universidade federal de santa...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE SANTA CATARINA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOQUÍMICA
Angélica Cavalett
INTERAÇÃO E CLIVAGEM DE DNA POR COMPLEXOS
METÁLICOS DE Cu(II) E Tb(III)
Dissertação submetida ao
Programa de Pós-Graduação em
Bioquímica da Universidade
Federal de Santa Catarina para a
obtenção do Grau de Mestre em
Bioquímica
Orientador: Prof. Dr. Hernán
Terenzi
Florianópolis
2011


Angélica Cavalett
INTERAÇÃO E CLIVAGEM DE DNA POR COMPLEXOS
METÁLICOS DE Cu(II) E Tb(III)
Esta Dissertação foi julgada adequada para obtenção do Título de
“Mestre em Bioquímica”, e aprovada em sua forma final pelo Programa
de Pós-Graduação em Bioquímica.
Florianópolis, 02 de dezembro de 2011.
________________________
Prof., Dr. Marcelo Farina
Coordenador do Curso
Banca Examinadora:
________________________
Prof., Dr. Hernán Terenzi,
Orientador
Universidade Federal de Santa Catarina
________________________
Prof.ª, Dr.ª Elene Cristina Pereira Mais,
Universidade Federal de Minas Gerais
________________________
Prof., Dr. Claus Tröger Pich,
Universidade Federal de Santa Catarina
________________________
Prof., Dr. Nelson H. Gabilan,
Universidade Federal de Santa Catarina
________________________
Prof., Dr. Marcelo Farina
Coordenador do Curso


Dedico este trabalho aos meus pais,
Alcidir e Ivete, ao meu irmão,
Gabriel, e à minha afilhada, Luiza.


i
AGRADECIMENTOS
Ao professor Hernán Terenzi pela oportunidade de estudo,
confiança e por disponibilizar todos os recursos para o desenvolvimento
deste trabalho.
Aos professores Adaílton Bortoluzzi e Hugo Gallardo pela
colaboração na realização deste trabalho. Ao professor Ademir Neves e
aos alunos do Laboratório de Bioinorgânica e Cristalografia do
departamento de Química da Universidade do Federal de Santa
Catarina, pela colaboração em outros projetos. Ao, agora mestre,
Everton Policarpi pela colaboração e ajuda neste trabalho.
Aos colegas do Centro de Biologia Molecular Estrutural: Angela,
Cami, Carol, Deise, Guilherme, Gabi Ecco, Gabi Müller, Gisa, Javier,
Jean, Louise, Manuel, Ricardo, Pri, Pati, Paty (minha IC favorita!) e
Tiago pela convivência, troca de conhecimento, ajuda nos experimentos,
pelas festinhas, comilanças e brincadeiras! Agradeço, em especial, o
Lab. 03, pela parceria e pelas risadas, o querido colega Tiago, por todos
os ensinamentos e todas as ajudinhas e a “crrrrazy” Gabrielle, por não
ser apenas uma colega de trabalho.
Aos meus pais, Alcidir e Ivete, pelo amor, zelo, paciência e
sabedoria em todos os momentos da minha vida. Por terem feito tudo
certo sempre! Pelo apoio e confiança sem cobranças! Muito, muito
obrigada!
Ao meu irmão Gabriel, minha amiga e cunhada e Cristiane, meu
segundo irmão Gabriel e minha amiga e concunhada Fernanda, à Stela e
ao Joaquim e à minha Best Friend Forever, Rafaela, por me aguentarem
nos meus dias insuportáveis e por serem responsáveis por momentos de
imensa alegria! À Luiza, por sempre ser um estímulo e me deixar
extremamente feliz simplesmente por existir!
Ao meu namorado Filipe, por todas as viagens nos finais de
semana, por toda a paciência e todo o amor nos meus melhores e piores
dias!
À UFSC e ao Programa de Pós-Graduação em Bioquímica.
À CAPES, pela concessão da bolsa de estudo e ao CNPQ, MCTI,
FINEP, FAPESC e Instituto de Ciência e Tecnologia em Biologia Estrutural e Bioimagem (INBEB).

ii

iii
"Não se pode contar com
nada. O único caminho viável é
viver e correr o sagrado risco do
acaso. E substituir o destino pela
probabilidade."
“Uma estrutura tão bonita
simplesmente tinha de existir.”
James Watson

iv

v
RESUMO
O ácido desoxirribonucléico (DNA) é alvo constante de estudo nas áreas
de bioquímica, biotecnologia e biologia molecular por sua importância
biológica, por isso a busca por novas ferramentas para sua manipulação
torna-se necessária. Com este objetivo foram estudadas a interação e
clivagem de DNA por complexos metálicos de Cu(II),
Cu(acac)dppz](ClO4), [Cu(bpma)dppz](ClO4)2.0,67H2O,
[Cu(diaza)dpq](ClO4).H2O e Tb(III), [Tb(tdzp)(acac)3]. Para avaliar a
capacidade de clivagem dos complexos foram realizados testes com
diferentes concentrações destas moléculas e na presença e ausência de
luz UV-A. O mecanismo de clivagem foi avaliado pelo aumento de
força iônica, adição de sequestradores de radicais livres e ligantes do
sulco maior e menor do DNA às reações. Testes na presença e ausência
de oxigênio também foram realizados. Os ensaios cinéticos ocorreram
no abrigo e na presença de luz UV-A. Os complexos de Cu(II) foram
avaliados quanto à sua interação ao CT-DNA, para tanto foram
realizadas titulações espectrofotométricas do complexo na presença e
ausência de CT-DNA, desnaturação térmica de CT-DNA na ausência e
presença dos complexos e avaliação de alterações na estrutura do DNA
na presença dos complexos por espectroscopia de dicroísmo circular. Os
quatro complexos foram capazes de clivar o DNA plasmidial em
concentrações micromolares em cinco minutos de reação quando
fotoativados por luz UV-A. O aumento de força iônica reduziu a
atividade dos complexos e o mecanismo de clivagem ocorre pela
formação de espécies reativas de oxigênio e carbono-centrados. Os
ensaios cinéticos revelaram tempos de meia-vida do DNA abaixo dos
cinco minutos quando as reações foram fotoativadas. Os complexos de
Cu(II) com ligantes dppz forneceram Kb de ~105 M
-1 e o Kb para o
complexo de Cu(II) com ligante dpq foi de ~104 M
-1. Os três complexos
de Cu(II) promoveram aumento da Tm do DNA e alterações na estrutura
do CT-DNA foram observadas pelos espectros de dicroísmo circular.
Palavras-chave: complexos de Cu(II), complexo de Tb(III),
fotoclivagem de DNA, interação com DNA.

vi

vii
ABSTRACT Deoxyribonucleic acid (DNA) is a constant target of study in
biochemistry, biotechnology and molecular biology, for this reason new
molecules capable of interaction and DNA cleavage are designed for
DNA manipulation. Herein we studied the DNA interaction and
cleavage by metal complexes of Cu(II), Cu(acac)dppz](ClO4),
[Cu(bpma)dppz](ClO4)2.0,67H2O , [Cu(diaza)dpq](ClO4).H2O, and
Tb(III), [Tb(tdzp)(acac)3]. To assess the ability of DNA cleavage by the
Cu(II) and Tb(III) complexes tests were performed at different
concentrations of the complexes and in the presence and absence of UV-
A light. The cleavage mechanism was studied by the increase of ionic
strength, addition of free radical scavengers and DNA groove binders to
the reactions. Tests in oxygen presence and absence were also
conducted. The kinetic assays were performed at dark and in UV-A light
presence. The Cu(II) complexes were evaluated for their interaction to
CT-DNA by spectrophotometric titrations of the complexes in DNA
presence and absence, CT-DNA thermal denaturation in the absence and
presence of complexes and changes evaluations in DNA structure by
circular dichroism technique. The four complexes were able to cleave
the plasmid DNA at micromolar concentrations in five minutes of
reaction under UV-A light. The complexes activity was reduced by ionic
strength increase and the cleavage mechanism occurs by the generation
of oxygen and carbon reactive species. The kinetics analysis revealed
DNA half-life times under five minutes when the reactions were
photoactive. The Cu(II) complexes with dppz ligands provided a Kb of
~105 M
-1 and the complex with dpq ligand provide a Kb of ~10
4 M
-1.
The three Cu(II) complexes increased the CT-DNA Tm and changes in
CT-DNA structure.
Keywords: Cu(II) complexes; Tb(III) complex; DNA photocleavage;
DNA interactions.

viii

LISTA DE FIGURAS
Figura 1. Formas de interação entre pequenas moléculas e a dupla hélice
de DNA…………………………………………………………………3
Figura 2. Mecanismo de clivagem de DNA por via hidrolítica. Etapa 1:
ataque nucleofílico do oxigênio de uma molécula de hidroxila ao fosfato
ligado à pentose do ácido nucléico, gerando um fosfato intermediário
penta-coordenado. Etapa 2: ruptura da ligação fosfodiéster em P-O5’ ou
P-O3’ (MANCIN et al., 2005) adaptado por Bortolotto (2011)……….4
Figura 3: Diagrama de Jablonski demonstrando os processos químicos
envolvidos na fotoexcitação de elétrons. Adaptado de Szaci łowski e
colaboradores (2005)…………………………………………………..5
Figura 4: Esquema de ação de um fármaco utilizado na terapia-
fotodinâmica. (A): Administração do fármaco; (B): concentração do
fármaco na região do tumor; (C): fotoativação do fármaco pela luz e
(D): destruição seletiva do tumor………………………………………6
Figura 5: Estrutura da fenantrolina (1) e seus derivados dpq (2) e dppz
(3) (ROY et al., 2010)………………………………………………….7
Figura 6: Estrutura do complexo [Cu(acac)(dpq)Cl] (1) e
[Cu(acac)(dppz)Cl] (2) (CHEN et al., 2011)…………………………..8
Figura 7: Exemplos de complexos de lantanídeos: [Tb(H2L)-
(NO3)(H2O)3]2+
(A), [Eu(H2L)(NO3)(H2O)3]2+
(B) e
[Gd(C34H41N6O8)]2+
(C) (CAMARGO et al., 2008; CAMARGO et al.,
2010b)………………………………………………………………….9
Figura 8: Ilustração esquemática da hibridização de oligonucleotídeos
dirigida pela complexação do íon e a sonda. A: sem a hibridização dos
oligonucleotídeos as sondas não são luminescentes. B: Formação do
complexo pela hibridização ao oligonucleotídeo alvo, o complexo é
excitado em 340 nm e emite luminescência de longa-vida em 615 nm,
modificado de Karhunem e colegas (2009)……………………………11
Figura 9: (A) Estrutura cristalina do complexo [Cu(bpma)(dppz)]
(ClO4).0,67H2O. (B) Estrutura cristalina do complexo

[Cu(acac)(dppz)](ClO4). (C) Estrutura cristalina do cátion do complexo,
[Cu(diaza)(dpq)]+. As figuras das estruturas foram gentilmente cedidas
pelo Mestre Everton de Britto Policarpi sob orientação do prof. Dr.
Adaílton J. Bortoluzzi………………………………………………….14
Figura 10: Estrutura cristalina do complexo [Tb(tdzp)(acac)3]
(GALLARDO et al., 2011)…………………………………………….15
Figura 11: Ilustração e micrografia das três formas do DNA plasmidial
(acima) e a sua diferenciação após eletroforese em gel de agarose
(abaixo) (BORTOLOTTO, 2011)……………………………………..16
Figura 12: Clivagem de DNA plasmidial pelo complexo 1 em diferentes
concentrações. Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-
HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 5 minutos em
luz UV-A………………………………………………………………23
Figura 13: Clivagem de DNA plasmidial pelo complexo 2 em diferentes
concentrações. Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-
HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 5 minutos em
luz UV-A………………………………………………………………24
Figura 14: Clivagem de DNA plasmidial pelo complexo 3 em diferentes
concentrações. Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-
HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 5 minutos em
luz UV-A………………………………………………………………25
Figura 15: Clivagem de DNA plasmidial pelo complexo 4 em diferentes
concentrações. Condições reacionais: [DNA] = 400 ng; 25 mM de
PIPES pH 7; NaCl 5 mM; [complexo]: 0 a 100 µM. Tempo de reação: 5
minutos em luz UV-A…………………………………………………26
Figura 16: Clivagem de DNA plasmidial pelo complexo 1 em diferentes
concentrações. Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-
HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 8h na ausência
de luz a 37°C…………………………………………………………27
Figura 17: Clivagem de DNA plasmidial pelo complexo 2 em diferentes
concentrações. Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-
HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 8h na ausência
de luz a 37°C…………………………………………………………..28

Figura 18: Clivagem de DNA plasmidial pelo complexo 3 em diferentes
concentrações. Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-
HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 8h na ausência
de luz a 37°C…………………………………………………………..29
Figura 19: Clivagem de DNA plasmidial pelo complexo 4 em diferentes
concentrações. Condições reacionais: [DNA] = 400 ng; 25 mM de
PIPES pH 7; NaCl 5 mM; [complexo]: 0 a 100 µM. Tempo de reação:
24h na ausência de luz a 37°C………………………………………...30
Figura 20: Atividade do CuCl2 nas mesmas condições testadas para os
complexos 1, 2 e 3. [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[CuCl2]: 2,5 a 20 µM. Tempo de reação: 5 minutos em luz UV-A…..32
Figura 21: Atividade do TbCl3 nas mesmas condições testadas para o
complexo 4. [DNA] = 400 ng; 25 mM de PIPES, pH ; [TbCl3]: 100 µM.
Tempo de reação: 5 minutos em luz UV-A…………………………..33
Figura 22: Atividade do ligante tdzp nas mesmas condições testadas
para o complexo 4. [DNA] = 400 ng; 25 mM de PIPES, pH ; [tdzp]: 50
e 100 µM. Tempo de reação: 5 minutos em luz UV-A e 24 horas no
escuro a 37°C………………………………………………………….34
Figura 23: Clivagem de DNA plasmidial pelo complexo 1 na ausência e
presença de diferentes concentrações de NaCl. Condições reacionais:
[DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexo]: 5 µM;
[NaCl]: 0 a 300 mM. Tempo de reação: 5 minutos em luz UV-A…..35
Figura 24: Clivagem de DNA plasmidial pelo complexo 2 na ausência e
presença de diferentes concentrações de NaCl. Condições reacionais:
[DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexo]: 20 µM;
[NaCl]: 0 a 300 mM. Tempo de reação: 5 minutos em luz UV-A……36
Figura 25: Clivagem de DNA plasmidial pelo complexo 3 na ausência e
presença de diferentes concentrações de NaCl. Condições reacionais:
[DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexo]: 7,5 µM;
[NaCl]: 0 a 300 mM. Tempo de reação: 5 minutos em luz UV-A……37
Figura 26: Clivagem de DNA plasmidial pelo complexo 4 na ausência e
presença de diferentes concentrações de NaCl. Condições reacionais:

[DNA] = 400 ng; 25 mM de PIPES, pH 7; [complexo]: 25 µM; [NaCl]:
0 a 300 mM. Tempo de reação: 5 minutos em luz UV-A……………38
Figura 27: Clivagem de DNA plasmidial pelo complexo 1 na ausência e
presença de diferentes sequestradores de radicais livres e D2O.
Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[complexo]: 5 µM; [DMSO]: 10% (v/v); [KI]: 10 mM; [SOD]: 15 U;
[D2O]: 70% (v/v); [NaN3]: 10 mM.Tempo de reação: 5 minutos em luz
UV-A………………………………………………………………….39
Figura 28: Clivagem de DNA plasmidial pelo complexo 2 na ausência e
presença de diferentes sequestradores de radicais livres e D2O.
Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[complexo]: 20 µM; [DMSO]: 10% (v/v); [KI]: 10 mM; [SOD]: 15 U;
[D2O]: 70% (v/v); [NaN3]: 10 mM Tempo de reação: 5 minutos em luz
UV-A…………………………………………………………………..40
Figura 29: Clivagem de DNA plasmidial pelo complexo 3 na ausência e
presença de diferentes sequestradores de radicais livres e D2O.
Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[complexo]: 7,5 µM; [DMSO]: 10% (v/v); [KI]: 10 mM; [SOD]: 15 U;
[D2O]: 70% (v/v); [NaN3]: 10 mM Tempo de reação: 5 minutos em luz
UV-A………………………………………………………………….41
Figura 30: Clivagem de DNA plasmidial pelos complexos 1, 2 e 3 na
ausência e presença de TEMPO. Condições reacionais: [DNA] = 400
ng; 10 mM de Tris-HCl, pH 7,4; [complexos]: 5; 20 e 7,5 µM; TEMPO:
500 µM. Tempo de reação: 5 minutos em luz UV-A…………………42
Figura 31: Clivagem de DNA plasmidial pelo complexo 4 na ausência e
presença de diferentes sequestradores de radicais livres. Condições
reacionais: [DNA] = 400 ng; 25 mM de PIPES, pH 7; [complexo]: 25
µM; [DMSO]: 10% (v/v); [KI]: 10 mM; [NaN3]: 10 mM; [TEMPO]:
500 µM Tempo de reação: 5 minutos em luz UV-A………………….44
Figura 32: Clivagem de DNA plasmidial por Fe-EDTA e pelos
complexos 1, 2 e 3 na ausência de oxigênio, em atmosfera de argônio.
Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[complexos]: 5; 20 e 7,5 µM. Tempo de reação: 5 minutos em luz UV-
A. ……………………………………………………………………..45

Figura 33: Clivagem de DNA plasmidial por Fe-EDTA e pelos
complexos 1, 2 e 3 na presença de oxigênio. Condições reacionais:
[DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexos]: 5; 20 e 7,5
µM. Tempo de reação: 5 minutos em luz UV-A………………………46
Figura 34: Clivagem de DNA plasmidial por CuCl2 + MPA e pelo
complexo 4 na ausência de oxigênio. Condições reacionais: [DNA] =
400 ng; 25 mM de PIPES, pH 7; NaCl 5 mM; [complexo]: 25 µM.
Tempo de reação: 5 minutos em luz UV-A……………………………47
Figura 35: Clivagem de DNA plasmidial por CuCl2 + MPA e pelo
complexo 4 na presença de oxigênio. Condições reacionais: [DNA] =
400 ng; 25 mM de PIPES, pH 7; NaCl 5 mM; [complexo]: 25 µM.
Tempo de reação: 5 minutos em luz UV-A……………………………48
Figura 36: Clivagem de DNA plasmidial pelo complexo 4 na ausência e
presenças dos ligantes dos sulcos de DNA. Condições reacionais:
[DNA] = 400 ng; 25 mM de PIPES, pH 7; NaCl 5 mM; [complexo]: 25
µM; [distamicina]: 50 µM; [verde de metila]: 50 µM. Tempo de reação:
5 minutos em luz UV-A……………………………………………….50
Figura 37: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 1. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 75 segundos
em luz UV-A………………………………………………………….51
Figura 38: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 2. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 75 segundos
em luz UV-A…………………………………………………………..52
Figura 39: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 3. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 75 segundos
em luz UV-A………………………………………………………….53
Figura 40: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento

com o complexo 4. Condições reacionais: [DNA] = 400 ng; 25 mM de
PIPES, pH 7; [complexo]: 100 µM. Tempo de reação: 5 minutos em luz
UV-A………………………………………………………………….54
Figura 41: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 1. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 9 horas no
escuro a 37°C………………………………………………………….55
Figura 42: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 2. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 9 horas no
escuro a 37°C………………………………………………………….56
Figura 43: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 3. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 9 horas no
escuro a 37°C………………………………………………………….57
Figura 44: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 4. Condições reacionais: [DNA] = 400 ng; 25 mM de
PIPES, pH 7; [complexo]: 100 µM. Tempo de reação: 48h no escuro a
37°C……………………………………………………………………58
Figura 45: Titulação espectrofotométrica do complexo 1 na presença de
diferentes concentrações de CT-DNA. Condições reacionais: [CT-
DNA]: 0,8 a 7,8 µM; 5 mM de Tris-HCl, pH 7,4; [complexo]: 10 µM;
temperatura: 37°C……………………………………………………..59
Figura 46: Titulação espectrofotométrica do complexo 2 na presença de
diferentes concentrações de CT-DNA. Condições reacionais: [CT-
DNA]: 1 a 9,8 µM; 5 mM de Tris-HCl, pH 7,4; [complexo]: 10 µM;
temperatura: 37°C……………………………………………………..60
Figura 47: Titulação espectrofotométrica do complexo 3 na presença de
diferentes concentrações de CT-DNA. Condições reacionais: [CT-

DNA]: 1,6 a 14 µM; 5 mM de Tris-HCl, pH 7,4; [complexo]: 25 µM;
temperatura: 37°C……………………………………………………..61
Figura 48: Desnaturação térmica de CT-DNA na ausência e presença
dos complexos 1, 2 e 3. Condições reacionais: [CT-DNA]: 100 µM; 1
mM tampão fosfato pH 7,2; [complexo]: 10 µM. Temperatura: 40 a
90°C, um grau por minuto……………………………………………63
Figura 49: Espectro de dicroísmo circular de CT-DNA na ausência e
presença do complexo 1. Condições reacionais: [CT-DNA]: 100 µM;
5mM de Tris-HCl pH 7,4; [complexo]: 5 a 16 µM……………………64
Figura 50: Espectro de dicroísmo circular de CT-DNA na ausência e
presença do complexo 2. Condições reacionais: [CT-DNA]: 100 µM;
5mM de Tris-HCl pH 7,4; [complexo]: 5 a 20 µM…………………..64
Figura 51: Espectro de dicroísmo circular de CT-DNA na ausência e
presença do complexo 3. Condições reacionais: [CT-DNA]: 100 µM;
5mM de Tris-HCl pH 7,4; [complexo]: 5 a 20 µM…………………..65


LISTA DE ABREVIATURAS E SIGLAS
acac: acetilacetonato
bpma: N-bis-(2-piridilmetil)-amina
CD: dicroísmo circular (do inglês, circular dichroism)
CT-DNA: DNA de timo de bezerro (do inglês, calf thymus
deoxyribonucleic acid)
diaza: 6-amino-6-metilperhidro-1,4-diazepina
DNA: ácido desoxirribonucléico (do inglês, deoxyribonucleic acid)
dppz: dipirido[3,2-a:2',3'-c]fenazina
dpq: dipirido[3,2-f:2',3'-h]quinoxalina
EDTA: ácido etilenodiamino tetra-acético (do inglês, ethylenediamine
tetraacetic acid)
ERO: espécie reativa de oxigênio
F I: forma superenovelada do DNA plasmidial, forma I
F II : forma circular aberta do DNA plasmidial, forma II
F III: forma linear do DNA plasmidial, forma III
pb: par de base
pH: potencial hidrogêniônico
phen: fenantrolina
PIPES: Piperazina-1,4-bis(ácido 2-etanosulfónico)
UV: ultravioleta
UV-Vis: ultravioleta - visível
tdzp: [1,2,5]thiadiazol[3,4-f][1,10]fenantrolina
Tris-HCl: hidrocloreto de tris(hidroximetil)aminometano


SUMÁRIO
AGRADECIMENTOS ............................................................ I
RESUMO ................................................................................ V
ABSTRACT ......................................................................... VII
LISTA DE FIGURAS ............................................................. 1
SUMÁRIO ................................................................................ 1
1. INTRODUÇÃO ................................................................... 1
1.1 ÁCIDO DESOXIRRIBONUCLÉICO ............................................ 1
1.2 COMPLEXOS METÁLICOS E A MOLÉCULA DE DNA ................ 2 1.2.2 Fotoclivagem de DNA por complexos metálicos ..... 5 1.2.3 Clivagem de DNA por complexos de cobre
derivados de fenantrolina .................................................. 6 1.2.4 Complexos de lantanídeos ........................................ 8
1.3 APLICAÇÕES DOS COMPLEXOS METÁLICOS ......................... 10
2. OBJETIVOS ...................................................................... 12
2.1. OBJETIVO GERAL ............................................................... 12 2.2. OBJETIVOS ESPECÍFICOS .................................................... 12
3. MATERIAL E MÉTODOS .............................................. 13
3.1. COMPLEXOS METÁLICOS ................................................... 13
3.2. EXTRAÇÃO DO DNA PLASMIDIAL PBSK II ........................ 15 3.3. CLIVAGEM DE DNA PLASMIDIAL ...................................... 16
3.3.1. Efeito da concentração do complexo .................... 17 3.3.2. Atividade do sal de cobre(II) e dos ligantes .......... 17
3.3.3. Atividade do sal de térbio(III) e dos ligantes ........ 18 3.3.4. Efeito da força iônica ............................................ 18
3.3.5. Efeito dos sequestradores de radicais livres e D2O
......................................................................................... 18 3.3.6. Efeito da presença e ausência de oxigênio ........... 18 3.3.7. Efeito dos ligantes dos sulcos do DNA .................. 19
3.3.8. Cinética de clivagem do DNA ............................... 19 3.4. INTERAÇÃO DOS COMPLEXOS COM CT-DNA .................... 20

3.4.1. Titulação espectrofotométrica (UV-Vis) ................ 20 3.4.2. Desnaturação térmica do DNA ............................. 21 3.4.3. Espectroscopia de dicroísmo circular (CD) .......... 22
4. RESULTADOS E DISCUSSÃO ....................................... 23
4.1. CLIVAGEM DE DNA PLASMIDIAL ....................................... 23
4.1.1. Efeito da concentração do complexo ..................... 23 4.1.2. Atividade do sal de cobre(II) e dos ligantes dpq e
dppz .................................................................................. 31
4.1.3 Atividade do sal de térbio(III) ................................ 32
4.1.3. Efeito da força iônica ............................................ 35
4.1.4. Efeito dos sequestradores de radicais livres e D2O
......................................................................................... 39 4.1.5. Efeito da presença e ausência de oxigênio ............ 45
4.1.6. Efeito dos ligantes dos sulcos do DNA .................. 49 4.3.7. Cinética de clivagem do DNA ................................ 51
4.2. INTERAÇÃO DOS COMPLEXOS COM CT-DNA .................... 59 4.2.1. Titulação espectrofotométrica (UV-Vis) ................ 59 4.2.2. Desnaturação térmica do DNA ............................. 62
4.2.3. Espectroscopia de dicroísmo circular (CD) .......... 63
5. CONCLUSÕES .................................................................. 66
6. PERSPECTIVAS ............................................................... 68
7. REFERÊNCIAS ................................................................. 69

1
1. INTRODUÇÃO
1.1 Ácido desoxirribonucléico
O ácido desoxirribonucléico (DNA) é formado por uma
sequência de nucleotídeos capaz de armazenar e transmitir a informação
genética de uma geração para outra. Os nucleotídeos são formados por
uma base nitrogenada (adenina, guanina, citosina ou timina), uma
desoxirribose e um fosfato, estes são ligados covalentemente por uma
ligação fosfodiéster, na qual o grupo 5´-fosfato de um nucleotídeo está
ligado ao grupo 3´-hidroxila do nucleotídeo seguinte. Em 1953, Watson
e Crick publicaram a estrutura tridimensional do DNA: duas cadeias
laterais de nucleotídeos torcidos na forma de uma dupla hélice, as duas
cadeias são unidas por associações fracas entre as bases, sendo duas
ligações de hidrogênio entre adenina e timina e três ligações de
hidrogênio entre guanina e citosina. O esqueleto hidrofílico dos grupos
fosfato e desoxirribose estão do lado de fora da dupla hélice
(GRIFFITHS et al., 2006; NELSON; COX, 2006).
A estrutura covalente das bases nitrogenadas da molécula de
DNA pode sofrer alterações espontâneas. Essas alterações ocorrem de
forma muito lenta, no entanto são extremamente importantes pela baixa
tolerância das células a estas modificações na informação genética. A
molécula de DNA pode sofrer desaminação dos grupos aminos
exocíclicos nas bases nucleotídicas, hidrólise da ligação N-ł-glicosídica
entre a base e a pentose e outras reações promovidas por radiação.
Químicos reativos desaminantes (ácido nitroso) e agentes alquilantes (S-
adenosilmetionina) também podem causar modificações no DNA, no
entanto a lesão oxidativa parece ser a maior causa de alterações na
molécula, sendo o radical hidroxila o principal responsável pelas lesões
oxidativas do DNA (NELSON; COX, 2006).
A constante de degradação espontânea do DNA em pH 7,0 a
25ºC é de ~10-11
s-1
(SREEDHARA; FREED; COWAN, 2000), a
estabilidade encontrada na molécula refere-se principalmente a
resistência à degradação das ligações fosfodiéster que fazem parte da
estrutura do DNA. A molécula de DNA é alvo de estudo em técnicas
bioquímicas, biotecnológicas e de biologia molecular e por isso torna-se importante o estudo de novas ferramentas de manipulação do DNA com
diferentes finalidades (COWAN, 2001; SREEDHARA; COWAN, 2001;
GRIFFITHS et al., 2006; NELSON; COX, 2006).

2
As nucleases catalisam a hidrólise do DNA e aceleram a reação
de 1012
a 1016
vezes em relação à hidrólise espontânea do DNA
(SIGMAN; MAZUMDER; PERRIN, 1993; KRÄMER, 1999;
NELSON; COX, 2006). O mecanismo mais aceito para hidrólise do
DNA catalisado pelas endonucleases ocorre pelo ataque nucleofílico do
oxigênio da água para formar um fosfato intermediário penta-
coordenado. Em seguida, a clivagem de P-O3´ ou P-O5´ fornece a cisão
da fita, liberando os terminais R-OH e R-O-PO3(H2), (LIU et al., 2004;
MANCIN et al., 2005). As nucleases promovem um ataque nucleofílico
à molécula de DNA preferencialmente pela coordenação de uma
molécula de água por um centro metálico da enzima. A presença de
metais nas nucleases está relacionada com o aumento de afinidade da
enzima pelos sítios de ligação e ao favorecimento cinético da reação
(DUPUREUR, 2008, XIE; et al, 2008). As nucleases são seletivas,
atuam em sequências específicas conservadas no DNA e estão presentes
em processos de reparação, degradação, recombinação e modificação da
estrutura de ácidos nucléicos (NICHOLSON, 1999; NELSON; COX,
2006).
1.2 Complexos metálicos e a molécula de DNA
Após a determinação da estrutura tridimensional do DNA e a sua
importância nos sistemas biológicos, o interesse no estudo da interação
entre pequenas moléculas e sequências específicas do DNA para o
controle da expressão gênica, sondas para diagnósticos e uso terapêutico
aumentou significativamente (PATRA, 2007). Pequenas moléculas,
como complexos metálicos, ligam-se ao DNA preferencialmente de três
formas identificáveis (DERVAN, 2001; ZEGLIS; PIERRE; BARTON,
2007) (figura 1). Uma das formas é interação eletrostática não específica
dos complexos com metais carregados positivamente e o esqueleto
carbono-fosfato do DNA carregado negativamente, esta forma de
ligação é a mais fraca. Outra forma é a ligação aos sulcos do DNA por
interações covalentes, eletrostáticas, ligações de hidrogênio, van der
Walls e hidrofóbicas. Os antibióticos antivirais distamicina e netropsina
são conhecidos como ligantes de sulco específicos em regiões A-T
(BAILLY; CHAIRS, 1998; NEIDLE, 2001). A terceira forma de
interação é a intercalação de moléculas planares entre os pares de base
do DNA. O brometo de etídio, a acridina e o azul de metileno são
agentes intercalantes de DNA devido às suas estruturas planares.

3
Figura 1: Formas de interação entre pequenas moléculas e a dupla
hélice de DNA.
O desenvolvimento de diversos complexos metálicos de transição
e de lantanídeos, baseados primeiramente em nucleases biológicas, com
capacidade de interação e/ou clivagem de DNA já foram reportados na
literatura (SIGMAN; CHEN, 1990; SIGMAN; MAZUMDER; PERRIN,
1993; HEGG; BURSTYN, 1998; ZHU et al., 1998; BRANUM; QUE
JR, 1999; KRÄMER, 1999; OTT; KRÄMER, 1999; BRANUM et al.,
2001a; COWAN, 2001; FRANKLIN, 2001; SREEDHARA; COWAN,
2001; LIU et al., 2004; JIANG et al., 2007; CAMARGO et al., 2008;
LIU; WANG, 2009).
A clivagem do DNA pode ocorrer pela fragmentação de seus
constituintes como as bases nitrogenadas, a desoxirribose ou as ligações
fosfodiéster. Os danos às estruturas do DNA ocorrem geralmente pela
oxidação da desoxirribose ou das bases, as quais possuem várias
posições susceptíveis a espécies reativas de oxigênio (EROs)(BREEN;
MURPHY, 1995; VALKO et al., 2007; CHEN et al., 2008a). A
hidrólise da ligação fosfodiéster, mecanismo pelo quais as nucleases
biológicas clivam o DNA, é outra forma de fragmentação da molécula.
De acordo com as características químicas dos complexos, estes
podem promover a clivagem do DNA por mecanismo hidrolítico,

4
oxidativo ou ambos. Quando a clivagem é hidrolítica (figura 2), os
centros metálicos podem coordenar moléculas de água facilitando a
interação com a ligação fosfodiéster do DNA e o ataque nucleofílico ao
átomo de fósforo (OTT; KRÄMER, 1999; LIU et al., 2004; LIU;
WANG, 2009). No mecanismo oxidativo ocorre a produção de espécies
radicalares a partir de uma reação de oxi-redução entre o núcleo
metálico e um co-reagente como o oxigênio ou o nitrogênio. As espécies
reativas produzidas podem se difundir e atacar preferencialmente as
bases nitrogenadas e a desoxirribose do DNA (SIGMAN;
MAZUMDER; PERRIN, 1993; ARMITAGE, 1998; POGOZELSKI;
TULLIUS, 1998), a quebra da fita do DNA ocorre pela abstração de um
átomo de hidrogênio da desoxirribose, a qual produz um radical
centrado no carbono do açúcar que pode sofrer rearranjo
(POGOZELSKI; TULLIUS, 1998).
Figura 2. Mecanismo de clivagem de DNA por via hidrolítica. Etapa 1:
ataque nucleofílico do oxigênio de uma molécula de hidroxila ao fosfato
ligado à pentose do ácido nucléico, gerando um fosfato intermediário
penta-coordenado. Etapa 2: ruptura da ligação fosfodiéster em P-O5’ ou
P-O3’ (MANCIN et al., 2005) adaptado por Bortolotto (2011).

5
1.2.2 Fotoclivagem de DNA por complexos metálicos
A fotoclivagem de DNA é realizada por complexos metálicos que
apresentem ligantes heterocíclicos capazes de absorver luz. Os
processos possíveis após a fotoativação de um agente estão
demonstrados no diagrama de Jablonski (figura 3) (SZACIłOWSKI et
al., 2005).
Figura 3: Diagrama de Jablonski demonstrando os processos químicos
envolvidos na fotoexcitação de elétrons. Adaptado de Szaci łowski e
colaboradores (2005).
Quando uma molécula absorve um fóton de luz, o elétron
presente nesta transição ganha energia e passa a ocupar um orbital mais
energético. A molécula neste momento encontra-se em um estado
excitado (singlete). Este estado tem uma meia-vida muito curta e pode
decair para o estado não ativado com a emissão de um fóton
(fluorescência). O estado singlete também pode sofrer uma conversão
interna inter-sistema para o estado triplete. O estado triplete possui uma
meia-vida maior e tende a ser importante em várias reações de
fosforescência (OCHSNER, 1997). O estado triplete pode reagir com o
oxigênio molecular para produzir EROs por duas reações diferentes.
Reação Tipo 1: O oxigênio molecular é reduzido e forma o radical
superóxido (O2ł-
), o qual dismuta para peróxido de hidrogênio (H2O2) e
pode formar o radical hidroxila (OHł) na presença de metais de
transição por reações de Fenton. Reação Tipo 2: envolve a excitação do
oxigênio molecular gerando o oxigênio singlete (1O2).
A fotoclivagem de ácidos nucléicos é vantajosa em relação à
hidrólise ou oxidação dependente de um co-fator, pois a reação só será
iniciada quando as amostras forem irradiadas. Esta característica é
essencial para moléculas utilizadas na terapia fotodinâmica (PDT, do

6
inglês, Photodynamic Therapy) (figura 4), a qual é utilizada contra
diversos tipos de tumores e apresenta menos efeitos colaterais por ter
ação apenas na região fotoativada e não ser funcional na ausência de luz
(CHAKRAVARTY, 2006; PATRA, 2007).
Figura 4: Esquema de ação de um fármaco utilizado na terapia-
fotodinâmica. (A): Administração do fármaco; (B): concentração do
fármaco na região do tumor; (C): fotoativação do fármaco pela luz e
(D): destruição seletiva do tumor.
1.2.3 Clivagem de DNA por complexos de cobre derivados de
fenantrolina
Diversos estudos relatam o uso de complexos de cobre(II) com
fenantrolina ou seus derivados (figura 5) como ligantes. A utilização de

7
bases heterocíclicas planares em complexos metálicos sempre foi
investigada por suas propriedades eletrônicas, reatividade química
diversificada e uma estrutura peculiar que resulta em interações não
covalentes com o DNA. A fenantrolina e suas bases são conhecidas por
apresentarem motivos de quinoxalina fotoativados por luz UV gerando
estados excitados na molécula que poderão reagir com o oxigênio
molecular e causar dano oxidativo ao DNA (TOSHIMA et al., 2002;
DHAR; NETHAJI; CHAKRAVARTY, 2005b; MUKHERJEE et al.,
2005; SASMAL et al., 2007; ROY et al., 2008a; ROY et al., 2010).
Figura 5: Estrutura da fenantrolina (1) e seus derivados dpq (2) e dppz
(3) (ROY et al., 2010).
O primeiro complexo de cobre, bis(phen)Cu(I), relatado como
“nuclease química” apresentou clivagem de DNA na presença de H2O2 e
um grupo tiol (SIGMAN et al., 1979). Complexos fotoativados possuem
a vantagem de não precisarem de co-reagentes redutores como H2O2
para sua ativação (CHAKRAVARTY, 2006), além desta característica
apresentam benefícios, já descritos no item anterior, em aplicações
terapêuticas. Neste sentindo, a busca por novos complexos de cobre (II)
com ligantes fotoativados é contínua. Roy e colaboradores (2010)
descreveram a fotoclivagem de DNA e atividade antitumoral por
complexos de cobre(II) com bases de fenantrolina. Chen e colegas
(2011) também demonstraram a clivagem de DNA fotoinduzida em luz
UV por dois complexos de cobre(II) com ligantes dpq e dppz:
[Cu(acac)(dpq)Cl] e [Cu acac)(dppz)Cl] (figura 6). Nosso grupo relatou
a fotoclivagem de DNA por quatro complexos de cobre(II) e
fenantrolina ou seus derivados em luz UV e laser vermelho ( ł = 635
nm) (SOUZA et al., 2010a).

8
Figura 6: Estrutura do complexo [Cu (acac) (dpq)Cl] (1) e [Cu (acac)
(dppz)Cl] (2) (CHEN et al., 2011).
1.2.4 Complexos de lantanídeos
Íons de lantanídeos e complexos dos mesmos são conhecidos por
sua capacidade de hidrólise de dinucleotídeos e DNA, algumas das
características que conferem esta atividade são: forte acidez de Lewis,
altas densidades de carga, altos números de coordenação, ausência de
química redox e fortes labilidades (FRANKLIN, 2001). Grandes
quantidades do íon de Ce(IV) hidrolisam de forma eficiente
didesoxirribonucleotídeos e promovem quebra simples do DNA
(KOMIYAMA et al., 1993; TAKASAKI; CHIN, 1994; RAMMO et al.,
1996; SUMAOKA et al., 1997; SUMAOKA; AZUMA; KOMIYAMA,
1998). Alguns complexos de Ce (IV), como o Ce2(HXTA) (HXTA = 5-
metil-2-hidroxi-1,3-xilano-ł,ł-diamino-N,N,N’,N’-ácido tetra-acético)
pode clivar a dupla fita de DNA na constante de aproximadamente 0,5h-
1, correspondendo a uma meia vida 1,4 horas para o DNA com apenas
10 µM do complexo (BRANUM et al., 2001b). Curiosamente, o íon de
Ce(III) promove a fragmentação do DNA por geração de radicais livres
na presença de H2O2, esse mecanismo diverge da característica
hidrolítica atribuída para o íon de Ce(IV)(HECKERT; SEAL; SELF,
2008).
Após esse estudo, vários outros reportam o uso de Ce(IV)
conjugado a oligonucleotídeos ou ácido nucléico peptídico (PNA) para
se obter a hidrólise específica do DNA (YAMAMOTO; TSUBOI;
KOMIYAMA, 2003; CHEN et al., 2004; YAMAMOTO et al., 2004;
CHEN; KOMIYAMA, 2005; KOMIYAMA et al., 2005; YAMAMOTO
et al., 2007; LÖNNBERG; SUZUKI; KOMIYAMA, 2008). Neste
contexto, foram realizados alguns estudos utilizando peptídeos com
sítios de ligação a íons de Eu(III) e Ce(IV) para se criar metalopeptídeos
com atividade de nuclease (SIRISH; FRANKLIN, 2002; KOVACIC;
9
WELCH; FRANKLIN, 2003; LIM; FRANKLIN, 2006; WONG-
DEYRUP; KIM; FRANKLIN, 2006). Complexos de outros lantanídeos
como Eu(III), La(III), Nd(III), Pr(III) e Gd(III) são capazes de hidrolisar
o DNA (RITTICH et al., 2004; CAMARGO et al., 2008; CAMARGO et
al., 2010a; CAMARGO et al., 2010b). No entanto novos aspectos do
mecanismo de clivagem dos lantanídeos foram recentemente
destacados. Novos complexos de La(III) e Gd(III) (HUSSAIN et al.,
2010) assim como de Eu(III) contendo derivados de fenantrolina
demonstraram fotoclivagem de DNA quando irradiados com luz UV-A
(CHEN et al., 2010). Nos dois estudos sequestradores de radicais livres
inibiram a atividade do complexo sugerindo a participação de EROs no
evento de clivagem do DNA. Esses estudos assinalam os complexos de
lantanídeos como agentes de clivagem oxidativa do DNA e não apenas
como agentes hidrolíticos.
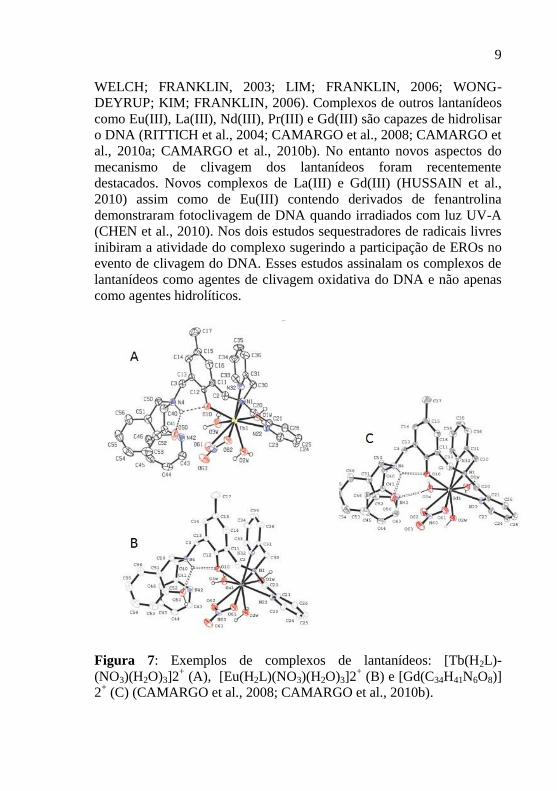
Figura 7: Exemplos de complexos de lantanídeos: [Tb(H2L)-
(NO3)(H2O)3]2+ (A), [Eu(H2L)(NO3)(H2O)3]2
+ (B) e [Gd(C34H41N6O8)]
2+ (C) (CAMARGO et al., 2008; CAMARGO et al., 2010b).

10
1.3 Aplicações dos complexos metálicos
Complexos metálicos que interagem e/ou clivam o DNA podem
ser utilizados como ferramentas em várias metodologias:
desenvolvimento de DNA recombinante, modelagem de enzimas de
restrição, aplicações terapêuticas, sequenciamento de DNA,
mapeamento genético e footprinting (BASHKIN, 1999; OTT;
KRÄMER, 1999; CHAKRAVARTY, 2006).
A técnica de footprinting ou photo-footprinting pode ser
melhorada pelo uso de complexos metálicos que produzem fragmentos
de clivagem muito menores e menos específicos que os gerados pela
enzima DNAse I utilizada tradicionalmente, o que promove um padrão
de DNA clivado de melhor resolução (ARMITAGE, 1998).
Complexos metálicos que apresentam mecanismo oxidativo
podem ser utilizados para investigar o processo de reparação do DNA
quando este é danificado por EROS (KIMURA; SAKAGUCHI, 2006;
SCHLACHER et al., 2006), assim como os que apresentam atividade
antimicrobiana podem ser aprimorados para utilização como fármacos
(EFTHIMIADOU et al., 2007; PATIL et al., 2011; RAMAN;
POTHIRAJ; BASKARAN, 2011).
Também são encontrados complexos com atividade antitumoral
(COLLINS et al., 2000; AFRASIABI et al., 2004), Fernandes e
colaboradores (2006), relatam a clivagem de DNA e ação antitumoral in
vitro para um complexo de cobre(II). Além disso, complexos com
atividade de fotoclivagem de DNA, atóxicos na ausência de luz, são
conhecidos por apresentarem efeitos localizados em aplicações
terapêuticas tornando-se úteis na terapia fotodinâmica (PDT)
(CHAKRAVARTY, 2006). Complexos de cobre(II) com atividade de
clivagem de DNA em luz vermelha, faixa de ação da PDT, também já
foram reportados (DHAR; NETHAJI; CHAKRAVARTY, 2005a;
PATRA et al., 2009; LAHIRI et al., 2010).
Complexos metálicos com capacidade de interação com o DNA,
porém sem atividade de clivagem, podem ser utilizados como sondas
(ZEGLIS; PIERRE; BARTON, 2007). Os mais cotados para essa
finalidade são os complexos de lantanídeos, devido à característica
luminescente pertencente aos íons desse grupo (MARUYAMA et al.,
2002). Karhunem e colegas (2009) descreveram a formação de um
complexo misto de Eu(III) com forte emissão de sinal luminescente
apenas quando os oligonucleotídeos eram hibridizados. Essa
metodologia pode ser empregada com eficiência para o estudo de DNA

11
complementar (figura 8). Complexos de Eu(III) e Tb(III) promoveram a
coloração seletiva de cromossomos em células mitóticas o que permite a
visualização dos cromossomos mitóticos de forma não invasiva. A baixa
toxicidade química e a capacidade de corar o DNA mitótico fazem
desses complexos excelentes sondas para estudos em células vivas
(LAW et al., 2010).
Figura 8: Ilustração esquemática da hibridização de oligonucleotídeos
dirigida pela complexação do íon e a sonda. A: sem a hibridização dos
oligonucleotídeos as sondas não são luminescentes. B: Formação do
complexo pela hibridização ao oligonucleotídeo alvo. O complexo é
excitado em 340 nm e emite luminescência de longa-vida em 615 nm,
modificado de Karhunem e colegas (2009).

12
2. OBJETIVOS
2.1. Objetivo geral
Analisar a interação de CT-DNA e clivagem de DNA
plasmidial por três complexos metálicos de cobre(II) e um
complexo de térbio(III).
2.2. Objetivos específicos
Analisar a clivagem de DNA pelos complexos de cobre(II) e
Tb(III) sob diferentes condições reacionais: concentração dos
complexos, presença e ausência de luz UV-A e influência da
força iônica.
Determinar o mecanismo de ação dos complexos na clivagem
de DNA por meio da adição de sequestradores de radicais livres
e D2O às reações e testes na ausência de oxigênio.
Analisar o modo de clivagem de DNA pelos complexos de
cobre(II) e térbio(III) na presença de inibidores específicos do
sulco menor e maior do DNA.
Determinar a cinética das reações de clivagem de DNA
plasmidial pelos complexos de cobre(II) e térbio(III).
Analisar a interação dos complexos de cobre(II) com DNA por
meio de titulação espectrofotométrica, desnaturação térmica de
DNA e espectroscopia de dicroísmo circular (CD).

13
3. MATERIAL E MÉTODOS
3.1. Complexos metálicos
Três complexos ternários mononucleares de cobre(II) foram
analisados para interação e clivagem de DNA: [Cu(acac)dppz](ClO4)
(1), publicado por Chen e colaboradores (2011),
[Cu(bpma)dppz](ClO4)2.0,67H2O (2) e [Cu(diaza)dpq](ClO4).H2O (3),
nos quais “bpma” corresponde ao ligante N-bis-(2-piridilmetil)-amina,
“acac” a acetilacetonato, “dppz” a dipirido[3,2-a:2',3'-c]fenazina,
“diaza” a 6-amino-6-metilperhidro-1,4-diazepina e “dpq” a dipirido[3,2-
f:2',3'-h]quinoxalina. Estes complexos foram sintetizados pelo Mestre
Everton de Britto Policarpi sob orientação do prof. Dr. Adaílton J.
Bortoluzzi do Laboratório de Bioinorgânica e Cristalografia do
departamento de Química da Universidade do Federal de Santa
Catarina. Um complexo mononuclear de térbio(III) também foi utilizado
para os testes de clivagem de DNA: [Tb(tdzp)(acac)3] (4), no qual tdzp:
[1,2,5]thiadiazol[3,4-f][1,10]fenantrolina e acac: acetilacetona
(GALLARDO et al., 2011). Esse complexo foi sintetizado por Hugo
Gallardo e Gilmar Conte no INCT-Catálise, Departamento de Química
da Universidade Federal de Santa Catarina.

14
Figura 9: (A) Estrutura cristalina do complexo [Cu(bpma)(dppz)]
(ClO4).0,67H2O. (B) Estrutura cristalina do complexo [Cu(acac)(dppz)](ClO4). (C) Estrutura cristalina do cátion do complexo,
[Cu(diaza)(dpq)]+. As figuras das estruturas foram gentilmente cedidas
pelo Mestre Everton de Britto Policarpi sob orientação do prof. Dr.
Adaílton J. Bortoluzzi.

15
Figura 10: Estrutura cristalina do complexo [Tb(tdzp)(acac)3]
(GALLARDO et al., 2011).
3.2. Extração do DNA plasmidial pBSK II
O substrato utilizado para os testes de clivagem de DNA
plasmidial foi o plasmídeo pBSK-II (Stratagene, USA) de 1961 pares de
base (pb). O DNA plasmidial pode apresentar três conformações
distintas em gel de agarose. A forma superenovelada do DNA
plasmidial (forma I, F I) é altamente tensionada, quando sofre uma
quebra simples (em apenas umas das fitas) ocorre o afrouxamento da
estrutura superenovelada e o DNA plasmidial assume a forma circular
aberta (forma II, F II). Se houver uma segunda quebra na fita oposta,
próxima à primeira quebra, há a formação de uma quebra dupla e o
DNA plasmidial fica na sua forma linear (forma III, F III). As três formas possuem mobilidade eletroforéticas diferentes (figura 3),
portanto podem ser separadas por eletroforese em gel de agarose e então
analisada a quantidade e a forma de DNA clivado.

16
Figura 11: Ilustração e micrografia das três formas do DNA plasmidial
(acima) e a sua diferenciação após eletroforese em gel de agarose
(abaixo) (BORTOLOTTO, 2011).
Para obtenção de quantidades suficiente do plasmídeo pBSK-II
para realização dos testes de clivagem de DNA este foi inserido em
Escherichia coli DH5ł por transformação utilizando o método de
CaCl2/choque térmico. Uma colônia transformante foi cultivada em 10
mL de meio LB suplementado com ampicilina (100 µg/mL) por 8 horas,
37°C a 200 rpm. Foram transferidos 500 µL deste pré-cultivo para 250
mL de meio LB suplementado com ampicilina, o qual foi cultivado por
16 horas, 37°C a 200 rpm. Após a multiplicação celular, a cultura foi
centrifugada a 3500 rpm por 15 minutos a 4°C e o DNA foi purificado
segundo o kit de extração e purificação de DNA plasmidial HiSpeed TM
Plasmid Maxi Kit (Qiagen). A quantificação de DNA plasmidial
extraído foi feita por espectrofotometria UV-Vis (A260 = 1 = 50 µg mL-1
de DNA).
3.3. Clivagem de DNA plasmidial
Neste trabalho foram realizados testes de clivagem de DNA em
reações de 20 µL contendo: 400 ng do plasmídeo pBSKII

17
superenovelado (~30 µM de DNA em pb); tampão Tris-HCl (10
mM, pH 7,4); e os complexos 1, 2 e 3 em diferentes concentrações. Para
o complexo 4 o tampão utilizado nas reações foi PIPES (pH 7 a 25 mM)
e NaCl (5 mM). A concentração final de acetonitrila (CH3CN), solvente
dos complexos, foi de 25%. O período de reação foi de 5 minutos sob
irradiação de luz UV-A ( ł = 365 nm) de um transluminador (UVP
USA). Essa condição foi determinada como padrão reacional após testes
em presença e ausência de luz UV-A.
As reações de clivagem do DNA plasmidial foram interrompidas
pela adição de tampão de corrida 6X concentrado (EDTA 0,25 M,
glicerol 50% e azul de bromofenol 0,01% - pH 8,0). As amostras foram
armazenadas a -20°C e posteriormente submetidas à eletroforese em gel
de agarose. (1%) contendo brometo de etídio (0,3 łg mL-1
) por 1 hora e
20 minutos a 90 V em tampão TBE 0,5x (Tris 44,5 mM, ácido bórico
44,5 mM, EDTA 1 mM - pH 8,0).
Os géis foram fotografados utilizando-se um sistema de foto-
documentação DigiDoc-It (UVP, USA) e as frações de cada forma de
DNA foram quantificadas por densitometria utilizando o software
KODAK Molecular Imaging Software 5.0 (Carestream Health,
USA). Como o intercalamento do brometo de etídio com a forma
superenovelada do DNA plasmidial é mais difícil, há uma menor
emissão de fluorescência pela forma I em relação às formas II e III. A
quantidade real de DNA superenovelado é corrigida multiplicando-se o
valor encontrado por 1,42, a proporcionalidade dos valores obtidos para
as demais formas também foi corrigido (BERNADOU et al., 1989).
3.3.1. Efeito da concentração do complexo
A atividade dos complexos foi analisada primeiramente quanto à
concentração dos mesmos nas reações, tanto na presença de luz UV-A
como no escuro. Reações controles, na ausência dos complexos, nas
mesmas condições reacionais também foram realizadas para observar a
degradação espontânea do DNA. A partir destes resultados foram
escolhidos os tempos reacionais e concentrações de complexos,
adequados para observação da clivagem do DNA, utilizados nos
próximos testes.
3.3.2. Atividade do sal de cobre(II) e dos ligantes
Ensaios de clivagem substituindo os complexos de Cu(II) por
seus ligantes isolados, dpq e dppz, e pelo sal de Cu(II) foram realizados
como controle para os experimentos padrão de clivagem. Os ensaios

