avaliação do efeito da gabapentina em modelo de dor ... · chronic musculoskeletal disorders are...
TRANSCRIPT

Alyne Santana Rosa
Avaliação do efeito da gabapentina em modelo de dor
muscular crônica em ratos
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Morfofuncionais do
Instituto de Ciências Biomédicas da
Universidade de São Paulo para obtenção do
título de Mestre em Ciências.
São Paulo
2018

Alyne Santana Rosa
Avaliação do efeito da gabapentina em modelo de dor
muscular crônica em ratos
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Morfofuncionais do
Instituto de Ciências Biomédicas da
Universidade de São Paulo para obtenção do
título de Mestre em Ciências.
Área de concentração: Ciências Morfofuncionais
Orientadora: Profa. Dra. Marucia Chacur
Versão original
São Paulo
2018


UNIVERSIDADE DE SÃO PAULO
INSTITUTO DE CIÊNCIAS BIOMÉDICAS
___________________________________________________________________________
Candidato (a): Alyne Santana Rosa
Título da Dissertação: Avaliação do efeito da gabapentina em modelo de dor muscular crônica
em ratos
Orientador (a): Marucia Chacur
A Comissão Julgadora dos trabalhos de Defesa da Dissertação de Mestrado, em sessão
pública realizada a ................./................./................., considerou
( ) Aprovado(a) ( ) Reprovado(a)
Examinador(a): Assinatura: ..............................................................................................
Nome: ......................................................................................................
Instituição: ..............................................................................................
Examinador(a): Assinatura: ..............................................................................................
Nome: ......................................................................................................
Instituição: ..............................................................................................
Examinador(a): Assinatura: ..............................................................................................
Nome: ......................................................................................................
Instituição: ..............................................................................................
Presidente: Assinatura: ..............................................................................................
Nome: ......................................................................................................
Instituição: ..............................................................................................


Dedico este trabalho a minha mãe, quem sempre me
inspirou e me deu forças para prosseguir, te amo muito!

AGRADECIMENTOS
Primeiramente e acima de tudo, agradeço a Deus por ter me capacitado e me dado
ânimo para concluir este trabalho.
Agradeço a minha família por todo incentivo, especialmente aos meus pais Sueli e
José, irmãos Felipe, Mayara, Tamires e Matheus, meu sobrinho Arthur, e ao meu esposo
Paulo. Vocês são minha base e quem eu sei que posso contar em todas as horas e em qualquer
situação. Amo vocês infinitamente! De nada valeria essa caminhada se eu não tivesse vocês!
Agradeço à minha orientadora, Profª Dra. Marucia Chacur, pela oportunidade e
paciência. Aos meus colegas de laboratório, Daniel, Igor, Rafael, Camilla, Lívia, Daniel
Marques, Cassio e Natalia, cada um de vocês contribuíram de alguma forma para que eu
chegasse até aqui, guardarei as boas lembranças! Também agradeço a Regina, Priscila, Aline
e Luana, pessoas que passaram pelo laboratório e ajudaram a construir nossa linha de
pesquisa!
Em especial, agradeço ao Fabio, sem o qual eu nem teria iniciado na pesquisa, serei
eternamente grata, por tudo! À Milena, quem me acompanhou desde o início e me ensinou
grande parte do que desenvolvi neste trabalho, muito obrigada! À Bianca e a Joyce, pessoas e
cientistas incríveis, por muito tempo foram meu braço direito aqui, obrigada, aprendi muito
com vocês!
Agradeço ainda a Mara e a Marina, amigas queridas que me apoiaram, enxugaram
minhas lágrimas e que não me deixaram desistir nas muitas vezes em que me abati e me
fizeram acreditar que não seria capaz. Meninas, eu amo vocês e desejo que vocês colham tudo
de bom que vocês plantaram aqui e que sejam reconhecidas pelo esforço e dedicação de
vocês, não se deixem vencer!
Aos professores e profissionais do Departamento de Anatomia e Programa de Pós-
graduação em Ciências Morfofuncionais do Instituto de Ciências Biomédicas da USP,
obrigada por todo suporte na execução deste projeto, bem como à FAPESP pelo
financiamento.
Sou muito grata a todos vocês!

Que os vossos esforços desafiem as impossibilidades, lembrai-vos de que as grandes coisas do
homem foram conquistadas do que parecia impossível.
Charles Chaplin

Resumo
Resumo
Rosa AS. Avaliação do efeito da gabapentina em modelo de dor muscular crônica em ratos.
[Dissertação (Mestrado em Ciências Morfofuncionais)] São Paulo: Instituto de Ciências
Biomédicas, Universidade de São Paulo; 2018.
Afecções musculoesqueléticas crônicas são um problema de saúde pública devido à sua alta
prevalência, seu alto custo econômico e por seu impacto negativo na qualidade de vida de
pacientes e seus familiares. Diante disso, pesquisadores têm estudado lesões musculares em
modelos animais para melhor compreensão dos mecanismos envolvidos na iniciação e
manutenção de distúrbios musculoesqueléticos. Na tentativa de esclarecer os mecanismos
nociceptivos envolvidos neste processo e encontrar terapias efetivas, nosso grupo de pesquisa
vem utilizando modelos experimentais de dor e potenciais tratamentos farmacológicos e não
farmacológicos como objeto de estudo. O presente estudo teve como finalidade avaliar se o
tratamento farmacológico com gabapentina é eficaz em reverter a dor muscular de ratos com
miosite crônica induzida pela injeção de Adjuvante de Freund Completo (CFA) no músculo
gastrocnêmio, e ainda, analisar a influência da gabapentina em células gliais e citocinas pró e
anti-inflamatórias no sistema nervoso central e periférico destes animais. Nossos resultados
demonstram o efeito da gabapentina em células gliais. Esta ação induz uma diminuição na
expressão de astrócitos e microglias, levando a um aumento de citocinas anti-inflamatórias e
inibição de citocinas pró-inflamatórias e consequentemente uma melhora do quadro
nociceptivo em nosso modelo experimental.
Palavras-chave: Dor muscular crônica. Gabapentina. Hiperalgesia. Alodinia. Astrócitos.
Microglia.

Abstract
Abstract
Rosa, AS. Evaluation of the gabapentin effect in model of chronic muscle pain in rats.
[Masters thesis (Morphofunctional Sciences)]. Sao Paulo: Biomedical Sciences Institute,
University of São Paulo; 2018.
Chronic musculoskeletal disorders are a public health problem due to its high prevalence,
high economic cost and negative impact on patients and their relatives quality of life. Due to
this, researchers have been studying muscle injury in animal models to better understand the
mechanisms involved in the initiation and maintenance of musculoskeletal disturbs. To
elucidate the nociceptive mechanisms involved in this process and seek for effective
therapies, our research group has been using experimental models of pain and potential
pharmacological and non-pharmacological treatments as object of study. The aim of this study
was to assess whether the treatment with gabapentin is effective in reversing the muscle pain
injury in rats with chronic myositis induced by the injection of Complete Freund's Adjuvant
(CFA) in the gastrocnemius muscle. Also to analyze the influence of gabapentin on glial cells
and pro-and anti-inflammatory cytokines in the central and peripheral nervous system of these
animals. Our results demonstrate the effect of gabapentin on glial cells. This action are able to
decrease expression of astrocytes and microglia, leadind to an increase in anti-inflammatory
cytokines and inhibition of proinflammatory cytokines and an improvement of the nociceptive
picture in our experimental mode,
Keywords: Chronic muscle pain. Gabapentin. Hyperalgesia. Allodynia. Astrocytes.
Microglia.

Lista de abreviaturas
LISTA DE ABREVIATURAS
AMPA ácido α-amino-3-hidroxi-5-metil-4-isoxazolepropionico
ANOVA Analysis of variance (Analise de Variância)
BSA Bovine Serum
CEUA Comissão de Ética em Uso Animal
CFA Complete Freund’s Adjuvant (Adjuvante Completo de Freund)
Cg Carragenina
CMD Concentração mínima detectada
COBEA Colégio Brasileiro de Experimentação Animal
CPME Corno/Coluna Posterior da Medula Espinal
DRG Dorsal Root Ganglia (Gânglio da Raiz Dorsal ou Gânglio da Coluna Posterior)
DTT Ditiotreitol
ECL Enhanced chemiluminescence
ELISA Enzyme-Linked Immunosorbent Assay (Ensaio de Imunoabsorção Enzimática)
e.p.m Erro padrão da média
EUA Estados Unidos da America
FMUSP Faculdade de Medicina da USP
GABA ácido γ-aminobutírico
GBP Gabapentina
GFAP Anti-Glial Fibrillary Acidic Protein
IASP International Association for the Study of Pain

Lista de abreviaturas
ICB Instituto de Ciências Biomédicas
IL-1 Interleucina 1
IL-1β Interleucina 1 beta
IL-4 Interleucina 4
IL-6 Interleucina 6
IL-10 Interleucina 10
i.m. Intramuscular
i.p. Intraperitoneal
i.t. Intratecal
L3 Segmento lombar 3 da medula espinal
L4 Segmento lombar 4 da medula espinal
L5 Segmento lombar 5 da medula espinal
M. Músculo
MI Medida Inicial
MP Medida Parcial
MF Medida Final
NMDA N-metil D-aspartato
PFA Paraformaldeído
Ph Potential of hydrogen (Potencial hidrogeniônico)
Sal Salina
SNC Sistema Nervoso Central
TNF-α Tumor Necrosis Factor alpha (Fator de Necrose Tumoral alfa)

Lista de abreviaturas
USP Universidade de Sao Paulo
vs. Versus
6º d Sexto dia
12º d Décimo segundo dia
13º d Décimo terceiro dia
14º d Décimo quarto dia
1 h Uma hora
3 h Três horas
5 h Cinco horas
7 h Sete horas
24 h Vinte e quatro horas
Unidades
cm Centímetros
oC Grau(s) Celsius
G Gauge
g Grama(s)
h Horas
log logaritmo
M Molar
mA Miliampère
mg Miligramas

Lista de abreviaturas
µg Microgramas
min Minutos
mL Mililitros
µL Microlitros
mM Milimolar
µm Micrometro
µM Micromolar
% Porcentagem
rpm Rotações por minuto
V Volts

Lista de figuras
LISTA DE FIGURAS
Figura 1 - Linha do tempo com as diferentes fases experimentais ao longo dos dias e horas.
Figura 2 - Ilustração do teste Von Frey eletrônico.
Figura 3 - Ilustração do teste de Hargreaves.
Figura 4 - Ilustração do teste Filamentos de Von Frey.
Figura 5 - Análise comportamental de animais com miosite crônica induzida por CFA após
tratamento com gabapentina.
Figura 6 - Análise comportamental da pata ipsilateral à lesão de animais com miosite crônica
induzida por CFA após tratamento com gabapentina.
Figura 7 - Análise comportamental da pata contralateral à lesão de animais com miosite
crônica induzida por CFA após tratamento com gabapentina.
Figura 8 - Dosagem de citocinas anti-inflamatórias no tálamo de animais com miosite crônica
induzida por CFA após tratamento com gabapentina.
Figura 9 - Dosagem de citocinas pró-inflamatórias no tálamo de animais com miosite crônica
induzida por CFA após tratamento com gabapentina.
Figura 10 - Dosagem de citocinas pró e anti-inflamatórias na medula espinal de animais com
miosite crônica induzida por CFA após tratamento com gabapentina.
Figura 11 - Dosagem de citocinas anti-inflamatórias no gânglio da coluna posterior de
animais com miosite crônica induzida por CFA após tratamento com gabapentina.
Figura 12 - Dosagem de citocinas pró-inflamatórias no gânglio da coluna posterior de
animais com miosite crônica induzida por CFA após tratamento com gabapentina
Figura 13 - Ilustração esquemática da região encefálica analisada.
Figura 14 - Imunorreatividade para astrócitos no tálamo de ratos com miosite crônica
induzida por CFA após tratamento com gabapentina.
Figura 15 - Imunorreatividade para microglia no tálamo de ratos com miosite crônica
induzida por CFA após tratamento com gabapentina.
Figura 16 - Expressão proteica de células gliais satélites de ratos com miosite crônica
induzida por CFA após tratamento com gabapentina.

Sumário
SUMÁRIO
1 INTRODUÇÃO E REVISÃO DA LITERATURA...........................................................18
1.1 DOR - CONSIDERAÇÕES GERAIS.................................................................................18
1.2 SENSIBILIZAÇÃO CENTRAL E PERIFÉRICA.............................................................20
1.3 DOR MUSCULOESQULÉTICA.......................................................................................22
1.4 DOR E CÉLULAS DA GLIA.............................................................................................23
1.5 GABAPENTINA................................................................................................................24
2 JUSTIFICATIVA E OBJETIVOS.....................................................................................26
2.1 OBJETIVOS ESPECÍFICOS..............................................................................................26
3 MATERIAL E MÉTODOS.................................................................................................27
3.1 ANIMAIS............................................................................................................................27
3.1.1 DELINEAMENTO EXPERIMENTAL..............................................................................27
3.2 INDUÇÃO DE MIOSITE CRÔNICA................................................................................27
3.3 AVALIAÇÃO DA SENSIBILIDADE NOCICEPTIVA....................................................28
3.3.1 TESTE DE HIPERALGESIA MECÂNICA - VON FREY ELETRÔNICO........................28
3.3.2 TESTE DE HIPERALGESIA TÉRMICA - HARGREAVES TEST ...................................29
3.3.3 ALODINIA MECÂNICA – FILAMENTOS DE VON FREY.............................................30
3.4 TRATAMENTO FARMACOLÓGICO.............................................................................30
3.5 ENSAIO PARA DOSAGEM DE CITOCINAS POR TECNOLOGIA MULTIPLEX......31
3.6 ENSAIO DE IMUNO-HISTOQUÍMICA...........................................................................31

Sumário
3.7 ENSAIO DE WESTERN BLOTTING...............................................................................32
3.8 ANÁLISE ESTATÍSTICA.................................................................................................33
4 RESULTADOS.....................................................................................................................34
4.1 PADRONIZAÇÃO DO TRATAMENTO FARMACOLÓGICO COM
GABAPENTINA......................................................................................................................34
4.2 EFEITO DO TRATAMENTO COM GABAPENTINA NA RESPOSTA NOCICEPTIVA
DE ANIMAIS COM MIOSITE CRÔNICA.............................................................................35
4.3 ANÁLISE DE CITOCINAS NO TÁLAMO DE RATOS COM MIOSITE
CRÔNICA.................................................................................................................................38
4.4 ANÁLISE DE CITOCINAS NA MEDULA ESPINAL DE RATOS COM MIOSITE
CRÔNICA.................................................................................................................................41
4.5 ANÁLISE DE CITOCINAS NO GÂNGLIO DA COLUNA POSTERIOR DE RATOS
COM MIOSITE CRÔNICA......................................................................................................43
4.6 ANÁLISE DE CÉLULAS GLIAIS NO TÁLAMO DE RATOS COM MIOSITE
CRÔNICA ................................................................................................................................46
4.7 ANÁLISE DE CÉLULAS GLIAIS SATÉLITES NO GÂNGLIO DA COLUNA
POSTERIOR DE RATOS COM MIOSITE CRÔNICA..........................................................49
5 DISCUSSÃO.........................................................................................................................50
6 CONCLUSÃO......................................................................................................................56
REFERÊNCIAS........................................................................................................................57

Introdução
18
1. INTRODUÇÃO E REVISÃO DA LITERATURA
1.1 DOR - CONSIDERAÇÕES GERAIS
Em 1994, a Associação Internacional para o Estudo da Dor (IASP) definiu a dor como
uma “experiência sensitiva e emocional desagradável associada a dano tecidual real ou
potencial ou à descrição desses danos” (Loeser; Treede, 2008; Merskey; Bogduk, 1994). Em
2016, Willians e Craig propuseram um novo conceito de dor: “experiência angustiante
associada a uma lesão tecidual atual ou potencial com componentes sensoriais, emocionais,
cognitivos e sociais” (Williams; Craig, 2016). Ainda, mais recentemente, Cohen e
colaboradores a definiram como “experiência somática mutuamente reconhecível que reflete a
apreensão de uma pessoa a ameaças à sua integridade corporal ou existencial” (Cohen et al.,
2018). Portanto, a dor é um complexo processo cognitivo subjetivo que depende de memória,
fatores sócio-culturais, crenças e emoções do indivíduo, e não apenas de seus aspectos
anatômicos e fisiológicos (Teixeira; Okada, 2010).
A experiência dolorosa é comum a todos os seres humanos e considerada o quinto
sinal vital (Casey, 2011; McCaffery; Pasero, 1997). A dor é uma das comorbidades mais
frequentes na atualidade e a principal queixa em unidades de emergência e pronto
atendimento (Cassell et al., 2018; McLean; Mercer, 2017; Oliveira et al., 2011). É
responsável por aproximadamente 80% da procura pelos serviços de saúde e está presente em
mais de 70% dos pacientes que buscam os consultórios brasileiros por motivos diversos
(Bottega; Fontana, 2010; Rocha et al., 2007). Embora seu tratamento seja um direito humano
fundamental, a dor ainda é um sintoma subtratado que gera sofrimento e prejuízo econômico
aos indivíduos e à sociedade (IASP, 2010).
A dor pode ser classificada em aguda e crônica. A dor aguda é um sintoma que alerta
para a ocorrência de lesões no organismo, sendo fundamental para a preservação da
integridade do indivíduo. Contudo a dor crônica não possui este papel biológico (Teixeira,
2001; Teixeira et al., 1999). A dor crônica perdura por mais de seis meses, é de difícil
tratamento e se constitui um problema de saúde pública por seu alto custo econômico e
impacto negativo na qualidade de vida de pacientes e seus familiares (Andrade, 2014;
Fernandes; Gomes, 2011). Este tipo de dor atinge 30 a 40% dos brasileiros e é a principal
causa de absenteísmo, licenças e aposentadorias por doença, baixa produtividade e
indenizações trabalhistas (Bottega; Fontana, 2010). Além disso, é predominante no sexo

