aspectos taxonÔmicos, alimentaÇÃo e reproduÇÃo da...
TRANSCRIPT

ASPECTOS TAXONÔMICOS, ALIMENTAÇÃO E
REPRODUÇÃO DA RAIA DE ÁGUA DOCE Potamotrygon
orbignyi (CASTELNAU)
(ELASMOBRANCHII:POTAMOTRYGONIDAE) NO RIO
PARANÃ-TOCANTINS
GETULIO RINCON FILHO
Tese apresentada ao Instituto de Biociências daUniversidade Estadual Paulista “Julio de Mesquita Filho”, Campus de Rio Claro, para a obtenção do título de Doutor em Ciências Biológicas (Área de Concentração: Zoologia).
Rio Claro Estado de São Paulo-Brasil
Março de 2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ASPECTOS TAXONÔMICOS, ALIMENTAÇÃO E
REPRODUÇÃO DA RAIA DE ÁGUA DOCE Potamotrygon
orbignyi (CASTELNAU)
(ELASMOBRANCHII:POTAMOTRYGONIDAE) NO RIO
PARANÃ-TOCANTINS
GETULIO RINCON FILHO
Orientador: Prof. Dr. MIGUEL PETRERE JR.
Tese apresentada ao Instituto de Biociências da Universidade Estadual Paulista “Julio de Mesquita Filho”, Campus de Rio Claro, para a obtenção do título de Doutor em Ciências Biológicas (Área de Concentração: Zoologia).
Rio Claro Estado de São Paulo-Brasil
Março de 2006


Agradecimentos
Agradeço aos meus pais, Getulio e Regina, pelo apoio, carinho e amor e aos meus
irmãos e irmãs, Regina, Lara, Rafael, Daniel, Estevão e Pepê.
À Claudia, por seu amor, carinho, paciência, força e por tudo mais. Sem você eu nunca
chegaria a esse ponto da minha vida. À minha família de coração, Dona Socorro, Seu
Hugo, Huguinho e Paulo, pelo apoio e carinho.
Ao meu orientador Prof. Dr. Miguel Petrere, pelo apoio, amizade, ensinamentos, puxões
de orelha e por me mostrar o caminho para me tornar um melhor pesquisador e ser
humano.
Aos membros da banca examinadora: Dr. Teodoro Vaske Jr., Dr. Otto B. F. Gadig, Dr.
Roberto Goitein e Dr. Paulo Emílio Vanzolini. Muito obrigado por seus comentários e
ajuda para o aprimoramento da tese.
Aos colegas de pós-graduação, ou não, que me apoiaram durante estes anos: Janice,
Marcelo, Kotas, Serguei, Saul, Lúcia, Jerry, Agostinho, Alonso, Yzel, Henrique,
Letizia, Tuca, Cinthia, Leandro, Rafael, Fábio, Sidney, Isabela, Simone, Valéria, Silvia,
Dona Maria, Beraldo, Denise e Andrea.
Aos amigos que não mediram informações e troca de experiências em conversas
infindáveis sobre as raias de água doce, Patricia Charvet-Almeida, Mauricio Almeida,
Lúcia Araújo e Ricardo Rosa. Em especial agradeço à Patricia e Maurício por sempre
me receberem com carinho em Belém e pela saída de campo na baía de Marajó em
2000.
Aos meus alunos de graduação do UniCEUB Ana, Francisco e Rodrigo, muito obrigado
pela convivência com vocês e pela ajuda nas análises dos animais e na morfometria.

Ao amigo Matthias Stehmann, pela ajuda na saída de campo, sugestões, idéias,
bibliografia, divulgação e inúmeras xícaras de chá tomadas na varanda.
Ao Prof. Dr. Hugo P. Castello, muito obrigado pela ajuda e atenção em enviar as
últimas separatas originais de seus trabalhos com raias de água doce na década de 70.
À Sharon Thorson, obrigado por enviar informações e livros de seu pai, Dr. Thomas
Thorson, sobre o seu trabalho com raias de água doce na América do Sul.
Ao meu irmão Pedro Paulo, Pepê, pela ajuda nas saídas de campo, noites mal dormidas
nas margens dos rios e por estar comigo no acidente de carro durante a última saída de
campo numa estrada de terra no meio do Tocantins. Muito obrigado.
Aos ladrões que roubaram meu computador e todos os meus cd’s de arquivos e
programas, agradeço por não terem levado meus cd’s de backup. A Deus, por não ter
deixado os ladrões levarem os cd’s de backup, escondendo-os debaixo de uma única
folha de papel sobre a mesa.
À Profa. Dra. Elineide E. Marques da Universidade Federal do Tocantins, que
compartilhou informações não publicadas sobre as raias de água doce do médio
Tocantins.
À Profa. Dra. Maria Julia Martins Silva da Universidade de Brasília, pelo apoio na
identificação de invertebrados bentônicos, principalmente os moluscos, encontrados na
alimentação das raias de água doce.
À CAPES, pela bolsa de doutorado.
À Tatiana Walter, pela amizade, ajuda e apoio. Brigadão Tati!
Ao Chicago Zoological Society, pelo financiamento do projeto de Conservação das
Raias de Água Doce do Rio Tocantins e que forneceu a oportunidade de colher maiores
informações sobre a gestação e comportamento em cativeiro destes animais.

Ao Monterey Bay Aquarium pelo apoio financeiro e material logístico de campo, em
especial à Dra. Ava Ferguson.
Ao IBAMA, pelo fornecimento das licenças de pesca, transporte e exposição de raias.
Ao Ray Troll, pelo apoio e ajuda nos projetos com as raias de água doce ao ceder uma
de suas obras para ilustrar os folders.
À ACEPOPA, pelo financiamento parcial do Programa de Monitoramento da Pesca
Ornamental de Raias de Água Doce, que ofereceu oportunidade de coleta de material
comparativo, estudo da dinâmica da pesca ornamental e infra-estrutura para futuras
coletas.
Ao UniCEUB, principalmente nas pessoas das Dra.’s Magda Carvalho Branco,
coordenadora do Labocien e Marta Rodrigues, coordenadora do curso de Biologia, pela
amizade, uso dos laboratórios e apoio logístico.
À IUCN Sharks Specialist Group-SSG, principalmente Sarah Fowler e Rachel
Cavanagh, que apoiaram a pesquisa e sempre abriram espaço para a sua divulgação no
boletim Shark News.
À Sociedade Brasileira para o Estudo de Elasmobrânquios-SBEEL, principalmente na
pessoa de sua ex-presidenta, Dra. Rosangela Lessa, que apoiou institucionalmente o
projeto de conservação e acompanha o monitoramento da pesca ornamental.
Ao Museu Paraense Emilio Goeldi-MPEG, Instituto Nacional de Pesquisas da
Amazônia-INPA e Universidade Federal da Paraíba-UFPB (em especial ao Prof. Dr.
Ricardo Rosa) por permitirem a análise de suas coleções de raias de água doce.
A Deus, muito obrigado.

ÍNDICE
Página
RESUMO...........................................................................................................................i
ABSTRACT.....................................................................................................................iii
INTRODUÇÃO GERAL..................................................................................................1
CAPÍTULO I – AS RAIAS DE ÁGUA DOCE DO RIO PARANÃ, ALTO
TOCANTINS. ASPECTOS TAXONÔMICOS E DISTRIBUIÇÃO DE
Potamotrygon orbignyi (CASTELNAU, 1855)...............................................................5
INTRODUÇÃO.................................................................................................................5
MATERIAL E MÉTODOS...............................................................................................7
RESULTADOS...............................................................................................................11
As Espécies......................................................................................................................11
Potamotrygon orbignyi....................................................................................................11
Potamotrygon sp. A…………………….........................................................................26
Paratrygon aiereba..........................................................................................................31
Identificação....................................................................................................................36
Distribuição de Potamotrygon orbignyi no rio Paranã....................................................38
Variabilidade do Colorido em Potamotrygon na Bacia Tocantins-Araguaia..................42
DISCUSSÃO...................................................................................................................45
LITERATURA CITADA................................................................................................53
Anexo 1............................................................................................................................60

CAPÍTULO II – ALIMENTAÇÃO DA RAIA DE ÁGUA DOCE Potamotrygon
orbignyi NO RIO PARANÃ, ALTO TOCANTINS....................................................62
INTRODUÇÃO...............................................................................................................62
MATERIAL E MÉTODOS.............................................................................................64
RESULTADOS...............................................................................................................69
Morfologia do Sistema Digestório..................................................................................69
A Captura.........................................................................................................................72
Dieta e Hábitos Alimentares............................................................................................73
Grau de Repleção e Digestão...........................................................................................79
Nível Trófico...................................................................................................................83
DISCUSSÃO...................................................................................................................86
LITERATURA CITADA................................................................................................93
CAPÍTULO III – REPRODUÇÃO DE Potamotrygon orbignyi NO RIO PARANÃ,
ALTO TOCANTINS...................................................................................................101
INTRODUÇÃO.............................................................................................................101
MATERIAL E MÉTODOS...........................................................................................103
RESULTADOS.............................................................................................................105
Tamanho de Maturação.................................................................................................105
O Sistema Reprodutor...................................................................................................109
Fecundidade Ovariana e Uterina...................................................................................114
Índice Hepatossomático.................................................................................................116
Ciclo Reprodutivo..........................................................................................................117
Idade de Maturação.......................................................................................................118
DISCUSSÃO.................................................................................................................119

LITERATURA CITADA..............................................................................................124
CONCLUSÃO GERAL................................................................................................129

i
RESUMO
A raia de água doce Potamotrygon orbignyi do rio Paranã, alto Tocantins, foi analisada
quanto a aspectos de sua taxonomia, identificação, alimentação e reprodução. Duas
espécies de raias de água doce do gênero Potamotrygon, P. orbignyi e Potamotrygon sp.
A (uma espécie ainda não descrita), e Paratrygon aiereba ocorreram nas capturas no rio
Paranã. P. orbignyi é identificada principalmente com base na sua morfologia e fórmula
dentária, colorido dorsal do disco, uma única fileira de espinhos caudais e presença de
sulco labial. Uma chave de identificação para as espécies do rio Tocantins é proposta
com base em caracteres externos. P. orbignyi e P. aiereba possuem ampla distribuição
na bacia Tocantins-Araguaia e são reportadas desde o médio Araguaia até o alto
Tocantins. O principal item alimentar de P. orbignyi foi ninfas de Ephemeroptera
(87,5% - IRI%), seguido por larvas de Diptera (10,6%) e Trichoptera (1,6%). Não foram
observadas diferenças significativas quali e quantitativamente na alimentação entre as
estações da seca e chuvas, entre machos e fêmeas, ou entre o dia e a noite. Contudo,
exemplares imaturos e adultos apresentaram diferença significativa, onde os jovens (até
180 mm LD) se alimentam principalmente de larvas de Diptera, enquanto que os
adultos de ninfas de Ephemeroptera. As raias Potamotrygonidae apresentaram um nível
trófico intermediário (NT = 3,76), predominantemente como consumidores secundários
(NT<4). O nível trófico apresentou correlação positiva significativa com a LD máxima.
A maturidade sexual (LD50) ocorreu aos 251 mm LD nos machos e 260 mm LD nas
fêmeas. A fecundidade uterina foi de um filhote por gestação e o parto da maior parte da
população adulta ocorreu nas chuvas, entre setembro e fevereiro. Os filhotes nascem
com aproximadamente 115 mm LD e pesos totais em torno dos 60 gramas, o que

ii
corresponde a um ganho de peso de 1605% em relação ao peso do ovo (3,8 g). A
variabilidade do índice hepatossomático ao longo do ano evidenciou uma queda nas
reservas hepáticas na estação da seca e acúmulo destas reservas durante as chuvas. Com
base na leitura de anéis etários em vértebras e a sua relação com o tamanho estimado de
maturidade sexual, a idade de maturação foi estimada em 5 anos e a longevidade
máxima de 10 anos em P. orbignyi.

iii
ABSTRACT
The smooth back river stingray Potamotrygon orbignyi of the Paranã river, upper
Tocantins river, was studied in relation to its taxonomy, identification, feeding and
reproduction aspects. Two Potamotrygon species, P. orbignyi and Potamotrygon sp. A
(undescribed species), and Paratrygon aiereba were also captured in the Paranã river.
P. orbignyi was identified based on the morphology of the tooth and its formula, dorsal
color pattern, one single row of spines on the tail and the presence of a labial fold. It is
proposed a key to the species of freshwater stingrays of the Tocantins river based
mainly on external characters. P. orbignyi and P. aiereba presented a broad distribution
in the Tocantins-Araguaia basin and records from the middle Araguaia to the upper
Tocantins river. Nymphs of Ephemeroptera (87,5% - IRI%) were the most important
feeding content of P. orbignyi, followed by Diptera (10,6%) and Trichoptera larvae
(1,6%). Comparisons between rainy and dry seasons, males and females, or day and
night periods have showed no evidence of difference in the specific composition or
amount of food ingested. However, immature specimens (especially those smaller than
180 mm disc width-DW) feed mainly on Diptera larvae, while adults feed on nymphs of
Ephemeroptera. The freshwater stingrays of the family Potamotrygonidae are in
intermediate trophic position (NT = 3,76) and they are largely second level consumers
(NT<4). Their disc width and trophic level presented a positive and highly significative
correlation. The sexual maturity (DW50) was estimated to occur in 251 mm DW for
males and 260 mm DW for females. The uterine fecundity per gestation was just one
embryo and the parturition occurred along the rainy season for most of the reproductive
population (September to February). The neonates are estimated to have 115 mm DW

iv
and 60 grams of total weight, which corresponds to a weight increase of 1605% in
relation to the initial ova weight (3,8 g). The hepatosomatic index variation along the
year indicates a lower hepatic reserve condition in the dry season and higher reserve
hepatic condition in the rainy season. Based on readings of vertebral rings, the sexual
maturity was estimated to occur at the age of 5 with a maximum longevity of 10 years.

1
Introdução Geral
As raias de água doce Neotropicais pertencem à família Potamotrygonidae
GARMAN e estão restritas a três gêneros reconhecidos, Potamotrygon, Paratrygon e
Plesiotrygon. Dentre as características em comum que determinam a inabilidade destas
raias em viver em ambiente marinho estão a redução da glândula retal e as baixas
concentrações de uréia no sangue (THORSON et al., 1983). A presença de um processo
pré-pélvico longo que se estende anteriormente até quase atingir a cintura peitoral
também é uma sinapomorfia deste grupo (THORSON et al., 1983, ROSA, 1985;
CARVALHO et al., 2003). Os gêneros Paratrygon e Plesiotrygon são monotípicos,
enquanto que Potamotrygon apresenta um número maior de espécies, podendo variar
segundo os autores de 16 (+3 a +6, segundo levantamento deste autor) a 18 (+2)
(ROSA, 1985; CARVALHO et al., 2003).
Embora as raias de água doce apresentem um longo registro histórico científico
datando do período colonial (q. v. CASTEX, 1963) e tenham sido tema de pesquisas
desenvolvidas entre as décadas de 60 e 70 (CASTEX, 1964, 1969; CASTEX et al.,
1963; CASTELLO, 1975; ACHENBACH & ACHENBACH, 1976), observa-se nestes
trabalhos uma análise pontual, baseada em poucos exemplares e sem preocupação com
o policromatismo expresso pelo grupo, o que gerou posteriormente um certo grau de
confusão. As principais questões taxonômicas da família Potamotrygonidae começaram
a ser elucidadas com a revisão de ROSA (1985a) e seus trabalhos subseqüentes (ROSA,
1985b, 1990; ROSA et al., 1987). Estudos filogenéticos procurando esclarecer a relação
das raias de água doce com as demais Myliobatiformes, e a sua possível origem no
Pacífico ou no Atlântico tomaram impulso na mesma época ou quase que

2
imediatamente após os primeiros resultados de ROSA (1985a, 1985b) e evidenciaram a
necessidade de uma melhor amostragem taxonômica e a escolha de caracteres
consistentes e com maior poder de inferência filogenética (BROOKS et al., 1981;
NISHIDA, 1990; LOVEJOY, 1996; LOVEJOY, 1997; LOVEJOY et al., 1998;
McEACHRAN et al., 1996; MARQUES, 2000; McEACHRAN & ASCHLIMAN,
2004; CARVALHO et al., 2004; GONZÁLEZ-ISÁIS & DOMINGUEZ, 2004). De
forma geral, estes estudos reiteram a monofilia da família Potamotrygonidae, e que o
evento invasivo no continente sul-americano ocorreu por um ancestral comum entre o
Eoceno e o Mioceno (54-5 milhões de anos atrás) (para maiores detalhes veja
CARVALHO et al., 2004). Contudo, o gênero Potamotrygon, o mais diverso da família
com aproximadamente 16-18 espécies, apresenta discordância quanto à sua suposta
origem comum (MARQUES, 2000) e ainda necessita melhor resolução filogenética.
Apesar do desenvolvimento recente dos estudos taxonômicos e filogenéticos do
grupo das raias de água doce, a biologia básica, ecologia e dinâmica populacional destes
animais ainda continuavam em sério descompasso com o desenvolvimento da
taxonomia. THORSON et al. (1983) foram os primeiros a descrever aspectos da
reprodução de P. motoro baseados em exemplares em cativeiro, ultrapassando o limite
da observação casual de poucos exemplares. Publicações sobre a biologia reprodutiva
ou alimentação continuaram durante os anos seguintes com P. magdalenae (TESHIMA
& TAKESHITA, 1992), P. orbignyi e Paratrygon aiereba (LASSO et al., 1996),
Potamotrygon sp. do rio Negro (ARAÚJO, 1998), Plesiotrygon iwamae (CHARVET-
ALMEIDA, 2001) e P. scobina (ALMEIDA, 2003) e com uma recente revisão sobre
aspectos da reprodução de cinco espécies, P. orbignyi, P. scobina, P. schroederi, P.

3
motoro, Potamotrygon sp. (raia cururu do rio Negro), P. aiereba e P. iwamae
(CHARVET-ALMEIDA et. al., 2005).
A raia de água doce Potamotrygon orbignyi apresenta ampla distribuição nas
bacias Amazônica e do Orinoco e em rios do Suriname e Guianas (CARVALHO et al.,
2003). LASSO et al., (1996) descreveram aspectos gerais da alimentação desta espécie
nos “llanos” Venezuelanos e identificaram esta espécie como predominantemente
insetívora, podendo se alimentar também de crustáceos. Estes autores relatam ainda
baixa fecundidade uterina e apontam evidências de atividade reprodutiva constante, com
maior incidência de partos ocorrendo na estação das chuvas. BRAGANÇA et al., (2004)
corroboram com a identificação de P. orbignyi como uma espécie insetívora e
CHARVET-ALMEIDA et al., (2005) apontam diferenças nos parâmetros reprodutivos
(fecundidade e tamanho de maturidade) desta espécie em exemplares capturados na baía
de Marajó. Embora tenha sido descrita para a bacia do Tocantins-Araguaia
(CASTELNAU, 1855), P. orbignyi não possui estudos quanto à sua taxonomia ou
variabilidade policromática, distribuição, alimentação e reprodução nesta bacia. A
lacuna de informações sobre as raias de água doce favorece um comportamento
corriqueiro por parte dos ictiólogos ao ignorar este grupo de animais e leva a engano nas
identificações em trabalhos bem intencionados e pioneiros realizados com as raias da
bacia Tocantins-Araguaia (PANTANO-NETO, 2001 possivelmente em parte do
material identificado).
Historicamente as raias de água doce não são consumidas pelas comunidades da
bacia do Tocantins-Araguaia, salvo por eventual uso medicinal do fígado e ferrão, que
são tidos como eficientes no tratamento de diversas enfermidades como asma, alergia,
dor de cabeça e outros. Contudo, recentemente uma pesca ornamental destas raias se

4
desenvolveu em certos pontos do baixo rio Tocantins, no Estado do Pará, e ao menos
duas espécies estão sendo capturadas para a comercialização no mercado internacional
(P. henlei e P. orbignyi). Desde de 2003, duas Portarias do IBAMA (Portaria No 36 de
25 de junho de 2003 e Portaria No 27 de 31 de agosto de 2005) autorizaram a pesca e
comercialização de raias de água doce e até o momento 2600 exemplares de P. henlei e
4600 exemplares de P. orbignyi foram capturados, de acordo com os números limites
estabelecidos pela portaria 36 e 27. O comércio destas duas espécies de raias equivale a
um montante aproximado entre US$88.000 a US$160.000, dependendo do mercado
final e do padrão de colorido da raia, e justifica a preocupação com a manutenção destes
estoques no baixo Tocantins, tendo em vista que a tendência atual é de expansão desta
pescaria, tanto pelo aumento das quotas de captura quanto pela busca de novos padrões
de colorido em outras regiões.
Desta forma, esta tese trata da identificação de P. orbignyi na bacia Tocantins-
Araguaia – com especial ênfase nos animais capturados no rio Paranã, alto Tocantins –
descrição de sua variabilidade de colorido, distribuição, análise de aspectos
taxonômicos, alimentação e reprodução, a fim de fornecer subsídios para uma
identificação de população/estoque com base em parâmetros biológicos para um correto
manejo frente ao recente crescimento da pesca ornamental ou possíveis impactos
ambientais.

5
CAPÍTULO I
As Raias de Água Doce do Rio Paranã, Alto Tocantins.
Aspectos Taxonômicos e Distribuição de Potamotrygon
orbignyi (CASTELNAU, 1855)
INTRODUÇÃO
Embora as raias de água doce sejam peixes de médio-grande porte (e.g.
Potamotrygon brachyura e Paratrygon aiereba, ROSA, 1985), e podendo pesar até 50
quilos, esses animais são geralmente ignorados ou fracamente considerados em
avaliações íctio-faunísticas em diversas bacias hidrográficas (SANTOS et al., 1984;
GÉRY, 1984; ORTEGA & VARI, 1986; PLANQUETTE, et al., 1996; GALVIS et al.,
1997; FERREIRA et al., 1998). Esta indiferença muito se deve à dificuldade de

6
identificação do material coletado, ao grande porte dos animais, o que dificulta sua
preservação e transporte para posterior análise e à falta de pesquisadores especialistas na
área para identificar corretamente as espécies encontradas. Da mesma forma, e pelos
mesmos motivos, não há estudos sistemáticos sobre a diversidade da chondrofauna nos
rios da bacia Tocantins-Araguaia (q. v. PAIVA, 1983; SANTOS et al., 1984).
Recentemente PANTANO-NETO (2001), em seu estudo de anatomia descritiva
funcional associada à alimentação em P. motoro e P. henlei, identificou diversas
espécies de raias de água doce para o rio Cristalino, médio Araguaia, incluindo P.
castexi, P. leopoldi e P. ocellata. No entanto, P. castexi pertence à bacia do rio Paraná e
seus registros na bacia Amazônica são questionados; P. leopoldi é considerada
endêmica do rio Xingu e P. ocellata possui registros esporádicos no baixo Amazonas,
sua validade taxonômica é duvidosa e não possui registro no rio Tocantins
(CASTELLO & YAGOLKOWSKI, 1969; CASTEX & CASTELLO, 1970; ROSA,
1985; CARVALHO et al., 2003). Estas identificações equivocadas evidenciam a notória
fragilidade de qualquer estudo biológico ou medida de manejo sem a identificação e a
descrição das espécies envolvidas.
Estudos recentes indicam que uma abordagem a partir de uma escala
microrregional pode ser o melhor caminho para uma correta estimativa da riqueza de
raias de água doce nas bacias hidrográficas onde ocorrem (ARAÚJO, 1998;
CHARVET-ALMEIDA, 2001 e ALMEIDA, 2003). O policromatismo intra-específico,
a dificuldade de identificação correta, amostras pequenas e sem boa representatividade e
a possibilidade de endemismos nas cabeceiras dos rios (ROSA, 1985) dificultam os
estudos biogeográficos e sistemáticos destes animais (NISHIDA, 1990; LOVEJOY,
1996; MARQUES, 2000). Somente com levantamentos faunísticos regionais a

7
verdadeira riqueza biológica poderá ser estimada, promovendo o desenvolvimento de
estudos sistemáticos envolvendo todas as espécies do grupo, corrigindo a validade dos
caracteres morfológicos e estimando sua variabilidade intra-específica, e
conseqüentemente, fornecendo maior poder de inferência filogenética às análises
(HILLIS, 1996).
Desta forma, este trabalho trata da identificação das raias de água doce do rio
Paranã, afluente do alto Tocantins e de sua distribuição na bacia Tocantins-Araguaia
com base em amostragens complementares realizadas desde o ano 2000. Esta
identificação será apresentada para fins práticos como um guia de campo, onde os
animais podem ser identificados sem dissecações para análise de suas estruturas
internas. Este tipo de identificação é crucial para o acompanhamento da pesca
ornamental, a fim de validar o manejo pesqueiro destes recursos, que atualmente é
baseado em quotas específicas de exportação.
MATERIAL E MÉTODOS
O material analisado consiste de 223 exemplares identificados previamente
segundo ROSA (1985) e pelo conjunto das seguintes características: colorido dorsal do
disco, colorido ventral da cauda, morfologia dentária, presença de sulco labial e número
de fileiras de espinhos na cauda. Todos os exemplares analisados foram coletados no
médio rio Paranã, afluente do alto Tocantins, às margens do município de Paranã-TO,
próximo à divisa com o Estado de Goiás (Fig. 1), com zagaia (arpão de mão) durante a
noite com o uso de facho luminoso, tarrafa e espinhel de fundo com 25 anzóis nos anos
de 2002 (janeiro-abril e agosto-dezembro) e 2003 (janeiro e agosto-setembro) e no alto

8
Paranã nas proximidades do município de Formosa-GO, distante 48 km do Distrito
Federal.
Figura 1. Identificação da área de coleta no rio Paranã às margens do município de
Paranã-TO e a distribuição da captura ao longo da área de coleta. Pontos pretos referem-
se a exemplares de P. orbignyi com cauda e os vermelhos aos exemplares sem cauda.
Os exemplares coletados foram ainda comparados a espécimes adicionais
depositados em museus e instituições de pesquisa (Museu Paraense Emílio Goeldi-
MPEG, Instituto Nacional de Pesquisas da Amazônia-INPA e Universidade Federal da
Paraíba-UFPB). A distribuição geográfica destas espécies ao longo da bacia Tocantins-
Araguaia foi inferida a partir de registros de captura dos animais depositados em
coleções e de cruzeiros adicionais efetuados no médio Araguaia (Município de Aruanã-

9
GO em junho de 2001) e baixo Tocantins (Municípios de Marabá e Tucuruí-PA em
julho e agosto de 2000, junho e julho de 2004 e maio de 2005) utilizando a mesma
metodologia de pesca descrita para os animais coletados no rio Paranã. Uma listagem
dos exemplares analisados e tombados em diferentes instituições encontra-se no Anexo
I. A terminologia do colorido segue ROSA (1985) e ALMEIDA (2003).
O colorido das espécies de Potamotrygon ocorrentes na bacia Tocantins-
Araguaia foi considerado para a inferência do padrão de colorido primitivo de
Potamotrygon para esta bacia hidrográfica. A análise do colorido para as espécies do rio
Tocantins segue a hipótese filogenética proposta por MARQUES (2000) com base em
seqüências genéticas do citocromo-b, que embora não inclua todas as espécies
ocorrentes na bacia Tocantins-Araguaia, atualmente é a única hipótese disponível que
considera o maior número possível de espécies do rio Tocantins para tal análise.
Somente parte do cladograma proposto por este autor, onde as espécies ocorrentes no
rio Tocantins estão agrupadas, foi considerada para esta análise (Figura 2.2-B, página
11). Os padrões de colorido considerados foram os observados como predominantes nas
capturas no rio Tocantins ou identificados por ROSA (1985) para cada espécie.
A morfometria segue ROSA (1985), modificada na inclusão das seguintes
medidas: 3-Largura do Disco aos Olhos (máxima largura do disco na linha
imediatamente posterior aos olhos); 4-Largura do Disco na Origem das Nadadeiras
Pélvicas (máxima largura do disco na linha por sobre a margem anterior das nadadeiras
pélvicas) e 22-Comprimento da Abertura Cloacal (comprimento máximo da abertura
cloacal). Todas as medidas foram tomadas com paquímetro em milímetros. A
morfometria do neurocranium também segue ROSA (1985), modificada com a inclusão
das seguintes medidas: 2-Comprimento Pré-Fontanela (comprimento desde a margem

10
anterior do crânio até a margem anterior da fontanela); 4-Largura do Crânio no Processo
Pós-Orbital (largura máxima do crânio através dos processos pós-orbitais); 6-
Comprimento da Fontanela Anterior (comprimento longitudinal da fontanela anterior);
8-Comprimento da Fontanela Posterior (comprimento longitudinal da fontanela
posterior); 9-Altura do Crânio nas Cápsulas Nasais (altura máxima do crânio na região
das cápsulas nasais); 10-Altura do Crânio na Cápsula Ótica (altura máxima do crânio na
região da cápsula ótica); 11-Largura do Forâmen do Nervo Óptico; 12-Largura do
Crânio no Processo Pré-Orbital (largura máxima do crânio através dos processos pré-
orbitais). Todas as medidas do neurocranium foram tomadas em centésimos de
milímetros com paquímetro digital Starrett.
A contagem de dentes e sua descrição foram feitas macroscopicamente ou
através de lupa estereoscópica (Nikon) e segue STEHMANN (1977). As contagens
foram expressas em fórmulas dentárias onde o numerador representa o número de
fileiras de dentes na arcada superior e o denominador representa o número de fileiras de
dentes na arcada inferior. A terminologia dentária segue HERMAN et al. (1999),
enquanto a terminologia para as estruturas do esqueleto segue COMPAGNO (1999) e
NISHIDA (1990) e ROSA (1985) para o clásper.
Todos os testes estatísticos foram executados pelos programas STATISTICA 5.5
da STATSOFT e Excel 2002 da Microsoft e consideraram o nível de significância de
5%.

11
RESULTADOS
As Espécies
Após a identificação preliminar, foram identificadas três espécies para o rio
Paranã. Duas espécies, Potamotrygon orbignyi e Paratrygon aiereba, apresentam ampla
distribuição na bacia Amazônica. Uma segunda espécie de Potamotrygon também foi
encontrada e identificada previamente como Potamotrygon sp. A, tendo em vista que
não se encaixa em nenhuma outra espécie já descrita e provavelmente se trata de uma
espécie nova. Ainda como resultado das amostragens complementares no baixo
Tocantins, na região de Marabá-PA, também foi identificada uma segunda espécie não
descrita e muito semelhante a P. henlei. Esta espécie está sendo descrita como parte de
uma revisão e redescrição de P. henlei e será denominada neste trabalho de
Potamotrygon sp. B.
A Tabela 1 apresenta a descrição das três espécies encontradas no rio Paranã em
termos percentuais de sua largura de disco (LD). A Tabela 2 apresenta a morfometria
dos neurocrania para as espécies P. orbignyi, P. henlei, P. scobina, Potamotrygon sp.A
e Potamotrygon sp. B em termos percentuais de seu comprimento.
Potamotrygon orbignyi (CASTELNAU, 1855)
Sinonímia - q. v. CARVALHO et al. (2003) para uma revisão. Localidade tipo no rio
Tocantins sem identificação do ponto de coleta. Holótipo MNHN 2333. Exemplar
preservado seco, com somente partes do corpo remanescentes (pele, cauda, olhos,
12
arcadas e nadadeiras pélvicas) (ROSA, 1985; Dr. Bernard Séret1, comunicação pessoal)
(Fig. 2).
Tabela 1. Descrição das espécies de potamotrygonídeos ocorrentes no rio Paranã.
Potamotrygon orbignyi Potamotrygon sp. A Paratrygon aierebaN Min Máx Média Var N Min Máx Média Var N Min Máx Média Var
1 Comprimento Total 64 230 880 433 15152 7 328 770 592 25736 16 501 838 637 103172 Largura do Disco 74 147 467 236 3928 8 264 452 364 5937 20 228 540 406 75873 Largura do Disco aos Olhos 74 125 400 198 2599 8 234 392 319 3922 20 218 506 375 6064
4Largura do Disco na Origem das Nadadeiras Pélvicas
73 121 385 200 2875 8 232 386 316 4568 19 187 491 342 5905
5 Comprimento do Disco 74 155 522 256 5103 8 283 489 397 7892 20 249 581 435 79056 Comprimento Interno do Disco 74 134 483 223 4085 8 247 439 351 6521 20 219 529 390 67707 Comprimento do Olho 74 7 17 11 4 8 10 14 12 2 20 5 8 7 18 Comprimento Espiracular 74 10 49 18 28 8 13 30 23 51 20 9 20 14 119 Largura Interocular 69 29 96 48 169 8 49 85 68 198 20 28 62 48 10610 Largura Interespiracular 73 25 76 40 110 8 50 80 66 142 20 27 67 49 11511 Comprimento Pré-Ocular 74 38 129 61 300 8 71 122 97 445 20 69 168 123 67812 Largura da Cauda na Base 74 15 66 29 105 8 30 61 47 141 20 12 38 26 3913 Largura da Cauda no Espinho 65 6 25 13 17 6 15 25 21 14 18 7 17 11 814 Altura da Cauda na Base 74 9 43 19 45 8 20 39 31 56 20 9 33 17 3215 Comprimento do Ferrão 61 34 96 57 221 6 73 98 86 110 12 19 46 30 8216 Boca Escapulocoracoide 74 44 184 78 571 7 80 139 114 579 20 50 110 82 31417 Boca Cloaca 72 93 367 163 2650 8 172 309 242 3056 20 121 291 215 1977
18 Boca 1o Fenda Branquial 69 18 71 33 106 7 43 67 55 120 19 19 50 37 72
19 Focinho 1o Fenda Branquial 69 37 156 77 459 7 103 172 135 849 19 93 236 164 1270
20 Cloaca ao Ferrão Caudal 63 63 264 117 1595 6 122 198 166 1041 17 45 134 89 54321 Comprimento Pré-Cloacal 72 125 451 208 3767 8 231 411 321 5177 20 189 488 344 564722 Comprimento da Abertura Cloacal 72 6 30 13 23 8 13 29 22 39 18 13 37 24 4323 Comprimento Pré-Oral 74 28 89 45 124 8 60 103 79 265 20 70 184 126 81524 Comprimento Pré-Narial 74 21 65 35 71 8 43 73 58 120 20 66 180 117 744
25Comprimento do Cesto Branquial (=1o a 5o Fendas Branquiais)
73 25 81 41 144 8 33 58 49 112 20 23 54 40 69
26Largura do Cesto Branquial (=Largura entre 1o' s Fendas Branquiais)
64 33 119 57 327 6 64 103 82 318 19 48 121 86 367
27 Largura da Boca 73 12 46 23 49 8 26 46 37 54 19 23 53 37 7428 Comprimento da Narina 74 5 28 13 18 8 12 23 19 19 20 2 7 4 229 Largura Internarial 73 12 36 19 25 8 22 33 29 17 20 19 45 33 6030 Comprimento da Cauda 63 8 410 213 3771 6 217 334 287 2366 17 109 413 265 11040
31Largura da Nadadeira Pélvica (=Largura da Margem Posterior da Pélvica)
74 30 154 69 518 8 69 128 104 484 20 42 119 83 427
32Comprimento da Nadadeira Pélvica (=Comprimento da Margem Anterior da Pélvica)
74 33 100 54 197 8 63 113 91 455 20 38 88 62 168
33Comprimento da Base da Nadadeira Pélvica
74 15 104 43 284 8 43 101 73 509 20 28 89 55 243
34 Comprimento do Clasper 36 10 71 30 384 4 25 113 75 1939 11 9 61 41 395
35Comprimento da Cauda Desde a MargemPosterior do Disco à Inserção do Espinho
64 45 251 94 1403 6 100 176 131 832 17 15 50 32 77
36 Comprimento Pré-Pélvico 35 139 422 205 3724 6 337 492 399 2911
37 Comprimento da Cauda (Cloaca) 34 145 415 223 3773 5 141 196 171 72438 Número de Espinhos na Cauda* 35 7 47 20 74 8 28 63 46 152 5 50 110 82 61739 Número de Fileiras de Espinhos* 35 1 1 1 0 8 1 3 2 0 5 4 6 5 1
40 Número de Dentes da Arcada Superior* 30 26 38 29 7 3 13 20 20 25 7 17 23 23
41 Número de Dentes da Arcada Inferior* 30 24 36 28 7 3 19 22 1 7 13 22 20
42 Número de Voltas da Válvula Espiral* 18 16 17 16 0 3 14 14 14 0 9 12 14 14 0
43 Número de Papillas Orais Desenvolvidas* 20 5 5 5 0 3 3 3 3 0 12 _ _ _ _
1 Dr. Bernard Séret – Museu Nacional de História Natural-MNHN, Paris-França.