18
foram realizados na presença e ausência de luz UV-A nas mesmas
condições reacionais padrão. Controles livres de ligantes ou CuCl2
também foram realizados.
3.3.3. Atividade do sal de térbio(III) e dos ligantes
Ensaios de clivagem substituindo o complexo de Tb(III) por seu
ligante isolado, tdzp, e pelo sal de Tb(III) também foram realizados
como controle para os experimentos padrão de clivagem. Os ensaios
foram realizados na presença e ausência de luz UV-A nas mesmas
condições reacionais padrão. Controles livres dos ligantes ou do sal de
Tb(III) também foram realizados.
3.3.4. Efeito da força iônica
O efeito da força iônica na clivagem de DNA plasmidial pelos
complexos foi verificado pela adição de concentrações crescentes (50,
100, 200 e 300 mM) de cloreto de sódio (NaCl) às misturas reacionais.
As reações foram testadas na presença de luz UV-A nas condições
padrão já citadas no item 3.3.
3.3.5. Efeito dos sequestradores de radicais livres e D2O
Para compreensão do mecanismo de clivagem do DNA (se por
via hidrolítica ou oxidativa), foram adicionados às reações uma série de
inibidores de radicais livres. Foram utilizados: 10% (v/v) de DMSO
(dimetil-sulfóxido), sequestrador do radical hidroxila (OHł); 10 mM de
KI (iodeto de potássio), inibidor da geração de peróxidos (R-O-OH); 10
U da enzima SOD (superóxido-dismutase), seqüestrador do radical
ânion superóxido (O2ł-
); 10mM de NaN3 (azida de sódio), sequestrador
de oxigênio singlete (1O2); 500 µM de TEMPO (N-oxil-2,2,6,6-
tetrametilpiperidina), um sequestrador de radicais oxigênio-, metal- e
carbono-centrados. Além dos sequestradores de radicais livres também
foram realizados testes com 70% (v/v) de D2O, a qual é conhecida por
aumentar o tempo de meia-vida do oxigênio singlete. Esses ensaios
foram realizados sob irradiação UV-A de acordo com as condições
reacionais padrão. Para o complexo 4 não foram realizados testes em
D2O.
3.3.6. Efeito da presença e ausência de oxigênio
Com o intuito de verificar qual a influência do oxigênio
molecular na clivagem do DNA pelos complexos, foram realizados

19
testes de clivagem, nas condições padrão, porém na ausência de
oxigênio, em atmosfera de argônio.
As soluções foram preparadas com água desoxigenada. O
preparo da água desoxigenada foi realizado da seguinte forma: a água
foi submetida à agitação sob vácuo, para eliminação das bolhas de ar e
em seguida foi borbulhada com argônio. Essa água permaneceu em
atmosfera de argônio até o momento do preparo das soluções e reações
de clivagem (FISCHER, 2010). As soluções e misturas reacionais foram
preparadas em uma “glove bag” (modelo X-17-17, 12R). As reações
foram então expostas a luz UV-A por cinco minutos. As mesmas
misturas reacionais também foram realizadas na presença de oxigênio.
Também foi realizado um controle de Fe (EDTA)2-
(100 µM) (NETTO;
STADTMAN, 1996) ou CuCl2 e ácido 3-mercaptopropriônico (MPA),
pois esses sistemas clivam o DNA somente na presença de oxigênio
molecular, sob irradiação UV-A por 5 minutos e 4 horas na ausência de
luz UV-A.
3.3.7. Efeito dos ligantes dos sulcos do DNA
A distamicina e o verde de metila são moléculas que se ligam
especificamente ao sulco menor e maior do DNA, respectivamente
(VAN DYKE; HERTZBERG; DERVAN, 1982; KIM; NORDEN,
1993). A inibição da clivagem demonstra por qual dos dois sulcos
ocorre a interação e consequente clivagem. Então, para verificar se os
complexos atuam preferencialmente por algum dos sulcos do DNA,
foram adicionados ao DNA, distamicina (50 łM) ou verde de metila
(50 łM), 30 minutos antes da adição dos complexos nas condições
reacionais padrão.
3.3.8. Cinética de clivagem do DNA
Para os testes cinéticos de clivagem de DNA plasmidial com os
complexos 1, 2, 3 as condições reacionais determinadas foram: [DNA] =
400 ng; 10 mM de Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de
reação: 75 segundos em luz UV-A e 9h no escuro a 37°C. Após o início
das reações uma alíquota foi retirada a cada 15 segundos para as reações
em luz UV-A e em 0 h, 2 h, 4 h, 6 h, 8 h e 9 h para as reações no escuro.
As condições reacionais para o complexo 4 foram: [DNA] = 400 ng; 25
mM de PIPES, pH 7; [complexo]: 100 µM. Tempo de reação: 5 minutos
em luz UV-A e 48 h no escuro a 37°C. Após o início das reações uma
alíquota foi retirada a cada um minuto para as reações em luz UV-A e
em 0 h, 6 h, 10 h, 24 h, 34 h e 48 h para as reações no escuro. Controles

20
reacionais, na ausência dos complexos também foram realizados para se
observar a degradação espontânea do DNA.
Como as condições reacionais testadas apresentaram excesso de
catalisador (complexo) em relação à concentração do substrato (DNA),
o perfil cinético de clivagem para essa reação foi considerado como de
pseudo-primeira-ordem. As constantes observadas de clivagem de DNA
(kobs) foram determinadas a partir da representação gráfica do plote do
logaritmo neperiano da fração intacta do DNA (F I) em função do tempo
reacional (BORTOLOTTO, 2011).
3.4. Interação dos complexos com CT-DNA
Para avaliação da interação do complexo com DNA, foram
realizadas análises espectrofotométricas. O DNA utilizado para esses
ensaios foi o CT-DNA (Calf Thymus DNA – DNA do timo de bezerro)
tipo XV (Sigma, USA). O CT-DNA é um DNA linear formado por uma
mistura de diferentes fragmentos de DNA genômico dupla-fita. Devido
as suas propriedades bioquímicas e físico-químicas já definidas essa é a
molécula de DNA mais utilizada em estudos de interação complexo-
DNA (BORTOLOTTO, 2011). Os testes a seguir foram realizados com
o complexo 1, 2 e 3.
3.4.1. Titulação espectrofotométrica (UV-Vis)
Para analisar a interação dos complexos com o CT-DNA foram
realizadas titulações espectrofotométricas. Essa técnica é utilizada, pois
quando o DNA interage com pequenas moléculas o espectro UV-Vis
das mesmas é modificado (BORTOLOTTO, 2011). O espectro UV-Vis
dos complexos e as titulações com o CT-DNA foram realizados em um
espectrofotômetro Ultrospec 2100 (Amersham Biosciences USA).
Para os complexos 1 e 2, a concentração dos complexos utilizada
para a titulação foi de 10 µM e para o complexo 3 foi de 25 łM. As
titulações ocorreram em 5 mM de Tris-HCl (pH 7,4), 25% (v/v) de
CH3CN com concentrações crescentes de CT-DNA (0,8 a 14 µM). As
varreduras foram na faixa de 200 a 600 nm, a 37°C logo após a adição
do CT-DNA. O controle foi realizado com uma amostra contendo
somente CT-DNA tamponado. Os espectros do controle foram
subtraídos dos espectros das titulações para eliminar o espectro de
absorção do DNA presente entre 200 e 300nm.
A constante intrínseca de ligação ao DNA (Kb) foi obtida pela
equação 1, abaixo, na qual: łf e łb são as absortividades molares do
complexo livre e do totalmente ligado ao DNA, respectivamente e ła é

21
a absortividade da amostra após cada adição de CT-DNA. O valor de
Kb foi obtido da razão entre o coeficiente angular e linear do gráfico de
[DNA] / ( ła - łf) por [DNA] (WOLFE; SHIMER; MEEHAN, 1987;
PYLE et al., 1989; BORTOLOTTO, 2011).
Equação 1:
)(
1
)(
[DNA]
)(
[DNA]
fbbfbfa K
3.4.2. Desnaturação térmica do DNA
A dupla-hélice do DNA pode ser desnaturada quando submetida
a extremos de pH ou temperaturas elevadas pelo rompimento das
ligações de hidrogênio entre as bases pareadas. Como resultado ocorre o
empilhamento das bases nitrogenadas e um desenovelamento da dupla
hélice formando duas fitas separadas ao longo da molécula. A interação
entre as bases nitrogenadas do DNA diminui a absorção de luz UV pela
molécula em relação a uma molécula de DNA simples fita. A
desnaturação do DNA pode então ser detectada pelo aumento da
absorção de luz (efeito hipercrômico) (NELSON; COX, 2006). A
temperatura necessária para que metade das moléculas de DNA seja
desnaturado é chamada de temperatura média de desnaturação (Tm),
quando outra molécula se liga ao DNA a Tm pode variar.
A partir desses conceitos as Tm do CT-DNA na ausência e na
presença dos complexos foram calculadas. Os ensaios foram realizados
no espectrofotômetro de UV-Vis Ultrospec 2100 (Amersham
Bioscienses, USA). 100 µM de CT-DNA não tratados e tratados com 10
µM dos complexos permaneceram 10 minutos na temperatura de início
do experimento (40 ºC). A temperatura foi então aumentada em
1°C/minuto até 90 °C. O tampão utilizado foi o tampão fosfato (1 mM)
pH 7,2 e 2 mM de NaCl (ARIAS et al., 2009; GARCÍA-GIMÉNEZ
et al., 2009; BORTOLOTTO, 2011). A Tm foi calculada pelo ponto
médio da progressão não linear sigmoidal de Boltzmann e confirmada pela primeira derivada da A260 nm pela temperatura.

22
3.4.3. Espectroscopia de dicroísmo circular (CD)
A técnica de dicroísmo circular (CD, do inglês, circular
dichroism) é muito utilizada para determinar a estrutura secundária de
proteínas e a conformação helicoidal do DNA (EFTHIMIADOU et al.,
2007; GHARAGOZLOU; BOGHAEI, 2008; SHENG et al., 2008) em
solução. O espectro de CD da dupla hélice direita na forma B do DNA
possui sinais característicos: uma banda positiva em 275 nm devido ao
empilhamento das bases do DNA e uma banda negativa em 245nm
referente à helipticidade direita do DNA. As mudanças observadas nas
duas bandas características de CD do DNA correspondem a mudanças
na estrutura do mesmo quando existe interação com outras moléculas
(CHEN et al., 2008b).
Os ensaios de CD foram realizados em um espectropolarímetro
de dicroísmo circular modelo J-815 (JASCO, USA). Os espectros de CD
foram obtidos a partir de uma amostra de 100 µM de CT-DNA em 5
mM de Tris-HCl (pH 7,4), esta foi titulada com concentrações
crescentes de complexos (5 a 25 µM). As varreduras foram realizadas
entre 220 e 400 nm, a 37°C, logo após a adição dos complexos.
23
4. RESULTADOS E DISCUSSÃO
4.1. Clivagem de DNA plasmidial
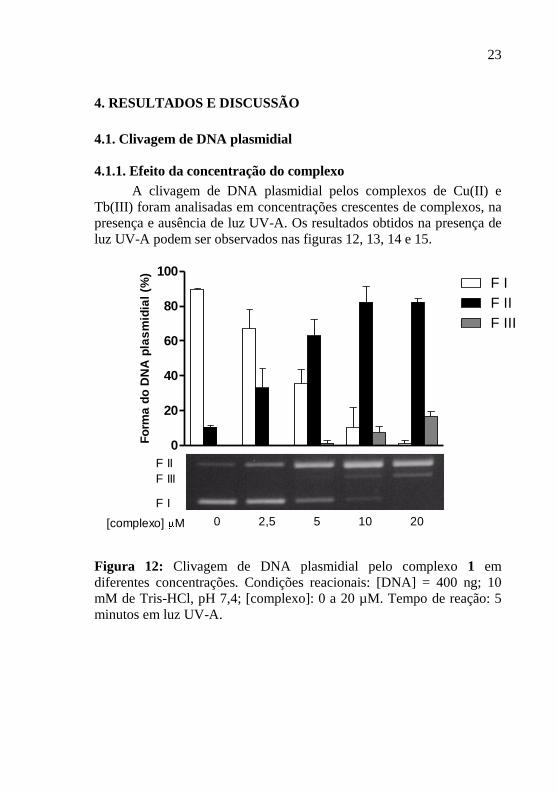
4.1.1. Efeito da concentração do complexo
A clivagem de DNA plasmidial pelos complexos de Cu(II) e
Tb(III) foram analisadas em concentrações crescentes de complexos, na
presença e ausência de luz UV-A. Os resultados obtidos na presença de
luz UV-A podem ser observados nas figuras 12, 13, 14 e 15.
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
[complexo] M 0 2,5 5 10 20
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 12: Clivagem de DNA plasmidial pelo complexo 1 em
diferentes concentrações. Condições reacionais: [DNA] = 400 ng; 10
mM de Tris-HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 5
minutos em luz UV-A.

24
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
[complexo] M 0 2,5 5 10 20
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 13: Clivagem de DNA plasmidial pelo complexo 2 em
diferentes concentrações. Condições reacionais: [DNA] = 400 ng; 10
mM de Tris-HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 5
minutos em luz UV-A.
25
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
[complexo] M 0 2,5 5 10 20
Fo
rma
do
DN
A p
las
mid
ial
(%)
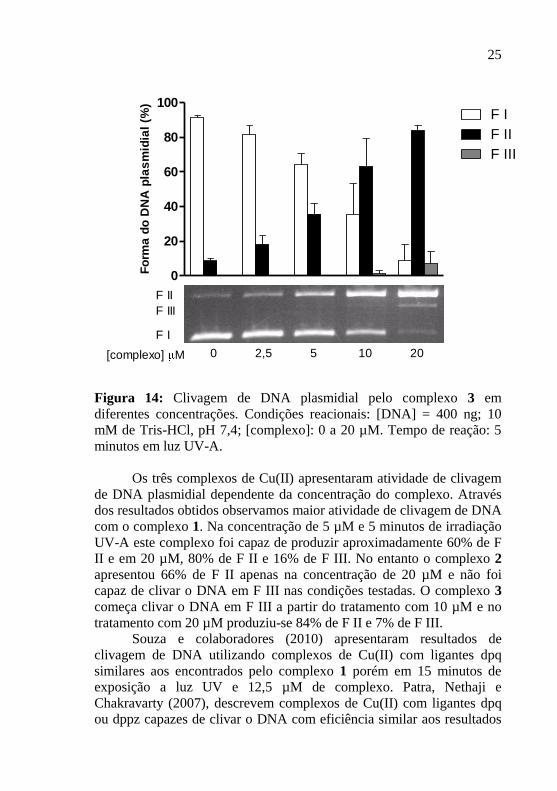
Figura 14: Clivagem de DNA plasmidial pelo complexo 3 em
diferentes concentrações. Condições reacionais: [DNA] = 400 ng; 10
mM de Tris-HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 5
minutos em luz UV-A.
Os três complexos de Cu(II) apresentaram atividade de clivagem
de DNA plasmidial dependente da concentração do complexo. Através
dos resultados obtidos observamos maior atividade de clivagem de DNA
com o complexo 1. Na concentração de 5 µM e 5 minutos de irradiação
UV-A este complexo foi capaz de produzir aproximadamente 60% de F
II e em 20 µM, 80% de F II e 16% de F III. No entanto o complexo 2
apresentou 66% de F II apenas na concentração de 20 µM e não foi
capaz de clivar o DNA em F III nas condições testadas. O complexo 3
começa clivar o DNA em F III a partir do tratamento com 10 µM e no
tratamento com 20 µM produziu-se 84% de F II e 7% de F III.
Souza e colaboradores (2010) apresentaram resultados de
clivagem de DNA utilizando complexos de Cu(II) com ligantes dpq
similares aos encontrados pelo complexo 1 porém em 15 minutos de
exposição a luz UV e 12,5 µM de complexo. Patra, Nethaji e
Chakravarty (2007), descrevem complexos de Cu(II) com ligantes dpq
ou dppz capazes de clivar o DNA com eficiência similar aos resultados

26
mostrados para os complexos 1 e 3, porém em maior tempo de
exposição a luz UV (1 hora) e maior concentração (50 µM).
A atividade de clivagem de DNA plasmidial pelo complexo 1 já
foi reportada na literatura, Chen e colaboradores (2011), descrevem a
formação de 92% de F II após 1 hora de exposição à luz UV-A com 0,2
µM do complexo. No trabalho mencionado não é citada a capacidade da
luz UV-A na qual os testes foram realizados, o tempo de exposição à luz
UV-A foi 12 vezes maior e a concentração do complexo foi 100 vezes
menor por isso. Uma comparação exata não pode ser realizada, já que as
três variáveis são importantes para a eficiência de clivagem dos
complexos.
0
20
40
60
80
100F I
F II
F III
F II
F I
[complexo] M 0 10 25 50 100
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 15: Clivagem de DNA plasmidial pelo complexo 4 em
diferentes concentrações. Condições reacionais: [DNA] = 400 ng; 25
mM de PIPES pH 7; NaCl 5 mM; [complexo]: 0 a 100 µM. Tempo de
reação: 5 minutos em luz UV-A.

27
O complexo 4 também apresentou clivagem de DNA plasmidial
dependente da concentração do complexo. Na presença de 50 µM do
complexo foram observadas 40% de F II e 5 % de F III. Estudos com
complexos de Gd(III) e La(III) registraram fotoclivagem de DNA em
luz UV-A (6W) durante 2 horas de reação (HUSSAIN et al., 2011).
Complexos de Eu(III) também demonstraram fotoclivagem de DNA,
porém de forma menos eficiente que a observada neste trabalho (CHEN
et al., 2010).
As figuras 16, 17, 18 e 19 mostram os resultados obtidos com
diferentes concentrações dos complexos 1, 2 e 3 na ausência de luz.
0
20
40
60
80
100F I
F II
F II
F I
[Complexo] M: 0 2,5 5 10 20
F III
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 16: Clivagem de DNA plasmidial pelo complexo 1 em
diferentes concentrações. Condições reacionais: [DNA] = 400 ng; 10
mM de Tris-HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 8h
na ausência de luz a 37°C.

28
0
20
40
60
80
100
F II
F I
[Complexo] M: 0 2,5 5 10 20
F I
F II
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 17: Clivagem de DNA plasmidial pelo complexo 2 em
diferentes concentrações. Condições reacionais: [DNA] = 400 ng; 10
mM de Tris-HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 8h
na ausência de luz a 37°C.

29
0
20
40
60
80
100F I
F II
F II
F I
[Complexo] M: 0 2,5 5 10 20
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 18: Clivagem de DNA plasmidial pelo complexo 3 em
diferentes concentrações. Condições reacionais: [DNA] = 400 ng; 10
mM de Tris-HCl, pH 7,4; [complexo]: 0 a 20 µM. Tempo de reação: 8h
na ausência de luz a 37°C.
A partir dos resultados obtidos podemos observar que a atividade
de clivagem na ausência de luz também é dependente da concentração
do complexo, porém a clivagem foi reduzida para os três complexos e
nenhum deles foi capaz de clivar o DNA em ambas as fitas (F III) nestas
condições.
Complexos de Cu(II) com ligantes dpq e dppz reportados na
literatura não apresentaram clivagem de DNA quando as reações foram
incubadas por 15 minutos com 12,5 µM de complexo (SOUZA et al.,
2010a), por 30 minutos e 50 µM de complexo (PATRA; NETHAJI;
CHAKRAVARTY, 2007) ou por 30 minutos e 5 µM de complexo
(PATRA et al., 2008) na ausência de fotoativação. Para o complexo 1, Chen e colaboradores (2011) apontam esse complexo como
praticamente inativo na ausência de luz.
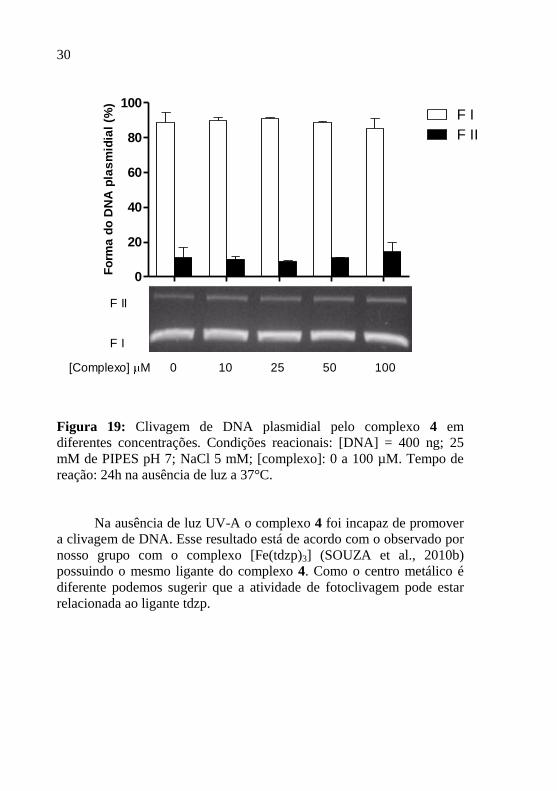
30
0
20
40
60
80
100F I
F II
F II
F I
[Complexo] M 0 10 25 50 100
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 19: Clivagem de DNA plasmidial pelo complexo 4 em
diferentes concentrações. Condições reacionais: [DNA] = 400 ng; 25
mM de PIPES pH 7; NaCl 5 mM; [complexo]: 0 a 100 µM. Tempo de
reação: 24h na ausência de luz a 37°C.
Na ausência de luz UV-A o complexo 4 foi incapaz de promover
a clivagem de DNA. Esse resultado está de acordo com o observado por
nosso grupo com o complexo [Fe(tdzp)3] (SOUZA et al., 2010b)
possuindo o mesmo ligante do complexo 4. Como o centro metálico é
diferente podemos sugerir que a atividade de fotoclivagem pode estar
relacionada ao ligante tdzp.

31
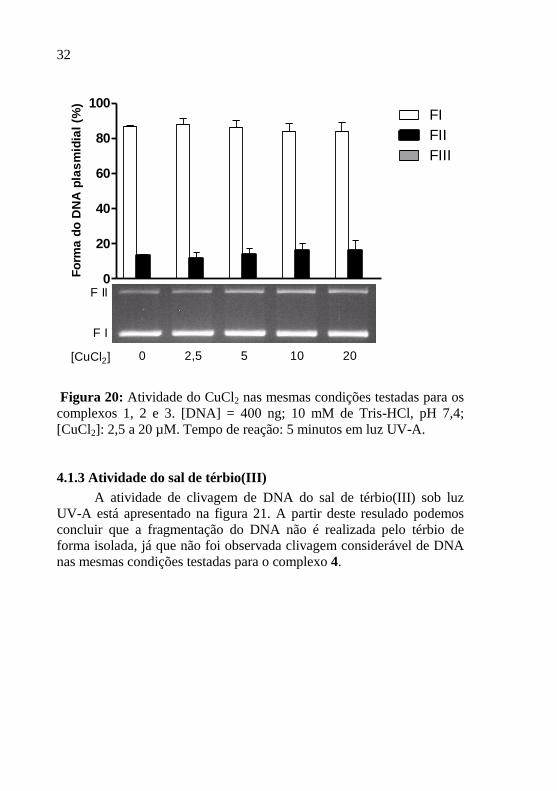
4.1.2. Atividade do sal de cobre(II) e dos ligantes dpq e dppz
Este ensaio foi realizado para descobrirmos o efeito do metal e
dos ligantes na clivagem de DNA plasmidial. A atividade de clivagem
de DNA do sal de cobre(II) sob luz UV-A está apresentado na figura 20.
Soluções homogêneas dos ligantes dos complexos não foram obtidas
devido à insolubilidade desses compostos e por esse motivo os testes
não foram realizados. A partir do resultado apresentado na figura 20
podemos assumir que a fragmentação do DNA não é realizada pelo
cobre de forma isolada, já que não se observou clivagem de DNA nos
testes com CuCl2 nas mesmas condições reacionais testadas para os
complexos. Diversos trabalhos relatam ausência de atividade dos
ligantes dppz e dpq, nas concentrações de 5 µM fotoirradiados por UV-
A durante trinta minutos (PATRA et al., 2008), 5 µM fotoirradiados por
UV-A durante uma hora (PATRA et al., 2009), 50 µM fotoirradiados
por UV-A durante uma hora (PATRA; NETHAJI; CHAKRAVARTY,
2007), 3,5 µM em luz UV-A por uma hora (CHEN et al., 2011). Os
ensaios dos trabalhos citados também foram realizados em tampão Tris-
HCl pH 7,2. Isso sugere que a clivagem de DNA observada pela
atividade dos complexos não é ocasionada pelo ligante dpq ou dppz
isolados dos complexos.
32
0
20
40
60
80
100FI
FII
FIII
F II
F I
[CuCl2] 0 2,5 5 10 20
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 20: Atividade do CuCl2 nas mesmas condições testadas para os
complexos 1, 2 e 3. [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[CuCl2]: 2,5 a 20 µM. Tempo de reação: 5 minutos em luz UV-A.
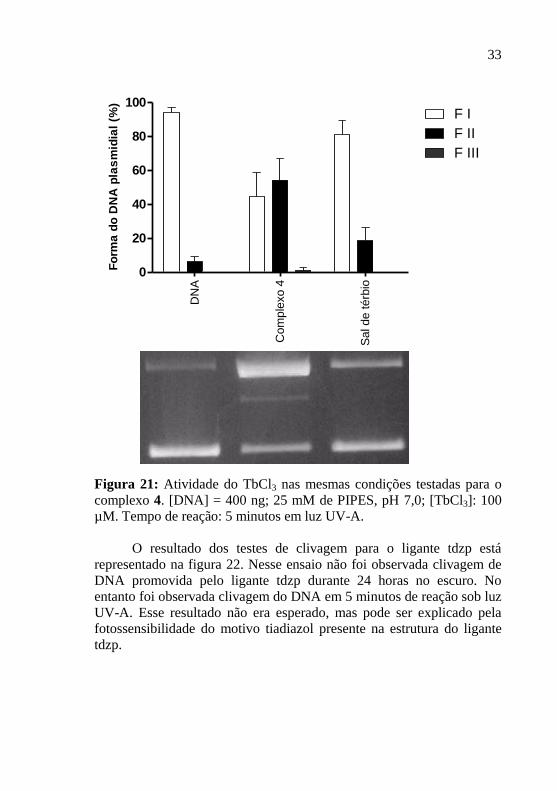
4.1.3 Atividade do sal de térbio(III)
A atividade de clivagem de DNA do sal de térbio(III) sob luz
UV-A está apresentado na figura 21. A partir deste resulado podemos
concluir que a fragmentação do DNA não é realizada pelo térbio de
forma isolada, já que não foi observada clivagem considerável de DNA
nas mesmas condições testadas para o complexo 4.
33
DN
A
Com
ple
xo 4
Sal de térb
io
0
20
40
60
80
100F I
F II
F III
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 21: Atividade do TbCl3 nas mesmas condições testadas para o
complexo 4. [DNA] = 400 ng; 25 mM de PIPES, pH 7,0; [TbCl3]: 100
µM. Tempo de reação: 5 minutos em luz UV-A.
O resultado dos testes de clivagem para o ligante tdzp está
representado na figura 22. Nesse ensaio não foi observada clivagem de
DNA promovida pelo ligante tdzp durante 24 horas no escuro. No
entanto foi observada clivagem do DNA em 5 minutos de reação sob luz
UV-A. Esse resultado não era esperado, mas pode ser explicado pela
fotossensibilidade do motivo tiadiazol presente na estrutura do ligante
tdzp.

34
0
20
40
60
80
100F I
F II
F III
F II
F II
[tdzp] M 0 50 100 0 50 100
Luz UV-A Escuro
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 22: Atividade do ligante tdzp nas mesmas condições testadas
para o complexo 4. [DNA] = 400 ng; 25 mM de PIPES, pH 7,0; [tdzp]:
50 e 100 µM. Tempo de reação: 5 minutos em luz UV-A e 24 horas no
escuro a 37°C.

35
4.1.3. Efeito da força iônica
O efeito da força iônica na clivagem de DNA pelos complexos
foi testado pelo aumento da concentração de NaCl nas misturas
reacionais. Como o DNA, em condições fisiológicas, apresenta carga
negativa devido ao grupo fosfato (DOMINGOS et al., 2003), moléculas
com comportamento de cátions teriam sua interação diminuída na
presença de NaCl. Nas figuras 23, 24, 25 e 26 podemos observar a
influência das interações eletrostáticas na clivagem de DNA pelos
complexos 1, 2, 3 e 4.
0
20
40
60
80
100FI
FII
FIII
[Complexo 1] (5 M)
NaCl
-
- -
+ ++++
50 100 200 300
Posição 1 2 3 4 5 6
FII
FI
FIII
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 23: Clivagem de DNA plasmidial pelo complexo 1 na ausência e
presença de diferentes concentrações de NaCl. Condições reacionais:
[DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexo]: 5 µM;
[NaCl]: 0 a 300 mM. Tempo de reação: 5 minutos em luz UV-A.

36
0
20
40
60
80
100FI
FII
FIII
[Complexo 2] (20 M)
NaCl
-
- -
+ ++++
50 100 200 300
FII
FIII
FI
Posição 1 2 3 4 5 6
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 24: Clivagem de DNA plasmidial pelo complexo 2 na ausência e
presença de diferentes concentrações de NaCl. Condições reacionais:
[DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexo]: 20 µM;
[NaCl]: 0 a 300 mM. Tempo de reação: 5 minutos em luz UV-A.
37
0
20
40
60
80
100FI
FII
FIII
[Complexo 3] (7,5 M)
NaCl
-
- -
+ ++++
50 100 200 300
FII
FIII
FI
Posição 1 2 3 4 5 6
Fo
rma
do
DN
A p
las
mid
ial
(%)
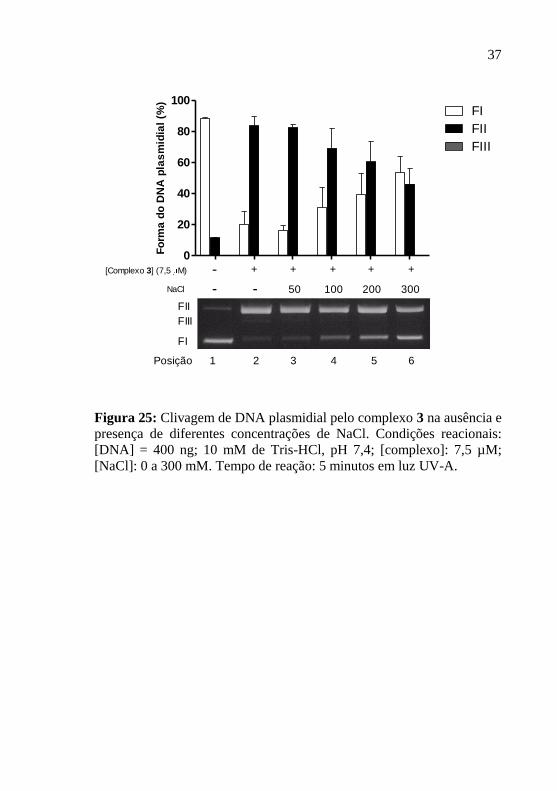
Figura 25: Clivagem de DNA plasmidial pelo complexo 3 na ausência e
presença de diferentes concentrações de NaCl. Condições reacionais:
[DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexo]: 7,5 µM;
[NaCl]: 0 a 300 mM. Tempo de reação: 5 minutos em luz UV-A.
38
0
20
40
60
80
100F I
F II
F II
F I
Posição 1 2 3 4 5 6
[Complexo 4] (25 M)
NaCl
-
- -
+ ++++
50 100 200 300
Fo
rma
do
DN
A p
las
mid
ial
(%)
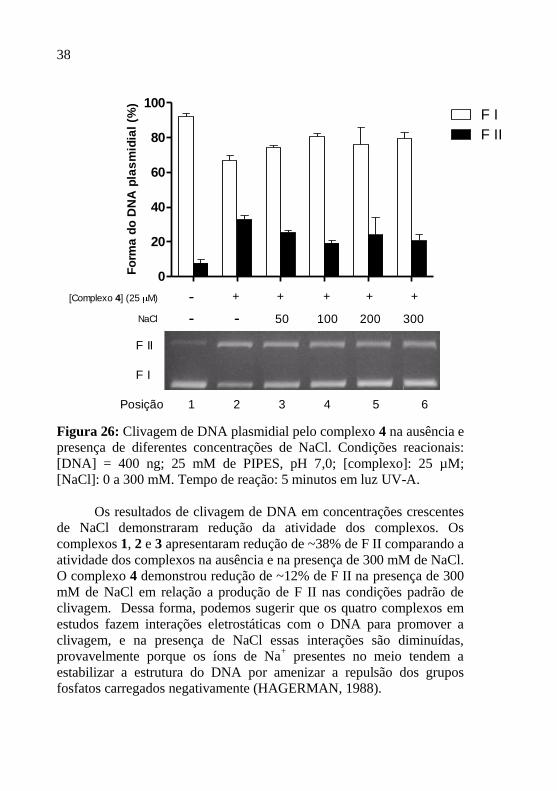
Figura 26: Clivagem de DNA plasmidial pelo complexo 4 na ausência e
presença de diferentes concentrações de NaCl. Condições reacionais:
[DNA] = 400 ng; 25 mM de PIPES, pH 7,0; [complexo]: 25 µM;
[NaCl]: 0 a 300 mM. Tempo de reação: 5 minutos em luz UV-A.
Os resultados de clivagem de DNA em concentrações crescentes
de NaCl demonstraram redução da atividade dos complexos. Os
complexos 1, 2 e 3 apresentaram redução de ~38% de F II comparando a
atividade dos complexos na ausência e na presença de 300 mM de NaCl.
O complexo 4 demonstrou redução de ~12% de F II na presença de 300
mM de NaCl em relação a produção de F II nas condições padrão de
clivagem. Dessa forma, podemos sugerir que os quatro complexos em
estudos fazem interações eletrostáticas com o DNA para promover a
clivagem, e na presença de NaCl essas interações são diminuídas, provavelmente porque os íons de Na
+ presentes no meio tendem a
estabilizar a estrutura do DNA por amenizar a repulsão dos grupos
fosfatos carregados negativamente (HAGERMAN, 1988).

39
4.1.4. Efeito dos sequestradores de radicais livres e D2O
Para avaliarmos o mecanismo de clivagem de DNA pelos
complexos foram realizados testes de clivagem na presença de
sequestradores de radicais livres e D2O. Os resultados obtidos estão
apresentados nas figuras 27, 28, 29, 30 e 31. D
NA
Co
mp
lexo
1
DM
SO KI
SO
D O2
D
3N
aN
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
Posição 1 2 3 4 5 6 7
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 27: Clivagem de DNA plasmidial pelo complexo 1 na ausência e
presença de diferentes sequestradores de radicais livres e D2O.
Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[complexo]: 5 µM; [DMSO]: 10% (v/v); [KI]: 10 mM; [SOD]: 15 U;
[D2O]: 70% (v/v); [NaN3]: 10 mM.Tempo de reação: 5 minutos em luz
UV-A.

40
DN
A
Com
ple
xo 2
DM
SO KI
SO
D O2
D
3N
aN
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
Posição 1 2 3 4 5 6 7
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 28: Clivagem de DNA plasmidial pelo complexo 2 na ausência e
presença de diferentes sequestradores de radicais livres e D2O.
Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[complexo]: 20 µM; [DMSO]: 10% (v/v); [KI]: 10 mM; [SOD]: 15 U;
[D2O]: 70% (v/v); [NaN3]: 10 mM Tempo de reação: 5 minutos em luz
UV-A.
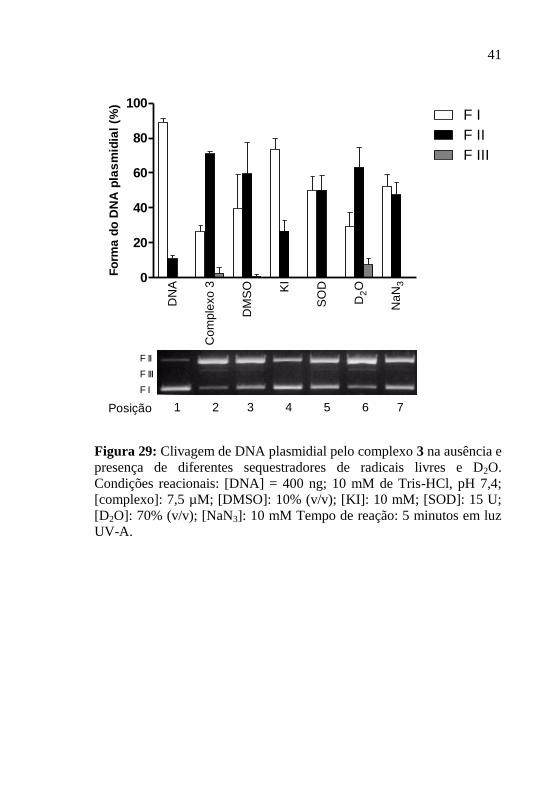
41
DN
A
Co
mple
xo
3
DM
SO KI
SO
D O2
D
3N
aN
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
Posição 1 2 3 4 5 6 7
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 29: Clivagem de DNA plasmidial pelo complexo 3 na ausência e
presença de diferentes sequestradores de radicais livres e D2O.
Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4;
[complexo]: 7,5 µM; [DMSO]: 10% (v/v); [KI]: 10 mM; [SOD]: 15 U;
[D2O]: 70% (v/v); [NaN3]: 10 mM Tempo de reação: 5 minutos em luz
UV-A.

