compartimento de remodelação Óssea - estudogeral.sib.uc.pt · concentração atingida por estes...
TRANSCRIPT

Imagem
João Filipe Brochado Martins
Compartimento de Remodelação Óssea
Dissertação de Mestrado em Patologia Experimental,
apresentada à Faculdade de Medicina da Universidade de Coimbra
Dezembro de 2017

Dissertação de candidatura ao grau de mestre
apresentada à Faculdade de Medicina da Universidade de Coimbra
A Faculdade de Medicina da Universidade de Coimbra não aceita qualquer responsabilidade em
relação à doutrina e a forma desta dissertação (Regimento da Faculdade de Medicina de
Coimbra,1931, Art. 108, & único).


Orientadores:
Professora Doutora Maria Helena Figueiredo
Professor Doutor Francisco do Vale


“Não andes apenas pelo caminho traçado,
pois ele conduz somente até onde os outros já foram.”
Alexander Graham Bell


Agradecimentos
__________________________________________________________________________
À Professora Doutora Helena Figueiredo, pela sua constante disponibilidade, pelo seu empenho
categórico apesar de todas as adversidades (que apenas aos grandes Homens são impostas), pela
motivação e entusiasmo contagiantes que demonstrou sempre no decorrer da elaboração deste
trabalho. A realização desta tese não teria sido possível sem a sua ajuda, orientação e partilha de
conhecimento científico. Agradeço também os enormes contributos que me deu ao longo da
realização deste trabalho, fomentando sempre o rigor e o raciocínio científico. Agradeço-lhe toda a
disponibilidade para me guiar no meu percurso académico, primeiro por acreditar em mim enquanto
aluno, e durante os anos seguintes como investigador e docente. As horas passadas ao microscópio,
nunca perdidas, mas sempre cheias de partilha de saber fizeram de mim, sem dúvida, um profissional
muito mais preenchido. Não é fácil agradecer a quem, além de Professora e orientadora, se tornou
acima de tudo família – a “mãe histológica”.
Ao senhor Prof. Francisco do Vale, coorientador deste trabalho, pela cedência dos materiais de
experiência, pelos conselhos dispensados e pela preciosa ajuda facultada, e ainda por todo o
incentivo e orientação científica, que só me fez crescer ao longo deste trabalho.
Ao Senhor Prof. Paulo Palma, que desde cedo me tem acompanhado na minha caminhada não só
pela investigação, mas também pela área clínica e cuja amizade, apoio e crítica construtiva permitiu a
abertura de caminhos na reflexão e aprofundamento das metas a que me propus. Agradeço também
a prontidão em ceder-me parte dos seus materiais da tese de doutoramento para que pudesse levar
a cabo o estudo a que me propus. Além disso, agradeço-lhe pela forma como, nos momentos chave,
me motivou, de forma entusiasmada e genuína, perante cada obstáculo ou sucesso alcançado.
À Senhora Prof. Ana Luísa Maló de Abreu, agradeço-lhe o apoio e constante disponibilidade para
a discussão científica, e não só, deste projeto. É com profunda gratidão e estima que lhe agradeço a
confiança em mim colocada, disponibilidade para discutir ideias e o cuidado depositado nas correções
para que esta tese tivesse a presente conclusão.
Á Dr. Catarina Cortez Vaz, pela amizade e pela preciosa disponibilidade com que nos auxiliou na
elaboração deste trabalho, sem a qual não teria sido possível levá-lo a bom termo.
Não poderia deixar de agradecer, numa nota mais pessoal, à minha família (em especial à minha
mãe), pilares inabaláveis da minha motivação, pelo apoio incondicional com o qual sei que posso
sempre contar.


Sumário
_________________________________________________________________________________
Agradecimentos
I – Introdução
1. Justificação do tema
1.2 Planificação do trabalho
2. Ciclo de Remodelação óssea (algumas considerações)
II – Compartimento de Remodelação Óssea
1. Objetivo
2. Materiais e Métodos
2.1 Animal de experimentação
2.2 Manutenção e bem-estar animal
2.3 Colheita do material
2.4 Processamento histológico de material descalcificado
2.5 Processamento histológico de material não descalcificado
3. Resultados
3.1 Deteção de compartimentos de remodelação óssea no tecido ósseo alveolar
3.2 Identificação de diferentes etapas do ciclo de remodelação óssea
3.3 Deteção de compartimentos de remodelação no cemento e dentina
4. Discussão e considerações finais
III – Resumo
IV – Abstract
V – Bibliografia
VI – Índice
VII- Anexos


Compartimento de Remodelação Óssea 2016
7
I. Introdução, justificação do tema e planificação do
trabalho
_______________________________________________________

Compartimento de Remodelação Óssea 2016
8
1.1 Introdução
Apesar de uma aparência inerte, quando considerados macroscopicamente, os ossos são, sob
um ponto de vista microscópico, estruturas plásticas, altamente dinâmicas, em permanente
remodelação1. A sua grande dinâmica funcional é conseguida sobretudo através de processos de
remodelação.
A remodelação óssea consiste num mecanismo de substituição ou reconstrução de áreas de
tecido ósseo de modo a preservar a sua integridade, otimizar a sua função e prevenir a sua
degradação, visando não só uma adaptação da estrutura óssea às solicitações mecânicas, como
também a remoção de áreas danificadas2. Por outro lado, a matriz óssea constitui um importante
reservatório de iões minerais (cálcio e fósforo), verificando-se um intercâmbio contínuo entre o
cálcio plasmático e o cálcio ósseo. Com efeito, a importância da homeostasia dos níveis séricos de
cálcio vem considerar também o processo de remodelação como um mecanismo de gestão do banco
de cálcio ósseo.
Deste modo, o objetivo da remodelação óssea parece incluir não só uma perspetiva mecânica,
dirigida para a reparação e adaptação da estrutura óssea ao meio, como uma perspetiva metabólica
relacionada com a homeostasia do cálcio e fosfato plasmático. É importante referir que, em
condições fisiológicas, as funções de suporte e de reserva metabólica apresentam-se num equilíbrio
estável3, 4. Porém, em situações de desequilíbrio, a função estrutural é sempre sacrificada em favor da
metabólica5-7.
Uma nota mais para sublinhar o facto de o tecido ósseo alojar e suportar, no seu interior, as
células constituintes da medula óssea, compartilhando, deste modo, um microambiente comum e
muitos dos seus fatores de regulação. Considerando a sua semelhança filogenética e proximidade
topográfica, não será difícil aceitar uma estreita comunicação e integração entre células ósseas,
células do sistema imunitário e células hematopoiéticas6, 8, 9,10. A osteoimunologia7, 11-18 assenta num
eixo de regulação que constitui uma ponte molecular entre o sistema imune e o metabolismo ósseo
e que muitas vezes resulta numa sobre-estimulação da atividade osteoclástica. Na verdade, um facto
que contribuiu enormemente para o desenvolvimento da osteoimunologia foi a constatação de uma
acelerada perda de massa óssea associada a muitos processos inflamatórios. Em situações patológicas
(doenças inflamatórias autoimunes e neoplásicas), o processo de remodelação óssea sofre, quase
sempre, um desvio para privilegiar a reabsorção óssea relativamente à formação4,11,19,20. A
remodelação óssea foi já, ela própria, considerada como um processo inflamatório ou uma forma
altamente controlada e especializada de autoimunidade sendo as áreas de tecido ósseo danificado
reconhecidas como non-self11-18.
A remodelação óssea engloba um processo pelo qual é eliminada uma área de tecido ósseo
que será substituída por outra, com pouca ou nenhuma alteração da massa óssea, constituindo a mais

Compartimento de Remodelação Óssea 2016
9
notável resposta do tecido ósseo ao seu meio envolvente. Qualquer ciclo de remodelação
compreende uma sequência ordenada e imutável de eventos que se inicia por uma fase de
reabsorção óssea e termina com uma etapa reparadora de formação óssea.
O conjunto de células responsáveis por estas diferentes atividades é coletivamente designado
por Unidades Multicelulares Básicas (Basic Multicellular Units - BMU)21-23,28,46,98,100 e reúne uma equipa
de células da linha osteoclástica (pré-osteoclastos e osteoclastos) e uma outra de células da linha
osteoblástica (pré-osteoblastos, osteoblastos, osteócitos e células de revestimento)8,9,10,25,38,46,98.
Desta unidade fazem também parte outras populações celulares que incluem células
mesenquimatosas indiferenciadas e células do tecido conjuntivo, muitas delas pertencentes ao
sistema imunitário11-18, bem como um suplemento vascular especializado para o efeito.
De facto, a remodelação óssea tem atrás de si uma complexa maquinaria celular que, por sua
vez, está sujeita à ação de numerosos fatores de regulação, que exigem um rigoroso e apertado
sistema de sinalização e comunicação, responsável pela ação concertada de todas estas células e pela
coordenação e transição equilibrada das diferentes etapas da remodelação1,98.
Era, pois, previsível que toda esta atividade celular tivesse que ser, de certo modo, “protegida”
de interferências estranhas, vindo em 2001 a ser descrita a existência de uma estrutura visando
proteger e abrigar este complexo e delicado conjunto celular responsável pelos processos de
remodelação. Com efeito, Hauge et al. (2001)24 foram os primeiros investigadores a detetar a
presença de um compartimento de remodelação óssea, como uma entidade real, ainda que
transitória, demonstrando histologicamente a sua existência. De facto, os trabalhos de Hauge et al
(2001) concluíram que as células constituintes das BMU (mesmo no caso do tecido ósseo esponjoso),
não estão em contacto direto com o tecido conjuntivo em seu redor, mas reunidas num espaço
fechado, separado do exterior por um conjunto de células que formam uma cúpula (habitualmente
conhecida por canopy), que constitui o limite externo de uma estrutura especializada24. Esta estrutura
foi então designada por Compartimento de Remodelação Óssea (Bone Remodeling Compartment -
BRC).
Se a remodelação óssea se processasse num espaço amplamente aberto, seria difícil o controlo
dos numerosos e importantes fatores de crescimento e de regulação, necessários aos processos de
diferenciação e maturação das células das linhas osteoclástica e osteoblástica. Com efeito, a
concentração atingida por estes fatores no interior de um espaço confinado irá facilitar a boa
prossecução dos mecanismos de remodelação óssea25.
Pode considerar-se, assim, que o BRC constitui um microambiente ideal para o desempenho
da intensa atividade celular que caracteriza uma BMU, permitindo encontrar no seu interior um
apropriado gradiente de citocinas e outros fatores moleculares capazes de recrutar as populações
celulares necessárias, permitir a sua diferenciação e, deste modo, modular a ligação entre a fase de
reabsorção e a fase de formação óssea.

Compartimento de Remodelação Óssea 2016
10
O aparecimento destes BRC, nos locais de remodelação óssea, coincide sempre com a fase de
ativação e início da fase de reabsorção do processo de remodelação24,26,99. Por outro lado, o seu
desaparecimento observa-se apenas quando já estiver completa a etapa de formação óssea82. Está
também confirmado que as situações de baixo turnover ósseo estão associadas a um decréscimo do
número de BRC, enquanto os casos de elevado turnover ósseo, são acompanhados por um aumento
do número de BRC21. De referir ainda que a perda de integridade de qualquer dos componentes
anatómicos constituintes destas estruturas, nomeadamente da canopy, está sempre relacionada com
uma paragem no processo de reconstrução óssea25-27,76,82. Estes factos reforçam bem a íntima relação
entre BRC e remodelação óssea, apontando mais uma vez para a importância crucial que esta
estrutura representa nos mecanismos de regulação coordenação e integração dos processos de
recrutamento e diferenciação das células da linha osteoclástica e osteoblástica.
Dos trabalhos de investigação realizados no âmbito da remodelação óssea, os estudos de
carácter histológico são escassos, com ilustrações de fraca qualidade e surgem quase sempre numa
posição secundária, constituindo uma base morfológica de apoio necessária à elaboração e
justificação de teorias de cariz bioquímico, conducentes ao desenvolvimento e otimização de
soluções terapêuticas relacionadas com o metabolismo ósseo. De facto, devido ao demasiado
enfoque dado à atividade das células da série osteoblástica ou osteoclástica de per si (assente em
técnicas de isolamento e culturas celulares cada vez mais sofisticadas) foi-se perdendo uma
perspetiva global da microanatomia e constituição histológica, no seu todo, da região onde se
desenrola o processo de remodelação óssea.
É realmente de admirar como só recentemente foi detetada a existência de um
compartimento de remodelação óssea e reconhecida a importância que a constituição e preservação
desta estrutura anatómica, e suas relações topográficas com o microambiente em seu redor,
nomeadamente os vasos sanguíneos, representa na funcionalidade do ciclo de remodelação óssea.
Todo o esforço que tem sido despendido visando aumentar a eficácia dos processos de
formação óssea com base em agentes anabólicos nunca será bem-sucedido se não se verificarem as
condições logísticas necessárias que assegurem o recrutamento e proteção das células
osteoprogenitoras e dos seus fatores de regulação, ou seja, a integridade do BRC.
1.2 Justificação e planificação do trabalho
O trabalho aqui apresentado pretende contribuir para um melhor conhecimento da
constituição histológica do compartimento de remodelação óssea, tendo por base uma análise em
microscopia de luz de material incluído em parafina e em metilmetacrilato.

Compartimento de Remodelação Óssea 2016
11
Uma vez que pretendemos integrar este estudo no âmbito da medicina dentária, e
considerando que a atividade de remodelação óssea atinge no osso alveolar uma frequência
notável1,28,101, incidimos a análise microscópica em cortes histológicos provenientes do tecido ósseo
esponjoso e do tecido ósseo compacto que formam o osso alveolar. Decidimos também estender as
observações para a dentina e para o cemento, uma vez que são tecidos mineralizados capazes de
sofrer processos de remodelação30-32.
Este trabalho encontra-se organizado em três partes distintas.
Na primeira parte (Parte A), após a introdução e justificação do tema, traçamos uma perspetiva
global do ciclo de remodelação óssea e dos elementos celulares que o constituem, dando particular
destaque à fase de ativação e à fase de reversão. Pretende-se com esta revisão proporcionar uma
base geral de apoio indispensável para uma melhor compreensão das imagens apresentadas nos
resultados.
Na segunda parte (Parte B), após uma descrição dos materiais utilizados e das metodologias
aplicadas, procedemos à apresentação pormenorizada das características histológicas dos
compartimentos de remodelação observados em trabéculas do tecido ósseo esponjoso, em cones de
reabsorção presentes no tecido ósseo compacto e em áreas de reabsorção de cemento e dentina.
Finalmente apresenta-se uma discussão e interpretação global dos resultados.
A última parte (Parte C) engloba toda a bibliografia consultada.

Compartimento de Remodelação Óssea 2016
12
1. Ciclo de remodelação óssea (algumas considerações)
A remodelação óssea consiste num mecanismo de substituição ou reconstrução de áreas de
tecido ósseo, de modo a otimizar a sua função ou prevenir a sua degradação34,99,100 seguindo uma
sequência imutável de etapas - ativação, reabsorção, inversão/reversão e formação (Figura 1 e 2).
Neste processo é eliminada uma área de tecido ósseo que será substituída por outra, com pouco ou
nenhuma alteração da massa óssea, englobando uma perfeita coordenação e integração de todas as
atividades celulares intervenientes22,27,33,104.
Uma característica importante da sequência de remodelação óssea é a sua especificidade de
localização. O processo de remodelação que ocorre numa determinada área não tem ligação
cronológica direta com a remodelação observada em qualquer outra área. Este facto leva a admitir
que a remodelação óssea está sobretudo sob controlo de fatores locais (com particular realce para
os estímulos mecânicos), gerados no microambiente de cada unidade ou área específica de
remodelação35,36. De facto, para além de fatores sistémicos (de natureza hormonal e não-hormonal)
as moléculas produzidas ou libertadas localmente, no compartimento de remodelação óssea,
parecem constituir os principais agentes moduladores do ciclo celular da remodelação18,25,35,26,38,46.
De registar ainda a variabilidade de duração das diferentes fases do ciclo de remodelação
óssea. Considerando o tecido ósseo esponjoso (que apresenta uma maior capacidade metabólica e
maior atividade de remodelação do que o tecido ósseo compacto), a fase de reabsorção estende-se
aproximadamente por 3 semanas101, enquanto a fase de formação demora cerca de 3-4 meses102.
Entre estas duas, observa-se uma fase intermédia mas igualmente crucial - fase de reversão - com
uma duração aproximada de 5 semanas101.
Figura 1 - Esquema representativo das diversas fases do ciclo de remodelação óssea. Adaptado de Henriksen e col. (2009)170

Compartimento de Remodelação Óssea 2016
13
A fase de ativação (A) compreende o reconhecimento de uma área precisa da superfície
óssea e a sua preparação para o processo de remodelação. O início desta fase parece ser da
responsabilidade de citocinas pró-inflamatórias produzidas pelos osteócitos em “sofrimento” ou em
apoptose, localizados na área que irá ser sujeita a remodelação2,22,37,46,47,117-119.
Será pertinente recordar aqui que à exceção da calcitonina, todas as hormonas e muitos
fatores de crescimento e citocinas que controlam a reabsorção do tecido ósseo, têm recetores nos
osteoblastos [Vitamina D3 – (1α,25(OH)2D3); paratormona – (PTH); peptídeo relacionado com a
paratormona - (parathyroid hormone related peptide PTHrP) e IL-11, entre outros] e não nos
osteoclastos18. Tudo indica, pois, que são as células da linha osteoblástica e não os osteoclastos que
desencadeiam o processo de remodelação óssea25,45-48.
Por outro lado, para que se inicie o seu processo de diferenciação, os osteoclastos necessitam
da presença de células da linha osteoblásticas, ou de células mesenquimatosas do estroma da medula
óssea, capazes de produzir fatores de diferenciação e ativação, que incluem, entre outros, o fator
estimulador de colónias de macrófagos (macrophage colony stimulating factor – M-CSF). Os
osteoblastos expressam também uma outra proteína transmembranar, o ligando para o recetor de
ativação do fator nuclear kappa-B (receptor activator of nuclear factor kappa-B ligand -
RANKL)25,46,47,104,105,109.
Para que possa dar-se a ativação das células pré-osteoclásticas é necessário que estas
desenvolvam entretanto também na sua superfície um recetor transmembranar designado por RANK
(receptor activator of nuclear factor kappa-B – RANK)4,39,45,49,50. A interação do RANKL (presente na
superfície dos osteoblastos) com o RANK (presente na superfície dos pré-osteoclastos) leva à
maturação dos osteoclastos, sendo um dos principais “motores de arranque” da osteoclastogénese.
Na verdade, está hoje bem estabelecido que o contacto célula-a-célula entre células da linha
osteoblástica e osteoclástica constitui um pré-requisito necessário para a maturação e ativação dos
osteoclastos. De facto, as células de revestimento ósseo, os osteoblastos e os osteócitos, através de
um mecanismo que envolve o contacto entre estas células e os precursores osteoclásticos,
constituem os principais indutores da osteoclastogénese116.
Os osteoblastos sintetizam ainda uma outra proteína, neste caso solúvel, a osteoprotegerina
(OPG), com uma alta afinidade para o RANKL, impedindo ou bloqueando a ligação do RANKL ao
seu recetor RANK51. Por este mecanismo, a OPG regula a população funcional dos osteoclastos,
atuando localmente como um “travão” à osteoclastogénese, reduzindo, deste modo, a reabsorção
óssea.
Com efeito, as células da linha osteoblástica podem controlar o desenvolvimento e a atividade
osteoclástica, ajustando os níveis de expressão do RANKL e de OPG44,113,116. A presença de altos
níveis de RANKL promove a osteoclastogénese, enquanto uma maior expressão de OPG resulta

Compartimento de Remodelação Óssea 2016
14
numa diminuição da osteoclastogénese, ou mesmo na apoptose dos osteoclastos52. Tradicionalmente,
o osteoblasto tem sido considerado como uma “master cell” na regulação do processo de
diferenciação, maturação e ativação dos osteoclastos (osteoclastogénese) e, deste modo, no
mecanismo de remodelação óssea. Porém, o aparecimento de um cada vez maior número de
informações leva-nos a repensar este conceito, já há muito estabelecido.
Na verdade, tem vindo a ser comprovado46,47,107,112,113 que é ao osteócito que cabe o papel
principal na orquestração de todo o mecanismo de remodelação óssea, incluindo o recrutamento e
maturação dos osteoclastos, através da produção de RANKL. A esclerostina, constituindo um
produto primário do osteócito, parece representar uma molécula chave na regulação da rácio
RANKL/OPG e, deste modo, no início, coordenação e transição equilibrada das várias etapas do
ciclo de remodelação45,46,110,111.
De notar, no entanto, que pré-osteoblastos, osteoblastos, osteócitos e células de revestimento
representam diferentes estádios de diferenciação da mesma célula. Assim sendo, é perfeitamente
compreensível que quanto mais diferenciada esteja a célula mais sofisticada e completa será a sua
atividade. Constituindo os osteócitos e as células de revestimento4,46,107,113 a etapa final do processo
de diferenciação não será de estranhar que sejam estas células as principais responsáveis pela
ativação e coordenação de todo o processo de remodelação óssea.
Devido ao seu elevado número (constituindo 90-95% do total de células óssea)105,114,115 e
complexa organização e disposição, facilmente se compreende que os osteócitos se encontrem numa
situação privilegiada para captar e transmitir quaisquer alterações existentes na matriz óssea,
formando uma rede interna capaz de “sentir” toda a estrutura óssea como uma única entidade46,47. É
já hoje, por demais evidente o papel dos osteócitos no desencadear dos processos de remodelação,
constituindo os principais mecanossensores e transdutores do tecido ósseo46,47,105,107,113,119.
O mecanismo pelo qual as forças mecânicas conseguem regular os processos celulares é
designado de uma forma global por mecanotransdução44,120-123.
A mecanotransdução pode ser entendida como a conversão de um sinal de natureza mecânica
(força) numa resposta celular de natureza bioquímica44,46,47. Por outras palavras, pode considerar-se o
mecanismo de transdução como um processo pelo qual certas células (células recetoras) detetam ou
“sentem” certos sinais mecânicos (forças ou tensões aplicadas) gerando uma resposta celular (de
natureza bioquímica) dirigida às células efetoras. Estas últimas células vão, por sua vez, ativar ou
modular os processos de remodelação. Os osteócitos constituem nestas circunstâncias as células
recetoras ou mecanossensoras (consideradas por isso como as “guardiães” do tecido ósseo), sendo
as células de revestimento ósseo, os osteoblastos e os osteoclastos considerados como células
efetoras.
A deformação da matriz óssea (que ocorre pela aplicação de uma determinada força) pode
causar diferenças de pressão no fluido que preenche os canalículos, alterando o seu fluxo, isto é, a

Compartimento de Remodelação Óssea 2016
15
sua velocidade. É nesta interação, entre o movimento do fluido e os osteócitos, que reside a chave
da mecanotransdução. As alterações do meio extracelular (variações de pressão e velocidade dos
fluidos) são, pois, captadas pelos osteócitos que as convertem em sinais e mensagens moleculares
que serão transmitidas às células efetoras quer através dos prolongamentos celulares e gap junctions
(ativando uma rede intracelular de comunicação), quer por difusão no fluido extracelular46,47,120-123. É
cada vez mais consensual que estes mecanismos não atuam de forma isolada, mas sim em sinergia,
observando-se diversas vias alternativas em simultâneo.
Importa acentuar ainda que as forças mecânicas aplicadas na matriz óssea são fundamentais
para assegurar a vitalidade das células ósseas117-119. Com efeito, a falta de estímulos mecânicos conduz
sempre e a curto prazo à apoptose dos osteócitos e à perda de massa óssea. Esta interação, entre o
movimento do fluido e os osteócitos, constitui um estímulo poderoso e imprescindível à manutenção
da sua atividade celular.
A presença de áreas de matriz danificada ou comprometida (pela presença de microfissuras) ao
provocar uma restrição na acessibilidade dos osteócitos aos nutrientes e oxigénio conduz à sua
apoptose117-119. A apoptose dos osteócitos desencadeia quase sempre processos de remodelação
óssea, verificando-se geralmente uma forte associação, no tempo e no espaço, entre microfissuras,
apoptose dos osteócitos e remodelação óssea119.
Por outro lado, foi também recentemente demonstrado que os osteócitos através dos seus
prolongamentos citoplasmáticos e do fluido tecidular, situado no interior do sistema lacuno-
canalicular, podem estabelecer contacto direto com os vasos sanguíneos da medula óssea (no tecido
ósseo esponjoso) ou com os capilares presentes no interior dos canais de Havers (no tecido ósseo
compacto)21, 46,47 ,constituindo um sistema de sinalização capaz de fornecer a localização e dimensão
das áreas a remodelar, estimulando em paralelo os processos de angiogénese e a quimiotaxia das
células osteoclásticas28. As inúmeras conexões dos osteócitos entre si, com com os osteoblastos e
osteoclastos, bem como as suas ligações vasculares, permitem um fácil acesso e uma rápida resposta
a estímulos mecânicos e a fatores sistémicos e hormonais (como aos estrogénios, a PTH e vitamina
D3)46,47.
A esclerostina parece ser considerada um mediador major na integração dos estímulos
mecânicos, hormonais e locais, detetados pelos osteócitos, controlando, desta forma, todo o
aparelho de remodelação. As alterações nos níveis de esclerostina regulam a expressão de outras
proteínas específicas nomeadamente o RANKL e a OPG, atrás referidas, bem como a expressão de
genes (PHEX, DMP1, MEPE, FGF-23) relacionados com os processos de mineralização46.
Pode afirmar-se portanto que, na fase de ativação, os osteócitos e as células de revestimento
presentes nas áreas a remodelar são determinantes para o processo de recrutamento das células
pré-osteoclásticas, para o reforço dos mecanismos de angiogénese e para a formação do
compartimento de remodelação óssea27,104-107,116.

