universidade federal da paraíba centro de ciências exatas e da … · 2018-09-05 · universidade...
TRANSCRIPT

Universidade Federal da Paraíba
Centro de Ciências Exatas e da Natureza
Departamento de Sistemática e Ecologia
Programa de Pós-Graduação em Ciências Biológicas - Zoologia
ESTRESSE OXIDATIVO EM HIPPOCAMPUS REIDI GINSBURG, 1933
(TELEOSTEI SYNGNATHIDAE): ESTUDO COMPORTAMENTAL
Kassiano dos Santos Sousa
.
João Pessoa, Paraíba
Junho de 2015

Universidade Federal da Paraíba
Centro de Ciências Exatas e da Natureza
Departamento de Sistemática e Ecologia
Programa de Pós-Graduação em Ciências Biológicas - Zoologia
ESTRESSE OXIDATIVO EM HIPPOCAMPUS REIDI GINSBURG, 1933
(TELEOSTEI SYNGNATHIDAE): ESTUDO COMPORTAMENTAL
KASSIANO DOS SANTOS SOUSA
Dissertação apresentada ao Programa
de Pós-Graduação em Ciências
Biológicas (Zoologia), da
Universidade Federal da Paraíba,
como requisito para obtenção do
título de Mestre em Ciências
Biológicas – Zoologia.
Orientador(a): Profª. Drª. Ierecê Maria de Lucena Rosa
João Pessoa, Paraíba
Junho de 2015

KASSIANO DOS SANTOS SOUSA
ESTRESSE OXIDATIVO EM HIPPOCAMPUS REIDI GINSBURG, 1933
(TELEOSTEI SYNGNATHIDAE): ESTUDO COMPORTAMENTAL
Dissertação apresentada ao Programa
de Pós-Graduação em Ciências
Biológicas (Zoologia), da
Universidade Federal da Paraíba,
como requisito para obtenção do
título de Mestre em Ciências
Biológicas – Zoologia.
.
Data:_________________________________
Resultado: ____________________________
BANCA EXAMINADORA:
Ierecê Maria de Lucena Rosa, Prof.ª Dr.ª, UFPB
Júlio Santos Rebouças, Prof. Dr., UfPB
Luiz Carlos Serrano Lopez, Dr., UFPB


À minha família, pelo respeito, amor,
carinho e incentivo que sempre recebi e
por compreenderem as tantas vezes que
fui ausente. Sem vocês eu nada seria.

5
AGRADECIMENTOS
Nunca fui bom com agradecimentos, mas isso não quer dizer que não tenho muito o que agradecer... tenho até demais. O problema é que traduzir sentimentos de minimamente dois anos, não é algo fácil.
Quero agradecer primeiramente a minha família, pais e irmãos (e um grupo no whatapp com TODA a variedade possível de parentesco da minha família que ignoro há quase dois anos), vocês são demasiadamente importantes para mim. Vocês que me aguentaram quando nem eu mesmo me suportei. Pai e mãe, exemplos de desapego, amo vocês. Kalina, Kardec e Kathiane, irmãos amados, Lívia, Cauã e Hélida, sobrinhos e cunhada, obrigado por tudo.
A Profª Ierecê, por ter me aceito no desafio de me tornar um mestre e por ter acreditado que eu seria capaz, junto com meus amigos do Lapec, de seguir adiante com tantas responsabilidades de se trabalhar com animais vivos, como também por todo o apoio financeiro sempre que necessário.
Ao profº. Júlio, por todos os ensinamentos, tarde e manhãs gastas tentando me fazer entender absorbância, molaridade, modelos multivariados e uma centena de outras coisas que não eram comuns à minha realidade acadêmica. Quero agradecer por toda paciência e me desculpar por tantas mensagens e e-mails. De fato, nada, nada disso seria possível sem sua ajuda também!
A Profª. Tacyana, que sempre me auxiliou e me ajudou quando precisei, por todos os bolos e cafés, por todas as conversas, científicas ou não! Muito, muito obrigado!
Ao profº Paulo, por todos os conselhos desde a graduação e por me ensinar o amor pela fisiologia, obrigado por tudo!
Ao profº. Wallace, que mesmo em meados de viajar ao exterior, aceitou o desafio de me auxiliar no que convém a testes estatísticos que não faziam parte da minha realidade, obrigado!
Ao pessoal do LAPEC (e é aqui que mais vou sentir falta), foram tantos momentos difíceis, momentos de que queríamos desistir de tudo e ir embora, coletas que deram errado, etc etc. No entanto, agora eu vejo o quanto crescemos quanto pessoas. Todos, TODOS são importantes para mim, obrigado.

6
Souto Neto, nem tenho palavras a agradecer a você, alguém que quero próximo a mim para sempre, você foi mais que um amigo, mais que um irmão... obrigado por tudo!
Lua e Ju, nosso rolo eterno durará mais que a eternidade viu (com redundância mesmo, porque sim!)? Não aceito menos que isso, porque como já foi dito certa vez, provavelmente sempre estaremos falando de dança.
Aline e Marianna, duas pessoas incríveis. Aline, obrigado por todas as conversas, dicas e desabafos e Mary, como te falei, não foi nós que nos conhecemos, foram nossas almas que se reconheceram (e pareciam ser velhas amigas). Obrigado por tudo!
Camys e Jessyca, Babys... NOOSSA, como sinto falta de vocês. Sinto falta até das nossas coletas! Sei que ainda nos veremos muito, muito MESMO. Finalmente “É HOJE (Ludmilla, 2015)”!
Lari, Lívia e Thamara... entendam que quando não respondo no Whatapp não é porque não gosto de vocês, tá? Nossa, sinto muita falta de todos nós juntos, das festas e das conversas na pioneira e, principalmente, de praticarmos bulliyng com Lívia. Obrigado mesmo!
Rebeca, Lais, Giovanna... não tenho como expressar meu agradecimento e apreço por vocês, sempre lembrarei de nossos momentos e sempre sentirei falta, mesmo dos mais difíceis. Obrigado por estarem presentes nessa minha jornada, obrigado.
E o que falar de vocês hein, Herta e Emmanuelle (Manu)? Minhas irmãs desde que o mundo é mundo, desde antes de eu me entender por gente. Nossa, crescemos juntos, enquanto pessoas e enquanto profissionais (apesar de Manu continuar com aquele apertão chato). E ainda cresceremos muito mais! Muito obrigado por tudo!
Aos Cavalos-Marinhos, que me mostraram uma nova paixão... ainda que eu venha a trabalhar com mil outros animais, sempre lembrarei de onde tudo começou e o que devo lutar para conservar. Obrigado.
A todos os amigos que tive a oportunidade de fazer no PPGCB e na EAD nesses últimos dois anos, a convivência durante as disciplinas, os aperreios durante os prazos da coordenação e as comemorações que tivemos, levareis essas emoções para sempre comigo.
No final, a única coisa que tenho a dizer é que amo vocês, meus amigos, definitivamente... AMO VOCÊS!

7
SUMÁRIO
INTRODUÇÃO GERAL ......................................................................................................... 16
Cavalo-marinho: O Modelo ..................................................................................................... 17
Desafios na Aquicultura .......................................................................................................... 19
Hipóteses ................................................................................................................................. 20
Contribuições para a Ciência e Industria ................................................................................. 20
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................................... 21
CAPÍTULO I - DEGRADAÇÃO DO PERÓXIDO DE HIDROGÊNIO EM CONDIÇÕES DE CULTIVO DE
HIPPOCAMPUS REIDI ......................................................................................................... 27
1. INTRODUÇÃO ................................................................................................................ 27
2. OBJETIVOS ..................................................................................................................... 29
2.1. Geral: ................................................................................................................................ 29
2.2. Específicos: ....................................................................................................................... 29
3. MATERIAIS E MÉTODOS ................................................................................................. 30
3.1. Qualidade da água ............................................................................................................ 30
3.2. Estudo da decomposição do H2O2 .................................................................................... 30
3.2.1. Procedimentos .......................................................................................................... 30
3.2.2. Medição Óptica ......................................................................................................... 31
3.3. Desenho Experimental ..................................................................................................... 32
3.4. Análises Estatísticas .......................................................................................................... 32
4. RESULTADOS ................................................................................................................. 34
4.1. Comparação entre os tratamentos .................................................................................. 36
5. DISCUSSÃO .................................................................................................................... 37
6. REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................................... 39
CAPÍTULO II - REAÇÕES COMPORTAMENTAIS PARA A AVALIAÇÃO DO ESTRESSE EM
HIPPOCAMPUS REIDI GINSBURG, 1933 (SYNGNATHIDAE) EXPOSTOS AO PERÓXIDO DE
HIDROGÊNIO EM TRATAMENTOS POR IMERSÃO (BANHO).................................................. 42
1. INTRODUÇÃO ................................................................................................................ 42
1.1. Comportamento ............................................................................................................... 43
2. OBJETIVOS ..................................................................................................................... 45
2.1. Geral: ................................................................................................................................ 45
2.2. Específicos: ....................................................................................................................... 45
3. MATERIAIS E MÉTODOS ................................................................................................. 46

8
3.1. Qualidade da água ............................................................................................................ 46
3.2. Desenho Experimental ..................................................................................................... 46
3.3. Análise comportamental .................................................................................................. 48
3.4. Medição Óptica ................................................................................................................ 51
3.5. Análises Estatísticas .......................................................................................................... 51
4. RESULTADOS ................................................................................................................. 52
4.1. Diferenças entre sexos ..................................................................................................... 52
4.2. Diferenças entre os tratamentos (0ml/L, 5ml/L, 12.5ml/L, 13.75ml/L e 15ml/L) ............ 54
5. DISCUSSÃO .................................................................................................................... 59
5.1. Diferenças entre sexos ..................................................................................................... 59
5.2. Diferenças entre os tratamentos (0ml/L, 5ml/L, 12.5ml/L, 13.75ml/L e 15ml/L) ............ 60
6. CONCLUSÕES ................................................................................................................. 65
7. REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................................... 66
CAPÍTULO III - ESTUDO INICIAL SOBRE A UTILIZAÇÃO DA PORFINATOMANGANÊS(III) (MNTE-
2-PYP5+) COMO REGULADOR REDOX CATALÍTICO EM HIPPOCAMPUS REIDI GINSBURG, 1933
(TELEOSTEI SYNGNATHIDAE) .............................................................................................. 73
1. INTRODUÇÃO ................................................................................................................ 73
1.1. O Estresse Oxidativo ......................................................................................................... 74
2. OBJETIVOS ..................................................................................................................... 76
2.1. Geral: ................................................................................................................................ 76
2.2. Específicos: ....................................................................................................................... 76
3. MATERIAIS E MÉTODOS ................................................................................................. 77
3.1. Qualidade da água ............................................................................................................ 77
3.2. Metodologia do Desenho Experimental: Matrizes Doehlert ........................................... 77
3.3. Desenho Experimental ..................................................................................................... 78
3.3. Análise comportamental .................................................................................................. 82
3.4. Medição Óptica ................................................................................................................ 85
3.5. Análises Estatísticas .......................................................................................................... 85
4. RESULTADOS ................................................................................................................. 86
5. DISCUSSÃO .................................................................................................................... 92
6. CONCLUSÃO .................................................................................................................. 94
7. REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................................... 95

9
ÍNDICE DE FIGURAS
INTRODUÇÃO GERAL
Figura 1: Foto de Hippocampus reidi in situ..................................................................18
CAPÍTULO I
Figura 1: Degradação Total (A) e Degradação Média (B) de peróxido de hidrogênio
(H2O2) em água do mar na presença e ausência de luminosidade em ambiente
controlado na ausência de H. reidi, LAPEC, Brasil, 2015. cl, Claro; es,
Escuro..............................................................................................................................34
Figura 2. Degradação Total (A) e Degradação Média (B) de peróxido de hidrogênio
(H2O2) em água do mar na presença e ausência de aeração em ambiente controlado na
ausência de H. reidi, LAPEC, Brasil, 2015. Ae, Aerção; Sae, Ausencia de
aeração.............................................................................................................................35
Figura 3. Degradação Total (A) e Degradação Média (B) de peróxido de hidrogênio
(H2O2) em água do mar em diferentes condições de área de contato da superfície da
água/ar: 5cm² e 20cm², em ambiente controlado na ausência de H. reidi, LAPEC, Brasil,
2015.................................................................................................................................35
Figura 4. Degradação Total (A) e Degradação Média (B) do peróxido de hidrogênio
(H2O2) em água do mar na presença e ausência de aeração em ambiente controlado e na
presença e ausência de H. reidi (3 indivíduos), LAPEC, Brasil, 2015. Ani, Presença de
Animais; SAni, Ausência de Animais.............................................................................35
Figura 5. Degradação Total (A) e Degradação Média (B) de peróxido de hidrogênio
(H2O2) em água do mar entre os tratamentos com condições Claro (cl)/Escuro (es),
Presença (Ae)/Ausência (Sae) de Aeração e Presença (Ani)/Ausência (SAni) de
Animais (n = 9) em ambiente controlado, LAPEC, Brasil,
2015.................................................................................................................................36

10
CAPITULO II
Figura 1. Sistema experimental com recirculação contínua de água, utilizada para
exposição de H. reidi a diferentes concentrações de peróxido de hidrogênio. 1: saída de
água do aquário; 2: entrada de água no aquário; 3: bomba de recirculação de água; 4:
substrato de apoio; 5: Mesa; 6: cadeira para apoio do aquário com bomba de
recirculação......................................................................................................................47
Figura 2. Gráficos dos comportamentos associados a locomoção, em ambos os sexos de
H. reidi (5 machos + 5 réplicas e 5 fêmeas + 5 réplicas), machos (1) e fêmeas (2), em
condições controladas, LAPEC, Brasil, 2015. A. Nados verticais; B. Movimentos
natatórios totais................................................................................................................54
Figura 3. Gráficos dos dados biométricos de H. reidi (n = 20) associados aos
tratamentos com diferentes concentrações de H2O2 em condições controladas, LAPEC,
Brasil, 2015. A. Ganho/perda de peso; B. Taxa de sobrevivência nas diferentes
concentrações...................................................................................................................55
Figura 4. Gráficos dos comportamentos de H. reidi (n = 20) associados a diferentes
concentrações de H2O2 em condições controladas, LAPEC, Brasil, 2015. A. Tempo em
repouso; B. Tempo aderido ao substrato de apoio...........................................................56
Figura 5. Gráficos dos comportamentos de H. reidi (n = 20) associados a diferentes concentrações de H2O2 em
condições controladas, LAPEC, Brasil, 2015. A. Frequência de Pipes; B. Frequência de
Clicks..................................................................................................................................56
Figura 6: Gráficos dos comportamentos de H. reidi (n = 20) associados a diferentes
concentrações de H2O2 em condições controladas, LAPEC, Brasil, 2015. A. Frequência
de vezes que o animal enrolou a cauda na cabeça; B. Frequência do batimento opercular
ao longo de 60min...........................................................................................................56

11
CAPITULO III
Figura 1. Cuboctaedro apresentando a localização dos pontos (13) experimentais para
três diferentes fatores codificados (X1, X2 e X3). Foram obtidos: ao longo do eixo X2,
em um trecho mediano, seis experimentos localizados nos vértices (2, 3, 5, 6, 8 e 11) e
um no centro de um hexágono, em ambos os lados; três experimentos (4, 12 e 13) estão
localizados no trecho superior e outros três experimentos (7, 9 e 10) em trecho
menor...............................................................................................................................78
Figura 2. Sistema experimental com recirculação contínua de água, utilizada para
exposição de H. reidi a diferentes concentrações de peróxido de hidrogênio e MnP. 1:
saída de água do aquário; 2: entrada de água no aquário; 3: bomba de recirculação de
água; 4: substrato de apoio; 5: Mesa; 6: cadeira para apoio do aquário com bomba de
recirculação; 7: Aquário menor para exposição ao MnP; 8: Aquário maior para
exposição ao
H2O2.................................................................................................................................81
Figura 3. Gráficos dos coeficientes de regressão para os experimentos utilizando H2O2
como estressor em H. reidi (n = 14) e a MnP como antioxidante em condições
controladas, LAPEC, Brasil, 2015.. A: Tempo de Nado; B: Tempo em Repouso; C:
Frequência dos comportamentos totais de
estresse.............................................................................................................................86
Figura 4. Superfície de resposta e valores de MnP vs. H2O2 no Ganho/Perda de peso de
H. reidi (n = 14) 30 dias após o experimento, em condições controladas. LAPEC, Brasil,
2015.................................................................................................................................91
Figura 5. Superfície de resposta e valores de MnP vs. H2O2 no tempo gasto por H. reidi
(n = 14) em repouso durante o experimento, em condições controladas. LAPEC, Brasil,
2015.................................................................................................................................91
Figura 6. Superfície de resposta e valores de MnP vs. H2O2 na taxa respiratória de H.
reidi durante o experimento, em condições controladas. LAPEC, Brasil,
2015.................................................................................................................................91

12
ÍNDICE DE TABELAS
CAPÍTULO I
Tabela 1: Sumário das condições experimentais: Claro/Escuro, Presença/Ausência de
Aeração, Presença/Ausência de Animais e Superfície de Contato
Água/Ar...........................................................................................................................33
Tabela 2: Efeito das variáveis na degradação do H2O2. ................................................36
CAPÍTULO II
Tabela 1: Etograma dos dados comportamentais de estresse em H. reidi......................49
Tabela 2: Etograma dos dados comportamentais de manutenção em H. reidi...............50
Tabela 3. Aspectos biométricos sob condições de estresse por H2O2, em ambos os sexos
de H. reidi (5 machos + 5 réplicas e 5 fêmeas + 5 réplicas) em condições controladas,
LAPEC, Brasil, 2015.......................................................................................................53
Tabela 4. Degradação do peróxido de hidrogênio em tratamentos experimentais, em
ambos os sexos de H. reidi (5 machos + 5 réplicas e 5 fêmeas + 5 réplicas) em
condições controladas, LAPEC, Brasil, 2015..................................................................53
Tabela 5. Dados comportamentais de manutenção, em ambos os sexos de H. reidi (5
machos + 5 réplicas e 5 fêmeas + 5 réplicas), sob condições de estresse por H2O2 em
condições controladas, LAPEC, Brasil, 2015..................................................................53
Tabela 6: Dados comportamentais de estresse, em ambos os sexos de H. reidi (5
machos + 5 réplicas e 5 fêmeas + 5 réplicas), sob condições de estresse por H2O2 em
condições controladas, LAPEC, Brasil, 2015..................................................................53
Tabela 7: Aspectos biométricos em H. reidi (n = 20), sob condições de estresse em
variadas concentrações de tratamentos por H2O2 em condições controladas, LAPEC,
Brasil, 2015......................................................................................................................57
Tabela 8: Degradação Total do peróxido de hidrogênio na presença de H. reidi (n = 20),
em variadas concentrações de tratamentos por H2O2 em condições controladas, LAPEC,
Brasil, 2015......................................................................................................................57
Tabela 9: Dados comportamentais de manutenção de H. reidi (n = 20), em variadas
concentrações de tratamentos por H2O2 em condições controladas, LAPEC, Brasil,
2015.................................................................................................................................57

13
Tabela 10: Dados comportamentais de estresse em H. reidi (n = 20), em variadas
concentrações de tratamentos por H2O2 em condições controladas, LAPEC, Brasil,
2015.................................................................................................................................58
CAPÍTULO III
Tabela 1: Valores originais e os valores correspondentes codificados para os
experimentos com H. reidi em condições controladas, LAPEC, Brasil,
2015.................................................................................................................................80
Tabela 2: Matriz experimental obtida pela metodologia de desenho experimental
Doehlert por três fatores. Valores experimentais H. reidi em condições controladas,
LAPEC, Brasil,
2015.................................................................................................................................80
Tabela 3: Etograma dos dados comportamentais de estresse em H.
reidi..................................................................................................................................83
Tabela 4: Etograma dos dados comportamentais de manutenção em H.
reidi..................................................................................................................................84
Tabela 5: Os coeficientes dos modelos obtidos para os dados biométricos em H. reidi
(n = 14) em condições controladas, LAPEC, Brasil,
2015.................................................................................................................................89
Tabela 6: Os coeficientes dos modelos obtidos para os Movimentos de estresse e de
cabeça para H. reidi (n = 14) em condições controladas, LAPEC, Brasil,
2015.................................................................................................................................89
Tabela 7: Os coeficientes dos modelos obtidos para os percentuais de posição e
locomoção de H.
reidi..................................................................................................................................90

14
RESUMO
A utilização do peróxido de hidrogênio (H2O2) como forma de prevenção e
controle de mortalidades associadas com infecções fúngicas, bacterianas, e de
infestações parasitárias em diferentes espécies e ciclos de vida dos peixes é comum na
aquicultura. Sabe-se que o H2O2 afeta a atividade de transcrição celular da base-redox, o
controle a respostas inflamatórias e as respostas imunológicas, estando diretamente
relacionados ao estresse. A grande vantagem do H2O2 é o impacto ambiental mínimo
relacionado com a ausência de resíduos tóxicos; nesse sentido, estudos direcionados ao
desenvolvimento de protocolos que se utilizem de produtos de baixo risco ambiental
para o tratamento de patologias se fazem necessários. Adicionalmente, estudos que
visam ampliar os conhecimentos etológicos como um método não-invasivo de avaliação
do bem-estar animal são de grande importância na avaliação do estresse. Neste trabalho,
foi realizado um estudo com Hippocampus reidi, visando ampliar o conhecimento a
respeito do seu comportamento em condições normais e de estresse, bem como avaliar o
impacto e implicações do uso de H2O2 no manejo/cultivo de cavalos-marinhos e
possível forma de remediação/mitigação desses efeitos. Foi utilizado um total de 43
espécimes de H. reidi, provenientes da população estoque para cultivo experimental do
LAPEC (Laboratório de Peixes – Ecologia e Conservação), Universidade Federal da
Paraíba, Brasil. Observou-se que a presença de animais parece ser um fator decisivo na
degradação do H2O2, por acréscimo de matéria orgânica e bactérias ao sistema, como
também pela própria absorção por parte dos animais. Foi constatado ainda que o H2O2
tem o potencial para ser usado em tratamento por imersão (banho) em H. reidi, tendo
em vista que concentrações abaixo de 545 µM mostraram-se seguras em um período de
60 min de exposição para a espécie. Por outro lado, o peróxido de hidrogênio é tóxico
em concentrações igual/superior a 599 µM e teve profundo impacto sobre o peso,
comportamento e frequência respiratória em H. reidi. Foi testado também se Mn-
porfirina MnTE-2-PyP5+
(MnP) é um potencial antioxidante catalítico para manutenção
e cultivo de H. reidi, minimizando os impactos causados por tratamentos de altas
concentrações de H2O2. Os resultados iniciais obtidos mostraram que o MnP foi
benéfico a H. reidi, uma vez que minimizou os impactos causados por tratamentos de
altas concentrações de H2O2 a longo prazo, apresentando assim um amplo potencial a
ser estudado nessa área.
Palavras-chave: Cavalo-marinho; Comportamento; Porfirina; Cultivo; Água
Oxigenada.

15
ABSTRACT
Hydrogen peroxide (H2O2) is widely used in aquaculture; in order to prevent and
control fish mortality, often associated with fungal, bacterial and others parasitic
infections. H2O2 affects cellular transcriptional activity of redox-based; additionally, it
controls inflammatory and immune responses, being directly related to stress. The main
advantage in H2O2’s usage is the reduced environmental impact related to a lack of toxic
waste. In this direction, research to develop protocols using products with low
environment risks used in pathologies treatment is much needed. Equally important,
studies aimed at expanding the ethological knowledge as a method non-invasive
assessment of animal welfare are central in the evaluation of stress behavior. This study
was conducted with Hippocampus reidi and aimed to understand it’s behavior under
normal and stressful conditions, to assess impact and implications of H2O2’s usage in
management/culture of seahorses, as well as to work with mitigation ways that may be
useful for developing protocols for culturing seahorses. It was used a total of 43
individuals of H. reidi from a population maintained under experimental culture at
LAPEC (Laboratório de peixes –Ecologia e Conservação), Universidade Federal da
Paraíba, Brasil. The presence of animals appeared to be a decisive factor in degrading
H2O2, by addition of organic matter and bacteria into the system, but also by the it’s
absorption by animals. It was also noticed that H2O2 has the potential to be used in
treatment by immersion (bath) in H. reidi, considering that concentrations below 545
µM were considered safe for a period of 60 min exposure of specimens. It should be
noted, however, that hydrogen peroxide was toxic at concentrations equal/greater than
599 µM and had a profound impact on weight, behavior and respiratory rate in H. reidi.
We also tested if Mn-porphyrin MnTE-2-PyP5+
(MnP) is a catalytic antioxidant
potential for maintenance and husbandry of H. reidi, minimizing impacts caused by
treatments of high concentrations of H2O2. Our initial results suggest that MnP has is
beneficial to H. reidi, as it minimized impacts caused by long-term high concentrations
of H2O2 treatment, thus presenting great potential to be studied in that context.
Keywords: Seahorse; Behavior; Porfiryn; Husbandry; Oxygenated water.