42
DN
A
Com
ple
xo 1
Com
ple
xo 2
Com
ple
xo 3
DN
A +
TE
MP
O
1 +
TE
MP
O
2 +
TE
MP
O
3 +
TE
MP
O
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
Posição 1 2 3 4 5 6 7 8
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 30: Clivagem de DNA plasmidial pelos complexos 1, 2 e 3 na
ausência e presença de TEMPO. Condições reacionais: [DNA] = 400
ng; 10 mM de Tris-HCl, pH 7,4; [complexos]: 5; 20 e 7,5 µM; TEMPO:
500 µM. Tempo de reação: 5 minutos em luz UV-A.
A fotoclivagem do DNA pelos complexos em estudo foi reduzida
quando adicionados os sequestradores de radicais livres (com exceção
da SOD para os complexos 1 e 2). O complexo 1 apresentou 71% de F
II na ausência de KI e 30% na presença de KI e quando o TEMPO foi
adicionado a F II diminui de 69% para 36%. A presença de DMSO e
NaN3 também inibiu parcialmente a clivagem de DNA pelo complexo
indicando a presença do radical hidroxila e oxigênio singlete. No
entanto a presença de D2O aumentou levemente a atividade do
complexo. O resultado encontrado por Chen e colaboradores (2011)

43
para o complexo 1 sugere o envolvimento de oxigênio singlete no
mecanismo de clivagem de DNA pois a adição de DMSO e KI não
reduziram a atividade do complexo. Os resultados do complexo 2
demonstraram maior redução de clivagem quando KI foi adicionado à
reação, 63% de F II para o tratamento com o complexo e 34% de F II na
presença de complexo e KI. Nas reações com TEMPO a F II diminui de
43% na presença apenas do complexo para 29%. Na presença de NaN3
foi observado aumento da atividade do complexo, esse evento pode ter
ocorrido pela formação do radical azidil como já descritos em outros
trabalhos. A formação do radical azidil pode ocorrer quando o cobre e
excesso de H2O2 oxidam a azida (YAMAMOTO; KAWANISHI, 1989;
LI et al., 1995; RAMIREZ; GOMEZ MEJIBA; MASON, 2005). Esse
resultado sugere formação de H2O2 para formação do radical azidil
aumentando assim a atividade do complexo e explicando a perda de
atividade na presença de KI. A redução de clivagem de DNA do
complexo 3 na presença de KI foi de 70% de F II (tratamento com
complexo) para 26% (adição de KI). A adição de DMSO, SOD e NaN3
também reduziram a clivagem de DNA mas de maneira menos evidente,
indicando a presença de radicais hidroxila, ânion superóxido e oxigênio
singlete. Na presença do TEMPO a redução de F II foi de 29%. Esses
resultados sugerem clivagem oxidativa e participação de várias espécies
radicalares na clivagem de DNA pelos complexos 1, 2 e 3.
Nas condições de fotoclivagem testadas podemos sugerir que a
formação de radicais livres é Cu(I)-dependente, a redução do Cu(II) para
Cu(I) pode ser explicada pela fotoativação do motivo
quinoxalina/fenazina dos ligantes dpq e dppz (TOSHIMA et al., 2002).
Após a formação de Cu(I), este pode participar de reações tipo-Fenton
nas quais ele reage com H2O2 formando OHł (HALLIWELL;
GUTTERIDGE, 2007).
Reação tipo-Fenton:
Cu(I) + H2O2 ł Cu(II) + OHł + OH
-
O Cu(I) também pode reagir com o oxigênio molecular gerando
O2ł-
que se dismuta em H2O2 e participa na geração de OHł pela reação
de Fenton (SOUZA et al., 2010a).

44
DN
A
Co
mple
xo
4
DM
SO KI 3
Na
N
TE
MP
O
0
20
40
60
80
100F I
F II
F II
F I
Posição 1 2 3 4 5 6
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 31: Clivagem de DNA plasmidial pelo complexo 4 na ausência e
presença de diferentes sequestradores de radicais livres. Condições
reacionais: [DNA] = 400 ng; 25 mM de PIPES, pH 7,0; [complexo]: 25
µM; [DMSO]: 10% (v/v); [KI]: 10 mM; [NaN3]: 10 mM; [TEMPO]:
500 µM Tempo de reação: 5 minutos em luz UV-A.
O complexo 4 apresentou redução de atividade na presença de
DMSO, KI, NaN3 e TEMPO (figura 31). Esses resultados sugerem
clivagem oxidativa do DNA pela formação de radicais livres de
oxigênio e carbono-centrados. O complexo [Fe(tdzp)3] possui o mesmo
ligante tdzp e também apresentou diminuição da sua atividade de
clivagem na presença do inibidor de radicais carbono-centrados
(SOUZA et al., 2010b).

45
4.1.5. Efeito da presença e ausência de oxigênio
Os testes em atmosfera de argônio foram realizados para que se
pudesse avaliar se o oxigênio molecular é necessário para clivagem de
DNA pelos complexos. Os resultados da atividade dos complexos na
ausência de oxigênio estão representados na figura 32 e 34. Nas figuras
33 e 35 estão os resultados das reações na presença de oxigênio
molecular, as quais foram realizadas em paralelo às reações na
atmosfera de argônio.
DN
A
Fe-E
DT
A
Com
ple
xo 1
Com
ple
xo 2
Com
ple
xo 3
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
Posição 1 2 3 4 5
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 32: Clivagem de DNA plasmidial por Fe-EDTA e pelos
complexos 1, 2 e 3 na ausência de oxigênio, em atmosfera de argônio.
Condições reacionais: [DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexos]: 5; 20 e 7,5 µM. Tempo de reação: 5 minutos em luz UV-
A.

46
DN
A
Fe
-ED
TA
Co
mple
xo
1
Co
mple
xo
2
Co
mple
xo
3
0
20
40
60
80
100F I
F II
F III
F II
F III
F I
Posição 1 2 3 4 5
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 33: Clivagem de DNA plasmidial por Fe-EDTA e pelos
complexos 1, 2 e 3 na presença de oxigênio. Condições reacionais:
[DNA] = 400 ng; 10 mM de Tris-HCl, pH 7,4; [complexos]: 5; 20 e 7,5
µM. Tempo de reação: 5 minutos em luz UV-A.
A fotoclivagem de DNA pelos complexos 1, 2 e 3 em atmosfera
de argônio foi aumentada em relação à fotoclivagem na presença de
oxigênio. Podemos observar que o complexo 3 clivou toda forma
superenovelada (F I) do DNA em F II e F III. A sugestão que pode ser
feita para esse comportamento seria a atuação do oxigênio molecular
como sequestrador dos radicais produzidos pelos complexos, dessa
forma estaria competindo com o DNA pelos radicais e por isso a
atividade dos complexos é aumentada na ausência do oxigênio
molecular.

47
DN
A
+ M
PA
2C
uC
l
Com
ple
xo 4
DN
A +
TE
MP
O
Com
ple
xo 4
+ T
EM
PO
0
20
40
60
80
100F I
F II
F II
F I
1 2 3 4 5Posição
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 34: Clivagem de DNA plasmidial por CuCl2 + MPA e pelo
complexo 4 na ausência de oxigênio. Condições reacionais: [DNA] =
400 ng; 25 mM de PIPES, pH 7,0; NaCl 5 mM; [complexo]: 25 µM.
Tempo de reação: 5 minutos em luz UV-A.

48
DN
A
+ M
PA
2C
uC
l
Com
ple
xo 4
DN
A +
TE
MP
O
Com
ple
xo 4
+ T
EM
PO
0
20
40
60
80
100F I
F II
F III
F II
F I
1 2 3 4 5Posição
F III
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 35: Clivagem de DNA plasmidial por CuCl2 + MPA e pelo
complexo 4 na presença de oxigênio. Condições reacionais: [DNA] =
400 ng; 25 mM de PIPES, pH 7; NaCl 5 mM; [complexo]: 25 µM.
Tempo de reação: 5 minutos em luz UV-A.
A fotoclivagem de DNA pelo complexo 4 não foi inibida em
atmosfera de argônio, no entanto na presença do sequestrador de radicais carbono-centrado, TEMPO, na ausência de oxigênio, a
clivagem foi totalmente inibida. Esse resultado corrobora o encontrado
por Souza e colaboradores (2010), com o complexo [Fe(tdzp)3] no qual
foi sugerido que o motivo tiadiazol no ligante tdzp forma um radical
tdzp-centrado após a exposição a luz UV-A, o qual oxidaria a estrutura

49
do DNA e consequentemente promove a quebra da fita. No entanto, o
complexo 4 parece agir também pela formação de oxi-radicais, os quais
poderiam ser gerados pela reação do radical tdpz-centrado com o
oxigênio molecular.
4.1.6. Efeito dos ligantes dos sulcos do DNA
Os dois ligantes dos sulcos do DNA, verde metila e distamicina,
utilizados nos ensaios promoveram clivagem de DNA nas condições
testadas para os complexos 1, 2 e 3 (10 mM de Tris-HCl pH7,4 e
fotoativação em luz UV-A por cinco minutos) e por esse motivo os
resultados foram desconsiderados. Os resultados encontrados para o
complexo 4 estão na figura 36 e demonstram que nenhum dos ligantes
de sulco reduziram a clivagem de DNA promovida pelo complexo 4, o
que sugere que a ligação específica à um dos sulcos não é necessária
para o evento de clivagem.

50
DN
A
DN
A +
Dis
tam
icin
a
DN
A +
Ve
rde
de m
etila
Com
ple
xo 4
Com
p 4
+ d
ista
mic
ina
Com
p 4
+ v
erd
e d
e m
etila
0
20
40
60
80
100F I
F II
F II
F I
Posição 1 2 3 4 5 6
Fo
rma
do
DN
A p
las
mid
ial
(%)
Figura 36: Clivagem de DNA plasmidial pelo complexo 4 na ausência e
presenças dos ligantes dos sulcos de DNA. Condições reacionais:
[DNA] = 400 ng; 25 mM de PIPES, pH 7,0; NaCl 5 mM; [complexo]:
25 µM; [distamicina]: 50 µM; [verde de metila]: 50 µM. Tempo de
reação: 5 minutos em luz UV-A.

51
4.3.7. Cinética de clivagem do DNA
Os perfis cinéticos dos complexos 1, 2, 3 e 4 foram analisados
tanto na presença de luz UV-A quanto no escuro. Os resultados dos
ensaios cinéticos sob luz UV-A para os complexos 1, 2, 3 e 4 estão
representados nas figuras 37, 38, 39 e 40.
0 20 40 603.0
3.5
4.0
4.5
5.0
kobs = 0,89 min-1
t1/2 = ~46 s
Tempo (s)
ln [
DN
A s
up
ern
ov
ela
do
%]
Figura 37: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 1. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 75 segundos
em luz UV-A.

52
0 20 40 603.8
4.0
4.2
4.4
4.6
4.8
kobs = 0,25 min-1
t1/2 = 2,8 min
Tempo (s)
ln [
DN
A s
up
ern
ov
ela
do
%]
Figura 38: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 2. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 75 segundos
em luz UV-A.

53
0 20 40 603.0
3.5
4.0
4.5
5.0
kobs = 0,92 min-1
t1/2 = ~44 s
Tempo (s)
ln [
DN
A s
up
ern
ov
ela
do
%]
Figura 39: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 3. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 75 segundos
em luz UV-A.
A constante observada de clivagem de DNA (Kobs) para o
complexo 1 na presença de luz UV-A foi de 0,89 min-1
, a qual resulta
em um t1/2 de ~46 segundos para o DNA superenovelado. Nas mesmas
condições o complexo 2 apresentou um kobs de 0,25 min-1
que representa
um t1/2 de ~2,8 minutos. O complexo 3 apresentou um resultado bastante
similar ao do complexo 1, kobs de 0,92 e um t1/2 de ~44 segundos para o
DNA superenovelado. Os complexos de Cu(II), Cu(dox)(phen) e
Cu(tc)(phen), quando fotoativados em luz UV-A apresentaram
resultados semelhantes com um t1/2 de ~ 30 segundos (BORTOLOTTO,
2011).

54
0 1 2 3 4 53.0
3.5
4.0
4.5
5.0
kobs = 0,17 min-1
t1/2 = ~4 min
Tempo (min)
ln [
DN
A s
up
ern
ov
ela
do
%]
Figura 40: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 4. Condições reacionais: [DNA] = 400 ng; 25 mM de
PIPES, pH 7,0; [complexo]: 100 µM. Tempo de reação: 5 minutos em
luz UV-A.
Na presença de 100 µM do complexo 4 o Kobs da reação foi
estimado em 0,17 min-1
resultando em um tempo de meia vida (t1/2) de
aproximadamente 4 minutos para o DNA nesta reação. O complexo
[Fe(tdzp)3] (o qual possui o mesmo ligante) apresentou um kcat (kobs no
nível de saturação) de 0,31 min-1
em condições reacionais similares
(SOUZA et al., 2010b).
Os resultados dos ensaios cinéticos no escuro para os complexos
1, 2, 3 e 4 estão representados nas figuras 41, 42, 43 e 44.

55
0 2 4 6 82.5
3.0
3.5
4.0
4.5
5.0
Kobs = 0,15 h-1
t1/2 = 4,4 h
Tempo (h)
ln [
DN
A s
up
ern
ov
ela
do
%]
Figura 41: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 1. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 9 horas no
escuro a 37°C.

56
0 2 4 6 83.5
4.0
4.5
5.0
Kobs = 0,08 h-1
t1/2 = 8,1h
Tempo (h)
ln [
DN
A s
up
ern
ov
ela
do
%]
Figura 42: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 2. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 9 horas no
escuro a 37°C.

57
0 2 4 6 83.8
4.0
4.2
4.4
4.6
4.8
Kobs = 0,035 h-1
t1/2 = 19,8 h
Tempo (h)
ln [
DN
A s
up
ern
ov
ela
do
%]
Figura 43: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 3. Condições reacionais: [DNA] = 400 ng; 10 mM de
Tris-HCl, pH 7,4; [complexos]: 100 µM. Tempo de reação: 9 horas no
escuro a 37°C.
Na ausência da luz UV-A os complexos apresentaram perfis
cinéticos inferiores como era esperado. Estes ensaios reforçam a
importância da fotoativação para uma melhor eficiência na atividade dos
complexos. No escuro o complexo 1 apresentou um Kobs de 0,15 h-1
e
um t1/2 de ~4,4 h, o Kobs para o complexo 2 foi de 0,08 h-1
gerando um
t1/2 de ~8 h. Entre os complexos de Cu(II), o complexo 3 é o que
apresenta maior redução de atividade na ausência da luz UV-A, Kobs de
0,035 h-1
e t1/2 de ~19 h. Bortolotto (2011) também descreve Kobs
menores para os complexos Cu(dox)(phen) e Cu(tc)(phen) na ausência
da fotoativação por luz UV-A.

58
0 10 20 30 40 504.0
4.2
4.4
4.6
4.8
kobs: 0,0032 h-1
t1/2: ~211 h
Tempo (h)
ln [
DN
A s
up
ern
ov
ela
do
%]
Figura 44: Gráfico do logaritmo neperiano do DNA plasmidial
superenovelado versus tempo (segundos) de reação após o tratamento
com o complexo 4. Condições reacionais: [DNA] = 400 ng; 25 mM de
PIPES, pH 7,0; [complexo]: 100 µM. Tempo de reação: 48h no escuro a
37°C.
A reação de clivagem pelo complexo 4 na ausência de luz UV-A
apresentou Kobs de 0,0032 h-1
e tempo de meia vida (t1/2) de
aproximadamente 211 horas para o DNA. Esse resultado demonstra a
importância da fotoativação para o aumento da velocidade da reação.
Quando a reação é fotoativada o tempo de meia vida do DNA diminui
aproximadamente três mil vezes.

59
4.2. Interação dos complexos com CT-DNA
4.2.1. Titulação espectrofotométrica (UV-Vis)
A realização deste experimento permitiu o cálculo da constante
intrínseca de ligação (Kb) dos complexos com o DNA e a determinação
da forma de ligação pelas mudanças observadas nos espectros. Os
resultados estão apresentados nas figuras 45, 46 e 47.
350 360 370 380 390 4000.00
0.05
0.10
0.15
0.20 Kb = 4,45 x 105 M
-1
(nm)
Ab
so
rbâ
nc
ia
Figura 45: Titulação espectrofotométrica do complexo 1 na presença de
diferentes concentrações de CT-DNA. Condições reacionais: [CT-
DNA]: 0,8 a 7,8 µM; 5 mM de Tris-HCl, pH 7,4; [complexo]: 10 µM;
temperatura: 37°C.

