universidade federal de mato grosso faculdade … · pelo carinho de ana maria, minha linda mãe,...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE MATO GROSSO FACULDADE DE AGRONOMIA E ZOOTECNIA
CURSO DE ZOOTECNIA
ANIELI LORENZET
MONITORAMENTO DE NINHOS DE JACARÉS – DO – PANTANAL (Caiman yacare, ALLIGATORIDAE) NA RPPN SESC – PANTANAL, POCONÉ - MT
CUIABA 2016

ANIELI LORENZET
MONITORAMENTO DE NINHOS DE JACARÉS – DO – PANTANAL (Caiman yacare, ALLIGATORIDAE) NA RPPN SESC – PANTANAL, POCONÉ - MT
Trabalho de Conclusão do Curso de Gradação em Zootecnia da Universidade Federal de Mato Grosso, apresentado como requisito parcial à obtenção do título de Bacharel em Zootecnia. Orientador: Prof. Dr. Felipe Franco Curcio
CUIABA 2016

ANIELI LORENZET
"MONITORAMENTO DE NINHOS DE JACARÉS-DO PANTANAL (Caiman yacare,
ALLlGATORIDAE"
Trabalho de curso aprovado como requisito parcial para obtenção do grau deBacharel em Zootecnia pela Universidade Federal de Mato Grosso .
BANCA EXAMINADORA
Prof. O . F L1PE FRANCO CURCIODe;;:p~~,.,;,I,....;.,<o de Biologia e Zoologia/lBUniversidade Federal de Mato Grosso
Presidente da Banca
Profa. Ora. THELMA MICHELLA SADDIDepartamento de Zootecn ia e Extensão Rural/FAMEVZ/UFMT
PfOt: Dr.kCIO AQUIO HOSHIBADepartamento de Zootecnia e Extensão RurallFAMEVZ/UFMT
Cuiabá2016

AGRADECIMENTOS
Primeiramente a Deus, pela graça de permitir ao homem desfrutar da beleza
de suas criações. Por não me deixar perder a fé e me conduzir até aqui.
A minha família, pelo respeito e amor mutuo. Meu amado paizinho Gilson,
meu orgulho, pelo trabalho e esforço dedicados a mim até hoje, pelos cabelos
brancos que lhe dei de preocupação e por tornar esse dia possível, o meu amor e
eterna gratidão. Pelo carinho de Ana Maria, minha linda mãe, pelas palavras nas
horas difíceis e aos meus irmãos, que tanto amo, Gilcieli, pela companhia, por me
aturar e sonhar meus sonhos comigo e Juliano, o orgulho da família.
Ao meu orientador e amigo, Felipe F. Curcio, excelente professor, exemplo de
humildade, por me aceitar como aluna e me proporcionar essa experiência única de
vivência em campo. Por acreditar sempre que seria capaz e por conhecer a
importância de cada palavra contida nesse trabalho. Pela atenção, bom humor,
preocupação, carinho e paciência, o meu respeito e gratidão.
Aos meus amigos Adrielle, Kellen, Wemerson e Jackeline, pelo privilégio de
vossas companhias durante o decorrer do curso, pelo afeto, risos e torcida.
Ao proprietário, por abrir as portas e aos funcionários da RPPN - SESC
Pantanal pelo apoio, dedicação e excelente trabalho.
E por fim, aos componentes da banca, Professores Márcio e Thelma, pela
alegria, disponibilidade, paciência e por aceitarem o convite de dividir comigo este
momento.
Obrigada.

“Nada te perturbe, nada te espante, tudo passa.
A paciência tudo alcança.
Quem a Deus tem nada lhe falta”
Santa Teresa de Ávila

LISTA DE ILUSTRAÇÕES
Figura 1. Localização da área de estudo: a elipse representa a abrangência da área amostrada; os triângulos representam as bases de apoio (1 – Posto Espírito Santo, 2 – Posto São Luiz); as texturas representam as formações do Pantanal (quadrículas), Cerrado (marrom) e matas secas transicionais (cinza sólido). ...................................................................... 12
Figura 2. Detalhe da área de estudo delimitada pela elipse da Figura 1, com os
pontos de busca demarcados em imagem de satélite. ........................... 12
Figura 3. Imagens de alguns pontos de busca que estão representados na Figura 2 entre as Bases de Apoio Espirito Santo e São Luiz. ............................... 13
Figura 4. Ninhadas encontradas nas áreas de monitoramento. ............................... 14 Figura 5. Período de transição entre a estação seca (A) e a cheia (B),
correspondendo ao período de deslocamento de populações em decorrência do início do período reprodutivo. .......................................... 15
Figura 6. Indivíduos enterrados na lama de uma lagoa localizada no posto de apoio
São Luiz.. ............................................................................................... 15 Figura 7. Uma das fêmeas encontradas montando guarda em seu ninho. ............... 16 Figura 8. Ninhos sendo cobertos pela água até completa submersão. ..................... 17 Figura 9. Ninho escolhido para avaliação do estado dos embriões e contagem de
ovos. ......................................................................................................... 17

LISTA DE TABELAS
Tabela 1. Coordenadas geográficas dos pontos de procura e monitoramento de ninhos de Caiman yacare na RPPN-SESC Pantanal............................ 11

LISTA DE ABREVIATURAS
CITES – Convention on International Trade in Endangered Species
CRC – Comprimento rostro-cloacal
LC – Menos Preocupante
RPPN – Reserva Particular do Patrimônio Natural
SBH – Sociedade Brasileira de Herpetologia
IUCN – União Internacional para a Conservação da Natureza

SUMÁRIO
1. INTRODUÇÃO ........................................................................................................ 1
2. OBJETIVO(S) .......................................................................................................... 3
3. REVISÃO ................................................................................................................ 4
3.1 Aspectos Gerais .................................................................................................... 4
3.2 Maturação Sexual ................................................................................................. 6
3.3 Ciclo Reprodutivo .................................................................................................. 7
3.4 Nidificação e Postura ............................................................................................ 7
4. MATERIAL E MÉTODOS ...................................................................................... 10
5. RESULTADOS E DISCUSSÃO ............................................................................ 14
6. CONCLUSÕES ..................................................................................................... 19
7. CONSIDERAÇÕES FINAIS .................................................................................. 20
REFERÊNCIAS ......................................................................................................... 21

RESUMO
O Pantanal mato-grossense é conhecido pela sua extensa área e por abrigar uma
grande diversidade de espécies. Dentre as mais conhecidas está o “jacaré-do-
pantanal” (Caiman yacare), muito abundante na região e que desperta interesse
econômico principalmente por sua pele e carne, embora as pesquisas sobre a
espécie no Pantanal do estado de Mato Grosso são escassas. Este estudo foi
desenvolvido na Reserva Particular SESC Pantanal entre os meses de agosto 2013
a abril de 2014 e teve como objetivo monitorar as perturbações sofridas pelos ninhos
de C. yacare decorrentes de intempéries naturais (e.g., o regime fluviométrico) e da
ação de predadores potencias. Apesar da Reserva apresentar uma vasta área de
mata fechada não inundável, com substrato e condições favoráveis ao processo de
nidificação conforme observado na literatura, foram encontrados 6 ninhos as
margens do Riozinho e 1 ninho as margens do Rio Cuiabá. Através do
acompanhamento dos mesmos, verificou-se que os sítios de nidificação escolhidos
pelas fêmeas reprodutoras dentro do local de foram determinantes para o insucesso
das ninhadas em questão. Além disso, corrobora-se que as chances de
sobrevivência individual de filhotes até a idade reprodutiva são baixas, mesmo com o
grande investimento no cuidado parental, já conhecido para a espécie. Dos ninhos
encontrados, todos foram submersos pelo aumento abrupto do nível do rio,
ocasionando a perda de todas as ninhadas monitoradas. Devido à falta de estudos
na região Pantaneira de Mato Grosso não foi possível a comparação dos resultados
apresentados, o que reforça o argumento de que estudos de longo prazo abordando
o tema serão fundamentais para avaliar o estado de conservação dos jacarés-do-
pantanal na área. Apenas com estes esforços será possível municiar a comunidade
científica de dados para subsidiar o manejo da espécie in situ e em cativeiro.
Palavras-chaves: crocodilianos, inundação, Mato Grosso, monitoramento, ovos,
Pantanal.

1
1. INTRODUÇÃO
Em sua grandiosa extensão, abrangendo uma área de 624.320 km², sendo
aproximadamente 62% no Brasil, 20% na Bolívia e 18% no Paraguai é tida como
berço de 4.700 espécies entre flora e fauna (WWF Brasil). Dentre elas, o “jacaré-do-
pantanal” (Caiman yacare) é uma das mais cobiçadas por ser muito abundante e
localmente explorada como recurso alimentar e econômico na região (COUTINHO &
CAMPOS, 1996), MOURÃO et al., 2002).
As populações de jacarés-do-pantanal foram intensamente caçadas nas
décadas de 1970 e 1980 e, após amparadas oficialmente por leis de proteção e
programas de manejo sustentável (THORBJARNARSON, 1992), se enquadram
atualmente na lista de categoria Menos Preocupante (LC), segundo a Avaliação
global da União Internacional para Conservação da Natureza (IUCN) de 2003.
Mesmo assim, sua abundancia local chama a atenção por seu potencial econômico,
instalado no apêndice II da Convention on International Trade in Endangered
Species (CITES), permitindo e regulamentando o comércio internacional. Sua
utilização por humanos se dá desde alimentação (ovos e carne), uso medicinal e
indústrial (LATHRAP, 1973). Também exercem papel fundamental na ecologia,
sendo considerados “controladores” de patógenos como a esquistossomose, devido
sua alimentação diversificada incluir caramujos transmissores da doença (MOLINA &
PAOLA, 2008). Adicionalmente, segundo os autores, suas fezes servem de alimento
para peixes e outros organismos aquáticos, contribuindo assim com a ciclagem de
nutrientes.
O sucesso reprodutivo é um dos parâmetros cruciais à estabilidade das
populações de crocodilianos uma vez que a maturação das gônadas inicia-se a
partir de 5 anos para machos enquanto que as fêmeas são mais tardias
apresentando ovários maduros aos 7 anos (COUTUNHO, 2001). Adicionalmente,
segundo Campos (1993), as inundações decorrentes dos ciclos plurianuais e a
predação de ninhos são as principais causas da mortalidade dos embriões, embora
a quantificação do reflexo destas pressões ambientais não seja precisa. No entanto,

2
para Campos et al. (2013), a maior ameaça à espécie não é decorrente de ações
climáticas, nem mesmo a ação de predadores como mamíferos, aves e outros
répteis, mas sim da ação antrópica que gera supressão dos hábitats naturais
(urbanização, desmatamentos, atividades agropecuárias e industriais, construções
de usinas e caça ilegal).
De acordo com o Livro Vermelho da Fauna Brasileira (2008), o maior entrave
à conservação dos répteis brasileiros se deve ao desconhecimento sobre sua
biologia e distribuição. Considerando que o número de ninhos presentes em uma
área equivale ao número de fêmeas reprodutoras naquele local e ano, os
levantamentos sobre nidificações representam uma forma eficiente de estimar o
sucesso reprodutivo e incremento populacional (CHABREK, 1966; BAYLISS, 1987).
Porém, o Livro Vermelho (2008), relata a inexistência de programas de
monitoramento de populações para a grande maioria das espécies o que
praticamente impossibilita avaliações seguras sobre seu estado de conservação.
Neste sentido, torna-se fundamental o estudo da biologia reprodutiva do
jacaré-do-pantanal através do acompanhamento dos ninhos ao longo do período de
nidificação postura e eclosão, não apenas registrando o sucesso e número de
neonatos, mas também eventuais perturbações, tais como a visitação de predadores
e a ação de intempéries naturais que podem afetar a dinâmica populacional.

3
2. OBJETIVO(S)
O presente estudo tem como objetivo estimar o sucesso de eclosão dos ovos
de C. yacare através de monitoramento de ninhos e registro de possíveis
perturbações.

4
3. REVISÃO
3.1 Aspectos Gerais
Os répteis, ao longo da evolução, desenvolveram diferentes especializações e
adaptações estruturais que os permitiram ocupar quase todos os ecossistemas
terrestres, nos mais variados ambientes (LEMA, 2002).
Atualmente são reconhecidas para o território brasileiro 808 espécies de
répteis, o que representa quase 8% das mais de 10.200 mil espécies conhecidas no
mundo (UETZ & HOSEK, 2016). Além desta enorme riqueza de espécies, mais de
um terço da nossa fauna é endêmica do Brasil.
Os crocodilianos apresentam o crânio e pelve especializados, pescoço curto e
rostro longo, esta última característica representando uma tendência morfológica
altamente associada à piscivoria (POUGH et al., 1998). Ao longo da evolução, os
crocodilianos desenvolveram um palato secundário que desloca a passagem de ar
para a porção caudal da boca, permitindo sua respiração ao mesmo tempo que se
alimentam, dilacerando suas presas sob a água (ANDRADE et al., 2006; MADER et
al., 2006); as mandíbulas são fortemente armadas com numerosos dentes (POUGH
et al., 1998), e uma dobra de tecido originária da base lingual, forma uma vedação
que impede a passagem de água entre a garganta e a boca, evitando a inalação
quando estiverem somente com as narinas expostas (POUGH et al., 2003). O corpo
é vigoroso e cilíndrico com uma cauda espessa e poderosa, usada para golpear e
impulsionar o corpo na água através de movimentos ondulatórios (TORRES, 2012).
Variam em tamanho, podendo na idade adulta atingir entre pouco mais de 1
até 7 m de comprimento, como o crocodilo de água salgada [Crocodylus porosus
(Crocodylidae)] (POUGH et.al., 2008). Segundo o mesmo autor, sua pele apresenta
osteodermos (Grego osteo = osso; dermis = pele) localizados sob as escamas,
cobertos por uma epiderme espessa e queratinosa, conferindo-lhes um revestimento
bastante rígido e protetor.

5
Os crocodilianos são eficazes predadores semiaquáticos de topo de cadeia
(MARTINS et al., 2001). Utilizam-se de emboscadas para capturar suas presas,
principalmente em águas rasas (POUGH et.al., 1998). Sua dieta inclui diversos itens,
podendo se alimentar de qualquer animal vivo passível de ser capturado incluindo
indivíduos de mesma espécie (SANTOS et al., 1993). A dieta varia de acordo com a
idade, habitat, estação do ano e área de ocorrência (WEBB et al., 1982). Filhotes
consomem principalmente insetos, crustáceos e moluscos, passando a predar
pequenos vertebrados conforme ganham massa corporal ao longo do crescimento
(SANTOS, 1997).
Estudos têm indicado o jacaré-do-pantanal, C. yacare, como uma das
espécies de crocodilianos mais vigorosas do mundo, com densidades superiores a
100 indivíduos/Km², distribuídos por toda planície Pantaneira (COUTINHO &
CAMPOS 1996, MOURÃO et al., 2002). Esse fato pode estar relacionado à sua
maior resiliência em resposta às alterações ambientais, quando comparado às
outras espécies de crocodilianos brasileiros (CAMPOS et al., 2010). São
normalmente encontrados ativos na água, ocorrendo também movimentos
migratórios em terra, nas regiões de clima sujeito a estiagens sazonais com estação
seca severa e de alta restrição hídrica (CAMPOS 2003, CAMPOS et al., 2006,
CAMPOS et al., 2008). Movimentam-se em grupos, fato relacionado à
disponibilidade de alimentos e comportamento reprodutivo, sendo comumente
encontrados em poços construídos ao lado das estradas, açudes para o gado e
outros reservatórios artificiais (CAMPOS et al., 2010).
Por se tratar de animais ectotérmicos, são altamente dependentes do
fotoperíodo, apresentando variações comportamentais ao longo do dia, com
alternâncias de sol e sombra e tempo de atividade (ROCHA & BERGALLO, 1990). A
ectotermia faz com que esses animais tenham a necessidade de poupar energia
metabólica para garantir a manutenção de seus processos vitais (PIANKA, 2003). A
temperatura média corporal dos jacarés varia de 25,7°C no inverno a 30,1 °C no
verão, com mínima de 16,9°C e máxima de 37,9°C (CAMPOS et al., 2005).
Coutinho (2000) relata que a temperatura corporal desses indivíduos tem
influência direta sobre o metabolismo, processos digestivos e condição corporal, que
por sua vez, tem impacto marcante sobre a reprodução, crescimento e dinâmica
populacional, que incluem número de fêmeas que irão se reproduzir anualmente e o
tamanho de cada ninhada.

6
3.2 Maturidade Sexual
Segundo Coutinho et al. (2001), C. yacare é uma espécie de vida
relativamente longa, cuja diferenciação gonadal ocorre no início da vida. Segundo
estes autores, machos com comprimento rostro-cloacal (CRC) de 40cm, equivalendo
a 2 ou 3 anos de idade, podem apresentar testículos púberes. Nesse mesmo estudo
foi identificado que, conforme o indivíduo cresce, ocorre o aumento da produção
espermática em função do aumento volumétrico dos tubos seminíferos. Os autores
supracitados observaram gônadas maduras em animais com aproximadamente 9 e
10 anos de idade, com um CRC maior que 90cm, indicando assim a idade
aproximada da maturidade sexual dos machos de jacarés-do-pantanal.
Já as fêmeas atingem a maturidade bastante tardiamente, apresentando
ovários maduros e folículos viáveis somente aos 7 anos de idade com CRC > 70cm
(COUTINHO, 2001). Entretanto, estudos de (CAMPOS, 1993; CAMPOS E
MAGNUSSON, 1995; COUTINHO, 2000) indicaram que o tamanho mínimo de
produção factual se torna ainda menor em animais de vida livre e, ainda que fêmeas
jovens (CRC < 75cm) sejam capazes de realizar a postura, há evidências de que a
maioria delas começam a nidificar com aproximadamente 9 e 10 anos de idade,
CRC de 80 cm e massa corpórea acima de 12Kg, idade em que ambos os sexos
atingem a maturação completa das gônadas.
Cabe ressaltar ainda que outros fatores como a qualidade do habitat e o
estado nutricional das fêmeas afetam a primeira reprodução e o número de ovos por
ninho (CAMPOS, 2003). Campos et al. (2008), identificaram que até 60% da
variação no tamanho das fêmeas está relacionada as características de postura,
indicando que os ninhos podem prover informações sobre a estrutura populacional.

7
3.3 Ciclo Reprodutivo
A sazonalidade reprodutiva de vertebrados tem sido registrada como uma
adaptação associada à otimização no tempo de nascimento, época em que mães e
filhotes encontram-se mais vulneráveis à mortalidade (SKINNER, 1971; DUVALL et
al., 1982). Para o jacaré-do-pantanal, o final da estação seca marca o início do
período reprodutivo da espécie. No Pantanal, o ciclo estral do C. yacare está
intimamente associado à temperatura e ao nível d’água, uma vez que estes são
fatores chaves relacionados à sobrevivência dos jovens e fêmeas, devido estarem
positivamente relacionados com o sucesso alimentar e a condição corporal dos
indivíduos (COUTINHO et al., 2005). Na Venezuela, o mesmo ocorre com C.
crocodilus, mas os períodos de nidificação correspondem aos meses de maio a
agosto (THORBJARNARSON, 1994). Isso indica que a mudança se deve
simplesmente a uma resposta quanto à diferenciação latitudinal (SILVEIRA et al.,
1997). Segundo o mesmo autor, na região da Amazônia central, jovens de C.
crocodilus são encontrados com frequência entre os meses de fevereiro a abril, onde
as características ambientais favorecem temperaturas altas e aumento dos níveis
d’água. Para o autor supracitado, os jacarés apresentam ciclo reprodutivo
sincronizado às variáveis ambientais reforçando a ideia de que o período de eclosão
é resultante da seleção natural.
3.4 Nidificação e Postura
Para C. yacare, observações feitas por diversos autores (CAMPOS, 1993;
CAMPOS & MAGNUSSON, 1995; COUTINHO, 2000; CAMPOS, et al., 2008)
indicam que altas temperaturas e o início das chuvas marcam o período de postura
dos ovos que ocorre entre final de dezembro e fevereiro, podendo ocorrer picos de
postura em meados de janeiro e inicio de fevereiro (COUTINHO, 2000; CAMPOS et
al., 2008).
De acordo com Campos (1993) e Mourão et al. (1994) após a postura dos
ovos, os ninhos são recobertos cuidadosamente com folhas e gravetos. Estes
autores, afirmam ainda que seu tamanho e composição variam de acordo com o
habitat e a oferta de material que, por sua vez, depende diretamente dos ciclos

8
plurianuais. Os fatores que determinam a escolha do local apropriado são
desconhecidos (CAMPOS 1993, CAMPOS & MAGNUSSON 1995).
Cintra (1988), Campos & Magnusson (1995) mencionam que, a partir da
postura até a eclosão, o desenvolvimento dos ovos depende das condições de
incubação e dos cuidados da fêmea, que permanece próxima ao ninho montando
sentinela contra possíveis predadores por um período de 70 a 90 dias. O ovo dos
répteis permite seu rápido desenvolvimento devido à presença e arranjo particular
das membranas embrionárias envoltas por uma casca rígida (calcária ou
pergaminácea) que oferece proteção mecânica, mas permite trocas gasosas com o
ambiente (CARROLL, 1988). Cintra (1993) e Campos & Magnusson (1995)
demonstram que o número de ovos depende do estado nutricional e do tamanho das
fêmeas, fatores variáveis de acordo com a idade.
A temperatura de incubação dos ovos determina o sexo dos embriões;
temperaturas baixas (< 31,5°C) produzem fêmeas e temperaturas altas (> 31,5°C)
produzem machos (CAMPOS, 1993). A temperatura de incubação também influencia
o tamanho dos recém-nascidos, sendo que ninhos mais aquecidos podem produzir
neonatos maiores quando comparados aos ninhos submetidos a temperaturas
menores (CAMPOS, 2003). Essa variação no ambiente interno do ninho ocorre em
resposta à insolação, à chuva e à temperatura do ar, em intervalos diferentes de
tempo (CAMPOS, 1993). Isso pode ser evidenciado ao comparar ninhos de mata,
dotados de alguma sombra conferida pela cobertura vegetal, com os de vegetação
flutuante, sujeitos à ação direta das intempéries, quase a céu aberto. Neste
contexto, os ninhos do interior de mata sofrem menos variações de temperatura.
O comportamento social dos crocodilianos é marcado pela vocalização dos
embriões ainda dentro do ovo, estimulando a eclosão coletiva (LANG, 1989). As
ninhadas variam entre 25 a 30 ovos (CAMPOS et al. 2010). Após o “aviso” (chamado
dos filhotes pela vocalização), a fêmea se aproxima do ninho e o desfaz, utilizando
os membros anteriores e posteriores, além do focinho (LANG & ANDREWS, 1994).
Os filhotes eclodem com cerca de 12 cm e peso aproximado de 50 g, podendo
alcançar 25 cm em um ano (COUTINHO, 2000). Após o nascimento os filhotes são
levados nas maxilas da mãe até a água, escolhendo regiões de possível
camuflagem, aumentando as chances de sobrevivência dos mesmos (MOLINA &
PAOLA, 2008). Mesmo assim a mortalidade dos juvenis é alta; apenas 7% dos
filhotes de cada ninhada atinge a idade reprodutiva, metade nem sequer chega a

9
eclodir e, do restante – a grande maioria – morre antes de completar um ano
(Oliveira, 2003). O autor argumenta que o cuidado parental das fêmeas por cerca de
três anos de acompanhamento dos filhotes é eficiente para que a taxa de
sobrevivência não seja ainda menor.

10
4. MATERIAL E MÉTODOS
Este estudo foi desenvolvido nas dependências da Reserva Particular do
Patrimônio Natural (RPPN) - SESC Pantanal, localizada na região de Poconé, às
margens do Rio Cuiabá. Parte importante da área é destinada ao turismo, contando
com a estrutura do Hotel SESC Porto Cerrado. A base administrativa localizada em
Várzea Grande está distante 145 km da base de apoio Porto Cercado que dá acesso
direto ao interior da Reserva. Foram realizadas 22 campanhas de campo; 19 com
duração de dois dias (finais de semana), 2 com duração aproximada de sete dias
(uma semana) e 1 com duração de três dias. Estas foram distribuídas da maneira
mais uniforme possível entre os meses de agosto de 2013 e abril de 2014. As
atividades de busca e monitoramento dos ninhos iniciavam-se as 07:30 da manhã e
se estendiam até aproximadamente as 17:30 horas.
Foram demarcados 22 pontos de possíveis nidificações, incluindo pontos
com ninhadas formadas para acompanhamento do desenvolvimento e contagem
dos juvenis. Todos os pontos foram devidamente georreferenciados e monitorados
(Tabela 1; Figuras 1, 2, e 3).
As bases de apoio utilizadas foram os Postos Espirito Santo (16º35’19” S;
56º16’51” W) e São Luiz (16º41’11” S; 56º10’29” W). Os critérios para escolha dos
pontos de possíveis nidificação seguiram as referências de Coutinho & Campos
(2005) que descreveram os locais de desova como ricos em matéria orgânica (e.g.
folhas, gravetos e terra) empregada na construção dos ninhos, que em geral se
encontram no interior da mata e próximos a corpos d'água.
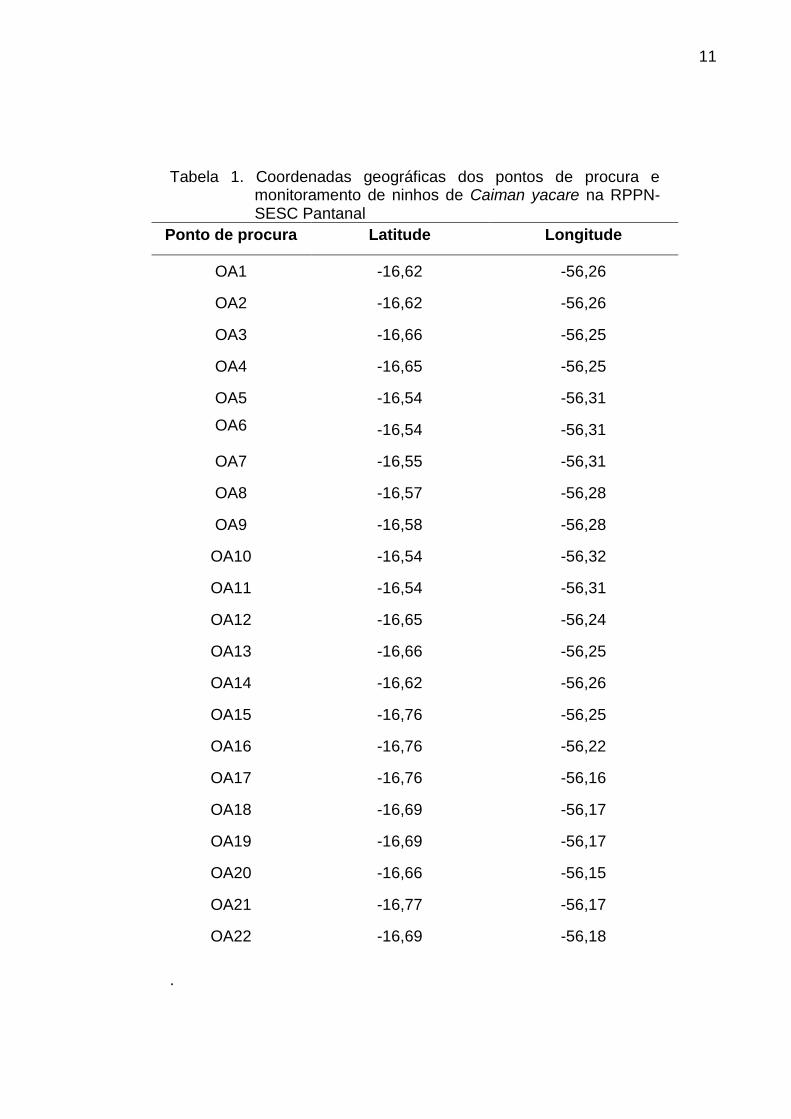
11
Tabela 1. Coordenadas geográficas dos pontos de procura e
monitoramento de ninhos de Caiman yacare na RPPN-SESC Pantanal
Ponto de procura Latitude Longitude
OA1 -16,62 -56,26
OA2 -16,62 -56,26
OA3 -16,66 -56,25
OA4 -16,65 -56,25
OA5 -16,54 -56,31
OA6 -16,54 -56,31
OA7 -16,55 -56,31
OA8 -16,57 -56,28
OA9 -16,58 -56,28
OA10 -16,54 -56,32
OA11 -16,54 -56,31
OA12 -16,65 -56,24
OA13 -16,66 -56,25
OA14 -16,62 -56,26
OA15 -16,76 -56,25
OA16 -16,76 -56,22
OA17 -16,76 -56,16
OA18 -16,69 -56,17
OA19 -16,69 -56,17
OA20 -16,66 -56,15
OA21 -16,77 -56,17
OA22 -16,69 -56,18
.
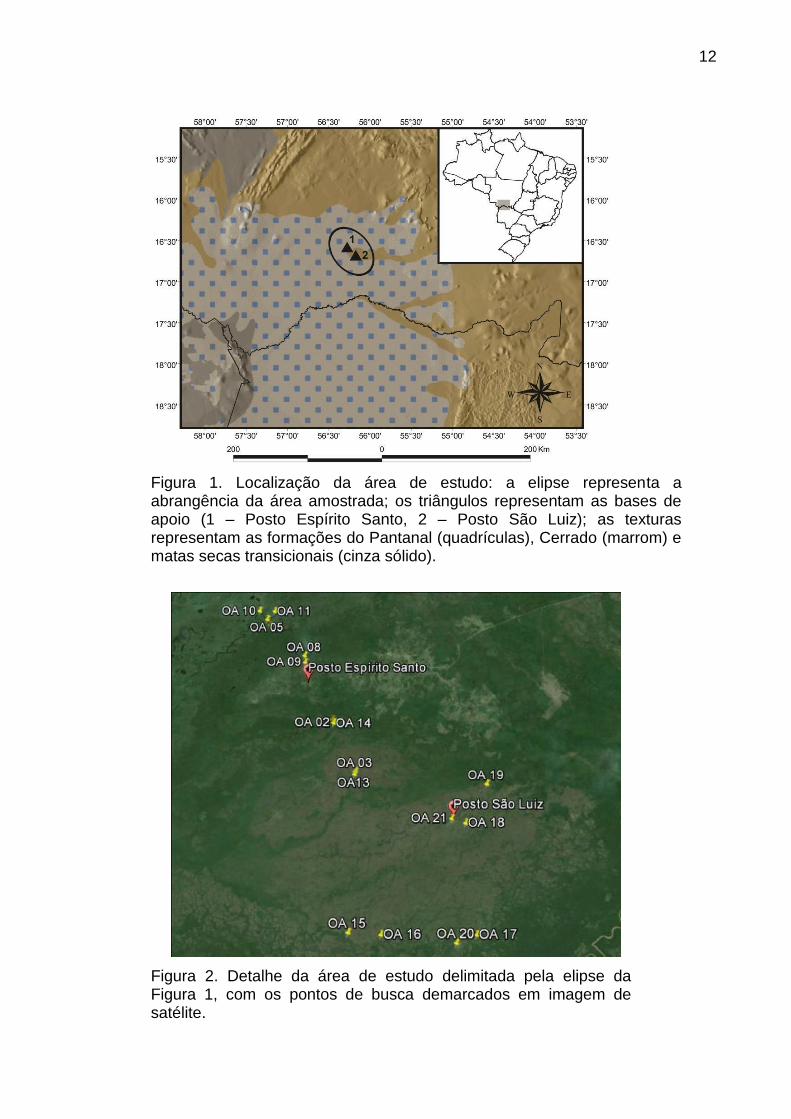
12
Figura 1. Localização da área de estudo: a elipse representa a abrangência da área amostrada; os triângulos representam as bases de apoio (1 – Posto Espírito Santo, 2 – Posto São Luiz); as texturas representam as formações do Pantanal (quadrículas), Cerrado (marrom) e matas secas transicionais (cinza sólido).
Figura 2. Detalhe da área de estudo delimitada pela elipse da Figura 1, com os pontos de busca demarcados em imagem de satélite.

13
Figura 3. Imagens de alguns pontos de busca que estão representados na figura 2 entre as Bases de Apoio Espirito Santo e São Luiz.

14
5. RESULTADOS E DISCUSSÃO
Entre os meses de agosto e setembro de 2013, foram registrados vestígios
de dois ninhos abandonados após eclosão de filhotes, além de seis ninhadas com
número de filhotes variando entre 05 e 27 (Figura 4).
Figura 4. Ninhadas encontradas nas áreas de monitoramento.
Segundo Coutinho et.al (2005) devido à sua vasta distribuição geográfica, o
período de eclosão é produto da seleção natural associada à otimização no tempo
de nascimento, época em que mãe e filhotes estão mais susceptíveis à mortalidade
natural (SKINNER, 1971; DUVALL et al., 1982).
O período de outubro a novembro compreende o período final da seca,
referente ao início da atividade reprodutiva da espécie. Observou-se que ao notar a
presença humana, os machos vocalizaram, refletindo possivelmente um
comportamento de demarcação de território provocando a reação de outros machos.
Fato semelhante foi descrito por Thorbjarnarson & Hernandez (1993), que sugeriu a
possibilidade de estar relacionado à corte dirigida às fêmeas próximas.

15
Neste período notou-se a diminuição abrupta em densidade de
determinadas populações que ocupavam lagoas isoladas, sugerindo migração em
busca de alimento ou locais apropriados para desova (Figura 5; ver também
CAMPOS, 2003; CAMPOS et al., 2006a, 2008).
Figura 5. Período de transição entre a estação seca (A) e a cheia (B), correspondendo ao período de deslocamento de populações em decorrência do início do período reprodutivo.
Foi registrado um exemplar com comportamento sugestivo de nidificação
em uma área de capinzal; o animal revolveu o substrato e permaneceu no local por
dois dias, evadindo-se em seguida sem qualquer prenúncio adicional de que
estivesse de fato em atitude reprodutiva. Ainda no mês de novembro, foram
registradas dezenas de espécimes enterrados na lama (Figura 6), descrito como
comportamento normal por Campos et al. (2006b).
Figura 6. Indivíduos enterrados na lama de uma lagoa localizada no posto de apoio São Luiz.
Os primeiros ninhos desta estação foram encontrados apenas na primeira
quinzena do mês de janeiro de 2014, corroborando as descrições de Coutinho
(2005), que registra picos de postura entre meados de janeiro e fevereiro. A partir
daí, foram localizados sete ninhos nos pontos meridionais do transecto (OA 05, OA
10 E OA 11), todos localizados às margens dos rios (Riozinho e Rio Cuiabá), longe
A B

16
da água; as fêmeas montavam guarda nas imediações de apenas três ninhos
(Figura 7).
Figura 7. Uma das fêmeas encontradas montando guarda em seu ninho.
No mês de março, o nível das águas começou a subir significativamente, os
tanques que abrigavam grandes populações de jacarés-do-pantanal no período da
seca foram invadidos pela cheia e os ninhos que aparentemente estavam distantes
da água, começaram a ficar progressivamente ilhados até serem totalmente
submersos na quinzena do mês de março (Figura 8). No final deste mesmo mês
quando o nível da água baixou, todos os ninhos estavam intactos e puderam ser
vistos pela equipe.

17
Figura 8. Ninhos sendo cobertos pela água até completa submersão.
Destes, um foi aleatoriamente escolhido e aberto para verificação. Foram
contados 26 ovos fecundos, com embriões mortos devido inundação como
postulado por Campos (2003) pois são dependentes das oscilações do nível da
água (Figura 9). As buscas por novos ninhos continuaram até o mês seguinte, sem
sucesso e novos filhotes começaram a surgir.
Figura 9. Ninho escolhido para avaliação do estado dos embriões e
contagem de ovos.
Na campanha mais extensa, foram monitorados três ninhos nos pontos OA 5
e OA 11 através de ciclos de observação com duração de 5 horas cada (das 9:00 às
14:00 hs), durante três dias (um ninho por dia). A observação a partir de uma

18
distância de aproximadamente 10m não rendeu dados diretos de tentativas de
predação, embora tenham sido avistados potenciais predadores como três lagartos
teiús (Tupinambis sp.), garças (Casmerodius albus) e gaviões (Falconiformes) nas
proximidades dos ninhos.

19
6. CONCLUSÕES
A seleção dos sítios de nidificação é um parâmetro fundamental no sucesso
reprodutivo da espécie, tendo em conta a variação dos ciclos pluviométricos anuais
de um ambiente sazonal como a planície inundável do Pantanal.
Considerando os ninhos construídos às margens dos rios, a inundação foi
um elemento decisivo na taxa de mortalidade das ninhadas. Todos os ninhos
monitorados foram inundados e nenhum apresentou sinal de perturbação por
predadores. Este resultado pode ser atípico devido ao regime de chuvas não ser
constante todos os anos.
Estudos de longa duração e com maior abrangência territorial contribuiriam
para encorpar os dados apresentados aqui, oferecendo base estatística sólida para
apontar a extensão do impacto causado pelas cheias e por pressão de predação
sobre as populações de C. yacare.

20
7. CONSIDERAÇÕES FINAIS
Estudos a respeito da espécie C. yacare no estado de Mato Grosso são de
fundamentais para averiguar do seu real estado de conservação no Pantanal norte.
Apesar dos esforços para regulamentar a comercialização dos produtos oriundos da
fauna silvestre, a caça ainda é registrada como uma das principais ameaças aos
estoques naturais, já que os órgãos ambientais não dispõem de infraestrutura para
fiscalização eficiente em ampla abrangência. Aliado a isso, a inexistência de
programas de monitoramento populacional contribui para a instabilidade de
populações de diversas espécies da fauna brasileira.
Por serem animais totalmente dependentes das condições ambientais e de
habitat, a preservação de áreas ambientais na região pantaneira é de suma
importância para a manutenção das espécies de jacarés-do-pantanal, visto que o
sucesso reprodutivo pode variar entre as estações em decorrência do regime hídrico
que, por sua vez, pode afetar o nascimento de novos indivíduos como relatado neste
trabalho. Estudos adicionais de maior extensão temporal devem ser conduzidos no
Pantanal de Mato Grosso para levantar dados de comparação com as informações
já existentes para a região de Mato Grosso do Sul.

21
REFERÊNCIAS
ANDRADE, M. B.; BERTINI, R. J.; PINHEIRO, A. E. P. Observation on the palate and choannae structures in Mesoeucrocodylia (Archosauria, Crocodylomorpha): Phylogenetic implication. Revista brasileira de Paleontologia, v. 9, p. 323-332, 2006. BAYLISS, P. Survey methods and monitoring within crocodile management programmes. In J. W. Webb, S. C. Manolis and P. J. Whiteheard (Eds.) Wildlife Managament: Crocodiles and Aligators, p. 157-175. Surrey Beatty and Sons Pty Limited. Canberra- Australia. CAMPOS, Z. Effect of habitat on survival of eggs and sex ratio of hatchlings of Caiman crocodilus yacare in the Pantanal, Brazil. Journal of Herpetology 27: 127-132, 1993. CAMPOS, Z. & MAGNUSSON, W. Relationship between rainfall, nesting habitat and fecundity of Caiman crocodilus yacare in the Pantanal, Brazil. Journal of Tropical Ecology, v.11: 351-358, 1995. CAMPOS, Z. Caiman crocodilus yacare (Pantanal Caiman). Food-related movement. Herpetological Review, 34(2): 140-141, 2003 CAMPOS, Z; Coutinho, M. & Magnusson, W.E. Field body temperatures of caimans in the Pantanal, Brazil. Herpetological Journal, 15: 97-106, 2005. CAMPOS, Z.; COUTINHO, M.; MOURÃO, G.; BAYLISS, P.; MAGNUSSON, W. Long distance movement by Caiman crocodilus yacare: implications for management of the species in the Brazilian Pantanal. Herpetological Journal,16: 123-132, 2006a CAMPOS, Z.; COUTINHO, M. & MAGNUSSON, W. Caiman crocodilus yacare (Pantanal Caiman). Aestivation. Herpetological Review, 37(3): 343-344, 2006b. CAMPOS, Z.; MAGNUSSON, W.; SANAIOTTI, T.; COUTINHO, M. Reproductive trade-effs in Caiman crocodilus crocodiles and C. c. yacare: implications for size-related management quotas. Herpetological Journal, 18: 91-96, 2008. CAMPOS, Z.; LLOBET, A.Q.; PIÑA, C.I. & MAGNUSSON, W. E. Caiman yacare (Yacare caiman). In: Manolis, S.C. & Stevenson, C. (eds.), Crocodiles. Status survey and conservation action plan. 3rd ed. IUCN SSC Crocodile Specialist Group, 2010. CARROLL, R. L. Vertebrate Paleontology and Evolution. New York. Freeman. 698p. 1988. CHABREK, R. H. Methods for determining the size and composition of alligator populations in Louisiana. Proceedings Annual Conference Southeastern Association Game and Fish Comm. 20: p.105-112, 1966.

22
CINTRA, R. Nesting ecology of the Paraguayan Caiman (Caiman yacare) in the Brasilian Pantanal. Journal of Herpetology, 22:p. 219-222, 1988. CITES. 2016. Convention on International Trade in Endangered Species, 2015. Disponivel em: https://cites.org/esp/app/appendices.php. Acessado em 26 de Fev. 2016. COUTINHO, M.; CAMPOS, Z. Effect of habitat and seasonality on the densities of caiman in southern Pantanal, Brazil. Journal of Tropical Ecology, 12: p.741-747, 1996. COUTINHO, M. Population ecology and the conservation and anagement of Caiman yacare in the Pantanal Brazil. Tese (Doutorado em Zoologia), University of Queensland, Austrália. 272p., 2000. COUTINHO, M.; CAMPOS, Z.; CARDOSO, F.; MARTINELLI, P.; CASTRO, A. Reproductive biology and its implication for management of caiman Caiman yacare in the Pantanal wetland, Brazil. In: GRIGG, G.; SEEBACHER, F. & FRANKLIN, C. (org.). Crocodilian Biology and Evolution. Surrey Beatty & Sons. Chipping Norton, p.229-342, 2001. COUTINHO, C.; CAMPOS, Z.; CARDSO, F.; MARTINELLI, P.; CASTRO, A. Ciclo reprodutivo de machos e fêmeas de jacaré-do-pantanal, cayman crocodilus yacare. Comunicado Técnico Embrapa, Corumbá, MS, 2005. CRAWSHAW, P. & SCHALLER, G. Nesting of Paraguayan caiman (Caiman yacare) in Brazil. Papéis Avulsos Zoologia,33: p.283-292, 1980. DUVAL, D.; GUILLETTE, L.; JONES, R. Environmental control of reptilian reproductive cycles. In: GANS, C. & POUGH, F. H. (org.), Biology of the Reptilia. Academic Press, New York, v. 13, p. 201-231, 1982. FARIAS, I. P.; MARIONI, B.; VERDADE, L. M.; BASSETTI, L.; COUTINHO, M. E.; MENDONÇA, S. H. S. T.; VIEIRA, T. Q.; MAGNUSSON, W. E.; CAMPOS, Z. Avaliação do risco de extinção do jacaré-do-pantanal Caiman yacare (Daudin, 1802) no Brasil. Instituto Chico Mendes de Conservação da Biodiversidade. Biodiversidade brasileira, 3(1), 21-30, 2013. IUCN - International Union for Conservation of Nature. Guidelines for application of IUCN red list criteria at regional levels: Version 3.0. IUCN Species Survival Commission. IUCN, Gland, Switzerland and Cambridge, 26p., 2003 LANG, J. W.; ANDREWS, H. V. Temperature-dependent sex determination in crocodilians. J. Exp. Zool., New York, v. 270, p. 28-44, 1994. LATHRAP, D. W. Gifts of the Caiman: some thoughts on the subsistence basis of Chavin. Lathrap, D. W.; Douglas, J. eds. Variation in Anthropology: Essays in Honor of John C. McGregor. Lllinois: lllinois Archaeological Survey, p. 91-105, 1973.

23
LEMA. T. Os répteis do Rio Grande do Sul: atuais e fósseis, biogeografia e ofidismo. Porto Alegre: EDIPUCRS, p 264, 2002. MADER, D. R. Reptile Medicine and Surgery. 2 ed. Saint Louis: Saunders Elsevier, 547p., 2006. MARTINS, M.; MOLINA, F. B.. Livro vermelho da fauna brasileira ameaçada de extinção. Panorama geral dos répteis ameaçados no Brasil. Brasilia,vol 2, p 326 – 377, 2008. MARTINS, M.; ARAUJO, M. S.; SAWAYA, R. J.; NUNES, R. Diversity and evolution of macrohabitat use, body size and morphology in a monophyletic group of Neotropical pitvipers (Bothrops). J. Zool. P. 529-538, 2001. MOLINA, F. B.; PAOLA, L. Jacaré-de-papo-amarelo. Zoológico de São Paulo, São Paulo, dez. 2008. Seção Animais: Répteis. Disponível em: http://www.zoologico.sp.gov.br/repteis/jacaredepapoamarelo.htm. Acesso em 24 de fev. 2016. MOURÃO, G.; CAMPOS, Z.; COUTINHO, M. Aerial surveys of caiman nests in wet savannas of Brazil. In: Proc. 12th working meeting of the crocodile specialist group/IUCN, 1994. MOURÃO, G.; COUTINHO, M.; MAURO, R.; CAMPOS, Z.; TOMÁS, W.; MAGNUSSON, W. Aerial surveys of caiman, marsh deer and pampas deer in the Pantanal Wetland of Brazil. Biological Conservation, 92: p.175-183, 2000. OLIVEIRA, P. M. A. Animais silvestres e exóticos na clínica particular. São Paulo, ed.Rocca, 375 p. 2003.
PIANKA, E. R; VITT, L. J. Lizards, windows to the evolution of diversity. University of California Press. Berkeley, California, 2003. POUGH, F. H.; HANDREWS, R. R.; CADLE, J. E.; CRUMP, M. L.; SAVITZKY, A. H.; WELLS, K. D. Herpetology. Prentice hall, New Jersey, USA, 577p., 1998. POUGH, F. H.; JANIS, C. M.; HEISER, J. B. A Vida dos Vertebrados. 3 ed. São Paulo: Atheneu, 699p., 2003. ROCHA, C. F.D.; BERGALLO H. G. Thermal biology and distance of Trapidurus oreadicus (Sauria, Iguanidae) in an area of Amazonian Brazil. Ethol. Ecol. Evol. 2(3): p. 263-268, 1990. SANTOS, S. A.; Dieta e nutrição de crocodilianos. Corumbá: EMBRAPA-CPAP, 59p., 1997. SANTOS, S. A.; PINHEIRO, M. S.; SILVA, R. A. Efeito de diferentes dietas naturais no desenvolvimento inicial de Caiman crocodilos yavare (Crocodilia Alligatoridae). Rev. Sociedade Brasileira de Zootecnia, Viçosa, v.22, n.3, p.406-412, 1993.

24
SKINNER, J. The effect of season on spermatogenesis in some ungulates. Journal Reproduction Fertilizacion, v.13: p. 29-37. 1971. THORBJARNARSON, J. & G. HERNANDEZ. Reproductive ecology of the Orinoco crocodile (Crocodylus Intermedius) in Venezuela. I. Nesting ecology and egg and clutch relationships. Journal of Herpetology 27(4): 363-370,1993. THORBJARNARSON, J. Reproductive ecology of the spectacle caiman (Caiman crocodilus) in the Venezuelan Llanos. Copeia 4: p. 907-919, 1994. THORBJARNARSON, J. Crocodiles: an action plan fortheir conservation. Gland: IUNC,132 p., 1992. TORRES, R. S. Adaptações evolutivas: Aspectos comportamentais, mecanismos de defesa e predação em répteis. Porto Alegre, RS, 2012. WEBB, G. J. W.; MANOLIS, S. C.; BUCKWORTH, R. Crocodilus johnstoni in the McKinlay River Area, N. T. I. Variation in diet, and a new method of assessing the relative importance of prey. Australian Journal Zoology, Melboume, v.3. p.887-899, 1982.
WWF Brasil. O Bioma. Disponível em: <http://www.wwf.org.br/natureza_brasileira/areas_prioritarias/pantanal/bioma_pantanal/> Acessado em 05 de agosto de 2016.