universidade federal de mato grosso§ões-teses/teses/2015... · localizado no centro de pesquisa...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E ZOOTECNIA
Programa de Pós-graduação em Agricultura Tropical
VARIAÇÃO DE ATRIBUTOS DO SOLO SOB SISTEMAS
AGROFLORESTAIS
LORENA DE SOUZA TAVARES
CUIABÁ- MT
2015

1
UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE AGRONOMIA, MEDICINA VETERINÁRIA E ZOOTECNIA
Programa de Pós-graduação em Agricultura Tropical
VARIAÇÃO DE ATRIBUTOS DO SOLO SOB SISTEMAS
AGROFLORESTAIS
LORENA DE SOUZA TAVARES
Engenheira Florestal
Orientadora: Profª. Dra. WALCYLENE LACERDA MATOS PEREIRA SCARAMUZZA
Tese apresentada à Faculdade de Agronomia, Medicina Veterinária e Zootecnia da Universidade Federal de Mato Grosso para obtenção do título de Doutora em Agricultura Tropical.
CUIABÁ – MT
2015

2

3

4
“Nada nos faltou, tudo Deus proveu
Nos deu coroa pra guardar
E fez tudo novo com amor glorioso
E nos deu vitórias ontem, hoje e nos dará no amanhã.”
Trecho do louvor: Já Chegamos Até Aqui!
Igreja Cristã Maranata

5
Aos meus pais, Alvinho Cândido Tavares
e Maria Lina de Souza Tavares e ao
meu irmão Romário Jonathas de Souza
Tavares que sempre me apoiaram e
incentivaram, sendo peças fundamentais
para a conclusão de mais essa etapa na
minha vida.
Dedico...

6
AGRADECIMENTO
Primeiramente, a Deus, que é a minha fortaleza e refúgio, que nunca me
desamparou, me sustentou e me proporcionou vitórias e experiências de
demonstração do seu amor;
Aos meus pais, Alvinho e Maria Lina pelas orações, pelo apoio e pela torcida, pois são
meus fiéis companheiros e minha inspiração para vencer cada obstáculo nessa vida;
Ao meu irmão, Romário, pelo incentivo e momentos de descontração ao telefone,
pois, mesmo de longe, me deu força e demonstrou amadurecimento de vida;
Ao Andherson pelo apoio, pelas longas conversas nos momentos difíceis, por
sempre me dizer para seguir em frente e acreditar mais em mim do que eu mesma;
por ter tido tanta paciência e por me incentivar a alcançar meu sonho;
À Profª. Dra. Walcylene L. M. P. Scaramuzza, pela orientação, confiança, força,
amizade e transmissão de seus conhecimentos fundamentais para a conclusão
deste trabalho;
À Profª. Dra. Oscarlina L. dos S. Weber, pela co-orientação, dedicação, paciência,
apoio, amizade e “injeções” de ânimo, que foram essenciais em muitos momentos;
Ao Prof. Dr. José Fernando Scaramuzza, pelas sinceras e válidas contribuições,
pelos “puxões” de orelha e por me fazer acreditar mais na minha capacidade;
À Professora Daniela T. da S. Campos e ao Professor Emilio Carlos de Azevedo por
terem cedido os Laboratórios de Microbiologia e Física do Solo, respectivamente,
para as análises.
À Professora Dra. Maria Aparecida P. Pirangeli e Selma B. Batista, pela participação
na banca de defesa e pelas valiosas contribuições.
À CEPLAC/Alta Floresta – MT, nas pessoas do Sr. Fernando pela liberação da
fazenda para pesquisa e pela disponibilidade dos recursos necessários; a
Eletissandra pelos contatos e toda assistência; ao Romildo e família pelo apoio,
pelas boas e descontraídas prosas após o trabalho;
Aos amigos, que participaram das coletas a campo, Gilmar, Diego, Túlio e Nelson;
Aos amigos de Pós-graduação: Ana, Daniel, Everton, Fabrício, Franciele, Kelly,
Liliane, Valdeir e Wininton pelo apoio e companheirismo;

7
Aos colegas dos laboratórios de Fertilidade do Solo, Física do Solo, Nutrição Mineral
de Plantas e Microbiologia do Solo: Dafne, Diogo, Eustáquio, Heriani, Lucas, Pedro
e Rayssa; e às funcionárias Maria Minervina, Berenice e Marcela pelo apoio e auxílio
nos laboratórios;
Aos amigos da Igreja Cristã Maranata pelas orações e companheirismo que me
impulsionaram a continuar;
À Universidade Federal de Mato Grosso, nas pessoas dos professores da
graduação, da pós-graduação e demais funcionários pelos conhecimentos
profissionais e experiências de aprendizado científico e humanitário durante toda a
vida acadêmica;
A CAPES, pela concessão da bolsa de estudo;
Ao Instituto Federal de Rondônia pelas liberações para dar seguimento ao curso e
aos amigos professores que trabalharam para que isso fosse possível,
especialmente à Elise, ao Raimundo e ao Wesley.
À D. Helena, aos meus amigos de infância e familiares que sempre torceram e
oraram muito por mim.
Bem, a todos aqueles que de alguma forma contribuíram para que eu pudesse
chegar, seja diretamente ou indiretamente, na realização deste trabalho.

8
VARIAÇÃO DE ATRIBUTOS DO SOLO SOB SISTEMAS AGROFLORESTAIS
RESUMO – As atividades agrícolas e agropecuárias quando desordenadas e intensivas podem causar a degradação das propriedades físicas, químicas e biológicas do solo. Nesse sentido, sistemas conservacionistas de exploração dos recursos naturais têm sido importantes por proporcionar a manutenção ou aumento da qualidade do solo, a exemplo dos sistemas agroflorestais, caracterizados principalmente pela combinação de espécies florestais com cultivos agrícolas e adicionados ou não às atividades pecuárias. Assim, objetivou-se neste trabalho avaliar as variações nos atributos químicos, físicos e biológicos do solo em diferentes sistemas florestais e épocas. Para isso, foram coletadas amostras de solo nas camadas de 0 a 0,10; 0,10 a 0,20 e 0,20 a 0,40 m em um experimento localizado no Centro de Pesquisa da Lavoura Cacaueira - CEPLAC de Alta Floresta – MT, em Argissolo Vermelho-Amarelo Eutrófico, textura franco-argilosa. Para este estudo, foi considerado um delineamento experimental em blocos casualizados em arranjo de parcelas subdivididas 5x2, com quatro repetições, sendo cinco sistemas florestais (floresta nativa; cupuaçu+mogno brasileiro; cupuaçu+teca; cupuaçu+cedro rosa; cupuaçu+paricá) e duas épocas de avaliação (período chuvoso e seco). Nas amostras de solo foram determinados: pH, componentes do complexo sortivo do solo, fósforo (P), teor de carbono orgânico (COT) e matéria orgânica (MOS), estoques de nutrientes, densidade, macro e microporosidade, porosidade total, estabilidade de agregados, carbono da biomassa microbiana, respiração basal, quociente metabólico e quociente microbiano. Quanto aos atributos químicos do solo, os sistemas florestais tiveram pouca influência sobre a acidez do solo, não sendo menos prejudiciais às culturas. Foi observada sazonalidade nos teores de COT e MOS na floresta nativa e nos sistemas consorciados de cupuaçu com mogno e cedro, sendo os maiores valores encontrados na época seca. Os maiores teores e estoques de bases e fósforo foram encontrados no período chuvoso devido ao efeito da decomposição da MOS neste período. Os sistemas consorciados tiveram menores teores de bases do solo em relação à floresta nativa, apesar disto, não houve alteração na saturação por bases do solo. O sistema de consórcio entre cupuaçu e paricá teve maior teor e estoque de P no solo em relação aos demais sistemas. Quanto aos atributos físicos, verificou-se que nenhum sistema consorciado manteve as propriedades semelhantes à floresta nativa, bem como, não houve diferença entre esses. Apesar disso, os atributos físicos dos sistemas consorciados não foram limitantes ao desenvolvimento das espécies. A floresta nativa e o consórcio entre cupuaçu e mogno se destacaram quanto aos atributos microbiológicos do solo sendo os mais eficientes em incorporar carbono à biomassa microbiana com menores taxas de quociente metabólico. Contrariamente, o sistema com cupuaçu e paricá foi o menos eficiente, com maior quociente metabólico. Para todos os sistemas, houve redução da atividade biológica no período seco. De forma geral, no período chuvoso foram obtidos maiores teores de nutrientes e atividade biológica. Os SAFs nas condições iniciais de avaliação, sete anos, apresentaram resultados satisfatórios, dando indícios de que uma avaliação temporal pelo mesmo período caberia resultados mais evidentes. Palavras-chave: consórcios florestais; estoque de carbono; física do solo; biomassa microbiana.

9
VARIATION OF SOIL ATTRIBUTES IN AGROFORESTRY SYSTEMS
ABSTRACT - The agricultural and livestock farming activities when disordered and intensive can cause degradation of the physical, chemical and biological soil properties. In this sense, conservationist systems of exploitation of natural resources have been important for providing the maintenance or increasing of soil quality, the example of agroforestry systems, characterized primarily by a combination of agricultural and forest species with agricultural crops and/or livestock activities. Thus, this study aimed to assess changes in chemical, physical and biological soil attributes in different periods and forest systems. For this, soil samples were collected in layers from 0 to 0.10; 0.10 to 0.20 and 0.20 to 0.40 m in an experiment located at the Research Center of Cocoa Farming – CEPLAC, in Alta Floresta – MT, in eutrophic Red-Yellow Utisol, clay loam texture. For this study, it was considered an experimental randomized block design in a subdivided plot arrangement 5x2, with four replications, with five forestry systems (native forest; Theobroma grandiflorum + Swietenia macrophylla; Theobroma grandiflorum + Tectona grandis; Theobroma grandiflorum + Cedrela fissilis; Theobroma grandiflorum + Schizolobium amazonicum) and two periods (rainy and dry season). In the soil samples were determined: pH, components of soil sorption complex, phosphorus (P), organic carbon (TOC) and organic matter (OMS), nutrient stocks, density, macro and microporosity, total porosity, aggregate stability, microbial biomass carbon, basal respiration, metabolic quotient and microbial quotient. In relation to the chemical attributes, the forest systems had little influence on soil acidity in which it was not harmful to crops. There was seasonality in TOC and OMS in the native forest and intercropping systems of Theobroma grandiflorum with Swietenia macrophylla and Cedrela fissilis, with the highest values found in the dry season. The highest levels and base and phosphorus stocks were found in the rainy season due to the effect of the decomposition of OMS in this period. The intercropping systems had lower soil base levels than the native forest, despite this, there was no change in the saturation of soil bases. The intercropping system between Theobroma grandiflorum + Schizolobium amazonicum had higher level and P stock in the soil in relation to other systems. Regarding the physical attributes, it was found that no intercropping system maintained the similar properties to the native forest, as well as, there was no difference between these. Nevertheless, the physical attributes of intercropping systems were not limiting to the development of the species. The native forest and the intercropping between Theobroma grandiflorum + Swietenia macrophylla stood out as the microbiological soil attributes, they were the most efficient in incorporating carbon to microbial biomass with lower metabolic quotient rates. In contrast, the system with Theobroma grandiflorum + Schizolobium amazonicum was the less efficient, with greater metabolic quotient. For all systems, there was reduction of biological activity in the dry season. In general, during the rainy season, highest values of nutrients were obtained. The SAF in the initial conditions of evaluation, seven years, showed satisfactory results, providing evidence that a temporal evaluation for the same period would fit more obvious results. Keywords: forest intercropping; carbon stock; soil physics; microbial biomass.

10
LISTA DE FIGURAS
Página
1 Croqui dos sistemas agroflorestais e floresta nativa.................................... 23 2 Dados meteorológicos de precipitação (Precip.), umidade (Umid.) e
temperatura (Temp.) do município de Alta Floresta – MT, do ano de 2012..............................................................................................................
24

11
LISTA DE TABELAS
Página
1 Caracterização textural do solo sob sistemas florestais, Alta Floresta - MT, Alta Floresta – MT, 2013...................................................................................
27
2 pH, acidez trocável (Al+3), acidez potencial (H+Al) e saturação por alumínio (m) no solo sob sistemas florestais, Alta Floresta - MT, 2013 .........................
31
3 Capacidade de troca catiônica (CTCpH7,0), capacidade de troca catiônica efetiva (CTCefetiva), carbono orgânico total (COT), matéria orgânica do solo (MOS) no solo sob sistemas florestais, Alta Floresta - MT, 2013 ....................
34 4 Teores de cálcio (Ca+2), magnésio (Mg+2), potássio (K+), saturação por base
(V%) e fósforo (P) no solo sob sistemas florestais, Alta Floresta - MT, 2013 ..
37 5 Estoques de carbono orgânico (Est-COT), de matéria orgânica (Est-MOS),
de fósforo (Est-P), de cálcio (Est-Ca), de magnésio (Est-Mg) e de potássio (Est-K) sob sistemas florestais, Alta Floresta - MT, 2013 ...............................
43 6 Densidade (Ds), macroporosidade (Mac), microporosidade (Mic), porosidade
total (PT), diâmetro médio ponderado (DMP), diâmetro médio geométrico (DMG) e estabilidade de agregados (EA) do solo sob sistemas florestais coletados no período chuvoso, Alta Floresta - MT, 2013..................................
46 7 Carbono da biomassa (C-BMS), respiração basal (C-CO2), quociente
metabólico (q-CO2) e quociente microbiano (q-MIC) do solo sob sistemas florestais, Alta Floresta - MT, 2013 .................................................................
49

12
SUMÁRIO
Página 1. INTRODUÇÃO................................................................................................... 13 2. REVISÃO DE LITERATURA ............................................................................ 15 2.1. Cenário do Uso do Solo na Amazônia Legal ................................................. 15 2.2. Benefícios dos SAFs Para os Atributos do Solo ............................................ 17 2.2.1. Benefícios dos SAFs para os atributos químicos e físicos do solo ............ 17 2.2.2. Benefícios dos SAFs para os atributos biológicos do solo .......................... 18 2.2.3. Características do solo com SAFs e períodos sazonais ............................. 20 3. MATERIAL E MÉTODOS ................................................................................. 22 3.1. Localização e Característica da Área de Estudo............................................ 22 3.2. Instalação Experimento .................................................................................. 23 3.3. Dados Meteorológicos .................................................................................... 25 3.4. Amostragem do Solo ...................................................................................... 25 3.5. Caracterização dos Atributos Químicos do Solo ............................................ 26 3.5.1. Estoque de carbono no solo e nutrientes .................................................... 27 3.6. Atributos Físicos do Solo ................................................................................ 27 3.7. Atributos Microbiológicos do Solo .................................................................. 29 3.8. Análise Estatística .......................................................................................... 30 4. RESULTADOS E DISCUSSÃO ........................................................................ 31 4.1. Atributos Químicos do Solo ............................................................................ 31 4.1.1. Componentes da acidez do solo ................................................................. 31 4.1.2. Trocas catiônicas, carbono orgânico e matéria orgânica do solo ............... 33 4.1.3. Componentes de bases e P no solo ........................................................... 37 4.2. Estoques de Nutrientes no Solo ..................................................................... 42 4.3. Variações dos Atributos Físicos do Solo......................................................... 46 4.4. Variação dos Atributos Microbiológicos.......................................................... 50 5. CONCLUSÕES ................................................................................................. 55 6. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................. 56

13
1. INTRODUÇÃO
A exploração excessiva dos recursos naturais pelas atividades agrícolas e
agropecuárias tem levado o ambiente à rápida degradação, contudo, sabendo da
importância desses sistemas para a população, é necessário que medidas sejam
tomadas para retardar esse processo. Nesse sentido, os sistemas consorciados de
culturas tradicionais com o componente arbóreo ou os chamados sistemas
agroflorestais (SAFs) surgem como alternativa sustentável.
Do ponto de vista ambiental, esses sistemas podem favorecer a redução da
degradação dos solos, mediante aporte de matéria orgânica e aumento da
biodiversidade do solo, beneficiando o aumento do estoque de nutrientes e da
capacidade retenção de água, além de diminuir a pressão sobre as florestas nativas.
Do ponto de vista econômico, esses sistemas têm maior rentabilidade para o
produtor, tendo em vista que numa mesma área sua produtividade multiplicaria pela
diversidade de produtos obtidos. Além disso, com maior diversidade de espécies
vegetais e a demanda de trabalho no campo, durante o ano todo, ocupa a mão-de-
obra familiar, proporcionando melhoria da qualidade de vida e contribuindo para
reduzir a taxa de êxodo rural.
Entretanto, todas essas qualidades promovidas pelos sistemas agroflorestais
podem diferenciar de um sistema para outro, tendo assim a necessidade de serem
quantificadas para poderem ser utilizadas como modelos mais precisos de êxito no
setor.
O presente trabalho partiu da hipótese de que os sistemas agroflorestais
alteram as características químicas, físicas e biológicas dos solos de acordo com as

14
espécies utilizadas, períodos sazonais e profundidade. Desse modo, reunir
informações que permitam inferir os ganhos obtidos da conversão da mata nativa,
torna esse estudo essencial, pois dessa forma auxiliará nas tomadas de decisões,
na implantação de metodologias e políticas mais efetivas para o setor. As
informações geradas orientarão novas práticas sustentáveis e servirão de subsídios
para incentivos fiscais.
Diante disso, objetivou-se neste trabalho quantificar os atributos químicos,
físicos e biológicos do solo em diferentes sistemas florestais e em épocas distintas
em uma região do estado de Mato Grosso, de modo que possa verificar os
benefícios dos sistemas agroflorestais para o solo.