60
350 360 370 380 390 4000.00
0.02
0.04
0.06
0.08
0.10Kb = 3,73 x 10
5 M
-1
(nm)
Ab
so
rbâ
nc
ia
Figura 46: Titulação espectrofotométrica do complexo 2 na presença de
diferentes concentrações de CT-DNA. Condições reacionais: [CT-
DNA]: 1 a 9,8 µM; 5 mM de Tris-HCl, pH 7,4; [complexo]: 10 µM;
temperatura: 37°C.
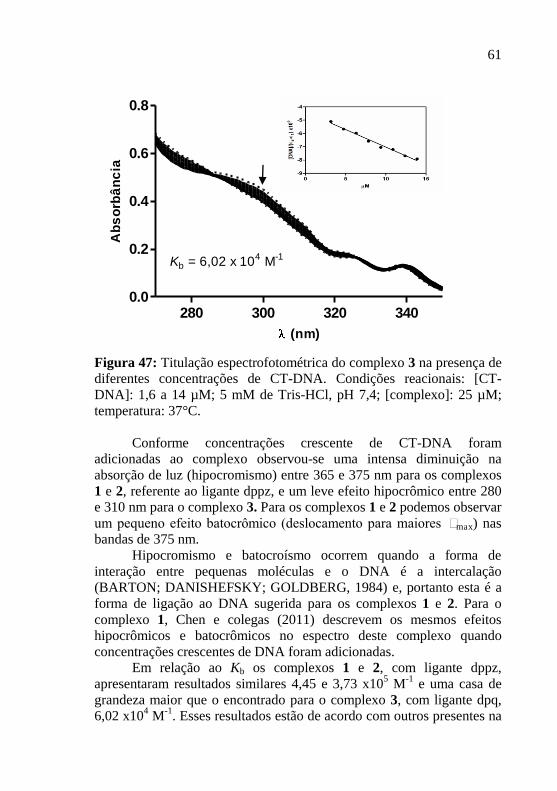
61
280 300 320 3400.0
0.2
0.4
0.6
0.8
Kb = 6,02 x 104 M
-1
(nm)
Ab
so
rbâ
nc
ia
Figura 47: Titulação espectrofotométrica do complexo 3 na presença de
diferentes concentrações de CT-DNA. Condições reacionais: [CT-
DNA]: 1,6 a 14 µM; 5 mM de Tris-HCl, pH 7,4; [complexo]: 25 µM;
temperatura: 37°C.
Conforme concentrações crescente de CT-DNA foram
adicionadas ao complexo observou-se uma intensa diminuição na
absorção de luz (hipocromismo) entre 365 e 375 nm para os complexos
1 e 2, referente ao ligante dppz, e um leve efeito hipocrômico entre 280
e 310 nm para o complexo 3. Para os complexos 1 e 2 podemos observar
um pequeno efeito batocrômico (deslocamento para maiores łmax) nas
bandas de 375 nm.
Hipocromismo e batocroísmo ocorrem quando a forma de
interação entre pequenas moléculas e o DNA é a intercalação
(BARTON; DANISHEFSKY; GOLDBERG, 1984) e, portanto esta é a
forma de ligação ao DNA sugerida para os complexos 1 e 2. Para o
complexo 1, Chen e colegas (2011) descrevem os mesmos efeitos
hipocrômicos e batocrômicos no espectro deste complexo quando
concentrações crescentes de DNA foram adicionadas. Em relação ao Kb os complexos 1 e 2, com ligante dppz,
apresentaram resultados similares 4,45 e 3,73 x105 M
-1 e uma casa de
grandeza maior que o encontrado para o complexo 3, com ligante dpq,
6,02 x104 M
-1. Esses resultados estão de acordo com outros presentes na

62
literatura, tanto nos valores encontrados de Kb como na ordem de maior
ligação para complexos de Cu com ligantes dppz em relação a
complexos de cobre com ligantes dpq (GUPTA et al., 2004; DHAR;
NETHAJI; CHAKRAVARTY, 2006; SELVAKUMAR et al., 2006;
PATRA et al., 2008; ROY et al., 2008b; LAHIRI et al., 2009; CHEN et
al., 2011).
4.2.2. Desnaturação térmica do DNA
Como a ligação de pequenas moléculas com o DNA pode alterar
a Tm do mesmo, nesse ensaio foi realizada a desnaturação térmica do
CT-DNA na ausência e presença dos complexos. O resultado pode ser
observado na figura 48.
A Tm do DNA livre dos complexos foi de 58,0°C. Na presença do
complexo 1 a Tm aumentou para 60,4°C, na presença do complexo 2 o
aumento foi para 60,2°C e na presença do complexo 3 para 59,8°C. O
aumento da Tm confirma a capacidade de ligação dos complexos com o
DNA e sugere um aumento da estabilidade da dupla-hélice dificultando
a desnaturação (BORTOLOTTO, 2011). Esses resultados concordam
com as constantes de ligação intrínseca calculadas para os complexos,
os complexos 1 e 2 com os ligantes dppz apresentaram maior Kb e maior
variação de Tm em relação ao complexo 3 com o ligante dpq.

63
40 50 60 70 80 900
20
40
60
80
100
DNA
Complexo 1
Complexo 2
Complexo 3
Temperatura (ºC)
Ab
so
rbâ
nc
ia r
ela
tiv
a
Figura 48: Desnaturação térmica de CT-DNA na ausência e presença
dos complexos 1, 2 e 3. Condições reacionais: [CT-DNA]: 100 µM; 1
mM tampão fosfato pH 7,2; [complexo]: 10 µM. Temperatura: 40 a
90°C, um grau por minuto.
4.2.3. Espectroscopia de dicroísmo circular (CD)
Através da técnica de CD podemos avaliar a estrutura secundária
do CT-DNA, empilhamento das bases e/ou helicidade direita da dupla
hélice, na presença e ausência dos complexos. Nas figuras 49, 50 e 51
estão os resultados obtidos.

64
220 240 260 280 300 320-4
-2
0
2
4
6CT-DNA
r=0,05
r=0,1
r=0,15
r=0,2
(nm)
CD
(m
deg
)
Figura 49: Espectro de dicroísmo circular de CT-DNA na ausência e
presença do complexo 1. Condições reacionais: [CT-DNA]: 100 µM;
5mM de Tris-HCl pH 7,4; [complexo]: 5 a 16 µM.
220 240 260 280 300 320-4
-2
0
2
4
6CT- DNA
r=0,1
r=0,15
r=0,2
r=0,25
(nm)
CD
(m
deg
)
Figura 50: Espectro de dicroísmo circular de CT-DNA na ausência e
presença do complexo 2. Condições reacionais: [CT-DNA]: 100 µM;
5mM de Tris-HCl pH 7,4; [complexo]: 5 a 20 µM.
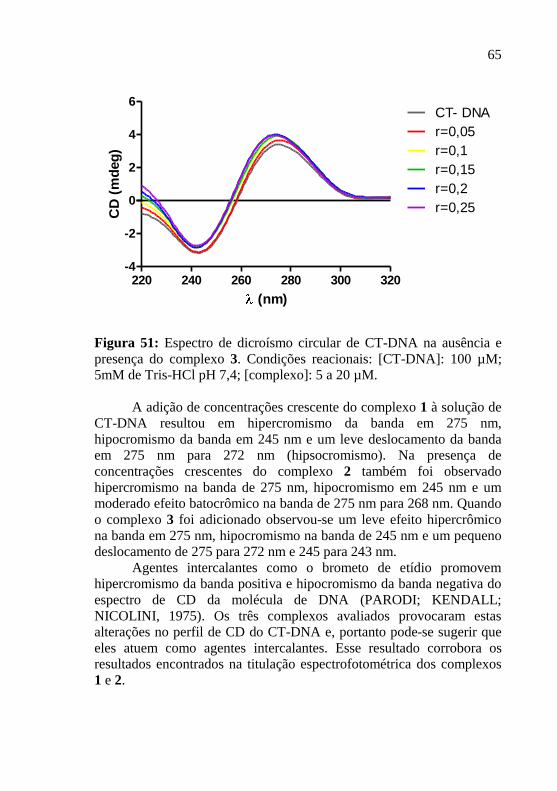
65
220 240 260 280 300 320-4
-2
0
2
4
6CT- DNA
r=0,05
r=0,1
r=0,15
r=0,2
r=0,25
(nm)
CD
(m
deg
)
Figura 51: Espectro de dicroísmo circular de CT-DNA na ausência e
presença do complexo 3. Condições reacionais: [CT-DNA]: 100 µM;
5mM de Tris-HCl pH 7,4; [complexo]: 5 a 20 µM.
A adição de concentrações crescente do complexo 1 à solução de
CT-DNA resultou em hipercromismo da banda em 275 nm,
hipocromismo da banda em 245 nm e um leve deslocamento da banda
em 275 nm para 272 nm (hipsocromismo). Na presença de
concentrações crescentes do complexo 2 também foi observado
hipercromismo na banda de 275 nm, hipocromismo em 245 nm e um
moderado efeito batocrômico na banda de 275 nm para 268 nm. Quando
o complexo 3 foi adicionado observou-se um leve efeito hipercrômico
na banda em 275 nm, hipocromismo na banda de 245 nm e um pequeno
deslocamento de 275 para 272 nm e 245 para 243 nm.
Agentes intercalantes como o brometo de etídio promovem
hipercromismo da banda positiva e hipocromismo da banda negativa do
espectro de CD da molécula de DNA (PARODI; KENDALL;
NICOLINI, 1975). Os três complexos avaliados provocaram estas
alterações no perfil de CD do CT-DNA e, portanto pode-se sugerir que
eles atuem como agentes intercalantes. Esse resultado corrobora os resultados encontrados na titulação espectrofotométrica dos complexos
1 e 2.

66
5. CONCLUSÕES
Os três complexos de cobre(II), [Cu(acac)dppz](ClO4),
[Cu(bpma)dppz](ClO4)2.0,67H2O e [Cu(diaza)dpq](ClO4).H2O foram
capazes de clivar o DNA plasmidial em concentrações micromolares e
pH próximo da neutralidade, na presença e ausência da luz UV-A. No
entanto, a atividade dos mesmos é melhorada quando ocorre a
fotoativação. O complexo [Tb(tdzp)(acac)3] também foi capaz de clivar
o DNA plasmidial em concentrações micromolares e em pH 7,0 quando
fotoativado, porém, em 24 horas de teste na ausência de luz UV-A não
foi observada a clivagem do DNA plasmidial. Os sais de cobre e térbio
não foram capazes de clivar o DNA plasmidial nas mesmas condições
testadas para os complexos. Os quatro complexos tiveram sua atividade
diminuída pelo aumento da força iônica, esse resultado indica a
participação de interações eletrostáticas entre os complexos e o DNA
para que ocorra a clivagem.
A utilização de sequestradores de radicais livres de oxigênio e
carbono-centrado reduziu a atividade dos complexos metálicos testados,
no entanto nenhum sequestrador de radical foi capaz de inibir totalmente
a atividade dos complexos. O mecanismo de clivagem é oxidativo e
sugere-se a formação de radical hidroxila, oxigênio singlete, radicais
peróxidos e carbono-centrado. Os ligantes dpq e dppz estão envolvidos
no processo de geração de radicais de oxigênio e carbono-centrado por
possuírem motivos fotossensíveis, assim como o motivo tiadiazol do
ligante tdzp gera o radical tdzp-centrado após exposição à luz UV-A. Na
presença da enzima superóxido dismutase o complexo 2 foi capaz de
gerar o radical azidil e aumentar seu potencial de clivagem. Em
atmosfera de argônio a atividade dos complexos 1, 2 e 3 não foi
diminuída provavelmente pela maior formação de radicais carbono-
centrados. O complexo 4 também não apresentou redução da atividade
em atmosfera de argônio, no entanto quando o sequestrador TEMPO foi
adicionado a clivagem foi totalmente inibida, confirmando a
participação do radical tdzp-centrado.
Os ensaios cinéticos com os quatro complexos demonstraram
diminuição do tempo de meia-vida do DNA superenovelado tanto na presença quanto na ausência de fotoativação por luz UV-A, no entanto a
fotoativação dos complexos acelera velocidade de clivagem do DNA.
Os complexos com ligantes dppz, 1 e 2, interagem de forma mais
forte com o CT-DNA em relação ao complexo 3, com ligante dpq. Os

67
três complexos foram capazes de aumentar a Tm do CT-DNA, as
maiores alterações foram observadas nos complexos 1 e 2. Os
complexos 1, 2 e 3 provocaram hipercromismo da banda positiva e
hipocromismo da banda negativa do espectro de CD da molécula de
DNA, o mesmo perfil encontrado para agentes intercalantes de DNA.

68
6. PERSPECTIVAS
Determinar os sítios de clivagem pelos quais os complexos
[Cu(acac)dppz](ClO4), [Cu(bpma)dppz](ClO4)2.0,67H2O ,
[Cu(diaza)dpq](ClO4).H2O e [Tb(tdzp)(acac)3] clivam o DNA
por espectrometria de massa e DNA footprinting.
Determinar as constantes de associação e dissociação dos quatro
complexos com o DNA por calorimetria de titulação isotérmica
(ITC, do inglês, Isothermal Titration Calorimetry).
Testar a atividade dos complexos em outros substratos de DNA
(linear dupla-fita e simples-fita).
Verificar os produtos de oxidação gerados pelos complexos por
cromatografia líquida acoplada à espectrometria de massas.

69
7. REFERÊNCIAS
AFRASIABI, Z. et al. Appended 1,2-naphthoquinones as anticancer agents 1:
synthesis, structural, spectral and antitumor activities of ortho-naphthaquinone
thiosemicarbazone and its transition metal complexes. Inorganica Chimica Acta, v.
357, n. 1, p. 271-278, 2004.
ARIAS, M. S. et al. Synthesis and copper-mediated nuclease activity of a
tetracationic tris(2,2 ł-bipyridine) ligand. Journal of Inorganic Biochemistry, v.
103, n. 7, p. 1067-1073, 2009.
ARMITAGE, B. Photocleavage of nucleic acids. Chemical Reviews, v. 98, n. 3, p.
1171-1200, 1998.
BAILLY, C.; CHAIRS, J. B. Sequence-Specific DNA Minor Groove Binders.
Design and Synthesis of Netropsin and Dystamicin analogues. Bioconjugate
Chemistry, v. 9, n. 5, 1998.
BARTON, J. K.; DANISHEFSKY, A.; GOLDBERG, J.
Tris(phenanthroline)ruthenium(II): stereoselectivity in binding to DNA. Journal of
the American Chemical Society, v. 106, n. 7, p. 2172-2176, 1984.
BASHKIN, J. K. Hydrolysis of phosphates, esters and related substrates by models
of biological catalysts. Current Opinion in Chemical Biology, v. 3, n. 6, p. 752-
758, 1999.
BERNADOU, J. et al. Potassium monopersulfate and a water-soluble manganese
porphyrin complex, [Mn(TMPyP)](OAc)5, as an efficient reagent for the oxidative
cleavage of DNA. Biochemistry, v. 28, n. 18, p. 7268-7275, 1989.
BORTOLOTTO, T. Interação e clivagem de DNA por novos complexos de
Cobre(II) derivados de tetraciclinas e 1,10-fenantrolina. Dissertação de
Mestrado. Universidade Federal de Santa Catarina. Florianópolis, 2011.
BRANUM, M. E.; QUE JR, L. Double-strand DNA hydrolysis by dilanthanide
complexes. Journal of Biological Inorganic Chemistry, v. 4, n. 5, p. 593-600,
1999.

70
BRANUM, M. E. et al. Double-Strand Hydrolysis of Plasmid DNA by Dicerium
Complexes at 37 °C. Journal of the American Chemical Society, v. 123, n. 9, p.
1898-1904, 2001a.
BRANUM, M. E. et al. Double-strand hydrolysis of plasmid DNA by dicerium
complexes at 37°C. Journal of the American Chemical Society, v. 123, n. 9, p.
1898-1904, 2001b.
BREEN, A. P.; MURPHY, J. A. Reactions of oxyl radicals with DNA. Free
Radical Biology and Medicine, v. 18, n. 6, p. 1033-1077, 1995.
CAMARGO, M. A. et al. Efficient phosphodiester hydrolysis by luminescent
terbium(III) and europium(III) complexes. Inorganic Chemistry, v. 49, n. 13, p.
6013-6025, 2010a.
CAMARGO, M. A. et al. New gadolinium complex with efficient hydrolase-like
activity: A 100-million-fold rate enhancement in diester hydrolysis. Inorganic
Chemistry, v. 47, n. 8, p. 2919-2921, 2008.
CAMARGO, M. A. et al. Synthesis, structure, and phosphatase-like activity of a
new trinuclear Gd complex with the unsymmetrical ligand H3L as a model for
nucleases. Inorganic Chemistry, v. 49, n. 6, p. 3057-3063, 2010b.
CHAKRAVARTY, A. R. Photocleavage of DNA by copper(II) complexes. Journal
of Chemical Sciences, v. 118, n. 6, p. 443-453, 2006.
CHEN, G.-J. et al. Synthesis, DNA binding, photo-induced DNA cleavage,
cytotoxicity and apoptosis studies of copper(II) complexes. Journal of Inorganic
Biochemistry, v. 105, n. 2, p. 119-126, 2011.
CHEN, G.-J. et al. Synthesis, DNA binding, photo-induced DNA cleavage and
cytotoxicity studies of europium(iii) complexes. Dalton Transactions, v. 39, n. 44,
p. 10637-10643, 2010.

71
CHEN, J. et al. Mechanistic studies on bleomycin-mediated DNA damage: multiple
binding modes can result in double-stranded DNA cleavage. Nucleic Acids Res, v.
36, n. 11, 2008a.
CHEN, L.-M. et al. Experimental and theoretical studies on the DNA-binding and
spectral properties of water-soluble complex [Ru(MeIm)4(dpq)]2+. Journal of
Molecular Structure, v. 881, n. 1-3, p. 156-166, 2008b.
CHEN, W. et al. Site-selective DNA hydrolysis by combining Ce(IV)/EDTA with
monophosphate-bearing oligonucleotides and enzymatic ligation of the scission
fragments. Journal of the American Chemical Society, v. 126, n. 33, p. 10285-
10291, 2004.
CHEN, W.; KOMIYAMA, M. Site-selective DNA hdrolysis by CeIV-EDTA with
the use of one oligonucleotide additive bearing two monophosphates.
ChemBioChem, v. 6, n. 10, p. 1825-1830, 2005.
COLLINS, M. et al. Metal complexes as anticancer agents: 2. Synthesis,
spectroscopy, magnetism, electrochemistry, X-ray crystal structure and
antimelanomal activity of the copper (II) complex of 5-amino-1-tolylimidazole-4-
carboxylate in B16F10 mouse melanoma cells. Inorganic Chemistry
Communications, v. 3, n. 9, p. 453-457, 2000.
COWAN, J. A. Chemical nucleases. Current Opinion in Chemical Biology, v. 5,
n. 6, p. 634-642, 2001.
DERVAN, P. B. Molecular recognition of DNA by small molecules. Bioorganic
and Medicinal Chemistry, v. 9, n. 9, p. 2215-2235, 2001.
DHAR, S.; NETHAJI, M.; CHAKRAVARTY, A. R. Designing molecules for PDT:
red light-induced DNA cleavage on disulfide bond activation in a dicopper(ii)
complex. Dalton Transactions, n. 2, p. 344-348, 2005a.
DHAR, S.; NETHAJI, M.; CHAKRAVARTY, A. R. Steric protection of a
photosensitizer in a N,N-bis[2-(2-pyridyl)ethyl]-2- phenylethylamine-copper(II)
bowl that enhances red light-induced DNA cleavage activity. Inorganic Chemistry,
v. 44, n. 24, p. 8876-8883, 2005b.

