abundancia relativa de insectos en un medio … · 2018-04-29 · abundancia relativa de insectos...
TRANSCRIPT

ABUNDANCIA RELATIVA DE INSECTOS EN UN MEDIO ESTACIONAL; SU INFLUENCIA EN LA HISTORIA DE
VIDA DE DOS IGUANIDOS SIMPATRICOS "
ALFREDO ORTEGA LUCINA HERNÁNDEZ Instituto de Ecologia Apartado Postal 18-845 11800 México, D. F. MEXICO
Folia Entomoldgica Mexicana No. 5.5: 129-144 (1983)
* Trahajo desarrollado dentro del proyecto "Reserva <le la Biosfera La Mictiilla, Ilgo.", como la contribiicidn No. 02 al proyecto "Ecologia y Comportamiento Animal" apoyado por la Direccidn Adjunta de Desarrrollo CientIfico del CONACYT (PCC-BCNA-O'L0119) . Así como, con el patrocinio de la Subsecretaría de Educación Superior e Investigacidn Cientlfica, SEP, el Gobierno del Estado de Durango y The Man and Biobphere l'rogram (UNESCO) .

La precipitación pluvial es el principal factor que determina la pro- ductividad de las comunidades de regiones que tienen una marcada esta- cionalidad de &pocas secas y húmedas, en estas zonas se encuentran drás- ticas diferencias en el número dc especies de insectos, de la cantidad de individuos de cada una de ellas y de su biomasa entre las dos épocas del año, entre años más secos y más húmedos y entre sitios más húmedos y más secos de la misma zona, sin embargo, la gran mayoría de este tipo de estudios han sido realizados en zonas tropicales (Janzen, 1973; sanzen and Pond, 1975; Janzen and Schoenner, 1968) o en zonas áridas (Ballinger, 1977; Davidson, 1977; Worthington, 1982), siendo relativamente poco lo que se sabe de este fenómeno en zonas montanas templadas.
Los efectos de la variación en la disponibilidad de alimento que estas diferencias en la abundancia de insectos representan para los organismos que se nutren de ellos son diversos y profundos. En el caso de las lagar- tijas insectívoras estas variaciones afectan aspectos tales como el tamaño y la frecuencia de las puestas (Ballinger, 1977, 1978; Goldberg, 1975), su crecimiento corporal (Ballinger and Congdon, 1980) el tamaño de su área de actividad (Simon, 1975), la edad y talla a la madurez sexual, el tama- ño de los huevos y su esfuerzo reproductivo (Martín, 1977).
Pretendemos analizar aquí qué niveles de fluctuación estaciona1 se pre- sentan en la abundancia de insertos de una zona montana, relacionarlos con la estacionalidad de la precipitación, con los cambios de biomasa de las plantas herbáceas y con la biología de las dos lagartijas más abundan- tes de la zona, contribuyendo de este modo a la integraci6n de los cono- cimientos obtenidos por diferentes equipos de especialistas que trabajan en una Reserva de la Biosfera.
El trabajo de campo se realizó en la Reserva de la Biosfera "La R~iichi- lia" la cual se localiza en la vertiente interna de la Sierra Madre Occidcii- tal en el Sureste del Estado de Durango, México. Sus límites convencio-

