a influência da heterogeneidade
estrutural de três áreas de uma floresta, em diferentes estágios sucessionais, sobre a
comunidade de besouros longicórneos. O principal objetivo foi responder se a riqueza, a
abundância e a dissimilaridade faunística dessa comunidade aumentam com o estágio
sucessional e com a heterogeneidade estrutural do habitat.

52
METODOLOGIA
Área de estudo. As coletas foram realizadas na Estação Ecológica do Tripuí
(EET), um fragmento de Mata Atlântica Estacional Semidecídua Montana, com uma
área de 337 ha, localizado na cidade de Ouro Preto, zona metalúrgica do estado de
Minas Gerais, Brasil (20°23’45” S e 43°34’33” W) (Fig. 2.1). Encontra-se na sub-bacia
do córrego Tripuí, e possui um gradiente altitudinal variando entre 1280 e 1450 m.
Segundo a classificação de Köppen, seu clima é Cwb, com verão temperado e o inverno
seco, com um índice pluviométrico anual próximo de 1.600 mm, e temperatura média
anual de 18ºC. Seus solos são litólicos, de natureza álica com textura areno-siltosa
(Alvares et al., 2013; Werneck et al., 2001, 2000).
Na EET foram escolhidas três áreas, já estudadas previamente por Werneck et
al. (2000), que sofreram diferentes impactos no passado e agora se encontram em
diferentes estágio sucessionais. Uma dessas áreas é conhecida como Repolheiro, que
corresponde ao trecho de floresta mais preservado da EET, onde não há relatos de
perturbação antrópica ou fogo nos últimos 45 anos; outra área é conhecida como
Macacos, que corresponde a um trecho intermediário de floresta na EET onde não há
relato de retirada de madeira ou fogo nos últimos 45 anos, mas que nos últimos anos
vem sofrendo perturbação sistêmica por equinos de moradores da EET e de fazendas
vizinhas; e uma área conhecida como Chá, onde havia uma plantação de chá preto
(Camellia sinensis (L.) Kuntze) que foi abandonada à aproximadamente 55 anos e
corresponde ao trecho da floresta em estágio sucessional mais jovem da EET (Werneck
et al., 2000).
Coleta de Besouros Longicórneos. Os besouros foram coletados através de
armadilhas luminosas nas três áreas de estudo durante os meses que compreenderam o
ápice da estação chuvosa, de outubro de 2014 a janeiro de 2015. Foram realizadas duas

53
campanhas de coleta por mês, sendo que cada campanha correspondeu a três noites de
coleta consecutivas, totalizando assim seis noites de coleta por mês. A armadilha
luminosa utilizada para a coleta foi a modelo Luiz de Queiroz, com fonte luminosa
fluorescente ultravioleta de 15 W e 100 V, fabricada pela empresa Biocontrole®. O
funcionamento das armadilhas é crepuscular noturno–vespertino, entre 18–06 hs. As
armadilhas foram posicionadas a 1,80 m de altura do solo para permitir uma maior
dispersão de luz (De Paula e Ferreira, 2000, 1998). Os insetos foram sacrificados com
acetato de etila ainda no campo. No laboratório, os espécimes coletados foram
montados e identificados até o menor nível taxonômico possível. Todos os espécimes
coletados estão depositados na coleção entomológica do Laboratório de Entomologia
Ecológica do Departamento de Biodiversidade, Evolução e Meio Ambiente da
Universidade Federal de Ouro Preto.
Estrutura da vegetação. Para caracterizar a estrutura da vegetação nas
diferentes áreas da floresta foram demarcadas 5 parcelas de 20x20 m em cada uma das
áreas descritas anteriormente, totalizando assim 2000 m² por área e 6000 m² no total.
Em cada parcela, todas as árvores vivas e mortas, troncos mortos caídos no solo, com
diâmetro a altura do peito (DAP) superior a 2,5 cm tiveram sua circunferência a altura
peito (CAP) medidos.
Ainda dentro de cada parcela uma foto hemisférica foi tomada com o intuído de
medir a abertura do dossel em cada área descrita anteriormente. Portanto, foram
tomadas cinco fotos hemisféricas por área, totalizando 15 fotos em todo o trabalho. As
fotos foram tiradas no mês de janeiro de 2015 (estação chuvosa), com o auxílio de um
tripé posicionado a 0,50 m do solo e a câmera fotográfica sempre posicionada para o
Norte, durante os momentos crepusculares do dia, evitando exposição direta de luz
solar. As lentes utilizadas foram as “olhos de peixe Panasonic® 8 mm” acoplada a uma

54
câmera modelo Panasonic® Lumix DMC-LX5. As fotografias foram analisadas com o
auxílio do Gap Light Analyzer software (Frazer et al., 1999).
Em cada área da EET foi mensurada a produção de serapilheira, para isso, um
coletor medindo de 1 m por 0,50 m foi instalado em cada parcela. As coletas foram
mensais ao longo da estação chuvosa, de outubro de 2014 à janeiro de 2015. A
serrapilheira coletada foi ensacada, etiquetada e levada ao laboratório. O material foi
separado em quatro categorias: (1) Material Reprodutivo, (2) Folha, (3) Galhos e (4)
não identificado, para posteriormente ser secado em estufa de ventilação forçada à
temperatura de 60ºC durante 5 dias e pesado em balança de precisão (Nassar et al.,
2008).
Análise Estatística. Para analisar a heterogeneidade ambiental, os parâmetros
fitofisionômicos, densidade de árvores vivas e mortas e seus respectivos CAP, troncos
mortos e seu respectivo CAP, abertura do dossel e produção de serapilheira foram
comparadas entre as áreas através da Análise de Variância (ANOVA). Para os
parâmetros que apresentaram variância significativa, foi realizado uma análise de
múltipla comparação (Teste de Tukey) para descobrir a real diferença dos parâmetros
entre as áreas. Todas as análises foram seguidas pela inspeção de resíduos para checar
as suposições dos modelos.
Para analisar a riqueza e a abundância de espécies de besouros longicórneos
entre as três áreas estudadas, os dias de coleta amostrados repetidamente foram
utilizados como unidades amostrais e são medidas repetidas no tempo dependentes entre
si. Nesse caso, como o desenho experimental é nitidamente longitudinal, foram
ajustados modelos mistos. A riqueza e abundância total de besouros longicórneos
foram as variáveis respostas, as três áreas foram considerados fatores fixos e os meses
como fator categórico aleatório. Primeiramente, foram ajustados modelos GLMM com

55
erros Poisson utilizando o pacote “lme4” no R (Bates et al., 2014). Esses modelos
apresentaram sobredispersão e como não há indicação de um padrão não linear, em um
segundo momento, foi aplicado um modelo GLMM com erros binomiais negativos que
pode ser utilizado para dados de contagem sobredispersos. A modelagem para GLMM
com efeitos aleatórios é limitada. Assim, foi utilizada a técnica de aproximação de
Laplace para otimização numérica disponível no pacote ‘glmmADMB’ (Fournier et al.,
2012). Após verificação dos dois modelos pelo valor de AIC, o modelo glmmADMB
com erros binomiais negativos obteve o melhor ajuste e foi escolhido para realizar as
análises.
Para analisar a composição faunística foi utilizado o índice de dissimilaridade de
Bray-Curtis (Legendre e Legendre, 1998; Quinn e Keough, 2002). Para cada região o
índice de dissimilaridade de Bray-Curtis foi usado para construir uma matriz de
dissimilaridade (matriz de distância ecológica) baseada na composição de espécies de
besouros longicórneos (Kindt e Coe, 2005; Legendre e Legendre, 1998). A distância de
Bray-Curtis foi calculada entre cada par de sítios de coleta. O índice de distâncias
euclidianas foi usado para construir uma matriz de dissimilaridade (matriz de distâncias
ambiental) baseada nas diferenças dos parâmetros ambientais analisados (densidade e
CAP de árvores vivas e mortas e dos troncos mortos, e a produção de serrapilheira). A
distância euclidiana foi calculada entre cada par de sítios de coleta. Para responder se a
dissimilaridade faunística de besouros longicórneos varia com a distância
ambiental entre as três áreas estudadas foi utilizado análises de correlação de Mantel,
seguindo a rotina proposta por Kindt e Coe (2005) no pacote ‘BiodiversityR’ (Kindt e
Coe, 2005) e ‘vegan’ (Oksanen et al., 2013). A correlação de Mantel (Kindt e Coe,
2005; Legendre e Legendre, 1998) foi calculada e sua significância obtida através da

