universidade estadual do norte fluminense darcy … · onde houver trevas, que eu leve a luz. ......
TRANSCRIPT

UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE DARCY RIBEIRO
CLÁUDIO FERNANDES SANTOS
LEUCOSE ENZOÓTICA BOVINA NA REGIÃO NORTE
FLUMINENSE: BIOQUÍMICA SÉRICA DE ANIMAIS SOROPOSITIVOS
E SORONEGATIVOS
Campos dos Goytacazes 2009

CLÁUDIO FERNANDES SANTOS
LEUCOSE ENZOÓTICA BOVINA NA REGIÃO NORTE
FLUMINENSE: BIOQUÍMICA SÉRICA DE ANIMAIS SOROPOSITIVOS
E SORONEGATIVOS
Tese apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense Darcy Ribeiro, como requisito parcial para a obtenção do grau de Doutor em Ciência Animal, na área de concentração de Sanidade Animal, linha de pesquisa Ensaios Farmacológicos, Afecções Clínicas e Cirúrgicas dos Animais.
ORIENTADOR: Prof. Dr. Claudio Baptista de Carvalho
Campos dos Goytacazes 2009

FICHA CATALOGRÁFICA
Preparada pela Biblioteca do CCTA / UENF 033/2009
Santos, Cláudio Fernandes
Leucose enzoótica bovina na região Norte Fluminense : bioquímica sérica de animais soropositivos e soronegativos / Cláudio Fernandes Santos. – 2009. 68 f. : il.
Orientador: Claudio Baptista de Carvalho Tese (Doutorado em Ciência Animal) – Universidade Estadual do Norte Fluminense Darcy Ribeiro, Centro de Ciências e Tecnologias Agropecuárias. Campos dos Goytacazes, RJ, 2009. Bibliografia: f. 57 – 68.
1. Leucose 2. Bovino 3. Bioquímica sérica 4. Histopatologia I. Universidade Estadual do Norte Fluminense Darcy Ribeiro. Centro de Ciências e Tecnologias Agropecuárias. II. Título.
CDD – 636.2089

CLÁUDIO FERNANDES SANTOS
LEUCOSE ENZOÓTICA BOVINA NA REGIÃO NORTE
FLUMINENSE: BIOQUÍMICA SÉRICA DE ANIMAIS SOROPOSITIVOS
E SORONEGATIVOS
Tese apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense Darcy Ribeiro, como requisito parcial para a obtenção do grau de Doutor em Ciência Animal, na área de concentração Sanidade Animal, e linha de pesquisa Afecções Clínicas e Cirúrgicas dos Animais.
Aprovada em 14 de abril de 2009
BANCA EXAMINADORA
Milton Masahiko Kanashiro (D.Sc., Biociências-Biotecnologia) - UENF
Bianca Brand Ederli (D.Sc., Produção Animal) – Médica Veterinária Autônoma
Prof. Carlos Eurico Pires Travassos (D. Sc., Microbiologia) – UENF
Prof. Claudio Baptista de Carvalho (D. Sc. Clínica Médica) - UENF (Orientador)

A meu avô, Claudio Fernandes Luiz, que, no início de minha
vida, ensinou-me a cuidar e amar os animais
A minha querida mãe Terezinha Fernandes Santos, pelo
exemplo de mulher, por todo apoio, incentivo e amor.
A Elenir Cruz Cabral, que nos deixou e de quem nunca
esquecerei.
A minha noiva Geanne Merys Cruz Cabral,
com todo o carinho.
DEDICO.

AGRADECIMENTOS
A Deus, por todas as bênçãos e graças que me fizeram chegar até aqui.
A minha família, pelo apoio, incentivo, compreensão e amor.
A meu orientador, Prof. Cláudio Baptista de Carvalho, pela confiança em mim
depositada, incentivo e amizade.
Aos colegas, Josias Alves Machado e Orlando Augusto Melo Júnior, pelo auxílio
sempre presente e amizade.
Ao Professor Carlos Eurico Pires Ferreira Travassos e Iliani Bianchi, pela concessão
do material de trabalho.
Ao Professor Antonio Peixoto Albernaz, pelo auxílio na elaboração do Projeto de
Pesquisa.
Ao Professor Eulógio Carlos Queiroz de Carvalho e Lio Moreira, pelo auxílio nas
análises histopatológicas.
À UENF/FAPERJ, pela concessão de bolsa de estudo.
À UENF, pela oportunidade de aprimoramento de meus conhecimentos.
Ao programa de Pós-Graduação em Produção Animal, que nos acolheu em seu
quadro, dando-nos a oportunidade do aperfeiçoamento. E ao Programa de Pós-
Graduação em Ciência Animal, que concretizou a nossa formação;
À Professora Célia Raquel Quirino e Professor Rogério Figueiredo Daher, pelo
auxílio no desenvolvimento das análises estatísticas.
Aos mestres que verdadeiramente contribuíram para a minha formação profissional
e pessoal.
À EMATER RIO/CENTERJ do município de Cordeiro, pela cessão dos animais
destinados ao estudo histopatológico.
Ao meu amor, pelo apoio e compreensão durante a realização deste trabalho.
Àqueles que contribuíram direta ou indiretamente para esta realização.
Meus sinceros agradecimentos.

Oração de São Francisco de Assis
Senhor, fazei de mim instrumento de vossa paz.
Onde houver ódio, que eu leve o amor;
Onde houver ofensa, que eu leve o perdão;
Onde houver discórdia, que eu leve a união;
Onde houver dúvida, que eu leve a fé;
Onde houver erro, que eu leve a verdade;
Onde houver desespero, que eu leve a esperança;
Onde houver tristeza, que eu leve a alegria;
Onde houver trevas, que eu leve a luz.
Ó Mestre, fazei que eu procure mais
consolar, que ser consolado;
compreender, que ser compreendido;
amar, que ser amado.
Pois, é dando que se recebe,
é perdoando que se é perdoado,
e é morrendo que se vive para a vida eterna.

RESUMO
SANTOS, Cláudio Fernandes, D.Sc., Universidade Estadual do Norte Fluminense
Darcy Ribeiro. Abril de 2009. Leucose Enzoótica Bovina na Região Norte
Fluminense: bioquímica sérica de animais soropositivos e soronegativos. Orientador:
Prof. Claudio Baptista de Carvalho.
Objetivou-se fornecer dados auxiliares ao estudo da fisiopatologia da leucose
enzoótica bovina (LEB). Foram examinadas amostras sorológicas de 240 fêmeas
bovinas, sendo 120 soropositivas para a LEB e 120 soronegativas, e necropsiadas
duas fêmeas da raça jersey, soropositivas que apresentavam lesões. No soro
sanguíneo, foram investigados os seguintes parâmetros: proteínas séricas totais
(PT), albumina (ALB), globulinas (GLO), razão albumina/globulina (A/G), aspartato
aminotransferase (AST), gamaglutamiltransferase (γGT), alanina aminotransferase
(ALT), ureia (URE), e creatinina (CRE), obtendo-se os seguintes valores para o
grupo de soropositivos: PT (7,81±1,32g/dl), ALB (2,29±0,48g/dl), GLO (5,52±1,1g/dl),
A/G (0,42±0,1), AST (46,37±17,82U/L), γGT (25,72±9,68U/L), ALT (11,57±4,93U/L),
URE (30,12±12,58mg/dl), CRE (1,11±0,29mg/dl); e para o grupo de soronegativos:
PT (7,14±0,82g/dl), ALB (2,45±0,38g/dl), GLO (4,69±0,68g/dl), A/G (0,53±0,1), AST
(33,38±13,96U/L), γGT (20,48±6,35U/L), URE (34,81±11,47 mg/dl), CRE
(1,13±0,39mg/dl). Concluiu-se que a LEB foi capaz de produzir alterações de
pequena magnitude na maioria dos parâmetros bioquímicos séricos avaliados,
quando comparados aos dos animais soronegativos. Os animais necropsiados
apresentaram características típicas de LEB nos linfonodos, fígado, cerebelo e dura-
máter da medula espinhal, associadas à forma leucêmica da doença.
Palavras-chave: Leucose, bovinos, bioquímica sérica, histopatologia.

ABSTRACT
SANTOS, Cláudio Fernandes, D.Sc., Universidade Estadual do Norte Fluminense
Darcy Ribeiro. April, 2009. Enzootic Bovine Leukosis in North Fluminense Region:
serum biochemical profile of animals serum positive and serum negatives. Advisor:
Prof. Claudio Baptista de Carvalho.
It was aimed to supply auxiliary data to the study of the fisiopathology of the enzootic
bovine leukosis (EBL). Serological samples of 240 bovine females were examined,
with 120 serum positives for EBL and 120 serum negatives, and additionally two
jersey breed females, serum positives presenting lesions were necropsied. From
blood serum, the following parameters were investigated: total serum protein (PT),
albumin (ALB), globulins (GLO), reason albumin / globulin (A/G), aspartate
aminotransferase (AST), gamma glutamyltransferase (γGT), alanine
aminotransferase (ALT), urea (URE), and creatine (CRE), being obtained the
following values for the serum positives group: PT (7,81 ± 1,32g / dl), ALB (2,29 ±
0,48g / dl), GLO (5,52 ± 1,1g / dl), A/G (0,42 ± 0,1), AST (46,37 ± 17,82U /L), γGT
(25,72 ± 9,68U /L), ALT (11,57 ± 4,93U /L), URE (30,12 ± 12,58mg / dl), CRE (1,11 ±
0,29mg / dl); and for the serum negatives group: PT (7,14 ± 0,82g / dl), ALB (2,45 ±
0,38g / dl), GLO (4,69 ± 0,68g / dl), A/G (0,53 ± 0,1), AST (33,38 ± 13,96U /L), γGT
(20,48 ± 6,35U /L), URE (34,81 ± 11,47 mg / dl), CRE (1,13 ± 0,39mg / dl).
Concluding, EBL was able to produce alterations of small magnitude in most of the
serum biochemical parameters evaluated when compared to serum negatives
animals. The necropsy data showed that animals presented typical characteristics of
EBL in the lymph nodes, liver, cerebellum and dura mater of spinal marrow
associated to the leukemic form of the disease.
Key-Words: leukosis, bovine, serum biochemistry, histopathology.

SUMÁRIO
1 INTRODUÇÃO.........................................................................................
10
2 REVISÃO DE LITERATURA................................................................... 12
2.1 Descrições da Leucose Enzoótica Bovina................................................ 12
2.2 Etiologia e propriedades gerais................................................................ 12
2.3 Epidemiologia........................................................................................... 13
2.4 Patogenia................................................................................................. 15
2.5 Manifestações clínicas............................................................................. 16
2.6 Diagnóstico clínico-laboratorial................................................................. 17
2.7 Controle e profilaxia..................................................................................
2.8 Importância econômica e sanitária...........................................................
19
20
2.9 Aspectos bioquímicos...............................................................................
2.9.1 Proteínas séricas totais (PT)....................................................................
2.9.2 Albumina...................................................................................................
2.9.3 Globulinas.................................................................................................
2.9.4 Aspartato aminotransferase – AST..........................................................
2.9.5 Gamaglutamiltransferase - γGT................................................................
2.9.6 Alanina Aminotransferase – ALT..............................................................
2.9.7 Uréia.........................................................................................................
2.9.8 Creatinina.................................................................................................
3 MATERIAL E MÉTODOS........................................................................
21
22
25
27
29
31
33
33
35 37
3.1 Animais..................................................................................................... 37
3.2 Análise laboratorial................................................................................... 37
3.3 Anatomopatologia..................................................................................... 38
3.3.1 Macroscopia............................................................................................
3.3.2 Histopatologia...........................................................................................
3.3.3 Imunoistoquímica.....................................................................................
3.4 Análise estatística.....................................................................................
39
39
39
40

4 RESULTADOS E DISCUSSÃO............................................................... 41
4.1 Análises estatísticas................................................................................. 41
4.2 Aspectos bioquímicos séricos.................................................................. 45
4.2.1 Proteínas séricas totais (PT)....................................................................
4.2.2 Albumina e razão albumina/globulina (A/G).............................................
4.2.3 Globulina..................................................................................................
4.2.4 Aspartato aminotranferase – AST............................................................
4.2.5 Gamaglutamiltranferase - γGT..................................................................
4.2.6 Alanina Aminotransferase – ALT..............................................................
4.2.7 Ureia.........................................................................................................
4.2.8 Creatinina.................................................................................................
4.3 Anatomopatologia.....................................................................................
45
46
47
48
48
49
49
50
51
5 CONCLUSÕES........................................................................................ 56
6 REFERÊNCIAS BIBLIOGÁFICAS.......................................................... 57

10
1 INTRODUÇÃO
A Leucose Enzoótica Bovina (LEB) é uma enfermidade de caráter
infectocontagioso causada por um vírus RNA, envelopado, medindo de 80 a 120 nm
de diâmetro, da família Retroviridae, subfamília Orthoretrovirinae, gênero
Deltaretrovirus.
A doença pode manifestar-se de duas formas, sendo elas: linfocitose
persistente e/ou linfossarcoma em bovinos adultos. Na primeira, o animal portador
da doença apresenta um aumento do número absoluto de linfócitos, o que ocorre em
29% dos casos. Já o linfossarcoma ocorre em torno de 5% dos animais infectados
pelo Vírus da Leucose Enzoótica Bovina (VLB), cujos sinais clínicos variam
dependendo da localização das massas tumorais no organismo dos animais, que
apresentam seus linfonodos superficiais, em geral, tumefeitos.
A LEB acomete, principalmente, animais de rebanho leiteiro com mais de
quatro anos de idade, fato este que, provavelmente, esteja relacionado ao manejo
neste tipo de exploração e à demorada soroconversão em alguns animais.
O VLB acomete, de forma natural, bovinos, ovinos, bubalinos e capivaras, e,
experimentalmente, caprinos, coelhos, suínos, gatos, cervídeos e chipanzés.
A infecção causada pelo VLB tem distribuição mundial. No Brasil, esta
patologia já foi descrita em várias regiões, tendo diferentes prevalências.
Em relação às vias de transmissão, a principal é a horizontal, principalmente
por exposição direta aos fluídos biológicos contaminados com linfócitos infectados,
como os que ocorrem com agulhas e instrumentos cirúrgicos contaminados com
sangue. A via de transmissão vertical ou transplacentária ocorre em menores
proporções. Um bovino, uma vez infectado, torna-se fonte de disseminação do VLB
por toda a sua vida.
Várias são as perdas econômicas produzidas pela Leucose Enzoótica
Bovina, dentre elas, podem-se citar: queda na produção leiteira, aumento no
intervalo entre partos e restrições ao comércio internacional.
O diagnóstico da LEB pode ser realizado por imunodifusão em gel de ágar
(IDGA), que é o teste recomendado pelo OIE (Office International des Epizooties), e
pelo enzime-linked immunosorbent assay (ELISA).
Devido à inexistência de vacinas e de um tratamento efetivo, os programas
de controle concentram-se em medidas de profilaxia sanitária que inibem a

11
disseminação do VLB, e, para tal é necessário o conhecimento da condição
sorológica do rebanho frente ao vírus. Entretanto, os criadores não têm acesso ao
diagnóstico.
Há varias décadas, as enzimas séricas têm sido mensuradas para
diagnosticar, monitorar e prognosticar os processos mórbidos. A despeito de nossa
familiaridade com o uso clínico empírico de enzimas séricas, as razões
patofisiológicas para as alterações observadas são muito pouco entendidas.
Algumas enzimas mitocondriais, normalmente, aparecem no sangue após uma lesão
severa, já que, aprisionadas, precisam atravessar duas ou mais membranas.
Quando há lesão tecidual, há aumento na liberação de algumas enzimas. A
importância do aumento depende de vários fatores: concentração tecidual da enzima
(que é variável com o tecido e a espécie), localização celular da enzima, quantidade
do tecido lesado, tipo de lesão tecidual e velocidade de remoção enzimática do soro.
Os objetivos do presente trabalho foram fornecer dados auxiliares ao estudo
da fisiopatologia da Leucose Enzoótica Bovina por meio da avaliação bioquímica
sérica dos animais soropositivos, devido ao fato de serem escassos os trabalhos
relativos a esse tema, bem como descrever os achados necroscópicos em dois
animais soropositivos.

12
2 REVISÃO DE LITERATURA
2.1 Descrições da Leucose Enzoótica Bovina
A primeira descrição da LEB foi publicada na Alemanha em 1871, sendo,
posteriormente, em 1969, descoberto o agente causal, utilizando o método de
isolamento e identificação por microscopia eletrônica. (MILLER et al., 1969;
LEISEREING, 1871 apud OLSON e MILLER, 1987).
No Brasil, a LEB foi citada inicialmente em 1943 (RANGEL; MACHADO,
1943 apud BIRGEL JUNIOR et al., 1995; LEITE et al., 2001), porém o primeiro
diagnóstico clínico da doença foi publicado em 1959 (MERKT et al., 1959, apud
LEITE et al., 2001).
A presença do vírus da LEB em células linfocitárias foi evidenciado em 1982,
mediante a microscopia eletrônica (ALENCAR FILHO et al.,1982 apud BIRGEL
JUNIOR et al., 1995), contudo, o isolamento viral só foi realizado, pela primeira vez
no Brasil, em 1985 (ANGELO et al., 1985).
O Vírus da Leucose Enzoótica Bovina (VLB) está mundialmente distribuído
(ONUMA, 1989; JONHSON; KANEENE, 1991). A provável introdução do VLB na
América do Sul foi proveniente da importação de bovinos contaminados vindos da
Europa e dos EUA (BRAGA et al., 1998b).
A prevalência do VLB varia tanto de um país a outro como também entre
regiões e/ou estados de um mesmo país (MODENA et al., 1984; ABREU et al., 1990;
BIRGEL JUNIOR et al., 1995; MORAES et al., 1996; BRAGA et al., 1998b).
2.2 Etiologia e propriedades gerais
Segundo o International Commitee on Taxonomy of Viruses (ICTV), o VLB
está classificado na família Retroviridae, subfamília Orthoretrovirinae, gênero
Deltaretrovirus.
O VLB é um vírus envelopado medindo de 80 a 120nm de diâmetro, com
genoma diploide formado por fitas idênticas de RNA simples, de polaridade positiva,
contidas em um capsídeo de simetria icosaédrica (MURPHY et al., 1999).