Introdução
19
feminino (56%), possui maior prevalência nas regiões sul e sudeste e 49% dos índividuos com
dor crônica estão instisfeitos com o manejo de sua dor (Souza et al., 2017).
Os termos dor e nocicepção se diferem. Nocicepção se refere aos sinais que chegam ao
sistema nervoso central (SNC) resultante da ativação de terminações nervosas livres de alto
limiar, denominadas nociceptores, que fornecem informações sobre a lesão tecidual
ocasionada por estímulos nocivos. Dor é a experiência emocional desagradável, que
geralmente acompanha a nocicepção. O processo nociceptivo é composto por quatro etapas:
transdução, transmissão, modulação e percepção (Fein, 2011).
Na transdução, os neurônios que transmitem a informação nociceptiva captam
estímulos nocivos da periferia por meio dos receptores sensoriais especializados,
nociceptores, e os transformam em potencial de ação. Na transmissão, as fibras nervosas
conduzem a informação nociceptiva ao sistema nervoso central por meio de fibras aferentes
primárias mielinizadas de grande calibre (fibras Aδ), com velocidade de condução de 5-30
m/s e resposta a estímulos térmicos e mecânicos e fibras não mielinizadas de pequeno calibre
(fibras C), com baixa velocidade de condução (0,5-2 m/s) e resposta a estímulos de origem
térmica, mecânica e química (Fernandes; Gomes, 2011; Julius; Basbaum, 2001).
Os corpos celulares dos neurônios aferentes primários localizam-se nos gânglios da
coluna posterior (DRG). As fibras desses neurônios são projetadas a partir da periferia em
direção à coluna posterior da medula espinal (CPME), chegando nas lâminas I e II de Rexed,
onde fazem sinapse com neurônios de segunda ordem. É no DRG e na CPME que o impulso
gerado é modulado. Então, os axônios dos neurônios de segunda ordem cruzam o plano
mediano da medula espinal pela comissura branca e ascendem cranialmente, constituindo as
principais vias de transmissão da informação nociceptiva (Costigan; Woolf, 2000; Millan,
1999).
Esses neurônios podem fazer conexões com os de terceira ordem em várias regiões da
formação reticular e com os de quarta ordem nos núcleos intralaminares do tálamo,
difundindo-se para diferentes regiões do córtex (trato espino-reticular) ou então conectar-se a
neurônios de terceira ordem no tálamo e projetar-se ao córtex somatossensorial (trato espino-
talâmico lateral). Suas projeções vão para áreas localizadas do córtex cerebral e correspondem
a diferentes regiões do corpo, ocorrendo assim, a percepção da dor (Costigan; Woolf, 2000;
Millan, 1999).

Introdução
20
Tecidos ilesos, ao sofrerem estímulos nocivos, ativam o reflexo de retirada, impedindo
a ocorrência de lesão tecidual. Essa dor é fisiológica e possui função protetora. Entretanto,
quando o tecido é inflamado ou de fato sofre uma lesão, ocorre a dor patológica com
sensibilização periférica e central. Esses mecanismos envolvem a liberação de mediadores
nociceptivos no local e em regiões do sistema nervoso central que reduz o limiar de ativação
da via nociceptiva e amplifica sua resposta, resultando em hiperalgesia (aumento da
sensibilidade à dor) e alodinia (resposta de dor a estímulos não nocivos) (Besson, 1999;
Kidd; Urban, 2001). Sabe-se, atualmente, que a alodinia ocorre em consequência ao
brotamento de fibras Aβ para a lâmina II da coluna posterior da medula espinal (Woolf et al.,
1992).
1.2 SENSIBILIZAÇÃO CENTRAL E PERIFÉRICA
Sensibilização é o fenômeno que corresponde à modificação no estado funcional dos
neurônios e das vias nociceptivas por todo o neuroeixo, sendo o aumento da atividade dessas
circuitarias muito importante na geração da dor crônica. Esse processo se desenvolve com a
presença de estímulos nocivos intensos ou repetitivos, resulta em redução do limiar de
ativação da via nociceptiva e amplificação de sua resposta. Assim, o indivíduo passa a
apresentar hiperalgesia e alodinia (Ashmawi; Freire, 2016; Latremoliere; Woolf, 2009).
A sensibilização pode ocorrer em nível periférico ou central. A sensibilização
periférica é aquela que se desenvolve no local da lesão tecidual e que ocorre devido a
mudanças nos limiares dos nociceptores locais. Enquanto que a sensibilização central pode ser
compredida como uma facilitação do sistema nervoso central à passagem de estímulos
nociceptivos. O processo de sensibilização pode se desenvolver por meio de alguns
mecanismos bioquímicos, como o aumento da excitabilidade da membrana, aumento da
eficácia sináptica e/ou pela redução da atividade da via inibitória nociceptiva (Ashmawi;
Freire, 2016; Latremoliere; Woolf, 2009).
Como elucidado, a ativação de nociceptores e canais iônicos na periferia geram
correntes de despolarização em resposta ao estímulo nocivo. Essas correntes são conduzidas
até a medula espinal e moduladas por meio de sinapses excitatórias mediadas pela ação de
glutamato em receptores de N-metil D-aspartato (NMDA) e de ácido α-amino-3-hidroxi-5-
metil-4-isoxazolepropionico (AMPA) e em canais iônicos kainato. Isto ocorre por meio da

Introdução
21
ativação de neurônios descendentes inibitórios, os quais liberam glicina e ácido γ-
aminobutírico (GABA). O impulso nervoso, projetado para o tálamo e para diversas outras
regiões encefálicas, segue até o córtex, onde a dor é percebida (Price, 2000).
No processo de sensibilização, há plasticidade do sistema nociceptivo, que se
manifesta por aumento na atividade deste sistema a estímulos repetitivos ou intensos. Os
nociceptores podem reduzir seu limiar aos estímulos nocivos devido a alterações na sua
conformação proteica ou devido à entrada de cálcio para o interior do neurônio. Além disso,
também ocorrem alterações plásticas de neurônios na coluna posterior da medula espinal, a
modulação da transmissão nociceptiva, desinibição e modificação neuronal. Estímulos de alta
frequência gerados por estímulos nociceptivos intensos ou sustentados resultam em liberação
de glutamato, gerando potenciais excitatórios pós-sinápticos (wind-up) (Thompson et al.,
1993; Woolf; Costigan, 1999).
A modulação da transmissão sináptica nociceptiva envolve ativação de uma cascata de
sinalização intracelular e resulta em facilitação sináptica excitatória e redução da sua inibição.
O envolvimento dos receptores de NMDA nessa modulação se dá por dois mecanismos.
Primeiro, a supressão do bloqueio dos receptores de NMDA pelo magnésio iônico gerada pela
somação de potenciais de ação com a ativação dos nociceptores. Segundo, o aumento da
atividade dos receptores de NMDA pela convergência dos sinais provindos de outros
receptores, como os receptores de tirosina quinase, que estão presentes na coluna posterior da
medula espinal. Receptores AMPA também são importantes na facilitação do processo de
transmissão sináptica. Esses receptores permitem o influxo de cálcio em quantidades
suficientes para produzirem uma facilitação da transmissão sináptica em nível medular. A
sensibilização não só envolve facilitação de sinapses excitatórias, mas também a falta da
inibição dessas sinapses. A inibição da transmissão sináptica nos neurônios da substância
gelatinosa pode ser desencadeada pela ativação de fibras Aδ. A desinibição requer a ativação
de receptores de NMDA e o aumento de cálcio na fenda sináptica (Thompson et al., 1993;
Woolf; Costigan, 1999).
Outro processo envolvido é a modificação dos neurônios nociceptivos.
Perifericamente, o aumento da liberação de substância P pelo primeiro neurônio nociceptivo
alteraria a atividade de neurônios na coluna posterior da medula espinal. E no sistema nervoso
central, haveria um aumento na expressão de receptores em neurônios da coluna posterior
com o aumento da ativação de neurônios sensitivos periféricos (Traub, 1996).

Introdução
22
1.3 DOR MUSCULOESQUELÉTICA
Pesquisas recentes e de caráter epidemiológico têm apontado a dor musculoesquelética
como um dos tipos de dor mais prevalente na população. Na Europa, 36% da população
referem apresentar dor musculoesquelética crônica (Cimas et al., 2018), nos Estados Unidos,
33% (Yelin et al., 2016). No Brasil, a dor muscular foi apontada em terceira posição entre as
dores mais prevalentes (Teixeira et al., 2001) e em primeira posição entre as dores mais
frequentes em Centros de Referencia em Dor no estado de São Paulo (dados FMUSP). Ainda,
a dor musculoesquelética esteve presente em 64% das brasileiras entrevistadas (dados
Mundipharma) e em 86% dos idosos (Miranda et al., 2012).
Distúrbios musculoesqueléticos representam um grupo de condições que afetam os
músculos, tendões, ligamentos, articulações, nervos periféricos e vasos sanguíneos de suporte
no corpo. A dor no tecido musculoesquelético pode resultar de uma variedade de desordens,
como inflamações dos tendões, distúrbios de compressão do nervo, osteoartroses, dores
miofasciais, fibromialgia, miosite ou distensões (Punnett; Wegman, 2004).
A dor no sistema musculoesquelético é frequentemente classificada como inflamatória
ou não inflamatória. A dor inflamatória pode ser localizada em um local, como a tendinite, ou
dispersa, como a artrite reumatóide. Em paralelo, a dor pode ser localizada no local da lesão
(hiperalgesia primária) ou pode se difundir para locais não lesionados (dor referida ou
hiperalgesia secundária), dependendo do local e da natureza da inflamação. As condições de
dor musculoesquelética não inflamatória são mais complexas e muito mais difíceis de
caracterizar ou diagnosticar devido à falta de dano tecidual observável ou inflamação
(Abdelhamid; Sluka, 2015). A lesão muscular resulta na liberação de substâncias pró-
inflamatórias e sensibilização de nociceptores na periferia e na coluna posterior da medula
espinal, transmitindo a informação nociceptiva para centros cerebrais superiores (Sluka,
1996).
Modelos animais de dor muscular são comumente usados para obter uma melhor
compreensão dos mecanismos relacionados ao processamento da dor. Geralmente, os modelos
de dor muscular são gerados por meio de inflamação muscular, injeção de soluções ácidas e
exercício físico intenso (Pratt et al., 2013). Ainda, pode-se gerar dor aguda ou crônica, como
nos modelos de inflamação muscular por Carragenina (Ren, Dubner, 1999; Radhakrishnan et
al., 2003) e Adjuvante Completo de Freund (Chacur et al., 2009; Rosa et al., 2017). O

Introdução
23
Adjuvante Completo de Freund (CFA) é uma solução de antígeno emulsionada em óleo
mineral utilizada como imunopotenciador e é composto por micobactérias inativadas. Após a
sua administração, este composto é capaz de atrair células do sistema imune e provocar uma
cascata de eventos ligados a este processo, induzindo um efeito inflamatório de longa duração
(Larson et al., 1986).
Estudos demonstraram que a inflamação crônica no músculo gastrocnêmio de ratos
induz alterações em neurônios da coluna posterior da medula espinal, sendo esta
sensibilização central a base para a dor espontânea, hiperalgesia e alodinia observada após a
lesão (Hoheisel et al., 2000, 2005; Tenschert et al., 2004). Estes estudos também
demonstraram que lesões no nervo ou inflamação muscular induzem alterações morfológicas
em neurônios algumas horas após lesão (Hoheisel et al., 1998) e que a miosite crônica
induzida no tecido musculoesquelético leva também a alterações morfológicas e funcionais de
astrócitos na medula espinal (Tenschert et al., 2004). Estudos eletrofisiológicos in vivo
também demonstraram que as células da glia participam da despolarização das fibras
nociceptivas no modelo de miosite crônica induzida pela injeção de CFA (Chacur et al.,
2009). Ainda, estudos demonstram um aumento de interleucinas (IL) dos tipos 1 e 6 após
indução de dor muscular aguda pela injeção de Carragenina no músculo gastrocnêmio de
animais (Loram et al., 2007).
1.4 DOR E CÉLULAS DA GLIA
Pesquisas têm demonstrado que as células da glia participam dos processos
nociceptivos, especialmente em condições crônicas, como em situações de inflamação
periférica e neuropatias (Milligan; Watkins, 2009; Tsuda et al., 2005), por meio da liberação
de substâncias que modulam a resposta nociceptiva, como prostaglandinas, glutamato, ácido
araquidônico, óxido nítrico e citocinas (Marchand et al., 2005), exercendo papel relevante na
transmissão da informação nociceptiva no SNC (Watkins; Maier, 2003; Watkins et al., 2003)
e geração de alodinia e hiperalgesia (Milligan et al., 2000; Sweitzer et al., 1999).
Modelos de dor crônica investigados até agora resultaram em mudanças fenotípicas da
glia, principalmente na medula espinal (Hains; Waxman, 2006) e no sistema nervoso
periférico (Ohtori et al., 2004; Rothermundt et al., 2007), mas também em estruturas
superiores, como o tálamo, onde ocorre a interpretação das informações transmitidas pela via
nociceptiva (LeBlanc et al., 2011). Isto sugere que a ativação da glia pode ser um mecanismo

Introdução
24
importante para a persistência de hipersensibilidade. Além disso, a inibição de células gliais
em modelos animais demonstrou atenuar dor inflamatória e neuropática (Bastos et al., 2013;
Chacur et al., 2004; Milligan et al., 2003).
Microglia e astrócitos parecem ser a fonte principal de citocinas no sistema nervoso.
Uma lesão neuronal induz a expressão de IL-1, primeiramente pela microglia, que
subsequentemente libera IL-1, IL-6 e TNFα. IL-1 formada a partir da microglia estimula
produção astrocítica de TNFα, IL-1 e IL-6. A ativação microglial aumenta a permeabilidade
da barreira hemato-encefálica, permitindo a invasão de mais células imunes, tornando a
inflamação persistente (Deleo, 2004).
Para que a resposta inflamatória ocorra, citocinas são necessárias. Elas podem ser
reconhecidas por neurônios e utilizadas para desencadear diversas reações celulares que
influenciam na atividade, proliferação e sobrevida da célula imune e na produção e atividade
de outras citocinas. Elas podem ser pró-inflamatórias, como por exemplo, IL-1 e TNF, ou
anti-inflamatórias, como a IL-10. Citocinas desempenham papel importante no processo
doloroso por meio de diferentes mecanismos ao longo das vias de transmissão da dor, desde a
condução da resposta inflamatória aos locais de lesão, cicatrização apropriada da ferida e são
essenciais para manutenção do processo nociceptivo (Kraychete et al., 2006; Marchand et al.,
2005; Oliveira CMBd et al., 2011).
1.5 GABAPENTINA
A gabapentina (Neurontin ®) foi originalmente desenvolvida como um análogo
químico do ácido gama - aminobutírico (GABA) para reduzir o reflexo da medula para
tratamento de espasticidade e mostrou ter efeito anticonvulsivo. Clinicamente, a gabapentina é
indicada como droga adjuvante para tratamento de crises convulsivas parciais e dor
neuropática.
Após seu efeito analgésico em pacientes com dor neuropática intratável, a gabapentina
passou a ter seu efeito antinociceptivo relatado em vários modelos de nocicepção animal, até
mesmo em modelos de dor inflamatória induzida e lesões de tecidos (Cheng; Chiou, 2006).
Um estudo realizado por Yang e colaboradores demonstrou que o uso de gabapentina foi
capaz de atenuar hiperalgesia térmica e ativação da microglia na medula espinal de ratos com
monoartrite (Yang et al., 2012). Ainda, estudos demonstraram que a gabapentina foi capaz de

Introdução
25
reverter alodinia mecânica em modelos de dor neuropática e artrite (Park et al., 2015;
Zvejniece et al., 2015).
Atualmente, para alívio da dor muscular crônica são muito utilizados opióides, como a
codeína e o tramadol, ou ainda, medicamentos tricíclicos como a ciclobenzaprina. Apesar de
seus diversos efeitos colaterais, como sedação, depressão respiratória, constipação e
dependência, os opióides e os tricíclicos ainda são os fármacos mais empregados para dor
crônica. Entretanto, a gabapentina pode ser útil na redução da dependência e da tolerância ao
opióide, pois em vários estados de dor o uso de opióides foi reduzido quando a gabapentina
foi coadministrada clinicamente. Além disso, a gabapentina é caracterizada por doses eficazes
toleráveis e poucos efeitos colaterais ou indesejados clinicamente, sendo que por via intratecal
(10-300μg) sua eficácia é maior (Cheng; Chiou, 2006).
Apesar de ser um aminoácido com a estrutura do neurotransmissor GABA, a
gabapentina não interage de modo significativo com esse ou outro neurotransmissor. Seu
mecanismo de ação consiste na redução da hiperexcitabilidade dos neurônios do corno dorsal
da medula espinal induzida pela sensibilização central. É possível que sua ação analgésica
ocorra por ligação pré-sináptica da gabapentina à subunidade α2δ de canais de cálcio
dependentes de voltagem do tipo N presentes nos neurônios, diminuindo a entrada de cálcio
nas terminações nervosas e reduzindo a liberação de neurotransmissores (Clivatti et al., 2009).
Estudos mais recentes sugerem que a gabapentina se liga especificamente na subunidade α2δ1
destes canais de cálcio (Morioka et al., 2015; Tsukumo et al., 2011; Yang et al., 2012;
Zhou; Luo, 2014).

