caracterizaÇÃo genÉtica do vÍrus influenza a … maria... · que me estimularam a realizar o...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS DA SAÚDE PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL
CARACTERIZAÇÃO GENÉTICA DO VÍRUS INFLUENZA A
(H1N1)pdm09 E DIAGNÓSTICO DIFERENCIAL DE CASOS SUSPEITOS DE INFLUENZA PANDÊMICA, NO ESTADO DE
PERNAMBUCO, NO PERÍODO DE MAIO DE 2009 A MAIO DE 2010
MARIA JOSÉ COUTO OLIVEIRA
RECIFE, PE
2012

MARIA JOSÉ COUTO OLIVEIRA
CARACTERIZAÇÃO GENÉTICA DO VÍRUS INFLUENZA A(H1N1)pdm09 E DIAGNÓSTICO DIFERENCIAL DE CASOS
SUSPEITOS DE INFLUENZA PANDÊMICA, NO ESTADO DE PERNAMBUCO, NO PERÍODO DE MAIO DE 2009 A MAIO DE 2010.
ORIENTADORA: PROFª. DRA. VERA MAGALHÃES DA SILVEIRA
CO-ORIENTADOR: DR. FERNANDO COUTO MOTTA
RECIFE, PE
2012
Tese apresentada a Banca Examinadora do Programa de Pós-Graduação em Medicina Tropical do Centro de Ciências da Saúde da Universidade Federal de Pernambuco, como parte dos requisitos para a obtenção do Título de Doutor em Medicina Tropical



UNIVERSIDADE FEDERAL DE PERNAMBUCO CENTRO DE CIÊNCIAS DA SAÚDE
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL REITOR
Anísio Brasileiro de Freitas Dourado
PRÓ-REITOR PARA ASSUNTOS DE PESQUISA E PÓS-GRADUAÇÃO
Francisco de Sousa Ramos
DIRETOR DO CENTRO DE CIÊNCIAS DA SAÚDE
José Tadeu Pinheiro
COORDENADORA DO PROGRAMA DE PÓS-GRADUAÇÃO
EM MEDICINA TROPICAL
Maria Rosângela Cunha Duarte Coelho
VICE-COORDENADORA DO PROGRAMA DE PÓS-GRADUAÇÃO
EM MEDICINA TROPICAL
Valdênia Maria Oliveira de Souza
CORPO DOCENTE
Ana Lúcia Coutinho Domingues
Célia Maria Machado Barbosa de Castro
Edmundo Pessoa de Almeida Lopes Neto
Fábio André dos Santos Brayner
Heloísa Ramos Lacerda de Melo
Maria Amélia Vieira Maciel
Maria de Fátima Pessoa Militão de Albuquerque
Maria do Amparo Andrade
Maria Rosângela Cunha Duarte Coelho
Marli Tenório Cordeiro
Ricardo Arraes de Alencar Ximenes
Valdênia Maria Oliveira de Souza
Vera Magalhães da Silveira
Vláudia Maria Assis Costa

DEDICATÓRIA
À minha querida e inesquecível
Mãe, Irene (In memorian).
Ao meu querido esposo, Marcelo pelo
companheirismo, renuncia, incentivo, compreensão e parceria.
Aos meus queridos filhos Flávio, Danielle,
Karina, Daniel e Mauro.
Às minhas netas Carolina e Mariana
que vieram para dar mais graça à minha vida.

AGRADECIMENTOS
Primeiro a Deus por me dar saúde e força para que eu pudesse concluir o
curso.
Aos meus familiares pelo constante apoio, especialmente ao meu esposo e
filhos por entenderem minhas constantes ausências.
A minha prezada orientadora Dra. Vera Magalhães da Silveira, pela
confiança, profissionalismo, paciência e incentivo nas horas de desânimo.
À Dra. Marilda Siqueira, chefe do Laboratório de Vírus Respiratórios e do
Sarampo da Fundação Oswaldo Cruz do Rio de Janeiro, grande pesquisadora e
amiga de longas datas que abriu as portas do Laboratório e disponibilizou seu corpo
técnico para realizarmos todos os ensaios. Sem a sua ajuda, não teria sido possível
a realização desse estudo.
Ao meu co-orientador Dr. Fernando Motta pela paciência e preciosas
orientações.
A amiga Marli Tenório pela indescritível ajuda e apoio na finalização do
estudo.
Aos colegas do Instituto Evandro Chagas, Dr Wyller Mello e Mirleide Santos
que me estimularam a realizar o projeto.
Aos colegas da Fiocruz, que estiveram ao meu lado na realização dos testes,
nas diversas vezes que estive no Rio de Janeiro, especialmente Ângela Pinhão,
Daniela Machado, Daniel Cohen, Maria de Lourdes Oliveira, Milena Mesquita, Paolla
Andrade, Priscilla Born, Sharon Carney e Thiago Moreno. Aos demais colegas que
não trabalharam na bancada, mas me acolheram como uma pessoa da casa.

A todos os colegas do Setor de Virologia do LACEN–PE, principalmente a
Ana Sinício e Risalva Travassos, pela compreensão nos momentos que precisei me
ausentar e que elas com muita eficiência executaram as atividades que eram de
minha responsabilidade e a Lílian Barros Lima responsável pelo diagnóstico de
influenza.
Aos colegas do Laboratório Municipal do Recife, principalmente os que
ajustaram seus períodos de férias, dando prioridade às minhas necessidades.
Aos colegas da Epidemiologia da Secretaria Estadual de Saúde que sempre
me atenderam quando precisei de informações, especialmente Lucilene Rafael
Aguiar, Ana Lúcia de Souza, Alice Rodovalho e Ana Antunes.
Aos meus superiores do LACEN-PE e do Laboratório Municipal do Recife por
terem compreendido a necessidade de me ausentar para a execução da parte de
bancada desse trabalho.
A todos os professores e membros do Programa de Pós-Graduação em
Medicina Tropical, especialmente a Walter pelas orientações ao longo do curso.

É graça divina começar bem.
Graça maior persistir na caminhada certa.
Mas graça das graças é não desistir nunca.
Dom Helder Câmara

RESUMO
OLIVEIRA, Maria José Couto, Caracterização genética do vírus influenza
A(H1N1)pdm09 e diagnóstico diferencial de casos suspeitos de influenza
pandêmica, no estado de Pernambuco, no período de maio de 2009 a maio de
2010. 158 folhas. (Tese de doutorado). Universidade Federal de Pernambuco -
Centro de Ciências da Saúde. Departamento de Medicina Tropical. Recife –
Pernambuco.
Durante a pandemia (2009-2010) com o vírus influenza A(H1N1)pdm09 foi
recomendado o tratamento com o oseltamivir ou zanamivir. Com o aumento da
detecção de vírus de Influenza A (H1N1) sazonal resistente ao oseltamivir houve a
preocupação de que o mesmo ocorresse com o novo vírus pandêmico. Nesta
pandemia, muitos pacientes com suspeita de infecção pelo vírus A(H1N1)pdm09
tiveram o teste negativo para influenza A, ficando sem uma definição do agente
etiológico. Testes moleculares podem detectar a presença de mutações
relacionadas à resistência ao oseltamivir, à virulência e antigenicidade do vírus,
assim como podem definir o diagnóstico etiológico por vírus respiratórios. Para
esclarecer essas questões dois estudos foram realizados. O primeiro foi a
“Caracterização genética dos vírus influenza A (H1N1)pdm09 detectados no Estado
de Pernambuco, Brasil, no período de maio de 2009 a maio de 2010”, com o objetivo
de verificar a resistência desse vírus ao oseltamivir e também avaliar a diversidade
genética dos vírus circulantes. Foram analisadas 118 amostras do vírus
A(H1N1)pdm09 através de pirosequenciamento, precedida da transcrição reversa e
reação em cadeia da polimerase em tempo real (rRT-PCR) para amplificação do
H1N1pdm-N1 fragmento C e posterior detecção da mutação H274Y, utilizando o
equipamento PyroMark Q-96 ID no modo SNP (single nucleotide polymorphism).
Foram sequenciados os genes da hemaglutinina de 31 amostras, pela técnica de
Sanger, de acordo com o Protocolo do CDC para Influenza. Foi utilizado o kit “Big
Dye® terminator Cycle Sequencing” (Applied Biosystem) e o produto submetido ao
método de precipitação X-terminator. A mutação H274Y não foi observada,
indicativo de que os vírus sequenciados eram sensíveis ao oseltamivir. As 31
amostras sequenciadas mostraram-se intimamente relacionadas com a cepa de
referência A/California/7/2009(H1N1), entretanto, foram detectados 14 tipos de
mutações, porém sem implicação no aumento da virulência. O segundo estudo

realizado: ”Aspectos epidemiológicos e virológicos da infecção por Influenza
A(H1N1)pdm09 e frequência de outros vírus respiratórios no Estado de
Pernambuco, Brasil: 2009 – 2010” teve como objetivo analisar a pandemia de
influenza no estado e identificar os vírus respiratórios responsáveis pelo quadro
clínico que levou à hipótese diagnóstica da influenza pandêmica. Foram analisados
espécimes de 705 casos para detecção do vírus da influenza A, utilizando-se a PCR
em tempo real, sistema TaqMan, de acordo com o Centers for Disease Control and
Prevention / Atlanta, das quais, 26,3% (186/705) foram positivas para o vírus
A(H1N1)pdm09 e 2,3% (16/705) positivas para influenza A sazonal. Para detecção
de outros vírus respiratórios foram analisadas 146 amostras negativas para o vírus A
(H1N1)pdm09 por RT-PCR multiplex, com o kit “FTD Respiratory21 PLUS”. Entre as
amostras negativas para o vírus A(H1N1)pdm09, 36,5% (53/146) foram positivas
para outros vírus respiratórios, com três casos de infecção viral múltipla. Foram
detectados: rhinovírus (41%), coronavírus 43 (14,3%), metapneumovírus humano
(14,3%), bocavírus (7,1%), vírus respiratório sincicial (5,3%), influenza B (3,6%),
parainfluenza 2 (3,6%), parainfluenza 3 (3,6%), adenovírus (1,8%), coronavírus HKU
(1,8%), enterovírus (1,8%) e parainfluenza 1 (1,8%). Estes resultados mostram a
circulação, além da Influenza A(H1N1)pdm09, de outros vírus respiratórios no
estado em 2009-2010; evidenciam a necessidade da análise laboratorial dos casos
suspeitos de influenza e a importância do monitoramento laboratorial das infecções
respiratórias, uma vez que o diagnóstico etiológico baseado apenas em critérios
clínicos nem sempre é acurado.
Palavras chave: Influenza; vírus influenza A(H1N1)pdm09; hemaglutinina;
oseltamivir; vírus respiratórios; infecção respiratória aguda; rhinovírus.

ABSTRACT
OLIVEIRA, Maria José Couto. Genetic characterization of Influenza A
(H1N1)pdm09 virus and diferential diagnostic of suspected pandemic influenza cases
in the State of Pernambuco, during the period of May 2009 to May 2010. Thesis
(Doctor of Tropical Medicine) - Universidade Federal de Pernambuco, Centro de
Ciências da Saúde, Departamento de Medicina Tropical. Recife, Pernambuco, 2012.
During the pandemic period (2009-2010) with Influenza A(H1N1)pdm09 virus it was
recommended the treatment using oseltamivir or zanamivir. With the increasing
detection of Influenza A (H1N1) virus seasonal resistant to oseltamivir there was the
preoccupation that the same could occur with the new pandemic virus. In this
pandemic, many patients suspected of Influenza A(H1N1)pdm09 virus infection had
a negative test result to influenza A, thus without a definition of the etiologic agent.
Molecular tests can detect the presence of mutations related to resistance to
oseltamivir, virulence and virus antigenicity, as well as they can define the etiologic
diagnose caused by respiratory virus. In order to clarify these questions two studies
were carried out. The first one was the “Genetic characterization of Influenza A
(H1N1)pdm09 viruses in Pernambuco State, Brazil, during the period of May 2009 to
May 2010”, with the objective of investigate the virus resistance to oseltamivir and
also to evaluate the genetic diversity of circulating viruses. It was tested 118 samples
of influenza A(H1N1)pdm09 virus by pyrosequencing, after the reverse transcription
and real time polymerase chain reaction (rRT-PCR), for H1N1pdm-N1 fragment C
amplification and detection of mutation H274Y using the PyroMark Q-96 ID in the
single nucleotide polymorphism (SNP) mode. It was sequenced the hemaglutinine
genes from 31 samples, through Sanger technique, according to CDC Influenza
Protocol. It was employed the kit Big Dye® terminator Cycle Sequencing” (Applied
Biosystem) and the product submitted to precipitation X-terminator method. It was
not observed the mutation H274Y, indicating that the sequenced virus were sensitive
to oseltamivir. The 31 samples sequenced reveled to be closed related to the
reference strain A/California/7/2009(H1N1), nevertheless, 14 types of mutation were
detected, but without implication in virulence increasing. The second study performed
was “Epidemiological and virological aspects of Influenza A(H1N1)pdm09 and
frequency other respiratory viruses in Pernambuco State, Brazil: 2009-2010” with the
objective of studying the influenza pandemic in the state and identify the respiratory

viruses responsible by the clinical symptoms that brought to the diagnose
hypotheses of pandemic influenza. It was tested 705 samples for influenza virus
detection by real time PCR (CDC protocol), from which 2.3% (16/705) were positive
for influenza A seasonal and 26.3% (186/705) were reactive for Influenza
A(H1N1)pdm09 virus. For presence of other respiratory viruses, 146 negative
samples for Influenza A(H1N1)pdm09 virus were tested by multiplex RT-PCR, using
the kit FTD Respiratory21 PLUS. Among the negative samples for Influenza
A(H1N1)pdm09 virus, 36.5% (53/146) were reactive for other respiratory viruses, and
three viral multiple-infection cases were found. It was detected: Rhinovirus (41.0%),
Coronavirus 43 (14.3%), human Metapneumovirus (14.3%), Bocavirus (7.1%),
Respiratory Syncytial Virus (5.3%), Influenza B (3.6%), Parainfluenza 2 (3.6%),
Parainfluenza 3 (3.6%), Adenovirus (1.8%), Coronavirus HKU (1.8%), Enterovirus
(1.8%) and Parainfluenza 1 (1.8%). These results show that besides Influenza
A(H1N1)pdm09 virus, other respiratory viruses were also circulating in Pernambuco
during 2009-2010; also show the necessity of laboratorial diagnoses and the
importance of monitoring the respiratory infections; taking in account that the
etiologic diagnose relying exclusively on clinical bases is not always accurate.
Key words: Influenza, virus influenza A(H1N1)pdm09; hemagglutinin; oseltamivir,
respiratory viruses, acute respiratory infection; rhinovirus.

LISTA DE FIGURAS
Figura 1 – Representação esquemática da partícula viral................................. 21
Figura 2 – Principais mecanismos evolutivos do vírus influenza ...................... 23
Figura 3 – Linha do tempo mostrando as pandemias provocadas pelo vírus
influenza A .........................................................................................................
28
Figura 4 – Fases de uma pandemia de influenza.............................................. 28
Figura 5 – Genótipos do vírus influenza A(H1N1) de linhagem suína america-
na e Influenza A(H1N1)pdm09 em casos recentes nos Estados Unidos...........
33
Figura 6 – Prevalência dos vírus influenza A (H1N1) sazonal resistentes ao
oseltamivir em julho de 2008..............................................................................
45
Figura 7– Fluxograma das etapas e técnicas utilizadas na realização do
estudo.................................................................................................................
64
ARTIGO 1
Figura 1– Árvore filogenética das sequências de HA dos vírus influenza A
(H1N1)pdm09 detectados em Pernambuco.....................................................
88
ARTIGO 2
Figura 1 – Distribuição dos casos de IRA e de infecção pelo vírus influenza
A(H1N1)pdm09 de acordo com as semanas epidemiológicas.........................
105

LISTA DE TABELAS
Tabela 1 – Proteínas do vírus Influenza A e respectivas funções.................... 22
ARTIGO 1
Tabela 1 – Mutações de aminoácidos no gene HA observados em pacientes
ambulatoriais e hospitalizados.......................................................................... 87
ARTIGO 2
Tabela 1 – Características clínico-epidemiológicas da população do estudo.. 103
Tabela 2 – Casos fatais por Influenza A(H1N1)pdm09 no Estado de
Pernambuco......................................................................................................
104
Tabela 3 – Frequência de outros patógenos virais entre os casos de IRA no
estado de Pernambuco.....................................................................................
104

LISTA DE ABREVIATURAS E SIGLAS
AdV Adenovírus
CDC Centers for Disease Control and Prevention
CT Ciclo no qual a fluorescência emitida atravessa o ponto de corte do teste de PCR em tempo real.
dNTP Desoxirribonucleotídeo trifosfato
EIA Ensaio imunoenzimático
EV Enterovírus
FLUA Influenza A
FLUB Influenza B
HA Hemaglutinina
HBoV Bocavírus humano
HCoV Coronavírus humano
HMPV Metapneumovírus humano
IEC Instituto Evandro Chagas
ILI Síndrome gripal
IOC Instituto Oswaldo Cruz
IRA Infecção respiratória aguda
LACEN Laboratório Central de Saúde Pública
LVRS Laboratório de Vírus Respiratórios e do Sarampo
M Proteína Matriz
MS Ministério da Saúde
NA Neuraminidase
NEP Proteína de exportação nuclear
NS Proteína não estrutural
OMS Organização Mundial de Saúde
One-Step- RT-PCR Transcrição reversa seguida de reação em cadeia da polimerase em uma única etapa.
PA Polimerase ácida
PB Polimerase básica
pb Pares de base
PCR Reação em cadeia da polimerase
Pdm Pandêmico
PeV Parechovírus

PIV Vírus parainfluenza
RIDTs Testes rápidos para o diagnóstico de influenza
RNA Ácido ribonucléico
RNP Ribonucleoproteína
rRT-PCR RT-PCR em tempo real
RT Transcrição reversa
RV Rhinovírus
SG Síndrome gripal
SRAG Síndrome respiratória aguda grave
SVO Serviço de verificação de óbitos
RSV Vírus respiratório sincicial

SUMÁRIO
RESUMO ABSTRACT 1 APRESENTAÇÃO............................................................................... 17
2 REVISÃO DA LITERATURA............................................................... 19 2.1 Vírus Influenza.................................................................................... 19 2.2 Caracterização genética do vírus influenza A (H1N1)pdm09 ............ 38
2.3 Outras viroses respiratórias ................................................................ 47 3 OBJETIVOS......................................................................................... 61 3.1 Geral.................................................................................................... 61 3.2 Específicos.......................................................................................... 61 4 MATERIAIS E MÉTODOS................................................................... 61
4.1 Desenho do estudo.............................................................................. 61 4.2 Local do estudo.................................................................................... 62 4.3 População do estudo............................................................................ 62 4.4 Variáveis............................................................................................... 62
4.4.1 Variáveis dependentes...................................................................... 62
4.4.2 Variáveis independentes................................................................. 62
4.5 Operacionalização da pesquisa.......................................................... 63 4.5.1 Detecção do vírus influenza A(H1N1)pdm09................................... 66 4.5.2 Sequenciamento do gene HA.......................................................... 67
4.5.3 Pirosequenciamento para detecção da mutação H274Y................ 70 4.5.4 Diagnóstico diferencial de influenza A............................................. 72 4.6 Considerações éticas.......................................................................... 74 4.7 Análise estatística................................................................................ 74
5 RESULTADOS...................................................................................... 75 5.1 ARTIGO 1 Caracterização genética dos vírus influenza A (H1N1)pdm09 detectados no estado de Pernambuco, no período de maio de
2009 a maio de 2010............................................................................. 75 5.2 ARTIGO 2
Aspectos epidemiológicos e virológicos da infecção por influenza A(H1N1)pdm 09 e frequência de outros vírus respiratórios no

Estado de Pernambuco, Brasil: 2009 – 2010......................................... 90
6 CONCLUSÕES...................................................................................... 106
7 CONSIDERAÇÕES FINAIS................................................................... 107 REFERÊNCIAS APÊNDICES
Apêndice A - Versão em inglês do Artigo 1 a ser submetido à Revista Memórias do Instituto Oswaldo Cruz
Apêndice B - Versão em inglês do Artigo 2 a ser submetido à Revista
Journal of Clinical Virology
ANEXOS
Anexo A – Aprovação do Comitê de Ética em Pesquisa da UFPE
Anexo B – Carta de anuência do LACEN-PE
Anexo C – Carta de anuência da Diretoria Geral de Vigilância
Epidemiológica e Ambiental /SES-PE.
Anexo D - Ficha de Investigação Influenza Humana por Novo
Subtipo (Pandêmico).

17
1 APRESENTAÇÃO
Em abril de 2009, a Organização Mundial da Saúde (OMS) notificou a
ocorrência de casos humanos de influenza que ocorreram no México e nos Estados
Unidos da América. Identificou-se um vírus influenza A subtipo H1N1 produto de
rearranjo que circulou pelo mundo, sendo responsável pela primeira pandemia do
século XXI e por mais de 18.000 mortes (ORGANIZAÇÃO MUNDIAL DA SAÚDE,
2010a). Os primeiros casos de infecção pelo vírus influenza A(H1N1)pdm09 no
Brasil foram confirmados em maio de 2009 e, no estado de Pernambuco, em junho
do mesmo ano.
Os vírus influenza são conhecidos por suas altas taxas de mutação,
principalmente, nos genes da hemaglutinina (HA) e da neuraminidase (NA). O gene
da hemaglutinina é o que apresenta a taxa de mutação mais elevada e a proteína
que ele codifica (HA) está diretamente associada ao sucesso da infecção viral, daí a
importância do seu monitoramento. Estudos com o vírus influenza A (H1N1)pdm09
têm detectado várias mutações no gene HA, sendo uma das principais a D222G,
que tem sido associada aos casos graves e fatais (LIU et al. 2010; LEDESMA 2011).
Mutações no gene NA podem levar à perda de sensibilidade aos inibidores da
atividade neuraminidásica viral ao oseltamivir (ABED et al. 2006, AOKI et al. 2007).
Estudos anteriores à pandemia de 2009 mostraram resistência do vírus influenza A
(H1N1) sazonal humano ao oseltamivir, superior a 10% em vários países (DHARAN
et al. 2009; ESHAGHI et al. 2009). No curto período de 8 de dezembro de 2008 a 24
de janeiro de 2009, trinta países notificaram à OMS a detecção de 1291 vírus
influenza A (H1N1) sazonal, resistentes ao oseltamivir (ORGANIZAÇÃO MUNDIAL
DA SAÚDE, 2009a).
Com a introdução do novo vírus influenza A(H1N1)pdm09 na população
mundial e do amplo uso do antiviral oseltamivir, principalmente nos países
desenvolvidos, surgiu a necessidade de monitoramento do referido vírus para a
identificação de variantes mais virulentas ou capazes de evadir da resposta imune
recém montada, assim como verificar se os vírus circulantes estão desenvolvendo
resistência às drogas antivirais aplicadas no momento.
Durante a pandemia, muitos casos de infecção respiratória aguda, analisados
laboratorialmente, foram negativos para o vírus Influenza A, permanecendo sem

18
diagnóstico etiológico. É sabido que além do vírus influenza, outros vírus podem
causar infecções respiratórias agudas, tais como o vírus respiratório sincicial,
parainfluenza, adenovírus, metapneumovírus (THOMAZELLI et al., 2007), rhinovírus,
coronavírus e enterovírus (WEISSENBACHER; ÁVILA, 1998). Esse fato faz com que
haja necessidade do exame laboratorial para determinar a etiologia das infecções
respiratórias agudas (IRAs) e assim evidenciar a necessidade de prevenção contra
outras viroses respiratórias. Para tanto, ensaios de multiplex RT-PCR com detecção
em tempo real constituem uma excelente ferramenta para detectar, além do vírus
pandêmico, outros patógenos associados aos casos de síndrome gripal (SG) ou
síndrome respiratória aguda grave (SRAG).
A ausência de dados sobre resistência ao oseltamivir e da análise genética
dos vírus influenza detectados durante a pandemia de 2009 no estado de
Pernambuco, assim como o grande número de casos suspeitos da infecção sem
diagnóstico etiológico, motivaram a realização deste estudo. Esta tese está
constituída de dois artigos para publicação em periódicos, e de uma parte
introdutória contendo uma breve revisão bibliográfica sobre o vírus influenza,
caracterização genética do vírus influenza A(H1N1)pdm09 e outros vírus
responsáveis pela etiologia de infecções respiratórias agudas.
No primeiro artigo, cujo título é “Caracterização genética dos vírus influenza A(H1N1)pdm09 detectados no estado de Pernambuco, Brasil, no período de maio de 2009 a maio de 2010” descrevem-se a frequência de
mutações detectadas no gene da hemaglutinina, relacionadas a antigenicidade e
virulência e da mutação H274Y no gene neuraminidase relacionada à resistência ao
oseltamivir. No segundo artigo, intitulado “Aspectos epidemiológicos e virológicos da infecção por Influenza A(H1N1)pdm09 e frequência de outros vírus respiratórios no estado de Pernambuco, Brasil: 2009-2010” foram descritos
os aspectos clínico-epidemiológicos dos casos confirmados por laboratório de
infecção com o vírus influenza A (H1N1)pdm09, assim como a etiologia viral de
alguns casos cujo resultado foi negativo para o vírus pandêmico.

19
2 REVISÃO DA LITERATURA 2.1 VÍRUS INFLUENZA
A influenza ou gripe é uma infecção viral aguda do sistema respiratório que
tem distribuição global e elevada transmissibilidade. Sua importância deve-se ao seu
caráter epidêmico, caracterizado pela disseminação rápida e marcada morbidade
nas populações atingidas (BRASIL, 2006). Epidemias de doença respiratória com
sintomas semelhantes aos ocasionados pelo vírus da Influenza já tinham sido
descritas por Hipócrates no ano 412 a.C. (NICHOLSON, 1998).
Em 1933, Wilson Smith e colaboradores do Instituto Nacional para Pesquisas
Médicas em Londres inocularam por via intranasal, lavados de nasofaringe de
pacientes com sintomas de gripe em furões. Observaram que a doença era
transmitida do homem para o furão e, que, após a inoculação, esses animais
apresentavam os mesmos sintomas dos pacientes e desenvolviam uma resposta
imunológica ao inóculo. Desse modo, foi descoberto o vírus denominado,
posteriormente, como vírus Influenza tipo A (SMITH; PARVIN; PALESE, 1986).
Em 1940, foi detectado o vírus influenza B, antigenicamente distinto do vírus
influenza A, mas com características estruturais semelhantes (FRANCIS, 1940) e em
1951 foi descoberto um novo gênero denominado Influenzavirus tipo C (TAYLOR,
1951).
O vírus Influenza pertence à família Ortomyxoviridae e de acordo com perfis
antigênicos característicos, são subdivididos em três gêneros: A, B e C, comumente
referidos como influenza “tipos” A, B e C (BUSH, 2007), entretanto, apenas os tipos
A e B têm relevância clínica em humanos (BEHRENS et al., 2006). Os principais
determinantes antigênicos dos vírus da Influenza A e da Influenza B são as
glicoproteínas de superfície, hemaglutinina (HA) e neuraminidase (NA) (PALESE;
SHAW, 2007). Representação esquemática da partícula viral pode ser vista na
Figura 1.
Os vírus Influenza A são classificados segundo as glicoproteínas (HA e NA).
Até o momento foram descritas 16 HA (H1 a H16) e 9 NA (N1 a N9). Sua completa
nomenclatura inclui o tipo do vírus, o hospedeiro de origem (exceto quando
humano), o local onde foi isolado, o número da cepa e o ano do isolamento, seguido
do subtipo antigênico de HA e NA entre parênteses. Como exemplo pode ser citado

20
o vírus Influenza A/Califórnia/4/2009 (H1N1). Subtipos antigênicos de Influenza B e
Influenza C não têm sido descritos (WRIGHT; WEBSTER, 2007).
As partículas virais são constituídas de 0,8 a 1% de RNA, 70% de proteína,
20% de lipídios, 5 a 8% de carboidratos. Seu genoma é constituído de RNA de fita
simples, polaridade negativa, segmentado, envolvido por um capsídeo protéico de
simetria helicoidal, composto pela proteína matriz e por um envelope lipoprotéico
(SANTOS; ROMANO; WIGG, 2008). Pela sua composição química é sensível ao
calor (56ºC durante 30 minutos), pH ácido (3,0) e solventes lipídicos (SANTOS;
ROMANO; WIGG, 2002).
Os vírus influenza são pleomórficos e, geralmente, se apresentam sob a
forma de partículas esféricas com um diâmetro de cerca de 100nm, mas partículas
filamentosas têm sido frequentemente observadas, com mais de 300nm (PALESE;
SHAW, 2007). As partículas do vírus influenza A possuem espículas denominadas
hemaglutinina (HA) e neuraminidase (NA) com comprimento em torno de 10 a 14 nm
respectivamente. Estas glicoproteínas são encontradas no vírion numa proporção de
quatro HA para cada NA. Embora a HA seja o principal alvo para anticorpos
neutralizantes, anticorpos contra a NA também podem reduzir a ocorrência e
gravidade da doença, e possivelmente, prevenir a infecção, se presente em altos
títulos (BUSH, 2007). A hemaglutinina é o principal antígeno viral que promove a
adsorção da partícula aos resíduos sializados das células do sistema respiratório e
após a endocitose da partícula, induz a fusão do envelope viral à membrana interna
do endossoma, permitindo a liberação do genoma viral no citoplasma celular. A
neuraminidase é outra glicoproteína de superfície e tem atividade de sialidase (cliva
o ácido siálico), sendo fundamental ao brotamento do vírus da superfície celular para
infectar novas células.
O genoma do vírus da Influenza A possui 10 genes em 8 segmentos (BUSH,
2007; BEHRENS et al., 2006). Cada segmento codifica importantes proteínas, cujas
funções encontram-se descritas na Tabela 1.
As proteínas básicas 1 e 2 ( PB1 e PB2) são componentes do complexo
polimerase, tendo como função o reconhecimento do iniciador 7-metil guanina do
RNA mensageiro celular, atividade endonucleásica e extensão da cadeia,
respectivamente. A proteína ácida (PA) completa o complexo polimerase. A
nucleoproteína (NP) tem função de proteção e importação do RNA para o núcleo
celular. A proteína de matriz (M1) tem função estrutural, interage com as RNPs e

21
glicoproteínas de superfície. M2 é uma proteína de membrana, tem função de canal
iônico e montagem. A NS1 é uma proteína não estrutural multifuncional e a
NEP/NS2 atua na exportação nuclear dos RNPs virais. Mais recentemente, outra
proteína foi descoberta, a PB1-F2 (PALESE; SHAW, 2007). Figura 1. Representação esquemática da partícula viral
Fonte: McHardy and Adams, 2009
A maioria das proteínas virais desempenha um papel direto na virulência.
Atualmente, as proteínas mais relacionadas ao potencial patogênico são a HA, PB1,
PB2 e NS1. As mais associadas com o desenvolvimento de resistência a drogas
antivirais são NA e M2 (ARIAS et al., 2009).

22
Tabela 1 – Proteínas do vírus Influenza A e respectivas funções
Segmento Proteínas Função 1 PB2 Componente da RNA polimerase, reconhecimento 2 PB1 Componente da RNA polimerase; atividade de endonuclease e alongamento 3 PA Componente da RNA polimerase 4 HA Glicoproteína de superfície, receptor de ligação, atividade de fusão e
principal antígeno 5 NP Ligação, síntese e importação nuclear de RNA 6 NA Glicoproteína de superfície; atividade de sialidase
M1 Proteína de matriz, interação com as RNPs virais e glicoproteínas de superfície
7
M2 Proteína de membrana, com atividade de canal iônico e montagem NS1 Proteína não estrurural multifuncional. Inibidora da resposta imune inata. 8
NEP/NS2 Proteína de exportação nuclear das RNPs virais do núcleo para o citoplasma celular.
Fonte: Adaptado de Palese e Shaw (2007). Variabilidade e evolução do vírus influenza
Dois aspectos do genoma do vírus da influenza permitem uma grande
flexibilidade de evolução: 1- o vírus tem um genoma segmentado, favorecendo o
rearranjo de segmentos; 2- o complexo polimerase viral não possui nenhum
mecanismo de revisão para corrigir erros de replicação (BUSH, 2007).
Os vírus influenza estão sujeitos a dois tipos de variações antigênicas:
a) Antigenic drift – durante a replicação viral, mutações decorrentes da baixa
fidelidade do complexo polimerase ocorrem gerando mutações pontuais no genoma
da progênie. Tais mutações pontuais podem resultar em mudanças nos aminoácidos
que compõem as glicoproteínas de superfície, particularmente da hemaglutinina.
Surgem então, novas variantes capazes de escapar da imunidade estimulada por
infecção ou vacinação prévia. b) Antigenic shift - são consequência de rearranjos
gênicos caracterizados pela completa substituição de um ou mais segmentos do
genoma viral. Essas alterações se devem ao reagrupamento entre vírus humanos e
vírus que infectam outras espécies animais. Quando ocorrem rearranjos antigênicos
entre amostras humanas e animais, a maioria da população não tem imunidade para
os novos vírus e a doença se dissemina rapidamente, podendo afetar indivíduos de
todas as faixas etárias. As grandes pandemias foram consequência de variações
antigênicas maiores (FORLEO-NETO et al., 2003). Demonstração de como pode
ocorrer variações antigênicas do vírus influenza pode ser observado na Figura 1.
Os vírus Influenza do tipo A infectam humanos, suínos, equinos, aves, focas,
baleias (SANTOS; ROMANO; WIGG, 2008) e caninos (DESHPANDE et al., 2009);
os do tipo B infectam humanos (HUNT, 2009) e focas (OSTERHAUS, 2000) e os do

23
tipo C infectam humanos, suínos (BRASIL, 2009a) e caninos (MANUGUERRA et al.,
1993). Apenas um subtipo de HA e um de NA são reconhecidos na Influenza B
(WRIGHT; WEBSTER, 2007).
O primeiro relato de transmissão interespécies envolvendo porcos ocorreu em
1938, quando Shope apresentou evidência sorológica de transmissão de um vírus
humano a porcos. Maior evidência de transmissão de vírus entre estas duas
espécies ocorreu em 1976, quando um vírus suíno (H1N1) foi isolado de um soldado
que tinha morrido de infecção por influenza em Nova Jersey. Posteriormente este
vírus foi isolado de outros cinco soldados e estudos sorológicos sugeriram que mais
de 500 pessoas foram infectadas (WRIGHT; WEBSTER, 2007).
Figura 2 – Principais mecanismos evolutivos do vírus influenza
Fonte: Adaptado de Glyco Word
Transmissão
A transmissão do vírus ocorre através de gotículas expelidas pelo indivíduo
doente através do espirro ou tosse, ou por contato direto ou indireto, com secreções
Geração de novos virus humanos (H3N2) que possuem hemaglutinina de vírus de influenza aviária
Antigenic shift (Rearranjo gênico)
Mutações pontuais vão se acumulando no genoma ocasionando a emergência de variantes virais

24
respiratórias de pessoas infectadas (NARAIN; KUMAR; BHATIA, 2009). Na influenza
sazonal o indivíduo infectado pode transmitir o vírus um dia antes, até cinco a sete
dias após o início dos sintomas (CENTERS FOR DISEASE CONTROL AND
PREVENTION, 2009a). Pessoas com elevado grau de imunodepressão podem
excretar vírus por semanas ou meses. As crianças, comparadas aos adultos,
também excretam vírus mais precocemente, por longos períodos e com maior carga
viral (BRASIL, 2009a). Na infecção pelo vírus A(H1N1)pdm09, observou-se que a
transmissão de adultos infectados ocorria um dia antes até o 7º dia do início dos
sintomas. Crianças menores de 12 anos de idade transmitiam o vírus um dia antes
até o 14º dia de início dos sintomas (BRASIL, 2009b).
Aspectos clínicos
O período de incubação é de um a cinco dias na influenza sazonal
(WRIGHT; WEBSTER, 2007), e de dois a três dias no caso do vírus da Influenza
A(H1N1)pdm09, podendo se estender até sete dias (ORGANIZAÇÃO MUNDIAL DA
SAÚDE, 2009b).
Os principais sintomas da infecção por Influenza são febre, tosse, coriza,
cefaléia, congestão nasal, calafrio, dor no corpo e, algumas vezes, diarréia e vômito
(CENTERS FOR DISEASE CONTROL AND PREVENTION, 2009b). A maioria dos
pacientes recupera-se em uma semana, sem exigir atenção médica, mas a gripe
pode causar doença grave ou morte (ORGANIZAÇÃO MUNDIAL DA SAÚDE,
2009c).
Complicações podem ocorrer em todas as faixas etárias e em indivíduos
saudáveis. Entretanto, idosos, particularmente os com doenças crônicas (asma,
diabetes, cardiopatia, enfisema), crianças de baixa faixa etária e gestante têm um
maior risco de desenvolvê-las (CENTERS FOR DISEASE CONTROL AND
PREVENTION, 2009b). Outras situações de risco de complicações incluem:
imunodepressão, doença renal crônica, doença pulmonar obstrutiva crônica e
hemoglobinopatias (BRASIL, 2006).
As complicações mais comuns são as pneumonias bacterianas secundárias
e, a mais grave, a pneumonia viral primária, podendo ocorrer as duas
simultaneamente. A gripe pode exacerbar doenças pulmonares, cardíacas e outras
doenças crônicas. Também tem sido associada com encefalopatia, mielite

25
transversa, miosite, miocardite, pericardite, síndrome de Reye (BEHRENS, G. et al.,
2006), e síndrome de Guillain-Barré (HUNT, 2009).
Diagnóstico laboratorial
Para o diagnóstico laboratorial é muito importante a qualidade da amostra que
deve ser coletada de preferência até o 4º ou 5º dia do início dos sintomas.
Dentre as amostras respiratórias, o aspirado de nasofarínge é normalmente mais
adequado do que amostras de swab combinado de orofaringe e nasofaringe. Em
pacientes entubados, secreção traqueal e lavado brônquico, podem ser utilizados
(VAN ZYL, 2006). Em caso de óbito, em situações especiais indicadas pela
vigilância epidemiológica, deverão ser colhidos fragmentos de brônquios e pulmões
(BRASIL, 2009c).
O diagnóstico laboratorial inclui o isolamento viral, ensaios imunoenzimáticos
(EIA) e testes rápidos para o diagnóstico de influenza (RIDTs) para detecção de
antígenos, reação em cadeia da polimerase (RT-PCR), imunofluorescência e
hemaglutinação/ inibição da hemaglutinação. A sensibilidade e a especificidade dos
testes variam entre os diferentes métodos, o tipo de amostra usada e os resultados
devem ser avaliados no contexto das informações clínicas e epidemiológicas. Testes
sorológicos com uma única amostra de sangue não são recomendáveis para o
diagnóstico de Influenza. Geralmente são realizados em pesquisas de saúde pública
utilizando amostras pareadas de sangue, uma na fase aguda da doença e outra na
fase convalescente. Os testes padrão para confirmação da infecção pelo vírus da
Influenza são a reação em cadeia da polimerase e o isolamento do vírus (CENTERS
FOR DISEASE CONTROL AND PREVENTION, 2009c; VAN ZYL, 2006,
ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2009d).
O vírus pode ser isolado em ovos embrionados de galinha e em células de
rim canino (MDCK). Os efeitos citopáticos são inespecíficos e vírus da influenza A
podem ser detectados e subtipados do sobrenadante de cultura de células através
das reações de RT-PCR, de imunofluorescência ou de inibição da hemaglutinação
(VAN ZYL, 2006).
Em 2009, foi desenvolvido um protocolo de RT-PCR em tempo real (rRT-
PCR) para detecção do vírus influenza A(H1N1)pdm09, pelo “Centers for Disease
Control and Prevention” (CDC) de Atlanta (ORGANIZAÇÃO MUNDIAL DA SAÚDE,
2009e h).

26
Reservatórios naturais
Aves são hospedeiros naturais de todos os subtipos conhecidos de vírus da
influenza A. Surtos de gripe aviária entre aves ocorrem no mundo inteiro. Apesar de
o vírus infectar predominantemente aves, eles podem e têm atravessado a barreira
das espécies e infectado seres humanos. Quando tal fato ocorre, pode desencadear
uma pandemia (CENTERS FOR DISEASE CONTROL AND PREVENTION, 2009d ).
Até 1997, acreditava-se que a transmissão direta de vírus das aves para o ser
humano não poderia acontecer, quando surgiram os primeiros casos de gripe aviária
em humanos em Hong Kong (ENGIN, 2007).
A exposição às aves infectadas, suas excreções e secreções (saliva,
secreção nasal e fezes) ou solo contaminado pode resultar em infecção humana.
Todas as aves são susceptíveis à infecção pelo vírus da influenza A, porém algumas
espécies são mais resistentes do que outras. O vírus tem, contudo, alta capacidade
de transmissibilidade e as aves migratórias contribuem para a disseminação
intercontinental (REVISTA DE SAÚDE PÚBLICA, 2006).
Todos os 16 subtipos de HA e os 9 subtipos de NA da Influenza A são
capazes de infectar aves aquáticas selvagens, resultando num imenso e
permanente reservatório. Nas aves, os vírus podem causar duas formas diferentes
de doença: uma suave que pode passar despercebida, e uma grave, de
disseminação rápida e taxa de mortalidade que pode aproximar-se de 100% dentro
de 48 horas. O vírus H5N1, altamente patogênico, pode sobreviver em fezes de
aves pelo menos 35 dias em baixa temperatura (4ºC) e em temperaturas mais altas
(37ºC) por seis dias (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2006).
Epidemiologia
O impacto das epidemias de influenza é reflexo da interação entre a variação
antigênica viral, o nível de proteção da população para as cepas circulantes e o grau
de virulência dos vírus (FORLEO-NETO et al., 2003). Anualmente, em período interpandêmico, cerca de três a cinco milhões de
pessoas infectadas pelo vírus da influenza apresentam um quadro grave da doença
e 250.000 a 500.000 pessoas evoluem para óbito (ORGANIZAÇÃO MUNDIAL DA
SAÚDE, 2009c). Os índices de hospitalização por doença grave são cerca de 3 por
1000 para crianças de 6 a 23 meses e de 9 por 1000, para crianças menores de 6
meses (GRIJALVA et al., 2006). A vacina é a melhor estratégia disponível para a

27
prevenção da gripe e suas consequências, proporcionando impacto indireto na
diminuição do absenteísmo no trabalho e dos gastos com medicamentos para
tratamento de infecções secundárias, das internações hospitalares e da mortalidade
evitável (BRASIL, 2009a).
A primeira vacina contra a gripe foi implementada em 1943, com vírus
inativados de influenza A e B. Como os vírus da influenza estão constantemente
apresentando mutações, as vacinas devem ser anualmente atualizadas, com base
nas cepas circulantes. (TURKINGTON; ASHBY, 1988). A vacina utilizada é
constituída de dois tipos de vírus da Influenza A e um tipo da Influenza B de acordo
com recomendação da Organização Mundial da Saúde (ORGANIZAÇÃO MUNDIAL
DA SAÚDE, 2011) e indicada para indivíduos com 60 anos de idade ou mais e é
oferecida por meio de campanhas anuais, cujo período deve ser anterior ao período
de maior circulação do vírus na população do país. Está disponível, também, nos
Centros de Referência para Imunobiológicos Especiais (CRIE) durante todo o ano,
para pessoas consideradas de maior risco para a doença e suas complicações e
familiares que estejam em contato com os referidos pacientes, além de profissionais
de saúde, trabalhadores de asilos e creches, de indígenas a partir de 6 meses de
idade e da população carcerária (BRASIL, 2009a). A despeito de ser o método mais
eficaz para minorar as consequências da infecção por influenza na comunidade, no
momento em que um novo subtipo ou cepa começa a ser transmitido na população
humana, dificilmente haverá uma vacina disponível para uso imediato.
Histórico das Pandemias de influenza
Pandemias de influenza têm sido documentadas pelo menos nos últimos
quatro séculos. No século XX ocorreram três pandemias e várias ameaças de
pandemias: 1918-1919 (gripe espanhola), 1957-1958 (asiática), 1968-1969 (Hong
Kong), 1976 (ameaça de gripe suína), 1977 e 1999 (ameaça de gripe russa), 1977
(ameaça de gripe aviária) (ORGANIZAÇÃO PANAMERICANA DA SAÚDE, 2009). A
Figura 3 mostra esquematicamente os períodos pandêmicos e os vírus que
circularam nos respectivos períodos.
Na gripe espanhola, considerada a mais grave de todos os tempos, calcula-se
que cerca de 20 a 40% da população mundial contraiu a infecção e mais de 50
milhões de pessoas morreram (ORGANIZAÇÃO PANAMERICANA DA SAÚDE,
2009). Muitas dessas mortes foram causadas por pneumonia bacteriana secundária,

28
mas a gripe espanhola também causou uma forma de pneumonia viral primária, com
extensa hemorragia dos pulmões, que matava em até 24 horas. A maioria das
mortes ocorreu em pessoas jovens e saudáveis de 15 a 35 anos e apenas 1% das
mortes ocorreu em indivíduos com 65 anos ou mais (BEHRENS et al., 2006). Figura 3. Linha do tempo mostrando as pandemias provocadas pelo vírus influenza A
De acordo com a OMS, existem seis fases que caracterizam uma pandemia
de influenza, conforme esquematicamente representado na Figura 4.
Figura 4 – Fases de uma pandemia de influenza Tempo
Infecção Transmissão Infecção Possibilidade Níveis de predominantemente entre humanos humana de eventos atividade animal; sustentada recorrentes sazonal algumas infecções da doença humanas
Fonte: Organização Mundial da Saúde, 2012.
Pós Pandemia
Fase 5-6 Pandemia Pós Pico
Fase 1 a 3
Fase 4

29
O agente etiológico da gripe espanhola foi o vírus Influenza A (H1N1). Este
vírus, de origem suína, espalhou-se em três ondas num período de 12 meses, em
1918-1919, cruzando a Europa, Ásia e América do Norte. A primeira onda, que
começou na primavera de 1918, foi altamente contagiosa, mas não particularmente
mortal. A segunda onda, que começou em setembro, espalhou-se de forma
altamente letal (BEHRENS et al., 2006). Uma terceira onda, de impacto semelhante
ao da segunda, ocorreu na primavera de 1919 (WRIGHT; WEBSTER, 2007).
Em fevereiro de 1957, a gripe asiática foi detectada no extremo oriente e em
maio do mesmo ano seu agente etiológico, o vírus da Influenza A do subtipo H2N2,
foi isolado no Japão (SCHOLTISSEK et al., 1978) e Singapura. Surtos eram
frequentemente explosivos, entretanto, o vírus quando comparado com o vírus da
pandemia de 1918 demonstrou ser responsável por um quadro clínico mais leve e
por um número de mortes bem menor (ORGANIZAÇÃO MUNDIAL DA SAÚDE,
2005a).
O mundo estava mais bem preparado para enfrentar uma pandemia, devido
a: 1- Maior conhecimento sobre os vírus da influenza em virtude do avanço da
virologia; 2- Vacinas para epidemias sazonais tinham sido desenvolvidas e
mostravam ser o método mais eficaz de prevenção, reduzindo a influenza sazonal
em dois terços ou mais, nos locais onde era aplicada; 3- Antibióticos estavam
disponíveis para tratamento das complicações, incluindo a pneumonia bacteriana; 4-
O monitoramento virológico da influenza vinha sendo realizado há mais de 10 anos,
através da Rede de Vigilância Global da Influenza (WHO Global Influenza
Surveillance Network) (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2005a). Em agosto
de 1957 já se dispunha de uma vacina, embora em quantidade limitada
(ORGANIZACÃO PANAMERICANA DA SAÚDE, 2009).
Pacientes portadores de doenças crônicas e gestantes eram mais
susceptíveis a desenvolver complicações pulmonares (LOURIA et al., 1957). A
mortalidade apresentou um padrão similar ao das epidemias sazonais, isto é, o
maior número de óbitos ocorreu em crianças e idosos. Durante a primeira onda, o
maior número ocorreu em crianças na idade escolar. Na maioria dos países, uma
segunda onda surgiu de um a três meses após o desaparecimento da primeira,
causando um maior número de pessoas infectadas e óbitos. Ao contrário da primeira
onda, a população mais atingida foi a de idosos, o que justifica o aumento da
mortalidade. Estima-se que mais de dois milhões de pessoas morreram. O maior

30
número de óbitos foi de indivíduos portadores de doença crônica e não de indivíduos
previamente saudáveis, como aconteceu durante a pandemia da gripe espanhola
(ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2005a).
Em 1968, onze anos depois da gripe asiática, o vírus influenza A (H2N2)
deixou de circular, sendo substituído pelo A (H3N2), dando origem à gripe de Hong
Kong. Os primeiros sinais de uma nova pandemia foram noticiados no sul da Ásia no
verão de 1968 e um vírus do subtipo H3N2 foi isolado em Hong Kong em julho de
1968. A população mais acometida pela doença foi o grupo de 10 a 14 anos de
idade (WRIGHT; WEBSTER, 2007).
Foi estimado cerca de um milhão de óbitos devido à gripe de Hong Kong.
Nos Estados Unidos, cerca de 50% de todos os óbitos por influenza ocorreram em
indivíduos com menos de 65 anos de idade (BEHRENS et al., 2006). Estudos
soroarqueológicos mostraram que a maioria dos indivíduos nascidos antes de 1893
possuía anticorpos para o subtipo A(H3), antes de terem sido expostos ao novo
vírus pandêmico de 1968 (DOWDLE, 1999), e que a pré-existência de anticorpos
para o subtipo H3 poderia ter protegido os idosos (>77 anos) durante a pandemia de
H3N2, em 1968 (BEHRENS et al., 2006).
Estudos utilizando técnicas de biologia molecular mostraram que o agente
etiológico da gripe espanhola foi um vírus aviário H1N1 que se adaptou ao homem e
foi capaz de se replicar eficientemente no mesmo. Ao contrário do vírus da
pandemia de 1918, os vírus da gripe asiática e da gripe de Hong Kong são produto
de rearranjos gênicos. O vírus da gripe asiática foi produto de rearranjo gênico entre
o subtipo A(H1N1) humano de 1918 e um vírus H2N2 aviário, resultando no novo
vírus H2N2 humano, composto por 3 segmentos do H2N2 aviário (HA, NA e PB1) e
5 segmentos do subtipo A (H1N1). O vírus da gripe de Hong Kong surgiu do
rearranjo gênico entre o subtipo H2N2 humano e um aviário (H3), resultando no
vírus humano subtipo A(H3N2) composto por dois segmentos do vírus aviário (HA e
PB1) e cinco segmentos do de 1918 (BELSHE, 2005).
Em 1997, centenas de pessoas se infectaram com o vírus da gripe aviária A
(H5N1) em Hong Kong. Dessas, 18 pessoas tiveram de ser hospitalizadas e seis
foram a óbito. Em 1999, foi detectado um novo vírus da gripe aviária, o A (H9N2),
que infectou duas crianças em Hong Kong. Apesar dos dois vírus não terem
causado uma pandemia, sua presença nas aves, sua habilidade para infectar os
seres humanos e, potencialmente, tornarem-se transmissíveis entre humanos, são

31
motivos de constante preocupação (ORGANIZAÇÃO PANAMERICANA DA SAÚDE,
2009). Do início de 2003 a 28 de janeiro de 2010 foram notificados à Organização
Mundial da Saúde 471 casos confirmados de Influenza A (H5N1) e 282 óbitos em 15
países (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2010b). Outras cepas de influenza
aviária têm causado infecções em humanos: H7N2, H7N3, H7N7 e H9N2.
Entretanto, o H5N1 é o que causa maior preocupação, pois tem sido responsável
pelo maior número de casos com doença grave em humanos e o maior número de
óbitos (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2006).
Na década de 1990, uma tripla recombinação de vírus influenza A (H1)
contendo segmentos de genes de influenza de origem aviária, humana e suína
emergiram e tornaram-se enzoóticos em rebanhos de porcos nos Estados Unidos na
década de 1990 (SHINDE et al., 2009).
Naquele momento sabia-se que a nova influenza, denominada inicialmente de
influenza suína, normalmente causava surtos de gripe entre os suínos e que em
geral, este vírus não infectava o homem, apesar de existirem registros de
transmissão pontual do vírus para os seres humanos, não havendo, entretanto
registro de transmissão deste novo subtipo da influenza suína para pessoas por
meio da ingestão de carne de porco e produtos derivados. É importante salientar
que no período de dezembro de 2005 a fevereiro de 2009, portanto, antes da atual
pandemia, o CDC notificou nos Estados Unidos, 11 casos esporádicos de infecção
em humanos pelo vírus Influenza A (H1N1) de origem suína, em indivíduos de 16
meses a 48 anos de idade. Destes, quatro foram hospitalizados, mas todos se
recuperaram, apesar de alguns deles apresentarem doença severa do aparelho
respiratório inferior e sintomas raros de gripe, como diarréia (SHINDE et al., 2009).
Diante da preocupação global da possibilidade de surgir um novo subtipo
pandêmico do vírus da influenza, os países reunidos na Assembléia Mundial da
Saúde aprovaram em 2003 uma resolução, incentivando a todos os países para a
elaboração de planos para enfrentamento a uma nova pandemia. Em novembro de
2005, o Brasil publicou a primeira versão do Plano de Preparação Brasileiro para
uma Pandemia de Influenza (BRASIL, 2005). O tempo de surgimento de uma nova
pandemia, o subtipo viral e sua patogenicidade são imprevisíveis. O planejamento
pandêmico tem por objetivo tornar os países mais preparados para reconhecer e
gerenciar uma pandemia de gripe. O nível de preparação para o enfrentamento de
uma pandemia de influenza pode ajudar a reduzir a transmissão do vírus, a diminuir

32
o número de casos, hospitalizações e óbitos, e manter os serviços essenciais,
reduzindo o impacto econômico e social da pandemia. Muitos países ganharam
experiência no planejamento e resposta à pandemia depois de lidar com a ameaça,
ou a realidade da síndrome respiratória aguda grave (SARS) e gripe aviária de alta
patogenicidade (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2005b). Pode-se afirmar
que uma das estratégias mais eficazes para o retardo da transmissão viral é a
vigilância virológica e epidemiológica, isto é, a detecção precoce dos primeiros
casos, o bloqueio da transmissão e a intervenção oportuna (DONALISIO, 2006).
Pandemia de 2009
A partir de 15 de março de 2009, o governo do México observou o aumento
não usual de infecção respiratória aguda grave. De 17 a 26 de abril foram notificados
1.455 casos prováveis de influenza com pneumonia grave, incluindo 84 óbitos, à
Organização Mundial da Saúde (OMS), que em 24 de abril de 2009 notificou aos
países membros a ocorrência de casos humanos de influenza suína que vinham
ocorrendo no México e nos Estados Unidos da América. Neste mesmo dia foi
identificado um novo subtipo do vírus de Influenza suína A(H1N1) classificada como
A/CALIFORNIA/04/2009 que não havia sido detectado previamente em humanos ou
suínos. Observou-se que sua transmissão ocorria de pessoa a pessoa,
principalmente por meio de tosse ou espirro e secreções respiratórias de pessoas
infectadas (BRASIL, 2009d ).
O novo subtipo viral de 2009 era composto por: dois genes (PA e PB2) da
linhagem aviária norte-americana; um gene (PB1) derivado da linhagem sazonal
H3N2; três genes (HA, NP e NS) da linhagem suína clássica norte-americana; e dois
genes (NA e M) da linhagem suína euro-asiática (DAWOOD et al., 2009), conforme
pode ser observado na Figura 3.
O primeiro grupo de triplo rearranjo suíno de vírus influenza A é encontrado
em suínos e pode ocasionalmente ser transmitido aos seres humanos, mas não se
transmite de forma eficiente de humano a humano. Em contraste, o vírus
A(H1N1)pdm09 não provoca epidemia em suínos (embora os porcos possam ser
infectados pela exposição aos seres humanos), mas a transmissão entre os seres
humanos ocorreu de maneira muito rápida e se espalhou por vários países
(BELSHE, 2009). O diferencial entre os subtipos A(H1N1)pdm09 e A (H1N1)
sazonal (circulante na população desde 1977) está especialmente localizado na HA

33
que apresenta distintos perfis de glicosilação e, por conseguinte, uma baixa
identificação antigênica entre as cepas (SETTEMBRE; DORMITZER; RAPPUOLI,
2010). Figura 5- Genótipos do vírus influenza A(H1N1) de linhagem suína americana e influenza A(H1N1)pdm09 em casos recentes nos Estados Unidos.
PB2 - proteína básica 2, PB1 – proteína básica 1, PA – proteína ácida, HA – hemaglutinina, NP –
nucleoproteína, NA – neuraminidase, M – proteína de matrix e NS – proteína não estrutural.
Fonte: Dawood, 2009
Em 25 de abril, seguindo o Regulamento Sanitário Internacional (RSI 2005), a
OMS declarou a pandemia de influenza como Emergência de Saúde Pública de
Importância Internacional (ESP II) e comunicou que o nível de alerta para
caracterizar uma pandemia de influenza estava na fase 3. No Brasil, a definição
inicial de caso suspeito era todo indivíduo que apresentasse febre alta de maneira
repentina (>38ºC) e tosse, podendo estar acompanhada de algum dos seguintes
sintomas: dor de cabeça, dores musculares e nas articulações, dificuldade
respiratória e ter apresentado sintomas até 10 dias após sair da área afetada pela
nova influenza (BRASIL, 2009d). Com a disseminação do vírus para vários países a

34
definição de caso foi se adequando à situação, tendo como base a situação
epidemiológica. Em 27 de abril de 2009, o nível de Emergência de Saúde Pública de
Importância Internacional foi elevado para a fase 4 (BRASIL, 2009e) e em 29 de
abril, para a fase 5. Em 30 de abril de 2009 a OMS adotou como denominação oficial
o termo Influenza A (H1N1), em substituição à denominação anterior de Influenza
Suína (BRASIL, 2009f).
Publicação da Organização Mundial da Saúde (2009f) em 22 de maio mostrou
através da análise de 10.243 casos confirmados por laboratório de infecção pelo
novo vírus influenza A(H1N1)pdm09 em 41 países, incluindo 80 óbitos, que os
principais sintomas da doença eram tosse, febre, dor de garganta, mal estar e
cefaléia; que a maioria dos pacientes apresentava uma síndrome gripal, sem
complicações, curando espontaneamente, mas que o estado clínico do paciente
podia variar de doença leve não febril do trato respiratório superior, à doença grave,
ou pneumonia fatal. De acordo com o “Centers for Disease Control and Prevention”
(2010a), dor no corpo, fadiga e algumas vezes vômito e diarréia também eram
sintomas da infecção pelo novo vírus pandêmico. No dia 7 de maio de 2009 foram
confirmados os primeiros casos do novo vírus Influenza A(H1N1)pdm09 no Brasil:
dois casos em São Paulo, um no Rio de Janeiro e um em Minas Gerais (BRASIL,
2009g) e no dia 9 de maio foi relatado o primeiro caso autóctone no Brasil. O
paciente apresentou sintomas”, após contato com um caso confirmado que havia
retornado do México (BRASIL, 2009h).
Em 11 de junho de 2009, o nível foi elevado da fase 5 para fase 6 (BRASIL,
2009i). Em 22 de junho de 2009 foi confirmado o primeiro caso de Pernambuco,
publicado no dia 24 de junho (BRASIL, 2009j). No dia 16 de julho de 2009, o
Ministério da Saúde declarou transmissão sustentada do vírus no Brasil (BOLETIM
ELETRÔNICO EPIDEMIOLÓGICO, 2009). A partir de 19 de julho de 2009 (Semana
Epidemiológica 29), após a declaração da transmissão sustentada, o Ministério da
Saúde passou a realizar o monitoramento apenas dos óbitos e casos de Síndrome
Respiratória Aguda Grave (SRAG), seguindo as orientações da OMS. Interrompeu a
investigação e notificação de casos leves suspeitos por Influenza A(H1N1), pois não
estava mais recomendada a identificação individual de cada caso suspeito de
influenza pelo novo vírus. No entanto, manteve o monitoramento de informações
sobre os grupos de risco para desenvolvimento de doença grave, assim como o
monitoramento da circulação do vírus no país (BRASIL, 2009c).

35
A coleta do material para o diagnóstico laboratorial passou a ser realizada
apenas dos óbitos, dos casos de SRAG e de surtos de síndrome gripal em
comunidades fechadas (BOLETIM ELETRÔNICO EPIDEMIOLÓGICO, 2010).
Até agosto de 2010, em todo o mundo, mais de 214 países e territórios ou
comunidades relataram casos laboratorialmente confirmados de infecção pelo vírus
influenza A(H1N1)pdm 2009, incluindo pelo menos 18.449 mortes (ORGANIZAÇÃO
MUNDIAL DA SAÚDE, 2010a). A taxa de letalidade no mundo foi estimada em 0,4%
(BAUMEISTER et al., 2010).
No Brasil, no período de 19 de abril a 22 de agosto de 2009 (Semanas
epidemiológicas 16 a 33) foram notificados 34.506 casos de infecção respiratória
com SRAG. Foram confirmados laboratorialmente 5.747 (16,7%) casos de infecção
pelo vírus influenza A(H1N1)pdm09, 917 (2,7%) casos de influenza sazonal e 4.176
(12,1%) casos descartados de influenza. Até o fechamento da coleta de dados dos
autores, restavam 23.668 (68,6%) de casos que ainda estavam sendo investigados.
Dos casos A(H1N1)pdm09 confirmados, 56,5% eram do sexo feminino e a faixa
etária mais atingida foi de 20 a 49 anos (56%). Fora os sintomas que faziam parte da
definição de SRAG, os mais frequentes foram mialgia (62,2%), coriza (54%) e
calafrio (53,4%). A incidência foi de 3.0/100.000 habitantes (OLIVEIRA et al., 2009).
No período de 19 de abril a 18 de julho de 2009 (Fase de contenção no
Brasil) foram notificados no país 12.919 casos de SG e SRAG e confirmados por
critério laboratorial ou clínico-epidemiológico, 4.434 casos, dos quais 1.556 (35%)
apresentavam SRAG. Dentre estes 14,5% evoluíram para óbito. Considerando os
casos confirmados por SG e SRAG a taxa de letalidade foi de 5,1% para o período
de contenção. Dos casos confirmados houve um predomínio do sexo masculino e a
faixa etária mais atingida foi a de 20 a 29 anos (29,3%) (BOLETIM ELETRÔNICO
EPIDEMIOLÓGICO, 2010).
No período de 19 de julho a 2 de janeiro de 2010 (Fase de mitigação no
Brasil) foram notificados 87.171 casos de SRAG e confirmados 44.544 (51,1%). Dos
casos confirmados houve predomínio do sexo feminino (57,2%) e a faixa etária mais
atingida também foi a de 20 a 29 anos (24,3%). A taxa de incidência nacional de
SRAG confirmada pela presença do vírus da influenza A (H1N1)pdm09 foi de 23,3
/100.000 habitantes e a maior incidência foi observada no grupo etário de menores
de 2 anos, seguido do grupo de 20 a 29 anos. A menor incidência ocorreu na faixa
etária acima de 60 anos. Entre os sintomas que definiam o caso de SRAG,

36
destacaram-se mialgia, calafrio, coriza e dor de garganta. Dos casos confirmados,
45,1% dos pacientes apresentavam algum tipo de condição crônica de saúde, sendo
o grupo das pneumopatias crônicas o mais frequente (19,1%), seguido de
cardiopatias, imunossupressão e doenças metabólicas (BOLETIM ELETRÔNICO
EPIDEMIOLÓGICO, 2010).
Em todo o ano de 2009, o Brasil registrou 2.051 óbitos por influenza
pandêmica e uma taxa de mortalidade de 1,1/100.000 habitantes, no sul a taxa de
mortalidade foi de 3,0 /100.000 habitantes e no nordeste 0,1/100.000 habitantes. A
letalidade dentre os casos de SRAG pelo vírus A (H1N1)pdm09 foi de 4,6%. Entre
os óbitos, houve uma predominância do sexo feminino (56,4%) e da faixa etária de
30 a 39 anos, seguida da faixa de 20 a 29 anos. Na distribuição etária das taxas de
mortalidade, a maior taxa ocorreu na faixa de 50 a 59 anos, seguida das faixas de
30 a 39 anos e menores de 2 anos de idade. Das mulheres que evoluíram para
óbito, 26,5% eram gestantes (BOLETIM ELETRÔNICO EPIDEMIOLÓGICO, 2010).
Possivelmente em decorrência do grande número de infectados, associado à
vacinação promovida pelo Ministério da Saúde, houve uma intensa queda no
número de casos. De janeiro a 4 de setembro de 2010, o Brasil notificou 8.366
casos, sendo até então confirmados 773 (9,2%) casos pelo vírus influenza
A(H1N1)pdm09. O nordeste notificou 614 casos sendo 103 (13,3%) confirmados.
Entre os casos confirmados do Brasil, a mediana foi de 24 anos e o sexo feminino foi
responsável por 61,1% dos casos, sendo que 68,9% estavam em idade fértil (10 a
49 anos) e destas 36% eram gestantes. Foram registrados 951 óbitos suspeitos de
influenza pandêmica, sendo 99 (10,4%) confirmados, 793 foram descartados e até o
dia 4 de setembro, 59 continuavam em investigação (BRASIL, 2010a).
No dia 10 de agosto de 2010, a OMS anunciou o fim da pandemia e início
da fase pós-pandêmica (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2010c).
Antivirais
Foram duas as drogas antivirais aprovadas e recomendadas para o
tratamento da nova influenza pandêmica: os inibidores de neuraminidase,
oseltamivir e zanamivir, mais comumente conhecidos pelos nomes comerciais
Tamiflu e Relenza respectivamente. Para os pacientes com sintomas de doença
grave, como dispnéia e febre alta, a OMS recomendou o tratamento com oseltamivir
que deveria começar imediatamente. Para indivíduos com maior risco de

37
desenvolver doença grave, como gestantes, crianças menores de 5 anos de idade,
portadores de doenças crônicas cardiovascular, respiratória ou doença hepática,
diabéticos ou indivíduos imunodeprimidos, transplantados e pacientes com câncer, a
OMS recomendou tratamento com oseltamivir ou zanamivir, logo no início dos
sintomas. Em todos os casos, onde o oseltamivir não estivesse disponível, ou não
pudesse ser usado por qualquer razão, o zanamivir poderia ser empregado
(ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2009f).
O oseltamivir liga-se à NA do vírus da influenza, impedindo-a de clivar o ácido
siálico da porção da membrana plasmática no momento do brotamento, bloqueando
o ciclo de infecção (IAMARINO, 2009).
Vacinas
A campanha de vacinação contra o vírus influenza A(H1N1)pdm09 no Brasil
foi realizada em cinco etapas e teve início em 8 de março de 2010. Foram
priorizados os seguintes grupos: trabalhadores dos serviços de saúde envolvidos na
resposta à pandemia, população indígena, gestantes, portadores de doenças
crônicas, crianças saudáveis maiores que seis meses e menores de dois anos de
vida, adultos saudáveis de 20 a 39 anos completos, e população com 60 anos e
mais de idade portadora de comorbidade (BRASIL, 2010b). Foi uma das maiores
campanhas de vacinação no Brasil, onde foram superadas diversidades regionais,
sociais, econômicas e culturais. Foram vacinadas mais de 89,5 milhões de pessoas,
contribuindo, certamente, de forma extremamente importante, para a redução da
incidência no país (BRASIL, 2010c).
Estudos relacionados à susceptibilidade viral ao oseltamivir vêm sendo
realizados. Até o momento, não foram identificadas variações antigênicas
importantes no vírus A(H1N1) pdm09 (Barr et al, (2010). Apesar de algumas
variantes genéticas terem sido relatadas, como a mutação E391K e a mutação
D222G que, segundo alguns autores, está ligada aos casos mais graves, nenhuma
variante tem predominado em um país ou região. De acordo com Adwan (2009), a
análise filogenética do gene HA de 54 cepas do vírus influenza A(H1N1)pdm09
isolados no Oriente Médio também mostraram que antigenicamente estão
relacionados a cepa protótipo da vacina A/California/7/2009(H1N1). Portanto, a cepa
A/California/7/2009-like recomendada para a formulação da vacina pela OMS desde

38
o início da pandemia, continuou a ser recomendada para uso no inverno de 2011, no
hemisfério sul, e também na estação de influenza de 2010-2011 no hemisfério norte
(ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2011).
2.2 CARACTERIZAÇÃO GENÉTICA DO VÍRUS INFLUENZA A(H1N1)pdm09
Mutações do vírus da influenza A (H1N1)pdm09 no gene HA
De acordo com Glinsky (2010), foi conduzida uma análise de grande volume
de dados clínicos, epidemiológicos e genômicos para a avaliação da evolução da
pandemia nos Estados Unidos, Canadá, Reino Unido, Austrália e Japão. A mutação
D222G foi frequentemente detectada, em vírus isolados de casos fatais e a análise
filogenética mostrou que 42,9% das amostras de autópsias de casos confirmados do
vírus A (H1N1)pdm09 possuíam a mutação Q310H. Entretanto, segundo Lee et al.
(2010) a mutação Q310H era comum na época, o que poderia facilmente explicar a
sua maior ocorrência entre os poucos casos fatais analisados, sem a necessidade
de, necessariamente, estar associada com a gravidade. Kawano et al. (2011)
detectaram 23 tipos de mutação no gene HA de amostras de vírus A(H1N1)pdm09
que circularam em Nagasaki, Japão, mas a D222G e a Q310H não foram
detectadas.
Na China, foi detectada a mutação D225G no gene da hemaglutinina do vírus
A(H1N1)pdm09. Foi observado que ela aumentava a virulência do novo vírus em
camundongos. Estudos anteriores realizados com o vírus H1N1 da pandemia de
1918 sugerem que essa mutação está associada à uma dupla especificidade de
ligação do vírus para os receptores de ácido siálico 2,3 e para o ácido siálico 2,6
do hospedeiro. Assim, o vírus com a mutação D225G poderia causar doença mais
grave devido à capacidade de adsorver nas células do trato respiratório inferior,
onde o ácido siálico 2,3 é mais predominante. O ácido siálico 2,6 está mais
presente nas células do trato respiratório superior (LIU et al., 2010, ZHENG et al.,
2010). Na Noruega, no período de maio de 2009 a janeiro de 2010, foram analisadas
61 amostras de casos graves e fatais e 205 amostras de casos leves através de
sequenciamento convencional e pirosequenciamento para detectar a mutação
D222G. Dentre os casos graves e fatais, foi detectado um percentual de 18% (11/61)

39
de amostras apresentando a mutação acima citada e, em contraste, nenhuma das
sequências obtidas a partir dos casos leves apresentaram esta mutação. Nesse
estudo, também foram detectadas as mutações D222E e D222N identificando-se a
co-circulação destes genótipos mutantes com o vírus selvagem 222D (KILANDER,
2010). No mesmo período, foram identificados vírus com a mutação D222G em dois
pacientes em Bordeaux, sudoeste da França, entre 24 internados em unidade de
terapia intensiva por complicações advindas da infecção pelo novo vírus pandêmico
(MALATO et al., 2011). Nesse estudo, foram detectadas também as mutações
S203T, D222E, Y230H, M257I, Q293H, I295V, K305R, V231I e V321F.
Ainda no período de maio de 2009 a janeiro de 2010, foram testadas 458
amostras respiratórias positivas para o vírus influenza A(H1N1)pdm09 que
circularam em Hong Kong, sendo 219 amostras provenientes de casos graves e 239
de casos leves. Nove dos casos graves (4,1%) apresentaram a mutação D222G,
enquanto nos casos leves nenhuma mutação D222G foi detectada. Quatro dos nove
pacientes cujas amostras apresentavam vírus com a referida mutação, foram a óbito,
havendo associação estatisticamente significativa entre a mutação D222G e a forma
grave da doença (p= 0,002). Outras substituições, como D222N (casos graves, n=3;
casos leves, n=1) e D222E (somente em casos leves, n=4) também foram
encontradas (MAK et al., 2010).
Foi realizado o sequenciamento de um segmento parcial de subunidade HA1
de vírus influenza A (H1N1)pdm09 detectados em amostras respiratórias de 273
casos graves e 533 casos leves de diferentes regiões espanholas para monitorar
substituições na posição 222. A mutação D222G foi detectada em vírus de 14 casos
graves (5,12%). A mutação D222E foi detectada em vírus de 47 casos graves
(17,21%) e de 52 casos leves (9,75%) e a mutação D222N ocorreu em vírus de três
casos graves (0,37%), sugerindo que as mutações D222G e D222E nos vírus
pandêmicos circulantes na Espanha estivessem relacionadas com doença
respiratória grave (LEDESMA, 2011).
Na Finlândia, foram analisados os genes HA e NA de vírus pandêmicos
detectados em 138 pacientes. Em dois indivíduos (1,44%) foram detectados vírus
com a mutação D222G e em outro indivíduo a D222Y, todos em estado grave. Não
foi detectada a mutação H274Y no gene da neuraminidase e todos os vírus foram
estreitamente relacionados aos vírus da vacina A/Califórnia/07/2009. Não foram

40
detectadas mudanças nos genes de HA e NA que pudessem levar à seleção de um
vírus com aumento de virulência e de potencial epidêmico (IKONEN et al., 2010).
Miller et al. (2010) sequenciaram a subunidade HA1 do gene HA de 58
amostras de pacientes do oeste da Escócia infectados com vírus influenza
A(H1N1)pdm09. As amostras foram subdivididas em dois grupos: o primeiro era
composto de 32 pacientes, dos quais nove tinham sido hospitalizados, mas haviam
se recuperado e 23 que tinham falecido, e o segundo grupo que era formado por 26
pacientes com quadro clínico leve. Foi detectado um aumento da incidência da
mutação D222G nos pacientes que faleceram (2/23 – 8,7%), quando comparados
com os pacientes que se restabeleceram (0/35 – 0%). Foi observado também um
aumento da incidência de D222N entre os pacientes do primeiro grupo (2/32 – 6,2%),
quando comparados com os pacientes do segundo grupo, formado de casos leves
(0/26 – 0%), não tendo sido detectada a mutação D222N nas amostras dos pacientes
que foram a óbito. Por fim, foram detectadas duas mutações D222E no primeiro
grupo, sendo uma em amostra de paciente que foi a óbito e outra na de um paciente
que foi hospitalizado, mas se recuperou, contra três que foram encontradas em
pacientes com quadro leve, não havendo portanto diferença significativa entre os dois
grupos.
Estudo realizado por Potdar et al. (2010) com vírus da influenza A
(H1N1)pdm09 isolados na Índia no período de maio a setembro de 2009, detectou no
gene HA as mutações S451N e I547T que só haviam sido detectadas na Índia e as
mutações K2E, Q310H, S220T, P100, I338V, D222G já descritas em outros países.
Análises moleculares de cepas circulantes no norte da Grécia revelaram uma série
de variações nas sequências de HA1, algumas das quais foram mais frequentes em
vírus que causaram infecções graves ou fatais. A mutação mais comum foi a D222G.
A análise filogenética confirmou a estreita correspondência da maioria das cepas
circulantes com a A/California/7/09. No entanto, também revelou uma tendência das
cepas de 2010 de acumular variações de aminoácidos e formar novos grupos
filogenéticos. Daí a importância da vigilância molecular constante para monitorar a
patogenicidade de cepas circulantes e avaliar a eficácia da vacina (MELIDOU et al.,
2010).
Em dezembro de 2009, de acordo com dados recebidos e repassados pela
OMS, a prevalência de substituição D222G era inferior a 1,8% (52 detecções entre
mais de 2.755 sequências de HA), mas levando-se em consideração apenas os

41
óbitos, a prevalência subiu para 7,1% (26 casos tinham a substituição D222G em
364 casos fatais analisados). Entretanto, a OMS salientou que a vigilância e o
laboratório vinham dando prioridade para amostras de pacientes hospitalizados e
gravemente doentes o que poderia levar a potenciais viezes (ORGANIZAÇÃO
MUNDIAL DA SAÚDE, 2009g). Análise filogenética dos genes HA e NA de vírus
influenza A(H1N1)pdm09 detectados de 253 pacientes no Japão, mostrou que no
início da pandemia as principais mutações encontradas foram a HA - S220T e NA -
N248D (MORLIGHEM et al., 2011).
Em Cingapura, foi detectada a mutação E391K que, de acordo com Maurer-
Stroh et al. (2010) pode alterar a estabilidade de uma região da HA importante para
a fusão do vírus à célula hospedeira. Vírus com a mutação E391K e N142D no gene
HA, foram detectados na Austrália, Nova Zelândia e Cingapura, no inverno de 2010.
Até o momento tais mutações não parecem representar uma mudança antigênica
significativa, no entanto, poderiam indicar o início de uma variação antigênica do
vírus influenza A(H1N1)pdm09 exigindo talvez uma atualização mais rápida da
vacina (BARR et al., 2010).
No período de abril de 2009 a junho de 2010, análises moleculares de 90
cepas de vírus influenza A(H1N1)pdm09 que circularam no Brasil, das quais 28
provenientes de pacientes que foram a óbito, revelaram uma série de substituições
nas sequências de HA. As cepas eram oriundas de cinco estados: São Paulo (76),
Goiás (7), Piaui (4), Mato Grosso (1), Mato Grosso do Sul (1) e Distrito Federal (1).
Foram encontradas as seguintes mutações: S220T (71%), D239G/N/S (20%),
Y247H (4,5%), E252K (3,3%), M274V (2,2%), Q310H (26,7%) e E391K (12%).
Casos fatais foram associados à mutação D239G (p<0,0001) (FERREIRA et al.,
2011).
De acordo com Mak (2011) em janeiro de 2011, época de inverno em Hong
Kong e tradicional estação da influenza, as mutações E391K e S202T cresceram
rapidamente e tornaram-se a cepa predominante no local, mas ainda não está claro
o seu significado.
Na Argentina, o sequenciamento do genoma completo de 26 cepas de vírus
de Influenza A(H1N1)pdm09 e de oito cepas sequenciadas parcialmente não
mostrou evidências de que a alta taxa de letalidade no país, que foi de 4,5%,
enquanto no resto do mundo foi estimada como menor que 1,0%, pudesse ser
atribuídas a mudanças no vírus. Nenhuma evidência de rearranjo, de mutações

42
associadas à resistência de drogas antivirais e mutações pontuais que pudessem
contribuir para aumentar a virulência do vírus foi observada. Apenas a mutação
S206T no gene de HA, já previamente detectada mundialmente, foi observada
(BAUMEISTER et al., 2010). Entretanto, Barrero et al. (2011) ao sequenciarem 28
cepas do vírus A (H1N1)pdm09, detectados na Argentina, observaram que todas
elas possuíam a mutação S220T no gene HA. Em 228 amostras detectaram cinco
cepas com a mutação H274Y no gene da NA.
No sul da China, foi realizada a caracterização genética do gene HA de 10
cepas do vírus A (H1N1)pdm09 e em 100% delas foram detectadas as mutações
P100S, T214A, S220T e I338V. Outras mutações presentes foram a I208L em 90%
das cepas, E391K e A409V (60%), S145P (40%), D144E (20%) e S340F e H416
(1%) (FAROOQUI et al., 2011).
Mutações do vírus influenza A (H1N1)pdm09 no gene NA
Estudos vêm sendo realizados em relação à resistência às drogas inibidoras
da neuraminidase (oseltamivir e zanamivir). Neste contexto, o pirosequenciamento
tem constituído uma ferramenta rápida e eficaz na vigilância de marcadores de
resistência (LACKENBY et al., 2008; DEYDE et al., 2009; DEYDE et al., 2010). A
mutação de resistência mais comumente encontrada tem sido a H274Y ou H275Y
que significa a substituição do aminoácido histidina (H) pela tirosina (Y) na posição
274 ou 275. Esta substituição altera a capacidade de interação da NA com o
oseltamivir, diminuindo a afinidade entre ambos (IAMARINO, 2009). Atualmente
utiliza-se H274Y para N1 e H275Y para N2 (KAWAY et al., 2009).
Um estudo realizado no Japão entre 2000–2001 com 43 crianças tratadas
com oseltamivir, detectou vírus influenza A (H1N1) com a mutação H274Y, em 16%
(7/43) das crianças (MOSCONA, 2005).
Em 2004, com o objetivo de investigar a resistência ao oseltamivir, foram
analisados vírus de influenza A (H3N2) isolados de 50 crianças no Japão, antes e
durante tratamento com oseltamivir. Foram detectadas mutações no gene de
neuraminidase em vírus de nove crianças (18%). Seis tinham a mutação R292K
(substituição da arginina na posição 292 pela lisina) e dois tinham a mutação E119V
(substituição do ácido glutâmico na posição 119 pela valina), que conferem

43
resistência a inibidores de neuraminidase. Também foi identificada numa criança a
mutação N294S (KISO et al., 2004).
Mutação subtipo específica de neuraminidase a inibidores de neuraminidase
tem sido notificada durante passagens in vitro e clínicas. Estudo realizado por Abed
et al. (2006) no Canadá, avaliou o impacto de várias mutações de NA (E119A/G/V,
H274Y, R292K e N294S) nos perfis de susceptibilidade a diferentes inibidores de NA
(oseltamivir, zanamivir e peramivir) usando proteínas de NA recombinante da
influenza A/WSN/33 (H1N1) e A/Sydney/5/97-like (H3N2). No subtipo N1, a mutação
E119V conferiu resistência cruzada ao oseltamivir, zanamivir e peramivir, ao passo
que resistência somente ao oseltamivir foi conferida pela mesma mutação no subtipo
N2. A mutação N294S conferiu resistência ao oseltamivir em ambos os subtipos, N1
e N2, ao passo que a mutação H274Y conferiu resistência ao oseltamivir e peramivir
só no subtipo N1.
Durante as estações de influenza de 2007 e 2008, a resistência entre os vírus
de influenza A (H1N1) ao oseltamivir aumentou, significativamente, pela primeira vez
no mundo. Amostras da Influenza A (H1N1) detectadas no período de 2007 a 2009
através da Vigilância da Influenza nos Estados Unidos foram testadas pelo CDC
para determinar a resistência ao oseltamivir através do ensaio de inibição da
atividade neuraminidásica e pelo método de pirosequenciamento. De 1.155 vírus da
influenza A (H1N1) testados, 142 (12,3%) eram resistentes ao oseltamivir. Nenhuma
exposição ao oseltamivir foi informada antes do diagnóstico de influenza nas
amostras coletadas, sugerindo que a resistência não teve ligação com o uso da
referida droga. Dados obtidos de 182 casos de vírus sensíveis ao oseltamivir,
quando comparados com os resistentes à mesma droga, não mostraram nenhuma
diferença significativa em relação às características demográficas, nem sintomas
clínicos (DHARAN, 2009).
Estudo realizado em Toronto no Canadá, também durante a estação de
influenza de 2007-2008, por Eshaghi et al. (2009), utilizando sequenciamento de
Sanger, demonstrou que 17% (82/474) dos vírus de influenza A (H1N1) isolados,
carregavam a mutação H274Y associada à resistência ao oseltamivir.
No dia 25 de janeiro 2008, a OMS foi notificada oficialmente por autoridades
norueguesas sobre um alto índice de resistência ao oseltamivir em vírus influenza A
(H1N1) de pacientes antes da estação de influenza em novembro e dezembro 2007

44
na Noruega. De 16 vírus isolados, 12 (75%) eram resistentes ao oseltamivir
(ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2008).
Na Oceania, sudeste da Ásia e sul da África, foram examinadas 264 amostras
de vírus da influenza A (H1N1) coletadas em 2008, para verificar a sensibilidade ao
oseltamivir, zanamivir e peramivir, utilizando um teste de fluorescência de inibição da
neuraminidase. Os vírus com reduzida sensibilidade ao oseltamivir foram analisados
por pirosequenciamento. A frequência da mutação H274Y, que confere resistência
ao oseltamivir, aumentou significativamente após maio de 2008, resultando em uma
proporção global de resistência entre as amostras de influenza A (H1N1) de 64%
(168/264), apesar de este subtipo representar apenas 11,6% de todos os isolados
entre os vírus identificados em 2008 (HURT et al., 2009).
No período de 2 de novembro a 27 de dezembro de 2008, a OMS recebeu a
notificação de alta prevalência de vírus de influenza A (H1N1) resistentes ao
oseltamivir detectada em diversos países: (19/19) no Canadá, (97/100) nos Estados
Unidos, (4/5) no México, (8/8) na Alemanha, (9/9) na Noruega, (5/5) na Espanha,
(36/37) no Reino Unido, (132/133) na Coréia e (34/35) no Japão. No total, 13 países
notificaram casos de influenza A (H1N1) resistentes ao oseltamivir, dando um
percentual de 92% (350/379) entre as amostras analisadas (ORGANIZAÇÃO
MUNDIAL DA SAÚDE, 2009h).
De outubro de 2008 a janeiro de 2009, 308 isolados de influenza, de 26
estados dos Estados Unidos, foram testados para resistência a medicamentos
antivirais no CDC. De 190 vírus de influenza A (H1N1) testados, 185 (97.4%) eram
resistentes ao oseltamivir e todos eram sensíveis a zanamivir. Todos 41 vírus de
influenza A (H3N2) e todos 77 vírus de influenza B testados eram sensíveis ao
oseltamivir e zanamivir (CENTERS FOR DISEASE CONTROL AND PREVENTION,
2009e). De dezembro de 2008 a 24 de janeiro de 2009, trinta países notificaram à
OMS a detecção de 1.291 isolados de influenza A (H1N1) resistentes ao oseltamivir
(ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2009a).
Estudo realizado na Noruega, no período de 2007-2008, com 265 pacientes
com vírus A (H1N1), 183 (67,3%) dos quais eram resistentes ao oseltamivir, mostrou
que a resistência não estava associada ao uso prévio de drogas antivirais, e que os
sintomas e índices de hospitalização e a transmissibilidade não eram diferentes
entre os vírus sensíveis e os resistentes ao oseltamivir (HAUGE et al., 2009).

45
Alguns estudos estão analisando a capacidade de replicação, transmissão e
infectividade dos vírus mutados resistentes ao oseltamivir em relação aos vírus que
não sofreram as respectivas mutações. Segundo Aoki; Boivin e Roberts (2007) vírus
com substituição na neuraminidase conferindo susceptibilidade reduzida ao
oseltamivir eram replicados com dificuldade in vitro, e estes mutantes geralmente
reduziam a infectividade e transmissibilidade, quando comparados com vírus
selvagem em modelos animais. Entretanto, posteriormente, Baz et al. (2010),
concluíram que a mutação H274Y não interfere aparentemente na aptidão viral, e
virulência de vírus de influenza A (H1N1), como o A/Brisbane/59/2007-like.
Figura 6 – Prevalência dos vírus influenza A (H1N1) sazonal resistentes ao oseltamivir.
Fonte: OMS - Julho de 2008
Com a instalação da pandemia em 2009, os inibidores da neuraminidase
foram largamente usados para o tratamento e quimioprofilaxia do vírus da influenza
A(H1N1)pdm09. Estudos vêm sendo realizados para verificar a resistência desse
vírus aos inibidores da neuraminidase e a maioria desses vírus tem mostrado ser
sensível a esse grupo de drogas (CHEN et al., 2011). No entanto, casos de
resistência ao oseltamivir já foram relatados na Dinamarca, Hong Kong, Japão e
Canadá foram notificados à OMS no final de julho de 2009. Todos os vírus
resistentes tinham uma mutação característica na posição 274/275 (ORGANIZAÇÃO
MUNDIAL DA SAÚDE, 2009i).

46
A possibilidade de resistência ao oseltamivir se disseminar tem sido uma
preocupação. Embora tenha sido focada no surgimento de resistência devido ao
tratamento, há relatos de resistência viral aos inibidores da neuraminidase entre
indivíduos sem história prévia de uso destas drogas. Anteriormente acreditava-se
que tais vírus teriam menos infectividade e transmissibilidade em seres humanos, o
que não se confirmou (MOSCONA, 2009). De acordo com Le et al. (2010) foi
observado no Vietnan que os vírus A(H1N1)pdm09 com a mutação H274Y,
resistentes ao oseltamivir podem replicar e causar doença em pessoas saudáveis,
mesmo na ausência de exposição prévia aos inibidores da neuraminidase.
Em agosto de 2009, o CDC detectou resistência ao oseltamivir em dois
pacientes imunossuprimidos infectados com o vírus da influenza A(H1N1)pdm09 em
Seattle, Washington. Os dois pacientes foram tratados com oseltamivir e tiveram
eliminação de vírus prolongada por, pelo menos, dois meses. Inicialmente o vírus
mostrou ser sensível ao oseltamivir, e a resistência se desenvolveu
subsequentemente, com o surgimento da mutação H274Y durante o tratamento
(CENTERS FOR DISEASE CONTROL AND PREVENTION, 2009f).
Também em agosto de 2009, o CDC identificou a mutação H274Y em
amostras de duas adolescentes que receberam oseltamivir como profilático na
Carolina do Norte, Estados Unidos. Na neuraminidase, uma segunda mutação I223V
também foi detectada. Este foi o primeiro relato de resistência ao oseltamivir em
casos de vírus influenza A (H1N1)pdm09 com vínculo epidemiológico (CENTERS
FOR DISEASE CONTROL AND PREVENTION, 2009g ).
No período de primeiro de setembro de 2009 até 13 de fevereiro de 2010,
algumas amostras de influenza foram testadas no CDC para resistência aos
inibidores de neuraminidase (oseltamivir e zanamivir). Os resultados da prova de
resistência antiviral mostraram que 1/1 da amostra de influenza sazonal A (H1N1) e
1,3% (48/3691) das amostras de Influenza A(H1N1)pdm09 eram resistentes ao
oseltamivir e que nenhuma das amostras testadas de Influenza A (H3N2) e Influenza
B apresentavam resistência a esse inibidor de neuraminidase. Quanto à resistência
ao zanamivir foram testadas 1.305 amostras de Influenza A(H1N1)pdm09 e todas
eram sensíveis à referida droga (CENTERS FOR DISEASE CONTROL AND
PREVENTION, 2010a).
Em outubro de 2009, foram detectados quatro casos infectados por vírus
resistentes de Influenza pandêmica em pacientes imunocomprometidos numa

47
enfermaria de oncologia hematológica num hospital da Carolina do Norte. Os
autores concluíram que a proximidade geográfica, a convivência por um
determinado período, a presença da mutação H274Y e a similaridade na sequência
viral fortemente sugeriam a transmissão nosocomial do vírus resistente e que o
imediato isolamento, poderia prevenir novas infecções (CHEN et al., 2011).
Até o dia 17 de fevereiro de 2010 foram notificados à OMS 248 casos de
infecção pelo vírus influenza A(H1N1)pdm09 resistentes ao oseltamivir. Todos
possuíam a mutação H274Y. De 248 casos, 23% estavam associados ao
tratamento, 6% associados à profilaxia, 24% eram pacientes imunodeprimidos, 6%
não tinham associação conhecida com tratamento ou provável transmissão pessoa a
pessoa e 41% dos casos tinham sido notificados preliminarmente, sem dados
completos dos pacientes (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2010d).
Estudo realizado por Souza et al. (2010) em pacientes oncológicos
hospitalizados no Rio de Janeiro, mostrou que os pacientes permaneciam
eliminando os vírus por um período prolongado, mas não detectou nenhum vírus
resistente. Corroborando esses achados ao analisarem amostras de 1.608 pacientes
hospitalizados com infecção pelo vírus influenza A(H1N1)pdm09, Harvala et al.
(2011) detectaram 10 (0,62%) amostras cujos vírus eram resistentes ao oseltamivir.
Todos os pacientes eram imunocomprometidos. Kawano et al. (2011) detectaram
pela primeira vez em Nagasaki, Japão, uma cepa de vírus A(H1N1)pdm09 com a
mutação H274Y entre 75 cepas isoladas.
A impossibilidade de uso do oseltamivir como uma opção de tratamento pode
ter consequências profundas. Para reduzir este risco, o uso de oseltamivir deve ser
restringido à profilaxia e tratamento de casos graves e pessoas sob maior risco de
complicações. As reservas de antivirais devem ser diversificadas, e dosagens ideais
e terapias de combinação devem ser urgentemente estudadas. Portanto, é essencial
monitorar e controlar a resistência aos inibidores de neuraminidase (LE et al., 2010),
para orientar o uso eficiente dessa droga.
A falta de estudos sobre os vírus influenza A(H1N1)pdm09 que circularam na
nossa região em relação à presença, ou ausência de mutações, implicadas na
infectividade ou na resistência ao oseltamivir motivaram a realização deste estudo.

48
2.3 OUTRAS VIROSES RESPIRATÓRIAS
As infecções respiratórias agudas (IRA) são responsáveis por considerável
morbidade e mortalidade no mundo inteiro, principalmente em crianças, idosos e
pacientes imunocomprometidos (RABONI et al., 2011), provocando
aproximadamente 30% de todos os óbitos infantis nos países em desenvolvimento,
(THOMAZELL et al., 2007). Frequentemente são causadas por vírus (COSTA et al.,
2006). Em adultos, os vírus respiratórios estão entre os principais agentes
causadores de pneumonia (De ROUX et al., 2004).
Os vírus influenza são uns dos patógenos mais importantes, pela alta taxa de
morbidade e mortalidade e pela sua capacidade de provocar epidemias e
pandemias. Entretanto, vários vírus podem ocasionar um quadro clínico semelhante,
denominado síndrome gripal ou “influenza-like illness” (ILI) (KELLY; BIRCH, 2004),
cuja definição inclui febre e pelo menos um sintoma respiratório (tosse e / ou dor de
garganta) e um sintoma constitucional (dor de cabeça, mal-estar, mialgia, calafrios
ou suor, ou fadiga) ( BELLEI et al., 2008). Dentre tais patógenos figuram vírus
respiratório sincicial (RSV), adenovírus (AdV), rhinovírus (RV), coronavírus (HCoV)
e enterovírus (EV) (TURKINGTON; ASHBY, 1988), metapneumovírus (HMPV),
(KELLY; BIRCH, 2004), bocavírus humano (HBoV) (PILGER et al., 2011) e o
parechovírus humano (PeV), descrito anteriormente como echovírus 22 e 23
(HARVALA; SIMMONDS, 2009) e vírus parainfluenza (PIV) (CENTERS FOR
DISEASE CONTROL AND PREVENTION, 2011).
Vírus Respiratório Sincicial (RSV)
O RSV pertence à família Paramyxoviridae e foi isolado pela primeira vez em
1956. São vírus envelopados, não segmentados, dotados de genoma composto por
RNA de fita simples, com polaridade negativa. Encontra-se classificado um único
sorotipo e dois subgrupos A e B (COLLINS; CROWER, 2007). Globalmente é uma
das principais causas de infecção do trato respiratório inferior em crianças (D’ELIA et
al., 2005). Em muitas regiões constitui a principal etiologia como causa de
pneumonia e bronquiolite em crianças menores de um ano. Portanto, é considerado
um dos patógenos mais importantes para o desenvolvimento de vacinas (COLLINS;
CROWER, 2007). Originalmente, era considerado como um patógeno apenas
pediátrico. Contudo, recentemente, tem se mostrado como uma relevante causa de

49
infecções respiratórias, em idosos e adultos sob maior risco (MURATA; FALSEY,
2007).
A transmissão viral ocorre através do contato direto com secreções de
indivíduos infectados, ou objetos contaminados. As pessoas infectadas normalmente
transmitem o vírus por um período de três a oito dias. Entretanto, algumas crianças
e imunocomprometidos podem transmitir o vírus por até quatro semanas. O período
de incubação é de quatro a seis dias, podendo variar de dois a oito dias. A maioria
das pessoas saudáveis se recupera da infecção por RSV em uma a duas semanas.
Os sintomas geralmente iniciam com coriza e diminuição do apetite. Tosse, espirros
e febre normalmente surgem de um a três dias depois. Sibilos também podem
ocorrer. Em crianças muito pequenas, irritabilidade, diminuição da atividade e
dificuldades respiratórias podem ser os únicos sintomas da infecção (CENTERS
FOR DISEASE CONTROL AND PREVENTION, 2010b). Entre os vírus respiratórios
é o mais frequente nos casos de otite média aguda (HEIKKINEN; THINT;
CHONMAITREE, 1999). Após a infecção natural, a proteção contra reinfecção é de
curta duração e estas são frequentemente observadas em crianças menores de dois
anos. Reinfecções em adultos também ocorrem, porém, clinicamente brandas
(SILVA et al., 2009).
Estudo realizado em crianças de baixa renda e menores de cinco anos,
atendidas em hospital pediátrico no Recife, evidenciou sua maior circulação no
período de abril a julho, durante a estação chuvosa, atingindo seu pico no mês de
maio (BEZERRA et al., 2011).
Evidências epidemiológicas indicam que o impacto da infecção por RSV em
idosos pode ser similar àquela ocasionada pela gripe (FALSEY; WALSH, 2005).
Até o momento não existe vacina licenciada (SLULLENDER, 2001). O uso de
imunoprofilaxia passiva com anticorpos monoclonais reduz o número de
hospitalizações em crianças sob risco (MEJIAS; RAMILLO, 2008).
Metapneumovírus
Os Metapneumovírus humanos (HPMV) também pertencem à família
Paramyxoviridae, subfamília Pneumovirinae, gênero Metapneumovirus, possuindo
similaridades com o RSV na sua organização genômica, estrutura viral,
antigenicidade e sintomas clínicos (SILVA et al., 2009). Foi descrito pela primeira
vez em 2001, quando foi isolado em 28 crianças holandesas, que apresentavam

50
desde doença do trato respiratório superior à bronquiolite grave e pneumonia (VAN
DER HOOGEN, 2001).
São vírus envelopados, cujo genoma é constituído de RNA de fita simples,
não segmentados, polaridade negativa (COLLINS; CROWER, 2007). Através da
análise genética foi caracterizado em dois grupos A e B e posteriormente, em dois
subgrupos cada: A1, A2 e B1, B2, respectivamente (BROOR; BHARAJ; CHAHAR,
2008). Estudo realizado na Austrália, no período de 2001 a 2004, mostrou a
circulação simultânea dos quatro subgrupos de HMPV, com um subgrupo diferente
predominando a cada ano. O subgrupo mais comum foi o B1 (MACKAY et al., 2008).
A infecção viral apresenta como característica o acometimento pulmonar,
associado, principalmente, aos casos de bronquiolite e pneumonia. (SILVA et al.,
2009). Tem distribuição mundial e suas manifestações clínicas podem variar de um
quadro leve do trato respiratório superior até doença grave. A infecção por
metapneumovírus humano ocorre em todas as idades (KELLY; BIRCH, 2004). Em
geral, os adultos apresentam sintomas brandos de resfriado, como tosse, rinorréia,
rouquidão, dor de garganta e, algumas vezes, febre. Entre os casos graves, são
comuns a bronquiolite, asma exarcebada e pneumonia, embora quadros de diarréia,
exantema, vômito, disfagia, conjuntivite e otite média já tenham sido descritos
(SANTOS; ROMANO; WIGG, 2008).
Em estudo realizado na cidade de Aracaju, abrangendo 111 crianças
menores de três anos e com infecção aguda do trato respiratório inferior foi
encontrada uma prevalência de 17% e, em oito (7%) a presença de infecção múltipla
com o RSV (co-infecção) (CUEVAS et al., 2003). Na cidade de Curitiba, foi
detectado o HPMV em 6,4% das 156 crianças hospitalizadas com IRA (DEBUR et
al., 2007). Achados semelhantes foram relatados em Campinas por Silva et al.
(2008). Na cidade do Recife a maior frequência ocorre em crianças menores de
cinco anos, nos meses de julho a outubro, atingindo seu pico no mês de agosto
(BEZERRA et al., 2011).
A infecção natural induz proteção parcial contra a doença. Não há proteção
cruzada contra as diferentes cepas do vírus (BROOR; BHARAJ; CHAHAR, 2008).
O desenvolvimento de vacinas eficientes para imunização contra o HSRV e HMPV
deve ser prioritário, bem como o aperfeiçoamento das terapias para recuperação de
pacientes infectados (SILVA et al., 2008).

51
Enterovírus
Os enterovírus humanos são membros da família Picornaviridae e estão entre
os vírus RNA mais simples. Constituem causa comum de doenças respiratórias,
embora as infecções sejam frequentemente subclínicas. As infecções respiratórias
por enterovírus ocorrem mais no trato respiratório superior, (como um resfriado
comum e crupe) do que no trato respiratório inferior (como a pneumonia)
(PALLANSCH; ROOS, 2007). Clinicamente, os quadros respiratórios causados por
enterovírus não diferem daqueles produzidos pelos vírus que causam infecção
respiratória primária, como os rhinovírus, RSV, influenza, parainfluenza, adenovírus.
Os principais sorotipos que causam infecções respiratórias são o Coxackie A21 e
A24, Coxsackie B4 e B5, Echovírus 4, 9, 11, 20, 25 e provavelmente 1-3, 6-8, 16, 19
e 22. Os enterovírus, 68 e 71 estão associados com quadros de pneumonia e
bronquiolite, respectivamente. Os casos graves ocorrem mais em crianças de faixa
etária baixa (GOMES et al., 1997). O enterovírus 68 foi isolado inicialmente em 1962
na Califórnia, em quatro crianças com doença respiratória (OBERSTE et al., 2004).
Rhinovírus
Os rhinovírus (RV) pertencem à família Picornaviridae, e foram agrupados
inicialmente em rhinovírus humano A (RV-A) e rhinovírus humano B (RV-B)
(TURNER; COUCH, 2007). Entretanto, durante o inverno de 2004, no estado de
Nova York, uma alta incidência de ILI com resultados negativos em testes
moleculares e em cultura de vírus, levou Lamson et al. (2006) a analisar através de
um novo ensaio multiplex, 79 amostras previamente negativas e esclareceu o
diagnóstico de 26 delas, sendo detectada uma grande frequência de rhinovírus,
metade dos quais pertenciam a um grupo genético que ainda não tinha sido
caracterizado, diferente dos RV-A e RV-B. Para avaliar se esse novo grupo circulava
fora do estado de Nova York foi realizado um estudo com amostras respiratórias da
África, Ásia, Austrália e Europa que tinham resultado negativo na avaliação
diagnóstica de rotina. Foi evidenciada a circulação global desse novo grupo (RV-C)
e sua associação com surtos de doença respiratória aguda e com infecção severa
do trato respiratório inferior em crianças (BRIESE et al., 2008).
Os rhinovírus são vírus não envelopados de RNA de fita simples, e de
polaridade positiva. Análises filogenéticas indicam a presença de 74 sorotipos do
grupo A e 25 sorotipos no grupo B (TURNER; COUCH, 2007). Tornaram-se

52
conhecidos como o vírus do resfriado comum porque estão implicados em
aproximadamente 50% das infecções do trato respiratório (BRIESE et al., 2008).
São associados a complicações do trato respiratório superior, como sinusite e otite
média. Estudos clínicos relatam que é o segundo agente associado à pneumonia e
bronquiolite em lactentes (HAYDEN, 2004).
Estudo realizado por Bellei et al. (2008), quando foram analisadas 420
amostras de lavados nasais de pacientes adultos com IRA e/ou ILI nos anos de
2001 a 2003, atendidos no ambulatório de um Hospital Universitário de São Paulo,
mostrou que dos vírus respiratórios detectados, 30,9% era influenza e 19,6% era
rhinovírus e que o pico de atividade dos dois vírus era concomitante. Esses
resultados destacam as implicações de outros vírus na etiologia das infecções
respiratórias na estação da influenza.
Swabs nasais de 47 idosos com sintomas de gripe ou resfriado que residiam
em Botucatu (São Paulo) foram colhidos entre 2002 e 2003 e testados para os vírus
da influenza A e B, rhinovírus, metapneumovírus e vírus respiratório sincicial. HPV
foi detectado em 28,6% das amostras (14/47) e HMPV em 2% (1/47). Os demais
vírus testados não foram detectados. Entre os RV, nove foram sequenciados, sendo
seis amostras pertencentes ao grupo A, uma ao grupo B e duas ao grupo C. De
acordo com os autores, a alta incidência de RV durante a estação da gripe,
necessita de estudos posteriores para avaliar o impacto desse vírus entre os idosos
(WATANABE et al., 2011b).
Coronavírus
Os coronavírus humanos (HCoV) pertencem à ordem Nidovirales, família
Coronaviridae e gênero Coronavírus. São vírus envelopados constituídos de RNA de
fita simples, de polaridade positiva e não segmentado. São os maiores vírus RNA
(SANTOS; ROMANOS; WIGG, 2008). Até o momento, cinco coronavírus (HCoV-
229E, HCoV-OC43, SARS-CoV, HCoV-NL63 e HCoV HKU-1) que infectam os seres
humanos têm sido identificados, dos quais, quatro (HCoV-229E, HCoV-OC43,
HCoV-NL63 e HCoV-HKU-1) circulam continuamente na população humana
(FIELDING, 2011). Os primeriros detectados, na década de 1960 foram os HCoV-
229E e HCoV-OC43 (VAN DER HOEK; PYRC; BERKHOUT, 2006) e o SARS-CoV,
HCoV-NL63 e HCoV HKU-1 foram descritos em 2003, 2004 e 2005, respectivamente
(VAN DER HOEK, 2007).

53
Os coronavírus causam doença do trato respiratório superior e
ocasionalmente, do trato respiratório inferior (CABEÇA; BELLEY, 2012). Até o
SARS-CoV ser identificado como o agente etiológico da síndrome respiratória aguda
grave (SARS), não foi dada muita importância aos dois coronavírus já conhecidos.
Acreditava-se que eles causavam apenas infecções leves do trato respiratório
superior em crianças, e que somente prematuros e crianças com doenças crônicas
poderiam desenvolver infecções graves do trato respiratório inferior. O isolamento do
SARS-CoV e a identificação de dois novos coronavírus HCoV-NL63 (FOUCHIER et
al., 2004) e HCoV-KHU1 levaram a estudos sobre o impacto clínico-epidemiológico
de infecções causadas por todos os coronavírus, que podem ser associadas com
manifestações respiratórias e extrarespiratórias, incluindo o envolvimento do sistema
nervoso central. Considerando a sua capacidade de recombinação, a circulação
desses vírus deve ser monitorada para detectar precocemente a identificação de
cepas particularmente virulentas e, dessa forma, facilitar o desenvolvimento de
medidas preventivas e terapêuticas adequadas (PRINCIPI; BOSIS; ESPOSITO,
2010).
Em Hong Kong, China, HCoV foram detectados em 26 (4,4%) de 587
crianças estudadas: 15 (2,6%) foram positivas para HCoV-NL63, 9 (1,5%) foram
positivas para HCoV-OC43 e 2 (0,3%) positivas para o HCoV-229E Além de causar
a doença do trato respiratório superior, o HCoV-NL63 pode provocar crupe,
exacerbação da asma, convulsão febril e febre alta. Ainda é um patógeno importante
que contribui para a hospitalização de crianças, e foi estimado ter causado a cada
ano, 224 admissões em hospital por 100.000 habitantes, com idade menor ou igual a
6 anos, em Hong Kong (CHIU et al., 2005). Dentre 4.181 aspirados pacientes com
infecção aguda do trato respiratório, 2,1% pacientes apresentaram infecção por
HCoV, sendo 13 (0,3%) positivos para HCoV-KHU-1, 17 (0,4%) positivos para
HCoV-NL63, 53 (1,3%) positivos para HCoV-OC43 e 4 (0,1%) positivos para HCoV-
229E. A infecção do trato respiratório superior foi a forma mais comum de
apresentação do HCoV-KHU-1, apesar de ter provocado também pneumonia,
bronquiolite e exarcebação asmática (LAU et al., 2006).
Nas estações inverno - primavera de 2003-2004 e 2004-2005, o HCoV foi
detectado em 5,7% pacientes italianos admitidos em Hospital com síndrome
respiratória aguda. As infecções foram causadas, respectivamente, por HCoV-OC43,
HCoV-229E e HCoV-NL63 em 25 (53,2%), 10 (21,3%) e 9 (19,1%) dos pacientes.

54
Além disso, 3 (6,4%) dos pacientes foram infectados por cepa não tipada. Co-
infecção com outros vírus respiratórios foi detectada em 14 (29,8%) dos 47
pacientes. Esses casos foram associados mais frequentemente com síndrome
respiratória grave, quando comparados com os casos de infecção única pelo HCoV
(p=0,002) (GERNA et al., 2006).
Parainfluenza
Os vírus parainfluenza (PIV) pertencem à família Paramyxoviridae, subfamília
Paramyxovirinae e gêneros Respirovírus (sorotipos 1 e 3) e Rubulavirus (sorotipo 2 e
subtipos 4a e 4b) (SANTOS; ROMANOS; WIGG, 2008). São constituídos de RNA de
fita simples e polaridade negativa. Cada um dos sorotipos tem características
clínicas e epidemiológicas diferentes. O vírion varia de tamanho (diâmetro médio
entre 150 e 200 nm) e forma, é instável no meio ambiente e facilmente inativado
com água e sabão.
Os quatro sorotipos virais podem causar infecções respiratórias e reinfecções,
particularmente o sorotipo 3 (HALL, 2001). O PIV-1 é a principal causa de crupe em
crianças, enquanto o PIV-2 é menos frequente. O PIV-3 é mais frequentemente
associado com bronquiolite e pneumonia. O PIV-4 é detectado com frequência, mas
normalmente causam doença mais branda (CENTERS FOR DISEASE CONTROL
AND PREVENTION, 2011).
Em lactentes, os vírus parainfluenza causam comumemente doença do
aparelho respiratório inferior. Podem causar doença grave do trato respiratório
inferior, como pneumonia, bronquite e bronqueolite, especialmente em idosos e
pacientes imunocomprometidos. Podem causar reinfecções por toda a vida. Estas
infecções, normalmente, ocorrem no trato respiratório superior provocando resfriado
e dor de garganta (CENTERS FOR DISEASE CONTROL AND PREVENTION,
2006). Em crianças, o tipo mais comum da doença consiste em faringite, rinite,
tosse e rouquidão, geralmente, com febre que dura de dois a três dias. Os piores
sintomas são observados mais frequentemente à noite (KARRON; COLLINS, 2007).
Como ocorre com outros vírus respiratórios, pode provocar otite média aguda
(HEIKKINEN; CHONMAITREE, 2003).
Inquéritos sorológicos mostraram que aproximadamente 75% das crianças a
partir dos cinco anos de idade têm anticorpos para o PIV-1 e PIV-2 e 90% a 100%
delas, tem anticorpos para o PIV-3 (CENTERS FOR DISEASE CONTROL AND

55
PREVENTION, 2011). Segundo Hall (2001), os parainfluenzavírus tipos 1 e 2
causam epidemias que ocorrem em ciclos a cada dois anos no outono, enquanto
que as infecções com o tipo 3 ocorrem anualmente, principalmente na primavera e
verão.
Bocavírus humano
O bocavírus humano (HBoV) pertence à família Parvoviridae, subfamília
Parvovirinae, gênero Bocavirus. Foi detectado pela primeira vez na Suécia, em
espécimes respiratórios (ALLANDER et al., 2005). Desde então, tem sido associado
a doenças do trato respiratório inferior e superior e doença gastrointestinal em
pacientes adultos e pediátricos, e apresenta distribuição global. (VALADARES,
2010). Entretanto, uma alta frequência de co-infecção tem sido descrita e a
patogenicidade viral ainda não se encontra totalmente esclarecida (CHOW; HUANG;
ESPER, 2008).
Trata-se de um vírus não envelopado, de simetria icosaédrica, medindo de 18
a 26 nanômetros de diâmetro. O genoma viral é composto por DNA de fita simples,
sentido positivo ou negativo com aproximadamente 5.3 Kb. Até o momento, estudos
mostram a existência de três tipos de HBoV (HBoV1, HBoV2 e HBoV3) sendo que
somente o HBoV1 está associado à infecção respiratória aguda (SILVA et al.,
2010). O HBoV-2 foi identificado pela primeira vez por Kapoor et al. (2009) e o
HBoV-3 por Arthur et al. (2009).
Na Espanha, Vicente et al. (2007), analisaram 520 amostras de aspirado de
nasofaringe de crianças com IRA e detectaram 40 amostras positivas (7,7%) para o
HBoV. Dessas, 25 (62,5%) mostraram co-infecção com outros vírus, sendo o mais
frequente o RSV (13). Amostras de 259 crianças internadas com sibilos expiratórios
agudos e média de idade de 1,6 anos, na Suécia, mostraram que 19% delas eram
positivas para o HBoV, mas esse vírus foi detectado isoladamente em apenas 5%
das crianças (ALLANDER, 2007). Estudo semelhante foi realizado por Valadares
(2010) em São Paulo, abrangendo crianças menores de dois anos, detectou 47
(4,7%) amostras positivas para HBoV e dessas, 27 (57,4%) amostras apresentaram
co-infecção com outros vírus respiratórios.
Foram analisadas 397 amostras de aspirado nasofaríngeo de pacientes com
diagnóstico de IRA atendidos ambulatorialmente no Pará, tendo sido detectado o
HBoV em três amostras de crianças menores de um ano. Uma delas apresentou co-

56
infecção com o RSV. O percentual de positividade obtido na investigação se revelou
inferior ao descrito na literatura, entretanto, vale ressaltar que os estudos já
publicados tinham sido realizados com pacientes hospitalizados (SILVA et al., 2010).
Segundo Pilger et al. (2011), a taxa de incidência do HBoV em amostras
respiratórias de crianças com infecção respiratória aguda é de 3 a 19%. Estudo
realizado pelo grupo com 455 amostras de aspirado de nasofaringe de 453 crianças
menores de dois anos, admitidas na emergência pediátrica em Porto Alegre,
detectou 60 (13,2%) amostras positivas para o HBoV.
Adenovírus
Os Adenovírus, na maioria das vezes, causam doenças respiratórias, no
entanto, dependendo do sorotipo infectante, podem causar várias outras doenças,
como gastroenterites, conjuntivite, cistite e doença exantemática. Os sintomas de
doença respiratória causada por adenovírus variam de síndrome do resfriado
comum à pneumonia, crupe e bronquite. Lactentes jovens, especialmente pacientes
com o sistema imunológico comprometido são mais suscetíveis a complicações
graves da infecção por adenovírus. Surtos de adenovírus associados a doenças
respiratórias têm sido mais comuns no final do inverno, primavera e início do verão,
no entanto, infecções por adenovírus podem ocorrer durante todo o ano. O período
de incubação no caso de infecções do trato respiratório é entre 2 e 14 dias
(CENTERS FOR DISEASE CONTROL AND PREVENTION, 2010c).
Os adenovírus pertencem à família Adenoviridae, gênero Mastadenovírus
(SANTOS; ROMANOS; WIGG, 2002), o genoma viral é constituído por DNA de
dupla fita e é protegido por capsídeo icosaédrico. Foram isolados e caracterizados
em 1953 (ROWE et al., 1953; HILLEMAN; WERNER, 1954). Um total de 51
sorotipos foram identificados. Esses 51 sorotipos foram classificados em 6
subgrupos (espécies de A a F) (BERCK, 2007). Alguns sorotipos, como o 3, 4, e 7
são mais comumente associados com doença respiratória aguda. Os profissionais
de saúde devem pensar em adenovírus como possível causa de casos graves de
pneumonia, e surtos de pneumonia de etiologia desconhecida (CENTERS DISEASE
CONTROL AND PREVENTION, 2010c).
Nos Estados Unidos, as doenças respiratórias agudas estão mais
frequentemente associadas aos adenovírus tipo 4 e 7 e mais recentemente ao
adenovírus 14. Surtos de adenovírus têm sido mais comuns no final do inverno,

57
primavera e início do verão; entretanto, a infecção pelo adenovírus pode ocorrer
durante todo o ano (CENTERS DISEASE CONTROL AND PREVENTION, 2010c). Parechovírus
Os parechovírus humanos pertencem à família Picornaviridae (VERBOON-
MACIOLEK et al., 2008), e não possuem envelope (DREXLER et al., 2009). Devido
às suas propriedades morfológicas e clínicas foram classificados inicialmente como
sendo do gênero enterovírus: o parechovírus humano 1 (PeV1) foi descrito como
Echovírus 22 e o parechovírus humano 2 (PeV2) como Echovírus 23 (HARVALA;
SIMMONDS, 2009). Entretanto, eles mostraram ser bem distintos dos demais
enterovírus no que concerne à organização do seu genoma e replicação e formaram
um novo gênero (JOKI–KORPELA; HYYPIÃ, 2001).
Com base em estudos genéticos e sorológicos, existem os seguintes tipos de
PeV: PeV-1, PeV-2, PeV-3, PeV-4 e PeV-5 (WATANABE et al., 2007) e PeV-6 (DE
VRIES et al, 2008). O PeV apresenta heterogeneidade genética e antigênica e um
número de tipos distintos circula amplamente nas populações humanas em todo o
mundo (HARVALA; SIMMONDS, 2009). De acordo com Chieochansin et al (2011),
há 16 genótipos reconhecidos, com distribuição global.
Estudos soroepidemiológicos têm demonstrado que a prevalência de
anticorpos PeV1 é surpreendentemente elevada, superior a 95% na população
adulta. De acordo com dados atuais, o PeV1 causa principalmente infecções
gastrointestinais e respiratórias, no entanto, podem causar doenças graves, como
miocardite e encefalite. Infecções por PeV2 parecem ser raras (JOKI–KORPELA;
HYYPIÃ, 2001). Estudo realizado por Harvala et al (2008), para investigar o
envolvimento dos parechovírus em doenças respiratórias foi realizado em
Edimburgo, na Escócia. Um total de 3.844 amostras respiratórias de 2.200
indivíduos foram coletadas de janeiro a dezembro de 2007. Foram detectadas 33
amostras positivas de 27 indivíduos (1,2%).
Diagnóstico diferencial
Apesar do diagnóstico de doenças virais ter melhorado significativamente nos
últimos 15 anos com o desenvolvimento das técnicas de PCR, a identificação do
agente etiológico das doenças respiratórias virais tem sido feita quase que
exclusivamente em crianças. Em idosos a frequência de identificação da etiologia

58
viral encontra-se abaixo de 40% (JARTTI et al., 2011). De acordo com Leung et al.,
(2009), o fato de grande parte de IRAs não ter seus agentes etiológicos identificados
pode ser devido às limitações dos ensaios de diagnóstico, ou devido a uma
proporção de IRAs ser causada por patógenos ainda desconhecidos.
Júven et al (2000) na Finlândia, pesquisaram 17 microorganismos entre vírus
e bactérias, em aspirado de nasofaringe de 254 crianças com pneumonia. O agente
etiológico foi detectado em 85% dos casos e destes 62% dos pacientes apresentava
infecção viral e 30% tinha evidência de infecção viral e bacteriana. Dentre os vírus
detectados o RSV (29%) e RV (24%) foram os agentes mais comuns associados
com a pneumonia. Dupla infecção viral foi detectada em 35 pacientes.
Na Índia, no período de abril de 2005 a março de 2007 foram coletados 301
aspirados de nasofaringe de crianças atendidas no ambulatório ou internadas em
hospitais de Nova Deli com problemas respiratórios. Um ensaio de PCR multiplex
detectou 130 vírus respiratórios em 106 (35,2%) amostras, sendo 61 (46,9%) RSV,
22 (16,9%) PIV3, 17 (13,1%) PIV2, 11 (8,5%) HMPV, 10 (7,7%) PIV1 e nove (6,9%)
FLUA (BHARAJ et al., 2009).
Utilizando um ensaio de RT-PCR multiplex, com cinco tubos de misturas de
primers capazes de detectar 17 diferentes vírus respiratórios, Wang et al (2010)
analisaram amostras de nasofaringe de 817 crianças menores de 9 anos com
infecção respiratória aguda, coletadas no período de outubro de 2006 a setembro de
2008, em Shangai, China. Foi detectado um único vírus em 373 amostras (45,7%) e
em 113 amostras (13,8%) foram detectados de dois a quatro vírus, perfazendo um
total de 486 amostras positivas. Ao todo, 618 vírus foram detectados, sendo o mais
frequente o RSV (19,4%), seguido do HBoV (19,3%), FLUA e FLUB (17,5%) PIV1, 3
ou 4 (12,5%), RV (11,8%), HMPV (7,1%), AdV (7%), HCoV (3,9%) e EV (1,8%).
Entre os 113 pacientes diagnosticados com múltipla infecção viral, os patógenos
mais presentes foram o HBoV, RSV, RV e AdV.
Aspirados de nasofaringe de 407 crianças menores de cinco anos com IRA
atendidas em Hospital Infantil na cidade do Recife, Pernambuco, no período de abril
de 2008 a março de 2009, foram analisadas por Bezerra et al. (2011). Foram
detectados os seguintes vírus: RSV (37,3% dos pacientes), AdV (24,8%), RV
(18,9%), HBoV (18,7%) e HMPV (10,3%). Coinfecções com um ou mais patógenos
foram detectadas em 161 amostras (39,6%). O RSV apresentou uma alta frequência

59
na estação chuvosa (abril a julho) e o HMPV de agosto a outubro, ambos mostrando
um padrão sazonal bem definido.
Depois da instalação da nova pandemia de influenza foi realizada a análise
de 58 aspirados de nasofaringe de crianças de 28 dias a 12 anos com doença
respiratória grave, internadas em Hospital de Porto Alegre, no período de 30 de julho
a 1 de setembro, utilizando-se a reação de imunofluorescência. O kit utilizado
detecta os vírus FLUA, FLUB, AdV, PIV-1, PIV-2, PIV-3 e RSV. Dos exames
realizados, 26 (44,8%) foram positivos para os vírus analisados. Destes, foram
detectados 18 (69,2%) casos de RSV, seis (23,2%) de FLUA, um (3,8%) de PIV-1 e
um (3,8%) de PIV-3. A associação de vírus foi verificada em cinco casos (19,6%)
para RSV e FLUA. Dos casos analisados, confirmou-se que três (5,2%) eram do
novo subtipo de Influenza H1N1 (CARNEIRO; KRUMMENAUER; MACHADO, 2010).
Também com o objetivo de determinar a etiologia viral durante a pandemia de
2009, estudo utilizando a técnica de PCR foi realizado em amostras coletadas de 98
crianças e 61 adultos internados num Hospital Sentinela de São Paulo, Brasil, com
suspeita de infecção pelo vírus pandêmico, no período de agosto a novembro de
2009. Foi detectado o vírus influenza A(H1N1)pdm09 em 19,5% (31/159), sendo
23% (14/61) em adultos e 18,4% (17/92) em crianças. As amostras negativas para
influenza A sazonal e pandêmica foram testadas para RV, HMPV, AdV e RSV.
Foram positivas para pelo menos um vírus, 61% (97/159) e 3,7% (6/159)
apresentaram coinfecção com múltiplos vírus. No total, 64,8% (103/159) de etiologia
viral foi determinada. A frequência de identificação viral nas crianças de até 12 anos
foi significativamente mais alta quando comparada com os adultos (p=0,0021). A
influenza A sazonal foi detectada em 6,2% (10/159) das amostras e dentre as
amostras que foram negativas para influenza A, 52,54% (62/118) foram positivas
para outros vírus respiratórios. Desses, 22,88% foram positivos para RV, 20,34%
para HMPV, 2,54% para AdV e 1,69% para RSV. Seis amostras (5,08%),
apresentaram coinfecção com dois vírus, sendo quatro (RV/ HMPV), um (AdV/RSV)
e um (AdV/RV) (WATANABE et al., 2011a).
Entre maio e agosto de 2009 foram testados em Buenos Aires, Argentina,
3.476 aspirados de nasofaringe para alguns vírus respiratórios, tendo sido positivas
para um dos vírus, 1.553 amostras (44,68%). Destas, 367 era influenza A, sendo
330 identificadas através de RT-PCR como vírus A(H1N1)pdm09, 30 amostras não
foram tipadas e duas foram identificadas como influenza A (H3N2) sazonal. Ao todo

60
foram detectados 1.158 vírus, uma vez que ocorreram três casos de infecções
duplas de FLUA–RSV no mês de junho. O RSV esteve presente em 1.142 amostras
(73,5%), PIV3 em 37 (2,4%) e AdV em 12 (0,7%). Não foram detectados IPV1, PIV2,
nem FLUB, apesar de terem sido testados (BARRERO et al., 2011).
No período de maio a dezembro de 2009 foram coletadas amostras
respiratórias de 171 pacientes internados com IRA no Hospital das Clínicas da
Universidade do Paraná. As amostras foram testadas para o vírus influenza A
(H1N1)pdm09 através da reação de rRT-PCR, de acordo com o protocolo CDC e
os outros vírus respiratórios através de uma reação RT-PCR multiplex, que permite a
detecção simultânea de vários agentes. Vírus respiratórios foram detectados de 129
amostras (75%), sendo 67 casos diagnosticados como vírus influenza
A(H1N1)pdm09 (39%) e 62 (36%) diagnosticados como outros vírus respiratórios.
O vírus da influenza A sazonal foi detectado em 15 amostras (9%). Considerando
apenas as amostras positivas, o maior percentual de detecção dos outros vírus
respiratórios foi: RSV (13,1%), RV (10%), HMPV (9,3%), PIV3 (5,4%), AdV (5,4%),
HCoV (3,8%), FLUB (0,7%), PIV1 (0,7%) e PIV2 (0,7%). Coinfecões virais foram
mais raras entre os casos do vírus influenza A (H1N1)pdm09 (1%) do que entre
outros vírus respiratórios (8%) (RABONI et al., 2011).
Zuccotii et al. (2011) estudaram 575 crianças menores de 15 anos,
hospitalizadas de dezembro de 2008 a dezembro de 2009, na cidade de Milão, com
infecção respiratória aguda. Amostras das referidas crianças foram testadas para
vírus respiratórios obtendo-se um percentual de positividade de 51%. A frequência
de detecção dos vírus foi: RSV 34,1%, HBoV 6,8%, HMPV 5%, influenza A sazonal
5%. O FLUB não foi detectado. No período de abril de 2009 até o final do estudo foi
detectado o vírus influenza A(H1N1)pdm09 em 11,6% das amostras.
Linde et al., 2009 ao analisar a súbita interrupção da propagação da influenza
pandêmica na Suécia levantou a hipótese de ter sido provocada por um aumento na
circulação de rhinovírus que teria competido com o vírus influenza A(H1N1)pdm09.
A falta de estudos sobre os vírus da influenza que circularam em Pernambuco
durante a última pandemia relacionados à resistência a drogas antivirais, aspectos
clínicos, epidemiológicos e moleculares, bem como a falta de dados sobre outros
vírus respiratórios que circularam no estado no mesmo período, levando os
profissionais de saúde à hipótese diagnóstica de infecção pelo vírus influenza
A(H1N1)pdm09, despertaram o interesse para a execução deste trabalho.

61
3 OBJETIVOS 3. 1 Objetivo Geral: Investigar aspectos clínicos, epidemiológicos e moleculares das infecções por
Influenza A(H1N1)pdm09, bem como a frequência de outros vírus respiratórios
circulantes no estado de Pernambuco, durante o período pandêmico (maio de 2009
a maio de 2010).
3. 2 Objetivos Específicos:
1. Determinar a frequência de vírus influenza A(H1N1)pdm09 no estado de
Pernambuco, no período de maio de 2009 a maio de 2010.
2. Estudar aspectos clínico-epidemiológicos apresentados pelos pacientes
infectados pelo vírus influenza A(H1N1)pdm09, no período de maio de
2009 e maio de 2010.
3. Investigar a frequência de outros vírus respiratórios em casos suspeitos
de infecção pelo vírus influenza A(H1N1)pdm09 com resultado
laboratorial negativo.
4. Caracterizar o gene hemaglutinina em amostras positivas para o vírus
pandêmico.
5. Investigar a prevalência do marcador de resistência ao oseltamivir,
H274Y no gene da Neuraminidase.
4 MATERIAIS E MÉTODOS
4.1 Desenho do estudo
Foi realizado um estudo descritivo, transversal, no período de maio de 2009 a
maio de 2010.

62
4.2 Local do estudo
A coleta dos swabs combinados de nasofaringe e orofaringe foi realizada em
hospitais do estado de Pernambuco que atenderam pacientes com suspeita clínica
de influenza pandêmica. No caso de pacientes que faleceram em decorrência da
doença, foram coletados fragmentos de brônquios e pulmões no Serviço de
Verificação de Óbito do Estado (SVO).
4.3 População de estudo
Indivíduos com síndrome gripal, com síndrome respiratória aguda grave e
indivíduos que evoluíram para óbito, atendidos nas Unidades Hospitalares do estado
de Pernambuco, com suspeita de infecção pelo vírus da Influenza A(H1N1)pdm09,
cujas amostras clínicas (swab combinado de nasofaringe e orofaringe ou tecidos de
brônquios e de pulmões) foram coletadas no período de maio de 2009 a maio de
2010. Os dados sócio-demográficos dos pacientes foram resgatados das Fichas de
Investigação: INFLUENZA HUMANA POR NOVO SUBTIPO (PANDÊMICO) e do
Sistema de Informação de Agravos de Notificação (SINAN). Do Ministério da Saúde.
4. 4 Variáveis 4. 4. 1 Variáveis dependentes
Infecção pelo vírus da Influenza A sazonal ou pelo vírus A(H1N1)pdm09.
Mutação no gene HA associadas à infectividade viral.
Mutação H274Y no gene NA associada à resistência ao oseltamivir.
Infecção por outros microrganismos.
4.4.2 Variáveis independentes
Dados sócio-demográficos e clínico-epidemiológicos:
Sexo: masculino e feminino
Faixa etária: - ≤ 2 anos, 3 a 5 anos, 6 a 9 anos, 10 a 19 anos, 20 a 29 anos, 30 a 39
anos, 40 a 49 anos, 50 a 59 anos e ≥ 60 anos.
Sintomas: Febre, tosse, mialgia, coriza, dor de garganta, calafrio, artralgia, dispnéia,
diarréia, conjuntivite, outros sintomas.

63
Grupos de risco: Gestantes, pneumopatias, cardiopatias, doenças metabólicas,
doenças renais, hemoglobinopatias e imunodeprimidos.
4.5 Operacionalização da Pesquisa
A coleta das amostras clínicas foi realizada em Unidades Hospitalares do
estado de Pernambuco que atendiam casos suspeitos de infecção pelo vírus
influenza A(H1N1)pdm09, e, de acordo com o Ministério da Saúde deveria,
preferencialmente, ser feita até três dias do início dos sintomas (fase aguda da
doença), tendo sido estendida até 7 dias após o início dos sintomas. Nos casos de
óbito, a coleta de fragmentos de brônquios e de pulmões foi realizada no Serviço de
verificação de óbito (SVO). Após a coleta, as amostras foram encaminhadas ao
Laboratório Central de Pernambuco (LACEN-PE), onde foram registradas e
permaneceram congeladas a -80ºC até posterior encaminhamento para o Instituto
Evandro Chagas (IEC) em Belém, Laboratório de Referência Regional e um dos três
Centros Nacionais para Influenza (OPAS/OMS) (BRASIL, 2008), cuja área de
abrangência é a região nordeste. Lá as amostras foram processadas para o
diagnóstico laboratorial do vírus influenza A(H1N1)pdm09 e vírus influenza A
sazonal, segundo o protocolo do CDC.
A caracterização genética e o diagnóstico diferencial para outros vírus
respiratórios foram realizados no Laboratório de Referência Nacional para Influenza,
o Laboratório de Vírus Respiratórios e do Sarampo da Fundação Oswaldo Cruz, Rio
de Janeiro.
Para execução deste trabalho, diferentes protocolos e técnicas foram
utilizados. O fluxograma contendo as etapas envolvidas no desenvolvimento do
presente estudo encontra-se ilustrado na Figura 5.
Os ensaios realizados seguiram protocolos mundialmente estabelecidos.
Para o diagnóstico do vírus A(H1N1)pdm09 e Influenza A sazonal foi realizada a
metodologia de PCR em tempo real (sistema TaqMan, seguindo o protocolo do
CDC/Atlanta). Foram utilizados o QIAamp® Viral RNA Mini Kit (Qiagen) para
extração do RNA, e o kit Primer and Probe Set da Invitrogen para a detecção do
RNA viral. Para o sequenciamento do gene HA foi utilizado o método de Sanger,
precedido de uma reação de RT-PCR, utilizando-se o kit SuperScript IIITM One-step
RT-PCR with Platinum Taq-DNA Polymerase (Invitrogen), de acordo com Protocolo
CDC para Influenza. As sequências foram analisadas e editadas utilizando-se o

64
programa Sequencher, e alinhadas com sequências de outros vírus disponíveis no
banco de dados do GenBank (http://www.ncbi.nlm.nih.gov), utilizando o programa
Mega 4.0 (TAMURA et al., 2011).
O pirosequenciamento para detecção da mutação H274Y no gene NA foi
precedido de uma reação de RT-PCR, utilizando-se o kit SuperScript IIITM One-step
RT-PCR with HiFi Taq system (Invitrogen), de acordo com Protocolo do CDC,
utilizando o método de análise SNP e o equipamento PyroMark Q-96 ID (DEYDE et
al., 2009)
Figura 7 – Fluxograma das etapas e técnicas utilizadas na realização do estudo

65
Técnicas utilizadas
Extração de RNA viral (RNAv) para o diagnóstico de Influenza (Protocolo CDC)
O RNAv foi extraído a partir de 140 µL do espécime biológico. Para obtenção
do ácido nucléico foi utilizado o QIAamp® Viral RNA Mini Kit (QIAGEN, Germany),
seguindo-se rigorosamente as orientações do fabricante. O método consiste em três
principais etapas: 1) extração - onde se utiliza o tampão AVL com RNA carreador
para inativar RNAses e liberar o RNAv. Condições tamponantes são, então,
ajustadas para permitir uma ligação do RNA à membrana de sílica em gel “QIAamp”;
2) lavagem - realizada em duas etapas sucessivas para eliminar os contaminantes,
utilizando-se os tampões AW1 e AW2, contendo etanol absoluto; 3) eluição - no final
do processo o RNAv foi eluído em tampão AVE (pH 8,0) num volume de 80 µL e
estocado a -80°C até o momento de uso. Foi utlizado na detecção do vírus de
influenza A, incluindo o A(H1N1)pdm09, no sequenciamento do gene HA e no
pirosequenciamento. Esta técnica baseia-se na capacidade que a sílica tem de
formar ligação com o RNAv em meios com alta concentração de sal combinada à
centrifugação em alta velocidade (8.000 a 14.000 rpm).
Para a posterior pesquisa de outros vírus respiratórios (vírus RNA e DNA),
uma nova extração foi realizada, utilizando o kit Magna Pure.
Extração de ácidos nucléicos para o diagnóstico diferencial
Foi realizada utilizando-se o sistema de extração de ácidos nucléicos e pós-
eluição com o kit Magna Pure LC Total Nucleic Acid Isolation Kit e o equipamento
MagNA Pure LC 2.0, da Roche, seguindo as instruções do fabricante. Este método
baseia-se na propriedade de ligação de moléculas de RNA e DNA à partículas
magnéticas. O DNA e RNA se ligam as partículas magnéticas de sílica (MGPs) na
presença de sal caotrópico com pH maior que 7.0. As MGPs apresentam uma
superfície vítrea (sílica) e um núcleo magnético.
A primeira etapa da extração foi realizada colocando em cada poço da placa
do equipamento MagNAPure, 300 µL do tampão de lise/ligação, 3 µL de controle
interno do kit Fast track (CI) e 200 µL das amostras respiratórias. Foi extraído um
controle negativo do Fast track para cada série de 12 amostras.

66
Em seguida foram colocados os descartáveis no MagNaPure e os reagentes
nos volumes descritos na tela de trabalho ao rack de reagentes e o equipamento foi
programado para fazer a extração. No final do processo, obtivemos o RNA e DNA
eluídos no volume de 100 µL.
4.5.1 Ensaio de RT-PCR em tempo real (rRT-PCR) para detecção do vírus
influenza A(H1N1)pdm09.
A técnica é composta das seguintes etapas: a) Extração do Ácido Nucléico; b)
Preparação das misturas de reação; c) Aplicação dos controles negativos; d)
Aplicação das amostras; e) Termociclagem – amplificação; f) Visualização, análise e
interpretação dos resultados.
Nas reações foram utilizados o Invitrogen SuperScriptTMIII Platinum One-
Step Quantitative RT-PCR System, kit com rox e o Kit Primer and Probe Set da
Invitrogen. Os iniciadores e as sondas do Kit Primer and Probe Set (Invitrogen)
utilizadas na reação, são específicos para detecção de: a) gene da matriz, que
permite a detecção de todos os subtipos de Influenza A; b) do gene da NP
específico para a variante A(H1N1)pdm09; c) da porção HA1 do gene HA também
específica da linhagem pandêmica e d) o gene housekeeping da RNase P, como
controle interno. As reações com cada conjunto de sondas e iniciadores foram feitas
em tubos separados nas seguintes condições de amplificação para o termociclador
7.500 da Applied, 50ºC/30min, 95ºC/2min, seguidos por 45 ciclos de 92ºC/15seg e
55ºC/35seg conforme recomendações do protocolo do Centers for Disease Control
and Prevention (CDC), disponível em
<http://www.who.int/csr/resources/publications/swineflu/CDCRealtimeRTPCR_Swine
H1Assay-2009_20090430.pdf.>. (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2009e).
O Kit Primer and Probe Set da Invitrogen é composto de painel com os
seguintes iniciadores e sondas - Sequência (5’>3’):
a) InfA que detecta todos Influenza A: InfA Forward GAC CRA TCC TGT CAC CTC TGA C 40 µM InfA Reverse AGG GCA TTY TGG ACA AAK CGT CTA 40 µM InfA Probe1 TGC AGT CCT CGC TCA CTG GGC ACG 10 µM

67
b) InfAsw que detecta especificamente todo Influenza A, linhagem suína SW InfA Forward GCA CGG TCA GCA CTT ATY CTR AG 40 µM SW InfA Reverse GTG RGC TGG GTT TTC ATT TGG TC 40 µM SW InfA Probe2 CYA CTG CAA GCC CA”T” ACA CAC AAG CAG GCA 10 µM
c) AH1sw que detecta Influenza A, subtipo H1 suíno SW H1 Forward GTG CTA TAA ACA CCA GCC TYC CA 40 µM SW H1 Reverse CGG GAT ATT CCT TAA TCC TGT RGC 40 40 µM SW H1 Probe2 CA GAA TAT ACA “T”CC RGT CAC AAT TGG ARA A 10 µM
d) RP detecta RNAse P humana – controle positivo da extração Rnase P Forward AGA TTT GGA CCT GCG AGC G 40 µM Rnase P Reverse GAG CGG CTG TCT CCA CAA GT 40 µM Rnase P Probe1 TTC TGA CCT GAA GGC TCT GCG CG 10 µM
As reações foram realizadas com um volume final de 25µL contendo, 5,5 µL de
H2O RNAse free, 0,5 µL de cada oligonucleotídeo iniciador específico, 0,5 µL da
sonda específica, 0.5 µL de SuperScriptTM III RT/Platinum® Taq Mix, 12,5µL de
tampão de reação 2X PCR Master Mix, e 5 µL de RNAv.
Em seguida foi feita a análise da reação usando o 7.500 Software v 2.0.5
(Applied Biosystems).
4.5.2 Sequenciamento do gene HA, utilizando o método de Sanger
A caracterização genética dos vírus foi realizada em três etapas principais: a)
extração do RNA viral (RNAv); b) amplificação do RNAv pela técnica de Reação em
Cadeia da Polimerase precedida de Transcrição Reversa (RT-PCR); e c) reação de
sequenciamento.
Amplificação do RNA através da Reação em Cadeia da Polimerase precedida de
transcrição reversa (RT-PCR)
A Reação em Cadeia da Polimerase precedida de transcrição reversa (RT-
PCR) em uma única etapa foi realizada utilizando-se o kit SuperScript IIITM One-step
RT-PCR with Platinum Taq-DNA Polymerase (Invitrogen) de acordo com Protocolo
CDC para Influenza (ORGANIZAÇÃO MUNDIAL DA SAÚDE, 2009j), disponível em
<http://www.who.int/csr/resources/publications/swineflu/GenomePrimers_20090512.
pdf.> As reações de RT-PCR foram feitas para um volume final de 25µL contendo,
12,5µL de tampão de reação 2X (contendo 0,4mM de cada dNTP, 2,4mM de
MgSO4), 2,0 µL de H2O RNAse free, 2,0 µL de cada oligonucleotídeo iniciador

68
específico (5 µM), 0,5 µL de inibidor de ribonuclease (RNAse OUT), 1,0 µL de mix
RT/Taq (SuperScriptTM III RT/Platinum Taq) e 5 µL de RNAv. Cada conjunto de
amostras submetido à amplificação foi acompanhado de um controle negativo.
Após a adição do RNAv a mistura foi para o termociclador, obedecendo a
seguinte ciclagem: 48°C por 45 min, 94°C por 2 minutos, 40 ciclos de 94°C por 20
segundos, 50°C por 30 segundos e 72°C por 1 minuto, finalmente 72°C por 7
minutos e 4°C até a retirada da placa do equipamento.
Iniciadores e sondas utilizados na reação:
Primers Forward pb 5’
HA1 460 tgt aaa acg acg gcc agt ata cga cta gca aaa gca ggg g
HA2 598 tgt aaa acg acg gcc agt acr tgt tac ccw ggr gat ttc a
HA3 865 TGT AAA ACG ACG GCC AGT ACR TGT TAC CCA GGR GAT TTC
HA4 604 tgt aaa acg acg gcc agt agr atg rac tat tac tgg ac
HA5 417 tgt aaa acg acg gcc agt tgg atg gta ygg tta yca yca
HA6 574 tgt aaa acg acg gcc agt aag atg aay acr car ttc aça g
Primers Reverse pb 5’
HA1 460 cag gaa aca gct atg acc tca tga ttg ggc cay ga
HA2 598 cag gaa aca gct atg acc gaa akg gga grc tgg tgt tta
HA3 865 CAG GAA ACA GCT ATG ACC TCT TTA CCY ACT RCT GTG AA
HA4 604 cag gaa aca gct atg acc ttc tkc att rta wgt cca aa
HA5 417 cag gaa aca gct atg acc tca taa gty cca ttt ytg a
HA6 574 cag gaa aca gct atg acc gtg tca gta gaa aça agg gtg ttt
Ao término da amplificação os produtos da RT-PCR foram analisados por meio
de eletroforese utilizando o E-Gel® 2% agarose (GP) da Invitrogen, conforme
instrução do fabricante (www.invitrogen.com/content/sfs/manuals/egel_qrc.pdf).
Os produtos que apresentaram a banda de interesse no E-gel, foram purificados
utilizando o kit comercial QIAquick PCR Purification spin columns (QIAGEN,
Canadá), de acordo com o protocolo fornecido pelo fabricante. Neste procedimento
o DNA é adsorvido à membrana de sílica da coluna na presença de altas
concentrações de sal, enquanto contaminantes passam através da coluna. Após

69
lavagens as impurezas são eliminadas e o DNA puro é eluído em tampão Tris. Após
a purificação o produto eluído foi armazenado a -20° C.
Reação de sequenciamento
Depois de purificados os produtos da RT-PCR foram sequenciados usando-
se o Kit Big Dye® terminator v 3.1 Cycle Sequencing (Applied Biosystem). Este
ensaio é baseado no sequenciamento de Sanger, onde ocorre uma reação de
polimerização contendo nucleotídeos sem o grupo hidroxil 3` terminal da pentose
(didesoxinucleotídeos) e quatro diferentes fluoróforos associados às bases Adenina
(A), Timina (T), Citosina (C) e Guanina (G), respectivamente. A presença das bases
modificadas bloqueia o processo de elongação, a partir da adição do nucleosídeo,
gerando produtos intermediários com a extremidade 5` fluorescente. No ensaio
foram utilizados os seguintes primers:
___________________________________________
Primer M13 F - tgt aas acg acg gcc agt
Primer M13 R - cag gaa aca gct atg acc____
Foram preparadas duas misturas de reação Mistura 1: composta de 11,5 µL
de água livre de RNAse, 2 µL de tampão de diluição e 4µL de Big Dye, perfazendo
um total de 17,5 µL. Mistura 2: preparados dois tubos, um com o primer M13
F(Forward) e outro com o primer M13 R (Reverse). Cada tubo foi composto de 17,5
µL da mistura 1, e 1 µL do primer respectivo. No final foi adicionado 1,5 µL do
fragmento purificado. Esta mistura foi processada em termociclador automático
(Máster Cycler, Eppendorf, Birkmann Instrument) onde foram realizados 30 ciclos a
96°C por 10 segundos, 50°C por 5 segundos e 60°C por 4 minutos.
Purificação do produto da reação de sequenciamento
Após a reação de sequenciamento os produtos marcados foram submetidos a
purificação pelo método de precipitação BigDye X-terminator (Applied Biosystems),
de acordo com orientação do fabricante, com a finalidade de retirar o excesso de
reagentes não incorporados na reação de (como íons de sal, ddNTPs
(didesoxinucleotídeos) não incorporados e dNTPs), além de garantir a linearidade
das sequências.

70
Sequenciador automático
Os produtos purificados foram analisados no sequenciador automático
ABIPrism 3130 XL (Applied Biosystem), onde o DNA purificado é injetado nos
capilares do sequenciador através de injeção eletrocinética. Esta técnica é baseada
no método de terminação de cadeia por didesoxirribonucleotídeos (Sanger et al.,
1977). Os contigs das sequências nucleotídicas obtidas para o gene da HA foram
montados com o auxílio do programa Sequencher (Gene codes).
Alinhamento das sequências e construção da árvore filogenética
Os eletroferogramas com a extensão *.ab1, gerados pelo sequenciador,
foram analisados com o uso do programa Sequencing Analises® (Applied
Biosystems), tendo sido avaliada a viabilidade das sequências obtidas. Para a
construção dos contigs das sequências, os arquivos com extensão *ab1 foram
inseridos em um projeto, criado com o programa Sequencher (Gene Codes, Ann
Arbor, USA). Este permite a comparação e montagem dos segmentos gênicos, com
base em sequências referência para o gene HA. As sequências foram alinhadas
utilizando o software MEGA v.5.0.5 (TAMURA et al., 2011) utilizando-se sequências
nucleotídicas de variantes virais depositadas no GenBank.
A análise filogenética foi realizada utilizando o programa MEGA v.5.0.5. As
árvores filogenéticas foram construídas com uso do método de agrupamento de
vizinhos- Neighbor-Joing, NJ (Saitou e Nei, 1987), com re-amostragem – “bootstrap”,
1000 repetições (Felsenstein, 1985). As distâncias evolucionárias foram calculadas
com base na distância p e de todas as posições que continham lacunas de
alinhamento (gaps) e dados perdidos foram eliminados apenas com a utilização da
opção Pairwise deletion. O aminoácido número 1 foi definido como o primeiro
aminoácido codificante. Os nucleotídeos foram numerados, utilizando-se a cepa
A/Califórnia/07/2009 como referência. A sequência inicia-se no primeiro codon
codificante e é composta de 566 aminoácidos.
4.5.3 Pirosequenciamento do gene NA para a detecção da mutação H274Y
Reação em Cadeia da Polimerase precedida de transcrição reversa (Pyro - RT-PCR)

71
Nesta reação foi utilizada a Superscript III HiFi Taq (Invitrogen) para
amplificação do H1N1pdm-N1 fragmento C, de acordo com o protocolo
disponibilizado pela OMS através do link
<http://www.who.int/csr/resources/publications/swineflu/NA_DetailedPyrosequencing
_20090513.pdf.> e (DEYDE et al., 2010).
Primers utilizados na Pyro- RT- PCR (20µM) que têm como gene alvo o vírus
(H1N1)pdm09 -N1 – fragmento C (marcador de resistência):
_________________________________________________________________ Uni-sw- N1-B-F780 5’- GGG GAA GAT TGT YAA ATC AGT YGA – 3’
Uni-sw-N1-B-R1273-biotinilado 5’- CWA CCCA GAA RCA AGG YCT TAT G - 3’
__________________________________________________________________________
À mistura contendo 25 µLde 2x Reaction Mix, 1 µL da enzima Super Script III
HiFi Taq, 1 µL do iniciador direto (20 µM), 1 µL do iniciador reverso biotinilado (20
µM), 0,5 µL do inibidor de RNAse e 16,5 µL de água livre de nucleases, foram
adicionados 5 µL do RNAv extraído, totalizando um volume final de 50 µL.
Em seguida a mistura com o RNAv foi para o termociclador, onde estava
programada a seguinte ciclagem: 50°C por 30 minutos para a transcrição reversa e a
PCR, 94°C por 2 minutos para desnaturação, 45 ciclos de (94°C por 15 segundos
para desnaturação, 55°C por 30 segundos para anelamento e 68°C por 60 segundos
para extensão), depois 68°C por 5 minutos para extensão final e por fim 4°C.
Purificação do produto de PCR e Reação de pirosequenciamento
Os produtos da RT-PCR foram analisados por meio de eletroforese utilizando-
se o E-Gel® 2% agarose (GP) da Invitrogen, conforme instrução do fabricante. Os
que apresentaram a banda de interesse, ou seja, os que presumidamente tinham a
fita biotinilada, foram purificados.
Reação de ligação - Foi colocado numa placa 60 µL de uma mistura de beads de
estreptavidina, com tampão de ligação (QIAGEN) e água e mais 20 µL de produto
biotinilado. A placa foi para o vortex por 10 minutos para que houvesse a ligação da
biotina do produto amplificado com a estreptavidina. Depois a placa foi colocada na
workstation e o suporte com as sondas foi introduzido na mesma, aspirando através

72
de vácuo, toda a mistura. Em seguida as sondas contendo as fitas biotiniladas foram
lavadas com álcool a 70% que foi aspirado por 20 segundos, com uma solução
desnaturante (NaOH) para separação das fitas não biotiniladas, aspirada por 20
segundos, e com um tampão de lavagem para terminar a purificação do produto que
foi aspirado também por 20 segundos.
Reação de anelamento – Numa placa do pirosequenciador foi colocado 40 µL do
Iniciador do Pirosequenciamento (100µM) diluído com tampão de anelamento
(QIAGEN), cujo alvo é o resíduo H274:
___________________________________________________________________
Uni-sw - N1-B-F804 seq, 5’ - GYT GAA TGC MCC TAA TT – 3’
Em seguida foram colocadas as beads de estreptavidina. A placa foi para o
vortex por 20 segundos e em seguida para o termobloco onde permaneceu por 2
minutos a 80°C. Depois ficou na temperatura ambiente por 5 minutos. Enquanto isso
foi preparado o cartucho do PyroMark Q96 ID, com os seguintes reagentes: guanina,
citosina, timina, substrato, adenina e enzima, de acordo com a posição
recomendada no protocolo. Para análise quantitativa da mutação H274Y, a alteração de codon CAC →
TAC foi investigada. A corrida foi realizada no modo SNP (single nucleotide
polymorphism) para detecção da alteração destacada em negrito. Através desta
técnica, qualquer amostra que apresente uma porcentagem menor do que 10 % de
T (mutante) é considerado selvagem, se apresentar um resultado maior que 10% é
considerado uma mistura verdadeira de variantes selvagem e mutantes, devendo
ser confirmado o ensaio (SOUZA et al., 2011).
4.5.4 Diagnóstico diferencial de influenza A através de PCR em tempo real (rRT – PCR multiplex)
A primeira etapa do diagnóstico diferencial consistiu na extração do ácido
nucléico que foi realizada por um sistema automatizado da Roche, seguida de uma
reação PCR em tempo real precedida de transcrição reversa (RT-PCR) em um único
tubo. Foi utilizado o kit Fast track21 plus®), capaz de detectar 23 patógenos
respiratórios, cujas informações podem ser obtidas através do link <

73
http://www.mikrogen.de/english/deutschland/products/testing-ystems/testsystem/respiratory-
21-plus-24.html>.
Reação de rRT- PCR multiplex utilizando o kit Fast track21 plus®
O kit Fast track21 plus® é constituído de seis misturas de iniciadores e
sondas para a detecção de patógenos respiratórios e dos seguintes controles:
interno, negativo, positivo para vírus e positivo para bactérias. Os demais reativos
para a reação de RT-PCR fazem parte do kit AgPath- ID ™ One Step RT-PCR
(Applied Biosystems).
Tubos Contém
Flu/ Rhino PP
Mistura de iniciador/sonda para FLUA,FLU B, FLUA (H1N1)pdm09 e RV
COR PP Mistura de iniciador/sonda para HCoV63, Cor43, Cor229 e CorKHU1
Para2/3/4 PP
Mistura de iniciador/sonda para Para 2, 3, 4 e IC (controle interno)
Bo/MP/Pf1 PP
Mistura de iniciador/sonda para HMPV A e B, HboV e Mycoplasma pneumoniae (Mpneu).
RS/ EPA PP Mistura de iniciador/sonda para RSVA, RSV A RSV B, AdV, EV e PEV
RespBac PP Mistura de iniciador/sonda para Chlamydia pneumoniae (Cpneu), Streptococcus pneumoniae (Spneu), Staphylococcus aureus (Saur) e Haemophilus influenzae (HiB)
PC Controle positivo (mistura de plasmídeo para Flu A, B, Flu A (H1N1)pdm09, RV, Cor63, Cor43, Cor229, CorKHU1, Para 1, 2, 3, 4, HMPV, HboV, Mpneu, RSV, ADV, EV e PEV)
PC Res Bac
Controle positivo: mistura de plasmídeo para Res Bac PP ( mistura de plasmídeos para Cpneu, Spneu, Saur e HiB)
NC Controle negativo (em agente desnaturante)
IC Controle interno (em agente desnaturante) Neste ensaio, o RNA viral é transcrito em cDNA usando um iniciador
específico mediado por uma etapa de transcrição reversa seguida imediatamente
por reação em cadeia da polimerase em tempo real. Inicialmente ocorre a
desnaturação do DNA, seguida da etapa de anelamento, onde os iniciadores e as
sondas hibridizam-se especificamente ao alvo. Na fase de extensão, as sondas
específicas para cada vírus e duplamente marcadas com fluoróforos são clivadas,
graças a atividade nuclease da enzima Taq polimerase e emitem sinal fluorescente.
Esse sinal fluorescente é revelado pelo termociclador em tempo real através do Ct
(ciclo no qual a fluorescência emitida atravessa o ponto de corte do teste). O ponto

74
de corte do teste é denominado threshold e deve ser ajustado na fase exponencial
da reação. As amostras positivas apresentam sinal de fluorescência emitido acima
do ponto de corte. Quanto menor o valor de Ct, maior a concentração das moléculas
alvo na amostra.
No procedimento técnico, inicialmente foi preparada a mistura de reação,
contendo1,5 µL de cada mistura de iniciadores e sondas, 12,5 µL de tampão 2X RT-
PCR e 1µL da mix de enzima 25X RT-PCR ( kit AgPath-ID™ One step RT-PCR).
Em seguida foram colocados 15 µL da mistura de reação nos devidos poços da
placa, e adicionados 10 µL de ácidos nucléicos. Foram utilizados controle positivo e
controle negativo do kit. A placa foi coberta com adesivo óptico e centrifugada por 1
minuto a 1.000 rpm. Em seguida foi colocada no equipamento de PCR em tempo
real ABI7500 e executado o programa Fast track contemplando os seguintes ciclos:
50°C por 15 minutos para a transcrição reversa, 95°C por 10 minutos para ativação
enzimática e 40 ciclos de 95°C por 8 segundos e 60°C por 34 segundos para
amplificação. A leitura da fluorescência é feita no passo 60ºC por 34 segundos de
cada ciclo. Para cada amostra, foi verificada a forma das curvas de amplificação
(logarítmica), para cada vírus. E o valor de cada Ct.
4.6 CONSIDERAÇÕES ÉTICAS
O estudo foi aprovado pelo Comitê de Ética do Centro de Ciências da Saúde
da Universidade Federal de Pernambuco, registro CEP/CCS/UFPE nº 219/10 (Anexo
A). Foram obtidas cartas de anuência da Diretoria Geral de Vigilância
Epidemiológica e Ambiental da Secretaria de Saúde de Pernambuco e do
Laboratório Central de Saúde Pública - LACEN-PE (Anexos B e C). Não foi
necessário o Termo de consentimento livre e esclarecido.
4.7 ANÁLISE ESTATÍSTICA
Na busca de associações entre as variáveis foi utilizado o teste Qui-quadrado
de Pearson com correção de Yates, e quando necessário o teste de Fisher. Para as
variáveis contínuas foi utilizado o teste T de Student. O nível de significância
adotado na análise dos testes estatísticos foi 5% (p<0,005) e o software utilizado foi
o STATA na versão 9.0.

75
5 RESULTADOS
5.1 ARTIGO 1
CARACTERIZAÇÃO GENÉTICA DOS VÍRUS INFLUENZA A (H1N1)pdm09 DETECTADOS NO ESTADO DE PERNAMBUCO, BRASIL, NO PERÍODO DE MAIO DE 2009 A MAIO DE 2010
Artigo a ser submetido à Revista Memórias do Instituto Oswaldo Cruz

76
CARACTERIZAÇÃO GENÉTICA DOS VÍRUS INFLUENZA A (H1N1)pdm09
DETECTADOS NO ESTADO DE PERNAMBUCO, BRASIL, NO PERÍODO DE MAIO DE 2009 A MAIO DE 2010
Maria José Couto Oliveira 1,2, Fernando do Couto Mota 3, Marilda M Siqueira 3, Maria
de Lourdes Aguiar Oliveira3, Thiago Moreno Lopes e Souza 3, Milena Mesquita 3,
Priscilla da Silva Born 3, Paola Cristina Resende Silva 3, Wyller Alencar de Mello 4,
Vera Magalhães da Silveira 2.
1 Laboratório Central de Pernambuco (LACEN-PE) Setor de Virologia, Recife,
Pernambuco, Brasil 2 Programa de Pós-graduação de Medicina Tropical, Universidade Federal de
Pernambuco, Recife, Pernambuco, Brasil. 3 Laboratório de Vírus Respiratórios e do Sarampo do Instituto Oswaldo Cruz,
Fiocruz, Rio de Janeiro, Brasil. 4 Instituto Evandro Chagas, Belém, Pará, Brasil.
Endereço para correspondência:
Maria José Couto Oliveira
Rua do Espinheiro, 360, Apto. 1801 – Espinheiro
Recife – PE, CEP 52020-020
e-mail: [email protected]

77
RESUMO
O vírus Influenza A(H1N1)pdm09 causou considerável morbidade e mortalidade no
mundo. Apesar do fim da pandemia de 2009, o vírus ainda circula na população
humana. O objetivo deste estudo foi investigar a frequência de resistência ao
oseltamivir, realizar a caracterização molecular dos vírus influenza A(H1N1)pdm09
circulantes no estado de Pernambuco e compará-los com outras amostras
representativas do Brasil e do mundo, coletadas durante o período da pandemia.
Foi realizado um estudo transversal entre casos de indivíduos com suspeita de
influenza pandêmica, atendidos nos hospitais de Pernambuco. Um total de 118
amostras foi investigado por pirosequenciamento para verificar a presença do
marcador de resistência antiviral H274Y, no gene da neuraminidase (NA). Além
disso, o gene da hemaglutinina (HA) de 31 amostras foi sequenciado pelo método
de Sanger et al. (1974) para verificar a presença de mutações. Os resultados do
pirosequenciamento demonstraram que nenhuma das amostras investigadas
possuía o marcador de resistência ao oseltamivir H274Y. Análises filogenéticas
mostraram alta similaridade entre as amostras de Pernambuco e aquelas que
circularam em outras regiões. Corroborando com outros achados, a cocirculação de
distintas linhagens de influenza A(H1N1)pdm09 foi demonstrada e todos os
agrupamentos (clusters) da árvore filogenética estavam estreitamente relacionados
à cepa protótipo da vacina A/Califórnia/07/09.
Palavras chaves: Influenza A(H1N1)pdm09, resistência ao oseltamivir, mutações,
hemaglutinina.

78
ABSTRACT
Worldwide, the influenza A(H1N1)pdm09 virus caused considerable morbidity and
mortality. Despite the end of 2009 pandemic, the virus still circulates within the
human population. The aim of this study was to investigate the frequency of
oseltamivir resistance and the molecular characterization of circulating influenza
A(H1N1)pdm09 viruses from Pernambuco State (PE), in comparison to other
Brazilian and worldwide representatives samples collected in the course of the
pandemic. A retrospective cross-sectional study was conducted among suspected
cases of pandemic influenza attended at PE reference hospitals. A total of 118
respiratory samples were submitted to the identification of the NA mutation H274Y, a
marker of antiviral resistance by pyrosequencing. Furthermore, HA gene of 31
samples was sequenced by the Sanger method in order to identify important
signature residues and unique mutations. Pyrosequencing results demonstrated that
none of the investigated samples bear the oseltamivir resistance mutation H274Y in
NA gene. Finally phylogenetic analysis showed a high similarity between PE samples
and those that circulated elsewhere. Corroborating previous findings, the co-
circulation of distinct lineages of influenza A(H1N1)pdm09 was demonstrated and all
clusters of phylogenetic tree were close related to the prototype vaccine strain
A/California/07/09.
Key words: Influenza A(H1N1)pdm09, oseltamivir resistance, hemagglutinin
mutations.

79
Em abril de 2009, a Organização Mundial da Saúde (OMS) notificou a
ocorrência de casos humanos de influenza que ocorreram no México e nos Estados
Unidos da América tendo sido identificado um vírus Influenza A subtipo H1N1 de
origem suína. Dentro de poucos meses o vírus circulou por todo o mundo, sendo
responsável pela primeira pandemia do século XXI e por mais de 18.000 mortes
(WHO 2010d). Os primeiros casos do vírus influenza A(H1N1)pdm09 no Brasil foram
confirmados em 7 de maio de 2009 e dois dias depois foi relatada a transmissão
autóctone desse vírus no país. No estado de Pernambuco, região nordeste do Brasil,
o primeiro caso foi confirmado em junho de 2009.
Os vírus influenza são conhecidos por suas altas taxas de mutação
principalmente nos genes da hemaglutinina (HA) e da neuraminidase (NA), gerando
cepas variantes com características antigênicas distintas em relação as
propriedades virais, tais como especificidade de ligação aos receptores sializados,
virulência ou patogenicidade (Arias et al. 2009). O gene HA apresenta a mais alta
taxa de mutação e a proteína que ele codifica está diretamente associada aos
primeiros passos do sucesso da infecção viral (Wagner et al. 2002). Muitos autores
têm monitorado as mutações no gene HA dos vírus influenza A(H1N1)pdm09, entre
as quais a mutação D222G que tem sido associada a casos graves ou fatais,
possivelmente devido a mudança na especificidade de ligação do vírus com o ácido
siálico α2-6 para o α2-3, mais predominantemente encontrado no trato respiratório
inferior de humanos (Liu et al. 2010, Zheng et al. 2010). Essa mutação foi detectada
em 18% dos casos graves e fatais na Noruega (Kilander 2010) em 8,7% na Escócia
(Miller et al. 2010), 5,12% na Espanha (Ledesma 2011), 4,1% em Hong Kong (Mak
et al. 2010), 1,4% na Finlândia (Ikonen et al. 2010).
Mutações no gene NA podem levar a perda de sensibilidade à terapia anti-
influenza utilizada no momento, baseada na inibição da atividade neuraminidásica
viral (Abed et al. 2006, Aoki et al. 2007). Estudos anteriores à pandemia de 2009
mostraram resistência do vírus influenza A (H1N1) sazonal humano ao oseltamivir
em vários países (Dharan et al. 2009; Eshaghi et al. 2009; Moscona et al. 2005).
Com a introdução do novo vírus influenza A(H1N1)pdm09 rearranjado na população
mundial, do amplo uso dos antivirais e os altos índices de resistência historicamente
detectados, tornou-se primordial a avaliação dos níveis de resistência nos vírus
emergentes. Consequentemente, muitos grupos iniciaram estudos a fim de

80
identificar mutações importantes, seguir a evolução dos principais genes e
determinar a antigenicidade e susceptibilidade do novo vírus ao oseltamivir.
Diante do impacto do vírus influenza A(H1N1)pdm09 e também das
peculiaridades da circulação de vírus respiratórios em diferentes regiões do país,
este estudo visa investigar a frequência de resistência ao oseltamivir, realizar a
caracterização molecular dos vírus influenza A(H1N1)pdm09 circulantes no estado
de Pernambuco e compará-los com outras amostras representativas do Brasil e do
mundo, coletadas durante o período da pandemia. Após a coleta, as amostras de
casos suspeitos de infecção pelo vírus influenza A(H1N1)pdm09 (triplo swabs
combinados e fragmentos de tecidos de pacientes que foram a óbito), foram
colocadas em meio de transporte, congeladas no Laboratório Central de
Pernambuco e posteriormente enviadas ao Instituto Evandro Chagas no estado do
Pará para realização do diagnóstico por RT-PCR em tempo real, de acordo com o
protocolo do CDC (Atlanta, USA), disponível em:
<http://www.who.int/csr/resources/publications/swineflu/CDCRealtimeRTPCR_Swine
H1Assay-2009_20090430.pdf>. As amostras positivas para o vírus influenza
A(H1N1)pdm09 foram encaminhadas para o Laboratório de Vírus Respiratórios e
Sarampo da Fiocruz, para realização deste estudo.
Um total de 118 amostras positivas de influenza A(H1N1)pdm09 foram
pirosequenciadas a fim de verificar a presença da mutação H274Y no gene NA, de
acordo com Deyde et al.(2010). O RNA foi extraído diretamente de 140 μL das
amostras clínicas, usando o kit QIAamp® Viral RNA Mini Kit (Qiagen, Germany), de
acordo com a orientação do fabricante. O volume final da eluição foi 80 μL. A
amplificação do fragmento C influenza A(H1N1)pdm09-N1 foi realizada usando a
SuperScript III One-step RT-PCR with HiFi Taq system (Invitrogen) de acordo com o
protocolo do CDC, disponível em <
http://www.who.int/csr/resources/publications/swineflu/NA_DetailedPyrosequencing_
20090513.pdf>. Após purificação, os amplicons foram analisados no Pyromark Q-96
ID (Qiagen, Germany), usando o modo SNP (single nucleotide polymorphism). Para
análise quantitativa da mutação H274Y no gene HA a alteração de codon CAC →
TAC foi investigada por pirosequenciamento. Através desta técnica, toda amostra
que apresente uma porcentagem menor do que 10 % de “T “(mutante) na primeira
posição do codon é considerada selvagem e consequentemente sensível ao
tratamento com oseltamivir”. Por outro lado, todos os valores acima de 10% são

81
considerados uma mistura de tipos selvagem e mutante, com uma sensibilidade
reduzida à droga (Souza et al. 2011).
Foi realizada a caracterização genética de 31 amostras representativas de
distintas fases da pandemia (início, meio e fim). O gene da HA, foi amplificado por
RT-PCR, utilizando-se SuperScript IIITM One-step RT-PCR with Platinum Taq-DNA
Polymerase (Invitrogen, USA) e foi sequenciado de acordo com protocolo da OMS
para influenza, disponibilizado em
http://www.who.int/csr/resources/publications/swineflu/GenomePrimers_20090512.p
df. Após purificação (Motta et al. 2006) os produtos foram sequenciados usando o
ABI Prism Big Dye Terminator Cycle Sequencing Ready Reaction (Applied
Biosystems, USA) com o ABI 3130 XL capillary automatic DNA analyzer (Applied
Biosystems). Todas as sequências do gene HA foram editadas usando o software
Sequencher v.4.10 (Gene Codes, Ann Arbor, USA) e alinhadas com o protótipo da
vacina A/California/07/2009 e outras sequências disponíveis no GenBank
(www.ncbi.nlm.nih.gov), usando o módulo Clustal W do pacote Molecular
Evolutionary Genetics Analyses (MEGA), version 5.0.5 (Tamura et al. 2011). As
sequências das 31 amostras obtidas neste trabalho estão disponíveis no GenBank,
códigos de acesso CY103911-CY103941.
O marcador de resistência da mutação H274Y não foi detectado em nenhuma
das 118 amostras avaliadas no estudo, estando de acordo com dados anteriormente
obtidos, sugerindo uma baixa frequência de resistência dos vírus influenza
A(H1N1)pdm09 ao oseltamivir, quando comparada com amostras sazonais (Souza
et al. 2010; Malato et al. 2011; Chen et al. 2011; Harvala et al. 2011 e Kawano et al.
2011).
Vinte e uma das 31 sequências de HA obtidas através do sequenciamento
automático foram de amostras de pacientes hospitalizados e dois deles evoluíram
para óbito. As sequências apresentaram várias mutações quando comparadas com
a A/California/07/2009 e estão listadas na Tabela I. No total, catorze mutações foram
identificadas das quais P100S e I338V foram encontradas em todas as sequências
avaliadas. A mutação S220T foi detectada em 23 amostras, sendo que a maioria (15
amostras) foi encontrada em pacientes hospitalizados, enquanto que de nove
mutações Q310H detectadas, seis pertenciam a esse subgrupo. Vale ressaltar que
enquanto a mutação D114N foi mais frequentemente encontrada em pacientes

82
ambulatoriais, as mutações N277K, K325E, Q310R, Y368F e I550L foram
detectadas apenas em pacientes que necessitaram de hospitalização.
Com o objetivo de detectar uma possível associação entre o quadro clínico do
paciente e um fundo genético, buscamos no gene HA, os marcadores de virulência
D222G (WHO, 2009) e Q310H (Glinsky et al. 2010). O primeiro não foi detectado
neste estudo e o segundo foi detectado em proporções similares nos dois grupos
(28,6% e 30,0% nos pacientes ambulatoriais e hospitalizados, respectivamente), não
sendo possível associar essa mutação com os casos graves e fatais. Estes
resultados corroboram com os resultados obtidos anteriormente com amostras
brasileiras, indianas e canadenses (Lee et al. 2010; Potdar et al. 2010; Graham et al.
2011). As mutações P100S e I338V foram identificadas em todas as amostras dos
dois grupos. Particularmente a mutação P100S tem sido associada com uma
excreção viral prolongada, sendo possível recuperar os vírus de pacientes durante
um período de até dois meses (Souza et al. 2010).
As relações filogenéticas entre as amostras de Pernambuco e outras
amostras brasileiras isoladas recentemente podem ser vistas na Figura I. Amostras
de Pernambuco foram agrupados em dois grandes “clusters” (um principal e um
menor), agrupadas junto com outras amostras brasileiras, coletadas no mesmo
período, de outros estados distantes de Pernambuco, como Santa Catarina ou Rio
Grande do Sul, no sul do Brasil. Os dois grupos são constituídos de alguns
subgrupos menores, a maioria deles caracterizada por mudança de um único
aminoácido. Todas as amostras são altamente similares ao protótipo vacinal, em
concordância com outras publicações (Ikonen et al. 2010; Melidou et al. 2010; Barr
et al. 2010). Notavelmente, a maioria das amostras de Pernambuco apresentou a
mutação S220T, mas somente sete amostras eram similares ao protótipo da vacina
para tal resíduo (Figura 1, grupamento inferior). Do contrário, todas as amostras do
grupo inferior apresentaram a mutação Q310 H/R, sendo que só duas amostras que
possuíam essa mutação faziam parte do grupo principal superior:
A/Pernambuco/175/09 e A/Pernambuco/414/09. Em relação aos subgrupos, aqueles
caracterizados pelas mutações D114N, P199Q e K325E foram construídos
basicamente com amostras de Pernambuco (incluindo a mais antiga amostra
enraizando cada subgrupo) as quais poderiam indicar pelo menos uma cadeia de
transmissão Figura 1).

83
Neste estudo não detectamos a mutação H274Y por pirosequenciamento,
indicando que todas as amostras eram sensíveis ao oseltamivir. Foi descrito o perfil
genético do vírus influenza A(H1N1)pdm09 coletados no estado de Pernambuco no
período de 2009-2010. Não houve diferença significativa entre as sequências do
gene HA das amostras de Pernambuco com sequências de amostras de outras
regiões brasileiras - amostras de diferentes regiões foram agrupadas juntas. Esses
subgrupos regionais formados por amostras coletadas entre julho/2009 e abril/2010,
são indicativos da cocirculação de linhagens, o que não só é esperado, mas foi
anteriormente observado (Galiano et al. 2011).
Apesar da declaração do final da pandemia pela OMS, o vírus continua
circulando globalmente, provocando, algumas vezes, óbitos. O monitoramento
contínuo da evolução viral e da resistência ao oseltamivir, são primordiais para a
vigilância da gripe em todo o mundo, e os resultados aqui descritos contribuem para
uma melhor compreensão das características genéticas dos vírus influenza
A(H1N1)pdm09 que circularam no nordeste do Brasil, durante o período pandêmico.
Agradecimentos:
À Dra. Marilda Siqueira por disponibilizar o Laboratório de Vírus Respiratórios
e do Sarampo do IOC-FIOCRUZ – Rio de Janeiro, para realização deste estudo.
Suporte Financeiro:
Instituto Oswaldo Cruz /Fundação Oswaldo Cruz.
REFERÊNCIAS: Abed Y, Baz M, Boivin G 2006. Impact of neuraminidase mutations conferring influenza resistance to neuraminidase inhibitors in the N1 and N2 genetic backgrounds. Antivir Ther 11: 971-976. Adwan G 2010. Molecular characterization and phylogenetic analysis of Middle East 2009 H1N1 pdm isolates. A PJTM: 624-628. Aoki FY, Boivin G, Roberts N 2007. Influenza virus susceptibility and resistance to oseltamivir. Antivir Ther 12: 603-616.

84
Arias CF, Escalera-Zammudio M, Soto-Del Rio, Cóbian-Güemes AG, Isa P, López S 2009. Molecular Anatomy of 2009 Influenza Vírus A (H1N1). Arch Med Res 40: 643-654. Barr IG, Cui L, Komandina N, Lee RT, Lin RT, Deng Y, Caldwell N, Shaw R, Maurer-Stroh S 2010. A new pandemic influenza A(H1N1) genetic variant predominated in the winter 2010 influenza season in Australia, New Zealand and Singapore. Euro Surveill 15 (42): pii = 19692. Chen LF, Dailey NJ, Rao AK, Fleischauer AT, Greenwald I, Deyde VM, Moore ZS, Anderson DJ, Duffy J, Gubareva LV, Sexton DJ, Fry AM, Srinivasan A, Wolfe CR 2011. Cluster of Oseltamivir-Resistant 2009 Pandemic Influenza A (H1N1) Vírus Infections on a Hospital Ward among Immunocompromised Patients-- North Carolina, 2009. J. Infect Dis 203: 838-846. Dharan NJ, Gubareva LV, Meyer JJ, Okomo-Adhiambo M, McClinton RC, Marshall SA , St George K, Epperson S, Drammer L, Klimov AI, Bresee JS, Fry AM 2009. Infections With Oseltamivir-Resistant Influenza A (H1N1) Virus in the United States. JAMA 301:1034-1041. Deyde VM, Okomo-Adhiambo M, Sheu TG, Wallis TR, Fry A, Dharan N, Klimov AI, Gubareva LV 2009. Pyrosequencing as a tool to detect molecular markers of resistance to neuraminidase inibitors in seasonal influenza A viruses. Antiviral Research 81: 16-24. Eshaghi A, Bolotin S, Burton L, Low DE, Mazzuli T, Drews SJ 2009. Genetic microheterogeneity of emerging H275Y influenza virus A (H1N1) in Toronto, Ontario, Canadá from 2007 – 2008 respiratory season. J Clin Virol 45: 142–145. Felsenstein J 1985. Confidence limits on phylogenies: Na approach using the bootstrap. Evolution 39: 783-791. Ferreira JLP, Borborema SET, Brígido LFM, Oliveira MI, Paiva TM, Santos CLS 2011. Sequence analysis of the 2009 pandemic influenza A H1N1 virus haemagglutinin gene from 2009-2010 Brazilian clinical samples. Mem Inst Oswaldo Cruz 106: 613-616. Galiano M, Agapow P-M, Thompson C, Platt S, Underwood A, Ellis J, Myers R, Green J, Zambon M 2011. Evolutionary Pathways of the Pandemic Influenza A (H1N1) 2009 in the UK. PLoS ONE 6(8): e23779 doi: 10.1371. Glinsky GV 2010. Genomic analysis of pandemic (H1N1) 2009 reveals association of increasing disease severity with emergence of novel hemagglutinin mutations. Cell Cycle 9: 958-970. Graham M, Liang B, Domselaar GV, Bastien N, Beaudoin C, Tyler S, Kaplen B, the National Influenza A/H1N1pdm Genomics Study Team (NIGST), Li Y. 2011. Nationwide Molecular Surveillance of Pandemic H1N1 Influenza A Virus Genomes: Canada, 2009. PLoS One 6: e16087 doi: 10.1371.

85
Harvala H, Gunson R, Simmonds P, Hardie A, Bennett S, Scott F, Roddie H, McKnight J, Walsh T, Rowney D, Clark A, Bremner J, Aitken C, Templeton K 2010. The Emergence Of Oseltamivir-Resistant Pandemic Influenza A(H1n1) 2009 Virus Amongst Hospitalised Immunocompromised Patients In Scotland, November-December, 2009. Euro Surveil 15: pii = 19536. Ikonen N, Haanpää M, Rönkkö E, Lyytikäinen O, Kuusi M, Ruutu P, Kallio-Kokko H, Mannonen L, Lappalainen M, Ziegler T, Julkunen I 2010. Genetic Diversity of the 2009 pandemic influenza A (H1N1) viruses in Finland. PLoS One 20: e13329 doi: 10.1371. Felsenstein J 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39:783-791.
Kawano H, Haruyama T, Hayashi Y, Sinoda Y, Sonoda M, Kobayashi N 2011. Genetic Analysis and Phylogenetic Caracterization of Pandemic (H1N1) 2009 Influenza Viruses that found in Nagasaki. Jpn J Infect Dis 63: 195-203. Kilander A, Rykkvin R, Dudman S. Hungnes O 2010. Observed association between the HA1 mutation D222G in the 2009 pandemic influenza a(H1N1) virus and severe clinical outcome, Norway 2009-2010. Euro Surveill 15 (9): pii = 19498. Ledesma J, Pozo F, Ruiz MP, Navarro JM, Piñero L, Montes M, Castro SP, Fernández JS, Costa JG, Fernández M, Galán JC, Cuevas MT, Casa I, Breña PP 2011. Substitutions in position 222 of haemagglutinin of pandemic influenza A (H1N1) 2009 viruses in Spain. J Clin Virol 51: 75-78. Lee RT, Santos CL, de Paiva TM, Cui L, Sirota FL, Eisenhaber F, Maurer-Stroh S 2010. All that glitters is not gold - founder effects complicate associations of flu mutations to disease severity. Virol J. 1: 297. Le QM, Wertheim HF, Tran ND, van Doorn HR, Nguyen TH, Horby P, Vietnam H1N1 Investigation Team 2010. A Community Cluster of Oseltamivir-Resistant Cases of 2009 H1N1 Influenza. N Engl J Med 362: 86-87. Liu Y, Childs RA, Matrosovich T, Wharton S, Palma AS, Chai W, Daniels R, Gregory V, Uhlendorff J, Kiso M, Klenk H-D, Hay A, Feizi T, Matrosovich M. Altered Receptor Specificity and Cell Tropismo f D222G Hemagglutinin Mutants Isolated from Fatal Cases of Pandemic A(H1N1) 2009 Influenza Vírus. J Virol 2010 84: 12069-74. Mak GC, Au KW, Tai LS, Chuang KC, Cheng KC, Shiu TC, Lim W. 2010. Association of D222G Substitution in Haemagglutinin of 2009 Pandemic Influenza A (H1N1) with Severe Disease. Euro Surveill 15 (14): pii =19534. Malato L, Llavador V, Marmier E, Youssef J, Balick Weber C, Rozé H, Bessede E, Fleury HJ 2011. Pandemic influenza A (H1N1)2009: molecular characterization and duration of viral shedding in intensive care patients in Bordeaux, south-west France, May 2009 to January 2010. Euro Surveill 16 (4) : pii = 19776.

86
Maurer-Stroh S, Lee RTC, Eisenhaber F, Cui L, Phuah SP, Lin RTP 2010. A new common mutation in the hemagglutinin of the 2009 (H1N1) influenza A virus. PLoS Curr 2: RRN1162. Melidou A, Gioula G, Exindari M, Chatzidimitriou D, Diza E, Malisiovas N 2010 Molecular and phylogenetic analysis of the haemagglutinin gene of pandemic influenza H1N1 2009 viruses associated with severe and fatal infections. Virus Res 151: 192-199. Miller RS, MacLean AR, Gunson RN, Carman WF 2010. Occurrence of Haemagglutinin Mutation D222G in Pandemic Influenza A(H1N1) Infected Patients in the West of Scotland, United Kingdom, 2009-10. Euro Surveill 15 (14) pii = 19546. Morlighem JE, Aoki S, Kishina M, Hanami M, Ogawa C, Jalloh A, Takahashi Y, Kaway Y, Saga S, Hayashi E, Ban T, Izumi S, Wada A, Mano M, Fukunaga M, Kijima Y, Shiomi M, Inoue K, Hata T, Koretsune Y, Kudo K, Himeno Y, Hirai A, Takahashi K, Sakai-Tagawa Y, Iwatsuki-Horimoto K, Kawaoka Y, Hayashizaki Y, Ishikawa T 2011. Mutation Analysis of 2009 Pandemic Influenza A (H1N1) Viruses Collected in Japan during the Peak Phase of the Pandemic. PLoS One Curr 6: e18956 PMCID: PMC3084724. Moscona A 2005. Global Transmission of Oseltamivir-Resistant Influenza. The N Engl J Med 360: 953-956.
Motta FC, Siqueira MM, Lugon AK, Straliotto SM, Fernandes SB, Krawczuk MM 2006. The reappearance of Victoria lineage influenza B virus in Brazil, antigenic and molecular analysis. J Clin Virol 36: 208-214.
Potdar VA, Chadha MS, Jadhav SM, Mullick J, Cherian SS, Mishra AC 2010. Genetic Characterization of the Influenza A Pandemic (H1N1) 2009 Virus Isolates from India. PLoS One Curr 5: e9693, PMCID: PMC 2837743. Saitou N, Nei M 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406-425. Sanger F, Donelson JE, Coulson AE, Kössel H, Fischer D 1974.Determination of a nucleotide sequence in bacteriophage f1 DNA by primed synthesis with DNA polymerase. J Mol Biol 90: 315-326.
Souza TML, Salluh JIF, Bozza FA, Mesquita M, Soares M, Motta FC, Pitrowsky MT, Oliveira ML, Mishin VP, Gubareva LV, Whitney A, Rocco AS, Gonçalves VMC, Marques VP, Velasco E, Siqueira MM 2010. H1N1pdm influenza Infection in Hospitalized Cancer Patients: Clinical Evolution and Viral Analysis. PLoS ONE 5: e14158 doi:10.1371. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S 2011. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood Evolutionary Distance and Maximum Parsimony Methods. Mol Biol Evol 28: 2731-2739.

87
Wagner R, Matrosovich M, Klenk HD 2002. Functional balance between haemagglutinin and neuraminidase in influenza virus infections. Rev Med Virol. 12:159-66. World Health Organization. Influenza A (H1N1) virus resistance to oseltamivir - 2008-2009 influenza season northern hemisphere. 18 March 2009. World Health Organization. Pandemic (H1N1) 2009 – update 112. 6 August 2010. Available from: <http://www.who.int/csr/don/2010_08_06/en/index.html>.
Zheng B, Chan KH, Zhang AJ, Zhou J, Chan CC, Poon VK, Zhang K, Leung VH, Jin DY, Woo PC, Chan JF, To KK, Chen H, Yuen KY 2010. D225G mutation in hemagglutinin of pandemic influenza H1N1 (2009) virus enhances virulence in mice. Exp Biol Med 235: 981-988. Tabela I - Mutações de aminoácidos no gene HA observadas em pacientes de ambulatório e hospitalizados.
Pacientes Resíduo Ambulatório
(n=10) Hospitalizados
(n=21)
Total (n=31)
S88F - 01 1* S91N 01 02 3
P100S 10 21 31 S101N 01 02 2 D114N 03 02 5 P199Q 01 06 7 S220T 08 15 23 N277K - 01 1* Q310H 03 06 9 Q310R - 01 1 K325E - 04 4 I338V 10 21 31 Y368F - 01 1 I550L - 01 1
* Óbito
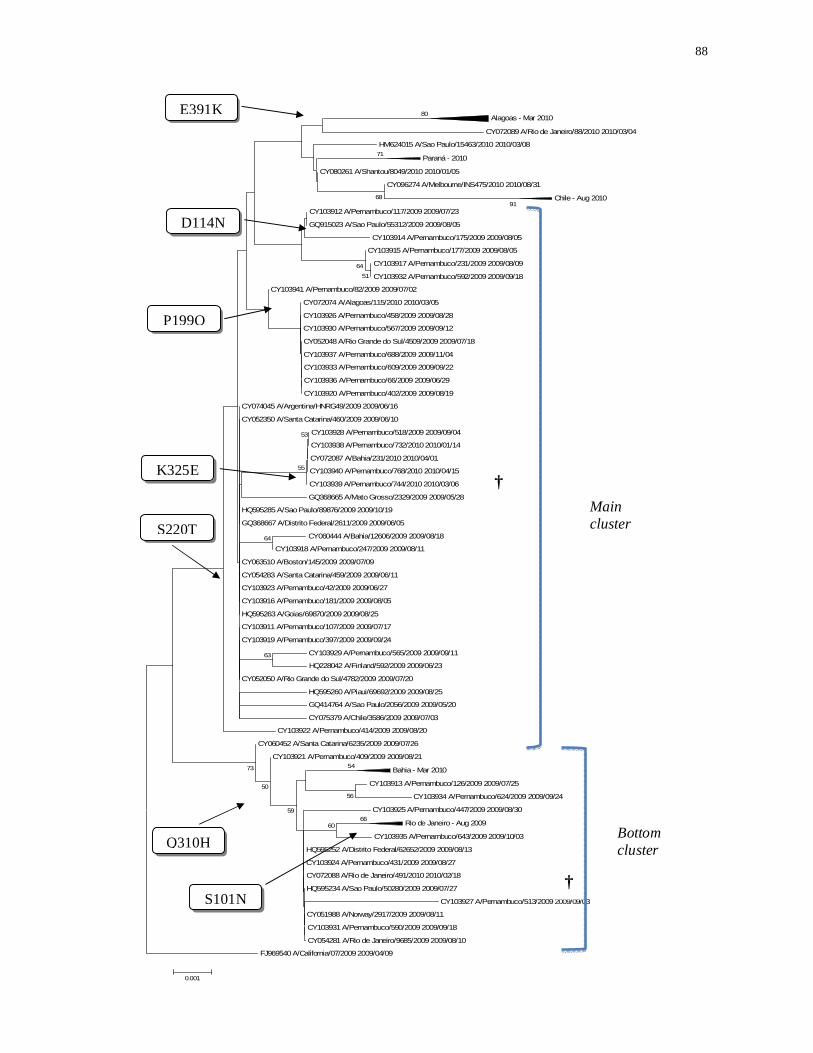
88
Alagoas - Mar 2010
CY072089 A/Rio de Janeiro/88/2010 2010/03/04
HM624015 A/Sao Paulo/15463/2010 2010/03/08
Paraná - 2010
CY080261 A/Shantou/8049/2010 2010/01/05
CY096274 A/Melbourne/INS475/2010 2010/08/31
Chile - Aug 2010
CY103912 A/Pernambuco/117/2009 2009/07/23
GQ915023 A/Sao Paulo/55312/2009 2009/08/05
CY103914 A/Pernambuco/175/2009 2009/08/05
CY103915 A/Pernambuco/177/2009 2009/08/05
CY103917 A/Pernambuco/231/2009 2009/08/09
CY103932 A/Pernambuco/592/2009 2009/09/18
CY103941 A/Pernambuco/82/2009 2009/07/02
CY072074 A/Alagoas/115/2010 2010/03/05
CY103926 A/Pernambuco/458/2009 2009/08/28
CY103930 A/Pernambuco/567/2009 2009/09/12
CY052048 A/Rio Grande do Sul/4509/2009 2009/07/18
CY103937 A/Pernambuco/688/2009 2009/11/04
CY103933 A/Pernambuco/609/2009 2009/09/22
CY103936 A/Pernambuco/66/2009 2009/06/29
CY103920 A/Pernambuco/402/2009 2009/08/19
CY074045 A/Argentina/HNRG49/2009 2009/06/16
CY052350 A/Santa Catarina/460/2009 2009/06/10
CY103928 A/Pernambuco/518/2009 2009/09/04
CY103938 A/Pernambuco/732/2010 2010/01/14
CY072087 A/Bahia/231/2010 2010/04/01
CY103940 A/Pernambuco/768/2010 2010/04/15
CY103939 A/Pernambuco/744/2010 2010/03/06
GQ368665 A/Mato Grosso/2329/2009 2009/05/28
HQ595285 A/Sao Paulo/89876/2009 2009/10/19
GQ368667 A/Distrito Federal/2611/2009 2009/06/05
CY060444 A/Bahia/12606/2009 2009/08/18
CY103918 A/Pernambuco/247/2009 2009/08/11
CY063510 A/Boston/145/2009 2009/07/09
CY054283 A/Santa Catarina/459/2009 2009/06/11
CY103923 A/Pernambuco/42/2009 2009/06/27
CY103916 A/Pernambuco/181/2009 2009/08/05
HQ595263 A/Goias/69870/2009 2009/08/25
CY103911 A/Pernambuco/107/2009 2009/07/17
CY103919 A/Pernambuco/397/2009 2009/09/24
CY103929 A/Pernambuco/565/2009 2009/09/11
HQ228042 A/Finland/592/2009 2009/06/23
CY052050 A/Rio Grande do Sul/4782/2009 2009/07/20
HQ595260 A/Piaui/69692/2009 2009/08/25
GQ414764 A/Sao Paulo/2056/2009 2009/05/20
CY075379 A/Chile/3586/2009 2009/07/03
CY103922 A/Pernambuco/414/2009 2009/08/20
CY060452 A/Santa Catarina/6235/2009 2009/07/26
CY103921 A/Pernambuco/409/2009 2009/08/21
Bahia - Mar 2010
CY103913 A/Pernambuco/126/2009 2009/07/25
CY103934 A/Pernambuco/624/2009 2009/09/24
CY103925 A/Pernambuco/447/2009 2009/08/30
Rio de Janeiro - Aug 2009
CY103935 A/Pernambuco/643/2009 2009/10/03
HQ595252 A/Distrito Federal/62652/2009 2009/08/13
CY103924 A/Pernambuco/431/2009 2009/08/27
CY072088 A/Rio de Janeiro/491/2010 2010/02/18
HQ595234 A/Sao Paulo/50280/2009 2009/07/27
CY103927 A/Pernambuco/513/2009 2009/09/03
CY051988 A/Norway/2917/2009 2009/08/11
CY103931 A/Pernambuco/590/2009 2009/09/18
CY054281 A/Rio de Janeiro/9685/2009 2009/08/10
FJ969540 A/California/07/2009 2009/04/09
91
80
66
54
56
60
71
68
51
64
59
50
73
53
55
64
63
0.001
S220T
Q310H
K325E
P199Q
D114N
E391K
S101N
†
†
Bottom cluster
Main cluster

89
Figura 1 - Árvore filogenética (Neighbor-Joing) de sequências completas do gene HA do vírus influenza A(H1N1) pdm09. Algumas sequências disponíveis no GenBank foram incluídas para representar grupos distintos que circularam no mundo durante o período de 2009 – 2010. A data de coleta está exibida após a identificação de cada amostra. A amostra protótipo A/Califórnia/07/2009 foi utilizada para a raiz da árvore. ✟: Paciente foi a óbito.

90
5.2 ARTIGO 2
ASPECTOS EPIDEMIOLÓGICOS E VIROLÓGICOS DA INFECÇÃO POR
INFLUENZA A (H1N1)PDM 09 E FREQUÊNCIA DE OUTROS VÍRUS
RESPIRATÓRIOS NO ESTADO DE PERNAMBUCO, BRASIL: 2009 – 2010
Artigo a ser submetido à Revista Journal of Clinical Virology

91
ASPECTOS EPIDEMIOLÓGICOS E VIROLÓGICOS DA INFECÇÃO POR
INFLUENZA A (H1N1)PDM 09 E FREQUÊNCIA DE OUTROS VÍRUS
RESPIRATÓRIOS NO ESTADO DE PERNAMBUCO, BRASIL: 2009 - 2010
Maria José Couto Oliveira1,2+, Marilda M Siqueira3, Maria de Lourdes Aguiar
Oliveira3, Fernando do Couto Motta3, Sharon Carney3, Marli Tenório Cordeiro1,2,
Wyller Alencar de Mello4, Vera Magalhães2.
1 Setor de Virologia, Laboratório Central de Saúde Pública (LACEN-PE), Secretaria
de Saúde do Estado de Pernambuco, Recife, PE, Brasil. 2 Programa de Pós-Graduação em Medicina Tropical de Universidade Federal de
Pernambuco, Recife, PE, Brasil. 3 Laboratório de Vírus Respiratórios e do Sarampo do Instituto Oswaldo Cruz,
Fiocruz, Rio de Janeiro, Brasil. 4 Instituto Evandro Chagas, Belém, Pará, Brasil.
+Endereço para correspondência:
Maria José Couto Oliveira
Rua do Espinheiro, 360, Apto. 1801 – Espinheiro
Recife – PE, CEP 52020-020
e-mail: [email protected]

92
RESUMO
As infecções respiratórias agudas podem ser causadas por diferentes patógenos e
relevantes padrões de morbidade e mortalidade podem ser observados
especialmente nos grupos de risco. Os padrões epidemiológicos da infecção por
influenza A(H1N1)pdm09 e a prevalência de outros vírus respiratórios foram
investigados no Estado de Pernambuco, durante o período pandêmico 2009-2010.
Neste estudo seccionado, amostras biológicas (swabs de orofaringe e nasofaringe
e/ou fragmentos de pulmão) de 705 casos suspeitos de influenza foram testadas por
reação em cadeia da polimerase em tempo real (rRT-PCR) para influenza
A(H1N1)pdm09 (Protocolo CDC), e uma sub-amostra (146 Influenza A negativa) foi
testada para a presença de outros vírus respiratórios. Dados clínicos e
epidemiológicos foram coletados usando formulário padronizado. Análises
descritivas/bivariadas (Qui-quadrado e/ou teste exato de Fisher e teste T de Student)
foram realizadas, e a significância foi considerada quando p<.005. A frequência de
infecção por influenza A(H1N1)pdm09 foi 26,4%; sendo significantemente superior
no sexo masculino (p=0.023) e na faixa etária entre 6 e 19 anos (p<0.001). Além
disso, 12 pacientes (6.4%) entre os 186 casos confirmados, evoluíram para o óbito.
Na subamostra Influenza A negativa, outros vírus foram detectados em 53 (36,3%):
Rhinovírus (41,0%), Coronavírus (16,1%), Metapneumovírus (14,3%), Parainfluenza
(8,9%), Bocavírus (7,1%), Vírus Respiratório Sincicial (5,4%), Influenza B (3,6%),
Adenovírus (1,8%) e Enterovírus (1,8%). Infecções múltiplas foram encontradas em
três casos. A magnitude da pandemia em Pernambuco foi menor do que no sul e
sudeste do Brasil. Além do vírus influenza A(H1N1)pdm09, outros patógenos
respiratórios circularam em 2009-2010 durante o período pandêmico, e parecem ser
responsáveis por cerca de um terço dos casos agudos de infecção respiratória com
sintomatologia de IRA. Apesar da importância do PCR em tempo real como
ferramenta diagnóstica, estes resultados devem ser interpretados no contexto
clínico-epidemiológico. Novos estudos estão sendo desenvolvidos para avaliar o
impacto desses outros patógenos em saúde pública.
Palavras chave: Influenza, vírus respiratórios, infecção respiratória aguda,
parainfluenza, coronavírus, metapneumovírus, bocavírus, adenovírus, rhinovírus,
vírus respiratório sincicial.

93
ABSTRACT
Acute respiratory infections can be caused by different pathogens and relevant
morbid-mortality patterns can be found especially some risk groups. The
epidemiological features of influenza A(H1N1)pdm09 and the prevalence of other
respiratory infections were investigated in Pernambuco state, during the 2009-2010
pandemic period. In this cross-sectional study, biological samples (nasopharyngeal
swab/lung fragments) from 705 Influenza suspected cases were tested by real time
PCR for influenza A(H1N1)pdm09 (CDC protocol) and a subsample (146 FluA-
negative) was tested for the presence of respiratory viruses. Epidemiological/clinical
data were collected using a standardized formulary. Descriptive/bivariate analyses
(Chi-square and/or Fisher exact test and T-test) were performed and significance
was considered as p<.005. The prevalence of influenza A(H1N1)pdm09 infection was
26.4%; being significantly higher among males (p=.023) and those aged between 6-
19 years (p>.001). Moreover 12 patients (6.4%) died. In the Influenza A - negative
subsample, other viruses could be detected in 53 (36.3%), as follows: Rhinovirus
(41,0%), Coronavirus (16.1%), Metapneumovirus (14.3%), Parainfluenza (8.9%).
Bocavirus (7.1%), Respiratory syncytial virus(5.4%), Influenza B (3.6%), Adenovirus
(1.8%), and Enterovirus (1.8%). Multiple infections were found in 3 cases. The
magnitude of Flu pandemic in Pernambuco was lower than in South/Southeast of
Brazil. Besides influenza A(H1N1)pdm09, other respiratory pathogens were
circulating during 2009-2010 Influenza season, and seemed to be responsible for
about one third of non-FluA clinical cases. Despite rRT-PCR importance as a
diagnostic tool, these results should be interpreted according to the
clinical/epidemiological contexts. New studies are under development to assess the
Public Health impact of these other pathogens.
Key-words: Influenza, Respiratory viruses, acute respiratory infection, Parainfluenza,
Coronavirus, Metapneumovirus, Bocavirus, Rhinovirus, Respiratory Syncytial Vírus.

94
INTRODUÇÃO
Mundialmente, as infecções respiratórias agudas (IRA) são uma importante
causa de morbimortalidade, principalmente entre alguns grupos, tais como idosos,
crianças e pacientes imunocomprometidos.1,2 A maioria dessas infecções é de
etiologia viral3 e clinicamente similar às infecções por influenza, sendo
conjuntamente denominadas “influenza-like illness” (ILI).4 Além do influenza (FLU), 5
outros vírus estão associados com ILI, como o vírus respiratório sincicial (RSV),6
rhinovírus (RV),7 coronavírus (CoV),8 parainfluenza (PIV) 1 a 4,9 adenovírus
(AdV),10 enterovírus (EV),11 parechovírus (PeV)12 e os mais recentemente
identificados metapneumovírus (HMPV)13 e bocavírus (HBoV).14
Recentemente, a reação em cadeia da polimerase em tempo real rRT-PCR)
tem sido empregada para detecção de múltiplos patógenos respiratórios.15-19
Entretanto, a relevância clínica dessas infecções no contexto da saúde pública ainda
se encontra sob investigação, especialmente no Brasil, onde os dados são muitos
escassos. 2
O objetivo deste estudo foi avaliar os aspectos virológicos e epidemiológicos
da infecção por influenza A(H1N1)pdm09 em Pernambuco, durante o período de
maio de 2009 a maio de 2010, assim como investigar a prevalência de outros
patógenos virais em casos clínicos de infecção respiratória aguda (IRA) negativos
para influenza A.
MÉTODOS
Desenho do estudo e População alvo
Neste estudo transversal, 705 pacientes com IRA foram investigados,
compreendendo o período de maio de 2009 a maio de 2010. Os indivíduos foram
atendidos em hospitais de referência do estado de Pernambuco para casos
suspeitos de influenza pandêmica. Até julho de 2009, o critério para a classificação
dos casos suspeitos consistiu na presença de febre >38ºC (ainda que apenas
referida) tosse e contato direto com pessoas infectadas ou viajantes para áreas com
casos confirmados de influenza A(H1N1)pdm09, nos últimos 10 dias. Após a
evidência da transmissão sustentada do vírus no Brasil (semana epidemiológica
29/09), o critério passou a contemplar apenas os casos com síndrome respiratória

95
aguda grave (SRAG), caracterizados pela presença de dispnéia, febre >38ºC e
tosse, assim como casos fatais.19 Durante a hospitalização, amostras clínicas
(swabs combinados de orofaringe e nasofaringe e/ou fragmentos de pulmão) foram
coletadas visando a realização de testes laboratoriais. Para a detecção de outros
patógenos virais, uma subamostra composta por 146 casos negativos para influenza
A (FLUA) foi aleatoriamente selecionada, sendo representativa das diversas
semanas epidemiológicas.
Detecção de Influenza A (H1N1)pdm09 e outros vírus respiratórios
Para a detecção do vírus influenza A (H1N1)pdm09, o RNA foi extraído
usando o kit QIAamp® Viral RNA Mini Kit (QIAGEN , Germany), e a reação em
cadeia da polimerase em tempo real (rRT-PCR) foi realizada, segundo o protocolo
desenvolvido pelo Centro de Controle e Prevenção de Doenças (CDC, EUA)21
(acessível em
<http://www.who.int/csr/resources/publications/swineflu/CDCRealtimeRTPCR_Swine
H1Assay-2009_20090430.pdf>). Estes ensaios foram realizados no Instituto
Evandro Chagas (Belém, Pará, Brasil).
Para a detecção de múltiplos patógenos respiratórios, a extração de ácido
nucléico foi realizada usando o kit Magna Pure LC Total Nucleic Acid Isolation
(Roche Diagnostics), seguido por RT- PCR em tempo real, onde foi usado o kit Fast
track21 plus® (Laboratoires Réunis – Luxemburgo). Este ensaio é capaz de detectar
23 patógenos respiratórios: FLUA, FLUB, FLUA(H1N1)pdm09, RV, HCoV229,
HCoV63, HCoV43 e HCoV KHU; PIV-2, PIV-3, PIV-4 HBoV, HMPV, PIV-1, AdV, EV,
PeV, RSV; Chlamydia pneumoniae, Haemophilus influenzae, Staphylococcus aureus
and Streptococcus pneumoniae. Somente vírus foram considerados nesta
investigação. Estes ensaios foram realizados no Laboratório de Vírus Respiratórios e
do Sarampo da Fundação Oswaldo Cruz (IOC, FIOCRUZ), Rio de Janeiro.
Análise Estatística
Análises descritivas/ bivariadas (Qui-quadrado e/ou teste exato de Fisher e
teste T para médias) foram realizadas usando o programa STATA, versão 9.0.
Possíveis associações entre as infecções pesquisadas e as variáveis demográficas,
clínicas e epidemiológicas foram analisadas e consideradas significativas, quando
p<0,005.

96
Aspectos Éticos
Este estudo foi aprovado pelo Comitê de Ética em Pesquisas do Centro de
Ciências da Saúde da UFPE. RESULTADOS
Aspectos demográficos, clínicos e epidemiológicos da infecção por Influenza
A(H1N1)pdm09
Os aspectos demográficos, clínicos e epidemiológicos dos casos de infecção
pelo vírus influenza A(H1N1)pdm09 encontram-se descritos na Tabela 1.
No período do estudo, 839 casos de influenza foram notificados e foram
coletadas amostras clínicas de 705 indivíduos (84,0%). Destes, 453 (64,3%) e 252
(35,7%) eram do sexo feminino e masculino, respectivamente, e a média de idade
foi 29,4 (±19) anos (3 meses a 89 anos). Em 67,4%, as amostras foram coletadas
em até 3 dias após o início dos sintomas e em 7,2% depois de 8 dias. Dos 705
indivíduos, 209 (29,6%) foram positivos para influenza A pelo teste do CDC e 186
(26,4%) foram positivos para influenza A(H1N1)pdm09. Destes, 23,4% (106/453)
eram do sexo feminino e 31,7% (80/252) do sexo masculino, com média de idade de
24,6 (±15,6) anos (faixa de um mês a 82 anos). Uma frequência significativamente
superior foi encontrada entre aqueles na faixa etária de 6 a 9 anos (p<0.001).
Os sintomas mais frequentes foram febre, tosse e dispnéia, de acordo com o
critério de classificação de casos do Ministério da Saúde. Dois picos no número de
casos de influenza A(H1N1)pdm09 foram observados: o primeiro na semana
epidemiológica 26 (13 casos) e o segundo entre as semanas 30 e 38 (113 casos)
(Figura 1). Durante a pandemia de influenza, as gestantes constituíram um dos principais
grupos de risco para a forma mais grave da doença.22 No Estado de Pernambuco,
entre 307 mulheres em idade fértil (entre 15 e 49 anos), 72 (23,5%) (IC: 18,7% a
28,2%) foram positivas para o vírus influenza A(H1N1)pdm09. De 79 gestantes, 22
(27,8%) foram positivas para influenza A (H1N1)pdm09 e três foram à óbito.
Óbitos por Influenza A(H1N1)pdm09
As características demográficas e epidemiológicas dos casos de óbitos por
influenza A(H1N1)pdm09 encontram-se descritas na Tabela 2.

97
Entre os casos fatais (n=35), influenza A(H1N1)pdm09 foi detectado em 12
indivíduos (34,3%; 8 em 2009 e 4 em 2010). A doença acometeu ambos os sexos,
na faixa etária entre 4 e 53 anos de idade. Nas três gestantes que faleceram (casos
4, 7 e 12 com 21, 28 e 23 anos, respectivamente) a gravidez foi o único fator de
risco apresentado.
Outras infecções respiratórias virais
Entre as 146 amostras negativas para influenza A(H1N1)pdm09 (Figura 2),
outros patógenos respiratórios foram detectados em 53 indivíduos (36,3%) (Tabela
3).
Os vírus mais prevalentes foram rhinovírus (n=23), metapneumovírus (n=8) e
Coronavírus OC43 (n=8). Entre os casos de infecção por rhinovírus, a média de
idade foi 23,1 (15,3) anos, com maior prevalência no sexo masculino (19,6%). O
pico dos casos ocorreu entre julho e setembro – o período de maior circulação de
influenza A(H1N1)pdm09 e notificação de casos de IRA. Em três amostras, infecção
dupla viral, Rhinovírus + Bocavírus; Rhinovírus + Metapneumovírus e Influenza B +
Coronavírus OC43 foram observadas. Ainda, entre os cinco casos fatais que foram
submetidos ao PCR multiplex, apenas RSV foi encontrado em uma única amostra de
uma criança de 2 anos.
DISCUSSÃO
A frequência de infecções por influenza A(H1N1)pdm09 em Pernambuco
durante 2009-2010, foi 26,4%, de modo que a magnitude da pandemia em nosso
estado foi inferior àquela observada no Sul e Sudeste do Brasil.22 Estes resultados
corroboram com outros achados em diferentes regiões do país.2 Segundo o Serviço
de Vigilância do Estado de Pernambuco, 846 casos suspeitos foram notificados
entre maio 2009 e maio 2010, e o coeficiente de mortalidade por 100.000 habitantes
foi da ordem de 0,1 em 2009 e 0,04 em 2010 (comunicação oral).
Em Pernambuco, a frequência de infecções foi significativamente mais alta
entre os indivíduos do sexo masculino do que no feminino (p=0,023), contrastando
com o perfil previamente descrito no Brasil.2,24 Achados similares também foram
observados em relação à idade. Em Pernambuco uma maior frequência foi

98
encontrada na faixa entre 6-19 anos (p<0,001). No Brasil os adultos jovens (entre 20
a 29 anos) foram os mais acometidos no período 2009-2010.24
De acordo com a literatura nacional e internacional,25 aqueles com idade ≥60
anos apresentaram frequência de infecção pelo vírus influenza A(H1N1)pdm09
inferior, quando comparados aos mais jovens. Como previamente publicado, este
evento parece ser devido à prévia exposição viral e imunidade cruzada entre os
vírus influenza, especialmente entre aqueles nascidos antes de 1957.26
Conforme observado nas demais regiões brasileiras,23 a presença de
doença respiratória crônica foi o principal fator de risco na casuística estudada,
sendo relatada por 40,4% dos indivíduos. Apesar da pequena amostra de
gestantes, cerca de 1/3 foi infectada por influenza A(H1N1)pdm09, incluindo três
casos fatais. Estes dados corroboram com a literatura, onde a gestação figura como
um importante fator de risco para a evolução clínica mais graves e óbito nesta
categoria de exposição, decorrentes da infecção por influenza.27, 28
Entre os casos suspeitos, somente 29,4% foram positivos para influenza A.
Os demais 496 casos permaneceram com etiologia desconhecida. No Brasil, o
menor índice de confirmação laboratorial de casos suspeitos foi observado na
Região Nordeste (32,3%).23 Muitos fatores podem ter contribuído para a baixa
frequência de detecção de influenza A(H1N1)pdm09 no estado: i) coleta tardia da
amostra – em alguns casos após o sétimo dia dos sintomas; ii) o material clínico
(swab de nasofaringe que é menos sensível do que aspirado para detecção por
PCR;29 iii) a temperatura inadequada durante o transporte, o que pode ter
comprometido a qualidade da amostra.
Para elucidar a etiologia dos casos de IRA, negativos para influenza A, uma
subamostra de 146 amostras foi investigada para a presença de outros 18
patógenos virais. O principal achado deste estudo foi que ao utilizar a abordagem de
PCR em tempo real multiplex, foi possível elucidar cerca de 40,0% destes casos,
onde o Rhinovírus foi o vírus mais prevalente (41,0%). Em estudo conduzido na
Suíça entre transplantados e paciente imunocomprometidos, o Rhinovírus foi o
segundo vírus respiratório mais frequente (22,6%) após o Coronavírus (32,2%).18
Resultados semelhantes foram também encontrados na Suécia onde os autores
levantaram a hipótese de que esta alta prevalência poderia ter contribuído para
reduzir a circulação de influenza A(H1N1)pdm09.30,31 Em nossa subamostra, 3
(2,0%) infecções virais múltiplas foram detectadas: RV + HMPV, RV + HBoV e FLUB

99
+ HCoV-OC43. A frequência foi mais baixa do que aquelas encontradas em outros
estudos, conduzidos em crianças,17,29,32,33 porém, similar ao encontrado em São
Paulo (3,77%).34 Devido ao pequeno tamanho da amostra e à baixa prevalência dos
patógenos pesquisados não fomos capazes de investigar possíveis associações
entre estas infecções e as variáveis clínico-epidemiológicas, a fim de conhecer
melhor o perfil destas infecções e a sua importância sob a perspectiva da saúde
pública.
O diagnóstico das doenças virais teve um grande impulso nos últimos 15 anos
com o advento da PCR. Entretanto, a maioria dos estudos têm sido desenvolvidos
em crianças.35 Apesar da praticicidade da rRT-PCR como ferramenta diagnóstica,
devido a sua alta sensibilidade, os resultados das infecções múltiplas devem ser
interpretados no contexto dos dados clínico-epidemiológicos.
Agradecimentos:
À Dra. Marilda Siqueira por disponibilizar o Laboratório de Vírus Respiratórios
e do Sarampo do IOC-FIOCRUZ – Rio de Janeiro, para realização deste estudo.
Suporte Financeiro:
Instituto Oswaldo Cruz /Fundação Oswaldo Cruz.
REFERÊNCIAS: 1. Boivin G, Côté S, Déry P, De Serres G, Bergeron MG. Multiplex Real-Time PCR
Assay for Detection of Influenza and Human Respiratory Syncytial Viruses. J Clin Microbiol 2004; 42: 45-51.
2. Raboni SM, Stella V, Cruz CR, França JB, Moreira S, Gonçalves L, et al.
Laboratory Diagnosis, Epidemiology, and Clinical Outcomes of Pandemic Influenza A and Community Respiratory Viral Infections in Southern Brazil. J Clin Microbiol 2011; 49: 1287-93.
3. Costa LF, Yokosawa J, Mantese O, Oliveira TFM, Silveira Hl, Nepomuceno Ll, et al. Respiratory viruses in children younger than five years old with acute respiratory disease from 2001 to 2004 in Uberlândia, MG, Brazil. Mem Inst Oswaldo Cruz Rio de Janeiro 2006; 101: 301-6.

100
4. Kelly H, Birch C. The causes and diagnosis of influenza-like illness. Australian Family Physician 2004; 33: 305-9.
5. Behrens G, Gottschalk R, Gürtler L, Harder TC, Hoffmann C, Kamps BS, et al.
Influenza Report 2006. Kamps-Hoffmann-Preiser Ed. Available from: www.InfluenzaReport.com>.
6. Murata Y, Falsey, AR. Respiratory syncytial virus infection in adults. Antiv Ther
2007; 12: 659-70. 7. Briese T, et al. Global Distribution of Novel Rhinovirus Genotype. Emerg Infect
Dis 2008; 14: 944-977. 8. Fielding BC. Human coronavirus NL63: a clinically important virus? Future
Microbiolog 2011; 6:153-159. 9. CDC.Human Parainfluenza Viruses. Available from:
<http://www.cdc.gov/ncidod/dvrd/revb/respiratory/PIVfeat.htm>. 10. Berk AJ. Adenoviridae: The Viruses and Their Replication. In: Knipe DM, Howley
PM editors. Fields Virology 2007. 5 th ed. CD-ROM. Lippincott Williams & Wilkins; 2: 2355-94.
11. Pallansch M, Roos R. Enteroviruses: Polioviruses, Coxsackieviruses,
Echoviruses, and Newer Enteroviruses. In: Knipe DM, Howley PM editors. Fields Virology 2007. 5 th ed. CD-ROM. Lippincott Williams & Wilkins; 2: 840-93.
12. Harvala H, Simmonds P. Human parechoviruses: biology, epidemiology and clinical significance. J Clinl Virol 2009; 45: 1-9.
13. Broor S, Bharaj P, Chahar HS. Human metapneumovirus: a new respiratory pathogen. J Biosci 2008; 33: 483-93.
14. Allander T, Jartti T, Gupta S, Niesters HGM, Lehtinen P, Osterback R, et al.
Human bocavirus and acute wheezing in children. Clin Infect Dis 2007; 44: 904-10.

101
15. Bellau-Pujol S, Vabret A, Legrand L, Dina J, Gouarin S, Petitjean-Lecherbonnier J, et al. Development of three multiplex RT-PCR assays for the detection of 12 respiratory RNA viruses. J Virol Methods 2005; 126: 53-63.
16. Freymuth F, Vabret A, Cuvillon-Nimal D, Simon S, Dina J, Legrand L, et al.
Comparison of multiplex PCR assays and conventional techniques for the diagnostic of respiratory virus infections in children admitted to hospital with an acute respiratory illness. J Med Virol 2006; 78: 1498-504.
17. Bonzel L, Tenenbaum T, Schroten H, Schildgen O, Schweitzer-Krantz S, Adams
O. Frequent detection of viral coinfection in children hospitalized with acute respiratory tract infection using a real-time polymerase chain reaction. Pediatric Infect Dis J 2008; 27: 589-94.
18. Garbino J, Soccal PM, Aubert JD, Rochat T, Meylan P, Thomas Y, et al. 2009. Respiratory viruses in bronchoalveolar lavage: a hospital-based cohort study in adults. Thorax 2009; 64: 339-404.
19. Vabret A, Dina J, Gouarin S, Petitjean J, Tripey V, Brouard J, et al. Human (non sever acute respiratory syndrome) coronavirus infections in hospitalisated children in France. J Paediatr Child Health 2008; 44: 176-81.
20. Oliveira WK, Carmo EH, Penna GO, Kuchenbecker RS, Santos HB, Araújo WN,
et al. Pandemic H1N1 influenza in Brazil: Analysis of the first 34.506 notified casesof influenza-like illness with severe acute respiratory infection (SARI). Euro Surveill 2009; 14: pii =19362.
21. WHO. CDC Protocol of real time RT-PCR for swine influenza A (H1N1)- revision 1
(30 April 2009). WHO 2009. Available from: <http://www.who.int/csr/resources/publications/swineflu/CDCRealtimeRTPCR_SwineH1Assay-2009_20090430.pdf.>.
22. BRASIL. Ministério da Saúde. Situação Epidemiológica da Nova influenza A
(H1N1) no Brasil, 2009.
23. BRASIL. Ministério da Saúde. Boletim Eletrônico Epidemiológico. CADERNO
ESPECIAL INFLUENZA. Influenza Pandêmica (H1N1) 2009. Análise da Situação Epidemiológica e da Resposta no Ano de 2009. Ano 10. Nº 2. Março de 2010a.
24. BRASIL. Ministério da Saúde. Informe técnico quinzenal de Influenza. Influenza
Pandêmica (H1N1) 2009 – Monitoramento da Síndrome Respiratória Aguda Grave. Ed. n. 9, 2010b.

102
25. WHO. Acute Respiratory Infections (Update Setember 2009). Available from: <http://www.who.int/vaccine_research/diseases/ari/en/index5.html>.
26. Hancock K, Veguilla V, Lu X, Zhong W, Butler EN, Sun Hong et al. Cross-
Reactive Antibody Responses to the 2009 Pandemic H1N1 Influenza Virus. N Engl J Med 2009; 361: 1945-52.
27. Louie JK, Acosta M, Jamieson DJ, Honein MA, California Pandemic (H1N1)
Working Group. Severe 2009 H1N1 Influenza in Pregnant and Postpartum Women in California. N Engl J Med 2010; 362: 27-35.
28. Mosby LG, Rasmussen SA, Jamieson DJ. 2009 pandemic influenza A (H1N1) in
pregnancy: a systematic review of the literature. Am J Obstet Gynecol 2011; 205:10-8.
29. Meerhoff TJ, Houben ML, Coenjaerts FE, Kimpen JL, Hofland RW, Schellevis F et
al. Detection of multiple respiratory pathogens during primary infection: nasal swab versus nasopharyngeal aspirate using real-time polymerase chain reaction. Eur J Clin Microbiol Infect Dis 29: 365-371.
30. Linde AM, Rotzén-Ostlund M, Zweygberg-Wirgart B, Rubinova S, Brytting M.
Does viral interference affect spread of influenza? Euro Surveill 2009; 8: pii=19354.
31. Casalegno JS, Ottmann, Duchamp MB, Escuret V, Billaud G, Frobert E et al.
Rhinoviruses delayed the circulation of the pandemic influenza A (H1N1) 2009 in France. Clin Microbiol Infect 2010; 16: 326-9.
32. Wang W, Cavailler P, Ren P, Zhang J, Dong W, Yan H, et al. Molecular
monitoring of causative viruses in child acute respiratory infection in endemo-epidemic situations in Shanghai. J Clin Virol 2010; 49: 211-8.
33. Bezerra PGM, Britto MCA, Correia JB, Duarte MCMB, Fonseca AM, Rose K et al.
Viral and Atypical Bacterial Detection in Acute Respiratory Infection in Children Under Five years. PLoS ONE 2011; 6: e18928.
34. Watanabe ASA, Carraro E, Moreira L, Camargo C, Sinohara J, Puerari D, et al.
Respiratory virus infection among hospitalized patients with suspected influenza A H1N1 2009 during the first pandemic wave in Brasil. Braz J Infect Dis 2011; 15: 220-24.
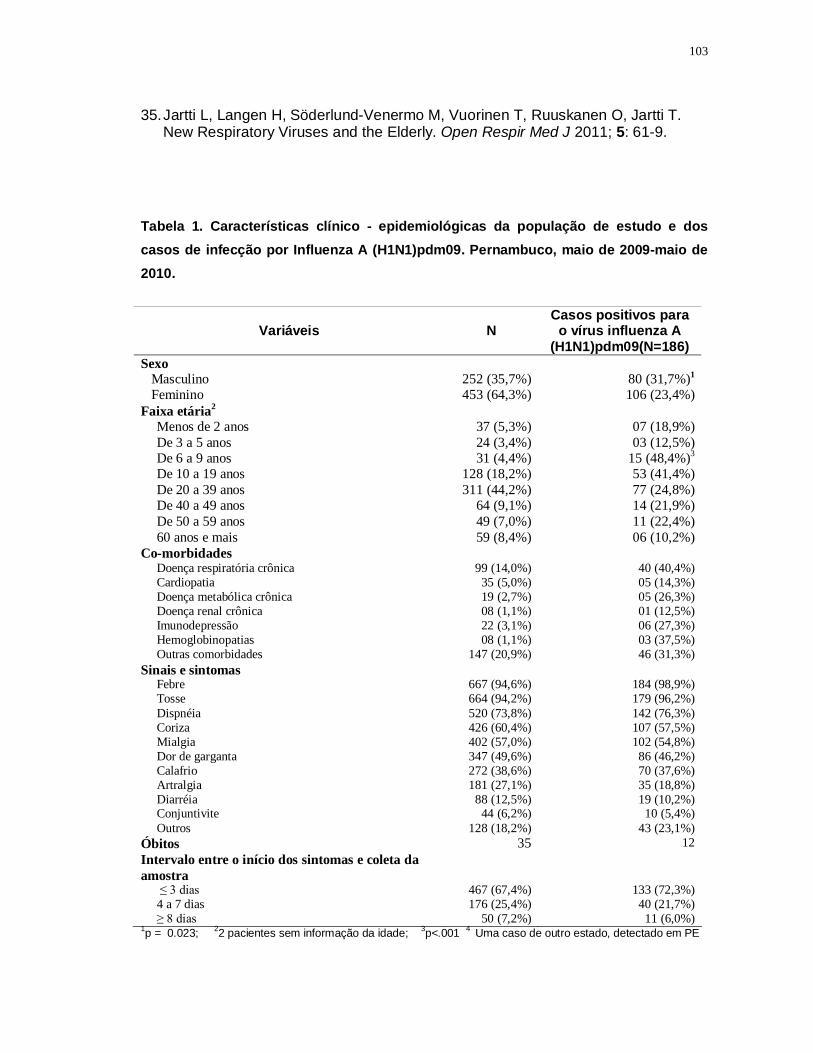
103
35. Jartti L, Langen H, Söderlund-Venermo M, Vuorinen T, Ruuskanen O, Jartti T. New Respiratory Viruses and the Elderly. Open Respir Med J 2011; 5: 61-9.
Tabela 1. Características clínico - epidemiológicas da população de estudo e dos casos de infecção por Influenza A (H1N1)pdm09. Pernambuco, maio de 2009-maio de 2010.
Variáveis
N Casos positivos para
o vírus influenza A (H1N1)pdm09(N=186)
Sexo Masculino 252 (35,7%) 80 (31,7%)1 Feminino 453 (64,3%) 106 (23,4%) Faixa etária2
Menos de 2 anos 37 (5,3%) 07 (18,9%) De 3 a 5 anos 24 (3,4%) 03 (12,5%) De 6 a 9 anos 31 (4,4%) 15 (48,4%)3
De 10 a 19 anos 128 (18,2%) 53 (41,4%) De 20 a 39 anos 311 (44,2%) 77 (24,8%) De 40 a 49 anos 64 (9,1%) 14 (21,9%) De 50 a 59 anos 49 (7,0%) 11 (22,4%) 60 anos e mais 59 (8,4%) 06 (10,2%)
Co-morbidades Doença respiratória crônica 99 (14,0%) 40 (40,4%) Cardiopatia 35 (5,0%) 05 (14,3%) Doença metabólica crônica 19 (2,7%) 05 (26,3%) Doença renal crônica 08 (1,1%) 01 (12,5%) Imunodepressão 22 (3,1%) 06 (27,3%) Hemoglobinopatias 08 (1,1%) 03 (37,5%) Outras comorbidades 147 (20,9%) 46 (31,3%) Sinais e sintomas
Febre 667 (94,6%) 184 (98,9%) Tosse 664 (94,2%) 179 (96,2%) Dispnéia 520 (73,8%) 142 (76,3%) Coriza 426 (60,4%) 107 (57,5%) Mialgia 402 (57,0%) 102 (54,8%) Dor de garganta 347 (49,6%) 86 (46,2%) Calafrio 272 (38,6%) 70 (37,6%) Artralgia 181 (27,1%) 35 (18,8%) Diarréia 88 (12,5%) 19 (10,2%) Conjuntivite 44 (6,2%) 10 (5,4%) Outros 128 (18,2%) 43 (23,1%)
Óbitos 35 12 Intervalo entre o início dos sintomas e coleta da amostra
≤ 3 dias 467 (67,4%) 133 (72,3%) 4 a 7 dias 176 (25,4%) 40 (21,7%) ≥ 8 dias 50 (7,2%) 11 (6,0%)
1p = 0.023; 22 pacientes sem informação da idade; 3p<.001 4 Uma caso de outro estado, detectado em PE

104
Tabela 2 – Casos fatais por Influenza A(H1N1)pdm09 no Estado de Pernambuco, Brasil. Maio 2009-Maio 2010. Amostra
# Idade (anos)
Sexo Mês/ Ano
Intervalo entre primeiros sintomas
e hospitalização (dias)
Intervalo entre
primeiros sintomas e óbito (dias)
Fator de Risco
1 27 Feminino 8/09 5 10 Desconhecido 2 50 Feminino 8/09 3 14 Desconhecido 3 53 Masculino 9/09 5 5 Desconhecido 4 28 Feminino 9/09 5 20 Gestação 5 40 Masculino 10/09 9 12 Desconhecido 6 4 Feminino 10/09 3 6 Desconhecido 7 21 Feminino 11/09 SI* 22 Gestação 8 24 Masculino 12/09 4 6 Obesidade, asma 9 29 Masculino 3/10 8 29 Obesidade
10 47 Masculino 4/10 7 7 Desconhecido 11 31 Masculino 4/10 5 5 Desconhecido 12 23 Feminino 4/10 2 6 Gestação
*SI – sem informação
Fonte: SINAN, Ministério da Saúde
Tabela 3 – Frequência de outros patógenos virais entre os casos de IRA no Estado de Pernambuco, Brasil. Maio 2009-Maio 2010.
N=146 Patógenos Virais N (%)
Negativo 93 (63,7%)
Positivo 53 (36,3%)
Outros patógenos virais 56 #
Rhinovírus 23 (41,0%)
Metapneumovírus 08 (14,3%)
Coronavírus 43 08 (14,3%)
Bocavírus humano 04 (7,1%)
Vírus Respiratório Sincicial 03 (5,4%)
Influenza B 02 (3,6%)
Parainfluenza 2 02 (3,6%)
Parainfluenza 3 02 (3,6%)
Parainfluenza 1 01 (1,8%)
Adenovírus 01 (1,8%)
Enterovírus 01 (1,8%)
Coronavírus HKU 01 (1,8%)
* 53 amostras e 56 vírus detectados (3 amostras com infeção múltipla)

105
Figura 1 – Distribuição de casos de IRA e de Influenza A(H1N1)pdm09, segundo a semana epidemiológica. Pernambuco, Brasil, Maio 2009- Maio 2010.
0
5
10
15
20
25
30
35
4045
50
55
60
65
70
75
80
85
17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49 51 1 3 5 7 9 11 13 15 17 19 21
Vírus A(H1N1)pdm09 Casos de IRA
Figura 2 – Fluxograma das amostras testadas para outros vírus respiratórios
Amostras testadas para o Vírus Influenza A
705
Amostras positivas
209
Amostras negativas
496
Ca s o s
Semanas epidemiológicas
146 amostras testadas
para outros vírus

106
6 CONCLUSÕES
- Na casuística estudada, não foram identificados vírus com a substituição
H274Y, corroborando achados de outros países sobre a baixa frequência de vírus
influenza A(H1N1)pdm09 resistentes ao oseltamivir. Entretanto, tal vigilância deve
ser encorajada.
- O pirosequenciamento constitui uma ferramenta importante para identificar
marcadores associados à resistência aos antivirais de uma forma rápida e em maior
escala.
- Uma alta similaridade entre as amostras investigadas e o protótipo vacinal
A/Califórnia/7/2009 foi encontrada, confirmando que, até o presente momento, não
foram observadas mudanças antigênicas importantes no vírus influenza
A(H1N1)pdm09.
- A frequência do vírus influenza A(H1N1)pdm09 em Pernambuco foi de
26,4%, inferior ao observado no Brasil (38,6%) e no nordeste (32,2%).
- Em Pernambuco, uma frequência significativamente superior de casos
positivos foi encontrada entre indivíduos do sexo masculino e na faixa etária de 6 a 9
anos, seguida da faixa de 10 a 19 anos.
- Com o uso de RT-PCR em tempo real, multiplex, foi possível identificar ao
menos um vírus respiratório em 36,3% dos casos clínicos, negativos para Influenza
A. Desta forma, tal ferramenta poderia ser empregada no esclarecimento diagnóstico
(diferencial), sobretudo nos casos de óbito. Ainda permitiria melhor conhecer a
importância clínico-epidemiológica destes vírus e seu respectivo impacto em Saúde
Pública.
- Excluindo os vírus influenza A, os rhinovírus foram os mais frequentes entre
os vírus detectados (41,0%), seguidos dos coronavírus (16,1%), e metapneumovírus
(14,3%). Esse achado é importante porque reforça a necessidade do diagnóstico

107
laboratorial para esclarecer a etiologia da IRA e também porque até então não havia
nenhum relato de outros vírus respiratórios detectados nos casos suspeitos de
influenza pandêmica no estado de Pernambuco.
- A maior circulação de rhinovírus ocorreu nos meses de julho, agosto e
setembro, coincidindo com a circulação do vírus influenza A(H1N1)pdm09. A de
coronavírus nos meses de julho e agosto.
7 CONSIDERAÇÕES FINAIS
- O monitoramento epidemiológico e virológico para conhecimento precoce
dos vírus respiratórios que estão circulando devem ser realizados rotineiramente,
utilizando-se técnicas moleculares mais sensíveis para nortear as medidas de
prevenção e controle a serem tomadas e mostrar a necessidade de produção de
vacinas para determinados patógenos.
- A caracterização genética dos vírus respiratórios, especialmente dos vírus
da influenza, deve ser repetida periodicamente, pois a vigilância genética permite a
detecção de mutações associadas à resistência aos antivirais e a detecção precoce
dos sítios antigênicos que são selecionados para os vírus escaparem das barreiras
imunológicas.

108
REFERÊNCIA
ABED, Y.; BAZ, M.; BOIVIN,G. Impact of neuraminidase mutations conferring influenza resistance to neuraminidase inhibitors in the N1 and N2 genetic backgrounds. Antiviral Therapy, v.11, n.8, p.971-976, 2006. ADWAN, G. Molecular characterization and phylogenetic analysis of Midle East 2009 H1N1 pdm isolates. Asian Pacific Journal of Tropical Medicine, p. 624-628, 2010. Disponível em <http://staff.najah.edu/sites/default/files/Molecular_characterization_and_phylogenetic_analysis_of_Middle_East_2009_H1N1_pdm_isolates.pdf.>. Acesso em 20 ago. 2011. ALLANDER T. et al. Cloning of a human parvovirus by molecular screening of respiratory tract samples. Proceedings of the National Academy of Sciences of the United States of America, v.102, n.36, p.12891-12896, 2005. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1200281/pdf/pnas-0504666102.pdf>. Acesso em 5 ago. 2011. ALLANDER, T. et al. Human bocavirus and acute wheezing in children. Clinical Infectious Diseases, v. 44, n. 7, p. 904-910, 2007. Disponível em < Disponível em < http://cid.oxfordjournals.org/content/44/7/904.full.pdf+html>. Acesso em 5 ago. 2011. AOKI, F.Y.; BOIVIN, G.; ROBERTS, N. Influenza virus susceptibility and resistance to oseltamivir. Antiviral Therapy, v.12, n.4, p. 603-616, 2007. ARIAS, C.F. et al. Molecular Anatomy of 2009 Influenza Vírus A (H1N1). Archives of Medical Research, v. 40, p. 643-654, 2009. ARTHUR, J.L. et al. A Novel Bocavirus Associated with Acute Gastroenteritis in Australian Children. PLoS Pathogens, v.5, n.4: e1000391. doi: 10.1371, 2009. BARRERO, P.R et al. Genetic and Phylogenetic Analysis of Influenza A H1N1pdm Vírus in Buenos Aires, Argentina. Journal of Virology. v. 85, n.2, p.1058 – 1066, 2011. Disponível em <http://jvi.asm.org/content/85/2/1058.full.pdf+html>. Acesso em 3 dez. 2011. BARRY, J.M. 1918 revisited: lessons and suggestions for further inquiry. In: Board on Global Health. The threat of Pandemic Influenza: Are We Ready? The National Academies Press 2004 apud BEHRENS, G. et al. Influenza Report 2006. Kamps-Hoffmann-Preiser Ed. Disponível em <http://influenzareport.com>. Acesso em 10 jul. 2009. BARR, I.G. et al. A new pandemic influenza A(H1N1) genetic variant predominated in the winter 2010 influenza season in Australia, New Zealand and Singapore. Eurosurveillance, v.15, n. 42, 2010. Disponível em < http://www.eurosurveillance.org/images/dynamic/EE/V15N42/art19692.pdf>. Acesso em 10 mar. 2011.

109
BAUMEISTER, E. et al. Molecular Characterization of Severe and Mild Cases of Influenza A (H1N1 2009 Strain from Argentina. Medicina (Buenos Aires), v.70, p. 518-523, 2010. BAZ, M. et al. Effect of the Neuraminidase Mutation H274Y Conferring Resistance to Oseltamivir on the Replicative Capacity and Virulence of Old and Recent Human Influenza A (H1N1) Viruses. The Journal of Infectious Diseases, v.201, n.5, p.740-745, 2010. BEHRENS, G. et al. Influenza Report 2006. Kamps-Hoffmann-Preiser Ed. Disponível em <http://influenzareport.com>. Acesso em: 10 jul.2009. BELLEI, N. et al. Acute respiratory infection and influenza-like illness viral etiologies in Brazilian adults. Journal of Medical Virology. v.80, p. 1824-1827, 2008. BELLAU- PUJOL, S. et al. Development of three multiplex RT-PCR assays for the detection of 12 respiratory RNA viruses. Journal of Virological Methods, v. 126, n. 1-2, p.53-63, 2005. BELSHE, R.B. The Origins of Pandemic Influenza: Lessons from the 1918 Virus. The New England Journal of Medicine, v.353, n.21, p.2209-11, 2005. BELSHE, R.B. Implications of the Emergence of a Novel H1 Influenza Virus. The New England Journal of Medicine, v. 360, p. 2667-2668, 2009. Disponível em <http://www.nejm.org/doi/full/10.1056/NEJMe0903995>. Acesso em 05 jan. 2010. BERK, A.J. Adenoviridae: The Viruses and Their Replication. In: KNIPE, D.M.; HOWLEY, P.M. editors. Fields Virology. 5 th ed. CD-ROM. Lippincott Williams & Wilkins, v. 2, p. 2355-2394, 2007. BEZERRA, P.G.M. et al. Viral and Atypical Bacterial Detection in Acute Respiratory Infection in Children Under Five Years. PLoS ONE , v.6 , e18928, 2011. BHARAJ, P. et al. Respiratory viral infections detected by multiplex PCR among pediatric patients with lower respiratory tract infections seen at an urban hospital in Delhi from 2005 to 2007. Virology Journal, v.6, n.89, 2009. Disponível em < http://www.virologyj.com/content/pdf/1743-422X-6-89.pdf>. Acesso em 10 out. 2011. BOLETIM ELETRÔNICO EPIDEMIOLÓGICO. Influenza - Edição Especial. Situação Epidemiológica da nova Influenza A (H1N1) no Brasil, 2009. Ano 09. n. 02. 2009. Disponível em < http://portal.saude.gov.br/portal/arquivos/pdf/boletim_epidemiologico_influenza_23_07_2009.pdf>. Acesso em 10 nov. 2009. BOLETIM ELETRÔNICO EPIDEMIOLÓGICO. Caderno Especial Influenza. Influenza Pandêmica (H1N1) 2009. Análise da Situação Epidemiológica e da Resposta no Ano de 2009. Ano 10. Nº 2. Março de 2010. <

110
http://portal.saude.gov.br/portal/arquivos/pdf/boletim_eletronico_influenza_25_03.pdf >. Acesso em 20 abr. 2010. BONZEL, L.et al. Frequent detection of viral coinfection in children hospitalized with acure respiratory tract infection using a real-time polymerase chain reaction. Pediatric Infect Disease Journal, v.27, n.7, p.589-594. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Plano de Contingência do Brasil para o Enfrentamento de uma Pandemia de Influenza. Brasília, 2005. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Guia de Vigilância Epidemiológia. 6 ed. Brasília. 2006, 815 p. BRASIL. Ministério da Saúde. Portaria SVS nº97 de 5 de outubro de 2008, 2008. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde/MS Guia de Vigilância Epidemiológica. 7.ed. Cad. 1. Brasília, 2009a, 24 p. Disponível < http://portal.saude.gov.br/portal/arquivos/pdf/gve_7ed_web_atual_influenza.pdf>. Acesso em 11 fev. 2010. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Vigilância Epidemiológica Integrada da Influenza: Vigilância de Doença Respiratória Aguda Grave. Brasília. 2009b. Disponível em <http://portal.saude.gov.br/portal/arquivos/pdf/guia_bolso_vigilancia_epidem_28jul09.pdf > . Acesso em 10 jan. 2010. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Informe Epidemiológio Influenza A (H1N1). Situação epidemiológica da nova influenza A (H1N1) no Brasil, 2009. Ed. n.3,2009c. Disponível em <http://portal.saude.gov.br/portal/arquivos/pdf/informe_influenza_se30_03_08_2009.pdf >. Acesso em 03 nov. 2009. BRASIL. Ministério da Saúde. Gabinete Permanente de Emergências de Saúde Pública. Emergência de Saúde Pública de Importância Internacional - ESPII. Ocorrências de casos humanos na América do Norte. Informe do dia 27.04.09, às 13h30. Brasília, 2009d, 7 p. BRASIL. Ministério da Saúde. Gabinete Permanente de Emergências de Saúde Pública. Emergência de Saúde Pública de Importância Internacional - ESPII. Ocorrências de casos humanos na América do Norte. Informe do dia 29.04.09, às 13h30. Brasília, 2009e. 7 p. BRASIL. Ministério da Saúde. Gabinete Permanente de Emergências de Saúde Pública. Emergência de Saúde Pública de Importância Internacional - ESPII. Ocorrências de casos humanos de infecção por Influenza A (H1N1). Informe do dia 01.05.09, às 16h30. Brasília, 2009f. 10 p.

111
BRASIL. Ministério da Saúde. Gabinete Permanente de Emergências de Saúde Pública. Emergência de Saúde Pública de Importância Internacional - ESPII. Ocorrências de casos humanos de infecção por Influenza A (H1N1). Informe do dia 08.05.09, às 13h. Brasília, 2009g. 14 p. BRASIL. Ministério da Saúde. Gabinete Permanente de Emergências de Saúde Pública. Emergência de Saúde Pública de Importância Internacional - ESPII. Ocorrências de casos humanos de infecção por Influenza A (H1N1). Informe do dia 09.05.09, às 13h. Brasília, 2009h. 12 p. BRASIL. Ministério da Saúde. Gabinete Permanente de Emergências de Saúde Pública. Emergência de Saúde Pública de Importância Internacional - ESPII. Ocorrências de casos humanos de infecção por Influenza A (H1N1). Informe do dia 12.06.09, às 14h. Brasília, 2009i. 13 p. BRASIL. Ministério da Saúde. Gabinete Permanente de Emergências de Saúde Pública. Emergência de Saúde Pública de Importância Internacional - ESPII. Ocorrências de casos humanos de infecção por Influenza A (H1N1). Informe do dia 24.06.09, às 13h. Brasília, 2009j. 8 p. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Informe técnico quinzenal de Influenza. Influenza Pandêmica (H1N1) 2009 – Monitoramento da Síndrome Respiratória Aguda Grave. Ed. n. 9, 2010a. Disponível em < http://portal.saude.gov.br/portal/arquivos/pdf/informe_influenza_8_agosto19_8_10.pdf.> Acesso em 5 nov. 2010. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Estratégia Nacional de Vacinação Contra o Vírus Influenza Pandêmico (H1N1) 2009. Brasília, 2010b. Disponível em < http://portal.saude.gov.br/portal/arquivos/pdf/informe_tecnico_vacina_18_03.pdf>.Acesso em 2 abr 2010. BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Informe técnico. Campanha Nacional de Vacinação contra a Influenza 2011. Brasília, 2010c. Disponível em < http://portal.saude.gov.br/portal/arquivos/pdf/informe_campanha_influenza_2011.pdf>. Acesso em 7 nov.2011. BRIESE, T. et al. Global Distribution of Novel Rhinovirus Genotype. Emerging Infectious Diseases, v. 14, n. 6, p. 944-977, 2008. Disponível em < http://wwwnc.cdc.gov/eid/article/14/6/pdfs/08-0271.pdf >. Acesso em 04 mar. 2011. BROOR, S.; BHARAJ, P.; CHAHAR, H.S. Human metapneumovirus: a new respiratory pathogen. Journal of Biosciences, v. 33, n. 4, p. 483-493, 2008. Disponível em < http://www.ias.ac.in/jbiosci/nov2008/483.pdf >. Acesso em 5 nov. 2011.

112
BUSH, R.M. Influenza Evolution. In: TIBAYRENC, M. Encyclopedia of Infectious Diseases – Modern Methodologies. John Wiley & Sons. New Jersey. 2007, p.199-208. CABECA, T.K.; BELLEI, N. Human coronavirus NL-63 infection in a Brazilian patiente suspect of H1N1 2009 influenza infection: Description of a fatal case. Journal of Clinical Virology, v.53, n.1, p.82-84, 2012. CANÉ, A. et al. First isolation of a oseltamivir-resistant Influenza A (H1N1) strain in Argentina. Pediatric Infectious Disease Journal, v.29, n.4, p. 384, 2010. CARNEIRO, M.; KRUMMENAUER, E.C.; MACHADO, J.A. Vigilância de vírus respiratórios em crianças hospitalizadas durante a pandemia de H1N1. Revista da Associação Médica do Rio Grande do Sul, Porto Alegre, v. 54, n. 2, p. 259-260, 2010. Disponível em http://www.amrigs.org.br/revista/54-02/24-571_vigilancia_carta_ao_editor.pdf > . Acesso em 8 out. 2011. CENTERS FOR DISEASE CONTROL AND PREVENTION. Human Parainfluenza Viruses (Common cold and croup), 2006. Disponível em <http://www.cdc.gov/ncidod/dvrd/revb/respiratory/PIVfeat.htm> Acesso em 10. Set. 2010. CENTERS FOR DISEASE CONTROL AND PREVENTION . Seasonal Influenza: Flu. How Flu Spreads 2009a. Disponível em < http://www.cdc.gov/flu/about/disease/spread.htm >. Página atualizada em 05.09.2009. Acesso em 04 jan. 2010. CENTERS FOR DISEASE CONTROL AND PREVENTION. Seasonal Influenza: Flu. Flu Symptoms et Severity. 2009b. Disponível em < http://www.cdc.gov/flu/about/disease/symptoms.htm >. Página atualizada em 07.12.2009. Acesso em 04 jan. 2010. CENTERS FOR DISEASE CONTROL AND PREVENTION. Role of Laboratory Diagnosis of Influenza. 2009c. Disponível em <http://www.cdc.gov/flu/professionals/diagnosis/labrole.htm>. Acesso em 10 mar. 2011. CENTERS FOR DISEASE CONTROL AND PREVENTION. Influenza (Flu) Research. 2009d. Disponível em < http://www.cdc.gov/flu/about/qa/research.htm>. Página atualizada em 08.09.2009. Acesso em 02 jan. 2010. CENTERS FOR DISEASE CONTROL AND PREVENTION. MMWR. Weekly Update: Influenza Activity -- United States, September 28, 2008 - January 31, 2009, v.58, n. 05, p.115-119, 2009e. Disponível em < http://www.cdc.gov/mmwr/preview/mmwrhtml/mm5805a4.htm>. Acesso em 15 dez. 2009. CENTERS FOR DISEASE CONTROL AND PREVENTION. MMWR. Oseltamivir-Resistant Novel Influenza A (H1N1) Virus Infection in Two

113
Immunosuppressed Patients - Seattle, Washington, 2009. v.58, p. 1 – 4, 2009f. Disponível em < http://www.cdc.gov/mmwr/preview/mmwrhtml/mm58d0814a1.htm,>. Acesso em 15 jan. 2010. CENTERS FOR DISEASE CONTROL AND PREVENTION. MMWR. Weekly. Oseltamivir-Resistant 2009 Pandemic Influenza A (H1N1) Virus Infection in Two Summer Campers Receiving Prophylaxis --- North Carolina, 2009, v. 58, n. 35, p.969 -972, 2009g. Disponível em < http://www.cdc.gov/mmwr/preview/mmwrhtml/mm5835a1.htm.>. Acesso em 10 fev. 2010. CENTERS FOR DISEASE CONTROL AND PREVENTION. MMWR. Safety of
Influenza A (H1N1) 2009 Monovalent Vaccines --- United States, October 1-- November 24, 2009. CENTERS FOR DISEASE CONTROL AND PREVENTION. 2009-2010 Influenza Season Week 6 ending February 13, 2010. 2010a. Disponível em < http://www.cdc.gov/flu/weekly/pdf/External_F1006.pdf >. Página revisada em 18.02.2010. Acesso em 20 fev. 2010. CENTERS FOR DISEASE CONTROL AND PREVENTION. Respiratory Syncytial Virus Infection (RSV), 2010b. Disponível em <http://www.cdc.gov/rsv/about/infection.html>. Acesso em 03 jun. 2011. CENTERS FOR DISEASE CONTROL AND PREVENTION. Adenoviruses. 2010c. Disponível em < http://www.cdc.gov/ncidod/dvrd/revb/respiratory/eadfeat.htm>. Acesso em 6 jun. 2011. CENTERS FOR DISEASE CONTROL AND PREVENTION. Human Parainfluenza Viruses. Disponível em < http://www.cdc.gov/ncidod/dvrd/revb/respiratory/PIVfeat.htm >. Página revisada em 25.03.2011. Acesso em 5 out. 2011. CHEN, L.F. et al. Cluster of Oseltamivir-Resistant 2009 Pandemic Influenza A (H1N1) Vírus Infections on a Hospital Ward among Immunocompromised Patients-- North Carolina, 2009. Journal of Infectious Diseases, v.203, n.6, p. 838-46. 2011. CHIEOCHANSIN, T. Molecular epidemiology, genome characterization, and recombination event of human parechovirus. Virology, 2011. CHIU, S.S. et al. Human coronavirus NL63 infection and other coronavirus infections in children hospitalized with acute respiratory disease in Hong Kong, China. Clinical Infectious Diseases, v. 40, n. 12, p. 1721-1729, 2005. CHOW, B.D.; HUANG, Y.T.; ESPER, F.P. Evidence of human bocavirus circulating in children and adults. Cleveland, Ohio. Journal of Clinical Virology, v.43, n.3, p.302-306, 2008.

114
COLLINS, P.L.; CROWER Jr, J.E. Respiratory Syncytial Virus and Metapneumovirus In: KNIPE, D.M. et al. Fields Virology. 5 ed. CD-ROM. Lippincott Williams & Wilkins, v.2, 2007. p. 1602-1646. COSTA, F.A. et al. Respiratory viruses in children younger than five years old with acute respiratory disease from 2001 to 2004 in Uberlândia, MG, Brazil. Memórias do Instituto Oswaldo Cruz, Rio de Janeiro. v 101, n.3, p. 301-306, 2006. Disponível em < http://www.scielo.br/pdf/mioc/v101n3/v101n3a14.pdf>. Acesso em 5 mai. 2010. CUEVAS, L.E. et al. Human metapneumovirus and respitatory syncytial virus, Brazil. Emerging Infectious Disease, v. 9, n. 12, p. 1626-1628, 2003. Disponível em < http://www.nc.cdc.gov/eid/article/9/12/pdfs/03-0522.pdf > Acesso em 3 mar. 2011. DAWOOD, F.S. et al. Emergence of a novel swine-origin influenza A (H1N1) virus in humans. The New England Journal of Medicine, v.360, p. 2605-15, 2009. DEBUR, M.C. et al. Acute respiratory infection by human metapneumovirus in children in southern Brazil. Journal of Clinical Virology, v. 39, n. 1, p. 59-62, 2007. D’ELIA, C. et al. Respiratory syncytial virus - associated lower respiratory tract infections in hospitalized infants. Revista da Sociedade Brasileira de Medicina Tropical, v. 38, n.1, p. 7- 10, 2005. De ROUX, A. et al. Viral community-acquired pneumonia in nonimmunocompromised adults. Chest, v. 125, n.4, p. 1343-1351, 2004. DESHPANDE, M.S. et al. Evaluation of the efficacy of a canine influenza virus (H3N8) vaccine in dogs following experimental challenge. Veterinary Therapeutic, v.10, n.3, p.103-112, 2009. DE VRIES, M. et al. Human parechovirus type 1, 3, 4, 5, and 6 detection in picornavirus cultures. Journal of Clinical Microbiology, v. 46, n. 2, p. 759 -762, 2008. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2238116/?tool=pubmed >. Acesso em 05 jun. 2011. DEYDE, V.M. et al. Pyrosequencing as a tool to detect molecular markers of resistance to neuraminidase inibitors in seasonal influenza A viruses. Antiviral Research, v81, p.16-24, 2009. DEYDE, V.M et al. Detection of molecular markers of drug resistance in 2009 pandemic influenza A(H1N1) viruses by pyrosequencing. Antimicrobial Agents Chemother, v. 54, n.3, p. 1102-1110, 2010.

115
DHARAN, N.J. et al. Infections With Oseltamivir-Resistant Influenza A (H1N1) Virus in the United States. The Journal of American Medical Association, v.301, n.10, p.1034-1041, 2009. DONALISIO, M.R. Influenza aviária: questões centrais. Revista Brasileira de Epidemiologia, v.9, n.1, 2006. Disponível em < http://www.scielosp.org/pdf/rbepid/v9n1/002.pdf> Acesso em 12 dez.2009. DOWDLE, W.R. Influenza A vírus recycling revisited. Bulletin of the World Health Organization, v.77, n.10, p. 820-8. 1999. Disponível em < http://whqlibdoc.who.int/bulletin/1999/Vol77-No10/bulletin_1999_77(10)_820-828.pdf> . Acesso em 11 dez. 2009. DREXLER, J.F. et al. Novel Human Parechovirus from Brazil. Emerging Infectious Diseases, v. 15, n. 2, 2009. Disponível em < http://wwwnc.cdc.gov/eid/article/15/2/08-1028_article.htm >. Acesso em 2 nov. 2011. ENGIN, A. Influenza type A (H5N1) virus infection. Mikrobiyoloji bulteni, 41, n.3, p.485-94. 2007. ESHAGHI, A. et al. Genetic microheterogeneity of emerging H275Y influenza virus A (H1N1) in Toronto, Ontario, Canadá from 2007 – 2008 respiratory season. Journal of Clinical Virology, v.45, n.2, p.142 – 145, 2009. FALSEY, A.R.; WALSH, E.E. Respiratory syncytial virus infection in elderly adults. Drugs Aging, v. 22, n.7, p.577-87, 2005. FAROOQUI, A. et al. Genetic and clinical assessment of 2009 pandemic influenza in southern China. The Journal of Infection in Developing Countries, v. 5, n.10, p. 700-710, 2011. Disponível em < http://www.jidc.org/index.php/journal/article/viewFile/21997938/611> Acesso em 30 nov. 2011. FELSENSTEIN, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, v.39, n. 4, p.783-791, 1985. FERREIRA, J.L.P et al. Sequence analysis of the 2009 pandemic influenza A H1N1 virus haemagglutinin gene from 2009-2010 Brazilian clinical samples. Memórias do Instituto Oswaldo Cruz. Rio de Janeiro, v.106, n. 5, p. 613-616, 2011. Disponível em < http://www.scielo.br/pdf/mioc/v106n5/14.pdf>. Acesso em 10 out. 2011. FIELDING, B.C. Human coronavirus NL63: a clinically important virus? Future Microbiology, v.6, n.2, p.153-159, 2011. FORLEO-NETO, E. et al. Influenza. Revista da Sociedade Brasileira de Medicina Tropical, v.36, n.2, p.267-274, 2003.

116
FOUCHIER, R.A.M. et al. A previously undescribed coronavirus associated with respiratory disease in humans. Proceedings of the National Academy of Sciences of the United States of America, v. 101, n. 16, p. 6212-6216, 2004. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC395948/?tool=pubmed >. Acesso em 5 nov. 2011. FRANCIS, T. Jr. A New Type of Virus from Epidemic Influenza. Science, v. 92, 405-408, 1940. GALIANO, M. et al. Evolutionary Pathways of the Pandemic Influenza A (H1N1) 2009 in the UK. PLoS One 6(8): e23779 doi: 10.1371, 2011. GARBINO, J. Et al. Respiratory viruses in bronchoalveolar lavage: a hospital-based cohort study in adults. Thorax, v., 64, n. 5, p. 339-404, 2009. GERNA, G. et al. Genetic variability of human coronavirus OC43-, 229E-, and NL63-like strains and their association with lower respiratory tract infections of hospitalized infants and immunocompromised patients. Journal of Medical Virology, v. 78, n. 7, p. 938-849, 2006. GLINSKY, G.V. Genomic analysis of pandemic (H1N1) 2009 reveals association of increasing disease severity with emergence of novel hemagglutinin mutations. Cell Cycle, v.9, n.5, p.958-970, 2010. GOMES, M.L.C et al. Enteroviroses. In : LEÃO, R.N.Q. et al. Doenças Infecciosas e Parasitárias - Enfoque Amazônico. Ed. CEJUP. Universidade do Estado do Pará. Instituto Evandro Chagas, p.243-252, 1997. GRAHAN, M. et al. Nationwide Molecular Surveillance of Pandemic H1N1 Influenza A Virus Genomes: Canada, 2009. PLoS One, 6(1): e16087. doi:10.1371, 2011. GRIJALVA, C.G. et al. Estimating influenza hospitalizations among children. Emerging Infectious Diseases, Atlanta, v.12, p.103-9, 2006. GRAHAN, M. et al. Nationwide Molecular Surveillance of Pandemic H1N1 Influenza A Virus Genomes: Canada, 2009. PLoS One 6: e16087 doi: 10.1371. HALL, C.B. Respiratory syncytial virus and parainfluenza virus, 2001 apud CARABALLO, L.P.; RAMIREZ, O.V. Vírus Parainfluenza Humanos, 2001. Disponível em: <http://www.paho.org/Spanish/AD/THS/EV/labs_ipk_6.pdf>. Acesso em: 04 nov. 2009. HARVALA, H. et al. Epidemiology and Clinical Associations of Human Parechovirus Respiratory infections. Journal of Clinical Microbiology, v.46, n.10, p. 3446 a 3453, 2008.

117
HARVALA, H; SIMMONDS, P. 2009- Human parechoviruses: biology, epidemiology and clinical significance. Journal of Clinical Virology, v.45, n. 1, p.1-9, 2009. HARVALA, H. et al. The Emergence of Oseltamivir-Resistant Pandemic Influenza A(H1N1) 2009 Virus Amongst Hospitalised Immunocompromised Patients in Scotland, November-December, 2009. Eurosurveillance, v. 15, n. 14, p. 1-3, 2010. HATTA, M.; GOTO, H.; KAWAOKA, Y. Influenza B Virus Requires BM2 Protein for Replication. Journal of Virology. v. 78, n. 11, p.5576-5583, 2004. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC415801/ >. Acesso em 08 abr. 2011. HAUGE, S.H.et al. Oseltamivir-resistant influenza viruses A (H1N1), Norway, 2007–08. Emerging Infectious Disease. v.15, n.2, p.155–62, 2009. HAYDEN, F.G. Rhinovirus and the lower respiratory tract. Reviews in Medical Virology, v. 14, n. 1, p. 17-31, 2004. HEIKKINEN, T.; THINT, M.; CHONMAITREE, T. Prevalence of various respiratory viruses in the middle ear during acute otitis media. The New England Journal of Medicine. , v. 340, n. 4, p. 260-264, 1999. HEIKKINEN, T.; CHONMAITREE, T. Importance of Respiratory Viruses in Acute Otitis Media. Clinical Microbiology Reviews, v. 16, n. 2, p. 230-241, 2003. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC153141/?tool=pubmed > . Acesso em 4 jun. 2011. HILLEMAN, M.R.; WERNER, J.H. Recovery of new agent from patients with acute respiratory illness. Proceedings of the Society for Experimental Biology and Medicine, v.85, n.1, p.183-188, 1954 apud CHINTAKUNTLAWAR, A.V; ASTLEY, R.; CHODOSH. Adenovirus Type 37 Keratitis in the C57BL/6J Mouse. Investigative Ophthalmology & Visual Science, v.48, n.2, p.781-788, 2007. HUNT, M. Virology. Influenza virus (Orthomyxovrus). In: Microbiology and Immunology On-line. University of South Carolina School of Medicine. Disponível em < http://pathmicro.med.sc.edu/mhunt/flu.htm >.Página revisada em 21.10.2009. Acesso em 12 dez. 2009. HURT, A.C. et al. Emergence and spread of oseltamivir-resistant A (H1N1) influenza viruses in Oceania, South East Asia and South Africa. Antiviral Research, v.83, n.1, p.90-93, 2009. Disponível em < http://download.thelancet.com/flatcontentassets/H1N1-flu/treatment/treatment-28.pdf > Acesso em 10 jan. 2010. IAMARINO, A. Influenza A (H1N1) Blog. Antivirais e resistência: inibidores de sialidase. Biblioteca Virtual em Saúde. 12/10/2009. Disponível em <

118
http://blog.h1n1.influenza.bvsalud.org/pt/2009/10/12/antivirais-e-resistencia-inibidores-de-sialidase/>. Acesso em 01 mar. 2010. IKONEN, N. et al. Genetic Diversity of the 2009 Pandemic Influenza A (H1N1) Viruses in Finland. PLoS One, v.5, n.10, 2010. Disponível em < http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0013329>. Acesso em 12 dez. 2010. JARTTI, L. et al. New Respiratory Viruses and the Elderly. The Open respiratory Medicine Journal, v.5, p. 65-69, 2011. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3134957/pdf/TORMJ-5-61.pdf >. Acesso em 7 jul. 2011. JOKI –KORPELA, P.; HYYPIÃ, T. Parechoviruses, a novel group of human picornaviruses. Annals of Medicine, v.33, n.7, p. 466-471, 2001. JUVÉN, T. et al. Etiology of community-acquired pneumonia in 254 hospitalized children. The Pediatric Infectious Disease Journal, v. 19, n. 4, p. 293-298, 2000. KAPOOR, A. et al. A newly identified bocavirus species in human stool. The Journal of Infectious Disease, v.199, n.2, p.196-200, 2009. KARRON, R.A.; COLLINS, P.L. Parainfluenza Viruses. In : KNIPE, D.M.; HOWLEY, P.M. editors. Fields Virology. 5 th ed. CD-ROM. Lippincott Williams & Wilkins, v. 2, p. 1498 -1526, 2007. KAWANO, H. et al. Genetic Analysis and Phylogenetic Caracterization of Pandemic (H1N1) 2009 Influenza Viruses that found in Nagasaki. Japanese Journal of Infectious Diseases. v.64, p. 195-203, 2011. Disponível em < http://www.nih.go.jp/JJID/64/195.pdf>. Acesso em 12 jun. 2011. KAWAY, N. et al. Clinical effectiveness of oseltamivir for influenza A (H1N1) vírus with H274Y neuraminidase mutation. Journal of Infection, v.59, p.207-212, 2009. Disponível em < http://download.thelancet.com/flatcontentassets/H1N1-flu/treatment/treatment-39.pdf>.. Acesso em 10 jan. 2010. KELLY, H.; BIRCH, C., The causes and diagnosis of influenza-like illness. Australian Family Physician, v.33, n.5, p.305-309, 2004. Disponível em <http://www.racgp.org.au/afp/200405/20040510kelly.pdf >. Acesso em 9 set.2010. KILANDER, A. et al. Observed association between the HA1 mutation D222G in the 2009 pandemic influenza a(H1N1) virus and severe clinical outcome, Norway 2009-2010. Eurosurveillance, v. 15, n. 9, 2010. Disponível em < http://www.eurosurveillance.org/images/dynamic/EE/V15N09/art19498.pdf> .Acesso em 14 mar. 2011.

119
KISO, M. et al. Resistant influenza A viruses in children treated with oseltamivir: descriptive study. The Lancet, v.364, p.759-765, 2004. LACKENBY, A. et al. Rapid quantitation of neuraminidase inhibitor drug resistance in influenza virus quasispecies. Antiviriral Therapy, v. 13, n.6, p. 809-820, 2008. LAU, S.K.P. et al. Coronavirus HKU1 and Other Coronavirus Infections in Hong Kong. Journal of Clinical Microbiology, v. 44, n. 6, p.2063-2071, 2006. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1489438/ >. Acesso em 4.nov. 2011. LEDESMA, J. et al. Substitutions in position 222 of haemagglutinin of pandemic influenza A (H1N1) 2009 viruses in Spain. Journal of Clinical Virology, v.51, n.1, p. 75-78, 2011. LEE, R.T.C et al. All that glitters is not gold - founder effects complicate associations of flu mutations to disease severity. Virology Journal, v.7, p. 297, 2010. Disponível em < http://www.virologyj.com/content/7/1/297> . Acesso em 30 abr. 2011. LE, Q.M. et al. A Community Cluster of Oseltamivir-Resistant Cases of 2009 H1N1 Influenza. The New England Journal of Medicine, v.362, n.1, p.86-87, 2010. Disponível em < http://content.nejm.org/cgi/content/full/362/1/86-a >. Acesso em 18 fev. 2010. LEUNG, T.F. et al. Epidemiology and Clinical Presentations of Human Coronavirus NL63 Infections in Hong Kong Children. Journal of Clinical Microbiology. V. 47, n. 11, p. 3486-3492, 2009. Disponível em < http://www.ncbi.nlm.nih.gov/pubmed/19759228 >. Acesso em 5 nov. 2011. LIU, Y et al. Altered Receptor Specificity and Cell Tropismo f D222G Hemagglutinin Mutants Isolated from Fatal Cases of Pandemic A(H1N1)2009 Influenza Virus. Journal of Virology, v. 84, n. 22, p. 12069-12074, 2010. LOURIA, D.B. et al. Studies on influenza in the pandemic of 1957-1958. II. Pulmonary complications of influenza. The Journal of Clinical Investigation, v.38, p.213-265, 1959. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC444127/>. Acesso em 10 jan. 2010. MACKAY, I.M et al. Genetic diversity of human metapneumovirus over 4 consecutive years in Australia. The Journal of Infectious Diseases, v.193, n.12, p.1630-1633, 2006. MAK, G.C. et al. Association of D222G Substitution in Haemagglutinin of 2009 Pandemic Influenza A (H1N1) with Severe Disease. Eurosurveillance, v. 15, n.14, 08 April 2010. Disponível em < http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19534>. Acesso em 6 jun. 2011.

120
MAK, G.C. et al. Evolution of the haemagglutinin gene of the influenza A (H1N1) 2009 virus isolated in Hong Kong, 2009–2011. Eurosurveillance, v.16, n.9, March 2011. Disponível em < http://www.eurosurveillance.org/images/dynamic/EE/V16N09/art19807.pdf>. Acesso em 12 jun. 2011. MALATO, L. et al. Pandemic influenza A (H1N1)2009: molecular characterization and duration of viral shedding in intensive care patients in Bordeaux, south-west France, May 2009 to January 2010. Euro Surveill, v.16, n.4 , pii=19776., January 2011. Disponível em < http://www.eurosurveillance.org/images/dynamic/EE/V16N04/art19776.pdf>. Acesso em 5 jun. 2001. MANUGUERRA, J.C. et al. Natural infection of dogs by influenza C virus: a serological survey in Spain. New Microbiology, v.16, n.4, p.367-71,1993. MAURER-STROH, S. et al. A new common mutation in the hemagglutinin of the 2009 (H1N1) influenza A virus. PLoS Currents, v.2. PMCID: PMC2880458, 2010. McHARD, A.C.; ADAMS, B. The Role of Genomics in Tracking the Evolution of Influenza A Virus. Plos Pathogens, 5(10): e1000566, doi: 10.1371, 2009. MEJÍAS, A.; RAMILO O. Review of palivizumab in the prophylaxis of respiratory syncytial vírus (RSV) in high-risk infants. Biologics, v. 2, n.3, p. 433-439, 2008. MELIDOU, A. et al. Molecular and phylogenetic analysis of the haemagglutinin gene of pandemic influenza H1N1 2009 viruses associated with severe and fatal infections. Virus Research, v. 151, n.2, p.192-199, 2010. MILLER, R.S. et al. Occurrence of Haemagglutinin Mutation D222G in Pandemic Influenza A(H1N1) Infected Patients in the West of Scotland, United Kingdom, 2009-10. Eurosurveillance, v.15, n.16, 2010. Disponível em < http://www.eurosurveillance.o rg/images/dynamic/EE/V15N16/art19546.pdf>. Acesso em 3 fev. 2011. MORLIGHEM, J.E. et al. Mutation Analysis of 2009 Pandemic Influenza A (H1N1) Viruses Collected in Japan during the Peak Phase of the Pandemic. PLoS One, v.6, n.4, Apr 29 2011. Disponível em <http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3084724/?tool=pubmed>. Acesso em 10 jun 2011. MOSCONA, A. Global Transmission of Oseltamivir-Resistant Influenza. . The New England Journal of Medicine, v.360, n.10, p.953-956, 2009. Disponível em < http://content.nejm.org/cgi/content/full/360/10/953 >. Acesso em 15 fev. 2010. MOSCONA, A. Oseltamir Resistance – Disabling Our Influenza Defenses. The England Journal of Medicine, v.355, n.25, p.2633-2636, 2005. Disponível em < http://content.nejm.org/cgi/content/full/353/25/2633 >. Acesso em 4 fev. 2010.

121
MOTTA et al. The reappearance of Victoria lineage influenza B virus in Brazil, antigenic and molecular analysis. Journal of Clinical Virology, v.36, p. 208-214, 2006. MURATA, Y.; FALSEY, A.R. Respiratory syncytial virus infection in adults. Antiviral Therapy, v.12, p. 659–670, 2007. NARAIN, J.P.; KUMAR, R.; BHATIA, R. Pandemic (H1N1) 2009: Epidemiological, clinical and prevention aspects. The National Medical Journal of Índia, v.22, n.5, 2009. Disponível em < http://www.searo.who.int/LinkFiles/CDS_Pandemic_influenza.pdf> Acesso em 05 jun. 2010. NICHOLSON, K.G. Textbook of Influenza. ed. K.G. Nicholson, R.G.Webster & A.J.Hay, Blackwell Science, London, 1998, 578p. OBERSTE, M.S et al. Enterovirus 68 is associated with respiratory illness and shares biological features with both the enteroviruses and the rhinoviruses. Journal of General Virology, v.85, p. 2577-2584, 2004. Disponível em < http://vir.sgmjournals.org/content/85/9/2577.full.pdf>. Acesso em 10 ago. 2011. OLIVEIRA, W.K. et al. Pandemic H1N1 influenza in Brazil: Analysis of the first 34.506 notified cases of influenza-like illness with severe acute respiratory infection (SARI). Euro Surveill, v.14 , pii = 19362, 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Avian influenza: assessing the pandemic threat. WHO/CDS/2005.29. 2005a. Disponível em < http://www.who.int/csr/disease/influenza/H5N1-9reduit.pdf >. Acesso em 12 dez. 2009. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Pandemic preparedness. January 2005b. Disponível em <http://www.who.int/csr/disease/influenza/pandemic/en/>. Acesso em 12 dez. 2009. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Avian Influenza (“bird flu”). February 2006. Disponível em < http://www.who.int/mediacentre/factsheets/avian_influenza/en/#history>. Acesso em 11 jan. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. WHO/ECDC frequently asked questions for Oseltamivir Resistance. February 2008. Disponível em < http://www.who.int/csr/disease/influenza/oseltamivir_faqs/en/index.html#whatare >. Acesso em 17 fev. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Influenza A (H1N1) virus resistance to oseltamivir - 2008-2009 influenza season northern hemisphere. 18 March 2009a. Disponível em < http://www.who.int/influenza/resources/documents/H1N1webupdate20090318_ed_ns.pdf >. Acesso em 25 dez. 2010.

122
ORGANIZAÇÃO MUNDIAL DA SAÚDE. Interim planning considerations for mass gatherings in the context of pandemic (H1N1) 2009 influenza. November 2009b. Disponível em< http://www.who.int/csr/resources/publications/swineflu/cp002_2009-0511_planning_considerations_for_mass_gatherings.pdf> . Acesso em 10 jan. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Influenza (Seasonal). Fact sheet N°211. April 2009c. Disponível em< http://www.who.int/mediacentre/factsheets/fs211/en/ >. Acesso em 10 jan. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Guidance to Influenza Laboratories. Diagnosing Swine Influenza A/H1N1 Infections of current concern. 25 April 2009. 2009d. Disponível em < http://www.who.int/csr/disease/swineflu/swineflu_guidance_labs_20090425.pdf>. Acesso em10 jan. 2011. ORGANIZAÇÃO MUNDIAL DA SAÚDE. CDC Protocol of realtime RT- PCR for Influenza A (H1N1). April 2009. 2009e Disponível em < http://www.who.int/csr/resources/publications/swineflu/CDCRealtimeRTPCR_SwineH1Assay-2009_20090430.pdf >. Acesso em 10 set. 2009. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Antiviral drugs for pandemic (H1N1) 2009: definitions and use. 2009f. Disponível em <http://www.who.int/csr/disease/swineflu/frequently_asked_questions/antivirals/definitions_use/en/index.html>. Acesso em 10 nov. 2009. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Preliminary review of D222G amino acid substitution in the haemagglutinin of pandemic influenza A (H1N1) 2009 viruses. 2009g. Disponível em <http://www.who.int/csr/resources/publications/swineflu/cp165_2009_2812_review_d222g_amino_acid_substitution_in_ha_h1n1_viruses.pdf. Acesso em 5 mar. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE Influenza A (H1N1) virus resistance to oseltamivir - 2008-2009 influenza season. 22 January 2009h. Disponível em <http://www.who.int/csr/disease/influenza/H1N120090122_final.pdf >. Acesso em 25 dez. 2009. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Pandemic (H1N1) 2009 - update 60. 2009i. Disponível em < http://www.who.int/csr/don/2009_08_04/en/index.html >. Acesso em 15 dez. 2009.
ORGANIZAÇÃO MUNDIAL DA SAÚDE. Sequencing primers and protocol. 12 May 2009 (first published 29 April 2009), 2009j. Disponível em < http://www.who.int/csr/resources/publications/swineflu/GenomePrimers_20090512.pdf>. Acesso em 5 jun 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Influenza A (H1N1) NA-H274 detailed

123
pyrosequencing protocol for antiviral susceptibility testing. 13 May 2009, 2009k. Disponível em < http://www.who.int/csr/resources/publications/swineflu/NA_DetailedPyrosequencing_20090513.pdf. Acesso em 2 set. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Pandemic (H1N1) 2009 – update 112. August 2010a. Disponível em < http://www.who.int/csr/don/2010_08_06/en/index.html >. Acesso em 08 ago. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Cumulative Number of Confirmed Human Cases of Avian Influenza A/(H5N1) Reported to WHO. January 2010b. Disponível em < http://www.who.int/csr/disease/avian_influenza/country/cases_table_2010_01_28/en/index.html>. Acesso em 01 fev. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. WHO recommendations for the post-pandemic period. Pandemic (H1N1) 2009 – briefing note 23. 10 August 2010c. Disponível em < http://www.who.int/csr/disease/swineflu/notes/briefing_20100810/en/index.html>. Acesso em 20 ago. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE . Weekly update on oseltamivir resistance to pandemic influenza A (H1N1) 2009 viruses. 2010d. Disponível em < http://www.who.int/csr/disease/swineflu/Oseltamivirresistant20100217.pdf >. Acesso em 20 fev. 2010. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Recommendations for influenza vaccines, 2011. Disponível em < http://www.who.int/csr/disease/influenza/vaccinerecommendations/en/index.html>.Acesso em 13 mar. 2011. ORGANIZAÇÃO MUNDIAL DA SAÚDE. Current WHO phase of pandemic alert for Pandemic (H1N1)2009, 2012. Disponível em < http://www.who.int/csr/disease/swineflu/phase/en/index.html>. Acesso em 5 jan. 2012. ORGANIZAÇÃO PANAMERICANA DA SAÚDE. Boletín de Inmunización. Vol. XXXI (3), 2009. OSTERHAUS, A.D. et al. Influenza B virus in seals. Science, v.288, p.1051–1053, 2000. PALLANSCH, M.; ROOS, R. Enteroviruses: Polioviruses, Coxsackieviruses, Echoviruses, and Newer Enteroviruses. In: KNIPE, D.M.; HOWLEY, P.M. editors. Fields Virology. 5 th ed. CD-ROM. Lippincott Williams & Wilkins, v. 2, p. 840-893, 2007.

124
PALESE, P.; SHAW, M.L. Orthomyxoviridae: The Viruses and Their Replication. In: KNIPE, D.M.; HOWLEY, P.M. editors. Fields Virology. 5th
ed. CD-ROM. Lippincott Williams & Wilkins, v. 2, p.1648-1689, 2007. PILGER, D.A. et al. Detection of human bocavirus and human metapneumovirus by real-time PCR from patients with respiratory symptoms in Southern Brazil. Memórias do Instituto Oswaldo Cruz, Rio de Janeiro, v. 106, n. 1, p. 56-60, 2011. POTDAR, V.A. et al. Genetic Characterization of the Influenza A Pandemic (H1N1) 2009 Virus Isolates from India. PLoS One, v.5, n.3, e9693, PMCID: PMC 2837743, 2010. Disponível em < http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2837743/pdf/pone.0009693.pdf)>. Acesso em 30 abr. 2011. PRINCIPI, N.; BOSIS, S.; ESPOSITO, S. Effects of coronavirus infections in children. Emerging Infectectious Diseases, v. 16, n. 2, 2010. Disponível em < http://wwwnc.cdc.gov/eid/article/16/2/09-0469.htm>. Acesso em 6 nov. 2011. RABONI, S.M. et al. Laboratory Diagnosis, Epidemiology, and Clinical Outcomes of Pandemic Influenza A and Community Respiratory Viral Infections in Southern Brazil. Journal of Clinical Microbiology, v. 49, n.4, p. 1287 – 1293, 2011. Disponível em < http://jcm.asm.org/content/49/4/1287.full>. Acesso em 5 jul. 2011. REVISTA DE SAÚDE PÚBLICA. Informes Técnicos Institucionais. Influenza aviária e casos em humanos, v. 40, n.1, p. 187-90. 2006. ROWE, W.P. et al. Isolation of a cytopathogenic agent from human adenoids undergoing spontaneous degeneration in tissue culture. Proceedings of the Society for Experimental Biology and Medicine, v.84, n.3, p. 570-573, 1953. apud CHINTAKUNTLAWAR, A.V; ASTLEY, R.; CHODOSH. Adenovirus Type 37 Keratitis in the C57BL/6J Mouse. Investigative Ophthalmology & Visual Science, v.48, n.2, p.781-788, 2007. RUIGROK, R.W.H. Structure of Influenza A, B and C viruses. In: NICHOLSON, K.G.; WEBSTER, R.G.; HAY, A.J. Textbook of Influenza. London, Blackwell Science. p. 29-42, 1998. SAITOU, N.; NEI M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, v.4, n.4, p.406-25, 1987. SANTOS, N. S. O.; ROMANOS, M. T. V.; WIGG, M. D. Introdução à Virologia Humana. Rio de Janeiro. Guanabara Koogan. 2002. 254p. SANTOS, N. S. O.; ROMANOS, M. T. V.; WIGG, M. D. Introdução à Virologia Humana. 2 ed. Rio de Janeiro. Guanabara Koogan. 2008. 532p.

125
SETTEMBRE, E.C.; DORMITZER, P.R.; RAPPUOLI, R. H1N1: can a pandemic cycle be broken? Science Translational Medicine, v. 2, n. 24, 24ps14, 2010. SHINDE, V. et al. Triple-Reassortant Swine Influenza A (H1) in Humans in the United States, 2005 – 2009. The New England Journal of Medicine, v.360, n. 25, p.2616-2625, 2009. Disponível em < http://content.nejm.org/cgi/content/full/NEJMoa0903812>. Acesso em 10/9/2009. SAITOU, N.; NEI, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution, v.4, n. 4, p.406-425, 1987. SANGER, F. et al. Determination of a nucleotide sequence in bacteriophage f1 DNA by primed synthesis with DNA polymerase. Journal of Molecular Biology, v. 90, n. 2, p. 315-326, 1974. SCHOLTISSEK, C.; ROHDE, W; VON HOYNINGEN, V.; ROTT, R. On the origin of the human influenza virus subtypes H2N2 and H3N2. Virology, v. 87, n.1, p. 13-20, 1978. SILVA, K.A. et al. Ocorrência de Bocavírus Humano associado às infecções respiratórias agudas em crianças de 0 a 2 anos de idade na Cidade de Belém, Pará, Brasil. Revista Pan-Amazônica de Saúde, v.1, n.1, p. 87-92, Ananindeua, 2010. Disponível em < http://scielolab.iec.pa.gov.br/pdf/rpas/v1n1/v1n1a13.pdf >. Acesso em 3 mai. 2011. SILVA, L.H.A. et al. Variant isolates of human metapneumovirus subgroup B genotype 1 in Campinas, Brazil. Journal of Clinical Virology, v. 42, n. 1, p. 78-81, 2008. SILVA, L.H.A. et al. Vírus Respiratório Sincicial Humano e Metapneumovírus Humano. Revista HCPA – Hospital de Clínicas de Porto Alegre, v.29, n.2, p.139-146, 2009. SOUZA, T.M.L. et al. H1N1pdm influenza Infection in Hospitalized Cancer Patients : Clinical Evolution and Viral Analysis. PLoS ONE, v.5, n. 11, e14158 doi:10.1371, 2010. SPEERS, D. J. et al. Oseltamivir-resistant pandemic (H1N1) 2009 influenza in a severely ill patient: the first Australian case. Medical Journal of Australia, v.192, n.3, p.166-168, 2010. SMITH, F.I.; PARVIN, J.D.; PALESE, P. Detection of single base substitutions in influenza virus RNA molecules by denaturing gradient gel electrophoresis of RNA-RNA or DNA-RNA heteroduplexes. Virology, v.150, p.55-64, 1986. SULLENDER, W.M. Respiratory syncycial virus genetic and antigenic diversity. Clinical Microbiology Rev, v.13, n.1, p.1-15, 2000.

126
TAMURA, K. et al. Molecular Evolutionary Genetics Analysis using Maximum Parsimony Methods. Molecular Biology and Evolution, v. 28, n.10, p.2731-2739, 2011. TAYLOR, R.M. A further note on 1233 influenza C virus. Arch Gesamte Virusforsch. v.4, p. 485-500, 1951. THOMAZELLI, L.M. et al. Surveillance of eight respiratory viruses in clinical samples of pediatric patients in Southeast Brazil. Jornal de Pediatria (Rio de Janeiro), v.83, n.5, p.422-8, 2007. Disponível em <http://www.scielo.br/pdf/jped/v83n5/v83n5a05.pdf>. Acesso em 3 mar.2009. TURKINGTON, C.; ASHBY, B. Encyclopedia of Infectious Diseases. New York, Facts on File Inc., 1988. 370p. TURNER, R.B.; COUCH, R.B. Rhinoviruses. In: KNIPE, D.M; HOWLEY, P.M. editors. Fields Virology. Philadelphia: Lippincott, Williams & Wilkins; 2007. p. 895–909. VABRET, A. et al. Human (non sever acute respiratory syndrome) coronavirus infections in hospitalisated children in France. Journal of Paedriatics Child and Health, v. 44, n.4, p. 176-181, 2008. VALADARES, M.P.O. Variabilidade genética de bocavírus humano isolado em crianças com doença respiratória aguda em São Paulo, Brasil. 2010. Tese (doutorado em Biotecnologia). Universidade de São Paulo, São Paulo, 2010. VAN DER HOOGEN, B.G. et al. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nature Medicine, v. 7, p.719 - 724 , 2001. VAN DER HOEK, L.; PYRC, K.; BERKHOUT, B. Human coronavirus NL63, a new respiratory virus. FEMS Microbiology Reviews, v. 30, n.5, p.760-773, 2006. VAN DER HOEK, L. Human coronaviruses: what do they cause? Antiviral Therapy, v.12, n.4, p.651-658, 2007. VAN ZYL, G. Laboratory Findings. In: BEHRENS, G. et al. Influenza Report 2006. Kamps-Hoffmann-Preiser Ed. Disponível em < http://influenzareport.com/ir/lab.htm>. Acesso em 10 mar. 2011. VERBOON-MACIOLEK, M.A. et al. Severe Neonatal Parechovirus Infection and Similarity With Enterovirus Infection. The Pediatric Infections Disease Journal, v. 27, n. 3, p.241-245, 2008. VICENTE, D. et al. Human Bocavirus, a Respiratory and Enteric Virus. CENTERS for Disease Control and Prevention. Emerging Infectious Diseases, v. 13, n.4, 2007.

127
WAGNER, R., MATROSOVICH, M., KLENK, H.D. Functional balance between haemagglutinin and neuraminidase in influenza virus infections. Reviews in Medical Virology, v. 12, n. 3, p.159-166. WANG, B. et. al. Detection of the rapid emergence of the H275Y mutation associated with oseltamivir resistance in severe pandemic influenza virus A/H1N1 09 infections. Antiviral Research, v. 87, p.16-21. 2010. WANG, W. et al. Molecular monitoring of causative viruses in child acute respiratory infection in endemo-epidemic situations in Shanghai. Journal of Clinical Virology, v. 49, p. 211-218, 2010. WATANABE, K. et al. Isolation and Characterization of Novel Human Parechovirus from Clinical Samples. CENTERSs for Disease Control and Prevention. Emerging Infectious Diseases, v. 13, n. 6, 2007. Disponível em < http://wwwnc.cdc.gov/eid/article/13/6/06-0896_article.htm >. Acesso em 20 set. 2011. WATANABE, A.S.A. et al. Respiratory virus infection among hospitalized patients with suspected influenza A H1N1 2009 during the first pandemic wave in Brasil. Brazilian Journal of Infectious Disease, v. 15, n. 3, p. 220-224, 2011a. Disponível em < http://www.scielo.br/pdf/bjid/v15n3/v15n3a07.pdf>. Acesso em 6 nov. 2011. WATANABE, A.S.A. et al. Viral etiology among the elderly presenting acute respiratory infection during the influenza season. Revista da Sociedade Brasileira de Medicina Tropical, v.44, n.1, p. 18-21, 2011b. Disponível em < http://www.scielo.br/pdf/rsbmt/v44n1/05.pdf >. Acesso em 5 jun. 2011. WEISSEMBACHER, M.C.; ÁVILA, M.M. Os vírus como causa de IRA alta e baixa em crianças: características gerais e diagnóstico. In: BENGUIGUI, Y.; SCHMUNIS, G.; YUNES, J. Infecções respiratórias em crianças, F.J.L.A. Washington: Organização Pan-Americana da Saúde, p. 91-103, 1998. Disponível em <www.paho.org/Portuguese/AD/DPC/CD/aiepi-1-5.pdf > Acesso em jul. 2011. WRIGHT, P.F; WEBSTER, R.G. Orthomyxoviruses. In: KNIPE, D.M. et al. Fields Virology. 5th. ed. CD-ROM. Lippincott Williams & Wilkins, Vol.II: 1693-1740, 2007. ZHENG, B. et al. D225G mutation in hemagglutinin of pandemic influenza H1N1 (2009) virus enhances virulence in mice. Experimental Biology and Medicine (Maywood, N.J), v. 235, n. 8, p.981-988, 2010. ZUCCOTTI, G et al. Epidemiological and clinical features of respiratory viral infections in hospitalizated children during the circulation of influenza virus A(H1N1) 2009. Influenza and Other Respiratory Viruses, v. 5, n. 6, p.528-534, 2011.

128
APÊNDICE A – VERSÃO DO ARTIGO 1 EM INGLÊS
A ser submetido à Revista: Memórias do Instituto Oswaldo Cruz
GENETIC CARACTERIZATION OF THE INFLUENZA A (H1N1)09 PDM VIRUSES
IN PERNAMBUCO STATE, BRAZIL, DURING THE PERIOD OF MAY 2009 TO
MAY 2010.
Maria José Couto Oliveira 1,2, Fernando do Couto Motta 3, Marilda M Siqueira 3,
Maria de Lourdes Aguiar Oliveira3 , Thiago Moreno Lopes e Souza 3, Milena
Mesquita 3, Priscilla da Silva Born 3, Paola Cristina Resende Silva3, Wyller Alencar
de Mello4, Vera Magalhães da Silveira2
1 Virology Sector, Central Public Health Laboratory of Pernambuco (LACEN-PE),
Recife, PE, Brasil 2 Postgraduate Program in Tropical Medicine, Federal University of Pernambuco,
Recife, PE, Brasil. 3 Respiratory Viruses and Measles Laboratory of Oswaldo Cruz Institute, Fiocruz,
Rio de Janeiro, Brazil. 4 Evandro Chagas Institute, Belém, Pará, Brasil.
Address correspondence to:
Maria José Couto Oliveira
Rua do Espinheiro, 360, Apto. 1801 – Espinheiro
Recife – PE, CEP 52020-020
e-mail: [email protected]

129
SUMMARY
The influenza A(H1N1)pdm09 virus caused considerable morbidity and mortality
worldwide. Despite the end of 2009 pandemic, the virus still circulates within the
human population. The aim of this study was to investigate the frequency of
oseltamivir resistance and the molecular characterization of circulating influenza
A(H1N1)pdm09 viruses from Pernambuco State (PE), in comparison to other
Brazilian and worldwide representatives samples collected in the course of the
pandemic. A retrospective cross-sectional study was conducted among suspected
cases of pandemic influenza attended at PE reference hospitals. A total of 118
respiratory samples were submitted to the identification of the NA mutation H274Y, a
marker of antiviral resistance by pyrosequencing. Furthermore HA gene of 31
samples was sequenced by the Sanger method in order to identify important
signature residues and unique mutations. Pyrosequencing results demonstrated that
none of the investigated samples bear the oseltamivir resistance mutation H274Y in
NA gene. Finally phylogenetic analysis showed a high similarity between PE samples
and those that circulated elsewhere. Corroborating previous findings, the co-
circulation of distinct lineages of influenza A(H1N1)pdm09 was demonstrated and all
clusters of phylogenetic tree were close related to the prototype vaccine strain
A/California/07/09.
Key words: Influenza A(H1N1)pdm09, oseltamivir resistance, haemagglutinin
mutations

130
In April 2009, the WHO notified the occurrence of a novel human influenza
cases in Mexico and in the United States, which was further identified as an influenza
A(H1N1) of a swine origin. Within few months, the virus had spread worldwide,
provoking the first pandemic of the 21st century, accounting for more than 18,000
enrolled deaths (WHO, 2010). In Brazil, the first cases were confirmed by real time
RT-PCR on the 7th of May 2009 and two days later the indigenous transmission of
A(H1N1)pdm09 influenza virus was noticed. In Pernambuco State (PE), Northeast
Brazil, the first case was confirmed in June.
Influenza viruses are well recognized by their high mutation rates, especially in
haemagglutinin (HA) and neuraminidase (NA) genes, generating variants
antigenically distinct in concern to viral properties, such as the specificity for sialic
acid receptors, virulence or pathogenicity (Arias et al. 2009). The HA gene presents
the highest mutation rates and the respective protein is directly associated to the first
steps of a successful infection (Wagner et al. 2002). Many authors has monitored
mutations in the HA gene of A(H1N1)pdm09 influenza viruses, among which, the
mutation D222G have been associated with severe or fatal cases, possibly as a
consequence of a shift on the viral binding specificity from alfa2-6 to alfa 2-3, the last
more frequently found in the lower respiratory tract of humans (Liu et al. 2010; Zheng
et al., 2010). This mutation was identified among 18.0% of severe/fatal cases in
Norway (Kilander 2010), 8.7% in Scotland (Miller et al. 2010), 5.2% in Spain
(Ledesma 2011), 4.1% in Hong Kong (Mak et al. 2010) and 1.4% in Finland (Ikonen
et al. 2010).
Mutations in NA can lead to loss of sensitivity to currently used anti-influenza
therapy, based on viral neuraminidase activity inhibition (Abed et al., 2006; Aoki et
al., 2007). The resistance rates of seasonal influenza A(H1N1) viruses were
identified in different countries by studies carried out previous to the pandemic
(Dharan et al. 2009; Eshaghi et al. 2009; Moscona et al. 2005). The worldwide
dissemination of the reassorted A(H1N1)pdm09 influenza virus in the human
population associated with the broad use of neuraminidase inhibitors and the high
levels of resistance historically observed in the H1N1 subtype, became crucial for the
evaluation of the levels of resistance in the emerging virus. In addition, many groups
initiated studies in order to identify important mutations, following the evolution of the
main genes and determining the antigenicity and susceptibility of the new virus to
oseltamivir.

131
Taking into account the impact of influenza A (H1N1)pdm09 as well as the
peculiarities of respiratory viruses circulation in the different regions of the country,
this study seeks to investigate the frequency of oseltamivir resistance and the
molecular characterization of circulating influenza A(H1N1)pdm09 viruses from
Pernambuco State (PE), in comparison to other Brazilian and worldwide
representatives samples collected in the course of the pandemic.
After collection, the suspected influenza A(H1N1)pdm09 specimens (triple
combined swabs and tissue fragment from deceased patients) were aliquoted and
frozen in transport medium at Pernambuco Central Laboratory (LACEN-PE) and then
sent to Evandro Chagas Institute in Para State for real time RT-PCR detection of
Influenza A(H1N1)pdm09 . The molecular diagnosis was carried out using the
detection protocol developed by CDC (Atlanta, USA), available at:
http://www.who.int/csr/resources/publications/swineflu/CDCRealtimeRTPCR_SwineH
1Assay-2009_20090430.pdf>. The influenza A(H1N1)pdm09 confirmed positive
samples were sent to the Respiratory and Measles Laboratory at Fiocruz for to
realize this study.
A total of 118 influenza A(H1N1)pdm09 positive samples were
pyrosequenced, in order to check for the presence of the H274Y mutation (NA gene),
according to Deyde et al. (2010). Total RNA was extracted directly from 140 μL of
clinical specimen using the Qiamp Viral RNA mini kit (Qiagen, Germany) as
described in the manufacture's manual. The final elution volume was 80 μL. The
influenza A(H1N1)pdm-N1 fragment C amplification were performed using the
SuperScript III One-step RT-PCR with HiFi Taq system (Invitrogen) protocol from
CDC, available from: <
http://www.who.int/csr/resources/publications/swineflu/NA_DetailedPyrosequencing_
20090513.pdf>. After purification, the amplicons were analyzed on the Pyromark Q-
96 ID (Qiagen, Germany), using the SNP (single nucleotide polymorphism) mode. To
carry out the quantitative analysis of H274Y mutation in the NA gene, we investigated
the codon change CAC → TAC by pyrosequencing. All samples presenting less than
10% of “T” at the first position of the codon is considered wild, and consequently,
sensitive to oseltamivir treatment. Conversely, all values above 10% are considered
as a mixture of wild and mutant types, with reduced sensitivity to the drug (Souza et
al. 2011).

132
The genetic characterizations from thirty-one samples, representatives of
distinct phases – beginning, middle and end of the pandemic, were performed. The
HA gene was amplified by RT-PCR using SuperScript III One-step RT-PCR with
Platinum Taq-DNA Polymerase (Invitrogen, USA) protocol from WHO, available at
http://www.who.int/csr/resources/publications/swineflu/GenomePrimers_20090512.p
df. After purification (Motta et al, 2006), the products were directly sequenced using
the ABI Prism Big Dye Terminator Cycle Sequencing Ready Reaction (Applied
Biosystems, USA) with an ABI 3130 XL capillary automatic DNA analyzer.
Sequences of total HA genes were edited using the Sequencher software, version
4.10 (Gene Codes, Ann Arbor, USA) and aligned with the vaccine prototype
A/California/07/2009 and other sequences available in GenBank
(www.ncbi.nlm.nih.gov) using the Molecular Evolutionary Genetics Analyses software
(MEGA), version 5.0.5 (Tamura et al. 2011). The phylogenetic analysis was
performed using the neighbor-joining method. The thirty-one sequences generated in
this study are available in the GenBank database under the accession numbers
CY103911-CY103941.
The resistance marker H274Y was not detected in any of 118 samples
evaluated in this study, which is in agreement with previous data, suggesting a very
low frequency of resistance in influenza A(H1N1)pdm09 in comparison to seasonal
viruses (Souza et al. 2010; Malato et al. 2011; Chen et al. 2011; Harvala et al. 2011
and Kawano et al. 2011).
Twenty-one out of thirty-one HA sequences obtained through automatic
sequencing came from hospitalized patients and two of then evolved to death. The
sequences presented several mutations in comparison to A/California/07/2009, which
are listed in Table I. In total, fourteen mutations were identified from which P100S
and I338V were found in all evaluated sequences. The mutation S220T was detected
in 23 samples, the majority (15 samples) from hospitalized patients, wheras Q310
was detected in six out of this subgroup. It is noteworthy that whereas mutation
D114N was most frequently found in the outpatient group, single mutations N277K,
K325E, Q310R, Y368F and I550L were detected only in patients requiring
hospitalization.
Aiming to identify any possible association between the clinical status of the
patient and genetic background, we sought for the putative virulence markers D222G
(WHO, 2009) and Q310H (Glinsky et al. 2010) in the HA gene. The former was not

133
detected in this study and the latter was detected in similar proportions in both patient
groups (28.6% and 30% for outpatient and hospitalized groups, respectively). We
could not associate this mutation with severe or fatal cases. These results
corroborate the results previously reported with Brazilian and Indian samples (Lee et
al. 2010; Potdar et al. 2010). The mutations P100S and I338V were identified in all
samples from both groups in this study, being characteristic of samples from PE.
Notably, mutation P100S was associated with extended viral shedding, with the
recovery of virus from patients being possible over a period of two months (Souza et
al. 2010).
Phylogenetic relationships between PE samples and other recent brazilian
isolates can be seen in Figure 1. PE samples were spread in two large clusters (main
and lower clusters), being grouped together with other Brazilian samples collected at
the same period from States further away from Pernambuco State, such as Santa
Catarina or Rio Grande do Sul, in the South of Brazil. The two clusters were
composed of some minor subclusters most of then characterized by single amino
acid changes. All samples were highly similar to the vaccine prototype, in agreement
with recent publications (Ikonen et al. 2010; Melidou et al. 2010; Barr et al. 2010).
Notably, the majority of PE samples presented the mutation S220T with only seven
samples being similar to the vaccine prototype for such a residue (Figure1, bottom
cluster). Otherwise, all samples in the bottom cluster presented the mutation
Q310H/R, with this mutation being found in only two samples in the main upper
cluster: A/Pernambuco/175/09 and A/Pernambuco/414/09. Regarding the
subclusters, those characterized by mutations D114N, P199Q and K325E were
constituted basically with samples from PE (including the oldest sample rooting each
subcluster), which could indicate at least two transmission and evolution chains
contained within the State (Figure1).
In this study, the genetic profile of influenza A(H1N1)pdm09 virus collected in
PE state during 2009-2010 has been described. There was no significant difference
in PE HA sequences when compared with those from other Brazilian regions -
samples from distinct regions were extensively grouped together. These regional
subclusters, formed by samples collected between July/2009 and April/2010, are
indicative of co-circulation of lineages, which is not only expected, as previously
observed (Galiano et al., 2011). In addition, we did not identify the mutation H274Y
by pyrosequencing , indicating that all samples were sensitive to oseltamivir.

134
Even after the WHO statement prompting the end of the pandemic, influenza
A(H1N1)pdm09 still circulates throughout the world, even provoking fatal cases. The
continuous monitoring of viral evolution and the emergence of oseltamivir resistance
are pivotal for worldwide influenza surveillance, and the findings depicted here
contribute to a better understanding of the genetic characteristics of influenza
A(H1N1)pdm09 virus that circulated in the Northeast of Brazil during the pandemic
period.
Acknowledgements
This work was realized in the Respiratory Viruses and Measles Laboratory and was
supported by IOC/FIOCRUZ.
REFERENCES: Abed Y, Baz M, Boivin G 2006. Impact of neuraminidase mutations conferring influenza resistance to neuraminidase inhibitors in the N1 and N2 genetic backgrounds. Antivir Ther 11: 971-976. Adwan G 2010. Molecular characterization and phylogenetic analysis of Middle East 2009 H1N1 pdm isolates. A PJTM: 624-628. Aoki FY, Boivin G, Roberts N 2007. Influenza virus susceptibility and resistance to oseltamivir. Antivir Ther 12: 603-616. Arias CF, Escalera-Zammudio M, Soto-Del Rio, Cóbian-Güemes AG, Isa P, López S 2009. Molecular Anatomy of 2009 Influenza Vírus A (H1N1). Arch Med Res 40: 643-654. Barr IG, Cui L, Komandina N, Lee RT, Lin RT, Deng Y, Caldwell N, Shaw R, Maurer-Stroh S 2010. A new pandemic influenza A(H1N1) genetic variant predominated in the winter 2010 influenza season in Australia, New Zealand and Singapore. Euro Surveill 15 (42): pii = 19692. Chen LF, Dailey NJ, Rao AK, Fleischauer AT, Greenwald I, Deyde VM, Moore ZS, Anderson DJ, Duffy J, Gubareva LV, Sexton DJ, Fry AM, Srinivasan A, Wolfe CR 2011. Cluster of Oseltamivir-Resistant 2009 Pandemic Influenza A (H1N1) Vírus Infections on a Hospital Ward among Immunocompromised Patients-- North Carolina, 2009. J. Infect Dis 203: 838-846. Dharan NJ, Gubareva LV, Meyer JJ, Okomo-Adhiambo M, McClinton RC, Marshall SA , St George K, Epperson S, Drammer L, Klimov AI, Bresee JS, Fry AM 2009.

135
Infections With Oseltamivir-Resistant Influenza A (H1N1) Virus in the United States. JAMA 301:1034-1041. Deyde VM, Okomo-Adhiambo M, Sheu TG, Wallis TR, Fry A, Dharan N, Klimov AI, Gubareva LV 2009. Pyrosequencing as a tool to detect molecular markers of resistance to neuraminidase inibitors in seasonal influenza A viruses. Antiviral Research 81: 16-24. Eshaghi A, Bolotin S, Burton L, Low DE, Mazzuli T, Drews SJ 2009. Genetic microheterogeneity of emerging H275Y influenza virus A (H1N1) in Toronto, Ontario, Canadá from 2007 – 2008 respiratory season. J Clin Virol 45:142–145. Felsenstein J 1985. Confidence limits on phylogenies: Na approach using the bootstrap. Evolution 39: 783-791. Ferreira JLP, Borborema SET, Brígido LFM, Oliveira MI, Paiva TM, Santos CLS 2011. Sequence analysis of the 2009 pandemic influenza A H1N1 virus haemagglutinin gene from 2009-2010 Brazilian clinical samples. Mem Inst Oswaldo Cruz 106: 613-616. Galiano M, Agapow P-M, Thompson C, Platt S, Underwood A, Ellis J, Myers R, Green J, Zambon M 2011. Evolutionary Pathways of the Pandemic Influenza A (H1N1) 2009 in the UK. PLoS ONE 6(8): e23779 doi: 10.1371. Glinsky GV 2010. Genomic analysis of pandemic (H1N1) 2009 reveals association of increasing disease severity with emergence of novel hemagglutinin mutations. Cell Cycle 9: 958-970. Graham M, Liang B, Domselaar GV, Bastien N, Beaudoin C, Tyler S, Kaplen B, the National Influenza A/H1N1pdm Genomics Study Team (NIGST), Li Y. 2011. Nationwide Molecular Surveillance of Pandemic H1N1 Influenza A Virus Genomes: Canada, 2009. PLoS One 6: e16087 doi: 10.1371. Harvala H, Gunson R, Simmonds P, Hardie A, Bennett S, Scott F, Roddie H, McKnight J, Walsh T, Rowney D, Clark A, Bremner J, Aitken C, Templeton K 2010. The Emergence Of Oseltamivir-Resistant Pandemic Influenza A(H1n1) 2009 Virus Amongst Hospitalised Immunocompromised Patients In Scotland, November-December, 2009. Euro Surveil 15: pii= 19536. Ikonen N, Haanpää M, Rönkkö E, Lyytikäinen O, Kuusi M, Ruutu P, Kallio-Kokko H, Mannonen L, Lappalainen M, Ziegler T, Julkunen I 2010. Genetic Diversity of the 2009 pandemic influenza A (H1N1) viruses in Finland. PLoS One 20: e13329 doi: 10.1371. Felsenstein J 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39:783-791.
Kawano H, Haruyama T, Hayashi Y, Sinoda Y, Sonoda M, Kobayashi N 2011. Genetic Analysis and Phylogenetic Caracterization of Pandemic (H1N1) 2009 Influenza Viruses that found in Nagasaki. Jpn J Infect Dis 63: 195-203.

136
Kilander A, Rykkvin R, Dudman S. Hungnes O 2010. Observed association between the HA1 mutation D222G in the 2009 pandemic influenza A(H1N1) virus and severe clinical outcome, Norway 2009-2010. Euro Surveill 15 (9): pii = 19498. Ledesma J, Pozo F, Ruiz MP, Navarro JM, Piñeiro L, Montes M, Castro SP, Fernández JS, Costa JG, Fernández M, Galán JC, Cuevas MT, Casas I, Breña PP 2011. Substitutions in position 222 of haemagglutinin of pandemic influenza A (H1N1) 2009 viruses in Spain. J Clin Virol 51: 75-78. Lee RT, Santos CL, de Paiva TM, Cui L, Sirota FL, Eisenhaber F, Maurer-Stroh S 2010. All that glitters is not gold - founder effects complicate associations of flu mutations to disease severity. Virol J. 1: 297. Le QM, Wertheim HF, Tran ND, van Doorn HR, Nguyen TH, Horby P, Vietnam H1N1 Investigation Team 2010. A Community Cluster of Oseltamivir-Resistant Cases of 2009 H1N1 Influenza. N Engl J Med 362: 86-87. Liu Y, Childs RA, Matrosovich T, Wharton S, Palma AS, Chai W, Daniels R, Gregory V, Uhlendorff J, Kiso M, Klenk H-D, Hay A, Feizi T, Matrosovich M. Altered Receptor Specificity and Cell Tropismo f D222G Hemagglutinin Mutants Isolated from Fatal Cases of Pandemic A(H1N1) 2009 Influenza Vírus. J Virol 2010; 84: 12069-74. Mak GC, Au KW, Tai LS, Chuang KC, Cheng KC, Shiu TC, Lim W. 2010. Association of D222G Substitution in Haemagglutinin of 2009 Pandemic Influenza A (H1N1) with Severe Disease. Euro Surveill 15 (14): pii =19534. Maurer-Stroh S, Lee RTC, Eisenhaber F, Cui L, Phuah SP, Lin RTP 2010. A new common mutation in the hemagglutinin of the 2009 (H1N1) influenza A virus. PLoS Curr 2: RRN1162. Melidou A, Gioula G, Exindari M, Chatzidimitriou D, Diza E, Malisiovas N 2010 Molecular and phylogenetic analysis of the haemagglutinin gene of pandemic influenza H1N1 2009 viruses associated with severe and fatal infections. Virus Res 151: 192-199. Miller RS, MacLean AR, Gunson RN, Carman WF 2010. Occurrence of Haemagglutinin Mutation D222G in Pandemic Influenza A(H1N1) Infected Patients in the West of Scotland, United Kingdom, 2009-10. Euro Surveill 15 (14) pii = 19546. Morlighem JE, Aoki S, Kishina M, Hanami M, Ogawa C, Jalloh A, Takahashi Y, Kaway Y, Saga S, Hayashi E, Ban T, Izumi S, Wada A, Mano M, Fukunaga M, Kijima Y, Shiomi M, Inoue K, Hata T, Koretsune Y, Kudo K, Himeno Y, Hirai A, Takahashi K, Sakai-Tagawa Y, Iwatsuki-Horimoto K, Kawaoka Y, Hayashizaki Y, Ishikawa T 2011. Mutation Analysis of 2009 Pandemic Influenza A (H1N1) Viruses Collected in Japan during the Peak Phase of the Pandemic. PLoS One Curr 6: e18956 PMCID: PMC3084724. Moscona A 2005. Global Transmission of Oseltamivir-Resistant Influenza. The N Engl J Med 360: 953-956.

137
Motta FC, Siqueira MM, Lugon AK, Straliotto SM, Fernandes SB, Krawczuk MM 2006. The reappearance of Victoria lineage influenza B virus in Brazil, antigenic and molecular analysis. J Clin Virol 36: 208-214.
Potdar VA, Chadha MS, Jadhav SM, Mullick J, Cherian SS, Mishra AC 2010. Genetic Characterization of the Influenza A Pandemic (H1N1) 2009 Virus Isolates from India. PLoS One Curr 5: e9693, PMCID: PMC 2837743. Saitou N, Nei M 1987. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406-425. Sanger F, Donelson JE, Coulson AE, Kössel H, Fischer D 1974.Determination of a nucleotide sequence in bacteriophage f1 DNA by primed synthesis with DNA polymerase. J Mol Biol 90: 315-326.
Souza TML, Salluh JIF, Bozza FA, Mesquita M, Soares M, Motta FC, Pitrowsky MT, Oliveira ML, Mishin VP, Gubareva LV, Whitney A, Rocco AS, Gonçalves VMC, Marques VP, Velasco E, Siqueira MM 2010. H1N1pdm influenza Infection in Hospitalized Cancer Patients: Clinical Evolution and Viral Analysis. PLoS ONE 5: e14158 doi:10.1371. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S 2011. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood Evolutionary Distance and Maximum Parsimony Methods. Mol Biol Evol 28: 2731-2739. Wagner R, Matrosovich M, Klenk HD 2002. Functional balance between haemagglutinin and neuraminidase in influenza virus infections. Rev Med Virol. 12:159-66. World Health Organization. Influenza A (H1N1) virus resistance to oseltamivir - 2008-2009 influenza season northern hemisphere. 18 March 2009. World Health Organization. Pandemic (H1N1) 2009 – update 112. 6 August 2010. Available from: http://www.who.int/csr/don/2010_08_06/en/index.html>.
Zheng B, Chan KH, Zhang AJ, Zhou J, Chan CC, Poon VK, Zhang K, Leung VH, Jin DY, Woo PC, Chan JF, To KK, Chen H, Yuen KY 2010. D225G mutation in hemagglutinin of pandemic influenza H1N1 (2009) virus enhances virulence in mice. Exp Biol Med 235: 981-988.

138
Table I – Influenza A(H1N1)pdm09 HA aminoacid substitutions among hospitalized and
outpatient patients attended at Pernambuco reference hospitals, Brazil, 2009-2010.
Patient Type Residue Outpatients
(n=10) Hospitalized
(n=21)
Total (n=31)
S88F 01 1 S91N 01 02 3
P100S 10 21 31 S101N 01 02 2 D114N 03 02 5 P199Q 01 06 7 S220T 08 15 23 N277K - 01 1 Q310H 03 06 9 Q310R - 01 1 K325E - 04 4 I338V 10 21 31 Y368F - 01 1 I550L - 01 1

139
Alagoas - Mar 2010
CY072089 A/Rio de Janeiro/88/2010 2010/03/04
HM624015 A/Sao Paulo/15463/2010 2010/03/08
Paraná - 2010
CY080261 A/Shantou/8049/2010 2010/01/05
CY096274 A/Melbourne/INS475/2010 2010/08/31
Chile - Aug 2010
CY103912 A/Pernambuco/117/2009 2009/07/23
GQ915023 A/Sao Paulo/55312/2009 2009/08/05
CY103914 A/Pernambuco/175/2009 2009/08/05
CY103915 A/Pernambuco/177/2009 2009/08/05
CY103917 A/Pernambuco/231/2009 2009/08/09
CY103932 A/Pernambuco/592/2009 2009/09/18
CY103941 A/Pernambuco/82/2009 2009/07/02
CY072074 A/Alagoas/115/2010 2010/03/05
CY103926 A/Pernambuco/458/2009 2009/08/28
CY103930 A/Pernambuco/567/2009 2009/09/12
CY052048 A/Rio Grande do Sul/4509/2009 2009/07/18
CY103937 A/Pernambuco/688/2009 2009/11/04
CY103933 A/Pernambuco/609/2009 2009/09/22
CY103936 A/Pernambuco/66/2009 2009/06/29
CY103920 A/Pernambuco/402/2009 2009/08/19
CY074045 A/Argentina/HNRG49/2009 2009/06/16
CY052350 A/Santa Catarina/460/2009 2009/06/10
CY103928 A/Pernambuco/518/2009 2009/09/04
CY103938 A/Pernambuco/732/2010 2010/01/14
CY072087 A/Bahia/231/2010 2010/04/01
CY103940 A/Pernambuco/768/2010 2010/04/15
CY103939 A/Pernambuco/744/2010 2010/03/06
GQ368665 A/Mato Grosso/2329/2009 2009/05/28
HQ595285 A/Sao Paulo/89876/2009 2009/10/19
GQ368667 A/Distrito Federal/2611/2009 2009/06/05
CY060444 A/Bahia/12606/2009 2009/08/18
CY103918 A/Pernambuco/247/2009 2009/08/11
CY063510 A/Boston/145/2009 2009/07/09
CY054283 A/Santa Catarina/459/2009 2009/06/11
CY103923 A/Pernambuco/42/2009 2009/06/27
CY103916 A/Pernambuco/181/2009 2009/08/05
HQ595263 A/Goias/69870/2009 2009/08/25
CY103911 A/Pernambuco/107/2009 2009/07/17
CY103919 A/Pernambuco/397/2009 2009/09/24
CY103929 A/Pernambuco/565/2009 2009/09/11
HQ228042 A/Finland/592/2009 2009/06/23
CY052050 A/Rio Grande do Sul/4782/2009 2009/07/20
HQ595260 A/Piaui/69692/2009 2009/08/25
GQ414764 A/Sao Paulo/2056/2009 2009/05/20
CY075379 A/Chile/3586/2009 2009/07/03
CY103922 A/Pernambuco/414/2009 2009/08/20
CY060452 A/Santa Catarina/6235/2009 2009/07/26
CY103921 A/Pernambuco/409/2009 2009/08/21
Bahia - Mar 2010
CY103913 A/Pernambuco/126/2009 2009/07/25
CY103934 A/Pernambuco/624/2009 2009/09/24
CY103925 A/Pernambuco/447/2009 2009/08/30
Rio de Janeiro - Aug 2009
CY103935 A/Pernambuco/643/2009 2009/10/03
HQ595252 A/Distrito Federal/62652/2009 2009/08/13
CY103924 A/Pernambuco/431/2009 2009/08/27
CY072088 A/Rio de Janeiro/491/2010 2010/02/18
HQ595234 A/Sao Paulo/50280/2009 2009/07/27
CY103927 A/Pernambuco/513/2009 2009/09/03
CY051988 A/Norway/2917/2009 2009/08/11
CY103931 A/Pernambuco/590/2009 2009/09/18
CY054281 A/Rio de Janeiro/9685/2009 2009/08/10
FJ969540 A/California/07/2009 2009/04/09
91
80
66
54
56
60
71
68
51
64
59
50
73
53
55
64
63
0.001
S220T
Q310H
K325E
P199Q
D114N
E391K
S101N
†
†
Bottom cluster
Main cluster

140
Figure1: Neighbor-Joining phylogenetic tree of complete HA gene sequences of A(H1N1)pdm09 viruses. Some sequences downloaded from GenBank were included to represent distinct clusters circulating in the world during 2009-2010 period. The collection date is displayed after the sample identification of each sample. The prototype sample A/California/07/2009 was used to root the tree. ✟: deceased patient.

141
APÊNDICE B – VERSÃO DO ARTIGO 2 EM INGLÊS
SUBMETIDO À REVISTA: Journal of Clinical Virology
EPIDEMIOLOGICAL AND VIROLOGICAL ASPECTS OF INFLUENZA A(H1N1)PDM09 INFECTIONS AND FREQUENCY OTHER RESPIRATORY VIRUSES IN PERNAMBUCO STATE, BRAZIL: 2009-2010
Maria José Couto Oliveira1,2+, Marilda M Siqueira3, Maria de Lourdes Aguiar
Oliveira3, Fernando do Couto Motta3, Marli Tenório Cordeiro1,2, Sharon Carney 3,
Wyller Alencar de Mello4, Vera Magalhães da Silveira2.
1 Virology Sector, Central Public Health Laboratory of Pernambuco (LACEN-PE),
Recife, PE, Brazil. 2 Postgraduate Program in Tropical Medicine, Federal University of Pernambuco,
Recife, PE, Brazil. 3 Respiratory Viruses and Measles Laboratory of Oswaldo Cruz Institute, Fiocruz,
Rio de Janeiro, Brazil. 3 Evandro Chagas Institute, Belém, Pará, Brasil.
+Address correspondence to:
Maria José Couto Oliveira
Rua do Espinheiro, 360, Apto. 1801 – Espinheiro
Recife – PE, CEP 52020-020
e-mail: [email protected]

142
ABSTRACT
Acute respiratory infections can be caused by different pathogens and relevant
morbid-mortality patterns can be found among some risk groups. The
epidemiological features of influenza A(H1N1)pdm09 and the prevalence of other
respiratory infections were investigated in Pernambuco state, during the 2009-2010
pandemic period. In this cross-sectional study, biological samples (nasopharyngeal
swab/lung fragments) from 705 Influenza suspected cases were tested by real time
PCR for influenza A(H1N1)pdm09 (CDC protocol) and a subsample (146 FluA-
negative) was tested for the presence of respiratory viruses. Epidemiological/clinical
data were collected using a standardized formulary. Descriptive/bivariate analyses
(Chi-square and/or Fisher exact test and T-test) were performed and significance
was considered as p<.005. The prevalence of influenza A(H1N1)pdm09 infection was
26.4%; being significantly higher among males (p=.023) and those aged between 6-
19 years (p>.001). Moreover 12 patients (6,4%) died. Among InfluenzaA-negative
subsample, other viruses could be detected in 53 (36.3%), as follows: Rhinovirus
(41,0%), Coronavirus (16.1%), Metapneumovirus (14.3%), Parainfluenza (8.9%).
Bocavirus (7.1%), Respiratory syncytial virus (5.4%), Influenza B (3.6%), Adenovirus
(1.8%) and Enterovirus (1.8%). Multiple infections were found in 3 cases. The
magnitude of Flu pandemic in Pernambuco was lower than in South/Southeast of
Brazil. Besides influenza A(H1N1)pdm09, other respiratory pathogens were
circulating during 2009-2010 Influenza season, and seemed to be responsible for
about one third of non-FluA clinical cases. Despite its importance as a diagnostic
tool, these results should be interpreted according to the clinical/epidemiological
contexts. New studies are under development to assess the Public Health impact of
these other pathogens.
Key-words: Influenza, Respiratory viruses, acute respiratory infection, Parainfluenza,
Coronavirus, Metapneumovirus, Bocavirus, Rhinovirus, Respiratory Syncytial Vírus.

143
INTRODUCTION
Globally, acute respiratory infections (ARI) are an important cause of morbidity
and mortality, especially among some groups, such as elderly, children and
immunocompromised patients.1,2 Most of these infections have a viral etiology3 and
are clinically similar to Influenza infections, being altogether referred as influenza-like
illness (ILI).4 Besides Influenza,5 other viruses are associated with ILI like respiratory
syncytial virus (RSV),6 Rhinovirus(RV),7 coronavirus (HCoV),8 parainfluenza (PIV)
viruses 1-4,9 adenovirus (AdV)10, enterovirus (EV),11 parechovirus (PeV)12 and the
lately identified metapneumovirus (HMPV)13 and bocavirus (HBoV).14 Recently, real
time PCR have been developed for detection of multiple respiratory pathogens.15-19
However, the clinical and public health relevance of these infections are still under
study, especially in Brazil, where data is very scarce. 2
The aim of this study was to explore the virological and epidemiological
aspects of influenza A(H1N1)pdm09 infections in Pernambuco during the 2009-2010
pandemic period as to investigate the prevalence of other viral pathogens in FluA-
negative clinical cases.
METHODS
Study design and population
This retrospective cross-sectional study, 705 ARI patients were investigated from
May 2009-May 2010. Subjects were attended at reference hospitals from
Pernambuco state. Until July, 2009 case classification criteria consisted of fever
>38ºC, cough and directed contact with travelers to areas with confirmed influenza
A(H1N1)pdm09 in the last 10 days. After sustained viral transmission in Brazil (epi
week 29/09), criteria included severe acute respiratory cases, characterized as the
presence of dyspnea, fever >38ºC and cough as well as fatal cases19. During
hospitalization, clinical samples (Nasopharyngeal swabs and/or lung fragments) were
collected for lab testing. For other viral pathogens detection, a subsample composed
of 146 FluA-negative cases was selected, representative of epidemiological weeks.

144
Influenza A (H1N1)pdm09 and other respiratory viruses detection
For Influenza A(H1N1)pdm09, RNA was extracted by using the kit QIAamp®
Viral RNA Mini Kit (QIAGEN Inc., CA) and real time RT-PCR was conduced
according to the CDC protocol 21 (accessible in
<http://www.who.int/csr/resources/publications/swineflu/CDCRealtimeRTPCR_
SwineH1Assay-2009_20090430.pdf>). These assays were performed at Evandro
Chagas Institute (Belém, Pará state, Brazil).
For real time detection of multiple respiratory viral pathogens, the nucleic acid
extraction was carried out using the Magna Pure LC Total Nucleic Acid Isolation Kit
(Roche Diagnostics), followed by Fast track21 plus® kit (Laboratoires Réunis –
Luxemburgo). This test is able to detect 23 respiratory pathogens as seasonal FLUA,
FLUB, FLUA(H1N1)pdm09, RV, HCoV229, HCoV63, HCoV43 e HCoV KHU; PIV-2,
PIV-3, PIV-4 HBoV, HMPV, PIV-1, AdV, EV, PeV, RSV; Chlamydia pneumoniae,
Haemophilus influenzae, Staphylococcus aureus and Streptococcus pneumoniae.
Only viruses were considered in this investigation. These assays were performed at
Respiratory Virus Laboratory from Oswaldo Cruz Foundation (IOC, FIOCRUZ), Rio
de Janeiro.
Statistical Analyses
Descriptive and bivariate (Chi-square, Fisher exact test and T-test) analyses
were performed using STATA software, version 9.0. Putative associations between
viral infections and demographic, clinical and epidemiological variables were
assessed and significance was established when p <.005.
Ethical aspects
This study was approved by the Health Sciences CENTERS from the Federal
Pernambuco University IRB.
RESULTS
Demographic, clinical and epidemiological features of Influenza A(H1N1)pdm09
infections

145
The demographic, clinical and epidemiological features of Influenza
A(H1N1)pdm09 infections are shown in Table I.
Within the studied period, 839 Flu cases were notified and samples were
collected from 705 subjects (84.2%). From these, 453 (64.3%) and 252 (35.7%) were
from female and male gender, respectively, and the mean age was 29,4 (±19) years
(range 3 months to 89 years). In 67.4%, samples were collected up to 3 days after
the first symptoms and in 7.2% after 8 days. From the 705 subjects, 209 were
influenza A positive at CDC test and 186 (26,4%) were positive for influenza
A(H1N1)pdm09. From these, 106 (23.4%) were females and 80 (31.7%) were males,
with mean age of 24,6 (±15.6) years (range 1 month to 82 years). A significantly
higher prevalence was found among those aged between 6-19 years (p<.001). The
most frequent symptoms were fever, cough, dyspnea, according to the case
classification criteria from the Brazilian MoH. Two peaks on the number of influenza
A(H1N1)pdm09 cases were observed: the first at the epidemiological week 26 (13
cases) and the second between weeks 30 and 38 (113 cases) (Figure 1).
During the Flu pandemics, pregnant women were one of the main risk groups
for severe outcomes.22 In Pernambuco state, among 307 women at reproducing
age, 23.5% (CI: 18.7% a 28.2%) were positive for influenza A(H1N1)pdm09. From
these, 22 (27.8%) were pregnant. From 79 pregnant, 22 (27.8%) were positive for
influenza A (H1N1)pdm09.
Influenza A(H1N1)pdm09 fatal cases
The demographic and epidemiological characteristics of influenza
A(H1N1)pdm09 fatal cases are described in Table II. Among fatal cases (35),
influenza A(H1N1)pdm09 were detected in 12 (34.3%) subjects (8 in 2009 e 4 in
2010). The diseases occurred in both genders and the age range was of 4-53 years.
In the two pregnant women who died (cases 4, 7 and 12, with 21, 28 and 23 years,
respectively) pregnancy was the only reported risk factor.
Other viral respiratory infections
Among the 146 influenza A(H1N1)pdm09-negative samples, other respiratory
pathogen was detected in 53 (36.3%) of subjects (Table III).

146
The most prevalent viruses were Rhinovirus (n=23), metapneumovirus (n=8) e
Coronavirus OC43 (n=8). Within the Rhinovirus cases mean age was 23.1 15.3
years, with major prevalence among males (19.6%). The peak of cases occurred
between July and September - the period of higher influenza A(H1N1)pdm09
circulation and notification of ARI cases. In three samples, double Rhinovirus +
Bocavirus; Rhinovirus + Metapneumovirus e Influenza B + Coronavirus OC43 were
observed. Furthermore, among the 5 fatal cases which were submitted to multiplex
testing, other viral pathogens were found in only one sample (RSV).
DISCUSSION
The prevalence of influenza A(H1N1)pdm09 infection during 2009-2010 in
Pernambuco was 26.4%, in such a way that the magnitude of Flu pandemics in our
state was significantly lower than those observed in the South and Southeast of
Brazil.22 The findings corroborate other reports on the different regional profiles in the
country.2 According to the Pernambuco Vigillance Surveillance Service, 846
suspected cases were notified between may 2009 – 2010. The mortality coefficient
per 100.000 habitants was 0.1 in 2009 and 0.04 in 2010 (oral communication ).
In Pernambuco, the frequency of infections was significantly higher among
males than in females (p=0,023), contrasting with the profile previously described in
Brazil.2,24 Similar figures were also found for age strata. Whereas in Pernambuco the
most affected subjects were aged between 6-19 years (p<.001), national data
identified the young adults ranged between 20 a 29 years, the main affected group
within the 2009-2010 period.24 According to the national 2 and international
literature,25 those aged ≥60 years showed lower prevalence of viral infection than
their counterparts. As previously published, these event seem to be due to previous
exposition and acquired cross-immunity between Flu viruses, especially among those
born before 1957.26
Chronic respiratory disease was the main risk factor, reported by 40.4% of
subjects, as also observed in national data.23 Despite the small sample composed of
pregnant women, about 1/3 was infected by influenza A(H1N1)pdm09, including
three fatal cases. These findings corroborate the literature, where pregnancy figures

147
as one of the most important risk groups for severe disease and outcomes from
Influenza infections.27, 28
Among the influenza A(H1N1)pdm09 suspected cases, only 29.4% were
positive for the pandemic Flu. The remaining 496 cases had their etiology unknown.
In Brazil, the lower rate of laboratory confirmation was seen in the Northeast region
(32.3%).23 Many factors can have contributed to the low frequency of influenza
A(H1N1)pdm09 detection: i) late sample collection - in some cases, even after the
7th Day after the symptoms; ii) the clinical material (nasopharyngeal swab, which is
less sensitive than aspirate for PCR detection;29 iii) inadequate temperature along
transportation, which could have compromised sample quality.
In order to elucidate the etiology of those FluA negative ARI cases, we tested
a subsample of 146 samples for the presence of other 18 viral pathogens. The main
finding of this study was that by using this real time RT-PCR multiplex approach, we
were able to elucidate about 40.0% of these cases, from which Rhinovirus was the
most prevalent viral pathogen (41.0%). In a Swiss study carried out among
transplanted and immunocompromised patients Rhinovirus was the second most
frequent respiratory virus (22.6%), after Coronavirus infections (32.2%).18 Similar
findings were also found in Sweden and the authors raised the hypothesis that this
high RV prevalence could have contributed to lowering influenza A(H1N1)pdm09
circulation.30,31 In our subsample, 3 (2,0%) multiple viral infections were detected: RV
+ HMPV, RV + HBoV e FLUB + HCoV-OC43. The frequency was lower than those
found in studies among children from elsewhere,17,29,32,33 but similar to the those
found in São Paulo (3.77%).34 Unfortunately, due to the small sample and low
prevalence, we were not able to explore putative associations between viral
infections and other demographic and epidemiological characteristics, in order to
better know the profile of these infections and their importance under the public
health perspective. Other study is under development in order to assess these
information.
In the PCR era, the diagnosis of viral diseases had improved significantly in
the last 15 years. However, most studies had been carried out among children.35 In
this context and with special regard to multiple infections, it is relevant to consider
that, despite the relevance of real time PCR as a very remarkable diagnostic tool,
results should be interpreted in the light of clinical data and epidemiological
plausibility, due to the high sensitivity of this technique.

148
REFERENCES: 1. Boivin G, Côté S, Déry P, De Serres G, Bergeron MG. Multiplex Real-Time PCR
Assay for Detection of Influenza and Human Respiratory Syncytial Viruses. J Clin Microbiol 2004; 42: 45-51.
2. Raboni SM, Stella V, Cruz CR, França JB, Moreira S, Gonçalves L, et al.
Laboratory Diagnosis, Epidemiology, and Clinical Outcomes of Pandemic Influenza A and Community Respiratory Viral Infections in Southern Brazil. J Clin Microbiol 2011; 49: 1287-93.
3. Costa LF, Yokosawa J, Mantese O, Oliveira TFM, Silveira Hl, Nepomuceno Ll, et al. Respiratory viruses in children younger than five years old with acute respiratory disease from 2001 to 2004 in Uberlândia, MG, Brazil. Mem Inst Oswaldo Cruz Rio de Janeiro 2006; 101: 301-6.
4. Kelly H, Birch C. The causes and diagnosis of influenza-like illness. Australian
Family Physician 2004; 33: 305-9. 5. Behrens G, Gottschalk R, Gürtler L, Harder TC, Hoffmann C, Kamps BS, et al.
Influenza Report 2006. Kamps-Hoffmann-Preiser Ed. Avaliable from: <www.InfluenzaReport.com>.
6. Murata Y, Falsey, AR. Respiratory syncytial virus infection in adults. Antiv
Ther 2007; 12: 659–70. 7. Briese T, et al. Global Distribution of Novel Rhinovirus Genotype. Emerg Infect
Dis 2008; 14: 944-977. 8. Fielding BC. Human coronavirus NL63: a clinically important virus? Future
Microbiolog 2011; 6:153-159. 9. CDC.Human Parainfluenza Viruses. Available from:
<http://www.cdc.gov/ncidod/dvrd/revb/respiratory/hpivfeat.htm> . 10. Berk AJ. Adenoviridae: The Viruses and Their Replication. In: Knipe DM, Howley
PM editors. Fields Virology 2007. 5 th ed. CD-ROM. Lippincott Williams & Wilkins; 2: 2355-94.

149
11. Pallansch M, Roos R. Enteroviruses: Polioviruses, Coxsackieviruses, Echoviruses, and Newer Enteroviruses. In: Knipe DM, Howley PM editors. Fields Virology 2007. 5 th ed. CD-ROM. Lippincott Williams & Wilkins; 2: 840-93.
12. Harvala H, Simmonds P. Human parechoviruses: biology, epidemiology and clinical significance. J Clinl Virol 2009; 45: 1-9.
13. Broor S, Bharaj P, Chahar HS. Human metapneumovirus: a new respiratory pathogen. J Biosci 2008; 33: 483-93.
14. Allander T, Jartti T, Gupta S, Niesters HGM, Lehtinen P, Osterback R, et al.
Human bocavirus and acute wheezing in children. Clin Infect Dis 2007; 44: 904-10.
15. Bellau-Pujol S, Vabret A, Legrand L, Dina J, Gouarin S, Petitjean-Lecherbonnier
J, et al. Development of three multiplex RT-PCR assays for the detection of 12 respiratory RNA viruses. J Virol Methods 2005; 126: 53-63.
16. Freymuth F, Vabret A, Cuvillon-Nimal D, Simon S, Dina J, Legrand L, et al.
Comparison of multiplex PCR assays and conventional techniques for the diagnostic of respiratory virus infections in children admitted to hospital with an acute respiratory illness. J Med Virol 2006; 78: 1498-504.
17. Bonzel L, Tenenbaum T, Schroten H, Schildgen O, Schweitzer-Krantz S, Adams
O. Frequent detection of viral coinfection in children hospitalized with acute respiratory tract infection using a real-time polymerase chain reaction. Pediatric Infect Dis J 2008; 27: 589-94.
18. Garbino J, Soccal PM, Aubert JD, Rochat T, Meylan P, Thomas Y, et al. Respiratory viruses in bronchoalveolar lavage: a hospital-based cohort study in adults. Thorax 2009; 64: 339-404.
19. Vabret A, Dina J, Gouarin S, Petitjean J, Tripey V, Brouard J, et al. Human (non sever acute respiratory syndrome) coronavirus infections in hospitalisated children in France. J Paediatr Child Health 2008; 44: 176-81.
20. Oliveira WK, Carmo EH, Penna GO, Kuchenbecker RS, Santos HB, Araújo WN,
et al. Pandemic H1N1 influenza in Brazil: Analysis of the first 34.506 notified casesof influenza-like illness with severe acute respiratory infection (SARI). Euro Surveill 2009; 14: pii=19362.

150
21. WHO. CDC Protocol of real time RT-PCR for swine influenza A (H1N1)- revision 1 (30 April 2009). WHO 2009. Available from: <http://www.who.int/csr/resources/publications/swineflu/CDCRealtimeRTPCR_SwineH1Assay-2009_20090430.pdf.>.
22. BRASIL. Ministério da Saúde. Situação Epidemiológica da Nova influenza A
(H1N1) no Brasil, 2009.
23. BRASIL. Ministério da Saúde. Boletim Eletrônico Epidemiológico. CADERNO
ESPECIAL INFLUENZA. Influenza Pandêmica (H1N1) 2009. Análise da Situação Epidemiológica e da Resposta no Ano de 2009. Ano 10. Nº 2. Março de 2010a.
24. BRASIL. Ministério da Saúde. Informe técnico quinzenal de Influenza. Influenza
Pandêmica (H1N1) 2009 – Monitoramento da Síndrome Respiratória Aguda Grave. Ed. n. 9, 2010b.
25. WHO. Acute Respiratory Infections (Update Setember 2009). Available from:
<http://www.who.int/vaccine_research/diseases/ari/en/index5.html>. 26. Hancock K, Veguilla V, Lu X, Zhong W, Butler EN, Sun Hong et al. Cross-
Reactive Antibody Responses to the 2009 Pandemic H1N1 Influenza Virus. N Engl J Med 2009; 361: 1945-52.
27. Louie JK, Acosta M, Jamieson DJ, Honein MA, California Pandemic (H1N1)
Working Group. Severe 2009 H1N1 Influenza in Pregnant and Postpartum Women in California. N Engl J Med 2010; 362: 27-35.
28. Mosby LG, Rasmussen SA, Jamieson DJ. 2009 pandemic influenza A (H1N1) in
pregnancy: a systematic review of the literature. Am J Obstet Gynecol 2011; 205:10-8.
29. Meerhoff TJ, Houben ML, Coenjaerts FE, Kimpen JL, Hofland RW, Schellevis F et
al. Detection of multiple respiratory pathogens during primary infection: nasal swab versus nasopharyngeal aspirate using real-time polymerase chain reaction. Eur J Clin Microbiol Infect Dis 29: 365-371.
30. Linde AM, Rotzén-Ostlund M, Zweygberg-Wirgart B, Rubinova S, Brytting M.
Does viral interference affect spread of influenza? Euro Surveill 2009; 8: pii=19354.

151
31. Casalegno JS, Ottmann, Duchamp MB, Escuret V, Billaud G, Frobert E et al. Rhinoviruses delayed the circulation of the pandemic influenza A (H1N1) 2009 in France. Clin Microbiol Infect 2010; 16: 326-9.
32. Wang W, Cavailler P, Ren P, Zhang J, Dong W, Yan H, et al. Molecular
monitoring of causative viruses in child acute respiratory infection in endemo-epidemic situations in Shanghai. J Clin Virol 2010; 49: 211-8.
33. Bezerra PGM, Britto MCA, Correia JB, Duarte MCMB, Fonseca AM, Rose K et al.
Viral and Atypical Bacterial Detection in Acute Respiratory Infection in Children Under Five years. PLoS ONE 2011; 6: e18928.
34. Watanabe ASA, Carraro E, Moreira L, Camargo C, Sinohara J, Puerari D, et al.
Respiratory virus infection among hospitalized patients with suspected influenza A H1N1 2009 during the first pandemic wave in Brasil. Braz J Infect Dis 2011; 15: 220-24.
35. Jartti L, Langen H, Söderlund-Venermo M, Vuorinen T, Ruuskanen O, Jartti T.
New Respiratory Viruses and the Elderly. Open Respir Med J 2011; 5: 61-9.
05
10152025303540455055606570758085
1718 192021 2223 242526 2728 2930 313233 3435 363738 3940 414243 4445 4647 484950 5152 1 2 3 4 5 6 7 8 9 10 1112 131415 1617 181920 2122
Vírus influenza A(H1N1)pdm09 IRA cases
Figure 1 – Distribution of ARI and Influenza A(H1N1)pdm09 cases according to the
epidemiological week. Pernambuco, Brazil, May 2009- May-2010.
Epidemiological Week
C a s e s

152
Table I. Demographic, clinical and epidemiological features of Influenza A
(H1N1)pdm09 infections in the state of Pernambuco, Brazil. May 2009- May 2010.
Variables
N
Positive cases for influenza A
(H1N1)pdm09 Gender
Males 252 (35,7%) 80 (31,7%)a Females 453 (64,3%) 106 (23,4%)
Age range (years)b ≤ 2 37 (5,3%) 07 (18,9%) 3 - 5 24 (3,4%) 03 (12,5%) 6 - 9 31 (4,4%) 15 (48,4%)c
10 - 19 128 (18,2%) 53 (41,4%) 20 - 39 311 (44,2%) 77 (24,8%) 40 - 49 64 (9,1%) 14 (21,9%) 50 - 59 49 (7,0%) 11 (22,4%)
≥60 59 (8,4%) 06 (10,2%) Comorbidities Chronic respiratory disease 99 (14,0%) 40 (40,4%) Cardiopathy 35 (5,0%) 05 (14,3%) Chronic metabolic disease 19 (2,7%) 05 (26,3%) Chronic renal disease 08 (1,1%) 01 (12,5%) Immunodeficiency 22 (3,1%) 06 (27,3%) Hemoglobinopathy 08 (1,1%) 03 (37,5%) Other comorbidities 147 (20,9%) 46 (31,3%) Signals and symptoms
Fever 667 (94,6%) 184 (98,9%) Cough 664 (94,2%) 179 (96,2%) Dyspnea 520 (73,8%) 142 (76,3%) Sore nose 426 (60,4%) 107 (57,5%) Myalgia 402 (57,0%) 102 (54,8%) Throat ache 347 (49,6%) 86 (46,2%) Calafrio 272 (38,6%) 70 (37,6%) Arthralgia 181 (27,1%) 35 (18,8%) Diarrea 88 (12,5%) 19 (10,2%) Conjunctivitis 44 (6,2%) 10 (5,4%) Others 128 (18,2%) 43 (23,1%)
Deaths 35 12 Interval between the first symptom and sample collection (days)
≤ 3 467 (67,4%) 133 (72,3%) 4 a 7 176 (25,4%) 40 (21,7%) ≥ 8 50 (7,2%) 11 (6,0%)
ap = 0.023; b2 patients have no age record ; cp<.001

153
Table II –Influenza A(H1N1)pdm09 fatal cases in the state of Pernambuco. May 2009-May 2010.
Sample
# Age Gender Month/
year Interval between
first symptom and hospitalization
(days)
Interval between first symptom and death (days)
Risk factor
1 27 Female 8/09 5 10 Unknown 2 50 Female 8/09 3 14 Unknown 3 53 Male 9/09 5 5 Unknown 4 28 Female 9/09 5 20 Pregnancy 5 40 Male 10/09 9 12 Unknown 6 4 Female 10/09 3 6 Unknown 7 21 Female 11/09 - 22 Pregnancy 8 24 Male 12/09 4 6 Obesity, asthma 9 29 Male 3/10 8 29 Obesity
10 47 Male 4/10 7 7 Unknown 11 31 Male 4/10 5 5 Unknown 12 23 Female 4/10 2 6 Pregnancy
Source: SINAN, MoH, Brazil
Table III – Frequency of other respiratory viral pathogens among ARI cases in the state of Pernanmbuco. May 2009-May 2010.
N=146 Viral pathogens N (%)
Negative 93 (63,7%)
Positive 53 (36,3%)
Any viral pathogen 56
Rhinovírus 23 (41,0%)
Metapneumovirus 08 (14,3%)
Coronavirus 43 08 (14,3%)
Human Bocavirus 04 (7,1%)
Respiratory syncytial virus 03 (5,4%)
Influenza B 02 (3,6%)
Parainfluenza 2 02 (3,6%)
Parainfluenza 3 02 (3,6%)
Parainfluenza 1 01 (1,8%)
Adenovirus 01 (1,8%)
Enterovirus 01 (1,8%)
Coronavirus HKU 01 (1,8%)

154
ANEXOS
ANEXO A – APROVAÇÃO DO COMITÊ DE ÉTICA

155
ANEXO B – CARTA DE ANUÊNCIA DO LACEN - PE

156
ANEXO C – CARTA DE ANUÊNCIA DA DIRETORIA GERAL DE VIGILÂNCIA EPIDEMIOLÓGICA E AMBIENTAL

157
ANEXO D – FICHA DE INVESTIGAÇÃO INFLUENZA HUMANA POR NOVO SUBTIPO (PANDÊMICO)

158