13
Figura 2. Fotografia de holótipo de P. orbignyi depositado no Museu Nacional de
História Natural (MNHN-2333). Foto enviada pela equipe do MNHN.
Distribuição no rio Paranã – registrada ao longo do médio e baixo Paranã; sem
registros no alto Paranã.
Nomes comuns - raia ou arraia branca ou arraia de fogo. Em todas as regiões
amostradas, o nome arraia é mais freqüentemente utilizado.
Diagnose - Esta raia caracteriza-se por possuir sulco labial desenvolvido, margeando os
cantos da boca e se estendendo de cada lado em direção à sínfise por aproximadamente
1/4 da largura da boca; musculatura débil da boca e a mesma pode ser aberta com a
mão, mesmo após a fixação; cinco papilas orais centrais no interior da boca; dentes
muito pequenos, com a coroa triangular e levemente monocúspide em machos e
trapezóide tricúspide em fêmeas; arranjados em 29-40 fileiras na arcada superior e 24-
42 fileiras na arcada inferior; uma única fileira de espinhos caudais, ocasionalmente
14
espinhos acessórios distribuídos aleatoriamente. P. orbignyi pode ser ainda confundida
com P. scobina, que se distribui no médio e baixo Tocantins e no rio Araguaia, mas P.
scobina não possui sulco labial acentuado, seu colorido ventral da cauda desde sua
origem não forma listras transversais, seus dentes são mono-cúspides e relativamente
maiores que os de P. orbignyi , e o colorido dorsal de adultos nunca forma retículas
hexagonais em fundo uniforme.
Tabela 2. Morfometria do neurocrania das espécies de potamotrygonídeos ocorrentes no
rio Tocantins. Os valores estão expressos em porcentagem do comprimento do
neurocranium
Medidas P. orbignyi P. orbignyi P. scobina P. sp. B P. henlei P. sp. ANúmero ES-15 ES-06 SN 31 SN 691005
Sexo Fêmea Fêmea Macho Fêmea Fêmea FêmeaComprimento do Neuro. (mm) 102.19 82.16 94.36 128.46 170 72.75Comprimento Pré-Fontanela 20.40 14.53 15.33 17.45 16.14 18.63Largura do Neuro. na Cápsula Nasal 56.41 55.61 66.25 60.42 64.25 64.18Largura do Neuro. no Processo Pós-Orbital 62.17 64.59 64.24 73.38 82.36 70.56Largura Interorbital 20.53 23.39 29.10 32.31 39.66 36.85Comprimento da Fontanela Anterior 25.09 25.06 19.92 23.74 24.96 23.11Largura da Fontanela Anterior 19.15 20.92 25.93 28.23 33.55 27.01Comprimento da Fontanela Posterior 42.20 41.91 43.24 42.77 45.03 39.70Altura do Neuro. nas Cápsulas Nasais 18.96 26.20 25.51 24.65 28.26 25.68Altura do Neuro. nas Cápsulas Óticas 26.92 24.50 22.13 23.88 25.34 25.10Largura do Forâmem do Nervo Ótico 16.04 13.78 6.17 8.87 7.89 7.42Largura do Neuro. no Processo Pré-Orbital 69.60 71.03 65.63 67.08 72.36 70.76
Variabilidade do colorido – Esta espécie apresenta policromatismo acentuado no
dorso do disco; o colorido dorsal de P. orbignyi apresentou seis padrões distintos
(reticulado, pontilhado, ocelado, spot, vermiculado e roseta-modificada) e seus
intermediários, onde os padrões se mesclam. O padrão mais conhecido é o reticulado
(Fig. 3c), sendo que este pode apresentar o retículo completo ou incompleto.
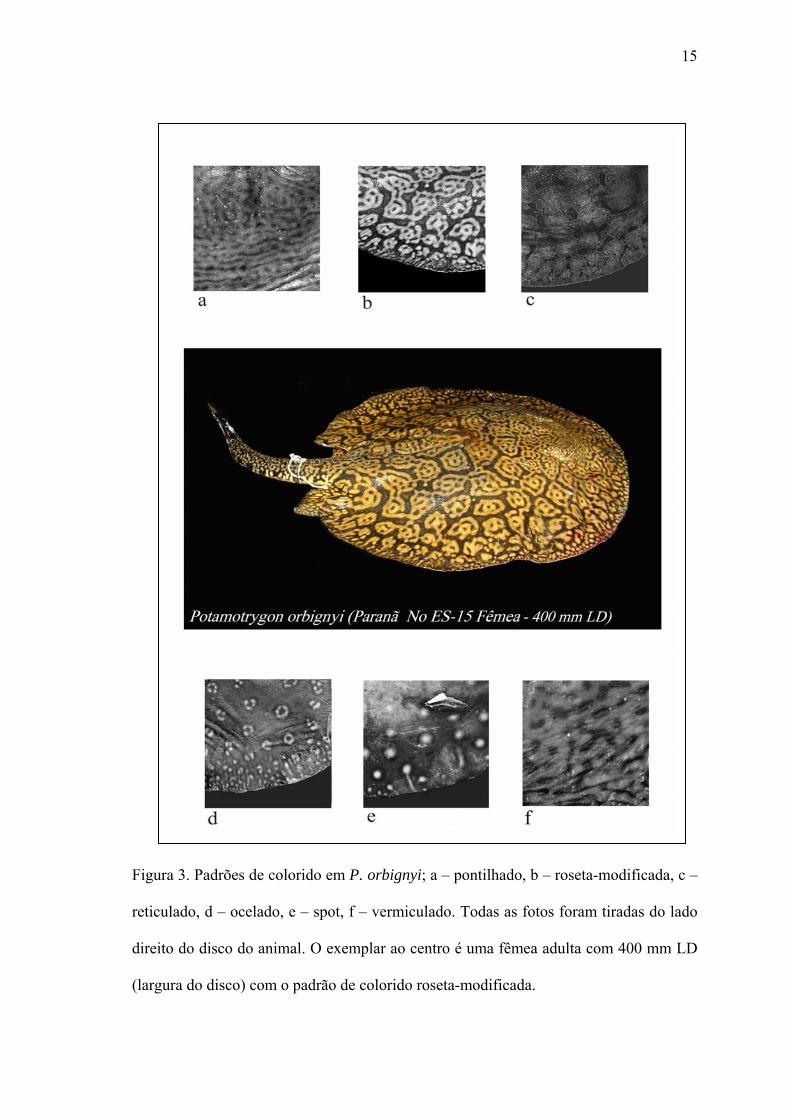
15
Figura 3. Padrões de colorido em P. orbignyi; a – pontilhado, b – roseta-modificada, c –
reticulado, d – ocelado, e – spot, f – vermiculado. Todas as fotos foram tiradas do lado
direito do disco do animal. O exemplar ao centro é uma fêmea adulta com 400 mm LD
(largura do disco) com o padrão de colorido roseta-modificada.

16
O padrão reticulado completo é menos freqüente nos exemplares do rio Paranã
(aproximadamente 27% dos exemplares analisados), mas aparece com alta freqüência
no baixo Tocantins, no reservatório da hidrelétrica de Tucuruí e baía de Marajó. No rio
Araguaia este padrão não foi encontrado, sendo mais freqüentes os padrões spot ou
ocelado, semelhante em alguns exemplares ao colorido de P. motoro (Fig. 3d). Quando
o padrão reticulado apresenta forte interrupção, este forma o padrão pontilhado, que
nada mais é que retículas extremamente quebradas (Fig. 3a). Este padrão nada tem em
semelhança com o padrão spot, pois os pontilhados não são sempre circulares e a sua
distribuição não apresenta padrão ou homogeneidade. Os padrões vermiculado e roseta-
modificada (Fig. 3b) são alterações do padrão ocelado, onde ocelos maiores encontram-
se margeados por pequenos spots que se fundem formando pequenas vermiculações e
conseqüentemente, formando as rosetas-modificadas; as vermiculações se formam com
o desmanche completo das rosetas e com a fusão de todos os spots e ocelos (Fig. 3f).
Neonatos apresentaram somente o padrão spot-ocelado, que logo após o nascimento
começa a modificar-se principalmente para os padrões reticulado ou pontilhado.
O colorido ventral é predominantemente branco ou creme claro, com as margens
do disco e margens posteriores das pélvicas podendo apresentar sombreado de
pontilhado amarelo claro ou castanho escuro a preto; o colorido ventral da cauda é
alternado em amarelo claro e preto, formando listras transversais. Geralmente ocorre um
ponto escuro por sobre a barra da cintura peitoral. Curiosamente este ponto localiza-se
exatamente onde a vesícula vitelínica está aderida ao ventre do animal durante a fase
embrionária.

17
Descrição morfológica do exemplar No 815180, macho adulto 230 mm LD - Disco
sub-circular, levemente oval, tão largo quanto longo e com proeminência anteromediana
na margem frontal do disco; largura do disco cabe 1,04 vezes dentro do seu
comprimento; margem do disco contínua, sem sinuosidades ou ponto de inflexão entre
as margens anterior ou posterior; disco relativamente baixo na região do neurocranium-
splanchnocranium, evidenciando débil musculatura associada. A largura interocular
cabe 1,26 vezes dentro do comprimento pré-ocular, equivale a 3,8 vezes o comprimento
do olho e cabe 5,4 vezes dentro da largura do disco. Olhos semi-ovais com leve
concavidade na face dorsal, relativamente pequenos e bem pedunculados em animais
fixados; operculum do olho em forma de “V” sem ornamentação aparente em animais
fixados.
Cauda ovalada em corte transversal, larga na origem e fortemente comprimida
dorso-ventralmente, afastada das bordas do disco na axila e relativamente curta,
dificilmente alcançando os espiráculos quando em posição de ataque; quilhas laterais
evidentes, uma a cada lado da cauda, desde a linha da extremidade das nadadeiras
peitorais à origem do ferrão caudal; quilha ventral desde a linha da origem do ferrão
caudal até o final da cauda; quilha dorsal desde a extremidade do ferrão até a ponta da
cauda; a cauda diminui de largura suavemente até a origem do ferrão, quando sofre
acentuada compressão lateral até a extremidade da cauda, formando o remo caudal.
Nadadeiras pélvicas musculosas, macias e com alto grau de mobilidade; margem
anterior ligeiramente convexa, com bordas espessas e arredondadas, seu comprimento
cabe 5,1 vezes na largura de disco; ápice sem ponto de inflexão distinto demarcando a
separação entre a margem anterior e posterior; margem posterior quase reta, com

18
endentações simples ou duplas (9/10 esquerda/direita) resultantes da disposição das
estruturas cartilaginosas de sustentação da nadadeira.
Comprimento pré-narial 6,8 vezes menor que o comprimento do disco, 2 vezes
maior que a largura internarial e 2,7 vezes maior que o comprimento da narina;
aberturas em forma de fenda, diagonalmente inclinadas com as margens anteriores
direcionadas ao focinho e posteriores às comissuras da boca; interior das narinas
levemente pigmentado de cinza; aberturas posteriores medialmente posicionadas em
relação às aberturas anteriores, lado a lado do frenum nasal; cortina nasal trapezóide,
com a margem posterior franjada não alcançando a boca, dividida em dois lóbulos pelo
frenum nasal e aproximadamente 1,8 vezes o comprimento da narina.
Boca localizada no primeiro 1/5 do comprimento do disco e sua largura cabendo
10,4 vezes na largura do disco; comprimento do cesto branquial igual a 1,25 vezes a
distância da boca à 1º fenda branquial; boca não recoberta pela cortina nasal, com
sulcos labiais evidentes, levemente arqueada quando fechada e com os dentes mais
externos visíveis em ambas as arcadas; cinco papilas orais desenvolvidas e centralizadas
internamente à arcada inferior. A válvula espiral é do tipo cônico-espiral com 16 voltas.
Cesto branquial com largura 3,8 vezes menor que a largura do disco e
comprimento 6 vezes menor que o comprimento do disco; fendas branquiais sinuosas,
dispostas lado a lado em “V” com o vértice sobre a barra coracóide e mergulhadas em
uma leve depressão do ventre.
Cloaca localizada entre as nadadeiras pélvicas e inserida na musculatura,
elipsóide e com uma fenda longilínea; a musculatura associada é longitudinal à fenda e
a mantém fechada.

19
Claspers robustos, relativamente curtos e cônicos, recobertos por tecido macio e
com todas as feições dispostas na face dorsal e lateral interna; a face ventral é lisa. O
canal do clasper estende-se obliquamente desde sua abertura anterior (apopyle), na face
dorsal próximo à sua base, e segue em direção à margem lateral externa até um ponto de
inflexão, que divide o comprimento do canal ao meio, e de onde segue levemente
oblíquo no sentido oposto em direção à face interna do clasper. A sua abertura posterior
(hypopyle) localiza-se na porção anterior do rhipidion, que segue paralelo ao
pseudosifão ventral. O pseudosifão dorsal é uma pequena fenda e localiza-se dorso-
lateralmente próximo à margem externa na altura do ponto de inflexão do canal do
clasper e se estende obliquamente em direção à face interna por um comprimento igual
à 1/3 da largura do clasper, na origem do pseudosifão.
Dentição arranjada em mosaico (Fig. 4ab), desde a sínfise até as comissuras.
Dentes relativamente pequenos (aprox. 1 mm largura) predominantemente triangulares,
mais largos que longos e de coroa tricúspide na face interna, ou monocúspide nos dentes
frontais mais desgastados; heterodontia monognástica acentuada entre os dentes
sinfisiais triangulares maiores e os comissurais ovalados menores; leve heterodontia
sexual, onde nos machos predominam dentes monocúspides ou com frágeis cúspides
laterais e nas fêmeas predominam dentes tricúspides. Raiz dupla com base triangular-
oval, curta e inclinada; cavidade com forâmen de nutrição do dente em posição mediana
entre as bases da raiz. Números dentários expressivos e que aumentam em proporção ao
crescimento do animal; exemplares adultos com mais de 40 dentes em cada arcada (Fig.
6; Tab. 3); fórmulas dentárias estão expressas na Tabela 1.
Dorso relativamente liso, recoberto por pequenos dentículos dérmicos dispersos
densamente sobre a região anterior do disco, sobre a cabeça, região mediana do corpo e

20
ao longo da cauda; demais margens do disco com fraca ou nenhuma espinulação, bem
como o dorso das nadadeiras pélvicas, onde somente escassos dentículos dérmicos são
encontrados aleatoriamente; dorso e laterais da cauda recobertos com dentículos
dérmicos.
Figura 4. Dentição das espécies de potamotrygonídeos ocorrentes no rio Tocantins; a, b
– P. orbignyi; c, d, e – Potamotrygon sp. A; f, g – Potamotrygon scobina; h – P. henlei;

21
i, j – Potamotrygon sp. B. Para cada espécie são apresentados primeiro os dentes da
arcada inferior direita e posteriormente os dentes da arcada superior esquerda.
Potamotrygon sp. A figura-d apresenta a linha divisória entre os dentes mais externos
em uso e os dentes mais internos ainda não utilizados da arcada inferior. Parte superior
da foto refere-se à face interna e a parte inferior à face externa. Todos os exemplares são
fêmeas.
Figura 5. Dentes inferiores próximos à sínfese de P. orbignyi (a, c, d, e – fêmea 155 mm
LD; b – macho 234 mm LD) e Paratrygon aiereba (f, g, h – fêmea 493 mm LD). a, h –
vista apical, b, c, g – vista da face interna, d, e – vista basal, f- vista lateral.
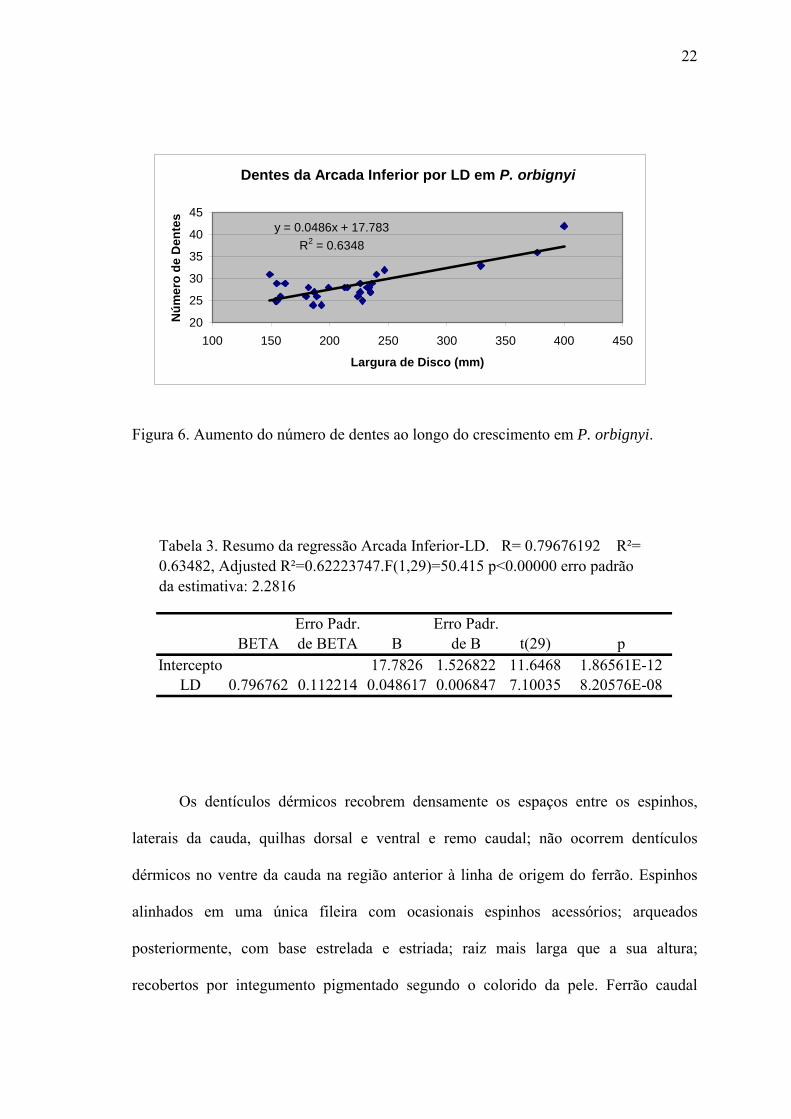
22
Dentes da Arcada Inferior por LD em P. orbignyi
y = 0.0486x + 17.783R2 = 0.6348
20
25
30
35
40
45
100 150 200 250 300 350 400 450
Largura de Disco (mm)
Núm
ero
de D
ente
s
Figura 6. Aumento do número de dentes ao longo do crescimento em P. orbignyi.
Tabela 3. Resumo da regressão Arcada Inferior-LD. R= 0.79676192 R²=0.63482, Adjusted R²=0.62223747.F(1,29)=50.415 p<0.00000 erro padrãoda estimativa: 2.2816
Erro Padr. Erro Padr.BETA de BETA B de B t(29) p
Intercepto 17.7826 1.526822 11.6468 1.86561E-12LD 0.796762 0.112214 0.048617 0.006847 7.10035 8.20576E-08
Os dentículos dérmicos recobrem densamente os espaços entre os espinhos,
laterais da cauda, quilhas dorsal e ventral e remo caudal; não ocorrem dentículos
dérmicos no ventre da cauda na região anterior à linha de origem do ferrão. Espinhos
alinhados em uma única fileira com ocasionais espinhos acessórios; arqueados
posteriormente, com base estrelada e estriada; raiz mais larga que a sua altura;
recobertos por integumento pigmentado segundo o colorido da pele. Ferrão caudal

23
posicionado após a linha de espinhos no dorso da cauda, em número variável (1-3) e
proporcional ao tamanho do animal; margens serrilhadas e direcionadas anteriormente.
Neurocranium pouco calcificado, mesmo em indivíduos adultos – descrição
baseada em dois exemplares fêmeas No ES-15 e No ES-06 com 400 e 319 mm LD,
respectivamente (Fig. 7abc)– mais longo que largo, cerca de 1,6 vezes (1,5;1,6) a sua
largura no processo pós-orbital e 1,7 (1,7;1,7) vezes a largura nas cápsulas nasais, que
são ovais e largas, inseridas no crânio com leve inclinação da lâmina basal (Fig. 8a) e
não ultrapassando a largura do crânio nos processos pré-orbitais; processos pré-orbitais
cônicos, relativamente longos e arqueados postero-ventralmente; processos pós orbitais
semi-transparentes, mais largos distalmente e levemente direcionados anteriormente.
Fossa parietal subdividida lateralmente com uma fenestra perilinfática e um forâmen
endolinfático em depressões individuais para cada lado. Forâmen do nervo óptico
relativamente maior e côndilo antorbital distintamente afastado da face ventral da
cápsula nasal. Cintura peitoral (Fig. 7de) com processo escapular triangular; forâmen
ântero-dorsal triangular menor que o forâmen ântero-ventral sub-retangular; forâmen
postero-ventral vestigial com pequena depressão em seu lugar. Cintura pélvica (Fig.
7fgh) com processo pré-púbico levemente cônico e longo; processos pré-púbicos
laterais oblíquos, direcionados para as laterais do disco; barra púbica fortemente
arqueada posteriormente e com processos ilíacos ornamentados na margem interna,
posteriormente ao processo isquial triangular; três foramina obturador posicionados
ventralmente ao côndilo radial e alinhados entre os processos pré-púbico lateral e
isquial.
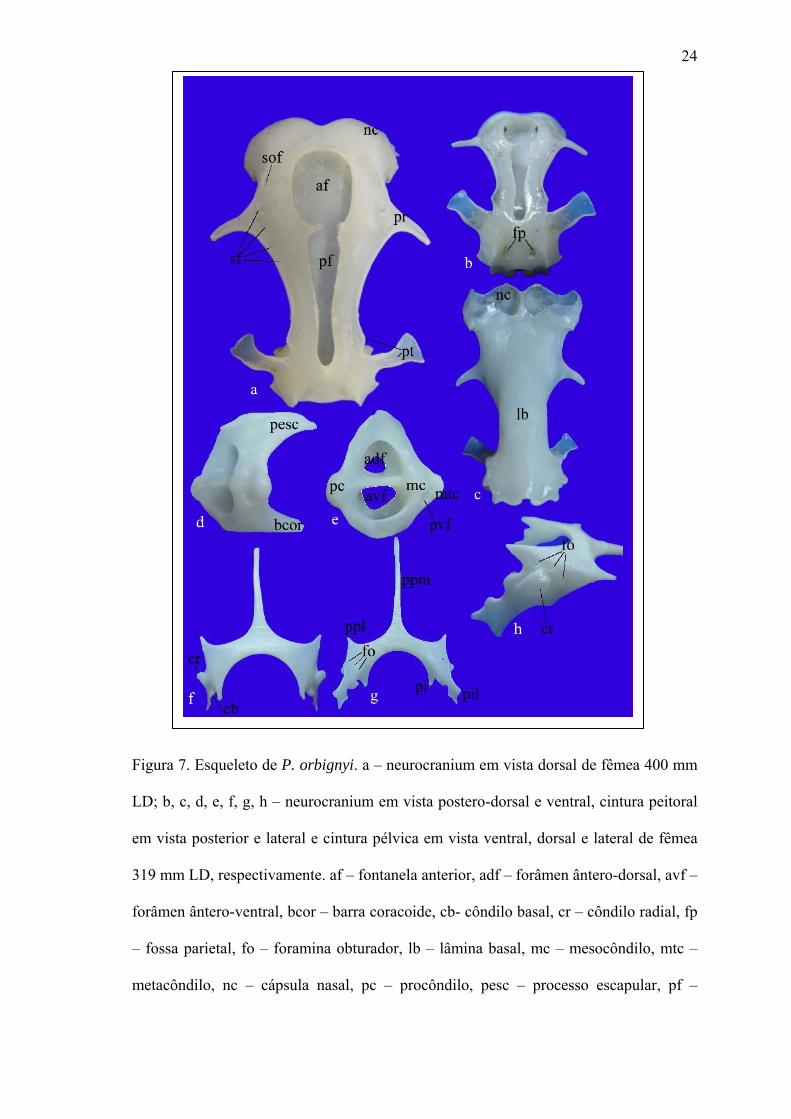
24
Figura 7. Esqueleto de P. orbignyi. a – neurocranium em vista dorsal de fêmea 400 mm
LD; b, c, d, e, f, g, h – neurocranium em vista postero-dorsal e ventral, cintura peitoral
em vista posterior e lateral e cintura pélvica em vista ventral, dorsal e lateral de fêmea
319 mm LD, respectivamente. af – fontanela anterior, adf – forâmen ântero-dorsal, avf –
forâmen ântero-ventral, bcor – barra coracoide, cb- côndilo basal, cr – côndilo radial, fp
– fossa parietal, fo – foramina obturador, lb – lâmina basal, mc – mesocôndilo, mtc –
metacôndilo, nc – cápsula nasal, pc – procôndilo, pesc – processo escapular, pf –
25
fontanela posterior, pi – processo isquial, pil- processo ilíaco, ppl – processo pré-púbico
lateral, ppm – processo pré-púbico medial, pr – processo pré-orbital, pt – processo pós-
orbital, pvf – forâmen postero-ventral, sf – foramina supraorbital, sof – forâmen
oftálmico superficial.
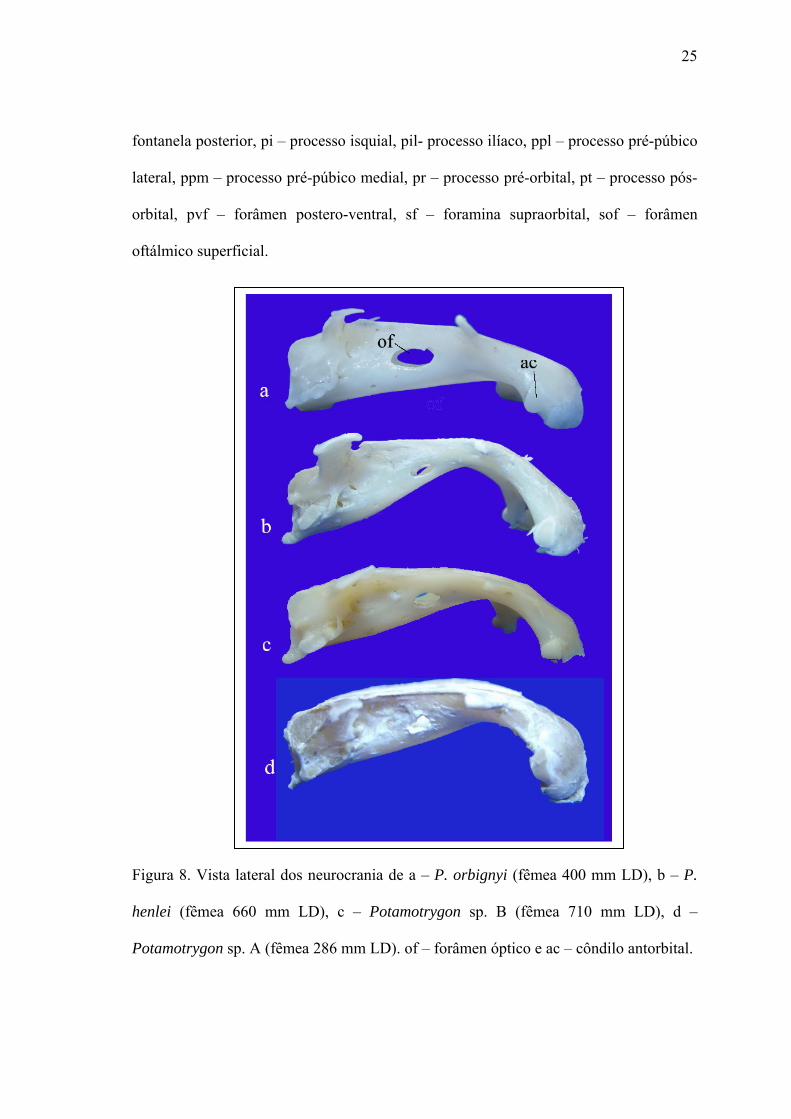
Figura 8. Vista lateral dos neurocrania de a – P. orbignyi (fêmea 400 mm LD), b – P.
henlei (fêmea 660 mm LD), c – Potamotrygon sp. B (fêmea 710 mm LD), d –
Potamotrygon sp. A (fêmea 286 mm LD). of – forâmen óptico e ac – côndilo antorbital.

26
Potamotrygon sp. A
Sinonímia – Não há.
Distribuição no rio Paranã - ao longo de todo o rio Paranã e ao menos até o médio
Tocantins. Aparentemente esta espécie é mais freqüente no médio Tocantins e seus
afluentes. Ocorre também no rio Maranhão (MARQUES2, material não publicado).
Nomes comuns - raia de fogo.
Diagnose - Esta raia caracteriza-se pela ausência de sulco labial; musculatura da boca
fortemente desenvolvida; três papilas orais centrais desenvolvidas e duas vestigiais
próximas às comissuras da boca; dentes hexagonais com coroa plana e sem cúspides,
arranjados em 13-20 fileiras na arcada superior e 19-22 fileiras na arcada inferior; duas
a três fileiras de espinhos no dorso da cauda; policromatismo pouco acentuado, variando
de spots a rosetas e vermiculado amarelo claro; colorido ventral é cinza escuro com as
regiões oral e branquial creme ou branco; a cauda é cinza escuro pontilhada por pontos
claros e três papilas orais desenvolvidas na boca. As duas únicas outras espécies da
bacia Tocantins-Araguaia com as quais esta espécie pode ser confundida por possuírem
mais de duas fileiras de espinhos sobre a cauda são P. henlei e Potamotrygon sp. B, que
se distribuem no baixo Tocantins e no rio Araguaia. No entanto, estas duas espécies
apresentam o dorso do disco coberto por ocelos em padrão circular que se expandem do
centro para as margens, enquanto que Potamotrygon sp. A possui um colorido uniforme
com spots menores que o diâmetro do olho e densamente distribuídos por sobre o dorso;
os dentes em Potamotrygon sp. B e são relativamente menores, com coroa em relevo e
um sulco transversal, losangulares a hexagonais e em maior número, enquanto que em
2 Dra. Elineide Eugênio Marques. Rua Paraguaçú 450. Setor Umuarama. 77500-000

27
P. henlei os dentes são relativamente maiores, losangulares, hexagonais e retangulares,
com coroa plana e sem sulco transversal e com profundas bases de fixação.
Variabilidade do colorido – O colorido dorsal de Potamotrygon sp. A apresentou três
padrões (spot, roseta e vermiculado) distintos ocorrendo dois a dois concomitantemente,
os padrões spot-roseta e spot-vermiculado (Fig. K). O padrão spot-roseta é caracterizado
por spots de pequeno tamanho distribuídos homogeneamente sobre a cabeça, focinho e
margem anterior do disco; no resto do disco surgem spots maiores margeados por spots
menores formando as rosetas, que estão intercaladas por spots pequenos. O padrão spot-
vermiculado aparentemente é formado quando os spots pequenos se fundem formando
vermículos que margeiam spots maiores distribuídos principalmente nas laterais do
disco e posterior à linha dos olhos.
Descrição do exemplar No 302, macho adulto, 428 mm LD - Disco sub-circular,
levemente oval, quase tão largo quanto longo e com proeminência anteromediana na
margem frontal do disco; largura do disco cabe 1,04 vezes dentro do seu comprimento;
margem do disco contínua, sem inflexões demarcando nitidamente margens anterior ou
posterior; disco alto na região do neurocranium-splanchnocranium, evidenciando forte
musculatura associada. A largura interocular cabe 1,3 vezes dentro do comprimento pré-
ocular, equivale a 5,5 vezes o comprimento do olho e cabe 5,4 vezes dentro da largura
do disco. Olhos ovais, relativamente pequenos e pouco pedunculados em animais
fixados; operculum do olho em forma de “V” sem ornamentação aparente em animais
fixados (Fig. 9).
Cauda ovalada em corte transversal, larga na origem e levemente comprimida
dorso-ventralmente, afastada das bordas do disco na axila e relativamente curta,
Porto Nacional, Tocantins. Fundação Universidade do Tocantins. Campus de Porto Nacional.

28
dificilmente alcançando os espiráculos em indivíduos adultos quando em posição de
ataque; quilhas laterais pronunciadas, uma a cada lado da cauda, que se estendem da
linha da extremidade das nadadeiras peitorais à origem do ferrão caudal; quilha ventral
desde a linha da origem do ferrão caudal até o final da cauda; quilha dorsal desde a
extremidade do ferrão até a ponta da cauda; a cauda diminui de largura suavemente até
a origem do ferrão, quando sofre acentuada compressão lateral até a extremidade da
cauda, formando o remo caudal.
Figura 9. Exemplar macho adulto de Potamotrygon sp. A (428 mm LD).
Nadadeiras pélvicas musculosas, macias e com alto grau de mobilidade; margem
anterior ligeiramente convexa com borda espessa e arredondada, cabendo 3,7 vezes na
largura de disco; ápice sem ponto de inflexão distinto demarcando a separação entre a

29
margem anterior e posterior; margem posterior quase reta, com endentações (10/11
esquerda/direita) resultantes da disposição das estruturas cartilaginosas de sustentação
da nadadeira.
Comprimento pré-narial 6,4 vezes menor que o comprimento do disco, 2,2 vezes
maior que a largura internarial e 3 vezes maior que a abertura anterior da narina;
aberturas em forma de fenda, diagonalmente inclinadas com as margens anteriores
direcionadas ao focinho e posteriores às comissuras da boca; interior das aberturas
anteriores fortemente pigmentado de preto; aberturas posteriores medialmente
posicionadas em relação às aberturas anteriores, lado a lado do frenum nasal; cortina
nasal trapezóide, com a margem posterior franjada não alcançando a boca, dividida em
dois lóbulos pelo frenum nasal.
Boca localizada no primeiro 1/5 do comprimento do disco e sua largura cabendo
11 vezes na largura do disco; distante do cesto branquial a uma distância
aproximadamente igual ao comprimento do cesto branquial; boca não recoberta pela
cortina nasal, sem sulcos labiais ou fossa oral, levemente arqueada quando fechada e
com os dentes mais externos visíveis em ambas as arcadas. Três papilas orais
centralizadas logo após a arcada inferior e outras duas severamente reduzidas, uma a
cada lado, próximas às comissuras da boca. A válvula espiral é do tipo cônico-espiral
com 14 voltas.
A largura do cesto branquial é 4,1 vezes menor que a largura do disco e seu
comprimento é 7,8 vezes menor que o comprimento do disco; as fendas são onduladas e
estão imersas em uma depressão em forma de arco que segue da primeira à quinta
fenda.

30
Cloaca posicionada entre as nadadeiras pélvicas, mergulhada na musculatura
onde apresenta uma forma elipsóide e uma fenda longilínea; a musculatura associada é
longitudinal à fenda e a mantém fechada.
Claspers robustos, curtos e cônicos, recobertos por tecido macio, sem feições
distintivas significantes que distinga Potamotrygon sp. A das demais espécies do
gênero; todas as feições do clásper estão na face dorsal e lateral; a face ventral é lisa e
não ornamentada.
Dentição arranjada em fileiras distintas ou em leve mosaico (Fig. 4cde), desde a
sínfise até as comissuras. Dentes predominantemente hexagonais, mais largos que
longos e de coroa plana. As fórmulas dentárias estão expressas na Tabela 1.
Dorso áspero, recoberto por pequenos dentículos dérmicos dispersos densamente
sobre a região mediana do corpo e ao longo da cauda; margens do disco com fraca
espinulação, bem como o dorso das nadadeiras pélvicas, onde somente escassos
dentículos dérmicos são encontrados aleatoriamente. Dorso e laterais da cauda
recobertos com dentículos dérmicos; espinhos arranjados em 1 a 2 fileiras com espinhos
acessórios ocasionais. Os dentículos dérmicos recobrem densamente os espaços entre os
espinhos, laterais da cauda, quilhas dorsal e ventral e remo caudal; ventre da cauda sem
dentículos dérmicos na região anterior à linha de origem do ferrão. Os espinhos são
arqueados posteriormente, com base estrelada e estriada, recobertos por integumento
pigmentado segundo o colorido da pele, dispersos em fileiras ligeiramente dispersas em
indivíduos jovens ou densamente arranjados em indivíduos adultos.