Justificativa e objetivos
26
2. JUSTIFICATIVA E OBJETIVOS
A dor musculoesquelética é um dos tipos mais recorrentes e persistentes de dor que
não responde satisfatoriamente às intervenções convencionais e sua fisiopatologia permanece
não totalmente esclarecida. Ainda, há poucos estudos com modelos de dor muscular. Portanto,
investigar os mecanismos envolvidos nesse quadro nociceptivo e novas alternativas
terapêuticas são necessárias.
Até o momento, nossas pesquisas vêm indicando que a indução de miosite crônica
pela administração de CFA é capaz de diminuir o limiar nociceptivo de ratos em testes
comportamentais e aumentar astrócitos e microglia no corno dorsal da medula espinal e
gânglio da coluna posterior desses animais, quando comparado aos animais controle (Rosa et
al., 2017).
Uma vez observado o envolvimento de células gliais em nosso modelo experimental,
optamos pelo uso da gabapentina como um tratamento potencial por seu efeito
antinociceptivo relatado em modelos experimentais (Cheng; Chiou, 2006; Quintao et al.,
2012) e por atenuar a ativação de células da glia na medula espinal de ratos (Yang et al.,
2012). Dessa forma, nosso objetivo neste trabalho é observar se a intervenção farmacológica
com gabapentina é capaz de reverter o processo nociceptivo dos animais com miosite crônica,
interferir em células gliais e na liberação de citocinas produzidas a nível central e periférico.
2.1 OBJETIVOS ESPECÍFICOS
Avaliação da sensibilidade nociceptiva dos animais com miosite crônica induzida após
tratamento farmacológico com gabapentina;
Analisar citocinas pró e anti-inflamatórias (IL-1β, IL-6, IL-10, TNFα) no gânglio da
coluna posterior, tálamo e porção lombar da medula espinal dos animais com miosite
crônica por meio de técnica de dosagem de citocinas Multiplex, após tratamento
farmacológico com gabapentina;
Analisar a imunorreatividade de células gliais (astrócitos e microglia) no tálamo dos
animais com miosite crônica por meio da técnica de imuno-histoquímica, após
tratamento farmacológico com gabapentina;
Analisar a expressão de astrócitos no gânglio da coluna posterior dos animais com
miosite crônica por meio da técnica western blotting, após tratamento farmacológico
com gabapentina;

Material e métodos
27
3. MATERIAL E MÉTODOS
3.1 ANIMAIS
Foram utilizados ratos da linhagem Wistar, machos, pesando entre 200-220 gramas,
fornecidos pelo Biotério Central do Instituto de Ciências Biomédicas e mantidos no Biotério
do Departamento de Anatomia, ambos da Universidade de São Paulo (ICB-USP). Os animais
foram mantidos em sala apropriada, com ciclo claro e escuro (12/12 h), isolamento acústico,
temperatura controlada (22 ºC 1) e acesso livre à água e ração. Todos os procedimentos
foram realizados de acordo com o protocolo aprovado pelos Princípios Éticos de
Experimentação Animal adotado pelo Colégio Brasileiro de Experimentação Animal
(COBEA) e pela Comissão de Ética no Uso de Animais do ICB (protocolo aprovado nº47, fls.
18, livro 03). Para a realização dos experimentos, os animais foram manipulados
considerando os princípios e o guia de uso de animais de laboratório envolvendo dor e
nocicepção (Zimmermann, 1983).
3.1.1 DELINEAMENTO EXPERIMENTAL:
Os animais foram alocados conforme divisão dos grupos abaixo:
Naive (animais controle) n=30
CFA + gabapentina (i.t., 200µg/50µl) n= 30
CFA (intramuscular - i.m., 150µg/300µl) + salina (veículo de diluição da gabapentina,
intratecal - i.t., 50µl) n=30
3.2 INDUÇÃO DE MIOSITE CRÔNICA
A miosite crônica foi induzida no músculo gastrocnêmio do membro posterior direito
de ratos, pela injeção de CFA (Sigma, St Louis, Mycobacterium tuberculosis) na dose de
150µg/300µl. Cada animal recebeu duas injeções de 150µL, uma na porção medial e outra na
porção lateral da cabeça do músculo gastrocnêmio. Animais com o músculo intacto foram
utilizados como controle (Chacur et al., 2009). Cabe mencionar que a injeção de CFA no
músculo foi realizada em um único dia. Os testes comportamentais foram aplicados antes de

Material e métodos
28
quaisquer procedimentos (medida inicial - MI), no 6º dia (medida parcial - MP), no 12º dia
após a injeção do irritante (medida final - MF) e em diferentes tempos (horas) após
administração da gabapentina.
Figura 1 - Linha do tempo com as diferentes fases experimentais ao longo dos dias e horas.
3.3 AVALIAÇÃO DA SENSIBILIDADE NOCICEPTIVA
Para a realização dos testes comportamentais, todos os ratos foram inicialmente
habituados por aproximadamente 30 minutos, em dois dias alternados que antecederam os
experimentos, na sala de experimentação e nos equipamentos utilizados. Realizamos um
estudo “duplo cego” e randomizado. Todos os resultados dos testes comportamentais foram
avaliados por meio de comparações das médias obtidas nos diferentes grupos experimentais.
Os testes foram aplicados nas patas experimental e contralateral à lesão.
3.3.1 TESTE DE HIPERALGESIA MECÂNICA - VON FREY ELETRÔNICO
Para determinação da hiperalgesia mecânica foi utilizado o teste de Von Frey
eletrônico (Anesthesiometer, IITC Inc. Life Science Instruments, Woodland Hills, Ca, USA),
ilustrado na figura 2, segundo método descrito por Guerrero et al. (Guerrero et al., 2006),
modificado. Neste teste, uma ponteira do próprio aparelho é pressionada e aplicada contra o
músculo gastrocnêmio do membro posterior dos animais. Neste teste, o limiar nociceptivo é
representado pela força em gramas necessária para a retirada do membro do animal.

Material e métodos
29
Figura 2 - Von Frey Eletrônico. Teste utilizado para aferir a hiperalgesia mecânica dos animais.
3.3.2 TESTE DE HIPERALGESIA TÉRMICA - HARGREAVES TEST
A determinação da hiperalgesia térmica foi feita por meio do teste plantar descrito por
Hargreaves (Hargreaves et al., 1988), conforme ilustrado na figura 3. Neste teste, os animais
são alocados em compartimentos de acrílico e posicionados sobre uma plataforma de vidro
que permite a passagem de luz e calor. Uma fonte de luz infravermelha com temperatura
constante acoplada a um monitor digital é colocada sob o vidro, sendo posicionada e acionada
sob o membro posterior direito do animal. Quando a fonte de luz é posicionada e acionada,
um cronômetro digital é deflagrado no monitor acoplado a fonte luminosa, até que o animal
retire a pata, quando, então, a fonte de luz cessa e o cronometro para automaticamente. Neste
teste, o limiar nociceptivo é representado pelo tempo em segundos necessário para a retirada
da pata.
Figura 3 - Teste de Hargreaves. Teste utilizado para aferir a hiperalgesia térmica dos animais.

Material e métodos
30
3.3.3 ALODINIA MECÂNICA - FILAMENTOS DE VON FREY
A determinação da alodinia mecânica foi avaliada por ensaio quantitativo, em resposta
a estímulo tátil aplicado à pata posterior do rato, segundo método descrito por Chaplan
(Chaplan et al., 1994), modificado, conforme ilustrado na figura 4. Neste teste, os ratos são
colocados, individualmente, em caixas de acrílico com base aberta. A caixa de acrílico fica
sobre base de arame fixada na parede, que mede 1 m de comprimento por 45 cm de largura,
com espaço entre os arames de 1 cm para permitir acesso às patas destes animais. Para o
ensaio de alodinia, é empregada uma série logarítmica de 10 filamentos de Von Frey
(Anesthesiometer Semmes-Weinstein, Stoelting Co., EUA). Os filamentos e seus valores
respectivos em gramas são: 3.61 (0.407g.); 3.84 (0.692g.); 4.08 (1.202g.); 4.17 (1.479g.); 4.31
(2.041g.); 4.56 (3.630g.); 4.74 (5.495g.); 4.93 (8.511g.); 5.07 (11.749g) e 5.18 (15.136g.). O
filamento capaz de induzir a retirada da pata, duas vezes consecutivas, foi considerado como a
força em gramas necessária para induzir a resposta (100% de resposta).
Figura 4 - Filamentos de Von Frey. Teste utilizado para aferir a sensibilidade tátil dos animais.
3.4 TRATAMENTO FARMACOLÓGICO
O tratamento farmacológico com gabapentina foi realizado por via intratecal, uma via
de administração que consiste na injeção de substâncias diretamente no líquido
cefalorraquidiano, no espaço subaracnóideo. Para padronização do tratamento, utilizamos as
doses de 100µg e 200μg de gabapentina diluídas em salina 0,9% (Takasusuki; Yaksh, 2011).
A gabapentina foi aplicada no 13º dia após a lesão induzida por CFA. Sua aplicação foi feita
sob anestesia inalatória com Isoflurano. Para análise da ação do fármaco, os testes
comportamentais foram repetidos 1, 3, 5, 7 e 24 horas após administração a fim de melhor
identificar o período de resposta a este medicamento.

Material e métodos
31
3.5 ENSAIO PARA DOSAGEM DE CITOCINAS POR TECNOLOGIA
MULTIPLEX
Após os ensaios comportamentais, os animais utilizados nesta metodologia foram
eutanasiados três horas após administração do fármaco para a retirada de tecido (gânglio da
coluna posterior, tálamo e porção lombar da medula espinal) de todos os grupos descritos
anteriormente e analisados para cada citocina específica. Os tecidos foram homogeneizados
com tampão extração de suplementado com inibidor de protease (Roche). Os homogenatos
foram centrifugados (12000RPM por 20 min a 4°C) e o sobrenadante recolhido. Para a
dosagem de citocinas IL-1β, IL-4, IL-6 e IL-10 e TNFα foram utilizados reagentes específicos
contidos no kit Milliplex®TM Map (Millipore Corporation, Darmstadt- Germany), conforme
protocolo especificado pelo fabricante. Foram adicionados 200μL de solução tampão por
poço (placa de 96 poços) e a placa foi fechada e homogeneizada por dez minutos à
temperatura ambiente. O sobrenadante foi retirado e 25μl da solução padrão/poço foram
adicionados. Em seguida, foram adicionados os controles, as amostras e as beads. A
incubação teve um total de tempo de 1 hora à temperatura ambiente e em seguida foi realizada
a incubação overnight à temperatura de 4ºC, sob agitação constante. Após a incubação
overnight a placa foi deixada por mais uma hora sob agitação constante à temperatura
ambiente. Em seguida, o conteúdo foi despejado e a placa lavada com tampão Wash buffer,
200µl em cada poço, por 3 vezes. Após a lavagem foram adicionados 25µl dos anticorpos de
detecção na placa que foi mantida sob agitação constante em temperatura ambiente.
Posteriormente foram adicionados 25µl do complexo estreptavidina-ficoeritrina por poço e a
placa permaneceu em agitação por 30 minutos em temperatura ambiente. Após o tempo
determinado de incubação a placa foi lavada novamente por 3 vezes com solução tampão
Wash buffer. Após as lavagens foi adicionada uma solução de parada (Sheat Fluid), 125μl por
poço. A placa foi levada para a leitura no aparelho Bioplex-200 System (Bio-Rad). A
concentração mínima detectada (CMD) foi calculada pelo aparelho em pg/ml e a dada curva
padrão comparada com cada amostra para as diferentes citocinas (Oliveira et al., 2017).
3.6 ENSAIO DE IMUNO-HISTOQUÍMICA
Após realização dos testes comportamentais e três horas após administração do
fármaco, os animais utilizados nesta metodologia foram eutanasiados com os grupos de
animais foram anestesiados (quetamina e xilazina 1:1, 100µL/100g de peso corpóreo) e

Material e métodos
32
submetidos à perfusão transcardíaca, com solução salina (300 ml), seguida de solução
fixadora (300 ml) constituída de paraformaldeído 4% dissolvido em tampão fosfato 0,1 M
(PB, pH 7,4). Após a perfusão, o tálamo dos animais foi coletado e armazenado em
paraformaldeído 4%, durante 4 horas e transferido para solução contendo sacarose a 30% em
PB para crioproteção. Após 24 horas, o tecido foi cortado por micrótomo de congelamento na
espessura de 40μm. Para análise de células gliais, os cortes foram incubados e submetidos à
metodologia de Imuno-histoquímica com anticorpo primário GFAP - marcador de astrócitos
(1:1000; Clone G-A-5 – Sigma/EUA) e OX-42 - marcador de microglia (1:1000; BD
Bioscience Pharmigen/EUA). A imunorreatividade foi analisada ao microscópio de luz. A
quantificação das células foi realizada por meio do software Image (NIH-EUA). Este método
consiste em análise por meio de microscopia de campo claro e contraste de interferência. A
densidade óptica encontrada no grupo controle será considerada 100% para a análise
comparativa entre o grupo controle e o experimental. Posteriormente, a média da diferença
porcentual de cada corte analisado foi calculada (média de 5 cortes para cada animal). A
determinação das regiões de interesse foi feita de acordo com o Atlas de Neuroanatomia “The
rat brain in stereotaxic coordinates” (Paxinos, Watson. 5. ed., 2005).
3.7 ENSAIO DE WESTERN BLOTTING
O conteúdo proteico do material isolado - gânglio da coluna posterior (L4-L6) foi
extraído e dosado pelo método de Bradford (Amresco, U.S.A) (Bradford, 1976). Após a
quantificação de proteína total pelo método de Bradford, os materiais foram diluídos em um
mesmo volume de tampão Laemmli, contendo 54 mg/ml de DTT, e o material foi aplicado em
gel de poliacrilamida a 12% e submetido a eletroforese com corrente contínua de 120V. Após
a separação eletroforética, as proteínas foram transferidas para uma membrana de
nitrocelulose (Millipore, 0,2 μm de diâmetro) de acordo com a técnica descrita por Towbin e
colaboradores (Towbin et al., 1979). Os antígenos presentes na membrana de nitrocelulose
foram submetidos à caracterização imunoenzimática. Após bloqueio com leite desnatado
(Molico, Nestlé, Brasil) 5% em tampão Tris-Salina (Tris 10 mM e NaCl 0,15 M, pH 7,5), por
2 horas, as membranas foram incubadas com anticorpos monoclonais específicos para
detecção de células gliais satélites, GFAP (1:1000 - Sigma Aldrich) em solução bloqueadora
(1% de BSA) por 18 horas a 4ºC. Em seguida, as membranas foram lavadas com Tris-Salina e
incubadas por 2 horas com o anticorpo secundário marcado com peroxidase de raiz forte

Material e métodos
33
(1:5000 AmershanBiosciences, NJ/EUA). O excesso de conjugado foi removido com mais de
um ciclo de lavagens. As membranas foram reveladas utilizando o Kit ECL
(AmershanBiosciences, NJ/EUA) de quimioluminescência e analisadas quanto à densidade
das bandas marcadas, com o programa ScionImage (Scion Corporation, Frederick, MD/EUA).
A análise dos dados foi feita pela média das diferenças percentual entre os diferentes grupos
experimentais. A correção foi realizada pela densidade óptica para a β-actina (1:10000 Sigma,
St. Louis, MO/USA), considerando as amostras dos animais controle como o padrão para a
normalização dos resultados.
3.8 ANÁLISE ESTATÍSTICA
Os dados foram representados como média ± e.p.m. e a análise estatística foi gerada
utilizando o programa GraphPad Prism 5.01 (GraphPad Software Inc., CA, USA). Para os
ensaios comportamentais, a comparação estatística entre os grupos e tempos foi realizada
usando a análise de variância de duas vias (Two-way ANOVA) e no grupo CFA diferentes
tempos usando a análise de variância de uma via (One-way ANOVA), seguidas pelo pós-teste
de Bonferroni. O índice de significância foi considerado de p < 0,05. Nos ensaios de
westernblotting, a comparação estatística entre os grupos foi realizada usando a análise de
variância de uma via (One-way ANOVA), seguidas pelo pós-teste de Bonferroni. O índice de
significância foi considerado de p < 0,05 (Sokal; Rohlf, 1981).
.

Resultados
34
4. RESULTADOS
4.1 PADRONIZAÇÃO DO TRATAMENTO FARMACOLÓGICO COM
GABAPENTINA
Para padronização do tratamento farmacológico, utilizamos o teste comportamental de
hiperalgesia mecânica - Von Frey Eletrônico, conforme detalhado anteriormente. Após
determinação do limiar nociceptivo dos animais (MI) por meio deste teste, realizamos a
injeção de CFA no músculo gastrocnêmio para induzir a miosite crônica, e então, repetimos o
teste comportamental no 6º (MP) e no 12º dia (MF - quando a miosite crônica está instalada)
após a lesão.
No 13º dia, o grupo experimental recebeu a aplicação intratecal de gabapentina (GBP)
sob anestesia inalatória com Isoflurano. Para avaliarmos o efeito do fármaco, o teste
comportamental foi repetido nas 1ª, 3ª, 5ª, 7ª e 24ª horas após administração do medicamento.
Para padronização do tratamento, testamos duas doses descritas na literatura, 100µg e 200µg
de gabapentina intratecal, diluídas em salina 0,9% (Takasusuki; Yaksh, 2011).
De acordo com nossos resultados (Figura 5), a dose de 200µg foi mais eficaz na
reversão do quadro nociceptivo em comparação a dose menor. Nota-se que o tratamento com
a dose mais alta de gabapentina (CFA + GBP 200µg) melhorou o quadro nociceptivo já na
primeira e terceira hora após a administração da droga (P<0,001 em comparação com o grupo
CFA). Após, na 5ª e 7ª hora, a ação analgésica do fármaco começou a diminuir. Na 24ª hora,
observou-se que já não havia qualquer efeito do medicamento.
Ainda, observamos que não houve alteração estatística após tratamento com
gabapentina na dose de 100µg (CFA + GBP 100µg) em nenhum dos tempos analisados. O
grupo CFA, sem tratamento farmacológico, permaneceu com nocicepção durante todo o
período analisado (P<0,05 em relação ao grupo controle), enquanto que o grupo naive,
utilizado como controle, manteve seu limiar estável ao longo do tempo. Portanto, elegemos a
dose de 200µg para aplicação da terapia farmacólogica com gabapentina intratecal.

Resultados
35
Figura 5 - Análise comportamental de animais com miosite crônica induzida por CFA após
tratamento com gabapentina. O limiar nociceptivo, avaliado pelo teste de Von Frey eletrônico e
expresso em gramas, foi determinado antes (tempo zero - MI), 6, 12 e 13 dias após a lesão com CFA -
i.m. Os animais foram tratados com gabapentina (GBP - nas doses de 100 ou 200μg) i.t., no 13º dia após
injeção do CFA. O teste comportamental foi novamente mensurado nos tempos 1, 3, 5, 7, e 24 horas após
administração do fármaco. Os resultados representam a média epm de 4 animais por grupo. #p<0.05
entre os grupos NAIVE e CFA. *p<0.05 entre os grupos CFA + GBP 200ug e CFA.
4.2 EFEITO DO TRATAMENTO COM GABAPENTINA NA RESPOSTA
NOCICEPTIVA DE ANIMAIS COM MIOSITE CRÔNICA
Após padronização do tratamento, foram realizados ensaios comportamentais para
avaliação de hiperalgesia térmica (painel A), alodinia tátil (painel B), e hiperalgesia mecânica
(painel C) nas patas ipsilateral (Figura 6) e contralateral à lesão (Figura 7). Novamente, os
testes foram aplicados na 1ª, 3ª, 5ª, 7ª e 24ª hora após administração do medicamento.
Nossos resultados demonstraram que a dose de 200μg de gabapentina aumentou o
limiar nociceptivo dos animais com miosite crônica nos testes de hiperalgesia térmica e
alodinia, no lado ipsilateral à lesão, durante cinco horas após sua aplicação, em comparação
ao grupo CFA + salina (Figura 6A e B, respectivamente). Em relação à hiperalgesia
mecânica, foi observado o aumento do limiar nociceptivo até a sétima hora após a
administração do fármaco, quando comparado ao grupo CFA + salina (Figura 6C).
MI
6 dia
s
12 d
ias
13d 1
h
13d 3
h
13d 5
h
13d 7
h
14d 2
4h
0
100
200
300
400
500
NAIVE
CFA
CFA+GBP 100ug
CFA+GBP 200ug
# ###
#
# #
**
Dias após injeção
Lim
iar
No
cic
ep
tivo
(g)
Resultados
36
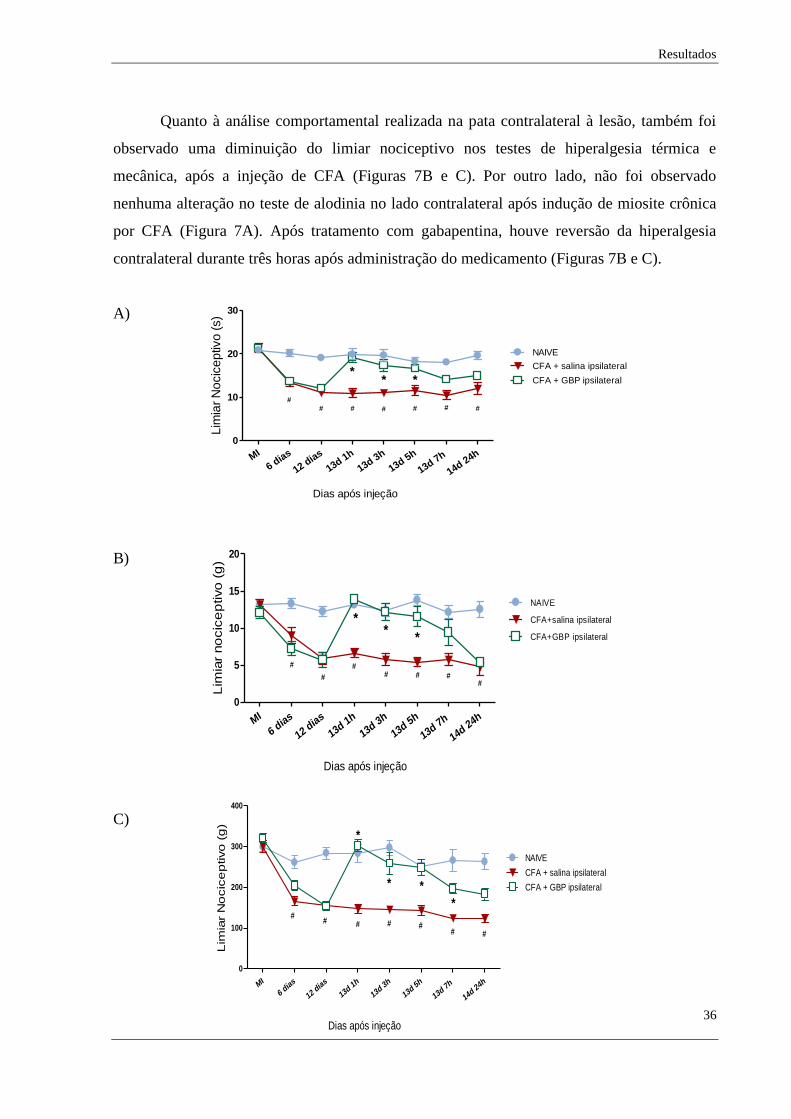
Quanto à análise comportamental realizada na pata contralateral à lesão, também foi
observado uma diminuição do limiar nociceptivo nos testes de hiperalgesia térmica e
mecânica, após a injeção de CFA (Figuras 7B e C). Por outro lado, não foi observado
nenhuma alteração no teste de alodinia no lado contralateral após indução de miosite crônica
por CFA (Figura 7A). Após tratamento com gabapentina, houve reversão da hiperalgesia
contralateral durante três horas após administração do medicamento (Figuras 7B e C).
A)
B)
C)
MI
6 dia
s
12 dia
s
13d 1h
13d 3h
13d 5h
13d 7h
14d 24h
0
10
20
30
*
#
# # # # ##
* *
CFA + salina ipsilateral
CFA + GBP ipsilateral
NAIVE
Dias após injeção
Lim
iar
No
cic
ep
tivo
(s)
MI
6 dia
s
12 dia
s
13d 1h
13d 3h
13d 5h
13d 7h
14d 24h
0
5
10
15
20
*
#
# ##
## #
**
NAIVE
CFA+salina ipsilateral
CFA+GBP ipsilateral
Dias após injeção
Lim
iar
no
cic
ep
tivo
(g
)
MI
6 dia
s
12 dia
s
13d 1h
13d 3h
13d 5h
13d 7h
14d 24h
0
100
200
300
400
*
*
*
*
# # ## #
##
NAIVE
CFA + salina ipsilateral
CFA + GBP ipsilateral
Dias após injeção
Lim
iar
No
cic
ep
tivo
(g
)

Resultados
37
Figura 6 - Análise comportamental da pata ipsilateral à lesão de animais com miosite crônica
induzida por CFA após tratamento com gabapentina. O limiar nociceptivo, avaliado por Teste de
Hargreaves (A - expresso em segundos), Filamentos de Von Frey (B - expresso em gramas) e Von
Frey Eletrônico (C - expresso em gramas), foi determinado antes (tempo zero - MI), 6 e 12 dias após a
lesão com CFA - i.m. No 13º dia após injeção de CFA, os animais foram tratados com gabapentina
200μg (GBP) por via intratecal e os testes comportamentais foram novamente mensurados nos tempos
1, 3, 5, 7, e 24 horas. Os resultados representam a média epm de 8 animais por grupo. # p<0,05 por
comparação entre os grupos NAIVE e CFA + salina ipsilateral. * p<0,05 por comparação entre os
grupos CFA + GBP ipsilateral e CFA + salina ipsilateral.
A)
B)
C)
MI
6 dia
s
12 dia
s
13d 1h
13d 3h
13d 5h
13d 7h
14d 24h
0
10
20
30
NAIVE
*
## # # # ##
CFA + salina contralateral
*
CFA + GBP contralateral
Dias após injeção
Lim
iar
No
cic
ep
tivo
(s)
MI
6 dia
s
12 dia
s
13d 1
h
13d 3
h
13d 5
h
13d 7
h
14d 2
4h
0
5
10
15
20
* * *
NAIVE
CFA + salina contralateral
CFA + GPB contralateral
####
# ###
Dias após injeção
Lim
iar
No
cic
ep
tivo
(g
)
MI
6 dia
s
12 dia
s
13d 1h
13d 3h
13d 5h
13d 7h
14d 24h
0
100
200
300
400
* *
#
## # #
##
NAIVE
CFA + salina contralateral
CFA + GBP contralateral
Dias após injeção
Lim
iar
No
cic
ep
tivo
(g)

Resultados
38
Figura 7 - Análise comportamental da pata contralateral à lesão de animais com miosite crônica
induzida por CFA após tratamento com gabapentina. O limiar nociceptivo, avaliado por Teste de
Hargreaves (A - expresso em segundos), Filamentos de Von Frey (B - expresso em gramas) e Von
Frey Eletrônico (C - expresso em gramas), foi determinado antes (tempo zero - MI), 6 e 12 dias após a
lesão com CFA - i.m. No 13º dia após injeção de CFA, os animais foram tratados com gabapentina
200μg (GBP) por via intratecal e os testes comportamentais foram novamente mensurados nos tempos
1, 3, 5, 7, e 24 horas. Os resultados representam a média epm de 8 animais por grupo. # p<0.05
entre os grupos NAIVE e CFA + salina contralateral. # p<0,05 entre os grupos NAIVE e CFA + salina
contralateral. * p<0,05 entre os grupos CFA + GBP contralateral e CFA + salina contralateral.
4.3 ANÁLISE DE CITOCINAS NO TÁLAMO DE RATOS COM MIOSITE
CRÔNICA
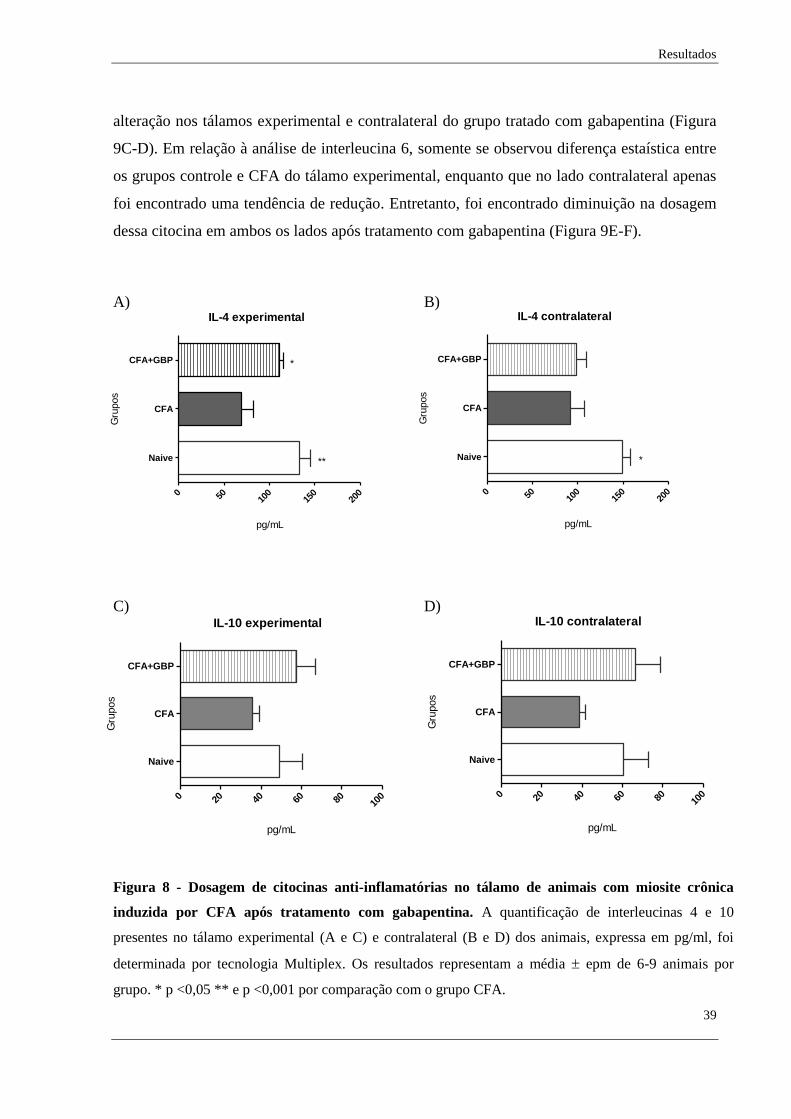
Para quantificação de citocinas pró e anti-inflamatórias no tálamo experimental e
contralateral dos animais, foi realizado ensaio para dosagem de citocinas pela tecnologia
Multiplex. As citocinas quantificadas foram IL-1β, IL-4, IL-6, IL-10 e TNF-α. Em relação à
análise de citocinas anti-inflamatórias, podemos observar que houve uma redução de
interleucina 4 nos tálamos experimental e contralateral do grupo em que a dor muscular
crônica foi induzida (CFA) e que esta alteração apenas foi revertida no tálamo experimental
após tratamento farmacológico com gabapentina (CFA+GBP), como demonstrado na Figura
8A-B.
Já em relação à interleucina 10, não foi observado alteração estatística entre os grupos
analisados, embora tenha sido observado uma tendência de diminuição dessa citocina nos
tálamos experimental e contralateral do grupo CFA (Figura 8C-D). Ainda, observa-se que o
grupo tratado apresentou uma tendência em ter maior nível dessa citocina, em comparação ao
grupo com miosite.
Quanto à dosagem de citocinas pró-inflamatórias, apenas foi encontrado diferença
estatística na dosagem de interleucina 1β entre os grupos controle e CFA no tálamo
experimental, contudo observou-se uma tendência de aumento dessa citocina após indução da
dor muscular crônica no lado contralateral. Além disso, foi demonstrado que o grupo tratado
com gabapentina obteve redução desta interleucina pró-inflamatória no tálamo experimental e
uma tendência de sua redução no tálamo contralateral (Figura 9A-B).
Ao quantificar a citocina TNFα, observamos que houve um aumento em sua dosagem
nos tálamos experimental e contralateral do grupo CFA, bem como houve reversão dessa
Resultados
39
alteração nos tálamos experimental e contralateral do grupo tratado com gabapentina (Figura
9C-D). Em relação à análise de interleucina 6, somente se observou diferença estaística entre
os grupos controle e CFA do tálamo experimental, enquanto que no lado contralateral apenas
foi encontrado uma tendência de redução. Entretanto, foi encontrado diminuição na dosagem
dessa citocina em ambos os lados após tratamento com gabapentina (Figura 9E-F).
A) B)
C) D)
Figura 8 - Dosagem de citocinas anti-inflamatórias no tálamo de animais com miosite crônica
induzida por CFA após tratamento com gabapentina. A quantificação de interleucinas 4 e 10
presentes no tálamo experimental (A e C) e contralateral (B e D) dos animais, expressa em pg/ml, foi
determinada por tecnologia Multiplex. Os resultados representam a média epm de 6-9 animais por
grupo. * p <0,05 ** e p <0,001 por comparação com o grupo CFA.
IL-4 contralateral
0 50 100
150
200
Naive
CFA
CFA+GBP
*
pg/mL
Gru
pos
IL-4 experimental
0 50 100
150
200
Naive
CFA
CFA+GBP
**
*
pg/mL
Gru
pos
IL-10 experimental
0 20 40 60 80 100
Naive
CFA
CFA+GBP
pg/mL
Gru
pos
IL-10 contralateral
0 20 40 60 80 100
Naive
CFA
CFA+GBP
pg/mL
Gru
pos

