universidade federal de goiÁs programa de pÓs … · nf - b fator nuclear kappa b oms...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE GOIÁS
PROGRAMA DE PÓS-GRADUAÇÃO EM MEDICINA TROPICAL
E SAÚDE PÚBLICA
ILDEFONSO ALVES DA SILVA JUNIOR
AVALIAÇÃO DA METACICLOGÊNESE IN VITRO DE
Leishmania (V.) braziliensis e Leishmania (L.) amazonensis
Goiânia
2013


ILDEFONSO ALVES DA SILVA JUNIOR
AVALIAÇÃO DA METACICLOGÊNESE IN VITRO DE
Leishmania (V.) braziliensis e Leishmania (L.) amazonensis
Dissertação de Mestrado apresentada ao
Programa de Pós-Graduação em Medicina Tropical
e Saúde Pública da Universidade Federal de
Goiás para obtenção do Título de Mestre em
Medicina Tropical e Saúde Pública.
Orientadora: Dra. Fátima Ribeiro Dias
Goiânia
2013


Programa de Pós-Graduação em Medicina Tropical e Saúde Pública
da Universidade Federal de Goiás
BANCA EXAMINADORA DA DISSERTAÇÃO DE MESTRADO
Aluno (a): ILDEFONSO ALVES DA SILVA JUNIOR
Orientador (a): Dra. FÁTIMA RIBEIRO DIAS
Membros:
1. Dra. Miriam Leandro Dorta
2. Dra. Maria de Fátima Horta
3. Dra. Fátima Ribeiro Dias
Data: 18 de fevereiro de 2013

edico este trabalho a todas as pessoas que caminham ao meu lado... transmitindo fé, amor, alegria, paciência, determinação e coragem, tornando os meus dias mais felizes e bonitos.

Agradecimentos
Ouvi dizer que agradecimentos acontecem quando todo o amor do mundo resolve se
exibir em palavras... então, espero que na simplicidade das palavras escritas abaixo eu consiga
expressar tamanha gratidão que sinto em ter compartilhado minha história com vocês. Assim,
agradeço...
Minha querida orientadora Profa. Dra. Fátima Ribeiro Dias, obrigado pela educação científica e
minha eterna gratidão pela confiança depositada em mim desde o princípio, da qual, muitas vezes,
nem eu mesmo acreditava que eu era capaz. A você devo minha formação, admiração e respeito. Foi
uma honra tê-la como minha orientadora por tantos anos, você com certeza será minha eterna
“mãe-científica”!
Aos meus queridos Familiares: Mãe, Pai, Priscilla, Patrícia, Jorge, Pedro, tias, tios, primas e primos...
obrigado por toda paciência e preocupação comigo durante todo esse período. Peço desculpa pela
ausência em alguns encontros familiares, tenham sempre em mente que sem o apoio de vocês, este
sonho que estou vivendo agora não seria possível.
À Dra. Miriam Leandro Dorta, obrigado por participar da minha banca de qualificação e do exame
de defesa, com certeza a senhora foi peça chave no meu desenvolvimento científico e pessoal durante
todos esses anos de convivência no laboratório...obrigado por todo apoio recebido.
Ao Dr. Milton, obrigado pela orientação e materiais cedidos ao laboratório. Certamente contribuístes
diretamente a este trabalho e a minha formação. Devo também deixar registrado um obrigado especial
pelo espírito crítico passado durante as discussões de artigos nas reuniões científicas, com certeza meu
“mundo imunológico” não seria o mesmo sem a sua contribuição.
Camila, minha querida amiga e companheira de laboratório, uma amizade fortalecida pelo trabalho
científico. Serei eternamente grato pelo tempo de convivência, pela cumplicidade e pela sempre boa
vontade em me ajudar.
Aos colegas, amigos e companheiros do Laboratório de Imunologia Natural e Imunobiologia das
Leishmanioses: Aline, Anna Karoline, Arissa, Barbara Pinheiro (Báh- minha querida visitante),
Delson, Eduarda, Grazzielle, Helio, Hermínio, João Pedro, Jose Vitor, Ledice, Livia, Lucas,
Nathalia, Priscila, Priscilla Castro, Rodrigo, Rosana, Stefanne, Valéria e Yasmim, obrigado pela

ajuda desde o preparo de materiais até a execução de alguns experimentos, com certeza sem a
convivência amistosa e as conversações “edificantes” que tivemos no lab, esse mestrado não teria o
“gostinho” que tem hoje.
Aos grandes amigos do Laboratório de Citocinas e da “parasito”: Clayson, Lucila, Lohane, Fernanda,
Juliana, Hânstter, Heloisa, Leticia, Tchesca, Camila, Carolina, Nathalia, Artur e aos demais colegas
da pós-graduação, que felicitaram meu dia-dia com gestos espontâneos de ajuda, conversas de
“corredor”, festas e “bafos” do departamento.
Aos caros colegas dos Laboratórios de Imunofarmacologia ICB/USP: Edson, Francisco, Mari,
Marlise, Mateusinho, Matheusão, Silvana e principalmente a Profa. Sônia Jancar, por me receberem
tão bem durante meu intercâmbio pelo PROCAD e também pela ajuda na execução dos experimentos.
Devo dizer que a estadia foi tão boa que agora estarei presente por mais três anos para o doutorado.
Aos colegas do Laboratório de Leishmanioses do ICB/USP: Adriano, Cris, Gê, Jordana, Juliana e a
Prof. Silvia Uliana, muito obrigado pela ajuda na caracterização dos isolados do Leishbank.
Aos inesquecíveis amigos de Rio Verde: Arcilene, Wiliam Pai, Laissa, Wiliam Jr., Ricardo e Eleni;
aos também recentes amigos de São Paulo: Douglas, Herminio, Marilu; Michelle, Miluska, Rayane e
aos eternos amigos de Goiânia: Eric, Graciana, Karen, Marcos, Michel, Rose, Thamiles, um
muitíssimo obrigado pela alegria e diversão em todos os momentos passados juntos.
Às professoras Ana Maria, Ledice, Ludmila e “Patiu” e ao professor Clecildo, por participarem da
minha banca de qualificação/defesa e pela correção da dissertação, do qual os apontamentos foram
cruciais para lapidar este trabalho.
A todos os professores do IPTSP, especialmente do departamento de imunologia, muito obrigado
pelos ensinamentos transmitidos durante o mestrado.
Aos funcionários da secretaria geral e da secretaria do curso de pós-graduação: Senhor Fernando,
Valeria, Divina, “Zezinho”, Kariny. Agradeço toda assistência.
Às agencias de fomento: CAPES, FAPEG e CNPq pelo financiamento do projeto e bolsa de estudo
do mestrado, certamente seria inviável realizar este trabalho sem estes apoios.
A todos os voluntários doadores de sangue.
A todos que contribuíram direta ou indiretamente para a realização deste trabalho.
Muito obrigado!


SUMÁRIO
TABELAS, FIGURAS E ANEXOS ................................................................................ x
ABREVIATURAS ......................................................................................................... xii
RESUMO....................................................................................................................... xiv
ABSTRACT ................................................................................................................... xv
1 INTRODUÇÃO / REVISÃO DA LITERATURA ..................................................... 16
1.1 A Leishmaniose Tegumentar Americana ............................................................. 16
1.2 O parasito Leishmania sp e o processo da metaciclogênese ................................ 19
1.3 Metodologias para identificar e quantificar formas metacíclicas ......................... 23
1.4 O parasito e a sua interação com os macrófagos do hospedeiro .......................... 27
2 JUSTIFICATIVA ........................................................................................................ 31
3 OBJETIVOS ................................................................................................................ 33
3.1 Objetivo Geral ...................................................................................................... 33
3.2 Objetivos Específicos ........................................................................................... 33
4 MÉTODOS .................................................................................................................. 34
4.1 Cultivo e manutenção dos parasitos ..................................................................... 34
4.2 Curvas de crescimento in vitro ............................................................................. 35
4.3 Análise da metaciclogênese ............................................................................. 35
4.3.1 Identificação das formas metacíclicas por análises morfométricas. ..........................36
4.3.2 Identificação e seleção de foras metacíclicas pelo método de aglutinação. .............36
4.3.3 Identificação das formas metacíclicas por citometria de fluxo ..................................36
4.3.4 Identificação das formas metacíclicas por ensaio de resistência ao Complemento ..37
4.4 Avaliação da infecção de macrófagos humanos com isolados de Leishmania sp. 37

4.4.1 Separação de monócitos do sangue periférico e derivação para macrófagos ...........37
4.4.2 Infecção dos macrófagos ...........................................................................................38
4.4.3 Avaliação do índice de infecção dos macrófagos ......................................................39
4.5 Considerações éticas ............................................................................................. 39
4.6 Análises estatísticas .............................................................................................. 39
5 RESULTADOS ........................................................................................................... 40
5.1 Análise da metaciclogênese das formas promastigotas de Leishmania spp in vitro
por morfometria .......................................................................................................... 40
5.2 Análise da metaciclogênese in vitro dos isolados por características de
aglutinação com lectina ou anticorpos........................................................................ 46
5.3 Análise da metaciclogênese dos isolados in vitro por citometria de fluxo ........... 51
5.4 Análise da metaciclogênese in vitro por teste de resistência ao Complemento. .. 57
5.5 Avaliação da infecção de macrófagos humanos por parasitos de diferentes fases
do crescimento ou formas metacíclicas selecionadas ................................................. 64
6 DISCUSSÃO ............................................................................................................... 77
7 CONCLUSÕES ........................................................................................................... 91
Conclusão geral .......................................................................................................... 91
Conclusões específicas ............................................................................................... 91
REFERÊNCIAS ............................................................................................................. 93
ANEXO 1 - PARECER DO COMITÊ DE ÉTICA ...................................................... 103
ANEXO 2 -TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO ............... 104
ANEXO 3 – ARTIGO DA DISSERTAÇÃO ............................................................... 106

x
TABELAS, FIGURAS E ANEXOS
Figura 1. Fases de desenvolvimento de Leishmania sp no trato gastrointestinal do
flebotomíneo ................................................................................................................... 21
Figura 2. Esquema da estrutura da molécula de LPG de formas procíclicas e
metacíclicas de L. major e L. (V.) braziliensis.. ............................................................. 25
Figura 3. Diferenças morfológicas existentes entre as formas promastigota procíclica e a
metacíclica em culturas in vitro.. .................................................................................... 27
Figura 4. Modelo de infecção in vitro de macrófagos humanos. ................................... 30
Figura 5. Curva de crescimento in vitro e morfologia dos isolados. .............................. 42
Figura 6. Morfometria dos isolados.. .............................................................................. 43
Figura 7. Metaciclogênese dos isolados avaliada por morfometria ................................ 44
Figura 8. Porcentagens de formas metacíclicas dos diferentes isolados identificadas por
morfometria.. .................................................................................................................. 45
Figura 9. Fotomicrografias dos parasitos PPS6m e PH8, antes e após purificação de
formas metacíclicas com lectina de Bauhnia purpurea (BPL) e anticorpo 3A1-La.. ..... 47
Figura 10. Metaciclogênese dos isolados avaliada por seleção negativa com lectina ou
anticorpo.. ....................................................................................................................... 49
Figura 11. Comparação entre as porcentagens de formas metacíclicas dos isolados,
detectadas após seleção com lectina de Bauhinia purpurea (L. (V.) braziliensis) ou
anticorpo 3A1-La (L. (L.) amazonensis). ....................................................................... 50
Figura 12. Identificação de formas metacíclicas de L. (V.) braziliensis IMG3 por
citometria de fluxo.. ........................................................................................................ 53
Figura 13. Identificação de formas metacíclicas de L. (L.) amazonensis MAB6 por
citometria de fluxo.. ........................................................................................................ 54
Figura 14. Metaciclogênese dos isolados avaliada por citometria de fluxo.. ................. 55

xi
Figura 15. Porcentagens de formas metacíclicas identificadas por citometria de fluxo. 56
Figura 16. Detecção de formas metacíclicas pelo ensaio de lise mediada pelo
Complemento.................................................................................................................. 58
Figura 17. Metaciclogênese dos isolados avaliada pelo ensaio do Complemento ......... 60
Figura 18. Porcentagens de formas metacíclicas de diferentes isolados avaliados por
resistência ao Complemento.. ......................................................................................... 61
Figura 19. Comparação entre as porcentagens de formas metacíclicas detectadas pelas
técnicas de morfometria, citometria de fluxo, seleção por lectina/anticorpo e ensaio de
resistência ao Complemento.. ......................................................................................... 63
Figura 20. Isolados de L. (V.) braziliensis ou L. (L.) amazonensis apresentam maior
capacidade de infecção quando estão na fase estacionária do crescimento.................... 65
Figura 21. Porcentagem de macrófagos infectados com isolados de L. (V.) braziliensis
ou L. (L.) amazonensis .................................................................................................... 66
Figura 22. Número de parasitos por macrófagos infectados com isolados de L. (V.)
braziliensis ou L. (L.) amazonensis ................................................................................ 67
Figura 23. Comparação entre os índices de infecção de macrófagos humanos após 24 h
ou 72 h de infecção com diferentes isolados.. ................................................................ 69
Figura 24. Seleção de formas metacíclicas das culturas de seis dias não altera o índice
de infecção de macrófagos humanos pelos isolados de L. (V.)b ou L. (L.)a. ................ 71
Figura 25. Isolados/cepas de diferentes dias de cultura apresentam diferenças na
suscetibilidade/resistência aos mecanismos microbicidas dos macrófagos (24 h) ......... 73
Figura 26. Isolados/cepas de diferentes dias de cultura apresentam diferenças na
suscetibilidade/resistência aos mecanismos microbicidas dos macrófagos (72 h). ........ 74
Figura 27. Infecção de macrófagos humanos com isolados de Leishmania spp antes e
após tratamento com IFN /LPS ...................................................................................... 76
Tabela 1. Descrição clínica epidemiológica dos isolados .............................................. 35

xii
ABREVIATURAS
3A1-La Anticorpo anti-LPG de L. (L.) amazonenses
BPL Bauhinia purpurea Lectin
CMN Células mononucleares
FSC Forward Scatter Cytometry
FSChigh
Forward Scatter Cytometry alto
FSClow
Forward Scatter Cytometrybaixo
GP63 glicoproteína de 63 KDaltons
IDRM Intradermoreação de Montenegro
IFN Interferon-gama
INGOH Instituto Goiano de Hematologia e Oncologia
IPTSP Instituto de Patologia Tropical e Saúde Pública
L. (L.) a Leishmania (Leishmania) amazonenses
L. (V.) b Leishmania (Viannia) braziliensis
LCD Leishmaniose Cutânea Difusa
LCDf Leishmaniose Cutânea Disseminada
LCL Leishmaniose Cutânea Localizada
LCM Leishmaniose Cutânea Mucosa
LM Leishmaniose Mucosa
LPG Lipofosfoglicana
LTA Leishmaniose Tegumentar Americana

xiii
NF- B Fator nuclear kappa B
OMS Organização Mundial de Saúde
PBS Salina tamponada com fosfato
PCR Reação da Polimerase em Cadeia
PNA Penaut Lectin of Arachis hypogaea
SFB Soro Fetal Bovino
SSC Side Scatter Cytometry
TLR2 Toll Like Receptor 2
TLR4 Toll Like Receptor 4
TNF Fator de Necrose Tumoral alfa
UFG Universidade Federal de Goiás
WHO World Health Organization

xiv
RESUMO
A Leishmaniose Tegumentar Americana (LTA), causada por protozoários
Leishmania, acomete a pele e/ou as mucosas nasal e orofaríngea. Durante o ciclo de
transmissão, as formas promastigotas procíclicas crescem no intestino do inseto vetor e
diferenciam-se em formas promastigotas metacíclicas infectantes, por um processo
chamado de metaciclogênese, que também ocorre em culturas axênicas in vitro. O
objetivo deste estudo foi avaliar a metaciclogênese in vitro de isolados L. (Viannia)
braziliensis e L. (L.) amazonensis, provenientes de pacientes com diferentes formas
clínicas de LTA, usando diferentes metodologias, e a capacidade destes isolados em
infectar macrófagos humanos. As curvas de crescimento (L. (V.) braziliensis: IMG3,
PPS6m, M2903 e L. (L.) amazonensis: MAB6 e PH8) foram realizadas em meio Grace
e após 2, 6 e 10 dias de cultivo, a quantidade de formas metacíclicas foi avaliada por
meio de morfometria, ensaio de seleção com a lectina de Bauhinia purpurea (BPL, para
L. (V.) braziliensis) ou anticorpo 3A1-La (para L. (L.) amazonensis), citometria de fluxo
e ensaio de resistência ao Complemento. Macrófagos humanos derivados de monócitos
de doadores sadios, ativados ou não com interferon gama (IFN e lipopolissacarídeo (LPS), foram infectados com os parasitos de diferentes dias de cultivo ou formas
metacíclicas selecionadas com BPL/3A1-La (1:1, parasito:macrófago) por 24 ou 72 h.
Os perfis das curvas de crescimento in vitro foram similares para todos os isolados.
Parasitos da fase estacionária do crescimento (6, 10 dias) apresentaram alterações
morfológicas, como a diminuição do tamanho do corpo celular e um aumento do
comprimento do flagelo, em relação aos parasitos do 2º dia. As formas promastigotas L.
(L.) amazonensis do 6º e do 10º dia de cultura apresentaram maiores comprimentos do
corpo celular e do flagelo do que os parasitos L. (V.) braziliensis. Na fase estacionária
houve um aumento significante da porcentagem de formas metacíclicas detectadas pelas
quatro técnicas (M2903: 71-86%; IMG3: 41-85%; PPS6m: 39-76%; PH8: 38-69% e
MAB6: 47-84%; 6º dia, valores min e max). Apesar das quatro técnicas mostrarem um
similar perfil de metaciclogênese para todos os isolados avaliados, as densidades de
formas metacíclicas variaram conforme a técnica utilizada, dependendo do dia de
cultura. A técnica de seleção com BPL/3A1-La foi a que detectou maiores frequências
de formas metacíclicas para todos os isolados no 6o dia. A metaciclogênese dos isolados
foi confirmada nos ensaios de infecção in vitro dos macrófagos, mostrando um aumento
significante no índice de infecção com parasitos do 6º e 10º dia em relação àqueles de 2o
dia (24 ou 72 h). Não foram atestadas diferenças entre os índices de infecção de
macrófagos incubados com parasitos totais ou formas metacíclicas selecionadas do 6º
dia. Os isolados mostraram diferentes perfis de suscetibilidade aos mecanismos
microbicidas dos macrófagos. As cepas M2903 e PH8 foram mais suscetíveis, enquanto
o isolado MAB6, mais resistente. Como as quantidades de formas metacíclicas nas
culturas, considerando a técnica de seleção BPL/3A1-La, foram elevadas para todos os
isolados, os resultados sugerem que a resistência/suscetibilidade aos mecanismos de
controle dos macrófagos seja uma característica intrínseca do isolado. Avaliar a
metaciclogênese de isolados clínicos, simultaneamente à capacidade dos parasitos em
infectar macrófagos humanos, define as condições necessárias para a obtenção de
parasitos eficientes em estabelecer uma infecção, para uma adequada avaliação da
patogenia e da resposta imune natural na leishmaniose.
Palavras-chave: Leishmania braziliensis, Leishmania amazonensis,
metaciclogênese, macrófago humano.

xv
ABSTRACT
The American Tegumentary Leishmaniasis (ATL), caused by Leishmania
protozoa, affects the skin and oropharyngeal/nasal mucosa. During transmission cycle,
the procyclic promastigote forms grow in the vector gut and differentiate into infective
metacyclic promastigotes by a process called metacyclogenesis, which also occurs in
axenic cultures. The aim of this study was to evaluate the in vitro metacyclogenesis of
L. (Viannia) braziliensis and L. (L.) amazonensis isolates obtained from patients with
different clinical forms of ATL, using different methodologies, and the ability of these
isolates to infect human macrophages. Parasites (L. (Viannia) braziliensis: IMG3,
PPS6m, M2903 and L. (L.) amazonensis: MAB6 and PH8) were cultured in Grace´s
insect medium and the amount of metacyclic forms was evaluated by using
morphometry, Bauhinia purpurea lectin (BPL, to L. (Viannia) braziliensis) or 3A1-La
antibody (to L. (L.) amazonensis) negative selection, flow cytometry and Complement-
resistance test after 2, 6 and 10 days of culture. Human monocyte-derived macrophages
from healthy donors, activated or not with interferon gamma (IFN ) and lipopolysaccharide (LPS), were infected with parasites from different days of culture or
with selected metacyclic forms with BPL/3A1-La (1:1 parasite: macrophage) for 24 or
72 h. Growth patterns were similar among the isolates. Stationary-phase promastigotes
(6, 10 days) showed morphological changes such as a reduction of the cell body and an
increase in the flagellum length, in comparison to the parasites of the 2nd day.
Leishmania. (L.) amazonensis promastigotes of the 6th and 10th day of culture
presented higher length of the cell body and flagellum than L. (Viannia) braziliensis
isolates. In stationary phase there was a significant increase in the percentage of
metacyclic forms detected by the four techniques used (M2903: 71-86%; IMG3: 41-
85%; PPS6m: 39-76%; PH8: 38-69% e MAB6: 47-84%; 6th day, medians of the
techniques used). Despite the four techniques demonstrated similar metacyclogenesis
profiles for all isolates evaluated, metacyclic forms densities varied with the technique
and the day of culture. The BLP/3A1-La selection consistently identified the highest
frequencies of metacyclic forms for all isolates (6th day) among the four techniques.
Metacyclogenesis of the isolates was confirmed in in vitro macrophage infection assays
which showed a significant increase of the index of infection with parasites of the 6th
and 10th day compared to those of the 2nd day (24 or 72 h). No significant differences
in capability to infect macrophages were detected between unfractionated or 6th day-
selected metacyclic forms. The isolates showed different profiles of susceptibility to
macrophage microbicidal mechanisms. The M2903 and PH8 strains were the most
susceptible whereas MAB6 isolate was the most resistant among the isolates. As high
amounts of metacyclic forms were present in all isolate cultures, considering BPL/3A1-
La selection, the results suggest that resistance/susceptibility to macrophage control
mechanisms is an intrinsic characteristic of the isolates. To evaluate the
metacyclogenesis of clinical isolates together with the ability of parasites to infect
human macrophages sets the conditions for obtaining efficient parasites to establish a
successful infection what is critical for the evaluation of the pathogenesis and natural
immune response in leishmaniasis.
Key words: Leishmania braziliensis, Leishmania amazonensis,
metacyclogenesis, human macrophage.

16
1 INTRODUÇÃO / REVISÃO DA LITERATURA
1.1 A Leishmaniose Tegumentar Americana
Os protozoários do gênero Leishmania sp são parasitos que causam a
leishmaniose em seres humanos e animais. Estudos epidemiológicos mostram que anualmente
mais de 1,5 milhão de pessoas estão infectadas e cerca de 350 milhões de pessoas correm o
risco de contrair a doença no mundo. As leishmanioses estão presentes em mais de 88 países e
por causa dos altos índices de morbidade e mortalidade, a Organização Mundial de Saúde
(OMS) incluiu as leishmanioses entre as seis doenças endêmicas de maior relevância no
mundo (WHO 2009). No Brasil, a doença é considerada de notificação compulsória, estando
presente em todos os estados brasileiros, principalmente nas regiões Norte, Nordeste e
Centro-Oeste, com incidência anual de mais de 23.000 novos casos (Brasil 2010).
Os parasitos causadores da leishmaniose foram descobertos provavelmente na
Índia por Cunningham (1885), em pacientes com a doença do Botão do Oriente. Em 1903,
Ross criou o gênero Leishmania e Wright identificou Leishmania tropica. Somente em 1909,
Lindenberg, assim como Carini e Paranhos, demonstraram, independentemente, a presença de
parasitos nas lesões de pacientes brasileiros. Em 1911, Gaspar Vianna descreveu o parasito
como causador da doença em nosso país, diferenciando-o dos protozoários isolados no Velho
Mundo e denominando-o Leishmania braziliensis (revisto por Momen & Cupolillo 2000,
Altamirano-Enciso et al. 2003).
Classificado dentro da ordem Kinetoplastida, família Trypanosomatidae, o gênero
Leishmania agrupa mais de 20 espécies de protozoários unicelulares responsáveis por causar
infecção. Dentre os hospedeiros têm-se os invertebrados e os vertebrados. Os hospedeiros
invertebrados, ou vetores de Leishmania, são insetos hematófagos conhecidos como
flebotomíneos, dos gêneros Phlebotomus e Lutzomyia. Já os hospedeiros vertebrados incluem
uma grande variedade de mamíferos, sendo os mais comuns os roedores e os canídeos, mas
também podem aparecer os endentados, marsupiais, procionídeos, ungulados e primatas,
incluindo o homem (Ashford 2000, Almeida et al. 2003, Amóra et al. 2009).