31
Paratrygon aiereba (MULLER & HENLE, 1841)
Sinonímia – q. v. ROSA (1991) e CARVALHO et al. (2003) para uma revisão.
Localidade tipo: Brasil. Exemplar tipo perdido segundo CASTEX & CASTELLO
(1969) e ROSA (1985).
Distribuição no rio Paranã - Ao longo de todo o rio. Esta espécie também é registrada
ao longo de todo o rio Tocantins e Araguaia.
Nomes comuns - arraia aramaçã ou aramaçá.
Diagnose - Esta raia caracteriza-se por possuir um grande porte (machos maturando em
torno dos 430 mm LD e fêmeas ainda imaturas aos 540 mm LD); comprimento pré-
ocular maior que em Potamotrygon e cerca de ⅓ da largura do disco; a margem anterior
do disco é distintamente côncava e não possui a proeminência anteromediana
característica de Potamotrygon; seus olhos são diminutos e não pedunculados, imersos
na musculatura do disco; cauda curta e fortemente armada com espinhos em indivíduos
adultos; freqüentemente possue pequeno ferrão; exemplares jovens possuem uma cauda
filiforme e longa, sem espinhos e sem quilhas; margem posterior do espiráculo com
projeção arredondada; boca relativamente pequena e sem sulcos labiais; papilas orais
extremamente vestigiais ou ausentes; dentes relativamente pequenos e arranjados em
17-23 dentes na arcada superior e 13-22 dentes na arcada inferior.
Variabilidade do colorido – O colorido dorsal desta espécie varia de um fundo marrom
claro com um malhado irregular marrom escuro ou preto sobre o dorso do disco; alguns
exemplares podem ainda ser castanho escuro a marrom escuro ou cinza sem o malhado
irregular; a cauda e nadadeiras pélvicas são da mesma cor do disco sem padrões
definidos (Fig. 10). Todos os exemplares analisados apresentavam manchas escuras
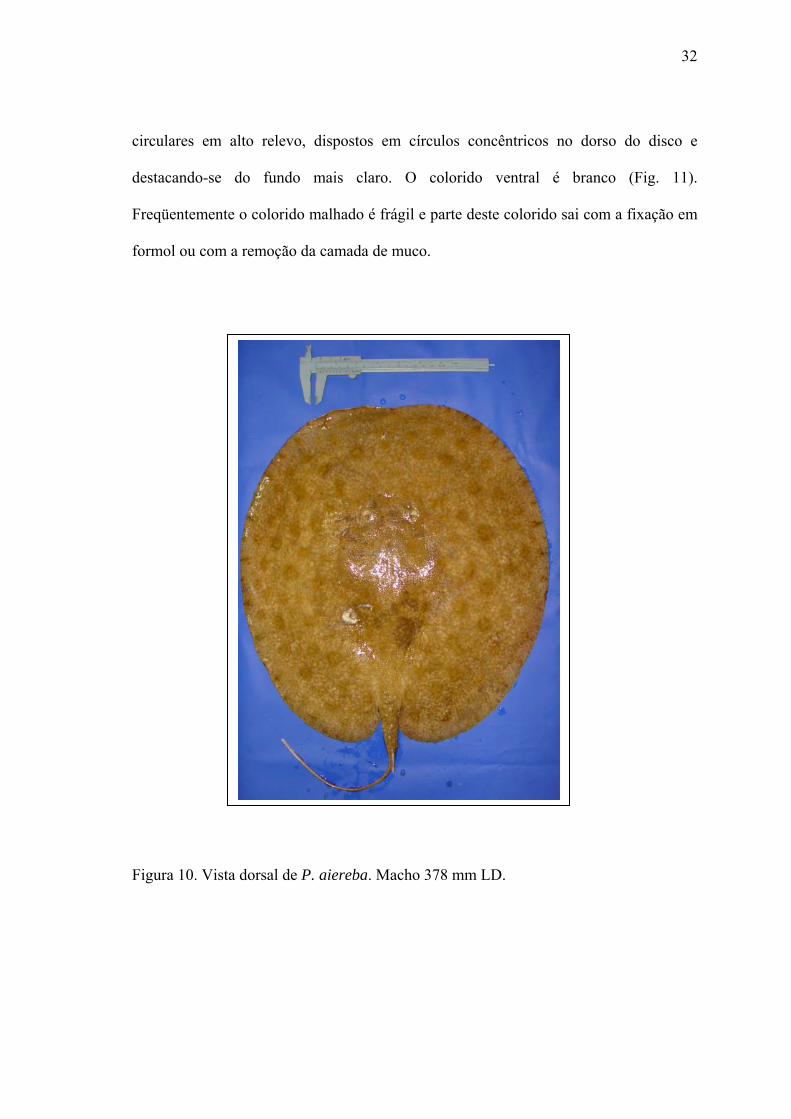
32
circulares em alto relevo, dispostos em círculos concêntricos no dorso do disco e
destacando-se do fundo mais claro. O colorido ventral é branco (Fig. 11).
Freqüentemente o colorido malhado é frágil e parte deste colorido sai com a fixação em
formol ou com a remoção da camada de muco.
Figura 10. Vista dorsal de P. aiereba. Macho 378 mm LD.

33
Figura 11. Vista ventral de P. aiereba. Macho 378 mm LD.
Descrição do exemplar No 691062, fêmea imatura, 327 mm LD - Disco oval, quase
tão largo quanto longo; margem anterior côncava e sem proeminência anteromediana;
largura do disco cabe 1,04 vezes dentro do seu comprimento; margem do disco
contínua, sem inflexões demarcando nitidamente margens anterior ou posterior; disco
alto na região do splanchnocranium, evidenciando forte musculatura associada. Órbitas
posicionadas posteriormente, comprimento pré-ocular cabe 3,9 vezes no comprimento
do disco; largura interocular cabe 2,3 vezes dentro do comprimento pré-ocular, equivale
a 6,1 vezes o comprimento do olho e cabe 8,8 vezes dentro da largura do disco. Olhos

34
semi-circulares com forte concavidade na face dorsal, extremamente reduzidos, não
pedunculados, mas posicionados dorso-lateralmente; operculum do olho em forma de
“V” sem ornamentação aparente em animais fixados e nadadeiras pélvicas totalmente
recobertas pelo disco.
Cauda ovalada, relativamente larga na origem, comprimida dorso-ventralmente e
afastada das bordas do disco na axila; comprimento variável com segmento filiforme
pós-ferrão geralmente ausente em indivíduos adultos; distintas quilhas laterais, uma a
cada lado da cauda, da linha da extremidade das nadadeiras peitorais até a origem do
ferrão caudal; quilhas ventral e dorsal ausentes; a cauda diminui de largura suavemente
até a origem do ferrão, quando sofre acentuada compressão lateral e forma um segmento
filiforme de comprimento variável, mas podendo ser igual à LD.
Nadadeiras pélvicas musculosas, relativamente curtas, macias e com alto grau de
mobilidade; margem anterior ligeiramente convexa com borda espessa e arredondada,
cabendo 6,1 vezes na largura de disco; ápice com ponto de inflexão distinto; margem
posterior reta, com endentações (17/16 esquerda/direita) resultantes da disposição das
estruturas cartilaginosas de sustentação da nadadeira.
Comprimento pré-narial 3,7 vezes menor que o comprimento do disco, 3,9 vezes
maior que a largura internarial e 30,3 vezes maior que a abertura anterior da narina;
aberturas circulares com interior claro; aberturas posteriores medialmente posicionadas
em relação às aberturas anteriores, lado a lado do frenum nasal; cortina nasal trapezóide,
com a margem posterior franjada, arqueada e alcançando a boca nas comissuras,
dividida em dois lóbulos pelo frenum nasal.
Boca localizada no primeiro 1/3 do comprimento do disco e sua largura cabendo
12,5 vezes na largura do disco; distante do cesto branquial a uma distância

35
aproximadamente igual a 4/5 do comprimento do cesto branquial; boca parcialmente
recoberta pela cortina nasal, sem sulcos labiais ou fossa oral, levemente arqueada
quando fechada e com os dentes não visíveis. Boca sem papilas orais desenvolvidas. A
válvula espiral é do tipo cônico-espiral com 14 voltas.
A largura do cesto branquial é 5,2 vezes menor que a largura do disco e seu
comprimento é 10,6 vezes menor que o comprimento do disco; as fendas são sinuosas e
estão distribuídas obliquamente em forma de arco, a cada lado, que segue da primeira à
quinta fenda.
A cloaca está entre as nadadeiras pélvicas, mergulhada na musculatura onde
apresenta uma forma elipsóide e uma fenda longilínea; a musculatura associada é
longitudinal à fenda e a mantém fechada.
Os cláspers são robustos, curtos e cônicos, recobertos por tecido macio, sem
feições distintivas significantes que distinga Paratrygon aiereba das demais espécies do
gênero; todas as feições do clásper estão na face dorsal e lateral; a face ventral é lisa e
não ornamentada.
Dentição arranjada em leve mosaico, desde a sínfise até as comissuras. Dentes
predominantemente losangulares (Fig. 5fgh), mais largos que longos e com cúspide
pronunciada. As fórmulas dentárias estão expressas na Tabela 1.
Dorso áspero, recoberto por pequenos dentículos dérmicos dispersos densamente
sobre a região mediana do corpo e ao longo da cauda; margens do disco com menor
espinulação, bem como o dorso das nadadeiras pélvicas, onde somente escassos
dentículos dérmicos são encontrados aleatoriamente. Dorso e laterais da cauda
recobertos com dentículos dérmicos; espinhos dispersos aleatoriamente sobre a cauda
até o ponto distal do ferrão; espinhos acessórios ocasionais. Os dentículos dérmicos

36
recobrem densamente os espaços entre os espinhos e laterais da cauda; não ocorrem
dentículos dérmicos no ventre da cauda antes da linha de origem do ferrão. Os espinhos
são arqueados posteriormente, recobertos por integumento pigmentado segundo o
colorido da pele, ligeiramente dispersos em indivíduos jovens ou densamente arranjados
em indivíduos adultos.
Identificação
Chave de identificação com base em caracteres externos para as espécies de
potamotrygonídeos do rio Tocantins (modificado a partir de ROSA (1985)).
1A. Distância da boca à margem anterior do disco (Comprimento Pré-Oral)
relativamente longa, 2,6 a 3,4 vezes na largura do disco; processo em forma de gota na
margem posterior dos espiráculos; olhos não pedunculados..............Paratrygon aiereba
MULLER & HENLE
1B. Distância da boca à margem anterior do disco (Comprimento Pré-Oral)
relativamente curta, 3,6 a 5,6 vezes na largura do disco; espiráculos sem processos em
forma de gota na margem posterior; olhos pedunculados.................................................2
2A. Sulco labial proeminente ou levemente demarcado; uma única fileira de espinhos na
cauda, podendo ocorrer espinhos acessórios*...................................................................3
2B. Sulco labial ausente; uma a três fileiras de espinhos na cauda*.................................4
3A. Sulco labial proeminente; dentes muito pequenos com menos de 1 mm de largura
da coroa em indivíduos adultos; forte policromatismo do colorido dorsal do disco,
variando entre reticulado, pontilhado, ocelado, vermiculado a roseta-modificada;

37
colorido ventral da cauda freqüentemente alternado entre amarelo e preto desde de sua
origem (Fig.X)...................................................................Potamotrygon orbignyi
CASTELNAU
3B. Sulco labial levemente demarcado; dentes relativamente maiores com mais de 2
mm de largura da coroa em indivíduos adultos; colorido dorsal com padrão spot;
colorido ventral da cauda freqüentemente com a base clara, com pigmentação após a
linha ventral referente ao ponto de origem do ferrão...Potamotrygon scobina GARMAN
4A. Sulco labial ausente; duas ou três fileiras de espinhos na cauda, podendo ocorrer
espinhos acessórios; colorido dorsal com ocelos ou spots e vermiculações claros (branco
ou laranja-avermelhado) em fundo preto ou cinza escuro; ocelos ou spots se estendendo
até a ponta da cauda; colorido ventral predominantemente cinza
escuro.................................................................................................................................5
4B. Sulco labial ausente; uma única fileira de espinhos na cauda, podendo ocorrer
espinhos acessórios; colorido com ocelos predominantemente laranja-avermelhado em
fundo marrom castanho ou marrom claro; ocelos não se estendem até a ponta da cauda,
restringindo-se à sua base; colorido ventral predominantemente claro........Potamotrygon
motoro NATTERER**
5A. Colorido dorsal com ocelos ou ocelos margeados por spots......................................6
5B. Colorido dorsal com spots, rosetas-modificadas ou vermiculações....Potamotrygon
sp. A.
6A. Dentes nas séries mais externas (dentes em uso) são brancos, pentagonais ou
hexagonais, mas predominantemente hexagonais e com coroa plana; até 27 dentes na
arcada superior e 33 dentes na arcada inferior (exemplar fêmea com 660 mm
LD)............................................................................Potamotrygon henlei CASTELNAU

38
6B. Dentes nas séries mais externas (dentes em uso) são caramelo, pentagonais ou
hexagonais, mas predominantemente pentagonais e com a coroa irregular e com uma
nítida crista no sentido da largura da boca; até 34 dentes na arcada superior e 39 dentes
na arcada inferior (exemplar fêmea com 710 mm
LD)......................................................................................................Potamotrygon sp. B
* As fileiras de espinhos na cauda aumentam com o tamanho do animal e desta forma,
neonatos e juvenis devem ser considerados com cuidado, pois todos possuem somente
uma única fileira de espinhos na cauda.
** A ocorrência de P. motoro para a porção do rio Tocantins à montante da barragem de
Tucuruí deve ser considerada com cuidado. Alguns exemplares de P. orbignyi
apresentam o padrão de colorido ocelado muito semelhante ao de P. motoro. Contudo,
P. motoro não possui sulco labial. Todos os exemplares analisados de P. motoro para
este estudo estavam em museus e foram capturados à jusante desta barragem e nenhum
exemplar de P. motoro foi capturado nas coletas. No entanto, esta espécie é considerada
na chave de identificação devido à possibilidade de ocorrência.
Distribuição de Potamotrygon orbignyi no rio Paranã
As raias foram coletadas em 33 locais diferentes do rio Paranã em 46 dias de
captura nas proximidades do município de Paranã dentro de um limite de 10 km à
montante e jusante deste município (Figura 1). A maior captura em um único dia foi de
17 raias em dois locais de captura, Barra do Genipapeiro e Ilha do Bonilha. Em 26 dias
de captura, ou 56% do esforço total, somente um ou dois indivíduos foram capturados,
enquanto que 76% das raias foram capturadas em apenas 16 dias, ou 34% do esforço

39
total. As maiores capturas diárias, acima de 5 indivíduos, ocorreram em um a quatro
pontos de pesca no mesmo dia. Em 81% (13 dias) dos dias de maiores capturas, apenas
um ou dois pontos foram responsáveis pela captura, enquanto que nos dias restantes (3
dias) quatro pontos registraram capturas. Embora o esforço de pesca tenha sido maior
nos pontos mais próximos do município, a saber: Arrozal, Auleria, Barra da Areia, Ilha
Bonita, Ilha do Bonilha, Micaela, Praia do Meio, Praião da Lagoa Verde, Praião do
Dominguinho, Praia do Retiro, Praia Três Bocas e São Vicente; somente as Praias do
Retiro, do Meio e Três Bocas e a Ilha do Bonilha apresentaram CPUE’s altas (8,3, 5,3, 4
e 5,7 indivíduos por dia, respectivamente). Nos demais pontos de captura a CPUE foi de
1 a 2,3 indivíduos por dia.
Tendo em vista o costume local de se amputar a cauda das raias de água doce
capturadas, antes de as devolverem ao rio, utilizou-se destes animais amputados como
exemplares marcados para uma estimativa de sua capacidade de dispersão. Desde que
não há comunidades ribeirinhas às margens do rio Paranã a pelo menos 50 km do
município de Paranã (montante e jusante), considerou-se que todas as raias amputadas
ocorrentes na região provinham da ação dos pescadores locais, já que em áreas distantes
de comunidades pesqueiras (alto Paranã e rio do Peixe no médio Araguaia) é rara a
ocorrência de animais amputados. As espécies encontradas com amputações foram P.
orbignyi e Potamotrygon sp. A, enquanto que nenhum exemplar de Paratrygon aiereba
apresentou evidências de amputações. De um lote de 147 exemplares de P. orbignyi
amostradas em três cruzeiros, 19 raias (13%) apresentavam a cauda amputada e
cicatrizada, mas não houve diferenças nas proporções de indivíduos amputados ao longo
dos três cruzeiros - 14%, 17%, 9% (n = 6, 7 e 6) de raias amputadas para o 1º, 2º e 3º
cruzeiros, respectivamente - (χ2 = 1,1678; p > 0,05). Dentre estas 19 raias amputadas, 8

40
raias (42%) foram capturadas na estação da seca, enquanto 11 raias (58%) foram
capturadas na estação das chuvas; não havendo diferenças significativas entre as
proporções de indivíduos amputados nestas duas estações (χ2 = 0,7647; p < 0,3818). A
grande maioria das raias amputadas encontrava-se dentro de um raio de 2 km do
município de Paranã, mas duas raias foram capturadas além deste perímetro, a
aproximadamente 3 e 4,5 km a jusante de Paraná, evidenciando portanto, sua restrita
mobilidade..
A distribuição das espécies de raia de água doce ao longo da bacia Tocantins-
Araguaia indica uma relação do número de espécies e o distanciamento da foz, onde nas
cabeceiras do rio Tocantins a riqueza de espécies é baixa – de uma a três espécies,
dependendo do afluente – em comparação com as porções médias e baixas com pelo
menos cinco espécies (Fig. 12). A espécie com maior distribuição geográfica na bacia
Tocantins-Araguaia é Paratrygon aiereba, ocorrendo inclusive nas cabeceiras do rio
Tocantins. A segunda espécie com maior distribuição geográfica é Potamotrygon
orbignyi, que aparentemente apresenta baixas densidades ou não ocorre em alguns
afluentes da cabeceira do Tocantins. Esta espécie apresenta a maior variabilidade de
coloridos no rio Tocantins e normalmente é confundida com outras espécies tais como
P. scobina, P. motoro e P. castexi. A raia de água doce P. scobina também apresenta
ampla distribuição, mas restringe-se ao rio Tocantins e à região da foz do rio Paranã
(MARQUES3, material não publicado), não tendo sido capturada no médio ou alto
Paranã. P. motoro apresenta ocorrência duvidosa na bacia Tocantins-Araguaia, e desta
forma, sua identificação e registro necessitam de confirmação. Potamotrygon sp. A é
uma espécie restrita ao alto e médio Tocantins, não sendo registrada para o baixo
3 Dra. Elineide Eugênio Marques. Rua Paraguaçú 450. Setor Umuarama. 77500-000 Porto Nacional, Tocantins. Fundação Universidade do Tocantins. Campus de Porto Nacional.
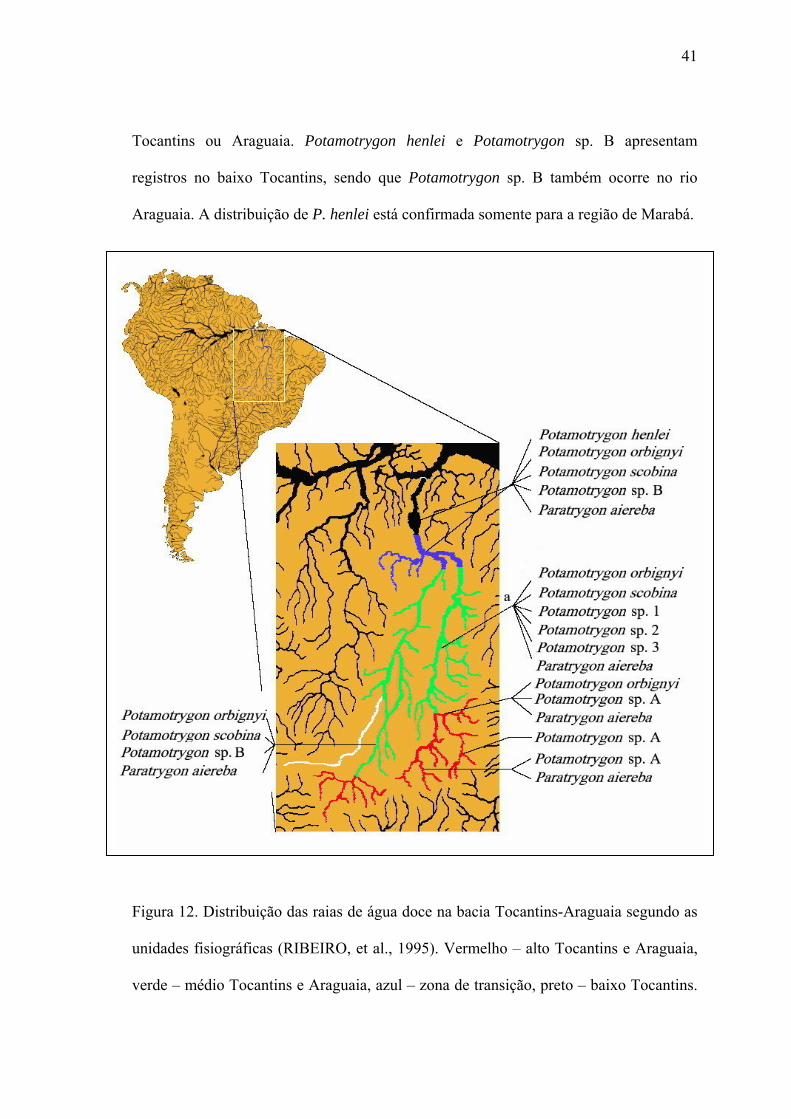
41
Tocantins ou Araguaia. Potamotrygon henlei e Potamotrygon sp. B apresentam
registros no baixo Tocantins, sendo que Potamotrygon sp. B também ocorre no rio
Araguaia. A distribuição de P. henlei está confirmada somente para a região de Marabá.
Figura 12. Distribuição das raias de água doce na bacia Tocantins-Araguaia segundo as
unidades fisiográficas (RIBEIRO, et al., 1995). Vermelho – alto Tocantins e Araguaia,
verde – médio Tocantins e Araguaia, azul – zona de transição, preto – baixo Tocantins.

42
“a” – Exemplares identificados pela equipe da Universidade do Tocantins-UNITINS;
Potamotrygon sp. 1 é sinônimo de P. orbignyi, Potamotrygon sp. 2 e 3 são sinônimos de
Potamotrygon sp. A.
Variabilidade do Colorido em Potamotrygon na Bacia Tocantins-Araguaia
Os padrões de colorido ocorreram freqüentemente mesclados e os animais foram
identificados segundo o seu padrão de colorido predominante. O padrão reticulado
correspondeu a 27% das raias amostradas, o padrão pontilhado a 14,8%, o padrão
vermiculado a 13,5%, o padrão roseta-modificada a 12,1% e os padrões ocelado e spot
corresponderam juntos a 32,4%. O colorido de uma mesma espécie de Potamotrygon
não é constante e varia tanto geograficamente quanto ao longo do crescimento do
animal. Enquanto embriões e neonatos de P. orbignyi apresentam o colorido
predominantemente nos padrões spot-ocelado (Fig. 3de), animais com mais de um mês
de nascimento apresentam padrões mesclados de ocelado-reticulado ou spot-reticulado.
Ao longo do crescimento esses padrões podem se alterar para padrões de colorido mais
complexos como o pontilhado → vermiculado → roseta-modificada. Esta variação é
confirmada com base na comparação das distribuições de larguras de disco (LD) para
cada padrão de colorido (Kruskal-Wallis-H = 18,21; p = 0,001), onde se observa que
indivíduos de menor porte apresentam os padrões ocelado (incluindo spot) e reticulado,
alternando ao longo do crescimento para os padrões pontilhado e vermiculado e
finalmente para o padrão roseta-vermiculado em indivíduos de maior porte (Fig. 13).
Em P. henlei e Potamotrygon sp. B observa-se um aumento da complexidade do
colorido ao longo do crescimento. Embriões e neonatos das duas espécies apresentam
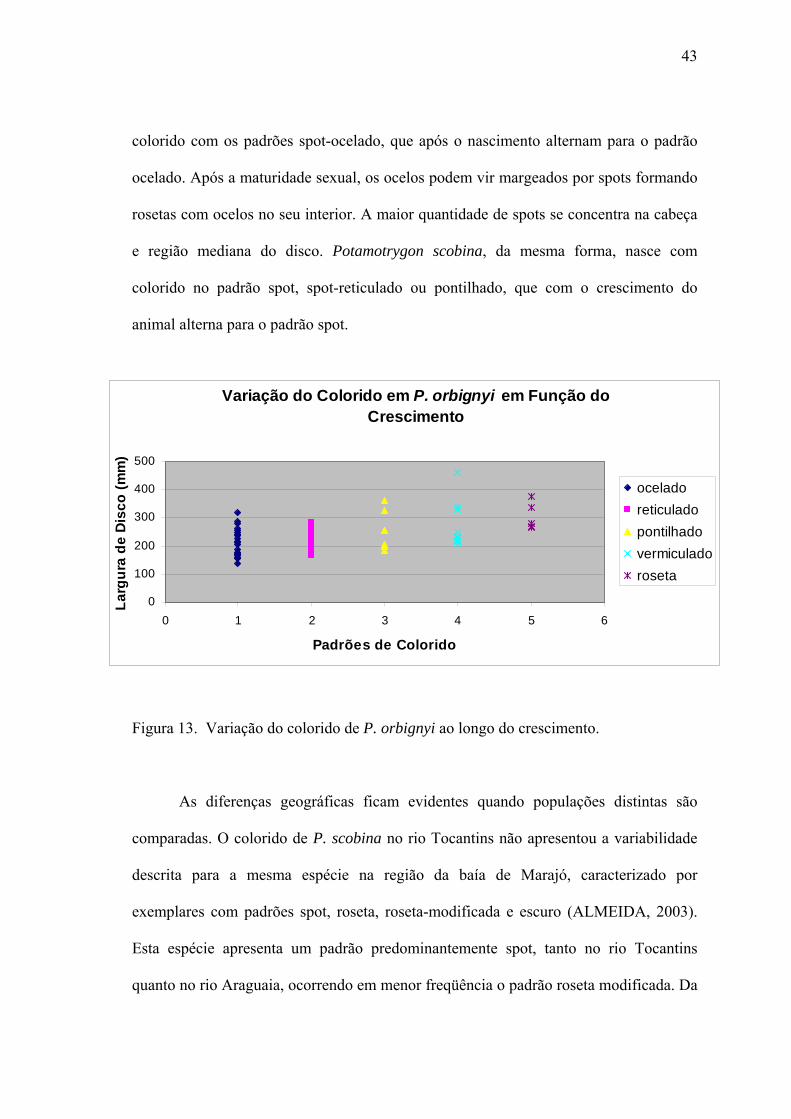
43
colorido com os padrões spot-ocelado, que após o nascimento alternam para o padrão
ocelado. Após a maturidade sexual, os ocelos podem vir margeados por spots formando
rosetas com ocelos no seu interior. A maior quantidade de spots se concentra na cabeça
e região mediana do disco. Potamotrygon scobina, da mesma forma, nasce com
colorido no padrão spot, spot-reticulado ou pontilhado, que com o crescimento do
animal alterna para o padrão spot.
Variação do Colorido em P. orbignyi em Função do Crescimento
0
100
200
300
400
500
0 1 2 3 4 5 6
Padrões de Colorido
Larg
ura
de D
isco
(mm
)
oceladoreticuladopontilhadovermiculadoroseta
Figura 13. Variação do colorido de P. orbignyi ao longo do crescimento.
As diferenças geográficas ficam evidentes quando populações distintas são
comparadas. O colorido de P. scobina no rio Tocantins não apresentou a variabilidade
descrita para a mesma espécie na região da baía de Marajó, caracterizado por
exemplares com padrões spot, roseta, roseta-modificada e escuro (ALMEIDA, 2003).
Esta espécie apresenta um padrão predominantemente spot, tanto no rio Tocantins
quanto no rio Araguaia, ocorrendo em menor freqüência o padrão roseta modificada. Da

44
mesma forma, P. orbignyi não apresenta o padrão vermiculado ou roseta-modificada na
baía de Marajó, o que a princípio gerou problemas de identificação dos exemplares
capturados no rio Paranã. Predominâncias de determinados padrões em afluentes ou
porções da bacia Tocantins-Araguaia foram observadas, mas o número de exemplares
amostrados ainda é insuficiente para uma identificação conclusiva dos padrões
predominantes por espécie e região nessa bacia hidrográfica.
O colorido predominante nas espécies ocorrentes na bacia Tocantins-Araguaia é
o ocelado, mas algumas espécies apresentam padrões mesclados principalmente de
ocelos e spots (P. henlei, P. orbignyi e Potamotrygon sp. B). Quando o padrão de
colorido predominante é plotado por espécie sobre a hipótese filogenética (Fig. 14),
observa-se um agrupamento das espécies com ocelos em um clado e das espécies com
colorido no padrão spot, ou derivado deste, em outro.
Figura 14. Padrões de coloridos predominantes das raias de água doce sobre a hipótese
filogenética de Marques (2000). Espécies com * ocorrem na bacia Tocantins-Araguaia.

45
Tendo em vista que o colorido ocelado provavelmente é uma derivação do
padrão spot, já que os embriões a termo e filhotes de espécies com padrão ocelado
apresentam o padrão spot antes de desenvolverem o padrão ocelado, é possível afirmar
que todas as espécies da hipótese filogenética em análise que ocorrem no rio Tocantins
apresentam o padrão spot ao menos em uma fase do seu desenvolvimento.
Ao considerarmos que o colorido dos filhotes provavelmente representa o
colorido mais geral (primitivo), enquanto o colorido dos adultos reflete os processos
adaptativos à caça, fuga de predadores e outros processos característicos da espécie
(ORTOLANI & CARO, 1996), e que o padrão spot é registrado em outras espécies não
considerados na hipótese filogenética ocorrentes na bacia Tocantins-Araguaia, é
possível inferir que o padrão spot é o padrão de colorido mais primitivo para as espécies
de Potamotrygon desta bacia hidrográfica.
DISCUSSÃO
Potamotrygon orbignyi foi identificada para o rio Paranã principalmente com
base na sua morfologia, tamanho e fórmula dentária, tendo em vista que esta espécie é a
única descrita para toda a bacia Tocantins-Araguaia com dentes levemente tricúspides,
larguras máximas da coroa em torno de 1 mm em indivíduos adultos e em números
superiores a 40 dentes por arcada. Estas características condizem com a descrição dos
dentes do holótipo de P. orbignyi (CASTELNAU, 1855; MNHN-2333) feitas por
ROSA (1985) em sua revisão da família Potamotrygonidae e considerados por este
autor como diagnósticas para a espécie. Embora o holótipo de P. orbignyi não possa
atualmente ser emprestado devido à fragilidade do material, é possível, através de

46
fotografias deste exemplar fornecidas por pesquisadores do Museu Nacional de História
Natural de Paris, comprovar a fileira única de espinhos na cauda e o colorido cinza
escuro. Análises de exemplares adicionais de P. orbignyi identificados por ROSA
(1985) e depositados em diversas instituições nacionais, e recentes avanços da pesquisa
em potamotrygonídeos desenvolvidos em conjunto com pesquisadores do Museu
Paraense Emilio Goeldi-MPEG e Universidade Federal da Paraíba-UFPB, identificaram
outras características potencialmente diagnósticas para esta espécie, tais como: presença
de fortes sulcos labiais, identificação dos padrões de colorido dorsal e ventral e fileira
única de espinhos caudais. Estas características, em conjunto, certamente identificam P.
orbignyi em qualquer fase da vida, ao contrário da chave de identificação anteriormente
proposta por ROSA (1985), que favorecia a identificação de exemplares adultos
somente com o padrão de colorido reticulado. A morfologia do endoesqueleto de P.
orbignyi também a difere das demais espécies do rio Tocantins no que tange a sua baixa
calcificação, permanecendo transparente em alguns pontos até mesmo em indivíduos
adultos, baixa inclinação da lâmina basal do crânio, maior largura relativa do forâmen
do nervo óptico, fossa parietal dupla e oblonga fontanela posterior. Embora LOVEJOY
(1996) tenha observado exemplares de P. orbignyi, nenhum dos caracteres citados
acima é abordado em sua análise sistemática da família Potamotrygonidae devido ao
fato deste autor não ter considerado mais intensivamente o neurocranium em seu estudo.
Um grande número de caracteres com marcante variabilidade interespecífica foi
observado no neurocranium, e desta forma, indicando sua potencialidade como fonte de
informações para análises filogenéticas. Na sua generalidade, as análises filogenéticas
da ordem Myliobatiformes não são totalmente conclusivas (ROSA, 1985; NISHIDA,
1990; LOVEJOY, 1996; McEACHRAN et al., 1996; MARQUES, 2000; DUNN et al.,

47
2003; McEACHRAN & ASCHLIMAN, 2004; ISÁIS & DOMINGUEZ, 2004,
CARVALHO et al., 2004), mas é consenso a identificação das Himanturas americanas
como grupo irmão das Potamotrygon e que estas deveriam ser incluídas na família
Potamotrygonidae (McEACHRAN et al., 1996). A validade dos caracteres escolhidos
nestes estudos e sua variabilidade intra e interespecíficas, necessitam confirmação
através de uma amostra maior e baseadas em identificações de campo fidedignas. De
outra forma, incorre-se no risco de utilizar caracteres com maior plasticidade e/ou
contaminação da amostra com espécies semelhantes, como por exemplo P. henlei e
Potamotrygon sp. B ou P. orbignyi e P. scobina.
O rio Paranã, que juntamente com o rio Maranhão formam os principais
afluentes da cabeceira do rio Tocantins (RIBEIRO et al., 1995), apresenta uma baixa
diversidade de espécies de raias de água doce quando comparado com as demais
porções do rio Tocantins. A baixa diversidade de espécies nas cabeceiras dos rios é um
padrão ecológico observado em rios tropicais (LOWE-McCONNELL, 1995) e muito se
deve a dificuldades de acesso a essas regiões e especificidade de habitats, com menor
variabilidade de nichos (RIBEIRO et al., 1995). No entanto, observa-se que as espécies
com maior distribuição geográfica na bacia Tocantins-Araguaia (Paratrygon aiereba e
Potamotrygon orbignyi) também são as únicas espécies registradas nestas cabeceiras,
evidenciando a sua alta adaptabilidade. Estas espécies também apresentam ampla
distribuição no restante da bacia Amazônica. Enquanto P. aiereba é registrada em quase
toda a bacia Amazônica, desde os seus principais afluentes (Ucayali, Solimões,
Amazonas, Negro, Branco e Madeira) até a bacia do rio Orinoco (CARVALHO et al.,
2003), P. orbignyi apresenta registros na bacia do rio Orinoco na Venezuela, na
Amazônia Colombiana, Guianas, Suriname e baixo Amazonas (ROSA, 1985;

48
CARVALHO et al., 2003). Segundo LOWE-McCONNELL (1995), é de se esperar
maior similaridade específica longitudinal ao longo da bacia Amazônica, o que se
reflete em maior semelhança entre as comunidades de raias de água doce entre as
Guianas, Suriname, foz do rio Amazonas e bacia Tocantins-Araguaia que entre rios
vizinhos como o Tapajós, Xingu e o Tocantins. Talvez isso explique o fato de haver ao
menos uma espécie endêmica em cada uma dessas bacias (CHARVET-ALMEIDA e
ARAÚJO4, com. pessoal) e um baixo número de espécies compartilhadas – somente P.
orbignyi e P. aiereba.
O registro de Potamotrygon sp. A., uma espécie nova na cabeceira do rio
Tocantins, evidencia a superficialidade do conhecimento acerca deste grupo de peixes e
a fragilidade das identificações destes animais em uma bacia supostamente bem
conhecida (COSTI et al., 1977; PAIVA, 1983; SANTOS, et al., 1984; SANTOS &
JEGU, 1989). A ocorrência de espécies novas nas porções médias e nas cabeceiras das
bacias hidrográficas é um padrão previsto (ROSA, 1985; RIBEIRO, et al., 1995) e
parece se confirmar também dentro dos elasmobrânquios de água doce. Contudo, a
identificação de uma nova espécie (Potamotrygon sp. B) em exemplares identificados
como P. henlei, uma espécie com grande importância econômica no mercado de peixes
ornamentais, evidencia não somente a fragilidade do conhecimento sobre a família
Potamotrygonidae, mas também o risco de depleção de um estoque antes mesmo de ser
identificado. O ordenamento da pesca ornamental por meio de quotas específicas no
Estado do Pará, que desde 2003 exportou aproximadamente 8000 exemplares de juvenis
e sub-adultos de três espécies diferentes de raias de água doce (P. henlei, P. leopoldi e
4 Patricia Charvet-Almeida: Universidade Federal da Paraíba-UFPB; [email protected]. Lúcia Góes de Araújo: Universidade Estadual do Amazonas; [email protected].