Compartimento de Remodelação Óssea 2016
16
Com efeito, na fase de ativação do ciclo de remodelação verifica-se uma retração e um
destacamento das células de revestimento, afastando-se da respetiva matriz óssea, dando início ao
aparecimento de uma canopy e ao desenvolvimento de um espaço fechado correspondente ao
BRC24,38. Importa salientar, ainda nesta fase, o desenvolvimento de uma rede de capilares sanguíneos
e a sua aproximação às células da canopy, formando no seu conjunto uma plataforma de comunicação
entre estruturas vasculares e compartimentos de remodelação39,40,108. A intervenção do componente
vascular, constituindo uma estrutura sempre presente, é determinante ao longo de todo o processo
de remodelação41-44, 108.
Na fase de reabsorção (R) os osteoclastos cavam, no tecido ósseo esponjoso, lacunas de
contornos muito irregulares denominadas lacunas de Howship e no tecido ósseo compacto
cavidades cilíndricas designadas por cones de reabsorção53,54. A dimensão das áreas reabsorvidas é
diretamente proporcional às áreas danificadas ou em sofrimento.
No final desta etapa observa-se uma inversão ou reversão nesta sequência, verificando-se
um conjunto de condições responsáveis pela chamada de células osteoprogenitoras e sua
diferenciação, dando início ao processo de reparação óssea54. Esta estimulação da atividade
osteoblástica, como resposta ao processo de reabsorção osteoclástica, é habitualmente conhecido
por coupling45,55.
Um aspeto curioso, e que assinala o início da fase de reversão (Rv), prende-se com o facto de
serem, mais uma vez, as células de revestimento da canopy as células responsáveis pela “limpeza” das
lacunas de Howship, no que respeita às moléculas de colagénio não digeridas e outros detritos
decorrentes da atividade osteoclástica48. Este processo constitui um pré-requisito obrigatório para o
desenvolvimento da fase de reversão, pois só em lacunas de reabsorção, bem preparadas e
limpas48,56, é possível a formação de uma linha cimentante e a deposição de uma camada de colagénio
fibrilar (colagénio III) e de osteocalcina, permitindo, deste modo, criar um ambiente osteogénico
capaz de levar a cabo um processo de maturação osteoblástica.
Ainda que o mecanismo de diferenciação e ativação das células da linha osteoclástica esteja,
como já foi referido, muito controlado pelas células da linha osteoblástica (pré-osteoblastos,
osteoblastos, osteócitos e células de revestimento), através do eixo de regulação
RANKL/RANK/OPG52, esta via não é de sentido único, uma vez que também os osteoclastos
condicionam e modulam muitas das atividades osteoblásticas. Nesta fase estabelece-se também uma
complexa comunicação bidireccional que vai inibir a atividade dos osteoclastos e promover a dos
osteoblastos.
Parecem verificar-se pelo menos quatro classes de sistemas de sinalização25 derivados dos
osteoclastos, ou da sua atividade, responsáveis pelo início e desenvolvimento da etapa de formação
óssea mas que começam a manifestar-se na fase de reversão: a) fatores de crescimento e citocina

Compartimento de Remodelação Óssea 2016
17
com efeito osteogénico contidos na matriz óssea e libertados durante o processo de reabsorção
osteoclástica; b) fatores de crescimento sintetizados e secretados pelos próprios osteoclastos,
exercendo um efeito parácrino nos osteoblastos; c) um contacto direto célula-célula entre
osteoclastos e osteoblastos; d) alterações deixadas pelos osteoclastos na nanotopografia da
superfície óssea reabsorvida com reconhecida ação osteoindutora25.
Com efeito, e de acordo com a alínea a), a matriz óssea representa um verdadeiro armazém
de citocinas e fatores de crescimento em fase latente (produzidos pelos osteoblastos) que são
libertados e ativados na sequência do processo de degradação osteoclástica com o principal objetivo
de estimular o recrutamento57-59, migração58,60-62 e diferenciação61,63,64 das células osteoprogenitoras.
Neste âmbito, podemos referir, entre muitos, o fator de transformação β (Transforming growth factor
β - TGFβ), os fatores de crescimento insulino-aparentados (Insulin growth factor - IGF I e II), o fator de
crescimento fibroblástico (Fibroblast growth factor - FGF), o fator de crescimento derivado das
plaquetas (Platelet derived growth factor - PDGF), fatores de crescimento das células endoteliais
(Vascular endothelial growth factor - VEGFs), as proteínas morfogenéticas ósseas (Bone morphogenic
proteins - BMP) e as metaloproteinases da matriz (Matrix metalloproteinases - MMP)65-70.
Entre os fatores de crescimento sintetizados diretamente pelos osteoclastos, e referidos na
alínea b), estão mais uma vez as TGF-βs e as PDGFs. Também o fator osteoclástico esfingosina 1-
fosfato (Sphingosine 1 phosphate - S1P) parece desempenhar um papel importante na fase de
reversão71. A síntese de S1P pelos osteoclastos estimula o recrutamento, migração e sobrevivência
das células da linha osteoblástica.
Por outro lado, e em concordância com a alínea c) está já confirmada uma sinalização direta
bidirecional entre osteoblastos e osteoclastos através de proteínas transmembranares presentes
nestas células, regulando simultaneamente a atividade de cada uma delas. Neste âmbito, encontramos
o sistema Ephrin-Eph como o principal mediador da interação entre osteoclastos e osteoblastos72, 73.
A interação entre as ephrin B2 (ligandos expressos nas membranas dos osteoclastos) e os seus
recetores EphB4 (presentes nos osteoblastos) requer um contacto físico direto célula-célula72, que
promove a diferenciação osteoblástica e, ao mesmo tempo, inibe a atividade osteoclástica. Pode
assim afirmar-se que os osteoclastos serão responsáveis, através da secreção de clastocinas, como a
S1P, pelo recrutamento das células osteoprogenitoras para os locais de reabsorção, estimulando ao
mesmo tempo a sua diferenciação através do sistema de sinalização Ehprina B2/Eph B472,73.
Compreende-se, pois, que a simples presença de osteoclastos, independentemente da sua
capacidade de reabsorção, constitua um fator essencial nos mecanismos de osteogénese, produzindo
fatores ativadores da função osteoblástica. A nova geração de agentes antirreabsorção (muito
utilizada nas situações de osteoporose) começa a ter em conta este facto, tentando não atingir a
sobrevivência dos osteoclastos, ainda que bloqueiem a sua atividade reabsortiva74.

Compartimento de Remodelação Óssea 2016
18
Por fim, as alterações na nanotopografia da superfície óssea, efetuadas pelos osteoclastos, e
referidas na alínea d), parecem ter uma importância determinante nos processos de adesão e
diferenciação das células da linha osteoblástica24,48. A este respeito, é impossível deixar de referir que
as concavidades escavadas pelos osteoclastos (lacunas de Howship) constituem locais estratégicos e
privilegiados para a formação da matriz osteóide e sua posterior mineralização. A geometria
conformacional, dimensão, composição, microarquitectura e ainda a nanoestrutura de superfície
destas cavidades criam um microambiente propício ao aparecimento de um fenótipo
osteoblástico75,98. Estas lacunas parecem favorecer, ou mesmo contribuir, para uma sequestração de
iões (nomeadamente cálcio) e de fatores de crescimento e outras proteínas com efeitos
osteoindutores, estimulando ao mesmo tempo os processos de angiogénese21,98.Todos estes fatores
terão uma influência determinante na coordenação e transição equilibrada entre a fase de reabsorção
e formação óssea.
Importa salientar, contudo, que as células que caracterizam a fase de reversão e que vão
beneficiar da ação osteoindutora dos fatores atrás mencionados, resultantes da atividade das células
osteoclásticas, não são osteoblastos maduros mas sim percursores de osteoblastos que colonizam as
superfícies ósseas após o desaparecimento dos osteoclastos76. As células da canopy, ainda que
derivadas das células de revestimento ósseo, parecem encontrar-se, no entanto, num estádio muito
precoce de desenvolvimento, podendo dar origem a grande parte das células que nesta fase vão
diferenciar-se em osteoblastos82,100. Para além disto, é também de considerar a forte presença de
células osteoprogenitoras que chegam, nesta altura, ao compartimento de remodelação provenientes
de outras origens, nomeadamente da corrente sanguínea e da parede vascular dos capilares
sanguíneos adjacentes77-81.
A fase de reversão parece constituir uma das etapas mais delicadas do ciclo de remodelação,
reunindo um conjunto de condições essenciais para a proliferação e diferenciação osteoblástica76.
Qualquer perturbação observada nesta etapa (nomeadamente a integridade da canopy) poderá
provocar uma paragem irreversível no processo de remodelação24-27,76,82,83,99.
Por último, o preenchimento e reparação da cavidade de reabsorção é naturalmente
decorrente da atividade secretora de osteoblastos maduros com origem nas células observadas
durante a fase de reversão e que, entretanto, beneficiaram de um processo de proliferação,
maturação e ativação27.
Como nota final gostaríamos de referir mais uma vez que, para além dos sistemas de
sinalização atrás descritos, a transição entre a fase de reabsorção e a fase de reversão/formação é
ainda modulada pelo aparecimento de novos estímulos mecânicos no interior do BRC (sendo
considerado um fenómeno “strain regulated”) e pelo aumento dos níveis extracelulares de fosfatos
inorgânicos libertados quer no interior do BRC quer para a circulação sanguínea46.

Compartimento de Remodelação Óssea 2016
19
Com efeito, estudos recentes27,46,47,117,118 têm vindo a chamar atenção para as alterações nas
forças de deformação verificadas no interior da lacuna de reabsorção que funcionariam como
importantes estímulos responsáveis pelo início da fase de reversão/formação. Assim, estes
investigadores defendem a ideia de que a transição da fase de reabsorção para a fase de reversão
seria também uma resposta adaptativa a alterações de estímulos mecânicos, ou seja, um fenómeno
regulado pelas forças de deformação levado a cabo pelos osteócitos, via esclerostina48. Assim, mais
uma vez, os osteócitos através do controlo dos níveis de esclerostina e sua repercussão no sistema
RANK/RANKL/OPG vão agora pôr termo à fase de reabsorção e desencadear o início de uma fase
osteogénica27,46,47. Esta noção faz todo o sentido tendo em conta o que referimos sobre os
fenómenos de mecanotransdução.
Em paralelo, estes trabalhos27,46,47,117,118 referem também que durante a fase de reabsorção os
cristais de hidroxiapatite são dissolvidos em iões Ca2+ e Pi (na forma de H2PO4- e HPO42-), causando
uma elevação transitória dos níveis de fosfatos quer no interior do BRC, quer na corrente sanguínea,
estando também esta situação associada ao início da fase de reversão, estimulando o
desenvolvimento dos processos de formação e mineralização da matriz óssea pela modulação da
expressão génica das células pré-osteoblásticas e osteoblásticas presentes no BRC46.
Neste contexto, o osteócito e a rede lacuno-canalicular funcionam como uma “unidade
externa” desempenhando, no entanto, e mais uma vez, um papel central no controlo da transição
adequada entre as diferentes fases do ciclo de remodelação óssea.
A fase de formação (F) consiste, pois, na síntese de matriz osteóide e na sua posterior
mineralização, tendo como resultado final a reconstrução da lacuna de Howship com novo tecido
ósseo ou de um cone de reabsorção, dando origem ao aparecimento de um novo sistema de Havers.
O término da sequência de remodelação parece ser também, tal como o seu início, da
responsabilidade dos osteócitos25,46,47,119,145. Foi já demonstrado que os osteócitos presentes na
matriz óssea recém-sintetizada e mineralizada estão também envolvidos no processo de regulação
desta fase, através da síntese de diversas moléculas como a esclerostina que determina a evolução
final dos osteoblastos secretores para células de revestimento ósseo, pondo fim ao ciclo de
remodelação46,48,65.

Compartimento de Remodelação Óssea 2016
20
Figura 2 – Representação esquemática das relações espaciais e funcionais entre capilares sanguíneos e compartimentos de remodelação
óssea observados no tecido ósseo esponjoso (topo) e no tecido ósseo compacto (em baixo). Esta proximidade permite um fácil acesso das
células da linha osteoclástica (provenientes da corrente sanguínea) para o interior do compartimento, bem como das células
osteoprogenitoras circulantes ou resultantes da diferenciação dos pericitos. (Adaptado de Lafage-Proust e col. 2015) 25.
Em síntese, pode afirmar-se que no processo de remodelação óssea intervêm duas atividades
opostas mas complementares, a reabsorção, a cargo das células da linha osteoclástica, e a formação
do tecido ósseo, a cargo das células da linha osteoblástica, funcionando em estreita coordenação e
integração. A reabsorção e a formação óssea são fenómenos que estão intimamente acoplados, visto
que o início da primeira estimula a atividade reparadora da segunda, de modo a que a reabsorção de
uma certa quantidade de tecido ósseo seja, em princípio, substituído pela mesma quantidade de novo
osso,25-27,41,50,55,76. O controlo da rácio RANKL/OPG verificado no interior do BRC parece ser
comandado pela atividade dos osteócitos, via esclerostina. Estas células determinam o “timing”, a
dimensão, sequência, velocidade e ritmo que marcam a coordenação e integração das diferentes fases
do ciclo de remodelação46.
O tecido ósseo constitui, pois, um notável material de construção, de natureza biológica, com
a singular capacidade de edificar estruturas muito resistentes que se remodelam e reparam a si
próprias.
***

Compartimento de Remodelação Óssea 2016
21
Ainda que a associação e alinhamento sequencial entre reabsorção (osteoclástica) e formação
(osteoblástica) raramente esteja afetada, o balanço final em termos de massa óssea pode variar
consideravelmente. No processo de remodelação óssea particularmente a fase de reabsorção pode
desenvolver-se, tendo como ponto de partida e base de sustentação o sistema imunológico7,11,12-18,35.
Muitos são os trabalhos que têm vindo a demonstrar que algumas citocinas pró-inflamatórias
(TNF- α e β; IL-1, IL-6, IL-11,IL-17)84-87 e os LPS (lipopolissacarídeos componentes da superfície da
parede de algumas bactérias Gram negativas) podem estar envolvidos na diferenciação terminal dos
osteoclastos, de uma forma independente dos osteoblastos (ou seja, sem a necessidade do contacto
direto com estas células). Tanto a IL-1, IL-6 como o TNF-α podem estimular por si só a
osteoclastogénese, sem recurso ao eixo RANKL/RANK18,88,89,90.
A maior parte das doenças metabólicas e inflamatórias que atingem o tecido ósseo
manifestam-se geralmente por perturbações na dinâmica da remodelação resultando, quase
invariavelmente, num decréscimo da massa óssea associada a processos de reabsorção osteoclástica.
Em casos patológicos como a artrite reumatóide observa-se uma forte participação de linfócitos T e
B4 e de fibroblastos sinoviais no processo de formação dos osteoclastos, traduzindo uma
contribuição determinante destas células (através da produção de RANKL) para a perda da massa
óssea e consequente deformação articular. Assim, a artrite reumatóide pode considerar-se
atualmente como uma RANKL disease. Neste contexto, a osteoporose é muitas vezes encarada como
uma “inflamm-aging”91.
Uma situação bastante semelhante parece verificar-se também com a doença periodontal140.
Os fibroblastos do ligamento periodontal podem expressar RANKL em resposta à estimulação por
bactérias periodontogénicas provocando severos processos de reabsorção no osso alveolar, bem
como lesões no cemento e na dentina88,92-94. A perda da radiopacidade típica da lâmina dura é muitas
vezes interpretada como uma imagem de marca da doença periodontal crónica, traduzindo processos
de reabsorção óssea sempre associados a uma resposta inflamatória, relacionados com uma alteração
da rácio RANKL/OPG140. É neste contexto que se insere a maior parte dos processos de reabsorção
do cemento e da dentina.

Compartimento de Remodelação Óssea 2016
22

Compartimento de Remodelação Óssea 2016
23
II. Constituição Histológica do Compartimento de
Remodelação Óssea
Alguns aspetos

Compartimento de Remodelação Óssea 2016
24

Compartimento de Remodelação Óssea 2016
25
Objetivo
Tendo em conta a participação e colaboração em diversos trabalhos experimentais
desenvolvidos na Área de Medicina Dentária da Faculdade de Medicina da Universidade de
Coimbra94,95,96 com particular incidência no tecido ósseo e dadas as limitações económicas
prementes para levar a cabo um novo estudo experimental, decidimos observar, com objetivos
completamente distintos, algumas das lâminas histológicas que estiveram na base dos trabalhos acima
referenciados e cujos resultados de índole morfológica estiveram à nossa responsabilidade. De facto,
a existência de compartimentos de remodelação óssea já tinha sido por nós detetada, aquando da
observação cuidadosa destes cortes histológicos de tecido ósseo, sem contudo ter sido descrita pois
encontrava-se fora do âmbito dos estudos experimentais em curso.
Assim, aproveitando as lâminas histológicas que constituíram os grupos controlo dos estudos
experimentais in vivo95,96 decidimos levar a cabo uma análise qualitativa em microscopia de luz no
âmbito da histologia básica, para deteção e identificação de compartimentos de remodelação óssea
(BRC) nas áreas de tecido ósseo alveolar mandibular, incidindo fundamentalmente na observação e
registo das suas características histomorfológicas. Decidimos ainda estender as observações para as
áreas de dentina e de cemento, uma vez que são também tecidos mineralizados com capacidade de
remodelação.
1. Materiais e Métodos
2.1. Animal de experimentação
Foram utilizados seis cães (Canis familiaris) da raça Beagle de origem pura, nascidos e criados
para fins de investigação, certificados pelo Colegio Official de Veterinarios da Cataluñya – Barcelona, e
adquiridos à Isoquimen, S.L. (Barcelona, Espanha), que constituíram os grupos controlo dos trabalhos
experimentais intitulados “Apexificação e revascularização em dentes permanentes imaturos –
estudo experimental in vivo”95 realizado pelo Prof. Paulo Palma e “Distração osteogénica dento-
ancorada – estudo experimental”96 realizada pelo Prof. Francisco do Vale no âmbito dos seus
respetivos trabalhos de doutoramentos.
Os animais tinham cerca de 14 meses de idade e pesos compreendidos entre os 15 e 18
quilogramas, apresentando todos uma identificação através de sistema de chips subcutâneos
localizados na zona do dorso.

Compartimento de Remodelação Óssea 2016
26
2.2. Manutenção e bem-estar animal
A receção, observação, manutenção, manipulação, cuidados e procedimentos cirúrgicos foram
efetuados no Hospital Veterinário da Universidade de Trás-os-Montes e Alto Douro, em Vila Real,
sob supervisão do Investigador Coordenador responsável pelo Bem-estar Animal.
O estado de saúde geral dos animais, à chegada à unidade hospitalar, foi avaliado por
veterinários com experiência prévia e qualificação na área de experimentação animal. Excluídas
quaisquer patologias agudas ou crónicas, os animais foram mantidos num período de quarentena,
com observação diária, durante um intervalo de tempo nunca inferior a duas semanas. Todos os
animais permaneceram numa fase de ambientação às novas instalações, aos cuidadores e ao sistema
de controlo clínico durante aproximadamente 12 semanas, antes de qualquer intervenção.
2.3. Colheita do material
Após a eutanásia, as mandíbulas foram retirados en bloc, após dissecção meticulosa dos tecidos
moles em planos e excisados com uma serra mecanizada refrigerada. De seguida foram separadas as
hemimandíbulas de cada animal, após osteotomia da sutura sinfisária e imersas em formol neutro
tamponado e posteriormente transportadas para o laboratório.
Todo o material utilizado nos estudos histológicos foi processado no Laboratório de Tecidos
Duros da Clínica Universitária de Medicina Dentária, da Faculdade de Medicina da Universidade de
Coimbra.
2.4. Processamento histológico de material descalcificado
Depois de colocados em cassetes “histológicas” individuais, devidamente identificadas, as peças
foram novamente colocadas em formalina tamponada a 10 % a +4º C durante uma semana. Após o
processo de fixação procedeu-se à descalcificação das amostras.
A descalcificação foi efetuada por imersão das amostras numa solução de Morse (ácido
fórmico a 50 % + citrato de sódio a 20 % em partes iguais), sob agitação permanente, numa unidade
de agitação (Jkika Labortechnik HS 501), durante um período mínimo de oito semanas.
Após a descalcificação das amostras, procedeu-se à sua desidratação por sucessivas passagens
em soluções crescentes de etanol, 50 %, 70 %, 90 % e 100 %, sob agitação permanente.
Posteriormente procedeu-se à fase de diafanização em xilol (Xileno PA Panreac®), impregnação e
inclusão em parafina (Paraplast® Regular), utilizando cassetes apropriadas (Kartell® Spa). Depois do
processo de inclusão, as amostras foram cortadas num micrótomo (Leica RM 2155), num plano
proximal, em cortes sequenciais de 7 μm de espessura.
Os cortes foram em seguida secos em estufa a 37º C, durante cerca de 16h, sendo
posteriormente selecionadas e coradas com Hematoxilina e Eosina (H.E.) ou com Tricrómio Masson
(T.M.). Durante todo o processamento foi assegurada uma rigorosa identificação das amostras.

