caracterizaÇÃo funcional de terpeno sintases de … · terpenos compreendem a maior e mais...
TRANSCRIPT

INSTITUTO AGRONÔMICO
CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
CARACTERIZAÇÃO FUNCIONAL DE TERPENO
SINTASES DE CITROS
CÉSAR AUGUSTO NASCIMENTO
Orientador: Dr. Marco Aurélio Takita
Dissertação submetida como requisito parcial
para obtenção do grau de Mestre em
Agricultura Tropical e Subtropical, Área de
Concentração em Genética, Melhoramento
Vegetal e Biotecnologia.
Campinas, SP
Abril 2016

Ficha elaborada pela bibliotecária do Núcleo de Informação e Documentação do Instituto Agronômico
N244c Nascimento, César Augusto
Caracterização funcional de terpeno sintases de citros /César Augusto Nascimento. Campinas, 2016. 49 fls.
Orientador: Marco Aurélio Takita Dissertação (Mestrado) Agricultura Tropical e Subtropical – Instituto Agronômico
1. Citros 2. Terpeno sintase. 3. Aroma 4.Metabolismo secundário, 5. Proteção vegetal 6. Sabor I. Takita, Marco Aurélio II. Título
CDD. 634.3


iii
DEDICATÓRIA
À minha família e amigos que, com
muito carinho е apoio, não mediram
esforços para que eu chegasse até
esta etapa de minha vida. Mãe, seu
cuidado е dedicação foi que deram, а
esperança e motivação para seguir.
Deus, sua presença significou
segurança е certeza de que eu não
estou sozinho nessa caminhada.
DEDICO

iv
AGRADECIMENTOS
Agradeço a Deus que possibilitou que essa jornada fosse possível, mesmo nas horas mais
incertas de grandes dúvidas, me mostrou o caminho certo a seguir;
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela concessão da
Bolsa, sem esse fomento não seria possível a conclusão deste trabalho;
A minha mãe, Luzia Aparecida Nascimento e ao meu padrasto, João Batista Fernandes pelo
suporte, pelos exemplos de vida, amor e fé que permeiam o que me tornei;
A minha irmã, Fernanda Aparecida Nascimento, pela amizade e carinho desde sempre;
Ao Dr. Marco Aurélio Takita, pela orientação em seu mais amplo significado e pela paciência,
amizade e confiança depositada nesses dois anos de trabalho;
À Dra. Márcia Ortiz Mayo Marques, pela disponibilidade da utilização do Laboratório de
Fitoquímica, e por me assessorar em análises de cromatografia e por compartilhar seus
conhecimentos;
Aos amigos do Laboratório de Biotecnologia do Centro APTA Citros “Sylvio Moreira”, pelos
cafés, risadas e muito trabalho;
Aos amigos do IAC, Acácia, Karina, Laura, Nicholas, Thais, Vanessa por tornarem esses dois
anos mais agradáveis;
Aos amigos da graduação Lilian, Patrícia, Mariele, Jonathan e Carlos pela amizade e
companheirismo desde os tempos de faculdade;
Aos amigos, Ricardo, Márcia, Rodrigo, Poliana e Claudio que mesmo indiretamente auxiliaram
nesse trabalho e em outras ocasiões sempre especiais e pela amizade sincera e acolhedora;
Aos amigos do alojamento Camila, Cristiano, Tiago, Simone, Bárbara, Inaiara, Laís e Romulo;
A toda equipe do IAC e do Centro APTA Citros “Sylvio Moreira”, pesquisadores, funcionários,
pós-graduandos e estagiários, que de alguma forma contribuíram para a conclusão de mais essa
etapa.

v
“Talvez não tenhamos conseguido fazer o
melhor, mas lutamos para que o melhor fosse
feito. Não somos o que deveríamos ser, não
somos o que iremos ser, mas Graças a Deus,
não somos o que éramos. ”
Martin Luther King

vi
SUMÁRIO
LISTA DE ABREVIAÇÕES E SÍMBOLOS ........................................................................... vii
LISTA DE TABELAS ............................................................................................................ viii
LISTA DE FIGURAS ............................................................................................................... ix
RESUMO .................................................................................................................................. xi
ABSTRACT ............................................................................................................................. xii
1 INTRODUÇÃO ....................................................................................................................... 1
2 REVISÃO DE LITERATURA ............................................................................................... 2
2.1 Aspectos Econômicos da Citricultura Brasileira .................................................................. 2
2.2 Terpenos ............................................................................................................................... 3
2.2.1 Classificação dos terpenos ................................................................................................. 4
2.2.2 Biossíntese de terpenos ...................................................................................................... 5
2.3 O Óleo essencial de citros .................................................................................................... 6
2.3.1 Composição dos óleos essenciais de citros ....................................................................... 9
2.4 Caracterização Funcional de Terpenos Sintases de Citros ................................................. 10
3 MATERIAL E MÉTODOS ................................................................................................... 12
3.1 Linhagens Bacterianas e Vetores. ....................................................................................... 12
3.2 Amplificação das Terpeno Sintases .................................................................................... 12
3.3 Clonagem Inicial ................................................................................................................ 14
3.3.1 Ligação em pJET 1.2 e pGEM-T..................................................................................... 14
3.3.2 Transformação bacteriana................................................................................................ 15
3.3.3 Confirmação da transformação ........................................................................................ 15
3.3.4 Extração de plasmídeo (Miniprep alcalina) ..................................................................... 16
3.4 Sequenciamento .................................................................................................................. 16
3.5 Clonagem em Vetor de Expressão...................................................................................... 17
3.6 Expressão de Proteína ......................................................................................................... 18
3.6.1 Indução da expressão de proteína .................................................................................... 18
3.6.2 Extração da proteína ........................................................................................................ 18
3.7 Ensaio de atividade enzimática .......................................................................................... 19
3.8 Identificação dos Produtos da Reação Enzimática ............................................................. 21
4 RESULTADOS E DISCUSSÃO .......................................................................................... 22
4.1 Amplificação das Terpeno Sintases .................................................................................... 22
4.2 Clonagem ............................................................................................................................ 23
4.3 Sequenciamento .................................................................................................................. 27
4.4 Construções Para Expressão Gênica................................................................................... 28
4.5 Extração de Proteína ........................................................................................................... 29
4.6 Identificação dos Produtos da Atividade Enzimática ......................................................... 32
5 CONCLUSÃO ....................................................................................................................... 41
6 REFERÊNCIAS .................................................................................................................... 43

vii
LISTA DE ABREVIAÇÕES E SÍMBOLOS
CCSM - Centro de Citricultura Sylvio Moreira
CG-EM - Cromatografia gasosa acoplado a Espectrometria de massas
DMAPP - Dimetilalil difosfato
DNA - Ácido desoxirribonucleico
DTT - Ditiotreitol
EST- Marcador de Sequência Expressa
ETDA - Ácido etilenodiaminotetracético
FAO - Food and Agriculture Organization
FCOJ - Suco Concentrado Congelado de Laranja
FPP - Farnesil difosfato
GGPP - Geranilgeranil difosfato
GPP - Geranil difosfato
IAC - Instituto Agronômico de Campinas
INCT - Institutos Nacionais de Ciência e Tecnologia
IPP - Isopentenil difosfato
IPTG - Isopropil-β-D-1-tiogalactopiranosídeo
LB - Meio Luria-Bertani
KCL - Cloreto de Potássio
MEP - 2C-metil-D- eritritol-4-fosfato
MgCl2 - Cloreto de Magnésio
MnCl - Cloreto de Manganês
MVA - Ácido Mevalônico
NaOH - Hidróxido de sódio
NaCl - Cloreto de sódio
NFC - Suco Não Concentrado
OE - Óleo Essencial
PCR - Reação de polimerase em cadeia
IR - Índice de retenção
SDS - Dodecil sulfato de sódio
SPME - Microextração por Fase Sólida
TI - Tempo de Retenção

viii
LISTA DE TABELAS
Tabela 1 – Genes de terpenos sintases caracterizados funcionalmente, até a presente data.
........................................................................................................................... 11
Tabela 2 – Lista de clones contendo sequências de terpeno sintases, resgatadas do CitEST.
........................................................................................................................... 12
Tabela 3 – Primers utilizados na amplificação dos genes de interesse contendo os sítios de
restrição SgfI e PmeI. As sequências de iniciação e terminação de transcrição do
gene de interesse estão em negrito, e os sítios de restrições estão sublinhados.
........................................................................................................................... 13
Tabela 4 – Tampões utilizados em reações de atividade enzimática. ................................. 20
Tabela 5 – Dados das sequências comparadas através da ferramenta BlastX em bancos de
dados de sequências. ......................................................................................... 28

ix
LISTA DE FIGURAS
Figura 1 – Biossíntese de terpenos em plantas superiores. MVA: Ácido mevalônico; MEP:
Metileritol Fosfato; IPP: isopentenil; DMAPP: dimetilalil difosfato; G3P:
Gliceroldeido-3-Fosfato; GPP: Geranil difosfato; FPP – Farnesil difosfato;
GGPP – Geranilgeranil difosfato. ....................................................................... 6
Figura 2 – Fluxograma do processamento da laranja. .......................................................... 7
Figura 3 – Mapa dos vetores utilizados para clonagem inicial. A – pJET 1.2; B – pGEM-T
........................................................................................................................... 14
Figura 4 – Mapa do vetor de expressão pF1K-T7 utilizado para expressão de proteína. .. 17
Figura 5 – Sistema de Captura dos voláteis utilizando a técnica de Microextração por Fase
Solida (SMPE). ................................................................................................. 19
Figura 6 – Cromatógrafo a gás acoplado a espectrômetro de massas (CG-EM). .............. 21
Figura 8 – Crescimento bacteriano em meio seletivo. Placas Meio de cultura LB contendo
ampicilina como agente seletivo foram usadas para crescimento de
transformantes, como representado para os genes CsTPS54, CrTPS15, CsTPS50
e CsTPS84. ........................................................................................................ 24
Figura 9 – Amplificação dos insertos contendo possíveis terpeno sintases de citros clonadas
em pJET 1.2 para confirmação positiva de clonagem. Os genes estão
identificados na parte superior do gel, MM – Marcador molecular 1 Kb Ladder.
........................................................................................................................... 25
Figura 10 – Análise de construções de pGEM-T. Amplificação dos genes CaTPS1,
CrTPS14, CrTPS15 e CsTPS84 de citros de modo a identificar-se aqueles que
apresentam inserto. Genes estão identificados na parte superior do gel, M –
Marcador molecular 1 Kb Ladder. .................................................................... 26
Figura 12 – Construção de plasmídeos para expressão gênica. Transformantes de E. coli
BL21 e Rosetta foram utilizados para amplificação dos genes de possíveis
terpeno sintases de citros clonadas em pF1K-T7 para confirmar aqueles que
apresentavam o inserto. Clones estão identificados na parte superior do gel, M –
Marcador molecular 1 Kb Ladder. .................................................................... 29
Figura 14 – SDS-PAGE (12% poliacrilamida). Ensaios de expressão a 37°C para a
construção CrTPS05, CrTPS26, CsTPS43 e CsTPS50 variando o tempo de
indução (0, 2 e 4 horas) As setas vermelhas indicam as bandas de expressão na
forma solúvel. ................................................................................................... 30
Figura 15 – SDS-PAGE (12% poliacrilamida). Ensaios de expressão a 37°C e 27°C para a
construção CsTPS51 variando o tempo de indução (4, 16) e a concentração de
IPTG (0,3; 0,5 e 1,0 mM). ................................................................................ 31

x
Figura 16 – Cromatograma de íons totais, utilizando solvente acetato de etila para captura
dos voláteis com GPP como substrato e tempo de reação de 1 hora a 30°C. A –
Controle (Proteínas totais de bactérias não transformadas). B – Gene CrTPS05.
........................................................................................................................... 32
Figura 17 – Cromatograma de íons totais, utilizando SPME para captura dos voláteis. GPP
como substrato e tempo de reação de 1 hora a 30°C A – Controle (Proteínas
totais de bactérias não transformadas). B – CrTPS05, C – CrTPS26, D –
CsTPS51, E – CsTPS54, F – CsTPS47. ........................................................... 33
Figura 18 – Cromatograma de íons totais, GPP como substrato e tempo de reação de 1 hora
a 30°C. A – Controle (Proteínas totais de bactérias não transformadas) B –
CsTPS51, a seta indica o pico relacionado a 1,8 cineol. ................................... 34
Figura 19 – Espectro de massa referênte ao pico com TR = 9,4 min do gene CsTPS51. A –
Espectro de massa do produto do CsTPS51, B – Espectro de massa do 1,8-cineol
obtido da biblioteca NIST62.LIB. .................................................................... 35
Figura 20 – Espectro de massa de 1,8 cineol segundo ADAMS, (2007). ............................ 35
Figura 21 – Cromatograma de íons totais, GPP como substrato e tempo de reação de 1 hora
a 30°C. A – Controle (Proteínas totais de bactérias não transformadas) B –
CrTPS26 (Pico 1 – mirceno, pico 3 - limoneno e pico 4 -1,8 cineol). .............. 36
Figura 22 – Espectros de massa dos produtos da reação do CrTPS26. A – Espectro de massa
do pico 1, C – Espectro de massas do produto do pico 3, E – Espectro de massa
do pico 4, B-D-F – Espectro de massa do mirceno, limoneno e 1,8-cineol obtido
da biblioteca NIST62.LIB. ................................................................................ 37
Figura 23 – Espectro de massa de A - Mirceno, B - limoneno C - 1,8 cineol, literatura
(ADAMS, 2007) ............................................................................................... 38

xi
Caracterização Funcional de Terpeno sintases de Citros
RESUMO
Terpenos compreendem a maior e mais diversa classe de produtos naturais obtidos de plantas.
São metabólitos secundários responsáveis por diversas funções como proteção térmica, atração
de polinizadores, estabilização de membrana, resistência contra herbívoros e microrganismos,
e sinalização planta-planta. Em Citrus spp, os terpenos estão presentes principalmente em
folhas, na epiderme dos frutos e na polpa dos frutos. O sequenciamento dos genomas de Citrus
sinensis e Citrus clementina permitiu a identificação de um grande número de possíveis terpeno
sintases entretanto, a caracterização funcional da maior parte dessas terpeno sintases ainda não
foi realizada. O presente estudo teve como objetivo a clonagem, expressão e caracterização
funcional de terpeno sintases de Citros para que, com isto, se obtivesse um maior conhecimento
da biologia/bioquímica de citros. Para isso, 18 genes foram primeiramente amplificados e
isolados a partir de clones existentes no banco do Centro de Citricultura - IAC e clonados em
pJET1.2 ou pGEM-T. Após sequenciamento, sete sequências codificadoras foram transferidas
para o vetor de expressão pF1K T7, e a indução de proteína em Escherichia coli se deu usando
IPTG. Proteínas solúveis foram extraídas e ensaios enzimáticos foram realizados com substrato
adequado e o produto sendo caracterizado por cromatografia gasosa acoplada a espectrometria
de massas. Foi possível identificar substâncias monoterpenicas, dentre elas 1,8 cineol, d-
limoneno e β-mirceno. Este estudo significou um grande passo no estudo de genômica
funcional de citros.
Palavras-chave: aroma, metabolismo secundário, proteção vegetal, sabor, terpeno sintase.