49
P. orbignyi), pode não estar sendo uma medida de manejo adequada devido a problemas
de identificação.
A distribuição de P. orbignyi nas coletas no rio Paranã, principalmente com base
nos indivíduos com a cauda amputada pelos pescadores e já cicatrizada, evidencia que
os indivíduos desta espécie não apresentam uma grande dispersão a partir do local onde
vivem e foram marcados. Embora a porcentagem de indivíduos marcados tenha variado
de 14 a 9% entre o primeiro e terceiro cruzeiros, esta variação não foi estatisticamente
significativa e provavelmente reflete muito mais o acaso do que um decréscimo de
indivíduos marcados devido à amostragem. O número de indivíduos marcados
capturados na estação das chuvas ou da seca foi muito próximo – 11 e 8,
respectivamente – indicando que estes animais possivelmente não efetuam migração ao
longo do rio, restando confirmar uma possível migração batimétrica ou de habitat.
Migrações batimétricas sazonais de elasmobrânquios para fins reprodutivos já foram
observadas em diversas espécies na costa brasileira tais como Squatina guggenheim,
Rhinobatos horkelii, Rhizoprionodon lalandii e Mustelus fasciatus (VOOREN, 1992,
1998; MOTTA, 2001; YOKOTA, 2005) e devido à variação do volume dos rios e
disponibilidade de alimentos, é possível que as raias de água doce troquem de habitat
entre as estações do ano. Os pescadores relatam que muitos exemplares de P. aiereba
são extremamente territorialistas e podem ser identificados ao longo do tempo por
preferirem certas bacias de pedras, raízes de árvores submersas ou entradas das margens
do rio, onde repousam longamente durante o dia. Segundo os pescadores, estes mesmos
animais são encontrados diversas vezes no mesmo local ao longo dos meses ou anos,
chegando a ponto de serem capturados, identificados, soltos e recapturados diversas
vezes. Este padrão de comportamento corrobora as evidências de baixa dispersão de P.

50
orbignyi e nos faz concluir que, ao menos em determinadas condições ambientais ou
trechos de rios, as raias de água doce parecem preferir permanecer em uma mesma
região ao longo de sua vida.
O colorido em elasmobrânquios tem gerado estudos sobre o seu mecanismo
fisiológico e suas implicações ecológicas (q. v. VISCONTI, et al., 1999 para uma
revisão dos processos fisiológicos; WILSON & DODD, 1973; GRUBER et al., 1975;
LOWE & GOODMAN-LOWE, 1996; KEMP, 1999). Particularmente nas raias de água
doce, o colorido tem sido historicamente utilizado como a principal característica
diagnóstica de muitas espécies (CASTELLO & YAGOLKOWSKI, 1969; ROSA, 1985)
e recentemente analisada e avaliada a sua variabilidade intra-específica (ALMEIDA,
2003). O forte policromatismo de P. orbignyi no rio Paranã corrobora os estudos de
ALMEIDA (2003), que também encontrou o mesmo padrão policromático em P.
scobina para a baía de Marajó e, desta forma, evidencia o policromatismo como uma
característica do gênero Potamotrygon. Esta característica se expressa na alternância de
padrões de colorido (spot, ocelado, pontilhado, reticulado, vermiculado e roseta-
modificada) ou no arranjo destes padrões sobre o dorso do animal. A alternância de
padrões de colorido ao longo do crescimento é sugerida por ALMEIDA (2003) para P.
scobina, que observou o padrão spot e roseta em indivíduos até 550 mm LD e o padrão
escuro como predominante em indivíduos maiores. Embora os fatores fisiológicos
envolvidos ainda não estejam totalmente esclarecidos, hormônios associados ao
crescimento e maturidade sexual (prolactina) podem estar relacionados a essa
alternância de padrões de colorido ao longo do crescimento do animal (VISCONTI et
al., 1999). Diversas outras espécies de elasmobrânquios também apresentam alternância
de padrão de colorido ao longo do crescimento (Galeocerdo cuvier, Chyloscyllium spp.,

51
Stegostoma fasciatum, Hemiscyllium spp. e outros) (COMPAGNO et al., 2005) e
acredita-se que a função ecológica desta alternância, principalmente em espécies
recifais, seja evitar o canibalismo intra-específico através de um aviso aos adultos ou
pela camuflagem (ALLEN, 1996). O surgimento do ocelo caudal em Cichla ocellaris, a
princípio como um spot, parece estar associado à comunicação intra-específica e ocorre
na fase juvenil mais vulnerável, logo após abandonar os pais (ZARET, 1977;
SCHROEDER & ZARET, 1979). A função ecológica desta alternância do colorido em
raias de água doce ainda requer maiores estudos tendo em vista que o canibalismo não
parece ocorrer nestes animais.
A análise de distribuição dos padrões de colorido sobre a hipótese filogenética
proposta por MARQUES (2000) indica uma predominância dos padrões spot-ocelado
no clado onde as espécies da bacia Tocantins-Araguaia se concentram. Embora P.
orbignyi, e P. scobina apresentem diversos padrões discutidos anteriormente, os
resultados apresentados neste estudo, ROSA (1985) e ALMEIDA (2003), evidenciam
que o padrão spot, ou variações diretas deste (roseta e ocelado), são predominantes
nestas espécies na bacia Tocantins-Araguaia. Desta forma, P. orbignyi e P. scobina
foram consideradas como spot-ocelado e spot, respectivamente. O padrão geral do
colorido em adultos, aliado ao colorido dos embriões e filhotes, que provavelmente
representa o colorido mais geral (primitivo) e não adaptado à caça, fuga de predadores
e/ou reprodução (ORTOLANI & CARO, 1996), evidenciam um colorido
predominantemente spot para as espécies do rio Tocantins e por conseqüência, o mesmo
padrão de colorido como o mais primitivo dentre as espécies do rio Tocantins. Contudo,
deve-se considerar a possibilidade de que o padrão spot em embriões e neonatos reflita
tão somente a incapacidade dos cromatóforos de se organizarem em padrões distintos ou

52
que o padrão spot esteja sendo determinado por uma relação filogenética maior que
englobe todas as espécies do gênero Potamotrygon - presumindo que o gênero seja
monofilético - e não somente a expressão da relação filogenética entre as espécies do rio
Tocantins. Ao analisar-se a hipótese filogenética proposta por MARQUES (2000)
baseada em seqüências genéticas do citocromo-b, deve-se considerar também a baixa
amostragem taxonômica perante a diversidade do gênero Potamotrygon e o método
utilizado. Tendo em vista o baixo número de espécies do gênero Potamotrygon, pelo
menos 16 espécies segundo avaliações recentes (CARVALHO et al., 2003), é
recomendável que uma análise filogenética seja feita com o maior número possível de
taxa a fim de se garantir a sua precisão (HILLIS, 1996).

53
LITERATURA CITADA
ALLEN, T. B. Shadows in the sea. The Sharks, skates and rays. Lyons & Burford
Publishers. 354p. 1996.
ALMEIDA, M. P. Pesca, policromatismo e aspectos sistemáticos de Potamotrygon
scobina Garman, 1913 (Chondrichthyes: Potamotrygonidae) da região da Ilha de
Colares – Baía de Marajó – Pará. Dissertação de Mestrado. Universidade Federal
do Pará e Museu Paraense Emílio Goeldi. 145p. 2003.
ARAÚJO, M. L. G. Biologia reprodutiva e pesca de Potamotrygon C (Chondrichthyes-
Potamotrygonidae), no médio rio Negro, Amazonas.171p. 1998.
BROOKS, D. R., THORSON, T. B. & MAYES, M. A. Freshwater stingrays
(Potamotrygonidae) and their helminth parasites: testing hypotheses of evolution
and coevolution. In: FUNK, V. A. & BROOKS, D. R. (eds.). Advances in
cladistics. Proceedings of the first meeting of the Willi Henning Society. New
York Botanical Garden, New York. p. 147-175. 1981.
CARVALHO, M.R., LOVEJOY, N.R. & ROSA, R.S. Potamotrigonidae. In: REIS,
R.E., KULLANDER, S. O. & FERRARIS, C. J. JR. (eds) Check List of the
Freshwater Fishes of South and Central America. Edipucrs, Pontifícia
Universidade Católica do Rio Grande do Sul. Porto Alegre. p. 22-28. 2003.
CASTELNAU, F. L. Animaux nouveaux ou rares recueillis pendant l’expédition dans
lês parties centrales de l’Amerique du Sud, de rio de Janeiro a Lima, et de Lima au
Para. Vol. 2. P. Bertrand, Paris. p. 101-103. Pl. 48. 1855.
CASTELLO, H. P. & YAGOLKOWSKI, D. R. Potamotrygon castexi n. sp., una nueva
especie de raya de agua dulce del rio Paraná. Acta Scientifica. ILAFIR. 21p. 1969.

54
CASTEX, M. N. Una nueva espécie de raya fluvial: P. labradori. Neotropica, La Plata,
9 (30). p. 117-121. 1963.
CASTEX, M. N. Estado actual de los estúdios sobre la raya fluvial neotropical. Ver.
Mus. Prov. Cs. Nat. Santa Fé. Número extraordinário del cincuentenario. p. 9-34.
1964.
CASTEX, M. N. Bases para el estúdio de las rayas de água dulce del sistema
Amazônico. Nuevas sinonímias de “P. motoro” (M. H., 1841). Atas do Simpósio
sobre a biota Amazônica. vol. 3 (Limnologia). P. 89-92. 1967.
CASTEX , M. N. & CASTELLO, H. P. Potamotrygon leopoldi, uma nueva espécie de
raya de água Dulce para el rio Xingu, Brasil (Chondrichthyes, Potamotrygonidae).
Acta Scientifica. ILAFIR. No 10. 16 p. 1970.
CHARVET-ALMEIDA, P. Ocorrência, biologia e uso das raias de água doce na Baía
de Marajó (Pará, Brasil), com ênfase na biologia de Plesiotrygon iwamae
(Chondrichthyes: Potamotrygonidae). Dissertação de mestrado, Programa de Pós-
graduação em Zoologia da Universidade federal do Pará e Museu Paraense Emílio
Goeldi. Departamento de Zoologia. 213p. 2001.
COMPAGNO, L. J. V. Systematics and body form. In: HAMLETT, W. C. (ed.) Sharks,
Skates and Rays.The Johns Hopkins University Press. Baltimore, p.02 – 42, 1999.
COMPAGNO, L. J. V., DANDO, M. & FOWLER, S. Sharks of the world. Princeton
Field Guides. Princeton University Press. 368p. 2005.
COSTI, C. G., RODRIGUES, C. C. B, PAIVA-FILHO, D. L. & RODRIGUES, J. B. R..
Estudo preliminar da ictiofauna do pólo Araguaia-Tocantins. Projeto de pesquisa
da Ictiofauna do pólo Araguaia-Tocantins, GO-Polamazônia. Superintendência do
Desenvolvimento da Pesca-SUDEPE. Brasília. 171 p. 1977.

55
DUNN, K. A., McEACHRAN, J. D. & HONEYCUTT, R. L. Molecular phylogenetics
of myliobatiform fishes (Chondrichthyes: Myliobatiformes), with comments on
the effects of missing data on parsimony and likelihood. Molecular Phylogenetics
and Evolution. 27, p. 259-270. 2003.
FERREIRA, E. J. G., ZUANON, J. A.S. & SANTOS, G. M. Peixes comerciais do
médio Amazonas. Região de Santarém-Pará. MMA-IBAMA. p. 17-27. 1998.
GALVIS, G., MOJICA, J. I. & CAMARGO, M. Peces Del Catatumbo. Asociación
Cravo Norte. 178p. 1997.
GARMAN, S. Plagiostomia – Sharks, skates and Rays. Benthic Press, Los Angeles,
California. 511 p. 1997.
GÉRY, J. The fishes of Amazonia. In: SIOLI, H. (ed.). The Amazon. Limnology and
landscape ecology of a mighty tropical river and its basin. Monographiae
Biologicae. Vol. 56. 353-370p. 1984.
GONZÁLES-ISÁIS, M. & DOMÍNGUEZ, H.M.M. Comparative Anatomy of the
Superfamily Myliobatoidea (Chondrichthyes) with some comments on phylogeny.
Journal of Morphology, 262(1): p. 517-535, 2004.
GRUBER, S. H., HAMASAKI, D. I. & DAVIS, B. L. Window to the epiphysis in
sharks. Copeia. p. 378-380. 1975.
HERMAN, J., HOVESTADT-EULER, M. HOVESTADT, D.C. & STEHMANN, M.
Contribution to the study of the comparative morphology of teeth and other
relevant ichthyodorulites in living supraespecific taxa of Chondrichthyan fishes.
Part B: Batomorphii 4b: Order Rajiformes – Suborder Myliobatoidei – Sperfamily
Dasyatoidea – Family Dasyatidae – Subfamily Dasyatinae – Genera: Taeniura,
Urogymnus, Urolophoides – Subfamily Potamotrygoninae – Genera: Disceus,

56
Plesiotrygon, and Potamotrygon (incl. supraespecific taxa of uncertain status and
validity), Family Urolophidae – Genera: Trygonoptera, Urolophus and Urotrygon
– Superfamily Myliobatidea – Family: Gymnuridae – Genus: Aetoplatea. Bulletin
de L’Institut Royal des Sciences Naturelles de Belgique. 69:161-200. 1999.
HILLIS, D. M. Inferring complex phylogenies. Nature, 383:130, 1996.
KEMP, N. E. Integumentary system and teeth. In: HAMLETT, W. C. (ed.) Sharks,
Skates and Rays.The Johns Hopkins University Press. Baltimore, p.02 – 42, 1999.
LOVEJOY, N. R. Systematics of myliobatoid elasmobranches: with emphasis on the
phylogeny and historical biogeography of neotropical freshwater stingrays
(Potamotrygonidae:Rajiformes). Zool. J. Linn. Soc. 117, p. 207-257. 1996.
LOVEJOY, N. R., BERMINGHAM, E., MARTIN, A. P. Marine incursion into South
America. Nature 396, p. 421-422. 1998.
LOWE, C., GOODMAN-LOWE, G. Suntanning in hammerhead sharks. Nature 383, p.
677. 1996.
LOWE-McCONNELL, R.H. Fish communities in tropical freshwaters. Longman Group
Limited. London. 337p. 1975.
LOWE-McCONNELL, R.H. Ecological studies in tropical fish communities. Terceira
Edição. Cambridge University Press. Cambridge. 1995.
MARQUES, F. P. L. Evolution of neotropical freshwater stingrays and their parasites:
taking into account space and time. Tese de doutorado. Department of Zoology.
University of Toronto, 325 p. 2000.
McEACHRAN, J. D., DUNN, K. & MIYAKE, T. Interrelationships of batoid fishes. In:
STIASNNY, M. L. J., JOHNSON, G. D., PARENTI, L. (eds.) Interrelationships
of fishes. Academic Press. San Diego. p. 63-84. 1996.

57
McEACHRAN, J. D. & ASCHLIMAN, N. Phylogeny of batoidea. In: CARRIER, J.
C., MUSICK, J. A. & HEITHAUS, M. R. Biology of sharks and their relatives.
CRC Press. Boca Raton. 79-113p. 2004.
MOTTA, F. S. A. A pesca artesanal e a reprodução de Rhizoprionodon lalandii
(Elasmobranchii-Carcharhinidae) no litoral sul do estado de São Paulo. Rio Claro,
2001. 88p. Dissertação de Mestrado em Zoologia. Universidade Estadual Paulista-
UNESP.
NISHIDA, K. Phylogeny of the suborder Myliobatoidei. Mem. Fac. Fish. Hok. Univ.
vol. 37. 108p. 1990.
ORTEGA, H. & VARI, R. P. Annotated checklist of the freshwater fishes of Peru.
Smithsonian Institution Press. Washington. 25p. 1986.
ORTOLANI, A. & CARO, T. M. The adaptive significance of colour patterns in
carnivores: phylogenetic tests of classic hypotheses. In: GITTLEMAN, J. L. (ed.).
Carnivore behavior, ecology and evolution. Vol. 2. Ithaca, Cornell University
Press: 132-188p. 1996.
PAIVA, M. P. Peixes e pescas de águas interiores do Brasil. Editerra, Brasília-DF.
158p. 1983.
PANTANO-NETO, J. Estudo preliminar da anatomia descritiva e funcional associada à
alimentação em raias de água doce (Potamotrygonidae, Myliobatiformes,
Elasmobranchii). Dissertação de mestrado, Instituto de biociências da
Universidade de São Paulo. Departamento de Zoologia. 93 p. 2001.
PLANQUETTE, P., KEITH, P. & LE BAIL, P. Y. Atlas des poisons d’eau douce de
Guyane. Muséum National D’Histoire Naturelle. Collection du Patrimoine
Naturel. vol. 22. p. 38-41. 1996.

58
RIBEIRO, M. C. L. B., PETRERE, M. & JURAS, A.A. Ecological integrity and
fisheries ecology of the Araguaia-Tocantins river basin, Brazil. Regulated Rivers:
Research and Management. v. 11, p. 325-350. 1995.
ROSA, R. S. A systematic revision of the south American freshwater stingrays
(Chondrichthyes: Potamotrygonidae). Dissertation for the degree of doctor of
philosophy, College of William and Mary Virginia. Faculty of the School of
Marine Science. 523 p. 1985.
ROSA, R. S., CASTELLO, H. P. & THORSON, T. B. Plesiotrygon iwamae, a new
genus and species of neotropical freshwater stingray (Chondrichthyes:
Potamotrygonidae). Copeia. No 2, p. 447-458. 1987.
SANTOS, G. M., JEGU, M. & MERONA, B. Catálogo de peixes comerciais do baixo
rio Tocantins. Projeto Tucuruí, ELETRONORTE/CNPq/INPA, Manaus, Brasil.
1984.
SANTOS, G. M. & JEGU, M. Inventário taxonômico e redescrição das espécies de
Anostomídeos (Characiformes: Anostomidae) do baixo rio Tocantins, PA. Brasil.
Acta Amazônica, 19(único), p. 159-213. 1989.
SANTOS, A.A. Morfologia externa, alimentação e reprodução de raias do alto e médio
rio Tocantins (Chondrichthyes, Potamotrygonidae). Relatório técnico-científico
final, Direção de Pesquisa e Extensão da Universidade do Tocantins. Curso de
Ciências Biológicas, (Material não publicado). 2000.
SCHROEDER, S. L. & ZARET, T. M. The adaptive significance of color patterns in
Cichla ocellaris. Copeia 1, p. 43-47. 1979.
SCHWARTZ, F. J. Biology of the clearnose skate, Raja eglanteria, from North
Carolina. Biol. Sci. (2). p. 82-95. 1996.

59
STEHMANN, M. Batoid fishes. In: FISHER, W., BIANCHI, G. & SCOTT, W. B.
(eds.) FAO Species identification sheets for fisheries purposes. Eastern Central
Atlantic. Fishing Area 31. p. 128-167. 1977.
VIEIRA, S. Bioestatística - Tópicos avançados. Teste não-paramétricos, tabelas de
contingência e análise de regressão. Editora Campus. Rio de Janeiro. 212 p. 2003.
VISCONTI, M. A., RAMANZINI, G. C. CAMARGO, C. R. & CASTRUCCI, A. M. L.
Elasmobranch color change: A short review and novel data on hormone
regulation. J. Exp. Zool. 284, p. 485-491. 1999.
VOOREN, C. M. Elasmobrânquios demersais. In: SEELIGER, U. & CASTELLO, J. P.
Os ecossistemas costeiro e marinho do extreme sul do Brasil. Editora Ecoscientia.
Rio Grande 157-162p. 1998.
VOOREN, C. M. Strategies reproductives compares de huit espèces de selaciens
vivipares d used du Brésil. Bull. Soc. Zool. Fr. 117(3), p. 303-312. 1992.
WILSON, J. F. & DODD, J. M. The role of the pineal complex and lateral eyes in the
colour change response of the dogfish. J. Endocrinol., 58, p. 591-598. 1973.
YOKOTA, L. Caiçara do Norte (RN): Um berçário de tubarões e raias? Recife. 89p.
2005. Dissertação de mestrado. Universidade Federal de Pernambuco-PE.
ZARET, T. M. Inhibition of cannibalism in Cichla ocellaris and hypothesis of predator
mimicry among South American fishes. Evolution 31, p. 421-437. 1977.

60
Anexo 1. Exemplares adicionais analisados.
Potamotrygon henlei – INPA-1762 (4 exemplares machos imaturos); 175 mm LD, 133
mm LD, 160 mm LD, 126 mm LD; Igarapé Valentim, Rio Tocantins-PA, 06/jul./82,
Det. Ricardo Rosa-1988. INPA-1748 fêmea com 172 mm LD; Jatobal, Rio Tocantins-
PA, 05/jul/82, Det. Ricardo Rosa. INPA-1749 (2 machos imaturos); 203 mm LD e 150
mm LD; Rio Tocantins (Igarapé Canoal), Breu Branco-PA, Nov./81, Det. Ricardo
Rosa-1988, Col. Equipe do INPA.
Potamotrygon scobina – INPA-20465 macho imaturo com 110 mm LD, Rio Xingu –
Altamira, Ilha do Sr. Izaltino (abaixo da ilha do Massaranduba) em 05/out./96, Col.
Zuanon. INPA-10157 fêmea imatura com 98 mm LD; Rio Amazonas, L. Grande em
Monte Alegre-PA, 5 a 12/ago./82, Col. Zuanon. INPA-1763 fêmea imatura com 129
mm LD; Rio Trombetas em Porto da Beira-PA, 08/out./85, Col. E. Ferreira e L. Py-
Daniel, Det. Ricardo Rosa-1988; exemplar identificado como P. orbignyi por Ricardo
Rosa, mas possui características de juvenil de P. scobina. INPA-15101 fêmea com
aproximadamente 270 mm LD; Rio Tefé – Jatuarana, 08/ago./79, Col. M. Gould.
Potamotrygon motoro – INPA-20478 Macho imaturo com 165 mm LD; Rio Araguaia,
Município de Caseara-TO, Lago de Volta Grande em 10/nov./00. INPA-15105 (2
machos, um adulto e o outro juvenil), 303 mm e 251 mm LD; Rio Tefé, Ressaca da
Paula em 03/ago./79, Col. W. M. Goulding, Det. Ricardo Rosa. INPA-1772 fêmea com
291 mm LD; Rio Tocantins a jusante da Barragem de Tucuruí-PA, V-85 Malha dura,
Det. Ricardo Rosa. INPA-15104 fêmea sem dados de coleta. Det. Ricardo Rosa.
MPEG-2466 fêmea imatura com 237 mm LD. MPEG-2468 macho adulto com 284

61
mm LD. UFPB-1474 fêmea com 172 mm LD; Volta Grande, Mun. Santo Antonio do
Leverger-MT, abr./1982, coletor J. B. Silva.
Potamotrygon sp. (Ilha de Marajó) – UFPB-3529 macho juvenil com 146 mm LD;
Praia Malhada, Cachoeira do Arari, rio Arari, Ilha de Marajó-PA, coletor C. S. Mendes.
Potamotrygon schroederi – F-0038, exemplar não tombado pela UFPB. Exemplar
fêmea com 293 mm LD.
Potamotrygon leopoldi – Sem numeração ou tombo pela UFPB. Exemplar fêmea com
297 mm LD.
Potamotrygon signata – UFPB-1518 macho juvenil com 155 mm LD; rio Sambito,
afluente do rio Poti, 22/ago./1986, coletor Jeremias P. S. Filho.
Potamotrygon magdalenae – UFPB-7638 macho adulto com 148 mm LD; coletor
Thomas B. Thorson. UFPB-7636 fêmea com 163 mm LD; coletor T. B. Thorson.
Potamotrygon yepezi – UFPB-1429 macho com 133 mm LD; rio Lemon, Venezuela,
6/dez./1982, coletor F. Mago-Leccia. UFPB-1427 macho adulto com 158 mm LD; lago
de Maracaibo, Venezuela, 1977, coletor T. B. Thorson.
Potamotrygon sp. (rio Negro) - UFPB-4867 macho adulto 150 mm LD; rio Itu, afluente
do rio Negro, Barcelos, out./2001, coletor M. L. G. de Araújo. UFPB-4866 fêmea com
160 mm LD; rio Itu, afluente do rio Negro, Barcelos, out./2001, coletor M. L. G. de
Araújo. UFPB-5217 fêmea com 233 mm LD; rio Itu, afluente do rio Negro, Barcelos,
3/jan./2001, coletor M. L. G. de Araújo.

62
CAPÍTULO II
Alimentação da Raia de Água Doce Potamotrygon
orbignyi no Rio Paranã – Alto Rio Tocantins
INTRODUÇÃO
As raias de água doce neotropicais pertencem à família Potamotrygonidae e
distribuem-se na maioria das bacias hidrográficas sul-americanas ao norte do rio da
Prata, com exceção daquelas que drenam para o Oceano Pacífico e da bacia do rio São
Francisco (ACHENBACH & ACHENBACH, 1976; BROOKS et al., 1981; ROSA,
1985; CARVALHO et al., 2003). Dezesseis espécies são reconhecidas em três gêneros,
sendo Paratrygon e Plesiotrygon gêneros monoespecíficos, e Potamotrygon com a
maior diversidade e 14 espécies (CARVALHO et al., 2003). Embora estas raias sejam
alvo de pesquisas desde o início do século XVIII (CASTEX, 1964), sua taxonomia e

63
biogeografia foram muito prejudicadas pelo forte policromatismo do grupo, descrições
superficiais, antigas, equivocadas e baseadas em poucos exemplares, perda de material
tipo, falta de informações precisas de coleta e revisões confusas (CASTELNAU, 1855;
GARMAN, 1913; CASTEX, 1963, 1964, 1967; ROSA, 1985).
A bacia Tocantins-Araguaia contém reconhecidamente seis espécies de raias de
água doce, a saber, Potamotrygon scobina, P. orbignyi, P. henlei, Potamotrygon sp. A
(alto e médio Tocantins) e Potamotrygon sp. B (baixo Tocantins e Araguaia) e
Paratrygon aiereba (ROSA, 1985; CARVALHO et al., 2003). Outras duas espécies
listadas para esta bacia, P. motoro e P. dumerilli, ainda não foram confirmadas ou, no
caso de P. dumerilli, foi sinonimizada como P. orbignyi (CARVALHO et al., 2003). O
rio Tocantins, principalmente na sua porção superior, é um rio que corre em canais, com
estreitas áreas de inundação, alta densidade de drenagem e gradiente íngreme (1400-600
m) (RIBEIRO et al., 1995). O rio Paranã é um dos dois principais afluentes formadores
do rio Tocantins e drena uma vasta área de inundação nas terras altas do Planalto
Central. Neste rio, apenas três espécies de raias de água doce foram registradas até o
momento, Paratrygon aiereba, Potamotrygon orbignyi e a espécie não descrita
Potamotrygon sp. A, o que confirma a previsão de ROSA (1985) de que as cabeceiras
das bacias hidrográficas poderiam conter espécies ainda não descritas.
A raia de água doce Potamotrygon orbignyi tem ampla distribuição geográfica
nas bacias dos rios Amazonas, Orinoco, Tocantins-Araguaia, Apure e em rios do
Suriname, Guiana e Guiana Francesa (ROSA, 1985; LASSO, 1985; LASSO et al.,
1996; CARVALHO et al., 2003). Embora apresente ampla distribuição em toda a bacia
Tocantins-Araguaia, pouco se sabe sobre a sua biologia reprodutiva, ecologia e relações
tróficas. LASSO et al., (1996), em estudos sobre a alimentação e reprodução de P.

64
orbignyi nos “llanos” venezuelanos da bacia do rio Apure, a identificam como
predadora de invertebrados aquáticos como larvas de insetos, crustáceos e Ostracoda.
PANTANO-NETO (2001), em estudo qualitativo da alimentação de P. motoro e P.
henlei em afluentes do rio Araguaia, corrobora com a identificação de P. orbignyi
(erroneamente identificada neste trabalho como P. castexi e possivelmente P. motoro)
como uma espécie basicamente insetívora. BRAGANÇA et al. (2004) também
identificam P. orbignyi como espécie predadora de invertebrados aquáticos tais como
isópodes, decápodes e larvas de díptera para a região da baía de Marajó (PA). Contudo,
surpreendentemente não há estudos quanto à cronologia da alimentação, câmbios
alimentares decorrentes do crescimento, estações do ano ou fases lunares e identificação
e avaliação do nível trófico desta espécie. Assim, o estudo da sua alimentação como
parte integrante da identificação de seu nicho é de fundamental importância para o
desenho das relações tróficas no ambiente onde vivem a fim de pautar a tomada de
decisões e medidas conservacionistas para todo o ecossistema.
Este capítulo tem por finalidade descrever a alimentação de Potamotrygon
orbignyi no rio Paranã, alto rio Tocantins, identificar sua posição trófica, variabilidade
ao longo do ano e possíveis câmbios de dietas decorrentes do crescimento e maturidade
do animal.
MATERIAL E MÉTODOS
O material analisado para o estudo da alimentação consiste de 83 exemplares de
Potamotrygon orbignyi coletados no médio rio Paranã, afluente do alto Tocantins, às
margens do município de Paranã-TO, próximo à divisa com o Estado de Goiás (Fig. 1),

65
com zagaia (arpão de mão) como arte de pesca ativa e espinhel de 25 anzóis como arte
de pesca passiva durante os anos de 2002 (janeiro-abril e agosto-dezembro) e 2003
(janeiro e agosto-setembro). No total, 181 indivíduos foram capturados em 38 dias de
coleta. Uma sub-amostra aleatória de 83 indivíduos foi escolhida para a análise da
alimentação. Desta sub-amostra, 67 indivíduos foram capturados em 26 dias de coleta.
Os demais 16 exemplares perderam suas etiquetas de campo no transporte ou manuseio.
As larguras de disco dos animais analisados variaram de 153 a 371 mm para machos e
de 150 a 460 mm LD para fêmeas (Fig. 2).
Figura 1. Mapa hidrográfico do Brasil e localização da região de coleta com imagem de
satélite (GoogleEarth). 1 – Rio Palmas e 2 – Rio Paranã. Escala = 2 km.
O alto Tocantins está compreendido na subregião de clima Xeroquimênico, que
é um clima tropical de monção, com um período seco no inverno e um período úmido
bastante acentuado, com chuvas torrenciais, no verão, sendo que a temperatura média
do mês mais frio é inferior a 20oC, o que condiciona a uma variação termoximênica
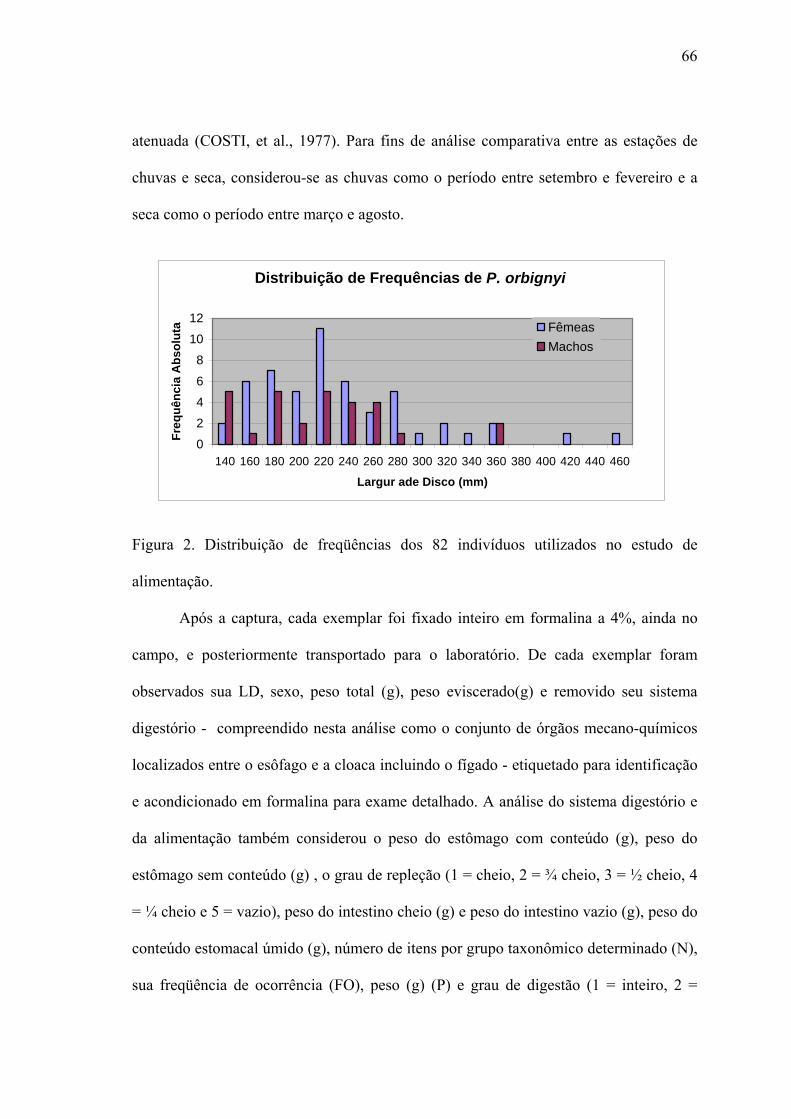
66
atenuada (COSTI, et al., 1977). Para fins de análise comparativa entre as estações de
chuvas e seca, considerou-se as chuvas como o período entre setembro e fevereiro e a
seca como o período entre março e agosto.
Distribuição de Frequências de P. orbignyi
0
2
4
6
8
10
12
140 160 180 200 220 240 260 280 300 320 340 360 380 400 420 440 460
Largur ade Disco (mm)
Freq
uênc
ia A
bsol
uta Fêmeas
Machos
Figura 2. Distribuição de freqüências dos 82 indivíduos utilizados no estudo de
alimentação.
Após a captura, cada exemplar foi fixado inteiro em formalina a 4%, ainda no
campo, e posteriormente transportado para o laboratório. De cada exemplar foram
observados sua LD, sexo, peso total (g), peso eviscerado(g) e removido seu sistema
digestório - compreendido nesta análise como o conjunto de órgãos mecano-químicos
localizados entre o esôfago e a cloaca incluindo o fígado - etiquetado para identificação
e acondicionado em formalina para exame detalhado. A análise do sistema digestório e
da alimentação também considerou o peso do estômago com conteúdo (g), peso do
estômago sem conteúdo (g) , o grau de repleção (1 = cheio, 2 = ¾ cheio, 3 = ½ cheio, 4
= ¼ cheio e 5 = vazio), peso do intestino cheio (g) e peso do intestino vazio (g), peso do
conteúdo estomacal úmido (g), número de itens por grupo taxonômico determinado (N),
sua freqüência de ocorrência (FO), peso (g) (P) e grau de digestão (1 = inteiro, 2 =

67
parcialmente digerido, 3 = digerido e identificado a grupo taxonômico superior e 4 =
digerido a resíduo). O Índice de Repleção Relativa (BM%) foi calculado como a razão
entre o peso úmido do conteúdo estomacal(g) e o peso total do animal (g).
Com base no número, freqüência de ocorrência e peso dos itens identificados,
foram calculados os seus respectivos N%, FO% e P%. O Índice de Importância Relativa
- IRI = FO% x (N% + P%) (PINKAS et al., 1971), modificado no uso do peso do item
ao invés de seu volume, e de seu respectivo IRI% (CORTÉS, 1997) e o Índice
Alimentar – IA = (FO% + P%)/∑( FO% + P%) (KAWAKAMI & VAZZOLER, 1980)
foram então calculados como índices de alimentação compostos para o cálculo do nível
trófico - NTk = 1 + ﴾∑ Pj x TLj ﴿, onde Pj é a proporção de um item alimentar e TLj é
o nível trófico deste item (CHRISTENSEN E PAULY, 1992; CORTÉS, 1999). Os
níveis tróficos dos itens alimentares invertebrados seguem VANDER ZANDEN et al.
(1997). Os níveis tróficos dos peixes identificados seguem o banco de dados do
FishBase para as espécies da Amazônia (www.fishbase.org) ou a média do nível trófico
da família identificada no conteúdo estomacal, caso a espécie não esteja listada no site.
Nos casos onde somente a categoria “peixes” é identificada, utilizou-se a média do nível
trófico para as espécies demersais do ecossistema amazônico listadas no FishBase
(NTmédio = 3,2; n = 157 espécies). Referências bibliográficas diversas como artigos
publicados em revistas científicas, resumos expandidos de congressos, anais de
congressos, relatórios, monografias e dissertações de mestrado também foram utilizados
como fonte de informação sobre a alimentação de diversas espécies de raias de água
doce para o cálculo de seus respectivos níveis tróficos a título de comparação com os
resultados obtidos neste trabalho.

68
A maturidade sexual (L50) nas fêmeas foi determinada por meio da presença de
folículos ovarianos com diâmetros maiores que 10 mm ou de fêmeas grávidas (L50 =
260 mm LD) e nos machos por meio da curva de crescimento do clásper (L50 = 241 mm
LD) e o grau de rigidez do mesmo (q. v. capítulo de reprodução). Com base nestes
valores de L50, os animais foram agrupados em juvenis e adultos para comparações
ontogenéticas de sua dieta.
Juvenis e adultos foram comparados entre si para cada sexo quanto aos
principais itens predados por meio do teste-χ2 em tabelas de contingência 2x2 com
correção de continuidade de Yates ou tabelas “rxs” (SOKAL & ROLF, 1969; VIEIRA,
2003). A alimentação para os principais itens alimentares foi comparada entre as
estações de seca e chuva e das fases lunares pelo teste-χ2 das freqüências destes itens
entre as estações e as fases lunares. As análises comparativas entre as estações de
chuvas e seca e entre os períodos diurno e noturno com base no BM% foram feitas por
meio do Teste-t, quando respeitadas suas premissas, ou pelo seu equivalente não
paramétrico Teste-U de Mann-Whitney. Para fins de análise circadiana da alimentação,
considerou-se o período diurno compreendido entre as 8:00 e 17:59 horas e o período
noturno entre as 18:00 e 7:59 horas. Todos os testes estatísticos foram executados pelos
programas STATISTICA 5.5 da STATSOFT e Excel 2002 da Microsoft e consideraram
o nível de significância de 5% (α = 0,05).
Todas as medidas foram tomadas com paquímetro em milímetros ou centésimos
de milímetros - paquímetro digital Starrett para o tamanho das presas, - e os pesos
medidos em balança digital Filizola modelo DC500 de precisão de 5 gramas para as
raias com mais de 500 gramas e com balança Plena Precision com precisão de 0,1
grama para raias com menos de 500 gramas. Os itens estomacais foram pesados em

69
balança Marte AL200c com precisão de milésimo de grama após secagem parcial em
papel toalha para a remoção do excesso de água.
Os itens estomacais foram identificados por meio de bibliografia especializada
(HYNES, 1970; McCAFFERTY, 1981; VOSHELL, 2003) e com auxílio da Dra. Maria
Julia Martins da Silva do Laboratório de Bentos Aquático da Universidade de Brasília-
UNB.
RESULTADOS
Morfologia do Sistema Digestório (Fig. 3)
Boca. Localizada ventralmente na porção anterior do disco, logo após as narinas e com
abertura que varia de 6,3 a 12% da largura do disco; parcialmente recoberta pela cortina
nasal. Fileiras centrais de dentes expostas com a boca fechada. Arcadas relativamente
frágeis e com musculatura associada pouco robusta. Sulcos labiais visíveis nos cantos
inferiores da boca, evidenciando uma relativa protractibilidade. A boca de indivíduos
formalizados se abre facilmente com a pressão dos dedos ou com um objeto introduzido
no interior da mesma.
Dentição. Arcadas armadas com fileiras de dentes arranjadas lado a lado e
acompanhando a arcada em diagonal. Dentes de machos com coroas monocúspides.
Fêmeas com coroas tricúspides; cúspide central maior e distinta e cúspides laterais
pouco evidentes. Dentes pequenos, cerca de 0,8 mm em indivíduos adultos, e com
distinto dimorfismo dignástico, monognástico e ginândrico; dimorfismo ontogenético
pouco evidente (maiores detalhes no capítulo 1).

70
Cortina oral. Desenvolvida e estendendo-se de uma comissura à outra; localizada
exatamente interior à arcada dentária, logo após a placa dentária; sua finalidade é
desconhecida, mas deve regular o fluxo de água para o interior da boca no momento da
ingestão dos alimentos.
Papilas orais. Localizadas em posição simétrica à da cortina oral, mas imediatamente
interior à cartilagem de Meckel. Apresenta cinco papilas orais desenvolvidas, próximas
entre si e centralizadas na boca.
Faringe. Cavidade trapezóide, ampla e livre de dentículos ou dentes para a mastigação;
não possui estruturas para a manipulação dos alimentos, que devem seguir diretamente
para o esôfago.
Esôfago. Estreitamento do sistema digestório a partir da faringe com desenvolvida
musculatura e evaginações do epitélio a fim de permitir a distensão do mesmo para a
entrada de alimentos maiores no estômago. Distensão limitada pela coluna vertebral e
barra coracóide da cintura peitoral.
Estômago. Órgão em forma de “J” com leve distinção entre o estômago cardíaco e
pilórico. Estômago cardíaco localizado imediatamente após o esfíncter fortemente
muscularizado e rico em evaginações do epitélio interno. Estômago pilórico sem
musculatura desenvolvida evidente e evaginações, semelhante a uma bolsa e terminando
em um esfíncter pilórico.
Intestino proximal. Logo após o esfíncter pilórico e anterior à válvula espiral existe
uma câmara relativamente ampla onde o alimento chega altamente hidratado.
Intestino. Órgão curto, cerca de duas vezes mais longo que largo; freqüentemente cheio
de areia e material em digestão; válvula espiral do tipo cônico-espiral, com 16
evoluções e segue até próximo ao final do intestino, que termina formando um reto com

71
um esfíncter. A glândula retal é visível na parede externa dorso-lateral esquerda do reto
e possui sua luz para dentro do mesmo. O bolo fecal chega ao reto e à cloaca
desidratado.
Figura 3. Sistema digestório de Potamotrygon orbignyi; a) Detalhe da boca em vista
ventral; b) Dentes superiores sinfisiais de fêmea No 815151 com 224 mm LD; c)
Sistema digestório em vista geral com boca e faringe à esquerda; d) Porção distal do
sistema digestório em vista ventral; e) Sistema digestório em vista ventral com o fígado
removido. a-cortina oral, b-arcada inferior, c-papilas orais, d-faringe, e-narinas, es-
esôfago, g-estômago, h-fígado, vb-vesícula biliar, i-intestino, r-glândula retal, ip-

72
intestino proximal, p-pâncreas, cg-estômago cardíaco, pg-estômago pilórico e cl-cloaca.
Escala em b é igual a 1 mm; escala nas demais figuras é igual a 20 mm.
A Captura
A pesca passiva com espinhel de fundo se mostrou pouco eficiente na captura de
qualquer espécie de raia de água doce, principalmente durante o dia, quando o
rendimento médio foi de 0,7 raias por dia. Desta forma, optou-se pela pesca ativa com a
zagaia, que apresentou um rendimento médio 2,57 indivíduos por dia. A proporção de
fêmeas e machos nos animais analisados foi de 1,7:1, indicando uma diferença
significativa em favor das fêmeas (χ2 = 5,90; p < 0,015). O rendimento diurno foi de 1,5
(18 animais em 12 dias) e noturno de 2,76 raias por dia (47 animais em 17 dias).
Embora as capturas diurnas e noturnas apresentem diferenças entre suas médias (1,5 vs.
2,76), essa diferença não se mostrou significativa (Teste-U de Mann-Whitney; Z = 1,87
e p = 0,06). A análise da captura por estação do ano revelou uma média para a seca de
2,33 e para as chuvas de 2,72 exemplares por dia, sem haver diferenças significativas
entre as estações (Teste-t; t = -0,54 e p = 0,59). A CPUE (captura por dia de pesca) por
fase lunar apresentou diferenças significativas (χ2 = 7,87; p = 0,019) entre as fases de
lua crescente, cheia e nova, evidenciando uma vulnerabilidade diferenciada de P.
orbignyi à pesca ativa. Na lua cheia a CPUE média foi a menor com 1,5 indivíduos por
noite, enquanto que nas luas crescente e nova a CPUE média foi respectivamente 4,5 e
2,3 indivíduos por noite. A fase de lua minguante não foi incluída na análise por ter sido
registrada em apenas dois dias de captura com média de 2 indivíduos por noite.
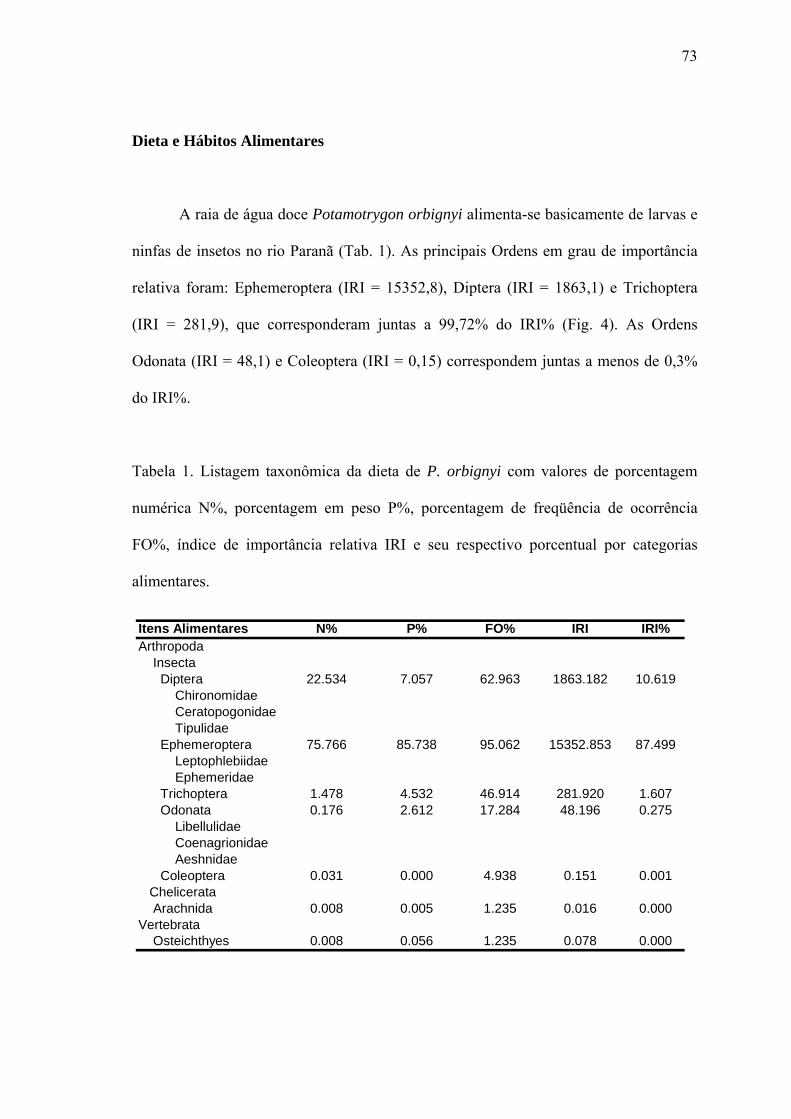
73
Dieta e Hábitos Alimentares
A raia de água doce Potamotrygon orbignyi alimenta-se basicamente de larvas e
ninfas de insetos no rio Paranã (Tab. 1). As principais Ordens em grau de importância
relativa foram: Ephemeroptera (IRI = 15352,8), Diptera (IRI = 1863,1) e Trichoptera
(IRI = 281,9), que corresponderam juntas a 99,72% do IRI% (Fig. 4). As Ordens
Odonata (IRI = 48,1) e Coleoptera (IRI = 0,15) correspondem juntas a menos de 0,3%
do IRI%.
Tabela 1. Listagem taxonômica da dieta de P. orbignyi com valores de porcentagem
numérica N%, porcentagem em peso P%, porcentagem de freqüência de ocorrência
FO%, índice de importância relativa IRI e seu respectivo porcentual por categorias
alimentares.
Itens Alimentares N% P% FO% IRI IRI%Arthropoda Insecta Diptera 22.534 7.057 62.963 1863.182 10.619 Chironomidae Ceratopogonidae Tipulidae Ephemeroptera 75.766 85.738 95.062 15352.853 87.499 Leptophlebiidae Ephemeridae Trichoptera 1.478 4.532 46.914 281.920 1.607 Odonata 0.176 2.612 17.284 48.196 0.275 Libellulidae Coenagrionidae Aeshnidae Coleoptera 0.031 0.000 4.938 0.151 0.001 Chelicerata Arachnida 0.008 0.005 1.235 0.016 0.000Vertebrata Osteichthyes 0.008 0.056 1.235 0.078 0.000

74
Figura 4. Diagrama IRI% com valores de FO%, N%, P% para os principais itens
alimentares de P. orbignyi (n = 83). Os três itens mais importantes corresponderam a
99,7% da dieta desta espécie.
Um único fragmento de peixe (IRI = 0,07) e uma aranha (IRI = 0,01) foram
registrados, mas correspondem a menos de 0,0005% de importância relativa percentual.
Itens digeridos a resíduo também foram encontrados e classificados como massa
amorfa, o que quase sempre correspondeu à massa digerida de larvas e ninfas de insetos,
evidenciado por uma grande quantidade de partes soltas destes animais, tais como
antenas, articulações e carapaças.
A massa amorfa, quando considerada no cálculo do IRI, alcançava o segundo
grau de importância relativa devido aos seus altos valores de peso (50,2%) e à sua
freqüência (82,7%), perdendo somente para Ephemeroptera. No entanto, como categoria

75
em si, não é efetivamente capturada pelo predador e não foi considerada para a análise
final do IRI.
Areia, pedras e fragmentos vegetais foram considerados itens não nutricionais,
provavelmente decorrentes de ingestão acidental ao revirar o fundo. Contudo, areia foi
um item muito pesado com uma média de 1,39 gramas em cada estômago analisado e
com freqüência de ocorrência em 30,8%. Embora pedras e fragmentos vegetais não
representem uma grande contribuição em peso, estes itens foram bem comuns com
45,6% e 27,1% de freqüência de ocorrência, respectivamente. Vermes nematóides
foram considerados parasitas por estarem sempre não digeridos, tanto no estômago
quanto no intestino.
Potamotrygon orbignyi apresentou distinta dominância de ninfas de
Ephemeroptera em sua dieta alimentar seguido de larvas de Diptera. Uma representação
gráfica tridimensional dos conteúdos estomacais em referência aos eixos FO%, N% e
P%, reiteram a dominância de Ephemeroptera com altos valores nestes eixos e
posicionamento desta categoria alimentar perto do vértice D (dominância) da Figura 5.
Categorias alimentares localizadas próximo ao vértice R, tais como peixes, Odonata,
Coleoptera e aracnida, indicam a raridade destas categorias, com baixos valores de
FO%, N% e P%. Diptera e Trichoptera, por outro lado, apesar de não apresentarem uma
representatividade em peso significativa, ocorrem freqüentemente e em grandes
números na dieta de P. orbignyi, evidenciando um alto grau de oportunismo para estas
categorias.

76
Figura 5. Representação Tridimensional da dieta de P. orbignyi com indicação dos
vértices de D-dominante e R-raro, em referência à presa, e S-especialista e G-
generalista, em referência ao hábito alimentar do predador.
A variabilidade percentual do número e peso dos principais itens encontrados
indica maior contribuição de Diptera nas menores classes de comprimento para ambos
os sexos (Fig. 6, 7, 8 e 9). Esta contribuição é gradativamente substituída por
Ephemeroptera, que a partir de 180 mm LD assume a maior importância relativa na
alimentação desta espécie. Em termos numéricos da participação de Diptera,
Ephemeroptera e Trichoptera na alimentação de P. orbignyi, houve diferença
significativa entre jovens e adultos (χ2 = 1682,5; p = 0,000), confirmando o câmbio
alimentar de Diptera para Ephemeroptera durante a fase juvenil. Exemplares de maior
porte – acima de 400 mm LD – foram escassos nas análises e estavam quase sempre
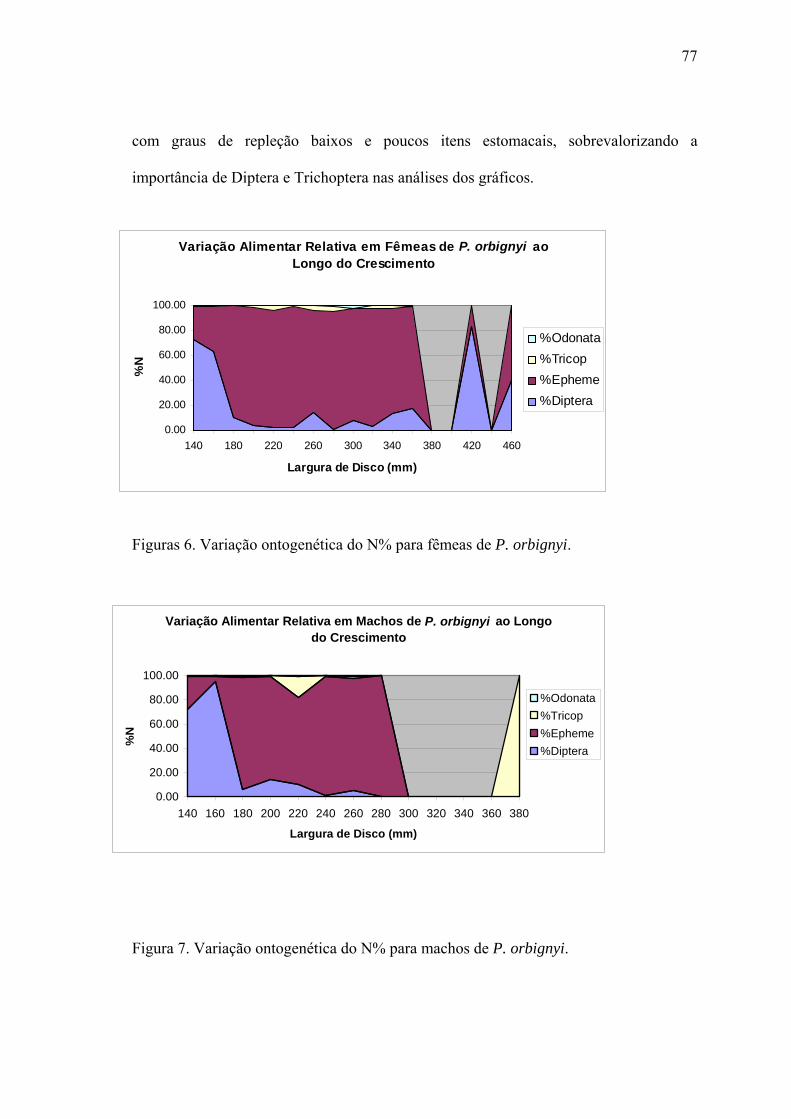
77
com graus de repleção baixos e poucos itens estomacais, sobrevalorizando a
importância de Diptera e Trichoptera nas análises dos gráficos.
Variação Alimentar Relativa em Fêmeas de P. orbignyi ao Longo do Crescimento
0.00
20.00
40.00
60.00
80.00
100.00
140 180 220 260 300 340 380 420 460
Largura de Disco (mm)
%N
%Odonata%Tricop%Epheme%Diptera
Figuras 6. Variação ontogenética do N% para fêmeas de P. orbignyi.
Variação Alimentar Relativa em Machos de P. orbignyi ao Longo do Crescimento
0.00
20.00
40.00
60.00
80.00
100.00
140 160 180 200 220 240 260 280 300 320 340 360 380
Largura de Disco (mm)
%N
%Odonata%Tricop%Epheme%Diptera
Figura 7. Variação ontogenética do N% para machos de P. orbignyi.
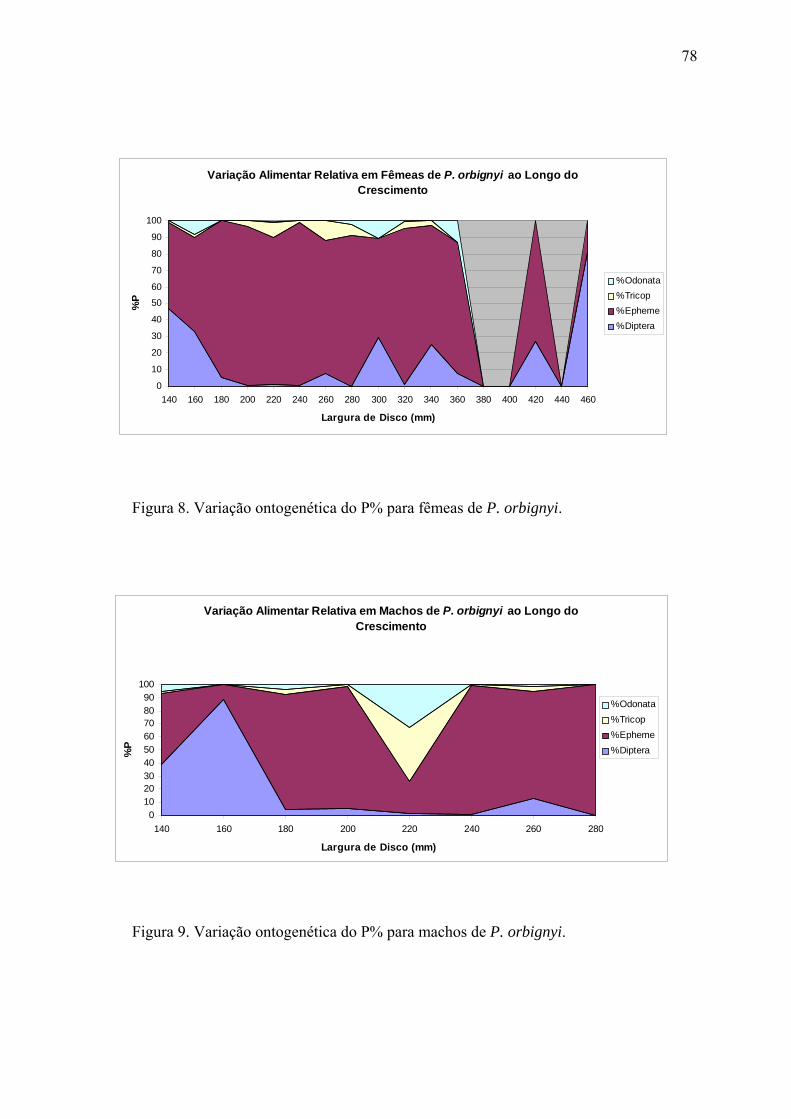
78
Variação Alimentar Relativa em Fêmeas de P. orbignyi ao Longo do Crescimento
0
10
20
30
40
50
60
70
80
90
100
140 160 180 200 220 240 260 280 300 320 340 360 380 400 420 440 460
Largura de Disco (mm)
%P
%Odonata%Tricop%Epheme%Diptera
Figura 8. Variação ontogenética do P% para fêmeas de P. orbignyi.
Variação Alimentar Relativa em Machos de P. orbignyi ao Longo do Crescimento
0102030405060708090
100
140 160 180 200 220 240 260 280
Largura de Disco (mm)
%P
%Odonata%Tricop%Epheme%Diptera
Figura 9. Variação ontogenética do P% para machos de P. orbignyi.

79
Os intestinos freqüentemente encontravam-se cheios de material em digestão –
carapaças de larvas e ninfas de insetos – e areia e pedras. A média em peso desse
conteúdo intestinal foi de 31,35 gramas (2,86-134,86 gramas), o que corresponde a
2,24% do peso total do animal e é aproximadamente 9 vezes o peso médio percentual
(0,25% e variância de 0,04) do conteúdo estomacal. Esta diferença evidencia uma baixa
taxa de evacuação gástrica em relação à freqüência alimentar de aproximadamente 1/9
vezes. Ou seja, se o intervalo entre o início de um período alimentar e o início do
próximo período alimentar for de três horas, o tempo para o início da evacuação gástrica
seria de aproximadamente 27 horas à temperatura ambiente média de 28oC, e sem a
interrupção de alimentação, pois o intestino necessariamente deve estar cheio, como
ocorre naturalmente.
A desidratação do conteúdo intestinal ocorreu na sua porção distal, indicando
maior absorção da água e nutrientes nesta porção do intestino. O peso médio da areia
encontrada no intestino foi aproximadamente 22 vezes maior que o peso médio da areia
encontrada no estômago.
Grau de Repleção e Digestão
Aproximadamente 96,3% (80 estômagos) das raias analisadas apresentaram
conteúdo estomacal. A distribuição de freqüências para o grau de repleção evidencia
que 51,2% dos estômagos analisados estavam com até ¼ do seu volume cheio de presas
e que 38,4% encontravam-se em categorias de ½ a ¾ do seu volume cheios (Fig. 10).
Cinco estômagos estavam cheios (6,4%) e somente três estômagos estavam vazios
(3,8%). A análise do Índice de Repleção Relativa (BM%) em função do período de
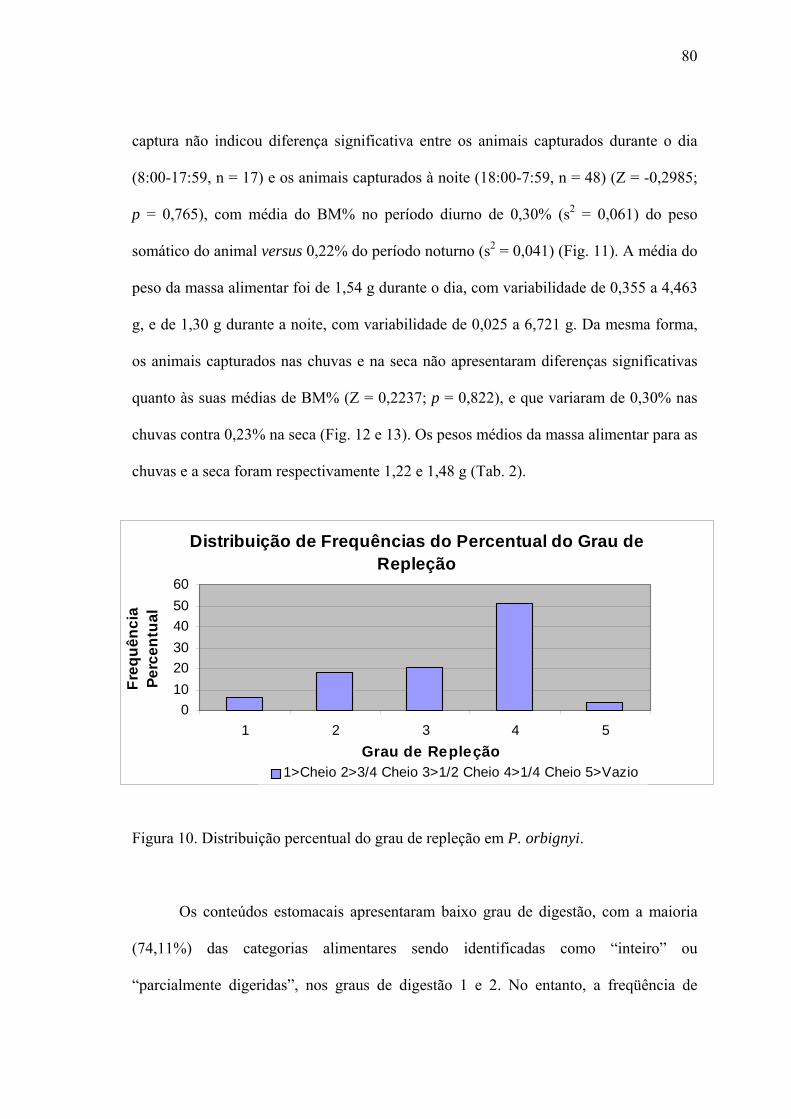
80
captura não indicou diferença significativa entre os animais capturados durante o dia
(8:00-17:59, n = 17) e os animais capturados à noite (18:00-7:59, n = 48) (Z = -0,2985;
p = 0,765), com média do BM% no período diurno de 0,30% (s2 = 0,061) do peso
somático do animal versus 0,22% do período noturno (s2 = 0,041) (Fig. 11). A média do
peso da massa alimentar foi de 1,54 g durante o dia, com variabilidade de 0,355 a 4,463
g, e de 1,30 g durante a noite, com variabilidade de 0,025 a 6,721 g. Da mesma forma,
os animais capturados nas chuvas e na seca não apresentaram diferenças significativas
quanto às suas médias de BM% (Z = 0,2237; p = 0,822), e que variaram de 0,30% nas
chuvas contra 0,23% na seca (Fig. 12 e 13). Os pesos médios da massa alimentar para as
chuvas e a seca foram respectivamente 1,22 e 1,48 g (Tab. 2).
Distribuição de Frequências do Percentual do Grau de Repleção
0102030405060
1 2 3 4 5Grau de Repleção
Freq
uênc
ia
Perc
entu
al
1>Cheio 2>3/4 Cheio 3>1/2 Cheio 4>1/4 Cheio 5>Vazio
Figura 10. Distribuição percentual do grau de repleção em P. orbignyi.
Os conteúdos estomacais apresentaram baixo grau de digestão, com a maioria
(74,11%) das categorias alimentares sendo identificadas como “inteiro” ou
“parcialmente digeridas”, nos graus de digestão 1 e 2. No entanto, a freqüência de
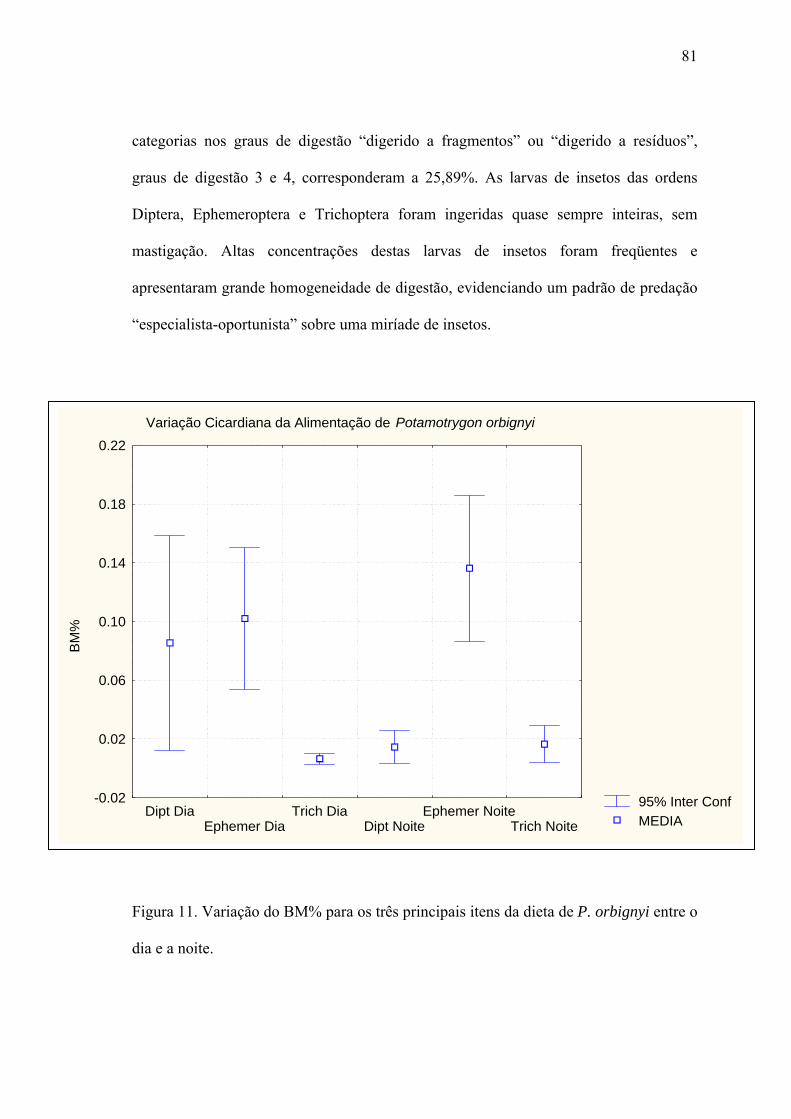
81
categorias nos graus de digestão “digerido a fragmentos” ou “digerido a resíduos”,
graus de digestão 3 e 4, corresponderam a 25,89%. As larvas de insetos das ordens
Diptera, Ephemeroptera e Trichoptera foram ingeridas quase sempre inteiras, sem
mastigação. Altas concentrações destas larvas de insetos foram freqüentes e
apresentaram grande homogeneidade de digestão, evidenciando um padrão de predação
“especialista-oportunista” sobre uma miríade de insetos.
95% Inter ConfMEDIA
Variação Cicardiana da Alimentação de Potamotrygon orbignyi
BM
%
-0.02
0.02
0.06
0.10
0.14
0.18
0.22
Dipt DiaEphemer Dia
Trich DiaDipt Noite
Ephemer NoiteTrich Noite
Figura 11. Variação do BM% para os três principais itens da dieta de P. orbignyi entre o
dia e a noite.
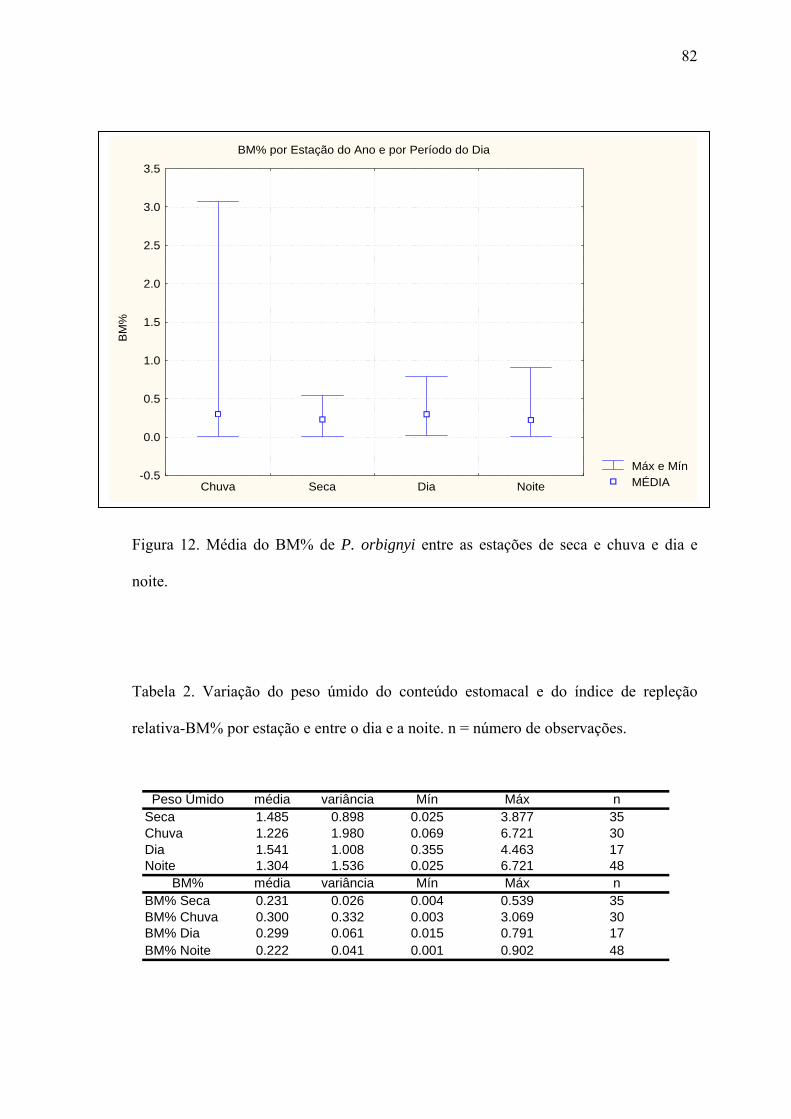
82
Máx e MínMÉDIA
BM% por Estação do Ano e por Período do Dia
BM
%
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
Chuva Seca Dia Noite
Figura 12. Média do BM% de P. orbignyi entre as estações de seca e chuva e dia e
noite.
Tabela 2. Variação do peso úmido do conteúdo estomacal e do índice de repleção
relativa-BM% por estação e entre o dia e a noite. n = número de observações.
Peso Úmido média variância Mín Máx nSeca 1.485 0.898 0.025 3.877 35Chuva 1.226 1.980 0.069 6.721 30Dia 1.541 1.008 0.355 4.463 17Noite 1.304 1.536 0.025 6.721 48
BM% média variância Mín Máx nBM% Seca 0.231 0.026 0.004 0.539 35BM% Chuva 0.300 0.332 0.003 3.069 30BM% Dia 0.299 0.061 0.015 0.791 17BM% Noite 0.222 0.041 0.001 0.902 48
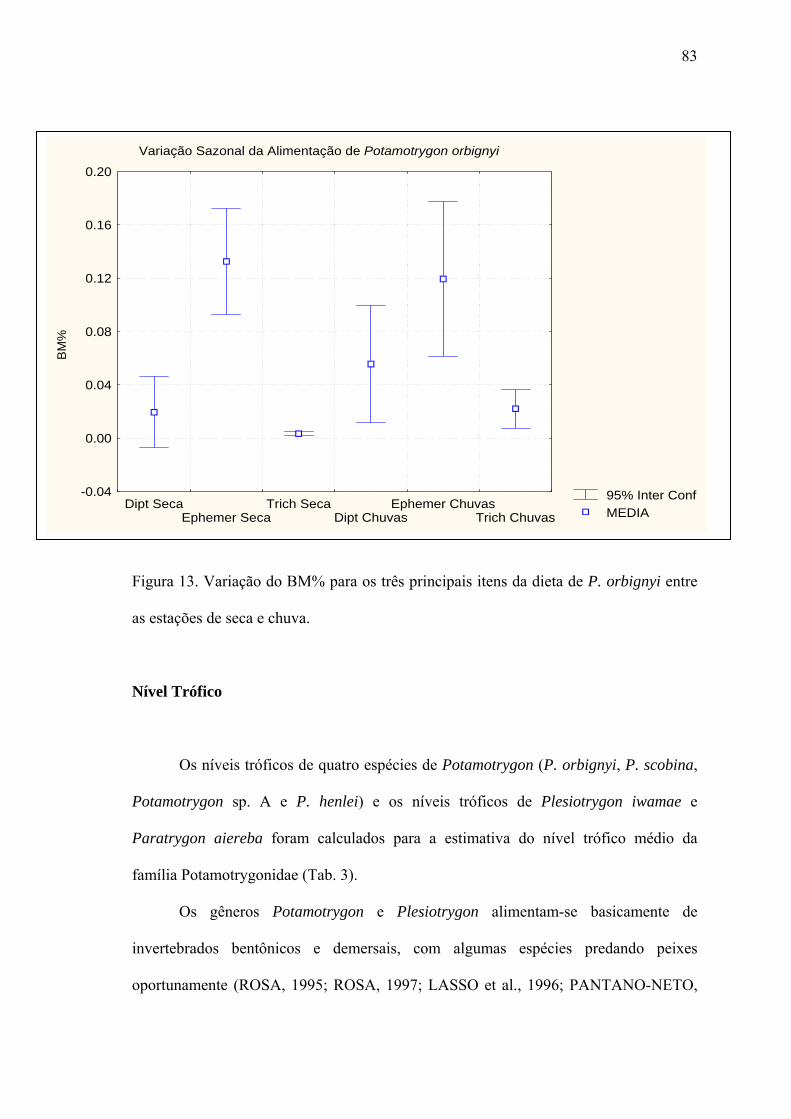
83
Figura 13. Variação do BM% para os três principais itens da dieta de P. orbignyi entre
as estações de seca e chuva.
Nível Trófico
Os níveis tróficos de quatro espécies de Potamotrygon (P. orbignyi, P. scobina,
Potamotrygon sp. A e P. henlei) e os níveis tróficos de Plesiotrygon iwamae e
Paratrygon aiereba foram calculados para a estimativa do nível trófico médio da
família Potamotrygonidae (Tab. 3).
Os gêneros Potamotrygon e Plesiotrygon alimentam-se basicamente de
invertebrados bentônicos e demersais, com algumas espécies predando peixes
oportunamente (ROSA, 1995; ROSA, 1997; LASSO et al., 1996; PANTANO-NETO,
95% Inter ConfMEDIA
Variação Sazonal da Alimentação de Potamotrygon orbignyi
BM%
-0.04
0.00
0.04
0.08
0.12
0.16
0.20
Dipt SecaEphemer Seca
Trich SecaDipt Chuvas
Ephemer ChuvasTrich Chuvas
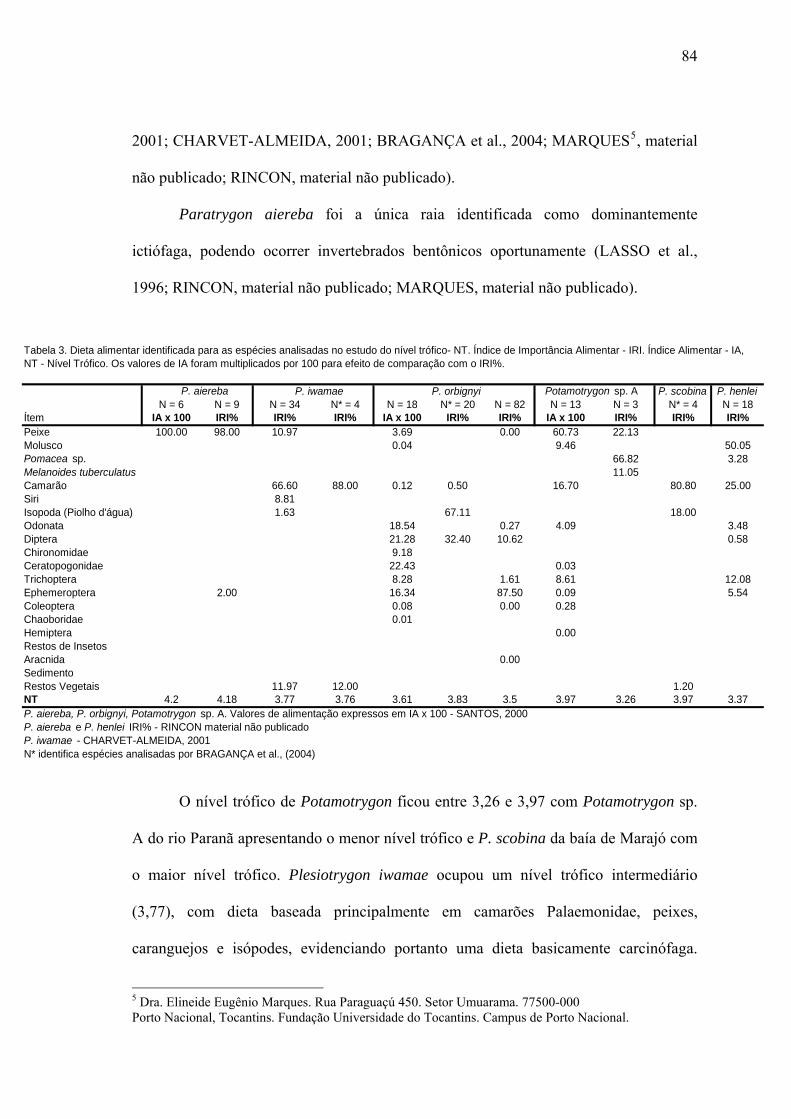
84
2001; CHARVET-ALMEIDA, 2001; BRAGANÇA et al., 2004; MARQUES5, material
não publicado; RINCON, material não publicado).
Paratrygon aiereba foi a única raia identificada como dominantemente
ictiófaga, podendo ocorrer invertebrados bentônicos oportunamente (LASSO et al.,
1996; RINCON, material não publicado; MARQUES, material não publicado).
Tabela 3. Dieta alimentar identificada para as espécies analisadas no estudo do nível trófico- NT. Índice de Importância Alimentar - IRI. Índice Alimentar - IA,NT - Nível Trófico. Os valores de IA foram multiplicados por 100 para efeito de comparação com o IRI%.
P. aiereba P. iwamae P. orbignyi Potamotrygon sp. A P. scobina P. henleiN = 6 N = 9 N = 34 N* = 4 N = 18 N* = 20 N = 82 N = 13 N = 3 N* = 4 N = 18
Ítem IA x 100 IRI% IRI% IRI% IA x 100 IRI% IRI% IA x 100 IRI% IRI% IRI%Peixe 100.00 98.00 10.97 3.69 0.00 60.73 22.13Molusco 0.04 9.46 50.05Pomacea sp. 66.82 3.28Melanoides tuberculatus 11.05Camarão 66.60 88.00 0.12 0.50 16.70 80.80 25.00Siri 8.81Isopoda (Piolho d'água) 1.63 67.11 18.00Odonata 18.54 0.27 4.09 3.48Diptera 21.28 32.40 10.62 0.58Chironomidae 9.18Ceratopogonidae 22.43 0.03Trichoptera 8.28 1.61 8.61 12.08Ephemeroptera 2.00 16.34 87.50 0.09 5.54Coleoptera 0.08 0.00 0.28Chaoboridae 0.01Hemiptera 0.00Restos de InsetosAracnida 0.00SedimentoRestos Vegetais 11.97 12.00 1.20NT 4.2 4.18 3.77 3.76 3.61 3.83 3.5 3.97 3.26 3.97 3.37P. aiereba, P. orbignyi, Potamotrygon sp. A. Valores de alimentação expressos em IA x 100 - SANTOS, 2000P. aiereba e P. henlei IRI% - RINCON material não publicadoP. iwamae - CHARVET-ALMEIDA, 2001N* identifica espécies analisadas por BRAGANÇA et al., (2004)
O nível trófico de Potamotrygon ficou entre 3,26 e 3,97 com Potamotrygon sp.
A do rio Paranã apresentando o menor nível trófico e P. scobina da baía de Marajó com
o maior nível trófico. Plesiotrygon iwamae ocupou um nível trófico intermediário
(3,77), com dieta baseada principalmente em camarões Palaemonidae, peixes,
caranguejos e isópodes, evidenciando portanto uma dieta basicamente carcinófaga.
5 Dra. Elineide Eugênio Marques. Rua Paraguaçú 450. Setor Umuarama. 77500-000 Porto Nacional, Tocantins. Fundação Universidade do Tocantins. Campus de Porto Nacional.
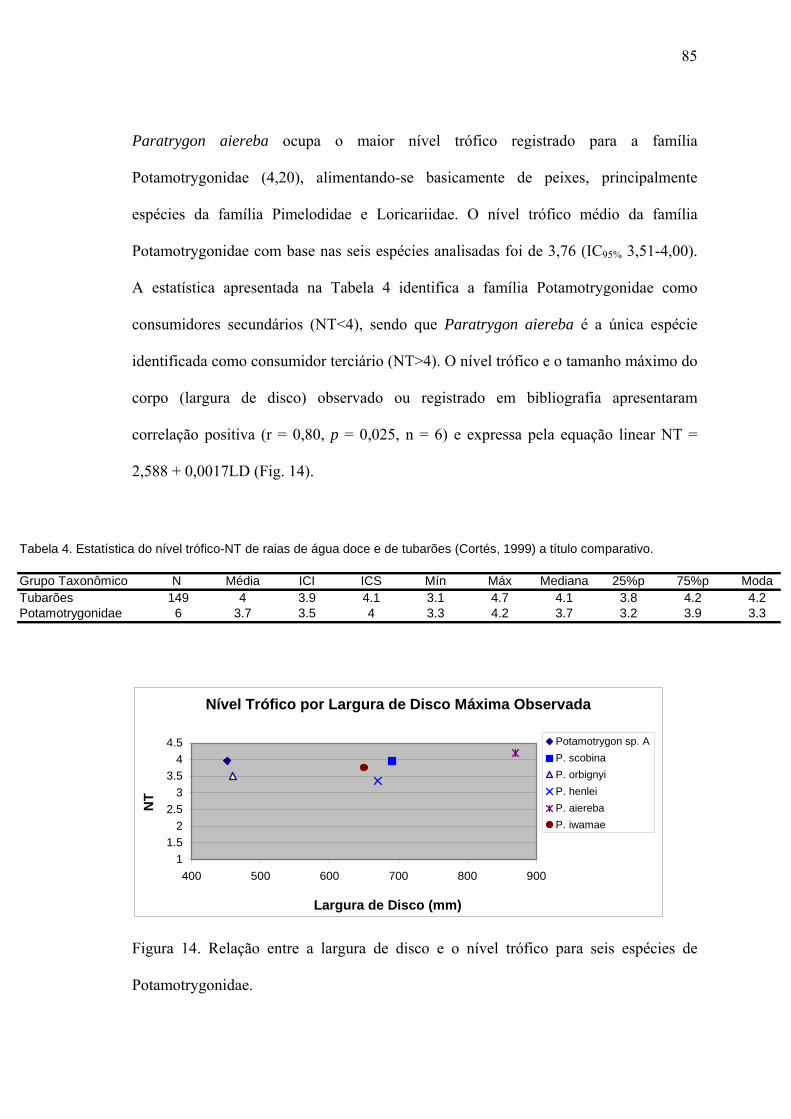
85
Paratrygon aiereba ocupa o maior nível trófico registrado para a família
Potamotrygonidae (4,20), alimentando-se basicamente de peixes, principalmente
espécies da família Pimelodidae e Loricariidae. O nível trófico médio da família
Potamotrygonidae com base nas seis espécies analisadas foi de 3,76 (IC95% 3,51-4,00).
A estatística apresentada na Tabela 4 identifica a família Potamotrygonidae como
consumidores secundários (NT<4), sendo que Paratrygon aiereba é a única espécie
identificada como consumidor terciário (NT>4). O nível trófico e o tamanho máximo do
corpo (largura de disco) observado ou registrado em bibliografia apresentaram
correlação positiva (r = 0,80, p = 0,025, n = 6) e expressa pela equação linear NT =
2,588 + 0,0017LD (Fig. 14).
Tabela 4. Estatística do nível trófico-NT de raias de água doce e de tubarões (Cortés, 1999) a título comparativo.
Grupo Taxonômico N Média ICI ICS Mín Máx Mediana 25%p 75%p ModaTubarões 149 4 3.9 4.1 3.1 4.7 4.1 3.8 4.2 4.2Potamotrygonidae 6 3.7 3.5 4 3.3 4.2 3.7 3.2 3.9 3.3
Nível Trófico por Largura de Disco Máxima Observada
11.5
22.5
33.5
44.5
400 500 600 700 800 900
Largura de Disco (mm)
NT
Potamotrygon sp. AP. scobinaP. orbignyiP. henleiP. aierebaP. iwamae
Figura 14. Relação entre a largura de disco e o nível trófico para seis espécies de
Potamotrygonidae.