56
permutação entre os sítios. Todas as análises foram realizadas no programa estatístico R
(R Core Team 2014).
57
Figura 2.1. Localização geográfica da Estação Ecológica do Tripuí, Ouro Preto, MG, Brasil, e a disposição dos três pontos amostrais.
58
RESULTADOS
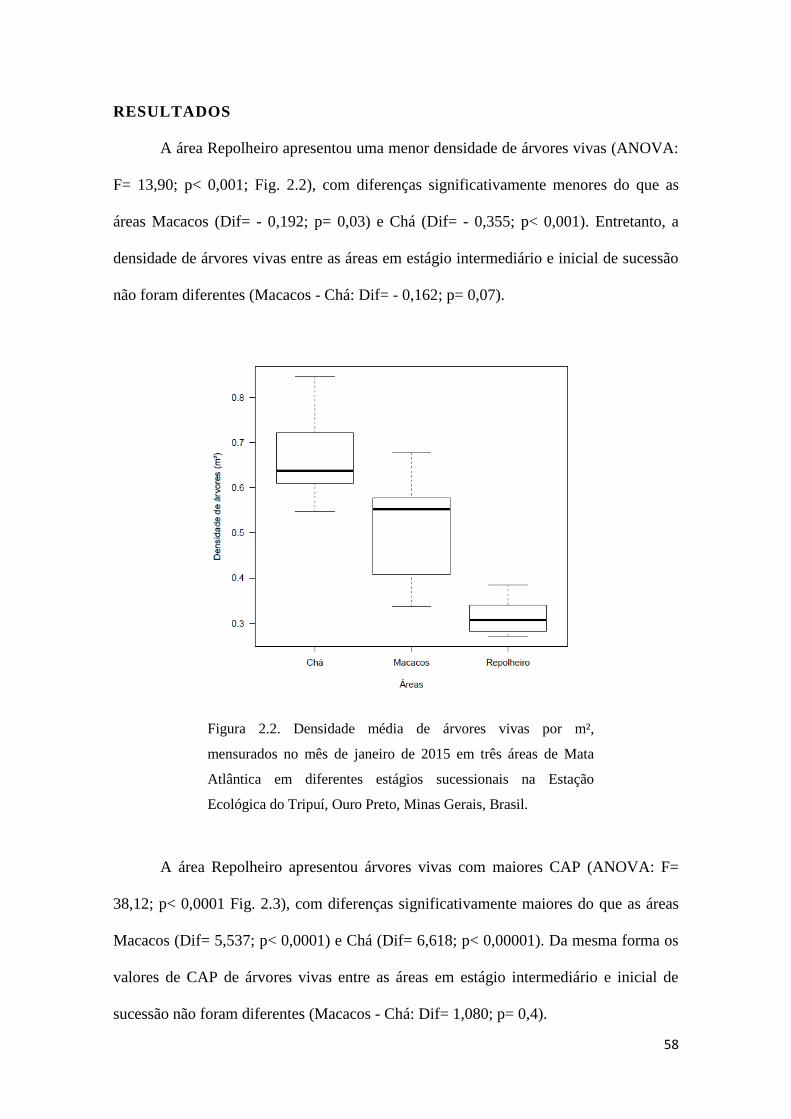
A área Repolheiro apresentou uma menor densidade de árvores vivas (ANOVA:
F= 13,90; p< 0,001; Fig. 2.2), com diferenças significativamente menores do que as
áreas Macacos (Dif= - 0,192; p= 0,03) e Chá (Dif= - 0,355; p< 0,001). Entretanto, a
densidade de árvores vivas entre as áreas em estágio intermediário e inicial de sucessão
não foram diferentes (Macacos - Chá: Dif= - 0,162; p= 0,07).
Figura 2.2. Densidade média de árvores vivas por m²,
mensurados no mês de janeiro de 2015 em três áreas de Mata
Atlântica em diferentes estágios sucessionais na Estação
Ecológica do Tripuí, Ouro Preto, Minas Gerais, Brasil.
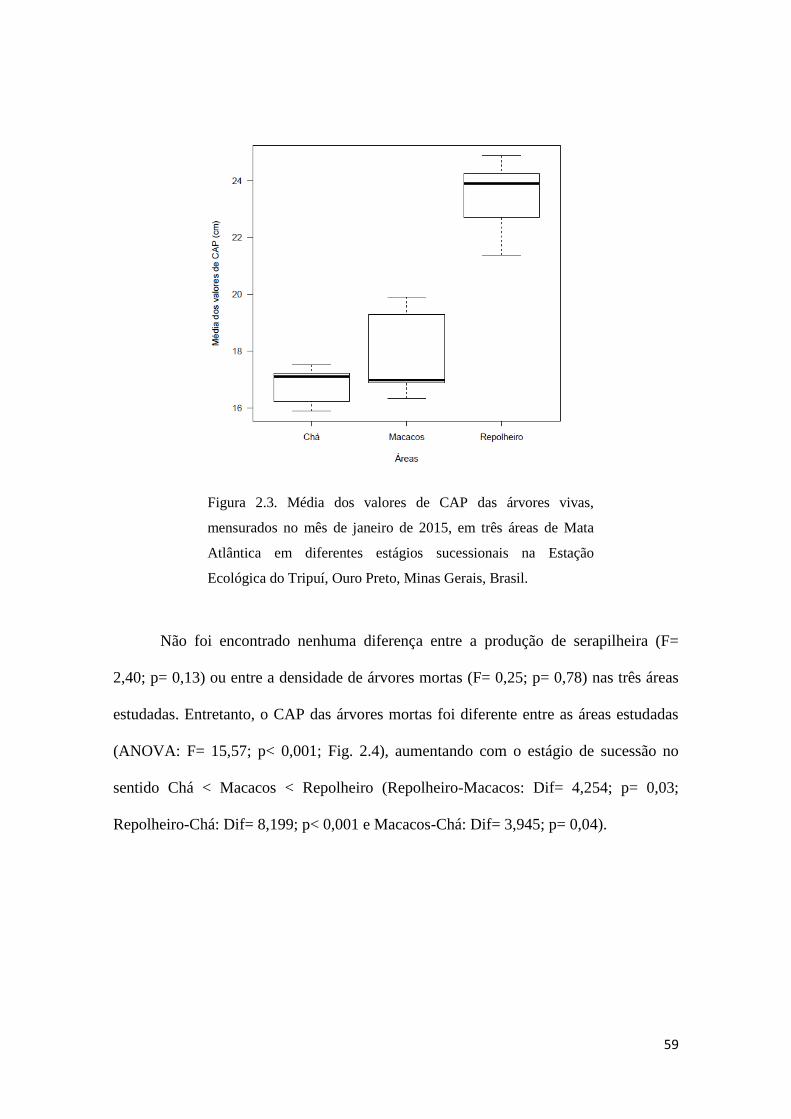
A área Repolheiro apresentou árvores vivas com maiores CAP (ANOVA: F=
38,12; p< 0,0001 Fig. 2.3), com diferenças significativamente maiores do que as áreas
Macacos (Dif= 5,537; p< 0,0001) e Chá (Dif= 6,618; p< 0,00001). Da mesma forma os
valores de CAP de árvores vivas entre as áreas em estágio intermediário e inicial de
sucessão não foram diferentes (Macacos - Chá: Dif= 1,080; p= 0,4).
59
Figura 2.3. Média dos valores de CAP das árvores vivas,
mensurados no mês de janeiro de 2015, em três áreas de Mata
Atlântica em diferentes estágios sucessionais na Estação
Ecológica do Tripuí, Ouro Preto, Minas Gerais, Brasil.
Não foi encontrado nenhuma diferença entre a produção de serapilheira (F=
2,40; p= 0,13) ou entre a densidade de árvores mortas (F= 0,25; p= 0,78) nas três áreas
estudadas. Entretanto, o CAP das árvores mortas foi diferente entre as áreas estudadas
(ANOVA: F= 15,57; p< 0,001; Fig. 2.4), aumentando com o estágio de sucessão no
sentido Chá < Macacos < Repolheiro (Repolheiro-Macacos: Dif= 4,254; p= 0,03;
Repolheiro-Chá: Dif= 8,199; p< 0,001 e Macacos-Chá: Dif= 3,945; p= 0,04).

60
Figura 2.4. Média dos valores de CAP das árvores mortas,
mensurados no mês de janeiro de 2015, em três áreas de Mata
Atlântica em diferentes estágios sucessionais na Estação
Ecológica do Tripuí, Ouro Preto, Minas Gerais, Brasil.
A densidade de troncos mortos caídos no solo (ANOVA: F= 8,068; p< 0,01; Fig.
2.5) foi maior na área Repolheiro do que nas áreas Macacos (Dif= 0,030; p= 0,01) e Chá
(Dif= 0,033; p< 0,01). Entretanto, não foi encontrada nenhuma variação entre as áreas
Macacos e Chá (Dif= 0,002; p= 0,9). O mesmo padrão foi encontrado para os valores de
CAP dos troncos caídos no solo (ANOVA: F= 8,541; p< 0,01; Fig. 2.6), ou seja, o
Repolheiro apresentou troncos caídos no solo mais grossos do que as outras áreas
(Repolheiro-Chá: Dif= 8,000; p< 0,01; Repolheiro-Macacos: Dif= 5,845; p= 0,03), e
esse parâmetro não variou entre as áreas Macacos e Chá (Dif= 2,155; p= 0,54).

61
Figura 2.5. Densidade média de troncos mortos por m²,
mensurados no mês de janeiro de 2015 em três áreas de Mata
Atlântica em diferentes estágios sucessionais na Estação
Ecológica do Tripuí, Ouro Preto, Minas Gerais, Brasil.

62
Figura 2.6. Média dos valores de CAP dos troncos mortos,
mensurados no mês de janeiro de 2015, em três áreas de Mata
Atlântica em diferentes estágios sucessionais na Estação
Ecológica do Tripuí, Ouro Preto, Minas Gerais, Brasil.
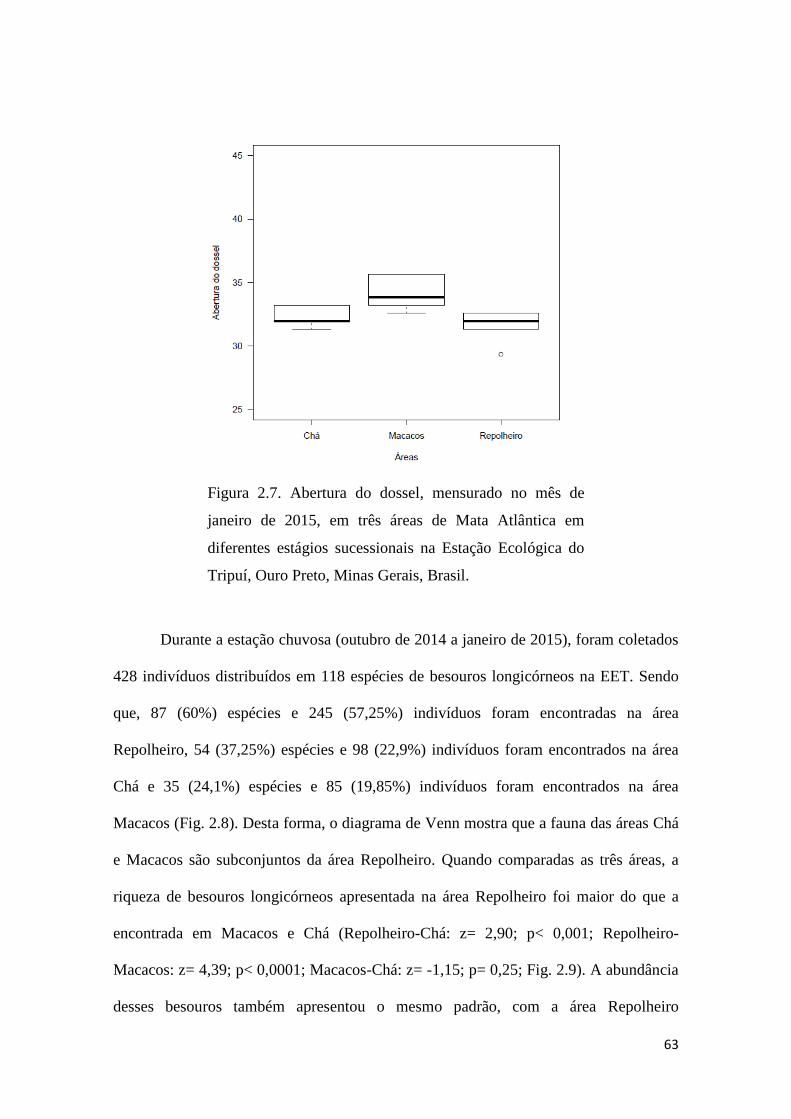
A abertura do dossel apresentou diferenças entre as três áreas estudadas
(ANOVA: F= 6,083; p< 0,01; Fig. 2.7). O dossel da área Repolheiro foi
significativamente mais fechado (Dif= - 2,642; p= 0,01) do que o dossel em Macacos,
mas não foi encontrada diferença entre as áreas Repolheiro e Chá (Dif= - 0,774; p=
0,59), ou entre a áreas Macacos e Chá (Dif= 1,867; p= 0,08).
63
Figura 2.7. Abertura do dossel, mensurado no mês de
janeiro de 2015, em três áreas de Mata Atlântica em
diferentes estágios sucessionais na Estação Ecológica do
Tripuí, Ouro Preto, Minas Gerais, Brasil.
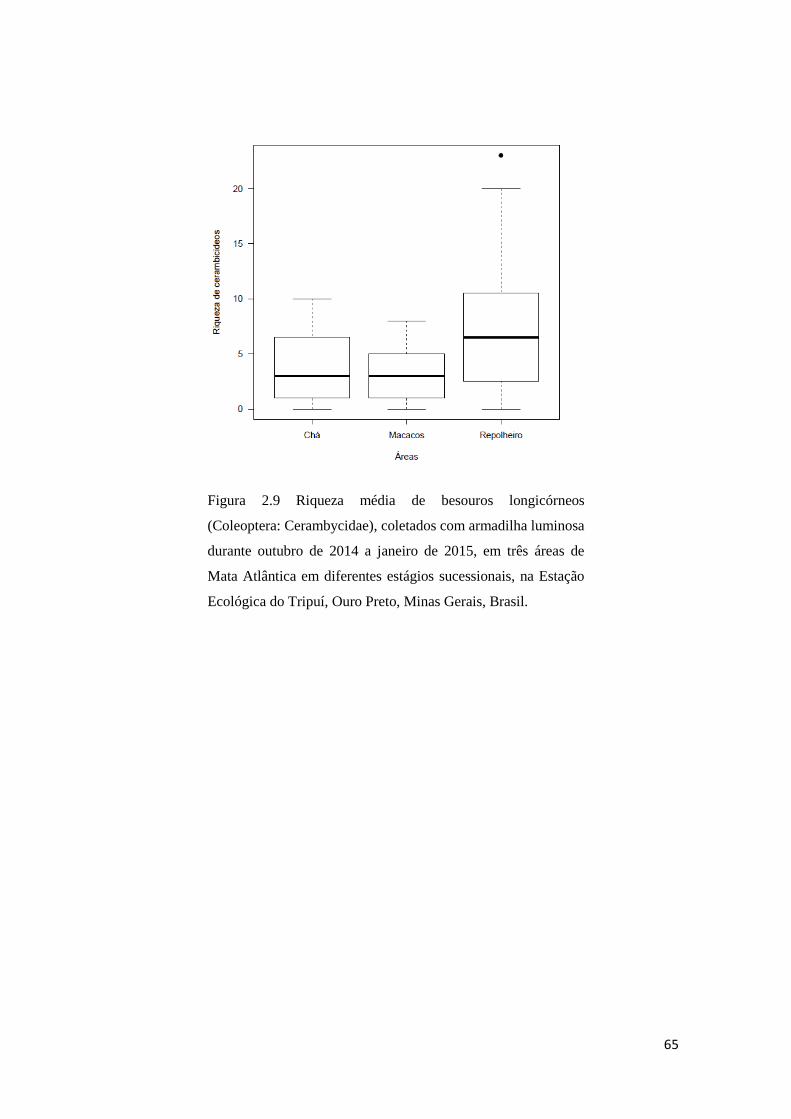
Durante a estação chuvosa (outubro de 2014 a janeiro de 2015), foram coletados
428 indivíduos distribuídos em 118 espécies de besouros longicórneos na EET. Sendo
que, 87 (60%) espécies e 245 (57,25%) indivíduos foram encontradas na área
Repolheiro, 54 (37,25%) espécies e 98 (22,9%) indivíduos foram encontrados na área
Chá e 35 (24,1%) espécies e 85 (19,85%) indivíduos foram encontrados na área
Macacos (Fig. 2.8). Desta forma, o diagrama de Venn mostra que a fauna das áreas Chá
e Macacos são subconjuntos da área Repolheiro. Quando comparadas as três áreas, a
riqueza de besouros longicórneos apresentada na área Repolheiro foi maior do que a
encontrada em Macacos e Chá (Repolheiro-Chá: z= 2,90; p< 0,001; Repolheiro-
Macacos: z= 4,39; p< 0,0001; Macacos-Chá: z= -1,15; p= 0,25; Fig. 2.9). A abundância
desses besouros também apresentou o mesmo padrão, com a área Repolheiro