13
O envelope viral possui projeções (glicoproteínas de superfície)
responsáveis pela interação do vírus com a superfície dos linfócitos (FAVA, 1995;
BRAGA et al., 1998a). A gp51 e p24 destacam-se entre as principais proteínas virais
(FAVA, 1995; BRAGA et al., 1998a; OLIVEIRA, 2000).
A inativação do VLB é possível com a utilização de detergentes e solventes
lipídicos, como álcool, clorofórmio e éter, sendo também inativados a 56ºC por 30
min, não resistindo também a sucessivos congelamentos e descongelamentos. Os
retrovírus possuem maior resistência aos raios ultravioleta e radiações X do que os
outros vírus, fato este que, provavelmente, seja devido ao seu genoma diploide
(FENNER et al.,1993).
2.3 Epidemiologia
A LEB já foi diagnosticada, com diferentes prevalências, em vários estados
do Brasil, sendo os de maior prevalência: Minas Gerais, Rio de Janeiro, Pará,
Paraná, Goiás, São Paulo, Rio Grande do Sul, Roraima (GARCIA LIMA, 1980;
KANTEK et al., 1982; MODENA et al., 1983; LEITE et al., 1984; BIRGEL JUNIOR et
al., 1995; SARDI et al., 2002; POLETTO et al., 2004; SPONCHIADO, 2008), como
demonstrado na Tabela 1.

14
Tabela 1. Prevalência do VLB em diferentes estados do Brasil, segundo diversos autores. Local Prevalência Autor
Rio de Janeiro 53,25% ROMERO ; ROWE, 1981
Paraná 18,33% KANTEK et al., 1982
Rio de Janeiro 26,94% CUNHA et al., 1982
Minas Gerais
Rondônia
Acre
Bahia
Goiás
70,87%
23,02%
9,72%
16,1%
35,67%
LEITE et al., 1984
ABREU et al., 1990
ABREU et al., 1990
TÁVORA, 1990
ANDRADE; ALMEIDA, 1991
Pernambuco 13,85 MELO, 1991
São Paulo 16,23% ARITA et al., 1992
Ceará 9,15% ABREU, 1993
Rio Grande do Sul 12,0% MORAES et al., 1996
Pará 49,8% MOLNÁR et al., 1999
Bahia 7,5% SARDI et al., 2002
Campos - RJ 17,30% BIANCHI, 2003
Serra do Botucatu - SP 51,80% MEGID et al., 2003
Amazonas 9,6% CARNEIRO et al., 2003
Passo Fundo - RS 23,52% POLETTO et al., 2004
Paraná 49,04% SPONCHIADO, 2008
Atribui-se a entrada do VLB à importação de animais infectados, o que
persiste até os dias de hoje (BRAGA et al., 1998b).
Demonstrou-se que o percentual de animais reagentes tende a aumentar
proporcionalmente com a idade, variando de 24,6% em animais de um a dois anos
para 86,2% em animais com mais de seis anos (BIRGEL JUNIOR et al., 1995) .
Em rebanhos leiteiros, uma maior prevalência da LEB tem sido observada,
principalmente naqueles estabelecimentos de criação intensiva e altamente
tecnificados, devido ao maior contato entre os animais e à maior utilização de
equipamentos susceptíveis à contaminação com sangue ou outro espécime
biológico que contenha linfócitos infectados, como o colostro e o leite, facilitando a
disseminação da doença. (LORENTZ; STRAUB, 1987 apud FAVA, 1995; OLIVEIRA
et al., 1999; VAN DER LAAN et al., 1999).

15
O desenvolvimento do linfossarcoma ocorre em animais com média de idade
de cinco anos, acometendo, devido ao tipo de manejo, mais ao gado leiteiro.
(OLIVEIRA et al., 1999).
2.4 Patogenia
O VLB pode ser transmitido de duas formas: vertical e horizontalmente
(BRAGA et al., 1998a), sendo que a transmissão vertical pode ocorrer em vacas
prenhes ou na passagem do feto pelo canal do parto (MOULTON,1990; JAIN, 1993).
O colostro e o leite também são vias de transmissão vertical (BIRGEL, 1982;
STRAUB et al., 1987 apud FAVA, 1995).
Os fômites e seringas, agulhas, instrumentos cirúrgicos, luvas, tatuadores e
material utilizado para a descorna contaminados com sangue são as principais
formas de transmissão horizontal (FAVA, 1995), que é a mais frequente (OLIVEIRA
et al., 1994; BRAGA et al., 1998a ).
A hipótese da contaminação pelo sêmen é aceita por alguns autores
(LUCAS et al., 1980). Tal contaminação é provável devido à presença de linfócitos
infectados no fluido seminal que ocorre com frequência nas doenças do trato genital
(THURMONT; BURRIDGE, 1982 apud BRAGA et al., 1998a). A disseminação do
VLB pode ocorrer em premunições contra anaplasma e babésia quando os animais
doadores estão infectados (FLORES et al.,1992; VAN DER LANN et al., 1999). A
transmissão via inseto hematófago também pode ocorrer (BIRGEL JUNIOR et al.,
1995; BRAGA et al., 1998b e VAN DER LANN et al., 1999).
A Leucose Enzoótica Bovina é uma doença infecciosa de caráter crônico e,
após a infecção do animal, este pode levar de três a 24 meses para que ocorra a
soroconversão e, uma vez infectado, o animal torna-se fonte de infecção por toda
sua vida (BIRGEL, 1982).
A principal célula sanguínea pela qual o VLB tem afinidade é o linfócito B, e,
uma vez tais células infectadas, infiltram-se nos tecidos linfoides do organismo do
animal, produzindo, assim, o linfossarcoma (BRAGA et al., 1998a).
O VLB tem também a capacidade de infectar, em menores proporções,
linfócitos T, monócitos e granulócitos (SCHWARTZ et al., 1999).
Há evidência de que linfócitos CD4 ativados, durante a infecção pelo VLB
promovem o desenvolvimento da Linfocitose Persistente (LP). Ainda não está

16
totalmente esclarecida a forma como os linfócitos CD4 são ativados pela infecção
pelo VLB. Foi observado que linfócitos T de bovinos com LP proliferam na presença
de células mononucleadas irradiadas e autólogas do sangue periférico. No entanto,
nenhuma proliferação foi observada em culturas de células de animais soropositivos
sem LP. A proliferação requereu contato direto entre as células, mas não foi
associada com a expressão de proteína viral ou inibida por anticorpos contra o VLB.
Estas observações e a magnitude da resposta proliferativa sugerem que a ativação
seja policlonal e envolva mecanismos diferentes da ativação antígeno-específica
causada pelo VLB (STONE et al., 2000).
Atualmente, existem evidências de que a expressão do VLB por um linfócito
infectado aumenta a sobrevivência desta célula por estacionar o ciclo celular em
G0/G1. Esta ação atrasa a apoptose fisiológica que, na ausência de estímulos
externos, ocorre na fase G2. O VLB pode ainda promover a proliferação de linfócitos
B não infectados devido à ação de proteínas virais em linfócitos T CD4+. Esta ação
aumenta os níveis de interleucina 2 (IL2), responsável pela proliferação dos linfócitos
B. Esta estratégia viral garante o sucesso da infecção por aumentar a produção de
partículas virais, prolongando a vida da célula infectada. Paralelamente, a
proliferação de linfócitos B não infectados aumenta a população de células-alvo. O
conjunto destes efeitos leva o animal a apresentar o quadro de LP, sendo esta a
principal via de transmissão do vírus aos animais não infectados (STONE et al.,
2000).
2.5 Manifestações clínicas
A maioria dos animais infectados não apresenta sinais clínicos
característicos, porém alguns podem desenvolver linfocitose persistente (VAN DER
LANN et al., 1999). Os sinais clínicos provenientes do linfossarcoma variam,
dependendo da sua localização, e a metástase pode comprometer vários órgãos
(OLIVEIRA et al., 1999).
O principal sinal clínico observado entre os animais clinicamente afetados é
o aumento no volume dos linfonodos periféricos. Outras observações relacionadas
com a leucose em rebanhos afetados são: anorexia, diminuição na produção de
leite, perda de peso e exoftalmia, dependendo do local de aparecimento dos
tumores e dos órgãos afetados (COELHO et al., 1998; BARROS, 2007), podendo

17
ocorrer, em alguns casos, pico febril, aumento dos linfonodos com o
comprometimento do coração, abomaso e intestino . À necropsia, são encontradas
formações tumorais em tecidos linfoides, no átrio direito do coração, na parede do
abomaso e útero, linfonodos aumentados com tecido neoplásico firme e branco,
muitas vezes, circundando um foco necrótico translúcido e amarelado e,
ocasionalmente, os rins e ureteres são afetados (OLIVEIRA et al., 1999). Observam-
se também aumento generalizado dos linfonodos superficiais e internos, superfície
de corte protrusa apresentando colorações variadas, entre branco-amarelado a
vermelho escuro (REIS et al., 2002; PEIXOTO et al., 2008).
São descritas também massas tumorais esbranquiçadas de consistência
firme a macia, com formações nodulares ou difusas nos diferentes órgãos, como
coração, rúmen, útero, pulmão, medula, cérebro, fígado, cadeia de linfonodos
mesentéricos, rins e tecido retrobulbar do globo ocular (YAMAMOTO, 1982;
REBHUN, 1984; SHERMANN, 1987; FIGHERA; BARROS, 2004; SILVA et al.,
2008).
O curso da doença é variável, podendo levar de algumas semanas a vários
meses, sendo que se tem, na insuficiência cardíaca, a principal causa-morte
(OLIVEIRA et al., 1999).
Uma das características dos animais infectados pelo VLB é a produção
persistente de anticorpos, sugerindo uma constante ou periódica estimulação do
sistema imunológico por antígenos virais (HEENEY et al., 1992 apud LEUZZI Jr. et
al., 2001).
2.6 Diagnóstico clínico-laboratorial
O diagnóstico da Leucose Enzoótica Bovina, inicialmente, era feito pela
observação das manifestações clínicas, baseando-se no aumento dos linfonodos e
na presença do linfossarcoma. Posteriormente, observou-se a ocorrência de
alterações hematológicas caracterizadas por linfocitose, ocorrendo tanto na
linfocitose persistente quanto no linfossarcoma. Visando ao diagnóstico precoce da
Leucose Enzoótica Bovina, foi elaborada uma chave leucométrica, buscando
combatê-la sistemicamente (GÖTZE et al., 1954 apud FAVA, 1995). Após o
isolamento viral em 1969, teve início a busca por metodologias diagnósticas mais
eficazes para a LEB (MILLER et al., 1969).

18
Atualmente, estão disponíveis vários métodos diagnósticos: leucograma
(GARCIA et al., 1991), histopatologia (COELHO et al., 1998), citologia (OLIVEIRA et
al., 1999), fixação de complemento (FC), radioimunoensaio (RIE), (MILLER; VAN
DER MAATEN, 1974 apud FAVA, 1995), imunofluorescência (IF) (GARCIA LIMA,
1980), ELISA (SIMARD et al., 2000a) e imunodifusão em gel de ágar (IDGA)
(SIMARD et al., 2000b).
A variação do leucograma, apesar de não muito utilizada, é capaz de
identificar um quadro de linfocitose persistente, precisando ser diferenciado de duas
outras linfocitoses: uma decorrente da passagem de linfócitos neoplásicos atípicos
para o sangue que ocorre em um linfossarcoma e as linfocitoses transitórias não
relacionadas ao VLB. Faz-se necessário, para diagnosticar a linfocitose persistente,
a realização de duas contagens de linfócitos em pelo menos duas amostras com
intervalo mínimo de três meses (OLIVEIRA et al., 1999).
Para exame citológico (método auxiliar para o diagnóstico da LEB), é
necessária a presença de uma massa neoplásica já instalada ou que os linfonodos
já estejam aumentados (locais da coleta do material), quando será possível
diferenciar um processo inflamatório de um processo neoplásico (OLIVEIRA et al.,
1999).
Ao exame histopatológico, observam-se infiltrações nodulares ou difusas de
células linfoides nos órgãos atingidos. Com base na morfologia celular, o
linfossarcoma é classificado como um tumor de células redondas. Neste grupo, as
células não possuem junções intercelulares, dispondo-se isoladamente. As células
neoplásicas malignas exibem uma ou mais das seguintes características: aumento
da relação núcleo/citoplasma, cromatina frouxa, presença de macronúcleo
(usualmente múltiplos), homogeneidade morfológica, pleomorfismo (presença de
células em um mesmo estágio de desenvolvimento, apresentando diferentes
formas), vacuolização intranuclear, anisocitose (presença de células em diferentes
tamanhos) e anisocariose (presença de células com núcleos de diferentes tamanhos
(CORDEIRO et al., 1994; OLIVEIRA et al., 1999). Deve-se coletar o material dos
órgãos que apresentarem tumorações e enviá-los em formalina a 10% para o
laboratório de histopatologia (LEITE et al.,2001). A histopatologia dos linfonodos
revela linfócitos com citoplasma escasso, fracamente eosinofílico, com contornos
irregulares e núcleos pleomórficos, por vezes, hipercromáticos com nucléolos

19
evidentes e frequentes figuras de mitose; há também alterações da arquitetura
normal do linfonodo e ocasionais áreas de necrose (PEIXOTO et al., 2008).
Simões (2007) e Tajima et al. (1998), ao estudarem bovinos com leucose
enzoótica, encontraram a expressão do p53 (proteína que se acumula devido a
mutações do DNA da célula neoplásca) em torno de 50% dos animais estudados.
O Radioimunoensaio é um teste mais sensível, que tem como fundamento
detectar anticorpos, porém é necessário a utilização de equipamentos altamente
especializados, o que torna este tipo de teste dispendioso (MILLER; VAN DER
MAATEN, 1974 apud FAVA, 1995).
As técnicas recomendadas pela OIE, para o diagnóstico da Leucose
Enzoótica Bovina, são o IDGA e o ELISA, o primeiro é menos sensível que o
segundo, porém devido à facilidade, praticidade, simplicidade de realização e
vantagens econômicas, tornou-se o teste-padrão a ser adotado, já tendo sido
utilizado com sucesso em programas de eliminação da LEB em alguns países
(OLIVEIRA et al., 1999).
O teste de IDGA atua detectando a presença de anticorpos para as
principais glicoproteínas do envelope viral, o que indica a infecção. Tal teste foi
desenvolvido utilizando como antígeno o polipeptídeo p24 que, depois, foi
substituído pela glicoproteína gp51, pois foi demonstrado que os animais portadores
da doença desenvolviam títulos de anticorpos mais elevados contra esta
glicoproteína (FAVA, 1995).
2.7 Controle e profilaxia
As medidas mundialmente adotadas no controle da Leucose Enzoótica
Bovina baseiam-se na identificação e eliminação de animais soropositivos. Todavia,
a conduta em um programa de controle dependerá da prevalência encontrada:
quando esta for baixa, a eliminação dos animais é recomendável, porém, quando a
infecção é encontrada em larga escala, torna-se mais viável separar os animais
soropositivos e mantê-los distante o suficiente para que não seja possível a
transmissão do vírus por meio de insetos hematófagos. Também é indispensável o
cuidado com as práticas de manejo adotadas na propriedade rural para o controle
e/ou eliminação do VLB no rebanho (BRAGA et al., 1998a).