Compartimento de Remodelação Óssea 2016
27
2.5. Processamento histológico de material não descalcificado
As amostras de material não descalcificado foram preparadas segundo o protocolo
desenvolvido por Karl Donath do Instituto de Patologia da Universidade de Hamburgo, na
Alemanha97.
As amostras individuais foram fixadas através de imersão numa solução de formaldeído a 10 %
em tampão fosfato (pH 7,4) imediatamente após a colheita e durante um período de 48 horas.
Posteriormente, e após lavagem com água corrente durante 30 minutos, iniciou-se o processo de
desidratação através do protocolo preconizado por Karl Donath. Numa primeira fase realizaram-se
passagens numa série crescente de concentrações de etanol (60 %, 80 %, 96 % e duas vezes a 100 %),
numa unidade de agitação e infiltração (Exakt® 510), com agitação contínua e com duração de 30 dias
para cada passo.
Posteriormente, procedeu-se à etapa de infiltração, utilizando-se soluções com graduações
crescentes de resina de metilmetacrilato (Technovit® 7200 VLC, Kulzer) em etanol (resina/álcool:
30/70 %, 50/50 %, 70/30 %, e duas vezes a 100 % de resina).
Finamente, incluíram-se as amostras na mesma resina de metilmetacrilato (Technovit® 7200
VLC, Kulzer) e foi realizada a fotopolimerização na unidade de polimerização (Exakt® 520), com
sistema de arrefecimento e em moldes plásticos.
As amostras incluídas nos blocos de resina sofreram de seguida um corte, na respetiva unidade
do sistema Exakt® (Exakt® 310 CP), consoante a orientação pretendida para subsequente preparação
da primeira lâmina acrílica.
De cada amostra, foram obtidos três cortes sagitais que envolviam o espaço compreendido
entre o terceiro e o quarto pré-molar, com a seguinte correspondência:
I. Corte sagital vestibular;
II. Corte sagital médio;
III. Corte sagital lingual.
A espessura final de 50 µm foi obtida através de polimento calibrado com discos de diamante
de granulometria decrescente (800, 1000, 1200, 2500 e 4000; Hermes®, Alemanha). Todas as
medições de controlo foram realizadas com micrómetro digital (Mitutoyo® 2093, Digimatic
Micrometer, Japão).
Concluiu-se a fase laboratorial efetuando uma técnica de coloração com azul de Toluidina
(A.T.) dos tecidos duros representados na lâmina final. Esta técnica, que utiliza um corante básico e
cora os componentes ácidos em diversas gradações de azul, permite estabelecer uma boa distinção
entre tecido osteóide e tecido ósseo mineralizado.

Compartimento de Remodelação Óssea 2016
28
A observação das lâminas foi efetuadas com recurso a um microscópio de luz de campo claro
(Nikon® Eclipse 600, Tóquio, Japão) e estereomicroscópio (Nikon® SMZ 1500, Tóquio, Japão). Estes
dispositivos têm capacidade para acoplar equipamento fotográfico convencional (Nikon® FDX 35,
com sensor multiponto Nikon® U-III, Tóquio, Japão), bem como câmaras fotográficas digitais
(Optronics® DEI 750D CE, Goleta, Califórnia, Estados Unidos da América e Nikon® Digital Camera
DXM-1200 C, Tóquio, Japão) ligadas a computadores PC (Intel®Pentium™ III e Intel®Core™ 2 Duo
Core™) com software para fotografia e análise de imagem (Nikon® ACT-1C, Tóquio, Japão).

Compartimento de Remodelação Óssea 2016
29
2. Resultados
__________________________________________________________

Compartimento de Remodelação Óssea 2016
30

Compartimento de Remodelação Óssea 2016
31
O trabalho aqui apresentado pretende contribuir para um melhor conhecimento da
constituição histológica de compartimentos de remodelação detetados no tecido ósseo alveolar e
ainda no cemento e dentina radicular, tecidos provenientes de mandíbulas de animais mantidos em
condições fisiológicas, tendo por base uma análise qualitativa realizada em microscopia de luz.
As imagens histológicas apresentadas compreendem cortes de material incluído em parafina,
corados com H.E. ou com o método T.M.. Apresentamos também cortes de material incluído em
metilmetacrilato, corados com A.T..
3.1 Deteção de compartimentos de remodelação óssea presentes no tecido ósseo
alveolar
O osso alveolar é constituído por áreas de tecido ósseo compacto (figuras 3 e 4) e por áreas
de tecido ósseo esponjoso (figuras 5 e 6), sendo considerado o local onde se verifica, em condições
fisiológicas, a mais alta taxa de remodelação de todo o esqueleto. Esta notável dinâmica de
remodelação está traduzida pela existência de inúmeras linhas de reversão (figuras 5 e 6). Estas linhas
de reversão são marcas indeléveis que definem uma nítida fronteira entre áreas de tecido ósseo
formadas por diferentes gerações de osteoblastos. De igual modo, e refletindo a sua intensa
capacidade de remodelação, foram também detetadas, no tecido ósseo que constitui o alvéolo
dentário, particularmente na apófise alveolar, numerosas áreas de remodelação em fase ativa.
Estas áreas em remodelação apresentam, no entanto, características bastante distintas,
considerando as regiões onde se encontram. De facto, no tecido ósseo compacto observam-se
verdadeiros túneis/cones de reabsorção (figura 7), alguns apresentando tamanho e profundidade
consideráveis. Nas trabéculas de tecido ósseo esponjoso os locais em remodelação ativa (figura 8 e
9) apresentam-se como escavações superficiais (figura 8), muitas vezes bastante discretas, observadas
junto à medula óssea ou já em fase de resolução (figura 9) apresentando uma razoável camada de
osteóide.
A identificação em microscopia de luz das regiões em remodelação baseou-se
maioritariamente na presença de nítidas lacunas de reabsorção com ou sem osteoclastos visíveis, ou
na observação de uma camada de células osteoblásticas quase sempre acompanhadas de osteóide.
Tendo em conta que os osteoclastos apresentam uma semivida bastante mais curta do que os
osteoblastos e a enorme diferença de duração das fases de reabsorção e de formação que compõem
o ciclo de remodelação óssea, foi possível encontrar a presença de osteoblastos num maior número
de vezes. Foram ainda consideradas como áreas em remodelação ativa, zonas onde as lacunas de
reabsorção estavam ocupadas por populações celulares que não apresentavam características típicas
de células osteoclásticas ou osteoblásticas, constituindo uma etapa intermédia e de ligação entre a
fase de reabsorção e a de formação, normalmente designada por fase de reversão.

Compartimento de Remodelação Óssea 2016
32
Uma observação mais atenta das áreas em remodelação, com recurso a maiores ampliações,
permite-nos distinguir a existência de estruturas que identificamos como compartimentos de
reabsorção óssea.
***
O compartimento de remodelação óssea apresenta-se no tecido ósseo esponjoso como um
espaço fechado (figuras 10-24) onde se encontram as células constituintes de uma BMU (nem sempre
fáceis de identificar em cortes de parafina), sendo, no entanto, de salientar a presença frequente de
osteoclastos e/ou osteoblastos localizados junto à matriz óssea. Este espaço encontra-se revestido
exteriormente por uma fina camada em forma de cúpula (canopy) formada por células extremamente
achatadas (figuras 13 e 14), muito semelhantes às células de revestimento ósseo, separando este
compartimento da medula óssea adjacente (figura 15). É também possível observar a continuidade
desta canopy com as restantes células de revestimento presentes na superfície da mesma trabécula
(figuras 15 e 16), mas em estado de repouso e que, por este motivo, permanecem em íntima ligação
com a matriz óssea.
De registar também que no caso do osso alveolar, observa-se entre as trabéculas de tecido
ósseo esponjoso a existência de medula óssea amarela, com nítido predomínio de células adiposas,
não sendo visíveis áreas de hematopoiese.
Será pertinente referir a existência de uma abundante rede capilar (figuras 17-24) localizada em
estreita associação de proximidade à canopy que reveste os compartimentos de remodelação, sendo
mesmo possível verificar a presença de inúmeros pontos de contacto (figuras 18,19) entre os
capilares e as células de revestimento que a compõem.
Esta rede capilar, particularmente desenvolvida junto às superfícies ósseas em remodelação,
apresenta quase sempre um trajeto paralelo à canopy (figuras 17,19,20), acompanhando-a em quase
toda a sua extensão. Por sua vez, nas superfícies ósseas em repouso, a densidade de capilares
presente é incomparavelmente menor.
***
No tecido ósseo compacto que forma o alvéolo dentário, o compartimento de remodelação
óssea (figuras 21-24) apresenta-se também como um espaço que recobre as zonas de matriz
mineralizada (que formam as paredes de um cone de reabsorção) e que estão a ser alvo de um
processo de remodelação. Também aqui é possível observar uma canopy, formando o teto do
compartimento em questão, constituída por uma camada de células achatadas, por vezes difíceis de
distinguir das células endoteliais dos vasos sanguíneos que as acompanham. De facto a proximidade
de vasos sanguíneos é sempre uma constante, mostrando em geral um percurso paralelo à canopy. A
população celular que ocupa o compartimento de remodelação vai variando consoante a fase do
ciclo em que se encontra.
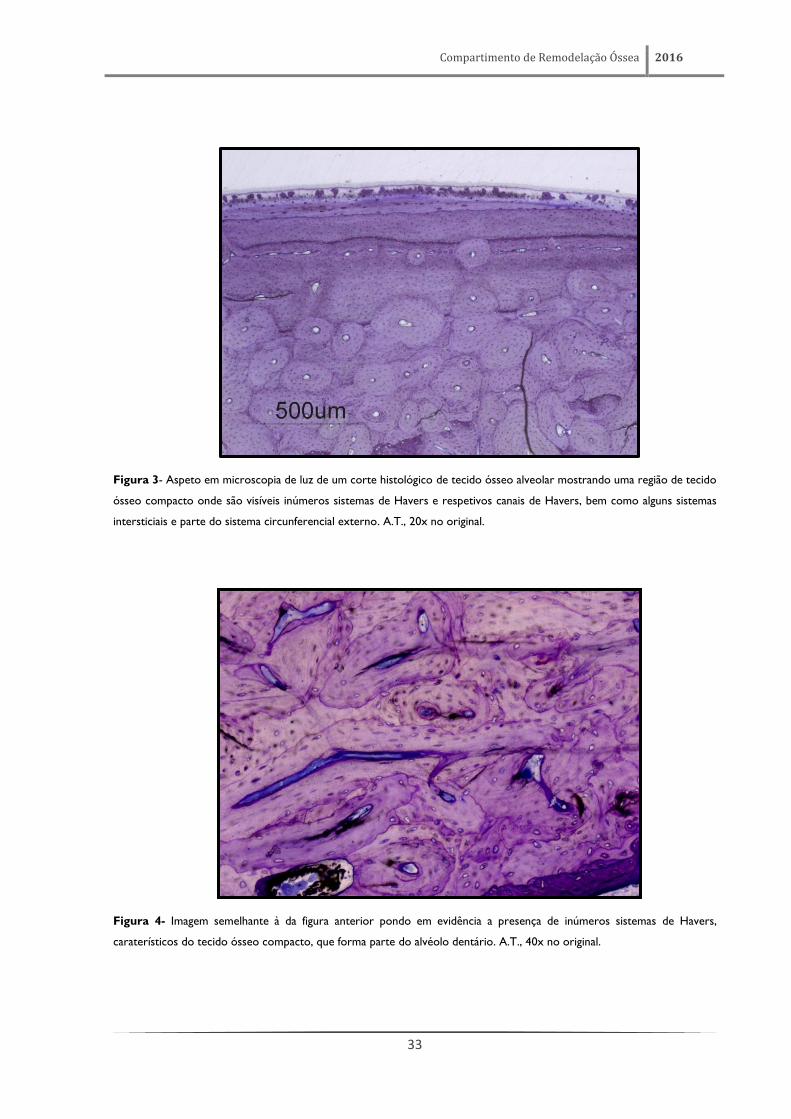
Compartimento de Remodelação Óssea 2016
33
Figura 3- Aspeto em microscopia de luz de um corte histológico de tecido ósseo alveolar mostrando uma região de tecido
ósseo compacto onde são visíveis inúmeros sistemas de Havers e respetivos canais de Havers, bem como alguns sistemas
intersticiais e parte do sistema circunferencial externo. A.T., 20x no original.
Figura 4- Imagem semelhante à da figura anterior pondo em evidência a presença de inúmeros sistemas de Havers,
caraterísticos do tecido ósseo compacto, que forma parte do alvéolo dentário. A.T., 40x no original.
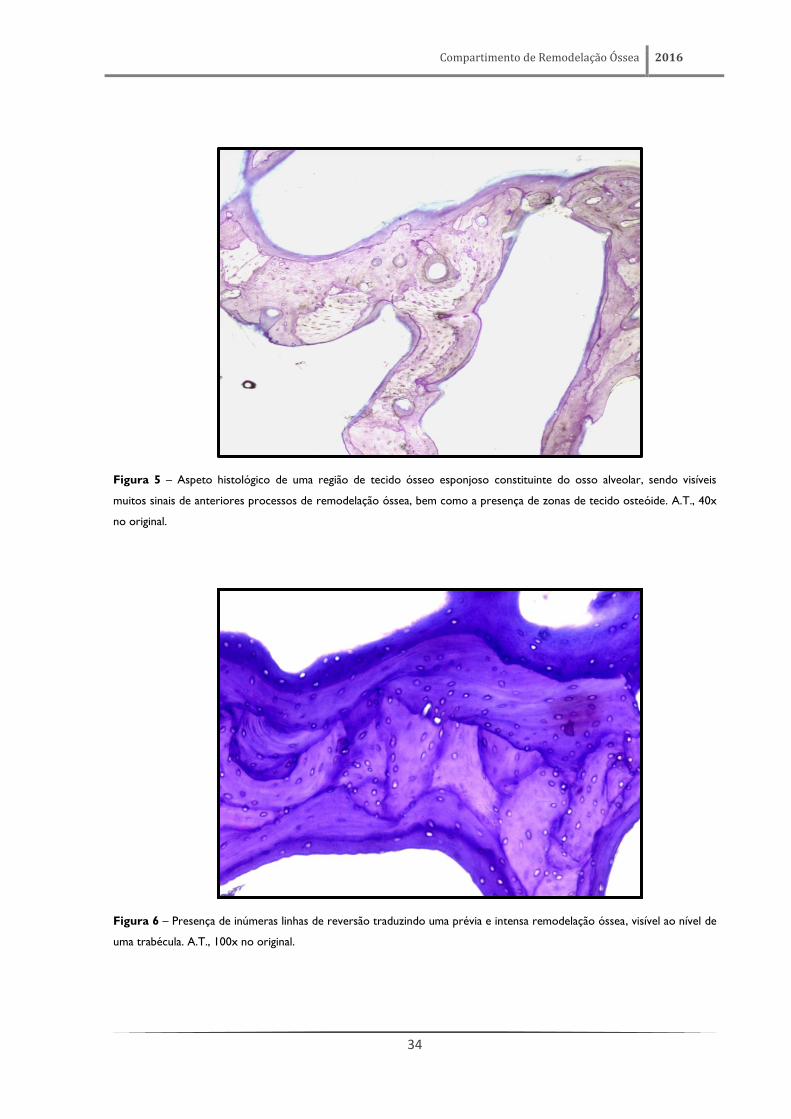
Compartimento de Remodelação Óssea 2016
34
Figura 5 – Aspeto histológico de uma região de tecido ósseo esponjoso constituinte do osso alveolar, sendo visíveis
muitos sinais de anteriores processos de remodelação óssea, bem como a presença de zonas de tecido osteóide. A.T., 40x
no original.
Figura 6 – Presença de inúmeras linhas de reversão traduzindo uma prévia e intensa remodelação óssea, visível ao nível de
uma trabécula. A.T., 100x no original.
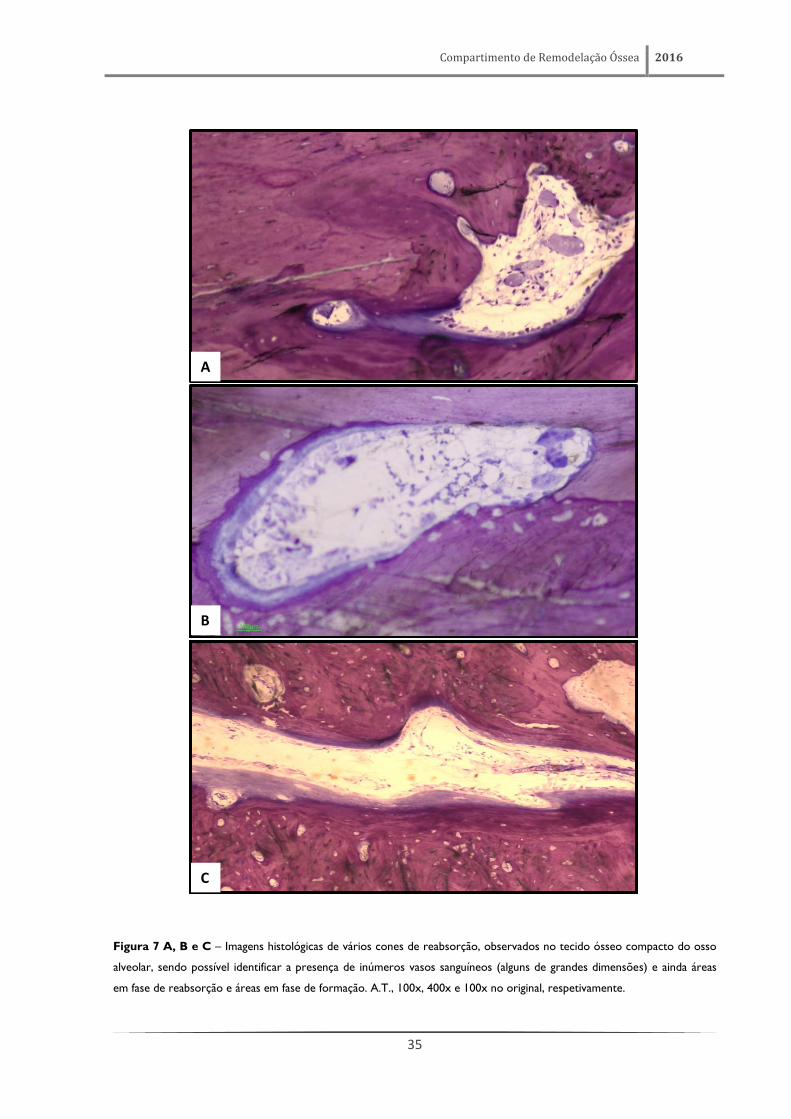
Compartimento de Remodelação Óssea 2016
35
Figura 7 A, B e C – Imagens histológicas de vários cones de reabsorção, observados no tecido ósseo compacto do osso
alveolar, sendo possível identificar a presença de inúmeros vasos sanguíneos (alguns de grandes dimensões) e ainda áreas
em fase de reabsorção e áreas em fase de formação. A.T., 100x, 400x e 100x no original, respetivamente.
A
B
C