17
São responsáveis pela leishmaniose tegumentar nas Américas principalmente as
espécies Leishmania (Viannia) braziliensis, L. (V.) panamensis, L. (V.) guyanensis, L. (V.)
peruviana, L. (Leishmania) amazonensis, L. (L.) mexicana, L. (L.) pifanoi, L. (L.)
venezuelensis e no Velho Mundo L. (L.) tropica, L. (L.) major e L. (L.) aethiopica. No Brasil,
foram identificadas sete espécies que são responsáveis pela leishmaniose tegumentar, sendo
seis do subgênero Viannia e uma do subgênero Leishmania. As três principais espécies são: L.
(V.) braziliensis, L. (V.) guyanensis e L. (L.) amazonensis e, mais recentemente, as espécies L.
(V.) lainsoni, L. (V.) naiffi , L. (V.) lindenberg e L. (V.) shawi foram identificadas nas regiões
Norte e Nordeste (Murray et al. 2005, Brasil 2006).
Existem diferentes manifestações clínicas nas leishmanioses e estas são
relacionadas com a espécie do parasito e a resposta imune do hospedeiro, podendo o
indivíduo apresentar desde lesões simples com cura clínica espontânea da doença até formas
graves, que levam à morte. Assim, na Leishmaniose Tegumentar Americana (LTA), a
infecção pode se manifestar de três formas comuns: a leishmaniose cutânea (localizada ou
disseminada), a leishmaniose mucocutânea e a leishmaniose cutânea difusa (Singh &
Sivakumar 2004, Murray et al. 2005, Brasil 2006).
A leishmaniose cutânea localizada (LCL) inicia-se com uma área de vermelhidão
e edema no sítio da inoculação do parasito pelo inseto vetor; a lesão inflamada aumenta de
tamanho e depois de três a quatro semanas, desenvolve-se uma única úlcera rasa, de
crescimento lento, na qual os parasitos são encontrados principalmente nas bordas da lesão. O
parasito pode permanecer no organismo por semanas ou mesmo meses antes do aparecimento
da lesão. A forma cutânea disseminada (LCD) acomete principalmente pacientes
imunossuprimidos, sendo caracterizada pela disseminação linfática a partir da lesão inicial
(adenomegalia satélite) e nódulos subcutâneos móveis, que mais tarde aderem à pele e
ulceram, formando múltiplas lesões (Silveira et al. 2009).
A leishmaniose mucocutânea (LM) ou cutâneo-mucosa (LCM) pode surgir
juntamente com a lesão cutânea ainda em atividade, ou anos após sua cicatrização. Há
frequentemente o aparecimento de lesões ulceradas destrutivas nas mucosas do nariz, boca e
faringe. A disseminação do parasito dá-se, provavelmente, por via hematogênica. Nódulos
com focos de macrófagos infiltrados podem surgir com raros parasitos e tendência à
ulceração, localizando-se na porção cartilaginosa do septo nasal. Essas úlceras costumam
progredir em extensão e profundidade, com destruição das cartilagens do nariz, da região
palatina ou do maciço facial. Progressivamente, ocorre a perfuração do septo ou do palato e o

18
processo inflamatório se estende à faringe e à laringe, sendo possível encontrar metástases
cutâneas. Esta forma de leishmaniose recebe também os nomes de espúndia, ferida brava,
nariz de tapir e úlcera de Bauru, sendo causada principalmente por L. (V.) braziliensis e L.
(V.) guyanensis (Amazônia brasileira) e raramente por L. (V.) panamensis (Naiff et al. 1988,
Osorio et al. 1998).
Na leishmaniose cutânea difusa (LCD), o processo tem início com o
desenvolvimento de lesão única no ponto de inoculação dos parasitos pelos flebotomíneos.
Segue-se uma disseminação metastática que dará origem a lesões múltiplas, não ulceradas,
disseminadas por toda a pele do paciente. Os nódulos são ricos em macrófagos repletos de
parasitos, mas a reação inflamatória local é muito reduzida ou mesmo ausente. São
responsáveis por essa forma de leishmaniose, L. (L.) pifanoi, L. (L.) mexicana e L. (L.)
amazonensis nas Américas; ou L. aethiopica, na África (Chaudhary et al. 2008, Vieira-
Gonçalves et al. 2008, Jafari et al. 2010, Machado et al. 2011).
O tratamento das diferentes formas clínicas da LTA é realizado com o uso de
antimoniais pentavalentes, com elevada porcentagem de cura clínica, quando usados
corretamente. Como fármaco de segunda escolha é usado o desoxicolato de anfotericina B,
que é um antibiótico poliênico com excelente atividade in vitro na destruição de Leishmania
intra e extracelular, porém pode provocar vários efeitos colaterais durante o uso in vivo (Brasil
2006).
O diagnóstico da LTA é baseado em exames moleculares, imunológicos e
parasitológicos. Os exames moleculares (reação da polimerase em cadeia, PCR) são muito
utilizados em pesquisas, e apesar de apresentarem alta sensibilidade e serem capazes de
identificar as espécies de Leishmania, são métodos ainda pouco utilizados na rotina de
diagnóstico (da Silva et al. 2012, Pita-Pereira et al. 2012, Khosravi et al. 2012). Os exames
imunológicos incluem a intradermorreação de Montenegro (IDRM) e teste sorológico
(imunofluorescência indireta), mas devido às reações cruzadas com outras doenças, esses
testes não devem ser realizados isoladamente. Juntamente com IFI e a IDRM, os exames
parasitológicos (demonstração do parasito) em fragmentos de biópsia, raspagem ou punção
das lesões e isolamento em cultivo ex vivo ou in vivo (inoculações em animais de
experimentação) são utilizados (Brasil 2006, Oliveira et al. 2010, Kobets et al. 2012).
A possibilidade de isolar o parasito in vitro, a partir das lesões e mantê-los com
relativa facilidade em ambientes artificiais de cultura, possibilitou um grande avanço no

19
estudo de várias características relacionadas à biologia da leishmânia. Em seguida, serão
descritos alguns aspectos importantes relacionados ao parasito, a fim de proporcionar um
melhor entendimento sobre os objetivos deste trabalho.
1.2 O parasito Leishmania sp e o processo da metaciclogênese
Leishmania sp é um parasito digenético (heteroxeno), que circula entre os
hospedeiros vertebrados e seus vetores, apresentando as formas amastigotas (ovais e com
flagelo rudimentar) dentro das células do sistema mononuclear fagocitário dos mamíferos e
várias formas flageladas no tubo digestivo dos flebotomíneos (promastigotas).
O ciclo de transmissão da leishmaniose começa quando fêmeas de fleblotomíneos
fazem seu repasto sanguíneo em hospedeiros infectados e ingerem células contendo os
parasitos. Dentro do intestino do vetor, os parasitos diferenciam-se em formas promastigotas
procíclicas flageladas que se replicam intensamente e passam por um processo chamado de
metaciclogênese, originando as formas promastigotas metacíclicas. Estas adquirem a
capacidade de infectar e uma vez na probócida do inseto vetor, podem, então, durante um
novo repasto sanguíneo, serem inoculadas em novo hospedeiro. As formas promastigotas, no
hospedeiro, são internalizadas pelas células do sistema fagocítico mononuclear, especialmente
os macrófagos. Nestas células, o parasito diferencia-se em forma amastigota, a qual se
multiplica continuamente, até que o macrófago se rompa e libere os parasitos, aptos a infectar
novas células (Reithinger et al. 2007, Naderer & McConville 2008).
A primeira evidência experimental da transmissão da leishmaniose pelo inseto
vetor foi demonstrada por Shortt et al. (1931) e Addler & Ber (1941). Esses pesquisadores
acreditavam que dentro do flebotomíneo existia uma forma promastigota do parasito que era
adaptada para viver dentro do hospedeiro vertebrado. Observações subsequentes mostraram
que existiam diferenças morfológicas entre as formas promastigotas que habitavam a parte
anterior e a posterior do intestino médio do vetor, aumentando as especulações sobre o
processo da metaciclogênese. Assim, em meados dos anos 80, vários autores começaram a
descrever as mudanças que ocorrem com o parasito dentro do inseto e em culturas axênicas in
vitro, introduzindo o conceito da metaciclogênese para espécies de leishmânia do Velho e do
Novo Mundo (revisto por Sacks 1989).
A metaciclogênese é um processo de diferenciação das formas promastigotas
procíclicas em formas promastigotas metacíclicas, altamente infectantes para o hospedeiro

20
vertebrado, consideradas o estágio final do desenvolvimento do parasito nos flebotomíneos. A
seguir será descrito o processo de metaciclogênese que ocorre in vivo (isto é, dentro do inseto
vetor) e em culturas axênicas in vitro.
O surgimento das formas promastigotas metacíclicas dentro do vetor foi descrito
primeiramente in vivo para as espécies L. major e L. mexicana por Sacks & Perkins (1984).
Estes autores demonstraram que clones de formas promastigotas destas duas espécies de
leishmânia, recuperadas do intestino médio de P. papatassi e L. longipalpis após três dias da
infecção, não eram infecciosas para camundongos BALB/c. Por outro lado, as formas
promastigotas intestinais recuperadas após 4 a 7 dias do repasto sanguíneo do inseto vetor
foram progressivamente mais virulentas, estabelecendo que era necessário um período de
“adaptação” do parasito dentro do inseto vetor para o surgimento de formas mais infecciosas,
que mais tarde, foram chamadas de formas metacíclicas. Após observar este fenômeno e com
o surgimento de estudos posteriores, utilizando outras espécies de leishmânia, ficou
esclarecido que quando o inseto vetor ingere o sangue com as formas amastigotas, estas, ao
chegarem ao intestino médio ou estômago do vetor, sofrem um primeiro ciclo replicativo,
transformando-se em formas promastigotas procíclicas flageladas, que também, por processos
sucessivos de divisão binária, multiplicam-se ainda no sangue ingerido, que é envolto por
uma membrana peritrófica, formada pela secreção das células do estômago do inseto. Após a
digestão do sangue, entre o 3º e o 4º dia, a membrana peritrófica se rompe e as formas
promastigotas ficam livres, sendo então, denominadas de promastigotas nectomônadas
(revisto por Kamhawi 2006). Estas, por sua vez, podem seguir por duas vias, dependendo da
espécie do parasito.
Na primeira via, as espécies pertencentes ao subgênero Viannia colonizam a
região do piloro e ílio (seção peripilária), onde são chamadas de promastigotas leptomônadas,
que permanecem aderidas pelo flagelo ao epitélio intestinal, por meio de junções
hemidesmossomas, usando moléculas de lipofosfoglicana (LPG). Nesta fase, os parasitos
sofrem sucessivas divisões celulares até atingir a fase metacíclica (término do ciclo
replicativo), quando ocorrem mudanças estruturais nas moléculas de LPG ancoradas na
membrana celular e o parasito deixa o intestino, seguindo em direção à faringe do inseto
(Gossage et al. 2003).
Na segunda via, as formas promastigotas leptomônadas, pertencentes ao
subgênero Leishmania, multiplicam-se livremente ou aderentes às paredes do estômago
(seção suprapilária). Em seguida, as formas flageladas vão para a região do esôfago e faringe

21
onde se diferenciam em formas metacíclicas, da mesma maneira que os parasitos do
subgênero Viannia. O tempo necessário para completar o processo de metaciclogênese varia
entre três e cinco dias para diferentes espécies, e geralmente coincide com o período de um
novo repasto sanguíneo do vetor, aumentando a probabilidade de um inóculo dos parasitos em
um novo hospedeiro vertebrado (Nieves & Pimenta 2000, Gossage et al. 2003, Kamhawi
2006, Bates 2008). A Figura 1 ilustra o processo de metaciclogênese dentro do intestino do
flebotomíneo.
Figura 1. Fases de desenvolvimento de Leishmania sp no trato gastrointestinal do flebotomíneo.
(Adaptado de Gossage et al. 2003).

22
A metaciclogênese também ocorre durante o cultivo dos parasitos em culturas
axênicas in vitro, onde as formas promastigotas procíclicas, da fase logarítmica do
crescimento, transformam-se em formas metacíclicas, durante a fase estacionária do
crescimento (Sacks & Perkins 1984, da Silva & Sacks 1987, Sacks 1989). As primeiras
evidências de que o processo de metaciclogênese dos parasitos também ocorria em culturas in
vitro foram observadas por Gianni (1974) e Keithly (1976), que utilizando cepas de L. (L.)
donovani, acharam uma correlação positiva entre o tempo de cultivo e a capacidade de
infecção de hamsters. Posteriormente, Sacks & Perkins (1984) observaram que os parasitos da
fase tardia da cultura (5 – 7 dias) da espécie L. tropica conseguiam infectar um maior número
de macrófagos de camundongos C57Bl/6 in vitro e geravam maiores lesões em patas de
camundongos BALB/c do que os parasitos da fase logarítmica da cultura (3 dias). Outros
autores também observaram este processo ocorrendo com isolados de diferentes espécies de
leishmânia, como L. major, L. mexicana, L. infantum, L. (L.) amazonensis, L. (V.) braziliensis
e L. (V.) peruviana (Sacks & da Silva 1987, Zakai et al. 1998, Louassini et al. 1998, Almeida
et al. 2003, Gamboa et al. 2008).
O mecanismo que estimula o aparecimento das formas metacíclicas ainda é pouco
entendido, mas sabe-se que in vitro, as formas promastigotas de Leishmania sp, cultivadas em
diferentes meios de cultura, crescem melhor em pH neutro ou ligeiramente alcalino (Sacks &
Perkins 1984). No entanto, é possível induzir maiores quantidades de formas metacíclicas de
L. (V.) braziliensis, L. donovani, L. major e L. mexicana in vitro, expondo as formas
promastigotas a pH baixo ou condições anaeróbicas de cultura (Bates & Tetley 1993, Zakai et
al. 1998, Louassini et al. 1998). A acidificação do meio de cultura e a carência de nutrientes
que se estabelece na fase estacionária do crescimento, parecem ser os principais estímulos que
determinam a diferenciação de formas promastigotas procíclicas em formas metacíclicas
(Bates 2008).
A metaciclogênese das formas promastigotas de leishmânia inclui uma série de
mudanças na expressão gênica, na morfologia e na estrutura da LPG presente na superfície do
parasito (Sacks & da Silva 1987). Genes que codificam uma proteína de choque térmico
chamada de HSP70 estão altamente expressos nas formas metacíclicas de L. major (Searle et
al. 1989, Coulson & Smith 1990). Além disso, alguns genes são expressos como proteínas
somente na forma metacíclica, como a proteína META e o fator de transcrição NAT-1, que
são considerados fatores de virulência do parasito. Outros fatores que estão alterados na forma
metacíclica, e que parecem exercer papel fundamental na infectividade e na proteção do

23
parasito durante a transmissão da doença, é a protease de superfície de leishmânia gp63
(glicoproteína de 63 KDaltons - aumento da expressão) e LPG (alterações estruturais) (Sacks
et al. 1990, Brodin et al. 1992, Marín et al. 2000, Ramos et al. 2004, 2011). As alterações que
ocorrem nestas moléculas serão descritas abaixo, pois permitiram o surgimento de
metodologias para caracterização e separação das formas metacíclicas nos estudos do
processo de metaciclogênese em culturas in vitro.
1.3 Metodologias para identificar e quantificar formas metacíclicas em culturas in vitro.
Existem diferentes métodos para identificar e separar formas metacíclicas e
procíclicas em cultura, estes métodos baseiam-se nas diferenças morfológicas e bioquímicas
existentes entre as diferentes formas evolutivas do parasito.
A primeira característica observada durante o processo de diferenciação dos
parasitos, dentro do inseto vetor ou em culturas in vitro, envolveu estudos de resistência das
formas promastigotas metacíclicas ao sistema Complemento do hospedeiro (Franke et al.
1985, Wozencraft & Blackwell 1987). A observação de que as formas metacíclicas,
provenientes da fase estacionária da cultura, são mais infecciosas do que as formas
promastigotas procíclicas da fase logarítmica (Sacks & da Silva 1987), levou a uma avaliação
de parasitos provenientes de diferentes fases de crescimento, considerando a sua
suscetibilidade ou resistência à lise mediada pelo Complemento. Parasitos da fase estacionária
de várias espécies de leishmânia (L. donovani, L. panamensis, L. major, L. mexicana, L.
amazonensis e L. braziliensis) são mais resistentes à lise mediada pelo Complemento do que
as promastigotas da fase logarítmica (Franke et al. 1985, Puentes et al. 1988, Louassini et al.
1998). Estes resultados foram relacionados ao alongamento na cadeia de LPG na superfície de
formas promastigotas metacíclicas, o qual parece proteger o parasito da lise mediada pelo
Complemento, agindo como uma barreira física, impedindo à inserção do complexo de ataque
à membrana (MAC) nos parasitos (Franke et al. 1985, Puentes et al. 1988). Outra modificação
que aparece na forma metacíclica e parece influenciar sua resistência à lise mediada pelo
Complemento é o aumento da expressão da metaloprotease de superfície gp63 (Brittingham et
al. 1995). O mecanismo desta resistência é dependente da atividade proteolítica desta enzima,
que cliva o componente do Complemento C3b em uma forma inativa (C3bi), impedindo,
assim, a continuidade da cascata do Complemento até a formação do MAC (Mosser &
Brittingham 1997). Assim, é possível separar as formas metacíclicas in vitro, por meio do

24
ensaio de lise mediada pelo Complemento, colocando os parasitos em contato com soro,
quando as formas suscetíveis (procíclicas) são lisadas e as formas metacíclicas (resistentes à
lise) continuam íntegras (Sacks & Melby 2001).
Outras técnicas para identificação e purificação de formas metacíclicas surgiram
baseadas nas mudanças que ocorrem na LPG exposta na membrana do parasito durante a
diferenciação destes para a forma metacíclica. Estudos com L. major e L. donovani mostraram
que mudanças na composição dos carboidratos que compõe a molécula de LPG, identificadas
por ligação diferencial à lectinas, são associadas ao processo de metaciclogênese dos parasitos
(da Silva & Sacks 1987, Sacks et al. 1990). Classicamente, o primeiro método de purificação
de formas metacíclicas, baseado nas alterações da LPG, foi descrito por Silva & Sacks (1987).
Estes autores utilizaram L. major para demonstrar que o alongamento da molécula de LPG
(aumento do número de unidades repetitivas de dissacarídeos fosforilados), juntamente com a
adição de resíduos de arabinose nesta molécula (Fig. 2A), fazia com que os parasitos da fase
estacionária perdessem a capacidade de se ligar a lectina PNA (Peanut Lectin of Arachis
hypogaea). Os parasitos PNA- (não aglutinados pela lectina) apresentaram maior capacidade
de infecção de macrófagos murinos do que os parasitos PNA+ (aglutinados pela lectina)
(McConville et al. 1992). O mesmo ocorreu para parasitos da espécie L. donovani, que
durante o processo de metaciclogênese apresentaram um aumento do tamanho da molécula de
LPG e mudança na conformação de seus açúcares, alterando a capacidade da lectina PNA de
se ligar na porção terminal da molécula (Sacks et al. 1995).
As formas metacíclicas de L. (V.) braziliensis também sofrem modificações na
molécula de LPG (Fig. 2B), adicionando resíduos de glicose e aumentando as unidades
repetidoras de Galactose - Manose (Gal-Man). Essas modificações também permitem que
estes parasitos possam ser purificados, por exemplo, pelo método de seleção negativa, usando
a lectina de Bauhinia purpurea (BPL). A BPL aglutina as formas procíclicas da cultura,
deixando as formas metacíclicas livres para serem coletadas (Pinto-da-Silva et al. 2002).
Protocolos similares para caracterização e purificação de formas promastigotas metacíclicas
tem sido desenvolvidos, utilizando anticorpos específicos para LPG de formas procíclicas de
L. (L.) amazonensis (Chaves et al. 2003, Pinto-da-Silva et al. 2005). O anticorpo 3A1-La é
capaz de aglutinar as formas procíclicas da cultura, impedindo sua ligação ao intestino do
vetor e sua posterior transmissão (Pinto-da-Silva et al. 2005). O método de purificação com o
anticorpo 3A1-La é útil para estudar infecções in vivo em modelos murinos, bem como de
macrófagos de diferentes espécies in vitro, usando as formas promastigotas metacíclicas

25
purificadas. Outros anticorpos contra LPG de leishmânia também já foram desenvolvidos para
purificar formas metacíclicas de L. tropica (Lira et al. 1998) e de L. donovani (Mahoney et al.
1999).
Figura 2. Esquema da estrutura da molécula de LPG de formas procíclicas e metacíclicas de L.
major e L. (V.) braziliensis.Durante a metaciclogênese, a molécula de LPG muda sua estrutura,
aumentando em comprimento, por adicionar unidades repetidoras de Gal-Man (Galactose - Manose) e
resíduos de arabinose (A: L. major) ou glicose (B: L. (V.) braziliensis), mudando sua especificidade
para a ligação com lectinas. Adaptado de Assis et al. 2012.
Uma abordagem alternativa para purificação de formas metacíclicas,
independente da molécula de LPG, tem sido utilizada, explorando o fato de que as formas
promastigotas procíclicas são morfologicamente distintas das formas metacíclicas. Spath e
Berveley (2001) propuseram um método de purificação de formas metacíclicas utilizando a
centrifugação dos parasitos em um gradiente de densidade. Nesta técnica, utilizando parasitos
da espécie L. major, os autores demonstraram que após centrifugação da cultura em gradiente
de densidade Ficoll 8-10%, foi possível obter na interface do gradiente, parasitos com

26
características de formas metacíclicas (PNA-) e que foram mais infecciosas para macrófagos
peritoneais de camundongos BALB/c do que os parasitos PNA+.
De maneira geral, as diferentes formas evolutivas do parasito podem ser
identificadas pela morfologia em culturas in vitro e dentro do inseto vetor, apesar de não
haver um consenso sobre a variedade de formas evolutivas existentes que podem ser
caracterizadas por este método (Gossage et al. 2003). Em culturas in vitro, o termo
promastigota procíclica é geralmente utilizado para determinar coletivamente aquelas formas
não-metacíclicas que estão em constante divisão celular (ou seja, na fase logarítmica do
crescimento) e o termo promastigota metacíclica é utilizado para definir aquelas formas
infectantes que aparecem durante a fase estacionária do crescimento (Sacks & da Silva 1987,
Pinto-da-Silva et al. 2002). Baseando-se nessa classificação, na Figura 3 são mostradas
microscopia eletrônica de varredura das formas evolutivas encontradas em culturas in vitro de
L. major (Besteiro et al. 2007). As formas metacíclicas exibem um corpo celular pequeno e
alongado (≤ 8,4 µm x 1,2 – 1,5 µm), com flagelo cerca de duas vezes ou mais alongado do
que o comprimento da célula. Em contraste, as formas procíclicas constituem uma população
heterogênea, com corpo celular arredondado (15 – 35 µm x 2 – 3 µm) e flagelo pequeno,
aproximadamente do mesmo tamanho da célula (Almeida et al 1993; Bates & Tetley et al.,
1993).
Análises morfométricas de culturas em fase estacionária (6º dia), cultivadas em
meio ácido (pH 5,0), mostraram que formas promastigotas de L. donovani, L. mexicana, L.
major e L. braziliensis que exibiam um corpo celular pequeno (≤ 8 m) e um longo flagelo,
eram formas metacíclicas, sendo que nesta fase, os parasitos apresentaram aumentada
resistência à lise mediada pelo Complemento e causaram maiores lesões em camundongos
BALB/c do que os parasitos cultivados em meio alcalino (Zakai et al. 1998).

