caps 11 a 20 livro imunologia
TRANSCRIPT

C A P Í T U L O 1 1
ConceitoCitocinas são um grupo de proteínas responsável pela comunicação entre as diversas células do sistema imunológico. Assim como os hormônios servem de mensageiros do sistema endócrino, as citocinas são as mensageiras do sis-tema imunológico. Entretanto, diferem dos hormônios endócrinos por não serem produzidas por glândulas especializadas, mas por células. Regulam a intensidade e a duração da resposta imunológica pela estimulação ou pela inibição da ativação/proliferação/diferenciação de diversas células. Também têm ação sobre a secreção de anticorpos e outras citocinas, regulam o cresci-mento de células, infl amação e reparo tecidual.
Linfocinas são citocinas secretadas na imunidade adaptativa por linfócitos ativados.
Interleucinas (IL) são citocinas secretadas por determinados leucócitos que têm ação sobre outros leucócitos. Foram identifi cadas e enumeradas, mas acredita-se que outras ainda possam ser descobertas.
Quimiocinas são interleucinas de baixo peso molecular, envolvidas na qui-miotaxia. Apresentam grande importância na resposta infl amatória.
PropriedadesAs citocinas se ligam a receptores específi cos na membrana das células, ge-rando transdução de sinais intracelulares, que levam à alteração na expres-são gênica. A suscetibilidade de uma célula a determinada citocina é causada pela presença desses receptores de superfície.
Uma determinada citocina pode ligar-se:– ao receptor da mesma célula que a secretou, exercendo ação autócrina;– às células próximas daquela que a secretou, exercendo ação parácrina;– às células em diversas regiões distantes do corpo, exercendo ação
endócrina.
Diferentemente dos hormônios, a maioria das citocinas apresenta ações autócrina e parácrina. Além disso, podem ser secretadas por muitos tipos de células e ligar-se aos receptores de diversas células. Dessa forma, linfócitos ativados por antígeno secretam citocinas que podem infl uenciar nas demais células da resposta imunológica. Por exemplo, citocinas secretadas por um linfócito TH ativado terão atividade sobre linfócitos B e T-citotóxico, células NK, macrófagos, granulócitos etc.
Júlia Berger Guimarães • Maria Lúcia Scroferneker
Citocinas

132 Imunologia Básica e Aplicada
FunçõesSua função está relacionada à mensagem inter-celular que provoca atividades biológicas par-ticulares, após sua ligação ao receptor de uma célula responsiva. Apesar de uma variedade de células secretarem citocinas, as principais são lin-fócitos TH e macrófagos.
O desenvolvimento das respostas imunes ce-lular e humoral, indução da resposta inflama-tória, regulação da hematopoiese e controle da proliferação e da diferenciação celulares estão entre as inúmeras respostas fisiológicas que en-volvem as citocinas.
As citocinas não são específicas a um antíge-no, podendo ligar-se a qualquer receptor para o qual apresentem afinidade. Entretanto, o siste-ma imunológico, de maneira global, apresenta especificidade a determinado antígeno.
É difícil de conciliar a não-especificidade das citocinas com a especificidade do sistema imu-ne. Uma das maneiras de conciliar é por meio da regulação da expressão de receptores para cito-cinas na superfície das células. Freqüentemente esse receptor só é expresso na superfície celular após o contato dessa célula com um antígeno. Dessa forma, a ativação de citocinas está limita-da a um linfócito ativado por um antígeno.
Outra maneira de manter a especificidade das citocinas é pela interação direta entre a célula produtora de citocina e a célula responsável pelo gatilho na secreção delas. No caso da célula TH, a maior produtora de citocinas, interações celu-lares fortes ocorrem quando o TCR reconhece o complexo MHC-antígeno na superfície da APC. Como exemplos de APCs, temos macrófagos, cé-lulas dendríticas e linfócitos B.
As citocinas secretadas na interação dessas células alcançam elevada concentração local, atingindo as APCs, mas não células mais distan-tes que não formem complexo com o linfócito TH. Além disso, a meia-vida das citocinas na cor-rente sangüínea ou em outros fluidos extracelu-lares é usualmente muito curta, permitindo que elas tenham ação por período limitado e em dis-tância curta.
Quando o organismo está sob o ataque de um patógeno específico, ocorre seu reconhecimen-to e a ativação da imunidade inata, que provoca mecanismos de inflamação. Além disso, a imuni-dade adaptativa também é estimulada. Ela age durante toda a vida da maioria dos vertebrados,
controlando a expressão de moléculas co-esti-mulatórias e induzindo a secreção de citocinas e quimiocinas adequadas. Essas vão direcionar os linfócitos da imunidade adaptativa para seus sí-tios de ação (Tabela 11.1).
Citocinas
Fator de necrose tumoral (TNF)Produzido por macrófagos, LT, NK, células endo-teliais e queratinócitos, pode ser de dois tipos: TNF-α e TNF-β.
Suas funções se equivalem às da IL-1 acrescida das seguintes:
– quimiotaxia para neutrófilos e aumento da desgranulação;
– aumento da adesão das células endoteliais, por meio da expressão de moléculas de adesão da membrana celular, quando em maior quantidade;
– estimulação de células endoteliais e da pro-dução de prostaglandinas, IL-6 e fator pró-coagulante, que tem a capacidade de iniciar a cascata de coagulação, quando presente em maior quantidade. Isso indica a capaci-dade do TNF de causar infartos e necroses hemorrágicas em tumores;
– reação de Schwartzman (hemorrágica) e a coagulação intravascular disseminada (CIVD) ocorrem após a exposição sistêmica ao TNF;
– aumenta a atividade fagocítica de macró-fagos e neutrófilos;
– efeito citotóxico;– estimula diferentes células envolvidas na
resposta inflamatória a secretarem outras citocinas.
Além disso, em geral o TNF tem a proprieda-de de aumentar expressão de MHC-I e está asso-ciado a quadros clínicos, como caquexia, artri-te reumatóide, linfomas, inflamação, infecção, choque séptico, esclerose múltipla, lúpus erite-matoso sistêmico, síndrome de Sjögren e tireoi-dite de Hashimoto.

133Citocinas
Tabela 11.1. Principais citocinas e suas respectivas funções
Funções biológicas
Citocinas Secretada por Células/tecidos Atividade
IL-1 Monócitos, macrófagos, células B, dendríticas e endoteliais, outros tipos
TH e células B Co-estimula ativação, promove maturação e expansão clonal
NK Aumenta atividade
Células endoteliais Aumenta expressão de ICAMs
Macrófagos e neutrófilos Atração quimiotática
Hepatócitos Induz síntese de proteínas de fase aguda
Hipotálamo Induz febre
IL-2 Células TH1 Células TH e T citotóxicas Induz proliferação
Células T Auxilia crescimento
Algumas células NK Aumenta atividade
IL-3 Células TH, NK e mastócitos Células hematopoiéticas e mastócitos
Auxilia crescimento e diferenciação, estimula secreção de histamina
IL-4 TH2, mastócitos e NK Células B ativadas Co-estimula ativação, estimula proliferação e diferenciação, induz switching de IgG e IgE
Células B em repouso Estimula a expressão de MHC-II
Timócitos e células T Induz proliferação
Macrófagos Inibe a expressão de MHC-II e aumenta atividade fagocítica
Mastócitos Estimula crescimento
IL-5 TH2 e mastócitos Células B ativadas Estimula proliferação e diferenciação, induz switching para IgA
Eosinófilos Promove crescimento e diferenciação
IL-6 Monócitos, macrófagos, TH2, células da medula óssea
Células B proliferadas Promove diferenciação terminal das células
Células plasmáticas Estimula secreção de anticorpo
Células tronco-hematopoiéticas, células mielóides
Auxilia na promoção da diferenciação
Hepatócitos Induz síntese de proteínas de fase aguda
IL-7 Medula óssea, células do estroma do timo
Células tronco-hematopoiéticas, células linfóides
Induz a diferenciação em células B e T
Células T em repouso Aumento da expressão de IL-2 e seus receptores
IL-8 Macrófagos e células endoteliais Neutrófilos Quimiocinas, atração quimiotática, indução de aderênciaao endotélio vascular e extravasamento para os tecidos
IL-9 TH Algumas TH Age como mitogênica e auxilia proliferação na ausência de antígeno
IL-10 TH2 Macrófagos Supressão da produção de citocinas, reduzindo assim indiretamente a produção de citocinas pelo TH1
APCs Down-regulation da expressão de MHC-II
IL-11 Células da medula óssea Progenitor de células B Promove diferenciação
Megacariócitos Promove diferenciação
Hepatócitos Induz síntese de proteínas de fase aguda
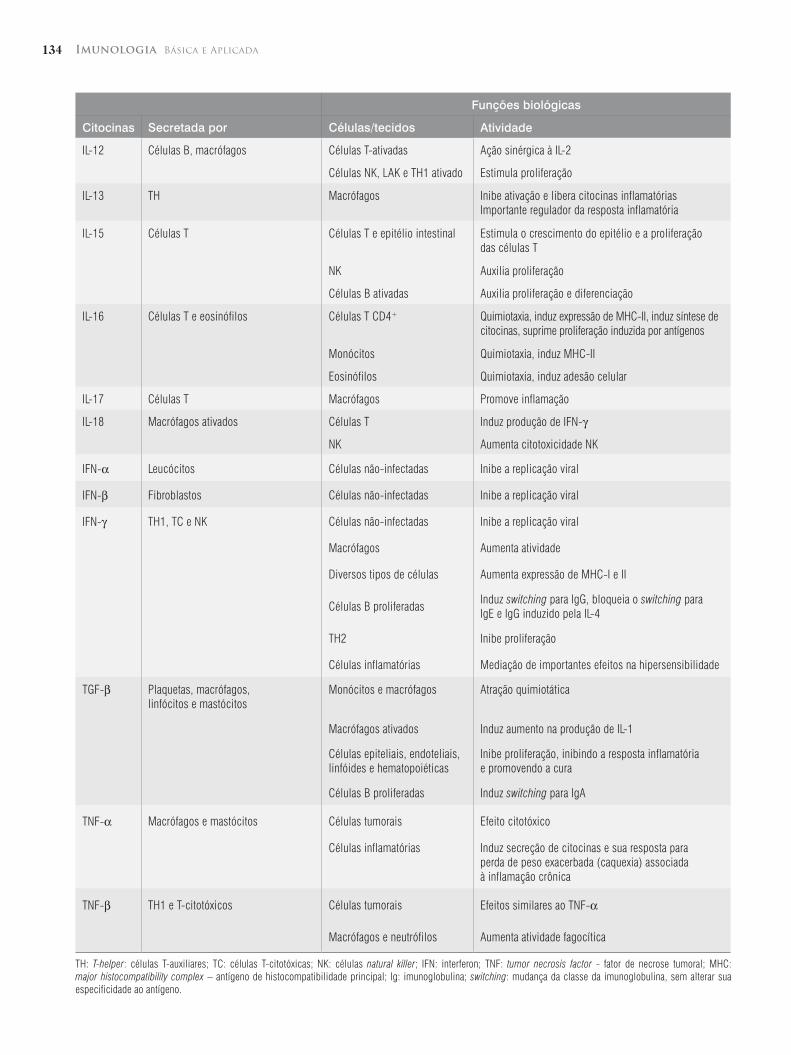
134 Imunologia Básica e Aplicada
Funções biológicas
Citocinas Secretada por Células/tecidos Atividade
IL-12 Células B, macrófagos Células T-ativadas Ação sinérgica à IL-2
Células NK, LAK e TH1 ativado Estimula proliferação
IL-13 TH Macrófagos Inibe ativação e libera citocinas inflamatóriasImportante regulador da resposta inflamatória
IL-15 Células T Células T e epitélio intestinal Estimula o crescimento do epitélio e a proliferação das células T
NK Auxilia proliferação
Células B ativadas Auxilia proliferação e diferenciação
IL-16 Células T e eosinófilos Células T CD4+ Quimiotaxia, induz expressão de MHC-II, induz síntese de citocinas, suprime proliferação induzida por antígenos
Monócitos Quimiotaxia, induz MHC-II
Eosinófilos Quimiotaxia, induz adesão celular
IL-17 Células T Macrófagos Promove inflamação
IL-18 Macrófagos ativados Células T Induz produção de IFN-γ
NK Aumenta citotoxicidade NK
IFN-α Leucócitos Células não-infectadas Inibe a replicação viral
IFN-β Fibroblastos Células não-infectadas Inibe a replicação viral
IFN-γ TH1, TC e NK Células não-infectadas Inibe a replicação viral
Macrófagos Aumenta atividade
Diversos tipos de células Aumenta expressão de MHC-I e II
Células B proliferadasInduz switching para IgG, bloqueia o switching para IgE e IgG induzido pela IL-4
TH2 Inibe proliferação
Células inflamatórias Mediação de importantes efeitos na hipersensibilidade
TGF-β Plaquetas, macrófagos, linfócitos e mastócitos
Monócitos e macrófagos Atração quimiotática
Macrófagos ativados Induz aumento na produção de IL-1
Células epiteliais, endoteliais, linfóides e hematopoiéticas
Inibe proliferação, inibindo a resposta inflamatória e promovendo a cura
Células B proliferadas Induz switching para IgA
TNF-α Macrófagos e mastócitos Células tumorais Efeito citotóxico
Células inflamatórias Induz secreção de citocinas e sua resposta para perda de peso exacerbada (caquexia) associada à inflamação crônica
TNF-β TH1 e T-citotóxicos Células tumorais Efeitos similares ao TNF-α
Macrófagos e neutrófilos Aumenta atividade fagocítica
TH: T-helper: células T-auxiliares; TC: células T-citotóxicas; NK: células natural killer; IFN: interferon; TNF: tumor necrosis factor - fator de necrose tumoral; MHC: major histocompatibility complex – antígeno de histocompatibilidade principal; Ig: imunoglobulina; switching: mudança da classe da imunoglobulina, sem alterar sua especificidade ao antígeno.

135Citocinas
Fator estimulador de colônias de granulócitos e macrófagos (GM-CSF)Liberado por neutrófilos e colônias de LB, esti-mula proliferação e diferenciação de progenito-res de granulócitos e macrófagos e aumenta a ativação e a sobrevida de granulócitos maduros. Está associado a quadros clínicos como mielodis-plasia, leucemia e osteossarcoma.
Transforming growth factor-β (TGF-β)É um grupo de fatores que possui papel funda-mental no crescimento e na diferenciação celu-lares. A maioria das células, inclusive linfócitos e macrófagos, tem capacidade de produzi-lo e quase todas as células apresentam receptores de alta afinidade para essa citocina. Por sua extensa ação biológica, é a única citocina secretada como complexo latente extracelular que deve ser con-vertida na sua forma ativa.
Seus efeitos sobre o sistema imunológico in-cluem inibição da proliferação e diferenciação de células, bem como efeitos estimulatórios, como a indução da proliferação de células CD8, ampliação da maturação de LT e células de me-mória. Além disso, tem a capacidade de inibir a citotoxicidade e a produção de anticorpos (com exceção da Ig-A)
Interleucina-4A IL-4 é uma citocina com múltiplas funções que modula a imunidade pela ação em diversas célu-las. Talvez a mais importante seja sua capacida-de de regular a diferenciação de linfócitos T e B. Além disso, também é reconhecida por modular a ativação de macrófagos. Estimula a expressão de MHC-II e induz ainda o switching de classes para IgE nos linfócitos B e plasmócitos.
Interleucina-10A IL-10 é uma das mais importantes citocinas imu-norreguladoras com efeitos antiinflamatórios e imunossupressivos, sendo macrófagos e monóci-tos as células mais afetadas. Nessas células, IL-10
inibe a produção de citocinas pró-inflamatórias e quimiocinas e inibe a expressão de MHC-II.
As IL-19, IL-20, IL-22, IL-24, IL-26, IL-28A/B e IL-29 foram descobertas e agrupadas na famí-lia da IL-10-interferon. Curiosamente, diferentes citocinas dessa família compartilham receptores. Apesar disso, parece que elas não executam efei-to biológico similar à IL-10. Em contraste com IL-10, cuja atividade foi extensamente estuda-da, pouco se sabe sobre essas citocinas. Supõe-se que tenham atividade pró-inflamatória. A IL-22 é a mais caracterizada do grupo.
Com relação à interação entre IL-10 e IL-22, foi visto que a presença de concentrações crescentes de IL-22 durante a estimulação de células por IL-10 não demonstrou efeito de modulação da IL-10. Além disso, foi possível comprovar a ação da IL-22 em receptores IL-10R2. Em contraparti-da, a presença da IL-10 também não demonstrou modulação nos efeitos da IL-22.
Interleucina-23Estudos utilizando ratos deficientes em IL-23 identificaram como essa citocina é importante na patogênese da doença auto-imune. Nesse caso, foi demonstrado que IL-23 promove o de-senvolvimento de uma população de linfócito T CD4+ que produz IL-6, IL-17 e TNF.
Estudos futuros deverão caracterizar os me-canismos moleculares pelos quais a IL-23 produz esse aumento de células T patogênicas e sugere-se que isso poderá proporcionar novas medidas terapêuticas para as doenças auto-imunes.
Receptores de citocinasPara exercerem suas finalidades biológicas, as ci-tocinas devem ligar-se a seus receptores específi-cos, expressos na membrana de células responsi-vas. Devido a esses receptores estarem presentes em variados tipos de células, as citocinas têm a capacidade de interagir com cada uma delas.
A clonagem de genes de receptores de citoci-nas tem permitido rápido avanço na identifica-ção e caracterização desses receptores.
Estrutura geral dos receptoresA maioria dos receptores pertence a uma das cin-co famílias:

136 Imunologia Básica e Aplicada
• receptor da superfamília das imunoglobulinas;• receptor da classe I de citocinas (também con-
hecido como receptor da hematopoetina);• receptor da classe II de citocinas (também conhe-
cido como receptor da família do interferon);• receptor de TNF;• receptor de quimiocinas.
A maior parte desses receptores, que perten-ce aos sistemas imune e hematopoiético, perten-ce ao grupo da classe I de citocinas.
Transdução de sinaisOs receptores das classes I e II de citocinas agem através da tirosina quinase. Estudos demonstra-ram que um dos primeiros eventos após a ligação da citocina com um desses receptores é uma su-cessão de fosforilações na proteína tirosina. Esses estudos utilizaram INF-γ, uma citocina da classe II. Essa molécula foi originalmente descoberta pela sua capacidade de bloquear ou inibir a replicação em uma variedade de vírus. Essa atividade antivi-ral é compartilhada com IFN-α e IFN-β. Entretan-to, ao contrário desses interferons, INF-γ participa do processo central da imunorregulação.
Além do INF-γ, inúmeros outros ligantes das classes I e II demonstraram causar a dimerização de seus receptores. Um elemento importante na especificidade da citocina vem da especificidade de sua ligação aos seus receptores. Além disso, cada citocina em particular induz a transcrição de genes específicos. Esse gene será responsável pela variedade de efeitos típicos daquela citocina.
Citocinas e linfócitos TCélulas T imaturas são células de vida longa que fazem parte do pool de linfócitos circulantes e migram continuamente do sangue para a linfa através de zonas especializadas do tecido lin-fóide secundário, do baço, dos linfonodos e das placas de Peyer. A sobrevivência dessas células requer contato contínuo com o MHC, combina-do com a exposição à IL-7. Acredita-se que esses dois ligantes induzem leve sinalização, que é su-ficiente para manter essas células vivas, mas não induzem a célula a entrar no ciclo celular.
Como os agentes infecciosos, geralmente, têm replicação muito rápida, o sistema imune primá-rio deve ter uma resposta tão intensa quanto possível. A divisão de células T antígeno-depen-dentes é muito rápida (três a quatro divisões ao
dia para células CD8+). Após a diferenciação em células efetoras, elas voltam à circulação através da linfa e disseminam-se pelo corpo. Pela expres-são de novas moléculas de superfície, as células efetoras adquirem a capacidade de penetrar nos vasos capilares e entrar em contato direto com células parenquimatosas infectadas, por exem-plo, epitélio respiratório infectado pelo vírus influenza. Essas células agem como alvo para as células citotóxicas e são mortas. No caso das in-fecções bacterianas, as células são mortas ou por células citotóxicas ou por citocinas, como IFN-γ, liberado por outras células efetoras.
Infecções e citocinasAssim como a atividade citolítica das células T-CD8+ e a produção de anticorpos por linfócitos B, a secreção de citocinas também contribui para o controle das infecções bacterianas.
A produção de citocinas inflamatórias como TNF-α, IL-1, IL-8 e IL-12 é crucial na resposta à infecção. Essas citocinas recrutam populações de células ao local da inflamação, induzem à ma-turação de células dendríticas e geram resposta específica ao microrganismo invasivo.
Estudos em humanos demonstraram que as células epiteliais respiratórias geram atividade antimicrobiana, mesmo na ausência de estruturas glandulares. A diferenciação da via aérea epitelial em células produtoras de muco é dependente de citocinas do linfócito TH2 (IL-4, 5, 9 e 13).
Assim como os vírus, algumas bactérias pato-gênicas desenvolveram mecanismos para modular a produção de citocinas, com modificação da resposta imunológica. A micobactéria é um bom exemplo da manipulação de citocinas. Ela pode in-duzir a formação de citocinas antiinflamatórias.
Macrófagos infectados produzem IL-6, que inibe a ativação das células T, assim como IL-10 e TGF-β. A IL-10 é imunossupressora de diversas maneiras, incluindo a inibição da ativação de macrófagos, a supressão da produção de citoci-nas inflamatórias e a produção de moléculas da imunidade específica (por exemplo, MHC-II).
A indução das citocinas imunossupressoras pela micobactéria também pode produzir a ativa-ção de células T-supressoras, contribuindo, dessa forma, para a inibição da atividade imunológica.

137Citocinas
A expressão de MHC-II em macrófagos inati-vos é muito baixa e INF-γ é um importante in-dutor de MHC-II nesses macrófagos. A micobac-téria tem a capacidade de inibir a expressão de MHC-II, interferindo na sinalização mediada pelo INF-γ, tornando-se um importante fator de dano à resposta das células TH a essa bactéria. Essa capacidade imunossupressora propicia meios de modular os mecanismos de defesa e o reconheci-mento imunológico.
A Chlamydia trachomatis, um patógeno trans-mitido sexualmente que causa doenças do trato urogenital e infecções oculares, também inibe a expressão das moléculas de MHC-II de superfície induzidas por INF-γ nas células infectadas. Além disso, ela também consegue inibir as moléculas MHC-I constitutivas e as induzidas por INF-γ.
QuimiocinasQuimiocinas são um grupo de citocinas pró-in-flamatórias com mecanismos de ação e especifi-cidade distintos. Suas funções são a promoção da quimiotaxia por meio de gradientes de concen-tração em direção ao sítio inflamatório, a ativa-ção celular através dos receptores de superfície, o auxílio na diferenciação das células da linhagem hematopoiética e também a opsonização. Como
exemplo de sua atividade, já foram descobertas 36 quimiocinas secretadas pelas células endo-teliais tímicas como: MCP-1 ou CCL2 (proteína quimioatrativa para monócitos), MCP-2 ou CCL8, MCP-3 ou CCL7, MCP-4 ou CCL13, CCL5, CCL21 e CCL25 (recrutamento de células T-precursoras do fígado para o timo fetal).
Existe uma mudança de nomenclatura des-se grupo especial de citocinas. Elas recebiam a denominação de família SCY (small cytokine), mas atualmente são chamadas de superfamília das quimiocinas. O subgrupo anteriormente cha-mado SCYC agora se chama C (C-chemokines), representado pela XCL1 e XCL2. O subgrupo anteriormente chamado de SCY-B agora se cha-ma de CXC (CX-chemokine). Seus principais re-presentantes são: MIP-2 (monocyte inflamatory protein), RTCK-1 (thymus chemokine-1), IL-8 ou CXCL8, que atuam ativando neutrófilos, e ainda CXCL9, CXCL10 até CXCL16. O subgrupo anterior-mente chamado de SCY-A agora é chamado de CC (CC chemokines). Seus representantes são a eotaxina (recrutamento de eosinófilos), a MCP-1 (monocyte chemoattractant protein-1), as MIP 1 e 3, a CHAK (chemokine-activate killer) e ainda CCL1 até CCL28. O último grupo é chamado de CXXXC (CXXXC chemokines) e é representado pela CX3CL1.
BibliografiaBenedict CA, Norris PS, Ware CF. To kill or be killed: viral evasion of apoptosis.
Nature Immunology 2002;3(11):1013-18.
Ganz T. Antimicrobial polypeptides in host defense of the respiratory tract (review). J Clin Invest 2002;109:693-7.
Goldsby RA, Kindt TJ, Osborne BA. Innate immune recognition. Ann Rev Immuno-logy 2002;20:197-216.
Hornef MW, Wick MJO, Rhen M et al. Bacterial strategies for overcoming host innate and adaptive immune responses. Nature Immunology 2002;3(11):1033-40.
Langrish CL, Chen Y, Blumenschein et al. IL-23 drives a pathogenic T cell population which induces autoimmune inflammation. J Exp Med 2005;201:233-40.
Major J, Fletcher JE, Hamilton TA. IL-4 Pretreatment selectively enhances cytokine and chemokine production in lipopolysaccharide-stimulated mouse perito-neal monocytes. J Immunology 2002;168:2456–63.
Orange JS, Fassett MS, Koopman LA et al. Viral evasion of natural killer cells. Nature Immunology 2002;3(11):1006-12.
Sprent J, Tough DF. T cell Death and Memory. Science, Vaccines and Immunity 2001;293:345-8.
Wolk K, Witte E, Reineke U et al. Is there an interaction between interleukin-10 and interleukin-22? Genes And Immunity 2005;6:8-18.


C A P Í T U L O 1 2
IntroduçãoO sistema imunológico desenvolveu-se com o objetivo de manter a homeos-tase e o adequado funcionamento dos sistemas biológicos, evitando distúr-bios causados pela presença de substâncias estranhas ao organismo. Após o contato com um antígeno, uma série de alterações no sistema imune ocorre com o objetivo de eliminá-lo e restabelecer a homeostase. Leucócitos poli-morfonucleares, sistema complemento, anticorpos, citocinas, linfócitos T e B, macrófagos, células dendríticas e células NK estão entre os componentes imunes que são ativados na resposta a um antígeno. Para que haja resposta imunológica adequada, é preciso que exista uma regulação de todos os com-ponentes do sistema imune. Isso é importante, pois as mesmas substâncias que eliminam os antígenos também têm o potencial de causar dano aos teci-dos e aos órgãos próprios. A seguir, os mecanismos de regulação da resposta imunológica referente a cada componente do sistema imune.
O antígenoO antígeno é o desencadeador da resposta imune, ativando os linfócitos T e B após ligação com seus receptores específi cos. Algumas características do antígeno, como sua natureza, dose e via de entrada, têm infl uência no tipo e na magnitude dessa resposta.
A principal diferença observada com relação à natureza dos antígenos é a sua constituição: eles podem ser protéicos ou constituídos por polissacarí-deos e lipídios. Os antígenos polissacarídicos e lipídicos, como as bactérias encapsuladas, não são apresentados aos linfócitos em associação ao MHC (major histocompatibility complex) e, por isso, a resposta celular contra eles será T-independente. Essa resposta consiste de anticorpos IgM e tem curta duração. Já os antígenos protéicos, como a maioria dos vírus, estimulam a imunidade humoral e celular, originando uma resposta celular T-dependen-te, com formação de memória imunológica de longa duração. Isso explica porque um indivíduo infectado ou vacinado contra um vírus permanece re-sistente à nova infecção por período longo ou até por toda a vida, o que não é tão freqüente com relação a bactérias encapsuladas.
A localização do antígeno no organismo infl uencia a resposta imune. Os vírus são microrganismos essencialmente intracelulares e utilizam tal recurso para escapar da resposta imune humoral específi ca exercida pelos anticor-pos presentes no plasma. Nesse caso, a resposta imune será em grande parte
Gustavo Brandão Fischer • Maria Lúcia Scroferneker
Regulação da resposta imune

140 Imunologia Básica e Aplicada
citotóxica, exercida pelos linfócitos TCD8+ e célu-las NK diretamente contra as células infectadas. Nesse caso, ocorre eliminação antigênica, porém também destruição das células próprias infecta-das. Isso pode gerar de certa forma comprometi-mento grave do tecido ou órgão afetado, como é o caso das hepatites virais. Os antígenos bacte-rianos são em grande parte extracelulares, o que por um lado facilita a resposta imune humoral específica, porém também pode gerar conseqü-ências negativas ao organismo. Por exemplo, an-tígenos provocam a formação de complexo antí-geno-anticorpo que pode se depositar em outros tecidos e causar dano tecidual, como os gloméru-los renais ou articulações. Da mesma forma, an-tígenos podem se ligar a moléculas de superfície de células do hospedeiro e ativar o sistema com-plemento, causando dano de estruturas do pró-prio hospedeiro.
A dose de antígeno a que o indivíduo é ex-posto influencia a amplitude da resposta imu-ne gerada. A dose ótima para que um antígeno seja imunogênico varia conforme o antígeno. Em geral, grandes doses de antígenos protéicos administrados repetidamente induzem tolerân-cia específica em linfócitos T e inibem a resposta imune, enquanto grandes doses de antígenos po-lissacarídicos induzem tolerância em linfócitos B específicos, impedindo a produção de anticorpos.
Com relação à porta de entrada do antígeno, sabe-se que antígenos administrados de forma subcutânea ou intradérmica costumam ser imu-nogênicos, o que não ocorre com antígenos ad-ministrados por via intravenosa ou oralmente, que induzem, dessa forma, tolerância imunoló-gica. Isso pode ser explicado pelo fato de que, nos tecidos subcutâneos e intradérmicos, estão localizadas as células especializadas na captação e na apresentação do antígeno para a ótima ati-vação do sistema imune, as chamadas APC pro-fissionais. Isso pôde ser observado em camun-dongos infectados com o vírus da coriomeningi-te linfocítica (LCMV) por via subcutânea, os quais desenvolveram imunidade.
Os leucócitos polimorfonuclearesApós a penetração do antígeno no hospedeiro, as primeiras células a chegarem no sítio de in-fecção são os leucócitos polimorfonucleares. São
células de vida curta que raramente excedem 24 horas e que morrem logo após liberarem os seus grânulos com toxinas. O processo que envolve a migração dessas células até o sítio infeccioso é complexo e requer o funcionamento de nume-rosas estruturas. Ao ocorrer uma infecção em determinada região do organismo, quimiocinas são liberadas a partir do local atingido e ativam células endoteliais próximas a apresentarem na sua superfície moléculas de adesão sinalizadoras. É assim que os polimorfonucleares que estão cir-culando na corrente sangüínea são capazes de perceber o local da injúria tecidual. No momen-to em que receptores de adesão na superfície dos polimorfonucleares se ligam às moléculas de adesão endoteliais, outros receptores são ex-pressos e o leucócito torna-se capaz, então, de realizar a diapedese e chegar aos tecidos peri-vasculares. Pelo estímulo de quimiocinas e de gradiente de toxinas e produtos inflamatórios, os microfilamentos de actina e miosina intrace-lulares são ativados e ocorre a migração da célu-la em direção ao sítio da infecção. Quando esses leucócitos reconhecem as partículas antigênicas por meio dos PAMPs (pathogen-associated mo-lecular patterns), são ativadas vias intracelulares que levam os grânulos de toxinas até a super-fície, liberando-as para o meio externo. Muitas dessas toxinas são formadas por radicais livres de oxigênio e enzimas de destruição tissular, como a elastase, mieloperoxidase, proteinases, cola-genases e outras. É importante observar que ao mesmo tempo que tais toxinas destroem o antí-geno, também causam dano tissular se ocorrer exposição prolongada ao antígeno ou a falta de mecanismos de controle. É o que acontece, por exemplo, na artrite reumatóide, em que a ati-vação crônica dos polimorfonucleares na cápsula articular causa destruição de todas as estruturas circunjacentes, levando às manifestações clínicas conhecidas. Da mesma forma, a SARA (síndrome da angústia respiratória do adulto) está relacio-nada com o estímulo inflamatório crônico e a liberação maciça de toxinas e citocinas dos poli-morfonucleares no pulmão. Sabe-se que indiví-duos neutropênicos apresentam manifestações pulmonares mais brandas do que indivíduos sem qualquer tipo de imunossupressão. A fisiopa-togenia do enfisema também está relacionada com a ação da elastase derivada dos leucócitos no pulmão.

141Regulação da resposta imune
Além dos riscos que envolvem a ativação leu-cocitária, como visto anteriormente, sabe-se que todo o processo de ativação, sinalização e qui-miotaxia é controlado por grande número de receptores e moléculas. É necessário um perfeito funcionamento de todas essas estruturas para uma resposta imune eficaz. Existem doenças li-gadas às deficiências de cada uma dessas etapas, como, por exemplo, Chediak-Higashi com anor-malidade na morfologia dos grânulos de toxinas, deficiência das moléculas de adesão integrinas e selectinas, deficiência de lactoferrina, deficiência de mieloperoxidase, deficiência de NADPH oxi-dase, deficiência de quimiocinas etc. Todas essas deficiências provocam infecções de repetição nos indivíduos afetados.
As células apresentadoras de antígenosAs células apresentadoras de antígenos (APCs) são indispensáveis para a indução da resposta imune dependente de linfócitos T. Elas têm o papel de apresentar o antígeno em associação com molé-culas do MHC ao linfócito, na presença de mo-léculas co-estimulatórias, aumentando sua proli-feração e diferenciação. Portanto, a presença de APCs competentes estimula a resposta imune, en-quanto sua ausência leva a respostas deficientes. A ausência de moléculas co-estimulatórias na su-perfície da APC pode levar à tolerância. Por outro lado, a apresentação por células dendríticas e ma-crófagos, ditas APC profissionais, que expressam grande quantidade de MHC-II e moléculas co-es-timulatórias, resulta em uma ativação altamente efetiva dos linfócitos T.
Uma vez estimulados, os linfócitos T produ-zem citocinas, entre elas o interferon-γ (IFN-γ), que provoca a ativação de macrófagos. Um resul-tado lento da liberação de IFN-γ é a aquisição de atividade citolítica pelos macrófagos, podendo destruir as células vizinhas, inclusive as próprias células T, que se ligam a eles através das molécu-las do MHC. Assim, os macrófagos ativados pas-sam a ter duas funções: ajudar a eliminar os antí-genos e limitar a duração da resposta imune por meio da lise das células antígeno-específicas.
Regulação genéticaAtualmente se sabe que a capacidade que um organismo tem de identificar antígenos estra-
nhos e defender-se é hereditária. Dessa forma, a capacidade da resposta imune varia de indivíduo para indivíduo.
Existem duas categorias de genes que são ca-pazes de controlar a resposta imunológica: ge-nes para receptores, que codificam os receptores das células B e T, determinando a existência de uma resposta a certo antígeno e os genes da res-posta imune, que influenciam na qualidade e na extensão da resposta.
Os genes da resposta imune se subdividem em genes que codificam o MHC ou não. Existem nu-merosos loci para esses genes, cada um influen-ciando em um nível a resposta a determinado antígeno.
Genes que codificam moléculas MHC-IIEstes genes influenciam nas interações entre as células do sistema imune e no seu processo de ativação. Com os genes para o MHC-I, determi-nam a responsividade da resposta imune. Tam-bém determinam a capacidade de ligação do antígeno (agretopo) ao seu local de ligação no MHC (desetopo). Se essa associação for fraca, o reconhecimento e a ativação da célula TH esta-rão comprometidos. Da mesma maneira, células TH incapazes de serem ativadas por uma ligação MHC + antígeno alterada não conseguirão ativar adequadamente as células T-citotóxicas. A fun-ção desses genes é antígeno-específica e, con-seqüentemente, o organismo estará preparado para responder melhor a determinados antíge-nos do que a outros.
Genes que codificam o MHC-IEstes genes regulam a ativação das células T cito-tóxicas e T supressoras.
As próprias moléculas do MHC-I podem ter função imunorregulatória. Essas moléculas exis-tem também na forma solúvel, além da forma ligada à membrana celular, e inibem a resposta imune pela interação com o TCR (T-cell receptor) e outros receptores. Nas células NK (natural killer cells), por exemplo, a ligação das moléculas do MHC-I provoca a inibição da função citotóxica.

142 Imunologia Básica e Aplicada
Genes não relacionados ao MHCNormalmente estes genes são menos polimorfos do que os genes para o MHC, tendo uma partici-pação menor na determinação da suscetibilidade do organismo aos antígenos. No entanto, altera-ções na sua codificação podem levar ao desenvol-vimento de doenças auto-imunes, como diabetes melito insulino-dependente, alergias e infecções. Estes genes afetam, primariamente, o metabolis-mo das APCs. No que diz respeito aos macrófa-gos, eles regulam a capacidade oxidativa dessas células, aumentam a atividade citotóxica tumoral e antimicrobiana e também são capazes de au-mentar a expressão do MHC-II na superfície dos macrófagos. Da mesma forma, a regulação feita nos eosinófilos determina o grau de resposta a in-vasores como parasitas (Tabela 12.1).
Rh (Rhogan) no puerpério imediato de uma mãe Rh-negativo previne a sensibilização primária ao Rh-positivo do feto, por inativar os antígenos estranhos provenientes do feto, presentes neste momento na circulação materna.
Os mecanismos pelos quais os anticorpos modulam a resposta imune não estão comple-tamente definidos. No entanto, duas interpre-tações são viáveis, no caso da regulação através da IgM: 1) os complexos imunes contendo IgM são capturados pela porção Fc do anticorpo ou receptores C3 nas APCs e são processados mais eficientemente do que o antígeno isolado; e 2) os complexos imunes contendo IgM estimulam a resposta antiidiotípica para a IgM, que, por sua vez, amplifica a resposta. A administração passi-va de IgM, juntamente com o antígeno, aumen-ta a resposta contra ele, enquanto a IgG suprime essa resposta.
O bloqueio antigênico e a reação cruzada en-tre receptores estão entre os mecanismos utiliza-dos para a supressão realizada pela imunoglobu-lina IgG. Por bloqueio antigênico (Figura 12.1.) se entende o mecanismo pelos quais anticorpos administrados passivamente se ligam ao antí-geno, competindo com as células B. O papel da IgG nesse fenômeno reside no fato de que a sua quantidade e capacidade de ligação ao antígeno estão diretamente relacionadas com sua afinida-de pelos receptores das células B. Esse mecanismo é independente da porção Fc do anticorpo. Ob-serva-se que altas doses do antígeno bloqueiam a interação do epítopo com o receptor da célula B, que se torna inapta a reconhecer esse antíge-no. Tal mecanismo de bloqueio também impede a multiplicação das células B.
Tabela 12.1. Genes que influenciam a resposta imune
Genes para receptores de superfície
Responsáveis pela ativação inicial das células e reconhecimento do antígeno
Genes para MHC-I Responsáveis pela ativação e expressão das células T citotóxicas e T supressoras
Genes para MHC-II Responsáveis pela ativação das células TH e potencialização das respostas humoral e celular, e também hipersensibilidade tardia
Genes não relacionados ao MHC
Afetam o metabolismo das APC, influenciando no processamento e apresentação do antígeno
Figura 12.1. Bloqueio antigênico. Altas doses de anticorpos bloqueiam a interação entre o determinante antigênico e o receptor da célula para aquele epítopo. A célula B torna-se inapta para re-conhecer aquele antígeno. Esse bloqueio também impede a duplicação das células B. Células B com epítopos diferentes não são afetadas.
MacrófagoReceptor Fc
Ac
Ag
Os anticorposOs anticorpos regulam o sistema imunológico por meio de um mecanismo de retroalimentação. A propriedade dos anticorpos de regular a resposta imune adquire importância em diversas situações práticas. As crianças menores de 1 ano de idade que são vacinadas precisam com freqüência de várias doses de reforços, uma vez que os níveis de IgG materna permanecem altos na criança pelo menos durante os 6 primeiros meses de vida. Esses anticorpos podem impedir o desenvolvimento de resposta protetora e duradoura na criança. Ou-tro exemplo é o da incompatibilidade do sistema Rh, no qual a administração de anticorpo anti-

143Regulação da resposta imune
A reação cruzada entre receptores (Figura 12.2) é o mecanismo pelo qual os anticorpos IgG, em uma ação Fc-dependente, inibem a diferenciação das células B por ligação cruzada entre o recep-tor do antígeno e a porção Fc da mesma célula. Doses de IgG que são insuficientes para inibir completamente a produção de anticorpos têm a ação de aumentar a afinidade destes, uma vez que somente células B de alta especificidade podem competir com anticorpos passivamente adquiridos pelo antígeno. Por essa razão, a re-troalimentação feita pelos anticorpos é um fator importante que determina o processo de matu-ração e afinidade dos linfócitos.
assumia que o sistema imunológico seria com-posto por um conjunto discreto de clones celu-lares originalmente em repouso e a atividade apenas existiria quando um estímulo externo se apresentasse no organismo. Porém, descobertas recentes mostraram que o conjunto discreto de clones celulares com imunoglobulinas de super-fície específicas é composto, na verdade, por um imenso grupo de células com uma gama infinita de especificidades. A idéia de que a especificida-de é um conjunto de receptores prontos e de-finidos na ontogenia e à espera dos antígenos foi substituída pelos conceitos de aleatoriedade nos processos de geração e maturação das mo-léculas de anticorpo. Essa aleatoriedade abran-ge os conceitos de imunoedição, maturação de afinidade e hipermutação somática, os quais são processos contínuos e independentes do contato com os antígenos. Essa aleatoriedade leva à idéia de que novas moléculas de anticorpos podem ser vistas como elementos invasores ao organismo, sendo tratadas como antígenos. Segundo Jerne: “O sistema imunológico é constituído por uma rede enorme e complexa de parátopos que re-conhecem conjuntos de idiótopos que são reco-nhecidos por conjuntos de parátopos, assim, cada elemento pode reconhecer e ser reconhecido”.
Idiótipo é o conjunto de epítopos encontra-do dentro ou próximo dos sítios de reconheci-mento de antígenos das imunoglobulinas e dos TCRs, sendo cada um desses epítopos chamado de idiótopo e a resposta do sistema imune con-tra cada idiótopo conhecida como antiidiotípi-ca. O parátopo representa a região variável de ligação da molécula de anticorpo. Os idiótopos podem se encontrar dentro ou fora do parátopo (Figura 12.3).
Os anticorpos antiidiotípicos são definidos como anticorpos que se ligam a conformações idiotípicas dentro ou próxima do sítio de com-binação do antígeno na imunoglobulina ou no TCR. Os anticorpos antiidiotípicos se ligam a idió-topos localizados no ou próximos ao parátopo.
O comportamento da rede idiotípica pode ser explicado do seguinte modo: quando um dado antígeno invade o nosso organismo, seu epítopo é reconhecido (com vários graus de es-pecificidade) por um conjunto de diferentes pa-rátopos, chamado p1. Esses parátopos p1 estão associados a um conjunto de idiótopos i1. O sím-bolo P1i1 denota o conjunto total de moléculas de anticorpo e linfócitos B capazes de reconhe-
Figura 12.2. Reação cruzada entre receptores. Baixas doses de anticorpos permitem a ligação cruzada pelo antígeno do receptor Fc da célula B e seu receptor para antígeno. Esse fenômeno permite a replicação celular, mas inibe a síntese de anticorpos. O efeito não é epítopo-específico.
Receptor Fc
Receptor LB
Antígeno
Complexo antígenoanticorpo
Rede idiotípicaEm vez de explicar os processos de sinalização celular e a interação de anticorpos, células e seus mecanismos efetores, a teoria da rede ini-cialmente proposta por Niels Kaj Jerne, em 1974 (Nobel em 1984), apresentava um novo ponto de vista sobre a atividade linfocitária, a produção de anticorpos, a seleção do repertório pré-imu-ne, a distinção do próprio e do não-próprio, a tolerância imunológica, a memória e a evolução do sistema imunológico. Foi sugerido que o sis-tema imunológico seria composto por uma rede regulada de células e moléculas que se reconhe-cem, mesmo na ausência de antígenos. Esse pon-to de vista estava em conflito com a teoria da seleção clonal já existente naquela época, que