16
INTRODUÇÃO GERAL
O manuseio dos peixes para diversos fins, como a medição, amostragem,
marcação e transporte na aquicultura é comum e frequentemente induzem uma resposta
primária de estresse, marcada pelo crescimento do cortisol plasmático e concentrações
de glicose, podendo levar a imunossupressão e ao retardo do crescimento (GRUTTER e
PANKURST, 2000; TREVES-BROWN, 2000; ROTLLANT et al, 2001; WEBER et al,
2009; HEO e SHIN, 2010). O estresse pode ser considerado como um estado que põe
em risco a homeostase e que pode se restabelecer através de um conjunto complexo de
respostas adaptativas (CHROUSOS, 1998). Barton et al. (1986) e Flos et al. (1988)
sugerem um efeito aditivo ou uma resposta fisiológica cumulativa ao estresse.
Para além dos fatores já citados, a utilização do peróxido de hidrogênio (H2O2)
como forma de prevenção e controle de mortalidades associadas com infecções fúngicas
(HOWE et al. 1999; RACH et al. 2004), bacterianas (SPEARE e ARSENAULT 1997;
LUMSDEN et al. 1998; RACH et al. 2000b, 2003; GAIKOWSKI et al. 2003), e de
infestações parasitárias (RACH et al. 2000a; MONTGOMERY-BROCK et al. 2001;
POWELL e CLARK 2004) em diferentes espécies e ciclos de vida dos peixes
(YANONG, 2008), pode estar relacionada ao estresse. Sabe-se, que o peróxido de
hidrogênio, juntamente com outras espécies reativas de oxigênio e/ou nitrogênio
(superóxido, radical hidroxila, peroxinitrito, óxido nítrico, dióxido de nitrogênio, entre
outras) são amplamente reconhecidos como sinalizadores, pelo fato de afetarem a
atividade de transcrição celular da base-redox, o controle a respostas inflamatórias e as
respostas imunológicas. Essas espécies estão diretamente relacionadas ao estresse
oxidativo primário e aumentam o estresse oxidativo secundário (BARTESAGHI et al,
2007; BONELLO et al, 2007; DU et al, 2008; HIDALFO e DONOSO, 2008; LI et al,
2007; RUBBO e RADI, 2008; SALVMINI et al, 2006; SOUZA et al, 2008;
WINTERBOURN, 2008; WINTERBOURN e HAMPTO, 2008). A U.S. Food and
Drug Administration (FDA - EUA) aprovou a utilização de produtos à base de peróxido
de hidrogênio para aquicultura (U.S. Food and Drug Administration, 2002),
aumentando assim o interesse em seu uso. Contudo, ainda não há estudos sobre o
impacto e implicações do estresse oxidativo associado ao uso de H2O2 no
manejo/cultivo de cavalos-marinhos – peixes mundialmente comercializados para fins
ornamentais e para os quais se busca desenvolver protocolo de cultivo (ROSA, 2005;

17
KOLDEWEY e MARTIN-SMITH, 2010; VINCENT et al, 2011; DIAS NETO, 2011) -
e possível forma de remediação/mitigação desses efeitos.
Nesse sentido, Mn-porfirinas (MnP) têm se destacado em estudos pré-clinicos
como uma das classes mais importantes classes de reguladores redox catalíticos
(Batinić-Haberle, 2010); estudos in vivo em diversos modelos animais de fisiopatologias
de caráter oxidativo têm mostrado excelentes resultados (ASLAN et al, 2001;
PIGANELLI et al, 2002; BENOV et al, 2005; CROW et al, 2005; CSONT et al, 2005;
WU et al, 2007; GAUTER-FLECKENSTEIN et al, 2008; REBOUÇAS et al, 2008b;
SOMPOL et al, 2008).
Cavalo-marinho: O Modelo
O gênero Hippocampus engloba peixes ósseos vulgarmente conhecidos como
cavalos-marinhos. O grupo tem distribuição mundial e, juntamente com gêneros de
dragões-marinhos, cavalos-cachimbo e peixes-cachimbo, constitui a família
Syngnathidae (KUITER, 2000; LOURIE et al., 1999; 2004).
Praticamente todas as espécies de cavalos-marinhos figuram na Lista Vermelha
da World Conservation Union (IUCN), o que reflete o status de ameaça global desses
animais, decorrente do comércio internacional, da degradação de seus ambientes e das
capturas diretas e incidentais a que se encontram submetidos (LOURIE et al., 2004).
Não menos relevante, todo o gênero Hippocampus figura no Apêndice II da CITES
(Convention on the International Trade in Endangered Species of Wild Flora and Fauna),
instrumento legal que requer a demonstração por parte dos países exportadores de
cavalos-marinhos de que o comércio não é prejudicial às populações naturais. O Brasil
participa ativamente do comércio internacional de cavalos-marinhos, sendo um dos
maiores exportadores de indivíduos vivos para fins de aquarismo no mundo, e o maior
da América Latina (BAUM; VINCENT, 2005), no qual o comércio destes animais
vivos incide sobre a espécie Hippocampus reidi (Fig. 1). Essa espécie de cavalo-
marinho tem suas densidades mais altas registradas, predominantemente, nas regiões
Norte e Nordeste do litoral brasileiro (ROSA et al., 2007).
Os maiores desafios para o cultivo de cavalos-marinhos nos países em
desenvolvimento são de ordem técnica, como o manejo e as doenças, uma vez que esse

18
cultivo enfrenta os mesmos problemas que a maioria das monoculturas intensivas de
peixes, como a manutenção de uma infraestrutura de cativeiro que diminua as altas
densidades de estocagem e os níveis elevados de estresse. A alimentação inadequada e
as doenças são as principais fontes de mortalidade desses animais em cativeiro,
(WILSON; VINCENT, 1998) que, em alguns casos, surgem em consequência de uma
nutrição deficiente (CASTRO et al., 2008) e ao estresse em que são submetidos. O
fornecimento de dietas apropriadas a uma equilibrada taxa de crescimento/custo viável e
questões ambientais (KOLDEWEY e MARTIN-SMITH, 2010; JENNINGS et al,
2001), como, por exemplo, o destino dos efluentes, requerem ainda estudos. Além
desses fatores, a indisponibilidade de substratos de apoio, o transporte e a circulação de
água inadequada, bem como a manutenção desses animais em aquários comunitários
com espécies mais ágeis e agressivas, levam à debilitação e/ou mortalidade de cavalos-
marinhos. Destaca-se a ocorrência de doenças, tais como infecções bacterianas,
fúngicas, parasitoses, embolia gasosa, entre outras (ALCAIDE et al, 2001; VINCENT;
CLIFTON-HADLEY, 1989; KOLDEWEY, 2005; VINCENT; KOLDEWAY, 2006;
ROSA et al, 2005, 2006); tais enfermidades estão entre os principais fatores que
impedem o pleno desenvolvimento da piscicultura. Tendo em vista a forte exploração
comercial e as dificuldades de manutenção ex-situ de cavalos-marinhos, estudos
voltados para os seus aspectos biológicos e fisiológicos podem contribuir diretamente
para o estabelecimento de protocolos de cultivo bem definidos que forneçam
alternativas à extração desses organismos no ambiente natural.
Fonte:
Acervo LAPEC.

19
Desafios na Aquicultura
A aquicultura é geralmente considerada como a exploração ou a criação de
organismos aquáticos para fins comerciais (LANDAU, 1992) e da aquicultura mundial
(peixes para alimentação e plantas aquáticas) tem crescido significativamente durante a
metade do século passado (KOLDEWEY e MARTIN-SMITH, 2010). Enquanto a maior
parte da produção aquícola no mundo inteiro é dedicada à produção de alimentos, a
produção de peixes ornamentais é um componente importante da aquicultura industrial
em várias nações, e este setor da indústria mostra um crescimento rápido (TLUSTY,
2002).
Em geral, os benefícios da aquicultura são considerados por ter aumentado a
produção mundial de alimentos, diminuído os impactos sobre as populações selvagens,
atingido uma produção mais eficiente, e pelo apoio econômico às comunidades costeiras
menores, através da produção de exemplares jovens e de tamanho comercial de uma
ampla variedade de espécies de peixes por ano (LANDAU, 1992; NEW, 1996; OLSEN,
1996; PILLAY, 1996; WABNITZ et al., 2003; ALVES et al, 2013). No entanto, as
monoculturas intensivas de peixes enfrentam problemas como: surtos de doenças
devido ao aumento da vulnerabilidade à infecção causada por altas densidades de
estocagem e níveis de estresse elevados; fornecimento de dietas apropriadas a uma
equilibrada taxa de crescimento/custo viável; e questões ambientais (WILSON;
VINCENT, 1998; KOLDEWEY e MARTIN-SMITH, 2010; JENNINGS et al, 2001),
como, por exemplo, o destino dos efluentes. Para além destes, o transporte de espécimes
vivos para sustentar o comércio ainda é um desafio. Por vezes, os animais ficam
expostos a resíduos de excreção, à diminuição do oxigênio dissolvido, a superlotação e
fome (IGUCHI et al, 2002; PATERSON et al., 2003; WRIGHT et al, 2007). Embora
geralmente não estudados, a maioria desses benefícios e os riscos se aplicam à
aquicultura de espécies ornamentais (KOLDEWEY e MARTIN-SMITH, 2010).
No Brasil, além desses, o uso indiscriminado de produtos considerados de
elevado risco ecológico no tratamento de doenças, e a escassez de estudos direcionados
à modificação dessa realidade, produzindo tratamentos de baixo risco ambiental ainda
compõem o cenário da aquicultura no país (CRUZ et al., 2005; MAXIMINIANO et al.,
2005; LUVIZOTTO-SANTOS et al., 2009; SILVA et al., 2009). De alguma forma, os
aquicultores devem equilibrar inúmeros fatores para propiciar um ambiente ideal para o

20
organismo do seu interesse (ANDERSON, 2009). Manter esses benefícios e, ao mesmo
tempo, minimizar todas essas problemáticas de cultivo é um dos principais desafios dos
estudos atuais voltados à aquicultura.
Hipóteses
O peróxido de hidrogênio, utilizado comumente na indústria da aquicultura,
demora quanto tempo para se degradar na água salgada? Aeração, luminosidade e
animais influenciam nessa degradação? Qual seria a quantidade ideal, não letal, para o
início de estudos utilizando tratamento de patologia por peróxido de hidrogênio em H.
reidi? Teria o H2O2 algum impacto na saúde dos peixes, comportamento e fisiologia?
Qual a viabilidade da utilização da Mn-porfirina - Mn(III) meso-tetraquis(N-
etilpiridinio-2-il)porfirina - na manutenção da estabilidade celular através de sua ação
antioxidante, utilizando o H2O2 como modelo ao estresse oxidativo?
Contribuições para a Ciência e Indústria
Essa dissertação aborda questões importantes para a etologia e aquicultura de
maneira geral, se mostrando necessário para o desenvolvimento de um protocolo de
manejo eficiente e exequível para criação em cativeiro de Cavalos-Marinho no Brasil.
Estabelecer um protocolo para tratamento de enfermidades e mitigar o estresse a que
esses animais são submetidos é um dos pontos mais delicados desse processo. Sendo
assim, antigas abordagens necessitam ser adaptadas e novas abordagens precisam ser
testadas.
Nesse contexto, estudos envolvendo o peróxido de hidrogênio, utilizado
amplamente na aquicultura para manter a estabilidade da água dos aquários
(combatendo principalmente fungos e cianobactérias), como também no tratamento de
algumas enfermidades que atingem peixes (bactérias e parasitas), associado a utilização
de um biomimético da enzima superóxido dismutase (SOD) que poderá vir a diminuir o
impacto associado ao estresse oxidativo a que esses animais são submetidos são
relevantes e necessários.

21
REFERÊNCIAS BIBLIOGRÁFICAS
ALCAIDE, E.; GIL-SANZ, C.; SANJUAN,E.; AMARO, C.; SILVEIRA, L. , 2001.
Vibrio harveyi causes disease in seahorse, Hippocampus sp. Journal of Fish Diseases,
24, 311-313;
ALVES, R. R. N., OLIVEIRA, T. P. R., ROSA, I. M. L., 2013. Wild animals used as
food and medicine in Brazil. Evidence-Based Complementary and Alternative
Medicine, v 2013, p. 1-12;
ANDERSON, P. A., 2009. The functions of sound production in the lined seahorse,
hippocampus erectus, and effects of loud ambient noise on its behavior and
physiology in captive environments. Dissertation to the graduate school, University of
Florida. 190pp;
ASLAN, M.; RYAN, T. M.; ADLER, B.; TOWNES, T. M.; PARKS, D. A.;
THOMPSON, J. A.; TOUSSON, A.; GLADWIN, M. T.; TARPEY, M. M.; PATEL, R.
P.; BATINIĆ-HABERLE, I.;WHITE, C. R.; FREEMAN, B. A., 2011. Oxygen radical
inhibition of nitric-oxide dependent vascular function in sickle cell disease. Proc.
Natl. Acad. Sci. U. S. A. 98:15215–15220;
BARTESAGHI, S.; FERRER-SUETA, G.; PELUFFO, G.; et al., 2007. Protein
tyrosine nitration in hydrophilic and hydrophobic environments. Amino Acids 32:
501–515;
BARTON, B.A.; SCHRECK, C. B.; SIGISMONDI, L.A, 1986. Multiple acute
disturbances evoque cumulative physiological responses in juvenile chinook
salmon. Trans. Am. Fish. Sot., 115: 245-251;
Batinić-Haberle, I.; Rebouças, J. S.; Spasojević, I., 2010. Superoxide Dismutase
Mimics: Chemistry, Pharmacology, and Therapeutic Potential. Antioxidants &
Redox Signaling. Jullho, 13(6): 877-918.
BAUM, J. K. VINCENT, A. C. J., 2005. Magnitude and inferred impacts of the
seahorse trade in Latin America. Environmental Conservation, v. 32, p. 305-319;
BENOV, L.; BATINIĆ-HABERLE, I. 2005. A manganese porphyrin SOD mimic
suppresses oxidative stress and extends the life span of streptozotocin-diabetic rats.
Free Radic. Res. 38:81–88;
BONELLO, S., ZAHRINGER, C., BELAIBA, R.S., et al., 2007. Reactive oxygen
species activate the HIF-1a promoter via a functional NFkB site. Arterioscl Thromb
Vasc Biol 27: 755–761;
BRASIL - MMA. Instrução normativa nº5, de 21 de maio de 2004. Diário Oficial da
União - seção 1, nº 102, sexta-feira, 28 de maio de 2004;
CASTRO, A.L.C; DINIZ, A.F.; MARTINS, I.Z.; VENDEL, A.L.; OLIVEIRA, T.P.R.;
ROSA, I.L. 2008. Assessing diet composition of seahorses in the wild using a non-
destructive method: Hippocampus reidi (Teleostei: Synhnathidae as a study-case.
Neotropical Ichthyology, v.6, n.4, p.637-644.

22
CHROUSOS, G. P. 1998. Stressors, stress, and neuroendocrine integration of the
adaptive response. Ann. N.Y. Acad. Sci. 851:311–335;
CROW, J. P. et al, 2005. Manganese Porphyrin Given at Symptom Onset Markedly
Extends Survival of ALS Mice. Ann Neurol;58:258–265;
CRUZ, C.; FUJIMOTO, R.Y.; LUZ, R.K.; et al., 2005. Toxicidade aguda e
histopatologia do fígado de larvas de trairão (Hoplias lacerdae) expostas à solução
aquosa de formaldeído a 10%. Pesticidas: r. Ecotoxicol. e meio ambiente. Curitiba.
15: 21-28;
CSONT, T.; VIAPPIANI, S.; SAWICKA, J.; SLEE, S.; ALTAREJOS, J. Y.;
BATINIĆ-HABERLE, I.; SCHULZ, R., 2005. The involvement of superoxide and
iNOS-derived NO in cardiac dysfunction induced by pro-inflammatory cytokines.
J. Mol. Cell. Cardiol. 39:833–840;
DIAS NETO, J. (org.), 2011. Proposta de Plano Nacional de Gestão para o uso
sustentável de cavalos-marinhos do Brasil. Brasília: Ibama. 104p.
DU, W.; ADAM, Z.; RANI, R.; et al., 2008. Oxidative stress in Fanconi anemia
hematopoiesis and disease progression. Antioxid Redox Signal 10: 1909–1921;
FLOS, R., REIG, L., TORRES, P. AND TORT, L. 1988. Primary and secondary
stress responses to grading and hauhng in rainbow trout, Salmo gairdneri.
Aquaculture. 71: 99-106;
GAIKOWSKI, M., A. JOHNSON, J. J. RACH, AND S. M. SCHLEIS, 2003. Efficacy
of hydrogen peroxide in controlling mortality associated with external columnaris
on walleye and channel catfish fingerlings. American Journal of Aquaculture 65:300–
305;
GAUTER-FLECKENSTEIN, B.; FLECKENSTEIN, K.; OWZAR, K.; JIAN, C.;
BATINIĆ-HABERLE, I.; VUJASKOVIĆ, Ž., 2008. Comparison of two Mn
porphyrin-based mimics of superoxidedismutase (SOD) in pulmonary
radioprotection. Free Radic. Biol. Med. 44:982–989;
GRUTTER, A. S.; PANKURST N. W. 2000. The effects of capture, handling,
confinement and ectoparasite on plasma levels of cortisol, glucose and lactate in
the coral reef fish Hemigymnus melapterus. J Fish Biol. 57, 391–401;
HOWE, G. E., W. H. GINGERICH, V. K. DAWSON, AND J. J. OLSON. 1999.
Efficacy of hydrogen peroxide for treating saprolegniasis in channel catfish.
Journal of Aquatic Animal Health 11:222–230;
HEO, G.J.; SHIN, G.2010. Efficacy of benzocaine as an anaesthetic for Crucian
carp (Carassius carassius). Vet. Anaesth. Analg., 37, pp. 132–135;
HIDALFO, C. e DONOSO P., 2008. Crosstalk between calcium and redox signaling:
from molecular mechanisms to health implications. Antioxid Redox Signal 10:
1275–1312;
IGUCHI, K., ITO, F., OGAWA, K., MATSUBARA, N., YODO, T. & YAMASAKI,
T., 2002. Reduction of transport stress of ayu by obligated schooling. Fisheries
Science 68, 849–853;

23
IUCN 2010. IUCN Red List of Threatened. Version 2010.4. Disponível em:
http://www.iucnredlist.org, acessado em 16 de Junho de 2015;
JENNINGS, S.; KAISER, M. J.; REYNOLDS, J. D., 2001. Marine Fisheries Ecology.
Blackwell Sciences, London;
KOLDEWEY, H. J., 2005. Seahorse Husbandry in Public Aquariums: 2005
Manual, with Chapter Contributed by Members of the Syngnathid Discussion
Group. ZSL, London, UK. 137 pp.;
KOLDEWEY, H. J.; MARTIN-SMITH K. M., 2010. A global review of seahorse
aquaculture. Aquaculture. Volume 302, Issues 3–4, Pages 131–152;
KUITER, R. H., 2000. Seahorses, Pipefishes and their Relatives: A Comprehensive
Guide to Syngnathiformes. Chorleywood: TMC Publishing, 240pp.;
LANDAU, M., 1992. Introduction to Aquaculture. Wiley, New York. 440 pp.;
LI, F.; SONVEAUX, P. P.; RABBANI, Z. N.; et al., 2007. Regulation of HIF-1a
stability through S-nitrosylation. Mol Cell 26: 63–74;
LOURIE, S. A.; FOSTER, S. J.; COOPER,E. W. T.; VINCENT, A C. J., 2004. A
Guide to the Identification of Seahorses. Project Seahorse and TRAFFIC North
America. Washington D.C.: University of British Columbia and World Wildlife Fund,
114p.;
LOURIE, S. A., VINCENT, A. C. J., HALL, H. J., 1999. Seahorses: an identification
guide to the world's species and their conservation. Project Seahorse, London, UK,
214pp.;
LUMSDEN, J. S., V. E. OSTLAND, AND H. W. FERGUSON, 1998. Use of hydrogen
peroxide to treat experimentally induced bacterial gill disease in rainbow trout.
Journal of Aquatic Animal Health 10:230–240;
LUVIZOTTO-SANTOS, R.; ELER, M.N.; ESPINDOLA, E.L.G; VIEIRA, E.M., 2009.
O uso de praguicidas nas pisciculturas e pesqueiros situados na bacia do rio Mogi-
Guaçu. B. Inst. Pesca, São Paulo. 35(3): 343 – 358;
MAXIMINIANO, A.A.; et al., 2005. Utilização de drogas veterinárias, agrotóxicos e
afins em ambientes hídricos: demandas, regulamentação e considerações sobre
riscos à saúde humana e ambiental. Ciência e Saúde Coletiva. 10(2): 483-491;
MONTGOMERY-BROCK, D., V. T. SATO, J. A. BROCK, AND C. S. TAMARU,
2001. The application of hydrogen peroxide as a treatment for the ectoparasite
Amyloodinium ocellatum (Brown 1931) on the Pacific threadfin Polydactylus
sexfilis. Journal of the World Aquaculture Society 32:250–254;
NEW, M., 1996. Sustainable global aquaculture. World Aquac. 27, 4–6;
OLSEN, J.H.T., 1996. Developing sustainable aquaculture. World Aquac. 27, 16–17;
PATERSON, B. D., RIMMER, M. A., MEIKLE, G. M. & SEMMENS, G. L., 2003.
Physiological responses of the Asian sea bass, Lates calcarifer, to water quality

24
deterioration during simulated live transport: acidosis, red-cell swelling and levels
of ions and ammonia in the plasma. Aquaculture 218, 717–728;
PIGANELLI, J. D.; FLORES, S. C.; CRUZ, C.; KOEPP, J.; YOUNG, R.; BRADLEY,
B.; KACHADOURIAN, R.; BATINIĆ-HABERLE, I.; HASKINS, K. A., 2002.
Metalloporphyrin superoxide dismutase mimetic (SOD mimetic) inhibits
autoimune diabetes. Diabetes 51:347–355;
PILLAY, T.V.R., 1996. Aquaculture and the Environment. Fishing News Books,
Oxford. 189 pp.;
POWELL, M. D., CLARK, G. A., 2004. Efficacy and toxicity of oxidative
disinfectants for the removal of gill amoebae from the gills of amoebic gill disease
affected Atlantic salmon (Salmo salar L.) in freshwater. Aquaculture Research
35:112–123;
RACH, J. J., M. P. GAIKOWSKI, AND R. T. RAMSAY. 2000a. Efficacy of hydrogen
peroxide to control mortalities associated with bacteria gill disease infections on
hatchery-reared salmonids. Journal of Aquatic Animal Health 12:119–127;
RACH, J. J., M. P. GAIKOWSKI, AND R. T. RAMSAY. 2000b. Efficacy of hydrogen
peroxide to control parasitic infections on hatchery-reared fish. Journal of Aquatic
Animal Health 12:267–273;
RACH, J. J., J. J. VALENTINE, T. M. SCREIER, M. P. GAIKOWSKI, AND T. G.
CRAWFORD. 2004. Efficacy of hydrogen peroxide to control saprolegniasis on
channel catfish (Ictalurus punctatus) eggs. Aquaculture 238:135–142;
REBOUÇAS, J.S.; DEFREITAS-SILVA, G.; SPASOJEVIC, I.; et al. , 2008. The
impact of electrostatics in redox modulation of oxidative stress by Mn porphyrins:
Protection of SOD-deficient Escherichia coli via alternative mechanism where Mn
porphyrin acts as a Mn-carrier. Free Radic. Biol. Med. 45: 201–210;
ROSA, I. L., ALVES, R. R. N., 2007. Pesca e comércio de cavalos-marinhos
(Syngnathidae: Hippocampus) no Norte e Nordeste do Brasil: subsídios para a
conservação e manejo. Povos e Paisagens. Recife: Núcleo de populações em ecologia
e etnobotânica aplicada, p. 115-134.
ROSA, I. L., 2005. National Report - Brazil. In: Bruckner, A.W.; Fields, J.D.;
Daves, N. (eds) The Proceedings of the International Workshop on CITES
Implementation for Seahorse Conservation and Trade. NOAA Techinical
Memorandum NMFS-OPR-35. 1 ed. vol 1. Silver Spring, p46-53.
ROSA, I. L.; ALVES, R. R. N.; BONIFÁCIO, K. M.; MOURÃO, J. S.; OSÓRIO, F.
M.; OLIVEIRA, T. P. R.; NOTTINGHAM, M. C. , 2005. Fishers’ knowledge and
seahorse conservation in Brasil. Journal of Ethnobiology and Ethnomedicene, v. 1,
n. 12;
ROSA, I. L.; SAMPAIO, C. L.; BARROS, A. T., 2006. Collaborative monitoring of
the ornamental trade of seahorses and pipefishes (Teleostei: Syngnathidae) in
Brazil: Bahia state as a case study. Neotropical Ichthyology. V. 4, n. 2, p. 247-252;

25
ROTLLANT, J.; BALM, P.H.M, PEREZ-SANCHEZ, J. et al. 2001. Pituitary and
internal function in gilthead sea bream (Sparus aurata L., Teleostei) after handling
and confinement stress. Gen Comp Endocrinol. 121, 333–342;
RUBBO, H. e RADI, R., 2008. Protein and lipid nitration: role in redox signaling
and injury. Biochim Biophys Acta 1780:1318–1324;
SALVEMINI, D.; DOYLE, T.M.; CUZZOCREA, S., 2006. Superoxide, peroxynitrite
and oxidative/nitrative stress in inflammation. Biochem Soc Trans 34: 965–970;
SILVA, A.L.; MARCASSI ALVES, F.C.; ANDRADE TALMELLI, E.F.; et al., 2009.
Utilização de cloreto de sódio, formalina e a associação destes produtos no controle
de ectoparasitas em larvas de tilápia (Oreochromis niloticus). B. Inst. Pesca, São
Paulo. 35(4): 597-608;
SOUZA, J.M.; PELUFFO, G.; RADI, R., 2008. Protein tyrosine nitration: functional
alteration or just a biomarker? Free Radic Biol Med 45: 357–366;
SOMPOL, P.; ITTARAT, W.; TANGPONG, J.; CHEN, Y.; DOUBINSKAIA, I.;
BATINIĆ-HABERLE, I.; ABDUL, H. M.; BUTTERFIELD, A.; ST. CLAIR, D. K.,
2008. A neuronal model of Alzheimer's disease: an insight into the mechanisms of
oxidative stress-mediated mitochondrial injury. Neuroscience 153:120–130;
SPEARE, D. J., AND G. J. ARSENAULT. 1997. Effects of intermitente hydrogen
peroxide exposure on growth and columnaris disease prevention of juvenile
rainbow trout (Oncorhynchus mykiss). Canadian Journal of Fisheries and Aquatic
Sciences 54:2653–2658;
TLUSTY, M., 2002. The benefits and risks of aquacultural production for the
aquarium trade. Aquaculture 205, 203–219;
TREVES-BROWN, K. M. Anaesthesia. In: Anaesthetics in Applied Fish
Pharmacology. Treves-BrownKM (ed.). Kluwer Academic Publishers, Dordrecht,
Netherlands. 2000. pp. 206–217;
VINCENT, A. C. J. e CLIFTON-HADLEY, R. S., 1989. Parasitic infection of the
seahorse (Hippocampus erectus) – a case report. Journal of Wildlife Diseases. 25,
404-406.
VINCENT, A. C. J.; FOSTER, S. J.; KOLDEWEY, H. J., 2011. Conservation and
management of seahorses and others Syngnathidae, Journal of Fish Biology, v. 78,
p. 1681-1724.
VINCENT, A. C. J; KOLDEWEY, H. J., 2005. An uncertain future for seahorse
aquaculture in conservation and economic contexts. Proceedings of the Regional
Technical Consulation on Stock Enhancement of Species Under International Concern.
Southeast Asian Fisheries Development Center. Iloilo, Panay, Philippines;
WABNITZ, C., TAYLOR, M., GREEN, E., RAZAK, T., 2003. From Ocean to
Aquarium. UNEP–WCMC,Cambridge, UK.
WEBER, R. A.; PELETEIRO, J.B.; GARCÍA MARTÍN, L. O.; ALDEGUNDE, M.
2009. The efficacy of 2-phenoxyethanol, metomidate, clove oil and MS-222 as

26
anaesthetic agents in the Senegalese sole (Solea senegalensis Kaup, 1858).
Aquaculture, 288, pp. 147–150;
WILSON, M. J.; VINCENT; A. C. J., 1998. Preliminary success in closing the life
cycle of exploited seahorse species, Hippocampus spp., in captivity. Aquarium Sci.
Cons. v. 2n. 4, p. 179-196;
WINTERBOURN, C.C., 2008. Reconciling the chemistry and biology of reactive
species. Nat Chem Biol 4: 278–286;
WINTERBOURN, C.C. e HAMPTON, M.B., 2008. Thiol chemistry and specificity in
redox signaling. Free Radic Biol Med 45: 549–561;
WRIGHT, K. A.; WOODS, C. M. C.; GRAY, B. E.; LOKMAN, P. M.; 2007. Recovery
from acute, chronic and transport stress in the pot-bellied seahorse Hippocampus
abdominalis. Journal of Fish Biology. 70, 1447–1457;
WU, T. J.; KHOO, N. H.; ZHOU, F.; DAY, B. J.; PARKS, D. A., 2007. Decreased
hepatic ischemiareperfusion injury by manganese-porphyrin complexes. Free
Radic. Res. 41:127–134;
YANONG, R. P. E., 2014. Use of Hydrogen Peroxide in Finfish Aquaculture.
Fisheries and Aquatic Sciences in the School of Forest Resources and Conservation,
UF/IFAS;
U.S. Food and Drug Administration. 2002. Title 21 Code of Federal Regulations Part
530 (21 CFR 530). Available: http://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/
CFRSearch.cfm?CFRPart=530 (Junho, 2015);

27
CAPÍTULO I - DEGRADAÇÃO DO PERÓXIDO DE HIDROGÊNIO EM
CONDIÇÕES DE CULTIVO DE HIPPOCAMPUS REIDI
1. INTRODUÇÃO
O peróxido de hidrogénio (H2O2) é um importante intermediário nos processos
redox aquáticos, reagindo com metais vestigiais e poluentes, agindo como um forte
agente oxidante em sistemas químicos e biológicos (PETASNE e ZIKA, 1997;
O’SULLIVAN et al, 2005; CLARK et al, 2008). Tem sido amplamente utilizado em
aquicultura como tratamento por imersão (banho) contra muitos organismos causadores
de doenças em diferentes espécies e fases da vida de peixes (YANONG, 2014). Muito
embora publicações que abordem metodologias que empregam o uso do peróxido no
tratamento de doenças e profilaxia em sistemas de cultivo sejam muito escassas, é
notável um consenso para o uso do mesmo pelos aquaristas e criadores de peixes.
O peróxido de hidrogênio é eficaz contra parasitas (RACH et al. 2000a;
MONTGOMERY-BROCK et al. 2001; POWELL e CLARK 2004), fungos (HOWE et
al. 1999; RACH et al. 2004), vírus e bactérias (SPEARE e ARSENAULT 1997;
LUMSDEN et al. 1998; RACH et al. 2000b, 2003; GAIKOWSKI et al. 2003),
apresentando geralmente, uma maior atividade contra as bactérias gram-negativas do
que contra as bactérias gram-positivas (RUSSO et al, 2007). O aumento dos níveis de
atividade de catalase, quando as bactérias são cultivadas sob determinadas condições,
tais como a presença de peróxido ou até à fase estacionária, tem sido relatado
(STORTZ, TARTAGLIA e AMES 1990; GONZALEZ-FLECHA e DEMPLE, 1997;
LOEWEN 1997). Adicionalmente, Angel et al (1999) demonstraram que mesmo níveis
baixos de peróxido de hidrogênio (100 nM) causam estresse oxidativo para bactérias em
águas costeiras, baseado no aumento da concentração de enzima catalase na coluna
d’água. No entanto, a sua toxicidade apresenta valores iniciais e sensibilidade espécie
específica, tanto para microrganismos como para peixes (RUSSO et al, 2007;
YANONG, 2014), sendo, portanto, inexistentes na literatura dados que demonstrem tais
valores para o gênero Hippocampus.

28
Uma grande vantagem do H2O2 é o impacto ambiental mínimo relacionado com
a ausência de resíduos tóxicos, representando uma alternativa mais amigável ao
ambiente (KIEMER e BLACK, 1997). Em água, o H2O2 se decompõe em H2O2(ℓ) →
H2O(ℓ) + ½ O2(g), e que também tem sido usado como uma fonte de oxigênio, durante
o transporte de peixe vivo ou em lagos ornamentais (TAYLOR e ROSS, 1988). Em
aquicultura ornamental de água doce, o uso de doses menores de medicamentos por
períodos de tempo mais longos pode se mostrar favoráveis em algumas situações. O
peróxido de hidrogênio em concentrações baixas como tratamento pode ser aplicado,
por exemplo, em tanques de criações e lagoas, onde uma troca de água pode não ser
uma opção (YANONG, 2014; RUSSO et al, 2007).
O peróxido de hidrogênio parece degradar-se relativamente rápido na presença
de matéria orgânica e de aeração e, na água do mar, esse processo de degradação tem
sido atribuído à decomposição biológica envolvendo enzimas peroxidase de plâncton e
bactérias marinhas (COOPER e ZEPP, 1990; MOFFETT e ZAFIRIOU, 1993;
PETASNE e ZIKA, 1997; YANONG, 2014). Outros dissipadores potenciais incluem
decomposição catalítica mediada por metais de transição, halogenetos e partículas,
decomposição fotoquímica e um mix de dinâmicas físicas (MOFFETT e ZAFIRIOU,
1993; PETASNE e ZIKA, 1997).
Nesse sentido, estudos para compreender a estabilidade do peróxido de
hidrogénio, em diferentes tratamentos e condições ambientais estáveis para o cultivo de
Hippocampus reidi, se fazem necessários para mensurar o tempo e a frequência dessa
degradação em um sistema controlado, uma vez que a exposição excessiva ao H2O2 e a
sua utilização de forma imprópria pode resultar em toxicidade para H. reidi.

29
2. OBJETIVOS
2.1. Geral:
Avaliar a taxa e o tempo de degradação de peróxido de hidrogênio em
condições controladas para o cultivo de H. reidi.
2.2. Específicos:
Mensurar a degradação total e média do peróxido de hidrogênio em água
salgada, ao longo de 24 horas, em um sistema controlado com diferentes
condições de luminosidade e aeração;
Avaliar a degradação total e média do peróxido de hidrogênio em água salgada,
ao longo de 24 horas, na presença e ausência de exemplares de H. reidi.
Identificar os fatores que regulam as taxas de decomposição do peróxido de
hidrogênio.

30
3. MATERIAIS E MÉTODOS
3.1. Qualidade da água
As variáveis físico-químicas dos aquários de populações estoque, bem como dos
demais aquários onde foram realizados os experimentos, foram controladas
constantemente. As variáveis de temperatura, salinidade e pH foram mantidas em 25°C, 27
ppm e 8,2 respectivamente. Os demais parâmetros químicos da água NH3, NH4+ foram
monitorados diariamente e não excederam os limites adequados para cavalos-marinhos
variando entre: 0 – 0,25mg/l (KOLDEWEY, 2005). Foi estabelecido também um
fotoperíodo de 10L : 14E, utilizando luz branca fria acima dos tanques, com intensidade
luminosa de cerca de 500lx na superfície da água.
3.2. Estudo da decomposição do H2O2
3.2.1. Procedimentos
Foram preparadas duas soluções (A e B) para o método I3-
de acordo com a
fórmula usual (Allen et al, 1952).
Solução “A” - Consiste de 33 g de KI, 1 g de NaOH, e 0,1 g de molibdato de
amônio tetrahidratado diluído com 500 mL com água. A solução foi agitada
durante ~10 min para a dissolução do molibdato sendo seu pH final de 12,7. A
solução foi armazenada no escuro total para inibir a oxidação do I-.
Solução “B” – É um tampão aquoso, contém 10g de KHP por 500mL de água, e
pH de 4,04;
O método I3- consiste em misturar volumes iguais das soluções A e B, seguido
pela adição em igual proporção da solução de H2O2. A absorbância da solução
resultante foi medida a 351.0 nm em uma cubeta, para se ter uma absorbância de ~0.9 (a
depender da concentração de H2O2 na solução).
Para se obter a absorbância do branco, foi calculado a absorbância esperada para
a diluição da mistura A/B com água pura e, posteriormente, foram misturados volumes

31
iguais da solução “A”, “B” e da água do mar (sem H2O2), assumindo que a diferença
entre as soluções foi devido a I3- fortuito.
Para os valores de max (I3-) a 351 nm foram assumidos os valores de Klassen et
al (1994).
A reação de H2O2 acidificada com KI para produzir I2, seguido da Iodometria
com tiossulfato de sódio padrão, é chamado de método de Kingzett (REICHERT et al,
1939; SCOTT, 1939; KINGZETT, 1880). Em altas concentrações de KI, o equilíbrio
entre I2, I- e I3
- é fortemente em favor de I3
-. A medição de I3
- pela sua absorção óptica
constitui a base do método I3- (KLASSEN et al, 1994).
H2O2 + 2I- + 2H
+ → I2 + 2H2O
I2 + I- ⇌ I3
-
3.2.2. Medição Óptica
Um espectrofotômetro BEL Photonics SP 1105, que mede a 0.001 unidade de
absorbância, foi utilizado. Os espectros apresentados no presente estudo tiveram o
Branco subtraído. As medições de absorbância foram feitas ~4 minutos após a
colocação da cubeta no espectrofotômetro, a fim de permitir que a temperatura das
soluções se igualem às do espectrofotômetro. Todas as medições de absorbância foram
feitas dentro da faixa de 24-25C.
A absorbância é proporcional à concentração de moléculas absorvedoras de luz
na amostra e ao caminho óptico da substância através da qual a luz viaja. Sendo assim, a
Lei de Beer expressa a dependência da absorbância (A) em relação ao caminho óptico
(b) e a concentração (c):
A = bc

32
3.3. Desenho Experimental
Foram selecionados nove animais com altura média 19.1 cm (±2.3) e peso
úmido 15,3 g (±0,3), respectivamente, provenientes da população estoque para cultivo
experimental do LAPEC (Laboratório de Peixes – Ecologia e Conservação),
Universidade Federal da Paraíba, Brasil. Os animais tiveram o comprimento estimado
com auxílio do software gratuito UTHSCSA Image Tool (WILCOX et al., 2002) e o
peso aferido com uma balança milesimal (Bel M-333).
Os experimentos foram feitos em quatro diferentes condições controladas
(Tabela 1). Para os ensaios envolvendo animais, utilizou-se um aquário de 21 x 36 x 30
cm (20 L), onde três animais, por vez, foram expostos a uma solução de H2O2 a 3% de
ingrediente ativo (Rioquímica) e concentração de 882 mM; os outros ensaios foram
realizados em beckers de 1 L e 2 L respectivamente, sem presença de animais. Para
cada condição experimental foram realizados três experimentos com uma réplica cada.
As concentrações testadas foram baseadas em informações empíricas e arbitradas a
partir da literatura (SAMUILOV et al, 2001; DRABKOVA et al, 2007; LARS-
FLEMMING et al, 2010; 2012). Foi definido o valor de 0,25 ml de H2O2 à 3% por 1 L
de água salgada, quantidade comumente utilizada na indústria para combate ao
surgimento e a proliferação de algas (DRABKOVA et al, 2007).
O H2O2 foi adicionado as 18:00 (h:m) de cada dia de experimento, sob as três
diferentes condições (claro/escuro, presença/ausencia de aeração e presença/ausencia de
animais). Foi retirada uma alíquota, por hora, de cada recipiente e submetido à
determinação espectrofotométrica de H2O2 por iodometria. Toda a água utilizada no
experimento passou por filtragem mecânica (Manta Acrílica – Perlon e Malha 150 µm)
e ultravioleta (Filtro UV Atman-36W), utilizado comumente para controle de
organismos planctônicos e microrganismos em sistemas de cultivo fechado.
3.4. Análises Estatísticas
Os resultados referentes às frequências e taxas de degradação do peróxido
obtidas por tratamento foram mensurados por meio da Análise de Variância (ANOVA),
sendo verificados para normalidade pelo Teste de Shapiro-Wilk (W-test). Diferenças
significativas apresentadas foram então analisadas com o teste a posteriori de Duncan,
diferenciando o(s) grupo(s) formados com as médias mais significativas. O Teste-t de

33
Student foi aplicado para verificar a existência de correlação entre as variáveis
mensuradas. A ferramenta estatística utilizada foi o STATISTICA 8 (Statsoft). Todos os
testes foram considerados significativos quando seus resultados demonstraram um nível
de p<0,05. Dados referentes às análises são apresentados na forma de porcentagem ±
erro padrão.
Condições Observações
Claro/Escuro Claro
Experimento submetido a condições normais de fotoperíodo
(10L : 14E) ao longo de 24 horas;
Escuro
Experimento submetido a condições de escuro constante, ao
longo de 24 horas;
Presença
Experimento submetido a condições normais de fotoperíodo
(10L : 14E) e aeração constante, ao longo de 24 horas. A
aeração consiste na utilização de uma bomba compressora
Boyu (D-200; 1,5 W) para aerar a água;
Presença/Ausência de
Aeração
Ausência
Experimento submetido a condições normais de fotoperíodo
(10L : 14E) e sem aeração, ao longo de 24 horas;
Presença/Ausência de
Animais Presença
Experimento submetido a condições normais de fotoperíodo
(10L : 14E) e aeração constante, com a presença de 3
cavalos-marinho ao longo de 24 horas;
Ausência
Experimento submetido a condições normais de fotoperíodo
(10L : 14E) e aeração constante, sem cavalos-marinho ao
longo de 24 horas;
Área de contato da
superfície da água/ar 5cm2
Experimento submetido a condições normais de fotoperíodo
(10L : 14E) e aeração constante ao longo de 24 horas, onde
a superfície de contato Água/Ar é de 5cm2;
20cm2
Experimento submetido a condições normais de fotoperíodo
(10L : 14E) e aeração constantee aeração constante ao
longo de 24 horas, onde a superfície de contato Água/Ar é
de 20cm2;
Tabela 1: Sumário das condições experimentais em ambiente controlado, LAPEC, Brasil, 2015.:
Claro/Escuro, Presença/Ausência de Aeração, Presença/Ausência de Animais e Superfície de Contato
Água/Ar

34
4. RESULTADOS
Neste experimento, os valores de concentração do H2O2 (~220,5 µM) se
mantiveram constantes na primeira hora de experimento para todos os tratamentos.
Após 15 horas, os tratamentos claro/escuro haviam perdido, respectivamente, 18,4 ±
0,5/15,3 ± 0,4 %; os tratamentos presença/ausência de aeração haviam perdido 4,2 ±
0,1/7 ± 0,6 % e os tratamentos presença/ausência de animais haviam perdido 79,2 ±
4,9/14,9 ± 1 % de sua concentração inicial de H2O2, respectivamente. Ao final das 24
horas, os tratamentos claro/escuro e presença/ausência de aeração ainda possuíam 6,8-
6,5 e 10,8-0,2 µM de H2O2, enquanto que ambos os tratamentos da condição
presença/ausência de animais já haviam degradado todo o H2O2.
A partir do Teste-t de Student realizado com todas as variáveis analisadas (taxa e
frequência de degradação) para cada situação experimental, não foi encontrada
diferença significativa entre os tratamentos para as condições claro/escuro (Fig. 1),
presença/ausência de aeração (Fig. 2) e condição área de contato da superfície da
água/ar (Fig. 3). Apenas para a condição presença/ausência de animais (Fig. 4) foi
observada diferença significativa a partir da análise, no que se refere apenas a taxa de
degradação (t = 2.74; p = 0.01).
Figura 1. Degradação Total (A) e Degradação Média (B) de peróxido de hidrogênio (H2O2) em água do
mar na presença e ausência de luminosidade em ambiente controlado na ausência de H. reidi, LAPEC,
Brasil, 2015. cl, Claro; es, Escuro.

35
Figura 2. Degradação Total (A) e Degradação Média (B) de peróxido de hidrogênio (H2O2) em água do
mar na presença e ausência de aeração em ambiente controlado na ausência de H. reidi, LAPEC, Brasil,
2015. Ae, Aerção; Sae, Ausencia de aeração.
Figura 3. Degradação Total (A) e Degradação Média (B) de peróxido de hidrogênio (H2O2) em água do
mar em diferentes condições de área de contato da superfície da água/ar: 5cm² e 20cm², em ambiente
controlado na ausência de H. reidi, LAPEC, Brasil, 2015.
Figura 4. Degradação Total (A) e Degradação Média (B) do peróxido de hidrogênio (H2O2) em água do
mar na presença e ausência de aeração em ambiente controlado e na presença e ausência de H. reidi (3
indivíduos), LAPEC, Brasil, 2015. Ani, Presença de Animais; SAni, Ausência de Animais.

36
4.1. Comparação entre os tratamentos
A taxa de degradação do H2O2 ao longo de 24 horas apresentou diferença
significativa entre os tratamentos (F5,289 = 6.1766, p < 0.01). A Frequência de
Degradação do peróxido não apresentou diferença significativa entre os tratamentos
(Tabela 2). A partir do teste a posteriori de Duncan foi observada a formação de dois
grupos distintos (G1 = Ani, G2 = demais tratamentos) (Fig. 5).
Claro/Escuro
(%)
Presença/ Ausência de
Aeração (%)
Presença/ Ausência de
Animais (%)
Variáveis Cl Es Ae Sae Ani SAni
Taxa de Degradação
(%) 33.2 ± 0.2 30.6 ± 0.8 20.2 ± 1.1 21.6 ± 1.6 47.2 ± 1.7* 29.3 ± 0.3
Freq. de degradação
(%) 8.9 ± 0.1 8.72 ± 0.1 8.62 ± 0.0 8.69 ± 0.0 12.19 ± 0.4 9.65 ± 0.0
Figura 5. Degradação Total (A) e Degradação Média (B) de peróxido de hidrogênio (H2O2) em água do
mar entre os tratamentos com condições Claro (cl)/Escuro (es), Presença (Ae)/Ausência (Sae) de Aeração
e Presença (Ani)/Ausência (SAni) de Animais (n = 9) em ambiente controlado, LAPEC, Brasil, 2015.
Tabela 2: Efeito das variáveis na degradação do H2O2.
Valores com asterisco (*) denotam diferença significativa.
Cl, Claro; Es, Escuro; Ae, Presença de Aeração; Sae, Ausência de Aeração; Ani, Presença de
Animais; SAni, Ausência de animais.

37
5. DISCUSSÃO
Os dados apresentados no presente estudo demonstram que as concentrações de
peróxido de hidrogênio podem sofrer alterações variadas ao longo do tempo, como
consequência de diversas condições abióticas e principalmente decorrente de fatores
bióticos, tais como a presença de microrganismos e de outros organismos, como peixes
ornamentais - cavalos-marinhos no caso deste trabalho. Os resultados obtidos
corroboram o que vem sendo apresentado na literatura: Russo e colaboradores (2007),
por exemplo, também observou que as concentrações de H2O2 se mantiveram constantes
durante a primeira hora de experimento para todas as condições testadas, e que
alterações nas taxas de degradação de peróxido foram mais elevadas e divergiram na
presença de cinco espécies de peixes ornamentais testadas.
Segundo Yanong (2014), certo número de diferentes elementos, enzimas e
compostos, bem como luz, calor e pH elevado, todos, aceleraram a degradação do
peróxido de hidrogênio. Tort et al. (2003) achou algo parecido em seus experimentos,
onde as concentrações de peróxido de hidrogênio iniciais de 10 e 100 mg/L em tanques
de cultivo não foram mensuráveis após 2-3 dias na presença de aeração e/ou de matéria
orgânica, no entanto, sob condições de água estática, sem aeração ou matéria orgânica,
as concentrações foram reduzidas para metade até 6 dias e se tornaram imensuráveis
após 10 dias. Os dados disponíveis na literatura são, de certa forma, ainda limitados
para fins comparativos, tendo em vista que, as diferentes condições e parâmetros
testados são bastante divergentes e abordam diversas espécies e/ou sua eficácia contra
diferentes patógenos.
Ao contrário do que se acreditava, o efeito da luminosidade e da aeração parece
estar, portanto, muito mais relacionado ao aumento da degradação do peróxido pela
presença de matéria orgânica e bactérias que se aproveitem dos efeitos benéficos destas
variáveis (Ex.: FUJIOKA et al, 1981) do que diretamente daqueles dois parâmetros.
. Experiências de filtragem indicam que partículas ou micro-organismos
desempenham o papel principal na determinação do tempo de vida do H2O2 na água do
mar (PETASNE e ZIKA, 1997). Spotte e Buck (1981) mostraram uma diminuição na
contagem de bactérias (> 75% de redução) em água do mar, após passagem por filtro de
irradiação UV. Neste sentido, o fato de não observarmos diferença significativa entre

38
nenhuma variável para os tratamentos Presença/Ausência de Aeração, Claro/escuro e
Área de Contato da Superfície da Água, pode estar relacionado ao tratamento dado à
água anteriormente aos experimentos, uma vez que a filtragem mecânica e ultravioleta
(comumente utilizadas em sistemas fechados para espécies ornamentais) elimina/retira
grande parte da matéria orgânica e possivelmente dos microrganismos presentes
anteriormente na água.
Spotte e Buck (1981) demonstraram que as fezes dos animais, e adicionalmente
porções de alimento ofertado (peixe cru), contribuem para contagens mais elevadas de
bactérias na água do mar em lagos artificiais, mesmo após a passagem pela irradiação
UV. No nosso caso, os animais não foram alimentados durante os experimentos, porém
podemos sugerir que a simples presenta de exemplares de H. reidi no aquário
experimental e a excreção de compostos nitrogenados com consequente aumento da
matéria orgânica dissolvida podem ter gerado um aumento na quantidade de
microrganismos que secundariamente degradem o peróxido.
Os resultados obtidos neste estudo mostraram que, para as variáveis testadas, a
presença de animais parece ser um fator decisivo na degradação do H2O2, seja por
acréscimo de matéria orgânica e bactérias ao sistema, como também pela possível
absorção por parte dos animais e, contrariamente ao esperado, a luminosidade, aeração e
área de superfície de contato água/ar possuem uma influência muito mais secundária, do
que direta nessa degradação.

39
6. REFERÊNCIAS BIBLIOGRÁFICAS
ALLEN, A. O.; DAVIS, T. W.; ELMORE, G.; GHORMLEY, J. A,; HAINES, B. M.;
HOCHANADEL, C. J. 1949.Oak Ridge Natl. Lab. Publ., ORNL 130;
ANGEL, D.L., FIEDLER, U., EDEN, N., KRESS, N., ADELUNG, D., HERUT, B.,
1999. Catalase activity in macro- and micro-organisms as an indicator of biotic
stress in coastal waters of the eastern Mediterranean Sea. Helgoland Mar. Res. 53,
209–218;
CLARK, C. D., DE BRUYN, W. J., JAKUBOWSKI, S. D., GRANT, S. B., 2008.
Hydrogen peroxide production in marine bathing waters: Implications for fecal
indicator bacteria mortality. Marine Pollution Bulletin 56, 397–401;
COOPER, W.J., ZEPP, R.G., 1990. Hydrogen peroxide decay in water with
suspended soils: evidence for biologically mediated processes. Can. J. Fish. Aquat.
Sci. 47, 888–893;
DRABKOVA, M.; ADMIRAAL, W.; MARSALEK, B., 2007. Combined exposure to
hydrogen peroxide and light : Selective effects on cyanobacteria, green algae, and
diatoms. American Chemical Society, Washington, DC, EUA. Vol. 41, no1, pp. 309-
314;
GAIKOWSKI, M., A. JOHNSON, J. J. RACH, AND S. M. SCHLEIS, 2003. Efficacy
of hydrogen peroxide in controlling mortality associated with external columnaris
on walleye and channel catfish fingerlings. American Journal of Aquaculture 65:300–
305.
GONZALEZ-FLECHA, B., DEMPLE, B., 1997. Homeostatic regulation of
intracellular hydrogen peroxide concentration in aerobically growing Escherichia
coli. J. Bacteriol. 47, 382–388;
HOWE, G. E., W. H. GINGERICH, V. K. DAWSON, AND J. J. OLSON. 1999.
Efficacy of hydrogen peroxide for treating saprolegniasis in channel catfish.
Journal of Aquatic Animal Health 11:222–230.
KIEMER, M.C.B., BLACK, K.D., 1997. The effects of hydrogen peroxide on the gill
tissues of Atlantic salmon, Salmo salar L. Aquaculture 153, 181–189;
KINGZETT, C. T. Journal of the Chemical Society. 1880, 37,792;
KLASSEN, N. V., MARCHINGTON, D., MCGOWANT, H. C. E., 1994. H2O2
Determination by the I3- Method and by KMnO4 Titration. Anal. Chem., 66, 2921-
2925;
KOLDEWEY, H. J., 2005. Syngnathid Husbandry in Public Aquariums: 2005
Manual, With Chapters Contributed by Members of the Syngnathidae Discussion
Group. Project Seahorse and Zoological Society of London. 137pp;
LOEWEN P.C., 1997. Bacterial catalases. In: Oxidative Stress and the Molecular
Biology of Antioxidant Defenses. Ed. by J.G. Scandalios, pp. 273–308. Cold Spring
Harbor Press, 5 Woodbury, NY, USA;

40
LUMSDEN, J. S., V. E. OSTLAND, AND H. W. FERGUSON, 1998. Use of hydrogen
peroxide to treat experimentally induced bacterial gill disease in rainbow trout.
Journal of Aquatic Animal Health 10:230–240;
MOFFETT, J.W., ZAFIRIOU, O.C., 1993. The photochemical decomposition of
hydrogen peroxide in surface waters of the Eastern Caribbean and Orinoco River.
J. Geophys. Res. 98, 2307–2313;
MONTGOMERY-BROCK, D., V. T. SATO, J. A. BROCK, AND C. S. TAMARU,
2001. The application of hydrogen peroxide as a treatment for the ectoparasite
Amyloodinium ocellatum (Brown 1931) on the Pacific threadfin Polydactylus
sexfilis. Journal of the World Aquaculture Society 32:250–254;
O’SULLIVAN, D.W., NEALE, P.J., COFFIN, R.B., BOYD, T.J., OSBURN, C.L.,
2005. Photochemical production of hydrogen peroxide and methylhydroperoxide
in coastal waters. Mar. Chem. 97, 14–33;
PETASNE, R.G., ZIKA, R.G., 1997. Hydrogen peroxide lifetimes in south Florida
coastal and offshore waters. Mar. Chem. 56, 215–225;
PEDERSEN, LARS-FLEMMING; GOOD, C. M.; PEDERSEN, P. B., 2012. Low-Dose
Hydrogen Peroxide Application in Closed Recirculating Aquaculture Systems.
North American Journal of Aquaculture 74:100–106;
PEDERSEN, LARS-FLEMMING; PEDERSEN, P. B., 2010. Hydrogen Peroxide:
Disinfectant For Recirculating Aquaculture Systems. Global Aquaculture Alliance;
POWELL, M. D., AND G. A. CLARK. 2004. Efficacy and toxicity of oxidative
disinfectants for the removal of gill amoebae from the gills of amoebic gill disease
affected Atlantic salmon (Salmo salar L.) in freshwater. Aquaculture Research
35:112–123;
RACH, J. J., M. P. GAIKOWSKI, AND R. T. RAMSAY. 2000a. Efficacy of hydrogen
peroxide to control parasitic infections on hatchery-reared fish. Journal of Aquatic
Animal Health 12:267–273;
RACH, J. J., M. P. GAIKOWSKI, AND R. T. RAMSAY. 2000b. Efficacy of hydrogen
peroxide to control mortalities associated with bacteria gill disease infections on
hatchery-reared salmonids. Journal of Aquatic Animal Health 12:119–127;
RACH, J. J., S. M. SCHLEIS, M. P. GAIKOWSKI, AND A. JOHNSON, 2003.
Efficacy of hydrogen peroxide in controlling mortality associated with external
columnaris on walleye and channel catfish fingerlings. North American Journal of
Aquaculture 4:300–305.
RACH, J. J., J. J. VALENTINE, T. M. SCREIER, M. P. GAIKOWSKI, AND T. G.
CRAWFORD. 2004. Efficacy of hydrogen peroxide to control saprolegniasis on
channel catfish (Ictalurus punctatus) eggs. Aquaculture 238:135–142;
REICHERT, J. S.; MCNEIGHT, S. A.; RUDEL, H. W., 1939. Ind. Eng. Chem. Anal.
Ed. 11, 194;

41
RUSSO, R., E. W. CURTIS, AND R. P. E. YANONG. 2007. Preliminary
investigations of hydrogen peroxide treatment of selected ornamental fishes and
efficacy against external bacteria and parasites in green swordtails. Journal of
Aquatic Animal Health 19:121–127;
SAMUILOV, V. D.; BEZRYADNOV, D. B.; GUSEV, M. V.; KITASHOV, A. V.;
FEDORENKO, T. A., 2001. Hydrogen Peroxide Inhibits Photosynthetic Electron
Transport in Cells of Cyanobacteria. Biochemistry, Moscow. Volume 66, Issue 6, pp
640-645;
SCOTT, W. W., 1939. In Standard Methods of Chemical Analysis, 5th ed.; Furman,
N. H., Ed.; Van Nostrand New York, Vol. 2, p 2180;
SPEARE, D. J., AND G. J. ARSENAULT. 1997. Effects of intermitente hydrogen
peroxide exposure on growth and columnaris disease prevention of juvenile
rainbow trout (Oncorhynchus mykiss). Canadian Journal of Fisheries and Aquatic
Sciences 54:2653–2658.
SPOTTE, S. E BUCK, J. D. 1981. The efficacy of UV irradiation in the microbial
disinfection of marine mammal water. Journal of Wildlife Diseases Vol. 17, No. 1,
January,
STORTZ G., TARTAGLIA L.A. & AMES B.N., 1990. Transcriptional regulation of
oxidative stress inducible genes: direct activation by oxidation. Science 248, 189–
194;
TAYLOR, N. I., AND L. G. ROSS., 1988. The use of hydrogen peroxide as a source
of oxygen for the transportation of live fish. Aquaculture 70:183–192;
TORT, J.M., WOOSTER, G.A., BOWSER, P.R., 2003. Effects of hydrogen peroxide
on hematology and blood chemistry parameters of walleye Stizostedion vitreum. J.
World Aquacult. Soc. 34 (2), 236–242;
YANONG, R. P. E., 2014. Use of Hydrogen Peroxide in Finfish Aquaculture.
Fisheries and Aquatic Sciences in the School of Forest Resources and Conservation,
UF/IFAS;
WILCOX, D.; DOVE, B.; MCDAVID, D. ; GREER, D, 2002. UTHSCSA ImageTool
Program, version 3.0. University of Texas Health Science Center, San Antonio, Texas.
Disponível em: <ftp:maxrad6.uthscsa.edu>