Compartimento de Remodelação Óssea 2016
36
Figura 8 – Aspeto histológico de uma região em fase de reabsorção presente numa trabécula óssea constituinte do osso
alveolar. A.T, 20x no original.
Figura 9 - Presença de grande quantidade de osteóide e de células osteoblásticas numa tentativa de reunião de duas
trabéculas ósseas. H.E, 20x no original.
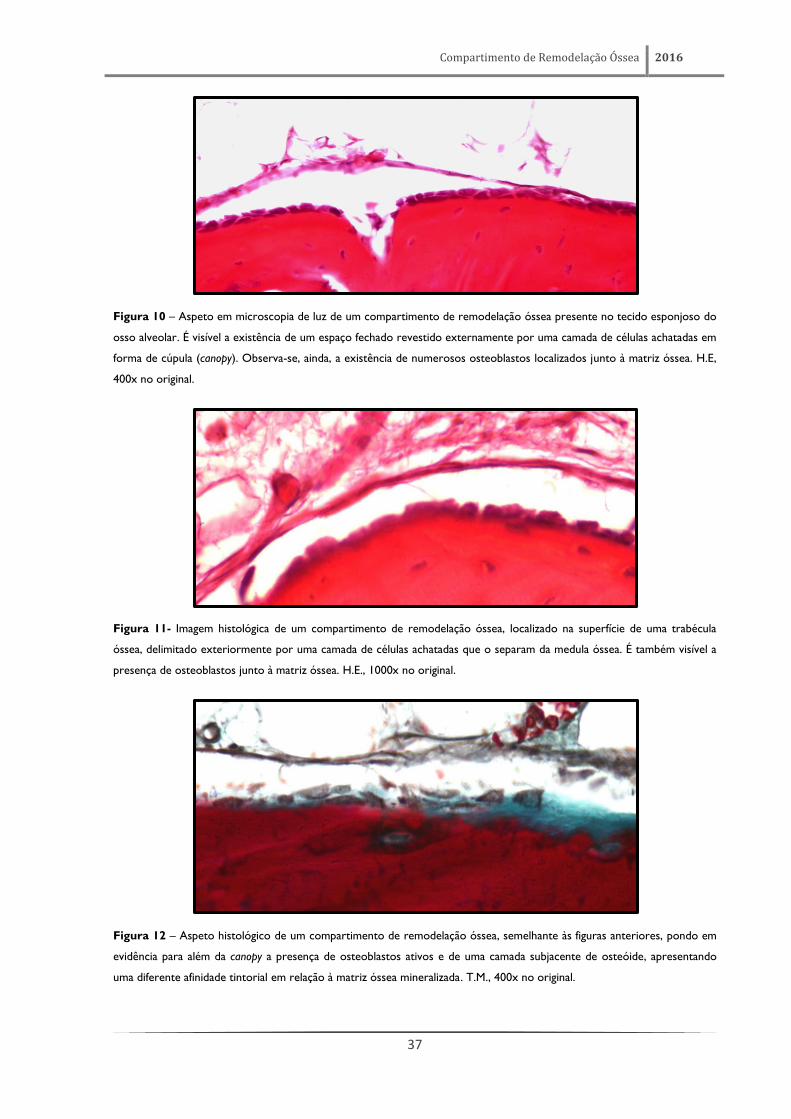
Compartimento de Remodelação Óssea 2016
37
Figura 10 – Aspeto em microscopia de luz de um compartimento de remodelação óssea presente no tecido esponjoso do
osso alveolar. É visível a existência de um espaço fechado revestido externamente por uma camada de células achatadas em
forma de cúpula (canopy). Observa-se, ainda, a existência de numerosos osteoblastos localizados junto à matriz óssea. H.E,
400x no original.
Figura 11- Imagem histológica de um compartimento de remodelação óssea, localizado na superfície de uma trabécula
óssea, delimitado exteriormente por uma camada de células achatadas que o separam da medula óssea. É também visível a
presença de osteoblastos junto à matriz óssea. H.E., 1000x no original.
Figura 12 – Aspeto histológico de um compartimento de remodelação óssea, semelhante às figuras anteriores, pondo em
evidência para além da canopy a presença de osteoblastos ativos e de uma camada subjacente de osteóide, apresentando
uma diferente afinidade tintorial em relação à matriz óssea mineralizada. T.M., 400x no original.
Compartimento de Remodelação Óssea 2016
38
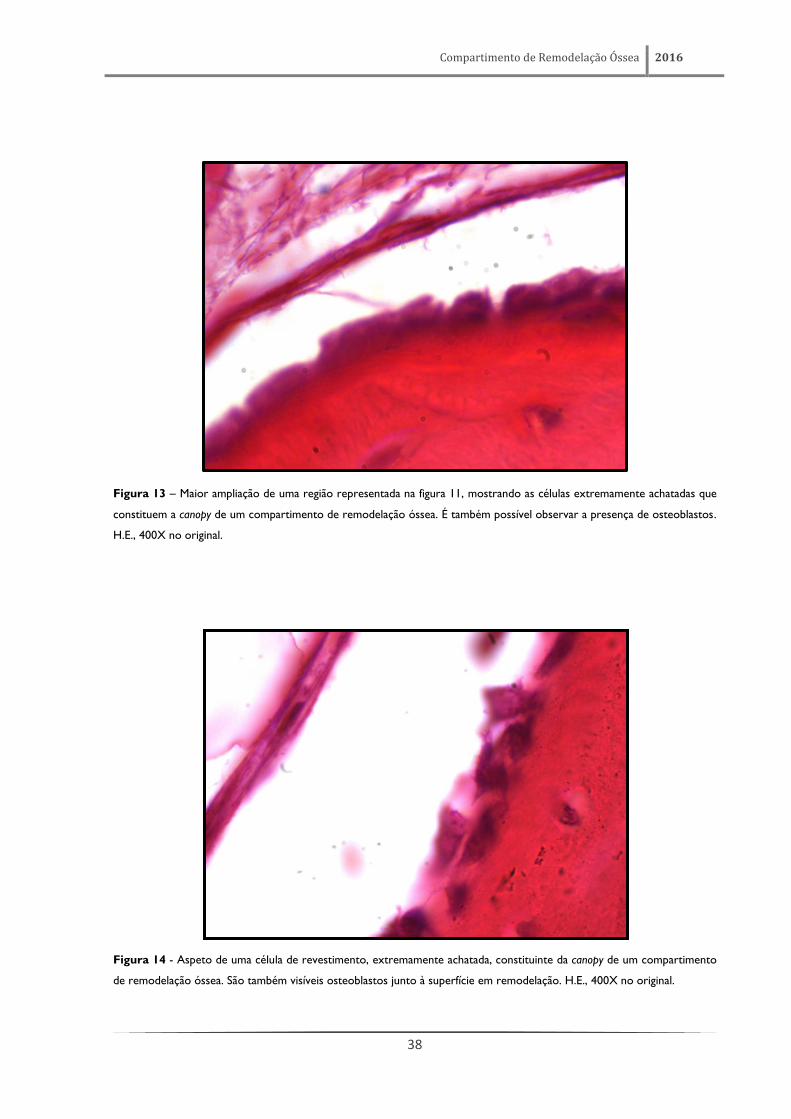
Figura 13 – Maior ampliação de uma região representada na figura 11, mostrando as células extremamente achatadas que
constituem a canopy de um compartimento de remodelação óssea. É também possível observar a presença de osteoblastos.
H.E., 400X no original.
Figura 14 - Aspeto de uma célula de revestimento, extremamente achatada, constituinte da canopy de um compartimento
de remodelação óssea. São também visíveis osteoblastos junto à superfície em remodelação. H.E., 400X no original.
Compartimento de Remodelação Óssea 2016
39
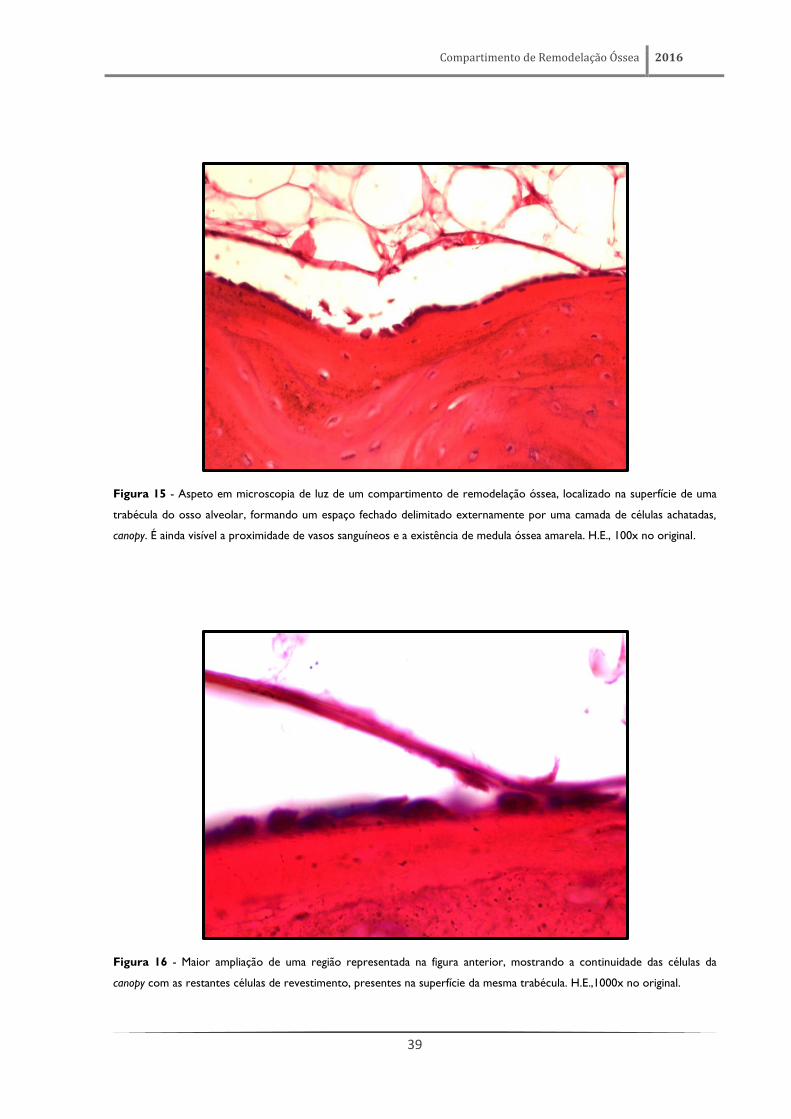
Figura 15 - Aspeto em microscopia de luz de um compartimento de remodelação óssea, localizado na superfície de uma
trabécula do osso alveolar, formando um espaço fechado delimitado externamente por uma camada de células achatadas,
canopy. É ainda visível a proximidade de vasos sanguíneos e a existência de medula óssea amarela. H.E., 100x no original.
Figura 16 - Maior ampliação de uma região representada na figura anterior, mostrando a continuidade das células da
canopy com as restantes células de revestimento, presentes na superfície da mesma trabécula. H.E.,1000x no original.
Compartimento de Remodelação Óssea 2016
40
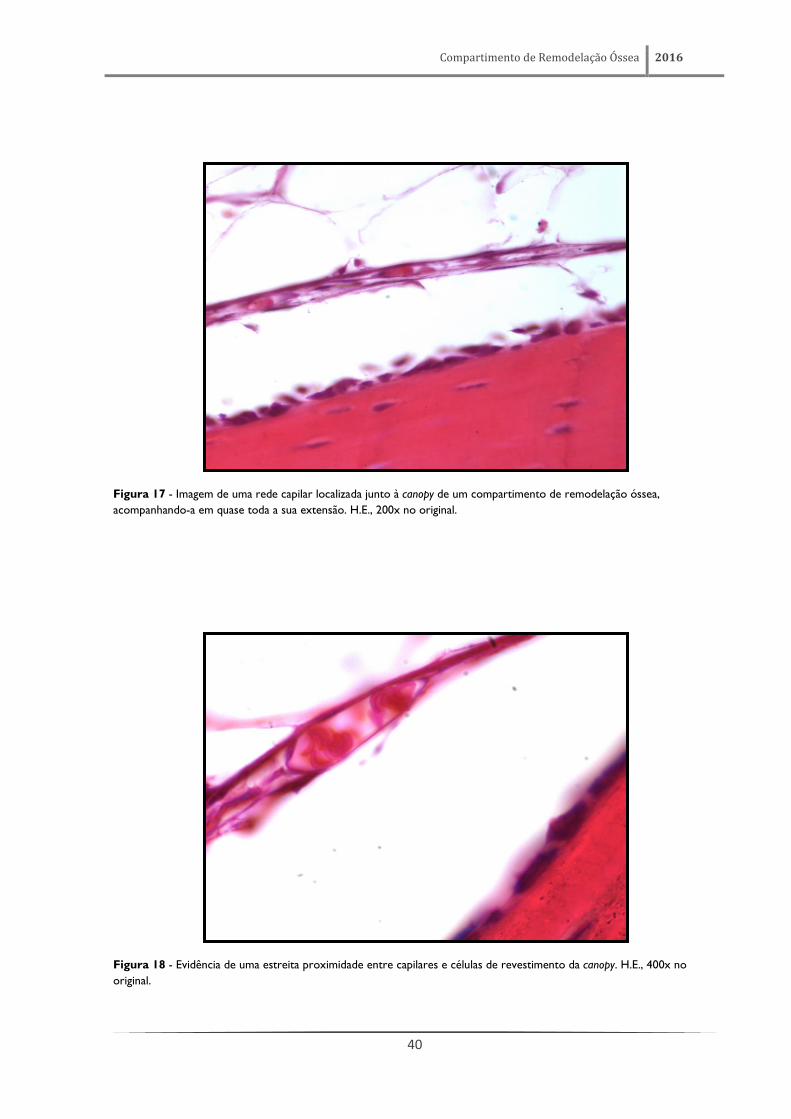
Figura 17 - Imagem de uma rede capilar localizada junto à canopy de um compartimento de remodelação óssea,
acompanhando-a em quase toda a sua extensão. H.E., 200x no original.
Figura 18 - Evidência de uma estreita proximidade entre capilares e células de revestimento da canopy. H.E., 400x no
original.
Compartimento de Remodelação Óssea 2016
41
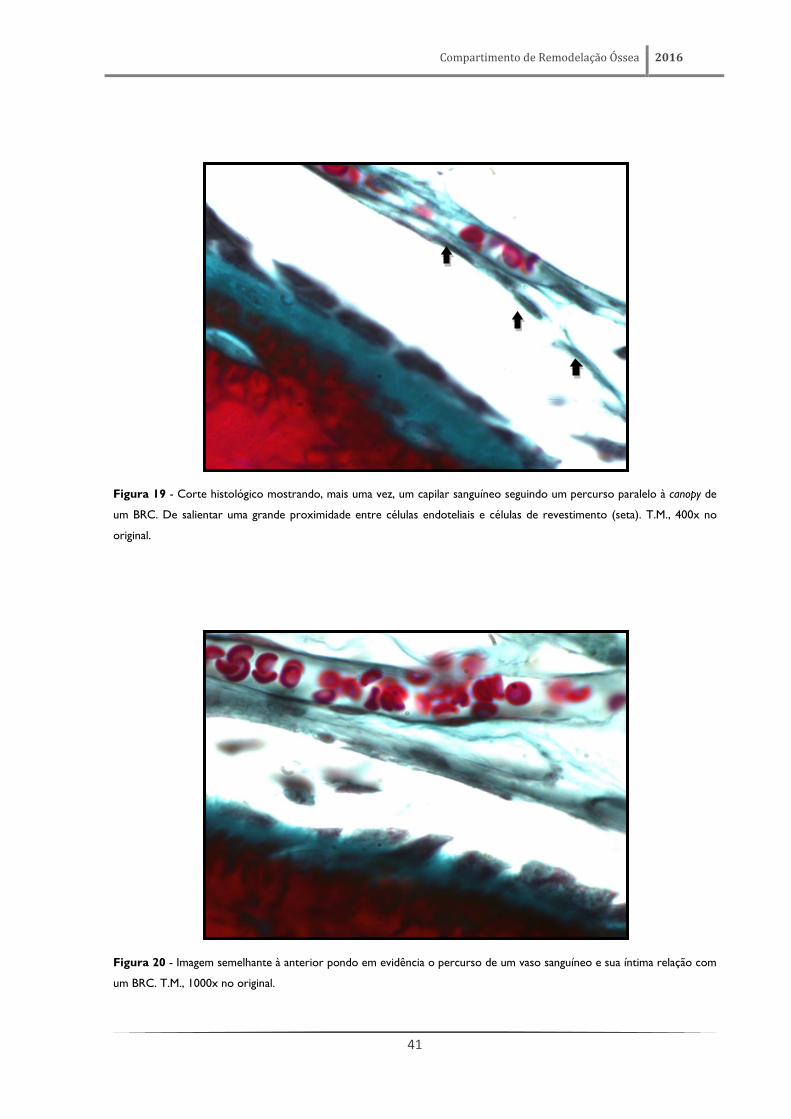
Figura 19 - Corte histológico mostrando, mais uma vez, um capilar sanguíneo seguindo um percurso paralelo à canopy de
um BRC. De salientar uma grande proximidade entre células endoteliais e células de revestimento (seta). T.M., 400x no
original.
Figura 20 - Imagem semelhante à anterior pondo em evidência o percurso de um vaso sanguíneo e sua íntima relação com
um BRC. T.M., 1000x no original.

Compartimento de Remodelação Óssea 2016
42
Figura 21 - Aspeto histológico de um compartimento de remodelação óssea, situado na parede de um cone de
reabsorção, presente no tecido ósseo compacto e a proximidade de um vaso sanguíneo. A.T., 100x no original.
Figura 22 - Imagem de um compartimento de remodelação óssea, em fase de reversão, presente no tecido ósseo
compacto. É possível observar a existência de um vaso sanguíneo particularmente desenvolvido situado em íntima
associação. A.T., 400x no original.
Compartimento de Remodelação Óssea 2016
43
Figura 23 - Corte histológico de material incluído em parafina mostrando um compartimento de remodelação óssea,
existente no tecido ósseo compacto do osso alveolar, e a sua relação com um vaso sanguíneo. T.M., 100x no original.
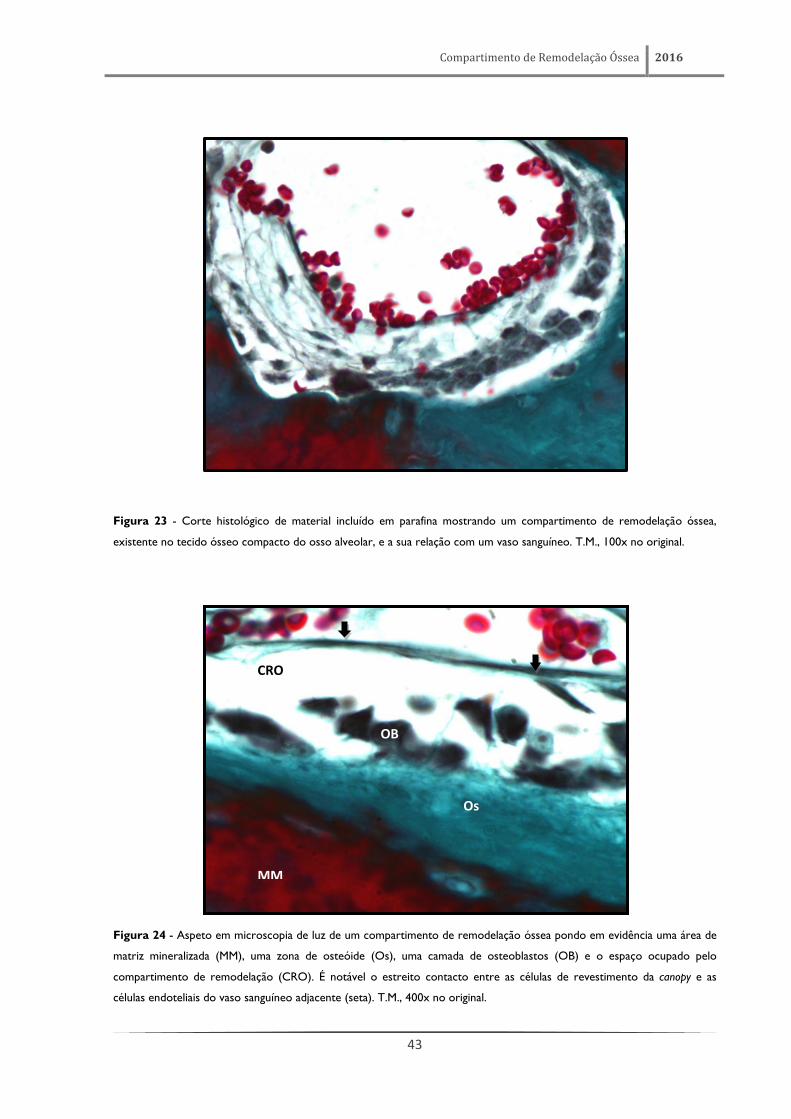
Figura 24 - Aspeto em microscopia de luz de um compartimento de remodelação óssea pondo em evidência uma área de
matriz mineralizada (MM), uma zona de osteóide (Os), uma camada de osteoblastos (OB) e o espaço ocupado pelo
compartimento de remodelação (CRO). É notável o estreito contacto entre as células de revestimento da canopy e as
células endoteliais do vaso sanguíneo adjacente (seta). T.M., 400x no original.
CRO
OB
Os
MM

Compartimento de Remodelação Óssea 2016
44

Compartimento de Remodelação Óssea 2016
45
3.2 Identificação de diferentes etapas do ciclo de remodelação óssea
Para além da deteção de numerosos compartimentos de remodelação no tecido ósseo que
compõe o alvéolo dentário, foi também possível identificar as diferentes etapas que constituem o
ciclo de remodelação óssea, tendo por base as características morfológicas das células localizadas
junto à matriz óssea em remodelação.
A fase de reversão, ainda que representando uma fase intermédia entre a reabsorção e a
formação óssea, parece assumir uma importância fundamental na integração e coordenação de todo
o processo de remodelação. Esta fase é entendida como uma etapa crucial, porém pouco esclarecida,
uma vez que é nesta fase que ocorre a criação de um microambiente osteogénico favorável para que
a transição entre a fase de reabsorção e a fase de formação, do ciclo de remodelação óssea, ocorra
sem perturbações. Assim, pareceu-nos vantajoso dar-lhe um relevo especial apresentando-a no final
e com maior número de imagens, de forma a permitir um melhor esclarecimento das características
histológicas do compartimento nesta fase.
A figura 25 mostra um BRC numa fase de reabsorção sendo possível reconhecer, na superfície
da matriz óssea que forma a parede de um cone de reabsorção, a existência de várias lacunas de
Howship, bem como a presença de vários osteoclastos. É também percetível a existência de outras
células, incluindo as células de revestimento da canopy e a proximidade de um vaso sanguíneo. Na
figura 26 observa-se, com mais detalhe, uma confluência de vários osteoclastos, formando células
gigantes multinucleadas, localizadas em cavidades por elas escavadas na superfície da matriz óssea. É
ainda possível identificar uma nítida interface, entre o osteoclasto e a matriz mineralizada,
demarcando o local onde irá surgir uma linha cimentante. Verifica-se também a existência de várias
zonas adjacentes à matriz mineralizada, contendo um material de aspeto fibrilar que deverá
corresponder a matriz orgânica (maioritariamente colagénio) ainda não completamente digerida.
***
Nas figuras 27 e 28 é possível observar em várias áreas de tecido ósseo compacto, a
diversidade celular e riqueza vascular existente num compartimento de remodelação em fase de
formação óssea. Com efeito, para além de uma zona de matriz osteóide, é visível uma extensa
camada de células osteoblásticas, uma complexa profusão celular e uma intrincada rede capilar que,
no seu conjunto, formam uma BMU.
Também as figuras 29-32 mostram, na superfície de trabéculas de tecido ósseo esponjoso, a
existência áreas de remodelação em fase de formação óssea caracterizadas pela presença de células
osteoblásticas, adjacentes à matriz osteóide por elas elaborada, recobertas por um compartimento
de remodelação. Neste compartimento, é possível observar as células de revestimento que formam a