86
DISCUSSÃO
A morfologia do sistema digestório de P. orbignyi confirma uma dieta baseada
em itens alimentares frágeis e de pouca resistência à predação, condizentes com o hábito
alimentar insetívoro encontrado nesta espécie. Todo o sistema digestório pré-esofágico
parece estar relacionado à captura e ingestão de presas frágeis, não havendo estruturas
auxiliares na digestão mecânica dos alimentos. O número de cinco papilas orais no
interior da boca parece ser freqüente em espécies com dieta dominante de invertebrados,
enquanto que espécies com durofagia ou peixes apresentam somente três papilas
centrais desenvolvidas e duas comissurais distintamente vestigiais (P. henlei,
Potamotrygon sp. A, Potamotrygon sp. B, P. aiereba e P. motoro). A relativa
protratibilidade da boca, evidenciada pelos sulcos labiais, permite brusca alteração do
volume buco-faringeano, provocando a sucção de itens alimentares aglomerados tais
como as larvas de insetos. Este mesmo padrão é observado em espécies de Myliobatis,
Rhinoptera e Aetobatus, que utilizam essa sucção para cavarem o fundo em busca de
moluscos bivalvos enterrados (HOWARD et al., 1977; GREGORY et al., 1979;
MOYLE & CECH, 1982).
A dentição de P. orbignyi é frágil, diminuta – com dentes menores que 1 mm de
largura máxima - e relativamente independente, onde cada dente pode agir
individualmente para a captura da presa. As pequenas cúspides e a fina cartilagem das
arcadas, permitem ao animal “pinçar” alguns de seus itens alimentares maiores tais
como os odonatas, coleopteras e possivelmente os trichopteras. Outras raias
microfágicas, do ponto de vista para elasmobrânquios, como Discopyge tschudii e
Narcine brasiliensis, também apresentam dentição de tamanho reduzido, o que parece

87
favorecer a manipulação da presa para a sua ingestão sem a mastigação (BIGELOW &
SCHROEDER, 1953; GARCIA, 1984). A ação mecânica do alimento ocorre no
interior da válvula espiral, onde, juntamente com uma grande quantidade de areia
acumulada e o deslocamento do alimento ao longo das evoluções, tritura as carapaças
das larvas de insetos e as separa para melhor absorção dos nutrientes. Esta absorção
ocorre na porção distal do intestino e é evidenciada pelo grau de desidratação do bolo
fecal.
A captura de raias de água doce durante o dia com o uso do espinhel de fundo
foi severamente prejudicada pela competição com outros peixes. Os baixos rendimentos
durante o dia induziram a troca de apetrecho e de comportamento de pesca. Segundo
BOWEN (1983), a pesca ativa no estudo da alimentação de peixes tem a vantagem de
capturar tanto os peixes que estão em busca de alimento quanto os saciados. Desta
forma, de uma arte de pesca tipicamente passiva, passou-se a uma pesca ativa, com o
uso da zagaia de três pontas, ou tridente. No entanto, o espinhel de fundo tem sido
utilizado com eficiência em outras regiões (CHARVET-ALMEIDA, 2001; PINTO DE
ALMEIDA, 2003). Esta diferença no rendimento do espinhel provavelmente se deve a
características regionais típicas na baía de Marajó como a presença de marés,
comportamento do pescador e diferença na composição específica da captura de raias de
água doce, que pode apresentar comportamentos alimentares diferenciados a presas
disponíveis no meio ambiente. A distribuição preferencial de P. orbignyi no rio Paranã é
próxima à margem do rio, em pontos de correnteza moderada a fraca onde encontra a
maior abundância de suas presas sobre fundo lodoso ou arenoso, intercalado por
vegetação subaquática e marginal. Desta forma, a sua distribuição sobrepõe estes pontos
ricos em alimento, onde a sua captura é favorecida. Da mesma forma, capturas em noite

88
de lua crescente e nova foram significativamente maiores que as capturas de lua cheia.
A diferença observada nas CPUE entre as fases lunares corrobora com o que os
pescadores identificam como comportamento tímido das raias de água doce nas noites
claras de lua cheia, dificultando sua captura com a zagaia. Estes animais são adaptados à
visão em condições de baixa intensidade luminosa através da reflexão da luz pelo
tapetum lucidum e pela seleção dos comprimentos de onda de luz por pigmentos
específicos dentro das células fotoreceptoras (HUETER et al., 2004). Desta forma, é
compreensível que o animal enxergue melhor em noites de lua cheia que em noites sem
lua ou mesmo de dia, quando teria dificuldades para lidar com o excesso de luz.
A raia de água doce Potamotrygon orbignyi é basicamente insetívora na região
do rio Paranã, alto Tocantins, e alimenta-se principalmente de ninfas de Ephemeroptera
e larvas de Diptera e Trichoptera ao longo de toda a sua vida. No entanto, esta espécie
parece adaptar-se facilmente à disponibilidade de recursos alimentares do ambiente,
alimentando-se de itens compatíveis com sua capacidade predatória e morfologia do
sistema digestório. O principal item alimentar desta espécie na baía de Marajó são
crustáceos isópodes (BRAGANÇA et al., 2004), os quais não foram encontrados em
exemplares da mesma espécie no alto Tocantins. Assim, LASSO et al. (1996) também
relatam uma variedade de invertebrados na dieta de P. orbignyi para os “llanos”
venezuelanos incluindo camarões, caranguejos e ostracodes, os quais também não
ocorreram na dieta dos exemplares analisados neste estudo. Este padrão não é incomum
em elasmobrânquios e já foi relatado para espécies de ampla distribuição geográfica
como Squalus acanthias, Raja radiata e Prionace glauca, dentre outras (HOLDEN,
1966; RAE, 1967; TRICAS, 1979; PEDERSEN, 1995; VASKE & RINCON, 1997).
Diferenças no tipo de habitat e profundidade foram identificadas como responsáveis por

89
influenciar a composição da dieta em elasmobrânquios (STILLWELL & KOHLER,
1982; CORTÉS et al., 1996). Assim, variações na dieta de P. orbignyi entre pontos
distintos geograficamente indica uma adaptabilidade da espécie às condições distintas
de habitat e às relações tróficas existentes.
A dominância de ninfas de Ephemeroptera deve ser cautelosamente interpretada
como uma especialização à predação desta presa. Embora P. orbignyi apresente
modificações morfológicas para uma dieta composta principalmente de invertebrados
aquáticos, populações distintas desta espécie apresentam diferentes itens dominantes em
suas dietas (LASSO et al., 1996; BRAGANÇA et al., 2004), indicando um oportunismo
de P. orbignyi para predar itens que apresentem um balanço compensatório entre
abundância, densidade e valor energético. A abundância e densidade necessárias
determinam um padrão comum em populações distintas de P. orbignyi, a preferência
por somente um ou poucos itens alimentares dominantes.
Câmbios alimentares devido à ontogenia e mesmo à maturidade sexual em
elasmobrânquios são freqüentes e relatados para diversas espécies (STILLWELL &
KOHLER, 1982, 1993; PEDERSEN, 1995; LOWE et al., 1996; ALONSO et al., 2001;
MUTO et al., 2001). Potamotrygon orbignyi apresenta dois câmbios alimentares na fase
juvenil; o primeiro entre 110-130 mm, quando troca a alimentação endógena pela
exógena ao exaurir as reservas vitelínicas; o segundo entre 160-180 mm LD, quando
troca as larvas de Diptera por ninfas de Ephemeroptera como seu principal item
alimentar. Nesta mesma fase do desenvolvimento - segundo câmbio alimentar - também
se observa um aumento da diversidade de itens (Odonata, Trichoptera e Coleoptera) na
dieta e de sua participação em número e peso. Este câmbio pode estar associado a um
aumento da experiência de captura das jovens raias, que começam a predar sobre ítens

90
alimentares com maior capacidade de locomoção, mas mais vantajosos energeticamente
(STEPHENS & KREBS, 1986). Este câmbio é ontogenético e não decorrente da
maturidade sexual, que ocorre somente em indivíduos maiores (240 mm LD para
machos e 260 mm LD para fêmeas). O mesmo padrão foi observado em Dipturus
chilensis (ALONSO et. al., 2001), que em seu primeiro câmbio alimentar ontogenético
troca uma dieta betônica, composta principalmente por crustáceos, por uma dieta
demersal-bentônica, composta principalmente por peixes.
A alta incidência de animais com conteúdo estomacal em todos os horários de
captura em um intervalo de 14 horas e a variedade de graus de digestão ocorrendo
concomitantemente evidencia contínua alimentação. De outra forma, seria de se esperar
períodos do dia onde os animais apresentassem estômagos vazios ou graus de
alimentação coincidentes, indicando sincronismo alimentar populacional. Este padrão é
ainda confirmado pela constância dos valores do índice de repleção relativa (BM%), os
quais não apresentam diferenças significativas entre o dia e a noite ou entre as estações
de seca e chuvas. O pequeno porte das suas presas corriqueiras e seu provável baixo
valor nutricional induzem o predador a uma constante busca por alimento a fim de
suprir suas necessidades nutricionais diárias. As ocorrências singulares de peixe e
aracnida indicam que P. orbignyi pode ter um comportamento alimentar oportunista. O
único peixe encontrado provavelmente foi ingerido morto, tendo em vista o fato de ser
somente um fragmento, o que contraria o comportamento usual da espécie de engolir
suas presas inteiras.
O valor médio do nível trófico da família Potamotrygonidae identifica as raias
de água doce como predadores intermediários na cadeia trófica dos rios, contrariando a
visão corriqueira de que estes animais sejam predadores de topo. Essa visão equivocada

91
do posicionamento trófico das raias de água doce provavelmente provém de uma
inferência a partir do posicionamento trófico do seu grupo irmão, os tubarões
(CORTÉS, 1999). Nos ambientes onde ocorrem, as raias de água doce provavelmente
utilizam recursos similares que a maioria dos peixes demersais da bacia Amazônica com
níveis tróficos estimados (n = 157), os quais apresentaram um nível trófico médio de 3,2
vs 3,6 das raias. No entanto, o impacto competitivo destas raias no rio Paranã somente
pode ser avaliado com base em estudos ecológicos mais complexos através de
levantamentos ictiofaunísticos regionais, avaliações de abundância bentônica e estudos
de alimentação e abundância de peixes.
A relação LD-NT apresentou boa correlação linear e cerca de 65% da variação
do nível trófico pode ser explicada pela reta de regressão (largura de disco e nível
trófico). No entanto, um número maior de espécies terá de ser incorporado neste modelo
para melhor avaliação de tendências. Com base neste modelo, espécies de grande porte
como P. brachyura, uma raia de água doce gigantesca das bacias dos rios Paraná,
Paraguay e Uruguay, devem possuir um nível trófico elevado (NT>4) e uma dieta
dominante de peixes. Por outro lado, espécies menores como a Potamotrygon sp. C
“cururu” do rio Negro, devem apresentar um nível trófico baixo, com dieta dominante
de invertebrados. No entanto, exceções podem ocorrer, como os grandes tubarões
planctófagos Cetorhinus maximus e Rhincodon typus, que apresentam NT muito abaixo
do esperado para seu grande porte (Cortés, 1999).
As fontes de variação e erro das estimativas dos níveis tróficos das raias de água
doce são diversas. O uso de diferentes índices de alimentação compostos tais como o
IRI e o IA, embora expressem a mesma relação de dominância alimentar, podem
apresentar variabilidade não estimada devido aos poucos trabalhos existentes sobre a

92
alimentação de raias de água doce e amostras pequenas. A padronização de todos os
trabalhos ao IRI, que incorpora as três dimensões alimentares (FO%, N% e P%), foi
impossibilitada pela falta de informações em alguns trabalhos (ausência do N%). A
possibilidade de padronização dos dados ao IA foi considerada restritiva com perda de
informação e, conseqüentemente, não utilizada. A categorização dos itens alimentares,
principalmente os peixes, pode ter afetado as estimativas de NT. Embora fosse
preferível a utilização do NT específico da presa, muitos trabalhos de alimentação de
raias apenas identificavam o item “peixes”. Tendo em vista que a maioria dos peixes
predados localizam-se no fundo ou perto dele, a utilização de um NT médio para peixes
demersais pareceu a mais acertada e exeqüível.
O aumento de pesquisas na área de alimentação das raias de água doce poderá
fornecer uma avaliação melhor do nível trófico médio da família e da largura da cadeia
trófica onde vivem, bem como estimativas da variabilidade intra-específica, correções
nas avaliações baseadas em poucos exemplares e comparações com estimativas por
isótopos estáveis.

93
LITERATURA CITADA
ACHENBACH, G. M. & ACHENBACH, S.V.M. Notas acerca de algunas especies de
raya fluvial (Batoidei, Potamotrygonidae) que frecuentan el sistema hidrográfico
del Paraná medio en el Departamento La Capital (Santa Fe - Argentina). Comun.
Mus. Prov. Ciencia. Nat. Florentino Ameghino, 8. p. 1-34, 1976.
ALMEIDA, M. P. Pesca, policromatismo e aspectos sistemáticos de Potamotrygon
scobina Garman, 1913 (Chondrichthyes: Potamotrygonidae) da região da Ilha de
Colares – Baía de Marajó – Pará. Dissertação de Mestrado. Universidade Federal
do Pará e Museu Paraense Emílio Goeldi. p. 145. 2003.
ALONSO, M. K., CRESPO, E. A., GARCIA, N. A., PEDRAZA, S. N., MARIOTTI, P.
A., BERÓN-VERA, B. & MORA, N. J. Food habits of Dipturus chilensis
(Pisces:Rajidae) off Patagonia, Argentina. ICES J. Mar. Sci. (58). p. 288-297.
2001.
BIGELOW, H. B. & SCHROEDER, W. C. Fishes of the Western North Atlantic. Part II
- Sawfishes, Guitarfishes, Skates and Rays. MEMOIR Sears Foundation for
Marine Research.1:588. 1953.
BOWEN, S. N. Quantitative description of the diet. In: NIELSEN, L. A., JOHNSON,
D. L. (ed.) Fisheries Techniques. American Fisheries Society. Bethesda,
Maryland. USA. p. 325-336. 1983.
BRAGANÇA, A. J. M.; CHARVET-ALMEIDA, P, & BARTHEM, R.B. Preliminary
obsevations on the feeding of the freshwater stingrays Potamotrygon orbignyi,
Potamotrygon scobina and Plesiotrygon iwamae (Chondrichthyes:
Potamotrygonidae) in the Cotijuba Island Region – Pará – Brasil. In: Biology and

94
Conservation of Freshwater Elasmobranchs: Symposium Proceedings.
International Congress on the Biology of Fish. Manaus, p. 49 – 60, 2004.
BROOKS, D. R., THORSON, T. B. & MAYES, M. A. Freshwater stingrays
(Potamotrygonidae) and their helminth parasites: testing hypotheses of evolution
and coevolution. In: FUNK, V. A., BROOKS, D. R. (eds.). Advances in cladistics.
Proceedings of the first meeting of the Willi Henning Society. New York Botanical
Garden, New York. p. 147-175. 1981.
CASTELNAU, F. L. Animaux nouveaux ou rares recueillis pendant l’expédition dans
lês parties centrales de l’Amerique du Sud, de rio de Janeiro a Lima, et de Lima au
Para. Vol. 2. P. Bertrand, Paris. p. 101-103. Pl. 48. 1855.
CASTEX, M. N. Una nueva espécie de raya fluvial: P. labradori. Neotropica, La Plata,
9 (30). p. 117-121. 1963.
CASTEX, M. N. Estado actual de los estúdios sobre la raya fluvial neotropical. Ver.
Mus. Prov. Cs. Nat. Santa Fé. Número extraordinário del cincuentenario. p. 9-34.
1964.
CASTEX, M. N. Bases para el estúdio de las rayas de água dulce del sistema
Amazônico. Nuevas sinonímias de “P. motoro” (M. H., 1841). Atas do Simpósio
sobre a biota Amazônica. vol. 3 (Limnologia). P. 89-92. 1967.
CARVALHO, M.R.; LOVEJOY, N.R. & ROSA, R.S. Potamotrigonidae. In: REIS, R.
E., S.O. KULLANDER, S. O. & FERRARIS, C. J. JR (Eds.) Check List of the
Freshwater Fishes of South and Central America. Edipucrs, Pontifícia
Universidade Católica do Rio Grande do Sul. Porto Alegre, 2003.
CHARVET-ALMEIDA, P. Ocorrência, biologia e uso das raias de água doce na Baía
de Marajó (Pará, Brasil), com ênfase na biologia de Plesiotrygon iwamae

95
(Chondrichthyes: Potamotrygonidae). Dissertação de mestrado, Programa de Pós-
graduação em Zoologia da Universidade federal do Pará e Museu Paraense Emílio
Goeldi. Departamento de zoologia, 2001.
CHRISTENSEN, V. & PAULY, D. The ECOPATH II – a software for balancing
steady-state models and calculating network characteristics. Ecological Modelling,
54: 726-738, 1997.
COMPAGNO, L. J. V. Systematics and body form. In: HAMLETT, W. C. (ed.) Sharks,
Skates and Rays.The Johns Hopkins University Press. Baltimore, p. 02 – 42, 1999.
CORTÉS, E. A critical review of methods of studying fish feeding based on analysis of
stomach contents: application to elasmobranch fishes. Can. J.l of Fish. and Aqua.
Sci., 54: p. 726-738, 1997.
CORTÉS, E. Standardized diet compositions and trophic levels of sharks. ICES J. of
Mar. Sci., 56: p. 707-717, 1999.
CORTÉS, E. & GRUBER, S. H. Diet, feeding habits and estimates of daily ration of
Young lemon sharks, Negaprion brevirostris (Poey). Copeia, (1). p. 204-218.
1990.
COSTI, C. G., RODRIGUES, C. C. B, PAIVA-FILHO, D. L. & RODRIGUES, J. B. R..
Estudo preliminar da ictiofauna do pólo Araguaia-Tocantins. Projeto de pesquisa
da Ictiofauna do pólo Araguaia-Tocantins, GO-Polamazônia. Superintendência do
Desenvolvimento da Pesca-SUDEPE. Brasília. 171 p. 1977.
GARCIA, M. L. Sobre la biologia de Discopyge tschudii (Chondrichthyes, Narcinidae).
Physis, Secc. A, 42(103): p. 101-112. 1984.
GARMAN, S. Plagiostomia – Sharks, skates and Rays. Benthic Press, Los Angeles,
California. 511 p. 1997.

96
GREGORY, M. R.; BALLANCE, P. F.; GIBSON, G. W. & AYLING, A. M. On how
some rays (Elasmobranchia) excavate feeding depressions by jetting water. J.
Sediment. Petrol. 49:1125-1129. 1979.
GONZÁLES-ISÁIS, M. & DOMÍNGUEZ, H.M.M. Comparative Anatomy of the
Superfamily Myliobatoidea (Chondrichthyes) with some comments on phylogeny.
J. of Morph., 262(1): p. 517-535, 2004.
HOWARD, J. D. MAYOU, T. V. & HEARD, R. W. Biogenic sedimentary structures
formed by rays. J. Sediment. Petrol. 47. p. 339-346. 1977.
HUETER, R. E., MANN, D. A., MARUSKA, K. P., SISNEROS, J. A. & DEMSKI, L.
S. Sensory biology of elasmobranches. In: CARRIER, J. C., MUSICK, J. A., &
HEITHAUS, M. R. Biology of sharks and their relatives. CRC Press, Boca Raton.
P. 325-368. 2004.
HYNES, H.B.N. The food of fresh-water stichbacks (Gasterosteus aculeatus and
Pugosteus pungitius), with a review of methods used in studies of the food of
fishes. J. of An. Ecol., 19(1). p. 36-58, 1950.
HYSLOP, E.J. Stomach contents analysis – a review of methods and their application.
Journal of Fish Biology, 17. p. 411-429, 1980.
KAWAKAMI, E. & VAZZOLER, G. Método gráfico e estimativa de índice alimentar
aplicado ao estudo da alimentação de peixes. Bolm. Inst. Oceanogr. 29(2). p. 205-
207. 1980.
LASSO, C. A., RIAL, A. B. & LASSO-ALCALÁ, O. Notes on the biology of the
freshwater stingrays Paratrygon aiereba (Müller & Henle, 1841) and
Potamotrygon orbignyi (Castelnau, 1855) (Chondrichthyes: Potamotrygonidae) in
the Venezuelan llanos. Aqua, v. 2, No 3, p. 39-52. 1996.

97
LOWE, C. G., WETHERBEE, B. M., CROW, G. L. & TESTER, A. L. Ontogenetic
dietary shifts and feeding behavior of the tiger shark, Galeocerdo cuvier, in
Hawaiian waters. Environ. Biol. Fish. 47. p. 203-211. 1996.
McCAFFERTY, W. P. Aquatic entomology. The fisherman’s and ecologists’ illustrated
guide to insects and their relatives. Science Books International, Boston,
Massachusetts. 448 p. 1981.
MOYLE, P. B. & CECH, JR. J. J. Fishes: An introduction to ichthyology. Prentice-Hall,
Inc. Englewood Cliffs, New Jersey. 593 p. 1982.
MUTO, E. Y., SOARES, L. S. H. & GOITEIN, R. Food resource utilization of the
skates Rioraja agassizi (Muller & Henle, 1841) and Psammobatis extenta
(Garman, 1913) on the continental shelf off Ubatuba, South-Eastern Brazil. Rev.
Brasil. Biol. 61(2). p. 217-238. 2001.
NELSON, G. A. & ROSS, M. R. Gastric evacuation in little skate. J. Fish Biol. (46). p.
977-986. 1995.
PANTANO-NETO, J. Estudo preliminar da anatomia descritiva e funcional associada à
alimentação em raias de água doce (Potamotrygonidae, Myliobatiformes,
Elasmobranchii). Dissertação de mestrado, Instituto de biociências da
Universidade de São Paulo. Departamento de Zoologia. 93 p. 2001.
PAULY, D.; CHRISTENSEN, V.; DALSGAARD, J.; FROESE, R. & TORRES, F.J.
Fishing down marine food webs. In: Science, 279. p. 860-863, 1998.
PEDERSEN, S. A. Feeding habits of starry ray (Raja radiata) in west Greenland
waters. ICES J. Mar. Sci. 52. p. 43-53. 1995.

98
PINKAS, L.; OLIPHANT, M.S. & IVERSON, I.L.K. Food habits of albacore, bluefin
tuna and bonito in California waters. In: California Fish and Game, 152. p. 1-105,
1971.
RIBEIRO, M. C. L. B., PETRERE, M. & JURAS, A.A. Ecological integrity and
fisheries ecology of the Araguaia-Tocantins river basin, Brazil. Regulated Rivers:
Research and Management. v. 11, p. 325-350. 1995.
ROSA, R. S. A systematic revision of the south American freshwater stingrays
(Chondrichthyes: Potamotrygonidae). Dissertation for the degree of doctor of
philosophy, College of William and Mary Virginia. Faculty of the School of
Marine Science. 523 p. 1985.
ROSA, R. S., CASTELLO, H. P. & THORSON, T. B. Plesiotrygon iwamae, a new
genus and species of neotropical freshwater stingray (Chondrichthyes:
Potamotrygonidae). Copeia. No 2, p. 447-458. 1987.
SANTOS, A.A. Morfologia externa, alimentação e reprodução de raias do alto e médio
rio Tocantins (Chondrichthyes, Potamotrygonidae). Relatório técnico-científico
final, Direção de Pesquisa e Extensão da Universidade do Tocantins. Curso de
Ciências Biológicas, (Material não publicado). 2000.
SCHWARTZ, F. J. Biology of the clearnose skate, Raja eglanteria, from North
Carolina. Biol. Sci. (2). p. 82-95. 1996.
SOARES, L. S. H., VAZZOLER, A. E. A. M. & CORREA, A. R. Diel feeding
chronology of the skate Raja agassizi (Muller & Henle) (Pisces, Elasmobranchii)
on the continental shelf off Ubatuba, southeastern Brazil. Revta bras. Zool. v.
16(1), p. 201-212. 1999.