15
2. REVISÃO DE LITERATURA
2.1. Cenário do Uso do Solo na Amazônia Legal
A cultura do corte e queima das florestas na Amazônia Legal, para dar lugar à
agricultura ou à pecuária é uma prática bem conhecida, no entanto, tem se
intensificado nos últimos anos.
De 1988 a 2013, foram desmatados nos nove Estados componentes da
Amazônia Legal o equivalente a 402.614 km2. Somente no ano de 2012 foram
desmatados 4.571 km2 e degradados 8.634 km2 de florestas nativas, no qual o
estado de Mato Grosso ficou em terceiro lugar no ranking do desmatamento com
757 km2 e liderou com 3.273 km2 de áreas degradadas, sendo que desse
quantitativo, ainda 171 km2 foram convertidos em corte raso para dar lugar à
agricultura ou pastagens (Instituto Nacional de Pesquisas Espaciais - INPE, 2014).
Essa prática consiste de uma atividade não sustentável, tendo em vista que
os solos se mantêm produtivo por apenas dois ou três anos, obrigando a migração
para outra área após esse período, dando início a um novo ciclo.
A transformação da floresta para cultivos agrícolas ou pecuária modifica as
propriedades químicas, físicas e biológicas do solo, cuja intensidade varia com as
condições do clima, uso e manejo adotado (Araújo et al., 2004). Essas perturbações
nas propriedades do solo afetam diretamente a microbiota do solo que é sensível às
alterações do pH, da umidade, da aeração, da temperatura e da disponibilidade de
nutrientes orgânicos e inorgânicos do solo (Melz e Tiago, 2009), pois se esses

16
microorganismos forem afetados, interferirão na capacidade de regeneração da
floresta.
Em algumas ocasiões, plantas consideradas invasoras se estabelecem
nessas áreas e acabam melhorando a fertilidade desse solo (Favero et al., 2000;
Meda et al., 2002), mas dependendo do estado desse solo, não há esse
estabelecimento e o que se acarreta é um avanço da degradação. Dessa forma, o
melhor uso do solo, com sistemas de manejo mais adequados que possibilitem
maior aporte de nutrientes e a redução de suas perdas, são fundamentais e
necessárias para a sustentabilidade agrícola.
Como alternativa de solução, os Sistemas Agroflorestais (SAFs) surgem como
uma importante ferramenta, capaz de controlar a erosão dos solos e manter a
capacidade produtiva da terra ao longo do tempo. Isso porque, a característica
desse sistema é implantar numa mesma área, combinações de espécies arbóreas
lenhosas (frutíferas e/ou madeireiras) com cultivos agrícolas e/ou animais, de forma
simultânea ou em sequência temporal e que interagem economicamente e
ecologicamente (Locatelli e Vieira, 2013).
O governo tem apostado nessa ideia e para incentivar o produtor rural lançou,
em 2010, o Plano Setorial de Mitigação e de Adaptação às Mudanças Climáticas
para a Consolidação de uma Economia de Baixa Emissão de Carbono na Agricultura
- Plano ABC. Esse plano tem por finalidade reduzir as emissões de gases do efeito
estufa – GEE no setor agropecuário do país com a adoção de produções
sustentáveis, através dos programas de incentivos fiscais. E de forma indireta,
também promover a manutenção da floresta em pé e/ou estimular reflorestamentos
nessas regiões.
Esses incentivos fiscais são fornecidos a quem implantar ou enquadrar em
uma das categorias do programa que inclusive tem os SAFs e a Integração Lavoura-
Pecuária-Floresta (ILPF) como uma das linhas de crédito do programa, assim como
a Recuperação de Pastagens Degradadas, o Sistema Plantio Direto (SPD), a
Fixação Biológica de Nitrogênio (FBN), as Florestas Plantadas, os Tratamentos de
Dejetos Animais e a Adaptação às Mudanças Climáticas. Estima-se que serão
disponibilizados recursos da ordem de R$197 bilhões durante todo o período de

17
vigência, financiados com fontes orçamentárias ou por meio de linhas de crédito
(Brasil, 2012).
Percebe-se então, que as vantagens desses sistemas são inúmeras, contudo
devido à diversidade de espécies utilizadas, torna-se necessário verificar se há
diferenças na contribuição da melhoria dos solos de acordo com as espécies
consorciadas.
2.2. Benefícios dos SAFs Para os Atributos do Solo
2.2.1. Benefícios dos SAFs para os atributos químicos e físicos do solo
Entre as vantagens desses sistemas, citam-se a variedade de espécies e
culturas diferentes, que trazem benefícios tanto para a fauna como para o solo da
região. Nesse último quesito, ressalta-se a importância da cobertura do solo pelas
copas das árvores em diferentes estratos, pois elas reduzem o impacto da gota da
chuva, insolação direta no solo, perda de solo e de nutrientes (Macedo et al., 2000).
Promove o acúmulo de material orgânico na superfície e contribuem com a melhoria
da fertilidade (Vital et al., 2004; Silva et al., 2012), sem depender de insumos
externos, além de reduzirem a supressão sobre as florestas nativas.
Essa deposição de matéria orgânica no solo, por meio da serapilheira,
beneficia as condições físicas do solo, resultando em maior retenção de água e
estabilidade dos agregados formados pela aproximação das partículas minerais
(Bayer e Mielniczuk, 1997) e ainda fornece energia para o crescimento microbiano, o
que reflete em maior ciclagem de nutrientes e aumento da capacidade de troca
catiônica (CTC) do solo (Lima et al., 2011).
Com relação às propriedades físicas, Carvalho et al. (2004) observaram que
os atributos físicos de um Latossolo Vermelho argiloso sob diferentes sistemas,
concluíram que nos sistemas agroflorestais a densidade aparente foi menor, com
maior porosidade, menor resistência à penetração e maior estabilidade de
agregados quando comparado ao solo sob sistema de plantio convencional.
Loss et al. (2009) avaliaram a influência de sistemas de manejo agroecológico
sobre os atributos físicos e químicos de um Argissolo Vermelho-Amarelo, sob área
de preparo convencional (milho/feijão), plantio direto (berinjela/milho), consórcio

18
maracujá/trevo de campo (Desmodium sp.), área cultivada com figo e sistema
agroflorestal (SAF) e verificaram que o SAF teve maiores percentuais de porosidade
total nas duas épocas avaliadas (verão de 2005 e inverno de 2006) na profundidade
de 0,05 a 0,10 m, porém não se observou alterações químicas.
2.2.2. Benefícios dos SAFs para os atributos biológicos do solo
Os microorganismos favorecem a incorporação da matéria orgânica e
melhoram o solo devido à mobilização de nutrientes, fragmentação dos resíduos
orgânicos e à mistura com o solo mineral (Ducatti, 2002), além de desempenhar
importante papel na aeração e permeabilidade do solo, através de galerias
construídas, facilitando a penetração das raízes (Silva et al., 2011b). Os
microorganismos são considerados bons indicadores biológicos, pois possuem
capacidade de fornecer informações rápidas sobre as alterações nas propriedades
do solo, causadas por cultivos e manejos ou devastações de florestas, ou seja, está
associada a alterações em função do ecossistema (Araújo et al., 2013).
A avaliação desses microorganismos pode ser estimada por métodos diretos
e indiretos que representam a densidade, diversidade e a atividade (Araújo e
Monteiro, 2007). No entanto, as técnicas consistem em isolar e avaliar o crescimento
desses microorganismos, porém representam apenas a comunidade cultivável do
solo e necessitam de outras variáveis, como o conhecimento do estado da biomassa
microbiana, atividade respiratória do solo e o metabolismo, que avaliados em
conjunto, podem fornecer informações da atividade biológica do solo (Sousa, 2014).
A biomassa microbiana do solo (BMS) representa o componente vivo da
matéria orgânica do solo, tirando a macrofauna e as raízes das plantas, e
geralmente compreende de 1 a 5% do carbono orgânico do solo (Araújo e Monteiro,
2007). Essa variável chamada de carbono da biomassa microbiana do solo (C-BMS)
é uma estimativa que permite aferir o acúmulo ou a perda de carbono liberado pela
morte dos microorganismos (proporção presente de células microbianas vivas
contendo carbono), em outras palavras é a quantidade de carbono que a biomassa
de microorganismos do solo imobiliza em suas células (Araújo e Monteiro, 2007;
Silva, 2008).

19
A respiração basal (C-CO2) representa a oxidação da matéria orgânica pelos
microorganismos de metabolismo aeróbio, ela indica a atividade metabólica e o
estádio fisiológico da célula microbiana que é influenciada por diversos fatores, em
outras palavras, expressa a qualidade biomassa microbiana (Sousa, 2014).
O quociente metabólico (q-CO2) representa a quantidade de CO2 liberado por
unidade de biomassa microbiana em determinado tempo, ou seja, indica a eficiência
da biomassa microbiana em utilizar o carbono disponível para biossíntese, sendo
sensível indicador para estimar a atividade biológica e qualidade do substrato
(Saviozzi et al., 2002). Pode ser usado como indicador de estresse, perturbação ou
estabilidade de um ecossistema.
Maiores valores de q-CO2 indicam maiores perdas de carbono no sistema na
forma de CO2 por unidade de carbono microbiano (Mercante et al., 2008). Uma
biomassa é mais eficiente quando menos carbono for perdido na forma de CO2,
devendo, então, uma fração significativa de carbono (C) estar incorporada ao tecido
microbiano (Mercante et al., 2008). Essa variação do q-CO2 pode ser causada por
desequilíbrio ambiental, podendo ser devido à matéria orgânica de baixa qualidade
ou comunidades microbianas submetidas a condições de estresse, como deficiência
de nutrientes, acidez e déficit hídrico (Ramos et al., 2010).
O quociente microbiano (q-MIC) expresso pela relação carbono
microbiano/carbono orgânico está diretamente relacionado com o acúmulo de C no
solo ao longo do tempo (Mercante et al., 2008).
O favorecimento do desenvolvimento da comunidade microbiana do solo é
afetado pela estabilidade de um sistema, que é influenciada pelo tempo de
adaptação. Tanto estabilidade quanto tempo de adaptação são dependentes do
incremento de matéria orgânica em quantidade e qualidade, favorecendo o
desenvolvimento da comunidade microbiana do solo (Pezarico et al., 2004).
Vasconcelos et al. (2005) ao avaliarem o carbono, nitrogênio e a atividade da
biomassa microbiana de um Latossolo Amarelo Concrecionário sob vegetações
secundárias de dois, seis e 14 anos de idade, nas estações seca e chuvosa,
verificaram que houve mudanças na estrutura da comunidade microbiana do solo,
indicando maior eficiência na biomassa microbiana em imobilizar um elevado teor de
carbono na vegetação secundária de 14 anos e durante a estação da seca, o

20
mesmo comportamento ocorreu para o q-MIC. Enquanto, a respiração basal (C-CO2)
e no quociente metabólico (q-CO2) foram superiores na estação chuvosa,
diferenciando no tempo de estabelecimento da vegetação secundária.
2.2.3. Características do solo com SAFs e períodos sazonais
Entende-se que nos sistemas cujo aporte de matéria orgânica é maior e não
há revolvimento do solo, as características deste tende a ser melhorada. Por isso,
devido à diversificação de espécies que os SAF promovem, há maiores chances de
se aproximarem de uma condição de equilíbrio e estabilidade, promovendo
resultados satisfatórios quanto à melhoria da qualidade do solo (Pezarico et al.,
2004).
Vários estudos com SAFs abordando as características do solo por meio da
aferição de indicadores físicos, químicos e biológicos comparativamente com outras
práticas de manejo e períodos sazonais foram realizados por Carvalho et al. (2004),
Pezarico et al. (2004), Vasconcelos et al. (2005), Barreto et al. (2006), Maia et al.
(2006), Loss et al. (2009), Collier e Araújo, (2010), Iwata et al. (2012), Lima et al.
(2011), Silva et al. (2011a), Silva Junior et al. (2012), entre outros.
Lima et al. (2011) avaliou diversos sistemas (SAF com 10 anos, SAF com seis
anos, sistema com base ecológica com três anos, agricultura de corte e queima e
floresta nativa) em um Argissolo Vermelho Amarelo no Piauí e verificaram que os
SAFs melhoraram a qualidade do solo, pois tiveram os maiores teores de fósforo (P),
estoque de carbono orgânico (Est-COT) e nitrogênio (Est-NT) que os demais
sistemas. Os teores de P nos SAFs foram maiores no período chuvoso e no período
seco, o SAF de 10 anos teve teores seis e sete vezes maior que a área de corte e
queima e floresta nativa, respectivamente. Efeito inverso ocorreu com o Est-COT e
Est-NT que teve maiores quantidades estocado no período seco. Isso demonstra a
eficiência do manejo com SAFs na ciclagem do P e armazenamento do COT e NT.
Iwata et al. (2012) ao avaliarem um SAF de seis anos, um SAF de 13 anos,
agricultura de corte e queima com seis anos de cultivo e floresta nativa, observaram
que os SAFs promoveram e garantiram melhoria da qualidade química do solo em
áreas sob Cerrado.

21
Collier e Araújo (2010) notaram diferença entre os períodos estacionais para
os atributos químicos do solo, e os maiores teores de nutrientes disponibilizados foi
no período chuvoso. No entanto, Silva Junior et al. (2012) ao avaliarem uma área de
pastagem de 20 anos, um SAF de oito anos de implantação, uma capoeira de 30
anos sem intervenção e uma área de cultivo de mandioca de dois anos sob
Latossolo Vermelho Distroférrico argissólico com textura média moderada,
verificaram que o SAF não proporcionou melhorias na fertilidade do solo, exceto no
teor de fósforo.
Enquanto, Silva et al. (2011a) ao avaliarem seis sistemas de SAFs, um cultivo
convencional e uma floresta nativa, verificaram que os indicadores químicos nos
sistemas agroflorestais tiveram melhoria na fertilidade do solo em relação ao sistema
nativo, que segundo os autores são indícios da necessidade de práticas corretivas e
de adubação, por considerarem que houve exportação de nutrientes por meio da
retirada dos frutos e grãos para comercialização, o que poderia ter diminuído as
reservas de nutrientes do solo. Já os atributos físicos do solo foram indicadores
favoráveis na avaliação da qualidade do solo, pois distinguiram os efeitos
proporcionados pelos SAFs em relação à floresta nativa.

22
3. MATERIAL E MÉTODOS
3.1. Localização e Característica da Área de Estudo
O município de Alta Floresta fica localizado no extremo Norte do estado de
Mato Grosso, a 757 km da capital Cuiabá, situado na Bacia Hidrográfica Amazônica,
constituída pelos rios Teles Pires, Cristalino, Santa Helena, Paranaíta e Apiacás,
tributários do rio Tapajós (Souza 2006).
O experimento está implantado na Fazenda experimental “Ariosto da Riva”
pertencente à Comissão Executiva do Plano da Lavoura Cacaueira – CEPLAC,
situado na Rodovia MT 208 a 25 km da cidade de Alta Floresta – MT, com
coordenadas (Latitude 9º 53’ 42” S e Longitude 56º 17’ 46” O). O solo da área é
classificado como Argissolo Vermelho-Amarelo Eutrófico típico de acordo com a
classificação do Sistema Brasileiro de Classificação do Solo (Embrapa, 2013).
O relevo da estação experimental se divide em três tipos, sendo o local da
coleta de solo, uma área plana com desnivelamentos muito pequeno, segundo a
classificação de Neves et al. (1981).
A vegetação nativa faz parte da região Amazônica e é classificada como
Floresta Ombrófila Aberta, composta de árvores de grande porte mais espaçadas,
com estrato arbustivo pouco denso e caracterizada pela presença de cipós (IBGE,
1997). Espécies de relevante importância comercial, como: a castanheira, cedro,
seringueira, carnaúba, copaíba, garapa, tatajuba, cambará, paricá entre outras,
foram encontradas nessa estação.