Compartimento de Remodelação Óssea 2016
46
canopy, para além de outras, de difícil identificação em microscopia de luz. É também possível
verificar a proximidade de capilares sanguíneos, mostrando um trajeto paralelo à canopy. Embora sem
possibilidades de recurso a métodos imunocitoquímicos parece-nos poder afirmar, com base nas suas
características morfológicas e localização, intimamente adjacentes às células endoteliais, a presença
de células (figuras 30-32) que consideramos como pericitos.
***
Ainda que o processo de reabsorção e de formação óssea sejam considerados fenómenos
intimamente ligados, visto que o mecanismo de reabsorção osteoclástica estimula a atividade
reparadora osteoblástica e que os osteoblastos, ou as suas células percursoras sejam recrutados
exatamente para os locais onde previamente se verificou um processo de reabsorção, estes dois
tipos de células raramente estão presentes em simultâneo na mesma superfície óssea. De facto,
verifica-se um intervalo de tempo de várias semanas entre o desaparecimento dos osteoclastos e o
aparecimento de osteoblastos no mesmo local. Este período intermediário, ainda mal esclarecido, é
geralmente classificado como uma fase de reversão e caracterizado pela presença de uma diferente
população celular. Assim, num compartimento de remodelação óssea em fase de reversão, podemos
identificar numerosas células alongadas com núcleo achatado, de certo modo semelhantes às células
de revestimento, porém apresentando-se mais volumosas e em maior número. As figuras 33-36
documentam bem o que acabámos de descrever, pondo em evidência vários compartimentos de
remodelação óssea em fase de reversão, muito bem definidos, contendo no seu interior inúmeras
células alongadas que nem sempre se diferenciam das células de revestimento da canopy. A
proximidade entre estas células e as células endoteliais dos vasos sanguíneos, que se situam em seu
redor, é notória (figuras 35 e 36) sendo problemática, nesta fase, e sem recurso a técnicas
imunocitoquímicas ou ultraestruturais, classificá-las como pericitos, células de revestimento da canopy
ou pré-osteoblastos.
Considerando que nesta etapa o BRC se apresenta como um importante reservatório de
citocinas, fatores de crescimento e maturação, facilmente se compreende a presença de uma grande
densidade de células, de difícil caracterização, por se encontrarem provavelmente em diferentes fases
de diferenciação e maturação osteoblástica. Uma vez que tanto os pericitos como as células da
canopy apresentam um forte potencial osteogénico, parece-nos perfeitamente aceitável a dificuldade
de identificação destas células tendo por base apenas as observações em microscopia de luz. Será
pois imprescindível avançar com outros métodos de caracterização no sentido de esclarecer a
identificação destas células.
No entanto a qualidade das imagens aqui apresentadas, decorrente do seu processamento em
metilmetacrilato, permitem, só por si, um avanço importante para um melhor conhecimento da
constituição histológica dos compartimentos de remodelação óssea em fase de reversão.
Compartimento de Remodelação Óssea 2016
47
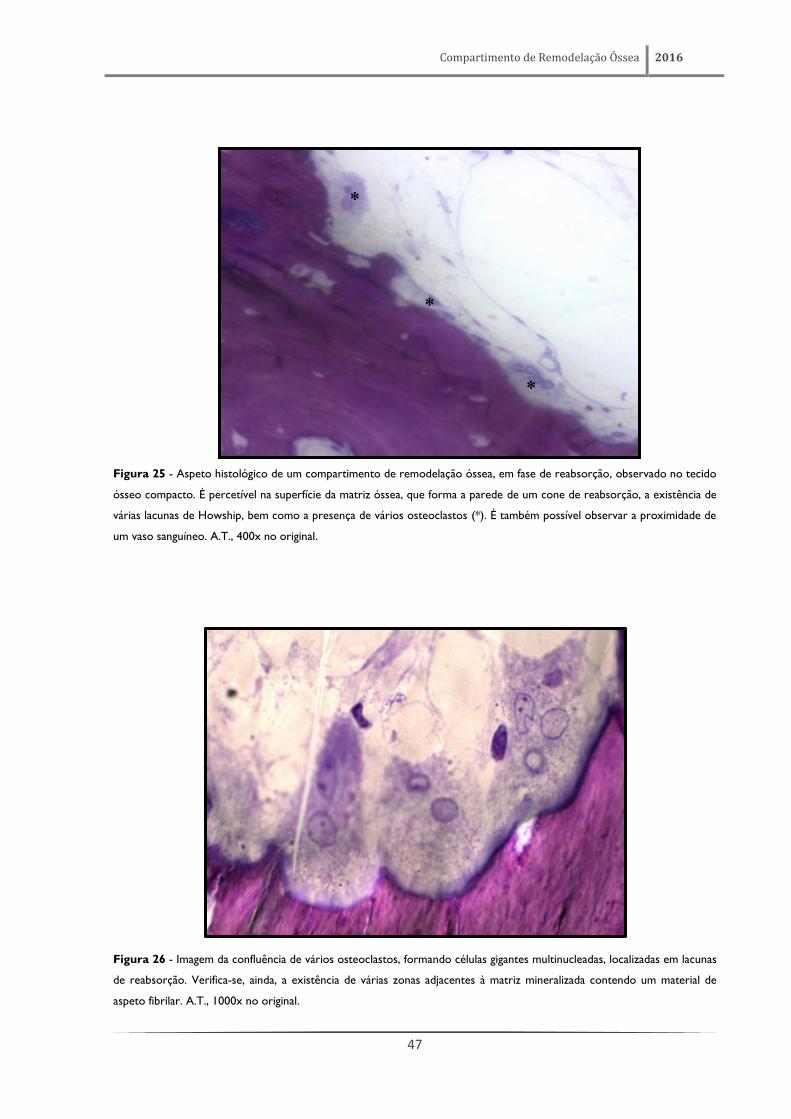
Figura 25 - Aspeto histológico de um compartimento de remodelação óssea, em fase de reabsorção, observado no tecido
ósseo compacto. É percetível na superfície da matriz óssea, que forma a parede de um cone de reabsorção, a existência de
várias lacunas de Howship, bem como a presença de vários osteoclastos (*). É também possível observar a proximidade de
um vaso sanguíneo. A.T., 400x no original.
Figura 26 - Imagem da confluência de vários osteoclastos, formando células gigantes multinucleadas, localizadas em lacunas
de reabsorção. Verifica-se, ainda, a existência de várias zonas adjacentes à matriz mineralizada contendo um material de
aspeto fibrilar. A.T., 1000x no original.
*
*
*
Compartimento de Remodelação Óssea 2016
48
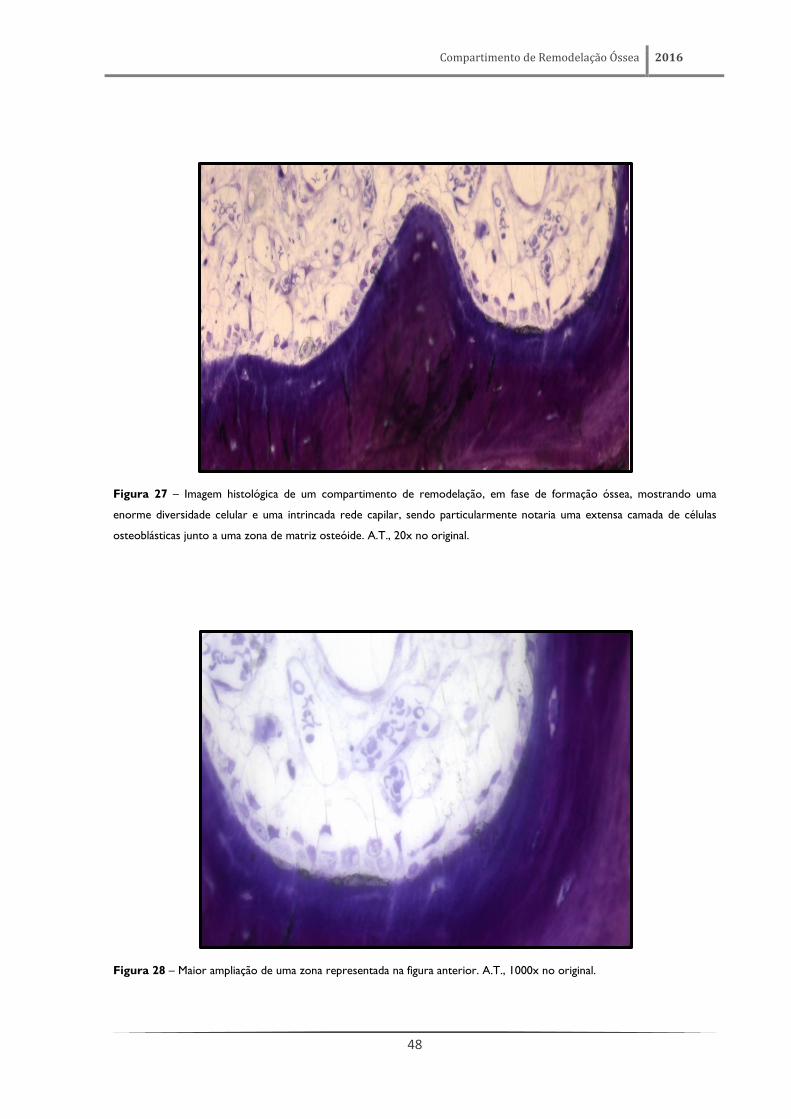
Figura 27 – Imagem histológica de um compartimento de remodelação, em fase de formação óssea, mostrando uma
enorme diversidade celular e uma intrincada rede capilar, sendo particularmente notaria uma extensa camada de células
osteoblásticas junto a uma zona de matriz osteóide. A.T., 20x no original.
Figura 28 – Maior ampliação de uma zona representada na figura anterior. A.T., 1000x no original.
Compartimento de Remodelação Óssea 2016
49
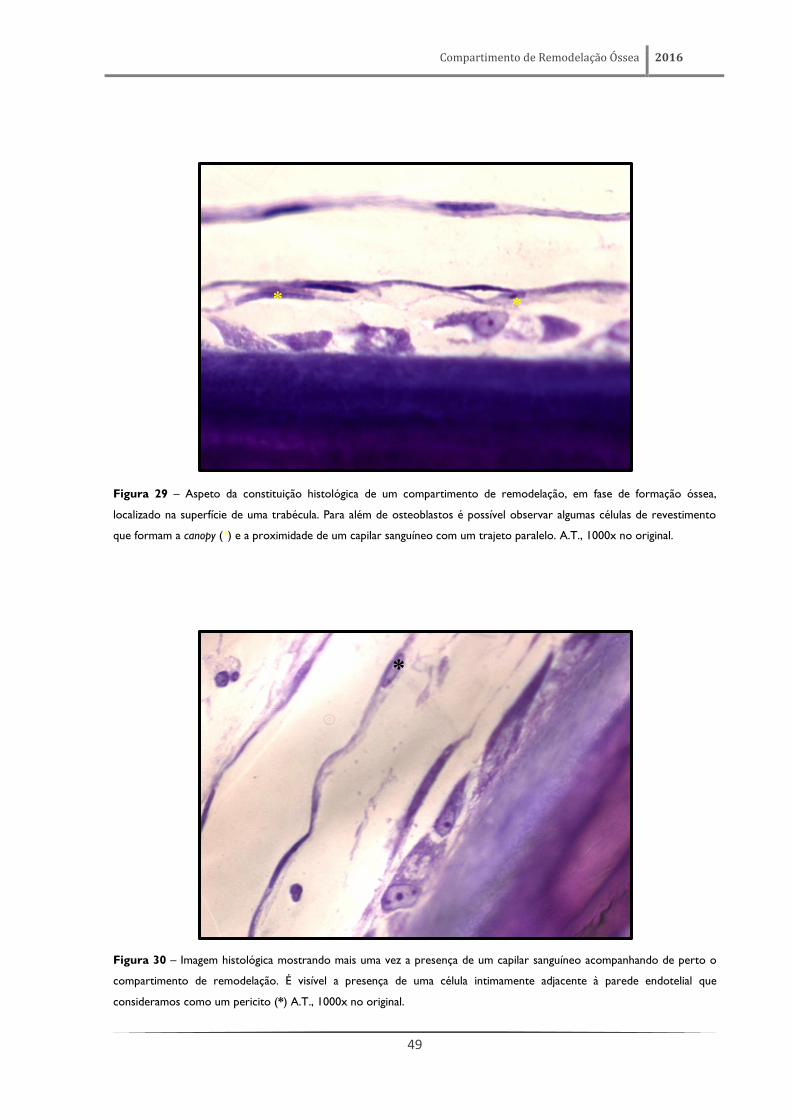
Figura 29 – Aspeto da constituição histológica de um compartimento de remodelação, em fase de formação óssea,
localizado na superfície de uma trabécula. Para além de osteoblastos é possível observar algumas células de revestimento
que formam a canopy (*) e a proximidade de um capilar sanguíneo com um trajeto paralelo. A.T., 1000x no original.
Figura 30 – Imagem histológica mostrando mais uma vez a presença de um capilar sanguíneo acompanhando de perto o
compartimento de remodelação. É visível a presença de uma célula intimamente adjacente à parede endotelial que
consideramos como um pericito (*) A.T., 1000x no original.
* *
*
Compartimento de Remodelação Óssea 2016
50
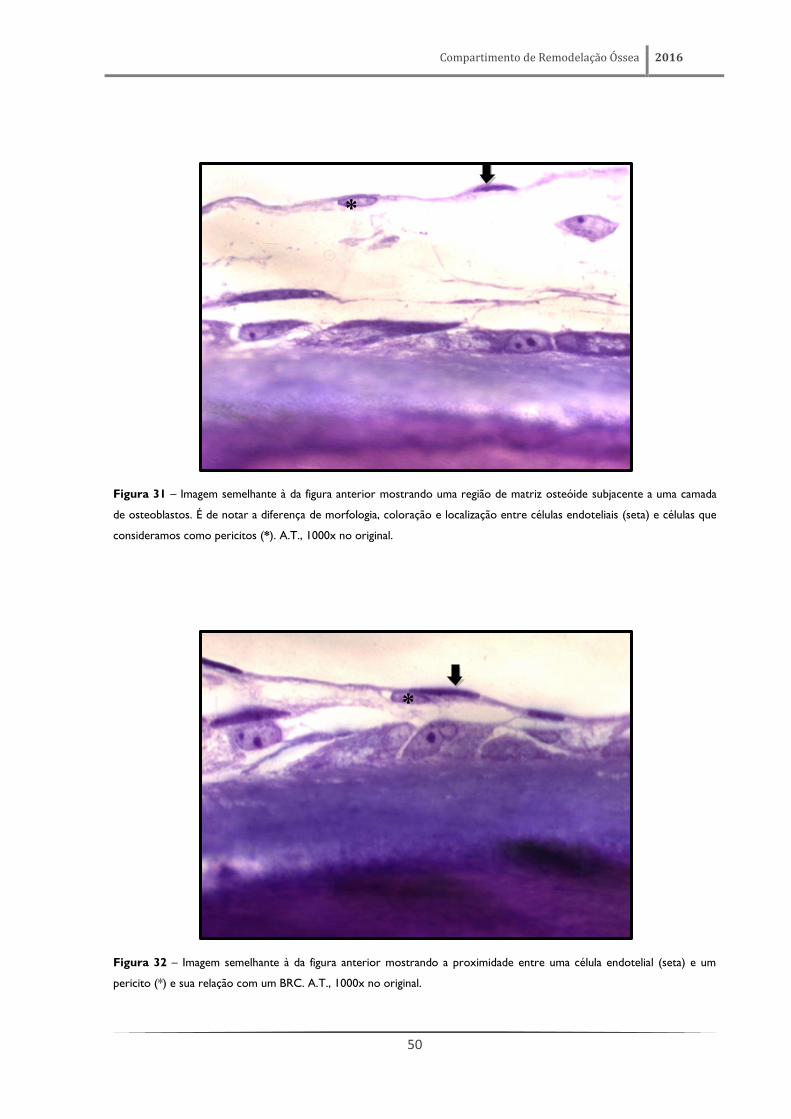
Figura 31 – Imagem semelhante à da figura anterior mostrando uma região de matriz osteóide subjacente a uma camada
de osteoblastos. É de notar a diferença de morfologia, coloração e localização entre células endoteliais (seta) e células que
consideramos como pericitos (*). A.T., 1000x no original.
Figura 32 – Imagem semelhante à da figura anterior mostrando a proximidade entre uma célula endotelial (seta) e um
pericito (*) e sua relação com um BRC. A.T., 1000x no original.
*
*

Compartimento de Remodelação Óssea 2016
51
Figura 33 – Aspeto de um compartimento de remodelação óssea, em fase de reversão, presente no tecido ósseo
compacto contendo no seu interior uma grande densidade de células alongadas, de difícil caracterização, não sendo possível
identificar células com morfologia de osteoblastos maduros. A.T., 400x no original.
Figura 34 – Imagem semelhante à da figura anterior mostrando um compartimento de remodelação óssea, em fase de
reversão, apresentando no seu interior células com diferente morfologia. É também de notar a existência de um vaso
sanguíneo em íntima aposição ao BRC. A.T., 400x no original.
*

Compartimento de Remodelação Óssea 2016
52
Figura 35 - Corte histológico mostrando um compartimento de remodelação óssea que contém uma população celular
difícil de definir, adjacente a uma vaso sanguíneo, sendo notável a proximidade entre células endoteliais (seta preta) e
pericitos ou células constituintes da canopy (seta vermelha). É também possível observar a presença do que consideramos
serem fibras de colagénio em redor dos vasos e no interior do compartimento. A.T., 200x no original.
Figura 36 - Aspeto semelhante ao da figura anterior pondo em evidência a morfologia e localização da população celular
presente num BRC durante uma fase de reversão. A.T., 400x no original.

Compartimento de Remodelação Óssea 2016
53
3.3 Deteção de compartimentos de remodelação presentes na dentina e
cemento
De modo semelhante ao que observámos nas regiões de tecido ósseo em remodelação,
também nas zonas de remodelação de cemento e dentina, foi possível identificar uma estrutura em
tudo idêntica ao compartimento de remodelação óssea. Porém, as áreas de reabsorção da dentina,
adjacentes ao cemento, são reparadas apenas com formação de novo cemento, e não de dentina.
Uma vez que a constituição histológica do cemento é muito idêntica à do tecido ósseo, seria
previsível que o seu processo de remodelação apresentasse também características muito
semelhantes. Com efeito, para além de zonas de reabsorção do cemento que podem atingir a
dentina, podemos identificar a existência de compartimentos de remodelação, revestidos por uma
canopy e em estreita proximidade de vasos sanguíneos.
Na figura 37 é possível observar uma zona de reabsorção de cemento que atinge a dentina,
formando um compartimento de remodelação, revestido por uma camada de células achatadas, que
consideramos como uma canopy, muito idêntico ao que se observa no tecido ósseo.
Uma fase correspondente à fase de formação óssea foi também reconhecida no processo de
remodelação do cemento. Com efeito, as figuras 38 e 39 põem em evidência a existência de um
compartimento de remodelação junto a uma área de cemento recoberta por cementóide e por
cementoblastos. A figura 40 permite observar, com mais pormenor, as células da canopy, o espaço
ocupado pelo compartimento e uma camada de cementoblastos (muito semelhantes a osteoblastos),
adjacente a uma zona de cemento não mineralizado. É também possível observar uma faixa de matriz
mineralizada de cemento e uma zona de dentina com túbulos dentinários.
A figura 40 mostra também uma área de reabsorção na dentina e no cemento, ocupada por
um espaço onde são visíveis diferentes tipos de células, bem como um vaso sanguíneo apresentando,
no seu conjunto, um aspeto muito semelhante ao que se verifica nos cones de reabsorção,
característicos do tecido ósseo compacto. Neste corte, é no entanto difícil distinguir as células de
revestimento da canopy das células endoteliais do vaso sanguíneo. A população celular presente no
compartimento de remodelação adjacente à superfície da dentina e do cemento, sem possibilidades
de métodos imunocitoquímicos, é de difícil caracterização. Todavia, por comparação com o tecido
ósseo, pode ser classificada como um compartimento de remodelação em fase de reversão.
A existência do que consideramos ser compartimentos de remodelação no cemento e na
dentina, com características semelhantes ao que se observa no tecido ósseo, não foi ainda
demonstrada nem publicada.
De referir, no entanto, que quando em comparação com o tecido ósseo alveolar encontramos
no cemento e dentina radicular compartimentos de remodelação em muito menor número e com

Compartimento de Remodelação Óssea 2016
54
uma constituição muito menos completa, apresentando com frequência perdas da canopy. Neste
contexto será ainda de assinalar que muitas vezes observamos áreas com uma excessiva reabsorção
e deficiente reparação.
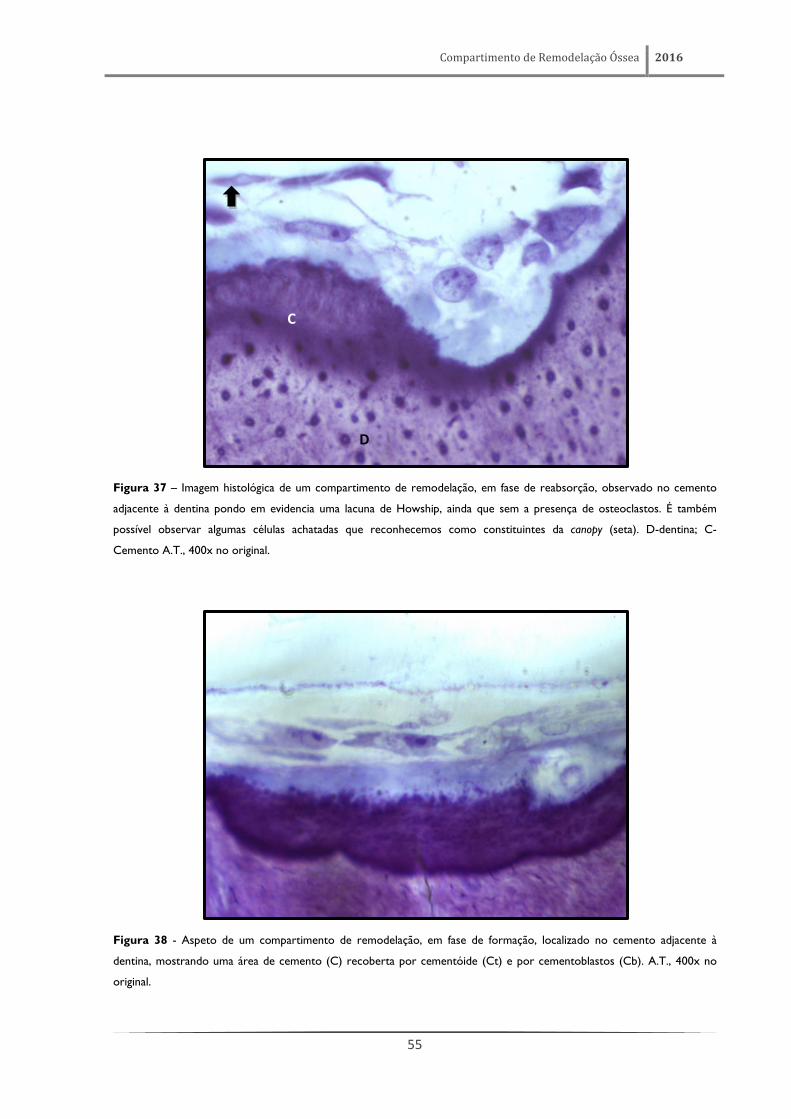
Compartimento de Remodelação Óssea 2016
55
Figura 37 – Imagem histológica de um compartimento de remodelação, em fase de reabsorção, observado no cemento
adjacente à dentina pondo em evidencia uma lacuna de Howship, ainda que sem a presença de osteoclastos. É também
possível observar algumas células achatadas que reconhecemos como constituintes da canopy (seta). D-dentina; C-
Cemento A.T., 400x no original.
Figura 38 - Aspeto de um compartimento de remodelação, em fase de formação, localizado no cemento adjacente à
dentina, mostrando uma área de cemento (C) recoberta por cementóide (Ct) e por cementoblastos (Cb). A.T., 400x no
original.
D
C
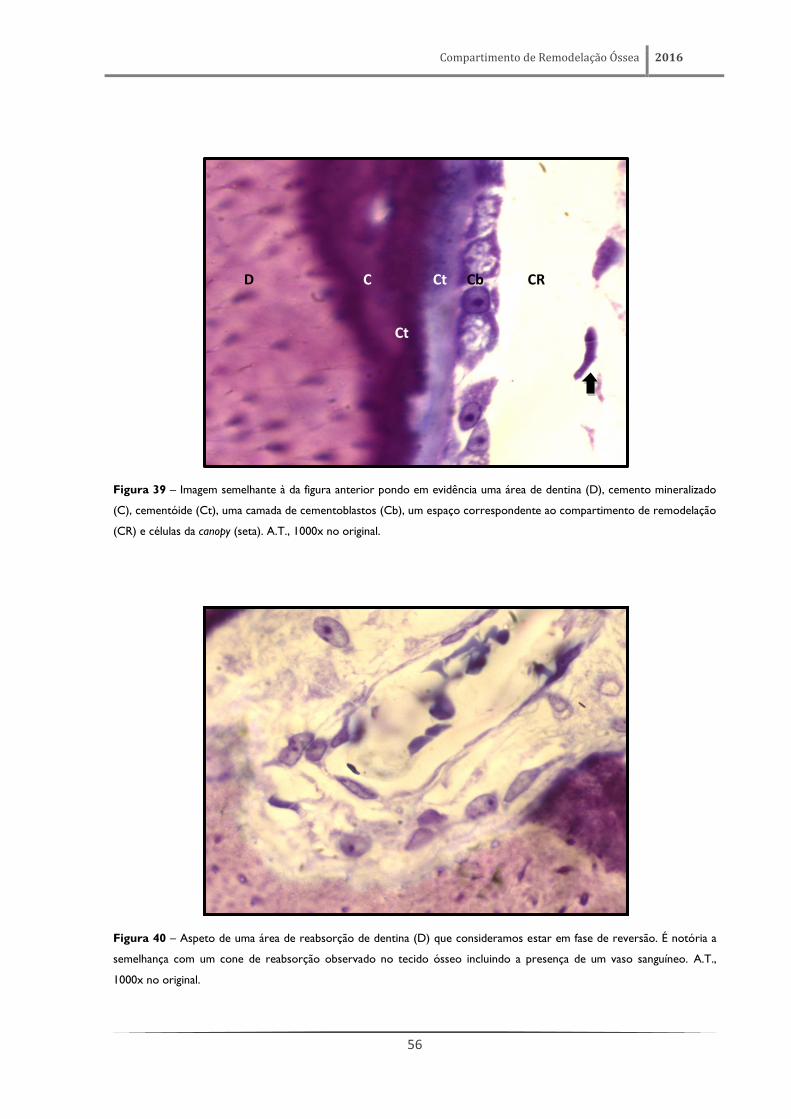
Compartimento de Remodelação Óssea 2016
56
Figura 39 – Imagem semelhante à da figura anterior pondo em evidência uma área de dentina (D), cemento mineralizado
(C), cementóide (Ct), uma camada de cementoblastos (Cb), um espaço correspondente ao compartimento de remodelação
(CR) e células da canopy (seta). A.T., 1000x no original.
Figura 40 – Aspeto de uma área de reabsorção de dentina (D) que consideramos estar em fase de reversão. É notória a
semelhança com um cone de reabsorção observado no tecido ósseo incluindo a presença de um vaso sanguíneo. A.T.,
1000x no original.
Ct
D C Ct Cb CR

Compartimento de Remodelação Óssea 2016
57
3. Discussão
__________________________________________________________

Compartimento de Remodelação Óssea 2016
58

Compartimento de Remodelação Óssea 2016
59
Com o presente trabalho pretendemos contribuir para uma maior compreensão dos
processos de remodelação óssea proporcionando um melhor conhecimento das características
histológicas dos compartimentos de remodelação óssea.
Considerando todo o esforço, tempo e elevado custo despendidos na preparação de cortes
histológicos realizados em tecido ósseo, no âmbito de vários estudos experimentais relacionados
com os processos de regeneração óssea95,96,97, realizados na Área de Medicina Dentária, pareceu-nos
manifestamente um desperdício não aproveitar este material para outros fins. Tendo em conta, ainda,
que muitos dos cortes histológicos efetuados eram provenientes de material incluído em
metilmetacrilato, efetuados com recurso ao sistema Exact®, que constitui um equipamento único no
país, seria imperdoável não rentabilizar este material. Para além de todas estas razões importa
sublinhar o facto de serem sobretudo pessoas com preparação em ciências morfológicas as mais
habilitadas para levar a cabo uma análise e interpretação abrangente e pormenorizada de toda a
informação presente num corte histológico. Assim, considerando toda a colaboração prestada no
desenvolvimento dos trabalhos atrás referidos, decidimos levar a cabo uma análise qualitativa em
microscopia de luz, no âmbito da histologia básica, para deteção e identificação de compartimentos
de remodelação óssea (BRC) nas áreas de tecido ósseo alveolar mandibular, incidindo
fundamentalmente na observação e registo das suas características histomorfológicas. Decidimos
ainda estender as observações para as áreas de dentina e do cemento, uma vez que são também
tecidos mineralizados com capacidade de remodelação.
Os resultados apresentados constituem, a nosso ver, um conjunto de observações únicas para
o esclarecimento da constituição histológica dos compartimentos de remodelação óssea, durante as
diferentes fases do ciclo de remodelação, que entendemos de grande utilidade didática e científica.
Ainda que não nos tivesse sido possível realizar estudos quantitativos, foi por demais evidente
que o número e variedade de imagens correspondentes a compartimentos de remodelação óssea,
que apresentamos nos resultados, demonstram bem a intensidade e frequência de aparecimento de
processos de remodelação que se verificam no osso alveolar. A dinâmica observada no osso alveolar
deve-se ao facto de apresentar uma constante e rápida formação e remodelação óssea resultante
inicialmente do crescimento radicular e erupção dentária e, posteriormente, das exigências funcionais
da mastigação1, 29, 101. Os pequenos movimentos que experimentam continuamente os dentes são as
principais causas de remodelação do osso alveolar.
A observação de um grande número de processos de remodelação verificados no osso
alveolar poderá estar ainda reforçada pela sua própria constituição estrutural. Com efeito, as áreas
de tecido ósseo adjacentes ao ligamento periodontal (osso alveolar propriamente dito) contêm