20
Algumas medidas podem e devem ser tomadas para evitar a disseminação
da Leucose em um rebanho, tais como: o controle de insetos vetores, o uso de luvas
individuais na palpação retal de vacas, atenção especial na utilização de
equipamentos para descorna, castração, utilização de agulhas e seringas
(RADOSTITS et al., 2002).
Atualmente, são grandes os esforços de vários grupos de pesquisas em
desenvolver uma vacina eficaz, capaz de proteger bovinos contra a infecção e
mesmo contra o desenvolvimento de Linfocitose Persistente e/ou linfossarcoma.
Entretanto, apesar de estarem sendo empregadas diferentes metodologias, tanto
tradicionais quanto moleculares, até o momento não há disponibilidade comercial de
uma vacina para o controle da LEB (LEUZZI Jr. et al., 2001).
2.8 Importância econômica e sanitária
A importância econômica da leucose vem aumentando nos últimos anos
(VAN DER LAAN et al., 1999). As perdas econômicas relacionadas à leucose em um
plantel afetado são elevadas, muito embora, frequentemente, afete os animais de
forma subclínica, causando: queda significativa na produção de leite (BRENNER et
al., 1989), restrições de comercialização no mercado internacional (JOHNSON;
KANEENE, 1991; BRAGA et al., 1997), aumento do intervalo entre partos
(BRENNER et al., 1989; HEALD et al., 1992) e mortalidade dos animais (BRAGA et
al., 1997). Em rebanhos leiteiros severamente afetados, a mortalidade anual chega a
2%, podendo atingir até 5% (RADOSTITIS et al, 2002).
Para importação dos animais, muitos países exigem o teste sorológico
negativo, inclusive o Brasil, porém não temos um programa oficial de controle desta
doença (BRAGA et al., 1997).
Em animais soropositivos, foi observada, no Brasil, uma diminuição na
produção leiteira da ordem de 11% (D’ANGELINO et al., 1998).
Foi estimada, nos EUA, uma perda financeira de 86 milhões de dólares por
ano em animais soropositivos que apresentavam linfocitose persistente por três anos
consecutivos, quando levado em conta fatores como: queda na produção leiteira e
diminuição nos teores de gordura no leite (DA et al., 1993).
Devido ao fato de já terem sido encontradas partículas virais no leite
(OLIVEIRA, 2000) - alimento básico na dieta de humanos e animais - têm-se a

21
preocupação que, além das perdas econômicas já citadas, outra questão vem sendo
objeto de estudo na virologia atual: o risco em potencial da transmissão do vírus a
humanos e outros animais nos quais a doença não ocorre naturalmente.
2.9 Aspectos bioquímicos
É fundamental a existência de parâmetros para avaliação dos resultados
obtidos num perfil bioquímico, o que só é possível diante de um número significativo
de amostras de uma população. Nesse sentido, teoricamente, 95% dos valores
populacionais saudáveis encontram-se situados ao redor de uma média, não
descartando a possibilidade de um outro valor fora deste intervalo ser absolutamente
normal. Indicam, ainda, que um teste nunca deve ser interpretado de forma isolada,
e sim avaliado de acordo com outros resultados obtidos e com as alterações clínicas
encontradas no animal em questão (ROUSSEL et al., 1997).
Inúmeros são os fatores, referidos na bibliografia especializada, que podem
influir sobre os parâmetros fisiológicos de um animal sadio, causando significativa
variabilidade. Entre estes, podemos destacar: a espécie animal; a raça; a idade; o
sexo; as condições de aclimatação; o manejo de criação e as normas de
alimentação. No caso específico dos bovinos, quer sejam taurinos ou zebuínos,
deve-se ressaltar a influência de doenças que determinam infecção latente ou
mesmo de enzootias, como o Vírus da Leucose dos Bovinos (GREGORY et al.,
2004).
Os médicos veterinários têm procurado meio auxiliares para avaliação e
diagnóstico que ofereçam informações cada vez mais precisas com menor
transtorno ao animal e seu proprietário. A enzimologia tem-se apresentado como
uma boa alternativa quando usada em conjunto com outros exames, ou mesmo
isoladamente (SCHEFFER, 2003).
As provas bioquímicas realizadas no soro sanguíneo dos animais
domésticos apresentam resultados que constituem excelente subsídio ao
diagnóstico clínico das enfermidades do fígado, sendo esses exames reunidos em
baterias de provas, referidas como avaliadoras da função hepática. Como rotina
clínica, recomenda-se, para essa avaliação, a realização das seguintes provas:

22
proteinograma, atividade enzimática da aspartato aminotransferase (AST) e
gamaglutamiltransferase (GGT) (SOUZA et al., 2004).
2.9.1 Proteínas séricas totais (PT)
A proteína é um composto de alto peso molecular, constituído primariamente
de cadeia de aminoácidos unidos por ligações peptídicas. A albumina é a menor das
proteínas plasmáticas, tem peso molecular de 69.000 dáltons; as alfaglobulinas, de
200.000 a 300.000 dáltons; as betaglobulinas de 150.000 a 350.000 dáltons; as
gamaglobulinas de 150.000 a 300.000 dáltons; e o fibrinogênio cerca de 400.000
daltons (COLES, 1984). O mesmo autor afirma ainda que, embora haja considerável
número de informações disponíveis sobre as propriedades químicas das proteínas
plasmáticas, somente nos últimos 20 anos essas propriedades e funções foram
associadas a um possível significado clínico-patológico.
O decréscimo na concentração das proteínas do sangue deve-se a:
requerimento insuficiente na dieta, má absorção proteica, deficiência na síntese de
albumina pelo fígado, evasão da albumina para o espaço tecidual, com o aumento
da permeabilidade capilar nos processos inflamatórios agudos, e nas enfermidades
crônicas como nas neoplasias, além do estado fisiológico do animal que influi na
variação do proteinograma (COLES 1984). Somente a determinação da proteína
total não reflete com precisão o estado do metabolismo proteico, sendo de particular
importância a determinação da albumina e da globulina (COLES 1984).
Ocasionalmente, uma destas proteínas pode apresentar valores diminuídos, como
hipoalbuminemia nas infestações parasitárias, e elevação da concentração de
globulinas nos processos infecciosos. Dados obtidos sobre o proteinograma de
vacas com diferentes enfermidades, inclusive a leucose, mostram alterações nas
concentrações proteicas (LIBERG, 1977; FERREIRA et al., 2001).
Com o conhecimento da razão albumina/globulina, os erros de interpretação
dos valores plasmáticos de proteínas são minimizados (HERZ; HOD 1969).
Analisando amostras de soro sanguíneo de 32 fêmeas zebuínas da raça
Haryana, criadas na Índia, com a finalidade de conhecer mais sobre as proteínas
séricas, encontraram-se valores variando de 6,46 a 6,74 g/dl, não sendo observadas
diferenças significativas entre resultados obtidos em novilhas e vacas, tanto no início
quanto no final da lactação (MURTUZA, 1977 apud BARROS FILHO, 1995). Porém,

23
outros autores afirmam que existem notáveis variações nas concentrações séricas
de proteínas totais de acordo com a idade, tipo de alimentação, condição fisiológica,
hereditariedade, raça e estados patológicos (BARROS FILHO, 1995).
Encontram-se, na literatura, trabalhos que citam diferenças significativas
entre os teores de proteínas séricas totais em novilhas taurinas (8,10 g/dl) e
zebuínas (7,00 g/dl), e que, em animais adequadamente alimentados, os níveis de
proteína sérica total eram maiores nos machos (SOUZA, 1997).
O primeiro relato no Brasil sobre os teores de proteína sérica total em
bovinos, mais especificamente no estado do Rio de Janeiro, informa que os valores
médios para as raças Nelore e Guzerá são, respectivamente, de 7,01± 1,23 g/dl e
7,23± 1,14 g/dl (VOGUEL et al., 1957 apud SOUZA, 1997).
Na década de 60, havia dificuldades em se estabelecer os padrões de
referência para proteína total em bovinos, devido à diversidade dos resultados
citados, ocasionados pela não consideração da influência de inúmeros fatores que
causavam variabilidade como: ambiente, sistema de criação, alimentação, idade e
sexo, bem como pelo método utilizado para as determinações (SOUZA, 1997).
Em sete novilhas soropositivas para a Leucose Enzoótica Bovina, obteve-se
para proteínas séricas totais, o valor médio de 8,7± 0,9 g/dl em três coletas
sucessivas realizadas em 2001, em diferentes fases reprodutivas, o que levou a
conclusão de que os animais permaneceram hiperproteinêmicos em todas as fases
(FERREIRA et al., 2001).
Em estudos com 30 fêmeas holandesas com idade entre três e 12 anos,
sendo 10 soronegativas para a LEB, 10 soropositivas e 10 soropositivas com
linfocitose persistente, obtiveram-se, respectivamente, os seguintes valores em g/dl
para proteínas séricas totais: 8,24, 6,75 e 6,69 (OLIVEIRA et al., 2001).
Em estudos com 60 fêmeas holandesas, sendo 20 soropositivas para a LEB
e com linfocitose, 20 soropositivas para a LEB sem linfocitose e 20 soronegativas,
obtiveram-se, respectivamente, os seguintes valores em g/dl para proteínas séricas
totais: 7,85 ± 0,44, 8,02 ± 0,55 e 8,01 ± 0,55, não sendo detectadas diferenças
significativas entre os três grupos (BIRGEL JUNIOR et al., 2001).
A seguir, na tabela 2, são apresentados valores normais de proteínas
séricas totais na espécie bovina, segundo diferentes autores.

24
Tabela 2 – Valores normais de proteínas séricas totais na espécie bovina, segundo diversos autores
Autores g/ dl
BRADISH et al., 1954 6,44 a 7,50
CARROL e KANEKO, 1967 6,92 a 7,28
AKIOSHI e GERSZTEIN, 1962 5,70 a 8,40
VIANA e CAMPOS, 1973 7,99 a 9,69
ALENCAR FILHO, 1976 5,70 a 8,40
IRFAN, 1982 7,16
MATOS et al., 1983 7,17 a 8,21
BLOOD et al., 1988 6,00 a 8,50
KANEKO, 1989 6,74 a 7,46
REKWOT et al., 1989 6,81
BIRGEL et al., 1991 5,97 a 7,30
MEYER et al., 1992 6,70 a 7,50
MUTURZA, 1977 apud BARROS FILHO, 1995
6,46 a 6,74
VILELA et al.,1971 apud BARROS FILHO, 1995
7,28 a 9,78
BRAUN et al., 1983 apud SOUZA, 1997 6,80 a 7,30
OLBRICH et al., 1971 apud SOUZA, 1997 7,00 a 8,10
GARNER, 1952 apud SOUZA, 1997 8,42 a 10,38
VOGEL et al., 1957 apud SOUZA, 1997 5,78 a 8,24
LOPES et al., 1996 6,76 a 7,46
NEIRA et.al.,1982 apud FERREIRA et al., 2001
7,6
FERREIRA et al., 2001 7,8 a 9,6
BIRGEL JUNIOR et al., 2001 7,46 a 8,56
SOUZA et al., 2004 5,66 a 7,52
CAMPOS et al., 2007 6,31 a 6,91

25
2.9.2 Albumina
Evidenciou-se que a taxa de albumina é maior em vacas quando comparada
com a de bezerras, recém natos e novilhas (MYLREA ; BAYFIELD, 1968).
Os principais contribuintes da pressão osmótica do plasma sanguíneo são
os íons sódio, potássio, cloro, bicarbonato e cálcio, havendo uma pequena
contribuição das proteínas plasmáticas (albumina e globulinas). Determinando a
quantidade de cada fração das citadas proteínas, obtemos a relação
Albumina/Globulina (A/G) que, na maioria das espécies domésticas, varia de 0,5 a
1,5. A albumina é sintetizada no fígado, portanto, qualquer processo que afete a
síntese de proteínas poderá afetar a relação A/G. Assim ocorre na fibrose hepática
ou mesmo na deficiência de aminoácidos (MEDWAY et al., 1973).
A albumina é a menor das proteínas plasmáticas, tem peso molecular de
69.000 dáltons e, no plasma dos animais sadios, constitui 40 a 60% da concentração
total das proteínas séricas. Sua concentração média varia de espécie para espécie,
atuando na manutenção da pressão osmótica, fonte primária de aminoácidos e
possuindo capacidade de ligação com uma série de substâncias (COLES, 1984).
A albumina constitui cerca de 35 a 50% do total de proteínas plasmáticas, é
sintetizada no fígado e catabolizada por todo os tecidos metabolicamente ativos e
sua deficiência causa o chamado edema hipoalbominêmico (KANEKO, 1989).
A relação Albumina/Globulina deve ser evidenciada em distúrbios variados,
como, por exemplo, nas desidratações e, em bovinos, esta relação varia de 0,8 a 0,9
(CARLSON, 1990).
As proteínas totais são responsáveis pela manutenção da homeostasia
corporal, participam da hemostasia, da resistência a infecções, do equilíbrio ácido-
base e carreiam constituintes plasmáticos. A albumina, globulinas (αglobulinas,
βglobulinas e γglobulinas) e o fibrinogênio constituem as frações que compõem as
proteínas totais. O mesmo autor informa que a desidratação é responsável pela
hiperproteinemia, citando, entre outras causas, a diarréia, indigestão vagal,
parasitismo e a pneumonia crônica. A hipoalbuminemia tem como principais causas
o decréscimo na sua produção e sua perda via aparelho digestivo e urinário
(JOHNSTON, 1990)
A hipoalbuminemia está associada a distúrbios hepáticos e renais, entre
outras causas, e a meia vida da albumina sérica em bovinos é de 16,5 dias. A

26
hipoalbuminemia é observada em forma conjunta com a perda de peso em animais
de grande porte (MAAS, 1990).
A fração albumina em animais mais velhos apresentou valores
significativamente inferiores aos obtidos nos animais mais jovens (BIRGEL et al.,
1991 apud BARROS FILHO, 1995).
No estado do Rio de Janeiro, foi descrito, para a albumina sérica, os valores
de 3,33 ± 0,38 g/dl em zebuínos da raça Nelore (VOGEL et al., 1957 apud BARROS
FILHO, 1995).
A albumina é a proteína primariamente responsável pela manutenção da
pressão oncótica no plasma e a hiperproteinemia pode ser causada por um aumento
de albumina, globulina ou de ambas, porém a única causa da hiperalbuminemia é a
desidratação, sendo que a hiperproteinemia, na ausência de desidratação, é
resultado de uma hiperglobulinemia. Em bovinos com doença hepática crônica, o
decréscimo na concentração de albumina pode ser severo. Os mesmos autores
afirmaram que a perda da albumina pode estar relacionada a uma doença renal,
particularmente, à doença glomerular, desordem gastrintestinal ou hemorragia
(ROUSSEL et al., 1997)
Em sete novilhas soropositivas para a Leucose Enzoótica Bovina, obteve-se,
para albumina, o valor médio de 4,02 ± 0,43 g/dl em três coletas sucessivas,
realizadas em diferentes fases reprodutivas. E, para a razão albumina/globulina, o
valor médio de 0,90 ± 0,24 (FERREIRA et al., 2001).
Em estudos com 30 fêmeas holandesas com idade entre três e 12 anos,
sendo 10 soronegativas para a LEB, 10 soropositivas e 10 soropositivas com
linfocitose persistente, obtiveram-se, respectivamente, os seguintes valores em g/dl
para albumina: 3,14, 2,76 e 3,37, não sendo detectadas diferenças significativas
entre os três grupos. E para razão albumina/globulina, respectivamente, 0,63, 0,67,
0,88 (OLIVEIRA et al., 2001).
Em estudos com 60 fêmeas holandesas, sendo 20 soropositivas para a LEB
e com linfocitose, 20 soropositivas para a LEB sem linfocitose e 20 soronegativas,
obtiveram-se, respectivamente, os seguintes valores em g/dl para albumina: 3,41 ±
0,39, 3,50 ± 0,30 e 3,46 ± 0,43, não sendo detectadas diferenças significativas entre
os três grupos. E, para a razão albumina/ globulina, respectivamente, 0,78 ± 0,16,
0,79 ± 0,14 e 0,78 ± 0,18 (BIRGEL JUNIOR et al., 2001).

27
Na tabela 3, são apresentados os valores normais da albumina sérica na
espécie bovina, segundo diferentes autores.
Tabela 3 – Valores normais de albumina sérica na espécie bovina, segundo diversos autores
Autores g/dl
BRADISCH et al., 1954 3,20
AKIOSHI e GERSZTEIN, 1962 3,50 a 3,71
VIANA; CAMPOS, 1973 2,79 a 3,63
ALENCAR FILHO, 1976 3,50 a 3,70
DIRKSEN et al., 1983 3,00 a 4,00
KOLB, 1984 2,78 a 4,00
BLOOD et al., 1988 2,10 a 3,60
FAGLIARI et al, 1988 2,00 a 2,07
KANEKO, 1989 3,03 a 3,55
CARLSON, 1990 3,03 a 3,55
JOHNSTON, 1990 3,03 a 3,55
MEYER et al., 1992 3,00 a 3,60
LOPES et al., 1996 3,03 a 3,55
BIRGEL JUNIOR et al., 2001 3,03 a 3,89
SOUZA et al., 2004 2,90 a 3,42
CAMPOS et al., 2007 2,16 a 2,98
2.9.3 Globulinas
As globulinas são subdivididas em três frações α, β e γ. As α globulinas
constituem a fração que mais rapidamente migra das globulinas, por isso recebe o
nome “alfa”, possuindo duas frações α1 e α2, atuando no transporte de um grande
número de substâncias. As β globulinas atuam no transporte de ferro e lipídios, as
frações C3 e C4 constituem fatores do complemento; e o fibrinogênio, cujo aumento

28
indica processo inflamatório agudo, é precursor da fibrina. As γ globulinas têm
função na imunidade humoral, compõem-se das imunoglobulinas e suas frações IgA,
IgG, IgM, IgE, (LOPES et al., 1996)
Raramente, há uma diminuição da concentração globulínica total, embora
possa ocorrer queda na concentração de frações individuais, que se deve ao fato de
que outras globulinas podem estar simultaneamente aumentadas para compensar a
queda de um componente atingido, ainda que existam casos de hipoglobulinemia
em animais privados de colostro. Raramente, são observadas reduções das α-
globulinas nos estados patológicos dos animais domésticos, entretanto, as
elevações nas concentrações plasmáticas são frequentes nas reações inflamatórias
(COLES, 1984). O componente α-2 das α-globulinas está significativamente
aumentado nas infecções bacterianas e virais (MEBUS; COLES 1977 apud COLES
1984).
Um aumento nas concentrações de β-globulinas pode estar associado a um
aumento das β-lipoproteínas, como ocorre em casos de hiperlipemia.
(DIMOPOULOS et al., 1968 apud COLES, 1984; MEBUS; COLES, 1977 apud
COLES 1984).
Não há variações significativas de algumas frações proteicas do soro de
bovinos que possam ser atribuídas à influência de fatores etários, pois a diferença
dos resultados obtidos em novilhas e vacas prenhes não diferem significativamente
dos valores das frações albumina, alfa e betaglobulinas. Comparando, entretanto, o
teor sérico de gamaglobulinas, os resultados são significativamente maiores nas
novilhas (MURTUZA, 1977 apud SOUZA, 1997).
Em sete novilhas soropositivas para a Leucose Enzoótica Bovina, obteve-se,
para globulina, o valor médio de 4,62 ± 0,96 g/dl em três coletas sucessivas,
realizadas em diferentes fases reprodutivas (FERREIRA et al., 2001).
Em estudos com 30 fêmeas holandesas com idade entre três e 12 anos,
sendo 10 soronegativas para a LEB, 10 soropositivas e 10 soropositivas com
linfocitose persistente, obtiveram-se, respectivamente, os seguintes valores em g/dl
para globulina: 4,97, 2,76 e 3,37 (OLIVEIRA et al., 2001).
Em estudos com 60 fêmeas holandesas, sendo 20 soropositivas para a LEB
e com linfocitose, 20 soropositivas para a LEB sem linfocitose e 20 soronegativas,
obtiveram-se, respectivamente, os seguintes valores em g/dl para globulina: 4,44 ±

29
0,29, 4,52 ± 0,30 e 4,55 ± 0,37, não sendo detectadas diferenças significativas entre
os três grupos. (BIRGEL JUNIOR et al., 2001).
Na tabela 4, são apresentados os valores normais de globulinas séricas
totais na espécie bovina, segundo diferentes autores.
Tabela 4. Valores normais de globulina sérica, segundo diversos autores
Autores g/dl
BRADISCH et al., 1954 3,77
VIANA E CAMPOS, 1973 5,63
ALENCAR FILHO, 1976 3,54
IRFAN, 1982 4,08
MATOS et al., 1983 4,40
FAGLIARI et al., 1988 3,22
KANEKO, 1989 3,00 a 3,48
MEYER et al., 1992 3,00 a 3,50
LOPES et al., 1996 3,00 a 3,48
FERREIRA et al., 2001 3,66 a 5,58
BIRGEL JUNIOR et al., 2001 4,18 a 4,92
SOUZA et al., 2004 2,53 a 4,33
CAMPOS et al., 2007 3,56 a 4,51
2.9.4 Aspartato aminotransferase – AST
A atividade da AST é mais intensa nos equinos, porém encontrou-se
atividade considerável em vários órgãos e nas mais variadas espécies; o fígado, o
coração, rins e músculo esquelético são os locais onde a enzima apresenta maior
atividade (MEDWAY et al., 1973).