23
3.2. Instalação do Experimento
A Fazenda experimental “Ariosto da Riva” possui um total de 505 ha e em
1981, parte da floresta primária foi derrubada para a implantação dos experimentos.
A área é ocupada por 95% de floresta nativa e o restante por experimentos em sua
maioria, de Sistemas Agroflorestais (SAFs).
Essa área ficou de três a quatro anos em pousio, para então instalar os
primeiros experimentos com SAFs e em 2005 foi implantado o experimento deste
estudo em delineamento em blocos casualizados com consórcios de espécies
arbóreas e cupuaçu (Theobroma grandiflorum).
A calagem e a adubação foram realizadas somente na implantação dos
sistemas consorciados, sendo aplicada nas covas o formulado de 00-25-20 de NPK
e mais 250 kg ha-1 de ureia aplicados via foliar.
Para este estudo, foi considerado o esquema de parcelas subdivididas 5x2,
sendo cinco sistemas florestais e duas épocas de avaliações (estação seca e
chuvosa), com quatro repetições. Nos sistemas florestais foi considerada a floresta
nativa - FN e quatro sistemas consorciados com cupuaçu, sendo: mogno brasileiro
(Switenia macrophyla) + cupuaçu – MBcp; teca (Tectona grandis) + cupuaçu –
TEcp; cedro rosa (Cedrela fissilis) + cupuaçu – CEcp; paricá (Schizolobium
amazonicum) + cupuaçu – PAcp.
Cada sistema consorciado era composto por sete fileiras de espécie arbórea
de grande porte e seis fileiras de cupuaçu no espaçamento 6x3 m para ambos os
plantios, arbóreos e cupuaçu. Para melhor entendimento, o desenho esquemático
dos consórcios florestais, em uma área de 1,61 ha está ilustrado na Figura 1.
A floresta nativa está adjacente aos sistemas cultivados e para efeito
estatístico, foi dividida em quatro blocos conforme blocos delimitados nos sistemas
consorciados (Figura 1).

24
6 m
3 m
P c P c P c P c P c P c P P c P c P c P c P c P c P P c P c P c P c P c P c P P c P c P c P c P c P c P P c P c P c P c P c P c P P c P c P c P c P c P c P
P c P c P c P c P c P c P
6 m
3 m M c M c M c M c M c M c M M c M c M c M c M c M c M M c M c M c M c M c M c M M c M c M c M c M c M c M M c M c M c M c M c M c M M c M c M c M c M c M c M
M c M c M c M c M c M c M
3 m
FIGURA 1. Croqui dos sistemas agroflorestais e floresta nativa.
Legenda: M: mogno brasileiro; T: teca; C: cedro rosa; P: paricá; c: cupuaçu

25
3.3. Dados Meteorológicos
O clima da região é tropical chuvoso do tipo Awi, segundo classificação de
Köppen e as estações são bem definidas, uma chuvosa e outra seca, com
precipitação anual de 2.750 mm, tendo os meses de Maio a Agosto com o menor
índice pluviométrico (Oliveira, 2006).
Durante o ano das coletas das amostras, em 2012, a temperatura máxima foi
de 32ºC e mínima de 21ºC, representada respectivamente nos meses de fevereiro e
julho, contudo a média anual foi 25ºC. Os meses de março e novembro foram os
mais chuvosos com precipitação mensal de aproximadamente 65 mm,
consequentemente com maior umidade relativa do ar (85%), segundo dados do
Instituto Nacional de Meteorologia - INMET (2014) (Figura 2).
Fonte: INMET (2014).
FIGURA 2. Dados meteorológicos de precipitação (Precip.), umidade (Umid.) e
temperatura (Temp.) do município de Alta Floresta – MT, do ano de
2012.
3.4. Amostragem do Solo

26
As coletas de solo foram realizadas em dois períodos distintos, considerando
um período chuvoso (abril/2012) e outro período de seca (agosto/2012) da região.
Os sistemas foram percorridos em zig-zag, amostrando-se três pontos em cada
sistema. Mediante a média aritmética simples dos três pontos, obtiveram-se os
resultados dos atributos do solo de forma a representar cada sistema. Em cada
ponto de amostragem, foi aberta uma mini-trincheira com enxadões e bocas de lobo
e coletadas amostras de terra nas profundidades 0,0 a 0,10; 0,10 a 0,20 e 0,20 a
0,40 m. Em cada profundidade foram coletadas três tipos de amostras:
indeformadas (para a análise física), semi-deformadas (agregados) e deformadas
(para a análise granulométrica, microbiológica e química). As amostras
indeformadas foram coletadas utilizando um trado amostrador de kopeck e as
demais com o auxílio de pás jardineiras.
As amostras para análise química e microbiológica foram acondicionadas em
sacos plásticos e colocadas em freezer para a preservação da biomassa microbiana;
as amostras para análise física foram armazenadas em vasilhames plásticos e em
anéis de Kopeck, os anéis foram colocados em estojos de madeira para preservação
da estrutura. Todas as amostras foram identificadas e encaminhadas ao Laboratório
de Solos da Faculdade de Agronomia, Medicina Veterinária e Zootecnia da
Universidade Federal de Mato Grosso para procedimento analítico.
Foram consideradas as camadas de 0,0 a 0,10; 0,10 a 0,20 e 0,20 a 0,40 m
para a análise dos atributos químicos, de 0,0 a 0,10 e 0,10 a 0,20 m para a análise
dos atributos físicos e de 0,0 a 0,10 m para análise dos atributos microbiológicos.
3.5. Caracterização dos Atributos Químicos do Solo
As amostras indeformadas foram destorroadas, secas e passadas em
peneiras com malha de 2 mm para análise dos atributos químicos. Foram
determinados o pH em CaCl2 0,01 mol L-¹ na proporção 1:2,5 por potenciometria; a
acidez potencial (H+Al) extraída pelo acetato de cálcio; o cálcio (Ca), magnésio (Mg)
e acidez trocável (Al3) extraídos por KCL 1 mol L-¹ e determinado por titulometria; o
fósforo (P) e o potássio (K) extraídos com solução de Mehlich 1, sendo o P

27
determinado por espectrocolorimetria e o K por fotometria de emissão de chama
(Embrapa, 2009).
Enquanto, a soma de bases (SB), a capacidade de troca catiônica (CTC), a
saturação por bases (V%), a saturação por alumínio (m%) e a matéria orgânica do
solo (MOS) foram obtidas por meio de cálculos, conforme Sousa e Lobato (2004).
O carbono orgânico total (COT) foi quantificado por oxidação da matéria
orgânica via úmida, utilizando a solução de dicromato de potássio em meio ácido
com fonte externa de calor e titulação com sulfato ferroso amoniacal, segundo
método de Yeomans e Bremner (1988).
3.5.1. Estoque do carbono do solo e dos nutrientes
Os estoques de COT para cada profundidade foram calculados utilizando as
seguintes expressões (Leite et al., 2003): Est-COT = (COT x Ds x E), em que: Est-
COT é o estoque de carbono orgânico total em determinada profundidade; COT o
teor de carbono orgânico total; Ds a densidade do solo, determinada pelo método do
anel volumétrico, segundo Embrapa (2009) em cada profundidade e E a espessura
da camada considerada.
Para o cálculo do estoque dos demais nutrientes, utilizou-se a mesma fórmula
supracitada, apenas substituindo o COT pelo teor do nutriente avaliado.
3.6. Atributos Físicos do Solo
As análises físicas do solo foram feitas de acordo com os métodos descritos
em Embrapa (1997). A análise textural foi realizada, nas amostras da profundidade
0,0 a 0,10; 0,10 a 0,20 e 0,20 a 0,40 m, com a finalidade de caracterização da área
para fins estatísticos, uma vez que sistemas com classificação textural diferentes
não poderiam ser comparados. Foi utilizado o método da pipeta, baseado no
princípio da velocidade de queda das partículas, conforme a Lei de Stokes,
utilizando-se como dispersante químico o NaOH a 0,1 mol L-1. Não foi constatada
diferença textural entre os sistemas, somente entre os blocos, o que permite a
comparação estatística entre sistemas. A caracterização textural da área com
médias e respectivo desvio-padrão encontra-se na Tabela 1.

28
TABELA 1. Caracterização textural do solo sob sistemas florestais, Alta Floresta -
MT, 2013
Sistemas Profund. Argila Silte Areia
(m) ----------------------- (g kg-1) -------------------------
FN
0,0 a 0,10
400,10±61,22 88,93±15,70 511,00±41,62
MBcp 303,00±40,91 96,72±46,80 600,30±42,10
TEcp 299,23±74,81 89,52±18,12 611,25±32,44
CEcp 305,00±27,45 76,42±5,62 618,60±45,72
PAcp 289,47±31,84 73,00±11,61 637,53±31,41
FN
0,10 a 0,20
528,27±29,58 88,93±25,67 382,80±23,76
MBcp 361,68±45,57 80,02±12,45 558,30±20,56
TEcp 400,62±41,67 86,73±19,67 512,65±65,89
CEcp 389,50±43,84 78,50±4,89 543,07±87,99
PAcp 360,28±25,22 41,08±5,45 598,63±37,12
FN
0,20 a 0,40
606,00±76,35 62,00±10,09 332,00±43,12
MBcp 515,88±151,15 61,92±12,09 422,20±9,44
TEcp 556,27±81,22 53,33±13,44 395,97±19,82
CEcp 518,62±45,12 61,97±7,86 419,42±11,34
PAcp 495,00±33,56 45,33±5,98 459,67±4,21
Floresta nativa – FN; mogno brasileiro + cupuaçu – MBcp; teca + cupuaçu – TEcp; cedro rosa + cupuaçu – CEcp; paricá + cupuaçu – PAcp.
Para a obtenção dos atributos físicos foram utilizadas as amostras das
camadas de 0,0 a 0,10 e de 0,10 a 0,20 m de profundidade.
A determinação da densidade do solo (Ds) foi pelo o método do anel
volumétrico (tipo Kopeck) com volume de 100 cm³. No laboratório, os anéis de
Kopeck após serem limpos e envolvidos com tecido tipo voal, foram saturados por
capilaridade para obtenção da macro e microporosidade e porosidade total pela
mesa de tensão (Mac, Mic e Pt em m³ m-³) conforme descrito em Embrapa (1997). A
microporosidade (Mic) foi obtida a partir da quantidade de água retida nas amostras
indeformadas de solo e submetidas à tensão de -0,006 MPa (60 cm de H2O). A
macroporosidade (Mac) foi calculada pela diferença entre o peso da amostra a
-0,006 MPa e a amostra seca em estufa a 105°C. A porosidade total foi obtida pela
soma entre macro e microporosidade.
As amostras semi-deformadas foram processadas manualmente de forma a
desmanchar os torrões e em seguida tamisados em peneiras de 4 a 2 mm para

29
obtenção das variáveis da estabilidade dos agregados do solo pelo método úmido.
Para obter a distribuição percentual das classes de tamanho dos agregados, os
retidos na peneira de 2 mm foram selecionados e colocados em agitação no
aparelho de Yoder por 4 minutos, em que foi aclopado um conjunto de peneiras com
malhas de 0,105; 0,25; 0,5; 1; e 2 mm de abertura (Embrapa, 1997). A partir dos
dados obtidos, foram calculados os índices: diâmetro médio geométrico (DMG),
diâmetro médio ponderado (DMP) e a estabilidade de agregados (EA) (Castro Filho
et al., 1998).
3.7. Atributos Microbiológicos do Solo
A quantificação do carbono da biomassa microbiana foi realizada pelo método
da fumigação-incubação adaptado por Jenkinson e Powlson (1976), no qual foram
utilizadas amostras fumigadas e incubadas e amostras incubadas sem fumigação.
Para a fumigação foram utilizados 20 mL de clorofórmio e deixadas às
amostras por 48 h no escuro e para a incubação foram utilizados frascos herméticos
de 280 mL, contendo 20 mL de NaOH 0,5 mol L-1, em que foram colocadas as
amostras por seis dias no escuro. Ao final do período de incubação, o NaOH dos
frascos das amostras foram titulados com HCl 0,3 mol L-1; para isso, em cada frasco
de 20 mL de NaOH foi adicionado 3 mL de solução saturada de BaCl2 e 3 gotas do
indicador (fenolftaleína a 1 %) sob agitação magnética.
Após a titulação do NaOH, calculou-se a quantidade de respiração basal (C-
CO2) liberada das amostras fumigadas e não fumigadas, em que o carbono da
biomassa microbiana foi calculado a partir da fórmula: C-BMS (μg C g-1) = (C
fumigado – C não-fumigado) / kc. Para a constante de mineralização (kc), foi
assumido o valor de 0,45, presumindo que apenas 45% da biomassa microbiana
presente no solo foi convertida a CO2 durante os 6 dias de incubação, após a
fumigação à 25ºC (Anderson e Domsch, 1978; Siqueira et al., 1994).
Os valores para a respiração basal foram obtidos por meio da seguinte
fórmula: C-CO2 (μg C-CO2 g-1 h-1) = (C amostra não fumigada – C branco) x
molaridade exata do HCl x 6 x proporção de NaOH/massa de solo seco (proporção
de NaOH 20 mL / 20 mL = 1), e depois foi dividido pela quantidade de horas dos dias

30
avaliados. Para a determinação do quociente metabólico, foi utilizada a fórmula: q-
CO2 (mg C-BMS h-1) = respiração basal/ carbono da biomassa microbiana (Anderson
e Domsch, 1990), e o quociente microbiano foi calculado pela fórmula: q-MIC (%) =
(carbono da biomassa microbiana/ carbono orgânico do solo).
3.8. Análise Estatística
Os resultados foram submetidos ao teste de normalidade dos resíduos pelo
teste Ryan Joiner (p>0,10) e homogeneidade de variância (p>0,10). Posteriormente,
foi realizada a análise de variância e quando o F foi significativo (p<0,05), realizou-se
o teste de média Scott-Knott (p<0,05) para comparação entre os sistemas florestais
e os períodos sazonais, exceto para as análises físicas que apenas os sistemas
florestais foram comparados. As análises estatísticas foram realizadas no programa
estatístico Sisvar (Ferreira, 2011).

31
4. RESULTADOS E DISCUSSÕES
4.1. Atributos Químicos do Solo
4.1.1. Componentes da acidez do solo
Os resultados do pH e a acidez potencial (H+Al) do solo (Tabela 2), não
diferiram em nenhum dos sistemas e épocas avaliadas. O pH variou de 4,38 a 5,31,
sendo classificada por Souza e Lobato (2004), como uma acidez de média a
adequada para o cultivo de culturas.
Em relação ao alumínio (Al+3), observou-se que na camada de 0,00 a 0,10 m
não houve diferença entre os sistemas quando a avaliação ocorreu no período
chuvoso (Tabela 2). Porém, quando a avaliação foi feita no período seco, verificou-
se que os sistemas Mogno Brasileiro + Cupuaçu (MBcp) e Paricá + Cupuaçu (PAcp)
tiveram maior teor de Al+3 em relação aos demais sistemas. Esses mesmos sistemas
não diferenciaram no teor de Al+3 em relação a épocas. Contrariamente, a Floresta
Nativa (FN) e a Teca + Cupuaçu (TEcp) tiveram menores valores de Al+3 e não
diferiram do Cedro Rosa + Cupuaçu (CEcp). Quando se compara a época de
avaliação, verificou-se que esses sistemas tiveram menores valores de Al+3 no
período seco.
Nas demais camadas, não houve diferença entre os sistemas independente
da época de avaliação, somente houve diferença nos teores de Al+3 no sistema
TEcp, na camada de 0,10 a 0,20 e 0,20 a 40 m e na FN de 0,20 a 0,40 m quando
comparados entre as épocas chuvosa e seca, com menores valores nessa última
época. A mesma tendência dos teores de Al+3 foi observada para a saturação por

32
alumínio (m%), ressaltando-se que os resultados da m% são considerados baixos
segundo a classificação de Sousa e Lobato (2004).
TABELA 2. pH, acidez trocável (Al+3), acidez potencial (H+Al) e saturação por
alumínio (m) no solo sob sistemas florestais, Alta Floresta - MT, 2013
CAMADA (m) ÉPOCA SISTEMAS
FN MBcp TEcp CEcp PAcp
pH (CaCl2)
0,00 a 0,10 Chuvoso 5,31 Aa 4,76 Aa 4,79 Aa 4,48 Aa 4,49 Aa
Seco 5,22 Aa 5,09 Aa 4,79 Aa 4,69 Aa 4,89 Aa
0,10 a 0,20 Chuvoso 5,17 Aa 4,55 Aa 4,48 Aa 4,39 Aa 4,55 Aa
Seco 5,09 Aa 5,01 Aa 4,52 Aa 4,63 Aa 4,93 Aa
0,20 a 0,40 Chuvoso 4,60 Aa 4,49 Aa 4,49 Aa 4,45 Aa 4,72 Aa
Seco 4,97 Aa 5,13 Aa 4,52 Aa 4,73 Aa 5,08 Aa
H+Al (cmolc dm-3
)
0,0 a 0,10 Chuvoso 5,70 Aa 4,01 Aa 3,72 Aa 4,14 Aa 4,35 Aa
Seco 3,82 Aa 3,97 Aa 5,31 Aa 4,01 Aa 3,52 Aa
0,10 a 0,20 Chuvoso 5,17 Aa 4,55 Aa 4,48 Aa 4,39 Aa 4,55 Aa
Seco 5,09 Aa 5,01 Aa 4,52 Aa 4,63 Aa 4,93 Aa
0,20 a 0,40 Chuvoso 4,76 Aa 2,97 Aa 3,42 Aa 3,21 Aa 3,05 Aa
Seco 3,95 Aa 3,07 Aa 3,90 Aa 3,15 Aa 2,38 Aa
Al+3
(cmolc dm-3
)
0,0 a 0,10 Chuvoso 0,13 Aa 0,12 Aa 0,13 Aa 0,16 Aa 0,15 Aa
Seco 0,01 Bb 0,12 Aa 0,03 Bb 0,05 Bb 0,13 Aa
0,10 a 0,20 Chuvoso 0,10 Aa 0,13 Aa 0,17 Aa 0,14 Aa 0,13 Aa
Seco 0,01 Aa 0,12 Aa 0,04 Ab 0,04 Aa 0,11 Aa
0,20 a 0,40 Chuvoso 0,24 Aa 0,12 Aa 0,18 Aa 0,13 Aa 0,15 Aa
Seco 0,01 Ab 0,10 Aa 0,05 Ab 0,06 Aa 0,11 Aa
m%
0,0 a 0,10 Chuvoso 1,27 Aa 1,70 Ab 2,19 Aa 2,67 Aa 2,14 Ab
Seco 0,40 Ba 6,45 Aa 1,39 Ba 3,58 Ba 6,92 Aa
0,10 a 0,20 Chuvoso 0,92 Aa 2,63 Aa 4,18 Aa 2,94 Aa 2,49 Aa
Seco 0,50 Aa 7,77 Aa 3,19 Aa 3,79 Aa 7,00 Aa
0,20 a 0,40 Chuvoso 2,35 Aa 2,98 Aa 4,60 Aa 3,11 Aa 3,45 Aa
Seco 0,50 Aa 7,18 Aa 3,82 Aa 5,03 Aa 8,55 Aa
*Médias seguidas por mesma letra, maiúscula na linha e minúscula na coluna, não diferem entre si pelo teste Scott-Knott (p<0,05). Floresta nativa – FN; mogno brasileiro + cupuaçu – MBcp; teca + cupuaçu – TEcp; cedro rosa + cupuaçu – CEcp; paricá + cupuaçu – PAcp.
Observou-se ainda, que os valores de acidez tenderam a aumentar em
profundidade, como era esperado. Entre os fatores que contribuem para o aumento
da acidez em profundidade está a mineralização da matéria orgânica do solo
acumulada em superfície que tende a consumir prótons elevando o pH nessa região

33
(Novais et al., 2007). Além disso, por meio da dissociação das hidroxilas com
formação de cargas negativas durante a mineralização conseguem complexar o H+ e
Al+3 livres em solução (Pavinato, 2008). Outro fator, que pode estar associado a
essa tendência é a mineralogia desse solo por se tratar de um Argissolo, que
naturalmente possui aumento de argila em profundidade associado ao aumento de
óxidos de Fe+2 e Al+3 (Embrapa, 2013).
Resultados contrários a este estudo foram observados por Iwata et al. (2012)
em um Argissolo Vermelho-Amarelo, em que os autores verificaram aumento da
acidez do solo em profundidade e encontraram menores teores de acidez nos
sistemas agroflorestais em relação à mata nativa e à área cultivada. Segundo os
autores, isso pode ter ocorrido porque o solo apresentava nas condições naturais
valores de pH mais baixos e teores de Al+3 e H+Al mais altos do que o do solo deste
estudo, estando mais propenso a modificações pelo manejo. Os autores não
constataram alterações na acidez do solo nos períodos chuvoso e seco,
demonstrando que essas características são pouco afetadas pela sazonalidade.
O aumento da acidez do solo em profundidade também foi observado por
Collier e Araújo (2010) quando estudaram sistemas de uso de solo em um Neossolo
Quartzarênico Órtico. Porém, os autores constataram que houve, em todas as
camadas para o período chuvoso, diferenças no pH e nos teores de Al+3 entre a
mata nativa, o sistema agroflorestal e lavoura de subsistência, sendo que no sistema
agroflorestal foram encontrados menores valores de pH e os maiores teores de Al+3
e H+Al. Maiores variações de pH devido ao uso do solo podem ser encontradas em
solos arenosos em relação aos argilosos, isso devido ao maior poder tampão dos
últimos em relação aos primeiros.
Portanto, todo o complexo de acidez do solo infere que os sistemas de uso e
as épocas de avaliação tiveram pouca influência sobre esses atributos químicos, de
forma que os mesmos não influenciaram negativamente no desenvolvimento das
espécies.
4.1.2. Trocas catiônicas, carbono orgânico e matéria orgânica do solo
Observou-se que no período chuvoso e em todas as camadas, a capacidade
de troca catiônica total (CTCpH7,0) do sistema FN teve maiores valores em relação

34
aos demais sistemas (Tabela 3). Enquanto no período seco, em todas as camadas,
não houve diferença entre os sistemas. No entanto, quando se avaliou a diferença
entre as épocas em cada camada, verificou-se que houve tendência de redução da
CTCpH7,0 no período seco em relação ao chuvoso. Do mesmo modo, Maia et al.
(2006), avaliando seis sistemas diferentes, dentre eles o agrossilvipastoril,
silvipastoril, cultivo convencional e matas, não verificaram diferenças da CTCpH7,0
entre os sistemas no período seco.
Os resultados da capacidade de troca catiônica efetiva (CTCefetiva) nos
sistemas e nas camadas, independente da época de avaliação, foram similares aos
resultados da CTCpH7,0, sempre com maiores valores para FN e no período chuvoso
(Tabela 3).
Quanto ao carbono orgânico (COT) e matéria orgânica do solo (MOS),
verificaram-se tendências semelhantes em relação aos sistemas e época de
avaliação em todas as camadas (Tabela 3). Na primeira camada (0,00 a 0,10 m),
especificamente na época chuvosa, não houve diferença entre os sistemas,
enquanto na estação seca a FN teve maior teor em relação aos demais. Conferiu-se
ainda, que nessa mesma camada, na FN o teor de COT e MOS foram maiores na
época seca em relação à época chuvosa. Para os demais sistemas não houve
diferença entre as épocas. Nessa camada a MOS foi classificada de baixa a
adequada, segundo Souza e Lobato (2004).
Na camada de 0,10 a 0,20 m no período chuvoso, os tratamentos FN e TEcp
se destacaram dos demais, em que apresentaram maiores teores de COT e MOS,
enquanto no período seco os sistemas não diferiram entre si. Na camada de 0,20 a
0,40 m não houve diferença entre os sistemas em nenhuma das épocas.
Observou-se que entre as camadas de 0,10 e 0,40 m, os sistemas MBcp e
CEcp tiveram seus teores de COT e MOS reduzidos na época chuvosa em relação à
época seca e que não foi constatado nos demais sistemas, pois não houve diferença
entre as épocas (Tabela 3). Na camada de 0,10 a 0,40 m a MOS foi classificada
como baixa segundo Souza e Lobato (2004).

35
TABELA 3. Capacidade de troca catiônica (CTCpH7,0), capacidade de troca catiônica
efetiva (CTCefetiva), carbono orgânico total (COT), matéria orgânica do
solo (MOS) no solo sob sistemas florestais, Alta Floresta-MT, 2013
*Médias seguidas por mesma letra, maiúscula na linha e minúscula na coluna, não diferem entre si pelo teste Scott-Knott (p<0,05). Floresta nativa – FN; mogno brasileiro + cupuaçu – MBcp; teca + cupuaçu – TEcp; cedro rosa + cupuaçu – CEcp; paricá + cupuaçu – PAcp.
Com relação ao aumento dos teores do COT e da MOS no período seco,
provavelmente se deve, segundo Salcedo e Sampaio (2008), à deposição de folhedo
e morte de raízes finas (diâmetro < 2 mm) que ocorrem durante este período e que
são as principais entradas de carbono no solo. Com o início das chuvas, a maior
umidade no solo determina um forte aumento da atividade biológica com
consequente mineralização de parte do carbono aportado, diminuindo os teores do
solo (Moreira e Siqueira, 2002). Na época seca, o efeito é o inverso do que acontece
CAMADA (m) ÉPOCA SISTEMAS
FN MBcp TEcp CEcp PAcp
CTCpH7,0 (cmolc dm-3
)
0,00 a 0,10 Chuvoso 17,15 Aa 11,08 Ba 9,91 Ba 10,11 Ba 11,22 Ba
Seco 6,29 Ab 5,75 Ab 7,00 Aa 5,38 Ab 5,30 Ab
0,10 a 0,20 Chuvoso 15,96 Aa 8,34 Ba 7,43 Ba 8,35 Ba 8,46 Ba
Seco 5,99 Ab 4,98 Aa 6,08 Aa 4,47 Ab 4,24 Ab
0,20 a 0,40 Chuvoso 13,94 Aa 7,03 Ba 7,34 Ba 7,37 Ba 7,37 Ba
Seco 5,92 Ab 4,43 Aa 5,07 Aa 4,24 Ab 3,59 Ab
CTCefetiva (cmolc dm-3
)
0,0 a 0,10 Chuvoso 11,59 Aa 7,19 Ba 6,32 Ba 6,12 Ba 7,01 Ba
Seco 2,48 Ab 1,90 Ab 1,73 Ab 1,42 Ab 1,91 Ab
0,10 a 0,20 Chuvoso 11,67 Aa 5,16 Ba 4,05 Ba 4,86 Ba 5,17 Ba
Seco 2,08 Ab 1,53 Ab 1,30 Ab 1,16 Ab 1,58 Ab
0,20 a 0,40 Chuvoso 9,42 Aa 4,18 Ba 4,10 Ba 4,28 Ba 4,47 Ba
Seco 1,98 Ab 1,46 Ab 1,23 Ab 1,15 Ab 1,31 Ab
COT (g kg-1
)
0,00 a 0,10 Chuvoso 16,22 Ab 13,41 Aa 15,70 Aa 13,71 Aa 13,05 Aa
Seco 25,24 Aa 13,38 Ba 13,14 Ba 13,87 Ba 14,20 Ba
0,10 a 0,20 Chuvoso 13,40 Aa 7,24 Bb 10,77 Aa 6,71 Bb 8,73 Ba
Seco 13,41 Aa 11,51 Aa 9,30 Aa 10,91 Aa 10,06 Aa
0,20 a 0,40 Chuvoso 10,28 Aa 5,78 Ab 7,57 Aa 6,10 Ab 8,13 Aa
Seco 10,77 Aa 10,52 Aa 7,62 Aa 9,83 Aa 6,93 Aa
MOS (g kg-1
)
0,00 a 0,10 Chuvoso 27,90 Ab 23,07 Aa 27,05 Aa 23,58 Aa 22,44 Aa
Seco 43,41 Aa 23,01 Ba 22,60 Ba 23,86 Ba 24,42 Ba
0,10 a 0,20 Chuvoso 23,04 Aa 12,45 Bb 19,52 Aa 11,54 Bb 15,01 Ba
Seco 23,06 Aa 20,79 Aa 17,00 Aa 18,76 Aa 17,30 Aa
0,20 a 0,40 Chuvoso 16,68 Aa 9,96 Ab 13,02 Aa 10,50 Ab 13,98 Aa
Seco 17,52 Aa 18,09 Aa 13,11 Aa 17,91 Aa 11,92 Aa

36
na época chuvosa, há maior aporte de resíduos vegetais sobre o solo devido às
características das espécies caducifólias que perdem as folhas no inverno, e menor
taxa de decomposição da matéria orgânica, pois as condições de umidade e
temperatura são limitadas ao processo.
Essa hipótese está respaldada nos resultados obtidos por Melz e Tiago (2009),
que verificaram redução de 20,61% dos teores de carbono orgânico e matéria orgânica
do solo no período chuvoso em relação ao período seco e conferiram aumento de
33,31% na população de bactérias e 33,96% na população de fungos do solo na época
chuvosa, quando o solo atingiu umidade 88,86% maior que a do período seco.
Vital et al. (2004), também, encontraram maior deposição e biomassa de
serapilheira no final do período seco em floresta semidecidual no Estado de São
Paulo, evidenciando as características sazonais, com pequena variação na
concentração dos nutrientes. Do mesmo modo, Silva Junior et al. (2012)
encontraram na camada de 0,00 a 0,10 m maior CTC na mata nativa e na camada
de 0,10 a 0,20 m, porém nesse caso se igualou ao sistema de capoeira, em um
Latossolo Vermelho Distroférrico argissólico, com textura média moderada.
Porém, esse efeito depende do tipo de resíduo depositado, pois foi observada
sazonalidade nos teores de COT e MOS para os sistemas TEcp e PAcp. Outra
relevância é que a sazonalidade de COT e MOS na FN ocorreu na superfície,
enquanto para MBcp e CEcp foi em subsuperfície.
O mogno e o cedro por pertencerem à mesma família e tem a mesma
característica das folhas e raízes, trata-se de materiais de mais fácil decomposição
que os demais consórcios, o que acelera sua decomposição na época chuvosa e
redução dos seus teores, assim o material depositado no solo, principalmente em
subsuperfície (raízes finas), não é suficiente para manter o equilíbrio entre a época
seca e chuvosa.
Bayer et al. (2003) e Ciotta et al. (2003) verificaram alta relação entre os
teores de MOS e a CTC, principalmente em solos de regiões tropicais e subtropicais,
em que a predominância da mineralogia nos componentes argilo minerais é a
caulinita que contribui apenas com 3 a 5 cmolc kg-1, enquanto os componentes
húmicos contribuem de 300 a 1000 cmolc kg-1 (Ernani, 2008).