Compartimento de Remodelação Óssea 2016
60
numerosas fibras de colagénio (osso fasciculado) provenientes do ligamento periodontal e que por
sua vez se inserem diretamente no cemento que reveste a dentina radicular1,6,28-32. Assim, qualquer
movimento dentário resultante da mastigação será fácil e rapidamente propagado a este tecido
ósseo, estimulando o desenvolvimento de processos de remodelação, visando uma constante
otimização e adaptação às suas exigências funcionais. Por outro lado, existe também um grande
número de comunicações entre o osso alveolar propriamente dito e o ligamento periodontal através
da presença de muitos canais de Volkman que transportam vasos sanguíneos estabelecendo uma
rápida e ampla ligação entre tecido ósseo e ligamento periodontal1,6,28. Qualquer variação fisiológica
verificada no órgão dentário ou qualquer situação fisiopatológica que ocorra no dente ou nas suas
estruturas de suporte e proteção (periodonto de suporte e de proteção) terão uma repercussão
direta no tecido ósseo que constitui o osso alveolar. A ortodontia aproveita toda a plasticidade que
lhe oferece o osso alveolar para, em conjunto com o ligamento periodontal, movimentar o dente
sem alterar as suas relações2, 28-32,121. O conhecimento deste jogo de forças, as suas consequências e
os mecanismos moleculares a elas subjacentes, abrem cada vez maiores perspetivas de intervenção e
eficácia na área de ortodontia e implantologia.
Uma nota mais para referir que os processos cariogénios e de periodontite101,169 são
acompanhados com grande frequência por fenómenos de remodelação óssea que na sua maioria
conduzem a uma perda de massa óssea por excesso de processos de reabsorção.
De modo semelhante ao que observamos nas regiões do tecido ósseo alveolar também nas
zonas de remodelação do cemento e dentina foi possível identificar estruturas com uma constituição
histológica em tudo idêntica ao compartimento de remodelação óssea, porém em muito menor
número e com uma constituição muito menos completa. Com efeito, detetamos com frequência
perdas da canopy e áreas com uma excessiva reabsorção e deficiente reparação. A este respeito
importa salientar que os processos de remodelação do cemento estão frequentemente associados a
processos inflamatórios e, embora partilhe grandes semelhanças morfológicas, funcionais e
bioquímicas com o tecido ósseo, é no entanto, um tecido avascular e bastante desestruturado28,124,125.
Assim, é facilmente compreensível que no cemento não estejam reunidas as condições ideais para
uma adequada formação de compartimentos de remodelação como se observam no tecido ósseo.
De qualquer modo, estes resultados, presença de compartimentos de remodelação no cemento e
dentina, tanto quanto é do nosso conhecimento, não foram ainda referidos nem publicados.
A existência de um compartimento de remodelação com características semelhantes (tanto no
tecido ósseo esponjoso como no tecido ósseo compacto como no cemento e dentina) e ainda a
manutenção do mesmo alinhamento ordenado de eventos pressupõe a existência de um mecanismo
comum a todos estes processos de remodelação. Assim, os parágrafos posteriores pretendem

Compartimento de Remodelação Óssea 2016
61
apresentar uma breve discussão sobre os mecanismos subjacentes e a problemática associada e aos
processos de remodelação dos tecidos mineralizados, tendo em conta os resultados apresentados.
***
O envolvimento direto das estruturas vasculares no desencadear dos fenómenos de
remodelação óssea e na sua manutenção não está ainda completamente esclarecido, mas são cada
vez mais as evidências que apontam nesta direção25,40,43,100,123-129. Este facto faz todo o sentido uma
vez que os mecanismos de angiogénese desempenham um papel crucial em todos os processos de
reabsorção, formação, crescimento, reparação de fraturas e regeneração óssea25,40,126,128. O
desenvolvimento de uma rede de vasos sanguíneos surgindo como uma estrutura sempre presente
ao longo de todas as fases do ciclo de remodelação óssea demonstra bem que a íntima relação entre
estruturas vasculares e compartimentos de remodelação terá uma importância crucial no início,
manutenção e coordenação de todo o processo de remodelação25,82,100,108.
Na verdade, um reforço dos mecanismos de angiogénese e a aproximação e/ou adesão de
capilares sanguíneos às células de revestimento de uma região da superfície óssea, que vai necessitar
de um processo de remodelação, parece constituir um dos primeiros sinais de ativação de um ciclo
de remodelação25,99,100,132. Este facto, que assinala a conversão de uma superfície óssea de um estado
quiescente para um estado ativo de remodelação, é acompanhado por alterações na morfologia e
comportamento das suas células de revestimento48,82. Com efeito, a fase de ativação do ciclo de
remodelação óssea é tradicionalmente caracterizada pela digestão da fina camada de endósteo, da
responsabilidade das células de revestimento, com recurso a MMP38,48, 65-70, conduzindo a uma mais
fácil exposição da matriz mineralizada. Ao mesmo tempo, as células de revestimento presentes nesta
região destacam-se da matriz mineralizada, localizando e confinando a área a ser remodelada, dando
início à formação de uma estrutura em forma de cúpula – canopy - e ao desenvolvimento de um
compartimento de remodelação óssea - BRC. As células que constituem a canopy são,
confirmadamente27,48,133-135, células da linha osteoblástica, que expressam a maioria dos marcadores
característicos do fenótipo osteoblástico (fosfatase alcalina, osteocalcina, osteonectina, IGF, TGFβ,
FGFβ, OPG e RANKL, entre outros)24,108,131,136, mas que parecem regredir para um estádio precoce
de diferenciação, mostrando inclusive um índice de proliferação superior a 10 %82,131.
Estes processos (formação da canopy e desenvolvimento e aproximação de estruturas
vasculares) são acompanhados por uma forte migração e recrutamento de células pré-osteoclásticas,
dirigindo-se exatamente para a região em causa, e por um reforço dos mecanismos de
osteoclastogénese. A superfície da matriz óssea, desnudada e exposta, atrai fortemente as células
percursoras dos osteoclastos contribuindo ainda para a sua maturação e ativação27,38,48,130. Com
efeito, só em contacto direto com a matriz óssea mineralizada, e sob a influência das células da linha
osteoblástica, é que os pré-osteoclastos se fundem formando células multinucleadas capazes de uma

Compartimento de Remodelação Óssea 2016
62
verdadeira atividade de reabsorção. As células pré-osteoclásticas podem alcançar o compartimento
de remodelação por diapedese, através das células da canopy, ou migrarem através de zonas livres
resultantes da retração de algumas das células da canopy38, 99,135,150. No caso das trabéculas do tecido
ósseo esponjoso, a proximidade da medula óssea deixa antever uma grande facilidade de acesso das
células percursoras dos osteoclastos e osteoblastos ao BRC. Porém, no tecido ósseo compacto
serão os capilares sanguíneos, presentes nos canais de Havers, as únicas vias capazes de conduzir
estas células para as áreas que delas necessitam.
Os pré-osteoclastos vão encontrar no BRC um ambiente ideal e uma concentração adequada
de citocinas, quimiocinas e fatores osteoclastogénicos, tendo sido atribuída às células de
revestimento da canopy (porque constituem células da linha osteoblástica) a principal
responsabilidade pela síntese de M-CSF e de RANKL27. A presença destas citocinas no interior do
BRC constitui, assim, um dos mais importantes fatores responsáveis pela diferenciação e ativação
osteoclástica.
Todavia e como já foi referido, os osteócitos não estão alheios a este processo. De facto, os
osteócitos comprometidos ou danificados119,145,166-168 podem estimular, a produção de RANKL nas
células de revestimento (através de gap junctions), ou mesmo nas células do estroma presentes em
territórios adjacentes, como é o caso da medula óssea no tecido ósseo esponjoso ou o endósteo nos
canais de Havers.
Por sua vez, os osteócitos em apoptose podem, eles próprios, para além de produzir citocinas
inflamatórias (PGE2, Il-1,4,6,11, TNF–α e TNF–β) ser também capazes de sintetizar RANKL, tendo
até sido recentemente considerados como a maior fonte de RANKL sempre que se inicia um
processo de remodelação46-48,117-119,141,142. De facto, os osteócitos em apoptose libertam corpos
apoptóticos que são apontados como os principais responsáveis pelos altos teores de RANKL
verificados nestas situações47,104,143,144. Neste contexto importa referir ainda que nas zonas que
requerem uma remodelação, ou reparação, foi detetado um aumento significativo na expressão de
VEGFs também da responsabilidade dos osteócitos, estimulando, deste modo, os mecanismos de
angiogénese nesta região109,129,139,146-148. Será também curioso registar que o RANKL não só promove
a formação, diferenciação e atividade de reabsorção osteoclástica, como estimula os processos de
angiogénese e a sobrevivência das células endoteliais109,129,139,146-149.
***
A grande densidade de capilares observada junto às canopies seguindo um percurso paralelo
representa uma ampla interface entre estas duas entidades25,40,43,100,123-129. Esta situação permite uma
grande superfície de contacto entre capilares e canopies, constituindo certamente um local
privilegiado de comunicação entre estruturas vasculares e compartimentos de remodelação. Este

Compartimento de Remodelação Óssea 2016
63
envolvimento vascular é muito mais do que uma resposta da circulação ao aumento das necessidades
locais. O aparecimento de novas células endoteliais e perivasculares (constituintes da sua parede)
parecem exercer um papel determinante no coupling entre a etapa de reabsorção e de formação
óssea151.
Na formação e desenvolvimento dos cones de reabsorção óssea, observados no tecido ósseo
compacto, o vaso sanguíneo cresce à mesma velocidade que a BMU avança (25 µm por dia),
situando-se usualmente a não mais de 100 µm de distância quer da zona de reabsorção, quer da zona
de formação óssea54.
De facto, o BRC tem sido frequentemente considerado como uma estrutura vascular, sendo
várias as representações esquemáticas em que se propõe a comunicação directa de um capilar
sanguíneo com o espaço do compartimento19,25,38,39,76,100,104,131. Todavia, um estudo recente, realizado
com recurso a microscopia electrónica100, revelou que capilares e células de revestimento da canopy
são estruturas fisicamente independentes, mostrando, no entanto, algumas regiões onde se verifica
uma muito estreita proximidade. Foi também demonstrado nesse estudo que os capilares adjacentes
às zonas de remodelação óssea, mesmo no caso de trabéculas ósseas do tecido ósseo esponjoso, são
constituídos por uma camada contínua de células endoteliais, não podendo, por isso, ser
considerados como sinusóides. Todavia estas estruturas apresentam geralmente muito maiores
dimensões do que a maioria dos capilares (50 µm de diâmetro)152. Estes trabalhos19,40,100,109,148,153
revelaram também que a parede dos capilares em questão apresentava, em redor da camada de
células endoteliais, uma bainha de células consideradas como pericitos, que durante as fases de
remodelação se encontram expostos a uma alta concentração de fatores de crescimento
provenientes, quer do compartimento de remodelação, quer sintetizados pelas células endoteliais.
Estas células (bastante indiferenciadas) apresentam inúmeras capacidades de diferenciação,
mostrando nomeadamente um forte potencial osteogénico. Os pericitos prestam uma forte
contribuição nas respostas osteogénicas, relacionadas com estimulações eléctricas, manipulações
ortodônticas, reparações da sutura palatina148 e sobretudo nos processos de distração
osteogénica148,155,156.
Com efeito, em estudos com timidina tritiada, realizados em várias das situações atrás citadas,
foi possível demonstrar o aparecimento de osteoblastos marcados provenientes de precursores
perivasculares. Por outro lado, os pericitos em cultura podem expressar muitas características do
fenótipo osteoblástico, incluindo a formação de nódulos mineralizados. Deste modo, será
compreensível que os pericitos vasculares possam representar uma importante fonte de precursores
osteoblásticos100,109,157,158.
Os pericitos parecem, pois, representar um importante reservatório de células
osteoprogenitoras que podem facilmente ser recrutadas para a canopy ou para o interior do

Compartimento de Remodelação Óssea 2016
64
compartimento e, consequentemente, colonizar as superfícies ósseas. Este facto poder-se-á verificar
sempre que se observe uma estreita proximidade entre capilares e canopies100,109,157,158.
Para além dos pericitos, as células endoteliais42,127,137,149 assumem também uma importância
crucial na ativação e manutenção dos processos de remodelação óssea. De facto, a íntima ligação
entre células endoteliais e osteoblastos levou mesmo à elaboração de uma teoria que afirmava que a
célula endotelial era, em si, um precursor osteoblástico46. De qualquer modo, e ainda que assim não
seja, o endotélio vascular presente no tecido ósseo parece apresentar características únicas46. De
facto, as células endoteliais vasculares são responsáveis pela secreção de numerosas moléculas de
regulação (fatores de crescimento, BMP, citocinas, endotelinas, radicais livres e prostanóides) que
vão exercer um efeito major no controlo da diferenciação, metabolismo, sobrevivência e função das
células da linha osteoblástica46.
Por sua vez, também os osteoblastos são capazes de sintetizar fatores parácrinos que atuam
sobre as células endoteliais. Durante esta “conversa” entre células endoteliais e osteoblastos, o VEGF,
que atua principalmente como um fator angiogénico, assume particular importância. Um estudo de
Crawford (2009)138 veio demonstrar ainda que o bloqueio da atividade endógena do VEGF tem como
consequência a inibição de todo o mecanismo de remodelação óssea (reabsorção e formação)138.
Pode, pois, inferir-se destes dados que o VEGF se revela como um elemento crucial nos mecanismos
de angiogénese, mas também indispensável para os processos de formação e remodelação óssea46,
109,129,139,146-148. Por outro lado, foi também demonstrado que as proteínas morfogénicas do osso 2 e 4
(BMP 2 e BMP 4) estimulam a síntese de VEGF (pelos osteoblastos), que por sua vez atuam ao nível
das células endoteliais, promovendo a síntese de mais BMP-2 e Prostaglandinas E139. Assim, o papel do
VEGF parece ser essencial, não apenas para a amplificação da estrutura vascular na área do tecido
ósseo danificado, mas também terá um desempenho fundamental na promoção do recrutamento e
diferenciação dos osteoblastos.
As alterações desta forte ligação estrutural e funcional entre tecido ósseo e vasos sanguíneos
parecem estar na base de uma forte associação entre osteoporose e ateroesclerose/calcificações
vasculares39,159,160,161. A escassez de vasos sanguíneos, redução da perfusão vascular e aumento da
medula óssea amarela, observada sobretudo em mulheres pós menopausa com idade avançada,
parece correlacionar-se diretamente com os problemas de osteoporose39,161. Compreende-se pois
que, para além de uma terapêutica de substituição de estrogénios, os vasos sanguíneos constituam
também um importante alvo terapêutico a atingir, no desenvolvimento e otimização de soluções
eficazes para muitas das doenças do metabolismo ósseo como a osteoporose.
***
Foi também detetado um alto índice de proliferação celular, acompanhando as regiões
adjacente às áreas de remodelação, ou seja, nos locais de convergência entre capilares e

Compartimento de Remodelação Óssea 2016
65
compartimentos25. Estas regiões podem, pois, ser consideradas como verdadeiras plataformas
angio/osteogénicas. De facto, uma vez iniciada a formação do compartimento de remodelação, as
células da canopy parecem encontrar-se numa posição ideal no que respeita à exposição de fatores
mitogénicos e osteogénicos provenientes, quer do interior do BRC, quer das regiões adjacentes. Por
sua vez, será durante a fase de reversão, ou seja, durante o tempo que medeia entre o final do
processo de reabsorção e o início do processo de formação óssea que o gradiente de citocinas,
fatores de crescimento e de diferenciação deverá atingir a sua concentração máxima no interior do
BCR54,76,82. Este facto irá facilitar o recrutamento de células osteoprogenitoras para o local deixado
pelas células osteoclásticas, promovendo ao mesmo tempo a sua proliferação e permitindo a sua
maturação no sentido de osteoblastos maduros. Parece ser absolutamente necessária também a
presença de uma alta densidade celular ao nível das superfícies reabsorvidas, de modo a atingir uma
“massa crítica” de células osteoblásticas capaz de dar início à síntese de matriz óssea25,131. Neste
âmbito foi também reconhecido que as células da canopy constituem um reservatório de
progenitores osteoblásticos, prontos a ser recrutados para as superfícies em reversão. Foi ainda
demonstrada a existência de um gradiente entre as células da canopy e as superfícies ósseas em
reparação com recurso a marcadores como Ki67, Osterix e P3NP25. Com efeito verificou-se um
aumento no seu estádio de diferenciação (associada a uma elevação dos níveis de marcação com
Osterix) e uma diminuição nos índices de proliferação (acompanhando o decréscimo dos níveis de
marcação com Ki67 e P3NP)25,76,87. Este recrutamento de células osteogénicas observado entre a
canopy e as superfícies em reversão é apontado como um pré-requisito necessário ao início do
processo de formação e reparação óssea.
Considerando os resultados por nós observados durante a fase de reversão e de acordo com
tudo o que atrás foi referido (uma alta densidade celular ao nível das superfícies reabsorvidas,
apresentando diferentes estádios de desenvolvimento), facilmente se justifica a presença de uma
grande densidade de células, mas difícil caracterização, uma vez que se encontram, provavelmente,
em diferentes fases de diferenciação e maturação osteoblástica.
A este respeito, ou seja, quanto à origem das células osteoprogenitoras considerava-se, até há
bem pouco tempo, a existência na circulação periférica de apenas um reduzido número de células
pré-osteoblásticas. Todavia, a possibilidade de identificação destas células com recurso a meios
tecnológicos mais sofisticados (citometria de fluxo e cell sorter analysis), permitiu encontrar e
caracterizar uma expressiva percentagem de células da linha osteoblástica no sangue periférico19. Este
facto vem reforçar a possibilidade de acesso destas células aos BRCs, sobretudo situados no tecido
ósseo cortical (geralmente bastante distantes da medula óssea) resultante da estreita relação entre
capilares e células da canopy100, 162-164.
Pode admitir-se, pois, que o desaparecimento ou perda da canopy numa fase de reversão, para
além de perturbar drasticamente o equilíbrio dentro do compartimento (considerado como um

Compartimento de Remodelação Óssea 2016
66
reservatório de fatores de crescimento e maturação), irá comprometer definitivamente a chamada
de células osteoprogenitoras e a sua diferenciação e, deste modo, parar o ciclo de remodelação24-
27,46,76,82,99,109,165. Nas situações em que se observa uma paragem no processo de coupling, a fase de
reversão apresenta uma baixa densidade de células (quando comparada com uma fase de reversão
ativa) que por sua vez se mostram muito mais achatadas, ou seja, muito semelhantes às células de
revestimento das superfícies em repouso27,46,82,99. As células da canopy assumem assim uma
importância crítica na reconstrução das áreas reabsorvidas. A este respeito importa referir que os
processos de reabsorção podem prosseguir mesmo sem a presença da canopy, justificando a
observação de muitas áreas onde se verifica uma excessiva reabsorção e uma deficiente ou ausência
de reparação.
Em jeito de conclusão, a figura 41 pretende ilustrar e fazer uma pequena sinopse do
envolvimento direto entre estruturas vasculares e compartimentos de remodelação óssea25,
mostrando a origem e localização das células osteogénicas e a proveniência de possíveis fatores
osteogénicos (derivados quer da corrente sanguínea e regiões perivasculares, quer inerentes aos
osteoclastos ou decorrentes da sua atividade, quer provenientes da matriz óssea a par de alterações
mecânicas - forças de deformação/estiramento geradas no interior do BRC).
Figura 41 – Representação esquemática da interligação de estruturas vasculares e perivasculares com o compartimento de
remodelação óssea e origem dos possíveis fatores de regulação das células com capacidade osteogénica. As células da
canopy encontram-se numa posição ideal para captar e responder aos inúmeros fatores osteogénicos (provenientes quer do
interior do BRC, quer das regiões adjacentes) constituindo verdadeiros reservatórios de células osteoprogenitoras. A
gradação dos tons de azul (claro para escuro) corresponde ao evoluir do processo de diferenciação das células
osteoblásticas. BLC – células de revestimento ósseo; OC – osteoclasto; Rv.Cs – células osteoblásticas características da
fase de reversão. Adaptado de JM Delaisse25

Compartimento de Remodelação Óssea 2016
67
Em síntese pode pois afirmar-se que vasos sanguíneos, regiões perivasculares e canopies
parecem realmente funcionar em conjunto, constituindo uma importante unidade de apoio, nutrição
e abastecimento celular e metabólico ao compartimento de remodelação óssea representando, como
já foi referido, uma verdadeira plataforma angio/osteogénica.
O desenvolvimento e otimização de soluções terapêuticas relacionadas com o metabolismo
ósseo nunca serão bem-sucedidos se não se verificarem as condições logísticas necessárias que
assegurem o recrutamento e proteção das células osteoprogenitoras e dos seus fatores de regulação,
ou seja, a integridade do BRC.
Como nota final sublinhamos, mais uma vez, como nos parece importante um conhecimento
pormenorizado da constituição histológica do compartimento de remodelação óssea, de modo a
permitir uma melhor interpretação da fisiologia e patologia do ciclo de remodelação óssea. A
preservação desta estrutura anatómica e suas relações topográficas com o microambiente em seu
redor, nomeadamente os vasos sanguíneos, é crucial para a vitalidade e eficácia dos processos de
remodelação óssea.
,

Compartimento de Remodelação Óssea 2016
68

Compartimento de Remodelação Óssea 2016
69
III. Resumo
__________________________________________________________

Compartimento de Remodelação Óssea 2016
70

Compartimento de Remodelação Óssea 2016
71
A remodelação óssea consiste num mecanismo de substituição ou reconstrução de áreas de
tecido ósseo, de modo a otimizar a sua função ou prevenir a sua degradação, seguindo uma
sequência imutável de etapas - ativação, reabsorção, inversão/reversão e formação.
O conjunto de células responsáveis por estas diferentes atividades reúne uma equipa de células
da linha osteoclástica (pré-osteoclastos e osteoclastos) e uma outra de células da linha osteoblástica
(pré-osteoblastos, osteoblastos, osteócitos e células de revestimento). Desta unidade fazem também
parte outras populações celulares que incluem células mesenquimatosas indiferenciadas e células do
tecido conjuntivo, muitas delas pertencentes ao sistema imunitário, bem como um suplemento
vascular especializado para o efeito.
Esta complexa maquinaria celular está sujeita à ação de numerosos fatores de regulação, que
exigem um rigoroso e apertado sistema de sinalização e comunicação, responsável pela ação
concertada de todas estas células e pela coordenação e transição equilibrada das diferentes etapas da
remodelação.
Toda a atividade celular do ciclo de remodelação óssea está protegida no interior de uma
estrutura anatómica, ainda que transitória, designada por Compartimento de Remodelação Óssea
(BRC), permitindo a criação de um microambiente osteogénico favorável para que a transição da fase
de reabsorção e a fase de formação possa ocorrer. Esta entidade só recentemente foi detetada e está
ainda mal caracterizada.
Deste modo, pretendemos com o presente trabalho contribuir para uma maior compreensão
dos processos de remodelação óssea, proporcionando um melhor conhecimento das características
histológicas dos compartimentos de remodelação. Decidimos ainda estender as observações para as
áreas de dentina e de cemento, uma vez que são também tecidos mineralizados com capacidade de
remodelação.
***
O início do processo de remodelação óssea (fase de ativação) é induzido por citocinas,
produzidas pelos osteócitos lesados, e caracterizado pelo recrutamento, maturação e ativação dos
osteoclastos. Este processo é controlado pelas células da linha osteoblástica, através do eixo de
regulação RANKL/RANK/OPG. Nesta fase, verifica-se uma retração e um destacamento das células
de revestimento da área em questão, afastando-se da respetiva matriz óssea, dando início ao
aparecimento de uma camada de células em forma de cúpula (canopy) e ao desenvolvimento de um
espaço fechado correspondente ao BRC. Importa salientar, ainda, o desenvolvimento de uma rede de
capilares sanguíneos e a sua aproximação às células da canopy, formando no seu conjunto uma
plataforma de comunicação entre estruturas vasculares e compartimentos de remodelação. A
intervenção do componente vascular, constituindo uma estrutura sempre presente, é determinante
ao longo de todo o processo de remodelação.