72
DHAR, S.; NETHAJI, M.; CHAKRAVARTY, A. R. DNA cleavage on
photoexposure at the d-d band in ternary copper(II) complexes using red-light laser.
Inorganic Chemistry, v. 45, n. 26, p. 11043-11050, 2006.
DOMINGOS, J. B. et al. A química dos ésteres de fosfato. Química Nova, v. 26, n.
5, 2003.
EFTHIMIADOU, E. K. et al. Metal complexes with the quinolone antibacterial
agent N-propyl-norfloxacin: Synthesis, structure and bioactivity. Journal of
Inorganic Biochemistry, v. 101, n. 3, p. 525-535, 2007.
FISCHER, F. L. Atividade de complexos metálicos sobre biomoléculas:
interação e clivagem. Dissertação de Mestrado. Universidade Federal de Santa
Catarina. Florianópolis, 2010.
FRANKLIN, S. J. Lanthanide-mediated DNA hydrolysis. Current Opinion in
Chemical Biology, v. 5, n. 2, p. 201-208, 2001.
GALLARDO, H. et al. Synthesis, structural characterization, and photo and
electroluminescence of a novel terbium(III) complex: {Tris(acetylacetonate)
[1,2,5]thiadiazolo[3,4-f][1,10]phenanthroline}terbium(III). Inorganica Chimica
Acta, v. 365, n. 1, p. 152-158, 2011.
GARCÍA-GIMÉNEZ, J. L. et al. Oxidative nuclease activity of ferromagnetically
coupled ł-hydroxo-ł-propionato copper(II) complexes [Cu3(L)2( ł-OH)2(ł-
propionato)2] (L = N-(pyrid-2-ylmethyl)R-sulfonamidato, R = benzene, toluene,
naphthalene). Journal of Inorganic Biochemistry, v. 103, n. 2, p. 243-255, 2009.
GHARAGOZLOU, M.; BOGHAEI, D. M. Interaction of water-soluble amino acid
Schiff base complexes with bovine serum albumin: Fluorescence and circular
dichroism studies. Spectrochimica Acta Part A: Molecular and Biomolecular
Spectroscopy, v. 71, n. 4, p. 1617-1622, 2008.
GRIFFITHS, A. J. F. et al. Introdução à Genética. Rio de Janeiro: Guanabara
Koogan. 2006.
GUPTA, T. et al. Bis(dipyridophenazine)copper(II) complex as major groove
directing synthetic hydrolase. Dalton Transactions, n. 12, p. 1896-1900, 2004.

73
HAGERMAN, P. J. Flexibility of DNA. Annual Review of Biophysics and
Biophysical Chemistry, v. 17, n. 1, 1988.
HALLIWELL, B.; GUTTERIDGE, J. M. C. Free Radicals in Biology and
Medicine. 4th, Oxford: Clarendon Press. 2007.
HECKERT, E. G.; SEAL, S.; SELF, W. T. Fenton-Like Reaction Catalyzed by the
Rare Earth Inner Transition Metal Cerium. Environmental Science & Technology,
v. 42, n. 13, p. 5014-5019, 2008.
HEGG, E. L.; BURSTYN, J. N. Toward the development of metal-based synthetic
nucleases and peptidases: A rationale and progress report in applying the principles
of coordination chemistry. Coordination Chemistry Reviews, v. 173, n. 1, p. 133-
165, 1998.
HUSSAIN, A. et al. Photocytotoxic Lanthanum(III) and gadolinium(III) complexes
of phenanthroline bases showing light-induced DNA cleavage activity. Inorganic
Chemistry, v. 49, n. 9, p. 4036-4045, 2010.
HUSSAIN, A. et al. Photocytotoxicity and DNA photocleavage activity of La(III)
and Gd(III) complexes of phenanthroline bases. Indian Journal of Chemistry -
Section A Inorganic, Physical, Theoretical and Analytical Chemistry, v. 50, n.
3-4, p. 519-530, 2011.
JIANG, Q. et al. Design of artificial metallonucleases with oxidative mechanism.
Coordination Chemistry Reviews, v. 251, n. 15-16, p. 1951-1972, 2007.
KIM, S. K.; NORDEN, B. Methyl green: A DNA major-groove binding drug. FEBS
Letters, v. 315, n. 1, p. 61-64, 1993.
KIMURA, S.; SAKAGUCHI, K. DNA Repair in Plants. Chemical Reviews, v. 106,
n. 2, p. 753-766, 2006.
KOMIYAMA, M. et al. Oligonucleotide bearing ethylenediamine-N,N,N ł-
triacetates for gap-selective DNA hydrolysis by Ce4+/EDTA. ChemBioChem, v. 6,
n. 1, p. 192-196, 2005.

74
KOMIYAMA, M. et al. Hydrolysis of oligoDNAs by lanthanide metal(III) chloride.
Polymer Journal, v. 25, n. 11, p. 1211-1214, 1993.
KOVACIC, R. T.; WELCH, J. T.; FRANKLIN, S. J. Sequence-Selective DNA
Cleavage by a Chimeric Metallopeptide. Journal of the American Chemical
Society, v. 125, n. 22, p. 6656-6662, 2003.
KRÄMER, R. Bioinorganic models for the catalytic cooperation of metal ions and
functional groups in nuclease and peptidase enzymes. Coordination Chemistry
Reviews, v. 182, n. 1, p. 243-261, 1999.
LAHIRI, D. et al. Anaerobic photocleavage of DNA in red light by dicopper(ll)
complexes of 3,3 ł-dithiodipropionic acid. Inorganic Chemistry, v. 48, n. 1, p.
339-349, 2009.
LAHIRI, D. et al. Anaerobic DNA cleavage activity in red light and
photocytotoxicity of (pyridine-2-thiol)cobalt(iii) complexes of phenanthroline bases.
Dalton Transactions, v. 39, n. 7, p. 1807-1816, 2010.
LAW, G.-L. et al. Observation of the selective staining of chromosomal DNA in
dividing cells using a luminescent terbium(iii) complex. Chemical
Communications, v. 46, n. 14, p. 2391-2393, 2010.
LI, Y. et al. ESR evidence for the generation of reactive oxygen species from the
copper-mediated oxidation of the benzene metabolite, hydroquinone: role in DNA
damage. Chemico-Biological Interactions, v. 94, n. 2, p. 101-120, 1995.
LIM, S.; FRANKLIN, S. J. Engineered lanthanide-binding metallohomeodomains:
Designing folded chimeras by modular turn substitution. Protein Science, v. 15, n.
9, p. 2159-2165, 2006.
LIU, C.; WANG, L. DNA hydrolytic cleavage catalyzed by synthetic multinuclear
metallonucleases. Dalton Transactions, n. 2, p. 227-239, 2009.
LIU, C. et al. DNA hydrolysis promoted by di- and multi-nuclear metal complexes.
Coordination Chemistry Reviews, v. 248, n. 1-2, p. 147-168, 2004.

75
LÖNNBERG, T.; SUZUKI, Y.; KOMIYAMA, M. Prompt site-selective DNA
hydrolysis by Ce(iv)-EDTA using oligonucleotide multiphosphonate conjugates.
Organic and Biomolecular Chemistry, v. 6, n. 19, p. 3580-3587, 2008.
MANCIN, F. et al. Artificial metallonucleases. Chemical Communications, n. 20,
p. 2540-2548, 2005.
MARUYAMA, K. et al. DNA Sensor with a Dipyridophenazine Complex of
Osmium(II) as an Electrochemical Probe. Analytical Chemistry, v. 74, n. 15, p.
3698-3703, 2002.
MUKHERJEE, A. et al. Ternary iron(II) complex with an emissive imidazopyridine
arm from Schiff base cyclizations and its oxidative DNA cleavage activity. Dalton
Transactions, n. 2, p. 349-353, 2005.
NEIDLE, S. DNA minor-groove recognition by small molecules. Natural Product
Reports, v. 18, n. 3, p. 291-309, 2001.
NELSON, D. L.; COX, M. M. Lehninger Princípios de Bioquímica. 4ª, São Paulo:
Sarvier. 2006.
NETTO, L. E. S.; STADTMAN, E. R. The Iron-Catalyzed Oxidation of
Dithiothreitol Is a Biphasic Process: Hydrogen Peroxide Is Involved in the Initiation
of a Free Radical Chain of Reactions. Archives of Biochemistry and Biophysics, v.
333, n. 1, p. 233-242, 1996.
NICHOLSON, A. W. Function, mechanism and regulation of bacterial
ribonucleases. FEMS Microbiology Reviews, v. 23, n. 3, p. 371-390, 1999.
OCHSNER, M. Photophysical and photobiological processes in the photodynamic
therapy of tumours. Journal of Photochemistry and Photobiology B: Biology, v.
39, n. 1, p. 1-18, 1997.
OTT, R.; KRÄMER, R. DNA Hydrolysis by Inorganic Catalysts. Applied
Microbiology and Biotechnology, v. 52, 1999.

76
PARODI, S.; KENDALL, F.; NICOLINI, C. A clarification of the comple spectrum
observed with ultraviolet circular dichroism of ethidium bromide bound to DNA.
Nucleic Acids Res, v. 4, n. 2, 1975.
PATIL, S. A. et al. Co(II), Ni(II) and Cu(II) complexes with coumarin-8-yl Schiff-
bases: Spectroscopic, in vitro antimicrobial, DNA cleavage and fluorescence studies.
Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, v. 79,
n. 5, p. 1128-1136, 2011.
PATRA, A. K. Studies on the photo-induced DNA cleavage activity of α-amino
acid copper complexes having phenantroline bases. Indian Institute of Science.
Bangalore, 2007.
PATRA, A. K. et al. DNA cleavage in red light promoted by copper(II) complexes
of ł-amino acids and photoactive phenanthroline bases. Dalton Transactions, n.
48, p. 6966-6976, 2008.
PATRA, A. K. et al. Copper(II) Complexes of l-Arginine as Netropsin Mimics
Showing DNA Cleavage Activity in Red Light. Inorganic Chemistry, v. 48, n. 7, p.
2932-2943, 2009.
PATRA, A. K.; NETHAJI, M.; CHAKRAVARTY, A. R. Synthesis, crystal
structure, DNA binding and photo-induced DNA cleavage activity of (S-methyl-L-
cysteine)copper(II) complexes of heterocyclic bases. Journal of Inorganic
Biochemistry, v. 101, n. 2, p. 233-244, 2007.
POGOZELSKI, W. K.; TULLIUS, T. D. Oxidative strand scission of nucleic acids:
Routes initiated by hydrogen abstraction from the sugar moiety. Chemical Reviews,
v. 98, n. 3, p. 1089-1107, 1998.
PYLE, A. M. et al. Mixed-ligand complexes of ruthenium(II): factors governing
binding to DNA. Journal of the American Chemical Society, v. 111, n. 8, p. 3051-
3058, 1989.
RAMAN, N.; POTHIRAJ, K.; BASKARAN, T. DNA interaction, antimicrobial,
electrochemical and spectroscopic studies of metal(II) complexes with tridentate
heterocyclic Schiff base derived from 2'-methylacetoacetanilide. Journal of
Molecular Structure, v. 1000, n. 1-3, p. 135-144, 2011.

77
RAMIREZ, D. C.; GOMEZ MEJIBA, S. E.; MASON, R. P. Mechanism of
hydrogen peroxide-induced Cu,Zn-superoxide dismutase-centered radical formation
as explored by immuno-spin trapping: the role of copper- and carbonate radical
anion-mediated oxidations. Free Radical Biology and Medicine, v. 38, n. 2, p. 201-
214, 2005.
RAMMO, J. et al. Catalysis of DNA cleavage by lanthanide complexes with
nucleophilic or intercalating ligands and their kinetic characterization. Chemical
Communications, n. 1, p. 105-107, 1996.
RITTICH, B. et al. Cleavage of double stranded plasmid DNA by lanthanide
complexes. Journal of chromatography. B, Biomedical sciences and
applications, v. 800, n. 1-2, p. 169-173, 2004.
ROY, M. et al. Double-strand DNA cleavage from photodecarboxylation of (ł-
oxo)diiron(III) L-histidine complex in visible light. Dalton Transactions, n. 27, p.
3542-3545, 2008a.
ROY, S. et al. Photosensitizer in a molecular bowl and its effect on the DNA-
binding and -cleavage activity of 3d-metal scorpionates. Inorganic Chemistry, v.
47, n. 13, p. 5625-5633, 2008b.
ROY, S. et al. DNA photocleavage and anticancer activity of terpyridine copper(II)
complexes having phenanthroline bases. Polyhedron, v. 29, n. 14, p. 2787-2794,
2010.
SASMAL, P. K. et al. DNA Cleavage by New Oxovanadium(IV) Complexes of N-
Salicylidene ł-Amino Acids and Phenanthroline Bases in the Photodynamic
Therapy Window. Inorganic Chemistry, v. 46, n. 26, p. 11112-11121, 2007.
SCHLACHER, K. et al. Roles of DNA Polymerase V and RecA Protein in SOS
Damage-Induced Mutation. Chemical Reviews, v. 106, n. 2, p. 406-419, 2006.
SELVAKUMAR, B. et al. Structures, spectra, and DNA-binding properties of
mixed ligand copper(II) complexes of iminodiacetic acid: The novel role of diimine
co-ligands on DNA conformation and hydrolytic and oxidative double strand DNA
cleavage. Journal of Inorganic Biochemistry, v. 100, n. 3, p. 316-330, 2006.

78
SHENG, X. et al. DNA binding, cleavage, and cytotoxic activity of the preorganized
dinuclear zinc(II) complex of triazacyclononane derivatives. Bioconjugate
Chemistry, v. 19, n. 2, p. 490-498, 2008.
SIGMAN, D. S.; CHEN, C. B. Chemical Nucleases: New Reagents in Molecular
Biology. Annual Review of Biochemistry, v. 59, 1990.
SIGMAN, D. S. et al. Oxygen-dependent cleavage of DNA by the 1,10-
phenanthroline cuprous complex. Inhibition of Escherichia coli DNA polymerase I. .
Journal of Biological Chemistry, v. 254, n. 24, 1979.
SIGMAN, D. S.; MAZUMDER, A.; PERRIN, D. M. Chemical nucleases. Chemical
Reviews, v. 93, n. 6, p. 2295-2316, 1993.
SIRISH, M.; FRANKLIN, S. J. Hydrolytically active Eu(III) and Ce(IV) EF-hand
peptides. Journal of Inorganic Biochemistry, v. 91, n. 1, p. 253-258, 2002.
SOUZA, B. et al. DNA photonuclease activity of four new copper(II) complexes
under UV and red light: Theoretical/experimental correlations with active species
generation. Dalton Transactions, v. 39, n. 8, p. 2027-2035, 2010a.
SOUZA, B. et al. Oxygen-independent photonuclease activity of a new iron(II)
complex. Chemical Communications, v. 46, n. 19, p. 3375-3377, 2010b.
SREEDHARA, A.; COWAN, J. A. Catalytic hydrolysis of DNA by metal ions and
complexes. Journal of Biological Inorganic Chemistry, v. 6, n. 4, p. 337-347,
2001.
SUMAOKA, J.; AZUMA, Y.; KOMIYAMA, M. Enzymatic manipulation of the
fragments obtained by cerium(IV)-induced DNA scission: Characterization of
hydrolytic termini. Chemistry - A European Journal, v. 4, n. 2, p. 205-209, 1998.
SUMAOKA, J. et al. Homogeneous DNA hydrolysis by
cerium(IV)/lanthanide(III)/dextran ternary complexes. Chemistry Letters, n. 6, p.
507-508, 1997.

79
SZACIłOWSKI, K. et al. Bioinorganic Photochemistry: Frontiers and
Mechanisms. Chemical Reviews, v. 105, n. 6, p. 2647-2694, 2005.
TAKASAKI, B. K.; CHIN, J. Cleavage of the phosphate diester backbone of DNA
with cerium(III) and molecular oxygen. Journal of the American Chemical
Society, v. 116, n. 3, p. 1121-1122, 1994.
TOSHIMA, K. et al. Molecular design and evaluation of quinoxaline-carbohydrate
hybrids as novel and efficient photo-induced GG-selective DNA cleaving agents.
Chemical Communications, n. 3, p. 212-213, 2002.
VALKO, M. et al. Free radicals and antioxidants in normal physiological functions
and human disease. International Journal of Biochemistry and Cell Biology, v.
39, n. 1, p. 44-84, 2007.
VAN DYKE, M. W.; HERTZBERG, R. P.; DERVAN, P. B. Map of distamycin,
netropsin, and actinomycin binding sites on heterogeneous DNA: DNA cleavage-
inhibition patterns with methidiumpropyl-EDTA.Fe(II). Proceedings of the
National Academy of Sciences, v. 79, n. 18, p. 5470-5474, September 1, 1982
1982.
WOLFE, A.; SHIMER, G. H.; MEEHAN, T. Polycyclic aromatic hydrocarbons
physically intercalate into duplex regions of denatured DNA. Biochemistry, v. 26,
n. 20, p. 6392-6396, 1987.
WONG-DEYRUP, S.; KIM, Y.; FRANKLIN, S. Sequence preference in DNA
binding: de novo designed helix–turn–helix metallopeptides recognize a family of
DNA target sites. Journal of Biological Inorganic Chemistry, v. 11, n. 1, p. 17-25,
2006.
YAMAMOTO, K.; KAWANISHI, S. Hydroxyl free radical is not the main active
species in site-specific DNA damage induced by copper (II) ion and hydrogen
peroxide. Journal of Biological Chemistry, v. 264, n. 26, p. 15435-15440,
September 15, 1989 1989.
YAMAMOTO, Y.; TSUBOI, W.; KOMIYAMA, M. Oligoamine-acridine
conjugates for promotion of gap-selective DNA hydrolysis by Ce(IV)/EDTA
complex. Nucleic Acids Research, v. 31, n. 15, p. 4497-4502, 2003.

80
YAMAMOTO, Y. et al. Development of artificial restriction DNA cutter composed
of Ce(Iv)/EDTA and PNA. Nucleosides, Nucleotides and Nucleic Acids, v. 26, n.
10-12, p. 1265-1268, 2007.
YAMAMOTO, Y. et al. Site-selective and hydrolytic two-strand scission of double-
stranded DNA using Ce(IV)/EDTA and pseudo-complementary PNA. Nucleic
Acids Research, v. 32, n. 19, 2004.
ZEGLIS, B. M.; PIERRE, V. C.; BARTON, J. K. Metallo-intercalators and metallo-
insertors. Chemical Communications, n. 44, p. 4565-4579, 2007.
ZHU, B. et al. Lanthanide binuclear macrocyclic complexes as synthetic enzymes
for the cleavage of DNA. Journal of Molecular Catalysis A: Chemical, v. 135, n.
1, p. 107-110, 1998.