37
Dessa forma, sistemas conservacionistas como os sistemas agroflorestais
que mantém ou aumentem significativamente os teores de MOS são importantes
para a manutenção da qualidade química desse solo.
4.1.3. Componentes de bases e de P no solo
Para o cálcio (Ca2+) foi observada diferença entre os sistemas apenas na
camada de 0,00 a 0,10 m no período chuvoso, tendo a FN como destaque com teor
de 46,03; 37,18; 49,03; 43,63% maior que os sistemas MBcp, TEcp, CEcp e PAcp,
respectivamente (Tabela 4). Entretanto, ao comparar as épocas avaliadas, verificou-
se que houve diferença em todas as camadas, sendo que nos períodos chuvosos
predominaram maiores teores de Ca2+ em relação aos períodos de seca, sendo
considerados adequados de acordo com a classificação de fertilidade do solo de
Souza e Lobato (2004).
Em relação ao magnésio (Mg2+), observou-se que, em todas as camadas
avaliadas, houve maior teor no sistema FN quando a avaliação ocorreu no período
chuvoso (Tabela 4). Nesse caso, na camada de 0,00 a 0,10 m, a FN resultou em teor
de Mg2+ de 35,59; 64,95; 49,29 e 46,62% maior que os sistemas MBcp, TEcp, CEcp e
PAcp, respectivamente. Porém, observou-se que nessa mesma camada, o sistema
TEcp teve menor teor de Mg2+ em relação à MBcp, CEcp e PAcp e nas demais
camadas não diferenciaram entre si.
No entanto, quando se compara as duas épocas em cada camada e sistema,
a tendência foi à mesma observada para o Ca2+, sendo os teores de Mg2+ sempre
maiores no período chuvoso (Tabela 4).
Para o potássio (K+) em todas as camadas de solo, independente da época
avaliada, não houve diferença entre os sistemas, demonstrando que os sistemas
consorciados não alteram os valores em relação à FN (Tabela 4). Em semelhança
ao Ca2+ e Mg2+, o K+ também teve maior teor no período chuvoso em relação ao
seco. O mesmo aconteceu com a saturação por bases (V%), exceto na camada de
0,20 a 0,40 m, em que os sistemas MBcp, CEcp e PAcp não tiveram diferenças
entre as épocas avaliadas, diferindo assim dos demais.
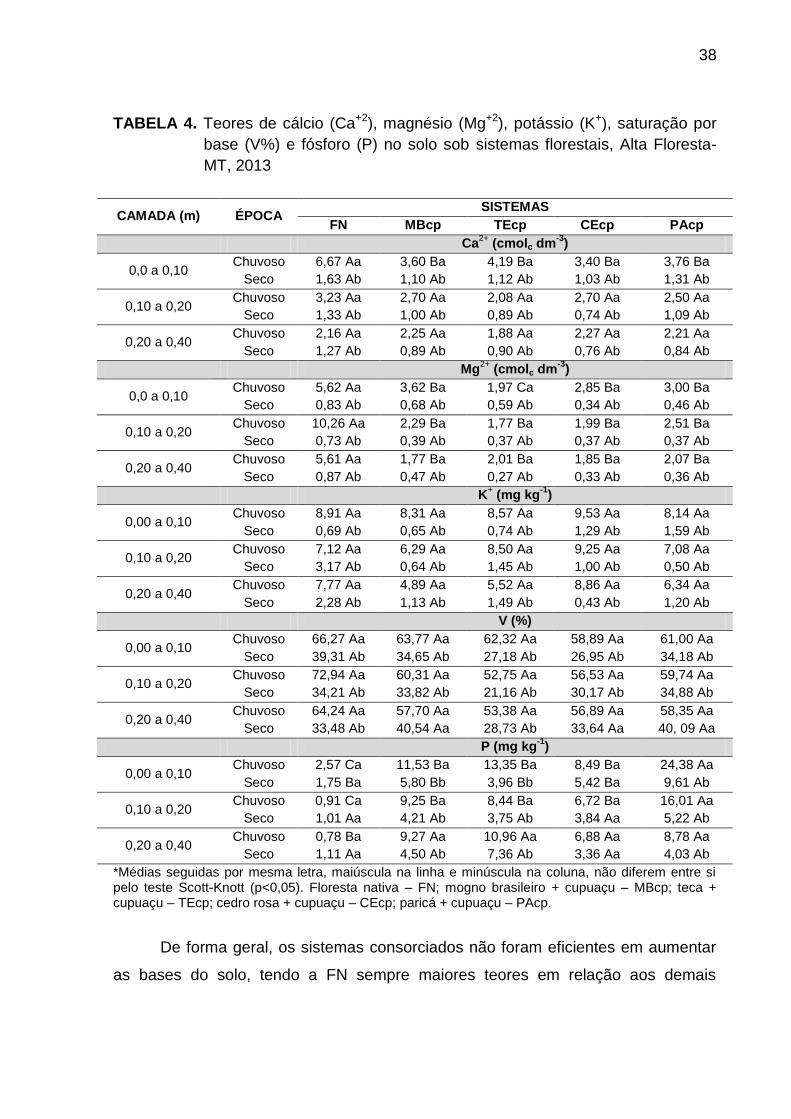
38
TABELA 4. Teores de cálcio (Ca+2), magnésio (Mg+2), potássio (K+), saturação por
base (V%) e fósforo (P) no solo sob sistemas florestais, Alta Floresta-
MT, 2013
CAMADA (m) ÉPOCA SISTEMAS
FN MBcp TEcp CEcp PAcp
Ca2+
(cmolc dm-3
)
0,0 a 0,10 Chuvoso 6,67 Aa 3,60 Ba 4,19 Ba 3,40 Ba 3,76 Ba
Seco 1,63 Ab 1,10 Ab 1,12 Ab 1,03 Ab 1,31 Ab
0,10 a 0,20 Chuvoso 3,23 Aa 2,70 Aa 2,08 Aa 2,70 Aa 2,50 Aa
Seco 1,33 Ab 1,00 Ab 0,89 Ab 0,74 Ab 1,09 Ab
0,20 a 0,40 Chuvoso 2,16 Aa 2,25 Aa 1,88 Aa 2,27 Aa 2,21 Aa
Seco 1,27 Ab 0,89 Ab 0,90 Ab 0,76 Ab 0,84 Ab
Mg2+
(cmolc dm-3
)
0,0 a 0,10 Chuvoso 5,62 Aa 3,62 Ba 1,97 Ca 2,85 Ba 3,00 Ba
Seco 0,83 Ab 0,68 Ab 0,59 Ab 0,34 Ab 0,46 Ab
0,10 a 0,20 Chuvoso 10,26 Aa 2,29 Ba 1,77 Ba 1,99 Ba 2,51 Ba
Seco 0,73 Ab 0,39 Ab 0,37 Ab 0,37 Ab 0,37 Ab
0,20 a 0,40 Chuvoso 5,61 Aa 1,77 Ba 2,01 Ba 1,85 Ba 2,07 Ba
Seco 0,87 Ab 0,47 Ab 0,27 Ab 0,33 Ab 0,36 Ab
K+ (mg kg
-1)
0,00 a 0,10 Chuvoso 8,91 Aa 8,31 Aa 8,57 Aa 9,53 Aa 8,14 Aa
Seco 0,69 Ab 0,65 Ab 0,74 Ab 1,29 Ab 1,59 Ab
0,10 a 0,20 Chuvoso 7,12 Aa 6,29 Aa 8,50 Aa 9,25 Aa 7,08 Aa
Seco 3,17 Ab 0,64 Ab 1,45 Ab 1,00 Ab 0,50 Ab
0,20 a 0,40 Chuvoso 7,77 Aa 4,89 Aa 5,52 Aa 8,86 Aa 6,34 Aa
Seco 2,28 Ab 1,13 Ab 1,49 Ab 0,43 Ab 1,20 Ab
V (%)
0,00 a 0,10 Chuvoso 66,27 Aa 63,77 Aa 62,32 Aa 58,89 Aa 61,00 Aa
Seco 39,31 Ab 34,65 Ab 27,18 Ab 26,95 Ab 34,18 Ab
0,10 a 0,20 Chuvoso 72,94 Aa 60,31 Aa 52,75 Aa 56,53 Aa 59,74 Aa
Seco 34,21 Ab 33,82 Ab 21,16 Ab 30,17 Ab 34,88 Ab
0,20 a 0,40 Chuvoso 64,24 Aa 57,70 Aa 53,38 Aa 56,89 Aa 58,35 Aa
Seco 33,48 Ab 40,54 Aa 28,73 Ab 33,64 Aa 40, 09 Aa
P (mg kg-1
)
0,00 a 0,10 Chuvoso 2,57 Ca 11,53 Ba 13,35 Ba 8,49 Ba 24,38 Aa
Seco 1,75 Ba 5,80 Bb 3,96 Bb 5,42 Ba 9,61 Ab
0,10 a 0,20 Chuvoso 0,91 Ca 9,25 Ba 8,44 Ba 6,72 Ba 16,01 Aa
Seco 1,01 Aa 4,21 Ab 3,75 Ab 3,84 Aa 5,22 Ab
0,20 a 0,40 Chuvoso 0,78 Ba 9,27 Aa 10,96 Aa 6,88 Aa 8,78 Aa
Seco 1,11 Aa 4,50 Ab 7,36 Ab 3,36 Aa 4,03 Ab
*Médias seguidas por mesma letra, maiúscula na linha e minúscula na coluna, não diferem entre si pelo teste Scott-Knott (p<0,05). Floresta nativa – FN; mogno brasileiro + cupuaçu – MBcp; teca + cupuaçu – TEcp; cedro rosa + cupuaçu – CEcp; paricá + cupuaçu – PAcp.
De forma geral, os sistemas consorciados não foram eficientes em aumentar
as bases do solo, tendo a FN sempre maiores teores em relação aos demais

39
sistemas e a época seca proporcionou redução dos teores na maioria dos sistemas.
Esses resultados foram diferentes daqueles obtidos por Silva et al. (2011) também
em Argissolo que encontraram maiores teores e porcentagem de bases nos
sistemas agroflorestais em relação à FN. Todavia, no trabalho feito por aqueles
autores houve correção da fertilidade do solo com calcário e adubos sintéticos na
camada de 0,0 a 0,20 m em todos os sistemas cultivados, o que garantiu aumento
de nutrientes em relação à FN.
Neste estudo, a correção da fertilidade mediante insumos externos ocorreu
apenas na implantação dos sistemas, sete anos atrás, não sendo suficiente para
aumentar os níveis de bases no solo e manter o equilíbrio entre absorção de
nutrientes e reposição pelos resíduos culturais. O cupuaçu utilizado nos sistemas
consorciados extrai grande quantidade de cátions básicos para a formação de frutos,
o que pode em princípio reduzir os teores no solo (Collier e Araujo, 2010). Como os
sistemas agroflorestais são recentes, pode ainda não ter dado tempo de ocorrer à
recuperação desses nutrientes, assim como observado por Bernardes et al. (2000),
que verificaram recuperação dos teores de nutrientes e matéria orgânica do solo
semelhantes da vegetação nativa após 16 anos de implantação dos sistemas
agroflorestais.
Devido a maior diversidade de espécies na FN é provável que a quantidade
de serapilheira desse sistema também seja maior que aquela depositada nos
sistemas consorciados, aumentando os teores de bases no solo. Silva et al. (2012)
verificaram que a FN acumulou aproximadamente 690 g m² de serapilheira, sendo
36% maior que o sistema agroflorestal 1 e 58% maior que o sistema agroflorestal 2,
que diferenciavam entre si nas espécies recuperadoras da fertilidade do solo. O
maior acúmulo de serapilheira na FN resultou na maior liberação de Ca e Mg em
relação aos sistemas agroflorestais.
Evidencia-se neste trabalho que, apesar dos sistemas agroflorestais terem
tido redução nos teores de Ca2+, Mg2+ e K+, isso não foi suficiente para alterar a
saturação de bases do solo. As diferenças entre a saturação por bases apenas
ocorreram entre épocas de avaliação e não entre sistemas (Tabela 3). Verifica-se
que o Argissolo estudado tem caráter eutrófico, o que de certa forma, evidencia o
equilíbrio dos sistemas.

40
Em relação à época de avaliação, foi observada, para as bases do solo,
tendência contrária do ocorrido com a MOS, pois houve redução na época seca,
evidenciando sazonalidade dos cátions básicos no solo, provavelmente devido ao
efeito da mineralização da MOS. Na época seca, com a menor atividade biológica há
redução na liberação de nutrientes, com isso, as culturas, passam a utilizar os
estoques de nutrientes do solo sem reposição pela decomposição da MOS, havendo
redução dos teores. Silva et al. (2012) também verificaram aumento nos teores de
bases no período chuvoso na FN e sistemas agroflorestais e atribuíram à
decomposição da matéria orgânica do solo.
Outro fator que pode corroborar com a redução de nutrientes no período seco, é
a ocorrência de chuvas eventuais nesse período, que podem ativar a liberação dos
nutrientes provenientes da MOS por parte da biomassa microbiana, que morre com
esse processo e estimula o crescimento de uma nova população de microorganismos,
essa por sua vez pode causar a imobilização de nutrientes que serão utilizados no seu
processo metabólico (Vasconcelos et al., 2005).
Por outro lado, Iwata et al. (2012) encontraram em Argissolo Vermelho-
Amarelo, menores teores de Ca2+, Mg2+ e K+ no período chuvoso e destacaram estar
relacionado à perda de bases pela enxurrada e demais ações da chuva, isso tem
sido evidenciado pela literatura, mesmo em sistemas de maior estabilização com os
sistemas agroflorestais e matas nativas. Nesse experimento, como a classe de
textura em superfície é franco-argilosa, que juntamente com a quantidade de MOS,
tenha oferecido resistência à perda de nutrientes via enxurrada.
A sazonalidade das bases do solo nos sistemas consorciados depende dos
tipos de consórcio, Collier e Araújo (2010) só encontraram diferenças entre as
épocas secas e chuvosas para vegetação nativa e sistemas de lavoura,
predominando os maiores teores na época chuvosa. No sistema agroflorestal
estudado pelos autores, que incluíam consórcio de árvores nativas, palmeira
babaçu, cupuaçu, limão e acerola, não houve diferença entre as épocas avaliadas,
isso porque a diversidade de espécies conduz maior reciclagem desses nutrientes,
no período seco, associada à menor exportação de nutrientes para os frutos.
Ao analisar o fósforo (P) no período chuvoso, nas camadas de 0,0 a 0,10 m e
0,10 a 0,20 m, observou-se que houve diferença entre os sistemas, em que o PAcp

41
resultou em maior teor e a FN no menor (Tabela 4). Na camada de 0,00 a 0,10 m, os
teores obtidos no PAcp foram 52,71; 45,24 e 65,18% maiores que os obtidos por
MBcp, TEcp e CEcp, respectivamente, e 89,46% daqueles obtidos por FN. Na
camada de 0,10 a 0,20 m, o PAcp proporcionou teores de P na ordem de 42,22;
47,28 e 58,02% maiores que o MBcp, TEcp e CEcp, respectivamente, e 94,32%
maior que o da FN, demonstrando a capacidade do PAcp em incorporar P ao solo
até 0,20 m de profundidade. Na camada de 0,20 a 0,40 m, o menor teor de P foi
observado na FN.
No período seco, os efeitos dos sistemas sobre os teores de P foram
observados apenas na camada de 0,00 a 0,10 m, sendo que o PAcp também
proporcionou maior teor em relação aos demais sistemas, porém, não houve
diferença de MBcp, TEcp e CEcp em relação à FN. Tanto na época chuvosa, quanto
na época seca, os teores de P no sistema PAcp, de acordo com a classificação de
Sousa e Lobato (2004), foram tidos como alto, enquanto os da FN como baixo.
Quando se compara os teores de P nas épocas em cada sistema, observa-se
em todas as camadas, que apenas os sistemas FN e CEcp não diferenciaram entre
as épocas chuvosa e seca. Todos os outros sistemas tiveram os teores de P
reduzidos na época seca em relação à chuvosa, resultado semelhante ao que
aconteceu com as bases do solo, podendo também ser atribuído à sazonalidade da
degradação da matéria orgânica do solo. A redução de P no período chuvoso
corrobora com os resultados de Loss et al. (2009), Collier e Araújo (2010), Lima et
al. (2011) e Iwata et al. (2012), sendo o primeiro e os dois últimos também em
Argissolo Vermelho-Amarelo.
Entre os atributos químicos do solo estudados nos sistemas agroflorestais, o
maior ganho foi em relação aos teores de P. Observa-se que o Argissolo deste
estudo, apesar de eutrófico, tem teores de P, originalmente classificados como
baixos. Têm sido observados, em sistemas agroflorestais, maiores teores de P em
relação à FN (Nogueira et al., 2008; Lima et al., 2011).
Neste estudo, verificou-se o ganho de P nos sistemas consorciados em
relação à FN até 0,40 m de profundidade no período chuvoso, principalmente no
consórcio entre cupuaçu e paricá. Diferentemente desses resultados, Lima et al.
(2011) verificaram na camada de 0,00 a 0,10 m de um Argissolo Vermelho-Amarelo

42
que os teores de P no solo de um sistema agroflorestal, com 10 anos no período
seco, foram sete vezes maiores que os da FN demostrando a eficiência do sistema
em reciclar P, considerando-se a baixa concentração desse elemento nos solos
tropicais. No período chuvoso houve aumento dos teores P e tanto o sistema com
seis quanto o com dez anos de agrofloresta teve seus teores maiores que a FN.
O paricá consegue extrair na parte aérea grande quantidade de P, sendo
esse o nutriente mais limitante para a cultura (Marques et al., 2004). Assim, deduz-
se que esta espécie acumule P em maiores quantidades na sua folha, permitindo na
decomposição de sua estrutura liberar grandes quantidades desse nutriente para o
solo.
Outro ponto relevante diz respeito à morfologia das folhas de paricá que são
menores que as demais espécies cultivadas, assim ao se acumularem no solo no
período seco, por ser espécie caducifólia, provavelmente no período chuvoso,
possuem maior taxa de decomposição, por isso, incorporando mais P ao solo em
relação às outras culturas. Oliveira (2006) encontrou maiores teores de P na
profundidade de 0,0 a 0,05 m para todos os tratamentos (gliricídia e cumaru;
cupuaçu, gliricídia e cumaru; monocultivo de café; monocultivo de pupunha e mata
nativa), e justificou a maior atividade biológica nos primeiros centímetros do solo.
Ao estudar o fracionamento de P em sistemas agroflorestais, Nogueira et al.
(2008) verificaram que o aporte e a manutenção de matéria orgânica nos sistemas
agroflorestais proporcionaram uma maior concentração de P no solo, além disso,
esses sistemas favoreceram a ciclagem desse elemento através do reservatório de
fósforo orgânico no solo.
4.2. Estoques de Nutrientes no Solo
Os estoques de COT (Est-COT) e de MOS (Est-MOS) seguiram as mesmas
tendências que seus teores para todos os fatores estudados (Tabela 5). Para Est-COT
só houve diferença da FN em comparação aos sistemas consorciados na camada de
0,10 a 0,20 m no período chuvoso. Nessa situação a FN pela sua diversidade teve os
mesmos estoques que TEcp e em maiores estoques que MBcp, CEcp e PAcp. Vale

43
destacar que TEcp proporcionou Est-CO 29,83; 37,99 e 22,19% maiores que MBcp,
CEcp e PAcp, respectivamente.
Para o Est-MOS, além das diferenças observadas na camada de 0,10 a 0,20
m semelhantes ao observado para o Est-COT, foi verificado também que na camada
de 0,00 a 0,10 m no período seco, a FN proporcionou maior estoque em relação aos
demais sistemas que não se diferenciaram entre si. Magalhães et al. (2013), em
Argissolo Vermelho-Amarelo, também verificaram maiores estoques de MOS em FN.
O Est-MOS confirmam a hipótese de que provavelmente o mogno e o cedro
forneçam resíduos de maior decomposição, pois no período seco, os Est-MOS são
altos, não diferindo da FN, porém, no período seco passam a ter os menores
estoques.
Para o estoque de fósforo (Est-P), na camada de 0,0 a 0,10 m na avaliação
do período chuvoso, observou-se que houve diferença entre os sistemas, sendo que
o PAcp proporcionou maior estoque e a FN o menor, o mesmo aconteceu na
camada de 0,0 a 0,20 m (Tabela 5).
No período seco, o estoque de fósforo diferiu na camada de 0,0 a 0,10 m,
sendo que o PAcp, MBcp e CEcp proporcionaram os maiores estoques no solo,
demonstrando a importância das culturas em incrementar P no solo e o Paricá como
cultura acumuladora de P.
Assim como observado para o teor de P, na época chuvosa foi constatado
para a FN o menor Est-P em todas as camadas. E com relação à avaliação entre
épocas da camada de 0,20 a 0,40 m, verificou-se que apenas os sistemas FN e
TEcp não diferenciaram entre os períodos de seca e chuva, os demais sistemas
também tiveram maiores estoques no período chuvoso. Como o P é um elemento
essencial para as plantas, é provável que na época seca seja necessária a entrada
do nutriente via fertilização para a manutenção do crescimento e produtividade do
sistema TEcp, pois este se mostrou mais próximo aos baixos valores da FN.
Magalhães et al. (2013) verificaram que os menores Est-P foram encontrados no
sistema com consórcio com teca, provavelmente pelo menor aporte de P em seus
resíduos.

44
TABELA 5. Estoques de carbono orgânico (Est-COT), de matéria orgânica (Est-MOS), de fósforo (Est-P), de cálcio (Est-Ca), de magnésio (Est-Mg) e de potássio (Est-K) sob sistemas florestais, Alta Floresta-MT, 2013
CAMADA (m) ÉPOCA SISTEMAS
FN MBcp TEcp CEcp PAcp
Est-COT (Mg ha-1
)
0,00 a 0,10 Chuvoso 17,86 Ab 20,46 Aa 22,92 Aa 20,36 Aa 18,60 Aa Seco 27,80 Aa 20,18 Aa 20,17 Aa 20,68 Aa 20,36 Aa
0,10 a 0,20 Chuvoso 17,83 Aa 11,36 Bb 16,19 Aa 10,04 Bb 12,50 Ba
Seco 17,91 Aa 18,08 Aa 13,42 Aa 16,79 Aa 14,53 Aa
0,20 a 0,40 Chuvoso 27,32 Aa 17,21 Ab 22,04 Aa 17,83 Ab 24,71 Aa
Seco 28,27 Aa 30,88 Aa 22,14 Aa 28,31 Aa 21,11 Aa
Est-MOS (Mg ha-1
)
0,00 a 0,10 Chuvoso 30,73 Ab 35,21 Aa 39,48 Aa 35,17 Aa 32,18 Aa Seco 47,81 Aa 35,12 Ba 32,98 Ba 35,59 Ba 35,02 Ba
0,10 a 0,20 Chuvoso 30,80 Aa 19,53 Bb 29,26 Aa 17,27 Bb 21,68 Ba
Seco 30,83 Aa 32,62 Aa 25,48 Aa 28,08 Aa 24,98 Aa
0,20 a 0,40 Chuvoso 44,25 Aa 29,32 Ab 37,78 Aa 30,78 Ab 42,72 Aa
Seco 46,47 Aa 53,68 Aa 38,02 Ba 52,51 Aa 36,43 Ba
Est-P (kg ha-1
)
0 a 0,10 Chuvoso 2,74 Ca 17,55 Ba 19,56 Ba 12,75 Ba 35,04 Aa
Seco 1,92 Ba 8,88 Ab 5,82 Bb 8,08 Aa 13,71 Ab
0,10 a 0,20 Chuvoso 1,21 Ca 14,60 Ba 12,70 Ba 10,01 Ba 23,07 Aa
Seco 1,35 Aa 6,60 Ab 5,63 Ab 5,74 Aa 7,54 Ab
0,20 a 0,40 Chuvoso 2,06 Ba 27,94 Aa 31,84 Aa 20,34 Aa 26,84 Aa
Seco 2,96 Ba 13,45 Ab 21,40 Aa 9,75 Ab 12,32 Ab
Est-Ca (kg ha-1
)
0,00 a 0,10 Chuvoso 1.439,97 Aa 1.101,21 Ba 1.209,91 Ba 1.007,09 Ba 1.077,61 Ba
Seco 357,70 Ab 336,28 Ab 326,36 Ab 306,68 Ab 371,81 Ab
0,10 a 0,20 Chuvoso 864,74 Aa 848,54 Aa 624,48 Ba 807,87 Aa 715,37 Ba
Seco 355,00 Ab 314,04 Ab 266,16 Ab 221,96 Ab 315,11 Ab
0,20 a 0,40 Chuvoso 1.149,54 Ba 1.333,39 Aa 1.092,30 Ba 1.338,52 Aa 1.347,98 Aa
Seco 675,96 Ab 517,66 Ab 522,39 Ab 446,42 Ab 510,89 Ab
Est-Mg (kg ha-1
)
0,00 a 0,10 Chuvoso 769,64 Aa 637,86 Aa 347,80 Ba 460,88 Ba 536,35 Ba
Seco 111,86 Ab 126,00 Ab 104,99 Aa 61,55 Ab 80,86 Ab
0,10 a 0,20 Chuvoso 1.664,96 Aa 435,74 Ba 322,94 Ba 365,11 Ba 441,11 Ba
Seco 118,49 Ab 74,13 Ab 67,84 Ab 67,69 Ab 65,75 Ab
0,20 a 0,40 Chuvoso 1.806,17 Aa 639,27 Ba 713,66 Ba 655,66 Ba 770,17 Ba
Seco 279,58 Ab 163,05 Ab 96,71 Ab 117,07 Ab 133,71 Ab
Est-K (kg ha-1
)
0,00 a 0,10 Chuvoso 8,65 Ba 27,91 Aa 12,51 Ba 14,23 Ba 11,77 Ba
Seco 1,50 Aa 1,98 Ab 2,23 Ab 3,86 Ab 4,35 Aa
0,10 a 0,20 Chuvoso 34,94 Aa 25,87 Aa 17,32 Ba 14,60 Ba 21,44 Ba
Seco 8,79 Ab 2,02 Ab 4,38 Ab 2,89 Ab 1,44 Ab
0,20 a 0,40 Chuvoso 33,87 Ba 44,07 Aa 27,72 Ba 28,89 Ba 43,79 Aa
Seco 3,01 Ab 1,69 Ab 2,13 Ab 0,62 Ab 1,81 Ab
*Médias seguidas por mesma letra, maiúscula na linha e minúscula na coluna, não diferem entre si pelo teste Scott-Knott (p<0,05). Floresta nativa – FN; mogno brasileiro + cupuaçu – MBcp; teca + cupuaçu – TEcp; cedro rosa + cupuaçu – CEcp; paricá + cupuaçu – PAcp.

45
O P foi o elemento menos estocado pelos sistemas, provavelmente devido a
sua dinâmica de se fixar aos colóides do solo, ficando menos disponível em solução
(Novais et al., 2007). Na época seca, os estoques variaram de 1,35 a 21,40 kg ha-1 e,
na época chuvosa, variaram de 2,74 a 35,04 kg ha-1.
Os sistemas tiveram a maior variação no estoque de cálcio (Est-Ca) nos
períodos chuvosos de cada camada avaliada (Tabela 5). Na camada de 0,0 a
0,10 m, o sistema FN teve maior Est-Ca em relação aos demais sistemas, já na
segunda camada (0,10 a 0,20 m), os sistemas FN, MBcp e CEcp estocaram mais
Ca, enquanto na camada 0,20 a 0,40 m foram os sistemas MBcp, CEcp e PAcp. Na
época seca não houve diferença entre os sistemas, proporcionando em todas as
camadas menores estoques em relação à época chuvosa. Na época seca, os
estoques variaram de 222 a 676 kg ha-1, enquanto na época chuvosa, a variação foi
de 624 a 1.440 kg ha-1 aproximadamente. Mas, independente da forma de uso do
solo, época de avaliação e profundidade, esse foi o nutriente mais estocado pelas
culturas.
Foi observado para o estoque de magnésio (Est-Mg) semelhança aos Est-Ca,
sendo o segundo nutriente mais estocado e que as maiores variações ocorreram na
época chuvosa, sem resultados significativos entre sistemas na época seca (Tabela
5). Na época chuvosa, a FN proporcionou maior Est-Mg em relação aos demais
sistemas, em todas as camadas.
Com relação ao estoque de potássio (Est-K) esse foi juntamente como P, um
dos nutrientes menos estocados. Do mesmo modo, não foi observada diferença entre
os sistemas na época seca e na época chuvosa (Tabela 5), sendo que o sistema
MBcp tendeu a ter maior acúmulo em relação aos demais sistemas no período
chuvoso. Porém, na camada de 0,10 a 0,20 m não se diferenciou da FN e na camada
de 0,20 a 0,40 m se comparou ao PAcp. Assim como o observado para os estoques
de Ca e Mg, a época seca proporcionou menores Est-K.
Os estoques de K ficaram abaixo dos obtidos por Silva (1995), que encontrou
em solos de floresta nativa estoques próximos de 66 kg ha-1 e Magalhães et al.
(2013) que encontraram valores médios de 40,03 kg ha-1 para sistemas
agroflorestais.

46
Maiores estoques de Ca e de Mg em relação ao K eram esperados, pois os
primeiros elementos apresentam preferência na série liotrópica de adsorção devido
ao complexo de carga e raio iônico (Novais et al., 2007), tornando o K mais
susceptível ao processo de lixiviação.
Verificou-se que, para todos os nutrientes em estudo, houve manutenção dos
estoques em profundidade até 0,40 m, provavelmente devido à textura do solo e
micorrizas, uma vez que a maior deposição de resíduos ocorre em superfície. Dentre
os fatores que podem ter contribuído para as respostas dos estoques no perfil,
destacam-se a preservação das características do solo nos sistemas de uso
estudados devido à ausência de revolvimento do solo e a provável existência de
canais contínuos, oriundos da decomposição de raízes e da atividade biológica
(Mokry 2003), possibilitando o aumento do estoque de nutrientes em profundidade
devido à translocação de nutrientes da camada superficial do solo (Magalhães et al.,
2013).
De forma geral, em termos de acúmulo nutricional em superfície do solo, foi
verificado que o sistema PAcp foi o que mais acumulou P; FN foi o que mais
acumulou Ca; FN e MBcp, os que mais acumularam Mg e MBcp, o que mais
acumulou K, demonstrando assim, a importância de cada sistema em cada
componente nutricional do solo.
4.3. Variações dos Atributos Físicos do Solo
A densidade do solo (Ds) teve menores valores no sistema FN para as duas
camadas avaliadas (Tabela 6). Com relação à macroporosidade do solo (Mac), foi
maior nos sistemas FN, na camada de 0,0 a 0,10 m e na camada de 0,10 a 0,20 m,
os resultados dos sistemas não apresentaram diferenças.
Respostas similares ao Mac, com relação aos sistemas nas duas camadas
avaliadas, foram verificadas para a porosidade total (PT), o diâmetro médio
ponderado (DMP), o diâmetro médio geométrico (DMG) e a estabilidade de
agregados (EA). Enquanto que para a microporosidade (Mic) não houve diferença
entre os sistemas nas duas camadas avaliadas (Tabela 6).

47
Segundo Silva et al. (2000) e Klein (2012), a densidade do solo é afetada pela
cobertura vegetal, teor de matéria orgânica, uso e manejo do solo, que de alguma
forma altere sua estrutura e por consequência arranjo e volume de poros.
Como a MOS do solo não variou consideravelmente nas camadas, pode-se
atribuir a diferença de Ds entre o sistema FN e MBcp, TEcp, CEcp e PAcp, ao
manejo do solo. Isso possivelmente se deve ao fato da FN não ter sido antropizada,
ao contrário das características solo nos consórcios florestais que foram alterados
na sua implantação, pelo revolvimento do solo, hipótese afirmada pela similaridade
dos resultados dos sistemas MBcp, TEcp, CEcp e Pacp.
TABELA 6. Densidade (Ds), macroporosidade (Mac), microporosidade (Mic),
porosidade total (PT), diâmetro médio ponderado (DMP), diâmetro
médio geométrico (DMG) e estabilidade de agregados (EA) do solo sob
sistemas florestais coletados no período chuvoso, Alta Floresta-MT,
2013
VARIÁVEL CAMADA (m) SISTEMAS
FN MBcp TEcp CEcp PAcp
Ds (Mg m-3
) 0 a 0,10 1,10 B 1,33 A 1,30 A 1,39 A 1,33 A
0,10 a 0,20 1,24 B 1,37 A 1,36 A 1,30 A 1,34 A
Mac (m-3
m-3
) 0 a 0,10 0,22 A 0,16 B 0,18 B 0,17 B 0,17 B
0,10a 0,20 0,19 A 0,17 A 0,18 A 0,17 A 0,18 A
Mic (m-3
m-3
) 0 a 0,10 0,38 A 0,32 A 0,32 A 0,33 A 0,32 A
0,10 a 0,20 0,32 A 0,31 A 0,31 A 0,32 A 0,31 A
PT (m-3
m-3
) 0 a 0,10 0,60 A 0,49 B 0,50 B 0,50 B 0,49 B
0,10 a 0,20 0,51 A 0,48 A 0,49 A 0,49 A 0,49 A
DMP 0 a 0,10 2,02 A 1,07 B 1,09 B 1,15 B 1,21 B
0,10 a 0,20 1,28 A 1,03 A 0,85 A 0,99 A 1,11 A
DMG 0 a 0,10 1,54 A 0,71 B 0,72 B 0,77 B 0,79 B
0,10 a 0,20 0,90 A 0,65 A 0,60 A 0,66 A 0,77 A
EA 0 a 0,10 95,10 A 87,67 B 88,49 B 89,32 B 88,46 B
0,10 a 0,20 89,26 A 86,07 A 86,75 A 87,05 A 87,61 A
*Médias seguidas por mesma letra, maiúscula na linha não diferem entre si pelo teste Scott-Knott (p<0,05). Floresta nativa – FN; mogno brasileiro + cupuaçu – MBcp; teca + cupuaçu – TEcp; cedro rosa + cupuaçu – CEcp; paricá + cupuaçu – PAcp.
Silva et al. (2011) encontraram diferença entre os sistemas, seis SAFs,
floresta nativa (FN) e cultivo convencional de mamão (CCM), em que a Ds e a PT,
tiveram menor e maior valor, respectivamente, para o sistema FN, contudo, não foi
observado diferença entre os sistemas para Mac e Mic em um Argissolo Amarelo
distrocoeso (PAdx).

48
A maior densidade do solo encontrada em profundidade pode ser explicada
pelas pressões exercidas das camadas superiores sobre as subjacentes, que
provocam a compactação, reduzindo a sua porosidade, bem como a movimentação
de material de menor granulometria dos horizontes superiores para os inferiores
(iluviação), que também concorre para a redução do espaço poroso e aumento da
densidade (Pauleto et al., 2006).
De modo geral, os maiores valores Ds proporcionaram os menores valores de
PT, concordando com os resultados de Loss et al. (2009), em Argissolo Vermelho-
Amarelo com textura franco-arenosa, que obtiveram o mesmo efeito em suas
avaliações de diferentes sistemas (milho/feijão, berinjela/milho, maracujá, figo e
SAF). Por outro lado, Barrreto et al. (2006) observaram que a Ds e a PT não
diferenciaram entre os sistemas mata Atlântica, cultura de cacau e pastagem nas
profundidades de 0,00 a 0,10 e 0,10 a 0,20 m, em um Latossolo Vermelho-Amarelo.
Nos sistemas avaliados, foi verificado que os resultados de Mac variaram
pouco e teve o mesmo efeito na PT, pois todos os valores ficaram acima de 0,10 m-3
m-3. Esses resultados estão de acordo com os encontrados por Ferreira (2010), não
indicando ter limitação para o crescimento e ao desenvolvimento radicular das
plantas.
De maneira geral, em todos os tratamentos, foram encontrados valores ideais
de porosidade total, pois segundo Camargo e Alleoni (1997), um solo ideal deve
apresentar 50% de volume de poros totais que, na capacidade de campo, teria
33,5% ocupado pela água e 16,5% ocupado pelo ar.
Os poros de menor diâmetro possuem maior resistência à deformação em
relação aos macroporos e maior capacidade de suporte de carga, conforme
observado por Giarola et al. (2009) e Bergamin et al. (2010), que ao estudarem o
efeito do uso do solo sobre a microporosidade, não encontraram alterações nesse
atributo. Araujo et al. (2004), também não verificaram diferenças em relação aos Mic
entre os sistemas de cultivo convencional e mata nativa, mas observaram maior
quantidade de Mac e PT na mata nativa, em Latossolo Vermelho Distrófico.
Com relação à Mic, a razão entre essa e a PT ficou entre 0,63 e 0,66, o que
indica que os sistemas não tiveram reflexos negativos na taxa de infiltração do solo
e crescimento radicular, pois os resultados ficaram próximos a 0,67 que é a relação

49
ideal, que permite adequada aeração do solo (Kiehl, 1979). A Mic é fortemente
influenciada pela textura, teor de COT e pouco afetada pela Ds (Araujo et al., 2004).
Os maiores diâmetros de agregados (DMP e DMG) foram encontrados na FN,
na camada de 0,00 a 0,10 m, em que também apresentou maior EA, sendo
observada ligeira diminuição de acordo com a profundidade. Essa redução nos
diâmetros e na EA nos sistemas cultivados em relação à FN, na camada de 0,00 a
0,10 m, pode estar relacionada à alteração na estrutura do solo com as práticas de
manejo e também à possibilidade de que o COT, presente no sistema FN, seja mais
estável que nos demais (Benjamim et al., 2008).
Valores, de DMG, maiores aos observados nesta pesquisa foram encontrados
por Silva et al. (2011) ao avaliar sistemas de SAFs, FN e cultivo convencional de
mamão, encontraram valores maiores, variando de 3,91 a 4,75 mm para os SAFs e
FN.
A maior estabilidade de agregados em água e agregados de maior tamanho
são propriedades desejáveis nos solos cultivados, uma vez que estão diretamente
ligadas ao processo erosivo, podendo refletir em maior resistência do solo à erosão,
em decorrência da maior resistência à desagregação e dispersão (Valadão et al.
(2011).
Oliveira (2006) quando avaliou as camadas de 0,00 a 0,05 e 0,05 a 0,20 m,
encontraram diferenças entre os sistemas (cupuaçu + gliricídia + cumaru, cacau +
gliricídia + cumaru, pupunha, café e mata) para DMP e DMG, somente no SAF
(cacau, gliricídia e cumaru) se destacou dos demais, também verificou que o
diâmetro dos agregados diminuiu de acordo com a profundidade.
Valadão et al. (2011), ao analisar os sistemas Cerrado nativo (S1), plantio
direto com adubação mineral (S2), plantio direto com adição de cama de frango crua
(S3) e plantio direto com cama de frango compostada (S4), verificaram que os
valores de PT, DMP, DMG e EA na camada de 0,00 a 0,20 m foram maiores para o
S1 que os demais sistemas e na EA, o sistema que mais se igualou à mata foi o S4.
Avaliando os atributos físicos estudados, exceto DS, verificou-se que os
valores observados na FN foram maiores em comparação aos SAFs, contudo
atributos dos SAFs não são consideradas limitantes ao desenvolvimento de culturas.

50
4.4 Variação dos Atributos Microbiológicos
Os dados, referentes aos componentes biológicos dos solos (Tabela 7),
respaldam a hipótese de maior atividade biológica no período chuvoso em relação
ao seco, conforme relatado para os componentes químicos e estoques de nutriente
do solo.
A sazonalidade dos componentes biológicos do solo tem sido verificada na
literatura (Mercante et al., 2008; Silva et al., 2012; Sousa, 2014), demonstrando que
esses atributos são altamente dependentes das condições climáticas.
TABELA 7. Carbono da biomassa (C-BMS), respiração basal (C-CO2), quociente
metabólico (q-CO2) e quociente microbiano (q-MIC) do solo sob
sistemas florestais, Alta Floresta-MT, 2013
CAMADA (m) ÉPOCA SISTEMAS
FN MBcp TEcp CEcp PAcp
C-BMS
(μg C g-1
)
Chuvoso 988,44 Aa 682,53 Ba 507,31 Ca 471,39 Ca 425,83 Ca
Seco 194,38 Ab 76,02 Bb 107,03 Bb 107,03 Bb 110,37 Bb
C-CO2
(μg C-CO2 g–1
h–1
)
Chuvoso 10,02 Aa 5,58 Ba 5,79 Ba 4,89 Ba 4,13 Ba
Seco 0,33 Ab 0,10 Ab 0,13 Ab 0,25 Ab 0,59 Ab
q-CO2
(mg C-BMS h–1
)
Chuvoso 9,21 Ba 8,73 Ba 12,38 Aa 12,60 Aa 12,00 Aa
Seco 1,72 Bb 1,01 Bb 1,22 Bb 3,36 Bb 6,39 Ab
q-MIC
%
Chuvoso 7,18 Aa 5,23 Ba 4,35 Ba 2,72 Ca 2,31 Ca
Seco 0,98 Ab 0,64 Ab 0,83 Ab 0,58 Ab 0,82 Ab
*Médias seguidas por mesma letra, maiúscula na linha e minúscula na coluna não diferem entre si pelo teste Scott-Knott (p<0,05). Floresta nativa – FN; mogno brasileiro + cupuaçu – MBcp; teca + cupuaçu – TEcp; cedro rosa + cupuaçu – CEcp; paricá + cupuaçu – PAcp.
Contudo, em algumas situações o COT e a MOS tenderam a reduzir no
período chuvoso, resultados contrários aos obtidos por Vasconcelos et al. (2005)
que encontraram maiores teores de C da biomassa microbiana do solo (C-BMS) na
época chuvosa, porém, essa tendência se manteve para o COT.
A redução de C-BMS, na época seca, pode estar relacionada à menor
disponibilidade hídrica no período. Essa relação inversa entre umidade e C-BMS
pode ser devido à ocorrência de nutrientes prontamente disponíveis no solo na
época chuvosa, o que pode ter contribuído para o crescimento microbiano, conforme
relatado por Vasconcelos et al. (2005).

51
Os teores de C-BMS variaram de 425,83 a 988,44 μg C g-1, no período
chuvoso, enquanto, no período seco, a variação foi de 76,02 a 194,38 μg C g-1
(Tabela 7). Nas duas épocas, a FN teve maior teor de C incorporado à biomassa
microbiana em relação aos demais sistemas, porém, na época chuvosa TEcp, Cecp
e PAcp tiveram menor aporte em relação à MBcp e na época seca, os sistemas
consorciados não se diferenciaram. O maior teor de C-BMS indica o maior fluxo de
resíduos na FN seguido do MBcp.
Os valores médios de C-BMS obtidos na FN, no período chuvoso, estiveram
acima dos limites de 412 a 822 μg C g-1, estabelecidos por Pfenning et al. (1992),
para o método de fumigação incubação em solos da Amazônia. Também foram
acima daqueles obtidos por Pezarico et al. (2004) em Nitossolo Vermelho de textura-
argilosa e mesmo tipo de vegetação primária deste estudo e foram compatíveis com
os dados de Vasconcelos et al. (2005) em Latossolo Amarelo da Amazônia
Ocidental.
O contínuo aporte de resíduos sobre o solo e a diversificação de espécie em
uma mesma área na FN, provavelmente proporcionaram diferentes fontes de
exsudatos liberados, bem como fontes de matéria orgânica com graus de
suscetibilidade à mineralização, diferenciados para serem metabolizados e
decompostos pela microbiota do solo. Com isso, aumentou a fonte de carbono, o
que pode ter favorecido o desenvolvimento da biomassa microbiana do solo (Souza,
2014).
Os maiores valores de respiração basal (C-CO2) também foram obtidos na FN
no período chuvoso, não havendo diferença entre os sistemas no período seco
(Tabela 7), evidenciando que a atividade microbiana foi estimulada pela
disponibilidade de água no solo. Resultados semelhantes, em relação a C-CO2,
foram obtidos por Pezarico et al. (2004), Mercante et al. (2008) e Silva et al. (2012),
o que pode ser explicado pela atividade biológica mais intensa em áreas com
elevada ciclagem de nutrientes devido ao maior aporte de resíduos orgânicos. Além
disso, a taxa de respiração basal do solo, para Tótola e Chaer (2002), também está
relacionada com a maior atividade biológica, a qual é diretamente proporcional à
fração de carbono lábil no solo.

52
Quanto ao quociente metabólico (q-CO2), verificou-se que os maiores valores
foram obtidos pelos sistemas TEcp, CEcp e PAcp no período chuvoso, porém, no
período seco, somente o PAcp passou a ter o maior q-CO2, mesmo assim, houve
redução em todos os sistemas do período chuvoso para o seco (Tabela 7).
Os baixos valores de q-CO2 durante a época seca confirmam a eficiência da
biomassa microbiana em imobilizar nutrientes nesse período, provavelmente devido
à produção estruturas de resistência, enquanto que na época chuvosa, os elevados
valores de q-CO2 sugerem que a biomassa microbiana está mineralizando mais
nutrientes do que imobilizando (Vasconcelos et al., 2005).
Maiores valores de q-CO2 indicam maiores perdas de C no sistema na forma
de CO2 por unidade de carbono microbiano e o maior requerimento energético da
população microbiana (Mercante et al., 2008). Nesse sentido, é considerado o
sistema mais eficiente àquele que incorpora mais C às suas células e proporcione
reduzidas perdas de C na respiração (Insam e Domsch, 1988).
Portanto, a FN e o MBcp se destacaram por resultarem nos menores q-CO2 e
foram os que mais incorporaram C à biomassa, demonstrando a capacidade dos
sistemas em manter o equilíbrio entre as taxas de entrada e saídas de C e manter
alto os estoques de MOS. Do mesmo modo, na época seca, o PAcp foi o menos
eficiente, apresentando o maior valor de q-CO2.
A qualidade do substrato disponível para decomposição é um fator a ser
considerado no valor de q-CO2, uma vez que, compostos muito lignificados, a sua
decomposição é mais lenta e desta forma a microbiota têm um elevado gasto
energético para degradar esse produto, até à sua incorporação (Santos, 2005).
Quanto ao quociente microbiano (q-MIC), verificou-se que a FN também
proporcionou maiores valores em relação aos demais sistemas e essa diferença só
ocorreu no período chuvoso (Tabela 7). Nesse período, CEcp e PAcp tiveram os
menores q-MIC demonstrando menores quantidades de C imobilizados como
biomassa. Por outro lado, observa-se que, em todos os sistemas, os valores de q-
MIC estiveram dentro ou maiores dos limites de 2 e 5% estabelecidos por Roscoe et
al. (2006) e acima dos valores encontrados por Mercante et al. (2008) em solos
cultivados com culturas anuais e vegetação nativa e Sousa (2014), em sistemas de
integração lavoura pecuária e vegetação nativa de Cerrado em transição

53
Amazônica, evidenciando grande imobilização de C como biomassa, o que pode
indicar que a biomassa microbiana não encontrava-se em condições de estresse,
com MOS de alta qualidade nutricional, tornando-se capaz de utilizar o C orgânico
do solo.
Segundo revisão de Roscoe et al. (2006), os dados sistematizados para solos
brasileiros ilustram a tendência de redução nos valores médios de q-MIC com o
aumento do estresse ao sistema (pH, condições nutricionais e disponibilidade
hídrica), sendo registrados valores decrescentes dos sistemas naturais (3,3%),
passando pela pastagem (1,4%) e chegando aos solos sob cultivo (1,2%). Segundo
esses autores, com o término de uma situação de estresse e/ou incremento de
matéria orgânica de boa qualidade ocorre um incremento de biomassa microbiana,
assim como no q-MIC, ainda que os teores de C permaneçam inalterados.
Os indicadores microbiológicos obtidos nesta pesquisa foram maiores quando
comparados com os de Silva et al. (2012), em sistemas agroflorestais no Rio de
Janeiro. Os valores elevados em relação aos componentes biológicos no período
chuvoso sugerem que os sistemas agroflorestais, mesmo que com valores inferiores
aos de FN, apresentam equilíbrio em suas propriedades orgânicas, sendo capazes
de manter a qualidade do sistema.
Ramos et al. (2010), quando estudaram indicadores da qualidade de um
Latossolo Vermelho-Amarelo textura média sob pastagem extensiva e floresta
nativa, encontraram que, no primeiro sistema, houve redução do C-BMS, C-CO2 e
agregação do solo comparativamente à floresta nativa e aumento do q-CO2. Os
autores classificaram o sistema de pastagem extensiva como um “sistema em
colapso” em comparação ao ambiente nativo. Isso não se verifica nos sistemas
agroflorestais estudados neste trabalho, com q-CO2 bem menores do que aqueles
observados por Ramos et al. (2010), demonstrando a sustentabilidade dos sistemas
florestais.
Considerando-se ainda, que os sistemas agroflorestais estudados são
sistemas relativamente jovens e diferem quanto à composição das espécies, pode-
se considerar que a sua estabilidade é influenciada pelo tempo de adaptação do
sistema, para que haja o incremento de matéria orgânica em quantidade e

54
qualidade, favorecendo o desenvolvimento da comunidade microbiana do solo e
propriedades físico-químicas adequadas às culturas.

55
5. CONCLUSÕES
1. Houve efeito da sazonalidade nos teores de COT e MOS na floresta nativa e nos
sistemas consorciados de cupuaçu com mogno e cedro, sendo os maiores valores
encontrados na época seca;
2. Os sistemas consorciados tiveram menores teores de bases do solo em relação à
floresta nativa; o sistema de consórcio entre cupuaçu e paricá proporcionou maior
teor e estoque de P no solo em relação aos demais sistemas;
3. Para os atributos físicos do solo, verificou-se que os valores observados na FN
foram maiores em comparação aos SAFs, exceto para densidade do solo;
4. A floresta nativa e o consórcio entre cupuaçu e mogno foram os mais eficientes
em incorporar carbono à biomassa microbiana com menores taxas de quociente
metabólico;
5. No período chuvoso, para os sistemas estudados, foram obtidos maiores teores
de nutrientes e atividade biológica.
6. A hipótese levantada foi aceita; os SAFs nas condições iniciais de avaliação, sete
anos, tiveram resultados satisfatórios, dando indícios de que uma avaliação
temporal, pelo mesmo período, caberia resultados mais evidentes.

56
7. REFERÊNCIAS BIBLIOGRÁFICAS
ANDERSON, J. P. E.; DOMSCH, K. H. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biology and Biochemistry, v.10, p. 215-221, 1978. ANDERSON, T.H.; DOMSCH, K. H. Application of ecophysiological quotients (q-CO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biology and Biochemistry, v. 22, n. 2, p. 251-255, 1990. ARAUJO, M. A; TORMENA, C. A; SILVA A. P. Propriedades físicas de um Latossolo Vermelho Distrófico cultivado e sob mata nativa. Revista Brasileira de Ciência do Solo, v. 28, n. 2, p. 337-345, 2004. ARAÚJO, A. S. F; MONTEIRO, R. T. R. Indicadores biológicos de qualidade do solo. Bioscience Journal, v. 23, n. 3, p. 66-75, 2007. ARAÚJO, A. S. F; et al. Soil microbial properties and temporal stability in degraded and restored lands of Northeast Brazil. Soil Biology & Biochemistry, v. 66, p. 175-181, 2013. BARRETO, A.C., et al. Características químicas e físicas de um solo sob floresta, sistema agroflorestal e pastagem no sul da Bahia. Revista Caatinga, v.19, n.4, p.415-425, 2006. BAYER, C. et al. Diminuição da humificação da matéria orgânica de um Cambissolo Húmico em plantio direto. Revista Brasileira Ciência do Solo, v.27, n. 3, p.537-544, 2003. BAYER, C.; MIELNICZUK, J. Características químicas do solo afetadas por métodos de preparo e sistemas de cultura. Revista Brasileira de Ciência do Solo, v. 21, n. 1, 105-112, 1997. BENJAMIN, J.G.; MIKHA, M.M.; VIGIL, M.F. Organic carbon effects on soil physical and hydraulic properties in a semiarid climate. Soil Science American Journal, v. 72, p.1357-1362, 2008.