Resultados
40
A) B)
C) D)
E) F)
Figura 9 - Dosagem de citocinas pró-inflamatórias no tálamo de animais com miosite crônica
induzida por CFA após tratamento com gabapentina. A quantificação de interleucina 1β, fator de
necrose tumoral α e interleucina 6 presentes no tálamo experimental (A, C e E) e contralateral (B, D e F)
dos animais, expressa em pg/ml, foi determinada por tecnologia Multiplex. Os resultados representam a
média epm de 4-9 animais por grupo. * p <0,05 e ** p <0,001 por comparação com o grupo CFA.
IL-1 experimental
0 10 20 30 40
NAIVE
CFA
CFA+GBP *
*
pg/mL
Gru
pos
IL-1 contralateral
0 10 20 30 40
NAIVE
CFA
CFA+GBP
pg/mL
Gru
pos
TNF-a contralateral
0 10 20 30
Naive
CFA
CFA+GBP
*
*
pg/mL
Gru
pos
TNF-a experimental
0 10 20 30
Naive
CFA
CFA+GBP
**
**
pg/mL
Gru
pos
IL-6 contralateral
050
010
0015
00
Naive
CFA
CFA+GBP *
pg/mL
Gru
pos
IL-6 experimental
050
010
0015
00
Naive
CFA
CFA+GBP
**
*
pg/mL
Gru
pos

Resultados
41
5.4 ANÁLISE DE CITOCINAS NA MEDULA ESPINAL DE RATOS COM
MIOSITE CRÔNICA
Igualmente, foi realizado quantificação das citocinas IL-1B, IL-4, IL-6, IL-10 e TNF-α
na medula espinal dos animais. Neste ensaio, foi observado menor dosagem de interleucina 10
na medula dos animais com miosite crônica (CFA), em comparação ao grupo naive, porém
não foi encontrado diferença estatística na interleucina 4 entre estes dois grupos (Figura 10A-
B).
Ainda em relação à dosagem de interleucina IL-10, não foi encontrado diferença
estatística no grupo tratado com gabapentina, somente foi observado uma tendência de seu
aumento após tratamento. Por outro lado, observa-se que houve aumento de interleucina IL-4
no grupo que recebeu tratamento farmacológico (Figura 10A-B).
Quanto à quantificação de citocinas pró-inflamatórias na medula espinal, foi
encontrado diferença estatística na dosagem de interleucina 1β entre os grupos naive e CFA.
No entanto, não foi observado alteração estatística desta citocina no grupo tratado com
gabapentina (Figura 10C).
Não encontramos diferença estatística na dosagem de TNFα entre os grupos controle e
CFA quando a medula espinal foi analisada. Entretanto, foi evidenciado a diminuição dessa
citocina nos animais tratados (Figura 10D).
Em relação à dosagem de interleucina 6, não foi encontrado diferença entre nenhum
dos grupos analisados, mas observa-se uma tendência de aumento dessa citocina após indução
da dor muscular crônica e uma tendência de sua diminuição no grupo tratado (Figura 10E).
Vale ressaltar que foi feita análise da medula completa, sem divisão de lado ipsi ou
contralateral, visto que nosso grupo já havia constatado alteração bilateral na morfologia da
medula espinal de ratos com dor muscular crônica induzida por CFA (Rosa et al., 2017).
A) B)
IL-10
0 50 100
150
Naive
CFA
CFA+GBP
*
pg/mL
Gru
pos
IL-4
010
020
030
040
050
0
Naive
CFA
CFA+GBP **
pg/mL
Gru
pos

Resultados
42
C) D)
E)
Figura 10 - Dosagem de citocinas pró e anti-inflamatórias na medula espinal de animais com
miosite crônica induzida por CFA após tratamento com gabapentina. A quantificação de interleucina
4, 10, 1β, fator de necrose tumoral α e interleucina 6 presentes na medula espinal (A, B, C, D e E,
respectivamente), expressa em pg/ml, foi determinada por tecnologia Multiplex. Os resultados
representam a média epm de 4-6 animais por grupo. * p <0,05 e ** p <0,001 por comparação com o
grupo CFA.
TNF-a
0 20 40 60
Naive
CFA
CFA+GBP *
pg/mL
Gru
pos
IL-6
050
010
0015
0020
0025
00
Naive
CFA
CFA+GBP
pg/mL
Gru
pos
IL-1
0 20 40 60 80
NAIVE
CFA
CFA+GBP
*
pg/mL
Gru
pos

Resultados
43
5.5 ANÁLISE DE CITOCINAS NO GÂNGLIO DA COLUNA POSTERIOR DE
RATOS COM MIOSITE CRÔNICA
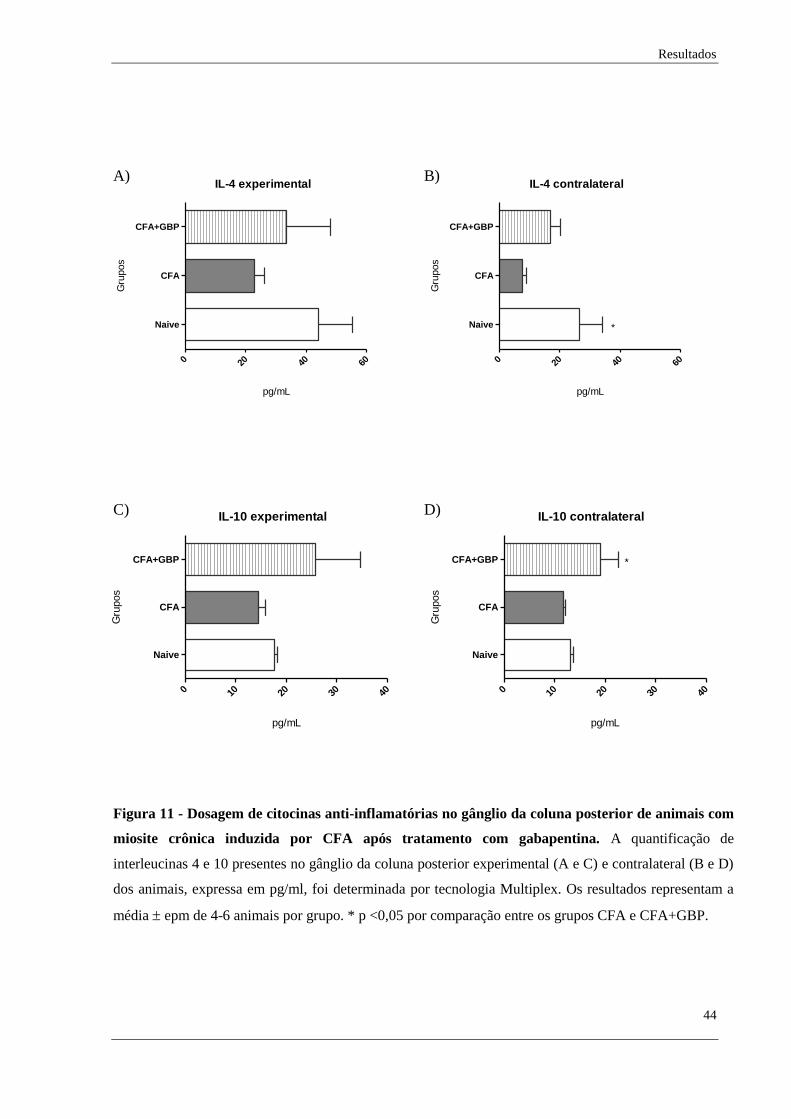
Também foi realizado quantificação de citocinas no gânglio da coluna posterior
experimental e contralateral dos animais. Novamente, as citocinas quantificadas foram IL-1B,
IL-4, IL-6, IL-10 e TNF-α.
Quanto à análise de interleucina 4 no gânglio experimental, não foi encontrado
diferença estatística entre os grupos analisados, mas uma tendência de diminuição dessa
citocina no grupo com dor muscular crônica e tendência de seu aumento após tratamento com
gabapentina foi obervado. (Figura 11A). Já no gânglio contralateral, encontrou-se diferença
estatítisca entre os grupos naive e CFA, mas não se observou alteração no grupo tratado
(Figura 11B).
Sobre a dosagem de interleucina 10 no gânglio experimental, não foi observado
diferença estatística entre os grupos analisados, somente uma tendência de aumento dessa
citocina no grupo tratado com gabapentina (Figura 11C). No gânglio contralateral, não foi
encontrado diferença entres os grupos controle e CFA, no entanto, observou-se o aumento de
IL-10 após tratamento (Figura 11D).
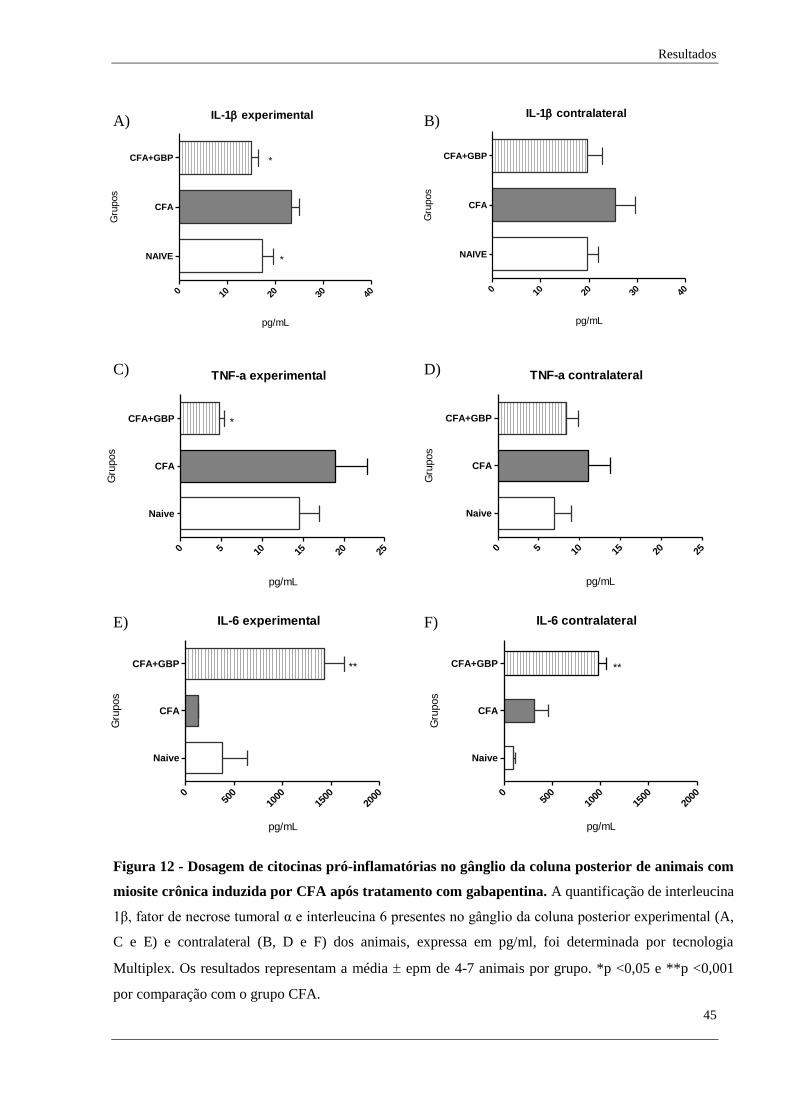
Quanto à quantificação de citocinas pró-inflamatórias no gânglio, foi encontrado
diferença estatística na dosagem de interleucina 1β entre os grupos naive e CFA, assim como
no grupo tratado com gabapentina, em comparação aos grupo CFA (Figura 12A). No lado
contralateral, apenas oberva-se a tendência de aumento dessa citocina após indução da dor
muscular crônica e a tendência de redução no grupo tratado com gabapentina (Figura 12B).
Ainda, não encontramos alteração estatística na dosagem de TNFα nos gânglios
experimental e contralateral de nenhum dos grupos analisados, apenas uma tendência de
aumento dessa citocina no grupo CFA (Figura 12C-D). Também, foi observado a diminuição
dessa citocina no gânglio experimental dos animais tratados (Figura 12C) e a tendência de
redução no gânglio contralateral (Figura 12D).
Em relação à dosagem de interleucina 6 nos gânglios experimental e contralateral, não
foi encontrado diferença entre os grupos controle e CFA, mas observou-se o aumento dessa
interleucina após tratamento farmacológico (Figura 12E-F).
Resultados
44
A) B)
C) D)
Figura 11 - Dosagem de citocinas anti-inflamatórias no gânglio da coluna posterior de animais com
miosite crônica induzida por CFA após tratamento com gabapentina. A quantificação de
interleucinas 4 e 10 presentes no gânglio da coluna posterior experimental (A e C) e contralateral (B e D)
dos animais, expressa em pg/ml, foi determinada por tecnologia Multiplex. Os resultados representam a
média epm de 4-6 animais por grupo. * p <0,05 por comparação entre os grupos CFA e CFA+GBP.
IL-4 experimental
0 20 40 60
Naive
CFA
CFA+GBP
pg/mL
Gru
pos
IL-4 contralateral
0 20 40 60
Naive
CFA
CFA+GBP
*
pg/mL
Gru
pos
IL-10 contralateral
0 10 20 30 40
Naive
CFA
CFA+GBP *
pg/mL
Gru
pos
IL-10 experimental
0 10 20 30 40
Naive
CFA
CFA+GBP
pg/mL
Gru
pos
Resultados
45
A) B)
C) D)
E) F)
Figura 12 - Dosagem de citocinas pró-inflamatórias no gânglio da coluna posterior de animais com
miosite crônica induzida por CFA após tratamento com gabapentina. A quantificação de interleucina
1β, fator de necrose tumoral α e interleucina 6 presentes no gânglio da coluna posterior experimental (A,
C e E) e contralateral (B, D e F) dos animais, expressa em pg/ml, foi determinada por tecnologia
Multiplex. Os resultados representam a média epm de 4-7 animais por grupo. *p <0,05 e **p <0,001
por comparação com o grupo CFA.
TNF-a experimental
0 5 10 15 20 25
Naive
CFA
CFA+GBP *
pg/mL
Gru
pos
TNF-a contralateral
0 5 10 15 20 25
Naive
CFA
CFA+GBP
pg/mL
Gru
pos
IL-6 contralateral
050
010
0015
0020
00
Naive
CFA
CFA+GBP **
pg/mL
Gru
pos
IL-6 experimental
050
010
0015
0020
00
Naive
CFA
CFA+GBP **
pg/mL
Gru
pos
IL-1 experimental
0 10 20 30 40
NAIVE
CFA
CFA+GBP *
*
pg/mL
Gru
pos
IL-1 contralateral
0 10 20 30 40
NAIVE
CFA
CFA+GBP
pg/mL
Gru
pos

Resultados
46
5.6 ANÁLISE DE CÉLULAS GLIAIS NO TÁLAMO DE RATOS COM MIOSITE
CRÔNICA
Para análise de células gliais no tálamo experimental e contralateral dos animais,
utilizamos à técnica de imuno-histoquímica para marcação de GFAP (marcador de astrócitos)
e Ox-42 (marcador de microglia). A área analisada foi referente ao núcleo ventral póstero-
lateral (VPL) e ao núcleo ventral póstero-medial (VPM), núcleos intralaminares do tálamo
que fazem parte do processamento da informação nociceptiva transmitida pelas vias aferentes,
como descrito anteriormente (Figura 13).
Pela imunorreatividade observada para estes marcadores, podemos observar que
houve aumento de astrócitos no tálamo experimental do grupo CFA, quando comparado ao
grupo controle. No entanto, não encontramos diferença estatística no tálamo contralateral,
apenas uma tendência de aumento de astrócitos nos animais com miosite crônica. Além disso,
notou-se que há inibição na imunorreatividade de astrócitos em ambos os lados analisados
após tratamento farmacológico com gabapentina (Figura 14).
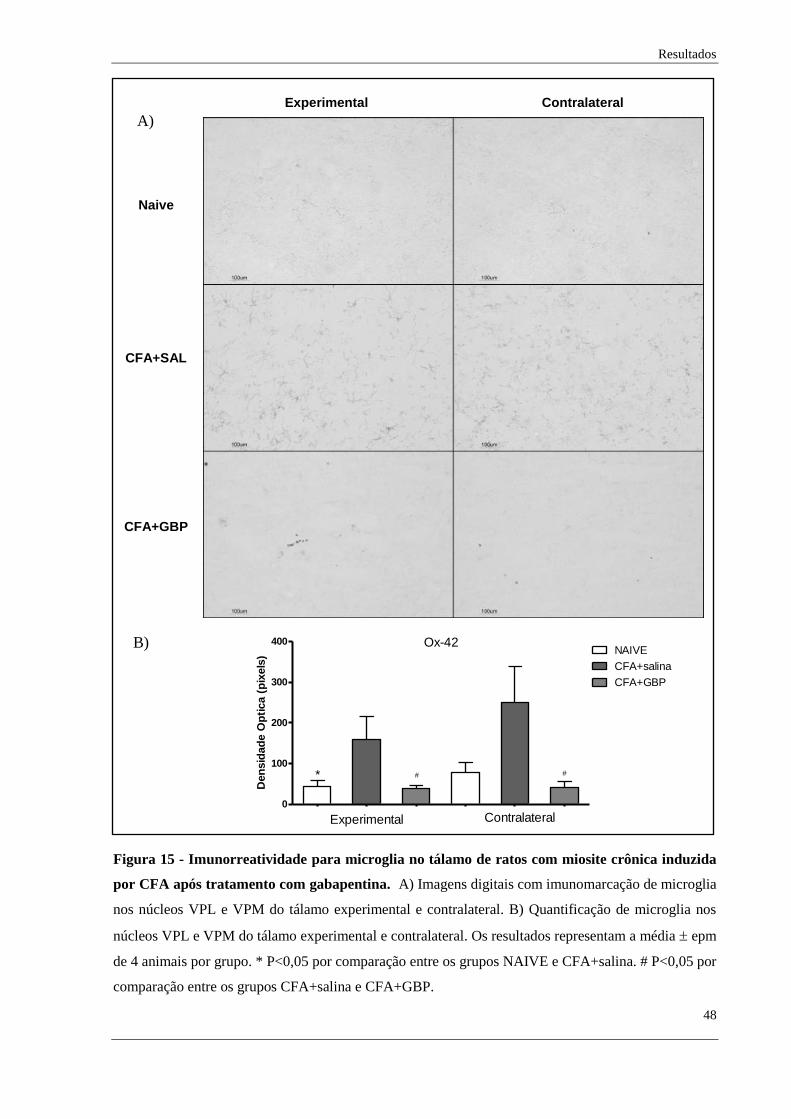
Ainda, observou-se que houve aumento de células microgliais nos tálamos
experimental do grupo com miosite crônica, porém não no tálamo contralateral, em
comparação aos animais naive. Da mesma forma, observamos que a imunorreatividade para
microglia foi reduzida após tratamento farmacológico nos tálamos experimental e
contralateral (Figura 15).
Figura 13 - Ilustração esquemática da região encefálica analisada. Imagens adaptadas do Atlas
The Rat Brain (Paxinos e Watson) que demonstra a região encefálica analisada durante o experimento,
os núcleos ventral póstero-lateral (VPL) e ventral póstero-medial (VPM) do tálamo.

Resultados
47
A)
B)
Figura 14 - Imunorreatividade para astrócitos no tálamo de ratos com miosite crônica induzida
por CFA após tratamento com gabapentina. A) Imagens digitais com imunomarcação de astrócitos
nos núcleos VPL e VPM do tálamo experimental e contralateral. B) Quantificação de astrócitos nos
núcleos VPL e VPM do tálamo experimental e contralateral. Os resultados representam a média epm
de 4 animais por grupo. * P<0,05 por comparação entre os grupos NAIVE e CFA+salina. # P<0,05
por comparação entre os grupos CFA+salina e CFA+GBP.
Experimental Contralateral
Naive
CFA+SAL
CFA+GBP
0
100
200
300
NAIVE
CFA+salina
CFA+GBP
Experimental Contralateral
#
*
#
GFAP
Den
sid
ad
e O
pti
ca (
pix
els
)
Resultados
48
A)
B)
Figura 15 - Imunorreatividade para microglia no tálamo de ratos com miosite crônica induzida
por CFA após tratamento com gabapentina. A) Imagens digitais com imunomarcação de microglia
nos núcleos VPL e VPM do tálamo experimental e contralateral. B) Quantificação de microglia nos
núcleos VPL e VPM do tálamo experimental e contralateral. Os resultados representam a média epm
de 4 animais por grupo. * P<0,05 por comparação entre os grupos NAIVE e CFA+salina. # P<0,05 por
comparação entre os grupos CFA+salina e CFA+GBP.
Experimental Contralateral
Naive
CFA+SAL
CFA+GBP
0
100
200
300
400NAIVE
CFA+salina
CFA+GBP
Experimental Contralateral
Ox-42
#* #
Den
sid
ad
e O
pti
ca (
pix
els
)

Resultados
49
5.7 ANÁLISE DE CÉLULAS GLIAIS SATÉLITES NO GÂNGLIO DA COLUNA
POSTERIOR DE RATOS COM MIOSITE CRÔNICA
Para análise de células gliais satélites no gânglio da coluna posterior, os tecidos foram
extraídos e submetidos à técnica de Western Blotting, sendo analisados os gânglios da porção
lombar da medula espinal (L4-L6). Como marcador de células gliais satélites, utilizamos o
anticorpo GFAP.
A partir da quantificação da proteína realizada, foi possível observar um aumento de
71% na expressão proteica de células gliais satélites no gânglio dos animais com miosite
crônica (CFA), em comparação ao grupo naive. Ainda, o grupo tratado com gabapentina
demonstrou uma diminuição de 62% na expressão dessas células gliais, em comparação ao
grupo CFA (Figura 16).
Figura 16 - Expressão proteica de células gliais satélites de ratos com miosite crônica induzida
por CFA após tratamento com gabapentina. Gráfico representativo para a quantificação proteica
das células glais satélites no gânglio da coluna posterior direito dos animais. Os resultados
representam a média epm de 5 animais por grupo. *P<0,05 por comparação entre os grupos CFA e
CFA+GBP.
GFAP
Naive CFA CFA + GBP0
50
100
150
200*
71%
62%Den
sid
ad
e ó
pti
ca
(%)

Discussão
50
5. DISCUSSÃO
O tecido musculoesquelético é um sítio muito comum de dor e, provavelmente, todos
irão vivenciar um episódio de dor muscular na vida. As afecções musculares crônicas
representam alto custo econômico e não respondem bem às intervenções convencionais,
afetando a qualidade de vida das pessoas. Deste modo, pesquisadores têm utilizado
substâncias algiogênicas, dentre elas o Adjuvante Completo de Freund (CFA), como modelo
experimental para estudo da dor muscular crônica (Chacur et al., 2009; Hoheisel et al., 1998;
Hoheisel et al., 2013; Reinert et al., 1998).
Neste trabalho, nós induzimos quadro de miosite crônica pela injeção de CFA no
músculo gastrocnêmico de ratos e avaliamos o efeito da gabapentina na dor muscular gerada.
Deste modo, este estudo tem como finalidade avaliar se o tratamento farmacológico com
gabapentina é eficaz em reverter a dor muscular crônica de animais induzida neste modelo
experimental de dor e analisar sua influência em células gliais, canais de cálcio dependentes
de voltagem e na produção e/ou liberação de citocinas a nível central e periférico.
Em nossos testes comportamentais, observamos que a dor muscular crônica induzida
pela injeção de CFA no músculo gastrocnêmio direito dos animais, não apenas diminui o
limiar nociceptivo em testes comportamentais realizados na pata experimental, como também
gera nocicepção na pata contralateral à lesão, o que corrobora com pesquisas anteriores. No
entanto, não observamos alodinia tátil na pata contralateral em nossos testes comportamentais.
Dessa forma, além de alodinia unilateral, foi observado que o quadro de miosite crônica
induzido produz hiperalgesia térmica e mecânica bilateral.
Um estudo de Sluka e seus colaboradores havia descrito anteriormente que ratos
injetados com salina ácida apresentavam quadro de hiperalgesia mecânica na pata
contralateral à lesão (Sluka et al., 2001). Este grupo também observou que ratos com dor
musculoesquelética aguda induzida por injeção de carragenina, além de apresentar
hiperalgesia mecânica na pata contralateral, também manifestavam hiperalgesia térmica
bilateral (Radhakrishnan et al., 2003). Por outro lado, outro grupo de pesquisa demonstrou
que a injeção de CFA no músculo masseter induz alodinia bilateral em ratos (Tariba Knezevic
et al., 2016).
Ainda, no teste Von Frey Eletrônico, observamos que o tratamento farmacológico com
gabapentina via intratecal na dose de 200μg foi mais eficaz em reverter a nocicepção dos
animais com miosite crônica, quando comparado com a dose de 100μg. Após padronização do

Discussão
51
tratamento, ao aplicarmos os testes Filamentos de Von Frey, Hargreaves e novamente o Von
Frey Eletrônico, foi demonstrado que o limiar nociceptivo dos animais aumentou em todos
eles, revertendo alodinia e hiperalgesia bilateral - térmica e mecânica (Rosa et al., 2017).
Desta forma, o efeito antinociceptivo da gabapentina em modelo de dor muscular crônica
induzida por CFA foi constatado. Estes resultados corroboram com um único estudo prévio de
dor musculoesquelética tratada com gabapentina que demonstrou seu efeito analgésico em
ratos com hiperalgesia mecânica após injeção de salina ácida no músculo gastrocnêmio (Kang
et al., 2014).
Nós escolhemos a gabapentina como um tratamento potencial para dor muscular
crônica por seu efeito antinociceptivo relatado na literatura (Allchorne et al., 2012;
Krzyzanowska et al., 2011; Mimenza Alvarado; Aguilar Navarro, 2016; Zhang et al., 2009;
Zhang et al., 2015), e também, devido à sua ampla eficácia em casos clínicos de dor crônica
(Atkinson et al., 2016; Beladi Mousavi et al., 2015; Cheng; Chiou, 2006; Dolgun et al.,
2014; Lewis et al., 2016; Robertson; Marshman, 2016; Rowbotham et al., 1998). Estudos
têm demonstrado que a gabapentina pode inibir respostas dolorosas por meio de sua ligação à
subunidade α2/δ-1 de canais de cálcio dependentes de voltagem presente em neurônios e
redução de substâncias como glutamato, substância P, NF-kB e CGRP (Fink et al., 2000;
Kukkar et al., 2013; Park et al., 2008; Quintero et al., 2011; Yaksh, 2006). Além disso,
sabe-se que a gabapentina apenas prejudica funções motoras em doses elevadas (Kang et al.,
2014).
As células da glia são células não condutoras que modulam a neurotransmissão a nível
sináptico e desempenham papel importante no desenvolvimento e manutenção da informação
nociceptiva. Após uma lesão, células microgliais são ativadas e liberam mediadores pró-
inflamatórios como prostaglandinas, óxido nítrico e citocinas, dando início ao processo
doloroso. A microglia propaga a neuroinflamação recrutando outras microglias e ativando
astrócitos, o que prolonga o estado inflamatório em condições de dor crônica (Milligan;
Watkins, 2009; Old et al., 2015; Tsuda et al., 2005; Vallejo et al., 2010).
Tendo em vista pesquisas anteriores que demonstraram que astrócitos e microglia da
medula espinal podem estar envolvidas na dor ipsilateral (Chacur et al., 2009; Tenschert et
al., 2004) e contralateral à lesão muscular crônica de ratos (Rosa et al., 2017), decidimos
avaliar estas células no tálamo experimental e contralateral dos animais após indução de
miosite e tratamento com gabapentina. Assim, durante análise de células gliais no tálamo

Discussão
52
experimental e contralateral dos animais, analisamos os núcleos ventral póstero-lateral e
ventral póstero-medial, que são relé das vias sensitivas e projetam suas fibras para o córtex do
giro pós-central (área somestésica primária).
Pela imunorreatividade observada para GFAP e Ox-42 na técnica de imuno-
histoquímica, pudemos observar que houve aumento de astrócitos no tálamo experimental do
dos animais com miosite crônica induzida, quando comparado aos animais controle. Contudo,
não encontramos diferença estatística no tálamo contralateral, apenas uma tendência de
aumento de astrócitos nos animais com miosite crônica. Além disso, observou-se que há
inibição na imunorreatividade de astrócitos em ambos os lados analisados do tálamo após
tratamento farmacológico com gabapentina. Ainda, observou-se que houve aumento de
células microgliais no tálamo experimental dos animais com dor muscular crônica, em
comparação aos animais brancos, porém não no tálamo contralateral, apesar da forte
tendência encontrada. Da mesma forma, observamos que a imunorreatividade de microglia foi
inibida nos tálamos experimental e contralateral após tratamento farmacológico.
Previamente, havia sido demonstrado na literatura que ratos com dor neuropática
expressam maior reatividade microglial no núcleo ventral póstero-lateral (LeBlanc et al.,
2011) e maior imunorreatividade de astrócitos e microglia no tálamo e mesencéfalo (Giardini
et al., 2017). Deste modo, sugerimos que há contribuição de astrócitos e microglia dos
núcleos talâmicos VPL e VPM na geração da hiperalgesia bilateral encontrada nos animais
com miosite crônica induzida por injeção de CFA. Além disso, sugerimos que a gabapentina
possui ação em núcleos intralaminares do tálamo, visto que pesquisadores encontraram
analgesia em casos de dor após acidente vascular encefálico em regiões talâmicas após
tratamento com gabapentina (Hesami et al., 2015; Yang et al., 2014) e que, neste trabalho,
encontramos inibição de microglia e astrócitos nos núcleos ventral póstero-lateral e ventral
póstero-medial de ratos com dor muscular crônica revertida após tratamento com gabapentina.
O gânglio da coluna posterior é um aglomerado de corpos de neurônios aferentes
primários localizado no SNP que modula de estímulos sensitivos, tais como a informação
nociceptiva, antes que estes cheguem ao SNC. Cada corpo celular existente no gânglio da
coluna posterior é envolvido por células gliais satélites, células que desempenham no SNP
atividade semelhante à dos astrócitos e têm sido relatadas por participar de processos
nociceptivos (Costa; Moreira Neto, 2015; Liu et al., 2012). Dessa forma, analisar a expressão
de células gliais no gânglio de animais foi mais um objeto de estudo neste trabalho.

Discussão
53
Pela quantificação da proteína marcadora de célula glial satélite no gânglio neste
trabalho, pudemos observar que os animais com miosite crônica induzida por CFA
apresentam maior expressão células gliais satélites, em comparação aos animais controle,
demonstrando modulação da dor muscular crônica por estas células. Além disso, foi
evidenciado neste estudo que o tratamento farmacológico com gabapentina é eficaz em inibir
a expressão destas células, contribuindo para o efeito antinociceptivo observado. Até o
momento, alguns estudos evidenciavam que animais com dor neuropática quando tratados
com gabapentina apresentavam redução da ativação dessas células (Park et al., 2008 ; Reda et
al., 2016; Suto et al., 2014).
Como mencionado, as células glias podem liberar citocinas pró-inflamatórias que
desempenham papel chave em desordens do SNC, como dor neuropática, epilepsia e doenças
neurodegenerativas. As citocinas são pequenas proteínas secretadas pelas células e têm um
efeito específico sobre as interações e comunicações celulares. Citocinas podem atuar sobre as
células que as secretam (ação autócrina), nas células próximas (ação parácrina) ou, em alguns
casos, em células distantes (ação endócrina). Há evidências significativas de que citocinas
estão envolvidas não apenas na iniciação, mas também na persistência de dor patológica por
ativação direta de neurônios sensoriais nociceptivos. Citocinas inflamatórias também estão
envolvidas na sensibilização central induzida por lesão nervosa e estão relacionadas com o
desenvolvimento de hiperalgesia e alodinia contralateral (Oliveira et al., 2017; Skaper; Facci,
2012; Zhang; An, 2007). Em razão disso, um de nossos objetivos foi quantificar algumas
citocinas pró-inflamatórias e anti-inflamatórias relacionadas à dor patológica.
A respeito do ensaio para dosagem de citocinas no tálamo, demonstrou-se que há
redução de interleucina IL-4 nos tálamos experimental e contralateral do grupo com dor
muscular crônica, no entanto não houve alteração dessa citocina após tratamento
farmacológico com gabapentina. Ainda, apesar de não ter sido observado alteração estatística
em interleucina IL-10 entre os grupos analisados, observa-se uma tendência de diminuição
desta citocina no tálamo contralateral do grupo CFA.
Já ao dosar citocinas pró-inflamatórias no tálamo, foi verificado maior marcação de
TNFα nos tálamos experimental e contralateral dos animais com dor muscular crônica, bem
como foi constatado a diminuição desse fator no tálamo experimental e tendência de redução
no tálamo contralateral dos animais após tratamento com gabapentina. Ao quantificar
interleucina IL-6, não foi encontrado alteração entre os grupos controle e CFA, todavia foi

Discussão
54
encontrado diminuição desta citocina após tratamento com gabapentina. Também não foi
verificado alteração estatística na interleucina 1β entre os grupos controle e CFA, mas
observou-se uma tendência de aumento dessa citocina após indução da dor muscular crônica e
redução desta interleucina no grupo tratado com gabapentina, tanto no tálamo experimental
quanto no tálamo contralateral.
Quando analizamos as mesmas interleucinas, porém no gânglio da coluna posterior
dos mesmos grupos de animais, observamos uma tendência de diminuição das interleucina IL-
4 no gânglio experimental do grupo CFA, bem como uma tendência de seu aumento após
tratamento com gabapentina. Apesar do pequeno número de animais por grupo, foi observado
uma tendência de aumento de interleucina IL-10 no lado experimental dos animais com dor
muscular, enquanto que no gânglio contralateral seu aumento foi constatado apenas após
tratamento. Ressalta-se que esses resultados podem sofrer alteração ao aumentar-se o número
de animais no experimento.
Já em relação à dosagem de citocinas pró-inflamatórias no gânglio da coluna posterior,
foi observado uma tendência de aumento na dosagem de interleucina 1β após indução de dor
muscular crônica, especialmente no lado contralateral. Ainda, foi encontrado redução desta
interleucina nos gânglios experimental e contralateral após tratamento com gabapentina. Não
foi observado alteração estatística na dosagem de TNFα nos gânglios experimental e
contralateral de nenhum dos grupos analisados, mas uma tendência de diminuição dessa
citocina no gânglio experimental dos animais após tratamento. Além disso, não foi encontrado
diferença na dosagem de interleucina IL-6 no gânglio experimental dos grupos analisados,
mas uma tendência de aumento dessa citocina no grupo tratado. Contudo, foi encontrado o
aumento dessa interleucina no gânglio contralateral dos animais tratados com gabapentina,
talvez em função do número pequeno de animais neste ensaio.
Em relação a análise da porção lombar da medula espinal, foi observado um aumento
estatisticamente significativo apenas na IL-4 quando tratados com a gabapentina. Já em
relação as citocinas pro-inflamatórias, observamos uma diminuição na dosagem de TNFα
após tratamento com gabapentina.
Desta forma, tais resultados sugerem que a dor muscular crônica interfere nas
interleucinas anti-inflamatórias IL-4 e IL-10 do tálamo e gânglio da coluna posterior dos
animais e apenas IL-4 em relação a avaliação da porção lombar da medula espinal. Ainda, há
participação das citocinas pró-inflamatórias IL-1β, IL-6 e TNF-α no tálamo, da IL-1β no

Discussão
55
gânglio da coluna posterior e do TNF-α na porção lombar da medula espinal durante este
processo nociceptivo.
Até o momento, estudos haviam encontrado aumento de IL-1β, IL-6 e TNF-α no
músculo gastrocnêmio de animais com dor muscular aguda (Loram et al., 2007; Manjavachi
et al., 2010) e apenas de IL-1β na medula espinal de ratos com dor muscular (Borghi et al.,
2014).
Além disso, sugere-se que a ação da gabapentina ocorre indiretamente por meio da
estimulação de citocinas anti-inflamatórias e, principalmente, pela inibição de citocinas pró-
inflamatórias. Estes dados complementam pesquisas que observaram que a gabapentina pode
reduzir IL-6, TNF-α e IL-1β e aumentar IL-10 na medula espinal de ratos com dor
neuropática (Hayashi et al., 2013; Kremer et al., 2016).
Portanto, concluímos que a gabapentina além de inibir a nocicepção bilateral induzida
em nosso modelo experimental também é capaz de reverter a reatividade glial e a
neuroinflamação existentes na dor muscular crônica. Diante da necessidade de inovação e
aperfeiçoamento das terapêuticas atuais, mais estudos são necessários, tanto para
compreensão da fisiopatologia da dor muscular crônica, como para conhecimento do efeito
benéfico da gabapentina.
* *
*
Em síntese, nossos resultados sugerem que o processo nociceptivo em nosso modelo
de dor muscular crônica interfere na produção e/ou liberação das interleucinas anti-
inflamatórias e pró-inflamatórias. Ainda, observamos que a este processo nociceptivo provoca
aumento de astrócitos e microglia no tálamo dos animais após indução de miosite crônica. No
que tange ao tratamento farmacológico, sugerimos que a gabapentina possui ação nos núcleos
intralaminares do tálamo e inibe astrócitos e microglia nos núcleos ventral póstero-lateral e
ventral póstero-medial em modelo de dor muscular crônica. Além disso, sugerimos que a ação
da gabapentina ocorre indiretamente por meio da estimulação de citocinas anti-inflamatórias
e, principalmente, pela inibição de citocinas pró-inflamatórias.

Conclusão
56
6. CONCLUSÃO
Nossos resultados em conjunto podem concluir que a modulação da dor muscular
crônica causada pelo aumento de neurotransmissores e citocinas pró-inflamatórias no tecido
musculoesquelético, como descrito na literatura, segue para o gânglio da coluna posterior e
ascende ao tálamo em terminais aferentes primários ipsilaterais à lesão, o que possibilita
ativação glial. As células gliais uma vez ativada, liberam mais mediadores que chegam aos
terminais do lado contralateral à lesão, levando também, à ativação de microglia e astrócitos
do lado contralateral. Os produtos gliais poderiam levar à uma liberação exacerbada de
neurotransmissores, sensibilização da membrana pós-sináptica, ativação de outras células
gliais e aumento da propagação glial de neuromediadores, facilitando o desenvolvimento de
hipersensibilidade neuronal, hiperalgesia e alodinia. Podemos concluir que, nosso tratamento
com a gabapentina, diminui a liberação de neurotransmissores que ativam células microgliais
e astrócitos no gânglio da coluna posterior e tálamo, inibindo a liberação de citocinas pró-
inflamatórias e estimulando a liberação de citocinas anti-inflamatórias; modulando assim a
resposta nociceptiva e por conseguinte, reversão do quadro doloroso.

Referências bibliográficas
57
REFERÊNCIAS
Abdelhamid RE, Sluka KA. ASICs Mediate Pain and Inflammation in Musculoskeletal
Diseases. Physiology. 2015; 30(6): 449-59.
Allchorne AJ, Gooding HL, Mitchell R, Fleetwood-Walker SM. A novel model of combined
neuropathic and inflammatory pain displaying long-lasting allodynia and spontaneous pain-
like behaviour. Neurosci Res. 2012; 74(3-4): 230-8.
Andrade RMQd. Dor crônica na atenção primária - um problema de saúde pública.
Universidade Federal de Minas Gerais. Faculdade de Medicina. Núcleo de Educação em
Saúde Coletiva . 2014; Monografia (Especialização em Atenção Básica em Saúde da
Família).
Ashmawi HA, Freire GMG. Peripheral and central sensitization. Revista Dor. 2016; 17: 31-4.
Atkinson JH, Slater MA, Capparelli EV, Patel SM, Wolfson T, Gamst A, Abramson IS,
Wallace MS, Funk SD, Rutledge TR, Wetherell JL, Matthews SC, Zisook S, Garfin SR. A
randomized controlled trial of gabapentin for chronic low back pain with and without a
radiating component. Pain. 2016; 8: 8.
Bastos LF, Godin AM, Zhang Y, Jarussophon S, Ferreira BC, Machado RR, Maier SF,
Konishi Y, de Freitas RP, Fiebich BL, Watkins LR, Coelho MM, Moraes MF. A minocycline
derivative reduces nerve injury-induced allodynia, LPS-induced prostaglandin E2 microglial
production and signaling via toll-like receptors 2 and 4. Neurosci Lett. 2013; 543: 157-62.
Beladi Mousavi SS, Zeraati A, Moradi S, Mousavi MB. The effect of gabapentin on muscle
cramps during hemodialysis: A double-blind clinical trial. Saudi J Kidney Dis Transpl. 2015;
26(6): 1142-8.
Besson JM. The neurobiology of pain. Lancet. 1999; 353(9164): 1610-5.
Borghi SM, Zarpelon AC, Pinho-Ribeiro FA, Cardoso RD, Cunha TM, Alves-Filho JC,
Ferreira SH, Cunha FQ, Casagrande R, Verri WA, Jr. Targeting interleukin-1beta reduces
intense acute swimming-induced muscle mechanical hyperalgesia in mice. J Pharm
Pharmacol. 2014; 66(7): 1009-20.
Bottega FH, Fontana RT. A dor como quinto sinal vital: utilização da escala de avaliação por
enfermeiros de um hospital geral. Texto & Contexto - Enfermagem. 2010; 19: 283-90.
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of
protein utilizing the principle of protein-dye binding. Anal Biochem. 1976; 72: 248-54.
Casey G. Pain--the fifth vital sign. Nurs N Z. 2011; 17(5): 24-9.
Cassell A, Edwards D, Harshfield A, Rhodes K, Brimicombe J, Payne R, Griffin S. The
epidemiology of multimorbidity in primary care: a retrospective cohort study. Br J Gen Pract.
2018; 68(669): e245-e51.

Referências bibliográficas
58
Chacur M, Gutierrez JM, Milligan ED, Wieseler-Frank J, Britto LR, Maier SF, Watkins LR,
Cury Y. Snake venom components enhance pain upon subcutaneous injection: an initial
examination of spinal cord mediators. Pain. 2004; 111(1-2): 65-76.
Chacur M, Lambertz D, Hoheisel U, Mense S. Role of spinal microglia in myositis-induced
central sensitisation: an immunohistochemical and behavioural study in rats. Eur J Pain. 2009;
13(9): 915-23.
Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile
allodynia in the rat paw. Journal of Neuroscience Methods. 1994; 53: 55-63.
Cheng JK, Chiou LC. Mechanisms of the antinociceptive action of gabapentin. J Pharmacol
Sci. 2006; 100(5): 471-86.
Cimas M, Ayala A, Sanz B, Agullo-Tomas MS, Escobar A, Forjaz MJ. Chronic
musculoskeletal pain in European older adults: Cross-national and gender differences. Eur J
Pain. 2018; 22(2): 333-45.
Clivatti J, Sakata RK, Issy AM. Revisão sobre o uso de gabapentina para controle da dor pós-
operatória. Revista Brasileira de Anestesiologia. 2009; 59: 87-98.
Cohen M, Quintner J, van Rysewyk S. Reconsidering the International Association for the
Study of Pain definition of pain. Pain Rep. 2018; 3(2).
Costa FA, Moreira Neto FL. [Satellite glial cells in sensory ganglia: its role in pain]. Rev Bras
Anestesiol. 2015; 65(1): 73-81.
Costigan M, Woolf CJ. Pain: molecular mechanisms. J Pain. 2000; 1(3 Suppl): 35-44.
Deleo JA, Ph.d.; Sorkin, Linda S., Ph.d.; Watkins, Linda R., Ph.d. Immune And Glial
Regulation of Pain, in 2004.
Dolgun H, Turkoglu E, Kertmen H, Gurer B, Yilmaz ER, Comoglu SS, Sekerci Z.
Gabapentin versus pregabalin in relieving early post-surgical neuropathic pain in patients after
lumbar disc herniation surgery: a prospective clinical trial. Neurol Res. 2014; 36(12): 1080-5.
Fein A. NOCICEPTORES: As células que sentem dor. 2011.
Fernandes BHP, Gomes CRdG. Mecanismos e Aspectos Anatômicos da Dor; 2011.
Fink K, Meder W, Dooley DJ, Göthert M. Inhibition of neuronal Ca2+ influx by gabapentin
and subsequent reduction of neurotransmitter release from rat neocortical slices. British
Journal of Pharmacology. 2000; 130(4): 900-6.
Giardini AC, Dos Santos FM, da Silva JT, de Oliveira ME, Martins DO, Chacur M. Neural
Mobilization Treatment Decreases Glial Cells and Brain-Derived Neurotrophic Factor

Referências bibliográficas
59
Expression in the Central Nervous System in Rats with Neuropathic Pain Induced by CCI in
Rats. Pain Res Manag. 2017; 7429761(10): 22.
Guerrero AT, Verri WA, Jr., Cunha TM, Silva TA, Rocha FA, Ferreira SH, Cunha FQ,
Parada CA. Hypernociception elicited by tibio-tarsal joint flexion in mice: a novel
experimental arthritis model for pharmacological screening. Pharmacol Biochem Behav.
2006; 84(2): 244-51.
Hains BC, Waxman SG. Activated microglia contribute to the maintenance of chronic pain
after spinal cord injury. J Neurosci. 2006; 26(16): 4308-17.
Hargreaves K, Dubner R, Brown F, Flores C, Joris J. A new and sensitive method for
measuring thermal nociception in cutaneous hyperalgesia. Pain. 1988; 32: 77-88.
Hayashi Y, Koyanagi S, Kusunose N, Okada R, Wu Z, Tozaki-Saitoh H, Ukai K, Kohsaka S,
Inoue K, Ohdo S, Nakanishi H. The intrinsic microglial molecular clock controls synaptic
strength via the circadian expression of cathepsin S. Sci Rep. 2013; 3(2744).
Hesami O, Gharagozli K, Beladimoghadam N, Assarzadegan F, Mansouri B, Sistanizad M.
The Efficacy of Gabapentin in Patients with Central Post-stroke Pain. Iran J Pharm Res. 2015;
14(Suppl): 95-101.
Hoheisel U, Kaske A, Mense S. Relationship between neuronal activity and substance P-
immunoreactivity in the rat spinal cord during acute and persistent myositis. Neurosci Lett.
1998; 257(1): 21-4.
Hoheisel U, Reuter R, de Freitas MF, Treede RD, Mense S. Injection of nerve growth factor
into a low back muscle induces long-lasting latent hypersensitivity in rat dorsal horn neurons.
Pain. 2013; 154(10): 1953-60.
Hoheisel U, Unger T, Mense S. A block of spinal nitric oxide synthesis leads to increased
background activity predominantly in nociceptive dorsal horn neurones in the rat. Pain. 2000;
88(3): 249-57.
Hoheisel U, Unger T, Mense S. The possible role of the NO-cGMP pathway in nociception:
different spinal and supraspinal action of enzyme blockers on rat dorsal horn neurones. Pain.
2005; 117(3): 358-67.
IASP. Guia para o Tratamento da Dor em Contextos de Poucos Recursos. IASP Press®. 2010;
Tradução por Andreas Kopf e Nilesh B. Patel.
Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature. 2001; 413(6852): 203-
10.
Kang TW, Sohn MK, Park NK, Ko SH, Cho KJ, Beom J, Kang S. Analgesic effect of
intrathecal gabapentin in a rat model of persistent muscle pain. Ann Rehabil Med. 2014;
38(5): 682-8.