99
SOKAL, R. R. & ROHLF, F. J. Biometry: The principles and practice of statistics in
biological research. First Edition. W. H. Freeman and Company, New York, 776
p., 1969.
STILLWELL, C. E. & KOHLER, N. E. Food, feeding habits, and estimatives of daily
ration of the shortfin mako (Isurus oxyrinchus) in the northwest Atlantic. Can. J.
Fish Aquat. Sci. v. 39, p. 407-414. 1982.
STILLWELL, C. E. & KOHLER, N. E. Food habits of the sandbar shark Carcharhinus
plumbeus off the U.S. northeast coast, with estimates of daily ration. Fish. Bull.
91.p. 138-150. 1993.
TRICAS, T. C. Relationships of the blue shark, Prionace glauca, and its prey species
near Santa Catalina Island, California. Fish. Bull. v. 77. p. 175-182. 1979.
VASKE, JR. T. & RINCON-FILHO, G. Conteúdo estomacal dos tubarões azul
(Prionace glauca) e anequim (Isurus oxyrinchus) em águas oceânicas no sul do
Brasil. Rev. Brasil. Biol. v. 58(3), p. 445-452. 1998.
VIEIRA, S. Bioestatística- Tópicos avançados. Teste não-paramétricos, tabelas de
contingência e análise de regressão. Editora Campus. Rio de Janeiro. 212 p. 2003.
VOSHELL, Jr. J. R. A guide to common freshawater invertebrates of North America.
The McDonald & Woodward Publishing Company. Blacksburg, Virginia. 442 p.
2002.
ZANDEN, M.J.V & RASMUSSEN, J.B. A trophic position model of pelagic food
webs: impact on contaminant biomagnification in lake trout. In: Ecological
Monographs, 66. p. 451-477, 1996.

100
ZANDEN, M.J.V.; CABANA, G. & RASMUSSEN, J.B. Comparing trophic position of
freshwater fish calculated using stable nitrogen isotope ratios ( )N15σ and literature
dietary data. Can. J. of Fish. and Aqua. Scie., 54. p. 1142-1158, 1997.
ZAVALA-CAMIN, L. A. Introdução os estudos sobre alimentação natural em peixes.
Paraná: Editora da Universidade Estadual de Maringá, 129 p. 1996.

101
CAPÍTULO III
Reprodução de Potamotrygon orbignyi
(CASTELNAU, 1855) no Rio Paranã, Alto
Tocantins.
INTRODUÇÃO
As raias de água doce apresentam um modo de reprodução identificado como
viviparidade aplacentária com trofonemata, o qual é seguido, de uma forma geral, por
todas as raias Myliobatiformes (HAMLETT & KOOB, 1999). Este padrão foi
evidenciado até mesmo em raias de água doce do Eoceno inferior da América do Norte
(CARVALHO et al., 2004), onde fêmeas grávidas e recém abortadas foram encontradas
em um depósito fossilífero de um sistema de paleolagos formados pela orogênese das
Montanhas Rochosas. O cenário descrito pelos fosseis é o mesmo encontrado nas

102
espécies atuais quando estas raias morrem grávidas, abortando seus filhotes na tentativa
de salvá-los se estiverem a termo.
Ao longo das últimas décadas, diversos trabalhos forneceram informações
superficiais e pontuais sobre aspectos da reprodução destes animais (CASTEX, 1963,
1964; ACHENBACH & ACHENBACH, 1976; YÉPEZ & ESPINOSA, 1970;
THORSON, et al., 1983), quase sempre baseados em encontros casuais, sem uma
hipótese clara e desenho amostral. A falta de estudos sistemáticos sobre a reprodução
das raias de água doce muito se deve à dificuldade em se obter coletas periódicas de
animais, extremamente necessárias devido aos longos períodos de gestação e ciclos
reprodutivos complexos. Estas características, por vezes, impossibilitam o desenho
esquemático de todas as etapas envolvidas na reprodução, desde a maturidade, cópula,
fecundação, gestação e parto, e conseqüente estimação de sua periodicidade.
Pesquisas recentes começaram a apresentar um melhor panorama do estudo da
reprodução em raias de água doce, fornecendo informações primordiais como a
morfologia do sistema reprodutor e sua variabilidade ao longo da gravidez, e estimando
parâmetros reprodutivos básicos como o tamanho de primeira maturação, fecundidade e
tempo de gestação (TESHIMA & TAKESHITA, 1992; LASSO et al., 1996; ARAÚJO,
1998; CHARVET-ALMEIDA, 2001; CHARVET-ALMEIDA et al., 2005). Somente
com base em informações biológicas mais detalhadas será possível estimar os melhores
parâmetros populacionais a serem utilizados em modelos para conservação e manejo de
estoques (CORTÉS, 2004).
Tendo em vista o desenvolvimento recente da pesca ornamental de raias de água
doce na bacia dos rios Tocantins-Araguaia e a necessidade de se estimar a fragilidade do
estoque perante esta atividade, faz-se necessário o estudo da reprodução destas raias,

103
tendo em vista que a maioria dos modelos de manejo pesqueiro se baseia em parâmetros
reprodutivos e de captura para uma avaliação do estoque. Este capítulo trata da
reprodução de Potamotrygon orbignyi no rio Paranã, afluente do alto rio Tocantins, e
fornece informações sobre a biologia básica reprodutiva e do aspecto morfológico dos
sistemas reprodutores, bem como estima parâmetros reprodutivos a serem utilizados nos
modelos de manejo pesqueiro.
MATERIAL E MÉTODOS
A amostra de animais dissecados para análise de seus sistemas reprodutores
consistiu de 47 fêmeas com larguras de disco-LD que variaram de 150 a 460 mm e de 37
machos com 153 a 371 mm LD. Todos os animais foram capturados no rio Paranã,
afluente do alto rio Tocantins na região do município de Paranã-TO com zagaia (arpão de
mão) durante a noite e com o uso de facho luminoso, tarrafa e espinhel de fundo com 25
anzóis nos anos de 2002 (janeiro-abril e agosto-dezembro) e 2003 (janeiro e agosto-
setembro). Em laboratório foi registrada a largura de Disco-LD (mm), peso total-PT (g) e
peso eviscerado-PE (g) de cada exemplar. Os órgãos reprodutores foram retirados e de
cada exemplar foram observados os seguintes aspectos: Fêmeas - forma e configuração do
sistema reprodutor, cor e diâmetro dos folículos ovarianos, conteúdo dos úteros,
classificados como vazios (fêmea não grávida) ou com ovos uterinos ou embriões (fêmea
grávida); os ovos uterinos foram contados, pesados (g) e medido o maior diâmetro (mm);
os embriões foram pesados (g), sexados e medido a LD (mm); Machos – foram medidos
os comprimentos dos claspers, desde sua axila até a sua extremidade posterior (mm);
largura e comprimento dos epidídimos, vaso deferente, vesículas seminais e testículos

104
(mm); os testículos também foram pesados (g). Todas as medidas foram tomadas com
paquímetro e os pesos com balança digital DIPSE PS-250 com precisão de décimo de
grama.
O critério de identificação de fêmeas adultas foi baseado nas seguintes
características em ordem de importância: presença de ovos ou embriões em seus úteros,
presença de folículos vitelogênicos com mais de 10 mm de diâmetro nos ovários,
morfologia dos seus úteros e largura e desenvolvimento das glândulas nidamentárias. Os
machos foram considerados adultos quando seus cláspers estão rigidamente calcificados e
existe conteúdo na vesícula seminal. O feto a termo é aquele com reduzida ou inexistente
vesícula vitelínica externa. Contudo, é importante ressaltar a capacidade que muitos
embriões possuem de sobreviver após o parto prematuro ou aborto induzido pela captura,
mesmo com vesículas vitelínicas relativamente grandes. A estimativa do comprimento
médio (L50) de primeira maturação foi calculada pelo método gráfico de VAZZOLER
(1996) e pelo método da regressão logística (ROA et al., 1999).
O índice hepatossomático em P. orbignyi foi calculado através da relação entre o
peso do fígado (PF) e o peso somático do animal (PT) pela equação: IHS = (PF/PT) x 100
e a sua variabilidade indicou o acúmulo de reservas energéticas hepáticas a serem
utilizadas na reprodução.
A leitura dos anéis nas vértebras se fez através da remoção das primeiras cinco
vértebras imediatamente posteriores ao sinarcual toracolumbar, emblocagem de uma
vértebra em resina epóxi e corte longitudinal, ou desgaste com serra manual de alta rotação
(11000-35000 RPM) com disco de lixa fina. Após o corte, a superfície era polida com
disco de lã. Os cortes foram lidos em lupa estereoscópica Nikon com luz incidente. A
terminologia e definições seguem CAILLIET et al., (1983).

105
A terminologia para a descrição do sistema reprodutor de P. orbignyi segue
BABEL (1967) e HAMLETT (1999) e a fecundidade foi interpretada como em PERES &
VOOREN (1991), na qual estes autores reconhecem a fecundidade uterina como o número
de filhotes e ovos nos úteros, e a fecundidade ovariana como o número de folículos
ovarianos em maturação (vitelogênese) no ovário.
Todos os testes estatísticos foram executados pelos programas STATISTICA 5.5
da STATSOFT, SYSTAT 10 da SPSS e Excel 2002 da Microsoft e consideraram o
nível de significância de 5%.
RESULTADOS
No total, 162 indivíduos (97 fêmeas e 65 machos) foram analisados para o estudo
da reprodução e 84 indivíduos foram dissecados (47 fêmeas e 37 machos) para análise do
sistema reprodutor. A proporção sexual encontrada foi 1,49:1 F:M, o que se mostrou
significativamente diferente da proporção esperada de 1:1 (χ2 = 6,32; p = 0,011).
Contudo, não foi encontrada diferença na razão sexual F:M entre adultos 1,71:1 (χ2 =
2,63; p = 0,10), mas ocorreu uma diferença significativa na proporção F:M entre jovens
1,44:1 (χ2 = 4,14; p = 0,04).
Tamanho de Maturação
As larguras de disco variaram de 153 a 371 mm LD nos machos (n = 37), onde o
menor macho maturo possuía 225 mm LD e o maior macho imaturo possuía 278 mm
LD. O comprimento médio de primeira maturação (LD50) foi estimado em 251 mm LD,
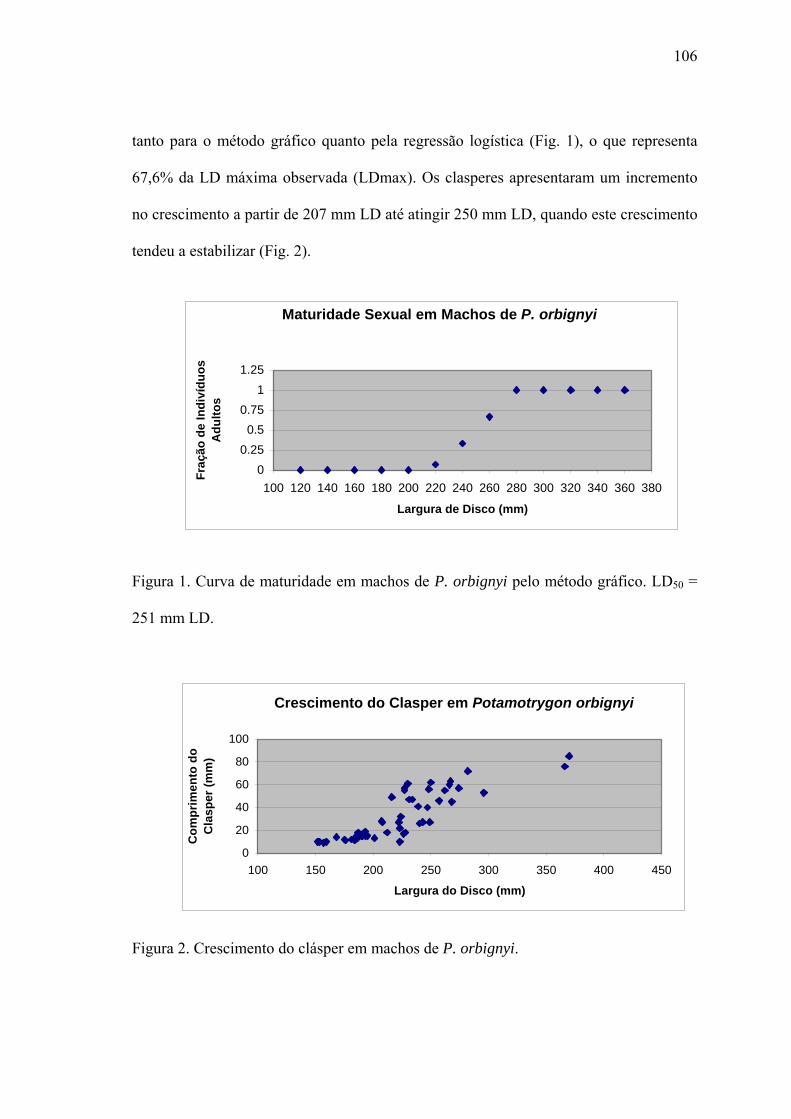
106
tanto para o método gráfico quanto pela regressão logística (Fig. 1), o que representa
67,6% da LD máxima observada (LDmax). Os clasperes apresentaram um incremento
no crescimento a partir de 207 mm LD até atingir 250 mm LD, quando este crescimento
tendeu a estabilizar (Fig. 2).
Maturidade Sexual em Machos de P. orbignyi
00.25
0.50.75
11.25
100 120 140 160 180 200 220 240 260 280 300 320 340 360 380
Largura de Disco (mm)
Fraç
ão d
e In
diví
duos
A
dulto
s
Figura 1. Curva de maturidade em machos de P. orbignyi pelo método gráfico. LD50 =
251 mm LD.
Crescimento do Clasper em Potamotrygon orbignyi
0
20
40
60
80
100
100 150 200 250 300 350 400 450
Largura do Disco (mm)
Com
prim
ento
do
Cla
sper
(mm
)
Figura 2. Crescimento do clásper em machos de P. orbignyi.
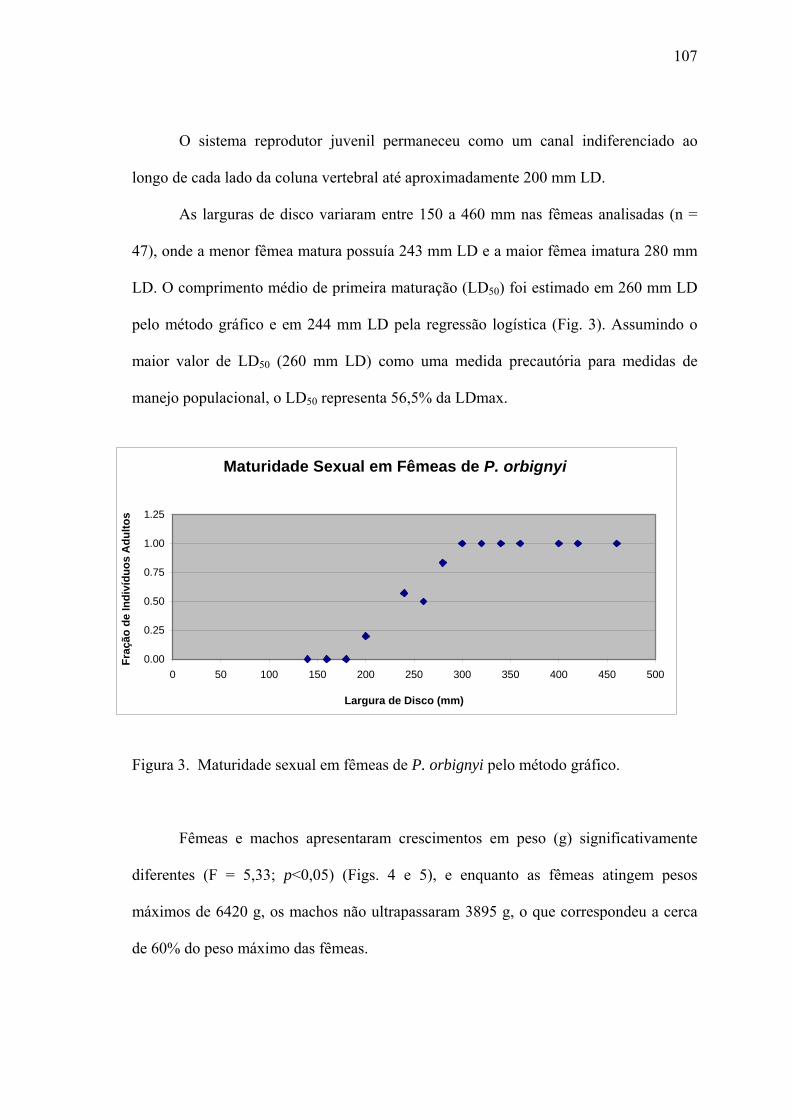
107
O sistema reprodutor juvenil permaneceu como um canal indiferenciado ao
longo de cada lado da coluna vertebral até aproximadamente 200 mm LD.
As larguras de disco variaram entre 150 a 460 mm nas fêmeas analisadas (n =
47), onde a menor fêmea matura possuía 243 mm LD e a maior fêmea imatura 280 mm
LD. O comprimento médio de primeira maturação (LD50) foi estimado em 260 mm LD
pelo método gráfico e em 244 mm LD pela regressão logística (Fig. 3). Assumindo o
maior valor de LD50 (260 mm LD) como uma medida precautória para medidas de
manejo populacional, o LD50 representa 56,5% da LDmax.
Maturidade Sexual em Fêmeas de P. orbignyi
0.00
0.25
0.50
0.75
1.00
1.25
0 50 100 150 200 250 300 350 400 450 500
Largura de Disco (mm)
Fraç
ão d
e In
diví
duos
Adu
ltos
Figura 3. Maturidade sexual em fêmeas de P. orbignyi pelo método gráfico.
Fêmeas e machos apresentaram crescimentos em peso (g) significativamente
diferentes (F = 5,33; p<0,05) (Figs. 4 e 5), e enquanto as fêmeas atingem pesos
máximos de 6420 g, os machos não ultrapassaram 3895 g, o que correspondeu a cerca
de 60% do peso máximo das fêmeas.
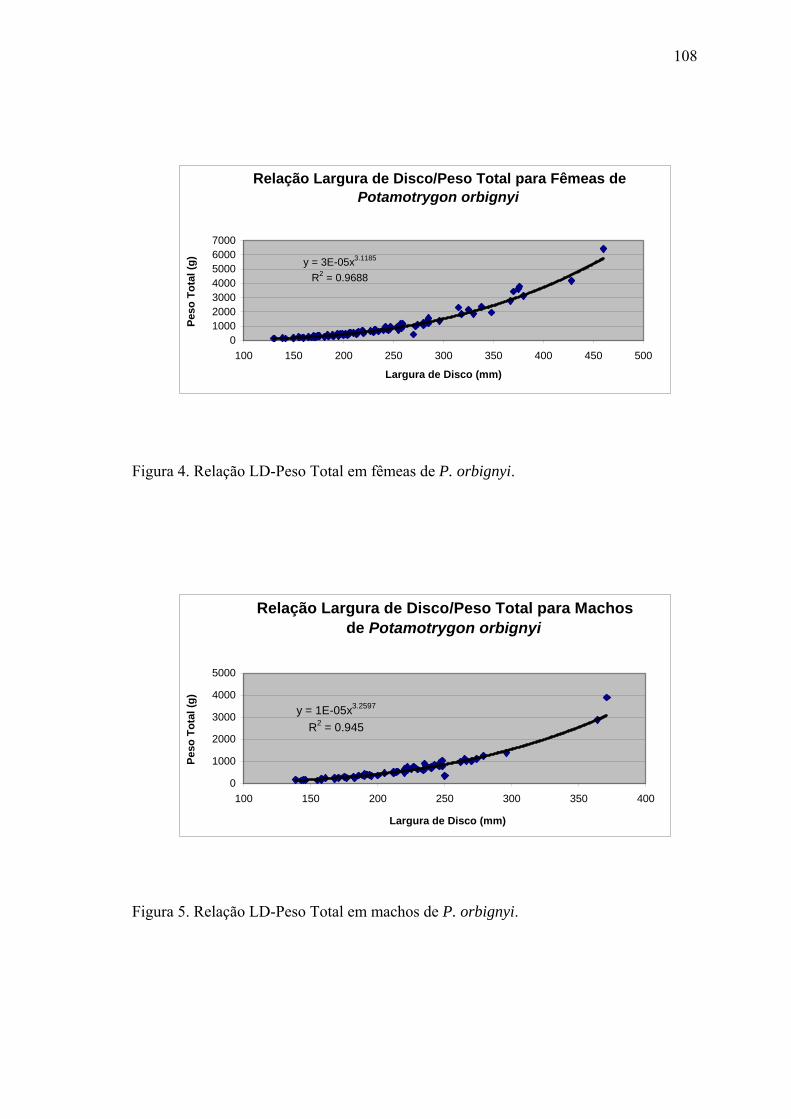
108
Relação Largura de Disco/Peso Total para Fêmeas de Potamotrygon orbignyi
y = 3E-05x3.1185
R2 = 0.9688
01000200030004000500060007000
100 150 200 250 300 350 400 450 500
Largura de Disco (mm)
Peso
Tot
al (g
)
Figura 4. Relação LD-Peso Total em fêmeas de P. orbignyi.
Relação Largura de Disco/Peso Total para Machos de Potamotrygon orbignyi
y = 1E-05x3.2597
R2 = 0.945
0
1000
2000
3000
4000
5000
100 150 200 250 300 350 400
Largura de Disco (mm)
Peso
Tot
al (g
)
Figura 5. Relação LD-Peso Total em machos de P. orbignyi.
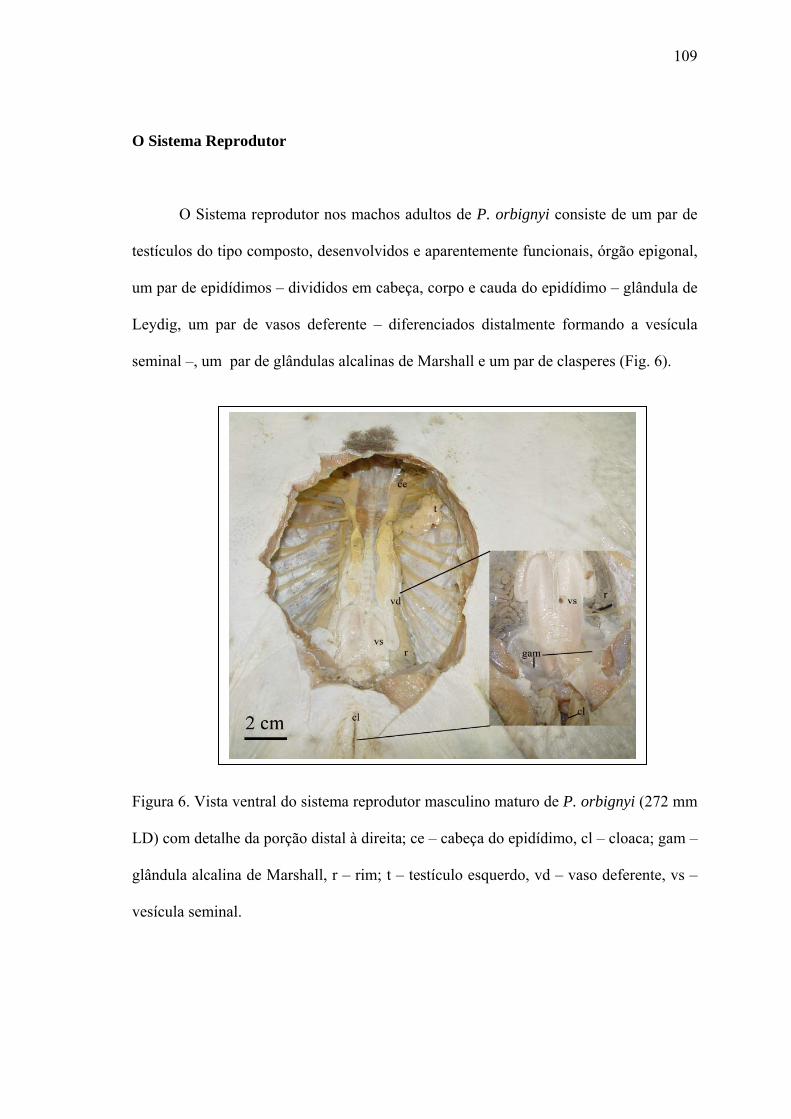
109
O Sistema Reprodutor
O Sistema reprodutor nos machos adultos de P. orbignyi consiste de um par de
testículos do tipo composto, desenvolvidos e aparentemente funcionais, órgão epigonal,
um par de epidídimos – divididos em cabeça, corpo e cauda do epidídimo – glândula de
Leydig, um par de vasos deferente – diferenciados distalmente formando a vesícula
seminal –, um par de glândulas alcalinas de Marshall e um par de clasperes (Fig. 6).
Figura 6. Vista ventral do sistema reprodutor masculino maturo de P. orbignyi (272 mm
LD) com detalhe da porção distal à direita; ce – cabeça do epidídimo, cl – cloaca; gam –
glândula alcalina de Marshall, r – rim; t – testículo esquerdo, vd – vaso deferente, vs –
vesícula seminal.

110
Em machos adultos, a cabeça do epidídimo está posicionada ventralmente em
relação à porção distal do sinarcual cervicotoráxico, lateralmente ao esôfago e entre os
testículos; os epidídimos seguem ventralmente inseridos na glândula de Leydig e
margeando a coluna vertebral até a cauda do epidídimo, demarcada por um
estreitamento do canal; o canal do vaso deferente apresenta um colorido esbranquiçado
e segue sinuosamente até a vesícula seminal, identificada por um distinto alargamento
do canal, seu posicionamento sobre a coluna vertebral e a sua abertura no sinus
urogenital. Estas vesículas apresentam compartimentações complexas, formadas por
prolongações das paredes dorsais da vesícula que se estendem até quase obstruí-la. Lado
a lado à porção distal das vesículas seminais estão as glândulas alcalinas de Marshall,
que são ovais, translúcidas e com um diâmetro igual à 1/3 do comprimento da vesícula.
O órgão epigonal estende-se medialmente desde os testículos até a vesícula seminal;
aderido dorsalmente na cavidade peritoneal como uma prega de tamanho e forma
variáveis.
O sistema reprodutor nas fêmeas consiste dos seguintes órgãos duplos: ovários,
ostia, ovidutos anteriores, glândulas nidamentárias e ovidutos posteriores ou úteros, que
terminam no sinus urogenital (Fig. 7). Os ovários são órgãos flácidos sem forma
definida e infiltrados no órgão epigonial, localizados na porção anterior da cavidade
peritoneal e aderidos à parede dorsal pelo mesovarium; cada oviduto anterior possui na
sua extremidade um afunilamento denominado ostium, por onde a ova entra e segue ao
longo do oviduto em direção ao útero; a glândula nidamentária diferencia-se do oviduto
anterior por um alargamento sinuoso da parede do oviduto e um estrangulamento
posterior demarcando o início do útero; o útero é mais largo que o oviduto anterior e
que a glândula nidamentária, mas a relação entre a sua largura e o comprimento são

111
variáveis com a maturidade e com a reprodução; a parede interna é altamente
vascularizada e rica em vilosidades denominadas trofonemata e apresenta grande
adaptabilidade ao desenvolvimento embrionário e ao crescimento do filhote.
Figura 7. Vista latero-ventral do sistema reprodutor de fêmea adulta de P. orbignyi (400
mm LD). A região anterior encontra-se à esquerda e a posterior à direita; fl – folículos
vitelogênicos, gn – glândula nidamentária, oa – oviduto anterior, ov – ovário, op –
órgão epigonal, os – óstium direito, ut – útero. O ovário esquerdo está mais
desenvolvido, enquanto o ovário da direita está reduzido com um pequeno folículo.
P. orbignyi apresentou simetria ovariana em 36% das fêmeas analisadas e na
maioria das vezes somente o ovário esquerdo é desenvolvido e funcional. Embora
menos freqüente, o ovário direito também desenvolveu folículos vitelogênicos e não
foram encontradas diferenças significativas na largura e comprimento médio entre os
ovários (Teste-t, t = 2,08 e p = 0,055 e t = 2,17 e p = 0,050). Os úteros de fêmeas
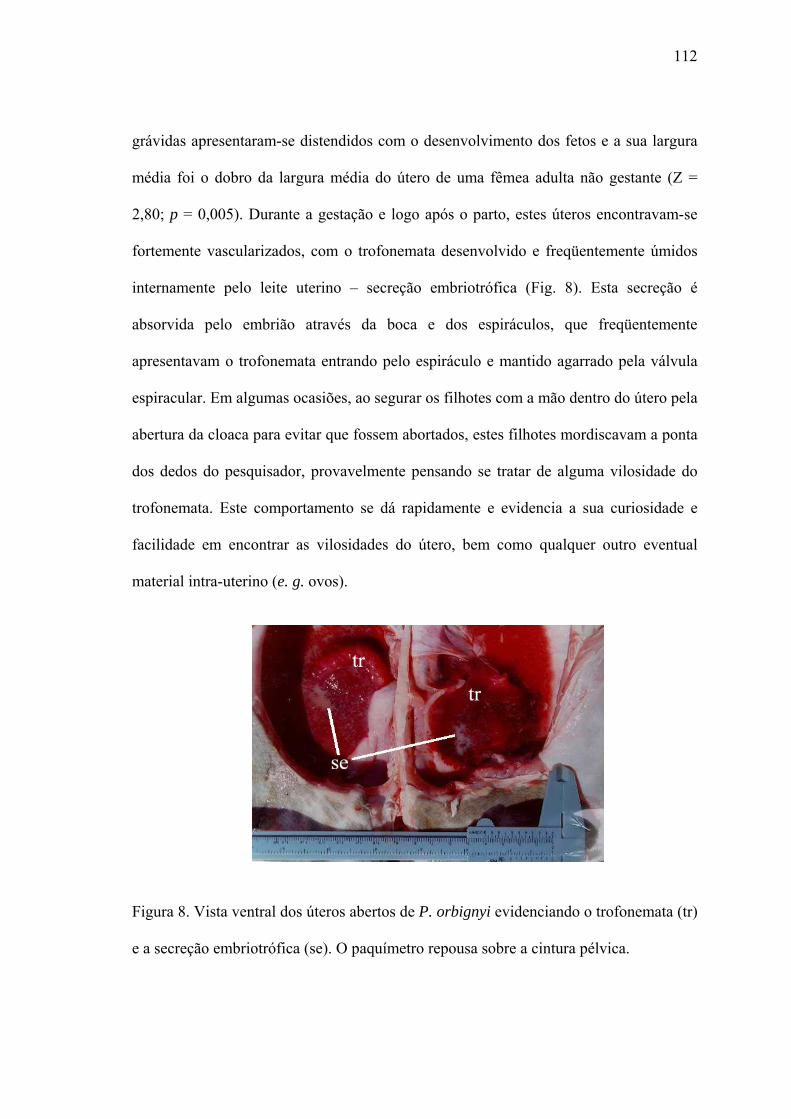
112
grávidas apresentaram-se distendidos com o desenvolvimento dos fetos e a sua largura
média foi o dobro da largura média do útero de uma fêmea adulta não gestante (Z =
2,80; p = 0,005). Durante a gestação e logo após o parto, estes úteros encontravam-se
fortemente vascularizados, com o trofonemata desenvolvido e freqüentemente úmidos
internamente pelo leite uterino – secreção embriotrófica (Fig. 8). Esta secreção é
absorvida pelo embrião através da boca e dos espiráculos, que freqüentemente
apresentavam o trofonemata entrando pelo espiráculo e mantido agarrado pela válvula
espiracular. Em algumas ocasiões, ao segurar os filhotes com a mão dentro do útero pela
abertura da cloaca para evitar que fossem abortados, estes filhotes mordiscavam a ponta
dos dedos do pesquisador, provavelmente pensando se tratar de alguma vilosidade do
trofonemata. Este comportamento se dá rapidamente e evidencia a sua curiosidade e
facilidade em encontrar as vilosidades do útero, bem como qualquer outro eventual
material intra-uterino (e. g. ovos).
Figura 8. Vista ventral dos úteros abertos de P. orbignyi evidenciando o trofonemata (tr)
e a secreção embriotrófica (se). O paquímetro repousa sobre a cintura pélvica.
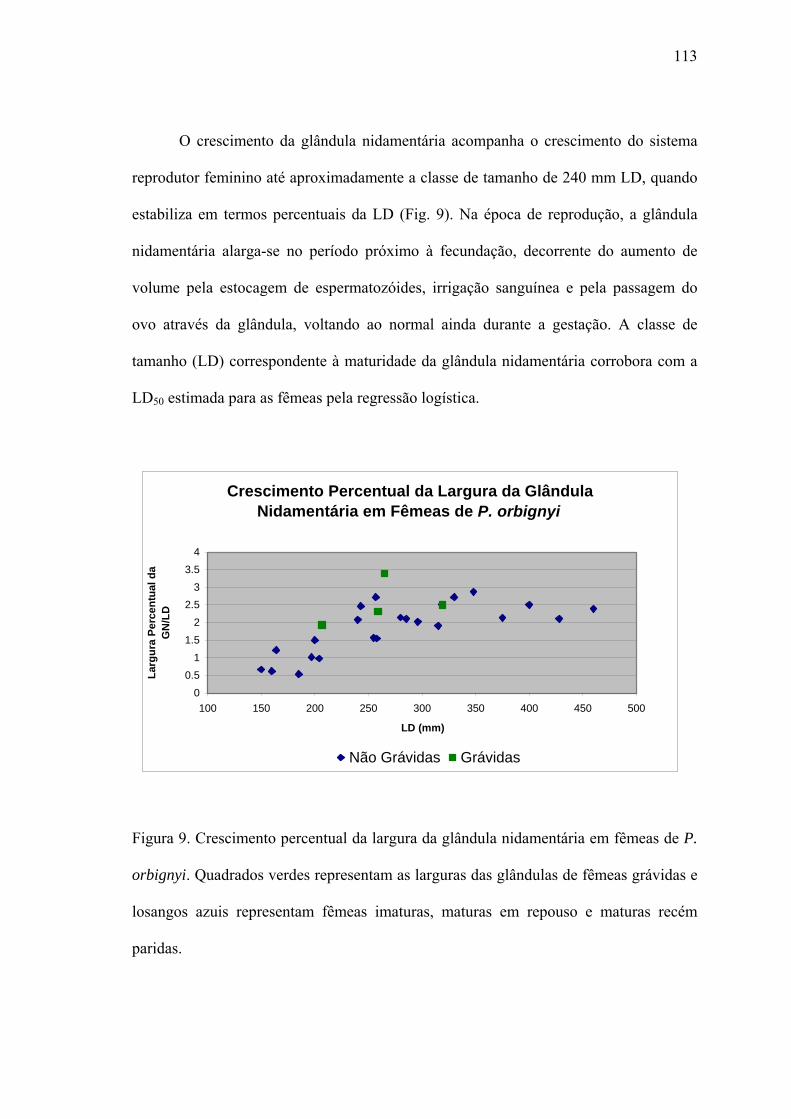
113
O crescimento da glândula nidamentária acompanha o crescimento do sistema
reprodutor feminino até aproximadamente a classe de tamanho de 240 mm LD, quando
estabiliza em termos percentuais da LD (Fig. 9). Na época de reprodução, a glândula
nidamentária alarga-se no período próximo à fecundação, decorrente do aumento de
volume pela estocagem de espermatozóides, irrigação sanguínea e pela passagem do
ovo através da glândula, voltando ao normal ainda durante a gestação. A classe de
tamanho (LD) correspondente à maturidade da glândula nidamentária corrobora com a
LD50 estimada para as fêmeas pela regressão logística.
Crescimento Percentual da Largura da Glândula Nidamentária em Fêmeas de P. orbignyi
00.5
11.5
22.5
33.5
4
100 150 200 250 300 350 400 450 500
LD (mm)
Larg
ura
Perc
entu
al d
a G
N/L
D
Não Grávidas Grávidas
Figura 9. Crescimento percentual da largura da glândula nidamentária em fêmeas de P.
orbignyi. Quadrados verdes representam as larguras das glândulas de fêmeas grávidas e
losangos azuis representam fêmeas imaturas, maturas em repouso e maturas recém
paridas.
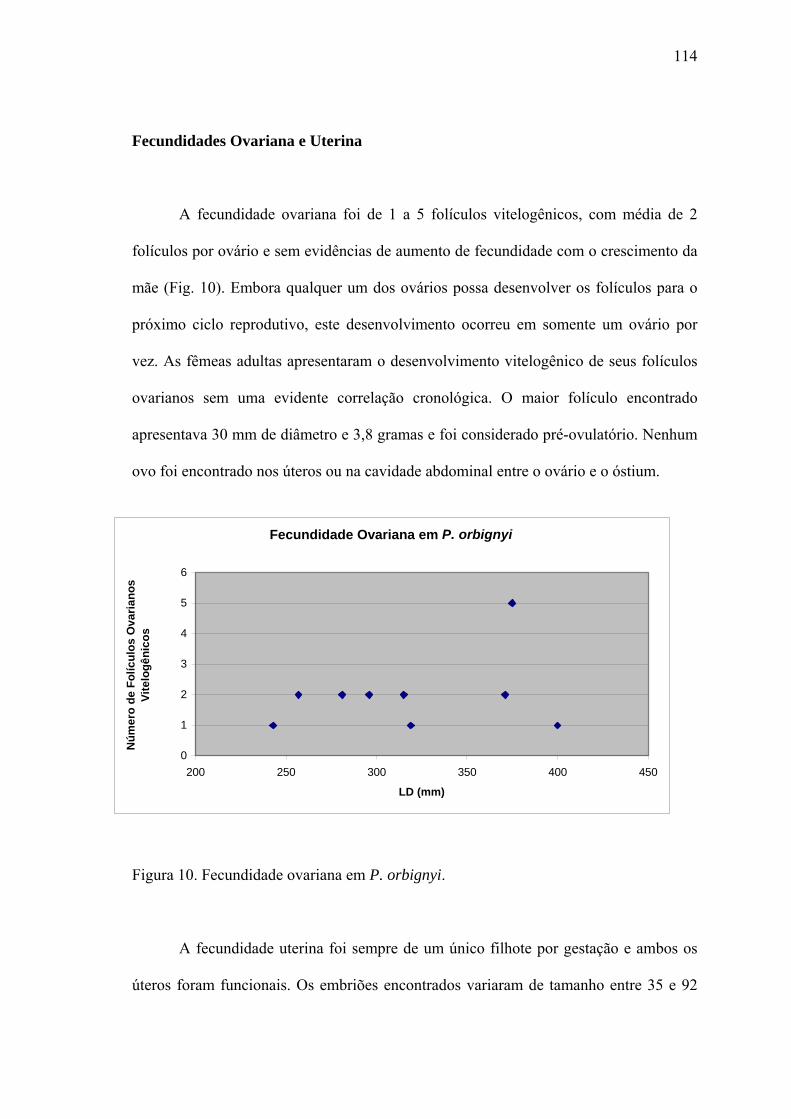
114
Fecundidades Ovariana e Uterina
A fecundidade ovariana foi de 1 a 5 folículos vitelogênicos, com média de 2
folículos por ovário e sem evidências de aumento de fecundidade com o crescimento da
mãe (Fig. 10). Embora qualquer um dos ovários possa desenvolver os folículos para o
próximo ciclo reprodutivo, este desenvolvimento ocorreu em somente um ovário por
vez. As fêmeas adultas apresentaram o desenvolvimento vitelogênico de seus folículos
ovarianos sem uma evidente correlação cronológica. O maior folículo encontrado
apresentava 30 mm de diâmetro e 3,8 gramas e foi considerado pré-ovulatório. Nenhum
ovo foi encontrado nos úteros ou na cavidade abdominal entre o ovário e o óstium.
Fecundidade Ovariana em P. orbignyi
0
1
2
3
4
5
6
200 250 300 350 400 450
LD (mm)
Núm
ero
de F
olíc
ulos
Ova
riano
s Vi
telo
gêni
cos
Figura 10. Fecundidade ovariana em P. orbignyi.
A fecundidade uterina foi sempre de um único filhote por gestação e ambos os
úteros foram funcionais. Os embriões encontrados variaram de tamanho entre 35 e 92

115
mm LD ou 6,6 e 42 g de PT, respectivamente. O menor exemplar livre natante
encontrado foi uma fêmea com 130 mm LD (130 g de PT) capturada no início de
agosto, durante a estação seca. Este exemplar já havia digerido todo o vitelo, o que
geralmente leva em torno de três a quatro semanas após o parto e que corresponde a um
acréscimo de 10 a 15 mm na LD durante este período (observação pessoal, Fig. 11).
Desta forma, o parto em P. orbignyi deve ocorrer quando os filhotes atingem de 100 a
115 mm LD e pesos entre 48 e 61 gramas, o que corresponde a um ganho em peso do
filhote de 1605% em relação ao peso inicial do ovo. Observações adicionais em
neonatos livre natantes capturados em Marabá pela pesca ornamental confirmam esses
valores.
Figura 11. Exemplar neonato macho de P. orbignyi mantido em aquário após o parto
(98 mm LD). A fita marrom clara ao lado é vitelo digerido defecado, indicando que este
exemplar ainda se nutre das reservas acumuladas durante a gestação.
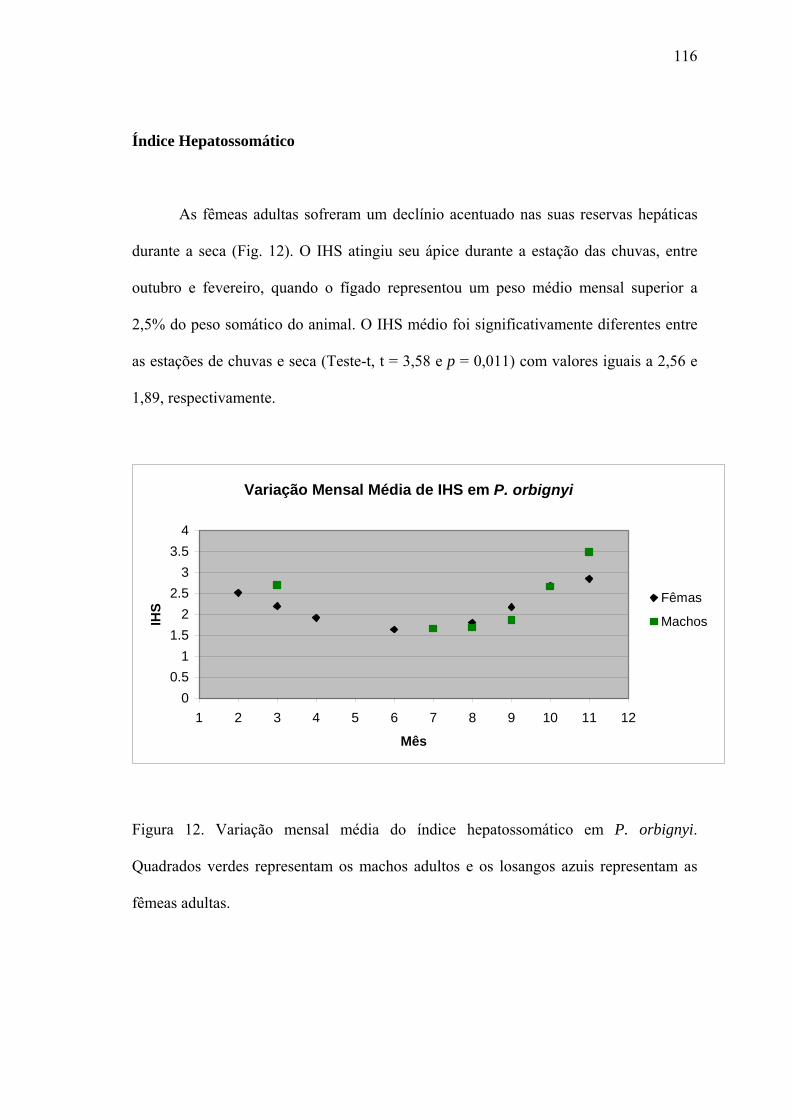
116
Índice Hepatossomático
As fêmeas adultas sofreram um declínio acentuado nas suas reservas hepáticas
durante a seca (Fig. 12). O IHS atingiu seu ápice durante a estação das chuvas, entre
outubro e fevereiro, quando o fígado representou um peso médio mensal superior a
2,5% do peso somático do animal. O IHS médio foi significativamente diferentes entre
as estações de chuvas e seca (Teste-t, t = 3,58 e p = 0,011) com valores iguais a 2,56 e
1,89, respectivamente.
Variação Mensal Média de IHS em P. orbignyi
00.5
11.5
22.5
33.5
4
1 2 3 4 5 6 7 8 9 10 11 12
Mês
IHS
Fêmas
Machos
Figura 12. Variação mensal média do índice hepatossomático em P. orbignyi.
Quadrados verdes representam os machos adultos e os losangos azuis representam as
fêmeas adultas.

117
Ciclo Reprodutivo
Todas as fêmeas grávidas foram capturadas nas chuvas (setembro-fevereiro),
representando 45% das fêmeas adultas capturadas nesta estação, e estavam em
adiantado estágio de desenvolvimento embrionário. O parto em P. orbignyi deve ocorrer
entre dezembro e fevereiro, quando os filhotes atingem o tamanho de nascimento e o rio
oferece maior abundância de alimentos, evidenciada pelos altos valores de reservas
hepáticas (IHS) acumuladas durante as chuvas. No entanto, pequenas raias (neonatos e
juvenis) com larguras de disco entre 130 e 140 mm também foram capturadas durante
os meses de seca. Tendo em vista que o desenvolvimento dos folículos ovarianos não
apresentou evidente correlação cronológica e a captura de fêmeas grávidas não foi
homogênea ao longo do ano, não foi possível estimar a época de cópula, fecundação e o
tempo de gestação. Contudo, o fato de que fêmeas grávidas apresentaram o
desenvolvimento de folículos vitelogênicos concomitantemente com a gestação é forte
evidência de que a cópula e o início de um próximo período gestacional ocorram logo
após o parto. Desta forma, com o parto ocorrendo nos meses das chuvas (dezembro-
fevereiro) para parte da população feminina adulta, a cópula e o início da nova gestação
devem ocorrer a partir de março.
Não foram identificadas áreas de berçário, entendidas como feições específicas
do rio (praias arenosas, igarapés, pedregal ou fundos lodosos) onde as raias preferissem
dar à luz e os neonatos e juvenis pudessem ocorrer em maiores densidades favorecidos
por maior proteção contra os predadores e disponibilidade de alimentos. Entretanto, esta
observação não foi considerada a fundo durante as capturas e requer futuros estudos.

118
Idade de Maturação
Uma análise comparativa do tamanho de maturação para P. orbignyi e de um
estudo preliminar das leituras dos anéis nas vértebras destes animais, revelou que, para
o mesmo intervalo de classe de largura de disco de maturação – 240-250 mm LD
(Fig.13) –, observa-se nas vértebras de fêmeas uma banda mais larga e distinta,
demarcando o que provavelmente seja o ano de maturidade sexual (Fig. 14) – o esforço
fisiológico para a formação de gametas, gestação e nutrição do embrião é maior nas
fêmeas e é melhor registrado nas suas vértebras que nos machos.
Figura 13. Vértebra de fêmea de P. orbignyi (460 mm LD) em corte longitudinal com as
bandas identificadas em ordem crescente da direita para a esquerda. A banda
correspondente ao 5º ano de vida identifica a maturidade sexual.
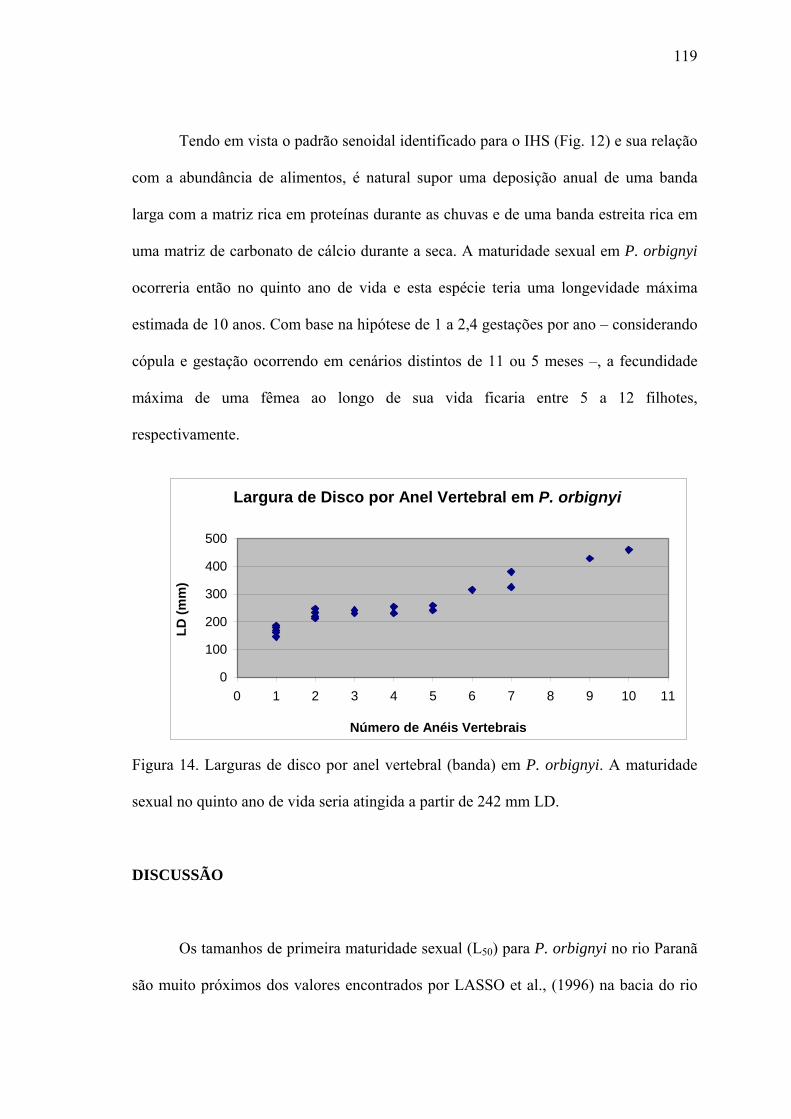
119
Tendo em vista o padrão senoidal identificado para o IHS (Fig. 12) e sua relação
com a abundância de alimentos, é natural supor uma deposição anual de uma banda
larga com a matriz rica em proteínas durante as chuvas e de uma banda estreita rica em
uma matriz de carbonato de cálcio durante a seca. A maturidade sexual em P. orbignyi
ocorreria então no quinto ano de vida e esta espécie teria uma longevidade máxima
estimada de 10 anos. Com base na hipótese de 1 a 2,4 gestações por ano – considerando
cópula e gestação ocorrendo em cenários distintos de 11 ou 5 meses –, a fecundidade
máxima de uma fêmea ao longo de sua vida ficaria entre 5 a 12 filhotes,
respectivamente.
Largura de Disco por Anel Vertebral em P. orbignyi
0
100
200
300
400
500
0 1 2 3 4 5 6 7 8 9 10 11
Número de Anéis Vertebrais
LD (m
m)
Figura 14. Larguras de disco por anel vertebral (banda) em P. orbignyi. A maturidade
sexual no quinto ano de vida seria atingida a partir de 242 mm LD.
DISCUSSÃO
Os tamanhos de primeira maturidade sexual (L50) para P. orbignyi no rio Paranã
são muito próximos dos valores encontrados por LASSO et al., (1996) na bacia do rio

120
Apure, Venezuela - 230 mm e 185-290 mm LD para machos e fêmeas, respectivamente.
No entanto, diferem drasticamente dos valores estimados para a mesma espécie na baía
de Marajó e rio Negro (CHARVET-ALMEIDA et al., 2005), que maturam com larguras
de disco muito superiores – 390 mm e 440 mm LD para machos e fêmeas,
respectivamente. Estas populações diferem ainda na fecundidade uterina (1 vs 1-5, para
Tocantins e Marajó-Negro, respectivamente) e no número de ovários funcionais (2 vs
1). Embora haja registros de variações intra-específicas nos parâmetros biológicos ou
populacionais em algumas espécies de elasmobrânquios (BRANSTETTER, et. al.,
1987; TANAKA et al., 1990; PARSONS, 1993; SMINKEY & MUSICK, 1995;
TANIUCHI & TACHIKAWA, 1999; CHIARAMONTE & PETTOVELLO, 2000), as
diferenças encontradas entre as populações de P. orbignyi do rio Tocantins e da baía de
Marajó-Negro são suficientes para questionar a validade da identificação específica.
Diferenças nos parâmetros reprodutivos de espécies simpátricas já foram utilizadas
como ferramentas taxonômicas na diagnose destas espécies (VOOREN & SILVA,
1991). Desta forma, baseando-se nestas evidências, faz-se necessária uma revisão
taxonômica comparativa de P. orbignyi entre os exemplares da baía de Marajó-Negro e
o rio Tocantins a fim de se confirmar a identificação.
As diferenças encontradas em alguns parâmetros reprodutivos entre as duas
populações poderiam ser explicadas por diferenças nas interações ecológicas, seja pela
ausência de predadores naturais na população Marajó-Negro ou por uma alimentação
qualitativamente mais rica e que fornecesse maior aporte nutricional. Segundo
BRAGANÇA et al., (2004), P. orbignyi teria larvas de Hemiptera (piolhos d’água)
como principal item alimentar na baía de Marajó, enquanto que Ephemeroptera é o
principal item alimentar desta raia no rio Tocantins (q. v. capítulo II). Assumindo que

121
larvas de Hemiptera sejam energeticamente mais compensatórias, essa diferença
alimentar justificaria um maior ganho de peso e tamanho nas raias de Marajó. No
entanto, se esta oferta nutricional beneficiasse P. orbignyi, necessariamente teria de
beneficiar outras espécies ecologicamente correlatas, tanto de teleósteos quanto
elasmobrânquios. Potamotrygon scobina é uma raia de água doce com hábitos
alimentares muito semelhantes a P. orbignyi, principalmente na baía de Marajó, onde
seu principal item alimentar também é larvas de Hemiptera (piolhos d’água)
(BRAGANÇA, et al., 2004). Contudo, P. scobina também é uma espécie comum no rio
Tocantins e o seu tamanho de maturidade sexual não difere do indicado para os
exemplares da baía de Marajó (350 e 400 mm LD para machos e fêmeas,
respectivamente; CHARVET-ALMEIDA et al., 2005), e os machos maturam com cerca
de 357 mm LD e as fêmeas com 430 mm LD (G. Rincon, material não publicado).
Desta forma, as diferenças de tamanho de maturidade sexual observadas em P. orbignyi
entre as populações do rio Tocantins e Marajó-Negro não podem ser explicadas com
base nas diferenças de hábitos alimentares ou na ausência de predadores naturais, haja
vista que a suposta ausência destes predadores na baía de Marajó também beneficiaria
P. scobina, o que não ocorre. Resta então a necessidade de uma revisão taxonômica de
P. orbignyi para a baía de Marajó e rio Negro e sua confirmação específica, o que está
em andamento pelos pesquisadores da Universidade Federal da Paraíba e Museu
Paraense Emílio Goeldi (Patricia Charvet-Almeida e Mauricio Almeida).
Potamotrygon magdalenae, uma espécie endêmica da Colômbia e distribuída
nos rios Magdalena, Cauca, San Jorge, Catatumbo e Atrato (GALVIS et al., 1997),
apresenta diversas similaridades reprodutivas com P. orbignyi do rio Tocantins, embora
seja ligeiramente menor que a espécie brasileira; são estas: dois ovários funcionais,

122
diâmetro do folículo ovariano maturo em aproximadamente 25 mm, tamanho de
maturidade sexual de machos e fêmeas – expresso como porcentagem da LDmax igual a
69% e 53%, respectivamente –, baixa fecundidade com apenas 2 filhotes por gestação,
tamanho ao nascer em 100 mm LD e período reprodutivo indefinido devido à presença
de fêmeas com vitelogênese e fêmeas grávidas em diversos estágios de
desenvolvimento embrionário concomitantemente (TESHIMA & TAKESHITA, 1992).
Da mesma forma, LASSO et al. (1996) também observaram fêmeas grávidas de P.
orbignyi, tanto na estação da seca quanto nas chuvas, e apesar de indicar um longo
período de atividade reprodutiva para a espécie, estes autores acreditam que a o parto
seja mais freqüente na época das chuvas, quando o rio está mais alto. Este padrão de
ciclo reprodutivo parece ser seguido, também, no rio Tocantins, onde parte da
população adulta está sincronizada e dá a luz nos meses da estação das chuvas
(aproximadamente 45% das fêmeas adultas). Da mesma forma, diversas outras espécies
de peixes reproduzem durante a estação das chuvas no rio Tocantins, quando a
produtividade é maior e a abundância de alimentos garante uma melhor sobrevivência
juvenil (LOWE-McCONNELL, 1975; COSTI et al., 1977; LOWE-McCONNELL,
1995).
Embora a análise de idade de maturação seja apresentada com base em
resultados preliminares, os valores encontrados de longevidade máxima das raias (10
anos), a baixa fecundidade (1 filhote por gestação) e a curta vida reprodutiva (5 anos) –
entendida como a máxima estimativa de longevidade em anos com atividade
reprodutiva – fornecem estimativas de fecundidade máxima (5 filhotes) – número de
filhotes produzidos ao longo da vida – muito baixas e são seriamente alarmantes para a
conservação desta espécie. Estes valores são mais próximos de exemplares de

123
Rajiformes, dos quais muitas espécies apresentam vidas reprodutivas de 1 a 13 anos
EBERT (2005). Este mesmo autor apresenta valores de estimativas de vida reprodutiva
para 17 espécies de Rajiformes, das quais 9 espécies possuem menos de 6 anos de vida
reprodutiva. Embora estas espécies sejam ovíparas e a sua fecundidade seja
significativamente maior que em potamotrygonídeos, a sua mortalidade juvenil também
é maior (HOLDEN, 1975).
A raia ticonha Rhinoptera bonasus apresenta uma fecundidade uterina de um
filhote por gestação, mas o seu filhote já nasce com uma largura de disco de 405 mm, o
que equivale a metade do tamanho de um adulto (HAMLETT et al., 1985). P. orbignyi
nasce com uma largura de disco equivalente a 45% da largura de disco de um adulto,
mas em termos absolutos é muito menor, com somente 115 mm LD e provavelmente
mais vulnerável a predadores (piranha, jacaré, alguns siluriformes, ariranha e boto). O
investimento energético em uma pequena prole deve ser o suficiente para garantir a
sobrevivência dos filhotes sem o completo esgotamento materno, o que impediria uma
nova gestação em curto prazo. Desta forma, embora P. orbignyi seja relativamente
pequena em relação às demais raias Myliobatiformes, sua mortalidade natural juvenil
deve ser baixa e a freqüência gestacional dentro do esperado para o grupo, o que
favorece a hipótese de uma gestação de 4 meses e pelo menos 2 ciclos reprodutivos por
ano (HAMLETT & KOOB, 1999) a fim de garantir a sobrevivência da espécie.
Contudo, esse equilíbrio é tênue, e diferentemente de espécies de elasmobrânquios mais
prolíficos como o tubarão-azul (Prionace glauca), que pode parir 130 filhotes em uma
única ninhada, estas raias dependem somente de uma baixa fecundidade para recuperar
a população de eventuais oscilações da mortalidade populacional, o que acarreta em um
maior tempo de recuperação, vulnerabilidade e muito baixa resiliência à pesca. Devido à

124
baixa fecundidade, o estoque de recrutamento é quase que imediatamente influenciado
pela pesca do estoque adulto, mesmo com a estimativa de 4 coortes sub-adultas. Embora
a pesca ornamental de P. orbignyi no rio Tocantins esteja baseada na captura de
indivíduos juvenis (Instrução Normativa No 27 de 6 de setembro de 2005), qualquer
mudança na arte ou comportamento de pesca deve ser imediatamente avaliado sob o
risco de uma sobre-pesca em curto prazo.
LITERATURA CITADA
ACHENBACH, G. M. & ACHENBACH, S.V.M. Notas acerca de algunas especies de
raya fluvial (Batoidei, Potamotrygonidae) que frecuentan el sistema hidrográfico
del Paraná medio en el Departamento La Capital (Santa Fe - Argentina). In:
Comun. Mus. Prov. Ciencia. Nat. Florentino Ameghino, 8. p. 1-34, 1976.
ARAÚJO, M. L. G. Biologia reprodutiva e pesca de Potamotrygon C (Chondrichthyes-
Potamotrygonidae), no médio rio Negro, Amazonas.171p. 1998.
BABEL, J. S. Reproduction, life history, and ecology of the round stingray, Urolophus
halleri Cooper. Fish Bull. No 137. 102p. 1967.
BRAGANÇA, A. J. M.; CHARVET-ALMEIDA, P, & BARTHEM, R.B. Preliminary
obsevations on the feeding of the freshwater stingrays Potamotrygon orbignyi,
Potamotrygon scobina and Plesiotrygon iwamae (Chondrichthyes:
Potamotrygonidae) in the Cotijuba Island Region – Pará – Brasil. In: Biology and
Conservation of Freshwater Elasmobranchs: Symposium Proceedings.
International Congress on the Biology of Fish. Manaus, p.49 – 60, 2004.

125
BRANSTETTER, S., MUSICK, J. A. & COLVOCORESSES, J. A. A comparison of
the age and growth of the tiger shark, Galeocerdo cuvieri, from off Virginia and
from the Northwestern Gulf of Mexico. Fish. Bull. v. 85, No 2, p. 268-279. 1987.
CAILLIET, G. M., MARTIN, L. K., KUSHER, D., WOLF, P., WELDEN, B. A.
Techniques for enhancing vertebral bands in age estimation of California
elasmobranches. In: PRINCE, E. D., PULOS, L. M. (Eds.). NOAA Technical
Report NMFS 8. Proceedings of the international workshop on age determination
of oceanic pelagic fishes: tunas, billfishes, and sharks. US Department of
Commerce. p. 157-165. 1983.
CARVALHO, M. R. DE., MAISEY, J. G. & GRANDE, L. Freshwater stingrays of the
Green River formation of Wyoming (Early Eocene), with the description of a new
genus and species and an analysis of its phylogenetic relationships
(Chondrichthyes: Myliobatiformes). Bull. Ame. Mus. Nat. Hist. No 284:136p.
2004.
CASTEX, M. N. Una nueva espécie de raya fluvial: P. labradori. Neotropica, La Plata,
9 (30). p. 117-121. 1963.
CASTEX, M. N. Estado actual de los estúdios sobre la raya fluvial neotropical. Ver.
Mus. Prov. Cs. Nat. Santa Fé. Número extraordinário del cincuentenario. p. 9-34.
1964.
CHARVET-ALMEIDA, P. Ocorrência, biologia e uso das raias de água doce na Baía
de Marajó (Pará, Brasil), com ênfase na biologia de Plesiotrygon iwamae
(Chondrichthyes: Potamotrygonidae). Dissertação de mestrado, Programa de Pós-
graduação em Zoologia da Universidade federal do Pará e Museu Paraense Emílio
Goeldi. Departamento de zoologia, 2001.

126
CHARVET-ALMEIDA, P., ARAÚJO, M. L. G. & ALMEIDA, M. P. Reproductive
aspects of freshwater stingrays (Chondrichthyes: Potamotrygonidae) in the
Brazilian Amazon basin. E-journal of the Northwest Atlantic Fishery Science, v.
35, art. 22. 2005.
CHIARAMONTE, G. E. & PETTOVELLO, A. D. The biology of Mustelus schimitti in
southern Patagonia, Argentina. J. Fish. Biol. v. 57, p. 930-942. 2000.
CORTÉS, E. Life history patterns, demography, and population dynamics. In:
CARRIER, J. C., MUSICK, J. A. & HEITHAUS, M. R. (Eds.). Biology of sharks
and their relatives. CRC Press. Boca Raton. 449-470p. 2004.
COSTI, C. G., RODRIGUES, C. C. B, PAIVA-FILHO, D. L. & RODRIGUES, J. B. R..
Estudo preliminar da ictiofauna do pólo Araguaia-Tocantins. Projeto de pesquisa
da Ictiofauna do pólo Araguaia-Tocantins, GO-Polamazônia. Superintendência do
Desenvolvimento da Pesca-SUDEPE. Brasília. 171 p. 1977.
EBERT, D. A. Reproductive biology of skates, Bathyraja (Ishiyama), along the eastern
Bering Sea continental slope. J. Fish Biol. v. 66, p. 618-649. 2005.
GALVIS, G., MOJICA, J. I. & CAMARGO, M. Peces Del Catatumbo. Asociación
Cravo Norte. 178p. 1997.
HAMLETT, W. C., WOURMS, J. P. & SMITH, J. W. Stingray placental analogues:
structure of trophonemata in Rhinoptera bonasus. J. Submicrosc. Cytol. v. 17, p.
541-550. 1985.
HAMLETT, W. C. & KOOB, T. J. Female reproductive system. In: HAMLETT, W. C.
(ed.) Sharks, Skates and Rays. The Johns Hopkins University Press. Baltimore,
p.398-443, 1999.

127
HOLDEN, M. J. The fecundity of Raja clavata in British waters. J. du Con. v. 36, p.
110-118. 1975.
LASSO, C. A., RIAL, A. B. & LASSO-ALCALÁ, O. Notes on the biology of the
freshwater stingrays Paratrygon aiereba (Müller & Henle, 1841) and
Potamotrygon orbignyi (Castelnau, 1855) (Chondrichthyes: Potamotrygonidae) in
the Venezuelan llanos. Aqua, v. 2, No 3, p. 39-52. 1996.
LOWE-McCONNELL, R.H. Fish communities in tropical freshwaters. Longman Group
Limited. London. 337p. 1975.
LOWE-McCONNELL, R.H. Ecological studies in tropical fish communities. Terceira
Edição. Cambridge University Press. Cambridge. 1995.
PARSONS, G. R. Geographic variation in reproduction between two populations of the
bonnethead shark, Sphyrna tiburo. Environ. Biol. Fish. 38, p. 25-35. 1993.
PERES, M. B. & VOOREN, C. M. Sexual development, reproductive cycle, and fecundity
of the school shark Galeorhinus galeus off Southern Brazil. Fish. Bull. v. 89, p.655-
667. 1991.
ROA, R., ERNST, B. & TAPIA, F. Estimation od size at sexual maturity: an evaluation of
analytical and resampling procedures. Fish. Bull. v. 97, p. 570-580. 1999.
SMINKEY, T. R. & MUSICK, J. A. Age and growth of the sandbar shark,
Carcharhinus plumbeus, before and after population depletion. Copeia, v. 4, p.
871-883. 1995.
TANAKA, S., CAILLIET, G. M. & YUDIN, K. G. Differences in growth of the blue
shark, Prionace glauca: Technique or population? In H. L. Pratt, Jr.; T. Taniuchi
and K. Matsuura (eds.). Elasmobranchs as living resources: advances in the

128
biology, ecology, systematics and the status of the fisheries. NOAA - Tech. Rep.
NMFS 90. p. 177-187. 1990.
TANIUCHI, T. & TACHIKAWA, H. Geographical variation in age and growth of
Squalus mitsukurii (Elasmobranchs: Squalidae) in the North Pacific. In: SÉRET,
B., SIRE, J. Y. (Eds.) Proceedings of the 5th Indo-Pacific Fish Conference.
Noumea, 1997. Society of French Ichthyologists, Paris. p. 321-328. 1999.
TESHIMA, K. & TAKESHITA, K. Reproduction of the freshawater stingray,
Potamotrygon magdalenae taken from the Magdalena river system in Colombia,
South América. Bull. Seikai Natl. Fish. Res. Inst. No 70, p. 11-27. 1992.
VAZZOLER, A. E. A. Biologia da reprodução de peixes teleósteos: teoria e prática.
Eduem – Editora da Universidade Estadual de Maringá. 169p. 1996.
VOOREN, C. M. & SILVA, K. G. On the taxonomy of the angel sharks from Southern
Brazil, with the description of Squatina occulta sp. n. Rev. Bras. Biol. v. 51, No 3,
p. 589-601. 1991.

129
Conclusão Geral
As raias de água doce identificadas no rio Paranã, afluente do alto rio Tocantins,
foram Potamotrygon orbignyi CASTELNAU, Potamotrygon sp. A e Paratrygon
aiereba MULLER & HENLE.
Potamotrygon orbignyi é uma espécie de médio porte, alcançando no máximo
460 mm LD e apresenta as maiores abundâncias nas capturas por pesca experimental no
rio Paranã. Esta espécie distingue-se das demais espécies ocorrentes no rio Tocantins
principalmente no reduzido tamanho dos dentes e no seu relativo maior número
dentário, colorido dorsal policromático, colorido ventral da cauda fortemente
pigmentada com faixas amarelas alternadas, uma única fileira de espinhos caudais e
presença de sulco labial desenvolvido. A sua distribuição é documentada desde o médio
Araguaia até o alto Tocantins, mas sem registros de capturas nas cabeceiras destes rios.
Potamotrygon sp. A é uma espécie de médio-grande porte, com fêmeas em
maturação com 452 mm LD. Esta espécie apresentou baixas abundâncias no rio Paranã
e distingue-se das demais espécies ocorrentes no rio Tocantins principalmente na
morfologia e fórmulas dentárias, colorido dorsal relativamente uniforme, duas ou mais
fileiras de espinhos na cauda e ausência de sulco labial. Os registros de captura
restringiram-se apenas para o alto e médio Tocantins.
Paratrygon aiereba é uma espécie de grande porte, com machos atingindo a
maturidade com aproximadamente 430 mm LD e fêmeas ainda imaturas aos 540 mm
LD. Esta espécie caracteriza-se pela morfologia dos dentes, concavidade acentuada na
margem anterior do disco, distância pré-ocular cerca de 1/3 do comprimento do disco,
olhos não pedunculados e relativamente pequenos e margem posterior dos espiráculos

130
com projeção arredondada. A raia aramaçã possui registros de captura ao longo de toda
a bacia Tocantins-Araguaia.
Nesta tese é proposta uma chave de identificação para as espécies de raias de
água doce ocorrentes na bacia Tocantins-Araguaia, com base em caracteres externos.
A análise da distribuição de P. orbignyi ao longo do rio Paranã, com base em
indivíduos amputados pela pesca local, indica baixa mobilidade, sem padrões de
migração evidentes.
O número de espécies registradas é menor nas cabeceiras e parece aumentar com
a proximidade da foz do rio Tocantins, onde ocorrem as raias com maior distribuição na
bacia. Tanto o alto quanto o baixo Tocantins registram espécies endêmicas e este padrão
sugere medidas específicas para o manejo destes estoques.
A raia P. orbignyi apresentou forte policromatismo, seu colorido predominante
foi o padrão mesclado de spot-ocelado (32,4%), seguido pelo padrão reticulado (27%),
o mais conhecido para esta espécie. Evidências sugerem que o colorido varie ao longo
do crescimento do animal, desde o padrão spot-ocelado, em indivíduos de menor porte,
até o padrão roseta-modificada nos indivíduos mais velhos. Variações populacionais de
colorido parecem ocorrer em P. orbignyi e P. scobina, com predominância de padrões
específicos para certas regiões.
A análise do colorido das raias de água doce da bacia Tocantins-Araguaia e de
seus filhotes, com base na hipótese filogenética de MARQUES (2000), evidencia o
padrão de colorido spot como o mais primitivo para estas raias.
P. orbignyi é uma espécie insetívora no rio Paranã e alimenta-se principalmente
de ninfas de Ephemeroptera, Diptera e Trichoptera. Não foram observadas diferenças na
alimentação (qualitativa e quantitativa) desta espécie entre o dia e a noite, entre as

131
estações de seca e chuvas e entre os sexos. No entanto, jovens alimentam-se mais
marcadamente de larvas de Díptera do que de ninfas de Ephemeroptera, mas esta
relação se inverte quando atingem 180 mm LD.
A análise do grau de repleção e de digestão em P. orbignyi evidencia
alimentação contínua de P. orbignyi, onde seus estômagos encontravam-se sempre com
conteúdos pouco digeridos, podendo ser identificados no nível de espécie. As larvas de
insetos foram predadas inteiras, sem maceração evidente e freqüentemente em grandes
números, evidenciando um padrão “especialista-oportunista”.
O nível trófico das raias de água doce é predominantemente intermediário –
consumidores secundários (NT<4) – devido ao seu hábito alimentar bentófago baseado
em invertebrados. A única espécie identificada como consumidora terciária foi P.
aiereba, por se alimentar basicamente de peixes. A largura máxima e o nível trófico das
raias de água doce apresentaram uma correlação positiva, correspondendo ao que foi
encontrado em tubarões, com exceção das espécies planctófagas.
Potamotrygon orbignyi atinge a maturidade com 251 mm LD nos machos e 260
mm LD nas fêmeas, o que corresponde também a uma diferença no seu crescimento em
peso, onde fêmeas alcançam um peso total maior que o dos machos para a mesma
largura de disco. Esta diferença é ainda mais evidente após o tamanho de maturidade
sexual e o peso máximo registrado para machos foi de 3895 g e de 6420 g para fêmeas.
A fecundidade ovariana variou de 1 a 5 folículos, mas a fecundidade uterina foi
sempre de 1 filhote por gestação. Os folículos estão maturos com diâmetros
aproximados de 30 mm e 3,8 g de peso. Os embriões estão à termo com 115 mm LD e
pesos totais entre 48 e 61 g, o que equivale a um ganho em peso de 1605% em relação
ao peso inicial do ovo.

132
O índice hepassomático apresentou um padrão senoidal com os menores valores
na estação da seca e os maiores valores durante a estação das chuvas. Este padrão
senoidal e a presença de fêmeas grávidas na estação das chuvas evidenciam que o parto
ocorra, para a maior parte da população, durante esta estação. Contudo, parte da
população dá a luz também durante a estação da seca, o que evidencia uma gestação de
4 meses e um mês de cópula.
A análise preliminar da deposição de bandas vertebrais identificou o 5º ano de
vida como a idade de maturação e longevidade máxima de 10 anos, o que corresponde a
cinco gestações após a idade de primeira maturação. Desta forma, a fecundidade total de
uma fêmea ao longo de sua vida é de 5 a 12 filhotes, dependendo do tempo de ciclo
reprodutivo considerado, 11 ou 5 meses.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo