a importância do conhecimento do potencial da água no ... · transporte da água no sistema...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO
ESCOLA SUPERIOR DE AGRICULTURA “LUIZ DE QUEIROZ”
DEPARTAMENTO DE ENGENHARIA RURAL
LCE – 5708 Processos de Transferência no Sistema Planta-Atmosfera
Professor: Luiz Roberto Angelocci
A importância do conhecimento do potencial
da água no sistema solo-planta-atmosfera
(É o potencial da água um bom indicador do estado hídrico da planta?)
Ralini Ferreira de Mélo
Tales Miler Soares
Piracicaba – SP
Novembro de 2004

2
INTRODUÇÃO
A disponibilidade de água e de energia radiante, junto com a temperatura,
são importantes fatores físicos do ambiente a governar a distribuição das espécies
vegetais na Terra. Na agricultura, a disponibilidade hídrica é um dos grandes
fatores que condicionam a produtividade e o sucesso de uma cultura. A
importância ecológica da água está associada ao fato de que todos os processos
fisiológicos das plantas são, direta ou indiretamente, influenciados pelo
suprimento hídrico (Kramer & Boyer, 1995). Sua importância fisiológica decorre
das funções que ela desempenha na planta, como principal constituinte dos
tecidos; como excelente solvente para nutrientes; como imprescindível reagente
em processos metabólicos; como mantenedora da turgescência e moderadora
térmica dos tecidos (Angelocci, 2002).
Conforme Kramer (1988), as várias razões para se avaliar o status da água
na planta podem ser sumarizadas em quatro categorias que são de interesse de
diferentes grupos de pessoas que requerem distintos tipos de informações:
1 - Determinar quando irrigar. De interesse dos agricultores.
2 - Selecionar populações de plantas. De interesse dos pesquisadores em
melhoramento.
3 - Estudar efeitos do estresse hídrico sobre o desenvolvimento e a
produtividade. De interesse dos agrônomos.
4 - Estudar efeitos do estresse hídrico sobre processos fisiológicos e
bioquímicos. De interesse dos fisiologistas e ecologistas.
O homem tem tentado de alguma forma estimar o status da água das plantas
desde que descobriu que poderia aumentar as produtividades com a irrigação. As
mais antigas observações científicas relatadas sobre o status da água nas plantas
foram provavelmente aquelas de Stephen Hales (1727). Hales mediu a seiva
ligando tubulações de vidro às extremidades cortadas de ramos e raízes, a partir
dos quais obteve observações suficientes para deduzir uma abundância de
informações sobre o fluxo da água nas plantas (Baker, 2003). Nos últimos 30
anos, muitas ferramentas têm sido desenvolvidas para medir o estado de

3
equilíbrio termodinâmico da água nos tecidos vegetais (Boyer, 1995), mas estas
não têm sido amplamente utilizadas em todos os campos com sentido diagnóstico
ou como base para se tomar decisões de manejo. O meio ambiente natural em
que operam as plantas é simplesmente não conducente às medidas de equilíbrio.
Insolação, temperatura, umidade atmosférica e velocidade do vento podem afetar
o status da água na planta ou as medidas do potencial da água, e são todos
inerentemente instáveis (Baker, 2003).
Mesmo se as medidas de potencial da água pudessem ser feitas de maneira
precisa e rotineira no campo, os valores dos dados seriam incertos. Como
mostrado por Sinclair & Ludlow (1985), muitos dos processos fisiológicos que
ocorrem dentro da planta, embora dependentes da água, não são correlacionados
com o potencial da água. Passioura (1988) discorreu sobre isto, notando o falso
conforto com que os fisiologistas têm abraçado o conceito aparentemente
unificador do potencial da água. Como notado por ele, uma das principais
atrações do potencial da água, isto é aquela que é a força motriz para o fluxo da
água, nem mesmo se aplica em plantas, onde o efeito de solutos e as
propriedades das membranas complicam o assunto. O que se mede então ?
Provavelmente não há uma resposta simples. Os fisiologistas estudando o
mecanismo de tolerância à seca precisam de informações diferentes das que
precisam os produtores que tentam otimizar a produção das culturas irrigadas.
Conseqüentemente, se reunidos os desejos, o requisitado pode ser uma série mais
ampla de ferramentas de medida (Baker, 2003).
ESTRESSE HÍDRICO
Em condições naturais e agricultáveis, as plantas estão freqüentemente
expostas ao estresse ambiental, seja por fatores bióticos ou abióticos. Alguns
fatores dessa natureza, como a temperatura do ar, por exemplo, podem se tornar
estressantes em poucos minutos; enquanto outros, como o conteúdo de água no
solo, podem levar dias ou até semanas e, ainda, fatores tais como as deficiências
minerais no solo necessitam de meses para se manifestar. Estima-se que, devido
ao estresse resultante de condições climáticas e edáficas (fatores abióticos), a

4
produtividade de culturas agrícolas nos EUA constitui apenas 22% da
produtividade potencial genética (Boyer, 1982). Em sua publicação, Taiz &
Zeiger (2004) dedicam um capítulo ao estudo da fisiologia do estresse,
destacando o estresse hídrico, dentre outros como os provocados pelo calor e
choque térmico, pelo resfriamento e congelamento, pela salinidade e pela
deficiência de oxigênio e nutrientes.
Além disso, o estresse desempenha um importante papel na determinação
de como o solo e o clima limitam a distribuição de espécies vegetais. Assim, a
compreensão dos processos fisiológicos subjacentes aos danos provocados por
estresse e dos mecanismos de adaptação e aclimatação de plantas a estresses
ambientais é de grande importância para a agricultura e o meio-ambiente. A
produtividade de plantas, limitada pela água, depende da quantidade disponível
deste recurso e da eficiência do seu uso pelo organismo. Uma planta capaz de
obter mais água porque tem maior eficiência no seu uso resistirá melhor à seca.
Algumas plantas possuem adaptação, como os tipos fotossintéticos C4 e CAM,
que lhes permitem explorar ambientes mais áridos. Além disso, as plantas
exibem mecanismos de aclimatação que são ativados em resposta ao estresse
hídrico (Taiz & Zeiger, 2004).
O déficit hídrico pode ser definido como todo conteúdo de água de um
tecido ou célula que está abaixo do conteúdo mais alto exibido no estado de
maior hidratação. Quando o déficit hídrico apresenta evolução suficientemente
lenta para permitir mudanças nos processos de desenvolvimento, o estresse
hídrico tem vários efeitos sobre o crescimento, um dos quais é a limitação da
expansão foliar. A área foliar é importante, porque em geral a fotossíntese é
proporcional a ela. No entanto, a expansão foliar rápida pode afetar
desfavoravelmente a disponibilidade de água (Taiz & Zeiger, 2004).
O estresse hídrico afeta praticamente todos os aspectos do crescimento da
parte aérea e da raiz, causando redução na expansão celular (Figura 1),
condutância estomática, fotossíntese e conseqüentemente na acumulação de
matéria seca (Hsiao, 1990). A Figura 2 mostra algumas mudanças fisiológicas
devido ao estresse hídrico.

5
Tax
a de crescim
ento
foliar
(mm
/h), G
R
Figura 1 – Dependência da expansão foliar em relação ao turgor. Indivíduos de
girassol (Helianthus annuus) foram submetidos a condições de crescimento
hídrico ou com água limitada no solo para produzir um estresse hídrico
moderado. Após re-hidratação, as plantas dos dois grupos de tratamentos foram
estressadas por falta de água, sendo as taxas de crescimento foliar (GR) e o
turgor (ψp) medidos periodicamente. A redução da extensibilidade (m) e o
aumento do limiar de turgor para crescimento (γ) limitam a capacidade de
crescimento da folha após exposição ao estresse (Matthews et al., 1984).
Os principais aspectos avaliados para análise da planta em tolerar déficit
hídrico elevado, principalmente sob baixa disponibilidade de água no solo,
referem-se à sensibilidade estomática, à troca gasosa e à capacidade de manter
alto teor de água na folha e suportar a desidratação imposta pelo ambiente
adverso. Tais aspectos resumem-se à medida da resistência oferecida pelos
estômatos a esta troca gasosa e pela análise do potencial da água na folha
(Brunini & Cardoso, 1998).

6
Figura 2 - Potencial da água de plantas sob várias condições de crescimento e
sensibilidade de vários processos fisiológicos ao potencial da água (Fonte: Taiz
& Zeiger, 2004).
Medidas do potencial da água na folha têm-se apresentado como importante
instrumento para quantificação do estresse hídrico a que os vegetais são
submetidos, e ainda permite observar e determinar a resistência ao transporte de
água no sistema solo-planta (Faiz & Weatherley, 1978; Taylor & Klepper, 1978;
Brunini, 1979).
SUPRIMENTO DE ÁGUA VIA IRRIGAÇÃO
A água é fator limitante para o desenvolvimento agrícola. Tanto a falta como o
excesso afeta o crescimento, a sanidade e a produção das plantas. A irrigação é
uma prática agrícola cujo propósito é manter adequado o estado hídrico da
planta, para assegurar o desenvolvimento, a produtividade e a rentabilidade
econômica. O objetivo da irrigação é maximizar a produção agrícola,
racionalizando o uso da mão-de-obra, energia, água e fertilizantes, e evitar a
ocorrência de problemas fitossanitários relacionados às aplicações excessivas ou

7
deficiências de água. Para este objetivo, deve-se adotar um manejo correto.
Portanto, para promover o uso racional da água, há necessidade do conhecimento
das variáveis de clima, solo e planta e de alguns aspectos relacionados ao
transporte da água no sistema solo-planta-atmosfera. Dentro de um contexto
amplo, o manejo da irrigação consiste na determinação do momento, da
quantidade e de como aplicar a água, levando-se em consideração os demais
aspectos do sistema produtivo, como a adubação, o controle fitossanitário, as
informações climatológicas e econômicas, e os tratos culturais. O manejo
racional da irrigação pode ser realizado, via solo (tensiômetros, sensores
eletrométricos e por dissipação térmica, sonda de Nêutrons, sonda enviroscan,
reflectometria no domínio do tempo – TDR, tomografia computadorizada,
atenuação de raios-gama, etc.), via clima (determinando-se a evapotranspiração
da cultura de interesse ou estimando-a mediante a evapotranspiração de
referência), via planta (avaliação visual ou outros métodos) ou pela associação
destes (Pires et al., 2001).
Cabe ressaltar que os métodos de avaliação do estado hídrico das plantas
são promissores, entretanto, devido à complexidade envolvida e também à falta
de informações mais específicas, eles ainda não têm sido utilizados em grande
escala. Se por um lado parece ser óbvio que o monitoramento da água
diretamente na planta seja a forma mais correta de se manejar e planejar
irrigações, por outro lado, a variabilidade encontrada entre plantas em função das
condições ambientais e a dificuldade em integrá-los tornam o seu uso pouco
prático para tal fim (Pires et al., 2001).
A avaliação visual é uma forma muito subjetiva para verificar o estresse
hídrico nas plantas, pois a sintomatologia é muito dependente da espécie vegetal.
Na maior parte dos casos, quando se visualiza o déficit hídrico os danos já estão
ocorrendo. A sintomatologia geral das plantas sob deficiência hídrica é a perda
de turgescência, mudança na posição e coloração das folhas, passando para um
verde azulado e redução de crescimento de suas partes mais expostas, aumento
da temperatura das folhas pelo fechamento dos estômatos, entre outros. A médio
e longo prazo, ocorrem morte das raízes superficiais, amarelecimento e

8
senescência das folhas inferiores. As plantas crescem menos, ficando com
estatura e enfolhamento menores em relação às áreas irrigadas. As gramíneas
apresentam enrolamento típico de folhas no sentido dos bordos para a nervura
central. Já as leguminosas apresentam movimentos de folíolos, num fechamento
típico para redução da carga de radiação recebida pelo sol (Pires et al., 2001)
(Figura 3).
Figura 3 – Aspecto característico de planta sob estresse hídrico.
Além da sintomatologia, o manejo das irrigações via planta pode ser
realizado por meio de avaliações do estado hídrico das plantas. Existem inúmeros
métodos para esse fim, que avaliam: temperatura foliar, resistência estomática,
grau de turgescência das plantas, diâmetro do caule, fluxo da seiva e, dentre
outros, potencial de água nas folhas.
Conforme Kramer (1988), parece que o status da água na planta pode ser melhor
estudado tanto em termos de conteúdo de água ou turgor celular quanto em
termos da energia livre ou potencial da água, sendo algumas características
desejáveis de um método para mensuração do status da água:

9
i) uma correlação razoavelmente boa entre o grau do estresse hídrico
medido e as taxas de importantes processos fisiológicos;
ii) um dado grau de estresse hídrico como medido pelo método deveria
ter similar efeito fisiológico em uma ampla gama de materiais
vegetais;
iii) as unidades usadas expressam o estresse hídrico e o método usado
para medi-lo deveria ser aplicável a uma ampla faixa de materiais
vegetais e ao solo;
iv) o método deveria ser tão simples e barato quanto possível;
v) deveria requerer uma quantidade mínima de tecidos vegetais para
cada medida, ou seja, deveria ser relativamente não destrutivo.
Infelizmente, ressalva Kramer (1988), nenhum método de medida do status
da água na planta encontra a maioria destas características.
O status hídrico das plantas usualmente é descrito em termos de potencial
da água, porque ele é fisicamente definido e permite que experimentos sejam
facilmente repetidos. Também, a água move-se através do sistema solo-planta,
por causa do potencial da água ou por forças incluídas no potencial da água.
Sinclair & Ludlow (1985) argumentam que o conteúdo relativo da água deveria
ser usado ao invés do potencial, pois ele é melhor correlacionado com processos
fisiológicos. As diminuições do conteúdo da água na célula podem concentrar
constituintes celulares e provavelmente causa mudanças na atividade enzimática.
Porém, o conteúdo de água freqüentemente falha como um indicador do status da
água na planta. O conteúdo relativo de água não detecta estas diferenças e,
enquanto, o conteúdo de água é importante, o potencial da água é preferível
(Kramer & Boyer, 1995).
Têm-se encontrado uma boa relação entre a evapotranspiração relativa
(razão da evapotranspiração real e a evapotranspiração máxima) de culturas e o
potencial da água foliar no pré-amanhecer, que parece ser dependente das
características do solo e estádio fenológico, portanto, o tratamento
termodinâmico é interessante para o manejo da irrigação (Itier et al., 1992).

10
São inúmeros os exemplos de averiguação do status da água na planta para
fins de manejo da agricultura irrigada. As Figuras 4 e 5 exemplificam alguns
estudos.
Figura 4 – Padrões sazonais do potencial da água foliar no pré-amanhecer e taxa
de assimilação líquida de videira medida ao meio-dia em 1992 (Fonte: Lopes,
1999).
Solo
Figura 5 – Potencial da água médio no solo e na folha de 5 genótipos de feijão,
cultivados nos meses de setembro a novembro de 1992, em casa de vegetação
(Fonte: Pimentel & Perez, 2000).

11
POTENCIAL DA ÁGUA NA PLANTA
As propriedades das soluções são descritas pelo conceito termodinâmico do
potencial, como primeiro exposto por J. Willard Gibbs (1875-1876) (Kramer &
Boyer, 1995). Todas as coisas vivas, incluindo as plantas, requerem uma adição
contínua de energia livre para aumentarem e repararem suas estruturas altamente
organizadas, assim como, para crescerem e se reproduzirem. Processos como
reações bioquímicas, acúmulos de solutos e transporte a longa distância são todos
movidos por um acréscimo de energia livre na planta. O potencial químico da
água é uma expressão quantitativa da energia livre a ela associada. Em
termodinâmica, energia livre representa potencial para realizar trabalho. Observa-
se que o potencial químico é uma grandeza relativa: ela é expressa como a
diferença entre o potencial de uma substância em um determinado estado e o
potencial químico da mesma substância em um estado padrão (Angelocci, 2002;
Libardi, 2004; Taiz & Zeiger, 2004).
Os principais fatores que influenciam o potencial da água em plantas são
concentração, pressão e gravidade. O potencial da água é simbolizado por ψw e o
potencial da água de soluções pode ser divido em componentes individuais,
sendo normalmente escrito pelos seguintes somatórios (Taiz & Zeiger, 2004):
ψw=ψs+ψp+ψg (1)
Os termos ψs, ψp, ψg expressam os efeitos de solutos, pressão e gravidade,
respectivamente, sobre a energia livre da água. O estado referencial utilizado
para definir potencial da água é a água pura sob pressão e temperatura ambientes.
O termo ψs, chamado de potencial de solutos ou potencial osmótico, representa o
efeito de solutos dissolvidos no potencial da água. Os solutos reduzem a energia
livre da água por diluição da mesma. Esse é primariamente um efeito de entropia;
ou seja, a mistura de solutos e água aumenta a desordem do sistema e, portanto,
reduz a energia livre. Isto significa que o potencial osmótico é independente da
natureza específica do soluto (Taiz & Zeiger, 2004). Para soluções diluídas de

12
substâncias indissociáveis o potencial osmótico pode ser estimado pela equação
de van´t Hoff:
ψσ= -RTCs (2)
onde R é a constante dos gases (8,32 J mol-1 K-1), T é a temperatura absoluta (em
K) e Cs é a concentração de solutos da solução, expressa como osmolalidade
(moles de solutos totais dissolvidos por litro de água) (mol L-1). O sinal negativo
indica que os solutos dissolvidos reduzem o potencial da água da solução em
relação ao estado de referência da água pura.
O termo ψp é a pressão hidrostática da solução. Pressões positivas
aumentam o potencial da água; pressões negativas reduzem-no. Às vezes, ψp é
chamado de potencial de pressão. A pressão hidrostática positiva dentro de
células é aquela que se refere à pressão de turgor. O valor de ψp também pode
ser negativo, tal como ocorre no xilema e nas paredes entre as células, onde uma
tensão ou pressão hidrostática negativa pode se desenvolver. Pressões negativas
fora da célula são muito importantes para o movimento de água de longas
distâncias na planta (Taiz & Zeiger, 2004).
A gravidade faz com que a água se mova para baixo, a não ser que à força
da gravidade se oponha uma força igual e oposta. O termo ψg depende da altura
da água acima dela no estado de referência, da densidade da água e da aceleração
da gravidade. Quando se lida com o transporte de água em nível celular, o
componente gravitacional é geralmente omitido porque ele é desprezível
comparado ao potencial osmótico e a pressão hidrostática (Taiz & Zeiger, 2004).
Assim, nestes casos a equação 3 pode ser simplificada como segue:
ψw=ψs+ψp (3)
Em discussões sobre solos secos, sementes e paredes celulares é comum
encontrar-se referência a um outro componente do potencial ψw, o potencial
mátrico. O conceito de potencial da água tem dois usos principais: primeiro, o

13
potencial da água governa o transporte através de membranas celulares,
conforme foi descrito. Segundo, o potencial da água é comumente utilizado como
uma medida do status hídrico de uma planta. Devido à perda de água
transpiratória para a atmosfera, as plantas raramente estão com plena hidratação.
Elas sofrem déficits hídricos que levam à inibição do crescimento vegetal e da
fotossíntese, assim como a outros efeitos prejudiciais. A Figura 2 lista algumas
das mudanças fisiológica as quais as plantas se expõem à medida que secam
(Taiz & Zeiger, 2004).
A Figura 2 também mostra valores representativos de ψw em vários
estágios de estresse hídrico. Em folhas de plantas bem aguadas, ψw varia de -0,2
até cerca de -1,0 MPa, mas as folhas de plantas em climas áridos podem ter
valores muito mais baixos, talvez de -2 a -5 MPa sob condições extremas.
Devido ao transporte de água ser um processo passivo, as plantas podem
absorver água somente quando ψw das plantas for menor que ψw do solo. À
medida que o solo torna-se mais seco, a planta, similarmente, torna-se menos
hidratada (atinge um menor ψw). Se esse não fosse o caso, o solo começaria a
extrair água da planta (Taiz & Zeiger, 2004).
O potencial total da água e seus principais componentes podem ser obtidos
por vários instrumentos e em diferentes partes da planta. No geral, a utilização do
potencial total é suficiente para uso em irrigação. Entretanto, há casos em que o
conhecimento do potencial de pressão é fundamental, visto ser este melhor
relacionado com processos biológicos, como o crescimento e expansão celular
(Pires et al., 2001). A medida de potencial da água como um indicador das
condições hídricas das plantas tem sido feita preferencialmente nas folhas,
embora nos últimos anos o desenvolvimento de técnicas apropriadas tenha
causado aumento do interesse de medidas na raiz e no caule (Angelocci, 1994).
O potencial total da água na folha é diretamente relacionado ao potencial da
água no solo na região das raízes e aos elementos meteorológicos que afetam o
processo de transpiração e abertura dos estômatos. Para cada cultura existe uma
relação em que potenciais da água do solo ou da planta podem ser críticos, a
partir do qual há redução da produção. A Figura 6 apresenta valores de potencial

14
da água no solo e na planta, durante um ciclo de secamento do perfil de solo. Em
um período ideal sem chuvas e de ambiente estável, o potencial da água do solo
se desenvolve paulatinamente, apresentando uma diminuição contínua em seu
valor. Por outro lado, o potencial da água nas folhas apresenta uma oscilação
diária e progressiva negativamente, até início dos períodos de fechamento de
estômato ao meio dia, seguido pela recuperação no final da tarde e à noite (Pires
et al., 2001).
Figura 6 - Alterações nos potenciais da água no solo, na superfície da raiz e nas
folhas, com o transcorrer da transpiração da cultura desde o molhamento
(ψraiz=0). As mesmas condições evaporativas são consideradas a cada dia
(Slatyer, 1967).
DETERMINAÇÃO DO POTENCIAL DA ÁGUA DA PLANTA
Há uma infinidade de métodos de medida do potencial da água e de seus
componentes desenvolvidos ao longo deste século, a maioria deles para tecidos
foliares. Os primeiros métodos para determinação de potencial osmótico
utilizando células, bastante tediosos e demorados, foram substituídos pela medida

15
na seiva extraída. Os métodos para determinação do potencial total da água na
folha evoluíram a partir da década de 1940, e podem ser classificados em três
grandes grupos, de acordo com o princípio: a) de compensação na fase líquida; b)
de equilíbrio na fase de vapor; c) equilíbrio de pressão (câmara de pressão). Nos
grupos a e b foram desenvolvidos métodos com técnicas diferentes de detecção
do ponto de compensação, mas de todos eles, somente o método do corante
(compensação na fase líquida) foi bastante utilizado, embora tenha perdido hoje
sua importância, enquanto que no grupo b tem boa aceitação o método baseado
na medida de pressão de vapor do ar em equilíbrio com a amostra, pela utilização
da técnica psicrométrica ou da higrométrica na medida, o qual permite também a
determinação do potencial osmótico da seiva extraída. O método de maior
utilização atualmente é o da câmara de pressão (Angelocci, 1994).
As determinações do potencial de água na planta devem ser realizadas com
muito cuidado para se obter resultados confiáveis. As modificações introduzidas
no ambiente durante as medições podem afetar a abertura dos estômatos e
conseqüentemente o potencial da água. Deve-se escolher folhas adultas, recém
expandidas, posicionadas no topo, horizontalmente bem exposta a radiação.
Devido ao dinamismo do sistema e a grande dependência do potencial às
condições do ambiente, dois horários têm sido utilizados para as medições no
campo: antes do amanhecer (pré-amanhecer) e no horário mais quente do dia.
Medições nas primeiras horas da manha expressam a integração da água no solo
e na planta. Após o período noturno de recuperação e antes do completo nascer
do sol, a planta expressa o seu maior valor de potencial da água nas folhas
(menor estresse). Boas correlações têm sido obtidas entre o potencial medido no
pré-amanhecer e a produção. Vários trabalhos têm sido desenvolvidos no sentido
de determinar um valor crítico de potencial da água na folha no pré-amanhecer
para estabelecer o momento de irrigação. As determinações no meio dia integram
a resposta da planta à umidade do perfil de solo e, além disso, respondem às
condições ambientais. É nesse horário do dia que a planta apresenta o menor
valor do potencial da água na folha (maior estresse). Em condições de clima de
baixa nebulosidade, o horário de medição é ao redor das 14:00 h (horário local).

16
Em regiões onde as manhãs se apresentam com céu limpo e à tarde com
nebulosidade crescente, as medições devem ser antecipadas. Medições feitas fora
desses horários podem não ter significado para uso na quantificação do estresse
para fins de irrigação, mas a obtenção de curvas de valores ao longo do dia são
de extrema importância para se conhecer a dinâmica da água na planta (Pires et
al., 2001).
Para auxiliar a interpretação de resultados, Cleary & Zaerr (1992) citam os
seguintes valores de potencial da água na folha: valores até -0,80 MPa são ideais
para o crescimento; de -0,90 a -1,20 MPa o estresse limita alguns processos de
crescimento resultando em menores altura e diâmetro; de -1,30 a -2,00 MPa
ocorre o fechamento acentuado dos estômatos, redução da fotossíntese, sendo
que a maioria das culturas sofre estresse e reduz a produção; de -2,00 a -4,00
MPa forte declínio do crescimento com o agravamento do déficit, há redução no
vigor e na capacidade da planta de recuperação quando re-hidratada; de -4,00 a -
5,00 MPa há um estresse muito acentuado e de ocorrência rara em cultivos
comerciais.
O estudo das relações hídricas de células, tecidos e órgãos é fundamental
para se entender a água em equilíbrio e em movimento nos vegetais. Um aspecto
importante a se considerar no estudo é a existência de compartimentagem da
água em simplasto (espaço interno a plasmalema) e apoplasto (paredes, vasos,
lúmen dos vasos, etc.). Caso se divida a planta nos seus “compartimentos”, o
potencial total da água no simplasto (ψsimp) pode ser representado pela equação
4, mediante a qual se computam os potenciais de pressão e osmótico. No
apoplasto, há um componente de solutos e um componente devido à tensão com
que a água fica submetida. O potencial de solutos (osmótico) do apoplasto é
menos negativo do que o do simplasto, devido à maior diluição da seiva
apoplástica (Angelocci, 2002). Alguns autores como Boyer (1995), reconhecem
o componente devido à tensão como sendo um potencial mátrico do apoplasto,
que será denominado de ψm (apo). O potencial total do apoplasto ψapo pode,
então, ser representado pela equação 5.

17
ψsimp=ψp(simp)+ψπ(simp) (4)
Em que:
ψp(simp)= potencial de pressão do simplasto;
ψπ(simp)= potencial osmótico do simplasto.
ψapo=ψs(apo)+ψm(apo) (5)
Em que:
ψs(apo)= potencial de solutos do apoplasto;
ψm(apo)= potencial mátrico do apoplasto.
POTENCIAL DA ÁGUA COMO INDICADOR DO ESTRESSE HÍDRICO
Há alguma dificuldade de interpretação do significado de valores do
potencial da água da folha como um indicador de deficiência hídrica, bem como,
na adoção de valores críticos de potencial na folha, que são muito variáveis de
espécie pára espécie. Um problema adicional é que o potencial total da folha é a
soma de componentes (pressão, osmótica, matricial) e as relações entre eles,
mostradas pelo diagrama de Höfler (Figura 7), são variáveis não somente de
espécie para espécie, mas de acordo com as condições de crescimento, podendo
ocorrer modificações, por ajustamento osmótico e modificação da elasticidade da
parede celular. Principalmente para estudos fisiológicos, muitas vezes torna-se
mais interessante a determinação em conjunto do potencial total e de seus
componentes (Angelocci, 2002).

18
Figura 7 – Diagrama de Hofler-Thoday ilustrando as relações entre o potencial da
água (Ψ), potencial de turgor (Ψp), potencial osmótico (Ψπ) e CRA (Θ) quando
uma célula ou tecido perde água a partir da turgidez plena. A linha tracejada
abaixo do turgor zero representa possível turgor negativo em células rígidas.
Muito da problemática em se encontrar um método ideal para se determinar
o status da água na planta é a falta de um entendimento do mecanismo ou
transdutor pelo qual a diminuição no potencial da água ou no conteúdo de água
pode afetar processos metabólicos mediados por enzimas (Kramer, 1988). A
atividade de enzimas sintéticas e hidrolíticas, tais como a amilase usualmente é
aumentada (Jacobsen, et al., 1986), mas a atividade da enzima pode ser
aumentada ou diminuída (Bradford & Hsiao, 1982). Por exemplo, prolina se
acumula em plantas em estresse hídrico porque: a) sua síntese da glutamina é
acelerada, b) sua oxidação é inibida e c) sua conversão em proteína é retardada
(Hanson & Hitz, 1982). Não está claro como uma diminuição no potencial da
água pode ser acompanhada pela diminuição na atividade de algumas enzimas e
aumento na atividade de outras.
A seguir, apresenta-se a transcrição das críticas de Sinclair & Ludlow
(1985) na qual eles argumentam pelo uso do conteúdo relativo de água, ao invés

19
do potencial da água, para avaliar o status da água na planta. Num terceiro
momento deste tópico, serão avaliadas as argumentações posteriores às críticas
destes autores.
“As propriedades termodinâmicas da água têm sido de interesse dos
fisiologistas vegetais desde que Dixon (1914), primeiro usou a câmara de
pressão para estudar a ‘tensão osmótica’ da seiva de uma árvore. Com o
desenvolvimento do psicrômetro (Spanner, 1951) e a moderna câmara de
pressão (Scholander et al., 1965) dados têm sido reportados regularmente sobre
o potencial químico da água e seus componentes. Além da relação entre o
potencial de turgor e a acumulação de ácido abscísico (Pierce & Raschke,
1980), nenhuma única relação fundamental tem sido estabelecida relatando o
estado termodinâmico da água para o comportamento fisiológico (Ritchie, 1981;
Turner et al., 1985; van Volkenburgh & Boyer, 1985).
De tempo em tempo, dúvidas têm sido levantadas sobre quais variáveis do
estado termodinâmico conduziriam às explicações do comportamento
fisiológico. Hsiao (1973) e Oertli (1976), ambos questionam fortemente que não
parece existir qualquer transdutor para explicar mudanças tanto no potencial da
água total (ψ) quanto na pressão osmótica (π) dentro das respostas fisiológicas.
Por exemplo, no tecido vegetal sofrendo estresse severo, o potencial químico da
água é reduzido somente em um pequeno percentual abaixo daquele da água
livre. Além disso, o único efeito da pressão osmótica parece ser via
concentração local de compostos deletérios em compartimentos específicos
dentro da célula (vacúolos).
Uma justificativa freqüentemente levantada para mensuração do potencial
da água é que ela influencia o fluxo da água. De fato, porém, muito do fluxo de
água no sistema solo-planta é governado pela pressão hidrostática (P). O fluxo
de água no solo, no xilema, no floema e no apoplasmo é dirigido pelos
gradientes de pressão hidrostática e não pelos gradientes de potencial total da
água (Nobel, 1974; Passioura, 1984). Os gradientes de potenciais da água são
provavelmente apenas de maior importância para o fluxo hídrico através de
membranas entre o apoplasma e o simplasma. Desde que o potencial da água no

20
equilíbrio ou próximo do equilíbrio é usualmente assumido sobre estas pequenas
distâncias e é, portanto, não mensurável, a caracterização do fluxo de água
pelos gradientes de potencial da água é de pequena relevância prática.
Não somente a pressão hidrostática mostra-se relevante ao problema de
fluxo de água, mas ela é também o componente termodinâmico na forma de
pressão de turgor, que recentemente tem recebido a maior atenção como a
variável de influência na fisiologia das plantas. Porém, para as plantas
transduzirem a pressão de turgor dentro de um sinal fisiológico, a energia do
potencial mecânico da pressão hidrostática deve ser determinada. Zimmerman
(1978) propôs que o plasmalema deve estar sujeito tanto à compressão quanto à
expansão em pressões de turgor maiores, resultando na alteração nas
propriedades da membrana. Em plantas superiores, há pouco suporte
experimental para esta hipótese. Além disso, os resultados de Lucas & Alexander
(1981), contradizem-no. Adicionalmente, só o plasmalema pode ser diretamente
responsivo a pressão de turgor, porque os gradientes de turgor através das
membranas das organelas e o tonoplasto não podem ser maiores ou as
membranas se romperiam (Nobel, 1974).
Parece que o mais significante transdutor da pressão de turgor em células
maduras está associado com mudanças volumétricas da célula íntegra. A
pressão hidrostática influencia o volume celular mediante mudanças na
elasticidade dimensional das paredes celulares (Tyree & Jarvis, 1982). As
mudanças elásticas no volume do tecido (V) podem ser descritas simplesmente
em termos de um módulo da elasticidade volumétrica (∈).
VdPdV∈
= (6)
A equação (6) ilustra um problema adicional na caracterização do estado
fisiológico do tecido em termos de P. A pressão de turgor não é uma variável
particularmente conservadora para o tecido como o das folhas, que têm volumes
pequenos e ∈ razoavelmente grande, porque pequenas mudanças no volume

21
causam grandes mudanças em P. Portanto, valores de P de folhas são
inerentemente instáveis porque pequenas mudanças nas variáveis ambientais
que alteram insignificantemente V causarão grandes mudanças em P, ainda que
o status da água em toda célula seja essencialmente não mudado. Esta
instabilidade não ocorre nos estômatos porque eles são regulados pela P, tanto
das células guarda quanto das células vizinhas (Cooke et al., 1976). As forças
contrárias dos dois tipos de células ajudam a estabilizar a abertura do complexo
estomático.
Há crescente evidência para sustentar a visão de que o volume da célula,
ou mais comumente o conteúdo relativo de água (CRA), é um importante, e
possivelmente um maior determinante da atividade metabólica e da
sobrevivência da folha. Diferenças na resposta da taxa fotossintética foliar ao
déficit hídrico entre várias espécies, e a salinidade nas mesmas espécies,
desaparecem quando as taxas são comparadas com base no relativo volume
osmótico do protoplasto (Kaiser 1982) ou no relativo volume osmótico (Kaiser,
1983). Esta evidência sugere uma forte relação entre os volumes e as taxas
fotossintéticas das folhas, protoplastos e cloroplastos (veja também Sharkey &
Badger, 1982). Respostas similares têm sido mostradas para a síntese de
proteína (Bewley, 1981), e para a atividade da redutase do nitrato (Sinha &
Nicholas, 1981). Além disso, recentes evidências (Flower & Ludlow, 1986)
sugerem que folhas de uma espécie particular ou um grupo de espécies morrem
em um específico CRA, mais do que em um específico potencial da água na folha
(ou pressão osmótica, porque a pressão de turgor é zero); por exemplo, folhas de
27 gramíneas C4 morreram em um CRA de 25± 1%, enquanto o potencial total
variou entre -7,5 e -13 MPa.
Uma alternativa para a visão termodinâmica do status da água na planta,
portanto, pode ser através do exame das mudanças do volume no tecido
diretamente ou ignorando as variáveis termodinâmicas.
Naturalmente, a equação (7) permanece válida e permite o intercâmbio entre as
duas visões se desejado. Experimentalmente, esta visão alternativa sugere que
maior atenção deveria ser dado ao volume relativo do tecido, ou CRA. A

22
mensuração do CRA foi originalmente proposta por Weatherley (1950) e
revisada em detalhes por Barrs (1968). A desaprovação principal do uso de CRA
tem se devido ás incertezas em sua determinação, mas Catský (1974) sugere que
os valores de CRA com erro absoluto de 1% poderiam ser obtidos com técnicas
cuidadosas.
Uma vantagem na caracterização do estresse hídrico vegetal pelo CRA é
que ele reflete o balanço dinâmico da água entre o fluxo de água dentro e fora
do tecido. Isto pode ser mais facilmente visualizado ao se considerar uma planta
muito simples com uma única raiz e uma única folha. O balanço hídrico da
planta, e finalmente o CRA são dados pela diferença entre a transpiração (T) e
absorção radicular (R):
dV/dT=T-R (7)
Em muitas situações, dV/dT deve ser próxima de zero ou, do contrário, a
planta explodiria prontamente de uma situação de muito maior conteúdo de
água ou secaria com a perda de água. Conseqüentemente, CRA também seria
uma variável razoavelmente estável, comparada à transpiração (T) e à absorção
radicular (R).
De fato, o balanço de água na planta e o status podem ser divididos dentro
de três preferenciais estágios distintos, com base na equação (X2). O estágio 1
ocorre quando a água do solo é abundante e a taxa de absorção radicular iguala
a taxa de transpiração. Desde que fossem esperadas as aberturas dos estômatos,
a taxa de transpiração é dominada pelo ambiente. O CRA, geralmente não cai
abaixo de 75-85% (Bradford & Hsiao, 1982). O estágio II inicia-se quando a
taxa de absorção radicular não pode se igualar à demanda da transpiração.
Neste estágio, a absorção radicular domina as taxas de perda de água, e os
estômatos são forçados a ajustar sua condutância para manter um balanço
hídrico mais ou menos estável nas folhas (Gardner, 1960; Cowan, 1965). Seria
esperado um CRA (assim como a pressão de turgor) mais baixo e diminuído
continuamente durante o estágio II, provavelmente na gama de 80-60%. O

23
estágio III começa quando o solo seca ainda mais, a absorção radicular de água
diminui, e os estômatos perdem a capacidade de manter a taxa de transpiração
aproximadamente igual à taxa de absorção. Neste estágio os estômatos estão
fechados e a taxa de transpiração é dependente da condutância de vapor da
epiderme. Quando a taxa de absorção radicular torna-se substancialmente
menor do que a taxa de transpiração da epiderme, o CRA cai e a folha morre
quando o CRA crítico é alcançado (Flower & Ludlow, 1986).
O potencial da água assim como o CRA diminui durante essas três fases.
Portanto, poderia ser argumentado que o CRA não é mais útil que o potencial
total para definir o status da água na planta quando ele influencia os processos
fisiológicos. Porém, a principal desvantagem do potencial total é que a relação
entre ele e o CRA não é única. Por exemplo, o ajuste osmótico causa alteração
nesta relação, de tal modo que potencial em que um específico CRA (ou pressão
de turgor) é alcançado diminui com o aumento do ajuste osmótico.
O conceito de estágios segregados da desidratação com base na regulação
tanto do ambiente atmosférico quanto das propriedades de transporte do solo
não é novidade. Gardner & Ehlij (1963) demonstraram uma fase inicial de
constante taxa de perda de água sob condições estáveis do meio ambiente,
seguido por uma taxa de diminuição que foi linearmente relatada ao conteúdo
volumétrico de água no solo. A confirmação para este padrão de perda de água
tem sido obtida sob condições controladas (Turner et al., 1985) e sob condições
de campo (Ritchie, 1974; Mason et al., 1980; Meyer & Green, 1981).
O uso do conceito de balanço hídrico para a visão do status da água na
planta e a resposta fisiológica parece ter várias vantagens sobre a aproximação
termodinâmica. Primeiro, a estrutura do balanço hídrico é consistente com a
substanciosa experiência de campo onde a taxa de perda de água não é regulada
pelas plantas sobre uma ampla variabilidade de água disponível no solo. Muitos
dos trabalhos sobre o uso da água pelas culturas durante a primeira parte deste
século (Veihmeyer & Hendrickson, 1950) concordam para esta conclusão, mas
ela não é popularmente prevalecente, porque estas pesquisas anteriores
confiavam em sintomas visuais, assim como o murchamento e o ponto de

24
murchamento permanente, para descrever o progresso da extração de água. Os
estágios como descritos acima são mais apropriadamente definidos pelo
comportamento estomático. Segundo, a aproximação do balanço hídrico oferece
uma nova perspectiva para análise das respostas fisiológicas ao secamento. Ao
invés de confiar em variáveis termodinâmicas relativamente instáveis que têm
falhado na revelação de relações universais às respostas fisiológicas, as taxas de
fluxo relativo de água dentro e entre tecidos vegetais e o CRA do tecido são
sugeridos como importantes variáveis independentes. A variação no fluxo de
água entre vários tecidos pode ser de considerável importância em influenciar a
destinação e as quantidades de compostos distribuídos na planta durante o
estresse hídrico. Assim como alterações nas quantidades de materiais chave
chegando nos tecidos, ligados com os efeitos diretos das mudanças de volume
(que estão vinculadas ao turgor), poderiam potencialmente ter efeitos profundos
nos processos fisiológicos. Terceiro, o uso do balanço hídrico e do CRA parece
conduzir a um estudo de variáveis que são inerentemente mais estáveis do que o
potencial químico da água ou seus componentes. Desde que o CRA é uma
propriedade dinâmica ainda estável que tende a integrar o balanço hídrico da
planta ou do tecido por dias e talvez semanas, ele poderia providenciar uma
ferramenta para estabelecer mais relações universais entre características
fisiológicas em níveis de estresse hídrico.”
Em qualquer evento, acreditam Sinclair & Ludlow (1985) que a
aproximação para a avaliação do status da água na planta precisa ser estendida
além dos pontos de referência da termodinâmica. Enquanto a visão
termodinâmica pode ser biofisicamente mais satisfatória, como um significado
prático ela não tem conduzido a qualquer profundo discernimento específico
sobre a resposta fisiológica ao déficit hídrico. Com sua proposta para explorar
novamente o conceito de balanço hídrico e a utilidade potencial da mensuração
do CRA, reabriram Sinclair & Ludlow (1985) o diálogo sobre as aproximações
experimentais apropriadas para o entendimento das relações da água na planta.
A integração das relações em cada compartimento aquoso (simplasto e
apoplasto) de um tecido leva a uma relação geral entre CRA e o potencial total da
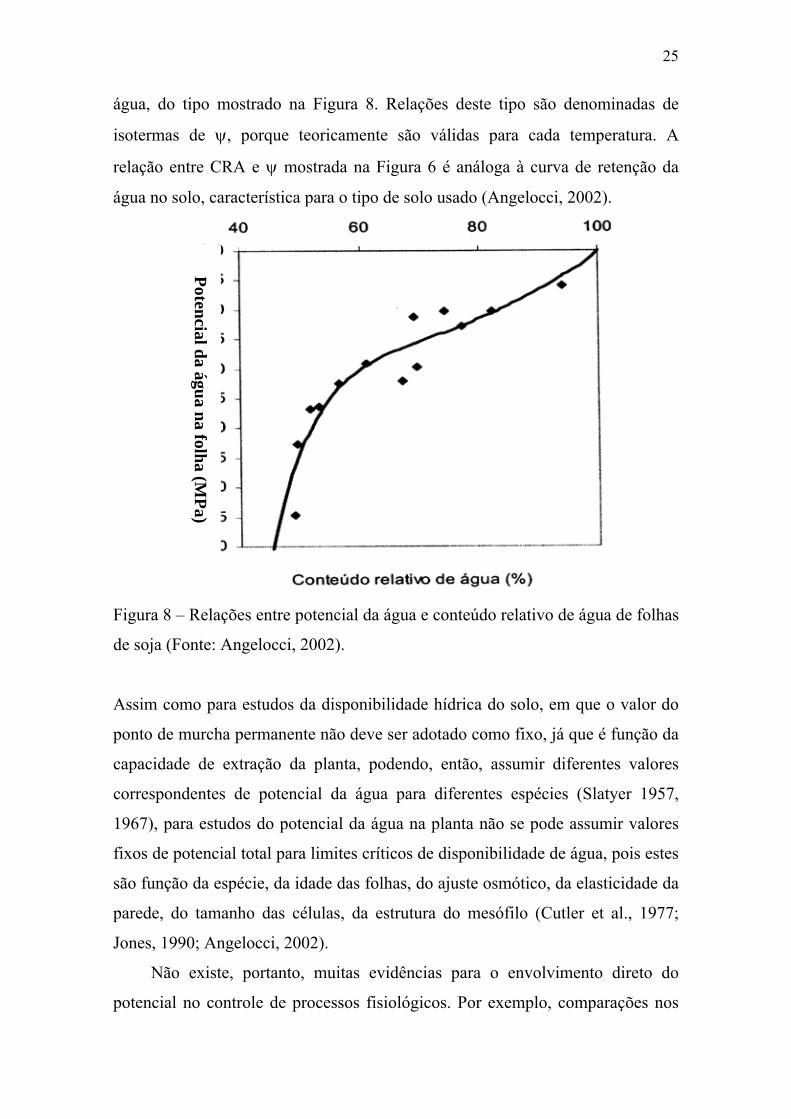
25
água, do tipo mostrado na Figura 8. Relações deste tipo são denominadas de
isotermas de ψ, porque teoricamente são válidas para cada temperatura. A
relação entre CRA e ψ mostrada na Figura 6 é análoga à curva de retenção da
água no solo, característica para o tipo de solo usado (Angelocci, 2002).
Potencial da água na folha (M
Pa) Figura 8 – Relações entre potencial da água e conteúdo relativo de água de folhas
de soja (Fonte: Angelocci, 2002).
Assim como para estudos da disponibilidade hídrica do solo, em que o valor do
ponto de murcha permanente não deve ser adotado como fixo, já que é função da
capacidade de extração da planta, podendo, então, assumir diferentes valores
correspondentes de potencial da água para diferentes espécies (Slatyer 1957,
1967), para estudos do potencial da água na planta não se pode assumir valores
fixos de potencial total para limites críticos de disponibilidade de água, pois estes
são função da espécie, da idade das folhas, do ajuste osmótico, da elasticidade da
parede, do tamanho das células, da estrutura do mesófilo (Cutler et al., 1977;
Jones, 1990; Angelocci, 2002).
Não existe, portanto, muitas evidências para o envolvimento direto do
potencial no controle de processos fisiológicos. Por exemplo, comparações nos

26
efeitos de fluxos osmóticos (que afetam tanto o potencial quanto o potencial
osmótico da célula) e fluxos não osmóticos (que também afetam o volume
celular e a pressão de turgor) mostraram que nem o potencial total nem o
potencial osmótico afetam processos fisiológicos como atividade fotossintética.
Outros estudos têm demonstrado claramente a importância do turgor para o
desenvolvimento celular, para a ação dos estômatos, e a produção de ácido
abscísico. Apesar de ser difícil distinguir entre o turgor e o volume celular (CRA)
como os principais fatores controladores, experimentos usando protoplastos
isolados têm demonstrado que o volume celular pode ser a variável importante
em alguns casos (Jones, 1990). As Figuras 9 e 10 mostram relações do potencial
da água com taxa fotossintética e expansão foliar, por exemplo, mas as relações
do potencial com estas variáveis não são universais.
Figura 9 – Efeitos do estresse hídrico sobre a fotossíntese e a expansão foliar do
girassol (Helianthus annus) (Adaptado de Taiz & Zeiger, 2004).
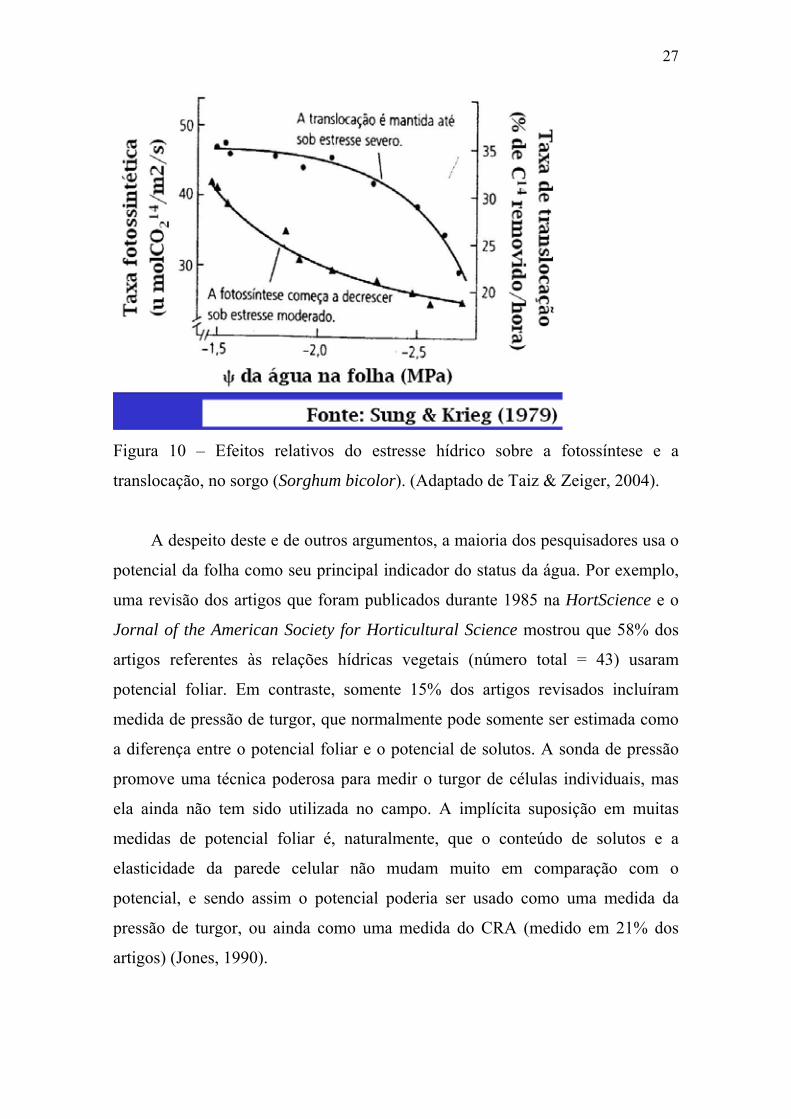
27
Figura 10 – Efeitos relativos do estresse hídrico sobre a fotossíntese e a
translocação, no sorgo (Sorghum bicolor). (Adaptado de Taiz & Zeiger, 2004).
A despeito deste e de outros argumentos, a maioria dos pesquisadores usa o
potencial da folha como seu principal indicador do status da água. Por exemplo,
uma revisão dos artigos que foram publicados durante 1985 na HortScience e o
Jornal of the American Society for Horticultural Science mostrou que 58% dos
artigos referentes às relações hídricas vegetais (número total = 43) usaram
potencial foliar. Em contraste, somente 15% dos artigos revisados incluíram
medida de pressão de turgor, que normalmente pode somente ser estimada como
a diferença entre o potencial foliar e o potencial de solutos. A sonda de pressão
promove uma técnica poderosa para medir o turgor de células individuais, mas
ela ainda não tem sido utilizada no campo. A implícita suposição em muitas
medidas de potencial foliar é, naturalmente, que o conteúdo de solutos e a
elasticidade da parede celular não mudam muito em comparação com o
potencial, e sendo assim o potencial poderia ser usado como uma medida da
pressão de turgor, ou ainda como uma medida do CRA (medido em 21% dos
artigos) (Jones, 1990).

28
Para qualquer medida do status da água usada, é claro que o ajuste osmótico
e mudanças na elasticidade do tecido ou no tamanho das células podem atuar
para estabilizar o turgor ou volume celular em resposta a diminuição do potencial
e são um importante fenômeno que deve ser considerado nos estudos das
respostas das plantas aos déficits hídricos. A adaptação, porém, é um processo
relativamente lento que é freqüentemente não completo quando as plantas são
rapidamente secas em experimentos em vasos, tanto quanto, em conseqüência,
muitos experimentos publicados usando plantas desenvolvidas em vasos dão
resultados enganosos que não podem ser aplicados a situações de campo. Além
disso as diferentes condições ambientais (especialmente radiação) em ambientes
controlados e no campo afetam substancialmente o desenvolvimento das plantas
e as relações hídricas (Davies, 1977; Davies et al., 1986; van Volkenberg &
Davies, 1977).
O potencial da água é mais comumente medido na folha, talvez porque
muitos dos trabalhos pioneiros tenham se preocupado com os processos foliares
tais como a fotossíntese e o comportamento estomático, apesar da facilidade
desta medida com a câmara de pressão ser provavelmente outro fator (Jones,
1990). Recentemente, há uma tendência crescente de estimar o potencial da água
no xilema do caule como o valor do potencial da água na folha para uma folha
envolta por um saco de alumínio por um período antes da medição na câmara de
pressão (Begg & Turner, 1970; Powell, 1974).
As principais desvantagens com mensurações no status da água na folha
podem ser sumarizadas, conforme Jones (1990) em:
a) O status da água na folha é extremamente variável como uma função das
condições ambientais e da taxa de transpiração, com a sensibilidade dependendo
das capacitâncias assim como das resistências do sistema. Mudanças rápidas do
potencial na folha em algodão em resposta às nuvens têm sido notadas (Stansell
et al., 1973), enquanto as conseqüências da variabilidade ambiental para
comparação do potencial da folha são bem conhecidas (Jones & Cumming,
1984). Taxas típicas e magnitudes das mudanças no potencial da folha em maçã e
morango em resposta às nuvens ou ao sombreamento são ilustradas na Figura 11.

29
Pelo menos em morangueiro, e provavelmente em outras plantas pequenas, o
potencial da folha pode variar em mais de 100% dentro de poucos minutos.
Assim uma pequena constante do tempo implica que tanto a resistência quanto a
capacitância são pequenos. Como grandes flutuações em curto prazo no potencial
da folha ocorrem continuamente em ambientes naturais, parece improvável que
elas tenham maior relevância no desenvolvimento a longo prazo, exceto talvez
em poucos casos especiais como nas clareiras em florestas (Woodward, 1981).
Apesar de teleológico, este argumento sugere que valores instantâneos do
potencial da folha não podem promover uma medida absoluta do status da água e
que eles podem somente ser utilmente empregados como medidas relativas
quando expressas como uma diferença do valor de referência obtido para um
tratamento ou cultivar padrão.
Figura 11 – Mudanças no potencial da folha em resposta ao sombreamento da
planta a luz solar direta para maçã. O tratamento sombreado diminui a irradiação
de ondas curtas nas superfícies foliares de mais de 650 W m-2 para < 80 W m-2.
Períodos durante os quais plantas tratadas foram sombreadas são indicados e
períodos de nuvens são mostrados hachurados. Valores de potencial foliar para

30
plantas controle (não sombreados) (•), para plantas tratadas durante o período de
sombra (ο), e durante a restauração após o período de sombra (∆) (Woodward,
1981).
b) Evidências adicionais para a irrelevância das flutuações em curto prazo
no potencial da folha como um fator controlador das funções da planta vem das
aparentemente anômalas observações onde plantas com um suprimento hídrico
restrito podem ter potencial da folha tão alto ou maior que plantas controle bem
supridas (Jones, 1985; Jones et al., 1983). Similarmente, onde o potencial da
folha controla a condutância foliar (gs), um gs esperado diminui à medida que o
potencial cai, mas, quando um considera mudanças diurnas normais nestas duas
variáveis, gs é freqüentemente relatada negativamente ao potencial da folha,
implicando um reverso do controle normal (Jones, 1983). Um controle reverso
similar com gs controlando o potencial também é observado às vezes em
experimentos de secagem (Jones, 1985), e tem sido visto em comparações de
cultivares [em uma comparação, por exemplo, de linhagens de gergelim tendo os
mais baixos valores de potencial da água onde se encontraram as maiores gs
(Hall & Yermanos, 1975)].
c) Os controles dominantes da adaptação das plantas ao estresse a longo prazo
são respostas de desenvolvimento (Jones, 1983; Turner, 1986) e estas são mais
provavelmente controladas nos meristemas, incluindo aqueles nas raízes. É
provável, portanto, que o status da água dos meristemas seja mais relevante do
que aquele da folha. Mesmo as propriedades foliares, tais como condutância
estomática, podem não ser diretamente controladas pelo valor local do potencial
da folha (ou mesmo pela pressão de turgor na folha). Gollan et al. (1985) e
Schulze (1986) variaram o potencial da folha de uma folha experimental
alterando a taxa de evaporação de outras folhas da planta, e mostraram que a
condutância foliar foi mais relacionada à água do solo do que à água da folha na
folha experimental em uma ampla faixa de umidade do solo neste sistema.

31
d) Muitas espécies vegetais podem manter o potencial da folha estável
sobre uma gama de potencial do solo (Bates & Hall, 1981; Jones, 1983b; Jones et
al., 1983; Lorrenzo-Minguez et al., 1985), ainda que o crescimento possa ser
significantemente alterado. Em tais casos, e mais convincentemente, onde o
potencial da folha é mantido mais alto em plantas secas do que em plantas
controle (Jones, 1983b), o potencial da folha não pode ser um bom mensurador
do estresse ambiental e provavelmente não mede um status da água relevante na
planta.
CRA X POTENCIAL DA ÁGUA
Conforme Kramer (1988), a medida do CRA promove um método bastante
conveniente e barato de mensuração das mudanças no conteúdo de água sem os
erros causados pelas mudanças em pesos fresco e seco. Ele também promove um
indicador de mudanças no volume celular e no turgor, e foi reivindicado por
Sinclair & Ludlow (1985) como melhor correlacionado aos processos
fisiológicos do que o potencial da água. Porém, a relação dos processos
metabólicos ao CRA não é mais evidente que sua relação com o potencial da
água. Além disso, o CRA varia com a idade e o habitat (Knipling, 1967), e entre
espécies (Slatyer, 1967) e não pode ser relacionado ao status da água no solo.
Entre as vantagens do potencial da água para avaliação do status da água em
plantas tem-se que ele é uma quantidade física amplamente reconhecida,
geralmente controla o movimento da água, e pode ser medido convenientemente
em unidades que descrevem o status da água na planta e no solo. Medidas
termodinâmicas que podem ser feitas no solo e na planta são mais úteis que
medidas de CRA, que não podem ser relacionadas facilmente ao status da água
no solo ou ao movimento no continuum solo-planta-atmosfera (Figura 12).
Também, medidas feitas com psicrômetros fornecem informações relacionadas
ao potencial da água total e ao potencial osmótico, e informações necessárias aos
estudos do movimento da água em tecidos de desenvolvimento (Boyer, 1985).

32
Figura 12 – Visão geral representativa do potencial da água e seus
componentes em vários pontos ao longo da rota de transporte do solo, por meio
da planta para a atmosfera. O potencial da água (Ψw) pode ser medido ao longo
deste continuum, mas os componentes variam. Na parte líquida do transporte,
pressão (Ψp), potencial osmótico (Ψs) e gravidade (Ψg) determinam Ψw. No ar,
somente a umidade relativa (RT/Vw x ln[RH]) é importante. Observe que,
embora o potencial da água seja o mesmo no vácuo de células do mesófilo e na
parede celular circundante, os componentes do Ψw podem variar bastante (por
exemplo, nesse caso Ψp é 0,2 MPa no interior da célula do mesófilo e –0,7 MPa
fora) (Nobel, 1999, apresentado por Taiz & Zeiger, 2004).
CONSIDERAÇÕES FINAIS
Na escolha de uma medida do status da água, é essencial notar o
mecanismo de controle que influencia o processo vegetal de interesse, que, para
muitos casos, será o desenvolvimento da planta (fisiologia).

33
É importante não rejeitar a relevância fisiológica pelo potencial
simplesmente porque ele é um termo baseado na termodinâmica.
Deve-se notar que a resposta inicial ao estresse hídrico pode ser muito
diferente das respostas a longo prazo quando processos adaptativos, como o
ajuste osmótico, ocorrem.
Para fins de manejo da irrigação, o uso do potencial da água na folha não
tem as restrições que sofre para fins de estudos fisiológicos.

34
REFERÊNCIAS BIBLIOGRÁFICAS
ANGELOCCI, L. R. Água na planta e trocas gasosas/energéticas com a
atmosfera: Introdução ao tratamento biofísico. Piracicaba: ESALQ/USP,
Departamento de Ciencias Exatas. Editora do autor. 272p. 2002.
ANGELOCCI, L. R. Determinação de potenciais da água em plantas.
Piracicaba: ESALQ/USP, 1994. 51p (apostila de curso de pós-graduação).
BAKER, J. M. Recalcitrant problems in environmental instrumentation.
Agronomy Journal, v.95, 2003. p.1404-1407.
BARRS, H. D. Determination of water deficits in plant tissues. In: Water deficits
and plant growth. T. T. KOZLOWSKI (Ed.), v. 1, p.235-368. Academic
Press: New York, 1968.
BATES LM, HALL AE. Stomatal closure with soil water depletion not
associated with changes in bulk leaf water status. Oecologia, v. 50, p. 62–65.
1981.
BEARDSELL, M. F.; COHEN, D. Relationships between leaf water status,
abscisic acid levels, and stomatal resistance in maize and sorghum. Plant
Physiol. v. 56, p. 207-212.
BEGG, J. E.; TURNER, N. C. Water potential gradients in field tobacco. Plant
Physiol., v.46, p.343-346, 1970.
BEWLEY, J. D. Protein synthesis. In: The physiology and biochemistry of
drought resistance in plants. L.G. PALEG; D. ASPINALL (Eds.). p.261-282.
Academic Press: Sydney. 1981.
BOYER, J. S. Plant productivity and envinroment. Science, v. 18. p. 443-448.
1982.
BOYER, J. S. Water transport. Annu. Rev. Plant Physiol., v.36, p.473, 1985.
BOYER, J. S. Measuring the water status of plant and soils. Academic Press.
1995.
BRADFORD, K.; HSIAO, T. C. Physiological response to moderate water stress.
In: Physiological plant ecology. II. Water relations and carbon assimilations.

35
O. L. LANG; P. S. NOBEL; C. B. OSMOND; H. ZIEGLER (Eds.). Encycl.
Plant Phyiology. New Ser., v. 12b, p.263-324. (Spring-Verlag, Berlin). 1982.
BRUNINI, O. New devices for in situ measurements of soil and root water
potentials and transport of water in the soil - plant - system. Guelph:
University of Guelph, 1979. 144p. Ph.D. Thesis.
BRUNINI, O.; CARDOSO, M. Efeito do déficit hídrico no solo sobre o
comportamento estomático e potencial da água em mudas de seringueira.
Pesq. Agropec. Bras., v.33, n.7, p.1053-1060. 1998.
CATSKÝ, J. Direct methods for water content determination. In: Methods of
studying plant water relations. B. SLAVIK (Ed.). p.121-156. Chapmann &
Hall Ltd: London. 1974.
CLEARY, B.; ZAERR, J. Guidelines for measuring plant moisture stress
with a pressure bomb. Corvalis: PMS Instruments Co., 1992. 22p.
COOKE, J. R.; DeBARDEMAEKER, J. G.; RAND, R. H.; MANG, H. A. A
finite element shell analysis of guard cell deformation. Trans. Am. Soc.
Agric. Eng., v.19, p.1107-1121. 1976.
COWAN, I. R. Transport of water in the soil-plant-atmosphere system. J. Appl.
Ecology, v.2, p.221-239. 1965.
CUTLER, J.M.; RAINS, D.W.; LOOMIS, R.S. On the importance of cell size in
the relations of plants. Physiologia Plantarum, v.40, p.256-259, 1977.
DAVIES, W. J.; WILKINSON, S.; LOVEYS, B. Stomatal control by chemical
signaling and the exploitation of this mechanism to increase water-use
efficiency in agriculture. New phytol. v. 153, p. 449-460. 2002.
DAVIES, W. J.; METCALFE; LODGE, T.A.; COSTA. A. R. plant growth
substances and the regulation of growth under drought. Australian Journal of
Plant Physiology, v. 13, p 105-125. 1986.
DAVIES, W. J. Stomatal response to water stress and light in plants grown in
cotrolled environments and in the field. Crop Science. v. 17, p. 735-740.
1977.
DENMEAD, O.T.; MILLER, B.O. Field studies of the conductance of wheat
leaves and transpiration. Agronomy Journal, v.6, n.8 p.307-311, 1976.

36
DIXON, H. H. Transpiration and the ascent of sap in plants (Mcmillan et al.:
London). 1914.
FAIZ, S.M.A.; WEATHERLEY, P.E. Further investigations into the location and
magnitude of the hydraulic resistances in the soil plant system. New
Phytologist, v.81, p.19-28, 1978.
FLOWER, D.; LUDLOW, M. M. Contribution of osmotic adjustment to the
dehidratation tolerance of water-stressed pigeon pea leaves. Plant, Cell
Environment, 1986.
GARDNER, W. R. Dinamics of aspects of water availability of plants. Soil
Science, v.2, p.63-73. 1960.
GARDNER, W. R.; EHLIJ, C. F. The influence of soil water transpiration by
plants. J. Geophys. Res., v. 68, p.5719-5724. 1963.
GOLLAN, T.; PASSIOURA, J. B.; MUNNS, R. Soil water status affects the
stomatal conductance of fully turgid wheat and sunflower leaves. Australian
Journal of Plant Physiology, v. 13, p.459-464, 1985.
HALES, S. Vegetable staticks. W. and J. Innys and T. Woodward, London.
1972.
HALL, A. E.; YERMANOS, D. M. Leaf conductance and leaf water status of
sesame strains in hot, dry climates. Crop Science, v.15, p.789-793, 1975.
HANSON, A. D.; HITZ. W. D. Metabolic responses of mesophytes to plant
water deficits. Annu. Rev. Plant Physiol., v. 33, p.163, 1982.
HSIAO, T. C. Measurements of plant water status. In: STEWART, B. A.;
NIELSEN, D. R (Eds.). Irrigation of agricultural crops. New York:
American Society of Agronomy, 1990. ch. 9, p. 244-280.
HSIAO, T. C. Plant responses to water stress. Ann. Rev. Plant Physiology, v.24,
p.519-570. 1973.
ITIER, B.; FLURA, D.; BELABBES, K.; KOSUTH, P.; RANA, G.
FIGUEIREDO, L. Relations between relative evapotranspiration and predawn
leaf water potential in soyabean grown in several locations. Irrigation
Sciense, v. 13, p. 109-114, 1992.

37
JACOBSEN, J. V.; HANSON, A. D; CHANDLER, P. C. Water stress enhances
expression of an α-amylase gene in barley leaves. Plant Physiol., v.80, p.350,
1986.
JONES, H. G. Estimation of effective soil water potential at the root surface of
transpiring plants. Plant, Cell and Environment, v.6, p. 671–674. 1983.
JONES, H. G. Physiological mechanisms involved in the control of leaf water
status: implications for the estimation of tree water status. Acta
Horticulturae, v. 171, 291–296. 1985.
JONES, H. G. Physilogical aspects of the control of water status in horticultural
crops. HortScience, v. 25. n.1, p. 19-26. 1990.
JONES, H. G.; LUTON, M.T.; HIGGS, K. H.; HAMER, P. J. C. Experimental
control of water status in apple orchard. Journal of Horticultural Science,
v.58, p.301-316, 1983.
TURNER, N. C; Cumming, I. G. Variation of the leaf conductance and leaf
water potential in apple orchads. Journal of Horticultural Science, v.59,
p.329-336, 1974.
KAISER, W. M. Correlation between changes in photosynthetic activity and
changes in total protoplast volume in leaf tissue from hygro-meso-and-
xerophytes under osmotic stress. Planta, v.154, p.538-545. 1982.
KAISER, W. M. Photosynthetic activity and osmotic volumes of isolated
chloroplasts and of cells in leaf tissue from various plants under osmotic
stress. In: Effects of stress on photosynthesis. R. MARCELLE; H.
CLIJSTERS; M. Van POUCKE (Eds.). p.55-63. (Martinus Nijhoff: The
Hague). 1983.
KNIPLING, E. B. Effect of leaf aging on water déficit-water potential
relationship of dogwood leaves growing in two environments. Physiol. Plant,
v. 20, p.65, 1967.
KRAMER, P. J. Measurement of plant water status: historical perspectives and
current concerns. Irrigation Science, v.9, p.275-287, 1988.
KRAMER, P. J.; BOYER, J. S. Water relations of plants and soils. Academic
Press: 1995. 495p.

38
LIBARDI, P. L. Dinâmica da água no solo. Piracicaba: O Autor. 2004. 327p.
LOPES, C. M. A. Relationships between leaf water potential and photosynthetic
activity of field-grown grapevines under a mediterranean environment. Acta
Horticulturae, 493, p.287-292, 1999.
LUCAS, W. J.; ALEXANDER, J. M. Influence of turgor pressure manipulation
on plasmalemma, transport of HCO3 e OH in Chara corallina. Plant
Physiology, v.68, p.553-559. 1981.
MASON, W.K.; CONSTABLE, G. A.; SMITH, R. C. G. Irrigation for crops in a
sub-humid environment. II. The water requirements of soybeans. Irrigation
Science, v. 2, p.13-22, 1980.
MATTHEWS, M. A.; VAN VOLKENBURGH, E.; BOYER, J. S. Acclimation
of leaf growth to low water potentials in sunflower. Plant Cell Environ. v. 7,
p. 199-206. 1984.
MEYER, W. S.; GREEN, G. C. Plant indicator of wheat and soybean crop water
stress. Irrigation Science, v.2, p. 167-176. 1981.
MILBORROW, B. V. The pathway of biosynthesis of abscisic acid in vascular
plants: A review of the present state of knowledge of ABA biosyntesis. J.
Exp. Bot. v. 52, p. 145-1164. 2001.
NOBEL, P. S. Introduction to biophysical plant physiology. (W. H. Freeman:
San Francisco). 1974.
OERTLI, J. J. The states of water in the plant. Theoretical consideration. In:
Water and Plant Life. Problems and modern approaches. O.L LANGE; L.
KAPPEN; E.D. SCHULZE (Eds). p. 19-31. (Springer-Verlag: New York).
1976.
PASSIOURA, J. B. Hydraulic resistance of plants. I. Constant or variable ?
Australian Journal of Plant Physiology, v. 11, p.333-339. 1984.
PASSIOURA, J. B. Response to Dr. P. J. Kramer’s article ‘Changing concepts
regarding plant water relations’. Plant Cell. Environ., v. 11, p.569-571,
1988.