144 Imunologia Básica e Aplicada
cer esse antígeno. Na teoria da rede imunoló-gica, cada parátopo do conjunto p1 reconhece um conjunto de idiótopos e todo o conjunto p1 reconhece um conjunto ainda maior de idióto-pos. Esse conjunto i2 de idiótopos é chamado de “imagem interna” do epítopo (ou antígeno), pois ele é reconhecido pelo mesmo conjunto p1 que reconhece o antígeno. O conjunto i2 está asso-ciado a um conjunto p2 de parátopos expresso por moléculas e receptores celulares do conjunto p2i2. Além disso, cada idiótopo do conjunto P1i1 é reconhecido por um conjunto de parátopos, de forma que todo o conjunto i1 é reconhecido por um conjunto p3 ainda maior de parátopos que estão associados aos idiótopos i3 de anticorpos e linfócitos pertencentes a um conjunto p3i3 cha-mado de conjunto antiidiotípico. Seguindo esse esquema, é possível chegar a conjuntos cada vez maiores de receptores que reconhecem e são re-conhecidos por conjuntos previamente definidos na rede. Além do conjunto P1i1, existe um con-junto paralelo P1i1 de moléculas e receptores
que apresentam idiótopos do conjunto i1 asso-ciados a parátopos que não reconhecem o antí-geno em questão. Ações dos anticorpos antiidio-típicos contra idiótopos que estão dentro de pa-rátopo podem desencadear uma reação cruzada, ativando a célula B. No entanto, se a porção Fab antiidiotípica estiver fora do parátopo, pode ha-ver interação do sinal entre a porção Fab e o re-ceptor Fc, resultando em regulação negativa da célula B (Figura 12.4).
Figura 12.3. Idiótipo. Um idiótipo consiste de epítopos associados com a região V da porção Fab do anticorpo. Cada epítopo é designado a um idiótopo. Estes, por sua vez, podem estar associados com o parátopo ou fora dele.
Porção Fab
Figura 12.4. Modelos de inibição e ativação segundo a localização da ligação entre anticorpos antiidiotípicos.
Porção Fab
Porção Fc
Idiótopos
IdiótipoParátopo
Linfócito B
Linfócito B
1. Ativação
2. Inibição
Receptor Fc
Ac antiidiótopo
Ac antiidiótopo
Exemplos do funcionamento da rede antiidio-típica envolvem, principalmente, a fisiopatogê-nese das doenças auto-imunes. Anticorpos an-tiinsulina (P1i1) causam a formação de anticorpos antiidiotípicos (P2i2) que apresentam o idiótopo semelhante à insulina, pois se ligam ao idiótopo que complementa a molécula de insulina, e isso é denominado de imagem em espelho ou imagem interna. Então, os anticorpos P2i2 ligam-se aos receptores de insulina, mimetizando a sua ação. Outro exemplo é o desenvolvimento de auto-an-ticorpos antiacetilcolina e sintomas miastênicos em coelhos imunizados com Bis-Q, uma substân-cia química orgânica que se liga à acetilcolina. Nesse caso, os coelhos desenvolveram anticorpos antiidiotípicos anti-Bis-Q (P1i1) e também uma população de anticorpos anti-receptor de acetil-colina (P2i2) com efeitos deletérios na placa mo-tora. Em outro exemplo, a imunização de ratos

145Regulação da resposta imune
com auto-anticorpos anti-DNA provocou o sur-gimento de vários outros auto-anticorpos com sinais patológicos de nefrite e depósitos renais de complexos antígeno-anticorpo.
Outra resposta antiidiotípica é a resposta hu-mana à imunização por toxóide tetânico. Indiví-duos previamente imunizados com toxóide te-tânico (TT) e toxóide diftérico receberam imuni-zações de reforço e foram monitorados para o desenvolvimento de anticorpos antitétano e an-ticorpos antiidiotípicos (contra os antitétano). Os anticorpos antiidiotípicos foram detectados em duas semanas após a imunização de reforço, per-manecendo detectáveis durante os quatro meses do estudo. O aparecimento dos anticorpos antii-diotípicos foi associado com a diminuição e pos-terior desaparecimento dos anticorpos antitétano (idiótopo). Mas o título total de anticorpos antité-tano só diminuiu parcialmente, o que indica que clones idiotípicos de anticorpos antitétano não foram afetados. A observação repetida da elimi-nação de um clone de anticorpos que apresentem idiótopo dominante pelos anticorpos antiidiotípi-co, sem haver diminuição significativa da popula-ção total de anticorpos contra o antígeno externo (em virtude da expansão clonal de linfócitos com idiótopos diferentes do dominante), tem levanta-do dúvidas sobre um papel principal do anticorpo antiidiotípico na regulação.
De qualquer forma, as aplicações clínicas dos conceitos de rede idiotípica são muitas. A pato-gênese de certas doenças auto-imunes poderia estar: 1) na falha da produção de anticorpos antiidiotípicos, permitindo a expansão descon-trolada de auto-anticorpos ou de um clone de linfócitos; 2) na formação persistente de anticor-pos antiidiotípicos, formando complexos com os respectivos idiótopos e provocando depósitos de imunocomplexos, já observado em glomérulos; ou 3) estimulação de receptores por anticorpos antiidiotípicos, mimetizando efeitos fisiológicos, como na doença de Graves, o que pode ocorrer quando o sistema idiotípico perde sua auto-re-gulação e deixa de ser auto-limitado, passando a induzir doenças imunomediadas.
Outros exemplos do uso terapêutico da rede antiidiotípica podem ser exemplificados com o uso das imunoglobulinas endovenosas polivalen-tes. Esses anticorpos antiidiotípicos podem ser usa-dos para bloquear auto-anticorpos patogênicos e assim tratar enfermidades imunes. A púrpura trombocitopênica imunológica e a sensibilização
Rh nas mulheres Rh-negativas são exemplos de como anticorpos introduzidos nos organismos de indivíduos com doenças causadas por anticorpos podem, por meio de suas interações idiotípicas, bloquear o seu efeito.
Portanto, o que ocorre é que, em alguns indi-víduos, existe quebra de tolerância que leva à fal-ta de regulação idiotípica com subseqüente pro-dução de múltiplos anticorpos antiidiotípicos em cadeia extensa. Tais indivíduos, provavelmente, têm fatores predisponentes, como fatores gené-ticos e hormonais, deficiências imunes ou exposi-ção a determinados agentes do meio ambiente.
Os linfócitosA ativação das células B e T arma o sistema imu-nológico para uma resposta adequada ao antí-geno invasor, realizando esse processo tanto por eliminação direta do antígeno por células efeto-ras (resposta celular) quanto por anticorpos espe-cíficos (resposta humoral). Nesse processo, uma série de mecanismos regulatórios são necessários para modular a ativação das células T efetoras e a produção de anticorpos pelos linfócitos B.
A expressão da função imunológica é o re-sultado de uma série complexa de eventos imu-norregulatórios que ocorrem em fases. Tan-to os linfócitos B como os T, por meio de sinais apropriados, são ativados, proliferam e se dife-renciam. Essa ativação pode ser o evento final, com a produção de anticorpos ou citotoxicidade, como pode servir de mecanismo regulador que modula outras funções.
Os linfócitos T chamados de “convencionais” são aqueles 5% de células T competentes TCD4+ ou TCD8+ que migraram do timo para os órgãos linfóides e estão aptas a exercer as suas funções de auxílio e citotoxicidade por meio de seus re-ceptores α/β de superfície. Porém, existem os lin-fócitos chamados não-convencionais no extenso repertório de células T. O primeiro grupo de lin-fócitos T não-convencionais foi descrito há vários anos e é constituído pelos linfócitos T γ/δ. Essas células não são selecionadas no timo e, portan-to, o seu repertório é independente da existên-cia desse órgão linfóide primário. Existem outras populações de linfócitos T recentemente desco-bertas que também não sofrem maturação tími-ca ou, porém, fazem-no de maneira diferente daquela dos linfócitos convencionais. Entre essas células, estão os linfócitos T-reg (T-reguladores),

146 Imunologia Básica e Aplicada
as NKT e os linfócitos TCD8 α/α, uma população que se origina a partir de timócitos duplo-positi-vos expostos a doses ótimas de antagonista.
Os linfócitos T-reg são importantes nos meca-nismos de tolerância periférica, atuando com su-pressão ativa direta. Diferentes tipos de células T-reg têm sido descritas, como as TH3, as Tr1 e, o exemplo mais conhecido, as células TCD4+CD25+, que expressam as moléculas CTLA-4, GITR e Foxp3. Essas células podem ser ativadas por antí-genos próprios e não-próprios e, uma vez ativa-das, podem suprimir células T de maneira antí-geno não-específica. Os efeitos supressivos não são restritos ao sistema imune adaptativo, mas podem também influenciar as células do sistema imune inato, como monócitos, macrófagos e cé-lulas dendríticas. Exercem sua ação pela secreção de citocinas inibitórias (como IL-10 e TGF-β) ou tolerizando APCs por interações célula-célula. Elas constituem apenas uma pequena fração das células TCD4 circulantes e são comumente iden-tificadas pela expressão de IL2Ra (CD25), assim como de Foxp3. O Foxp3 foi identificado como marcador celular característico das T-reg e sua expressão é essencial para o desenvolvimento e a função dessa célula. A expressão de Foxp3 em linfócitos T convencionais pode conferir ativida-de supressora e esses também passam a expressar CD25. O Foxp3 age estabelecendo e mantendo o programa genético da T-reg e funciona como regulador negativo da ativação de células T e tal-vez como efetor transcricional de programas de citocinas antiinflamatórias.
As células Tr1 produzem IL-10 e IL-4, enquanto as células TH3 produzem IL-10 e TGF-β.
Os linfócitos NKT parecem ser originados dos mesmos precursores duplo-positivos dos linfócitos T convencionais. Essas células podem apresentar moléculas CD4 ou CD8 na superfície, apresentam um TCR com repertório limitado e possuem re-ceptores característicos como os NK1.1 (CD161) e receptores da família Ly-49 e NKG2. Distribuem-se no fígado, no baço, no timo e na medula óssea, sendo menos abundantes nos linfonodos. Repre-sentam menos de 1% da população de linfócitos. Secretam IFN-γ, TNF-α, IL-4 e IL-13. Possuem uma molécula de CD1d ligada ao TCR e tal fato possibi-lita que sejam reconhecidos alguns antígenos que não o são pelos linfócitos T convencionais. Apre-sentam receptores do tipo toll e são capazes de reconhecer glicosilceramidas das bactérias Gram-negativas pelos seus CD1d. Tal via de sinalização
possibilita o reconhecimento de patógenos que não apresentam ligantes para TLR em sua pare-de celular. Estão envolvidas nas respostas alérgi-cas, inflamatórias, tumorais e na auto-imunidade, além de participarem na regulação da resposta imune. A ativação dessa célula parece ser benéfica nos modelos murinos de diabetes e artrite induzi-da pelo colágeno, além de mediar a tolerância ao enxerto pela supressão da resposta celular através da secreção de IL-10 e IL-13.
Os linfócitos CD8 α/α apresentam a molécula CD8 homodimérica ao contrário da forma hete-rodimérica α/β dos linfócitos T convencionais. A forma homodimérica não parece ser um co-re-ceptor eficiente do TCR. Células CD8 α/α são am-plamente presentes nos linfócitos intra-epiteliais do intestino. A sua origem é controversa, porém alguns autores demonstraram a sua origem tí-mica. Esses linfócitos parecem expressar genes típicos do sistema imune inato, como recepto-res NK. Alguns linfócitos TCD8 convencionais podem expressar CD8 α/α, que estão envolvidas na memória imunológica celular. Mais estudos são necessários para o completo entendimento do comportamento dessas populações de linfó-citos T não-convencionais. Apesar disso, existe a tendência crescente em acreditar que as doenças auto-imunes, apesar de multifatoriais, são causa-das primariamente por deficiência na regulação imune efetuada pelas células com atividade su-pressora em vez da simples ação de células auto-reativas nos tecidos-alvos (Figura 12.5).
Figura 12.5. Esquema do mecanismo imunológico causal nas doenças auto-imunes, envolvendo a re-gulação do grupo de células com atividade supressora.
Célulaauto-reativa
Célularegulatória
Auto-imunidade

147Regulação da resposta imune
Os linfócitos TCD4+ convencionais são subdi-vididos em duas populações distintas conforme a produção de citocinas. Assim, encontramos a subpopulação TH1, produtora de INF-γ e IL-2, e os TH2, produtores de IL-4, IL-5, IL-6 e IL-10. O primeiro subgrupo ativa as células T citotóxicas e leva à resposta do tipo hipersensibilidade tardia, enquanto os TH2 regulam negativamente a inten-sidade da resposta imune através da secreção de IL-10, inibidora da produção das demais citocinas. Os TH2 induzem à produção de grande quantida-de de imunoglobulinas específicas pelos linfócitos B, respondendo a antígenos que requerem altos títulos de anticorpos para sua eliminação.
A regulação cruzada das subpopulações de TH demonstra que citocinas como o INF-γ, secreta-das pelas TH1, podem inibir a ativação das TH2, assim como a secreção de IL-10 pelas TH2 impede a expressão da molécula B7 nos linfócitos B e a expressão de IL-12 nas APC, inibindo a ativação de TH1. A ativação preferencial de um subgrupo resulta na regulação da resposta em torno de um efetor particular, tornando-a mais específica. Es-tudos recentes observaram que a resposta imune é totalmente desregulada quando se inativa a subpopulação de TH2 (mas não de TH1), suge-rindo que o processo mediado por essas células seja diretamente responsável, ao menos em par-te, pela regulação fisiológica normal da resposta imunológica.
Grande parte dos efeitos reguladores dos linfócitos e também dos monócitos ocorre por meio de seus mediadores solúveis, que regulam a maturação das células B e T. A presença de ma-crófagos é necessária para a ativação ótima dos linfócitos. A interação macrófago-antígeno leva à produção de IL-1, IL-6 e TNF (tumor necrosis factor), que induzem à produção de receptores de IL-2 nos linfócitos e à secreção de citocinas, resultando na ativação e expansão das células efetoras e reguladoras.
Em um indivíduo normal, as respostas longa e elevada das células NK podem levar ao desen-volvimento de uma série de reações auto-imu-nes, necessitando, portanto, de um sistema que regule sua atividade. A existência de equilíbrio dinâmico entre as populações de células NK e de linfócitos T-citotóxicos parece ser essencial para os mecanismos de defesa do hospedeiro. Se os precursores T não reconhecem o antígeno que lhes está sendo apresentado, as células NK po-dem suprir essa falha com uma resposta menos
específica, mas eficaz. No entanto, se os linfócitos T-citotóxicos reconhecem o antígeno e são ati-vados, as células NK imediatamente limitam sua atividade, permitindo que sejam os T citotóxicos os principais responsáveis pela defesa mediada por células. A partir dessas observações, verifi-cou-se que os linfócitos são capazes de controlar a ativação e a resposta das células NK por meio de vários mediadores solúveis, entre eles, IL-4 e TGF-β, ambos produzidos pelos linfócitos TH2, responsáveis pelos efeitos supressores. As células T citotóxicas CD8+, ativadas durante uma infec-ção viral, também são capazes de competir com as NK por fatores tróficos, como a IL-2, processo esse que termina por controlar a proliferação de ambas as populações.
A depleção de células NK em animais de la-boratório resulta em uma imunidade suprimida não somente contra uma série de patógenos, mas também de tumores, bem como da resposta vírus-específica realizada pelas células T. As cé-lulas NK determinam quando um linfócito CD8+ ativado vai apenas proliferar, mas não se dife-renciar (na ausência de células NK), ou prolife-rar e sofrer diferenciação em um efetor citolítico (na presença das células NK). A maturação nessas condições requer o contato direto entre as célu-las, sugerindo que a expressão do CD56 seja um fator importante, modulando o reconhecimen-to aloantigênico e proporcionando ativação NK adequada, além de servir como molécula aces-sória para o linfócito T citotóxico. É interessante salientar que as células T de memória requerem igualmente a presença das células NK para que se diferenciem em linfócitos efetores.
Estudos em linhagens tumorais mostraram que a supressão de células NK impede a dife-renciação dos linfócitos em T citotóxicos tumo-res-específicos, abolindo a imunidade protetora contra o tumor.
As células NK também exercem atividade aces-sória/reguladora para os linfócitos auxiliares CD4+, tanto na sua interação com APCs como na sua ativação em células efetoras, fazendo parte, jun-tamente com a IL-12, da cascata que leva à dife-renciação preferencial dos TH0 em TH1. O auxílio prestado pelas células NK aos linfócitos auxiliares CD4+ também facilita a diferenciação dos precur-sores T citotóxicos CD8+ em células efetoras.
Recentemente, foi identificada uma popula-ção de linfócitos TH produtores de IL-17 e dife-rentes das populações tradicionais TH1 e TH2.

148 Imunologia Básica e Aplicada
Além disso, a formação dessa população não é dependente das citocinas que medeiam o desen-volvimento dos linfócitos TH1 e TH2. Ainda não está claro se essas células denominadas de TH17 são linfócitos TH0 diferenciados diretamente em células produtoras de IL-17 ou se são células TH modificadas pela presença de IL-23, importan-te citocina envolvida no desenvolvimento dos LTH17. A IL-17 (também conhecida como IL-17 A) é membro da família da IL-17, a qual tem ainda como membros a IL-17B até a IL-17F. O receptor de IL-17 está presente em todos os tecidos, e a sua ligação provoca a ativação das vias intrace-lulares NF-κβ (nuclear factor-kappa beta) e JNK (JUN amino-terminal kinase). A IL-17 possui ação pró-inflamatória, induzindo a produção de IL-6, GM-CSF e de outras quimiocinas por fibroblas-tos e células epiteliais. Portanto, a IL-17 exerce importante papel na defesa contra agentes in-fecciosos. Porém, sua superexpressão resultou em dano inflamatório ao hospedeiro, incluindo artrite reumatóide, lúpus eritematoso sistêmico, esclerose múltipla e asma.
Interações neuroimunoendócrinasEntre as diversas evidências da interação funcio-nal entre o sistema imune e o neuroendócrino, um dos achados mais importantes é o fato de que os dois sistemas compartilham uma diversi-dade de mediadores endógenos e exógenos que tem por objetivo final a homeostase do organis-mo. Existem vários níveis de interação entre os dois sistemas: hormônios clássicos e neurotrans-missores têm receptores específicos nas células imunes e modulam sua atividade; as citocinas po-dem agir em células do sistema neuroendócrino, modificando sua função e o estímulo imune; os hormônios liberados pelo hipotálamo induzem as células linfóides a sintetizar neuropeptídeos que, por sua vez, podem influenciar a atividade do sistema neuroendócrino, assim como as cito-cinas e outras moléculas citocina-like, que poten-cialmente estão aptas a modular a atividade do sistema imune e são produzidas por células do sistema nervoso.
Entre as células do sistema imune, os linfóci-tos são aqueles que mais se assemelham a uma célula neuroendócrina, apresentando em sua su-perfície receptores para peptídeos neuroendócri-nos e hormônios hipotalâmicos, assim como tam-
bém possuem a capacidade de produção desses neuropeptídeos.
Pode-se verificar a presença e a atividade de peptídeos derivados da pró-opiomelanocortina (POMC), como o ACTH, β-endorfinas e hormônio α-melanócito estimulante (α-MSH), bem como citocinas e óxido nítrico (NO) em diversas célu-las imunológicas, entre elas, fagócitos e macró-fagos. A migração dessas células para os locais de inflamação, como sua ativação, são processos básicos afetados e modulados por hormônios, opióides e citocinas.
Dentro da regulação neuroimunológica, está incluída a presença de células imunes como mo-nócitos, macrófagos e linfócitos, com as mesmas características encontradas no restante do orga-nismo, fazendo parte da microglia e secretando, sob condições adversas, citocinas e mediadores inflamatórios, o que pode provocar desde leves alterações de comportamento até coma e des-truição neuronal permanente. Entre os mediado-res mais potentes de neurotoxicidade liberados por monócitos/macrófagos, pode-se observar o NO (óxido nítrico), que age como uma exotoxina. Por outro lado, um número grande de citocinas também é sintetizado pelas células da glia, prin-cipalmente IL-1, IL-2, IL-4 e TNF-α que, além das funções de defesa, também agem como fatores neurotróficos. Sabe-se que a IL-1 inibe a secre-ção neuronal de acetilcolina, interferindo direta-mente no processo da memória, enquanto a IL-6 pode estimular a expressão do HIV nos monócitos infectados, o que sugere que processos inflama-tórios, não somente cerebrais, mas em qualquer local do corpo, podem precipitar a encefalopatia da síndrome da imunodeficiência adquirida.
Influências do sistema nervoso regulam dire-tamente o sistema imune. Estudos clássicos com indução de imunossupressão e lesões de hipo-tálamo em animais mostram redução importante das respostas celular e humoral a antígenos exó-genos. Da mesma forma, a secreção de neuro-transmissores modifica o processo inflamatório.
Alterações induzidas experimentalmente no sistema nervoso alteram a função imune sistêmi-ca, sugerindo que a imunocompetência seja regu-lada centralmente por uma série de mecanismos: cada hormônio secretado pela hipófise anterior está sob controle neuroendócrino do hipotálamo, e sua secreção é alterada por diversos estímulos, como sinais do ambiente externo, ritmo de sono e estresse físico e emocional. Por sua vez, cada

149Regulação da resposta imune
um desses hormônios tem ação direta ou indireta na resposta imune. O exemplo mais convincente dessa modulação é a ativação da secreção adre-nocortical em resposta a estímulos inflamatórios ou antigênicos. As células imunocompetentes são também afetadas diretamente pela ação do hor-mônio do crescimento (GH) e da prolactina.
A capacidade das células dos órgãos linfóides primários e secundários depende do contato com o hormônio do crescimento e com a prolactina. Os sinais que regulam o crescimento e a diferen-ciação linfocitária podem ser divididos em três categorias: sinais hormonais (GH e prolactina), sinais antigênicos e de moléculas de adesão, e sinais liberados por citocinas.
O eixo hipotálamo-hipófise-adrenal medeia a imunossupressão e serve como antagonista endó-crino dos efeitos imunoestimulatórios do GH e da prolactina. Outros hormônios, incluindo tiroxina e hormônios sexuais, têm efeitos imunorregula-tórios. Um controle particularmente importante é mediado pelos corticosteróides, endorfinas e ence-falinas, todos sendo liberados durante situações de estresse, tendo ação imunossupressiva. Os próprios linfócitos podem responder ao fator liberador de corticotrofina e gerar seu próprio ACTH que, por sua vez, induz a liberação de corticosteróide.
Há muito se estabeleceu o papel imunossu-pressor dos corticosteróides, ignorando o fato de que, nas concentrações fisiológicas, eles podem aumentar ou suprimir as funções imunológicas. Quando em situações de estresse físico ou emo-cional, por exemplo, os corticosteróides suprimem a produção de citocinas, mas em concentrações fi-siológicas podem promover a produção e a libera-ção de citocinas, como IL-6 e TNF-α. Outro efeito observado é a indução do aumento de receptores para as citocinas.
Na resposta de fase aguda, reação sistêmica à lesão tecidual e/ou à infecção, ocorrem produção e liberação de citocinas e corticosteróides e a in-teração entre IL-6 e corticosteróides (endógenos) teria a função de proteger o indivíduo das con-seqüências potencialmente letais do processo de septicemia, induzindo a produção de proteínas de fase aguda. Contrariamente, a administração exógena de corticosteróides durante a sepse tem efeito supressor.
Acredita-se que os corticosteróides tenham também outras funções: seleção das células T du-rante seu desenvolvimento, influência na apre-sentação de antígenos, modulando a expressão
de moléculas do MHC, e modificação na produção de citocinas, induzindo alteração no balanço TH1/TH2, favorecendo respostas baseadas na função TH2. As células imunocompetentes contêm quan-tidade importante de peptídeos, hormônios e receptores classicamente encontrados nas células nervosas ou do tecido endócrino (Tabela 12.2).
Tabela 12.2. Hormônios e neuropeptídeos encontrados nas células imunocompetentes
Hormônio Meio encontrado
Corticotropina Linfócito B
Hormônio de crescimento Linfócito T
Tireotrofina Células T
Prolactina Células mononucleares
Gonadotrofina coriônica Células T
Encefalinas Linfócitos B
Peptídeo intestinal vasoativo Leucócitos mononucleares, mastócitos
Somatostatina Leucócitos mono e polimorfonucleares
Vasopressina Timo
Ocitocina Timo
Neurofisina Timo
Os linfócitos dos órgãos linfóides periféricos são inervados pelos sistemas simpático e paras-simpático, como os da placa de Peyer do intesti-no que, quando estimulados, secretam peptídeo vasoativo intestinal (VIP). O VIP regula o tráfico de células imunocompetentes através do intesti-no delgado. Essas células também contêm RNAm da POMC que codifica corticotrofina e endorfi-nas, responsáveis pela sua imunorreatividade. A expressão genética e a liberação hormonal pelos linfócitos, assim como na hipófise, estão condicio-nadas ao eixo hipófise-adrenal, aumentando com a secreção de ACTH e diminuindo com o aumento dos glicocorticóides séricos. Esse fenômeno tam-bém é observado nos macrófagos. A secreção de hormônio da tireóide e de prolactina está sob a mesma regulação descrita anteriormente.
O correto funcionamento da reação imune depende do perfeito funcionamento da resposta hipófise-adrenal. Estudos em animais mostram que defeitos hormonais podem levar, em situa-ções de estresse, ao desenvolvimento de doen-ças auto-imunes, como artrite reumatóide, o que não se observa em animais com o funcionamen-to íntegro do eixo.

150 Imunologia Básica e Aplicada
BibliografiaChen Y, Ma Y. Roles of cytotoxic T-lymphocyte-associated antigen-4 in the induc-
tive phase of oral tolerance. Immunology 2002;105(2):171-80.
Dong C. Diversification of T-helper-cell lineages: finding the family root of IL-17 producing cells. Nature Immunology 2006;6:329-33.
Furtado GC, De Lafaille MA, et al. Interleukin 2 signaling is required for CD4(+) regulatory T cell function. J Exp Med 2002;196(6):851-7.
Gangadharan D, Cheroutre H. The CD8 isoform CD8aa is not a functional homologue of the TCR co-receptor CD8ab. Curr Op Immunology 2004; 16(3):264-70.
Itoh M, Takahashi T, Sakaguchi N, Kuniyasu Y, Otsuka F, Sahaguchi S. Thymus and autoimmunity production of CD25+CD4+ naturally anergic and supressive T cells as a key fynction of the thymus in maintaining immunologic self-tolerance. J Immunology 2004;162:5317-26.
Jiang H, Chess L. Regulation of immune responses by T cells. N Engl J Med 2006;354:1166-76.
Kaneko M, Mizunuma T, Takimoto H, Kumasawa Y. Development of TCRab. CD8aa intestinal intraepithelial lymphocytes promoted by interleukin-15-producing epithelial cells constitutively stimulated by gram negative bacteria via TLR 4. Bio. Pharm Bull 2004;27(6):883-9.
Kenny E, Mason D, Saoudi A, Pombo A, Ramirez F. CDalfa is an activation mar ker for a subset of a peripheral CD4 T cells. European J Immunology 2004;34(5):1262-71.
Kim SV, Flavell RA. CD8alfa/alfa and T cell memory. Science 2004; 304(5670):529-30.
Mchugh RS, Whitters MJ et al. CD4(+)CD25(+) immunoregulatory T cells: gene expression analysis reveals a functional role for the glucocorticoid-induced TNF receptor. Immunity 2002;16(2):311-23.
Piccirillo CA, Letterio JJ et al. CD4(+)CD25(+) regulatory T cells can mediate suppressor function in the absence of transforming growth factor beta1 pro-duction and responsiveness. J Exp Med 2002;196(2):237-46.
Poussier P, Ning T, Banerjee D, Julius M. A unique subset of self-especific intraintes-tinal T cells maintains gut integrity. J Exp Medicine 2002;193(11):1491-7.
Sakaguchi S. Naturally arising Foxp3-expressing CD25+CD4+ regulatory T cells in immunological tolerance to self and non-self. Nature Immunology 2005;6(4):345-52.
Schwartz RH. Natural regulatory T cells and self-tolerance. Nature Immunology 2005;6(4):327-30.
Shimizu J, Yamazaki JS et al. Stimulation of CD25(+)CD4(+) regulatory T cells through GITR breaks immunological self-tolerance. Nat Immunol 2002;3(2):135-42.

C A P Í T U L O 1 3
IntroduçãoHistoricamente o estudo da tolerância iniciou em 1905, quando Ehrlich e Morgenroth publicaram os primeiros experimentos sugerindo que os indi-víduos não poderiam se tornar facilmente imunizados contra seus próprios antígenos. A seguir, Owen sugeriu que a tolerância aos antígenos próprios era adquirida durante o desenvolvimento do sistema imune. Em 1953, Billin-gaham, Brent e Medawar, em um trabalho reconhecido com o prêmio Nobel, injetaram tecidos alogênicos (diferentes indivíduos da mesma espécie) em fetos de camundongos durante a vida intra-uterina, os quais desenvolveram tolerância aos mesmos antígenos teciduais durante a vida adulta, evidencia-da por transplantes cutâneos do mesmo camundongo doador. Esses expe-rimentos foram a base para os estudos subseqüentes na área da tolerância imunológica.
O sistema imunológico tem como princípio básico desenvolver mecanis-mos capazes de destruir potenciais microrganismos invasores em prol da de-fesa do hospedeiro. Desde o surgimento dos elementos mais primitivos do sistema imune, a capacidade de diferenciar os antígenos próprios dos estra-nhos ao organismo se fez essencial, processo que se tornou cada vez mais específi co com o desenvolvimento da imunidade adaptativa.
O sistema imune inato diferencia o próprio (self) do não-próprio (non-self) pela identifi cação de padrões moleculares comuns presentes em com-ponentes microbianos. Já a resposta imune adaptativa é mediada essencial-mente por linfócitos T e B, os quais expressam altas concentrações de recep-tores antigênicos específi cos, gerados de modo aleatório por recombinação gênica. Teoricamente acredita-se que esse processo possa gerar cerca de 109 receptores de células T (TCR) diferentes, muitos deles capazes de reagir con-tra antígenos próprios do organismo, além de inúmeras células B capazes de produzir anticorpos auto-reativos. A tolerância imunológica compreen-de os processos de eliminação ou neutralização de linfócitos com potencial de auto-reatividade, preservando uma resposta efi caz contra os antígenos externos. Neste capítulo, estudaremos os mecanismos de geração e manu-tenção de tolerância das células envolvidas com a imunidade adaptativa, essencialmente os linfócitos do tipo T e B.
Conforme já estudado no capítulo de regulação da resposta imune, todos os processos imunológicos são modulados por diversos reguladores humo-rais e celulares, os quais participam tanto de mecanismos efetores diretos
Angela Zanonato • Fábio Muradás Girardi • Ricardo Machado Xavier
Tolerância imunológica

152 Imunologia Básica e Aplicada
quanto da geração de tolerância a determinados antígenos. Dessa forma, os estudos da regulação e da tolerância se sobrepõem em diversos aspec-tos e sua dissociação nem sempre é possível.
Tolerância central e periférica – relação com a ontogenia do sistema imuneOs mecanismos de tolerância obedecem à on-togenia das células do sistema imune, sendo divididos em centrais, quando ocorrem na fase do desenvolvimento celular no nível dos órgãos linfóides primários (medula óssea e timo), e pe-riféricos, em relação à fase de desenvolvimento ocorrida dentro dos órgãos linfóides secundários (linfonodos, baço e tecidos linfóides associados à mucosa).
Embora os mecanismos de tolerância centrais sejam eficientes, eles não são capazes de elimi-nar todos os linfócitos auto-reativos, em parte porque nem todos os auto-antígenos são expres-sos nos órgãos linfóides primários. Os mecanis-mos de tolerância periférica permitem o “refina-mento” do processo de tolerância de linfócitos potencialmente auto-reativos fora do seu sítio primário de desenvolvimento.
Tolerância de células T e de células B
Tolerância T
Tolerância central de células T
A marca registrada da tolerância central das cé-lulas T é a deleção clonal, por meio da indução de apoptose de progenitores de células T que te-nham alta afinidade por auto-antígenos. Outros processos incluem a negligência e a edição do re-ceptor de células T. Esses três processos reduzem ou eliminam a maturação de células auto-reati-vas, sendo considerados mecanismos de seleção negativa. Essa é a principal forma de eliminação de linfócitos T auto-reativos em nível central, permitindo a sobrevivência de apenas 3% a 5% dos linfócitos que saem do timo em estágio ma-duro para a periferia.
O grau de afinidade na ligação de timócitos com os complexos apresentadores de antígenos define o destino dessas células. Aqueles que não
se ligam ao MHC, ou se ligam com muito baixa afinidade, sofrem morte induzida por negligên-cia, já que não recebem sinais provindos da inte-ração com TCR que permitam a continuação de sua maturação (seleção positiva). Por outro lado, células com alta afinidade pelo complexo MHC/auto-antígeno sofrem apoptose (deleção clonal, principal forma de seleção negativa). Esse proces-so ocorre ainda na fase em que os timócitos são duplamente positivos (CD4+ e CD8+). As remanes-centes com afinidade intermediária terminam o processo de maturação e migram para os tecidos linfóides periféricos/secundários. Os processos de seleção se desenvolvem principalmente na medu-la tímica, zona rica em moléculas co-estimulató-rias (CD80 e CD86), células dendríticas e epiteliais medulares, cruciais na apresentação de antígenos característicos de tecidos periféricos.
A edição do receptor é um mecanismo de to-lerância no qual a ligação de um auto-antígeno ao TCR, durante o desenvolvimento de um timó-cito, promove rearranjo secundário do gene do receptor de antígeno, na tentativa de gerar um novo TCR que não seja auto-reativo. Esse é um dos principais mecanismos de tolerância no de-senvolvimento de um linfócito B, mas é de me-nor importância para os processos de tolerância central de células T (Tabela 13.1).
Tabela 13.1. Relação entre afinidade de ligação MHC-TCR e tolerância imunológica
Apresentação de antígenos pelas células epiteliais tímicas
Alta afinidade Deleção clonal
Moderada afinidade Seleção positiva
Baixa afinidade Negligência
Tolerância periférica de células T (fine tune – ajuste fino)
Apesar da alta eficiência dos mecanismos centrais de tolerância, linfócitos T com algum potencial de auto-reatividade acabam escapando para a peri-feria, já que nem todos auto-antígenos se encon-tram à disposição no timo. Ao receberem estímulo antigênico adequado, esses linfócitos podem pro-vocar quebra da tolerância, o que exige a necessi-dade de mecanismos periféricos de controle.
Algumas dessas células auto-reativas, após in-teração de alta afinidade com o complexo MHC/auto-antígeno, escapam da indução de apopto-

153Tolerância imunológica
se, recebendo sinais estimulatórios e diferencian-do-se em células T regulatórias. Essas se dirigirão à periferia, onde posteriormente participarão de mecanismos de tolerância periférica, restringin-do a resposta imune pela interação com contato direto com outros linfócitos ou da síntese de ci-tocinas inibitórias, como IL-10 e TGF-β, mecanis-mo conhecido como supressão. Detalhes desses processos ainda não são muito bem conhecidos, mas atualmente são objeto de intensa investiga-ção. Três principais tipos celulares têm sido con-siderados como potenciais fenótipos de células T regulatórias: célula T regulatória CD4+ CD25+; linfócitos epiteliais intestinais CD8αα+; e células T natural killer (NKT). Alguns estudos demons-traram que os corpúsculos de Hassal, agrupados concêntricos de células reticulares epiteliais ex-clusivos do timo, podem estar envolvidos nesse processo, produzindo citocinas capazes de esti-mular células dendríticas para a geração de célu-las T regulatórias.
Células T não suficientemente estimuladas ou ineficientemente co-estimuladas podem ser leva-das à irresponsividade (anergia) frente aos antí-genos específicos para seu TCR. Algumas das cé-lulas T anérgicas produzem IL-10, o que suprime a ativação de outras células T. Outras expressam CD152 (também conhecido por CTLA-4 – proteína associada a linfócito T citotóxico), o qual se liga com alta afinidade às moléculas co-estimulatórias CD80 (B7-1) e CD86 (B7-2) das células apresentado-ras de antígeno, impedindo a ligação dessas com seu receptor nos linfócitos T (CD28) e inibindo sua ativação.
Outras células T auto-reativas simplesmente ignoram seus peptídeos-alvo. Vários mecanismos podem estar envolvidos na ignorância, como ní-veis baixos de expressão antigênica; presença de separação física entre as células T e os antígenos (barreira hematoencefálica); ausência de molé-culas co-estimulatórias na interação do complexo peptídeo-MHC com o TCR; ou ausência de co-es-timulação das células T-auxiliares quando da ati-vação de células T CD8.
Entretanto, a apresentação de antígenos na ausência de co-estimulação pode não apenas ge-rar ignorância, mas também deleção por apop-tose das células T auto-reativas. Isso também pode ocorrer em virtude da carência de fatores de crescimento pelos quais todas as células T ati-vadas competem ou pelo fato de alguns tecidos apresentarem normalmente ligante para Fas (via
indutora de apoptose), como, por exemplo, as câ-maras oculares, levando à morte de células T que entrarem nesse tecido.
Estudos com transplantes alogênicos de cé-lulas tumorais mostraram que alguns sítios do organismo são protegidos da resposta imunoló-gica, já que o desenvolvimento de reação imune nesses locais poderia ser potencialmente lesivo. Exemplos de sítios imunoprivilegiados (santuá-rios imunológicos) são: olhos (córnea, câmara anterior, cavidade vítrea e espaço subretiniano), cérebro (ventrículos e corpo estriado), útero gra-vídico e placenta, testículos, córtex adrenal e folículos pilosos, bem como certos tumores po-dem apresentar essas características que favore-cem o seu desenvolvimento. O mesmo princípio pode ser observado em relação a alguns órgãos ou tecidos que, quando transplantados a sítios convencionais, duram por longo tempo ou até indefinidamente, sem sofrer ataque do sistema imune e conseqüente rejeição. O fenômeno do sítio imunoprivilegiado dá-se pela inibição de efetores dos sistemas imune inato e adaptativo por meio de diversos mecanismos geradores de tolerância: falta de drenagem linfática na câ-mara anterior do olho, cavidade vítrea e espa-ço subretinal; ausência de expressão de MHC-II e baixa expressão de MHC-I de superfície, apre-sentando de forma ineficaz os antígenos intra-citoplasmáticos; presença da barreira anatômica sangue-olhos, similar à hematoencefálica; presen-ça de fatores imunomodulatórios solúveis e de su-perfície celular, inibindo a ativação de células efe-toras do sistema imune em qualquer fase de sua ativação; menor imunogenicidade dos antígenos localizados na câmara anterior do olho, mediada por células e moléculas incapazes de amplificar a resposta imune (células TCD8+ e anticorpos não-fixadores de complemento).
Tolerância de células BDurante o desenvolvimento das células B, existe delicado balanço entre apoptose e proliferação para moldar um repertório diversificado de anti-corpos. À semelhança dos linfócitos T, o regulador central desse processo seletivo é o receptor de células B (B cell receptor – BCR). Para sobreviver até a maturidade, um linfócito B imaturo necessi-ta ultrapassar diversos check-points no interior da medula e dos órgãos linfóides periféricos.

154 Imunologia Básica e Aplicada
Tolerância central de células B
Uma vez que as células B imaturas passem a ex-pressar um receptor de superfície, seu destino é guiado de acordo com a natureza do ligante. Antígenos próprios guiam os linfócitos imaturos para morte celular por apoptose (deleção); pro-dução de novo receptor por edição; ou indução de estado permanente de insensibilidade, tam-bém denominado ignorância imunológica.
Uma célula ignorante é definida como uma célula que possui alta afinidade para um auto-antígeno, mas não o percebe. Isso ocorre porque o antígeno está em baixa concentração ou não faz ligação cruzada entre receptores de células B (não proporciona o sinal necessário para a ativa-ção de células B independente de linfócitos T).
Algumas células passam por um intervalo antes da morte celular, durante o qual a célula B auto-reativa pode ser resgatada, sofrer rear-ranjos gênicos e trocar de um receptor por ou-tro que não mais seja auto-reativo (edição de receptores). Isso só ocorre porque as proteínas RAG (recombination activating gene), que regu-lam o rearranjo gênico dos receptores de células B, somente deixam de ser expressas quando os linfócitos atingem seu completo estado de ma-turidade, já na periferia.
Tolerância periférica de células B
A tolerância central dos linfócitos B é menos rí-gida que dos linfócitos T. Cerca de metade das células pré-B precoces que entram na medula é perdida nos processos de indução de tolerância, tanto por falhas no processo de rearranjo gênico do receptor quanto por deleção clonal de linfóci-tos B imaturos. Das células liberadas para a peri-feria, mais de 90% são de longa sobrevida, e uma fração menor morre a cada dia, sendo substituí-da por novos pools de linfócitos. A especificida-de do receptor influi na sobrevivência de células B periféricas. Algumas morrem porque se ligam a antígenos próprios, apesar dos mecanismos de tolerância central. A sucessiva perda de células B garante que receptores diferentes sejam produ-zidos continuamente por síntese de novas células na medula óssea, visando ao encontro de novos desafios antigênicos, enquanto a persistência da progênie que sobreviveu é retida para assegurar o combate às infecções.
A teoria mais aceita para explicar a tolerância periférica de células B maduras é baseada na hipó-
tese do duplo sinal, na qual a ativação de células B por antígenos protéicos depende de dois sinais estimulatórios: um proveniente do próprio antí-geno e outro derivado da ação co-estimulatória das células T (ativação T-dependente de células B). Em alguns casos as células B podem ser ativadas por antígenos multivalentes capazes de prover, por reação cruzada, os dois sinais estimulatórios sem a presença das células T (ativação B por via T-independente). No entanto, essas considerações não explicam como as células B distinguem os an-tígenos T-dependentes de antígenos multiméricos próprios. Na tentativa de resolver essas questões, foi recentemente proposta a hipótese de que as células B usariam um sistema semelhante ao das células NK para discriminar o próprio do não-pró-prio. Frações dos antígenos próprios são capazes de estimular receptores inibitórios das células B, impedindo o desencadeamento de resposta imu-ne a esses determinados antígenos. Alguns auto-res acreditam em uma via evolutiva comum entre uma célula primordial inata que se diferencia em células NK e células B e T, pois essas não expres-sam receptores inatos, mas mantêm uma gama de receptores inibitórios. O descobrimento de células NKT reforçou essa idéia, na medida em que es-sas preservam características tanto das células NK quanto das células T.
A noção de que as células B expressam recep-tores capazes de reconhecer ligantes próprios amplamente distribuídos no nosso corpo tem várias implicações na tolerância imunológica e na auto-imunidade. Por exemplo, os tecidos são protegidos dos efeitos deletérios do siste-ma complemento por meio da expressão de vá-rias moléculas reguladoras, como CD46, CD55 e CD35, que trabalham suprimindo a ativação do complemento e, portanto, a fase inicial da res-posta imune mediada por anticorpos. Entretan-to, os tecidos diferem na expressão de marcado-res próprios capazes de inibir a resposta imune mediada por células B. Essa heterogeneidade te-cidual pode explicar a maior vulnerabilidade de determinados tecidos à quebra da tolerância.
Os mecanismos de tolerância T e B são alvos de intensas especulações e ganham ou perdem cre-dibilidade conforme novas evidências e achados ocorrem. Certamente vários aspectos discutidos an-teriormente não são definitivos, e sua importância e aceitação variam de enfoque conforme as crenças de cada autor ou o surgimento de novas evidências experimentais (Figura 13.1 e Tabela 13.2).

155Tolerância imunológica
Tabela 13.2. Quadro comparativo da tolerância B e T, central e periférica
CentralMedula óssea e timo (células T)
Periférica (refinamento da tolerância)Órgãos Linfóides secundários
Linfócitos T Seleção negativa Ignorância
Deleção clonal Deleção clonal
Negligência Anergia
Edição de receptoresSítios e órgãos imunoprivilegiados
Seleção positiva
Supressão
Linfócitos B Deleção clonal Hipótese do duplo sinal
Edição de receptores (receptores inibitórios)
Ignorância
Figura 13.1. Ilustração do mecanismo de tolerância B pe-riférica, conhecido como “hipótese do duplo sinal”, em que sinais inibitórios provindos de ligantes de células próprias inibiriam a ativação de células B, à semelhança do pro-cesso ocorrido com as células NK.
Receptores inibitórios Ligantes inibitóriosLinfócito B
Peptídeos própriosCélula própria
O denominado tecido linfóide associado ao in-testino (gut-associated lymphoid tissue – GALT) con-siste em um sistema de linfócitos espalhados pela lâmina própria e pelo epitélio da mucosa intestinal, além de coleções de grandes folículos de células B entremeados por áreas de células T, compondo as placas de Peyer e os linfonodos mesentéricos.
Acredita-se que bactérias comensais intestinais e antígenos alimentares, por estarem em constan-te contato com os enterócitos do epitélio intestinal, seriam captados por esses, sendo expressos nas suas superfícies juntamente com moléculas do MHC-II. Entretanto, como os enterócitos não produzem moléculas co-estimulatórias, a interação de um lin-fócito T CD4+ com um complexo antígeno-MHC-II do enterócito levaria a estado de tolerância.
Por outro lado, bactérias patogênicas seriam captadas por células M, células intestinais espe-cializadas, e entregues a células dendríticas, que, nas placas de Peyer e linfonodos mesentéricos, gerariam linfócitos reativos a esses patógenos com a participação de moléculas co-estimulató-rias, dando origem a células efetoras.
Outro modelo que explica a diferenciação de resposta a antígenos patogênicos e inócuos tem colocado as células dendríticas como protago-nistas principais desse processo. Originárias da medula óssea, essas células são encontradas na periferia em órgãos linfóides e não-linfóides, e livres nas circulações sangüínea e linfática. Uma vez em contato com o antígeno, as células den-dríticas usam diversas vias para facilitar a cap-tação de antígenos. Embora essas vias pareçam ser comprometidas com a captação de antígenos patogênicos, elas também podem estar relacio-nadas com peptídeos próprios, seguindo um ca-minho de desenvolvimento tolerogênico.
Proteínas alimentares e produtos de bactérias comensais seriam captados por células dendríticas e, na ausência de inflamação e na presença de prostaglandina E (produzida construtivamente por células mesenquimais e macrófagos), leva-riam à maturação parcial de células dendríticas na placa de Peyer ou lâmina própria. Os antígenos seriam então apresentados a células T nos linfo-nodos mesentéricos ou nas placas de Peyer, as quais se diferenciariam até as células T regulató-rias, produtoras de IL-10, ou células T-auxiliares do tipo 3, produtoras de TGF-β, gerando homeostase imune em âmbito local. Quando surgem patóge-nos no intestino, a inflamação local é induzida pe-los efeitos dos produtos patogênicos em contato
Tolerância a antígenos intestinaisO sistema imune intestinal é a maior e mais complexa parte do sistema imune. Não somen-te encontra mais antígenos que qualquer outra parte do corpo, mas também deve discriminar claramente entre organismos invasivos e antíge-nos inofensivos, como as proteínas alimentares e bactérias comensais. A imunidade ativa contra materiais não-patogênicos deve ser evitada, e respostas de hipersensibilidade contra antígenos dietéticos ou bactérias comensais podem levar a distúrbios inflamatórios como a doença celíaca e a doença de Crohn, respectivamente.

156 Imunologia Básica e Aplicada
com receptores do tipo toll, expressos pelas célu-las mesenquimais, macrófagos e células epiteliais. Como resultado, as células dendríticas na placa de Peyer ou lâmina própria maturam completamen-te depois de captar o antígeno e produzem IL-12.
Depois de migrar para os linfonodos mesentéri-cos, ativam as células T, porém induzem sua dife-renciação em células T-auxiliares do tipo 1, produ-toras de IFN-γ, causando maior infl amação local (Figura 13.2.a e 13.2.b).
Figura 13.2.a e 13.2.b. Ilustração dos mecanismos de tolerância a antígenos intestinais. Antígenos não-patogênicos (inertes) são captados pelos enterócitos, os quais são incapazes de estimular a resposta imune. Os antígenos patogênicos são captados pelas células M, intercaladas entre os enterócitos, as quais interagem com células dendríticas e estimulam uma via que culminará com a diferenciação de células T-auxiliares em subtipos inflamatórios (T-auxiliar do tipo 1). As próprias células dendríticas são capazes de captar diretamente os antígenos patogênicos, levando a mesma via de ativação. Quando as células dendríticas captam antígenos inertes à resposta tende a diferenciação de células T-auxiliares em formas inibitórias (T-reg e T-auxiliar do tipo 3).
EnterócitoEnterócito
Membrana basalCélula dendríticaCapilar
Sem resposta
Célula M
Antígenos intestinais inertes Antígenos intestinais
patogênicos
Antígenos intestinais inertes
Antígenos intestinais patogênicos
Célula dendrítica
IL-10TGF-β
IL-12INF-γ
T-regT-auxiliar 3
T-auxiliar 1
Homeostase
Infl amação
BibliografiaAluvihare VR, Kallikourdis M, Betz AG. Regulatory T cells mediate maternal tole-
rance to the fetus. Nature Immunol 2004;3:266-71.
Gavin A, Aït-Azzouzene D, Märtensson A et al. Peripheral B lymphocyte tolerance. Keio J Med 2004; 53(3):151-8.
Hogquist KA, Baldwin TA, Jameson SC. Central tolerance: learning self-control in the thymus. Nature Reviews Immunology 2005;5:772-82.
Jiang H, Chess L. Regulation of immune responses by T cells. N Engl J Med 2006;354:1166-76.
Kamradt T, Mitchison NA. Tolerance and autoimmunity. N Engl J Med 2001; 344(9):655-64.
Kanellopoulos-Langevin C, Caucheteux SM, Verbeke P, Ojcius DM. Tolerance of the fetus by the maternal immune system: role of inflammatory mediators at the feto-maternal interface. Reproductive Biology and Endocrinology 2003;1:121.
Janeway CA, Travers P, Walport M, Shlomchik M. O desenvolvimento e a sobrevivência dos linfócitos. In: Janeway CA, Travers P, Walport M, Shlomchik M. Imunobiolo-gia: O sistema imune na saúde a na doença. 5a ed. Porto Alegre, Artmed, 2002.
Makrigiannakis A, Zoumakis E, Kalantaridou S et al. Corticotropin-releasing hor-mone promotes blastocyst implantation and early maternal tolerance. Nature Immunol 2001;2:1018-23.
Mowat AM. Anatomical basis of tolerance and immunity to intestinal antigens. Nature Reviews Immunology 2003;3:331-41.
Mowat AM. Anatomical basis of tolerance and immunity to intestinal antigens. Nature Reviews Immunology 2003;3:331-41.
Palmer E. Cell death and immunity: Negative selection – clearing out the bad apples from the T-cell repertoire. Nature Reviews Immunology 2003; 3:383-91.
Quah BJC, O’Neill HC. Maturation of function in dendritic cells for tolerance and immunity. J Cell Mol Med 2005;9(3):643-54.
Schwartz RH. Natural regulatory T cells and self-tolerance. Nat Immunol 2005;6(4):327-30.
Streilein JW. Ocular immune privilege: therapeutic opportunities from an experi-ment of nature. Nature Reviews Immunology 2003;3:879-89.
Takahama Y. Journey through the thymus: stromal guides for T-cell development and selection. Nature Reviews Immunology 2006;6:127-35.
Tolerância do feto pelo sistema imune maternoO sistema imune tolera a persistência de aloan-tígenos fetais durante a gestação, permitindo a sobrevivência do feto na vida intra-uterina. O imunoprivilégio fetal está envolvido com diversos mecanismos indutores de tolerância. Logo após a implantação do embrião no útero materno, a liberação de CRH (hormônio liberador de cortico-
trofi na) induz a expressão de ligante Fas (CD95L) e HLA-G pelas células trofoblásticas, levando, res-pectivamente, à apoptose de linfócitos maternos e à inibição da ativação de células NK na interface materno-fetal. Acredita-se que a expansão da po-pulação de células T regulatórias também esteja envolvida no processo de tolerância fetal, o que poderia explicar a taxa de remissão de algumas doenças auto-imunes e o aumento de tolerância a alguns enxertos do pai durante a gestação.

C A P Í T U L O 1 4
IntroduçãoUm dos maiores desafi os da medicina moderna é a tradução dos avanços da imunoquímica e imunobiologia em procedimentos diagnósticos e terapêuti-cos que serão de grande utilidade na prática da clínica médica. No laborató-rio, os testes que utilizam os princípios da imunologia, que estão sendo gra-dativamente elucidados e confi rmados, permitem a análise e o diagnóstico de várias doenças em diversos tipos de indivíduos. Os resultados desses pro-cedimentos, tanto quantitativos quanto qualitativos, são utilizados pelos clí-nicos para diagnóstico, estadiamento, prognóstico e possível tratamento de várias doenças, assim como seu levantamento epidemiológico. No entanto, é sempre importante o entendimento de que esses raramente oferecerão cem por cento de exatidão, ocorrendo muitas vezes a positividade em indivíduos não portadores da doença que está sendo investigada e casos negativos na-queles em que, realmente, o processo patológico está em desenvolvimento.
Ao se analisar uma metodologia imunológica, algumas variáveis se sobres-saem, o que permite verifi car a qualidade do teste e a resposta que poderá oferecer. Dessa, algumas combinações serão importantes para a avaliação da reação quanto a sua qualidade, envolvendo os resultados positivos e negativos e suas complicações, que passam a ser analisadas a seguir.
Em primeiro lugar, um teste poderá sempre levar às seguintes combina-ções segundo o quadro abaixo:
Teste Doença presente Doença ausente
Positivo Verdadeiro-positivos A Falso-positivos B
Negativo Falso-negativos C Verdadeiro-negativos D
SensibilidadeRefere-se à porcentagem de pacientes doentes com testes positivos detec-tados em população sabidamente infectada. A sensibilidade de um teste é estabelecida pela seguinte relação:
Paulo Jaconi Saraiva
Reações antígeno-anticorpo in vitro
Sensibilidade: A A + C

158 Imunologia Básica e Aplicada
Assim, um teste bastante sensível é aquele que diagnostica o maior número possível de pa-cientes que tenham determinada doença.
EspecificidadeRefere-se à percentagem de indivíduos sem doen-ça com teste negativo em população sabidamen-te não-infectada. É estabelecida pela seguinte relação:
estudada. Deve ser diferenciada da prevalência sorológica, que é aquela definida pelo teste, ou seja, os verdadeiros mais os falso-positivos sobre a população estudada.
Valor preditivo positivo (VPP)
Refere-se à probabilidade de ocorrência de uma do-ença se o resultado do teste for positivo. Ou seja:
Dessa forma, o teste específico é aquele que diagnostica como “normais”, ou seja, sem doença, o maior número possível de indivíduos realmente não doentes. Esse resultado pode ser influenciado por vários fatores que levam a falsos resultados positivos, como, por exemplo, indivíduos poliin-fectados por parasitas intestinais que apresentam uma série de determinantes antigênicas que aca-bam reagindo cruzadamente com os antígenos alvos dos testes sorológicos. Outras doenças pre-sentes que não as pesquisadas também podem causar o mesmo efeito.
Muitas vezes é difícil para um laboratório ga-rantir a confiabilidade de um teste. Assim, na in-terpretação de valores, deve-se sempre levar em conta a sensibilidade e especificidade do teste utilizado.
EficiênciaO teste eficiente é aquele que diagnostica como positivo os verdadeiros positivos e como negati-vo os verdadeiros negativos. Em outras palavras, refere-se à relação entre os verdadeiros resulta-dos positivos e negativos sobre toda população estudada. Veja:
Sensibilidade: A + D A + B + C + D
Sensibilidade: D D + B
VP: A A + B
Valor preditivo negativo (VPN)
Refere-se à probabilidade de não ocorrência de doenças se o resultado do teste for negativo:
VPN: D C + D
Assim, quanto maior a prevalência da doença, maior o valor preditivo positivo do teste, e quan-to menor a prevalência, maior o valor preditivo negativo do teste.
O objetivo da medicina laboratorial é melhorar cada vez mais a avaliação, a acurácia e a precisão desses testes, garantindo interpretação correta, a fim de facilitar a análise dos dados. Um entendi-mento melhor dos métodos usados na imunolo-gia laboratorial proporcionaria ao estudante e ao médico guia útil para a aplicação e a interpreta-ção correta dessa fonte de informações.
A combinação do anticorpo (Ac) com o antí-geno (Ag) é a reação fundamental em imunolo-gia e se processa em dois estágios: no primeiro, há combinação específica entre o determinan-te antigênico e o anticorpo correspondente e, no segundo, dá-se o fenômeno visível, como a precipitação ou a aglutinação, por exemplo. O antígeno, quando colocado em presença do an-ticorpo correspondente, interage formando um complexo (Ag-Ac) reversível, no qual não há in-terferência de ligações não-covalentes.
Essa interação Ag-Ac é dependente da con-centração dos reagentes (Ag e Ac), da concentra-ção dos íons na solução, do pH, da temperatura e do tempo de reação. A estabilidade do imuno-complexo formado é dependente da afinidade do anticorpo para com o antígeno. Do ponto de
PrevalênciaÉ a porcentagem de indivíduos infectados em uma população, ou seja, os verdadeiros positi-vos mais os falso-negativos sobre a população

159Reações antígeno-anticorpo in vitro
vista físico-químico, a afinidade é a capacidade de ligação que se exprime matematicamente por um número dado pela equação derivada da lei de ação das massas, mas, do ponto de vista conceitual, a afinidade representa determinada configuração da molécula de anticorpo mono-valente a único sítio de união com o antígeno. Outro fator importante se refere à avidez que é caracterizada pela força total de ligação de um anticorpo a um antígeno ou partícula. Para anti-corpos IgG, a ligação bivalente pode aumentar a avidez de modo significativo e, com o passar do tempo, essa adesão aos epítopos fica mais forte.
Outro ponto importante a ressaltar são as forças de ligação, compostas pelo somatório das forças coulombianas, envolvendo os grupos ami-na (NH3+) e carboxila (COO.)-, as pontes de hi-drogênio, as forças de Van der Waals (atração de elétrons do antígeno pelos elétrons da molécula do anticorpo), bem como as forças hidrofóbicas, que expelem a água intramolecular, provocando reações estáveis em meio líquido.
Tipos de reações imunológicas
Reações de precipitaçãoÉ uma modalidade de reação Ag-Ac em meio líquido ou semi-sólido, onde antígenos multir-reativos solúveis, em presença de seus anticor-pos correspondentes, formam um precipitado. Para que ocorra a formação do precipitado, a concentração dos reagentes deve ser adequada a ponto de permitir a constituição de complexos em concentração suficiente. Quando antígenos são colocados junto a anticorpos, três coisas po-dem ocorrer:
• Como um antígeno é portador de vários epítopos, existe a possibilidade de vários anticorpos ligar-se a uma só molécula de antígeno, representando um imunocom-plexo com excesso de anticorpos. Isso é chamado fenômeno de pró-zona, que se refere à reação de precipitação subótima. Para que se obtenha precipitação máxima, é necessária a diluição do anti-soro. O fenômeno de pró-zona é a causa de erros de interpretação em alguns testes quando há grandes quantidades de anticorpos;
• Pode haver também excesso de antígeno e o anticorpo, no caso IgG, monomérico, só pode ligar-se a dois antígenos e suas valên-cias serão ocupadas rapidamente, não ha-vendo possibilidade de formar uma rede e, conseqüentemente, não havendo for-mação de precipitado. Esse é o chamado fenômeno pós-zona;
• Por fim, pode haver concentração ótima de antígenos e anticorpos, favorecendo a precipitação do imunocomplexo, chamado zona de equivalência. A precipitação máxi-ma ocorre nessa área. Assim, a formação de linhas de precipitação em qualquer tipo de imunodifusão é dependente das concentra-ções de antígeno e anticorpo (Figura 14.1).
Figura 14.1. Curva de precipitação.
Aumento da concentração de antígenos
Pró-zona
Zona de excesso de anticorpos
Zona de equivalência
Zona de excesso de antígenos
Pós-zona
Con
cent
raçã
o d
o co
mp
lexo
an
tígen
o-an
ticor
po
Reações de imunodifusão em meio gelificadoAs reações de imunodifusão podem ser classifica-das em simples e duplas. Na imunodifusão sim-ples, o antígeno ou o anticorpo permanece fixo e o reagente que estiver livre move-se e forma um complexo com o reagente fixo. Na imunodi-fusão dupla, tanto o antígeno quanto o anticor-po estão livres, a fim de se mover um em direção ao outro e precipitar.
Reação de imunodifusão radial ou simples
Mistura-se o anticorpo a um gel de ágar e colo-ca-se em uma placa de vidro ou plástico. Após a solidificação do ágar/anticorpo, faz-se orifícios

160 Imunologia Básica e Aplicada
de três milímetros separados por uma distância de 15 milímetros. Nesses orifícios, coloca-se o antígeno e a placa é mantida em repouso por 48 horas em câmara úmida. Onde houver reação do antígeno com o anticorpo, irão se formar halos de precipitação, cujo diâmetro informará sobre a concentração do antígeno. Assim, essa reação parte do princípio de que existe relação quan-titativa entre a quantia de antígenos colocada no orifício da placa de ágar contendo anticorpos e o diâmetro do anel de precipitação resultan-te, ou seja, quanto maior o diâmetro do halo, maior a concentração do antígeno. Uma impor-tante aplicação clínica dessa técnica é a medida de proteínas séricas, por exemplo, a concentra-ção de imunoglobulinas (Igs), de C3, de C4, de α-feto proteína, de transferrina, e também in-fecções fúngicas. Um anti-soro contra a porção Fc ou a cadeia pesada da imunoglobulina pode ser colocado na placa de ágar, a fim de determinar a concentração das imunoglobulinas séricas. Na prática, apenas os níveis de IgG, IgA, IgM e IgE são medidos. Isso porque anormalidades na IgD sérica ainda não foram associadas com nenhuma doença específica. Já a medida da IgE é útil no diagnóstico de alergias e parasitoses. A medida da IgG sérica é particularmente importante no diagnóstico e na monitorização dos pacientes com hipogamaglobulinemia (Figura 14.2).
imunocomplexos estáveis que podem ser anali-sados visualmente. O ágar é colocado em placas de Petri e as amostras contendo antígeno e an-ticorpo são introduzidas em orifícios em lados opostos da placa, sendo deixados por 48 horas em uma câmara úmida. Após, se houver preci-pitação, as linhas resultantes desta, que repre-sentam os imunocomplexos, serão analisadas confirmando ou não um diagnóstico. Se houver maior concentração de antígeno ou anticorpo, as linhas se formarão próximas ao que estiver em menor concentração. Pode ser utilizada para o diagnóstico das parasitoses, micoses e doenças auto-imunes (Figura 14.3).
Figura 14.2. Reação de imunodifusão radial ou simples.
P
A 1/2
1/41/8
Figura 14.3. Reação de Ouchterlony.
Ag1 Ag2
Ac1
Ag1 Ag2
Ac1/2
Ag1
Ac1/3
Ag1/3
Reação de dupla difusão ou Ouchterlony
É baseada no princípio de que, quando o antí-geno e o anticorpo se difundem através de um meio sólido (ágar, por exemplo), eles formam
Reação de eletroforese
Nesta reação, não imunológica, as proteínas são separadas de acordo com sua carga de superfície. O meio aonde a reação ocorre é teoricamente inerte, não interferindo no fluxo das moléculas no campo elétrico. Nesta técnica, soro ou outros fluidos biológicos são separados pela eletrofo-rese por tempo determinado, precipitados com corante, fixados, sendo após as tiras eluídas ou analisadas por densitometria. Quando colocadas

161Reações antígeno-anticorpo in vitro
em densitômetro, são detectadas absorções va-riáveis em virtude de diferentes concentrações protéicas, por uma célula fotoelétrica, e repro-duzidas por traçado que converte essa concen-tração em picos. O soro humano normal é sepa-rado em cinco principais regiões eletroforéticas por esse método, isto é, albumina, α1-globulina, α2-globulina, β-globulina e γ-globulina. A eletro-forese é útil no diagnóstico de paraproteinemias humanas, como, por exemplo, o mieloma múlti-plo. Nesse distúrbio um pico isolado geralmente ocorre na região das γ-globulinas (Figura 14.4).
cipitação. Este método permite a comparação de misturas complexas de antígenos, como são en-contradas no soro.
No diagnóstico de paraproteinemias, essa si-tuação deve ser associada à imunoeletroforese. A presença de pico na região de γ-globulinas na imunoeletroforese sugere, fortemente, a presen-ça de paraproteína monoclonal, porém a imuno-eletroforese deve ser realizada para determinar qual o tipo exato de cadeias leve e pesada desta imunoglobulina. A imunoeletroforese distingue aumentos policlonais dos monoclonais das γ-glo-bulinas. Além disso, a diminuição ou a ausência de imunoglobulinas (Igs) observadas nas imuno-deficiências pode ser analisada com essa técnica. Porém, os níveis dessas imunoglobulinas devem ser analisados por outras técnicas (imunodifusão radial, por exemplo). Como com essa técnica, é possível identificar cadeias leves de imunoglobuli-nas na urina; ela é importante para a detecção da proteína de Bence Jones, encontrada na urina de pacientes com mieloma múltiplo. Também pode ser utilizada para o diagnóstico de doenças parasi-tárias, como hidatidose e amebíase (Figura 14.5).
Figura 14.4. Perfil eletroforético obtido por densitometria.
Pro
teín
as d
o so
ro
Globulinas
Albumina
Motilidade eletroforética
γ β α2 α1
-
Reação de imunoeletroforese
Em conseqüência de sua insensibilidade reativa, a imunoeletroforese é quase sempre um teste de rastreamento para anormalidades nas proteínas séricas. Testes imunológicos mais específicos e quantitativos devem ser realizados para confir-mar o diagnóstico.
A heterogeneidade de proteínas séricas hu-manas pode ser analisada por eletroforese, téc-nica que separa as proteínas de acordo com seu campo elétrico. A imunoeletroforese combina essa técnica com a imunodifusão na mesma placa de ágar. A identificação e a quantificação apro-ximada podem ser aplicadas para cada proteína individualmente, presente no soro, na urina ou em outros fluidos biológicos. É realizada em dois tempos: primeiro, os antígenos que compõem o soro são separados, em uma placa de gel, de acordo com suas cargas elétricas por meio de ele-troforese. Após, uma depressão entre os orifícios contendo antígenos é preenchida com anticor-pos. O material é então deixado em repouso por determinado tempo a fim de propiciar a difusão. Os antígenos e anticorpos formam arcos de pre-
Figura 14.5. Reação de imunoeletroforese.
Teste
Anti-soroPadrão
Teste
Padrão
Anti-soro
Reações de eletroimunodifusãoNas técnicas de imunodifusão descritas anterior-mente, o antígeno e o anticorpo entram em con-tato e precipitam no ágar puramente por difu-são. Entretanto, a chance de este encontro ocor-rer, bem como a velocidade do desenvolvimento da linha de precipitação, pode ser aumentada na presença de corrente elétrica. Essa técnica de eletroimunodifusão é útil no diagnóstico soroló-gico de doenças infecciosas pela detecção do an-tígeno sérico. Apesar de muitas variantes terem sido descritas, apenas as duas citadas a seguir têm aplicabilidade.
+

162 Imunologia Básica e Aplicada
Figura 14.7. Reação de imunoeletroforese de Laurell ou em fuso.
Figura 14.6. Reação de contra-imunoeletroforese.
Corrente eletroforética
Eletroendosmose
Ag Ac
e outras componentes em suspensão são agluti-nadas com freqüência quando colocadas com o anti-soro. O princípio das reações de precipita-ção e aglutinação é fundamentalmente o mes-mo, sendo os resultados obtidos é que tornam diversas umas das outras. Enquanto as reações de precipitação são qualitativas, semiquantitati-vas ou quantitativas, as de aglutinação são quali-tativas ou semiquantitativas, sendo, porém, mais rápidas e sensíveis. A aglutinação é a formação de agregados suficientemente grandes de célu-las interligadas por pontes moleculares de anti-corpos que se combinam simultaneamente com dois epítopos iguais, porém situados à superfície de dois elementos diferentes.
No fenômeno de aglutinação, a visualização depende somente do tamanho dos agregados formados e, conseqüentemente, da relação en-tre anticorpo e partículas, pois, havendo distri-buição eqüitativa, se couber apenas uma mo-lécula de anticorpo por partícula, não se dará aglutinação; a célula ficará com o anticorpo na sua superfície, constituindo o que se chama de molécula sensibilizada.
É um método simples e rápido para identifi-car várias bactérias, fungos e grupos sangüíneos (ABO, Rh), além disso, se dispusermos do antíge-no, é possível determinar e quantificar anticor-pos contra ele. As reações de aglutinação podem ser classificadas diretas ou ativas, indiretas ou passivas, reversas ou de inibição.
Reação de aglutinação direta ou ativa
Nesta técnica, o antígeno insolúvel particulado é aglutinado diretamente por um anticorpo. Eri-trócitos, bactérias, fungos e uma variedade de
Reação de contra-imunoeletroforese
O princípio básico deste método envolve a ele-troforese do antígeno e do anticorpo, presentes em orifícios separados em meio gelificado, em direções opostas simultaneamente, com preci-pitação resultante em ponto intermediário en-tre suas origens. É semelhante à imunodifusão dupla, porém usa corrente elétrica para tornar a reação mais rápida (20 minutos em vez de 48 horas), sendo dez vezes mais sensível que esta. Porém, é uma técnica apenas semiquantitativa. Sua aplicação clínica é na detecção de Crypto-coccus, Meningococcus e Haemophilus no liquor, e aspergilose, candidíase, histoplasmose e para-coccidiodomicose, no soro (Figura 14.6).
Reação de imunoeletroforese de Laurell ou em fuso
Consiste na combinação da imunodifusão radial e da eletroforese. O soro contendo uma quanti-dade desconhecida de antígeno é colocado em um pequeno orifício, em placa de ágar com an-ticorpo específico para determinado antígeno (por exemplo, albumina). A eletroforese des-te antígeno é realizada e, a partir dos orifícios com antígeno, formam-se cones de precipitação, cujas áreas variam de acordo com a concentra-ção de albumina em cada orifício. Há proporcio-nalidade entre as áreas projetadas desses cones e as concentrações do antígeno. Com essa técnica, é possível quantificar imunoglobulinas e compo-nentes do complemento (Figura 14.7).
Reações de aglutinaçãoAglutinação é o acúmulo observável de partículas que ocorre como resultado da interação especí-fica entre uma aglutinina, usualmente o anticor-po, e o aglutinógeno, ou o antígeno. Bactérias

163Reações antígeno-anticorpo in vitro
outras espécies microbianas podem ser agluti-nados diretamente pelo anticorpo. A pesquisa de anticorpo específico no soro, dirigido contra determinado antígeno, é feita com diferentes diluições do soro (contendo anticorpos) contra a mesma quantidade de antígenos. Após alguns minutos ou horas de incubação, a aglutinação é completada e as partículas são examinadas dire-tamente ou microscopicamente para a evidência de aglutinação. Os resultados são expressos de forma qualitativa ou em titulação de anticorpos, isto é, pela maior diluição na qual a aglutinação ocorreu. Este título deve ser diferente em pelo menos duas vezes de um padrão normal, dois tubos de ensaio, para ser considerado significati-vo. Por exemplo, se a reação é iniciada em dilui-ção de 1/2 (uma medida de Ac para duas de Ag), deve-se ter aglutinação pelo menos em diluição de 1/8 (uma medida de Ac para oito de Ag, ou seja, uma quantidade duas vezes menor de anti-corpo ainda é capaz de aglutinar a mesma quan-tidade de antígeno) para que seja considerado significativo (Figura 14.8).
executados em lâminas, microplacas e tubos, e o tempo, a temperatura e o material de diluição da amostra biológica são fatores que devem ser ob-servados em decorrência da sensibilidade do mé-todo e das características do antígeno e da reação (Figura 14.9).
Figura 14.8. Reação de aglutinação direta ou ativa.
Reação de aglutinação indireta ou passiva
Aglutinação passiva ou indireta é o teste que não usa o antígeno isolado, mas sim acoplado a outra partícula. A quantidade de antígenos que passaram a ser passivamente adsorvidos ou quimi-camente conjugados a eritrócitos ou outras par-tículas inertes, como o látex, a bentonita, a gela-tina e ao carvão ativado, provocou aumento do uso das reações de aglutinação. Muitos antígenos se conjugam espontaneamente com eritrócitos, formando reagentes estáveis para a detecção de anticorpos no soro ou outro fluido biológico que contenha a proteína. Os testes são normalmente
Figura 14.9. Reação de aglutinação indireta ou passiva.
Partículas de poliestireno de látex
Moléculas de proteína
Moléculas de proteína ligadas covalentemente às partículas de poliestireno de látex
Anticorpos IgM com especificidade para as proteínas ligadas
Aglutinação passiva reversa
Consiste na aderência do anticorpo a partículas inertes (látex) que podem então ser usadas para detectar a presença de um antígeno. Em virtude da natureza protéica do anticorpo, a absorção pode ocorrer espontaneamente ou pode precisar de manipulação para que ocorra. É usada para a de-tecção de uma série de antígenos, como: Haemo-philus influenzae, Neisseria meningitidis, rotavírus, estreptococos do grupo B e várias drogas de uso terapêutico (Figura 14.10).
Figura 14.10. Reação de aglutinação passiva reversa.
Poliestireno de látex
Moléculas de anticorpo com especificidade para antígeno a ser selecionado
Antígeno
1
2

164 Imunologia Básica e Aplicada
Reação de inibição da aglutinação
É uma reação que envolve interferência pelo an-tígeno ou anticorpo, com reação Ag-Ac que te-ria resultado em aglutinação, caso esta inibição não ocorresse. A técnica é chamada inibição da hemaglutinação quando a partícula inerte utili-zada na reação é um eritrócito. Há duas técnicas de inibição da aglutinação utilizadas rotineira-mente:
1. É usada para detectar antígenos solúveis (HBsAg, gonadotrofi na coriônica, fator VII). Neste procedimento a amostra bio-lógica (fonte de antígenos) é adicionada a uma quantidade de anticorpos neutrali-zantes. Se a amostra do paciente não pos-suir antígenos, a adição de uma partícula ligada a um antígeno (látex, eritrócitos) re-sultará em aglutinação, pois os anticorpos livres ligar-se-ão a ela. Por outro lado, se houver antígenos na amostra do paciente, nenhuma aglutinação será obser vada, pois os anticorpos estarão li gados aos antíge-nos, não se ligando às partículas adiciona-das (Figura 14.11).
aglutinação será evidente. A utilização clás-sica dessa técnica é a detecção de anticorpos contra o vírus da rubéola (Figura 14.12).
Figura 14.11. Reação de inibição da aglutinação.
Anticorpo aglutinante IgM Amostra do paciente
com antígeno solúvel
Não ocorre ligação
Antígeno
Figura 14.12. Reação de inibição da hemaglutinação utilizando partículas virais.
Controle
Amostra do paciente com anticorpo antiviral
da hemaglutinaçãoInibição
Hemácia Ligação do vírus às hemácias
Aglutinação
Vírus não pode se ligar às hemácias
Não há aglutinação
1
2
1
2
Vírus
2. Uma série de vírus tem a capacidade de in-duzir hemaglutinação quando combinados, in vitro, com eritrócitos tripsinizados. Esse fenômeno é a base para a segunda técnica de aplicação da inibição da aglutinação. An-ticorpo para um antígeno hemaglutinante pode ser detectado no soro de pessoas que tenham o agente etio lógico. Se anticorpos contra o vírus estiverem presentes no soro do paciente, ao se adicionar eritrócitos não haverá aglutinação, pois o agente hemaglu-tinante estará bloqueado por esses anticor-pos. Se os anticorpos estiverem ausentes, a
Reações mediadas pelo sistema complementoO sistema complemento é um dos principais me-canismos efetores humorais do sistema imune capaz de induzir dano tecidual. A utilização ou fi xação do complemento, que ocorre como con-seqüência de reações Ag-Ac, proporciona um meio sensível e útil para detectar antígenos e anticorpos no soro. Veja a seguir como o sistema complemento pode ser utilizado.
Reação de citotoxicidade
É aquela que ocorre quando os anticorpos se fi xam a antígenos presentes na superfície das células e, adicionando-se complemento (soro fresco de cobaias, coelho, humano) ocorrerá a lise dessas células. É uma reação muito usada para a tipifi cação dos antígenos do sistema de histocompatibilidade. Tendo-se os anti-soros específi cos, colocando-se os linfócitos a serem pesquisados (do doador e receptor) em presen-ça de complemento (de coelho), e deixando-se incubar a 37oC, haverá lesão da membrana dos linfócitos que possuam os mesmos antígenos de histocompatibilidade contra os quais o anti-soro foi preparado. A verifi cação da lesão é realizada
às hemácias
pode se ligar às hemácias
Ligação do vírus
da hemaglutinação

165Reações antígeno-anticorpo in vitro
Reação de fixação do complemento
A fi xação do complemento ocorre durante a in-teração de antígenos e anticorpos. Dessa forma, o consumo de complemento in vitro pode ser uti-lizado como fator para detectar e medir anticor-pos, antígenos ou ambos. Esse teste é uma reação bifásica. Na primeira fase, deixa-se o antígeno re-agir com o soro a ser testado e, se anticorpos pro-curados estiverem presentes, imunocomplexos formar-se-ão. O complemento é então adiciona-do à mistura e, se os imunocomplexos estiverem presentes, o complemento se ligará a eles, sendo consumido, fi xado.
Na segunda fase, a atividade hemolítica do complemento é medida, a fi m de determinar a quantidade de anticorpo ou antígeno presente na mistura inicial. Células indicadoras (eritróci-tos) são sensibilizadas com anticorpos antieritró-citos (hemolisina), formando um complexo EA
Reações com antígenos e anticorpos marcados
Os imunoensaios são nomeados de acordo com o marcador que é utilizado para evidenciar a re-ação Ag-Ac. Assim, a imunofl uorescência utiliza um marcador fl uorescente, a técnica de enzima imunoensaio utiliza um marcador enzimático, a técnica de radioimunoensaio utiliza um marca-dor radioativo e as técnicas de quimiolumines-cência e imunofl uorimétricas utilizam marcado-res que emitem luz e podem ser medidas quanti-tativamente em luminômetro.
Figura 14.13. Reação de citotoxidade.
Células + anti-soro
+ complemento + corante Reagente
Não-reagente
adicionando-se corante à suspensão, o azul de tripan ou eosina Y, já que as células mortas fi cam permeáveis ao corante. Os linfócitos são consi-derados da mesma espécie do anti-soro quando pelo menos 20% deles são lisados. Assim, tipifi -cando linfócitos do doador e do receptor, quan-to maior a mortalidade dos linfócitos de ambos, com os mesmos anti-soros, maior a coincidência de antígenos da histocompatibilidade. As provas cruzadas (cross-match) pré-transplante são reali-zadas por citotoxicidade, cruzando o soro do re-ceptor com as células do doador (Figura 14.13).
(eritrócito – anticorpo antieritrócito) que será adicionado ao complexo inicialmente formado e, após período de incubação, observa-se se houve ou não lise do eritrócito sensibilizado.
Quando se pesquisa a presença de anticorpo em fl uido biológico, a ausência de lise do siste-ma hemolítico indica a presença de anticorpo no fl uido em estudo, pois, como o complemento foi consumido pelo imunocomplexo, não estará dis-ponível para lisar o sistema EA. Quando há lise das hemácias, diz-se que não houve consumo do complemento e que este foi desviado para a he-mácia sensibilizada e a reação é negativa. A rea-ção pode ser negativa se não houver anticorpos no soro testado ou se houver anticorpos, mas não houver antígenos, não havendo formação de imunocomplexos. Essa reação é usada para o diagnóstico da doença de Chagas (Machado Guerreiro), sífi lis (Wasserman), herpes e citome-galovírus, além de outros agentes etiológicos (Figura 14.14).
Figura 14.14. Reação de fixação do complemento.
Anticorpo anti-hemácia de carneiro
Anticorpo
Antígeno
Reação positiva
Complemento
Reação negativa
Hemácia de carneiro
Não há hemólise
Hemólise

166 Imunologia Básica e Aplicada
Reação de imunofluorescênciaSão reações que utilizam marcadores fluores-centes para indicar reações entre antígenos e anticorpos. O fenômeno conhecido como fluo-rescência consiste na capacidade de certas molé-culas ou compostos (fluorocromos, fluoroforos) de absorver energia, em geral, de uma fonte de luz incidente e converter esta energia em fótons de luz com diferentes características e compri-mentos de onda. A maioria desses compostos é de partículas orgânicas e a mais freqüentemente usada é a fluoresceína em virtude de suas carac-terísticas, que incluem alta intensidade de fluo-rescência, boa fotoestabilidade, eficiência de-senvolvida e emissão de comprimento de onda, usualmente distinguível dos demais. Na prática, a fluoresceína é geralmente preparada como iso-tiocianato, a fim de facilitar sua ligação ao antí-geno ou anticorpo, assim, fluoresceína isotiocia-nato (FITC) se combina ao antígeno ou anticorpo sendo chamado “conjugado”.
As reações de imunofluorescência podem ser divididas em diretas e indiretas. As diretas são reações de única fase, nas quais, normalmente, são pesquisados os antígenos com o uso de con-jugado específico marcado com a fluorescência. As reações indiretas são aquelas em que são pes-quisados anticorpos contra antígenos já prede-terminados, obedecendo à seqüência de incuba-ções e separações, na qual o conjugado utilizado é um antianticorpo marcado com fluoresceína.
Imunofluorescência direta
É utilizada para detectar um antígeno qualquer. O antígeno reagirá com anticorpo conhecido e marcado. O teste é incubado, as moléculas não-reagentes são dispensadas e a positividade é indicada pela presença de fluorescência, visua-lizada em microscópio específico, que indicará a reação entre o anticorpo conhecido e o antí-geno presente. Essa reação é, na prática, utili-zada para identificar marcadores de superfície (CDs nos linfócitos, por exemplo), estreptococos β-hemolíticos do grupo A, Chlamydia trachoma-tis. É também chamada de técnica de fase única (Figura 14.15).
Imunofluorescência indireta
É utilizada para pesquisar a presença de classes de anticorpos no soro do paciente. Um antígeno
conhecido, contra o qual se está pesquisando an-ticorpos, é colocado no meio sólido (lâmina) para reagir com o soro do paciente. Deixa-se incuban-do por determinado tempo e após lava-se a re-ação a fim de retirar partículas não-reagentes. Uma imunoglobulina anti-Ig humana marcada com fluorocromo é subseqüentemente incubada com os reagentes. Estes são novamente lavados e após, em microscópio específico, a observação da fluorescência indica que o anticorpo, especí-fico para o antígeno usado, estava presente na amostra do paciente. Utiliza-se essa técnica na pesquisa de anticorpos circulantes na toxoplas-mose, doença de Chagas, sífilis (FTA-ABS), anti-corpos antinucleares e outros. O reagente tam-bém pode ser usado para detectar antígenos celulares pela incubação dessas células com an-ticorpo específico para o marcador a ser identi-ficado, seguido pela adição de anticorpo anti-Ig marcado com fluorocromo (Figura 14.16).
Figura 14.15. Reação de imunofluorescência direta.
Método direto
Anticorpo marcado
Antígeno
LavagemFluorescência
F
F
F
F
Figura 14.16. Reação de imunofluorescência indireta.
Método indireto
Antígeno
LavagemFluorescência
Lavagem
Anticorpo
ConjugadoF
F
F
Reações de radioimunoensaio (RIA)É o método no qual um radionuclídeo marcado é utilizado para detectar a concentração de molécu-

167Reações antígeno-anticorpo in vitro
las biológicas. É uma das técnicas disponível mais difundidas, úteis e precisas. Sua extrema sensibili-dade é superada apenas por técnicas novas de imu-nofluorimetria e quimioluminescência. Antígenos e anticorpos marcados com isótopos radioativos são muito úteis em imunologia, pois proteínas as-sim marcadas podem ser detectadas em quantida-des muito pequenas e, desta forma, a sensibilidade do teste é aumentada. Os marcadores utilizados podem ser trítio (3H) ou iodos radioativos (131I ou 125I ). O 125I é o marcador de escolha pela facilida-de de marcação e por seu metabolismo restrito. A vantagem dessa técnica é poder determinar quan-tidades menores que um picograma por mililitro e a desvantagem é meia-vida curta do radionuclídeo utilizado ( I125– 57 dias).
Sendo um ensaio com ligantes, o radioimuno-ensaio apresenta muitas variações, mas o princípio básico envolve a quantidade de reagente marcado (antígeno ou anticorpo) para quantificar antígeno ou anticorpo na amostra. Sendo um teste quanti-tativo, a construção de uma curva de calibração com padrões e calibradores de concentrações co-nhecidas é imperiosa para a posterior interpolação dos resultados obtidos com as amostras. Apesar das inúmeras variáveis que a metodologia ofere-ce, na prática laboratorial, os ensaios competitivos ou com excesso de reagentes utilizando fase sóli-da são os mais empregados. O uso desses ensaios da fase sólida facilita a retirada de componentes pelas lavagens, principalmente o material radioa-tivo não-ligado, sendo esta também mais rápida e com controle de qualidade mais eficiente.
Reação imunorradiométrica (IRMA)
No ensaio imunorradiométrico, o anticorpo deve estar em excesso na fase sólida, utilizando siste-ma de dois sítios ou duplo anticorpo (“sanduí-che”) com quantidade fixa de anticorpo espe-cífico para o antígeno aderido à fase sólida. Os padrões para a construção da curva e as soluções teste são então adicionados. Após incubação, remove-se o antígeno não-ligado por lavagens sucessivas e incorpora-se o conjugado radioiso-tópico específico contra o antígeno. O anticorpo marcado (conjugado) não-ligado é removido por lavagem e faz-se a medida da radioatividade na fase sólida. Sendo uma reação cujo resultado é diretamente proporcional à quantidade de ana-lito na amostra, maior será a radioatividade me-dida na fase sólida (Figura 14.17).
Reação de competição com antígeno marcado
Nesta variante, uma quantidade fixa de anticor-po é ligada ao suporte. Adiciona-se quantidade padronizada de antígeno marcado com radio-isótopo em conjunto com as amostras e os pa-drões que determinarão a curva de calibração e, posteriormente, a interpolação. Após incubação, remove-se por lavagem o material não-ligado e mede-se a radioatividade. Os antígenos da amostra inibem, por competição, a ligação do antígeno marcado ao anticorpo aderido na fase sólida. Dessa forma, quanto menor a radioativi-dade medida, maior a concentração de analito na amostra biológica (Figura 14.18).
Figura 14.17. Reação imunorradiométrica.
Medir radioatividade
Anticorpo Amostra Conjugado radioativo
Figura 14.18. Reação de competição com antígeno marcado.
Medir radioatividade
Fase sólida anticorpo
Antígeno marcado
+ antígeno amostra
Reação + separação
Reação de competição com anticorpo marcado
Uma quantidade fixa do antígeno é imobiliza-da em suporte inerte, adicionando-se quantida-de também padronizada de anticorpo marcado com o radioisótopo, além do material biológico e padrões contendo antígeno. Após incubação, o anticorpo que não se ligou ao antígeno na fase sólida e formou imunocomplexo solúvel é retirado do meio por lavagem e faz-se a medida da radioatividade da fase sólida. O antígeno da amostra ou solução padrão inibe, por competi-

168 Imunologia Básica e Aplicada
ção, a ligação do anticorpo marcado ao antígeno imobilizado na fase sólida, de modo que, de for-ma inversamente proporcional, quanto maior a concentração de antígeno na amostra em teste, menor será a quantidade de anticorpo marcado ligado à fase sólida (Figura 14.19).
Figura 14.19. Reação de competição com anticorpo marcado.
capacidade do anticorpo em mediar mudança na atividade de uma enzima ligada a seu antígeno específico.
Nesta técnica, o antígeno no material bioló-gico do paciente a ser detectado competirá com o antígeno ligado à enzima (AgE) por pequena quantidade de anticorpos. Quem estiver em maior quantidade reagirá com o anticorpo. Quando o AgE liga-se ao anticorpo, a atividade enzimática diminui detectavelmente, ou seja, há menor co-loração, por exemplo. Essa atividade enzimática é diretamente proporcional à quantidade de antí-geno pesquisado, ou seja, quanto mais antígenos livres, mais anticorpos ligar-se-ão a eles, não se ligando ao AgE e não inibindo sua atividade. Na prática laboratorial, essa técnica é utilizada para detecção de antígenos de baixo peso molecular, como drogas terapêuticas, hormônios e alguns antimicrobianos. É considerada simples, mas que perde pouco da sensibilidade das outras reações imunoenzimáticas (Figura 14.20).
Medir radioatividade
Fase sólida antígeno
Fase sólida (Ag) + amostra (Ag) +
anticorpo marcado
Reação + separação
Reações de enzima imunoensaio A capacidade inerente das enzimas de agir sobre múltiplos substratos moleculares, sem serem con-sumidas na reação, resulta em amplificação, ou seja, a presença de quantidade relativamente pe-quena pode reagir com grandes quantidades de substrato, produzindo níveis detectáveis de pro-dutos vindos de sua quebra. Na prática laborato-rial, o antígeno ou o anticorpo pode ser ligado a enzimas de maneira que, ao adicionar o substrato da enzima à reação, é gerado um produto colorido que poderá ser medido por espectrofotometria.
A enzima imunoensaio (EIE) é um método quantitativo ou qualitativo em que a reação Ag-Ac é monitorada por medida da atividade enzi-mática. Essa técnica constitui alternativa para a detecção de reações Ag-Ac que suplanta muitas das dificuldades da técnica de radioimunoensaio, como a meia-vida curta dos agentes radioativos, o custo e o dano potencial à saúde. O método de EIE é de baixo custo, as enzimas utilizadas têm longa duração (a peroxidase, por exemplo, pode durar até nove anos), é simples e fácil de usar, e não necessita de aparelhagem sofisticada. Pode ser dividido em duas grandes categorias, basea-do no local em que a reação ocorrer.
Reação homogênea
A reação ocorre de forma homogênea na solução, sem separação das partes. O princípio baseia-se na
Reação heterogênea
Esta variante de reação imunológica necessita de diferentes etapas, de modo semelhante às téc-nicas de radioimunoensaio. Na reação heterogê-nea, a separação dos componentes se torna ne-cessária, pois a atividade da enzima não é altera-da pela reação antígeno-anticorpo, como ocorre na reação homogênea. Os reagentes (anticorpos ou antígenos) serão absorvidos em sua fase só-lida, sendo as placas e os tubos de poliestireno os mais comuns. Conjugados compostos por anti-corpos antiantígenos ou antianticorpos ligados a uma enzima constituem a peça fundamental da especificidade do ensaio, que será complementa-do pelo uso de substratos condizentes com cada enzima. O termo Elisa (enzyme-linked immuno-sorbent assay) identifica o ensaio imunoenzimá-
Figura 14.20. Reação de enzimaimunoensaio homogênea.
Droga
Anticorpo
Substrato
Reação
Droga marcada com enzima

169Reações antígeno-anticorpo in vitro
tico heterogêneo que apresenta várias alternati-vas, sendo que as mais utilizadas em nosso meio estão descritas a seguir.
Enzimaimunoensaio indiretoÉ utilizado para a pesquisa de anticorpos. Nes-te caso, o antígeno é absorvido à placa, sendo adicionado posteriormente o material biológico que, supostamente, contém anticorpos especí-ficos para os antígenos fixados ao meio sólido. Após incubação e lavagem, adiciona-se o conju-gado enzimático específico que se ligará ao an-ticorpo aderido ao antígeno. Após incubação e lavagem para retirar elementos não-aderidos ao complexo principal, adiciona-se o substrato. O desenvolvimento de cor indica a presença de an-ticorpos contra o antígeno definido na fase sóli-da. A variação de cor está relacionada à quanti-dade de anticorpo no material biológico analisa-do. Para um mesmo antígeno, pode-se pesquisar classes de anticorpos diversos (principalmente IgG, IgA e IgM), possibilitando diversidade enor-me de caracterizações, desde doenças infecciosas a doenças auto-imunes (Figura 14.21).
to antígeno que se quer identificar, é colocada para incubação com o conjugado antígeno-enzi-ma na fase sólida. Depois de obtido o equilíbrio da reação, ocorre a lavagem para a retirada dos elementos não-aderidos e adiciona-se o substra-to. Se houver concentração elevada do analito pesquisado no material biológico, ele ocupa-rá os espaços do anticorpo, impedindo reação do antígeno marcado com a enzima. Portanto, quanto menos cor na reação, maior a concen-tração do analito na amostra, visto o antígeno marcado não-aderido ter sido retirado pelas lavagens sucessivas. Portanto, há relação inver-samente proporcional entre a concentração de antígenos na amostra e as densidades óticas ob-tidas (Figura 14.22).
Figura 14.21. Reação de enzimaimunoensaio indireto.
Antígeno Antígeno +amostra
Conjugação Substrato (cor)
D.O.
Concentração
Enzimaimunoensaio competitivo
Com antígeno marcadoA fase sólida é sensibilizada com anticorpo es-pecífico. A amostra biológica, contendo o supos-
Figura 14.22. Reação de enzimaimunoensaio competitivo com antígeno marcado.
Anticorpo Antígeno marcado +
amostra
Reação + separação
Substrato (cor)
D.O.
Concentração
Com anticorpo marcadoO antígeno ligado à fase sólida e a ligação do anticorpo marcado com enzima são, competi-tivamente, inibidos pela adição do antígeno presente no material biológico que está sendo analisado. As etapas são iguais às do método an-teriormente descrito, quando utilizado antíge-no marcado. As densidades ópticas obtidas são inversamente proporcionais à concentração do antígeno na amostra (Figura 14.23).

170 Imunologia Básica e Aplicada
Figura 14.23. Reação de enzima imunoensaio competitivo com anticorpo marcado. do um anticorpo antiantígeno específico marca-
do com enzima que, em contato com substrato, formará cor medida em espectrofotômetro. A taxa de degradação do substrato é proporcional à concentração do antígeno (Figura 14.25).
D.O.
Concentração
Fase sólida
(antígeno)
Amostra (Ag) + Ac marcado
Reação + separação
Figura 14.24. Reação de enzimaimunoensaio de captura para anticorpos IgM.
Figura 14.25. Reação de enzimaimunoensaio de captura de antígenos.
Anti-IgM Amostra Antígeno Substrato (cor)
D.O.
Concentração
Anticorpo marcado
Anticorpo Amostra Substrato (cor)
D.O.
Concentração
Anticorpo marcado
Substrato
Enzimaimunoensaio de captura para anticorpos IgMA detecção de IgM pelo método indireto pode sofrer interferências principalmente pela presen-ça do fator reumatóide, gerando resultados fal-so-positivos. Também a presença de excesso de IgG pode competir pelo antígeno imobilizado na fase sólida e provocar resultado falso-negativo. Para evitar esses problemas, utilizam-se anticor-pos anti-IgM fixados à fase sólida. Adiciona-se ao material biológico no qual se quer pesquisar o IgM específico para determinado antígeno. Após lavagem do material, adiciona-se o antígeno es-pecífico para o qual se está pesquisando a IgM. A seguir, após separação, adiciona-se o conjugado enzimático antiantígeno específico e, posterior-mente, observadas todas as etapas necessárias para o bom funcionamento da técnica, adicio-na-se o substrato. Este teste de captação de IgM é indicado para caracterizar fase aguda de um processo infeccioso (Figura 14.24).
Enzimaimunoensaio de captura de antígenosVariante de técnica imunoenzimática, muito uti-lizada para identificação de hormônio e marca-dores tumorais. A fase sólida é sensibilizada com anticorpo monoclonal antiantígeno específico. A amostra contendo o possível antígeno é incuba-da com a fase sólida. Na seqüência, é incorpora-
Reação de quimioluminescênciaÉ uma técnica que utiliza marcadores lumines-centes, diferentes de outros fenômenos, como a fluorescência e a fosforescência, pois o even-to excitante é causado por reação química e não

171Reações antígeno-anticorpo in vitro
por fotoiluminação. A quimioluminescência en-volve a oxidação de um componente orgânico como o luminol, ou ésteres de acridina, ou por um oxidante, havendo a emissão de luz do pro-duto excitado formado na reação. Essas reações ocorrem na presença de enzimas, íons metálicos ou complexos metálicos que catalisam o proces-so. A conseqüência é a diminuição do limiar de detecção de antígenos pela ampliação do sinal. A sensibilidade das reações de quimiolumines-cência, com limiar de detecção do atomol à zep-tomol, faz deste tipo de reação importante ins-trumento na detecção de marcadores de baixa concentração em fluidos biológicos por técnicas de imunoensaio.
O princípio básico de reação consiste na de-tecção do complexo antígeno-anticorpo utili-zando uma enzima, uma molécula sintetizada (quimioluminescente) ou a mistura de molécu-las que atuará como substrato para a enzima e será o emissor de luz. O luminol emite energia luminosa em meios alcalinos, na presença de um agente catalítico ou co-oxidante. Como a enzima (peroxidase, fosfatase alcalina) pode servir como um catalítico, os ensaios usam como marcador acoplado ao anticorpo antiantígeno ou antianti-corpo. O luminol, por sua vez, produzirá luz que será diretamente proporcional à quantidade da enzima presente. Quimioluminômetros especial-mente desenvolvidos são utilizados para a quan-tificação da resposta, tornando o teste totalmen-te automatizado e com programas fechados para cada fabricante (Figura 14.26).
Reação de imunofluorimetriaMétodo comumente utilizado para a determi-nação de agentes, que desencadeiam doenças infecciosas, determinação de marcadores tumo-rais, hormônios, bem como a identificação de alérgenos específicos.
Os ensaios imunofluorimétricos seguem o mesmo padrão das reações de enzimaimunoen-saio heterogênea, utilizando enzimas como a fosfatase alcalina ou ß-galactosidase, como ele-mentos associados ao anticorpo específico para antígenos ou outros anticorpos. O que muda em relação ao enzimaimunoensaio tradicional é o substrato utilizado, sendo empregado o 4-metil-umbeliferil fosfato, que, em contato com a en-zima, fornece composto fluorescente, o 4-metil-umbeliferilferona, medida na faixa de 450 nm.
Esses ensaios apresentam variantes produzi-das por fabricantes tradicionais. A Biolab Merieux desenvolveu o sistema Vidas, que utiliza a téc-nica Elfa (enzime linked fluorescent immunoas-say), para a pesquisa de antígenos e anticorpos com alta sensibilidade (99,3%) e alta especifici-dade (98%). Da mesma forma, com pequenas variações a Abbott desenvolveu o sistema Meia (microparticle enzyme imunoassay), utilizando os mesmos conjugados e substratos, além dos índices de sensibilidade, especificidade e repro-dutividade serem praticamente os mesmos. A determinação de hormônios, marcadores tumo-rais, doenças infecciosas, tanto na pesquisa do antígeno como do anticorpo, é utilizada rotinei-ramente empregando esse sistema imunológico de ensaio in vitro.
O sistema Unicap da Pharmacia, também imu-nofluorimétrico enzimático, mede a concentra-ção de IgE total e IgE específico contra mais de 300 alérgenos. A técnica segue o mesmo princí-pio das outras já comentadas, na qual um anti-corpo anti-IgE estará ligado à fase sólida para a determinação do IgE total na amostra biológica ou então quando se quer determinar qual alér-geno está produzindo reação de hipersensibili-dade no indivíduo, adsolve-se antígeno especí-fico na fase sólida. Para identificar um ou outro adiciona-se conjugado específico marcado com β-galactosidase. Quando se adiciona o substrato metil-umbeliferil-β-D-galactosídeo, obtém-se um composto fluorescente 4-metil-umbeliferilfero-na, com emissão de luz medida em fluorímetro.Figura 14.26. Reação de quimioluminescência.
Incubar
Conjugado luminescente
Decantar
Incubar
Detecção da luminescência
Padrão Soro
Fase sólida

172 Imunologia Básica e Aplicada
nado à reação e, se o antígeno estiver presente, já ligado ao anticorpo da fase sólida, tem-se um complexo formado. A reação se completa quan-do é adicionada uma reação facilitadora que pro-duz quelatos de Európio, que, no fenômeno de tempo resolvido, são excitados com luz incidente. O comprimento de onda da luz excitadora fica na faixa dos 340 nm, permitindo que o decaimento da luz emitida fique na faixa de 1 milissegundo, enquanto a faixa de decaimento da fluorescência convencional é de 5 a 20 nanossegundos.
Nos fenômenos de tempo resolvido, os quela-tos de Európio ou Samário são excitados por fa-chos de luz rápidos, com duração de menos de 0,5 microssegundo e, durante os 400 microssegundos a cada facho de luz, o fotodetector fica fechado, enquanto a fluorescência inespecífica é emitida. Nos próximos 400 microssegundos, a fluorescência do quelato é medida, na faixa de 613 nm e, a cada 1 milissegundo, um novo ciclo se inicia. Os ciclos são repetidos mil vezes por segundo, fornecendo resultados de alta sensibilidade, especificidade e, principalmente, reprodutibilidade (Figura 14.28).
Figura 14.27. Reação de imunofluorimetria.
Fase sólida
Antígeno
Conjugado
Substrato
MU
Fluorescência
P
Figura 14.28. Reação de imunofluorimetria em tempo resolvido.
AnticorpoAntígeno
Conjugado
EUFluorescência
EU
EUSolução facilitadora
Reação de imunofluorimetria em tempo resolvidoO ensaio fluorimétrico de tempo resolvido em-prega equipamentos e reagentes especiais a fim de aumentar a sensibilidade do ensaio, dosando concentrações na faixa do nano e do picomolar, também utilizando materiais biológicos diversos dos convencionais, como o sangue total impreg-nado em papel filtro.
Os quelatos de lantanídeos, como o marcador não-isotópico Európio, em combinação com os fluorômetros de tempo controlado eliminam rea-ções de fundo que os marcadores fluorescentes convencionais produzem e conseqüentemente interferem na especificidade da reação.
A técnica em tempo resolvido permite medi-da cinética da reação antígeno-anticorpo, mi-nimizando a interferência da reação de fundo, aumentando a sensibilidade do ensaio, já que a substância fluorescente possui meia-vida longa e a fluorescência é medida após transcorrido perío-do de excitação.
A reação utiliza normalmente um anticorpo ligado a uma fase sólida, a qual é adicionado o material biológico previamente preparado. Um lantanídeo (Európio ou Samário) aderido ao anti-corpo para formar o conjunto indicador é adicio-
Essas metodologias exigem equipamentos es-pecíficos, que utilizam programas individualiza-dos, nos quais os protocolos são inseridos e não se adaptam a outros aparelhos a não ser aqueles para os quais foram programados (Figura 14.27).
Reação de fluorescência polarizadaÉ outro ensaio que utiliza o marcador fluorescen-te. Esse imunoensaio homogêneo utiliza luz pola-rizada para ativar o marcador fluorescente. Essa luz polarizada consiste de ondas de luz paralelas orientadas em plano e é criada quando a luz passa através de filtros especiais. Quando a luz polariza-da é utilizada para ativar um marcador fluores-cente, a luz emitida por este pode ser polarizada ou despolarizada, dependendo do tamanho do marcador fluorescente. Moléculas grandes, como as formadas quando o marcador fluorescente se
MU

173Reações antígeno-anticorpo in vitro
liga ao anticorpo, por si só giram lentamente e emitem luz polarizada paralela à luz polarizada de ativação. Pequenas moléculas, como haptenos fluorescentes livres (que não se ligaram aos anti-corpos), giram mais rapidamente, emitindo assim luz em várias direções, produzindo luz despola-rizada. A luz polarizada é medida em ângulo de 90o e comparada com o “caminho” da luz de ati-vação. No ensaio de fluorescência polarizada, um marcador fluorescente é ligado a um pequeno hapteno e, após a ativação com luz polarizada, o hapteno marcado livre emitirá principalmente luz despolarizada e pouca luz polarizada. Por exem-plo, em um ensaio competitivo, haptenos mar-cados (do laboratório) e haptenos não-marcados (dos antígenos no soro do paciente) competem por sítios na molécula de anticorpo. Quando ne-nhum antígeno ou hapteno não-marcado estiver presente no soro do paciente, o hapteno marcado não sofrerá competição, ligando-se totalmente ao anticorpo, gerando grandes imunocomplexos que girarão lentamente, demonstrando alto nível de polarização. Quando haptenos não-marcados (Ags) estiverem presentes no soro do paciente, haverá competição com haptenos marcados pelos sítios de ligação no anticorpo, resultando em mais haptenos marcados livres. Estes girarão rapida-mente, resultando em polarização diminuída. O grau de polarização é inversamente proporcional à quantidade de antígenos presentes no soro do paciente (Figura 14.29).
Figura 14.29. Reação de fluorescência polarizada.
AntígenoAnticorpo
Antígeno marcado
Antígeno ligado ao anticorpo – aumento da luz polarizada
Antígeno não-ligado ao anticorpo – diminuição da luz polarizada
Reações de turbidimetria e nefelometriaQuando um feixe de luz passa através de uma solução, alterações na natureza do feixe ocor-rerão como resultado de sua colisão com partí-culas suspensas na solução, de vários tamanhos e formas. Ondas de luz dentro do feixe podem ser refletidas, absorvidas ou dispersadas quando em contato com as partículas. Na prática, muito pode ser reduzido sobre a natureza dessas partí-culas solúveis pela observação do grau de refle-xão, absorção ou dispersão da luz incidente.
Turbidimetria
É a medida de luz transmitida através de suspen-são de partículas. Dependendo da concentração dos analitos e do complexo imune formado, tem-se a diminuição ou não da intensidade de luz transmitida em virtude da reflexão, do absor-ção ou do espalhamento. As leituras são defini-das em unidades de absorbância, que refletem a relação entre a luz incidente e a transmitida. Es-pectrofotômetros convencionais ou equipamen-tos específicos, que utilizem grade de difração monocromática e tenham capacidade de absor-bância na faixa de 290 a 430 nm, são os ideais. Os fotodetectores dos aparelhos devem medir a luz transmitida em ângulo de 0o com luz incidente, o que reduz o efeito do espalhamento da luz na medida de transmitância.
É uma reação antígeno-anticorpo convencio-nal, na qual os complexos formados são medi-dos conforme vão sendo produzidos em tempo pré-fixado ou na medida cinética da mudança de absorbância.
O emprego de substâncias, como partículas de poliestireno ou polímeros como o polieti-lenoglicol, aumenta a sensibilidade do ensaio. É uma técnica que apresenta várias vantagens como a de não necessitar de separação das par-tes integrantes do teste, poder ser ensaiada diretamente, ser rápida, sensível, econômica e pode ser realizada em equipamentos conven-cionais. É empregada, principalmente, para a determinação de antígenos e anticorpos espe-cíficos, proteína plasmática, além de outros me-tabólitos (Figura 14.30).

174 Imunologia Básica e Aplicada
Nefelometria
É um método direto para a medida de luz disper-sada por partículas suspensas na solução. Um equi-pamento específico, que utiliza como fonte de luz lâmpadas de mercúrio ou helioneônio (laser) e mede o espalhamento da luz incidente, alterada pela formação de imunocomplexos. Com o aumen-to de precipitação do complexo antígeno-anticorpo formado, aumenta o espalhamento e a reflexão de luz e diminui a transmitância. Uma vez que a esta-bilização da intensidade da luz incidente, o compri-mento de onda e o ângulo de detecção de medida (entre 30o e 70o) são estabelecidos, a dispersão da luz torna-se uma medida exata, correspondente ao complexo antígeno-anticorpo formado, desenvolvi-do na amostra do paciente, se a concentração do anticorpo reagente for controlada (Figura 14.31).
zar o efeito de turvação de fundo. Exige um tem-po maior de incubação dos reagentes e analitos e a diferença entre os valores é correlacionada à curva de referências. Mede principalmente pro-teínas plasmáticas.
Nefelometria cinéticaNesta reação não há necessidade da formação do platô, medindo a taxa de aumento do espalha-mento em curto espaço de tempo. A relação entre a quantidade do espalhamento e o tempo, após o início da reação, é sigmoidal, sendo que no iní-cio do teste os complexos pequenos não espalham luz. Após limiar de tempo e reação, o espalhamen-to aumenta proporcionalmente, até o esgotamen-to dos reagentes. A taxa máxima de aumento para cada reação é convertida em concentração, em-pregando-se curvas já preestabelecidas.
Esta variante de técnica é utilizada para a dosa-gem de proteínas plasmáticas, drogas e hormônios.
Reação de imunoeletrotransferênciaVários tipos de membranas sintéticas absorvem tão intensamente proteínas que podem ser usa-das como suportes para meios sólidos de imuno-ensaios. As proteínas ligadas mantêm sua antige-nicidade e são prontamente acessíveis para provas com anticorpos. Isto levou a uma série de ensaios altamente sensíveis e específicos coletivamente conhecidos como immunoblots. Um experimen-to típico de imunoeletrotransferência consiste de seis passos inter-relacionados:
1. As proteínas são separadas por eletro-forese em gel de poliacrilamida, contendo dodecil sulfato de sódio (SDS-PAGE);
2. As proteínas são transferidas do gel para uma membrana de nitrocelulose, onde se tornam imobilizadas, como uma réplica da banda padrão no gel. Inúmeras técnicas foram tentadas a fim de transferir proteí-nas do gel para a membrana. A mais efi-ciente é o uso de campos elétricos para levar as proteínas do gel à superfície das membranas. A eletrotransferência produz réplica fiel na membrana, mantendo a boa resolução do gel;
3. Os sítios de ligação das proteínas na mem-brana que não estão ligados são saturados
Figura 14.30. Reação de turbidimetria.
Fonte de luz
Sistema de detecção
Solução com imunocomplexo
Figura 14.31. Reação de nefelometria.
Fonte de luz
Sistema de detecção
Solução com imunocomplexo
Sistema de detecção
Há duas variações no ensaio da nefelometria, utili-zando equipamentos próprios e bem caracterizados.
Nefelometria em ponto finalA leitura é feita na região do platô da precipita-ção, necessitando de correções a fim de minimi-
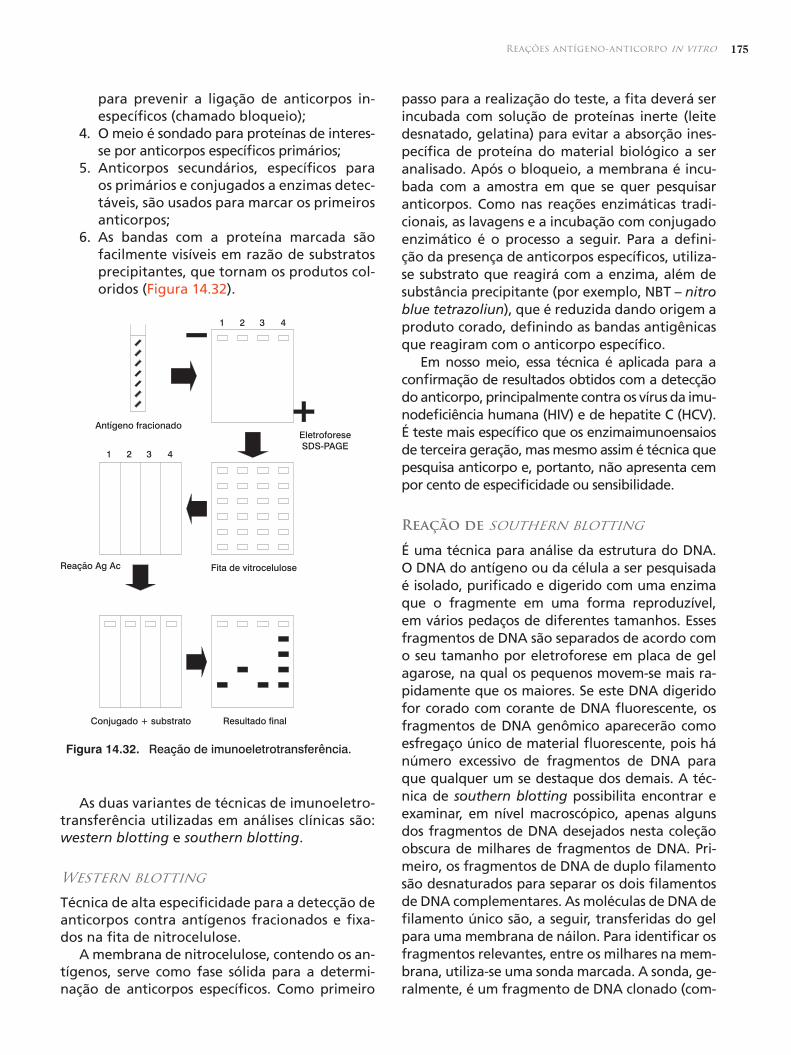
175Reações antígeno-anticorpo in vitro
para prevenir a ligação de anticorpos in-específicos (chamado bloqueio);
4. O meio é sondado para proteínas de interes-se por anticorpos específicos primários;
5. Anticorpos secundários, específicos para os primários e conjugados a enzimas detec-táveis, são usados para marcar os primeiros anticorpos;
6. As bandas com a proteína marcada são facilmente visíveis em razão de substratos precipitantes, que tornam os produtos col-oridos (Figura 14.32).
passo para a realização do teste, a fita deverá ser incubada com solução de proteínas inerte (leite desnatado, gelatina) para evitar a absorção ines-pecífica de proteína do material biológico a ser analisado. Após o bloqueio, a membrana é incu-bada com a amostra em que se quer pesquisar anticorpos. Como nas reações enzimáticas tradi-cionais, as lavagens e a incubação com conjugado enzimático é o processo a seguir. Para a defini-ção da presença de anticorpos específicos, utiliza-se substrato que reagirá com a enzima, além de substância precipitante (por exemplo, NBT – nitro blue tetrazoliun), que é reduzida dando origem a produto corado, definindo as bandas antigênicas que reagiram com o anticorpo específico.
Em nosso meio, essa técnica é aplicada para a confirmação de resultados obtidos com a detecção do anticorpo, principalmente contra os vírus da imu-nodeficiência humana (HIV) e de hepatite C (HCV). É teste mais específico que os enzimaimunoensaios de terceira geração, mas mesmo assim é técnica que pesquisa anticorpo e, portanto, não apresenta cem por cento de especificidade ou sensibilidade.
Reação de southern blotting
É uma técnica para análise da estrutura do DNA. O DNA do antígeno ou da célula a ser pesquisada é isolado, purificado e digerido com uma enzima que o fragmente em uma forma reproduzível, em vários pedaços de diferentes tamanhos. Esses fragmentos de DNA são separados de acordo com o seu tamanho por eletroforese em placa de gel agarose, na qual os pequenos movem-se mais ra-pidamente que os maiores. Se este DNA digerido for corado com corante de DNA fluorescente, os fragmentos de DNA genômico aparecerão como esfregaço único de material fluorescente, pois há número excessivo de fragmentos de DNA para que qualquer um se destaque dos demais. A téc-nica de southern blotting possibilita encontrar e examinar, em nível macroscópico, apenas alguns dos fragmentos de DNA desejados nesta coleção obscura de milhares de fragmentos de DNA. Pri-meiro, os fragmentos de DNA de duplo filamento são desnaturados para separar os dois filamentos de DNA complementares. As moléculas de DNA de filamento único são, a seguir, transferidas do gel para uma membrana de náilon. Para identificar os fragmentos relevantes, entre os milhares na mem-brana, utiliza-se uma sonda marcada. A sonda, ge-ralmente, é um fragmento de DNA clonado (com-
Figura 14.32. Reação de imunoeletrotransferência.
Antígeno fracionadoEletroforese SDS-PAGE
Reação Ag Ac Fita de vitrocelulose
Conjugado + substrato Resultado final
1 2 3 4
1 2 3 4
As duas variantes de técnicas de imunoeletro-transferência utilizadas em análises clínicas são: western blotting e southern blotting.
Western blotting
Técnica de alta especificidade para a detecção de anticorpos contra antígenos fracionados e fixa-dos na fita de nitrocelulose.
A membrana de nitrocelulose, contendo os an-tígenos, serve como fase sólida para a determi-nação de anticorpos específicos. Como primeiro

176 Imunologia Básica e Aplicada
plementar ao antígeno ou à célula a ser pesquisa-da) que foi marcado radiativamente e desnatura-do até a condição de filamento único. Essa sonda marcada e a membrana de náilon são incubadas juntas em solução sob condições que favoreçam a formação de moléculas de DNA de duplo filamen-to. Graças à especificidade apurada do código do DNA, a sonda associa-se novamente apenas ao seu filamento complementar na membrana e ignora todos os outros filamentos de DNA. Após lavar a membrana e remover a sonda não-ligada, expõe-se a membrana a um filme de radiografia, para revelar a posição dos fragmentos com os quais a sonda se ligou, hibridizou.
Existem várias aplicações dessa técnica na prática laboratorial, como a análise de rearran-jos genéticos de linfócitos T e B em linfomas e leucemias que não são bem definidos quanto à linhagem celular, tipagem do complexo de histo-compatibilidade principal (MHC), doenças auto-imunes, detecção do HIV1.
Reação de imunodot
Método que utiliza fita de nitrocelulose rígida, também denominada de dot-Elisa, tem como ca-racterística a fixação de antígenos ou anticorpos no suporte sólido sob a forma de gotas.
Como as membranas de nitrocelulose absor-vem proteínas com grande eficiência, pequenas concentrações de antígenos ou anticorpos po-dem ser adicionadas à fita, permitindo que uma reação imunoenzimática, idêntica em passos ao western blotting, seja realizada. As enzimas são utilizadas como marcadores associados a anti-corpos específicos e o substrato contendo subs-tâncias precipitantes é utilizado para quando o teste for reagente e desenvolver cor.
Esse teste é muito utilizado como confirma-tório de outras reações, principalmente aquelas que utilizam os métodos de imunofluorescência mediante o enzimaimunoensaio qualitativo. É aplicado principalmente para identificação de antígenos ou anticorpos presentes nas doenças infecciosas (Figura 14.33).
Reação imunocromatográfica
Utilizados como testes de triagem, os testes imu-nocromatográficos permitem a determinação de antígenos e anticorpos. São qualitativos simples, não exigindo equipamentos ou instrumentos para a leitura e a interpretação.
O método utiliza membrana de nitrocelulose como fase sólida na qual são fixados os antíge-nos ou anticorpos e os correspondentes contro-les do teste. O reagente de detecção pode ser marcado com enzima, corante coloidal ou ouro coloidal. Após a aplicação da amostra biológica (soro, urina, fezes) por processo cromatográfico, as proteínas difundem e, quando encontram as suas identidades, precipitam, sendo reveladas e fornecendo os resultados qualitativos.
A determinação de anticorpos contra agentes etiológicos que produzem doenças infecciosas, a pesquisa de alguns hormônios e os marcadores tumorais são situações em que a técnica é em-pregada (Figura 14.34).
Figura 14.33. Reação de imunodot.
Controle
Controle
Controle
Controle
Teste reagente
Teste não-reagente
Teste invalidado
Teste
Teste
Teste
Teste
Figura 14.34. Reação imunocromatográfica.
Área de teste
Amostra
Teste
Controle
Teste
Controle
Teste
Controle
Teste não-reagente
Teste reagente Teste invalidado

177Reações antígeno-anticorpo in vitro
orocromos permite a identifi cação de antígenos de superfície diversos e a técnica poderá ser dire-ta ou indireta.
A citometria de fl uxo direta utiliza anticorpos monoclonais conjugados com marcador fl uores-cente e fotômetro para medir a energia lumino-sa emitida pelas células. A análise simultânea de marcadores de superfície, volume e tamanho ce-lular é então permitida. Nesta técnica, a amostra de sangue do paciente é incubada com um ou dois tipos de anticorpos monoclonais, a fi m de detec-tar diferentes marcadores de superfície celular.
Na forma indireta, os anticorpos monoclonais podem ser ou não conjugados a um marcador. Após a incubação com amostra contendo linfócitos, por exemplo, um anticorpo marcado contra o anticorpo monoclonal é adicionado à solução, tornando esse método mais sensível que o método direto.
Uma vez que as células são marcadas, elas são postas em uma câmara de fl uxo rodeada por fl uido isotônico pressurizado. As diferenças en-tre as pressões deste fl uido e da amostra criam fl uxo laminar ou corrente de células fl uindo em um sentido único, permitindo a análise individu-al de partículas. Cada célula cruzará com um fei-xe de luz originado de uma lâmpada de laser ou mercúrio. Quando a célula interromper o raio de luz, esta será dispersada e medida em ângulo de aproximadamente 90o através de um detector. A dispersão da luz, detectada a 0o ou 360o da fonte de luz, é proporcional ao tamanho da célula, en-quanto a medida de dispersão em ângulo de 90o
é proporcional a certas propriedades da estrutura interna da célula, como a granularidade. Assim, esses ângulos podem ser usados para caracterizar tipos individuais de células e estão relacionados como: eritrócitos < linfócitos < monócitos < gra-nulócitos polimorfonucleares. Baseando-se nesses parâmetros de dispersão da luz, registros eletrô-nicos, chamados histogramas, podem ser dese-nhados para a população celular a ser analisada. Cada tipo celular estará em determinada parte do histograma, dependendo de seu tamanho.
Citometria de fluxoTécnica que permite medir partículas ou células enquanto elas passam uma a uma através de um sensor, sendo medidas uma partícula ou célula de cada vez e não como valor médio. O citômetro de fl uxo é o aparelho empregado para a análi-se, que avalia eletronicamente os sinais gerados pelas células ou partículas em suspensão, quando são interceptadas por um feixe de luz que forne-ça comprimento de onda específi co. As células, ao passar pelo feixe de luz, promovem o espalha-mento desta em todas as direções em padrão que depende do tamanho, da forma e da estrutura da partícula ou célula. Já aqueles cujas estruturas estão marcadas com fl uorocromos são excitados e produzem fl uorescência. Os parâmetros de sig-nifi cado biológico, avaliados pela citometria de fl uxo, são o espalhamento frontal e lateral de luz, medindo o tamanho e diferenciando as partículas e a emissão de fl uorescência (Figura 14.35).
Figura 14.35. Reação de citometria de fluxo.
Fluxo contínuo
Laser
Placas de deflexão do fluxo celular
Células marcadascom anticorpos fluorescentes
Analisador
Vídeo/computador
Como a principal função de um citômetro de fl uxo é a detecção de fl uorescência, a utilização de vários conjugados marcados com diversos fl u-
BibliografiaAbbas AK, Lichtman AH, Pober JS. Imunologia celular e molecular. 4a ed. Revinter.
2002.
Carla Burtis C, Edward R, Ashwdos ER. Tietz textbook of clinical chemistry. 3a ed. W.B. Saunders, 1999.
Chapel H, Haeney M, Misbah S, Snowden N. Imunologia para o clínico. 4a ed. Revinter, 2003.
Ferreira AW, Ávila SLM. Diagnóstico laboratorial das principais doenças infeccio-sas e auto-imunes 2a ed. Guanabara Koogan, 2001.
Goldsby RA, Kindt TJ, Osborne BA. Imunologia – Kuby. 4a ed. Revinter, 2002.
Gorczynski R, Stanley J. Imunologia clínica. 1a ed. Reichmann & Affonso, 2001.
Henry JB. Diagnósticos clínicos e tratamento por métodos laboratoriais 19a ed. Ma-nole, 1999
Miller LE, Ludke HR, Peacock JE, Tomar RH. Manual of laboratory immunology 2a ed. Lea & Febiger, 1991.

178 Imunologia Básica e Aplicada
Rich RR, Fleisher TA, Schwartz BD, Shearer WT, Strober W. Clinical immunology – principles and practice. 1a ed. Mosby – Year Book, Inc. 1996.
Rose NR, de Macário EC, Folds JD, Lane HC, Nakamura RM. Manual of clinical laboratory immunology, 6a ed. American Society for Microbiology, 2001.
Scroferneker ML, Pohlmann PR. Imunologia básica e aplicada. 1a ed. Sagra-Luzzaro, 1998.
Sheehan C. Clinical immunology – principles and laboratory 2a ed. Diagnosis. 1997.
Stites DP, Terr AI, Parslow TG. Medical immunology. 9a ed. Appleton & Lange, 1997.

C A P Í T U L O 1 5
Reações de hipersensibilidadeAs reações de hipersensibilidade (RH) constituem-se em respostas imunes que se apresentam exageradas ou inapropriadas, causando muitas vezes infl amação e lesão tecidual. Muitas substâncias podem estar envolvidas no desencadeamento de uma reação de hipersensibilidade, de pendendo, entre outras coisas, de uma suscetibilidade inerente aos indivíduos. Foram descri-tos quatro tipos de reações de hipersensibilidade. Cada uma dessas reações tem causas e mecanismos fi siopatogênicos distintos, implicando manejos clí-nicos diferentes. As três primeiras são mediadas por anticorpos, enquanto a quarta está relacionada à ativação de células T e macrófagos.
A reação de hipersensibilidade tipo I, também conhecida como anafi la-xia, ocorre quando mastócitos sensibilizados por imunoglobulinas da classe E (IgEs) liberam mediadores químicos gerando uma resposta infl amatória aguda. Como exemplos clínicos, podemos citar a asma e a rinite alérgica. A hipersensibilidade tipo II ocorre quando anticorpos se ligam a auto-an-tígenos ou antígenos externos, estimulando a fagocitose, a atividade das células NK (natural killers) ou a lise mediada pelo complemento. Chamamos ADCC (antibody dependent cellular citotoxicity – citotoxicidade celular de-pendente de anticorpos), sendo seu melhor exemplo a reação de hemólise por incompatibilidade ABO, que ocorre quando é transfundido inadverti-damente sangue não-compatível. A reação de hipersensibilidade tipo III é mediada por complexos antígeno-anticorpo que são produzidos em grande quantidade ou que não são retirados adequadamente de circulação pelo sistema mononuclear fagocítico (SFM). O dano tecidual ocorre pelo depósi-to dos imunocomplexos, associados à ativação da via alternativa do sistema complemento. São exemplos desse tipo de reação as glomerulonefrites indu-zidas por drogas, vasculites em geral e reações subagudas e crônicas de pele. A hipersensibilidade tipo IV, ou celular, é a mais lenta de todas as reações e é mediada por linfócitos T e macrófagos. Os exemplos clínicos mais importan-tes são as doenças granulomatosas, como a tuberculose e a sífi lis.
HistóricoO conceito alergia foi empregado pela primeira vez em 1906 por Clemens von Pirquet como “uma capacidade alterada do corpo de reagir a uma subs-tância estranha”, uma defi nição bastante ampla. Hoje, defi ne-se alergia como reação imunológica específi ca (= hiperegia), provocadora de doença,
Sócrates Salvador • Maria Lúcia Scroferneker
Reações de hipersensibilidade tipo I

180 Imunologia Básica e Aplicada
contra substâncias inócuas. Coombs e Gell, em 1963, propuseram a classificação das alergias nos tipos I até IV, ainda hoje empregada. As graves reações alérgicas imediatas do tipo I são também chamadas de anafilaxias. Richet e Postier utiliza-ram esse conceito pela primeira vez em 1902, para uma reação alérgica aguda contra o soro do coelho. O conceito de atopia foi introduzido por Coca e Cooke em 1923. Ele se refere à tendência hereditária de desenvolver alergias do tipo I con-tra antígenos por inalação e eczemas. A atopia inclui, por conseguinte, eczema atópico, rinite alérgica e asma.
EpidemiologiaSegundo dados epidemiológicos, 25% a 40% da população de países industrializados manifesta sintomas alérgicos. A prevalência de uma rini-te alérgica fica em torno de 10% a 15%; a de dermatite atópica, em 10%; a de uma urticária alérgica, em 10%; e a de uma asma brônquica alérgica, em 5% a 10%. A distribuição da fre-qüência das doenças alérgicas é dependente da idade. Na lactação e na infância, dominam as dermatites atópicas e alergias a alimentos. Na adolescência e na idade adulta, encontram-se a rinite alérgica e a asma brônquica alérgica. Na terceira idade, diminui a prevalência das doen-ças atópicas. Além disso, encontram-se diferen-ças regionais. Nesse sentido, a taxa das doenças atópicas em áreas urbanas é nitidamente mais alta do que em zonas rurais. As pessoas em paí-ses industrializados são mais freqüentemente acometidas do que a população de países em desenvolvimento. Na infância, afetam mais os meninos e, na idade adulta, mais as mulheres. Alterações na dieta também são responsáveis por elevada incidência em pessoas suscetíveis. No todo nas últimas décadas, as doenças alérgi-cas tiveram um nítido aumento.
As mudanças na exposição aos patógenos mi-crobianos até o momento são a explicação mais plausível para o aumento da atopia. Ela está as-sociada negativamente com história de infecção com vírus do sarampo e hepatite A e com testes cutâneos de tuberculinas positivos. Estudos têm mostrado que a infecção por esses patógenos induziria produção de INF-γ, aumentando a ex-pressão de linfócitos TH1 que inibiriam os TH2. Por outro lado, há evidências de que crianças que tiveram crises de bronquiolite por vírus sincicial
respiratório (VSR) são suscetíveis ao desenvolvi-mento de asma. Crianças hospitalizadas com essa doença apresentam um desvio de produção de citocinas como o INF-γ para IL-4, a citocina que induz resposta TH2. É possível que a infecção por um organismo que evoca uma resposta imune TH1 precocemente possa reduzir a probabilidade de respostas de TH2 tardiamente e vice-versa.
Como vimos, fatores não-genéticos também desempenham papel importante na gênese da alergia, tais como a sensibilização prévia (uma pessoa só pode ser alérgica a uma substância com a qual já teve contato), a quantidade da ex-posição, a via de administração, a reatividade do órgão-alvo, o estado nutricional do indivíduo, a presença de infecções concomitantes agudas ou crônicas, a temperatura ambiente ou o estado emocional do indivíduo. Há, portanto, um con-junto de fatores que conduzem a uma predispo-sição individual em produzir uma reação hiper-sensível diante de um estímulo imunogênico.
DefiniçõesOs requisitos básicos para que ocorra uma rea-ção de hipersensibilidade tipo I são:
a) a exposição prévia a um alérgeno capaz de gerar produção suficiente de IgE em res-posta ao estímulo em indivíduo genetica-mente suscetível;
b) a presença do mastócito;c) a sensibilização;d) a reexposição ao mesmo antígeno;e) a ligação cruzada com conseqüente libe-
ração de mediadores químicos.
AlérgenosA maioria dos alérgenos são proteínas, pois so-mente as proteínas induzem respostas de células T, fundamentais para a produção da IgE. Além de serem proteínas, os alérgenos são enzimati-camente ativos, apresentados em baixas doses e possuem baixo peso molecular, alta solubilidade e estabilidade, qualidades necessárias para que possam se difundir pela mucosa e não alterar sua composição quando forem dissecados.

181Reações de hipersensibilidade tipo I
Células presentes na reação de hipersensibilidade do tipo I
Linfócitos TAs células T presentes nas lesões alérgicas in-cluem duas variedades: TH1 e TH2. O tipo TH2 predomina. É conhecido que as células TH2 se-cretam IL-4 e IL-5, mas não INF-γ, enquanto as células TH1 secretam INF-γ e IL-2, mas não IL-4 e IL-5. A IL-4 estimula os linfócitos B a produzirem mais anticorpos IgE do que os outros tipos, e o INF-γ, por sua vez, retarda tal síntese. Esses resul-tados sugerem que a quantidade individual de células TH1 e TH2 determina se os linfócitos B da-rão origem a moléculas de IgE ou outros tipos de anticorpos. Além disso, drogas capazes de inter-ferir na síntese ou na atividade da IL-4 poderão ajudar a reduzir os níveis de IgE e assim prevenir as reações alérgicas. Sabe-se, então, que para a produção de IgE é necessário células TH2.
O destino das células TH virgens, responden-do a um peptídeo apresentado por uma célula dentrítica, é determinado pelas citocinas expos-tas antes e durante essa resposta e pelas proprie-dades intrínsecas de antígeno, dose de antígeno e via de apresentação.
Um fator bastante importante na diferenciação das células TH em seus diferentes subgrupos efe-tores é a quantidade e a seqüência exata do peptí-deo específico utilizado para iniciar uma resposta. Grandes quantidades de peptídeos que alcançam
elevada densidade na superfície das células apre-sentadoras de antígeno tendem a estimular uma resposta de células TH1, enquanto apresentações de baixa densidade levam ao estímulo de respos-tas de células TH2. Além disso, peptídeos que in-teragem fortemente com o receptor de célula T tendem ao estímulo de respostas semelhantes às do TH1, ao passo que peptídeos que se unem fra-camente estimulam respostas do TH2. Essa dife-rença é muito bem exemplificada pela alergia. Os antígenos que induzem alergia mediada por IgE geralmente são liberados em doses diminutas e fazem com que as células TH2 secretem IL-4, mas não INF-γ (Figura 15.1).
Linfócito B e IgEO linfócito B , após o estímulo antigênico, como é caso da apresentação de um alérgeno por uma APC, começa a se proliferar e diferenciar em plas-mócitos. Essas células possuem capacidade de expressar vários isótopos de imunoglobulinas, le-vando à síntese de diferentes classes de anticor-pos com a mesma especificidade antigênica. Esse fenômeno é denominado switching de classes. A mudança de classe das células B para a produção de IgE é induzida por dois sinais separados, e am-bos fornecidos pelas células TH2. O primeiro dos sinais é proporcionado pelas citocinas IL-4 e IL-13, interagindo com os receptores de superfície das células, estimulando com isso o switching de clas-ses para IgE. O segundo sinal para a mudança de classe para IgE é a interação co-estimuladora en-
Figura 15.1. Diferenças das fases efetoras nas respostas TH1 e TH2.
Linfócito T-auxiliarativado
Ativação de macrófagos
Linfócito B PlasmócitoTH2
TH1
IgE
IFN-γIL-2, 4, 5
IL-4, 5, 6, 9, 10, 13
IgE

182 Imunologia Básica e Aplicada
tre o ligante CD 40 na superfície da célula T com o CD40 da superfície da célula B.
A resposta de IgE, uma vez iniciada, pode ser amplificada ainda pelos basófilos, mastócitos e eosinófilos, que também podem estimular a pro-dução de IgE. Quando essas células especializadas são ativadas pela ligação cruzada do antígeno com IgE ligada ao FcεRI, elas podem expressar o CD 40L na superfície celular e secretar IL-4, assim como as células TH2. Portanto, elas podem esti-mular a mudança de classe e a produção de IgE pelas células B. A IgE, um anticorpo monomérico com peso molecular de 190 mil dáltons, liga-se às células efetoras por intermédio de sua porção Fc e tem papel-chave na mediação de reações alér-gicas e citotóxicas contra parasitos após a ligação com antígeno específico.
Uma das características mais importantes da IgE é a sua capacidade de se ligar aos mastócitos e basófilos com alta afinidade por meio da sua porção Fc. Com essa ligação, será possível o de-sencadeamento da reação de hipersensibilidade tipo I. A produção de IgE pelos linfócitos B de-pende da apresentação do antígeno pelas APCs e da cooperação entre as células B e os linfócitos TH2. As IgEs produzidas sensibilizam primeiro os mastócitos locais e depois o excedente entra na circulação, sensibilizando basófilos circulantes e mastócitos teciduais em todo o organismo.
A grande afinidade da IgE pelo receptor FcεRI de mastócitos e basófilos permite que, combina-da a essas células, a imunoglobulina esteja pro-tegida da ação de proteases, aumentando sua meia-vida sérica de poucos dias, quando livre, para vários meses. A IgE encontra-se em concen-trações mínimas no sangue, contribuindo com menos de 0,001% do total de imunoglobulinas. Está normalmente presente na superfície de ba-sófilos e mastócitos de todos os indivíduos.
Os níveis de IgE costumam estar elevados ao longo de infestações parasitárias, sendo impor-tantes no recrutamento de eosinófilos durante a imunidade a parasitos. Especula-se, inclusive, que essa imunoglobulina tenha se desenvolvido como uma defesa contra esse tipo de infecção. Embora fatores genéticos e ambientais devam ser levados em consideração na análise de um paciente, níveis elevados de IgE na ausência de parasitas sugerem fortemente a presença de atopia. A realização de testes de sensibilidade demonstra que 30% da população tem respos-ta positiva a alérgenos comuns. Como apenas
10% da população realmente manifesta sinais clínicos, acredita-se que o restante dos indiví-duos, embora produza IgE, não possua nenhum fator que precipitaria o aparecimento das rea-ções alérgicas.
Enquanto as concentrações de anticorpos não-IgE no sangue tipicamente variam pouco de pessoa para pessoa, os anticorpos IgE podem va-riar consideravelmente, nunca ultrapassando os níveis de IgM e IgG.
SuscetibilidadeA sensibilização poderá causar doença localizada na mucosa nasal, na mucosa brônquica, na pele, no trato gastrintestinal ou em uma combinação de dois ou mais desses sítios. Isso é particular-mente verdadeiro na atopia, situação na qual existe uma propensão hereditária em responder imunologicamente a vários antígenos comuns da natureza por meio da produção contínua de anticorpos da classe IgE. Alguns indivíduos rea-gem ao mesmo antígeno com produção de IgE, enquanto outros o fazem produzindo IgG. Em-bora fatores como dose, via de administração e número de exposições possam estar implicados, há fortes evidências de que o fator principal é o genético. Até 40% das pessoas na população ocidental mostram tendência exagerada a pro-duzir respostas IgE a uma ampla variedade de alérgenos ambientais mais comuns. Esse estado é denominado de atopia. Os indivíduos atópicos têm níveis mais elevados de IgE na circulação e níveis mais elevados de eosinófilos que pessoas normais. Estudos de famílias atópicas sugerem loci nos cromossomos 11q e 5q, que podem ser importantes para determinar presença de ato-pia. O gene candidato 11q codifica a subunidade β do receptor de IgE de alta afinidade, ao passo que no cromossomo 5q existe um grupo de ge-nes intimamente ligados, que inclui aqueles para a formação da IL-3, IL-4, IL-5, IL-9, IL-12, IL-13 e GM-CSF. Essas citocinas são importantes na mu-dança de isótopo para IgE, na sobrevivência dos eosinófilos e na proliferação de mastócitos.
Um segundo tipo de variação hereditária nas respostas IgE está ligado à região MHC de classe II e afeta as respostas a alérgenos específicos. Mui-tos estudos mostraram que a produção específica de IgE está associada a certos tipos de alelos de HLA, que, em combinações particulares de MHC/peptídeo, favorecem uma forte resposta TH2.

183Reações de hipersensibilidade tipo I
A incidência de alergia nos membros de uma família tende a ser maior do que naquelas sem esse tipo de doença, muitas vezes com expres-sões clínicas variadas (asma, rinite alérgica etc.). As crianças herdam uma predisposição à atopia com prevalência de história familiar positiva três vezes maior nos pacientes atópicos. Pais alérgi-cos têm chance maior do que 50% de ter filhos alérgicos. Quando apenas um dos pais é alérgico, o risco é de cerca de 30%. Apesar disso, alguns estudos sugerem que pessoas atópicas e não-ató-picas têm o mesmo risco de desenvolver anafila-xia sistêmica ao entrar em contato com drogas ou picadas de insetos.
Mastócitos e basófilos
Ambos se originam da medula óssea. Enquan-to os basófilos estão distribuídos em pequeno número no sangue circulante, os mastócitos en-contram-se no tecido conjuntivo dos diferen-tes órgãos, mais próximos de vasos sangüíneos, vasos linfáticos e nervos. Os dois tipos celulares apresentam receptores de alta afinidade para a fração Fc de IgE (FcεRI).
Os mastócitos e os basófilos possuem em seu citoplasma grânulos que contêm substâncias químicas como heparina e histamina (Tabela 15.1). Essas substâncias desencadeiam várias al-terações características da reação de hipersen-sibilidade tipo I (Tabela 15.2). Após a ligação cruzada de FcεRI por imunoglobulinas da classe IgE, esses grânulos são liberados. Esse processo é denominado desgranulação (Figura 15.2).
Tabela 15.2. Conseqüências da reação anafilática
Ação Mediador
Infiltração celular (quimiotaxia)
• Leucotrieno B4• PAF• TNF-α• Fatores quimiotáticos para eosinófilos e neutrófilos
Vasodilatação e aumento da permeabilidade vascular
• Histamina • PAF• Leucotrienos C4, D4 e E4• Prostaglandina D2
Espasmo da musculatura lisa brônquica
• Leucotrienos C4 e D4• Histamina• Prostaglandina D2• PAF
Tabela 15.1. Mediadores derivados dos mastócitos
Mediadores pré-formados e liberados rapidamente em condições fisiológicas
• Histamina • Fatores quimiotáticos para eosinófilos e neutrófilos• Cininogenase• Fator de necrose tumoral (TNF-α)• Interleucina-6• Endotelina-1
Mediadores produzidos durante o processo de desgranulação
• Superóxidos • Leucotrienos C4, D4, E4 (antigo SRS-A)• Adenosina• Bradicinina• Fator ativador de plaquetas (PAF)• Fator gerador de prostaglandinas da anafilaxia• Prostaglandina D2
Mediadores intimamente associados aos grânulos • Heparina, sulfato de condroitina E• Triptase• Catepsina G• Carboxipeptidase• Peroxidase• Arilsulfatase• Fatores inflamatórios• Citocinas geradas após a ativação• IL-1, 2, 3, 4, 5, 6• Fator estimulante de colônias de granulócitos e macrófagos• Proteína inflamatória de macrófagos 1a e 1b• Fator quimiotático e ativador de macrófagos• Fator de necrose tumoral (TNF-α)• Endotelina-1

184 Imunologia Básica e Aplicada
inflamatórios que são capazes de exacerbar as reações atópicas.
Pacientes com eczema atópico (um tipo de rea-ção cutânea) costumam apresentar células de Lan-gerhans cobertas por IgE em sua pele, o que po-deria estar envolvido na apresentação do alérgeno e na ativação das células T específicas. Essa com-binação, célula de Langerhans/IgE, não é vista nos pacientes atópicos livres de reações eczematosas.
FisiopatogeniaNas reações anafiláticas, alguns componentes do sistema imune reagem a substâncias estranhas que normalmente são consideradas inofensivas. Os alérgenos que costumam induzir essas rea-ções são geralmente proteínas ou polipeptíde-os grandes. Algumas moléculas menores, como produtos da degradação da penicilina (grupos peniciloil), podem se combinar com proteínas do paciente e tornarem-se imunogênicas, funcio-nando como haptenos. Para que se inicie uma reação de hipersensibilidade tipo I, é necessário que haja exposição a um antígeno apropriado e que o indivíduo responda a esse estímulo com uma produção aumentada de IgE. O alérgeno induz, então, uma resposta imune denominada sensibilização em vez da tradicional imunização. A sensibilização acontece na primeira vez em
Figura 15.2. Sensibilização e desgranulação mastocitária.
A desgranulação dos mastócitos pode ocorrer por causa de vários estímulos. As lectinas (presen-tes em grande quantidade no morango) podem gerar ligação cruzada entre IgEs, ligando-se a sua porção Fc, prescindindo da especificidade da por-ção Fab. Os componentes do complemento C5a e C3a (anafilatoxinas) podem ativar os mastócitos e os basófilos diretamente ao se ligarem em recep-tores específicos na superfície celular. Além deles, outras substâncias também dispensam a presença de IgE, como citocinas derivadas de mastócitos (IL-8), algumas drogas (codeína, morfina e hormônio adrenocorticotrófico sintético), a melitina (pre-sente no veneno da abelha) e estímulos físicos e químicos (calor, frio e luz solar): pacientes atópicos costumam perceber no cotidiano a importância desses últimos três estímulos no agravamento de suas crises alérgicas. Essas reações não desencadea-das por IgE são denominadas de anafilactóides.
Outras células do sistema imune também po-dem se ligar à IgE. Eosinófilos, plaquetas, macró-fagos e células de Langerhans expressam recep-tores de alta e baixa afinidade para essa imuno-globulina. Eosinófilos e plaquetas, quando sen-sibilizados, têm aumentada sua citotoxicidade contra certos parasitos, como o esquistossoma. Esses dois tipos celulares têm também papel im-portante na alergia, já que contêm mediadores
IL-4
IgEPlasmócito
T CD4
MHC-II
APC
LB
Antígenos no pólen

185Reações de hipersensibilidade tipo I
Figura 15.3. Ativação do mastócito pela porção Fc da IgE com conseqüente liberação de aminas vasoativas.
Mastócito
Grânulos
IgE
HistaminaPGDLTC4
VasodilataçãoConstrição brônquica
Secreção de mucoEstímulos nervosos
pidas nos níveis celulares de AMPc. Essa elevação transitória ativa proteínas quinases responsáveis pela fosforilação das membranas dos grânulos dos mastócitos. A fosforilação das membranas as tor-na permeáveis ao cálcio e à água, resultando em edema dos grânulos. Ao mesmo tempo, enzimas ativadas pelo cálcio liberam energia para micro-túbulos e microfilamentos que transportam esses grânulos para a superfície celular, na qual eles se fundem à membrana e liberam seu conteúdo para o exterior. Vários estímulos físicos e químicos (além da ligação cruzada de IgEs) podem levar à desgranulação de mastócitos e basófilos, como as anafilatoxinas C3a e C5a, morfina e codeína, a luz solar, traumatismos, o calor e o frio.
Entre os mediadores liberados pelos mastóci-tos, alguns estão bem estudados e merecem es-pecial atenção. A histamina pode estimular a pro-dução de muco pelo epitélio das vias aéreas con-tribuindo para sua congestão. Causa contração dos músculos lisos dos brônquios e do intestino. Pode também dilatar pequenos vasos sangüíneos e aumentar a sua permeabilidade, o que leva ao extravasamento de líquido para os tecidos. As alterações vasculares dão origem a hiperemia e edema. Entre os mediadores secundários, me-recem atenção os lipídios, principalmente pros-taglandinas e leucotrienos. Ambos se formam a partir do ácido araquidônico pelas vias da cicloxi-genase e 5-lipoxigenase, respectivamente, e cau-sam constrição dos brônquios e dilatação vascu-lar. O fator ativador de plaquetas (PAF) é também um mediador secundário. Ele age indiretamente pela liberação de histamina e diretamente cau-sando broncoconstrição, vasodilatação e aumen-to da permeabilidade vascular (Figura 15.3).
que o alérgeno entra no organismo e ocorre da seguinte maneira: as células apresentadoras de antígeno (APCs) englobam a substância estra-nha, fragmentando-a e expondo os epítopos na superfície celular. Os linfócitos TH reconhecem alguns desses fragmentos e se ligam a eles. Essa ligação estimula as células T a secretarem IL-4 e outras citocinas que, por sua vez, estimulam os linfócitos B vizinhos a se diferenciarem até plas-mócitos. A partir da IgM, esses plasmócitos, in-fluenciados pelos tipos de linfocinas do ambien-te, produzem quantidades aumentadas de IgE. Os anticorpos IgE formados possuem uma ten-dência muito forte de se ligar a receptores para sua porção Fc localizados em mastócitos e basó-filos. Essas células cobertas por anticorpos se tor-nam sensibilizadas. Uma vez que a sensibilização ocorra, o indivíduo permanecerá assintomático até que haja uma nova exposição ao alérgeno.
Em exposições subseqüentes, antígenos mul-tivalentes do alérgeno se ligam prontamente a mais de uma molécula de IgE de uma mesma célu-la, formando uma ligação cruzada de anticorpos IgE adjacentes, induzindo mobilidade nas molé-culas de IgE e provocando alterações na mem-brana plasmática de mastócitos e basófilos. Com isso, ocorre o desencadeamento de dois processos paralelos e interdependentes: (1) a desgranulação dos mastócitos com liberação de mediadores quí-micos pré-formados e a (2) síntese com posterior liberação de mediadores secundários. Esses me-diadores, liberados pelos mastócitos nos tecidos circunjacentes, geram a resposta alérgica, e a na-tureza dos sintomas vai depender do sítio envolvi-do e da quantidade de mediadores liberados.
A desgranulação mastocitária se dá por um processo ativo envolvendo ascensão e queda rá-

186 Imunologia Básica e Aplicada
As substâncias liberadas pelos mastócitos ati-vados podem também induzir basófilos, eosi-nófilos e outras células a migrarem através dos vasos sangüíneos para dentro dos tecidos. Essas células recrutadas também secretam substâncias que podem sustentar a resposta imune e causar lesão tecidual (Figura 15.4). A Tabela 15.1 apre-senta uma visão geral das alterações básicas da reação de hipersensibilidade tipo I e seus respec-tivos mediadores.
TratamentoAs terapias utilizadas podem atuar em dois ní-veis: (1) atenuar ou bloquear os sintomas e (2) diminuir a quantidade de IgE no indivíduo por
modulação da resposta imune (imunoterapia). Em alguns pacientes, evitar o contato com o alér-geno é a melhor forma de impedir o desencadea-mento da reação de hipersensibilidade. Em ou-tros casos, isso é impraticável, exigindo controle farmacológico (Tabela 15.3).
A modulação da resposta imune pode ter de 30% a 70% de eficácia. Não foi descrito o meca-nismo de ação desse tratamento sobre o sistema imune, porém há quatro hipóteses:
1. Hipossensibilização: produção de anticor-pos IgG bloqueadores da reação de hiper-sensibilidade tipo I. Exposições repetidas ao alérgeno por uma via diferente da usual (injeções, por exemplo), em doses maiores do que as naturais, repetidamente, podem
Tabela 15.3. Agentes farmacológicos e suas ações sobre mastócitos
Classe Exemplo Mecanismo de ação Efeito
Anti-histamínicos Difenidramina (H1)Cimetidina (H2)
Bloqueio da ação da histamina em seus receptores H1 e H2
Bloqueio dos efeitos da histamina nos diversos tecidos (útil especialmente antes do início dos sintomas)
β-adrenérgicos SalbutamolIsoproterenol
Aumenta o AMPc pela ativação da adenilatociclase
Diminuição da liberação de mediadores e contraposição aos seus efeitos nos órgãos-alvo
Corticosteróides BeclometasonaPrednisona
Estímulo da síntese de lipocortina, proteína inibidora da produção defosfolipase A2; aumento do número de receptores β
Inibição da síntese e liberação de leucotrienos e prostaglandinas; inibição da quimiotaxia e adesão celulares
Metilxantinas TeofilinaAminofilina
Aumento do AMPc pela diminuição da fosfodiesterase
Diminuição da liberação de mediadores e contraposição aos seus efeitos nos órgãos-alvo
Outras Cromolin Inibe a fosfodiesterase; diminui o fluxo de cálcio pela membrana plasmática dos mastócitos
Inibe liberação de mediadores por mastócitos, macrófagos e eosinófilos
Figura 15.4. Ativação do mastócito pela porção Fc da IgE com conseqüente liberação de citocinas.
Mastócito
Grânulos
IgE
Citoninas
Síntese de IgEAtivação de leucócitos
Inflamação crônicaAcúmulo de leucócitosAdesão de leucócitos

187Reações de hipersensibilidade tipo I
causar desvio da resposta imune de TH2 para TH1. Com a queda da produção de IL-4 e IL-13, haveria um bloqueio na diferen-ciação de plasmócitos produtores de IgE. A IgG formada em maiores quantidades tem a mesma especificidade da IgE, competin-do com ela pela ligação aos alérgenos;
2. Dessensibilização: é produzida por inocu-lação repetida de pequenas doses do an-tígeno, suficientes para que se liguem às IgEs e impeçam sua ligação aos mastócitos
teciduais. Com a suspensão do tratamento, os sintomas tendem a reaparecer;
3. Tolerância: perda de responsividade de lin-fócitos T e B específicos, por deleção. Pa-rece estar relacionada à ação de linfócitos T-supressores;
4. Supressão: aumento na população de linfóci-tos T-supressores específicos para o alérgeno (o indivíduo alérgico parece ter uma diminu-ição da função supressora). Isso pode resul-tar na suspensão da produção de IgE e do recrutamento de mastócitos.
BibliografiaAbbas AK, Lichtman AH, Pober JS. Immediate hypersensitivity. In: Abbas AK, Licht-
man AH, Pober JS (eds.). Cellular and Molecular Immunol. 5.ed. New York: W. B. Saunders, 2003. pp. 432-52.
Benn CS, Melbye M, Wohlfahrt J, Björkstén B, Aaby P. Cohort study of sibling effect, infectious diseases, and risk of atopic dermatitis during first 18 months of life. BMJ, doi:10.1136/bmj.38069.512245.FE (publicado em 30 de abril de 2004).
Bochner B. Anaphylaxis. The New England Journal of Medicine 1991; 324(25): 1785-90.
Brostoff J, Hall T. Hypersensitivity – Type I. In: Roitt I, Brostoff J, Male D. Immuno-logy. 4a ed. St. Louis, Mosby, 1996. pp. 1-17.
Buentke E, Heffler LC, Wallin RPA, Fman CLO, Ljunggren HG, Scheynius A. The allergenic yeast Malassezia furfur induces maturation of human dendritic cells. Clinical and Experimental Allergy 2001; 31:1583-9.
Charlesworth EN. The skin as a model to study the pathogenesis of IgE-mediated acute and late-phase responses. Journal of allergy and clinical immunology 1994; 6(94):1240-50.
Hawrylowicz M, O’Garra A. Potential role of interleukin-10-secreting regulatory T cells in allergy and asthma c. nature reviews. Immunology 2005; (5):271-83.
Kay AB. Allergic and allergic diseases. New England J Med 2001; 344(1):30-7.
Marone G. The clinical relevance of basophil releasability. Journal of allergy and clinical immunology 1994; (6)94:1293-303.
Mori A, Suko M, Tsuruoka N et al. Allergen-specific human T cell clones produce interleukin-5 upon stimulation with the Th1 cytokine interleukin-2. Interna-tional Archives of Allergy and Immunology 1995; 1-3(107):220-2.
Reisman RE. Insect Stings. The New England Journal of Medicine 1994; 8(331):523-7.
Tanus T, Mines D. Serum tryptase in idiopatic anaphylaxis: a case report and a review of the literature. Annals of Emergency Medicine 1994; (1)24:104-6.


C A P Í T U L O 1 6
IntroduçãoA reação de hipersensibilidade tipo II também é denominada reação citolí-tica mediada por anticorpo. Consiste na formação de anticorpos das classes IgG e IgM que reagem contra antígenos presentes na membrana das células ou fi xos em tecidos, ativando mecanismos de injúria efetuados pela ativação do complemento e fagocitose.
FisiopatogeniaA reação de hipersensibilidade tipo II é ativada, na maioria dos casos, por anticorpos formados contra antígenos próprios, ou seja, auto-anticorpos. Em alguns casos, ocorre a formação de anticorpos contra antígenos estranhos que acabam reagindo contra componente tecidual próprio, o que também é denominado de reação cruzada.
Geralmente os anticorpos são das classes IgG e IgM. Ambas apresentam grande efi ciência em ativar o complemento e células infl amatórias.
Ao ocorrer a ligação dos anticorpos com os antígenos de membrana ou tecidos, ativam-se mecanismos efetores da hipersensibilidade. Freqüente-mente, os diferentes mecanismos efetores são ativados conjuntamente, le-vando à liberação de espécies reativas (enzimas, metabólitos de O2 etc.) e à injúria tecidual. São eles:
1. Fagocitose da célula recoberta por anticorpo (Figura 16.1):a) a célula-alvo opsonizada pelo anticorpo se liga à porção Fc do receptor
dos macrófagos presentes principalmente no fígado e no baço;b) essa ligação promove a sua fagocitose, sendo eliminada no interior do
fagolisossoma por metabólitos de O2, radicais livres, íons e enzimas;c) por fi m, ocorre depleção do tipo celular acometido. Por exemplo, na
anemia hemolítica auto-imune, os eritrócitos é que são o alvo dos auto-anticorpos, provocando anemia.
2. Ativação do complemento (Figura 16.2):a) ocorre quando as imunoglobulinas da classe IgM e algumas IgG se li-
gam aos seus antígenos específi cos, ativando o complemento pela via clássica e formando o MAC (membrane attack complex – complexo de ataque a membrana). Com isso, promove a lise celular;
Renata Farinon • Maria Lúcia Scroferneker
Reações de hipersensibilidade tipo II

190 Imunologia Básica e Aplicada
b) ocorre a formação de outros componentes do complemento como o C5a e C3a, que apre-sentam atividade quimiotática e de anafilaxia, atraindo neutrófilos e macrófagos para o lo-cal, e C3b, que facilita a ação de fagócitos.
3. Recrutamento e ativação de células inflamatórias: a) ocorre por meio de dois mecanismos:
Lesão tecidual
A ligação de anticorpo + antígeno ativa a cascata do complemento, pro-duzindo C5a, que é qui-miotático para leucócitos (Figura 16.2)
Porção Fc do fagócito se liga ao antígeno + anticor-po, estimulando a fagoci-tose sem ativação do com-plemento (Figura 16.1)
Atração de neutrófilospara o local
Liberaçãode enzimas, radicais livres etc.
Figura 16.1. Promoção da fagocitose por meio da ligação entre receptor Fc e complexo antígeno-anticorpo.
Figura 16.2. Ataque pelo sistema complemento.
4. Citotoxicidade não-fagocítica (Figura 16.3):a) também chamada de ADCC (antibody de-
pendent cellular citotoxicity – citotoxidade celular dependente de anticorpo);
b) depende da ligação dos anticorpos com a fração Fc dos receptores das células NK ou dos fagócitos, provocando ataque da cé-lula com o antígeno na superfície por um mecanismo diferente da fagocitose.
Célula-alvo
MacrófagoReceptor Fc
Ag
Ac
Célula-alvo
Ag
Ac
Ativação da via clássica do complemento
C5b 6 7 8 9 = MAC Lise
C3a, C5a
C3aOpsonização/
fagocitose
Quimiotaxia e anafilaxia
Figura 16.3. Citotoxicidade dependente de anticorpos mediada por células NK.
Linfócito NK
Grânulos de perfurina e granzimas
Célula-alvo
Ag
Ac
Ag
Célula-alvo Linfócito NK
O receptor da célula NK interage com a porção Fc do anticorpo, mudando sua configuração para célula K (killer). Esta destrói a célula-alvo não por fagocitose, mas sim por um mecanismo semelhante ao usado pelo linfócito CD8, ou seja, por meio de grânulos de perfurina que se aderem à membrana da célula-alvo. Ocorrem a formação de poros e o conseqüente desequilíbrio hidroeletrolítico.
Nos casos em que a célula é muito grande para ser fagocitada, o fagócito também atua por esse mecanismo, liberando enzimas lisossômicas para o meio, danificando não só a célula-alvo como também as células vizinhas.
Aspectos clínicosAs reações citotóxicas mediadas por anticorpos provocam repercussão clínica, acarretando o sur-gimento de diversas doenças. O tipo de célula envolvida, as características dos anticorpos e o nú-mero de sítios antigênicos são alguns fatores que interferem no tipo de resposta clínica a ser obser-

191Reações de hipersensibilidade tipo II
vada. As células mais freqüentemente acometidas são as sangüíneas: eritrócitos (anemia hemolítica auto-imune), leucócitos (agranulocitose) e plaque-tas (trombocitopenia auto-imune). Além das célu-las, outras estruturas também são afetadas. Como exemplo, temos o ataque aos receptores de acetil-colina da placa motora no caso da Miastenia grave, o que caracteriza o acometimento tecidual.
Os anticorpos produzidos contra a membra-na celular e os tecidos, conforme já mencionado anteriormente, são geralmente auto-anticorpos. O mecanismo por meio do qual são formados ainda é desconhecido. Contudo, sabe-se que são capazes de promover o surgimento de doenças tecido-específicas ou órgão-específicas. Algumas das doenças auto-imunes surgem por esse meca-nismo (Tabela 16.1).
As reações de hipersensibilidade tipo II também aparecem nas reações transfusionais, eritroblasto-se fetal e reações hemolíticas a drogas, e nestas não é comum a formação de auto-anticorpos, e sim anticorpos contra antígeno externo estranho ao organismo (aloanticorpos) (Tabela 16.2).
Doenças órgão/tecido-específicas causadas por auto-anticorpos
Síndrome de Goodpasture
É caracterizada pela presença de auto-anticor-pos contra o colágeno tipo IV presente tanto na membrana basal glomerular quanto na membrana basal dos alvéolos (anticorpo anti-MBG). Por isso, essa doença promove não só acometimento re-nal (glomerulonefrite grave), mas também pul-monar (hemorragias). A maioria dos casos inicia com sintomas respiratórios como hemoptise.
Os auto-anticorpos são IgGs e parecem fixar bem o complemento (Figura 16.4). Contudo, o fator de-sencadeante para a sua formação ainda é desconhe-cido. A ativação do complemento e a deposição de fibrina promovem a destruição da membrana basal.
Para o diagnóstico, emprega-se a imunoflores-cência marcada com IgG, em que se percebe pa-drão linear difuso e homogêneo de deposição dos anticorpos ao longo da membrana basal.
Tabela 16.1. Alguns exemplos de doenças órgão ou tecido-específicas causadas por auto-anticorpos
Doença Característica clínica Auto-anticorpo (auto-Ac)
Anemia hemolítica auto-imune Presença de anemia em virtude da hemóliseContra antígeno presente na superfície do eritrócito
Pênfigo vulgar Formação de bolhas em pele e mucosasContra moléculas de adesão (desmossomas) presentes na pele e mucosas
Síndrome de GoodpastureCaracterizada por alteração renal (glomerulonefrite) e alteração pulmonar (hemorragia)
Contra colágeno tipo IV presente na membrana basal do glomérulo renal e alvéolo pulmonar
Miastenia grave Fraqueza muscular progressiva Contra receptor de acetilcolina da placa motora
Púrpura trombocitopênica auto-imune Presença de trombocitopenia e sangramentos Contra proteínas de membrana das plaquetas
Neutrocitopenia auto-imune Presença de neutropeniaContra antígeno presente na membrana de neutrófilos
Doença de Graves HipertireoidismoContra o receptor de TSH das células da tireóide, promovendo sua estimulação
Tabela 16.2. Alguns exemplos de doenças órgão ou tecido-específicas causadas por aloanticorpos
Doença Característica clínica Aloanticorpo (Ac)
Doença hemolítica do recém-nascido =eritroblastose fetal
Lise de eritrócitos fetais, ocorrendo anemia e icterícia
Ac anti-Rh materno atravessa a placenta e chega ao feto
Reação tranfusional Lise de eritrócitos no receptor da doação Ac do receptor reage com eritrócitos de doador incompatível
Reação induzida por drogas Anemia e/ou trombocitopeniaAc contra a droga forma imunocomplexo que é adsorvido à membrana do eritrócito

192 Imunologia Básica e Aplicada
Púrpura trombocitopênica auto-imune
Ocorre formação de auto-anticorpos IgG contra antígenos próprios das plaquetas. A destruição das plaquetas promove hemorragias e hemato-mas. Pode haver esplenomegalia por causa da atividade aumentada do baço.
O mecanismo imunológico é parecido com o da anemia hemolítica auto-imune: fagocitose de célula opsonizada e ativação do complemento, com seqüestro da plaqueta ligada ao anticorpo por macrófagos esplênicos, promovendo sua re-moção acelerada na circulação.
A diferenciação da trombocitopenia auto-imune e das outras formas de trombocitopenia depende da detecção de anticorpo antiplaquetário.
Neutrocitopenia auto-imune
Doença em que há formação de anticorpos alta-mente específicos contra antígenos presentes na superfície de neutrófilos. É dividida em:
1. Primária: em crianças de até 3 anos sem as-sociação com doença. Em neonatos, pode ocorrer leucopenia devido ao ataque de anticorpos maternos aos leucócitos fetais (sensibilização da mãe com leucócitos do filho que atravessaram a placenta);
2. Secundária: neutrocitopenia associada com outras doenças, como lúpus eritematoso sistêmico, trombocitopenia auto-imune e linfomas, tendo remissão ao tratar doença de base.
Miastenia grave
É uma doença caracterizada pela formação de anticorpos da classe IgG contra receptores de acetilcolina (ACh) presentes na junção neuro-muscular da placa motora. O paciente se apre-senta com fraqueza muscular progressiva.
Para a transmissão do impulso nervoso nor-mal, é necessário que haja liberação e ligação da ACh com seus receptores presentes na fibra mus-cular. O paciente miastênico não possui problema na produção de ACh, porém apresenta número reduzido de receptores. Isso porque a ligação do auto-anticorpo ao receptor promove sua endo-citose ou degradação celular (pelo sistema com-plemento ativado pela presença de anticorpos). Dessa forma, ocorre interferência na transmissão
Figura 16.4. Anticorpos IgG fixam o complemento e ativam a cascata, causando dano tecidual na síndrome de Goodpasture.
Ativação do complemento
Ac
Ac
Ac
Lise
Anemia hemolítica auto-imune
Ocorre por causa da formação de auto-anticor-pos contra antígenos próprios dos eritrócitos. Os anticorpos podem ser: IgG (denominados quen-tes por agir somente em temperatura superior a 37oC, reagem mais com antígenos Rh e pro-vocam destruição dos eritrócitos por macrófa-gos do baço) ou IgM (chamados de frios pois só agem abaixo dos 37oC, reagem com precursores dos antígenos ABO e provocam destruição por ativação do complemento).
A reação contra os eritrócitos ocorre na circu-lação periférica, promovendo a ativação do com-plemento e a fagocitose da célula opsonizada, ou seja, a ligação da IgG ao eritrócito estimula o seqüestro dele pelo fagócito, promovendo lise celular e anemia.
A diferenciação da anemia auto-imune e das outras formas de anemia hemolítica depende da demonstração de anticorpos anti-eritrocitários. Nos casos de anemia congênita, não se detectam esses anticorpos, visto que os eritrócitos é que são defeituosos.
O principal teste diagnóstico da anemia he-molítica auto-imune é o teste de Coombs:
1. Direto: pesquisa a presença de anticorpos ligados aos eritrócitos. É positivo em 95% das vezes;
2. Indireto: pesquisa a presença de anticor-pos livres no plasma. Possui baixa sensibi-lidade, visto que os anticorpos estão mais presentes nos receptores dos eritrócitos e não livres na circulação.