xii
Functional Characterization of Terpene synthases of Citrus
ABSTRACT
Terpenes constitute the largest class of natural products in plants, they are secondary
metabolites that have different roles like thermal protection, pollinator attraction, membrane
stabilization, resistance against herbivorous and microorganisms, plant-plant signalization. In
Citrus spp, terpenes are mainly present in leaves, fruit endocarp and epidermis. The genome
sequencing of Citrus sinensis and Citrus clementina allowed the identification of a high number
of putative terpene synthases however, the characterization of most of these terpene synthases
has not been performed. This study aimed the cloning, expression and functional
characterization of terpene synthases of Citrus, to get a better understanding of citrus
biology/biochemistry. For this, 18 genes were first PCR amplified from clones available at the
bank of the Citrus Center - IAC and cloned into pJET1.2 or pGEM-T. After sequencing to
confirm absence of errors, seven coding sequences were cloned into the pF1K T7 expression
vector, and the protein induction in Escherichia coli was attained with IPTG. Soluble proteins
were extracted and enzymatic assays were performed with appropriate substrate and the
products being characterized by gas chromatography coupled to mass spectrometry. It was
possible to identify different monoterpenes, among them 1,8 cineole, d-limonene e β-mircene.
This study represents a big step in functional genomics of citrus.
Key words: aroma, flavor, plant protection, secondary metabolism, terpene synthesis

1
1 INTRODUÇÃO
A citricultura apresenta um papel fundamental no agronegócio brasileiro, tendo a
produção de suco concentrado congelado como a principal atividade, e o Estado de São Paulo
como o principal produtor, processador e exportador (FAO, 2016).
Além do suco concentrado, outros produtos como óleo essencial, d-limoneno, e farelo
de polpa cítrica compõem uma importante parcela nas exportações brasileiras. Dentre estes, os
óleos essenciais de citros, em especial o de laranja, destacaram-se por movimentar mais de 422
milhões de dólares em exportações entre janeiro de 2012 e dezembro de 2015, constituindo o
Brasil como maior produtor mundial deste segmento de óleo essencial (CITRUSBR, 2016).
Devido à importância dos citros para o agronegócio brasileiro, o Laboratório de
Biotecnologia de Citros do Centro de Citricultura "Sylvio Moreira" do Instituto Agronômico
de Campinas viu a necessidade de investir em estudos genômicos, e assim, coordenou um
projeto de sequenciamento de ESTs (CitEST - Programa Institutos do Milênio), onde foram
obtidas aproximadamente 300 mil sequências de laranja doce, tangerina e de outras plantas de
espécies diferentes de citros ou pertencentes a gêneros correlatos (REIS et al., 2007),
constituindo-se no maior banco de sequências de citros do mundo gerado em um único
laboratório, até aquele momento. Esta base genômica possibilitou o desenvolvimento de
diversos trabalhos de bioinformática como os realizados com o banco de ESTs, que
possibilitaram a identificação de 49 possíveis terpeno sintases em Citros, bem como outros
componentes das vias de síntese de Isopentenil difosfato (IPP) e Dimetilalil difosfato (DMAPP)
(DORNELAS & MAZZAFERA, 2007; TAKITA et al., 2007). Posteriormente, o mesmo centro
de pesquisa coordenou o projeto INCT-Citros do programa Institutos Nacionais de Ciência e
Tecnologia (INCTs) do Ministério de Ciência e Tecnologia/CNPq onde, trabalhando com
sequenciamento de nova geração, focou em análises de transcriptômas (RNA-seq) e genomas
de diferentes espécies de citros e gêneros correlatos, os quais auxiliaram o Consórcio
Internacional para o Sequenciamento do Genoma de Citros, resultando em recente publicação
dos genomas de laranja doce (Citrus sinensis) e clementina (Citrus clementina) (WU et al.,
2015). A disponibilização destes genomas contribuiu para identificação de um número maior
de terpeno sintases putativas neste grupo, 120 no total, distribuídos nas duas espécies.
Entretanto a caracterização funcional da maioria destas terpeno sintases ainda não foi realizada
e, portanto, seus produtos ainda são desconhecidos. Para plantas do gênero Citrus, foram

2
caracterizadas funcionalmente algumas proteínas desta classe, como por exemplo, duas d-
limoneno sintases, duas γ-terpineno sintases, uma β-pineno síntase, uma (E)-β-ocimeno sintase
e 1,8 cineol sintase de Citrus unshiu (SHIMADA et al., 2004, 2005b; SUZUKI et al., 2004),
uma valenceno sintase de Citrus sinensis (SHARON-ASA et al., 2003), duas d-limoneno
sintases, uma β-pineno sintase e uma γ-terpineno sintase de Citrus limon (LÜCKER et al.,
2002), uma β-farneseno sintase de Citrus junos (MARUYAMA et al., 2001), e duas sabineno
sintases, uma d-limoneno sintase, e um geraniol sintase de Citrus jambhiri: (YAMASAKI &
AKIMITSU, 2007; KOHZAKI et al., 2009; SHISHIDO et al., 2012) entre outras. As proteínas
ainda não caracterizadas apresentam indicações de função baseado em análises de similaridade
com outras sequências disponíveis em bancos públicos. E, neste caso, a propagação de erros é
muito grande, o que reforça a ideia da necessidade de caracterização funcional. Além disso,
somente o conhecimento destas proteínas, com suas atividades e produtos gerados possibilita o
que se conhece hoje como engenharia metabólica "preditiva" (DIXON, 2005). Nesta, pode-se
modificar o metabolismo das plantas levando ao aumento da produção de determinada
substância envolvida na qualidade dos frutos de citros ou que tenha valor comercial. O presente
estudo teve como objetivo caracterizar funcionalmente atividades de possíveis terpeno sintases
de citros, empregando técnicas de biologia molecular ampliando assim, os conhecimento da
biologia/bioquímica de citros.
2 REVISÃO DE LITERATURA
2.1 Aspectos Econômicos da Citricultura Brasileira
A citricultura desempenha um papel importante na agroindústria brasileira, sendo o
Brasil o maior produtor mundial de laranjas, seguido pelos Estados Unidos (EUA), China,
Índia, México, Espanha e Egito. Estes países produzem 70% de toda a laranja disponível, sendo
que somente o Brasil é responsável por cerca de 30% da produção mundial, com
aproximadamente 18 milhões de toneladas anuais (FAO, 2016). No cenário nacional, a região
sudeste é responsável por 83% da produção de laranja e o Estado de São Paulo detém a
hegemonia de produção com 75% do volume nacional, sendo há vários anos o maior produtor
deste fruto (IBGE, 2015).

3
Os citros são as frutíferas mais extensamente produzidas no mundo, principalmente
laranjas, tangerinas, limas e limões, provavelmente devido ao sabor, valor nutritivo, várias
formas de consumo (in natura, sucos, licores, doces etc.), facilidade de industrialização, relativa
resistência a danos resultantes da manipulação pós-colheita e do transporte de frutos, e ampla
adaptação das plantas a diferentes condições de clima e de solo. No cultivo de citros existentes
no Brasil, os pomares de laranja doce (Citros sinensis L. Osbeck) são os mais importantes em
área e volume de frutos produzidos, tanto para consumo in natura ou processada na forma de
suco, possuindo uma área superior a 800 mil hectares (ha), distribuídos em todos os estados
brasileiros, o que representa um rendimento de 24 t ha-1. Em São Paulo, alguns pomares
produzem volumes superiores, resultado da adoção de inovações tecnológicas como produção
de mudas de alta qualidade genética e sanitária, adensamento de plantios, adubação e irrigação
(MATTOS JR. et al., 2012).
O Brasil destina 71% das laranjas que produz para a indústria, que processa e exporta
mais de 90% do suco produzido. Apenas 1% é exportado como fruto in natura, ocupando
apenas a 13ª posição mundial na exportação do fruto, e os 28% restantes são consumidos no
mercado interno. A principal característica que influi no destino da produção citrícola é o
rendimento da fruta, que vai determinar se as laranjas possuem a quantidade de sólidos solúveis
adequada para o uso industrial. A indústria utiliza 15% das laranjas na produção de NFC (“Not
From Concentrate”) e 85% na produção de FCOJ (“Frozen Concentrated Orange Juice”). O
processamento de laranja apresentou um movimento ascendente nos últimos anos, confirmando
a tendência da citricultura brasileira para a produção de suco (CITRUSBR, 2016).
Além do suco concentrado, outros produtos como óleo essencial, d-limoneno, e farelo
de polpa cítrica compõem uma importante parcela nas exportações brasileiras. Dentre estes, os
óleos essenciais de citros atingiram mais de 422 milhões de dólares em exportações entre
janeiro de 2012 e dezembro de 2015. O óleo essencial de laranja é um subproduto da produção
de suco, o que torna o Brasil seu maior produtor mundial (CITRUSBR, 2016).
2.2 Terpenos
Terpenos e terpenoides, também conhecidos como isoprenoides, além de constituírem
a maior e mais diversa classe de produtos naturais obtidos de plantas, são os que apresentam a

4
maior variedade estrutural e funcional. Apesar de alguns terpenos estarem envolvidos no
metabolismo primário de plantas em processos como respiração e desenvolvimento, como por
exemplo na produção de hormônios giberelina (HEDDEN & KAMIYA, 1997) e ácido abscísico
(SCHWARTZ et al., 1997), pigmentos fotossintéticos como carotenoides (FURUBAYASHI et
al., 2014), a vasta maioria são classificados como metabólitos secundários, tendo como função
intermediar a relação planta x ambiente. Apresentam funções ecológicas, oferecendo defesa
contra herbívoros ou patógenos, atraindo animais que dispersam pólen e sementes, ou inibindo
a germinação de plantas vizinhas, etc. (LANGENHEIM, 1994; BOUWMEESTER et al., 1999;
DICKE, 1999; PICHERSKY & GERSHENZON, 2002). A maioria destes compostos
apresentam baixa massa molecular, natureza lipofílica, grande variedade de estruturas e alta
pressão de vapor à temperatura ambiente (BAKKALI et al., 2008), sendo estimados mais de 40
mil terpenoides diferentes na natureza (CROTEAU et al., 2000; BOHLMANN & KEELING,
2008).
Os terpenos compõem a essência volátil de flores e dos óleos essenciais de ervas e
especiarias, pertencendo a uma classe de compostos naturais com propriedades biológicas
funcionais e desejáveis. Por suas características, os terpenos também são muito utilizados como
agentes de sabores e fragrâncias adicionados a alimentos, bebidas, perfumes, sabões, creme
dental e outros produtos, além de apresentarem importância farmacológica (CROWELL et al.,
1992; VAN GELDRE et al., 1997).
2.2.1 Classificação dos terpenos
Os terpenos são classificados pelo número de carbonos que eles contêm. São derivados
das moléculas precursoras de cinco carbonos, o isopentenil difosfato (IPP) e seu isômero
dimetilalil difosfato (DMAPP). Assim, a unidade básica contém cinco carbonos e é chamada
isopreno. Os menores terpenos que existem apresentam um único isopreno e são denominados
hemiterpenos. Terpenos que contém duas unidades de isopreno são chamados de monoterpenos
(C10), os quais são constituintes de componentes de essências voláteis de flores e óleos
essenciais extraídos de plantas medicinais e ervas aromáticas, justificando sua importância para
a indústria de perfumes e aromatizantes (CROTEAU et al., 2000).

5
Os sesquiterpenos (C15) são os terpenoides que derivam de três unidades de isopreno e,
portanto, contém quinze carbonos. Estes também são encontrados em óleos essenciais e vários
deles atuam como fitoalexinas, compostos antimicrobianos produzidos em resposta a ataque de
micro-organismos e compostos anti-herbivoria que desencorajam ataques de herbívoros. O
grupo dos diterpenos (C20) contém 20 carbonos e incluem resinas ácidas apresentadas por
coníferas e leguminosas, os hormônios giberelinas, fitoalexinas, e metabólitos secundários
farmacologicamente importantes (como Taxol®, quimioterápico, e forscolina, usada no
tratamento de glaucoma) (WILDUNG & CROTEAU, 1996). Os triterpenos (C30) apresentam
30 átomos de carbono e são sintetizados a partir da condensação cauda-cauda de duas unidades
de sesquiterpeno (pela extremidade fosfato). Os brassinoesteroides, fitoesteroides de
membrana, algumas fitoalexinas, toxinas e componentes de ceras cuticulares compõem esta
ampla classe de compostos químicos (CROTEAU et al., 2000).
Os terpenos que contêm oito unidades de isopreno são chamados tetraterpenos, sendo o
principal deles os carotenoides, pigmentos intimamente ligados aos processos fotossintéticos.
Terpenos que apresentam mais de oito unidades de isopreno são denominados politerpenos,
onde se encontram compostos como ubiquinonas, poliprenoides e polímeros longos como a
borracha encontrada no látex (CROTEAU et al., 2000).
2.2.2 Biossíntese de terpenos
Nas plantas superiores, existem duas vias distintas capazes de sintetizar os precursores
universais dos isoprenoides, isopentenil difosfato (IPP) e seu isômero dimetilalil difosfato
(DMAPP) (Figura 1). No citosol, IPP é gerado pela via do ácido mevalônico (MVA), e através
da ação da enzima IPP-isomerase, pode originar DMAPP; ao passo que a síntese nos
cloroplastos ocorre pela via do 2C-metil-D-eritritol-4-fosfato (MEP), sendo esta presente
também em alguns protozoários e na maioria das eubactérias (CHENG et al., 2007; PHILLIPS
et al., 2008).
DMAPP é condensado com uma, duas ou três unidades de IPP, pela ação de
preniltransferases gerando geranil difosfato (GPP), farnesil difosfato (FPP) e geranilgeranil
difosfato (GGPP), respectivamente (RAMOS-VALDIVIA et al., 1997; OGURA & KOYAMA,
1998; KOYAMA & OGURA, 1999), os quais servem como precursores para a síntese dos
monoterpenos (C10), sesquiterpenos (C15) e diterpenos (C20) por enzimas conhecidas como

6
terpeno sintases ou terpeno ciclases (CANE, 1990, 1999; WISE & CROTEAU, 1999;
MACMILLAN & BEALE, 2000).
As primeiras pesquisas indicavam que o IPP sintetizado no citosol seria precursor do
farnesil difosfato (FPP) para a síntese de sesquiterpenos e triterpenos, da mesma forma que o
IPP sintetizado nos plastídios seria precursor para o geranil difosfato (GPP), utilizado na síntese
de monoterpenos, e geranilgeranil difosfato (GGPP), utilizado na síntese de diterpenos e
tetraterpenos. Porém, através do bloqueio das vias MVA e MEP por inibidores, foi demonstrado
que a separação entre essas duas rotas é quase inexistente, uma vez IPP e DMAPP sintetizados
no citosol podem ser desviados para o metabolismo nos cloroplastos, e vice-versa, como
esquematizado na Figura 1 (BISWAS et al., 2009).
Figura 1 – Biossíntese de terpenos em plantas superiores. MVA: Ácido mevalônico; MEP:
Metileritol Fosfato; IPP: isopentenil; DMAPP: dimetilalil difosfato; G3P: Gliceroldeido-3-
Fosfato; GPP: Geranil difosfato; FPP – Farnesil difosfato; GGPP – Geranilgeranil difosfato.
Fonte: Adaptado de CHENG et al. (2007); SCHMIDT et al. (2010).
2.3 O Óleo essencial de citros
Os frutos cítricos contêm bolsões em sua casca ricos em óleos essenciais, que são
substâncias aromáticas e flavorizantes, muito utilizada por diversos setores industriais, na