30
O aumento da AST sérica em bovinos ocorre na gestação adiantada, no
dano degenerativo e necrótico agudo do parênquima hepático e nas lesões agudas
do miocárdio e músculo esquelético (DIRKSEN et al., 1983).
Os valores da AST em fêmeas bovinas não sofrem diferenças estatísticas a
partir dos 24 meses de idade (GREGORY et al., 1999).
Os níveis séricos da AST podem estar aumentados nas hepatopatias,
praticamente, em todas as espécies, entretanto, não é um teste específico para
lesões hepáticas. A AST ocorre em concentrações elevadíssimas no músculo
esquelético e cardíaco e sua mensuração sérica é valiosa na confirmação do
diagnóstico de degeneração muscular, o que propicia o extravasamento de
quantidades significativas dessa enzima para a circulação. Em casos de necrose
hepatocelular, seus níveis podem estar extremamente aumentados (COLES, 1984).
A presença da AST em poucos tecidos faz com que ela seja um bom
indicador para danos teciduais específicos, observados na mensuração de seus
níveis séricos (KANEKO, 1989).
Aproximadamente 60 a 80% da AST estão no interior do hepatócito
associada à mitocôndria, enquanto o restante fica na forma solúvel no citoplasma;
em doenças hepáticas severas em estágio avançado, por exemplo, cirrose, os níveis
séricos da AST podem estar normais ou apenas pouco aumentados, provavelmente,
devido a uma diminuição da massa hepática viável e pela alteração da arquitetura
hepática (MEYER et al., 1992).
Em 47 bovinos da raça Jersey, soropositivos para a LEB, criados no estado
de São Paulo, obtiveram-se os seguintes valores para AST: 36,95 ± 10,18 U/l, que
não diferiram do grupo dos animais soronegativos, no qual se obteve o valor, para
AST, de 33,89 ± 5,51 U/l (GREGORY et al., 1999).
Em estudos com 118 fêmeas bovinas soropositivas para LEB, obtiveram-se
os valores para AST que variaram entre 26 a 40U/L (HAGIWARA et al., 1986 apud
GREGORY et al., 1999).
Nas condições brasileiras, o valor da atividade sérica da AST, em bovinos
sadios, não deveriam exceder 50U/l (GREGORY et al., 1999).
Na tabela 5, os valores normais de AST sérica na espécie bovina, segundo
diferentes autores, são apresentados.

31
Tabela 5 – Valores normais de AST sérica na espécie bovina, segundo diversos autores
Autores U/L
MEDWAY, et al., 1973 42 a 70
DIRKSEN et al., 1983 10 a 50
BLOOD et al., 1988 50 a 150
KANEKO, 1989 78 a 132
CARLSON, 1990 78 a 132
MEYER et al., 1992 78 a 132
LOPES et al., 1996 20 a 34
GREGORY et al., 1999 23 a 45
CAMPOS et al., 2007 39 a 73
2.9.5 Gamaglutamiltransferase - γγγγGT
É uma carboxipeptidase que cliva os grupos glutamil terminal C e os
transfere para peptídeos (COLES, 1984).
As maiores quantidades de γGT celular estão nos rins e no epitélio do ducto
biliar. A γGT renal é a fonte da γGT urinária e é um indicador de nefrotoxicidade,
juntamente com a fosfatase alcalina urinária. Desordens colestáticas, em todas as
espécies examinadas, resultaram em um aumento sérico da γGT (BRAUN et al.,
1987).
Com exceção do músculo, todas as células têm alguma participação da γGT
em suas atividades de membrana ou no citosol. Estima-se que seu peso molecular
varie de 90.000 a 350. 000 dáltons (KANEKO, 1989).
Não houve diferença significativa entre os níveis séricos de γGT obtidos em
machos e fêmeas, em estudo com bezerros das raças Frísia Piednoir, Garonnaise e
Pardo Suíço, criados na França, com idade variando entre quatro e seis meses de
idade (BRAUN et al., 1977 apud BARROS FILHO, 1995).
A mensuração da γGT representa um dos testes mais específicos para a
avaliação da função hepática na veterinária (ROUSSEL et al., 1997). Afirmaram
ainda que o nível de γGT aumenta exageradamente em doenças tubulares renais.
Os mesmos autores informaram que, após o consumo do colostro, que é rico em
γGT, os bezerros apresentaram um rápido aumento dos níveis séricos dessa

32
enzima, chegando a ser 50 a 100 vezes maior quando comparados aos níveis de
bezerros que não tiveram contato com colostro.
Avaliando bovinos recém natos das raças Nelore (Bos taurus indicus) e
Holandesa (Bos taurus taurus), e de bubalinos (Bubalus bubalis) da raça Murrah, foi
verificado que a atividade de γGT foi maior no dia do nascimento, com acentuado
decréscimo com o avanço da idade. Afirmaram, ainda, que há uma queda da
atividade enzimática ao longo do tempo (FAGLIARI et al., 1998).
Os valores da γGT em fêmeas bovinas não sofrem diferenças estatísticas a
partir dos 24 meses de idade (GREGORY et al., 1999).
Em 47 bovinos da raça Jersey, soropositivos para a LEB, criados no estado
de São Paulo, obtiveram-se os seguintes valores para γGT: 12,56 ± 7,78 U/l, que
não diferiram grupo dos animais soronegativos, no qual se obteve o valor para γGT
de 11,32 ± 4,20 U/l (GREGORY et al., 1999).
Em estudos com 118 fêmeas bovinas soropositivas para LEB, obtiveram-se
os valores para γGT que variaram entre 6 a 17U/L (HAGIWARA et al., 1986 apud
GREGORY et al., 1999).
Nas condições brasileiras, o valor da atividade sérica da γGT em bovinos
sadios não deveriam exceder 25 U/l (GREGORY et al., 1999).
Na tabela 6, são apresentados os valores séricos de γGT, considerados
normais em bovinos, segundo diversos autores.
Tabela 6 – Valores normais de γGT sérica na espécie bovina, segundo diversos autores
Autores U/L
SIMENSEN; NANSEN, 1974 17,47
BRAUN et al., 1977 11,6 a 19,0
MÁRQUEZ et al., 1977 12,0 a 22,7
BOTELHO et al., 1980 12,025
DIRKSEN et al., 1983 10,0 a 20,0
KANEKO, 1989 6,1 a 17,4
MEYER et al., 1992 11,0 a 24,0
BARROS FILHO, 1995 7,83 a 15,23
LOPES et al., 1996 6,1 a 17,4
GREGORY et al., 1999 13,21 ± 12,72
CAMPOS et al., 2007 7,43 a 17,12

33
2.9.6 Alanina Aminotransferase - ALT
O aumento da ALT ocorre em lesões do músculo cardíaco e esquelético e
necrose grave de hepatócitos (DIRKSEN et al., 1983).
O citoplasma do hepatócito é rico em ALT no cão, gato e primatas; os
equinos e ruminantes são notáveis exceções. Uma agressão (toxina, hipoxia) à
membrana hepatocelular resulta em um aumento da ALT sérica. De forma aguda
(horas ou dias), a magnitude do aumento torna-se mais ou menos paralela ao
número de hepatócitos com a permeabilidade da membrana alterada. Segundo os
mesmos autores, a atividade marcadamente aumentada das enzimas ALT e AST
circulante sugere um insulto hepático severo e difuso, especialmente se a icterícia
estiver presente. A meia vida plasmática da ALT é de aproximadamente 60 horas;
caso a injúria permaneça por alguns dias, os níveis séricos da ALT decrescerão
(MEYER et al.,1992).
A atividade da ALT requer o cofator piridoxal 5’-fosfato, o metabólito ativo da
vitamina B6, uma deficiência desta vitamina resulta na diminuição da atividade
enzimática (MEYER et al.,1992).
O valor médio para bovinos adultos saudáveis é de 10 U/l, podendo variar de
7 a 14 U/l (DIRKSEN et al., 1983). Segundo Kaneko (1989), o valor da ALT em
bovinos pode variar de 11 a 40 U/l (27±14); de 14 a 38 U/l (LOPES et al, 1996); de
18 a 47 UI/l (CAMPOS et al., 2007).
2.9.7 Ureia
A ureia é um composto orgânico relativamente simples produzido pelos
mamíferos. No fígado, como produto final do catabolismo de proteínas, é uma das
substâncias mais difusíveis pelo corpo e se encontra em todos os líquidos orgânicos
e é eliminada principalmente pelos rins. Ao se avaliarem os níveis séricos de ureia,
observam-se possíveis transtornos renais. Por exemplo, as tetraciclinas produzem
aumento nos níveis de ureia. Em ruminantes, a determinação de ureia sérica não
tem tanta importância na avaliação do funcionamento renal, pois há reciclagem da
ureia sanguínea no rúmen. Há variação que ocorre praticamente em todos os

34
herbívoros que demonstra que os níveis de ureia tornam-se mais elevados na
primavera (MEDWAY et al, 1973).
Existem variações dos teores de ureia na dependência do tipo de
alimentação, bem como diferenças significativas, conforme categorização etária dos
grupos experimentais ou de acordo com o estado fisiológico dos bovinos (BOSE,
1983; BOTELHO et al., 1984 ).
A ureia é proveniente das proteínas, hidrolisadas no intestino, em
aminoácidos e estes absorvidos pelas células da mucosa intestinal. Bactérias
intestinais podem degradar os aminoácidos em amônia. Os aminoácidos e a amônia
são transportados para o fígado via circulação portal. Aproximadamente 75% dos
aminoácidos (ou amônia) são transportados através da membrana celular hepática
até as mitocôndrias. Nestas, existem mecanismos de desaminação, um processo
cíclico chamado de ciclo da ureia. A excreção da ureia pelos rins representa a maior
rota de excreção de nitrogênio. A ureia foi introduzida como aditivo alimentar para
ruminantes na forma de um substituto barato de proteína (BLOOD et al., 1988).
Ruminantes intoxicados por ureia morrem rapidamente. Animais com doença
hepática difusa têm reduzida capacidade de sintetizar a ureia. Nesses casos, os
níveis séricos dos precursores da ureia, como aminoácidos e amônia, são
geralmente elevados (KANEKO, 1989).
O ciclo da ureia fica completamente alterado diante da cirrose hepática.
Nesses casos, a ureia sérica pode apresentar-se em baixas concentrações e passa
a ser excretada pela urina também em concentrações mais baixas. (KANEKO,
1989).
Com o avançar da idade, há tendência de aumento da ureia sanguínea,
além de outros fatores que poderiam influenciar as variações da taxa de ureia no
organismo animal: o manejo, a dieta, a raça e o sexo (GREATOREX, 1955 apud
BARROS FILHO, 1995).
Com o avançar da idade, há pequena diminuição da ureia sanguínea e, em
estudos realizados no estado do Rio Grande do Sul, em 1976, foram observadas
diferenças significativas nos níveis obtidos nos machos e fêmeas bovinas (LANE et
al., 1977 apud BARROS FILHO, 1995).
Os valores de ureia em fêmeas bovinas não sofrem diferenças estatísticas a
partir dos 24 meses de idade (GREGORY et al., 2004).

35
Em estudos com fêmeas bovinas da raça Jersey soropositivas para a LEB,
criadas no estado de São Paulo obtiveram-se os seguintes valores para a ureia de
29,39 ± 11,76 mg/dl, que não diferiram do grupo dos animais soronegativos, no qual
se obteve o valor para uréia de 28,34 ± 11,16 mg/dl (GREGORY et al., 2004).
Na Tabela 7, os valores normais de ureia sérica na espécie bovina são
apresentados, segundo diversos autores.
Tabela 7 - Valores normais de ureia sérica na espécie bovina, segundo diversos autores
Autores mg/dl
DIRKSEN et al., 1983 10 a 40
COLES, 1984 6 a 27
KOLB, 1984 20 a 40
BLOOD et al., 1988 6 a 27
KANEKO, 1989 20 a 30
MEYER et al., 1992 19,89 a 29,98
LOPES et al., 1996 42,8 a 64,2
GREGORY et al., 2004 17,41 a 39,29
CAMPOS et al., 2007 16,05 a 29,21
2.9.8 Creatinina
A creatinina é formada durante o metabolismo da creatina e fosfocreatina
muscular. Como ocorre com a ureia, a creatinina é um índice grosseiro da filtração
glomerular. A creatinina sérica não é marcadamente influenciada pela dieta ou por
hemorragias intestinais. Uma severa perda muscular poderá reduzir a quantidade de
creatinina formada. O seu aumento na circulação sanguínea ocorre mais
tardiamente nos estados de insuficiência renal, quando comparado com a uréia
sanguínea, por ser facilmente eliminada (MEYER et al., 1992; LOPES et al., 1996).
De forma semelhante à ureia, uma redução na taxa de filtração glomerular
aumenta a concentração sérica de creatinina. Os mesmos fatores pré-renal, renal e
pós-renal, que influenciam a uréia, influenciam também a creatinina sérica
(desidratação, doença cardiovascular, choque (séptico ou traumático), doença renal,

36
causando destruição dos néfrons, obstrução ou ruptura do trato urinário) (MEYER et
al., 1992).
O valor médio normal da creatinina no soro de bovinos adultos é de 1,19 (1-
1,5) mg/dl, (touros: maior ou igual a 2,82 mg/dl), e em bezerros é de 1,47 (1,24 a
2,03) mg/dl (DIRKSEN et al., 1983). O valor médio da creatinina em bovinos é de 1,0
a 2,0 mg/dl (KANEKO, 1989), 1,0 a 2,0 mg/dl (LOPES et al., 1996) e 1,14 a 1,56
mg/dl (GREGORY et al., 2004), 0,79 a 1,67 mg/dl (CAMPOS et al., 2007).
Os valores de creatinina, em fêmeas bovinas, não sofrem diferenças
estatísticas a partir dos 24 meses de idade (GREGORY et al., 2004).
Em estudos com fêmeas bovinas da raça Jersey soropositivas para a LEB,
criadas no estado de São Paulo, obtiveram-se os valores para creatinina de 1,42 ±
0,18 mg/dl, que não diferiram dos valores do grupo dos animais soronegativos, no
qual se obteve o valor para creatinina de 1,36 ± 0,15 mg/dl (GREGORY et al.,
2004).

37
3 MATERIAL E MÉTODOS
3.1 Animais
Foram utilizados soros de 120 fêmeas bovinas soropositivas para a Leucose
Enzoótica Bovina e soros de 120 fêmeas soronegativas (grupo controle). Em ambos
os grupos, as fêmeas eram mestiças com idade superior a quatro anos, provenientes
de 18 propriedades localizadas na região Norte Fluminense, criadas a campo, cujas
amostras foram coletadas por punção da veia jugular em tubos “vacutainer” de 10
ml. No Laboratório de Sanidade Animal (LSA-CCTA/UENF), as amostras de sangue
foram mantidas à temperatura ambiente até a dessoragem completa; em seguida,
centrifugadas a 600 g por 5 min. Os soros obtidos foram fracionados em duas
alíquotas e armazenados em microtubos tipo “Eppendorf” e congelados a -20 ºC, até
a realização do teste sorológico de IDGA. Para a verificação de anticorpos no soro,
foi utilizado um kit comercial proveniente da Sanofi/França, que consta de soro-
controle positivo e antígeno gp51. Todos os animais foram protocolados em fichas
individuais, com informações sobre: idade, sexo, raça e manejo realizado na
propriedade.
3.2 Análise laboratorial
Foi realizada análise espectrofotométrica com o Labmax Plenno®, um
analisador bioquímico automático, multiparamétrico de acesso aleatório da marca
Labtest Diagnóstica S/A em todas as 240 amostras; resultando em um total de 1680
análises. Nas referidas análises, foram utilizados kits comerciais específicos para
cada analito, e os controles de qualidade Qualitrol 1 e Qualitrol 2 foram empregados
antes do início de cada bateria de testes, todos da marca Labtest Diagnóstica S/A,
seguindo os protocolos de automação do equipamento.
Os seguintes analitos foram mensurados:
-PROTEÍNAS SÉRICAS TOTAIS (PT);
-ALBUMINA;
-ASPARTATO AMINOTRANSFERASE (AST);
-GAMAGLUTAMILTRANSFERASE (γGT);

38
-ALANINA AMINOTRANSFERASE (ALT);
-UREIA;
-CREATININA.
As técnicas seguidas foram disponibilizadas nos próprios kits reagentes,
seguindo os protocolos de automação para o Labmax Plenno®.
O valor da Globulina foi obtido pela diferença entre a proteína sérica total e a
albumina de cada amostra.
O valor da razão albumina/globulina (A/G) foi obtido dividindo-se o valor
obtido da albumina pelo da globulina de cada amostra.
3.3 Anatomopatologia
Realizou-se exame clínico de rotina para grandes animais preconizado por
Dirksen et al. (1983).
Paralelamente ao exame clínico, foram coletados 5 ml de sangue total da
veia jugular, que foram acondicionados em tubos plásticos contendo EDTA 10%.
Tais tubos foram transportados em caixa isotérmica, contendo gelo reciclável, para
posterior realização do hemograma.
As amostras de sangue total para a realização do hemograma foram
processadas utilizando-se o contador hematológico de células MS4® (Melet
Schloesing Laboratories, France), sendo avaliados: hematócrito, hematimetria,
hemoglobinometria, volume corpuscular médio (VCM), concentração de
hemoglobina corpuscular média (CHCM), hemoglobina corpuscular média (HCM),
leucometria global e plaquetometria. A determinação da leucometria específica foi
realizada no esfregaço sanguíneo confeccionado com a amostra de sangue total
colhido.
Foram eutanasiadas duas fêmeas da raça Jersey soropositivas ao VLB, com
18 meses de idade, pertencentes à EMATER RIO, CENTERJ do município de
Cordeiro, utilizando-se sedação com xilazina, seguida de superdosagem de
barbitúricos (tiopental sódico) e, após obtenção da anestesia, foi aplicada a solução
saturada de cloreto de potássio, atendendo ao recomendado pelo Conselho Federal
de Medicina Veterinária.

39
3.3.1 Macroscopia
Procedeu-se à necropsia de rotina, quando se observaram os órgãos do
sistema linfático (baço, linfonodos, timo), do sistema digestório (esôfago, pré-
estômagos, estômago e fígado), além dos rins, coração, pulmões, traqueia e sistema
nervoso central (encéfalo e medula).
3.3.2 Histopatologia
Foram coletadas amostras com cerca de 5 cm de comprimento e 0,5 cm de
diâmetro dos órgãos citados no item 3.3.1., subsequentemente fixados em formol
neutro tamponado a 10% e submetidos a processamento tecidual por inclusão em
parafina e coloração pela hematoxilina e eosina (com exceção do decalque de
superfície de corte do linfonodo pré-escapular que foi fixado em álcool a 70% e,
posteriormente, corado pelo HE), no setor de morfologia e anatomia patológica da
UENF.
As amostras foram microtomizadas com 5 µm de espessura e coletadas em
banho-Maria, a 38ºC, em lâminas histológicas para microscopia óptica de luz branca.
3.3.3 Imunoistoquímica
Seções em parafina de quatro µm foram obtidas das amostras selecionadas
e colhidas em lâminas histológicas silanizadas (Silano, Sigma-Aldrich, MO, USA),
submetidas à desparafinização em cinco banhos em xilol (10 min) e reidratação em
cinco banhos de alcoóis graduados (5 min) e três banhos em água deionizada (5
min). A seguir, os espécimes foram tratados com solução aquosa de peróxido de
hidrogênio 30V (Vetec, RJ, Brasil) a 30% por 30 min para bloqueio da peroxidase
endógena. Posteriormente, eram colocadas em banho-maria a 98oC em tampão
citrato (Vetec, RJ, Brasil), por 30 min para recuperação antigênica, e mantidas em
temperatura ambiente por 20 min. Subsequentemente, as lâminas foram
cuidadosamente enxutas com papel toalha, os cortes envolvidos circularmente com

40
Dako pen® (Dako, CA, USA), e os espécimes incubados por 1 hora com solução
para bloqueio de ligações inespecíficas Trisma NaCl (Vetec, RJ, Brasil) com 1% de
soroalbumina sérica bovina livre de ácidos graxos (BSA – Sigma-Aldrich, MO, USA)
e 1% de leite em pó desnatado (MolicoTM – Nestlé, SP, Brasil). Depois de descartada
a solução de bloqueio, as seções foram incubadas com anticorpo monoclonal
anticitoqueratinas (Dako, CA, USA, clone AE1/AE3) (fígado) e anti-p53 (Calbiochem,
USA, clone Ab-3, mutant) para todas as amostras citadas no item 3.3.1. Os
anticorpos foram diluídos 1:100 e incubadas “overnight” a 4oC em câmara úmida. No
dia seguinte, as lâminas foram lavadas com solução salina de Trisma com tween 20-
TBS (Vetec, RJ, Brasil) por 5 min e tratadas com kit LSAB-HRP+ (Dako, CA, USA),
lavadas por 5 min com TBS e reveladas com Kit cromógeno DAB (Dako, CA, USA);
contra-coradas com hematoxilina de Harris, desidratadas em banhos de alcoóis (1
min) e montadas com Permount® (Sigma-Aldrich, MO, USA) e, posteriormente,
analisadas em microscópio óptico visando observar as imunomarcações.
3.4 Análise estatística
As análises estatísticas foram realizadas por meio de intervalo de confiança
para os valores das variáveis (COCHRAN, 1955), com auxílio do programa
computacional GENES, versão 2006.4.1 (CRUZ, 2006), considerando o nível de
significância α igual a 5%, admitindo-se uma amostra representativa de uma
população infinita de bovinos da região Norte Fluminense, RJ.

41
4 RESULTADOS E DISCUSSÃO
4.1 Análises estatísticas
Realizou-se a análise descritiva contendo média e desvio-padrão para os
grupos de bovinos soropositivos e soronegativos (tabela 8). Subsequentemente as
comparações entre as médias foram feitas pela sobreposição ou não dos intervalos
de confiança (probabilidade de conter a média da população igual a 95%). Caso
houvesse diferença significativa, os intervalos de confiança não se sobreporiam
(figuras 1 a 6).
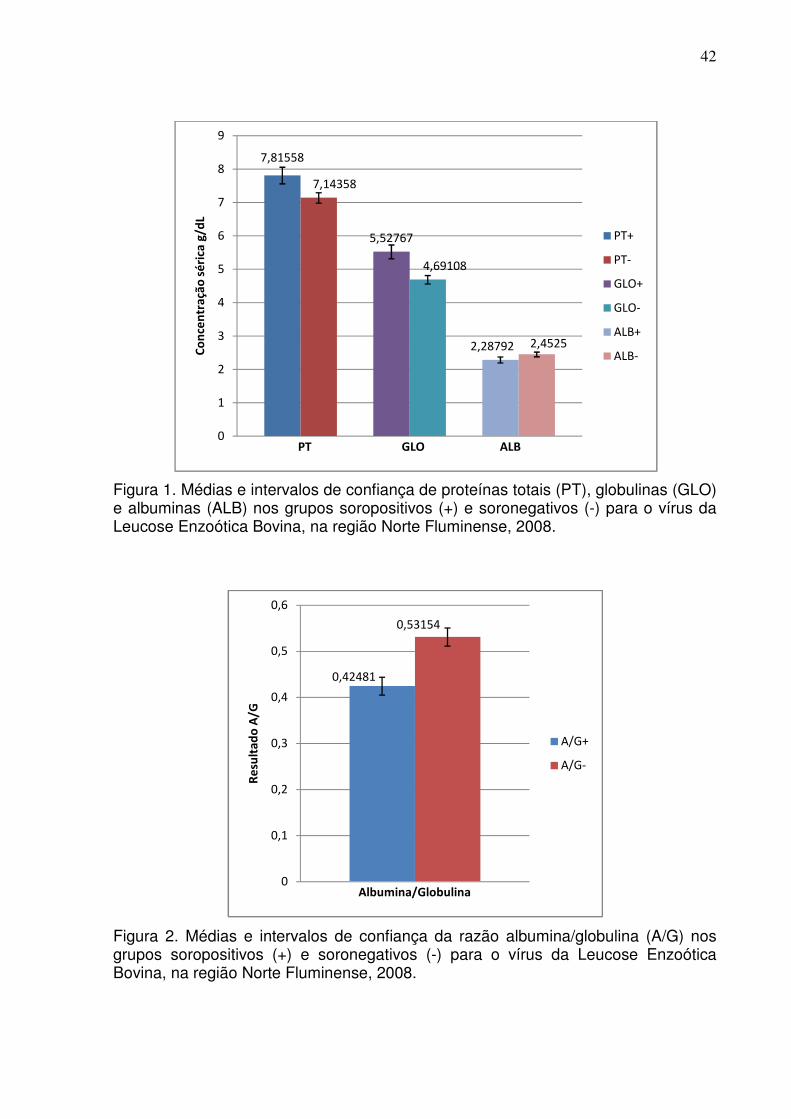
Tabela 8 – Bioquímica sérica de bovinos da região Norte Fluminense soropositivos e soronegativos para o vírus da Leucose Enzoótica Bovina, 2008. Número de amostras Média Desvio-Padrão
+ - + - + -
Proteínas Totais (g/dl)
120
120
7,81
7,14
1,32
0,82
Albumina (g/dl) 120 120 2,29 2.45 0,48 0,38
Globulinas (g/dl) 120 120 5,52 4,69 1,1 0,68
AST (U/L) 1 120 120 46,37 33,38 17,82 13,96
ALT(U/L) 2 120 120 11,57 9,87 4,93 3,44
γγγγGT (U/L) 3 120 120 25,72 20,48 9,68 6,35
Creatinina (mg/dl) 120 120 1,11 1,13 0,29 0,39
Ureia (mg/dl) 120 120 30,12 34,81 12,58 11,47
Razão A/G 120 120 0,42 0,53 0,10 0,10
1. Aspartato aminotransferase
2. Alanina aminotransferase
3.Gamaglutamiltranferase
+. Soropositivos
-. Soronegativos
Se utilizarmos, para comparação, a amplitude de variação de um desvio-
padrão para cada parâmetro mensurado no grupo soropositivo e soronegativo,
verificaremos que, em nenhum dos parâmetros mensurados, será possível observar
diferenças significativas, pois sempre observaremos sobreposição dos valores
quando consideramos média ± desvio-padrão como amplitude de variação.
A seguir, as figuras 1, 2, 3, 4, 5 e 6 mostram a média e os intervalos de
confiança obtidos para os parâmetros mensurados.
42
Figura 1. Médias e intervalos de confiança de proteínas totais (PT), globulinas (GLO) e albuminas (ALB) nos grupos soropositivos (+) e soronegativos (-) para o vírus da Leucose Enzoótica Bovina, na região Norte Fluminense, 2008.
Figura 2. Médias e intervalos de confiança da razão albumina/globulina (A/G) nos grupos soropositivos (+) e soronegativos (-) para o vírus da Leucose Enzoótica Bovina, na região Norte Fluminense, 2008.
7,81558
7,14358
5,52767
4,69108
2,28792 2,4525
0
1
2
3
4
5
6
7
8
9
Co
nce
ntr
açã
o s
éri
ca g
/dL
PT GLO ALB
PT+
PT-
GLO+
GLO-
ALB+
ALB-
0,42481
0,53154
0
0,1
0,2
0,3
0,4
0,5
0,6
Re
sult
ad
o A
/G
Albumina/Globulina
A/G+
A/G-
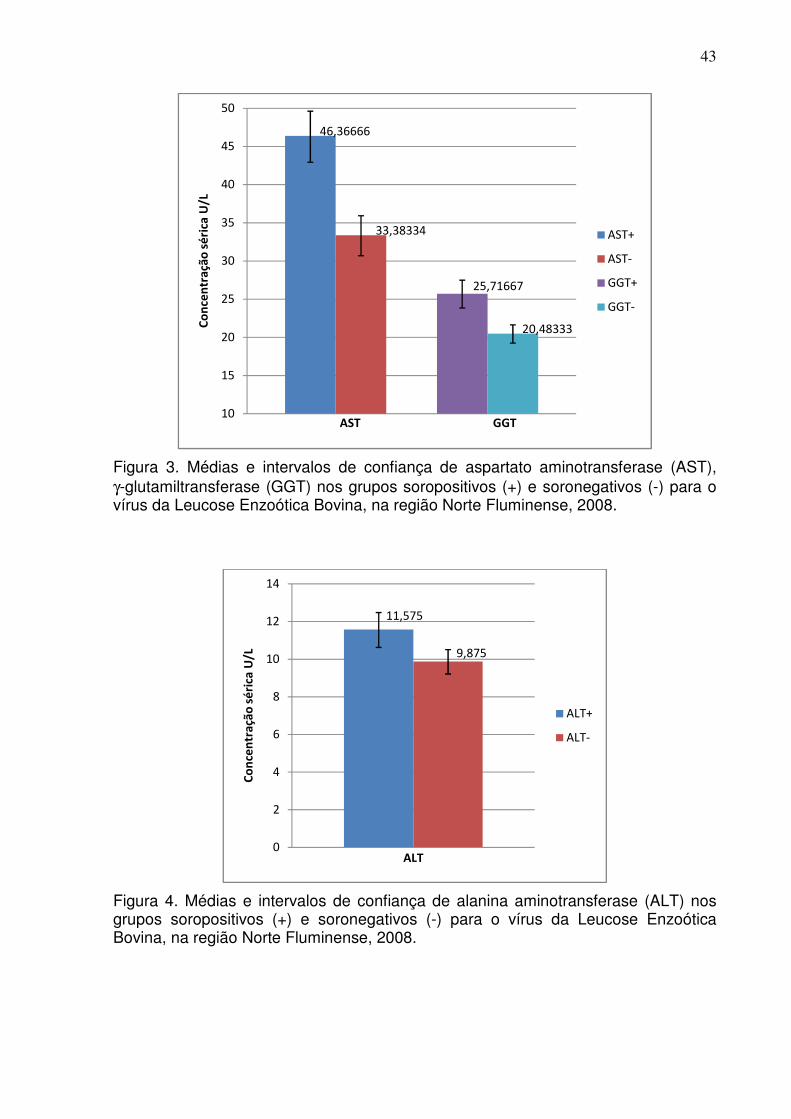
43
Figura 3. Médias e intervalos de confiança de aspartato aminotransferase (AST), γ-glutamiltransferase (GGT) nos grupos soropositivos (+) e soronegativos (-) para o vírus da Leucose Enzoótica Bovina, na região Norte Fluminense, 2008.
Figura 4. Médias e intervalos de confiança de alanina aminotransferase (ALT) nos grupos soropositivos (+) e soronegativos (-) para o vírus da Leucose Enzoótica Bovina, na região Norte Fluminense, 2008.
46,36666
33,38334
25,71667
20,48333
10
15
20
25
30
35
40
45
50
Co
nce
ntr
açã
o s
éri
ca U
/L
AST GGT
AST+
AST-
GGT+
GGT-
11,575
9,875
0
2
4
6
8
10
12
14
Co
nce
ntr
açã
o s
éri
ca U
/L
ALT
ALT+
ALT-

44
Figura 5. Médias e intervalos de confiança de ureia (URE) nos grupos soropositivos (+) e soronegativos (-) para o vírus da Leucose Enzoótica Bovina, na região Norte Fluminense, 2008.
Figura 6. Médias e intervalos de confiança de creatinina (CRE) nos grupos soropositivos (+) e soronegativos (-) (há sobreposição dos intervalos de confiança), para o vírus da Leucose Enzoótica Bovina, na região Norte Fluminense, 2008.
30,11667
34,80833
0
5
10
15
20
25
30
35
40
Co
nce
ntr
açã
o s
éri
ca m
g/d
L
Ureia
URE+
URE-
1,1111,13317
0,6
0,7
0,8
0,9
1
1,1
1,2
1,3
Co
nce
ntr
açã
o m
g/d
L
Creatinina
CRE+
CRE-

45
4.2 Aspectos bioquímicos séricos
4.2.1 Proteínas séricas totais (PT)
Os valores médios e desvios-padrão obtidos na mensuração das proteínas
séricas totais foram de 7,81 ± 1,32 g/dl para o grupo de soropositivos e de 7,14 ±
0,82 g/dl para o grupo de soronegativos. Resultados semelhantes foram obtidos por
Ferreira et al. (2001) e Neira et al. (1982) apud Ferreira et al. (2001), que também
encontraram valores de proteínas séricas totais maiores em animais soropositivos
quando comparados aos dos soronegativos, atribuindo a esse resultado o fato de
ocorrer produção de paraproteínas pelos animais leucóticos.
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio padrão (entre 6,49 e 9,13 g/dl), é considerado
normal, segundo Bradisch et al. (1954); Carroll, Kaneko (1967); Akioshi, Gersztein
(1962); Viana, Campos (1973); Alencar Filho (1976); Irfan (1982); Matos et al.
(1983); Blood et al. (1988); Kaneko (1989); Rekwot et al. (1989); Birgel et al. (1991);
Meyer et al. (1992); Muturza (1977) apud Barros Filho (1995); Vilela et al. (1971)
apud Barros Filho (1995); Braun et al. (1983) apud Souza (1997); Olbrich et al.
(1971) apud Souza (1997); Garner (1952) apud Souza (1997); Vogel et al (1957)
apud Souza (1997); Lopes et al. (1996); Neira et al. (1982) apud Ferreira et al.
(2001); Ferreira et al. (2001); Birgel Junior et al. (2001) e Campos et al. (2007).
Considerando os estudos realizados por outros autores que também
trabalharam com animais soropositivos para LEB, os valores obtidos, no presente
estudo para o grupo soropositivo, foram considerados normais, segundo os dados
de Oliveira et al. (2001), Ferreira et al. (2001), Birgel Junior et al. (2001).
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 6,32 e 7,96 g/dl), está dentro da
normalidade, segundo Bradisch et al. (1954); Carroll, Kaneko (1967); Akioshi,
Gersztein (1962); Alencar Filho (1976); Irfan (1982); Matos et al. (1983); Blood et al.
(1988); Kaneko (1989); Rekwot et al. (1989); Birgel et al. (1991); Meyer et al. (1992);
Muturza (1977) apud Barros Filho (1995); Vilela et al. (1971) apud Barros Filho
(1995); Braun et al. (1983) apud Souza (1997); Olbrich et al.(1971) apud Souza
(1997); Vogel et al. (1957) apud Souza (1997); Lopes et al. (1996); Neira (1982)

46
apud Ferreira et al. (2001); Ferreira et al. (2001); Birgel Junior et al. (2001) e
Campos et al. (2007); e abaixo do normal segundo Viana, Campos (1973) e Garner
(1952) apud Souza (1997).
4.2.2 Albumina e razão albumina/globulina (A/G)
Os valores médios e desvios-padrão obtidos na mensuração da albumina
sérica foram de 2,29 ± 0,48 g/dl para o grupo de soropositivos, e de 2,45 ± 0,38 g/dl,
para o grupo de soronegativos.
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 1,81 e 2,77 g/dl), é considerado
normal, segundo Blood et al. (1988); Fagliari et al. (1988); Campos et al. (2007); e
abaixo do normal segundo Bradisch et al. (1954); Akioshi, Gersztein (1962); Viana,
Campos (1973); Alencar Filho (1976); Dirksen et al. (1983); Kolb (1984); Kaneko
(1989); Carlson (1990); Johnston (1990); Meyer et al. (1992); Lopes et al. (1996);
Birgel Junior et al. (2001) e Souza et al. (2004).
Considerando os estudos realizados por outros autores que também
trabalharam com animais soropositivos para LEB, os valores obtidos, no presente
estudo, para o grupo soropositivo foram considerados normais, segundo os dados
de Oliveira et al. (2001); e abaixo do normal segundo, Ferreira et al. (2001) e Birgel
Junior et al. (2001).
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 2,07 e 2,83 g/dl), é considerado
normal segundo Viana, Campos (1973); Kolb (1984); Blood et al.(1988); Fagliari et
al. (1988); Campos et al. (2007); e abaixo do normal, segundo Bradisch et al. (1954);
Akioshi, Gersztein (1962); Alencar Filho (1976); Dirksen et al. (1983); Kolb (1984);
Kaneko (1989); Carlson (1990); Johnston (1990); Meyer et al. (1992); Lopes et al.
(1996); Birgel Junior et al. (2001) e Souza et al. (2004).
Os valores médios e desvios-padrão obtidos na mensuração da razão
albumina/globulina foram de 0,42 ± 0,10 para o grupo de soropositivos, e de 0,53 ±
0,10, para o grupo de soronegativos.
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 0,32 e 0,52), é considerado

47
normal, segundo Medway et al. (1973); e abaixo do normal, segundo Carlson (1990);
Birgel Junior et al. (2001); Oliveira et al. (2001) e Ferreira et al. (2001).
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 0,43 e 0,63), é considerado
normal, segundo Medway et al. (1973); Oliveira et al. (2001); Birgel Junior et al.
(2001); e abaixo do normal, segundo Carlson (1990) e Ferreira et al. (2001).
4.2.3 Globulina
Os valores médios e desvios-padrão obtidos na mensuração da globulina
sérica foram de 5,52 ± 1,1 g/dl para o grupo de soropositivos e, de 4,69 ± 0,68 g/dl,
para o grupo de soronegativos.
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 4,42 e 6,62 g/dl), é considerado
normal, segundo Viana, Campos (1973); Ferreira et al. (2001); Birgel Junior et al.
(2001); e Campos et al. (2007); e acima do normal segundo Bradisch et al. (1954);
Alencar Filho (1976); Irfan (1982); Matos et al. (1983); Fagliari et al.(1988); Kaneko
(1989); Meyer et al. (1992); Lopes et al. (1996), Birgel Junior et al. (2001) e Souza et
al. (2004).
Considerando os estudos realizados por outros autores que também
trabalharam com animais soropositivos para LEB, os valores obtidos, no presente
estudo, para o grupo soropositivo, foram considerados normais, segundo os relatos
de Ferreira et al. (2001) e Birgel Junior et al. (2001); e acima do normal, segundo
Oliveira et al. (2001).
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio padrão (entre 4,01 e 5,37 g/dl), é considerado
normal, segundo Irfan (1982); Matos et al. (1983); Ferreira et al. (2001); Birgel Junior
et al. (2001); Souza et al. (2004) e Campos et al. (2007); e acima do normal,
segundo Bradisch et al. (1954); Alencar Filho (1976); Fagliari et al.(1988); Kaneko
(1989); Meyer et al. (1992); Lopes et al. (1996); e considerado abaixo do normal
segundo Viana, Campos (1973).

48
4.2.4 Aspartato aminotransferase – AST
Os valores médios e desvios-padrão obtidos na mensuração da aspartato
aminotransferase foram de 46,37 ± 17,82 U/L para o grupo de soropositivos e, de
33,38 ± 13,96 U/L, para o grupo de soronegativos.
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 28,55 e 64,19 U/L), é considerado
normal, segundo Medway et al. (1973); Dirksen et al. (1983); Blood et al. (1988);
Lopes et al. (1996); Gregory et al. (1999); Campos et al. (2007); e considerado
abaixo do normal, segundo Kaneko (1989); Carlson (1990) e Meyer et al. (1992).
Considerando os estudos realizados por outros autores que também
trabalharam com animais soropositivos para LEB, os valores obtidos no presente
estudo para o grupo soropositivo foram considerados normais, segundo as
pulicações de Hagiwara et al. (1986) apud Gregory et al. (1999); e Gregory et al.
(1999).
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 19,42 e 47,34 U/L), é considerado
normal, segundo Medway et al. (1973); Dirksen et al. (1983); Lopes et al. (1996);
Gregory et al. (1999); Campos et al. (2007); e considerado abaixo do normal,
segundo Blood et al. (1988); Kaneko (1989); Carlson (1990) e Meyer et al. (1992).
4.2.5 Gamaglutamiltransferase - γγγγGT
Os valores médios e desvios-padrão obtidos na mensuração da
gamaglutamiltransferase foram de 25,72 ± 9,68 U/L para o grupo de soropositivos e,
de 20,48 ± 6,35 U/L, para o grupo de soronegativos.
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 16,04 e 35,40 U/L), é considerado
normal, segundo Simensen, Nansen (1974); Braun et al. (1977); Márquez et al.
(1977); Dirksen et al. (1983); Kaneko (1989); Meyer et al. (1992); Lopes et al. (1996);
Gregory et al. (1999); Campos et al. (2007); e acima do normal, segundo Botelho et
al. (1980); e Barros Filho (1995).

49
Considerando os estudos realizados por outros autores que também
trabalharam com animais soropositivos para LEB, os valores obtidos, no presente
estudo, para o grupo soropositivo, foram considerados normais, segundo
informações de Hagiwara et al. (1986) apud Gregory et al. (1999); e Gregory et al.
(1999).
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 14,13 e 26,83 U/L), é considerado
normal, segundo Simensen, Nansen (1974); Braun et al. (1977); Márquez et al.
(1977); Dirksen et al. (1983); Kaneko (1989); Meyer et al. (1992); Barros Filho
(1995); Lopes et al. (1996); Gregory et al. (1999); Campos et al. (2007); e acima do
normal, segundo Botelho et al. (1980).
4.2.6 Alanina aminotransferase - ALT
Os valores médios e desvios-padrão obtidos na mensuração da alanina
aminotransferase foram de 11,57 ± 4,93 U/L para o grupo soropositivo e, de 9,87 ±
3,44 U/L, para o grupo soronegativo.
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 6,64 e 16,5 U/L), é considerado
normal, segundo Dirksen et al. (1983); Kaneko (1989); Lopes et al. (1996); e abaixo
do normal, segundo Campos et al. (2007); não sendo encontrado estudos na
literatura que mensurem ALT em animais soropositivos para LEB.
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 6,43 e 13,31 U/L), é considerado
normal, segundo Dirksen et al. (1983); Kaneko (1989); e abaixo do normal, segundo
Lopes et al. (1996) e Campos et al. (2007).
4.2.7 Ureia
Os valores médios e desvios-padrão obtidos na mensuração da ureia sérica
foram de 30,12 ± 12,58 mg/dl para o grupo soropositivo e, de 34,81 ± 11,47 mg/dl,
para o grupo soronegativo.

50
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 17,54 e 42,70 mg/dl), é
considerado normal, segundo Dirksen et al. (1983); Coles (1984); Kolb (1984); Blood
et al. (1988); Kaneko (1989); Meyer et al. (1992); Gregory et al. (2004); Campos et
al. (2007); e abaixo do normal, segundo Lopes et al. (1996).
Considerando os estudos realizados por outros autores que também
trabalharam com animais soropositivos para LEB, os valores obtidos, no presente
estudo, para o grupo soropositivo, foram considerados normais segundo os estudos
de Gregory et al. (2004).
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 23,34 e 46,28 mg/dl), é
considerado normal, segundo Dirksen et al. (1983); Coles (1984); Kolb (1984); Blood
et al. (1988); Kaneko (1989); Meyer et al. (1992); Gregory et al. (2004); Campos et
al. (2007) e Lopes et al. (1996).
4.2.8 Creatinina
Os valores médios e desvios-padrão obtidos na mensuração da creatinina
sérica foram de 1,11 ± 0,29 mg/dl para o grupo soropositivo e, de 1,13 ± 0,39 mg/dl,
para o grupo soronegativo.
O valor médio obtido para o grupo soropositivo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 0,82 e 1,40 mg/dl), é considerado
normal, segundo Dirksen et al. (1983); Kaneko (1989); Lopes et al. (1996); Gregory
et al. (2004); Campos et al. (2007).
Considerando os estudos realizados por outros autores que também
trabalharam com animais soropositivos para LEB, os valores obtidos, no presente
estudo, para o grupo soropositivo, foram considerados normais, segundo os relatos
de Gregory et al (2004).
O valor médio obtido para o grupo soronegativo, por estar dentro da
amplitude de variação de um desvio-padrão (entre 0,74 e 1,52 mg/dl), é considerado
normal, segundo Dirksen et al. (1983); Kaneko (1989); Lopes et al. (1996); Gregory
et al. (2004); Campos et al. (2007).

51
4.3 Anatomopatologia
Os linfonodos pré-escapulares, pré-crurais, submandibulares e mesentérios
exibiram megalia e consistência firme. Os linfonodos pré-escapulares (Prancha 1A),
ao corte longitudinal, revelaram protusão do parênquima para superfície de corte,
nodulações em seu interior de coloração que variou de branco a amarelo-cinza, com
perda da arquitetura tecidual e do limite córtico-medular (Prancha 1B). Estes
achados estão de acordo com aqueles encontrados por Oliveira et al. (1999); Reis et
al. (2002); Barros et al. (2007); Silva et al. (2008). Contudo, Peixoto et al. (2008)
também encontraram coloração branca e amarelada, mas também áreas vermelho-
escuras intercaladas.
O exame macroscópico revelou aumento de volume discreto do baço em
ambos os animais (Prancha1C), fato que, possivelmente, tenha ocorrido devido ao
método de eutanásia empregado no presente estudo.
À microscopia dos linfonodos aqui estudados, observou-se população celular
representada por linfócitos anisocarióticos que, em algumas vezes, revelavam
hipercromatismo e mitoses bizarras (Prancha 1D). Essas observações coincidem
com as de Oliveira et al. (1999), Peixoto et al. (2008) e Cordeiro et al. (1994).
Seções teciduais hepáticas, desta investigação, mostraram moderada
população de linfócitos anisocarióticos, polimórficos e hipercromáticos no tecido
conjuntivo e interior dos vasos do espaço-porta, e entre os sinusoides hepáticos
(Prancha 1E).
Yamamoto (1982) afirma que a ocorrência de linfócitos neoplásicos no
espaço-porta hepático é compatível com a manifestação leucêmica desta
enfermidade em bovinos. Essa afirmação corrobora os achados do presente
experimento, que apresentou linfócitos atípicos no sangue periférico, com
leucocitose e linfocitose absoluta, observadas no hemograma deste animal
necropsiado (Leucócitos: 20.900/µl; linfócitos: 13.617/µl). Comparados aos valores
de referência, segundo Lopes et al. (1996): Leucócitos: 4.000 a 12.000/µl; Linfócitos:
2.500 a 7.500/µl).
Yamamoto (1982) ainda descreve que a lesão hepática é caracterizada pela
ploriferação neoplásica nas tríades do espaço-porta, o que também foi observado
nesta investigação (Prancha 1E).

52
Os dutos biliares da tríade portal foram melhores evidenciados pela
imunomarcação com a pancitoqueratina, o que permitiu uma visualização plena dos
linfócitos proliferados e entremeados no tecido conjuntivo do espaço-porta e entre os
sinusoides hepáticos aqui investigados (Prancha 1F).
A citopatologia, em decalque da superfície de corte do linfonodo pré-
escapular, revelou linfócitos anisocarióticos e pleomórficos, compatíveis com a
leucose bovina (Prancha 2 A).
Os vasos sanguíneos da dura-máter da medula espinhal apresentaram, à
microscopia em HE, manguitos de células linfoides, que exibiam atipias e
hipecromasia, compatíveis com aquelas encontradas na leucose enzoótica bovina
(Prancha 2B). Ao passo o que, em seções teciduais cerebelares, foram mostrados
manguitos perivasculares, representados por células compatíveis com aquelas
acima descritas (Prancha 2C/D).
Os infiltrados de linfócitos neoplásicos na membrana dura-máter da medula
espinhal e cerebelares explicam os sinais percebidos no exame clínico, e estes
incluíam incoordenação e ataxia dos membros posteriores. Esse animal tinha, na
leucometria global, um total de 26.700 leucócitos/µl de sangue e contagem de
linfócitos de 17.889/µl, revelando um quadro de leucocitose com linfocitose absoluta,
segundo Lopes et al. (1996).
A ocorrência de linfossarcomas intracerebrais e no canal vertebral
comprimindo a medula espinhal causa quadro clínico de disfunção medular
(REBHUN, 1984; SHERMANN, 1987; FIGHERA, 2004; BARROS, 2004). A
localização intracraniana de linfossarcomas é relatada como sendo incomum por
Summers et al. (1995). A compressão da medula espinhal por linfossarcomas
localizados no canal vertebral, causando sinais clínicos, também foi descrita por Valli
(1993). Apesar de não terem sido observadas massas tumorais pronunciadas no
cerebelo e medula espinhal, nos cortes realizados no animal eutanasiado neste
experimento, que apresentava sintomatologia nervosa, acredita-se que este quadro
tenha sido desencadeado pelo infiltrado linfocitário.
Não houve imunomarcação para o antígeno p53 nas seções teciduais dos
animais necropsiados no presente trabalho. Simões (2007) e Tajima et al. (1998), ao
estudarem bovinos com leucose enzoótica, a expressão do p53 foi detectada em
torno de 50% dos animais estudados. Portanto, é provável que nem em todos os
casos de leucose enzoótica bovina ocorra a expressão e o acúmulo da proteína p53

53
estável, o que pode ter ocorrido nesta investigação, o que explica os valores
negativos encontrados nas amostras analisadas. Além do que, a leucose enzoótica
bovina tem progressão crônica, caracterizada pelo aparecimento lento de massas
tumorais, que somente são percebidas nos animais mais velhos. Os infiltrados
linfocitários, observados em amostras dos animais aqui estudados, eram discretos e
incipientes, talvez fosse interessante realizar análise do p53 em massas tumorais
mais evoluídas.
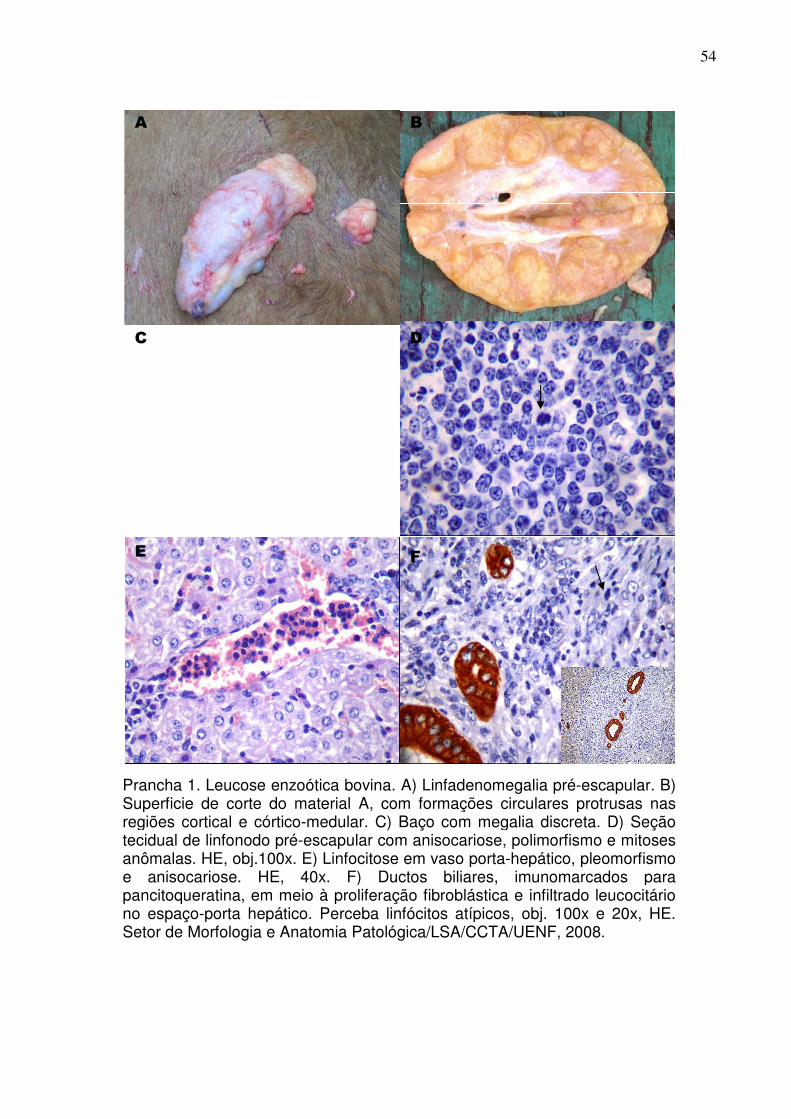
Prancha 1. Leucose enzoótica bovina. A) Linfadenomegalia préSuperficie de corte do material Aregiões cortical e córticotecidual de linfonodo préanômalas. HE, obj.100x. E) Linfocitose em vaso portae anisocariose. HE, 40x. F) Ductos biliares, imunomarcados para pancitoqueratina, em meio àno espaço-porta hepático. Perceba linfócitos atípicos, obj. 100x e 20x, HE.Setor de Morfologia e Anatomia Patológica/LSA/CCTA/UENF, 2008.
A
C
E
Leucose enzoótica bovina. A) Linfadenomegalia préuperficie de corte do material A, com formações circulares protrusas nas
regiões cortical e córtico-medular. C) Baço com megalia discreta. D) Stecidual de linfonodo pré-escapular com anisocariose, polimorfismo e mitoses anômalas. HE, obj.100x. E) Linfocitose em vaso porta-hepático, pleomoe anisocariose. HE, 40x. F) Ductos biliares, imunomarcados para
oqueratina, em meio à proliferação fibroblástica e infiltrado leucocitário porta hepático. Perceba linfócitos atípicos, obj. 100x e 20x, HE.
Anatomia Patológica/LSA/CCTA/UENF, 2008.
B
D
F
54
Leucose enzoótica bovina. A) Linfadenomegalia pré-escapular. B) formações circulares protrusas nas o com megalia discreta. D) Seção
escapular com anisocariose, polimorfismo e mitoses hepático, pleomorfismo
e anisocariose. HE, 40x. F) Ductos biliares, imunomarcados para filtrado leucocitário
porta hepático. Perceba linfócitos atípicos, obj. 100x e 20x, HE. Anatomia Patológica/LSA/CCTA/UENF, 2008.

55
Prancha 2. Leucose enzoótica bovina, HE. A) Decalque de superfície de corte de linfonodo pré-escapular, exibindo linfócitos anisocarióticos e polimórficos, obj.100x. B) Dura-máter da medula espinhal infiltrada por linfócitos neoplásicos (com atipias), obj. 20x. C/D) Vaso cerebelar com manguito de linfócitos neoplásicos (anisocarióticos, hipercromáticos e polimórficos, objetivas 40x e 100x. Setor de Morfologia e Anatomia Patológica/LSA/CCTA/UENF, 2008.
A B
C D

56
5 CONCLUSÕES
Não houve imunomarcação para o antígeno p53 nas lesões dos animais
soropositivos necropsiados.
A LEB foi capaz de produzir alterações de pequena magnitude na maioria
dos parâmetros bioquímicos séricos avaliados, incluindo a ALT ainda não estudada.
Os animais necropsiados apresentaram características típicas da leucose
enzoótica bovina nos linfonodos, fígado, cerebelo e dura-máter da medula espinhal
associados à forma leucêmica da doença.
Estudos da LEB devem ser incentivados.
Deverá ser estimulado o desenvolvimento de um programa de controle e
erradicação da Leucose Enzoótica Bovina no rebanho brasileiro.
Há necessidade de se estabelecer valores bioquímicos séricos regionais.

57
6 REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, J.M.G. Leucose enzoótica dos bovinos. Prevalência de anticorpos séricos anti-vírus da leucose bovina em animais criados na bacia leiteira de Fortaleza, Estado do Ceará. Tese (Mestrado em Clínica Veterinária) – São Paulo- SP, Faculdade de Medicina Veterinária e Zootecnia – USP, 75p, 1993. ABREU, V.L.V.; SILVA, J.A.; MODENA, C.M.; MOREIRA, E.C. Prevalência da Leucose Enzoótica Bovina nos Estados de Rondônia e Acre. Arquivo Brasileiro Medicina Veterinária. v.42, n. 3, p. 203-210, 1990. AKIOSHI, H.T.; GERSZTEIN, A. Atlas de diagramas eletroforéticos. Buenos Aires. Ed. Artecnica, 102 p, 1962. ALENCAR FILHO, R.A. Considerações sobre exames hematológicos nas enfermidades animais. Bioquímica do sangue – Indicações clínicas. O Biológico, São Paulo, v.42, n.6, p.140-147, 1976. ANDRADE, J.R.A.; ALMEIDA, M.M.R. Prevalência da leucose enzoótica bovina na bacia leiteira de Goiânia, Goiás. Hora Veterinária, Porto Alegre, v. 10, n. 60, p. 49-53. 1991. ANGELO, N.J.O.; BIRGEL, E.H.; HAGIWARA, M.K.; BENESI, F.J.; D’ANGELINO, J.L.; DAHME, H.W.P.F.; CARVALHO, R.P.S. Isolamento do vírus da leucose bovina de animais com leucocitose persistente. In: 13º Congresso Brasileiro de Microbiologia, Anais...São Paulo, p.284, 1985. ARITA, G.M.M.; GONÇALVES, C.D.; SABER, A.F.; GERMANO, P.M.L.; DEAK, J.G.; KOTAIT, I. Estudo Epidemiológico da Leucose Enzoótica dos Bovinos no Vale do Paraíba, São Paulo, Brasil. In: Reunião Anual do Instituo Biológico de São Paulo, 5, Anais..., São Paulo: Secretaria da Agricultura e Abastecimento do Estado de São Paulo, p.30, 1992.. BARROS, C.S.L. Leucose Bovina. In: RIET-CORREA, F.; SCHILD, A.L.; LEMOS, R.A.A.; BORGES, J.R.J.(eds) Doenças de ruminantes e eqüídeos. Fenovi, p.159-169, 2007. BARROS FILHO, I.R. Contribuição ao estudo da bioquímica clínica em zebuínos da raça Nelore (Bos Indicus, Linnaeus, 1758) criados no estado de São Paulo. Tese (Mestrado em Clínica Veterinária) – São Paulo – SP. Universidade de São Paulo – USP, 133p, 1995.

58
BIANCHI, I. Prevalência da Leucose Enzoótica Bovina na Região Norte Fluminense. Tese (Mestrado em Produção Animal) – Campos dos Goytacazes – RJ, Universidade Estadual do Norte Fluminense – UENF, 31p, 2003. BIRGEL, E.H. Leucose linfática enzoótica dos bovinos adultos. Aspectos clínicos e diagnósticos. In: BIRGEL, H.E.H., BANESI, F.J. Patologia clínica veterinária. São Paulo: Sociedade Paulista de Medicina Veterinária, p. 249-260, 1982. BIRGEL, E.H.; BIRGEL JR., E.H.; D’ANGELINO, J.L.; TAKAHIRA, R.K.; SILVA, M. M. Determinação de valores padrões bioquímicos do soro sanguíneo de bovinos sadios da raça Jersey, criados no Estado de São Paulo. Anais da Conferência Anual da Sociedade Paulista de Medicina Veterinária, 46, São Paulo, v. 46. p. 37, 1991. BIRGEL JUNIOR, E.H.; DÂNGELO, J.; BENESI, J.; BIRGEL, E.H. Prevalência da infecção da leucose dos bovinos, em animais da raça Jersey, criados no Estado de São Paulo. Pesquisa Veterinária Brasileira, v.15, n. 4, p. 93-99, 1995. BIRGEL JUNIOR, E.H.; SALVATORE, L.C.A.; NEVES, F.S.; MIRANDOLA, R.M.S.; SOUZA, P.M.; BIRGEL, E.H. Dinâmica das proteínas séricas de fêmeas bovinas da raça holandesa naturalmente infectadas pelo vírus da leucose dos bovinos. Ciência Rural, v. 31, n.4, p. 615-619, 2001. BLOOD, D.C.; HENDERSON, J.A.; RADOSTITS, O.M. Clínica Veterinária. 5a ed. Rio de Janeiro. Ed. Guanabara Koogan S. A . 1121 p, 1988. BOSE, M.L.V. Contribuição para a determinação do nível normal de componentes do sangue bovino Canchim. Anais da Escola Superior de Agricultura Luiz de Queiroz,1, Piracicaba, SP, v. 1, p. 119-36, 1983. BOTELHO, G.G.; NASCIMENTO, S.C.A.G.; SILVEIRA, J.M.; NASCIMENTO, F.G. Gama-glutamil-transferase em bovinos de abate. Resumos do Congresso Brasileiro de Medicina Veterinária, 17, Fortaleza, p. 30, 1980. BOTELHO, G.G.; OLIVEIRA, A.R.; PACHECO, R.G. Uréia, creatinina e ácido úrico em bovinos da raça Canchim. Revista Brasileira de Medicina Veterinária. v. 6, n. 2, p. 31-32, 1984. BRADISCH, C.S.; HENDERSON, W.M.; BROOKSBY, J.B. Electrophoretic studies of ox serum. The sera of normal cattle. Biochem. v.56, p. 329-333, 1954.

59
BRAGA, F.M.; VAN DER LAAN, C.W.; HALFEN, D.C.; VIDOR, T. Avaliação de métodos de controle da infecção pelo vírus da leucose enzoótica bovina. Ciência Rural, Santa Maria, v.27, n. 4, p.635-640, 1997. BRAGA, F.M.; VAN DER LAAN, C.W.; SCHUCH, L.F.; HALFEN, D.C. Infecção pelo Vírus da Leucose Enzoótica Bovina (BLV). Ciência Rural, Rio Grande do Sul, v. 28 n. 1, p. 163-172, 1998a. BRAGA, A.C.; ROSA, J.C.A.; OLIVEIRA, L.G.; RECKZIEGEL, P.E.; TEIXEIRA, J.C.F.; LISBOA, C.S. Anticorpos contra o vírus da leucose bovina em animas da raça leiteira importados do Uruguai. Pesquisa Agropecuária Gaúcha, v.4, n.1, p. 35-38, 1998b. BRAUN, J.P.; RICO, A.G.; BERNARD, P.; THOUVENOT, J.P; BONNEFIS, M.J. Blood and tissue distribuition of gama glutamyl transferase in calves. Journal of Dairy Science. v.61, n. 5, p.596-599, 1977. BRAUN, J.P.; SIEST, G.; RICO, A.G. Uses of gamma-glutamyltransferase in experimental toxicology. Adv. Vet. Sci. Comp. Med. v.31, p.151-172, 1987. BRENNER, J.; VAN-HAAM, M.; SAVIR, D.; TRAININ, Z. The implications of BLV infection in the productivity, reproductive capacity and survival rate of dairy cow. Veterinary Immunology Imunopatology. v.22, p. 299-305, 1989. CAMPOS, R.; CUBILLOS, C.; RODAS, A.G. Indicadores metabólicos em razas lecheras especializadas em condiciones tropicales em Colombia. Acta Agon. (Colombia). v.56, n. 2, p. 85-92, 2007. CARLSON G.P. Clinical Chemistry Tests. In: SMITH, B. P. Large Animal Internal Medicine. Toronto. The C. V. Mosby Company, 22 cap., p 386-413, 1990. CARNEIRO, P.A.M.; ARAÚJO, W.P; BIRGEL, E.H; SOUZA, K.W. Prevalência da infecção pelo vírus da leucose dos bovinos em rebanhos leiteiros criados no Estado do Amazonas, Brasil. Acta Amazônia. v.33, n. 1, p. 111-125, 2003. CARROLL, E.J.; KANEKO, J.J. The clinical significance of serum protein fractionation by electroforesis. The California veterinariam. v. 21, n. 1, p. 22-25, 1967. COCHRAN, W.G. Técnicas de amostragem. Rio de Janeiro. Fundo de Cultura, 555p,1955.

60
COELHO, H.E.; MERHI, S.A.; REIS, D. Linfossarcoma em bovinos de Uberlândia – MG e região durante 20 anos (1976-1995). Higiene Alimentar, v.12, n. 58, p. 27-32, 1998.
COLES, E.H. Patologia Clínica Veterinária. São Paulo. Ed. Manole. 566 p, 1984. CORDEIRO, J.L.F.; DESCHAMPS, F.C.; MARTINS, E.; MARTINS, V.M.V. Identificação e controle da Leucose enzoótica bovina (LEB) em um rebanho leiteiro. Pesq. Agropec. Bras., v. 29, n.8, p. 1287-1292, 1994. CRUZ, C.D. Programa Genes: Estatística Experimental e Matrizes. Editora UFV. Viçosa, MG. 285p., 2006. CUNHA, R.G.; TEIXEIRA, A.C.; SOUZA, D.M. Antígenos do vírus da leucose bovina e anticorpos precipitantes em soros de bovinos. Pesquisa Agropecuária Brasileira, v.17, n. 9, p. 1363-1370, 1982. DA, Y.; SHANKS, R.D.; STEWART, J.A.; LEWIN, H.A. Milk and fat decline in bovine leukemia virus-infected Holstein cattle with persistent lymphocytosis. Proceedings National Academy Science USA, v. 90, n. 14 p. 6538-6541, 1993. D’ANGELINO, J.L.; GARCIA, M.; BIRGEL, E.H. Productive and reproductive performance in cattle infected with bovine leucosis vírus. Journal Dairy Research, v.65, n.4, p. 693-695, 1998. DIRKSEN, G.; GRÜNDER, H.; STÖBER, M. Rosenberger Exame Clínico dos Bovinos.3.ed.Guanabara Koogan, 419p.,1983. FAGLIARI, J.J.; PASSIPIERI, M.; CURI, P.R.; FERREIRA NETO, J.M; LUCAS, A. Valores padrões das proteínas séricas de bovinos da raça Guzerá II. Proteinograma sérico de bezerros recém-nascidos. Arqs. Veterinária.v.4, n. 2, p. 225-232, 1988. FAGLIARI, J.J.; SANTANA, A.E.; LUCAS, F.A.; CAMPOS FILHO, E.; CURI, P.R.; Constituintes sanguíneos de bovinos recém-nascidos das raças Nelore (Bos indicus) e Holandesa (Bos taurus) e de bubalinos (Bubalus bubalis) da raça Murrah. Arq. Bras. Med. Vet. Zootec. v.50, n. 3, p. 253-262, 1998. FAVA, C.D. Leucose Enzoótica Bovina em Búfalos. Tese (Mestrado em Medicina Veterinária) – Jaboticabal-SP, Universidade Estadual Paulista – UNESP, 79p., 1995.

61
FENNER, J.F.; GIBBS, E.P.J.; MURPHY, F.A. Retroviridae. In: Veterinary Virology. San Diego: Academic Press, cap. 33, p. 561-595, 1993. FERREIRA, M,M.; MELO, M.M.; MARQUES JÚNIOR, A.P. Concentração de proteína sérica total, albumina e globulinas em novilhas holandesas soro-reagentes para leucose bovina a vírus durante a gestação e pós-parto. Rev. Bras. Saúde Prod. An. v.1, n.1, p.1-7, 2001. FIGHERA, R.A.; BARROS, C.S.L. Linfossarcoma intracerebral em bovino. Ciência Rural, v. 34, n. 3, p. 943-945, 2004. FLORES, E.F.; WEIBLEN, R.; OLIVEIRA, C.; KREUTZ, L.C. Anticorpos contra o vírus da leucose bovina (VLB) em soro de bovinos provenientes da República Ocidental do Uruguai. A Hora Veterinária, v.12, n.68, p.5-8, 1992. GARCIA LIMA, E. Contribuição para o estudo diagnóstico da leucemia bovina. Revista de Patologia Tropical, Goiânia, v.9, n. 1-2, p. 1-12, 1980. GARCIA, M.; D’ANGELINO, J.L.; BIRGEL, E.H.; MARÇAL, W.S. Avaliação do leucograma de fêmeas da raça holandesa naturalmente infectadas pelo vírus da leucose bovina. Pesquisa Veterinária Brasileira. v.11, n. 3-4, p. 61-64, 1991. GREGORY, L.; BIRGEL JUNIOR, E.H.; MIRANDOLA, R.M.S.; ARAÚJO, W.P.; BIRGEL, E.H. Valores de Referência da atividade enzimática da aspartato-aminotransferase e da gama-glutamiltranferase em bovinos da raça Jersey. Influência dos fatores etários, sexuais e da infecção pelo vírus da leucose dos bovinos. Arq. Bras. Med. Vet. Zootec. v.51, n. 6, p.515-522, 1999. GREGORY, L.; BIRGEL JUNIOR, J.L.; D´ANGELINO, F.J.; BENESI, W.P.A.; BIRGEL, E.H. Valores de referência os teores séricos da uréia e creatinina em bovinos da raça Jersey criados no Estado de São Paulo. Influência dos fatores etários, sexuais e da infecção pelo vírus da leucose dos bovinos. Arq. Inst. Biol. v.71, n. 3, p. 339-345, 2004. HEALD, M.T.S.; WALTNER-TOEWS, D.; JACOBS, R.M.; McNAB, W.B. The Prevalence of anti-bovine Leukemia Virus Antibodies in Dairy Cows and Associations with farm Management Pratices, Production and Culling in Ontário. Preventive Veterinary Medicine, v.14, p. 45-55, 1992.

62
HERZ, A.; HOD, I. The albumin alpha-globulin ration invarious physiological states in cattle. Br. Vet. J., v.125, n.7, p. 326-329, 1969. IRFAN, M. The electroforetic pattern of serum proteins in normal animals. Research in Veterinary Science. v.8, p.137-142, 1982. JAIN, C.N. Essentials of Veterinary Hematology. Philadelphia: Lea & Febiger, p.416, 1993. JOHNSTON, J.K. Alterations in Blood Proteins, In: SMITH, B.P., Large Animal Internal Medicine. Toronto. The C. V. Mosby Company, 26 cap. p. 435-443, 1990. JOHNSON, R., KANEENE, J.B. Bovine leukemia virus. Part I. Descriptive epidemiology, clinical manifestations, and diagnostic tests. In: Compendium on continuing education of the practicing veterinarian. p. 315-319, 1991. KANEKO, J. Clinical Biochemistry of Domestic Animals. 4a ed., New York. Academic Press. 725 p., 1989. KANTEK, C.E.; KRUGER, E.R.; WELTE, V.R. Infecção com o vírus da leucose enzoótica bovina em um lote de vacas produtoras de leite importadas do Uruguai. Pesquisa Veterinária Brasileira, Rio de Janeiro, v.3, n. 4, p. 843-845, 1982. KOLB, E. Fisiologia dos líquidos orgânicos. In: Kolb, E. Fisiologia Veterinária. 4a ed., Rio de Janeiro. Ed. Guanabara Koogan S. A. p. 208-248, 1984. LEITE, R.C.; MODENA, C.M.; MOREIRA, E.C.; ABREU, J.J. Evolução Clínica da Leucose Enzoótica Bovina. Arquivo Brasileiro Medicina Veterinária e Zootecnia, Belo Horizonte, v. 36, n. 1, p.47-57, 1984. LEITE, R.C.; LOBATO, Z.I.P.; CAMARGOS, M.F. Leucose Enzoótica Bovina. Revista do Conselho Federal de Medicina Veterinária, Brasília, Ano VII, v.24 p. 20-28, 2001. LEUZZI Jr, L.A.; ALFIERI, A.F.; ALFIERI, A.A. Leucose enzoótica bovina e vírus da leucemia bovina. Semina: Ci Agrárias, Londrina, v. 22, n. 2, p. 211-221, 2001. LIBERG, P. Agarose gel eletrophoretic fractionation of serum proteins in adult cattle. A study Of cows with diferents diseases. Acta Vet. Scand., v.18, n.3, p.335-348, 1977.

63
LOPES, S.T.A.; CUNHA, C.M.A.; BIONDO, A.; FAN, L.C.R. Patologia Clínica Veterinária. Santa Maria. 166p, 1996. LUCAS, M.H.; DAWSON, M.; CHASEY, D. Enzootic bovine leucosis virus in semem. Veterinary Record, v.106, p.128, 1980. MAAS, J. Alterations in body weight or size. In: Smith B. P. Large Animal Internal Medicine. Toronto. The C. V. Mosby Company. 9 cap. p 171-194, 1990. MÁRQUEZ, A.G.; FRATTINI, J.F.; GRIMOLD, R.J.; FERNÁNDEZ, G.; TAMAMES, F.A.; WILLIAMS, M.B. Perfil enzimático em sueros de ruminantes: lactico desidrogenase; gamma glutamil transpeptidase; aldolasa; leucin aminopeptidasa; colinesterasa. Gaceta Veterinaria. v. 39, n. 37, p. 33-42, 1977. MATOS, M.S.; GARGUR, M.B.; GUIMARÃES, J.E.; ARAÚJO, C.F. Perfil eletroforético de bovinos com brucelose. Arquivos da Escola de Medicina Veterinária da Universidade Federal da Bahia, Salvador. v. 8, n.1, p.80- 87, 1983. MEDWAY, W.; PRIER J. E.; WILKINSON, J. S. Patologia Clinica Veterinaria. Mexico. Union Tipografica Editorial Hispano-Americana. 532 p., 1973. MEGID, J.; NOZAKI, C.N.; KURODA, R.B.S.; CRUZ, T.F.; LIMA, K.C. Ocorrência de leucose enzoótica bovina na microrregião da Serra de Botucatu. Arq. Bras. Med. Vet. Zootec., v.55, n. 5, p.645-646, 2003. MELO, L.E.H. Leucose enzoótica dos bovinos: Prevalência da infecção em rebanhos leiteiros criados no agreste meridional do Estado de Pernambuco. Tese (Mestrado em Patologia Bovina) – São Paulo – SP, Faculdade de Medicina Veterinária e Zootecnia – USP, 104p.,1991. MEYER, D.J., COLES, E.H., RICH, L.J. Veterinary Laboratory Medicine – Interpretation and diagnosis. Philadelphia. W. B. Saunders Company. 349 p., 1992. MILLER, J.N., MILLER, L.D., OLSON, C., GILLETTE, K.G. Virus-like particles in phytohemagglutinn-stimulated lymphocyte cultures with reference to bovine linfossarcoma, J. Naltl. Cancer Inst., Washington, v. 43, n. 6, p. 1297-1305, 1969. MODENA, C.M.; ABREU, V.L.V.; SILVA, J.A.; MOREIRA, E.C.; AZEVEDO, N.A.; REHFED, O.A.M. Ocorrência de infecção pelo virus da leucose enzoótica bovina em

64
animais importados. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, Belo Horizonte, v. 35, n. 4, p. 565-573, 1983. MODENA, C.M.; SILVA, J.A.; GOUVEIA, A.M.G.; VIANA, F.C.; AZEVEDO, N.A.; REHFED, O.A.M. Leucose enzoótica bovina: Prevalência em rebanhos de alta linhagem no Estado de Minas Gerais. Arquivo Brasileiro de Medicina Veterinária e Zootecnia. v.36, n.1, p.39-45, 1984. MOLNÁR, E.; MOLNAR, L.; DIAS, H.T.; SILVA, A.O.A.; VALE, W.G. Ocorrência da Leucose Enzoótica dos Bovinos no Estado do Pará, Brasil. Revista Brasileira de Medicina veterinária, v. 21, n.4, p. 171-175, 1999. MOULTON, J.E. Tumors in Domestic Animals. 3. ed. London: University of California Press, p. 672, 1990. MORAES, M.P.; WEIBLEN, R.; FLORES, E.F.; OLIVEIRA, J.C.D. Levantamento sorológico da infecção pelo vírus da leucose bovina nos rebanhos leiteiros do Estado do Rio Grande do Sul, Brasil. Ciência Rural, Sant Maria, v.26, n. 2, p. 257-262, 1996. MURPHY, F.A.; GIBBS, E.P.J.; HARZINEK, M.C.; STUDDRT, M.J. Retroviridae. In: Veterinary Virology. Academic Press, p. 363-383, 1999. MYLREA, P.J.; BAYFIELD, R.F. Concentrations of some components in the blood and serum of apparently healthy dairy cattle. Electrolyte and minerais. Aust. Vet. J. v. 44, n. 12, p. 565-569, 1968. OLIVEIRA, A.R.; MACHADO, P.E.A.; KROLL, L.B.; CORREIA, C.M. Leucose Bovina: Caracterização do proteinograma eletroforético em dois rebanhos identificados como soropositivos e com linfocitose persistente. Arq. Inst. Biol., São Paulo, v.67, n.1, p.31-36, 2001. OLIVEIRA, K.B. (2000) Leucose enzoótica bovina – Uma revisão bibliográfica e a importância do seu diagnóstico. Dissertação de Graduação PUC-Paraná, 17p., 2000. OLIVEIRA, S.R.; GUEDES, R.M.C.; NOGUEIRA, R.H.G; Exame citológico como método auxiliar no diagnóstico da leucose enzoótica bovina. Veterinária Notícia, Minas Gerais, v. 5, n.2, p. 103-109, 1999. OLIVEIRA, A.R.; IKUNO, A.A.; MUELLER, S.B.K.; BARRETO, C.S.F.; SAAD, V.M. Leucose bovina: ocorrência de anticorpos em bovinos de várias faixas etárias. Revista Brasileira de Medicina Veterinária, v.16, n. 2, p. 46-50, 1994.

65
OLSON, C.; MILLER, J. History and terminology of enzootic bovine leucosis. In: BURNY, A., MAMMERICKX, M. (Eds.) Enzootic bovine leucosis and bovine leukemia virus. Boston: Martinus Nijhoff, p. 3-11, 1987. ONUMA, M. (1989) Diagnosis of bovine leucosis using monoclonal antibody against tumor-associated antigen of bovine leukemia cells. Folia Veterinária, v. 33, n. 1, p. 9-21, 1989. PEIXOTO, T.C.; MARTINI-SANTOS, B.J.; YAMAZAKI, E.M.; GALVÃO, A.; SALLES, S.P.X. Leucose juvenil bovina em uma bezerra no Rio de Janeiro, Brasil. In. CONGRESSO BRASILEIRO DE MEDICINA VETERINÀRIA, 35., 2008, Gramado – RS, 2008. CD-ROM. POLETTO, R.; KREUTZ, L.C.; GONZALES, J.C.; BARCELLOS, L.J.G. Prevalência de tuberculose, brucelose e infecções víricas em bovinos leiteiros do município de Passo Fundo, RS. Ciência Rural, Santa Maria, v.34, n. 2, p.595-598, 2004. RADOSTITIS, O.M. GAY, C.C.; BLOOD, D.C., Clínica Veterinária 9ed, Guanabara-Koogan, Rio de Janeiro, p. 940-951, 2002. REBHUN, W.C. Compressive neoplasms affectingh the bovine spinal cord. Comp. Comt. Educ. Pract. Vet., v.6, p.396-440, 1984 REIS, D.O.; HORTÊNCIO, M.C.; ALMEIDA, L.P.; MOREIRA, M.D. Linfoma em bovinos abatidos sob inspeção federal: aspectos morfológicos e linfonodos mais acometidos. Uberlândia, MG. Biosci J., v. 18, n. 2, p. 109-112, 2002. REKWOT, P.I.; OYEDIPE, E.O.; AKEREJOLA, O.O.; VOH JR., A.A.; DAWUDA, P.M. Serum biochemistry of zebu and their fresian crosses fed two planes of protein. British Veterinary Journal, v.145, n.1, p. 85-88, 1989. ROMERO, C.H.; ROWE, C.A. Enzootic bovine leucosis virus in Brazil. Tropical. Animal. Health. Production., Edinburgh, v.13, n. 2, p. 107-111, 1981. ROUSSEL, A.J.; WHITNEY, M.S.; COLE, D.J; Interpreting a bovine serum chemistry profile: Part 1. Food-Animal practice, Veterinary Medicine. 6ed: p.553-566, 1997. SARDI, S.I.; CAMPOS, G.S.; BARROS, S.B.; EDELWEISS, L.G.; MARTINS, D.T. Detecção de anticorpos contra o vírus da parainfluenza bovina tipo 3 (pi-3) e o vírus

66
da leucose bovina (vlb) em bovinos de diferentes municípios do Estado da Bahia, Brazil. Rev. Ci. Med. Biol., Salvador, v.1, n.1, p. 61-65, 2002. SCHEFFER, J.F. Enzimologia clínica em medicina veterinária. Trabalho de conclusão de curso, Universidade Federal do Rio Grande do Sul – UFRGS, 21p., 2003. SCHWARTZ, I.; BENSAID, A.; POLACK, B.; PERRIN, B.; BERTHELEMY, M.; LEVY, D. (1999) In vivo Leucocyte Tropism of Bovine Leukemia Virus in Sheep and cattle. Journal of Virology, v.68, n. 7, p. 4589-4596, 1999. SHERMAN, D.M. Localized diseases of the bovine brain and spinal cord. Vet. Clin. North Am. Food Anim. Pract., v.3, p.179-191, 1987. SILVA, R.C.; FONTANA, I.; MEIRELLES, F.C.; RUGGIERO, A.P.M.; BENATO, N.; BORGES, J.R.J. Ocorrência de Leucose Enzoótica Bovina na forma de linfossarcomas no Distrito Federal: Relato de caso. Arq. Inst. Biol., São Paulo, v. 75, n.4, p. 507-512, 2008. SIMARD, C.; RICHARDSON, S.; DIXON, P.; BÉLANGER, C.; MAXWELL, P. Enzime-linked immunosorbent assay for diagnosis of bovine leucosis: comparison with the agar gel immunodiffusion test approved by the Canadian Food Inspection Agency. The Canadian Journal of Veterinary Research, v.67, n.2, p.101-106, 2000a. SIMARD, C.; RICHARDSON, S.; DIXON, P.; KOMAL, J.; Agar gel immunodiffusion test for the detection of bovine leukemia virus antibodies: lack of trans-Atlantic standardization. The Canadian Journal of Veterinary Research, v. 64, n. 2, p. 96-100, 2000b. SIMENSEN, M.G.; NANSEN, P. Serum gama glutamiltranspeptidase (GGT) and aspartate-aminotransferase (AST), activities in adult cattle with chronic Fasciola hepatica infection. Acta Veterinaria Scandianavia. v. 15, n. 2, p. 239-43, 1974. SIMÕES, C.G.C. Expressão imunoistoquímica de p53 e bcl-2 em casos de leucose enzoótica bovina. Dissertação (Mestrado em Ciências – Patologia Experimental e Comparada) – São Paulo – USP, 88p, 2007. SOUZA, R.M.; BIRGEL, E.H.; AYRES, M.C.C. Influência dos fatores raciais na função hepática de bovinos da raça holandesa e Jersey. Braz. J. Vet. Res. Anim. Sci. v. 41, n. 5, p. 306-312, 2004.

67
SOUZA, P.M. Perfil bioquímico sérico de bovinos das raças Gir, Holandesa e Girolanda, criados no estado de São Paulo – influência de fatores de variabilidade etários e sexuais. Tese (Doutorado em Clínica Veterinária) – São Paulo – SP., Universidade de São Paulo – USP, 168 p., 1997. SPONCHIADO, D; Prevalência de anticorpos séricos anti-Vírus da Leucose Enzoótica Bovina em rebanhos da raça Holandesa Preta e Branca, criados no Estado do Paraná. Dissertação (Mestrado em Ciências Veterinárias) UFPR – Curitiba, 101p, 2008 STONE, D.M.; NORTON, L.K.; DAVIS, W.C. Spontaneously proliferating lymphocytes from bovine leukaemia virus infected, lymphocytotic cattle are not the virus-expressing lymphocytes, as these cells are delayed in G0/G1 of the cell cycle and are spared from apoptosis. Journal of General Virology, v. 81, n.4, p. 971-981, 2000. SUMMERS, B.A.; CUMMINGS, J.F.; DE LAHUNTA, A. Veterinary neuropathology. Baltimore : Mosby, 527p, 1995. TAJIMA, S.; ZHUANG, W.Z.; KATO, M.V.; OKADA, K.; IKAWA, Y.; AIDA, Y. Function and Conformation of Wild-type p53 Protein are Influenced by Mutations in Bovine Leukemia Virus-Induced B-Cell Lymphosarcoma. Virology, v. 243, p. 235-246, 1998. TAVORA, J.P.F. Prevalência da Infecção pelo Vírus da Leucose Bovina em Rebanhos Leiteiros Criados na Região do Pólo Itabuna, Estado da Bahia. Tese (Mestrado em Patologia Bovina) – São Paulo-SP, Faculdade de Medicina Veterinária e Zootecnia, USP, 106p., 1990. VALLI, V.E.O. The hematopoietic system. In: JUBB, K.V.F., KENNEDY, P.C., PALMER, N. Pathology of domestic animals. 4. ed. San Diego : Academic, 653p. v.3, cap. 2, p.101-265, 1993. VAN DER LAAN, C.W.; VIDOR, T., BRAGA, F.M., HALFEN, D., HÜBNER, D.E.O. Leucose enzoótica bovina em bovinos produtores de leite importados do Uruguay. Pesquisa Agropecuária Gaúcha, v. 5, n. 1, p. 139-141, 1999. VIANA, E.S.; CAMPOS, J.M. Eletroforese do soro, proteína total e relação albumina/globulina (A/G) em bezerros com verminose gastrintestinal naturalmente adquirida. Arquivos da Escola de Veterinária da Universidade Federal de Minas Gerais. v. 25, n. 3, p. 279-85, 1973.

68
YAMAMOTO, H. Histopathological definition of the adult and calf types of bovine leukosis. Nati. Inst. Anim. Health. Tokyo, v. 22, n, 3, p. 115-129, 1982.