Compartimento de Remodelação Óssea 2016
72
Na fase de reabsorção os osteoclastos cavam, no tecido ósseo esponjoso, lacunas de contornos
muito irregulares denominadas por lacunas de Howship e no tecido ósseo compacto cavidades
cilíndricas designadas por cones de reabsorção. A dimensão das áreas reabsorvidas é diretamente
proporcional às áreas danificadas ou em sofrimento. No final desta etapa observa-se uma inversão ou
reversão nesta sequência, verificando-se um conjunto de condições responsáveis pela chamada de
células osteoprogenitoras e sua diferenciação, dando início ao processo de reparação óssea. Esta
estimulação da atividade osteoblástica, como resposta ao processo de reabsorção osteoclástica, é
habitualmente conhecido por coupling.
A fase de reversão parece constituir uma das etapas mais delicadas do ciclo de remodelação,
reunindo um conjunto de condições essenciais para a proliferação e diferenciação osteoblástica.
Qualquer perturbação observada nesta etapa (nomeadamente a integridade na canopy) poderá
provocar uma paragem irreversível no processo de remodelação. O preenchimento e reparação da
cavidade de reabsorção é naturalmente decorrente da atividade secretora de osteoblastos maduros
com origem nas células observadas durante a fase de reversão e que, entretanto, beneficiaram de um
processo de proliferação, maturação e ativação.
A fase de formação consiste, pois, na síntese de matriz osteóide e na sua posterior
mineralização, tendo como resultado final a reconstrução da lacuna de Howship com novo tecido
ósseo ou de um cone de reabsorção, dando origem ao aparecimento de um novo sistema de Havers.
***
Os resultados apresentados neste estudo comprovam que o compartimento de remodelação
óssea se apresenta como um espaço fechado que recobre as zonas de matriz mineralizada que estão
a ser alvo de um processo de remodelação. Este espaço encontra-se revestido por uma canopy
formada por células extremamente achatadas. De referir ainda a existência de uma abundante rede
capilar localizada em estreita associação de proximidade à canopy, que reveste os compartimentos de
remodelação, sendo mesmo possível verificar a presença de inúmeros pontos de contacto entre os
capilares e as células de revestimento que a compõem.
Ainda que não nos tivesse sido possível realizar estudos quantitativos, foi por demais evidente
que o número e variedade de imagens correspondentes a compartimentos de remodelação óssea,
que apresentamos nos resultados, demonstram bem a intensidade e frequência de aparecimento
destes processos no osso alveolar. De modo semelhante ao que observamos nas regiões do tecido
ósseo alveolar também nas zonas de remodelação do cemento e dentina foi possível identificar
estruturas com uma constituição histológica em tudo idêntica ao compartimento de remodelação
óssea, porém em muito menor número e com uma constituição muito menos completa.

Compartimento de Remodelação Óssea 2016
73
***
A grande densidade de capilares observada junto às canopies, seguindo um percurso paralelo,
representa uma ampla interface entre estas duas entidades. Esta situação permite uma grande
superfície de contacto entre capilares e canopies, constituindo certamente um local privilegiado de
comunicação entre estruturas vasculares e compartimentos de remodelação.
A parede dos capilares apresenta, em redor da camada de células endoteliais, uma bainha de
células consideradas como pericitos, que durante as fases de remodelação se encontram expostos a
uma alta concentração de fatores de crescimento provenientes, quer do compartimento de
remodelação, quer sintetizados pelas células endoteliais. Estas células apresentam inúmeras
capacidades de diferenciação, mostrando nomeadamente um forte potencial osteogénico. Os
pericitos parecem, assim, representar um importante reservatório de células osteoprogenitoras que
podem facilmente ser recrutadas para a canopy ou para o interior do compartimento e,
consequentemente, colonizar as superfícies ósseas em remodelação.
Para além dos pericitos, as células endoteliais assumem também uma importância crucial na
ativação e manutenção dos processos de remodelação óssea.
Foi também detetado um alto índice de proliferação celular, acompanhando as regiões
adjacente às áreas de remodelação, ou seja, nos locais de convergência entre capilares e
compartimentos. De facto, uma vez iniciada a formação do compartimento de remodelação, as
células da canopy parecem encontrar-se numa posição ideal, no que respeita à exposição de fatores
mitogénicos e osteogénicos provenientes, quer do interior do BRC, quer das regiões adjacentes. Por
outro lado, as células da canopy, ainda que derivadas das células de revestimento ósseo, parecem ter
regredido para um estádio precoce de diferenciação, mostrando inclusive um índice de proliferação
superior a 10 %, constituindo também verdadeiros reservatórios de células osteoprogenitoras. O
BRC irá fornecer um ambiente propício para o recrutamento destas células pré-osteoblásticas para o
local deixado pelas células osteoclásticas, promovendo, ao mesmo tempo, a sua proliferação e
maturação no sentido de osteoblastos maduros.
Recentemente, a possibilidade de identificação de célula osteoprogenitoras na circulação
periférica, com recurso a meios tecnológicos mais sofisticados (citometria de fluxo e cell sorter analysis),
permitiu encontrar e caracterizar uma expressiva percentagem de células da linha osteoblástica no
sangue periférico. Este facto vem reforçar a possibilidade de acesso destas células aos BRCs,
resultante da estreita relação entre capilares e células da canopy.
Em síntese pode pois afirmar-se que vasos sanguíneos, regiões perivasculares e canopies
parecem realmente funcionar em conjunto, constituindo uma importante unidade de apoio, nutrição
e abastecimento celular e metabólico ao compartimento de remodelação óssea.
O desenvolvimento e otimização de soluções terapêuticas relacionadas com o metabolismo
ósseo nunca serão bem-sucedidos se não se verificarem as condições logísticas necessárias que

Compartimento de Remodelação Óssea 2016
74
assegurem o recrutamento e proteção das células osteoprogenitoras e dos seus fatores de regulação,
ou seja, a integridade do BRC.
Como nota final sublinhamos, mais uma ve,z como nos parece importante um conhecimento
pormenorizado da constituição histológica do compartimento de remodelação óssea, de modo a
permitir uma melhor interpretação da fisiologia e patologia do ciclo de remodelação óssea. A
preservação desta estrutura anatómica e suas relações topográficas com o microambiente em seu
redor, nomeadamente os vasos sanguíneos, é crucial para a vitalidade e eficácia dos processos de
remodelação óssea.

Compartimento de Remodelação Óssea 2016
75
IV. Abstract
__________________________________________________________

Compartimento de Remodelação Óssea 2016
76

Compartimento de Remodelação Óssea 2016
77
Bone remodelling represents the most notable response of the bone tissue, as it is supported by
complex cellular machinery which is under the influence of numerous regulation factors.
Furthermore, bone remodeling is a strictly localized process.
Bone remodeling is a continuous process during adulthood, which preserves the bone strength
through a constant renewal of the bone. This process is conducted by numerous tightly regulated
bone remodeling units (BRUs), which include four sequential events taking place on the bone surface:
activation of the precise area of bone which needs to be repair, bone resorption by osteoclasts
(OCs), a reversal phase, and bone formation by osteoblasts (OBs).
This way, it is plausible to state that bone resorption and formation are events which are closely
attached, as the beginning of the first one stimulates the repairing activity of the second, in the way
that the resorption of a certain amount os bone tissue shall be replaced by the same amount of new
bone.
Truthfully, considering the fact that in the sequence of bone remodeling the formation phase
(performed by osteoblastos) is always preceded by a resorption phase (performed by osteoclastos),
the way in which the transition ou reversion takes placed is still widely unknown. Can osteoclastos
trigger osteoblast differentiation, representingan essential factor for osteoblástica differentiation
There are multiple possibilities of answers to this question, namely: a) the presence of growth
factors produced by osteoclastos will have a paracrine effect on osteoblastos; b) the action of growth
factors and cytokines with osteogenic potential present within the bone matrix are released with the
beginning os osteoclástica resorption and c) a direct cell-to-cell contact between osteoclastos and
osteoblastos.
Despite the alignment and association between bone resorption and formation is rarely affected,
the final balance concerning the level of bone mass can vary considerably. As it is conventionally
accepted, with the aging process and in situation of bone disease, such as osteoporosis, the final
balance between bone resorption and formation tends to be negative.
Furthermore, mosto of inflammatory and metabolic diseases that hit bone tissue usually manifest
themselves by disturbances in the dynamics of the remodeling process, resulting almos invariably in a
decrease in bone mass, associated with osteoclástic resorption.
***
It is also important to stress out that all cellular activity within the bone remodeling cycle has to
be protected by a specific structure created for that matter.
Hauge et al (2001) were the first investigators to point out the existence os a bone remodeling
compartment as a real and temporary entity, demonstrating its existence histologically.
The cells which forme the canopy expresse the majority os the markers characteristic of the
osteoblástica phenotype (alkaline phosphatase, osteocalcine and osteonectine), most likely being

Compartimento de Remodelação Óssea 2016
78
bone linning cells. These investigations also raised the hypothesis that this compartment may very
well form a specialized vascular structure
Later on, numerous investigations demonstrated as well that there is a strict association between
blood capillaries and the bone remodeling compartment’s canopy, concluding that the integrity of the
BRC itself is also a pre-requirement fundamental for bone remodeling and formation.
The marrow microenviroment surrounding the BRC is one of the main sources of cells that form
the BMU. In spite of this, it is important to state that the bone marrow is not the only origin of these
cells. In fact, multiple blood capillaries penetrate the BRC canopy and serve as a method of
transportation of the cells needed to the functioning of the BMU.
The blood vessel is a structure strategically present e located in all BMU’s, once the formations of
new blood vessels in intimately related with the processes of osteogenesis and ossification.
In another perspective, it is ever more evident the existence os a common lineage between
vascular endothelial cells and osteoblástica cells. Furthermore, the osteoblasts are always located
near endothelial cells and vascular invasion is a pre-requirement for bone formation, remodeling and
regeneration.
Concerning the osteoblástica cells, merely a low number of cells from this lineage has been
detected in the peripheral circulation until recently. However, with the possibility of identifying these
cells through technologically more sophisticated methods (flow citometry and cell sorter analysis)
allowed investigators to find a describe an expressive percentage of osteoblástica cells on the
peripheral blood.
***
In summary, these recent studies demonstrated that bone remodeling (both in cortical and
trabecular bone tissue) takes place in specialized vascular structures – the bone remodeling
compartment. According to this model, osteoblastic precursor cells arrive in the BRC not only from
the bone marrow, but also via blood capillaries which invade the BRC. Furthermore, osteoblastic
lineage cells may enter the BRC through the bloog circulation (afferent capillary), they may be
originated from perivascular structures (perycites) and also from the endothelial cells themselves.
This way, the possibility of performing the identification e harvesting of these cells and the
subsequent expansion through culture represents a promising tool for the formulation of future
therapeutics of diseases related to bone tissue metabolism.

Compartimento de Remodelação Óssea 2016
79
V. Bibliografia _________________________________________________________________________________

Compartimento de Remodelação Óssea 2016
80

Compartimento de Remodelação Óssea 2016
81
1. Garant PR. Oral Cells and Tissues: Quintessence Publishing (IL) 2003.p. 195-238.
2. Martin, T.J. and E. Seeman, Bone remodelling: its local regulation and the emergence of bone fragility. Best Pract
Res Clin Endocrinol Metab, 2008. 22(5): p. 701-22.
3. Schmitt JM, Buck DC, Joh SP, Lynch SE, Hollinger JO. Comparison of porous bone mineral and biologically
active glass in critical-sized defects. J Periodontol 1997;68(11):1043-53.
4. Sims, N.A. and J.H. Gooi, Bone remodeling: Multiple cellular interactions required for coupling of bone
formation and resorption. Semin Cell Dev Biol, 2008. 19(5): p. 444-51.
5. Junqueira LCCJ. Tecido ósseo. In: Koogan G, editor. Histologia Básica. 11ª edição ed; 2008. p. 108-26.
6. Kierszenbaum AL. Histologia e Biologia Celular. In: Kierszenbaum AL, Editor. Tradução da 2º Edição ed:
Mosby-Elsevier; 2008. p. 136-59.
7. Datta HK, Ng WF, Walker JA, Tuck SP, Varanasi SS. The cell biology of bone metabolism. J Clin Pathol
2008;61(5):577-87.
8. Standring S. Gray's Anatomy, 40th Edition - The Anatomical Basis of Clinical Practice, Expert Consult Churchill
Livingstone 2009. p. 27-59.
9. Geddes AC. The Origin of the Osteoblast and of the Osteoclast. J Anat Physiol 1913;47(Pt 2):159-76.
10. Parfitt, A.M., Targeted and nontargeted bone remodeling: relationship to basic multicellular unit origination and
progression. Bone, 2002. 30(1): p. 5-7.
11. Galliera E, Locati M, Mantovani A, Corsi MM. Chemokines and bone remodeling. Int J Immunopathol
Pharmacol 2008;21(3):485-91.
12. Baud'huin M, Lamoureux F, Duplomb L, Redini F, Heymann D. RANKL, RANK, osteoprotegerin: key partners of
osteoimmunology and vascular diseases. Cell Mol Life Sci 2007;64(18):2334-50.
13. Lee SH, Kim TS, Choi Y, Lorenzo J. Osteoimmunology: cytokines and the skeletal system. BMB Rep
2008;41(7):495-510.
14. J CL, H C, J EF. Osteoimmunology--the hidden immune regulation of bone. Autoimmun Ver 2009;8(3):250-5.
15. Targonska M, Kochanowska I, Ostrowski K, Gorski A. [Osteoimmunology: new area of research on the
associations between the immune and bone systems]. Pol Arch Med Wewn 2001;105(5):435-40.
16. David JP. Osteoimmunology: a view from the bone. Adv Immunol 2007;95:149-65.
17. Rauner M, Sipos W, Pietschmann P. Osteoimmunology. Int Arch Allergy Immunol 2007;143(1):31-48.
18. Cohen MM, Jr. The new bone biology: pathologic, molecular, and clinical correlates. Am J Med Genet A
2006;140(23):2646-706.
19. Eghbali-Fatourechi, G.Z., et al., Characterization of circulating osteoblast lineage cells in humans. Bone, 2007.
40(5): p. 1370-7.
20. Robling, A.G., A.B. Castillo, and C.H. Turner, Biomechanical and molecular regulation of bone remodeling. Annu
Rev Biomed Eng, 2006. 8: p. 455-98.
21. Epstein, S., Serum and urinary markers of bone remodeling: assessment of bone turnover. Endocr Rev, 1988.
9(4): p. 437-49.
22. Parfitt AM. Bone remodeling, normal and abnormal: a biological basis for the understanding of cancer-related
bone disease and its treatment. Can J Oncol 1995;5 Suppl 1:1-10.

Compartimento de Remodelação Óssea 2016
82
23. Doblare M, Garcia JM. Anisotropic bone remodelling model based on a continuum damage-repair theory. J
Biomech 2002;35(1):1-17.
24. Hauge, E.M., et al., Cancellous bone remodeling occurs in specialized compartments lined by cells expressing
osteoblastic markers. J Bone Miner Res, 2001. 16(9): p. 1575-82.
25. Delaisse, JM The reversal phase of the bone-remodeling cycle: cellular prerequisites for coupling resorption and
formation. BoneKEy Reports 3, Article number: 561 (2014) | doi:10.1038/bonekey.2014.56 & 2014
International Bone & Mineral Society 2047-6396/14
26. Andersen TL, Sondergaard TE, Skorzynska KE, Dagnaes-Hansen F, Plesner TL, Hauge EM, Plesner T, Delaissé
JM: A physical mechanism for coupling bone resorption and formation in adult human bone. Am J Pathol 2009,
174:239e247
27. Sims, N., Martin, T.J, Coupling signals between the osteoclast and osteoblast: how are messages transmitted
between these temporary visitors to the bone surface?. Frontiers in Endocrinology | Bone Research, 2015, Vol.
6, Art 41, (1-4)| doi:10.3389/fendo.2015.00041
28. Nanci A, Ten Cate AR. Ten Cate´s - Oral Histology - Development, Structure, and Function In: Nanci A, editor.
Sixth Edition ed; 2003. p.34-49.
29. Sodek, J. and McKee, M. D. (2000), Molecular and cellular biology of alveolar bone. Periodontology 2000, 24:
99–126. doi:10.1034/j.1600 0757.2000.2240106.x
30. Roberts WE, Jeffery AR, Bruce NE, David BB, James KH. Remodeling of Mineralized Tissues, Part I: The Frost
Legacy. Seminars in orthodontics 2006;12(4):216-37.
31. Roberts WE, Bruce NE, David BB, James KH, Jeffery AR. Remodeling of Mineralized Tissues, Part II: Control
and Pathophysiology. Seminars in orthodontics 2006;12(4):238-53.
32. Roberts WE. Bone physiology of tooth movement, ankylosis, and osseointegration. Seminars inorthodontics
2000;6(3):173-82.
33. Roberts WE, Simmons KE, Garetto LP, DeCastro RA. Bone physiology and metabolism in dental implantology:
risk factors for osteoporosis and other metabolic bone diseases. Implant Dent 1992;1(1):11-21.
34. Parfitt, A.M., The cellular basis of bone remodeling: the quantum concept reexamined in light of recent
advances in the cell biology of bone. Calcif Tissue Int, 1984. 36 Suppl 1: p. S37-45.
35. Mensah KA, Li J, Schwarz EM. The emerging field of osteoimmunology. Immunol Res 2009.
36. Cohen MM, Jr. The new bone biology: pathologic, molecular, and clinical correlates. Am J Med Genet A
2006;140(23):2646-706.
37. Roberts WE, Simmons KE, Garetto LP, DeCastro RA. Bone physiology and metabolism in dental implantology:
risk factors for osteoporosis and other metabolic bone diseases. Implant Dent1992;1(1):11-21.
38. Perez-Amodio S, Beertsen W, Everts V., (Pre-)Osteoclasts Induce Retraction of Osteoblasts Before Their Fusion
to Osteoclasts., J Bone Miner Res, 2004; 19:1722-1731; doi:10.1359/JBMR.040509
39. Lafage-Proust MH., et al., Assessment of bone vascularization and its role in bone remodeling. BoneKEy
Reports 4, Article number: 662 (2015) | doi:10.1038/bonekey.2015.29
40. Brandi ML et al. Vascular biology and the skeleton; J Bone Miner Res. 2006;21:183-192
41. Parfitt AM, The mechanism of coupling: a role for the vasculature. Bone 2000, 26:319-323

Compartimento de Remodelação Óssea 2016
83
42. Burkhardt R, Bartl R, Frisch B, Jager K, Mahl C, Hill W, Kettner G: The structural relationship of bone forming
and endothelial cells of the bone marrow. Bone Circulation. Edited by Arlet J, Ficat RP, Hungerford DS.
Baltimore, Williams and Wilkins, 1984, pp 2-14
43. Laroche M, Barbier A, Ludot I, Vernhet C, Thiechart M, Viguier G, Mazieres B: Effect of ovariectomy on
intraosseous vascularization and bone remodeling in rats: action of tiludronate. Osteoporos Int 1996, 6:127-
129
44. Chow, J.W., et al., Mechanical loading stimulates bone formation by reactivation of bone lining cells in 13-week-
old rats. J Bone Miner Res, 1998. 13(11): p. 1760-7.
45. Lemaire V, Tobin FL, Greller LD, Cho CR, Suva LJ. Modeling the interactions between osteoblast and osteoclast
activities in bone remodeling. J Theor Biol 2004;229(3):293-309.
46. Sapir-Koren, R., Livshits G., Osteocyte control of bone remodeling: is sclerostin a key molecular coordinator of
the balanced bone resorption-formation cycles?,Osteoporos Int 2014, 25: 2685-2700. Doi:10.1007/s00198-
014-2808-0
47. Bellido, T., Osteocyte-Driven Bone Remodeling. Calcif Tissue Int. 2014; 94(1): 25-34. Doi:10.1007/s00223-
013-9774-y
48. Everts, V., et al., The bone lining cell: Its Role in Cleaning Howship´s Lacunae and Initiating Bone Formation.
Journal of Bone and Mineral Reser, 2002, 17(1): 77-90.
49. Matsuo K, Irie N. Osteoclast-osteoblast communication. Arch Biochem Biophys 2008;473(2):201-9.
50. Martin T, Gooi JH, Sims NA. Molecular mechanisms in coupling of bone formation to resorption. Crit Rev
Eukaryot Gene Expr 2009;19(1):73-88.
51. Boyce BF, Xing L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch Biochem, Biophys
2008;473(2):139-46
52. Khosla, S., Minireview: the OPG/RANKL/RANK system. Endocrinology, 2001. 142(12): p. 5050-5.
53. Agerbaek MO, Eriksen EF, Kragstrup J, Mosekilde L, Melsen F. A reconstruction of the remodelling cycle in
normal human cortical iliac bone. Bone Miner. 1991;12:101–12
54. Eriksen E., Cellular mechanisms of boné remodeling. Rev Endocr Metab Disord (2010) 11:219–227, DOI
10.1007/s11154-010-9153-1
55. Martin, T.J. and N.A. Sims, Osteoclast-derived activity in the coupling of bone formation to resorption. Trends
Mol Med, 2005. 11(2): p. 76-81.
56. Chambers, TJ, Darby JA, Fuller K. Mammalian collagenase predisposes bone surfaces to osteoclastic resorption.
Cell Tissue Res 1985, 241: 671-675
57. Hock JM, Canalis E. Platelet-derived growth factor enhances bone cell replication, but not differentiated
function of osteoblasts. Endocrinology (1994) 134:1423–8.doi:10.1210/endo.134.3.8119182
58. Fiedler J, Roderer G, Gunther KP, Brenner RE. BMP-2, BMP-4, and PDGF-b stimulate chemotactic migration of
primary human mesenchymal progenitor cells. J Cell Biochem (2002) 87:305–12. doi:10.1002/jcb.10309
59. Xian L, Wu X, Pang L, Lou M, Rosen CJ, Qiu T, et al. Matrix IGF-1maintains bone mass by activation of mTOR
in mesenchymal stem cells. Nat Med (2012) 18:1095–101.doi:10.1038/nm. 2793
60. Sanchez-Fernandez MA, Gallois A, Riedl T, Jurdic P, Hoflack B. Osteoclasts control osteoblast chemotaxis via
PDGF-BB/PDGF receptor beta signaling. PLoSOne (2008) 3:e3537.doi:10.1371/ journal.pone.0003537

Compartimento de Remodelação Óssea 2016
84
61. Tang Y, Wu X, Lei W, Pang L, Wan C, Shi Z, et al. TGF-beta1-induced migration of bone mesenchymal stem
cells couples bone resorption with formation. Nat Med (2009) 15:757–65.doi:10.1038/ nm.1979
62. Kreja L, Brenner RE, Tautzenberger A, Liedert A, Friemert B, Ehrnthaller C, et al. Non-resorbing osteoclasts
induce migration and osteogenic differ- entiation of mesenchymal stem cells. J Cell Biochem (2010) 109:347–
55.doi:10.1002/jcb.22406
63. Rickard DJ, Sullivan TA, Shenker BJ, Leboy PS, KazhdanI. Induction of rapid osteoblast differentiation in rat
bone marrow stromal cell cultures by dexamethasone and BMP-2. Dev Biol (1994) 161:218–
28.doi:10.1006/dbio.1994.1022
64. Mitlak BH, Finkelman RD, Hill EL, Li J, Martin B, Smith T, et al. The effect of systemically administered PDGF-
BB on the rodent skeleton. J Bone Miner Res (1996) 11:238–47.doi:10.1002/ jbmr.5650110213
65. Hanamura H, Higuchi Y, Nakagawa M, Iwata H, Nogami H, Urist MR. Solubilized bone morphogenetic
protein (BMP) from mouse osteosarcoma and rat demineralized bone matrix. Clin Orthop Relat Res (1980)
148:281–90.
66. Centrella M, Canalis E. Local regulators of skeletal growth: a perspective. Endocr Rev (1985) 6:544–51.
doi:10.1210/edrv-6-4-544
67. Pfeilschifter J, Mundy GR. Modulation of type beta transforming growth factor activity in bone cultures by
osteotropic hormones. Proc Natl Acad Sci U SA (1987) 84:2024–8.doi:10.1073/pnas.84.7. 2024
68. Oreffo RO, Mundy GR, Seyedin SM, Bonewald LF. Activation of the bone-derived latent TGF beta complex by
isolated osteoclasts. Biochem Biophys Res Commun (1989) 158:817–23.doi:10.1016/ 0006-291X(89)92795-
2.
69. Hock JM, Canalis E, Centrella M. Transforming growth factor-beta stimulates bone matrix apposition and bone
cell replication in cultured fetal rat calvariae. Endocrinology (1990) 126:421–6.doi:10. 1210/endo-126-1-421
70. Dallas SL, Rosser JL, Mundy GR, Bonewald LF. Proteolysis of latent transforming growth factor-beta (TGF-beta)-
binding protein-1 by osteoclasts. A cellular mechanism for release of TGF-beta from bone matrix. J Biol Chem
(2002) 277:21352–60. doi:10.1074/jbc.M111663200
71. Lu Q, Sun EE, Klein RS et al. Ephrin-B reverse signaling is mediated by a novel PDZ-RGS protein and selectively
inhibits G protein-coupled chemoattraction. Cell 2001; 105(1): 69–79.
72. Zhao, C., et al., Bidirectional ephrinB2-EphB4 signaling controls bone homeostasis. Cell Metab, 2006. 4(2): p.
111-21.
73. Edwards CM, Mundy GR. Eph Receptors and Ephrin Signaling Pathways: A Role in Bone Homeostasis. Int. J.
Med. Sci. 2008,5(5): 263-279
74. Canalis, E., et al., Glucocorticoid-induced osteoporosis: pathophysiology and therapy.Osteoporos Int, 2007.
18(10): p. 1319-28.
75. Everts, V., et al., The bone lining cell: its role in cleaning Howship's lacunae and initiating bone formation. J
Bone Miner Res, 2002. 17(1): p. 77-90.
76. AndersenTL, Abdelgawad ME, Kristensen HB, Hauge EM, Rolighed L, Bollerslev J,et al. Understanding coupling
between bone resorption and formation: are reversal cells the missing link? Am J Pathol (2013) 183:235–
46.doi:10.1016/j.ajpath. 2013.03.006
77. Karsdal MA, Martin TJ, Bollerslev J, Christiansen C, Henriksen K: Are nonresorbing osteoclasts sources of bone
anabolic activity? J Bone Miner Res 2007, 22:487e494

Compartimento de Remodelação Óssea 2016
85
78. Kousteni S,, Bilezikian J: The cell biology of parathyroid hormone in osteoblasts. Curr Osteoporos Rep 2008,
6:72e7
79. Colnot C, Zhang X, Tate MLK: Current insights on the regenerative potential of the periosteum: molecular,
cellular, and endogenous engineering approaches. J Orthop Res 2012, 30:1869e1878
80. Diaz-Flores L, Gutierrez R, Madrid JF, Varela H, Valladares F, Acosta E,Martin-Vasallo P,Diaz-FloresLJr.:
Pericytes:morphofunction, interactions and pathology in a quiescent and activated mesenchymal cell niche.
Histol Histopathol 2009, 24:909e969
81. Menton DN, Simmons DJ, Orr BY, Plurad SB: A cellular investment of bone marrow. Anat Rec 1982,
203:157e164
82. Andersen TL et al., Correlation between absence of bone remodeling compartment canopies, reversal phase
arrest, and deficient bone formation in post-menopausal osteoporosis. 2014, The American journal of
pathology, vol.184,no.4
83. Baron R, et al., Estimation of trabecular bone resorption by histomorphometry: evidence for a prolonged
reversal phase with normal resorption in post-menopausal osteoporosis and couple increase in primary
hyperparathyroidism. Clin Disorders Bone Miner Metab 1983:191-195
84. Aaron JR, Choi Y. Osteoimmunology: bone versus immune system. Nature 2000;408:535-6.
85. Walsh MC, Kim N, Kadono Y, Rho J, Lee SY, Lorenzo J et al. Osteoimmunology: interplay between the
immune system and bone metabolism. Ann Rev Immunol 2006;24:33-63.
86. Takayanagi H. New developments in osteoimmunology. Nat Rev Rheumatol 2012;8:684-9.
87. Arboleya L, Castañeda S, Osteoimmunology: The Study of the Relationship Between the Immune System and
Bone Tissue, Reumatol Clin 2013;9:303-15 - Vol. 9 Num.5 DOI: 10.1016/j.reumae.2013.02.004
88. Theoleyre S, Wittrant Y, Tat SK, Fortun Y, Redini F, Heymann D. The molecular triad OPG/RANK/RANKL:
involvement in the orchestration of pathophysiological bone remodeling. Cytokine Growth Factor Rev
2004;15(6):457-75
89. Deng ZL, Sharff KA, Tang N, Song WX, Luo J, Luo X, et al. Regulation of osteogenic differentiation during
skeletal development. Front Biosci 2008;13:2001-21.
90. Theoleyre S, Wittrant Y, Couillaud S, Vusio P, Berreur M, Dunstan C, et al. Cellular activity and signaling
induced by osteoprotegerin in osteoclasts: involvement of receptor activator of nuclear factor kappaB ligand
and MAPK. Biochim Biophys Acta 2004;1644(1):1-7.
91. Franceschi C, Campisi J, Chronic Inflammation (Inflammaging) and Its Potential Contribution to Age-Associated
Diseases,J Gerontol A Biol Sci Med Sci 2014 June;69(S1):S4–S9/doi:10.1093/gerona/glu057
92. Kwan Tat S, Padrines M, Theoleyre S, Heymann D, Fortun Y. IL-6, RANKL, TNF-alpha/IL-1: interrelations in
bone resorption pathophysiology. Cytokine Growth Factor Rev 2004;15(1):49-60.
93. Ukai T, Yumoto H, Gibson FC, 3rd, Genco CA. Macrophage-elicited osteoclastogenesis in response to bacterial
stimulation requires Toll-like receptor 2-dependent tumor necrosis factor-alpha production. Infect Immun
2008;76(2):812-9.
94. Lu HK, Chen YL, Chang HC, Li CL, Kuo MY. Identification of the osteoprotegerin/receptor activator of uclear
factor-kappa B ligand system in gingival crevicular fluid and tissue of patients with chronic periodontitis. J
Periodontal Res 2006;41(4):354-60.

Compartimento de Remodelação Óssea 2016
86
95. Rocha, P., et al, “Apexificação e Revascularizaçção em dentes permanentes imaturos – estudo experimental In
vivo”, tese de doutoramento apresentada à Faculdade de Medicina da Universidade de Coimbra, Coimbra,
2013
96. Vale, F., et al, “Distração Osteogénica Mandibular dento-ancorada – estudo experimental”, tese de
doutoramento apresentada à Faculdade de Medicina da Universidade de Coimbra, Coimbra, 2014
97. Palma, P., “Estudo de novas formulações para regeneração óssea em defeitos de dimensão crítica”, tese de
mestrado apresentada à Faculdade de Medicina da Universidade de Coimbra
98. Judas, F; Palma, P; Falacho, RI; Figueiredo, MH., Estrutura e Dinâmica do tecido ósseo., Texto de apoio para os
alunos de medicina – disciplina de ortopedia; 2012, Coimbra
99. Jensen PR, Andersen TL, Søe K, Hauge EM, Bollerslev J, Amling M, Barvencik F, Delaissé JM: Premature loss of
bone remodeling compartment canopies is associated with deficient bone formation: a study of healthy
individuals and patients with Cushing’s syndrome. J Bone Miner Res 2012, 27:770e780
100. Kristensen HB, Andersen TL, Marcussen N, Rolighed L, Delaissé JM: Increased presence of capillaries next to
remodeling sites in adult human cancellous bone. J Bone Miner Res 2013, 28:574e585
101. Jonasson, G, Rythén, M.: Alveolar bone loss in osteoporosis: a loaded and cellular affair?. Clinical, Cosmetic and
Investigational Dentistry 2016:8 95-103
102. Eriksen EF, Melsen F, Mosekilde L. Reconstruction of the resorptive site in iliac trabecular bone: a kinetic
model for bone resorption in 20 normal individuals. Metab Bone Dis Relat Res (1984) 5:235–
42.doi:10.1016/0221-8747(84)90065-1
103. Eriksen EF, Gundersen HJ, Melsen F, Mosekilde L. Reconstruction of the formative site in iliac trabecular bone
in 20 normal individuals employing a kinetic model for matrix and mineral apposition. Metab Bone Dis Relat
Res (1984) 5:243–52.doi:10. 1016/0221-8747(84)90065-1
104. Matsomoto,M, et al., Bone Tissue Healing Dynamics: From Damage to Reconstruction. Journal of Molecular
Signaling Updates, 2016,1,33-40
105. Bonewald LF. The amazing osteocyte. J Bone Miner Res. 2011; 26(2): 229-38.
http://dx.doi.org/10.1002/jbmr.320
106. Capulli M, Paone R, Rucci N. Osteoblast and osteocyte: games without frontiers. Arch Biochem Biophys. 2014;
561: 3-12. http://dx.doi.org/10.1016/j.abb.2014.05.003
107. Xiong J, Onal M, Jilka RL, Weinstein RS, Manolagas SC, O'Brien CA. Matrix-embedded cells control osteoclast
formation. Nat Med. 2011; 17(10): 1235-41. http://dx.doi.org/10.1038/nm.2448
108. Parfitt, A.M., The bone remodeling compartment: a circulatory function for bone lining cells. J Bone Miner Res,
2001. 16(9): p. 1583-5.
109. Dirckx, N, Van Hul, M, Maes, C., Osteoblast Recruiment to sites of bone formation in skeletal development,
homeostasis and regeneration. Birth Defects Research (Part C) 99:170-191(2013)
110. Li X, Ominsky MS, Warmington KS, Morony S, Gong J, Cao J, Gao Y, Shalhoub V, Tipton B, Haldankar R,
Chen Q, Winters A, Boone T, Geng Z, Niu QT, Ke HZ, Kostenuik PJ, Simonet WS, Lacey DL, Paszty C (2009)
Sclerostin antibody treatment increases bone formation, bone mass, and bone strength in a rat model of
postmenopausal osteoporosis. J Bone Miner Res 24:578–588.

Compartimento de Remodelação Óssea 2016
87
111. Tian X, Setterberg RB, Li X, Paszty C, Ke HZ, Jee WS (2010) Treatment with a sclerostin antibody increases
cancellous bone formation and bone mass regardless of marrow composition in adult female rats. Bone
47:529–533.
112. Xu F., Teitelbaum S., Osteoclasts: new insights. Bone research (2013)1: 11-26
113. Nakashima T. et al., Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat
Med. 2011; 17:1231-1234.
114. Franz-Odendaal TA, Hall BK, Witten PE (2006) Buried alive: how osteoblasts become osteocytes. Dev Dyn
235:176–190
115. Bonewald LF (2007) Osteocytes as dynamic multifunctional cells. Ann N YAcad Sci 1116:281–290
116. Boyle, W.J., W.S. Simonet, and D.L. Lacey, Osteoclast differentiation and activation. Nature, 2003. 423(6937):
p. 337-42.
117. Lin C, Jiang X, Dai Z, Guo X, Weng T, Wang J, Li Y, et al. Sclerostin mediates bone response to mechanical
unloading through antagonizing Wnt/beta-catenin signaling. J Bone Miner Res. 2009; 24:1651–1661.
[PubMed: 19419300]
118. Li X, Ominsky MS, Niu QT, Sun N, Daugherty B, D'Agostin D, Kurahara C, et al. Targeted deletion of the
sclerostin gene in mice results in increased bone formation and bone strength. J Bone Miner Res. 2008;
23:860–869. [PubMed: 18269310]
119. Aguirre JI, Plotkin LI, Stewart SA, Weinstein RS, Parfitt AM, Manolagas SC, Bellido T. Osteocyte apoptosis is
induced by weightlessness in mice and precedes osteoclast recruitment and bone loss. J Bone Min Res. 2006;
21:605–615.
120. Huiskes, R., et al., Effects of mechanical forces on maintenance and adaptation of form in trabecular bone.
Nature, 2000. 405(6787): p. 704-6.
121. Robling, A.G., A.B. Castillo, and C.H. Turner, Biomechanical and molecular regulation of bone remodeling. Annu
Rev Biomed Eng, 2006. 8: p. 455-98.
122. Bonewald, L.F., Mechanosensation and Transduction in Osteocytes. Bonekey Osteovision, 2006. 3(10): p. 7-
15.
123. Huiskes, R., et al., Effects of mechanical forces on maintenance and adaptation of form in trabecular bone.
Nature, 2000. 405(6787): p. 704-6.
124. Bosshardt DD, Schroeder HE. Cementogenesis reviewed: a comparison between human premolars and rodent
molars. Anat Rec 1996, 245(2): 267-292.
125. Bosshardt DD, Nanci A. Hertwig’s epithelial root sheath, enamel matrix proteins, and initiation of
cementogenesis in porcine teeth. J Clin Periodontol 2004, 31(3): 184-192.
126. Eriksen E. et al., Perspective Remodeling and Vascular Spaces in Bone. Journal of bone and mineral research,
Volume 22, Number 1, 2007; Published online on October 2, 2006; doi: 10.1359/JBMR.060910
127. Collin-Osdoby, P., Role of vascular endothelial cells in bone biology. J Cell Biochem, 1994. 55(3): p. 304-9.
128. Carlevaro, M.F., et al., Vascular endothelial growth factor (VEGF) in cartilage neovascularization and
chondrocyte differentiation: auto-paracrine role during endochondral bone formation. J Cell Sci, 2000. 113 ( Pt
1): p. 59-69.
129. Street, J., et al., Vascular endothelial growth factor stimulates bone repair by promoting angiogenesis and bone
turnover. Proc Natl Acad Sci U S A, 2002. 99(15): p. 9656-61.

Compartimento de Remodelação Óssea 2016
88
130. Chambers, TJ, Darby JA, Fuller K. Mammalian collagenase predisposes bone surfaces to osteoclastic
resorption. Cell Tissue Res 1985, 241: 671-675
131. Kristensen HB., et al., Osteoblast recruitment routes in human cancellous bone remodeling. Am J Pathol 2014.
184:778-789
132. Khosla, S., J.J. Westendorf, and U.I. Modder, Concise review: Insights from normal bone remodeling and stem
cell-based therapies for bone repair. Stem Cells, 2010. 28(12): p. 2124
133. Barou O, Laroche N, Palle S, Alexandre C, Lafage-Proust MH: Preosteoblastic proliferation assessed with BrdU
in undecalcified, Eponembedded adult rat trabecular bone. J Histochem Cytochem 1997, 45:1189e1195
134. Maehata Y, Takamizawa S, Ozawa S, Izukuri K, Kato Y, Sato S,Lee MC, Kimura A, Hata RI: Type III collagen
is essential for growth acceleration of human osteoblastic cells by ascorbic acid 2- phosphate, a long-acting
vitamin C derivative. Matrix Biol 2007, 26:371e381
135. Ferrier J, Xia SL, Lagan E, Aubin JE, Heersche JNM: Displacement and translocation of osteoblast-like cells by
osteoclasts. J Bone Miner Res 1994, 9:1397e1405
136. Wegrzyn J., et al.: Role of trabecular micro-architecture and its heterogeneity parameters in the mechanical
behaviour of ex vivo human L3 vertebrae. J Bone Miner Res 2010,25:2324-2331
137. Villars, F., et al., Effect of human endothelial cells on human bone marrow stromal cell phenotype: role of
VEGF? J Cell Biochem, 2000. 79(4): p. 672-85.
138. Crawford, Y. and N. Ferrara, VEGF inhibition: insights from preclinical and clinical studies. Cell Tissue Res,
2009. 335(1): p. 261-9.
139. Kempen, D.H., et al., Effect of local sequential VEGF and BMP-2 delivery on ectopic and orthotopic bone
regeneration. Biomaterials, 2009. 30(14): p. 2816-25.
140. Taubman, M.A. and T. Kawai, Involvement of T-lymphocytes in periodontal disease and in direct and indirect
induction of bone resorption. Crit Rev Oral Biol Med, 2001. 12(2): p. 125-35.
141. Kurata, K., et. al., Bone marrow cell differentiation induced by mechanically damaged osteocytes in 3D gel-
embedded culture. J Bone Miner Res. 2006;21:616-25
142. Bonewald, LF., Osteocyte messages from a bony tomb. Cell Metab. 2007;5:410-411[PubMed:17550775].
143. Kogianni G, Mann V, Noble BS. Apoptotic bodies convey activity capable of initiating osteoclastogenesis and
localised bone destruction. J Bone Miner Res. 2008; 23:915–927. [PubMed: 18435576]
144. Kennedy OD, Herman BC, Laudier DM, Majeska RJ, Sun HB, Schaffler MB. Activation of resorption in fatigue-
loaded bone involves both apoptosis and active pro-osteoclastogenic signaling by distinct osteocyte populations.
Bone. 2012; 50:1115–1122. [PubMed: 22342796]
145. Bellido T. Osteocyte apoptosis induces bone resorption and impairs the skeletal response to weightlessness.
BoneKEy-osteovision. 2007; 4:252–256.
146. Maes, C., et al., Hypoxia-driven pathways in bone development, regeneration and disease. Nat Rev
Rheumatol. 2012b;8:358-366
147. Riddle, RC., et al., Role of hypoxia-inducible factor-1alpha in angiogenic-osteogenic coupling. J Mol Med
2009;87:583-590.
148. Schipani, E., et al., Regulation of osteogenesis-angiogenesis coupling by HIFs and VEGF. J Bone Miner Res
2009; 24:1347-1353

Compartimento de Remodelação Óssea 2016
89
149. McGonigle JS., et al., Osteoprotegerin and RANKL differentially regulate angiogenesis and endothelial cell
function. Angiogenesis (2009) 12:35–46 DOI 10.1007/s10456-008-9127-z
150. Saltel F, Chabadel A, Zhao Y, Lafage-Proust MH, Clezardin P, Jurdic P et al. Transmigration: a new property of
mature multinucleated osteoclasts. J Bone Miner Res 006;21:1913–1923.
151. Mundy, G.R., Cytokines and growth factors in the regulation of bone remodeling. J Bone Miner Res, 1993. 8
Suppl 2: p. S505-10.
152. Roodman, G.D.; Mechanisms of bone metastasis. New England J Med 2004;350:1655-1664
153. Towler DA. The osteogenic-angiogenic interface: novel insights into the biology of bone formation and fracture
repair. Curr Osteoporos Rep 2008;6:67–71.
154. Zelzer E, Olsen BR. Multiple roles of vascular endothelial growth factor (VEGF) in skeletal development,
growth, and repair. Curr Top Dev Biol. 2005;65:169–87.
155. Glowacki J. Angiogenesis in fracture repair. Clin Orthop Relat Res. 1998 Oct; (355 Suppl); S82–9. 6.
156. Pacicca DM, Patel N, Lee C, Salisbury K, Lehmann W, Carvalho R, Gerstenfeld LC, Einhorn TA. Expression of
angiogenic factors during distraction osteogenesis. Bone. 2003 Dec; 33(6):889–98.
157. Ribatti, D., B. Nico, and E. Crivellato, The role of pericytes in angiogenesis. Int J Dev Biol, 2011. 55(3): p. 261-
8.
158. Edelman, D.A., et al., Pericytes and their role in microvasculature homeostasis. J SurgRes, 2006. 135(2): p.
305-11.
159. Laroche M. Intraosseous circulation from physiology to disease. Joint Bone Spine. 2002 May; 69(3):262–9.
160. Anagnostis P, Karagiannis A, Kakafika A, Tziomalos K, Athyros V, Mikhailidis D. Atherosclerosis and
osteoporosis: age-dependent degenerative processes or related entities?. Osteoporos Int. 2009; Feb 1;
20(2):197–207.
161. Thompson B, Towler DA. Arterial calcification and bone physiology: role of the bone-vascular axis. Nat Rev
Endocrinol 2012; 8: 529–543.
162. Baltadjiev G. Ultrastructural features in the structure of blood capillaries of the Haversian canal. Folia Med
(Plovdiv). 1994;36(2):47–53.
163. Marotti G, Zambonin Zallone A. Changes in the vascular network during the formation of Haversian systems.
Cells Tissues Organs. 1980;106(1):84–100. 10.
164. Parfitt AM. Osteonal and hemi-osteonal remodeling: the spatial and temporal framework for signal traffic in
adult human bone. J Cell Biochem. 1994 Jul; 55(3):273–86.
165. Baron R, Vignery A, Lang R: Reversal phase and osteopenia: defective coupling of resorption to formation in
the pathogenesis of osteoporosis. Edited by Deluca HF, Frost HM, Jee WSS, Johnston CC, Parfitt AM.
Osteoporosis: Recent Advances in Pathogenesis and Treatment. Baltimore, University Park Press, 1980, pp
311e320
166. Noble BS, Peet N, Stevens HY, Brabbs A, Mosley JR, Reilly GC, Reeve J, et al. Mechanical loading: biphasic
osteocyte survival and the targeting of osteoclasts for bone destruction in rat cortical bone. Am J Physiol Cell
Physiol. 2003; 284:C934–C943. [PubMed: 12477665]
167. Bellido T, Plotkin LI. Novel actions of bisphosphonates in bone: Preservation of osteoblast and osteocyte
viability. Bone. 2011; 49:50–55. [PubMed: 20727997]

Compartimento de Remodelação Óssea 2016
90
168. Jilka, RL.; Bellido, T.; Almeida, M.; Plotkin, LI.; O'Brien, CA.; Weinstein, RS.; Manolagas, SC Apoptosis in bone
cells. In: Bilezikian, JP.; Raisz, LG.; Martin, TJ., editors. Principles of Bone Biology. Academic Press; San Diego,
San Francisco, New York, London, Sydney, Tokyo: 2008.p. 237-261
169. Makris GP, Saffar JL. Disturbances in bone remodeling during the progress of hamster periodontitis. A
morphological and quantitative study. J Periodontal Res 1985;20:411–420.
170. Henriksen K. et al. Local communication on and within bone controls bone remodelling. 2009; Elsevier Bone;
44:1026-10.

Compartimento de Remodelação Óssea 2016
91
VI. Índice
__________________________________________________________
Agradecimentos---------------------------------------- ---- --------------------- 4
Sumário--------------------------------------------------------- ----------------- ---5
I – Introdução
1. Justificação do tema 7
1.2 Planificação do trabalho 10
2. Ciclo de Remodelação óssea (alguns aspetos) 12
II – Compartimento de Remodelação Óssea (BRC)
1. Objetivo 25
2. Materiais e Métodos 25
2.1 Animal de experimentação
2.2 Manutenção e bem-estar animal
2.3 Colheita do material
2.4 Processamento histológico de material descalcificado
2.5 Processamento histológico de material não descalcificado
3. Resultados
3.1 Deteção de BRC no tecido ósseo alveolar 31
3.2Identificação de diferentes etapas do ciclo de remodelação óssea 45
3.3 Deteção de compartimentos de remodelação no cemento e dentina 53
4. Discussão 59
III – Resumo 71
IV – Abstract 77
V – Bibliografia 83
VII- Anexos 95

Compartimento de Remodelação Óssea 2016
92
VII. Anexos
__________________________________________________________
Listagem de Acrónimos
BMU – Basic Multicellular Unit
BRC – Bone Remodelling Compartment
LPS – Lipopolissacarídeos
RANKL – Receptor Activated of Nuclear Factor Kappa-B Ligand
RANK – Receptor Activated of Nuclear Factor Kappa-B
OPG – Osteoprotegerin
TGFβ- Tumoral Growth Factor β
PDGF – Platelet Derived Growth Factor
IGF-I – Insulin-like Growth Factor I
BMP – Bone Morphogenic Protein
S1P – Sphingosin-1-phosphate
ET-I – Endothelin I
VEGF – Vascular Endothelial Growth Factor