avaliação das propriedades antioxidantes e antimutagénicas ... · dissertação para obtenção...
TRANSCRIPT

Susana Margarida Vieira Carreira
Licenciada em Ensino de Física e Química
Avaliação das propriedades antioxidantes e
antimutagénicas de diferentes cultivares Vaccinium spp.:
Partiot, Bluecrop, Duke, Earlyblue e O’Neal, produzidas na
Sub-região do Baixo Vouga
Dissertação para obtenção do Grau de Mestre em
Tecnologia e Segurança Alimentar
Orientadora: Professora Doutora Maria Paula Amaro de Castilho Duarte,
Professora Auxiliar, FCT/UNL
Júri:
Presidente: Prof. Doutora Ana Lucia Monteiro Durão Leitão
Arguente: Prof. Doutora Ana Sofia Gregório Fernandes
Vogal: Doutora Maria Paula Amaro de Castilho Duarte
Setembro 2012

i
Susana Margarida Vieira Carreira
Licenciada em Ensino de Física e Química
Avaliação das propriedades antioxidantes e
antimutagénicas de diferentes cultivares Vaccinium spp.:
Partiot, Bluecrop, Duke, Earlyblue e O’Neal, produzidas na
Sub-região do Baixo Vouga
Dissertação para obtenção do Grau de Mestre em
Tecnologia e Segurança Alimentar
Orientadora: Professora Doutora Maria Paula Amaro de Castilho Duarte,
Professora Auxiliar, FCT/UNL
Setembro 2012

ii
Copyright – Susana Margarida Vieira Carreira, UNL, FCT
A Faculdade de Ciências e Tecnologia e a Universidade Nova de Lisboa têm o direito, perpétuo e
sem limites geográficos, de arquivar e publicar esta dissertação através de exemplares impressos
reproduzidos em papel ou de forma digital, ou por qualquer outro meio conhecido ou que venha a ser
inventado, e de a divulgar através de repositórios científicos e de admitir a sua cópia e distribuição
com objectivos educacionais ou de investigação, não comerciais, desde que seja dado crédito ao autor
e editor.

iii
Agradecimentos
Gostaria de agradecer:
À Professora Doutora Benilde Mendes, coordenadora deste Mestrado, por ter proporcionado todas as
condições para a realização deste trabalho.
À Professora Doutora Maria Paula Duarte pela excelente orientação científica, pelos conhecimentos
que me transmitiu, pela pronta colaboração face a todas as minhas solicitações e total disponibilidade
em todo o trabalho desenvolvido e ainda pela amizade, pois sem este apoio incondicional este tabalho
não teria sido possível, o meu eterno agradecimento.
Ao Professor Doutor José Rueff, Diretor do Departamento de Genética da Faculdade de Ciências
Médicas da Universidade Nova de Lisboa, por ter disponibilizado as instalações e os equipamentos
necessários à realização do ensaio ORAC.
À minha colega e amiga Eugénia Rasteiro, pelo apoio e colaboração neste trabalho, bem como pelo
incentivo, encorajamento nos momentos mais difíceis ao longo deste percurso. Obrigada pelo que
passamos por muito de bom e de mau, pois ficarás sempre no meu coração.
Ao Pedro Oliveira, pelo apoio, colaboração neste trabalho e pela sua total disponibilidade. Um
obrigado pela tua descontração, boa disposição e optimismo que teve um papel curcial ao longo deste
percurso e não só... E ainda pelo amor, carinho e pela compreensão. Um obrigado por existires na
minha vida, pois mesmo as pessoas mais fortes têm momentos de fraqueza e felizmente que têm
alguém em quem se apoiar.
À minha família e amigos por acreditarem em mim, pelas palavras de incentivo e encorajamento.
À Mirtilusa, Sociedade de Produtores Hortofrutícolas, Sever do Vouga, por me terem recebido e
disponibilizou as amostras das cultivares de mirtilo, sem as quais não teria sido possível a realização
deste trabalho.
À empresa Mirtisul – Produção de Mirtilos Lda., localizada na Aldeia do Pico – Grândola, que
generosamente me recebeu e disponibilizaram amostras de cultivares que, embora não sejam objeto de
estudo neste trabalho, foram utilizadas no desenvolvimento das teses dos meus colegas Eugénia
Rasteiro e Pedro Oliveira.

iv
Resumo
Estudaram-se as atividades antioxidante (ensaios FRAP, CUPRAC, ORAC, quelação de ferro
e sequestro de peróxido de hidrogénio e do radical anião superóxido) e antimutagénica (teste de Ames)
em cinco diferentes variedades de mirtilo (Partiot, Bluecrop, Duke, Earlyblue e O’Neal) produzidas no
ano de 2010, em Portugal, na sub-região do Baixo-Vouga.
Todas as variedades analisadas apresentaram atividade antioxidante, detetada através da
avaliação da atividade redutora e de sequestro de espécies reativas de oxigénio, bem como atividade
antimutagénica, não tendo demonstrado capacidade de quelação do Fe(II).
Das várias variedades estudadas, a Duke foi a que apresentou um teor mais elevado em
polifenóis, tendo apresentado a maior capacidade antioxidante em todos os ensaios realizados, com
excepção do sequestro do peróxido de hidrogénio. Por outro lado, a variedade O’Neal foi a que
apresentou o mais baixo conteúdo em compostos fenólicos, tendo sido aquela que apresentou mais
elevada capacidade de sequestro do peróxido de hidrogénio. Assim, com excepção do sequestro de
peróxido de hidrogénio e da quelação de Fe(II), todas as restantes atividades antioxidantes
determinadas parecem estar relacionadas com os compostos fenólicos e, particularmente, com as
antocianinas presentes nas diferentes variedades de mirtilos estudadas.
Palavras-chave: Mirtilo, Atividade antioxidante, Atividade antimutagénica, Compostos fenólicos

v
Abstract
In this study it was investigated the antioxidant activity (FRAP, CUPRAC, ORAC, iron
chelating activity, hydrogen peroxide scavenging and superoxide anion radical scavenging) and the
antimutagenic activitiy (Ames test) of five blueberry varieties (Partiot, Bluecrop, Duke, EarlyBlue and
O'Neal) produced in 2010, in Portugal, in the sub-region of Baixo-Vouga.
All analyzed blueberry varieties presented antioxidant activity, detected by the evaluation of
the reductive activity and the scavenging of reactive oxygen species, and antimutagenic activity. The
ferrous ions chelating activity of the samples were not demonstrated.
Of the several varieties studied, Duke showed the highest level of total phenolic coumpounds,
and showed the highest antioxidant capacity in all methods except in the hydrogen peroxide
scavenging assay. The variety O'Neal showed the lowest content of phenolic compounds, and the
higher capacity for scavenging the hydrogen peroxide. Thus, with the exception of hydrogen peroxide
scavenging assay and Fe(II) chelation assay, all other antioxidant activities appear to be related to
certain phenolic compounds, and particularly with anthocyanins present in the different blueberry
varieties studied.
Keywords: Blueberries, antioxidant activity, antimutagenic activity, phenolic compounds

vi
Índice de Matérias
Introdução ..........................................................................................................................................1
1. Breve caraterização do mirtilo.........................................................................................................3
1.1. Variedades de mirtilo.................................................................................................................3
1.2. Produção e consumo de mirtilo em Portugal .............................................................................5
1.3. Caraterísticas físico-químicas e nutricionais do Mirtilo ...............................................................7
2. Mirtilo e importância na saúde...................................................................................................... 10
2.1. Mirtilo alimento funcional ...................................................................................................... 10
2.2. Os compostos fenólicos nos mirtilos ....................................................................................... 11
2.3. Espécies reativas de oxigénio ................................................................................................. 14
2.4. Sistemas de Defesas Antioxidantes ......................................................................................... 19
2.5. Mutagéneos e Antimutagéneos nos alimentos ....................................................................... 22
2.5.1. Mutagéneos alimentares ................................................................................................. 22
2.5.2. Antimutagéneos alimentares ........................................................................................... 25
2.6. Efeitos dos polifenóis na saúde ............................................................................................... 27
3. Materiais e Métodos ..................................................................................................................... 30
3.1. Caraterização das amostras de mirtilo .................................................................................... 30
3.2. Reagentes e meios de cultura ................................................................................................. 30
3.3. Preparação dos extratos de mirtilo ......................................................................................... 31
3.4. Determinação dos fenóis totais pelo Método de Folin-Ciocalteu ............................................ 31
3.5. Quantificação das antocianinas monoméricas totais pelo método do pH diferencial .............. 32
3.6. Determinação do teor de sólidos solúveis (grau Brix) .............................................................. 34
3.7. Avaliação da Capacidade Antioxidante ................................................................................... 35
3.7.1. Determinação da capacidade de quelação de Fe(II) ......................................................... 35
3.7.2 Determinação da atividade de redução Fe(III) a Fe(II) pelo ensaio FRAP (“Ferric Reduction
Antioxidant Power”) .................................................................................................................. 36
3.7.3. Determinação da redução do Cu(II) pelo ensaio CUPRAC (“Cupric Reducing Antioxidant
Capacity”) ................................................................................................................................. 37
3.7.4. Determinação do sequestro do peróxido de hidrogénio ................................................... 38
3.7.5. Determinação do sequestro do radical anião superóxido ................................................. 39

vii
3.7.6. Determinação do resgate do radical peroxilo pelo ensaio ORAC (“Oxygen Radical
Absorbance Capacity”) .............................................................................................................. 40
3.8. Avaliação da atividade mutagénica e antimutagénica através do Teste de Ames .................... 42
3.8.1. Caraterização da estirpe de S. typhimurium utilizada ....................................................... 43
3.8.2. Realização dos testes de Ames......................................................................................... 44
3.9. Análise estatística dos resultados ........................................................................................... 45
4. Resultados e Discussão ................................................................................................................. 46
4.1. Determinação dos fenóis totais .............................................................................................. 46
4.2. Quantificação das antocianinas monoméricas totais............................................................... 47
4.3. Determinação do teor de sólidos solúveis (grau Brix) .............................................................. 49
4.4. Determinação da atividade antioxidante das diferentes variedades de mirtilo........................ 49
4.4.1. Determinação da atividade antioxidante por capacidade redutora da amostra ................ 49
4.4.1.1. Determinação da atividade de redução do Fe(III) a Fe(II) pelo ensaio FRAP ................... 50
4.4.1.2. Determinação da atividade de redução do Cu(II) a Cu(I) pelo ensaio CUPRAC ............... 51
4.4.2. Determinação da capacidade de quelação de ferro .......................................................... 52
4.4.3. Determinação da atividade antioxidante por sequestro de espécies reativas de oxigénio 52
4.4.3.1. Determinação da capacidade de sequestro do peróxido de hidrogénio por medição da
absorção no UV ......................................................................................................................... 53
4.4.3.2. Determinação da capacidade de sequestro do radical anião superóxido detetada pela
diminuição da formação de formazano ..................................................................................... 53
4.4.3.3. Determinação da atividade antioxidante através da análise da capacidade de resgate do
radical peroxilo pelo ensaio ORAC ............................................................................................. 54
4.5. Avaliação da atividade mutagénica e antimutagénica através do teste de Ames ..................... 55
4.6. Análise global dos resultados.................................................................................................. 58
5. Conclusão ..................................................................................................................................... 64
6. Referências Bibliográficas.............................................................................................................. 67

viii
Índice de Figuras
Figura 1.1 – Aspeto da planta de mirtilo Vaccinium spp. ....................................................................3
Figura 1.2 – Baga de mirtilo ...............................................................................................................4
Figura 1.3 – Mapa da Sub-região Baixo Vouga ..................................................................................7
Figura 2.1 - Compostos fitoquímicos presentes em alimentos de origem vegetal. .............................. 12
Figura 3.1 - Formas estruturais predominantes de antocianinas a diferentes valores de pH. ............... 33
Figura 3.2 - Caraterísticas espetrais das antocianinas em soluções a pH 1,0 e a pH 4,5..................... 33
Figura 3.3 - Formação do complexo (Fe2+
-TPTZ) após redução do Fe3+
por um antioxidante. .......... 36
Figura 3.4 - Redução do complexo Cu(II)–neocuproína a Cu(I)–neocuproína, por ação de uma
molécula antioxidante (HA) originando uma molécula de antioxidante oxidada (A+). ....................... 37
Figura 3.5 - Redução do NBT2+
(A) pelo radical anião superóxido, dando origem ao azul de
formazano (B). .................................................................................................................................. 39
Figura 3.6 - Formação do radical anião superóxido através do sistema PMS/NADH ......................... 39
Figura 3.7 - Esquema para a decomposição do AAPH originando radicais peroxilo .......................... 40
Figura 3.8 – Mecanismos propostos para a oxidação da fluoresceína pelo radical peroxilo ................ 41
Figura 4.1 – Curva de dose-resposta do t-BHP com a estirpe TA100. ............................................... 55
Figura 4.2 - Curvas de dose-resposta dos extratos de mirtilo das diferentes variedades (A) Patriot, (B)
Bluecrop, (C) Duke, (D) Earlyblue (E) O’Neal na presença e na ausência de t-BHP, na estirpe TA100.
......................................................................................................................................................... 57
Figura 4.3 - Percentagem máxima de inibição da mutagenicidade do t-BHP exercida pelos extratos
das variedades de mirtilo Patriot, Bluecrop, Duke, Earlyblue e O’Neal. ............................................. 58

ix
Índice de Tabelas
Tabela 1.1 – Composição química e nutricional média de frutos de mirtilo do género Vaccinium
(grupos Highbush (V. corymbosum e V. ashei) e Lowbush (V. augustifolium)) por 100 g de parte
edível ..................................................................................................................................................8
Tabela 2.1- Principais classes de polifenóis ...................................................................................... 13
Tabela 2.2 - Conteúdo em flavónoides de mirtilos cultivados. ........................................................... 14
Tabela 2.3 – Caraterização das principais Espécies Reativas de Oxigénio formadas in vivo............... 17
Tabela 3.1 - Composição da gelose de superfícies por litro. .............................................................. 44
Tabela 3.2 - Composição do meio VB (por litro). ............................................................................. 45
Tabela 4.1 - Teor de compostos fenólicos totais nas amostras testadas .............................................. 46
Tabela 4.2 – Teor de antocianinas monoméricas nas amostras testadas ............................................. 47
Tabela 4.3 – Teor de sólidos solúveis (grau Brix) nas amostras testadas............................................ 49
Tabela 4.4 – Valores do ensaio FRAP nas diferentes amostras .......................................................... 50
Tabela 4.5 – Valores do ensaio CUPRAC nas diferentes amostras .................................................... 51
Tabela 4.6 – Percentagem de sequestro do H2O2 nas diferentes variedades de mirtilo ........................ 53
Tabela 4.7 – Valores de inibição do superóxido obtidos nas diferentes variedades de mirtilo. ........... 53
Tabela 4.8 - Valores obtidos nas diferentes variedades de mirtilo no ensaio ORAC........................... 54
Tabela 4.9 - Classificação da correlação em função do valor do coeficiente de correlação de Pearson.
......................................................................................................................................................... 59
Tabela 4.10 - Coeficiente de Pearson para as correlações entre a composição em fenóis totais e
antocianinas monoméricas das cinco amostras estudadas. .................................................................. 59
Tabela 4.11 - Coeficiente de Pearson para as correlações entre a composição em fenóis totais e
antocianinas monoméricas nas amostras das variedades Bluecrop, Duke, Earlyblue e O’Neal. ........... 60
Tabela 4.12 - Coeficiente de Pearson para as correlações entre composição química e atividade
antioxidante das cinco variedades em estudo. .................................................................................... 61
Tabela 4.13 - Coeficiente de Pearson para as correlações entre composição química e atividade
antioxidante das variedades Bluecrop, Duke, Earlyblue e O’Neal. ..................................................... 61

x
Simbologia e Notações
AAPH - 2,2´- Azobis(2-metilpropionamidina) dihidrocloreto
AH- Aminas heterocíclicas
CAT - Catalase
CUPRAC - Cupric Reducing antioxidant capacity
DNA - Ácido desoxirribonucleico
EAT- Equivalentes de -Tocoferol
EDTA – Ácido etilenodiaminotetracético
eq – Equivalente
ERO - Espécies reativas de oxigénio
FRAP - Ferric Reduction Antioxidant Power
GPX - Glutationa peroxidase
GSH - Glutationa reduzida
GSSG - Glutationa oxidada
HAPs - Hidrocarbonetos aromáticos policíclicos
HAT - Hydrogen atom transfer (transferência de um átomo de hidrogénio)
LDL - Lipoproteínas de baixa densidade
nm - nanómetro
NADH- Dinucleotideo de adenina e nicotinamida na forma reduzida
NADPH- Fosfato de dinucleotideo de adenina e nicotinamida na forma reduzida
NB - Meio de cultura Nutrien Broth
NBT2+
- Azul de nitrotetrazólio
NHB - Northern Highbush Blueberry
ORAC - Oxygen Radical Absorbance Capacity
OTA - Ocratoxina A
PhIP - 2-amino-1-metil-6-fenilimidazole[4,5-b]piridina
PMS – Metossulfato de fenazina
r – Coeficiente do correlação de Pearson

xi
RNA - Ácido ribonucleico
rpm – rotações por minuto
SET - Single electron transfer (transferência de um eletrão)
SHB - Southern Highbush Blueberry
SOD - Superóxido dismutase
spp - Espécie
t-BHP - tert-Butil-hidroperóxido
TPTZ - 2,4,6-Tris(2-piridil)-s-triazina
UV – Radiação ultravioleta
VB – Meio de cultura Vogel-Bonner
λ - Comprimento de onda
m- micrómetro

1
Introdução
O progressivo crescimento e desenvolvimento do sector alimentar implicou novos hábitos
alimentares, associados a uma alimentação mais saudável, que incluem um maior consumo de
pequenos frutos entre os quais se destaca o mirtilo (Vaccinium spp.). Estes novos hábitos surgiram nos
países desenvolvidos e rapidamente se espalharam pelo mundo, observando-se um segmento de
adeptos em constante crescimento. Para além de garantir o suprimento das necessidades nutricionais, a
seleção dos produtos alimentares visa, igualmente, o fornecimento de alimentos que possam implicar
melhorias na saúde, bem-estar e longevidade dos consumidores. Na lista dos alimentos preferenciais
estão as frutas por serem altamente nutritivas, sendo importantes fontes de vitaminas e minerais, fibras
alimentares e flavonoides com capacidades antioxidantes que podem proteger o organismo contra os
efeitos prejudiciais do stress oxidativo. As frutas fornecem ainda energia rápida, em função do seu
teor em açúcar natural, além de serem de fácil digestão.
O mirtilo está incluído no grupo das pequenas frutas, sendo reconhecido o seu conteúdo rico
em antioxidantes, tanto na pelicula como na polpa (Sousa et al., 2007). Diversos trabalhos têm
demonstrado a existência nos mirtilos de compostos com atividade antioxidante, anti-inflamatória,
antibacteriana e anticancerígena (Neto, 2007; Zafra-Stone et al., 2007), podendo, no entanto, o teor
nestes compostos bioativos variar em função de fatores genéticos e de fatores ambientais, tais como o
tipo de solo ou a temperatura.
Em Portugal o mirtilo é conhecido vulgarmente como arando, erva-escovinha ou uva-monte.
As plantas nativas encontram-se nas regiões de floresta, matos e pinhais de montanha entre 400 a 2000
metros de altitude, no Alto Minho, Marão e Serra da Estrela. Atualmente, as variedades cultivadas são
híbridos que permitem uma melhor adaptação a várias zonas geográficas. O mirtilo é uma baga cujas
características sensoriais o tornam apetecível para ser consumido em fresco, como conserva, em
gelados, doces, licores e aguardentes, podendo ser também utilizado como corante graças à sua
riqueza em antocianinas. A cultura do mirtilo em Portugal apresenta um elevado potencial devido às
condições climáticas existentes (Lavadinho et al., 2001).
Tendo em consideração a aceitação, o aumento do consumo de mirtilos por parte dos
consumidores, o elevado potencial económico e os potenciais efeitos benéficos na saúde e sabendo
que as atividades biológicas dos mirtilos podem variar consoante as características de solo e do clima
da região em que são produzidos, o objetivo deste trabalho foi avaliar a capacidade antioxidante e
antimutagénica de cinco variedades de mirtilo (Vaccinium spp.) produzidas nas plantações da região
de Sever do Vouga e gentilmente cedidas pela Mirtilusa (Sociedade de Produtores Hortofrutícolas),

2
nomeadamente as variedades Earlyblue, Patriot, Bluecrop, Duke e O`Neal. Tentou-se avaliar o
conteúdo em compostos fenólicos totais e, em particular em antocianinas monoméricas totais, das
diferentes cultivares. Para avaliar a capacidade antioxidante recorreu-se a diferentes ensaios,
nomeadamente, ensaios de avaliação da sua capacidade redutora total, da sua capacidade para
sequestrar várias espécies reativas de oxigénio e capacidade de quelação de ferro. Foi ainda avaliado o
potencial antimutagénico destas cultivares através do estudo da sua capacidade para inibir a
mutagenicidade do mutagéneo oxidativo tert-butil-hidroperóxido (t-BHP). Tentou-se ainda estabelecer
uma correlação entre o conteúdo em compostos fenólicos, em particular em antocianinas, e a atividade
antioxidante e antimutagénica das diferentes variedades.
Esta dissertação inicia-se com dois capítulos teóricos, sendo que no primeiro capítulo é feita
uma pequena descrição das cultivares de mirtilo em estudo, sua importância económica na Sub-região
do Baixo Vouga, sua composição química e propriedades funcionais, e no segundo uma revisão
bibliográfica sobre a problemática do stress oxidativo e o papel protetor dos antioxidantes, bem como
sobre os mutagéneos e antimutagéneos alimentares. No terceiro capítulo descrevem-se os
procedimentos experimentais seguidos e, no quarto apresentam-se os resultados obtidos e a sua
respetiva discussão. Por fim, no capítulo cinco, resumem-se as principais conclusões da presente
dissertação.

3
1. Breve caraterização do mirtilo
1.1. Variedades de mirtilo
Os mirtilos pertencem à família das Ericaceae, subfamília das Vaccinoiodae, género
Vaccinium spp. (Spiers et al., 2004). Os géneros são muito diversos, contendo de 150 a 450 espécies,
sendo, na maioria dos casos arbustos (Figura 1.1) de tamanho e formas variadas (Riegger, 2006).
Figura 1.1 – Aspeto da planta de mirtilo Vaccinium spp.
O mirtilo é uma baga (Figura 1.2) de aproximadamente um centímetro de diâmetro e com um
peso médio de 1,5 g, sendo o tamanho exato do fruto uma característica da variedade (Eck, 1988). O
fruto baciforme e globular é sumarento e tem um sabor agridoce. Amadurece dois a três meses após a
floração. A baga pequena com sementes apresenta em geral cor azul com tonalidades variando de mais
claro a mais escuro e intenso. A cor do mirtilo é influenciada pela presença de pruína, cera
epicuticular, que produz o efeito glauco responsável pela cor azul típica dos mirtilos. Esta camada
cerosa constitui uma barreira importante à perda de água, impedindo o murchar do fruto (Albrigo et
al., 1980). A baga apresenta uma cicatriz, diametralmente oposta ao ápice, de dimensão e formato
variáveis, segundo a espécie e a cultivar. Os mirtilos apresentam um grande número de sementes
(Makus e Morris, 1987; Westwood, 1982) que contribuem para um sabor menos acentuado e um certo
grau de arenosidade conferido pela presença de escleritos (Sousa et al., 2007). O fruto pode ser
consumido in natura ou após processamento (congelamento, desidratação, conservas, fabrico de
geleias ou licores) (Makus e Morris, 1987).

4
Figura 1.2 – Baga de mirtilo
As variedades de mirtilos comerciais podem ser divididas em cinco grupos de acordo
características que incluem, entre outras, o genótipo, necessidades de crescimento ou o tipo de fruto
produzido (Galletta e Ballington, 1996):
a) Northern highbush (arbusto alto), plantas com mais de dois metros de altura e necessidade de
650 a 850 horas de frio hibernal;
b) Half high (arbusto médio), plantas que atingem 0,5 m a 1,0 metro de altura e apresentam menor
exigência em frio que as highbush. Este grupo envolve híbridos de V. angustifolium e V.
corymbosum;
c) Southern highbush, plantas de porte alto, conhecidas como highbush de baixa exigência em frio,
com maior resistência à doença e que suportam solos pobres em matéria orgânica, sendo mais
exigentes em água, drenagem e matéria orgânica que rabbiteye;
d) Rabbiteye (“olho-de-coelho”), plantas alcançam 2 a 4 m de altura. São características da espécie
V. ashei o vigor, a longevidade, produtividade, tolerância ao calor e à seca e baixa necessidade
em frio. São produtoras de frutos ácidos, firmes e de longa conservação. As limitações dessa
espécie são a tendência de rachar a película em períodos húmidos, desenvolver a cor completa
das frutas antes do ponto ideal de colheita e longo período para alcançar o máximo de
produtividade;
e) Lowbush, são plantas com porte menor que 0,5 m. A maioria pertence à espécie V.
angustifolium, neste grupo encontra-se o mirtilo do Canadá (V. myrtilloides e V.boreale).
As espécies de mirtilos com expressão comercial mais elevada pertencem aos grupos
highbush, rabbiteye e lowbush, sendo as práticas de manejo diferenciadas para cada um dos grupos,
desde a produção de mudas até a colheita e utilização dos frutos (Fachinello, 2008).

5
Ao longo do tempo, o género Vaccinium spp. evoluiu principalmente em climas temperados,
nomeadamente no norte do continente Americano, Europa e Ásia, sendo que a maior parte das atuais
cultivares foram obtidas por hibridação de diferentes espécies norte-Americanas (Sousa et al., 2007).
Em Portugal existem ainda espécies silvestres do género Vaccinium: o Vaccinium myrtillus (mirtilo
Europeu) na Serra do Gerês, o Vaccinium padifloium na Ilha da Madeira e o Vaccinium cylindraceum
(uva-da-serra) no arquipélago dos Açores (Sousa et al., 2006).
Por ser uma espécie frutífera nativa dos Estados Unidos da América e do Canadá são nestes
países que se encontram as maiores produções. Segundo a FAO (2010), os Estados Unidos da América
produziram 157 451,20 toneladas anuais no período entre 2004 e 2008, ocupando uma superfície de 21
946,80 hectares, tendo este valor representado 50% da produção mundial. O Canadá, segundo maior
produtor mundial, produziu, no mesmo período, 81 240,20 toneladas em 30 655,20 hectares, valor que
representou cerca de 33% da produção mundial. Os Estados Unidos da América e o Canadá são
também os maiores consumidores e exportadores deste fruto.
1.2. Produção e consumo de mirtilo em Portugal
Em Portugal, o progressivo crescimento e desenvolvimento do sector alimentar e a influência
dos restantes países da Europa, introduziram novos hábitos alimentares, como o consumo de pequenos
frutos, nos quais se inclui o mirtilo. Por outro lado, o nosso país, devido às suas condições edafo-
climáticas, apresenta um elevado potencial para a cultura do mirtilo. Nos últimos anos assistiu-se a um
aumento da área de produção, que passou de zero para cerca de quarenta e cinco hectares, com
tendência para aumentar. O Alentejo e a zona do Sever do Vouga são as principais regiões de
produção de mirtilo. A introdução no país das diferentes cultivares de mirtilo tem conduzido à
implementação de tecnologias de produção e concomitantemente à necessidade de se determinar a
qualidade dos frutos obtidos, de forma a elegerem-se as cultivares com maior interesse económico
(Sousa et al., 2007).
As primeiras plantações de mirtilo surgiam em Portugal, na década de 90, no concelho de
Sever do Vouga, transformando-se o produto da cultura do mirtilo num acrescento à economia
familiar, que foi ganhando maior relevância com o passar dos anos. Atualmente cerca de 95% da
produção é exportada, com a inerente mais-valia dos produtores do mirtilo de Sever, colocarem o fruto
no mercado europeu, nomeadamente nos Países do norte da Europa, nos meses de Maio/Junho de cada
ano, ou seja, antes dos demais concorrentes.

6
As primeiras experiências de produção em Portugal foram efetuadas em 2007, pela Fundação
Lockorn, da Holanda, nos concelhos de Sever do Vouga e Trancoso, e tentavam perceber se haveria
condições para a produção precoce de mirtilo. Depois das várias experiências realizadas pela
Fundação e a Cooperativa Agrícola de Sanfins, conclui-se que Sever de Vouga reunia condições ideais
de caraterísticas do solo e do micro clima para a produção precoce de mirtilo. A primeira experiência
piloto ocorreu na Quinta do Linheiro, localizada na freguesia de Rocas do Vouga. Desde então, a
produção deste fruto no concelho tem vindo a crescer em média 15% ao ano, o que tem sido uma
mais-valia para a região e tem permitido diversificar o sector agrícola e dinamizar a economia local,
tornando-se uma importante fonte de rendimento para as famílias e uma verdadeira alternativa à
agricultura tradicional (Serrada et al, 2008).
A produção de mirtilo em Sever do Vouga ronda as 120 toneladas, mais de 80% são
exportadas, nomeadamente para França, Bélgica e Holanda. A produção nacional é escassa face à
procura. O consumo nacional é irrisório apesar de ter duplicado de 2007 para 2008. Em 2010 o preço
do mirtilo, foi de cerca de 3,5 euros o quilograma pago ao produtor, sendo posteriormente
comercializado a 25/30 euros o quilograma (Costa, 2010). No Alentejo os mirtilos são produzidos por
uma empresa designada por Mirtisul, situada na região de Grândola, sendo esta o maior produtor de
mirtilo em Portugal. Em 2007 a Mirtisul atingiu uma produção de 140 toneladas em 14 hectares
(Sousa et al., 2007).
Os grupos que apresentam maior interesse para as condições climáticas de Portugal são os
Northern Highbush Blueberry (NHB) e Southern Highbush Blueberry (SHB) e eventualmente
algumas cultivares do grupo Rabbiteye. Na zona centro/norte de Portugal as cultivares que melhor
desempenho apresentam, pertencem principalmente ao grupo NHB, ao passo que a sul do Tejo têm-se
revelado interessantes os híbridos dos grupos SHB e Rabbiteye, por necessitarem de menos horas de
frio (Sousa et al., 2007).
De um modo geral, as cultivares do grupo Northern Higbush, por necessitarem de um maior
número de horas de frio durante o repouso invernal, são as mais adequadas para o norte e centro de
Portugal e as cultivares do grupo Southern Highbush para o sul. Em Sever do Vouga as cultivares mais
utilizadas são de Vaccinium Corymbosum. Esta escolha deveu-se a vários fatores: clima, possibilidade
de ocorrência de geadas tardias, a resistência a doenças e pragas e ainda, por se tratar de uma
plantação comercial, à produção e à qualidade.
As cultivares pertencentes ao grupo Northern Highbush são as mais plantadas em todo o
mundo, necessitando as plantas de muitas horas de frio para quebrar dormência durante o Inverno e
entrar em produção. O intervalo de horas de frio vai de 800 a 1000 horas abaixo dos 7ºC. As cultivares
Southern Highbush necessitam menos horas de frio durante o Inverno para quebra de dormência da

7
planta, diferindo este número de horas de cultivar para cultivar. O intervalo varia entre 150 a 600
horas de frio abaixo dos 7ºC (Sousa et al., 2006).
As cultivares estudadas neste trabalho pertencem maioritariamente ao grupo Northern
Highbush (variedades Earlyblue, Patriot, Bluecrop e Duke) e apenas uma pertence ao grupo Southern
Highbush (variedade O`Neal).
As amostras das cultivares em estudo foram gentilmente cedidas pela Empresa Mirtilusa,
Sociedade de Produtores Hortofrutícolas, sediada em Sever do Vouga. A área geográfica de produção
de mirtilo abrangida por esta empresa engloba alguns concelhos da Sub-região do Baixo Vouga
(Figura 1.3) tais como Viseu, Sever do Vouga (mais apto), Águeda e Vale de Cambra.
Figura 1.3 – Mapa da Sub-região Baixo Vouga
1.3. Caraterísticas físico-químicas e nutricionais do Mirtilo
A qualidade das frutas e vegetais é caraterizada com base em atributos como aparência, sabor,
textura e valor nutritivo (Chitarra e Chitarra, 2005). Esta caraterização físico-química e nutricional é
importante para o controle e melhoria da qualidade dos frutos, para a comercialização no mercado

8
interno e externo, bem como para o desenvolvimento de técnicas de armazenamento e de manipulação
pós-colheita adequadas.
A composição nutricional do mirtilo varia em função de diversos fatores que incluem a
variedade, práticas de cultivo, fertilidade do solo, época do ano ou grau de maturação. A tabela 1.1
apresenta a composição química e nutricional média de frutos de mirtilo do género Vaccinium.
Tabela 1.1 – Composição química e nutricional média de frutos de mirtilo do género Vaccinium
(grupos Highbush (V. corymbosum e V. ashei) e Lowbush (V. augustifolium)) por 100 g de parte
edível (adaptado USDA, 2011).
Valor nutricional por 100 g edível de mirtilo
Energia (kcal) 57,00
Proteínas (g) 0,74
Gorduras (g) 0,33
Açúcares (g) 14,49
Fibra (g) 2,40
Água (g) 84,21
Minerais
Cálcio (mg) 6,00
Ferro (mg) 0,28
Magnésio (mg) 6,00
Manganésio (mg) 0,34
Fósforo (mg) 12,00
Potássio (mg) 77,00
Selénio (µg) 0,10
Sódio (mg) 1,00
Zinco (mg) 0,16
Vitaminas
Vitamina C (mg) 9,70
Tiamina (mg) 0,04
Riboflavina (mg) 0,04
Niacina (mg) 0,42
Ácido pantoténico (mg) 0,12
Vitamina B6 (mg) 0,05
Folato (µg) 6,00
Vitamina A (UI) 54,00
Vitamina E (mg EAT) 0,57
Lípidos
Ácidos gordos saturados, total (g) 0,028
Ácidos gordos monoinsaturados, total (g) 0,047
Ácidos gordos polinsaturados, total (g) 0,146
(EAT- Equivalentes de Alfa-tocoferol; UI - Unidades Internacionais)

9
De acordo com a tabela 1.1 pode observar-se que o mirtilo apresenta um teor de água bastante
elevado, sendo superior a 80%. O conteúdo de água nos tecidos depende, entre outros fatores, da
disponibilidade hídrica do solo no momento da colheita (Sousa et al., 2007). A perda de água nos
frutos conduz à redução de volume e perda de massa, enquanto o elevado teor de água os torna,
geralmente, mais suscetíveis à deterioração, aumentando a possibilidade de contaminação
microbiológica. O mirtilo apresenta um baixo valor calórico e um baixo teor em lípidos e em
proteínas. Os componentes de maior quantidade são os sólidos solúveis constituídos pela glucose e
frutose, que representam cerca de 80% da matéria seca, e os sólidos insolúveis constituídos pelas
fibras e cinzas. Os frutos produzidos em regiões de verões quentes e secos têm, em relação aos que
crescem em regiões mais amenas e húmidas, uma concentração mais elevada de açúcares e um aroma
e coloração mais intensos (Rieger, 2006).
Os mirtilos apresentam ainda na sua composição uma variedade de vitaminas (A, B, C, K,
ácido fólico), minerais (potássio, magnésio, cálcio, fósforo, ferro, manganésio), açúcares, pectinas e
taninos. Os ácidos orgânicos com teores mais elevados no mirtilo são o quínico, o málico e o cítrico.
No mirtilo o ácido quínico representa 40% dos ácidos orgânicos, o ácido málico apresenta valores
entre 0,06 a 0,14 g/100 g de fruto maduro (Sousa et al., 2007) e o ácido cítrico 0,4 a 0,5 g/100 g de
fruto (Raseira e Antunes, 2004).
Os mirtilos são muito ricos em fito nutrientes. A baga contém quantidades significativas de
antocianidinas, compostos antioxidantes, pertencentes ao grupo dos flavonoides, que dão cores azul,
roxo e vermelho a frutas e legumes. Incluídas nas antocianinas dos mirtilo encontram-se a malvidina,
delfinidina, petunidina, peonidina e cianidina. Além das antocianinas, os mirtilos também contêm
ácidos fenólicos, estilbenos e outros flavonoides como flavanóis e proantocianidinas (Prior et al.,
1998, Sellappan et al., 2002).
O aumento do interesse pelo consumo de mirtilos deriva não só dos seus sabores exóticos mas
também das suas alegações terapêuticas (Raseira e Antunes, 2004). Com efeito, nas últimas décadas,
muitos estudos têm vindo a associar as antocianinas e outros compostos presentes em algumas
espécies de Vaccinium, com diversas propriedades biológicas que incluem atividade antioxidante
(Wang et al, 1997; Mazza et al., 2002; Lohachoompol et al., 2004), anti-inflamatória (Youdin et al.,
2002) e atividade antibacteriana (Zafra-Stone et al., 2007).

10
2. Mirtilo e importância na saúde
2.1. Mirtilo alimento funcional
A definição mais comum para alimento funcional é a de um alimento para o qual se
demonstrou capacidade para influenciar positivamente uma ou mais funções alvo no organismo, além
de possuir efeitos nutricionais adequados, desta forma, estes alimentos podem ser relevantes para o
bem-estar e a para a saúde, uma vez que o seu consumo está associado com a redução do risco de
doenças (Roberfroid, 2002). É ainda assumido que os alimentos funcionais são todos os alimentos ou
bebidas que, consumidos na alimentação quotidiana, podem trazer benefícios fisiológicos específicos,
graças à presença de compostos fisiologicamente ativos (Candido e Campos, 2005).
Os compostos bioativos podem ser provenientes de produtos de origem animal, como, por
exemplo, os ácidos gordos da família ómega 3 ou ácidos gordos conjugados, ou de origem vegetal, tais
como, por exemplo, os carotenoides, fitoesteróis, terpenos ou compostos fenólicos. Os compostos
bioativos de vegetais (fitoquímicos) compreendem uma grande variedade de classes de compostos
químicos com diferentes propriedades físico-químicas (polaridade, solubilidade, capacidade de formar
pontes de hidrogénio, potencial de oxidação-redução) que irão determinar tanto o tipo como a
eficiência de atividade, assim como o meio e a estrutura celular em que podem atuar (Oliveira e
Bastos, 2011).
Os frutos do género Vaccinium podem ser considerados alimentos funcionais, sendo utilizados
há centenas de anos na medicina tradicional no tratamento de problemas urinários, prevenção de
escorbuto e aplicação tópica em ferimentos (Beattie et al., 2005). Estudos mais recentes têm sugerido
que os mirtilos possam conferir proteção contra doenças cardiovasculares, ter propriedades
antidiabéticas (Neto, 2007; Zafra-Stone et al., 2007) e contribuir para melhorar a acuidade visual
(Burdulis et al., 2007; Zafra-Stone et al., 2007). Os mirtilos, ou alguns dos seus compostos
isoladamente, mostraram ainda ter atividade antimutagénica contra diversos mutagéneos alimentares
(Kong et al., 2003), capacidade de induzir a apoptose ou de inibir a proliferação de células neoplásicas
in vitro (Katsube et al., 2003; Neto, 2007; Wang e Stoner, 2008) e in vivo em animais de laboratório
(Wang e Stoner, 2008), prevenir problemas inflamatórios como, por exemplo, a artrite reumatoide
(Törronen et al., 2000), reduzir a fragilidade capilar (Kong et al., 2003) e o aparecimento de doenças
degenerativas (Alves et al., 2008). O mirtilo parece, igualmente, impedir a fixação e o
desenvolvimento da Escherichia coli, bactéria causadora de infeções no trato urinário (Smith et al.,
2000). Os extratos de mirtilo conseguiram inibir, in vitro, o crescimento da bactéria Helicobacter

11
pylori, conhecida como um fator de risco associado ao surgimento de algumas doenças
gastrointestinais, incluindo úlcera duodenal e cancro de estômago (Zafra-Stone et al., 2007).
Ensaios realizados em ratos alimentados com extratos de mirtilos mostraram que estes
levavam a um aumento das defesas antioxidantes do cérebro e músculo-esquelético, retardando os
efeitos do envelhecimento, melhorando as funções cognitivas (Carlsen et al., 2003) e os distúrbios da
memória (Ramirez et al., 2005). Estudos em ratos velhos demonstram que administração por 8
semanas de mirtilo promovia uma melhoria da atividade de locomoção (equilíbrio e coordenação)
(Joseph et al., 1999).
Todos estes efeitos benéficos na saúde parecem estar, pelo menos em parte, associados com o
elevado teor em polifenóis que estes frutos apresentam. Com efeito, numerosos estudos têm vindo a
considerar os compostos fenólicos como sendo um dos principais grupos de compostos responsáveis
pelos efeitos benéficos das dietas ricas em frutas e vegetais (Horst e Lajolo, 2007).
2.2. Os compostos fenólicos nos mirtilos
Os compostos fenólicos são um dos maiores grupos de substâncias, não essenciais à dieta,
encontrados nos alimentos de origem vegetal, tipicamente, em pequenas quantidades, e englobam mais
de 8000 moléculas diferentes (Kris–Etherton et al., 2002). A capacidade dessas moléculas para quelar
metais, inibir a ação da lipoxigenase e desativar os radicais livres tem aumentado o interesse dos
investigadores pelo estudo destes compostos (Martinez-Valverde et al., 2000). A figura 2.1 representa
as principais classes de compostos fitoquímicos ou compostos bioativos presentes em alimentos de
origem vegetal.
Os compostos fenólicos contêm anéis aromáticos com um ou mais substituintes hidroxilos,
variando desde moléculas simples, como os ácidos fenólicos com uma única estrutura aromática,
bifenóis, como ácido elágico, flavonoides contendo 2 a 3 anéis aromáticos e polifenóis mais
complexos, contendo de 12 a 16 anéis (Ribeiro e Servalli, 2004). De acordo com a sua estrutura, os
compostos fenólicos podem ser divididos em diversas categorias que incluem os ácidos fenólicos
(derivados de ácido hidroxibenzóico e hidroxicinâmico), flavonoides, estilbenos (resveratrol)
cumarinas e taninos (Peterson e Dwyer, 1998; Ribeiro e Servalli, 2004). Nos alimentos os fenóis
pertencem geralmente à classe dos ácidos fenólicos, flavonoides, lignanas, estilbenos, cumarinas e
taninos (Figura 2.1) (Shahidi e Naczk, 1995).

12
Fitoquímicos
Carotenóides Fenólicos Alcalóides Compostos contendo
azoto
Compostos Organossulfurados
Ácidos
fenólicos
Flavonóides Estilbenos Cumarinas Taninos
Flavonóis Flavonas Antiocianidinas IsoflavonasFlavanóisÁcidos hidroxibenzóicos
Ácidos
hidroxicianâmicos
-Caroteno
-Caroteno
Luteína
ZeaxantinaAstaxantina
Licopeno
Isotiocianato
Indóis
Compostos alílicos
sulfurados
Gálico
Vanílico
Seríngico
p-cumáricoCafeico
Sinápico
Ferúlico
Quercetina
Caempferol
Miricertina
Galangina
ApigeninaCrisina
Luteolina
Catequina
Epicatequina
Epigalocatequina
Flavanonas
Eriodictiol
Hesperitina
Naringenina
CianidinaDelfinidina
Peonidina
Malvidina
GenisteínaDaidzeína
Gliciteína
Fitoquímicos
Carotenóides Fenólicos Alcalóides Compostos contendo
azoto
Compostos Organossulfurados
Ácidos
fenólicos
Flavonóides Estilbenos Cumarinas Taninos
Flavonóis Flavonas Antiocianidinas IsoflavonasFlavanóisÁcidos hidroxibenzóicos
Ácidos
hidroxicianâmicos
-Caroteno
-Caroteno
Luteína
ZeaxantinaAstaxantina
Licopeno
Isotiocianato
Indóis
Compostos alílicos
sulfurados
Gálico
Vanílico
Seríngico
p-cumáricoCafeico
Sinápico
Ferúlico
Quercetina
Caempferol
Miricertina
Galangina
ApigeninaCrisina
Luteolina
Catequina
Epicatequina
Epigalocatequina
Flavanonas
Eriodictiol
Hesperitina
Naringenina
CianidinaDelfinidina
Peonidina
Malvidina
GenisteínaDaidzeína
Gliciteína
Figura 2.1 - Compostos fitoquímicos presentes em alimentos de origem vegetal (adaptado de Liu,
2004).
De todos as categorias de compostos fenólicos enumeradas, a dos flavonoides é, talvez, aquela
que é mais bem estudada. Esta categoria é composta por mais de 4000 compostos diferentes, que têm
em comum a estrutura C6-C3-C6, constituída por dois anéis aromáticos ligados por um heterocíclico
oxigenado (Liu, 2004). Os flavonoides dividem-se em diversas subclasses, que incluem as subclasses
dos flavonóis, flavonas, flavanóis, flavanonas, anticianidinas e isoflavonas (Figura 2.1). Nestas classes
existe uma grande variação de estruturas, que dependem do nível de hidrogenação, hidroxilação e
metilação das moléculas. De um modo geral, estes compostos, não se encontram na natureza na forma
livre mas sim complexados, principalmente, com açúcares (Lopes et al., 2000). A Tabela 2.1 mostra a
estrutura básica das diferentes classes de polifenóis, bem como os principais alimentos onde estes
podem ser encontrados.
As frutas vermelhas estão entre as fontes alimentares mais importantes em compostos
fenólicos (Häkkinen e Törrönen, 2000). Em particular, o mirtilo (Vaccinium spp.) apresenta na sua
composição uma elevada concentração em compostos fenólicos, sendo superior à mesma concentração
na maioria das frutas (Wolfe et al., 2008).
Nestes últimos anos muitos tem sido os estudos sobre os compostos fenólicos dos mirtilos,
Zadernowski e seus colaboradores (2005) identificaram 17 ácidos fenólicos presentes em algumas das
cultivares de mirtilo por cromatografia gasosa acoplada à espetrometria de massa. Assim, foram

13
identificados os ácidos hidroxibenzóicos gentísico, gálico, o-pirocatecuico, protocatecuico, salicílico,
siríngico, vanílico e verátrico. Como representantes dos ácidos hidroxicinâmicos, identificaram-se os
ácidos cafeico, m-cumárico, o-cumárico, p-cumárico, 3,4- dimetoxicinâmico, ferúlico, hidroxicafeico
e sinápico.
Tabela 2.1- Principais classes de polifenóis (adaptado de Ferguson, 2001).
Classe Esqueleto básico Exemplo Fonte principal
Ácidos
hidroxibenzóicos
C6-C1 Ácidos gálico, vanílico, siríngico,
protocatéquico e tânico
Comum nas plantas superiores e
fetos
Ácidos
hidroxicinâmicos
C6-C3 Ácidos ferúlico, p-cumárico,
cafeico e sinápico.
Comum nas plantas superiores,
muitas vezes como componentes
da parede celular
Cumarinas,
isocumarinas
C6-C3 Umbeliferona, esculetina,
escopoletina
-
Estilbenos C6-C2-C6 Resveratrol Especialmente na pele das uvas
Antraquinonas C6-C2-C6 - -
Flavonóides C6-C3-C6 Apigenina, genisteína,
caempferol, epigalocatequina
galato, miricetina,quercetina
Difundidos por todo o reino
vegetal.
Diarilheptanos C6-C7-C6 Curcumina, iaquichinona A e B Presentes no gengibre
Lignanas (C6-C3)2 - Comum nas sementes de linho
Ligninas (C6-C3)n - Componentes das paredes
celulares de diversas plantas
(Fibra dietética)
As antocianinas (antocianidinas glicosiladas) identificadas nos mirtilos foram a delfinidina-3-
galactose, delfinidina-3-glucose, delfinidina-3-arabinose, cianidina-3-galatoctose, cianidina-3-glucose,
petunidina-3-galactose, cianidina-3-arabinose, petunidina-3-glucose, peonidina-3-galactose,
petunidina-3-arabinose, peonidina-3-glucose, malvidina-3-galactose, peonidina-3-arabinose,
malvidina-3-glucose e malvidina-3-arabinose (Blumenthak et al., 2000; Faria et al., 2005;
Zadernowski et al., 2005). Além das antocianinas, os mirtilos também contêm ácidos fenólicos,
estilbenos e outros flavonoides como flavonóis, flavanóis e proantocianidinas (tabela 2.2). O perfil
exato destes compostos, bem como as quantidades em que se encontram, variam, não só, com a
cultivar, como também com aspetos climáticos, podendo algumas diferenças resultar de diferentes

14
graus de stress hídrico, da maior ou menor disponibilidade de nutrientes e da intensidade da radiação
ultravioleta (Giovanelli e Buratti, 2009).
Tabela 2.2 - Conteúdo em flavónoides de mirtilos cultivados (Adaptado de USDA, 2011).
Conteúdo em flavónoides mg/100 g de edível de mirtilo
Classe Flavonóide Valores médios
Antocianidinas Cianidina 7,10
Delfinidina 30,91
Malvidina 59,64
Peonidina 15,36
Petunidina 28,02
Flavanóis (-) -Epicatequina 0,62
(-) –Epigalocatequina 0,66
(+) -Catequina 5,29
(+) -Galocatequina 0,12
Flavonas Luteolina 0,20
Flavonóis Caempferol 1,66
Miricetina 1,26
Quercetina 7,67
Proantocianidinas
Monómeros
Dímeros
Trímeros
4-6 monómeros 7-10 monómeros
Polímeros
3.46
5,71
4,16
19,57 14,55
129,05
Os compostos fenólicos interferem em alvos fisiológicos específicos, parecendo modelar a
defesa contra processos oxidativos, inflamatórios e mutagénicos, os quais estão relacionados com o
desenvolvimento de diversas patologias, podendo, desta forma, os compostos fenólicos ser
considerados como essenciais para a manutenção da saúde (Oliveira e Bastos, 2011).
2.3. Espécies reativas de oxigénio
Espécies reativas de oxigénio (ERO) é nome genérico dado a uma variedade de moléculas e
radicais livres derivados do oxigénio. Os radicais livres são moléculas reativas que apresentam um
eletrão desemparelhado. São produzidos continuamente nas células, podendo ser gerados no
citoplasma, nas mitocôndrias ou na membrana celular (Holmqvist et al., 1994), no decurso de
processos metabólicos oxidativos como a respiração aeróbia ou processos que envolvam a atuação das
enzimas NADPH oxidase, xantina oxidase, ciclo-oxigenases, lipoxigenases ou monoxigenases como
os citocromos P450. Para além destes processos endógenos existem ainda fontes exogénas capazes de

15
induzir um aumento da produção de ERO. Nestas fontes exógenas encontram-se a exposição ao ozono,
à radiações gama e ultravioleta, a diversos compostos presentes na dieta, fármacos ou tabaco
(Holmqvist et al., 1994).
Diversas evidências científicas têm comprovado que as ERO podem desempenhar papeis
fisiológicos importantes ao funcionarem como mensageiros secundários nas células de mamíferos,
regulando vias de transdução de sinal, que controlam a expressão de genes e a indução de alterações
pós-traducionais de proteínas, estando, assim, implicadas no controlo dos processos de divisão e
diferenciação celular (Al-Gubory et al., 2010). Além disso as ERO são de extrema utilidade quando há
necessidade de ativação do sistema imunológico e na produção do fator relaxante derivado do
endotélio, o óxido nítrico, extremamente importante nos processos que desencadeiam o relaxamento
dos vasos sanguíneos (Schneider e Oliveira, 2004).
A formação das ERO deve-se ao fato do oxigénio ter uma elevada tendência para receber um
eletrão de cada vez. A adição de um eletrão a uma molécula de oxigénio no estado fundamental
origina a formação de um radical anião superóxido (O2●-
) (Schneider e Oliveira, 2004) (reação 1).
Reação 1:
O radical anião superóxido, pelo processo de dismutação, pode receber mais um eletrão
(originando o ião peróxido) e dois iões hidrogénios formando o peróxido de hidrogénio. Esta reação é
catalisada pela enzima superóxido dismutase (SOD) que se encontra em quantidades elevadas nas
células dos mamíferos e acelera 104
vezes a frequência do processo de dismutação espontânea a pH
fisiológico (Yu, 1994) (reação 2).
Reação 2: + O2
Enquanto que a dismutação do radical anião superóxido produz o peróxido de hidrogénio, que
por sua vez pode ser reduzido a água e O2, a sua protonação conduz ao aparecimento do radical
peroxilo (HOO•).
Quando o ião peróxido recebe mais um eletrão e um ião de hidrogénio é formado radical
hidroxilo (OH●), que é um intermediário muito reativo, podendo reagir e alterar qualquer estrutura
celular que esteja próxima e assim influenciar enzimas, membranas ou ácido nucleicos (Jenkins,
1988). O radical hidroxilo pode ser formado quando o peróxido de hidrogénio reage com os iões ferro
e cobre, esta reação é conhecida pela reação Fenton (reação 3) (Schneider e Oliveira, 2004).

16
Reação 3:
Os iões dos metais de transição de ferro e cobre podem funcionar como catalisadores na
reação entre o peróxido de hidrogénio e o radical anião superóxido, conduzindo, igualmente, à
formação do radical hidroxilo. Esta reação é conhecida por reação de Haber-Weiss (reação 4)
(Schneider e Oliveira, 2004).
Reação 4:
Os radicais anião superóxido e hidroxilo são designados por radicais livres porque apresentam
eletrões desemparelhados na sua orbital mais externa. O peróxido de hidrogénio não é um radical
livre, mas é metabolito de oxigénio parcialmente reduzido. Outra espécie reativa de oxigénio é o
singleto de oxigénio que é uma forma de oxigénio com spin alterado.
O radical anião superóxido é o precursor da maioria das ERO (Covarrubias et al., 2008). Para
além das já citadas as ERO incluem ainda outras espécies, tais como, os radicais alcoxilo (RO●) e
hidroperoxilo (HOO●), o ácido hipocloroso, o ozono e o peroxinitrito (Dalle-Done et al., 2003). Todas
as ERO têm as suas caraterísticas, mostrando diferentes reatividades e tempos de semi-vida (Yu, 1994)
(tabela 2.3).
As espécies radicalares têm um tempo de semi-vida curto, reagindo rapidamente com outras
moléculas. Por outro lado, as espécies não radicalares podem possuir tempos de semi-vida mais
prolongados, podendo variar de partes de segundos até horas, dependendo da espécies em causa e
também de outros fatores, como o pH ou a presença de outros compostos. De uma forma geral,
espécies com maior tempo de semi-vida podem difundir-se e ir interagir e causar danos em alvos
biológicos mais afastados do seu local de produção. Por exemplo, o radical anião superóxido, apesar
de ser uma espécie radicalar, possui um tempo de semi-vida suficientemente longo que lhe permite
mover-se e interagir com alvos biológicos mais distantes do seu local de produção. Pelo contrário, os

17
possíveis danos causados pelo radical hidroxilo estão mais confinados aos alvos biológicos que se
encontram imediatamente adjacentes ao seu local de produção (Kohen & Nyska, 2002).
Tabela 2.3 – Caraterização das principais Espécies Reativas de Oxigénio formadas in vivo (Ribeiro et
al., 2005).
Intermediário Comentário Semi-Vida Sítios de formação
Radical anião
superóxido
(O2●-
)
Formado a partir da redução
parcial do oxigénio
molecular por eletrão
Decomposição
enzimática
Reação de autoxidação
envolvendo flavoproteínas e
ciclo redox
Peróxido de
hidrogénio
(H2O2)
Formado a partir da redução
parcial do oxigénio
molecular por dois eletrões
Decomposição
enzimática
Vias catalisadas por oxidases, e
pela superóxido dismutase.
Radical
Hidroxilo
(OH●)
Formado a partir da redução
do oxigénio molecular, por
três eletrões nas reações de
Fenton e Haber-Weiss,
catalisadas por metais
10-9 s Locais adjacentes à formação de
anião superóxido/ peróxido de
hidrogénio na presença de
metais, principalmente do ferro;
produto da reação do óxido
nítrico com o radical
superóxido.
Radical alcoxilo
(RO●)
Radical orgânico centrado
no oxigénio
10-6 s Intermediário na peroxidação de
lípidos de membrana.
Radical
Peroxilo(ROO●)
Formado a partir de hidroperóxidos orgânicos
7 s Intermediário na peroxidação de lípidos de membrana.
Oxigénio
Singleto
Primeiro estado excitado do
oxigénio molecular
10-5 s Sem sítios metabólicos
definidos
Aumentos descontrolados nas concentrações dos oxidantes, situação que se designa por stress
oxidativo, levam a uma reação em cadeia, mediada pelas ERO, que atacam, indiscriminadamente
proteínas (Stadtman e Levine, 2003), lípidos (Rubbo et al., 1994) e DNA (Richter et al., 1988),
comprometendo as funções biológicas normais. O alvo principal do stress oxidativo pode variar,
dependendo da célula, do tipo de exposição e severidade do stress (Fridovich, 1998). As lesões
oxidativas no DNA e em outras estruturas celulares, estão provavelmente relacionadas com o processo
de envelhecimento celular. A eventual degenerescência celular, deve-se à senescência do DNA,
devido ao acumular de lesões e desempenho menos eficiente da sua função informativa (Fridovich,
1998).

18
Muitas patologias estão associadas a um aumento na formação de radicais livres ou a uma
redução da capacidade de defesa antioxidante, ou simultaneamente a ambas as situações. O equilíbrio
entre oxidantes e antioxidantes pode ser então o limiar entre a saúde e o aparecimento de doenças
crónicas não transmissíveis e de outras condições patológicas, como doenças neurológicas
inflamatórias, aterosclerose, cancro e diabetes (Halliwell, 2007a e b, Rahman et al., 2006, Vatten e
Shetty, 2005).
A oxidação de proteínas pode acarretar danos em recetores e transportadores celulares,
enzimas, vias de transdução de sinais, pode ainda ativar o sistema imunitário, induzir a formação de
anticorpos e originar o aparecimento de doenças autoimunes (Galloway e Handy, 2003). As lesões
oxidativas em proteínas podem ainda causar danos indiretos em outras biomoléculas, por exemplo, as
lesões nos sistemas enzimáticos de reparação do DNA podem elevar os níveis de lesões não reparadas
nesta molécula e, deste modo, contribuir para o aumento da frequência de mutações. As lesões nas
proteínas envolvidas na manutenção essencial do gradiente iónico entre os fluídos do exterior e do
interior das células pode comprometer a manutenção destes gradientes (Halliwell e Gutteridge, 1989).
A susceptibilidade das proteínas depende da sua composição em aminoácidos e da importância
que os aminoácidos mais susceptíveis têm na sua estrutura tridimensional e atividade catalítica. Os
produtos mais comuns, resultantes da oxidação proteica são carbonilos (Kohen & Nyska, 2002).
Algumas patologias, como por exemplo artrite reumatoide, doença de Alzheimer, síndroma de
distúrbio respiratório, doença de Parkinson e aterosclerose, estão associadas ao aumento destes
produtos de oxidação proteica (Zwart et al., 1999).
Existem evidências do envolvimento das ERO no desenvolvimento do cancro, quer por efeitos
diretos, por meio ataque químico às bases do DNA ou à desoxirribose, quer indireto, pela alteração
dos processos de transdução de sinal, proliferação e morte e celular ou ainda da comunicação
intercelular (Halliwell e Gutteridge, 1989). A modificação das bases do DNA pode levar ao
aparecimento de mutações (Beckman e Ames, 1997).
A peroxidação lipídica é um processo bioquímico que envolve a oxidação dos ácidos gordos
polinsaturados como os componentes das membranas celulares. Os danos que a peroxidação lipídica
pode causar nas membranas biológicas podem conduzir à lise celular. Por outro lado, no decurso das
reações de lipoperoxidação podem formar-se compostos capazes de reagir com diversas biomoléculas,
incluindo o DNA, causando danos celulares (Chung et al., 1996). Para além da lipoperoxidação de
ácidos gordos polinsaturados, foi igualmente descrito que as ERO podem modificar as lipoproteínas de
baixa densidade (LDL) e, desta forma, contribuir para a patogénese da aterosclerose (Ryan e Aust,
1992).

19
As doenças cardiovasculares representam uma das principais causas de morte em todo o
mundo (WHO, 2011). O stress oxidativo está associado a diversas destas doenças, tais como, por
exemplo, a doença arterial coronária, hipertensão ou enfarte do miocárdio. Também a diabetes
mellitus não dependente de insulina parece estar associada ao stress oxidativo. Este tipo de patologias
é multifatorial e inclui inflamação vascular e aumento da geração vascular do radical anião superóxido
e de outras ERO (Kojda e Harrison, 1999).
Para combater as espécies reativas de oxigénio os organismos aeróbios possuem um sistema
de defesa antioxidantes endógenos, o qual inclui várias enzimas e moléculas antioxidantes de elevada
e baixa massa molecular (Kaur e Kapoor, 2001). Para além dos antioxidantes endógenos existem ainda
os antioxidantes exógenos, que são aqueles que têm de ser obtidos através da dieta e que incluem o
ácido ascórbico (vitamina C), o alfa-tocoferol (vitamina E), carotenóides e os compostos fenólicos que
englobam diversos dos antioxidantes mais ativos presentes nas frutas e nos vegetais (Soares, 2002).
2.4. Sistemas de Defesas Antioxidantes
A manutenção da vida aeróbica na terra só é possível porque os organismos aeróbios
evoluíram no sentido de desenvolver mecanismos de defesa capazes de minimizar os efeitos oxidantes
do oxigénio. A definição, em termos biológicos, de antioxidante é a de “qualquer substância que,
quando presente em baixas concentrações comparadas àquelas de um substrato oxidável,
significativamente previne a oxidação deste substrato” (Halliwell e Gutteridge, 2000) O termo
“substrato oxidável” refere-se a todo o tipo de molécula encontrada in vivo. Assim, a principal função
dos sistemas de defesa antioxidante dos organismos é o de inibir ou reduzir os danos celulares
causados pelas espécies reativas de oxigénio.
De acordo com o seu modo de ação, os antioxidantes podem classificar-se como antioxidantes
de prevenção (impedem a formação de EROs), eliminação (impedem o ataque das EROs às células) e
de reparação (Alderton et al., 2001). Nos mecanismos de reparação de danos causados pelas EROs
encontram-se os sistemas enzimáticos que favorecem a reconstituição das membranas celulares
danificadas e os sistemas enzimáticos de reparação do DNA, capazes de reparar as lesões oxidativas
que ocorrem nesta biomolécula. As lesões oxidativas podem ser reparadas através de diferentes
mecanismos: reparação por excisão de bases, reparação por excisão de nucleótidos e reparação de
erros de emparelhamento (Lunec et al., 2002).
Exemplos de sistemas de ação antioxidante por prevenção de EROs, são a restrição de spin do
oxigénio que diminui a sua reatividade face às biomoléculas, o transporte do oxigénio na forma ligada

20
e não livre e a quelação de metais durante o transporte e armazenamento, evitando a ocorrência da
reação de Fenton (Sies, 1993, citado por Ribeiro et al., 2005).
A eliminação dos EROs formadas pode ser efetuada através de mecanismos enzimáticos e não
enzimáticos. Dentro dos sistemas de defesa antioxidante enzimáticos encontram-se as enzimas
superóxido dismutase (SOD), a catalase (CAT) e a glutationa peroxidase (GPX). Estas enzimas
desativam diversas EROs inativando, assim, os seus efeitos negativos sobre o organismo (Ribeiro et
al., 2005).
A SOD catalisa a dismutação do radical anião superóxido a peróxido de hidrogénio (H2O2)
(Yu, 1994).
A CAT desempenha importante papel na eliminação do H2O2, promovendo a sua catálise até
água. A CAT atua apenas nas porções aquosas da célula, portanto, as zonas lipídicas, como a
membrana celular, permanecem desprotegidas e suscetíveis a ação do peróxido de hidrogénio (Ribeiro
et al., 2005).
A GPX também funciona como mecanismo de proteção contra o stress oxidativo, removendo
o peróxido de hidrogénio ao mesmo tempo que converte a glutationa reduzida (GSH) em glutationa
oxidada (GSSG) (Ferrari et al., 1985).
Desta forma, a SOD, a CAT e a GPX evitam a acumulação tanto do radical anião superóxido
como do peróxido de hidrogénio, contribuindo, desta forma, para a diminuição da produção do radical
hidroxilo, contra o qual não existe sistema enzimático de defesa (Yu, 1994).
A presença zinco (Zn), cobre (Cu), mangnésio (Mn) e selénio (Se), além de quantidades
suficientes de vitaminas é essencial para a produção das enzimas antioxidantes. Tanto o Cu como o Zn
e o Mn são particularmente importantes para a produção da superóxido dismutase, sendo o Mn
particularmente importante para a síntese da SOD mitocondrial, onde a maior parte dos radicais livres
são produzidos. O Se é essencial para a formação da GPX e a vitaminas C e do complexo B são

21
necessárias para a produção da CAT em grandes quantidades e da GPX. A vitamina B6 (piridoxina),
por exemplo, parece desempenhar um papel fundamental na biossíntese da GPX. Assim, o consumo
regular de determinados minerais e vitaminas é indispensável para o correto funcionamento do sistema
antioxidante do organismo (Ribeiro et al., 2005).
O sistema de defesa antioxidante não enzimático inclui compostos sintetizados pelo organismo
humano como, por exemplo, a bilirrubina, ceruloplasmina, melatonina, coenzima Q, ácido úrico e, de
forma muito importante, a glutationa reduzida (GSH) (Regulski e Tully, 1995; Schneider e Oliveira,
2004) e compostos não sintetizados pelo organismo e que têm que ser ingeridos através da dieta
regular, ou via suplementação, como ácido ascórbico (vitamina C), -tocoferol (vitamina E), -
caroteno (precursor de vitamina A) e compostos fenólicos (Schneider e Oliveira, 2004). Da interação
entre estas moléculas antioxidantes e as EROs pode resultar o aparecimento de espécies radicalares
derivadas da molécula antioxidante, sendo que estes são por definição pouco reativos. As vitaminas
atuam de forma conjunta em diferentes compartimentos intracelulares. Com caraterísticas
lipossolúveis, as vitaminas A e E atuam essencialmente ao nível das membranas celulares enquanto a
vitamina C, por ser hidrossolúvel, atua no citosol, participando ainda na reoxidação das vitaminas
lipossolúveis (Regulski e Tully, 1995).
A maioria das defesas antioxidantes não-enzimáticas não são sintetizadas endogenamente pelo
ser humano, devem ser obtidas através da dieta. O reino vegetal dispõe de uma enorme gama de
compostos com potencial antioxidante in vivo (Hermes-Lima, 2004). Daqui a importância da relação
entre a nutrição e as defesas antioxidantes. Com efeito, existem evidências epidemiológicas
significativas que o consumo de alimentos ricos em antioxidantes está associado a um envelhecimento
saudável e à longevidade funcional (Rice-Evans e Miller, 1997). Dentro dos compostos antioxidantes
produzidos pelos vegetais a categoria dos polifenóis é a mais diversificada, compreendendo uma
ampla variedade de compostos e de fontes alimentares de origem vegetal (Shahidi e Naczk, 1995),
entre os quais se encontram os mirtilos.
Os polifenóis podem apresentar diferentes propriedades protetoras e agir em diversas etapas
do processo oxidativo, funcionando por diferentes mecanismos e são, portanto, classificadas em duas
categorias principais: antioxidantes primários e secundários. São considerados primários os compostos
de ação antioxidante capazes de inibir ou retardar a oxidação por inativação de EROs graças à doação
de átomos de hidrogénio (mecanismo HAT de hydrogen atom transfer) ou atividade redutora ou de
transferência eletrónica (mecanismo SET de single electron transfer), o que transforma os radicais em
substâncias estáveis. Os antioxidantes secundários apresentam uma grande variedade de modos de
ação: Capacidade de quelação de iões metálicos, em especial o Fe2+
e o Cu+, evitando, deste modo, a
formação de radicais hidroxilo através das reações de Fenton; Indução das enzimas antioxidantes

22
(catalase, superóxido dismutase ou glutationa peroxidase); Inibição de enzimas oxidativas como a
xantina oxidase ou as ciclooxigenases; Absorção de radiação ultravioleta (Maisuthisakul et al., 2007;
Magalhães et al., 2008).
2.5. Mutagéneos e Antimutagéneos nos alimentos
2.5.1. Mutagéneos alimentares
A ingestão de alimentos constitui uma das principais vias de exposição do homem a diferentes
compostos químicos. Algumas desses compostos podem ter atividade mutagénica/cancerígena ou
antimutagénica; isto é, podem induzir mutações no ácido desoxirribonucleico (DNA) e/ou podem
favorecer o desenvolvimento de tumores, enquanto outros podem atenuar ou anular os efeitos destes.
Muitos destes compostos encontram-se presentes nos alimentos de forma natural enquanto que outros
são adicionados ou produzidos durante o seu processamento e o armazenamento (Antunes e Araújo,
2000).
A macromolécula do DNA possui uma estrutura complexa, passível de sofrer modificações
por exposição a agentes potencialmente mutagénicos (Wogan et al., 2004). O DNA, ao longo da vida,
vai sofrendo alterações que se denominam por mutações, podendo estas ser causadas por erros que
ocorrem durante o processo de replicação, pela exposição a mutagéneos endógenos, como, por
exemplo, as espécies reativas de oxigénio, ou induzidas por exposição a agentes externos. Estes
agentes externos podem ser divididos em três classes:
1) Agentes físicos, como as radiações ultravioleta e ionizantes;
2) Agentes biológicos, como alguns vírus, que se integram ao genoma celular interrompendo
as sequências génicas ou promovendo rearranjos génicos;
3) Agentes químicos, como os hidrocarbonetos aromáticos policíclicos ou as N-nitrosaminas.
Os compostos químicos capazes de lesar o DNA são, normalmente, compostos de natureza
electrofílica capazes de reagir com esta molécula causando-lhe alterações, que podem envolver apenas
um par de bases ou podem envolver deleções, inserções ou rearranjos de segmentos de nucleótidos
(Miller e Miller, 1986). Dependendo dos genes em que ocorrem e do tipo de alteração que provocam,
as mutações podem causar maior ou menor perturbação ao funcionamento celular. Assim, enquanto
que umas mutações podem não implicar mudanças detetáveis na atividade metabólica celular, outras
ser letais para célula. Em particular, quando ocorrem nos genes envolvidos no processo de divisão

23
celular ou nos genes responsáveis pela manutenção da integridade do DNA, as mutações podem
conduzir ao aparecimento de tumores (Bishop, 1991).
Os compostos mutagénicos, como já foi referido anteriormente, podem estar presentes em
produtos alimentares de forma natural, acidental (caso dos contaminantes ambientais) ou ser gerados
no decurso do seu processamento ou conservação (Ferguson e Philpott, 2008). Alguns agentes
mutagénicos alimentares são a aflatoxina B1, ocratoxina A, vários alcalóides pirrolizidínicos, aminas
heterocíclicas (AH), como, por exemplo, a 2-amino-1-metil-6-fenilimidazole[4,5-b]piridina (PhIP),
hidrocarbonetos aromáticos policíclicos (HAPs), como, por exemplo, o benzo(a)pireno, as N-
nitrosaminas e produtos das reações de Maillard de que é exemplo a acrilamida.
A aflatoxina B1 é um metabolito produzido pelo fungo Aspergillus flavus, que pode crescer em
cereais, como o milho ou o arroz, e frutos secos, como o amendoim, mal armazenados (Busby e
Wogan, 1984). As populações com uma exposição alimentar elevada a esta micotoxina apresentam um
aumento da incidência de hepatocarcinomas, especialmente quando em associação a infeção pelo vírus
da hepatite B (Hussain et al., 2007). A ocratoxina A (OTA) é também uma micotoxina produzida por
fungos através de contaminação de produtos alimentares mal armazenados. Esta micotoxina é
nefrotóxica e tem sido associada ao desenvolvimento de tumores do trato urinário. O mecanismo de
genotoxicidade da OTA parece envolver a formação de adutos de DNA e a formação de espécies
reativas de oxigénio (Pfohl-Leszkowicz e Manderville, 2007).
Os alcalóides pirrolizidínicos são comuns em plantas alimentares, especialmente as utilizadas
como medicamentos tradicionais. Os seres humanos podem ser expostos a este grupo de compostos de
uma forma direta, por ingestão das ervas ou dos chás com elas preparados, ou de forma indireta, por
ingestão de produtos derivados destas plantas, como o mel, ou de produtos obtidos a partir de animais
com elas alimentados, como, por exemplo, o leite. Estes alcaloides só se tornam reativos face ao DNA
após sofrerem metabolização pelas enzimas de biotransformação (Fu et al., 2004).
As aminas heterocíclicas são substâncias indesejadas produzidas durante a exposição de
alimentos a temperaturas elevadas. Muitas AH são formadas ao assar, fritar ou grelhar alimentos ricos
em proteínas, como as carnes e o pescado (Delfino et al., 2000). As AH encontram-se entre as
substâncias mutagénicas conhecidas mais potentes, causando tumores das glândulas mamárias,
próstata, pulmão, cólon, pele, pâncreas, fígado e principalmente bexiga em animais de laboratório.
Existem ainda evidências de que AH afetam o sistema vascular, as glândulas salivares e de que levam
à degeneração do miocárdio (Gross e Grüter, 1992; Felton et al., 2007, Garcia-Closas et al., 2007; Ahn
et al., 2008). As aminas heterocíclicas também requerem metabolização para se tornarem reativas face
ao DNA (Felton et al., 2007). Dados epidemiológicos recentes apontam no sentido da existência de
uma associação entre a exposição alimentar às AH e o aparecimento de cancro da bexiga em humanos,

24
sendo esta associação mais significativa no grupo da população com polimorfismos genéticos
considerados mais desfavoráveis nas enzimas de biotransformação das aminas hetrocíclicas (Lin et al.,
2012).
Os hidrocarbonetos aromáticos policíclicos (HAP) são produtos da combustão incompleta de
material orgânico (Phillips, 1999), podendo aparecer nos alimentos por se formarem no decurso da sua
confeção a temperaturas elevadas (peixe e carne grelhados) ou da sua conservação (alimentos
fumados), ou por ocorrem naturalmente no ambiente como contaminantes (cereais, legumes, pescado)
(Fergusun et al., 1996). Segundo a Organização Mundial da Saúde, dentro do grupo dos HAP, há 13
compostos que são claramente carcinogénicos e genotóxicos (WHO, 2005). O mecanismo de
toxicidade dos HAP dá-se pela formação de produtos intermediários reativos, responsáveis pelos
efeitos mutagénicos/carcinogénicos (Falahatpsisheh et al., 2004; Varlet et al., 2007). Estudos
epidemiológicos associam a exposição de misturas de HAP contendo benzo(a)pireno ao risco
aumentado de desenvolver cancro de pulmão e outros tumores. Também foi observado maior
frequência de cancro de estômago em populações que consomem tradicionalmente grandes
quantidades de produtos cárneos fumados (Kazerouni et al., 2001; Falahatpisheh et al., 2004).
As N-nitriosaminas são compostos que têm sido reconhecidos como compostos mutagénicos e
potencialmente cancerígenos. Estes compostos podem ser encontrados numa ampla variedade de
alimentos, incluindo carnes processadas, como bacon, presunto e enchidos, peixe fumado e certos
queijos (Ferguson e Philpott, 2008). Estes compostos podem, igualmente, ser sintetizados
endogenamente no estômago a partir de nitrito e de aminas secundárias e terciárias existentes nos
alimentos (AICR-WCRF, 2007). As N-nitrosaminas requerem ativação metabólica para exercerem sua
ação mutagénica e carcinogénica. A etapa inicial da biotransformação envolve uma hidroxilação do
carbono do grupo alquilo, catalisada pelos citocromos P450 formando um aldeído ou cetona e uma
nitrosamina primária, instável, a qual, por sua vez, acaba alquilando sítios núcleofilicos do DNA e do
ácido ribonucleico (RNA). Esta etapa é considerada fundamental no processo de iniciação do cancro,
(Tricker e Preussmann, 1991) sendo que o fígado é o principal órgão de biotransformação das
nitrosaminas, mas outros tecidos humanos também as podem biotransformar (Järgestad e Skog, 2005).
A acrilamida pode formar-se naturalmente em alimentos ricos em açúcares e cozinhados a
temperaturas elevadas, como resultado das reações de Maillard, ou seja, como resultado das reações
não enzimáticas que ocorrem entre açúcares e aminoácidos a temperaturas elevadas. Embora seja
claramente um carcinogénico animal e uma neurotoxina, a extrapolação dos efeitos em sistemas
celulares e em animais para os efeitos em humanos tem sido controversa (Exon, 2006). Diversos
estudos apontam no sentido da acrilamida poder interferir com os mecanismos de reparação do DNA
(AICR-WCRF, 2007).

25
2.5.2. Antimutagéneos alimentares
O modo de ação dos agentes antimutagénicos podem dividir-se em dois mecanismos
principais: atuação direta sobre os compostos que induzem mutações no DNA, inativando-os química
ou enzimaticamente, inibindo a sua ativação metabólica ou sequestrando moléculas reativas, e os que
atuam sobre o processo que leva a indução de mutações ou sobre a reparação das lesões causadas no
DNA (Kada et al., 1978).
Os estudos com os agentes antimutagénicos foram iniciados nos anos cinquentas, estes estudos
centram-se na identificação de antimutagéneos, principalmente os de origem natural. A identificação
de agentes antimutagénicos e/ou anticarcinogénicos em alimentos é indispensável e extremamente
importante na busca de estratégias para a prevenção do cancro, por meio de modificações dos hábitos
alimentares (Wargovich, 1997). Grandes progressos foram feitos desde meados da década de 1990 na
compreensão do processo de cancerigénese, havendo crescentes evidências que mostram ou sugerem
que a alimentação esteja profundamente relacionada com este processo (AICR- WCRF, 2007).
As mutações podem ser relacionadas com a ausência de alguns componentes alimentares,
especialmente micronutrientes (Fenech e Ferguson, 2001). A dose diária recomendada (recommended
dietary allowances- RDA), indica a quantidade de um nutriente por dia que é necessário para a maioria
das pessoas se manter saudável, estes valores são atualizado periodicamente para refletir novos
conhecimentos. Muitos compostos antimutagénicos encontrados nos alimentos são agentes
antioxidantes e atuam sequestrando EROs, quando administrados com o agente que induz as mutações
no DNA.
A vitamina E aparece nos alimentos predominantemente como -tocoferol. Os tocoferóis são
encontrados em grandes concentrações no germe de trigo, amêndoas e avelãs, óleos vegetais como os
extraídos do germe de trigo, girassol, caroço de algodão, dendê, amendoim, milho e soja. Uma das
principais funções atribuídas à vitamina E é a proteção que confere às membranas celulares contra a
destruição oxidativa, talvez atuando em conjunto com pequenas moléculas e enzimas para defender as
células contra os danos causados pelos radicais de oxigénio. Assim, a vitamina E possui propriedades
antioxidantes (Mccall et al., 1989).
O selénio é um elemento essencial para o ser humano pois foi demonstrada a sua incorporação
na glutationa peroxidase (GPX), enzima vital na proteção de membranas celulares e subcelulares
contra a agressão por peróxidos solúveis. Do ponto de vista biológico, o selénio é um elemento
necessário para a produção de enzimas fundamentais na neutralização de radicais livres e na proteção
contra a peroxidação lipídica de membranas celulares e subcelulares. Age em sinergismo com a
vitamina E na sua função antioxidante. Também está envolvido nas funções da tiroide, além das

26
funções vitais. O selénio está presente na maioria dos alimentos com grande quantidade de proteínas
como: aipo, atum, brócolos, cebola, cereais integrais, fígado, frango, frutos do mar, gema de ovo,
germe de trigo, leite pepino e repolho (Olken, 1994).
Os carotenóides são corantes naturais presentes nas frutas e vegetais (cenouras, tomates,
espinafre, laranjas, pêssegos, etc.), sendo que sua estrutura química é composta por ligações duplas
conjugadas, que são responsáveis pela sua cor e por algumas de suas funções biológicas. Diversos
estudos revelam a existência de uma relação entre o aumento do consumo de alimentos ricos em
carotenóides e a diminuição no risco de várias doenças. Estes revelam que os carotenóides sequestram
o oxigénio singleto, desativam espécies radicalares, modelam o metabolismo dos cancerígenos, inibem
a proliferação celular, estimulam a comunicação entre as células e elevam a resposta imunitária.
(Olson, 2004). Os carotenoides como o betacaroteno, licopeno, zeaxantina e luteína, exercem funções
antioxidantes em fases lipídicas, bloqueando os radicais livres que danificam as membranas
lipoprotéicas (Hickey et al., 1997).
Os folatos são micronutrientes que existem essencialmente em vegetais de folhas verdes e tem
sido sugerido que o seu consumo possa diminuir o risco de desenvolvimento de doenças
cardiovasculares e de vários tipos de cancro (Stahl e Sies, 1997). Existem estudos que mostram que a
deficiente ingestão deste micronutriente provoca um nível de danos no DNA comparável ao que é
causado por uma dose elevada de radiação (Courtemanche et al., 2004). Os mecanismos pelos quais a
deficiência de folato pode influenciar a carcinogénese parecem envolver o aumento dos erros na
replicação de DNA e a hipermetilação de genes de reparação de DNA ou de genes supressores de
tumor (Shrubsole et al., 2001).
As fibras alimentares podem ser consideradas antimutagéneos alimentares uma vez que
reduzem a concentração de compostos mutagénicos no intestino, bem como o seu tempo de
permanência, diminuindo o tempo de exposição da mucosa intestinal a esses compostos e reduzindo a
possibilidade da sua absorção. Mais ainda, estas fibras podem adsorver compostos mutagénicos, como,
por exemplo, as aminas heterocíclicas, impedindo, também desta forma, a sua absorção (Ferguson,
2001).
Diversos trabalhos têm demonstrado que os polifenóis apresentam capacidade antimutagénica
que pode manifestar-se através de diferentes mecanismos. Assim, diversos polifenóis têm mostrado
inibir a absorção de mutagéneos como o benzo(a)pireno ou inibir a formação gástrica de N-
nitrosaminas, podendo, por isso, o estômago ser o principal local onde os polifenóis previnem a
cancerigénese. Mais ainda, certos polifenóis parecem influenciar a atividade de enzimas de reparação
do DNA através da modelação da expressão dos seus genes, inibir a ação de enzimas oxidativas e
desativar espécies oxidativas. Além do já referido, alguns flavonoides, e outros compostos fenólicos,

27
obtidos através da dieta podem inibir citocromos P450 específicos envolvidos na bioativação de
cancerígenos e proteger contra mutagénese através deste mecanismo (Ferguson, 2001).
2.6. Efeitos dos polifenóis na saúde
Numerosos são os estudos analíticos realizados na determinação, distribuição e quantificação
de compostos polifenóis em frutas, leguminosas e bebidas. As concentrações destes compostos variam
de acordo com as numerosas variedades, os fatores ambientais e tecnológicos, alguns dos quais podem
ser controlados para otimizar o conteúdo de polifenóis nos alimentos. É de todo o interesse a
identificação das variedades de plantas mais ricas em polifenóis, melhorar os seus métodos de cultivo
e limitar as perdas durante o decurso do seu processamento industrial e na cozinha (Manach et al.,
2004).
A importância de ingerir alimentos com elevado teor em polifenóis prende-se com as
evidências epidemiológicas que apontam no sentido da ingestão insuficiente destes compostos
bioativos poder constituir uma componente de risco para o desenvolvimento de doenças crónicas não
transmissíveis, entre as quais se encontram as doenças cardiovasculares e o cancro (Holst e
Williamson, 2008; Bastos et al., 2009). Com efeito, os polifenóis presentes na dieta têm sido
associados com a promoção da saúde através de efeitos que compreendem a diminuição dos níveis de
açúcar no sangue, a redução o peso corporal, atividade antimutagénica/anti carcinogénica, anti-
inflamatória, anti envelhecimento, antitrombótica, antimicrobiana e, principalmente, atividade
antioxidante (Ferguson, 2001; Scalbert et al., 2005; Bernal et al., 2011).
Diversos trabalhos têm demonstrado a capacidade de diferentes polifenóis para inibirem as
enzimas envolvidas na síntese de prostaglandinas e leucotrienos, potentes mediadores da resposta
inflamatória (Anjo, 2004), contribuindo, assim, para a diminuição desta resposta e conferindo, deste
modo, proteção contra uma diversidade de patologias nas quais a inflamação pode desempenhar um
papel importante, tais como, por exemplo, a aterosclerose e o cancro (Leighton e Urquiaga, 2007;
Moraes e Locatelli, 2010).
Diversos estudos sugerem que os compostos fenólicos sugerem que os efeitos benéficos dos
polifenóis sobre a saúde possa resultar de outros mecanismos diferentes das suas capacidades
antioxidantes e anti-inflamatórias. Dentro desses outros mecanismos encontram-se a modelação da
atividade de diferentes enzimas como as telomerases ou as enzimas de biotransformação, interações
com recetores celulares e com vias de transdução de sinal e ainda a regulação do ciclo celular,
mecanismos essenciais para a manutenção da homeostase dos organismos vivos (D’ Archivio et al.,

28
2007). Os polifenóis estão ainda envolvidos na regeneração das vitaminas antioxidantes, como, por
exemplo, a vitamina E, a vitamina C e a vitamina A (Araújo et al., 2005).
Um estudo recente, e o primeiro destes ensaios realizado in vivo, teve o propósito de avaliar o
possível efeito pré-biótico dos polifenóis na flora intestinal humana (Queipo-Ortuño et al., 2012).
Neste estudo, que envolveu a participação de 10 homens saudáveis, foi possível observar mudanças
significativas na microbiota fecal de todos os participantes, tendo os polifenóis revelado capacidade
para inibir o crescimento de bactérias prejudiciais, como algumas estirpes de Clostridium, e
potenciado o crescimento de bactérias pró bióticas, como a Bifidobacteria, implicadas na redução de
bio marcadores inflamatórios (proteína C reativa) e da concentração de colesterol, promovendo deste
modo benefícios para o hospedeiro (Queipo-Ortuño et al., 2012).
As mesmas propriedades que determinam a atividade antioxidante dos compostos fenólicos
podem ser responsáveis por eventuais efeitos nefastos. Assim, em determinadas condições, como, por
exemplo, na presença do óxido nítrico (NO), ou quando em doses elevadas, e em determinados
tecidos, os polifenóis podem oferecer mais riscos oxidativos do que benefícios antioxidantes
(Ferguson, 2001; Behling et al., 2004). Com efeito, os polifenóis podem reduzir o ferro (III) a ferro
(II) e, assim, gerar radicais hidroxilo através da reação de Fenton. Os polifenóis podem ainda ser
reduzidos a quinonas e entrar em ciclos redox originando a produção de ERO ou podem, no decurso
da sua regeneração, gerar radicais secundários reativos capazes de oxidar proteínas e lípidos
(Ferguson, 2001). Embora alguns ensaios in vivo, realizados em animais de laboratório, atribuam
efeitos pró-carcinogénicos a alguns flavonoides, a maioria destes ensaios apontam no sentido destes
compostos terem uma atividade anticancerígena. É possível que as enzimas de biotransformação
possam desempenhar um papel importante na limitação dos efeitos mutagénicos dos polifenóis in vivo
(Scalbert et al., 2005).
Outro efeito negativo que os compostos fenólicos podem exercer no organismo reside no fato
destes poderem ser potencialmente anti nutricionais. Esta propriedade pode dever-se à sua capacidade
de quelação de metais, como, por exemplo, o ferro ou o cálcio, e de ligação a aminoácidos e proteínas
no trato gastrointestinal. O consumo elevado destes compostos pode estar, assim, relacionado com
uma redução da biodisponibilidade destes nutrientes e consequente perda de valor biológico dos
alimentos (Ferguson, 2001; Ross e Kasum, 2002).
As atividades biológicas dos compostos fenólicos são influenciadas pelas suas caraterísticas
estruturais, pela natureza do alvo biológico, pelas condições ambientais, como por exemplo o pH dos
tecidos, pela dosagem ingerida, bem como pela biodisponibilidade destes compostos no organismo
(Behling et al., 2004; Manach et al., 2005). Existem evidências, embora indiretas, da absorção do áci-
do clorogénico e de outros compostos fenólicos, uma vez que existem ensaios em humanos em que se

29
observara um aumento na capacidade antioxidante do plasma após o consumo de alimentos ricos
nestes compostos (Manach et al., 2005).
Um melhor conhecimento da biodisponibilidade é essencial para a investigação
os efeitos na saúde dos polifenóis, qualquer que seja a abordagem
usada. Vários fatores alteram a biodisponibilidade de compostos bioativos presentes nos alimentos: a
complexidade da matriz alimentar, a forma química do composto de interesse, a estrutura e quantidade
de outros compostos ingeridos simultaneamente (fatores exógenos) e ainda o tempo de trânsito
intestinal, esvaziamento gástrico, metabolismo do composto e grau de conjugação, possíveis
interações com proteínas na circulação sanguínea e tecidos, composição da microflora intestinal e o
perfil genético do indivíduo (fatores endógenos) (Scholz e Williamson, 2007; Crozier et al., 2009).
Avaliados em conjunto, estes fatores respondem pelas variações intra e inter-individuais na
biodisponibilidade de uma determinada substância (Holst e Williamson, 2008).
Os polifenóis têm biodisponibilidade limitada e podem ser extensamente metabolizados. No
entanto, apesar desta limitada disponibilidade, estudos efetuados em humanos sugerem que mesmo
essa pequena quantidade que é absorvida possa ser suficiente para exercer efeitos benéficos na saúde
(Ferguson, 2001; Williamson e Manach, 2005).
A biodisponibilidade difere grandemente entre os vários tipos de polifenóis sendo o ácido
gálico e as isoflavonas os que parecem ser mais bem absorvidos, seguidos pelas catequinas, flavanonas
e glicosídeos de quercetina, sendo as proantocianidinas, as catequinas galato do chá e as antocianinas
os polifenóis de mais difícil absorção (Manach et al., 2005). No entanto, a determinação da taxa de
absorção destes compostos, e, em particular, da taxa de absorção das antocianinas, pode vir
subestimada dada a possibilidade deles serem rapidamente convertidos a metabolitos que ainda não
tenham sido identificados (Manach et al., 2005).
Apesar das antocianinas terem uma limitada absorção em mamíferos, ensaios realizados com
ratos, mostram se conseguem obter concentrações plasmáticas destes compostos dentro da gama em
que estes exercem as suas atividades antioxidantes in vitro (Passamonti et al., 2003).
A metabolização pela flora intestinal pode, igualmente, ter um efeito preponderante na
biodisponibilidade dos polifenóis. Com efeito, os polifenóis que não forem absorvidos no trato
gastrointestinal, podem ser metabolizados por estas bactérias, originando metabolitos passíveis de ser
absorvidos no epitélio do cólon e, desta forma, chegar à corrente sanguínea. Por exemplo, as
proantocianidinas têm demonstrado alguns efeitos em ensaios epidemiológicos embora se saiba que a
sua absorção intestinal seja muito reduzida. Desta forma, é possível que os efeitos biológicos
associados a estes compostos resultem de algum metabolito formado no decurso da sua metabolização
pelas bactérias da flora intestinal (Williamson e Manach, 2005).

30
3. Materiais e Métodos
3.1. Caraterização das amostras de mirtilo
Na realização deste trabalho foram analisadas as variedades de mirtilo Northen Highbush
Earlyblue, Patriot, Bluecrop e Duke e a variedade Southern highbush O`Neal, produzidas na zona
geográfica abrangida pela cooperativa de Sever do Vouga, Mirtilusa (Sociedade de Produtores
Hortofrutícolas). As variedades foram fornecidas pela Mirtilusa no estado de maturação em que as
bagas são, normalmente, comercializadas, ou seja, no estado de plena maturação quando os frutos se
apresentam com a totalidade da coloração desenvolvida mas sem perda de turgescência.
A colheita foi sempre efetuada de manhã tendo cada amostra sido composta por
aproximadamente 300 g de cada cultivar, de entre três a seis plantas de cada genótipo. Depois de
colhidas, as amostras foram transportadas em saco térmico até à Faculdade de Ciências e Tecnologia
(Campus da Caparica) no escuro, sob refrigeração, tendo sido preparadas para análise (realização dos
extratos) num no prazo máximo de duas horas.
3.2. Reagentes e meios de cultura
Na realização do presente trabalho foram utilizados os seguintes reagentes e meios de cultura:
acetato de amónio (Riedel-de Haën, 98%); acetato de sódio tri-hidratado (Panreac, 99%); ácido L (+)
ascórbico (Panreac, 99%); ácido cítrico mono-hidratado (JMGS, 99,8%); ácido clorídrico (Panreac,
37%); ácido gálico (Merck, 99,5%); 2,2’-azobis(2-metilpropionamidina)dihidrocloreto (AAPH)
(Sigma-Aldrich, 97%); azul de nitrotetrazólio (NBT2+
) (Sigma); Bacto agar (Becton Dickinson and
Company); biotina (Sigma-Aldrich, 99%); carbonato de sódio (VReis); cloreto de cobre (II)
bihidratado (Riedel-de Haën, 99%); cloreto de ferro (III) hexa-hidratado (Merck, 99%); cloreto de
potássio (Panreac, 99%); cloreto de sódio (Panreac, 99,5%); dihidrogenofosfato de sódio (Panreac,
98%); dinucleótido de nicotinamida e adenina na forma reduzida (NADH) (Sigma, 97%); etanol
(Riedel-de Haën, 99%); fluoresceína (Sigma-Aldrich); D-glucose monohidratada (Panreac);
hidrogenofosfato de dipotássio (Becton Dickinson and Company, 97,5%); hidróxido de sódio (Merck,
99%); histidina (Merck, 99%); meio de cultura Nutrien Broth (NB) nº2 (Oxoid); metanol (Riedel-de
Haën, 99,8%); metossulfato de fenazina (PMS) (Sigma, 90%); neocuproína (2,9–dimetil–1,10–
fenantrolina) (Sigma); peróxido de hidrogénio (Panreac, 30%); reagente de Folin-Ciocalteu (Panreac);
sal dissódico de ácido etilenodiaminatetracético (EDTA) (Panreac, 99%); sal dissódico de ácido 3-(2-
piridil)-5,6-difenil1,2,4-triazina4’,4’’-dissulfónico (ferrozina) (Sigma-Aldrich); sulfato de magnésio
heptahidratado (Merck, 99,5%); sulfato ferroso hepta-hidratado (Riedel-de Haën, 99%); tert-butil
hidroperóxido (t-BHP) (Sigma-Aldrich, 70% em H2O); 2,4,6-tri(2-piridil)-s-triazina (TPTZ) (Fluka,
99%); hidrogenofosfato de amónio e sódio tetrahidratado (NaNH4HPO4.4H2O (Sigma, 99%) e trolox

31
(Acros Organics, 97%). Na preparação de todas as soluções, diluições e meios de cultura, utilizou-se
água ultrapura, captada a partir de um sistema de purificação Milli-Q (Millipore, Molsheim, França).
3.3. Preparação dos extratos de mirtilo
De acordo com Smith e colaboradores (2000), a melhor estratégia de armazenamento dos
extratos é mantê-los no escuro a baixas temperaturas, separados em alíquotas, de forma a retirar
apenas a quantidade necessária a cada ensaio. Assim, em todos os passos realizados até à obtenção dos
extratos, separação em alíquotas e realização dos ensaios, foi minimizado o contacto com a luz,
oxigénio e temperaturas elevadas.
Os extratos foram preparados de acordo com o procedimento descrito por Connor e
colaboradores (2002), com pequenas adaptações. Assim, aos mirtilos previamente triturados foi
adicionado metanol acidificado com ácido clorídrico (0,1% volume/volume) e arrefecido em gelo,
numa proporção de 10 g de fruto por cada 20 mL de solvente. A mistura foi agitada em gelo durante
20 minutos e filtrada, por gravidade, através de um filtro de papel (Watman – Lote EW0287-1; Ref. Nº
10311612). Repetiu-se a extração, a partir dos resíduos recuperados do papel de filtro, usando igual
volume de metanol acidificado, nas mesmas condições. O filtrado total recuperado foi diluído a 50
mL, num balão volumétrico, com o solvente da extração. Os extratos assim obtidos foram evaporados
em rotavapor (Büchi R-200), utilizando-se um banho termo estatizado a 37 °C (Büchi heating bath B-
490), até à evaporação total do metanol. Os extratos secos foram ressuspensos em 50 mL de água
ultrapura, em balão volumétrico, ficando com uma concentração final de 0,20 g/mL. Os extratos foram
filtrados em ambiente asséptico (câmara de fluxo laminar) através de membranas estéreis Millipore
Millex GP com poro de 0,22 μm. Depois de esterilizados, os extratos foram distribuídos em alíquotas
de 1,5 mL em tubos de polipropileno estéreis e congelados a -50 °C (UltraCongelador New Brunswick
Scientific U410).
3.4. Determinação dos fenóis totais pelo Método de Folin-Ciocalteu
O ensaio Folin-Ciocalteu baseia-se na redução a pH alcalino do heteropolianião
molibdotungnesteniofosfórico do reagente de Folin-Ciocalteu, efetuada pelos compostos fenólicos, na
forma de ião fenolato, da amostra e que origina o aparecimento de um produto azul com um máximo
de absorção a 765 nm, sendo a intensidade da coloração desenvolvida proporcional ao teor em
compostos fenólicos (Singleton et al., 1999; Huang et al., 2005). Este método é simples, sensível e
preciso mas pode sofrer interferências de outros componentes da amostra que possam, igualmente,
reagir com o reagente de Folin-Ciocalteu como, por exemplo, adenina, adenosina, alanina, ácido

32
aminobenzóico, ácido ascórbico, benzaldeído, cisteína, creatinina, histamina, histidina, índole, frutose,
sacarose, sulfito de sódio, sulfato de ferro e outros (Prior et al., 2005).
Para a determinação dos compostos fenólicos totais utilizou-se o método descrito por Kosar e
colaboradores (2008) com algumas modificações. Assim, em balões volumétricos de 10 mL, contendo
cerca de 6 mL de água ultrapura, adicionou-se 100 μL de cada extrato, ou das suas diluições, e 500 μL
de reagente de Folin-Ciocalteu sem diluição. Deixou-se um minuto à temperatura ambiente,
adicionou-se 1,5 mL de uma solução de carbonato de sódio a 20% (peso/volume) e perfez-se o volume
a 10 mL com água ultrapura. As misturas assim preparadas foram incubadas durante 2 horas, a 25 ºC e
no escuro. Findo este tempo, procedeu-se então à medição da absorvância das amostras num
espectrofotómetro (SPEKOL 1500) a 765 nm, contra um branco preparado da mesma forma mas
substituindo a amostra por água ultrapura.
Todas as amostras foram analisadas em quadruplicado, tendo a concentração em fenóis sido
determinada por interpolação de uma reta de calibração, preparada da forma já descrita mas
substituindo as amostras por soluções de ácido gálico com concentrações entre os 50 e os 500 mg/L.
Os resultados obtidos foram expressos em mg de equivalentes de ácido gálico por 100 grama
de peso fresco de amostra (por 100 grama de fruto).
3.5. Quantificação das antocianinas monoméricas totais pelo método do pH
diferencial
Este método baseia-se na transformação estrutural reversível em função do pH que as
antocianinas monoméricas sofrem (Figura 3.1) que leva à obtenção de diferentes espectros de absorção
(Figura 3.2). Assim, a pH 1, a forma predominante é a de oxónio colorido (λmax-vis 520 nm) enquanto a
pH 4,5 a forma predominante é a de hemiacetal sem coloração (sem absorção a 520 nm).

33
Figura 3.1 - Formas estruturais predominantes de antocianinas a diferentes valores de pH (Lee et al.,
2005).
Figura 3.2 - Caraterísticas espetrais das antocianinas em soluções a pH 1,0 e a pH 4,5 (Lee et al.,
2005).
O método do pH diferencial permite medir, com rapidez e precisão, o total de antocianinas,
mesmo quando em presença de pigmentos polimerizados e de outros compostos interferentes, uma vez
- H+
pH=7, coloração azul
pH=1, coloração laranja a púrpura
+H2O / - H+
pH=4,5, incolor
pH=4,5, incolor
(nm)
Ab
sorv
ânci
a

34
que estes não sofrem a mesma alteração de absorção em função do pH. A diferença de absorção a 520
nm de um extrato a pH 1 e a pH 4,5 é então proporcional à quantidade de antocianinas totais.
A determinação das antocianinas totais foi efetuada de acordo com o método descrito por Lee
e colaboradores (2005). Assim, os extratos foram diluídos numa solução de cloreto de potássio
0,025M, acidificada com ácido clorídrico até pH 1, até que a absorvância a 520 nm das soluções
ficasse dentro da zona linear do espectrofotómetro (SPEKOL 1500) (entre 0,2 e 1), para que não fosse
excedida a capacidade tampão dos reagentes, a percentagem de amostra na solução final nunca foi
superior a 20%. Procedeu-se à mesma diluição dos extratos utilizando uma solução tampão de acetato
de sódio 0,4 M a pH 4,5. Deixaram-se estabilizar as soluções durante 20 minutos e leu-se a
absorvância no referido espectrofotómetro a 520 nm e a 700 nm, utilizando água ultrapura como
branco. A leitura a 700 nm foi feita com o objetivo de eliminar possíveis interferências de alguns
materiais uma vez que a este comprimento de onda as antocianinas não absorvem.
As amostras foram analisadas em quadruplicado tendo o teor em antocianinas no extrato,
expresso em mg equivalentes de cianidina-3-glucósido, sido determinado recorrendo à seguinte
expressão:
mg equivalentes/L = (A x MM x FD x 103) / ε x d
Sendo que: A corresponde a [(A520-A700)pH1-(A520-A700)pH4,5]; MM é a massa molar da
cianidina-3-glucósido (449,2 g/mol); FD é o fator de diluição determinado; 103 o fator de conversão de
g para mg; d o percurso ótico (1 cm) e ε o coeficiente de absortividade molar da cianidina-3-glucósido
(26900 Lmol-1
cm-1
) (Lee et al., 2005).
Os resultados finais foram expressos em mg de equivalentes de cianidina-3-glucósido por 100
gramas de peso fresco de amostra.
3.6. Determinação do teor de sólidos solúveis (grau Brix)
O grau Brix indica a percentagem de sólidos solúveis no sumo do fruto. Nos mirtilos, este
parâmetro pode ser influenciado por vários fatores nos quais se incluem, a variedade, região de
cultivo, fatores climatéricos e estado de maturação (Türkmen e Eksi, 2011). Cada grau Brix
corresponde a 1g de sólidos solúveis/açúcares por 100 g de sumo. A determinação do grau Brix é
usada como referência de ponto de colheita e consumo para a maioria das frutas, especialmente para as
não-climatéricas.

35
O sumo extraído de 5 bagas de cada cultivar foi utilizado para determinar a concentração de
sólidos solúveis, através de um refratómetro manual, com correção de temperatura, Modelo
EUROMEX – 5532 (0-32%), sendo o resultado expresso em percentagem (grau Brix).
3.7. Avaliação da Capacidade Antioxidante
3.7.1. Determinação da capacidade de quelação de Fe(II)
A ferrozina (3-(2-piridil)-5,6-difenil1,2,4-triazina4’,4’’- sal dissulfónico de sódio) pode formar
complexos de cor vermelha com os iões Fe2+
, [Fe(Ferrozina)3]4-, apresentando estes complexos um
máximo de absorção a 562nm. Na presença de agentes quelantes, a formação desses complexos é
dificultada, resultando numa diminuição da coloração vermelha característica. As medições da
diminuição da coloração, através da medição da diminuição da intensidade da absorção a 562 nm,
permitem efetuar uma estimativa da capacidade de quelação do ferro da amostra (Geckil et al., 2005).
A capacidade de quelação de ferro foi avaliada de acordo com a técnica descrita por Geckil e
colaboradores (2005). Assim, as misturas reacionais foram preparadas misturando diferentes volumes
de amostra, ou de suas diluições, com 50 µL de uma solução FeSO4.7H2O (0,2 mM). Após uma
incubação de cinco minutos à temperatura ambiente adicionaram-se 200µL de ferrozina (5 mM),
ajustou-se o volume a 2000 µL com água, agitou-se vigorosamente a mistura obtida e incubou-se
durante 10 minutos, também à temperatura ambiente. Terminada esta segunda incubação, leu-se a
absorvância a 562 nm no espectrofotómetro (SPEKOL 1500). Realizaram-se igualmente um controlo
negativo, substituindo a amostra por igual volume de água ultrapura (solvente da amostra), e um
controlo de interferência da cor da amostra, utilizando igual volume de amostra e completando a 2000
µL com água ultrapura.
As amostras foram analisadas em quadruplicado tendo a percentagem de inibição da formação
dos complexos [Fe(Ferrozina)3]4-
sido calculada através da seguinte expressão:
%Inibição = [(Abscontrolo negativo)–(Absamostra com ferrozina–Absamostra sem ferrozina)/(Abscontrolo negativo)]x100

36
3.7.2 Determinação da atividade de redução Fe(III) a Fe(II) pelo ensaio FRAP (“Ferric
Reduction Antioxidant Power”)
O ensaio FRAP baseia-se na determinação da capacidade da amostra para reduzir o Fe(III) a
Fe(II), medindo assim a sua capacidade redutora. Quando o Fe(III) do complexo de Fe(III)-
tripiriditriazina (Fe3+
-TPTZ) é reduzido a Fe(II) forma-se, em meio ácido, uma intensa coloração azul
suscetível de ser quantificada espectrofotometricamente a 593 nm e que é proporcional à quantidade
de espécies redutoras (mecanismo antioxidante SET) presentes na amostra (Figura 3.3) (Benzie e
Strain, 1996).
Figura 3.3 - Formação do complexo (Fe2+
-TPTZ) após redução do Fe3+
por um antioxidante
(adaptado de Moon e Shibamoto, 2009).
Para a realização do ensaio FRAP seguiu-se o método descrito por Ramful e colaboradores
(2010) com algumas modificações. O reagente FRAP foi preparado no momento da utilização,
adicionando 25 mL de tampão acetato 0,25 M (pH 3,6), 2,5 mL de TPTZ (10 mM) em HCl (40 mM) e
2,5 mL de cloreto de ferro (III) (FeCl3.6H2O, 20 mM). O ensaio iniciou-se com a adição, num tubo de
ensaio, de 3,0 mL do reagente FRAP, preparado de fresco e pré-aquecido a 37ºC, com 300 µL de água
e 100 µL de cada um dos extratos. As misturas foram incubadas no escuro durante 4 minutos a 37°C.
Durante esta incubação foi possível observar o aparecimento de uma coloração azul, cuja intensidade
variava entre as várias amostras, e que foi quantificada por leitura da absorvância a 593 nm num
espectrofotómetro (SPEKOL 1500) utilizando como branco a absorvância do reagente FRAP.
A diferença entre a absorvância da amostra e a absorvância do branco foi determinada e usada
para calcular o valor de FRAP. As amostras foram analisadas em quadruplicado, tendo a atividade
antioxidante sido determinada por interpolação de uma reta de calibração, preparada da forma já
Complexo Fe2+ -TPTZ,
coloração azul intensa Complexo Fe3+ -TPTZ
Antioxidante
(+ e-)

37
descrita para as amostras, mas substituindo as amostras por soluções de sulfato ferroso com
concentrações entre os 0 e os 1,25 mM. Os resultados foram expressos em mmol de Fe2+
/por 100
grama de peso fresco de mirtilo.
3.7.3. Determinação da redução do Cu(II) pelo ensaio CUPRAC (“Cupric Reducing
Antioxidant Capacity”)
O ensaio CUPRAC consiste na redução do Cu(II) para Cu(I) por ação de redutores
(antioxidantes) presentes numa amostra. Ao contrário da forma oxidada (Cu(II)-neocuproína), a forma
reduzida do complexo Cu-neocuproína (Cu(I)-neocuproína) apresenta uma coloração intensa com um
máximo de absorção a 450 nm (Figura 3.4). Assim, a redução da forma oxidada deste complexo
(Cu(II)-neocuproína) pelos compostos da amostra pode ser determinada espectrofotometricamente
através da leitura da absorvância a 450 nm (Apak et al., 2004).
Figura 3.4 - Redução do complexo Cu(II)–neocuproína a Cu(I)–neocuproína, por ação de uma
molécula antioxidante (HA) originando uma molécula de antioxidante oxidada (A+). Os protões
libertados na reação são neutralizados numa solução tampão de acetato de amónio (Gülçin, 2012).
A realização do ensaio CUPRAC foi efetuada de acordo com o método descrito por Apak e
colaboradores (2004) com algumas adaptações. Assim, num tubo de ensaio juntou-se 1 mL de cada
uma das seguintes soluções: cloreto de cobre (II) bihidratado 10 mM, acetato de amónio 1M,
neocuproína 7,5 mM em etanol. Em seguida, adicionou-se 50 µL de amostra e completou-se a 4100
µL com água. Os tubos foram incubados ao abrigo da luz durante 1 hora à temperatura ambiente
procedendo-se então à leitura da absorvância (espectrofotómetro SPEKOL 1500) a 450 nm, utilizando
como branco a mesma mistura com 1,1 mL de água ultrapura em vez da amostra. As amostras foram
analisadas em quadruplicado tendo a atividade antioxidante sido determinada por interpolação de uma
reta de calibração, preparada da forma já descrita mas substituindo as amostras por 1,1 mL das

38
soluções padrão de ácido ascórbico (15,625; 31,25; 62,5; 125; e 250 µM). Os resultados foram
expressos em mmol equivalentes de ácido ascórbico por 100 grama de peso fresco de mirtilo.
Em relação aos ensaios de redução do ferro, o CUPRAC tem a vantagem de conseguir detetar
antioxidantes do tipo tiol (glutationa, cisteína, etc.). Por outro lado o facto do potencial redox para o
par Cu(II)-neocuproína /Cu(I)-neocuproína ser inferior ao do Fe(III)/Fe(II) com os respetivos ligandos
do FRAP faz com que determinados compostos, tais como açúcares simples ou o ácido cítrico não
possam ser reduzidos no CUPRAC podendo-o ser no FRAP. O método é rápido e não necessita de
aparelhos muito sofisticados, sendo a reação relativamente insensível ao ar, luz, humidade ou pH
(Apak et al., 2004).
3.7.4. Determinação do sequestro do peróxido de hidrogénio
O peróxido de hidrogénio apresenta um máximo de absorção a 230 nm. Desta forma, a
percentagem de sequestro deste composto pode ser determinada pela diminuição da absorvância a 230
nm de uma solução de peróxido de hidrogénio na presença de amostra, por comparação com um
branco (Magalhães et al., 2008; Esmaeili e Sonboli, 2010).
A percentagem de sequestro do peróxido de hidrogénio foi determinada de acordo com o
procedimento descrito por Esmaeili e Sonboli (2010). Assim, em tubos de ensaio, misturaram-se 400
µL de extrato e 4,6 mL de solução 10 mM de H2O2 preparada em tampão fosfato (50 mM, pH 7,4). A
mistura foi incubada, no escuro, durante 30 minutos, a 25ºC. Findo o período de incubação, leu-se a
absorvância das soluções num espectrofotómetro (SPEKOL 1500) a 230 nm, utilizando cuvetes de
quartzo. Para cada uma das amostras foi preparado um branco com o mesmo volume de cada um dos
extratos e 4,6 mL de tampão fosfato (50 mM, pH 7,4), sem peróxido de hidrogénio. Foi ainda
preparado um controlo para o ensaio, com 400 µL de água ultrapura e 4,6 mL da solução de H2O2 em
tampão fosfato (50 mM, pH 7,4). As amostras foram analisadas em quadruplicado tendo a
percentagem de sequestro de H2O2 sido calculada utilizando a seguinte expressão:
% sequestro = [(Abscontrolo – (Absamostra – Absbranco da amostra)/(Abscontrolo)] x 100
O peróxido de hidrogénio forma-se in vivo, no decurso de diversos processos metabólicos,
nomeadamente pela ação de diversas enzimas como a glucose oxidase ou a superóxido dismutase.
Desta forma uma das vantagens deste ensaio é a de se estudar a capacidade de sequestro de um
oxidante com relevância fisiológica.

39
3.7.5. Determinação do sequestro do radical anião superóxido
O azul de nitrotetrazólio (NBT2+
) reage com o radical anião superóxido (O2●-
) dando origem
ao azul de formazano (Figura 3.5) que possui um máximo de absorção a 560 nm. Desta forma, se à
mistura de reação se adicionar um composto com capacidade para captar o O2●-
, passarão a existir, no
meio reacional, dois compostos a competir pelo radical anião superóxido, o que se traduz por uma
diminuição da extensão da redução do NBT2+
, com uma consequente diminuição da taxa de aumento
da absorvância a 560 nm (Valentão et al., 2001; Magalhães et al., 2008).
Figura 3.5 - Redução do NBT2+
(A) pelo radical anião superóxido, dando origem ao azul de
formazano (B).
Neste ensaio o radical anião superóxido é gerado utilizando o sistema metossulfato de
fenazina (PMS)/NADH em condições aeróbias (Figura 3.6) (Nakamura et al., 1992).
NADH + H+ + PMS NAD
+ + PMSH2
PMSH2 + O2 2 O2●-
+ PMS
Figura 3.6 - Formação do radical anião superóxido através do sistema PMS/NADH (Nakamura et al.,
1992).
A capacidade das amostras para sequestrar o radical anião superóxido foi avaliada de acordo
com o método descrito por Valentão e colaboradores (2001) com pequenas alterações. Assim, em
cuvetes do espectrofotómetro adicionaram-se 200 µL de cada uma das amostras. Em seguida
adicionaram-se 300 µL de NADH 1,66 mM em tampão fosfato (19 mM, pH 7,4), 300 µL de NBT2+
430 µM em tampão fosfato (19 mM, pH 7,4) e por fim tampão fosfato (19 mM, pH 7,4) para
completar a 2 950 µL. A reação iniciou-se com a adição de 50 µL de PMS 162 µM em tampão fosfato
(19 mM, pH 7,4) tendo-se acompanhado a variação da absorvância a 560 nm, durante dois minutos à
A B
O2●-

40
temperatura ambiente. A estabilidade do NBT2+
em presença das amostras foi verificada, através da
observação da estabilidade da absorvância a 560 nm antes da adição do PMS, como forma de garantir
que as amostras em si não tinham capacidade de reduzir o NBT2+
a formazano. Realizou-se,
igualmente, um ensaio controlo substituindo a amostra por igual volume de solvente da amostra
(água). As determinações foram realizadas em quadruplicado, no espectrofotómetro (SPEKOL 1500).
A percentagem de inibição da redução do NBT2+
de cada uma das amostras foi calculada em relação
ao controlo utilizando a seguinte expressão:
% Inibição = [(Variaçãoabsorvância do controlo – Variaçãoabsorvância da amostra)/(Variaçãoabsorvância do controlo) x 100]
O radical anião superóxido resulta da redução do oxigénio molecular e pode surgir no decurso
de diversos processos metabólicos. Desta forma, uma das vantagens deste ensaio é a de se utilizar um
oxidante com relevância fisiológica. Este ensaio pode dar resultados erróneos quando a amostra
interfere com o sistema gerador do radical anião superóxido ou quando a amostra reduz diretamente o
NBT2+
, embora esta última situação possa ser despistada pela observação da estabilidade da
absorvância antes da adição do PMS.
3.7.6. Determinação do resgate do radical peroxilo pelo ensaio ORAC (“Oxygen Radical
Absorbance Capacity”)
Este ensaio permite determinar a capacidade da amostra em estudo para sequestrar radicais
peroxilo (ROO•) gerados pela decomposição térmica (a 37 ºC) do 2,2′-Azobis(2-metilpropionamidina)
dihidrocloreto (AAPH) (Figura 3.7), através da determinação da inibição da oxidação da fluoresceína
induzida por estes radicais.
Figura 3.7 - Esquema para a decomposição do AAPH originando radicais peroxilo. Da
termodecomposição do AAPH resulta o radical R• que, ao reagir com O2, dá origem ao radical ROO
•
(Dunlap et al, 2003).

41
O radical peroxilo oxida a fluoresceína originando produtos não fluorescentes (Figura 3.8).
Desta forma a incubação da fluoresceína na presença de uma fonte de radicais peroxilo leva a uma
diminuição imediata da fluorescência. Quando esta incubação se processa na presença de um
antioxidante verifica-se um período de latência antes de se iniciar essa diminuição, sendo que esse
período de latência é tanto maior quanto maior for a capacidade antioxidante da amostra (Ou et al.,
2001).
A realização deste ensaio com diferentes concentrações de Trolox permite a construção de
uma curva de calibração que relaciona a concentração deste antioxidante com a área sob a curva de
decaimento da fluoresceína. Desta forma, sabendo o valor desta área em presença da amostra, é
possível avaliar a capacidade de sequestro do radical peroxilo pela mesma, expressa em equivalentes
de Trolox, por interpolação da respetiva reta de calibração (Ou et al., 2001).
.
Figura 3.8 – Mecanismos propostos para a oxidação da fluoresceína pelo radical peroxilo (Ou et al.,
2001).
Para a realização do ensaio ORAC seguiu-se o método descrito por Ou e colaboradores (2001)
com algumas modificações, tendo-se efetuado todas as determinações em triplicado. As misturas
reacionais compostas por 20 μL das diferentes amostras diluídas em tampão fosfatos (75 mM, pH 7,4),
20 μL de diluições de trolox (entre 0,781 e 50 μM em tampão fosfatos 75 mM, pH 7,4) ou, no caso do
Produto final FL4 (perda de fluorescência)

42
branco, 20 μL de tampão fosfatos (75 mM, pH 7,4), e 160 μL de fluoresceína (0,075 μM em tampão
fosfatos 75 mM, pH 7,4) foram preparadas em microplacas de 96 poços. Após as placas terem sido
pré-incubadas durante cinco minutos a 37 ºC, iniciou-se a reação com a adição a cada poço de 20 μL
de uma solução de AAPH (35,7 g/L em tampão fosfatos 75 mM, pH 7,4) pré-aquecida a 37 ºC. As
placas foram colocadas num leitor de microplacas (Anthos Zenyth 3100) e incubadas a 37 ºC, com
agitação durante 30 minutos, tendo a fluorescência (λ de excitação = 485 nm e λ de emissão = 535 nm)
sido medida em intervalos de 1 minuto.
A área sob a curva (AUC) de decaimento da fluorescência da fluoresceína ao longo do tempo
foi calculada recorrendo à seguinte expressão (Ou et al., 2001):
AUC = 1+f1/f0+f2/f0+f3/f0+………+f30/f0
Sendo f0 a f30 a fluorescência registada no minuto zero (f0) e nos outros minutos até ao minuto 30.
A atividade antioxidante, expressa em μmol de equivalentes de trolox por grama de peso
fresco de mirtilos, foi quantificada por interpolação de uma reta de regressão linear entre os valores
das concentrações de trolox e a área sob a curva de decaimento da fluoresceína obtida com as diversas
concentrações de trolox testadas, após a subtração da área obtida no ensaio em branco.
3.8. Avaliação da atividade mutagénica e antimutagénica através do Teste
de Ames
O teste de Ames ou “bacterial reverse mutagenicity test”, desenvolvido por Bruce Ames em
1975, deteta o potencial mutagénico de substâncias através da indução da mutação reversa no operão
histidina em mutantes de Salmonella typhimurium LT2. Este teste é um dos testes de curto termo mais
utilizados em toxicologia genética para determinar a mutagenicidade de compostos puros ou de
misturas complexas, como, por exemplo, extratos obtidos a partir de alimentos (Maron e Ames, 1983).
O teste de Ames utiliza estirpes de Salmonella typhimurium com diferentes mutações em
vários genes do operão histidina, e que, por isso, se encontram impedidas de sintetizar este aminoácido
(his-). Quando estas estirpes crescem num meio de cultura sem histidina (meio mínimo contendo
apenas sais minerais e uma fonte de carbono e azoto), apenas uma pequena percentagem de células,
característica de cada estirpe, é capaz de reverter espontaneamente a mutação e tornar-se prototrófica
para a histidina (his+), conseguindo, por isso, crescer e formar colónias. A exposição destas estirpes a
agentes mutagénicos conduz a um aumento da taxa de reversão da mutação e, consequentemente, a um
aumento do número de células prototróficas para a histidina. Assim, quando um mutagéneo é

43
adicionado ao meio, o número de revertentes aumenta proporcionalmente à dose de mutagéneo
utilizado (revisto em Mortelmans e Zeiger, 2000).
A realização do teste consiste em adicionar uma quantidade fixa de bactérias (cerca de 108)
com quantidades variáveis do composto a testar e em plaquear estas misturas em meio de cultura
contendo uma quantidade mínima e limitante de histidina (geralmente 0,05 mM). As placas são
incubadas a 37 °C, durante 48 horas, sendo, então, contado o número de colónias de revertentes. A
baixa concentração de histidina no meio permite que as bactérias se dividam um número limitado de
vezes, de modo a que as lesões que tenham ocorrido no DNA possam ser fixadas em mutações.
Durante o crescimento surge na placa uma turvação característica (“background”) visível a olho nu,
que pode funcionar como um indicador de citotoxicidade, uma vez que a sua ausência indica que o
composto em estudo levou a uma morte celular generalizada. Quando a histidina do meio se esgota,
apenas as bactérias que reverteram a mutação, bactérias que passaram a his+, conseguem continuar a
crescer formando colónias isoladas (revisto em Mortelmans e Zeiger, 2000).
Os resultados são expressos em termos de número de revertentes/placa ou número de
revertentes/quantidade de substância testada (revisto em Mortelmans e Zeiger, 2000). Um composto é
considerado positivo no teste de Ames quando se obtém um efeito consistente de dose-resposta, isto é,
quando se observa um aumento do número de revertentes com o aumento da concentração e,
simultaneamente, quando para uma das doses estudadas se observa uma duplicação do número de
revertentes espontâneos. A atividade mutagénica específica pode ser calculada pelo declive da parte
linear da curva de dose-resposta.
3.8.1. Caraterização da estirpe de S. typhimurium utilizada
No presente trabalho utilizou-se a estirpe TA100 de S. typhimurium gentilmente cedida pelo
Departamento de Genética da Faculdade de Ciências Médicas da Universidade Nova de Lisboa. Trata-
se de uma estirpe com uma mutação por substituição de base, na qual o códão CTC da estirpe
selvagem foi substituída por CCC no alelo hisG46. Para além desta mutação no operão da histidina, a
estirpe TA100 exibe outras características genotípicas adicionais que aumentam a sua sensibilidade
para a deteção de mutagéneos. Estas mutações adicionais incluem a mutação rfa, que causa a perda
parcial da barreira lipopolissacárida aumentando a permeabilidade da parede celular bacteriana a
moléculas de grande dimensão; A deleção uvrB, que corresponde a uma deleção do gene que codifica
para uma das enzimas envolvidas no sistema de reparação de DNA por excisão; E a inserção do
plasmídeo pKM101, que confere resistência à ampicilina, e que contém os genes de um sistema de
reparação errónea, o que permite aumentar a frequência de mutação. A deleção uvrB abrange parte do
gene que codifica para a biotina, o que torna esta estirpe incapaz de crescer na ausência desta

44
vitamina. Esta estirpe apresenta um número de revertentes espontâneos que varia entre 150 e 210
(Maron e Ames, 1983; Mortelmans e Zeiger, 2000).
3.8.2. Realização dos testes de Ames
Para avaliar a atividade mutagénica e antimutagénica dos diferentes extratos de mirtilo estudados,
procedeu-se à realização do teste de Ames, na variante de ensaio com pré-incubação, conforme o
descrito por Maron e Ames (1983). Foi preferida esta variante, pois ela permite aumentar a
sensibilidade do teste, tendo em conta que as bactérias ficam expostas aos compostos em estudo
durante algum tempo antes de efetuar a sua diluição na placa de Petri (Maron e Ames, 1983). Todos os
ensaios foram efetuados em triplicado.
A estirpe TA100 foi inoculada em 5 mL de meio NB, incubada durante 12 a 16 horas, a 37 ºC,
com agitação (210 rpm) e ao abrigo da luz, numa incubadora orbital, tendo sido mantida ao abrigo da
luz e à temperatura de 4 °C até à realização do ensaio.
Para a realização dos ensaios de mutagenicidade, os diferentes volumes dos extratos analisados,
entre 0 e 1000 μL, foram misturados com 250 μL de tampão de fosfatos de sódio (0,2 M, pH 7,4) e
100 μL da cultura bacteriana. Para que o volume final em todas as misturas de pré-incubação fosse o
mesmo, este foi sempre ajustado com água estéril até ao volume final de 1350 μL. As misturas assim
preparadas foram incubadas a 37 ºC e 210 rpm, durante 20 minutos numa incubadora orbital. Após a
incubação adicionou-se a cada tubo 2 mL de gelose de superfície (“top-agar”) (Tabela 3.1) fundida a
100 ºC e arrefecida até 45 º C. Esta mistura foi então agitada e plaqueada em placas de Petri com meio
VB (Tabela 3.2). As placas foram incubadas invertidas em estufa a 37 ºC durante 48 horas,
procedendo-se então à contagem manual das colónias de revertentes his+ para cada concentração.
Tabela 3.1 - Composição da gelose de superfícies por litro (Maron e Ames, 1983).
Reagentes Quantidade / Concentração
Agar 6 g
NaCl 5 g
Histidina 0,05 mM
Biotina 0,05 mM
Na preparação da gelose de superfície a histidina e a biotina foram adicionadas após esterilização da
mistura de NaCl e agar, a partir de uma solução estéril e equimolar de histidina e de biotina (0,5 mM).
Após esta suplementação a gelose de superfície foi imediatamente distribuída, em alíquotas de 2 mL, por
tubos de vidro esterilizados.

45
Tabela 3.2 - Composição do meio VB (por litro) (Maron e Ames, 1983).
Reagentes Quantidade (g)
Glucose 20
Agar 15
K2HPO4 10
NaHNH4PO4.4H2O 3,5
Ácido cítrico monohidratado 2
MgSO4.7H2O 0,2
O meio VB resulta da mistura, após esterilização, de soluções de agar (1,5%), glucose (40%) e de uma solução
de sais composta por MgSO4.7H2O (10g/L), ácido cítrico monohidratado (100g/L), K2HPO4 (500g/L) e
NaHNH4PO4.4H2O (175g/L).
Nos ensaios de atividade antimutagénica procedeu-se da mesma forma descrita para os ensaios
de mutagenicidade, com a diferença que as misturas de pré incubação continham, simultaneamente,
um volume fixo (10 µL) de uma solução de t-BHP (10 µg/µL,) preparada de fresco, por diluição em
água estéril, a partir da respetiva solução comercial. Esta quantidade corresponde a uma concentração
final de 1,11 µmol de t-BHP/placa. A atividade antimutagénica, expressa em percentagem de inibição
da mutagenicidade, foi calculada através da seguinte expressão (Yen e Chen, 1995):
%Inibição = [(RI-RE)-(RIM-RE)] x 100/(RI-RE)
Sendo RI o número de revertentes induzidos pelo mutagéneo, RE o número de revertentes
espontâneos e RIM o número de revertentes induzidos pelo mutagéneo na presença dos extratos em
estudo.
3.9. Análise estatística dos resultados
O tratamento estatístico dos dados foi efetuado utilizando o software “Microsoft Office Exel 2007”
(Microsoft Corporation, Washington). Nesse sentido foram realizados o teste t, regressões lineares,
análises de correlação e análise ANOVA.

46
4. Resultados e Discussão
A capacidade antioxidante é influenciada pelo solvente e pela técnica de extração utilizada,
bem como pelo binómio tempo-temperatura. No que se refere aos solventes orgânicos, o metanol, por
conseguir extrair elevada quantidade de compostos bioativos, tem sido apontado como o mais eficaz
(Naczk e Shahidi, 2004). Assim, neste trabalho, foi utilizada uma solução de metanol acidificada como
solvente extrator. No entanto, tendo em conta que era também objetivo do estudo avaliar a atividade
antimutagénica dos extratos, através da realização do teste de Ames, foi necessário proceder à
evaporação do solvente extrator, para posteriormente se ressuspender o resíduo em água ultrapura e
esterilizar o extrato aquoso por filtração, em câmara de fluxo laminar, usando membranas estéreis com
poro de 0.22 μm.
4.1. Determinação dos fenóis totais
A composição em compostos fenólicos das amostras de mirtilo, preparadas de acordo com o
método descrito anteriormente, é apresentada na tabela 4.1.
Tabela 4.1 - Teor de compostos fenólicos totais nas amostras testadas
Amostras Compostos fenólicos totais
(mg eq. ác. gálico/100g de mirtilo)
Patriot 153,4a ± 3,0
Bluecrop 155,7a ± 1,7
Duke 201,1b ± 2,3
Earlyblue 161,0c ± 3,4
O’Neal 147,2d ± 1,5
Médias com letras diferentes são significativamente diferentes de acordo com o teste t (p<0,05).
Os teores em compostos fenólicos variaram entre 147,2 e 201,1 mg equivalentes de ácido
gálico por 100 g de mirtilo. A variedade que apresentou menor teor em compostos fenólicos foi a
O’Neal, seguindo-se as variedades Patriot e Bluecrop, que apresentaram valores semelhantes de
acordo com o teste t (p<0,05), de seguida foi a variedade Earlyblue e finalmente, com maior teor em
compostos fenólico totais, a variedade Duke. Todas as amostras apresentaram valores que foram
significativamente diferentes de acordo com o teste t, excetuando as amostras das variedades Patriot e

47
Bluecrop. O fato da variedade O’Neal ter sido aquela com mais baixo teor em polifenóis pode estar
relacionado com esta ser a única pertencente ao grupo Southern Higbush mais adaptada às condições
climatográficas da região sul de Portugal.
Os valores obtidos estão de acordo com os valores apresentados por Koca e Karadeniz (2009) para
mirtilos de outras variedades, cultivados na região do mar negro, na Turquia, e com os valores
reportados por Ehlenfeldt e Prior (2001) para as mesmas variedades de mirtilos, cultivadas em Nova
Jersey, nos Estados Unidos da América. Neste último estudo os valores variaram entre 139 para a
variedade O’Neal e 216 mg equivalentes de ácido gálico/100 g de peso fresco, para a variedade Duke.
Outros autores descrevem teores de fenóis totais superiores aos obtidos neste trabalho, chegando a
ser superiores a 300 mg ácido gálico/100 g de fruto, em cultivares de V. corymbosum, e superiores a
500 mg ácido gálico/100 g de fruto em mirtilos selvagens (Prior et al., 1998; Ehlenfeldt e Prior, 2001;
Giovanelli e Buratti, 2009; Koca e Karadeniz, 2009; Bunea et al., 2011; You et al., 2011). Esta
variação dos resultados pode ser justificada com a variação das espécies e variedades analisadas, graus
de maturação, tipo de solo, condições climáticas e práticas de cultivo (Moyer et al., 2002). Prior e
colaboradores (1998), verificaram que com o teor polifenóis totais dos mirtilos aumentava entre 113%
e 169% com o aumento do grau de maturação da baga.
4.2. Quantificação das antocianinas monoméricas totais
Os resultados do teor das antocianinas monoméricas totais, expressos em mg de equivalente de
cianidina por 100 g de mirtilo, encontram-se na tabela 4.2.
Tabela 4.2 – Teor de antocianinas monoméricas nas amostras testadas
Amostras Antocianinas monoméricas
(mg eq. cianidina/100 g de mirtilo)
Patriot 53,28a ± 0,58
Bluecrop 46,49b ± 0,45
Duke 52,49a ± 0,14
Earlyblue 46,96b± 0,37
O’Neal 46,14b± 0,70
Médias com letras diferentes são significativamente diferentes de acordo com o teste t (p<0,05).

48
Pela análise dos resultados apresentados na tabela 4.2, pode constatar-se que o teor em
antocianinas monoméricas das amostras de mirtilos das variedades Bluecrop, Earlyblue e O’Neal são
significativamente semelhantes de acordo com o teste t (p<0,05), sendo as variedades Patriot e Duke
significativamente diferentes das anteriores, mas não significativamente diferentes entre si de acordo
com o teste t (p<0,05). Os resultados mostram que as variedades com maior teor em antocianinas
monoméricas são a Patriot e a Duke com cerca de 53 mg equivalentes de cianidina por 100 g de
mirtilo, seguindo-se as restantes variedades com aproximadamente 46 mg equivalentes de cianidina
por 100 g de mirtilo.
Efetuando a razão entre as antocianinas (expressas em mg eq. cianidina/100 g de mirtilo) e os
compostos fenólicos (expressos em mg eq. ác. gálico/100g de mirtilo) totais verifica-se que as
antocianinas representam entre 26% (Duke) a 35% (Patriot) da concentração de compostos fenólicos
presentes nas variedades avaliadas. Os níveis de antocianinas em compostos de fenóis totais estão de
acordo com os resultados obtidos entre 23% e 37% no estudo efetuado por Bunea e os seus
colaboradores (2011) e por Dragovic-Uzelac e seus colaboradores (2010). Esta análise aponta no
sentido das antocianinas contribuírem mais para o total de compostos fenólicos na variedade Patroit,
seguido das variedades O’Neal, Bluecrop e Earlyblue e terem o menor peso na variedade Duke.
Os valores de antocianinas obtidos estão de acordo com os valores obtidos, por outros autores,
para as variedades Earlyblue e Bluecrop cultivadas em Nova Jersey, nos Estados Unidos da América
(Ehlenfeldt e Prior, 2001), ou para a variedade Duke cultivada na Roménia (Bunea et al. 2011), sendo,
no entanto, os valores obtidos para as restantes variedades analisadas inferiores aos valores reportados
por esses mesmos autores. Procurando na literatura verifica-se que o teor em antocianinas apresenta
uma grande variação entre as várias variedades de mirtilos colhidas em diferentes zonas e em
diferentes condições. Assim este valor varia entre 12,7 e 235,4 mg/100 g fruto nas variedades de V.
corymbosum (Prior et al., 1998; Sellappan et al., 2002; Giovanelli e Buratti, 2009; Bunea et al., 2011),
atingindo valores bastante superiores (cerca de 350 mg eq. cianidina/100 g de mirtilo) no caso dos
mirtilos selvagens, em que a própria polpa tem uma coloração avermelhada (Giovanelli e Buratti,
2009; Bunea et al., 2011). Tal como já anteriormente referido em relação às variações encontradas no
teor em compostos fenólicos totais também a variação do teor em antocianinas pode dever-se às
diferenças genéticas das cultivares, a fatores climáticos e ambientais, tais como, a temperatura, a
radiação UV, o tipo de solo ou as técnicas agrícolas aplicadas (Koca e Karadeniz, 2009).

49
4.3. Determinação do teor de sólidos solúveis (grau Brix)
Os resultados do teor de sólidos solúveis, expressos em percentagem, obtidos nas diferentes
amostras analisadas encontram-se na tabela 4.3.
Tabela 4.3 – Teor de sólidos solúveis (grau Brix) nas amostras testadas
Amostras Grau Brix
(%)
Patriot 10,9a ± 0,7
Bluecrop 10,4a ± 0,6
Duke 11,4a,b,c ± 1,0
Earlyblue 11,9b ± 0,4
O’Neal 12,2b,c ± 0,6
Médias com letras diferentes são significativamente diferentes de acordo com o teste t (p<0,05)
Os valores do teor de sólidos solúveis variaram entre 10,4% para a variedade Bluecrop e 12,2%
para a variedade O’Neal. De acordo com o teste t (p<0,05), a variedade Duke não difere
significativamente das outras amostras e as amostras das variedades Patriot e Bluecrop diferem
significativamente das amostras das variedades Earlyblue e O’Neal. Os teores em sólidos solúveis
obtidos encontram-se na gama de valores obtidos por outros autores (entre 10 e 19%) para diversas
cultivares de V. corymbosum na altura da plena maturação (Prior et al., 1998; Saftner et al., 2008;
Giovanelli e Buratti, 2009).
4.4. Determinação da atividade antioxidante das diferentes variedades de
mirtilo
Para a caraterização da atividade antioxidante das diferentes variedades de mirtilo em estudo
procedeu-se à avaliação da sua capacidade redutora com os ensaios FRAP, CUPRAC, à avaliação da
sua capacidade de quelação de metais (capacidade de quelação do ferro (II)), e, por fim, à avaliação da
sua capacidade de sequestro de espécies reativas de oxigénio, nomeadamente do radical anião
superóxido, do peróxido de hidrogénio e do radical peroxilo (ensaio ORAC).
4.4.1. Determinação da atividade antioxidante por capacidade redutora da amostra
Os ensaios FRAP e CUPRAC são ensaios que empregam mecanismos de transferência de eletrões
(SET). O CUPRAC é um ensaio que se baseia na redução de iões complexos Cu2+
para Cu+ por ação

50
de redutores (antioxidantes) presentes numa amostra. O FRAP é um ensaio que se baseia na redução
de iões Fe(III) a Fe(II), medindo assim a capacidade redutora da amostra.
4.4.1.1. Determinação da atividade de redução do Fe(III) a Fe(II) pelo ensaio FRAP
Os resultados do ensaio FRAP, expressos em mmol de Fe
2+ por 100 g de mirtilo, para as várias
amostras estudadas encontram-se na tabela 4.4.
Tabela 4.4 – Valores do ensaio FRAP nas diferentes amostras
Amostras Valores de FRAP
(mmol Fe2+
/100 g de mirtilo) Patriot 1,620a ± 0,042
Bluecrop 1,287b ± 0,012
Duke 1,719c ± 0,046
Earlyblue 1,277b ± 0,040
O’Neal 1,167d ± 0,022
Médias com letras diferentes são significativamente diferentes de acordo com o teste t (p < 0,05)
Os valores de FRAP obtidos para as variedades analisadas são significativamente diferentes de
acordo com o teste t (p<0,05), com exceção das variedades Bluecrop e Earlyblue que são
significativamente semelhantes. Em relação à variedade Earlyblue foi possível observar que o valor de
FRAP encontrado foi aproximadamente o dobro do valor encontrado para a mesma variedade quando
cultivada no estado brasileiro do Rio Grande do Sul (0,699 mmol Fe2+
/100 g) (Rodrigues et al., 2011).
Esta diferença pode refletir as diferenças de clima, solo e técnica de cultivo verificadas entre a zona de
Sever do Vouga e o estado do Rio Grande do Sul.
Os valores obtidos variaram entre 1,167 mmol Fe2+
/100 g de mirtilo para a variedade O’Neal e
1,719 mmol Fe2+
/100 g de mirtilo para a variedade Duke. Assim, nas variedades em estudo, o maior
valor de FRAP foi encontrado na variedade Duke, que foi aquela que apresentou maior valor em
compostos fenólicos, e o menor na variedade O’Neal, que foi a que apresentou menor teor em fenóis.
Esta tendência está de acordo com os resultados publicados por outros autores, que reportam que os
valores de poder redutor vão aumentando com concentrações crescentes de compostos fenólicos nos
extratos de mirtilo (Dragovic-Uzelac et al, 2010).
Analisando a literatura verifica-se a existência de uma grande amplitude nos valores de FRAP
reportados, variando estes entre os 0,699 e o 2,64 mmol Fe2+
/100 g (Koca e Karadeniz, 2009;

51
Giovaneli e Buratti, 2009; Rodrigues et al., 2011) para diversas variedades de mirtilos cultivados,
apresentando os mirtilos selvagens valores substancialmente mais elevados (entre 3,4 e 5,7) (Giovaneli
e Buratti, 2009; Koca e Karadeniz, 2009). Assim, os valores de FRAP encontrados para as variedades
de mirtilos em estudo encontram-se dentro da gama de valores obtida por outros autores para os
mirtilos cultivados.
4.4.1.2. Determinação da atividade de redução do Cu(II) a Cu(I) pelo ensaio CUPRAC
A tabela 4.5 mostra os resultados obtidos na determinação da redução do Cu2+
pelo ensaio de
CUPRAC, expressos em mmol equivalentes de ácido ascórbico por 100 g de mirtilo.
Tabela 4.5 – Valores do ensaio CUPRAC nas diferentes amostras
Amostras Valores de CUPRAC
(mmol eq. de ácido ascórbico/ 100g de mirtilo)
Patriot 1,082a ± 0,029
Bluecrop 1,094a ± 0,015
Duke 1,384b ± 0,021
Earlyblue 1,110a ± 0,033
O’Neal 1,114a ± 0,014
Médias com letras diferentes são significativamente diferentes de acordo com o teste t (p<0,05).
A observação dos resultados obtidos permite concluir que os valores das diferentes variedades
são significativamente semelhantes de acordo com o teste t (p<0,05) à exceção da variedade Duke que
apresenta um valor significativamente diferente. O valor do CUPRAC para a maioria das variedades
analisadas é de aproximadamente 1,1 mmol eq. de ácido ascórbico/ 100 g de mirtilo e para a variedade
Duke é de 1,384 mmol eq. de ácido ascórbico/100 g de mirtilo. Não foi possível encontrar qualquer
estudo que reportasse valores de CUPRAC (utilizando o ácido ascórbico como padrão) relativamente a
amostras de mirtilo, de forma a serem comparados com os obtidos neste ensaio.
Analisando de forma conjunta os valores obtidos, para as diferentes variedades de mirtilo em
estudo, nos ensaios FRAP e CUPRAC é possível verificar que é a variedade Duke a que apresenta os
valores mais elevados em ambos os ensaios.

52
4.4.2. Determinação da capacidade de quelação de ferro
A capacidade de quelação do ferro pode ser encarada como um mecanismo de atividade
antioxidante (Geckil et al., 2005), uma vez que, nas reações de Fenton, os iões Fe2+
podem gerar
espécies reativas de oxigénio ao reagirem com o peróxido de hidrogénio ou com hidroperóxidos
lipídicos.
Pela análise dos ensaios dos vários extratos de mirtilo não foi possível detetar nenhuma
quelação do Fe(II), uma vez que não se verificou inibição da formação dos complexos entre o Fe(II) e
a ferrozina em nenhuma das amostras testadas até uma concentração de 50 mg/mL. Este resultado
indica que os compostos fenólicos presentes nos extratos de mirtilos, ou não têm capacidade de quelar
o Fe(II) ou têm uma afinidade para com este ião inferior à da ferrozina.
Conforme a estrutura dos compostos fenólicos, estes apresentam maior ou menor afinidade
química para com os catiões dos metais de transição, tendo, assim, capacidade para os complexar,
tornando-os indisponíveis para participar nas reações de Fenton. Esta caraterística dos polifenóis faz
com que extratos preparados a partir de alimentos com elevada concentração destes compostos
possam, igualmente, apresentar capacidade de quelação de iões metálicos. Com efeito, extratos de
plantas aromáticas (Dorman et al., 2003 e 2004), de própolis (Geckil et al., 2005) ou de bagas de
zimbro (Micelli et al., 2009) têm mostrado capacidade de quelação de Fe(II). No entanto esta mesma
caraterística de quelação de metais, pode, igualmente, acarretar um lado menos positivo quando
contribui para a redução da absorção digestiva de diversos elementos minerais essenciais (Ferguson,
2001).
4.4.3. Determinação da atividade antioxidante por sequestro de espécies reativas de
oxigénio
A avaliação da capacidade de sequestro de espécies reativas de oxigénio foi realizada
recorrendo a ensaios que detestam a capacidade das amostras para efetuar o sequestro de três
importantes espécies reativas de oxigénio: o peróxido de hidrogénio, o radical anião superóxido e o
radical peroxilo (ensaio ORAC). O facto de todas estas espécies reativas de oxigénio se poderem
formar in vivo confere, a todos estes ensaios, importante relevância fisiológica.

53
4.4.3.1. Determinação da capacidade de sequestro do peróxido de hidrogénio por
medição da absorção no UV
A tabela 4.6 mostra os resultados obtidos na avaliação da capacidade que as diferentes
variedades de mirtilo apresentaram para sequestrar o peróxido de hidrogénio.
Tabela 4.6 – Percentagem de sequestro do H2O2 nas diferentes variedades de mirtilo
Amostras Valores da percentagem de sequestro de H2O2
(% de sequestro do H2O2) Patriot 18,1a,b± 1,0
Bluecrop 19,3b,c ± 1,0
Duke 17,7a ± 0,5
Earlyblue 19,5c ± 0,8
O’Neal 22,4d ± 0,8
Médias com letras diferentes são significativamente diferentes de acordo com o teste t (p<0,05)
As percentagens de sequestro variam entre 17,7%, para a variedade Duke, e 22,4%, para a
O’Neal. Assim, os valores obtidos neste ensaio mostram que a variedade O’Neal é a que apresenta a
maior percentagem de sequestro do peróxido de hidrogénio com 22,4%, valor que é significativamente
diferente das outras variedades, de acordo com o teste t (p<0,05).
4.4.3.2. Determinação da capacidade de sequestro do radical anião superóxido detetada
pela diminuição da formação de formazano
A tabela 4.7 mostra os valores das percentagens de sequestro do radical anião superóxido nas
diferentes variedades de mirtilo em estudo.
Tabela 4.7 – Valores de inibição do superóxido obtidos nas diferentes variedades de mirtilo.
Amostras %
sequestro do anião superóxido
Patriot 93,3a ± 1,9
Bluecrop 93,8a ± 1,1
Duke 93,9a ± 2,4
Earlyblue 93,6a ± 0,0
O’Neal 93,6a ± 0,0
Médias com letras diferentes são significativamente diferentes de acordo com o teste t (p<0,05)

54
Os valores das percentagens de inibição são muito próximos, variando entre 92,99% e 93,88%
de inibição. Não se verificaram, nas condições experimentais utilizadas, diferenças significativas entre
essas percentagens diferenças significativas de acordo com o teste t (p<0,05), entre nenhuma das
amostras, sendo as percentagens de sequestro do radical anião superóxido muito elevadas. Este fato
pode ter-se devido à utilização de uma quantidade de amostra demasiado elevada para permitir detetar
diferenças entre as amostras.
4.4.3.3. Determinação da atividade antioxidante através da análise da capacidade de
resgate do radical peroxilo pelo ensaio ORAC
A tabela 4.8 mostra os valores obtidos na atividade antioxidante por sequestro do radical
peroxilo, expressa em mol de equivalente de trolox por g de mirtilo, nas diferentes variedades
estudadas. Os valores obtidos mostram que todas as amostras apresentaram capacidade de inibir a
oxidação da fluoresceína induzida pelo radical peroxilo, ou seja, em todas as amostras existem
compostos capazes de doar um átomo de hidrogénio ao radical promovendo a sua desativação.
Tabela 4.8 - Valores obtidos nas diferentes variedades de mirtilo no ensaio ORAC.
Amostras ORAC
(μmol eq trolox/g de mirtilo)
Patriot 13,51a,b ± 0,87
Bluecrop 12,74b ± 0,30
Duke 15,51a± 0,17
Earlyblue 14,52a ± 0,78
O’Neal 12,81a,b ± 1,16
Médias com letras diferentes são significativamente diferentes de acordo com o teste t (p<0,05).
Os valores de ORAC obtidos variaram entre os 12,74 μmol eq trolox/g de mirtilo, para a
variedade Bluecrop, e 15,51 μmol eq trolox/g de mirtilo para a variedade Duke. De um modo geral os
valores de ORAC obtidos aproximaram-se bastante dos valores reportados por Ehlenfeldt e Prior
(2001) para as mesmas variedades de mirtilos cultivadas em Nova Jersey nos Estados Unidos da
América. Contudo, esta comparação pode estar afetada de algum erro uma vez que no ensaio realizado
por Ehlenfeldt e Prior (2001) o composto fluorescente utilizado foi a R-ficoeritrina enquanto que no
presente ensaio foi a fluoresceína. Outros autores, no entanto, reportam valores mais elevados para as
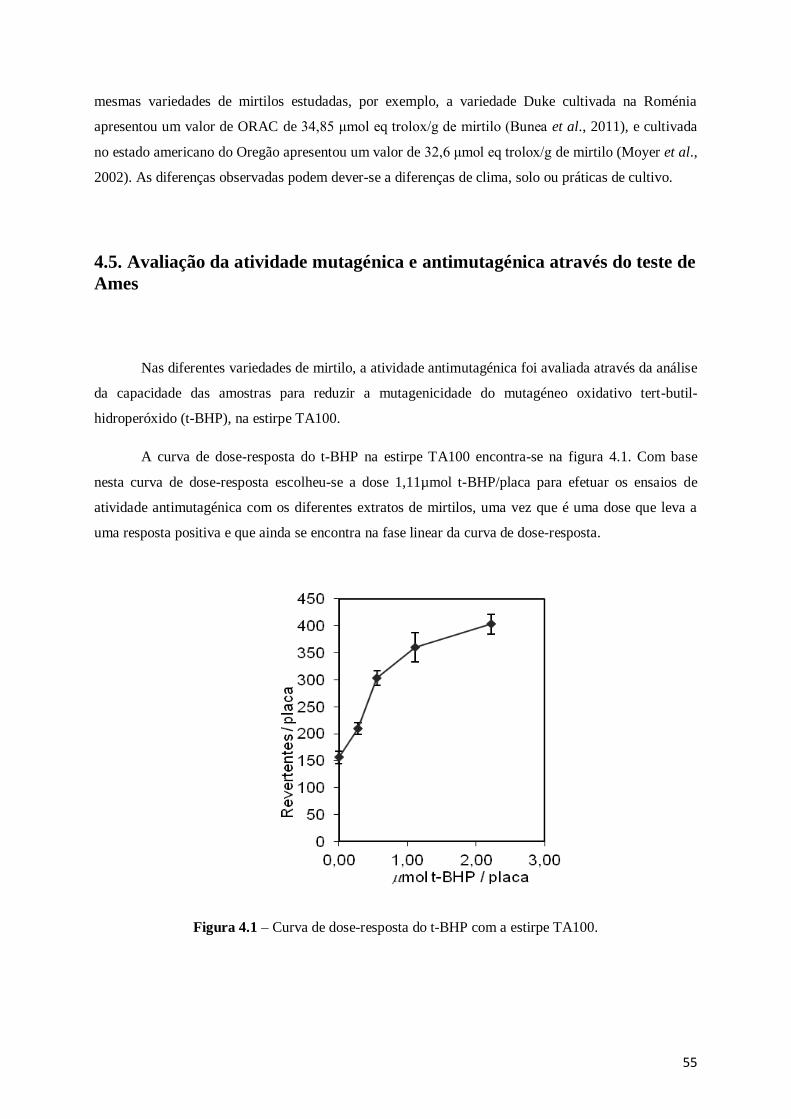
55
mesmas variedades de mirtilos estudadas, por exemplo, a variedade Duke cultivada na Roménia
apresentou um valor de ORAC de 34,85 μmol eq trolox/g de mirtilo (Bunea et al., 2011), e cultivada
no estado americano do Oregão apresentou um valor de 32,6 μmol eq trolox/g de mirtilo (Moyer et al.,
2002). As diferenças observadas podem dever-se a diferenças de clima, solo ou práticas de cultivo.
4.5. Avaliação da atividade mutagénica e antimutagénica através do teste de
Ames
Nas diferentes variedades de mirtilo, a atividade antimutagénica foi avaliada através da análise
da capacidade das amostras para reduzir a mutagenicidade do mutagéneo oxidativo tert-butil-
hidroperóxido (t-BHP), na estirpe TA100.
A curva de dose-resposta do t-BHP na estirpe TA100 encontra-se na figura 4.1. Com base
nesta curva de dose-resposta escolheu-se a dose 1,11µmol t-BHP/placa para efetuar os ensaios de
atividade antimutagénica com os diferentes extratos de mirtilos, uma vez que é uma dose que leva a
uma resposta positiva e que ainda se encontra na fase linear da curva de dose-resposta.
Figura 4.1 – Curva de dose-resposta do t-BHP com a estirpe TA100.

56
A figura 4.2 apresenta as curvas dose-resposta das diferentes variedades de mirtilo Patriot,
Bluecrop, Duke, Earlyblue e O’Neal na presença e na ausência do mutagéneo t-BHP, na estirpe
TA100. Observando a figura 4.2 (A a E), os extratos das diferentes variedades de mirtilo, quando
testadas sem o mutagéneo t-BHP (curvas a cinzento), não apresentam mutagenicidade para a estirpe
TA100, uma vez que não se verificou uma duplicação do número de revertentes espontâneos em
nenhuma das doses ensaiadas. A figura 4.2 permite verificar que todos os extratos das diferentes
variedades causaram uma diminuição no número de revertentes induzidos pelo t-BHP e que essa
diminuição foi consistente com o aumento da quantidade de amostra (curvas a preto nos gráficos A, B,
C, D e E).
Em todas as amostras, o facto do número de revertentes nunca ter descido para valores
inferiores aos revertentes espontâneos, em paralelo com o declive pouco acentuado das curvas de
atividade antimutagénica e com a existência em algumas das amostras de um patamar no final dessas
curvas, prova a ideia de que a diminuição de número de revertentes por placa é resultado da
diminuição do número de revertentes induzidos pelo t-BHP, não sendo, por isso, resultado da morte
das bactérias devida a fenómenos de toxicidade aguda.

57
A B C
D E
Figura 4.2 - Curvas de dose-resposta dos extratos de mirtilo das diferentes variedades (A) Patriot, (B)
Bluecrop, (C) Duke, (D) Earlyblue (E) O’Neal na presença ( ) e na ausência ( ) de
t-BHP, na estirpe TA100.
O cálculo da percentagem de máxima de inibição da atividade do mutagéneo t-BHP, para as
cinco variedades de mirtilo (Figura 4.3) que todas as variedades apresentaram atividade
antimutagénica moderada, uma vez que as percentagens de inibição nunca chegaram aos 50%. Não se
verificaram diferenças significativas entre as percentagens de inibição obtidas com as diferentes
variedades de mirtilo do estudo.

58
Amostras com letras diferentes significam percentagens de inibição significativamente diferentes de acordo com o teste t (p<0.05).
Figura 4.3 - Percentagem máxima de inibição da mutagenicidade do t-BHP exercida pelos extratos
das variedades de mirtilo Patriot, Bluecrop, Duke, Earlyblue e O’Neal.
4.6. Análise global dos resultados
Com o objetivo de tentar identificar classes de compostos presentes nas variedades de mirtilo
que possam ser responsáveis pelas atividades antioxidantes identificadas, bem como de estabelecer
relações entre diversos constituintes das amostras, foram efetuadas várias correlações lineares. Assim,
foram determinadas as equações das retas que melhor se ajustavam aos vários pontos experimentais
obtidos e foi determinado o respetivo coeficiente de correlação linear de Pearson (r). Este coeficiente
mede a intensidade da associação linear existente entre as variáveis e pode tomar valores entre -1 e 1
(Tabela 4.9).

59
Tabela 4.9 - Classificação da correlação em função do valor do coeficiente de correlação de Pearson
(Santos, 2007).
Coeficiente de Pearson (r) Tipo de correlação
r =1 Perfeitamente positiva
0,8 ≤ r < 1 Fortemente positiva
0,5 ≤ r < 0,8 Moderadamente positiva
0,1 ≤ r < 0,5 Fracamente positiva
0 < r < 0,1 Infimamente positiva
r = 0 Nula
-0,1 < r < 0 Infimamente negativa
-0,5 < r ≤ -0,1 Fracamente negativa
-0,8 < r ≤ -0,5 Moderadamente negativa
-1 < r ≤ -0,8 Fortemente negativa
r = -1 Perfeitamente negativa
Na tabela 4.10 apresentam-se os valores do coeficiente de Pearson para as correlações entre o
teor em fenóis totais, o teor em antocianinas monoméricas totais e grau Brix para as cinco variedades
de mirtilos analisadas (Patriot, Bluecrop, Duke, Earlyblue e O’Neal).
Tabela 4.10 - Coeficiente de Pearson para as correlações entre a composição em fenóis totais e
antocianinas monoméricas das cinco amostras estudadas.
a) mg eq ác.gálico/100 g de mirtilo; b) mg eq. cianidina /100g de mirtilo; c) %.
Analisando os valores do coeficiente de Pearson para as correlações entre o teor em fenóis
totais, o teor em antocianinas monoméricas e grau Brix para as cinco variedades de mirtilos estudadas
(Patriot, Bluecrop, Duke, Earlyblue e O’Neal) (Tabela 4.10), verifica-se a existência de uma
correlação moderadamente positiva (r = 0,5285) entre o teor em fenóis totais e o teor de antocianinas
monoméricas, enquanto que o grau Brix parece variar de uma forma independente, uma vez que não se
verifica a existência de associações significativas entre estes dois parâmetros e o valor do grau Brix.
Analisando os resultados individualmente verificou-se que uma das variedades, a variedade Patriot,
Fenóisa
Antocianinas monoméricasb Brix
c
Fenóisa 1
Antocianinas monoméricasb 0,5285 1
Brixc 0,0224 -0,2857 1

60
apresentava um comportamento diferente das restantes uma vez que apesar de ter sido uma das
variedades com menor teor em fenóis totais apresentou um teor em antocianinas monoméricas mais
elevado. Assim, efetuou-se igualmente a análise de correlação excluindo esta variedade (Tabela 4.11).
Tabela 4.11 - Coeficiente de Pearson para as correlações entre a composição em fenóis totais e
antocianinas monoméricas nas amostras das variedades Bluecrop, Duke, Earlyblue e O’Neal.
a) mg eq ác.gálico/100 g de mirtilo; b) mg eq. cianidina /100g de mirtilo; c) %.
Quando se exclui a variedade Patriot, verifica-se que a correlação entre o teor em fenóis totais
e antocianinas monoméricas totais altera-se para fortemente positiva, continuando, no entanto, a não se
verificar nenhuma associação entre estes dois parâmetros e o valor do grau Brix (Tabela 4.11). Desta
forma, os resultados sugerem que, com exceção da variedade Patriot, quanto maior o teor em fenóis
das amostras maior o seu teor em antocianinas monoméricas. A variedade Patroit contraria esta
correlação, uma vez que foi a que apresentou o teor mais elevado em antocianinas monoméricas
(Tabela 4.2), ao mesmo tempo que foi uma das que apresentou um menor teor em fenóis totais (Tabela
4.1). Desta forma, as antocianinas monoméricas parecem ter um maior peso nos fenóis totais na
variedade Patriot do que nas restantes variedades.
Na tabela 4.12 e 4.13 apresentam-se os valores de coeficiente de Pearson para as correlações
entre o teor em fenóis, antocianinas e grau Brix e a atividade antioxidante para as cinco variedades de
mirtilos estudadas (Tabela 4.12) e apenas para as variedades Bluecrop, Duke, Earlyblue e O’Neal
(Tabela 4.13).
Fenóisa
Antocianinas monoméricasb Brix
c
Fenóisa 1
Antocianinas monoméricasb 0,9912 1
Brixc -0,1292 -0,0710 1

61
Tabela 4.12 - Coeficiente de Pearson para as correlações entre composição química e atividade
antioxidante das cinco variedades em estudo.
Fenóisa
Antocianinas
monoméricasb
Brixc FRAP
d CUPRAC
e Sequestro
do H2O2f
Sequestro do
superóxidof
ORACg
Fenóisa 1
Antocianinas
monoméricasb 0,5285 1
Brixc -0,0224 -0,2857 1
FRAPd
0,7131 0,9644 -0,3447 1
CUPRACe 0,9660 0,4741 0,1142 0,6420 1
Sequestro do
H2O2f -0,6254 -0,7750 0,5943 -0,8616 -0,4416 1
Sequestro do
superóxidof 0,6423 -0,2335 -0,0625 0,0194 0,6639 -0,0824 1
ORACg 0,8805 0,5259 0,2242 0,6505 0,8015 -0,6083 0,3519 1
a) mg eq ác.gálico/100 g de mirtilo; b) mg eq. cianidina /100 g mirtilo; c) %; d) mmol Fe2+
/100 g de mirtilo; e) mmol eq. de ácido
ascórbico/ 100g de mirtilo; f) %de inibição; g) mol eq. Trolox/g.
Tabela 4.13 - Coeficiente de Pearson para as correlações entre composição química e atividade
antioxidante das variedades Bluecrop, Duke, Earlyblue e O’Neal.
Fenóisa
Antocianinas
monoméricasb
Brixc FRAP
d CUPRAC
e Sequestro
do H2O2f
Sequestro do
superóxidof
ORACg
Fenóisa
1
Antocianinas
monoméricasb 0,9912 1
Brixc -0,1292 -0,0710 1
FRAPd 0,9942 0,9881 -0,2143 1
CUPRACe 0,9648 0,9911 -0,0014 0,9631 1
Sequestro do
H2O2f -0,8252 -0,7493 0,5295 -0,8339 -0,6573 1
Sequestro do
superóxidof 0,7753 0,7670 -0,6689 0,8372 0,7380 -0,7864 1
ORACg 0,8829 0,8500 0,1869 0,8289 0,8065 -0,7318 0,4157 1
a)mg eq ác.gálico/100 g de mirtilo; b) mg eq. cianidina /100 g mirtilo; c) %; d) mmol Fe2+
/100 g de mirtilo; e) mmol eq. de ácido
ascórbico/ 100g de mirtilo; f) %de inibição; g) mol eq. Trolox/g.
A análise das Tabelas 4.12 e 4.13 mostra a inexistência de associações significativas entre o
grau Brix e os ensaio FRAP e CUPRAC e a existência de correlações fortemente positivas entre o teor
em fenóis totais e a atividade antioxidante determinada no ensaio CUPRAC e entre o teor em
antocianinas e a atividade antioxidante determinada pelo ensaio FRAP, verificando-se a existência de
correlações moderadamente positivas entre o teor em fenóis totais e o ensaio FRAP e fracamente
positivas entre o teor em antocianinas e o ensaio CUPRAC. Contudo, quando se elimina a amostra
Patriot, que, conforme anteriormente referido, parece ser diferente de todas as restantes amostras, as

62
correlações entre fenóis, antocianinas e estes dois ensaios de atividade antioxidante passam a ser todas
fortemente positivas.
Assim, estes resultados indicam que as variedades com maior teor de fenóis totais, e, em
particular, em antocianinas, foram as que apresentaram maiores valores de FRAP e de CUPRAC,
indicando que os polifenóis contribuem significativamente para a atividade antioxidante detetada por
estes ensaios, ou seja, os resultados obtidos suportam a hipótese dos polifenóis e, em particular, as
antocianinas, estarem envolvidos nos mecanismos de atividade antioxidante por transferência
eletrónica. No entanto, esta relação entre o teor em antocianinas e o ensaio CUPRAC não se verifica
com a variedade Patriot que, apesar de ser das variedades estudadas a que apresentou um teor mais
elevado em antocianinas foi a variedades que apresentou o menor valor de CUPRAC.
A existência de uma correlação fortemente positiva entre os ensaios FRAP e CUPRAC (r =
0,9631) também aponta no sentido dos compostos responsáveis por estas duas atividades serem, na sua
maioria, os mesmos. A existência de correlações fortemente positivas entre o ensaio FRAP e os teores
em fenóis e em antocianinas foi igualmente verificada por outros autores (Giovaneli e Buratti, 2009;
Koca e Karadeniz, 2009; Bunea et al., 2011). No estudo de Bunea e colaboradores (2011) esta
correlação foi encontrada em várias variedades de mirtilo cultivados e selvagens da Roménia, entre as
quais se encontravam as variedades Duke e Bluecrop igualmente estudadas no presente trabalho.
Em relação aos ensaios de sequestro das espécies reativas de oxigénio (peróxido de
hidrogénio, radical anião superóxido e radical peroxilo) o que melhor se correlacionou com o teor em
fenóis totais foi o sequestro do radical peroxilo, que apresentou uma correlação fortemente positiva,
seguido do radical anião superóxido, que apresentou uma correlação moderadamente positiva. Entre o
teor de antocianinas monoméricas e os valores de ORAC verificou-se uma correlação moderadamente
positiva, contudo esta correlação passa a fortemente positiva quando se elimina a amostra Patriot
(Tabela 4.13). Estes resultados sugerem o envolvimento dos compostos fenólicos, e em particular das
antocianinas, nestas duas atividades de sequestro. Em relação às associações verificadas entre a
composição fenólica das amostras e o sequestro do peróxido de hidrogénio, verificaram-se sempre
correlações negativas, sugerindo que os compostos fenólicos em geral, e as antocianinas em particular,
não constituam o principal grupo de compostos envolvido nesta atividade de sequestro.
Comparando os valores para os diferentes ensaios de atividade antioxidante, obtiveram-se
correlações fortemente ou moderadamente positivas entre os ensaios FRAP e CUPRAC e os ensaios
de sequestro do radical anião superóxido e ORAC, enquanto que se obtiveram correlações negativas
entre os ensaios de atividade redutora e o sequestro de peróxido de hidrogénio. Entre os ensaios de
sequestro do radical anião superóxido e ORAC a correlação obtida foi fracamente positivas, sugerindo
que possam ser compostos diferentes, particularmente fenóis diferentes, que mais contribuem para as
atividades detetadas.

63
O envolvimento dos fenóis nas atividades de sequestro já tinha sido sugerido por outros
autores. Assim, Wang e seus colaboradores (2012) mostraram que a polpa de mirtilo apresenta a
existência de uma correlação fortemente positiva entre o teor de fenóis e o sequestro de radical
peroxilo e uma correlação moderadamente positiva no sequestro do peróxido de hidrogénio e do
radical anião superóxido, mas os extratos das amostra da película do mirtilo apresentaram uma
correlação fortemente positiva no sequestro do peróxido de hidrogénio, do radical anião superóxido e
do radical peroxilo. Outros estudos (Bunea et al., 2011) mostraram a existência de uma correlação
fortemente positiva entre o teor de fenóis totais e os valores de ORAC.
A discrepância que se verifica entre os resultados publicados por outros autores e os resultados
obtidos com as amostras em estudo, em relação ao envolvimento dos fenóis na atividade de sequestro
do peróxido de hidrogénio, pode resultar do pequeno número de amostras estudadas ou do fato de, em
muitos casos, as diferenças obtidas não terem sido sequer significativas. Além disso, esta análise
baseada unicamente em ferramentas estatísticas apresenta diversas limitações. A análise de correlações
que se efetuou pretende apenas sugerir possíveis associações entre as classes de compostos presentes
nas amostras de mirtilo e as atividades antioxidantes detetadas. Assim, as propriedades antioxidantes
dos compostos isolados dentro de um grupo podem variar consideravelmente de modo que os mesmos
níveis de compostos fenólicos não correspondam necessariamente às mesmas respostas antioxidantes.
O fato das amostras de mirtilo serem misturas complexas, com múltiplos compostos diferentes, faz
com que possam coexistir compostos com atividades opostas, por exemplo, compostos com atividade
antioxidante juntamente com compostos com atividade pró-oxidante, ou ainda que possam ocorrer
fenómenos de sinergia e/ou potenciação entre os vários compostos, que acabam por, de alguma forma,
mascarar os resultados obtidos na análise de correlação.

64
5. Conclusão
O género Vaccinium spp. (Ericaceae) compreende cerca de 200 espécies, algumas das quais
produzem frutos comestíveis. Nas últimas décadas tem-se verificado um interesse crescente em
algumas espécies de Vaccinium spp., devido às propriedades farmacológicas e de promoção da saúde
que têm vindo a ser associadas a estes frutos. Estas propriedades incluem atividade antioxidante
(Wang et al., 1997; Mazza et al., 2002; Lohachoompol et al., 2004), anti-inflamatória (Youdim et al.,
2002), de proteção cardiovascular, antidiabéticas e de melhoria da visão (Katsube et al., 2003).
Muitas destas propriedades são atribuídas ao elevado teor em polifenóis dos mirtilos. O teor
das bagas nestes compostos, bem como o perfil exato em que estes aparecem, varia em função de
fatores intrínsecos (variedade, estádio de maturação, etc.) e extrínsecos (condições de clima, solo e
práticas de cultura), assim as frutas apresentam, em termos quantitativos e qualitativos, composição
variada desses compostos. Por sua vez, a eficácia da ação antioxidante depende da estrutura química e
da concentração destes fitoquímicos nos alimentos (Melo et al., 2008).
Assim, este trabalho teve por objetivo fazer um estudo comparativo do teor de fenóis totais,
antocianinas e da atividade antioxidante e antimutagénica em cinco variedades de mirtilo produzidas
em Portugal (Partiot, Bluecrop, Duke, Earlyblue e O’Neal) mais concretamente na Sub-região do
Baixo-Vouga. O fato de todas as plantas terem estado sujeitas aos mesmos fatores de cultivo (mesma
localização e ano de produção) elimina os efeitos de diferenças climatéricas, níveis de radiação UV,
temperatura ambiente, stress hídrico, composição do solo e disponibilidade de fatores nutricionais.
Todas as amostras foram, igualmente, colhidas com o grau de maturação adequado à sua
comercialização, transportadas para o laboratório e analisadas em iguais condições. Assim, as
diferenças observadas devem-se principalmente a diferenças genéticas entre as diferentes cultivares.
A capacidade para inibir o efeito do mutagéneo oxidativo do tert-butil-hidroperóxido foi
utilizada para avaliar a atividade antimutagénica, tendo a atividade antioxidante sido avaliada com
recurso a diferentes tipos de ensaios com o objetivo cobrir os vários dos mecanismos pelos quais esta
atividade pode ser exercida. Assim, foram então realizados ensaios de avaliação da atividade redutora
(FRAP e CUPRAC), da capacidade de sequestro de três espécies reativas de oxigénio passíveis de se
formarem in vivo, nomeadamente, do peróxido de hidrogénio e dos radicais anião superóxido e
peroxilo, e ainda a capacidade de quelação de Fe (II).
Os resultados mostram que todas a variedades de mirtilo estudadas apresentaram atividade
antioxidante e atividade antimutagénica. A variedade Duke foi a que apresentou o teor mais elevado
em compostos fenólicos totais e a que mais se distinguiu na atividade redutora, apresentando os
valores de FRAP e CUPRAC mais elevados. Esta variedade também mostrou maior capacidade de

65
desativação do radical peroxilo, detetada pelo ensaio ORAC. Por outro lado, a variedade O’Neal, foi a
que apresentou uma maior capacidade de sequestro do peróxido de hidrogénio. Com exceção da
atividade de sequestro do peróxido de hidrogénio, todas as outras atividades antioxidantes parecem
relacionar-se com o teor em compostos fenólicos e, em particular, em antocianinas.
Concluindo, neste trabalho foi possível detetar in vitro a existência de atividade antioxidante e
de atividade antimutagénica nas cinco variedades de mirtilos estudadas. No entanto, a extrapolação
dos resultados obtidos em ensaio in vitro para a situação in vivo, tem que ser cuidadosamente
ponderada, uma vez que a manutenção das propriedades verificadas in vitro depende de vários fatores,
como, por exemplo, do fato dos compostos por elas responsáveis resistirem ao processo de digestão
gastrointestinal, serem absorvidos pelas células do intestino e não serem rapidamente inativados nem
pelas bactérias da flora intestinal, nem pelas enzimas de biotransformação humanas. Para se poder
ponderar a extrapolação dos resultados seriam necessários estudos mais aprofundados sobre a
biodisponibilidade dos compostos bioativos dos mirtilos estudados.
A absorção digestiva dos polifenóis parece variar dependendo da sua estrutura, o que faz com
que nem sempre sejam os polifenóis mais abundantes nos alimentos aqueles que podem atingir
maiores concentrações no interior das células humanas. Estudos relatam que as proanticianidinas são
pouco absorvidas e a sua ação é limitada ao intestino, já a ingestão de flavonoides monoméricos,
flavonas e flavonoides são relativamente baixos e as concentrações plasmáticas raramente excedem
1mol/L por causa da absorção limitada e eliminação rápida. As flavanonas e isoflavonas são os
flavonoides com os perfis de melhor biodisponibilidade e as concentrações plasmáticas podem chegar
a 5 mol/L (Manach et al., 2004). Apesar da absorção dos fenóis poder ser limitada, alguns estudos,
efetuados em voluntários humanos saudáveis, mostram que após a ingestão de mirtilos a capacidade
antioxidante do plasma aumenta (Serafini et al., 2009).
Um importante contributo para possibilitar a extrapolação dos resultados obtidos para a
situação real in vivo seria a realização de ensaios sobre a resistência ao processo de digestão, a
possibilidade de absorção e sobre a biotransformação dos compostos bioativos presentes nestes
mirtilos. Seria igualmente interessante o estudo da atividade antimutagénica utilizando diferentes
mutagéneos alimentares, por exemplo, a aflatoxina B1, o benzo(a)pireno ou aminas aromáticas
heterocíclicas. Todos estes compostos são mutagéneos indiretos, ou seja requerem ativação metabólica
para exercerem os efeitos lesivos sobre o DNA. Por outro lado, para melhor poder compreender as
diferenças encontradas entre as diversas cultivares em estudo seria importante a identificação e
caracterização dos compostos fenólicos presentes nas várias cultivares de modo a tentar estabelecer
correlações entre as atividades biológicas e a presença ou ausência de compostos específicos. Este tipo
de análise permitiria identificar os principais compostos bioativos presentes nas diferentes cultivares
de mirtilo.

66
Uma vez que existem espécies silvestres do género Vaccinium em Portugal na Serra do Gerês,
na Ilha da Madeira e no arquipélago dos Açores, seria interessante quantificar e caraterizar o perfil dos
polifenóis presentes nestas variedades assim como as suas capacidades antioxidantes e
antimutagénicas e compará-las com as variedades de mirtilo cultivadas e comercializadas.

67
6. Referências Bibliográficas
Ahn J.H., Kim Y.P., Seo E.M., Choi Y.K. e Kim H.S., (2008). Antioxidant effect of natural plant extracts on the microencapsulated high oleic sun ower oil. Journal of Food Engineering, 84(2), 327-34.
AICR-WCRF American Institute for Cancer Research/World Cancer Research Fund, (2007).
Food, Nutrition, Physical Activity and the Prevention of Cancer: a Global Perspective.
Washington, DC.
Alderton W.K., Cooper C.E. e Knowles R.G., (2001). Nitric oxide synthases: structure,
function and inhibition. Journal Biochemistry, 1(357), 593-615.
Al-Gubory K.H., Fowler P.A. e Garrel C., (2010). The roles of cellular reative oxygen species, oxidative stress and antioxidants in pregnancy outcomes. The International Journal of Biochemistry & Cell Biology, 42(10), 1634–1650.
Alves R.E., Brito E.A., Rufino M.S. M. e Sampaio C. G., (2008). Antioxidant activity
measurement in tropical fruits: A case study acerola. Act Horticulturae, 773, 299-305.
Anjo D.L.C., (2004). Alimentos funcionais em angiologia e cirurgia vascular. Jornal Vascular
Brasileiro, 3, 145-154.
Antunes L.M.G. e Araújo M.C.P., (2000). Mutagenicidade e Antimutagenicidade dos
Principais Corantes para Alimentos. Revista Nutrição, 13 (2), 81-88.
Apak R., Güglü K., Özyürek M. e Karademir S.E., (2004). Novel total antioxidant capacity
index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC Method. Journal of Agricultural and Food
Chemistry, 52, 7970–7981.
Araújo P.W.B., Júnior L.J.Q., Vasconcelos H.D. e Almeida J.R.G.S., (2005). Flavonóides e
Hipertensão. Revista Brasileira de Hipertensão, 12(3), 188-189.
Bastos D. H. M., Rogero M. M. e Arêas J. A. G., (2009). Mecanismos de ação de compostos
bioativos dos alimentos no contexto de processos inflamatórios relacionados à obesidade.
Arquivos Brasileiros de Endocrinologia e Metabologia, 53, 646.
Beattie J., Crozier, A. e Duthie, G. G., (2005). Potencial Haelth Benefits of Berries. Current
Nutrition e Food Science, 1, 71-86.
Beckman K.B. e Ames B. N., (1997). Oxidative decay of DNA. The Journal of Biological
Chemistry, 272 (32), 19633-19636.
Behling E.B., Sendão M.C., Francescato H.D.C., Antunes L.M.G. e Bianchi M.L.P., (2004).
Flavonóide Quercetina: Aspectos gerais e ações biológicas. Alimentos e Nutrição, 15(3), 285-292.
Benzie I.F.F. e Strain J.J., (1996). The Ferric Reducing Ability of Plasma (FRAP) as a
measure of ‘‘antioxidant power’’: The FRAP Assay. Analytical Biochemistry, 239, 70–76.
Bernal J., Mendiola J.A., Ibáñez E. e Cifuentes A., (2011). Advanced analysis of
nutraceuticals. Journal of Pharmaceutical and Biomedical Analysis, 55(4), 758-774.

68
Bishop J.M., (1991). Molecular themes in oncogenesis. Cell, 64, 235-248.
Blumenthak M., Goldberg A. e Brinckmann J., (2000). Herbal Medicine: expanded
commission and monographs. Newton (USA): American Botanical Council, 519.
Bunea A., Rugina D. O., Pintea A.M., Sconta Z., Bunea C.I. e Socaciu C., (2011).
Comparative Polyphenolic Content and Antioxidant Activities of Some Wild and Cultivated
Blueberries from Romania. Notulae Botanicae Horti Agrobotanici, 39(2), 70-76.
Burdulis D., Ivanauskas L., Dirsė V., Kazlauskas S. e Ražukas A., (2007). Study of diversity
of anthocyanin composition in bilberry (Vaccinium myrtillus L.) fruits. Medicina (Kaunas); 43(12), 971–977.
Busby W.F.J. e Wogan G.N., (1984). Aflatoxins. Chemical Carcinogens, ed. C Searle, 945–
1136.
Candido L.M.B. e Campos A. M., (2005). Alimentos funcionais. Uma revisão. Boletim da
SBCRA, 29, 193-203.
Carlsen H., Myhrstad M.C.W., Thoresen M., Moskaug J.Ø. e Blomhoff R, (2003). Berry
intake increases the activity of the glutamylcysteine synthetase promoter in transgenic reporter
mice. The Journal of Nutrition, 133, 2137–2140.
Chitarra M.I.F. e Chitarra A.B., (2005). Pós-colheita de futas e hortaliças: fisiologia e
manuseio. 2º ed. Lavras: UFLA, 783.
Chung, F.-L., Chen, H.-J. C., e Nath, R. G., (1996). Lipid peroxidation as a potential
endogenous source for the formation of exocyclic DNA adducts. Carcinogenesis, 17(10), 2105-2111.
Connor A.M., Luby J.J., Tong C.B.S., Finn C.E. e Hancock J.F., (2002). Genotypic and
environmental variation in antioxidant activity, total phenolic content, and anthocyanin
content among Blueberry Cultivars. Journal of the American Society for Horticultural Science, 127(1), 89–97.
Costa J. P., (2010). Jornal de Noticias – “Capital do mirtilo" pretende quadruplicar produção
até 2014 - Rentabilidade e garantia de escoamento fez disparar o número de produtores -2010-06-25 consultado em 7 de julho 2011.
Courtemanche C., Huang A.C., Elson-Schwab I., Kerry N., Ng B.Y. e Ames B.N., (2004).
Folate deficiency and ionizing radiation cause DNA breaks in primary human lymphocytes: a comparison. FASEB Journal, 18, 209–11.
Covarrubias L., Hernández-García D., Schnabel D., Salas-Vidal E. e Castro–Obregón S.,
(2008). Function of reactive oxygen species during animal development: Passive or active? Developmental Biology, 320, 1-11.
Crozier A., Jaganath I. B. e Clifford M. N., (2009). Dietary phenolics: chemistry,
bioavailability and effects on health. Natural Product Reports, 26, 1001.
D’Archivio M., Filesi C., Di Benedetto R., Gargiulio R., Giovannini C. e Masella, R., (2007).
Polyphenols, dietary sources and bioavailability. Annali dell'Istituto Superiore di Sanità, 43, 348.
Dall-Done R., Giustarini D., Milzani A. e Colombo R., (2003). Protein carbonyl groups as
biomarkers of oxidative stress. Clinica Chimica Acta, 329, .23-38.

69
Delfino R.J., Sinha R., Smith C., West J., White E. e Lin H.J., (2000). Breast cancer, heterocyclic aromatic amines from meat and N-acetyltransferase 2 genotype. Carcinogenesis, 21(4), 607-15.
Dorman H.J.D., Bachmayer O., Koşar M. e Hiltunen R., (2004). Antioxidant properties of
aqueous extracts from selected Lamiaceae species grown in Turkey. Journal of Agriculture
and Food Chemistry, 52, 762–770.
Dorman H.J.D., Koşar M.¸ Kahlos K., Holm Y. e Hiltunen R., (2003). Antioxidant properties
and composition of aqueous extracts from Mentha species, hybrids, varieties, and cultivars.
Journal of Agriculture and Food Chemistry, 51, 4563–4569
Dragovic-Uzelac D., Savic Z., Brala A., Levaj B.,Bursac D., Kovacevic B. D. e Bisko A., (2010). Evaluation of Phenolic Content and Antioxidant Capacity of Blueberry Cultivars
(Vaccinium corymbosum L.) Grown in the Northwest Croatia. Food Technology and
Biotechnology, 48 (2), 214–221.
Dunlap W., Llewellyn L., Doyle J. e Yamamoto Y., (2003). A microtiter plate assay for
screening antioxidant activity in extracts of marine organisms. Marine Biotechnology, 5(3),
294–301.
Eck P., (1988). Blueberry Science. Rutgers University Press, New Brunswick and London.
Ehlenfeldt M.K. e Prior R.L., (2001. Oxygen radical absorbance capacity (ORAC) and
phenolic and anthocyanin concentrations in fruit and leaf tissues of highbush blueberry.
Journal of Agricultural and Food Chemistry, 49, 2222–2227.
Esmaeili M.A. e Sonboli A., (2010). Antioxidant, free radical scavenging activities of Salvia
brachyantha and its protective effect against oxidative cardiac cell injury. Food and Chemical
Toxicology, 48, 846–853
Exon J.H., (2006). A review of the toxicology of acrylamide. Journal of Toxicology and
Environmental Health Part B: Critical Reviews, 9, 397–412.
Fachinello J.C., (2008). Mirtilo. Revista Brasileira de Fruticultura, 30(2).
Falahatpisheh M.H., Kerzee J.K., Metz R.P., Donnelly K.C. e Ramos K.S., (2004). Inducible cytochrome P450 activities in renal glomerular mesangial cells: biochemical basis for antagonistic interactions among nephrocarcinogenic polycyclic aromatic hydrocarbons. Journal Carcinogenesis, 3(1),12.
FAO, (2010). Production Crops. Disponível em http://faostat.fao.org. Acesso em mar.2010.
Faria A., Oliveira J., Neves P., Gameiro P., Santos-Buelga C., Freitas V. e Mateus N., (2005).
Antioxidant properties of prepared Bluberry (Vaccinium myrtilus) extracts. Jornal of
Agricultural and Food Chemistry, 53(17), 6896-6902.
Felton J.S., Knize M.G., Wu R.W., Colvin M.E., Hatch F.T. e Malfatti M.A., (2007). Mutagenic potency of food-derived heterocyclic amines. Mutation Research, 616(1-2), 90-4.
Fenech M. e Ferguson L.R., (2001). Vitamins/minerals and genomic stability in humans.
Mutation Research, 475, 1–6.
Ferguson L. R. e Philpott M., (2008). Nutrition and Mutagenesis. Annu. Rev. Nutr., 28, 313-
32.

70
Ferguson L.R., (2001). Role of plant polyphenols in genomic stability. Mutation Research,
475, 89-111.
Ferguson L.R., Gregory T.J., Pearson A.E., Hay J.E. e Lewis G.D., (1996). Mutagenicity tests
as a monitoring tool for potential mutagens and carcinogens in shellfish gathering areas of New Zealand. Marine and Freshwater Research, 30, 413–21.
Ferrari R., Ceconi C., Curello S., Guarnieri C., Caldarera M., Albertini A. e Visioli O., (1985). Oxygen-mediated myocardial damage during ischaemia and reperfusion: role of the cellular defenses against oxygen toxicity. Journal of Molecular and Cellular Cardiology, 17, 937-45.
Fridovich I., (1998). Oxygen toxicity: radical explanation. The Journal of Experimental
Biology, 201, 1203-1209.
Fu P.P., Xia Q., Lin G. e Chou M.W., (2004). Pyrrolizidine alkaloids—genotoxicity,
metabolism enzymes, metabolic activation, and mechanisms. Drug Metabolism Reviews, 36,
1–55.
Galletta G.J. e Ballington J.R., (1996). Blueberry, cranberries, and lingonberries In: Janick J.e
Moore J.N., (1995). Fruit breeding: vine and small fruits. New York: Willey & Sons, 1-108.
Galloway T. e Handy R., (2003). Immunotoxicity of organophosphorous pesticides.
Ecotoxicology, 12, 345-363.
García-Closas R., García-Closas M., Kogevinas M., Malats N., Silverman D., Serra C., (2007). Food, nutrient and heterocyclic amine intake and the risk of bladder cancer. European Journal of Cancer, 43(11), 1731-40.
Geckil H., Ates B., Durmaz G., Erdogan S. e Yilmaz I., (2005). Antioxidant, free radical
scavenging and metal chelating characteristics of propolis. American Journal of Biochemistry
and Biotechnology, 1, 27–31.
Giovanelli G. e Buratti S., (2009). Comparison of polyphenolic composition and antioxidant
activity of wild Italian blueberries and some cultivated varieties. Food Chemistry, 112, 903–
908.
Gross G.A e Grüter A., (1992). Quantitation of mutagenic/ carcinogenic heterocyclic aromatic amines in food products. Journal of Chromatography A, 592(1-2), 271-8.
Gülçin I., (2012). Antioxidant activity of food constituents: an overview. Archives of
Toxicology, 86(3), 345-91.
Häkkinen S. e Törrönen A.R., (2000). Contento f flavonois and selected phenolic acids in
strawberries and Vaccinium species: Influence of cultivar, cultivation site and technique. Food
Recearoh International, 33, 517-524.
Halliwell B., (2007). Dietary polyphenols: good, bad, or indifferent for your health?
Cardiovascular Research. 73, 341– 47.
Halliwell B., (2007). Oxidative stress and cancer: have we moved forward? Journal of
Biochemistry, 401, 1-11.
Halliwell B. e Gutteridge J. M. C., (1989). Lipid peroxidation, a radical chain reaction. In:
Halliwell B., Gutteridge J.M.C. Free Radicals in Biology and Medicine, 3 rd ed. Clarendon,
Oxford, 900.

71
Halliwell B. e Gutteridge J.M.C., (2000). Free Radicals in Biology and Medicine. 3ª ed.,
Oxford, University Press.
Hermes-Lima M., (2004). in Functional Metabolism: Regulation and Adaptation. Storey K. B.
Jonh Wiley and Sons, New Jersey.
Hickey M.J., Sharkey K.A., Sihota E.G., Reinhardt P.H., Macmicking J.D., Nathan C. e Kubes
P., (1997). Inducible Nitric oxide synthase-deficient mice have enhanced leukocyte-
endothelium interactions in endotoxemia, Faseb Journal, 11, 955-964.
Holmqvist B.I., Ostholm T., Alm P. e Ekstrom P., (1994). Nitric oxide synthase in the brain of
a teleost. Neurological letters, 25, 171(1-2), 205-7.
Holst B. e Williamson G., (2008). Dietary reference intake (DRI) value for dietary
polyphenols: are we heading in the right direction? British Journal of Nutrition, 99(3), S55–S58.
Horst M.A. e Lajolo F.M., (2007). Biodisponibilidade de compostos bioativos de alimentos In:
Silvia M.F.C., Biodisponibilidade de Nutrientes. 2 ed. São Paulo: Manole, 1, 697-731.
Huang D., Ou B. e Prior R.L., (2005). The Chemistry behind Antioxidant Capacity Assays
Journal of Agricultural and Food Chemistry, 53, 1841-1856.
Hussain S.P., Schwank J., Staib F., Wang X.W. e Harris C.C., (2007). TP53 mutations and
hepatocellular carcinoma: insights into the etiology and pathogenesis of liver cancer.
Oncogene, 26, 2166–76.
Järgestad M. e Skog, K., (2005). Review: genotoxicity of heat-processed foods. Mutation
Research, 574, 156-172.
Jenkins R.R., (1988). Free radical chemistry relationship to exercise. Sports Medicine, 5, 156-
70.
Joseph J.A., Shukitt-Hale B., Denisova N.A., Bielinski D., Martin A., Mcewen J.J. e Bickford
P.C., (1999). Reversals of Age-Related Declines in Neuronal Signal Transduction, Cognitive, and Motor Behavioral Deficits with Blueberry, Spinach, or Strawberry Dietary
Supplementation. Journal of Neuroscience, 19, 8114-8121.
Kada T., Morita K. e Inoue T., (1978). Anti-mutagenic action of vegetable factor(s) on the
mutagenic principle of tryptophan pyrolysate. Mutation Research, Amsterdam, 53(3), 351-
353.
Katsube N., Iwashita K., Tsushida T., Yamaki K. e Kobori M., (2003). Induction of apoptosis
in cancer cells by bilberry (Vaccinium myrtillus) and the anthocyanins. Journal of
Agricultural and Food Chemistry 51, 68-75.
Kaur C. e Kapoor H. C., (2001). Antioxidants in fruits and vegetables – the millennium’s health. International Journal of Food Science e Technology, 36, 703-725.
Kazerouni N., Sinhá R., Hsu C.H., Greenberg A. e Rothman N., (2001). Analysis of 200 food items benzo[a] pyrene and estimation of its intake in an epidemiologic study. Food Chem Toxicol., 39(5), 423-36.
Koca I. e Karadeniz B., (2009). Antioxidant properties of blackberry and blueberry fruits grown in the Black Sea Region of Turkey. Scientia Horticulturae, 121, 447–450.

72
Kohen R. e Nyska A. (2002). Oxidation of Biological Systems: Oxidative Stress Phenomena, Antioxidants, Redox Reactions, and Methods for Their Quantification. Toxicologic Pathology, 30(6), 620-650.
Kojda G. e Harrison D.G., (1999). Interactions between NO and reactive oxygen species:
pathophysiological importante in atherosclerosi, hypertension, diabetes and heart failure.
Cardiovascular Research, 43, 562-571.
Kosar M., Fatih G. e Baser K.H.C., (2008). In vitro antioxidant properties and phenolic
composition of Salvia virgata Jacq. from Turkey. Journal of Agricultural and Food
Chemistry, 56, 2369–2374.
Kris-Etherton P. M., Hecker K.D., Bonanome A., Coval, S.M., Binkoski A. E., Hilpert K.F., Griel A.E. e Etherton T.D., (2002). Bioactive Compounds in Food: Their Role in the Prevetion
of Cardiovascular Disease and Cancer. The American Journal of Medicine, 113 (9b), 718-883.
Lavadinho C., Sousa M. B. e Moldão-Martins M., (2001). Influência da data de colheita na qualidade do mirtilo. Atas do 5º Encontro de Química de Alimentos-Qualidade, Segurança e Inovação, Porto.
Lee J., Durst R.W. e Wrolstad R.E., (2005). Determination of total monomeric anthocyanin
pigment content of fruit juices, beverages, natural colorants, and wines by the pH Differential Method: Collaborative Study. The Journal of AOAC International, 88, 269–278.
Leighton F. e Urquiaga I., (2007). Changes in cardiovascular risk factors associated with wine
consumption in intervention studies in humans. Annals of Epidemiology, 17(5), S32-S36.
Lin J., Forman M.R., Wang J., Grossman H.B., Chen M., Dinney C.P., Hawk E.T. e Wu X.,
(2012). Intake of red meat and heterocyclic amines, metabolic pathway genes and bladder
cancer risk International Journal of Cancer, 131, 1892–1903.
Lohachoompol V., Srzednicki G. e Craske J., (2004). The change of total anthocyanins in
blueberries and their antioxidant effect after drying and freezing. Journal of Biomedicine and
Biotechnology, 5, 248-252.
Lopes R. M., Oliveira T.T., De Nagem T.J., Pinto A.S., (2000). Flavonóides. Biotecnologia
ciência & desenvolvimento, 17, 18-22.
Lui R.H., (2004). Potential synergy of phytochemicals in cancer prevention: mechanism of
action. The Journal of Nutrition, 134(12), 3479S-3485S.
Lunec J., Holloway K.A., Cook M.S., Faux S., Griffiths H.R. e Evans M.D., (2002). Urinary
8-oxo-2-deoxyguanasina: redox regulation of DNA repair in vivo? Free Radical Biology and
Medicine, 33(7), 875-885
Magalhães L.M., Segundo M.A., Reis S. e Lima J.L.F.C., (2008) Methodological aspects
about in vitro evaluation of antioxidant properties. Analytica Chimica Ata, 613, 1–19.
Maisuthisakul P., Suttajit M. e Pongsawatmanit R., (2007). Assessment of phenolic content and free radical-scavenging capacity of some Thai indigenous plants. Food Chemistry, 100, 1409-1418.
Makus D.J. e Morris J.R., (1987). Highbush vs. Rabbiteye blueberry: a comparison of fruit
quality. Arkansas Farm Research, 36(3), 5.

73
Manach C., Scalbert A., Morand C., Rémésy C. e Jiménez L., (2004). Polyphenols: food
sources and bioavailability. American Society for Clinical Nutrition, 79, 727–47.
Manach C., Williamson G., Morand C., Scalbert A. e Rémésy C., (2005). Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. The American Journal of Clinical Nutrition, 81, 230S.
Maron D.M. e Ames B.N., (1983). Revised methods for the Salmonella mutagenicity test.
Mutation Research, 113(3-4), 173-215.
Martinez-Valverde I., Periogo M.J. e Ros G., (2000). Significado nutricional de los
compuestos fenólicos de la dieta. Arohivos Latinoamericanos de Nutrition, 50 (1), 5-18.
Mazza G., Kay C.D., Cottrell T. e Holub B.J., (2002). Absorption of anthocyanins from
blueberries and serum antioxidant status in human subjects. Journal of Agriculture and Food
Chemistry, 50, 7731-7737.
Mccall T.B., Boughton-Smith N.K., Palmer R.M., Whittle B.J. e Moncada S., (1989).
Synthesis of Nitric oxide from L-arginine by neutrophils. Release and interaction with
superoxide anion. Journal Biochemistry, 261, 293-296
Melo E. A., Maciel M.I.S., Lima, V.L.A.G. e Nascimento R.J., (2008). Capacidade
antioxidante de frutas. Brazilian Journal of Pharmaceutical Sciences, 44(2), 193-201.
Micelli N., Trovato A., Dugo P., Cacciola F., Donato P., Marino A., Bellinghieri V., La
Barbera T.M., Güvenç A. e Taviano M.F., (2009). Comparative analysis of flavonoid profile,
antioxidant and antimicrobial activity of the berries of Juniperus communis L. var. communis
and Juniperus communis L. var. saxatilis Pall. from Turkey. Journal of Agricultural and Food Chemistry, 57, 6570–6577.
Miller E.C. e Miller J.A., (1986). Carcinogens and mutagens that may occur in foods. Cancer,
58(8), 1795-1803.
Moon J-K. e Shibamoto T., (2009). Antioxidant Assays for Plant and Food Components.
Journal of Agricultural and Food Chemistry, 57, 1655–1666.
Moraes V. e Locatelli C., (2010). Vinho: uma revisão sobre a composição química e
benefícios à saúde. Evidência, 10(1-2), 57-68.
Mortelmans K. e Zeiger E. (2000). The Ames Salmonella/microsome mutagenicity assay.
Moyer R.A., Hummer K.E., Finn C.E., Frei B. e Wrolstad R.E., (2002). Anthocyanins,
phenolics and antioxidant capacity in diverse small fruits: Vaccinium, Rubus and Ribes.
Journal of Agricultural and Food Chemistry, 50 (3), 519–525
Mutation Research, 455, 29–60.
Naczk M. e Shahidi F., (2004). Extraction and analysis of phenolics in food – Review, Journal
of Chromatography A, 1054(1-2), 95-111
Nakamura S, Kato A. e Kobayaahit K., (1992) Enhanced antioxidative effect of ovalbumin
due to covalent binding of polysaccharides. Journal of Agricultural and Food Chemistry, 40, 2033–2037.
Neto C. (2007) Cranberry and blueberry: Evidence for protective effects against cancer and
vascular diseases. Molecular Nutrition & Food Research, 51, 652–664.

74
Oliveira D.M. e Bastos D. H. M., (2011). Biodisponibilidade de ácidos fenólicos Quimíca
Nova, 34(6), 1051-1056.
Olken N.M., Osawa, Y. e Marletta, M.A., (1994). Characterization of the inactivation of Nitric
oxide synthase by NG-methyl-L-arginine: evidence for heme loss. Biochemistry, 33, 14784-14791.
Ou B., Hampsch-Woodill M. e Prior R.L., (2001). Development and validation of an improved
Oxygen Radical Absorbance Capacity assay using fluorescein as the fluorescent probe.
Journal of Agricultural and Food Chemistry, 49, 4619–4626.
Passamonti A. S., Vrhovsekb U., Vanzoa A. e Mattivib F., (2003). The stomach as a site for
anthocyanins absorption from food. FEBS letters, 544, 210-213.
Peterson J.E. e Dwyer J., (1998). Flavonoids: dietary occurrence and biochemical activity.
Nutrition Research. 18, 1995-2018.
Pfohl-Leszkowicz A. e Manderville R.A., (2007). Ochratoxin A: an overview on toxicity and
carcinogenicityn in animals and humans. Molecular Nutrition and Food Research, 51, 61–99.
Phillips D.H., (1999). Polycyclic aromatic hydrocarbons in the diet. Mutation Research, 443,
139-47.
Prior R.L, Wu X. e Schaich K., (2005). Standardized methods for the determination of
antioxidant capacity and phenolics in foods and dietary supplements. Journal of Agricultural
and Food Chemistry, 53, 4290–4302.
Prior R.L., Cao G., Martin A., Sofic E., McEwen J. e O’Brien C., (1998). Antioxidant capacity
as influenced by total phenolic and anthocyanin content, maturity, and variety of Vaccinium
species. Journal of Agricultural and Food Chemistry, 46(7), 2686-2693.
Queipo-Ortuño M.I., Boto-Ordóñez M., Murri M., Gomez-Zumaquero J.M., Clemente-Postigo
M., Estruch R., Diaz F.C., Andrés-Lacueva C. e Tinahones F., (2012). Influence of red wine polyphenols and ethanol on the gut microbiota ecology and biochemical biomarkers.
American Journal of Clinical Nutrition, 95, 1323-1334.
Rahman I., Biswas S.K. e Kirkham P.A., (2006). Regulation of inflammation and redox
signaling by dietary Polypehenols. Biochemical Pharmacology, 72, 1439-1452.
Ramful D., Bahorun T., Bourdon E., Tarnus E. e Aruoma O.I., (2010). Bioactive phenolics
and antioxidant propensity of flavedo extracts of Mauritian citrus fruits: Potential prophylactic ingredients for functional foods application. Toxicology, 278, 75-87.
Ramirez M.R., Izquierdi I., Raseira M.D.C.B., Zuanazzi J.A., Barros D. e Henriques A.T.,
(2005). Effect of lyophilised vaccium Berries on memory, anxiety and locomotion in aldult rats. Pharmacological Research, 52, 457-462.
Raseira R.J. e Antunes L.E.C., (2004). A cultura do mirtilo (Vasccinium myrtillus). Pelotas:
Embrapa Clima Temperado, 69.
Regulski M. e Tully T., (1995). Molecular and biochemical characterization of dNOS: a
Drosophila Ca2+
/calmodulin-dependent Nitric oxide synthase. Proceedings of the National
Academy of Sciences, 26, 9072-6.
Ribeiro E.P. e Seravalli E.A.G., (2004). Química de alimentos. São Paulo: Editora Edgard
Blucher: Instituto Mauá de Tecnologia.

75
Ribeiro S.M.R., Queiroz J.H., Pelúzo M.C. G., Costa N.M.B., Matta S.L.P. e Queiroz M.
E.L.R., (2005). A formação e os efeitos das espécies reativas de oxigénio no meio biológico.
Bioscience Journal, 21(3), 133-149.
Rice-Evans C.A. e Miller N.J.E., (1997). Paganga, G. Antioxidant properties of phenolic
compounds. Trends in Plant Sience, 2, 152-159.
Richter C., Park J.W. e Ames B.N., (1988). Normal oxidative damage to mitochondrial and
nuclear DNA is extensive. Proceedings of the National. Academy of Science. USA, 85, 6465-
6467.
Riegger M., (2006). Introduction Fruit Crops. Binghamton: Food Products Press, an Imprint of
the Haworth Press, Inc., 462.
Roberfroid M., (2002). Functional food concept and its application to prebiotics. Digestive and
Liver Disease, 34, 105-10.
Rodrigues E., Poerner N., Rockenbach I.I., Gonzaga L.V., Mendes C. R. e Fett R., (2011)
Phenolic compounds and antioxidant activity of blueberry cultivars grown in Brazil. Ciências. Tecnologia Alimentares, Campinas, 31(4), 911-917.
Ross J.A. e Kasum C.M., (2002). Dietary flavonoids: bioavailability, metabolic effects, and
safety. Annual Review of Nutrition, 22, 19-34.
Rubbo H., Radi R., Trujillo M., Telleri R., Kalyanaraman B., Barnes S., Kirk M. e Freeman
B.A., (1994). Nitric oxide regulation of superoxide and peroxynitrite-dependent lipid
peroxidation. Formation of novel nitrogen-containing oxidized lipid derivatives. Journal of Biological of Chemistry, 269, 26066-26075.
Ryan T.P. e Aust S.D., (1992). The role of iron in oxygen-mediated toxicities. Critical Reviews in Toxicology, 22(1), 119-141.
Saftner R., Polashock J., Ehlenfeldt M. e Vinyard B., (2008) Instrumental and sensory quality
characteristics of blueberry fruit from twelve cultivars. Postharvest Biology and Technology,
49, 19–26.
Santos C., (2007). Estatística Descritiva - Manual de Autoaprendizagem, Edições Sílabo,
Lisboa.
Scalbert A., Manach C., Morand C., Remesy C. e Jimenez L., (2005). Dietary polyphenols and
the prevention of diseases. Critical Reviews in Food Science and Nutrition, 45(4), 287–306.
Schneider C.D. e Oliveira A.R., (2004). Radicais livres de oxigênio e exercício: mecanismos
de formação e adaptação ao treinamento físico. Artigo de revisão. Revista Brasileira de Medicina do Esporte, 10(4), 308-313.
Scholz S. e Williamson G., (2007). Interactions affecting the bioavailability of dietary polyphenols in vivo. International Journal for Vitamin and Nutrition Research, 77, 224.
Sellappan S., Akoh C.C. e Krewer G., (2002). Phenolic compounds and antioxidant capacity
of Georgia-grown blueberries and blackberries. . Journal of Agricultural and Food Chemistry, 50, 2432-2438.
Serafini M., Testa M.F., Villaño D., Pecorari M., Wieren K., Azzini E., Brambilla A. e Maiani
G., (2009). Antioxidant activity of blueberry fruit is impaired by association with milk. Free Radical Biology e Medicine, 46, 769–774.

76
Serrada F., Pereira M., Fretas S., Martins S. e Dias T., (2008). Mirtilo guia de boas práticas
para produção, promoção e comercialização. ADRIMAG.
Shahidi, F. e Naczk M., (1995). Food phenolics: sources, chemistry, effects and applications.
Ed Lancaster: Technomic Publishing, 331.
Shrubsole M.J., Jin F., Dai Q., Shu X., Potter J.D. e Hebert J.R., (2001). Dietary folate intake
and breast cancer risk: results from the Shanghai Breast Cancer Study. Cancer Research,
61(19), 7136-41.
Sies H., (1993). Strategies of antioxidant defence. Review. European Journal of Biochemistry,
215(2), 213-219. In: Ribeiro S.M.R., Queiroz J.H., Pelúzo M.C.G., Costa N.M.B., Matta
S.L.P. e Queiroz M.E.L.R., (2005). A formação e os efeitos das espécies reativas de oxigénio
no meio biológico. Bioscience Journal, 21(3), 133-149.
Singleton V.L., Orthofer R. e Lamuela-Raventos R.M., (1999). Analysis of total phenols and
other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods in
Enzymology, 299, 152–178.
Smith M., Marley K.A., Seigler D., Singletary K.W. e Meline B., (2000). Bioactive properties
of wild blueberry fruits. Journal of food science – Sensory and Nutritive Qualities of Food,
65(2), 352–356.
Soares, S. E., (2002). Ácidos fenólicos como antioxidantes. Revista de Nutrição, 15, 71-81.
Sousa M.B., Curado, T., Vasconcellos F.N. e Trigo M.J., (2007). Mirtilo: qualidade pós-
colheita. Agro divulgação, 556(8), disponível em http://www.inrb.pt/fotos/editor2/inia/manuais/8_mirtilo_qualidade_pos_colheita.pdf.
Sousa M.B., Curado T., Lavadinho C. e Moldão-Martins M., (2006). A survey of Quality
Factors in Highbush and Rabbiteye Blueberry cultivars in Portugal. Acta Horticulturae, 715,
567-572.
Spiers J.M., Marshall D.A., Smith B.J. e Braswell J.H., (2004). Method to determine chilling
requirement in Blueberries. Acta Horticulturae, 715, 105-110.
Stadtman E. R. e Levine R.L., (2003). Free radical-mediated oxidation of free amino acids and
amino acid residues in proteins. Amino Acids, 25, 207-218.
Stahl W e Sies H., (1997).Antioxidant defense: vitamins E, C and carotenoids. Diabete, 46,
14-8.
Törronen R.S., Hänninnen O., Kaartinen K., Rauma A.-L., Nenonen M., Häkkinen S.,
Adlercreutz H. e Laakso J., (2000). Antioxidants in vegan diet and rheumatic disorders.
Toxicology, 155, 45-53.
Tricker A. R. e Preussmann R., (1991). Carcinogenic Nnitrosamines in diet: occurrence,
formation, mechanism and carcinogenic potential. Mutation Research, 259, 277- 289.
Turkmen I. e Eks A., (2011). Brix degree and sorbitol/xylitol level of authentic pomegranate
(Punica granatum) juice. Food Chemistry, 12, 118
USDA (2011) USDA National Nutrient Database for Standard Reference, Release 24,
http://ndb.nal.usda.gov/ndb/foods/show/2266?fg=Fruits+and+Fruit+Juices&man=&lfacet=&f
ormat=&count=&max=25&offset=25&sort=&qlookup, acedido em 14 de abril 2012.

77
Valentão P., Fernandes E., Carvalho F., Andrade P.B., Seabra R.M. e Bastos M.L., (2001).
Antioxidant activity of Centaurium erythraea infusion evidenced by its superoxide radical
scavenging and xanthine oxidase inhibitory activity. Journal of Agricultural and Food
Chemistry, 49, 3476–3479.
Varlet V., Prost C. e Serot T., (2007). Analitycal, nutritional and clinical methods volatile
aldehydes in smoked fish: analysis methods, occurence and mechanisms of formation. Food
Chemistry, 105, 1536-56.
Vattem D.A. e Shetty K., (2005). Biological functionality of ellagic acid: a review. Jornual of
Food Biochemistry, 29, 234-266.
Wang H., Cao G. e Prior R.L., (1997). Oxygen radical absorbing capacity of anthocyanins.
Journal of Agricultural and Food Chemistry, 45, 304-309.
Wang L.S. e Stoner G.D., (2008). Anthocyanins and their role in cancer prevention. Cancer
Letters, 269, 281-290.
Wang S.Y., Camp M.J. e Ehlenfeldt M.K., (2012). Antioxidant capacity and a-glucosidase
inhibitory activity in peel and flesh of blueberry (Vaccinium spp.) cultivars. Food Chemistry
132, 1759–1768.
Wargovich M.J., (1997). Experimental evidence for cancer preventive elements in foods.
Cancer Letters, 114(1), 11-17.
Westwood M.N., (1982). Fruticultura de zonas templadas. Barcelona: Mundi-Prensa, 461.
WHO, World Health Organization, (2011). 10 facts on the global burden of disease.
Cardiovascular diseases are the leading causes of death in the world. Disponivel:
http://www.who.int/features/factfiles/glogal_burden/facts/en/index1.html. Acedido em 12
dezembro 2011
WHO, World Health Organization, (2005). Summary and conclusions of the sixty-fourth
meeting of the joint FAO/WHO Expert Committee on Food Additives.
Williamson G. e Manach C., (2005). Bioavailability and bioefficacy of polyphenols in
humans. II. Review of 93 intervention studies. The American Journal of Clinical Nutrition, 81 (suppl), 243S–245S.
Wogan G.N., Hecht S. S., Felton S. J., Conney A. H. e Loebe L.A., (2004). Environmental and
chemical carcinogenesis. Seminars in Cancer Biology, 14, 473–486
Wolfe K.L., Kang X., He, X., Dong M., Zhang, Q. e Liu R.H., (2008). Cellular antioxidant
activity of common fruits. Journal of Agricultural and Food Chemistry, 56, 8418-8426.
Yen G.C. e Chen H.Y., (1995) Antioxidant activity of various tea extracts in relation to their
antimutagenicity. Journal of Agricultural and Food Chemistry, 43(1), 27–32.
You Q., Wang B., Chen F., Huang Z., Wang X. e Luo P.G., (2011). Comparison of
anthocyanins and phenolics in organically and conventionally grown blueberries in selected cultivars. Food Chemistry, 125, 201–208.
Youdim K.A., McDonald J., Kalt W. e Joseph J.A., (2002). Potential role of dietary flavonoids
in reducing microvascular endothelium vulnerability to oxidative and inflammatory insults.
Journal of Nutritional Biochemistry, 13, 282-288.

78
Yu B.P., (1994). Cellular defenses against damage from reactive oxygen species.
Physiological Reviews, 74, 139-62.
Zafra-Stone S., Yasmin T., Bagchi M., Chatterjee A., Vinson J.A. e Bagchi D., (2007). Berry
anthocyanins as novel antioxidants in human health and disease prevention. Molecular Nutrition & Food Research, 51, 675–683.
Zardernowski R., Naczk M. e Nesterowicz J., (2005). Phenolic Acid profiles in some small
berries. Journal of Agricultural and Food Chemistry, 53, 2118-2124.
Zwart L.L., Meerman J.H.W., Commandeur J.N.M., Vermeulen N.P.E., (1999). Biomarkers of
radical damage. Applications in experimental animals and in humans. Free Radical Biology
Medicine, 26, 202-206.