39
PIERCE, M.; RASCHKE, K. Correlation between los s of turgor and
accumulation of abscisic acid in detached leaves. Planta, v. 148, p.174-182.
1980.
PIMENTEL, C.; PEREZ, A. J. C. Estabelecimento de parâmetros para avaliação
de tolerância à seca, em genótipos de feijoeiro. Pesquisa Agropecuária
Brasileira, v.35, n.1, p.31-39, 2000.
PIRES, R. C. M.; SAKAI, E.; ARRUDA, F. B.; FOLEGATTI, M. V.
Necessidades Hídricas das culturas e manejo da irrigação. In: Miranda, J.
H.; Pires, R. C. M. (eds.). Irrigação. Piracicaba: FUNEP, 2001. cap.1, p. 121-
194.
POWELL, D. B. B. Some effects of water stress in late spring on apple trees.
Journal of Horticultural Science, v. 49, p.257-272, 1974.
RITCHIE, J. T. Atmospheric and soil water influences on the plant wtaer
balance. Agric. Meteorol., v.14, p.183-198. 1974.
SAUTER, A.; DAVIES, W. J.; HARTUNG, W. The long distance abscisic acid
signal in the droughted plant: The fate of the hormone on its way from the
root to the shoot. J. Exp. Bot. v. 52, p. 1-7. 2001.
SCHOLANDER, P. F.; HAMMEL, H. T.; BRADSTREET, E. D.;
HEMMINGSEN, E.A. Sap pressure in vascular plants. Science, v. 148,
p.339-346, 1965.
SCHULZE, E. –D. Whole-plant responses to drought. Australian Journal of
Plant Physiology, v.13, p.127-141, 1986.
SHARKEY, T. D.; BADGER, M. R. Effects of water stress on photosythetic
electron transport, photophosphorylation and metabolite levels of Xanthium
strumarium mesophyll cells. Planta, v.156, p. 199-206. 1982.
SINCLAIR, T. R.; LUDLOW, M. M. Who taught plants thermodynamics? The
unfulfilled potential of plant water potential. Australian Journal of Plant
Physiology, v. 12, p.213-217, 1985.
SLATYER, R.O. Plant water relationships. London and New York: Academic
Press. 1967.

40
SLATYER, R.O. Significance of the permanent wilting percentage in studies of
plant and soil water relations. Bot. Rev. , v.23, p.585-636, 1957.
SINHA, S.K.; NICHOLAS, D. J. D. Nitrate reductase. In: The Physiology and
Biochemistry of Drought Resistance in plants. L. G. PALEG; D. ASPINALL
(Eds.). p.145-169. Academic Press: Sydney. 1981.
SPANNER, D. C.The Peltier effect and its use in the measurements of suction
pressure. Journal of Experimental Botanny, v. 2, p. 145-168. 1951.
STANSELL, J. R.; KLEPPER, B.; BROWNING, V. D.; TAYLOR, H. M. Plant
water status in relation of clouds. Agronomy Journal, v.65, p.677-678, 1973.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. Porto Alegre: Artmed. 2004. 719p.
TAYLOR, H.M.; KLEPPER, R.B. The role of rooting characteristics in the
supply of water to plants. Advances in Agronomy, v.30, p.90-128, 1978.
TURNER, N. C.; SCHULZE, E. D.; GOLLAN, T. The response of stomata and
leaf gas exchange to vapour pressure deficits and soil water content. II. In the
mesophytic herbaceous species Helliantus annus. Oecologia, v. 65, p.348-
355. 1985.
TURNER, N. C. Adaptation to water deficits: a changing perspective.
Australian Journal of Plant Physiology, v.13, p.175-190, 1986.
TYREE, M. T.; JARVIS, P. G. Water and tissues and cells. In: Physiological
plant ecology. II. Water relations and carbon assimilation. O. L. LANGE; P.
S. NOBEL; C. B. OSMOND; H. ZIEGLER (Eds). Encycl. Plant Physiology.
New Ser., v.12b, p.35-77. (Springer-Verlag: New York). 1972.
VAN VOLKENBERG, DAVIES, W. J. Leaf anatomy and water relations of
plants grown in controlled environments and in the field. Crop Science. V. 17,
p. 353-358. 1977.
VEIHMEYER, F. J.; HENDRICKSON, A. H. Soil moisture in relation to plant
growth. Annu. Rev. Plant Physiology, v.2, p.285-304. 1950.
WEATHERLEY, P. E. Studies in the water relations of the cotton plant. I. The
field measurements of water deficits in leaves. New Phytol., v.49, p.81-97,
1950.

41
WOODWARD, F. I. Shoot extension and water relations of Circaea lutetiana in
sunflecks. In: GRACE, J.; FORD, E. D.; JARVIS, P. G. (Eds.). Plant and
their atmospheric environment. Blackwell, Oxford, UK. P. 83-91. 1981.
ZIMMERMAN, U. Physics of turgor and osmoregulation. Annu. Rev. Plant
Physiology, v.29, p.121-148. 1978.

42
ANEXO
Acumulação de ácido abscísico
O ácido abscísico é o hormônio vegetal encontrado em todas as plantas
vasculares. Foi também detectado nos musgos, mas parece estar ausente nas
hepáticas. Vários musgos secundários produzem o ABA como metabólito
secundário. Nas plantas, ele tem sido detectado na maioria dos órgãos ou tecidos
vivos, desde a coifa até a gema apical, sendo sintetizado em quase todas as
células que possuem cloroplastos ou amiloplastos (Milborrow, 2001).
A perda de solutos a partir das células-guarda pode ser desencadeada por
um decréscimo no conteúdo de água da folha, e o ácido abscísico (ABA)
desempenha um importante papel nesse processo. O ácido abscísico é sintetizado
continuamente em taxas baixas nas células do mesófilo e tende a se acumular nos
cloroplastos. Quando o mesófilo torna-se moderadamente desidratado acontece o
seguinte: a) parte do ABA armazenado nos cloroplastos é liberada para o
apoplasto (o espaço entre as paredes celulares) do mesófilo (Hartung et al., 1998
citados por Taiz & Zeiger, 2004). A redistribuição de ABA depende de
gradientes de pH dentro da folha, de propriedades fracamente ácidas da molécula
de ácido abscísico e das propriedades de permeabilidade de membranas celulares.
A redistribuição de ABA possibilita ao fluxo da transpiração transportar parte
dele para as células-guarda; b) o ABA é sintetizado em taxas mais altas e a maior
quantidade dele se acumula no apoplasto foliar. As concentrações mais altas do
ABA resultantes de taxas mais altas de sua síntese parecem acentuar ou
prolongar o efeito inicial de fechamento por este ácido armazenado.
O papel do ABA nas respostas ao estresse hídrico, ao frio e à salinidade levou à
sua caracterização como um hormônio do estresse. Conforme, observada
inicialmente a concentração do ABA nas folhas pode aumentar até 50 vezes sob
condições de seca – a alteração de concentração mais severa descrita para um
hormônio em resposta a um sinal ambiental. A redistribuição ou a biossíntese do
ABA é muito eficaz no fechamento estomático e seu acúmulo em folhas

43
estressadas exerce um importante na redução da perda de água, pela transpiração,
sob condições de estresse hídrico (Figura 1).
O fechamento estomático pode também ser causado pelo transporte do
ABA sintetizado nas raízes para a parte aérea. Os mutantes que perderam a
capacidade para produzir o ABA exibem uma murcha permanente chamados de
mutantes wilty, devido à sua incapacidade de fechar os estômatos. A aplicação
exógena do ABA nesses mutantes leva ao fechamento estomático e à restauração
da pressão de turgor (Beardsell & Cohen, 1975).
As respostas estomáticas à desidratação foliar podem apresentar ampla
variação, intra e interespecífica. Os estômatos de algumas espécies com
desidratação retardada, como feijão-de-corda (Vigna unguiculata) e a mndioca
(Manihot esculenta) são incomumente responsáveis pelo decréscimo na
disponibilidade de água; a condutância estomática e a transpiração decrescem
tanto que o potencial da água da folha (ψw) pode permanecer quase constante
durante a seca.
Sinais químicos das raízes podem afetar as respostas estomáticas ao estresse
hídrico (Davies et al., 2002). A condutância estomática freqüentemente está
muito mais intimamente relacionada com o status hídrico do solo do que com o
status hídrico da folha. A única parte da planta que pode ser diretamente afetada
pelo status hídrico do solo é o sistema subterrâneo. De fato, a desidratação de
apenas parte das raízes pode causar fechamento estomático, mesmo que uma
porção bem hidratada do sistema subterrâneo ainda libere grande quantidade de
água para a parte aérea.

44
FIGURA 1 - Alterações no potencial da água, na resistência estomática (o
inverso da condutância estomática) e no conteúdo de ABA em milho, em
resposta ao estresse hídrico. À medida que o solo perde água, o potencial da água
das folhas decresce e aumenta tanto o conteúdo de ABA quanto à resistência
estomática. O processo foi revertido pela rega (Beardesell & Cohen, 1975).
Quando indivíduos de milho (Zea mays) crescem com as raízes orientadas
em dois vasos separados e um dos vasos não foi suprido de água, os estômatos
fecham parcialmente e o potencial da água da folha aumenta, justamente como
nos casos de retardo da desidratação descritos. Tais resultados mostram que os
estômatos podem responder às condições percebidas nas raízes. Além do ABA
(Sauter et al., 2001), outros sinais, como o pH e a redistribuição iônica
inorgânica, parecem desempenhar um papel na sinalização de longa distância
entre as raízes e as partes aéreas (Davies et al., 2002).