27
Figura 3. Diferenças morfológicas existentes entre as formas promastigota procíclica e a
metacíclica em culturas in vitro.Microscopia eletrônica de varredura adaptado de Besteiro et al.
2010.
Devido às diferenças morfológicas existentes entre as diferentes formas
evolutivas, também é possível identificar as populações de formas promastigotas procíclicas e
metacíclicas, de diferentes espécies de Leishmania spp, por meio de citometria de fluxo,
avaliando propriedades como tamanho celular e granulosidade interna das células (Saraiva et
al. 2005). A citometria de parasitos, como a leishmânia, constitui o método mais simples para
avaliar se ocorreu ou não o processo de metaciclogênese in vitro. A identificação de formas
metacíclicas de Leishmania spp, utilizando citometria de fluxo, foi primeiramente realizada
por Saraiva et al. (2005), que analisando os parasitos por tamanho (FSC) e granulosidade
(SSC), identificaram duas populações, uma constituída de formas de tamanho grande
(FSChigh
) e outra de tamanho pequeno (FSClow
). Essas formas de tamanho pequeno
apresentam morfologia similar à das formas metacíclicas, aumentam em número de acordo
com o tempo de cultura e possuem três vezes maior capacidade de infectar macrófagos
murinos do que as formas de tamanho maior (FSChigh
), comprovando que as formas pequenas,
são as formas metacíclicas nas culturas.
Assim, é possível identificar e quantificar as formas metacíclicas da cultura,
utilizando a citometria de fluxo, as análises morfométricas e o teste de resistência ao
Complemento. Para obter populações homogêneas de formas metacíclicas, porém, é
necessário utilizar um dos métodos de purificação baseado nas alterações da molécula de
LPG, isto é, na utilização de lectinas/anticorpos, ou na densidade dos parasitos, usando
gradientes de densidade.
1.4 O parasito e a sua interação com os macrófagos do hospedeiro
Quando os parasitos são introduzidos no hospedeiro mamífero, as formas
promastigotas metacíclicas encontram um ambiente constituído por componentes do sangue
(células e plasma), extravasados para o tecido durante o repasto sanguíneo do inseto.
Imediatamente após este contato, há o início da resposta imune natural contra os parasitos. Foi
demonstrado, no entanto, que as formas metacíclicas inoculadas no hospedeiro são hábeis
para evadir da resposta imune natural. Vários mecanismos foram descritos, tais como (1)
inativação do sistema Complemento, (2) modulação da produção de citocinas e quimiocinas,
(3) interferência nos processos de migração e apoptose celulares e (4) modificação do micro-

28
ambiente intracelular para diminuir os efeitos microbicidas dos macrófagos (revisto por
Awasthi et al. 2004). As formas metacíclicas são, portanto, as formas que, após a fagocitose
por macrófagos, resistem aos mecanismos microbicidas destas células, estando envolvidas
diretamente na patogênese da doença.
O contato do parasito com os componentes do sistema imune inato do hospedeiro
mamífero representa um passo crucial na resistência ou suscetibilidade do hospedeiro à
leishmaniose. Como referido anteriormente, diferentemente das formas procíclicas, as formas
metacíclicas, por meio do alongamento da molécula de LPG e da superexpressão da enzima
gp63 na membrana, são capazes de se proteger da lise pelo sistema Complemento
(Brittingham et al. 1995, Sacks & Noben-Trauth 2002, Sacks & Sher 2002). Por outro lado, a
opsonização das formas metacíclicas com componentes C3b ou C3bi (considerados
opsoninas) facilita a ligação do parasito com os receptores do Complemento CR1 e CR3,
respectivamente, mediando sua adesão ao macrófago hospedeiro e conseqüentemente, a
internalização dos parasitos. Outros receptores no macrófago do hospedeiro vertebrado, que
reconhecem padrões moleculares dos parasitos ou opsoninas, têm sido identificados, tais
como o receptor para fucose-manose, o receptor para fibronectina, o receptor para a porção Fc
dos anticorpos e o receptor para proteína C reativa, que facilitam a entrada da forma
promastigota metacíclica, geralmente de forma silenciosa, na célula hospedeira (Sacks & Sher
2002, Akilov et al. 2007, Naderer & McConville 2008, Ueno et al. 2009).
No processo de fagocitose do parasito pelos macrófagos, as leishmânias
localizam-se inicialmente em vacúolo parasitóforo, circundado pela membrana da célula
hospedeira, que, a seguir, funde-se com lisossomos, que contém enzimas proteolíticas capazes
de lisar os parasitos, formando o fagolisossomo (Lippuner et al. 2009, Millington et al. 2010,
Hsiao et al. 2011). No entanto, as formas metacíclicas podem retardar a formação do
fagolisossomo e a enzima gp63 pode atuar degradando as enzimas lisossomais. Assim, as
formas metacíclicas, atuam inicialmente, modificando o ambiente intracelular, para
transformar-se com sucesso na forma amastigota e promover sua replicação (Burchmore &
Barrett 2001, Lodge & Descoteaux 2005, Real et al. 2010). Além disso, dependendo da
espécie de leishmânia fagocitada, o vacúolo parasitóforo exibe características distintas. Os
fagolisossomos contendo parasitos do subgênero Leishmania são grandes (cerca de um terço
da célula), contendo várias amastigotas aderidas à membrana. Já os fagolisossomos contendo

29
parasitos do subgênero Viannia são pequenos e apertados, contendo somente uma amastigota
por vacúolo (revisto por Rittig & Bogdan 2000).
Apesar dos vários mecanismos de evasão que os parasitos utilizam, as células do
sistema imune inato possuem receptores que reconhecem padrões moleculares do parasito e
podem ativar vários fatores de transcrição, desencadeando a produção de citocinas e
quimiocinas, que culminam na formação da resposta inflamatória. Na infecção por
Leishmania, o receptor do tipo TLR2 (Toll Like Receptor 2), presente em macrófagos, células
dendríticas e células NK (Natural Killer), pode ser acionado pelas moléculas LPG (Becker et
al. 2003, Veer et al. 2003, Kavoosi et al. 2009, 2010). O TLR2 pode sinalizar para a ativação
do fator de transcrição NF-kB nessas células e induzir a produção de diversas citocinas
proinflamatórias, que, entre várias funções, ativam os macrófagos (Kawai & Akira 2010).
O interferon-gama (IFN ) e o fator de necrose tumoral alfa (TNF) são citocinas
proinflamatórias que atuam sinergicamente nos macrófagos para a eliminação dos parasitos,
induzindo a produção de importantes moléculas microbicidas, como óxido nítrico, peróxido
de hidrogênio e peroxinitrito (Liew et al. 1990, Vouldoukis et al. 1995). Estudos in vitro de
infecção de macrófagos humanos por diferentes espécies de leishmânia (Fig. 4) mostram que
estas células são permissivas à infecção por estes parasitos, e quando ativadas por citocinas,
como o IFN e por produtos bacterianos, tais como o lipopolissacarídeo (LPS), um agonista
de TLR4, são capazes de eliminar os parasitos intracelulares (Vouldoukis et al. 1995, Bosque
et al. 1998, Panaro et al. 1999). Os estudos de interação entre macrófagos humanos e
leishmânias são escassos, considerando a infecção por L. (V.) braziliensis ou por L. (L.)
amazonensis (Rey et al. 1990, Calegari-Silva et al. 2009, Giudice et al. 2012).
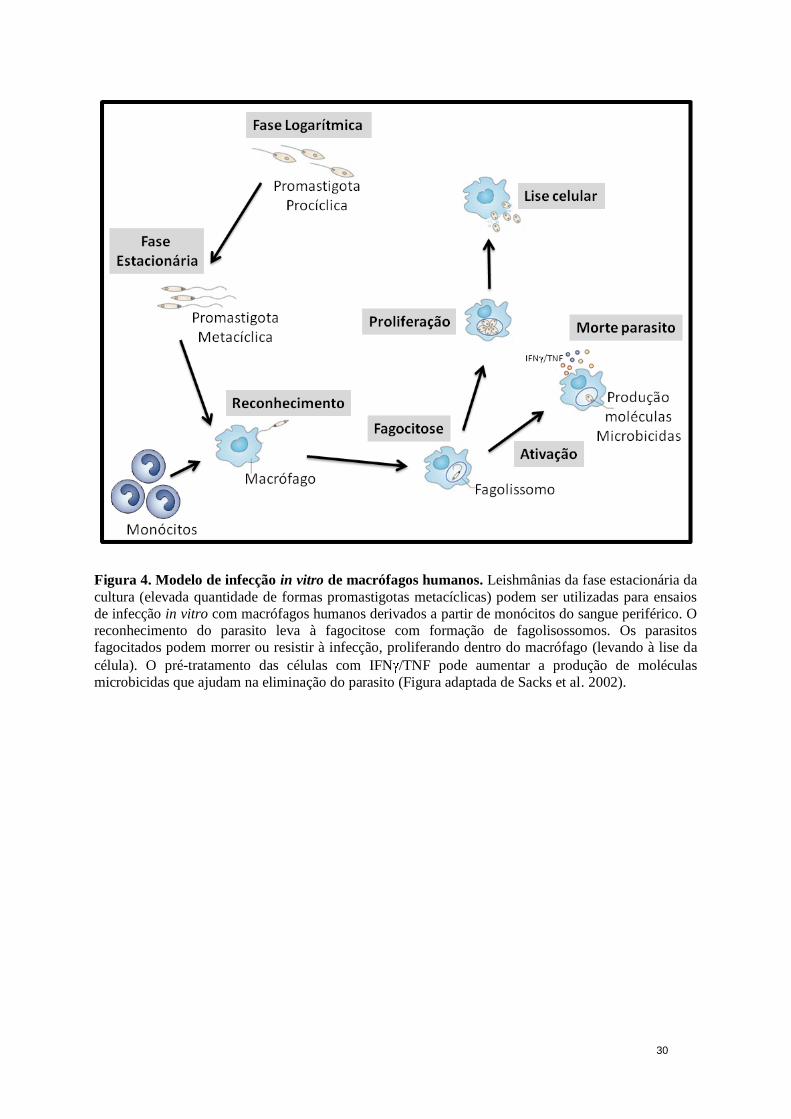
30
Figura 4. Modelo de infecção in vitro de macrófagos humanos. Leishmânias da fase estacionária da
cultura (elevada quantidade de formas promastigotas metacíclicas) podem ser utilizadas para ensaios
de infecção in vitro com macrófagos humanos derivados a partir de monócitos do sangue periférico. O
reconhecimento do parasito leva à fagocitose com formação de fagolisossomos. Os parasitos
fagocitados podem morrer ou resistir à infecção, proliferando dentro do macrófago (levando à lise da
célula). O pré-tratamento das células com IFN /TNF pode aumentar a produção de moléculas
microbicidas que ajudam na eliminação do parasito (Figura adaptada de Sacks et al. 2002).

31
2 JUSTIFICATIVA
A LTA é uma doença que apresenta diferentes formas clínicas, sendo que L. (V.)
braziliensis e L. (L.) amazonensis causam formas graves, como a leishmaniose mucosa e a
cutânea difusa, respectivamente. O estabelecimento das diferentes formas clínicas depende da
relação parasito-hospedeiro. Portanto, é necessário avaliar características dos parasitos, bem
como do hospedeiro, que determinam as formas clínicas da doença e sua evolução. Neste
sentido, a possibilidade de isolar o parasito in vitro, a partir das lesões de pacientes, e mantê-
los com relativa facilidade em ambientes artificiais de cultura, tem possibilitado um grande
avanço no estudo de várias características relacionadas à biologia de Leishmania. No entanto,
ainda há grandes lacunas no conhecimento da imunobiologia das espécies do Novo Mundo,
tais como L. (V.) braziliensis e L. (L.) amazonensis, que são as duas principais espécies
causadoras da LTA no Brasil.
Os parasitos Leishmania sp são inoculados no hospedeiro na forma promastigota
metacíclica, a qual origina-se da forma promastigota procíclica, por um processo conhecido
como metaciclogênese. A importância de avaliar a metaciclogênese de diferentes isolados de
Leishmania reside no fato de que as formas promastigotas metacíclicas são hábeis em evadir
da resposta imune e assim, estabelecer a infecção. Pouco ainda foi descrito sobre a
metaciclogênese in vitro das espécies L. (V.) braziliensis e L. (L.) amazonensis e estudar este
processo pode levar ao conhecimento das características da metaciclogênese de diferentes
isolados e à determinação das melhores condições experimentais para avaliação de infecções
in vivo e in vitro. Tais condições são de fundamental importância para estudos sobre a
patogênese da leishmaniose, uma vez que as formas procíclicas e metacíclicas, por
apresentarem diferenças nas moléculas de superfície, podem levar à diferentes consequências
na infecção in vivo ou in vitro.
É necessário estudar o parasito e sua capacidade de infectar macrófagos,
identificando as variáveis que possam estar relacionadas com o eficiente estabelecimento da
infecção, bem como com a eliminação do parasito. O macrófago é a principal célula
hospedeira deste parasito e pouco é conhecido sobre a infecção de macrófagos humanos pelas
espécies L. (V.) braziliensis e L. (L.) amazonensis. Estabelecer modelos de infecção de

32
macrófagos humanos com isolados oriundos de pacientes com diferentes formas clínicas da
doença pode revelar características dos parasitos importantes para o sucesso ou não da
infecção de macrófagos. Neste sentido, avaliar a metaciclogênese de isolados clínicos de L.
(V.) braziliensis e L. (L.) amazonensis, utilizando diferentes metodologias, baseadas na
morfologia e nas alterações das moléculas de superfície dos parasitos, em um mesmo estudo,
seguido da avaliação da capacidade dos parasitos em infectar macrófagos humanos, deve
definir as condições necessárias para a obtenção de parasitos que estabeleçam uma infecção
com sucesso. Posteriormente, a detecção de diferenças na metaciclogênese de diferentes
isolados clínicos pode levar à detecção de moléculas responsáveis pela melhor adaptação das
formas metacíclicas para sobreviver dentro dos macrófagos, transformando-se em formas
amastigotas eficientes para estabelecer uma infecção. Tais moléculas podem ser alvos para o
desenvolvimento de futuras terapias ou imunização.

33
3 OBJETIVOS
3.1 Objetivo Geral
Avaliar a metaciclogênese in vitro de isolados L. (V.) braziliensis e L. (L.)
amazonensis, provenientes de pacientes com diferentes formas clínicas de Leishmaniose
Tegumentar Americana, usando diferentes metodologias, e a capacidade destes isolados em
infectar macrófagos humanos.
3.2 Objetivos Específicos
1. Comparar os perfis das curvas de crescimento in vitro de diferentes isolados de L. (V.)
braziliensis e L. (L.) amazonensis, definindo as fases logarítmica e estacionária do
crescimento.
2. Identificar as mudanças morfológicas que ocorrem nos parasitos durante a
metaciclogênese in vitro, por meio da morfometria dos isolados.
3. Avaliar o curso temporal da metaciclogênese in vitro e comparar as porcentagens de
formas metacíclicas, obtidas para cada isolado nos diferentes dias de cultura, pelos métodos
morfometria, seleção negativa com lectina de Bauhinia purpurea (L. (V.) braziliensis) ou o
anticorpo 3A1-La (L. (L.) amazonensis), citometria de fluxo e ensaio de resistência ao
Complemento.
4. Avaliar os índices de infecção de macrófagos humanos com preparações de parasitos
totais do 6o dia de cultivo ou formas metacíclicas obtidas pela técnica de seleção negativa com
lectina de Bauhinia purpurea (L. (V.) braziliensis) ou o anticorpo 3A1-La (L. (L.)
amazonensis).
5. Avaliar a infecção de macrófagos humanos, não ativados ou ativados, pelas formas
promastigotas dos diferentes isolados, em fases logarítmica (2 dias) ou estacionária (6 e 10
dias) do crescimento.

34
4 MÉTODOS
4.1 Cultivo e manutenção dos parasitos
Foram estudadas as cepas de referência da Organização Mundial da Saúde
(OMS), L. (V.) braziliensis (MHOM/BR/1975/M2903), referida no trabalho como leishmânia
M2903 e L. (L.) amazonensis (IFLA/BR/67/PH8), referida no trabalho como leishmânia PH8
(gentilmente doadas pelo Dr. Luis Carlos Crocco Afonso, da Universidade Federal de Ouro
Preto, MG). Essas leishmanias têm sido extensivamente estudadas, servindo de controle para
as nossas condições experimentais.
Neste trabalho, também foram objetos de estudo, dois isolados de L. (V.)
braziliensis: MHOM/BR/2003/IMG, referida como IMG3, isolado de um paciente com
leishmaniose cutânea e o isolado MHOM/BR/2006/PPS, referido no trabalho como PPS6m,
isolado de paciente com leishmaniose mucosa. Um isolado de L. (L.) amazonensis,
MHOM/BR/2006/MAB, referido como MAB6, obtido de paciente com leishmaniose cutânea
difusa, também foi avaliado. Todos os isolados encontram-se estocados, congelados em
nitrogênio líquido na forma promastigota procíclica, no Leishbank (Banco de leishmânias do
Laboratório de Imunobiologia das Leishmanioses,no Instituto de Patologia Tropical e Saúde
Pública, Universidade Federal de Goiás). Os dados clínicos-epidemiológicos dos pacientes
encontram-se na Tabela 1.
Os parasitos foram mantidos em meio Grace completo (Grace’s Insect Medium,
Sigma Chemical Co., St Louis, MD, EUA), pH 5,8, suplementado com 20% de Soro Fetal
Bovino (SFB, Cripion, Andradina, SP, Brasil) inativado, 2 mM de L-glutamina (Sigma), 100
U/mL de penicilina (Sigma) e 100 μg/mL de estreptomicina (Sigma), à temperatura de 26 ºC.
As leishmânias foram repicadas de três em três dias, sempre iniciando a cultura com 5 x 105
promastigotas/mL, sendo que os parasitos foram utilizados para os experimentos entre a 3ª e a
6ª passagem. A quantificação dos parasitos foi realizada após diluição de uma pequena
alíquota da cultura em solução fixadora (salina tamponada com fosfato – PBS, contendo 0,2%
de paraformaldeído), realizando a contagem em hematocitômetro, sob microscopia de luz

35
(400x). Para a estocagem, formas promastigotas procíclicas dos parasitos foram preservadas
em nitrogênio líquido, em SFB contendo 10% de dimetilsulfóxido (DMSO, Sigma).
Tabela 1. Descrição clínica epidemiológica dos isolados
ISOLADO
ANO/LOCALa
ESPÉCIEb
FORMA
CLÍNICAc
DESCRIÇÃO DAS LESÕES
M2903 L.b - -
IMG3/GO L.b LCL 2 lesões ulceradas (3 meses)
PPS6/BA L.b LM Lesões ulceradas na região nasal, palato e úvula.
PH8 L.a - -
MAB6/TO L.a LCDf Varias lesões disseminadas ulceradas, verrucosas
e nodulares.
aÉ apresentado o nome do isolado (iniciais do nome do paciente), seguido do ano do isolamento (3:
2003; 6: 2006), o provável local de infecção (GO: Goiás; BA: Bahia; TO: Tocantins). bL.b Isolados de L. (V.) braziliensis, L.a Isolados de L. (L.) amazonensis.
cLCL= Leishmaniose Cutânea Localizada; LM = Leishmaniose Mucosa; LCDf = Leishmaniose
Cutânea Difusa.
4.2 Curvas de crescimento in vitro
Os parasitos foram repicados em triplicatas (5 x 105
parasitos/mL) em placa de 24
poços (Costar, Cambridge, MA, EUA), contendo meio Grace completo e mantidos à
temperatura de 26ºC. Os parasitos foram quantificados, diariamente, durante 12 dias, por
meio de contagem em hematocitômetro, como descrito acima. Os resultados foram expressos
como número de parasitos/mL.
4.3 Análise da metaciclogênese
Para cada isolado, os ensaios para avaliar morfometria, separação com lectina ou
anticorpo e resistência ao complemento foram realizados simultaneamente.

36
4.3.1 Identificação das formas metacíclicas por análises morfométricas.
Após 2, 6 ou 10 dias de cultivo, os parasitos foram coletados e centrifugados
sobre lâminas (2000 rpm, 30 segundos, citocentrífuga), fixados em metanol, corados com kit
Instant Prov (Laborclin, Pinhais, PR, Brasil) e fotografados digitalmente, sob microscopia de
luz (aumento 1000x - 5 fotos/lâmina). As imagens digitalizadas foram analisadas com auxílio
do programa Image J (National Institute of Health), onde foram mensurados o comprimento e
a largura do corpo celular e o comprimento do flagelo de 300 parasitos/dia e determinadas as
porcentagens de formas metacíclicas presentes na cultura. Foram consideradas formas
metacíclicas os parasitos cuja relação do comprimento do flagelo/corpo celular era maior do
que dois.
4.3.2 Identificação e separação das formas promastigotas metacíclicas pelo método de
aglutinação com lectina ou anticorpo.
Para seleção negativa foi utilizada a técnica descrita por Saraiva et al. (2005). Os
parasitos foram coletados após 2, 6 ou 10 dias de cultivo, lavados três vezes em PBS (1.000 g,
4ºC, 15 min) e ressuspendidos para 108 parasitos/mL de PBS (300 L). Os isolados da espécie
L. (V.) braziliensis foram incubados com 100 g/mL de lectina de Bauhinia purpurea (BLP,
Vector Laboratórios, Burlingame, CA, EUA) enquanto os da espécie L. (L.) amazonensis,
com 100 g/mL de anticorpo 3A1-La (gentilmente doados pelo Dr. Milton Adriano Pelli de
Oliveira, Laboratório de Citocinas, UFG). Após 30 minutos de incubação à temperatura
ambiente (T. a), a suspensão foi centrifugada à 40 g, 5 min. As formas promastigotas
metacíclicas, não aglutinadas, foram coletadas cuidadosamente do sobrenadante, lavadas três
vezes com PBS, quantificadas e utilizadas para análises por citometria de fluxo ou infecção de
macrófagos. Neste método, a porcentagem e o número absoluto de formas metacíclicas
presentes na cultura, nos dias avaliados, foram obtidos quantificando-se os parasitos antes e
após a separação das formas metacíclicas.
4.3.3 Identificação das formas metacíclicas por citometria de fluxo
Parasitos totais e parasitos enriquecidos em formas metacíclicas como descrito no
item 4.3.2, do 2º, 6º ou 10º dia de cultura (cultivados de maneira a ter no mesmo dia da
aquisição no citômetro, parasitos com 2, 6 e 10 dias de cultivo, a partir de uma mesma

37
cultura), fixados em solução de paraformaldeído 1% em PBS, foram adquiridos (FSC vs.
SSC, 20.000 eventos) em Citômetro de Fluxo (FACScan FlowCytometer; Becton Dickinson
Immunocytometry Systems, San Jose, CA, EUA). Os parasitos foram analisados em relação
ao tamanho (FSC) e complexidade celular (SSC) para determinar as formas promastigotas
procíclicas e metacíclicas de acordo com Saraiva et al (2005), usando o software Cyflogic
(Turcu, Finlândia). Os resultados são expressos em porcentagens de formas metacíclicas
encontradas nas culturas.
4.3.4 Identificação das formas metacíclicas por ensaio de resistência ao Complemento
Para o ensaio de resistência ao Complemento foi utilizada a técnica descrita por
Louassini et al. (1998). Soro de coelho liofilizado (CEDARLANE, Burlington, Canadá), ricos
em proteínas do sistema Complemento, foi ressuspendido em HBSS (do inglês, Hank’s
Balanced Salt Solution, Sigma, CO, EUA) e mantido no gelo até o uso. Parasitos do 2º, 6º ou
10º dia de cultura foram coletados, lavados três vezes em PBS (1.000 g, 4ºC, 15 min),
ressuspendidos em HBSS e incubados (5 x 105 parasitos/200 L/tubo) na ausência ou
presença de diferentes diluições do soro (1%, 5%, 10%, 20%, 40% e 80%), à 37ºC. Após 1 h,
os tubos foram colocados no gelo e o número de parasitos vivos (com estrutura morfológica
mantida) foi determinado por microscopia de luz, usando hematocitômetro. Os resultados
foram expressos como porcentagem de parasitos sobreviventes comparados com os controles
que não foram expostos ao soro.
4.4 Avaliação da infecção de macrófagos humanos com isolados de Leishmania sp.
4.4.1 Separação de monócitos do sangue periférico e derivação para macrófagos
Do sangue periférico (12 mL) de indivíduos doadores sadios (n = 18), do Banco
de Sangue do Hospital das Clínicas da UFG ou do Instituto Goiano de Hematologia e
Oncologia, foram separados os monócitos. As amostras de sangue venoso foram colhidas em
tubos vacutainer (BD Vacutainer, Juiz de Fora, MG, Brasil), contendo solução de EDTA
como anticoagulante, bem como em tubos sem anticoagulante (5 mL), para retirar o soro. O
sangue com EDTA foi aplicado sobre gradiente de densidade Ficoll (volume/volume) e
centrifugado a 1.000 g, por 20 min, T.a. O anel, contendo as células mononucleares (CMN)
foi colhido da interface e as células foram lavadas em solução de PBS-EDTA duas vezes: 1ª
lavagem: 600 g por 10 min, a 4°C; 2º lavagem: 200 g por 10 min, a 4°C. As CMN foram

38
ressuspendidas em 1 mL de meio RPMI 1640 (Sigma) sem SFB. As células foram
quantificadas, em hematocitômetro, após a diluição de uma pequena alíquota da suspensão
celular em solução de azul de tripano 0,1% em PBS. A concentração foi ajustada para 2,5 x
106 células/mL.
Em seguida, as CMN foram distribuídas em placas de 24 poços (Costar), sobre
lamínulas de vidro de 13 mm de diâmetro (Knitel, Alemanha), na concentração de 5 x 105
células/300 L. Primeiro fazendo uma gota de 200 L sobre a lamínula e deixando
sedimentar por 30 min, depois completando, com meio RPMI 1640 contendo 10% de SFB
(Gibco) e 5% de soro autólogo, para 300 L.
As CMN foram derivadas para macrófagos durante sete dias de cultivo, sendo que
o meio completo de cultura foi trocado a cada 48 h. No 6° dia de derivação, as CMN foram
incubadas na ausência ou na presença de 100 U/mL de rIFN humano (Sigma), por 24 h, a
36ºC, em atmosfera úmida, contendo 5% de CO2 e, após este período, os macrófagos foram
infectados com os parasitos.
4.4.2 Infecção dos macrófagos
A fim de avaliar se a porcentagem de formas metacíclicas presentes na cultura
influenciam no processo de infecção dos macrófagos, os parasitos totais, em diferentes dias de
cultivo (2, 6 ou 10 dias) ou formas metacíclicas do 6º dia de cultura, selecionadas como
descrito no item 4.4.2, foram lavados três vezes com PBS ou salina, a 10oC, 1.000 g, 15 min.
Em seguida, os parasitos foram ressuspendidos em meio RPMI 1640 contendo 5% de SFB e
quantificados como descrito no item 4.1. Os parasitos foram adicionados na proporção de um
parasito por célula (5 x 105 parasitos/poço) aos macrófagos cultivados na ausência ou
presença de IFN por 24 h, sendo os poços tratados com IFN , simultaneamente ativados com
50 ng/mL de lipopolissacarídeo (LPS, E. coli 0111:B4, Sigma). As placas foram incubadas a
36 ºC, em atmosfera úmida, contendo 5% de CO2 por 24 ou 72 h. Após este período, as
lamínulas foram colhidas, sendo as células fixadas em metanol absoluto e coradas com
corante panótico, utilizando o kit comercial Instant Prov (Newprov).

39
4.4.3 Avaliação do índice de infecção dos macrófagos
As lamínulas foram fixadas sobre lâminas para microscopia, usando Entellan
Novo (Merck KgaA, Damstadt, Germany) e as células foram analisadas sob microscopia de
luz (aumento de 1.000 vezes), sendo determinadas a frequência de células infectadas (300 a
500 células analisadas) e o número médio de parasitos por célula infectada. O índice de
infecção foi obtido multiplicando-se a porcentagem de células infectadas pelo número médio
de parasitos por células infectadas.
4.5 Considerações éticas
O presente trabalho foi aprovado pelo Comitê de Ética em Pesquisa, do Hospital
das Clínicas da Universidade Federal de Goiás, sob protocolo nº 192/2011 (Anexo 1). Os
doadores de sangue leram e assinaram o Termo de Consentimento Livre e Eclarecido (Anexo
2) antes de doarem o sangue.
4.6 Análises estatísticas
Para variáveis paramétricas, os dados foram apresentados como média ± EPM,
sendo os resultados analisados por teste t de Student ou ANOVA/Bonferoni. Para variáveis
não paramétricas, os dados foram apresentados como medianas e valores mínimos e máximos.
Os resultados foram avaliados pelos testes pareado de Wilcoxon (para amostras pareadas),
Mann Whitney (para amostras não pareadas) ou por ANOVA Kruskal-Wallis, seguida de
teste de Dunn. O nível de significância foi estabelecido em p < 0,05. As análises foram
realizadas utilizando o GraphPad Prism 4.0 Software (San Diego, CA, EUA).

40
5 RESULTADOS
5.1 Análise da metaciclogênese das formas promastigotas de Leishmania spp in vitro por
morfometria
A primeira etapa para determinar o perfil de metaciclogênese dos isolados foi
estabelecer a curva de crescimento in vitro dos parasitos e acompanhar as alterações
morfológicas que ocorrem na cultura para identificação das formas metacíclicas. Os parasitos
da espécie L. (V.) braziliensis M2903, IMG3 e PPS6m, bem como os da espécie L. (L.)
amazonensis PH8 e MAB6 foram cultivados a 26ºC durante 12 dias e quantificados
diariamente. A Figura 5A mostra que foi detectada proliferação dos parasitos entre o 1º e 4º
dia de cultura, sendo esta fase denominada de fase logarítmica do crescimento. A partir do 5º
dia de cultura, a proliferação dos parasitos diminuiu, iniciando a fase estacionária do
crescimento. O número máximo de parasitos nas culturas variou de ~4 a 7 x 107 parasitos/mL.
Não houve diferença significante entre os perfis das curvas de crescimento ou entre as
quantidades de parasitos obtidos nas culturas.
Na Figura 5B são mostradas as diferentes formas dos parasitos, analisados sob
microscopia de luz, durante a fase logarítmica (2º dia) e a estacionária (6o e 10º dia). Como
pode ser observado, o aspecto morfológico das formas promastigotas de todos os isolados
variou conforme o tempo de cultura, indicando que durante o surgimento das formas
metacíclicas, que aparecem na fase estacionária (6º ou 10º dia), os parasitos sofrem uma
diminuição significante no tamanho do comprimento do corpo celular e um aumento
progressivo no tamanho do flagelo, em relação às formas promastigotas procíclicas, presentes
no 2º dia de cultura, mas não há diferenças significantes nestas medidas entre o 6º e o 10º dia
de cultura (Fig. 6A).
As análises da morfometria mostraram que existem diferenças significantes entre
o tamanho das formas promastigotas em fase estacionária do crescimento dos isolados de
diferentes espécies, sendo que os isolados da espécie L. (L.) amazonensis apresentaram maior
comprimento do corpo celular (Fig. 6B e 6D) e do tamanho do flagelo (Fig. 6C e 6E, parasitos
do 6º e 10º dia, respectivamente) do que os parasitos pertencentes à espécie L. (V.)

41
braziliensis. Não foram detectadas diferenças significantes entre os resultados da morfometria
dos isolados de uma mesma espécie.
Por meio da morfometria dos parasitos, foi possível determinar a porcentagem de
formas metacíclicas presentes nas culturas dos isolados, nos três dias avaliados (2, 6 ou 10
dias). Foram consideradas formas metacíclicas aquelas que apresentavam o comprimento do
flagelo duas vezes maior do que o comprimento do corpo celular. Houve um aumento
significante na quantidade de formas metacíclicas detectadas na fase estacionária do
crescimento (6º ou 10º dia) em relação aos parasitos do 2º dia de cultura, para todos os
isolados avaliados (p < 0,05, Fig. 7A a 7E). Os parasitos L. (L.) amazonensis (PH8 e MAB6)
apresentaram menores porcentagens de formas metacíclicas no 6º e 10º dia em relação aos
demais isolados L. (V.) braziliensis (p < 0,05, Fig. 8B e 8C, respectivamente).

42
Figura 5. Curva de crescimento in vitro e morfologia dos isolados. Os parasitos, em fase
logarítmica do crescimento, foram cultivados na concentração inicial de 5 x 105 parasitos/mL em meio
Grace, à 26°C: (A) Os parasitos foram quantificados diariamente em hematocitômetro e cada ponto
representa a média ± EPM de 3 experimentos independentes, realizados em triplicatas. As linhas
pontilhadas verticais representam os dias em fase logarítmica, estacionária inicial e estacionária tardia,
escolhidos para análises do processo de metaciclogênese in vitro. (B) Após 2, 6 ou 10 dias de cultivo,
os parasitos foram fixados sobre lâmina, corados e analisados sob microscopia de luz (aumento
1.000x). A barra de escala representa 10 m. M2903, IMG3, PPS6m (L. (V.) braziliensis); PH8,
MAB6 (L. (L.) amazonensis)

43
Figura 6. Morfometria dos isolados. Após 2, 6 ou 10 dias de cultivo em meio Grace, os parasitos
foram fixados sobre lâmina, corados e analisados sob microscopia de luz (aumento 1.000x). As
medidas do comprimento, largura do corpo celular e tamanho do flagelo dos parasitos foram obtidas
com auxílio do programa Image J. Em (A) são mostradas as análises morfométricas (médias ± EPM)
dos isolados (*p < 0,05, 2º dia vs 6º dia ou 2º vs 10º dia, n = 300 parasitos avaliados/dia). Em (B) e (D)
são mostradas as medidas do comprimento do corpo celular e em (C) e (E) as medidas do
comprimento do flagelo (parasitos do 6º e 10º dia, respectivamente) dos diferentes isolados agrupados
por espécie (M2903, IMG3 e PPS6m – L. (V.) braziliensis e PH8 e MAB6 – L. (L.) amazonensis). Os
dados representam a média ± EPM (3 experimentos realizados em duplicatas para cada isolado/cepa).
*p < 0,05.
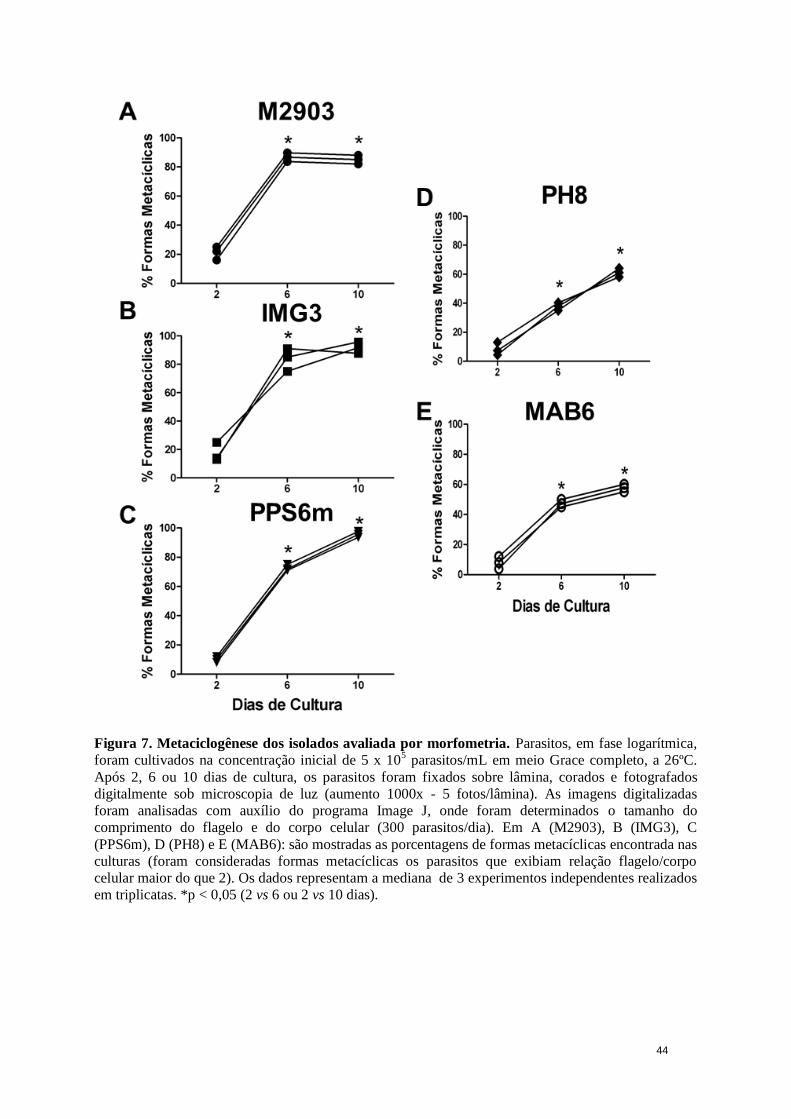
44
Figura 7. Metaciclogênese dos isolados avaliada por morfometria. Parasitos, em fase logarítmica,
foram cultivados na concentração inicial de 5 x 105 parasitos/mL em meio Grace completo, a 26ºC.
Após 2, 6 ou 10 dias de cultura, os parasitos foram fixados sobre lâmina, corados e fotografados
digitalmente sob microscopia de luz (aumento 1000x - 5 fotos/lâmina). As imagens digitalizadas
foram analisadas com auxílio do programa Image J, onde foram determinados o tamanho do
comprimento do flagelo e do corpo celular (300 parasitos/dia). Em A (M2903), B (IMG3), C
(PPS6m), D (PH8) e E (MAB6): são mostradas as porcentagens de formas metacíclicas encontrada nas
culturas (foram consideradas formas metacíclicas os parasitos que exibiam relação flagelo/corpo
celular maior do que 2). Os dados representam a mediana de 3 experimentos independentes realizados
em triplicatas. *p < 0,05 (2 vs 6 ou 2 vs 10 dias).

45
Figura 8. Porcentagens de formas metacíclicas dos diferentes isolados identificadas por
morfometria. Os parasitos, em fase logarítmica, foram cultivados na concentração inicial de 5 x 105
parasitos/mL em meio Grace completo, a 26ºC. Após 2 (A), 6 (B) ou 10 (C) dias de cultura, a
porcentagem de formas metacíclicas foi determinada por morfometria. Os dados representam a
mediana e os valores mínimos e máximos. *p < 0,05 (PH8 e MAB6 vs M2903, PPS6m e IMG), n = 3.

46
5.2 Análise da metaciclogênese in vitro dos isolados por características de aglutinação
com lectina ou anticorpos
Ao sofrer metaciclogênese, os parasitos mostram, além das alterações
morfológicas, modificações bioquímicas que permitem sua identificação e purificação
(Nieves & Pimenta 2000, Pinto-da-Silva et al. 2002). Dentre essas mudanças, aquelas que
ocorrem em carboidratos presentes na superfície do parasito foram utilizadas no presente
estudo para determinar o perfil de metaciclogênese in vitro dos isolados. Nesta técnica, foi
utilizada a BPL (para isolados da espécie L. (V.) braziliensis) ou o anticorpo 3A1-La (para
isolados da espécie L. (L.) amazonensis), os quais aglutinam as formas procíclicas da cultura,
pois se ligam a carboidratos presentes somente na superfície destes parasitos, deixando as
formas metacíclicas livres para serem purificadas e quantificadas. Assim, parasitos BPL- ou
3A1-La- foram consideradas como as formas metacíclicas da cultura.
Na Figura 9 são visualizadas fotomicrografias do isolado PPS6m (Fig. 9A e 9B) e
da cepa PH8 (Fig. 9C e 9D) do 6º dia de cultura, antes (Fig. 9A e 9C) e após (Fig. 9B e 9D) o
processo da seleção de formas metacíclicas, utilizando BPL ou anticorpo 3A1-La. Notam-se
que os parasitos foram enriquecidos em formas que possuem as características morfológicas
das formas metacíclicas (Fig. 9B e 9D), como flagelo alongado e diminuição do tamanho do
corpo celular, em relação aos parasitos antes da separação (Fig. 9A e 9C). Também é
mostrada, na Figura 9E (aumento de 200x) e na Figura 9F (aumento de 1.500x), a aglutinação
das formas procíclicas do 6º dia de cultura (BPL+) do isolado PPS6m.

47
Figura 9. Fotomicrografias dos parasitos PPS6m e PH8, antes e após purificação de formas
metacíclicas com lectina de Bauhnia purpurea (BPL) e anticorpo 3A1-La. Os parasitos, em fase
logarítmica do crescimento, foram cultivados na concentração inicial de 5 x 105 parasitos/mL em meio
Grace, à 26°C. No 6º dia de cultura, as formas metacíclicas foram purificadas pelo método de seleção
negativa com BPL (L. (V.) braziliensis, PPS6m) ou anticorpo 3A1-La (L. (L.) amazonensis, PH8) e
fixados sobre lâmina, corados e analisados sob microscopia de luz (aumento 1.000x). Em A e C são
mostrados os parasitos antes da purificação e em B e D, após a seleção (B: BPL-; D: 3A1-La
-). Em E
(aumento 200x) e F (aumento 1.500x), sendo mostrados os parasitos do isolado PPS6m aglutinados
com BPL (BPL+).

48
Padronizado o processo de seleção das forma metacíclicas, foram determinadas as
porcentagens de formas metacíclicas presentes nas culturas dos isolados, nos três dias
avaliados (2, 6 ou 10 dias). Houve um aumento significante na quantidade de formas
metacíclicas detectadas na fase estacionária do crescimento (6º ou 10º dia) em relação aos
parasitos do 2º dia de cultura, para todos os isolados avaliados (p < 0,05, Fig. 10A a 10E).
Não foram atestadas diferenças significantes entre as porcentagens de formas metacíclicas
encontradas nas culturas do 6º e 10º dia (Fig. 10A a 10E). Entre os isolados, M2903 e PPS6m
apresentaram maiores porcentagens de formas metacíclicas no 2º dia em relação aos outros
isolados neste mesmo período (p < 0,05, Fig. 11A). No 6º dia de cultura, o isolado MAB6
apresentou maiores porcentagens de formas metacíclicas do que os isolados IMG3 e PPS6m
(Fig. 11B) e no 10º dia de cultura, as culturas de todos os isolados apresentaram porcentagens
similares de formas metacíclicas (Fig. 11C).

49
Figura 10. Metaciclogênese dos isolados avaliada por seleção negativa com lectina ou anticorpo.
Parasitos em fase logarítmica foram cultivados na concentração inicial de 5 x 105parasitos/mL em
meio Grace completo, a 26ºC. Em A (M2903), B (IMG3), C (PPS6m), D (PH8) e E (MAB6) são
mostradas as porcentagens de formas metacíclicas encontradas nas culturas após seleção utilizando a
lectina de Bauhinia purpurea (para isolados L.(V.) braziliensis A, B, C) ou anticorpo 3A1-La (para
isolados de L. (L.) amazonensis D, E) no 2º, 6º ou 10º dia de cultura. As linhas representam os
resultados de um experimento realizado em duplicata (n = 3 experimentos independentes). *p < 0,05, 2
vs 6 ou 2 vs 10 dias.

50
Figura 11. Comparação entre as porcentagens de formas metacíclicas dos isolados, detectadas
após seleção com lectina de Bauhinia purpurea (L. (V.) braziliensis) ou anticorpo 3A1-La (L. (L.)
amazonensis). Parasitos em fase logarítmica do crescimento foram cultivados na concentração inicial
de 5 x 105parasitos/mL em meio Grace completo, a 26ºC. Após 2 (A), 6 (B) ou 10 (C) dias de cultura,
as formas metacíclicas foram selecionadas e quantificadas. Os dados representam a mediana e os
valores mínimo e máximo. *p < 0,05, M2903 e PPS6m vs os demais isolados (A) e MAB6 vs IMG3 e
PPS6m (B), n = 3 experimentos realizados em duplicatas.

51
5.3 Análise da metaciclogênese dos isolados in vitro por citometria de fluxo
Foi demonstrado que, baseado no tamanho dos parasitos, é possível identificar
formas promastigotas procíclicas e metacíclicas usando a citometria de fluxo (Saraiva et al.
2005). Por esta técnica, as formas promastigotas metacíclicas são as que exibem tamanho
celular menor (FSClow
) do que o das formas promastigotas procíclicas (FSChigh
).
Os parasitos foram colhidos em diferentes dias de cultura (2, 6 ou 10) e analisados
por citometria de fluxo, antes e após a seleção das formas metacíclicas da cultura, utilizando
lectina BPL ou o anticorpo 3A1-La. As análises foram realizadas de acordo com Saraiva et al.
(2005). As Figuras 12 e 13 mostram análises de citometria de fluxo de um experimento
representativo do isolado IMG3 (L. (V). braziliensis) e um do isolado MAB6 (L. (L.)
amazonensis), respectivamente. As análises foram realizadas após 2 (A e D), 6 (B e E) ou 10
(C e F) dias de cultivo. Em A, B e C são mostradas as análises dos parasitos totais e em D, E e
F são mostradas as análises após a purificação das formas metacíclicas com a lectina ou o
anticorpo. Podem ser visualizadas duas diferentes populações de parasitos bem definidas, uma
de tamanho menor (FSClow
, janela à esquerda) e uma de tamanho maior (FSChigh
, janela à
direita). As formas de tamanho pequeno (FSClow
) são as metacíclicas, cuja porcentagem
aumenta de acordo com o tempo de cultivo (2, 6 e 10 dias: 25%, 34% e 68%, Fig. 12A, 12B,
12C, IMG3; 30%, 66% e 92%, Fig. 13A, 13B, 13C, MAB6, parasitos totais).
A aglutinação das formas procíclicas das culturas, utilizando BPL ou o anticorpo
3A1-La mostrou que os parasitos de maior tamanho foram eliminados, enriquecendo as
suspensões em formas metacíclicas (2, 6 e 10 dias: 96%, 86% e 95%, Fig. 12D, 12E, 12F,
IMG3; 92%, 90%, 96%, Fig. 13D, 13E, 13F, MAB6). Estes dados também confirmaram os
achados visualizados nas fotomicrografias (Fig. 9B e 9D), nas quais os parasitos, após
separação com BPL ou anticorpo, apresentam corpos celulares menores do que os dos
parasitos totais (antes da purificação). Em todas as análises foi observada uma população de
tamanho intermediário, que não foi eliminada totalmente durante a seleção das metacíclicas.
Nas análises dos perfis de metaciclogênese dos diferentes isolados, a citometria de
fluxo revelou que as maiores porcentagens de formas metacíclicas nas culturas foram obtidas
no 6º ou 10º dia de cultivo para todos os isolados quando comparadas às porcentagens de
formas metacíclicas encontradas no 2º dia (p < 0,05, Fig. 14). A análise por citometria
mostrou que independente do dia de cultivo, para todos os isolados, a técnica de seleção com

52
BPL/Anticorpo seleciona ~75-97% de formas metacíclicas das culturas (Fig. 14). Não houve
diferenças significantes entre as porcentagens de formas metacíclicas detectadas pela
citometria de fluxo, entre os isolados, nos três dias avaliados (Fig. 15).

53
Figura 12. Identificação de formas metacíclicas de L. (V.) braziliensis IMG3 por citometria de
fluxo. Parasitos totais (A, B e C) e formas metacíclicas selecionadas, utilizando a lectina BPL (D, E e
F), de diferentes dias de cultura (2: A e D; 6: B e E; 10: C e F) foram analisados por citometria de
fluxo. Análise do tamanho (Eixo x: FSC) e granulosidade (Eixo y: SSC) das subpopulações de
parasitos. FCSlow
, consideradas formas metacíclicas (janela à esquerda) e FCShigh
, consideradas formas
procíclicas (janela à direita).

54
Figura 13. Identificação de formas metacíclicas de L. (L.) amazonensis MAB6 por citometria de
fluxo. Parasitos totais (A, B e C) e formas metacíclicas selecionadas, utilizando o anticorpo 3A1-La
(D, E e F), de diferentes dias de cultura (2: A e D; 6: B e E; 10: C e F) foram analisados por citometria
de fluxo. Análise do tamanho (Eixo x: FSC) e granulosidade (Eixo y: SSC) das subpopulações de
parasitos. FCSlow
, consideradas formas metacíclicas (janela à esquerda) e FCShigh
, consideradas formas
procíclicas (janela à direita).
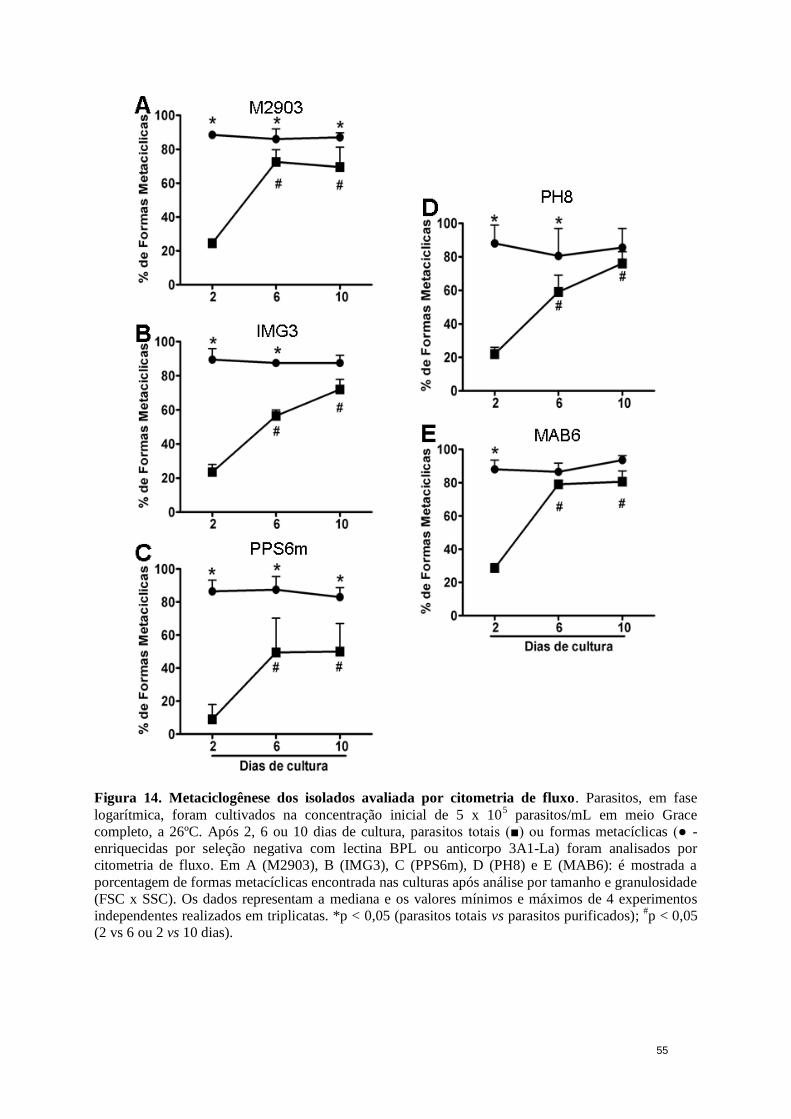
55
Figura 14. Metaciclogênese dos isolados avaliada por citometria de fluxo. Parasitos, em fase
logarítmica, foram cultivados na concentração inicial de 5 x 105 parasitos/mL em meio Grace
completo, a 26ºC. Após 2, 6 ou 10 dias de cultura, parasitos totais (■) ou formas metacíclicas (● -
enriquecidas por seleção negativa com lectina BPL ou anticorpo 3A1-La) foram analisados por
citometria de fluxo. Em A (M2903), B (IMG3), C (PPS6m), D (PH8) e E (MAB6): é mostrada a
porcentagem de formas metacíclicas encontrada nas culturas após análise por tamanho e granulosidade
(FSC x SSC). Os dados representam a mediana e os valores mínimos e máximos de 4 experimentos
independentes realizados em triplicatas. *p < 0,05 (parasitos totais vs parasitos purificados); #p < 0,05
(2 vs 6 ou 2 vs 10 dias).

56
Figura 15. Porcentagens de formas metacíclicas dos diferentes isolados identificadas por
citometria de fluxo. Os parasitos, em fase logarítmica, foram cultivados na concentração inicial de 5
x 105
parasitos/mL em meio Grace completo, a 26ºC. Após 2 (A), 6 (B) ou 10 (C) dias de cultura, os
parasitos foram analisados por tamanho e granulosidade por citometria de fluxo. Os dados representam
a mediana e valores mínimos e máximos das porcentagens de formas metacíclicas nas culturas. n = 4
(experimentos independentes).

57
5.4 Análise da metaciclogênese in vitro por teste de resistência ao Complemento.
Durante a passagem da fase logarítmica para a fase estacionária do crescimento, o
surgimento das formas metacíclicas é acompanhado por um aumento na capacidade de
resistência à lise mediada pelo sistema do Complemento (Sacks, 1984). Na Figura 16 são
mostradas as curvas de sobrevivência dos parasitos de diferentes dias de cultura (2, 6 ou 10
dias) da cepa M2903 (A) e do isolado IMG3 (B), após incubação com diferentes
concentrações de soro de coelho rico em proteínas do sistema Complemento. As formas
promastigotas da fase logarítmica (2º dia) foram as mais sensíveis à lise mediada pelo
Complemento (M2903 e IMG3 50% lise, 2º dia: 0,6% e 0,7 % de soro, Fig. 16A e 16B). Os
parasitos da fase estacionária inicial (6º dia) e tardia (10º dia) apresentaram similar resistência
ao complemento (M2903 50% lise, 6º e 10º dia: 19,7% e 20% de soro; IMG3 50% lise, 6º e
10º dia: 10% e 19% de soro).

58
Figura 16. Detecção de formas metacíclicas pelo ensaio de lise mediada pelo Complemento. Os
parasitos (A: M2903 e B: IMG3) de diferentes dias de cultura (2, 6 ou 10) foram incubados com
diferentes concentrações de soro de coelho e a viabilidade das formas promastigotas foi avaliada após
1 hora de incubação à 37ºC. As formas promastigotas incubadas na ausência de soro foram usadas
como controle. Os dados representam a média ± EPM de três experimentos independentes realizados
em triplicata.

59
A fim de avaliar as porcentagens de formas metacíclicas presentes nas culturas,
por meio da capacidade de resistência ao Complemento, os isolados de diferentes fases de
crescimento (2, 6 ou 10 dias) foram incubados com 10% de soro e após incubação, foram
determinadas as frequências de parasitos sobreviventes (de acordo com Sacks & Melby
2001). Como já demonstrado na figura anterior (Fig. 16), os parasitos da fase estacionária
foram mais resistentes à lise mediada pelo Complemento, de modo que as culturas de todos os
isolados apresentaram maiores porcentagens de formas metacíclicas no 6º ou 10º dia de
cultivo (p < 0,05, Fig. 17). Quando comparamos as porcentagens de formas metacíclicas
resistentes à lise pelo Complemento, entre os isolados, observamos que no 2º dia de cultura,
as culturas do isolado PPS6m apresentaram menores porcentagens de formas metacíclicas do
que as dos outros isolados (p < 0,05, Fig. 18A). No 6º dia de cultura, as culturas das cepas de
referência M2903 e PH8 apresentaram significantemente maiores porcentagens de formas
metacíclicas do que as do isolado PPS6m (Fig. 18B). Já no 10º dia, as culturas de todos os
isolados apresentaram porcentagens similares de formas metacíclicas (Fig. 18C).

60
Figura 17. Metaciclogênese dos isolados avaliada pelo ensaio do Complemento. Parasitos de
diferentes dias de cultura (2, 6 ou 10) foram incubados com 10% de soro de coelho e a viabilidade dos
parasitos foi avaliada após 1 hora de incubação à 37ºC. Em A (M2903), B (IMG3), C (PPS6m), D
(PH8) e E (MAB6) são mostradas as porcentagens de formas metacíclicas encontradas nas culturas
após o ensaio de resistência ao Complemento. As linhas representam os resultados de um experimento
realizado em triplicata (n = 3 experimentos independentes). *p < 0,05 (2 vs 6 ou 2 vs 10 dias).

61
Figura 18. Porcentagens de formas metacíclicas de diferentes isolados avaliados por resistência
ao Complemento. Os parasitos em fase logarítmica foram cultivados na concentração inicial de 5 x
105
parasitos/mL em meio Grace completo, a 26ºC. Após 2 (A), 6 (B) ou 10 (C) dias de cultura, os
parasitos foram incubados com 10% de soro de coelho e a viabilidade dos parasitos foi avaliada após 1
hora de incubação à 37ºC. Os dados representam a mediana e os valores mínimos e máximos (n = 3).
*p < 0,05, (PPS6m vs todos os isolados, A); #p < 0,05, (M2903 e PH8 vs PPS6m, B).

62
A Figura 19 mostra um resumo comparativo das porcentagens de formas
metacíclicas encontradas nas culturas dos isolados no 2º, 6º e 10º dia de cultura, avaliados
pelas diferentes técnicas (morfometria, citometria de fluxo, seleção por BPL/anticorpo e
resistência ao Complemento). Um mesmo isolado, avaliado por diferentes técnicas, apresenta
perfis de metaciclogênese similares, mas a porcentagem de formas metacíclicas podem ser
diferentes. Para os isolados clínicos IMG3 e PPS6m maiores porcentagens de formas
metacíclicas foram detectadas por morfometria e seleção com BPL, enquanto a citometria e o
ensaio de resistência ao Complemento detectaram menores porcentagens de metacíclicas,
especialmente no 6o dia de cultura, quando houve a maior variação das porcentagens de
formas metacíclicas detectadas pelas diferentes técnicas (Fig. 19B, 19C, p < 0,05). Para os
isolados L. (L.) amazonensis, a análise morfométrica indicou uma menor porcentagem de
formas metacíclicas, seguida pelo ensaio de resistência ao Complemento, no caso do isolado
MAB6 (Fig. 19D, 19E, p < 0,05).

63
Figura 19. Comparação entre as porcentagens de formas metacíclicas detectadas pelas técnicas
de morfometria, citometria de fluxo, seleção por lectina/anticorpo e ensaio de resistência ao
Complemento. Os parasitos, em fase logarítmica, foram cultivados na concentração inicial de 5 x 105
parasitos/mL em meio Grace completo, a 26ºC. São mostradas as medianas das porcentagens de
formas metacíclicas detectadas pela morfometria ( parasitos que exibiam relação tamanho do
flagelo/corpo celular maior do que 2), citometria ( Populações de parasitos FCSlow
), seleção com
lectina/anticorpo ( formas metacíclicas selecionadas negativamente por lectina de Bauhinia
purpurea ou anticorpo 3A1-La) e ensaio de resistência ao Complemento ( parasitos viáveis após 1h
de incubação com 10% de soro de coelho); após 2, 6 ou 10 dias de cultura dos isolados M2903 (A),
IMG3 (B), PPS6m (C), PH8 (D) e MAB6 (E). Os ensaios de morfometria, seleção e Complemento
foram realizados simultaneamente com parasitos provenientes das mesmas culturas (n = 3); para
citometria foram utilizados parasitos provenientes de experimentos separados (n = 4). *p < 0,05
(Citometria vs todos os outros ensaios); #p < 0,05, (morfometria vs todos os outros ensaios); ςp < 0,05,
(seleção vs todos os outros ensaios); θp < 0,05, (Lise pelo Complemento vs todos os outros ensaios).

64
5.5 Avaliação da infecção de macrófagos humanos por parasitos de diferentes fases do
crescimento ou formas metacíclicas selecionadas pelo ensaio de lectina/anticorpo
As formas promastigotas metacíclicas são mais infecciosas e resistentes aos
mecanismos microbicidas dos macrófagos (Awasthi et al. 2004), portanto, os isolados, em
diferentes dias de cultivo (2, 6 ou 10 dias), foram utilizados para ensaios de infecção de
macrófagos humanos derivados de monócitos in vitro, a fim de determinar as condições ideais
para infecção de macrófagos com os diferentes isolados.
Como mostrado na Figura 20, o índice de infecção dos macrófagos, após 24 horas
de incubação, foi maior naquelas culturas infectadas com parasitos em fase estacionária (6º ou
10º dia de cultura) do que com parasitos em fase logarítmica (2º dia), devido a um aumento na
porcentagem de células infectadas (Fig. 21) e/ou no número de parasitos por célula (Fig. 22).
Analisando os perfis de infecção dos diferentes isolados, detectamos que para a cepa de
referência de L. (V.) braziliensis M2903 e o isolado IMG3, os maiores índices de infecção (6o
e 10o dia) foram devidos a um maior número de parasitos dentro das células (Fig. 22A e 22B,
respectivamente) e não à porcentagem de células infectadas (Fig. 21A e 21B,
respectivamente). Com os isolados PPS6m, PH8 e MAB6, o aumento significante do índice
de infecção foi devido tanto ao aumento na porcentagem de células infectadas (Fig. 21C, 21D
e 21E) quanto a um maior número de parasitos por célula (Fig. 22C, 22D e 22E). Em paralelo,
as formas metacíclicas foram identificadas por citometria de fluxo nas culturas utilizadas para
infecção, como descrito no item 5.3, sendo apresentadas as medianas das porcentagens de
formas metacíclicas junto a cada resultado do índice de infecção (Fig. 20), porcentagens de
células infectadas (Fig. 21) e número de parasitos por macrófagos (Fig. 22).
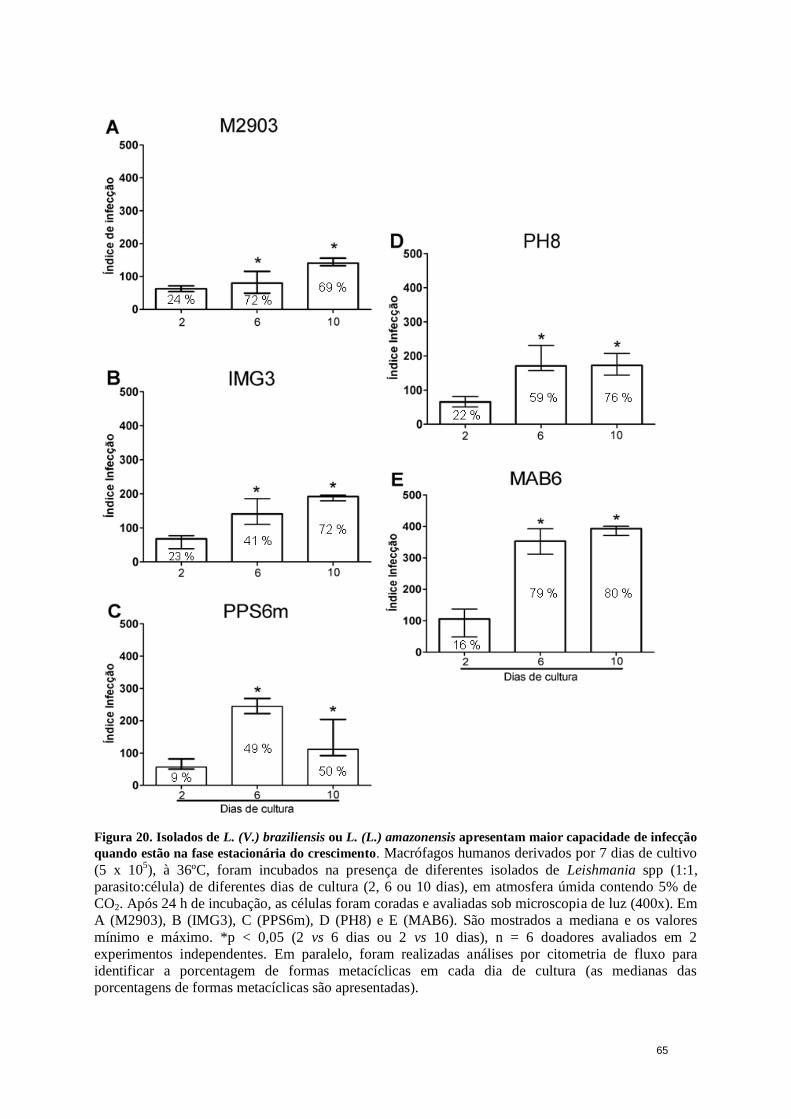
65
Figura 20. Isolados de L. (V.) braziliensis ou L. (L.) amazonensis apresentam maior capacidade de infecção
quando estão na fase estacionária do crescimento. Macrófagos humanos derivados por 7 dias de cultivo
(5 x 105), à 36ºC, foram incubados na presença de diferentes isolados de Leishmania spp (1:1,
parasito:célula) de diferentes dias de cultura (2, 6 ou 10 dias), em atmosfera úmida contendo 5% de
CO2. Após 24 h de incubação, as células foram coradas e avaliadas sob microscopia de luz (400x). Em
A (M2903), B (IMG3), C (PPS6m), D (PH8) e E (MAB6). São mostrados a mediana e os valores
mínimo e máximo. *p < 0,05 (2 vs 6 dias ou 2 vs 10 dias), n = 6 doadores avaliados em 2
experimentos independentes. Em paralelo, foram realizadas análises por citometria de fluxo para
identificar a porcentagem de formas metacíclicas em cada dia de cultura (as medianas das
porcentagens de formas metacíclicas são apresentadas).
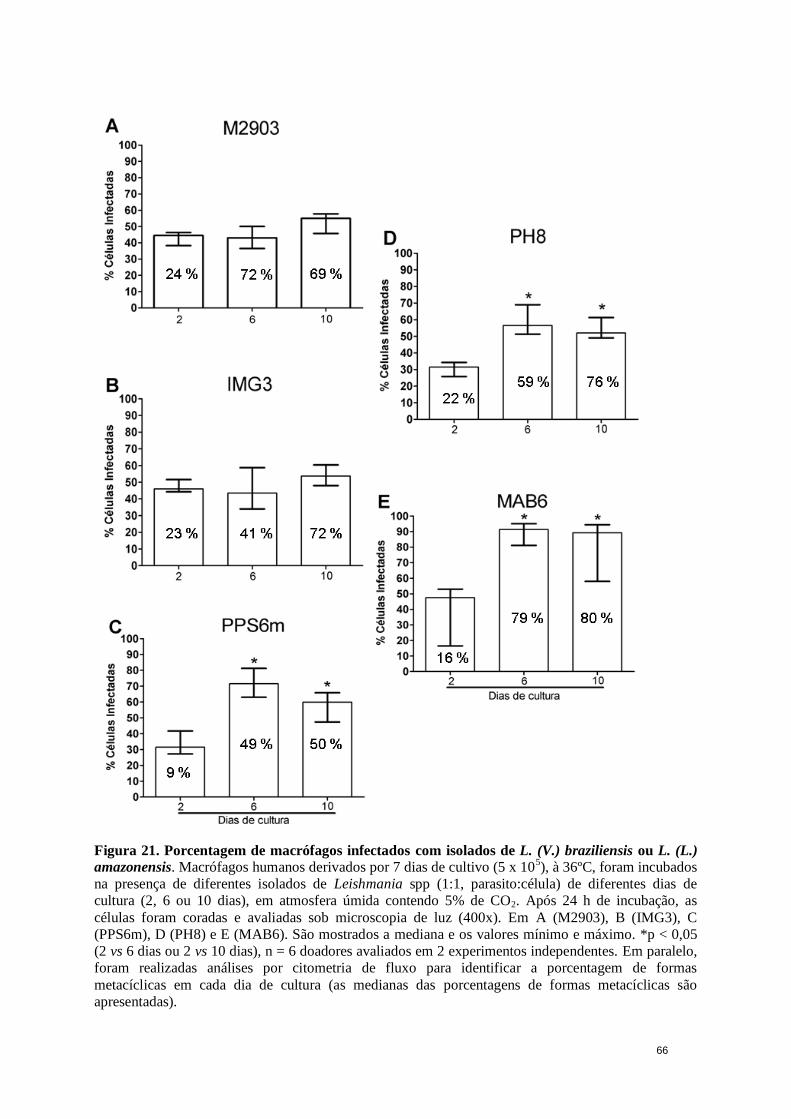
66
Figura 21. Porcentagem de macrófagos infectados com isolados de L. (V.) braziliensis ou L. (L.)
amazonensis. Macrófagos humanos derivados por 7 dias de cultivo (5 x 105), à 36ºC, foram incubados
na presença de diferentes isolados de Leishmania spp (1:1, parasito:célula) de diferentes dias de
cultura (2, 6 ou 10 dias), em atmosfera úmida contendo 5% de CO2. Após 24 h de incubação, as
células foram coradas e avaliadas sob microscopia de luz (400x). Em A (M2903), B (IMG3), C
(PPS6m), D (PH8) e E (MAB6). São mostrados a mediana e os valores mínimo e máximo. *p < 0,05
(2 vs 6 dias ou 2 vs 10 dias), n = 6 doadores avaliados em 2 experimentos independentes. Em paralelo,
foram realizadas análises por citometria de fluxo para identificar a porcentagem de formas
metacíclicas em cada dia de cultura (as medianas das porcentagens de formas metacíclicas são
apresentadas).

67
Figura 22. Número de parasitos por macrófagos infectados com isolados de L. (V.) braziliensis ou
L. (L.) amazonensis. Macrófagos humanos derivados por 7 dias de cultivo (5 x 105), à 36ºC, foram
incubados na presença de diferentes isolados de Leishmania spp (1:1, parasito:célula) de diferentes
dias de cultura (2, 6 ou 10 dias), em atmosfera úmida contendo 5% de CO2. Após 24 h de incubação,
as células foram coradas e avaliadas sob microscopia de luz (400x). Em A (M2903), B (IMG3), C
(PPS6m), D (PH8) e E (MAB6). São mostrados a mediana e os valores mínimo e máximo. *p < 0,05
(2 vs 6 dias ou 2 vs 10 dias), n = 6 doadores avaliados em 2 experimentos independentes. Em paralelo,
foram realizadas análises por citometria de fluxo para identificar a porcentagem de formas
metacíclicas em cada dia de cultura (as medianas das porcentagens de formas metacíclicas são
apresentadas).

68
Para avaliar se o perfil de infecção dos macrófagos com parasitos de diferentes
dias de cultivo seria alterado após um período maior de incubação, as culturas foram avaliadas
após 72 horas de incubação com os parasitos e os resultados foram comparados com aqueles
das culturas de 24 horas. O mesmo perfil de infecção dos macrófagos, observado inicialmente
nas primeiras 24 h de incubação, foi mantido após 72 h de infecção, pois para todos os
isolados, o índice de infecção dos macrófagos foi maior quando estes foram incubados com
parasitos em fase estacionária (6º ou 10º dia) do que com parasitos em fase logarítmica (2º
dia, p < 0,05, Fig. 23).
Avaliando M2903 e IMG3, foi detectada uma diminuição significante no índice
de infecção após 72 h de incubação, quando comparados com o índice obtido após 24 h de
infecção, especialmente quando os macrófagos foram infectados com parasitos do 2º ou 10º
dia de cultivo (Fig. 23A e 23B). Para o isolado PPS6m, apesar de haver uma tendência para
diminuição do índice de infecção de 24 h para 72 h (parasitos do 2º, 6º ou 10º dia), não foram
encontradas diferenças significantes entre os índices de infecção nestes períodos de incubação
(Fig. 23C). Como pode ser obervado nas Figuras 23D e 23E, para os isolados L. (L.)
amazonensis PH8 e MAB6, similares índices de infecção foram obtidos após 24 h ou 72 h de
incubação (parasitos do 2º, 6º ou 10º dia de cultura).
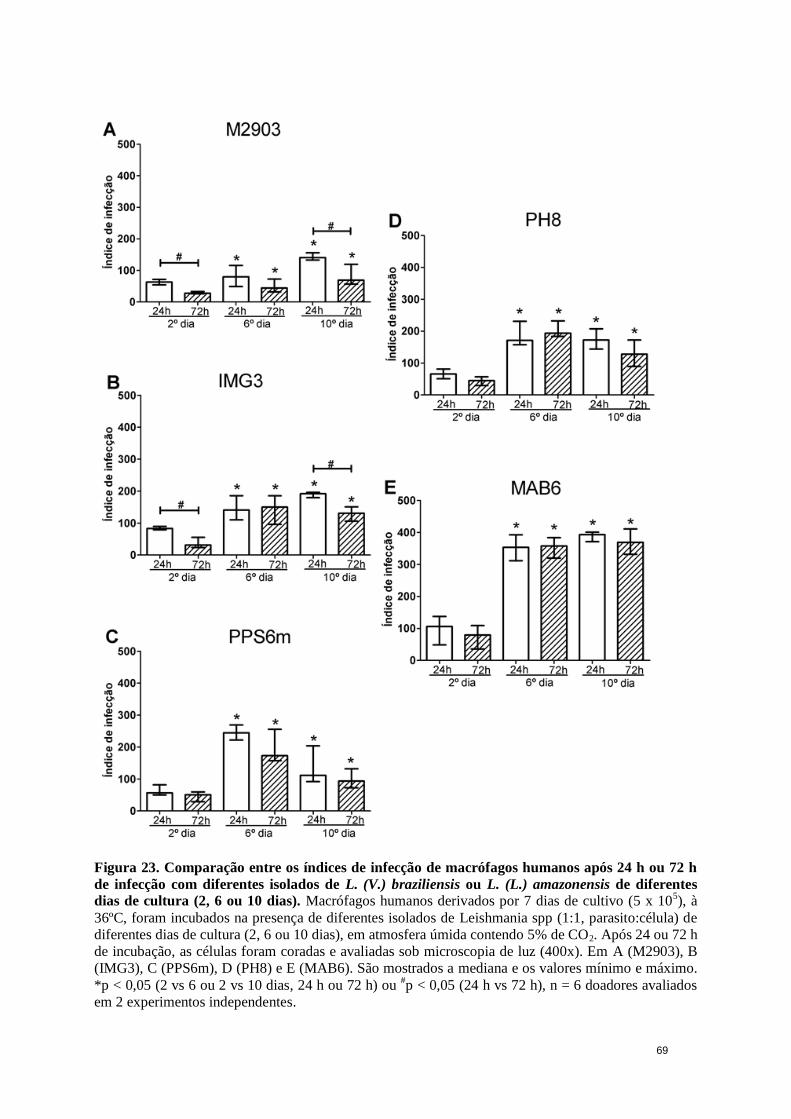
69
Figura 23. Comparação entre os índices de infecção de macrófagos humanos após 24 h ou 72 h
de infecção com diferentes isolados de L. (V.) braziliensis ou L. (L.) amazonensis de diferentes
dias de cultura (2, 6 ou 10 dias). Macrófagos humanos derivados por 7 dias de cultivo (5 x 105), à
36ºC, foram incubados na presença de diferentes isolados de Leishmania spp (1:1, parasito:célula) de
diferentes dias de cultura (2, 6 ou 10 dias), em atmosfera úmida contendo 5% de CO2. Após 24 ou 72 h
de incubação, as células foram coradas e avaliadas sob microscopia de luz (400x). Em A (M2903), B
(IMG3), C (PPS6m), D (PH8) e E (MAB6). São mostrados a mediana e os valores mínimo e máximo.
*p < 0,05 (2 vs 6 ou 2 vs 10 dias, 24 h ou 72 h) ou #p < 0,05 (24 h vs 72 h), n = 6 doadores avaliados
em 2 experimentos independentes.

70
Os resultados obtidos das infecções dos macrófagos com parasitos de diferentes
dias de cultivo (contendo formas metacíclicas e procíclicas) mostrou que o índice de infecção
dos macrófagos aumenta quando estas células são infectadas com parasitos da fase
estacionária da cultura, sendo que foi nesta fase que foram detectadas as maiores quantidades
de formas metacíclicas. Assim, para avaliar se havia necessidade de selecionar as formas
metacíclicas para obter melhores índices de infecção, preparações de macrófagos foram
cultivadas com estes parasitos, totais ou metacíclicas selecionadas pela técnica de aglutinação
com lectina BPL ou anticorpo, e os índices de infecção foram comparados. Como mostrado
na Figura 24, não houve diferenças significantes entre as infecções realizadas com parasitos
totais do 6º dia de cultura ou as preparações contendo formas metacíclicas selecionadas, após
24 ou 72 h de incubação.
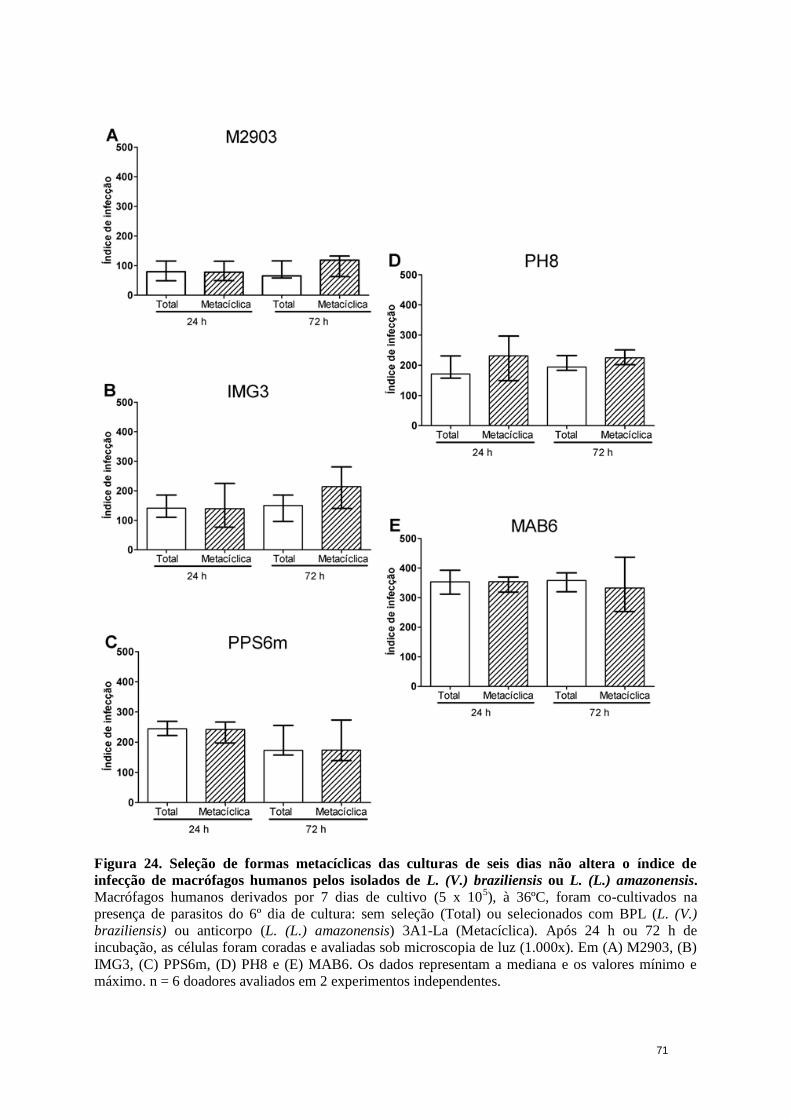
71
Figura 24. Seleção de formas metacíclicas das culturas de seis dias não altera o índice de
infecção de macrófagos humanos pelos isolados de L. (V.) braziliensis ou L. (L.) amazonensis.
Macrófagos humanos derivados por 7 dias de cultivo (5 x 105), à 36ºC, foram co-cultivados na
presença de parasitos do 6º dia de cultura: sem seleção (Total) ou selecionados com BPL (L. (V.)
braziliensis) ou anticorpo (L. (L.) amazonensis) 3A1-La (Metacíclica). Após 24 h ou 72 h de
incubação, as células foram coradas e avaliadas sob microscopia de luz (1.000x). Em (A) M2903, (B)
IMG3, (C) PPS6m, (D) PH8 e (E) MAB6. Os dados representam a mediana e os valores mínimo e
máximo. n = 6 doadores avaliados em 2 experimentos independentes.

72
A detecção de similares índices de infecção entre 24 h e 72 h de cultivo indica que
os parasitos podem resistir aos mecanismos microbicidas dos macrófagos, persistindo dentro
das células. Para posteriormente avaliar a resistência dos isolados de diferentes fases de
cultura aos mecanismos leishmanicidas dos macrófagos humanos, as células foram pré-
tratadas com rIFN por 24 h e, em seguida, co-cultivadas com os parasitos na presença de
LPS. Da mesma forma que observado nas culturas não tratadas (Fig. 23), o índice de infecção
foi maior naquelas culturas tratadas e infectadas com parasitos em fase estacionária do
crescimento, tanto no período inicial de 24 h, quanto após 72 h de infecção (p < 0,05, Fig. 25).
No entanto, observamos que o tratamento com IFN /LPS diminuiu significantemente o índice
de infecção inicial (após 24 h de incubação) naquelas culturas infectadas com parasitos
M2903, IMG3 e MAB6 do 2º dia de cultivo (Fig. 26A, 26B e 26E, respectivamente).
Também houve uma redução significante no índice de infecção nas culturas tratadas e
infectadas com parasitos M2903, PPS6m e PH8 do 6º dia de cultura em relação às culturas
não tratadas (Fig. 25A, 25C e 25D, respectivamente). Nas infecções realizadas com parasitos
do 10º dia, somente para a cepa PH8 (Fig. 25E) foi detectada uma redução no índice de
infecção quando as culturas foram ativadas. De um modo geral, resultados similares foram
obtidos após 72 h de incubação (Fig. 26), à exceção de que com M2903, IMG3 e PPS6m a
ativação com IFN /LPS não causou alterações significantes nos índices de infecção após 72 h
de incubação (Fig. 26A, 26B e 26C).

73
Figura 25. Isolados/cepas de diferentes dias de cultura apresentam diferenças na
suscetibilidade/resistência aos mecanismos microbicidas dos macrófagos (24 h de infecção).
Macrófagos humanos derivados por 6 dias de cultivo (5 x 105) à 36ºC, foram pré-tratados com rIFN
(100 U/mL) por 24 h e co-cultivados na presença de diferentes isolados de Leishmania spp (1:1
parasito:célula) de diferentes dias de cultura (2, 6 ou 10 dias) e LPS (50 ng/mL), em atmosfera úmida,
contendo 5% de CO2. Após 24 h de incubação, as células foram coradas e avaliadas sob microscopia
de luz (1.000x). Os dados representam à mediana e os valores mínimo e máximo.#p < 0,05 (tratados vs
não tratados) ou *p < 0,05 (2 vs 6 ou 2 vs 10 dias tratados com IFN /LPS), n = 6 doadores avaliados
em 2 experimentos independentes.

74
Figura 26. Isolados/cepas de diferentes dias de cultura apresentam diferenças na
suscetibilidade/resistência aos mecanismos microbicidas dos macrófagos (72 h de infecção).
Macrófagos humanos derivados por 6 dias de cultivo (5 x 105) à 36ºC, foram tratados com rIFN (100
U/mL) por 24 h e co-cultivados na presença de diferentes isolados de Leishmania spp (1:1
parasito:célula) de diferentes dias de cultura (2, 6 ou 10 dias) e LPS (50 ng/mL), em atmosfera úmida,
contendo 5% de CO2. Após 72 h de incubação, as células foram coradas e avaliadas sob microscopia
de luz (1.000x). Os dados representam a mediana e os valores mínimo e máximo. #p < 0,05 (tratados
vs não tratados) ou *p < 0,05 (2 vs 6 ou 2 vs 10 dias tratados com IFN /LPS), n = 6 doadores avaliados
em 2 experimentos independentes.

75
Para visualização da infecção dos macrófagos, na Figura 27 são mostradas
fotomicrografias (aumento de 1.000x) dos macrófagos humanos infectados pelos diferentes
isolados, não tratados (A, B e C) ou tratadas (D, E, F) com IFN /LPS. A infecção com
isolados da espécie L. (V.) braziliensis (IMG3, Fig. 27A e 27D; PPS6m, Fig. 27B e 27E,
respectivamente) exibe parasitos fagocitados dentro de pequenos e apertados vacúolos
parasitóforos, contendo somente uma amastigota livre por vacúolo. Um aspecto importante a
ser notado na infecção dos macrófagos com o isolado PPS6m (Fig. 27B) do 6º dia de cultura é
que algumas células estavam intensamente parasitadas. Os vacúolos parasitóforos contendo
parasitos da espécie L. (L.) amazonensis (Fig. 27C) são diferentes, pois são grandes (cerca de
um terço da célula), contendo uma ou várias amastigotas aderidas à membrana do vacúolo. O
tratamento dos macrófagos com IFN /LPS causou uma maior vacuolização das células,
aspecto típico de ativação do macrófago, e dentro da maioria destes vacúolos estavam
presentes fragmentos de parasitos mortos (Fig. 27D, 27E e 27F).

76
Figura 27. Infecção de macrófagos humanos com isolados de Leishmania spp antes e após
tratamento com IFN /LPS. Macrófagos humanos (5 x 105 células) foram derivados por 6 dias a 36
ºC, em atmosfera úmida contendo 5% de CO2. Em seguida, as células foram cultivados (24 h) na
ausência (A, B, C) ou na presença (D, E, F) de IFN e depois co-cultivadas com formas promastigotas
de 6o dia de cultivo por mais 24 h, na presença de LPS. Em A e D: IMG3; B e E: PPS6m e C e F:
MAB6. As setas retilíneas indicam formas amastigotas e as setas tracejadas indicam vacúolos.
Aumento 1.000 x.

77
6 DISCUSSÃO
No presente estudo, foi avaliada a metaciclogênese in vitro de três isolados
clínicos de leishmânia, dois isolados (IMG3 e PPS6m) caracterizados como L. (V.)
braziliensis e um isolado (MAB6), pertencente à espécie L. (L.) amazonensis. Estes isolados
foram selecionados do Leishbank/IPTSP/UFG para este estudo, por serem provenientes de
pacientes com diferentes formas clínicas da doença (LCL, LM e LCDf). Também foram
estudadas no trabalho, as cepas de referência da OMS, M2903 (L. (V.) braziliensis) e PH8 (L.
(L.) amazonensis), que por serem amplamente conhecidas foram utilizadas como controles
experimentais, para comparação com nossos isolados. A avaliação da metaciclogênese foi
realizada para cada isolado/cepa por três metodologias simultaneamente: análise
morfométrica, aglutinação com lectina/anticorpo e resistência ao Complemento. Análises por
citometria de fluxo e infecção de macrófagos humanos foram realizadas, simultaneamente, em
outra série de experimentos, pois não havia viabilidade técnica para a realização de todas as
técnicas com todos os isolados ao mesmo tempo. Como o processo de metaciclogênese pode
sofrer alterações no seu padrão, dependendo do tempo de adaptação dos parasitos ao meio de
cultura, o tipo de meio de cultura, seu pH e sua suplementação (Celeste & Guimarães 1988,
Lemesre et al. 1988, O’Daly & Rodriguez 1988, Bates & Tetley 1993, Zakai et al. 1998,
Louassini et al. 1998), de tal forma que o aparecimento da fase estacionária do crescimento,
juntamente com as formas metacíclicas dos parasitos, pode demorar mais ou ser mais rápida
para se estabelecer, em algumas condições de cultura, para alguns isolados, no presente
trabalho todos os isolados já estavam adaptados às mesmas condições de cultura e foram
utilizados após descongelamento e repiques por até no máximo seis vezes.
A primeira etapa do estudo foi identificar as fases logarítmica e estacionária do
crescimento dos isolados in vitro, uma vez que durante estas fases ocorre a metaciclogênese
(Sacks & Perkins 1984, Franke et al. 1985). A análise das curvas de crescimento dos parasitos
in vitro mostrou similaridade no curso temporal do crescimento entre todos os isolados e
cepas de referência, não havendo também diferenças significantes entre as quantidades de
parasitos obtidos nas culturas, mesmo comparando culturas de parasitos de espécies
diferentes. Foi estabelecido que a fase logarítmica do crescimento ocorre até o 4º dia da

78
cultura (fase de intensa proliferação dos parasitos), iniciando a fase estacionária a partir deste
dia de cultivo. O perfil de crescimento dos isolados, obtido nas condições do presente estudo,
foi similar àquele descrito anteriormente por outros autores, mesmo com uso de diferentes
meios de cultura, usando a cepa L. (V.) braziliensis M2903 e também outros isolados da
mesma espécie (Rey et al. 1990, Costa et al. 2011). Diferentes perfis de crescimento podem
ser obtidos in vitro, variando as condições de cultura. Alguns autores variam estas condições,
alterando a composição de nutrientes ou o pH do meio, temperatura da cultura, a fim de obter
culturas mais ricas em formas metacíclicas (Celeste & Guimarães 1988, Lemesre et al. 1988,
O’Daly & Rodriguez 1988, Torrico et al. 1999, Gamboa et al. 2007, da Luz et al. 2009). Em
todos estes trabalhos, parasitos L. (V.) braziliensis de 4º ao 8º dia de cultivo são considerados
em fase estacionária inicial e nestes dias têm sido condicionados para experimentos de
infecção in vitro ou in vivo. Embora tenham sido descritas diferenças nos padrões de
crescimento de L. (V.) braziliensis e L. (L.) amazonensis, dependendo do meio de cultura
(Lemesre et al. 1988, Niño & Camacho 2005), em acordo com estudos prévios, no presente
trabalho foram estabelecidos o 2o dia para análise de parasitos em fase logarítmica, o 6
o dia
para parasitos em fase estacionária inicial e o 10º dia para parasitos em fase estacionária
tardia, tanto para L. (V.) braziliensis quanto L. (L.) amazonensis.
Em relação às alterações morfológicas que ocorrem com a leishmânia durante a
fase promastigota, os primeiros estudos utilizaram modelos in vivo de infecção de
flebotomíneos com L. major (Killick-Kendrick 1978, Sacks & Perkins 1984, Franke et al.
1985). Posteriormente, outros autores relataram experimentos similares (Nieves & Pimenta
2000, Gossage et al. 2003), utilizando parasitos das espécies L. (V.) braziliensis, L. (L.)
amazonensis, L. mexicana e L. infantum. Nestes trabalhos, os autores identificaram quatro
formas promastigotas morfologicamente diferentes (procíclica, nectomônadas, leptomônadas
e metacíclica), que existem em locais diferentes do trato gastrointestinal do inseto vetor.
Porém, em culturas in vitro, como neste trabalho, as mudanças morfológicas vistas no inseto
vetor não são relatadas da mesma maneira. Como vem sendo descrito, para as culturas in vitro
os parasitos são classificados apenas como parasitos em fase logarítmica, contendo
especialmente formas procíclicas, ou em fase estacionária do crescimento, contendo
especialmente metacíclicas (Pinto-da-Silva et al.; Soares et al. 2002).
Na análise morfológica dos parasitos neste trabalho, durante o crescimento, foram
visualizados na fase logarítmica principalmente parasitos com corpo celular grande e flagelo
pequeno. Ao entrar na fase estacionária (6º dia), os parasitos apresentaram diminuição do

79
tamanho do corpo celular e aumento do comprimento do flagelo. Essas mudanças coincidem
com aquelas descritas durante o processo de metaciclogênese por Almeida et al. (1993) para
L. (V.) braziliensis, onde na fase estacionária do crescimento há formas metacíclicas de corpo
pequeno em forma de seta, bem como com corpo oval/arredondado e longos flagelos,
similares aos encontrados no presente trabalho. Para os parasitos da espécie L. (L.)
amazonensis, as mudanças morfológicas ocorridas na cultura foram mais homogêneas,
podendo ser observados parasitos com o corpo celular mais alongado e afilado, de acordo
com o aumento do tempo de cultura, corroborando os dados de Courret et al. (1999) e
Felizardo et al. (2007).
Mensurar o tamanho do corpo celular e do flagelo dos parasitos é necessário para
confirmar a metaciclogênese, uma vez que, dependendo das condições da cultura, populações
heterogêneas podem ser encontradas e as alterações podem não ser óbvias (Sacks, 1989;
Almeida et al. 1993; Bates & Tetley, 1993). Usando análises morfométricas, relatamos pela
primeira vez, que as formas metacíclicas do 6º e 10º dia de cultura dos isolados pertencentes à
espécie L. (L.) amazonensis (LLa) apresentam maior comprimento do corpo celular e do
flagelo do que os parasitos pertencentes à espécie L.(V.) braziliensis (LVb) destes mesmos
dias de cultura (corpo - LLa: 7-10 m x LVb: ~4 m; flagelo - LLa: ~15-20 m x LVb: ~12
m). Considerando como formas metacíclicas aquelas que possuem a relação tamanho do
flagelo/corpo ≥ 2, foi demonstrado aqui que um perfil similar de metaciclogênese é observado
para todos os isolados/cepas avaliados, com um aumento de formas metacíclicas nos 6o e 10
o
dia das culturas. No entanto, a porcentagem de formas metacíclicas detectadas nestes dias de
cultura dos parasitos L. (L.) amazonensis (~40-60%) foram significantemente menores do que
aquelas dos parasitos L.(V.) braziliensis (~70-90%). Estes resultados podem ser explicados
pelo fato de que os comprimentos do corpo celular e do flagelo de L. (L.) amazonensis são
grandes e o aumento detectado no tamanho do flagelo na fase estacionária do crescimento não
seja suficiente para alcançar uma relação comprimento do flagelo/corpo maior do que 2 e
atestar a presença de formas metacíclicas.
Ao sofrer metaciclogênese, os parasitos mostram, além das alterações
morfológicas, modificações bioquímicas na molécula de LPG que recobre a membrana
externa do parasito, permitindo sua identificação e purificação por meio da aglutinação com
lectinas e anticorpos (Sacks et al. 1990, Pinto-da-Silva et al. 2002, 2005). No presente estudo,
foi realizada a seleção das formas metacíclicas da cultura, utilizando a BPL ou anticorpo 3A1-
La. Os resultados mostraram que os diferentes isolados apresentam um perfil de

80
metaciclogênese semelhante nos dias avaliados, pois houve aumento significante na
quantidade de formas metacíclicas detectadas na fase estacionária do crescimento (6º ou 10º
dia), em relação aos parasitos do 2º dia de cultura, para todos os isolados avaliados,
coincidindo com o período de mudanças morfológicas descritas acima. A cepa de referência
M2903 e o isolado PPS6m apresentaram maiores quantidades de formas metacíclicas no 2º
dia de cultura do que os outros isolados. O fato de formas metacíclicas estarem presentes
durante a fase logarítmica da cultura pode ocorrer em função da manutenção dos parasitos in
vitro, pois quantidades variadas de formas metacíclicas podem ser passadas entre um repique
e outro da cultura, havendo necessidade de controlar o número de repiques utilizados durante
os experimentos (Rebello et al. 2010), que no caso deste estudo, foram de no máximo seis
após o descongelamento das formas promastigotas. Além disso, os parasitos podem
apresentar diferentes afinidades dos carboidratos presentes na membrana externa pelas
lectinas/anticorpos devido a polimorfismos estruturais na molécula de LPG, como visto por
Coelho-Finamore et al. (2011), que analisaram LPG de isolados de L. infantum e Mahoney et
al. (1999), analisando LPG de parasitos da espécie L. donovani. Outra característica que pode
influenciar na capacidade de aglutinação das formas procíclicas da cultura com
lectinas/anticorpos, durante o crescimento in vitro dos parasitos, diz respeito ao padrão
temporal de mudanças da molécula de LPG, que pode ser mais rápido para alguns isolados do
que para outros. Alguns autores demonstraram que, dentro do vetor, os parasitos podem
expressar enzimas que irão controlar o tamanho da molécula de LPG durante o processo de
metaciclogênese, podendo acelerar este processo. Tal fato poderia ocorrer in vitro também,
dependendo das condições de cultura dos parasitos que poderiam influenciar na expressão
destas enzimas nas leishmânias (Sacks et al. 1990, McConville et al. 1992, Soares et al. 2005,
Nikolaev & Sizova 2011).
No presente estudo, apesar de haver uma maior porcentagem de formas
metacíclicas nas culturas dos isolados M2903 e PPS6m no 2o dia de cultivo, usando a técnica
de aglutinação com BPL/anticorpo, similares porcentagens de formas metacíclicas foram
detectadas no 6o dia para todos os isolados, exceto para o isolado MAB6. No 10
o dia de
cultura todos os isolados apresentaram quantidades similares de formas metacíclicas nas
culturas. Estes dados indicam que, utilizando a técnica de identificação de formas
metacíclicas baseado em alterações da LPG com lectina ou anticorpo, uma maior variação nas
porcentagens de formas metacíclicas nos primeiros dias de cultura pode ser encontrada entre
os isolados, mas à medida que passa o tempo, similares porcentagens serão obtidas,

81
independentemente da espécie dos parasitos ou de isolados de uma mesma espécie. Nos 6o e
10o dias de cultura, cerca de 70 a 90% dos parasitos foram classificados como formas
metacíclicas para isolados L. (V.) braziliensis e cerca de 50 a 90%, para isolados L. (L.)
amazonensis. Pinto-da-Silva et al. (2002) relataram ~50% de formas metacíclicas de L. (V.)
braziliensis, em culturas em fase estacionária do crescimento, usando técnica similar à
utilizada no presente estudo. As diferenças entre os nossos resultados e os destes autores
podem ser devidas ao tipo de meio de cultura (Schneider´s insect medium), o isolado (H3456)
e a concentração da lectina BPL (32 g/mL) utilizados. No presente trabalho, utilizamos uma
concentração mais elevada de BPL (100 g/mL), fazendo uma seleção mais rigorosa das
formas metacíclicas. Alguns autores já utilizaram outras lectinas para purificar populações de
formas metacíclicas de L. (V.) braziliensis, tais como as lectinas PNA e LL (Almeida et al.
1993, Muskus et al. 1997), que também já foram utilizadas para purificação de formas
promastigotas metacíclicas de L. major e L. donovani (McConville et al. 1992, Sacks et al.
1995). Nestes trabalhos e naqueles que usaram BPL ou anticorpo 3A1-La (Pinto-da-Silva et
al. 2002, Pinto-da-Silva et al. 2005), não foi objetivo comparar isolados de diferentes espécies
ou da mesma espécie de Leishmania.
No presente trabalho, utilizando a mesma metodologia, foi demonstrado que é
possível haver diferença entre as porcentagens de formas metacíclicas de isolados de
diferentes espécies, no 6o dia de cultivo (MAB6 vs PPS6m e IMG3, Fig. 11B). A maior
porcentagem de metacíclicas detectada nas culturas de MAB6 no 6o dia, assim como as
elevadas porcentagens destas formas nas culturas de 6o e 10
o dias de leishmânias PH8,
ilustram diferenças entre os resultados das análises morfométricas e do uso da técnica de
seleção de metacíclicas com o anticorpo 3A1-La. É possível que as alterações bioquímicas
dos parasitos L. (L.) amazonensis sejam mais expressivas do que as alterações morfológicas
durante o processo de metaciclogênese (6o e 10
o dias). Nas condições experimentais aqui
apresentadas, parece que as alterações morfológicas e bioquímicas ocorrem mais rapidamente
em parasitos L. (V.) braziliensis do que em L. (L.) amazonensis, mas é possível obter similares
porcentagens de formas metacíclicas de parasitos das duas espécies no 6o dia de cultura
usando a técnica de aglutinação com BPL/anticorpo. Devido às diferenças morfológicas dos
parasitos encontrados nas culturas, entre a fase logarítmica e a estacionária, a técnica
desenvolvida por Saraiva et al. (2005) foi utilizada, no presente trabalho, para identificar e
quantificar as formas metacíclicas dos diferentes isolados, por citometria de fluxo. Esta é uma
técnica sensível para detecção de alterações morfológicas e de fácil execução. Em paralelo,

82
para confirmar a determinação das populações de formas metacíclicas e procíclicas da cultura,
foram realizadas análises das populações de formas metacíclicas selecionadas com
BPL/anticorpo. O presente trabalho é o primeiro a usar a técnica de citometria de fluxo
descrita por Saraiva et al. (2005) para comparar diferentes isolados clínicos de uma mesma
espécie. A citometria de fluxo revelou que não existem diferenças entre os perfis de
metaciclogênese dos isolados da mesma espécie ou de espécies diferentes. Apesar do ensaio
de citometria não ter sido realizado simultaneamente com os ensaios de morfometria e seleção
com BPL/anticorpo, os perfis de metaciclogênese foram similares nas três técnicas. Em
termos de porcentagens de formas metacíclicas comparadas entre os diferentes isolados, a
técnica de citometria não mostrou diferenças significantes entre isolados da mesma espécie ou
de espécies diferentes. Porém, houve uma tendência a menor porcentagem de formas
metacíclicas do isolado PPS6m nos três dias avaliados e do isolado IMG3 no 6o dia. Os
resultados da citometria de fluxo mostraram elevada variabilidade interexperimental,
especialmente nas análises dos dias 6 e 10 de cultivo para isolados L. (V.) braziliensis.
As porcentagens de formas metacíclicas, identificadas pela citometria no 2º (16-
26%) e 6º (47-80%) dia de cultura para a cepa de referência M2903 foram similares àquelas
obtidas por Saraiva et al. (2005): 16% (2o dia) e 75% (5
o dia), utilizando L. (V.) braziliensis
H3456. Para os isolados clínicos IMG3 (2o vs 6
o dia: 21-28% vs 30-60%) e PPS6m (7-21% vs
40-77%), as porcentagens foram similares às de Saraiva et al. (2005), mas com tendência a
uma menor detecção de formas metacíclicas de IMG3 no 6o dia e de PPS6m no 2
o dia. Em
relação a parasitos L. (L.) amazonensis, tanto para a cepa de referência PH8 (18-26% vs 54-
69%) quanto para o isolado MAB6 (8-29% vs 68-80%), maiores porcentagens de formas
metacíclicas foram detectadas no presente trabalho, comparando com os resultados de Saraiva
et al. (2005) que utilizaram L. (L.) amazonensis Josefa (3% no 2o dia vs 37% no 5
o dia). Neste
caso, como estes autores avaliaram as culturas no 5o dia, talvez a diferença de 24 h em relação
ao presente estudo (6o dia) seja suficiente para aumentar a quantidade de formas metacíclicas
nas culturas de L. (L.) amazonensis.
Em relação às análises dos parasitos por citometria após seleção/enriquecimento
com BPL/anticorpo, as porcentagens de formas metacíclicas purificadas encontradas para
todos os isolados no 6º dia de cultura (variando de 67% a 87% para L. (V.) braziliensis e de
48% a 92% para L. (L.) amazonensis) foram similares às porcentagens de formas metacíclicas
encontradas por Saraiva et al. (2005) que também utilizaram as mesmas técnicas
(BPL/anticorpo/citometria: L. (V.) braziliensis 93%; L. (L.) amazonensis 81%). Existe uma

83
heterogeneidade na morfologia das formas promastigotas durante os dias de cultivo in vitro,
que poderia ser detectada pela citometria de fluxo. Essa heterogeneidade de diferenças
morfológicas observada nas culturas foi confirmada pela seleção com BPL/anticorpo, que
mostrou que numa população BPL-/3A1
- (somente de formas metacíclicas) há uma variação
de cerca de 75% a 97% de formas metacíclicas que se encaixam no tamanho específico da
análise das formas metacíclicas por citometria de fluxo (FSClow
, de acordo com os parâmetros
descritos por Saraiva et al., 2005). Portanto, nas análises da citometria é observada uma
pequena população de tamanho intermediário que não é eliminada totalmente durante a
seleção com BPL/Anticorpo, sendo, portanto, considerada metacíclica pelo critério de
alteração bioquímica da LPG, mas não pela citometria. Estes dados estão de acordo com
aqueles de Saraiva et al. (2005) que encontraram heterogeneidade entre as formas da fase
estacionária do crescimento de diferentes espécies, as quais estão associadas ao processo de
metaciclogênese que representa um contínuo de diferenciação celular e não uma passagem
abrupta de formas procíclicas para metacíclicas. Apesar desta heterogeneidade, a maioria das
formas metacíclicas são detectadas em FSClow
. A análise por citometria mostrou-se útil para
avaliar se após a utilização da técnica de purificação BPL/anticorpo houve realmente seleção
de formas metacíclicas (eliminação das formas procíclicas de maior tamanho).
Outra característica que acompanha o processo de metaciclogênese dos parasitos
em culturas in vitro é o aumento da resistência à lise mediada pelo sistema Complemento
(Sacks & Perkins 1984). No presente estudo, após análise da resistência ao Complemento de
uma cepa de referência e um isolado clínico foi estabelecido que 10% de soro seria uma
concentração que poderia causar ≥50% de lise dos parasitos da fase estacionária e ~90%
daqueles da fase logarítmica. Os parasitos da fase estacionária de isolados de ambas as
espécies foram mais resistentes à lise mediada pelo Complemento do que os parasitos do 2º
dia (Fase Log.). Estes dados corroboram os achados de Almeida et al. (1993) e Pinto-da-Silva
et al. (2005) que, utilizando cepas de L. (V.) braziliensis e L. (L.) amazonensis, identificaram
que as formas metacíclicas purificadas da fase estacionária da cultura foram mais resistentes à
lise mediada pelo Complemento do que os parasitos da fase logarítmica. No presente trabalho,
os resultados da avaliação do perfil de metaciclogênese pelo ensaio de resistência ao
Complemento mostraram que os perfis de todos os isolados são similares aos obtidos com as
análises morfométricas, seleção com BPL/anticorpo e citometria de fluxo, identificando o
aumento significante de formas metacíclicas a partir do 6o dia de cultura. Porém,
considerando que os ensaios de seleção com BPL/anticorpo e de resistência ao Complemento,

84
ambos avaliam alterações bioquímicas da LPG, seria esperado que as porcentagens de formas
metacíclicas encontradas nas culturas fossem similares ao utilizar simultaneamente estas duas
técnicas para identificar estas formas. No entanto, isto não ocorreu para o isolado L. (V.)
braziliensis PPS6m no 2o dia de cultivo (BPL vs Complemento: 26-36% vs 10-20%), nem
para os isolados L. (L.) amazonensis PH8 e MAB6, para os quais o ensaio do Complemento
detectou maiores porcentagens de formas metacíclicas do que o ensaio de seleção com
anticorpo. No 6o dia de cultura, quando aumentam as porcentagens das formas metacíclicas, a
seleção com BPL detectou maior porcentagem destas formas do que o ensaio do
Complemento nas culturas de PPS6m, IMG3 e MAB6 (p < 0,05). Com 10 dias, os resultados
foram similares com as duas técnicas. Estes resultados sugerem que os isolados PPS6m,
IMG3 e MAB6, apesar de sofrerem alterações bioquímicas da LPG, demoram mais para
tornarem-se resistentes ao Complemento. Estas alterações bioquímicas da LPG, ocorridas
durante a metaciclogênese, parecem ser necessárias à resistência ao Complemento, o que é
importante para a virulência do parasito (Muskus et al. 1997, Naderer & McConville 2008,
McConville & Naderer 2011, Franco et al. 2012), mas para alguns isolados clínicos este
processo pode requerer um tempo maior.
Um dos objetivos de avaliar o processo de metaciclogênese in vitro de diferentes
leishmânias, além de avaliar características dos parasitos, é determinar as condições ideais
para infecção de macrófagos humanos, para estudos da interação entre estas células e os
parasitos. Os macrófagos são a célula alvo no processo de infecção por Leishmania sp, sendo
uma das primeiras células a entrar em contato com as formas metacíclicas inoculadas pelo
inseto vetor, tornando-se parasitada (Aderem & Underhill 1999, Kaye & Scott 2011). No
presente trabalho, foram realizados ensaios de infecção in vitro de macrófagos humanos com
parasitos provenientes de diferentes dias de cultura e com aqueles antes e após seleção de
formas metacíclicas. Além de determinar as condições para a adequada infecção dos
macrófagos humanos, os experimentos de infecção devem confirmar que houve o processo de
metaciclogênese in vitro, sendo esperado que culturas de parasitos ricas em formas
metacíclicas ou formas metacíclicas selecionadas sejam mais infectantes do que aquelas
pobres nestas formas. De fato, os resultados mostraram que o índice de infecção dos
macrófagos, após 24 ou 72 horas de incubação com os parasitos, foi maior naquelas culturas
infectadas com formas promastigotas da fase estacionária do 6º ou 10º dia de cultura, devido a
um aumento na porcentagem de células infectadas e/ou no número de parasitos por célula. O
aumento no índice de infecção coincidiu com o aumento das porcentagens de formas

85
metacíclicas, detectadas por todas as técnicas aqui utilizadas. A avaliação das culturas dos
parasitos, utilizados para os ensaios de infecção dos macrófagos, por citometria de fluxo,
simultaneamente, confirmaram a metaciclogênese dos parasitos nas culturas.
Em macrófagos murinos, tem sido demonstrado que as formas promastigotas
metacíclicas de L. (V.) braziliensis e L. (L.) amazonensis infectam estas células (Pinto-da-
Silva et al. 2002, Felizardo et al. 2007, Gamboa et al. 2007, da Luz et al. 2009, Zauli-
Nascimento et al. 2010), mas muito pouco é conhecido sobre a infecção de macrófagos
humanos (Rey et al. 1990, Calegari-Silva et al. 2009, Giudice et al. 2012). Zauli-Nascimento
et al. (2010) mostraram que a cepa de referência M2903 causa um baixo índice de infecção
em linhagem monocítica humana (THP-1). De acordo com nossos resultados, os menores
índices de infecção foram obtidos com esta cepa, apesar de, com as quatro técnicas utilizadas,
uma elevada porcentagem de formas metacíclicas (~70%) terem sido detectadas já no 6o dia
de cultivo. Giudice et al. (2012) mostraram que a cepa LTCP15344 (L. (V.) braziliensis)
infecta macrófagos humanos derivados de monócitos (6 dias de derivação; 2:1 relação
parasito:macrófago) de indivíduos sadios, causando índices de infecção ~200, similares aos
índices encontrados no presente trabalho com os isolados IMG3 e PPS6m.
No presente trabalho, apesar de poder ser detectada uma maior porcentagem de
formas metacíclicas no 10o dia de cultura de alguns isolados, sendo este um período com
menor variação entre as porcentagens de metacíclicas dos isolados avaliados, os índices de
infecção com estes parasitos não foram alterados significantemente em relação àqueles
atingidos com parasitos do 6o dia de cultivo. Em alguns experimentos, especialmente com o
isolado PPS6m, o índice de infecção com parasitos do 10o dia chegaram a diminuir. Tais
resultados podem ser atribuídos à morte dos parasitos no 10o dia de cultura, quando foram
observados vários parasitos rompidos ou imóveis. Estes resultados indicam que não é
adequado utilizar parasitos em fase estacionária tardia para ensaios de infecção dos
macrófagos. Dependendo das condições de cultura, a morte de parasitos pode ser detectada
após o início da fase estacionária e vai se acentuando com o tempo (Celeste & Guimarães
1988, Lemesre et al. 1988, Niño & Camacho 2005). Embora tenha sido descrito por
Wanderley et al. (2009) que parasitos em apoptose juntamente com os parasitos viáveis
contribuem para o sucesso da infecção, 10 dias de cultura podem gerar parasitos
enfraquecidos que morram facilmente dentro dos macrófagos.
No presente trabalho, observamos que para a cepa M2903 e o isolado IMG3,
houve uma diminuição significante no índice de infecção de 24 h para 72 h de incubação,

86
quando os macrófagos foram infectados com parasitos do 2º ou 10º dia. Uma tendência a
resultados similares foram encontrados com PPS6m, PH8 e MAB6 (2o dia; 10
o dia, exceto
para MAB6). Estes resultados sugerem que as formas amastigotas oriundas dos parasitos de
2o (ainda não adaptados para sobreviver dentro dos macrófagos) ou 10
o dia (enfraquecidos por
falta de nutrientes) podem ser mais suscetíveis aos mecanismos microbicidas dos macrófagos
ou que estes parasitos ativam mais os macrófagos nas primeiras horas de infecção, induzindo
um melhor controle do crescimento do parasito. Em contrapartida, aquelas formas
amastigotas oriundas de parasitos do 6o dia mantiveram níveis similares de infecção entre 24
h e 72 h, sendo, portanto, mais resistentes aos mecanismos microbicidas dos macrófagos.
Dentre os parasitos avaliados, IMG3, PH8 e MAB6 parecem ser os mais resistentes à
atividade microbicida dos macrófagos, quando estes são infectados com parasitos do 6o dia de
cultura. Essas diferenças entre os isolados podem ser resultado da capacidade intrínseca dos
parasitos de sobreviver dentro dos macrófagos. Hsiao et al. (2011) demonstraram que a
infecção de macrófagos humanos por L. infantum chagasi leva à ativação celular, com
formação de fagolisossomos, onde são produzidas moléculas microbicidas que levam ao
controle da infecção, induzindo a morte ou inibindo a replicação dos parasitos numa fase mais
tardia da infecção (72 h). De acordo com nossos resultados, a infecção por L. (V.) braziliensis
ou L. (L.) amazonensis pode ser controlada pelos macrófagos humanos até o período avaliado,
72 h.
Os resultados dos experimentos de infecção dos macrófagos demonstraram que,
quando estão presentes na cultura maiores quantidades de formas metacíclicas (como
naquelas infecções realizadas com parasitos da fase estacionária inicial ou tardia) os isolados
causam maiores índices de infecção e podem resistir aos mecanismos microbicidas dos
macrófagos no período de 72 h de incubação. Alterações bioquímicas que ocorrem nos
parasitos durante o processo de metaciclogênese, como alterações na LPG, facilitam a evasão
dos parasitos (Handman et al. 1986, Sacks et al. 1990). A LPG inibe a fusão do fagossomo ao
lisossomo, favorecendo possivelmente a transformação da forma promastigota para
amastigota (Desjardins & Descoteaux 1997), e ainda, a LPG seqüestra radicais hidroxila e
ânion superóxido, diminuindo os efeitos microbicidas dos macrófagos (revisto por Franco et
al. 2012). Outra molécula abundante na superfície da forma metacíclica é a enzima gp63, que
possui papel protetor no parasito contra a citólise e a degradação no interior do fagolisossomo
(Seay et al. 1996). Assim, os diferentes isolados podem apresentar diferenças em relação a
estas moléculas, evadindo ou não aos mecanismos microbicidas dos macrófagos ativados

87
durante as primeiras horas de infecção, o que pode contribuir para a persistência do parasito
nos macrófagos (Depledge et al. 2010, de Souza Carmo et al. 2010, Matta et al. 2010).
Se a ativação ou não dos macrófagos nas primeiras horas de infecção são
determinantes no sucesso da geração das formas amastigotas e resistência destas aos
mecanismos microbicidas dos macrófagos, é possível que a presença de formas procíclicas e
metacíclicas em diferentes estágios de diferenciação nas preparações dos parasistos, possa
alterar a capacidade dos macrófagos em eliminar os parasitos. As formas metacíclicas de L.
major são menos ativadoras do que as procíclicas (Sartori et al. 1997), mas uma avaliação
deste tipo para infecção de macrófagos com L. (V.) braziliensis ou L. (L.) amazonensis ainda
estava faltando. No presente trabalho, das quatro técnicas avaliadas, a seleção negativa de
formas metacíclicas pela técnica de aglutinação com lectina BPL ou anticorpo foi escolhida
para obter formas metacíclicas. As formas metacíclica de 6o dia foram selecionadas e os
macrófagos foram infectados com parasitos totais (contendo uma mistura de formas dos
parasitos) ou com as formas metacíclicas. Surpreendentemente, não houve diferenças entre o
índice de infecção dos macrófagos (24 h ou 72 h) quando estas células foram infectadas com
parasitos totais ou formas metacíclicas selecionadas. Mas se considerarmos que a técnica de
seleção com BPL/Anticorpo foi a que detectou maiores frequências de formas metacíclicas
para todos os isolados do 6o dia de cultura (Fig. 19, medianas de 66 a 84%; valores abaixo de
50% foram obtidos somente com a cepa PH8 e as análises morfométricas mostraram maiores
frequências de metacíclicas nas culturas de M2903 e IMG3, ~85%), elevadas porcentagens de
metacíclicas estavam presentes nas culturas e o enriquecimento obtido com a técnica
BPL/Anticorpo pode, por isto, não ter resultado em aumento da infecção dos macrófagos com
as formas metacíclicas selecionadas. Assim, demonstrando que não há necessidade de
utilizar-se de métodos de purificação de formas metacíclicas para ensaios de infecção de
macrófagos humanos quando estão presentes grandes quantidades de formas metacíclicas nas
culturas. Os resultados sugerem que as formas procíclicas ou intermediárias entre procíclicas
e metacíclicas de L. (V.) braziliensis ou L. (L.) amazonensis presentes na fase estacionária
inicial não estariam ativando os macrófagos para que estes conseguissem controlar mais a
infecção pelas formas metacíclicas, não interferindo no sucesso da infecção. Ainda, é possível
que o ensaio de macrófagos estabelecido não tenha sensibilidade para mostrar estas
diferenças.
Diante dos resultados discutidos acima, a avaliação da capacidade dos isolados em
resistir aos mecanismos microbicidas dos macrófagos ativados com IFN /LPS foram

88
avaliadas em culturas de macrófagos infectados com parasitos não selecionados, dos dias 2, 6
ou 10 de cultura. A atividade microbicida dos macrófagos pode ser aumentada por ativação
clássica dos macrófagos (IFN /LPS) ( Hayes et al. 1995, Gordon 2012), que induz a produção
de moléculas microbicidas, tais como derivados de ROI/RNI, que conhecidamente matam
leishmânias (Liew et al. 1997, Panaro et al. 1999, Mukbel et al. 2007). Portanto, nós pré-
tratamos os macrófagos com IFN e adicionamos LPS no momento da infecção com os
parasitos. A sensibilização dos macrófagos com IFN causa um aumento da expressão de
TLR4, o receptor para o LPS, aumentando a capacidade dos macrófagos humanos de
responder ao LPS (Bosisio et al. 2002). No presente trabalho, similares resultados foram
obtidos com macrófagos não ativados ou ativados com IFN /LPS, ou seja, ao avaliar os
índices de infecção os parasitos de 6o e 10
o dia causaram os maiores índices de infecção. No
entanto, os macrófagos ativados conseguiram diminuir significantemente os índices de
infecção (24 h) causados pelos parasitos M2903 (de 2 e 6 dias), IMG3 (de 2 dias), PPS6m (de
6 dias) e PH8 (de 6 e 10 dias), mas não o do isolado MAB6. Estes resultados se repetiram
apenas para M2903 e PH8, que continuaram sendo mais sensíveis aos mecanismos de
controle dos macrófagos, e MAB6, que continuou apresentando alto índice de infecção após
72 h de incubação. Os dados sugerem que M2903 e PH8 foram mais suscetíveis aos
mecanismos microbicidas dos macrófagos, enquanto o isolado MAB6 parece ser o mais
resistente. Os isolados PPS6m e IMG3 também mostraram um certo grau de resistência, uma
vez que após 72 h de cultivo, a ativação dos macrófagos não resultou em diminuição
significante dos índices de infecção. Como as culturas dos parasitos de 6o dia de cultivo são
ricas em formas metacíclicas, com densidades similares destas formas nas culturas (M2903:
76-87%; IMG3: 67-72%; PPS6m: 75-80%; PH8: 48-85% e MAB6: 80-92%; técnica
BPL/3A1-La), é possível que a maior ou menor resistência dos isolados aos mecanismos
microbicidas dos macrófagos sejam características intrínsecas de cada isolado. Em
fortalecimento desta ideia, a cepa de referência M2903, que causou baixos índices de infecção
e foi mais sensível aos mecanismos de controle dos macrófagos independente da ativação
destes, vem sendo usada em experimentos de infecção de camundongos mostrando um baixo
grau de patogenicidade. Em geral, causa lesões pequenas que curam espontaneamente
rapidamente, mesmo quando são utilizadas formas metacíclicas selecionadas (Maioli et al.
2004; Souza-Neto et al. 2004; Pereira et al. 2009). In vitro, nas nossas culturas, M2903 gerou
elevadas porcentagens de formas metacíclicas (~78%, BPL/anticorpo). Além disto, o isolado
MAB6, que causou maiores índices de infecção nos experimentos e mostrou-se mais

89
resistente à atividade microbicida dos macrófagos, é oriundo de um paciente com LCDf que
recebeu vários tratamentos sem sucesso. Somente após forte ativação da imunidade natural
com BCG e Leishvacin, juntamente com a quimioterapia, o paciente apresentou uma remissão
transitória das lesões (Pereira et al. 2009b). Experimentos para avaliar a patogenicidade dos
isolados clínicos utilizados no presente estudo estão em andamento.
Este é o primeiro trabalho a avaliar o processo de metaciclogênese in vitro de
parasitos das espécies L. (V.) braziliensis e L. (L.) amazonensis, utilizando, simultaneamente,
diferentes métodos já estabelecidos. Os resultados revelam que, avaliando critérios de
alterações morfológicas e bioquímicas (alteração de LPG), por meio de análises
morfométricas, citometria de fluxo, aglutinação com lectinas/anticorpos e ensaio de
resistência ao Complemento, não há diferenças nos perfis de metaciclogênese dos isolados
selecionados para os experimentos. Os isolados L. (V.) braziliensis M2903, IMG3 e PPS6m,
bem como os L. (L.) amazonensis PH8 e MAB6, apresentam significantes porcentagens de
formas metacíclicas no 6o dia de cultivo que aumentam, mas não significantemente, no 10
o
dia. Este perfil de metaciclogênese foi confirmado nos ensaios de infecção de macrófagos
humanos, que mostraram que com parasitos de culturas ricas em formas metacíclicas maiores
índices de infecção são alcançados.
A densidade de formas metacíclicas obtidas em cada dia de cultivo, no entanto,
pode ser diferente entre os isolados, dependendo da técnica utilizada. Ao comparar as
porcentagens de formas metacíclicas identificadas pelas quatro técnicas, é aparente que para
os isolados L. (V.) braziliensis, as técnicas que detectam maiores porcentagens de formas
metacíclicas são a morfometria e a seleção com lectina BPL. Já para os isolados L. (L.)
amazonensis, a seleção com anticorpo 3A1-La e citometria de fluxo detectaram elevadas
porcentagens de formas metacíclicas. Considerando que os resultados dos ensaios de infecção
dos macrófagos com parasitos não selecionados ou selecionados, pela técnica de aglutinação
com BPL/anticorpo, foram similares, acreditamos que as culturas de 6 e 10 dias apresentam
elevadas porcentagens de formas metacíclicas e que as formas procíclicas ou intermediárias
entre procíclicas e metacíclicas não interferem no estabelecimento da infecção, dispensando a
necessidade de purificação de formas metacíclicas. Apesar de não ter sido demonstrada
diferença entre os índices de infecção dos macrófagos incubados com parasitos totais ou
selecionados, é recomendável que a cada novo isolado clínico obtido seja realizada a análise
da metaciclogênese in vitro, usando a técnica que quantifica as formas metacíclicas por meio
de alterações bioquímicas dos parasitos (lectina/anticorpo), para avaliar se há ou não a

90
necessidade de uma purificação, no caso de baixa densidade de formas metacíclicas no 6o dia
de cultivo, antes de infectar os macrófagos.
As técnicas de seleção com BPL/anticorpo e citometria de fluxo podem ser
realizadas conjuntamente para a avaliação da metaciclogênese, avaliando duas características
importantes: alteração bioquímica da LPG e morfologia, respectivamente, e para
quantificação das formas metacíclicas nas culturas, a fim de padronizar as quantidades destas
formas a serem usadas em experimentos de infecção. Utilizar um mesmo número de formas
metacíclicas é ideal para a comparação da virulência de diferentes isolados clínicos,
especialmente quando houver diferenças nas porcentagens de metacíclicas na fase
estacionária do crescimento.

91
7 CONCLUSÕES
Conclusão geral
Isolados clínicos de L. (V.) braziliensis e L. (L.) amazonenses apresentam
similares perfis de metaciclogênese, com maiores porcentagens de formas metacíclicas
detectadas após seis dias de cultura. A técnica de identificação de formas metacíclicas usando
lectina/anticorpo para identificar alterações da LPG é ideal para a quantificação e seleção das
formas metacíclicas dos isolados clínicos. Os parasitos em fase estacionária do crescimento
(6º dia) infectam macrófagos humanos similarmente às formas metacíclicas selecionadas
quando elevadas porcentagens de formas metacíclicas são detectadas. Os resultados sugerem
que diferentes isolados clínicos podem apresentar diferenças na capacidade de infecção de
macrófagos humanos.
Conclusões específicas
1. O curso temporal de crescimento in vitro dos diferentes isolados, avaliado pela contagem
diária dos parasitos, foi similar para todos os isolados, sendo detectada a fase estacionária do
crescimento a partir do 5º dia de cultivo.
2. Os parasitos sofrem uma diminuição significante no tamanho do comprimento do corpo
celular e um aumento progressivo no tamanho do flagelo na fase estacionária do crescimento
(6o ou 10
o dia), em relação às formas promastigotas presentes no 2º dia de cultura. As formas
promastigotas dos isolados de L. (L.) amazonensis do 6º e 10º dia possuem o comprimento do
corpo celular e do flagelo maiores do que os parasitos pertencentes à espécie L. (V.)
braziliensis.
3. Os perfis de metaciclogênese dos isolados foram similares, sendo detectado um aumento na
porcentagem de formas metacíclicas durante a fase estacionária da cultura (6º e 10º dia). No

92
entanto, apesar das quatro técnicas mostrarem um similar perfil de metaciclogênese para todos
os isolados avaliados, as densidades de formas metacíclicas variaram conforme a técnica
utilizada, dependendo do dia de cultura. A técnica de seleção com BPL/3A1-La foi a que
detectou maiores frequências de formas metacíclicas para todos os isolados no 6o dia.
4. Não houve diferença entre o índice de infecção de macrófagos infectados com parasitos
totais ou com formas metacíclicas purificadas do 6º dia de cultura.
5. Os parasitos em fase estacionária (6º ou 10º dia de cultivo) causaram maiores índices de
infecção dos macrófagos humanos do que os parasitos de fase logarítmica. Os isolados
apresentaram diferenças quanto à suscetibilidade aos mecanismos microbicidas dos
macrófagos ativados, sendo que as cepas M2903 e PH8 foram mais suscetíveis, enquanto o
isolado MAB6, mais resistente.

93
REFERÊNCIAS
Aderem A, Underhill DM 1999. Mechanisms of phagocytosis in macrophages. Annu. Rev.
Immunol. 17: 593–623.
Akilov OE, Kasuboski RE, Carter CR, McDowell MA 2007. The role of mannose receptor
during experimental leishmaniasis. J. Leukoc. Biol. 81: 1188–1196.
Almeida MC, Cuba CA, Sa CM, Pharoah MM, Howard KM, Miles MA 1993.
Metacyclogenesis of Leishmania (Viannia) braziliensis in vitro: evidence that lentil lectin is a
marker of complement resistance and enhanced infectivity. Trans. R. Soc. Trop. Med. Hyg.
87: 325–329.
Almeida MC, Vilhena V, Barral A, Barral-Netto M 2003. Leishmanial infection: analysis of
its first steps: A review. Mem. Inst. Oswaldo Cruz 98: 861–870.
Altamirano-Enciso AJ, Marzochi MCA, Moreira JS, Schubach AO, Marzochi KBF 2003. On
the origin and spread of cutaneous and mucosal leishmaniasis, based on pre- and post-
colombian historical source. História, ciências, saúde-Manguinhos 10: 852–882.
Amóra SSA, Bevilaqua CML, Feijó FMC, Alves N, Maciel M 2009. Control of phlebotomine
(Diptera: Psychodidae) leishmaniasis vectors. Neo. Entomol 38: 303–310.
Ashford RW 2000. The leishmaniases as emerging and reemerging zoonoses. Int J Parasitol.
30: 1269–1281.
Assis RR, Ibraim IC, Noronha FS, Turco SJ, Soares RP (2012) Glycoinositolphospholipids
from Leishmania braziliensis and L. infantum: Modulation of Innate Immune System and
Variations in Carbohydrate Structure. PLoS Negl Trop Dis 6(2): e1543.
doi:10.1371/journal.pntd.0001543.
Awasthi A, Mathur RK, Saha B 2004. Immune response to Leishmania infection. Indian J.
Med. Res. 119: 238–258.
Bates PA 2008. Leishmania sand fly interaction: progress and challenges. Cur. Opin in
Microbiol. 11: 340–344.
Bates PA, Tetley L 1993. Leishmania mexicana: induction of metacyclogenesis by cultivation
of promastigotes at acidic pH. Exp. Parasitol. 76: 412–423.
Becker I, Salaiza N, Aguirre M, Delgado J, Carrillo-Carrasco N, Kobeh LG, Ruiz A,
Cervantes R, Torres AP, Cabrera N, González A, Maldonado C, Isibasi A 2003. Leishmania
lipophosphoglycan (LPG) activates NK cells through toll-like receptor-2. Mol. Bioch.
Parasitol. 130: 65–74.

94
Besteiro S, Williams RAM, Coombs GH, Mottram JC 2007. Protein turnover and
differentiation in Leishmania. Int J Parasitol. 37: 1063–1075.
Bosisio D, Polentarutti N, Sironi M, Bernasconi S, Miyake K, Webb GR, Martin MU,
Mantovani A, Muzio M 2002. Stimulation of toll-like receptor 4 expression in human
mononuclear phagocytes by interferon-gamma: a molecular basis for priming and synergism
with bacterial lipopolysaccharide. Blood 99: 3427–3431.
Bosque F, Milon G, Valderrama L, Saravia NG 1998. Permissiveness of human monocytes
and monocyte-derived macrophages to infection by promastigotes of Leishmania (Viannia)
panamensis. J. Parasitol. 84: 1250–1256.
Brasil 2006. Atlas de Leishmaniose tegumentar Americana: Diagnóstico clinico e diferencial.
Ministério da Saúde. Manual de vigilância da Leishmaniose Tegumentar Americana.
Brasil 2010. Boletim Eletrônico Epidemiológico. Ministério da Saúde. Secretaria de
vigilância sanitária. Brasília
Brittingham A, Morrison CJ, McMaster WR, McGwire BS, Chang KP, Mosser DM 1995.
Role of the Leishmania surface protease gp63 in complement fixation, cell adhesion, and
resistance to complement-mediated lysis. J. of Immunol. 155: 3102–3111.
Brodin TN, Heath S, Sacks DL 1992. Genes selectively expressed in the infectious
(metacyclic) stage of Leishmania major promastigotes encode a potential basic-zipper
structural motif. Mo. Bioche. Parasitol. 52: 241–250.
Burchmore RJ, Barrett MP 2001. Life in vacuoles--nutrient acquisition by Leishmania
amastigotes. Int J Parasitol. 31: 1311–1320.
Calegari-Silva TC, Pereira RMS, Melo LDB, Saraiva EM, Soares DC, Bellio M, Lopes UG
2009. NF-kappaB-mediated repression of iNOS expression in Leishmania amazonensis
macrophage infection. Immunol. Letters 127: 19–26.
Celeste BJ, Guimarães MC 1988. Growth curves of Leishmania braziliensis promastigotes
and surface antigen expression before and after adaptation to Schneider’s Drosophila medium
as assessed by anti-Leishmania human sera. Rev. Inst. Med. Trop. de São Paulo 30: 63–67.
Chaudhary RG, Bilimoria FE, Katare SK 2008. Diffuse cutaneous leishmaniasis: co-infection
with human immunodeficiency virus (HIV). Ind. J. Dermatol. Venereol. Leprology 74: 641–
643.
Chaves CS, Soares DC, Silva RP, Saraiva EM 2003. Characterization of the species- and
stage-specificity of two monoclonal antibodies against Leishmania amazonensis. Exp.
Parasitol. 103: 152–159.
Coelho-Finamore JM, Freitas VC, Assis RR, Melo MN, Novozhilova N, Secundino NF,
Pimenta PF, Turco SJ, Soares RP 2011. Leishmania infantum: Lipophosphoglycan
intraspecific variation and interaction with vertebrate and invertebrate hosts. Int J Parasitol.
41: 333–342.

95
Costa TL, Ribeiro-Dias F, Oliveira MAP, Bezerra JCB, Vinaud MC 2011. Energetic
metabolism of axenic promastigotes of Leishmania (Viannia) braziliensis. Exp. Parasitol.
128: 438–443.
Coulson RM, Smith DF 1990. Isolation of genes showing increased or unique expression in
the infective promastigotes of Leishmania major. Mol. Bioche. Parasitol. 40: 63–75.
Souza-Neto SM, Carneiro CM, Vieira LQ, Afonso LC 2004. Leishmania braziliensis: partial
control of experimental infection by interleukin-12 p40 deficient mice. Mem. Inst. Oswaldo
Cruz 99:289-94.
Depledge DP, MacLean LM, Hodgkinson MR, Smith BA, Jackson AP, Ma S, Uliana SRB,
Smith DF 2010. Leishmania-specific surface antigens show sub-genus sequence variation and
immune recognition. PLoS Neglect. Trop. Diseases 4: 829.
Desjardins M, Descoteaux A 1997. Inhibition of phagolysosomal biogenesis by the
Leishmania lipophosphoglycan. J. Exp. Med. 185: 2061–2068.
Felizardo TC, Toma LS, Borges NB, Lima GMC, Abrahamsohn I 2007. Leishmania
(Leishmania) amazonensis infection and dissemination in mice inoculated with stationary-
phase or with purified metacyclic promastigotes. Parasitol. 134: 1699–1707
Franco LH, Beverley SM, Zamboni DS 2012. Innate immune activation and subversion of
Mammalian functions by leishmania lipophosphoglycan. J. Parasitol. Res. 12: 165-226.
Franke ED, McGreevy PB, Katz SP, Sacks DL 1985. Growth cycle-dependent generation of
complement-resistant Leishmania promastigotes. J. Immunol. 134: 2713–2718.
Gamboa D, Eys GV, Victoir K, Torres K, Adaui V, Arevalo J, Dujardin JC 2007. Putative
markers of infective life stages in Leishmania (Viannia) braziliensis. Parasitol. 134: 1689–
1698.
Gamboa D, Torres K, Doncker S De, Zimic M, Arevalo J, Dujardin J-C 2008. Evaluation of
an in vitro and in vivo model for experimental infection with Leishmania (Viannia)
braziliensis and L. (V.) peruviana. Parasitol. 135: 319–326.
Giudice A, Vendrame C, Bezerra C, Carvalho LP, Delavechia T, Carvalho EM, Bacellar O
2012. Macrophages participate in host protection and the disease pathology associated with
Leishmania braziliensis infection. BMC Inf. Diseases 12: 75.
Gordon S 2012. Innate Immune Functions of Macrophages in Different Tissue Environments.
J. Innate Immunity. 1:35-46
Gossage SM, Rogers ME, Bates PA 2003. Two separate growth phases during the
development of Leishmania in sand flies: implications for understanding the life cycle. Int J
Parasitol. 33: 1027–1034.
Handman E, Schnur LF, Spithill TW, Mitchell GF 1986. Passive transfer of Leishmania
lipopolysaccharide confers parasite survival in macrophages. J. Immunol 137: 3608–3613.

96
Hayes MP, Freeman SL, Donnelly RP 1995. IFN-gamma priming of monocytes enhances
LPS-induced TNF production by augmenting both transcription and mRNA stability.
Cytokine 7: 427–435.
Hsiao CHC, Ueno N, Shao JQ, Schroeder KR, Moore KC, Donelson JE, Wilson ME 2011.
The effects of macrophage source on the mechanism of phagocytosis and intracellular
survival of Leishmania. Microbes and infection13: 1033–1044.
Jafari S, Hajiabdolbaghi M, Mohebali M, Hajjaran H, Hashemian H 2010. Disseminated
leishmaniasis caused by Leishmania tropica in HIV-positive patients in the Islamic Republic
of Iran. Eastern Mediterranean Health Journal 16: 340–343.
Kamhawi S 2006. Phlebotomine sand flies and Leishmania parasites: friends or foes? Trends
in Parasitol. 22: 439–445.
Kavoosi G, Ardestani SK, Kariminia A 2009. The involvement of TLR2 in cytokine and
reactive oxygen species (ROS) production by PBMCs in response to Leishmania major
phosphoglycans (PGs). Parasitol. 136: 1193–1199.
Kavoosi G, Ardestani SK, Kariminia A, Alimohammadian MH 2010. Leishmania major
lipophosphoglycan: discrepancy in Toll-like receptor signaling. Exp. Parasitol. 124: 214–218.
Kawai T, Akira S 2010. The role of pattern-recognition receptors in innate immunity: update
on Toll-like receptors. Nature Immunol. 11: 373–384.
Kaye P, Scott P 2011. Leishmaniasis: complexity at the host-pathogen interface. Nature Rev.
Microbiol. 9: 604–615.
Khosravi S, Hejazi SH, Hashemzadeh M, Eslami G, Darani HY 2012. Molecular diagnosis of
Old World leishmaniasis: Real-time PCR based on tryparedoxin peroxidase gene for the
detection and identification of Leishmania spp. Journal of Vector Borne Diseases 49: 15–18.
Killick-Kendrick R 1978. Recent advances and outstanding problems in the biology of
phlebotomine sandflies. A review. Acta Tropica 35: 297–313.
Kobets T, Grekov I, Lipoldova M 2012. Leishmaniasis: prevention, parasite detection and
treatment. Current. Med. Chemistry 19: 1443–1474.
Lemesre JL, Darcy F, Kweider M, Capron A, Santoro F 1988. Requirements of defined
cultivation conditions for standard growth of Leishmania promastigotes in vitro. Acta Tropica
45: 99–108.
Liew FY, Parkinson C, Millott S, Severn A, Carrier M 1990. Tumour necrosis factor (TNF
alpha) in leishmaniasis. I. TNF alpha mediates host protection against cutaneous
leishmaniasis. Immunol. 69: 570–573.
Liew FY, Wei XQ, Proudfoot L 1997. Cytokines and nitric oxide as effector molecules
against parasitic infections. Biological Sciences 352: 1311–1315.

97
Lippuner C, Paape D, Paterou A, Brand J, Richardson M, Smith AJ, Hoffmann K, Brinkmann
V, Blackburn C, Aebischer T 2009. Real-time imaging of Leishmania mexicana-infected
early phagosomes: a study using primary macrophages generated from green fluorescent
protein-Rab5 transgenic mice. Exp. Biology 23: 483–491.
Lira R, Méndez S, Carrera L, Jaffe C, Neva F, Sacks D 1998. Leishmania tropica: the
identification and purification of metacyclic promastigotes and use in establishing mouse and
hamster models of cutaneous and visceral disease. Exp. Parasitol. 89: 331–342.
Lodge R, Descoteaux A 2005. Modulation of phagolysosome biogenesis by the
lipophosphoglycan of Leishmania. Clinical Immunology 114: 256–265.
Louassini M, Adroher FJ, Foulquié MR, Benítez R 1998. Investigations on the in vitro
metacyclogenesis of a visceral and a cutaneous human strain of Leishmania infantum. Acta
Tropica 70: 355–368.
da Luz RI da, Vermeersch M, Dujardin JC, Cos P, Maes L 2009. In vitro sensitivity testing of
Leishmania clinical field isolates: preconditioning of promastigotes enhances infectivity for
macrophage host cells. Antimicrobial Agents and Chemotherapy 53: 5197–5203.
Machado PR, Rosa MEA, Costa D, Mignac M, Silva JS, Schriefer A, Teixeira MM, Bacellar
O, Carvalho EM 2011. Reappraisal of the immunopathogenesis of disseminated
leishmaniasis: in situ and systemic immune response. Trans. R. Soc. Trop. Med. Hyg. 105:
438–444.
Mahoney AB, Sacks DL, Saraiva E, Modi G, Turco SJ 1999. Intra-species and stage-specific
polymorphisms in lipophosphoglycan structure control Leishmania donovani-sand fly
interactions. Biochemistry 38: 9813–9823.
Maioli TU, Takane E, Arantes RM, Fietto JL, Afonso LC 2004. Immune response induced by
New World Leishmania species in C57BL/6 mice. Parasitol Res 94:207-12.
Marín M, Muskus C, Ramírez JR, Arbelaez LF, Alzate JF, Berberich C 2000. The gene
encoding the metacyclogenesis-associated transcript Mat-1 is conserved in the genus
Leishmania and shows a tendency to form dimers upon protein expression. Parasitol. Res. 86:
431–435.
Matta NE, Cysne-Finkelstein L, Machado GMC, Da-Cruz AM, Leon L 2010. Differences in
the antigenic profile and infectivity of murine macrophages of Leishmania (Viannia)
parasites. J. Parasitol. 96: 509–515.
McConville MJ, Naderer T 2011. Metabolic pathways required for the intracellular survival
of Leishmania. Annual Rev. Microbiol. 65: 543–561.
McConville MJ, Turco SJ, Ferguson MA, Sacks DL 1992. Developmental modification of
lipophosphoglycan during the differentiation of Leishmania major promastigotes to an
infectious stage. The EMBO Journal 11: 3593–3600.
Millington OR, Myburgh E, Mottram JC, Alexander J 2010. Imaging of the host/parasite
interplay in cutaneous leishmaniasis. Exp. Parasitol. 126: 310–317.

98
Momen H, Cupolillo E 2000. Speculations on the origin and evolution of the genus
Leishmania. Mem. Inst. Oswaldo Cruz 95: 583–588.
Mosser DM, Brittingham A 1997. Leishmania, macrophages and complement: a tale of
subversion and exploitation. Parasitol. 115: 9–23.
Mukbel RM, Patten C, Gibson K, Ghosh M, Petersen C, Jones DE 2007. Macrophage killing
of Leishmania amazonensis amastigotes requires both nitric oxide and superoxide. The
American J. Trop. Med. Hyg. 76: 669–675.
Murray HW, Berman JD, Davies CR, Saravia NG 2005. Advances in leishmaniasis. Lancet
366: 1561–1577.
Muskus C, Segura I, Oddone R, Turco SJ, Leiby DA, Toro L, Robledo S, Saravia NG 1997.
Carbohydrate and LPG expression in Leishmania viannia subgenus. J. Parasitol. 83: 671–678.
Naderer T, McConville MJ 2008. The Leishmania-macrophage interaction: a metabolic
perspective. Cellular Microbiology 10: 301–308.
Naiff RD, Talhari S, Barrett TV 1988. Isolation of Leishmania guyanensis from lesions of the
nasal mucosa. Mem. Inst. Oswaldo Cruz 83: 529–530.
Nieves E, Pimenta PF 2000. Development of Leishmania (Viannia) braziliensis and
Leishmania (Leishmania) amazonensis in the sand fly Lutzomyia migonei (Diptera:
Psychodidae). J. Med. Entomol. 37: 134–140.
Nikolaev AV, Sizova OV 2011. Synthetic neoglycoconjugates of cell-surface phosphoglycans
of Leishmania as potential anti-parasite carbohydrate vaccines. Biochemistry. 76: 761–773.
Niño A, Camacho M 2005. Leishmania (Viannia) braziliensis growth in vitro culture relies
more on folic acid availability than Leihsmania (Leishmania) amazonensis. Mem. Inst.
Oswaldo Cruz 100: 309–310.
Oliveira MAP, Pires A, Bastos RP, Lima GMCA, Pinto SA, Pereira LIA, Pereira AJCS,
Abrahamsohn IA, Dorta ML, Ribeiro-Dias F 2010. Leishmania spp. parasite isolation through
inoculation of patient biopsy macerates in interferon gamma knockout mice. Ver. Inst. Med.
Trop. de São Paulo 52: 83–88.
Osorio LE, Castillo CM, Ochoa MT 1998. Mucosal leishmaniasis due to Leishmania
(Viannia) panamensis in Colombia: clinical characteristics. The American J. Trop. Med. Hyg.
59: 49–52.
O’Daly JA, Rodriguez MB 1988. Differential growth requirements of several Leishmania spp.
in chemically defined culture media. Acta Tropica 45: 109–126.
Panaro MA, Acquafredda A, Lisi S, Lofrumento DD, Trotta T, Satalino R, Saccia M, Mitolo
V, Brandonisio O 1999. Inducible nitric oxide synthase and nitric oxide production in
Leishmania infantum-infected human macrophages stimulated with interferon-gamma and
bacterial lipopolysaccharide. Int. J. Clin. & Lab. Res. 29: 122–127.

99
Pereira LIA, Dorta ML, Pereira AJCS, Bastos RP, Oliveira MAP, Pinto SA, Galdino H,
Mayrink W, Barcelos W, Toledo VPCP, Lima GMCA, Ribeiro-Dias F 2009. Increase of NK
cells and proinflammatory monocytes are associated with the clinical improvement of diffuse
cutaneous leishmaniasis after immunochemotherapy with BCG/Leishmania antigens. The
American J. of Trop. Med. Hyg. 81: 378–383.
Pereira CG, Silva AL, de Castilhos P, Mastrantonio EC, Souza RA, Romão RP, Rezende RJ,
Pena JD, Beletti ME, Souza MA 2009b. Different isolates from Leishmania braziliensis
complex induce distinct histopathological features in a murine model of infection. Vet
Parasitol 12:231-40.
Pinto-da-Silva LH, Camurate M, Costa K, Oliveira SMP, Cunha-e-Silva NL, Saraiva EMB
2002. Leishmania (Viannia) braziliensis metacyclic promastigotes purified using Bauhinia
purpurea lectin are complement resistant and highly infective for macrophages in vitro and
hamsters in vivo. Int J Parasitol. 32: 1371–1377.
Pinto-da-Silva LH, Fampa P, Soares DC, Oliveira SMP, Souto-Padron T, Saraiva EM 2005.
The 3A1-La monoclonal antibody reveals key features of Leishmania (L) amazonensis
metacyclic promastigotes and inhibits procyclics attachment to the sand fly midgut. Int J
Parasitol. 35: 757–764.
Pita-Pereira D, Lins R, Oliveira MP, Lima RB, Pereira BAS, Moreira OC, Brazil RP, Britto C
2012. SYBR Green-based real-time PCR targeting kinetoplast DNA can be used to
discriminate between the main etiologic agents of Brazilian cutaneous and visceral
leishmaniases. Parasites & vectors 5: 15.
Puentes SM, Sacks DL, Silva RP, Joiner KA 1988. Complement binding by two
developmental stages of Leishmania major promastigotes varying in expression of a surface
lipophosphoglycan. J. Exp. Med. 167: 887–902.
Ramos CS, Franco FAL, Smith DF, Uliana SRB 2004. Characterisation of a new Leishmania
META gene and genomic analysis of the META cluster. FEMS Microbiology Letters 238:
213–219.
Ramos CS, Yokoyama-Yasunaka JKU, Guerra-Giraldez C, Price HP, Mortara R, Smith DF,
Uliana SRB 2011. Leishmania amazonensis META2 protein confers protection against heat
shock and oxidative stress. Exp. Parasitol. 127: 228–237.
Real F, Mortara RA, Rabinovitch M 2010. Fusion between Leishmania amazonensis and
Leishmania major parasitophorous vacuoles: live imaging of coinfected macrophages. PLoS
Neglec. Trop. Diseases 4: 905.
Rebello KM, Britto C, Pereira BAS, Pita-Pereira D de, Moraes MO, Ferreira ABR, Cysne-
Finkelstein L, Otto TD, Côrtes LM de C, Da-Silva GG, Alves CR 2010. Leishmania
(Viannia) braziliensis: influence of successive in vitro cultivation on the expression of
promastigote proteinases. Exp. Parasitol. 126: 570–576.
Reithinger R, Dujardin JC, Louzir H, Pirmez C, Alexander B, Brooker S 2007. Cutaneous
leishmaniasis. The Lancet 7: 581–596.

100
Rey JA, Travi BL, Valencia AZ, Saravia NG 1990. Infectivity of the subspecies of the
Leishmania braziliensis complex in vivo and in vitro. The American J. Trop. Med. Hyg. 43:
623–631.
Rittig MG, Bogdan C 2000. Leishmania-host-cell interaction: complexities and alternative
views. Parasitology Today 16: 292–297.
Sacks DL 1989. Metacyclogenesis in Leishmania promastigotes. Exp. Parasitol. 69: 100–103.
Sacks DL, Brodin TN, Turco SJ 1990. Developmental modification of the lipophosphoglycan
from Leishmania major promastigotes during metacyclogenesis. Mo. Bioche. Parasitol. 42:
225–233.
Sacks DL, Melby PC 2001. Animal models for the analysis of immune responses to
leishmaniasis. Cur. Prot. in Imunol. 19: 19.2-6
Sacks D, Noben-Trauth N 2002. The immunology of susceptibility and resistance to
Leishmania major in mice. Nat. Rev. Immunol. 2: 845–858.
Sacks DL, Perkins PV 1984. Identification of an infective stage of Leishmania promastigotes.
Science 223: 1417–1419.
Sacks DL, Pimenta PF, McConville MJ, Schneider P, Turco SJ 1995. Stage-specific binding
of Leishmania donovani to the sand fly vector midgut is regulated by conformational changes
in the abundant surface lipophosphoglycan. J. Exp. Med. 181: 685–697.
Sacks D, Sher A 2002. Evasion of innate immunity by parasitic protozoa. Nat. Immunol. 3:
1041–1047.
Sacks DL, Silva RP 1989. The generation of infective stage Leishmania major promastigotes
is associated with the cell-surface expression and release of a developmentally regulated
glycolipid. J. Immunol. 139: 3099–3106.
Saraiva EM, Pimenta PF, Brodin TN, Rowton E, Modi GB, Sacks DL 1995. Changes in
lipophosphoglycan and gene expression associated with the development of Leishmania
major in Phlebotomus papatasi. Parasitol. 111: 275–287.
Saraiva EM, Pinto-da-Silva LH, Wanderley JLM, Bonomo AC, Barcinski M a, Moreira MEC
2005. Flow cytometric assessment of Leishmania spp metacyclic differentiation: validation by
morphological features and specific markers. Exp. Parasitol. 110: 39–47.
Sartori A, Oliveira MA, Scott P, Trinchieri G 1997. Metacyclogenesis modulates the ability
of Leishmania promastigotes to induce IL-12 production in human mononuclear cells. J.
Immunol. 159: 2849–2857.
Searle S, Campos AJ, Coulson RM, Spithill TW, Smith DF 1989. A family of heat shock
protein 70-related genes are expressed in the promastigotes of Leishmania major. Nucleic
Acids Res. 17: 5081–5095.

101
Seay MB, Heard PL, Chaudhuri G 1996. Surface Zn-proteinase as a molecule for defense of
Leishmania mexicana amazonensis promastigotes against cytolysis inside macrophage
phagolysosomes. Infect. and Immunity 64: 5129–5137.
da Silva R, Sacks DL 1987. Metacyclogenesis is a major determinant of Leishmania
promastigote virulence and attenuation. Infect. and Immunity 55: 2802–2806.
da Silva JGL, Silva TM, Peloso EF, Machado-Coelho GLL, Mayrink W, Ariosa MCF, Faria,
Silva PM, Marques MJ 2012. Comparison among three polymerase chain reaction assays on
detection of DNA from Leishmania in biological samples from patients with American
cutaneous leishmaniasis. Rev. Soc. Bras. de Med. Trop. 45: 257–259.
Silveira FT, Lainson R, Castro GCM, Laurenti MD, Corbett CEP 2009. Immunopathogenic
competences of Leishmania (V.) braziliensis and L. (L.) amazonensis in American cutaneous
leishmaniasis. Parasite Immunol. 31: 423–431.
Singh S, Sivakumar R 2004. Challenges and new discoveries in the treatment of
leishmaniasis. J. Inf. Chemot. 10: 307–315.
Soares RPP, Cardoso TL, Barron T, Araújo MSS, Pimenta PFP, Turco SJ 2005. Leishmania
braziliensis: a novel mechanism in the lipophosphoglycan regulation during
metacyclogenesis. Int J Parasitol. 35: 245–253.
Späth, G.F. and S. M. Beverley (2001), “An LPG-independent method for isolation of
infective Leishmania metacyclic promastigotes by density gradient centrifugation”, Exptl.
Parasitol. 99: 97-103.
da Souza Carmo, Katz S, Barbiéri CL 2010. Neutrophils reduce the parasite burden in
Leishmania (Leishmania) amazonensis-infected macrophages. PloS one 5: e13815.
T. Naderer & McConville., 2008. The Leishmania–macrophage interaction: a metabolic
perspective. Cellular Microbiology, nº 10(2), pág. 301–308.
Torrico MC, Doncker S, Arevalo J, Ray DL, Dujardin JC 1999. In vitro promastigote fitness
of putative Leishmania (Viannia) braziliensis/Leishmania (Viannia) peruviana hybrids. Acta
Tropica 72: 99–110.
Ueno N, Bratt CL, Rodriguez NE, Wilson ME 2009. Differences in human macrophage
receptor usage, lysosomal fusion kinetics and survival between logarithmic and metacyclic
Leishmania infantum chagasi promastigotes. Cellular Microbiol. 11: 1827–1841.
Veer MJD, Curtis JM, Baldwin TM, DiDonato JA, Sexton A, Mcconville MJ, Handman E,
Schofield L, Veer MJ de 2003. MyD88 is essential for clearance of Leishmania major:
possible role for lipophosphoglycan and Toll-like receptor 2 signaling. Eur. J. of Immunol.
33: 2822–2831.
Vieira-Gonçalves R, Pirmez C, Jorge ME, Souza WJS, Oliveira MP, Rutowitsch MS, Da-
Cruz AM 2008. Clinical features of cutaneous and disseminated cutaneous leishmaniasis
caused by Leishmania (Viannia) braziliensis in Paraty, Rio de Janeiro. Int. J. of Dermatol. 47:
926–932.

102
Vouldoukis I, Riveros-Moreno V, Dugas B, Ouaaz F, Bécherel P, Debré P, Moncada S,
Mossalayi MD 1995. The killing of Leishmania major by human macrophages is mediated by
nitric oxide induced after ligation of the Fc epsilon RII/CD23 surface antigen. Proc. of the
Nat. Acad. of Sci. of EUA 92: 7804–7808.
Wanderley JLM, da Silva LHP, Deolindo P, Soong L, Borges VM, Prates DB, Souza APA,
Barral A, Balanço JMF, Nascimento MTC, Saraiva EM, Barcinski MA 2009. Cooperation
between Apoptotic and Viable Metacyclics Enhances the Pathogenesis of Leishmaniasis.
PLoS ONE 4(5):5733.
World Health Organization WHO 2009. Leishmaniasis. Disponível em
<http://www.who.int/topics/leishmaniasis/en/ >. Acesso em: 21 fevereiro 2010.
Wozencraft AO, Blackwell JM 1987. Increased infectivity of stationary-phase promastigotes
of Leishmania donovani: correlation with enhanced C3 binding capacity and CR3-mediated
attachment to host macrophages. Immunol. 60: 559–563.
Zakai HA, Chance ML, Bates PA 1998. In vitro stimulation of metacyclogenesis in
Leishmania braziliensis, L. donovani, L. major and L. mexicana. Parasitol. 116: 305–309.
Zauli-Nascimento RC, Miguel DC, Yokoyama-Yasunaka JKU, Pereira LI, Oliveira MAP,
Ribeiro-Dias F, Dorta ML, Uliana SRB 2010. In vitro sensitivity of Leishmania (Viannia)
braziliensis and Leishmania (Leishmania) amazonensis Brazilian isolates to meglumine
antimoniate and amphotericin B. Trop. Med. & Int. Health 15: 68–76.

103
ANEXO 1 - PARECER DO COMITÊ DE ÉTICA

104
ANEXO 2 -TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO
Você está sendo convidado(a) para participar, como voluntário(a), de uma pesquisa. Após
receber os esclarecimentos e as informações a seguir, no caso de aceitar fazer parte do estudo,
assine ao final deste documento, que está em duas vias. Uma delas é sua e a outra é do
pesquisador responsável. Em caso de recusa, você não será penalizado(a) de forma alguma.
Em caso de dúvida, você pode procurar o Comitê de Ética em Pesquisa do Hospital das
Clínicas da UFG (CEP/HC-UFG), pelo telefone: (62) 3269-8338 e 3269-8426.
INFORMAÇÕES SOBRE A PESQUISA:
TÍTULO DO PROJETO: Avaliação da metaciclogênese in vitro de isolados de Leishmania
(Associado ao Banco imunológico de leishmanias (Leishbank) da região Centro-Oeste).
PESQUISADOR RESPONSÁVEL: Fátima Ribeiro Dias
TELEFONES PARA CONTATO (inclusive ligações a cobrar): Laboratório de Imunidade
Natural / IPTSP: 062-3209 6116 – Dra. Fátima Ribeiro Dias: 62 3225 3751
PESQUISA COM RISCO MÍNIMO.
Neste estudo pretendemos entender melhor como as pessoas que têm leishmaniose se
defendem da doença. Uma das formas que podemos fazer isso é estudando as células e seus
produtos que participam da resposta contra o parasito. A sua colaboração nesse sentido, seria
a doação de cerca de 10 mL de sangue. Esclarecemos que esta quantidade será obtida
seguindo a colheita da bolsa de sangue, não sendo necessária uma nova punção. O material
será colhido uma única vez e será usado para pesquisar as células. Os resultados obtidos
poderão acrescentar informações importantes no conhecimento da doença, que poderá
beneficiar no futuro todas as pessoas que estão expostas à leishmaniose. Você poderá pedir
qualquer informação sobre os procedimentos, riscos e dúvidas relacionadas à pesquisa,
utilizando os telefones para contato. Finalmente, gostaríamos de dizer que os resultados
obtidos serão publicados em congressos e revistas especializadas, onde não terá nenhuma
informação capaz de fazer a sua identificação como componente do grupo de pesquisa.
Assinatura do pesquisador _____________________________________________

105
CONSENTIMENTO DA PARTICIPAÇÃO DA PESSOA COMO SUJEITO DA PESQUISA
Eu, _______________________________________________________________________,
RG_________________________ CPF_______________________________, abaixo
assinado, concordo em participar do estudo Avaliação da metaciclogênese in vitro de isolados
de Leishmania sp, sob a responsabilidade da pesquisadora Dra. Fátima Ribeiro Dias, como
sujeito voluntário. Fui devidamente informado e esclarecido pelo pesquisador responsável
sobre a pesquisa, os procedimentos nela envolvidos, assim como os possíveis riscos e
benefícios decorrentes de minha participação. Foi me garantido que posso retirar meu
consentimento a qualquer momento, sem que isto leve à qualquer penalidade.
Local e data _______________________________________________________
Nome e Assinatura do sujeito ou responsável:_______________________________
Assinatura Dactiloscópica:
Nome e assinatura do Pesquisador Responsável_____________________________
Presenciamos a solicitação de consentimento, esclarecimento sobre a pesquisa e aceite do
sujeito em participar.
Testemunhas (não ligadas à equipe de pesquisadores):
Nome: _____________________Assinatura: ____________________________
Nome: _____________________Assinatura: ____________________________
• Observações complementares:

106
ANEXO 3 – ARTIGO DA DISSERTAÇÃO