análise citogenética das espécies do gênero hypophthalmus...
TRANSCRIPT

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA CONSERVAÇÃO E
BIOLOGIA EVOLUTIVA
Análise citogenética das espécies do gênero Hypophthalmus (Siluriformes, Pimelodidae) da região
do Lago Catalão, Amazonas, Brasil
LEILA BRAGA RIBEIRO
MANAUS – AM 2009

ii
LEILA BRAGA RIBEIRO
Análise citogenética das espécies do gênero Hypophthalmus (Siluriformes, Pimelodidae) da região
do Lago Catalão, Amazonas, Brasil ORIENTADOR(A): Eliana Feldberg, Dra. CO-ORIENTADOR: Alberto Akama, Dr.
Dissertação apresentada ao Programa de Pós-
Graduação em Genética, Conservação e
Biologia Evolutiva, como parte dos requisitos
para obtenção do título de Mestre em
Genética, Conservação e Biologia Evolutiva.
MANAUS – AM 2009

iii
FICHA CATALOGRÁFICA
SINOPSE
As espécies do gênero Hypophthalmus que ocorrem na região do lago Catalão foram caracterizadas citogeneticamente e o número diplóide foi igual a 56 cromossomos para todas. Porém, a fórmula cariotípica, o padrão de distribuição de heterocromatina constitutiva e a localização das regiões organizadoras de nucléolo foram espécie-específicas. Os dados cromossômicos obtidos evidenciam que o grupo estudado possui várias características comuns à família Pimelodidae, o que sugere uma macroestrutura cariotípica conservada para as espécies de Hypophthalmus. Palavras-chave:
Cariótipo, Mapará, Bacia amazônica.
R484 Ribeiro, Leila Braga Análise citogenética das espécies do gênero Hypophthalmus (Siluriformes, Pimelodidae) da região do Lago Catalão, Amazonas, Brasil / Leila Braga Ribeiro.--- Manaus : [s.n.], 2009. xiii, 72f. : il. color. Dissertação (mestrado)-- INPA/UFAM, Manaus, 2009 Orientador : Eliana Feldberg Co-orientador : Alberto Akama Área de concentração : Genética, Conservação e Biologia Evolutiva
1. Mapará. 2. Hypophthalmus. 3. Cromossomos. 4. Citogenética. I. Título.
CDD 19. ed. 597.520415

iv
À minha família, pelo amor e incentivo a cada desafio.

v
“O mundo não deve ser restringido até que seja compreendido...
mas a compreensão deve ser expandida até que possa assimilar o mundo.” Francis Bacon
“Aprender é a única coisa de que a mente nunca se cansa,
nunca tem medo e nunca se arrepende.” Leonardo da Vinci

vi
Apoio
Programa de Pós-Graduação em Genética, Conservação e Biologia Evolutiva.
Laboratório de Genética Animal INPA/CPBA.
MCT/INPA/PPI - Caracterização genética (cromossomos, proteínas e DNA) de
vertebrados amazônicos.
MCT/CNPq/CT-Amazônia/CT-Energ nº 13/2006, Proc. 554057/2006-9 -
Biotecnologia aplicada ao estudo das populações de peixes de importância
econômica para a Amazônia sob coordenação da prof. Dra. Izeni Pires Farias.
Convênio do Conselho de Aperfeiçoamento de Pessoal de Nível Superior
(CAPES) e da Fundação de Amparo a Pesquisa do Estado do Amazonas (FAPEAM),
pela concessão da bolsa de estudo.

vii
Agradecimentos
À minha orientadora, Dra. Eliana Feldberg, por me receber de braços abertos
em Manaus, pela confiança depositada, pela atenção e dedicação na realização
deste trabalho. Obrigada também pela compreensão, pela paciência, pela amizade e
exemplo profissional.
Ao meu co-orientador, Dr. Alberto Akama, pelas sugetões, ajuda na escolha do
objeto de estudo e identificação dos espécimes.
Ao Dr. Jorge Porto, pelo carinho e apoio no dia-a-dia. A todos os companheiros
de laboratório, Brenda, Carlos, Claudia, Eduardo, Érica, Leandra e Rodrigo, pela
ajuda e companheirismo durante o mestrado.
A Carlos Sotero e Mário Picanço, que me acompanharam nos trabalhos de
campo e foram fundamentais na busca dos maparás.
Ao Dr. Roberto Ferreira Artoni, por me apresentar a citogenética e despertar
meu interesse pela pesquisa. A Dr. Mara Cristina de Almeida e ao Dr. Marcelo
Ricardo Vicari, pelas aulas de genética e ensinamentos na bancada. A Miguel Airton
de Carvalho, Américo Moraes Neto, e demais companheiros de laboratório da UEPG.
Ao Programa de Pós-Graduação em curso de Genética, Conservação e Biologia
Evolutiva. Especialmente à secretária Alessandra, sempre disposta a atender nossas
solicitações.
Aos colegas de mestrado Arlisson, Alessandra, Elizabete, Fabíola, Rodrigo e
Suzana, pelas boas conversas na hora do almoço e companhia nas disciplinas. A
turma do mestrado de 2008, que me “adotou” e me acompanhou em muitos
momentos de diversão. À Bárbara, Mariana e Suzana, pela amizade e carinho

viii
dedicados.
Aos amigos Carlos Schneider, Maria Claudia Gross e Maria Leandra Terencio,
pela ajuda nas coletas, no laboratório, pelos ensinamentos e pelas risadas no dia-a-
dia. Vocês foram essenciais na execução deste trabalho e na minha adaptação em
Manaus. Obrigada à Maria Claudia pelas sugestões, pelo apoio, pelo esclarecimento
de dúvidas e por atuar como “orientadora” diversas vezes.
Aos amigos Edson e Eduardo, pelas risadas, pelas conversas, pela paciência
que tiveram comigo e por cuidarem de mim quando precisei. Minha estadia em
Manaus se tornou mais agradável depois que os conheci. Adoro vocês meninos!
Às velhas amigas de Iguape, Deborah, Edinah e Glauce, que apesar da
distância continuam presentes em minha vida e compartilham comigo cada conquista.
Aos colegas da universidade, com os quais dividi quatro anos de muito esforço e
diversão, em especial ao Neto, amigo e companheiro de citogenética.
Aos meus pais Dália e Antônio (In memorian), que além de me concederem a
vida deram sentido a ela. Obrigada pelos ensinamentos, pelo amor e pela estrutura
familiar que me deram. Às minhas “irmães” Cátia e Leidi, pela dedicação, pelo carinho
e incentivo, vocês foram fundamentais na minha formação. Ao meu irmão Pedro, ao
meu cunhado Paulo, aos meus primos Cristiane, João Carlos e Nilson, por desejarem
o meu bem e torcerem pelo meu sucesso a cada desafio. Aos meus sobrinhos Miguel
e Raquel, que tornam minha vida mais feliz simplesmente por existirem. Amo todos
vocês!
Agradeço a todos, que de alguma maneira, contribuiram para a realização deste
trabalho.

ix
Resumo
O gênero Hypophthalmus é constituído por peixes de couro e de hábito
pelágico, incluindo quatro espécies válidas, três das quais (H. edentatus, H.fimbriatus
e H. marginatus) ocorrem na bacia amazônica. Esse grupo apresenta uma taxonomia
baseada apenas em dados morfológicos que é ainda bastante confusa. Dados
citogenéticos do gênero são escassos, não existindo nenhuma informação sobre as
espécies amazônicas. Com base nisso os objetivos desse trabalho foram otimizar
técnicas para obtenção de cromossomos mitóticos em Hypophthalmus spp.,
caracterizar citogeneticamente as espécies do gênero e seus morfotipos que ocorrem
no lago Catalão e buscar marcadores citogenéticos espécie-específicos que
auxiliassem na elucidação taxonômica do grupo. Para isso foram capturados 167
indivíduos, pertencentes às três espécies e testadas três metodologias de obtenção
de preparações cromossômicas. Desse total foi possível obter preparações
cromossômicas de boa qualidade de 43 exemplares, totalizando 1192 metáfases
analisadas. O número diplóide obtido para todos os indivíduos foi 56 cromossomos,
sem heteromorfismo de cromossomos sexuais e sem diferença entre as espécies e
seus morfotipos. Entretanto, diferenças interespecíficas foram vistas com relação à
fórmula cariotípica, número fundamental e localização das RONs. As RONs foram
simples e sempre localizadas na porção terminal de braços curtos de cromossomos
submetacêntricos, que variaram quanto à posição no cariótipo, mostrando-se um bom
marcador. Blocos heterocromáticos foram observados nas regiões centroméricas e
teloméricas de vários cromossomos do complemento, porém, de uma maneira geral,
foram difusos e fracamente corados com apenas alguns blocos mais conspícuos,
sendo que as RONs foram positivas para banda C. A caracterização citogenética
realizada nesse trabalho permite afirmar que as espécies do gênero Hypophthalmus
apresentam uma macroestrutura cariotípica conservada dentro da família
Pimelodidae, porém a presença de rearranjos não robertsonianos foram evidentes na
diferenciação das espécies. Pode-se sugerir também que houve uma
heterocromatinização, uma vez que alguns cromossomos apresentaram os braços
curtos totalmente heterocromáticos.

x
Abstract
The genus Hypophthalmus includes pelagic catfishes and is composed by four
valid species, with three of them (H. edentatus, H. fimbriatus e H. marginatus)
distributed throughout Amazon basin. The species of the group are not well
established and present a taxonomy based only in morphological features. The
objectives of this work are: to improve cytogenetic analyses on Hypophthalmus spp.
testing some methods; and to describe the karyotypic results of the genus. To
undertake the study, 167 specimens from Catalão Lake representing the three species
were collected and three methods were applied to obtain karyotypes. From these, 43
specimens were analyzed cytogenetically. The diploid number 2n=56 was observed in
all the individuals, without evidences of sex chromosome heteromorphism or
supernumerary chromosomes. Inter-specific differences were detected in karyotype
formula, fundamental number and NOR localization. The constitutive heterochromatin
was detected at telomeric and centromeric positions in several chromosomal pairs in
all the species. The nucleolus organizer regions (Ag-NORs) were located at terminal
position on short arms of the submetacentric chromosomal pairs, suggesting a
positive association with heterochromatic regions. The karyotypic analysis of
Hypophthalmus spp. allowed the identification of various characters common to other
members of Pimelodidae, indicating conserved characters in the group.

xi
Sumário
Lista de Figuras ........................................................................................................ xii
Lista de Tabelas ....................................................................................................... xiii
1. Introdução ............................................................................................................... 1 1.1 A bacia amazônica .............................................................................................. 1 1.2 Aspectos ecológicos e taxonômicos dos maparás .............................................. 2 1.3 Citogenética de Pimelodídeos ............................................................................. 4 1.4 Objetivos ............................................................................................................ 13
1.4.1 Objetivo geral ............................................................................................... 13 1.4.2 Objetivos específicos ................................................................................... 13
2. Material e métodos ............................................................................................... 14 2.1 Material .............................................................................................................. 14 2.2 Métodos ............................................................................................................. 17
2.2.1 Obtenção de cromossomos mitóticos .......................................................... 17 2.2.2 Preparação das lâminas para visualização dos cromossomos mitóticos .... 19 2.2.3 Detecção das regiões organizadoras de nucléolos (RONs) ........................ 20 2.2.4 Detecção da heterocromatina constitutiva (banda C) .................................. 21 2.2.5 Análises cromossômicas ............................................................................. 21
3. Resultados ............................................................................................................ 23 3.1 Obtenção de cromossomos mitóticos ................................................................ 23 3.2 Caracterização cariotípica de Hypophthalmus edentatus .................................. 26 3.3 Caracterização cromossômica de Hypophthalmus fimbriatus ........................... 28 3.4 Caracterização cromossômica de Hypophthalmus cf. fimbriatus ...................... 30 3.5 Caracterização cromossômica de Hypophthalmus marginatus ......................... 32
4. Discussão .............................................................................................................. 34 4.1 Considerações sobre a obtenção de cromossomos metafásicos das espécies de Hypophthalmus spp. ................................................................................................ 34 4.2 Características cromossômicas de Hypophthalmus spp. .................................. 35 4.3 Padrão de bandeamento C e regiões organizadoras de nucléolo ..................... 37 4.4 Evolução cariotípica de Hypophthalmus spp. ................................................... 40
5. Conclusões ........................................................................................................... 44
6. Perspectivas futuras ............................................................................................ 44
7. Referências Bibliográficas ................................................................................... 45

xii
Lista de Figuras Figura 1. Local de coleta, região do lago Catalão....................................................... 15 Figura 2. Exemplares de Hypophthalmus edentatus. a) H. edentatus tipo 1; b) H. edentatus tipo 2. As setas indicam a diferença no local de inserção das nadadeiras entre os dois tipos........................................................................................................ 15 Figura 3. Exemplares de Hypophthalmus marginatus. a) H. marginatus tipo 1; b) H. marginatus tipo 2. Setas indicando diferentes locais de inserção das nadadeiras nos dois tipos...................................................................................................................... 16 Figura 4. Exemplar de Hypophthalmus fimbriatus...................................................... 16 Figura 5. Exemplar classificado como H. cf. fimbriatus. Seta indicando os longos barbilhões.....................................................................................................................16 Figura 6. Gráfico mostrando a eficiência das técnicas empregadas para obtenção de cromossomos mitóticos em Hypophthalmus spp. Técnica 1: Moreira-Filho & Bertollo, 1990; Técnica 2: Bertollo et al. 1978; Técnica 3: Barreto-Netto et al. 2007.................24 Figura 7. Metáfases de Hypophthalmus spp., mostrando diferenças na qualidade das preparações celulares: a) Técnica de Moreira-Filho & Bertollo, 1990; b) Técnica de Barreto-Netto et al. 2007.............................................................................................. 24 Figura 8. Cariótipo de H. edentatus: a) em coloração convencional; b) região organizadora de nucléolo evidenciada pela impregnação com nitrato de prata; c) coloração para banda C............................................................................................... 27 Figura 9. Cariótipo de H. fimbriatus: a) em coloração convencional; b) região organizadora de nucléolo evidenciada pela impregnação com nitrato de prata; c) metáfase em coloração para banda C. As setas indicam blocos heterocromáticos terminais.......................................................................................................................29 Figura 10. Cariótipo de Hypophthalmus cf. fimbriatus: a) em coloração convencional; b) região organizadora de nucléolo evidenciada por impregnação com nitrato de prata; c) Coloração para banda C.......................................................................................... 31 Figura 11. Cariótipo de H. marginatus: a) em coloração convencional; b) região organizadora de nucléolo evidenciada pela impregnação com nitrato de prata; c) coloração para banda C............................................................................................... 33 Figura 12. Hipótese filogenética proposta para a família Pimelodidae, em destaque o gênero Hypophthalmus................................................................................................ 42

xiii
Lista de Tabelas Tabela 1. Dados cariotípicos da família Pimelodidae compilados da literatura (2n=número diplóide; FC=fórmula cariotípica; NF=número fundamental ou número de braços; m=metacêntrico;sm=submetacêntrico; st=subtelocêntrico; a=acrocêntrico). ... 7 Tabela 2. Dados citogenéticos de Hypophthalmus spp.............................................. 25

1
1. Introdução
1.1 A bacia amazônica
A bacia amazônica é o maior sistema fluvial já conhecido, com cerca de
7.000.000 km² de área de drenagem e 23.000 km de rios navegáveis (Santos &
Ferreira, 1999). A drenagem amazônica apresenta habitats bastante heterogêneos,
pois além de sua grande extensão, a bacia amazônica ainda é formada por diversas
sub-bacias altamente influenciadas pelos períodos de seca e cheia, comuns na região
(Goulding et al., 2003). Essa heterogeneidade de ambientes é um dos fatores
responsáveis pelo grande número de peixes de água doce da região Neotropical, que
pode variar de 6.025 a 8.000 espécies (Vari & Malabarba, 1998; Reis et al., 2003).
A região amazônica apresenta a mais diversificada ictiofauna de água doce do
mundo, que mesmo não estando totalmente conhecida abrange cerca de 54 famílias
(43% Characiformes, 39% Siluriformes e 3% Gymnotiformes), as quais não se
distribuem uniformemente ao longo da bacia (Santos & Ferreira, 1999).
Avalia-se que 50% da ictiofauna do território brasileiro se encontra no sistema
amazônico, uma vez que apenas para o rio Negro são estimadas de 700 a 950
espécies (Chao, 2001; Goulding et al., 2003). Em média, 200 espécies oriundas da
bacia amazônica são direcionadas à exportação, constituindo um importante item no
mercado externo, quer na forma de pescado semi-industrializado para consumo
humano, quer na forma de peixes ornamentais (Santos et al., 1991; Batista, 2003).
Com relação à pesca, a Amazônia assume uma posição de destaque quando
comparada às demais regiões brasileiras, tanto pela diversidade de espécies
envolvidas quanto pela quantidade de pescado capturado, contribuindo com 25% da

2
produção do pescado nacional. Nos últimos anos houve uma expansão da pesca de
espécies de Siluriformes na Amazônia Central, com perspectivas de ampliação
(Barthem & Fabré, 2004). Este aumento se deve ao fato de algumas espécies de
peixes lisos como Brachyplatystoma vaillantii (piramutaba), Pseudoplaystoma tigrinum
(surubim) e Hypophthalmus spp. (maparás) dominarem o mercado internacional
(Cabral Jr. & Almeida, 2004).
Atualmente os representantes do gênero Hypophthalmus, conhecidos
popularmente como maparás, vêm sendo objeto de estudos para a aplicação de
novas técnicas, tais como “minced-fish”, também conhecida como carne
mecanicamente separada (CMS), que têm como objetivo aumentar o valor agregado
do pescado, visto que o mercado externo valoriza mais estes produtos, como filés,
hambúrguer, entre outros (Almeida & Androczevecz, 2004; Cabral Jr. & Almeida,
2004). No estado do Amazonas, a produção pesqueira do mapará apresenta-se maior
que de outras espécies amplamente comercializadas, como por exemplo: matrinchã
(Brycon spp.), pescada (Plagioscion spp.), piramutaba (Brachyplatystoma vaillantii),
surubim (Pseudoplatystoma tigrinum), tambaqui (Colossoma macropomum) e
pirarucu (Arapaima gigas) (Ruffino et al., 2006).
1.2 Aspectos ecológicos e taxonômicos dos maparás
O gênero Hypophthalmus foi recentemente enquadrado na família Pimelodidae
(Lunberg et al., 1991a; de Pinna, 1993) e compreende quatro espécies válidas até o
momento: H. edentatus Spix & Agassiz, 1829; H. fimbriatus Kner, 1857; H. marginatus
Valenciennes, 1840; e H. oremaculatus Nani & Fuster, 1947. Distribui-se na América

3
do Sul, sendo que três das quatro espécies descritas ocorrem na bacia amazônica. H.
edentatus e H. marginatus ocorrem nas bacias do Orinoco e Amazonas, e em alguns
rios da Guiana, Guiana Francesa e Suriname, H. fimbriatus distribui-se apenas na
bacia do rio Negro e áreas de influência desse rio, do Brasil até a Venezuela e H.
oremaculatus distribui-se apenas na bacia do Paraná (Lundberg & Littmann, 2003;
Ferraris Jr., 2007).
Os maparás são peixes de couro, de corpo alongado e comprimido lateralmente
(Ferreira et al., 1998), com três pares de barbilhões, coloração acinzentada e porte
grande, chegando a atingir 50 cm (López-Fernández & Winemiller, 2000; Santos et
al., 2006). Diferem-se dos demais pimelodídeos quanto ao hábito alimentar e sua
localização na coluna d’água, pois enquanto a maioria dos indivíduos da família
possui hábito carnívoro e demersal, os maparás mostram-se planctófagos e pelágicos
(Cutrim & Batista, 2005). Formam cardumes e realizam migrações verticais diárias,
relacionadas à alimentação e fuga de predadores, concentrando-se na superfície
durante a noite (Barthem & Goulding, 1997; Abujanra & Agostinho, 2002).
A taxonomia do grupo é baseada em caracteres morfológicos e ainda não está
completamente estabelecida. Variações morfológicas encontradas entre exemplares
de H. fimbriatus que ocorrem no rio Negro e na Venezuela mostram a necessidade de
uma revisão detalhada para essa espécie, com descrição de nova entidade
taxonômica para os indivíduos classificados como Hypophthalmus cf. fimbriatus, ou
inclusão desses em H. fimbriatus (López-Fernández & Winemiller, 2000). Além disso,
análises morfológicas de H. edentatus e H. marginatus sugerem que essas duas
espécies sejam subdivididas em dois grupos cada uma, e que a espécie H.

4
oremaculatus, de ocorrência restrita à bacia do Paraná, seria na verdade um dos
subgrupos de H. edentatus (Michael Littmann com. pess.).
Essa subdivisão de H. edentatus e H. marginatus baseia-se em diferenças
morfológicas observadas em exemplares da mesma espécie, principalmente com
relação ao local de inserção da nadadeira dorsal, ao comprimento dos barbilhões e
ao número de vértebras. Assim sendo, registra-se para a bacia amazônica a
ocorrência de seis grupos: H. edentatus tipo 1, H. edentatus tipo 2, H. fimbriatus, H.
cf. fimbriatus, H. marginatus tipo 1 e H. marginatus tipo 2 (Michael Littmann com.
pess.).
O emprego de técnicas genéticas é de fundamental importância para elucidar
problemas taxonômicos, como os encontrados no gênero Hypophthalmus,
principalmente por meio da citotaxonomia.
1.3 Citogenética de Pimelodídeos
A citotaxonomia pode ser definida como a nomeação e designação de
organismos a táxons, utilizando a citogenética como ferramenta auxiliar, a qual
permite individualizar espécies por meio de características cromossômicas. A
citotaxonomia está intimamente ligada à citogenética, que é a ciência que estuda os
cromossomos, seja de forma isolada ou direcionada ao conjunto cromossômico de
uma espécie, abrangendo aspectos morfológicos, funcionais e evolutivos (Guerra,
1988). Os estudos citotaxonômicos realizados com peixes têm como principais
ferramentas para a compreensão da estrutura organizacional dos cromossomos a
determinação do número diplóide e a fórmula cariotípica. Outros marcadores eficazes

5
na caracterização de espécies são a localização das regiões organizadoras de
nucléolo (RONs) no cromossomo e no cariótipo e o padrão de distribuição da
heterocromatina constitutiva no cariótipo. As regiões organizadoras de nucléolo são
regiões cromossômicas envolvidas na transcrição de RNA ribossomal, enquanto o
padrão de heterocromatina indica como o genoma eucariótico está organizado
(Feldberg et al. 1993; Almeida-Toledo, 1998; Margarido & Galetti Jr., 1999).
A citogenética tem contribuído significativamente no que diz respeito ao
conhecimento da biodiversidade genética e da evolução dos peixes amazônicos,
principalmente na identificação de espécies crípticas (Nakayama et al., 2001; Teixeira
et al., 2006; Nakayama et al., 2008), na detecção de polimorfismos intra-específicos
(Nakayama et al., 2000; Centofante et al., 2002; Mesquita et al., 2008), na detecção
de cromossomos supranumerários (Feldberg et al., 2004), na identificação de híbridos
(Alves-Brinn et al., 2004), na determinação de sistemas de cromossomos sexuais (de
Oliveira et al., 2007; 2008; Terencio et al., 2008) e evolução cromossômica
(Benzaquem et al., 2008).
A família Pimelodidae inclui 93 espécies, distribuídas em 29 gêneros (Ferraris
Jr., 2007), e do ponto de vista citogenético apenas 28 espécies válidas foram
estudadas. O número diplóide predominante nesta família é 56 cromossomos,
entretanto: 2n=50 (Pinirampus pirinampu e Callophysus macropterus); 2n=54
(Pimelodus sp., P. fur e Megalonema platanum) e 2n=58 (Pimelodus blochii e P. cf.
maculatus) também foram observados (Tabela 1). Por outro lado, os pimelodídeos
mostram variabilidade quanto à estrutura cariotípica, ou seja, no número fundamental,
sugerindo a presença de rearranjos do tipo inversão pericêntrica e/ou translocações,
os quais modificam o tipo cromossômico, mas não o número diplóide (Swarça et al.,

6
2006). Além disso, cromossomos B foram encontrados em seis espécies e um caso
de sistema sexual do tipo XX/XY foi descrito para Steindachneridion melanodermatum
(Tabela 1).
Em relação às espécies amazônicas observa-se uma carência de dados
citogenéticos, principalmente no que se refere aos maparás (Tabela 1), visto que para
o gênero Hypophthalmus só existe a descrição do número diplóide para H. edentatus
da bacia do Paraná (Martins-Santos et al., 1990). Entretanto, essa espécie pode ser
na verdade a espécie descrita como H. oremaculatus, que tem ocorrência restrita à
bacia do Paraná, e que assim como H. edentatus possui taxonomia confusa.
Considerando a crescente pressão de pesca sobre os estoques pesqueiros de
espécies do gênero Hypophthalmus (Cutrim & Batista, 2005) e a taxonomia incerta do
grupo, como mencionado anteriormente, uma caracterização cromossômica é de
fundamental importância para a correta determinação das espécies desse grupo.
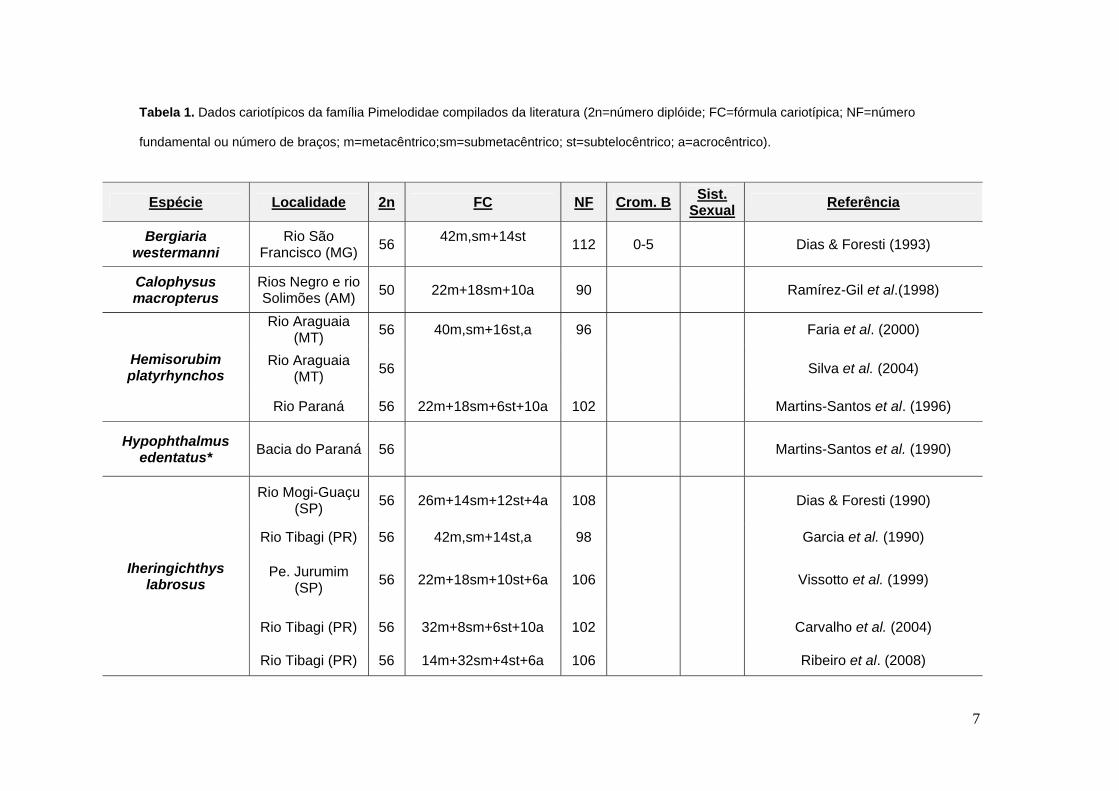
7
Tabela 1. Dados cariotípicos da família Pimelodidae compilados da literatura (2n=número diplóide; FC=fórmula cariotípica; NF=número
fundamental ou número de braços; m=metacêntrico;sm=submetacêntrico; st=subtelocêntrico; a=acrocêntrico).
Espécie Localidade 2n FC NF Crom. B Sist. Sexual Referência
Bergiaria westermanni
Rio São Francisco (MG) 56 42m,sm+14st
112 0-5 Dias & Foresti (1993)
Calophysus macropterus
Rios Negro e rio Solimões (AM) 50 22m+18sm+10a 90 Ramírez-Gil et al.(1998)
Hemisorubim platyrhynchos
Rio Araguaia (MT) 56 40m,sm+16st,a 96 Faria et al. (2000)
Rio Araguaia (MT) 56 Silva et al. (2004)
Rio Paraná 56 22m+18sm+6st+10a 102 Martins-Santos et al. (1996)
Hypophthalmus edentatus* Bacia do Paraná 56 Martins-Santos et al. (1990)
Iheringichthys labrosus
Rio Mogi-Guaçu (SP) 56 26m+14sm+12st+4a 108 Dias & Foresti (1990)
Rio Tibagi (PR) 56 42m,sm+14st,a 98 Garcia et al. (1990)
Pe. Jurumim (SP) 56 22m+18sm+10st+6a 106 Vissotto et al. (1999)
Rio Tibagi (PR) 56 32m+8sm+6st+10a 102 Carvalho et al. (2004)
Rio Tibagi (PR) 56 14m+32sm+4st+6a 106 Ribeiro et al. (2008)

8
Espécie Localidade 2n FC NF Crom. B Sist. Sexual Referência
Megalonema platanum
Rio Paraná (Argentina) 54 14m+18sm+12st+10a 98 Sanches (2006)
Parapimelodus valenciennis Rio Guaíba (RS) 56 Costa & Reggi (1986)
Phractocephalus hemioliopterus Piscicultura 56 Castilho-Almeida et al. (1997)
Pimelodus absconditus Rio Paraná (PR) 56 24m+18sm+8st+6a
106 Borin & Martins-Santos (2002)
P. argenteus Rio Paraguai (MS) 56 24m+16sm+12st+4a
108 Souza et al. (2003)
P. blochii
Rio Araguaia (MT) 56 36m,sm+20st,a 92 Faria & Venere (2000)
Rio Araguaia (MT) 56 36m,sm+20st,a 92 Silva et al. (2004)
Rio Solimões (AM) 58 Della-Rosa et al. (1980)
P. clarias Argentina 56 Fenocchio et al. (1994)
Uruguai 56 Gonzales (1994)
P. fur Não definida 56 30m+14sm+12a 100 Toledo & Ferrari (1976a)
Rio São Francisco (MG) 54 32m+8sm+6st+8a 100 Garcia & Moreira-Filho (2005)

9
Espécie Localidade 2n FC NF Crom. B Sist. Sexual Referência
P. heraldoi Rio Tibagi (PR) 56 22m+22sm+6st+6a 106 Souza et al. (2004)
Rio Piquiri (PR) 56 18m+24sm+6st+8a 104 Treco & Dias (2009)
Não definida 56 30m+14sm+12a 100 Toledo & Ferrari (1976b) Rio Guaíba (RS) 56 Costa & Reggi (1986)
P. maculatus
Araquari (MG) 56 56m,sm,st,a Moreira et al. (2004) Rio São
Francisco (MG) 56 40m,sm+16st,a 96 Dias & Foresti (1993)
Rio Mogi-Guaçu (SP) 56 40m,sm+16st,a
96 Dias & Foresti (1993)
Rio Sapucaí (MG) 56 40m,sm+16st,a 96 Marques et al. (1998)
Rio Paraná (PR) 56 Vasconcelos & Martins-Santos (2000)
Pe.Jurumirim 56 Vissotto et al. (1999)
Rio Paraná (PR) 56 20m+20sm+10st+6a 106 Swarça et al. (2001a) Rio Paraná (PR) 56 20m+20sm+10st+6a 106 Borin & Martins-Santos (2002) D.Paranaense 56 24m+14sm+12st+6a 106 Heras & Mendoza (2002)
Rio Tejuco (MG) 56 56m,sm,st,a Moreira et al. (2004)
Rio Paraguai (MS) 56 22m+16sm+10st+8a 104 Souza et al. (2003)
São Caetanos 56m,sm,st,a Moreira et al. (2004)
Rio São Francisco (MG) 56 32m+12sm+12st
112
Garcia & Moreira-Filho (2005)
Rio Paranapanema 56 20m+20sm+10st+6a 106 Mazzuchelli et al. (2007)

10
Espécie Localidade 2n FC NF Crom. B Sist. Sexual Referência
P. cf. maculatus Rio Jari (PA) 58 30m,sm+28st,a 116 Souza et al. (2000)
P. mysteriosus Rio Paraguai (MS) 56 26m+20sm+2st+8a 104 Souza et al. (2003)
P. ornatus Rio Paraná (PR) 56 18m+22sm+6st+10a
102 Abucarma & Martins-Santos (1996)
Rio Paraná (PR) 56 20m+18sm+8st+10a 102 Borin & Martins-Santos (2002)
P. ortmanni
Rio Iguaçu (PR) 56 24m+18sm+8st+6a 106 0-4 Terencio et al. (2001)
Rio Iguaçu (PR) 56 24m+18sm+8st+6a 106 Borin & Martins-Santos (2004)
P. paranaensis Rio Piquiri (PR) 56 22m+22sm+4st+8a 104 Treco & Dias (2009)
Pimelodus sp.
Não definida 30m+14sm+12a 100 Toledo & Ferrari (1976b)
Rio Iguaçu (PR) 56 22m+22sm+4st+8a 104 Dias & Foresti (1993)
Rio Iguaçu (PR) 56 24m+26sm+4st+2a 110 Souza et al. (2004)
Rio Iguaçu (PR) 56 30m+14sm+8st+4a 108 0-4 Borin & Martins-Santos (2004)
Rio São Francisco (MG) 56 32m+12sm+6st+6a
106 Garcia & Moreira-Filho (2005)
Rio Tejuco (MG) 54 54m,sm,st,a Moreira & Morelli (2001)
Rio Araguari (MG) 54 54m,sm,st,a
Moreira & Morelli (2001)

11
Espécie Localidade 2n FC NF Crom. B Sist. Sexual Referência
Pinirampus pirinampu
Rio Tibagi (PR)
50
26m+12sm+2st+10a
90
Swarça et al. (1999)
Rio Paraná (PR) 50 22m+12sm+4st+12a 88 Vasconcelos & Martins-Santos (2000)
Araguari (MG) 50 Molina & Morelli (2004)
Pseudoplatystoma corruscans
Coxim (MS) 56
42m,sm+14st,a
98 Souza et al. (1992)
Rio Mogi-Guaçu 56 18m+18sm+10st+10a 102 Bigoni et al. (1992)
Três Marias (MG) 56
20m+12sm+12st+12a
100 Fenocchio (1993)
Porto Rico (PR) 56 18m+16sm+10st+12a
100 Martins-Santos et al. (1996)
Rio Mogi-Guaçu 56 18m+18sm+10st+10a 102 Borin & Martins-Santos (2002)
Rio Paraná (PR) 56 26m+10sm+6st+14a 98 Swarça et al. (2005)
Rio Paraguai (MS) 56 20m+16sm+8st+12a
100 Swarça et al. (2005)
Rio Miranda (MT) 56 56m,sm
112 Porto-Foresti et al. (2003)

12
Espécie Localidade 2n FC NF Crom. B Sist. Sexual Referência
P. corruscans
Rio Aquidauana (MG) 56 56m,sm
112 Porto-Foresti et al. (2003)
Rio Paraguai (MS) 56 56m,sm
112 Porto-Foresti et al. (2003)
P. fasciatum Rio Solimões (AM) 56 18m+14sm+10st+14a
98 Fenocchio & Bertollo (1992)
P. tigrinum Rio Solimões (AM) 56 18m+16sm+8st+14a
98 Fenocchio & Bertollo (1992)
Sorubim lima
Rio Solimões (AM) 56 18m+12s+14st+12a
100 Fenocchio & Bertollo (1992)
Porto Rico (PR) 56 20m+14sm+10st+12a 100 Martins-Santos et al. (1996)
Steindachneridion scriptum
Rio Paranapanema
(PR) 56 24m+20sm+4st+8a
104 Swarça et al. (2005)
Rio Tibagi (PR) 56 24m+20sm+4st+8a 104 Swarça et al. (2005)
Steindachneridion melanodermatum Rio Iguaçu (PR) 56
20m+24sm+2st+10a 21m+23sm+2st+10a
102 XX/
XY Swarça et al. (2006)
Zungaro zungaro
Foz do Iguaçu (PR) 56 26m+10sm+6st+14a
98
Martins-Santos et al. (1996)
Jupiá (SP) 56 32m+6sm+8st+10a
102 Swarça et al. (2001b)
* Esta espécie pode ser a descrita como H. oremaculatus, de ocorrência restrita à bacia do Paraná.

13
1.4 Objetivos
1.4.1 Objetivo geral
Fazer um estudo cromossômico das espécies de mapará que ocorrem na região
do Catalão (Hypophthalmus edentatus tipo 1, H. edentatus tipo 2, H. fimbriatus, H. cf.
fimbriatus, H. marginatus tipo 1 e H. marginatus tipo 2) para se detectar possíveis
marcadores que possam auxiliar na taxonomia destes peixes.
1.4.2 Objetivos específicos
a) Determinar o cariótipo das espécies do gênero Hypophthalmus que
ocorrem na região do Catalão.
b) Estabelecer o padrão de distribuição de heterocromatina constitutiva e
das regiões organizadoras de nucléolo das espécies estudadas.
c) Verificar a ocorrência de variabilidade cariotípica intra-específica.
d) Identificar marcadores cromossômicos espécie-específicos.

14
2. Material e métodos
2.1 Material
A coleta dos exemplares ocorreu na região do lago Catalão (3º09’57’’S,
59º54’44’W), a qual está situada próximo à confluência dos rios Negro e Solimões
(Figura 1). Tal lago conecta-se ao rio Negro de forma quase permanente e ao rio
Solimões por um curto canal após os primeiros meses de subida das águas. Sendo
assim, o nível de água do lago Catalão varia de acordo com o nível dos rios
adjacentes, oferecendo abrigo e alimento a diversas espécies e servindo de local de
desova para muitos peixes (Leite et al., 2006).
As atividades de campo foram realizadas em períodos de cinco dias nos meses
de setembro/2007, fevereiro/2008, junho/2008 e novembro/2008, com o objetivo de
encontrar os animais em período de migração, que ocorre nos períodos de enchente
e vazante, visto que os mesmos utilizam o lago como local de desova. Os indivíduos
foram coletados no período noturno, quando se concentram na superfície d’água,
com o auxílio de malhadeiras. As medidas das malhadeiras foram de 2,80m de altura,
55,0m de comprimento e com distância entrenós de 25x30mm e 25x 5mm.
Capturou-se um total de 167 indivíduos, distribuídos entre as três espécies do
gênero: Hypophthalmus edentatus, H. fimbriatus e H. marginatus. No caso das
espécies H. edentatus e H. marginatus que apresentam dois subtipos, foram obtidos
exemplares de cada subtipo (Figuras 2 e 3). Foram capturados também alguns
indivíduos que apresentavam longos barbilhões maxilares, diferentes dos observados
em H. marginatus, mas menos espessos do que os presentes em H. fimbriatus
(Figura 4). Tais espécimes foram tratados como H.cf. fimbriatus, como sugerido em

15
López-Fernández & Winemiller (2000) (Figura 5). Os peixes coletados foram levados
vivos ao Laboratório do Flutuante do Instituto Nacional de Pesquisa da Amazônia –
INPA, no Catalão, onde foram obtidas as suspensões celulares.
Figura 1. Local de coleta, região do lago Catalão (Modificado de Teixeira et al., 2002).
Figura 2. Exemplares de Hypophthalmus edentatus. a) H. edentatus tipo 1; b) H. edentatus tipo 2. As setas indicam a diferença no local de inserção das nadadeiras entre os dois tipos.

16
Figura 3. Exemplares de Hypophthalmus marginatus. a) H. marginatus tipo 1; b) H. marginatus tipo 2. Setas indicando diferentes locais de inserção das nadadeiras nos dois tipos.
Figura 4. Exemplar de Hypophthalmus fimbriatus.
Figura 5. Exemplar classificado como H. cf. fimbriatus. Seta indicando os longos barbilhões.

17
2.2 Métodos
2.2.1 Obtenção de cromossomos mitóticos
Para obtenção dos cromossomos mitóticos os animais foram anestesiados em
água gelada e sacrificados em seguida. Foi retirada a porção anterior e/ou posterior
do rim, órgão hematopoiético dos peixes. Em virtude dos maparás morrerem com
muita facilidade em situação de estresse, três técnicas foram testadas e adaptadas,
visando otimizar a obtenção de cromossomos em metáfase. Após a preparação
cromossômica todos os exemplares foram fixados em formol 10%, lavados e
guardados em álcool 70%, para posteriormente serem depositados na coleção
ictiológica do Instituto Nacional de Pesquisas da Amazônia.
A primeira técnica testada foi descrita por Moreira-Filho & Bertollo (1990), que
consiste na adição de colchicina “in vitro”. Nessa metodologia a porção anterior do rim
foi colocada em 15mL de solução hipotônica de KCL 0,075M e dissociada com pinças
de dissecação e seringas de vidro desprovidas de agulha. Em seguida colocou-se 0,5
mL de colchicina na concentração de 0,0062% e a suspensão celular foi mantida em
banho-maria a 37° C por 30 minutos. Após esse tempo, o material foi ressuspendido
e transferido para um tubo de centrífuga, utilizando-se uma pipeta Pasteur. Adicionou-
se 0,3 mL de fixador Carnoy (metanol: ácido acético 3:1) à suspensão celular, que foi
centrifugada durante 10 minutos a 900 rpm e teve seu sobrenadante posteriormente
descartado. Ressuspendeu-se o material novamente com o auxílio de uma pipeta
Pasteur, e adicionou-se 8 mL de fixador Carnoy à suspensão antes de nova
centrifugação a 900 rpm por 10 minutos. Esse procedimento foi repetido por mais
duas vezes. Após a última centrifugação e a eliminação do sobrenadante, 1,5 mL de

18
fixador foram adicionados e o material ressuspendido com cuidado. Essa suspensão
celular foi então estocada em tubo tipo “eppendorf” e mantida em freezer (-10º C).
A segunda metodologia (Bertollo et al., 1978), que consiste na aplicação da
colchicina “in vivo”, sofreu modificações e foi testada apenas nos indivíduos que
apresentaram maior resistência, sobrevivendo por cerca de uma hora em aquário
bem aerado. Injetou-se intraperitonealmente uma solução de colchicina 0,0062% na
proporção de 1mL para cada 100 g de peso do animal. Os peixes foram mantidos em
aquários por cerca de 50 minutos. Em seguida os indivíduos foram sacrificados e
tiveram a porção anterior do rim retirada, a qual foi colocada em uma solução
hipotônica de KCL 0,075M e divulsionada com o auxílio de uma seringa de vidro. Em
seguida, retirou-se o sobrenadante com o auxílio de uma pipeta Pasteur, que foi
colocado em tubo de centrífuga. A suspensão celular foi incubada a 37 °C por 30
minutos. Esse material foi pré-fixado com seis gotas de metanol:ácido acético (3:1) e
ressuspendido. Essa suspensão ficou em repouso por 5 minutos e centrifugado por
10 minutos a 900 rpm. O sobrenadante foi desprezado e completou-se para 6mL com
fixador, ressuspendendo o material novamente. A suspensão foi centrifugada por 10
minutos a 900 rpm, descartando o sobrenadante e completando para 6mL de fixador.
Esse processo foi repetido por mais duas vezes. Após a última centrifugação e a
eliminação do sobrenadante, 1,5 mL de fixador foram adicionados e o material
ressuspendido. Essa suspensão celular foi estocada em tubo “eppendorf” e mantida
em freezer para posterior análise.
A terceira metodologia testada foi descrita por Barreto Netto et al. (2007), que
consiste na utilização de meio de cultura RPMI 1640. Uma porção pequena do rim foi
retirada, lavada em meio RPMI 1640 gelado e dissociada em 5mL de meio RPMI

19
1640 gelado, com auxílio de pinças de dissecação e seringas sem agulha. A
suspensão foi transferida para um tubo de centrífuga, completando para 10 mL com
meio RPMI 1640. Essa suspensão foi mantida refrigerada por um período de 12
horas. Passado esse tempo adicionou-se 10 gotas de colchicina a 0,0062% e o
material foi mantido em temperatura ambiente por 30 minutos. O material foi
centrifugado a 900 rpm durante 10 minutos e o sobrenadante foi descartado.
Acrescentou-se 10 mL de solução hipotônica KCL 0,075M e após a homogeneização
a suspensão foi mantida em temperatura ambiente por 20 minutos. Cinco gotas de
fixador Carnoy recém preparado na proporção 3:1 (metanol: ácido acético) foram
adicionadas à suspensão e o material foi novamente centrifugado a 900 rpm por 10
minutos. Após o descarte do sobrenadante foram adicionados 6mL de fixador. O
material foi ressuspendido e submetido à nova centrifugação a 900 rpm durante 10
minutos, sendo o sobrenadante posteriormente descartado. Tal procedimento foi
repetido mais duas vezes. Após a última centrifugação o sobrenadante foi descartado
e adicionou-se 1,5 mL de fixador Carnoy, previamente preparado na proporção 1:1
(metanol: ácido acético). Tal suspensão foi armazenada em tubo “eppendorf” e
mantida em freezer (-10º C) para análise posterior.
2.2.2 Preparação das lâminas para visualização dos cromossomos mitóticos
Para a visualização dos cromossomos mitóticos, as lâminas foram lavadas,
secas ao ar e posteriormente imersas em água destilada a 50º C, em banho-maria.
Após cinco minutos, as lâminas foram retiradas do banho-maria de forma a manter
uma película de água sobre a sua superfície, sobre a qual foi gotejada a suspensão

20
celular em diferentes regiões. As lâminas foram secas diretamente ao ar, sendo
posteriormente coradas com Giemsa 5% diluído em tampão fosfato 0,06M e pH 6,8
por 15 minutos, lavadas em água destilada, secas ao ar e analisadas ao microscópio
óptico.
2.2.3 Detecção das regiões organizadoras de nucléolos (RONs)
Para a caracterização das regiões organizadoras de nucléolos (RON) foi
utilizada a técnica de precipitação de cristais de Prata (Ag-RON), descrita por Howell
& Black (1980), com pequenas modificações. A técnica consiste em pingar sobre a
lâmina com a preparação cromossômica uma gota de uma solução coloidal, obtida
com 1g de gelatina comercial sem sabor dissolvida em 50 mL de água destilada e
acrescida de 0,5 mL de ácido fórmico. Em seguida, adicionou-se sobre a solução
coloidal duas gotas de solução aquosa de AgNO3 (Nitrato de Prata) a 50%. A lâmina
foi levemente agitada e coberta com lamínula. A lâmina foi mantida sobre chapa
aquecedora a 60 ºC por cerca de 5 minutos. Em seguida foi lavada em água destilada
de forma a permitir que a lamínula fosse retirada com o próprio jato de água. As
lâminas secaram diretamente ao ar. Para facilitar a visualização das metáfases
algumas lâminas foram posteriormente coradas com Giemsa 5% em tampão fosfato
0,06M e pH 6,8 por 2 minutos, lavadas em água corrente e secas ao ar.

21
2.2.4 Detecção da heterocromatina constitutiva (banda C)
Para a caracterização da heterocromatina constitutiva utilizou-se a técnica de
banda C descrita por Sumner (1972), com adaptações.
As lâminas contendo as preparações cromossômicas foram tratadas durante 2
minutos com HCl 0,2N a 46 ºC, lavadas em água destilada à temperatura ambiente e
secas ao ar. Em seguida, as lâminas foram incubadas em solução de hidróxido de
bário a 5%, recém preparada e filtrada, a 46 ºC, por 2 minuto e 30 segundos.
Interrompeu-se a ação do hidróxido de bário imergindo-se a lâmina em solução de
HCl 0,2N (46 ºC) por cerca de 30 segundos, sendo posteriormente lavada em água
destilada. Após secas, as lâminas foram incubadas em solução 2xSSC (Cloreto de
Sódio 0,3M e Citrato Trisódico 0,03M, pH 6,8) em banho-maria a 60 ºC, por um
período de 15 minutos, lavadas e secas ao ar. Posteriormente foram coradas com
Giemsa 10% em tampão fosfato 0,06M e pH 6,8 por 20 minutos, lavadas em água
corrente e secas novamente ao ar.
2.2.5 Análises cromossômicas
As preparações cromossômicas foram analisadas em microscópio óptico
comum com objetiva de imersão e as células selecionadas foram fotografadas, com
máquina digital Sony Cybershot 7.2MP ou em fotomicroscópio Olympus Bx-51. A
montagem dos cariótipos se deu com auxílio do programa Adobe Photoshop, versão
CS3, a partir de cromossomos metafásicos mitóticos, os quais foram recortados e
tentativamente emparelhados. Os cromossomos foram medidos pelo programa livre
ImageJ e colocados em ordem decrescente de tamanho. A morfologia cromossômica

22
foi determinada levando em consideração a posição do centrômero, segundo a
proposta de Levan et al. (1964). Os cromossomos mitóticos foram agrupados com
base no índice de relação de braços (RB=comprimento do braço maior/comprimento
do braço menor), de acordo com a seguinte classificação: metacêntricos (m) (RB=
1,0-1,70), submetacêntricos (sm) (RB=1,71-3,00), subtelocêntricos (st) (RB=3,01-
7,00) e acrocêntricos (a) (RB>7,00). O número fundamental (NF) foi determinado de
acordo com o número de braços cromossômicos, considerando-se os metacêntricos,
submetacêntricos e subtelocêntricos como tendo dois braços e os cromossomos
acrocêntricos como tendo apenas um braço.

23
3. Resultados
3.1 Obtenção de cromossomos mitóticos
Realizou-se análise citogenética de 167 exemplares pertencentes ao gênero
Hypophthalmus (7 H. edentatus tipo 1, 13 H. edentatus tipo 2, 3 H. fimbriatus, 8 H. cf.
fimbriatus, 107 H. marginatus tipo 1 e 29 H. marginatus tipo 2). Entretanto,
cromossomos mitóticos, em condições de análise, só foram obtidos em 43 indivíduos:
dois H. edentatus tipo 1 (1 fêmea, 1 macho), sete H. edentatus tipo 2 (3 fêmeas, 4
machos), três H. fimbriatus (2 fêmeas, 1 macho), cinco H. cf. fimbriatus (2 fêmeas, 3
machos), 24 H. marginatus tipo 1 (10 fêmeas, 9 machos, 5 com sexo indeterminado)
e dois H. marginatus tipo 2 (1 fêmea, 1 com sexo indeterminado), o que representa
apenas 25,74% de sucesso na obtenção de cromossomos mitóticos destes peixes.
As três técnicas foram utilizadas de início como descritas pelos autores, mas
ainda passaram por algumas modificações, como por exemplo, a alteração da
concentração de colchicina para 0,0062% e tempo de hipotonização. Comparando as
três técnicas utilizadas neste trabalho, a que permitiu obter cromossomos com melhor
qualidade para análise citogenética em Hypophthalmus spp. foi a descrita por Barreto
Netto et al. (2007). Tanto a metodologia descrita por Moreira-Filho & Bertollo (1990),
quanto a descrita por Bertollo et al. (1978), apresentaram um índice baixo de
metáfases e a qualidade das mesmas não foi satisfatória para análise dos
cromossomos (Figura 6). Na metodologia com meio de cultura a longo prazo foi
possível observar maior número de metáfases e cromossomos de melhor qualidade,
principalmente no que diz respeito ao nível de compactação dos mesmos (Figura 7).

24
Figura 6. Gráfico mostrando a eficiência das técnicas empregadas para obtenção de cromossomos mitóticos em Hypophthalmus spp. Técnica 1: Moreira-Filho & Bertollo, 1990; Técnica 2: Bertollo et al. 1978; Técnica 3: Barreto-Netto et al. 2007.
Figura 7. Metáfases de Hypophthalmus spp., mostrando diferenças na qualidade das preparações celulares: a) Técnica de Moreira-Filho & Bertollo, 1990; b) Técnica de Barreto-Netto et al. 2007.

25
Para a determinação do número diplóide das espécies foi analisado um total de
1192 metáfases, sendo que todos os exemplares apresentaram 2n=56 cromossomos
e ausência de heteromorfismo cromossômico sexual. Porém, variações na fórmula
cariotípica e localização da região organizadora de nucléolo foram verificadas entre
as espécies (Tabela 2).
Tabela 2. Dados citogenéticos de Hypophthalmus spp (presente trabalho).
Espécie Número Diplóide Fórmula Cariotípica Número
Fundamental Localização
da RON H. edentatus 2n=56 18m+20sm+6st+12a NF=100 Par nº 13 (sm)
H. fimbriatus 2n=56 18m+16sm+6st+16a NF=96 Par nº 11 (sm)
H. cf. fimbriatus 2n=56 10m+22sm+10st+14a NF=98 Par nº 10 (sm)
H. marginatus 2n=56 12m+22sm+8st+14a NF=98 Par nº 16 (sm)

26
3.2 Caracterização cariotípica de Hypophthalmus edentatus
Os indivíduos pertencentes a H. edentatus apresentaram um número diplóide
modal de 56 cromossomos sem heteromorfismo de cromossomos sexuais. A fórmula
cariotípica foi igual a 18m+20sm+6st+12a e NF=100 (Figura 8a). A impregnação por
nitrato de Prata (Ag-RON) evidenciou a região organizadora de nucléolo em posição
terminal, nos braços curtos de um par submetacêntrico, provavelmente o par 13,
coincidente com a constrição secundária visível em coloração convencional (Figura
8b). A distribuição da heterocromatina constitutiva se deu em porções terminais e
centroméricas de vários cromossomos do complemento. Os pares cromossômicos 1
e 3 apresentaram blocos heterocromáticos centroméricos e biteloméricos, e cerca de
cinco pares submetacêntricos mostraram os braços curtos totalmente
heterocromáticos. A RON também foi positiva para banda C (Figura 8c).
Apesar de H. edentatus ser morfologicamente diferenciada em tipo 1 e tipo 2,
nenhuma diferença citogenética foi verificada entre os subtipos, seja em fórmula
cromossômica, posição da região organizadora de nucléolo ou padrão de distribuição
de heterocromatina constitutiva.

27
Figura 8. Cariótipo de H. edentatus: a) em coloração convencional; b) região organizadora de nucléolo evidenciada pela impregnação com nitrato de prata; c) coloração para banda C.

28
3.3 Caracterização cromossômica de Hypophthalmus fimbriatus
Os indivíduos pertencentes a H. fimbriatus apresentaram um número diplóide
modal de 56 cromossomos sem heteromorfismo de cromossomos sexuais. A fórmula
cariotípica foi igual a 18m+16sm+6st+16a e NF=96 (Figura 9a). A região organizadora
de nucléolo foi observada em posição terminal, nos braços curtos de um par
submetacêntrico, provavelmente o de número 11 (Figura 9b). Nessa espécie o
resultado de banda C não foi satisfatório para se determinar o padrão de distribuição
da heterocromatina constitutiva, devido principalmente à qualidade das metáfases.
Porém, pode-se observar uma quantidade menor de blocos heterocromáticos,
comparada à H. edentatus e estes foram observados, principalmente em porções
teloméricas (Figura 9c).
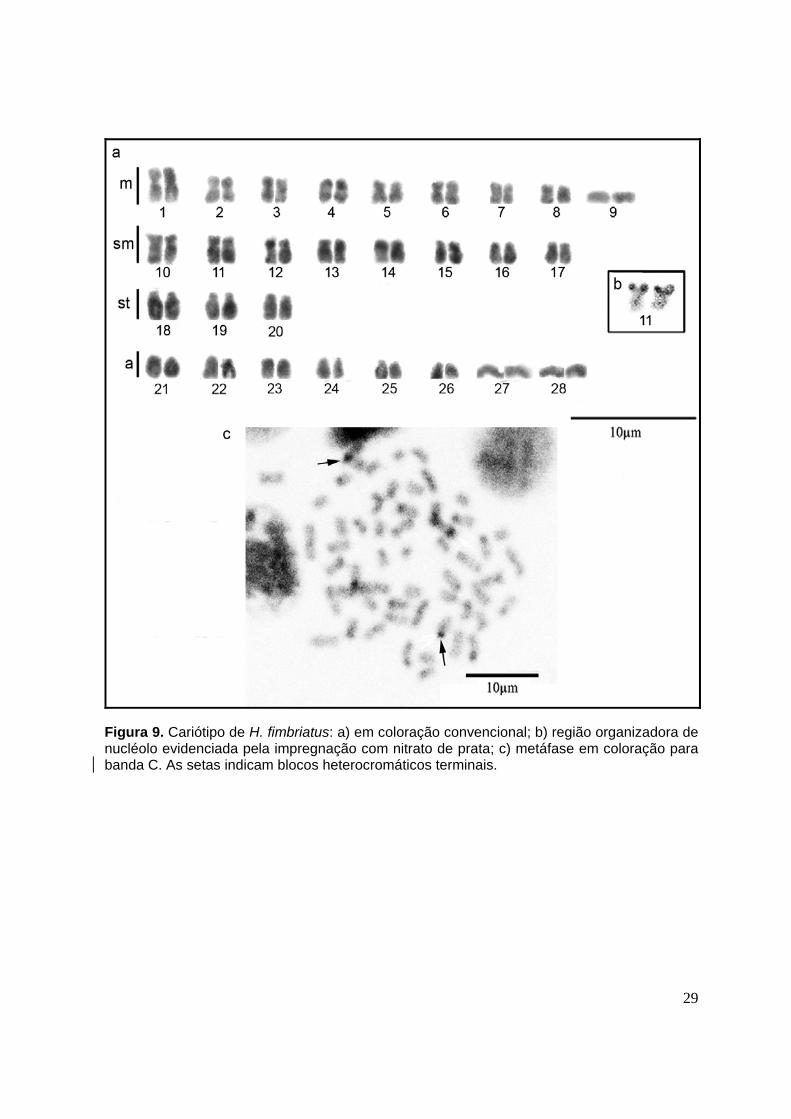
29
Figura 9. Cariótipo de H. fimbriatus: a) em coloração convencional; b) região organizadora de nucléolo evidenciada pela impregnação com nitrato de prata; c) metáfase em coloração para banda C. As setas indicam blocos heterocromáticos terminais.

30
3.4 Caracterização cromossômica de Hypophthalmus cf. fimbriatus
Os indivíduos pertencentes a H.cf. fimbriatus apresentaram um número diplóide
modal de 56 cromossomos, sem heteromorfismo de cromossomos sexuais. A fórmula
cariotípica foi igual a 10m+22sm+10st+14a e NF=98 (Figura 10a). A localização da
região organizadora de nucléolo foi detectada no par submetacêntrico número 10, ora
marcando apenas um dos homólogos e ora apresentando comportamento de
associação de RONs (Figura 10b), visível inclusive com coloração convencional. Nas
metáfases em que a associação não foi observada apenas um dos cromossomos do
par subtelocêntrico apresentou marcação positiva para o nitrato de prata, com
heteromorfismo de tamanho das RONs. Esse padrão foi visto apenas nessa espécie
e mostrou-se positivo para a banda C. Essa espécie apresentou também poucos
blocos heterocromáticos no complemento cromossômico, os quais estão distribuídos
em regiões terminais e centroméricas, porém marcações biteloméricas foram
evidenciadas apenas no par subtelocêntrico número 17 (Figura 10c).

31
Figura 10. Cariótipo de Hypophthalmus cf. fimbriatus: a) em coloração convencional; b) região organizadora de nucléolo em associação evidenciada por impregnação com nitrato de prata; c) Coloração para banda C (par nucleolar em associação).

32
3.5 Caracterização cromossômica de Hypophthalmus marginatus
Os indivíduos pertencentes a H. marginatus apresentaram um número diplóide
modal de 56 cromossomos, sem heteromorfismo de cromossomos sexuais. A fórmula
cariotípica foi igual a 12m+22sm+8st+14a e NF=98 (Figura 11a). A localização da
região organizadora de nucléolo foi detectada no par submetacêntrico número 16
(Figura 11b), coincidente com a constrição secundária. Essa espécie apresentou
blocos heterocromáticos distribuídos nas porções teloméricas e centroméricas em
vários cromossomos do complemento, inclusive alguns biteloméricos. Pelo menos em
seis pares (4, 8,10, 12, 20,21) os braços curtos foram totalmente heterocromáticos. A
região organizadora de nucléolo foi positiva para a banda C (Figura 11c).
Apesar de H. marginatus ser morfologicamente diferenciado em tipo 1 e tipo 2,
nenhuma diferença citogenética foi verificada entre os subtipos, seja em fórmula
cromossômica ou padrões de bandeamento.

33
Figura 11. Cariótipo de H. marginatus: a) em coloração convencional; b) região organizadora de nucléolo evidenciada pela impregnação com nitrato de prata; c) coloração para banda C.

34
4. Discussão
4.1 Considerações sobre a obtenção de cromossomos metafásicos das
espécies de Hypophthalmus spp.
A extrema fragilidade dos maparás dificultou a obtenção de preparações
cromossômicas de boa qualidade, pois muitos exemplares já se encontravam mortos
antes de serem retirados das malhadeiras. Os indivíduos retirados com vida não
sobreviviam tempo suficiente para receberem tratamento adequado, como por
exemplo, a indução de mitoses por uso de fermento biológico, que necessitaria da
sobrevivência por no mínimo 24 horas. Devido a isso, tanto a técnica de colchicina “in
vitro” (Moreira-Filho & Bertollo, 1990) quanto a técnica de colchicina “in vivo” (Bertollo
et al., 1978) mostraram-se pouco eficazes na obtenção de cromossomos mitóticos,
provavelmente pelo fato da divisão celular não ser estimulada anteriormente. Tal fato
resultou em suspensões com baixo índice metafásico e cromossomos muito
condensados, dificultando a análise citogenética.
De forma geral, a obtenção de cromossomos metafásicos em pimelodídeos
não é tida como fácil, o que pode ser percebido nos resultados compilados na Tabela
1, onde não foi possível determinar a fórmula cariotípica de várias espécies e em
algumas delas os cromossomos foram classificados em apenas dois grupos (m-sm;
st-a), o que evidencia a dificuldade em se determinar a morfologia cromossômica.
A metodologia que mostrou maior eficiência na obtenção de cromossomos
mitóticos em Hypophthalmus spp. foi a descrita por Barreto Netto et al. (2007), que
consiste em manter o órgão hematopoiético dos peixes em meio de cultura RPMI
1640 por um período mínimo de 12 horas. Isso permitiu a indução da atividade

35
mitótica nas células incubadas e favoreceu a sincronização da divisão celular,
resultando em maior número de metáfases.
Também foi observado que os indivíduos apresentavam cromossomos
pequenos e de difícil visualização, sendo assim, realizou-se testes para encontrar
uma concentração de colchicina adequada. Utilizou-se colchicina a 0,0062% pois
nessa concentração o grau de condensação cromossômica diminuiu, tornando
possível a determinação da fórmula cariotípica e de bandeamentos cromossômicos,
que até o momento não haviam sido efetuados em espécies desse gênero. Essa
técnica foi utilizada após vários testes, nos quais diversos indivíduos foram
sacrificados sem sucesso na obtenção de cromossomos mitóticos, e devido a isso os
resultados aqui apresentados são de apenas 25,74% do total de exemplares
analisados. Entretanto, o número diplóide e a fórmula cromossômica puderam ser
determinados com segurança.
4.2 Características cromossômicas de Hypophthalmus spp.
A maioria das espécies da família Pimelodidae apresenta um número diplóide
igual a 56 cromossomos, como pode ser observado na Tabela 1. Das 29 espécies
descritas citogeneticamente, 27 apresentaram 2n=56 cromossomos, sendo que três
dessas ainda mostraram uma variação no número diplóide em algumas populações
(Pimelodus blochii – Della-Rosa et al. (1980); Pimelodus sp. – Moreira & Morelli
(2001); P. fur – Garcia & Moreira-Filho (2005).
Considerando os números fundamentais (NF) descritos na Tabela 1, pode-se
observar que há uma variação de 92 a 112. Portanto, apesar de compartilharem o
mesmo número diplóide, o NF, as fórmulas cariotípicas e a localização das RONs no

36
cariótipo variam de forma considerável. Também se pode observar na Tabela 1 que
uma mesma espécie quando coletada em mais de uma localidade pode possuir até
oito fórmulas cariotípicas, como por exemplo, Pimelodus maculatus e
Pseudoplatystoma corruscans.
As três espécies de Hypophthalmus aqui analisadas (H.edentatus, H. fimbriatus e
H. marginatus), mais H. cf. fimbriatus, também apresentaram 2n=56 cromossomos,
número também encontrado para H. edentatus da bacia do Paraná por Martins-
Santos et al. (1990), e considerado ancestral para a ordem Siluriformes (Oliveira &
Gosztonyi, 2000). Apesar de compartilharem o número diplóide as fórmulas
cariotípicas entre os quatro grupos variaram, o que evidencia a presença de
rearranjos não robertsonianos.
Rearranjos cromossômicos não robertsonianos têm sido considerados por alguns
autores como os mais importantes na diferenciação cariotípica da família Pimelodidae
(Garcia & Moreira-Filho, 2005; Mazzuchelli et al., 2007), uma vez que a morfologia
cromossômica é alterada e o número diplóide mantido. Segundo Andreata et al.
(2006), variações no número fundamental em peixes podem estar relacionadas com a
presença de segmentos positivos para banda C, já que o acúmulo desses segmentos
pode mudar a morfologia cromossômica.
Em Hypophthalmus spp. as diferenças nas fórmulas cariotípicas indicam a
ocorrência de rearranjos cromossômicos, como por exemplo inversões pericêntricas.
Entretanto, no caso específico dessas espécies, essa variação também poderia estar
relacionada a rearranjos do tipo duplicação in tandem devido à presença de braços
cromossômicos inteiros heterocromáticos, vistos em alguns cromossomos
submetacêntricos e subtelocêntricos.

37
As espécies H. edentatus e H. marginatus, que podem ser separadas em dois
subtipos cada uma com base em caracteres morfológicos, conforme mencionado
anteriormente, não apresentaram diferenças cariotípicas entre os subtipos. Acredita-
se que essa diferença morfológica intraespecífica seja um polimorfismo relacionado a
fatores ambientais, ou esteja em nível gênico, num estágio inicial de especiação.
Assim sendo, tem-se a necessidade de uma revisão taxonômica com estudos
pormenorizados a respeito do comportamento, fisiologia, e reprodução dessas
espécies, aliados a análises moleculares.
4.3 Padrão de bandeamento C e regiões organizadoras de nucléolo
A heterocromatina, detectada pela técnica de bandeamento C, constitui parte
importante do genoma de eucariotos por possuir funções como segregação
cromossômica, organização nuclear e regulação da expressão gênica (Grewal & Jia,
2007). O padrão de distribuição da heterocromatina visto nas espécies de
Hypophthalmus é o mesmo previamente descrito para espécies da família
Pimelodidae e para a maioria dos Siluriformes, ou seja, blocos heterocromáticos
localizados em regiões teloméricas e centroméricas, e muitos cromossomos do
complemento com ausência de heterocromatina ou presença de blocos pálidos e
difusos. Um aspecto interessante nas espécies desse gênero são os cromossomos
com braços curtos totalmente heterocromáticos. Este padrão foi identificado em
outras espécies da ordem Siluriformes por diferentes autores (LeGrande, 1981;
Fenocchio & Bertollo, 1992; Swarça et al., 2001b; Treco et al., 2008), que chamaram
atenção para a dificuldade em se detectar a heterocromatina constitutiva devido a

38
características peculiares da cromatina nesses peixes quanto ao grau de
condensação.
Variações na quantidade e na distribuição da heterocromatina constitutiva têm
sido consideradas características importantes na evolução de alguns grupos de
peixes e permitido a diferenciação de espécies e de populações, além de ser uma
ferramenta útil em relações filogenéticas (Andreata et al., 2006; Treco et al., 2008).
Essas diferenças são resultantes de mecanismos epigenéticos como remodelamento
da cromatina e metilação do DNA, que auxiliam na compactação e organização do
genoma em domínios cromáticos. Isso é importante devido ao fato de diversas
atividades gênicas serem dependentes do estado de condensação da cromatina, que
pode mudar em resposta a sinais celulares e atividade gênica, respondendo inclusive
a mudanças ambientais (Grewal & Jia, 2007).
Tais fenômenos epigenéticos podem explicar o padrão diferenciado de
distribuição da heterocromatina, onde alguns blocos heterocromáticos podem ser
marcadores de gênero de peixes, como observado para as espécies de piranha do
gênero Serrasalmus, as quais possuem um grande bloco proximal no braço longo de
um cromossomo relacionado com o DNAr 5S (Nakayama et al, 2008) ou de
populações como a diferença no padrão de distribuição de banda C vista em
Geophagus brasiliensis (Vicari et al. 2006). Para as espécies de Hypophthalmus o
padrão de banda C foi muito similar, embora pôde-se perceber uma diferença
interespecífica com relação à quantidade da heterocromatina. Apesar da dificuldade
na obtenção dos bandeamentos verificou-se que a distribuição da heterocromatina
coincide com o padrão previamente descrito na família Pimelodidae (Swarça et al.,
2001a; Carvalho et al., 2004; Treco et al., 2008), onde os indivíduos apresentam

39
pouca quantidade de heterocromatina, distribuída principalmente em blocos
centroméricos e terminais.
Segundo Grewal & Jia (2007), as regiões heterocromáticas apresentam alta
densidade de DNA repetitivo, o qual contém “clusters” de DNA satélite, RNA
ribossomal e elementos transponíveis e essa associação da heterocromatina com
seqüências repetitivas é crucial para organização funcional de estruturas
cromossômicas como centrômeros e telômeros, bem como para a regulação da
atividade de sítios ribossomais (Boisvert et al., 2007). Quanto a este aspecto, ficou
evidente a relação de heterocromatina com as RONs, uma vez que as quatro
espécies apresentaram a RON positiva para a banda C, confirmando a presença de
heterocromatina associada à cístrons ribossomais (Swarça et al., 2005).
A associação das RONs com as regiões de heterocromatina constitutiva promove
um controle epigenético do genoma, uma vez que pode ocorrer tanto a manutenção
de cópias de DNA ribossomal expressas para as células, quanto o silenciamento
desses genes. Considerando que a região de DNA ribossomal é instável e responde
a danos causados por diversos fatores, incluindo fatores ambientais, existe a
necessidade de se ter um mecanismo de regulação da expressão gênica eficiente
nessas regiões (Grewal & Jia, 2007). Além disso, associações de RONs com blocos
heterocromáticos têm promovido a mudança de posição dos organizadores
nucleolares no cariótipo (Tabela 2) por meio de rearranjos, sendo esses mecanismos
já relatados em diferentes grupos de peixes, e são mais evidentes em famílias que
apresentam um número diplóide conservado, como por exemplo Curimatidae,
Anostomidae e Pimelodidae (Galetti et al., 1991; Feldberg et al., 1992; 1993; presente
trabalho).

40
Com relação ao heteromorfismo de tamanho da RON descrito em H. cf. fimbriatus
pode-se dizer que é bastante comum em peixes, e já foi relatado em outras espécies
da mesma família, como por exemplo, em Iheringichthys labrosus (Vissotto et al.,
1999; Carvalho et al., 2004; Ribeiro et al., 2008) e Pimelodus maculatus (Vissotto et
al., 1999; Souza et al., 2003; 2004; Treco et al., 2008). Esta variação pode ser
oriunda de mecanismos como “crossing-over” desigual, transposições ou outros
rearranjos incluindo deleções e/ou duplicações, freqüentemente atribuídos a
mecanismos de modificações estruturais das RONs (Carvalho et al., 2004; Ribeiro et
al., 2008). O comportamento de associação das RONs também pode ser o
responsável pela presença de tal heteromorfismo, uma vez que envolve segmentos
de cromossomos homólogos. Essa característica da RON em H. cf. fimbriatus pode
representar um marcador citogenético para esta espécie, uma vez que tal
característica não foi observada nas demais espécies analisadas.
Assim, o que se observa no presente estudo e nos dados da literatura é que as
espécies da família Pimelodidae, bem como a ordem Siluriformes, apresentam uma
compactação da cromatina muito diferente das ordens Characiformes e Perciformes,
o que pode ser observado pelo pequeno tamanho dos cromossomos e pelo padrão
de banda C.
4.4 Evolução cariotípica de Hypophthalmus spp.
Entre os Siluriformes observa-se uma variabilidade considerável de cariótipos,
envolvendo principalmente o número diplóide e a fórmula cariotípica. Os números
diplóides encontrados nessa ordem variam de 2n=22 a 2n=132 cromossomos, com

41
um número modal de 58 (±2) cromossomos (Oliveira et al., 1988). Tal diversidade é
considerada uma característica distinta nos Siluriformes, principalmente entre as
espécies das famílias Heptapteridae, Pseudopimelodidae e Pimelodidae.
Para a família Pimelodidae a variação ocorre principalmente no que diz respeito
aos números fundamentais descritos, que foi de 92 a 112, mas o número diplóide é
mantido (Swarça et al., 2007). A fixação de mecanismos de manutenção do número
diplóide reforça a idéia proposta por Oliveira & Gosztonyi (2000), de que 2n=56
cromossomos seria o número diplóide ancestral nos Siluriformes.
As espécies da família Pimelodidae que apresentam número diplóide igual a 56
cromossomos foram subdivididas em dois grupos: “Pimelodus” e “Sorubiminae”, por
apresentarem características citogenéticas peculiares, como por exemplo a
localização das RONs. No grupo “Pimelodus” as RONs localizam-se em porções
terminais em braços longos, em contrapartida, os “Sorubiminae” apresentam RONs
em porções terminais de braços curtos de cromossomos submetacêntricos ou
subtelocêntricos (Swarça et al., 2007). Considerando a árvore filogenética proposta
por Lundberg et al.(1991b) e tendo em vista que dados cromossômicos da família
Pimelodidae foram sobrepostos a esta árvore (Swarça et al., 2007), é possível
confirmar a inclusão de Hypophthalmus no grupo “Sorubiminae” com base nos
resultados citogenéticos aqui apresentados (Figura 12), principalmente no que diz
respeito às características das regiões organizadoras de nucléolo.

42
Figura 12. Hipótese filogenética proposta para a família Pimelodidae, em destaque o gênero Hypophthalmus (Modificado de Swarça et al., 2007).
Apesar da dificuldade de se fazer comparações entre os cariótipos dos
Siluriformes devido ao alto nível de condensação cromossômica (LeGrande, 1981),
que dificulta a determinação do tipo cromossômico e a organização do cariótipo, é
possível observar que dentro do gênero Hypophthalmus a variação das fórmulas
cariotípicas entre as espécies se deve a rearranjos cromossômicos associados ao
processo de especiação e não decorrentes de eventos de poliploidia, como sugerido
para a origem de outros gêneros pertencentes à mesma ordem (Fenerich et al.,
2004). Tais rearranjos podem ser considerados não robertsonianos, sendo os

43
principais: inversões pericêntricas, visto pelo número de pares cromossômicos por
categoria (m, sm, st, a), translocações ou transposições, visto pela diferença na
posição das RONs nos cariótipos e duplicações “in tandem” que resultaram na
heterocromatinização de braços inteiros.
A diversidade cariotípica encontrada nos Siluriformes é determinada por
diferenças no número diplóide, na fórmula cariotípica e cromossomos sexuais em
alguns gêneros, como por exemplo, em Ancistrus (Loricariidae), para o qual foram
descritos quatro sistemas de cromossomos sexuais (de Oliveira et al., 2007; 2008),
indicando que essa ordem constitui um modelo interessante para estudos de
mecanismos envolvidos no processo de especiação dos peixes neotropicais.
Entretanto, a revisão sistemática dos diferentes táxons dessa ordem faz-se
necessária, e os dados citogenéticos podem ser de grande valia, uma vez que novas
espécies podem apresentar características citogenéticas particulares. Como visto em
H. cf. fimbriatus, analisada nesse trabalho, que possui características cromossômicas
mais semelhantes a H. marginatus do que a H. fimbriatus.

44
5. Conclusões
a) A caracterização cariotípica descrita mostra que o gênero Hypophthalmus
apresenta uma macroestrutura cariotípica conservada entre os pimelodídeos.
b) Os bandeamentos cromossômicos, juntamente com a fórmula cariotípica e
localização das RONs permitem distinção entre as espécies do gênero.
c) A divisão das espécies H. edentatus e H. marginatus em dois tipos cada uma,
não é confirmada citogeneticamente, podendo estar associada a fatores ambientais
e/ou comportamento reprodutivo, que podem resultar em diferenças morfológicas
intraespecíficas.
6. Perspectivas futuras
No decorrer desse trabalho surgiram novos questionamentos, os quais poderão
ser explorados em trabalhos posteriores, o que contribuirá para a compreensão da
evolução do gênero Hypophthalmus. Todos os exemplares coletados no presente
estudo tiveram tecido muscular armazenado, que deverá ser utilizado na produção de
sondas para mapeamento físico cromossômico de genes ribossomais e elementos
transponíveis, bem como para análises moleculares. Tais análises associadas aos
dados citogenéticos aqui obtidos permitirão maior esclarecimento acerca da evolução
cariotípica e da diferenciação de espécies no gênero em questão.

45
7. Referências Bibliográficas
Abucarma, M.; Martins-Santos, I.C. 1996. Caracterização cromossômica de duas
espécies da família Pimelodidae (Pisces, Siluriformes). In: Anais do VI Simpósio
de Citogenética Evolutiva Aplicada a Peixes Neotropicais. São Carlos, SP. p.73.
Abujanra, F.; Agostinho, A.A. 2002. Dieta de Hypophthalmus edentatus (Spix, 1829)
(Osteichthyes, Hypophthalmidae) e variações de seu estoque no reservatório de
Itaipu. Acta Scientiarum, 24(2): 401-410.
Almeida, O.T.; Androczevecz, S. 2004. Novas espécies comerciais e novos produtos
de pescado na Amazônia: as instituições de pesquisa e as indústrias. In: Almeida,
O.T. A indústria pesqueira na Amazônia. IBAMA/Pró-Várzea, Manaus, Amazonas.
p. 41-66.
Almeida-Toledo, L.F. 1998. Cytogenetic markers in Neotropical freshwater fishes. In:
Malabarba, L.R.; Reis, R.E.; Vari, R.P.; Lucena, R.P.; Lucena, Z.M.S.; Lucena,
C.A.S. 1998. Phylogeny and classification of neotropical fishes. Editora da
Pontifícia Universidade Católica, Porto Alegre, Brasil. p. 583-588.
Alves-Brinn, M.N.; Porto, J.I.R.; Feldberg, E. 2004. Karyological evidence for
interspecific hybridization between Cichla monoculus and C. temensis
(Perciformes, Cichlidae) in the Amazon. Hereditas, 141 (1): 1-6.
Andreata, A.A.; Oliveira, C.; Foresti, F. 2006. Karyological characterization of four
Neotropical fish species of the genus Hisonotus (Teleostei, Loricariidae,
Hypoptopomatinae) from distinct Brazilian river basins. Genetics and Molecular
Biology, 29(1): 62-66.

46
Barreto Neto, M.R.C.; Pauls, A.; Affonso, P.R.A.M. 2007. A standart protocol for
obtaining fish chromosomes under post-mortem condictions. Micron, 38: 214-217.
Barthem, R.; Goulding, M. 1997. Os bagres balizadores: ecologia, migração e
conservação de peixes amazônicos. Instituto de Proteção Ambiental do Estado do
Amazonas / Sociedade Civil de Mamirauá, Brasília, Distrito Federal. 129 pp.
Barthem, R.B.; Fabré, N.N. 2004. Biologia e diversidade dos recursos pesqueiros da
Amazônia. In: Rufino, M.L. A pesca e os recursos pesqueiros na Amazônia
brasileira. IBAMA/Pró-Várzea, Manaus, Amazonas. p. 17-62.
Batista, V.S. 2003 A pesca na Amazônia Central. In: Rufino, M.L. A pesca e os
recursos pesqueiros na Amazônia brasileira. IBAMA/Pró-Várzea, Manaus,
Amazonas. p. 213-244.
Benzaquem, D.C.; Feldberg, E.; Porto, J.I.R.; Gross, M.C.; Zuanon, J.A.S. 2008.
Cytotaxonomy and karyoevolution of the genus Crenicichla (Perciformes,
Cichlidae). Genetics and Molecular Biology, 31(1): 259-255.
Bertollo, L.A.C.; Takahashi, C.S.; Moreira-Filho, O. 1978. Cytotaxonomic
considerations on Hoplias lacerdae (Pisces, Erytrinidae). Revista Brasileira de
Genética, 1: 103-120.
Bigoni, A.P.V.; Almeida-Toledo, L.F.; Toledo Filho, S.A. 1992. Estudos citogenéticos
em Pseudoplatystoma corruscans (Pimelodidae, Sorubiminae) do rio Mogi-Guaçú,
SP. In: Anais do IX Simpósio de Citogenética Evolutiva Aplicada a Peixes
Neotropicais. p.32.
Boisvert, F.M.; Koningsbruggen, S.; Navascués, J.; Lamond, A. 2007. The
multifunctional nucleolus. Molecular Cell Biology, 8: 574-585.

47
Borin, L.A.; Martins-Santos, I.C. 2002. Cytogenetic aspects in species of the genus
Pimelodus (Pisces, Siluriformes) of the river Paraná Basin. Cytologia, 67: 199-
204.
Borin, L.A.; Martins-Santos, I.C. 2004. Study on karyotype and occurrence of B
chromosomes in two endemic species Pimelodus (Siluriformes, Pimelodidae) from
the Iguaçu River. Hereditas, 140: 201-209.
Cabral Jr, W.; Almeida, O.T. 2004. Avaliação do mercado e indústria pesqueira na
Amazônia. In: Almeida; O.T. A indústria pesqueira na Amazônia. IBAMA/Pró-
Várzea, Manaus, Amazonas. p. 17-39.
Carvalho, R.A.; Giuliano–Caetano, L.; Dias, A.L. 2004. Cytogenetic analisis of A- and
B- chromosomes of Iheringichthys labrosus from Tibagi River, Paraná, Brazil.
Cytologia, 60(4): 381-385.
Castilho-Almeida, R.B.; Porto-Foresti, F.; Oliveira, C.; Foresti, F. 1997. Análise
citogenética de Pirarara (Phractocephalus hemeliopterus) reproduzida em
piscicultura na região de Terenus, MS. In: 43º Congresso Brasileiro de Genética,
Goiânia, GO. Revista Brasileira de Genética, 20 (3-suplemento): 103.
Centofante, L.; Porto, J.I.R.; Feldberg, E. 2002. Chromosomal polymorphism in
Serrasalmus spilopleura, Kner, 1858 (Characidae, Serrasalminae) from Central
Amazon Basin. Caryologia, 55 (1): 37-45.
Chao, N.L. 2001. The fishery, diversity and conservation of ornamental fishes in the
Rio Negro Basin, Brazil - A Review of Project Piaba (1989-1999). In: Chao, N.L.;
Petry, P.; Prang, G.; Sonneschien, L.; Tlusty, M. (Eds). Conservation and
management of ornamental fish resources of the rio Negro Basin. Amazonia,

48
Brazil - Project Piaba. Editora da Universidade do Amazonas, Manaus,
Amazonas. p. 161-204.
Costa, L.J.; Reggi, R. 1986. Estudos preliminares de duas espécies da família
Pimelodidae: Pimelodus maculatus e P. valenciennes do rio Guaíba – RS. Anais
do I Simpósio de citogenética evolutiva aplicada de peixes Neotropicais. São
Carlos – SP, p. 34.
Cutrim, L.; Batista, V.S. 2005. Determinação de idade e crescimento do mapará
(Hypophthalmus marginatus) na Amazônia Central. Acta Amazonica, 35 (1):85-
92.
Della-Rosa, V.A. Bertollo, L.A.C.; Ferrari, I.; Takahashi, C.S.; Moreira-Filho, O.;
Foresti, F. 1980. Estudos Citogenéticos de Peixes da Amazônia. II. Ordem
Siluriformes. Ciência e Cultura, 32:735.
Dias, A.L.; Foresti, F. 1990. Algumas considerações a respeito do cariótipo de
Iheringichthys labrosus (Siluriformes, Pimelodidae) do rio Mogi-Guaçú. In: Anais
do III Simpósio de Citogenética Evolutiva Aplicada em Peixes Neotropicais.
Botucatu, SP. p. 32.
Dias, A.L.; Foresti, F. 1993. Cytogenetic studies on fishes of the family Pimelodidae
(Siluroidei). Brazilian Journal of Genetics, 16(3): 585-600.
Faria, A.A.; Venere, P.C. 2000. Caracterização cariotípica de Pimelodus blochii
(Pisces, Siluriformes). Anais do XXIII Congresso Brasileiro de Ictiologia, Cuiabá.
p. 353.
Faria, A.A.; Brito, J.G.; Venere, P.C. 2000. Citogenética de Pimelodidae:
caracterização cromossômica de Pimelodus blochii, Pimelodella cristata e

49
Hemisorubim platyrhynchos (Siluriformes) do médio Araguaia. Anais do VIII
Simpósio de Citogenética e Genética de Peixes, Manaus. p. 92.
Feldberg, E. Porto, J.I.R.; Bertollo, L.A.C. 1992. Karyotype evolution in Curimatidade
(Teleostei, Characiformes) from the Amazon region. I. Studies on the genera
Curimata, Psectrogaster, Steindachnerina and Curimatella. Revista Brasileira de
Genética, 15 (2): 369-383.
Feldberg, E.; Porto, J.I.R.; Nakayama, C.M. 1993. Karyotype evolution in
Curimatidade (Teleostei, Characiformes) from the Amazon region. II. Centric
fissions in the genus Potamorhina. Genome, 36: 372-376.
Feldberg, E.; Porto, J.I.R.; Alves-Brinn, M.N.; Mendonça, M.N.C.; Benzaquem, D.C.
2004. B Chromosomes in Amazonian cichlid species. Cytogenetic and Genome
Research, 106: 195-198.
Fenerich, P.C.; Foresti, F. Oliveira, C. 2004. Nuclear DNA content in 20 species of
Siluriformes (Teleostei: Ostariophysi) from the Neotropical region. Genetics and
Molecular Biology, 27(3): 350-354.
Fenocchio, A.S.; Bertollo, L.A.C. 1992. Karyotype similarities among Pimelodidae
(Pisces, Siluriformes) from the Brazilian Amazon region. Cytobios, 69: 41-46.
Fenocchio, A.S. 1993. Cromossomos supranumerários no gênero Rhamdia (Pisces).
Caracterização cromossômica e considerações sobre a evolução cariotípica nos
Siluroidei. PhD. Tese. Faculdade de Medicina de Ribeirão Preto, Universidade de
São Paulo.
Fenocchio, A.S.; Pastori, M.C.; Lopéz, P.A.; Sanchéz, S.; Alberdi, A.J.; Bordenave, S.;
Dib, M.C. 1994. Levantamento citogenético em peixes de água doce da

50
Argentina: resumo das espécies estudadas. In: Anais do V Simpósio de
Citogenética Evolutiva Aplicada em Peixes Neotropicais. Botucatu, SP. p. 8.
Ferraris Jr., C.J. 2007. Checklist of catfishes, recent and fossil (Osteichthyes:
Siluriformes), and catalogue of siluriform primary types. Zootaxa,1418: 1-628.
Ferreira, E.J.G.; Zuanon, J.A.S.; Santos, G.M. 1998. Peixes comerciais do médio
Amazonas: Região de Santarém-PA. IBAMA/Pró-Várzea, Brasília, Distrito
Federal. 211 pp.
Galetti Jr, P.M.; Mestriner, C.A.; Venere, P.C. Foresti, F. 1991. Heterochromatin and
karyotype reorganization in fish of the family Anostomidae (Characiformes).
Cytogenetic Cell Genetic, 56: 116-121.
Garcia, R.M.G.; Sachete, S.; Martins-Santos, I.C. 1990. Aspectos citogenéticos de
Iheringichthys labrosus (Pisces, Pimelodidae) do rio Paraná, região de Porto Rico,
PR. In: Anais do III Simpósio de Citogenética Evolutiva Aplicada em Peixes
Neotropicais. Botucatu, SP. p. 32.
Garcia, C; Moreira–Filho, O. 2005. Cytogenetical analyses in three fish species of the
genus Pimelodus (Siluriformes: Pimelodidae) from rio São Francisco:
considerations about the karyotypical evolution in the genus. Neotropical
Ichthyology, 3: 285-290.
Gonzales, A. 1994. Estudio preliminar de tres species pertenecientes al ordem
Siluriformes (Pisces). In:Anais do V Simpósio de Citogenética Evolutiva Aplicada
em Peixes Neotropicais. p.21.
Goulding, M.; Barthem, R.B.; Ferreira, E.J.G. 2003. The Negro and the Trombetas –
black and clear waters from ancient lands. In: The Smithsonian Atlas of the
Amazon. Princeton Editorial Associates, Inc. Hong Kong. p. 211-226.

51
Grewal S.I.S.; Jia S. 2007. Heterochromatin revised. Nature Review, 8: 35-46.
Guerra, M. 1988. Introdução à citogenética geral. Editora Guanabara, Rio de Janeiro.
142 pp.
Heras, M.P.; Mendoza, N.R. 2002. Analísis cariotípico de Pimelodus maculatus
(LACÉPÈDE, 1803) (Pisces, Siluriformes, Pimelodidae) provenientes del delta
Paranaense. In: Proceedings of XXXI Congreso Argentino de Genetica, La Plata,
Argentina. p. 95
Howell, W.M.; Black, D.A. 1980. Controlled silver staining of nucleolus organizer
regions with a protective colloidal developer: a 1-step method. Experientia, 36:
1014-1915.
LeGrande, W.H. 1981. Chromosomal evolution in North American catfishes
(Siluriformes: Ictaluridae) with particular emphasis on the madtoms, Noturus.
Copeia 1: 33-52.
Leite, R.G.; da Silva, J.V.V.; Freitas, C.E. 2006. Abundância e distribuição das larvas
de peixes no lago Catalão e no encontros dos rios Solimões e Negro, Amazonas,
Brasil. Acta Amazonica, 36 (4): 557-562.
Levan, A., Fredga, K.; Sandberg, A.A. 1964. Nomenclature for centromeric position on
chromosomes. Hereditas, 52: 201-220.
López-Fernández, H.; Winemiller, K.O. 2000. A review of Venezuelan species of
Hypophthalmus (Siluriformes: Pimelodidae). Ichthyology Explorer Freshwaters,
11(1): 35-46.
Lundberg, J.G.; Mago Leccia, F; Nass, P. 1991a. Exallodontus aguanai, a new genus
and species of Pimelodidae (Pisces, Siluriformes) from deep river channels of

52
South America and delimitation of the subfamily Pimelodidae. Proc. Biol. Soc.
Washington, 104: 840-869.
Lundberg, J.G. Bornbush, A.H. Mago Leccia, F. 1991b. Gladioglanis consquistator N.
Sp. From Ecuador with diagnoses of the subfamilies Rhamdiinae Bleeker and
Pseudopimelodidae N. Subf. (Siluriformes, Pimelodidae). Copeia, 1: 190-209.
Lundberg, J.G; Littmann M.W. 2003. Family Pimelodidae (Long-whiskered catfishes).
In: Reis, R.E.; Kullander, S.O.; Ferraris Jr., C.J. Check list of the freshwater fishes
of South and Central America. Editora da Pontifícia Universidade Católica, Porto
Alegre, Brasil. p. 432-446.
Margarido, V.P.; Galetti Jr., P.M. 1999. Heterochromatin patterns and karyotype
relationships within and between the genera Brycon and Salminus (Pisces,
Characidae). Genetics and Molecular Biology, 22: 357-362.
Marques, S.; Maistro, E.L.; Oliveira, C.; Foresti, F. 1998. Estudos cariotípicos na
espécie Pimelodus maculatus (Pisces, Pimelodidae) coletada no rio Sapucaí,
represa de Furnas, MG. In: Anais do 44º Congresso Brasileiro de Genética.
Águas de Lindóia, SP. p. 89.
Martins-Santos, I.C.; Santos, I.C.M.; Portela, A.L.B.; Julio-Jr, H.F. 1990. Análise
cromossômica de espécies da ordem Siluriformes do Rio Paraná. In: Anais do III
Simpósio de Citogenética Evolutiva Aplicada de Peixes Neotropicais, Botucatu. p.
24.
Martins-Santos, I.C.M.; Julio, H.F.; Burin, I. 1996. Karyotypic studies of the subfamily
Surubiminae (Pisces, Siluriformes). Caryologia, 49: 73-80.
Mazzuchelli, J.A.; Swarça, A.C.; Dias, A.L. 2007. Structural chromosome
polymorphism in a Pimelodus maculates La Cepède, 1803 population

53
(Siluriformes, Pimelodidae) from the Paranapanema river basin, PR, Brazil.
Brazilian Jounal of Biology, 67 (4): 935-937.
Mesquita, D.R.; Porto, J.I.R.; Feldberg, E. 2008. Chromosomal variability in the
ornamental fish Symphysodon spp. (Perciformes, Cichlidae) from Amazon.
Neotropical Ichthyology 6(2): 181-190.
Molina, R.A.S.; Morelli, S. 2004. Análise citogenética de uma população de
Pinirampus pirinampu (Pisces, Siluriformes) da bacia do rio Araguari (Uberlândia,
MG). In: Anais do X Simpósio de Citogenética e Genética de Peixes. Natal, RN.
p.143.
Moreira, R.G.; Morelli, S. 2001. Citogenética do gênero Pimellodus sp. (Pisces,
Pimelodidae) In: Anais da III Mostra científica de Pós-Graduação em genética e
bioquímica, Uberlândia. p. 103.
Moreira, R.G.; Morelli-Shimizu, L.; Morelli, S. 2004. Aspectos citogenéticos de duas
populações do mandi amarelo Pimelodus maculatus (Siluriformes, Pimelodidae).
In: Anais do X Simpósio de Citogenética e Genética de Peixes. Natal, RN. p. 145.
Moreira-Filho, O.; Bertollo, L.A.C. 1990. Astyanax scabripinis (Pisces, Characidae): A
species complex. Revista Brasileira de Genética, 14: 331-347.
Nakayama, C.M.; Porto, J.I.R.; Feldberg, E. 2000. Ocorrência de dois citótipos de
Serrasalmus spilopleura Kner, 1958 (Characiformes, Serrasalmidae) da região de
confluência dos rios Negro e Solimões, Amazonas, Brasil. Acta Amazonica, 30
(1): 149-154.
Nakayama, C.M.; Jégu, M. Porto, J.I.R.; Feldberg, E. 2001. Karyological evidence for
a cryptic species of piranha within Serrasalmus rhombeus (Characidae,
Serrasalmidae) in the Amazon. Copeia, 3: 866-869.

54
Nakayama, CM; Feldberg, E.; Bertollo, L.A.C. 2008. Mapping of ribosomal genes and
chromosomal markers in three piranha species of the genus Serrasalmus
(Characidae, Serrasalminae) from the Amazon basin. Genetics and Molecular
Biology 31 (4):868-873.
Oliveira, C.; Almeida-Toledo, L.F.; Foresti, F.; Britski, H.A.; Toledo Filho, S.A. 1988.
Chomosome formulae of Neotropical freshwater fishes. Revista Brasileira de
Genética, 11 (3): 577-624.
Oliveira, C.; Gosztonyi, A.E. 2000. A cytogenetic study of Diplomystes mesembrinus
(Teleostei, Siluriformes, Diplomystidae) with a discussion of chromosome
evolution in siluriforms. Caryologia, 53: 31-37.
de Oliveira, R.R.; Feldberg, E.; dos Anjos, M.B.; Zuanon, J. 2007. Karyotype
characterization and ZZ/ZW sex chromosome heteromorphism in two species of
the catfish genus Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the
Amazon basin. Neotropical Ichthyology, 5(3): 301-306.
de Oliveira, R.R.; Feldberg, E.; dos Anjos, M.B.; Zuanon, J. 2008. Occurrence of
multiple sexual chromosomes (XX/XY1Y2 and Z1Z1Z2Z2/Z1Z2W1W2) in catfishes of
the genus Ancistrus (Siluriformes: Loricariidae) from the Amazon basin. Genética,
134: 243-249.
de Pinna, M.C. 1993. Higher-level phylogeny of Siluriformes, with a new classification
of the order (Teleostei, Ostariophysi). PhD. Tese. City University of New York,
New York.
Porto-Foresti, F.; Silva, M.F.Z.D.; Senhorini, J.A.; Cecarelli, P.C. Almeida-Toledo, L.F.;
Oliveira, C.; Foresti, F. 2003. Pesquisas genéticas em recursos pesqueiros da
bacia do alto rio Paraguai. I Pintado (Pseudoplatystoma corruscans), curimbatá

55
(Prochilodus lineatus) e piauçu (Leporinus macrocephalus). Anais do 49º
Congresso Nacional de Genética. Águas de Lindóia. CD-room.
Ramírez-Gil, H.; Feldberg, E.; Almeida-Val, V.M.F.; Val, A.L. 1998. Karyological,
biochemical, and physiological aspects of Callophysus macropterus (Siluriformes,
Pimelodidae) from the Solimões and Negro Rivers. Braz. J. Med. Biol. Res.
31(11): 1449-1458.
Reis, R.E.; Kullander, S.O; Ferraris Jr., C.J. 2003. Check list of the freshwater fishes
of South and Central America. Editora da Pontifícia Universidade Católica, Porto
Alegre, Brasil. 742 pp.
Ribeiro, L.B; Matoso, D.A.; Almeida, M.C; Vicari, M.R.; Moraes-Neto, A.; Svidinicki,
M.C.C.M.; Artoni, R.F. 2008. Karyotypic variability in Iheringichthys labrosus
(Teleostei, Pimelodidae) from the Tibagi River basin (Paraná State, Brazil).
Genetics and Molecular Research, 7 (3): 718-724.
Ruffino, M.L.; Soares-Silva, E.C.; Silva, C.O.; Barthem, R.B.; Silva, V.B.; Estupinan,
G.; Pinto, W. 2006. Estatística pesqueira do Amazonas e Pará/2003. IBAMA/Pró-
Várzea, Manaus, Amazonas. 76 pp.
Sanches, S. 2006. Estudios citogeneticos em peces de la família Pimelodidae
(Pisces, Siluriformes) de la cuenca del rio Paraná, Argentina. Ph.D. Dissertation.
Universidad Nacional de Córdoba, Argentina.
Santos, G.M.; Ferreira, E.J.G.; Zuanon, J.A.S. 1991. Ecologia de Peixes da
Amazônia. In: Val, L.A.; Figliuolo, R.; Feldberg, E. (eds) Bases Científicas de
Preservação e Desenvolvimento da Amazônia: Fatos e Perspectivas, vol.I.
Instituto Nacional de Pesquisas da Amazônia, Manaus, Amazonas. p. 263-280.

56
Santos, G.M.; Ferreira, E.J.G.1999. Peixes da Bacia Amazônica. In: Lowe-McConnell,
R.H. Estudos ecológicos de comunidades de peixes tropicais. Editora da
Universidade de São Paulo, São Paulo, São Paulo. p.345-373.
Santos, G.M.; Ferreira, E.J.G.; Zuanon, J. 2006. Peixes comerciais de Manaus.
IBAMA/Pró-Várzea, Manaus, Amazonas. 141 pp.
Silva, L.V.B.; Abreu, M.F.; Souza, I.L. Venere, P.C. 2004. Caracterização
cromossômica de Hemissorubin platyrhynchos, Sorubim lima e Pimelodus blochii
(Pisces, Siluriformes, Pimelodidae) do Médio Araguaia. In: Anais do X Simpósio
de Citogenética e Genética de Peixes, Natal, RN. 132pp.
Souza, A.B.; Fonseca, C.G.; Pinehiro, L.E.L.; Ribeiro, L.P. 1992. Estudos
citogenéticos preliminares em Pseudoplatystoma corruscans (Siluriformes,
Pimelodidae) da bacia do rio Paraguai. In: Anais do IV Simpósio de Citogenética
Evolutiva Aplicada a Peixes Neotropicais. Rio de Janeiro, RJ. p.28.
Souza, A.C.P.; Nagamachi, C.Y. Rissino, J.D. Junior, J.R.C. Barros, R.M.S.;
Pieczarka, J.C. 2000. Descrição cariotípica de Pimelodus cf. maculatus
(Siluriformes, Pimelodidae). In: Anais do VIII Simpósio de Citogenética e Genética
de Peixes. Manaus, AM. p. 90.
Souza, L.; Giuliano-Caetano, L.; Dias, A.L. 2003. Karyotypic study of three species of
Pimelodus (Pisces, Pimelodidae), from the Paraguai River Basin. Cytologia, 68(4):
345-350.
Souza, L.; Giuliano-Caetano, L.; Dias, A.L. 2004. Banding chromosome pattern of two
species of Pimelodus (Siluriformes, Pimelodidae) from Paraná river basin of
Brazil. Folia Biologica, 52(3-4): 165-169.

57
Sumner, A.T. 1972. A simple technique for demonstrating centromeric
heterochromatin. Experimental Cell Research, 75: 304-306.
Swarça, A.C.; Giuliano-Caetano, L. Dias, A.L. 1999. Cytogenetic characterization
through chromosomic banding of Pinirampus pirinampu from the Tibagi River
Basin, PR/Brazil. Caryologia, 52: 31-35.
Swarça, A.C.; Giuliano-Caetano, L.; Dias, A.L. 2001a. Analyses of nucleolus organizer
regions and heterochromatin of Pimelodus maculatus. Genetica, 1(110): 97-100.
Swarça, A.C.; Cestari, M.M.; Giuliano-Caetano, L.; Dias, A.L. 2001b. Cytogenetic
characterization of the large South American Siluriform fish species Zungaro
zungaro (Pisces, Pimelodidae). Chromosome Science, 5: 51-55.
Swarça, A.C.; Fenocchio, A.S. Cestari, M.M. Dias, A.L. 2005. Chromosomal
divergence among populations of Pseudoplatystoma corruscans (Pisces,
Pimelodidae): I. Evidences obtained from conventional giemsa staining, C and G
banding. Ichtholyological Exploration of Freshwaters, 16(4): 325-330.
Swarça, A.C.; Fenocchio, A.S.; Cestari, M.M.; Dias, A.L. 2006. First report of a sex
chromosome system in a Sorubiminae fish, Steindachneridion sp. (Pimelodidae),
with an uncommon large Y chromosome. Mitotic and meiotic analysis. Cytogenetic
and Genome Research, 112(3-4): 325-328.
Swarça. A.C.; Fenocchio, A.S.; Dias, A.L. 2007. An update cytogenetic review for
species of the families Pseudopimelodidae, Pimelodidae and Heptapteridae
(Pisces, Siluriformes). Suggestion of a cytotaxonomical classification. Caryologia,
60(4): 338-348.

58
Teixeira, A.S.; Jamielson, A.; Raposo, J.C.P. 2002. Transferrin polymorphism in
Central Amazon populations of pescada, Plagioscion squamosissimus. Genetics
and Molecular Research, 1 (3): 216-226.
Teixeira, A.S.; Nakayama, C.M.; Porto, J.I.R.; Feldberg, E. 2006. Esterase-D and
chromosome patterns in Central Amazon piranha (Serrasalmus rhombeus Linnaeus,
1766) from lake Catalão. Genetics and Molecular Biology, 29(3): 498-502.
Terencio, M.L.; Almeida, M.C. Artoni, R.F. 2001. Citogenética de Pimelodus ortmanni
uma espécie de mandi endêmica ao rio Iguaçu. Anais do 47º Congresso Nacional
de Genética, Águas de Lindóia. CD-room.
Terencio, M.L.; Schneider, C.H.; Gross, M.C.; Silva, A.M.; Feldberg, E.; Porto, J.I.R.
2008. Comparative cytogenetics of Carnegiella marthae and Carnegiella strigata
(Characiformes, Gasteropelecidae) and description of a ZZ/ZW sex chromosome
system. Genetics and Molecular Biology, 31(1): 231-234.
Toledo, V.; Ferrari, I. 1976a. Estudo citogenético de Pimelodella sp. e Rhamdia hilarii
(Pimelodinae, Pimelodidae, Pisces): Cromossomo marcador. Científica, 4: 120-
123.
Toledo, V.; Ferrari, I. 1976b. Estudos citogenéticos de três espécies do gênero
Pimelodus (Pisces, Pimelodidae). Científica, 4: 101-106.
Treco, F.R.; Malabarba, L.R.; Giuliano-Caetano, L.; Dias, A.L. 2008. Cytogenetic
study of two species of the family Pimelodidae (Siluriformes) collected in lago
Guaíba, Rio Grande do Sul, Brazil. Neotropical Ichthyology, 6(1): 87-92.
Treco, F.R.; Dias, A.L. 2009. Karyotypes of two species of the genus Pimelodus
(Siluriformes, Pimelodidae). Folia Biologica-Krakow, 57(2): 43-48.

59
Vari, R.P.; Malabarba, L.R. 1998. Neotropical ichthyology: An overview In: Malabarba,
L.R.; Reis, R.E.; Vari, R.P.; Lucena, Z.M.S.; Lucena, C.A.S. (Eds). Phylogeny and
classification of Neotropical fishes. Editora da Pontifícia Universidade Católica,
Porto Alegre, Brasil. p. 1-11.
Vasconcelos, C.; Martins–Santos, I.C. 2000. Chromosome polymorphism in species of
the Pimelodidae family (Pisces, Siluriformes). Hereditas, 132: 103-109.
Vicari, M.R.; Moreira-Filho, O.; Artoni, R.F.; Bertollo, L.A.C. 2006. Basic and molecular
cytogenetics in freshwater Cichlidae (Osteteichthyes, Perciformes). Karyotic and
conservationism and divergence. Caryologia, 59: 260-266.
Vissotto, P.C.; Foresti, F.; Oliveira, C. 1999. Supernumerary chromosomes in two
species of the family Pimelodidae (Teleostei, Siluriformes). Chromosome Science,
3: 9-13.