Referências bibliográficas
60
Kidd BL, Urban LA. Mechanisms of inflammatory pain. Br J Anaesth. 2001; 87(1): 3-11.
Kraychete DC, Calasans MTdA, Valente CML. Citocinas pró-inflamatórias e dor. Revista
Brasileira de Reumatologia. 2006; 46: 199-206.
Kremer M, Salvat E, Muller A, Yalcin I, Barrot M. Antidepressants and gabapentinoids in
neuropathic pain: Mechanistic insights. Neuroscience. 2016; 338: 183-206.
Krzyzanowska A, Pittolo S, Cabrerizo M, Sanchez-Lopez J, Krishnasamy S, Venero C,
Avendano C. Assessing nociceptive sensitivity in mouse models of inflammatory and
neuropathic trigeminal pain. J Neurosci Methods. 2011; 201(1): 46-54.
Kukkar A, Bali A, Singh N, Jaggi AS. Implications and mechanism of action of gabapentin in
neuropathic pain. Archives of Pharmacal Research. 2013; 36(3): 237-51.
Latremoliere A, Woolf CJ. Central sensitization: a generator of pain hypersensitivity by
central neural plasticity. J Pain. 2009; 10(9): 895-926.
LeBlanc BW, Zerah ML, Kadasi LM, Chai N, Saab CY. Minocycline injection in the ventral
posterolateral thalamus reverses microglial reactivity and thermal hyperalgesia secondary to
sciatic neuropathy. Neurosci Lett. 2011; 498(2): 138-42.
Lewis SC, Bhattacharya S, Wu O, Vincent K, Jack SA, Critchley HO, Porter MA, Cranley D,
Wilson JA, Horne AW. Gabapentin for the Management of Chronic Pelvic Pain in Women
(GaPP1): A Pilot Randomised Controlled Trial. PLoS One. 2016; 11(4).
Liu FY, Sun YN, Wang FT, Li Q, Su L, Zhao ZF, Meng XL, Zhao H, Wu X, Sun Q, Xing
GG, Wan Y. Activation of satellite glial cells in lumbar dorsal root ganglia contributes to
neuropathic pain after spinal nerve ligation. Brain Res. 2012; 3: 65-77.
Loeser JD, Treede RD. The Kyoto protocol of IASP Basic Pain Terminology. Pain. 2008;
137(3): 473-7.
Loram LC, Fuller A, Fick LG, Cartmell T, Poole S, Mitchell D. Cytokine profiles during
carrageenan-induced inflammatory hyperalgesia in rat muscle and hind paw. J Pain. 2007;
8(2): 127-36.
Manjavachi MN, Motta EM, Marotta DM, Leite DF, Calixto JB. Mechanisms involved in IL-
6-induced muscular mechanical hyperalgesia in mice. Pain. 2010; 151(2): 345-55.
Marchand F, Perretti M, McMahon SB. Role of the immune system in chronic pain. Nat Rev
Neurosci. 2005; 6(7): 521-32.
McCaffery M, Pasero CL. Pain ratings: the fifth vital sign. Am J Nurs. 1997; 97(2): 15-6.
McLean G, Mercer SW. Chronic migraine, comorbidity, and socioeconomic deprivation:
cross-sectional analysis of a large nationally representative primary care database. J Comorb.
2017; 7(1): 89-95.

Referências bibliográficas
61
Merskey H, Bogduk N. Part III: Pain Terms, A Current List with Definitions and Notes on
Usage. IASP Press. 1994.
Millan MJ. The induction of pain: an integrative review. Prog Neurobiol. 1999; 57(1): 1-164.
Milligan ED, Mehmert KK, Hinde JL, Harvey LO, Martin D, Tracey KJ, Maier SF, Watkins
LR. Thermal hyperalgesia and mechanical allodynia produced by intrathecal administration of
the human immunodeficiency virus-1 (HIV-1) envelope glycoprotein, gp120. Brain Res.
2000; 861(1): 105-16.
Milligan ED, Twining C, Chacur M, Biedenkapp J, O'Connor K, Poole S, Tracey K, Martin
D, Maier SF, Watkins LR. Spinal glia and proinflammatory cytokines mediate mirror-image
neuropathic pain in rats. Journal of Neuroscience. 2003; 23(3): 1026-40.
Milligan ED, Watkins LR. Pathological and protective roles of glia in chronic pain. Nat Rev
Neurosci. 2009; 10(1): 23-36.
Mimenza Alvarado A, Aguilar Navarro S. Clinical Trial Assessing the Efficacy of Gabapentin
Plus B Complex (B1/B12) versus Pregabalin for Treating Painful Diabetic Neuropathy. J
Diabetes Res. 2016; 4078695(10): 17.
Miranda VS, Decarvalho VB, Machado LA, Dias JM. Prevalence of chronic musculoskeletal
disorders in elderly Brazilians: a systematic review of the literature. BMC Musculoskelet
Disord. 2012; 13(82): 1471-2474.
Morioka N, Zhang FF, Nakamura Y, Kitamura T, Hisaoka-Nakashima K, Nakata Y. Tumor
necrosis factor-mediated downregulation of spinal astrocytic connexin43 leads to increased
glutamatergic neurotransmission and neuropathic pain in mice. Brain Behav Immun. 2015;
49: 293-310.
Ohtori S, Takahashi K, Moriya H, Myers RR. TNF-alpha and TNF-alpha receptor type 1
upregulation in glia and neurons after peripheral nerve injury: studies in murine DRG and
spinal cord. Spine. 2004; 29(10): 1082-8.
Old EA, Clark AK, Malcangio M. The role of glia in the spinal cord in neuropathic and
inflammatory pain. Handb Exp Pharmacol. 2015; 227: 145-70.
Oliveira, Silva MdFN, Araujo IEM, Carvalho-Filho MA. Profile of the Population Cared for
in a Referral Emergency Unit. Revista Latino-Americana de Enfermagem. 2011; 19: 548-56.
Oliveira CMBd, Sakata RK, Issy AM, Gerola LR, Salomão R. Citocinas e dor. Revista
Brasileira de Anestesiologia. 2011; 61: 260-5.
Oliveira ME, Santos FM, Bonifacio RP, Freitas MF, Martins DO, Chacur M. Low level laser
therapy alters satellite glial cell expression and reverses nociceptive behavior in rats with
neuropathic pain. Photochem Photobiol Sci. 2017; 16(4): 547-54.

Referências bibliográficas
62
Park HJ, Sandor K, McQueen J, Woller SA, Svensson CI, Corr M, Yaksh TL. The effect of
gabapentin and ketorolac on allodynia and conditioned place preference in antibody-induced
inflammation. Eur J Pain. 2015; 30(10): 816.
Park S, Ahn ES, Han DW, Lee JH, Min KT, Kim H, Hong YW. Pregabalin and gabapentin
inhibit substance P-induced NF-kappaB activation in neuroblastoma and glioma cells. J Cell
Biochem. 2008; 105(2): 414-23.
Pratt D, Fuchs PN, Sluka KA. Assessment of avoidance behaviors in mouse models of muscle
pain. Neuroscience. 2013; 248: 54-60.
Price DD. Psychological and neural mechanisms of the affective dimension of pain. Science.
2000; 288(5472): 1769-72.
Punnett L, Wegman DH. Work-related musculoskeletal disorders: the epidemiologic evidence
and the debate. J Electromyogr Kinesiol. 2004; 14(1): 13-23.
Quintao NL, Antonialli CS, da Silva GF, Rocha LW, de Souza MM, Malheiros A, Meyre-
Silva C, Lucinda-Silva RM, Bresolin TM, Filho VC. Aleurites moluccana and its main active
ingredient, the flavonoid 2''-O-rhamnosylswertisin, have promising antinociceptive effects in
experimental models of hypersensitivity in mice. Pharmacol Biochem Behav. 2012; 102(2):
302-11.
Quintero JE, Dooley DJ, Pomerleau F, Huettl P, Gerhardt GA. Amperometric measurement of
glutamate release modulation by gabapentin and pregabalin in rat neocortical slices: role of
voltage-sensitive Ca2+ alpha2delta-1 subunit. J Pharmacol Exp Ther. 2011; 338(1): 240-5.
Radhakrishnan R, Moore SA, Sluka KA. Unilateral carrageenan injection into muscle or joint
induces chronic bilateral hyperalgesia in rats. Pain. 2003; 104(3): 567-77.
Reda HM, Zaitone SA, Moustafa YM. Effect of levetiracetam versus gabapentin on peripheral
neuropathy and sciatic degeneration in streptozotocin-diabetic mice: Influence on spinal
microglia and astrocytes. Eur J Pharmacol. 2016; 771: 162-72.
Reinert A, Kaske A, Mense S. Inflammation-induced increase in the density of neuropeptide-
immunoreactive nerve endings in rat skeletal muscle. Exp Brain Res. 1998; 121(2): 174-80.
Robertson KL, Marshman LA. Gabapentin Superadded to a Pre-Existent Regime Containing
Amytriptyline for Chronic Sciatica. Pain Med. 2016; 3.
Rocha AP, Kraychete DC, Lemonica L, de Carvalho LR, de Barros GA, Garcia JB, Sakata
RK. Pain: current aspects on peripheral and central sensitization. Rev Bras Anestesiol. 2007;
57(1): 94-105.
Rosa AS, Freitas MF, Rocha IR, Chacur M. Gabapentin decreases microglial cells and
reverses bilateral hyperalgesia and allodynia in rats with chronic myositis. Eur J Pharmacol.
2017; 10(17): 012.

Referências bibliográficas
63
Rothermundt M, Ohrmann P, Abel S, Siegmund A, Pedersen A, Ponath G, Suslow T, Peters
M, Kaestner F, Heindel W, Arolt V, Pfleiderer B. Glial cell activation in a subgroup of
patients with schizophrenia indicated by increased S100B serum concentrations and elevated
myo-inositol. Prog Neuropsychopharmacol Biol Psychiatry. 2007; 31(2): 361-4.
Rowbotham M, Harden N, Stacey B, Bernstein P, Magnus-Miller L. Gabapentin for the
treatment of postherpetic neuralgia: a randomized controlled trial. Jama. 1998; 280(21): 1837-
42.
Skaper SD, Facci L. Mast cell–glia axis in neuroinflammation and therapeutic potential of the
anandamide congener palmitoylethanolamide. Philosophical Transactions of the Royal
Society B: Biological Sciences. 2012; 367(1607): 3312-25.
Sluka KA. Pain mechanisms involved in musculoskeletal disorders. J Orthop Sports Phys
Ther. 1996; 24(4): 240-54.
Sluka KA, Kalra A, Moore SA. Unilateral intramuscular injections of acidic saline produce a
bilateral, long-lasting hyperalgesia. Muscle Nerve. 2001; 24(1): 37-46.
Sokal RR, Rohlf FJ. Biometry. New York; 1981. p. 859.
Souza JB, Grossmann E, Perissinotti DMN, de Oliveira Junior JO, da Fonseca PRB, Posso IP.
Prevalence of Chronic Pain, Treatments, Perception, and Interference on Life Activities:
Brazilian Population-Based Survey. Pain Res Manag. 2017; 4643830(10): 26.
Suto T, Severino AL, Eisenach JC, Hayashida K. Gabapentin increases extracellular
glutamatergic level in the locus coeruleus via astroglial glutamate transporter-dependent
mechanisms. Neuropharmacology. 2014; 81: 95-100.
Sweitzer SM, Colburn RW, Rutkowski M, DeLeo JA. Acute peripheral inflammation induces
moderate glial activation and spinal IL-1beta expression that correlates with pain behavior in
the rat. Brain Res. 1999; 829(1-2): 209-21.
Takasusuki T, Yaksh TL. The effects of intrathecal and systemic gabapentin on spinal
substance P release. Anesth Analg. 2011; 112(4): 971-6.
Tariba Knezevic P, Vukman R, Antonic R, Kovac Z, Uhac I, Simonic-Kocijan S. The role of
P2X3 receptors in bilateral masseter muscle allodynia in rats. Croat Med J. 2016; 57(6): 530-
9.
Teixeira M, Okada M. História da Dor; 2010.
Teixeira MJ. Fisiopatologia da nocicepção e da supressão da dor. In: Dor orofacial :
diagnóstico, terapêutica e qualidade de vida.Editora Maio; 2001.
Teixeira MJ, SOUZA ECD, YENG LT, PEREIRA WC. A lesão do trato de Lissauer e do
corno posterior da substância cinzenta da medula espinal e a estimulação elétrica do sistema

Referências bibliográficas
64
nervoso central para o tratamento da dor por avulsão de raízes do plexo braquial. Arquivos de
Neuro-Psiquiatria. 1999; 57: 56-62.
Teixeira MJ, Teixeira WGJ, Santos FPS, Andrade DCA, Bezerra SL, Figueiró JB, Okada M.
Clinical epidemiology of muscleskeletal pain. 2001. 2001; 80: 21.
Tenschert S, Reinert A, Hoheisel U, Mense S. Effects of a chronic myositis on structural and
functional features of spinal astrocytes in the rat. Neurosci Lett. 2004; 361(1-3): 196-9.
Thompson SW, Woolf CJ, Sivilotti LG. Small-caliber afferent inputs produce a
heterosynaptic facilitation of the synaptic responses evoked by primary afferent A-fibers in
the neonatal rat spinal cord in vitro. J Neurophysiol. 1993; 69(6): 2116-28.
Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins from polyacrylamide
gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A.
1979; 76(9): 4350-4.
Traub RJ. The spinal contribution of substance P to the generation and maintenance of
inflammatory hyperalgesia in the rat. Pain. 1996; 67(1): 151-61.
Tsuda M, Inoue K, Salter MW. Neuropathic pain and spinal microglia: a big problem from
molecules in "small" glia. Trends Neurosci. 2005; 28(2): 101-7.
Tsukumo Y, Matsumoto Y, Miura H, Yano H, Manabe H. Gabapentin and pregabalin inhibit
the itch-associated response induced by the repeated application of oxazolone in mice. J
Pharmacol Sci. 2011; 115(1): 27-35.
Vallejo R, Tilley DM, Vogel L, Benyamin R. The role of glia and the immune system in the
development and maintenance of neuropathic pain. Pain Pract. 2010; 10(3): 167-84.
Watkins LR, Maier SF. GLIA: A novel drug discovery target for clinical pain. Nat Rev Drug
Discov. 2003; 2(12): 973-85.
Watkins LR, Milligan ED, Maier SF. Glial proinflammatory cytokines mediate exaggerated
pain states: implications for clinical pain. Adv Exp Med Biol. 2003; 521: 1-21.
Williams AC, Craig KD. Updating the definition of pain. Pain. 2016; 157(11): 2420-3.
Woolf CJ, Costigan M. Transcriptional and posttranslational plasticity and the generation of
inflammatory pain. Proc Natl Acad Sci U S A. 1999; 96(14): 7723-30.
Woolf CJ, Shortland P, Coggeshall RE. Peripheral nerve injury triggers central sprouting of
myelinated afferents. Nature. 1992; 355(6355): 75-8.
Yaksh TL. Calcium channels as therapeutic targets in neuropathic pain. J Pain. 2006; 7(1
Suppl 1): S13-30.

Referências bibliográficas
65
Yang F, Fu H, Lu YF, Wang XL, Yang Y, Yu YQ, Sun W, Wang JS, Costigan M, Chen J.
Post-stroke pain hypersensitivity induced by experimental thalamic hemorrhage in rats is
region-specific and demonstrates limited efficacy of gabapentin. Neurosci Bull. 2014; 30(6):
887-902.
Yang JL, Xu B, Li SS, Zhang WS, Xu H, Deng XM, Zhang YQ. Gabapentin reduces
CX3CL1 signaling and blocks spinal microglial activation in monoarthritic rats. Mol Brain.
2012; 5(18): 1756-6606.
Yelin E, Weinstein S, King T. The burden of musculoskeletal diseases in the United
StatesSemin Arthritis Rheum. 2016 Dec;46(3):259-260. doi:
10.1016/j.semarthrit.2016.07.013. Epub 2016 Jul 26.; 2016.
Zhang J-M, An J. Cytokines, Inflammation and Pain. International anesthesiology clinics.
2007; 45(2): 27-37.
Zhang WS, Xu H, Xu B, Sun S, Deng XM, Zhang YQ. Antihyperalgesic effect of systemic
dexmedetomidine and gabapentin in a rat model of monoarthritis. Brain Res. 2009; 6: 57-66.
Zhang YB, Guo ZD, Li MY, Fong P, Zhang JG, Zhang CW, Gong KR, Yang MF, Niu JZ, Ji
XM, Lv GW. Gabapentin Effects on PKC-ERK1/2 Signaling in the Spinal Cord of Rats with
Formalin-Induced Visceral Inflammatory Pain. PLoS One. 2015; 10(10).
Zhou C, Luo ZD. Electrophysiological characterization of spinal neuron sensitization by
elevated calcium channel alpha-2-delta-1 subunit protein. Eur J Pain. 2014; 18(5): 649-58.
Zimmermann M. Ethical guidelines for investigations of experimental pain in conscious
animals. Pain. 1983; 16(2): 109-10.
Zvejniece L, Vavers E, Svalbe B, Veinberg G, Rizhanova K, Liepins V, Kalvinsh I,
Dambrova M. R-phenibut binds to the alpha2-delta subunit of voltage-dependent calcium
channels and exerts gabapentin-like anti-nociceptive effects. Pharmacol Biochem Behav.
2015; 137: 23-9.