64
apresentando maior abundância do que Macacos e Chá (Repolheiro-Chá: z= 3,50; p<
0,001; Repolheiro-Macacos: z= 4,04; p< 0,001; Macacos-Chá: z= -0,81; p= 0,4; Fig.
2.10), sendo que entre essas duas últimas áreas não houve diferença significativa.
Figura 2.8. Diagrama de Venn mostrando o número de espécies
de besouros longicórneos (Coleoptera: Cerambycidae) totais,
exclusivas e compartilhadas entre as três áreas estudadas,
coletados entre outubro de 2014 e janeiro de 2015 na Estação
Ecológica do Tripuí, Ouro Preto, Brasil.
65
Figura 2.9 Riqueza média de besouros longicórneos
(Coleoptera: Cerambycidae), coletados com armadilha luminosa
durante outubro de 2014 a janeiro de 2015, em três áreas de
Mata Atlântica em diferentes estágios sucessionais, na Estação
Ecológica do Tripuí, Ouro Preto, Minas Gerais, Brasil.

66
Figura 2.10. Abundância média de besouros longicórneos
(Coleoptera: Cerambycidae), coletados com armadilha luminosa
durante outubro de 2014 a janeiro de 2015, em três áreas de
Mata Atlântica em diferentes estágios sucessionais, na Estação
Ecológica do Tripuí, Ouro Preto, Minas Gerais, Brasil.
A composição de espécies foi diferente entre as áreas estudadas, com valores dos
índices de Bray-Curtis superiores a 0,60 (Chá-Macacos= 0,60; Chá-Repolheiro = 0,67;
Macacos-Repolheiro= 0,72). A área Repolheiro apresentou 55 e 68 espécies exclusivas
em relação às áreas Chá e Macacos, respectivamente (Fig. 2.8). Entretanto, a
dissimilaridade não variou com a distância ambiental entre os pares de áreas estudados
(Mantel: r= 1,00; p= 0,17).

67
DISCUSSÃO
Os parâmetros fitofisionômicos mostraram que a área Repolheiro apresentou
características de florestas em estágio de sucessão mais avançado em relação as áreas
Macacos e Chá. A menor densidade de árvores (Fig. 2.2), porém, sendo essas mais
grossas (Fig. 2.3 e 2.4), é um forte indicativo de que a floresta na área Repolheiro está
em um estágio mais maduro de desenvolvimento (Bauhus et al. 2009; DeWalt et al.,
2003; Finegam, 1996; Guariguata e Ostertag, 2001; Pascarella et al. 2004; Quesada et
al. 2009). O Repolheiro também apresentou um maior aporte de madeira morta caída no
solo (Fig. 2.5 e 2.6). Em ecossistemas florestais naturais, a biomassa de madeira morta é
considerada uma característica estrutural importante, mantida naturalmente através dos
processos de sucessão e distúrbios naturais, que ajuda a criar uma diversidade de
habitats proporcionando condições adequadas de vida para muitos organismos, entre
eles os besouros longicórneos (Bauhus et al., 2009; Burrascano et al., 2008; Grove,
2002a, 2002b, 2002c; Hjältén et al., 2012; Lachat et al., 2006; Siitonen, 2001;
Stenbacka et al., 2010).
Apesar das diferenças entre as metodologias limitarem as comparações,
Werneck et al. (2000) avaliando a fitofisionomia das mesmas áreas aqui estudadas
encontraram resultados que suportavam a proximidade entre as áreas Repolheiro e
Macacos, distanciando-as da área Chá, que se encontrava em um estágio inicial de
sucessão. Entretanto, passados 15 anos desse estudo, os parâmetros fitofisionômicos da
área Macacos, com exceção da produção de serapilheira e a densidade de árvores
mortas, foram significativamente diferentes dos parâmetros da área Repolheiro, e iguais,
com exceção do CAP das árvores mortas, aos da área Chá. Esses resultados sugerem
que desde o estudo de Werneck et al., (2000) a área Macacos vem sofrendo algum tipo
de impacto que está retardando o seu desenvolvimento sucessional. Essa região

68
específica da EET (ver Fig. 2.1) é invadida sistematicamente por equinos de fazendas
vizinhas e de moradores que ainda ocupam a EET. As atividades de grandes herbívoros
causam impactos negativos muito fortes sobre o ambiente. Eles alteram a
heterogeneidade espacial da vegetação, influenciando assim os processos
ecossistêmicos e a biodiversidade (Adler et al., 2001; Olff e Ritchie, 1998). O pastejo
aumenta o número de espécies dominantes, diminuindo assim a diversidade de plantas.
Em áreas que apresentam árvores e arbustos, ele impede o desenvolvimento do sub-
bosque, pois limita o crescimento da biomassa aérea da planta (Milchunas e Lauenroth,
1993), diminuindo também a produção de material reprodutivo por essas plantas
(Gomez e Zamora, 2000). Esses grandes herbívoros também comem e pisoteiam
plântulas e brotos, impedindo o seu desenvolvimento natural. Todos esses impactos
causados pelos equinos, provavelmente, estão contribuindo para retardar o
desenvolvimento sucessional da área (Gibson e Brown, 1992).
A assembleia de besouros longicórneos do Repolheiro representa cerca de 60%
das espécies da EET e, conforme indicado no diagrama de Venn (Fig. 2.8), as demais
áreas estudadas são subconjuntos dessa assembleia. A maior riqueza e abundância de
besouros encontrada no Repolheiro (Fig. 2.9 e 2.10) pode ser explicada pela
heterogeneidade e disponibilidade de recursos, nesse caso a madeira. A madeira é um
recurso fundamental para os besouros longicórneos, usada tanto para alimentação
quanto para a nidificação. Os besouros adultos escolhem minuciosamente o local de
nidificação, levando em consideração a sobrevivência, saúde e disponibilidade da
madeira que vai ser hospedeira de suas larvas, pois essas são completamente
dependentes desse recurso para se desenvolverem (Hanks, 1999). O trabalho realizado
por Maeto et al. (2002), mostrou que a riqueza de besouros longicórneos aumenta com o
tamanho máximo do diâmetro das árvores, e que normalmente essas árvores estão

