alteraÇÕes das fibras musculares estriadas …livros01.livrosgratis.com.br/cp007566.pdf · 5...
TRANSCRIPT

USC
UNIVERSIDADE DO SAGRADO CORAÇÃO
ALTERAÇÕES DAS FIBRAS MUSCULARES ESTRIADAS ESQUELÉTICAS DESNERVADAS E
REINERVADAS EM DIFERENTES PERÍODOS
MARCELO MORGUETI
Bauru-2005

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

1
Dissertação de Mestrado – Morgueti, M.
USC UNIVERSIDADE DO SAGRADO CORAÇÃO
ALTERAÇÕES DAS FIBRAS MUSCULARES ESTRIADAS ESQUELÉTICAS DESNERVADAS E
REINERVADAS EM DIFERENTES PERÍODOS
MARCELO MORGUETI
Dissertação apresentada à Pró-reitoria de Pesquisa e Pós-graduação da USC - Universidade do Sagrado Coração, como parte dos requisitos para obtenção do título de Mestre, no Programa de Pós-graduação em Odontologia. Área de Concentração: Biologia Oral Orientador: Prof. Dr. Jesus Carlos Andreo
Bauru- 2005

2
Dissertação de Mestrado – Morgueti, M.
USC
UNIVERSIDADE DO SAGRADO CORAÇÃO
NOTAS DA BANCA EXAMINADORA DA DEFESA DO MESTRADO
MARCELO MORGUETI
ALTERAÇÕES DAS FIBRAS MUSCULARES ESTRIADAS ESQUELÉTICAS DESNERVADAS E REINERVADAS EM
DIFERENTES PERÍODOS
DATA DA DEFESA: ____/_____/_____
PROF. DR. JESUS CARLOS ANDREO
AVALIAÇÃO: _________________ ASSINATURA________________
PROF. DR. ANTONIO DE CASTRO RODRIGUES
AVALIAÇÃO: _________________ ASSINATURA________________
PROFª. DRª. ANGELA M. O. KINOSHITA
AVALIAÇÃO: _________________ ASSINATURA________________

3
Dissertação de Mestrado – Morgueti, M.
DEDICATÓRIAS
Á DEUS,
Pela presença constante em minha vida, sempre iluminando os melhores caminhos
a seguir. Tudo o que conquistei até hoje, foi pela força de SUA luz divina.
“SEM ELE NADA É POSSÍVEL”;
AOS MEUS PAIS,
DORIVAL E SÔNIA, pelos ensinamentos dados durante todos esses anos, que
auxiliaram na construção do meu alicerce de vida e são meus grandes exemplos e
apoio em todos os momentos;
AO MEU IRMÃO
LUCIANO, por todos esses anos de convivência e amizade entre nós como
alicerce de grande união;
Á FLÁVIA, minha companheira de todas as horas, pelo apoio, incentivo,
companheirismo, carinho e paciência. Sem sua ajuda este trabalho não se
realizaria, obrigada.
AO MARCOS E BRUNA, pelo carinho e compreensão. Que este trabalho lhes
sirva de exemplo para seus futuros.

4
Dissertação de Mestrado – Morgueti, M.
AGRADECIMENTOS
AO PROF. DR. JESUS CARLOS ANDREO, orientador que sempre me recebeu
com atenção e carinho, transmitindo-me valiosos ensinamentos;
AO PROF. DR. ANTÔNIO CASTRO RODRIGUES, que juntamente com meu
orientador, colaborou na elaboração deste trabalho com dedicação e carinho;
Á PROF. DRA. ANGELA MITIE OTTA KINOSHITA, professora de Técnicas Físicas
Aplicadas ao Complexo Bucomaxilofacial da Universidade do Sagrado Coração –
Bauru, PARTICIPANTE DA MINHA BANCA DE DEFESA DE MESTRADO, o meu
muito obrigado pela participação ativa;
A UNIVERSIDADE DO SAGRADO CORAÇÃO, na pessoa do Sr. Pró-Reitor de
Pesquisa e Pós-Graduação Dr. JOSÉ JOBSON DE ANDRADE ARRUDA, o meu
muito obrigada pela realização de mais um sonho;
A COORDENADORIA DO CURSO DE PÓS-GRADUAÇÃO EM BIOLOGIA ORAL
PROF. DR. SÉRGIO CATANZARO, pela sua amizade, dedicação, ensinamentos e
orientação;
A SECRETARIA DO CURSO DE PÓS-GRADUAÇÃO, na pessoa da Srta. ANGELA
MORAES, pela dedicação durante o decorrer dos anos de curso;
À PROF.ª MARIA BEATRIZ DE BARROS MORAES TRAZZI, PRÓ-REITORA DE
AÇÃO COMUNITÁRIA E UNIMAR - UNIVERSIDADE DE MARÍLIA, pela dedicação
e presteza na correção deste trabalho;

5
Dissertação de Mestrado – Morgueti, M.
AO PROF. DR. PADOVANI, Prof. da disciplina de Bioestatística da USC –
Universidade do Sagrado Coração – Bauru, pela presteza e dedicação na
confecção da parte estatística deste trabalho, o meu muito obrigado
AOS COLEGAS DO LABORATÓRIO DE HISTOLOGIA DA UNIVERSIDADE DO
SAGRADO CORAÇÃO – BAURU, pela elaboração da parte histológica deste
trabalho;
AOS MEUS COLEGAS DO MESTRADO, EM ESPECIAL À DANIELA VIEIRA
BUCHAIM E FÁBIO FREIRIA BARBOSA, pelo companheirismo e amizade durante
o curso;
AOS FUNCIONÁRIOS DO BIOTÉRIO, DA UNIVERSIDADE DE MARÍLIA,
UNIMAR, ROBERTO E NADIR, pela atenção dispensada durante a realização
deste trabalho;
A COORDENADORA DO CURSO DE FISIOTERAPIA DA UNIVERSIDADE DE
MARÍLIA, UNIMAR, PROF. DRA. CECÍLIA BARROS, pela compreensão durante
minhas ausências;
AO DIRETOR DO CURSO DE CIÊNCIAS DA SAÚDE DA UNIVERSIDADE DE
MARÍLIA – UNIMAR, PROF. DR. ARMANDO CASTELLO BRANCO JUNIOR, pela
compreensão durante o curso de mestrado;
Ao amigo SALVIANO OLIVEIRA, secretário da Pró-Reitoria da Graduação da
UNIMAR – Universidade de Marília, pela dedicação e desempenho na confecção
dos gráficos deste trabalho.
A TODOS QUE DE ALGUMA FORMA CONTRIBUÍRAM PARA A ELABORAÇÃO E
CONCLUSÃO DESTE TRABALHO, O MEU MUITO OBRIGADO.

6
Dissertação de Mestrado – Morgueti, M.
“EMBORA NINGUEM POSSA
VOLTAR ATRÁS E FAZER UM
NOVO COMEÇO, QUALQUER UM
PODE COMEÇAR AGORA E
FAZER UM NOVO FIM”.
(CHICO XAVIER)

7
Dissertação de Mestrado – Morgueti, M.
RESUMO
O objetivo deste trabalho foi realizar um estudo experimental sobre a desnervação
e a reinervação do músculo sóleo de rato e observar sob critérios morfológicos e
morfométricos, em qual período pós reinervação pode se obter um melhor padrão
do referido músculo. Para isso foram utilizados vinte e sete animais, divididos
aleatoriamente em três grupos de nove cada um deles. No grupo G1(grupo zero
dia), foi realizada uma incisão entre o túber isquiático e o trocanter maior, onde o
nervo ciático foi dissecado e seccionado na altura do terço médio da coxa e
imediatamente submetido à técnica de reparo através da neurorrafia término
terminal. No grupo G2(grupo dez dias), o mesmo procedimento foi realizado, mas
após o seccionamento do nervo, foi colocado no coto proximal um tubo de
polietileno de fundo cego, fixado através de fio 6-0. Transcorridos dez dias os
animais foram novamente anestesiados para serem submetidos à reinervação
através da técnica descrita no grupo 1. No grupo G3 (grupo 30 dias), o mesmo
procedimento foi realizado, somente diferenciando o tempo e espera de trinta dias,
para se proceder a técnica de reparo. A sutura da pele foi realizada com o fio de
nylon 5-0. Os animais foram sacrificados com superdosagem de anestésicos
sempre decorridos 10 dias de reparo para todos os grupos. O músculo retirado foi
primeiramente colocado por alguns minutos em formol tamponado a 10% para dar
forma à estrutura e em seguida foi acondicionado em recipiente apropriado com
formol 10% por vinte e quatro horas. Após este período, os músculos foram
lavados em água corrente por mais vinte e quatro horas e novamente colocados
nos mesmos recipientes contendo álcool a 70%, até serem levados para
procedimentos laboratoriais de rotina. Os resultados obtidos mostraram o sucesso
da técnica de neurorrafia término terminal na recuperação do nervo periférico sem
perda tecidual, onde em todos os grupos, as fibras musculares se apresentaram
reinervadas e os fusos neuromusculares se mantiveram com seus números de
fibras intrafusais normais, compreendendo de 5 a 6 fibras em cada um fuso.
Palavras Chaves: desnervação, reinervação, músculo sóleo, neurorrafia término
terminal.

8
Dissertação de Mestrado – Morgueti, M.
ABSTRACT
The objective of this work was to carry out an experimental study on the
desnervation and the reinervation of the rat muscle soleus and also observe under
morphologic and morphometric criteria, in which period after reinervation you can
get a better standard of the related muscle. In order for that, twenty seven animals
were used, aleatory divided in three groups of nine. In the group G1 (group zero
day), it was carried out an incision between the tuber ischiatic and the trochanter
bigger, where the sciatic nerve was dissected and sectioned in the height part of
mid-thigh and immediately submitted to the technique of repair through the
neurorhaphy terminus terminal. In the group G2 (group 10 days), the same
procedure was carried out, but after the nervous section it was placed in the stump
proximalis a polyethylene pipe of the blind fundus, fixed through wire 6-0. After ten
days, the animals were again anesthetized to be submitted the reinervation through
the described technique in group 1. In the group G3 (group 30 days), the same
procedure was carried out, only differentiating the time and the wait of 30 days, to
proceed the repair technique. The suture of the skin was made with the nylon wire
5-0. The animals had been sacrificed with superdosage of anesthetic, always after
10 days of repair to all groups. The removed muscle was first placed for some
minutes in formaldehyde at 10% , to give form to the structure and after it was
conditioned in appropriate container with formaldehyde 10% for twenty and four
hours. After this period, the muscles were washed in running water for twenty and
four hours and placed again in the same containers with alcohol 70%, until being
taken to laboratorial procedures of routine. The gotten results had shown the
success of the neurorrhaphy terminus terminalis in the recovery of the peripheral
nervous without tissue loss, where in all groups the muscle fiber presented
reinervation and the neuromuscular spindles, had kept with its normal intrafusal
fiber numbers, understanding of 5 to 6 fiber in each one spindle.
Key words: desnervation, reinervation, muscle soleum, neurorhaphy terminus
terminalis.

9
Dissertação de Mestrado – Morgueti, M.
ÍNDICE
RESUMO ...................................................................................................................................7
ABSTRACT ...............................................................................................................................8
1. INTRODUÇÃO ....................................................................................................................10
2. REVISÃO DE LITERATURA...............................................................................................11
3. PROPOSIÇÃO......................................................................................................................18
4-MATERIAL E MÉTODOS ....................................................................................................19
4.1-AMOSTRA ........................................................................................................................19 4.2-PROCEDIMENTOS............................................................................................................19 Figura 1 - Depilação por tração manual dos pêlos da face lateral da coxa esquerda. ....................21 Figura 2 - Incisão entre o túber isquiático e o trocanter maior.....................................................21 Figura 3 - Nervo ciático dissecado na altura do terço médio da coxa. ..........................................22 Figura 4 - Injúria causada no nervo ciático. ...............................................................................22 Figura 5 - No coto proximal foi colocado um tubo de plástico rijo de fundo cego para prevenir a reinervação...............................................................................................................................23 Figura 6 - Técnica de reinervação através da neurorrafia término terminal..................................23 Figura. 7- Sutura local com o fio de nylon 5-0. ...........................................................................24
5. RESULTADOS.....................................................................................................................24
5.1 ANÁLISE MORFOLÓGICA ......................................................................................................25 Figura 8 – Corte transversal das fibras musculares estriadas esqueléticas do músculo sóleo dos animais do grupo G1.................................................................................................................26 Figura 9 - Corte transversal das fibras musculares estriadas esqueléticas do músculo sóleo dos animais do grupo G2.................................................................................................................27 Figura 10 – Corte transversal das fibras musculares estriadas esqueléticas do músculo sóleo dos animais do grupo G3.................................................................................................................28 5.2 ANÁLISE MORFOMÉTRICA ....................................................................................................29 Figura 11 – Gráfico de barras referentes às áreas de todos os animais pesquisado do grupo G1....30 Figura 12 - Gráfico de barras referentes às áreas de todos os animais pesquisado do grupo G2. ...30 Figura 13 - Gráfico de barras referentes às áreas de todos os animais pesquisado do grupo G3....31 Figura 14 – Gráfico das médias dos animais de todos os grupos pesquisados.. .............................31
6. DISCUSSÃO ........................................................................................................................32
7. CONCLUSÕES.....................................................................................................................38
8. REFERÊNCIAS BIBLIOGRÁFICAS.....................................................................................39

10
Dissertação de Mestrado – Morgueti, M.
1. INTRODUÇÃO
As fibras musculares estriadas esqueléticas apresentam a forma de um
longo cilindro multinucleado quando examinadas em nível de microscopia óptica.
Tais fibras agrupadas paralelamente constituem os chamados fascículos de fibras
musculares, visíveis inclusive a olho nu em músculos de pouca espessura, por
exemplo, os músculos do bulbo ocular. Esses fascículos organizam-se de forma
variada em relação a seus tendões de tal forma que os músculos estriados
esqueléticos podem ser classificados de várias maneiras segundo este critério.
Os fascículos de fibras musculares juntos constituem o chamado ventre
muscular, estrutura de cor vermelha escuro, com capacidade contrátil que é
envolta por tecido conjuntivo que tem por função proteger, individualizar e permitir
um melhor deslizamento de um ventre muscular contra outro durante as contrações
de vários grupamentos musculares na realização de determinado movimento. Os
tecidos conjuntivos presentes nos músculos estriados esqueléticos constituem seu
estroma e são designados de diferentes maneiras para facilitar seu estudo.
Assim, a camada mais superficial, quando observada em corte
transversal é denominada epimísio; o septo que divide o ventre em fascículos é
denominado perimísio e finalmente há uma delicada trama de fibras reticulares que
envolvem cada fibra muscular denominada endomísio.
Conforme descrito os núcleos das fibras estriadas esqueléticas são
numerosos, podendo em certas fibras de alguns centímetros de comprimento,
conter centenas de mionúcleos. Os mionúcleos apresentam-se de forma alongada
sempre com seu maior eixo na direção do maior eixo da fibra muscular.
A posição das fibras musculares esqueléticas pode variar dependendo do
tipo de músculo e espécie de animal, muito embora na maioria dos mamíferos
adultos elas se localizem na periferia das fibras imediatamente abaixo do
sarcolema.
Com isso pareceu-nos de grande interesse avaliar os resultados da
desnervação e reinervação do músculo sóleo de ratos, de acordo com a literatura
especificada para podermos concluir qual é a melhor época de se realizar a
tentativa de reinervação, ou seja, imediatamente após a cirurgia de seu nervo
motor ou em um período posterior a tal lesão.

11
Dissertação de Mestrado – Morgueti, M.
2. REVISÃO DE LITERATURA
De acordo com MAURO (1964), as células satélites ou mioblastos
apresentam a capacidade de se dividirem e de se fundirem na ausência de
inervação em meio de cultura e em vivo, porém a inervação é necessária para a
maturação e a manutenção das novas fibras (BISCHOFF, 1974; MUSSINI et al.,
1987; GULATI, 1988; WHALEN et al., 1990; SKETELJ et al., 1991; SESODIA &
CULLEN, 1991).
As fibras presentes em uma unidade motora pertencem sempre do mesmo
tipo, lembrando ainda que a modulação de tais fibras musculares esteja a cargo da
fibra nervosa. Vários experimentos têm comprovado tal fato, especialmente
experimentos envolvendo a inervação cruzada, “cross inervation” (BULLER et al.,
1965; MAMMAERTS et al., 1969 e BARANY & CLOSE, 1971). Assim, para alguns
autores o número de fibras musculares num determinado músculo é estabelecido
ao nascimento (CHYAKULAS & PAULY, 1965; MAXWEL et al., 1974) enquanto
que para outros, em condições de exercício programado, pode haver um discreto
aumento do número de fibras musculares em animais adultos (GONYEA et al.,
1986).
Com relação às características contráteis e o tipo de metabolismo, as
fibras musculares estriadas esqueléticas podem ser classificadas em três tipos já
consagrados pela literatura: FG (fast-twitch-glicolytic); FOG (fast-twitch-oxidative-
glycolitic) e SO (slow-twitch-oxidative) (PETER et al., 1972).
A porcentagem de células satélites em relação aos mionúcleos, em
músculos normais é de cerca de 8% (SCHMALBRUCH & HELLHAMER, 1977),
dobram dentro dos primeiros dez dias após desnervação e então diminuem para
menos de ¼ do normal.
O papel da inervação, todavia, não deve estar limitado a estes eventos
durante a regeneração muscular.
A capacidade proliferativa dos mioblastos em músculos jovens é nervo
dependente (BONNER, 1978, 1980, 1987), e a proliferação dos mioblastos em
meio de cultura é estimulada por fatores tróficos produzidos pelos nervos e
contidos em extratos embrionários normalmente adicionados a tal meio (OH &
MARKELONIS, 1980; BONNER & ADAMS, 1982; POPIELA et al. 1984).

12
Dissertação de Mestrado – Morgueti, M.
Já no período senil e em determinadas patologias (DUBOWITZ, 1985;
ADACHI & SHER, 1990; DAL PAI et al., 1991) pode haver redução do número de
fibras musculares.
Em músculos de rãs as fibras musculares desnervadas por quase quatro
anos desaparecem totalmente (ANZIL & WERNIG, 1989).
As fibras musculares estriadas esqueléticas desnervadas atrofiam,
degeneram e eventualmente morrem sendo substituídas por novas fibras as quais
novamente degeneram. A força do músculo sóleo de rato desnervado declina em
escala logarítmica durante os quatro meses iniciais de desnervação ficando
constante a partir deste tempo até por seis meses. Isto sugere que a atrofia,
degeneração e perda de fibras de um lado e a formação de novas fibras do outro
lado estão balanceadas após quatro meses (AL-AMOOD et al., 1991;
SCHMALBRUCH et al., 1991).
Estas observações são importantes e relevantes para o entendimento das
alterações patológicas nas doenças musculares em humanos. Músculos
esqueléticos em pacientes com Distrofia de Duchenne não apresentam somente
necrose, mas também uma extensiva regeneração a qual, contudo não é
compensada pelo desgaste muscular, pois aquelas novas fibras que não se
estendem através da zona da placa motora nunca serão inervadas e desaparecem
(SCHMALBRUCH, 1992).
Por outro lado condições neuropatológicas crônicas podem causar quebra
e regeneração de fibras submetidas a carga de trabalho excessiva o que leva às
alterações miopatológicas secundárias (JENNEKENS, 1992).
SCHMALBRUCH & LEWIS (1994), estudaram as propriedades
morfológicas e fisiológicas no músculo sóleo do rato e compararam-nas com
aquelas de músculos desnervados experimentalmente. Assim o papel
desempenhado pelas células satélites na recuperação do músculo está
diretamente relacionado à sua capacidade proliferativa. Isto também explica a
diminuição no poder de contração após a desnervação que nos primeiros três
meses decresce logaritmicamente para se manter estável até aproximadamente
vinte e quatro semanas pós desnervação. Este processo tem sido atribuído à
exaustão do pool de células satélites no processo de regeneração (RODRIGUES &
SCHMALBRUCH, 1995).

13
Dissertação de Mestrado – Morgueti, M.
MARQUES et al., (1996), estudaram o tempo de regeneração motora do
músculo EDL (extensor longo dos dedos), depois do nervo periférico ser
transeccionado e entubado. Em ratos adultos após transeccionado o nervo ciático
o coto proximal e o distal, foram suturados com um tubo de polietileno longo e no
espaço de duas a quarenta semanas, o músculo EDL, foi retirado e submetido ao
processo histoquímico para visualizar os locais ricos em colinesterase. Concluíram
que as fibras musculares apresentaram um aumento da concentração de colágeno
e fibroblastos nos locais onde ocorreram as sinapses. O processo de reinervação
começou após quatro semanas da implantação do tubo, e doze semanas após a
realização da cirurgia, toda junção neuromuscular voltou ao normal e de doze a
quarenta semanas toda a musculatura motora voltou a ser inervada.
YOSHIMURA & HARII (1997), estudaram os eventos celulares e
moleculares coincidentes com os músculos desnervados, principalmente a
mudança regenerativa, observando o papel das células satélites e a possibilidade
de apoptose como um mecanismo de perda durante a atrofia do músculo
desnervado. Concluíram que a maioria das células satélite se fundiu às fibras
desnervadas com o propósito de arrumá-las, ao invés de se fundirem e formarem
novas fibras como um mecanismo compensatório. Em relação a apoptose concluiu-
se que ela contribui para a perda de mionúcleos associados com a atrofia pela
desnervação.
De acordo com RODRIGUES (1998), a porcentagem de células satélites
rapidamente diminui no músculo sóleo de rato em regeneração desprovido de
inervação, como também que, a desnervação de músculos intactos causa a
degeneração e a perda de fibras, enquanto que o comportamento das células
satélites nestas condições não foi conhecido.
DEVOR & FAULKNER (1999), estudaram a hipótese específica após um
protocolo de prolongamento das contrações, onde as fibras recém regeneradas
foram tratadas com bupivacaina injetada nos músculos EDL de ratos jovens e
adultos. Os resultados demonstraram um déficit menor de força e também uma
menor quantidade de fibras danificadas nos músculos que não foram tratados com
a bupivacaina e ainda, que a regeneração das fibras musculares tanto nos animais
jovens como nos velhos requerem a mesma resistência para o dano causado.

14
Dissertação de Mestrado – Morgueti, M.
DESGRANGES et al., (1999), estudaram um polímero sintético chamado
de RGTA (agente regenerador) que pode estimular tecidos quando aplicado no
local do dano por agentes diversos. Em músculos esmagados, o RGTA foi
utilizado para acelerar a regeneração e a reinervação e tanto a isquemia quanto a
desnervação o músculo EDL tratados com o RGTA diferiu em vários aspectos: o
pós-operatório foi diminuído e a área de tecido de fibrose no meio das fibras foi
reduzida; o número de fibras de músculos intactos foi maior que o dobro. Estes
resultados sugerem que o RGTA protege as fibras dos músculos degenerados e
preserva o estado diferenciado das fibras sobreviventes. Com isso ficou
demonstrado que o polímero pode prevenir algum tipo de dano resultante de
músculo em isquemia.
Recentemente, alguns pesquisadores têm proposto uma classificação
mais ampla levando-se em conta critérios mais específicos, e desta forma
classificando as fibras em unidades morfofuncionais (BEGO et al., 1996).
RODRIGUES et al., (1999), relataram que diferentes tipos de fibras
estriadas esqueléticas encontram-se na maioria das vezes espalhadas pelo ventre
muscular a chamada distribuição em mosaico, embora em alguns músculos de
alguns mamíferos, como o músculo sóleo do porco, por exemplo, encontramos
fibras do tipo SO agrupadas constituindo os chamados grupos metabólicos.
WANEK & SNOW (2000), realizaram uma pesquisa para entender porque
a atrofia muscular é limitada quando ocorre a imobilização do membro. O trabalho
foi realizado em ratos adultos no músculo sóleo, no qual o animal era imobilizado
com gesso durante duas a dez semanas. Após esse período os animais eram
sacrificados e o músculo sóleo examinado para evidenciar as fibras em
degeneração ou regeneração e quantificar os núcleos das células satélites.
Detectou a ocorrência de dano na fibra muscular secundária, inclusive uma atrofia
miofibular, com rompimento do disco e funções. O número de células satélites
diminuiu significantemente de 6,4% para 3,3% entre oito a dez semanas de
imobilização. Somente 1% das fibras parecia estar regenerado.
IJKEMA-PAASSEN et al., (2001), estudaram a morfologia do músculo e a
distribuição dos tipos de fibras depois do nervo ciático ter sido transeccionado no
membro de ratos.

15
Dissertação de Mestrado – Morgueti, M.
Eles realizaram seccionamento do nervo ciático em 12 mm próximo a
bifurcação do nervo tibial e fibular, depois esse nervo era invertido e reimplantado
como um enxerto de nervo autólogo. Após um período de sete, quinze e vinte e
uma semanas, os músculos gastrocnêmio lateral, o tibial anterior e o músculo sóleo
foram dissecados e a distribuição dos tipos de fibras foi estudada. O estudo
demonstrou que depois de sete semanas as áreas seccionadas foram diminuídas
em todos os músculos. Nos músculos gastrocnêmios, tibiais anteriores e músculo
sóleo o número de fibras permaneceram inalterados. Por volta de sete a quinze
semanas o número de fibras tipo II diminuiu consideravelmente voltando ao normal
na 20ª-21ª semanas. Como resultados finais concluíram que a reinervação não
seletiva e os reajustes posteriores por regressão de inervação de vários nervos
servem em parte para explicar as mudanças nas distribuições da vários tipos de
fibras.
THIERRY LAUNAY et al., (2001), realizaram o estudo para demonstrar o
padrão de expressão do myf-5, o myod e o mrf4 (proteínas) durante a regeneração
do músculo sóleo em ratos depois de causar danos com o cardiotoxin, utilizando a
hibridação in situ e também para investigar a influência da inervação durante o
processo de degeneração e regeneração completa. Concluíram que nas
experiências de regeneração com o músculo sóleo desnervado o myf-5, o myod e
o mrf4 (proteínas) apareceram em células satélites derivadas de mioblastos
resultantes da primeira regeneração do músculo analisado.
MINAMOTO et al., (2001), avaliaram os aspectos e as mudanças
morfológicas na área e a incidência dos tipos de fibras musculares, quando
provocadas contusões periódicas no músculo tibial anterior de ratos. Foram oito
traumas sucessivos, um por semana, durante oito semanas, sendo avaliados
depois de quatro meses após a última contusão. Os resultados demonstraram que
o peso muscular era diminuído quando comparado ao músculo contralateral intacto
e mostrava presença abundante de fibras em “splitting” com núcleos centralizados,
principalmente, na porção profunda. Os músculos danificados apresentavam uma
incidência maior de fibras semidiferenciadas quando comparada aos músculos
intactos.

16
Dissertação de Mestrado – Morgueti, M.
O estudo demonstrou que as fibras musculares eram principalmente
danificadas na porção profunda, provavelmente devido à compressão feita com a
tíbia; as contusões periódicas não mudaram a porcentagem do tipo I e II das fibras
musculares e as áreas que sofreram as contusões levaram quatro meses para se
regenerarem completamente.
MORACZEWSKI et al., (2002), estudaram através de um modelo de
regeneração induzido por regeneração à implicação de CPK (proteína quinase) em
conserto de músculo após um dano qualquer. Foi utilizado o músculo sóleo e o
músculo EDL que possuem regeneração muscular diferente. Após o
esmagamento, foi utilizada a atividade de CPK durante quatorze dias para se
analisar a regeneração muscular. A degeneração ocorreu logo após o
esmagamento e estavam associados à enzima em ambos os músculos lesados. A
reconstrução das fibras do músculo EDL era associada com um aumento da
atividade de CPK, onde por volta do sétimo dia o músculo se regenerava. Ficou
demonstrado que o CPK é utilizado no processo regenerativo em diferentes
tempos e locais e sugere que possua funções distintas. A eventual exaustão do
pool de células satélites pode ser responsável posteriormente pela ocorrência do
declíneo da força muscular como é visto na poliomielite.
A área da fibra (média da área seccionada transversalmente) em músculos
regenerados desnervados foi de 139 µm2 depois de dez dias; a força específica
nesses músculos foi normal. Mais tarde, com o início da degeneração e atrofia, a
força específica diminuiu. O tamanho médio da fibra foi somente de 26µm2 depois
de setenta dias.
Nesse período, sinais de quebra e fibras em regeneração imaturas foram
encontradas em ambos os modelos experimentais. Os músculos desnervados já
continham poucos miotubos aos dez dias e aos quarenta dias as fibras musculares
em regeneração já eram distintas.
Por outro lado a desnervação de alguns músculos após setenta dias,
triplica a porcentagem de células satélites enquanto que em outros diminuem para
a metade do normal. Isto vem demonstrar que a regeneração de músculos
desprovidos de inervação alcança seu alto grau de diferenciação estrutural e então
atrofiam, degeneram e desaparecem.

17
Dissertação de Mestrado – Morgueti, M.
Tais fibras são substituídas por novas fibras, as quais não alcançam o
tamanho das originais, presumivelmente porque o pool de células satélites sem
inervação se torna exaurido.
A lenta e não sincronizada quebra das fibras musculares desnervadas do
músculo sóleo induzem a proliferação de células satélites como em músculos
necróticos; aqueles músculos que apresentem menos células satélites do que os
normais após setenta dias possam já estar entrando em processo de exaustão.
Investigações preliminares de músculos sóleos desnervados por sete
meses sugerem que o número de células satélites normalmente cai para menos de
quinze, isto é, para cerca de 10% do normal. Estes resultados indicam que a
proliferação de células satélites e a habilidade das fibras musculares para se
regenerarem é nervo dependente.
Assim, o fato de que músculos desnervados por longo período perderem a
sua capacidade de regeneração depois de reinervado pode ser devido à exaustão
do pool de células satélites muito mais do que por sua fibrose ou substituição das
fibras musculares por adipócitos.
Segundo DOLFINI et al., (2002), até dezesseis semanas após a
desnervação o músculo comporta-se de maneira oscilatória entre tentativas de
regeneração e degeneração, com o auxílio das células satélites e conseqüente
diminuição na quebra do DNA. Reafirmou-se a idéia de que neste período a perda
de fibras por um lado e a tentativa de regeneração de outro, estão equilibradas.
Estes autores sugerem frente a esses resultados que o melhor período para se
obter uma melhor recuperação de um músculo desnervado deve ser no período
onde é observada a menor quebra de DNA, ou seja, entre o décimo segundo e a
décima sexta semana pós desnervação.
De acordo com DAHLIN (2004), um dano causado no nervo periférico, tem
um impacto profundo na vida. Este dano é considerado complexo porque envolve
ambos componentes periféricos e centrais do sistema nervoso. Este dano pode
comprometer as células de Schwann em condicionamento regenerativo e
proliferativo respectivamente.

18
Dissertação de Mestrado – Morgueti, M.
3. PROPOSIÇÃO
Foi propósito deste trabalho estudar experimentalmente o processo de
desnervação e reinervação do músculo sóleo de ratos e observar sob critérios
morfológicos e morfométricos, em qual período pós lesão ( zero, dez e trinta dias)
se pode obter um melhor padrão de recuperação desse músculo.

19
Dissertação de Mestrado – Morgueti, M.
4-MATERIAL E MÉTODOS
4.1-AMOSTRA
Para desenvolvimento deste trabalho, foram utilizados vinte e sete ratos
(Rattus novergicus albinus, Wistar) machos, com peso variando entre cento e
oitenta a duzentos gramas, procedentes do Biotério da Universidade de Marilia
(UNIMAR) SP.
Os animais foram mantidos em gaiolas apropriadas e distribuídas
aleatoriamente em três grupos de nove animais cada, denominados de G1 (grupo
zero dia), G2 (grupo 10 dias) e G3 (grupo 30 dias).
4.2-PROCEDIMENTOS CIRÚRGICOS
O experimento foi realizado no Biotério da Universidade de Marilia
(UNIMAR), onde os animais receberam água e ração* livremente e foram mantidos
em ambiente controlado à temperatura de 20ºC e ciclos de doze horas de claro e
escuro.
Os animais foram anestesiados com ZOLETIL 50** na proporção de 0,1
ml para cada 100g.
Aplicada à anestesia intramuscular na coxa direita do animal foi realizada
a depilação por tração manual dos pêlos da face lateral da coxa esquerda (fig. 1).
A seguir, através de uma incisão entre o túber isquiático e o trocanter
maior, o nervo ciático foi dissecado e seccionado na altura do terço médio da coxa
(fig. 2 e 3).
No coto proximal foi colocado um tubo de plástico rijo de fundo cego nos
grupos G2 e G3 para prevenir a reinervação espontânea. Após o ato cirúrgico
todos os animais foram suturados com fio monofilamentar, não absorvível, 5-0***.
O primeiro grupo foi formado por nove animais denominados de G1(grupo
zero dias), que após terem seus nervos ciáticos seccionados, foram submetidos

20
Dissertação de Mestrado – Morgueti, M.
imediatamente à reparação através da neurorrafia término terminal com o fio 6-0 e
a seguir foi realizada a sutura da pele utilizando o fio 5-0*** (fig. 4).
O segundo grupo foi formado por nove animais denominados de G2
(grupo 10 dias), que após terem seus nervos seccionados, tiveram seus cotos
proximais colocados em um tubo de polietileno de fundo cego, fixado através de fio
6.0***. Após este período (dez dias), os animais foram submetidos novamente à
anestesia e em seguida foi provocada à reparação através da técnica de
neurorrafia termino terminal (fig.5).
O terceiro grupo foi formado por nove animais denominados de G3
(grupo 30 dias), onde foi realizado o mesmo procedimento do grupo anterior,
somente diferenciando o tempo de permanência do tubo de polietileno de dez para
trinta dias.
Após a cirurgia de reparação do grupo G2(10 dias) e do G3(30 dias),
onde foi respeitado o período de 10 e 30 dias respectivamente, os animais foram
sacrificados oito semanas após, por superdosagem do anestésico Zoletil 50**
sendo que o grupo G1(grupo zero), no qual a reparação foi realizada
imediatamente, também foi respeitado este mesmo período (fig. 6).
A pele dos animais foi suturada com o fio de nylon 5-0 após os
procedimentos cirúrgicos (fig. 7).
Após o sacrifico, o músculo sóleo dos animais foram coletados e
preparados para processamento para microscopia de luz.
O músculo retirado foi primeiramente colocado por alguns minutos em
formol tamponado a 10% para dar forma à estrutura, e em seguida colocada em
recipiente apropriado com o mesmo fixador por vinte e quatro horas.
Após este período, os músculos foram lavados em água corrente por
mais vinte e quatro horas e novamente colocados em recipiente apropriado com
álcool a 70%, até ser levado para procedimentos laboratoriais de rotina.
*Nuvilab. Nutrimental
**Zoletil 50. Virbac
***Nylon 5-0. Shalon
***Nylon 6-0. Shalon

21
Dissertação de Mestrado – Morgueti, M.
Figura 1 - Depilação por tração manual dos pêlos da face lateral da coxa esquerda.
Figura 2 - Incisão entre o túber isquiático e o trocanter maior.

22
Dissertação de Mestrado – Morgueti, M.
Figura 3 - Nervo ciático dissecado na altura do terço médio da coxa.
Figura 4 - Injúria causada no nervo ciático.

23
Dissertação de Mestrado – Morgueti, M.
Figura 5 - No coto proximal foi colocado um tubo de plástico rijo de fundo cego para prevenir a reinervação.
Figura 6 - Técnica de reinervação através da neurorrafia término terminal.

24
Dissertação de Mestrado – Morgueti, M.
Figura. 7- Sutura local com o fio de nylon 5-0.

25
Dissertação de Mestrado – Morgueti, M.
5- RESULTADOS
5.1 Análise Morfológica
Em todos os animais operados, foi observado macroscopicamente
resultado de sucesso no reparo do nervo ciático através da técnica de neurorrafia
termino terminal. Cabo de regeneração pôde ser encontrado em todos os animais
operados e uma leve fibrose, como era esperado foi constatado. Não foi observado
em nenhum dos animais operados qualquer evidência de necrose ou processo
inflamatório no local da cirurgia.
Em relação ao músculo sóleo, no grupo G1 (zero dia), suas fibras
apresentaram maiores áreas se comparadas às áreas das fibras dos demais
grupos. Outro fato que merece destaque em relação a esse grupo é a observação
de seus fusos neuromusculares. Todos os fusos dos músculos desse grupo
apresentaram em média 5 fibras intrafusais (fig. 8). Finalmente, pode ser
encontradas fibras com características poliédricas e com pouco tecido conjuntivo
entre seus fascículos.
Nos animais do grupo G2 (10 dias), as fibras musculares apresentaram-
se mais heterogêneas em relação a suas áreas. Um aumento bastante acentuado
de tecido conjuntivo, fibras colágenos também foram observadas. As fibras de
menor área encontravam-se preferencialmente localizadas na periferia do ventre
muscular (fig.9). Foram encontradas nesse grupo algumas fibras com mionúcleos
de posição central.
Finalmente, os animais do grupo G3 (30 dias), atrofia das fibras mais
intensa pode ser observada. Os fascículos apresentaram-se em maior número,
embora contendo menor número de fibras musculares em seu interior. A
heterogeneidade das fibras com relação às suas áreas foi mais intensa nesse
grupo. A invasão de tecido conjuntivo, especialmente tecido adiposo, pode ser
encontrada nesse grupo. Raramente fibras com áreas de aspecto normal puderam
ser encontradas (fig.10).

26
Dissertação de Mestrado – Morgueti, M.
Figura 8 – Corte transversal das fibras musculares estriadas esqueléticas do músculo sóleo dos animais do grupo G1. Note fuso com 5 fibras intrafusais. Fibras intrafusais poliédricas com mionúcleos em posição subsarcolemal.

27
Dissertação de Mestrado – Morgueti, M.
Figura 9 - Corte transversal das fibras musculares estriadas esqueléticas do músculo sóleo dos animais do grupo G2. Note o aparecimento de fibras de menores áreas agrupadas na periferia do músculo. Observe ainda um maior aumento de tecido conjuntivo entre os fascículos.

28
Dissertação de Mestrado – Morgueti, M.
Figura 10 – Corte transversal das fibras musculares estriadas esqueléticas do músculo sóleo dos animais do grupo G3. Note a invasão de grande quantidade de tecido conjuntivo para o interior do músculo.

29
Dissertação de Mestrado – Morgueti, M.
5.2 Análise Morfométrica
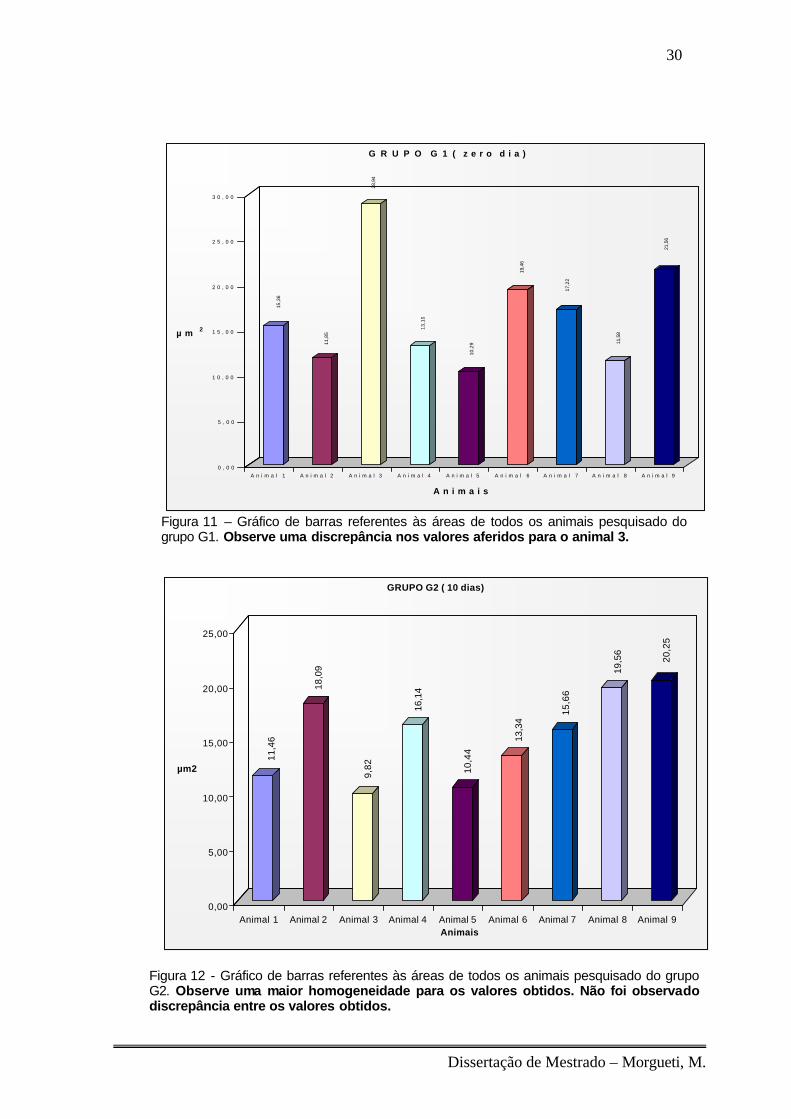
O grupo (G1) apresentou heterogeneidade na área das fibras,
principalmente no animal número três no qual se mostrou extremamente
aumentado em sua média. Para esse grupo foi encontrado um valor de 16,61
µm2 para a média geral de suas fibras (fig.11).
No grupo (G2) foi aferido valor médio da área de suas fibras de 14,99 µm2.
Este grupo apresentou-se mais homogêneo se comparado ao grupo anterior
(fig.12).
Para os animais do grupo (G3), foi constatado valor médio de suas fibras
menor do que dos grupos anteriores. Nesse grupo a média da área de suas fibras
foi de 5,63µm2. .Por outro lado foi observada uma heterogeneidade de suas fibras,
principalmente nos animais 07 e 08 do referido grupo (fig.13).
Tomando-se todos os animais de todos os grupos pesquisados,
observamos que a tendência das médias das áreas das fibras musculares é de
diminuição, sendo que nos períodos iniciais essa queda é mais intensa (fig.14).
30
Dissertação de Mestrado – Morgueti, M.
11,4
6
18,0
9
9,82
16,1
4
10,4
4
13,3
4
15,6
6
19,5
6
20,2
5
0,00
5,00
10,00
15,00
20,00
25,00
µm2
Animal 1 Animal 2 Animal 3 Animal 4 Animal 5 Animal 6 Animal 7 Animal 8 Animal 9 Animais
GRUPO G2 ( 10 dias)
15,3
6
11,8
5
28,9
4
13,1
9
10,2
9
19,4
6
17,2
2
11,5
8
21,5
6
0 , 0 0
5 , 0 0
1 0 , 0 0
1 5 , 0 0
2 0 , 0 0
2 5 , 0 0
3 0 , 0 0
µ m 2
A n i m a l 1 A n i m a l 2 A n i m a l 3 A n i m a l 4 A n i m a l 5 A n i m a l 6 A n i m a l 7 A n i m a l 8 A n i m a l 9
A n i m a i s
G R U P O G 1 ( z e r o d i a )
Figura 11 – Gráfico de barras referentes às áreas de todos os animais pesquisado do grupo G1. Observe uma discrepância nos valores aferidos para o animal 3.
Figura 12 - Gráfico de barras referentes às áreas de todos os animais pesquisado do grupo G2. Observe uma maior homogeneidade para os valores obtidos. Não foi observado discrepância entre os valores obtidos.

31
Dissertação de Mestrado – Morgueti, M.
4,59
4,24
2,75
2,15
1,50
3,34
13,8
5
13,0
9
5,17
0 , 0 0
2 , 0 0
4 , 0 0
6 , 0 0
8 , 0 0
1 0 , 0 0
1 2 , 0 0
1 4 , 0 0
µ m 2
An ima l 1 An ima l 2 An ima l 3 An ima l 4 An ima l 5 An ima l 6 An ima l 7 An ima l 8 An ima l 9
A n i m a i s
G R U P O G 3 ( 3 0 d i a s )
T a b e la Y = 1 0 1 , 2 6 2 0 5 - 0 , 0 1 9 7 6 X
1 6 , 6 1
1 4 , 9 9
5 , 6 3
0
2
4
6
8
10
12
14
16
18
0 5 10 15 2 0 2 5 3 0 3 5
D i a s P ó s C i r ú r g i a
µ m 2
Y e x p
Figura 13 - Gráfico de barras referentes às áreas de todos os animais pesquisado do grupo G3. Note uma homogeneidade para os animais 1, 2, 3, 4, 5 e 6 e uma discrepância para os valores obtidos para os animais 7,8 e 9.
Figura 14 – Gráfico das médias dos animais de todos os grupos pesquisados. Observe uma queda nos valores obtidos principalmente para os animais do grupo G3.

32
Dissertação de Mestrado – Morgueti, M.
6. DISCUSSÃO
Quando se discute o melhor momento para se tentar a recuperação de
um nervo e o músculo por ele inervado, deve-se levar em consideração
evidentemente, as alterações que ocorrem em ambos para se tentar estabelecer
em que momento o nervo e o músculo estão mais suscetíveis a um melhor padrão
de regeneração. Assim, relacionados aos nervos, é sabido que em suas fibras que
sofreram axotomia, a produção de NGF (neuro growth factor) atinge nível máximo
24 horas após e mantém níveis elevados de NGF pelo menos até duas semanas
após a lesão os músculos, ou melhor, as fibras musculares desnervadas sofrem
uma atrofia perdendo progressivamente massa tecidual. (RODRIGUES &
SCHMALBRUCH, 1995).
Há tempos vários pesquisadores discutem o melhor momento para se
tentar a reinervação em músculos que tiveram seus nervos motores seccionados.
Para alguns os músculos estariam mais aptos a receber novo contato do nervo
motor entre 12 e 16 semanas pós-lesão (DOLFINI, 2002) enquanto que para
outros, quando não há perda tecidual, quanto mais rápida for a tentativa de reparo
nervoso, melhor será o resultado de sua regeneração (RODRIGUES, 1998).
Um músculo desnervado por longo tempo, acima de 25 semanas,
apresenta redução significativa no número de células satélites (RODRIGUES &
SCHMALBRUCH, 1995), fato este importante para a recuperação do músculo
como um todo. Portanto músculos com tempos longos de desnervação podem
estar fadados a não apresentarem condições para uma possível reinervação.

33
Dissertação de Mestrado – Morgueti, M.
A favor dessa idéia podemos encontrar um trabalho realizado por ANZIL
& WERNIG (1989) que procedeu a desnervação do músculo peitoral de rã e
observou, após quase 4 anos, a completa ausência de fibras musculares. No local
foi observada apenas e tão somente uma lâmina conjuntiva.
A idade do indivíduo evidentemente também é fator a ser considerado
nessa situação, lembrando que animais recém-nascidos apresentam uma queda
no número de células satélites muito mais pronunciada que nos animais adultos
(RODRIGUES et al., 2002).
Classicamente, a tentativa de reinervação de músculo, deve ser
realizada o mais rápido possível. No entanto, ao se analisar músculos
desnervados, podem perceber que morfologicamente tais músculos apresentam
fibras musculares com características de fibras jovens após 12 semanas de
desnervação. É sabido que os músculos estriados esqueléticos existem como
estruturas anatômicas muito antes de sua inervação. Os nervos desempenham
papel importante na modulação de tais fibras, alterando sua constituição, ou
melhor, modulando os diferentes tipos de fibras para melhor atender à demanda do
músculo. Se o raciocínio for feito levando-se em conta o desenvolvimento natural
dos músculos estriados esqueléticos, um pequeno retardo de algumas semanas
parece-nos mais acertado do que a tentativa imediata de reinervação.
Em nossa pesquisa realizamos experimentalmente lesão no nervo ciático
do rato (sem perda tecidual) com recuperação, através da técnica da neurorrafia
término terminal, em três tempos: imediatamente após a lesão; 10 dias após e 30
dias após. O músculo escolhido para análise morfológica de sua recuperação foi o
músculo sóleo.

34
Dissertação de Mestrado – Morgueti, M.
A justificativa para tal escolha foi o fato do músculo sóleo ser
considerado um músculo postural, rico em fibras vermelhas (tipo SO e FOG),
portanto mais homogêneo se comparado a um músculo branco como o músculo
EDL (extensor longo dos dedos). O músculo sóleo recebe inervação exclusiva do
nervo ciático que foi seccionado conforme descrito anteriormente.
Os resultados apontam para o tempo imediato após lesão do nervo
(primeiro grupo), padrão de recuperação muscular mais próximo do normal. A
explicação para tal resultado deve estar no tempo necessário para que as fibras
nervosas em regeneração estabeleçam contato com as fibras musculares. Assim,
levando-se em conta que as fibras nervosas crescem em média de 1 a 3 mm por
dia e, que a distância entre o local da secção nervosa e o músculo sóleo foi de
aproximadamente 5 cm (ou 50 mm), os novos neuritos devem ter atingido as fibras
do músculo sóleo após 40 ou 50 dias (6 a 7 semanas) pós desnervado.
Esse resultado vem de encontro aos resultados obtidos por outros
pesquisadores que puderam observar um padrão para as fibras musculares
estriadas esqueléticas desnervadas, semelhantes àqueles observados por ocasião
da primeira inervação sofrida por elas.
Assim, fibras com núcleos centrais denominadas miotubos e uma baixa
ocorrência do fenômeno de apoptose nesse período viria a favorecer uma
reinervação mais facilitada.
Seguindo o mesmo raciocínio poderíamos então sugerir que na tentativa
de se reinervar um determinado músculo que fora alvo de uma desnervação e
onde não houve perda tecidual, ou seja, o nervo em questão tenha sofrido tão
somente sua secção (neurotomia), a neurorrafia término terminal deverá ser
realizada considerando-se em conta a distância da secção ao músculo alvo.

35
Dissertação de Mestrado – Morgueti, M.
Como a fibra nervosa varia em sua velocidade de crescimento entre 1 a
3 mm2 por dia, seria razoável adotar-se em média um crescimento de 2 mm2 por
dia e assim estabelecer o melhor momento para se fazer o reparo nervoso.
Com relação ao músculo sóleo observado neste experimento, em nível
de microscopia óptica, foi observada a presença de duas áreas musculares
distintas; uma apresentando fibras musculares organizadas, de diâmetros
regulares entre si, e outra com fibras heterogêneas. Provavelmente estas
diferenças sejam devidas às características metabólicas diferentes de cada tipo de
fibra muscular. Essa desorganização vista ao microscópio óptico reflete as
alterações nucleares e citoplasmáticas que através da técnica de microscopia
eletrônica certamente seriam melhor observáveis.
Conforme foi mencionado anteriormente, o fenômeno de apoptose deve
ser levado em conta em pesquisas de regeneração tecidual. No caso de músculo
estriado esquelético normal, a quebra de DNA, que é um indicativo de apoptose,
pode ser devido ao próprio “turnover” do tecido. Assim, é comum observar-se tal
fenômeno mesmo em músculo normal.
Já o músculo lesado recorre a sua via de reparação, ou seja, às células
satélites que funcionam como verdadeiros mioblastos. Para a maioria dos
pesquisadores, a própria injúria muscular atua como estímulo que ativa as células
satélites. Essa ativação em nosso caso, muito provavelmente, foi desencadeada
direta ou indiretamente, pela desnervação, o que também foi observado em outras
pesquisas com músculo desnervado (ONTELL, 1975; SNOW, 1983; MCGEACHIE,
1989; BORISOV & CARLSON, 2000).

36
Dissertação de Mestrado – Morgueti, M.
Assim, por um período a partir de aproximadamente 10 semanas pós
desnervação, o que certamente aconteceu, foram oscilações do músculo sóleo na
tentativa de regeneração, com o auxílio das células satélites e conseqüentes
diminuição na quebra do DNA.
LEWIS & SCHMALBRUCH (1994) observaram a diminuição do poder de
contração de músculo estriada esquelético após desnervação, encontrando que
nos primeiros três meses ocorre um decréscimo em escala logarítmica para depois
se estabilizar até aproximadamente 24 semanas pós desnervação, reafirmando a
idéia de que neste período a perda de fibras por um lado, e a tentativa de
regeneração por outro, estão equilibradas.
Em face dos resultados obtidos nesta pesquisa, e as informações
referentes à literatura supracitada, parece-nos pertinente inferir que a melhor época
para a reinervação de um músculo desenervado também seja, dentre outros
fatores, o período onde o fenômeno de apoptose, ou seja, onde ocorra a menor
quebra da cadeia de DNA. Quanto maior for o tempo de desnervação, maior o
declínio da quantidade de células satélites, sendo que após 30 semanas há uma
diminuição na porcentagem de tais células para menos de 1/5 de um músculo
normal. (Rodrigues & Schmalbruch, 1995).
Essa observação é coincidente com o aumento rápido da quebra do
DNA, que a partir de 16 semanas pós desnervação, passa a ser ascendente
(DOLFINI et al., 2002). Levando-se em consideração o fato de não ter sido
constatado em nenhum dos animais operados (desnervados) sinais de necrose; a
distância entre o local onde o nervo ciático foi seccionado experimentalmente e o
músculo pesquisado; a velocidade de regeneração das fibras nervosas; e os

37
Dissertação de Mestrado – Morgueti, M.
resultados dos trabalhos consultados na literatura podem concluir que no protocolo
proposto por esta investigação o melhor período para o emprego da técnica de
neurorrafia término terminal do nervo ciático foi obtido nos animais do primeiro
grupo, ou seja, imediatamente após a secção do nervo.

38
Dissertação de Mestrado – Morgueti, M.
7. CONCLUSÕES
1- Na recuperação de um músculo estriado esquelético que tenha sofrido
transecção total de seu nervo motor, é de fundamental importância
correlacionar à distância da lesão nervosa e o referido músculo.
2- O melhor momento para se restabelecer o contato entre o nervo em
regeneração e seu músculo é entre 6 e 7 semanas pós desnevação.
3- No modelo proposto, os melhores resultados foram obtidos nos animais
que sofreram reparo do nervo ciático imediatamente após a lesão.

39
Dissertação de Mestrado – Morgueti, M.
8. REFERÊNCIAS BIBLIOGRÁFICAS
ADACHI, M., J. H. SHER. Neuromuscular disease. Igaku-Shoin Ltda.,
New York, p.347, 1990.
AL-AMOOD, W. S., D. M. LEWIS & H. SCHMALBRUCH. Effects of chronic
electrical stimulation on contractile properties of long-term denervated
rat skeletal muscle. J. Physiol., v.441, p.243-256, 1991.
ANZIL, A. P. & A. WERNIG. Muscle fibre loss and reinnervation after long-
term denervation. J. Neurocytol., v.18, p.833-845, 1989.
BARANY, M. & R. I. CLOSE. The transformation of myosin in cross-
innervated rat muscle. J. Physiol., v.213, p.455-474, 1971.
BEGO, U., V. MITIN & M. ZOBUNDZIJA. New approach to the functional
classification of horse diaphragm muscle fibers. Praxis veterinaria, v. 44,
p.159-175, 1996.
BISCHOFF, R. Enzymatic liberation of myogenic cells from adult rat
muscle. Anat. Rec., v.180, p.645-662, 1974.
BONNER, P. H. Nerve-dependent changes in clonable myoblast
populations. Dev. Biol., v.66, p.207-219, 1978.
BONNER, P. H. Differentiation of chick embryo myoblasts is transiently
sensitive to functional denervation. Dev. Biol., v.76, p.79-86, 1980.
BONNER, P. H. Myoblast differentiation is induced by nerve transplanted
to chick embryo legs. J. Exp. Zool., v.243, p.63-70, 1987.

40
Dissertação de Mestrado – Morgueti, M.
BONNER, P. H. & T. R. ADAMS. Neural induction of chick myoblast
differentiation in culture. Dev. Biol., v. 90, p.175-184, 1982.
BORISOV, A. B., CARLSON, B. M. Cell death in denervated skeletal
muscle is distinct from classical apoptosis. Anat. Rec ., v. 258(3), p. 305-
318, 2000.
BULLER, A. J. & D. M. LEWIS. Further observations on mammalian cross-
innervated skeletal muscle. J. Physiol., v.178, p.343-358, 1965.
CHIAKULAS, J. J., J. E. PAULY. A study of postnatal growth of skeletal
muscle in the rat. Anat. Rec., v. 152, p. 55-62, 1965.
DAHLIN B. LARS. The biology of nerve injury and repair. Department of
hand surgery, Malmö University Hospital, Malmö, Sweden. Available online 15
September 2004.
DALL PAI, V., L. C. T. CARMELLO & DAL PAI, M. Morfologia e tipos de
fibras do músculo extensor longo do hálux do rato Wistar. Acta Biol.
Leopol., v. 1, p. 5-16, 1991.
DESGRANGES, P et al. A substituted dextran enhances muscle fiber
survial and regeneration in ischemic and denervated rat EDL muscle,
v.13, n.6, p.761-766, April 1999.
DOLFINI, M.I. Apoptose e células satélites em músculo desnervado por
longo período, 2002. p.98. Dissertação de Mestrado-
Medicina/Unesp/Botucatu.
DUBOWITZ, V. Muscle biopsy. . Bailliere Tindall, London, 2ª ed., p.403,
1985.

41
Dissertação de Mestrado – Morgueti, M.
GU YU HUAI, et al.Survival, regeneration and functional recovery of
motoneurons after delayed reimplantation of avulsed spinal root in adult
rat. Department of Anatomy, Faculty of Medicine of Hong Kong, 21Sassoon
Road, Hong Kong SAR, China. Available online 19 January 2005.
GULATI, A. K. Long-term retention of regenerative capability after
denervation of skeletal muscle, and dependency of late differentiation on
inervation. Anat. Rec., v. 220, p. 429-434, 1988.
GONYEA, W. J., D. G. SALE, F. G. GONYEA. Exercise induced increases
in muscle fiber number. Eur. J. Appl. Physiol., v. 55, p. 137-141, 1986.
IJKEMA-PAASSEN, J.; MEEK, M.F.; GRAMSBERGEN, A. Muscle
differentiation after sciatic nerve transaction and reinnervation in adult
rats, Ann Anat; 183(4), p.369-77, jul. 2001.
JENNEKENS, F. G. I. Neurogenic disorders of muscle. In: Skeletal Muscle
Pathology Edinburgh and London: Churchill Livingstone. (edited by
MASTAGLIA, F. L. and LORD WALTON) 2ª ed.; p.563-597, 1992.
LAUNAY, T et al. Expression and Neural Control of Myogenic Regulatory
Factor Genes during Regeneration of Mouse Soleus. Journal of
histochemistry and cytochemistry, v. 49, n.7, p.887-899, 2001.
LEWIS, D. M. & H. SCHMALBRUCH. Contractile properties of aneurally
regerated compared with denervated muscles of rat. J. Muscle Res. Cell
Motil, v. 15, p. 267-277, 1994.
MARQUES, M.J.; DA-SILVA, C.F.; SANTOS NETO, H.An Ultra structural
Study Of The Temporal Progress Of Neuromuscular Junction
Regeneration After Sciatic Nerve Transection And Tubulization Repair.
Braz. J. morphol. Sci., v.13, n.2, p. 181-187, 1996.

42
Dissertação de Mestrado – Morgueti, M.
MAXWEEL, L. C., Y. A. FAULKNER, G. Y. HYAT. Estimation of number of
fibers in guinea pig skeletal muscle. J. Appl. Physiol., v. 37, p. 259-264,
1974.
MAMMAERTS, W. F. H., A. J. Buller and K. Seraydarian. The modification
of some biochemical properties of muscle by cross-inervation. Proc. Nat.
Acad. Sci., v. 64, p. 128-133, 1969.
MAURO, A. Satellite cell of skeletal muscle fibers. J. Biopsy. Biochem.
Cytol., v.9, p. 493-495, 1961.
MCGEACHIE, J.K. Sustained cell proliferation in denervated skeletal
muscle of mice. Cell Tissue Res ., v. 257(2), p. 455-457, 1989.
MINAMOTO, V.B; BUNHO, S.R; SALVINI, T.F. Regenerated rat skeletal
muscle after periodic contusions. Brazilian Journal of Medical and
Biological Research, v.34, n. 11, p. 1447-1452, 2001.
MORACZEWSKI, J et al. Differential changes in protein kinase C
associated with regeneration of rat extensor digit rum longs and soleus
muscle. The International Journal of Biochemistry & Cell Biology, v.34, p. 938-
949, 2002.
MUSSINI, I., D. FAVARO & U. CARRARO. Maturation dystrophic changes
and the continuous production of fibers in skeletal muscles regenerating
in the absence of nerve. J. Neuropath. Exp. Neural, v. 46, p. 315-331, 1987.
OH, T. H. & G. J. MARKELONIS. Dependence of in vitro myogenesis on a
tropic protein present in chicken embryo extract. Proc. Natl. Acad. Sci., v.
77, p. 6922-6925, 1980.
ONTELL, M. Evidence for myoblast potential of satellite cells in
denervated muscle. Cell Tissue Research, v. 160(3), p. 345-53, 1975.

43
Dissertação de Mestrado – Morgueti, M.
PETTER, J. B., P. J. BARNARD., V. R. Edgerton. Metabolic profiles of
three types of skeletal muscle fibers in guinea pigs and rabbits.
Biochemistry, v. 11, p. 2627-2633, 1972.
POPIELA, H et al. Regulation of mitotic activity and the cell cycle in
primary chick muscle cells by neurotransferin. J. Cell Physiol., v. 119,
p.234-240, 1984.
RODRIGUES, A. C. & H. SCHMALBRUCH. Satellite cells and myonuclei in
long-term desnervated rat muscle. Anat. Rec., v. 243, p.430-437, 1995.
RODRIGUES, A.C. et al. C.R. Comparative study between the fiber-type
composition in the flexor and extensor muscles in the pig (Sus scrofa).
Rev. Chil. Anat, v. 17(1), p. 53-60, 1999.
RODRIGUES, A.C.; et al. Satellite cells and myonuclei in neonatallt
denervated rat muscle. Italian Journal Anatomy and Embriology. Roma, v.
107(1), p.51-56, 2002.
RODRIGUES, A.C. Estudo das alterações das fibras musculares
estriadas esqueléticas desnervadas por longo período. 1998.p. 54. Tese
apresentada a Universidade Paulista Julio de Mesquita Filho¨, para obtenção
do grau de Livre Docente. Botucatu.
SCHMALBRUCH, H. THE MUSCULAR DYSTROPHIES. In: Skeletal Muscle
Pathology (edited by MASTAGLIA F. L. AND LORD WALTON) 2ª ed., p. 283-
318. Edinburgh and London: Churchill Livingstone, 1992.
SCHMALBRUCH, H. & U. HELLHAMMER. The number of nuclei in adult rat
muscles with special reference to satellite cells. Anat. Rec., v.189, p. 169-
176, 1977.

44
Dissertação de Mestrado – Morgueti, M.
SCHMALBRUCH, H. & D. M. LEWIS. A comparison of the morphology of
desnervated with aneurally regenerated soleus muscle of the rat. J.
Muscle Res. Cell Motil, v. 15, p. 256-266, 1993.
SCHMALBRUCH, H. & D. M. LEWIS. A comparison of the morphology of
denervated with aneurally regenerated soleus muscle of rat. J. Muscle
Res. Cell. Motil. v.15, p. 256-266, 1994.
SCHMALBRUCH, H. W. S. AL-AMOOD & D. M. LEWIS. Morphology of
long-term denervated rat soleus muscle and the effect of chronic
electrical stimulation. J. Physiol., v.441, p. 233-241, 1991.
SCHMALBRUCH, H et al. A new mouse mutant with progressive motor
neuronopathy. J. Neuropathol. Exp. Neurol., v.50, p. 192-204, 1991.
SESODIA, S. & M. J. CULLEN. The effect of denervation on the
morphology of regenerating rat soleus muscles. Acta Neuropathol, v.82, p.
21-32, 1991.
SKETELJ, J et al. Interactions between intrinsic regulation and neural
modulation of acetyl cholinesterase in fast and slow skeletal muscles.
Cell. Molec. Neurobiol, v. 11, p. 35-54, 1991.
SNOW, M. H. A quantitative ultra structural analysis of satellite cells in
denervated fast and slow muscles of the mouse. Anat. Rec., v. 207, p.
593-604, 1983.
STEVEN, T; DAVOR; FAULKER, J.A. Regeneration of new fibers in
muscles of old rats reduces contraction-induced injury. Journal of applid
physiology, v.87, n.2, p.750-756, 1999.
WANEK, L.J; SNOW, M.H. Activy-induced Fiber Regeneration in Rat
Soleus Muscle. The Anatomical Record, v. 258, p. 176-185, 2000.

45
Dissertação de Mestrado – Morgueti, M.
WHALEN, R. G et al. Expression of myosin isoforms during notexin-
induced regeneration of rat soleus muscles. Dev. Biol., v.141, p. 24-40,
1990.
YOSHIMURA, K.; HARII, K. A Regenerative Change during Muscle
Adaptation to Denervation in Rats. Journal of Surgical Research, v. 81, p.
139-146, 1999.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo