allane maria lacerda ferreira de oliveira …dspace.bc.uepb.edu.br/jspui/bitstream/123456789/671...o...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DA PARAÍBA
CENTO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE - CCBS
DEPARTAMENTO DE BIOLOGIA
LICENCIATURA PLENA E BACHARELADO EM CIÊNCIAS BIOLÓGICAS
ALLANE MARIA LACERDA FERREIRA DE OLIVEIRA
CONSTRUÇÃO DO MAPA DE RESTRIÇÃO DO GENE DA TIREOPEROXIDASE
(TPO) A PARTIR DE MUTAÇÕES QUE CAUSAM HIPOTIREOIDISMO
CONGÊNITO
Campina Grande, PB
2012

ALLANE MARIA LACERDA FERREIRA DE OLIVEIRA
CONSTRUÇÃO DO MAPA DE RESTRIÇÃO DO GENE DA TIREOPEROXIDASE
(TPO) A PARTIR DE MUTAÇÕES QUE CAUSAM HIPOTIREOIDISMO
CONGÊNITO
Trabalho de conclusão de Curso apresentado ao
Curso de Licenciatura e Bacharelado em
Ciências Biológicas da Universidade Estadual da
Paraíba em cumprimento às exigências para a
obtenção do grau de Licenciada e Bacharel em
Ciências Biológicas
Orientadora: Prof. Drª. Simone Silva dos Santos Lopes
Campina Grande, PB
2012

F ICHA CATALOGRÁFICA ELABORADA PELA BIBLIOTECA CENTRAL – UEPB
O48c Oliveira, Allane Maria Lacerda Ferreira de.
Construção do mapa de restrição do gene da
tireoperoxidase (TPO) a partir de mutação que causam
hipotireoidismo congênito [manuscrito] / Allane Maria
Lacerda Ferreira de Oliveira. – 2012.
65 f. : il. color.
Digitado.
Trabalho de Conclusão de Curso (Graduação em Ciências
Biológicas) – Universidade Estadual da Paraíba, Centro de
Ciências Biológicas e da Saúde, 2012.
“Orientação: Profa. Dra. Simone Silva dos Santos Lopes,
Departamento de Biologia”.
1. Mutação. 2. Genética humana. 3. Hipotireoidismo.
I. Título.
CDD 21. ed. 576

ALLANE MARIA LACERDA FERREIRA DE OLIVEIRA
CONSTRUÇÃO DO MAPA DE RESTRIÇÃO DO GENE DA TIREOPEROXIDASE
(TPO) A PARTIR DE MUTAÇÕES QUE CAUSAM HIPOTIREOIDISMO
CONGÊNITO
Orientadora: Prof. Drª. Simone da Silva dos Santos Lopes
MONOGRAFIA APROVADA EM: 21/08/2012

A Deus,
toda honra e toda glória seja dada a Ti,
que sempre esteve comigo.

AGRADECIMENTOS
Agradeço a Deus, hoje mais do que nunca eu sei que se não fosse pela força que Senhor
esteve me dando todos esses anos eu não estaria aqui, não teria força pra prosseguir.
Agradeço-te infinitamente meu Deus. Tu és o motivo por qual ainda estou aqui!
As duas pessoas que mais contribuíram, incentivaram e acreditaram que eu seria capaz...
Mainha (Kleber Ferreira) e Vovó (Eliraci Oliveira), o que seria de mim se não fosse por todo
o apoio que sempre me deram, por toda dedicação e amor. Pela paciência que demonstravam
quando muitas vezes eu ligava chorando dizendo que não aguentava mais e queria ir pra
casa. Como sou grata a Deus por ter me presenteado com duas jóias raras que são vocês na
minha vida. Obrigada por tudo. Espero poder retribuir de alguma forma TUDO que sempre
fizeram por mim. Amo incondicionalmente!
Ao meu Pai (Antonio Silva) e meus irmãos (Klethonio e José Neto) por todo amor, carinho e
contribuição na minha jornada. Pela paciência comigo nos meus momentos de “estresse”.
Apesar das nossas brigas de irmãos, amo vocês, são uma parte de mim.
Ao meu namorado Jefferson Honório pelo amor e companheirismo, por estar sempre
presente na minha vida, me apoiando e incentivando a conquistar meus ideias, nossos ideais.
Cada dia tenho mais certeza que foi Deus quem te colocou na minha vida, pra me ajudar a
superar todos os obstáculos que a vida nos impõe, pra me dizer você precisa tentar, só assim
vai conseguir. Quantas vezes me irritei por você me mandar ir estudar, mais hoje diante de
tantas graças recebidas entendo que era Deus te usando para não permitir que eu desistisse.
Á toda minha família, Tios, primos, obrigada pela confiança e apoio.
A professora Simone Silva dos Santos Lopes pela oportunidade de trabalhar com aquilo que
eu sempre almejei, por todos os ensinamentos, todas as orientações, pela paciência nos
momentos das falhas. Só tenho a agradecer por ter compartilhado comigo tantos
ensinamento... de Biologia Molecular, humildade e dedicação.
A todos do Laboratório de Biologia Molecular e Genética-UEPB, especialmente à Rayssa,
Mayara1, Mayara2, Thuany, Tafnys, Bárbara e Renata por toda ajuda no desenvolvimento
do meu trabalho e principalmente pela amizade construída. E também as amizades que foram
cultivadas esses 4 anos, Àkyla, Suziane, Raquel, Rayssa, Daniella.
A banca examinadora pela gentileza e disponibilidade em aceitar o convite.
A Universidade Estadual da Paraíba e ao Conselho Nacional de Desenvolvimento Científico
e Tecnológico – CNPq, pelo apoio financeiro.

"Deus nos concede, a cada dia, uma página de vida nova no livro do tempo. Aquilo que
colocarmos nela, corre por nossa conta."
(Emmanuel, psicografado por Chico Xavier)
“Existe um tempo certo para cada coisa, momento oportuno para cada propósito debaixo do
Céu: Tempo de nascer, tempo de morrer; tempo de plantar, tempo de colher”.
(Eclesiastes 3:4)

RESUMO
O Hipotireoidismo Congênito (HC) é a causa mais comum de retardo mental prevenível em crianças, e se
deve a ausência ou deficiência na produção dos hormônios tireoidianos (HTs), sua incidência varia entre 1:
3.000 a 4.000. A enzima Tireoperoxidase (TPO) é uma enzima chave na biossíntese dos HTs, sendo responsável
por catalizar importantes reações, como a organificação do iodo para formação dos hormônios tireoidianos.
Vários estudos moleculares do gene TPO revelaram que existe uma grande heterogeneidade de defeitos ao
nível desta proteína. Diante disto, foi realizada uma análise “in sílico” no gene da tireoperoxidase para
identificar os sítios de restrição presentes nos 17 éxons e assim criar um Mapa de Restrição desse gene, além de
realizar um levantamento bibliográfico para a construção de uma tabela contendo todas as mutações descritas no
gene TPO até o momento. A Construção do Mapa de Restrição do gene TPO foi realizada utilizando como base
a sequência completa do RNA mensageiro (NCBI Ref. Seq: NM_000547.5) posteriormente, utilizou-se o
programa NEBcutter para conhecer as enzimas que possuem sítios de restrição e suas posições nos éxons da
sequência do DNA. Também foi realizada uma pesquisa bibliográfica para selecionar todos os artigos que
possuem mutações descritas no gene TPO para elaboração de uma tabela revisada e atualizada, além de elaborar
uma tabela contendo as mutações e as enzimas que possuem sítios no local de cada mutação descrita no gene
TPO, como contribuição científica para posteriores estudos com esse gene. Construi-se um Mapa de Restrição
do gene TPO e uma tabela com as enzimas que possuem sítios de restrição correlacionados com as mutações que
causam HC no gene TPO. Durante o levantamento bibliográfico das mutações descritas no gene TPO,
encontramos disparidades entre as posições descritas por cada autor e a sequência depositada no banco de dados
do GenBank que foi utilizado como base. Também foi construída uma tabela com as enzimas que podem ser
utilizadas na detecção de mutações no gene TPO, pois essas enzimas possuem sítios nesse gene, ou em alguns
casos as mutações eliminam sítios. A partir desse estudo conclui-se que é possível construir um Mapa de
Restrição do gene TPO com base na sequência analisada. No entanto, a investigação da literatura permitiu
verificar que as mutações descritas nos artigos referenciados podem não apresentar concordância com a
sequência do gene TPO presente no GenBank.
Palavras-chave: Mutações, TPO, Mapa de Restrição

ABSTRACT
The Congenital Hypothyroidism (CH) is the most common cause of preventable mental retardation in children,
and is due to absence or deficiency in the production of thyroid hormones (TH), his incidence varies between 1:
3,000 to 4,000. The enzyme thyroperoxidase (TPO) is a key enzyme in the biosynthesis of CH, being responsible
for catalyzing important reactions, such as organification of iodine for the formation of thyroid hormones.
Several molecular studies of the TPO gene revealed that there is a great variety of defects at the level of this
protein. Given this, an analysis was performed "in silico" of the mutation in the thyroperoxidase to identify the
restriction sites present in exons 17 and thus creating a restriction map of this gene, in addition to conducting a
literature review to build a table containing all TPO gene mutations described so far. Construction of restriction
map of the TPO gene was performed based on the complete sequence of the messenger RNA (NCBI Seq Ref:
NM_000547.5) then, the software used to meet enzymes was the NEBcutter which have restriction sites and
their positions exons in the sequence of DNA. We also performed a literature search to select all items that have
in the TPO gene mutations described for the preparation of a revised and updated the table, and draw up a table
containing the mutations and the enzymes that have sites on the site of each mutation described in the TPO gene
, as a scientific contribution to further studies with this gene. Build up a restriction map of the TPO gene and a
table with the enzymes which have restriction sites correlated with mutations in the TPO gene causing CH .
During the literature of the mutations described in the TPO gene, we found differences between the positions
described by each author and the sequence deposited in the GenBank database that was used as a base. Was also
constructed a table with the enzymes which may be used in detecting mutations in the TPO gene, as these
enzymes have sites in this gene, or some cases eliminate the mutation sites. From this study it is concluded that it
is possible to construct a restriction map of the TPO gene based on the sequence analyzed. However, research
literature has shown that the mutations described in the referenced articles may not show agreement with the
TPO gene sequence present in GenBank.
Keywords: Mutations, TPO, Restriction Map.

LISTA DE ABREVIAÇÕES
DT – Disgenesia tireoidiana
HC – Hipotireoidismo congênito
HP – Hipotireoidismo primário
HTs – Hormônios tireoideanos
TG – Tireoglobulina
TPO – Tireoperoxidase
NIS- Simportador de sódio e iodeto
PIOD – Defeito parcial na organificação do iodeto
TIOD – Defeito total na organificação do iodeto
TRH – Hormônio liberador de tirotrofina
TSH – Hormônio tireo-estimulante
PCR – Reação em Cadeia da Polimerase
RFLP – Polimorfismo de comprimento de fragmentos de restrição
SSCP- Polimorfismo de Conformação de fita simples
DGGE- Eletroforese em gel com gradiente de desnaturação
BsrFI- Bacillus stearothermophilu
MseI- Micrococcus species
NarI - Norcardia argentinensis
HinP1I - Haemophilus influenzae
HpyCH4V - Helicobacter pylori
PhoI - Pyrococcos horikoshii OT3
XhoI - Xanthomonas holcicola
PaeR7I - Pseudomonas aeruginosa
TliI - Thermococcus litoralis

BssSI - Bacillus stearothermophilus
AscI - Arthrobacter species
Bfa I - Bacteroides fragilis
CviQ-I - Chlorella virus
HaeIII - Haemophilus aegypticus
AluI - Arthrobacter luteus
BglII - Bacillus globigii
PciI gene from Planococcus citreus
BsrGI - Bacillus stearothermophilus
RsaI - Rhodopseudomonas sphaeroides
HhaI - Haemophilus influenzae
SfoI - Serratia fonticola
KasI - Kluyvera ascorbata
MspI - Moraxella species
HpaII - Haemophilus parainfluenzae
NgoMIV- Neisseria gonorrhoeae
PvuII - Proteus vulgaris
MluI - Micrococcus luteus
FseI - Frankia species
NaeI - Nocardia aerocolonigenes
BamHI - Bacillus amyloliquefaciens
MluCI - Micrococcus luteus.
ClaI - Caryophanon latum

MspI - Moraxella species
Tsp509I - Thermus species
BmgBI - Bacillus megaterium
PspOMI - Pseudomonas species
NgoMIV - Neisseria gonorrhoeae
SacI - Streptomyces achromogenes
Eco53kI - Escherichia coli 53k
ApaLI - Acetobacter pasteurian
NlaIII - Neisseria lactamica
CviAII - Chlorella virus
FatI - Flavobacterium aquatile
PstI - Providencia stuartii
BsiWI - Bacillus species

LISTA DE TABELAS
Tabela 1 - Éxon 1: Éxon com enzimas de restrição......................................................... 30
Tabela 2- Éxon 2: Éxon com enzimas de restrição........................................................... 31
Tabela 3 - Éxon 3: Éxon com enzimas de restrição........................................................... 31
Tabela 4- Éxon 4: Éxon com enzimas de restrição........................................................... 32
Tabela 5- Éxon 5: Éxon com enzimas de restrição........................................................... 32
Tabela 6- Éxon 6: Éxon com enzimas de restrição........................................................... 33
Tabela 7- Éxon 7: Éxon com enzimas de restrição........................................................... 34
Tabela 8- Éxon 8: Éxon com enzimas de restrição........................................................... 34
Tabela 9- Éxon 9: Éxon com enzimas de restrição........................................................... 35
Tabela 10- Éxon 10: Éxon com enzimas de restrição......................................................... 36
Tabela 11- Éxon 11: Éxon com enzimas de restrição......................................................... 36
Tabela 12- Éxon 12: Éxon com enzimas de restrição......................................................... 37
Tabela 13- Éxon 13: Éxon com enzimas de restrição......................................................... 38
Tabela 14- Éxon 14: Éxon com enzimas de restrição......................................................... 38
Tabela 15- Éxon 15: Éxon com enzimas de restrição......................................................... 39
Tabela 16- Éxon 17: Éxon com enzimas de restrição......................................................... 39
Tabela 17- Tabela contendo mutações e enzimas de restrição.......................................... 41

LISTA DE GRÁFICOS
Gráfico 1- Comparação entre o número de mutações e o número de enzimas de restrição
disponíveis em cada
éxon...................................................................................30

LISTA DE FIGURAS
Figura 1- Glândula Tireóide.........................................................................................13
Figura 2- Eixo hipotálamo-hipófise-tireóide.............................................................14
Figura 3- Desenho esquemático com mutações descritas no Gene
Tireoperoxidase............................................................................................21
Figura 4- Desenho esquemático com enzimas disponíveis no Gene
Tireoperoxidase............................................................................................28

SUMÁRIO
1. INTRODUÇÃO................................................................................................................. 15
2. REFERENCIAL TEÓRICO............................................................................................ 17
2.1 Tireóide .............................................................................................................................. 17
2.2 Hipotireoidismo congênito...................................................................................................... 18
2.3 Tireoperoxidase .................................................................................................................. 20
2.4 Mutações no gene da Tireoperoxidase................................................................................ 22
2.5 Análise de Mutação............................................................................................................. 23
2.5.1 Enzima de restrição ............................................................................................................ 23
2.5.2 Polimorfismo de comprimento de fragmento de restrição.................................................. 24
2.5.3 Mapa de Restrição .............................................................................................................. 25
3. OBJETIVOS...................................................................................................................... 26
3.1 Objetivo Geral .............................................................................................................. 26
3.2 Objetivo Específico....................................................................................................... 26
4. METODOLOGIA.............................................................................................................. 27
4.1 Identificação das mutações descritas na literatura no gene TPO .................................. 27
4.2 Construção do Mapa de Restrição do gene TPO.............................................................. 27
4.3 Mutações identificadas e analisadas por RFLP na literatura ........................................ 28
5. RESULTADOS ................................................................................................................ 29
5.1 Mapa de restrição do gene TPO.......................................................................................... 29
5.2 Levantamento das enzimas de restrição e mutações no gene TPO............................... 40
6. DISCUSSÃO...................................................................................................................... 43
7. CONCLUSÃO .................................................................................................................. 46
8. PERSPECTIVAS.............................................................................................................. 47
REFERÊNCIAS BIBLIOGRÁFICAS............................................................................ 48
APÊNDICES .................................................................................................................... 53
APÊNDICE A ................................................................................................................... 59
APÊNDICE B ................................................................................................................... 58

15
1. INTRODUÇÃO
A glândula tireóide é a primeira a se formar durante a embriogênese. Responsável pela
síntese dos hormônios tireoidianos (HTs), que atuam regulando uma variedade de processos
metabólicos. A redução ou ausência na produção desses hormônios pode ocasionar retardo no
desenvolvimento cognitivo e motor da criança, ocasionando uma desordem endócrina
denominada Hipotireoidismo Congênito (HC).
O HC é a causa mais comum, prevenível, de retardo mental em crianças, sua
incidência varia entre 1: 3.000 a 4.000 nascidos (RAMOS et al., 2008).
Vários estudos realizados permitiram a caracterização dos genes que são essenciais
para síntese e desenvolvimento normal dos hormônios tireoidianos. Dentre eles, um grande
número de mutações no gene da tireoperoxidase (TPO) foram identificadas em pacientes com
HC por disormonogênese, evidenciando que defeitos nesse gene são a causa mais comum no
HC (RODRIGUES, 2004).
Os estudos moleculares do gene TPO revelaram que existe uma grande variação
alélica e que são visíveis ao nível da proteína. Foram descritas nesse gene mutações do tipo
missense (o que leva uma alteração no códon, alterando a proteína), nonsense (mutação de
ponto em uma sequência de DNA que resulta em um códon de parada), de frameshift
(mutação causada por inserções ou deleções onde o número de pares de bases não é múltiplo
de três, causando alteração na fase de leitura da proteína) e de splicing (excisão inapropriada
de um íntron ou deleção parcial ou até total de um éxon) ao longo de todo o gene
(NOGUEIRA et al., 2010).
Diante da grande heterogeneidade das mutações do gene TPO é necessário a
realização de estudos de caracterização molecular nesse gene para estabelecer uma relação
entre o genótipo e fenótipo apresentado pelo paciente. Na genética humana, as técnicas de
Biologia Molecular tornaram-se fundamentais no diagnóstico de doenças, permitindo não só a
identificação do gene afetado, mas também da mutação responsável pela doença. A
disponibilidade da análise molecular tem possibilitado o diagnóstico pré-natal e a triagem
populacional para tais doenças.
Os estudos genéticos moleculares utilizam várias técnicas para analisar mutações no
gene. A primeira descoberta que permitiu o avanço da tecnologia do DNA recombinante foi a
descoberta das enzimas de restrição, elas são fabricadas naturalmente por microorganismos

16
para destruir o DNA de vírus invasores, reconhecem pontos específicos - sítios de restrição
no DNA duplo, local em que cortam as sequências de nucleotídeos, originando os fragmentos
conhecidos como extremidades coesivas, que podem se ligar a outras pontas de moléculas de
DNA que tenham sido cortadas com a mesma enzima. Cada extremidade pode se unir a outra,
desde que seja um palíndromo, isto é, uma região de DNA com repetições invertidas.
Algumas enzimas geram extremidades cegas, isto é, sem bases despareadas, nesse caso o
DNA não é capaz de se ligar a outra sequência. As enzimas de restrição são utilizadas para
realização da técnica RFLP- Polimorfismo do Comprimento de Fragmento de Restrição
(Restriction Fragment Length Polymorfism) elas digerem o DNA em sítios específicos do
fragmento amplificado pela PCR e os padrões são detectados pela mobilidade eletroforética,
essa técnica é útil na elaboração de mapas de restrição.
No Mapa de Restrição estão contidas todas as enzimas disponíveis para um segmento
de DNA e essas enzimas são utilizados no momento da análise de uma mutação.
Normalmente elabora-se um mapa de restrição para uma região de interesse de um
cromossomo. Eles são utilizados na comparação de alelos normais e afetados para uma
determinada mutação estudada.
Outras técnicas utilizadas para detecção de mutações são: Polimorfismo de
conformação de fita simples (Single-Stranded Conformational Polymorfism - SSCP) a qual
possibilita determinar os polimorfismos em DNA de fita simples; Técnicas de amplificação de
alvos-específicos- a PCR Real-time; e o Sequenciamento- o sequenciamento de ácidos
nucléicos permite determinar a ordem das bases nucleotídicas na molécula do DNA, a análise
de mutações no seqüenciamento automático se dá pela identificação de picos diferentes na
leitura. Como possuímos dois alelos, quando há alteração na seqüência, o que observamos são
dois picos na leitura no fragmento de DNA (SAEZ et al., 2008). Os resultados destas técnicas
podem ser favoráveis no diagnóstico e na avaliação do tipo de mutação observado nos
pacientes.
No presente estudo foi realizada uma análise “in sílico” no gene da tireoperoxidase
para identificar os sítios de restrição presentes nos 17 éxons e assim criar um Mapa de
Restrição desse gene, como contribuição científica para análise das mutações que estão
relacionadas ao Hipotireoidismo Congênito no gene da Tireoperoxidase. Além de realizar um
levantamento bibliográfico para construção de uma tabela atualizada das mutações descritas
no gene TPO.
17
2. REVISÃO DE LITERATURA
2.1 Tireóide
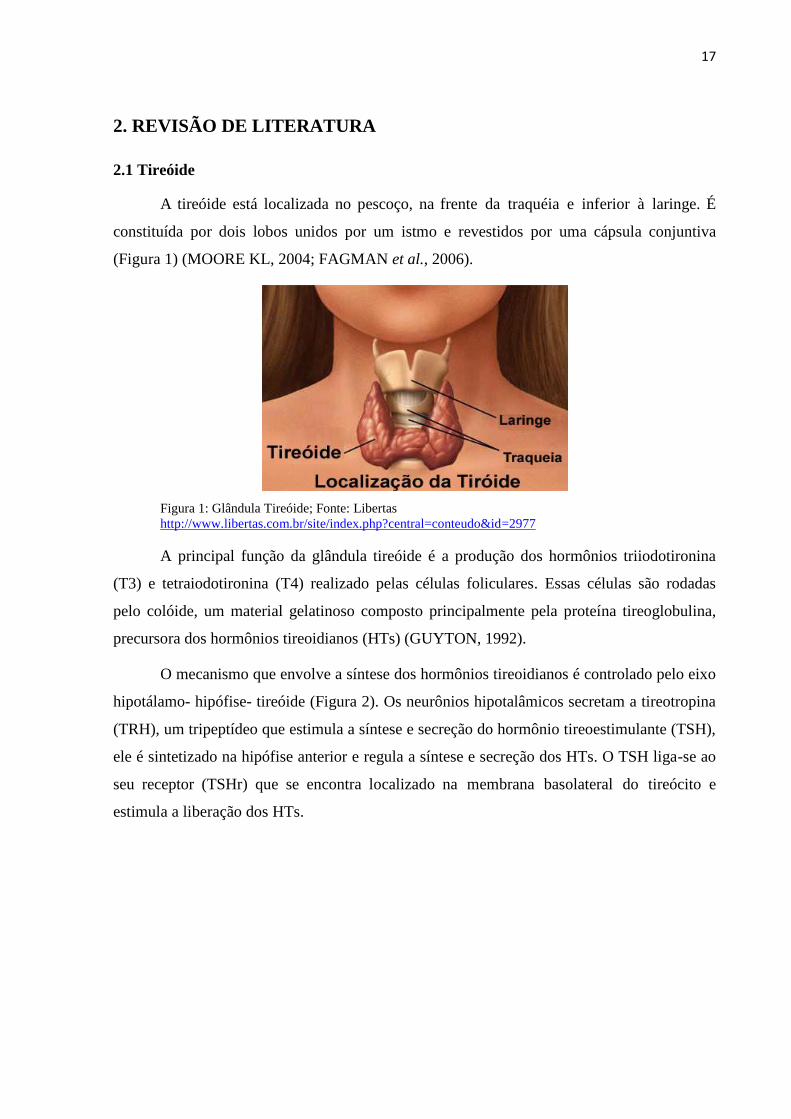
A tireóide está localizada no pescoço, na frente da traquéia e inferior à laringe. É
constituída por dois lobos unidos por um istmo e revestidos por uma cápsula conjuntiva
(Figura 1) (MOORE KL, 2004; FAGMAN et al., 2006).
Figura 1: Glândula Tireóide; Fonte: Libertas
http://www.libertas.com.br/site/index.php?central=conteudo&id=2977
A principal função da glândula tireóide é a produção dos hormônios triiodotironina
(T3) e tetraiodotironina (T4) realizado pelas células foliculares. Essas células são rodadas
pelo colóide, um material gelatinoso composto principalmente pela proteína tireoglobulina,
precursora dos hormônios tireoidianos (HTs) (GUYTON, 1992).
O mecanismo que envolve a síntese dos hormônios tireoidianos é controlado pelo eixo
hipotálamo- hipófise- tireóide (Figura 2). Os neurônios hipotalâmicos secretam a tireotropina
(TRH), um tripeptídeo que estimula a síntese e secreção do hormônio tireoestimulante (TSH),
ele é sintetizado na hipófise anterior e regula a síntese e secreção dos HTs. O TSH liga-se ao
seu receptor (TSHr) que se encontra localizado na membrana basolateral do tireócito e
estimula a liberação dos HTs.

18
Figura 2: Regulação da síntese e secreção dos hormônios da tireóide - Eixo hipotalâmico-hipofisário-tiróide
(Fonte: GREENSPAN, 2004. In RODRIGUES, 2004).
Segundo Rodrigues (2004), o TSH atua aumentando a transcrição dos genes essenciais
na hormonogênese coma a Tireoglobulina (TG), a Tireoperoxidase (TPO) e o Transportador
N a+ / I - (NIS), além de aumentar a produção de peróxido de hidrogênio, composto
indispensável na atuação da enzima TPO.
A produção dos hormônios TRH e TSH é controlada pelos níveis de T3 e T4 no
sangue, mecanismo de feedback negativo. Níveis elevados de T3 e T4 inibem a produção
de TSH e TRH que deixam de estimular os tireócitos diminuindo a síntese e secreção das
HTs. Outro fator que interfere na produção de HTs é a quantidade de iodo presente no
citoplasma, o excesso de iodo inibe o processo de iodização inibindo a síntese. Esse
mecanismo é conhecido como efeito Wolff-Chaikoff (VAISMAN, ROSENTHAL &
CARVALHO, 2004)
De acordo com Vaisman, Rosenthal & Carvalho (2003), a adequada produção dos
HTs depende do normal funcionamento das proteínas que são necessárias para a captação de
iodeto através da membrana dos tireócitos e sua incorporação à proteína aceptora, a
tireoglobulina, esta etapa é denominada organificação e depende da ação da enzima
Tireoperoxidade (TPO), principal enzima envolvida nesse processo.
2.2 Hipotireoidismo Congênito
O Hipotireoidismo Congênito (HC) é a uma doença endócrina que, afeta cerca

19
de 1: 3000 a 4000 recém-nascidos, causando retardo mental se tratamento ocorrer
tardiamente. ( GILLAM ;KOPP, 2001).
A deficiência é causada quando ocorre uma diminuição ou ausência na produção dos
hormônios tireoidianos. Os HTs são indispensáveis nos primeiros dias de desenvolvimento
da criança, confirmando a importância do diagnóstico precoce, para que o tratamento seja
iniciado mais rápido e possa evitar danos irreversíveis. Um recém-nascido não apresenta
sintomas específicos e a doença começa a se tornar grave no tempo em que a criança
permanece com déficit na produção HTs (AVBELJ et al, 2007).
A ausência dos HTs no início do desenvolvimento da criança pode ocasionar alguns
danos neurológicos, como deficiências nas funções cognitivas e coordenação motora
(NOGUEIRA et al., 2010).
O Programa de Triagem Neonatal é utilizado para detecção de algumas doenças
como Fenilcetonúria, Hemoglobinopatias, Fibrose Cística e Hipotireoidismo Congênito,
com o intuito de se estabelecer o diagnóstico precoce de tais doenças (BRASIL, 2003).
Gruters (1992) afirma que uma vez diagnosticado o HC, a terapia de reposição dos
hormônios tireoidianos deve ser iniciada até o primeiro mês de vida, pois durante esse
período, maiores são as possibilidades de evitar danos mentais e motores aos afetados.
De acordo com Foley (2000) o HC se classifica em quatro grupos principais:
1. Permanente - quando a ausência dos HTs ocorre durante toda vida do paciente.
2. Transitório - é diagnosticado precocemente (triagem neonatal) e desaparece
espontaneamente.
3. Primário - a tiróide não é capaz de produzir a quantidade suficiente dos hormônios
T3 e T4.
4. Secundário - Causado por doenças hipofisiárias ou hipotalâmicas (hipotireoidismo
hipotalâmico-pituitário), (RODRIGUES, 2004).
No Hipotireoidismo permanente esporádico os defeitos na organogênese da tireóide
são classificados como Disgenesia tireoidiana e ocorrem em aproximadamente 85% dos
casos de HC. A disgenesia tireoideana pode se apresentar com agenesia glandular (25% a
35% dos casos), definida como ausência de tecido tireóideo detectável; ectopia (40% a
60%), com tecido tireóideo encontrado desde a base da língua até o mediastino ou
hipoplasia (aproximadamente 5%), onde a glândula de tamanho reduzido se situa em

20
posição cervical normal e ainda ateriose que são defeitos na diferenciação inicial ou a
sobrevivência de células foliculares da tireóide (KNOBEL, NOGUEIRA & MEDEIROS-
NETO, 2001).
Os 10% dos casos de HC estão relacionados a defeitos da síntese hormonal causados
por disormonogênese e geralmente criam condições para fenótipo com presença do bócio e
grau variável de hipotireoidismo (PERONE et al., 2003). Esses defeitos são transmitidos na
maioria das vezes de forma autossômica recessiva, com exceção dos defeitos do receptor do
hormônio tireoidiano, que são autossômicos dominantes (KNOBEL; MEDEIROS, 2003). Os
5% restantes resultam da transferência de anticorpos maternos para o recém-nascido
(PERONE et al., 2003).
Os defeitos na síntese dos hormônios podem ser causados por mutações no gene
codificador para a proteína transportadora de iodeto (NIS), no gene pendrina (síndrome de
Pendred), no gene da tireoglobulina (TG), além de mutações que afetam o receptor de
hormônio tireóideo (resistência genética ao HT) ou aos vários defeitos no transporte de HT na
circulação periférica (RUBIO et al., 2008). No entanto, a forma mais comum resulta de
defeitos na organificação do iodo causado por mutações no gene da tireoperoxidase (TPO)
(RODRIGUES, 2004).
A causa mais comum de disormonogênese ocorre devido a alterações na enzima TPO.
De acordo com Menzalira (2010) três mecanismos podem alterar a atividade enzimática da
TPO:
1. Alterações na sequência correta de DNA, resultando em um códon de parada que
produzirá uma proteína incompleta conhecida como proteína truncada, essa proteína apresenta
expressão deficitária.
2. A troca de aminoácidos pode alterar a estrutura terciária da proteína, impedindo que
ela saía do retículo endoplasmático e perde sua função pela localização incorreta.
3. A substituição de um aminoácido que não altera a estrutura nem a localização da
proteína, mas a atividade da proteína permanece danificada.
2.3Tireoperoxidase
O gene da TPO humana (GenBank Acession number: NM_000547), localiza-se no
braço curto do cromossomo 2p2.5, está clonado e a sequência completa do RNAm possui
3146 nucleotídeos, composto de 17 éxons e 16 íntrons que se estendem por uma região de

21
aproximadamente 150 kb (ENDO et al., 1995). No banco de dados do GenBank estão
depositadas 7 isoformas do gene TPO, a mais atualizada foi a utilizada no nosso estudo
(NM_000547).
A expressão do gene da TPO ocorre principalmente na tireóide, traquéia e glândulas
salivares (AZA BLANC et al., 1993). Sua expressão é controlada pelo TSH através de
um sistema dependente de 3’5’-adenosina monofosfato cíclico (AMPc)/proteína quinase A
(GERARD et al., 1989) e também pelos fatores de transcrição, NKX2.1, FOXE1 e PAX8
que se ligam em sítios específicos do promotor, regiões TATAbox localizadas na posição -
25 do ponto de início da transcrição (ABRAMOWICZ et al., 1992).
A tireoperoxidase é sintetizada no retículo endoplasmático rugoso da célula e,
transferido para a membrana apical através do complexo de Golgi, onde irá catalisar os
processos de iodação e posteriormente o acoplamento de resíduos de tirosina na tiroglobulina
para formação dos hormônios da tireóide. É uma proteína glicosilada, com um grupo
prostético heme ligado à membrana, permanece localizada na membrana apical dos
tireócitos com o sítio catalítico exposto para o colóide (VAISMAN, ROSENTHAL &
CARVALHO, 2004).
A TPO possui 933 aminoácidos e peso molecular de 103 kDa é uma da principais
enzima envolvida na síntese dos hormônios tireoidianos (BAKKER et al., 2000), sendo
responsável por três reações:
A oxidação de íons iodeto (I-),
Iodação das tirosinas da TG e o
Acoplamento das tirosinas iodinizadas (acoplamento) para a formação dos hormônios
T4 e T3 (TAUROG A, 2000; LARSEN ET AL., 2003; VAISMAN; ROSENTHAL;
CARVALHO, 2004).
Já foram descritas outras isoformas de TPO encontrada em tireóides normais, chamada
TPO 2, apresenta 876 aminoácidos, e a TPO zanelli presente em doença de Graves que possui
929 aminoácidos. O papel dessas TPO ainda não foi esclarecido.
Várias mutações que atuam inativando ambos os alelos do gene da TPO, foram
identificadas em pacientes com HC, a grande da maioria localizada nos éxons 8, 9 e 10,
que contêm os resíduos de histidina de ligação heme, distais e proximais, que

22
correspondem à região catalítica da enzima. Alterações de seqüências nestes importantes
sítios podem, provavelmente, levar a inativação da TPO (PANNAIN et al., 1999).
2 .4 Mutações descritas no gene da Tireoperoxidase
A primeira mutação no gene TPO foi relatada por Abramowicz (1992). No estudo
realizado por Ris-Stalpers & Bikker (2010) foram descritas 61 mutações neste gene (como é
apresentado na Figura 3), essas mutações se disseminam por quase todo o gene variando entre
mutações missense, nosense, mutações no splicing e frameshift o que comprova sua elevada
heterogeneidade.
Figura 3: Mutações no gene TPO humano identificadas em pacientes com Hipotireoidismo Congênito.
(Fonte: RIS-STALPERS & BIKKER, 2010).

23
2.5 Análises de mutação
Para detecção de alterações genéticas envolvendo uma única base, ou grupos de bases,
são necessárias técnicas que comparem a forma mutante com a comum, dentre estas técnicas
podem destacar:
Eletroforese em gel com gradiente de desnaturação (DGGE) - esse método
utiliza uma eletroforese com gel de poliacrilamida desnaturante, ou seja, com uréia ou
formamida (substâncias desnaturantes). Nestas condições, ocorre a separação das moléculas
de DNA de acordo com sua temperatura de desnaturação. Os produtos são marcados por
coloração com brometo de etídio ou nitrato de prata para observar fitas homoduplex e
heteroduplex (ARIAS et al., 2003).
Polimorfismo de conformação de fita simples (SSCP) - essa técnica
possibilita a diferenciação de fragmentos de DNA de mesmo tamanho, a diferença de um
nucleotídeo na sequência do DNA altera o padrão de migração eletroforética (MOLINA;
TOBO, 2004). Na análise de SSCP, o DNA de fita dupla é desnaturado por calor na presença
de formamida e as fitas simples de DNA são analisadas por eletroforese em gel não
desnaturante de poliacrilamida (ARIAS et al., 2003).
Sequenciamento - O sequenciamento é realizado pela PCR utilizando, além
dos deoxinucleotídeos normais (dNTPs), os dideoxinucleotídeos (ddNTPs) marcados com
material fluorescente, uma cor para cada base, permitindo a detecção de diferentes fragmentos
durante a migração pela eletroforese capilar. Os fragmentos são lidos automaticamente por
lasers, com o auxílio de softwares para leitura dos dados (SAEZ, et al. ,2008)
2.5.1 Enzimas de restrição
As atuais técnicas utilizadas em Biologia Molecular tornaram-se possível graças à
identificação e purificação de enzimas nucleares. Entre elas destacam-se as enzimas de
restrição que reconhecem uma sequência específica de pares de bases de nucleotídeos e
clivam o DNA nesse sítio, gerando fragmentos de vários tamanhos que são separados e
analisados por eletroforese.
As enzimas são específicas e cada uma reconhece apenas o seu sítio de restrição
(HIRATA et al., 2006). Esses sítios são seqüências específicas de 4 a 8 pares de base (pb) na
molécula de DNA.

24
As enzimas são isoladas de bactérias e sua função, descoberta nos anos 70, é destruir
vírus que invadem a bactéria, os bacteriófagos. Uma vez cortados, os fagos tornam-se
inofensivos. As bactérias se protegem de suas próprias enzimas de restrição através da
metilação do seu DNA. Este sistema, pelo qual as endonucleases de restrição reconhecem o
DNA exógeno pode ser considerado um sistema imune primitivo da bactéria. Como a enzima
evoluiu até obter este grau de especificidade, ainda é desconhecido. Em organismos mais
evoluídos, incluindo humanos, a metilação está associada à capacidade da célula de ligar e
desligar genes.
Já foram identificadas mais de 1000 enzimas de restrição. E elas são nomeadas de
acordo com o nome da bactéria da qual a enzima foi isolada. A primeira letra representa o
gênero e as outras duas à espécie, seguido de um algarismo romano (ou outra letra) que indica
a ordem da descoberta ou a linhagem da qual ela foi isolada. Por exemplo, a enzima de
restrição denominada de EcoRI é purificada de uma Escherichia coli que carrega um fator de
transferência de resistência RI (NASCIMENTO et al., 2003).
2.5.2 Polimorfismo dos Comprimentos de Fragmento de Restrição (RFLP)
Em muitas doenças genéticas, o alelo causador da doença pode estar muito próximo a
um sítio de reconhecimento de uma enzima de restrição particular. Sabe-se que variações nas
sequências de DNA ocorrem aleatoriamente através do genoma inteiro e podem está
associadas a uma doença específica. Estas alterações de bases resultam na perda de um sítio
de restrição existente ou de aquisição de um novo sítio (HIRATA; HIRATA, 1999)
Este é o princípio em que se baseia a técnica de RFLP (Polimorfismo dos
Comprimentos de Fragmento de Restrição). Estes polimorfismos de fragmentos de restrição
(RFLP) podem ser facilmente testados em indivíduos, facilitando estudos com grandes
populações para identificação de polimorfismos (BOTSTEIN et al., 1980).
Alguns polimorfismos no genoma podem ser detectados pela comparação de mapas de
restrição de diferentes indivíduos, observado a mudança na mobilidade eletroforética do
padrão de fragmento produzido pela clivagem com a enzima de restrição (LEWIN, 2001). A
base para o polimorfismo entre os alelos são as diferentes mutações que podem ocorrer na
sequência de DNA. Os Polimorfismos de Comprimento de Fragmentos de Restrição (RFLP)
representam a ausência ou presença de sítios de reconhecimentos de enzimas de restrição em
diferentes indivíduos (GRIFFITHS, et al., 2006).

25
Muitas vezes uma mutação responsável por uma doença específica remove um sítio de
restrição que está normalmente presente no gene. Ou, ao contrário uma mutação ocasional
associada a uma doença altera a sequência normal, criando um sítio de restrição para uma
determinada enzima. É o que ocorre no íntron 3 do gene TPO, mutações nesse lócus criam um
novo local de corte para a enzima AluI, diferindo no padrão de bandeamento de um indivíduo
que não apresenta essa mutação. Também ocorre no éxon 8 que cria um local de corte para a
enzima DdeI.
Os RFLP são herdados como características genéticas simples obedecendo às leis
mendelianas de herança. Se for demonstrado em estudos que um gene produtor de uma
doença está ligado a um RFLP, este pode, então, fornecer um meio de detectar o gene
defeituoso sem efetivamente conhecê-lo (HIRATA; HIRATA, 1999).
2.5.3 Mapas de restrição
Os mapas de restrição são construídos para fornecer uma caracterização do genoma,
como por exemplo, o tamanho completo do genoma, número de sítios de restrição por enzima,
posição relativa desses sítios, e em comparação com outras espécies, a determinação de
regiões mais ou menos conservadas desse genoma. A posição de sítios de restrição no mapa
serve como um marcador que pode auxiliar na definição de espécies e na determinação das
relações filogenéticas entre espécies próximas (ARIAS et al., 2003).
O mapa de restrição construído durante o trabalho apresentado poderá ser utilizado na
análise de mutação dos pacientes que possuem HC por disormonogênese tratados no Hospital
Universitário Alcides Carneiro (HU).

26
3. OBJETIVOS
3.1 Objetivo Geral
Construir um mapa de restrição do gene da Tireoperoxidase (TPO) para correlacionar os
sítios de restrição e as mutações descritas que causam Hipotireoidismo Congênito (HC).
3.2 Objetivos específicos
Buscar na literatura as principais mutações no gene TPO que causam HC;
Identificar sítios de restrição nos 17 éxons do gene TPO para construção de um Mapa
de Restrição;
Investigar as enzimas descritas na literatura utilizadas na identificação de mutações no
gene TPO presente na literatura;
Correlacionar as mutações descritas na literatura com a sequência do GenBank e criar
uma tabela com as enzimas atualizada.

27
4. METODOLOGIA
4.1 Identificação das mutações descritas na literatura no gene TPO e construção de um
Mapa de Restrição
Foi realizado um levantamento bibliográfico, com dados obtidos através de uma busca
na literatura nos bancos de dados como Portal Capes que possui livre acesso as revistas
conceituadas como: Science, Nature, Clinical Endocrinology. Foram selecionados todos os
artigos que possuem mutações descritas no gene TPO para elaboração de uma tabela. Essa
tabela foi comparada com a tabela apresentada na dissertação de Menzalira (2010) e o artigo
publicado por Ris-Stalpers (2010), para contrapor as mutações com os respectivos artigos,
além de verificar a veracidade das tabelas apresentadas.
Com base nesse estudo foi elaborado um mapa de restrição com a utilização do
programa NEBcutter (http://tools.neb.com/NEBcutter2/), esse programa esta disponível no
site da empresa comercial New England BioLabs, responsável por comercializar enzimas para
a realização de pesquisas. As mutações descritas foram confirmadas na sequência do gene
TPO disponível do GenBank (NCBI Ref. Seq: NM_000547.5) e também utilizada pelos
autores que descreveram as mutações.
4.2 Construção do Mapa de Restrição do gene TPO
Inicialmente utilizou-se a sequência completa do RNAm (NCBI Ref. Seq:
NM_000547.5) do gene TPO para conferir e separar a sequência referente a cada éxon, para
posterior utilização desse sequência na construção do mapa de restrição, essa sequência foi
utilizada por ser a sequência de maior comprimento depositada no GenBank. A sequência
referente a cada éxon (1 a 17) do gene TPO foi copiada e colada na caixa de texto do
programa NEBcutter disponível no site BioLabs (http://tools.neb.com/NEBcutter2/index.php),
este programa é uma ferramenta utilizada para reconhecer os sítios de restrição de enzimas e
suas posições nos éxons da sequência do DNA. Selecionou-se as opções para obtenção de um
mapa no formato linear que apresentasse todas as enzimas com sítios no éxon, independente
de serem comercializadas ou não. As imagens com as sequências específicas foram
arquivadas para análise no momento da construção do mapa de restrição. Também utilizamos
o programa WEBcutter (http://rna.lundberg.gu.se/cutter2/) para confirmar as enzimas que o
programa NEBcutter apresentou para cada éxon.

28
Na construção do mapa foram excluídas as enzimas que possuem sítios degenerados, o
que significa que eles contêm um ou mais pares de bases que não são especificamente
definidos. Um exemplo é a enzima BsrFI que reconhece o sítio RCCGGY, onde na posição
representada por R pode está uma base A (Adenina) ou G (Guanina) e na posição Y pode está
uma base C (Citosina) ou T (Timina). Essas enzimas foram excluídas, pois caso ocorresse
uma alteração na posição degenerada a mesma não seria detectada pela enzima no momento
da análise de restrição.
4.3 Mutações identificadas e analisadas por RFLP na literatura
Durante a revisão descrita no tópico 3.2 os artigos que utilizaram a técnica RFLP
foram selecionados para posterior elaboração de uma tabela contendo as mutações e as
enzimas que possuem sítios no local da mutação ou em regiões próximas, sítios que são
criados ou eliminados na presença de uma alteração na sequência do gene TPO.

29
5. RESULTADOS
5.1 Mapa de Restrição do gene TPO
A figura 4 apresenta as enzimas que estão disponíveis nos respectivos éxons, essas
enzimas poderão ser utilizadas para analisar e/ou confirmar a presença de mutações no gene
TPO, uma vez que as mutações poderão criar ou eliminar os sítios de restrição presentes nos
éxons. Essa é uma maneira de facilitar a identificação e confirmação de mutações no gene
TPO. Esse mapa de restrição poderá ser utilizado como uma ferramenta no momento da
análise do gene TPO em pesquisas posteriores.
Figura 4: Desenho esquemático com enzimas disponíveis no Gene Tireoperoxidase. O número de enzimas disponíveis nos éxons
está representado abaixo.
A comparação entre o número de mutações e o número de enzimas de restrição
disponíveis em cada éxon foi realizada e está apresentada no gráfico 1.

30
Gráfico 1: Comparação entre o número de mutações e o número de enzimas de restrição disponíveis em
cada éxon.
As tabelas abaixo apresentam a descrição dos 17 éxons presentes no gene TPO, estão
apresentadas as enzimas e seus sítios de restrição e as posições em que ocorrem os cortes no
gene.
O éxon 1 do gene TPO apresenta 90 nucleotídeos e sete enzimas possuem sítios de
restrição nesse segmento. Até o momento nenhuma mutação foi descrita nesse éxon.
Tabela 1- Éxon 1:
Enzima Sítios de restrição Posição no éxon
MseI 5'... T T A A ... 3'
3'... A A T T ... 5'
8
KasI 5'... G G C G C C ... 3'
3'... C C G C G G ... 5'
12
NarI 5'...G G C G C C... 3'
3'...C C G C G G... 5'
13
HinP1I 5'... G C G C ... 3'
3'... C G C G ... 5'
13
SfoI 5'... G G C G C C ... 3'
3'... C C G C G G ... 5'
14
HhaI 5'... G C G C ... 3'
3'... C G C G ... 5'
15
HpyCH4V 5'... T G C A ... 3'
3'... A C G T ... 5'
58

31
O éxon 2 possui 94 nucleotídeos, representados da posição 91...185 no gene. Uma
mutação foi descrita nesse éxon, uma duplicação da posição 31 á posição 50, resultando em
uma mutação do tipo frameshift (Bikker et al., 1994). Nove enzimas possuem sítios de
restrição nesse segmento.
Tabela 2 - Tabela Éxon 2:
Enzima Sítio de restrição Posição no éxon
HinP1I 5'... G C G C ... 3'
3'... C G C G ... 5'
1
HhaI 5'... G C G C ... 3'
3'... C G C G ... 5'
3
PhoI 5'... G G C C ... 3'
3'... C C G G ... 5'
34
HaeIII 5'... G G C C ... 3'
3'... C C G G ... 5'
34
HpyCH4V 5'...T G C A... 3'
3'...A C G T... 5'
38
XhoI 5'...C T C G A G... 3'
3'...G A G C T C... 5'
63
TliI 5'... C T C G A G ... 3'
3'... G A G C T C ... 5'
63
PaeR7I 5'...C T C G A G... 3'
3'...G A G C T C... 5'
63
TaqI 5'... T C G A ... 3'
3'... A G C T ... 5'
64
O éxon 3 possui 84 nucleotídeos, está representado na posição 186...270, sete enzimas
possuem sítios de restrição e apenas uma mutação foi descrita nesse éxon. A mutação descrita
na posição c.157G>C, está apresentada no artigo como ocorrendo no éxon 3 , no entanto essa
posição encontra-se no éxon 2 (NIU, et al. 2002).
Tabela 3 - Éxon 3:
Enzima Sítio de restrição Posição no éxon

32
BssSI 5'... C A C G A G ... 3'
3'... G T G C T C ... 5'
9
BfaI 5'...C T A G... 3'
3'...G A T C... 5'
18
HinP1I 5'... G C G C ... 3'
3'... C G C G ... 5'
40
HhaI 5'... G C G C ... 3'
3'... C G C G ... 5'
42
AciI 5'... C C G C ... 3'
3'... G G C G ... 5'
54
CviAII 5'...C A T G... 3'
3'...G T A C... 5'
58
CviQI 5'... G T A C ... 3'
3'... C A T G ... 5'
61
HpyCH4V 5'... T G C A ... 3'
3'... A C G T ... 5'
73
O éxon 4 possui 169 nucleotídeos iniciando na posição 271...440, apenas uma enzima
possui sítio de restrição nesse segmento e duas mutações foram descritas. A mutação
c.292C>A descrita por Neves (2008) causa uma substituição da Leucina por uma Isoleucina
causando defeito total na incorporação do iodeto (DIIT).
Tabela 4- Éxon 4:
Enzima Sítio de restrição Posição no éxon
AciI 5'... C C G C ... 3'
3'... G G C G ... 5'
65
O éxon 5 possui 132 nucleotídeos e está representado na posição 441...573, cinco
enzimas possuem sítios de restrição, apenas duas mutações foram descritas nesse éxon. A
mutação c.387delC descrita por (Rivolta et al., 2003) é do tipo frameshift e resulta em um
códon de parada originando uma proteína truncada. Essa mutação está descrita na posição do
éxon 4.
Tabela 5- Éxon 5:
Enzima Sítio de restrição Posição no éxon

33
BglII 5'... A G A T C T ... 3'
3'... T C T A G A ... 5'
7
DpnI 5'...G A T C... 3'
3'...C T A G... 5'
9
PciI 5'...A C A T G T... 3'
3'...T G T A C A... 5'
30
HaeIII 5'... G G C C ... 3'
3'... C C G G ... 5'
100
AluI 5'... A G C T ... 3'
3'... T C G A ... 5'
113
O éxon 6 possui 129 nucleotídeos, está representado na posição 574...703, este
possui o maior número de enzimas com sítios disponíveis (10), mas apenas uma mutação foi
descrita. A mutação 613C>T dá origem a um códon de parada na proteína (R175X), causando
a perda de uma parte catalítica da enzima. Além de eliminar um sítio de restrição para a
endonuclease BsaXI , que está presente na ausência da mutação, a técnica de RFLP foi usada
para identificar essa mutação (AVBELJ et al., 2007).
Tabela 6- Éxon 6:
Enzima Sítio de restrição Posição no éxon
KasI 5'... G G C G C C ... 3'
3'... C C G C G G ... 5'
10
NarI 5'... G G C G C C ... 3'
3'... C C G C G G ... 5'
11
HinP1I 5'... G C G C ... 3'
3'... C G C G ... 5'
11
SfoI 5'...G G C G C C... 3'
3'...C C G C G G... 5'
12
HhaI 5'... G C G C ... 3'
3'... C G C G ... 5'
13
PhoI 5'... G G C C ... 3'
3'... C C G G ... 5'
25
HaeIII 5'... G G C C ... 3'
3'... C C G G ... 5'
25
BsrGI 5'... T G T A C A ... 3'
3'... A C A T G T ... 5'
98
CviQI 5'... G T A C ... 3'
3'... C A T G ... 5'
99
RsaI 5'... G T A C ... 3'
3'... C A T G ... 5'
100

34
O éxon 7 possui 206 nucleotídeos e está representado na posição 704...910, possui
sete enzimas com sítios disponíveis, e três mutações já foram descritas nesse éxon. A mutação
c.796C> T [p.Q266X], gera uma proteína truncada de 265 aminoácidos, que provoca a perda
da parte catalítica da enzima. Assim, grande parte de um dos principais domínios funcionais
da TPO é eliminado (BELFORT et al., 2012).
Tabela 7- Éxon 7:
Enzima Sítio de restrição Posição no éxon
PciI 5'... A C A T G T ... 3'
3'... T G T A C A ... 5'
11
AciI 5'... C C G C ... 3'
3'... G G C G ... 5'
53
TaqI 5'...T C G A... 3'
3'...A G C T... 5'
91
BstUI 5'... C G C G ... 3'
3'... G C G C ... 5'
105
AluI 5'... A G C T ... 3'
3'... T C G A ... 5'
132
MspI 5'... C C G G ... 3'
3'... G G C C ... 5'
204
HpaII 5'... C C G G ... 3'
3'... G G C C ... 5'
204
O maior comprimento esta presente no éxon 8, este possui 518 nucleotídeos e está
representado na posição 911...1429, possui sete enzimas com sítios disponíveis, e também
apresenta o maior número de mutações descritas, apresentando 17 ao longo do éxon. Os éxons
8, 9 e 10 codificam para a parte catalítica da enzima e são os locais mais freqüentes de
inativação causados por mutações (AVBELJ et al., 2007). Uma análise “in sílico” realizada
por Neves, 2008 detectou que a estrutura secundária da proteína pode ser alterada pela
presença do ácido glutâmico na posição 319 localizada em regiões de ligação de cálcio
alterando a conformação da proteína e função da enzima, ocasionada pela mutação 1046G>A
presente no éxon 8.
Tabela 8- Éxon 8:

35
Enzima Sítio de restrição Posição no éxon
NgoMIV 5'... G C C G G C ... 3'
3'... C G G C C G ... 5'
11
NaeI 5'... G C C G G C ... 3'
3'... C G G C C G ... 5'
13
FseI 5'... G G C C G G C C ... 3'
3'... C C G G C C G G ... 5'
15
BsrBI 5'... C C G C T C ... 3'
3'... G G C G A G ... 5'
46
MluI 5'... A C G C G T ... 3'
3'... T G C G C A ... 5'
148
BfaI 5'... C T A G ... 3'
3'... G A T C ... 5'
180
PvuII 5'... C A G C T G ... 3'
3'... G T C G A C ... 5'
191
O éxon 9 possui 258 nucleotídeos e está representado na posição 1430...1688, possui
nove enzimas com sítios disponíveis e 10 mutações já foram descritas. O éxon também
codifica uma parte catalítica da enzima. A mutação 1496delC origina um códon de parada
resultando em uma proteína TPO truncada, essa proteína não apresenta uma parte da
transmembrana e não pode ser adequadamente inserido na membrana apical (DELADOEY et
al., 2008).
Tabela 9- Éxon 9:
Enzima Sítio de restrição Posição no éxon
BamHI 5'... G G A T C C ... 3'
3'... C C T A G G ... 5'
22
StuI 5'... A G G C C T ... 3'
3'... T C C G G A ... 5'
39
CviQI 5'...G T A C... 3'
3'...C A T G... 5'
50
RsaI 5'... G T A C ... 3'
3'... C A T G ... 5'
51
AluI 5'... A G C T ... 3'
3'... T C G A ... 5'
175
HpaII 5'... C C G G ... 3'
3'... G G C C ... 5'
199
SmaI 5'... C C C G G G ... 3'
3'... G G G C C C ... 5'
200

36
HpyCH4V 5'... T G C A ... 3'
3'... A C G T ... 5'
212
NcoI 5'... C C A T G G ... 3'
3'... G G T A C C ... 5'
231
O éxon 10 possui 170 nucleotídeos e está representado na posição 1689...1859,
possui seis enzimas com sítios disponíveis e 10 mutações já foram descritas nesse éxon. A
mutação 1842 G>A (Arg584Gln) está presente no sítio catalítico da enzima TPO, essa região
é altamente conservada nas diferentes espécies, essas mutações alteram a conformação da
proteína e a função da enzima (NEVES, 2008)
Tabela 10- Éxon 10:
Enzima Sítio de restrição Posição no éxon
StuI 5'... A G G C C T ... 3'
3'... T C C G G A ... 5'
18
PvuII 5'...C A G C T G... 3'
3'...G T C G A C... 5'
58
Tsp509I 5'... A A T T ... 3'
3'... T T A A ... 5'
100
MluCI 5'... A A T T ... 3'
3'... T T A A ... 5'
100
HpaII 5'... C C G G ... 3'
3'... G G C C ... 5'
145
MspI 5'... C C G G ... 3'
3'... G G C C ... 5'
145
O éxon 11 possui 237 nucleotídeos, está representado na posição 1860... 2097
possui seis enzimas com sítios disponíveis, e oito mutações já foram descritas nesse éxon. A
substituição 1978 G>C no éxon 11 resulta na substituição de ácido glutâmico por glutamina
na posição 660 (Gln660Glu). Esta mutação já foi descrita em seis famílias portuguesas e duas
brasileiras (DELODEY et al., 2008).
Tabela 11- Éxon 11:
Enzima Sítio de restrição Posição no éxon

37
ApaI 5'... G G G C C C ... 3'
3'... C C C G G G ... 5'
177
ClaI 5'... A T C G A T ... 3'
3'... T A G C T A ... 5'
120
BspDI 5'... A T C G A T ... 3'
3'... T A G C T A ... 5'
120
BseYI 5'... C C C A G C ... 3'
3'... G G G T C G ... 5'
130
AluI 5'...A G C T... 3'
3'...T C G A... 5'
143
PspOMI 5'... G G G C C C ... 3'
3'... C C C G G G ... 5'
173
O éxon 12 possui 208 nucleotídeos e está representado na posição 2098...2306, possui
sete enzimas com sítios disponíveis, mas apenas duas mutações foram descritas. A delT na
posição 2243/2244 é causada por uma mutação Frameshift, que gera um códon de parada na
posição 718 originando uma proteína truncada, essa mutação foi descrita pela primeira vez em
pacientes holandeses (BAKKER et al.,2000).
Tabela 12- Éxon 12:
Enzima Sítio de restrição Posição no éxon
HpyCH4IV 5'... A C G T ... 3'
3'... T G C A ... 5'
28
BmgBI 5'... C A C G T C ... 3'
3'... G T G C A G ... 5'
15
AluI 5'... A G C T ... 3'
3'... T C G A ... 5'
42
NcoI 5'... C C A T G G ... 3'
3'... G G T A C C ... 5'
101
Tsp509I 5'... A A T T ... 3'
3'... T T A A ... 5'
124
MluCI 5'... A A T T ... 3'
3'... T T A A ... 5'
124
StuI 5'... A G G C C T ... 3'
3'... T C C G G A ... 5'
178
O éxon 13 possui 170 nucleotídeos, está representado na posição 2307...2477,
também possui dez enzimas com sítios disponíveis, e cinco mutações descritas. A mutação
tipo frameshift localizada na posição 2243insT foi primeiro relatada em pacientes de Taiwan
com defeito total na organificação do iodo (NIU et al.,2002).

38
Tabela 13- Éxon 13:
Enzima Sítio de restrição Posição no éxon
ApaLI 5'...G T G C A C... 3'
3'...C A C G T G... 5'
38
HinP1I 5'... G C G C ... 3'
3'... C G C G ... 5'
61
BstUI 5'... C G C G ... 3'
3'... G C G C ... 5'
63
HhaI 5'... G C G C ... 3'
3'... C G C G ... 5'
63
NgoMIV 5'... G C C G G C ... 3'
3'... C G G C C G ... 5'
81
NaeI 5'... G C C G G C ... 3'
3'... C G G C C G ... 5'
83
Eco53kI 5'... G A G C T C ... 3'
3'... C T C G A G ... 5'
97
SacI 5'... G A G C T C ... 3'
3'... C T C G A G ... 5'
99
HaeIII 5'...G G C C... 3'
3'...C C G G... 5'
105
PhoI 5'... G G C C ... 3'
3'... C C G G ... 5'
105
O éxon 14 possui 131 nucleotídeos e está representado na posição 2478...2609,
possui cinco enzimas com sítios disponíveis, e nove mutações descritas. Uma substituição na
posição 2512 leva a troca de um aminoácido cistina por uma serina, esta mutação foi descrita
em pacientes portugueses (RODRIGUES et al, 2005).
Tabela 14- Éxon 14:
Enzima Sítio de restrição Posição no éxon
HinP1I 5'...G C G C... 3'
3'...C G C G... 5'
81
BstUI 5'... C G C G ... 3'
3'... G C G C ... 5'
83
HhaI 5'...G C G C... 3'
3'...C G C G... 5'
83
MspI 5'... C C G G ... 3'
3'... G G C C ... 5'
129

39
HpaII 5'...C C G G... 3'
3'...G G C C... 5'
129
O éxon 15 possui 99 nucleotídeos e está representado na posição 2610...2709, este
possui quatro enzimas com sítios disponíveis, apenas três mutações descritas. A mutação de
ponto presente no éxon 15 (2669G>A (G860R)) é responsável por criar um novo sítio de
restrição para a enzima BclI. Essa alteração é primeira que resulta em uma substituição de
aminoácido na região transmembranar da enzima TPO (AVBELJ et al., 2007).
Tabela 15- Éxon 15:
Enzima Sítio de restrição Posição no éxon
FatI 5'... C A T G ... 3'
3'... G T A C ... 5'
27
CviAII 5'... C A T G ... 3'
3'... G T A C ... 5'
28
NlaIII 5'... C A T G ... 3'
3'... G T A C ... 5'
31
TaqI 5'... T C G A ... 3'
3'... A G C T ... 5'
74
O éxon 16 possui 129 nucleotídeos, está representado na posição 2710..2839 e não
apresenta enzimas disponíveis. Cinco mutações já foram descritas nesse éxon. A mutação
2738 C>T foi descrita pela primeira vez em pacientes brasileiros (Neves, 2008), esses
pacientes apresentam defeito total na organificação do iodo.
Éxon 16
Não há sítios de restrição com enzimas disponíveis. E 4 mutações já foram descrtitas
nesse éxon.
O éxon 17 possui o segundo maior cumprimento com 306 nucleotídeos e esta
representado na posição 2840...3146, possui cinco enzimas com sítios disponíveis, e nenhuma
mutações foi descrita nesse éxon.
Tabela 16- Éxon 17:
Enzima Sítio de restrição Posição no éxon

40
MspI 5'... C C G G ... 3'
3'... G G C C ... 5'
14
HpaII 5'...C C G G... 3'
3'...G G C C... 5'
14
PstI 5'... C T G C A G ... 3'
3'... G A C G T C ... 5'
70
BsiWI 5'... C G T A C G ... 3'
3'... G C A T G C ... 5'
97
BfaI 5'... C T A G ... 3'
3'... G A T C ... 5'
168
5.2 Levantamento das enzimas de restrição e mutações no gene TPO
Muitos autores (AMBRUGGER et al., 2001; AVBELJ et al., 2007; RIVOLTA et al.,
2007; RODRIGUES et al., 2005; WU et al., 2002;) utilizaram as enzimas de restrição como
uma maneira de confirmar a presença de uma mutação presente no gene TPO.
Estudos realizados com pacientes na Eslovênia, Bósnia e Eslováquia observaram
mutações no gene TPO que eliminavam os sítios de restrição para a endonuclease BsaXI, essa
análise foi realizada através da técnica de RFPL, e também indentificou mutações no éxon 8
(P280P) que eliminou o sítio de restrição para endonucleases NaeI e a mutação no éxon 11
(G667S) que eliminou o sítio de restrição da enzima HphI (AVBELJ et al, 2007).
Outro estudo realizado com pacientes Chineses utilizou a análise com enzimas de
restrição para confirmar a presença de mutações na sequência do gene TPO. Em um dos
pacientes foi detectado a presença de uma mobilidade eletroforética anormal no éxon 13, essa
mutação foi confirmada com a utilização de enzimas de restrição (WU et al, 2002).
Com base na revisão realizada, os artigos que utilizaram a técnica RFLP foram
selecionados e usados na elaboração da Tabela 17. Na tabela estão presentes os éxons com a
mutação e a enzima utilizadas na verificação. A mutação c.180-47A>C está descrita no éxon
3 (Rodrigues et al.,2004), no entanto a posição 180 está presente no éxon 2 e não condiz com
a sequência do GenBank.

41
Tabela 17- Tabela apresenta todas as mutações e respectivas enzimas utilizadas na comprovação ou
detecção descritas na literatura, reunidas após uma análise “in sílico”.
Éxon Mutação Enzima Referência
3 c.180-47A>C AluI (C) Rodrigues et al., 2004
6 c.613C>T BsaXI (E) Avbelj at al, 2007
8 1183_1186dupGGCC DdeI (C) Rodrigues et al., 2005
8 c.1159G>A Apa I (C) Rivolta et al,2007
8 1227delGGCC NaeI (C) Bakker et al, 2000
8 c.930 G>A NaeI (E) Avbelj et al, 2007
8 1273_1276dupGGCC NaeI (C) Avbelj et al, 2007
9 c.1463T>C MspI (C) Ambrugger, et al. 2001
9 1519_1539del BsaXI (E) Avbelj et al, 2007
9 c.1477G>A HaeIII(C) Wu et al, 2002
10 C1708C>T MslI (C) Tenebrum et al,2007
11 2033G>A DdeI (C) Pannain et al,1998
11 c.2089G>A HphI (E) Avbelj et al, 2007
13 c.2386G>T NaeI (C) Wu et al, 2002
14 2585G>A TaqI Pannain et al, 1999
15 c.2669G>A BclI (C) Avbelj et al, 2007
13 2268insT HincII (C) Niu et al., 2005
Legenda: C (Cria sítio de restrição); E (Elimina sítio de restrição)
A primeira mutação descrita no gene TPO foi uma duplicação do segmento CCGG no
éxon 8 apresentada por Abramowicz et al (1992), essa mutação encontra-se descrita em várias
populações apresentando uma elevada incidência. Analisando os dados que apresentam
mutações descritas no gene TPO, observou-se que até o momento já foram descritas 74
mutações que ocorrem ao logo de todo o gene, e tem maior incidência nos éxons 8, 9, 10
(responsáveis por codificar o sítio catalítico da enzima TPO).
A tabela apresentada no Apêndice A foi construída utilizando como base as tabelas
apresentadas na dissertação de Menzalira (2010) e no artigo científico publicado por Ris-
Stalpers (2010), a averiguação nos 38 artigos referenciados apresentou discordância com as
tabelas apresentadas nos trabalhos supracitados, pois nos respectivos trabalhos as posições em
que as mutações estão apresentadas não estão em concordância com os artigos de origem,
diferindo na posição da maioria das mutações, apesar de citar como base para descrição a
mesma sequência do GenBank (NCBI Ref. Seq: NM_000547.5) utilizada nos artigos.

42
No Apêndice A, apresenta-se a tabela revisada e atualizada, na qual se encontram 43
mutações missense (ocasionadas pela substituição de apenas uma base), 7 mutações nonsense
(que originam um códon de parada, formando uma proteína truncada), 5 mutações que afetam
o mecanismo de splicing, 16 mutações frameshift, com duas sendo deleções e 3 mutações
neutras que não ocasionam dano a nível de proteína.
As mutações missense, nonsense e as deleções revisadas foram verificadas na
sequência do gene TPO e a maioria apresentou discordância na posição da base, pois onde
eles indicam conter um tipo de base apresenta outra. Das 52 mutações revisadas apenas 22
estão em concordância com a sequência do gene TPO. A partir dessas 22 mutações foi
elaborada uma tabela presente no apêndice B com as enzimas que possuem sítios de restrição
na sequência normal do gene, essas enzimas possuem sítios em locais próximos da mutação.
Na presença das mutações esses sítios podem ser eliminados e não haverá corte no fragmento
o que possibilita uma verificação da ocorrência de uma mutação nessa posição. A mutação
também pode criar um local de corte para uma nova enzima de restrição. A tabela apresentada
no Apêndice B possui os sítios de restrição presentes na sequência normal do gene TPO e
sítios formados ou eliminados na presença da mutação.

43
6. DISCUSSÃO
Para a construção do Mapa de Restrição do gene TPO utilizamos como sequência base
a mesma descrita nos artigos encontrados durante o levantamento bibliográfico (GenBank
Acession number: NM_000547), os resultados apresentaram muitas enzimas disponíveis em
cada éxon, as que não são comercializadas ou possuem sítios degenerados foram excluídas no
momento da construção. Esse mapa será muito importante para futuras pesquisas com o gene
TPO, pois a utilização de enzima de restrição é uma ferramenta acessível e rápida na detecção
de mutações, muitos autores utilizam as enzimas como uma forma de confirmar uma mutação.
Em estudos realizados em Taiwan um paciente foi caracterizado por SSCP e apresentou uma
migração eletroforética anormal no éxon 9, com o sequenciamento observou-se que esse
paciente apresentava uma substituição de guanina por adenina na posição 1477, e essa
alteração elemina o sítio de restrição da enzima HaeIII, esse estudo utilizou a enzima HaeIII
para determinar a segregação do alelo na família e sua frequência de alelos na população em
geral (Wu et al., 2002), por se tratar de uma ferramenta mais acessível que o sequenciamento.
Dessa forma, evidencia-se a importância da construção de mapas de restrição para outras
doenças genéticas, pois conhecendo as enzimas que podem ser utilizadas em uma análise de
RFLP o produto dos fragmentos digeridos pode ser comparado com o de pacientes normais e
que possuem a doença, facilitando a descoberta da natureza da mutação, correlação genótipo-
fenótipo e utilização no diagnóstico de doenças genéticas.
Em outro estudo realizado uma mutação cria um sítio de restrição para a enzima MspI
no éxon 9, uma análise de restrição foi realizada na família desse paciente para observar se
algum parente também apresentava a referida mutação (AMBRUGGER et al., 2001). A
grande heterogeneidade alélica do gene TPO e o fato das características das mutações serem
específicas para diferentes populações permitem a utilização do mapa de restrição para a
realização de triagem das mutações nas famílias analisadas.
A princípio o Mapa de Restrição seria construído para as mutações descritas no gene
TPO, no entanto a posição de nucleotídeos dessas mutações diferiram da sequência disponível
no GenBank o que dificultou a construção do mapa para as mutações encontradas. Das
mutações que as sequências coincidiram construímos uma tabela com as enzimas que
possuem sítios próximos ou dentro do sítio da mutação, essas enzimas poderão ser utilizadas
diretamente na análise da respectiva mutação, facilitando a detecção da mutação.

44
A partir desse levantamento também foi elaborada uma tabela com todas as enzimas
que já foram confirmadas como ferramenta na análise de restrição do gene TPO, essas
enzimas estão confirmadas em experimentos descritos na literatura. No caso dos exemplos
supracitados.
Também foi construída uma Tabela atualizada contendo todas as mutações descritas
no gene TPO relacionadas ao HC.
De acordo com o nosso estudo a maioria das mutações apresentaram disparidade entre
a sua posição descrita e a sequência do GenBank indicada por cada autor, fato que dificultou a
construção do Mapa de Restrição para essas mutações, das 52 mutações (missense, nonsense e
deleções) descritas até o momento apenas 22 estavam em concordância com a sequência do
GenBank.
No artigo de Rivolta (2007) uma deleção descrita na posição 215 (c.215delA) está
descrita como presente no éxon 4, o autor citado afirma utilizar como base a mesma sequência
do GenBank (NM_000547) na qual foi realizada a análise “in sílico” do trabalho apresentado,
nossa análise na sequência de gene TPO indica que o nucleotídeo presente na posição 215
pertence ao éxon 3, essa mutação é do tipo frameshift e resulta em um códon de parada no
aminoácido 86 da proteína (p.Q72fsX86). Segundo Belfort (2012) a mutação do tipo
missense na posição 1784(c.1784G>A) esta presente no éxon 11, embora utilize a mesma
sequência do GenBank usada na análise apresentada.
Para a construção dessa tabela (Apêndice A) tomamos como base as tabelas
apresentados nos artigos de Ris-Stalpers (2010) e a dissertação de Menzalira (2010),
realizamos uma verificação entre as mutações apresentadas por esses autores e os artigos
referenciados e constatamos que muitas mutações apresentadas não estavam em concordância
com o autor citado, como a mutação c.1994G>A presente no éxon 11 citada na dissertação de
Menzalira (2010) como descrita por Nascimento (2003), durante a constatação no respectivo
artigo observamos que não há descrição para tal mutação. Outro fato observado foi que
algumas mutações estão descritas pelo autor citado em outras posições, mas com o mesmo
efeito a nível da proteína é o que ocorre na mutação apresenta da no artigo de Ris-Stalpers
(2010) ele indica a mutação na posição 1597 (c.1597G>T) que causa uma mudança de
aminoácido na posição 533, essa troca de aminoácido é apresentada no artigo como ocorrendo
devido a uma mutação na posição 1687 e causada pela mesma substituição de nucleotídeo.

45
As divergências supracitadas foram encontradas mesmo a maioria dos autores
afirmando ter utilizado a mesma sequência do gene TPO, essas discordâncias podem ter
ocorrido devido a atualizações na sequência apresentado no Genbank, pois com a realização
de novas pesquisas no gene TPO, novos sequenciamento são realizados em pacientes e
controles e os dados desses resultados são depositados no banco de dados do Genbank. Ou de
alguma forma, pode ter ocorrido erros no momento das anotações das mutações ou
consolidação dos resultados.

46
7. CONCLUSÃO
A partir dessa sequência foi possível construir um mapa de restrição que poderá ser
utilizado em estudos posteriores e uma tabela com as enzimas de restrição utilizadas na
detecção de mutações no gene TPO. Esse mapa é fundamental em futuras pesquisas, pois
permitem analisar o tipo de mutação de forma mais rápido e acessível, tendo em vista que as
enzimas possuem um baixo custo comparado ao do sequenciamento, à utilização também é
essencial para o aconselhamento genético das famílias investigadas.
A investigação de literatura realizada nesse estudo permitiu verificar que as mutações
descritas nos artigos referenciados por Menzalira (2010) e Ris-Stalpers (2010) não apresentam
na sua totalidade concordância com a sequência do gene TPO presente no GenBank, sendo a
mesma que a maioria dos autores (alguns autores não descrevem a sequência que utilizaram
como base) afirmam ter utilizado para nomear as mutações encontradas por cada um.
O presente trabalho realizou um levantamento bibliográfico para construção de um
Mapa de Restrição para os 17 éxons presentes no gene TPO com a finalidade de auxiliar em
futuras pesquisas com esse gene.

47
8. PERSPECTIVAS
O mapa de restrição do gene TPO e as enzimas desenvolvidas nesse trabalho poderão
ser utilizadas em estudos posteriores com enzimas de restrição para análise de mutação no
gene TPO e a partir da sua publicação poderão ser utilizadas em futuros estudos para a
triagem das mutações e o uso no aconselhamento genético.

48
REFERÊNCIAS BIBLIOGRÁFICAS
ABRAMOWICZ, Marc J. et al. Identification of a Mutation in the Coding Sequence of the
Human Thyroid Peroxidase Gene Causing Congenital Goiter. Journal of Clinical
Investigation. 90 (4): 1200-1204, 1992.
AMBRUGGER, P. et al. Novel mutations of the thyroid peroxidase gene in patients with
permanent congenital hypothyroidism", Eur.J.Endocrinol., 145 (1): 19-24, 2001.
ARIAS M. C., FRANCISCO F. O., SILVESTRE D. O DNA mitocondrial em estudos
populacionais e evolutivos de meliponíneo. In G. A. R. Melo & I. Alves-dos-Santos, Apoidea
Neotropica: Homenagem aos 90 anos de Jesus Santiago Moure. Editora UNESC,
Criciúma, 2009.
AVBELJ, Magdalena. et al. High prevalence of thyroid peroxidase gene mutations in patients
with thyroid dyshormonogenesis. European Journal of Endocrinology. 156( 5):511–519,
2007.
AZA-BLANC P., DI LAURO R., SANTISTEBAN P. Identification of a cis-regulatory
element and a thyroid-specific nuclear factor mediating the hormonal regulation of rat
thyroid peroxidase promoter activity. Mol. Endocrinol, 1993.
BAKKER, B. et al. Two decades of screening for congenital hypothyroidism in the
Netherlands: TPO gene mutations in total iodide organification defects (an update). J Clin
Endocrinol Metab. 85, 3708-3712, 2000.
BELFORT F. S. et al. Congenital goitrous hypothyroidism: mutation analysis in the thyroid
peroxidase gene. Clinical Endocrinology 76, 568–576, 2012
BIKKER, H. Et al. A 20-basepair duplication in the human thyroid peroxidase gene results in
a total iodide organification defect and congenital hypothyroidism",
J.Clin.Endocrinol.Metab, 79 (1): 248-252, 1994.
BIKKER, H. Et al. "Identification of five novel inactivating mutations in the human thyroid
peroxidase gene by denaturing gradient gel electrophoresis", Hum.Mutat., 6 (1): 9-16, 1995.
BIKKER, H., BAAS, F., DE VIJLDER, J. J. "Molecular analysis of mutated thyroid
peroxidase detected in patients with total iodide organification defects",
J.Clin.Endocrinol.Metab, 82 (2): 649-653, 1997.
BRASIL. Ministério da Saúde. Secretaria de Atenção à Saúde. Coordenação-Geral de Média
Complexidade Ambulatorial. Manual de normas técnicas e rotinas operacionais do programa
nacional de triagem neonatal. – 1. ed., Brasília: Ministério da Saúde, 2003.
BOTSTEIN, D. et al. Construction of a genetic linkage map in man using restriction fragment
length polymorphisms. Am. J. Hum. Genet. 32, 314-331. 1980.
DELADOEY, J. et al. "Pseudodominant inheritance of goitrous congenital hypothyroidism
caused by TPO mutations: molecular and in silico studies", J.Clin.Endocrinol.Metab, 93(2):
627-633, 2008.

49
ENDO Y. et al. Regional localization of the gene for thyroid peroxidase to human
chromosome 2p25 and mouse chromosome 12C. Genomics. 25, 760–761. 1995.
FAGMAN H., ANDERSSON L., NILSSON M. The developing mouse thyroid: embryonic
vessel contacts and parenchymal growth pattern during specification, budding, migration,
and lobulation. Dev Dyn. 235(2):444-45, 2006.
FOLEY T.P. Congenital hypothyroidism. In: Werner and Ingbar’s The Thyroid, edn 8, part
1, ch 4, pp 977-983. Eds LE Braverman & RD Utiger, Philadelphia: Lippincott Williams
& Wilkins, 2000.
FUCHS, O. et al. "Congenital hypothyroidism caused by a novel homozygous mutation in the
thyroid peroxidase gene", J.Pediatr.Endocrinol.Metab, 21(11): 1093-1097, 2008.
GÉRARD C.M.,et al. Control of thyroperoxidase and thyroglobulin transcription by
cAMP: evidence for distinct regulatory mechanisms. Mol Endocrinol. 3(12): 2110-2118,
1989.
GILLAM M.P., KOPP P. Genetic defects in thyroid hormone synthesis. Curr
OpiPediatr.13:364-72, 2001.
GRIFFITHS, A. J. F., et al. Introdução à genética. 8ª ed. Rio de Janeiro: Guanabara Koogan,
2005.
GRUTERS A. Congenital hypothyroidism. Pediatric Annals. 21(1): 15-21; 1992.
GUYTON, Arthur C. Tratado de Fisiologia Médica. 8. ed. Rio de Janeiro: Guanabara
Koogan, p. 731-738, 1992.
HIRATA, M. H. et al. Da biologia molecular à medicina: métodos comumente utilizados em
farmacogenética. Medicina (Ribeirão Preto); 39 (4): 522-34, 2006.
KOTANI, T. et al."A novel mutation in the human thyroid peroxidase gene resulting in a total
iodide organification defect", J.Endocrinol. 160 (2): 267-273, 1999.
KOTANI, T. et al. "Partial iodide organification defect caused by a novel mutation of the
thyroid peroxidase gene in three siblings", Clin.Endocrinol.(Oxf). 59 (2): 198-206, 2003.
KNOBEL, M. et al. Genética Molecular do Hipotireoidismo Congênito. Arquivo Brasileiro
de Endocrinologia & Metabologia. 45 (1). 2001.
KNOBEL M.; MEDEIROS-NETO G. An outline of inherited disorders of the thyroid
hormone generating system. Thyroid. 13(8), 771-801. 2003.
LARSEN P. R. et al. Thyroid physiology and diagnostic evaluation of patients with
thyroid disorders. In: Larsen PR, Kronenberg HM, Melmed S, Polonsky KS, editors.
Williams’ textbook of endocrinology. 10th ed. Philadelphia: W.B.Saunders, p.331-73;
2003.

50
Lewin B., Genes VII. 7ª. Ed Artemed, 2001. 960 p.
PANNAIN, S. et al. "Two different mutations in the thyroid peroxidase gene of a large inbred
Amish kindred: power and limits of homozygosity mapping", J.Clin.Endocrinol.Metab, 84
(3): 1061-1071, 1999.
PFARR, N. et al. "Goitrous congenital hypothyroidism and hearing impairment associated
with mutations in the TPO and SLC26A4/PDS genes", J.Clin.Endocrinol.Metab, 91 (7):
2678-2681, 2006a.
PFARR, N. et al. "Congenital primary hypothyroidism with subsequent adenomatous goiter in
a Turkish patient caused by a homozygous 10-bp deletion in the thyroid peroxidase (TPO)
gene", Clin.Endocrinol.(Oxf), 64(5): 514-518, 2006b.
PERONE D et al. Aspectos Genéticos do Hipotireoidismo Congênito. Arquivo Brasileiro
Endocrinologia & Metabolismo. vol 48 nº 1, 2004.
MEZALIRA, P. R. Rastreamento e estudo funcional de alterações no gene da
tireoperoxidase associadas ao hipotireoidismo congênito com defeito parcial de
incorporação de iodeto. Dissertação (Mestrado em Endocrinologia) - Faculdade de
Medicina, Universidade de São Paulo, SP, 2010.
MOLINA, A. L., TOBO, P. R. Uso das técnicas de biologia molecular para diagnóstico.
Einstein. 2(2):139, 2004.
MOORE, K. L. Fundamentos de anatomia clínica. 2ª ed. Rio de Janeiro: Guanabara Koogan,
p. 501. 2004.
NASCIMENTO, A. C. et al. "Thyroperoxidase gene mutations in congenital goitrous
hypothyroidism with total and partial iodide organification defect", Thyroid. 13 (12): 1145-
1151, 2003.
NEVES, S. C. Hipotireoidismo congênito: rastreamento e identificação de mutações no
gene TPO em pacientes com defeito parcial ou total de incorporação do iodeto. 131 f.
Dissertação [Mestrado em Ciências com concentração em endocrinologia] – Faculdade de
Medicina da Universidade de São Paulo, 2008.
NIU, D. M. et al. "High prevalence of a novel mutation (2268 insT) of the thyroid peroxidase
gene in Taiwanese patients with total iodide organification defect, and evidence for a founder
effect", J.Clin.Endocrinol.Metab. 87( 9): 4208-4212, 2002.
NIU, D. M. et al. "Contribution of genetic factors to neonatal transient hypothyroidism",
Arch.Dis.Child Fetal Neonatal. 90(1): F69-F72, 2005.
NOGUEIRA C. et al. Caracterização molecular do gene TPO em crianças Portuguesas com
hipotireoidismo congênito causado por disormonogénese. Acta Pediátrica Portuguesa,
2010.

51
RAMOS H. E. et al. Novos aspectos da genética e dos mecanismos moleculares da
morfogênese da tiróide para o entendimento da Disgenesia Tiroidiana. Arq Bras Endocrinol
Metab, 52/9.2008.
RIS-STALPERS, C.; BIKKER, H. Genetics and phenomics of hypothyroidism and goiter due
to TPO mutations. Molecular and Cellular Endocrinology. 322, 38–43. 2010.
RIVOLTA, C. M. et al. "Five novel inactivating mutations in the thyroid peroxidase gene
responsible for congenital goiter and iodide organification defect", Hum.Mutat. 22 (3):,
2003.
RIVOLTA, C. M. et al. "Two compound heterozygous mutations (c.215delA/c.2422T-->C
and c.387delC/c.1159G-->A) in the thyroid peroxidase gene responsible for congenital goitre
and iodide organification defect", Clin.Endocrinol.(Oxf), 67(2): 238-246, 2007.
RODRIGUES, C. et al. "Mutation screening of the thyroid peroxidase gene in a cohort of 55
Portuguese patients with congenital hypothyroidism", Eur.J.Endocrinol. 152 (2): 193-198,
2005.
RODRIGUES, C. F. Rastreio molecular do gene da peroxidase da tireóide em doentes
com hipotiroidismo congênito. 2004. 105f. Dissertação [Mestrado em Genética Molecular] -
Universidade do Minho. Escola de Ciências. Departamento de Biologia, 2004.
RUBIO, G. et al. Hipotireoidismo Congênito: Recentes Avanços em Genética Molecular.
Arquivo Brasileiro de Endocrinologia & Metabolismo. 46 (4): 391-401, 2002.
SAEZ, C. R. N. et al. Seqüenciamento automático de parte da região 3’ do gene c3-22 de
rhynchosciara americana (diptera: sciaridae). IV Mostra Interna de Trabalhos de Iniciação
Científica do Cesumar. 2008.
SANTOS, C. L. et al. "A novel mutation in the TPO gene in goitrous hypothyroid patients
with iodide organification defect", Clin.Endocrinol.(Oxf), 51 (2): 165-172, 1999.
SIMM, D. et al. "Two novel mutations in the human thyroid peroxidase (TPO) gene: genetics
and clinical findings in four children", Acta Paediatr., 98 (6): 1057-1061, 2009.
TAJIMA, T. et al. "Two novel mutations in the thyroid peroxidase gene with goitrous
hypothyroidism", Endocr.J., 52 (5): 643-645, 2005.
TAUROG A. Hormone synthesis: thyroid iodine metabolism. In: Braverman LE, Utiger
RD, editors. Werner and Ingbar’s The Thyroid, 8th ed, part 1, ch 4. Philadelphia:
Lippincott Williams & Wilkins, 2000.
TENENBAUM-RAKOVER, Y. et al. "Clinical and genetic characteristics of congenital
hypothyroidism due to mutations in the thyroid peroxidase (TPO) gene in Israelis",
Clin.Endocrinol.(Oxf), 66(5): 695-702, 2007.
UMEKI, K. et al. "Two novel missense mutations in the thyroid peroxidase gene, R665W and
G771R, result in a localization defect and cause congenital hypothyroidism",
Eur.J.Endocrinol. 146(4):491-498, 2002.

52
UMEKI, K. et al. "Congenital hypothyroidism caused by a unique thyroid peroxidase allele
containing two mutations, C1708T and C2737T", J.Pediatr.Endocrinol.Metab, 17(2): 231-
234, 2004.
VAISMAN, M.; ROSENTHAL, D.; CARVALHO, D. P. Enzimas Envolvidas na Organificação
Tireoideana do Iodo. Arq Bras Endocrinol Metab. 48(1). 2004.
Wu, J. Y. et al. "Mutation analysis of thyroid peroxidase gene in Chinese patients with total
iodide organification defect: identification of five novel mutations", J.Endocrinol., 172(3):
627-635, 2002
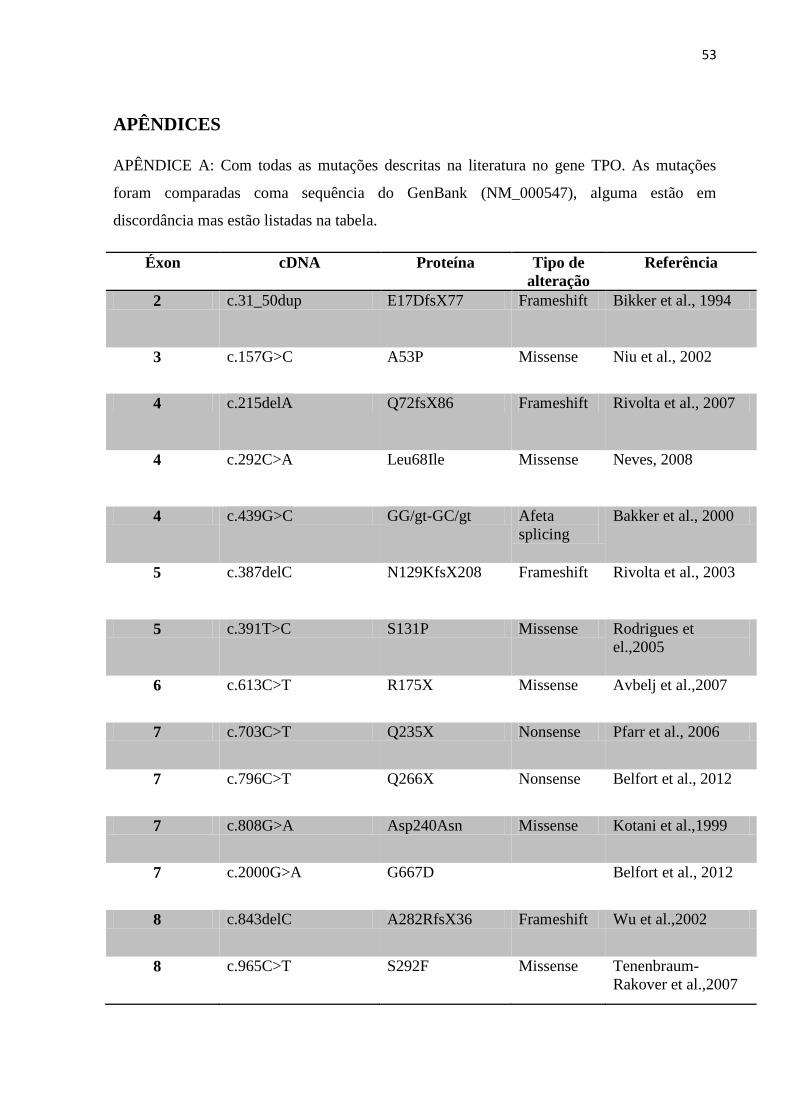
53
APÊNDICES
APÊNDICE A: Com todas as mutações descritas na literatura no gene TPO. As mutações
foram comparadas coma sequência do GenBank (NM_000547), alguma estão em
discordância mas estão listadas na tabela.
Éxon
cDNA Proteína Tipo de
alteração
Referência
2 c.31_50dup E17DfsX77 Frameshift Bikker et al., 1994
3 c.157G>C A53P Missense Niu et al., 2002
4 c.215delA Q72fsX86 Frameshift Rivolta et al., 2007
4 c.292C>A Leu68Ile Missense Neves, 2008
4 c.439G>C GG/gt-GC/gt Afeta
splicing
Bakker et al., 2000
5 c.387delC N129KfsX208 Frameshift Rivolta et al., 2003
5 c.391T>C S131P Missense Rodrigues et
el.,2005
6 c.613C>T R175X Missense Avbelj et al.,2007
7 c.703C>T Q235X Nonsense Pfarr et al., 2006
7 c.796C>T Q266X Nonsense Belfort et al., 2012
7 c.808G>A Asp240Asn Missense Kotani et al.,1999
7 c.2000G>A G667D Belfort et al., 2012
8 c.843delC A282RfsX36 Frameshift Wu et al.,2002
8 c.965C>T S292F Missense Tenenbraum-
Rakover et al.,2007

54
8 c.920A>C N307T Missense Rivolta et al., 2003
8 c.930G>A P280P Neutra Avbelj et al., 2007
8 c.940C>T R314W Missense Fuchs et al., 2008
8 c.943C>T Q315X Nonsense Alper et al., 2010
8 c.1066G>A Ala326Thr Missense Bakker et al., 2000
8 c.1046G>A Gly319Glu Missense Neves, 2008
8 c.1159G>A G387R Missense Rivolta et al., 2007
8 c.1222G>A E378K Missense Tajima et al., 2005
8 c.1274A>G N425S Missense Rodrigues et al.,
2005
8 c.1297G>A V433M Missense Rivolta et al., 2003
8
1338G>C Q446H
Missense Simm et al., 2009
8 c.1367C>G Ala426Gly Missense Neves, 2008
8 c.1425delC Q446RfsX5 Frameshift Bakker et al., 2000
9 c.1429A>T I447F Missense Bikker et al., 1997
9 c.1447T>G Tyr453Asp Missense Bikker et al., 1997
9 c.1463T>C Leu458Pro Missense Ambrugger et al.,
2001
9 c.1477G>A G493S Missense Wu et al., 2002
9 c.1496C>T P499L Missense Rivolta et al., 2003

55
9 c.1496delC Pro499Argfs2X Frameshift Deladoey et al.,2008
9 c.1519_1539del A477_N483del Frameshift Avbelj et al.,2007
9 c.1562G>A Arg491His Missense Ambrugger et al.,
2001
9 1597G>T G533C
c.1687G>T G533C Missense Kotani et al.,2003
9 c.1671 G>T Trp527Cys Missense Bakker et al., 2000
10 c.1780C>A L564I Missense Nascimento et
al.,2003
10 c.1768+1G>A Sítio de
splicing
Bakker et al., 2000
10 +1 intron 10 AG/gt3AG/at Afeta
splicing
Bakker et al., 2000
10 c.1708C>T Arg540stop Nonsense Bikker et al., 1997
10 1808–13del Asp574/Leu575d
el
Frameshift Kotani et al.,2003
10 c.1842G>A Arg584Gln Missense Neves et al., 2008
10 c.1858G>A Gly590Ser Afeta
splicing
Bikker et al., 1997
11 c.1942G>A Val618Mat Missense Neves et al., 2008
11 c.1784G>A R595K Missense Belfort et al., 2012
11 c.1955insT Phe653Valfs15X Frameshift Deladoe y et al.,
2008
11 c.2084G>A R665Q Missense Nascimento et al.,
2003

56
11 c.2033 G >A R648Q Missense Pannain et al., 1999
11 c. 2068C>G Q66OE Missense Santos et al., 1999
11 c.2083C>T R665W Missense Umeki et al.,2002
11 c.2089G>A G667S Missense Avbelj et al., 2007
12 2243_2244delT F718X Nonsense Bakker et al., 2000
12 c.2167C>T Arg693Trp Missense Bakker et al., 2000
13 c.2268insT E756X Nonsense Niu et al., 2002
13 c.2268-2269insT Asp796Tyr Frameshift Wu et al., 2002
13 c.2386G>T Asp796Tyr Missense Wu et al., 2002
13 c. 2401G>A G771R Missense Umeki et al., 2002
13 c.2243delT V748GfsX50 Frameshift Niu et al., 2002
14 c.2413delC H805WfsX27 Frameshift Wu et al., 2002
14 c.2421dupC C808LfsX72 Frameshift Bikker et al., 1995
14 c.2422T>C C808R Missense Rivolta et al., 2003
14 c. 2565C>T C825C Neutra Avbelj et al., 2007
14 c.2485G>A Glu 799 Lys Missense Bikker et al., 1997

57
14 c.2512delT C838AfsX1 Frameshift Bakker et al., 2000
14 c.2512T>A C838S Missense Rodrigues et al.,
2005
14 c.2505insC Frameshift Bikker et al.,1995
VS15-
exon16
c.2619_2622del ? Deleção Tajima et al,2005
15 2619G>A W873X Nonsense Simm et al., 2009
15 c.2669G>A G860R Missense Avbelj et al.,1997
16 c.2738C>T Pro883Leu Missense Neves, 2008
16 c.2748G>A Q916Q Neutra/Sítio
splicing
Rodrigues et
al.,2005
16 c.2815G>A Ala909Thr Missense Neves, 2008
16 c.2863dup10pb A909fsX49 Missense Neves, 2008

58
APÊNDICE B- Tabela com mutações confirmadas na sequência do gene TPO com sítios próximos
da região da mutação.
Mutação Enzima Sítio
215 del A X X
292C>A X X
613C>T
(S)
(C)
CviKI-1
A mutação elimina
esse sitio e não cria
nenhum
5'... R G C Y
... 3'
3'... Y C G R
... 5'
808G>A
(S)
(C)
BstUI
Elimina o sítio da
enzima acima e
cria para:
HpyCH4IV
5'...C GC G... 3'
3'...G CG C... 5'
5'... A CGT ... 3'
3'... TGC A ... 5'
843delC X X
613C>T
(S)
(C)
CviKI-1
A mutação elimina
o sítio de restrição
da enzima acima e
não cria sítio.
5'... R G C Y
... 3'
3'... Y C G R
... 5'

59
930G>A
(S)
(C)
BstUI
AciI
SacII
Na presença da
mutação, os sítios
de restrição são
eliminados, criando
um sitio de
restrição que não
está
comercialmente
disponível
RdeGBI
5'... C C G C ... 3'
3'... G G C G ... 5'
5'...C C G C A G... 3'
3'...G G C G T C... 5'
5'... C G C G
... 3'
3'... G C G C
... 5'
5'... C C G C G G ... 3'
3'... G G C G C C ... 5'
c.943C>T Não há enzimas
disponíveis
c.965C>T
(S)
EaeI
EagI
CviKI-1
5'...Y G G C C R... 3'
3'...R C C G G Y... 5'
5'...R G C Y... 3'
3'...Y C G R... 5'
5'... C GGCC G ... 3'
3'... GC CGG C ... 5'

60
PhoI
HaeIII
AspCNI
AciI
Na presença da
mutação todos os
sítios de restrição
apresentados acima
são eliminados
5'... GG CC ... 3'
3'... CC GG ... 5'
5’...GGCC...3’
3’...CCGG...5’
5'... GCCGC ... 3'
3'... CGGCG ... 5'
5'...C C G C... 3'
3'...G G C G... 5'
c.1297G>A Não há enzimas
disponíveis
c.1425delC
(S)
Sau3AI
MboI
DpnII
BfuCI
DpnI
Na presença da
mutação todos os
5'...
G A T C
... 3'
3'...
C T A G
... 5'
5'...
G A T C
... 3'
3'...
C T A G
... 5'
5'...
G A T C
... 3'
3'...
C T A G
... 5'
5'...
G A T C
... 3'
3'...
C T A G
... 5'
5'...
G A T C
... 3'
3'...
C T A G
... 5'

61
(C)
sítios de restrição
apresentados acima
são eliminados,
criando um sitio
para a enzima
EcoRV
5'... G A T A T C
... 3'
3'... C T A T A G
... 5'
c.1429A>T Não há enzimas
disponíveis
c.1562G>A
(S)
(C)
EaeI
GdiII
CviKI-1
PhoI
HaeIII
Elimina todos os
sítios de restrição
acima e cria um
novo:
TaqI
5'...
Y G G C C R
... 3'
3'...
R C C G G Y
... 5'
5'...
C G G C C R
... 3'
3'...
G C C G G Y
... 5'
5'...
R G C Y
... 3'
3'...
Y C G R
... 5'
5'...
G G C C
... 3'
3'...
C C G G
... 5'
5'...G G C C... 3'
3'...C C G G... 5'
5'...
T C G A
... 3'
3'...
A G C T
... 5'
c.1942G>A Não há enzimas
disponíveis

62
c.2033 G >A
(S)
PspOMI
Sau96I
PhoI
HaeIII
Na presença da
mutação todos os
sítios de restrição
apresentados acima
são eliminados.
5'... G G G C C C
... 3'
3'... C C C G G G
... 5'
5'...
G G N C C
... 3'
3'...
C C N G G
... 5'
5'...
G G C C
... 3'
3'...
C C G G
... 5'
5'...G G C C... 3'
3'...C C G G... 5'
c.2084G>A
(S)
HpyCH4III
Na presença da
mutação o sítio de
restrição
apresentado acima
é eliminado.
5'...A C N G T... 3'
3'...T G N C A... 5'
c. 2401G>A Não há enzimas
disponíveis
c.2413delC
(S)
PhoI
5'...
G G C C
... 3'
3'...
C C G G
... 5'

63
BsskI
HaeIII
MspI
HpaII
Na presença da
mutação todos os
sítios de restrição
apresentados acima
são eliminados.
5'...
C C N G G
... 3'
3'...
G G N C C
... 5'
5'...
G G C C
... 3'
3'...
C C G G
... 5'
5'...
C C G G
... 3'
3'...
G G C C
... 5'
5'...
C C G G
... 3'
3'...
G G C C
... 5'
c.2485G>A
(S)
HpyCH4III
Na presença da
mutação o sítio de
restrição
apresentado acima
é eliminado,
criando sítios para
as enzimas:
BsrGI
CviQI
RsaI
5'...
T G C A
... 3'
3'...
A C G T
... 5'
5'...
T G T A C A
... 3'
3'...
A C A T G T
... 5'
5'... G TAC ... 3'
3'... CAT G ... 5'
5'... G T A C ... 3'
3'... C A T G ... 5'

64
c.2738C>T
(S)
Hpy188I
Na presença da
mutação o sítio de
restrição
apresentado acima
é eliminado.
5'... T C N G A
... 3'
3'... A G N C T
... 5'
Legenda: C- com mutação; S- Sem mutação