69
presentes em florestas mais antigas. Outros trabalhos sugerem que a disponibilidade de
madeira morta pode aumentar a diversidade desses besouros nas florestas (Grove,
2002b; Lachat et al., 2006; Raje et al., 2012; Stenbacka et al., 2010). Como vimos, a
área Repolheiro apresentou tanto árvores com maiores diâmetros (Fig. 2.3 e 2.4) quanto
maior disponibilidade de madeira morta (Fig. 2.4, 2.5 e 2.6) do que as outras áreas.
Além disso, os estágios sucessionais mais avançados podem apresentar um
maior número de espécies de plantas arbóreas hospedeiras, enquanto o número de
espécies de plantas arbustivas e de lianas diminuem (Capers et al., 2005; González-
Iturbe et al., 2002; Guariguata et al., 1997; Madeira et al., 2009; Peña-Claros e de Boo,
2002; Weaver, 2000). Por exemplo, Meng et al. (2013) mostrou que a riqueza e
abundância de besouros longicórnoes foram positivamente relacionadas com a maior
riqueza de árvores encontradas em florestas mais antigas. A riqueza de plantas é um
importante componente da heterogeneidade do habitat para insetos (Strong et al., 1984),
e aumenta per se o número de nichos disponíveis para espécies especialistas, e cria um
ambiente físico estruturalmente mais heterogêneo em função das diferentes partes
aéreas das espécies de plantas hospedeiras (Strong et al. 1984). Desta forma, é esperado
uma relação positiva entre a riqueza desses besouros e a riqueza de plantas arbóreas
hospedeiras.
Apesar da composição entre as espécies de besouros longicórneos ter variado
entre as áreas (Bray-Curtis, Chá-Macacos= 0,60; Chá-Repolheiros = 0,67; Macacos-
Repolheiro= 0,72), essa dissimilaridade não foi explicada pela distância ambiental. Esse
fato pode ser explicado pelo pequeno número de áreas estudadas, dificultando a
obtenção de uma correlação significativa.
Esses resultados mostraram que a heterogeneidade estrutural do habitat foi
determinante para a composição da comunidade de besouros longicórneos na Mata

70
Atlântica. Mostrando que remanescentes florestais mais antigos são elementos-chave
para a manutenção da diversidade desses besouros, pois aportam maior riqueza e
abundância dessa família de insetos. E justamente por estarem intimamente associados
aos componentes estruturais de florestas mais antigas e serem fáceis de amostrar, a
diversidade de besouros longicórneos pode ser um valioso indicador do estado de
conservação dos remanescentes florestais (Maeto et al., 2002; Meng et al., 2013) e da
comunidade de outros organismos (Buse et al., 2008; Calderón-Cortés et al., 2011).

71
Conclusão
Os resultados do primeiro capítulo mostraram que a EET apresenta uma grande
diversidade de besouros longicórneos. Quando comparada com trabalhos similares, essa
diversidade foi maior que a encontrada em Viçosa, Ilha Grande e Porto Velho. Entretanto, regiões
onde trabalhos foram realizados a longo prazo apresentaram riqueza muito maior que a observada
na EET. A riqueza e a abundância de besouros longicórneos apresentaram grandes flutuações
sazonais, atingindo seu máximo amostral nos meses de novembro e dezembro de 2014, período
que corresponde ao ápice da estação chuvosa e a primavera no hemisfério sul. Este trabalho
mostrou que inventários faunísticos de grupos hiperdiversos, realizados com apenas um ano de
coleta, podem subestimar a riqueza de espécies de uma região. Assim, séries temporais maiores
associadas a mais métodos de coletas são essenciais para uma amostragem mais precisa. No segundo capítulo, os resultados mostraram que a área Repolheiro apresentou
características fitofisionômicas de uma área em estágio mais avançado de sucessão em
relação às demais áreas estudadas. Esse fato refletiu na comunidade de besouros
longicórneos, pois, nessa área se observou maior riqueza e abundância desses besouros.
Este trabalho mostrou a importância de florestas mais preservadas para a manutenção da
diversidade dos besouros longicórneos, mostrando também que essa íntima relação com
a estrutura da floresta torna o grupo um bom indicador para a avaliação da
biodiversidade e qualidade do habitat.

72
REFERÊNCIAS
Adler, P., Raff, D., Lauenroth, W., 2001. The effect of grazing on the spatial
heterogeneity of vegetation. Oecologia 128, 465-479.
Alvares, C.A., Stape, J.L., Sentelhas, P.C., de Moraes, G., Leonardo, J., Sparovek, G.,
2013. Köppen's climate classification map for Brazil. Meteorol. Z. 22, 711-728.
Barreto, M.R., Machiner, R., Smiderle, E.C., 2013. Cerambycinae (Coleoptera,
Cerambycidae) in Mato Grosso, Brazil. Biota Neotrop. 13, 331-335.
Basset, Y., Novotny, V., 1999. Species richness of insects herbivores communities on
Ficus in Papua New Guinea. J. Linn. Soc. 67, 477-499.
Bates, D., Maechler, M., Bolker, B.M., Walker, S., 2014. “lme4: Linear mixed-effects
models using Eigen and S4.” ArXiv e-print; submitted to _Journal of Statistical
Software, URL: http://arxiv.org/abs/1406.5823.
Bauhus, J., Puettmann, K., Messier, C., 2009. Silviculture for old-growth attributes. For.
Ecol. Manag. 258, 525-537.
Baur, B., Coray, A., Minoretti, N., Zschokke, S., 2005. Dispersal of the endangered
flightless beetle (Coleoptera: Cerambycidae) in spatially realistic landscapes. Biol.
Conser. 124, 49-61.
Baur, B., Zschokke, S., Coray, A., Schlapfer, M., Erhardt, A., 2002. Habitat
characteristics of the endangered flightless beetle Dorcadion fuliginator
(Coleoptera: Cerambycidae): implications for conservation. Biol. Conser. 105, 133-
142.
Beeson, C.F.C., Bhatia, B.M., 1939. On the Biology of the Cerambycidae (Coleopt.).
Ind. For. Rec. 5, 1-235.

73
Brown, K.S., 1997a. Insetos como rápidos e sensíveis indicadores de uso sustentável de
recursos naturais. In: Martos, H.L., Maia, N.B. (Eds.), Indicadores Ambientais.
Sorocaba, PUC/SP, pp. 143-155.
Brown, K.S., 1997b. Diversity, disturbance, and sustainable use of Neotropical forests:
insects as indicators for conservation monitoring. J. Insect Conserv. 1, 25-42.
Bullock, S.H., Solís-Magallanes, A., 1990. Phenology of canopy trees of a tropical
deciduous forest in Mexico. Biotropica 22, 22-35.
Burrascano, S., Lombardi, F., Marchetti, M., 2008. Old-growth forest structure and
deadwood: Are they indicators of plant species composition? A case study from
central Italy. Plant Biosyst. 142, 313-323.
Buse, J., Ranius, T., Assmann, T., 2008. An endangered longhorn beetle associated with
old oaks and its possible role as an ecosystem engineer. Conserv. Biol. 22, 329-337.
Calderón-Cortés, N., Quesada, M., Escalera-Vázquez, L.H., 2011. Insects as Stem
Engineers: Interactions Mediated by the Twig-Girdler Oncideres albomarginata
chamela Enhance Arthropod Diversity. PLoS ONE 6, e19083.
Capers, R.S., Chazdon, R.L., Brenes, A.R., Alvarado, B.V., 2005. Successional
dynamics of woody seedling communities in wet tropical secondary forests. J. Ecol.
93, 1071-1084.
Chao, A., 1984. Non-parametric estimation of the number of classes in a population.
Scand. J. Stat. 11, 265-270.
Chao, A., 1987. Estimating the population size for capture–recapture data with unequal
catchability. Biometrics 43, 783-791.
Chao, A., Lee, S.M., 1992. Estimating the number of classes via sample coverage. J.
Am. Stat. Assoc. 87, 210-217.