42
CAPÍTULO II - REAÇÕES COMPORTAMENTAIS PARA A AVALIAÇÃO DO
ESTRESSE EM HIPPOCAMPUS REIDI GINSBURG, 1933 (SYNGNATHIDAE)
EXPOSTOS AO PERÓXIDO DE HIDROGÊNIO EM TRATAMENTOS POR
IMERSÃO (BANHO)
1. INTRODUÇÃO
O peróxido de hidrogênio (H2O2), comumente utilizado como um desinfetante
para assepsia de ferimentos de maneira geral, tem sido amplamente utilizado em
aquicultura como tratamento por imersão (banho) contra muitos organismos causadores
de doenças e como forma de prevenção e controle de mortalidades associadas com
infecções fúngicas (HOWE et al. 1999; RACH et al. 2004), bacterianas (SPEARE e
ARSENAULT 1997; LUMSDEN et al. 1998; RACH et al. 2000b, 2003; GAIKOWSKI
et al. 2003), e de infestações parasitárias (RACH et al. 2000a; MONTGOMERY-
BROCK et al. 2001; POWELL e CLARK 2004) em diferentes espécies e ciclos de vida
dos peixes (YANONG, 2008). Muitos agentes patogênicos (incluindo bactérias
anaeróbias) não produzem catalase e superóxido dismutase, o que os tornam mais
sensíveis ao composto, já que não podem quebrar os componentes do peróxido
(KEYER e IMLAY, 1996; IMLAY 2002, 2003). No Brasil, essas enfermidades estão
entre os principais fatores que impedem o pleno desenvolvimento da piscicultura, uma
vez que, no tratamento de doenças, o uso indiscriminado de produtos considerados de
elevado risco ecológico, e a escassez de estudos direcionados a produtos de baixo risco
ambiental ainda compõem o cenário da aquicultura no país (CRUZ et al., 2005;
MAXIMINIANO et al., 2005; LUVIZOTTO-SANTOS et al., 2009; SILVA et al.,
2009).
Uma grande vantagem do H2O2 é o impacto ambiental mínimo relacionado com
a ausência de resíduos tóxicos, representando uma alternativa mais amigável ao
ambiente (KIEMER e BLACK, 1997). No entanto, fatores como a concentração do
produto, o tempo de exposição e a temperatura da água pode levar o peróxido de
hidrogênio a se tornar tóxico, resultando em danos como morbidade (por exemplo,
danos nas brânquias) ou mesmo mortalidade, dependendo do tamanho do peixe
(JOHNSON et al. 1993; THOMASSEN 1993; CLAYTON e SUMMERFELT 1996;
KIEMER e BLACK 1997; RACH et al. 1997; GAIKOWSKI et al. 1999; POWELL e

43
CLARK 2004). Além disso, sua toxicidade apresenta valores iniciais de exposição e
sensibilidade espécie específica, tanto para microrganismos como para peixes (RUSSO
et al, 2007; YANONG, 2014).
Inexiste na literatura, dados que demonstrem tais valores para o gênero
Hippocampus. Uma vez que as doenças estão entre uma das principais fontes de
mortalidade desses animais em cativeiro (WILSON e VINCENT, 1998), estudos
direcionados ao desenvolvimento de protocolos que se utilizem de produtos de baixo
risco ambiental (como o H2O2) para o tratamento de patologias se fazem necessários.
1.1. Comportamento
O peróxido de hidrogênio, juntamente com outras espécies reativas de oxigênio
e/ou nitrogênio (superóxido, radical hidroxila, peroxinitrito, óxido nítrico, dióxido de
nitrogênio, entre outras) são amplamente reconhecidos como sinalizadores, pelo fato de
afetarem a atividade de transcrição celular da base-redox, o controle a respostas
inflamatórias e as respostas imunológicas (BARTESAGHI et al, 2007; BONELLO et al,
2007; DU et al, 2008; HIDALFO e DONOSO, 2008; LI et al, 2007; RUBBO e RADI,
2008; SALVEMINI et al, 2006; SOUZA et al, 2008; WINTERBOURN, 2008;
WINTERBOURN e HAMPTO, 2008; ).
O H2O2, bem como a manipulação, pode gerar uma resposta primária de
estresse, marcada pelo crescimento do cortisol plasmático e concentrações de glicose
(HEO e SHIN, 2010). Dependendo da intensidade e duração do stress, podem ocorrer
ainda respostas secundárias de estresse, que incluem alterações no plasma, nos níveis
dos metabólitos, nos íons dos tecidos, nas características hematológicas e nas proteínas
de choque térmico ou de tensão (PCT), todas relacionadas com os ajustes fisiológicos,
como o metabolismo, a respiração, o estado ácido-base, equilíbrio hidromineral, função
imune e respostas celulares (PICKERING, 1981; IWAMA et al., 1997, 1998;
MOMMSEN et al., 1999) e respostas terciárias como alterações no crescimento,
condição resistência total à doença, o escopo para a atividade metabólica,
comportamento, e em última análise, a sobrevivência, resumidamente, aspectos do
desempenho de todo o animal (WEDEMEYER e MCLEAY, 1981; WEDEMEYER et

44
al., 1990). Essas respostas afetam os peixes em todos os níveis de organização, desde
molecular e bioquímica à população e comunidade (ADAMS, 1990).
Após a exposição a um estressor, a adrenalina aumenta rapidamente
(WENDELAAR BONGA, 1997) e eleva as catecolaminas em circulação
(principalmente noradrenalina e adrenalina) que estão associadas tanto com a
hipoventilação como com a hiperventilação em uma ampla variedade de vertebrados,
quer através de mecanismos reflexos periféricos ou centrais (PEYRAUD-
WAITZENEGGER, 1979; FOLGERING, 1980; FOLGERING et aL, 1982;
ELDRIDGE et aL, 1985; JANSEN et ai, 1986; TAYLOR e RANDALL, 1990;
RANDALL e TAYLOR, 1991). As injeções de catecolaminas na corrente sanguínea do
peixe podem resultar num aumento da ventilação branquial em peixes (PEYRAUD-
WAITZENAGGER, 1979; TAYLOR e WILSON, 1989; MCKENZIE, 1990;
RANDALL e TAYLOR. 1991).
A frequência respiratória tem sido empregada como indicador indireto da
condição de estresse oxidativo em peixes (BARRETO e VOLPATO, 2006; AURÉLIO,
2012), bem como a outros vertebrados, a exemplo dos roedores (GAUTER-
FLECKENSTEIN, 2010). O comportamento é tanto uma sequência de ações
quantificáveis, operando através dos sistemas nervosos central e periférico, como uma
manifestação cumulativa de processos genéticos, bioquímicos e fisiológicos essenciais à
vida (alimentação, reprodução e fuga de predadores) (DUBE e HOSETTI, 2010),
oferecendo uma perspectiva única que liga a fisiologia e ecologia de um organismo e
seu ambiente, servindo, portanto, como biomarcadores de estresse (BEITINGER, 1990;
LITTLE e BREWER, 2001). Apesar de métodos comportamentais possuírem seus
inconvenientes, como a inconsistência de resposta, períodos de formação
excessivamente longos, e estímulos estressantes (KENYON et al., 1998), a frequência
ventilatória vem se mostrando uma forma eficaz para avaliar o estresse, principalmente
porque é um método não-invasivo que concorda, em certo grau, com as medições de
cortisol, evitando assim técnicas dolorosas ou estressantes para serem usadas na
avaliação de estresse, tais como colheita de sangue (BARRETO e VOLPATO, 2004;
2006). Além disso, é um parâmetro barato e facilmente mensurável, já que podem ser
contadas visualmente, de modo que não requer equipamento sofisticado (BARRETO e
VOLPATO, 2006).

45
2. OBJETIVOS
2.1. Geral:
Determinar a toxicidade aguda do peróxido de hidrogênio, com referência aos
seus efeitos sobre o peso, comportamento e a disfunção do consumo de oxigénio
para Hippocampus reidi.
2.2. Específicos:
Avaliar a segurança da utilização do H2O2 como um tratamento químico externo
em H. reidi;
Descrever comportamentos que possam vir a ser bons sinalizadores de estresse
em H. reidi;
Verificar se a frequência respiratória de H. reidi é alterada em função das
concentrações de peróxido de hidrogênio (estressor);
Observar se há diferenças no comportamento entre machos e fêmeas de H. reidi,
quando em situação de estresse agudo;

46
3. MATERIAIS E MÉTODOS
3.1. Qualidade da água
As variáveis físico-químicas dos aquários de populações estoque, bem como dos
demais aquários onde foram realizados os experimentos, foram controladas
constantemente. As variáveis de temperatura, salinidade e pH foram mantidas em 25°C, 27
ppm e 8,2 respectivamente. Os demais parâmetros químicos da água NH3, NH4+ foram
monitorados diariamente e não excederam os limites adequados para cavalos-marinhos
variando entre: 0 – 0,25mg/l (KOLDEWEY, 2005). Foi estabelecido também um
fotoperíodo de 10L : 14E, utilizando luz branca fria acima dos tanques, com intensidade
luminosa de cerca de 500lx na superfície da água.
3.2. Desenho Experimental
Foram selecionados 20 animais (10 machos e 10 fêmeas) com altura média e
peso úmido equivalente a 18,5 g (±2.6) e 14,1 g (±3,6), respectivamente, provenientes
da população estoque para cultivo experimental do LAPEC (Laboratório de Peixes –
Ecologia e Conservação), Universidade Federal da Paraíba, Brasil. Os animais tiveram o
comprimento estimado com auxílio do software gratuito UTHSCSA Image Tool
(WILCOX et al., 2002) e o peso aferido com uma balança milesimal (Bel M-333).
Os tratamentos e durações foram concebidos com base em protocolos que
podem ser facilmente aplicadas em explorações agrícolas comerciais (Gaikowski et al.
1999; A U.S. Food and Drug Administration – EUA, 2002; Yanong, 2014): 60 minutos
de imersão em altas concentrações de H2O2. Este tipo de tratamento, com uma
concentração mais elevada de H2O2, pode ser adaptado a reservatórios de interiores, em
que todo o volume de água pode ser facilmente alterado.
Foi preparado um sistema de exposição que consiste em 1 aquários de 20L (21 x
36 x 30 cm) com recirculação contínua da água (Submersible Pump SP-2500, Boyu),
para homogeneização do peróxido de hidrogênio e constante oxigenação, e um substrato
de apoio (Fig. 1). Todos os experimentos ocorreram entre as 11:00 am e 18:00 pm, uma

47
vez que sabe-se que a frequência respiratória em peixes se alteram ao longo do dia
(BARRETO e VOLPATO, 2006).
Os animais foram expostos, um por vez, a quatro concentrações de tratamento
por H2O2 (218 µM, 545 µM, 599 µM, 654 µM) e um controle sem qualquer
interferência química (0 µM), com uma repetição/concentração, para cada sexo testado,
durante 60min de exposição. A mais alta concentração testada foi determinada com base
em métodos padrão de toxicidade aguda, utilizado para estimar dose críticas em ensaios
de dose-resposta (BLISS, 1935; PELTIER e WEBER, 1985, VALADARES, 2006).
Observações de aspectos biométricos, saúde e comportamento dos animais foram
observados. Foi retirada uma alíquota de água, no início e no final de cada experimento
e medido a sua absorbância em um período máximo de 40 minutos, seguindo o método
I3- de acordo com a fórmula e procedimento usual (ALLEN et al, 1952; KLASSEN et al,
1994).
Foi utilizada uma solução de H2O2 a 3% de ingrediente ativo (Rioquímica) e
concentração de 882 mM. Toda a água utilizada no experimento passou por filtragem
mecânica (Manta Acrílica – Perlon e Malha 150 µm) e ultravioleta (Filtro UV Atman-
36W), utilizado comumente para controle de organismos planctônicos e
microrganismos em sistemas de cultivo fechado, antes de cada experimento.
Figura 1. Sistema experimental com recirculação contínua de água, utilizada para exposição de H. reidi a
diferentes concentrações de peróxido de hidrogênio. 1: saída de água do aquário; 2: entrada de água no
aquário; 3: bomba de recirculação de água; 4: substrato de apoio; 5: Mesa; 6: cadeira para apoio do
aquário com bomba de recirculação.
4

48
3.3. Análise comportamental
Os experimentos foram realizados com uma população de H. reidi clinicamente
saudável (ou seja, sem parasitas externos na pele, nadadeiras e brânquias; sem lesões
externas; com comportamento e alimentação normais; nenhuma mortalidade ou
morbidade conhecida ou observada). Os animais foram filmados (HDR-XR160, Sony) e
monitorados em todo o experimento para análise comportamental e posterior contagem
dos batimentos operculares por minuto (BARRETO e VOLPATO, 2006;
TANTARPALE et al, 2012; AURÉLIO, 2012), utilizando-se de métodos não invasivos
para mensurar o estresse oxidativo. Além disso, a cada 10 min durante o período do
experimento (60min), o número total do tempo que os peixes passaram no fundo, em
natação, em repouso, em exaustão e preso no substrato foi mensurado, bem como a
frequência total de movimentos natatórios horizontais, no fundo, verticais e totais, a
frequência de clicks e gapes (em ambos, a cabeça do animal se inclina para cima e o
hióide se sobressai, no click é instantâneo e no gape é feito de forma lenta) e a
frequência de batimentos operculares/min foram aferidos. Movimentos típicos de
estresse em peixes também foram observados.
As medidas comportamentais foram escolhidas para análise com base nos
trabalhos de Øverli e colaboradores (2002), Barreto e Volpato (2006), Faleiro e
colaboradores (2008), Anderson e colaboradores (2011); são elas:
A proporção do tempo gasto por cada animal, na parte inferior do tanque;
A proporção do tempo gasto por cada animal nadando;
A proporção de tempo gasto por cada animal em repouso (estacionário);
A proporção de tempo gasto pelo animal aderido ao substrato (segurando/preso
sobre apoio) contra o tempo total que o animal estava em repouso;
A proporção do tempo gasto por cada animal em exaustão;
A frequência de movimentos de natação (horizontal, vertical e no fundo do
aquário);
A frequência de comportamentos característicos de estresse em peixes;
Número de Clicks;
Número de Pipes (quando o focinho do animal rompe a superfície da água);

49
Número de Gapes;
Número de Batimentos Operculares por minuto.
Comportamentos que ocorreram tanto na ausência de qualquer estímulo externo
(adição de H2O2) como na presença, foram classificados como comportamento de
manutenção (Tab. 2), são eles: presença dos animais na parte inferior do aquário,
comportamentos de nado (horizontal, vertical e no fundo), repouso (estacionário) do
animal no fundo do aquário e/ou aderido ao substrato (segurando/preso sobre apoio),
Clicks e Gapes. Para este caso, foi medida a alteração do tempo gasto realizando estas
ações e/ou sua frequência em relação aos animais controle. A posição é um artifício de
confinamento e não é traduzível para o comportamento do cavalo-marinho em estado
selvagem; porém, em um ambiente de aquário, cavalos-marinhos podem posicionar-se
em zonas mais calmas (ANDERSON et al, 2011). Por essa razão, foi incluída,
quantificada e analisada.
Os comportamentos que só ocorreram na presença do peróxido de hidrogênio,
foram categorizados como comportamentos de estresse (Tab. 1) para H. reidi, com base
na literatura de comportamentos semelhantes descrito para outras espécies de peixes
(FRANCIS-FLOYD, 1988; BARRETO e VOLPATO, 2006; DUBE e HOSETTI, 2010;
ANDERSON et al, 2011).
Tabela 1: Etograma dos dados comportamentais de estresse em H. reidi.
Etograma de cavalo-marinho Hippocampus reidi
Comportamento de estresse
Posição Posição do animal em relação ao tanque
Exaustão
Quando ocorre à perda da capacidade de realização de ações, onde o animal fica a
deriva no aquário.
Locomoção
Posturas enquanto em locomoção
Nados irregulares
Os animais demonstram movimentos de natação irregulares, erráticos e
dardejantes, seguidos de perda de equilíbrio, podendo ou não serrem
acompanhados do peixe ficar "pendurado" verticalmente na água.
Movimentos da
cabeça
Movimentos da cabeça e suas partes
Pipe
O focinho do animal rompe a superfície da água enquanto os movimentos
operculares continuam.
Cauda enrolada
na cabeça
O animal enrola, insistentemente, a cauda na cabeça e/ou partes da mesma.

50
Tabela 2: Etograma dos dados comportamentais de manutenção em H. reidi.
Etograma de cavalo-marinho Hippocampus reidi
Comportamento manutenção
Posição Posição do animal em relação ao tanque
Topo
Quando o corpo do animal localiza-se em sua maior perção, na metade superior da
altura vertical do tanque;
Fundo
Quando o animal está localizado, em sua maior porção, na metade inferior da altura
vertical do tanque;
Estacionário
Posturas enquanto estiver parado
Empoleirado
O animal está em repouso no tanque, aderido ao substrato.
Sentado
O animal está em repouso no fundo do tanque, livre do substrato. Cabeça e tronco
estão em uma posição vertical.
Locomoção
Posturas enquanto em locomoção
Nado
horizontal
Animal se desloca na horizontal, lateralmente, através da água em uma posição vertical
ou horizontal, nadadeira peitoral e barbatanas dorsais em movimento ondulatório. A
cauda pode ser ondulado ou para a frente, estendendo-se abaixo ou atrás do animal.
Nado
vertical
Animal se desloca na vertical, através da coluna d'água em uma posição vertical,
nadadeira peitoral e barbatanas dorsais em movimento ondulatório. A cauda pode ser
ondulado ou para a frente, estendendo-se abaixo ou atrás do animal.
Nado no
fundo
O animal atravessa o fundo do tanque com o tronco em posição vertical ou
obliquamente na posição vertical, nadadeira peitoral e barbatanas dorsais em
movimento ondulatório. A cauda arrasta na parte inferior, estendida por trás do animal.
Movimentos
da cabeça
Movimentos da cabeça e suas partes
Click
A cabeça se inclina para cima instantaneamente, o hióide se sobressai e a boca está
aberta. Associado a um som de clique.
Gape
O animal abre a boca lentamente, com a cabeça um pouco inclina para cima e o hióide
se projeta. Não há som de clique.

51
3.4. Medição Óptica
Um espectrofotômetro BEL Photonics SP 1105, que mede a 0.001 unidade de
absorbância, foi utilizado. Os espectros apresentados no presente estudo tiveram o
Branco subtraído. As medições de absorbância foram feitas ~4 minutos após a
colocação da cubeta no espectrofotômetro, a fim de permitir que a temperatura das
soluções se igualem as do espectrofotômetro. Todas as medições de absorbância foram
feitas dentro da faixa de 24-25C.
A absorbância é proporcional à concentração de moléculas absorvedoras de luz
na amostra e ao caminho óptico da substância através da qual a luz viaja. Sendo assim, a
Lei de Beer expressa a dependência da absorbância (A) em relação ao caminho óptico
(b) e a concentração (c):
A = bc
3.5. Análises Estatísticas
Os resultados referentes ao Peso Inicial, Peso Inicial/Final, Sobrevivência,
Degradação do H2O2 (Deg. H2O2), Tempos no Fundo, Nadando, em Exaustão, Repouso
e Substrato, Frequência de nados Horizontais (FNH), no fundo (FNF), Verticais (FNV)
e totais (FNT), bem como a frequência de movimentos típicos de estresse como Focinho
para Fora da água, Cauda enrolada na cabeça, Nados Irregulares e Movimento totais de
estresse, Frequência de Clicks e Gapes e Batimento Opercular, obtidas por tratamento
foram mensurados por meio da Análise de Variância (ANOVA), sendo verificados para
normalidade pelo Teste de Shapiro-Wilk (W-test). A fim de normalizar os dados
(quando necessário) os valores foram elevados a raiz quadrada. Diferenças
significativas apresentadas foram então analisadas com o teste a posteriori de Duncan,
diferenciando o(s) grupo(s) formado(s) com as médias significativas. O Teste-t de
Student foi aplicado para verificar a existência de correlação entre as variáveis
mensuradas. A ferramenta estatística utilizada foi o STATISTICA 8 (Statsoft).
Todos os testes foram considerados significativos quando seus resultados
demonstraram um nível de p<0,05. Dados referentes às análises são apresentados na
forma de média ± erro ou porcentagem ± erro padrão.

52
4. RESULTADOS
A toxicidade aguda do peróxido de hidrogênio para a espécie Hippocampus
reidi, durante um período de imersão (banho) de 60 minutos, foi verificada em 15ml/L,
ou 12781 80 µM, onde houve mortalidade de 100%. As concentrações de 11606 73
µM (13,75 ml/L) apresentaram nível subletal, com mortalidade de 50%. Para
concentrações iguais/inferiores a 10474 54 µM (12,5 ml/L), não houve mortalidade
em nenhum dos experimentos, mostrando-se seguras nas condições experimentais.
Entre os comportamentos observados, 4 deles só ocorreram na presença do H2O2
(t = -2.84, p = 0,01) (Tab. 2), são eles: exaustão, nados irregulares, pipe e cauda
enrolada na cabeça. Este último, não descrito até o momento para o gênero
Hippocampus, foi expresso principalmente quando os indivíduos foram expostos a
concentrações letais e subletais do peróxido de hidrogênio (Tab. 10). H. reidi enrolava
a cauda preênsil no focinho/cabeça, variando o tempo de duração do movimento (de 0 a
3 minutos), onde o animal parecia inferir alguma força no movimento.
4.1. Diferenças entre sexos
O Teste-t de Student foi realizado com as variáveis peso inicial/final,
sobrevivência (Tab. 3), nadando, em exaustão, repouso e substrato, frequência de nados
no fundo (FNF), verticais (FNV) e totais (FNT), bem como a frequência de movimentos
típicos de estresse como cauda enrolada na cabeça, nados Irregulares e movimento
totais de estresse, frequência de clicks, não mostrando diferença significativa em
nenhum dos casos (Tab. 5). Para as variáveis peso inicial (Tab. 3), degradação do H2O2
(Deg. H2O2) (Tab. 4), tempos no fundo, frequência de nados horizontais (FNH),
verticais (FNV) e totais (FNT), frequência de gapes e batimento opercular (Tab. 5) e
frequência de movimentos típicos de estresse como frequência de pipes (Tab. 6), foi
realizado o teste de Wilcoxon-Mann-Whitney, onde apenas as variáveis frequência dos
nados verticais (U = 23.5, p < 0.05) (Fig. 2 - A) e nados totais (U = 20.5, p = 0.03) (Fig.
2 - B) apresentaram diferença significativa.

53
Tabela 3: Aspectos biométricos sob condições de estresse por H2O2, em ambos os sexos de H. reidi (5 machos + 5
réplicas e 5 fêmeas + 5 réplicas) em condições controladas, LAPEC, Brasil, 2015.
Aspectos biométricos
Sexo
Variável Macho Fêmea Teste Estatístico P
Peso Inic. - Final (mg) -209.1 ± 191.6 -473.7 ± 322.1 t-value = 0.65 0.53
Sobrevivência (%) 70 ± 14.5 70 ± 15.5 t-value = 0 1.00
Tabela 4: Degradação do peróxido de hidrogênio em tratamentos experimentais, em ambos os sexos de H. reidi
(5 machos + 5 réplicas e 5 fêmeas + 5 réplicas) em condições controladas, LAPEC, Brasil, 2015.
Degradação do H2O2
Sexo
Variável Macho Fêmea Teste Estatístico P
Deg. H2O2 (%) 2 ± 0.5 3 ± 0.6 U = 37.0 0.33
Tabela 5: Dados comportamentais de manutenção, em ambos os sexos de H. reidi (5 machos + 5 réplicas e 5
fêmeas + 5 réplicas), sob condições de estresse por H2O2 em condições controladas, LAPEC, Brasil, 2015.
Dados comportamentais de manutenção
Sexo
Variável Macho Fêmea Teste Estatístico P
T. no Fundo (%) 76.7 ± 6.3 66 ± 6.3 U = 36.5 0,31
T. de Nado (%) 46.8 ± 9 51.9 ± 7.9 t-value = -1.41 0,69
T. de Repouso (%) 53.2 ± 9 34.8 ± 8 t-value = 1.45 0,16
T. no Substrato (%) 17.4 ± 7.9 14.6 ± 9.2 t-value = 0.23 0,82
FNH 9 ± 2.5 10.9 ± 2.4 U = 41.5 0,52
FNF 6.7 ± 2.1 12.5 ± 2.9 t-value = -1.56 0,14
FNV 18.8 ± 3.6 34.3 ± 5.5 U = 23.5 0,05
FNT 34.5 ± 6.1 57.8 ± 9.8 U = 20.5 0,03
Click 3.2 ± 0.8 2.5 ± 0.6 t-value = 0.59 0,57
Gape 0 0.5 ± 0.1 U = 21.0 0,65
Btm. Opercular 49.4 ± 2.4 44.8 ± 2.2 U = 32 0,17
Tabela 6: Dados comportamentais de estresse, em ambos os sexos de H. reidi (5 machos + 5 réplicas e 5 fêmeas + 5
réplicas), sob condições de estresse por H2O2 em condições controladas, LAPEC, Brasil, 2015.
Dados comportamentais de estresse
Sexo
Variável Macho Fêmea Teste Estatístico P
T. de Exaustão (%) 0 13.3 ± 6.5 t-value = -1.94 0,07
Nados irregulares 3.5 ± 1.2 3 ± 1.3 t-value = 0.27 0,79
Pipe 0.9 ± 0.3 2.5 ± 0.7 U = 28.5 0,10
Cauda enrolada cabeça 1.7 ± 0.7 3.2 ± 1.3 t-value = -0.96 0,35
Mov. Estresse totais 9.8 ± 2.5 8.7 ± 2.7 t-value = 0.30 0,77 Deg. H2O2, Degradação do peróxido em durante os 60 min de experimento; FNH, Frequência de Nado na
Horizontal; FNF, Frequência de Nados no Fundo; FNV, Frequência de Nados na Vertical; FNT, Frequência de
Movimentos Natatórios Totais.
Deg. H2O2, Degradação do peróxido em durante os 60 min de experimento; FNH, Frequência de Nado na
Horizontal; FNF, Frequência de Nados no Fundo; FNV, Frequência de Nados na Vertical; FNT, Frequência de
Movimentos Natatórios Totais.

54
Figura 2. Gráficos dos comportamentos associados a locomoção, em ambos os sexos de H. reidi (5
machos + 5 réplicas e 5 fêmeas + 5 réplicas), machos (1) e fêmeas (2), em condições controladas,
LAPEC, Brasil, 2015. A. Nados verticais; B. Movimentos natatórios totais.
4.2. Diferenças entre os tratamentos com H2O2 (0 µM, 218 µM, 545 µM, 599 µM e
654 µM)
H. reidi expostos ao H2O2 tiveram perda de peso (-656,9 ± 190,2 mg) em relação
aos animais que não foram expostos (447,3 ± 40,1 mg), sendo a perda de peso maior
para as maiores concentrações de H2O2 (F4, 15 = 11.7, p < 0.00) (Fig. 3 – A). A
sobrevivência apresentou também diferença significativa entre os tratamentos (F4, 15 =
19.0, p < 0.00) (Tab. 7). A partir do teste a posteriori de Duncan foi observada a
formação de dois grupos distintos (G1 = 599 µM e 654 µM, G2 = demais tratamentos)
(Fig. 3 - A) para a variável peso inicial/final e de três grupos distintos (G1 = 599 µM,
G2 = 654 µM e G3 = demais tratamentos) para a sobrevivência (Fig. 3 - B).
A Degradação do peróxido durante o experimento não apresentou diferença
significativa entre tratamentos (Tab. 8).
Para os dados de comportamento, foi observado que animais em aquários
contendo H2O2 passavam um tempo muito maior nadando (58,6 ± 4,8 %) do que os
animais em ausência total do peróxido (12,3 ± 10,1 %, F4, 15 = 4.99, p < 0.01), e
consequentemente, menos tempo em repouso (F4, 15 = 6.48, p = 0.003). Foi encontrada
diferença significativa também entre frequência de nados horizontais (F4, 15 = 3.6, p =
0.03), verticais (F4, 15 = 3.68, p < 0.03) e totais (F4, 15 = 4.44, p = 0.01), as frequências de
A
A
B
B
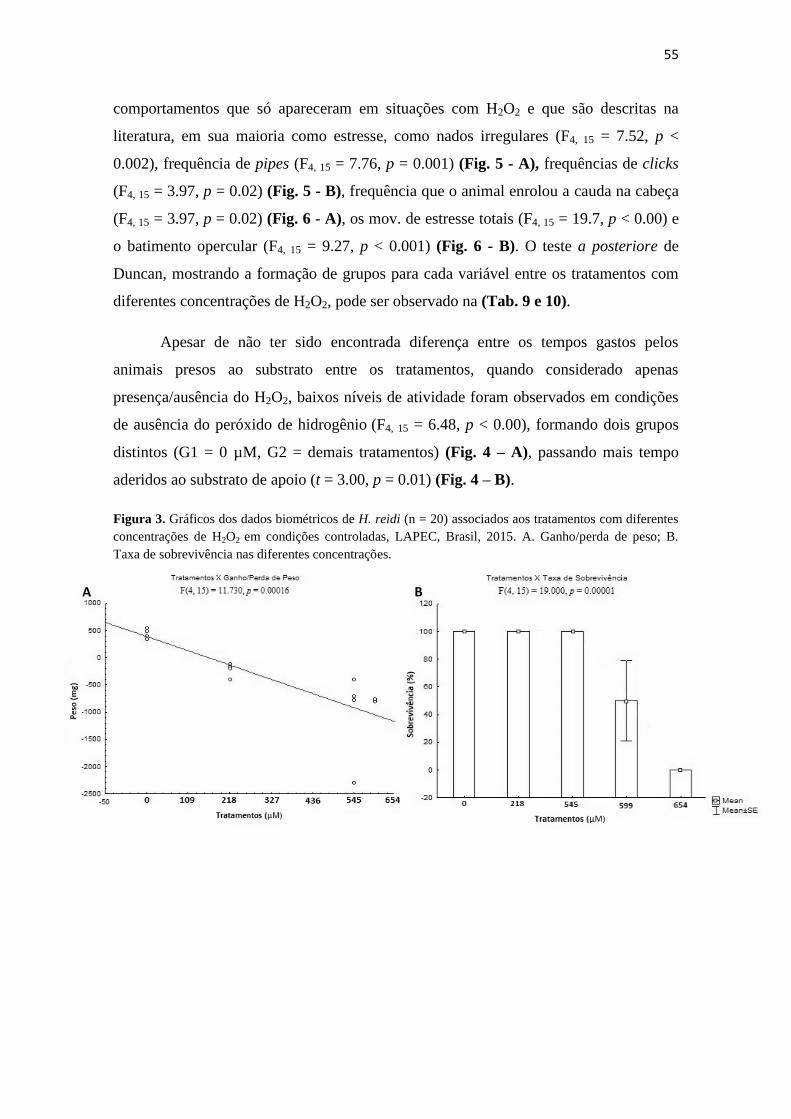
55
comportamentos que só apareceram em situações com H2O2 e que são descritas na
literatura, em sua maioria como estresse, como nados irregulares (F4, 15 = 7.52, p <
0.002), frequência de pipes (F4, 15 = 7.76, p = 0.001) (Fig. 5 - A), frequências de clicks
(F4, 15 = 3.97, p = 0.02) (Fig. 5 - B), frequência que o animal enrolou a cauda na cabeça
(F4, 15 = 3.97, p = 0.02) (Fig. 6 - A), os mov. de estresse totais (F4, 15 = 19.7, p < 0.00) e
o batimento opercular (F4, 15 = 9.27, p < 0.001) (Fig. 6 - B). O teste a posteriore de
Duncan, mostrando a formação de grupos para cada variável entre os tratamentos com
diferentes concentrações de H2O2, pode ser observado na (Tab. 9 e 10).
Apesar de não ter sido encontrada diferença entre os tempos gastos pelos
animais presos ao substrato entre os tratamentos, quando considerado apenas
presença/ausência do H2O2, baixos níveis de atividade foram observados em condições
de ausência do peróxido de hidrogênio (F4, 15 = 6.48, p < 0.00), formando dois grupos
distintos (G1 = 0 µM, G2 = demais tratamentos) (Fig. 4 – A), passando mais tempo
aderidos ao substrato de apoio (t = 3.00, p = 0.01) (Fig. 4 – B).
Figura 3. Gráficos dos dados biométricos de H. reidi (n = 20) associados aos tratamentos com diferentes
concentrações de H2O2 em condições controladas, LAPEC, Brasil, 2015. A. Ganho/perda de peso; B.
Taxa de sobrevivência nas diferentes concentrações.
A
A
B
B

56
Figura 4. Gráficos dos comportamentos de H. reidi (n = 20) associados a diferentes concentrações de H2O2 em
condições controladas, LAPEC, Brasil, 2015. A. Tempo em repouso; B. Tempo aderido ao substrato de apoio.
Figura 5. Gráficos dos comportamentos de H. reidi (n = 20) associados a diferentes concentrações de H2O2 em
condições controladas, LAPEC, Brasil, 2015. A. Frequência de Pipes; B. Frequência de Clicks.
Figura 6: Gráficos dos comportamentos de H. reidi (n = 20) associados a diferentes concentrações de H2O2 em
condições controladas, LAPEC, Brasil, 2015. A. Frequência de vezes que o animal enrolou a cauda na cabeça;
B. Frequência do batimento opercular ao longo de 60min.
A
A
B
B
A
A
B
B
B
B
A
A

57
Tabela 7: Aspectos biométricos em H. reidi (n = 20), sob condições de estresse em variadas
concentrações de tratamentos por H2O2 em condições controladas, LAPEC, Brasil, 2015.
Biometria
Variável 0 µM
(0 ml/L)
218 µM
(5 ml/L)
545 µM
(12,5 ml/L)
599 µM
(13,7 ml/L)
654 µM
(15 ml/L) P
Peso Inic./Final (mg) 447.3 ±
40
-214.25 ±
55
-1040 ±
368.3a
-776 ±
13.4a - 0,01
Sobrevivência (%) 100 100 100 50 ± 25a 0
b 0,00
Tabela 8: Degradação Total do peróxido de hidrogênio na presença de H. reidi (n = 20), em variadas
concentrações de tratamentos por H2O2 em condições controladas, LAPEC, Brasil, 2015.
Degradação do H2O2
Variável 0 µM
(0 ml/L)
218 µM
(5 ml/L)
545 µM
(12,5 ml/L)
599 µM
(13,7 ml/L)
654 µM
(15 ml/L) p
Deg. H2O2 (%) - 2.5 ± 0.2 2.7 ± 0.8 3.5 ± 1.1 3.8 ± 0.2 0,60
Tabela 9: Dados comportamentais de manutenção de H. reidi (n = 20), em variadas concentrações de
tratamentos por H2O2 em condições controladas, LAPEC, Brasil, 2015.
Comportamento de manutenção
Variável 0 µM
(0 ml/L)
218 µM
(5 ml/L)
545 µM
(12,5 ml/L)
599 µM
(13,7 ml/L)
654 µM
(15 ml/L) p
T. no Fundo (%) 96.9 ± 2.5 70.3 ± 5.2 55.7 ± 11.8 67.4 ± 6.3 66.5 ± 9.1 0,08
T. de Nado (%) 12.3 ± 10.1 68 ± 8.2a 43.7 ± 13.7
a 58.6 ± 5
a 64.2 ± 2.2
a 0,01
T. de Repouso (%) 87.7 ± 10.1 32 ± 8.2a 35.7 ± 11.4
a 41.4 ± 5
a 23.2 ± 6.8
a 0,00
T. no Substrato (%) 47.2 ± 21.2 13.8 ± 9.9 11.7 ± 5 6.4 ± 2.7 0.8 ± 0.4 0,13
FNH 2.4 ± 2 16.3 ± 3.9a 8.9 ± 4
a 13.2 ± 2.6
a 9.1 ± 2.6
a 0,03
FNF 3.9 ± 3.1 15.7 ± 4.4 5.9 ± 2.3 9.6 ± 4.4 13 ± 3.3 0,30
FNV 11.4 ± 9.6 35.2 ± 5a 18.4 ± 5 38.3 ± 7.8
a 29.5 ± 4
a 0,03
FNT 17.7 ± 14.7 67.2 ± 12.5a 33.3 ± 7.2 61 ± 11.9
a 51.6 ± 3.8
a 0,01
Click 0.7 ± 0.5d 1.2 ± 0.6
cd 3.3 ± 0.5
bcd 3.8 ± 1
ac 5.2 ± 1.2
ab 0,02
Gape 0.2 ± 0.1 0,0 0,0 0,0 0.2 ± 0.1 0,26
Btm. Opercular 38.5 ± 1.3 40.9 ± 2.0 52.3 ± 2.6a 54.6 ± 2.8
a 49.3 ± 1.3
a 0,00
Valores com letras iguais denotam diferença não significativa entre elas.
Deg. H2O2, Degradação do peróxido em durante os 60 min de experimento; FNH, Frequência de Nado na
Horizontal; FNF, Frequência de Nados no Fundo; FNV, Frequência de Nados na Vertical; FNT,
Frequência de Movimentos Natatórios Totais.
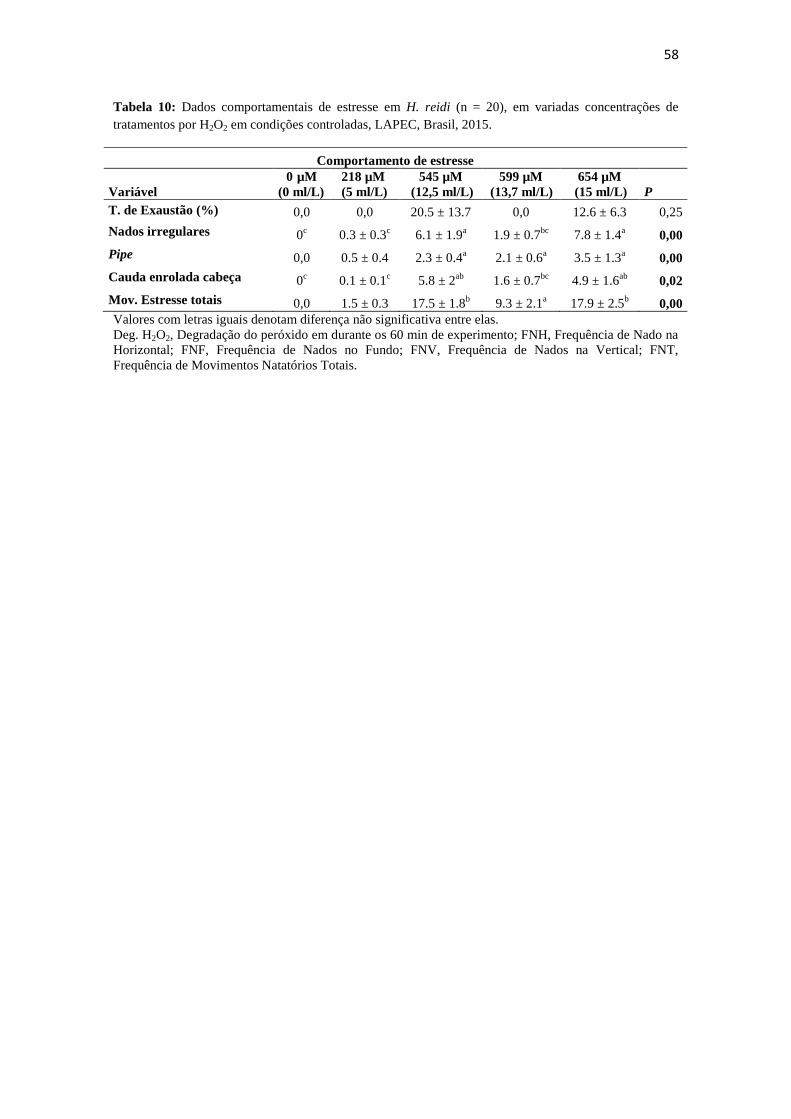
58
Tabela 10: Dados comportamentais de estresse em H. reidi (n = 20), em variadas concentrações de
tratamentos por H2O2 em condições controladas, LAPEC, Brasil, 2015.
Comportamento de estresse
Variável
0 µM
(0 ml/L)
218 µM
(5 ml/L)
545 µM
(12,5 ml/L)
599 µM
(13,7 ml/L)
654 µM
(15 ml/L) P
T. de Exaustão (%) 0,0 0,0 20.5 ± 13.7 0,0 12.6 ± 6.3 0,25
Nados irregulares 0c 0.3 ± 0.3
c 6.1 ± 1.9
a 1.9 ± 0.7
bc 7.8 ± 1.4
a 0,00
Pipe 0,0 0.5 ± 0.4 2.3 ± 0.4a 2.1 ± 0.6
a 3.5 ± 1.3
a 0,00
Cauda enrolada cabeça 0c 0.1 ± 0.1
c 5.8 ± 2
ab 1.6 ± 0.7
bc 4.9 ± 1.6
ab 0,02
Mov. Estresse totais 0,0 1.5 ± 0.3 17.5 ± 1.8b 9.3 ± 2.1
a 17.9 ± 2.5
b 0,00
Valores com letras iguais denotam diferença não significativa entre elas.
Deg. H2O2, Degradação do peróxido em durante os 60 min de experimento; FNH, Frequência de Nado na
Horizontal; FNF, Frequência de Nados no Fundo; FNV, Frequência de Nados na Vertical; FNT,
Frequência de Movimentos Natatórios Totais.

59
5. DISCUSSÃO
No Brasil, Hippocampus reidi é a espécie de cavalo-marinho que tem suas
densidades mais altas registradas, predominantemente, nas regiões Norte e Nordeste do
litoral brasileiro (ROSA et al., 2007). Uma proposta de Plano Nacional de Gestão para o
Uso Sustentável dos Cavalos-Marinhos foi publicada em 2011 (DIAS NETO, 2011),
devido a sua inclusão na Lista Nacional de Espécies de Invertebrados Aquáticos e
Peixes Sobre-Explotadas ou Ameaçadas de Sobre-Explotação, de acordo com a
Instrução Normativa nº 05, de 21 de maio de 2004, do Ministério do Meio Ambiente,
instrumento legal que prevê a elaboração de um Plano de Gestão. Sendo assim, é
necessário evoluir em direção à construção do Plano propriamente dito.
O estudo atual é o primeiro a observar respostas comportamentais de estresse
agudo sob diferentes concentrações de H2O2 em H. reidi, para a utilização em
tratamento por imersão (banho) (YANONG, 2014).
5.1. Diferenças entre sexos
Ambos os sexos apresentaram comportamentos parecidos sob as variadas
situações experimentais, no entanto, os machos mostraram significativamente menos
atividade locomotora que as fêmeas. Não se sabe, até então, o mecanismo fisiológico
responsável pela diferença sexual observada na resposta locomotora ao estresse. Øverli
et al (2006) encontraram um comportamento semelhante em Truta-arco-íris
(Oncorhynchus mykiss) em seus experimentos, onde também houve uma diferença
significativa na atividade locomotora entre os sexos, porém, foram as fêmeas que
apresentaram uma menor atividade locomotora. No mesmo estudo, não foi encontrada
diferença para os níveis de cortisol entre os sexos, o que pode indicar que, isoladamente,
a variável não é representativa para esse parâmetro. Para peixes salmonídeos, sabe-se
que o cortisol, hormônios de crescimento e outras substâncias de sinal central e
periférico, como o fator de liberação da corticotropina (CRF), 5-hidroxitriptamina (5-
HT, ou serotonina), podem afetar a atividade locomotora (WINBERG et al, 1993;
ØVERLI et al, 2002; CLEMENTS et al, 2003; JOHANSSON et al, 2004;
JOHANSSON et al, 2005). Entretanto, para muitas espécies de peixes ornamentais,

60
incluindo as do gênero Hippocampus, ainda há uma lacuna em relação ao
funcionamento de tais mecanismos e respostas fisiológicas ao estresse entre indivíduos
de sexo diferentes.
É, portanto, de grande interesse notar que, entre os indivíduos machos e fêmeas
e, consequentemente, na população como um todo, não houve distinção dos
comportamentos associados restritamente ao estresse, nem tampouco aos níveis de
tolerância ao H2O2.
5.2. Diferenças entre os tratamentos com H2O2 (0 µM, 218 µM, 545 µM, 599 µM e
654 µM)
O cortisol é conhecido por diminuir a ingestão de alimentos em animais
expostos, reduzindo assim seu peso (STEVENS, 1972; GREGORY e WOOD, 1999). H.
reidi tiveram perda de peso quando expostos ao H2O2 e ganho de peso nos tratamentos
controle, o ganho de peso é inversamente proporcional à concentração do peróxido de
hidrogênio ao qual foram expostos (Fig. 3 - A). Muito embora, utilizando um estressor
diferente em seu trabalho (estresse sonoro), Anderson e colaboradores (2011)
encontraram resultados semelhantes aos nossos para a espécie H. erectus onde animais
expostos a tanques com ruído (estressor), tiveram perda de peso (ΔWt = −2.23 ± 0.30 g)
e em tanques sem ruído a perda de peso foi significantemente menor (ΔWt = −1.44 ±
0.28 g).
Sabendo que a ingestão de toxinas, aglomeração, cor do tanque e confinamento
são fatores estressantes que tem provocado uma diminuição nas taxas de crescimento
entre peixes expostos (PICKERING e POTTINGER, 1987; LUMLERTDACHA et al.,
1995; PROCARIONE et al., 1999; POTTINGER et al., 2002; COTTER et al., 2005) e
que neste experimento, todos os animais, após os três primeiros dias, já se alimentavam
normalmente, equivalente ao apresentado por Anderson e colaboradores (2011). Isso
sugere que, provavelmente, a perda de peso não ocorreu pela não ingestão de alimentos,
mas sim por um maior gasto energético do animal para recuperar a homeostase.
Os comportamentos de manutenção observados foram similares aos encontrados
para H. guttulatus (FALEIRO et al, 2008) e H. erectus (ANDERSON et al, 2011).
Faleiro e colaboradores (2008) observaram que os peixes submetidos a um estresse

61
social, em tanques com maior densidade populacional, passavam menos tempo presos
ao substrato, em repouso (U = 3935.00, N1 = 120, N2 = 80, p < 0.05). Anderson e
colaboradores (2011) submeteram espécimes de H. erectus a um estresse sonoro e não
identificaram diferença para o tempo gasto pelos animais em repouso nos tanques com
ruído e sem ruído (F3,88 = 3.96, p = 0.011); em ambos os casos, os animais passavam 94
± 1 % em repouso, porém, na primeira semana, o número de ajustes no substrato nos
tanques com ruído (69±18 %) foi significativamente maior que nos tanques sem ruído
(27±7 %, F1,120 = 7.40, p < 0.01), sugerindo assim algum desconforto no animal. H.
reidi apresentou comportamento semelhante a H. guttulatus, onde animais em aquários
contendo H2O2 passaram um tempo muito maior nadando (58.6 ± 4.8 %) do que os
animais em ausência total do peróxido (12.3 ± 10.1 %); ou seja, consequentemente,
animais expostos ao H2O2 passaram muito menos tempo em repouso (Fig. 4 - A). Øverli
e colaboradores (2006), em um estudo com Oncorhynchus mykiss, observaram uma
forte correlação entre atividade locomotora em testes de estresse agudo e estresse
induzido pelo cortisol. Especificamente, o tratamento agudo de cortisol (1h) aumentou
natação e agressividade em peixe submetido a um teste de intrusão territorial, mas
nenhum efeito foi observado no comportamento de peixes sem tratamento (ØVERLI et
al, 2002). Tais dados nos mostram que mesmo as espécies agrupadas no gênero
Hippocampus sejam consideradas espécies com baixa motilidade natatória em relação
às demais espécies de peixe (FOSTER e VINCENT, 2004), tal comportamento merece
uma atenção relevante num contexto experimental ex-situ.
Clicks em cavalos-marinhos já foram descritos em vários contextos, incluindo
agressão/competição (VINCENT, 1994; OLIVEIRA et al, 2014) e alimentação
(COLSON et al., 1998; OLIVEIRA et al, 2014). Anderson e colaboradores (2011)
observaram um aumento no número de clicks em tanques com ruídos altos, que vai ao
encontro dos índices fisiológicos de aumento do estado de estresse. Esse
comportamento já havia sido relatado por Fish (1953), que observou clicks de H.
erectus em resposta a transferências para novos recipientes de água do mar e a uma
transferência rápida entre duas massas de água que diferiam na temperatura de 12 °C, o
que pode ter representado estressores agudos. H. reidi mostrou um comportamento
similar quando em contato com H2O2, onde os tratamentos com altas concentrações
apresentaram significativamente um aumento no número de clicks, indicando assim um
maior nível de estresse (Fig. 5 - B). No contexto ambiental, os clicks podem servir

62
como uma vocalização de aflição (distress) em um ambiente não favorável (Anderson et
al, 2011). Em mamíferos e aves, esse tipo de vocalização pode ser percebido em
momentos de predação, onde os animais encontram-se nas garras/boca de um predador
(HOGSTEDT, 1983), quando animais sociais são separados de seus pares ou mães e
para anunciar sua localização para indivíduos da mesma espécie, foi associado como a
resposta ao estresse fisiológico (NORCROSS e NEWMAN, 1999; STAHL et al., 2002;
FELTENSTEIN et al., 2003). A maioria das espécies de cavalos-marinhos (incluindo H.
reidi) apresenta comportamentos de animais sociais; pode ser monogâmica (VINCENT
e SADLER, 1995; KVARNEMO et al.,2000; VINCENT et al., 2005) ou poligâmica
(LOURIE et al., 1999; WOODS, 2000; KVARNEMO et al., 2000; FOSTER e
VINCENT, 2004; SILVEIRA, 2005), ou seja, estes animais podem manter o mesmo
parceiro ao longo de pelo menos uma estação reprodutiva (FOSTER e VINCENT,
2004); uma vocalização de angústia poderia servir como alerta e ser reforçada sob
estresse (ANDERSON et al, 2011).
Os comportamentos que ocorreram apenas na presença do estressor (H2O2) são
similares aos encontrados para outros peixes em estudos de estresse agudo. Foi
observada a presença de nados irregulares, semelhante ao registrado em Labeo rohita
por Dube e Hosetti (2010), onde foi testada a toxicidade do cianeto de sódio. Assim
como para L. rohita, H reidi apresentou momentos de letargia, chegando à exaustão nos
níveis de concentração mais altos de H2O2 (12781 80 µM e 11606 73 µM) (Tab.
10), seguidos intermitentemente por hiperanimação em alguns exemplares, resultando
em movimentos erráticos. Selye (1973) descreveu um estágio de alarme inicial, a fase
seguinte de resistência e fase final de exaustão, onde esta última pode ser alcançada se
as condições estressantes são graves e/ou suficientemente prolongadas. Nos mamíferos,
esta fase é caracterizada por diminuição dos níveis de cortisol, deleção de glicogênio no
fígado, imunossupressão, e outras alterações que diminuam a sobrevivência dos
organismos (HONTELA et al, 1992).
Outro comportamento é o que Anderson e colaboradores (2011) descrevem
como Pipe, que é o ato do animal pôr o focinho para fora d’água, este comportamento é
provavelmente similar em função ao que, pra outros peixes, é descrito como Gulping.
Ambos podem ajudar a evitar o contato direto com o meio tóxico e também melhorar a

63
hipóxia, fazendo uso de uma fina camada de água rica em oxigênio na superfície,
aliviando o stress respiratório (FRANCIS-FLOYD, 1988; DUBE e HOSETTI, 2010).
O Pipe foi observado para H. reidi em todos os tratamentos onde houve presença
de peróxido de hidrogênio, aumentando gradativamente à medida que aumentou sua
concentração (Fig. 5 - A), o que pode indicar um estresse/desconforto respiratório nos
níveis de maior concentração do H2O2. Em consonância a este, registrou-se o aumento
da frequência respiratória que, similar ao ocorrido para o número de pipes, também teve
um aumento de sua frequência em concentrações mais altas de H2O2 (Fig. 6 - B) em H.
reidi, esse aumento pode ser atribuído ao desconforto respiratório, como consequência
do comprometimento do metabolismo oxidativo (DUBE e HOSETTI, 2010). A
atividade respiratória de um peixe é muitas vezes a primeira resposta fisiológica a ser
afetada pela presença de contaminantes no ambiente aquático (DUBE e HOSETTI,
2010), uma vez que as brânquias são os principais órgãos respiratórios dos peixes e
todas as vias metabólicas dependem de sua eficiência para o fornecimento de energia,
nesse contexto, danos a estes órgãos vitais causam uma cadeia de eventos que, em
última análise, levam ao estresse respiratório (MAGARE e PATIL, 2000). Barreto e
Volpato (2006) em seus estudos com Oreochromis niloticus, mostraram um contexto
onde a frequência respiratória pode distinguir entre os efeitos estressores
adequadamente pela quantificação não invasiva.
O movimento de enrolar a cauda na cabeça (Fig. 6 – A), possibilitado pelas
características peculiares dos representantes inclusos no gênero Hippocampus, como a
posição da cabeça formando um ângulo reto em relação ao eixo do corpo, uma cauda
preênsil, utilizada geralmente para se prender a substratos de apoio como algas, corais e
plantas de mangue (ROBINS; RAY, 1986; SMITH, 1997; LOURIE et al., 2004;
KUITER, 2009), ainda não havia sido descrito para os cavalos-marinhos. Similarmente,
quando exposto a uma situação de manuseio por um período prolongado, H. reidi
expressa o mesmo comportamento de enrolar a cauda na cabeça (observação pessoal), o
que pode representar um comportamento agonístico.
Comportamentos de natação irregulares, hiperexcitação, disfunção respiratória,
Piping e natação na superfície da água foram observados durante a exposição ao H2O2.
Trabalhos anteriores sobre outras espécies de peixe indicam a recuperação total entre 24
horas e 7 dias (TORT et al., 2003), enquanto no presente trabalho o tempo de

64
recuperação foi de três dias. Em Salmo salar (Salmão do Atlântico) exposto ao H2O2, o
cortisol permaneceu elevado entre 6 e 12 horas pós-tratamento e voltou para estado de
normal após 24 horas (BOWERS et al., 2002). No entanto, para Dicentrarchus labrax,
após esse período os níveis de cortisol continuavam altos (ROQUE et al, 2010). Essa
diferença pode ser explicada pelo tempo de exposição a que Salmo salar (20 min) e
Dicentrarchus labrax (60 min) a que foram expostos ao H2O2, este último com tempo
de exposição igual a este estudo. Mansell e colaboradores (2005) sugerem ainda que os
efeitos agudos do banho contendo peróxido de hidrogênio são menos prejudiciais para o
peixe do que os efeitos de infecção crônica persistente.

65
6. CONCLUSÕES
Os resultados deste estudo utilizando como modelo cavalos-marinhos indicaram
que H2O2 tem potencial para ser usado em tratamento por imersão (banho) em H. reidi,
tendo em vista que concentrações abaixo de 12.5ml/L mostraram-se seguras em um
período de 60 min de exposição para a espécie. Trabalhos com outras espécies de peixes
ornamentais mostraram que concentrações de H2O2 acima de 3,1ml/L podem ser
utilizadas para tratar bactérias externas e concentrações superiores a 6,5ml/L, podem ser
utilizadas no tratamento de Ichthyobodo spp. (parasita externo). Entretanto, os efeitos
tóxicos do H2O2, para H. reidi, podem ter causado as mortalidades observadas nas
concentrações iguais/superiores a 13.75ml/L nos tratamentos. Essas concentrações
devem ser evitadas, embora mais testes sejam justificados.
Para além dos resultados específicos obtidos neste estudo, destaca-se a
importância do comportamento animal como ferramenta relevante e útil para o
aprimoramento da aquicultura. As alterações registradas no presente trabalho mostraram
que o peróxido de hidrogênio é tóxico e que teve profundo impacto sobre o peso,
comportamento e frequência respiratória em H. reidi, nas concentrações letais e
subletais.
Nessa mesma direção, a disfunção registrada de comportamento e respiração é
um sinal importante e visualmente observável, como método não invasivo, do índice de
toxicidade por H2O2. Assim, estudos etológicos mostram-se como importantes aliados
para ampliar a compreensão acerca do estresse oxidativo provocado pelo H2O2 no
metabolismo dos peixes.

66
7. REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, S. M., 1990. Biological indicators of stress in fish. Am. Fish. Sot. Symp. Ser.
ANDERSON, P. A., BERZINS, I. K., FOGARTY, F., HAMLIN, H. J., GUILLETTE JR, L. J.,
2011. Sound, stress, and seahorses: The consequences of a noisy environment to animal
health. Aquaculture 311, 129–138;
ALLEN, A. O.; DAVIS, T. W.; ELMORE, G.; GHORMLEY, J. A,; HAINES, B. M.;
HOCHANADEL, C. J. Oak Ridge Natl. Lab. Publ. 1949, ORNL 130;
AURÉLIO, M. L., 2012. Physiological and behavioral responses of temperate seahorses
(Hippocampus guttulatus) to environmental warming. Departamento de Biologia Animal,
Universidade de Lisboa;
BARRETO, R. E.; VOLPATO, G. L., 2004. Caution for using ventilatory frequency as an
indicator of stress in fish. Behavioural Processes 66 (2004) 43–51;
BARRETO, R. E.; VOLPATO, G. L., 2006. Ventilatory frequency of Nile tilapia subjected
to different stressors. Journal of Experimental Animal Science 43, 189–196;
BARTESAGHI, S.; FERRER-SUETA, G.; PELUFFO, G.; et al., 2007. Protein tyrosine
nitration in hydrophilic and hydrophobic environments. Amino Acids 32: 501–515;
BEITINGER, T. L., 1990. Behavioral reactions for the assessment of stress in fishes. J. Great
Lakes Res. 16(4):495-528
BONELLO, S., ZAHRINGER, C., BELAIBA, R.S., ET AL., 2007. Reactive oxygen species
activate the HIF-1a promoter via a functional NFkB site. Arterioscl Thromb Vasc Biol 27:
755–761;
Bowers, J. M., Speare, D. J., Burka, J. F., 2002. The effects of hydrogen peroxide on the
stress response of Atlantic Salmon (Salmo salar). J. Vet. Pharmacol. Therap. 25, 311–313;
BLISS, C. I. 1935. The calculator of the dosage-mortality curve. Annals of Applied Biology.
v.22, p. 134-167.
COLSON, D. J., PATEK, S. N., BRAINERD, E. L., LEWIS, S. M., 1998. Sound production
during feeding in Hippocampus seahorses (Syngnathidae). Environ. Biol. Fish. 51, 221–229;
COTTER, P. A., HARRIS, J. O., MCLEAN, E., CRAIG, S., SCHWARZ, M., RASMUSSEN,
M. R., 2005. The effect of tank color upon growth performance and stress response of
summer flounder Paralichthys dentatus. Poster. Aquaculture America 2005, 17–21 Jan 2005,
New Orleans, LA;
CLAYTON, R. D., AE SUMMERFELT, R. C., 1996. Toxicity of hydrogen peroxide to
fingerling walleyes. Journal of Applied Aquaculture 6:39–49;
CLEMENTS, S., MOORE, F. L., SCHRECK, C. B., 2003. Evidence that acute serotonergic
activation potentiates the locomotor-stimulating effects of corticotropinreleasing hormone
in juvenile chinook salmon (Oncorhynchus tshawytscha). Horm Behav, 43:214–21;
CRUZ, C.; FUJIMOTO, R.Y.; LUZ, R.K.; et al., 2005. Toxicidade aguda e histopatologia do
fígado de larvas de trairão (Hoplias lacerdae) expostas à solução aquosa de formaldeído a
10%. Pesticidas: r. Ecotoxicol. e meio ambiente. Curitiba, 15: 21-28;

67
DIAS NETO, J. (org.) 2011. Proposta de Plano Nacional de Gestão para o uso sustentável
de cavalos-marinhos do Brasil. Brasília: Ibama. 104p;
DU, W.; ADAM, Z.; RANI, R.; et al., 2008. Oxidative stress in Fanconi anemia
hematopoiesis and disease progression. Antioxid Redox Signal 10: 1909–1921;
DUBE, P.N.; HOSETTI, B.B., 2010. Behaviour surveillance and oxygen consumption in the
fresh water fish labes rohita exposed to sodium cyanide. Biotechnology in Animal
husbandary, 26(1-2), p. 91-103.
FALEIRO, F., NARCISO, L. E VICENTE, L., 2008. Seahorse behaviour and aquaculture:
How to improve Hippocampus guttulatus husbandry and reproduction? Aquaculture 282,
33–40;
FELTENSTEIN, M. W., LAMBDIN, L. C., WEBB, H. E., WARNICK, J. E., KHAN, S. I.,
KHAN, I. A., ACEVEDO, E. O., SUFKA, K. J., 2003. Corticosterone response in the chick
separation-stress paradigm. Physiol. Behav. 78, 489–493.
FRANCIS-FLOYD, R., 1988. Behavioral diagnosis. Vet. Clin. N. Am.-Small Anim. Pract. 18
(2), 305–316;
FISH, M. P., 1953. The production of underwater sound by the northern seahorse,
Hippocampus hudsonius. Copeia 1953, 98–99;
FOSTER, S. J. E VINCENT, A. C. J., 2004. Life history and ecology of seahorses:
implications for conservation and management. Journal of Fish Biology, v.65, p. 1-61;
GAIKOWSKI, M., A. JOHNSON, J. J. RACH, AND S. M. SCHLEIS, 2003. Efficacy of
hydrogen peroxide in controlling mortality associated with external columnaris on walleye
and channel catfish fingerlings. American Journal of Aquaculture 65:300–305;
GAIKOWSKI, M. P., J. J. RACH, AND R. T. RAMSAY, 1999. Acute toxicity of hydrogen
peroxide treatments to selected lifestages of cold-, cool-, and warmwater fish. Aquaculture,
178:191–207;
GAUTER-FLECKENSTEIN, B.; FLECKENSTEIN, K.; OWZAR, K.; et al., 2010. Early and
late administration of MnTE-2-PyP5+
in mitigation and treatment of radiation-induced
lung damage. Free Radic. Biol. Med. 48: 1034-1043;
GREGORY, T. R., WOOD, C. M., 1999. The effects of chronic plasma cortisol elevation on
the feeding behaviour, growth, competitive ability, and swimming performance of juvenile
rainbow trout. Physiol. Biochem. Zool. 72 (3), 286–295;
HEO, G.J.; SHIN, G., 2010. Efficacy of benzocaine as an anaesthetic for Crucian carp
(Carassius carassius). Vet. Anaesth. Analg., 37, pp. 132–135;
HIDALFO, C. e DONOSO P., 2008. Crosstalk between calcium and redox signaling: from
molecular mechanisms to health implications. Antioxid Redox Signal 10: 1275–1312;
HÖGSTEDT, G., 1983. Adaptation unto death: function of fear screams. Am. Nat. 121 (4),
562–570;
HONTELA, A., RASMUSSEN, J. B., AUDET, C. E CHEVALIER, G., 1992. Impaired
cortisol stress response in fish from evironments polluted by PAHs, PCBs, and mercury.
Archives of Environmental Contaminantion and Toxicology. 22:278-285;

68
HOWE, G. E., W. H. GINGERICH, V. K. DAWSON, AND J. J. OLSON. 1999. Efficacy of
hydrogen peroxide for treating saprolegniasis in channel catfish. Journal of Aquatic Animal
Health 11:222–230;
IMLAY, J. A., 2002. How oxygen damages microbes: oxygen tolerance and obligate
anaerobiosis. Advances in Microbial Physiology 46:111–153;
IMLAY, J. A., 2003. Pathways of oxidative damage. Annual Review of Microbiology
57:395–418.
IWAMA, G.K., PICKERING, A.D., SUMPTER, J.P., SCHRECK, C.B. (Eds.), 1997. Fish
stress and health in aquaculture. Society for Experimental Biology Seminar Series 62,
Cambridge University Press, Cambridge, 278pp.
IWAMA, G. K.; VIJAYAN, M. M.; FORSYTH, R. B.; ACKERMAN, P. A., 1998. Heat shock
proteins and physiological stress in fish. American Zoologist, 39: 901-909.
JOHNSON, S. C., J. M. CONSTIBLE, AND J. RICHARD; 1993. Laboratory investigations
on the efficacy of hydrogen peroxide against the salmon louse Lepeophtheirus salmonis and
its toxicological and histopathological effects on Atlantic salmon Salmo salar and Chinook
salmon Oncorhynchus tshawytscha. Diseases of Aquatic Organisms 17:197–204.
JOHANSSON V., WINBERG S., BJÖRNSSON B. T., 2005. Growth hormone-induced
stimulation of swimming and feeding behaviour of rainbow trout is abolished by the D1
dopamine antagonist SCH23390. Gen Comp Endocrinol, 141:58–65;
JOHANSSON, V., WINBERG, S., JONSSON, E., HALL, D., BJÖRNSSON, B. T., 2004.
Peripherally administered growth hormone increases brain dopaminergic activity and
swimming in rainbow trout. Horm Behav, 46:436– 43;
KENYON, T.N., LADICH, F., YAN, H.Y., 1998. A comparative study of hearing ability in
fishes: the auditory brainstem response approach. J. Comp. Physiol. A 182, 307-318
KEYER, K., AND J. A. IMLAY, 1996. Superoxide accelerates DNA damage by elevating
free-iron levels. Proceedings of the National Academy of Sciences of the USA 93:13635–
13640.
KIEMER, M.C.B., BLACK, K.D., 1997. The effects of hydrogen peroxide on the gill tissues
of Atlantic salmon, Salmo salar L. Aquaculture 153, 181–189;
KLASSEN, N. V., MARCHINGTON, D., MCGOWANT, H. C. E., 1994. H2O2 Determination
by the I3- Method and by KMnO4 Titration. Institute for National Measurement Standards,
Ionizing Radiation Standards, National Research Council, Canada. Anal. Chem., 66, 2921-2925;
KOLDEWEY, H. J., 2005. Syngnathid Husbandry in Public Aquariums: 2005 Manual,
With Chapters Contributed by Members of the Syngnathidae Discussion Group. Project
Seahorse and Zoological Society of London. 137pp.
KUITER, R. H., 2009. Seahorses and their relatives. Seaford: Aqua Photographics. 334 p.
KVARNEMO, C., MOORE, G. I., JONES, A. G., NELSON, W. S. E AVISE J. C., 2000.
Monogamous pair bonds and mate switching in the western Australian seahorse
Hippocampus subelongatus. J. Evol. Biol., v. 13, p. 882- 888;
LORIE, S. A., VINCENT, A. C. J. E HALL, H. J., 1999. Seahorses: an identification guide to
the word’s species and their conservation. Project Seahorse, London, UK. 224 p;

69
LOURIE, S. A. et al., 2004 .A Guide to the Identification of Seahorses. Project Seahorse and
TRAFFIC North America. Washington D.C.: University of British Columbia and World
Wildlife Fund;
LI, F.; SONVEAUX, P. P.; RABBANI, Z. N.; et al., 2007. Regulation of HIF-1a stability
through S-nitrosylation. Mol Cell 26: 63–74;
LITTLE E.E., BREWER S.K., 2001. Neurobehavioral toxicity in fish. Schlenk, D. and
Benson, W.H. (Ed.), Target Organ Toxicity in Marine and Freshwater Teleost New
Perspectives: Toxicology and the Environment, Taylor and Francis, London and New York. 2,
139-174;
LUMLERTDACHA, S., LOVELL, R. T., SHELBY, R. A., LENZ, S. D., KEMPPAINEN, B.
W., 1995. Growth, hematology, and histopathology of channel catfish, Ictalurus punctatus,
fed toxins from Fusarium moniliforme. Aquaculture 130, 201–218;
LUMSDEN, J. S., V. E. OSTLAND, AND H. W. FERGUSON, 1998. Use of hydrogen
peroxide to treat experimentally induced bacterial gill disease in rainbow trout. Journal of
Aquatic Animal Health 10:230–240;
LUVIZOTTO-SANTOS, R.; ELER, M.N.; ESPINDOLA, E.L.G; VIEIRA, E.M. , 2009. O uso
de praguicidas nas pisciculturas e pesqueiros situados na bacia do rio Mogi-Guaçu. B. Inst.
Pesca, São Paulo, 35(3): 343 – 358;
MAGARE S. R., PATIL H. T., 2000. Effect of pesticides on oxygen consumption, red blood
cell count and metabolites of a fish, Puntius ticto. Environ. Ecology. 18, 891-894;
MANSELL, B., POWELL, M. D., ERNST, I., NOWAK, B. F., 2005. Effects of the gill
monogenean Zeuxapta seriolae (Merserve 1938) and treatment with hydrogen peroxide on
pathophysiology of kingfish, Seriola lalandi Valenciennes, 1833. J. Fish Dis. 28, 253–262;
MAXIMINIANO, A.A.; et al. , 2005. Utilização de drogas veterinárias, agrotóxicos e afins
em ambientes hídricos: demandas, regulamentação e considerações sobre riscos à saúde
humana e ambiental. Ciência e Saúde Coletiva, 10(2): 483-491;
MOMMSEN, T. P., VIJAYAN, M. M., MOON, T. W., 1999. Cortisol in teleosts: dynamics,
mechanisms of action, and metabolic regulation. Reviews on Fish Biology and Fisheries, 9:
211–268;
MONTGOMERY-BROCK, D., V. T. SATO, J. A. BROCK, AND C. S. TAMARU, 2001. The
application of hydrogen peroxide as a treatment for the ectoparasite Amyloodinium
ocellatum (Brown 1931) on the Pacific threadfin Polydactylus sexfilis. Journal of the World
Aquaculture Society 32:250–254;
NORCROSS, J. L., NEWMAN, J. D., 1999. Effects of separation and novelty on distress
vocalizations and cortisol in the common marmoset (Callithrix jacchus). Am. J. Primatol.
47, 209–222;
OLIVEIRA, T. P. R., LADICH, F., ABED-NAVANDI, D., SOUTO, A. S., ROSA, I. L., 2014.
Sounds produced by the longsnout seahorse: a study of their structure and functions. Journal of Zoology, The Zoological Society of London.
ØVERLI Ø., SØRENSEN, C., NILSSON, G. E., 2006. Behavioral indicators of stress-coping
style in rainbow trout: Do males and females react differently to novelty? Physiology &
Behavior 87, 506 – 512;

70
ØVERLI Ø., KOTZIAN S, WINBERG S., 2002. Effects of cortisol on aggression and
locomotor activity in rainbow trout. Horm Behav, 42:53–61;
PELTIER, W.H., WEBER, C.I., 1985. Methods for measuring the acute toxicity of effluents
to freshwater and marine organisms. US Environmental Protection Agency EPA 600r4-85-
013, Cincinnati, OH, 216 pp;
PICKERIN. A.D., 1981. Stress and fish. Academic P&s, London, 367 pp.
POWELL, M. D., AND G. A. CLARK. 2004. Efficacy and toxicity of oxidative disinfectants
for the removal of gill amoebae from the gills of amoebic gill disease affected Atlantic
salmon (Salmo salar L.) in freshwater. Aquaculture Research 35:112–123;
PICKERING, A. D., POTTINGER, T. G., 1987. Crowding causes prolonged leucopenia in
salmonid fish, despite interrenal acclimation. J. Fish Biol. 30, 701–712;
POTTINGER, T. G., CARRICK, T. R., YEOMANS, W. E., 2002. The three-spined
stickleback as an environmental sentinel: effects of stressors on whole-body physiological
indices. J. Fish Biol. 61, 207–229;
PROCARIONE, L. S., BARRY, T. P., MALISON, J. A., 1999. Effects of high rearing
densities and loading rates on the growth and stress responses of juvenile rainbow trout. N. Am. J. Aquac. 61, 91–96;
RACH, J. J., M. P. GAIKOWSKI, AND R. T. RAMSAY. 2000a. Efficacy of hydrogen
peroxide to control mortalities associated with bacteria gill disease infections on hatchery-
reared salmonids. Journal of Aquatic Animal Health 12:119–127;
RACH, J. J., M. P. GAIKOWSKI, AND R. T. RAMSAY. 2000b. Efficacy of hydrogen
peroxide to control parasitic infections on hatchery-reared fish. Journal of Aquatic Animal
Health 12:267–273;
RACH, J. J., T. M. SCHREIER, G. E. HOWE, AND S. D. REDMAN, 1997. Effect of species,
life stage, and water temperature on the toxicity of hydrogen peroxide to fish. Progressive
Fish-Culturist 59:41–46;
RACH, J. J., S. M. SCHLEIS, M. P. GAIKOWSKI, AND A. JOHNSON, 2003. Efficacy of
hydrogen peroxide in controlling mortality associated with external columnaris on walleye
and channel catfish fingerlings. North American Journal of Aquaculture 4:300–305;
RACH, J. J., J. J. VALENTINE, T. M. SCREIER, M. P. GAIKOWSKI, AND T. G.
CRAWFORD. 2004. Efficacy of hydrogen peroxide to control saprolegniasis on channel
catfish (Ictalurus punctatus) eggs. Aquaculture 238:135–142;
ROBINS, C. R; RAY, G. CARLETON. 1986. A field guide to Atlantic coast fishes. Boston:
Houghton Miffin Company, 354p;
ROQUE, A., YILDIZ, H. Y., CARAZO, I., DUNCAN, N., 2010. Physiological stress
responses of sea bass (Dicentrarchus labrax) to hydrogen peroxide (H2O2) exposure. Aquaculture 304, 104–107;
ROSA, I. L., ALVES, R. R. N., 2007. Pesca e comércio de cavalos-marinhos (Syngnathidae:
Hippocampus) no Norte e Nordeste do Brasil: subsídios para a conservação e manejo. In:
ALBUQUERQUE, U. P. de; ALVES, A. G. C.; ARAÚJO, T. A. de S. (Org.). Povos e
Paisagens. Recife: Núcleo de populações em ecologia e etnobotânica aplicada, p. 115-134;
RUBBO, H. e RADI, R., 2008. Protein and lipid nitration: role in redox signaling and
injury. Biochim Biophys Acta 1780:1318–1324;

71
Russo, R., E. W. Curtis, and R. P. E. Yanong. 2007. Preliminary investigations of hydrogen
peroxide treatment of selected ornamental fishes and efficacy against external bacteria
and parasites in green swordtails. Journal of Aquatic Animal Health 19:121–127;
SALVEMINI, D.; DOYLE, T.M.; CUZZOCREA, S., 2006. Superoxide, peroxynitrite and
oxidative/nitrative stress in inflammation. Biochem Soc Trans 34: 965–970;
Selye, H., 1973. The evolution of stress concept. Am. Sci. 61, 692–699;
SILVA, A.L.; MARCASSI ALVES, F.C.; ANDRADE TALMELLI, E.F.; et al. , 2009.
Utilização de cloreto de sódio, formalina e a associação destes produtos no controle de
ectoparasitas em larvas de tilápia (Oreochromis niloticus). B. Inst. Pesca, São Paulo, 35(4):
597-608;
SILVEIRA, R. B., 2005. Dinâmica populacional do cavalo-marinho Hippocampus reidi
(Syngnathidae) no manguezal de Maracaípe, Ipojuca, PE. Tese. (Doutorado em Zoologia).
Porto Alegre, 2005. 127 f. Departamento de Zoologia, Pontifícia Universidade Católica do Rio
Grande do Sul;
SMITH, C. L., 1997. National Audubon Society Field Guide to Tropical Marine Fishes of
the Caribbean, the Gulf of Mexico, Florida, the Bahamas, and Bermuda. 2ed. New York:
Alfred A. Knopf, Inc. 718p;
SOUZA, J.M.; PELUFFO, G.; RADI, R., 2008. Protein tyrosine nitration: functional
alteration or just a biomarker? Free Radic Biol Med 45: 357–366;
SPEARE, D. J., AND G. J. ARSENAULT. 1997. Effects of intermitente hydrogen peroxide
exposure on growth and columnaris disease prevention of juvenile rainbow trout
(Oncorhynchus mykiss). Canadian Journal of Fisheries and Aquatic Sciences 54:2653–2658;
STAHL, C. E., REDEI, E., WANG, Y., BORLONGAN, C. V., 2002. Behavioral, hormonal
and histological stress markers of anxiety-separation in postnatal rats are reduced by
prepro-thyrotropin-releasing hormone. Neurosci. Lett. 178–199. 321, 85–89;
STEVENS, E. D., 1972. Change in body weight caused by handling and exercise in fish. J.
Fish. Res. BoardC an. 29:202-203;
TANTARPALE, V.T.; RATHOD, S.H.; KAPIL, S. Temperature Stress on Opercular Beats
and Respiratory Rate of Freshwater Fish Channa Punctatus. International Journal of
Scientific and Research Publications, Volume 2, Issue 12, December 2012;
THOMASSEN, J. M., 1993. Hydrogen peroxide as a delousing agent for Atlantic salmon.
Pages 290–295 in G. A. Boxhall and D. Defaye, editors. Pathogens of wild farmed fish: sea lice.
Ellis Horwood, London;
TORT, J. M., WOOSTER, G. A., BOWSER, P. R., 2003. Effects of hydrogen peroxide on
hematology and blood chemistry parameters of walleye Stizostedion vitreum. J. World
Aquacult. Soc. 34 (2), 236–242;
VALADARES, M. C. Avaliação de toxicidade aguda: estratégias após a “Era do Teste
DL50”. Revista Eletrônica de Farmácia, 2006. Vol. 3(2), 93-98;
VINCENT, A.C.J., 1994. Seahorses exhibit conventional sex roles in mating competition,
despite male pregnancy. Behaviour 128 (1–2), 135–151;
VINCENT, A. C. J., EVANS, K. L., MARSDEN, A. D., 2005. Home range behavior on the
monogamous Australian seahorse, Hippocampus whitei. Environmental Biology of Fishes, v.
72, p. 1-12;

72
VINCENT, A. C. J., SADLER, R. M., 1995. Faithful pair bonds in wild seahorses,
Hippocampus whitei. Animal Behavior, v. 50, p. 1557-1569;
YANONG, R. P. E., 2014. Use of Hydrogen Peroxide in Finfish Aquaculture. Fisheries and
Aquatic Sciences in the School of Forest Resources and Conservation, UF/IFAS;
WENDELAAR BONGA, S.E., 1997. The stress response in fish. Physiol, Rev. 77(3), 591-
626;
WEDEMEYER, G. A., AND D. J. MCLEAY., 1981. Methods for determining the tolerance
of fishes to environmental stressors. A.D. Pickering, editor. Stress and fish. Academic Press,
London, Pages 247-275;
WEDEMEYER, G. A., B. A. BARTON, and D. J. MCLEAY., 1990. Stress and acclimation.
In C. B. Schreck and P. B. Moyle (eds.), Methods for fish biology pp. 451–489. American
Fisheries Society, Bethesda, Maryland;
WINBERG, S., NILSSON, G. E., SPRUIJT, B. M., HÖGLUND, U., 1993. Spontaneous
locomotor activity in Arctic charr measured by a computerized imaging technique: role of
brain serotonergic activity. J Exp Biol, 179:213– 32;
WILCOX, D.; DOVE, B.; MCDAVID, D. ; GREER, D, 2002. UTHSCSA ImageTool
Program, version 3.0. University of Texas Health Science Center, San Antonio, Texas.
Disponível em: <ftp:maxrad6.uthscsa.edu>;
WILSON, M. J.; VINCENT; A. C. J., 1998. Preliminary success in closing the life cycle of
exploited seahorse species, Hippocampus spp., in captivity. Aquarium Sci. Cons. v. 2n. 4, p.
179-196;
WINTERBOURN, C.C., 2008. Reconciling the chemistry and biology of reactive species.
Nat Chem Biol 4: 278–286;
WINTERBOURN, C.C. e HAMPTON, M.B., 2008. Thiol chemistry and specificity in redox
signaling. Free Radic Biol Med 45: 549–561;
WOODS, C. M. C., 2000. Preliminary observations on breeding and rearing the seahorse
Hippocampus abdominalis (Teleostei: Syngnathidae) in captivity. New Zeland Journal of
Marine and Freshwater Research, v.34, p. 475-485;
U.S. FOOD AND DRUG ADMINISTRATION. 2002. Title 21 Code of Federal Regulations
Part 530 (21 CFR 530). Available: http://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/
CFRSearch.cfm?CFRPart=530 (Junho, 2015);

73
CAPÍTULO III - ESTUDO INICIAL SOBRE A UTILIZAÇÃO DA Mn-PORFIRINA
(MnTE-2-PYP5+
) COMO REGULADOR REDOX CATALÍTICO EM
HIPPOCAMPUS REIDI GINSBURG, 1933 (TELEOSTEI SYNGNATHIDAE)
1. INTRODUÇÃO
Na aquicultura de modo geral, é comum o manuseio dos peixes para diversos
fins, como a medição, amostragem, marcação, transporte, entre muitos outros. Essas
manipulações frequentemente induzem uma resposta primária de estresse, marcada pelo
crescimento do cortisol plasmático e concentrações de glicose, podendo levar a
resultados indesejáveis como imunossupressão e retardo do crescimento (GRUTTER e
PANKURST, 2000; TREVES-BROWN, 2000; ROTLLANT et al, 2001; WEBER et al,
2009; HEO e SHIN, 2010). Respostas secundárias de estresse, que incluem alterações
no plasma, nos níveis dos metabólitos, nos íons dos tecidos, nas características
hematológicas e nas proteínas de choque térmico ou de tensão (PCT), todas
relacionadas com os ajustes fisiológicos, como o metabolismo, a respiração, o estado
ácido-base, equilíbrio hidromineral, função imune e respostas celulares (PICKERING,
1981; IWAMA et al., 1997, 1998; MOMMSEN et al., 1999) e respostas terciárias como
alterações no crescimento, condição resistência total à doença, o escopo para a atividade
metabólica, comportamento, e em última análise, a sobrevivência, resumidamente,
aspectos do desempenho de todo o animal (WEDEMEYER e MCLEAY, 1981;
WEDEMEYER et al., 1990) ainda podem ocorrer, dependendo da intensidade e duração
do stress. Essas respostas afetam os peixes em todos os níveis de organização, desde
molecular e bioquímica à população e comunidade (ADAMS, 1990). Dhabhar e
Mcewen (1997) definem stress como uma constelação de eventos, que começa com um
estímulo (estressor), que precipita uma reação no cérebro (percepção de stress), que
subsequentemente resulta na ativação de certos sistemas fisiológicos no corpo (resposta
ao stress).
O estresse também pode resultar como uma consequência de outros fatores
ambientais, incluindo condições adversas de temperatura, salinidade, pH ou oxigênio
dissolvido (FLOS et al., 1988). Esse estado põe em risco a homeostase, que pode se
restabelecer através de um conjunto complexo de respostas adaptativas (CHROUSOS,

74
1998). Barton et al. (1986) e Flos et al. (1988) sugerem um efeito aditivo ou uma
respostas fisiológicas cumulativa ao estresses. Se a intensidade do estressor é
excessivamente severa ou duradoura, os mecanismos de resposta fisiológica podem ser
comprometidos e tornar-se prejudiciais para a saúde e bem-estar dos peixes. Minimizar
fatores que levem esses animais a essa má adaptação são de grande interesse dos
gestores e aquicultores (SELYE, 1974; BARTON e IWAMA, 1991; BARTON, 2002).
1.1. O Estresse Oxidativo
A produção de ATP (Adenosina Trifosfato) nas mitocôndrias está ligada
diretamente ao O2 que é absorvido pelos organismos, bem como está envolvida no
grupo heme e na biossíntese dos centros de ferro-enxofre, de aminoácidos e do
metabolismo do nitrogênio, e na modulação da homeostase do cálcio (STOREY, 1996;
SCHEFFLER, 1999; MURPHY e SMITH, 2000; NICHOLLS e FERGUSON, 2002).
Mais de 90% de todo o oxigênio absorvido pelo corpo humano é usado na citocromo
oxidase que acrescenta quatro elétrons em cada molécula de O2 para formar água (O2 +
4H+ + 4e
- → 2 H2O) (HALLIWELL, 1992).
A cadeia respiratória mitocondrial é a maior fonte de superóxido (O2-), podendo
formar outras espécies reativas de oxigênio (ROS), como peróxido de hidrogênio
(H2O2), o radical hidroxila (OH.), e peroxinitrito (ONOO
-). Outros componentes
enzimáticos dentro da mitocôndria podem formar ROS (FINKEL, 2011; MURPHY,
2009). ROS são muitas vezes gerados na fase aquosa de plasma, por exemplo, O2 e
H2O2 pode surgir a partir de enzimas, compostos oxidantes e células fagocíticas ativadas
(HALLIWELL e GUTTERIDGE, 1989). Todos os componentes celulares são
susceptíveis ao ataque por ROS (O2, H2O2 e .OH), particularmente por
.OH (STOREY,
1996). Danos oxidativos patológicos para as mitocôndrias em doenças e envelhecimento
são consequências da produção de ROS, embora haja muitas respostas protetoras e
adaptativas para prevenir e reparar os danos oxidativos (WALLACE, 1999;
SHIGENAGA et al, 1994).
O ânion superóxido é desintoxicado pela Mn superóxido dismutase mitocondrial
(MnSOD) ou pela SOD citoplasmática (Cu,ZnSOD) para o peróxido de hidrogénio
(H2O2), e H2O2 é convertido em H2O pela glutationa peroxidase (GPx) (WALLACE,
1999):

75
O2 + e- → O2
- superoxido
O2- + e
- + 2 H
+ → H2O2 peróxido de hidrogênio
H2O2 + e- + H
+ → H2O +
. OH
radical hidroxila
.OH + e
- + H
+ → H2O
A redução de H2O2 por superóxido (a reação de Haber-Weiss) produz a maior
parte dos radicais hidroxila in vivo. O H2O2, na presença de metais de transição
reduzidos (Fe2+, algumas vezes Cu+), pode ser convertido também no radical hidroxila
(OH.), que é altamente reativo, pela reação de Fenton.
Fe3+
+ O2- → Fe
2+ + O2
Fe2+
+ H2O2 → Fe3+
+ OH- +
. OH Reação de Fento
O2- + H2O2 → O2 + OH
- +
. OH Reação de Haber-Weiss
O estresse oxidativo é caracterizado por um desequilíbrio celular no qual os
níveis normais de oxidantes (espécies reativas) predominam sobre os níveis normais de
antioxidantes (endógenos), ocasionando, eventualmente, danos oxidativos. Este
desequilíbrio tem implicação em uma série de doenças e estados fisiopatológicos que
compreendem desde processos inflamatórios simples, a doenças neurodegenerativas e
até mesmo o câncer (BATINIĆ-HABERLE et al, 2010).
Mn-porfirinas têm se destacado em estudos pré-clinicos como uma das classes
mais importantes classes de reguladores redox catalíticos e encontram-se hoje entre uns
dos melhores modelos biomiméticos das enzimas SOD (TSE et al, 2004; MOELLER et
al, 2004; ZHAO et al, 2005; REBOUÇAS et al, 2008a). Estudos in vivo com Mn-
porfirinas apareceram há apenas alguns anos, mas os resultados em diversos modelos
animais de fisiopatologias de caráter oxidativo têm sido excelentes (ASLAN et al, 2001;
PIGANELLI et al, 2002; BENOV et al, 2005; CROW et al, 2005; CSONT et al, 2005;
WU et al, 2007; GAUTER-FLECKENSTEIN et al, 2008; REBOUÇAS et al, 2008b;
SOMPOL et al, 2008). Entre as Mn-porfirinas que se destacam nesses estudos,
encontram-se a meso-tetraquis (N-etilpiridinio-2-il)porfinatomanganês(III) (MnTE-2-
PyP5+
) e a meso-tetraquis(N-hexilpiridinio-2-il)porfinatomanganês(III) (MnTnHex-2-
PyP5+
) (BATINIĆ-HABERLE et al, 2010).

76
2. OBJETIVOS
2.1. Geral:
Verificar se a Mn-porfirina MnTE-2-PyP5+
(MnP) é um antioxidante catalítico
em potencial para manutenção e cultivo de H. reidi;
2.2. Específicos:
Avaliar se MnP é capaz de minimizar os impactos causados por tratamentos de
altas concentrações de H2O2 em H. reidi;
Mensurar, de forma preliminar, tempo de exposições e concentrações ideais e/ou
mínimas para que MnP traga benefícios a saúde de H. reidi, através de um
estudo etológico;

77
3. MATERIAIS E MÉTODOS
3.1. Qualidade da água
As variáveis físico-químicas dos aquários de populações estoque, bem como dos
demais aquários onde foram realizados os experimentos, foram controladas
constantemente. As variáveis de temperatura, salinidade e pH foram mantidas em 25°C,
27ppm e 8.2 respectivamente. Os demais parâmetros químicos da água NH3, NO2 foram
monitorados diariamente e não excederam os limites adequados para cavalos-marinhos
variando entre: 0 – 0,25 (KOLDEWEY, 2005). Foi estabelecido também um fotoperíodo
de 10L : 14E, utilizando luz branca fria acima dos tanques, com intensidade luminosa de
cerca de 500lx na superfície da água.
3.2. Metodologia do Desenho Experimental: Matrizes Doehlert
Um desenho experimental é o meio mais eficiente para chegar a conclusões com
um mínimo de ensaios e a utilização de técnicas de desenho experimental multivariados
vem sendo cada vez mais difundido em vários campos de pesquisa (BENSALAH et al,
2010). Neste experimento, foi usado um design Doehlert. Matrizes Doehlert
(DOEHLERT, 1970) apresentam a vantagem de, através de poucos experimentos, varrer
de forma mais eficiente todo o domínio experimental, considerando a interação entre as
variáveis (QUIGNON et al., 1997). O design Doehlert também tem vantagens sobre
outros modelos (como Central Composite (CCD), Box–Behnken e Plackett-Burman),
devido à possibilidade de aumentar o domínio experimental por adição de outro fator,
sendo mais eficientes e podendo mover-se através do domínio experimental (KIRAN et
al, 2010; SAUTOUR et al, 2001), considerando também o efeito das interações entre as
variáveis (KIRAN et al, 2010). Outras vantagens das Matrizes Doehlert são: (1)
apresenta um domínio experimental esférico com uma uniformidade no espaço
experimental, (2) a sua capacidade de explorar a totalidade do domínio, e (3) o seu
potencial de reutilização consecutiva, onde os experimentos podem ser reutilizados
quando os limites não foram bem escolhidos inicialmente (BENSALAH et al., 2010).

78
Este desenho experimental permitiu não só a avaliação da influência relativa dos
parâmetros testados a baixo custo (necessidade de 13 combinações no lugar de 81 para
um design clássico completo, onde seria usado 3 posições (valores) para cada uma das
três variáveis testadas), mas também para determinar as interações inesperadas entre
alguns destes parâmetros.
3.3. Desenho Experimental
A matriz Doehlert foi aplicada para encontrar uma tendência a uma possível
diminuição do estresse oxidativo em H. reidi. O número de experimentos necessários
(N) é dado por N = n + n2 + n0, onde “n” é o número de variáveis e “n0” é o número de
pontos centrais. Neste experimento, o valor “n0” foi fixada em 2, assim, com três fatores
(concentração de MnTE-2-PyP5+
, concentração de peróxido de hidrogênio e tempo de
exposição MnTE-2-PyP5+
), o total de pontos na matriz foi 13 (Fig. 1). As réplicas
Doehlert foram realizadas no nível central das variáveis a fim de validar o modelo por
meio de uma estimativa de variância experimental.
Figura 1. Cuboctaedro apresentando a localização dos pontos (13) experimentais para três diferentes
fatores codificados (X1, X2 e X3). Foram obtidos: ao longo do eixo X2, em um trecho mediano, seis
experimentos localizados nos vértices (2, 3, 5, 6, 8 e 11) e um no centro de um hexágono, em ambos os
lados; três experimentos (4, 12 e 13) estão localizados no trecho superior e outros três experimentos (7, 9
e 10) em trecho menor.
(X2)
(X2)
(X3)
(X3)

79
Os experimentos foram feitos utilizando soluções estoque de H2O2 a 3% de
ingrediente ativo (Rioquímica) e concentração de 882 mM e de Mn-porfirina - Mn(III)
meso-tetraquis(N-etilpiridinio-2-il)porfirina - de concentração igual a 1,99 mM.
A Mn-porfirina (MnP) foi estudado em cinco níveis, o peróxido de hidrogênio
em sete e o tempo de exposição ao MnP em 3, com seus domínios experimentais
definidos entre: 0-10 µM (MnP), 0-654 µM (H2O2) e 0-60 min (exposição ao MnP). As
concentrações de H2O2 testadas foram determinadas com base nos experimentos do
Capítulo 2.
O desenho experimental Doehlert é a distribuição uniforme dos experimentos
em um espaço tridimensional (SAUTOUR et al., 2001). Assim, 12 experimentos são
equidistantes a partir de um experimento central, tendo os valores codificados (0, 0, 0) e
são distribuídos sobre uma esfera com um raio de 1 (Fig. 1). As variáveis experimentais
Xi foram codificados como xi de acordo com o trabalho de Sena et al (2012), uma vez
que para configurar ambos, modelo e os experimentos, é necessário utilizar os valores
iniciais e os valores codificados correspondentes a estes (Tabela 1), através da seguinte
equação:
xi = ((Xi – X0i) / ΔXi)*αi
Onde “xi” é o valor codificado da variável “i”, Xi é o valor natural, X0i é o valor
no ponto central, ΔXi é o valor mudança, e αi é o valor máximo da variável codificada
(ou seja, 1,0, 0,866 e 0,816 por cinco, sete e três níveis, respectivamente) (KANARAM
e MEDICHERLA, 2010) (Tabela 2).
Foram selecionados 14 animais com altura média e peso úmido equivalente a
15.7 g (±1.9) e 12,2 g (±1,2), respectivamente, provenientes da população estoque para
cultivo experimental do LAPEC (Laboratório de Peixes – Ecologia e Conservação),
Universidade Federal da Paraíba, Brasil. Os animais tiveram o comprimento estimado
com auxílio do software gratuito UTHSCSA Image Tool (WILCOX et al., 2002) e o
peso aferido com uma balança milesimal (Bel M-333).
Os tratamentos e durações foram concebidos com base na Matriz Doehlert
(Tabela 2), seguindo o mesmo princípio dos experimentos do Capítulo 2 desta
dissertação, que foram baseados em protocolos que podem ser facilmente aplicadas em
explorações agrícolas comerciais (GAIKOWSKI et al. 1999; A U. S. Food and Drug

80
Administration – EUA, 2002; YANONG, 2014): 60 minutos de imersão em diferentes
concentrações de H2O2, precedido por exposição de 0,35 ou 60 minutos de MnP..
Foi preparado um sistema de exposição que consiste em 1 aquários aquário
pequeno de 140ml (7 x 4 x 8cm) com oxigenação constante para o tratamento com MnP
e um aquário maior, de 20L (21 x 36 x 30cm) com recirculação contínua da água
(Submersible Pump SP-2500, Boyu), para homogeneização do peróxido de hidrogênio e
constante oxigenação, e um substrato de apoio (Fig. 2). Todos os experimentos
ocorreram entre as 11:00am e 18:00pm, uma vez que sabe-se que a frequência
respiratória em peixes se alteram ao longo do dia (BARRETO e VOLPATO, 2006).
Tabela 1: Valores originais e os valores correspondentes codificados para os experimentos com H. reidi
em condições controladas, LAPEC, Brasil, 2015.
Variáveis
independentes
Valores codificados e naturais
-1 -0.866 -0.816 -0.577 -0.5 -0.289 0 0.289 0.5 0.577 0.816 0.866 1
V. MnP (μl/mL) X1 0
177
352.00
528
703
V. H2O2 (ml/L) X2
0
50
100 150.00 200
250
300
TEMnP (min) X3
10
35
60
Tabela 2: Matriz experimental obtida pela metodologia de desenho experimental Doehlert por três
fatores. Valores experimentais H. reidi em condições controladas, LAPEC, Brasil, 2015.
Experimentos
Valores codificados Valores naturais Concentrações
utilizadas
X1 X2 X3
V. MnP
(μl)
V. H2O2
(ml/L)
TEMnP
(min)
[MnP]
(Mol/L)
[H2O2]
(mM)
1 0 0 0
352 150 35
0.00199 872
2 0 -0.577 -0.816
352 50 10
0.00199 872
3 +1 0 0
704 150 35
0.00199 872
4 0 +0.577 +0.816
352 250 60
0.00199 872
5 -1 0 0
0 150 35
- 872
6 -0.5 -0.866 0
176 0 35
0.00199 -
7 +0.5 -0.866 0
528 0 35
0.00199 -
8 +0.5 -0.289 +0.816
528 100 60
0.00199 872
9 -0.5 -0.289 +0.816
176 100 60
0.00199 872
10 -0.5 +0.289 -0.816
176 200 10
0.00199 872
11 +0.5 +0.289 -0.816
528 200 10
0.00199 872
12 +0.5 +0.866 0
528 300 35
0.00199 872
13 -0.5 +0.866 0 176 300 35 0.00199 872

81
Os animais foram expostos, um por vez, a diferentes concentrações de MnP
(0μl/ml, 1.26μl/ml, 2.51μl/ml, 3.77μl/ml e 5.03μl/ml) no aquário menor por um
determinado tempo (10, 35 e 60 minutos) e, em seguida, transferidos para o aquário
maior contendo diferentes concentrações de H2O2 (0 µM, 218 µM, 545 µM, 599 µM,
654 µM) (Tabela 2), onde permaneceram por 60 minutos. A transferência entre o
aquário menor e o aquário maior não excedeu 10 segundos, não houve contato algum
com os animais no processo.
Observações de aspectos biométricos, saúde e comportamento dos animais
foram realizadas. Para quantificação da concentração de H2O2 nos aquários, foi retirada
uma alíquota de água, no início e no final de cada experimento e medido a sua
absorbância em um período máximo de 40 minutos, seguindo o método I3-
de acordo
com a fórmula e procedimento usual (ALLEN et al, 1952; KLASSEN et al, 1994).
Foi utilizada uma solução de H2O2 a 3% de ingrediente ativo (Rioquímica). Toda
a água utilizada no experimento passou por filtragem mecânica (Manta Acrílica –
Perlon e Malha 150µm) e ultravioleta (Filtro UV Atman-36W), utilizado comumente
para controle de organismos planctônicos e microrganismos em sistemas de cultivo
fechado, antes de cada experimento.
Figura 2. Sistema experimental com recirculação contínua de água, utilizada para exposição de H. reidi a
diferentes concentrações de peróxido de hidrogênio e MnP. 1: saída de água do aquário; 2: entrada de
água no aquário; 3: bomba de recirculação de água; 4: substrato de apoio; 5: Mesa; 6: cadeira para apoio
do aquário com bomba de recirculação; 7: Aquário menor para exposição ao MnP; 8: Aquário maior para
exposição ao H2O2.
8
8

82
3.3. Análise comportamental
Os experimentos foram realizados com uma população de H. reidi clinicamente
saudável (ou seja, não haviam parasitas externos observados na pele, nadadeiras e
brânquias; sem lesões externas; comportamento e alimentação normal; nenhuma
mortalidade ou morbidade conhecida ou observada). Os animais foram filmados (HDR-
XR160, Sony) e monitorados em todo o período de 60 minutos no aquário maior para
análise comportamental e posterior contagem dos batimentos operculares por minuto
(BARRETO e VOLPATO, 2006; TANTARPALE et al, 2012; AURÉLIO, 2012),
utilizando-se de métodos não invasivos para mensurar o estresse oxidativo. Além disso,
a cada 10 min durante o período do experimento (60min), o número total do tempo que
os peixes passaram no fundo, em natação, em repouso, em exaustão e preso no substrato
foi mensurado, bem como a frequência total de movimentos natatórios horizontais, no
fundo, verticais e totais, a frequência de clicks e gapes e a frequência de batimentos
operculares/min foram aferidos. Movimentos típicos de estresse em peixes também
foram observados.
As medidas comportamentais foram escolhidas para análise com base nos
experimentos do capítulo anterior, são elas:
A proporção do tempo gasto por cada animal, na parte inferior do tanque;
A proporção do tempo gasto por cada animal nadando;
A proporção de tempo gasto por cada animal em repouso (estacionário);
A proporção de tempo que o animal gasto pelo animal aderido ao substrato
(segurando/preso sobre apoio) contra o tempo total que o animal estava em
repouso;
A proporção do tempo gasto por cada animal em exaustão;
A frequência de movimentos de natação (horizontal, vertical e no fundo do
aquário);
A frequência de comportamentos característicos de estresse em peixes;
Número de Clicks;
Número de Pipes;
Número de Gapes;
Número de Batimentos Operculares por minuto.

83
Os comportamentos foram descritos seguindo os resultados do Capítulo 2, onde
foi possível descrever quais seriam os comportamentos de manutenção e de estresse
para H. reidi. Nele, os comportamentos que ocorreram tanto na ausência de qualquer
estímulo externo (adição de H2O2) como na presença, foram classificados como
comportamento de manutenção (Tab. 3), são eles: presença dos animais na parte
inferior do aquário, comportamentos de nado (horizontal, vertical e no fundo), repouso
(estacionário) do animal no fundo do aquário e/ou aderido ao substrato
(segurando/preso sobre apoio), Clicks e Gapes. Para este caso, foi medida a alteração do
tempo gasto realizando estas ações e/ou sua frequência em relação aos animais controle.
A posição é um artifício de confinamento e não são traduzíveis para o comportamento
do cavalo-marinho em estado selvagem, porém, em um ambiente de aquário, cavalos-
marinhos podem posicionar-se em zonas mais calmas do aquário (ANDERSON et al,
2011), assim foi incluída, quantificada e analisada. Nesse mesmo sentido, os
comportamentos que só ocorreram na presença do peróxido de hidrogênio, foram
categorizados como comportamentos de estresse (Tab. 4) para H. reidi, com base na
literatura de comportamentos semelhantes descrito para outras espécies de peixes
(FRANCIS-FLOYD, 1988; BARRETO e VOLPATO, 2006; DUBE e HOSETTI, 2010;
ANDERSON et al, 2011).
Tabela 3: Etograma dos dados comportamentais de estresse em H. reidi.
Etograma de cavalo-marinho Hippocampus reidi
Comportamento de estresse
Posição Posição do animal em relação ao tanque
Exaustão
Quando ocorre à perda da capacidade de realização de ações, onde o animal fica a
deriva no aquário.
Locomoção
Posturas enquanto em locomoção
Nados
irregulares
Os animais demonstram movimentos de natação irregulares, erráticos e
dardejantes, seguidos de perda de equilíbrio, podendo ou não serrem
acompanhados do peixe ficar "pendurado" verticalmente na água.
Movimentos da
cabeça
Movimentos da cabeça e suas partes
Pipe
O focinho do animal rompe a superfície da água enquanto os movimentos
operculares continuam.
Cauda
enrolada na
cabeça
O animal enrola, insistentemente, a cauda na cabeça e/ou partes da mesma.

84
Tabela 4: Etograma dos dados comportamentais de manutenção em H. reidi.
Etograma de cavalo-marinho Hippocampus reidi
Comportamento manutenção
Posição Posição do animal em relação ao tanque
Topo
Quando o corpo do animal se localiza em sua maior perção, na metade superior da
altura vertical do tanque;
Fundo
Quando o animal está localizado, em sua maior porção, na metade inferior da altura
vertical do tanque;
Estacionário
Posturas enquanto estiver parado
Empoleirado
O animal está em repouso no tanque, aderido ao substrato.
Sentado
O animal está em repouso no fundo do tanque, livre do substrato. Cabeça e tronco
estão em uma posição vertical.
Locomoção
Posturas enquanto em locomoção
Nado
horizontal
Animal se desloca na horizontal, lateralmente, através da água em uma posição vertical
ou horizontal, nadadeira peitoral e barbatanas dorsais em movimento ondulatório. A
cauda pode ser ondulada ou para a frente, estendendo-se abaixo ou atrás do animal.
Nado
vertical
Animal se desloca na vertical, através da coluna d'água em uma posição vertical,
nadadeira peitoral e barbatanas dorsais em movimento ondulatório. A cauda pode ser
ondulada ou para a frente, estendendo-se abaixo ou atrás do animal.
Nado no
fundo
O animal atravessa o fundo do tanque com o tronco em posição vertical ou
obliquamente na posição vertical, nadadeira peitoral e barbatanas dorsais em
movimento ondulatório. A cauda arrasta na parte inferior, estendida por trás do animal.
Movimentos
da cabeça
Movimentos da cabeça e suas partes
Click
A cabeça se inclina para cima instantaneamente, o hioide se sobressai e a boca está
aberta. Associado a um som de clique.
Gape
O animal abre a boca lentamente, com a cabeça um pouco inclina para cima e o hioide
se projeta. Não há som de clique.

85
3.4. Medição Óptica
Um espectrofotômetro BEL Photonics SP 1105, que mede a 0.001 unidade de
absorbância, foi utilizado. Os espectros apresentados no presente estudo tiveram o
Branco subtraído. As medições de absorbância foram feitas ~4 minutos após a
colocação da cubeta no espectrofotômetro, a fim de permitir que a temperatura das
soluções se igualem as do espectrofotômetro. Todas as medições de absorbância foram
feitas dentro da faixa de 24-25 C.
A absorbância é proporcional à concentração de moléculas absorvedoras de luz
na amostra e ao caminho óptico da substância através da qual a luz viaja. Sendo assim, a
Lei de Beer expressa a dependência da absorbância (A) em relação ao caminho óptico
(b) e a concentração (c):
A = bc
3.5. Análises Estatísticas
Os resultados foram processados em planilhas eletrônicas, utilizando-se do
software Excel® da Microsoft para realizar cálculos de planejamentos fatoriais e
otimização empregando Metodologias de Superfície de Resposta (RSM), desenvolvidos
por Teófilo e Ferreira (2006), que tem seus resultados e cálculos validados através dos
softwares Matlab 6.5 e Statistica 6.0.
Todos os testes foram considerados significativos quando seus resultados
demonstraram um nível de p ≤ 0,05. Dados referentes às análises são apresentados na
forma de média ± erro ou porcentagem ± erro padrão.

86
4. RESULTADOS
Os resultados das análises de regressão múltiplas que forneceu as estimativas
dos coeficientes do modelo estão listadas na Tabela 3. Os coeficientes de regressão, r2,
foram iguais a 0.903, 0.901 e 0.895 (Fig. 3), respectivamente, para o tempo gasto pelo
animal nadando, em repouso e a frequência de comportamentos típico de estresse;
portanto, cerca de 90% da fração da variação em torno da média, para estes parâmetros,
pode ser explicada pelos modelos (BOX e DRAPER, 1987). A variável ganho/perda de
peso do animal e as variáveis comportamentais de cauda enrolada, nados irregulares,
click e batimento opercular, apresentaram r2
com valores de 0.800, 0.771, 0.753, 0.800,
0.672. Isso claramente pode ser comprovado uma vez que o coeficiente de resposta, b0,
apresenta os mesmos valores experimentais obtidos quando todos os fatores são
ponderados com o nível codificada em 0, ou seja, b0 = Condição Experimental 1 da
Tabela 2. Nesse sentido, quanto maior for o valor absoluto dos coeficientes lineares, b1,
b2 e b3, mais importante foi a variável que influenciou a resposta.
Figura 3. Gráficos dos coeficientes de regressão para os experimentos utilizando H2O2 como estressor
em H. reidi (n = 14) e a MnP como antioxidante em condições controladas, LAPEC, Brasil, 2015.. A:
Tempo de Nado; B: Tempo em Repouso; C: Frequência dos comportamentos totais de estresse.
Figura 3. Gráficos dos coeficientes de regressão para os experimentos utilizando H2O2 como estressor em H. reidi
e a MnP como antioxidante. A: Tempo de Nado; B: Tempo em Repouso; C: Frequência dos comportamentos totais
de estresse.

87
Similar aos observado nos experimentos do Capítulo 2, o peróxido de
hidrogênio (b2) foi a substância que causou maior impacto sobre as variáveis estudadas
(Tabelas 5, 6 e 7), influenciando de forma negativa ao bem-estar do animal em todos os
parâmetros testados. A segunda variável que teve maior influência foi o tempo de
exposição do animal ao MnP (b3), seguido da concentração de MnP (b1). O animal
perde peso com o aumento da concentração de H2O2 (0-654 µM) e tende a ganhar com o
aumento da concentração de MnP (0-10 µM) a que será exposto e com o aumento do
tempo de exposição ao mesmo (0-60min) (Tabela 5). Todas as variáveis para esse
parâmetro apresentaram um efeito quadrático (Tabela 5). Este resultado sugere que,
dependendo dos valores dos parâmetros, o valor ótimo para essas variáveis está dentro
do desenho amostral estudado. Foi demonstrada também uma interação significativa
entre [MnP] e [H2O2] (b12), [MnP] e tempo de exposição ao mesmo (b13), [H2O2] e
tempo de exposição ao MnP (b23) (Fig. 4). Esses valores sugerem uma influência
sinérgica de ambas as variáveis sobre ganho/perda de peso nos animais, com destaque
para b13 que proporciona um ganho de peso em até 923.1±55.1 mg (t = 16.75, p <
0.004) (Tabela 5).
Para o parâmetro locomoção, temos que aumento da concentração de H2O2 (0-
654 µM) e do tempo de exposição ao MnP (0-60min) tendem a tornar o animal mais
agitado, aumentando em até 28 e 16% o tempo que o animal está nadando e,
consequentemente, diminui o tempo do animal em repouso em -29 e -16%,
respectivamente (Tabela 7). O aumento da concentração de MnP (0-10 µM) não se
demonstrou significativa, no entanto, foi observado um efeito quadrático para esse
parâmetro (Tabela 7).
Os comportamentos descritos como comportamentos de estresse no Capítulo 2,
apresentaram-se em menor frequência que no capítulo anterior, uma vez que os valores
aqui achados, em alguns casos, como os da frequência de pipe e exaustão não se
mostraram suficiente para os testes. Quanto aos demais comportamentos, como a
frequência de Nados Irregulares e frequência de vezes que o animal enrolou a cauda na
cabeça, é interessante notar o efeito sinérgico presente, onde em ambos os casos,
novamente b13 mostrou uma mitigação desses comportamentos, chegando a diminuir a
frequência em até -2.9 ±0.1 e -4.5 ±0.5, respectivamente (Tabela 6). Esse efeito se torna
ainda mais claro quando observamos a frequência de movimentos totais de estresse,

88
uma vez que b13 apresentou uma diminuição nesse parâmetro ainda maior, de -8.7±2
(Tabela 6).
Os clicks, diferente do que se esperava, mostraram resultados semelhantes entre
as variáveis, onde b1, b2 e b3 proporcionaram um aumento na sua frequência em 1.2
±0.1, 1.9±0.1 e 1.5 ±0.1, respectivamente (Tabela 6). Com um efeito quadrático em b22
e b33. Também apresentou interação significativa em b23, com um aumento em até
4.1±0.3 na frequência de clicks (Tabela 6). No entanto, vale notar que por se tratar de
um estudo inicial, mesmo não tendo a significância estipulada, as sinergias entre b12 e
b13 não devem ser ignoradas, uma vez que estão próximos a esta, onde b12 apresentou
uma diminuição na quantidade de clicks (-0.7 ±0.3, t = 3,46, p = 0,07) e b13 um aumento
de 1 ±0.3 (t = 3,97, p < 0,06) (Tabela 6).
Os batimentos operculares aumentaram sua frequência em b2 (8.06±0.82) e b3
(4.83±0.82), uma vez que para esses valores, os animais também ficavam mais agitados
(Tabela 6). Todas as variáveis para esse parâmetro apresentaram um efeito quadrático,
com destaque para b2, onde o efeito quadrático apresentou um valor negativo (-
7.11±1.49), contrário ao seu valor quando consideramos só a variável linear (Tabela 6).
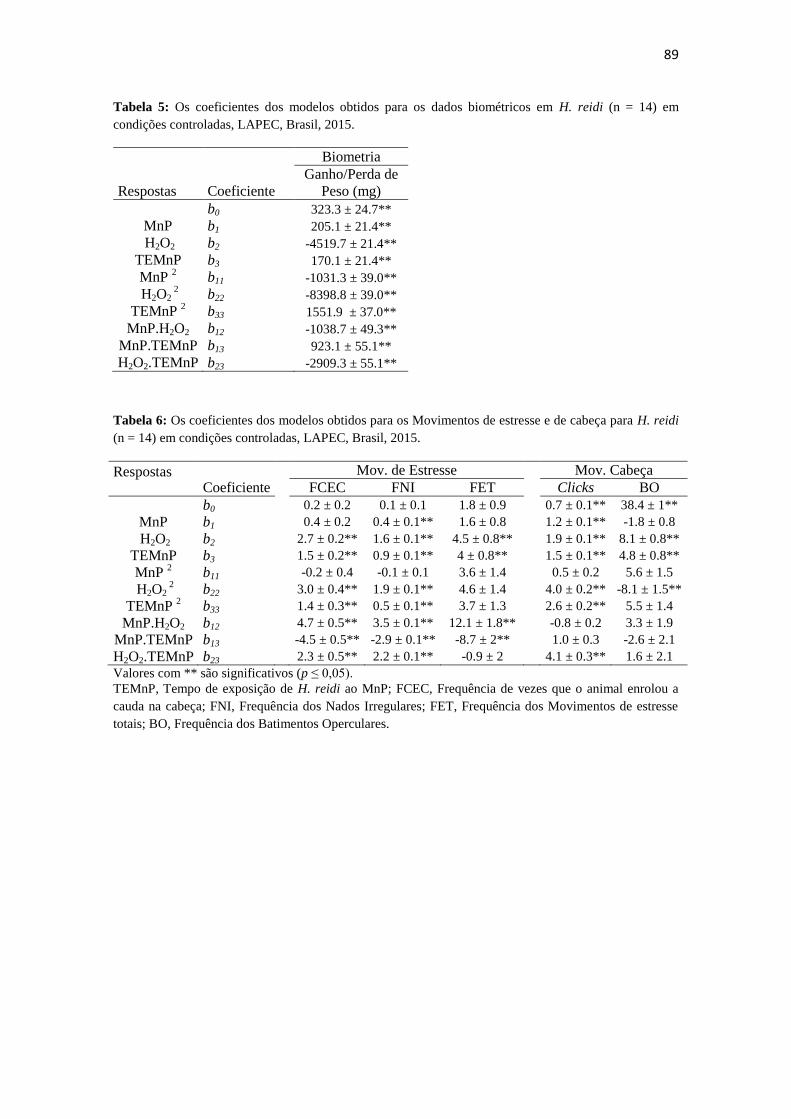
89
Tabela 5: Os coeficientes dos modelos obtidos para os dados biométricos em H. reidi (n = 14) em
condições controladas, LAPEC, Brasil, 2015.
Respostas Coeficiente
Biometria
Ganho/Perda de
Peso (mg)
b0 323.3 ± 24.7**
MnP b1 205.1 ± 21.4**
H2O2 b2 -4519.7 ± 21.4**
TEMnP b3 170.1 ± 21.4**
MnP 2 b11 -1031.3 ± 39.0**
H2O2 2 b22 -8398.8 ± 39.0**
TEMnP 2 b33 1551.9 ± 37.0**
MnP.H2O2 b12 -1038.7 ± 49.3**
MnP.TEMnP b13 923.1 ± 55.1**
H2O2.TEMnP b23 -2909.3 ± 55.1**
Tabela 6: Os coeficientes dos modelos obtidos para os Movimentos de estresse e de cabeça para H. reidi
(n = 14) em condições controladas, LAPEC, Brasil, 2015.
Respostas Coeficiente
Mov. de Estresse Mov. Cabeça
FCEC FNI FET
Clicks BO
b0 0.2 ± 0.2 0.1 ± 0.1 1.8 ± 0.9
0.7 ± 0.1** 38.4 ± 1**
MnP b1 0.4 ± 0.2 0.4 ± 0.1** 1.6 ± 0.8
1.2 ± 0.1** -1.8 ± 0.8
H2O2 b2 2.7 ± 0.2** 1.6 ± 0.1** 4.5 ± 0.8**
1.9 ± 0.1** 8.1 ± 0.8**
TEMnP b3 1.5 ± 0.2** 0.9 ± 0.1** 4 ± 0.8**
1.5 ± 0.1** 4.8 ± 0.8**
MnP 2 b11
-0.2 ± 0.4 -0.1 ± 0.1 3.6 ± 1.4
0.5 ± 0.2 5.6 ± 1.5
H2O2 2 b22
3.0 ± 0.4** 1.9 ± 0.1** 4.6 ± 1.4
4.0 ± 0.2** -8.1 ± 1.5**
TEMnP 2 b33
1.4 ± 0.3** 0.5 ± 0.1** 3.7 ± 1.3
2.6 ± 0.2** 5.5 ± 1.4
MnP.H2O2 b12 4.7 ± 0.5** 3.5 ± 0.1** 12.1 ± 1.8**
-0.8 ± 0.2 3.3 ± 1.9
MnP.TEMnP b13 -4.5 ± 0.5** -2.9 ± 0.1** -8.7 ± 2**
1.0 ± 0.3 -2.6 ± 2.1
H2O2.TEMnP b23 2.3 ± 0.5** 2.2 ± 0.1** -0.9 ± 2 4.1 ± 0.3** 1.6 ± 2.1
Valores com ** são significativos (p ≤ 0,05).
TEMnP, Tempo de exposição de H. reidi ao MnP; FCEC, Frequência de vezes que o animal enrolou a
cauda na cabeça; FNI, Frequência dos Nados Irregulares; FET, Frequência dos Movimentos de estresse
totais; BO, Frequência dos Batimentos Operculares.
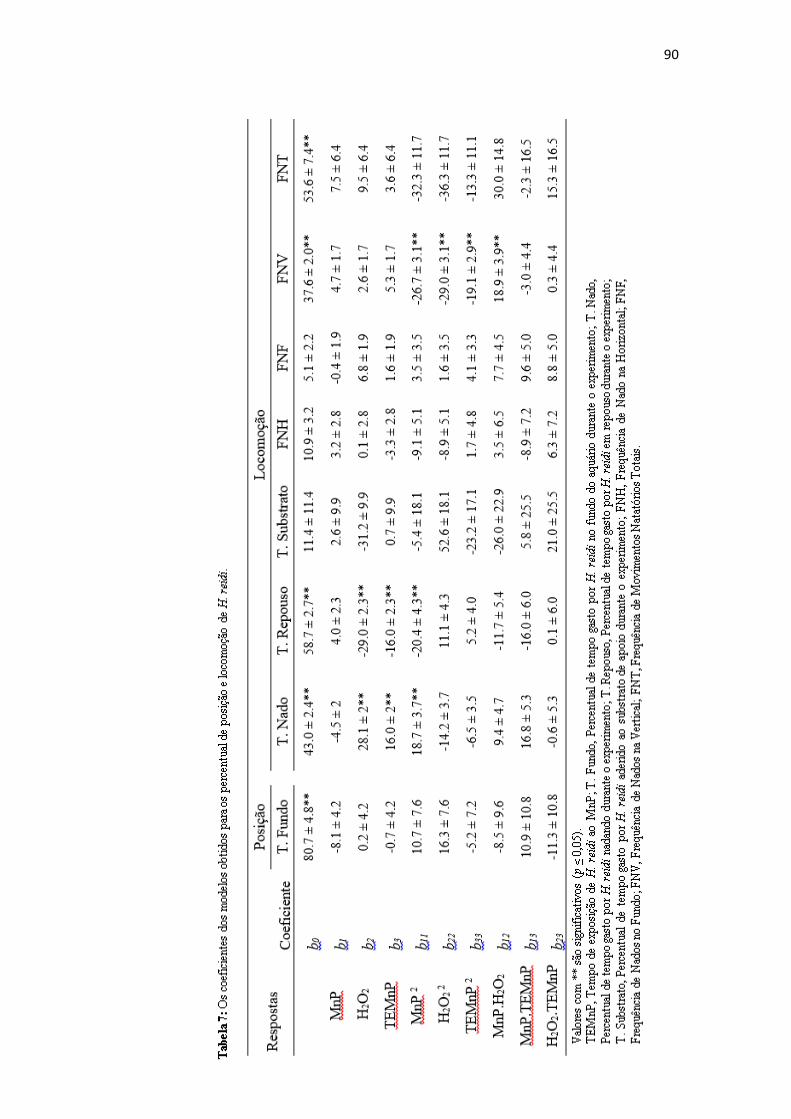
90

91
Figura 4. Superfície de resposta e valores de MnP vs. H2O2 no Ganho/Perda de peso de H. reidi (n = 14)
30 dias após o experimento, em condições controladas. LAPEC, Brasil, 2015.
Figura 5. Superfície de resposta e valores de MnP vs. H2O2 no tempo gasto por H. reidi (n = 14) em
repouso durante o experimento, em condições controladas. LAPEC, Brasil, 2015.
Figura 6. Superfície de resposta e valores de MnP vs. H2O2 na taxa respiratória de H. reidi durante o
experimento, em condições controladas. LAPEC, Brasil, 2015.

92
5. DISCUSSÃO
As análises e parâmetros comportamentais foram selecionadas e realizadas com
base nos resultados dos estudos já apresentados anteriormente no Capítulo 2 e na
bibliografia (ØVERLI et al, 2002; BARRETO e VOLPATO, 2006; FALEIRO et al,
2008; ANDERSON et al, 2011). Por outro lado, no presente estudo, um desenho
experimental foi usado para avaliar a influência das variáveis independentes sobre o
bem-estar dos animais, sob uma nova abordagem para mensurar o efeito combinado de
diferentes concentrações de MnP (b1) e H2O2 (b2), e variados tempos de exposição de H.
reidi ao MnP (b3), por meio de um estudo comportamental associado.
O cortisol é, sabidamente, conhecido por diminuir a ingestão de alimentos em
animais expostos, reduzindo seu peso (STEVENS, 1972; GREGORY e WOOD, 1999).
Os experimentos realizados neste capítulo reforçam os resultados do Capítulo 2, uma
vez que o desenho experimental demonstrou que o H2O2 em altas concentrações possui
um efeito nocivo ao animal (Tabelas 5, 6 e 7). É interessante notar também que b1 e b3,
de forma isolada, ocasionaram um ganho no peso do animal e sua interação (b13) em
condições máximas de concentração e tempo de exposição ao MnP apresentou um sinal
de ganho de peso ainda maior (Tabela 5), indicando que para esse parâmetro, o MnP se
mostrou benéfico. Sua interação com H2O2 também demonstrou resultados satisfatórios,
uma vez que tanto b12 quanto b23 apresentaram uma perda de peso menor quando em
condições máximas do que se esperaria da variável H2O2 isoladamente (Fig. 4).
Um padrão semelhante também foi observado para outros parâmetros quando
consideramos as variáveis isoladas. Parâmetros como o tempo gasto nadando e em
repouso (Fig. 5), e nas frequências de movimentos típicos de estresse como enrolar a
cauda na cabeça, os nados irregulares e a soma dos movimentos totais de estresse, bem
como na frequência dos batimentos operculares (Fig. 6), mostraram que um aumento
em suas frequências (e diminuição, no caso do repouso) quando o H2O2 é avaliado de
forma isolada (b2) e uma diminuição em relação a b2 desses valores em b1 e b3 e, quando
observamos a interação entre as variáveis, os movimentos típicos de estresse se
destacam, por praticamente inexistirem em b13 (Tabelas 6 e 7). Em nenhum dos casos
anteriores, b12 e b23 se mostraram benéficos, o que pode indicar que as concentrações
utilizadas de MnP ou o tempo de exposição ao mesmo, não foram suficientes para

93
antagonizar os efeitos nocivos dessas altas concentrações de H2O2 em um primeiro
momento, porém, o ganho de peso dos animais após 30 dias de experimento pode
demonstrar que houve uma minimização desses efeitos nocivos a longo prazo. Vale
mencionar que nos experimentos do capítulo anterior, após 30 dias, não foi observado
ganho de peso em nenhum animal exposto ao H2O2.
O número de Clicks não pareceu se diferenciar quando consideramos as
variáveis b1, b2 e b3 isoladas, em todos os casos houve um aumento no número de clicks.
Esse aumento pode estar associado não necessariamente ao estresse, cerca de 90% dos
clicks ocorreram nos primeiros minutos após a passagem do animal do aquário menor
ao maior. Nesses primeiros minutos, os animais pareciam “clicar” repetidamente numa
tentativa de limpar o focinho, uma vez que no aquário menor, eles estavam totalmente
imersos em concentrações de MnP. Não obstante, a interação b12 apresentou um sinal de
diminuição de clicks, apesar de seu valor não ser significativo (t = 3,46, p = 0,07), por
este ser um trabalho de caráter exploratório, vale ser considerado.
Apesar de não se poder fazer uma comparação direta entre os experimentos
deste capítulo com os do Capítulo 2 por possuírem metodologias exploratórias
diferentes, é válido notar que para todos os casos, os exemplares de H. reidi deste
capítulo reduziram os comportamentos associados ao estresse (quando na presença de
MnP) do que os animais do capítulo anterior, expostos apenas ao H2O2. Também houve
uma diminuição, visual e numérica, entre os valores de todos os parâmetros, onde
parâmetros como Exaustão e frequência de Pipes não puderam participar das análises
devido a uma limitação dos testes a valores muito baixos.
Por não haver trabalhos envolvendo MnP em estudos com animais marinhos,
tampouco havendo estudos comportamentais, as informações existentes são difíceis de
comparar devido aos distintos objetivos e metodologias. No entanto, trabalhos
demonstrando o valor da MnP enquanto antioxidante catalítico in vivo são unanimidade.
Benov et al (2005) demonstraram uma supressão da diabetes induzida por estresse
oxidativo em ratos; Crow et al (2005) mostraram que MnP é um agente promissor para
o tratamento de esclerose lateral amiotrófica em camundongos, quando tratada no
início; Gauter-Fleckenstein et al (2008), através de medidas de frequência respiratória
e avaliações histopatológicas em ratos Fisher expostos a radiação, mostraram uma
capacidade de melhorar danos pulmonares e reduzir consideravelmente a expressão de

94
fatores pulmonares chave que desempenham um importante papel nas lesões
pulmonares induzidas por radiação, entre outros (ASLAN et al, 2001; PIGANELLI et
al, 2002; CSONT et al, 2005; WU et al, 2007; REBOUÇAS et al, 2008b; SOMPOL et
al, 2008).
6. CONCLUSÃO
O uso de um desenho experimental não permitiu apenas estimar a importância
relativa (coeficientes padronizados) dos diferentes efeitos sobre a resposta
comportamental de H. reidi, mas também revelou interações específicas que podem
facilmente não serem detectadas utilizando a metodologia clássica de “modificar um
fator a cada vez”, como por exemplo, o tempo de exposição ao MnP versus a sua
concentração, e a minimização dos efeitos do H2O2 em função do MnP.
Em conclusão, parece prudente julgar que os resultados do presente estudo
mostraram, de forma preliminar e exploratória, que o MnP se demonstrou benéfico a H.
reidi, uma vez que minimizou os impactos causados por tratamentos de altas
concentrações de H2O2 a longo prazo. Um tempo mais prolongado de exposição ao
MnP pelos cavalos-marinhos também tiveram um maior impacto sobre esses efeitos do
que uma maior concentração da mesma em um menor tempo de exposição. No entanto,
por se tratar de um primeiro estudo envolvendo o MnP para minimização de estresse
oxidativo em animais marinhos, mais estudos são necessários para refinar esses
resultados, utilizando outros valores de concentração e diferentes técnicas de exposição
ao MnP.

95
7. REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, S.M. Biological indicators of stress in fish. Am. Fish. Sot. Symp. Ser. 1990
ALLEN, A. O.; Davis, T. W.; Elmore, G.; Ghormley, J. A,; Haines, B. M.; Hochanadel, C. J.
Oak Ridge Natl. Lab. Publ. 1949, ORNL 130;
ANDERSON, P. A., BERZINS, I. K., FOGARTY, F., HAMLIN, H. J., GUILLETTE JR, L. J.,
2011. Sound, stress, and seahorses: The consequences of a noisy environment to animal
health. Aquaculture 311, 129–138;
ASLAN, M.; RYAN, T. M.; ADLER, B.; TOWNES, T. M.; PARKS, D. A.; THOMPSON, J.
A.; TOUSSON, A.; GLADWIN, M. T.; TARPEY, M. M.; PATEL, R. P.; BATINIĆ-
HABERLE, I.;WHITE, C. R.; FREEMAN, B. A. Oxygen radical inhibition of nitric-oxide
dependent vascular function in sickle cell disease. Proc. Natl. Acad. Sci. U. S. A. 98:15215–
15220; 2001
AURÉLIO, M. L., 2012. Physiological and behavioral responses of temperate seahorses
(Hippocampus guttulatus) to environmental warming. Departamento de Biologia Animal,
Universidade de Lisboa;
BARTON, B. A. Stress in Fishes: A Diversity of Responses with Particular Reference to
Changes in Circulating Corticosteroids. Integ. and comp. biol., 2002. 42:517–525
BARRETO, R. E.; VOLPATO, G. L., 2006. Ventilatory frequency of Nile tilapia subjected
to different stressors. Journal of Experimental Animal Science 43, 189–196
BARTON, B. A. AND G. K. IWAMA. Physiological changes in fish from stress in
aquaculture with emphasis on the response and effects of corticosteroids. Ann. Rev. Fish
Dis., 1991. 1:3–26.
BARTON, B.A., SCHRECK, C.B. AND SIGISMONDI, L.A. Multiple acute disturbances
evoque cumulative physiological responses in juvenile chinook salmon. Trans. Am. Fish.
Sot., 1986. 115: 245-251
BATINIĆ-HABERLE; REBOUÇAS, J.S.; SPASOJEVIĆ, I. Superoxide Dismutase Mimics:
Chemistry, Pharmacology and Therapeutic Potential. Antioxid Redox Signal, 2010. 13,
877–918;
BENOV, L.; BATINIĆ-HABERLE, I. A manganese porphyrin SOD mimic suppresses
oxidative stress and extends the life span of streptozotocin-diabetic rats. Free Radic. Res.
38:81–88; 2005;
BENSALAH, W. et al, 2010. Thick and Dense Anodic Oxide Layers Formed on Aluminum
in Sulphuric Acid Bath. Journal of Materials Science and Technology, v. 26, n. 2, p. 113-118;
BOX, G. E. P.; DRAPER, N. R., 1987. Empirical Model-Building and Response Surfaces.
Wiley: New York.
CHROUSOS, G. P. Stressors, stress, and neuroendocrine integration of the adaptive
response. Ann. N.Y. Acad. Sci., 1998. 851:311–335
CROW, J. P. et al, 2005. Manganese Porphyrin Given at Symptom Onset Markedly
Extends Survival of ALS Mice. Ann Neurol 2005;58:258–265;

96
CSONT, T.; VIAPPIANI, S.; SAWICKA, J.; SLEE, S.; ALTAREJOS, J. Y.; BATINIĆ-
HABERLE, I.; SCHULZ, R. The involvement of superoxide and iNOS-derived NO in cardiac
dysfunction induced by pro-inflammatory cytokines. J. Mol. Cell. Cardiol. 39:833–840; 2005
DHABHAR, F. S., MCEWEN, B. S., & SPENCER, R. L. Adaptation to prolonged or
repeated stress — Comparison between rat strains showing intrinsic differences in
reactivity to acute stress. Neuroendocrinology, 1997. 65, 360–368
DOEHLERT, D. H., 1970. Appl. Statist., 19, 231.
DUBE, P.N.; HOSETTI, B.B., 2010. Behaviour surveillance and oxygen consumption in the
fresh water fish labes rohita exposed to sodium cyanide. Biotechnology in Animal
husbandary, 26(1-2), p. 91-103
FALEIRO, F., NARCISO, L. E VICENTE, L., 2008. Seahorse behaviour and aquaculture:
How to improve Hippocampus guttulatus husbandry and reproduction? Aquaculture 282,
33–40;
FINKEL, T. Signal transduction by reactive oxygen species. J. Cell Biol, 2011. 194, 7-15
FLOS, R., REIG, L., TORRES, P. AND TORT, L. Primary and secondary stress responses
to grading and hauhng in rainbow trout, Salmo gairdneri. Aquaculture, 1988. 71: 99-106
FRANCIS-FLOYD, R., 1988. Behavioral diagnosis. Vet. Clin. N. Am.-Small Anim. Pract. 18
(2), 305–316;
GAIKOWSKI, M. P., J. J. RACH, AND R. T. RAMSAY, 1999. Acute toxicity of hydrogen
peroxide treatments to selected lifestages of cold-, cool-, and warmwater fish. Aquaculture,
178:191–207;
GAUTER-FLECKENSTEIN, B.; FLECKENSTEIN, K.; OWZAR, K.; JIAN, C.; BATINIĆ-
HABERLE, I.; VUJASKOVIĆ, Ž. Comparison of two Mn porphyrin-based mimics of
superoxidedismutase (SOD) in pulmonary radioprotection. Free Radic. Biol. Med. 44:982–
989; 2008
GREGORY, T. R., WOOD, C. M., 1999. The effects of chronic plasma cortisol elevation on
the feeding behaviour, growth, competitive ability, and swimming performance of juvenile
rainbow trout. Physiol. Biochem. Zool. 72 (3), 286–295;
GRUTTER, A. S.; PANKURST N. W. The effects of capture, handling, confinement and
ectoparasite on plasma levels of cortisol, glucose and lactate in the coral reef
fish Hemigymnus melapterus. J Fish Biol, 2000. 57, 391–401
HALLIWELL B. and GUTTERIDGE, J.M.C. Free Radicals in Biology and Medicine, 2nd
edition. Clarendon Press; U.K, 1989
HALLIWELL, B.; 1992. Reactive oxygen species and the central nervous system. J
Neurochem. Nov, 59(5):1609-23.
HEO, G.J.; SHIN, G. Efficacy of benzocaine as an anaesthetic for Crucian carp (Carassius
carassius). Vet. Anaesth. Analg., 37 (2010), pp. 132–135
http://www.microsoft.com, acessada em julho 2015
IWAMA, G.K., PICKERING, A.D., SUMPTER, J.P., SCHRECK, C.B. (Eds.). Fish stress and
health in aquaculture. Society for Experimental Biology Seminar Series 62, Cambridge
University Press, Cambridge, 1997. 278pp

97
MOELLER, B.J.; CAO, Y.; LI, C.Y.; DEWHIRST, M.W., 2004. Radiation activates HIF-1 to
regulate vascular radiosensitivity in tumors: role of reoxygenation, free radicals, and
stress granules. Cancer Cell, 5, 429
KANARAM, S. K.; MEDICHERLA, N. R., 2010. Application of Doehlert experimental
design for the optimization of medium constituents for the production of L-asparaginase
from Palm Kernal cake (Elaeis guineensis). Microbial & Biochemical Technology, v. 2, n. 1,
p. 7-12;
KIRAN, R. R. S. et al, 2010. Statistical optimization of endo-polygalacturonase production
by overproducing mutants of Aspergillus niger in solid-state fermentation. Journal of
Biochemical Technology, v. 2, n. 2, p. 154-157;
KLASSEN, N. V., MARCHINGTON, D., MCGOWANT, H. C. E., 1994. H2O2 Determination
by the I3- Method and by KMnO4 Titration. Institute for National Measurement Standards,
Ionizing Radiation Standards, National Research Council, Canada. Anal. Chem., 66, 2921-2925;
KOLDEWEY, H. J., 2005. Syngnathid Husbandry in Public Aquariums: 2005 Manual,
With Chapters Contributed by Members of the Syngnathidae Discussion Group. Project
Seahorse and Zoological Society of London. 137pp;
Matlab, The Language of Technical Computing; Natick, USA, 2003
MOMMSEN, T. P., VIJAYAN, M. M., MOON, T. W. Cortisol in teleosts: dynamics,
mechanisms of action, and metabolic regulation. Reviews on Fish Biology and Fisheries,
1999. 9: 211–268
MURPHY, M. P. 2009. How mitochondria produce reactive oxygen species. Biochem. J..
417, 1-13
MURPHY, M. P., SMITH, R.A.J., 2000. Drug delivery to mitochondria: the key to
mitochondrial medicine. Adv. Drug Deliv. Rev. 41, 235–250;
ØVERLI Ø., KOTZIAN S, WINBERG S., 2002. Effects of cortisol on aggression and
locomotor activity in rainbow trout. Horm Behav, 42:53–61;
NICHOLLS, D. G., FERGUSON, S. J., 2002. Bioenergetics 3. Academic Press, London;
POWELL, M. D., CLARK, G. A., 2004. Efficacy and toxicity of oxidative disinfectants for
the removal of gill amoebae from the gills of amoebic gill disease affected Atlantic salmon
(Salmo salar L.) in freshwater. Aquaculture Research 35:112–123;
PICKERING. A.D. Stress and fish. Academic P&s, London, 1981. 367 pp.;
PIGANELLI, J. D.; FLORES, S. C.; CRUZ, C.; KOEPP, J.; YOUNG, R.; BRADLEY, B.;
KACHADOURIAN, R.; BATINIĆ-HABERLE, I.; HASKINS, K. A., 2002. Metalloporphyrin
superoxide dismutase mimetic (SOD mimetic) inhibits autoimune diabetes. Diabetes
51:347–355;
QUIGNON, F., HUYARD, A., SCHWARTZBROD, L., THOMAS, F.,1997. Use of Doehlert
matrices for study of poliovirus-1 absorption. Journal of Virological Methods 68, 33±44;
REBOUÇAS, J.S.; SPASOJEVIĆ, I.; TJAHJONO, D.H.; et al., 2008a. Redox modulation of
oxidative stress by Mn porphyrin-based therapeutics: The effect of charge distribution. Dalton Trans., 1233–1242;

98
REBOUÇAS, J.S.; DEFREITAS-SILVA, G.; SPASOJEVIC, I.; et al., 2008b. The impact of
electrostatics in redox modulation of oxidative stress by Mn porphyrins: Protection of
SOD-deficient Escherichia coli via alternative mechanism where Mn porphyrin acts as a
Mn-carrier. Free Radic. Biol. Med., 45: 201–210
ROTLLANT, J.; BALM, P.H.M, PEREZ-SANCHEZ, J. et al. 2001. Pituitary and internal
function in gilthead sea bream (Sparus aurata L., Teleostei) after handling and
confinement stress. Gen Comp Endocrinol,. 121, 333–342;
SAUTOUR, M. et al., 2001. Application of Doehlert design to determine the combined
effects of temperature, water activity and pH on conidial germination of Penicillium
chrysogenum. Journal of Applied Microbiology, v. 91, p. 900-906;
SCHEFFLER, I. E., 1999. Mitochondria. Wiley-Liss, New York;
SELYE, H., 1974. Stress without distress. McClelland Stewart, Toronto;
SENA, A. R. et al, 2011. Production, purification and characterization of a thermostable β-
1,3-glucanase (laminarinase) produced by Moniliophthora perniciosa. Anais da Academia
Brasileira de Ciências, v. 83, p. 599-609;
SHIGENAGA, M. K., HAGEN, T. M., and AMES, B. N. Proc. Natl. Acad. Sci. U. S. A., 1994.
91, 19771–19778;
SOMPOL, P.; ITTARAT, W.; TANGPONG, J.; CHEN, Y.; DOUBINSKAIA, I.; BATINIĆ-
HABERLE, I.; ABDUL, H. M.; BUTTERFIELD, A.; ST. CLAIR, D. K., 2008. A neuronal
model of Alzheimer's disease: an insight into the mechanisms of oxidative stress-mediated
mitochondrial injury. Neuroscience 153:120–130;
Statistica, Data Analysis Software System; Tulsa, USA, 2001;
STEVENS, E. D., 1972. Change in body weight caused by handling and exercise in fish. J.
Fish. Res. BoardC an. 29:202-203;
STOREY, K. B., 1996. Oxidative stress: animal adaptations in nature. Braz. J. Med. Biol. Res.
29, 1715-1733;
TANTARPALE, V.T.; RATHOD, S.H.; KAPIL, S. 2012. Temperature Stress on Opercular
Beats and Respiratory Rate of Freshwater Fish Channa Punctatus. International Journal of
Scientific and Research Publications, Volume 2, Issue 12;
TEÓFILO, R.F., FERREIRA, M.M.C., 2006. Quimiometria II:planilhas eletrônicas para
cálculos de planejamentos experimentais, um tutorial. Quím. Nova, Abr, vol.29, no.2, p.338-
350;
TREVES-BROWN, K. M., 2000. Anaesthesia. In: Anaesthetics in Applied Fish
Pharmacology. Treves-BrownKM (ed.). Kluwer Academic Publishers, Dordrecht, Netherlands.
pp. 206–217;
TSE, H.M.; MILTON, M.J.; PIGANELLI, J.D., 2004. Mechanistic analysis of the
immunomodulatory effects of a catalytic antioxidant on antigen-presenting cells:
implication for their use in targeting oxidation–reduction reactions in innate immunity.
Free Radical Biol. Med., 36, 233
U.S. Food and Drug Administration. 2002. Title 21 Code of Federal Regulations Part 530 (21
CFR 530). Available: http://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/
CFRSearch.cfm?CFRPart=530 (Junho, 2015);

99
YANONG, R. P. E., 2014. Use of Hydrogen Peroxide in Finfish Aquaculture. Fisheries and
Aquatic Sciences in the School of Forest Resources and Conservation, UF/IFAS
WALLACE, D. C. Science, 1999. 283, 1482–1488;
WEBER, R. A.; PELETEIRO, J.B.; GARCÍA MARTÍN, L. O.; ALDEGUNDE, M., 2009. The
efficacy of 2-phenoxyethanol, metomidate, clove oil and MS-222 as anaesthetic agents in
the Senegalese sole (Solea senegalensis Kaup, 1858). Aquaculture, 288, pp. 147–150;
WEDEMEYER, G. A., AND D. J. MCLEAY., 1981. Methods for determining the tolerance
of fishes to environmental stressors. A.D. Pickering, editor. Stress and fish. Academic Press,
London. Pages 247-275;
WEDEMEYER, G. A., B. A. BARTON, and D. J. MCLEAY., 1990. Stress and acclimation.
In C. B. Schreck and P. B. Moyle (eds.), Methods for fish biology. pp. 451–489. American
Fisheries Society, Bethesda, Maryland;
WILCOX, D.; DOVE, B.; MCDAVID, D.; GREER, D, 2002. UTHSCSA ImageTool
Program, version 3.0. University of Texas Health Science Center, San Antonio, Texas.
Disponível em: <ftp:maxrad6.uthscsa.edu>;
WU, T. J.; KHOO, N. H.; ZHOU, F.; DAY, B. J.; PARKS, D. A., 2007. Decreased hepatic
ischemiareperfusion injury by manganese-porphyrin complexes. Free Radic. Res. 41:127–134;
ZHAO, Y.; CHAISWING, L.; OBERLEY, T.D.; BATINIĆ-HABERLE, I.; ST. CLAIR, W.;
EPSTEIN, C.J.; ST. CLAIR, D., 2005. A Mechanism-Based Antioxidant Approach for the
Reduction of Skin Carcinogenesis. Cancer Res., 65, 1401;