aditivos probiÓticos bacterianos na alimentaÇÃo … · 2011b) o termo aditivo refere se aos...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE GOIÁS
ESCOLA DE VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
Disciplina: SEMINÁRIOS APLICADOS
ADITIVOS PROBIÓTICOS BACTERIANOS NA ALIMENTAÇÃO DE
BOVINOS LEITEIROS
Evando Alves Filgueiras
Orientador: Cláudio Ulhôa Magnabosco
GOIÂNIA
2011

ii
EVANDO ALVES FILGUEIRAS
ADITIVOS PROBIÓTICOS BACTERIANOS NA ALIMENTAÇÃO DE BOVINOS
LEITEIROS
Seminário apresentado à Disciplina
Seminários Aplicados do Programa de Pós-
Graduação em Ciência Animal da Escola de
Veterinária da Universidade Federal de
Goiás.
Nível: Mestrado
Área de Concentração:
Produção Animal
Linha de pesquisa:
Fatores genéticos e ambientais que influenciam o desempenho dos animais
Orientador:
Dr. Cláudio Ulhôa Magnabosco – EMBRAPA CERRADOS
Comitê de orientação:
Dr. Roberto Daniel Sainz Gonzalez – EMBRAPA SGE
Dr. Carlos Frederico Martins – EMBRAPA CERRADOS
GOIÂNIA
2011

iii
SUMÁRIO
LISTA DE TABELAS .............................................................................................. iv
LISTA DE FIGURAS ............................................................................................... v
1 INTRODUÇÃO .................................................................................................... 1
2 REVISÃO DA LITERATURA ............................................................................... 3
2.1 Histórico e conceituação de probióticos ........................................................... 3
2.2 Legislação brasileira sobre aditivos .................................................................. 5
2.3 Mecanismos de ação dos probióticos .............................................................. 6
2.3.1 Exclusão competitiva ..................................................................................... 6
2.3.2 Síntese de bacteriocinas ............................................................................... 7
2.3.3 Prevenção da acidose ruminal ...................................................................... 7
2.3.4 Ativação do sistema imune ............................................................................ 8
2.4 Efeitos dos Aditivos Probióticos para Bovinos Leiteiros ................................... 9
2.4.1 Redução da incidência de diarréia em bezerros ........................................... 9
2.4.2 Ganho em peso ............................................................................................. 9
2.4.3 Consumo de matéria seca ........................................................................... 11
2.4.4 Parâmetros ruminais ................................................................................... 12
2.4.5 Produção de leite ........................................................................................ 14
2.4.6 Composição do leite .................................................................................... 15
2.4.7 Qualidade do leite ....................................................................................... 17
2.4.8 Ausência de resultados com o uso de probióticos ...................................... 18
3 CONSIDERAÇÕES FINAIS .............................................................................. 20
REFERÊNCIAS .................................................................................................... 21

iv
LISTA DE TABELAS
TABELA 1 Microrganismos com propriedades probióticas......................... 4
TABELA 2 Efeito de Prevotella bryantii 25A e tempo de amostragem
sobre as concentrações de ácidos graxos de cadeia curta
(AGCC) e de cadeia ramificada (AGCR) (mM), lactato(mM),
NH3-N (mg/L) e pH ruminal antes e 2 e 3 h após a
alimentação................................................................................
13

v
LISTA DE FIGURAS
FIGURA 1 Consumo de matéria seca do feno ........................................... 11
FIGURA 2 Consumo de matéria seca do concentrado .............................. 11
FIGURA 3 Aumento na produção de leite ajustado para 4% de gordura
em vacas multíparas e primíparas alimentadas com
Propionibacterium P169 ...........................................................
14
FIGURA 4 Aumento nas concentrações de lactose no leite de vacas
multíparas e primíparas alimentados com Propionibacterium
estirpe P169 .............................................................................
16
FIGURA 5 Aumento nas concentrações de sólidos não gordurosos no
leite de vacas multíparas e primíparas alimentados com
Propionibacterium estirpe P169 ...............................................
16

1 INTRODUÇÃO
Nas últimas décadas tem se percebido uma grande preocupação com
a segurança alimentar, principalmente no que se refere aos resíduos de
antibióticos nos alimentos. Neste contexto, os produtos agropecuários,
principalmente a carne e o leite, tem sido alvo de críticas e embargos comerciais,
já que a busca por uma maior eficiência de produção levou pecuarista e técnicos
a utilizarem uma gama muito grande de antibióticos, antiparasitários e até mesmo
hormônios como ferramentas para viabilizem tal anseio.
Todavia, existe a possibilidade de que, se utilizado de forma incorreta,
tais princípios deixam resíduos nos produtos de origem animal, o que poderia
ocasionar a seleção de bactérias patogênicas resistentes a antibióticos e
quimioterápicos, diminuindo ou anulando o efeito benéfico destes produtos nos
tratamentos de doenças em humanos.
Estas preocupações com um produto final de qualidade, livre do
resíduo de antibióticos tem sido notada em todos os elos da cadeia de lácteos,
por isso o uso de fontes alternativas que proporcionem incremento de produção,
melhoria da saúde dos animais e melhoria da qualidade do leite, sem oferecer
risco à saúde humana ou animal tem tomado espaço no mercado atual.
Uma destas alternativas tem sido o uso de aditivos probióticos na dieta
de vacas em lactação ou animais em fase de aleitamento. Isto ocorre porque de
acordo com KREHBIEL et al. (2003) os probióticos tem a capacidade de prevenir
a colonização de patógenos indesejáveis no intestino, melhorando a saúde e o
desempenho animal, diminuindo ainda o risco de acidose ruminal em vacas
leiteiras, melhorando também a resposta imune em vacas e bezerros.
Todavia, apesar de vários autores relatarem os benefícios com o uso
destes produtos (SAINZ et al. 2011, MAGNABOSCO et al. 2010, STEIN et al.
2006, FRANCISCO et al. 2002) os mecanismos de ação que levam a tais
benefícios, principalmente dos probióticos a base de bactérias, ainda não estão
devidamente elucidados, o que se percebe de uma forma geral é que os
probióticos melhoram a saúde do hospedeiro, resultando em melhores índices
produtivos, melhorando ainda a qualidade do produto final.

2
Objetivou-se com esta revisão apresentar conhecimentos científicos
relacionados à suplementação dietética de bovinos leiteiros com probióticos
bacterianos, os mecanismos de ação conhecidos até o momento, seus reflexos
na saúde e produção destes animais, bem como sua influência na composição e
qualidade do leite.

3
2 REVISÃO DA LITERATURA
2.1 Histórico e conceituação de probióticos
O consumo de microrganismos através de leites fermentados para
melhoria da saúde humana é relatado desde tempos imemoriais, porém, estudos
em animais são relatados a partir do ano de 1980 (GONZALES 2011). Em 1989,
FULLER (1989) definiu o termo probiótico como sendo um suplemento alimentar
composto por microorganismos vivos que beneficiam a saúde do hospedeiro,
através do equilíbrio da microflora intestinal.
Em trabalhos norte-americanos com aditivos probióticos nota-se o
emprego do termo DFM (Direct-fed microbials). Esta nomenclatura foi adotada por
que não existia um consenso referente á inclusão de culturas de leveduras e
bactérias nas dietas de ruminantes, por isso, em 1989 o FDA (Food and Drug
Administration) entidade responsável pelo registro e liberação de drogas,
antibióticos, aditivos ou qualquer outra substância de uso em humanos e animais
dos EUA, exigiu que os fabricantes adotassem o termo DFM (Direct-fed
microbials), em vez de probiótico (MARTIN & NISBET, 1992). O FDA define DFM
como uma fonte de microorganismos vivos (viável) de ocorrência natural no
organismo animal (KREHBIEL et al. 2003).
No Brasil utiliza-se ainda o termo probiótico, que pode ser definido
como cepas de microrganismos vivos e viáveis, que agem como auxiliares na
recomposição da microbiota do trato digestivo dos animais, diminuindo o número
dos microrganismos patogênicos ou indesejáveis (MAPA, 2011a).
De acordo com COPPOLA & TURNES (2004) os microrganismos
classificados como probióticos devem apresentar as seguintes características:
não ser patogênicos aos humanos e animais, ser resistentes aos ácidos e
enzimas do trato digestivo, não transportar genes de resistência a antibióticos,
possuir propriedades antimutagênicas e anticarcinogênicas, permanecer viável no
alimento até o momento da ingestão e gerar benefícios ao hospedeiro.
HOLZAPFEL (2001) subdivide os microrganismos probióticos em dois
grupos: bactérias do acido láctico e bactérias não ácidas lácticas conforme

4
descrito na Tabela 1. É importante ressaltar que os microorganismos
Saccharomyces cerevisiae e boulardii não são bactérias, mas sim fungos
conforme descrito por BITENCOURT et al. (2011).
TABELA 1 – Microrganismos com propriedades probióticas
Bactérias ácido lácticas Bactérias não ácido lácticas
Lactobacillus Bifidobacterium Outras Bactérias
L. acidophilus B. adolescentis Enterococcus
faecalis Bacillus cereus var. toyoi
L. amylovorus B. animalis Enterococcus
faecium Bacillus subtilis
L. casei B. bifidum Lactococcus lactis Escherichia coli cepa nissle
L. crispatus B. breve Leuconstoc
mesenteroides
Propionibacterium
freudenreichii
L. bugaricus B. infantis Pediococcus
acidilactici Saccharomyces cerevisiae
L. galinarum B. lactis Sporolactobacillus
inulinus Saccharomyces boulardii
L. gasseri B. longum Streptococcus
thermophilus
L. johmssnii
L. paracasei
L. plantarum
L. reuteri
L. rhamnous
Fonte: Adaptado de HOLZAPFEL (2001)
Os principais aditivos probióticos comerciais para ruminantes incluem
bactérias do gênero Lactobacilli, como o Lactobacillus acidophilus e várias
espécies de Bifidobacterium, Enterococcus e Bacillus e ainda leveduras como
Saccharomyces cerevisiae e boulardii. No entanto, trabalhos científicos têm sido
conduzidos com outros microrganismos potencialmente viáveis para a nutrição de
ruminantes, como por exemplo, a bactéria Prevotella bryantii estirpe 25 A
(CHIQUETTE et al., 2008) e Megasphaera elsdenii (HASHIZUME et al., 2003) e
Propionibactérias (STEIN et al., 2006).

5
2.2 Legislação brasileira sobre aditivos
Conforme redação dada pela Instrução Normativa 15/2009 (MAPA,
2011b) o termo aditivo refere se aos produtos destinados à alimentação animal,
sendo substância, microorganismo ou produto formulado, adicionado
intencionalmente aos produtos, que não é utilizado normalmente como
ingrediente, tenha ou não valor nutritivo e que melhore as características dos
produtos destinados à alimentação animal ou dos produtos animais, melhore o
desempenho dos animais sadios e atenda às necessidades nutricionais ou tenha
efeito anticoccidiano.
O Regulamento técnico sobre aditivos para produtos destinados à
alimentação animal (IN 12/2004) (MAPA, 2011a) classifica os aditivos de acordo
com suas funções e propriedades nas seguintes categorias: aditivos tecnológicos
compreendem qualquer substância adicionada ao produto destinado à
alimentação animal com fins tecnológicos; aditivos sensoriais melhoram ou
modificam as propriedades organolépticas ou as características visuais dos
produtos; aditivos nutricionais correspondem a toda substância utilizada para
manter ou melhorar as propriedades nutricionais do produto; aditivos zootécnicos
influem positivamente na melhoria do desempenho dos animais; aditivos
anticoccidianos eliminam ou inibem protozoários.
Os aditivos zootécnicos incluem os seguintes grupos funcionais:
digestivo, substância que facilita a digestão dos alimentos ingeridos, atuando
sobre determinadas matérias primas destinadas à fabricação de produtos para a
alimentação animal; equilibradores da flora, microorganismos que formam
colônias ou outras substâncias definidas quimicamente que têm um efeito positivo
sobre a microbiota do trato digestório; e melhoradores de desempenho,
substâncias que melhoram os parâmetros de produtividade (MAPA, 2011a).
Os probióticos enquadram-se como aditivos zootécnicos equilibradores
da microbiota do trato digestório e são definidos como cepas de microrganismos
vivos (viáveis), que agem como auxiliares na recomposição da microbiota do trato
digestivo dos animais, diminuindo o número dos microrganismos patogênicos ou
indesejáveis (MAPA, 2011a).

6
2.3 Mecanismos de Ação dos Probióticos
De acordo com GARCIA (2008) os mecanismos de ação dos
probióticos no organismo animal ainda não estão bem definidos, porém acredita-
se que os mesmos apresentem pelo menos quatro tipos de mecanismos de ação
que ocasionam benefícios ao hospedeiro. São eles: a exclusão competitiva seja
por nutrientes ou por sítios de ativação; síntese de bacteriocinas que agem como
antibióticos naturais; prevenção da acidose ruminal; e ativação do sistema imune.
2.3.1 Exclusão competitiva
SALMINEN et al. (1996) relatou que os microrganismos presentes no
trato gastrointestinal (TGI) precisam aderir à mucosa intestinal para produzir suas
toxinas, uma vez que a adesão evita que os mesmos sejam eliminados pelos
movimentos peristálticos. FULLER & BROOKER (1974) afirmam que a aderência
dos microrganismos no intestino depende da interação entre os
mucopolissacarídeos da parede celular e da camada de mucopolissacarídeos
semelhante na mucosa intestinal, sendo assim os vários gêneros de
microrganismos presentes no TGI possuem sítios de aderência específicos na
mucosa intestinal.
CROSS (2002) e KREHBIEL et al. (2003) relatam que um dos
principais efeitos dos probióticos no intestino seria a exclusão competitiva dos
microrganismos patogênicos, uma vez que os mesmo ocupariam os mesmo sítios
de ativação de tais microrganismos. KREHBIEL et al. (2003) relata ainda que
outra forma de competição seria por alimentos, uma vez que microrganismos
benéficos colonizaram a mucosa intestinal, a disponibilidade de nutrientes para os
microrganismos patogênicos tendem a ser menor.
Dessa forma fica evidenciada a importância de se fornecer de forma
contínua doses diárias de probióticos, para que se maximize seu efeito benéfico
ao hospedeiro (COPPOLA & TURNES, 2004).

7
2.3.2 Síntese de bacteriocinas
Algumas cepas probióticas possuem ainda a capacidade de inibir a
multiplicação de microrganismos patogênicos, através da síntese de
bacteriocinas, influenciando diretamente a saúde do hospedeiro (JIN et al., 2000;
OGAWA et al., 2001).
De acordo com RUSSEL & MANTOVANI (2002) bacteriocinas são
peptídeos antimicrobianos de baixo peso molecular, sendo produzidas por
bactérias gram-positivas e gram-negativas, que apresentam espectro de ação
variado. A atuação das bacteriocinas ocorre primariamente sobre a membrana
plasmática bacteriana, formando poros que possibilitam a perda de componentes
citoplasmáticos, ocasionando a morte celular (MOLL et al., 1999).
MATOVANI et al. (2001) identificaram e caracterizaram algumas
bacteriocinas em bactérias do rúmen, sendo estas substâncias mais efetivas
contra bactérias gram-positivas. A ação antimicrobiana desses peptídeos
assemelha se à dos ionóforos (RUSSEL & MANTOVANI, 2002).
LEE et al. (2002) demonstraram que a bateriocina ruminal Bovicina
HC5 produzida por Streptococcus bovis HC5 inibiu a produção de amônia por
culturas puras de Clostridium aminophilum e a produção de metano in vitro, por
culturas mistas de microrganismos do rúmen. CALLAWAY et al. (1997)
verificaram que o Lactococcus lactis produzia uma bacteriocina conhecida como
Nisina, que possuía efeito inibitório sobre a produção de amônia por algumas
bactérias.
2.3.3 Prevenção da acidose ruminal
Existem trabalhos que demonstram que os probióticos bacterianos
possuem efeitos benéficos no ambiente ruminal, principalmente na prevenção de
acidose. Um dos mecanismos propostos seria de que a produção de lactato pelas
cepas probióticas induziria a seleção de bactérias consumidoras deste produto
(GHORBANI et al., 2002). Outro mecanismo seria que as próprias cepas
probióticas utilizariam o lactato, reduzindo assim suas concentrações no ambiente

8
ruminal, como por exemplo, as Propionibacterium, que utilizam o lactato para
produzir propionato e outros ácidos graxos de cadeia curta (KREHBIEL et al.,
2003)
Todavia, a maioria dos relatos sobre ação dos probióticos na
prevenção de acidose ruminal é atribuído as leveduras, que podem atuar
controlando os níveis de oxigênio no ambiente ruminal e estimulando o
crescimento de bactérias desejáveis no rúmen através de pequenos grupos de
peptídeos não identificados ainda, que estimulam o crescimento desses
microrganismos (BARBOSA et al., 2004).
2.3.4 Ativação do sistema imune
Além de seu papel na digestão e absorção dos nutrientes, o TGI
fornece ao animal proteção contra a presença de antígenos presentes nos
alimentos e microrganismos patogênicos que colonizam o lúmen intestinal. Estas
respostas podem ser inespecíficas (inatas) ou específicas (adaptativas)
(KREHBIEL et al., 2003).
De acordo com SAXELIN et al. (2005) os probióticos podem atuar no
intestino ativando o sistema imune, reforçando a barreira da mucosa e suprimindo
as inflamações intestinais. Esses efeitos podem estar relacionados à capacidade
de os microrganismos do probiótico interagirem com as placas de Peyer e as
células epiteliais intestinais, estimulando as células B produtoras de IgA e a
migração de células T para a mucosa intestinal (PERDIGÓN et al. 2003).
Também tem sido demonstrado que os probióticos favorecem a atividade
fagocítica inespecífica dos macrófagos alveolares, sugerindo uma ação sistêmica
por secreção de mediadores que estimulariam o sistema imune (CROSS, 2002).
ISOLARI et al. (2001) relatam que a administração oral de Lactobacillos
sp resultou em um aumento da resposta imune inata, ou seja, aumento da
fagocitose, bem como uma elevada produção de imunoglobulina (IGA) e uma
diminuição da produção de IgE em humanos e animais.

9
2.4 Efeitos dos Aditivos Probióticos para Bovinos Leiteiros
2.4.1 Redução da incidência de diarréia em bezerros
TIMMERMAN et al. (2005) utilizando um composto com cepas de
Lactobacillus, Lactococcus e Enterococcus na alimentação de bezerros recém
nascidos verificaram redução nos casos de diarréia, e também no número de dias
que os animais permaneciam com diarréia no lote de animais que consumiam o
probiótico (P<0,05).
De acordo com BATISTA et al. (2008) a adição de probiótico
(Lactobacillus acidophillus, Bifidobacterium bifidum e Enterococus faecium) ao
leite sem resíduo de antimicrobiano reduziu o número de dias com diarréia de
bezerras holandesas, de um a 60 dias de idade (P<0,05).
SIGINORINI et al. (2011) realizou uma meta-análise com trabalhos
referentes ao uso de probióticos em bezerros e constatou que em 70% dos
experimentos o uso destes aditivos reduziu a incidência de casos de diarréia
(P<0,05).
TIMMERMAN et al. (2005) relatam que bezerros geralmente são
submetidos a estresses como transporte, mudanças alimentares e criação em
bezerreiros que não oferecem boas condições sanitárias, o que resulta em
constante exposição à patógenos, agravando os casos de diarréia.
Desta forma, a redução da incidência de diarréia em bezerros com uso
de probióticos pode ser explicada pelos relatos de KREHBIEL et al. (2003) e
CHIQUETTE (2009) que afirmam que algumas cepas probióticas podem auxiliar
na eliminação de bactérias patogênicas do intestino, seja por ação das
bacteriocinas, ou por exclusão competitiva, impedindo que tais bactérias
colonizem a mucosa intestinal e produzam suas toxinas.
2.4.2 Ganho em peso
GARCIA (2008) avaliou o ganho em peso de 38 animais da raça
holandesa recém nascidos recebendo sucedâneo do leite (controle) e sucedâneo

10
do leite mais 1, 2 ou 3 gramas de um probiótico comercial a base de Bacillus
subtilis, e verificou um maior ganho em peso diário nos animais que consumiam 4
gramas do probiótico (P<0,05).
MEYER et al. (2001) utilizando um probiótico à base de Lactobacillus
acidophilus, Enterococcus faecium e Saccharomyces cerevisiae, também
verificaram um aumento no ganho em peso de 79 bezerros recém nascidos da
raça holandesa.
TIMMERMAN et al. (2005) utilizando um composto com cepas de
Lactobacillus, Lactococcus e Enterococcus na alimentação de bezerros recém
nascidos verificaram uma diferença de 46% no ganho em peso dos animais
tratados quando comparados com o lote controle, o autor enfatiza ainda que o
ganho em peso dos animais foi potencializado pelas condições precárias de
higiene, expressando desta forma o efeito imunoprotetor dos probióticos.
Antes do nascimento o trato digestivo de bezerros geralmente é estéril,
ocorrendo à colonização por vários microrganismos somente após o nascimento.
Em condições saudáveis e não estressantes, uma microflora benéfica coloniza o
rúmen e o intestino em uma relação de simbiose em que os microrganismos
suprem nutrientes para o hospedeiro (KUNG JR, 2011).
Porém, conforme relatado por TIMMERMAN et al. (2005), bezerros
oriundos de propriedades leiteiras são geralmente submetidos a estresses,
mudanças alimentares e criação em bezerreiros que não oferecem boas
condições sanitárias, o que resulta em constantes exposições à patógenos,
elevando assim o risco do estabelecimento de uma microbiota indesejável,
resultando em comprometimento da eficiência alimentar e incidência de diarréias.
Dessa forma, o melhor desempenho em ganho de peso obtido pelos
animais que receberam o probiótico na alimentação pode ser explicado pela
função de exclusão competitiva e colonização da mucosa intestinal (KREHBIEL et
al. 2003), o que pode auxiliar no estabelecimento de uma microbiota benéfica,
ocasionando assim um melhor desempenho desses animais.
Este fato pode ser reforçado pelo maior consumo de matéria seca
observado por GARCIA (2008) no lote que consumiu o probiótico, indicando um
11
melhor desenvolvimento do trato digestivo destes animais (LESMEISTER et al.,
2004). MEYER et al. (2001) não verificaram aumento no consumo de matéria
seca, porém foi observada uma melhoria de 18,2% (P<0,05) na conversão
alimentar.
2.4.3 Consumo de matéria seca
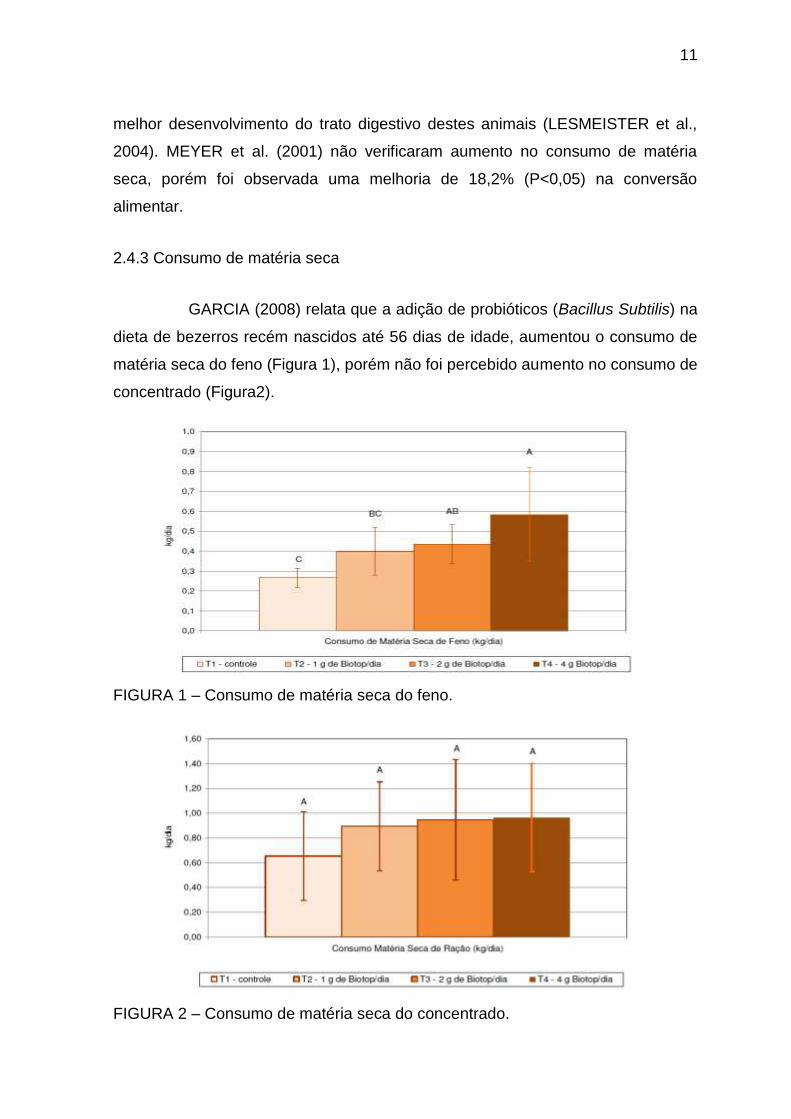
GARCIA (2008) relata que a adição de probióticos (Bacillus Subtilis) na
dieta de bezerros recém nascidos até 56 dias de idade, aumentou o consumo de
matéria seca do feno (Figura 1), porém não foi percebido aumento no consumo de
concentrado (Figura2).
FIGURA 1 – Consumo de matéria seca do feno.
FIGURA 2 – Consumo de matéria seca do concentrado.

12
A influência dos probióticos no consumo de matéria seca em bovinos
na fase de aleitamento não foi explicada por nenhum dos autores, porém acredita-
se que a eliminação de bactérias patogênicas ocasionando o aumento da flora
benéfica, devido aos probióticos no TGI, conforme descrito por KREHBIEL et al.
(2003), possa de alguma forma estar associada ao evento observado. Já a
ausência de diferença no consumo de matéria seca do concentrado pode ser
explicada pela interrupção do fornecimento do sucedâneo do leite, o que
provocou um aumento no consumo de concentrado (GARCIA, 2008).
Em contrapartida, FRANCISCO et al. (2002) verificaram uma redução
no consumo de matéria seca ao adicionar 17 gramas de Propionibacterium estirpe
P169 na dieta de vacas da raça holandesa avaliadas desde duas semanas pré
parto até 12 semanas pós parto, 35,3 e 38,9 kg de MS/animal/dia para os lotes
tratamento e controle, respectivamente.
Conforme relatado pelo autor, a redução no consumo de matéria seca
no lote de animais que consumiram a Propionibacterium pode ter sido provocada
pelo aumento nos níveis plasmáticos de leptina. Hormônio este que parece atuar
como um modulador no consumo de alimento (INGVARTSEN & ANDERSEN,
2000). Já o aumento deste hormônio no sangue pode ter sido provocado pela
tendência (P<0,10) observada de melhoria no balanço energético dos animais que
receberam o probiótico, aumentado a gordura corporal destes animais.
De acordo com DELAVAUD et al. (2000) a leptina é produzida pelos
adipócitos ou células gordurosas, dessa forma a melhoria do balanço energético
pode ocasionar o aumento das concentrações de leptina no sangue (EHRHARDT
et al. 2000)
2.4.4 Parâmetros ruminais
STEIN et al. (2006) avaliaram a suplementação de vacas holandesas
primíparas e multíparas com Propionibacterium estirpe P169 e constataram que
os níveis de propionato ruminal apresentaram maior porcentagem molar no lote
tratamento quando comparado ao lote controle.
13
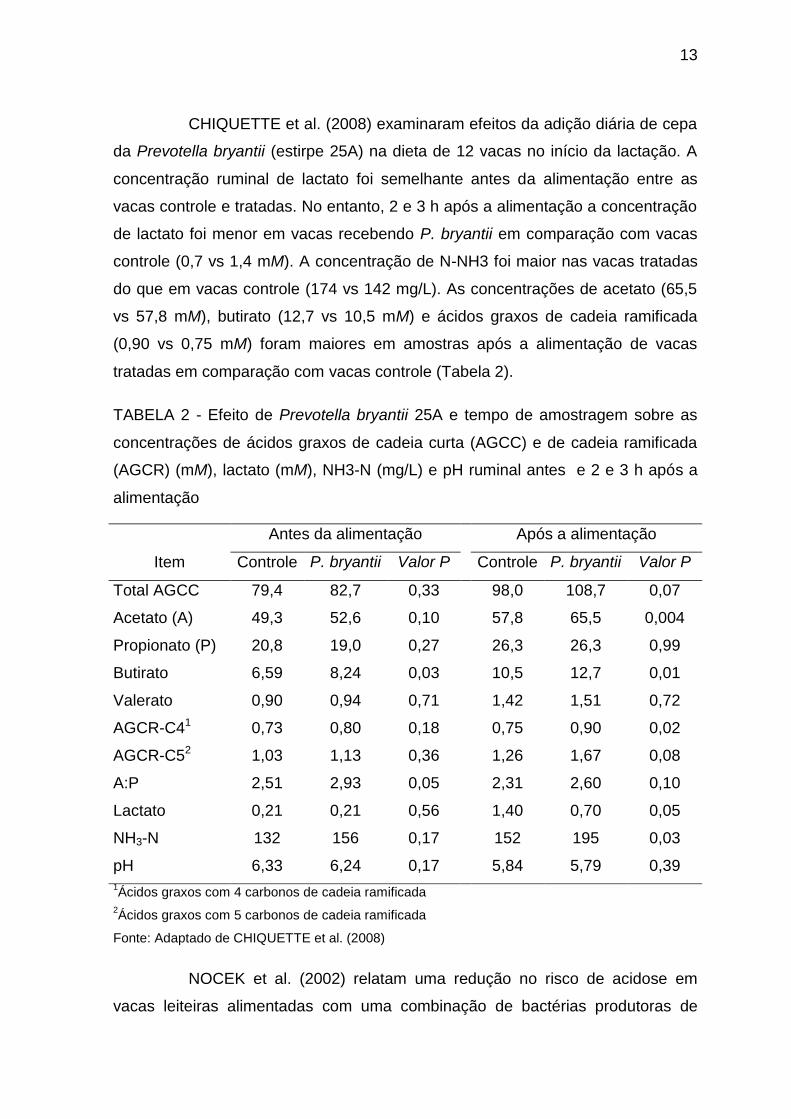
CHIQUETTE et al. (2008) examinaram efeitos da adição diária de cepa
da Prevotella bryantii (estirpe 25A) na dieta de 12 vacas no início da lactação. A
concentração ruminal de lactato foi semelhante antes da alimentação entre as
vacas controle e tratadas. No entanto, 2 e 3 h após a alimentação a concentração
de lactato foi menor em vacas recebendo P. bryantii em comparação com vacas
controle (0,7 vs 1,4 mM). A concentração de N-NH3 foi maior nas vacas tratadas
do que em vacas controle (174 vs 142 mg/L). As concentrações de acetato (65,5
vs 57,8 mM), butirato (12,7 vs 10,5 mM) e ácidos graxos de cadeia ramificada
(0,90 vs 0,75 mM) foram maiores em amostras após a alimentação de vacas
tratadas em comparação com vacas controle (Tabela 2).
TABELA 2 - Efeito de Prevotella bryantii 25A e tempo de amostragem sobre as
concentrações de ácidos graxos de cadeia curta (AGCC) e de cadeia ramificada
(AGCR) (mM), lactato (mM), NH3-N (mg/L) e pH ruminal antes e 2 e 3 h após a
alimentação
Antes da alimentação
Após a alimentação
Item Controle P. bryantii Valor P
Controle P. bryantii Valor P
Total AGCC 79,4 82,7 0,33
98,0 108,7 0,07
Acetato (A) 49,3 52,6 0,10
57,8 65,5 0,004
Propionato (P) 20,8 19,0 0,27
26,3 26,3 0,99
Butirato 6,59 8,24 0,03
10,5 12,7 0,01
Valerato 0,90 0,94 0,71
1,42 1,51 0,72
AGCR-C41 0,73 0,80 0,18
0,75 0,90 0,02
AGCR-C52 1,03 1,13 0,36
1,26 1,67 0,08
A:P 2,51 2,93 0,05
2,31 2,60 0,10
Lactato 0,21 0,21 0,56
1,40 0,70 0,05
NH3-N 132 156 0,17
152 195 0,03
pH 6,33 6,24 0,17
5,84 5,79 0,39
1Ácidos graxos com 4 carbonos de cadeia ramificada
2Ácidos graxos com 5 carbonos de cadeia ramificada
Fonte: Adaptado de CHIQUETTE et al. (2008)
NOCEK et al. (2002) relatam uma redução no risco de acidose em
vacas leiteiras alimentadas com uma combinação de bactérias produtoras de

14
lactato (Lactobacillus e Enterococcus). Provavelmente porque a produção de
lactato por estas bactérias acabou aumentando as populações de bactérias
consumidoras de lactato, prevenindo assim o acúmulo deste acido no rúmen
(GHORBANI et al., 2002).
2.4.5 Produção de leite
STEIN et al. (2006) trabalhando com 38 vacas primíparas e multíparas
da raça Holandesa, divididas em 3 tratamentos contendo ou não
Propionibacterium estirpe P169 (Controle, Baixa dose de P169 – 6 x 1010, e alta
P169 – 6 x 1011) constataram aumento na produção de leite corrigido para 4% de
gordura, 7,1 e 8,5% (Figura 1) nos tratamentos alta e baixa dose de P169 em
relação ao lote controle (P<0,003).
FIGURA 3 – Aumento na produção de leite ajustado para 4% de gordura em
vacas multíparas e primíparas alimentadas com Propionibacterium P169.

15
MAGNABOSCO et al. (2010) utilizando um simbiótico à base de
Lactobacillus acidophilus, Enterococcus faecium, Bacillus subtilis e
Saccharomyces cerevisiae, com mais três enzimas digestivas (celulase,
hemicelulase e xilanase) verificaram um aumento de 10% na produção de animais
da raça Girolando (P<0,05).
NOCEK & KAUTZ (2006) relatam um aumento de 6% (P<0,01) na
produção de leite de vacas da raça Holandesa quando utilizou um probiótico a
base de Enterococcus faecium e Saccharomyces cerevisiae. Resultados
semelhantes foram encontrados por NOCEK et al. (2003).
Vários podem ser os mecanismos pelos quais o uso de probióticos
estimula a produção de leite, pois os efeitos benéficos dos mesmos conforme
descrito por KREHBIEL et al. (2003) como ativação do sistema imune e
prevenção de acidose ruminal, podem melhorar a saúde dos animais como um
todo, refletindo em maiores produções.
É importante ressaltar também que os resultados encontrados por
SAINZ et al. (2011) e MAGNABOSCO et al. (2010) indicando uma redução na
contagem de células somáticas (CCS) de animais em lactação, podem também
contribuir para o aumento na produção de leite, uma vez que a contagem de
células somáticas exerce efeito negativo na produção de leite (COLDEBELLA et
al. 2004)
2.4.6 Composição do leite
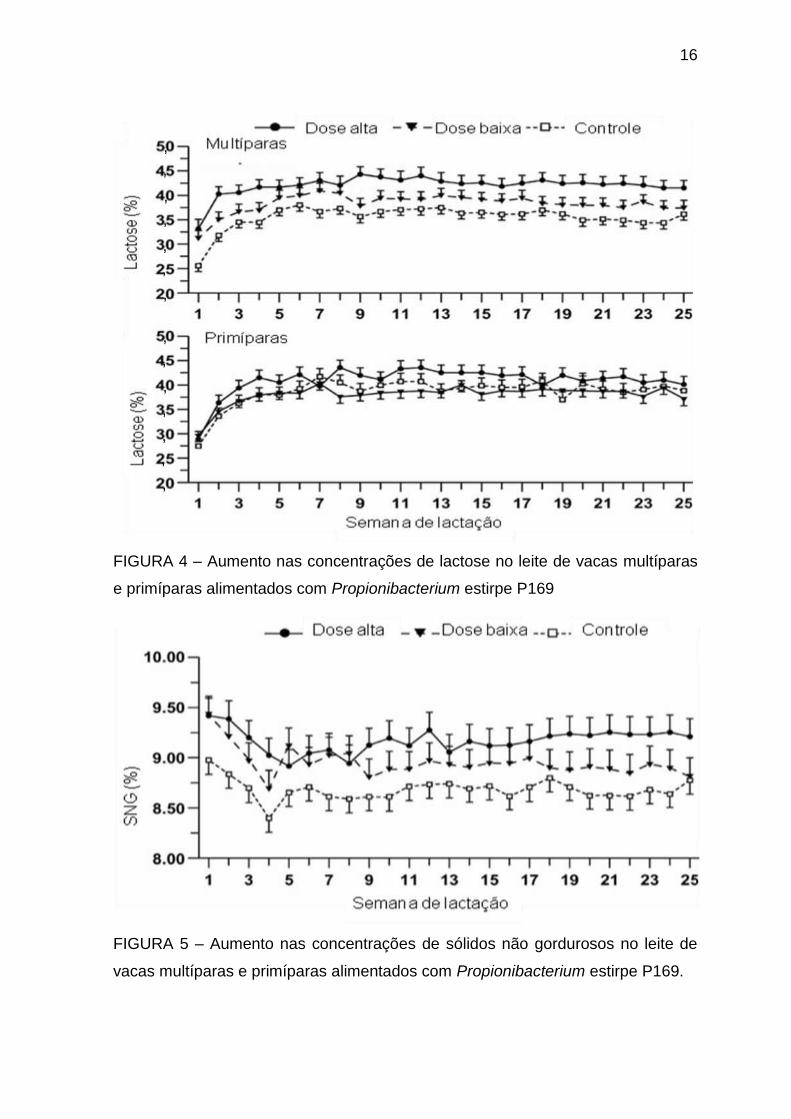
STEIN et al. (2006) trabalhando com 38 vacas holandesas primíparas e
multíparas, divididas em 3 tratamentos contendo ou não Propionibacterium estirpe
P169 (Controle, Baixa dose de P169 – 6 x 1010, e alta P169 – 6 x 1011)
constataram aumento na concentração de lactose (Figura 2) e sólidos
desengordurados (Figura 3) nos lotes tratamentos em relação ao lote controle.
16
FIGURA 4 – Aumento nas concentrações de lactose no leite de vacas multíparas
e primíparas alimentados com Propionibacterium estirpe P169
FIGURA 5 – Aumento nas concentrações de sólidos não gordurosos no leite de
vacas multíparas e primíparas alimentados com Propionibacterium estirpe P169.
,
,
,
,
,
,
,
,
,
,
,
,
,
,

17
NOCEK & KAUTZ (2006) relatam um aumento na concentração de
gordura do leite de vacas da raça Holandesa, 4,65 vs 4,59%, quando utilizou um
probiótico a base de Enterococcus faecium e Saccharomyces cerevisiae em
comparação com o lote controle. Resultados semelhantes foram encontrados por
NOCEK et al. (2003).
FRANCISCO et al. (2002) não observou aumento nas concentrações
de lactose e gordura quando adicionaram 17 gramas de Propionibacterium estirpe
P169 na dieta de vacas da raça holandesa avaliadas desde 2 semanas pré parto
até 12 semanas pós parto, porém os mesmos observaram aumento na proteína
na primeira semana pós parto (P<0,05).
O aumento nos teores sólidos do leite pode estar diretamente
relacionados com as mudanças ocorridas na fermentação ruminal, resultando em
mudanças nas concentrações de ácidos graxos de cadeia curta, como acetato e
propionato (STEIN et al., 2006, CHIQUETTE et al., 2008), estes ácidos são de
grande importância na síntese da gordura do leite, sendo ainda o propionato o
principal precursor da glicose (ANTUNES et al., 2011) percussor da lactose no
leite.
De acordo com RULQUIN et al. (2004) a síntese de propionato
economiza aminoácidos glicogênicos na gliconeogênese, o que por sua vez pode
resultar em aumento da disponibilidade de aminoácidos para síntese de proteína
no leite (STEIN et al., 2006).
Outro fato que pode estar diretamente relacionado com o aumento dos
teores de sólidos do leite seria a redução de células somáticas ocasionada pelos
probióticos conforme descrito por SAINZ et al. (2011) e MAGNABOSCO et al.
(2010), uma vez que autos índices de CCS podem influenciar de forma negativa
na composição do leite (SCHAELLIBAUM, 2000).
2.4.7 Qualidade do leite
SAINZ et al. (2011) utilizando um simbiótico à base de Lactobacillus
acidophilus, Enterococcus faecium, Bacillus subtillis e Sacharomices cerevisiae
associado a três enzimas digestivas (celulase, hemicelulase e xilanase)

18
verificaram uma redução média de 53% na contagem de células somáticas (CCS)
em vacas da raça holandesa e girolando. MAGNABOSCO et al. (2010)
trabalhando com a mesma composição verificaram redução de 34% na CCS de
vacas girolando.
Como ao aumento da CCS geralmente é ocasionado pela presença de
células de defesa do organismo para combater uma inflamação na glândula
mamária, o efeito dos probióticos favorecendo a atividade fagocitária de forma
sistêmica, conforme descrito por CROSS (2002) poderia prevenir o aumento
destas células no leite. A fagocitose é responsável pela ativação precoce da
resposta inflamatória antes da produção de anticorpos, produzindo agentes
tóxicos como reativos de oxigênio e enzimas líticas (ISOULARI, 2001)
2.4.8 Ausência de resultados com o uso de probióticos
Existem relatos na literatura de ausência de resultados positivos com o
uso de probióticos, como no ganho em peso (GONÇALVES et al. 2000,
GORGULU et al. 2003), redução da incidência de diarréia (GARCIA, 2008,
GORGULU et al. 2003) consumo de matéria seca (RAETH-KINIG et al. 2007,
STEIN, 2006), produção de leite (SAINZ et al., 2011, FRANCISCO et al., 2002) e
composição do leite (SAINZ et al., 2011, MAGNABOSCO et al., 2010).
KREHBIEL et al. (2003) relatam que os casos de ausência de efeito da
adição de probióticos na dieta de bezerros estão geralmente associados à falta de
desafio sanitário, dado este reforçado por TIMMERMAN et al. (2005) que
relataram uma potencialização no ganho em peso de bezerros criados em
condições sanitárias desfiadoras recebendo dietas a base de probióticos.
Nos trabalhos de SAINZ et al. (2011) também percebe-se que houve
um aumento na contagem de células somáticas das vacas que não recebiam a
suplementação com culturas probióticas quando foram desafiadas, principalmente
pela entrada no período chuvoso, que por sua vez agrava as condições sanitárias
do ambiente, contudo não foi verificado aumento na CCS de vacas recebendo a
cultura probiótica.

19
Estes relatos reforçam os mecanismos de ação propostos por ISOLARI
et al. (2001) e KREHBIEL et al. (2003) em que os probióticos exercem a função
de proteção do organismo animal, seja pela exclusão competitiva, pela produção
de bacteriocinas ou pela indução de ativação do sistema imune, podendo ainda
melhorar de forma sensível o aproveitamento dos nutrientes.

20
3 CONSIDERAÇÕES FINAIS
Os probióticos têm apresentado potencial para melhorar a saúde e
desempenho de bovinos leiteiros, porem os mecanismos de ação propostos,
muitas vezes parece não ser devidamente identificados em alguns experimentos,
sendo necessário um maior detalhamento para sua compreensão.
A variação de resultados sejam elas no desempenho produtivo ou na
saúde dos animais pode estar relacionadas à ausência de um consenso quanto à
dosagem, tipo de microorganismo e forma de fornecimento, o que muitas vezes
pode gerar resposta equivocadas.
Nos trabalhos onde o uso de aditivos probióticos refletiu em benefícios
para a produção ou saúde animal, geralmente não são apresentadas discussões
sobre a viabilidade econômica destes produtos, o que por sua vez é de
fundamental importância para a disseminação desta tecnologia no campo.

21
REFERÊNCIAS
1. ANTUNES, R. C.; RODRIGUEZ, N. M.; SALIBA, E. O. S. Metabolismo dos carboidratos não estruturais. In: BERCHIELLI, T. T.; PIRES, A. V.; OLIVEIRA, S. G. Nutrição de Ruminantes. 2. Ed. Jaboticabal; Funep, 2011. cap. 8, p. 239 – 260.
2. BARBOSA, F. A.; FARIA, G. A.; VILELA, H. Leveduras vivas na nutrição de bovinos – Uma revisão. Bioscience Journal, Uberlandia, v. 20, n. 1, p. 143-150, 2004.
3. BATISTA, C. G.; COELHO, S. G.; RABELO, E.; LANA, A. M. Q.; CARVALHO, A. U.; REIS, R. B.; SATURNINO, H. M. Desempenho e saúde de bezerras alimentadas com leite sem resíduo de drogas antimicrobianas ou leite de vacas tratadas contra mastite adicionado ou não de probiótico. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, Belo Horizonte, v. 60, n. 1, p. 185 - 191, 2008.
4. BITENCOURT, L. L.; MARTINS SILVA, J. R.; OLIVEIRA, B. M. L.; DIAS JÚNIOR, G. S.; LOPES, F.; SIÉCOLA JÚNIOR, S.; ZACARONI, O. F.; PEREIRA, M. N. Diet digestibility and performance of dairy cows supplemented with live yeast. Scientia Agricola, Piracicaba, v. 68, n. 3, p. 301 - 307, 2011.
5. CALLAWAY, T.R.; CARNEIRO DE MELLO, A.M. S.;RUSSEL, J.B. The effect of Nisin and Monensin on ruminal fermentations in vitro. Current Microbiology, New York, v. 35, p. 90 - 96, 1997.
6. CHIQUETTE, J. Evaluation of the protective effect of probiotics fed to dairy cows during a subacute ruminal acidosis challenge. Animal Feed Science and Technology, v. 153 p. 278 – 291, 2009.
7. CHIQUETTE, J.; ALLISON, M. J.; RASMUSSEN, M. A. Prevotella bryantii 25A used as a probiotic in early-lactation dairy cows: effect on ruminal fermentation characteristics, milk production, and milk composition. Journal of Dairy Science, Champaign, v. 91, n. 9, p. 3536 - 3543, 2008.
8. COLDEBELLA, A.; MACHADO, P. F.; DEMETRIO, P. C. G. B.; RIBEIRO JUNIOR, P. J.; MEYER, P. M.; CORASSIN, C. H.; CASSOLI, L. D. Contagem de Células Somáticas e Produção de Leite em Vacas Holandesas Confinadas. Revista Brasileira de Zootecnia, Viçosa, v. 33, n. 3, p. 623 - 634, 2004.
9. COPPOLA, M. M.; TURNES, C. G. Probióticos e a resposta imune. Ciência Rural, Santa Maria, v. 34, p. 1297 - 1303, 2004.

22
10. CROSS, M. L. Microbes vs. Microbes: imune signals generated by probiótico lactobacilli and their role in protection against microbial pathogens. FEMS Immunology and Medical Microbiology, Amsterdam, v. 34, n. 4, p. 545 - 553, 2002.
11. DELAVAUD, C.; BOCQUIER, F.; CHILLIARD, Y.; KEISLER, D. H.; GERTLER, A.; KANN, G., Plasma leptin determination in ruminants: effect of nutritional status and body fatness on plasma leptin concentration assessed by a specific RIA in sheep. Journal of Endocrinology, Bristol, v. 165, p. 519 – 526, 2000.
12. EHRHARDT, R. A.; SLEPETIS, R. M.; SIEGAL-WILLOTT, J.; VAN AMBURGH, M. E.; BELL, A. W.; BOISCLAIR, R. Development of a specific radioimmunoassay to measure physiological changes of circulating leptin in cattle and sheep. Journal of Endocrinology, Bristol, v. 166, p. 519 – 528, 2000.
13. FRANCISCO, C. C.; CHAMBERLAIN, C. S.; WALDNER, D. N.; WETTEMANN, R. P.; SPICER, L. J. Propionibacteria fed to dairy cows: effects on energy balance, plasma metabolites and hormones, and reproduction. Journal of Dairy Science, Champaign, v. 85, n. 7, p. 1738 - 1751, 2002.
14. FULLER, R. Probiotic in man and animals – a review. Journal Applied Bacteriology, Malden, v. 66, p. 365-378, 1989.
15. FULLER, R.; BROOKER, B. E. Lactobacilli which attach to the crop epithelium of the fowl American Journal of Clinical Nutrition, New York, v. 27, p. 1305 – 1312, 1974.
16. GARCIA, G. R. Caracterização microbiológica e avaliação de uma cepa de Bacillus subtilis no desempenho de bezerros da raça holandesa. 2008. 68f. Tese (Doutorado em Microbiologia Agropecuária) - Faculdade de Ciências Agrárias e Veterinária – Universidade Estadual Paulista Julio de Mesquita, Jaboticabal.
17. GHORBANI, G.R.; MORGAVI, D. P.; BEAUCHEMIN, K. A.; LEEDLE, J. A. Z. Effects of bacterial direct-fed microbials on ruminal fermentation, blood variables, and the microbial populations of feedlot cattle. Journal of Animal Science, Champaign, v. 80, p.1977-1985, 2002.
18. GONÇALVES, G. D.; SANTOS, G. T.; RIGOLON, L. P.; DAMASCENO, J. C.; RIBAS, N. P.; VEIGA, D. R.; MARTINS, E. N. Influencia da adição de probióticos na dieta sobre o estado sanitário e desempenho de bezerros da raça Holandesa. Brazilian Journal of Veterinary Research and Animal Science, São Paulo, v. 37, n. 1, 10 p, 2000.

23
19. GONÇALVES, M.; MALUTA, R. P.; DAHLKE, F.; et al. Avaliação da capacidade imunoestimulante e da estabilidade de um probiótico empregado para cães. Archives of Veterinary Science, Curitiba, v.12, n.2, p.25-30, 2007.
20. GONZALES, E. Aditivos para rações de aves e suínos. Botucatu: Faculdade de Medicina Veterinária e Zootecnia – Universidade Estadual Paulista Julio de Mesquita, 2011. 183 p. [Apostila]
21. GORGULU, M.; SIUTA, A.; ONGEL, E.; et al. Effect of probiotic on growing performance and health of calves. Pakistan Journal of Biological Science, v.6, n.7, p.651-654, 2003.
22. HASHIZUME, K.; TSUKAHARA, T.; YAMADA, K.; KOYAMA, H.; USHIDA, K. Megasphaera elsdenii JCM1772T normalizes hyperlactate production in the large intestine of fructooligosaccharide-fed rats by stimulating butyrate production. Journal of Nutrition, Philadelphia, v. 133, n. 10, p. 3187 - 3190, 2003.
23. HOLZAPFEL, W.; HABERER, P.; GEISEN, R.; BJÖRKROTH, J.; SCHILLINGER, U. Taxonomy and important features of probiotic microorganisms in food and nutrition, American Journal of Clinical Nutrition, New York, v. 73(suppl), p. 365S–373S, 2001
24. INGVARTSEN, K. L.; ANDERSEN, J. B. Integration of Metabolism and Intake Regulation: A Review Focusing on Periparturient Animals. Journal of Dairy Science, Champaing, v. 83, p. 1573–1597, 2000.
25. ISOLAURI, E.; SÜTAS, Y.; KANKAANPÄÄ, P.; ARVILOMMI, H.; SALMINEN, S. Probiotics: effects on immunity, American Journal of Clinical Nutrition, New York, v. 73 (suppl), p. 444S-450S, 2001.
26. JIN, L. Z.; MARQUARDT, R. R.; BAIDOO, S. K. Inhibition of enterotoxigenic Escherichia coli K88, K99 and 987P by the Lactobacillus isolates from porcine intestine. Journal of the Science of Food and Agriculture, London, v.80, n.5, p.619-624, 2000.
27. KREHBIEL, C. R.; RUST, S. R.; ZHANG, G.; GILLILAND, S. E. Bacterial direct-fed microbials in ruminant diets: Performance response and mode of action. Journal of Animal Science, Champaign, v. 81, (Suplemento especial 2), p.120 - 132, 2003.
28. KUNG Jr., L. Direct fed microbials and enzymes for ruminants. In: ANNUAL FLORIDA RUMINANT NUTRITION SYMPOSIUM, 12., 2001. Gainesville. Proceedings… Gainesville: University of Florida, 2001. p. 11 – 12.

24
Disponível em: http://dairy.ifas.ufl.edu/rns/2001/Kung.pdf. Acesso em 24 de novembro de 2011.
29. LEE, S. S.; HSU, J. T.; MANTOVANI, H. C.; RUSSEL, J. B. The effect of bovicin HC5, a bacteriocin from Streptococcus bovis HC5, on ruminal methane production in vitro. FEMS Microbiology Letters, Amsterdam, v.217, p. 51-55, 2002.
30. LESMEISTER, K. E.; HEINRICHS, A. J.; GABLER, M. T. Effects of feeding yeast culture and propionibacteria on milk yield and milk components on Holstein cows. Journal of Dairy Science, Champaing, v.88, p.1832-1839, 2004.
31. MAGNABOSCO, C. U.; CARNEVALLI, R. A.; SAINZ, R. D.; FILGUEIRAS, E. A.; MAMEDE, M. M. S.; CASTRO, L. M. Probióticos melhoram a qualidade do leite de vacas Girolando. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 47, 2010, Salvador. Anais eletrônico... [CD-ROM], Salvador: SBZ, 2010.
32. MANTOVANI, H. C.; KAM, D. K.; HA, J. K.; RUSSEL, J. B. The antibacterial activity and sensitivity of Streptococcus bovis strains isolated from the rumen of cattle. FEMS Microbiology Ecology, Amsterdam, v.37, p.223-239, 2001.
33. MAPA, Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa Nº 13, de 30 de novembro de 2004. Disponível em: http://extranet.agricultura.gov.br/sislegis/action/detalhaAto.do?method=consultarLegislacaoFederal. Acesso em 24 de novembro de 2011a.
34. MAPA, Ministério da Agricultura, Pecuária e Abastecimento. Instrução Normativa Nº 15, de 26 de maio de 2009. Disponível em: http://extranet.agricultura.gov.br/sislegis/action/detalhaAto.do?method=consultarLegislacaoFederal. Acesso em 24 de novembro de 2011b.
35. MARTIN, S. A.; NISBET, D. J. Effect of direct-fed microbials on rumen microbial fermentation. Journal of Dairy Science, Champaign, v. 75, n. 6, p. 1736 – 1744,1992.
36. MEYER, P. M.; PIRES, A. V.; BAGALDO, A. R.; SIMAS, J. M. C.; SUSIN,I. Adição de probiótico ao leite integral ou sucedâneo e desempenho de bezerros da raça holandesa. Scientia Agricola, Piracicaba, v.58, n. 2, p.215-221, 2001.
37. MOLL, G. N.; KONINGS, W. N.; DRIESSEN, A. J. Bacteriocins: mechanism of membrane insertion and pore formation. Antonie van Leeuwenhoek, Amsterdam, v.76, p.185-198, 1999.

25
38. NOCEK, J. E.; KAUTZ, W. P.; LEEDLE, J. A. Z.; ALLMAN, J. G. Ruminal Supplementation of Direct-Fed Microbials on Diurnal pH Variation and In Situ Digestion in Dairy Cattle. Journal of Dairy Science, Champaing, v. 85, p. 429–433, 2002.
39. NOCEK, J. E.; KAUTZ, W. P.; LEEDLE, J. A. Z.; BLOCK, E. Direct-Fed Microbial Supplementation on the Performance of Dairy Cattle During the Transition Period. Journal of Dairy Science, Champaing, v. 86, p. 331–335, 2003.
40. NOCEK, J.E.; KAUTZ, W. P. Direct-Fed Microbial Supplementation on Ruminal Digestion, Health, and Performance of Pre- and Postpartum. Journal of Dairy Science, Champaing, v. 89, p. 260–266, 2006.
41. OGAWA, M.; SHIMIZU, K.; NOMOTO, K.; TANAKA, R.; HAMABATA, T.; YAMASAKI, S.; TAKEDA, T.; TAKEDA, Y. Inhibition of in vitro growth of Shiga toxinproducing Escherichia coli O157:H7 by probiotic Lactobacillus strains due to production of lactic acid. International Journal of Food Microbiology, Amsterdam, v.68, n.1-2, p.135-140, 2001.
42. RAETH-KNIGHT, M. L.; LINN, J. G.; JUNG, H. G. Effect of Direct-Fed Microbials on performance, diet digestibility, and rumen characteristics of Holstein dairy cows. Journal of Dairy Science, Champaing, v. 90, p. 1802-1809, 2007.
43. RULQUIN, H.; RIGHOUT, S.; LEMOSQUET, S.; BACH, A. Infusion of glucose directs circulating amino acids to the mammary gland in well fed dairy cows. Journal of Dairy Science, Champaing, v .87, p. 340–349, 2004.
44. RUSSEL, J. B.; MANTOVANI, H. C. The bacteriocins of ruminal bacteria and their potential as an alternative antibiotics. Journal of Molecular Microbiology and Biotechnology, La Jolla, v. 4, p.347-355, 2002.
45. SAINZ, R. D.; MAGNABOSCO, C. U.; FILGUEIRAS, E. A.; GUIMARÃES, R.; FREITAS, F. M. C.; MATTOS, L. R. Effects of a direct-fed microbial and fibrolytic enzyme product on somatic cell counts in milk produced by crossbred dairy cows in the Brazilian Cerrado. Journal of Dairy Science, Champaing. v. 94, E-Suppl. 1: p. 126. 2011.
46. SALMINEN, S.; ISOLAURI, E.; SALIMEN, E. Clinical uses of probiotics for stabilizing the gut mucosal barrier: successful strains and future challenges. Antonie van Leeuwenhoek, Amsterdam, v. 70, p. 347– 358. 1996.

26
47. SAXELIN, M.; TYNKKYNEN, S.; MATTILA-SANDHOLM, T.; VOS, W. Probiotic and other functional microbes: from markets to mechanisms. Current Opinion in Biotechnology, London, v.16, p.1-8, 2005.
48. SCHAELLIBAUM, M. Efeitos de altas contagens de células somáticas sobre a produção e qualidade de queijos. In: Simpósio Internacional sobre Qualidade do Leite, 2, 2000, Curitiba. Anais... Curitiba: CIETEP/FIEP, 2000. p.21-26.
49. SIGNORINI, M.L.; SOTO, L. P.; ZBRUM, M. V; SIQUEIRA, G. J.; ROSMINI, M. R.; FRIZZO, L. R. Impact of probiotic administration on the health and fecal microbiota of young calves: A meta-analysis of randomized controlled trials of lactic acid bacteria. Research in Veterinary Science, London, Online, 2011.
50. STEIN, D. R.; ALLEN, D. T.; PERRY, E. B.; BRUNER, J. C.; GATES, K. W.; REHBERGER, T. G.; MERTZ, K.; JONES, D.; SPICER L. J. Effects of feeding propionibacteria to dairy cows on milk yield, milk components, and reproduction. Journal of Dairy Science, Champaign, v. 89, n. 1, p. 111 - 125, 2006.
51. TIMMERMAN, H.M.; MULDER, L.; EVERST, H. et al. Health and grow of veal calves fed milk replacers with or without probiotics. Journal of Dairy Science, Champaing, v. 88, n. 6, p. 2154 - 2165, 2005.