74
Chao, A., Yang, M.C., 1993. Stopping rules and estimation for recapture debugging
with unequal failure rates. Biometrika 80, 193-201.
Cobb, T.P., Hannam, K.D., Kishchuk, B.E., Langor, D.W., Quideau, S.A., Spence, J.R.,
2010. Wood-feeding beetles and soil nutrient cycling in burned forests:
implications of post-fire salvage logging. Agr. Forest Entomol. 12, 9-18.
Collinge, S.K., Holyoak, M., Barr, C.B., Marty J, T., 2001. Riparian habitat
fragmentation and population persistence of the threatened valley elderberry
longhorn beetle in central California. Biol. Conser. 100, 103-113.
Costa Lima, A., 1955. Insetos do Brasil. 3ª Parte. Vol. 9. Escola Nacional de
Agronomia, Rio de Janeiro.
Costa, C., 2000. Estado de conocimiento de los Coleoptera neotropicales. In: Hacia un
proyecto para el inventario y estimación de la diversidad entomológica en
iberoamérica: Pribes 2000. Sociedad Entomológica Aragonesa, pp. 99-114.
De Paula, A.S., Ferreira, P.S.F., 1998. Fauna de Heteroptera de la “Mata do Córrego do
Paraíso”, Viçosa, Minas Gerais, Brasil. I. Riqueza y diversidade específicas. Anales
Inst. Biol. Univ. Nac. Auton. México, Ser. Zool. 69, 39-51.
De Paula, A.S., Ferreira, P.S.F., 2000. Fauna de Heteroptera de la “Mata do Córrego do
Paraíso”, Viçosa, Minas Gerais, Brasil. II. Patrones temporales. Distribuición anual
y estacionalidad. Anales Inst. Biol. Univ. Nac. Autón. México, Ser. Zool. 71, 7-19.
DeWalt, S.J., Maliakal, S.K., Denslow, J.S., 2003. Changes in vegetation structure and
composition along a tropical forest chronosequence: implications for wildlife.
Forest Ecol. Manag. 182, 139-151.
Duval, B.D., Whitford, W.G., 2008. Resource regulation by a twig-girdling beetle has
implications for desertification. Eco. Entomol. 33, 161-166.
Fahrig, L., 2001. How much habitat is enough? Biol. Conserv. 100, 65-74.

75
Fahrig, L., 2003. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol.
Evol. Syst. 34, 487-515.
Feinsinger, P., 2001. Designing field studies for biodiversity conservation. The Nature
Conservancy. Island Press, Washington DC.
Feller, I.C., 2002. The role of herbivory by wood-boring insects in mangrove
ecosystems in Belize. Oikos 97, 167-176.
Finegan, B., 1996. Pattern and process in Neotropical secondary rain forests: The first
100 years of succession. Trends Ecol. Evol. 11, 119–124.
Fournier, D.A., Skaug, H.J., Ancheta, J., Ianelli, J., Magnusson, A., Maunder, M.,
Nielsen, A., Sibert, J., 2012. AD Model Builder: using automatic differentiation for
statistical inference of highly parameterized complex nonlinear models. Optim.
Methods Softw, 27, 233-249.
Fundação SOS Mata Atlântica, INPE, 2011. Atlas dos remanescentes florestais da Mata
Atlântica período 2008–2010. Fundação SOS Mata Atlântica/Instituto Nacional de
Pesquisas Espaciais, São Paulo.
Galileo, M.H.M., Martins, U.R., 2010. New species of Cerambycinae (Coleoptera,
Cerambycidae) from South America. Insecta Mundi 0115, 1-9.
Galileo, M.H.M.; Martins, U.R., Francisco, E.D.L., 2014. Cerambycidae (Coleoptera)
do Parque Nacional da Serra das Confusões, Piauí, Brasil: novas espécies e novos
registros. Iheringia Sér. Zool. 103, 393-397.
Gaston, K.J., 1996. Species richness: measure and measurement. In: Gaston K.J. (Ed.),
Biodiversity: a biology of numbers and difference. Blackwell Science, Cambridge,
UK, pp. 77-113.

76
Gibson, C.W.D., Brown, V.K., 1992. Grazing and vegetation change: deflected or
modified succession? J. Appl. Ecol. 29, 120-131.
Golfari, L., 1975. Zoneamento ecológico do Estado de Minas Gerais para
reflorestamento. Centro de pesquisa florestal da Região do Cerrado,
PNUD/FAO/IBDF, Belo Horizonte.
Gomez, J.M., Zamora, R., 2000. Differential impact of vertebrate and invertebrate
herbivores on the reproductive output of Hormathophylla spinosa. EcoScience 7,
299-306.
Gómez-Pompa, A., Wiechers, B.L., 1979. Regeneracion de los ecossistemas tropicales
y subtropicales. In: Gómez-Pompa, A., Amo, R.S. (Eds.), Investigaciones sobre la
regeneración de las selvas altas en Vera Cruz, México. Companhia Editorial
Continental, México, pp.11-30.
González-Iturbe, J., Olmsted, I., Tun-Dzul, F., 2002. Tropical dry forest recovery after
long term Henequen (sisal, Agave fourcroydes Lem.) plantation in northern
Yucatan, Mexico. For. Ecol. Manage. 167, 67-82.
Grove, S.J., 2002a. Saproxylic insect ecology and the sustainable management of
forests. Annu. Rev. Ecol. Syst. 23, 1-23.
Grove, S.J., 2002b. The influence of forest management history on the integrity of the
saproxylic beetle fauna in an Australian lowland tropical rainforest. Biol. Conser.
104, 149-171.
Grove, S.J. 2002c. Tree basal area and dead wood as surrogate indicators of saproxylic
insect faunal integrity: a case study from the Australian lowland tropics. Ecol.
Indic. 1, 171-188.
Guariguata, M.R., Chazdon, R.L., Denslow, J.S., Dupuy, J.M., Anderson, L., 1997.
Structure and foristics of secondary and old-growth forest stands in lowland Costa
Rica. Plant Ecol. 132, 107-120.

77
Guariguata, M.R., Ostertag, R., 2001. Neotropical secondary forest succession: changes
in structural and functional characteristics. Forest Ecol. Manag. 148, 185-206.
Hanks, L.M., 1999. Influence of the larval host plant on reproductive strategies of
cerambycid beetles. Annu. Rev. Entomol. 44, 483–505.
Hjältén, J., Stenbacka, F., Pettersson, R.B., Gibb, H., Johansson, T., Danell, K., Ball,
J.P., Hilszczański, J., 2012. Micro and macro-habitat associations in saproxylic
beetles: implications for biodiversity management. PLoS ONE 7, e41100.
Holland, J.D., 2007. Sensitivity of Cerambycid biodiversity indicators to definition of
high diversity. Biodivers. Conserv 16, 2599-2609.
Jansson, N., Bergman, K.O., Jonsell, M., Milberg, P., 2009. An indicator system for
identification of sites of high conservation value for saproxylic oak (Quercus spp.)
beetles in southern Sweden. J. Insect Conserv. 13, 399-412.
Kindt, R., Coe, R., 2005. Tree diversity analysis. A manual and software for common
statistical methods for ecological and biodiversity studies. World Agroforestry
Centre (ICRAF), Nairobi. ISBN 92-9059-179-X.
Lachat, T., Nagel, P., Cakpo, Y., Attignon, S., Goergen, G., Sinsin, B., Peveling, R.,
2006. Dead wood and saproxylic beetle assemblages in a semi-deciduous forest in
Southern Benin. Forest Ecol. Manag. 225, 27-38.
Legendre, P.L., Legendre, L.L., 1998. Numerical ecology. Second English Edition,
Amsterdam Elsevier Science.
Lieutier, F., Day K.R., Battisti A., Grégoire J.C., Evans H.F., 2004. Bark and wood
boring insects in living trees in Europe, a synthesis. Springer.
Linsley, E.G., 1959. Ecology of cerambycidae. Annu. Rev. Entomol. 4, 99-138.

