universidade luterana do brasil - livros grátislivros01.livrosgratis.com.br/cp064952.pdf · das...
TRANSCRIPT

UNIVERSIDADE LUTERANA DO BRASIL
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA E TOXICOLOGIA
APLICADA
ANÁLISE DO PERFIL FITOQUÍMICO E AVALIAÇÃO DE
EFEITOS BIOLÓGICOS DE Solidago chilensis MEYEN
Dissertação para obtenção do Título de
Mestre em Genética e Toxicologia
Aplicada
FRANCIELI TRÊS GRIZA
Orientador: Prof. Dr. ALEXANDRE DE BARROS FALCÃO FERRAZ
CANOAS
2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

2
Este trabalho foi desenvolvido nas instalações dos Laboratórios de
Farmacognosia, Farmacocinética e Genética Toxicológica da Universidade Luterana do
Brasil (ULBRA), e no Laboratório de Microbiologia da Universidade de Passo Fundo
(UPF), sendo financiado pela ULBRA.

3
AGRADECIMENTOS
Ao meu orientador Prof. Dr. Alexandre B. F. Ferraz pelas orientações,
contribuições, sugestões, disponibilidade e por todas as experiências vividas durante este
período de trabalho, e, principalmente, pela amizade em mais uma etapa de minha vida.
Aos professores Dr. Flávio H. Reginatto, Dr. Marc F. Richter, Dr.ª Jaqueline N.
Picada, Dr.ª Jenifer Saffi, Dr. Sérgio A. L. Bordignon e demais professores que de alguma
forma contribuíram para o desenvolvimento desta dissertação.
À Universidade Luterana do Brasil (ULBRA) pelo apoio financeiro na pesquisa e
à Universidade de Passo Fundo (UPF) pela disponibilidade dos laboratórios.
A todos os colegas de mestrado pelo convívio e companheirismo, em especial,
Giovanni Cignachi e Thaís B. Longo pela amizade, auxílio técnico, disponibilidade e
colaboração.
À Dinara J. Moura, Jacqueline H. Dias, Ângela Castoldi, Kely C. Fulanetto,
Shandale E. Cappelari, Carlos H. Silva, Aline Parisi e a todos os estagiários e bolsistas pela
amizade e colaboração na realização desta pesquisa.
Às funcionárias dos laboratórios pelo auxílio técnico nos trabalhos.
Aos meus amigos Gabriel e Gibran Braga pela amizade e acolhida em São
Leopoldo, e a minha prima Jocasta por me hospedar em Passo Fundo.
A toda minha família, meus pais, Elio e Marlene, e minhas irmãs, Francesca e
Taynara, pela dedicação, preocupação, carinho, apoio e estímulo.
Ao meu amor, Juliano, pelo carinho, compreensão, paciência durante este período
e principalmente pelo incentivo na realização deste mestrado.
A todas as pessoas que, de alguma maneira, colaboraram nesta etapa de
crescimento profissional e pessoal.
E, especialmente, a Deus por ter me iluminado para finalizar este trabalho.
Muito obrigada!

4
ÍNDICE GERAL
1 - INTRODUÇÃO ...................................................................................................... 13
1.1 Família Asteraceae ................................................................................................... 14
1.2 Gênero Solidago ...................................................................................................... 15
1.3 Aspectos fitoquímicos ............................................................................................. 15
1.3.1 Aspectos botânicos ............................................................................................... 15
1.3.2 Aspectos químicos ................................................................................................ 17
1.3.2.1 Constituição química ......................................................................................... 17
1.3.2.1.1 Ácidos fenólicos e flavonóides ...................................................................... 17
1.3.2.1.2 Óleos essenciais .............................................................................................. 19
1.3.2.1.3 Terpenos ......................................................................................................... 21
1.3.2.1.4 Saponinas ........................................................................................................ 23
1.3.2.1.5 Outros constituintes ........................................................................................ 24
1.4 Aspectos biológicos ................................................................................................ 25
1.4.1 Usos populares de espécies de Solidago ............................................................... 25
1.4.2 Atividade antimicrobiana ......................................................................................26
1.4.3 Atividade antioxidante .......................................................................................... 28
1.4.4 Atividade antigenotóxica/genotóxica ................................................................... 33
1.4.5 Outras atividades .................................................................................................. 36
2 - OBJETIVOS ........................................................................................................... 40
2.1 Objetivo geral .......................................................................................................... 40
2.2 Objetivos específicos ............................................................................................... 40
3 - MATERIAIS E MÉTODOS .................................................................................. 42
3.1 Material vegetal ....................................................................................................... 42
3.2 Preparação dos extratos e frações de Solidago chilensis ......................................... 42
3.3 Análise fitoquímica .................................................................................................. 43
3.3.1 Caracterização fitoquímica ................................................................................... 43
3.3.1.1 Alcalóides .......................................................................................................... 44
3.3.1.2 Antraquinonas .................................................................................................... 44
3.3.1.3 Cardiotônicos .................................................................................................... 44

5
3.3.1.4 Cumarinas .......................................................................................................... 45
3.3.1.5 Flavonóides ........................................................................................................ 45
3.3.1.6 Saponinas ........................................................................................................... 46
3.3.1.7 Taninos .............................................................................................................. 46
3.3.1.8 Triterpenos e esteróides .................................................................................... 46
3.3.2 Determinação de flavonóides totais ...................................................................... 47
3.3.3 Análise cromatográfica ......................................................................................... 48
3.4 Avaliações biológicas .............................................................................................. 50
3.4.1 Avaliação da atividade antibacteriana .................................................................. 50
3.4.2 Avaliação da atividade antioxidante ..................................................................... 51
3.4.2.1 Ensaio antioxidante in vivo ............................................................................... 52
3.4.2.2 Ensaio antioxidante in vitro ............................................................................... 53
3.4.3 Avaliação da atividade antigenotóxica/genotóxica .............................................. 55
3.5 Análise estatística .................................................................................................... 57
4 - RESULTADOS ...................................................................................................... 59
4.1 Rendimentos dos extratos e frações de Solidago chilensis ..................................... 59
4.2 Análise fitoquímica ................................................................................................. 59
4.2.1 Análise da constituição química ........................................................................... 59
4.2.2 Avaliação do teor de flavonóides totais ................................................................ 60
4.2.3 Análise cromatográfica ......................................................................................... 61
4.3 Avaliações biológicas............................................................................................... 62
4.3.1 Atividade antibacteriana ....................................................................................... 62
4.3.2 Atividade antioxidante .......................................................................................... 62
4.3.2.1 Ensaio antioxidante in vivo ............................................................................... 62
4.3.2.2 Ensaio antioxidante in vitro ............................................................................... 66
4.3.3 Atividade antigenotóxica/genotóxica.................................................................... 67
5 - DISCUSSÃO ........................................................................................................... 71
6 - CONCLUSÕES ...................................................................................................... 82
7 - REFERÊNCIAS ..................................................................................................... 84

6
LISTA DE FIGURAS
Figuras Descrição Página
1 Partes aéreas da Solidago chilensis Meyen .......................................... 16
2 Estruturas químicas dos principais flavonóides do gênero Solidago ... 17
3 Estrutura química do ácido caféico (a) e do ácido clorogênico (b) .... 18
4 Estrutura química do pumilóxido ......................................................... 20
5 Estrutura química do germacreno D .................................................... 21
6 Estrutura química da solidagenona ...................................................... 21
7 Estrutura química do lupeol ................................................................. 22
8 Estrutura química da virgaureasaponina 1 ........................................... 23
9 Estrutura do núcleo básico de poliacetilenos ....................................... 24
10 Estrutura do núcleo básico de benzilbenzoatos ................................... 24
11 Vias endógenas de destruição enzimática das espécies reativas de
oxigênio (ERO) e localização subcelular das principais vias ..............
29
12 Esquema da preparação dos extratos e frações da S. chilensis ............ 43
13 Esquema do ensaio para medir atividades antioxidantes utilizando o
sistema da xantina oxidase ...................................................................
54
14 Reação dos radicais HO• com o ácido salicílico, levando a formação
dos compostos estáveis 2,3- e 2,5-diidroxi-ácido-benzóico (DHBA)..
55
15 Ensaio de sobrevivência com células de S. cerevisiae na fase
estacionária após tratamento com extratos clorofórmico (EC), bruto
(EB) e fração butanólica (FB) das flores de S. chilensis ....................
63
16 Ensaio de sobrevivência com células de S. cerevisiae na fase
estacionária após tratamento com extratos clorofórmico (EC), bruto
(EB) e fração butanólica (FB) das folhas de S. chilensis .....................
64
17 Ensaio de sobrevivência com células de S. cerevisiae na fase
estacionária após tratamento com extratos clorofórmico (EC), bruto
(EB) e fração butanólica (FB) das raízes de S. chilensis .....................
65
18 Inibição da geração de espécies reativas de oxigênio por S. chilensis
no sistema hipoxantina/xantina oxidase ...............................................
66
19 Imagens de lâminas com as diferentes classes de cometas ................. 68

7
20 Classificação dos cometas na avaliação genotóxica do EB das flores
nas doses de 5, 7,5 e 10 mg/mL em comparação ao controle negativo
69

8
LISTA DE TABELAS
Tabelas Descrição Página
1 Susceptibilidade antimicrobiana frente aos controles positivos ........ 51
2 Linhagens de Saccharomyces cerevisiae utilizadas neste estudo ...... 52
3 Rendimentos dos diferentes extratos e frações da S. chilensis .......... 59
4 Metabólitos secundários presentes nas flores, folhas e raízes da S.
chilensis ..............................................................................................
60
5 Perda de água por dessecação e teor de flavonóides das diferentes
partes da S. chilensis ..........................................................................
60
6 Avaliação da genotoxicidade/antigenotoxicidade dos extratos
brutos das flores, folhas e raízes da S. chilensis ................................
67

9
ABREVIATURAS
DL50 Dose letal de uma substância capaz de matar 50% da população em estudo
EDTA Ácido etilenodiamino tetra-acético
H2SO4 Ácido sulfúrico
HCl Ácido clorídrico
IC50 Concentração inibitória que reduz o efeito em 50%
K2HPO4 Fosfato monoácido de potássio
KH2PO4 Fosfato monopotássico
KOH Hidróxido de potássio
M Molar
mA Miliamperes
mM Milimolar
NaCl Cloreto de sódio
NaOH Hidróxido de sódio
NADPH Fosfato dinucleotídeo adenina-nicotinamida
(NH4)2SO4 Sulfato de amônio
nm Nanômetro
PBS “Phosphate-buffered saline”, solução tampão salina fosfatada
UFC Unidade formadora de colônias
UI Unidade Internacional
V Volts

10
RESUMO
Solidago chilensis Meyen, conhecida no Brasil principalmente como “erva-
lanceta”, é utilizada na medicina tradicional como diurética, cicatrizante, analgésica,
antiinflamatória e para tratar distúrbios gastrointestinais. O uso popular de S. chilensis e a
escassez de estudos biológicos e toxicológicos motivaram analisar o perfil fitoquímico e
avaliar as propriedades antibacteriana, antioxidante, antigenotóxica e genotóxica dos
diferentes extratos e frações das flores, folhas e raízes desta espécie. A análise fitoquímica,
determinação de flavonóides totais e caracterização por métodos cromatográficos destes
produtos foram realizadas para as três partes da planta. A atividade antibacteriana contra
Staphylococcus aureus, Enterococcus faecalis, Enterococcus faecium, Escherichia coli e
Klebsiella pneumoniae foi investigada através do método da difusão de disco. Para a
atividade antioxidante, extratos e frações foram analisados pelo ensaio de sobrevivência in
vivo com linhagens de Saccharomyces cerevisiae proficiente e deficientes em defesas
antioxidantes, assim como pelo método in vitro da hipoxantina/xantina oxidase. Avaliou-se
o efeito protetor dos extratos brutos frente a danos oxidativos ao DNA induzidos por H2O2
através do teste cometa alcalino in vitro. A partir da análise fitoquímica, detectou-se na
planta a presença de substâncias com núcleos esteroidal e triterpênico, saponinas e
flavonóides, sendo os teores de flavonóides totais maiores nas flores e folhas do que nas
raízes. Nos extratos brutos (EB) e frações butanólicas (FB) das flores e folhas foram
detectados, por cromatografia de camada delgada, ácidos fenólicos e flavonóides, já nas
raízes não foram detectadas estas substâncias. Os extratos e frações testados não
demonstraram atividade antibacteriana, mas apresentaram significativa ação antioxidante
in vivo e in vitro, tendo os EB e as FB das flores e folhas os maiores efeitos de proteção
das linhagens de levedura contra danos oxidativos induzidos por H2O2. No teste cometa, o
EB das flores apresentou efeito antigenotóxico em todas as doses testadas, mas também
efeito genotóxico nas maiores concentrações, no entanto, os EB das folhas e raízes não
apresentaram tais efeitos. A partir destes resultados e dos dados da literatura, sugere-se que
os flavonóides sejam os responsáveis pelo efeito antioxidante e pela proteção contra danos
no DNA induzidos por H2O2, por outro lado, terpenos, saponinas e até mesmo os
flavonóides em maiores concentrações presentes no extrato, podem ser os responsáveis
pela ação genotóxica.

11
ABSTRACT
Solidago chilensis Meyen, known in Brazil mainly as “erva-lanceta”, is used in
folk medicine as diuretic, healing, analgesic, antiinflammatory and to treat gastrointestinals
disorders. The popular use of S. chilensis and the few biological and toxicological studies
had motivated to analyse the phytochemical profile and to evaluate the antibacterial,
antioxidant, antigenotoxic and genotoxic properties of the different extracts and fractions
of the flowers, leaves and roots of this species. The phytochemical screening, total
flavonoids determinations and characterization by chromatographic methods of these
products had been carried for the three parts of the plant. Antibacterial activity against
Staphylococcus aureus, Enterococcus faecalis, Enterococcus faecium, Escherichia coli e
Klebsiella pneumoniae was investigated through the disk diffusion method. For the
antioxidant activity, extracts and fractions had been analyzed in vivo, by Saccharomyces
cerevisiae survival assay, with strains proficient and deficient in antioxidant defenses, and
in vitro, by hypoxanthine/xanthine oxidase method. The protective effect in vitro of crude
extracts front the oxidative damage to the DNA induced for H2O2 was evaluateded by the
alkaline comet assay. The phytochemical analyzis detected, in the plant, the presence of
substances with nucleus steroidal and triterpenes, saponins and flavonoids, being the total
flavonoid contents in flowers and leaves higher that in the roots. By thin layer
chromatography, in crude extracts (EB) and butanol fractions (FB) of flowers and leaves
were detected phenolic acids and flavonoids, but in root extracts these substances had not
been detected. The extracts and fractions assayed not demonstrate antibacterial activity.
But EB and FB of flowers and leaves presented significant antioxidant activity at in vivo
and in vitro test, displaying higher protection effect with S. cerevisiae strains against
oxidatives damages induced for H2O2. In the comet assay, EB of flowers presented
antigenotoxic effect in all the tested doses, but also showed genotoxic effect in higher
concentration, however, the EB of leaves and roots presented none effect. According these
results and literature data, we can suggest that the flavonoids are the responsible by
antioxidant effect and protection against damages to DNA induced for H2O2, and the other
hand, terpenes, saponins and even though the flavonoids in higher concentrations presents
in the extract, can be causing genotoxic effect.

12
_________________________________________________________________________
INTRODUÇÃO

13
1 INTRODUÇÃO
Desde o início dos tempos, o homem tem utilizado espécies vegetais como
alimento e agente terapêutico para fins de tratamento e cura de doenças. Como exemplos
clássicos desta prática farmacológica podemos citar a medicina milenar chinesa e indiana
(ayurvédica). Nos países em desenvolvimento e subdesenvolvidos, devido à pobreza e à
falta de acesso à medicina moderna, faz com que a maior parte da população dependa
essencialmente do conhecimento sobre plantas medicinais para seus cuidados de saúde,
pois, muitas vezes, é o único recurso terapêutico de muitas comunidades. Deste modo, as
informações terapêuticas acumuladas durante séculos têm sido utilizadas como guia para
pesquisas farmacológicas, comprovando a importância da etnofarmacologia. Estes fatos
estimularam o interesse médico pelas plantas e algumas destas já têm sido cientificamente
estudadas para avaliar a sua qualidade, segurança e eficácia (MACIEL et al., 2002;
CALIXTO, 2005; MICHELIN et al., 2005; NOLDIN et al., 2006). Além disso, em 1978 a
Organização Mundial de Saúde (OMS), reconheceu os medicamentos fitoterápicos como
recurso terapêutico, e rapidamente os produtos naturais voltaram a ser vistos como fonte de
medicamentos levando ao desenvolvimento de muitos fármacos importantes como a
artemisina (antimalárico), paclitaxel, topotecan (quimioterápicos) e extratos de Ginkgo
biloba (tratamento de problemas vasculares), entre outros (YUNES & CECHINEL FILHO,
2001; LORENZI & MATOS, 2002).
Para a utilização segura dos fitoterápicos, a padronização é necessária para
garantir a autenticidade da planta e seu conteúdo de substâncias ativas de acordo com os
parâmetros utilizados como critérios de qualidade. Métodos cromatográficos são aplicados
para o controle de qualidade de plantas medicinais devido às muitas vantagens, como alta
eficiência, rapidez e a possibilidade de sua utilização em sistemas automatizados
(CELEGHINI et al., 2001). O isolamento de substâncias ativas de plantas tem contribuído
para o fortalecimento da indústria farmacêutica, cuja produção e caracterização na forma
sintética de muitas estruturas químicas complexas seriam inviáveis técnica e
economicamente. Por possuírem alta diversidade molecular, tais substâncias podem ser
utilizadas em processo de semi-síntese ou para modificações estruturais, propiciando o
desenvolvimento de medicamentos mais eficazes ou menos tóxicos (MONKS et al.,

14
2002b; NOLDIN et al., 2006). Neste contexto, espécies de Solidago utilizadas na medicina
popular da Europa, América do Norte e na América do Sul, inclusive nos Estados do Sul
do Brasil, com fins profiláticos e curativos de infecções e inflamações compreendem um
grande grupo de plantas medicinais com reconhecidas aplicações terapêuticas, dentre estas
37 já foram estudadas sob aspectos botânicos e fitoquímicos, como S. virgaurea L., S.
gigantea Ait. e S. canadensis L. (SIMÕES et al., 1998; VILA et al., 2002; APÁTI et al.,
2003; WEBER & JAKOBS, 2005). Desta forma, surgiu o interesse em estudar as
características químicas e atividades biológicas da espécie S. chilensis.
1.1 Família Asteraceae
Asteraceae é considerada uma das famílias mais numerosa e evoluída do reino
vegetal, com aproximadamente 1.500 gêneros e 25.000 espécies já identificadas, sendo no
Brasil representados cerca de 180 gêneros (BARROSO et al., 1991; DILLON & ALVA,
2001). O nome Asteraceae deriva da estrutura característica da inflorescência, em capítulos
florais, da qual fazem parte plantas de hábitos variados (ervas, arbustos, árvores e
trepadeiras), embora predomine no gênero plantas de pequeno porte. Esta família é
cosmopolita, com grande concentração de espécies em regiões subtropicais, tropicais,
temperadas e montanhosas, principalmente na América do Sul (DILLON & ALVA, 2001;
HEEMANN et al., 2004).
Plantas da família Asteraceae têm sido muito utilizadas na medicina popular,
como exemplo podemos citar arnica (Arnica montana L.), marcela (Achyrocline
satureioides (Lam.) DC.), alcachofra (Cynara scolymos L.) e guaco (Mikania glomerata
Spreng.), amplamente empregadas, respectivamente, como cicatrizante, antiinflamatória,
para insuficiência hepática e para problemas respiratórios (SIMÕES et al., 2000). Várias
espécies dessa família já tiveram a composição química e atividades biológicas estudadas,
sendo isoladas uma grande variedade de classes de metabólitos como: alcalóides,
benzofuranos, cumarinas, flavonóides, monoterpenos, diterpenos, triterpenos,
poliacetilenos, saponinas, sesquiterpenos e lactonas sesquiterpênicas. Cabe salientar que
esta última classe, além de colaborar como importante marcador quimiotaxonômico para

15
Asteraceae, tem fornecido substâncias ativas para estudos de desenvolvimento de novos
fármacos (LIMA, 2001; VERDI et al., 2005).
1.2 Gênero Solidago
O gênero Solidago é um dos maiores da família Asteraceae, tribo Asterae,
subtribo Solidaginae, com cerca de 120 representantes (WEBER & JAKOBS, 2005), sendo
a maioria de suas espécies nativas da América do Norte (VILA et al., 2002) e conhecidas
na literatura internacional pelo nome de “Goldenrod” (CORREIA, 1998). Algumas
espécies como S. altissima L., S. gigantea Ait. e S. rugosa Mill. são nativas da América do
Norte e hoje estão distribuídas em várias partes do mundo (LU et al., 1995; TORI et al.,
1999; INOGUCHI et al., 2003; WEBER & JAKOBS, 2005). Outra espécie freqüentemente
estudada é S. virgaurea L., nativa da Europa, Ásia e Norte da África, e, também, ocorre na
América do Norte, encontrando-se em terrenos arenosos e pedregosos (CUNHA et al.,
2003; WEBER & JAKOBS, 2005).
1.3 Aspectos fitoquímicos
1.3.1 Aspectos botânicos
Solidago chilensis Meyen é conhecida popularmente como erva-lanceta, arnica-
do-Brasil, arnica-silvestre, cordão-de-ouro, espiga-de-ouro, lanceta, rabo-de-rojão e sapé-
macho (Figura 1). Esta espécie é comum em campos, beira de estradas e lavouras
abandonadas, sendo nativa do Centro e Sul do Brasil, Argentina, Chile, Paraguai, Peru e
Uruguai (ALICE et al., 1995; CORREIA, 1998; SIMÕES et al., 1998; DILLON & ALVA,
2001; VILA et al., 2002).
Solidago chilensis é uma erva anual, com ramos eretos, delgados, sem
ramificações, glabra ou levemente pilosa, densamente coberta de folhas até a
inflorescência, que pode atingir até 1,7 m de altura. Apresenta folhas simples, alternas,
sésseis, lanceoladas a linear-lanceoladas, trinervadas, ápice e base agudos, margem inteira
ou pouco dentada, glabras ou pilosas em ambas as faces, com pêlos curtos e curvos na

16
margem, com até 10 cm de comprimento e 1 cm de largura. As flores são amarelas,
reunidas em inflorescências do tipo capítulo, sendo estes agrupados em panículas terminais
compostos de ramos secundários curvos e unilaterais de 16-30 cm, dispostos
perpendicularmente ao eixo principal. O caule é ereto, geralmente simples, cilíndrico,
piloso, rizomatoso, de consistência fibro-lenhosa, medindo 80-120 cm de altura. Depois do
florescimento e da frutificação, o caule seca e a planta morre, sendo reproduzida por
sementes e raízes. Apresenta sistema radicular com raiz principal axial, atingindo até 30
cm de comprimento e raízes atingindo até 50 cm de comprimento por 4 mm de diâmetro, e
superfície lisa de coloração castanho-claro com manchas arroxeadas (ALICE et al., 1995;
CORREIA, 1998; SIMÕES et al., 1998).
Figura 1 – Partes aéreas da Solidago chilensis Meyen (www.geocities.com).

17
1.3.2 Aspectos químicos
1.3.2.1 Constituição química
Investigações químicas com espécies de Solidago demonstraram que nas partes
aéreas das plantas há mono e sesquiterpenos (KASALI et al., 2002; GRESSLER et al.,
2003), diterpenos (LU et al., 1995; VILA et al., 2002), flavonóides (APÁTI et al., 2002;
WANG et al., 2006a; WU et al., 2007a), saponinas (REZNICEK et al., 1991; REZNICEK
et al., 1996; SOVOVÁ et al., 1998), entre outras substâncias; enquanto das raízes já foram
isolados poliacetilenos (INOGUCHI et al., 2003) e diterpenos (solidagenona), este último
na espécie S. chilensis (SCHMEDA-HIRSCHMANN et al., 2002).
1.3.2.1.1 Ácidos fenólicos e flavonóides
Os ácidos fenólicos e os flavonóides são algumas das substâncias que constituem
o grupo dos compostos fenólicos, presente no gênero Solidago (Figura 2).
Figura 2 - Estruturas químicas dos principais flavonóides do gênero Solidago.