193Reações de hipersensibilidade tipo II
Pênfigo vulgar
Doença bolhosa auto-imune em que há formação de auto-anticorpos IgG contra os desmossomas da junção intercelular da epiderme. Geralmente, ini-cia-se com comprometimento de mucosa oral que persiste por vários meses antes de afetar a pele. Há formação de bolhas que podem sofrer erosão.
O pênfigo do tipo vulgar é o mais comum, havendo outros três tipos de pênfigo: bolhoso, violáceo e eritematoso.
O diagnóstico é feito por imunofluorescência direta da pele (mostrando depósito de imuno-globulina ao longo da membrana das células da epiderme) ou indireta do soro.
Doença de Graves
Doença na glândula tireóide causada por auto-an-ticorpos contra o receptor TSH presente na superfí-cie das células epiteliais. É caracterizada por hiper-tireoidismo, ou seja, aumento da função da glân-dula, que passa a produzir maior quantidade de hormônios T3 e T4. O TSH é um hormônio produzi-do na hipófise e é responsável pela estimulação da produção de hormônios T3 e T4 pela tireóide.
A ligação do auto-anticorpo com um recep-tor pode provocar seu estímulo ou inibição. Na doença de Graves, o auto-anticorpo, ao se ligar ao receptor de TSH, estimula a produção exces-siva dos hormônios T3 e T4. Ou seja, age como o próprio TSH (o auto-anticorpo mimetiza-o), esti-mulando as células epiteliais da tireóide a exer-cerem sua função (Figura 16.6).
Doenças órgão/tecido-específicas causadas por aloanticorpos
Reações transfusionais
A reação imune transfusional ocorre quando há reação entre:
Figura 16.5. Anticorpos anti-receptores de acetilcolina na Miastenia grave.
Figura 16.6. Ligação do auto-anticorpo aos receptores de células tireoidianas, provocando estímulo da secreção de hormônios tireoidianos como o T3 e T4.
Neurônio
Músculo
Ac
Vesículasde ACh
Receptorde ACh
Ac
Ac
Célula do folículo tireoidiano
T4 e T3 elevados
neuromuscular, não ocorrendo despolarização do músculo de forma efetiva (Figura 16.5).
A maioria dos antígenos implicados nessas re-ações são fortes imunogênicos e pertencem ao grupo sangüíneo ABO e Rh (Figura 16.7).
Anticorpos naturaisdo receptor
Antígenos de umdoador incompatível
Figura 16.7. Diferenças entre as reações de hipersensi-bilidade tipo II nas incompatibilidades Rh e ABO com relação ao anticorpo.
Eritrócito do doador
Ac IgM
Aglutinação
Antígeno do sistema ABO
Ativação do complemento
Hemólise intravascular
Eritrócito do doador
Ac IgG
Antígeno do sistema Rh
Estimula fagocitose

194 Imunologia Básica e Aplicada
Durante uma transfusão entre grupos incom-patíveis, o receptor produz anticorpos contra os antígenos presentes na membrana dos eritró-citos do sangue recebido do doador, causando destruição dessas células.
Os anticorpos contra antígenos do grupo ABO são, geralmente, da classe IgM. A incom-patibilidade promove aglutinação, ativação do complemento (ocasiona fagocitose de hemácias opsonizadas por suas frações ou por anticorpo) e reação hemolítica das hemácias transfundidas (hemólise intravascular).
Já os anticorpos contra antígenos do grupo Rh são IgG. Promovem a destruição da célula-alvo por meio dos fagócitos presentes no fígado e baço (hemólise extravascular).
Doença hemolítica do recém-nascido
Caracterizada pela formação de anticorpos ma-ternos da classe IgG contra antígenos das hemá-cias do feto. É também conhecida como eritro-blastose fetal. A incompatibilidade Rh constitui-se um exemplo conhecido das incompatibilida-des materno-fetais.
A incompatibilidade Rh só ocorre em mães Rh negativas que tenham sido sensibilizadas com sangue Rh positivo (seja por parto prévio, trans-fusão etc.). É caracterizada pela formação de an-ticorpos maternos da classe IgG contra antígeno do grupo Rh.
No caso da sensibilização ter ocorrido por causa da gestação, é necessário que o feto seja Rh posi-tivo para que seus eritrócitos com tal antígeno, ao entrar em contato com o sangue materno na hora do parto, provoquem a produção de anticorpos anti-Rh na mãe, sensibilizando-a. No momento do parto, o grande sangramento é que garante que as células Rh positivas da criança entrem em con-tato com a circulação materna, formando os an-ticorpos. Podem ocorrer micro-hemorragias trans-placentárias durante a gestação, mas, na maioria dos casos, o volume transfundido é insuficiente para provocar uma resposta imune. Portanto, o risco de sensibilização da mãe aumenta durante a gestação, sendo máximo durante o parto.
Assim, a doença hemolítica raramente afeta o primeiro filho (a não ser que a mãe já esteja sen-sibilizada por causa de contato prévio com san-gue Rh positivo), pois a imunização ocorre em virtude da troca de sangue nesse primeiro parto. A doença só vai realmente ocorrer numa futura
gestação, desde que o feto seja novamente Rh positivo. Nesse caso, os anticorpos anti-Rh IgG já prontos serão capazes de cruzar a barreira pla-centária, promovendo a lise de eritrócitos fetais (Figura 16.8).
Figura 16.8. Produção de anticorpos anti-Rh causando lise das hemácias fetais na eritroblastose fetal.
1a gestação
HemóliseSensibilização da mãe e produção
de Ac anti-Rh
2a gestação
Mãe Rh-
Feto Rh+
O recém-nascido com eritroblastose fetal se apresenta com anemia, icterícia progressiva e hepatoesplenomegalia, além de reticulocitose no hemograma.
A profilaxia da doença hemolítica deve ser realizada em mães Rh negativas ainda não-sen-sibilizadas que estejam grávidas de feto Rh po-sitivo. Consiste em administrar anticorpos anti-RhD (Rhogan) próximo ao parto ou até 72 horas depois para que os eritrócitos do feto, ao entrar em contato com o sangue da mãe, sofram blo-queio antigênico, não permitindo a formação de anticorpos maternos. Os anticorpos administra-dos devem aparecer no soro da mãe entre 12 e 60 horas, não durando mais de cinco meses. Se durarem por mais de seis meses, podem indicar sensibilização e falha do anti-RhD em bloquear a reação. O Rhogan previne a imunização da mãe, mas não a reverte. Logo, não será efetivo se a paciente já estiver sensibilizada.
A profilaxia também está recomendada em caso de aborto, hemorragia, gravidez ectópica e procedimentos obstétricos (amniocentese, bióp-sia de vilosidades coriônicas etc.), visto que au-mentam o risco de hemorragia transplacentária. A administração de imunoglobulina deve ser fei-ta logo após esses eventos.
A realização de exames como tipagem ABO, Rh e anticorpos irregulares (dirigidos contra ou-

195Reações de hipersensibilidade tipo II
tros grupos sangüíneos) deve ocorrer na primei-ra consulta pré-natal, a fim de detectar possível incompatibilidade. O teste de Coombs indireto é um exame importante em gestantes Rh negati-vas para verificar a presença e a titulação de an-ticorpos anti-RhD. No caso de o teste de Coombs indireto ser negativo, deve-se repetir na 28a, 32a e 36a semana de gestação para averiguar possí-vel sensibilização nesse período.
Doença induzida por drogas
Algumas drogas são capazes de provocar reação de hipersensibilidade contra células sanguíneas, principalmente eritrócitos e plaquetas. Com isso, provocam anemia hemolítica e trombocitopenia.
A reação induzida por drogas é dividida em três grupos de acordo com o agente:
1. Sulfonamida, fenotiazina: o organismo pas-sa a produzir anticorpos IgG contra as dro-gas e/ou metabólitos já que os reconhece como estranhos, induzindo a formação de imunocomplexos que se depositam na mem-brana eritrocitária. Ocorre ativação do com-plemento e destruição do eritrócito;
2. Penicilina, cefalotina: ao administrar grande quantidade dessas drogas, uma porção de-las e/ou seus metabólitos aderem-se direta-mente na membrana do eritrócito, induzindo a formação de anticorpos IgG contra esse eri-trócito. Ocorre ativação do complemento e fagocitose, causando anemia;
3. Metildopa, levodopa: induz a formação de anticorpos IgG contra antígenos Rh dos er-itrócitos, levando à hemólise.
BibliografiaAbbas AK, Lichtman AH, Pober JS. Diseases caused by immune responses: Hyper-
sensivity and autoimmunity. In: Abbas AK, Lichtman, Abbas AK, Lichtman AH, Pober JS (eds.) Cellular and Molecular Immunology. 5.ed. New York: W. B. Saunders, 2003. pp. 411-31.
Abbas AK, Lichtman AH, Pober JS. Immediate hypersensivity. In: Abbas AK, Lichtman AH, Pober JS (eds.). Cellular and Molecular Immunology. 5.ed. New York: Saunders, 2003. pp. 432-52
Chapel H, Haeney M, Misbah S, Snowden N. Autoimmunity. In: Essentials of clini-cal immunology. 4a ed. Blackwell Science, 1999, pp.96-97.
Chilcott J, Jones ML, Wight J, Forman K, Wray K, Beverly C, Tappenden P. A review of the clinical effectiveness and cost-effectiveness of routine anti-D prophyla-xis for pregnant women who are rhesus-negative. Health Technology Asses-ment 2003; (4)7.
Coleman RM, Lombard MF, Sicard RE, Rencricca NJ. Fundamental immunology. 5a ed. Dubrique, Wm.C.Brown, 1989, pp. 360-5.
Hudson BG. The molecular basis of Goodpasture and Alport syndromes: beacons for the discovery of the collagen IV family. Journal of the American Society of Nephrology 2004; 15:2514-27.
Kay AB, Ravetch J, van de Winkel JGJ. Allergy and hypersensivity. In: Janeway CA, Travers P, Walport M, Shlomchik M. Immuno Biology. 4.ed. Edimbourgh: Churchill Livingstone, 1999. pp. 461-88.
Maayan-Metzger A, Schwartz T, Sulkes J, Merlob P. Maternal anti-D profilaxis du-ring pregnancy does not cause neonatal hemolysis. Archives of disease in childhood. Fetal and neonatal edition 2001; 84:60-2.
Magalhães JA, Dias RSP. Rotinas em obstetrícia. In: Freitas F, Martins-Costa SH, Ramos JGL, Magalhães JA. Doença Hemolítica Perinatal. 5.ed. Porto Alegre: Artmed, 2006. pp.105-114.Cidade Porto Alegre.
Male D. Hypersensivity-type 2. In: Roitt I, Brostoff J, Male D. Immunology. 6.ed. Filadélfia: Mosby, 2001. pp. 345-56.
Parham P. Reações excessivas do sistema imune. In: O sistema imune. 1.ed. Porto Alegre: Artmed, 2001. pp. 269-98.
Robbins SL, Cotran RS, Kumar V, Collins T. Patologia estrutural e funcional. 6.ed. Rio de Janeiro: Guanabara & Koogan, 2000. pp. 123-4, 864-5, 1152
Salama AD, Levy JB. Tolerance and autoimmunity in anti-GBM disease. Journal of the American Society of Nephrology 2003; 14:2.988-9.
Scroferneker ML, Pohlmann PR. Hipersensibilidade tipo II. In: Imunologia básica e aplicada. 2a ed. Sagra Luzzato, 1998, pp. 278-89.
Vicent A. Immunology of disorders of neuromuscular transmission. Acta Neurolo-gica Scandinavica Supplementum 2006; 183:1-7


C A P Í T U L O 1 7
IntroduçãoA hipersensibilidade tipo III pode ser defi nida como uma reação infl amatória induzida pela deposição de complexos antígeno-anticorpo em determinados tecidos do organismo. A reação tem início quando antígenos se combinam com anticorpos na circulação (imunocomplexos circulantes) ou quando os complexos são formados em regiões extracelulares, onde o antígeno pode ter sido depositado previamente (imunocomplexos in situ). A deposição des-ses complexos antígeno-anticorpo provoca lesões teciduais resultantes da ativação de mediadores séricos, principalmente o sistema complemento.
A simples formação de imunocomplexos na circulação não signifi ca a pre-sença de doença, pois esses são formados durante muitas respostas imunes e representam um mecanismo normal de remoção de antígenos. Os fatores que determinam se os imunocomplexos serão patogênicos ou não ainda são pouco compreendidos. Porém, seu papel na patogênese de doenças sistê-micas, como a doença do soro e as doenças auto-imunes como a reação de Arthus, está bem estabelecido.
Patogenia das reações de hipersensibilidade tipo IIIA patogênese da reação de hipersensibilidade mediada por imunocomple-xos pode ser dividida em três fases:
Formação do complexo antígeno-anticorpoA formação do imunocomplexo é iniciada pela introdução do antígeno no orga-nismo, o qual geralmente se constitui de uma proteína. Quando esse antígeno entra em contato com os linfócitos B específi cos, induz sua diferenciação em plasmócitos. Ocorre, então, o processo de seleção clonal, formando-se vários lin-fócitos que passam a sintetizar e secretar anticorpos específi cos para esse mesmo antígeno. A formação dos anticorpos que constituem, com o antígeno, o com-plexo imune circulante ocorre aproximadamente uma semana após a penetra-ção do antígeno no organismo.
Samuel Uez • Maria Lúcia Scroferneker
Reações de hipersensibilidade tipo III

198 Imunologia Básica e Aplicada
Deposição de imunocomplexos nos tecidosOs fatores que determinam se os imunocomple-xos formados irão se depositar em diversos te-cidos e provocar doença ainda são pouco com-preendidos. Existem alguns fatores que possivel-mente estão envolvidos nesse processo, que é responsável pelo início da reação.
Tamanho e constituição do imunocomplexoOs complexos grandes são praticamente ino-fensivos, pois são formados com grande excesso de antígenos, fixando o complemento de uma maneira mais efetiva. Esses complexos são rapi-damente removidos da circulação pelo sistema fagocítico mononuclear. Os complexos mais pa-togênicos são os de tamanhos pequenos e inter-mediários, pois são formados com excesso de an-tígenos e não conseguem fixar adequadamente o complemento. Dessa forma, a propriedade de imunoaderência do complemento fica prejudi-cada e os complexos, por não conseguirem es-tabelecer uma ligação eficaz com os eritrócitos, ficam mais tempo circulando.
Estado funcional do sistema fagocitário mononuclear (SFM)O SFM é localizado particularmente no fígado e no baço, tem a função de filtrar e retirar de circulação os imunocomplexos circulantes. Esse processo de eliminação ocorre a partir da fixa-ção do C3b do complemento na porção Fc dos anticorpos dos imunocomplexos, seguida pela li-gação do C3b aos receptores CR1 dos eritrócitos (Figura 17.1). Estes carregam os imunocomple-xos até o SFM para que sejam destruídos. Assim, qualquer interferência nesse processo, seja por sobrecarga ou por disfunção intrínseca, eleva o tempo de permanência desses complexos antíge-no-anticorpo na circulação e a chance de deposi-ção nos tecidos.
Classe da imunoglobulinaOs imunocomplexos podem ser formados com IgM, IgG e IgA, sendo a classe da imunoglobulina um fator determinante na eliminação circulatória. A IgG ligada ao antígeno é capaz de ativar o com-plemento pela via clássica e provocar sua redução gradual da circulação. Porém, os imunocomplexos formados com IgA não ativam o complemento pela via clássica e apresentam uma maior predis-posição ao depósito tecidual.
Outros fatoresExistem ainda outros fatores que influenciam na deposição dos complexos nos tecidos: a carga nos imunocomplexos (aniônica versus catiônica); a valência do antígeno; a afinidade do antígeno em relação a vários componentes teciduais; a ca-pacidade de ativação do sistema complemento; a configuração tridimensional dos complexos e fatores hemodinâmicos.
Sítios de deposiçãoOs glomérulos estão entre os tecidos-alvo mais estudados quanto às influências que determinam a deposição dos imunocomplexos. No entanto, existem vários outros sítios propensos ao depósito dos complexos: articulações, pele, coração, zonas de turbulência em que há alta pressão sangüínea (plexo coróide e bifurcação de artérias), superfí-cies serosas e pequenos vasos sangüíneos.
Para que os imunocomplexos formados na circulação deixem o meio vascular e depositem-se nas paredes dos vasos, é necessário que haja
Figura 17.1. Ligação do complexo antígeno-anticorpo com o receptor de C3b na hemácia, caracterizando o processo de imunoaderência.
Complexo Ag/Ac
Receptor para C3b
C3b

199Reações de hipersensibilidade tipo III
um aumento da permeabilidade vascular. Essa alteração vascular ocorre em virtude da ação dos fragmentos do sistema complemento, que, de-pois de formados na circulação, induzem a libe-ração de mediadores vasoativos.
Reação inflamatória causada por imunocomplexosUma vez depositados nos tecidos, os imunocom-plexos desencadeiam uma resposta inflamatória aguda. O evento principal dessa reação é a ativa-ção da cascata do sistema complemento e conse-qüente liberação de substâncias biológicas ativas. A importância do complemento na patogênese da lesão tecidual é sustentada pela observação do de-créscimo de seus níveis séricos durante a fase ativa da hipersensibilidade tipo III.
A ativação do sistema complemento promo-ve inflamação principalmente pela produção de fatores quimiotáticos, que induzem a migração dos leucócitos polimorfonucleares e monócitos (C5a), e pela liberação de anafilatoxinas (C3a e C5a), que aumentam a permeabilidade vascu-lar quando se ligam aos mastócitos e basófilos, provocando sua desgranulação. Nesse processo, ocorre a liberação de aminas vasoativas e fator ativador de plaquetas (FAP), que induz a agrega-ção plaquetária.
Os imunocomplexos também podem intera-gir diretamente com as plaquetas por meio de receptores para Fc e C3b. Essa interação estimu-la a liberação de aminas vasoativas bem como a agregação plaquetária, resultando na formação de microtrombos sobre a superfície exposta do colágeno da membrana basal do endotélio, exa-cerbando a resposta inflamatória.
Os leucócitos que são atraídos para o sítio de in-flamação pelos fatores quimiotáticos são ativados pelo acoplamento de seus receptores de C3b e Fc por imunocomplexos, resultando na liberação de uma série de mediadores inflamatórios, incluindo prostaglandinas e peptídeos vasodilatadores, assim como diversas enzimas lisossômicas (proteases, co-lagenases e elastases) capazes de digerir a membra-na basal, o colágeno, a elastina e a cartilagem dos tecidos (Figura 17.2). Os radicais livres de oxigênio produzidos por neutrófilos e macrófagos também atuam no dano tecidual. A lesão conseqüente à ação desses mediadores é denominada vasculite, se ocorrer em vasos sangüíneos; glomerulonefrite, caso ocorra nos glomérulos renais; artrite, se ocor-rer nas articulações; e assim sucessivamente.
Situações clínicas de reações de hipersensibilidade tipo IIIOs exemplos clássicos de reações de hipersensibi-lidade tipo III são a doença do soro, a reação de Arthus e as doenças auto-imunes (Tabela 17.1).
Figura 17.2. Patogênese da reação inflamatória mediada por imunocomplexos. 1. Reconhecimento antigênico pelo linfócito B. 2. Transformação do linfócito B em plasmócito. 3. Secreção de anticorpos pelo plasmócito. 4. Ligação de antígeno e anticorpo formando imunocomplexos. 5. Atração de polimorfonucleares. 6. Liberação de substâncias vasoativas e lise pelo complemento no sítio inflamatório com lesão tecidual.
Neutrófilos
Plasmócito
4
2 3
15 6
Anticorpos
Antígenos Linfócito B
Imunocomplexos
Complemento
Agregação plaquetária
Necrose fibrinóide

200 Imunologia Básica e Aplicada
Doença do soroA doença do soro foi descrita inicialmente em pacientes submetidos à soroterapia (soro hete-rólogo geralmente eqüino) para tratamento de doenças infecciosas. Atualmente, tornou-se uma doença rara em virtude do advento da antibioti-coterapia e da restrição do uso de soro heterólo-go a poucas doenças.
A doença do soro é uma patologia autolimita-da aguda causada por uma reação inflamatória a imunocomplexos formados por antígenos exó-genos (como os antígenos eqüinos), constituin-do-se no protótipo das doenças sistêmicas causa-das por imunocomplexos.
Depois de transcorridos seis a oito dias da in-jeção do soro heterólogo, surge o quadro clínico que caracteriza a doença: febre, artralgias, mial-gias, vasculite e glomerulonefrite aguda. Ocorre inflamação do sítio de injeção do soro e pode haver angioedema. Ocasionalmente, há cefaléia, náuseas, vômitos e comprometimento neurológi-co (mononeurite). O acometimento cardíaco ou renal grave é incomum. É importante salientar que a dose de antígeno administrada é preditora da gravidade da doença do soro.
Os antígenos estranhos induzem a produção de anticorpos, levando à formação de imunocom-plexos circulantes. Esses complexos são formados com excesso de antígeno, pois a quantidade de anticorpos formados é muito inferior aos antí-genos, dificultando a fixação do complemento. Além disso, o tamanho pequeno ou intermediário dos complexos circulantes dificulta a sua elimina-
ção pelas células do SFM. Como conseqüência, há forte tendência desses imunocomplexos à depo-sição tecidual. Entretanto, uma vez depositados na membrana basal de vasos, os complexos acu-mulam-se formando agregados de anticorpos e adquirem a capacidade de ativar o sistema com-plemento, desencadeando a reação inflamatória (Tabela 17.2). Esse processo resulta na queda dos níveis totais de complemento sérico.
Os achados laboratoriais são: leve leucocitose; aumento do número de plasmócitos na medula óssea (podendo ser encontrados no sangue); eo-sinofilia; aumento da velocidade de eritrossedi-mentação ou de sedimentação globular (VSG); presença de imunocomplexos circulantes e dimi-nuição do complemento.
O diagnóstico é clínico, com história compatível e sinais e sintomas típicos. A dosagem do comple-mento e a detecção de anticorpos séricos IgG e IgE por imunofluorescência são ferramentas auxiliares utilizadas na confirmação diagnóstica.
A doença do soro é benigna e tem bom prog-nóstico, pois a quantidade de anticorpos formados aumenta e facilita a eliminação dos imunocomple-xos pelo sistema fagocitário (Figura 17.3). Raramen-te ocorrem complicações, sendo a principal delas a obstrução respiratória por edema de laringe.
Pode ocorrer uma forma crônica de doença do soro mediante administração repetida de baixas doses de antígenos. Sua cronicidade se deve à ele-vada concentração antigênica que estimula conti-nuamente a formação de anticorpos.
Tabela 17.1. Exemplos de doenças de hipersensibilidade tipo III
Doença Composição do imunocomplexo Tipo de deposição Local de deposição
Doença do soro Excesso de antígeno Generalizada Arteríolas, glomérulos renais e articulações
Reação de Arthus Excesso de anticorpos Localizada Arteríolas da derme
Doenças auto-imunes Auto-anticorpos e antígenos endógenos Generalizada Arteríolas, glomérulos renais e articulações
Tabela 17.2. Reação inflamatória da doença do soro
Reação inicial Reação tardia
Ag e Ac combinam-se formando complexos imunes Complexos Ag-Ac depositam-se na parede dos vasos sangüíneos
Complexos Ag-Ac ativam o complemento e promovem a liberação de histamina por basófilos e plaquetas
Complexos Ag-Ac ativam o complemento, estimulam plaquetas a formar microtrombos, atraem neutrófilos e estimulam a liberação de enzimas lisossômicas
Aumenta a permeabilidade vascular, e a resposta inflamatória se inicia Dano sustentado na parede dos vasos sangüíneos

201Reações de hipersensibilidade tipo III
da ligação antígeno-anticorpo, surgem os fenô-menos inflamatórios característicos da hipersen-sibilidade tipo III. A lesão vascular resultante é mediada pela ativação do complemento, segui-da de recrutamento e estimulação celular.
Na reação de Arthus, a ativação de neutrófi-los é mais efetiva, uma vez que a liberação de enzimas lisossômicas e radicais livres de oxigê-nio por essas células leva à necrose fibrinóide da parede dos vasos, podendo produzir hemorra-gia. A ativação plaquetária que ocorre por meio da liberação de FAP pelos mastócitos e basófilos ou diretamente por imunocomplexos também é mais pronunciada, causando trombos plaque-tários com neutrófilos nos vasos da derme e de-terminando uma vasculite seguida de isquemia e necrose tecidual. Ocorre, assim, um desequilí-brio na coagulação com uma concentração de plaquetas nos trombos dos pequenos vasos. Es-sas alterações da hemostasia, com o aumento da permeabilidade vascular, causam o edema e a hemorragia (Figura 17.4).
As manifestações clínicas da reação de Arthus são os sinais típicos de inflamação: edema inten-so, eritema, calor e dor à palpação, somados à necrose e hemorragia, surgindo em poucas horas, com pico entre quatro e dez horas após a inocu-lação do antígeno e localizando-se no seu sítio de injeção na pele. Além dessa forma clássica, pode ocorrer uma forma leve de reação de Arthus ca-racterizada por leve eritema e endurecimento no local da injeção. Essa forma freqüentemente está associada à terapia de dessensibilização a alérge-nos, na qual os níveis de anticorpos são insuficien-tes para causar a forma clássica.
A ocorrência de necrose hemorrágica no sítio de injeção de uma droga ou no local da picada de um inseto pode sugerir uma reação de Arthus, mas a distinção com uma reação tóxica ou uma infecção secundária requer evidências laboratoriais.
O prognóstico da reação de Arthus é bom, havendo regressão da resposta inflamatória em poucos dias. Essa limitação inflamatória é resul-tante da migração dos macrófagos ao sítio teci-dual. Estes substituem os neutrófilos em 12 a 24 horas e são capazes de eliminar os imunocom-plexos circulantes.
Assim como a reação de Arthus, a anafilaxia (reação de hipersensibilidade tipo I) também pode ocorrer após a inoculação intradérmica de um an-tígeno. Por isso, é importante destacar as diferen-ças entre essas duas doenças (Tabela 17.3).
Figura 17.3. Correlação temporal entre as lesões e os níveis de Ag, Ac e complemento na doença do soro.
Complemento
Antígenos
Complexos imunes
Lesões teciduais
Anticorpo livre
2 4 6 8 10 12 14 16 18 20 22 24Dias após injeção do antígeno
Nív
el s
éric
o d
e re
agen
tes
(Esc
ala
LOG
)
A prática atual de utilização de antibioticote-rapia em altas doses propiciou o surgimento de uma doença iatrogênica clinicamente semelhante à doença do soro clássica. Todavia, tal patologia caracteriza-se por uma apresentação clínica mais branda e não se acompanha de diminuição dos níveis séricos de complemento. Trata-se de uma doença imunológica reacional às drogas admi-nistradas, na qual ocorre a formação de anticor-pos contra haptenos efetivos, como antibióticos β-lactâmicos e sulfonamidas, após a conjugação espontânea destes com proteínas carreadoras.
Reação de Arthus A reação de Arthus refere-se a um modelo expe-rimental em que uma lesão localizada é induzi-da por imunocomplexos. Essa reação foi descrita por Maurice Arthus, em 1903, quando observou que injeções intradérmicas de um antígeno pro-téico em coelhos hiperimunizados levava a um processo inflamatório local que progredia para uma lesão necrótica hemorrágica e ulceração. Essa reação caracterizada por inflamação cutâ-nea localizada se dava em resposta à deposição de imunocomplexos em vasos sangüíneos da der-me com conseqüente vasculite aguda e necrose tecidual. Tal reação é rara em humanos e ocorre apenas quando há altos níveis séricos de anticor-pos, principalmente IgG.
À medida que o antígeno injetado na derme migra em direção à parede vascular, ele se liga aos anticorpos, e imunocomplexos grandes são formados na parede dos vasos dérmicos. A partir

202 Imunologia Básica e Aplicada
Figura 17.4. Etapas da reação de Arthus. 1. Injeção do antígeno. 2. Reação antígeno-anticorpo. 3. Anafilaxia/quimiotaxia. 4. Liberação de enzimas lisossômicas. 5. Necrose fibrinóide. 6. Cicatrização ou hemorragia ou resolução.
Tabela 17.3. Diferenças entre reação anafilática e reação de Arthus
Reação anafilática Reação de Arthus
Reação imediata e fugaz Reação lenta, requer horas (4-10h) para alcançar seu máximo e dias para desaparecer
Apresenta anticorpo fixo a células e a reação pode ocorrer na ausência de anticorpos circulantes
Não necessita de anticorpos fixados em tecidos e a reação não se processa na ausência de anticorpos circulantes
Não necessita de anticorpos precipitantes Necessita de anticorpos precipitantes
Não requer complemento Requer complemento
As alterações vasculares são causadas por mediadores solúveis, não havendo formação e precipitação de imunocomplexos na parede dos vasos acometidos
As alterações vasculares são desencadeadas pela formação e precipitação de imunocomplexos na parede dos vasos comprometidos
O fenômeno principal é o aumento da permeabilidade capilar, com intensidade moderada
Apresenta edema intenso, hemorragia, infiltrado celular, principalmente neutrófilos e, nos casos mais graves, isquemia com necrose e perda de tecido na área afetada
Necessita de sensibilização prévia Necessita de imunização, mas não sensibilização prévia
Reação antígeno-anticorpo
Injeção de antígeno Resolução
Anafilaxia e quimiotaxia
Hemorragia
Necrose fibrinóide
Cicatrização (endarterite)
Pele
Liberação de enzimas lisossômicas pelos neutrófilos
+ Complemento
1
2
3
4
5
Doenças auto-imunesNas patologias auto-imunes, ocorre a produção contínua de auto-anticorpos contra antígenos próprios do organismo, gerando uma prolongada formação de imunocomplexos. Isso ocasiona uma
sobrecarga do sistema fagocítico mononuclear,
eritrócitos e sistema complemento, responsáveis
pela remoção dos complexos. São exemplos o lú-
pus eritematoso sistêmico, a artrite reumatóide, a
poliarterite nodosa e outras vasculites graves.
6

203Reações de hipersensibilidade tipo III
Glomerulonefrite pós-infecciosa aguda
A glomerulonefrite pós-infecciosa aguda é uma complicação que ocorre, com maior freqüên-cia, por determinadas cepas de estreptococo β-hemolítico do grupo A denominadas nefri-togênicas por causarem síndrome nefrítica. Em alguns casos, pode estar relacionada a infecções estafilocócicas (por exemplo, endocardite estafi-locócica aguda) e, mais raramente, é resultante de infecções virais (por exemplo, hepatite B) ou parasitárias (por exemplo, malária).
O pico de incidência dessa patologia ocor-re na infância, dos 6 aos 10 anos de idade, mas pode acometer indivíduos de qualquer faixa etá-ria, estando os casos mais graves entre os adul-tos. No inverno, é freqüentemente observada após faringite estreptocócica, embora possa ser conseqüente a infecções em outros sítios, como a pele (impetigo). Nos países desenvolvidos, sua incidência vem diminuindo, porém ainda é uma das doenças renais mais comuns da infância.
A glomerulonefrite pós-infecciosa é uma doen-ça mediada por imunocomplexos e caracteriza-se por uma reação inflamatória do parênquima renal desencadeada pela deposição de comple-xos imunes formados com antígenos do micror-ganismo (geralmente estreptocócico). Existe um período de latência de 6 a 21 dias entre a infec-ção e o surgimento dos sintomas de síndrome nefrítica, sendo esse período correspondente ao intervalo de tempo necessário para a produção de anticorpos específicos.
Os complexos imunológicos depositados nos glomérulos renais ativam o sistema complemento e iniciam uma resposta inflamatória. A ativação é tão intensa que mais de 90% dos pacientes desenvolvem hipocomplementemia devido ao consumo dentro dos glomérulos. A microscopia óptica revela glomérulos muito aumentados, com espaço de Bowman comprimido, infiltração difusa de monócitos e neutrófilos e proliferação de células endoteliais, mesangiais e epiteliais, o que determina a oclusão da luz dos capilares. Na imunofluorescência, são observados depósi-tos granulares focais e esparsos de IgG, IgM e C3 no mesângio e ao longo da membrana basal. Os aglomerados de imunocomplexos podem ser vis-tos à microscopia eletrônica como depósitos dis-cretos, amorfos e densos, com aparência seme-
lhante a corcovas de camelo (humps) e situados na superfície epitelial da membrana basal.
O quadro clínico da glomerulonefrite pós-in-fecciosa caracteriza-se por mal-estar, febre, náu-seas, oligúria, hematúria, edema periorbitário e hipertensão leve à moderada. O exame qualita-tivo de urina revela proteinúria e presença de cilindros hemáticos e leucocitários. Há aumento dos títulos séricos de anticorpos específicos para antígenos estreptocócicos como antiestreptolisi-na, ASLO (antistreptolysin O – antiestreptolisina O), anti-hialuronidase (AHT), antiestreptoquina-se e anti-DNAse. A diminuição do C3 sérico du-rante a síndrome aguda, retornando ao normal com uma a duas semanas, é típica.
O diagnóstico é realizado na presença de uma história clínica clássica e achados labora-toriais compatíveis. O prognóstico é bom, ocor-rendo resolução espontânea da nefrite em 95% dos casos. Contudo, a hematúria e a proteinúria podem persistir durante alguns meses. Alguns pacientes podem apresentar alterações no se-dimento urinário durante anos após a infecção. Nos outros 5%, desenvolve-se a glomerulonefri-te rapidamente progressiva.
Pneumonite por hipersensibilidade
A pneumonite por hipersensibilidade, também chamada de alveolite alérgica extrínseca, com-preende um grupo de distúrbios pulmonares que se desenvolvem devido a uma deposição lo-calizada de complexos imunes. A hipersensibili-dade é uma conseqüência, na maioria das vezes, da inalação de poeiras orgânicas contendo antí-genos formados por esporos de bactérias termo-fílicas, fungos verdadeiros, proteínas animais ou produtos bacterianos.
A doença pulmonar do fazendeiro, por exem-plo, é uma pneumonite causada por inalação de poeiras geradas pelo feno mofado, que permite a rápida proliferação de actinomicetos termofíli-cos. Nos indivíduos sensibilizados por exposição repetida a esses antígenos, a presença de grande quantidade de anticorpos específicos leva à for-mação de imunocomplexos, que se depositam nos capilares dos alvéolos. A partir do depósito, tem início uma reação inflamatória local. Os pacientes acometidos apresentam tosse e dispnéia intensa algumas horas após a inalação do antígeno.
A doença do criador de pombos (também de-nominada de doença do criador de aves), pro-

204 Imunologia Básica e Aplicada
vocada por proteínas do soro, excretas ou penas de aves, tem uma patogenia semelhante. Esses exemplos de doenças patologicamente similares não esgotam o conjunto chamado pneumonite de hipersensibilidade. É importante ressaltar que a resposta imunológica aos antígenos extrínsecos envolve tanto reações de imunocomplexos como reações de hipersensibilidade do tipo IV.
As manifestações clínicas são variadas e carac-terizam-se por ataques agudos e recorrentes de febre, dispnéia, tosse e leucocitose. A radiografi a de tórax com infi ltrados nodulares e difusos e os testes de função pulmonar demonstrando um dis-túrbio restritivo agudo podem auxiliar no diag-nóstico. O reconhecimento precoce dessas do-enças no início do curso clínico é extremamente importante, uma vez que a progressão para uma doença pulmonar fi brótica crônica pode ser evita-da por meio da remoção do agente ambiental.
Doença de Berger
A doença de Berger ou nefropatia por IgA é uma glomerulonefrite causada pelo acúmulo de imunocomplexos compostos predominan-temente de IgA. Os mecanismos precisos de acúmulo e os antígenos envolvidos ainda não são conhecidos. Entretanto foram detectados
níveis sangüíneos elevados e complexos imunes formados por IgA nos pacientes acometidos por essa doença descrita primeiramente por Jean Berger. Acredita-se que esses indivíduos produ-zam alta quantidade de IgA, que poderia não ser tão efi ciente quando expostos a uma infec-ção dos tratos respiratório ou gastrintestinal. Assim, os anticorpos se combinariam com os mi-crorganismos infectantes (antígenos) e dariam origem à grande quantidade de complexos que se depositam nos glomérulos. Estudos têm su-gerido que os imunocomplexos contendo IgA dentro do mesângio ativam o complemento por meio da via alternativa.
A doença de Berger é a forma mais comum de glomerulonefrite no mundo e apresenta maior incidência em países asiáticos. É mais comum em homens jovens, com um pico de idade entre 15 e 30 anos. As manifestações clínicas são variáveis e incluem hematúria microscópica assintomática, hematúria macroscópica intermitente, síndrome nefrótica e insufi ciência renal.
A resolução completa é rara, sendo mais fre-qüente um curso clínico marcado por perío dos de remissão agudizados por infecções do trato respiratório. Cerca de 20% dos pacientes evo-luem lentamente para a insufi ciência renal ter-minal após dez anos.
BibliografiaAbbas AK, Lichtman AH. Diseases caused by immune responses: hypersensitivity.
In: Abbas AK, Lichtman AH. Cellular and Molecular Immunology. 5.ed. New York: W. B. Saunders, 2003. pp. 411-31.
Burtis C, Ashwood ER. Tietz textbook of clinical chemistry. 3a ed. W.B.- Saunders Company, 1999, pp. 94-112 e pp. 205-24.
Chapel H, Haeney M, Misbah S, Snowden N. Imunologia para o clínico. 4a ed. Revinter, 2003, pp. 314-32.
Cotran RS, Kumar V, Robbins SL. O Rim. In: Bases patológicas das doenças. 7.ed. Rio de Janeiro: Guanabara Koogan, 2005. pp. 999-1066.
Fernández N, Jancar S, Sánchez CM. Blood and endothelium in immune com-plex-mediated tissue injury. In: Trends in Pharmacological Sciences 2004; (10)25:512-17.
Ferreira AW, Ávila SLM. Diagnóstico laboratorial das principais doenças infeccio-sas e auto-imunes. 2ª ed. Guanabara Koogan, 2001, pp. 1-8 e 9-48.
Goldsby RA, Kindt TJ, Osborne BA. Imunologia – Kuby. 4a ed. Revinter, 2002, pp. 149-68.
Gorczynski R, Stanley J. Imunologia clínica. 1a ed. Reichmann & Affonso Editores, 2001, pp. 186-201.
Henry JB. Diagnósticos clínicos e tratamento por métodos laboratoriais. 19ª ed. São Paulo, Manole Ltda., 1999, pp.851-76.
Miller LE, Ludke HR, Peacock JE, Tomar RH. Manual of laboratory immunology. 2ª ed. Lea & Febiger, 1991, pp. 31-70.
Rich RR, Fleisher TA, Schwartz BD, Shearer WT, Strober W. Clinical immu-nology – Principles and Practice. 1a ed. Mosby – Year Book, Inc, 1996, pp. 2083-219
Rose NR, Macário EC, Folds JD, Lane HC, Nakamura RM. Manual of clinical labora-tory immunology. 6ª ed. American Society for Microbiology, 2001, pp. 1-74.
Scroferneker ML, Pohlmann PR. Imunologia Básica e Aplicada. 1a ed. Sagra- Luz-zaro, 1998, pp. 238-64.
Sheehan C. Clinical immunology – Principles and laboratory diagnosis. 2a ed. 1997, pp. 115-202.
Stites DP, Terr AI, Parslow TG. Medical immunology. 9a ed. Appleton & Lange, 1997, pp. 211-320.

C A P Í T U L O 1 8
Introdução A reação de hipersensibilidade tipo IV é também conhecida como reação de hipersensibilidade tardia (RHT) e caracteriza-se pela utilização da imunidade celular (linfócitos T) como via efetora. Assim, ela não pode ser transferida de uma pessoa sensibilizada para outra não-sensibilizada através de anticorpos séricos. Representa uma imunidade protetora contra patógenos intracelula-res, entretanto, se exagerada, pode gerar efeitos deletérios, atacando célu-las do próprio organismo.
Existem dois tipos básicos de linfócitos: os linfócitos B, responsáveis pela imunidade humoral por meio da produção de anticorpos, e os linfócitos T, responsáveis pela imunidade celular. Estes últimos subdividem-se em linfóci-tos TH CD4+, que podem ser TH0, 1 ou 2, linfócitos TC-CD8+ (ou citotóxicos) e TS-CD8- (ou supressores).
A reação de hipersensibilidade tardia é dividida em três fases: sensibiliza-ção, fase efetora (ou infl amatória) e resolução. Para que ocorra o desfecho clínico, é necessária uma sensibilização prévia dos linfócitos TH.
O passo central para o desenvolvimento da hipersensibilidade tipo IV é a apresentação antigênica na qual os LTH0 se transformam em TH1. O IFN-γ é o mediador da RHT, aumentando a ativação fagocítica dos macrófagos e a expressão de MHC-II, secretando ainda TNF, IL-1 e IL-12. O TNF aumenta a secreção de prostaciclina (vasodilatador), IL-8 e PE selectina (moléculas de ade-são de linfócitos e monócitos). A IL-12 é produzida pelos macrófagos e células dendríticas durante a apresentação antigênica, induzindo a diferenciação de LTH0 em LTH1, além de induzir a secreção de IFN-γ pelos LT e células NK.
Existem três tipos básicos de reações de hipersensibilidade tardia: de contato (48 a 72 horas), tuberculínica (48 a 72 horas) e granulomatosa (21 a 28 dias). As reações iniciam-se quando antígenos são aplicados diretamente na pele ou inje-tados via intradérmica, ativando de formas diferentes o sistema imunológico.
Dermatite de contatoA dermatite de contato é uma reação geralmente eczematosa e pruriginosa que pode ocorrer em qualquer região do corpo, embora seja mais freqüente em mãos, pés e face – áreas associadas com exposição ocupacional, uso de sapatos e maquiagens, respectivamente. A exposição ocupacional destaca-se entre donas-de-casa que utilizam detergentes e sabões em pó, trabalhadores que manejam comidas cruas (especialmente frutos-do-mar), látex e couro.
Júlia Rostirolla Schmitt • Maria Lúcia Scroferneker
Reações de hipersensibilidade tipo IV

206 Imunologia Básica e Aplicada
A reação de hipersensibilidade tardia tipo der-matite de contato se inicia quando haptenos pe-netram na pele e ligam-se a proteínas plasmáti-cas presentes nos capilares da derme, que atuam como carreadores. Ocorre a seguir a internaliza-ção do complexo hapteno-proteína pelas células dendríticas, as quais expressam CD1 e MHC-II, atuando como células apresentadoras de antígeno (APC), e possuem receptores para a fração Fc dos anticorpos e complemento. Estas migram para a região paracortical dos linfonodos regionais e ali apresentam o complexo hapteno-proteína para o linfócito T CD4+, formando linfócitos T CD4+ de memória. Durante esse processo, os queratinóci-tos induzidos por haptenos liberam citocinas in-flamatórias como IL-1, fator de necrose tumoral α (TNF-α) e fator estimulante de colônias de macró-fagos e granulócitos (GM-CSF), ativando as células dendríticas que produzem IL-1ß, IL-6, IL-12 e indu-zindo sua migração juntamente com o complexo hapteno-carreador aos linfonodos, aumentando a expressão do MHC-I e II, moléculas de adesão, e amplificando a apresentação antigênica.
Essa primeira fase é conhecida como sensibi-lização e possui duração média de 10 a 14 dias (Figura 18.1).
Apesar de a hipersensibilidade de contato (HC) fazer parte do grupo das RHT, estudos recentes
apontam diferenças nas suas fisiopatogenias. A RHT é mediada por MHC-II e LTH, mas dados re-centes apontam para a HC mediada por MHC-I e LT-citotóxicos, tendo os LTH efeito supressor na HC, e o contrário se sucedendo na RHT. Um fato que suporta a hipótese é o de que pacientes si-déticos em estágio terminal podem desenvolver dermatite de contato, mas não RHT tipo tuber-culínica. Sabe-se também que as células dendrí-ticas não são as únicas APCs que podem realizar a apresentação antigênica, há também os que-ratinócitos, os mastócitos da derme e as células endoteliais, embora esses o façam via MHC-I e linfócitos T-citotóxicos.
Ainda sobre a fase de sensibilização, experi-mentos mostram a capacidade dos haptenos de induzir a produção de LT-citotóxicos e supressor e que altas doses de hapteno produzem ambos os tipos celulares, suprimindo a HC, enquanto baixas doses deste estimulam apenas LT-citotóxicos.
A fase efetora é a fase de recrutamento de células inflamatórias para o local de aplicação do hapteno na pele. Ocorre quando o indiví-duo previamente sensibilizado entra novamen-te em contato com o antígeno. Forma-se nova-mente o complexo hapteno-proteína, o qual é internalizado pelas células dendríticas que saem da epiderme e levam o complexo até a derme,
Figura 18.1. Fases da RHT: o complexo hapteno-carreador é processado pelas células de Langerhans e apresentado para os linfócitos TH que, por sua vez, ativarão os mecanismos efetores da resposta imune.
Hapteno Carreador
Tempo
Células de Langerhans
IFN-γ Citocinas pró-inflamatórias
Arteríola
Vênula
Linfático

207Reações de hipersensibilidade tipo IV
onde é apresentado aos LTH de memória. Esses liberam interferon γ (IFN-γ) que induz a expres-são de ICAM-1 e MHC-II na membrana dos que-ratinócitos e células endoteliais dos capilares e induz a produção de IL-1, IL-6 e GM-CSF pelos queratinócitos. Além disso, também liberam his-tamina, serotonina, óxido nítrico e prostaglan-dinas (principalmente PGE), os quais ativam o endotélio local e atraem leucócitos. Desse modo, amplifica-se a resposta inflamatória e, em 4 a 8 horas, células mononucleares (linfócitos TH não-específicos e poucos LT-citotóxicos) infiltram ca-pilares e a epiderme. Os LTH de memória ainda, através da secreção de IFN-γ, ativam macrófagos aumentando a expressão de moléculas co-esti-mulatórias e secretam IL-2 que realiza a expan-são clonal de LT específicos. Em 48 horas, che-gam os macrófagos e, de 48 a 72 horas, inicia-se a fase de resolução com as respostas inibitórias através da produção de prostaglandina E (PGE), TGF-β pelos mastócitos da derme e a degradação do complexo hapteno-proteína-LTH. O peptídeo associado ao gene da calcitonina e o hormônio melanócito-estimulante também participam da fase de resolução.
A IL-1 inibe o metabolismo dos macrófagos e a IL-10 diminui a expressão de MHC-II, a produ-ção de citocinas e a proliferação clonal de LTH1. Células dendríticas podem, ainda, ser inativadas pelos raios ultravioleta, razão pela qual as der-matites de contato costumam melhorar após a exposição solar. Parece que a IL-4 possui efeito supressor na dermatite de contato com vias efe-toras não bem estabelecidas.
A heterogeneidade dos resultados experimen-tais na dermatite de contato pode-se dever aos diferentes tipos de respostas (LTH, LT-citotóxico ou LT-supressor) de acordo com os diferentes ti-pos de imunógenos. Além disso, o contato com alérgenos nem sempre resulta em dermatite de contato em indivíduos sensibilizados e determi-nadas áreas da pele não são afetadas apesar do contato, o que sugere que outros fatores, além do alérgeno específico, são necessários para o desencadeamento da resposta efetora na HC.
TuberculínicaO exemplo clássico de RHT tuberculínica é o teste de Mantoux ou PPD.
A tuberculose é uma das principais causas de morte por infecção no mundo, e o número de pa-
cientes com a doença ativa aumentou significati-vamente nos últimos anos devido à concomitância da infecção HIV-tuberculose e ao desenvolvimento de microrganismos resistentes a múltiplas drogas. A doença é causada pelo Mycobacterium tubercu-losis e a estimativa é de que 20% a 43% da popu-lação mundial seja portadora assintomática.
A vacina BCG (extrato do Mycobacterium bo-vis) protege contra as formas graves da doença, mas não previne a primo-infecção.
Noventa por cento dos indivíduos infectados permanecem com infecção latente e não desen-volvem sintomas da doença, entretanto situa-ções que perturbem o status imunológico do in-divíduo podem propiciar a reativação, que ocor-re em menos de 10% dos casos, como: infecção pelo HIV, desnutrição, idade avançada, uso de medicações imunossupressoras (corticosteróides, terapia anti-TNF), gastrectomia, silicose e diabe-tes melito.
O screening para indivíduos de baixo risco para adquirir a infecção não é recomendado. O teste de Mantoux é o método de escolha para o rastrea-mento, porém não faz distinção entre doença ativa e passada. Consiste num teste cutâneo com 0,1 ml de derivado protéico purificado (PPD), contendo cinco unidades de tuberculina injetadas intrader-micamente na face anterior do antebraço. A me-dida transversa da induração (indica sensibilização prévia de LT de memória) na pele deve ser mensu-rada após 48 a 72 horas por um técnico experiente. Em geral, leva de 2 a 10 semanas após a infecção para que haja a conversão cutânea do teste. Falso-positivos no Mantoux ocorrem em indivíduos pre-viamente vacinados e naqueles com infecção por outras micobactérias. Falso-negativos resultam de má técnica, infecções concomitantes, desnutrição, idade avançada, desordens imunológicas, vacinas feitas com vírus, neoplasias de tecido linfóide, te-rapia com corticosteróides, estresse agudo, insufi-ciência renal crônica, infecção pelo HIV e tubercu-lose fulminante (Tabela18.1).
A resposta imune inicia quando o M. tubercu-losis chega ao espaço alveolar e encontra macrófa-gos, sendo internalizado e levado aos linfonodos regionais, onde é apresentado aos linfócitos TH e T-citotóxicos. Há uma hipótese alternativa na qual o Mycobacterium causaria erosão direta do epitélio pulmonar com disseminação para o interstício e ca-pilares, tendo os macrófagos papel de carreadores do bacilo até o tecido linfático associado aos brôn-quios (Balt). Na hipótese clássica, há a liberação de

208 Imunologia Básica e Aplicada
Tabela 18.1. Classificação do CDC para teste tuberculínico positivo
Tamanho da induração
5 mm1. Indivíduos HIV-positivos2. Contato recente com indivíduos com tuberculose ativa3. Pacientes com alterações fibróticas à radiografia sugestivas
de tuberculose prévia4. Pacientes transplantados e com outras imunossupressões
(recebendo o equivalente a > 15 mg/d de prednisona por um mês ou mais)
10 mm1. Imigrantes recentes (< 5 anos) de países com alta prevalência
de tuberculose (como Ásia, África, América Latina)2. Usuários de drogas endovenosas HIV-negativos3. Trabalhadores de laboratórios de micobacteriologia4. Moradores e trabalhadores de instituições penitenciárias; asilos;
hospitais e outras instituições de saúde; casas de abrigo para pacientes com SIDA e indigentes
5. Pessoas com as seguintes condições médicas que aumentam o risco para tuberculose: gastrectomia, 10% abaixo do peso ideal, bypass jejunoileal, diabetes melito, silicose, insuficiência renal crônica, algumas doenças hematológicas (como leucemias, linfomas), e outras neoplasias malignas (como carcinoma de cabeça ou pescoço e pulmão)
6. Crianças < 4 anos ou crianças e adolescentes expostos a adultos de grande risco
15 mm1. Pessoas sem fatores de risco para tuberculose
Observação 1: uma reação tuberculínica é considerada positiva se o diâmetro transverso da área indurada alcança o tamanho requerido para o grupo específico. Todas as outras reações são consideradas negativasObservação 2: pessoas que estão sob alto risco e são testadas ao ingressar num emprego, uma induração > 15 mm é considerada positiva
Adaptado de: Screening for tuberculosis and tuberculosis infection in high-risk populations: recommendations of the Advisory Council for the Elimination of Tuberculosis. MMWR Morb Mortal Wkly Rep 1995; 44(RR-11):19.
linfócitos T e B, células dendríticas, macrófagos e fibroblastos migram para a região pulmonar in-fectada, culminando na posterior formação do granuloma. Sabe-se que o TNF, produzido por ma-crófagos ativados, tem atividade antimicrobiana essencial (reguladora da expressão de quimiocinas e seus receptores, moléculas de adesão, modulação da migração celular), mantendo a resposta granu-lomatosa em sua forma latente. Monócitos são a maioria do infiltrado celular (80% a 90%) local, e a proporção de LTH e LT-citotóxicos é de 2:1.
Na RHT tuberculínica, os macrófagos são a principal APC, embora também se encontrem cé-lulas dendríticas. Os primeiros têm papel central, pois são as células efetoras responsáveis pela morte das bactérias intracelulares, mas também o hábitat onde reside o bacilo resistente. O ma-crófago, após a ativação por citocinas, reduz o pH intrafagossomal e expõe o bacilo ao oxigênio e seus radicais livres.
A lesão tuberculínica dura geralmente 5 a 7 dias, mas, se houver persistência da micobacté-ria, pode-se desenvolver uma reação granuloma-tosa. Há diversos mecanismos imunológicos para a persistência bacilífera:
a) atenuação do processamento antigênico;b) impedimento da maturação do fagossomo;c) seqüestro intracelular de dímeros do MHC-II;d) inibição da expressão do gene indutor de IFN e do ativador do MHC-II;e) transformação dos radicais livres de O2 em metabólitos menos tóxicos;f) impedimento da fusão dos macrófagos con-
tendo o Mycobacterium com os lisossomos.
Experimentos em animais sugerem que o au-mento da IL-10 (principalmente produzida pelos macrófagos) no sítio da infecção conjuntamente com a diminuição do TNF sistêmico pode ser o evento-chave na reativação da doença.
Parece que a produção precoce de IFN-γ pelo LTH pode determinar o desfecho da infecção, e que os LTH possuem outras funções protetoras contra o M.tuberculosis como a apoptose de macrófagos infectados via Fas-Fas ligante. Em indivíduos HIV positivos, a depleção de LTH diminui a produção de IFN-γ com diminuição da atividade macrofágica e reativação da infecção. Caso haja excessiva multi-plicação bacilífera, há perda da estrutura granulo-matosa, necrose e cavitação (Figura 18.2).
IL-1 pelos macrófagos, a qual induz a produção de IL-2 nos LT, realizando a expansão clonal de linfó-citos específicos e a liberação de IL-12 pelos macró-fagos, estimulando a secreção de IFN-γ pelos LTH, amplificando a atividade macrofágica para a elimi-nação do Mycobacterium intracelular e produzin-do TNF-α e mais IL-1. Citocinas pró-inflamatórias agem nas células endoteliais dos vasos sangüíneos da derme e induzem a expressão de moléculas de adesão E-selectina, ICAM-1 e VCAM-1. Estas ligam-se em receptores nos leucócitos (integrinas LFA-1 e VFA-1), recrutando-os para o local da inflama-ção. Em quatro horas, chegam os neutrófilos, em 12 horas monócitos e LT. Esse infiltrado rompe as fibras de colágeno e tem seu pico em 48 horas com a expressão de MHC-II nos queratinócitos. Assim,

209Reações de hipersensibilidade tipo IV
GranulomatosaA RHT granulomatosa ocorre devido à persistência de microrganismos ou partículas não destruídas no interior dos macrófagos após uma reação tipo tuberculínica. Como resultado, o infiltrado peri-vascular linfocítico é substituído por macrófagos, os quais sofrem transformação morfológica e for-mam os granulomas imunológicos ou de corpo estranho. O primeiro é formado por células epite-lióides características (macrófagos ativados croni-camente, com abundante retículo endoplasmático e secretores de TNF, amplificando a resposta), as quais podem fundir-se e formar células gigantes multinucleadas (células de Langhans), cercadas, ainda, por linfócitos e fibroblastos.
Granulomas de corpo estranho podem ser de-sencadeados por materiais como a sílica, o talco (associado a abuso de drogas endovenosas) e os fios de suturas e diferem do tipo imunológico pela ausência de linfócitos.
Granulomas imunológicos estão presentes na tuberculose, esquistossomose, histoplasmose, goma sifilítica, doença de Crohn, reações a zircônio, berí-lio, de Kveim na sarcoidose e a de Mitsuda para han-seníase, entre outros. A principal doença que exem-plifica a reação granulomatosa é a hanseníase.
A hanseníase é uma infecção crônica causada pelo Mycobacterium leprae, parasita intracelular obrigatório, de multiplicação lenta e sem cultu-ra in vitro reproduzível. Permanece viável vários
dias ex vivo. A doença é mais comum em países pobres, de clima tropical, no sexo masculino. A prevalência mundial é de 1,25:10.000 habitantes e países como Índia, Brasil, Burma, Indonésia, Madagáscar e Nepal são os mais afetados, con-centrando 83% dos casos registrados.
Reservatórios não humanos são ratos, tatus e macacos. A transmissão ocorre principalmente pelo aerossol da secreção nasal que penetra na mucosa respiratória, não atravessando pele intacta. O prin-cipal fator de risco determinante da transmissão é a proximidade de pacientes com a doença – con-tactantes domésticos têm risco bastante aumenta-do de adquirir a doença e devem receber profilaxia com 2 doses da vacina BCG. Sabe-se que o período de incubação varia de alguns meses até 30 anos. O M. leprae não secreta toxinas, sua virulência está associada às propriedades da parede celular. O an-ticorpo contra o ácido glicolipídico-fenólico I (an-tígeno da parede celular) reflete carga bacilífera, mas não confere imunidade.
A apresentação clínica varia conforme a res-posta imunológica do paciente de acordo com um espectro de doença que vai da forma tuberculói-de até a lepromatosa. O tipo tuberculóide possui máculas hipopigmentadas ou lesões eritematosas anestésicas com margens elevadas ou placas com centro tendendo a cura. Já o tipo lepromatoso, conhecido como hanseníase anérgica devido à irresponsividade do sistema imune, apresenta-se
Figura 18.2. Esquema das etapas do teste tuberculínico.
E-selectina
ICAM-I
VCAM-I
Capilar
TNF
Tuberculina
Epiderme
Derme
0 4 h 12 h 48 h
Inflamação e edema
Expressão de MHC-II

210 Imunologia Básica e Aplicada
com máculas, pápulas e nódulos eritematosos dis-seminados, sem granulomas organizados, com es-pessamento da epiderme, tendência a cronicidade e complicações em longo prazo. A resposta imune celular é fundamental na defesa contra o patóge-no e determina a extensão da doença. A resposta imune celular vigorosa contra o Mycobacterium na forma tuberculóide limita a doença a poucas lesões (granulomas bem delimitados) com ativa-ção da via LTH1, secreção de IL-2 e INF-γ. O subtipo lepromatoso possui níveis baixos de IL-12 ou uma irresponsividade dos LT a esses níveis, reduzindo a resposta LTH1, assim predominando a via LTH2 com produção de IL-4, IL-5 e IL-10 e aumentando a resposta imune humoral. Outras vias da imuni-dade que contribuem no combate ao patógeno são a apresentação antigênica via CD1 e a da gra-nulolisina (proteína antimicrobiana expressa por linfócitos T na resposta imune tuberculóide).
Segundo a OMS, a hanseníase é classifi cada em paucibacilífera (cinco ou menos lesões na pele, sem bacilos nas amostras coletadas) e mul-tibacilífera (seis ou mais lesões com positividade ou não nas amostras).
O bacilo tem predileção pelas células de Schwamm dos nervos periféricos, mais especifi -camente pela laminina, onde se replica e forma imunocomplexos que são reconhecidos por LT. Assim, ocorre aumento da resposta infl amató-ria local e conseqüente mononeurite múltipla (neuropatia autonômica motora e sensorial). Os nervos mais acometidos são, em ordem de fre-qüência, os dos pés, mãos e rosto. A neurite, se não tratada, pode causar paralisias irreversíveis. A prevalência de neuropatia clinicamente detec-tável é de 10% na doença paucibacilífera e 40% na multibacilífera. Podem, ainda, ser produzi-dos anticorpos, não protetores, os quais formam imunocomplexos com antígenos livres, podendo
desencadear o eritema nodoso, irite, orquite, dactilite, artrite, vasculite e glomerulonefrite.
O diagnóstico é clínico e baseado na positivida-de de um ou mais sinais cardinais da doença:
a) lesões hipopigmentadas ou eritematosas com sensibilidade diminuída;
b) espessamento de nervo periférico acom-panhado de alteração de sensibilidade;
c) bacilo álcool-ácido resistente em amostras de pele ou biópsia. Entretanto, o padrão-ouro para diagnóstico é a histologia.
A prevenção parcial da doença foi adquirida com a vacinação BCG (M. tuberculosis e leprae possuem antígenos comuns), entretanto, uma vacina DNA específi co custo-efetiva ainda não está disponível.
A reação de Mitsuda determina a reatividade de pacientes sensibilizados, estando associada à alta re-sistência à infecção quando positiva (Tabela 18.2).
BibliografiaAbulafia J, Vignale RA. Leprosy: accessory immune system as effector of infec-
tious, metabolic, and immunologic reactions. Internal Journal of Dermatology 2001; 40:673-87.
Brasil. Ministério da Saúde. Doenças infecciosas e parasitárias: Guia de bolso. Brasília: FUNASA, 2000.
Friedmann PS. Contact sensitisation and allergic contact dermatitis: immunobiological mechanisms. Toxicol Lett 2006; 162(1):49-54. Epub 2005.
Kumar V, Abbas AK, Fausto N. Robbins and Cotran: Pathologic Basis of Disease. 7a ed. Elsevier Saunders, Philadelphia, 2005.
Leprosy. The Lancet 2004; 363:1209-19.
Mark BJ, Slavin RG. Allergic contact dermatitis. Med Clin North Am 2006; 90(1):169-85.
McFadden J. Contact allergic reactions in patients with atopic eczema.Acta Derm Venereol Suppl (Stockh) 2005;(215):28-32.
Orme IM, Cooper AM. Cytokine/chemokine cascades in immunity to tuberculosis. Immunology Today 1999; 20:307-12.
Roitt I, Brostoff J, Male D. Immunology. 6a ed. Mosby, 2001.
Sasaki S, Takeshita F, Okuda K et al. Mycobacterium leprae and Leprosy: a Com-pendium. Microbiology Immunology 2001; (11)45:729-36.
Scroferneker ML, Pohlmann PR. Imunologia básica e aplicada. Sagra Luzzatto, Porto Alegre, 1998.
Seabrook TJ, Borron PJ, Dudler L, Hay JB, Young AJ. A novel mechanism of im-mune regulation: interferon-gamma regulates retention of CD4 T cells du-ring delayed type hypersensitivity. Immunology 2005; 116(2):184-92.
Tufariello JM, Chan J, Flynn JL. Latent tuberculosis: mechanisms of host and bacillus that contribute to persistent infection. The Lancet Infectious Diseases 2003; (9)3.
Tabela 18.2. Reação de Mitsuda
Estágio 1 (24 a 48 horas) ou reação de Fernandez (hipersensibilidade tardia tuberculínica por injeção do antígeno)Negativa: ausência de reação ou eritema sem edema ou com edema de diâmetro < 5 mmDuvidosa: eritema/edema de diâmetro entre 5 mm e 10 mmPositiva fraca: eritema/edema de diâmetro > 10 mm e < 15 mmPositiva moderada: eritema/edema de diâmetro entre 15 mm e < 20 mmPositiva forte: eritema/edema de diâmetro > 20 mm
Estágio 2 (2 a 4 semanas) ou reação de Mitsuda (mensuração do granuloma)Negativa: ausência eritema ou edemaDuvidosa: eritema/edema < 3 mmPositiva fraca: infiltrado com diâmetro ente 3 mm e 5 mmPositiva: infiltrado com diâmetro > 5 mmPositiva forte: ulceração

C A P Í T U L O 1 9
IntroduçãoO desenvolvimento das infecções virais, em função das características pró-prias de cada vírus e da resposta imune do hospedeiro, pode desencadear di-ferentes situações. Podem ocorrer a eliminação viral, a permanência do vírus no organismo nas infecções latentes ou, ainda, a persistência, com replicação viral contínua, mesmo na presença da resposta imune.
Os estudos da resposta imune a vírus têm permitido um avanço conside-rável no estudo da imunologia em geral, mas principalmente no desenvolvi-mento de vacinas e nas metodologias aplicadas no diagnóstico das infecções virais, como é o caso do vírus da imunodefi ciência humana, o HIV (human immunodefi ciency virus – vírus da imunodefi ciência humana).
Para melhor compreensão dos mecanismos envolvidos na resposta imune a vírus, abordaremos inicialmente alguns aspectos desse microrganismo que conferem certas particularidades à sua interação com o hospedeiro.
Características gerais das infecções viraisOs vírus são microrganismos obrigatoriamente intracelulares, pois necessi-tam da maquinaria enzimática e bioquímica da célula hospedeira, a fi m de produzir novas partículas virais.
Os vírus apresentam uma grande diversidade quanto à sua estrutura e complexidade genética. De maneira geral, podem apresentar-se como enve-lopados ou não, com simetria simples ou complexa, com DNA ou RNA como material genético, cujo tamanho pode variar de 1,7 kb (circovírus) a 235 kb (herpes-vírus).
A partícula viral completa apresenta o ácido nucléico (DNA ou RNA) en-volto em uma camada protéica morfologicamente defi nida, o capsídeo. Juntos, o ácido nucléico e o capsídeo formam o nucleocapsídeo, os quais podem estar recobertos pelo envelope formado por uma camada lipopro-téica (Figura 19.1). Projeções a partir do capsídeo ou do envelope são de-nominadas peplômeros ou espículas e estão relacionadas com a ligação do vírus ao receptor celular, com as características antigênicas ou mesmo com a capacidade patogênica do vírus.
Ubirajara Maciel da Costa • Ana Paula Ravazzolo
Imunidade a vírus

212 Imunologia Básica e Aplicada
Padrões de infecção viral
Infecção aguda Ocorre a infecção primária com altas taxas de re-plicação viral em um curto espaço de tempo (por exemplo, rotavírus). A evolução da doença é defi-nida pela morte do hospedeiro ou eliminação do vírus através da ação do sistema imune.
Infecção latente Após a entrada no organismo e uma primeira fase de replicação (aguda), o vírus persiste sob uma forma não-infecciosa que, de tempos em tempos, sofre reativação com liberação de partí-culas infecciosas (por exemplo, herpes-vírus).
Infecção crônicaQuando o vírus mantém uma contínua liberação de partículas virais ou está presente em tecidos-alvo do hospedeiro (por exemplo, vírus da hepa-tite C – HCV). A infecção crônica pode se estabe-lecer caso o sistema imune não consiga eliminar os vírus gerados durante a infecção aguda. Nas infecções crônicas, o quadro é semelhante ao observado na infecção aguda, mas parece haver aumento na escala de tempo. O vírus tenta man-ter seu ciclo de replicação enquanto o sistema imune mantém seus mecanismos de defesa cons-tantemente ativados.
Infecção lenta e crônica Nesse tipo de infecção, não ocorre a fase aguda ou esta passa despercebida. O vírus entra no or-ganismo, infecta as células-alvo e permanece em um estágio de “hibernação” (incubação) longo, até que se inicie a replicação com maior inten-sidade, levando às manifestações clínicas (por exemplo, HIV).
Fases da replicação viral
Adsorção É a ligação de uma proteína específica externa do vírus a uma porção constituinte da superfície celular ou receptor. Um receptor viral é definido como um componente da superfície celular em que o vírus se liga especificamente, e esta inte-ração (vírus-receptor) resulta na penetração do vírus na célula hospedeira. A ligação do vírus a receptores específicos determina o tropismo por um hospedeiro ou tipo celular (Tabela 19.1).
PenetraçãoAcontece quase instantaneamente após a adsor-ção e pode ocorrer pela translocação, na qual o vírus completo passa pela membrana da célula; endocitose, em que as partículas virais ficam dentro de vesículas no citoplasma celular (ade-novírus); ou pela fusão do envelope viral com a membrana celular (herpes simplex virus – HSV).
DesnudamentoÉ a liberação do genoma viral que ocorre antes do início da sua replicação. Para alguns vírus, essa etapa ocorre durante a penetração na célu-la, enquanto para outros é necessário que ocor-ra proteólise do envoltório viral para liberação do genoma.
Replicação gênica e expressão das proteínas viraisEm virtude da grande variedade na forma de apresentação do genoma (DNA/RNA, fita dupla ou simples, segmentado ou não-segmentado), diferentes vírus desenvolveram diferentes es-tratégias de replicação do material genômico
Figura 19.1. Representação esquemática das principais es-truturas de um vírus, neste caso envelopado.
Envelope
Espículas ou peplômeros
Ácido nucléico
Núcleo capsídeo
Capsídeo

213Imunidade a vírus
Figura 19.2. Representação da replicação viral, neste caso um vírus não envelopado: a) adsorção; b) penetração; c) desnudamento; d) replicação do genoma e expressão de proteínas virais; e) empacotamento e liberação das partículas virais.
Antígenos virais
EnvelopeB
C
D
E
e síntese de proteínas. De maneira geral, nessa fase, ocorrem a síntese de RNA mensageiro viral a síntese das proteínas virais e a produção das cópias do genoma viral por meio dos processos de transcrição e tradução.
Empacotamento e liberaçãoO empacotamento ocorre à medida que as pro-teínas estruturais vão sendo sintetizadas, asso-ciando-se e formando os capsômeros. Estes, por sua vez, vão rearranjando-se e gerando o cap-sídeo dentro do qual o genoma viral é empa-cotado. Nesse processo, podem estar envolvidas etapas de clivagem de algumas proteínas virais. Após a montagem da partícula viral, ocorre a liberação por brotamento (vírus envelopados), exocitose ou lise celular (Figura 19.2).
Mecanismos efetores antivirais
Resposta imune inataA importância da resposta inata ou inespecífica no controle das infecções virais vem sendo de-monstrada em modelos animais, como, por exem-plo, em camundongos transgênicos cuja atividade
do interferon-α esteja comprometida. Em virtude dessa deficiência, os animais tornam-se mais sus-cetíveis às manifestações clínicas induzidas pelo vírus influenza. Por outro lado, a administração de interferon tem sido empregada no tratamento de algumas infecções virais. É o caso de pacientes infectados pelo vírus da hepatite C.
Interferons (IFNs)
Uma das primeiras reações de um organismo contra a infecção viral é a síntese de citocinas
Tabela 19.1. Exemplo de receptores virais
Receptor Vírus (família) Células-alvo Classe do receptor
CD4 HIV, SIV (Retroviridae); HHV-7 (Herpesviridae) Monócitos, macrófagos, linfócitos T
Proteínas de adesão celular e de contato entre células
ICAM-1 HRV, coxsackievírus (Picornaviridae) Várias células ativadas
MHC-ISV 40 (Papovaviridae); adenovírus 5 (Adenoviridae)
Várias
CD46Vírus do sarampo (Paramyxoviridae); HHV-6 (Herpesviridae)
VáriasProteína envolvida na regulação do complemento
Heparan sulfatoHSV, HCMV (Herpesviridae); HIV (Retroviridae); vírus da dengue (Flaviviridae)
VáriasProteínas componentes da matriz extracelular
Ácido siálicoInfluenza (Orthomyxoviridae), coronavírus humano (Coronaviridae)
Várias
HIV – vírus da imunodeficiência humana; SIV – vírus da imunodeficiência símia; HHV-7 – herpes-vírus humano tipo 7; HRV – rinovírus humano; SV40 – vírus símio 40; HHV-6 – herpes-vírus humano tipo 6; HSV – vírus herpes simples; HCMV – citomegalovírus humano.
A

214 Imunologia Básica e Aplicada
antivirais, como os interferons do tipo I (IFN-α e -β). Os IFNs geram um ambiente intracelular que restringe a replicação viral e sinalizam a pre-sença de vírus para a resposta imune adaptativa ou específica. Uma série de reações em cascata ocorre na célula infectada por vírus (transdução ou sinalização celular), a qual leva à síntese de mediadores que regulam positivamente a secre-ção de IFNs.
Após serem transcritos e traduzidos, os IFNs do tipo I, ao ligarem-se aos seus receptores (li-gantes) presentes nas superfícies celulares, indu-zem a expressão de genes em células vizinhas, que levam a um estado antiviral. Os fatores in-tracelulares reguladores de IFN (IRFs) são ativa-dos diretamente por componentes virais como o capsídeo e o RNA dupla fita. A cooperação de vários fatores de transcrição leva à síntese de três inibidores principais: uma quinase (PKR), uma RNAse (2’-5’ oligoadenylate synthase) e as proteínas Mx. Essas proteínas atuam inibindo os processos de transcrição e tradução virais.
Além dos interferons do tipo I, o IFN-γ ou tipo II atua na resposta antiviral. Sua síntese, diferen-te dos IFNs do tipo I, ocorre em linfócitos T ati-vados e células natural-killer (NK). Embora tenha sido inicialmente descrito como uma citocina antiviral, o IFN-γ apresenta um leque de ações muito mais amplo: a ativação da atividade bac-tericida dos fagócitos promove a apresentação de antígeno via MHC classes I e II, coordena as interações entre leucócitos e células endoteliais e tem efeitos na proliferação e na apoptose celu-lares. Mais de 200 genes são regulados pela ação do IFN-γ.
Os IFNs de tipo II atuam por meio de recepto-res diferentes, mas podem desencadear ações si-nérgicas por mediadores comuns na transdução de sinais intracelulares como a STAT1. O IFN-γ também ativa as proteínas PKR e a 2’-5’ oligoa-denylate synthase.
Células NK (natural-killer)
São descritas como grandes linfócitos granulares com capacidade citotóxica, atuando como pri-meiras células imunitárias na infecção viral. Além da lise de células infectadas por vírus, as células
são responsáveis pela síntese de IFN-γ que irá in-fluenciar a expansão clonal dos linfócitos T.
O principal mecanismo de lise da célula-alvo é por meio da exocitose dos grânulos citoplas-máticos, embora a apoptose induzida pelas mo-léculas FasL (Fas ligand) e TNF (tumor necrosis factor – fator de necrose tumoral) também ocor-ra. Os grânulos citoplasmáticos contêm várias proteínas, como a perforina e serina proteases (granzimas). A perforina, a mesma proteína se-cretada pelos linfócitos T-citotóxicos, atua por meio da formação de um poro na membrana da célula infectada, levando à lise por choque osmótico.
O reconhecimento das células infectadas por vírus e a ativação do processo de citotoxicidade se dão pela ligação de receptores presentes na superfície das células NK (por exemplo, CD16). O processo de inibição se dá por meio de recepto-res que reconhecem as moléculas de MHC clas-se I (KIR, para killer Ig-like receptors) presentes em todas as células nucleadas do organismo. A diminuição da expressão de proteínas celulares, induzida pelo processo de infecção viral (que uti-liza a maquinaria celular em benefício próprio, para produzir suas proteínas), faz com que haja diminuição no número de moléculas MHC classe I na superfície da célula infectada, que se torna, então, alvo para as células NK. As células NK par-ticipam igualmente no mecanismo de ADCC (an-tibody dependent cellular cytotoxicity – citotoxi-dade celular dependente de anticorpos).
Outras substâncias
Há várias décadas, pesquisadores descreveram substâncias presentes no sangue, mucosas e se-creções que não se enquadram na categoria dos interferons. Essas substâncias vêm sendo carac-terizadas em diferentes espécies, sendo possível detectá-las desde humanos até crustáceos.
Como exemplo, podemos citar a glicoprote-ína UTIβ e a lipoproteína HDL (high-density li-poprotein – lipoproteína de alta densidade), en-contradas no soro humano e de outras espécies animais. A atividade antiviral é exercida pela ini-bição da adsorção (UTIβ) e/ou de outras etapas subseqüentes da replicação viral. Essas substân-cias parecem desempenhar papel importante no controle da viremia.
Outros inibidores foram igualmente descritos em cultivos celulares, urina, no trato gastrintestinal, no

215Imunidade a vírus
sistema nervoso central e na saliva. A maioria deles é formada por glicoproteínas e lipoproteínas que, em grande parte das vezes, conservam sua ativida-de inibitória, mesmo sob a ação de enzimas e outros agentes desnaturantes.
Resposta imune adquirida
Anticorpos
Um dos principais mecanismos de ação dos anti-corpos nas infecções virais é o processo de neu-tralização. A definição do termo compreende a anulação do poder infeccioso do vírus por meio da ligação de anticorpos neutralizantes à superfí-cie viral. O requisito mínimo para a neutralização é a associação dos parátopos dos anticorpos com os epítopos dos antígenos virais, os quais devem estar expostos na superfície da partícula viral. A título ilustrativo, pode-se citar o número de mo-léculas IgG ou IgM necessárias para neutralizar o vírus da raiva: 130-150 e 40-50, respectivamen-te. Demonstrou-se que existe uma relação linear entre a área da superfície da partícula viral e o número de anticorpos neutralizantes necessários para inibir a infecção.
A neutralização ocorre, principalmente, pelo bloqueio das proteínas virais responsáveis pelo reconhecimento do receptor celular na primeira etapa da replicação viral, a adsorção. Altos títu-los de anticorpos levam à neutralização, enquan-to baixos títulos levariam à facilitação da infec-ção, segundo alguns estudos in vitro.
Além do bloqueio direto, mudanças confor-macionais das proteínas externas e a agregação das partículas virais têm sido citadas como meca-nismos mediados por anticorpos que levariam à inibição da replicação viral, inclusive em etapas posteriores à adsorção, como a internalização por endocitose.
Em células infectadas, que expressam as pro-teínas virais na superfície, os anticorpos podem ser responsáveis por opsonização para macrófa-gos, neutrófilos e células NK. Ocorre a lise celu-lar, mediada por anticorpos via receptor Fc.
Linfócitos T
Os linfócitos T apresentam várias funções na imunidade contra os vírus, mas, sem dúvida, sua maior atividade diz respeito à eliminação de cé-
lulas já infectadas. Por apenas alguns dias, entre a queda da titulação das células NK e o aumento quantitativo dos linfócitos T, estes poderão atuar em conjunto com as células NK.
Os mecanismos de atuação dos linfócitos TH são os já descritos no capítulo referente à respos-ta celular. Basicamente, atuam sobre as células imunitárias por meio da produção de citocinas – desde a ativação e a proliferação até a síntese de anticorpos pelos linfócitos B, atuando também na troca de classes das imunoglobulinas. Ativam monócitos, macrófagos, células NK, neutrófilos, além de agirem de forma autócrina. Entretanto, são os linfócitos T-citotóxicos que têm a capacida-de de reconhecer os antígenos virais sintetizados na célula hospedeira e expressos em associação com o MHC classe I, presente em praticamente todas as células do organismo.
Linfócitos TH CD4+: duas características bási-cas diferem os linfócitos TH dos TC, a sua função e o mecanismo de reconhecimento dos antíge-nos virais pelos seus receptores de superfície (TCR – T-cell receptor). Os linfócitos TH reco-nhecem as proteínas virais em associação com o MHC classe II na superfície das células apresen-tadoras de antígeno (APCs). Esse tipo de apre-sentação é denominado exógeno, pois o vírus sofre um processamento após ter sido fagoci-tado ou endocitado pelas APCs (macrófagos, células dendríticas, linfócitos B). Sua atuação é ampla, ativando praticamente todas as células imunitárias efetoras.
Linfócitos T-citotóxicos: esse grupo de linfóci-tos T reconhece os antígenos virais quando estes são apresentados pela molécula MHC-I, expressa em praticamente todas as células nucleadas. A apresentação dos antígenos virais aos linfócitos T-citotóxicos é denominada endógena. As proteí-nas virais são sintetizadas na mesma célula em que ocorre a associação com o MHC classe I. Após a ação de proteases celulares sobre as proteínas virais, os peptídeos gerados são transportados ao retículo endoplasmático por um complexo de proteínas transportadoras (TAPs). No retículo endoplasmático, irão se associar ao MHC classe I para finalmente ser expressos na superfície celu-lar e reconhecidos pelos linfócitos TC por meio do TCR. A resposta imune de linfócitos T-citotóxi-cos é mais tardia e relativamente maior quando comparada com a resposta das NK nas infecções virais (Figura 19.3).

216 Imunologia Básica e Aplicada
dos herpes-vírus e poxvírus, por exemplo, os ge-nes destinados a esse tipo de controle chegam a representar mais de 50% do genoma total.
As proteínas virais podem agir em vários me-canismos da resposta imune (Tabela 19.2). Os principais são:
Tabela 19.2. Mecanismos de escape virais e moléculas associadas
Vírus Gene/proteína Mecanismo
HCMV US3 Retenção do MHC no RE
HSV ICP-47 Inibe a ligação de peptídeos ao TAP
EBV EBNA-1 Inibe a degradação dentro do proteassoma
HCMV UL-18 MHC-I homólogo (efeito sobre as NK)
EBV EBNA-2 Diminui a transcrição de IFN
Vírus do sarampo Hemaglutinina Liga a CD46 e inibe a produção de IL-12 pelos macrófagos
EBV, EHV BCRF-1, IL-10 gene Atividade de IL-10,diminui a resposta de Th1
Adenovírus 14.7 K Inibidor de caspases
HHV-8 ORF-16 Homólogo da proteína Bcl-2 (antiapoptótica)
HCMV – citomegalovírus humano; HSV – vírus herpes simplex; EBV – vírus Epstein-Barr; EHV – herpes-vírus eqüino; HHV-8 – herpes-vírus humano tipo 8.
Figura 19.3. Comparação entre a intensidade e o início de resposta imune das células NK e linfócitos T-citotóxicos
Dias pós-infecção
Res
pos
ta r
elat
iva
NK
LTc
2 3 4 5 6 7 8
Mecanismos de escape viralPara se manterem na natureza, é essencial que os vírus possam replicar e infectar novos hospe-deiros. Entretanto, no processo de replicação, ocorre a produção de proteínas antigênicas que tornam o vírus vulnerável ao sistema imune. Des-sa forma, os vírus desenvolveram mecanismos de escape do sistema imune.
Variabilidade antigênica É a forma mais efetiva para despistar as defesas do organismo. O vírus da imunodeficiência hu-mana (HIV) e o influenza apresentam claramen-te essa propriedade. Este último possui em sua superfície proteínas como a hemaglutinina e a neuraminidase, que interagem com anticorpos e podem sofrer mutações. Pequenas mudanças nes-sas moléculas são devidas a mutações pontuais no genoma viral (antigen drift), enquanto mutações maiores podem ser formadas quando ocorre tro-ca entre material genético de diferentes vírus, isto é, recombinação genética (antigen shift). Durante uma infecção por influenza, são gerados anticor-pos neutralizantes contra a hemaglutinina viral, e alterações na estrutura dessa molécula tornam os anticorpos preexistentes ineficazes.
Interações com moléculas do sistema imuneAlguns vírus contam com genes destinados ao controle do sistema imune do hospedeiro. No caso
1. Interferência com MHC e a apresentação do antígeno: a diminuição da expressão das moléculas do MHC-I na superfície da célula infectada a torna temporariamente invisível aos linfócitos T-citotóxicos, dando ao vírus um tempo extra para sua replica-ção. Essa menor expressão pode ser feita pela retenção das moléculas do MHC-I no citoplasma da célula (adenovírus, citomega-lovírus humano – HCMV), pelo controle da transcrição dos genes de MHC-I (herpes-vírus humano-8 ou HHV-8; gamaherpes-vírus murino 68 ou MHV-68) e até mesmo pela destruição de moléculas do MHC-I (proteínas U2 e U11 do HCMV). Entretan-to, a diminuição de moléculas de MHC-I da superfície leva ao ataque da célula in-fectada pelas células NK, que reconhecem

217Imunidade a vírus
células com deficiente expressão de MHC-I. Mesmo assim, alguns vírus como HCMV e MCMV (citomegalovírus murino) podem gerar moléculas homólogas ao MHC-I que serviriam para enganar as células NK.
Outra via que pode servir para fuga viral do sistema imune é o transporte de peptídeos pro-duzidos no citosol até o retículo endoplasmático (ER) pelo complexo TAP (transporter associated with antigen processing). O HSV-1 e o HSV-2 (herpes-vírus simplex, tipos 1 e 2) codificam poli-peptídeos inibidores de TAP, prejudicando a for-mação do complexo com MHC.
Uma das primeiras etapas para geração de peptídeos antigênicos ocorre por meio de pro-teólise dentro do proteassoma no citoplasma da célula. Alguns vírus como o HCMV e o EBV (vírus Epstein-Barr) podem interferir nessa eta-pa, inibindo a geração de epítopos específicos e afetando diretamente a proteólise dentro do proteassoma.
2. Inibição da resposta humoral: a alteração na resposta imune humoral pode ocorrer não só pela variação antigênica, mas tam-bém pela interferência de proteínas virais como, por exemplo, fatores do sistema de complemento. O vírus vaccinia, por exem-plo, codifica uma proteína (VCP – vaccinia complement-control protein) que está relacionada com a clivagem de C3b e C4b, interferindo tanto na via clássica como na via alternativa do complemento.
3. Interferência com interferon (IFN): alguns vírus, como os poxvírus, podem codificar proteínas com homologia aos receptores de IFN e, dessa forma, bloquear a ação deste. Além disso, vários vírus inibem a atividade do IFN-γ, que é um importante ativador da imunidade celular, pelo bloqueio de sua síntese ou de fatores necessários para sua produção, como IL-12 ou IL-18.
4. Produção de homólogos de citocinas e seus receptores: um dos mais interessantes mecanismos identificados nos últimos anos é a produção, por agentes virais (herpes-vírus e poxvírus), de proteínas virais seme-lhantes aos receptores celulares de citocinas e até mesmo de proteínas semelhantes às
próprias citocinas do hospedeiro. Como as citocinas podem regular a resposta imune tanto positiva como negativamente, alguns vírus codificam suas próprias citocinas. O EBV, por exemplo, codifica uma proteína homóloga à IL-10, que é uma reguladora negativa de IL-12.
5. Inibição da apoptose: a indução da apop-tose de células infectadas por vírus, feita pe-los linfócitos T-citotóxicos e NKs, através de citocinas como TNF, também pode ser alvo de proteínas virais. Agentes virais podem inibir a ativação das caspases e codificar homólogos às proteínas anti-apoptóticas, como a Bcl-2 (Tabela 19.2).
Imunopatologia
Células dendríticasEmbora as células dendríticas venham sendo apontadas como as principais responsáveis pela apresentação de antígeno aos linfócitos T que desencadeiam uma efetiva ativação, elas tam-bém podem impedir a eliminação viral, desem-penhando o papel de “reservatórios”. A maioria das infecções virais se inicia por mucosas e pele, sítios onde se encontram as células dendríticas que capturam os antígenos por fagocitose e pi-nocitose. Após a captura, as células dendríticas sofrem maturação funcional enquanto dirigem-se aos linfonodos regionais.
A infecção das células dendríticas já foi de-monstrada para o vírus HIV, vírus da hepatite C (HCV), citomegalovírus (CMV) e vírus do saram-po. Pelo contato com linfócitos T, estas pode-riam ser infectadas. Ou seja, as células dendrí-ticas seriam os “reservatórios” de vírus para a infecção de linfócitos T. Na maioria das vezes, a replicação viral é bloqueada ou mantida em bai-xos níveis nas células dendríticas, embora estas últimas continuem sendo fonte de infecção para os linfócitos T.
Situações clínicas
SarampoO vírus do sarampo é altamente contagioso e é responsável pela morte de muitas crianças em todo o mundo (mais de 1 milhão de mortes por ano em crianças do terceiro mundo). É transmitido

218 Imunologia Básica e Aplicada
por secreções orais e respiratórias. Após uma repli-cação inicial, o vírus pode se disseminar para vários órgãos. De particular importância é a infecção do sistema nervoso, que pode levar a uma panence-falite esclerótica subaguda anos após a infecção inicial. O organismo desenvolve uma forte respos-ta imunológica contra o vírus, mas a imunidade contra outros patógenos parece fi car comprome-tida. Essa imunossupressão permite o aparecimen-to de infecções oportunísticas e está relacionada às complicações associadas ao sarampo.
Vírus respiratório sincicial (VSR)O VSR é o principal agente de bronquiolite e pneumonia em crianças e jovens, causando apro-ximadamente 4.500 mortes e 91 mil hospitali-zações anualmente nos Estados Unidos. O vírus
também está associado a 3,3 milhões de casos de doença do trato respiratório em idosos. A imuni-dade protetora contra o VSR é efetivada por meio da produção de anticorpos neutralizantes contra as proteínas virais de fusão e ligação à célula.
Vírus da hepatite C (HCV)O HCV é o mais importante agente infeccioso re-lacionado com doença hepática crônica, cirrose e câncer hepatocelular. Dos indivíduos infectados, menos de 30% recuperam-se da hepatite aguda; os restantes desenvolvem uma infecção crônica. Estudos em chimpanzés demonstraram que a re-solução da doença está relacionada com uma for-te resposta de linfócitos T-citotóxicos e uma baixa resposta humoral. Por outro lado, animais que ma-nifestaram evolução crônica da doença apresenta-ram fraca resposta de linfócitos T-citotóxicos.
BibliografiaAlcami A, Koszinowski UH. Viral mechanisms of immune evasion. Trends in Mi-
crobiology 2000; (9)8:410-8.
Alcami A. Viral mimicry of cytokines, chemokines and their receptors. Nature Re-views 2003; 3:36-50.
Baron S, Singh I, Chopra A, Coppenhaver D, Pan J. Innate antiviral defenses in body fluids and tissues. Antiviral Research 2000; 48:71-89.
Bernet J, Mullick J, Singh AK, Sahu A. Viral mimicry of the complement system. Journal of Bioscience 2003; (3)28:249-64.
Boehm U, Klamp T, Groot M, Howard JC. Cellular responses to interferon-γ. Annu-al Reviews in Immunology 1997; 15:749-95.
Brander C, Walker BD. Modulation of host immune responses by clinically rele-vant human DNA and RNA viruses. Current Opinion in Microbiology 2000; 3:379-86.
Burton DR, Williamson RA, Parren PWHI. Antibody and virus: binding and neutra-lization. Virology 2000; 270:1-3.
Gewurz BE, Gaudet R, Tortorella D, Wang EW, Ploegh HL. Virus subversion of immuni-ty: a structural perspective. Current Opinion in Immunology 2001; 13:442-50.
Grandvaux N, Tenoever BR, Servant MJ, Hiscott J. The interferon antiviral response: from viral invasion to evasion. Current Opinion in Infectious Diseases 2002; 15:259-67.
Kaiserlian D, Dubois B. Dendritic cells and viral immunity: friends or foes? Sem-minars in Immunology 2001; 13:303-10.
Klasse PJ, Sattentau QJ. Occupancy and mechanism in antibody-mediated neutraliza-tion of animal viruses. Journal of General Virology 2002; 83:2091-108.
Mahalingam S, Meanger J, Foster PS, Lidbury BA. The viral manipulation of the host cellular and imune enviroments to enhance propagation and survival: a focus on RNA viruses. Journal of Leukocyte Biology 2002; 72:429-39.
Warren HS, Smyth MJ. NK cells and apoptosis. Immunology and Cell Biology 1999; 77:64-75.

C A P Í T U L O 2 0
IntroduçãoAs bactérias são células procarióticas desprovidas de núcleos e retículo endo-plasmático. As espécies Gram-negativas apresentam paredes celulares rela-tivamente rígidas, compostas de duas bicamadas fosfolipídicas, com uma ca-mada de peptideoglicana intercalada. As bactérias Gram-positivas têm uma única bicamada coberta por peptideoglicana. As bactérias sintetizam seus próprios DNA, RNA e proteínas, mas dependem do hospedeiro para obter condições favoráveis ao crescimento. As bactérias podem ser intra ou extra-celulares em nosso organismo.
Nossos mecanismos de defesa podem ser inatos ou adaptativos. Caso os me-canismos não sejam efetivos, a infecção bacteriana pode conduzir a quadros graves como sepses e choque séptico, cujo desenvolvimento também depende da ação do sistema imunológico. Para se sobressair ao sistema imunológico, al-gumas bactérias desenvolvem mecanismos de resistência contra nossas defesas.
Bactérias extracelularesSão as bactérias que têm capacidade de replicar-se fora da célula do hospe-deiro, como na circulação sangüínea, vias aéreas, tecido conjuntivo, trato urinário, cavidade intestinal, seios paranasais.
As bactérias extracelulares patogênicas podem causar doença por infl ama-ção, o que resulta em destruição tecidual no sítio da infecção. Exemplos são as bactérias piogênicas, que causam infecções supurativas, como S. aureus. Além disso, essas bactérias podem produzir toxinas, que causam efeitos patológi-cos diversos. Algumas têm endotoxinas, componentes da parede da bactéria, como lipopolissacarídeos (LPS) das bactérias Gram-negativas, como a E. coli. Outras secretam exotoxinas ativamente, como a C. tetani, podendo ser cito-tóxicas, matando a célula por mecanismos diversos, interferindo na função celular normal ou estimulando a produção de citocinas (Tabela 20.1).
Bactérias intracelularesSão aquelas que têm capacidade de sobreviver e replicar-se dentro de células fagocitárias, como os macrófagos em diferentes tecidos (alvéolo, pele, intestino etc.) e neutrófi los. Dessa forma, elas tornam-se inacessíveis a qualquer proteí-na ou célula do sistema imune na circulação. Freqüentemente, o resultado da interação dessas bactérias intracelulares com o sistema imune gera conseqüên-cias patológicas em virtude da resposta do hospedeiro a esses microrganismos e também pela maior permanência do patógeno no hospedeiro (Tabela 20.2).
Laura Prates Vitório • Maria Lúcia Scroferneker
Imunidade a bactérias

220 Imunologia Básica e Aplicada
Tabela 20.1. Exemplos de bactérias extracelulares
Bactéria Mecanismo patogênico Doenças
Staphylococcus aureus Pele: inflamação aguda e morte celular induzida por toxinas.Sistêmica: indução de produção de citocinas
Infecções de pele e tecidos moles, abscesso pulmonarSíndrome do choque tóxico, envenenamento alimentar
Streptococcus pyogenes Inflamação aguda induzida por toxinas FaringiteImpetigo, erisipela, celuliteEscarlatina
Streptococcus pneumoniae Inflamação aguda induzida por constituintes de parede PneumoniaMeningite
Escherichia coli Ação da toxina no epitélio intestinal, aumentando a secreção de cloro e água. Estímulo de secreção de citocinas
Infecções do trato urinárioGastroenteriteChoque séptico
Clostridium tetani Sua toxina se liga a estruturas da junção neuromuscular, causando contração muscular irreversível
Tétano
Helicobacter pyloriAltera as propriedades normais do epitélio gástrico, induz alterações mutagênicas nas células epiteliais e perda do controle inibitório normal da secreção gástrica
Gastrite crônicaÚlcera pépticaNeoplasia gástrica
Leptospira spp. Vasculite infecciosa, que danifica as células endoteliais capilares, com aumento da permeabilidade intravascular, hemorragia e choque
LeptospiroseDoença de Weil
Klebsiella pneumoniae Infecção piogênica hemorrágica ou necrotizante PneumoniaInfecções de vias urinárias
Tabela 20.2. Exemplos de bactérias intracelulares
Bactéria Mecanismo patogênico Doenças
Mycobacterium tuberculosis
Macrófagos ativados formam inflamação granulomatosa e destruição tecidual Tuberculose
Mycobacterium leprae Macrófagos ativados formam inflamação granulomatosa e destruição tecidual Hanseníase
Neisseria gonorreae Produção de fatores citotóxicos que danificam as células ciliadas das mucosas, supuração aguda seguida de inflamação crônica e fibrose
Gonorréia ou blenorragia
Neisseria meningitidis Inflamação aguda, com doença sistêmica causada por endotoxina Meningite
Mecanismos da imunidade a bactérias
Barreiras físicas
As bactérias são excluídas dos tecidos do hospedei-ro por barreiras anatômicas, pele e mucosas.
A integridade de superfícies mucosas é mantida pela remoção ativa de bactérias, como o movimen-to ciliar no trato respiratório superior, o contínuo fluxo de urina no trato urinário baixo, a renovação das células epiteliais, as enzimas produzidas por algumas células e o pH de alguns tecidos. Todos esses mecanismos são formas de dificultar a entra-

221Imunidade a bactérias
da e a sobrevivência das bactérias em nosso orga-nismo. Um dos melhores exemplos desse sistema é a mucosa intestinal que, com seus mais de 400 m2
de superfície, é uma grande porta de entrada para microrganismos patogênicos. Entretanto, a secre-ção de enzimas por parte dessa mucosa mantém os níveis de pH da flora microbiana normal, além de apresentar defesas específicas, como a imunoglo-bulina A secretora (IgAs), que controla a penetra-ção das bactérias nas células. Outro exemplo seria o epitélio traqueal, que, com o movimento de seus cílios e a secreção de muco, elimina bactérias que entram por via inalatória. Lágrimas e saliva tam-bém dificultam a entrada das bactérias.
Flora microbiana normalÉ constituída por bactérias que possuem uma re-lação comensal ou simbiótica com o hospedeiro e que auxiliam no controle do desenvolvimento de bactérias patogênicas. Essa flora é influencia-da por condições ambientais como a dieta, o sa-neamento básico, a poluição do ar e os hábitos de higiene. As condições hormonais do indivíduo também exercem influência sobre essa flora, as-sim como o uso de fármacos como corticóides e antimicrobianos.
Bactérias extracelulares
Imunidade inataOs principais mecanismos são: ativação do siste-ma complemento e fagocitose.
O complemento pode ser ativado pela via alternativa através dos peptideoglicanos da pa-rede celular das bactérias Gram-positivas e li-popolissacarídeos (LPS) da parede celular das Gram-negativas. Isso pode ocorrer mesmo na ausência de anticorpos, ainda enquanto a imu-nidade adaptativa não foi ativada. Essa ativação pode levar a opsonização, lise da bactéria pelo MAC (membrane attack complex – complexo de ataque à membrana), quimiotaxia, anafilaxia e ativação de leucócitos.
Os fagócitos ligam-se às bactérias por meio de vários receptores de membrana, além da li-gação pela opsonização (por anticorpos, C3b do complemento etc.). A partir daí, eles se ativam, adquirindo atividade microbicida, secretando citocinas e causando infiltração leucocitária no
sítio da infecção. Conseqüências dessa resposta são a injúria ao tecido normal do hospedeiro e a indução de manifestações sistêmicas, pelas cito-cinas, como febre.
ReconhecimentoMesmo após esse esclarecimento, fica uma dú-vida interessante: como o sistema imune inato diferencia o próprio do não-próprio, se ele não tem especificidade? Ou seja, como vai haver ati-vação do sistema complemento, por via alter-nativa, e como os fagócitos vão reconhecer os microrganismos patogênicos, e tudo isso ainda induzir inflamação?
Os componentes do sistema imune inato que discriminam entre microrganismos e células pró-prias são capazes de reconhecer estruturas en-contradas nos microrganismos. Quando essas estruturas estão presentes nas células, elas são chamadas de padrão de reconhecimento por receptores. Ou seja, possuímos receptores para algumas estruturas comuns encontradas em al-guns microrganismos patogênicos. Esses recep-tores, em fagócitos, fatores do complemento ou outros leucócitos, podem induzir fagocitose, quimiotaxia ou secreção de mediadores pró-in-flamatórios.
Uma importante estrutura conhecida é o pep-tideoglicano, como comentado anteriormente, que é um componente único e essencial da parede celular de virtualmente todas as bactérias, similar em Gram-positivas (mais abundante) e em Gram-negativas. Envolvendo a membrana citoplasmáti-ca bacteriana, é responsável pela manutenção da forma e da pressão osmolar da bactéria. Também é um alvo conhecido para quase todos os antibi-óticos clinicamente úteis, que inibem a síntese de parede bacteriana. Além disso, peptideoglicano não está presente em eucariontes, sendo assim um excelente alvo para o sistema imune inato.
Imunidade adaptativaA imunidade humoral é o principal mecanismo imune protetor contra bactérias extracelulares, sua eliminação e neutralização de suas toxinas.
Os anticorpos, produzidos pelos linfócitos B ativados ou linfócitos B de memória, são especí-ficos contra antígenos da parede celular e antí-genos associados às toxinas dessas bactérias, que podem ser polissacarídeos ou proteínas.

222 Imunologia Básica e Aplicada
A maior função da imunidade humoral contra bactérias extracelulares é a defesa contra bacté-rias encapsuladas ricas em polissacarídeos (que é um artifício utilizado por elas para sobrevivência dentro do hospedeiro). Os polissacarídeos são os antígenos protótipos T-independentes, ou seja, que não podem ser fagocitados, degradados e acoplados ao MHC dentro da APC para ser apre-sentados ao linfócito T, e assim contar com seu auxílio por meio da secreção de citocinas estimu-ladoras e ativadoras de diversas funções do siste-ma imune. Sendo assim, os mecanismos efetores utilizados pelos anticorpos para o combate a es-sas bactérias incluem:
- neutralização;- opsonização e fagocitose;- ativação do complemento pela via clássica
(Figura 20.1).
júrias como inflamação e choque séptico (mais comumente secundário à infecção por bactérias Gram-negativas). Essas situações ocorrem em virtude da secreção de citocinas, principalmen-te por macrófagos ativados, o que acontece com muito mais intensidade na resposta T-dependen-te (Figura 20.2).
Figura 20.1. Imunidade adaptativa a antígenos T-indepen-dentes de bactérias extracelulares.
Neutralização
Ativação do complemento
OpsonizaçãoPlasmócito
Figura 20.2. Imunidade adaptativa a antígenos T-depen-dentes de bactérias extracelulares.
Bactérias
MHC-II
APC
TCR
LTH
TNF
Outras citocinas
IFN-γ
Apresentação antigênica para células TH. Há produção de interferon-γ, que ativa macrófagos para fagocitose e eliminação da bactéria; TNF, que promove inflamação local; e várias outras citocinas que ativam a resposta humoral; APC, célula apre-sentadora de antígeno; APC, célula apresentadora de antígeno; MHC-II, complexo de histocompatibi-lidade principal; TCR, receptor de célula T; LTH, lin-fócito T-auxiliar; TNF, fator de necrose tumoral.
Uma complicação tardia da resposta imune humoral a infecções bacterianas pode ser a gê-nese de doenças secundárias à produção de anti-corpos. Um exemplo importante é a infecção por Streptococcus da faringe ou da pele, em que as conseqüências manifestam-se de semanas a me-ses após a infecção. Uma seqüela da infecção fa-ríngea por alguns tipos sorológicos de Streptococ-cus β-hemolítico é a febre reumática. Nesse caso, a infecção leva à produção de anticorpos contra uma proteína constituinte da parede microbiana (proteína M). Alguns desses anticorpos podem fa-zer reação cruzada com proteínas constituintes do endocárdio e depositar-se no coração, causando inflamação (cardite) e, além disso, causando ar-trite, nódulos subcutâneos, coréia de Sydenham. Outra seqüela em virtude da infecção da faringe ou da pele por outros sorotipos de Streptococcus β-hemolítico é a glomerulonefrite pós-estrepto-cócica. Nesse caso, os anticorpos contra as bacté-rias formam complexos com o antígeno bacteria-no, podendo depositar-se nos glomérulos renais e causar nefrite. Essa situação é conhecida como reação de hipersensibilidade do tipo III.
Produção de anticorpos por linfócitos B ativa-dos (plasmócitos) após o reconhecimento de an-tígenos T-independentes. Os anticorpos são res-ponsáveis por neutralizar e eliminar as bactérias e suas toxinas pelos mecanismos já apresentados.
Antígenos protéicos das bactérias extracelu-lares ativam células TCD4+,, são T-dependentes e induzem produção de importantes citocinas, o que diferencia daquela resposta na ausência de estímulo por linfócito T. Essas citocinas estimu-lam a produção de anticorpos, inflamação local, e ativam fagocitose e atividade microbicida dos macrófagos. A citocina responsável pela ativação dos macrófagos é o interferon-γ (IFN-γ), liberada por linfócitos T, além de TNF (tumor necrosis fac-tor – fator de necrose tumoral) e outras.
As principais conseqüências da resposta do hospedeiro às bactérias extracelulares são in-

223Imunidade a bactérias
Bactérias intracelulares
Imunidade inata
A resposta imune inata a bactérias intracelulares consiste principalmente de macrófagos e células natural-killers (NK).
Hoje se sabe que o sistema imune inato não apenas serve como barreira contra microrganis-mos como também determina a resposta adap-tativa subseqüente. Isso se deve à capacidade do sistema imune inato de distinguir microrganis-mos invasores do self, o que já foi comentado anteriormente neste capítulo, e da capacidade de induzir o tipo de resposta mais adequada para o patógeno.
Os fagócitos, inicialmente neutrófilos e após macrófagos, fagocitam e tentam destruir esses microrganismos, mas bactérias intracelulares pato-gênicas são resistentes a essa degradação.
O hospedeiro desenvolve uma série de me-canismos de defesa contra os microrganismos. Dentro de macrófagos, a defesa contra essas bactérias se deve a alguns mecanismos antibac-terianos. Sabe-se que intermediários reativos de nitrogênio (IRN) e intermediários reativos de oxigênio (IRO) têm papel bioquímico impor-tante como defesa. Tem sido sugerido que esses dois mecanismos possam compensar a ausência um do outro, o que chama a atenção para a sua importância.
Os fagócitos eliminam o microrganismo intra-celular durante a fase inicial oxidativa dependen-te de NADPH (nicotina adenina dinucleotídeo fosfato hidrogenada) – oxidase, seguida de uma fase nitrosativa prolongada, quando o crescimen-to bacteriano é inibido por NO sintase (NOS ou sintase do óxido nítrico). Além disso, sabe-se que os níveis de IRN dentro do macrófago determi-nam se o microrganismo será eliminado por via enzimática ou por apoptose. Nesse último caso, ainda não está claro, pois também pode ser um mecanismo da bactéria exterminar células da pri-meira linha de defesa. Além da ação direta mi-crobicida, IRN têm efeitos imunorregulatórios relevantes para o controle de infecção.
Em infecção por M. tuberculosis, tem sido en-contrada forte correlação entre expressão de NOS em sítios da doença e subseqüente resolução da infecção. Isso sugere que NO possa eliminar essa
bactéria, o que já foi observado pela administra-ção de NO em doses seguras em pacientes com doença pulmonar. Pode ser que também exista a produção de NOS em monócitos. Entretanto, M. tuberculosis pode inibir a resposta de IFN-γ. Estudos recentes sugerem que espécies de Sal-monella sp. são mais sensíveis a NADPH-oxidase fagocitária (phox) do que a NOS.
Essas bactérias intracelulares também ativam células NK, tanto diretamente quanto pela esti-mulação da produção de IL-2 por macrófagos, a citocina mais importante na ativação de NK. As células NK produzem IFN-γ, o qual por sua vez ativa macrófagos e promove a morte da bactéria fagocitada. Além disso, células NK pro-movem uma defesa rápida contra esses micror-ganismos, ainda antes do desenvolvimento da imunidade adaptativa.
Isso foi comprovado por meio de experimentos com ratos, em que aqueles com imunodeficiência grave de linfócitos B e T são capazes de contro-lar a infecção da bactéria intracelular Listeria mo-nocytogenes através das células NK.
A imunidade inata pode limitar o crescimen-to bacteriano por algum tempo, mas geralmente não consegue erradicar essas infecções. A erradi-cação requer a imunidade adaptativa mediada por células (Figura 20.3).
Figura 20.3. Imunidade inata e adaptativa a bactérias intracelulares.
Controle de infecção
Núm
ero
de
bac
téria
s vi
ávei
s
NK LTH
LTCMacrófago
Neutrófilo
Imunidade inata
Dias após a infecção
Imunidade adquirida
Macrófago
Macrófago ativado
0 7 14
A resposta imune inata a bactérias intracelula-res é realizada por fagócitos (macrófagos e neu-trófilos), interagindo com células NK por meio de citocinas. Os macrófagos estimulam as células NK através da IL-2, e as células NK estimulam ma-

224 Imunologia Básica e Aplicada
crófagos através de IFN-γ. Inicialmente, essa res-posta inata ocorre por neutrófilos e pelas célu-las NK. A resposta imune adaptativa típica para bactérias intracelulares é a mediada por células, havendo ativação de macrófagos por linfócitos T. A imunidade inata controla o crescimento bacte-riano, mas a imunidade adaptativa elimina esses microrganismos. NK, célula natural-killer; LTC, linfócito T-citotóxico; LTH, linfócito T-auxiliar.
As principais e mais estudadas bactérias intra-celulares causadoras de doenças no homem são Mycobacterium tuberculosis e Salmonella typhi, causadoras de tuberculose e febre tifóide, res-pectivamente.
Tuberculose e febre tifóide, embora sejam doenças com fisiopatologias muito diferentes, apresentam encontro inicial com o hospedeiro similar. Ambas as bactérias residem dentro de macrófagos.
Os macrófagos alveolares são as primeiras cé-lulas com que as micobactérias entram em con-tato. No caso da salmonela, são os macrófagos presentes nas placas de Peyer do intestino delga-do. Esse primeiro contato do microrganismo com células do hospedeiro determina três possíveis desfechos:
- a efetividade do fagócito e a eliminação do organismo ingerido;
- indução de morte da célula pela bactéria;- adaptação do microrganismo ao ambiente
hostil dentro do fagócito, que se torna sua célula hospedeira, estabelecendo a infecção.
Abrigada dentro do macrófago, a bactéria pode se disseminar para linfonodos, fígado e baço. Ela se replica em sítios sistêmicos antes de atingir a circulação novamente e chegar ao seu sítio de entrada. A partir desse ponto, tuberculo-se e febre tifóide desenvolvem diferentes rotas.
A infecção por salmonela pode resultar em bacteremia, o que pode ser fatal em caso de dis-seminação irrestrita. Pode também resultar em microabscessos (ricos em neutrófilos e células mononucleares), o que leva à resolução da infec-ção, quando controlada com sucesso. O micror-ganismo também pode persistir na vesícula biliar de portadores assintomáticos, contribuindo para a disseminação da doença.
Em contraste, a infecção por M. tuberculosis é latente em um terço da população mundial. Entretanto, menos de 10% desses indivíduos de-
senvolvem a doença ativa, enquanto os outros geram uma resposta imune efetiva, contendo o bacilo dentro de lesões granulomatosas. Nesse estado, a bactéria persiste por longos períodos, mas não é capaz de transmitir a infecção. Para isso, é necessário reativação da infecção. O que mantém essa latência é o mecanismo imune, pre-venindo a reativação e os fatores de virulência, por meio dos quais o bacilo adapta-se e aguarda para a reemergência.
Imunidade adaptativaA resposta imunológica mediada por células é a mais importante contra bactérias intracelulares. Prova disso é que indivíduos com deficiência na imunidade mediada por células, como pacientes com síndrome da imunodeficiência adquirida (SIDA), são extremamente suscetíveis a infecções por bactérias e vírus intracelulares.
A imunidade mediada por células consiste em dois tipos de reação:
- ativação de macrófagos infectados, por sinali-zação de células T (ligação a CD40 e IFN-γ), resultando em morte dos microrganismos fagocitados, ainda dentro do fagócito;
- lise da célula infectada por linfócito T-citotóxi-co (LTC), resultando em morte do microrganis-mo e da célula infectada (Figura 20.4).
Figura 20.4. Reações de imunidade adaptativa a bactérias intracelulares mediada por células.
APC
LTH
TCRMHC-II
IFN-γ
Bactéria no fagossomo
APC
MHC-II
MHC-I
Perfurinas e granzimas
Bactéria no citosol
TCRMHC-I
LTC
Morte da célula infectada
Ativação dos macrófagos e morte das bactérias dentro do fagossomo
Bactérias intracelulares são fagocitadas por ma-crófagos e podem sobreviver dentro do fagossoma ou escapar dentro do citoplasma. Os LTH respon-dem a antígenos ligados ao MHC-II, derivados das bactérias intravesiculares, secretando IFN-γ, que ativa os macrófagos para eliminar as bactérias in-trafagossomais. Os LTC respondem a antígenos li-

225Imunidade a bactérias
gados ao MHC-I, derivados das bactérias do citosol, liberando perfurinas e granzimas para eliminar a célula macrofágica infectada. MHC, complexo de histocompatibilidade; APC, célula apresentadora de antígeno; TCR, receptor de célula T, LTH, linfóci-to T-auxiliar; LTC, linfócito T-citotóxico.
Células T CD4+ e T CD8+ respondem a antíge-nos protéicos dos microrganismos fagocitados apresentados na membrana através do acopla-mento a MHC-I e II, respectivamente. Células T CD4+ diferenciam-se em efetores TH1, importan-te estimulador da imunidade celular e ativador de macrófagos, sob estímulo de macrófagos e células dendríticas.
Por meio da ligação a CD40, expresso na mem-brana de células T, e do estímulo de IFN-γ, macró-fagos produzem substâncias microbicidas, incluin-do IRO, NO e enzimas lisossômicas. Além disso, IFN-γ também estimula linfócitos B a produzirem anticorpos, que ativam complemento e opsoni-zam as bactérias (nesse caso, aquelas que ainda não foram fagocitadas), aumentando as funções efetoras dos macrófagos.
Já se sabe que ratos com deficiência em cito-cinas como IFN-γ são extremamente suscetíveis a infecções por bactérias intracelulares, como M. tuberculosis e L. monocytogenes.
Após a fagocitose de uma bactéria, seus antí-genos podem ser transportados dos fagossomos para dentro do citosol ou a bactéria escapa dos fagossomos, entrando no citoplasma das células infectadas. Em ambos os casos, há estímulo de resposta de linfócitos T CD8+.
Microrganismos que residem predominan-temente em fagossomos, como micobactéria, salmonela, brucela, clamidia, leishmania e plas-módio, ativam células CD4+, restritas a MHC-II, pois seus antígenos são processados por enzimas lisossômicas e o caminho de apresentação é o endossômico. Nesse caso, o estímulo da resposta de linfócitos T CD8+ é por outro mecanismo, por meio de vesículas apoptóticas, que será explica-do posteriormente.
Ao permanecerem livres no citoplasma, como L. monocytogenes, Rickettsia sp. ou T. cruzi, os mi-crorganismos não estão muito suscetíveis aos me-canismos microbicidas dos fagócitos, que ocorrem dentro dos fagossomos, pela digestão enzimática. Nesse caso, a infecção só é erradicada pela elimina-ção das células infectadas, que, conseqüentemen-te, elimina os microrganismos que viviam no seu citosol. Essa eliminação se faz pelos LT-citotóxicos,
por meio do reconhecimento das células infecta-das, que é estabelecido com a sua ligação ao MHC-I, expondo antígeno bacteriano, com conseqüente liberação de grânulos de perfurinas e citocinas pe-los linfócitos TC.
Além da destruição da célula infectada por cé-lulas T, uma série de estímulos diferentes pode in-duzir apoptose celular, desde estímulos do pató-geno, estímulos da própria célula infectada (auto-indução de apoptose) e fatores imunológicos do hospedeiro, como citocinas pró-inflamatórias ou mediação por células NK.
Ativação de linfócitos T CD8+
por bactérias em fagossomosBactérias intracelulares, primariamente infectam macrófagos, em que o patógeno é confinado a fagossomas, e seus antígenos são apresentados por meio de MHC-II. Esse mecanismo de apresen-tação antigênica por via endossômica nem sem-pre é possível.
Sugere-se que haja outro caminho para a ati-vação de células T CD8+ contra bactérias intracelu-lares, através de vesículas apoptóticas de macró-fagos infectados. Assim, antígenos derivados dos patógenos de células infectadas chegam às célu-las dendríticas não-infectadas. Estas, sendo APCs profissionais, apresentam antígenos microbianos através de MHC-I a células T. Ou seja, as vesícu-las apoptóticas têm papel importante na função imunológica intrínseca, possibilitando a ativação das células TCD8+ quando esse mecanismo está impedido na célula infectada. Muitos caminhos já haviam sido propostos para explicar o enigma de como microrganismos intrafagossomos podem in-duzir ativação de células CD8+, restritas a MHC-I.
Essa nova função da morte celular na imu-nidade a bactérias requer consideração para desenvolvimento de vacinas contra infecções como tuberculose, febre tifóide e malária. Para aumentar a apoptose nas células infectadas, va-cinas poderiam ser equipadas com propriedades de indução de apoptose. Essa estratégia ativaria células CD8+, restritas a MHC-I, e também poten-cializaria o estímulo de células CD4+, restritas a MHC-II.
Esse processo também foi considerado impor-tante na infecção por tuberculose, pois a APC in-fectada pela micobactéria rapidamente perde sua capacidade de apresentação antigênica.

226 Imunologia Básica e Aplicada
Injúria tecidualA ativação dos macrófagos que ocorre em respos-ta a microrganismos intracelulares também é ca-paz de causar injúria tecidual. Essa injúria pode ser manifestada como reações de hipersensibilidade tipo tardia a antígenos protéicos microbianos.
Em função de as bactérias intracelulares terem desenvolvido resistência à morte, dentro dos fa-gócitos, elas freqüentemente persistem por lon-gos períodos e causam persistência do estímulo antigênico e ativação de células T e macrófagos, o que pode resultar na formação de granulomas ao redor dos microrganismos. A marca histoló-gica de algumas infecções por bactérias intrace-lulares é a formação de inflamação granuloma-tosa. Esse tipo de reação inflamatória pode ser-vir para localizar e prevenir a disseminação dos microrganismos, mas isso também está associado com grave comprometimento funcional causado por necrose e fibrose tecidual.
As diferenças entre os indivíduos nos padrões de resposta das células T a microrganismos intra-celulares são importantes determinantes da pro-gressão da doença e do desfecho clínico. Como exemplo, a infecção por M. leprae.
Uma possível razão para as diferenças nessas duas formas de doença causada pelo mesmo mi-crorganismo pode ser a existência de diferentes padrões individuais de diferenciação de células T e produção de citocinas.
Mecanismos de escape das bactériasMicrorganismos desenvolveram maneiras de vencer as barreiras de defesa do hospedeiro e, em alguns casos, até se utilizar de suas respostas inatas. Muitos microrganismos patogênicos evi-tam o reconhecimento ou prejudicam a ativação imune subseqüente por meio de sofisticadas in-terações com as defesas do hospedeiro. Alguns patógenos beneficiam-se da estimulação de rea-ção inflamatória.
As bactérias patogênicas são capazes de inva-dir sítios corporais íntegros, proliferar-se e causar dano tecidual substancial ou inflamação sistêmi-ca, como na infecção por Shigella dysenteriae ou Mycobacterium tuberculosis.
O sucesso de muitos patógenos está na sua habilidade de contornar, resistir, contra-atacar ou manipular o sistema imune do hospedeiro. Além disso, algumas bactérias provocam a ati-
vação do sistema imune, o que, algumas vezes, leva ao rompimento da barreira epitelial e inva-são da bactéria.
Ataque e colonização da superfície corporalEm função dos mecanismos de barreiras anatômi-cas (pele e mucosas), comentados anteriormen-te, motilidade e fatores de adesão (chamados adesinas) encontrados na maioria das bactérias patogênicas são essenciais para a abordagem de superfícies celulares e resistência aos mecanis-mos de remoção.
Além disso, a secreção de toxinas bacterianas impede as funções de proteção e facilita a coloni-zação. A produção de matriz extracelular protetora também pode facilitar a resistência contra os meca-nismos de proteção da superfície do hospedeiro.
A formação de biofilme, como na Pseudomo-nas aeruginosa, é um tópico de grande interesse. Isso ocorre pela habilidade de muitas espécies de bactérias de crescer nessas estruturas multicelu-lares, chamadas de biofilmes. O crescimento em biofilmes leva a um metabolismo microbiano al-terado, produção de fatores de virulência extra-celular e diminuição da suscetibilidade a biocidas, a agentes antimicrobianos e a defesa molecular e celular do hospedeiro.
Outra defesa do hospedeiro é a extensa micro-flora residente. Nesses sítios, para haver coloni-zação efetiva de um patógeno, este necessita de estratégias para haver sucesso na competição por recursos nutricionais e espaciais. Como exemplo, podemos citar as exotoxinas de leveduras, que são capazes de induzir efeitos letais em células sensíveis da mesma espécie ou gênero e assim são designadas por toxinas killer. Várias dessas toxi-nas já foram caracterizadas fisiológico-bioquími-co e geneticamente.
Escape do reconhecimento imune pelas superfícies mucosasApesar de funcionarem como barreiras mecâni-cas, as membranas mucosas são cobertas de fa-tores de opsonização solúveis, como anticorpos. Algumas bactérias fazem degradação proteolíti-ca dessas imunoglobulinas. Esse método de esca-pe é utilizado principalmente por bactérias que

227Imunidade a bactérias
colonizam o trato respiratório superior, como Haemophilus influenzae.
Muitas células epiteliais têm a habilidade de reconhecimento específico de algumas estrutu-ras dos patógenos (PAMPs). As estratégias mais efetivas para evitar o reconhecimento seriam proteção externa, como cápsulas, ou modifica-ção das estruturas reconhecidas.
Em algumas situações, a inflamação durante o início do curso da infecção pode ser vantajosa, como no caso da infecção por Shigella, em que a ativação celular é importante para induzir in-filtrado leucocitário intestinal, que se segue com rompimento da camada mucosa intestinal, facili-tando a invasão da bactéria.
Em contraste, microrganismos de colonização crônica requerem escape do estímulo e ativação das defesas do hospedeiro. Um exemplo é o Heli-cobacter pylori, que coloniza a mucosa gástrica hu-mana e causa infecção crônica em grande parte da população. A bactéria destrói um gene codificador da principal proteína reconhecida pelas células epi-teliais, reduzindo a resposta inflamatória.
Mais uma vez, a formação de biofilme pode pro-teger a bactéria mesmo em inflamação vigente.
A inibição da ativação celular por microrganis-mos comensais ou patogênicos pode representar uma estratégia para a tolerância da mucosa gas-trintestinal a estímulos pró-inflamatórios.
Resistência a efetores antibacterianos nas superfícies epiteliaisAlém da habilidade de atração de células imunes profissionais, os epitélios de superfície corporal por si só desencadeiam uma resposta antimicro-biana efetiva.
Uma grande variedade de peptídeos protege as superfícies internas e externas contra patóge-nos microbianos da maior parte dos organismos multicelulares. Esses peptídeos antimicrobianos ligam-se e integram-se na superfície do invasor, induzindo distúrbio da sua membrana celular e morte. Embora esses peptídeos sejam considera-dos relativamente resistentes à digestão enzimá-tica, já foi encontrada degradação de alguns por proteases bacterianas. Algumas bactérias, como Staphylococcus aureus e Gram-negativas, em cer-tas circunstâncias, podem modificar sua estrutura externa para tornar-se menos suscetíveis a eles.
Entretanto, não existe resistência completa a pep-tídeos antimicrobianos.
Estratégias para invadir e atravessar o epitélioA invasão de barreiras epiteliais permite prote-ção contra as moléculas de defesa da superfície, já que a maior parte dos microrganismos que chegam diretamente à pele intacta está destina-da a morrer, pois a sobrevivência na pele ou em folículos pilosos requer resistência a ácidos gra-xos, baixo pH e outros fatores antimicrobianos da pele.
Algumas bactérias induzem sinalização para as células do hospedeiro para haver seu englo-bamento e conseqüente invasão celular pela bactéria. Um exemplo é a E. coli, que, ao invadir o epitélio do trato urinário, escapa da limpeza da superfície, que é um mecanismo de defesa do hospedeiro.
A bactéria Shigella flexneri, quando internaliza-da, libera produtos que lisam o fagossomo e per-mitem o escape da bactéria dentro do citoplasma.
Já a penetração direta da pele é encontrada em patógenos transmitidos por vetores, como no caso da doença de Lyme, em que a barreira pro-tetora da pele é rompida pelo vetor. Pelo mesmo mecanismo, a Borrelia burgdorferi alcança dire-tamente o espaço subepitelial, de onde inicia a infecção sistêmica. Leptospira sp. é um exemplo de migração transcutânea ativa. Essa bactéria tem a habilidade excepcional de penetrar a pele ativamente, sem o auxílio de um vetor.
Outras bactérias, como Streptococcus pyogenes ou Clostridium perfringens, ambos causadores de infecção de tecidos moles, atravessam o epitélio via injúrias preexistentes e degradam enzimatica-mente a matriz extracelular do hospedeiro.
Outras bactérias que têm maneira importan-te de entrada através da barreira epitelial intes-tinal são Salmonella e Shygella, que se utilizam de um tipo especial de célula, as células M, que estão nas placas de Peyer no intestino delgado e podem translocar pequenos antígenos luminais (e até mesmo uma bactéria inteira, como neste caso) para o lado basolateral do epitélio e evitar o reconhecimento pelas células imunes da região (Figura 20.5).

228 Imunologia Básica e Aplicada
Escape da fagocitoseAo alcançar o espaço subepitelial, a bactéria en-contra as células locais e as células fagocitárias pro-fissionais recentemente infiltradas, atraídas pelas quimiocinas liberadas pelas células epiteliais.
Os fagócitos são equipados com uma varie-dade de receptores que detectam a presença de microrganismos invasores e se ligam a superfícies opsonizadas destes.
Conseqüentemente, as bactérias utilizam vá-rias estratégias para evitar o englobamento e a degradação pelos fagócitos e facilitar a prolife-ração e disseminação pelos tecidos do hospedei-ro. Exemplos são a inibição da fagocitose pela formação de cápsula ou destruição e necrose ce-lular mediadas por toxinas.
Em contraste, a indução de apoptose evita a liberação de sinais pró-inflamatórios. A indução de apoptose das células do epitélio pulmonar do hospedeiro durante a infecção por Pseudo-monas aeruginosa tem papel importante na re-dução da infiltração leucocitária e manutenção da função essencial do pulmão: a oxigenação do sangue. A Salmonella e a Shigella ativam cami-nhos pró-apoptóticos com a intenção de parali-sar a defesa fagocítica.
Algumas bactérias ainda possuem mecanis-mos para manipular o tráfico endossômico intra-celular, escapar da ação enzimática e interferir na sinalização intracelular, mimetizando alguns sinais (Figura 20.6).
Resistência a mecanismos de defesa humoraisO sucesso do escape dos microrganismos da fagoci-tose abre caminho para a disseminação sistêmica no hospedeiro por via sangüínea ou linfática.
Entretanto, o suprimento limitado de nutrientes essenciais, como ferro, requer alto grau de adap-tação a esse ambiente. Um exemplo é a Yersinia, uma bactéria que carrega genes codificadores de um sistema de alta afinidade ao ferro.
Além disso, as bactérias encontram a defesa hu-moral, como opsoninas e fixadores do complemen-to. S. pyogenes e S. pneumoniae possuem estru-turas de superfície que se ligam a um componen-te regulador do sistema complemento, inibindo a quimiotaxia e formação do MAC.
Interferência bacteriana com a secreção de citocinasAlgumas bactérias patogênicas desenvolveram mecanismos para modular a produção de citoci-nas pelo hospedeiro, modificando sua resposta imune. Micobactéria pode induzir a produção de
Figura 20.5. Algumas estratégias de escape das bacté-rias dos mecanismos epiteliais de defesa. Prevenção da opsonização (1), utilizada para facilitar a colonização de superfícies do hospedeiro (2). Secreção de citocinas (3) pode alterar as defesas do hospedeiro. A produção de citocinas (4) pode interferir com o reconhecimento dos antígenos e a resposta do hospedeiro. Modificação do pa-drão de apresentação molecular (5) pode alterar a sinalização intracelular. Escape dos fagossomos com inibição do reconhecimento celular (6) impede a eliminação pelos meca-nismos de defesa do hospedeiro.
Figura 20.6. Algumas estratégias de escape das bac-térias dos mecanismos de resposta imune adaptativa. Interferência com a fagocitose bacteriana (1). Indução de citocinas imunos-supressoras, como IL-10, IL-6 e TGF-β (2). Interferência com maturação no fagossomo (3) e com processamento antigênico (4).
Células epiteliais
Fagócito
IL-8
1
2
3
4
5
6
1
2 3
4MHC-II
TGF-βIL-10lL-6

229Imunidade a bactérias
citocinas antiinflamatórias. Os macrófagos infec-tados são induzidos a produzir citocinas imunos-supressivas, como IL6, TGF-β e IL10. Algumas bac-térias, como Yersinia e L. monocytogenes, tam-bém desenvolveram mecanismos de interferência nos caminhos de transdução do sinal para regula-ção de expressão de citocinas e outras proteínas pró-inflamatórias.
Interferência bacteriana com a apresentação antigênicaEssa é outra estratégia usada pelas bactérias pato-gênicas para prevenir a ativação da resposta imu-ne adaptativa. Exemplos são H. pylori, que diminui a degradação dos antígenos internalizados, e M.
tuberculosis, que inibe a maturação do fagossomo e regula a expressão de MHC-II extracelular. Chla-mydia trachomatis, um patógeno transmitido se-xualmente, também inibe a expressão de molécu-las de MHC-II de superfície.
Inibição da função efetora de células T e BUm exemplo é H. pylori que é capaz de enviar fatores que sinalizam apoptose em células T. A Neisseria gonorrheae sinaliza a inibição de ati-vação e proliferação de células CD4+, por meio de um fator que se liga a um receptor da célula T, o que contribui para a baixa especificidade da resposta imune contra essa bactéria.
BibliografiaAbbas AK, Lichtman AH. Immunity to microbes. 5ª ed. In: Cellular and Molecular
Immunology. Saunders, Philadelphia, 2003. pp. 345-366
American College of Chest Physicians; Society of Critical Care Medicine Con-sensus Conference Committee. Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. Critical Care Medi-cine 1992; 6(20):864-74.
Bernard GR, Artigas A, Brigham KL et al. The American Consensus Conference on ARDS: definitions, mechanisms, relevant outcomes, and clinical trial coordination. American Journal Of Respiratory and Critical Care Medicine 1994; 149:818-24.
Dziarski R. Recognition of bacterial peptidoglycanby the innate immune system. Cellular and Molecular Life Sciences 2003; 60:1793-804.
Flynn JL, Chan J. Immunology of tuberculosis. Annual Review of Immunology 2001; 19:93-129.
Goosney DL, Celli J, Kenny B et al. Enteropathogenic Escherichia coli inhibits phagocytosis. Infection and Immunity 1999; 67:490-5.
Hajjar AN, Ernst RK, Tsai JH et al. Human Toll-like recptor 4 recognizes host-spe-cific LPS modifications. Nature Immunol 2002; 3:354-9.
Harding CV, Ramachandra L, Wick MJ. Intercation of bacteria with antigen pre-senting cells: influences on antigen presentation and antibacteria immunity. Current Opinion in Immunology 2003; 15:112-9.
Hornef MW, et al. Bacterial strategies for overcoming host innate and adaptive immune responses. Nature Immunology 2002; (11)3:1033-9.
Janeaway Jr CA, Medzhitov R. Innate immune recognition. Annual Review of Im-munology 2002; 20:197-216.
Maganin S, Maguilnick I. Mecanismos imunogênicos da infecção por Helicobac-ter Pylori. Revista HCPA 1997; (2)17:76-82.
Mastroeni P, Vazquez-Torres A, Fang FC et al. Antimicrobial actions of the NADPH phagocyte oxidase and inducible nitric oxide synthase in experimental sal-monellosis. II. Effects on microbial proliferation and host survival in vivo. The Journal of Experimental Medicine 2000; 192:237-48.
Nathan C, Shiloh MU. Reactive oxygen and nitrogen intermediates in the relation-ship between mammalian hosts and microbial pathogens. Proc Nati Acad Sci USA 2000; 97:8841-8.
Navarre WW, Zychlinsky A. Pathogen-induced apoptosis of macrophages: a com-mom end for different pathogenic strategies. Cellular Microbiology 2000; 2:265-73.
Pieters J. Evasion of host cell defense mechanisms by pathogenic bacteria. Cur-rent Opinion in Immunology 2001, 13:37-44.
Philpott DJ, Girardin SE, Sansonetti PJ. Innate immune responses of ephitelial cells following infection with bacterial pathogens. Current Opinion in Immu-nology 2001; 13:410-6.
Raupach B, Kaufmann SHE. Immune reponses to intracellular bactéria. Current Opinion in Immunology 2001; 13:417-28.
Scroferneker M L, Pohlmann PR. Imunidade a bactérias. In: Imunologia Básica e Aplicada. 1ª ed. Sagra Luzzato, Porto Alegre, 1998. pp. 327-346.
Schaible UE, Collins HL, Kaufmann SHE. Confrontation between intracellular bacteria and the immune system. Advances in Immunology 1999, 71:267-377.
Shiloh MU, Nathan CF. Reactive nitrogen intermediates and the pathogenesis of Sal-monella and mycobacteria. Current Opinion in Microbiology 2000; 3:35-42.
Vazquez-Torres A, Jones-Carson J, Mastroeni P. et al. Antimicrobial actions of the NADPH phagocyte oxidase and inducible nitric oxide syntase in experimental salmonellosis. I. Effects on microbial killing by activated peritoneal macro-phages in vitro. The Journal of Experimental Medicine 2000, 192:227-36.
Webwe JR, Moreillon P, Tuomanen EI. Innate sensors for Gram-positive bacteria. Current Opinion in Immunology 2003; 15:408-15.
Winau F, Kaufmann SHE, Schaible UE. Apoptosis paves the detour path for CD8T cell activation against intracellular bacteria. Cellular Microbiology 2004; 7(6):599-607.
Weinrauch Y, Zychlinsky A. The induction of apoptosis by bacterial pathogens. Annual Review of Microbiology 1999; 53:155-87.
Zasloff M. Antimicrobial peptides of multicellular organisms. Nature 2002; 415:389-94.