78
Longino, J.T., Coddington, J., Colwell, R.K., 2002. The ant fauna of a tropical rain
forest: estimating species richness three different ways. Ecology 83, 689–702.
Madeira, B.G., Espírito-Santo, M.M., D’Ângelo Neto, S., Nunes, Y.R.F., Sánchez-
Azofeifa, A.S., Fernandes, G.W., Quesada, M., 2009. Changes in tree and lianas
communities along a successional gradient in a tropical dry forest in southeastern
Brazil. Plant Ecol. 201, 291-304.
Maeto, K., Sato, S., Miyata, H., 2002. Species diversity of longicorn beetles in humid
warm-temperate forests: the impact of forest management practices on old-growth
forest species in southwestern Japan. Biodivers. Conserv. 11, 1919–1937.
Makino, S., Goto, H., Hasegawa, M., Kimiko Okabe, K., Tanakah, H., Inoue, T.,
Okochi, I., 2007. Degradation of longicorn beetle (Coleoptera, Cerambycidae,
Disteniidae) fauna caused by conversion from broad-leaved to man-made conifer
stands of Cryopomeria japonica (Taxodiaceae) in central Japan. Ecol. Res. 22, 134-
156.
Martínez-Yrízar, A. 1995. Biomass distribution and primary productivity of tropical dry
forests. In: Bullock, S.H., Mooney, H.A., Medina, E. (Eds.), Seasonally dry tropical
forests. Cambridge University Press, New York, pp. 326-245.
Martins, U.R., 1997. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 1.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 1998. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 2.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 1999a. Cerambycidae. In: Joly, C.A., Bicudo, C.E.M. (Eds.)
Biodiversidade do Estado de São Paulo. FAPESP , São Paulo, pp. 279.
Martins, U.R., 1999b. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 3.
Sociedade Brasileira de Entomologia, São Paulo.

79
Martins, U.R., 2002. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 4.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 2003a. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 5.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 2003b. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 6.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 2005. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 7.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 2006. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 8.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 2007. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 9.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 2008. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 10.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 2010. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 11.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., 2011. Cerambycidae Sul-Americanos (Coleoptera). Taxonomia. Vol. 12.
Sociedade Brasileira de Entomologia, São Paulo.
Martins, U.R., Galileo, M.H.M., Limeira-de-Oliveira, F., 2009a. Cerambycidae
(Coleoptera) do estado do Maranhão, Brasil. I. Pap. Avulsos Zool. 49, 229-247.
Martins, U.R., Galileo, M.H.M., Limeira-de-Oliveira, F., 2009b. Cerambycidae
(Coleoptera) do estado do Maranhão, Brasil. II. Pap. Avulsos Zool. 49, 503-527.

80
Martins, U.R., Galileo, M.H.M., Limeira-de-Oliveira, F., 2011. Cerambycidae
(Coleoptera) do estado do Maranhão, Brasil: III. Pap. Avulsos Zool. 51, 275-293.
Martins, U.R., Galileo, M.H.M., Santos-Silva, A., Rafael, J.A., 2006. Cerambycidae
(Coleoptera) coletados à luz a 45 metros de altura, no dossel da floresta amazônica,
e a descrição de quatro espécies novas. Acta Amaz. 36, 265-272.
Meng, L-Z., Martin, K., Weigel, A., Yang, X.D., 2013. Tree Diversity Mediates the
Distribution of Longhorn Beetles (Coleoptera: Cerambycidae) in a Changing
Tropical Landscape (Southern Yunnan, SW China). PLoS ONE 8: 10.
Milchunas, D.G., Lauenroth, W.K., 1993. Quantitative effects of grazing on vegetation
and soils over a global range of environments. Ecol. Monogr. 63, 327-366.
Monné, M.A. 2005. Catalogue of the Cerambycidae (Coleoptera) of the Neotropical
region. Part I. Subfamily Cerambycinae. Zootaxa 946,1‑765.
Monné, M.A., Bezark, L., 2009. Checklist of the Cerambycidae, or longhorned beetles
(Coleoptera) of the Western Hemisphere. Rancho Dominguez, BioQuip
Publications.
Monné, M.L., Monné, M.A., Aragão, A.C., Quintino, H.Y., Botero, J.P., Machado,
V.S., 2010. Inventário das espécies de Lepturinae, Parandrinae e Prioninae (Insecta,
Coleoptera, Cerambycidae) do Parque Nacional do Itatiaia, RJ, Brasil. Biota
Neotrop. 10, 325-335.
Monné, M.L., Monné, M.A., Quintino, H.Y., Botero, J.P., Machado, V.S., Aragão,
A.C., Simões, M.V.P., Cupello, M., 2012. Inventory of the Lamiinae species
(Insecta, Coleoptera, Cerambycidae) of the Parque Nacional do Itatiaia, RJ, Brazil.
Biota Neotrop. 12, 39-76.
Monné, M.L., Monné, M.A., Mermudes, J.R.M., 2009. Inventário das espécies de
Cerambycinae (Insecta, Coleoptera, Cerambycidae) do Parque Nacional do Itatiaia,
RJ, Brasil. Biota Neotrop. 9, 1-30.

81
Morillo, S.I.E., 2007. Biodiversidade e análise faunística de Cerambycidae (Insecta:
Coleoptera) em Reserva de Mata Atlântica, Viçosa, Minas Gerais. Dissertação
(Mestrado Entomologia) – Universidade Federal de Viçosa, Viçosa, Minas Gerais.
Muona, J., Rutanen, I., 1994. The short-term impact of fire on the beetle fauna in boreal
coniferous forest. Ann. Zool. Fenn. 31, 109-109.
Napp, D.S., 1994. Phylogenetic relationships among the subfamilies of Cerambycidae
(Coleoptera, Chrysomeloidea). Rev. Bras. Entomol. 38, 265-419.
Nassar, J., Rodríguez, J.P., Sánchez-Azofeifa, A., Garvin, T., Quesada, M., 2008.
Manual of methods: Human, Ecological and Biophysical Dimensions of Tropical
Dry Forests. IVIC. Caracas, Venezuela.
New, T.R., 1995. Introduction to invertebrate conservation biology. Oxford University
Press.
Novotný, V., Basset, Y., 2000. Rare species in communities of tropical insect
herbivores: pondering the mystery of singletons. Oikos 89, 564-572.
O'hara, R.B., 2005. Species richness estimators: how many species can dance on the
head of a pin? J. Anim. Ecol. 74, 375-386.
Olff, H., Ritchie, M.E., 1998. Effects of herbivores on grassland plant diversity. Tr.
Ecol. Evol. 13, 261-265.
Oliveira-Filho, A.T., Machado, J.N.M., 1993. Composição florística de uma floresta
semidecídua montana, na Serra de São José, Tiradentes, MG. Acta Bot. Bras. 7,71-
88.
Oksanen, J.F., Blanchet, G., Kindt, R.; Legendre, P., Minchin, P.R., O'Hara, R.B.,
Simpson, G.L., Solymos, P., Stevens M.H.H., Helene Wagner, H., 2015. vegan:

82
Community Ecology Package. R package version 2.2-1. http://CRAN.R-
project.org/package=vegan.
Orozco-Santos, M., García-Mariscal, K., Vázquez-Jiménez, J.L., Robles-González, M.,
Velázquez-Monreal, J.J., Manzo-Sánchez, G., Nieto-Ángel, D., 2011. The Long-
jawed Longhorn Beetle (Coleoptera: Cerambycidae) in Tamarid Trees in the Dry
Tropic of Mexico-A Brief Revision. Southwest. Entomol. 36, 197-202.
Paniago, M.D.C., 1983. Evolução histórica e tendências de mudanças sócio-culturais na
comunidade de Viçosa-MG. Dissertação de Mestrado em Educação. Universidade
Federal de Viçosa, Viçosa, MG.
Pascarella, J., Aide, T., Zimmerman, J., 2004. Short-term response of secondary forests
to hurricane disturbance in Puerto Rico, USA. For. Ecol. Manage. 199, 379–393.
Paula, A.D., Silva, A.D., Souza, A.D., Santos, F. D., 2002. Alterações florísticas
ocorridas num período de quatorze anos na vegetação arbórea de uma floresta
estacional semidecidual em Viçosa-MG. Rev. Árvore 26, 743-749.
Pearson, D.L., 1994. Selecting indicator taxa for the quantitative assessment of
biodiversity. Phil. Trans. R. Soc. B. 345, 75-79.
Pires, E.M., Moreira, I., Soares, M.A., Marinho, J.A., Pinto, R., Zanuncio, J.C., 2011.
Oxymerus aculeatus (Coleoptera: Cerambycidae) causing damage on corn plants
(Zea mays) in Brazil. Rev. Colomb. Entomol. 37, 82-83.
Peña-Claros, M., de Boo. H., 2002. The effect of forest successional stage on seed
removal of tropical rain forest tree species. J. Trop. Ecol. 18, 261-274.
Quesada, M., Sanchez-Azofeifa, G.A., Alvarez-Anorve, M., Stoner, K.E.,
Avila-Cabadilla, L., Calvo-Alvarado, J., Espirito-Santo, M.M., Fagundes, M.,
Fernandes, G.W., Gamon, J., Lopezaraiza-Mikel, M., Lawrence, D., Cerdeira
Morellato, L.P., Powers, J.S., Neves, F.S., Rosas-Guerrero, V., Sayago, R.,
Sanchez-Montoya, G., 2009. Succession and management of tropical dry

83
forests in the Americas: review and new perspectives. Forest Ecol. Manag. 258,
1014-1024.
Quinn, G.P., Keough, M.J., 2002. Experimental design and data analysis for biologists.
Cambridge University Press.
R Development Core Team (2012). R: A language and environment for statistical
computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-
900051-07-0, URL http://www.R-project.org/.
Raje, K.R., Abdel-Moniem, H.E.M., Farlee, L., Ferris, V.R., Holland, J.D., 2011.
Abundance of pest and benign Cerambycidae both increase with decreasing forest
productivity. Agr. Forest Entomol. 14, 165-169.
Rodrigues, J.M.S., Monné, M.A., Mermudes, J.R.M., 2010. Inventário das espécies de
Cerambycidae (Coleoptera) de Vila Dois Rios (Ilha Grande, Angra dos Reis, Rio de
Janeiro, Brasil). Biota Neotrop. 10, 311-321.
Saint-Germain, M., Drapeau, P., Hébert, C., 2004. Comparison of Coleoptera
assemblages from a recently burned and unburned black spruce forests of
northeastern North America. Biol. Conserv. 118, 583-592.
Schilling, A.C., Batista, J.L.F., 2008. Curva de acumulação de espécies e suficiência
amostral em florestas tropicais. Rev. Bras. Bot. 31, 179-187.
Schoener, T.W., 1987. The geographical distribution of rarity. Oecologia 74, 161-173.
Siitonen, J. 2001. Forest management, coarse woody debris and saproxylic organisms:
Fennoscandian boreal forests as an example. Ecol. Bull. 49,11-42.
Silveira Neto, S., Monteiro, R.C., Zucchi, R.A., de Moraes, R.C.B., 1995. Uso da
análise faunística de insetos na avaliação do impacto ambiental. Sci. Agric. 52, 9-
15.

84
Souza, D.D.S., Silva, A.D.A., 2012. Cerambycidae (Insecta: Coleoptera) of the Parque
Natural Municipal de Porto Velho, Rondônia, Western Amazon, Brazil. Biota
Neotrop. 12, 237-240.
Speight, M.C.D., 1989. Saproxylic invertebrates and their conservation. Nature and
Environment Series 46, Council of Europe, Strasbourg.
Stenbacka, F., Hjältén, J., Hilszczanski, J., Dynesius, M., 2010. Saproxylic and non-
saproxylic beetle assemblages in boreal spruce forests of different age and forestry
intensity. Ecol. Appl. 20, 2310-2321.
Stokland, J.N., Siitonen, J., Jonsson, B.G., 2012. Biodiversity in dead wood. Cambridge
University Press.
Strong, D.R., Lawton, J.H., Southwood, S.R., 1984. Insects on plants. Community
patterns and mechanisms. Blackwell Scientific Publications, Oxford.
Sugiura, S., Tsuru, T., Yamaura, Y., Makihara, H., 2009a. Small off-shore islands can
serve as important refuges for endemic beetle conservation. J. Insect Conserv 13,
377-385.
Sugiura, S., Yamaura, Y., Tsuru, T., Goto, H., Hasegawa, M., Makihara, H., Makino,
S.I., 2009b. Beetle responses to artificial gaps in an oceanic island forest:
implications for invasive tree management to conserve endemic species diversity.
Biodivers. Conserv. 18, 2101-2118.
Taki, H., Inoue, T., Tanaka, H., Makihara, H., Sueyoshi, M., Isono, M., Okabe, K.,
2010. Responses of community structure, diversity, and abundance of understory
plants and insect assemblages to thinning in plantations. Forest Ecol. Manag. 259,
607-613.
Teixeira, A.M.G., Soares-Filho, B.S., Freitas, S.R., Metzger, J.P., 2009. Modeling
landscape dynamics in an Atlantic Rainforest region: implications for conservation.
Forest Ecol. Manag. 257, 1219-1230.

85
Toledo, V.H., Noguera, F.A., Chemsak, J.A., Hovore, F.T., Giesbert, E.F., 2002. The
cerambycid fauna of the tropical dry forest of “El Aguacero,” Chiapas, México
(Coleoptera: Cerambycidae). Coleopts. Bull. 56, 515-532.
Vives, E., 2000. Coleoptera, Cerambycidae. Fauna Ibérica, Vol. 12. Museo Nacional de
Ciencias Naturales, CSIC Press, Madrid, España.
Vodka, S., Konvicka, M., Cizek, L., 2009. Habitat preferences of oak-feeding
xylophagous beetles in a temperate woodland: implications for forest history and
management. J. Insect Conserv. 13, 553-562.
Wagner, M.R., Cobbinah, J.R., Bosu, P.P., 2008. Forest entomology in west tropical
Africa: forest insects of Ghana. Springer.
Weaver, P.L., 2000. Elfin woodland recovery 30 years after a plane wreck in Puerto
Rico’s Luquillo Mountains. Carib. J. Sci. 36, 1-9.
Werneck, M.S., Pedralli, G., Koenig, R., Gieseke, L.F., 2000. Florística e estrutura de
três trechos de uma floresta semidecídua na Estação Ecológica do Tripuí, Ouro
Preto, MG. Rev. Bras. Bot. 23, 97-106.
Werneck, M.S., Pedralli, G., Gieseke, L.F., 2001. Produção de serapilheira em três
trecho de uma floresta semidecídua com diferentes graus de perturbação na Estação
Ecológica do Tripuí, Ouro Preto, MG. Rev. Bras. Bot. 24, 195-198.
Wolda, H., 1978a. Fluctuations in abundance of tropical insects. Am. Nat. 112, 1017-
1045.
Wolda, H., 1978b. Seasonal fluctuations in rainfall, food and abundance of tropical
insects. J. Anim. Ecol. 47, 369-381.
Wolda, H., 1988. Insect seasonality: why? Annu. Rev. Ecol Syst. 19, 1-18.

86
Zajciw, D. 1967. Contribuição para o estudo da fauna dos longicórneos do Estado do
Maranhão. Bol. Soc. Cear. Agron. 8, 1‑12.
Zanúncio, J.C., Pires, E.M., de Paula Almado, R., Zanetti, R., Monné, M.A., Milagres
Pereira, J., Serrão, J., 2009. Damage assessment and host plant records of
Oxymerus basalis (Dalman, 1823) (Cerambycidae: Cerambycinae: Trachyderini) in
Brazil. Coleopts. Bull. 63, 179-181.