18
Solidago chilensis contém grande quantidade de flavonóides, os quais já foram
identificados nas partes aéreas da planta, entre estes o campferol, a quercetina (GÜNTNER
et al., 1999) e a quercitrina, que foi extraída com 35% de rendimento (CORREIA, 1998).
No Chile foram detectados nas raízes de S. chilensis, por cromatografia líquida de alta
eficiência (CLAE), rutina (0,4%) e ácido clorogênico (0,5%), sendo os maiores teores
encontrados na planta durante o mês de maio (SCHMEDA-HIRSCHMANN et al., 2005).
As características fitoquímicas dos extratos (em infusão, decocção e maceração) e
das tinturas (40, 70, 96% v/v em etanol) de S. canadensis foram determinadas por
cromatografia líquida de alta eficiência (CLAE). Nesta espécie, identificaram ácido
caféico, ácido clorogênico (Figura 3), quercetina, quercitrina, isoquercitrina, rutina,
hiperosídeo, afzelina e nicotiflorina (APÁTI et al., 2002). Além dessas substâncias,
também foram identificados ácido cafeoilquínico, campferol, campferol-3-/6"-O-acetil-/-β-
glicopiranosídeo, astragalina, isoramnetina, isoramnetin-3-/6"-O-acetil-/-β-
glicopiranosídeo, narcissina e quercetin-3-/6"-O-acetil-/-β-glicopiranosídeo (APÁTI et al.,
2004). A infusão e a tintura 70% mostraram ser as melhores fontes de rutina, enquanto
ácido clorogênico foi mensurado em altas concentrações na infusão, na decocção e nas
tinturas 40% e 70% (APÁTI et al., 2003). A composição do extrato hidroalcoólico da S.
canadensis foi determinada por CLAE, sendo rutina, quercitrina e quercetina os compostos
majoritários do extrato (APÁTI et al., 2006). Em outro estudo, foi identificado um novo
glicosídeo fenólico, 2′-hidroxi-4′,6′-di-O-β-D-glicopiranosil-butirrofenona, das partes
aéreas de S. canadensis (ZHANG et al., 2007).
Figura 3 - Estrutura química do ácido caféico (a) e do ácido clorogênico (b) (BONGARTZ
& HESSE, 1995).

19
As partes aéreas de S. virgaurea contêm vários compostos fenólicos como
quercitrina (WAGNER & BLADT, 1996; EUROPEAN PHARMACOPOEIA, 2006),
ácido ferrúlico, ácido clorogênico, hiperosídeo, rutina, isoquercitrina, apigenin-7-O-
glicosídeo (IVANCHEVA & VITKOVA, 1997), ácido cafeoilquínico, campferol,
nicotiflorina (CHOI et al., 2004), ácido caféico, ramnetina, astragalina e, também, taninos
(CUNHA et al., 2003), além de fenólicos biglicosídeos, como leiocarposídeo e
virgaureosídeo A (SOVOVÁ et al., 1998). Extratos aquosos e metanólicos de S. gigantea
contêm flavonóides como rutina, quercitrina, isoquercitrina e ácidos fenólicos, como o
ácido clorogênico, os quais estão relacionados com as propriedades espasmolíticas e
diuréticas da planta (KRISTÓ et al., 2002; YARNELL, 2002; WEBER & JACOBS, 2005).
Numa pesquisa mais recente, dois novos flavonóides, campferol-3-O-β-D-
apiofuranosil-(1→6)-β-D-glicopiranosídeo e quercetin-3-O-β-D-apiofuranosil-(1→6)-β-D-
glicopiranosídeo, juntamente com seis já conhecidos, isoquercitrina, rutina, astragalina,
nicotiflorina, campferol-3-O-β-D-apiofuranosil-(1→2)-β-D-glicopiranosídeo e campferol-
3-O-β-D-apiofuranosil-(1→2)-O-[α-L-ramnopiranosil-(1→6)]-β-D-glicopiranosídeo,
foram isolados da fração butanólica das folhas de S. altissima L. (WU et al., 2007a).
1.3.2.1.2 Óleos essenciais
Pesquisas com óleos essenciais de espécies de Solidago reportam monoterpenos e
sesquiterpenos como os maiores componentes. No óleo essencial das folhas de S. chilensis
identificou-se 36 compostos, compreendendo 45% do total do óleo, dentre estes os
sesquiterpenos (23,6 %) e diterpenos (15,8 %) constituiram-se nos principais grupos
detectados. O componente majoritário foi identificado como pumilóxido (Figura 4), um
diterpeno labdano com um anel-furano 2,3-dissubstituído. Ainda neste estudo, no óleo
essencial das flores, foram identificados 24 constituintes, representando cerca de 52 % do
total do óleo, sendo detectados sesquiterpenos (18,9 %), monoterpenos (14,7 %)
hidrocarbonados e diterpenos (12,7 %) como os maiores grupos, e entre os principais
compostos foram caracterizados limoneno, pumilóxido, γ-cadineno, óxido cariofileno, β-
elemeno e germacreno-D (VILA et al., 2002).

20
Figura 4 - Estrutura química do pumilóxido.
Os índios Americanos utilizam S. odora Ait., há muito tempo, como erva
medicinal, no tratamento de desordens digestivas e urinárias (http://botanical.com). No
óleo essencial das flores de duas subespécies, S. odora fo. odora, original de Maryland, e
S. odora fo. inodora, de Delaware (Estados Unidos), foram caracterizados em maior
quantidade estragol (70,81 %) e mirceno (12,45 %) na primeira, enquanto mirceno (31,28
%), limoneno (27,09 %) e metilisoeugenol (12,89 %) na segunda subespécie (TUCKER et
al., 1999).
Na determinação dos constituintes majoritários do óleo essencial das folhas de S.
microglossa DC. foram encontrados vários sesquiterpenos hidrocarbonados (β-cubebeno,
germacreno B, cariofileno, β-elemeno e β-farneseno) e um sesquiterpeno oxigenado
(espatulenol). Além destes, houve a identificação de outros compostos, como os
monoterpenos hidrocarbonados (limoneno, α-felandreno e β-ocimeno) (GRESSLER et al.,
2003; MOREL et al., 2006).
Solidago canadensis teve seu óleo essencial das folhas analisado por vários
estudos, entre estes o de SCHMIDT e colaboradores (1999), que encontraram os
sesquiterpenos germacreno D (Figura 5), ciclocolorenona, α-gurgujeno e γ-gurgujeno
como os principais produtos. PROSSER e colaboradores (2002) também identificaram o
germacreno D como o constituinte majoritário do óleo (71%) e outros sesquiterpenos
hidrocarbonados, como germacreno A, α-humuleno e β-cariofileno. Enquanto KASALI e
colaboradores (2002) detectaram, por cromatografia gasosa, além dos sesquiterpenos

21
hidrocarbonados (como germacreno D, germacreno B, γ-cadineno e β-elemeno), os
monoterpenos hidrocarbonados (como mirceno, α-pineno e limoneno) e um monoterpeno
oxigenado (acetato de bornil).
Figura 5 – Estrutura química do germacreno D.
1.3.2.1.3 Terpenos
O estudo químico da raiz de S. chilensis mostrou a predominância de diterpenos
entre os produtos naturais elaborados pela planta, sendo os do tipo labdano em maior
quantidade, ao contrário do que ocorre na maioria das outras espécies estudadas, onde
predominam os clerodanos (CORREIA, 1998). Das raízes desta espécie foram isolados
solidagenona (diterpeno labdano) e outros diterpenos secundários (CORREIA, 1998;
SCHMEDA-HIRSCHMANN et al., 2002). O teor de solidagenona (Figura 6) em nove
amostras de raízes de S. chilensis, cultivadas no Chile, variou de 0,5-3,5%, sendo a maior
concentração encontrada na planta durante os meses de março e abril (2,2-3,5%)
(SCHMEDA-HIRSCHMANN et al., 2005).
Figura 6 - Estrutura química da solidagenona.

22
Os diterpenos são comuns também em outras espécies de Solidago, sendo estes
constituintes já investigados nas raízes e partes aéreas de S. rugosa, das quais foram
isolados diterpenos de diferentes classes como clerodano (colavenol); labdano ((+)-18-
tigloiloximanool; abietano (ácido ent-abiético) e diterpeno ácido caurenóico (LU et al.,
1995). Também, foram caracterizados no extrato metanólico das raízes de S. altissima
nove novos diterpenos (TORI et al., 1999), assim como na S. gigantea são relatados
diterpenobutenolídeos (WEBER & JAKOBS, 2005).
Em outro estudo, CHATURVEDULA e colaboradores (2004) isolaram do extrato
metil etil cetona de S. canadensis sete compostos conhecidos como lupeol (Figura 7),
acetato de lupeol, ácido ursólico, cicloartenol, cicloartenil palmitato, α-amirina e
estigmasterol, e ainda quatro novos triterpenóides lupano.
Figura 7 - Estrutura química do lupeol
Três compostos citotóxicos, eritrodiol-3-acetato, α-tocoferol-quinona e trans-fitol,
foram isolados da fração hexano da S. virgaurea (SUNG et al., 1999). Da mesma planta,
foram obtidos do extrato metanólico das partes aéreas seis terpenóides: trans-fitol, ent-
germacra-4(15),5,10(14)-trien-1α-ol, acetato de β-amirina, ent-germacra-4(15),5,10(14)-
trien-1β-ol, β-dictiopterol e ácido oleanólico (CHOI et al., 2004).

23
1.3.2.1.4 Saponinas
As saponinas também estão presentes em várias espécies de Solidago, como é
relatado grande teor destas substâncias em S. chilensis (VILA et al., 2002) e em S.
gigantea (WEBER & JAKOBS, 2005). No estudo realizado com o extrato metanol:água
(80:20) de S. gigantea foram detectadas por CLAE quatro principais saponinas
(giganteasaponinas 1-4) e o teor destas variou de 0,8 a 1,9% (REZNICEK et al., 1996). Já
na pesquisa com a espécie S. canadensis, REZNICEK e colaboradores (1991) isolaram
quatro novas saponinas bidesmosídicas, sendo denominadas canadensissaponinas 1 a 4.
Assim como S. gigantea e S. canadensis, a S. virgaurea contém saponinas bidesmosídicas
como as virgauresaponinas 1 a 3 (Figura 8) e solidagosaponinas (I a XXIX) (SOVOVÁ et
al., 1998).
Figura 8 – Estrutura química da virgaureasaponina 1 (http://pubchem.ncbi.nlm.nih.gov).
Na espécie S. virgaurea foi isolado, das partes aéreas, um triterpenóide glicosídeo
3,28-bidesmosídico determinado como ácido 3-O-β-D-glicopiranosil-(1→3)-β-D-
glicopiranosilpoligalácico (BADER et al., 1992). Em outro estudo, com a mesma espécie,
foram identificados, nas partes aéreas e nas raízes, dois novos e dois conhecidos

24
triterpenóides glicosídeos bidesmosídicos do ácido poligalácico, sendo as estruturas
elucidadas como compostos 4-O-fucopiranosil-acilados (BADER et al., 1995).
1.3.2.1.5 Outros constituintes
Embora cumarinas sejam freqüentemente encontradas em plantas da família
Asteraceae, no gênero Solidago há poucos relatos da presença destas substâncias, ficando
restrito aos relatos nas espécies S. gigantea (WEBER & JACOBS, 2005) e S. virgaurea
(CUNHA et al., 2003).
Poliacetilenos (Figura 9) são conhecidos por serem produzidos
predominantemente nas raízes de Asteraceae (INOGUCHI et al., 2003; WEBER &
JACOBS, 2005). O composto C-10 poliacetileno, cis-deidromatricaria éster (cis-DME), foi
extraído e identificado nas raízes de S. altissima, e por apresentar efeitos inibitórios de
crescimento sobre outras plantas foi considerado um composto aleloquímico (ITO et al.,
1998; INOGUCHI et al., 2003).
Figura 9 - Estrutura do núcleo básico de poliacetilenos.
Ao pesquisar a fração hexano das partes aéreas da S. virgaurea, CHOI e
colaboradores (2005) isolaram um novo benzilbenzoato (2-metoxibenzil-2-
hidroxibenzoato) juntamente com quatro conhecidos benzilbenzoatos (Figura 10). Já as
raízes de S. gigantea contêm ácido solidagóico (WEBER & JAKOBS, 2005), enquanto nas
raízes de S. microglossa há acetofenona e 3-metoxibenzaldeído (CORREIA, 1998).
Figura 10 - Estrutura do núcleo básico de benzilbenzoatos.

25
1.4 Aspectos biológicos
1.4.1 Usos populares de espécies de Solidago
Há mais de 700 anos, espécies do gênero Solidago têm sido utilizadas na medicina
popular de vários países, como diuréticas (YARNELL, 2002), espasmolíticas
(BORCHERT et al., 2004), mas são especialmente indicadas no tratamento de infecções e
inflamações (APÁTI et al., 2003). Na América Latina, S. chilensis é um representante
deste gênero, utilizado na medicina popular como diurético e anti-helmíntico (MONKS et
al., 2002b; VILA et al., 2002), no tratamento de inflamações e úlceras (SCHMEDA-
HIRSCHMANN et al., 2002), em feridas, escoriações, contusões e hematomas. A partir
das partes aéreas desta planta, são produzidas preparações na forma de extrato fluído,
tintura, cataplasma ou compressa (CORREIA, 1998), que são empregadas em distúrbios
gastrointestinais, como adstringentes, analgésicas, emolientes e cicatrizantes, enquanto a
raiz é usada no tratamento de cefaléias (ALICE et al., 1995; NOELLI, 1996; CORREIA,
1998; SIMÕES et al., 1998; MONKS et al., 2002b). Além dessas propriedades
farmacológicas é importante ressaltar que S. chilensis foi considerada tóxica para o gado,
devido as suas propriedades fotosensibilizantes e seu alto teor de saponinas (VILA et al.,
2002).
Solidago microglossa é uma espécie que possui propriedades semelhantes a S.
chilensis, sendo popularmente utilizada como estomáquica, laxante (NOELLI, 1996),
estimulante gastrointestinal, cicatrizante e antiinflamatória (MOREL et al., 2006), e a
tintura é empregada em escoriações, traumatismos, contusões, varizes e reumatismo
(GRESSLER et al., 2003). Diferentemente, S. odora é utilizada para tratar disenteria e
ulceração intestinal, seu óleo essencial é indicado para flatulência e vômitos, a infusão das
flores é benéfica ao trato urinário, além de serem tônicas e adstringentes
(http://botanical.com). Na Europa, a espécie S. canadensis é indicada para o tratamento de
cistite, nefrite crônica, urolitíase e reumatismo (APÁTI et al., 2003), de maneira
semelhante, as partes aéreas de S. gigantea são indicadas em desordens renais como
diurético e antiinflamatório para a bexiga (VILA et al., 2002).

26
Solidago virgaurea é, popularmente, indicada para tratar inflamações, infecções
renais, cálculos renais, cistite, afecções prostáticas, reumatismo, gota, hipertensão arterial,
diarréia, colite, infecções cutâneas, fadiga e também em dismenorréia e amenorréia. Em
sua constituição química é descrita a presença de cumarinas, flavonóides e saponinas, a
esta última são atribuídas as ações antifúngica e antiinflamatória (FEROW & AVILA,
2000; THIEM & GOSLINSKA, 2002; CUNHA et al., 2003) e o seu óleo essencial atua
como repelente de insetos (THIEM & GOSLINSKA, 2002).
1.4.2 Atividade antimicrobiana
Desde a antiguidade, o homem tem utilizado as plantas como fonte para a
elaboração de remédios com fins profiláticos e curativos de infecções (RANGEL et al.,
2001). Contudo, os trabalhos relacionados à atividade antimicrobiana de plantas são
recentes, tendo início na década de 1940, após a descoberta da penicilina (LIMA, 2001).
Devido aos crescentes problemas associados aos antimicrobianos, como a resistência
microbiana aos fármacos tradicionais, e considerando-se o limitado espectro de ação e
efeitos colaterais destes medicamentos, entre outros fatores, os estudos de substâncias
oriundas de plantas adquiriram novas perspectivas, pelo fato de possuírem alto percentual
de diversidade de estruturas, essencial para a descoberta e produção de novos fármacos
seguros, estáveis e eficazes pela indústria farmacêutica (LIMA, 2001; RANGEL et al.,
2001; CUSHNIE & LAMB, 2005; KUMAR et al., 2005; MICHELIN et al., 2005;
OLIVEIRA et al., 2005).
Trabalhos recentes sobre a atividade antimicrobiana de extratos e óleos essenciais
mostram o grande potencial de aplicação de plantas nativas de diversas regiões do mundo,
com o objetivo de isolar e elucidar substâncias químicas com potencial antibacteriano
frente às bactérias resistentes de importância médica. No Brasil, país com uma rica
biodiversidade, estudos com a mesma finalidade são de grande importância, uma vez que
plantas medicinais são utilizadas em várias áreas da saúde como forma alternativa de
tratamento (DUARTE et al., 2004).

27
Um grande número de plantas medicinais tem atividade antimicrobiana
(YARNELL, 2002), e, entre elas, espécies do gênero Solidago têm sido utilizadas pela
população para esta finalidade (CORREIA, 1998; SIMÕES et al., 1998). Para investigar
esta atividade antimicrobiana, o método da difusão de disco é um dos mais empregados,
pois o ensaio se baseia no uso de discos como reservatórios contendo soluções dos extratos
das plantas a serem avaliados e após a incubação é estabelecida a sensibilidade do
microorganismo (OPLUSTIL et al., 2000; KUMAR et al., 2005; OLIVEIRA et al., 2005).
Em algumas espécies de Solidago foi observada a atividade antibacteriana, como
verificado por DUARTE e colaboradores (2004), com o extrato hidroalcoólico de S.
chilensis, o qual apresentou uma forte inibição (concentração inibitória mínima -CIM- de
0,1mg/mL) frente a Staphylococcus aureus, utilizando o método de microdiluição. Em
outro estudo, pelo método da difusão de disco, os extratos etanólico (E) e metanólico (M)
da S. virgaurea mostraram uma moderada atividade bactericida (concentração bactericida
mínima, mg/mL) contra Bacillus subtilis (1,8 mg/mL - E; 3,9 mg/mL - M), Bacillus
pumilis (15,6 mg/mL - E; 31,2 mg/mL - M), Proteus mirabilis (3,9 mg/mL - E; 7,8 mg/mL
- M), Proteus vulgaris (31,2 mg/mL - E; 62,5 mg/mL - M), Micrococcus luteus 7,8 mg/mL
- E; 15,6 mg/mL - M), Pseudomonas aeruginosa (31,2 mg/mL - E; 62,5 mg/mL - M),
Staphylococcus aureus (62,5 mg/mL - E; 125 mg/mL - M) Staphylococcus epidermidis
(31,2 mg/mL – E; 31,2 mg/mL - M) e Escherichia coli (31,2 mg/mL - E; 62,5 mg/mL - M)
(THIEM & GOSLINSKA, 2002).
Em outro trabalho, o extrato metanólico das raízes de S. microglossa e os
compostos isolados (quercitrina, α-spinasterol e solidagenona) mostraram atividade
moderada (CIM>1 mg/mL) contra os microorganismos testados (Staphylococcus
epidermidis, Staphylococcus aureus, Klebsiella pneumoniae, Escherichia coli, Salmonella
setubal, Bacillus subtilis, Pseudomonas aeruginosa, Saccharomyces cerevisiae e Candida
albicans) pelo método de microdiluição. Já o óleo essencial das folhas da mesma planta
inibiu efetivamente o crescimento de todas as bactérias através de ambos os métodos de
microdiluição (CIM>1 mg/mL) e da difusão de disco (dose de 500 µg) (MOREL et al.,
2006). Da mesma forma, o extrato de S. arguta Ait., na concentração de 100 µg/mL inibiu
100% o crescimento de Mycobacterium tuberculosis H37Rv (CANTRELL et al., 1998).

28
As plantas medicinais utilizadas por moradores da cidade de Governador
Valadares, MG, Brasil, foram avaliadas quanto às atividades antimicrobiana e citotóxica. O
extrato etanólico das partes aéreas de S. microglossa apresentou toxicidade frente às larvas
de Artemia salina (DL50<1000 ppm). Por outro lado, este mesmo extrato não apresentou
atividade antibacteriana contra Escherichia coli, mas se mostrou ativo contra
Staphylococcus aureus nas concentrações testadas (5, 10 e 20 mg/mL) através do método
de difusão do disco (BRASILEIRO et al., 2006).
A atividade antifúngica do óleo essencial das folhas de S. chilensis foi avaliada e
demonstrou uma eficiente ação contra Microsporum gypseum e Trichophyton
mentagrophytes nas doses de 5µL e 10µL, e Cryptococcus neoformans apenas na maior
dose de óleo essencial testada (10 µL) (VILA et al., 2002). A atividade contra os
dermatófitos Trichophyton menthagrophytes, Microsporum canum e Microsporum
gypseum foi reportada para o extrato etanólico das partes aéreas de S. virgaurea, enquanto
os de S. gigantea mostraram atividade contra Candida pseudotropicalis (PEPELJNJAK et
al., 1998). Ademais, o óleo essencial de S. canadensis apresentou atividade antifúngica,
com significante inibição no crescimento de Thanatephorus cucumeris e Rhizoctonia
solani e efeito moderado contra Botrytis cinerea, mas uma fraca atividade contra
Alternaria solani, Colletotrichum lindemuthianum e Glanerlla cingulata (WANG et al.,
2006b).
1.4.3 Atividade antioxidante
Numerosos processos fisiológicos produzem radicais livres ou espécies reativas
de oxigênio (ERO), em quantidades controladas por uma variedade de mecanismos de
defesas celulares (PIETTA, 2000; RASPOR et al., 2005). Um desequilíbrio entre a
produção de radicais livres e remoção destes pelas defesas antioxidantes leva a uma
situação de estresse oxidativo, causando danos nas células, processo que pode ser
decorrente da diminuição dos níveis das enzimas antioxidantes, aumento da produção de
ERO, ou por ambos os processos. A alteração deste equilíbrio está associado a uma série
de eventos fisiológicos envolvendo danos aos componentes celulares ou, em alguns casos,
morte celular, ao mesmo tempo que estão diretamente envolvidos em vários processos

29
patológicos, como asma, câncer, catarata, diabetes, desordens neurodegenerativas, doenças
cardiovasculares, reações inflamatórias, entre outras, e não patológicos, como o
envelhecimento (PIETTA, 2000; CAI et al., 2004; TEPE et al., 2005; VALKO et al.,
2006).
Para proteger o organismo do ataque de ERO existem mecanismos de defesas
antioxidantes, sendo os principais: (1) supressão da formação de ERO pela inibição de
enzimas ou quelação de elementos envolvidos no estresse oxidativo; (2) captação de ERO
pelos sistemas enzimáticos ou não enzimáticos, que impedem a continuação das reações
em cadeia; (3) reparação de biomoléculas oxidadas por ERO, especialmente o ácido
desoxirribonucléico (DNA) (HALLIWELL & GUTTERIDGE, 1999; PIETTA, 2000;
LUSHCHAK & GOSPODARYOV, 2005). Algumas das defesas antioxidantes celulares
estão representadas na Figura 11.
Figura 11 - Vias endógenas de destruição enzimática das espécies reativas de oxigênio
(ERO) e localização subcelular das principais vias (Adaptado de ENGELHARDT, 1999).