nales están dados por los paralelos 23O25' y 23O30' de Iatitud norte y por los meridianos 104015' y 104O21' de longitud oeste, la altitud del área varía entre los 2350 y los 2850 m.s.n.m., su clima es de templado semi- seco a templado subhúmedo fluctuando la precipitación entrc los 500 y 700 mm anuales y la temperatura media anual entre 17.4 y 20.4OC. Geo- 10gicamente la región está constituída por rocas ígneas de extrusiones vol- cánicas terciarias, riolitas y basaltos, y ticne una vegetacicín sumamente diversificada (hlartíncz y Saldivar, 1978) encontrándose once unidades f i - sonómico-llorísticas distintas.
Las especies de reptiles más abundantes de la zona son los iguánidos Sceloporus granzn7.iczr.s microlepido t us Wiegmann y Sceloporus .scaTaris Wiegmann, las cuales son lagartijas diurnas insectívoras y más bien pe- queñas (de 50 a 62 mm cuando adultas). Para detcrminar más específi- camente qué tipo de insectos consumen estas dos especies, individuos adul- tos de ellas fueron colectados entre las 10 y las 18 horas en septiembre de 1979 y abril de 1980, una vez capturados fueron transladados al labo- ratorio de la estación "Piedra Herrada" del Instituto de Ecología en La - Michilía, donde se proccs;tron dentro de las 6 hrs postcriores a su captura, anotando su longitud corporal (de la punta del hocico a la abertura cloa- cal), la longitud de la cola (de la abertura cloaca1 a la punta de la misma) y el peso del organismo, a continuación fueron sacrificados con una sobre- dosis de nembutal, se les extrajo el estbmago, se fijó en alcohol al 70% y se mandó a la Ciudad dc Mcxico en dondc fuc cuiclnclosniil<!iit~~ :il>icr.to. Todos los artrópodos encontrados eii su interior fueron identilicacios hasta nivel de orden y medidos con aproximación de 0.01 mm con un ocular micrométrico en un microscopio estereoscópico. Los cuerpos de las lagar- tijas están depositados en la colección de vertebrados del Instituto de Ecología en la Ciudad de México.
Una vez conocidos cl tipo y la talla de los inscctos que consumcn en mayor cantidad estas dos especies, sc procedió a detcrminar si existen diferencias en su abuiidancia en diferentes estaciones del año, para lo cual, durante los 5 primeros días de mayo, junio, agosto y octubrc dc 1981 y dentro de los transectos utilizados para estudios de marca-rccap- tura de las dos especies de lagartijas se colocaron en el suelo distribuidos aleatoriamentc 10 cuadros de madera de 100 cm2 cubiertos de material nd- hesivo, estas trampas eran colocadas diariamente a las 10 hrs y se recogían a las 18 hrs, los insectos capturados eran entonces fijados en alcohol al 70%. Durante los mismos T> días y al inicio de cada hora entre las 10 y las 17 hrs se aislaron en el suelo, mediante un marco metálico, Arcas de 2500 cm2 escogidas también aleatoriamente recogiendo de manera directa y con una pequeña red o con aspiradores bucales los insectos encontrados

dentro del cuadro fijándolos inmediatamente en alcohol al 70%. En total se recogieron 200 trampas y se efectuaron 160 muestreos en el suelo.
La Tabla 1 resume el porcentaje de los órdenes de insectos niás consu- midos por Sc. grammicu.~ y Sc. scalaris así como el porcentaje que tienen en talla la mayoría de las presas que consumen. Como se observa cerca del 80% de la dieta de estas dos especies la constituyen 3 6rdenes de in- sectos y cerca del 75% de todas las presas que ingieren miden entre 2.1 y 8.0 mm.
Tabla No. l.- Porcentaje de los principales ordenes de insectos consumidos por las dos especies de lagartijas más abundantes de la reserva de la biosfera .LA MICHI LIAD, y porcentaje del total de presas que consumen y que miden entre 2.1 y 8.0 mni.
'A Ortópteros
% Himenópteros
'4 Coleópteros
% de talla entre 2.1 y 8.0 mm
Sc. grammicus
32
28
19
72
Sc. scalaris -
3 1
29
22
74


En base a estos datos, el análisis de la abundancia relativa dc insectos por estación, fue restringido al estudio de los ortóptcros, himenópteros y cole0pteros cuyo tamaño estuviera entre los 2.1 y los 8.0 mm. La Tabla 2 resume los rcsult,idos encontrados para el iiíiincro dc individuos de estos insectos colectados por loi dos métodos y en los cuatro meses de trabajo de campo. A pesar de que conio se observa en la Fig. 1 la proporcihn de los diicrentes íxdcncs se modifica a lo largo del año, la tendencia dentro cle cad'i orden (Iiig. 2) cs de que aumenta el número de individuos co- lectados tambibn a lo 1.iigo dcl año, por lo cual es 1í)gico esperar (Janzen and Poncl, 197.5) q ~ i c t'irnbién su diversidad específica y su bioinasa se incremeriten a lo largo del niismo.
Para la zona se cuenta con datos climatológicos a partir de septiembrc de 1980, sin embargo, analizando los datos de 1976 a 1980 de 2 estaciones -
climatol6gicas cerc'inas (Fig. 3) se observa con claridad que aproximada- rriente cl 90% de la precipitación total anual se concentra en los mcscs de junio, julio, agosto y septienibre, lo cual establece un patrím anual <lc aburid,irite Iiulned'ld de junio a septiembre y de sequía de octubre a mayo.
Al compara1 los datos dcl año de 1981 de precipitación pluvial (Tabla 3) de la estación climatológica instalada en el laboiatorio "Piedra He- rrada", con cl número de insectos de las categorias previamente definidas inuestreado en los .1 mese5 (Fig. 4) observamos que no existe una cstreclia rcl;iciOn entre la cantidad promedio de lluvia por día en cada mes y cl númeio cle hectos atrapado (r = 0.03) -donde r es cl coeficiente <lc corrclacicín de Pearson (Sokal and Rohlf, 1969)- así aunque la precipi- tacicín ;iunienta bruscamente de mayo a junio (alcanando en este mes el niáxi~rio) declinando un poco, pero permaneciendo alta, hasta septiem- brc para después cacr abriiptamente en octubre, el níimero de insectos comien~a a increnicntarse significativanicnte de mayo a junio aumentando signilicativamentc de este mcs a agosto y alcaii7ando el máximo en octu- bre (toda\ diferencias significativas P < 0.01, para x L 7.38).
El efecto de la cantidad de lluvia tampoco es automático sobrc la pio- ductivida<l cle la vegetación, así Carrillo (1982) tomando 134 puntos de muestico eri varias lonas de la Reserva para estimar la l>iomasa cle las herbáceas y su productividad por los métodos de Pecliancc y Pickford (1937) y por el de Wiegert y Evans (1964), encuentra que mayo y octubre
son los meses con los valores máximo y minimo de bioinasa respectivamente (en gramos de materia seca por m2) observándose de marzo a mayo dismi-

Tabla No.3; Precipitaciori pluvial promedio diaria durante 1981 eii la Reserva de la Biosfera -LA MICHILIA..
nuciones significativas en la cantidad de biomasa y de mayo a agosto un incremento tambiéri significativo que se continúa hasta octubre (Fig. 5). En esta misma figura se comparan estos resultados con el patríin de preci- pitación en la zona para 1981, coino se observa tambikn existe poca relación entre estos valores (r = 0.32) sobre todo porque a pesar de que las lluvias van en declinación desde junio (primero lentamente liasta septiembre) hasta noviembre (con un brusco descenso de septiembre a octubre), la bio- masa de las lierbáceas va en incremeiito constante de mayo a octubre.
Carrillo (1982) indica que este patrón es debido a que las herbáceas anuales incrementan su biomasa principalmente de junio a agosto pero las pcrennes lo Iiaccn principalmente de agosto a octubre, ya que estas tienen sus raíces inás profundas y son capaces de aprovechar el agua que está a mayor distancia de la superficie y por lo tanto menos accesibles para las anuales; este patrón de incremento de la biomasa de las herbáceas de la zona se mantiene con poca variación interanual, alcanzando siempre el máximo en octubre (Sonia Gallina com. pers.).
Los datos de abundancia de insectos tienen una muy estrecha corres- pondencia (r = 0.95) con los valores de la biomasa de herbáceas: los dos tienen sus valores máximo y mínimo en octubre y mayo respectivamente y tienen además, incrementos semejantes (Fig. 6).
Asi el hecho de que no se encuentre una clara relación entre la canti- dad de lluvia y el número de insectos en el área, pero si exista una estre- cha correspondencia entre la biomasa de herbáceas y la cantidad de insec- tos nos indica sin lugar a diidas que el efecto de la precipitación pluvial no es directo ni inmediato sobre la abundancia de insectos sino que está mediado a través de la productividad de las plantas, conclusión que ha sido reiteradamente sugerida en otros trabajos, la mayoría de los cuales,

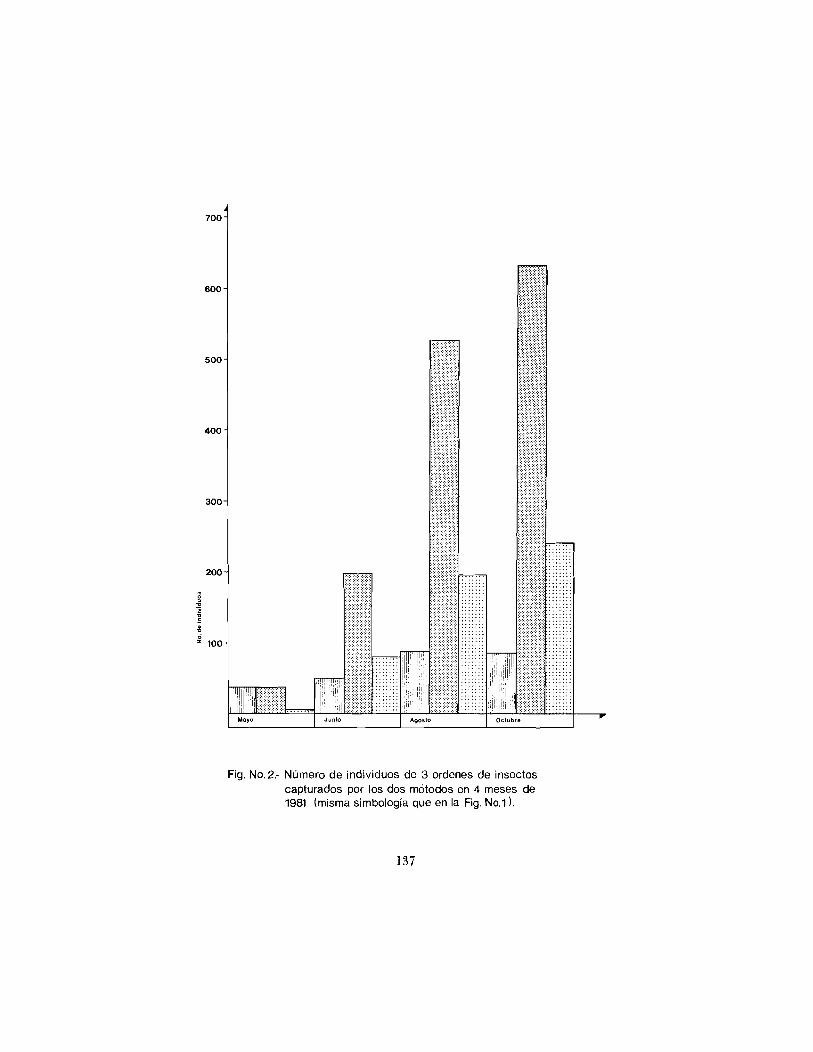
Fig. No.2. Número de individuos de 3 ordenes de insectos capturados por los dos métodos en 4 meses de 1981 (misma simbologia que en la Fig. No.1).

f - : ; ! , ,k E F M A M J J A S O N D
5 5 0 Eslacio11 C H A L C H I H U I T E S
I'romcdio: 1976 1 9 8 0
1 7 . 0 6 " C . 517.4 mni
o Y-+#aLJ E F M A M J J A S O N D
Estacion M E S Q U I T A L
Promedio: 1976 1980
1 9 . 2 6 ' C . 5 3 4 . 8 m m
Fig. No. 3.- Climogramas de las estaciones Meteorológicas de Chalchihui tes (~acatecas) y Mesquital (~urango), para los,anos 1976 - 1980 (tomado de ~ a r r i I lo, 1982).

Fig. lVo. 4. Precipitación pluvial promedio mensual en rnilirnetros por dia y abundancia de insectos zapturados en número de individ~ios.
sin embargo, carecen de datos acerca de la productividad de las plantas (Martin, 1977; Mayhew 1965, 1966 y 1967).
El periodo de mAxima disponibilidad de alimento en la zona para Sc. g r a m m i c ~ ~ s y Sc. scalaris va de junio a octubre, y sus ciclos reproductivos están siilcronizados con este, así la ovulación, el cortejo y la cópula en Sc. scalaris ocurren en mayo y las hembras depositan los huevos en junio eclosionando estos y apareciendo los juveniles cn agosto (Ortega, en pre- paracihn), una segunda época de cortejo y apareamiento ocurre en julio, 10s huevos fecundados se depositan en agosto y estos eclosionan en octu- bre. Se. grammicus (Ortega y Rarbault, en prensa) se aparca en noviem- bre y los embriones son mantenidos por las hembras hasta su completo desarrollo y nacimiento lo cual ocurre en mayo.

lp,ccll,,l3t,~,,, ,>1,,",31
G-O biomasa de hcrbacenc
Fig. No. 5.- Precipitacion pluvial promedio mensual en milimetros por dia y biomasa de herbaceas en gramos de peso seco por rnetro cuadrado.
Entonces el periodo de mayor abundancia de alimento es aprovechado por las liembras cle Sceloporus scalnri.~ para transferir mayor cantidad de nutrientes a los Iiuevos (le la primera puesta y para acceder a una segunda puesta, y por sus juveniles para crecer lo más posible antes del invierno; en el caso dc Sc. grarnmictl.~ sus juveniles aprovechan este periodo de gran cantidad de alimento para crecer rápidamente y así a fines de la estación estar en condiciones de aparearse.
Otros autores han sugerido la existcncia de una relacicin positiva entre la precipitacibn y el tiempo de reproduccií,n en lagartijas (Hoddenbach In, Turner, 1Yti8; Mayhew, 1!)661>; Paikci-, 1973; Pianka, 1970; Zweifel and Lowe, 1966), sin embargo, en este trabajo se evidencía el cómo la energía obtenida en la época de mayor abundancia de alimento no necesariamente

biomasa de herbaceas
0--0 abundancia de inseclos
Fig. No. 6.- Biomasa de Herbaceas en gramos de peso seco por metro cuadrado y abcindancia de insectos en número de individuos capturados.
tiene que ser transferida por las hembras a los juveniles de la nueva gene- racicín (ya sea a travCs de vitelo o a través de la placenta), sino que esta energía puede ser asignada diferencialmente en especies simpátricas como en este caso, en una sí directamente en reproducciOn, pero en la otra para el crecimiento corporal, lo cual influencía 1;i sincronía de sus ciclos repro- ductivos y por eiidc sil liistoria (le vida.
Aunque cstá claro que existen diferencias estacionales muy marcadas en la Reserva <le 1;t Riosfera de 1.a Micliilia en 1;i abundancia de los insectos, sc liace prioritario electuar estudios expcrimentalcs mrjs cspecíficos para determinar la causalidacl cntre la productivi<l;id rle 1;is plantas cle la zona y el número y diversidacl de los mismos así como el establecer de forma inás precisa la dinámica sucesional dc sus poblaciones a lo largo dcl aíío (por ejeniplo, el chino varíari las poblaciones de Litófagos, cariiívorob y piirrísitos), para acercarlos m3s a la corriprensitin de la estructura y turi-

cióil de un ecosistema que ocupa decenas de miles de hectáreas en la Rc- pública klexicana.
Estudiando la dieta de las dos lagartijas insectívoras más abundantes de la Reserva de Ida hlicliilía en el Estado de Ilurango, Mcxico, Scc1oporu.s gramrnicus y Sceloporu .~ .scaluri.s, se encucntra que aproxiniadaniente cl 80y, de su alimento lo constituyen insectos de los Ordenes Ortlioptera, EIy- menoptera y Coleoptera, y que aproxiiriadamente cl 75y,, cle todas sus prc- sas miden entre 2.1 y 8.0 mm en ambas especies. 1)urarite 4 mescs de 1981 se estudia la abundancia de este tipo de insectos por dos métodos clistintos (trampas dc adhcsivo y colectas directas en áreas de 0.25 m') encontrando que estos datos tienen una pobre correlación (r = 0.03) con los de preci- pitación pluvial, pero una alta (r = 0.95) con los datos de biomasa de las herbáceas de la zona. El período de máxima abundancia de alimento para Sc. grammicus y Sc. scaluris es de junio a octubre lo cual influencia la sincronía de sus ciclos reproductivos aprovechando las dos especies de forma diferencial estas variaciones en la disponibilidad de este recurso en su historia de vida.
Studying diets of the two more abundant insectivorous lizards of the "La Micliilía" Biosphere Kescrve at tlie State ol Durango, Mexico, it is found that approxiniately 80% of its nourisli~nent is composed by threc insect's orders: C)rthoptera, Elyiiierioptcra antl Coleoptera and nearly 75% of al1 its prcys llave a leng~li between 2.1 arid 8.0 mni in both spccies. During 4 months of the year 1981 the abundance of these insects is studied by two different methods (sticky traps and direct collects in 0.25 m%rcas). 'I'hese data liave ;I poor corrclation (r = 0.03) witli rainlall, but a higli correlation (r = 0.95) witli grass I>ioiiiass of tlie site. Tlie period of ma- ximuni abiindancc of iood lor Sr. gra~nrr~ icu , and Sc. ~ c a l a r i ~ is between June and Octobcr. 'l'liis Eact influcnces their reproductive cycles synch- rony, uhing eacli specie, in a tlilrccre~it I'oiiri tIic v~triationb in iood a\a- ilability in thcir life histories.

BALLINGER, R. E. 1977. Reproductivc strategies: Cood avai1ai)ility as a source oi prox- iinal variatioii in a lizarcl. Ecology, 5R(S) :628-635. --- 1978. Variation iti ancl eiolution o£ clutch and litter sizc. 111: R. E. Joncs (ed.),
T h e verlebrate ovary. Plenum llublisliing Corp. U.S.A. Cap. 23:789-825. BALLINCER, R. E., anrl J. D. C o ~ c n o ~ . 1980. Food resource limitation oi I)ody growth
ratcs in Sceloporiis sculuris (Sauria: Igiiaiiidac). COPEIA, 1980 (4) :921-923. CARICILLO, A. S. 1982. Produccicin priirraricz neta aérea del eslralo herbáceo y efecto del
ganado sobre su composicidn flori~tica en la Reserva d e la Biosfera "L,a Michilta". Tesis proEcsiona1 UNAM. Pac. (le Ciencias.
I )AVIDS~N, D. W. 1977. Species diversity and commiinity orgaiiization in desert seed- eating ants. Ecoloby 58 (4) :711-724.
Go~osenc, R. S. 1975. Yearly variations in tlie ovarian cycle o£ the lizard Sceloporil~ occidentalis. J . Herp . 9(?) :187-189.
IIODDENBACH, A. G . and F. B. TURNIR. 1968. Clutch size o£ the lizard Uta stunsburiana in southern Nevada. Amer. Miti. Nat . 80:262-265.
JANZEN, D. H. 1973. Sweep samplcs o£ tropical Coliage insects: effects of seasons, vege- tation types, elevation, time o£ day, and insiilarity. Ecology 54:687-708.
JANZEN, D.-H. and C. M. POND. 1975. A comparison, by sweet sampling, o£ the arthopod fauna o£ secondary vcgetation iii Micliigan, England and Costa Rica. Trans. R . Ent . Soc. Lond . 127 (1) :33-50.
JANZEN, D. H. and T. W. SCHOENER. 1968. Differences in insect abundante and diversity between wettcr anrl drier sites during a tropical dry season. Ecology 49:69-110.
MARTIN, R. F. 1977. Variation in reprotluctive procluctivity of range margin tree lirards (Urosaurus ornaliis). COPEIA, 1977 (1) :82-95.
MART~NEZ, E. O. y hl. C. SALDIVAR. 1978. Unidades de vegetacibn en la Reserva de la Biosfera La Michilia, Durango. En: G. Halffter (ed.). Reservas d e la Uiosfera, Instituto de Ecología, México. Publ. 4:134-198.
MAYHKW, W. \Y. 1965. Reprodriction in the sanrl-dwelling lizard U~rra inornaln. Herpe- lologica 21 :39-55. - 1966a. Reprodiiction in the psamophilous lizard Urna scoparia. COPEIA, 1966:
114-122. --- 19661,. Reprodiiction in tlie arenicolous lizard Urna notata. Ecology 47:9-18. ---- 1967. Comparatice rcproduction in three species of the genus Unta. In: W. W .
Milstead (cd.). I,izar(l ecology: a syrnposiz~m. Univ. Missouri Press, Columbia: 45-61. O r r e c ~ , A. Reprodiictive cycles in thc lizard Sreloporus scalaris at tlic Biosphere Rcscr-
ve "La Michilia" Mexico. I n prcp. O ~ r e c a , A. and R. BARUAULT. Rcproductioir in tlic incz<liiitc lizard Scelol>orzis grarn~riicus
(Sauria: Iguanidae). In press. PAKKER. S. W. 1973. Natural history notes on thc iguanid lizartl Urosaz~rus ornatus. J.
Herp . 7:21-26. l l ~ c r r ~ ~ ~ c , J. F. anrl G. 1) . I>ICKPOKU. 1937. A weight estimatc method for the detcrm-
ination o£ rangc or püsture prodiiction. J. Amer Soc. Agron. 29594-904. I>IANl<A, E. R. 1970. Comparative autoccology oC tlic lizard Cizeitritioi)hor~is Ligris in dif-
ferent parts o£ its geograpliic range. E c o l o g , 51:703-720. SIMON, C. A. 1975. T h c influence of food abiindancc on tcrritory siae in tlie iguaiiitl
lizarrl Sceloporiis jarrovi. Ecology, 56:993-998. SOKAL, R. R. aiid F. J . KOHI.P. 1969. Iliomelry. W. H. Freeman anrl Co. (Ed.). San Fran-
cisco.

WIEGERT, R. G. and 1:. C. EVANS. 1964. Piiniary production aiitl tlic (lisappearailce o f <lea<i vcgetation on an olcl ficld in southeasterri Micliigan. Ecoloby, 45 (1) :49-69.
W ~ K ~ I I I N G T O N , R. 1). 1982. ilry ancl wct year cornparisons of cliilch antl acliilt body sizcs o f Uta stan,sbt~riciilu stejtcegeri. J . Hevpelol. 16(3): 332-994.
ZWEIFKL, R. G . and C . H. LOWE. 1966. 'Tlie ecology o f a populatiori o l Xarltusia uigilis, the desert riight lizarcl. A ~ n e r . A t r ~ s . Novi t . 2247:l-57.