aspectos docrescimentoe atividade da redutasedo nitrato
TRANSCRIPT

Hoehnea 33( I): 89-97, 4 fig., 2006
Aspectos do crescimento e atividade da redutase do nitrato em plantasde Vernonia herbacea (Yell.) Rusby submetidas a
diferentes fontes de nitrogenio
Patricia Gaya de Carvalho', Marcos Pereira Marinho Aidar l, Lilian Beatriz Penteado Zaidan l e
Maria Angela Machado de Carvalho 1,2
Recebido: 05.07.2005; aceito: 05.01.2006
ABSTRACT - (Growth and activity of nitrate reductase in plants of Vernonia herbacea (VeIl.) Rusby submited to differentnitrogen sources). Vernonia herbacea (Veil.) Rusby, Asteraceae from the Cerrado, bears underground organs (rhizophores)which accumulate inulin-type fructans and nitrate. Previous studies showed that 10 mM nitrate was the most efficientconcentration for total plant biomass and fructan accumulation in the rhizophores. However, other nitrogen sources werenot used. Thus, the aim of this project was to analyse and compare the effect of different nitrogen sources on growth of Vherbacea. The plants were treated with nutrient solutions containing ammonium «NH4hS04)' nitrate (KN03) and a mixtureof nitrate and ammonium (NH4N03), both at a final concentration of 10 mM. No preference for the nitrogen source wasobserved when growth parameters were analysed. Therefore, the results suggest that plants of V herbacea can be cultivatedunder any of the nitrogen sources used in this study.Key words: ammonium, cerrado, nitrate, plant growth
RESUMO - (Aspectos do crescimento e atividade da redutase do nitrato em plantas de Vernonia herbacea (VeIl.) Rusbysubmetidas a diferentes fontes de nitrogenio). Vernonia herbacea (VeIl.) Rusby, Asteraceae do Cerrado, possui orgaossubterraneos denominados rizOforos que armazenam frutanos do tipo inulina e nitrato em seus vacuolos. Experimentosanteriores mostraram que plantas tratadas com 10 mM de nitrato apresentaram melhor eficiencia no acumulo de biomassatotal e no acumulo de frutanos nos rizOforos. Entretanto, outras fontes de nitrogenio nao foram testadas. Assim, estetrabalho teve 0 objetivo de verificar a melhor fonte de nitrogenio no cultivo dessa especie. As plantas foram submetidas aocultivo com nitrato (KN03), amonio «NH4)2S04) e a mistura de nitrato e amonio(NH4N03), ambas na concentrayao de 10mM. Os resultados mostraram que plantas dessa especie nao apresentaram preferencia pela fonte nitrogenada indicandoque 0 seu cultivo pode ser realizado sob qualquer uma das fontes nitrogenadas estudadas.Palavras-chaves: amonio, cerrado, crescimento, nitrato
Introdu~ao
°nitrogenio (N) eum elemento fundamental paraas plantas (Pereira et al. 198 I) e de baixadisponibilidade nos solos, principal mente nas areastropicais, onde e lixiviado, volatilizado e utilizado pormicroorganismos em maior velocidade do que naszonas temperadas (Pimentel 1998). °N molecular(N2)' apesar de constituir mais de 78 % da atmosfera,so e utilizado por microorganismos fixadores de N2. Amaioria dos ecossistemas naturais e agnirios apresentaum ganho expressivo da produtividade apos afertilizal):ao com nitrogenio inorganico, 0 quedemonstra a importancia desse elemento para asplantas (Malavolta 198 I).
As principais fontes de N disponiveis para asplantas terrestres sao 0 nitrato (N03-) e 0 amonio(NH/). °N03-, absorvido pelas raizes, pode serassimilado nesses orgaos ou nos orgaos aereos,dependendo da sua disponibilidade e da especievegetal. No processo de assimilal):ao, 0 N03- e reduzidoa nitrito (N02-) no citosol pela enzima redutase donitrato (RN); posteriormente, 0 N02- e reduzido aNH4+ nos plastideos da raiz ou nos cloroplastos pelaenzima redutase do nitrito (Tischiner 2000). °NH4+
absorvido pela raiz ou produzido por assimilal):ao doN03-, ou ainda originario da fotorrespiral):ao, econvertido a glutamina e glutamato pel a al):aosequencial das enzimas glutamina sintetase eglutamato sintase, localizadas no citosol enos plastideos
I. Instituto de Botiinica de Sao Paulo, Se~ao de Fisiologia e Bioquimica de Plantas, Caixa Postal 4005, 0 I061-970 Sao Paulo, Brasil2. Autor para correspondencia: [email protected]

90 Hoehnea 33( I), 2006
das raizes ou nos cloroplastos (Bloom et al. 1992,Becker et al. 1993).
Os ions NH/ e N03-, liberados pOl'decomposiyao da materia organica do solo, tomam-seobjeto de intensa competiyao entre microorganismose plantas (Havlin et af. 1999), as quais desenvolverammecanismos para capturar rapidamente esses ions apartir da soluyao do solo.
Sob concentrayoes elevadas de N no solo, queocorrem ap6s a fertilizayao, a absoryao do NH/ e do
03- pelas raizes pode exceder a capacidade de umaplanta em assimilar esses ions, levando ao acumulo donitrato, mas nao do amonio, que e t6xico e nao podeser acumulado. Niveis altos de N03-podem ser acumulacos no vacuolo para posterior utilizayao ou podemser translocados atraves dos tecidos sem efeitos prejudiciais (Lea et al. 1992). A maior parte do NH/ naplanta deve ser incorporada em N organico ja na raiz,pois nao pode permanecer livre na celula enos tecidos.
Embora a maioria das plantas assimilepreferencialmente 0 N03-ao NH4+ (Forde 2000), emalguns casos observa-se 0 oposto. Portanto, enecessario encontrar 0 balanyo adequado entre N03e NH4+, para obter-se um 6timo crescimento edesenvolvimento. Na planta, a utilizayao do N edeterminada pOl' algumas variaveis, como eficienciada absoryao de N03- OU NH4+, atividade da redutasedo nitrato, estoque de N03-, habilidade de mobilizare translocar N para os 6rgaos dreno e adaptayao abaixa disponibilidade de N no meio (Duncan &Baligar 1991).°conhecimento sobre 0 comportamento vegetalquanta ao usa de nutrientes permite manusear oumodificar 0 sistema de cultivo para melhorar aeficiencia na utilizayao de N. Alguns gen6tipos, pOl'exemplo, tem mel hoI' capacidade de absoryao de NN03-, e outros, de N-NH/ e produzem mais materiaseca pOl' unidade de N aplicada; outros apresentammaior capacidade de mobilizayao de N das folhas ede outros 6rgaos para 0 grao; outros, ainda, sao maisadaptados ao alto suprimento de N, enquanto outrossao adaptados a baixos suprimentos (Vose 1990).
Estudos sobre nutriyao mineral em especiesnativas do Cerrado sao escassos e foram realizadosprincipalmente com arb6reas, como Qualeagrandi/lora Mart. (Felippe & Dale 1990) eDalbergia miscolobium Benth. (Paulilo & Felippe1995, Sassaki & Felippe 1998) e, em menor numero,com herbaceas, como Bidens gardneri Baker(Felippe & Dale 1990). Entretanto, a preferencia
pela fonte de nitrogenio nao foi abordada nessestrabalhos.
Estudos realizados com Vernonia herbacea(VeIl.) Rusby, uma Asteraceae perene de Cerrado,mostraram que as plantas acumulam mais materia secaaerea e subterranea quando recebem soluyoesenriquecidas com nitrato (Cuzzuol et al. 2005,Carvalho 2005). Entretanto, a fonte preferencial denitrogenio dessa especie nao foi investigada ate 0momento. Plantas de V herbacea apresentam 6rgaossubterraneos espessados de estrutura caulinar,denominados riz6foros, pOl' meio dos quais a planta sereproduz vegetativamente. Os riz6foros atuam como6rgaos de reserva para a planta, armazenando emseus tecidos grandes quantidades de frutanos(Carvalho & Dietrich 1993).
Os frutanos sao polimeros de frutose originadosda sacarose e consistem de series hom610gas de oligoe polissacarideos nao redutores, em que cada membroda serie contem urn residuo a mais de frutose que 0membro anterior (Edelman & lefford 1968, Pollocket al. 1996). Alem da sua funyao de reserva, osfrutanos parecem estar associ ados a resistencia dasplantas a seca e ao frio (Hendry & Wallace 1993,Livingston & Henson 1998 e Pilon-Smits et al. 1995,1999), possivelmente devido a sua atuayao naestabilizayao de membranas (Hincha et al. 2000,Vereyken et al. 2003).
Os frutanos sao de grande interesse economicodevido as suas propriedades beneficas a saudehumana (Tomomatsu 1994). Segundo Coussement &Franck (1998), esses carboidratos tem sidoempregados com sucesso nos casos de constipayao ena prevenyao de doenyas do intestino, pOl' estimular aproliferayao das bifidobacterias, e no controle daosteoporose, pOl' estimular a absoryao de dlcio. Alemdisso, os frutanos sao utilizados como adoyantesalternativos de baixo teor cal6rico, pois nao saoabsorvidos pelo organismo humano (Hidaka &Hirayama 1991).
Estudos de nutriyao mineral para a adequayaode tratamentos de adubayao e fertilizayao visando aprodutividade de plantas de interesse economico saode extrema importancia. Tendo em vista 0 teor elevadode frutanos em V herbacea, este trabalho tern pOl'objetivo analisar os efeitos do N03-, NH/ e da misturadessas duas fontes de nitrogenio no crescimento e naalocayao de biomassa, na atividade da redutase donitrato nos riz6foros e folhas e na concentrayao denitrogenio total nas folhas dessas plantas.

P.G. Carvalho et at.: Crescimento e redutase do nitrato em V. herbacea 91
Material e metodos
Material vegetal e condic;:oes de crescimento - foramutilizadas plantas de Vernonia herbacea (Yell.) Rusby(Asteraceae) obtidas por propagac;:ao vegetativa apartir de fragmentos de riz6foros de plantas coletadasna Reserva Biologica e Estac;:ao Experimental de MojiGuac;:u, Sao Paulo, em julho de 2004. Plantas detamanho uniforme (cerca de 3 cm de altura) foramselecionadas do is meses apos a brotac;:ao dosfragmentos de riz6foros cultivados em bandejascontendo quartzo umedecido com agua destilada. Asplantas foram transferidas para vasos plasticos (3 L),contendo 0 mesmo substrato, e submetidas ascondic;:oes de luz, temperatura e fotoperiodo naturais,em casa de vegetac;:ao. As plantas foram separadasem tres lotes, cada urn com 38 plantas, e receberamsemanalmente 150 mL de soluc;:ao nutritiva contendo0,5 mM K2HP04, 1,3 mM NaS04, 2,3 mM KCL,2,5 mM CaCI2.2H20 e 1,7 mM MgS04·7H20. °primeiro lote foi enriquecido com 10 mM de KN03, 0
segundo, com 5 mM de (NH4)2S04' e 0 terceiro com5 mM de NH4N03 . Todos os tratamentos foramsuplementados com os sais de micronutrientes dasoluc;:ao de Hoagland (Hoagland & Amon 1938) e 0
pH ajustado para 6,0-6,5 com KOH. Quandonecessario, as plantas foram regadas com aguadestilada para evitar 0 dessecamento.
A primeira coleta de dados (TO) foi realizada emsetembro, quando as plantas foram transferidas dasbandejas para vasos individuais e passaram a receberas soluc;:oes nutritivas. As coletas seguintes foramrealizadas mensalmente, 30 (outubro), 60 (novembro),90 (dezembro) e 120 Uaneiro) dias apos 0 inicio dotratamento. As analises de crescimento, quantificac;:aode nitrato e determinac;:ao da atividade da redutase donitrato foram realizadas em seis plantas, coletadasaleatoriamente, constituindo cada urna, uma replicata.As plantas foram analisadas quanta a altura do caule,numero de folhas, area foliar e massa de materiafresca e seca aerea e subterranea. Para determinac;:aoda massa de materia seca, os ramos aereos e amostrasde riz6foros foram colocados em estufa a 60°C atemassa constante. A area foliar foi estimada pelaequac;:ao da curva de regressao linear y = 0,2737 +0,7158x, onde x equivale ao produto do comprimentopela largura das folhas, com urn coeficiente decorrelac;:ao de 0,9658 (Carvalho et al. 1997).
Os parametros derivados de crescimento, taxade crescimento relativo (TCR), razao de area foliar(RAF) e taxa de assimilac;:ao liquida (TAL) foram
calculados de acordo com Hunt (1982) e com umaferramenta de software (Hunt et al. 2002). A forc;:ade dreno aparente foi calculada em func;:ao da taxa deincremento de massa seca dos riz6foros (massa finaldos riz6foros - massa inicial dos riz6foros) / (t2 - t I),conforme Schubert & Fuerle (1997).
Ensaio da atividade da redutase do nitrato (RN) - aatividade da redutase do nitrato foi determinada pelometodo de Stewart et al. (1986) em folhas e riz6foros.Amostras dos tecidos frescos (0,1 g) foram colocadasem tubos de ensaio com tampao fosfato de potassio50 mM pH 7,5 contendo 100 mM de nitrato depotassio, 1,5% de I-propanol e triton para analise dasfolhas, e contendo 50 mM de nitrato de potassio e1,0% de I-propanol para analise dos riz6foros. Emseguida, os tecidos foram submetidos a infiltrac;:ao avacuo para retirada do ar. As amostras foram entaoincubadas a 25°C por I h, no escuro. Da mistura deincuba9ao, retirou-se I mL, sendo adicionados I mLde acido sulfanilico 1% e I mL de a-naphtil etilenodiamida (NED) 0,05%, para colorac;:ao do produtoformado, 0 nitrito. Apos repouso de 30 min emtemperatura ambiente, a absorbancia foi lida emespectrofotometro a 540 nm.
Determinac;:ao do nitrogenio total (NT) - 0 NT foideterminado pelo metodo de Malavolta et al. (1987)e realizado no Departamento de Solos e Nutri9ao daESALQ-USP. Amostras de 0, I g de massa seca defolhas foram colocadas em urn tuba digestor com 5 mLde uma mistura de acido sulfUrico e sais catalisadores.A temperatura foi elevada de 50°C a cada 30 min ate350°C. Apos a digestao, foi adicionado urn volume deNaOH 15 N, suficiente para neutralizar 0 acido edeixar 0 meio alcalino. Em seguida, foi adicionado acidoborico como soluc;:ao indicadora. A absorbancia damistura resultante foi lida em espectrofotometro a410 nm.
Analise estatistica - os efeitos dos tratamentos foramanalisados por ANOYA e as medias comparadas peloteste de Duncan a 5%, no software Winstat:
Resultados e Discussao
Os resultados apresentados na figura I mostramurn aumento nos parametros de crescimento avaliadosnas plantas cultivadas com nitrato (KN03), amonio((NH4)2S04) e a mistura de nitrato e amonio(NH4N03), ao longo do periodo de observac;:oes.
Durante os meses de tratamento, tanto a areafoliar quanta a massa seca aerea nao apresentaram

92 Hoehnea 33( I), 2006
diferenc;:as significativas entre os tratamentos.Entretanto, foi possivel observar que as plantastratadas com nitrato mostraram maior massa secaaerea e de riz6foros no terceiro mes do periodoexperimental. Em relac;:ao a area foliar, ostratamentos com amonio e a mistura nitrogenadapropiciaram maior area foliar em relayao aotratamento com nitrato.
Em cultura de batata, Cao & Tibbitts (1992),usando uma mistura de nitrato e amonio, constataramque esse tratamento aumentou a massa seca e a areafoliar, quando comparado aos tratamentos contendosomente nitrato ou amonio. Na presenc;:a de nitrato asplantas cresceram melhor do que sob amonio, mas ao
final do experimento, nao apresentaram massa secatotal superior a daquelas que receberam a misturanitrogenada.
Outros estudos realizados com plantas cultivadasrevelaram diferenyas quanta apreferencia pela fontenitrogenada. A batata-doce, por exemplo, apresentoumaior numero de tuberculos quando tratada compequenas concentrac;:oes de N03' (Kim et al. 2002).Duas cultivares de arroz (IAC-4440 e IAC-l20),tratadas com NH/, produziram maior numero de graospor planta (Silveira & Machado 1990). Por outro lado,a alfafa apresentou maior crescimento e maior numerode folhas quando recebeu a mistura de N03' e NH/(Meuriot et al. 2003).
Area foliar
90
-;'60'"C
'"0..N
30Eu
0TO nitrato amonio nitrato + amonio
Massa seca aerea1,2
-;'
'"C0,8'"0..
bf)
0,4
0,0TO nitrato amonio nitrate + amonio
Massa seca dos rizOforos2,0
-;'1,5
~1,0c
'"0..bf) 0,5
0,0TO nitrato am6nio nitrato + amonio
Fontes de nitrogenio
Figura I. Medidas de crescimento de plantas de V. herbacea tratadas com 10 mM de KNO) (nitrato), 5 mM de (NH4)zS04 (am6nio) e5 mM de NH4NO) (nitrato + am6nio). Determina~aorealizada no material inicial TO 0 e I mes~, 2 meses., 3 meses IIIID e 4 meses iIIap6s 0 inicio do tratamento. Os dados representam medias de seis repeti~oes (n = 6) e as barras representam 0 erro padrao. Nao houvediferen~as significativas entre os tratamentos dentro de cada tempo.

P.G. Carvalho el 01.: Crescimento e redutase do nitrato em V. herboceo 93
De acordo com Poorter (2002), a TCR reflete 0
incremento de produyao de massa de materia secaem urn determinado intervalo de tempo. Dessa forma,foi possivel observar que as plantas que receberamnitrato como fonte de nitrogenio apresentaraminicialmente maior ganho de materia seca. Entretanto,a produyao de materia seca foi decrescendo apos 0
segundo mes, ate se igualar aos outros doistratamentos. A queda gradual da TCR, verificada emtodos os tratamentos, apresentou padrao semelhanteao que e detectado na maioria das plantas, ou seja, 0
incremento de massa tornou-se proporcionalmentedecrescente em relayao a massa total do individuo.Portanto, quanta maior a sua massa total, menor ataxa de crescimento (Beadle 1993).
Segundo Lambers et al. (1998), as respostas decrescimento a adiyao de nutrientes em plantas nativasrefletem uma pequena elevayao da TCR, indicio deque as plantas possuem baixo potencial de TCR. BaixaTCR e considerada por Grime & Hunt (1975) umaestrategia de sobrevivencia de plantas de ambientesmenos ferteis. Segundo Chapin (1980, 1988), taisplantas, quando cultivadas em solos mais ferteis,acumulam os nutrientes ao inves de aproveita-Iosimediatamente, vindo a utiliza-Ios mais tarde, quandosua disponibilidade for limitante, 0 que corrobora osresultados encontrados em plantas de V. herbacea.
Os tratamentos com nitrogenio causaramvariayoes na RAF (figura 2), que e uma medida daeficiencia da planta como produtora de areaassimiladora. As plantas tratadas com nitratoapresentaram urn valor de RAF significativamentemaior no inicio do experimento, quando comparadasas que receberam outras fontes de nitrogenio. Asplantas tratadas com a mistura nitrogenada mostraramincremento significativo no segundo periodo decrescimento, mas ao final do periodo analisado, asplantas sob os tres tratamentos nao diferiramsignificativamente entre si.
A TAL (figura 2), que mede a eficiencia da plantacomo produtora de materia seca, foi mais elevada nasplantas tratadas com nitrato e apenas no inicio doexperimento. Entretanto, ao final, plantas de todos ostratamentos apresentaram valores semelhantes.
Valores mais elevados de TAL, encontrados noinicio do experimento para as plantas dos trestratamentos de nitrogenio, sugerem que nesta fase asplantas utilizam recursos originados do rizOforo. Defato, no inicio do crescimento de plantas deV. herbacea, 0 rizOforo atua como fonte de carbono,
90-120del - jan
90-120del - jan
90-120dez - jan
60-90nov - del
60-90nov - del
60-90nov - dez
RAF
TAL
30-60out - nov
a
30-60oul - nov
30-60oul - nov
Periodo (dias/meses)
b
b
0-30set - oul
a
0-30set - out
0-30set - out
40
30
10-l----~-----.----~---__,
20
0.004
o.oo+----~----~---__,.---~
0.01
0.05
0,000-1---------.-----,--------.,----,
0.06
TCRom
Com relayao aos parametros derivados de crescimento, TCR, RAF e TAL (figura 2), observou-se que,no primeiro periodo analisado (0 a 30 dias), as plantastratadas com nitrato apresentaram a maior TCR,seguindo-se uma queda continua ate 0 final doexperimento. As plantas tratadas com soluyoesnutritivas con tendo apenas amonio ou a misturanitrogenada apresentaram valores simi lares entre si emais baixos, ao fongo do periodo analisado.
a
'OJ) 0.04N8 0,Q3
0.02 b
Figura 2. Taxa de crescimento relativo (TCR), razao de area foliar(RAF) e taxa de assimila'Yao liquida (TAL) em plantas deV. herbacea tratadas com 10 mM de KN03, 5 mM de (NH4)2S04
e 5 mM de NH4N0
3.• nitrato, • am6nio, e _ nitrato + am6nio.
Os dados representam medias de seis repeti'Yoes (n = 6).Diferen'Yas significativas entre os tratamentos, quando existentes,sao indicadas por letras diferentes (p < 0,05).
'OJ)
N
E()
~ 0.003
N
E() 0.002
'OJ) 0001
94 Hoehnea 33( I), 2006
Periodo (dias/meses)
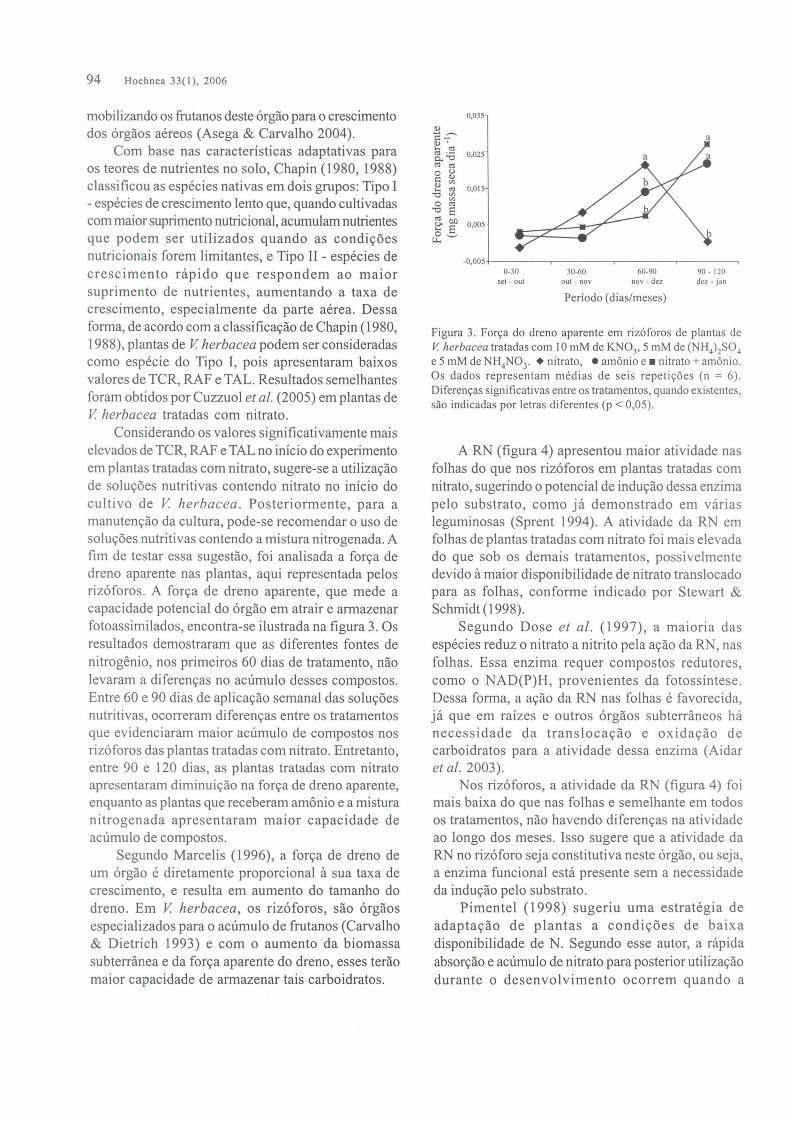
Figura 3. Fon;:a do dreno aparente em rizOforos de plantas deV. herbacea tratadas com 10 mM de KN03, 5 mM de (NH4)2S04e 5 mM de NH4N03.• nitrato, • amonio e _ nitrato + amonio.Os dados representam medias de seis repeti~5es (n = 6).Diferen~as significativas entre os tratamentos, quando existentes,sao indicadas por letras diferentes (p < 0,05).
A RN (figura 4) apresentou maior atividade nasfolhas do que nos riz6foros em plantas tratadas comnitrato, sugerindo 0 potencial de induc;;ao dessa enzimapelo substrato, como ja demonstrado em variasleguminosas (Sprent 1994). A atividade da RN emfolhas de plantas tratadas com nitrato foi mais elevadado que sob os demais tratamentos, possivelmentedevido amaior disponibilidade de nitrato translocadopara as folhas, conforme indicado por Stewart &Schmidt (1998).
Segundo Dose et al. (1997), a maioria dasespecies reduz 0 nitrato a nitrito pela ac;;ao da RN, nasfolhas. Essa enzima requer compostos redutores,como 0 NAD(P)H, provenientes da fotossintese.Dessa forma, a ac;;ao da RN nas folhas e favorecida,ja que em raizes e outros 6rgaos subterraneos hanecessidade da translocac;;ao e oxidayao decarboidratos para a atividade dessa enzima (Aidaret al. 2003).
Nos riz6foros, a atividade da RN (figura 4) foirna is baixa do que nas folhas e semelhante em todosos tratamentos, nao havendo diferenyas na atividadeao longo dos meses. Isso sugere que a atividade daRN no riz6foro seja constitutiva neste 6rgao, ou seja,a enzima funcional esta presente sem a necessidadeda induyao pelo substrato.
Pimentel (1998) sugeriu uma estrategia deadaptac;;ao de plantas a condiyoes de baixadisponibilidade de N. Segundo esse autor, a rapidaabsoryao e acumulo de nitrato para posterior utilizayaodurante 0 desenvolvimento ocorrem quando a
60-90 90 . 120nov - dez del - jan
0.Q35
4)
E~4).... ellell .- 0.0250.-0ell ell0 uI:: 4)
4)Vl
.... ell 0.015-0 Vl
Vl0 ell-0 Eell 0/)
0.005~E~~
-0.0050-30 30·60
sct - QUI out - nov
mobilizando os frutanos deste 6rgao para 0 crescimentodos 6rgaos aereos (Asega & Carvalho 2004).
Com base nas caracteristicas adaptativas paraos teores de nutrientes no solo, Chapin (1980, 1988)classificou as especies nativas em dois grupos: Tipo 1- especies de crescimento lento que, quando cultivadascom maior suprimento nutricional, acumulam nutrientesque podem ser utilizados quando as condic;;oesnutricionais forem limitantes, e Tipo II - especies decrescimento Hipido que respondem ao maiorsuprimento de nutrientes, aumentando a taxa decrescimento, especialmente da parte aerea. Dessaforma, de acordo com a classificac;;ao de Chapin (1980,1988), plantas de V herbacea podem ser consideradascomo especie do Tipo I, pois apresentaram baixosvalores de TCR, RAF e TAL. Resultados semelhantesforam obtidos por Cuzzuol et al. (2005) em plantas deV. herbacea tratadas com nitrato.
Considerando os valores significativamente maiselevados de TCR, RAF e TAL no inicio do experimentoem plantas tratadas com nitrato, sugere-se a utilizac;;aode soluc;;oes nutritivas contendo nitrato no inicio docultivo de V herbacea. Posteriormente, para amanutenc;;ao da cultura, pode-se recomendar 0 usa desoluc;;oes nutritivas contendo a mistura nitrogenada. Afim de testar essa sugestao, foi analisada a forc;;a dedreno aparente nas plantas, aqui representada pelosriz6foros. A forc;;a de dreno aparente, que mede acapacidade potencial do 6rgao em atrair e armazenarfotoassimilados, encontra-se ilustrada na figura 3. asresultados demostraram que as diferentes fontes denitrogenio, nos primeiros 60 dias de tratamento, naolevaram a diferenc;;as no acumulo desses compostos.Entre 60 e 90 dias de aplicac;;ao semanal das soluc;;oesnutritivas, ocorreram diferenc;;as entre os tratamentosque evidenciaram maior acumulo de compostos nosriz6foros das plantas tratadas com nitrato. Entretanto,entre 90 e 120 dias, as plantas tratadas com nitratoapresentaram diminuic;;ao na forc;;a de dreno aparente,enquanto as plantas que receberam amonio e a misturanitrogenada apresentaram maior capacidade deacumulo de compostos.
Segundo Marcelis (1996), a forc;;a de dreno deurn 6rgao e diretamente proporcional a sua taxa decrescimento, e resulta em aumento do tamanho dodreno. Em V herbacea, os riz6foros, sao 6rgaosespecializados para 0 acumulo de frutanos (Carvalho& Dietrich 1993) e com 0 aumento da biomassasubterranea e da forc;;a aparente do dreno, esses teraomaior capacidade de armazenar tais carboidratos.

P.G. Carvalho el at.: Crescimeillo e redulase do nitrato em V. herbacea 95
Folhas
10
8
6
nitrate amonio
RizQforosRlzoforos
nitrato + amonio
TO nitrato amonio nitrato + amonio
Fontes de nitrogenio
Figura 4. Atividade da redutase do nitrato em folhas e rizOforos de plantas de V herbacea tratadas com 10 mM de KNO) (nitrato), 5 mMde (NH4)2S04 (am6nio) e 10 mM de NH 4N03(nitrato + am6nio). Detemlinayao realizada no material inicial TO 0 e 1 mes ~, 2 meses.,3 meses fiI!II e 4 meses mI ap6s 0 inicio do tratamento. as dados representam medias de seis repetiyoes (n = 6) e as barras representamo erro padrao. Diferenyas significativas entre os tratamentos, quando existentes, sao indicadas por letras diferentes (p < 0,05).
disponibilidade do nutriente diminui. 0 Cerrado, adiminui y30 da disponibilidade de inorgftnico podeocorrer pOl' lixiviayao pelas chuvas e pelo usocrescente do nutriente pelos microorganismos do solo,portanto e razoavel assumir que a estocagem, parauso posterior, ocorre quando a oferta do nutriente paraa planta e elevada.
A analise do conteudo de nitrogenio total nasfolhas nao mostrou diferenyas significativas entre ostratamentos. Os valores encontrados foram de 1,78%,1,91 % e 1,95% em plantas tratadas respectivamentecom nitrato, am6nio e a mistura nitrogenada e estaode acordo com os dados relatados para a maioria dasespecies cultivadas, entre 1,35 e 6% da massa secatotal da planta (Marschner 1995). Entretanto, nopresente estudo considerou-se apenas a massa secafoliar. Como os riz6foros apresentam concentrayoesmais elevadas de nitrato (cerca de 2,5 vezes) do queas folhas (Carvalho 2005), e provavel que aconcentrayao de nitrogenio total em funyao da massaseca total da planta seja mais elevada.
Cuzzuol et af. (2005) observaram que a forya dedreno em plantas de Vernonia herbacea foi estimuladapela concentrayao mais elevada de N (10,7 mmol L- I ),
que promoveu tambern 0 aumento da massa seca do
riz6foro. 0 autor verificou que as plantas quereceberam mais N tambem apresentaram concentrayao inferior de frutanos nos riz6foros, quando comparadas as que receberam menos N (1,3 mMol L- I ).
Contudo, resultados obtidos pOI' Carvalho (2005)revelaram que plantas submetidas a concentrayao de10 mM de nitrato apresentaram resultados semelhantesaos de Cuzzuol et al. (2005), exceto no que se refereao teo I' de frutanos. Assim, estudos futuros deveraoconsiderar, alem do controle das condiyoesexperimentais, a fase de desenvolvimento das plantas.
Os resultados obtidos no presente trabalho,referentes ao crescimento, incremento de biomassade riz6foros e forya de dreno, permitem concluirque as plantas de Vernonia herbacea podem sercultivadas sob as tres fontes de nitrogenio.Entretanto, considerando 0 mel hoI' crescimento dasplantas tratadas com nitrato no inicio do perfodo,recomenda-se inicialmente 0 uso do N na formadesse composto e, posteriormente, a aplicayao damistura nitrogenada de am6nio e nitrato.
Agradecimentos
A FAPESP, pelo apoio financeiro (BIOTASPprocesso 98/05124-8) e pela bolsa concedida a Patricia

96 Hoehnea 33( I), 2006
G. Carvalho (processo 02/11226-5) e ao CNPq, pelabalsa de Produtividade em Pesquisa a Lilian B.P.Zaidan.
Literatura citada
Aidar, M.P.M., Schmidt, S., Moss, G, Stewart, GR & Joly,C.A. 2003. Nitrogen use strategies of neotropicalrainforest trees in threatened Atlantic Forest. Plant, Celland Environment 26: 389-399.
Asega,A.F. & Carvalho, M.A.M. 2004. Fructan metabolizingenzymes in rhizophores of Vernonia herbacea (VeIl.)Rusby upon excision ofaerial organs. Plant Physiologyand Biochemistry 42: 313-319.
Beadle, c.L. 1993. Growth analysis. In: D.O. Hall, lM.O.Scurlock, H.R. Bolhar-Nordenkampf & R.C. Leegood(eds.). Photosynthesis and production in a changingenvironment: a field and laboratory manual. Chapmanand Hall, London, pp. 36-46.
Becker, T.W., Perrot-Rechenmann, c., Suzuki,A. & Hirel,B. 1993. Subcellular and immunocytochemicallocalization of the enzymes involved in ammoniaassimilation in mesophyll and bundle-sheath cells ofmaize leaves. Planta 191: 129-136.
Bloom, A.J., Sukapanna, S.S. & Warner, RL. 1992. Rootrespiration associated with ammonium and nitrateabsorption and assimilation by barley. Plant Physiology99: 1294-1301.
Cao, W. & Tibbits, T.W. 1992. NH/N03 mixtures enhancegrowth in potatoes. HortScience, Alexandria, v. 27, 177 p.
Carvalho, M.A.M. & Dietrich, S.M. 1993. Variation infructan content in the underground organs of Vernoniaherbacea (VeIl.) Rusby at different phenological phases.New Phytologist 123: 735-740.
Carvalho, M.A.M., Dietrich, S.M.C. & Zaidan, L.B.P. 1997.Growth and fructan content of plants of Vernoniaherbacea (Asteraceae) regenerated from rhizophores.New Phytologist 136: 153-161.
Carvalho, P.G 2005. Efeito do nitrogenio no crescimento emetabolismo de frutanos em plantas de Vernoniaherbacea (VeIl.) Rusby. Disserta9ao de Mestrado,Escola Superior de Agricultura "Luiz de Queiroz",Piracicaba, 103 p.
Chapin, F.S. III. 1980. The mineral nutrition ofwild plants.Annual Review ofEcology Systematics II: 233-260.
Chapin, F.S. III. 1988. Ecological aspects ofplant mineralnutrition. In: B. Tinker & A. Uiuchli (eds.). Advances inPlant Nutrition. Praeger, New York, v. 3, pp. 161-191.
Coussement, P. & Franck, A. 1998. New food applicationfor inulin. Agro-food-industry Hi-Tech 9: 26-28.
Cuzzuol, GRF., Carvalho, M.A.M. & Zaidan, L.B.P. 2005.Growth, photosynthase partitioning and fructanaccumulation in plants of Vernonia herbacea (VeIl.)Rusby under two nitrogen levels. Brazilian Journal ofPlant Physiology 17:273-281.
Dose, M.M., Hirasawa, M., Kleis-Sanfrancisco, S., Lew,E.L. & Knaff, D.B. 1997. The ferredoxin-binding site offerredoxin: nitrite oxidoreductase. Plant Physiology 114:1047-1053.
Duncan, R.R. & Baligar, V.C. 1991. Genetic andphysiological basis of nutrient uptake and useefficiency: an overview. In: R.R. Duncan & Vc. Baligar(eds.). Crops as enhancers of nutrient use. AcademicPress, New York, pp. 238-282.
Edelman, J. & Jefford, T.G 1968. The mechanism offructosan metabolism in higher plants as exemplified inHelianlhus tuberosus. New Phytologist 67: 517-531.
Felippe, GM. & Dale, J.E. 1990. The effects ofphosphatesupply on growth of plants from Brazilian Cerrado:experiments with seedlings of the annual weed, Bidensgardneri Baker (Compositeae) and the tree, Qualeagrandiflora Mart. (Vochysiaceae). Oecologia 82: 81-86.
Forde, B.G 2000. Nitrate transporters in plants: structure,function and regulation. Biochimica et Biophysica Acta1465:219-235.
Grime, J.P. & Hunt, R 1975. Relative growth-rate: its rangeand adaptive significance in a local flora. Journal ofEcology 63: 393-422.
Havlin, J.L., Beaton, J.D., Tisdale, S.L. & Nelson, W.L.1999. Soil fertility and fertilizers. 6 ed. Prentice Hall, NewJersey, 499 p.
Hendry, G.A.F. & Wallace, R.K. 1993. The origin,distribution and evolutionary significance offructans.In: M. Suzuki & J.N. Chatterton (eds.). Science andtechnology of fructans. CRC Press, Boca Raton,pp.119-139.
Hidaka, H. & Hirayama, M. 1991. Useful characteristicsand commercial applications offructo-oligosaccharides.Biochemical Society Transactions 19: 561-565.
Hincha, D.K., Hellwege, E.M., Heyer, A.G & Crowe, J.H.2000. Plant fructans stabilize phosphatidylcholineliposomes during freeze-drying. European Journal ofBiochemistry 267: 535-534.
Hoagland, D.R. & Arnon, D.1. 1938. The water-culturemethod for growing plants without soil. CaliforniaAgricultural Experimental Station. Circ. No. 347.
Hunt, R. 1982.Plant growth curves: the functional approachto plant growth analysis. Edward Arnold Publishers,London, 80 p.
Hunt, R, Causton, D.R, Shipley, B. & Askew, A.P. 2002. Amodern tool for classical plant growth analysis. AnnalsofBotany 90: 485-488.
Kim, S-H., Mizuno, K., Sawada, S. & Fujimara, T. 2002.Regulation oftuber and ADP-glucose pyrophosphorylase(AGPase) in sweet potato (Ipomoea balalas (L.)Lam.) by nitrate. Plant Growth Regulation 37: 207213.

P.G. Carvalho ef al.: Crcscimento e redutase do nitrato em V. Izerbacea 97
Lambers, J., Chapin, F.S. & Pons, T.L. 1998. PlantPhysiological Ecology. Springer-Verlag, New York,540p.
Livingston, D.P. & Henson, C.A. 1998. Apoplastic sugars,fructan exohidrolase, and invertase in winter oat:response to second-phase cold hardening. PlantPhysiology 116: 403-408.
Malavolta, K, Vitti, Gc. & De Oliveira, S.A. 1997. Avaliac;:aodo estado nutricional das plantas: principios eaplicac;:oes. 2 ed. Potafos, Piracicaba, 319 p.
Malavolta, K 1981. Manual de quimica agricola - Adubos eadubac;:ao. Ceres, Sao Paulo, 596 p.
Marcelis, L.F.M. 1996. Sink strength as a determinant ofdry matter partitioning in the whole plant. Journal ofExperimental Botany 47: 1281-1291.
Marschner, H. 1995. Mineral nutrition of higher plants.Academic Press, Orlando, 649 p.
Meuriot, F., Avice, J-C., Decau, M-L., Simon, J-C, Laine,P., Volenec, J.J. & Ourry, A. 2003. Accumulation ofNreserves and vegetative storage protein (YSP) intaproots of non-nodulated alfafa (Medicago sativa L.)are affected by mineral N availability. Plant Science 165:709-718.
Paulilo, M.T.S. & Felippe, GM. 1995. Resposta de plantulasde Qualea grandiflora Mart., uma especie arb6rea decerrado, a adic;:ao de nutrientes minerais. RevistaBrasileira de Botanica 18: 109-112.
Pereira, P.A.A., Baldani, J.I., Blana, RA.G & Neyra, C.A.1981. Assimilac;:ao e translocac;:ao de nitrogenio emrelac;:ao aproduc;:ao de graos e proteinas em milho (Zeamays L.). Revista Brasileira de Ciencias do Solo 5: 28-31.
Pilon-Smits, KA.H., Ebskamp, M.J.M., Paul, M.J., Jeuken,M.J.W., Weisbeek, P.J. & Smeekens, S.C.M. 1995.Improved performance of transgenic fructanaccumulating tobacco under drought stress. PlantPhysiology 107: 125-130.
Pilon-Smits, KA.H., Terry, N., Sears, T. & Van Dun, K.1999. Enhanced drought resistance in fructanproducing sugar beet. Plant Physiology andBiochemistry 37: 313-317.
Pimentel, C. 1998. Metabolismo de carbono na agriculturatropical. EDUR, Rio de Janeiro, 159 p.
Pollock, C.J., Cairns, A.J., Sims, I.M. & Housley, T.L.1996. Fructans as reserve carbohydrates in crop plants.In: E. Zamski & A.A. Shaffer, (eds.). Photoassimilatedistribution in plants and crops: source-sinkrelationships. Marcel Dekker Inc, New York, pp. 97-113.
Poorter, H. 2002. Plant growth and carbon economy.Encyclopedia of Life Sciences, pp. 1-6. http://www.els.net(acessoem20.11.2004).
Sassaki, RM. & Felippe, GM. 1998. Response ofDalbergiamiscolobium Benth. seedlings, a cerrado tree species,to mineral nutrients supply. Revista Brasileira deBotanica 21: 65-72.
Schubert, S. & Fuerle, R 1997. Fructan storage in tubersofJerusalem artichoke: characterization ofsink strength.New Phytologist 136: 115-122.
Silveira, J.A.G. & Machado, KC. 1990. Mobilizac;:ao denitrogenio e de carboidratos durante 0 desenvolvimentode paniculas de duas cultivares de arroz. RevistaBrasileira de Fisiologia Vegetal 2: 37-46.
Sprent, J.I. 1994. Nitrogen acquisition systems in theLeguminosae. In: J.I. Sprent & D. McKey (eds.).Advances in Legume Systematics. The nitrogen factor.Royal Botanic Gardens, Kew, v.5, pp.I-16.
Stewart, GR, Popp, M., Holzapfel, I., Stewart, J.I. & DickieEskew, A. 1986. Localization ofnitrate reduction in fernsand its relationship to environmental and physiologicalcharacteristics. New Phytologist 104: 373-384.
Stewart, GR & Schmidt, S. 1998. Evolution and ecologyof plant mineral nutrition. In: M.e. Press, J.D. Scholes& M.G. Barker (eds.). Physiological plant ecology.Blackwell Science, Oxford, pp. 91-113.
Tischner, R 2000. Nitrate uptake and reduction in higherand lower plants. Plant, Cell and Environment 23:1005-1024.
Tomomatsu, H. 1994. Health effects ofoligosaccharides.Food Technology 48: 61-65.
Vereyken, I.J., Van Kuik, A.J., Evers, T.R., Rijken, P.J. &Kruijff, B. 2003. Structural requirements of the fructanlipid interaction. Biophysical Journal 84: 3147-3154.
Vose, P.B. 1990. Plant nutrition relationships at the wholeplant level. In: v.e. Baligar & R.R. Duncan (eds.). Cropsas enhancers of nutrient use. Academic Press, SanDiego, pp. 65-80.