30
Nas células eucarióticas, os principais sistemas antioxidantes enzimáticos incluem
a superóxido dismutase (SOD), a catalase (CAT) e a glutationa peroxidase (GSH-Px), as
quais catalisam a redução dos oxidantes, prevenindo a formação de ERO (PIETTA, 2000;
VALKO et al., 2006). A SOD catalisa a dismutação do ânion superóxido (O2•−) a peróxido
de hidrogênio (H2O2) e oxigênio molecular (O2) (LUSHCHAK, 2006), sendo encontrada
em diferentes isoformas, diferindo na natureza do metal ativo e nos diferentes papéis na
proteção dos organismos do estresse oxidativo. Entre as SOD mais importantes estão a
SODCuZn, que possui cobre (Cu2+) e zinco (Zn2+), codificada pelo gene SOD1, e está
presente principalmente no citosol das células; a SODMn com um íon de manganês (Mn3+)
em cada uma de suas subunidades, que é codificada pelo gene SOD2 e está localizada
primariamente na mitocôndria, protegendo-a de ERO gerados durante respiração e
exposição ao etanol (PEREIRA et al., 2003; VALKO et al., 2006); e a SODEC, que
contém Cu2+ e Zn2+ e atua no espaço extracelular. As catalases citosólicas e peroxissomais
convertem H2O2 à água (H2O) e O2 (PICADA et al., 2003; WILMSEN et al., 2005). A
glutationa peroxidase também reduz o H2O2 à H2O e seus peróxidos orgânicos a seus
respectivos álcoois, através da conversão da glutationa reduzida ao dissulfeto
correspondente glutationa oxidada (GRANT et al., 1998; PICADA et al., 2003).
Os antioxidantes não enzimáticos podem prevenir a formação de ERO através de
mecanismos como seqüestro de radicais livres (glutationa, urato, carotenóides, vitamina C
e vitamina E), estabilização de elétrons (carotenóides, licopenos e ubiquinona) e quelação
de íons ferro e cobre (urato) (PIETTA, 2000; PICADA et al., 2003; VALKO et al., 2006).
Além desses, nas plantas há uma grande variedade de classes químicas como os compostos
fenólicos, substâncias nitrogenadas, terpenóides, vitaminas e outros metabólitos endógenos
que apresentam atividade antioxidante. Por isso, a introdução dessas substâncias na dieta é
necessária para diminuir os efeitos cumulativos do dano oxidativo no organismo (PIETTA,
2000; CAI et al., 2004; VALKO et al., 2006).
A capacidade antioxidante dos extratos de plantas pode ser avaliada utilizando
vários métodos como DPPH (radical 1,1-difenil-2-picrilidrazila), ABTS [radical
2,2’azinobis-(3-etilbenzotiazolin-6-ácido sulfônico], hipoxantina/xantina oxidase,
atividade de superóxido dismutase, catalase, glutationa, ensaio de sobrevivência com a

31
Saccharomyces cerevisiae, entre outros. Dentre estes, destacamos o método químico in
vitro da hipoxantina/xantina oxidase e o método de avaliação da atividade in vivo com a
levedura Saccharomyces cerevisiae, por estarem ligados aos objetivos do presente estudo.
A xantina oxidase catalisa as reações em que a hipoxantina é oxidada à xantina e
esta em ácido úrico (VALKO et al., 2006). Durante este processo, a hipoxantina/xantina
oxidase produz também O2•−, a partir do oxigênio, e H2O2, a partir da água. Na presença de
ferro (Fe3+) e de EDTA, o O2•− reduz o Fe3+ a Fe2+, formando numa segunda etapa o H2O2,
o qual gera radicais hidroxilas (HO•), que são espécies altamente reativas. A concentração
destes HO• no teste é baseada na sua reação com o ácido salicílico, formando, assim, dois
compostos estáveis 2,3- e 2,5-diidróxi-ácido-benzóico, fáceis de quantificá-los via CLAE
(OWEN et al., 1996 e 2000).
Os ensaios utilizando leveduras são muito úteis nos testes de várias substâncias
naturais e sintéticas, sendo rápidos, sensíveis, reprodutíveis, bem como produzem
resultados em termos da identificação da atividade biológica e antioxidante (WILMSEN et
al., 2005). A levedura Saccharomyces cerevisiae é um organismo eucarioto
extensivamente estudado, com seu genoma completamente seqüenciado (LUSHCHAK,
2006), e notavelmente semelhante às células de mamíferos no que se refere às
macromoléculas, organelas e proteínas com homologia a proteínas humanas (COSTA &
FERREIRA, 2001). Esta levedura possui uma variedade de defesas antioxidantes
enzimáticas, incluindo a glutationa redutase, a glicose-6-fosfato desidrogenase (ZHANG et
al., 2003; BAYLIAK et al., 2006), as catalases citoplasmáticas e peroxissomais
(WILMSEN et al., 2005), as superóxido dismutases SODCuZn e SODMn, as quais
representam 90% e 5%-15% do total de atividade da SOD durante o crescimento aeróbico,
respectivamente (DELLA CROCE et al., 2003; LUSHCHAK et al., 2005).
Com as linhagens deficientes nestas defesas antioxidantes é possível investigar a
ação de agentes físicos ou químicos que interferem com o estado redox celular
(LUSHCHAK & GOSPODARYOV, 2005), reforçando a utilidade de S. cerevisiae como
modelo nas pesquisas sobre mutagênese, reparo de DNA e mecanismos que respondem ao
estresse oxidativo (COSTA & FERREIRA, 2001). Assim, no ensaio de sobrevivência, o

32
aumento da viabilidade celular ao tratamento sugere uma atividade protetora (antioxidante)
e a diminuição da viabilidade a um efeito deletério (pró-oxidante) (MARIS et al., 2000).
A seguir serão descritos relatos de estudos sobre a atividade antioxidante em
algumas espécies de Solidago. O estudo de GÜNTNER e colaboradores (1999) monitorou
espectrofotometricamente a capacidade antioxidante total do extrato das flores de S.
chilensis frente a soluções de β-caroteno na presença de H2O2 0,1% e comparou ao
antioxidante padrão hidroxitolueno butilado (BHT). O extrato que inibiu 40% mais a
oxidação do β-caroteno do que o BHT era constituído por 50% de quercetina.
MCCUNE & JOHNS (2002) demonstraram que a atividade in vitro do extrato
metanólico das raízes de S. canadensis em sequestrar radicais peroxila no ensaio
DCF/AAPH [2,7-diclorofluorescina-diacetato/2,2’-diazobis(2-amidinopropano)
diidrocloreto] foi maior quando comparado ao chá verde, ácido ascórbico e Trolox,
enquanto no ensaio NBT/XO (Nitroblue tetrazolium/hipoxantina-xantina oxidase) a
atividade de captar O2•− foi semelhante ao ácido ascórbico e maior que o Trolox. Para os
autores, a atividade antioxidante desta planta pode ser devida à presença de vitaminas
antioxidantes, compostos fenólicos ou taninos. Em outro estudo com essa espécie, foram
correlacionadas características fitoquímicas e propriedades antioxidantes na forma de
infusão, decocção, maceração e tinturas preparadas em várias concentrações de etanol (40,
70, 96% v/v). Baseados na taxa de dissolução, variedade de flavonóides, doação de
hidrogênio e equivalentes de ácido ascórbico, os autores concluíram que a tintura (70%) e
a infusão são as preparações que apresentaram melhor capacidade antioxidante in vitro
(APÁTI et al., 2003).
Devido ao alto teor de flavonóides presentes em S. canadensis, o efeito do extrato
hidroalcoólico na atividade de glutationa-S-transferase (GST) utilizando células HepG2 foi
comparado com quercetina e seus glicosídeos quercitrina e rutina, constituintes
majoritários da planta. Na mesma concentração, rutina e quercitrina aumentaram a
atividade de GST em 50% e 24,5%, respectivamente, enquanto a aglicona quercetina inibiu
a enzima em 30% (APÁTI et al., 2006).

33
A atividade antioxidante de nove flavonóides isolados de S. canadensis foram
avaliados pelo método in vitro DPPH. Neste ensaio, quercetina apresentou maior atividade,
comparada ao controle positivo vitamina C. Os demais flavonóides 3-metóxi-quercetina e
isoquercitrina mostraram atividade moderada, enquanto quercitrina, rutina, campferol,
astragalina, afzelina e nicotiflorina exibiram atividade mais fraca do que vitamina C
(WANG et al., 2006a). Em outro estudo, segundo os autores, o extrato hidroalcoólico de S.
virgaurea também apresentou propriedades antioxidantes, pois foi capaz de inibir reações
bioquímicas (xantina oxidase, NADPH-diaforase, lipoxigenase) que ocorrem em situações
de inflamação e levam à formação de ERO (MEYER et al., 1995).
1.4.4 Atividade antigenotóxica/genotóxica
Ensaios de genotoxicidade com células de mamíferos são freqüentemente
utilizados na análise do potencial genotóxico de agentes químicos e físicos que podem
resultar em mutações. O teste de eletroforese para célula única em gel (SCGE - single cell
gel electrophoresis) ou teste cometa é um ensaio de genotoxicidade que determina a
extensão do dano ao DNA em células individuais, que podem levar a doenças como o
câncer (FAIRBAIRN et al., 1995; VILLELA et al., 2003; HARTMANN et al., 2004;
BRENDLER-SCHWAAB et al., 2005). Para a execução do teste cometa, há dois
protocolos principais relacionados com o pH. A versão neutra detecta quebras duplas no
DNA e ligações cruzadas, enquanto a versão alcalina detecta quebras de fita simples e
dupla, danos álcali-lábeis e ligações cruzadas (FAIRBAIRN et al., 1995; VILLELA et al.,
2003; CAVALCANTI et al., 2006). Lesões indiretas ao DNA promovem efeitos como
metilação e aductos (dímeros de pirimidinas), que, sendo álcali-lábeis, se expressam como
quebras simples frente ao tratamento alcalino. As quebras resultantes de reparo por excisão
não concluído podem ser detectadas utilizando enzimas de reparo (FAIRBAIRN et al.,
1995; VILLELA et al., 2003).
O teste cometa é sensível para detectar baixos níveis de dano ao DNA e avaliar o
seu reparo. Este ensaio é de custo relativamente baixo, simples e rápido pois requer um
pequeno número de células analisadas, utilizado para testes de genotoxicidade in vitro e in
vivo, incluindo estudos de biomonitoramento humano e ambiental, sendo aplicado para

34
várias células de eucariotos (SILVA et al., 2000; MALUF & ERDTMANN, 2003;
VILLELA et al., 2003; BRENDLER-SCHWAAB et al., 2005). Como o teste cometa é
muito sensível, necessita de controles bem estabelecidos, como a curta duração de tempo
(24 h no máximo) entre exposição à genotoxina e a preparação das lâminas até a lise, pois,
como é um teste de genotoxicidade, o dano pode ser reparado. A citotoxicidade pode
potencializar resultados tanto falsos positivos como falsos negativos devido à
fragmentação do DNA como uma conseqüência da necrose ou apoptose. Desse modo, este
ensaio deve ser usado apenas como indicativo de risco (MALUF & ERDTMANN, 2003;
VILLELA et al., 2003; HARTMANN et al., 2004).
Este teste consiste em dispor as células embebidas em gel de agarose sobre
lâminas de microscopia, submetendo as mesmas a uma solução com alta concentração de
sais e detergentes a fim de lisar as células. As lâminas, depois de imersas em um tampão de
pH variável (neutro ou alcalino) (FAIRBAIRN et al., 1995), são submetidas à eletroforese
de modo a induzir a migração de fragmentos de DNA para fora do núcleo (SILVA et al.,
2000; VILLELA et al., 2003; PEREIRA et al., 2005). Após a eletroforese, as lâminas são
coradas e, na visualização ao microscópio, as células normais contêm um núcleo redondo,
sem danos ao DNA, enquanto as células lesadas apresentam forma similar a um “cometa”
(célula com cauda), devido à migração de fragmentos de DNA, que se apresentam em
diferentes tamanhos e podem estar ligados ao núcleo por uma cadeia simples
(FAIRBAIRN et al., 1995; VILLELA et al., 2003). A extensão do dano é medida pelo
comprimento do DNA que migrou e pela avaliação visual das células em diferentes classes
(0-4) através de análise microscópica, sendo possível determinar se todas as células de uma
população demonstram o mesmo nível de dano (FAIRBAIRN et al., 1995).
Devido aos poucos relatos sobre danos ao DNA com o gênero Solidago, a revisão
da literatura apresentada em seguida inclui dados de mutagenicidade e de genotoxicidade
em plantas da família Asteraceae.
O efeito mutagênico de S. microglossa foi avaliado pelo teste de Ames, usando
linhagens de Salmonella typhimurium (TA97a, TA98, TA100 e TA104), sendo que a maior
taxa de mutagenicidade foi obtida com o extrato aquoso, quando a linhagem TA98 foi

35
utilizada com ativação metabólica (S9) e com TA97a com ou sem a adição de S9
(RIVERA et al., 1994). Devido ao efeito mutagênico e por ser utilizada nas contusões via
absorção da pele, há a necessidade de mais estudos de lesões no DNA sobre esta e outras
espécies do gênero Solidago (CORREIA, 1998).
Extratos de várias plantas medicinais utilizadas na medicina popular da Turquia
foram avaliados no teste cometa alcalino. Arctium minus Bernh., assim como as demais
plantas investigadas, apresentaram mais lesões no DNA do que o controle negativo
(BASARAN et al., 1996).
O efeito genotóxico do extrato aquoso de Baccharis genistelloides Pers. (EABg),
foi avaliado pelo teste cometa, em células hepáticas e renais de camundongos de diferentes
grupos (saudáveis, com artrite induzida por colágeno e com EABg). Grupos de
camundongos saudáveis e com artrite tratados com EABg em ambas as doses (4,2 mg/kg e
42 mg/kg) não mostraram efeitos genotóxicos nas células analisadas, quando comparados
aos controles. A ausência de hepatotoxicidade e nefrotoxicidade aguda pelo EABg foi
confirmada pela não detecção de alterações nos órgãos e nos parâmetros bioquímicos dos
animais em estudo (COELHO et al., 2004).
Extratos etanólicos das plantas Crepis vesicaria L. e Scolymus hispanicus L.
foram analisados pelo teste cometa alcalino quanto à capacidade de proteger danos
oxidativos no DNA induzidos por H2O2 em linfócitos humanos pré-tratados com os
extratos das plantas nas concentrações de 0,1, 1 e 10 µg/mL. Na concentração de 10
µg/mL, o extrato de S. hispanicus mostrou capacidade de proteção ao DNA acima de 20%,
enquanto a proteção pelo extrato de C. vesicaria foi menor que 10%. Quando C. vesicaria
foi utilizada em baixas concentrações (0,1 e 1 µg/mL), a planta apresentou significativa
proteção (acima de 20%) (KAPISZEWSKA et al., 2005).
Extratos metanólicos e aquosos de 13 plantas do Sul da África foram investigados
quanto às propriedades genotóxicas. Os resultados do teste cometa utilizando linfócitos
humanos mostraram que os extratos (0,01 g/mL) de Callilepis laureola DC. não
protegeram contra os danos no DNA na presença de H2O2. O extrato metanólico resultou

36
em danos semelhantes aos do controle positivo (H2O2), podendo esta indução ser um dos
mecanismos dos efeitos hepatotóxicos e renais desta planta (STEENKAMP et al., 2005).
1.4.5 Outras atividades
Solidago chilensis vem sendo estudada mostrando diversas atividades, como na
pesquisa de GOULART e colaboradores (2007), em que os extratos aquosos das raízes,
folhas e flores desta planta demonstraram um significativo efeito antiinflamatório em
modelo in vivo inibindo células (leucócitos e neutrófilos) na inflamação induzida por
carragenina. Entretanto, os principais efeitos observados foram nas raízes, em que seu
extrato aquoso (EA), fração butanólica (BuOH) e fração residual aquosa (RA) inibiram a
migração das células e a atividade das enzimas mieloperoxidase, adenosina-deaminase e do
fator de necrose tumoral alfa (TNF- α). Também foi observada a inibição da exudação, do
óxido nítrico e da interleucina-1 pelo EA e fração BuOH das raízes, demonstrando uma
evidente ação antiinflamatória. Ainda, o EA das raízes e suas frações reduziram
significativamente os leucócitos na inflamação induzida por bradicinina e histamina e os
neutrófilos na inflamação induzida por histamina. Contudo, apenas a fração RA inibiu
neutrófilos induzidos por bradicinina.
A ação antinociceptiva do extrato hidroalcoólico das partes aéreas de S. chilensis
foi verificada em ratos utilizando o teste de formalina na pata. Nos 5 primeiros minutos os
animais controles apresentaram índice de nocicepção (IN) entre 2-2,5, indicando dor
moderada, os animais tratados com o extrato (3 mg/kg) apresentaram dor leve (IN entre 1-
1,5), enquanto com os extratos de 10 mg/kg e 30 mg/kg não apresentaram dor. Na fase
inflamatória, os animais tratados com 3 mg/kg mostraram um IN máximo de 2,0, com 10
mg/kg mostraram IN de 1,5 e com 30 mg/kg sem dor até o minuto 50. Os controles a partir
dos minutos 20-25 chegaram a um IN de 3,0 (dor intensa) mantido até o minuto 50
(MOLSKA et al., 2003b). O mesmo extrato hidroalcoólico (10 e 30 mg/kg) inibiu
significativamente a hipernocicepção mecânica aguda, induzida em ratos pela injeção
subcutânea de prostaglandina E2 na pata, de maneira dose-dependente depois de 30 min da
sua administração (1,8±1,7 e -1,3±1,0 respectivamente) quando comparados com o
controle (23,9±2,8), sendo que este efeito permaneceu durante todo o tempo (até o minuto

37
90). Na dose de 3 mg/kg, o extrato reduziu a hipernocicepção somente 90 min após a sua
administração (6,3±1,1) (MOLSKA et al., 2003a).
Numa triagem de plantas brasileiras, MONKS e colaboradores (2002a)
verificaram que os extratos orgânicos das folhas, caules e raízes de S. chilensis na
concentração de 100 mg/mL exibiram, no ensaio in vitro, atividade inibitória do
crescimento em ambas linhagens celulares tumorais humanas, HT29 (adenocarcinoma de
cólon) e NCI-H460 (carcinoma de pulmão).
Em outro estudo, foi demonstrado que a solidagenona, isolada das raízes de S.
chilensis, apresentou efeito gastroprotetor em lesões gástricas induzidas por etanol e
aspirina em ratos na dose de 600 mg/kg (RODRÍGUEZ et al., 2002). O efeito da
solidagenona e seus derivados semi-sintéticos, também, foi avaliado em lesões gástricas
induzidas por HCl e etanol. O efeito gastroprotetor da solidagenona nas doses de 100 e 200
mg/kg foi similar, sendo tão ativo quanto o lansoprazol 20 mg/kg; já na dose de 100
mg/kg, os derivados 3α-hidroxisolidagenona e solidagen-6β-ol apresentaram efeito maior
que a solidagenona, reduzindo a lesão em 72,4% e 75,6%, respectivamente (SCHMEDA-
HIRSCHMANN et al., 2002).
Solidagenona e seus derivados semi-sintéticos (2-10) foram avaliados quanto ao
efeito gastroprotetor e anti-úlcera utilizando células epiteliais gástricas humanas (AGS) e
fibroblastos de pulmão humano (MRC-5). Uma redução significativa no dano celular após
incubação com taurocolato de sódio (NaT) foi observada nas células AGS pré-tratadas com
os compostos 2 e 6. O tratamento com os compostos 4-6, 8 e 9 estimulou o conteúdo de
glutationa em culturas de células AGS e o composto 5 aumentou o conteúdo de
prostaglandina E2. O efeito estimulante de proliferação foi observado para os compostos 2,
8 e 9 nas células AGS e para 5, 7-9 nos fibroblastos MRC-5. A solidagenona apresentou
maior toxicidade, enquanto os compostos 4 e 7 foram os menos tóxicos (RODRÍGUEZ et
al., 2005).
Folhas e flores de S. virgaurea, S. gigantea e S. canadensis exibem propriedades
espasmolíticas, e, além de aumentar a diurese, diminuem a excreção de potássio e sódio
(CHODERA et al., 1991; YARNELL, 2002). Estas propriedades podem estar relacionadas

38
com flavonóides, saponinas e outras substâncias químicas, como o ácido 3,5 di-O-cafeoil,
presente em várias espécies de Solidago que apresentam propriedades antiinflamatórias
(APÁTI et al., 2004; WEBER & JAKOBS, 2005). Um estudo em ratos com uma forma
comercial de S. gigantea (extrato etanólico 60%) mostrou atividade antiinflamatória
semelhante ao diclofenaco de sódio, com redução do edema induzido pela carragenina,
além de moderados efeitos espasmolíticos e diuréticos (LEUSCHNER, 1995). Extratos de
Solidago são utilizados como um adjuvante na terapia em pacientes com infecções do trato
urinário, para prevenir cálculos renais e ajudar a remover cristais urinários, sendo a dose
usual da infusão da planta de 2-4 g/xícara três vezes ao dia (YARNELL, 2002).
O fracionamento do extrato das folhas de S. virgaurea rendeu a fração G-100 que
exibiu significativa atividade citotóxica in vitro em várias linhagens de células tumorais
humanas, incluindo células da próstata (PC3), mama (MDA435), melanoma (C8161) e
carcinoma de pulmão (H520). A atividade citotóxica em tumor de próstata em
camundongos também foi avaliada, sendo a fração G-100 (5 mg/kg) administrada, por 3
dias, via intraperitoneal ou subcutânea, e, depois de 25 dias, o crescimento do tumor foi
significativamente suprimido pela fração G-100 (GROSS et al., 2002). Os compostos ent-
germacra-4(15),5,10(14)-trien-1β-ol, β-dictiopterol e ácido 3,5-di-O-cafeoilquínico,
isolados do extrato metanólico das partes aéreas de S. virgaurea, mostraram moderada
citotoxicidade in vitro contra culturas de cinco linhagens de células de tumor humano
(CHOI et al., 2004). Efeitos biológicos de benzilbenzoatos extraídos da fração hexano do
extrato metanólico das partes aéreas de S. virgaurea foram investigados in vitro utilizando
macrófagos de camundongos. Neste estudo, verificou-se que os benzilbenzoatos podem
servir como agentes imunoterapêuticos por estimular as funções dos macrófagos, com
potencial uso no tratamento de doenças infecciosas (SANG et al., 2005). Em outro estudo,
detectou-se que o extrato aquoso das folhas desta mesma espécie inibiu contrações da
bexiga de ratos e de humanos (amostras de pacientes cistectomizados) quando comparados
ao carbacol, por inibir o receptor muscarínico indicando um efeito espasmolítico
(BORCHERT et al., 2004).

39
_________________________________________________________________________
OBJETIVOS

40
2 OBJETIVOS
2.1 Objetivo geral
Estudar as propriedades químicas, biológicas e toxicológicas de diferentes extratos
de Solidago chilensis Meyen.
2. Objetivos específicos
� Caracterizar os principais grupos de metabólitos secundários nas flores, folhas
e raízes de S. chilensis, através de análise fitoquímica.
� Determinar o conteúdo de flavonóides totais e caracterizá-los nas diferentes
partes de S. chilensis.
� Verificar a atividade antibacteriana dos extratos clorofórmicos e brutos, frações
butanólicas e aquosas obtidos das flores, folhas e raízes da espécie S. chilensis, através do
método de difusão em disco.
� Determinar a atividade antioxidante in vivo dos extratos clorofórmicos, brutos
e butanólicos obtidos das diferentes partes de S. chilensis, utilizando linhagens de
Saccharomyces cerevisiae proficiente e deficientes em proteção antioxidante.
� Determinar a atividade antioxidante in vitro dos extratos brutos e frações
butanólicas obtidos das diferentes partes de S. chilensis, através do método da
hipoxantina/xantina oxidase.
� Avaliar a ação antigenotóxica/genotóxica dos extratos brutos das flores, folhas
e raízes de S. chilensis, através do teste cometa in vitro em células sanguíneas de
camundongos.

41
_________________________________________________________________________
MATERIAIS E MÉTODOS

42
3 MATERIAIS E MÉTODOS
Os procedimentos de preparação do material vegetal e obtenção dos extratos, bem
como a caracterização de metabólitos secundários, foram realizados no Laboratório de
Farmacognosia da ULBRA, sendo os solventes e reagentes empregados no
desenvolvimento deste trabalho de procedência LabSynth (Diadema, SP, Brasil), Merck
(Rio de Janeiro, RJ, Brasil) e Nuclear (Diadema, SP, Brasil).
3.1 Material vegetal
A planta Solidago chilensis Meyen foi coletada em Canoas, Estado do Rio Grande
do Sul, Brasil, em abril de 2006. A identificação botânica da espécie vegetal foi realizada
pelo Prof. Dr. Sérgio A. L. Bordignon e uma exsicata desta espécie foi depositada no
Herbário da Universidade Luterana do Brasil (HERULBRA nº 3779).
3.2 Preparação dos extratos e frações de Solidago chilensis
O material vegetal foi separado (em flores, folhas e raízes) e submetido à secagem
a temperatura ambiente e ao abrigo da luz. A seguir, a matéria-prima vegetal foi moída, e
50 g de cada parte da planta foi macerada com 500 mL de clorofórmio a temperatura
ambiente, em recipientes fechados e ao abrigo da luz, até a exaustão. Durante 10 dias, a
cada 48 h o extrato foi filtrado do recipiente contendo a amostra com clorofórmio, e
concentrado em evaporador rotativo a uma temperatura inferior a 50ºC, para obtenção do
extrato clorofórmico (EC). Após este procedimento, o material vegetal utilizado foi
colocado sobre papel filtro para secagem sendo reutilizado na extração por infusão com
água destilada (1/10 m/v) a 80ºC por 30 min. Depois de resfriar, o material foi filtrado e
separado em duas alíquotas de 250 mL cada, sendo a primeira submetida à evaporação
para obtenção do extrato bruto aquoso (EB). A segunda alíquota foi particionada com n-
butanol (5 x 50 mL) para obtenção da fração butanólica (FB) e resíduo aquoso (RA), as
quais foram submetidas à evaporação (Figura 12).