57
BERGAMIN, A.C. et al. Compactação em um Latossolo Vermelho distroférrico e suas relações com o crescimento radicular do milho. Revista Brasileira de Ciência do Solo, v. 34, n. 4, 681-691, 2010. BERNARDES, M.S. et al. Recuperação de solo degradado com sistema agroflorestal no extremo Sul da Bahia. In: Resumos Expandidos da XII Reunião Brasileira de Manejo e Conservação do Solo e da Água [CD-ROM] Ilhéus. Ilhéus: CEPLAC/CEPEC; 2000. BRASIL. Plano setorial de mitigação e de adaptação às mudanças climáticas para a consolidação de uma economia de baixa emissão de carbono na agricultura: plano ABC (Agricultura de Baixa Emissão de Carbono). Ministério da Agricultura, Pecuária e Abastecimento, Ministério do Desenvolvimento Agrário, coordenação da Casa Civil da Presidência da República. Brasília: MAPA/ACS, 2012. 173 p. CAMARGO, O. A.; ALLEONI, L. R. F. Compactação do solo e o desenvolvimento das plantas. São Paulo: Divisão de biblioteca e documentação - ESALQ/USP, 1997. 132 p. CASTRO FILHO, C.; MUZILLI, O.; PODANOSCHI, A.L. Estabilidade dos agregados e sua relação com o teor de carbono orgânico num Latossolo Roxo distrófico, em função de sistema de plantio, rotações de culturas e métodos de preparo das amostras. Revista Brasileira de Ciência do Solo, v.22, n.3, p.527-38, 1998. CARVALHO, R.; GOEDERT, W.J.; ARMANDO, M.S. Atributos físicos da qualidade de um solo sob sistema agroflorestal. Pesquisa Agropecuária Brasileira, v. 39, n. 11, p.1153-1155, 2004. CIOTTA, M. N. et al. Matéria orgânica e aumento da capacidade de troca de cátions em solo com argila de atividade baixa sob plantio direto. Ciência Rural, v. 33, n. 6, p. 1161-1164, 2003. COLLIER, L. S.; ARAÚJO, G. P. Fertilidade do Solo sob Sistemas de Produção de Subsistência, Agrofloresta e Vegetação Remanescente em Esperantina – Tocantins. Floresta e Ambiente, v. 17, n.1, p. 12-22, 2010. DUCATTI, F. Fauna edáfica em fragmentos florestais e em áreas reflorestadas com espécies da mata atlântica. Dissertação de mestrado, Escola Superior de Agricultura Luiz de Queiroz, Piracicaba, 2002. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA - EMBRAPA. Manual de métodos análises de solo. 2 ed. rev. atual.- Rio de Janeiro, 1997. 212 p. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA – EMBRAPA. Manual de análises químicas de solos, plantas e fertilizantes. 2. ed. rev. amp. Brasília: Embrapa Informação Tecnológica, 2009. 627p.

58
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA – EMBRAPA. Sistema Brasileiro de Classificação de Solos. 3 ed. ver. amp. Brasília: Embrapa, 2013. 353 p. ERNANI, P.R. Química do solo e disponibilidade de nutrientes. Lages-SC: O autor, 2008. 230 p. FAVERO, C. et al. Crescimento e acúmulo de nutrientes por plantas espontâneas e por leguminosas utilizadas para adubação verde. Revista Brasileira de Ciência do Solo, v. 24, n. 1, p. 171-177, 2000. FERREIRA, D.F. Sisvar: A computer statistical analysis system. Ciência e Agrotecnologia, v.35, n.6, p.1039-1042, 2011. FERREIRA, M.M. Caracterização física do solo. In: Física do solo. Quirijn de Jong van Lier. Viçosa: Sociedade Brasileira de Ciência do Solo, p. 23, 2010. 298 p. GIAROLA, N.F.B. et al. Cultivares de soja sob plantio direto em Latossolo Vermelho compactado. Acta Scientia Agronomica, v. 31, n. 3, p. 641-646, 2009. IBGE – Instituto Brasileiro de Geografia e Estatística. Recursos Naturais e Meio Ambiente: uma visão do Brasil. 2ª edição. Ministério do Planejamento e Orçamento, Rio de Janeiro, 1997. INPE – Instituto Nacional de Pesquisas Espaciais. Projeto PRODES monitoramento da Floresta Amazônica brasileira por satélite: Taxas anuais. Disponível em: http://www.obt.inpe.br/prodes/prodes_1988_2014.htm Acesso em: 02 Ago 2014. INMET – Instituto Nacional de Meteorologia. Estações automáticas: Alta Floresta. Ministério da Agricultura, Pecuária e Abastecimento. 2014. Disponível em: http://www.inmet.gov.br/portal/index.php?r=home/page&page=rede_estacoes_auto_graf Acesso em: 02 Ago 2014. INSAM, H.; DOMSCH, K. H. Relationship between soil organic carbon and microbial biomass on chronosequences of reelamation sites. Microbial Ecology, v.15, n.1, p.177-188, 1988. IWATA, B. F. et.al., Sistemas agroflorestais e seus efeitos sobre os atributos químicos em Argissolo Vermelho-Amarelo do cerrado piauiense. Revista Brasileira de Engenharia Agrícola e Ambiental, v.16, n. 7, p. 730-738, 2012. JENKINSON, D.S. Determination of microbial biomass carbon and nitrogen in soils. In: WILSON, J.R., ed. Advances in nitrogen cycling in agricultural systems. Wallingford, CAB International, p.368-386, 1988.

59
JENKINSON, D.S. POWLSON, D.S. The effects of biocidal treatments on metabolism in soil. I. Fumigation with chloroform. Soil Biologic Biochemica, v. 8, p.167-177, 1976. KIEHL, E. J. Manual de edafologia. São Paulo: Ceres, 1979. 264 p KLEIN, V. A. Física do solo. 2ª edição. Passo Fundo: UPF, 2012, 240p. LEITE, L.F.C. et.al., Estoques totais de carbono e seus compartimentos em Argissolo sob floresta e sob milho cultivado com adubação mineral e orgânica. Revista Brasileira de Ciência do Solo, v. 27, n. 5, p. 821-832, 2003. LIMA, S. S. et.al., Atributos químicos e estoques de carbono e nitrogênio em Argissolo vermelho-amarelo sob sistemas agroflorestais e agricultura de corte e queima no norte do Piauí. Revista Árvore, v.35, n.1, p.51-60, 2011. LOCATELLI, M.; VIEIRA, A. H. Sistemas agroflorestais e a conservação do solo. Referência Florestal, v. 15, n. 144, p. 54-57, 2013. LOSS, A. et. al., Atributos químicos e físicos de um Argissolo vermelho-amarelo em sistema integrado de produção agroecológica. Pesquisa agropecuária brasileira, v.44, n.1, p.68-75, 2009. MACEDO, R.L.G. Princípios básicos para o manejo sustentável de sistemas agroflorestais. Lavras: UFLA: FAEPE, 2000. 157p. MAGALHÃES, S.S.A; et.al., Estoque de nutrientes sob diferentes sistemas de uso do solo de Colorado do Oeste-RO. Acta Amazônica, v. 43, n. 1, p. 63-72, 2013. MAIA, S.M.F. et a. Impactos de sistemas agroflorestais e convencional sobre a qualidade do solo no semi-árido cearense. Revista Árvore, v. 30, p.837-848, 2006. MARQUES, T.C.L.L.S.M. et.al. Crescimento inicial do paricá (schizolobium amazonicum) sob omissão de nutrientes e de sódio em solução nutritiva. Cerne, v. 10, n. 2, p. 184-195, 2004. MEDA, A. R. et al. Plantas invasoras para melhorar a eficiência da calagem na correção da acidez subsuperficial do solo. Revista Brasileira de Ciência do Solo, v. 26, v. 1, p. 647-654, 2002. MELZ, E. M; TIAGO P. V. Propriedades físico-químicas e microbiológicas do solo de um Parque em Tangará da Serra, MT, uma área de transição entre Amazônia e Cerrado. Acta Amazônica, v. 39, n. 4, p. 829-834, 2009. MERCANTE, F. M. et.al. Biomassa microbiana, em um Argissolo vermelho, em diferentes coberturas vegetais, em área cultivada com mandioca. Acta Science Agronomy, v. 34, n. 4, p. 479-485, 2008.

60
MOKRY, M. Makroporen Transport Von Phosphor Gulleausbringung auf Loss-und Tonboden. 2003. Disponível em http:/www.gumpenstein.at/publikationen/lysime ter2003/mokry.pdf. Acesso em 20/04/2014. MOREIRA, F.M.S.; SIQUEIRA, J.O. Microbiologia e bioquímica do solo. UFLA/FAEPE, Lavras. 2002. 626 p. NEVES, A.D.S; DIAS, A.C.C.P; BARBOSA, R.C.M. Levantamento detalhado dos solos da Estação Experimental de Alta Floresta. Bahia: CEPLAC, 1981, 29 p. (boletim técnico nº 83). NOGUEIRA, R. S; et al. Formas de fósforo em Luvissolo Crómico Órtico sob sistemas agroflorestais no município de Sobral-CE. Revista Ciência Agronômica, v. 39, n. 4, p. 494-502, 2008. NOVAIS, R.F. et al. Fertilidade do solo. Viçosa-MG; Sociedade Brasileira de Ciência do Solo, 2007. p.471-550. OLIVEIRA, A.D. Qualidade do solo em sistemas agroflorestais em Alta Floresta – MT. 59f. Dissertação (Programa de Pós-graduação em Solos e Nutrição de Plantas). Universidade Federal de Viçosa, Viçosa, 59p. 2006. PAULETO, E. A, et.al., Avaliação da densidade e da porosidade de um gleissolo submetido a diferentes sistemas de cultivo e diferentes culturas. Revista Brasileira Agrociência, v. 11, n. 2, p. 207-2010, 2006. PAVINATO, P.S. Disponibilidade de nutrientes no solo – decomposição e liberação de compostos orgânicos de resíduos vegetais. Revista Brasileira de Ciência do Solo, v. 32, n. 3, p 911-920, 2008. PEZARICO, C. R, et.al., Indicadores de qualidade do solo em sistemas agroflorestais. Revista de Ciências Agrárias, v. 56, n. 1, p. 40–47, 2004. PFENNING, L; EDUARDO, B. P.; CERRI, C.C. Os métodos da fumigação incubação e fumigação-extração na estimativa da biomassa microbiana de solos da Amazônia. Revista Brasileira de Ciência do Solo, v. 16, n. 1, p. 31-37, 1992. RAMOS, F.T.; et al. Indicadores de qualidade em um Latossolo Vermelho-Amarelo sob pastagem extensiva no Pantanal Matrogrossense. Revista Caatinga, v. 23, n. 1, p. 112-120, 2010. ROSCOE, R; MERCANTE, F. M; SALTON, J. C. Dinâmica da Matéria Orgânica do Solo em Sistemas Conservacionistas – Modelagem Matemática e Métodos Auxiliares. 1ª Ed. Dourados: Embrapa Agropecuária Oeste, 2006, 304 p.

61
SALCEDO, I.H.; SAMPAIO, E.V.S.B. Matéria orgânica do solo no bioma caatinga. In: SANTOS, G.A. et al., eds. Fundamentos da matéria orgânica do solo: Ecossistemas tropicais e subtropicais. 2.ed. Porto Alegre, Metrópole, 2008. p.419-441. SANTOS, M. N. Método de controle de plantas daninhas na cultura do cafeeiro e seus efeitos na agregação e em frações da matéria orgânica do solo. 2005. 74f. Tese (Doutorado em Solos e Nutrição de Plantas) Universidade Federal de Lavras, Lavras - MG, 2005. SAVIOZZI, A.; et al. Biochemical activities in a degraded soil restored by two amendments: a laboratory study. Biology and Fertility of Soils, v. 35, n. 1, p. 96-101, 2002. SILVA, S. C. Condições edafoclimáticas para a produção de Panicum sp. In: SIMPÓSIO SOBRE MANEJO DE PASTAGEM, 12, Piracicaba, 1995. Anais... Piracicaba, Escola Superior de Agricultura “Luiz de Queiroz”, p.147-196, 1995. SILVA, V. R. et al. Densidade do solo, atributos químicos e sistema radicular do milho afetados pelo pastejo e manejo do solo. Revista Brasileira de Ciência do Solo, v.24, n.1, p.191-199, 2000. SILVA, L. G. Uso e monitoramento de indicadores microbiológicos para avaliação da qualidade dos solos de Cerrado sob diferentes agroecossistemas. 2008. 117f. Dissertação (Mestrado em Agronomia) Universidade de Brasília: Faculdade de Agronomia e Medicina Veterinária, 2008. SILVA, D. C. et.al. Atributos do solo em sistemas agroflorestais cultivo convencional e floresta nativa, Revista de Estudos Ambientais, v. 13, n. 1, p. 77-86, 2011a. SILVA, R. C. S. et al. Os indicadores físicos, químicos e biológicos da qualidade do solo e da sustentabilidade dos ambientes naturais. Repositório Eletrônico Ciências Agrárias, Coleção Ciências Ambientais, p. 1-13, 2011b. SILVA, M. S. C. et.al., Estoque de serapilheira e atividade microbiana em solo sob sistemas agroflorestais. Floresta e Ambiente, v. 19, n. 4, p 431-441, 2012. SILVA JUNIOR, C. A.; BOECHAT, C. L.; CARVALHO, L. A. Atributos químicos do solo sob conversão de floresta amazônica para diferentes sistemas na região norte do Pará, Brasil. Bioscience Journal, v. 28, n. 4, p. 566-572, 2012. SIQUEIRA, J. O.; et al., S. Microorganismos e processos biológicos do solo: perspectiva ambiental. Brasília: EMBRAPA-SPI, 1994. 142p. (EMBRAPA-CNPAF. Documentos, 45). SOUSA, D.M.G.; LOBATO, E. Cerrado: correção do solo e adubação. 2 ed. Brasília, DF: Embrapa Informação Tecnológica, 2004. 416 p.

62
SOUSA, H. M. Atributos microbiológicos do solo em sistemas de integração lavoura-pecuária no ecótono cerrado – Amazônia. 70 f. Dissertação (Pós Graduação em Agricultura Tropical). Faculdade de Agronomia, Medicina Veterinária e Zootecnia da Universidade Federal de Mato Grosso, 70 f. 2014. SOUZA, S. C. Desmatamento e clima em Alta Floresta – Amazônia Mato-Grossense. 2006. 94f. Dissertação (Mestrado em Geografia). Universidade Federal de Mato Grosso, Instituto de Ciências Humanas e Sociais, Pós-graduação em Geografia, 2006. 94p. TÓTOLA, M.R.; CHAER, G.M. Microorganismos e processos microbiológicos como indicadores da qualidade dos solos. In: ALVAREZ V.,V.H. et al. eds. Tópicos em ciência do solo. Viçosa, MG, Sociedade Brasileira de Ciência do Solo, 2002. v.2. p.195-276. VALADÃO, F.C.A. et al. Variação nos atributos do solo em sistemas de manejo com adição de cama de frango. Revista Brasileira de Ciência do Solo, v. 35, n. 6, p. 2073 – 2082, 2011. VASCONCELOS, L.G.T.R et al., Carbono, Nitrogênio e Atividade da Biomassa Microbiana de um solo sob vegetação secundária de diferentes idades na Amazônia Oriental. Revista Ciência Agrária, v.2, n. 44, p. 49-63, 2005. VITAL, A. R. T. et al. Produção de serrapilheira e ciclagem de nutrientes de uma floresta estacional semidecidual em zona ripária. Revista Árvore, v.28, n.6, p.793-800, 2004. YEOMANS, J.C.; BREMNER, J.M. A rapid and precise method for routine determination of organic carbon in soil. Communications Soil Science Plant Analysis, v. 19, n. 6, 1467-1476, 1988.