7
aromatização de produtos. O óleo essencial de laranja em especial, é extraído das células do
pericarpo do fruto, e pode ser considerado um dos mais importantes subprodutos no
processamento das laranjas pela indústria de suco, apresentando crescente uso nas indústrias de
aromas graças à sua delicada e persistente fragrância, altamente estimada como adjuvante em
inúmeras composições para aromas e sabores (STUART et al., 2000).
Os processos de extração do suco e do óleo essencial são realizados pelas próprias
unidades produtoras de suco em uma única operação através de prensagem a frio (cold pressed),
método mais empregado para extração de óleos de frutos cítricos. A laranja é esmagada em
uma prensa hidráulica, e o óleo essencial contido na casca é liberado mediante esta ação
mecânica. Óleo essencial é então lavado, separado e coletado por um conjunto de técnicas que
visa sua purificação (BAUER et al., 2006).
A Figura 2 apresenta o fluxograma descrevendo as etapas do processamento da laranja
pelas unidades produtoras de suco, e a representatividade do óleo essencial de laranja nesse
processo.
Figura 2 – Fluxograma do processamento da laranja.
Fonte: TETRA PAK (1998).
Frutas
Pesagem, Seleção, Lavagem
Extração
Suco + Oleo
Finisher
Centrifuga Concentradora
Suco
Tanque pulmão
Concentração
Suco Concentrado
Recuperação Aromas
Fases Oleosas, Aquosa
Emulsão Oleosa
Centrifuga Polidora
Óleo
Deceramento
Óleo CP

8
A grande produção de laranjas no Brasil e no mundo faz com que os subprodutos
resultantes de seu processamento também tenham grande importância econômica (BENETI,
2009).
Além da obtenção do óleo essencial, diversas unidades citrícolas também fazem a sua
desterpenização, em virtude do elevado teor de hidrocarbonetos terpênicos, insolúveis em água
e quimicamente instáveis, submete-se frequentemente o óleo a um processo que reduz a fração
terpênica do óleo, que se degrada com facilidade, e concentra, ao mesmo tempo, a fração
oxigenada, responsável pelo seu sabor e aroma. Como resultado, tem-se um óleo com melhor
estabilidade, solubilidade e aroma (BENETI, 2009; AZAMBUJA, 2016).
Conforme o tipo de fruta e técnica utilizada, o rendimento máximo de extração de óleos
cítricos varia entre 0,2% e 0,5%, ou seja, para cada 1000 kg de matéria processada são obtidos
2 até 5 kg de óleo essencial na forma de emulsão (BIZZO et al., 2009).
Ao longo dos últimos vinte anos, o óleo essencial de laranja respondeu por 80% das
exportações do segmento das indústrias produtoras de óleos essenciais, sendo o primeiro item
na pauta nacional de produção, comercialização e exportação desse setor. Além de influenciar
diretamente a qualidade dos frutos por sua relação direta com aroma e sabor, os terpenos são
crescentemente utilizados na indústria para os mais diversos fins. Além da indústria alimentícia,
essa cadeia produtiva atende a diferentes segmentos de consumidores industriais, como
cosmética, perfumaria e indústrias farmacêutica, fazendo uso dos óleos essenciais, seus
constituintes e derivados, e a pectina cítrica (SANTOS & SANTOS, 2011). Grande parte desta
demanda ocorre pela importância do d-limoneno, sendo o principal terpeno no óleo essencial
de citros.
Alguns genes responsáveis pela síntese de terpenos já foram caracterizados em citros
(MARUYAMA et al., 2001; LÜCKER et al., 2002; SHARON-ASA et al., 2003; SHIMADA et
al., 2004, 2005a; YAMASAKI & AKIMITSU, 2007; KOHZAKI et al., 2009; SHISHIDO et
al., 2012). Todavia, há ainda muitos outros genes que codificam proteínas desta família,
presentes no genoma de laranja doce, que ainda não foram caracterizados funcionalmente. A
caracterização da atividade destas proteínas possibilitaria a identificação de substâncias com
possível potencial econômico que poderiam ser utilizadas em diversos processos industriais,
inclusive abrindo perspectivas novas para a economia como, por exemplo, a produção de
terpenos para a utilização como biocombustível.

9
2.3.1 Composição dos óleos essenciais de citros
Os óleos essenciais são constituídos normalmente de uma mistura complexa de
terpenos, álcoois, ésteres, éteres, aldeídos, cetonas, lactonas, fenóis, e uma pequena quantidade
de resíduos viscosos ou sólidos como parafinas, ceras, etc. Os monoterpenos e sesquiterpenos
são representados pelo d-limoneno, β-pineno, γ-terpineno, valenceno, β-Farneseno, δ-elemeno.
Estes têm efeito antiviral, antisséptico, bactericida e anti-inflamatório.
Mais de 40 compostos voláteis são encontrados nos óleos cítricos, entre os componentes
principais estão: limoneno (67.90–90.95%), 1,8-cineol (14.72%) em laranja azeda (Citrus
aurantium); limoneno (37.63–69.71%), β-pineno (0.63–31.49%), γ-terpineno (0.04–9.96%) em
limão (Citrus limon); e no caso das tangerinas (Citrus reticulata), o limoneno (51.81–69.00%),
1,8-cineol (0.01–26.43%), e γ-terpineno (2.53–14.06%) são os mais representativos, os
resultados mostram que todos os óleos essenciais de citros apresentam principalmente o
limoneno. (MOUFIDA & MARZOUK, 2003; BOURGOU et al., 2012). Segundo BAKKALI
et al. (2008), os óleos essenciais podem conter entre 2-60 componentes em concentrações muito
diferentes, e são caracterizados por dois ou três componentes principais presentes em
concentrações bastante elevadas (20-70%, em comparação com os outros componentes
presentes em pequenas quantidades), e que geralmente determinam as propriedades biológicas
dos óleos essenciais.
A composição química dos óleos essenciais de laranja doce comum (C. sinensis)
prensada a frio foram analisados por CG e CG-EM e um total de 51 e 55 componentes químicos
voláteis foram identificados respectivamente. Os principais grupos químicos foram os
hidrocarbonetos monoterpenos (94,4 e 97,3%), álcoois (1,4 e 1,0%), aldeídos alifáticos (1,6 e
0,8%) e traços de outros aldeídos (1,4%). Os principais compostos foram: limoneno; 87,9 e
92,5%, mirceno; 2,4 e 2,0%, α-pineno; 0,5 e 2,4%, linalol; 1,2 e 0,9%, octanol; 1,3 e 0,6% e
decanal, 0,2%. (NJOROGE et al., 2009).
A composição dos óleos essenciais de citros pode variar amplamente, mesmo dentro de
um mesmo gênero, bem como em tecidos diferentes (VEKIARI et al., 2002). DELLACASSA
et al. (2011) examinaram a composição química do óleo essencial extraído de três cultivares de
mandarim e dois híbridos de mandarim cultivadas no Uruguai, alguns componentes chegaram
a variar de 75 – 96% entre os óleos essenciais.

10
Os frutos cítricos contêm mais de 30 terpenoides, com casca e folhas possuindo
composições diferentes (VEKIARI et al., 2002). O óleo essencial de 22 cultivares de limão e
21 de limas foram analisadas nas mesmas condições de cultivos e clima para investigar a
composição e variação química entre eles sem interferências externas. Os resultados mostraram
a variabilidade química dos óleos essenciais de limão extraídos de folhas e casca dos frutos.
Óleos essenciais extraídos da casca da maioria das variedades de limões apresentam
principalmente o d-limoneno, β-pineno e γ-terpineno, enquanto para a maioria das variedades
de óleos das folhas de limão apresentam o d-limoneno, β-pineno, geraniol, neral (LOTA et al.,
2002).
TEIXEIRA et al (2013) avaliaram o teor e a composição dos óleos essenciais de 15
genótipos de limão (Citrus limon Burm.) pertencentes ao Banco de Germoplasma de Citros –
BAG CITROS, do Centro de Citricultura do Instituto Agronômico (IAC), em dois períodos de
colheita distintos, por cromatografia a gás e espectrometria de massas. As três principais
substâncias presentes nos óleos essenciais dos genótipos estudados foram d-limoneno (46,20 –
67,80%), γ-terpineno (8,46 -13,45%) e β-pineno (7,98 -17,81%). Chegando a variância total
dos dados de composição dos óleos essenciais de 81% entre os genótipos avaliados e as safras
estudadas. Tais variações podem ser atribuídas a dois fatores principais: genética e meio
ambiente (solo, práticas de cultura e clima).
Existem diversos outros trabalhos mostrando a composição de óleos essenciais de citros
(VERZERA et al., 1998, 2000, 2003, 2005; FRIZZO et al., 2004; SAWAMURA et al., 2004;
STATTI et al., 2004), porém há uma carência de estudos sobre os mecanismos fisiológicos,
bioquímicos e moleculares da síntese desses compostos.
2.4 Caracterização Funcional de Terpenos Sintases de Citros
A riqueza de esqueletos de carbono de terpeno pode ser atribuída a uma classe de
enzimas conhecidas como terpeno sintases. Estes catalisadores convertem os prenil difosfatos
acíclicos e esqualeno em uma infinidade de formas cíclicas e acíclicas. As principais causas da
diversidade de terpenos são o grande número de diferentes terpeno sintases existentes e pelo
fato de que algumas terpeno sintases produzirem diversos produtos (DEGENHARDT et al.,
2009).

11
Muitos genes relacionados à atividade de terpeno síntase já foram caracterizados em
diversas espécies descritas. Para plantas do gênero Citrus, foi relatado até o presente momento,
a caracterização funcional de 21 terpenos (18 monoterpenos e 3 sesquiterpenos), entretanto,
esses genes provêm de apenas cinco espécies do gênero, que por sua vez é muito diverso,
podendo apresentar uma riqueza maior desses produtos (Tabela 1).
Tabela 1 – Genes de terpenos sintases caracterizados funcionalmente, até a presente data.
Acesso Produto Descrição Espécie Referencias
AAK54279 (E)-β-Farneseno CjFS C. junos MARUYAMA et al. (2001)
AAQ04608 Valenceno CsTPS1 C. sinensis SHARON-ASA et al. (2003)
AAM53945 (-)-β-Pineno Cl(-)βPINS C. limon LÜCKER et al. (2002)
AAM53944 (+)-(4R)-Limoneno Cl(+)LIMS1 C. limon LÜCKER et al. (2002)
AAM53946 (+)-(4R)-Limoneno Cl(+)LIMS2 C. limon LÜCKER et al. (2002)
AAM53943 γ -Terpineno ClγTS C. limon LÜCKER et al. (2002)
BAD91045 1,8-Cineol CitMTSL1 C. unshiu SHIMADA et al. (2005)
BAD91046 (E)-β-Ocimeno CitMTSL4 C. unshiu SHIMADA et al. (2005)
BAD27260 β-Pineno CitMTS62 C. unshiu SHIMADA et al. (2004)
BAD27256 (+)-(4R)-Limoneno CitMTSE1 C. unshiu SHIMADA et al. (2004)
BAD27257 (+)-(4R)-Limoneno CitMTSE2 C. unshiu SHIMADA et al. (2005)
BAD27258 γ -Terpineno CitMTSL3 C. unshiu SUZUKI et al. (2004)
BAD27259 γ -Terpineno CitMTS61 C. unshiu SHIMADA et al. (2004)
BAF73932 limoneno RlemTPS1 C. jambhiri YAMASAKI et al. (2007)
BAF73933 β-Pineno RlemTPS2 C. jambhiri YAMASAKI et al. (2007)
- Sabineno RlemTPS2 C. jambhiri KOHZAKI et al. (2009)
BAM29049 Geraniol RlemTPS3 C. jambhiri SHISHIDO et al. (2012)
BAP74389 δ-elemeno RlemTPS4 C. jambhiri UJI et al. (2015)
BAP75559 Linalool CuSTS3-1 C. unshiu SHIMADA et al. (2014)
BAP75560 Linalool CuSTS3-2 C. unshiu SHIMADA et al. (2014)
BAP75561 Linalool CuSTS4 C. unshiu SHIMADA et al. (2014)

12
3 MATERIAL E MÉTODOS
3.1 Linhagens Bacterianas e Vetores.
As linhagens de Escherichia coli utilizadas foram:
- DH5α [ΔlacZ ΔM15 Δ (lacZYA-argF) U169 recA1 endA1 hsdR17(rK-mK+) supE44
thi-1 gyrA96 relA1],
- BL21 (DE3) [F– ompT gal dcm lon hsdSB(rB- mB-) λ [DE3 (lacI lacUV5-T7 gene
1ind1 sam7 nin5)] e
- ROSETTA (DE3). [F– ompT hsdSB (rB– mB–) gal dcm (DE3) pRARE (CamR)].
Os vetores utilizados foram pF1K T7 (Promega) como vetor de expressão e os vetores
pJET 1.2 (Thermo Fisher Scientific) e pGEM-T (Promega) para clonagem de amplicons.
3.2 Amplificação das Terpeno Sintases
Os clones contendo as sequências que codificam terpeno sintases foram resgatadas do
CitEST (Tabela 2), estando os genes que codificam estas proteínas clonados no vetor pSport1
(Invitrogen).
Tabela 2 – Lista de clones contendo sequências de terpeno sintases, resgatadas do CitEST.
Gene Nome CitEST Organismo
CaTPS01 CA26-C1-002-067-D10 Citrus aurantium
CrTPS05 CR05-C1-100-017-G04 Citrus reticulata
CrTPS11 CR05-C1-100-051-H03 Citrus reticulata
CrTPS14 CR05-C1-100-071-B06 Citrus reticulata
CrTPS15 CR05-C1-100-090-E06 Citrus reticulata
CrTPS26 CR05-C1-700-109-C04 Citrus reticulata
CsTPS43 CS00-C1-100-017-B09 Citrus sinensis
CsTPS47 CS00-C1-100-077-A08 Citrus sinensis
CsTPS50 CS00-C1-101-041-G09 Citrus sinensis
CsTPS51 CS00-C1-102-079-C06 Citrus sinensis
CsTPS54 CS00-C3-700-044-G09 Citrus sinensis
CsTPS64 CS00-C3-702-064-A10 Citrus sinensis
CsTPS66 CS00-C3-702-076-D05 Citrus sinensis
CsTPS67 CS00-C3-703-002-B05 Citrus sinensis
CsTPS81 CS00-C3-704-014-A01 Citrus sinensis
CsTPS84 CS00-C3-705-022-D05 Citrus sinensis
CsTPS88 CS00-C3-705-074-A07 Citrus sinensis
ClTPS92 LT33-C1-003-037-F04 Citrus latifolia

13
Para isolamento destes genes do vetor, foram desenhados primers específicos (contendo
os sítios de restrição SgfI e PmeI), de forma a amplificarmos a sequência completa da região
codificadora (Tabela 3).
Tabela 3 – Primers utilizados na amplificação dos genes de interesse contendo os sítios de
restrição SgfI e PmeI. Os códons de iniciação e terminação estão marcados em negrito, e os
sítios de restrições estão sublinhados. As sequências Forward e Reverse são referentes às
extremidades amino e carboxi das proteínas codificadas, respectivamente.
Primer Sequência (5’ > 3’)
SEQ10-C-FORWARD TGCGATCGCCATGGAAGTTTCAGCCTCTTCTGC
SEQ10-C- REVERSE AGTTTAAACTGCTGATATCGGCACAGGATT
SEQ15-C- FORWARD TGCGATCGCCATGTCTTGCATTAATCCCTCAACC
SEQ15-C- REVERSE AGTTTAAACGCCAGGAGATGCTGTGAAAG
SEQ17-C- FORWARD TGCGATCGCCATGTACTTGCAAAGACTTGGATTGG
SEQ17-C- REVERSE AGTTTAAACTCCTTTGGTGCCAGGTGATGAT
SEQ19-C- FORWARD TGCGATCGCCATGGCGGCATGCAATTTCACCAGAT
SEQ19-C- REVERSE AGTTTAAACAGGAATGGGATCAATAAATA
SEQ25-C- FORWARD TGCGATCGCCATGGATCTTAAGAGTCTTCCATCTTC
SEQ25-C- REVERSE AGTTTAAACCATGGGAAGAGGATCAACA
SEQ29-C- FORWARD TGCGATCGCCATGTCTCTTCAAGTTTCAGCCTCTCC
SEQ29-C- REVERSE AGTTTAAACCGGCACAGGATTAATAAGCAA
SEQ33-C- FORWARD TGCGATCGCCATGTCTGATGCCGATAAACCTG
SEQ33-C- REVERSE AGTTTAAACGAGCTTAATGGGGTCCCTCAG
SEQ34-C- FORWARD TGCGATCGCCATGCTAGCAACGGTTTCCAGTTCA
SEQ34-C- REVERSE AGTTTAAACAGTGACAGGGTCTCTGAGCA
SEQ40-C- FORWARD TGCGATCGCCATGTCGTCCTACTGCAGATTTCC
SEQ40-C- REVERSE AGTTTAAACAAATGGAACGTGGTCTCC
SEQ43-C- FORWARD TGCGATCGCCATGCTGAAAGCTGCTAACAAGA
SEQ43-C- REVERSE AGTTTAAACTGGGCACAGGATTAATAAGCA
SEQ52-C- FORWARD TGCGATCGCCATGTCGTCTGATTCATCCCACA
SEQ52-C- REVERSE AGTTTAAACACACTCCTTGAGTAGGCAAA
SEQ56-C- FORWARD TGCGATCGCCATGTTGATGCAGTACACCAATACGG
SEQ56-C- REVERSE AGTTTAAACTGGGAAGAGGATCAACAAGC
Estes primers foram utilizados em reações de polimerização em cadeia, PCR,
utilizando-se a GoTaq DNA polimerase (Promega). A amplificação foi feita com 1x tampão,
0,2mM de cada dNTP, 1ng de plasmídeo, 250ng de cada primer, 1U de enzima em um volume
total de 50L. As condições de ciclização foram: um passo inicial de 45 segundos a 95oC, 25

14
ciclos de 45 segundos a 95oC, 45 segundos a 55oC, 1 minutos a 72oC, e um passo final de
incubação a 72oC por 10 min. A amplificação foi verificada em gel de Agarose 0.7%.
3.3 Clonagem Inicial
3.3.1 Ligação em pJET 1.2 e pGEM-T
Os fragmentos contendo os genes de terpeno sintase previamente descritos foram
purificados utilizando-se o Illustra GFX PCR DNA and Gel Band Purification kit (GE
Healthcare), seguindo as orientações do fabricante. Os fragmentos foram clonados inicialmente
em pJET 1.2 e pGEM-T. Esses vetores foram escolhidos pois são de fácil ligação,
transformação e seleção. O pGEM-T possui um sistema de seleção por coloração (branco/azul),
onde a enzima β-galactosidase é inativada pelo inserção na região de clonagem permitindo a
identificação de recombinantes com o uso do substrato cromogênico X-Gal (Sigma), que
normalmente é quebrado pela ação da enzima, liberando 4-chloro-3-brom-indigo, que precipita
formando uma coloração azul intensa. Assim sendo, colônias que apresentaram a coloração
azul foram descartadas e as que apresentaram coloração branca foram selecionadas.
O pJET 1.2 possui o gene letal eco47IR que é interrompido pela ligação do inserto no
local de clonagem, como resultado apenas células com plasmídeos recombinantes são capazes
de se propagar. Ambos vetores possuem genes de resistência a ampicilina, facilitando assim a
seleção dos transformantes (Figura 3).
Figura 3 – Mapa dos vetores utilizados para clonagem inicial. A – pJET 1.2; B – pGEM-T
Fonte: https://www.thermofisher.com/; https://www.promega.com/
Para a ligação do amplicon no vetor pJET 1.2 utilizou-se 10μL de tampão de ligação
A B

15
2x, 100ng do amplicon, 1μL DNA Blunting Enzyme e água até o volume de 18μL, a reação foi
ligeiramente vortexada e incubada por 5 minutos a 70oC e resfriada em gelo. Posteriormente
foram adicionados 50ng de pJET1.2/blunt Cloning Vector e 1μL de T4 DNA ligase totalizando
um volume final de 20μL. Após 5 minutos em temperatura ambiente o DNA foi utilizado para
transformação. Para a ligação em pGEM-T utilizou-se 5μL de tampão de ligação 2x, 50ng do
vetor pGEM-T, 100ng do amplicon, 3U de T4 DNA ligase, e água para um volume final de
10μL. Após incubação em temperatura ambiente por 1 hora, a reação de ligação foi utilizada
para transformação de E. coli.
3.3.2 Transformação bacteriana
Células competentes das linhagens bacterianas foram preparadas com 50mM cloreto de
cálcio e armazenadas a -80oC. Cinco microlitros das ligações foram usados para transformação
misturando-se a 100L de suspensão de células da linhagem DH5 ou BL21 DE3 ou
ROSETTA. O tubo foi incubado em gelo por 30 minutos e submetido a um choque térmico a
42oC por 45 segundos. As células voltaram para o gelo por 2 minutos, adicionando-se 1mL de
meio SOC (2% triptona, 0,5% extrato de levedura, 0,05% Na Cl; 10mM MgCl2, e 20mM glicose
adicionados após autoclavação) pré-aquecido. Os tubos foram incubados a 37oC por 1 hora com
agitação (250 rpm) e 200L de células foram plaqueados em meio LB sólido (1% Na Cl, 1%
triptona, 0,5% extrato de levedura, pH 7,0, 2% ágar (0.5mM IPTG e 80µg/mL X-Gal foram
adicionados ao meio LB, apenas para os genes clonados em pGEMT-T)) contendo 100g/mL
ampicilina ou 30g/mL kanamicina. O restante das células foi concentrado por centrifugação e
também plaqueado em meio LB sólido. As placas foram incubadas durante a noite em estufa a
37oC para crescimento.
3.3.3 Confirmação da transformação
As colônias que apresentaram crescimento em meio de cultura com antibiótico
apropriado após a transformação, foram selecionadas e transferidas para uma nova placa com
meio LB sólido, PCR de colônia foi realizado para confirmação da transformação, utilizando
colônias isoladas de células transformadas como molde. A amplificação foi realizada utilizando
a enzima TopTaq Master Mix (Qiagen). Para tanto, foram usados 12,5μL do mix, 0,2μM de
cada primer, 3μL de DNA, e água para volume final de 25μL. O programa usado foi 2 minutos

16
a 95oC, 30 ciclos de 20 segundos a 94oC, 20 segundos a 58oC e 1 minuto e 30 segundos a 72oC,
seguido de um período de extensão de 10 minutos a 72oC. A amplificação foi verificada em gel
de Agarose 0.7%.
3.3.4 Extração de plasmídeo (Miniprep alcalina)
Os transformantes foram inoculados em 1mL de meio LB Liquido e crescidos durante
a noite. Foram feitas mini preparações alcalinas para extração dos plasmídeos. As células foram
coletadas por centrifugação e ressuspensão em 100L de Solução I (50mM glicose, 25mM Tris-
HCl pH 8.0, 10mM EDTA, pH 8.0). Foram adicionados 200L de Solução II (0.2N NaOH, 1%
SDS), invertendo-se o tubo e adicionando-se 150L de Solução III (60mL de 5M acetato de
potássio, 11,5mL de ácido acético glacial, em um volume total de 100mL), misturando
delicadamente. O DNA foi recuperado na fração solúvel por centrifugação e purificado através
de tratamento com igual volume de fenol:clorofórmio:álcool isoamílico (25:24:1), transferindo-
se a fase aquosa para novo tubo e precipitando-se com 1/10 de volume de Acetato de Sódio 3M
pH 5,2 e 1 volume de isopropanol, com incubação a -80˚C por 30 minutos seguido de
centrifugação por 10 minutos a 13362 g. O DNA foi lavado com 500μL de 70% Etanol gelado
e ressuspensas em 50μL de TE (10mM Tris-HCl pH 8,0, 1mM EDTA pH 8,0). Os plasmídeos
extraídos foram confirmados por PCR e sequenciados para verificação de erros na sequência,
sendo posteriormente utilizados para a clonagem em vetor de expressão.
3.4 Sequenciamento
O sequenciamento foi realizado após a clonagem dos amplicons nos vetores pJET1.2 e
pGEM-T, de forma a confirmar-se a identidade das sequências. Os primers utilizados foram
pJET Forward e pJET Reverse para genes clonados a pJET 1.2 e T7 e SP6 para genes clonados
em pGEM-T, além de primers internos, visando sequenciar a região codificante completa.
Assim, para o sequenciamento, as células contendo os plasmídeos foram crescidas em 1mL de
meio CircleGrow (BIO101) por 16 horas. Foi feito uma mini preparação alcalina seguida de
purificação com sílica (BIO101). Este DNA foi utilizado para sequenciamento, o que foi feito
utilizando-se 2,5 pmol de primer, 0,4μL de BigDye 3.1 (Applied Biosystems), 2μL de tampão
save money (2,5μL MgCL2 2M, 200μL Tris-HCl pH 9,0 1M em 1mL), 3μL de DNA em um
volume final de 10μL. As reações foram incubadas em termoclicador Veriti (Applied

17
Biosystems) com o seguinte protocolo: desnaturação inicial a 96oC por 2 minutos, 25 ciclos de
96oC por 45 segundos, 50oC por 30 segundos, 60oC por 4 minutos. As reações foram
precipitadas com 80μL isopropanol 65% e lavadas por duas vezes com 200μL etanol 60%. Os
DNAs foram secados e ressuspendidos em 10μL HiDi formamida (Applied Biosystems). As
corridas foram feitas em um sequenciador automático ABI3730 (Applied Biosystems).
3.5 Clonagem em Vetor de Expressão.
Para clonagem em vetor de expressão, foi utilizado o Flexi System (Promega), com o
vetor pF1K-T7, seguindo as instruções do fabricante. Neste sistema, vetor e fragmentos são
cortados com as enzimas de restrição SgfI e PmeI, possibilitando clonagem direcional do
fragmento, no caso, a região codificadora dos genes de terpeno sintase. O vetor contém um
promotor T7 para expressão bacteriana ou expressão de proteínas in vitro. O vetor também
contém o gene letal “barnase” para seleção positiva da inserção, e um gene de resistência à
Kanamicina para seleção do plasmídeo (Figura 4).
Figura 4 – Mapa do vetor de expressão pF1K-T7 utilizado para expressão de proteína.
Fonte: https://www.promega.com/
A digestão dos vetores contendo os genes de possíveis terpeno sintases foi realizada
utilizando 4μL do tampão de digestão, 1μg do plasmídeo, 4μL do composto de enzimas (SgfI e
PmeI) e água para 20μL. Para a digestão do vetor de expressão utilizou-se 12μL de água, 4μL
do tampão de digestão, 200ng de pF1K T7, 2μL do composto de enzimas (SgfI e PmeI).

18
As reações foram incubadas por 30 minutos a 37oC, e posteriormente aquecidas a 65oC
por 20 minutos para inativar as enzimas de restrição. Para reação de ligação utilizou-se 10μL
de tampão de ligação, 50ng do vetor digerido, 100ng do fragmento, 20 U de T4 DNA ligase,
em um volume de 20μL. Após incubação em temperatura ambiente por 1 hora, o DNA foi
utilizado para transformação de E. coli, da mesma forma como descrito no item 3.4.2
3.6 Expressão de Proteína
3.6.1 Indução da expressão de proteína
Ensaios iniciais de expressão foram realizados em E. coli BL21(DE3) e
ROSETTA(DE3) variando as concentrações de IPTG (0,25; 0,5 e 1mM) e o tempo de indução
(0, 2 e 4 horas). Os experimentos de indução foram realizados a 37°C. Para cada construção,
foram utilizados como pré-inoculo 5mL de meio LB contendo antibiótico apropriado (30g/mL
kanamicina), inoculados com amostras de estoque em glicerol e incubados a 37°C sob agitação
de 225 rpm por 16 horas. O pré-inoculo foi diluído 1:100 em 100mL de meio LB com
antibiótico apropriado e incubado a 37°C sob agitação de 225 rpm até atingir uma DO600 entre
0,5 e 0,6. Neste ponto, foi retirado uma alíquota da cultura de 30mL, que foi usada como
controle sendo esse o tempo 0, não induzida (sem adição de IPTG). A cultura restante submetida
ao experimento de indução, por meio da incubação das mesmas a 37°C sob agitação de 225
rpm. Alíquotas das colônias foram removidas após 2 horas e 4 horas de indução (em um volume
de 20 e 10 mL respectivamente). Ao final do experimento, o restante da cultura
(aproximadamente 40mL), foi separado e posteriormente utilizado para ensaio de atividade
enzimática. As alíquotas foram centrifugadas a 2.000 g por 20 minutos a 4°C, o precipitado
com as bactérias foi armazenado no freezer (-20°C), descartando-se o sobrenadante.
3.6.2 Extração da proteína
O precipitado com as bactérias foram ressuspendidas em 1000 L de tampão Tris-HCL
50mM, pH 7,5, NaCl 300mM, sonicadas 3 vezes com potência 70% por 10 segundos, com
intervalos de 30 segundos, e centrifugadas a 15.700 g por 30 minutos a 4°C. O sobrenadante
foi transferido para uma novo tubo, e o precipitado foi ressuspendido em 400 L de tampão
Tris-HCL 50mM, pH 7,5, NaCl 300mM. O extrato de proteínas totais (solúveis e insolúveis)

19
foi removido (aproximadamente 20L) aos quais foram adicionados outros 20L de tampão de
amostra (100mM Tris-HCl pH 6,5, 4% SDS, 0,2% azul de bromofenol, 20% glicerol). As
amostras foram fervidas a 95oC por 5 minutos, aplicadas e analisadas em gel de Poliacrilamida
(SDS-PAGE 12%) para verificar o rendimento e a solubilidade da proteína. As proteínas foram
visualizadas após coloração do gel com uma solução de coomassie brilliant blue R-250 (2,5g
em 400mL Metanol, 100mL ácido acético glacial e água para 1 litro) e descoloração com uma
solução de metanol: ácido acético (300mL:400mL para 4 litros).
3.7 Ensaio de atividade enzimática
Para avaliar a atividade das proteínas, ensaios enzimáticos foram realizados em reações
contendo 200μL de extrato de proteínas totais obtido após indução com IPTG, 10μg de substrato
(geranil difosfato – GPP, farnesil difosfato - FPP), em 800μL de tampão para o volume final de
1mL e incubadas e mantidas a 30oC por 60 Minutos em Vial de 15mL.
Após o período de incubação as amostras foram agitados em vortex e os voláteis foram
capturados por meio da técnica de Microextração por Fase Sólida (SPME), empregando-se a
fibra Carboxen/Polidimetilsiloxano (75μm CAR/PDMS, Supelco, Bellefonte, PA, USA), A
fibra foi pré-condicionada seguindo as orientações do fabricante e exposta por 30 minutos no
vial para captura dos voláteis, cujo tempo de captura foi otimizado após a realização de pré-
testes (Figura 5).
Figura 5 – Sistema de Captura dos voláteis utilizando a técnica de Microextração por Fase
Solida (SMPE).

20
Para otimização do tampão utilizado dos ensaios enzimáticos, foram montados diversas
soluções com diferentes tampões e sais, alterando pH e adicionando glicerol ou ditiotreitol
(DTT). As reações foram realizadas utilizando o substrato geranil difosfato e ao diversos
tampões descritos na tabela 4.
Tabela 4 – Tampões utilizados em reações de atividade enzimática.

21
Extrato de proteínas totais de bactérias não transformadas (sem genes de terpeno sintase)
foi obtido, seguindo as mesmas condições de tratamento e tempo de incubação, e utilizado como
controle para comparação com as amostras que havia a presença de terpeno sintases, para cada
tratamento utilizado nas reações de atividade enzimática.
Após a captura, a fibra SPME foi diretamente inserida no injetor do cromatógrafo a gás
acoplado a espectrômetro de massas (CG-EM), para dessorção das substâncias e a análise da
composição química.
3.8 Identificação dos Produtos da Reação Enzimática
Os produtos foram identificados através de cromatografia gasosa acoplada a
espectrometria de massas (CG-EM Shimadzu, modelo QP-5000), operando por impacto de
elétrons a 70eV, coluna capilar de sílica fundida DB-5 (30m de comprimento, 0,25 cm de
diâmetro interno e 0,25μm de espessura de filme líquido). A temperatura do injetor foi de
240ºC, detector a 230oC, hélio como gás de arraste (1mL/min-1) (Figura 6).
Figura 6 – Cromatógrafo a gás acoplado a espectrômetro de massas (CG-EM).
A temperatura inicial da coluna cromatográfica foi de 60ºC com elevação de
temperatura na razão de 3ºC por minuto até 170ºC; 170ºC - 280ºC elevação de temperatura na

22
razão de 15ºC por minuto; permanecendo por 2 minutos a 280ºC, Split: 1/20, com tempo total
de corrida de 46 minutos. Todas as amostras obtidas das reações de atividade enzimáticas
inclusive os controles foram analisados nas condições cromatográficas citadas.
A identificação dos constituintes químicos foi efetuada através da análise comparativa
dos espectros de massas das substâncias analisadas com o banco de dados do sistema CG-EM
(NIST12.LIB, NIST62.LIB, WILEY139.LIB); índice de retenção (IR) e literatura (ADAMS,
2007).
Os índices de Retenção (IR) foram obtidos pela injeção de uma série homologa de
padrões de referência de n-alcanos (C9-C25) no CG-EM com o seguinte programa de
temperatura 60°C - 240°C, elevando a temperatura por 3°C/min-1. Os índices foram calculados
empregando-se a equação:
IR = 100n + 100 (𝑡𝑅(𝑖) − 𝑡𝑅(𝑛)
𝑡𝑅(𝑛+1) − 𝑡𝑅(𝑛))
Onde 𝑡𝑅 é o tempo de retenção
𝑖 – analito
𝑛 – número de carbonos do padrão adjacente menos retido, e 𝑛 + 11 é o número de
carbonos do padrão adjacente mais retido. (VAN DEN DOOL & DEC. KRATZ, 1963; ZINI &
MÜHLEN, 2009).
4 RESULTADOS E DISCUSSÃO
4.1 Amplificação das Terpeno Sintases
Os genes utilizados nesse trabalho foram identificados anteriormente em trabalhos
efetuados pelo Centro de Citricultura Sylvio Moreira (CCSM) do Instituto Agronômico em
Cordeirópolis, onde inicialmente foram feitas buscas por sequências que codificam terpeno
sintases no site do Phytozome (phytozome.org), tanto para C. sinensis quanto para C.
clementina, as duas espécies com genomas completos sequenciados. Estas buscas foram feitas
através de palavra-chave (“terpene”) e resultou na identificação de 123 sequências no total, que
foram usadas para buscas por similaridade com a base de dados de proteína do NCBI, para
confirmação de que codificavam proteínas similares a terpeno sintases. Algumas sequências
curtas ou redundantes foram eliminadas nesta análise, resultando em 55 diferentes terpenos

23
sintases de citros que foram usadas para desenho dos primers com o programa PrimerSelect do
LASERGENE99, obtendo-se 50 primers diretos e 55 primers reversos, que foram sintetizados
(Tabela 3). Posteriormente, foram feitas buscas de similaridade com sequências existentes no
banco de clones do CitEST, para identificar-se aqueles que haviam apresentado expressão
nestes organismos e, uma vez que a base do CitEST são ESTs, facilitaria o trabalho de
amplificação e clonagem destes genes.
Os genes que codificam estas proteínas foram amplificados por PCR, sendo a
amplificação verificada em gel de agarose 0.7%. Assim sendo, das 55 sequências, foram
amplificadas 18 a partir de clones do CitEST, codificando possíveis terpeno sintases. Dentre
estes 16 clones resultaram em amplificação de intensidade alta e 2 com baixa intensidade
(CrTPS15 e CsTPS88), sendo todas selecionados e utilizados posteriormente para clonagem
inicial nos vetores pJET 1.2 e pGEM-T (Figura 7).
Figura 7 – Gel de agarose 0,7% (m/v) Amplificação de genes de possíveis terpeno sintases de
citros. Na parte superior do gel estão identificados os clones utilizados, MM- Marcador
molecular 1 Kb Ladder.
4.2 Clonagem
Os fragmentos purificados contendo os genes de terpeno sintase foram clonados
inicialmente em pJET 1.2, sendo utilizado para transformação de E. coli.
Esta etapa inicial resultou na clonagem da maioria dos amplicons. Todavia, para os genes
CaTPS01, CrTPS14, CrTPS15 e CsTPS84 não foi possível obter-se colônias utilizando o vetor
pJET 1.2. Deste modo, outro vetor, pGEM-T, foi utilizado como alternativa de clonagem. As
MM
CA
TP
S01
CR
TP
S05
CR
TP
S11
CR
TP
S14
CR
TP
S15
CR
TP
S26
CS
TP
S43
CS
TP
S47
CS
TP
S50
CS
TP
S51
CS
TP
S54
CS
TP
S64
CS
TP
S66
CS
TP
S67
CS
TP
S81
CS
TP
S84
CS
TP
S88
ClT
PS
92
1,5 Kb

24
colônias que cresceram em meio seletivo foram passadas para novas placas com meio LB +
ampicilina, como exemplificado na Figura 8.
Figura 8 – Crescimento bacteriano em meio seletivo. Placas Meio de cultura LB contendo
ampicilina como agente seletivo foram usadas para crescimento de transformantes, como
representado para os genes CsTPS54, CrTPS15, CsTPS50 e CsTPS84.
As clonagens utilizando o vetor pJET 1.2 em E. coli DH5α apresentaram alta eficiência,
sendo que para maioria das colônias selecionadas foi possível confirmar sua transformação com
amplificação do fragmento esperado, de aproximadamente 1,5 Kb (Figura 9).
54 15 50 84
MM CrTPS05 CrTPS11 CrTPS26
MM CsTPS43
1,5Kb
1,5Kb

25
Figura 9 – Amplificação dos insertos contendo possíveis terpeno sintases de citros clonadas
em pJET 1.2 para confirmação positiva de clonagem. Os genes estão identificados na parte
superior do gel, MM – Marcador molecular 1 Kb Ladder.
Apenas o gene 81 não mostrou amplificação de banda de tamanho esperado, sendo
menor que 1Kb, por isso esse gene foi descartado do ensaio. Para todos os outros genes foi
MM CsTPS47 CsTPS50
MM CsTPS51 CsTPS54 CsTPS64
MM CsTPS67 CsTPS81
MM CsTPS88 ClTPS92
1,5Kb
1,5Kb
1,5Kb
1,5Kb

26
possível obter um número alto de colônias transformadas, com exceção dos genes CsTPS43 e
ClTPS92, dos quais foi possível obter-se 2 e 4 colônias transformadas, respectivamente.
Os genes CaTPS01, CrTPS14, CrTPS15 e CsTPS84 foram clonados no vetor pGEM-T,
vetor que possui seleção por coloração para varredura das colônias positivas. Apenas uma
colônia do gene CrTPS15 mostrou amplificação de uma banda de tamanho esperado. Para os
outros genes clonados nesse vetor, foi possível obter várias colônias positivas (Figura 10).
Figura 10 – Análise de construções de pGEM-T. Amplificação dos genes CaTPS1, CrTPS14,
CrTPS15 e CsTPS84 de citros de modo a identificar-se aqueles que apresentam inserto. Genes
estão identificados na parte superior do gel, M – Marcador molecular 1 Kb Ladder.
MM CaTPS01 CrTPS14
MM CrTPS15
MM CsTPS84
1,5Kb
1,5Kb
1,5Kb

27
4.3 Sequenciamento
O sequenciamento foi realizado após a clonagem dos amplicons nos vetores pJET1.2 e
pGEM-T.
As sequencias obtidas, foram devidamente montadas e analisadas, através do programa
Seqman do pacote LASERGENE 99 (DNASTAR, Inc.). Após a montagem dos contigs, as
sequências foram comparadas com as disponíveis nos bancos dados do phytozone e do NCBI
através da ferramenta BlastX afim de verificar se essas sequências codificariam proteínas
similares terpeno sintases, se elas estavam apenas anotadas ou já caracterizadas funcionalmente,
e ainda verificar os dados de anotações referente a essas sequências.
As sequências mostraram variações típicas de comparação de diferentes organismos
mas sem que houvesse problema em relação à criação de stop códons, como exemplificado na
figura 11.
Figura 11 – Análise de busca por similaridade dos genes CrTPS26 (A) e CsTPS51 (B) através
de Blastx.
Fonte: www.ncbi.nlm.nih.gov
A
B

28
Interessantemente, a sequência obtida do gene CsTPS51 mostrou similaridade com uma
região localizada à jusante do códon de início, onde fora desenhado o primer. Isto mostra que
a amplificação se deu a partir de um anelamento inespecífico, resultando em uma perda de
aproximadamente 40 aminoácidos da extremidade amino desta proteína.
Os dados de sequências obtidos foram analisados quanto à similaridade com proteínas
no banco de dados e os resultados estão demonstrados na tabela 5, evidenciando os possíveis
produtos gênicos que essas sequências codificariam.
Tabela 5 – Dados das sequências comparadas através da ferramenta BlastX em bancos de dados
de sequências.
Sequência Anotação Organismo Descrição
CrTPS05 Predito Citrus sinensis (-)-germacreno D sintase
CrTPS11 Predito Citrus sinensis γ-terpineno sintase
CrTPS15 Predito Citrus sinensis (-)-germacreno D sintase
CrTPS26 RlemTPS4 Citrus jambhiri δ-elemeno sintase
CsTPS43 Predito Citrus jambhiri δ-elemeno sintase
CsTPS47 Predito Citrus sinensis (-)-germacreno D sintase
CsTPS50 Predito Citrus sinensis (E)-β-farneseno sintase
CsTPS51 Predito Citrus sinensis (-)-germacreno D sintase
CsTPS54 Predito Citrus sinensis (E)-β-farneseno sintase
CsTPS64 CitMTSE2 Citrus unshiu d-limoneno sintase
CsTPS66 Predito Citrus sinensis (-)-germacreno D sintase
CsTPS67 CitMTSE2 Citrus unshiu d-limoneno sintase
CsTPS84 Predito Citrus sinensis (R)-limoneno sintase
CsTPS88 Predito Citrus sinensis nerolidol sintase
ClTPS92 Predito Citrus sinensis (E)-β-farneseno sintase
4.4 Construções Para Expressão Gênica
Uma vez que todas as etapas de clonagem inicial foram concluídas, foi feita a
transferência dos genes para um vetor de expressão, no caso o pF1K T7.

29
O DNA plasmidial extraído foi quantificado no espectrofotômetro NanoDrop 8000
(Thermo Scientific), onde a razão 260/280 ficou próxima a 1,8 e a 260/230 próxima a 2,2.
A clonagem das sequências codificadoras em pF1K T7 não foi tão eficiente quanto o
observado anteriormente para pJET 1.2 e pGEM-T, ocasionando uma diminuição de 18 para 7
genes a serem levados para análise de atividade. Estas construções foram usadas para
transformação de E. coli BL21 e Rosetta (Figura 12).
Figura 12 – Construção de plasmídeos para expressão gênica. Transformantes de E. coli BL21
e Rosetta foram utilizados para amplificação dos genes de possíveis terpeno sintases de citros
clonadas em pF1K-T7 para confirmar aqueles que apresentavam o inserto. Clones estão
identificados na parte superior do gel, M – Marcador molecular 1 Kb Ladder.
Com exceção do gene CsTPS66, todos os outros genes foram transferidos para os
vetores de expressão com sucesso.
4.5 Extração de Proteína
Os ensaios de expressão foram realizados em E. coli BL21(DE3). Inicialmente foram
realizados testes de expressão com a construção com o gene CrTPS05 utilizando a temperatura
de 37°C variando-se o tempo de indução, no intuito de verificar o rendimento e a solubilidade
da proteína. Exemplo do perfil de expressão está representado para os resultados obtidos depois
de 2 e 4 horas de indução com a concentração de IPTG de 1mM e o tempo 0 como controle. É
possível verificar a presença de bandas intensas nos tempos 2 e 4 na porção insolúvel, que
indicam maior expressão próximo a região esperada do peso molecular esperada para essa
MM
C
rTP
S0
5
BL
21
CrT
PS
26
BL
21
CsT
PS
43
BL
21
CsT
PS
50
BL
21
CsT
PS
66
Bl2
1
CrT
PS
26
Ro
sett
a
CsT
PS
47
Ro
sett
a
CsT
PS
51
Ro
sett
a
1,5Kb

30
proteína aproximadamente 60 kDa. Essa mesma banda aparece também na fração solúvel
porém com menor intensidade (Figura 13).
Figura 13 – SDS-PAGE (12% poliacrilamida). Ensaios de expressão a 37°C para a construção
CrTPS05 nos tempo de indução (0, 2 e 4 horas) O retângulo indicam as bandas de expressão na
forma solúvel e insolúvel.
Outras corridas foram efetuadas, e utilizando um visualizador de imagens com luz
branca, para melhor analisar os dados do gel de SDS, é possível verificar bandas de alta
intensidade na fração solúvel que não aparece ou aparece em baixa intensidade no controle T0
(Figura 14).
Figura 14 – SDS-PAGE (12% poliacrilamida). Ensaios de expressão a 37°C para a construção
CrTPS05, CrTPS26, CsTPS43 e CsTPS50 variando o tempo de indução (0, 2 e 4 horas) As
setas vermelhas indicam as bandas de expressão na forma solúvel.
kDa
70,0 -
60,0 -
50,0 -
40,0 -
30,0 -
25,0 -
20,0 -
15,0 -
10,0 -
CsTPS43 CrTPS05 CrTPS26 CsTPS50

31
Outro ensaio foi realizado modificando outros parâmetros, como temperatura e
concentração do indutor (IPTG). As concentrações de IPTG utilizadas foram de 0,3mM, 0,5mM
e 1mM em duas temperaturas (27°C e 37°C) e o tempo de indução foi de 4 e 16 horas, apenas
a fração solúvel foi aplicada no gel (Figura 15).
Figura 15 – SDS-PAGE (12% poliacrilamida). Ensaios de expressão a 37°C e 27°C para a
construção CsTPS51 variando o tempo de indução (4, 16) e a concentração de IPTG (0,3; 0,5 e
1,0 mM).
Nesse ensaio foi possível verificar que o tempo de indução de 16 horas se mostrou a
condição mais eficiente, sendo que é possível verificar algumas bandas expressas nessas
condições que não aparecem com 4 horas de indução mesmo entre as 3 concentrações de IPTG
testadas nesse ensaio (Figura 15). As concentração de IPTG de acima de 0,5mM parece ter
influência tanto a 27°C quanto 37oC quando induzidas por 16 horas. Nesse ensaio foi possível
determinar que a melhor condição para expressão dessas proteínas foi utilizando a concentração
1mM de indutor de expressão (IPTG), por 16 horas a 37°C. Todas os ensaios de expressão
seguintes foram utilizando essas condições de indução.
O gene CsTPS51 foi aquele que apresentou tamanho de amplificação menor que o
esperado, com consequente diminuição da proteína. Isto se confirma neste gel uma vez que a
indução observada é de uma proteína realmente menor, com tamanho entre 55 e 58 KDa.
kDa
116,0-
66,2 -
45,0 -
35,0 -
25,0 -
18,0 -
~58 KDa.
16 horas

32
4.6 Identificação dos Produtos da Atividade Enzimática
Inicialmente, os voláteis foram extraídos do meio reacional utilizando o solvente acetato
de etila de acordo com o procedimento descrito na literatura (SHIMADA et al., 2004;
SHIMADA et al., 2005), porém as amostram apresentaram um grande número de substâncias
não relacionadas à classe de terpeno, além deste fato, observou-se que o perfil químico das
amostras controle (ensaios enzimáticos realizados com proteínas totais de bactérias não
transformadas) eram similares comparadas com as amostras, realizados com proteínas totais de
bactérias transformadas com gene de terpeno sintase (Figura 16).
Tempo de Retenção (Min.)
Figura 16 – Cromatograma de íons totais, utilizando solvente acetato de etila para captura dos
voláteis com GPP como substrato e tempo de reação de 1 hora a 30°C. A – Controle (Proteínas
totais de bactérias não transformadas). B – Gene CrTPS05.
Deste modo, foi utilizada uma outra estratégia para a captura dos voláteis, o que foi feito
pela técnica de Microextração por Fase Solida (SPME). Na literatura vários trabalhos utilizaram
essa técnica para captura de monoterpenos e sesquiterpenos com sucesso, KOHZAKI et al
(2009) caracterizou sabineno sintase utilizando SPME para captura. Mais recentemente
A
B

33
SHIMADA et al (2014) identificou três genes, dois codificam linalol sintase e um codificava
nerolidol/linalol sintase em C. unshiu utilizando essa técnica. Para soja também utilizou-se
dessa técnica para captura e caracterização de nerol sintase com sucesso (ZHANG et al., 2013).
Utilizando esta técnica de captura, foi possível observar diferenças entre o controle e as
outras amostras que continham os genes de terpeno sintase clonados. Na amostra controle
observou-se a presença de quatro substâncias no intervalo de tempo de retenção de 10 a 20
minutos (Figura 17A).
Tempo de Retenção (Min.)
Figura 17 – Cromatograma de íons totais, utilizando SPME para captura dos voláteis. GPP
como substrato e tempo de reação de 1 hora a 30°C A – Controle (Proteínas totais de bactérias
não transformadas). B – CrTPS05, C – CrTPS26, D – CsTPS51, E – CsTPS54, F – CsTPS47.
As amostras que continham os genes apresentaram as mesmas substâncias com
proporção relativa (intensidade) diferente em relação à amostra controle e entre si. Apesar
destes resultados, não foi observada a formação dos produtos desejados ou que nos garantia
concluir que as substâncias presentes na amostra estavam relacionadas com a atividade de uma
enzima de terpeno síntase (Figura 17 B-F).
A B
C D
E F

34
Para que a atividade enzimática ocorra é necessário que a enzima esteja em condições
ótimas para seu melhor funcionamento, para isso é necessário que vários fatores estejam em
condições ideais. Os fatores que mais influenciam na funcionalidade de uma enzima são pH e
temperatura, que em níveis adequados levarão a conversão do substrato no produto final. Para
isso foram feitos outros ensaios alterando o tampão entre TRIS-HCl, MOPS e HEPES, variando
a temperatura entre 23°C e 31°C, pH variando entre 7 e 8, e tempo de reação entre 1 e 24 horas.
A melhor condição avaliada para os ensaios enzimáticos foram utilizando o tampão
15mM MOPS pH 7,4, 25mM MgCl2, 2mM DTT, em um tempo de reação de 1 hora com
temperatura de 30°C. Nessas condições e utilizando como substrato o geranil difosfato foi
possível observar atividade de duas enzimas codificadas pelos genes CsTPS51 e CrTPS26.
O cromatograma de íons totais do produto do gene CsTPS51 de C. sinensis, apresentou
uma substância com tempo de retenção (TR) de 9.4 minutos, correspondendo a 91,3% da
amostra (Figura 18B).
Tempo de Retenção (Min.)
Figura 18 – Cromatograma de íons totais, GPP como substrato e tempo de reação de 1 hora a
30°C. A – Controle (Proteínas totais de bactérias não transformadas) B – CsTPS51, a seta indica
o pico relacionado a 1,8 cineol.
A
B

35
A comparação do espectro de massas da substância (Figura 19A) com o banco de dados
do Equipamento CG-EM (nist62.lib, wiley139.lib) (Figura 19B), em conjunto com o índice de
retenção da substância (IR = 1030), e o espectro de massas da literatura (ADAMS, 2007)(Figura
20), permitiu identificar a substância como 1,8-cineol.
m/z
Figura 19 – Espectro de massa referênte ao pico com TR = 9,4 min do gene CsTPS51. A –
Espectro de massa do produto do CsTPS51, B – Espectro de massa do 1,8-cineol obtido da
biblioteca NIST62.LIB.
m/z
Figura 20 – Espectro de massa de 1,8 cineol segundo ADAMS, (2007).
Fonte: ADAMS, (2007).
Assim, pode-se observar que mesmo a proteína tendo uma deleção de aminoácido em
uma de suas extremidades, a atividade enzimática se manteve, mostrando que o sítio catalítico
da enzima não foi afetado.
A análise da composição química das substâncias voláteis da reação gerado pelo produto
do gene CrTPS26 de C. reticulata também apresentou um resultado interessante, na análise do
A B
IR = 1031
IR = 1030

36
cromatograma de íons totais foi possível identificar três picos com a proposta de três
monoterpenos, sendo o primeiro pico, com tempo de retenção de 8 minutos, identificado como
mirceno (8.5%), o terceiro pico com TR = 9,3 minutos como limoneno (2,5%), e o quarto pico,
em maior abundância na amostra com TR = 9,4 minutos como 1,8 cineol (57,3%). (Figura 21).
Tempo de Retenção (Min.)
Figura 21 – Cromatograma de íons totais, GPP como substrato e tempo de reação de 1 hora a
30°C. A – Controle (Proteínas totais de bactérias não transformadas) B – CrTPS26 (Pico 1 –
mirceno, pico 3 - limoneno e pico 4 -1,8 cineol).
Por meio da equação sugerida por VAN DEN DOOL & DEC. KRATZ (1963) foi possível obter
os seguintes índices de retenção para as substâncias: IR = 993 para o pico 1, IR = 1027 para o
pico 3 e IR = 1029 para o pico 4. Esses índices e os espectros de massa foram comparados com
os espectro de massas disponíveis na literatura (ADAMS, 2007), permitindo a confirmação dos
produtos de reação como: mirceno, limoneno e 1,8 cineol (Figura 23A-C).
A
B
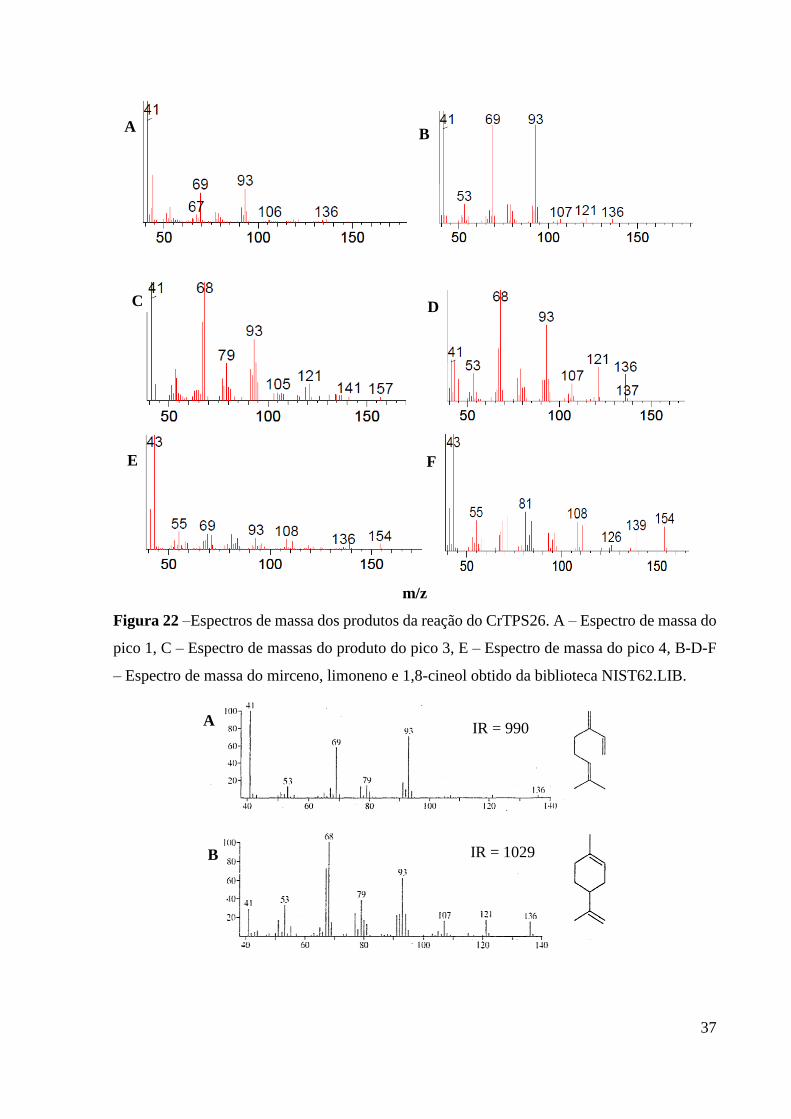
37
m/z
Figura 22 –Espectros de massa dos produtos da reação do CrTPS26. A – Espectro de massa do
pico 1, C – Espectro de massas do produto do pico 3, E – Espectro de massa do pico 4, B-D-F
– Espectro de massa do mirceno, limoneno e 1,8-cineol obtido da biblioteca NIST62.LIB.
A B
C D
E F
A
B
IR = 990
IR = 1029

38
m/z
Figura 23 – Espectro de massa de A - Mirceno, B - limoneno C - 1,8 cineol, literatura
(ADAMS, 2007)
As reações enzimáticas utilizando como substrato o farnesil difosfato (FPP) foram
analisadas e não foi observada a formação de substâncias sesquiterpênicas. Porém, as reações
com este substrato não foram otimizadas, assim como efetuado com o GPP.
Terpenos sintases são conhecidas por pertencerem a uma família de enzimas que
possuem alta similaridade funcional entre elas, capazes de produzir vários tipos de terpenos
(BOHLMANN et al., 1998). Todos os genótipos de citros contêm uma ampla diversidade de
terpenos voláteis, como hemiterpenos, monoterpenos e sesquiterpenos, os quais são abundantes
em folhas, frutos e flores. Mais de 49 genes de possíveis terpenos sintases foram identificados
in silico entre espécies de citros (DORNELAS & MAZZAFERA, 2007), porém poucos genes
foram clonados e analisados para suas funções (Tabela 1).
Um fato interessante sobre as terpeno sintases é a habilidade dessas enzimas produzirem
um ou mais produtos, por exemplo, pineno sintase de muitas plantas produzem α- e β-pineno
(CROTEAU et al., 2000). Porem essa característica não é observada para todos genes de
terpeno sintase, em Citrus limon foram identificadas quatro genes que codificam monoterpeno
sintases e a caracterização do produto de dois destes genes através de expressão em E. coli,
confirmaram que as enzimas codificadas para ambos os genes produziram especificamente (+)-
limoneno como produto principal, as outras duas monoterpeno sintases produziram γ-terpineno
e (-)β-pineno respectivamente, porém observou-se a formação de produtos secundários em
menor quantidade (LÜCKER et al., 2002).
No presente estudo, as análises por CG-EM possibilitaram identificar a presença de
algumas substâncias monoterpenicas nas amostras de dois genes avaliados. Na amostra do gene
CrTPS26, pertencente a Citrus reticulata, foram identificados três monoterpenos, sendo que a
substância mais abundante na amostra foi o 1,8 cineol e os produtos com menor abundância
C IR = 1031

39
foram d-limoneno e β-mirceno. Como mencionado anteriormente, é uma característica comum
de muitas monoterpeno sintases serem capazes de formar múltiplos produtos a partir de geranil
difosfato como substrato. Resultados semelhantes, quanto à formação de diversas produtos em
uma amostra foi observado por KOHZAKI et al (2009), aonde foi confirmada a presença de
sabineno como o produto principal e α-pineno, β-pineno e β-felandreno como produtos com
menor quantidade na amostra dos genes de limão rugoso (C. x jambhiri). UJI et al (2015),
também isolaram e avaliaram a expressão de uma terpeno sintase e identificaram para o mesmo
gene as seguintes substâncias sesquiterpenicas: δ-elemeno como principal produto e β-elemeno,
δ-selineno, γ-elemeno e germacreno D como produtos secundários.
A representatividade destes compostos no óleo essencial de laranja doce é muito
elevada, monoterpenos é a principal classe química entre os voláteis, os quais em algumas
variedades corresponderam a até 96,9% da composição total dos óleos essenciais avaliados.
Limoneno foi o composto mais abundante (aproximadamente 94,6 %) de acordo com relatórios
publicados sobre diversas variedades de óleos essenciais extraídos de laranja doce de diferentes
origens (COLEMAN & SHAW, 1971; WOLFORD et al., 1971; MOSHONAS & SHAW, 1974;
SHAW, 1979; CHOI & SAWAMURA, 2000; MITIKU et al., 2000; TU et al., 2002). O
monoterpenos 1,8-cineol estava presente em níveis residuais em cada óleo essencial. Mirceno
foi classificado como um componente importante de vários tipos de óleos essenciais de laranja
doce, apesar de corresponder a menos de 0,05% das amostras avaliadas (MOSHONAS &
SHAW, 1974; SHAW, 1979). Os outros terpenos proeminentes nos óleos essenciais descritos
na literatura foram α-terpineno (1.5-1,7%), α-pineno (0,3-0,5%), e sabineno (0,1-0,2%). β-
pineno e terpinoleno correspondem menos de 0,05% em cada óleo essencial (NJOROGE et al.,
2009).
Na amostra utilizando o produto do gene CsTPS51 de Citrus sinensis (Figura 18), foi
possível verificar a presença majoritária de 1,8 cineol, correspondendo a mais de 90% da
composição química da amostra avaliada, apesar de se tratar do mesmo produto, a proteína
codificada por esse gene é mais especifica para síntese desse monoterpeno, quando comparada
ao produto do gene CrTPS26 de Citrus reticulata que apresentou produção de outras
substâncias monoterpenicas secundárias (Figura 21).
O 1,8-cineol foi o principal componente de óleos essenciais obtidos de diferentes partes
(caules, folhas, flores adultos imaturos e frutos) de Eucalyptus oleosa, avaliados quanto as suas
propriedades antioxidantes e antimicrobianas e a sua composição química. Os óleos essenciais
mostraram atividade antibacteriana contra bactérias gram-positivas e gram-negativas, e
significativa ação antifúngica (MARZOUG et al., 2011).

40
Em tangerina Satsuma (C. unshiu) foi demostrado que 1,8 cineol e (E)- β-ocimeno foram
encontrados em níveis abundantes em flores, entretanto, em frutos foi verificado o decréscimo
no níveis destas substâncias, com o decorrer do amadurecimento. SHIMADA et al. (2005)
demonstraram que a regulação da expressão de genes relacionados a terpenos pode ser
controlada durante o desenvolvimento da planta, de acordo com as suas necessidades, e que os
diferentes perfis de voláteis nos tecidos pode depender da expressão dos genes e da competição
das enzimas pelo substrato. Foi descrito que 1,8 cineol é emitido por flores como mecanismo
de atração de polinizadores, como abelhas, mariposas e morcegos. No entanto, em folhas, foi
verificado que 1,8 cineol atua inibindo a alimentação foliar de grandes herbívoros, como lebres
e veados, e também fornece vantagem competitiva para várias espécies de angiospermas, como
agente inibidor da germinação de sementes de outras espécies (CROTEAU et al., 2000). Além
de estar envolvido em interações planta-planta ele também possui ação antifúngica
(PATTNAIK et al., 1997).
A sequência de aminoácidos do gene CrTPS26 mostrou alta identidade com uma
sesquiterpeno sintase, reportado por Uji et al. (2015), identificado como δ-elemeno sintase em
limão rugoso (Citrus jambhiri)(Tabela 5), resultado obtido utilizando-se FPP como substrato.
UJI et al. (2015), também avaliou a atividade de monoterpeno sintase para este gene utilizando
GPP como substrato e a principal substância encontrado foi β-mirceno, porém os dados não
foram publicados. Esse resultado indica que a atividade das terpeno sintases nas células pode
depender da regulação a nível de transcrição e da competição de enzimas correlacionadas pelo
substrato.
Outro trabalho mostra a identificação de dois produtos de diferentes classes de terpenos
gerados pela mesma enzima, utilizando substratos diferentes, convertendo GPP em linalol e
FPP em nerolidol (SHIMADA et al., 2014), demonstrando a capacidade de uma enzima de
produzir ambos, monoterpenos e sesquiterpenos.
O gene CsTPS51 também mostrou alto nível de identidade com sequências preditas de
(-) germacrene D sintases de Citrus sinensis, porém essa predição é equivalente a sesquiterpeno
sintases, todavia, a nossas análises resultaram em síntese de monoterpenos, denotando uma
possível falha no processo de anotação por similaridade, de sequências de terpeno sintases
(Tabela 5).
Sequências de aminoácidos de terpeno sintases em citros possuem grande semelhança
entre si, evidenciado em análises filogenéticas, maior do que outros grupos de genes com a
mesma função em outras espécies. Na literatura os genes que codificam proteínas que
sintetizam monoterpenos em citros foram fortemente agrupadas, sugerindo que estes genes

41
podem ter sido duplicados e especializados após a especiação de citros. Além disso, o acúmulo
de variações continuas nos aminoácidos, deleções, inserções resultaram na diversidade
molecular para a família de genes de monoterpeno sintases e criaram uma alta diversidade em
Citrus spp. Assim estas características de evolução dificultam a predição da função do gene
através de domínios específicos ou identidade de sequências. (SHIMADA et al., 2004; 2005).
Segundo UJI et al. (2015), em geral, os produtos gerados por atividades de terpeno
sintases não podem ser preditos apenas por análises de homologia e identidade de sequências.
O monoterpeno sintase RlemTPS2, por exemplo, que foi isolada de limão rugoso e
caracterizada como sabineno sintase (KOHZAKI et al., 2009), tem 95% e 97% de identidade
de sequência com β-pineno sintases anteriormente identificadas em Citrus limon e Citrus
unshiu (LÜCKER et al., 2002; SHIMADA et al., 2004), evidenciando a necessidade da
realização de testes in vitro para caracterizar a atividade destas enzimas. Nossos resultados
confirmam esta dificuldade, uma vez que através da análise de homologia de sequências dos
genes CrTPS26 de C. reticulata e CsTPS51 de C. sinensis, verificou-se alta similaridade com
δ-elemeno sintase e (-)-germacreno D sintase, respectivamente, todavia foram encontrados
monoterpenos como produtos em nossas análises.
5 CONCLUSÃO
Muitos terpenos ainda serão descobertos, possibilitando novas aplicações. Para que isto
aconteça, é muito importante que haja a caracterização funcional das proteínas. Neste sentido,
o presente estudo teve como objetivo a clonagem, expressão e caracterização funcional de
terpeno sintases de Citros para que, com isto, se obtivesse um maior conhecimento da
biologia/bioquímica de citros. Foi possível identificar substâncias pertencentes da classe de
monoterpenos, por cromatografia gasosa acoplada a espectrometria de massas, dentre os quais
1,8 cineol, d-limoneno e β-mirceno, a partir da utilização de geranil difosfato como substrato
por duas terpenos sintases de citros, caracterizando-as funcionalmente.
Análises de similaridade indicaram que estas proteínas seriam sesquiterpeno sintases
entretanto, a análise funcional provou que são, na verdade, monoterpeno sintases, reforçando a
idéia de que há a necessidade de testar-se a funcionalidade das enzimas in vitro é fundamental
para que se caracterize sua atividade.
Este trabalho demonstrou também a capacidade destas enzimas de citros produzirem
mais de um produto a partir de um mesmo substrato, característica comum de enzimas desta
classe.

42
O sucesso obtido com duas terpenos sintases de citros estimula a caracterizar as outras
enzimas clonadas.
Finalmente, o desenvolvimento do projeto abre possibilidades de utilização
biotecnológica destes genes para produção de 1,8 cineol, por exemplo, o que poderia levar a
um produto mais barato que o comercializado hoje (U$ 120,00 por 25 mL).
.

43
6 REFERÊNCIAS
ADAMS, R. Identification of Essential Oil Components by Gas Chromatography/Mass
Spectrometry. In: STREAM, C. (Ed.). 4th. ed. [s.l.] Allured Publishing Corporation, 2007. p.
804.
AZAMBUJA, W. Oleosessencial.org. Disponível em:
<http://www.oleosessenciais.org/metodos-de-extracao-de-oleos-
essenciais/#Prensagem_Frio>. Acesso em: 7 fev. 2016.
BAKKALI, F.; AVERBECK, S.; AVERBECK, D.; IDAOMAR, M. Biological effects of
essential oils - A review. Food and chemical toxicology : an international journal published
for the British Industrial Biological Research Association, v. 46, n. 2, p. 446–75, 2008.
Disponível em:
<http://linkinghub.elsevier.com/retrieve/pii/S0278691507004541\nhttp://www.ncbi.nlm.nih.g
ov/pubmed/17996351>.
BAUER, K.; GARBE, D.; SURBURG, H. Common Fragrance and Flavor Materials:
Preparation, Properties and Uses. Common Fragrance and Flavor Materials: Preparation,
Properties and Uses, v. 4, p. 1–318, 20 fev. 2006. Disponível em:
<http://doi.wiley.com/10.1002/3527608214>.
BENETI, S. C. Fracionamento de óleos essenciais em coluna de destilação a vácuo em modo
batelada. 2009. 105f Dissertação ( Mestrado em Engenharia de Alimentos) - Universidade
Regional Integrada do Alto Uruguai e das Missões - URI, 2009.
BISWAS, K. K.; FOSTER, A. J.; AUNG, T.; MAHMOUD, S. S. Essential oil production:
Relationship with abundance of glandular trichomes in aerial surface of plants. Acta
Physiologiae Plantarum, 31, 13-19, 2009.
BIZZO, H. R.; ANA MARIA, C. H.; REZENDE, C. M. Oleos essenciais no brasil: aspectos
gerais, desenvolvimento e perspectivas. Quimica Nova, v. 32, n. 3, p. 588–594, 2009.
BOHLMANN, J.; KEELING, C. I. Terpenoid biomaterials. Plant Journal, v. 54, n. 4, p. 656–
669, 2008.
BOHLMANN, J.; MEYER-GAUEN, G.; CROTEAU, R. Plant terpenoid synthases: molecular
biology and phylogenetic analysis. Proceedings of the National Academy of Sciences of the
United States of America, v. 95, n. 8, p. 4126–4133, 1998.
BOURGOU, S.; RAHALI, F. Z.; OURGHEMMI, I.; SAÏDANI TOUNSI, M. Changes of Peel
Essential Oil Composition of Four Tunisian Citrus during Fruit Maturation. The Scientific
World Journal, v. 2012, p. 1–10, 2012.
BOUWMEESTER, H. J.; VERSTAPPEN, F. W. A.; POSTHUMUS, M. A.; DICKE, M. Spider
mite-induced (3S)-(E)-nerolidol synthase activity in cucumber and lima bean. The first
dedicated step in acyclic C11-homoterpene biosynthesis. Plant physiology, v. 121, n. 1, p. 173–
180, 1999.
CANE, D. E. Enzymic formation of sesquiterpenes. Chemical Reviews (Washington, DC,

44
United States), v. 90, n. 7, p. 1089–1103, 1990.
CANE, D. E. Sesquiterpene biosynthesis: Cyclization mechanisms. Compr. Nat. Prod.
Chem., v. 2, p. 155–200, 1999.
CHENG; AI-XIA, L.; YONG-GEN, M.; YING-BO; LU, SHAN, W.; LING-JIAN.; CHEN, X.-
Y. Plant Terpenoids:Biosynthesis and ecological functions. Journal of Integrative Plant
Biology, v. 49, n. 2, p. 179–186, 2007.
CHOI, H.-S.; SAWAMURA, M. Composition of the Essential Oil of Citrus t amurana Hort. ex
Tanaka (Hyuganatsu). Journal of Agricultural and Food Chemistry, v. 48, n. 10, p. 4868–
4873, out. 2000. Disponível em: <http://pubs.acs.org/doi/abs/10.1021/jf000651e>.
CITRUSBR. ASSOCIAÇÃO NACIONAL DOS EXPORTADORES DE SUCOS
CÍTRICOS. Disponível em: <http://www.citrusbr.com/>. Acesso em: 26 jan. 2016.
COLEMAN, E. C.; SHAW, P. E. Analysis of Valencia orange essence and aroma oils. Journal
of Agricultural and Food Chemistry, v. 19, n. 3, p. 520–523, 1971. Disponível em:
<\\Robsrv-05\reference manager\Articles\2889.pdf>.
CROTEAU, R.; KUTCHAN, T. M.; LEWIS, N. G. Secondary Metabolites. Biochemistry
Molecular Biology of Plants, v. 7, n. 7, p. 1250–1318, 2000. Disponível em:
<http://www.ncbi.nlm.nih.gov/pubmed/21671613>.
CROWELL, P. L.; LIN, S.; VEDEJS, E.; GOULD, M. N. Identification of metabolites of the
antitumor agentd-limonene capable of inhibiting protein isoprenylation and cell growth.
Cancer Chemotherapy and Pharmacology, v. 31, n. 3, p. 205–212, 1992. Disponível em:
<http://link.springer.com/10.1007/BF00685549>.
DEGENHARDT, J.; KÖLLNER, T. G.; GERSHENZON, J. Monoterpene and sesquiterpene
synthases and the origin of terpene skeletal diversity in plants. Phytochemistry, v. 70, n. 15-
16, p. 1621–1637, 2009. Disponível em:
<http://dx.doi.org/10.1016/j.phytochem.2009.07.030>.
DELLACASSA, E.; ROSSINI, C.; MENENDEZ, P.; MOYNA, P.; VERZERA, A.; TROZZI,
A.; DUGO, G. Citrus essential oils of Uruguay. Part 1. Composition of oils of some varieties
of mandarin. Journal of Essential Oil Research, v. 4, n. 3, p. 265–272, 1992. Disponível em:
<\\Robsrv-05\reference manager\Articles\10215.pdf>.
DICKE, M. Specificity of herbivore-induced plant defences. Novartis Foundation
symposium, v. 223, n. FEBRUARY 1999, p. 43–54; discussion 54–59, 160–165, 1999.
DIXON, R. a. Engineering of plant natural product pathways. Current Opinion in Plant
Biology, v. 8, n. 3 SPEC. ISS., p. 329–336, 2005.
DORNELAS, M. C.; MAZZAFERA, P. A genomic approach to characterization of the Citrus
terpene synthase gene family. Genetics and Molecular Biology, v. 30, n. 3, p. 832–840, 2007.
Disponível em: <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1415-
47572007000500011&lng=en&nrm=iso&tlng=en>.
FAO. Food and Agriculture Organization of United Nations. Disponível em:
<http://faostat.fao.org>. Acesso em: 26 jan. 2016.

45
FRIZZO, C. D.; LORENZO, D.; DELLACASSA, E. Composition and Seasonal Variation of
the Essential Oils from Two Mandarin Cultivars of Southern Brazil. Journal of Agricultural
and Food Chemistry, v. 52, n. 10, p. 3036–3041, 2004.
FURUBAYASHI, M.; IKEZUMI, M.; KAJIWARA, J.; IWASAKI, M.; FUJII, A.; LI, L.;
SAITO, K.; UMENO, D. A high-throughput colorimetric screening assay for terpene synthase
activity based on substrate consumption. PloS one, v. 9, n. 3, p. e93317, 2014. Disponível em:
<http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3969365&tool=pmcentrez&rend
ertype=abstract>.
HEDDEN, P.; KAMIYA, Y. GIBBERELLIN BIOSYNTHESIS: Enzymes, Genes and Their
Regulation. Annual Review of Plant Physiology and Plant Molecular Biology, v. 48, p. 431–
460, 1997.
IBGE. Estatística da Produção Agrícola. Disponível em:
<ftp://ftp.ibge.gov.br/Producao_Agricola/Fasciculo_Indicadores_IBGE/estProdAgr_201512.p
df>. Acesso em: 26 jan. 2016.
KOHZAKI, K.; GOMI, K.; YAMASAKI-KOKUDO, Y.; OZAWA, R.; TAKABAYASHI, J.;
AKIMITSU, K. Characterization of a sabinene synthase gene from rough lemon (Citrus
jambhiri). Journal of Plant Physiology, v. 166, n. 15, p. 1700–1704, 2009. Disponível em:
<http://dx.doi.org/10.1016/j.jplph.2009.04.003>.
KOYAMA, T.; OGURA, K. Isopentenyl diphosphate isomerase and prenyltransferases.
Comprehensive Natural Product Chemistry. Isoprenoids Including Carotenoids and
Steroids, Vol 2. Pergamon Press, Oxford, pp 69–96, p. 93, 1999.
LANGENHEIM, J. H. Higher plant terpenoids: A phytocentric overview of their ecological
roles. Journal of Chemical Ecology, v. 20, n. 6, p. 1223–1280, 1994.
LOTA, M. L.; DE ROCCA SERRA, D.; TOMI, F.; JACQUEMOND, C.; CASANOVA, J.
Volatile components of peel and leaf oils of lemon and lime species. Journal of Agricultural
and Food Chemistry, v. 50, n. 4, p. 796–805, 2002.
LÜCKER, J.; EL TAMER, M. K.; SCHWAB, W.; VERSTAPPEN, F. W. a.; VAN DER PLAS,
L. H. W.; BOUWMEESTER, H. J.; VERHOEVEN, H. a. Monoterpene biosynthesis in lemon
( Citrus limon ). European Journal of Biochemistry, v. 269, n. 13, p. 3160–3171, 2002.
Disponível em: <http://doi.wiley.com/10.1046/j.1432-1033.2002.02985.x>.
MACMILLAN, J.; BEALE, M. H. Diterpene Biosynthesis. Comp. Nat. Prod. Chem., v. 2, p.
217, 2000.
MARUYAMA, T.; ITO, M.; HONDA, G. Molecular cloning, functional expression and
characterization of (E)-beta farnesene synthase from Citrus junos. Biological &
pharmaceutical bulletin, v. 24, n. 10, p. 1171–1175, 2001.
MARZOUG, H. N. Ben; ROMDHANE, M.; LEBRIHI, A.; LEBRIHI, F.; COUDERC, F.;
ABDERRABA, M.; ABDERRABA, M. L.; BOUAJILA, J. Eucalyptus oleosa essential oils:
Chemical composition and antimicrobial and antioxidant activities of the oils from different
plant parts (stems, leaves, flowers and fruits). Molecules, v. 16, n. 2, p. 1695–1709, 2011.
MATTOS JR., D. F. C. B.; ZAMBROSI, F. C. B. .; BOARETTO, R. M. .; QUAGGIO, J. A. .;

46
CANTARELLA, H. Adubação fosfatada em pomares de citros: avanços na pesquisa.
Informações Agronômicas, n. 139, p. 1–8, 2012.
MITIKU, S. B.; SAWAMURA, M.; ITOH, T.; UKEDA, H. Volatile components of peel cold-
pressed oils of two cultivars of sweet orange (Citrus sinensis (L.) Osbeck) from Ethiopia.
Flavour and Fragrance Journal, v. 15, n. 4, p. 240–244, 2000.
MOSHONAS, M. G.; SHAW, P. E. Quantitative and qualitative analysis of tangerine peel oil.
Journal of Agricultural and Food Chemistry, v. 22, n. 2, p. 282–284, 1974. Disponível em:
<\\Robsrv-05\reference manager\Articles\10110.pdf>.
MOUFIDA, S.; MARZOUK, B. Biochemical characterization of blood orange, sweet orange,
lemon, bergamot and bitter orange. Phytochemistry, v. 62, n. 8, p. 1283–1289, 2003.
NJOROGE, S. M.; PHI, N. T. L.; SAWAMURA, M. Chemical composition of peel essential
oils of sweet oranges (Citrus sinensis) from uganda and Rwanda. J.Essent.Oil Bearing Plants,
v. 12, n. 1, p. 26–33, 2009. Disponível em: <\\Robsrv-05\reference
manager\Articles\10789.pdf>.
OGURA, K.; KOYAMA, T. Enzymatic Aspects of Isoprenoid Chain Elongation. Chemical
reviews, v. 98, n. 4, p. 1263–1276, 1998. Disponível em:
<http://www.ncbi.nlm.nih.gov/pubmed/11848932>.
PAK, T.; SYSTEMS, P.; BOOK, T. O.; COMMUNICATION, P.; PRINTER, A. B.; PRINTED,
R. P. The orange book. [s.l: s.n.]
PATTNAIK, S.; SUBRAMANYAM, V. R.; BAPAJI, M.; KOLE, C. R. Antibacterial and
antifungal activity of aromatic constituents of essential oils.Microbios, 1997. .
PHILLIPS, M. A.; LE??N, P.; BORONAT, A.; RODR??GUEZ-CONCEPCI??N, M. The
plastidial MEP pathway: unified nomenclature and resources. Trends in Plant Science, v. 13,
n. 12, p. 619–623, 2008.
PICHERSKY, E.; GERSHENZON, J. The formation and function of plant volatiles: Perfumes
for pollinator attraction and defense. Current Opinion in Plant Biology, v. 5, n. 3, p. 237–
243, 2002.
RAMOS-VALDIVIA, A. C.; VAN DER HEIJDEN, R.; VERPOORTE, R. Isopentenyl
diphosphate isomerase: a core enzyme in isoprenoid biosynthesis. A review of its biochemistry
and function. Natural Product Reports, v. 14, n. 6, p. 591, 1997. Disponível em:
<http://xlink.rsc.org/?DOI=np9971400591>.
REIS, M. S.; TAKITA, M. a.; PALMIERI, D. a.; MACHADO, M. A. Bioinformatics for the
Citrus EST Project (CitEST). Genetics and Molecular Biology, v. 30, n. 3 SUPPL., p. 1024–
1029, 2007.
SANTOS, A. D. S.; SANTOS, L. C. D. S. Aplicação das classificações do sistema de
informação estatística brasileiro à cadeia produtiva óleo-suco-citrícola nacional. Ciência
Rural, v. 41, n. 4, p. 731–737, 2011.
SAWAMURA, M.; THI MINH TU, N.; ONISHI, Y.; OGAWA, E.; CHOI, H.-S. Characteristic
Odor Components of Citrus reticulata Blanco (Ponkan) Cold-pressed Oil. Bioscience,

47
Biotechnology and Biochemistry, v. 68, n. 8, p. 1690–1697, 2004. Disponível em:
<http://www.tandfonline.com/doi/abs/10.1271/bbb.68.1690>.
SCHMIDT, A.; WÄCHTLER, B.; TEMP, U.; KREKLING, T.; SÉGUIN, A.; GERSHENZON,
J. A bifunctional geranyl and geranylgeranyl diphosphate synthase is involved in terpene
oleoresin formation in Picea abies. Plant physiology, v. 152, n. 2, p. 639–655, 2010.
SCHWARTZ, S. H.; TAN, B. C.; GAGE, D. a; ZEEVAART, J. a; MCCARTY, D. R. Specific
oxidative cleavage of carotenoids by VP14 of maize. Science (New York, N.Y.), v. 276, n.
5320, p. 1872–1874, 1997.
SHARON-ASA, L.; SHALIT, M.; FRYDMAN, A.; BAR, E.; HOLLAND, D.; OR, E.; LAVI,
U.; LEWINSOHN, E.; EYAL, Y. Citrus fruit flavor and aroma biosynthesis: Isolation,
functional characterization, and developmental regulation of Cstps1, a key gene in the
production of the sesquiterpene aroma compound valencene. Plant Journal, v. 36, n. 5, p. 664–
674, 2003.
SHAW, P. E. Review of Quantitative Analyses of Citrus Essential Oils. Quantitative Analysis
of Essential Oils, v. 27, n. 2, p. 246–257, 1979.
SHIMADA, T.; ENDO, T.; FUJII, H.; HARA, M.; OMURA, M. Isolation and characterization
of (E)-beta-ocimene and 1,8 cineole synthases in Citrus unshiu Marc. Plant Science, v. 168, n.
4, p. 987–995, 2005a.
SHIMADA, T.; ENDO, T.; FUJII, H.; HARA, M.; UEDA, T.; KITA, M.; OMURA, M.
Molecular cloning and functional characterization of four monoterpene synthase genes from
Citrus unshiu Marc. Plant Science, v. 166, n. 1, p. 49–58, 2004.
SHIMADA, T.; ENDO, T.; FUJII, H.; OMURA, M. Isolation and characterization of a new d-
limonene synthase gene with a different expression pattern in Citrus unshiu Marc. Scientia
Horticulturae, v. 105, n. 4, p. 507–512, 2005b. Disponível em:
<http://www.sciencedirect.com/science/article/pii/S0304423805000737\nhttp://linkinghub.els
evier.com/retrieve/pii/S0304423805000737>.
SHIMADA, T.; ENDO, T.; FUJII, H.; RODRÍGUEZ, A.; PEÑA, L.; OMURA, M.
Characterization of three linalool synthase genes from Citrus unshiu Marc. and analysis of
linalool-mediated resistance against Xanthomonas citri subsp. citri and Penicilium italicum in
citrus leaves and fruits. Plant Science, v. 229, p. 154–166, 2014. Disponível em:
<http://www.ncbi.nlm.nih.gov/pubmed/25443842>.
SHISHIDO, H.; MIYAMOTO, Y.; OZAWA, R.; TANIGUCHI, S.; TAKABAYASHI, J.;
AKIMITSU, K.; GOMI, K. Geraniol synthase whose mRNA is induced by host-selective ACT-
toxin in the ACT-toxin-insensitive rough lemon (Citrus jambhiri). Journal of Plant
Physiology, v. 169, n. 14, p. 1401–1407, 2012. Disponível em:
<http://dx.doi.org/10.1016/j.jplph.2012.05.003>.
STATTI, G. A.; CONFORTI, F.; SACCHETTI, G.; MUZZOLI, M.; AGRIMONTI, C.;
MENICHINI, F. Chemical and biological diversity of Bergamot (Citrus bergamia) in relation
to environmental factors. Fitoterapia, v. 75, n. 2, p. 212–216, 2004.
STUART, G. R.; DARIVA, C.; OLIVEIRA, J. V. High-pressure vapor-liquid equilibrium data
for CO2-orange peel oil. Brazilian Journal of Chemical Engineering, v. 17, n. 2, p. 181–189,

48
jun. 2000. Disponível em: <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0104-
66322000000200006&lng=en&nrm=iso&tlng=en>. Acesso em: 24 fev. 2016.
SUZUKI, Y.; SAKAI, H.; SHIMADA, T.; OMURA, M.; KUMAZAWA, S.; NAKAYAMA,
T. Characterization of gamma-terpinene synthase from Citrus unshiu (Satsuma mandarin).
BioFactors (Oxford, England), v. 21, n. 1-4, p. 79–82, 2004. Disponível em:
<http://www.ncbi.nlm.nih.gov/pubmed/15630174>.
TAKITA, M. A.; BERGER, I. J.; BASÍLIO-PALMIERI, A. C.; BORGES, K. M.; DE SOUZA,
J. M.; TARGON, M. L. N. P. Terpene production in the peel of sweet orange fruits. Genetics
and Molecular Biology, v. 30, n. 3 SUPPL., p. 841–847, 2007.
TEIXEIRA, J. P. F.; MARQUES, M. O. M.; FIGUEIREDO, J. O. Composição química de
óleos essenciais de quinze genótipos de limão em duas épocas de colheita. Citrus Research &
Technology, v. 34, n. 2, 2013. Disponível em: <http://www.gnresearch.org/doi/10.5935/2236-
3122.20130008>.
TU, N. T. M.; THANH, L. X.; UNE, A.; UKEDA, H.; SAWAMURA, M. Volatile constituents
of Vietnamese pummelo, orange, tangerine and lime peel oils. Flavour and Fragrance
Journal, v. 17, n. 3, p. 169–174, 2002.
UJI, Y.; OZAWA, R.; SHISHIDO, H.; TANIGUCHI, S.; TAKABAYASHI, J.; AKIMITSU,
K.; GOMI, K. Isolation of a sesquiterpene synthase expressing in specialized epithelial cells
surrounding the secretory cavities in rough lemon (Citrus jambhiri). Journal of Plant
Physiology, v. 180, p. 67–71, 2015. Disponível em:
<http://linkinghub.elsevier.com/retrieve/pii/S0176161715000826>.
VAN DEN DOOL, H.; DEC. KRATZ, P. A generalization of the retention index system
including linear temperature programmed gas—liquid partition chromatography. Journal of
Chromatography A, v. 11, n. 3, p. 463–71, jan. 1963. Disponível em:
<http://linkinghub.elsevier.com/retrieve/pii/S002196730180947X>.
VAN GELDRE, E.; VERGAUWE, A.; VAN DEN EECKHOUT, E. State of the art of the
production of the antimalarial compound artemisinin in plants. Plant Molecular Biology, v.
33, n. 2, p. 199–209, 1997. Disponível em:
<http://link.springer.com/10.1023/A:1005716600612>.
VEKIARI, S. A.; PROTOPAPADAKIS, E. E.; PAPADOPOULOU, P.; PAPANICOLAOU,
D.; PANOU, C.; VAMVAKIAS, M. Composition and seasonal variation of the essential oil
from leaves and peel of a cretan lemon variety. Journal of Agricultural and Food Chemistry,
v. 50, n. 1, p. 147–153, 2002.
VERZERA, A.; TROZZI, A.; COTRONEO, A.; LORENZO, D.; DELLACASSA, E.
Uruguayan essential oil. 12. Composition of Nova and Satsuma mandarin oils. Journal of
Agricultural and Food Chemistry, v. 48, n. 7, p. 2903–2909, 2000.
VERZERA, A.; TROZZI, A.; GAZEA, F.; CICCIARELLO, G.; COTRONEO, A. Effects of
rootstock on the composition of bergamot (Citrus bergamia risso et poiteau) essential oil.
Journal of Agricultural and Food Chemistry, v. 51, n. 1, p. 206–210, 2003.
VERZERA, A.; TROZZI, A.; MONDELLO, L.; DELLACASSA, E.; LORENZO, D.
Uruguayan essential oils. Part X. Composition of the oil of Citrus clementine Hort. Flavour

49
and Fragrance Journal, v. 13, n. 3, p. 189–195, 1998.
VERZERA, A.; TROZZI, A.; ZAPPALÁ, M.; CONDURSO, C.; COTRONEO, A. Essential
oil composition of Citrus meyerii Y. Tan. And Citrus medica L. cv. Diamante and their lemon
hybrids. Journal of Agricultural and Food Chemistry, v. 53, n. 12, p. 4890–4894, 2005.
WILDUNG, M. R.; CROTEAU, R. A cDNA clone for taxadiene synthase, the diterpene cyclase
that catalyzes the committed step of taxol biosynthesis. Journal of Biological Chemistry, v.
271, n. 16, p. 9201–9204, 1996.
WISE, M. L.; CROTEAU, R. 2.05 - Monoterpene Biosynthesis. Comprehensive Natural
Products Chemistry, p. 97–153, 1999. Disponível em:
<http://www.sciencedirect.com/science/article/pii/B9780080912837000382>.
WOLFORD, R. W.; KESTERSON, J. W.; ATTAWAY, J. A. Physicochemical properties of
citrus essential oils from Florida. Journal of Agricultural and Food Chemistry, v. 19, n. 6,
p. 1097–1105, 1971. Disponível em:
<http://dx.doi.org/10.1021/jf60178a006\nhttp://pubs.acs.org/doi/abs/10.1021/jf60178a006\nht
tp://pubs.acs.org/doi/pdf/10.1021/jf60178a006>.
WU, G. A.; PROCHNIK, S.; JENKINS, J.; SALSE, J.; HELLSTEN, U.; MURAT, F.;
PERRIER, X.; RUIZ, M.; SCALABRIN, S.; TEROL, J.; TAKITA, M. A.; LABADIE, K.;
POULAIN, J.; COULOUX, A.; JABBARI, K.; C, F. C.; ROKHSAR, D. Sequencing of diverse
mandarin, pummelo and orange genomes reveals complex history of admixture during citrus
domestication. Nat bitechnol., v. 32, n. 7, p. 656–662, 2015.
YAMASAKI, Y.; AKIMITSU, K. In situ localization of gene transcriptions for monoterpene
synthesis in irregular parenchymic cells surrounding the secretory cavities in rough lemon
(Citrus jambhiri). Journal of plant physiology, v. 164, n. 11, p. 1436–48, 2007. Disponível
em: <http://www.scopus.com/inward/record.url?eid=2-s2.0-
35348885429&partnerID=tZOtx3y1>.
ZHANG, M.; LIU, J.; LI, K.; YU, D. Identification and Characterization of a Novel
Monoterpene Synthase from Soybean Restricted to Neryl Diphosphate Precursor. PLoS ONE,
v. 8, n. 10, p. e75972, 2013. Disponível em:
<http://dx.plos.org/10.1371/journal.pone.0075972>.
ZINI, C.; MÜHLEN, C. VON. Índices De Retenção Em Cromatografia Gasosa Bidimensional
Abrangente. Scientia Chromatographica, v. 1, n. 3, p. 21–29, 2009.