43
Figura 12 - Esquema da preparação dos extratos e frações da S. chilensis.
MPV (Matéria-prima vegetal), EC (Extrato Clorofórmico), EB (Extrato Bruto Aquoso), FB (Fração Butanólica), RA (Resíduo Aquoso).
3.3 Análise fitoquímica
3.3.1 Caracterização fitoquímica
A caracterização fitoquímica das flores, folhas e raízes da S. chilensis, moídas e
secas, foi realizada de acordo com a metodologia proposta por HARBORNE (1984), com
modificações (ALICE et al., 1995; WAGNER & BLADT, 1996; FALKENBERG et al.,
2000; BARBOSA et al., 2004). Visando caracterizar os principais grupos de metabólitos
secundários na espécie em estudo, utilizou-se os seguintes métodos: alcalóides (Bertrand,
Bouchardat, Dragendorff e Mayer), antraquinonas (reação de Bornträger), cardiotônicos
(Baljet, Keller-Killiani, Salkowsky), cumarinas (fluorescência em papel alcalinizado e
comparação com padrão por CCD), flavonóides (reação da cianidina), saponinas (teste de
formação de espuma e comparação com plantas de referência por CCD), taninos (teste da
MPV - Flores, folhas e raízes → secas e trituradas
1ª Extração: 50 g MPV + 500 mL clorofórmio
Maceração (5 x 48 h)
Levar à secura → ECFiltrar
Alíquota de 250 mL
Levar à secura - EB 5 x 50 mL butanolLevar à secura - FB
Resíduo aquoso Levar à secura - RA
2ª Extração: 50 g MPV reutilizada + 500 mL água destilada
Infusão a 80ºC por 30’
Filtrar
Alíquota de 250 mL

44
gelatina e reação com cloreto férrico) e triterpenos/esteróides (reação de Liebermann-
Burchard).
3.3.1.1 Alcalóides
Na técnica para os alcalóides, 1 g da amostra foi extraído com 50 mL de HCl 10%
por 15 min em ebulição. Após esfriar, o material foi filtrado com algodão e separado 4 mL
para os testes de identificação, o qual foi dividido em cinco vidros de relógio (reações e
branco). A seguir, acrescentou-se sob a solução no vidro de relógio, três gotas dos
reagentes de Mayer, Bouchardat, Dragendorff e Bertrand. O aparecimento de precipitado
indica provável presença de alcalóides (ALICE et al., 1995).
3.3.1.2 Antraquinonas
Para detectar as agliconas antraquinônicas, cerca de 0,5 g da amostra foi extraído
com 10 mL de HCl 10% em ebulição por 15 min. Depois de filtrado o material foi extraído
com acetato de etila (2 x 10 mL) em funil de separação. Posteriormente, adicionou-se 4 mL
da fração acetato de etila em um tubo de ensaio e acrescentou-se 3 mL de KOH 5% (reação
de Bornträger). O desenvolvimento de coloração rósea avermelhada na fase aquosa (KOH)
sugere a presença de antraquinonas livres (ALICE et al., 1995).
3.3.1.3 Cardiotônicos
Para verificar a presença de cardiotônicos é necessário avaliar as três partes que
compõem essas substâncias (anel lactônico, núcleo esteróide e desóxi-oses). Para estes
ensaios 3 g da amostra foram extraídos a quente com 20 mL de etanol 50% e 10 mL de
acetato de chumbo a 10% por 30 min. Após este período, a amostra resfriada e filtrada, foi
extraída com clorofórmio (3 x 15 mL), sendo esta fase clorofórmica distribuída em
cápsulas de porcelana (5 mL). Na primeira cápsula, foi evaporado o solvente e adicionado
ao resíduo algumas gotas do reagente de Baljet (ácido pícrico 1% em etanol 50% + NaOH
10%, 1:1) para detectar a presença de anel lactônico caracterizado pelo desenvolvimento
de cor que varia do alaranjado ao vermelho-escuro. À segunda cápsula foi realizada a

45
reação de Salkowsky adicionando algumas gotas de H2SO4 concentrado, que na presença
de núcleo esteróide, leva ao desenvolvimento de coloração vermelha. Na terceira cápsula,
foi realizada a reação de Keller-Killiani adicionando ao resíduo 3 mL de ácido acético
glacial e 1 gota de cloreto férrico 5% em metanol, sendo este material vertido a um tubo de
ensaio contendo 2 mL de H2SO4 concentrado de modo que não se misturassem. O
desenvolvimento de anel marrom avermelhado na interface e a progressiva coloração azul
esverdeada na fase acética indicam a presença de 2-desóxi-oses (ALICE et al., 1995;
FALKENBERG et al., 2000).
3.3.1.4 Cumarinas
Para detecção de cumarinas, foi aquecido em banho-maria (100ºC) 0,5 g da
amostra colocada em tubo de ensaio, o qual foi coberto com papel de filtro seco
previamente impregnado com solução de KOH 5% em metanol. Após 10 min, o papel foi
exposto à luz ultravioleta (UV), sendo a fluorescência amarela ou azul indicativa da
presença de cumarinas voláteis. Amostras dos extratos e frações da planta também foram
comparadas com padrão de cumarina (Merck, Darmstadt, Alemanha) por CCD, utilizando
fase móvel hexano:éter etílico (1:1) seguida de revelação com KOH 5% (ALICE et al.,
1995; FALKENBERG et al., 2000; MARINOFF et al., 2005).
3.3.1.5 Flavonóides
A ocorrência de flavonóides foi investigada utilizando 2 g da amostra extraídos
com 20 mL de água destilada fervente por 10 min. Após esfriar, o material filtrado foi
extraído com n-butanol (2 x 10 mL) em funil de separação. Em seguida, evaporou-se a
fração butanólica à secura em cápsula de porcelana em banho-maria, acrescentando ao
resíduo 10 mL de metanol. A solução obtida foi dividida em dois tubos de ensaio, branco e
reação da cianidina, na qual adicionou-se uma gota de HCl concentrado e uma pitada de
magnésio em pó. Os derivados flavônicos, de cor amarela, são reduzidos a antocianidinas,
de coloração vermelha (flavonóis), laranja (flavonas) e violeta (flavanonas), indicando a
presença das diferentes estruturas dos flavonóides (ALICE et al., 1995; BARBOSA et al.,
2004).

46
3.3.1.6 Saponinas
No método para detectar saponinas, a amostra (1 g) foi extraída com 20 mL de
água destilada por 10 min em ebulição. Depois de resfriada e filtrada, 10 mL da amostra
foram agitados vigorosamente em tubo de ensaio durante 30 segundos. A seguir, mediu-se
a altura do anel de espuma formado logo após a agitação, 10 min depois da agitação e após
adição de 1 mL de HCl 10%. O desenvolvimento de espuma com altura superior a 10 mm
e persistência após adição de 1 mL de HCl 10% sugerem a presença de saponinas. Para
confirmação deste teste, extratos brutos e frações butanólicas da planta também foram
comparados com extratos butanólicos das plantas de referência (Aesculus hippocastanum
L. e Sapindus saponaria L.) por CCD, utilizando fase móvel clorofórmio:metanol:ácido
acético glacial (80:40:6) seguida de revelação com anisaldeído sulfúrico e aquecimento da
placa (ALICE et al., 1995; WAGNER & BLADT, 1996; BARBOSA et al., 2004).
3.3.1.7 Taninos
Para caracterizar taninos, 2 g da amostra foram extraídos com 20 mL de água
destilada fervente por 30 min. Posteriormente, foram adicionados 6 mL da amostra filtrada
em dois tubos de ensaio (reação e branco). Para a reação da gelatina adicionou-se 5 mL de
solução de NaCl 2% e 5 mL de solução de gelatina 1%, sendo a reação positiva quando
ocorre a formação de turvação ou precipitado, comparado ao branco. Para a reação com
sais de ferro, adicionou-se quatro gotas de cloreto férrico 1% (solução aquosa) em um tubo
de ensaio com 6 mL da amostra filtrada. O desenvolvimento de substâncias de coloração
azulada sugere a presença de taninos hidrolisáveis e a cor verde taninos condensados
(ALICE et al., 1995; FALKENBERG et al., 2000).
3.3.1.8 Triterpenos e esteróides
Na reação para a detecção de triterpenos e esteróides, a amostra (1 g) foi
macerada com 25 mL de ciclohexano por 20 min, filtrada e dividida em duas alíquotas de 5
mL em cápsulas de porcelana, levando ao banho-maria uma das alíquotas até obtenção do
resíduo. Para a reação de Liebermann-Burchard, adicionou-se ao resíduo 3 mL da solução

47
10 mL de anidrido acético e 2 gotas de H2SO4 concentrado. Após alguns minutos, verifica-
se o desenvolvimento de cores, que vão do azul ou verde indicando núcleo esteróide; e
vermelha, rósea ou violeta indicando núcleo triterpênico. Para a confirmação do resultado,
concentrou-se a alíquota restante e realizou-se cromatografia em camada delgada (CCD)
com fase móvel ciclohexano:acetato de etila (70:30), utilizando revelador Liebermann-
Burchard e após 10 min, aquecimento da placa em estufa a 125ºC. A presença desta classe
é sugerida pela presença de manchas azul ou rosa após revelação (ALICE et al., 1995).
3.3.2 Determinação de flavonóides totais
Para cada parte da planta (flor, folha e raiz) foi preparada uma solução-mãe,
adicionando a um balão de fundo redondo de 100 mL, 1,25 g da amostra moída e 15 mL de
metanol, sendo aquecido em banho-maria, sob refluxo, por 30 min entre 45-50ºC. A
mistura foi filtrada em algodão para balão volumétrico de 25 mL e o resíduo insolúvel no
algodão foi recolocado ao mesmo balão de fundo redondo e adicionado 10 mL de metanol.
A amostra foi aquecida sob refluxo, por mais 10 min, e filtrada para o mesmo balão
volumétrico de 25 mL. Após resfriamento à temperatura ambiente, completou-se o volume
para 25 mL com metanol. Para a determinação dos flavonóides totais, foram preparadas em
balões volumétricos de 25 mL, a solução-amostra (1 mL da solução-mãe, 0,5 mL de
solução de cloreto de alumínio a 2% em metanol e completou-se o volume com metanol) e
a solução-branco (1 mL da solução-mãe e completou-se o volume com metanol). A
absorbância (A) da solução-amostra foi medida em 425 nm 30 min após seu preparo,
utilizando a solução-branco para ajuste do zero. O ensaio foi realizado em duplicata e
calculou-se o teor de flavonóides totais segundo a expressão (FARMACOPÉIA
BRASILEIRA, 2003):
H = A x 62500 .
500 x m x (100 – Pd)
Em que H= hiperosídeo, A= absorbância medida, m= massa do vegetal (g) e Pd=
determinação de água (%).

48
A determinação da perda de água foi realizada por dessecação – método
gravimétrico. Foram pesados 2 g da amostra moída e transferida para pesa-filtro chato
previamente dessecado durante 30 min nas mesmas condições a serem empregadas na
determinação. O pesa-filtro contendo a amostra foi pesado e colocado na estufa por 5 h a
105ºC. Após, a amostra esfriar à temperatura ambiente em dessecador, foi pesada e
calculada a porcentagem de água em relação ao vegetal seco ao ar. As operações de
pesagem foram repetidas até atingir peso constante. A porcentagem de perda por
dessecação é dada pela equação (FARMACOPÉIA BRASILEIRA, 1988):
Pu – Ps x 100
Pa
Em que Pu = peso do pesa-filtro contendo a amostra antes da dessecação, Ps = peso
do pesa-filtro contendo a amostra após a dessecação e Pa = peso da amostra.
3.3.3 Análise cromatográfica
Os extratos e frações foram submetidos à análise cromatográfica em camada
delgada (CCD) e cromatografia em coluna (CC), utilizando diferentes sistemas eluentes,
visando conhecer sua constituição química e o isolamento de novos compostos.
Para os procedimentos, utilizou-se colunas cromatográficas de vidro cilíndricas
com dimensões variando de acordo com a quantidade de amostra a ser cromatografada. A
cromatografia em camada delgada foi empregada para análise e reunião das frações obtidas
por cromatografia em coluna. As substâncias analisadas por CCD foram evidenciadas pelo
uso de luz UV sob os comprimentos de onda de 254 e 366 nm, e quando necessário através
de revelação com revelador natural (1% em metanol) e polietilenoglicol (3% em etanol).
Para os processos cromatográficos utilizou-se cromatofolhas de sílica gel 60 F254, 20x20
cm e sílica gel 60 (0,063-0,200 mm, 70-230 mesh) da Merck (Darmstadt, Alemanha),
além dos padrões de ácido clorogênico, campferol, hiperosídeo, isoquercitrina, luteolina,
quercetina e rutina da Sigma (St. Louis, MO, Estados Unidos).

49
Todos os extratos de S. chilensis foram analisados por CCD com diferentes
sistemas de fase móvel. A fração butanólica das flores de S. chilensis foi selecionada
devido a baixa concentração de clorofila e o destacado efeito antioxidante apresentado. A
partir disso, FB foi filtrada com carvão ativo, concentrada e fracionada (2 g) por
cromatografia em coluna “flash”. Utilizando como eluente misturas de acetato de
etila:metanol combinados em ordem crescente de polaridade (100:0, 70:30, 50:50, 30:70,
0:100), obteve-se cinco frações (F1-F5). As frações F1 (425,2 mg), F2 (550,7 mg), F3
(472,3 mg), F4 (265,4 mg) e F5 (181,8 mg) foram analisadas por CCD, utilizando como
fase móvel acetato de etila:metanol (90:10), sob luz UV após nebulização com revelador
natural e polietilenoglicol.
As frações F1 e F2 foram reunidas por apresentarem perfil semelhante e
submetidas a cromatografia em coluna, utilizando-se solventes de polaridades crescentes
clorofórmio:acetato de etila (100-0 a 0:100) e acetato de etila:metanol (100:0 a 0:100). Um
total de 52 frações de 100 mL foi coletado, monitorando-as por CCD reuniu-se estas em 7
subfrações: Fr1 (F1-7) 151 mg, Fr2 (F8-11) 123 mg, Fr3 (F12-20) 105 mg, Fr4 (F21-30)
109 mg, Fr5 (F31-38) 95 mg, Fr6 (F39-46) 136 mg e Fr7 (F47-52) 90 mg. Posteriormente,
comparou-se as subfrações com os padrões por CCD utilizando como fase móvel, para
agliconas, o sistema acetato de etila:clorofórmio (70:30) e, para heterosídeos
flavonoídicos, o sistema acetato de etila:ácido fórmico:ácido acético glacial:água
(100:11:11:26), seguidos de revelação com revelador natural e polietilenoglicol e
visualização sob luz UV. Seguindo os sistemas cromatográficos anteriormente
mencionados, analisou-se o perfil de flavonóides presentes no EB das flores e EB e FB das
folhas e raízes.

50
3.4 Avaliações biológicas
3.4.1 Avaliação da atividade antibacteriana
Esta parte do estudo foi desenvolvida no Laboratório de Microbiologia da
Faculdade de Farmácia, na Universidade de Passo Fundo (UPF), Passo Fundo-RS, sendo
os microrganismos testados fornecidos pelo mesmo Laboratório. Os meios de cultura
empregados no desenvolvimento deste trabalho foram de procedência Biobrás
Diagnósticos (Montes Claros, MG, Brasil), discos de antimicrobianos da Sensidisc
(Araçatuba, SP, Brasil) e solventes da Sigma (St. Louis, MO, Estados Unidos).
Os extratos clorofórmicos (EC), extratos brutos (EB), frações butanólicas (FB) e
resíduos aquosos (RA) das flores, folhas e raízes da S. chilensis, foram submetidos à
avaliação da atividade antibacteriana empregando-se o método da difusão de disco
conforme as normas preconizadas pelo National Committee for Clinical Laboratory
Standard (NCCLS, 2004).
Uma triagem da atividade antibacteriana foi realizada frente a linhagens padrão da
“American Type Culture Collection” (ATCC) de bactérias Gram positivas, Staphylococcus
aureus (25923), Enterococcus faecalis (29212), Enterococcus faecium (10541), e bactérias
Gram negativas, Escherichia coli (25922), e Klebsiella pneumoniae (cepa clínica fornecida
pelo Hospital Municipal de Passo Fundo). As suspensões bacterianas foram preparadas
pelo método direto, inoculando-se colônias de repiques de 24 h de incubação em Ágar
Nutriente (desenvolvidas previamente em caldo BHI, “Brain-heart infusion”), em solução
de NaCl 0,9% esterilizada até atingir a concentração de 1,5 x 108 UFC/mL. O ajuste desta
concentração foi feito através de leitura espectrofotométrica, no comprimento de onda de
635 nm e absorbância de 0,08 a 0,10. O meio de cultura utilizado para o ensaio
antibacteriano foi Ágar Müeller-Hinton recém-preparado e submetido previamente ao teste
de esterilidade (OPLUSTIL et al., 2000; OLIVEIRA et al., 2005).

51
A semeadura nas placas foi realizada através da técnica de oito direções. Após a
semeadura, as placas ficaram entreabertas, em zona asséptica, por aproximadamente 3 min
para absorção uniforme do inóculo (OPLUSTIL et al., 2000). Em seguida foram aplicados
os discos de papel de filtro (6 mm de diâmetro) impregnados com 20 µL dos extratos
vegetais na concentração de 50 mg/mL em DMSO, os controles negativos (discos de papel
de filtro sem nenhuma substância e discos de papel de filtro com 20 µL de DMSO) e
controles positivos (discos de antimicrobianos) (OLIVEIRA et al., 2005), os quais foram
selecionados de acordo com a sensibilidade da bactéria testada (Tabela 1). Todos os testes
foram realizados em triplicata.
Tabela 1 - Susceptibilidade antimicrobiana frente aos controles positivos.
Bactéria Antimicrobiano Halo Padrão (mm) Enterococcus faecalis ATCC 29212 Ampicilina (10 µg) 27-35 Enterococcus faecium ATCC 10541 Ampicilina (10 µg) 27-35 Staphylococcus aureus ATCC 25923 Ampicilina (10 µg) 27-35 Escherichia coli ATCC 25922 Cloranfenicol (30 µg) 21-27 Klebsiella pneumoniae Cloranfenicol (30 µg) 21-27
Posteriormente, as placas foram incubadas em estufa bacteriológica à 37 ºC (±1
ºC) por 18 h de incubação. Depois deste tempo, foram medidos os diâmetros dos halos de
inibição dos controles positivos e dos extratos, sendo considerado como resultado
negativo, quando houve crescimento bacteriano ao redor do disco igual aos controles
negativos (OPLUSTIL et al., 2000).
3.4.2 Avaliação da atividade antioxidante
Os ensaios antioxidantes foram realizados nos Laboratórios de Genética
Toxicológica e de Farmacocinética do Centro de Pesquisa em Ciências Médicas da
ULBRA. Extrato de levedura, Bacto Peptona e Bacto Ágar empregados no
desenvolvimento dos ensaios foram de procedência Difco Laboratories (Detroit, MI,
Estados Unidos), os solventes e reagentes foram de procedência Nuclear (Diadema, SP,
Brasil), Sigma (St. Louis, MO, Estados Unidos) e Merck (Rio de Janeiro, RJ, Brasil).

52
3.4.2.1 Ensaio antioxidante in vivo
Linhagens da levedura Saccharomyces cerevisiae proficiente (SOD) e deficientes
(sod1, sod2 e sod1sod2) no sistema de defesa antioxidante (Tabela 2) foram analisadas
pelo Ensaio de Sobrevivência. Os extratos (EC, EB, FB) das flores, folhas e raízes foram
primeiramente dissolvidos em DMSO 5% e em seguida em água destilada para
concentração final de 1 mg/mL. A concentração final de DMSO na mistura não excedeu
0,2%.
Tabela 2 - Linhagens de Saccharomyces cerevisiae utilizadas neste estudo.
Meios e soluções foram preparados previamente como descrito por BURKE e
colaboradores (2000). O inóculo das diferentes linhagens foi preparado em meio líquido
YEL (“Yeast Extract Líquid”, 1% extrato de levedura, 2% Bacto Peptona e 2% glicose) e
mantido em estufa à 30 ºC por 48 h para crescimento celular. Meio YEPD (“Yeast Extract-
Peptone-Dextrose”) contendo 1% extrato de levedura, 2% Bacto Peptona, 2% Bacto Ágar e
2% glicose foi utilizado para as placas na rotina de crescimento celular das leveduras.
Uma solução de NaCl 0,9% foi empregada para diluição das suspensões celulares
e água destilada estéril para diluição do H2O2 utilizado nos experimentos para testar a
atividade antioxidante. Células tratadas com DMSO e com H2O2 (1 mM) foram utilizadas
como controles negativo e positivo, respectivamente (BOEIRA et al., 2002; DELLA
CROCE et al., 2003; LOPES et al., 2004; ROSA et al., 2006).
Linhagens Genótipo Deficiência do sistema enzimático
Fonte
SOD+ Matα his3∆1 leu2∆0 trp1-289 ura3-52
Nenhuma E.B.Gralla, Los Angeles
sod1∆ mesmo que SOD+ exceto sod1::URA3
Cu-Zn superóxido dismutase
E.B.Gralla, Los Angeles
sod2∆ mesmo que SOD+ exceto sod2::TRP1
Mn superóxido dismutase
E.B.Gralla, Los Angeles
sod1∆sod2∆ mesmo que SOD+ exceto sod1::URA3 sod2::TRP1
Todas superóxido dismutases
E.B.Gralla, Los Angeles

53
A atividade antioxidante de S. chilensis foi avaliada pela sobrevivência das
leveduras de S. cerevisiae na fase estacionária de crescimento tratadas com H2O2. Culturas
em fase estacionária foram obtidas pela inoculação de uma colônia isolada em meio
líquido YEL. Após 48 h de incubação a 30 ºC, as células foram agitadas, centrifugadas (5
min, 2500 rpm) e ressuspendidas com 5 mL de solução NaCl 0,9%. O número de células
foi determinado utilizando uma câmara de Neubauer. As linhagens foram avaliadas por
inoculação de 2x107 células/mL por 1 h a 30 ºC, contendo NaCl 0,9%, as diferentes doses
do extrato (50, 100, 250 e 500 µL/mL) e o agente oxidante (100 µL H2O2 1 mM). Para a
determinação das células sobreviventes, alíquotas das células tratadas foram diluídas
apropriadamente e plaqueadas (100 µL) em meio YEPD. Células sobreviventes foram
contadas após incubação a 30 ºC por 48 h (BOEIRA et al., 2002; LOPES et al., 2004). O
percentual de sobrevivência foi calculado em relação ao número de colônias sobreviventes
no controle negativo. Os ensaios foram realizados em triplicata e os dados representam a
média das contagens dos experimentos.
3.4.2.2 Ensaio antioxidante in vitro
O método da hipoxantina/xantina oxidase, empregado para a análise em CLAE,
foi baseado no método descrito por OWEN e colaboradores (1996) (Figura 13). Neste
ensaio foram utilizados os extratos brutos e as frações butanólicas das flores, folhas e
raízes da S. chilensis. Para cada amostra utilizada foi preparada uma solução-mãe na
concentração de 8,0 mg/mL e apropriadamente diluídas (em triplicata) no mix
hipoxantina/ácido salicílico em um volume final de 1,0 mL, resultando em concentrações
de 0,1, 0,25, 0,5, 1,0, e 2,0 mg/mL. As amostras foram dissolvidas no Mix
hipoxantina/ácido salicílico (19,05 mL K2HPO4 1M, 30,95 mL KH2PO4 1M, 0,146
mg/mL EDTA, 0,065 mg/mL Fe3+, 0,041 mg/mL hipoxantina e 0,25 mg/mL ácido
salicílico, água destilada – q.s.p. 500 mL, protegido da luz) e uma alíquota de 5 µL da
enzima xantina oxidase dissolvida em 3,2 M (NH4)2SO4 foi adicionada às amostras para
iniciar a reação. Em seguida, as amostras foram incubadas em um termomixer
(Eppendorff, Alemanha) por 3 h a 37 °C com agitação para completar a reação, que após
este tempo foi interrompida com 10 µL de HCl puro. As amostras foram mantidas
refrigeradas até o momento da análise de uma alíquota de 30 µL da amostra no aparelho de

54
CLAE, utilizando condições cromatográficas como descrito por OWEN e colaboradores
(2000).
Figura 13 - Esquema do ensaio para medir atividades antioxidantes utilizando o sistema da
xantina oxidase (OWEN et al., 1996).
Na análise cromatográfica foi utilizado um gradiente a base dos solventes
metanol:água:ácido acético (100:95:5), com fluxo de 1 mL/min durante 30 min, uma
coluna de fase reversa µBondapack C18 (300 x 3,9 mm – 4 µm) (Waters) e detecção a 325
nm. O equipamento de CLAE Alliance (Waters) é constituído pelo módulo de separação
2695 (Waters) com autoinjetor e um detector UV-VIS 2487 dual λ (Waters). Os ácidos
diidróxi-benzóicos e o ácido úrico foram monitorados na A= 325 nm e A= 278 nm,
respectivamente.

55
A quantidade de diidroxifenóis, 2,5-diidroxi-ácido-benzóico e 2,3-diidroxi-ácido-
benzóico (DHBAs) (Figura 14), produzidos pela reação do ácido salicílico com produção
de radicais hidroxilas (HO•) foi determinada pela curva padrão dos respectivos
diidroxifenóis.
Figura 14 - Reação dos radicais HO• com o ácido salicílico, levando a formação dos
compostos estáveis 2,3- e 2,5-diidroxi-ácido-benzóico (DHBA) (OWEN et al., 1996).
3.4.3 Avaliação da atividade antigenotóxica/genotóxica
Este experimento foi realizado no Laboratório de Genética Toxicológica da
ULBRA. A agarose normal 1,5% e a agarose “low melting” 0,75% empregadas no
desenvolvimento deste ensaio foram de procedência Promega Corporation (Madison, WI,
USA) e Gibco BRL (Grand Island, NY, USA), respectivamente. Os solventes e reagentes
foram de procedência Acros Organics (New Jersey, Estados Unidos), Invitrogen
Corporation (Carlsbad, CA, Escócia), LabSynth (Diadema, SP, Brasil), Merck
(Darmstadt, Alemanha), Nuclear (Diadema, SP, Brasil), Plat-Lab (Guarulhos, SP,
Brasil) e Sigma (St. Louis, MO, Estados Unidos).
O teste cometa alcalino in vitro foi realizado conforme descrito por SINGH e
colaboradores (1988) e TICE e colaboradores (2000), com modificações (HARTMANN et
al., 2004). Para o teste coletaram-se amostras de sangue periférico de seis camundongos
machos SR-1, utilizando tubos com heparina sódica 25.000 UI/5mL (Liquemine,
Roche, Basiléia, Suíça). As amostras coletadas no mesmo dia do experimento foram
mantidas refrigeradas e protegidas da luz até o processamento. O sangue periférico (150
µL) foi tratado com 20 µL dos extratos brutos das flores, folhas e raízes de S. chilensis nas

56
concentrações de 5, 7,5 e 10 mg/mL (TICE et al., 2000) ou com PBS (controle negativo)
por 1 h a 37 ºC. Lâminas de microscópio pré-cobertas com agarose normal 1,5%
(preparada em PBS a 60 °C), foram secas à temperatura ambiente por 24 h e, após,
estocadas a 4 °C até o uso (SILVA et al., 2000).
A avaliação do efeito genotóxico foi realizada adicionando, isoladamente, 7,5 µL
de cada concentração tratada a 95 µL de agarose “low melting” 0,75% (mantida a 37 °C
em banho-maria). Posteriormente transferiu-se esta solução para quatro lâminas pré-
cobertas com agarose normal a 1,5% (denominadas A, B, C e D) sendo cobertas com
lamínulas (SILVA et al., 2000). Depois da remoção das lamínulas, duas das lâminas (A e
B) foram mergulhadas em uma solução de lise (2,5 M NaCl; 100 mM EDTA; 10 mM Tris,
pH 10-10,5; 1% Triton X-100 e 10% DMSO) por no mínimo 1 h a no máximo 72 h a 4 oC,
para o rompimento das membranas celulares (HARTMANN et al., 2004). Para avaliação
da atividade antigenotóxica no tratamento in vitro, colocou-se as outras duas lâminas (C e
D) por 5 min na solução de H2O2 0,25 mM (controle positivo) a 4 ºC, e, depois de lavadas
três vezes com solução de NaCl, foram mergulhadas em solução de lise por no mínimo 1 h
a no máximo 72 h a 4 oC.
Após permanência na solução de lise, removeu-se todas as lâminas, sendo estas
colocadas em uma cuba de eletroforese contendo tampão alcalino gelado (300 mM NaOH
e 1 mM EDTA, pH > 13, 4 °C) por 20 min. A corrida eletroforética foi realizada por 15
min, a 25 V (0,90 V/cm) e 300 mA. Todos os procedimentos (preparação das lâminas, lise
e eletroforese) foram realizados no escuro com luz vermelha para prevenir adicionais
danos ao DNA (HARTMANN et al., 2004). Após a eletroforese, colocou-se as lâminas em
posição horizontal lavando-as duas vezes (5 min cada) com tampão Tris 0,4 M, pH 7,5, e
mais duas vezes com água destilada, para neutralizar a alcalinidade, sendo secadas a
temperatura ambiente por no mínimo 5 h. Depois deste período, fixou-se as lâminas por 10
min com uma solução de fixação (ácido tricloroacético 15%, sulfato de zinco 5% e glicerol
5%), e estas foram lavadas três vezes com água destilada e secadas a temperatura ambiente
(5 h). Reidratou-se as mesmas com água destilada por 5 min para serem coradas com
solução de prata (5% carbonato de sódio, mais solução de 0,1% nitrato de amônia, 0,1%
nitrato de prata, 0,25% ácido tungstosilícico e 0,15% formaldeído) em banho-maria a 37

57
°C. A seguir colocou-se as lâminas em solução de parada (ácido acético 1%) por 5 min,
sendo lavadas três vezes com água destilada e secadas a temperatura ambiente (5 h)
(SILVA et al., 2000; NADIN et al., 2001; SILVA et al., 2007).
Analisou-se as lâminas em microscópio óptico, sendo observados os parâmetros
tamanho e formato de cauda para 100 células/lâmina. Os resultados foram expressos em
índice de danos (ID) e freqüência de danos (FD). O ID foi obtido pela avaliação visual das
classes de dano (de 0–4), extraindo-se um índice que expressa o dano geral sofrido por
uma população de células (100 células por lâmina em duplicata), os núcleos contendo o
DNA intacto aparecem redondos (classe 0 – sem dano), já nas células lesadas, o DNA livre
migra em direção ao ânodo, durante a eletroforese, mostrando uma “cauda” de fragmentos
sedimentados, semelhante a um cometa e são classificados entre classes um (dano mínimo)
a quatro (dano máximo). Assim, o ID de cada extrato estudado variou de zero (100X0; 100
células observadas completamente sem danos) a 400 (100X4; 100 células observadas com
dano máximo) (SILVA et al., 2007). A FD foi calculada baseada no número de células
com cauda versus aquelas sem cauda. A percentagem de redução do ID foi calculada de
acordo com a equação: I%: = [ID do peróxido – ID do extrato com o peróxido] / [ID do
peróxido – ID do controle negativo] x 100 (KAPISZEWSKA et al., 2005).
3.5 Análise estatística
Os resultados dos ensaios antioxidantes foram submetidos à análise da variância
(ANOVA) de uma via e, após, teste de Tukey. Para análise dos dados do teste cometa
utilizou-se ANOVA e teste de Dunett, comparando-se os tratamentos com os extratos e o
controle (dose zero de extrato). Os valores calculados estão expressos como média ±
desvio padrão e os considerados estatisticamente significativos destacados com *P<0,05 ou
**P<0,01.

58
_________________________________________________________________________
RESULTADOS

59
4 RESULTADOS
4.1 Rendimentos dos extratos e frações de Solidago chilensis
O rendimento dos diferentes extratos e frações das flores, folhas e raízes de
Solidago chilensis utilizados nos ensaios biológicos deste estudo estão apresentados na
Tabela 3.
Tabela 3 – Rendimentos dos diferentes extratos e frações da S. chilensis.
Amostra Massa (g) Extrato/ Fração Rendimento (g) Rendimento (%)
Flores 50 g EC 7,69 15,38 EB 2,31 4,62 FB 2,65 5,30 RA 4,32 8,64 Folhas 50 g EC 6,85 13,70 EB 6,89 13,78 FB 2,32 4,64 RA 4,59 9,18 Raízes 50 g EC 1,13 2,26 EB 9,07 18,14 FB 0,46 0,92 RA 8,53 17,06
EC (Extrato Clorofórmico), EB (Extrato Bruto Aquoso), FB (Fração Butanólica), RA (Resíduo Aquoso).
4.2 Análise fitoquímica
4.2.1 Análise da constituição química
Na análise da constituição química das diferentes partes da S. chilensis verificou-
se a presença de substâncias com núcleo esteróide (vermelho) e núcleo triterpênico
(violeta), saponinas (persistência da espuma após adição de HCl e manchas violáceas na
CCD após revelação) e flavonóides (avermelhado), sendo que nas raízes o último
metabólito citado apresentou uma coloração menos intensa do que nas flores e folhas. O
resultado da análise dos metabólitos secundários detectados nas flores, folhas e raízes da
planta em estudo encontra-se na Tabela 4.

60
Tabela 4 – Metabólitos secundários presentes nas flores, folhas e raízes da S. chilensis.
Metabólito secundário
Caracterização fitoquímica
Flores Folhas Raízes
Alcalóides Reação de Bertrand Reação de Bouchardat Reação de Dragendorff Reação de Mayer
- - - -
- - - -
- - - -
Antraquinonas Reação de Bornträger - - - Cardiotônicos Reação de Baljet (anel lactônico)
Reação de Keller-Killiani (desóxi-oses) Reação de Salkowsky (núcleo esteróide)
- -
+
- -
+
- -
+ Cumarinas Fluorescência em papel alcalinizado
Comparação com padrão por CCD - -
- -
- -
Flavonóides Reação da cianidina + + + Saponinas Teste de formação de espuma
Comparação com plantas de referência por CCD
+
+
+
+
+
+ Taninos Teste da gelatina
Reação com cloreto férrico -
verde -
verde -
verde Triterpenos Reação de Liebermann-Burchard + + +
No ensaio de caracterização de taninos, a reação de cloreto férrico, das três partes
da planta, apenas indica a presença de compostos fenólicos, pois não houve turvação no
teste com a gelatina.
4.2.2 Avaliação do teor de flavonóides totais
Os teores de flavonóides totais encontrados nas flores, folhas e raízes de S.
chilensis estão apresentados na Tabela 5, o qual foi expresso em hiperosídeo de acordo
com técnica descrita na FARMACOPÉIA BRASILEIRA, 4 ed. (2003).
Tabela 5 - Perda de água por dessecação e teor de flavonóides totais das diferentes partes
da S. chilensis.
Amostra Massa (g)
Perda de água por dessecação (%)
Massa (g)
Teor de flavonóides (%)
Flores 2 g 9,520 1,25 g 2,0601 Folhas 2 g 8,655 1,25 g 1,9377 Raízes 2 g 10,045 1,25 g 0,0278

61
4.2.3 Análise cromatográfica
Os extratos clorofórmicos das flores, folhas e raízes foram submetidos a diversas
CCD em diferentes sistemas cromatográficos. Nestas amostras verificou-se uma grande
variedade de produtos, em pequenas concentrações, que muitas vezes ficavam sobrepostos
formando uma mancha única, dificultando a separação e isolamento dos constituintes
destes extratos. Por outro lado, FB mostrava-se propicia a análise cromatográfica. Após os
fracionamentos realizados em coluna cromatográfica com FB das flores, previamente
descrito no item Materiais e Métodos, as sete frações reunidas (Fr1-Fr7) foram analisadas
por cromatografia de camada delgada e comparadas a padrões de ácidos fenólicos e
flavonóides, quanto à coloração, fator de retenção (Rf) e dados da literatura (WAGNER &
BLADT, 1996). A partir da análise destes dados pôde-se caracterizar a presença de
produtos com o comportamento cromatográfico de campferol (Rf 0,76) e quercetina (Rf
0,59) em Fr5, quercitrina (Rf 0,80), hiperosídeo (Rf 0,62) e rutina (Rf 0,41) em Fr6 e ácido
caféico (Rf 0,92) em Fr7. Dentre estes, a quercitrina pode ser sugerida como a substância
majoritária na FB devido ao tamanho da mancha correspondente.
No EB das flores, as manchas detectadas por CCD apresentaram perfil
cromatográfico do campferol, quercetina, quercitrina, hiperosídeo, rutina e ácido
clorogênico (Rf 0,48). Desta mesma maneira, nas folhas foram caracterizadas quercetina
na FB, e quercitrina, rutina e ácido clorogênico em ambos EB e FB. No entanto nas raízes
não foram detectados ácidos fenólicos e flavonóides nos sistemas utilizados.

62
4.3 Avaliações biológicas
4.3.1 Atividade antibacteriana
Na avaliação pelo método de difusão do disco, cinco diferentes bactérias
(Staphylococcus aureus, Enterococcus faecalis, Enterococcus faecium, Escherichia coli e
Klebsiella pneumoniae) foram utilizadas para avaliar a possível atividade antimicrobiana
dos extratos e frações (EC, EB, FB e RA) das flores, folhas e raízes de S. chilensis. O
efeito dos extratos e frações foi comparado aos antibióticos utilizados como controle
positivo (ampicilina e cloranfenicol). De acordo com os resultados obtidos, não foi
observada inibição significativa dos extratos e frações na concentração de 50 mg/mL,
frente às bactérias testadas, quando comparadas aos controles positivos.
4.3.2 Atividade antioxidante
4.3.2.1 Ensaio antioxidante in vivo
Com base nos resultados encontrados foi possível determinar a sobrevivência das
linhagens de S. cerevisiae proficiente (SOD) e deficientes em defesas antioxidantes (sod1,
sod2 e sod1sod2), incubadas com diferentes doses (50 a 500 µL/mL) dos extratos e frações
(EC, EB e FB) das flores, folhas e raízes de S. chilensis e expostas ao peróxido de
hidrogênio (1mM) em fase estacionária de crescimento. Nesta fase as leveduras podem
sobreviver por semanas ou meses na ausência de nutrientes e a atividade das enzimas
antioxidantes está aumentada, para proteger a célula das ERO formadas devido ao
metabolismo respiratório (LUSHCHAK & GOSPODARYOV, 2005). Os resultados
obtidos (Figuras 15, 16 e 17) mostram claramente que a incubação das linhagens com os
extratos e frações resultou em um aumento da sobrevivência das linhagens de S. cerevisiae
quando expostas ao H2O2, principalmente com os extratos brutos e frações butanólicas das
flores e folhas. Em quase todos estes extratos, o aumento da sobrevivência ocorreu de
forma dose-dependente e foi semelhante nas linhagens utilizadas, independente do tipo de
perda de defesa enzimática.

63
Figura 15 - Ensaio de sobrevivência com células de S. cerevisiae na fase estacionária após
tratamento com extratos clorofórmico (EC), bruto (EB) e fração butanólica (FB) das flores
de S. chilensis. Os resultados representam a média dos experimentos realizados em
triplicata, sendo considerados estatisticamente significativos *P<0,05 e **P<0,01.
Entretanto, verifica-se, na linhagem sod1sod2 uma menor sobrevivência em
relação às demais linhagens. Nos tratamentos com extratos brutos e frações butanólicas, na
maioria das doses, as linhagens da levedura apresentaram atividade antioxidante
estatisticamente significativa. A atividade da FB das raízes se mostrou semelhante à FB
das folhas. Porém, a linhagem sod1sod2 tratada com FB das folhas e a linhagem sod2
tratada com EB das raízes não apresentaram, em nenhuma das doses testadas, ação
antioxidante contra danos causados pelo H2O2. Os extratos clorofórmicos (EC), também
SOD sod1 sod2 sod1sod20
25
50
75
100
******
*****
***
Flores - EB
Linhagens
Sob
revi
vênc
ia (
%)
****
**** **
Flores - EC
SOD sod1 sod2 sod1sod20
25
50
75
100
****
******
**
**
**
**
Linhagens
Sob
revi
vênc
ia (
%)
**
H2O2 (100 µL, 1mM)
50 µL extrato + H2O2
100 µL extrato + H2O2
250 µL extrato + H2O2
500 µL extrato + H2O2
Flores - FB
SOD sod1 sod2 sod1sod20
25
50
75
100
** ******
*
Linhagens
Sob
revi
vênc
ia (
%)
******
******
****
****
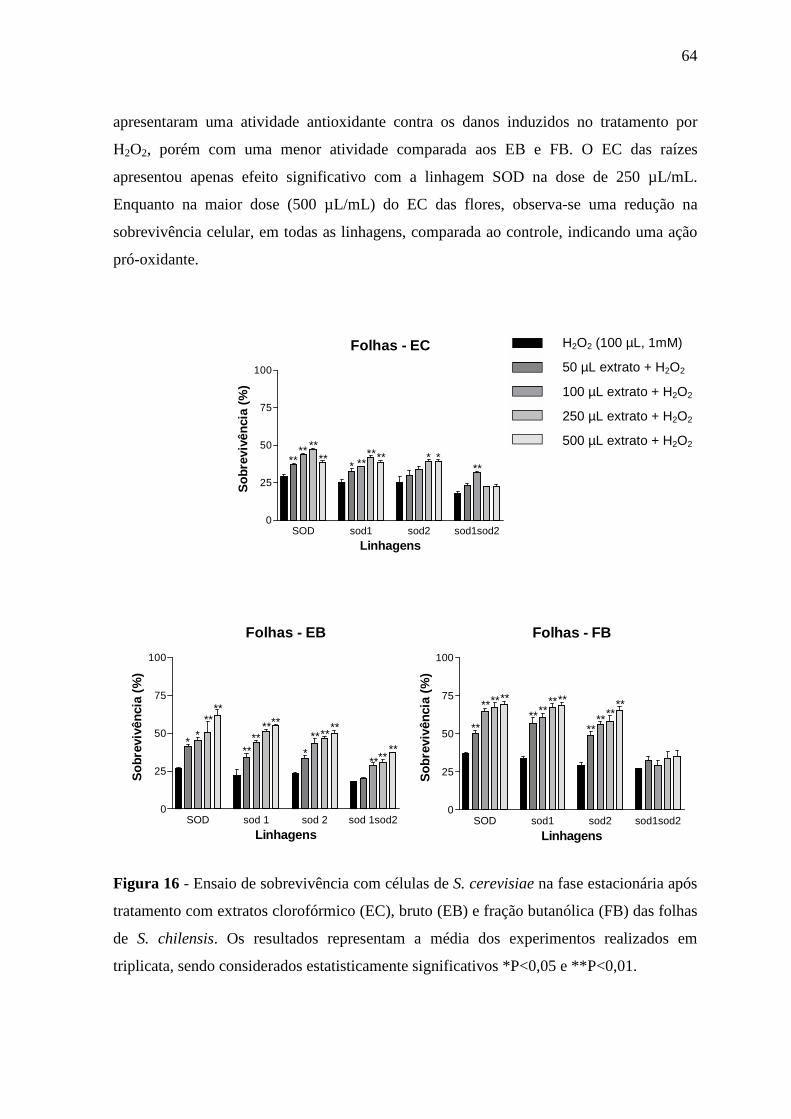
64
apresentaram uma atividade antioxidante contra os danos induzidos no tratamento por
H2O2, porém com uma menor atividade comparada aos EB e FB. O EC das raízes
apresentou apenas efeito significativo com a linhagem SOD na dose de 250 µL/mL.
Enquanto na maior dose (500 µL/mL) do EC das flores, observa-se uma redução na
sobrevivência celular, em todas as linhagens, comparada ao controle, indicando uma ação
pró-oxidante.
Figura 16 - Ensaio de sobrevivência com células de S. cerevisiae na fase estacionária após
tratamento com extratos clorofórmico (EC), bruto (EB) e fração butanólica (FB) das folhas
de S. chilensis. Os resultados representam a média dos experimentos realizados em
triplicata, sendo considerados estatisticamente significativos *P<0,05 e **P<0,01.
Folhas - EB
SOD sod 1 sod 2 sod 1sod20
25
50
75
100
****
*
*
****
****
****
Linhagens
Sob
revi
vênc
ia (
%)
******
***
Folhas - EC
SOD sod1 sod2 sod1sod20
25
50
75
100
** ******
**
Linhagens
Sob
revi
vênc
ia (
%)
** ** ** **
*
H2O2 (100 µL, 1mM)
50 µL extrato + H2O2
100 µL extrato + H2O2
250 µL extrato + H2O2
500 µL extrato + H2O2
Folhas - FB
SOD sod1 sod2 sod1sod20
25
50
75
100
** ******
**
**
Linhagens
Sob
revi
vênc
ia (
%)
**
**
********

65
Figura 17 - Ensaio de sobrevivência com células de S. cerevisiae na fase estacionária após
tratamento com extratos clorofórmico (EC), bruto (EB) e fração butanólica (FB) das raízes
de S. chilensis. Os resultados representam a média dos experimentos realizados em
triplicata, sendo considerados estatisticamente significativos *P<0,05 e **P<0,01.
Raízes - EB
SOD sod 1 sod 2 sod1sod20
25
50
75
100
****
**
Linhagens
Sob
revi
vênc
ia (
%)
** ****
Raízes - EC
SOD sod1 sod2 sod1sod20
25
50
75
100
*
Linhagens
Sob
revi
vênc
ia (
%)
H2O2 (100 µL, 1mM)
50 µL extrato + H2O2
100 µL extrato + H2O2
250 µL extrato + H2O2
500 µL extrato + H2O2
Raízes - FB
SOD sod1 sod2 sod1sod20
25
50
75
100
****
** ****
Linhagens
Sob
revi
vênc
ia (
%)
*
**********

66
4.3.2.2 Ensaio antioxidante in vitro
Uma vez que o maior efeito antioxidante, in vivo, foi demonstrado nos extratos
brutos e frações butanólicas de S. chilensis, decidiu-se avaliar a atividade antioxidante in
vitro destes extratos e frações utilizando o ensaio hipoxantina/xantina oxidase. Através
deste método é realizado um monitoramento da produção de ácidos diidroxifenóis (DHBA)
devido ao ataque de ERO ao ácido salicílico (OWEN et al., 1996). A figura 18 mostra uma
significativa atividade antioxidante dos extratos e frações de forma dose-dependente. Com
base neste gráfico, nota-se que todos os extratos e frações testados apresentaram efeito
inibitório na geração de DHBA. Dentre estes, observa-se a maior atividade na FB-folha
(IC50 = 0,728 mg/mL), que reduziu em 78,04 % a formação de ambas as espécies 2,3-
DHBA e 2,5-DHBA na maior concentração (2 mg/mL), seguidos pela FB-flor (IC50 =
0,889 mg/mL), EB-flor (IC50 = 1,87 mg/mL), FB-raiz (IC50 = 1,88 mg/mL), EB-folha (IC50
= 2,42 mg/mL) e EB-raiz (IC50 = 3,15 mg/mL), que na maior concentração reduziram em
73,07 %, 52,34 %, 50,10 %, 40,54 % e 30,77 % a formação de DHBA, respectivamente.
Figura 18 - Inibição da geração de espécies reativas de oxigênio por S. chilensis no
sistema hipoxantina/xantina oxidase.
EB (Extrato Bruto), FB (Fração Butanólica).
0.0 0.5 1.0 1.5 2.0 2.50
25
50
75
100 EB raizEB folhaEB flor
FB folhaFB raiz
FB flor
concentração (mg/mL)
% D
HB
A

67
4.3.3 Atividade antigenotóxica/genotóxica
Devida à freqüente utilização de S. chilensis na medicina popular, ausência de
estudos quanto a sua toxicidade, e pelo extrato aquoso da espécie S. microglossa ter
apresentado atividade mutagênica (RIVERA et al., 1994), surgiu o interesse em investigar
o possível efeito antigenotóxico/genotóxico, in vitro, dos extratos brutos das flores, folhas
e raízes de S. chilensis, já que uma das formas de utilização desta planta pela população é a
infusão. Os resultados referentes ao índice de dano (ID) e freqüência de dano (FD) e
respectivos desvios padrões para os extratos avaliados estão apresentados na Tabela 6.
Tabela 6 - Avaliação da genotoxicidade/antigenotoxicidade dos extratos brutos das flores,
folhas e raízes da S. chilensis.
Tratamento EB-Flores EB-Folhas EB-Raízes Genotoxicidade ID c ID c ID c
CN ª 6,50 ± 2,38 10,75 ± 3,77 8,00 ± 2,94 5 mg/mL 24,50 ± 9,85 7,50 ± 5,74 12,00 ± 3,74 7,5 mg/mL 26,00 ± 18,67 6,00 ± 2,45 13,00 ± 7,75 10 mg/mL 36,50 ± 10,60 * 14,50 ± 8,10 9,50 ± 1,91 FD d FD d FD d CN ª 2,75 ± 0,96 5,75 ± 1,50 5,25 ± 1,50 5 mg/mL 8,50 ± 2,88 4,25 ± 1,89 9,50 ± 4,93 7,5 mg/mL 8,75 ± 5,06 * 3,25 ± 0,96 7,75 ± 4,57 10 mg/mL 11,00 ± 2,16 ** 6,50 ± 4,04 4,00 ± 1,83 Antigenotoxicidade ID c (I%) e ID c ID c H2O2
b 197,75 ± 38,00 226,00 ± 14,00 228,25 ± 10,81 5 mg/mL + H2O2 119,25 ± 29,65 ** (41 %) 218,50 ± 36,39 231,25 ± 10,31 7,5 mg/mL+ H2O2 117,75 ± 17,63 ** (41,8 %) 233,00 ± 31,08 219,50 ± 16,42 10 mg/mL+ H2O2 140,25 ± 17,65 * (30 %) 217,00 ± 26,28 248,50 ± 3,53 FD d FD d FD d H2O2
b 61,25 ± 6,40 63,50 ± 6,86 61,50 ± 4,43 5 mg/mL+ H2O2 41,25 ± 12,28 * 67,75 ± 4,65 61,75 ± 1,71 7,5 mg/mL+ H2O2 33,00 ± 8,28 ** 67,25 ± 8,62 58,00 ± 3,16 10 mg/mL+ H2O2 43,25 ± 3,30 * 58,00 ± 10,54 68,50 ± 0,71
a CN: Controle negativo (solvente PBS); b Controle positivo (H2O2, 0,25 mM). c ID: Índice de dano – de zero (sem dano, 100 células X 0) a 400 (com máximo de dano, 100 X 4). d FD: Freqüência de dano (%), calculada com base no número de células com cauda versos aquelas sem cauda. e I%: percentagem de inibição do ID = [ID do peróxido – ID do extrato com o peróxido] / [ID do peróxido – ID do controle negativo] x 100. Média e desvio padrão obtidos por análise de 100 células por tratamento - total de 2 experimentos para cada extrato. Dados significativos em relação aos controles *P<0,05 e **P<0,01, Teste ANOVA- Dunett.

68
Como pode ser visualizado, os EB das folhas e raízes não são capazes de causar
danos significativos ao DNA das células em estudo, nem apresentaram efeito
antigenotóxico nas concentrações utilizadas neste ensaio. Entretanto, uma significante ação
antigenotóxica do EB das flores é verificada contra danos induzidos por H2O2 em todas as
concentrações testadas (5, 7,5 e 10 mg/mL), sendo os índices e as freqüências de danos
menores do que o controle. Por outro lado, este mesmo extrato demonstrou ação
genotóxica, com valores aumentados no ID (36,50 ± 10,60) e FD (11,00 ± 2,16%) na maior
concentração testada (10 mg/mL), e na dose de 7,5 mg/mL somente na FD (8,75 ± 5,06%).
Com base na tabela apresentada, observa-se um efeito similar do EB das flores na inibição
do ID nas concentrações de 5 e 7,5 mg/mL, produzindo uma redução de dano em relação
ao controle positivo (41 e 41,8 %, respectivamente). Para a maior concentração (10
mg/mL) verificou-se uma menor inibição no ID (30 %) devido aos indícios de
genotoxicidade. Os efeitos não foram doses-dependentes, no entanto, com o aumento da
concentração do extrato, a proteção diminuiu, evidenciando os danos ao DNA. Imagens
das lâminas com as classes de cometas obtidas neste ensaio são apresentadas na figura 19 e
a classificação dos cometas na avaliação genotóxica do EB das flores nas doses testadas
pode ser visualizada na figura 20.
(a) (b)
(c) (d)
Figura 19 – Imagens de lâminas com as diferentes classes de cometas. (a) EB-Folha 5
mg/mL, (b) EB-Folha 5 mg/mL + H2O2, (c) EB-Flor (10 mg/mL) e (d) EB-Flor 10 mg/mL
+ H2O2.

69
CN5
mg/mL 7,5mg/mL 10
mg/mL
Classe 0Classe 1Classe 2
Classe 3Classe 4
0
10
20
30
40
50
60
70
80
90
100
Figura 20 – Classificação dos cometas na avaliação genotóxica do EB das flores nas doses
de 5, 7,5 e 10 mg/mL em comparação ao controle negativo.

70
_________________________________________________________________________
DISCUSSÃO

71
5 DISCUSSÃO
Espécies de Solidago são utilizadas na medicina popular de vários países, no
entanto, há poucos estudos farmacológicos e toxicológicos dos extratos dessas espécies.
Para o gênero Solidago são reportadas atividades antifúngica (PEPELJNJAK et al., 1998;
WANG et al., 2006b), antibacteriana (THIEM & GOSLINSKA, 2002; MOREL et al.,
2006), antioxidante (MEYER et al., 1995; MCCUNE & JOHNS, 2002; APÁTI et al.,
2003; WANG et al., 2006a), mutagênica (RIVERA et al., 1994), antiinflamatória,
espasmolítica e diurética (LEUSCHNER, 1995). Estudos prévios com S. chilensis
demonstraram propriedades antimicrobianas contra alguns tipos de fungos como
Microsporum gypseum, Trichophyton mentagrophytes e Cryptococcus neoformans (VILA
et al., 2002) e contra a bactéria Staphylococcus aureus (DUARTE et al., 2004), além de
atividade antinociceptiva (MOLSKA et al., 2003b), antiinflamatória (GOULART et al.,
2007) e antioxidante in vitro (GÜNTNER et al., 1999). A partir das raízes desta espécie foi
isolado um diterpeno, denominado solidagenona, que apresenta efeito gastroprotetor
(RODRÍGUEZ et al., 2002; SCHMEDA-HIRSCHMANN et al., 2002). Com base nestas
informações, neste estudo realizou-se a análise fitoquímica e avaliou-se as atividades
antibacteriana, antioxidante e antigenotóxica/genotóxica de diferentes extratos e frações
das flores, folhas e raízes de S. chilensis.
Nas flores e folhas os rendimentos dos extratos clorofórmicos são maiores do que
nas raízes por extraírem substâncias mais lipofílicas como ceras, terpenos e clorofila.
Também as frações butanólicas das flores e folhas apresentaram um rendimento maior em
relação às raízes, refletindo a concentração de compostos fenólicos.
Através da análise fitoquímica das diferentes partes de S. chilensis detectou-se a
presença de substâncias com núcleo esteróide e triterpênico, saponinas e flavonóides
(Tabela 4). Além disso, verificou-se que o último metabólito citado apresentou uma
coloração menos intensa nas raízes do que em flores e folhas, sendo que esta observação se
encontra em concordância com os resultados da avaliação do teor de flavonóides totais
(Tabela 5). Comparando os teores de flavonóides totais das flores de S. virgaurea,
determinados pela EUROPEAN PHARMACOPOEIA (2006) e expressos em hiperosídeo

72
com mínimo de 0,5% e máximo de 1,5%, a concentração de flavonóides em S. chilensis foi
superior, entretanto, menor em relação aos teores das flores de S. gigantea e S. canadensis,
que apresentam teor mínimo de 2,5% de flavonóides expressos em hiperosídeo
(EUROPEAN PHARMACOPOEIA, 2006). Cabe salientar que os métodos de extração
para as espécies citadas são diferentes ao utilizado em nosso estudo, e, por isso, podem
interferir nos resultados dos teores totais de flavonóides.
A partir dos processos cromatográficos empregados na fração butanólica das
flores de S. chilensis, foi possível detectar um ácido fenólico (ácido caféico) e cinco
flavonóides conhecidos (campferol, quercetina, quercitrina, hiperosídeo e rutina),
caracterizados por comparação com padrões e pelo fator de retenção (Rf) disponível na
literatura (WAGNER & BLADT, 1996). Acredita-se que a quercitrina seja o flavonóide
majoritário, devido ao tamanho da mancha correspondente. No extrato bruto das flores
também foram detectados por CCD os mesmos flavonóides citados e o ácido clorogênico.
Já nas folhas, a quercetina estava presente na FB e quercitrina, rutina e ácido clorogênico
em ambos os EB e FB, com maior intensidade de cor nas FB. O perfil cromatográfico de
flavonóides de S. chilensis revelou uma composição semelhante àqueles encontrados em
outras espécies de Solidago (IVANCHEVA & VITKOVA, 1997; APÁTI et al., 2002;
KRISTÓ et al., 2002; WU et al., 2007a).
As pesquisas sobre as propriedades antibacterianas de produtos obtidos de plantas
têm sido promissoras quanto ao potencial bactericida que extratos, frações, óleos essenciais
e outros produtos exercem sobre as mais variadas espécies de bactérias (LIMA, 2001). Em
alguns países, certas espécies de Solidago são utilizadas, na medicina popular, na forma de
infusões para o tratamento de infecções (APÁTI et al., 2003; CUNHA et al., 2003). Para a
avaliação da atividade antibacteriana dos diferentes extratos e frações de S. chilensis foram
utilizadas cepas ATCC de Staphylococcus aureus, Enterococcus faecalis, Enterococcus
faecium, Escherichia coli e Klebsiella pneumoniae (amostra clínica). Os resultados obtidos
apontaram que os referidos extratos e frações não foram capazes de inibir o crescimento
bacteriano nas condições empregadas. Em estudo prévio, a literatura relata ação
antibacteriana do extrato hidroalcoólico nas concentrações entre 2,0 - 0,031 mg/mL de S.
chilensis apenas frente à bactéria Staphylococcus aureus tendo apresentado forte inibição

73
(CIM de 0,1mg/mL) através do método de microdiluição, mas não teve efeito contra
Bacillus subtilis, Micrococcus luteus, Staphylococcus epidermides, Rhodococcus equi,
Salmonella choterasuis, Pseudomonas aeruginosa, Streptococcus faecium, Enterococcus
faecium e Escherichia coli (DUARTE et al., 2004), as duas últimas também utilizadas em
nosso trabalho. Em pesquisa realizada por BRASILEIRO e colaboradores (2006),
utilizando o método de difusão do disco, o extrato etanólico das partes aéreas de S.
microglossa também demonstrou atividade contra Staphylococcus aureus nas
concentrações (5, 10 e 20 mg/mL) menores que as utilizadas no presente estudo, mas não
inibiu o crescimento de Escherichia coli. Em outro trabalho, utilizando o método de
microdiluição, o extrato metanólico das raízes desta mesma espécie foi moderadamente
ativo (CIM> 1mg/mL) contra sete bactérias (Staphylococcus epidermidis, Staphylococcus
aureus, Klebsiella pneumoniae, Escherichia coli, Salmonella setubal, Bacillus subtilis e
Pseudomonas aeruginosa) (MOREL et al., 2006). O estudo relatado por THIEM &
GOSLINSKA (2002), empregando o método da difusão do disco, reporta que os extratos
etanólicos e metanólicos de S. virgaurea foram ativos frente Staphylococcus aureus e
Escherichia coli, diferentemente dos resultados encontrados na nossa pesquisa. Desta
maneira, ao contrário do que se verificou para as outras espécies do gênero, o resultado
reportado para S. chilensis foi negativo neste trabalho. Cabe salientar, que apesar de
algumas pesquisas empregarem a mesma metodologia, as cepas, a preparação do extrato,
as concentrações e as plantas são diferentes às utilizadas no nosso estudo.
No presente trabalho, a capacidade antioxidante dos extratos e frações de S.
chilensis foi avaliada por métodos in vitro, técnica da hipoxantina/xantina oxidase, e in
vivo, através do ensaio de sobrevivência com a levedura S. cerevisiae. Esta levedura
produz enzimas antioxidantes para detoxificar ERO, entre as quais superóxido dismutases
(SODCuZn e SODMn), que protegem a célula contra danos oxidativos por catalisar a
dismutação do O2•− para H2O2 (WILMSEN et al., 2005; BAYLIAK et al., 2006). Através
do ensaio de sobrevivência, utilizando linhagens da S. cerevisiae, em fase estacionária de
crescimento, proficiente e deficientes no sistema de defesa antioxidante da enzima
superóxido dismutase (SOD, sod1, sod2, sod1sod2), foram verificadas propriedades
antioxidantes dos extratos e frações de S. chilensis. Esta propriedade foi demonstrada pelo
aumento na sobrevivência da levedura S. cerevisiae (Figuras 15, 16 e 17), independente

74
das linhagens testadas. Os resultados deste ensaio mostraram proteção das células contra
H2O2 quando tratadas, principalmente, com extratos brutos e frações butanólicas das flores
e folhas. A FB das raízes, embora tenha baixo teor de flavonóides, apresenta atividade
antioxidante devido aos outros constituintes presentes na amostra, como saponinas e outros
compostos fenólicos não detectados neste estudo. A partir destes dados, sugere-se que esta
proteção ocorra devido às substâncias antioxidantes presentes nas amostras que
detoxificam ERO, como os ácidos fenólicos e flavonóides, os quais possuem propriedades
seqüestradoras de radicais livres (ARNOUS et al., 2002; RIBEIRO et al., 2005; CAILLET
et al., 2007).
Os extratos clorofórmicos também apresentaram atividade antioxidante contra os
danos induzidos pelo H2O2 (Figuras 15, 16 e 17), verificados principalmente nas linhagens
SOD e sod1, porém, com uma menor atividade comparada aos EB e FB. O EC das raízes
apresentou efeito significativo com a linhagem SOD apenas na dose de 250 µL/mL. No
entanto, é importante notar que, na maior dose do EC das flores foi verificado um efeito
pró-oxidante em todas as linhagens, o que pode indicar que os compostos presentes no
extrato estejam induzindo citotoxicidade e formação de ERO (VALKO et al., 2006).
Considerando que não foi verificada ação pró-oxidante, mesmo nas altas doses dos EB e
FB utilizadas, acredita-se que a ação pró-oxidante observada no EC possa estar relacionada
à presença de compostos com características apolares como algumas classes de terpenos já
identificados em outras espécies de Solidago, os quais são relatados como tóxicos a várias
linhagens celulares (SUNG et al., 1999; CHOI et al., 2004; CAVALCANTI et al., 2006).
Alguns estudos têm indicado que os carotenóides, responsáveis pela coloração das flores,
podem atuar tanto como antioxidantes como pró-oxidantes. O β-caroteno, precursor de
vitamina A, age como antioxidante sob pressões normais de oxigênio, porém, sob pressões
mais altas de oxigênio, perde sua capacidade antioxidante e passa a exercer um efeito pró-
oxidante (PALOZZA et al., 1997). Ácidos fenólicos, como o ácido caféico e ácido
clorogênico, também são reportados como antioxidantes ou pró-oxidantes, pois, sob certas
condições, podem inibir ou acelerar a oxidação de LDL (lipoproteína de baixa densidade)
(YAMANAKA et al., 1997).

75
Verificou-se, neste trabalho, que as linhagens mutantes de S. cerevisiae foram
mais sensíveis ao H2O2 do que a linhagem selvagem (SOD) (Figuras 15, 16 e 17),
indicando que as superóxido dismutases são importantes para proteção celular contra
estresse oxidativo causado por oxidantes exógenos, como o H2O2 (DELLA CROCE et al.,
2003; BAYLIAK et al., 2006). Na linhagem mutante sod1∆sod2∆, a sobrevivência celular
foi menor em relação às outras linhagens, pois, sendo deficiente em ambas as enzimas
SODCuZn e SODMn, esta linhagem está em um estado crônico de estresse oxidativo e,
assim, é mais sensível a ERO, mesmo em baixas concentrações (DELLA CROCE et al.,
2003). No estudo de LUSHCHAK (2006), a SOD da levedura demonstrou a máxima
atividade seguida das mutantes sod2∆ e sod1∆, contudo, a duplo mutante sod1∆sod2∆
também apresentou uma pequena, mas reprodutível atividade. Isto sugere que o efeito de
proteção da sod1∆sod2∆, demonstrado em alguns dos extratos e frações testadas, provém
de outras enzimas presentes na levedura, como as catalases (WILMSEN et al., 2005) e a
glutationa redutase (BAYLIAK et al., 2006), ou da atividade dos compostos dos extratos
da planta.
Nos extratos brutos e frações butanólicas das flores e folhas foram detectados
ácidos fenólicos e flavonóides conhecidos e já relatados em outras espécies de Solidago.
Quanto aos flavonóides detectados por CCD neste trabalho, vários estudos já avaliaram
suas atividades biológicas. A capacidade dos ácidos fenólicos e flavonóides para atuar
como antioxidantes in vitro também tem sido muito estudada nos últimos anos, como a do
ácido caféico (MEYER et al., 1998; OWEN et al., 2000; SOARES, 2002; WU et al.,
2007b), campferol (COS et al., 2001), quercetina, rutina (RATTY & DAS, 1988; COOK &
SAMMAN, 1996; CHOI et a., 2002; CAILLET et al., 2007), quercitrina (RATTY & DAS,
1988; COOK & SAMMAN, 1996; RIBEIRO et al., 2005; YAMAZAKI et al., 2007) e
hiperosídeo (YAMAZAKI et al., 2007).
Os EB e as FB de S. chilensis demonstraram a capacidade de inibir a formação de
espécies reativas, a qual foi avaliada no ensaio in vitro da hipoxantina/xantina oxidase. A
xantina oxidase catalisa a hidroxilação de hipoxantina a xantina e de xantina a ácido úrico,
e em ambas as fases produz também o O2•− a partir do O2, e H2O2 da água. Na presença de
íons ferro, o H2O2 pode originar um dos intermediários mais reativos do oxigênio, o radical

76
OH•, através da reação de Haber-Weiss/Fenton (WILMSEN et al., 2005). Na figura 18, os
resultados mostraram que houve uma significativa redução na formação de ácidos
diidroxibenzóicos (DHBA), produtos da ação do radical OH• com o ácido salicílico, sendo
a atividade antioxidante maior nas FB das folhas e flores do que nos EB e nas raízes. É
interessante comentar que a única FB menos ativa do que o EB foi a das raízes, sugerindo
que sua menor concentração de flavonóides esteja relacionado com o menor efeito
antioxidante, pois nos EB há uma menor concentração destes por estarem diluídos em uma
grande gama de outros constituintes. A partir disso, o efeito antioxidante dos EB e das FB
da S. chilensis, tanto nos ensaios in vivo como in vitro, pode ser atribuído aos compostos
presentes na planta, como ácidos fenólicos e flavonóides que atuam inibindo a formação de
radicais livres. Com base nos resultados obtidos, constata-se a propriedade antioxidante de
S. chilensis, a qual encontra-se em concordância com estudos anteriormente realizados
nesta espécie (GÚNTNER et al., 1999) e em outras espécies de Solidago, como S.
virgaurea (MEYER et al., 1995) e S. canadensis (MCCUNE & JOHNS, 2002; APÁTI et
al., 2003).
Nas plantas, os ácidos fenólicos e os flavonóides participam nos processos
fisiológicos e bioquímicos, durante o desenvolvimento normal e em resposta às condições
de estresse, além de atraírem polinizadores, contribuírem para a pigmentação do vegetal e
atuarem como fitoalexinas, antioxidantes, repelentes, fotoprotetores, entre outros (NACZK
& SHAHIDI, 2004). Os flavonóides apresentam uma significativa atividade antioxidante e
são capazes de reduzir a formação de radicais livres causadores do estresse oxidativo, em
ambientes aquosos e orgânicos, agindo como inibidores de enzimas, quelantes de metais,
seqüestradores e estabilizantes de ERO (ARNOUS et al., 2002; CHOI et al., 2002; APÁTI
et al., 2003). Uma das explicações para a proteção de ERO é a capacidade do flavonóide
de doar átomo de hidrogênio ou elétron aos radicais, finalizando a reação em cadeia
(RICE-EVANS et al., 1997; YANG et al., 2001; VALKO et al., 2006). Esta capacidade de
doar hidrogênio foi demonstrada, como concentração dependente, em várias preparações
investigadas da S. canadensis na presença de DPPH• (APÁTI et al., 2003).
A ingestão regular de compostos fenólicos está associada à baixa incidência de
câncer, doença cardiovascular (VALKO et al., 2006), diabetes (MCCUNE & JOHNS,

77
2002), doenças inflamatórias (SHAPIRO et al., 2007) e outras patologias associadas à
exposição ao estresse oxidativo. Muitas destas ações são atribuídas às propriedades
antioxidantes dos flavonóides e sua possível influência do estado redox intracelular (RICE-
EVANS et al., 1997; PIETTA, 2000; WILMSEN et al., 2005). A revisão de MIDDLETON
JR. e colaboradores (2000) abordou os efeitos dos flavonóides nas células de mamíferos,
que se referem aos efeitos sobre uma variedade de enzimas, na modulação da inflamação,
nas doenças cardiovasculares, nas atividades hepatoprotetora, antiviral, antioxidante,
anticarcinogênica e mutagênica. Em outros estudos, também têm sido relatadas atividades
estrogênica (LEE et al., 1998), antibacteriana, antifúngica (CUSHNIE & LAMB, 2005;
MOREL et al., 2006), antiviral (SEMPLE et al., 1999), antiinflamatória (GARCÍA-
MEDIAVILLA et al., 2007; SHAPIRO et al., 2007), inibidora da lipoxigenase in vitro
(VALKO et al., 2006), anticarcinogênica (YANG et al., 2001; KUO et al., 2004) e efeitos
de proteção contra isquemia (CHANG et al., 2007).
Atualmente, a avaliação de compostos com potencial farmacológico ou
nutracêutico se inicia com um teste de genotoxicidade, pois certas substâncias podem tanto
prevenir quanto induzir lesões no DNA, favorecendo o desenvolvimento de câncer
(KAPISZEWSKA et al., 2005). Radicais livres podem produzir lesões no DNA,
danificando bases, causando quebras simples de cadeia, criando sítios apurínicos e
pirimidínicos (sítios AP) e ligações cruzadas entre DNA e proteínas (LOPES et al., 2004).
Devido a isto e ao evidente efeito antioxidante observado nos EB e FB, avaliou-se neste
trabalho, através do teste cometa alcalino in vitro, uma possível ação antigenotóxica ou
genotóxica dos EB das flores, folhas e raízes. A partir deste ensaio, pôde-se observar que
os extratos das folhas e das raízes, na ausência ou na presença de H2O2, não foram capazes
de induzir atividades genotóxica ou antigenotóxica, pois não houve alteração significativa
no índice e na freqüência de danos nas doses utilizadas (Tabela 6). Além disso, nota-se que
apenas o EB das flores da S. chilensis apresentou significativa ação antigenotóxica, em
todas as doses testadas, contra danos oxidativos induzidos pelo H2O2 nas células em
estudo. Pelo menos em parte, o efeito protetor do extrato contra quebras induzidas por
H2O2 no DNA pode ser atribuído à presença de flavonóides e outros compostos fenólicos,
detectados na análise comatográfica, os quais têm capacidade de seqüestrar ERO gerados

78
na fase aquosa e aumentar a resistência de lipídeos contra peroxidação (VALKO et al.,
2006).
Com base no cálculo de inibição de ID encontrado para EB das flores (30-41%)
(Tabela 6), verifica-se que a capacidade de proteção ao DNA de S. chilensis foi similar a
extratos de plantas da mesma família como C. vesicaria e S. hispanicus (menor de 30%)
(KAPISZEWSKA et al., 2005). Além disso, é possível relacionar a menor inibição de ID
apresentada na maior concentração (10 mg/mL) com o efeito genotóxico anteriormente
verificado para esta concentração, conforme demonstrado na figura 20 em que há um
aumento de cometas de classe 4, com dano máximo ao DNA, e assim, elevam os valores
do índice e freqüência de dano comparados ao controle (Tabela 6). A partir deste ensaio,
verifica-se uma ação direta dos compostos sobre as células, ou seja, apresentam efeito
genotóxico sem necessidade de metabolização.
Embora as flores e folhas apresentem um perfil semelhante de flavonóides, nas
flores uma maior concentração de alguns flavonóides, ácidos fenólicos, ou outros
compostos fenólicos, não detectados neste estudo, podem estar relacionados também às
propriedades pró-oxidantes e genotóxicas observadas nos ensaios com a levedura e no
teste cometa, respectivamente, causado pela autooxidação dos compostos fenólicos que
seriam capazes de formar HO• acumulando danos intermediários no DNA (YAMANAKA
et al., 1997; SILVA et al., 2002; GARCÍA-MEDIAVILLA et al., 2007). Tais
propriedades dependem de fatores como o potencial de redução de metal, pH e das
características de solubilidade (DECKER, 1997; VALKO et al., 2006). Todavia, há
controvérsias sobre os efeitos dos flavonóides sobre o DNA, pois, no estudo de BROWN
& DIETRICH (1979), quercetina e campferol exibiram efeitos mutagênicos, ao passo que
quercitrina e rutina não apresentaram tal efeito. SILVA e colaboradores (2002)
demonstraram, através do teste cometa alcalino in vivo, que a quercetina, em altas doses
(2 x 2500, 2 x 1250 e 2 x 625 mg/kg), apresentou efeitos genotóxicos, e a rutina também
aumentou danos ao DNA na dose 2 x 1250 mg/kg. Já na pesquisa in vitro de
KAPISZEWSKA e colaboradores (2005), quercetina protegeu o DNA contra danos
oxidativos induzidos pelo H2O2 e aumentou a capacidade seqüestradora de ERO de forma

79
dose-dependente, provando que a quercetina exerce proteção em um sítio específico, o
qual está relacionado a sua capacidade de remover ERO.
Como na análise fitoquímica, realizada neste estudo, foi detectada a presença de
saponinas em todas as partes da planta, não se pode desconsiderar a participação desta
classe no efeito genotóxico observado no EB das flores pelo teste cometa. As saponinas
apresentam um grau de toxicidade devido a sua elevada concentração como afirmam VILA
e colaboradores (2002). Além deste relato, estudos prévios com saponinas isoladas de
espécies deste gênero, como giganteasaponinas demonstraram atividade hemolítica in vitro
(REZNICEK et al., 1996), e virgaureasaponinas apresentaram efeito citotóxico in vitro
(BADER et al., 1998) e antitumoral in vivo (PLOHMANN et al., 1997). Assim como as
saponinas, outra classe que pode estar ocasionando efeitos tóxicos no DNA é a dos
diterpenos, comuns em espécies de Solidago, que apresentam efeitos repelentes e
citotóxicos (GONZÁLEZ-COLOMA et al., 2005; RODRÍGUEZ et al., 2005). O diterpeno
ácido caurenóico, presente na S. rugosa (LU et al., 1995), apresentou potencial genotóxico
contra linhagem de células de pulmão de hamster chinês (V79) nas maiores concentrações
testadas no ensaio cometa in vitro (CAVALCANTI et al., 2006). Outros terpenos como o
ent-germacra-4(15),5,10(14)-trien-1β-ol e o β-dictiopterol isolados do extrato metanólico
das partes aéreas de S. virgaurea mostraram moderada citotoxicidade in vitro contra
culturas de linhagens de células de tumor humano (CHOI et al., 2004). Ademais, as folhas,
caules e raízes de S. chilensis e as folhas de S. virgaurea também mostraram atividade
inibitória do crescimento de linhagens de células tumorais humanas nos estudos in vitro
realizados por MONKS e colaboradores (2002a) e GROSS e colaboradores (2002),
respectivamente.
Portanto, os dados deste trabalho indicam que, nas condições testadas, S. chilensis
não apresenta atividade antibacteriana, mas os extratos das flores e folhas possuem
significativa propriedade antioxidante in vivo e in vitro, que parece estar relacionada aos
efeitos antigenotóxicos observados no teste cometa. Com isso, supõe-se que os compostos
fenólicos sejam os responsáveis pela proteção da levedura S. cerevisiae e do DNA à
exposição dos efeitos tóxicos induzidos pelo H2O2. Ainda, verifica-se que compostos mais
lipofílicos estão relacionados ao efeito pró-oxidante; e acredita-se que terpenos, saponinas,

80
outros compostos não caracterizados e até mesmo os flavonóides possam estar
relacionados ao efeito genotóxico. Por outro lado, fatores como interação entre os
compostos dos extratos da planta e a célula, a variada composição dos polifenóis e
compostos terpênicos, e as diferenças nas concentrações de cada composto podem ter um
papel importante nos efeitos pró-oxidantes e genotóxicos das flores. Enfim, este é o
primeiro relato da atividade antioxidante in vivo e antigenotóxica/genotóxica in vitro de S.
chilensis, e considerando que substâncias naturais podem ser responsáveis pela ação
protetora contra os riscos de muitas patologias em que os radicais livres estão implicados,
os resultados descritos neste trabalho estimulam a continuidade dos estudos a fim de
avaliar a ação antioxidante e antigenotóxica de substâncias a serem isoladas da espécie S.
chilensis.

81
_________________________________________________________________________
CONCLUSÕES

82
6 CONCLUSÕES
Nas flores, folhas e raízes de S. chilensis foram detectadas a presença de
substâncias contendo núcleo esteroidal e núcleo triterpênico, além de saponinas e
flavonóides. Os maiores teores de flavonóides foram encontrados nas flores (2,060%) e
folhas (1,938%) em relação às raízes (0,028%).
Na análise cromatográfica da FB das flores foram detectados compostos com
comportamento cromatográfico dos flavonóides campferol, quercetina, quercitrina,
hiperosídeo e rutina, e o ácido fenólico, ácido caféico. No EB das flores também foram
identificados os mesmos flavonóides, além do ácido clorogênico. Na CCD dos EB e FB
das folhas foram detectados quercitrina, rutina e ácido clorogênico, e na FB quercetina.
Nos EB e FB das raízes não foram detectadas estas substâncias.
Extratos e frações (50 mg/mL) de S. chilensis não apresentaram atividade
antibacteriana contra S. aureus, E. faecalis, E. faecium, E. coli e K. pneumoniae através do
método de difusão do disco.
As maiores doses dos EB e FB das flores e folhas mostraram um maior efeito
antioxidante in vivo, do que os EC e as raízes, frente aos danos induzidos por H2O2 em
linhagens de S. cerevisiae. Na maior dose (500 µL) do EC das flores foi verificado um
efeito pró-oxidante em todas as linhagens da levedura.
No ensaio antioxidante in vitro, na maior concentração de 2 mg/mL, a atividade
de redução na formação de DHBA foi maior nas FB das folhas e flores do que nos EB e
nas raízes.
No teste cometa, os EB das folhas e raízes não induziram efeitos genotóxicos ou
antigenotóxicos. O EB das flores apresentou significativa ação antigenotóxica, em todas as
doses testadas (5, 7,5 e 10 mg/mL) e o mesmo extrato induziu genotoxicidade nas maiores
doses testadas.

83
_________________________________________________________________________
7 REFERÊNCIAS

84
7 REFERÊNCIAS
ALICE, C.B., SIQUEIRA, N.C.S., MENTZ, L.A., SILVA, G.A.A.B., JOSÉ, K.F.D. Plantas medicinais de uso popular: atlas farmacognóstico. Canoas (RS): ULBRA, 1995. 205 p. APÁTI, P., HOUGHTON, P.J., GEOFFREY, K., STEVENTON, G.B., KÉRY, A. In-vitro effect of flavonoids from Solidago canadensis extract on glutathione S-transferase. Journal of Pharmacy and Pharmacology, v. 58, p. 251-256, 2006. APÁTI, P., HOUGHTON, P.J., KÉRY, A. HPLC investigation of antioxidant components in Solidago herba. Acta Pharmaceutica Hungarica, v. 74, p. 223-231, 2004. APÁTI, P., SZENTMIHÁLYI, K., BALÁZS, A., BAUMANN, D., HAMBURGER, M., KRISTO, T.S., SZOKE E., KERY, A. HPLC analysis of the flavonoids in pharmaceutical preparations from canadian Goldenrod (Solidago canadensis). Chromatographia, v. 56, p. 65-68, 2002. APÁTI, P., SZENTMIHÁLYI, K., KRISTO, S.T., PAPP, I., VINKLER, P., SZOKE E., KERY, A. Herbal remedies of Solidago - correlation of phytochemical characteristics and antioxidative properties. Journal of Pharmaceutical and Biomedical Analysis, v. 32, p. 1045-1053, 2003. ARNOUS, A., MAKRIS, D.P., KEFALAS, P. Correlation of pigment and flavanol content with antioxidant properties in selected aged regional wines from Greece. Journal of Food Composition and Analysis, v.15, p. 655-665, 2002. BADER, G., WRAY, V., HILLER, K. The main saponins from the aerial parts and the roots of Solidago virgaurea subsp. virgaurea. Planta Medica, v. 61, p. 158-161, 1995. BADER, G., WRAY, V., HILLER, K. Virgaureasaponin 3, a 3,28-bisdesmosidic triterpenoid saponin from Solidago virgaurea. Phytochemistry, v. 31, p. 621-623, 1992. BADER, G., WRAY, V., JUST, U., HILLER, K. Enzymatic hydrolysis of the cytotoxic triterpenoid glycoside virgaureasaponin 1. Phytochemistry, v. 49, p. 153-156, 1998. BARBOSA, W.L.R., QUIGNARD, E., TAVARES, I.C.C., PINTO, L.N., OLIVEIRA, F.Q., OLIVEIRA, R.M. Manual para análise fitoquímica e cromatográfica de extratos vegetais. Revista Científica da UFPA, v. 4, 2004. Disponível em http://www.ufpa.br/rcientifica: Acesso em: 10/09/2006. BARROSO, G.M., PEIXOTO, A.L., COSTA, C.G., ICHASO, C.L.F., GUIMARÃES, E.F., LIMA, H.C. Ordem 9 – Asterales. In: BARROSO, G.M. (Ed.). Sistemática de Angiospermas do Brasil. Viçosa (MG): Imprensa Universitária, 1991. v.3. BASARAN, A.A., YU, T.W., PLEWA, M.J., ANDERSON, D. An investigation of some Turkish herbal medicines in Salmonella typhimurium and in the Comet assay in human lymphocytes. Teratogenesis Carcinogenesis and Mutagenesis, v.16, p. 125-138, 1996.

85
BAYLIAK, M., SEMCHYSHYN, H., LUSHCHAK, V. Effect of hydrogen peroxide on antioxidant enzyme activities in Saccharomyces cerevisiae is strain-specific. Biochemistry, v. 71, p. 1013-1020, 2006. BOEIRA, J.M., VIANA, A.F., PICADA, J.N., HENRIQUES, J.A.P. Genotoxic and recombinogenic activities of the two β-carboline alkaloids harman and harmine in Saccharomyces cerevisiae. Mutation Research, v. 500, p. 39-48, 2002. BONGARTZ, D., HESSE, A. Selective extraction of quercetrin in vegetable drugs and urine by off-line coupling of boronic acid affinity chromatography and high-performance liquid chromatography. Journal of Chromatography B, v. 673, p. 223-230, 1995. BORCHERT, V.E., CZYBORRA P., FETSCHER, C., GOEPEL M., MICHEL, M.C. Extracts from Rhois aromatica and Solidaginis virgaurea inhibit rat and human bladder contraction. Naunyn-Schmiedeberg’s Archives Pharmacology, v. 369, p. 281-286, 2004. BRASILEIRO, B.G., PIZZIOLO, V.R., RASLAN, D.S., JAMAL, C.M., SILVEIRA, D. Antimicrobial and cytotoxic activities screening of some Brazilian medicinal plants used in Governador Valadares district. Revista Brasileira de Ciências Farmacêuticas, v. 42, p. 195-202, 2006. BRENDLER-SCHWAAB, S., HARTMANN, A., PFUHLER, S., SPEIT, G. The in vivo comet assay: use and status in genotoxicity testing. Mutagenesis, v. 20, p. 245-254, 2005. BROWN, J.P., DIETRICH, P.S. Mutagenicity of plant flavonols in the Salmonella/mammalian microsome test Activation of flavonol glycosides by mixed glycosidases from rat cecal bacteria and other sources. Mutation Research/Genetic Toxicology, v. 66, p. 223-240, 1979. BURKE, D., DAWSON, D., STEARNS, T. Methods in yeast genetics: A Cold Spring Harbor Laboratory Course Manual. New York: CSH Laboratory Press, p. 171-205, 2000. CAI, Y., LUO, Q., SUN, M., CORKE, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Science, v. 74, p. 2157-2184, 2004. CAILLET, S., YU, H., LESSARD, S., LAMOUREUX, G., AJDUKOVIC, D., LACROIX, M. Fenton reaction applied for screening natural antioxidants. Food Chemistry, v. 100, p. 542-552, 2007. CALIXTO, J.B. Twenty-five years of research on medicinal plants in Latin America: A personal view. Journal of Ethnopharmacology, v. 100, p. 131–134, 2005. CANTRELL, C.L., FISCHER, N.H., URBATSCH, L., MCGUIRE, M.S., FRANZBLAU, S.G. Antimycobacterial crude plant extracts from South, Central, and North America. Phytomedicine, v. 5, p. 137-145, 1998.

86
CAVALCANTI, B.C., COSTA-LOTUFO, L.V., MORAES, M.O., BURBANO, R.R., SILVEIRA, E.R., CUNHA, K.M.A., RAO, V.S.N., MOURA, D.J., ROSA, R.M., HENRIQUES, J.A.P., PESSOA, C. Genotoxicity evaluation of kaurenoic acid, a bioactive diterpenoid present in Copaiba oil. Food and Chemical Toxicology, v. 44, p. 388-392, 2006. CELEGHINI, R.M.S., VILEGAS, J.H.Y., LANÇAS, F.M. Extraction and Quantitative HPLC analysis of coumarin in hydroalcoholic extracts of Mikania glomerata Spreng: ("guaco") leaves. Journal of the Brazilian Chemical Society, v. 12, p.706-709, 2001. CHANG, W.T., SHAO, Z.H., YIN, J.J., MEHENDALE, S., WANG, C.Z., QIN, Y., LI, J., CHEN, W.J., CHIEN, C.T., BECKER, L.B., HOEK, T.LV., YUAN, C.S. Comparative effects of flavonoids on oxidant scavenging and ischemia-reperfusion injury in cardiomyocytes. European Journal of Pharmacology, v. 566, p. 58-66, 2007. CHATURVEDULA, V.S., ZHOU, B.N., GAO, Z., THOMAS, S.J., HECHT, S.M., KINGSTON, D.G. New lupane triterpenoids from Solidago canadensis that inhibit the lyase activity of DNA polymerase beta. Bioorganic & Medicinal Chemistry, v. 12, p. 6271-6275, 2004.
CHODERA, A., DABROWSKA, K., SLODERBACH, A., SKRZYPCZAK, L., BUDZIANOWSKI, J. Effect of flavonoid fractions of Solidago virgaurea L on diuresis and levels of electrolytes. Acta Poloniae Pharmaceutica, v. 48, p. 35-7, 1991. CHOI, C.W., KIM, S.C., HWANG, S.S., CHOI, B.K., AHN, H.J., LEE, M.Y., PARK, S.H., KIM, S.K. Antioxidant activity and free radical scavenging capacity between Korean medicinal plants and flavonoids by assay-guided comparison. Plant Science, v. 163, p. 1161-1168, 2002. CHOI , S.Z., CHOI, S.U., BAE, S.Y, PYO, S., LEE, K.R. Immunobiological activity of a new benzyl benzoate from the aerial parts of Solidago virga-aurea var. gigantea. Archives of Pharmacal Research, v. 28, p. 49-54, 2005. CHOI, S.Z., CHOI, S.U., LEE, K.R. Phytochemical constituents of the aerial parts from Solidago virga-aurea var. gigantea. Archives of Pharmacology Research, v. 27, p. 164-168, 2004. COELHO, M.G.P., REIS, P.A., GAVA, V.B., MARQUES, P.R., GAYER, C.R., LARANJA, G.A.T., FELZENSWALB, I., SABINO, K.C.C. Anti-arthritic effect and subacute toxicological evaluation of Baccharis genistelloides aqueous extract. Toxicology Letters, v. 154, p. 69-80, 2004. COOK, N.C., SAMMAN, S. Flavonoids - Chemistry, metabolism, cardioprotective effects, and dietary sources. Nutritional Biochemistry, v. 7, p. 66-76, 1996. CORREIA, E. Aspectos da propagação sexuada e vegetativa da arnica brasileira (Solidago chilensis Meyen, Asteraceae). In: MING, L.C., SCHEFFER, M.C., CORRÊA JÚNIOR, C., BARROS, I.B.I., MATTOS, J.K.A. (Eds.). Plantas medicinais aromáticas e

87
condimentares: avanços na pesquisa agronômica. Botucatu (SP): UNESP, p. 193-195, 1998. v. 2. COS, P., CALOMME, M., SINDAMBIWE, J.B., DE BRUYNE, T., CIMANGA, K., PIETERS, L., VLIETINCK, A.J., BERGHE, D.V. Cytotoxicity and lipid peroxidation-inhibiting activity of flavonoids. Planta Medica, v. 67, p. 515-519, 2001. COSTA, V., FERREIRA, P.M. Oxidative stress and signal transduction in Saccharomyces cerevisiae: insights into ageing, apoptosis and diseases. Molecular Aspects of Medicine, v. 22, p. 217-246, 2001. CUNHA, A.P., SILVA, A.P., ROQUE, O.R. Plantas e produtos vegetais em fitoterapia. Lisboa: Fundação Calouste Gulbenkian, 2003. 702 p. CUSHNIE, T.P.T., LAMB, A.J. Antimicrobial activity of flavonoids. International Journal of Antimicrobial Agents, v. 26, p. 343-356, 2005. DECKER, E.A. Phenolics: prooxidants or antioxidants? Nutrition Review, v. 55, p. 396-407, 1997. DELLA CROCE, C., BRONZETTI, G., CINI, M., CALTAVUTURO, L., POI, G. Protective effect of lipoic acid against hydrogen peroxide in yeast cells. Toxicology in Vitro, v. 17, p. 753-759, 2003. DILLON, M.O., ALVA, A.S. Tribal classification and diversity in the Asteraceae of Peru. Arnaldoa, v. 8, p. 25-44, 2001. Disponível em http://www.sacha.org/Peru_Asteraceae/Peru_Tribes.htm: Acesso em: 07/04/2006. DUARTE, M.C.T., FIGUEIRA, G.M., PEREIRA, B., MAGALHÃES, P.M. DELARMELINA, C. Atividade antimicrobiana de extratos hidroalcólicos de espécies da coleção de plantas medicinais CPQBA/UNICAMP. Revista Brasileira de Farmacognosia, v. 14, p. 06-08, 2004. ENGELHARDT, J.F. Redox-mediated gene therapies for environmental injury: approaches and concepts. Antioxidants & Redox Signaling, v. 1, p. 5-27, 1999. EUROPEAN PHARMACOPOEIA. 5 ed. Strasbourg: Council of Europe, 2006. FAIRBAIRN, D.W., OLIVE, P.L., O'NEILL, K.L. The comet assay: a comprehensive review. Mutation Research, v. 339, p. 37-59, 1995. FALKENBERG, M.B., SANTOS, R.I., SIMÕES, C.M.O. Introdução à análise fitoquímica. In: SIMÕES, C.M.O., SCHENKEL, E.P., GOSMANN, G., MELLO, J.C.P., MENTZ, L.A., PETROVICK, P.R. (Eds.). Farmacognosia: da planta ao medicamento. 2.ed. Porto Alegre/Florianópolis: Ed. Universidade UFRGS/ Ed. UFSC, p. 163-179, 2000. FARMACOPÉIA BRASILEIRA. Parte I. 4 ed. São Paulo: Atheneu Editora, 1988.

88
FARMACOPÉIA BRASILEIRA. Parte II. 4º fascículo. 4 ed. São Paulo: Atheneu Editora, 2003. FEROW, C.W., AVILA, J.R. The complete guide to herbal medicines. Springhouse (PA): Springhouse Corporation, 2000. 618 p. GARCÍA-MEDIAVILLA, V., CRESPO, I., COLLADO, P.S., ESTELLER, A., SÁNCHEZ-CAMPOS, S., TUÑÓN, M.J., GONZÁLEZ-GALLEGO, J. The anti-inflammatory flavones quercetin and kaempferol cause inhibition of inducible nitric oxide synthase, cyclooxygenase-2 and reactive C-protein, and down-regulation of the nuclear factor kappa B pathway in Chang Liver cells. European Journal of Pharmacology, v. 557, p. 221-229, 2007. GONZÁLEZ-COLOMA, A., GUADAÑO, A., TONN, C.E., SOSA, M.E. Antifeedant/insecticidal terpenes from Asteraceae and Labiatae species native to Argentinean semi-arid lands. Zeitschrift Für Naturforschung, v. 60, p. 855-861, 2005. GOULART, S., MORITZ, M.I.G., LANG, K.L., LIZ, R., SCHENKEL, E.P., FRÖDE, T.S. Anti-inflammatory evaluation of Solidago chilensis Meyen in a murine model of pleurisy. Journal of Ethnopharmacology, v. 113, p. 346–353, 2007. GRANT, C.M., PERRONE, G., DAWES, I.W. Glutathione and catalase provide overlapping defenses for protection against hydrogen peroxide in the yeast Saccharomyces cerevisiae. Biochemical and Biophysical Research Communications, v. 253, p. 893-898, 1998. GRESSLER, V., DIAS, G.O.C., SIMIONATTO, E., DESSOY, E.C.S.M., MOREL, A.F. Isolamento de sesquiterpeno a partir do óleo volátil de Solidago microglossa D.C. In: 11 ENCONTRO DE QUÍMICA DA REGIÃO SUL, 2003, Pelotas. Resumos dos trabalhos apresentados no 11 Encontro de Química da Região Sul. Pelotas: SBQSUL, 2003. p. 100. GROSS, S.C., GOODARZI, G., WATABE, M., BANDYOPADHYAY, S., PAI, S.K., WATABE ,, K. Antineoplastic activity of Solidago virgaurea on prostatic tumor cells in an SCID mouse model. Nutrition and Cancer, v. 43, p. 76-81, 2002. GÜNTNER, C., BARRA, C., CESIO, M.V., DELLACASSA, E., FERRANDO, L., FERREIRA, F., GARCÍA, C., GONZÁLEZ, G., HEINZEN, H., LLORET, A., LORENZO, D., MENÉNDEZ, P., PAZ, D., SOULE, S., VÁZQUEZ, A., MOYNA, P. Antioxidant properties of Solidago chilensis L. flavonoids. In: II WOCMAP CONGRESS MEDICINAL AND AROMATIC PLANTS, 1999, Mendoza. Resumos dos trabalhos II WOCMAP Congress Medicinal and Aromatic Plants. Mendoza: Acta Horticulturae (ISHS) 501, 1999. 159-164. Disponível em http://www.actahort.org/books/501/501.23.htm: Acesso em: 07/04/2006. HALLIWELL, B., GUTTERIDGE, J.M.C. Free radicals in biology and medicine. 3. ed. New York: Oxford University Press, 1999. 936 p.

89
HARBORNE, J.B. Phytochemical Methods: a guide to modern techniques of plant analysis. 2. ed. London: Chapman and Hall, 1984. 288 p. HARTMANN, A., SCHUMACHER, M., PLAPPERT-HELBIG, U., LOWE, P., SUTER, W., MUELLER, L. Use of the alkaline in vivo Comet assay for mechanistic genotoxicity investigations. Mutagenesis, v. 19, p. 51-59, 2004. HEEMANN, A.C., MIGUEL, O.G., MIGUEL, M.D. Revisão do gênero Pterocaulon - Aspectos fitoquímicos e atividades biológicas. Visão Acadêmica, v. 5, p. 53-60, 2004. http://botanical.com/botanical/mgmh/g/golrod26.html: Acesso em: 18/07/2006. http:// pubchem.ncbi.nlm.nih.gov/summary.cgi?cid=441955: Acesso em: 28/08/2007. INOGUCHI, M., OGAWA, S., FURUKAWA, S., KONDO, H. Production of an allelopathic polyacetilene in hairy root cultures of goldenrod (Solidago altissima L.). Bioscience Biotechnology Biochemistry, v. 67, p. 863-868, 2003. ITO, I., KOBAYASHI, K., YONEYAMA, T. Fate of dehydromatricaria ester added to soil and its implications for the allelopathic effect of Solidago altissima L. Annals of Botany, v. 82, p. 625-630, 1998. IVANCHEVA, S., VITKOVA, A. Flavonoids from Solidago virgaurea L. Pharmacia, v. 44, p. 17-19, 1997. KAPISZEWSKA, M., SOŁTYS, E., VISIOLI, F., CIERNIAK, A., ZAJAC, G. The protective ability of the Mediterranean plant extracts against the oxidative DNA damage. The role of the radical oxygen species and the polyphenol content. Journal of Physiology and Pharmacology, v. 56, p. 183-197, 2005. KASALI, A.A., EKUNDAYO, O., PAUL, C., KONIG, W.A. Epi-Cubebanes from Solidago canadensis. Phytochemistry, v. 59, p. 805-810, 2002. KRISTÓ, S.T., GANZLER, K., APÁTI, P., SZOKE, É., KÉRY, Á. Analysis of antioxidant flavonoids from Asteraceae and Moraceae plants by capillary electrophoresis. Chromatographia, v. 56, p. 121-126, 2002. KUMAR, R.S., SIVAKUMAR, T., SUNDERAM, R.S., GUPTA, M., MAZUMDAR, U.K., GOMATHI, P., RAJESHWAR, Y., SARAVANAN, S., KUMAR, M.S., MURUGESH, K., KUMAR, K.A. Antioxidant and antimicrobial activities of Bauhinia racemosa L. stem bark. Brazilian Journal of Medical and Biological Research, v. 38, p. 1015-1024, 2005. KUO, P.C., LIU, H.F., CHAO, J.I. Survivin and p53 modulate quercetin-induced cell growth inhibition and apoptosis in human lung carcinoma cells. The Journal of Biological Chemistry, v. 279, p. 55875-55885, 2004.

90
LEE, J.S., CHUNG, H.Y., MAIER, C.G.A., WOOD, A.R., DIXON, R.A., MABRY, T.J. Estrogenic flavonoids from Artemisia vulgaris L. Journal of Agricultural and Food Chemistry, v. 46, p. 3325-3329, 1998. LEUSCHNER, J. Anti-inflammatory, spasmolytic and diuretic effects of a commercially available Solidago gigantea herb extract. Arzneimittel Forschung Drug Research, v. 45, p. 165-168, 1995. LIMA, E.O. Plantas e suas propriedades antimicrobianas: uma breve análise histórica. In: YUNES, R.A., CALIXTO, J.B. (Eds.). Plantas medicinais sob a ótica da moderna química. Chapecó (SC): Argos, p. 482-501, 2001. LOPES, M.I.L., SAFFI, J., ECHEVERRIGARAY, S., HENRIQUES, J.A.P., SALVADOR, M. Mutagenic and antioxidant activities of Croton lechleri sap in biological systems. Journal of Ethnopharmacology, v. 95, p. 437-445, 2004. LORENZI, H., MATOS, F.J.A. Plantas Medicinais no Brasil: nativas e exóticas cultivadas. Nova Odessa, SP: Instituto Plantarum, 2002. 512 p. LU, T., VARGAS, D., FRANZBLAU, S.G., FISCHER, N.H. Diterpenes from Solidago rugosa. Phytochemistry, v. 38, p. 451-456, 1995. LUSHCHAK, V.I. Budding yeast Saccharomyces cerevisiae as a model to study oxidative modification of proteins in eukaryotes. Acta Biochimica Polonica, v. 53, 679-684, 2006. LUSHCHAK, V.I., GOSPODARYOV, D.V. Catalases protect cellular proteins from oxidative modification in Saccharomyces cerevisiae. Cell Biology International, v. 29, p. 187-192, 2005. LUSHCHAK, V.I., SEMCHYSHYN, H., LUSHCHAK, O., MANDRYK, S. Diethyldithiocarbamate inhibits in vivo Cu,Zn-superoxide dismutase and perturbs free radical processes in the yeast Saccharomyces cerevisiae cells. Biochemical and Biophysical Research Communications, v. 338, p.1739–1744, 2005. MACIEL, M.A.M., PINTO, A.C., VEIGA JR., V.F. Plantas medicinais: a necessidade de estudos multidisciplinares. Química Nova, v. 15, p. 429-438, 2002. MALUF, S.W., ERDTMANN, B. Biomonitorização do dano genético em humanos. In: SILVA, J., ERDTMANN, B., HENRIQUES, J.A.P. (Eds.). Genética Toxicológica. Porto Alegre: Alcance, p. 193-194, 2003. MARINOFF, M.A., ZAGO, G.L., PZOCIK, H.J., CHIFA, C., GIMÉNEZ, M.C. Contribucion al conocimiento de la actividad farmacologica de Medicago sativa L. (Fabaceae), “alfalfa”. Comunicaciones Científicas y Tecnológicas, Universidad Nacional del Nordeste, 2005. Disponível em http://www.unne.edu.ar: Acesso em: 20/09/2006. MARIS, A.F., KERN, A.L., PICADA, J.N., BOCCARDI, F., BRENDEL, M., HENRIQUES, J.A.P. Glutathione, but not transcription factor Yap1, is required for carbon

91
source-dependent resistance to oxidative stress in Saccharomyces cerevisiae.Current Genetics, v. 37, p. 175-182, 2000. MCCUNE, L.M., JOHNS, T. Antioxidant activity in medicinal plants associated with the symptoms of diabetes mellitus used by the Indigenous Peoples of the North American boreal forest. Journal of Ethnopharmacology, v. 82, p. 197-205, 2002. MEYER, B., SCHNEIDER, W., ELSTNER, E. F. Antioxidative properties of alcoholic extracts from Fraxinus excelsior, Populus tremula and Solidago virgaurea Arzneimittel Forschung Drug, v. 45, p. 174-176, 1995. MEYER, A.S., HEINONEN, M., FRANKE, E.N. Antioxidant interactions of catechin, cyanidin, caffeic acid, quercetin, and ellagic acid on human LDL oxidation. Food Chemistry, v. 61, p. 71-75, 1998. MICHELIN, D.C., MORESCHI, P.E., LIMA, A.C., NASCIMENTO, G.G.F., PAGANELLI, M.O., CHAUD, M.V. Avaliação da atividade antimicrobiana de extratos vegetais. Revista Brasileira de Farmacognosia, v. 15, p. 316-320, 2005. MIDDLETON JR., E., KANDASWAMI, C., THEOHARIDES, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacological Reviews, v. 52, p. 673-751, 2000. MOLSKA, G.R., ZANONI, C.I.S., PARADA, C.A., MALPEZZI-MARINHO, E.L.A., MARINHO, E.A.V. Efeito do extrato hidroalcóolico de Solidago chilensis (EHSc) sobre a hipernocicepção mecânica aguda induzida por prostaglandina E2. In: XXXV CONGRESSO BRASILEIRO DE FARMACOLOGIA- SBFTE, 2003, Águas de Lindóia. Resumos de trabalhos apresentados no XXXV Congresso Brasileiro de Farmacologia- SBFTE, Águas de Lindóia, 2003, p. 28. (a). Disponível em http://www.sbfte.org.br/Setor08.pdf: Acesso em: 18/06/2007. MOLSKA, G.R., ZANONI, C.I.S., PAULA-FREIRE, L.I.G., JIMENEZ, R.S., CATENACI, V.K., GRACIA, V.P., MALPEZZI-MARINHO, E.L.A., MARINHO, E.A.V. Atividade antinociceptiva de extrato hidroalcoólico de Solidago chilensis Meyen (Compositae). In: XXXV CONGRESSO BRASILEIRO DE FARMACOLOGIA- SBFTE, 2003, Águas de Lindóia. Resumos de trabalhos apresentados no XXXV Congresso Brasileiro de Farmacologia- SBFTE, Águas de Lindóia, 2003, p. 49. (b). Disponível em http://www.sbfte.org.br/Setor08.pdf: Acesso em: 18/06/2007. MONKS, N.R., BORDIGNON, S., FERRAZ, A., MACHADO, K.R., FARIA, D.H., LOPES, R.M., MONDIN, C.A., SOUZA, I.C.C., LIMA, M.F.S., ROCHA, A.B., SCHWARTSMANN, G. Anti-tumour Screening of Brazilian Plants. Pharmaceutical Biology, v. 40, p 1-14, 2002. (a) MONKS, N.R., FERRAZ, A., BORDIGNON, S., MACHADO, K.R., LIMA, M.F.S., ROCHA, A.B., SCHWARTSMANN, G. In vitro cytotoxicity of extracts from Brazilian Asteraceae. Pharmaceutical Biology, v. 40, p 1-6, 2002. (b)

92
MOREL, A.F., DIAS, G.O., PORTO, C., SIMIONATTO, E., STUKER, C.Z., DALCOL, I.I. Antimicrobial activity of extractives of Solidago microglossa. Fitoterapia, v. 77, p. 453-455, 2006. NACZK, M., SHAHIDI, F. Extraction and analysis of phenolics in food. Journal of Chromatography A, v. 1054, p. 95-111, 2004. NADIN, S.B., VARGAS-ROIG, M.L., CIOCCA, D.R. A silver staining for single-cell gel assay. Journal of Histochemistry Cytochemistry, v. 49, p. 1183-1186, 2001. NCCLS. Performance Standards of Antimicrobial Disk Susceptibility Testing. Fourteenth edition. NCCLS document M100-514. NCCLS, Wayne, PA, USA, 2004.
NOELLI, F.S. Múltiplos usos de espécies vegetais pela farmacologia Guarani através de informações históricas. In: 1 SIMPÓSIO DE ETNOBIOLOGIA E ETNOECOLOGIA, 1996, Feira de Santana. Resumos de trabalhos apresentados no 1 Simpósio de Etnobiologia e Etnoecologia. Feira de Santana: Universidade Estadual de Feira de Santana, 1996. Disponível em http://www.dhi.uem.br/publicacoesdhi/dialogos/volume01/Revista%20Dialogos/DI%C1LOGOS10.doc: Acesso em: 18/05/2006. NOLDIN, V.F., ISAIAS, D.B., CECHINEL FILHO, V. Gênero Calophyllum: Importância química e farmacológica. Quimica Nova, v. 29, p. 549-554, 2006. OLIVEIRA, S.Q., TRENTIN, V.H., KAPPEL, V.D., BARELLI, C., GOSMANN, G., REGINATTO, F.H. Screening of antibacterial activity of South Brazilian Baccharis species. Pharmaceutical Biology, v. 43, p. 434-438, 2005. OPLUSTIL, C.P., ZOCCOLI, C.M., TOBOUTI, N.R., SINTO, S.I. Procedimentos básicos em microbiologia clínica. São Paulo: Sarvier, 2000. 254 p. OWEN, R.W., MIER, W., GIACOSA, A., HULL, W.E., SPIEGELHALDER, B., BARTSCH, H. Phenolic compounds and squalene in olive oils: the concentration and antioxidant potencial of total phenols, simple phenols, secoiridoids, lignans and squalene. Food and Chemical Toxicology, v. 38, p. 647-659, 2000. OWEN, R.W., WIMONWATWATEE, T., SPIEGELHALDER, B., BARTSCH, H. A high performance liquid chromatography method for quantification of hydroxyl radical formation by determination of dihydroxy benzoic acids. European Journal of Cancer Prevention, v. 5, p. 233-240, 1996. PALOZZA, P., LUBERTO, C., CALVIELLO, G., RICCI, P., BARTOLI, G.M. Antioxidant and prooxidant role of beta-carotene in murine normal and tumor thymocyte: Effects of oxygen partial pressure. Free Radical Biology and Medicine, v. 22, p. 1065-1073, 1997.

93
PEPELJNJAK, S., KUSTRAK, D., VUKUSIC, I. Investigation of the antimycotic activity of Solidago virgaurea and Solidago gigantea extracts. Pharmaceutical and Pharmacological Letters, v. 8, p. 85-86, 1998. PEREIRA, M.D., HERDEIRO, R.S., FERNANDES, P.N., ELEUTHERIO, E.C.A., PANEK, A.D. Targets of oxidative stress in yeast sod mutants. Biochimica et Biophysica Acta, v. 1620, p. 245-251, 2003. PEREIRA, B.K., PEREIRA, A.B., GROFF, A.A., SCHRÖDER-PFEIFER, N.T., SILVA, J. Avaliação do efeito fotoprotetivo de Polytrichum juniperinum Hedw., Colobanthus quitensis (Kunth.) bartl. e Deschampsia antarctica Desv. através de ensaio cometa em Helix aspersa (Müller, 1774). Revista de Iniciação Científica da ULBRA, v. 3, p. 73-78, 2005. PICADA, J.N., KERN, A.L., RAMOS, A.L.L.P, SAFFI, J. O estresse oxidativo e as defesas antioxidantes. In: SILVA, J., ERDTMANN, B., HENRIQUES, J.A.P. (Eds.). Genética Toxicológica. Porto Alegre: Alcance, p. 251-268, 2003. PIETTA, P.G. Flavonoids as Antioxidants. Journal of Natural Products, v. 63, p. 1035-1042, 2000. PLOHMANN, B., BADER, G., HILLER, K., FRANZ, G. Immunomodulatory and antitumoral effects of triterpenoid saponins. Pharmazie, v. 52, p. 953-957, 1997. PROSSER, I., PHILLIPS, A.L., GITTINGS, S., LEWIS, M.J., HOOPER, A.M., PICKETT, J.A., BEALE, M.H. (+)-(10R)-Germacrene A synthase from goldenrod, Solidago canadensis; cDNA isolation, bacterial expression and functional analysis. Phytochemistry, v. 60, p. 691-702, 2002. RANGEL, D., GARCÍA, I., VELASCO. J., BUITRAGRO, D., VELAZCO, E. Actividad antimicrobiana de los extractos etanólico, acetónico y acuoso de Baccharis nitida (Ruiz et Pavon) Pers. Revista de la Facultad de Farmácia, v. 42, p. 43-46, 2001. RASPOR, P., PLESNICAR, S., GAZDAG, Z., PESTI, M., MIKLAVCIC, M., LAH, B., LOGAR-MARINSEK, R., POLJSAK, B. Prevention of intracellular oxidation in yeast: the role of vitamin E analogue, Trolox (6-hydroxy-2,5,7,8-tetramethylkroman-2-carboxyl acid). Cell Biology International, v. 29, p. 57-63, 2005. RATTY, A.K., DAS, N.P. Effects of flavonoids on nonenzymatic lipid peroxidation: Structure-activity relationship. Biochemical Medicine and Metabolic Biology, v. 39, p. 69-79, 1988. REZNICEK, G., FREILER, M., SCHADER, M., SCHMIDT, U. Determination of the content and the composition of the main saponins from Solidago gigantea AIT. using high-performance liquid chromatography. Journal of Chromatography A, v. 755, p. 133-137, 1996.

94
REZNICEK, G., JURENITSCH, J., PLASUN, M., KORHAMMER, S., HASLINGER, E., HILLER, K., KUBELKA, W. Four major saponins from Solidago canadensis. Phytochemistry, v. 30, p. 1629-1633, 1991. RIBEIRO, A.B., BOLZANI, V.S., YOSHIDAB, M., SANTOS, L.S., EBERLIN, M.N., SILVA, D.H.S. A new neolignan and antioxidant phenols from Nectandra grandiflora. Journal of the Brazilian Chemical Society, v. 16, p. 526-530, 2005. RICE-EVANS, C.A., MILLER, N.J., PAGANGA, G. Antioxidant properties of phenolic compounds. Trends in Plant Science, v. 2, p. 152-159, 1997. RIVERA, I.G., MARTINS, M.T., SANCHEZ, P.S., SATO, M.I.Z., COELHO, M.C.L., AKISUE, M., AKISUE, G. Genotoxicity assessment through the Ames test of medicinal plants commonly used in Brazil. Environmental Toxicology and Water Quality, v. 9, p. 87-93, 1994. RODRÍGUEZ, J.A., BUSTAMANTE C., ASTUDILLO, L., SCHMEDA-HIRSCHMANN, G. Gastroprotective activity of solidagenone on experimentally induced gastric lesions in rats. Journal of Pharmacy and Pharmacology, v. 54, p. 399-404, 2002. RODRÍGUEZ, J.A., THEODULOZA, C., SÁNCHEZ, M., RAZMILICB, I., SCHMEDA-HIRSCHMANN, G. Gastroprotective and ulcer-healing effect of new solidagenone derivatives in human cell cultures. Life Sciences, v. 77, p. 2193–2205, 2005. ROSA, R.M., MELECCHI, M.I.S., HALMENSCHLAGER, R.C., ABAD, F.C., SIMONI, C.R., CARAMÃO, E.B., HENRIQUES, J.A.P., SAFFI, J., RAMOS, A.L.L.P. Antioxidant and antimutagenic properties of Hibiscus tiliaceus L. methanolic extract. Journal of Agricultural and Food Chemistry, v. 54, p. 7324-7330, 2006. SANG, Z.C., SANG, U.C., SEONG, Y.B., SUHK, N.P., KANG, R.L. Immunobiological activity of a new benzyl benzoate from the aerial parts of Solidago virga-aurea var. gigantea. Archives of Pharmacal Research, v. 28, p. 49-54, 2005. SCHMEDA-HIRSCHMANN, G., JORDAN, M., GERTH, A., WILKEN, D. Secondary metabolite content in rhizomes, callus cultures and in vitro regenerated plantlets of Solidago chilensis. Zeitschrift Für Naturforschung, v. 60, p. 5-10, 2005. SCHMEDA-HIRSCHMANN, G., RODRÍGUEZ, J., ASTUDILLO, L. Gastroprotective activity of the diterpene solidagenone and its derivatives on experimentally induced gastric lesions in mice. Journal of Ethnopharmacology, v. 81, p. 111-115, 2002. SCHMIDT, C.O., BOUWMEESTER, H.J., BÜLOW, N., KÖNIG, W.A. Isolation, characterization, and mechanistic studies of (-)-α-gurjunene synthase from Solidago canadensis. Archives of Biochemistry and Biophysics, v. 364, p. 167-177, 1999. SEMPLE, S.J., NOBBS, S.F., PYKE, S.M., REYNOLDS, G.D., FLOWER, R.L.P. Antiviral flavonoid from Pterocaulon sphacelatum, an Australian Aboriginal medicine. Journal of Ethnopharmacology, v. 68, p. 283-288, 1999.

95
SHAPIRO, H., SINGER, P., HALPERN Z., BRUCK, R. Polyphenols in the treatment of inflammatory disease and acute pancreatitis. Gut, v. 56, p. 426-436, 2007. SIMÕES, C.M.O., MENTZ, L.A., SCHENKEL, E.P., IRGANG, B.E., STEHMANN, J.R. Plantas da medicina popular no Rio Grande do Sul. 5. ed. Porto Alegre: Ed. Universidade/UFRGS, 1998. 173 p. SIMÕES, C.M.O., SCHENKEL, E.P., GOSMANN, G., MELO, J.C.P., MENTZ, L.A., PETROVICK, P.R. Farmacognosia: da planta ao medicamento. 2 ed. Porto Alegre/Florianópolis: Ed. Universidade/UFRGS/ Ed. da UFSC, 2000. 821 p. SINGH, N.P., MCCOY, M.T., TICE, R.R., SCHNEIDER, E.L. A simple technique for quantification of low levels of DNA damage in individual cells. Experimental Cell Research, v. 175, p. 184-191, 1988. SILVA, J., FREITAS, T.R.O., MARINHO, J.R., SPEIT, G., ERDTMANN, B. An alkaline single-cell gel electrophoresis (comet) assay for environmental biomonitoring with native rodents. Genetics and Molecular Biology, v. 23, p. 241-245, 2000. SILVA, J., HERMANN, S.M., HEUSER, V., PERES, W., MARRONI, N., GONZÁLEZ-GALLEGO, J., ERDTMANN, H. Evaluation of the genotoxic effect of rutin and quercetin by comet assay and micronucleus test. Food and Chemical Toxicology, v. 40, p. 941-947, 2002. SILVA, J., SIMEONI, N., GROFF, A.A., IANISTCKI, M., BENVEGNÚ, V., SCHRÖDER, N.T. O Ensaio cometa na avaliação da genotoxicidade induzida por poluentes atmosféricos utilizando como biomonitor o molusco Cantareus aspersus (Müller, 1774). Journal of the Brazilian Society of Ecotoxicoogy, v. 2, p. 45-51, 2007. SOARES, S.E. Ácidos fenólicos como antioxidantes. Revista de Nutrição, v. 15, p. 71-81, 2002. SOVOVÁ, M., FISEROVÁ, J., SOVA, J., OPLETAL, L. Pharmaceutical significance of the species of the genus Solidago 1. The glycosides of Solidago virgaurea L. Ceska a Slovenska Farmacie, v. 47, p. 103-109, 1998. STEENKAMP, V., GRIMMER, H., SEMANO, M., GULUMIAN, M. Antioxidant and genotoxic properties of South African herbal extracts. Mutation Research, v. 581, p. 35-42, 2005. SUNG, J.H., LEE, J.O., SON, J.K., PARK, N.S., KIM, M.R., KIM, J.G., MOON, D.C. Cytotoxic constituents from Solidago virga-aurea var. gigantea MIQ. Archives of Pharmacal Research, v. 22, p. 633-637, 1999. TEPE, B., SOKMEN, M., AKPULAT, H.A., SOKMEN, A. In vitro antioxidant activities of the methanol extracts of four Helichrysum species from Turkey. Food Chemistry, v. 90, p. 685-689, 2005.

96
THIEM, B., GOSLINSKA, O. Antimicrobial activity of Solidago virgaurea L. from in vitro cultures. Fitoterapia, v. 73, p. 514–516, 2002. TICE, R.R., AGURELL, E., ANDERSON, D., BURLINSON, B., HARTMANN, A., KOBAYASHI, Y. et al. Single cell gel/comet assay: guidelines for in vitro and in vivo genetic toxicology testing. Environmental and Molecular Mutagenenesis, v. 35, p. 206-221, 2000. TORI, M., KATTO, A., SONO, M. Nine new clerodane diterpenoids from rhizomes of Solidago altissima. Phytochemistry, v. 52, p. 487-493, 1999. TUCKER, A.O., MACIARELLO, M.J., CLANCY, K. Sweet goldenrod (Solidago odora, Asteraceae): A medicine, tea, and state herb. Economic Botany, v. 53, p. 281-284, 1999. VALKO, M., RHODES, C.J., MONCOL, J., IZAKOVIC, M., MAZUR, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chemico-Biological Interactions, v. 160, p. 1-40, 2006. VERDI, L.G., BRIGHENTE, I.M.C., PIZZOLATTI, M.G. Gênero Baccharis (Asteraceae): aspectos químicos, econômicos e biológicos. Química Nova, v. 28, p. 85-94, 2005. VILA, R., MUNDINA, M., TOMI, F., FURLÁN, R., ZACCHINO, S., CASANOVA, J., CAÑIGUERAL, S. Composition and antifungal activity of the essencial oil of Solidago chilensis. Planta Medica, v. 68, p. 164-167, 2002. VILLELA, I.V., LAU, A., SILVEIRA, J., PRÁ, D., ROLLA, H.C., SILVEIRA, J.D. Bioensaios para o monitoramento de genotoxicidade ambiental. In: SILVA, J., ERDTMANN, B., HENRIQUES, J.A.P. (Eds.). Genética Toxicológica. Porto Alegre: Alcance, p. 160-161, 2003. WAGNER, H., BLADT, S. Plant drug analysis a thin layer chromatography atlas. 2a ed. Berlin: Springer, 1996. 384 p. WANG, K.J., CHEN, L.Z., LI, N. , YU, X.P. Antioxidant and radical-scavenging activity of flavonoids from Solidago canadensis. Chinese Pharmaceutical Journal, v. 41, p. 493-497, 2006. (a) WANG, K.J., LI, N., CHEN, L.Z., YU, X.P. Chemical constituents and antifungal activity of essential oil from Solidago canadensis. Journal of Plant Resources and Environment, v. 15, p. 34-36, 2006. (b) WEBER, E., JAKOBS, G. Biological flora of central Europe: Solidago gigantea Aiton. Flora, v. 200, p. 109–118, 2005. WILMSEN, P.K., SPADA, D.S., SALVADOR, M. Antioxidant activity of the flavonoid hesperidin in chemical and biological systems. Journal of Agricultural and Food Chemistry, v. 53, p. 4757-4761, 2005.

97
WU, B., TAKAHASHI, T., KASHIWAGI, T., TEBAYASHI, S.I., KIM, C.S. New flavonoid glycosides from the leaves of Solidago altissima. Chemical and Pharmaceutical Bulletin, v. 55, p. 815-816, 2007. (a) WU, W.M., LU, L., LONG, Y., WANG, T., LIU, L., CHEN, Q., WANG, R. Free radical scavenging and antioxidative activities of caffeic acid phenethyl ester (CAPE) and its related compounds in solution and membranes: A structure–activity insight. Food Chemistry, v. 105, p.107-115, 2007. (b) www.geocities.com: Acesso em: 20/07/2007. YAMANAKA, N., ODA, O., NAGAO, S. Prooxidant activity of caffeic acid, dietary non-flavonoid phenolic acid, on Cu+2 –induced low density lipoprotein oxidation. FEBS Letters, v. 405, p. 186-190, 1997. YAMAZAKI, E., INAGAKI, M., KURITA, O., INOUE, T. Antioxidant activity of Japanese pepper (Zanthoxylum piperitum DC.) fruit. Food Chemistry, v. 100, p. 171-177, 2007. YANG, C.S., LANDAU, J.M., HUANG, M.T., NEWMARK, H.L. Inhibition of carcinogenesis by dietary polyphenolic compounds. Annual Reviews of Nutrition, v. 21, p. 381-406, 2001. YARNELL, E. Botanical medicines for the urinary tract. World Journal of Urology, v. 20, p. 285-293, 2002. YUNES, R.A., CECHINEL FILHO, V. Breve análise histórica da química de plantas medicinais: sua importância na atual concepção de fármacos segundo os paradigmas ocidental e oriental. In: YUNES, R.A., CALIXTO, J.B. (Eds.). Plantas medicinais sob a ótica da moderna química. Chapecó (SC): Argos, p. 18-42, 2001. ZHANG, L., ONDA, K., IMAI, R., FUKUDA, R., HORIUCHI, H., OHTA, A. Growth temperature downshift induces antioxidant response in Saccharomyces cerevisiae. Biochemical and Biophysical Research Communications, v. 307, p. 308-314, 2003. ZHANG, J., ZHANG, X., LEI, G., LI, B., CHEN, J., ZHOU, T. A new phenolic glycoside from the aerial parts of Solidago canadensis. Fitoterapia, v. 78, p. 69-71, 2007.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo