universidade estadual do cearÁ - uece.br · revisão de literatura figura 1. mapa do genoma do...
TRANSCRIPT

1
UNIVERSIDADE ESTADUAL DO CEARÁ
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
FACULDADE DE VETERINÁRIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS VETERINÁRIAS
TESE DE DOUTORADO
CARACTERIZAÇÃO MOLECULAR DE LENTIVÍRUS DE
PEQUENOS RUMINANTES ISOLADOS NO BRASIL
Doutoranda: Aryana Lushese Vasconcelos Lima Feitosa
Orientadora: Dra. Maria Fátima da Silva Teixeira
FORTALEZA – CE
2011

2
ARYANA LUSHESE VASCONCELOS LIMA FEITOSA
CARACTERIZAÇÃO MOLECULAR DE LENTIVÍRUS DE
PEQUENOS RUMINANTES ISOLADOS NO BRASIL
Tese apresentada ao Programa de Pós-Graduação
em Ciências Veterinária da Faculdade de Veterinárias da
Universidade Estadual do Ceará, como requisito parcial
para a obtenção do título de Doutor em Ciências
Veterinárias.
Área de Concentração: Reprodução e Sanidade de
Pequenos Ruminantes
Orientadora: Profa. Dra. Maria Fátima da Silva Teixeira
FORTALEZA – CEARÁ
2011

3
ARYANA LUSHESE VASCONCELOS LIMA FEITOSA
F311c Feitosa, Aryana Lushese Vasconcelos Lima Caracterização molecular de lentivírus de pequenos
ruminantes isolados no Brasil / Aryana Lushese Vasconcelos Lima Feitosa. – 2011.
175 f. : il. color., enc. ; 30 cm. Tese (Doutorado) – Universidade Estadual do Ceará,
Faculdade de Veterinária, Programa de Pós-Graduação em Ciências Veterinárias, Fortaleza, 2011.
Área de Concentração: Reprodução e Sanidade Animal. Orientação: Profª. Drª. Maria Fátima da Silva Teixeira . Co-orientação: Prof. Dr. Raymundo Rizaldo Pinheiro. 1. Lentivírus. 2. Filogenia. 3. gag. 4. pol. 5. CAEV. 6. MVV. . I.
Título.
CDD: 636.089

4
CARACTERIZAÇÃO MOLECULAR DE LENTIVÍRUS DE PEQUENOS
RUMINANTES ISOLADOS NO BRASIL
Tese apresentada ao Programa de Pós-Graduação
em Ciências Veterinária da Faculdade de Veterinária da
Universidade Estadual do Ceará, como requisito parcial
para a obtenção do título de Doutor em Ciências
Veterinárias.
Aprovada em: 05/12/2011
BANCA EXAMINADORA

5
LINHA DE PESQUISA
Reprodução e Sanidade de pequenos ruminantes
LOCAL DE EXECUÇÃO
1. Laboratório de Virologia (LABOVIR) do Programa de Pós-Graduação em
Ciências Veterinárias (PPGCV) – Faculdade de Veterinária (FAVET) –
Universidade Estadual do Ceará (UECE);
2. Empresa Brasileira de Pesquisa Agropecuária - Embrapa Caprinos e Ovinos –
Sobral-CE
3. Núcleo de Biotecnologia de Sobral – NUBIS em parceria com Universidade
Estadual Vale do Acaraú (UVA); Universidade Federal do Ceará – Faculdade de
Medicina (UFC) e Embrapa Caprinos e Ovinos.
4. Laboratório Aplicado à Biologia Molecular e Sorologia - LABMAS ––
Faculdade de Medicina Veterinária e Zootecnia – FMVZ – VPS – Universidade
de São Paulo – USP.
EQUIPE EXECUTORA
1. Profa. Dra. Maria Fátima da Silva Teixeira – Orientadora
PPGCV/FAVET/UECE, Bolsista de Produtividade em Pesquisa do CNPq;
2. Dr. Raymundo Rizaldo Pinheiro – Coorientador, Pesquisador Embrapa – CNPC;
3. Dra. Alice Andrioli – Colaboradora; Pesquisadora Embrapa – CNPC;
4. MSc. Aryana Lushese Vasconcelos Lima Feitosa – Doutoranda–PPGCV/UECE;
5. Dra. Lúcia Sider – Colaboradora; Pesquisadora Embrapa – CNPC;
6. Prof. Rodrigo Maranguape S. da Cunha – Pesquisador e Coordenador do
NUBIS;
7. Prof. Dr. João Paulo de Matos – Pesquisador da Universidade Federal do Rio
Grande do Norte – UFRN;
8. Prof. Dr. Paulo Eduardo Brandão – FMVZ-VPS-USP;

6
9. Dalva Alana Aragão de Azevedo - Bolsistas IC/Embrapa (Curso de Ciências
Biológicas - UEVA)
10. Bolsistas IC/CNPq (Curso de Medicina Veterinária – FAVET/UECE)

7
Aos meus pais,
Aos meus irmãos,
Ao meu noivo Leonardo E. Marinho,
Dedico.

8
“Nada em biologia faz sentido a não ser à luz da evolução”.
Theodosius Dobzhansky (1900- 1975)

9
AGRADECIMENTOS
À UECE e ao Programa de Pós-Graduação em Ciências Veterinárias que propiciou a
realização deste projeto.
À Embrapa e ao Centro Nacional de Pesquisa em Caprinos que disponibilizou as
condições técnicas e pessoal para a realização deste trabalho.
Ao Núcleo de Biotecnologia de Sobral, NUBIS-UEVA, que disponibilizou
infraestrutura, reagentes, equipamentos e pessoal.
Ao Laboratório Aplicado à Biologia Molecular e Sorologia - LABMAS –– Faculdade
de Medicina Veterinária e Zootecnia – FMVZ – VPS – Universidade de São Paulo –
USP, que muito gentilmente disponibilizou infraestrutura, reagentes, equipamentos e
pessoal.
À FUNCAP e ao Banco do Nordeste pelo auxílio financeiro que possibilitou a
realização deste trabalho.

10
AGRADECIMENTOS
A Deus pela força nos momentos difíceis.
Aos meus pais Jauneval e Irlene pelo amor, incentivo e exemplo de luta.
À Profa. Maria Fátima Teixeira pela orientação, confiança e exemplo profissional.
Ao Dr. Raymundo Rizaldo Pinheiro por ter me iniciado na pesquisa e pelos
ensinamentos e à Dra. Alice Andrioli que juntos me propiciaram incalculável apoio
neste trabalho.
Ao Professor Rodrigo Maranguape – NUBIS-UEVA, que disponibilizou infraestrutura,
reagentes e equipamentos, além de transmitir todos os seus conhecimentos de biologia
molecular. Ao Dr. João Paulo Mattos, por me ajudar com as sequências do capítulo III.
Ao Professor Paulo Eduardo Brandão – LABMAS-USP, que me acolheu de forma tão
generosa, disponibilizando infraestrutura, livre acesso a reagentes e equipamentos, pela
contribuição imensurável à realização deste trabalho. Ao Prof. Fábio Gregori, por sua
disponibilidade em ajudar, tirando dúvidas, dando sugestões, por auxiliar na edição das
sequências do capítulo IV, meu muito obrigada!
Aos colegas do LABOVIR: Tânia (agora orgulhosamente presente na banca como
Pesquisadora da Embrapa). Além Neilson, Valeska, Edmara, Dávila, Esmaile, Cinthia,
Gabrielle, Igor, Alfredo, Rosivaldo Júnior, Ronaldo, Carlos, Luís Antônio, pelo
convívio e cooperação na realização deste trabalho.
Aos colegas de laboratório da Embrapa: Dra. Lúcia Sider, por me disponibilizar
reagentes; Dalva Azevedo, sua ajuda foi essencial no cultivo, obrigada por me
acompanhar sempre que precisei. Samilly, Kelma, Osmarilda e Ricardo, pelo auxílio
nas horas precisas.

11
Às colegas do LABMAS: Haila, Karen, Carol, Cinthia Baldin, Cíntia Favero, Camila,
Sueli, Giselle, Katarina, obrigada pelas sugestões, pelo convívio, pelas horas de
descontração. Em especial à Dra. Alessandra Marnie, pela ajuda, sugestões e dicas
valiosas e à Sheila de Oliveira pelos sequenciamentos e pelas sugestões.
Aos meus irmãos por acreditarem em mim, pelo carinho.
Ao meu noivo Leonardo Ellery Marinho pelo amor, apoio e exemplo.
Aos meus sogros que me acolheram em Fortaleza, Helenira Ellery e João Batista
Marinho; à Francisca por cuidar de mim.
Aos meus amigos de Sobral, Isana Mara, Juracy e Bartolomeu e Daniele Val, por me
acolherem e pela estimável amizade.
A todos aqueles que, direta ou indiretamente, contribuíram para a realização deste
trabalho, os meus mais sinceros agradecimentos!

12
RESUMO
Os Lentivírus de Pequenos Ruminantes incluem os Vírus da Artrite Encefalite Caprina
(CAEV) e o Vírus Maedi-Visna (MVV), estes têm sido considerados geneticamente
distintos, mas antigenicamente relacionados como patógenos de caprinos e ovinos,
respectivamente. Além disso, foi demonstrado que estes vírus estão constantes e
facilmente transgredindo a barreira entre caprinos e ovinos, com CAEV infectando
ovinos e MVV infectando caprinos. Assim, como todos os lentivírus, o genoma do
CAEV tem uma alta taxa de mutação e o seu grau de heterogeneidade pode estar
relacionado com a baixa fidelidade da transcriptase reversa, que carece da atividade de
leitura da enzima exonuclease (proof reading). A análise genética de Lentivírus de
Pequenos Ruminantes (LVPRs) pode ajudar a compreender a genética, proteínas e
antigenicidade destes vírus; sua patogênese, epidemiologia, relações filogenéticas e,
assim, a sua inclusão nos subgrupos de LVPRs recentemente criados. Também pode ser
relevante para o desenvolvimento de testes de diagnósticos específicos às estirpes
locais. Neste sentido, os principais objetivos deste trabalho consistem em isolar cepas
virais circulantes e analisar a filogenia por meio de sequências gag e pol; bem como,
implementar uma biblioteca genômica de LVPRs isolados em alguns estados
brasileiros. Para isso, inicialmente, amostras sanguíneas de caprinos e ovinos foram
analisados utilizando a técnica de Imunodifusão em Gel de Agarose (IDGA),
confirmados por Western Blotting com consequente isolamento dos vírus em Mem
brana Sinovial Caprina (MSC) a partir de amostras positivas. Em seguida, foi realizada
Reação em Cadeia da Polimerase (Nested-PCR) para confirmação da presença de vírus,
assim como para a amplificação e sequenciamento das regiões de interesse. Também
foram realizadas comparações através de alinhamentos entre as regiões gag e pol das
cepas isoladas, além da montagem da biblioteca genômica de LVPRs isolados. Foram
isoladas cepas virais circulantes de animais naturalmente infectados nos estados do Rio
Grande do Norte, Ceará, Piauí, Bahia e Minas Gerais. A cepa isolada, denominada
BrRN-CNPC.G1 foi considerada o primeiro isolamento do CAEV no Estado do Rio
Grande do Norte. Todas as cepas isoladas mostraram-se ser fenotipicamente líticas, do
tipo rápido/alto. Foram amplificadas e sequenciadas cepas virais isoladas do Ceará e
Minas Gerais e Rio Grande do Norte, tanto para o gene gag quanto para o gene pol. Pela
genealogia proposta com base nas sequências nucleotídicas dos genes gag e pol, a sua

13
topologia mostrou-se compatível com os genótipos do subtipo B de CAEV. O
alinhamento das sequências de aminoácidos da região gag indicaram divergências entre
as linhagens deste estudo com a cepa padrão CAEV-Cork. Essa divergência de
aminoácidos foi ainda mais acentuada quando comparas sequências de aminoácidos do
gene pol..
Palavras-chave: Lentivírus, Filogenia, gag, pol, CAEV, MVV.

14
ABSTRACT
CAEV and MVV have been considered genetically distinct but antigenically related as
pathogens of goats and sheep, respectively. Moreover, it was demonstrated that these
viruses are constantly and easily breaking the barrier between sheep and goats, sheep
being infected with CAEV and MVV infecting goats. Like all lentiviruses, the genome
of CAEV has a high mutation rate and degree of heterogeneity that may be related to the
low fidelity of reverse transcriptase, which lacks proof reading activity of the
exonuclease subunit. Genetic analysis of Small Ruminants Lentiviruses (SRLVs) may
help to understand the genetics, the proteins and the antigenicity of these viruses, their
pathogenesis, epidemiology, phylogenetic relationships and thus their inclusion in the
newly created subgroups of SRLVs. It may also be relevant to the development of
diagnostic tests specific to the local strains. In this sense, the aim of this work was to
isolate circulating viral strains and to study the phylogeny with gag and pol sequences,
as well as to implement a genomic library of SRLVs isolated in some Brazilian states.
Initially, goats and sheep were screened using the AGID technique, confirmed by
Western blotting with subsequent isolation of virus in goat synovial membrane (GSM)
from positive samples. Nested-PCR was performed to confirm the presence of the virus
and for amplification of regions of interest, followed by sequencing of individual
samples. Comparisons were made through alignments of the gag and pol regions of the
strains, besides assembling the genomic library of isolated SRLVs. The results were the
isolation of circulating virus strains of naturally infected animals at Rio Grande do
Norte, Ceará, Piauí, Bahia and Minas Gerais states. The strain isolate, called BrRN-
CNPC.G1 were considered the first isolation of small ruminant lentiviruses
from naturally infected goat in flock of Rio Grande do Norte, Brazil. The
strains demonstrated phenotypically litic, type rapid/high. In the present
work, virus strains isolated from Ceará, Minas Gerais and Rio Grande do
Norte were amplified and sequenced for gag and pol genes. The genealogy
proposed for gag and pol gene was compatible with subtypes B from
CAEV. The alignment of amino acid sequences from gag gene was
divergent between the strains this study and the standart strain CAEV-
Cork. The amino acids were more divergent in sequences of the pol gene.
Key-words: Lentiviruses, Phylogeny, gag, pol, CAEV, MVV

15
LISTA DE FIGURAS
Revisão de Literatura
Figura 1. Mapa do genoma do lentivírus.
Figura 2. Um esboço do ciclo infeccioso de retrovírus. Painel A: passos da primeira fase
do ciclo de replicação. Fixação do vírus ao se ligar aos receptores de membrana
presentes nas células hospedeiras suscetíveis ocorre através das proteínas virais Env.
Esta interação leva a alterações conformacionais nas proteínas Env que facilitam a
fusão das membranas celular e viral, entregando o capsídeo viral para o
citoplasma. Painel B: A transcrição reversa ocorre no citoplasma e produz
uma molécula de DNA de fita dupla que se integra no genoma do hospedeiro, gerando
um provírus. Últimos estágios do ciclo de vida incluem a expressão do RNA viral a
partir do provírus. Alguns desses RNAs são emendados e exportados para o
citoplasma juntamente com RNAs unspliced. O RNA unspliced serve tanto
como RNA genômico como para a síntese das poliproteínas Gag e Gag-Pol.
Formas unidas são usadas para fazer proteínas de Env e complexo de
proteínas reguladoras do retrovírus. Painel C: Montagem das proteínas
virais e encapsidação dos RNAs genômicos são representados, levando à formação da
progênie viral. Adaptada do Livro Viral Genome Replication (2009).
Figura 3. DNA províral resultante da retrotranscrição.

16
Capítulo II
Figura 1. Isolamento a partir de cocultivo de monócitos/macrófagos em MSC
apresentando efeito citopático característico do CAEV com presença de células gigantes
multinucleadas.
Capítulo III
Figura 1. In the phylogenetic tree a group identified by the prototype VISNA was
clearly distinguished from the CAEVs with a high bootstrap value of 100.
Figura 2. Alignment of nucleotide sequences from the gag region of SRLV strains.
Asterisks indicate identity among the strains.
Figura 3. Alignment of deduced amino acid sequences from the capsid protein of SRLV
strains. Asterisks indicate identity between the strains.
Capítulo IV
Figura 1. Eletroferograma mostrando a sobreposição de bases nas cepas 229CE e
Pan12CE para o gene gag.
Figura 2. Alinhamento do fragmento parcial de 620 nucleotídeos do gene codificador da
proteína p28 do capsídeo gerados no presente estudo comparadas com sequências
padrões depositadas no Genbank.
Figura 3. Alinhamento do fragmento parcial de 472 nucleotídeos do gene gerados no
presente estudo comparadas com sequências padrões depositadas no Genbank.

17
Figura 4. Árvore filogenética diferenciando os grupos de CAEV e MVV com alto valor
de bootstrap.
Figura 5. Alinhamento do fragmento parcial de 472 nucleotídeos do gene pol gerados
no presente estudo comparadas com sequências padrões depositadas no Genbank.
Figura 6. Alinhamento das sequências de aminoácidos do gene pol correspondente a
uma região intergência de dUTPase e integrase.
Figura 7. Árvore filogenética do gene pol diferenciando os grupos de CAEV e MVV
com alto valor de bootstrap.

18
LISTA DE TABELAS
Capítulo II
Tabela 1 - Sequências de primers utilizados nas reações de Nested-PCR e seus
respectivos tamanhos de fragmentos amplificados.
Tabela 2. Correlação: dia do experimento, coleta de sobrenadante e resultado de Nested-
PCR.
Capítulo IV
Tabela 1 - Sequências de primers utilizados nas reações de PCR e seus respectivos
tamanhos de fragmentos amplificados.
Tabela 2 - Sequências e nucleotídeos analisados neste estudo.
Tabela 3 – Relação da origem das amostras e tamanho do fragmento dos respectivos
genes sequenciados.
Tabela 4. Matriz de identidade com os valores de similaridade nucleotídica do
fragmento parcial do gene gag comparadas com as demais cepas depositadas do
Genbank.
Tabela 5. Matriz de identidade com os valores de similaridade nucleotídica do
fragmento parcial do gene pol comparadas com as demais cepas depositadas do
Genbank.

19
LISTA DE ABREVIATURAS E SIGLAS
% - Porcentagem
3'OH – hidroxila na porção 3´do DNA
A25 – garrafa de cultivo celular com capacidade de 25 cm3
AGID – sigla em inglês para Imunodifusão em gel de agarose
AIE – Anemia infecciosa equina
AP-1 - Ativador de proteína-1
AP2 – Ativador de proteína-1
BDV – Borna Disease Virus
BIV – Vírus da imunodeficiência bovina
bp – sigla do inglês para pares de bases
BR/CNPC – BR, Brasil; CNPC, Centro Nacional de Pesquisa em Caprinos
BR/MG – Cepa brasileira de LVPRs isolada em Minas Gerais
G - goat
BrRN-CNPC.G1 – BR, Brasil; RN, Rio Grande do Norte; CNPC, Centro Nacional de
Pesquisa em Caprinos
CA – Proteína do Capsídeo
CAE – Artrite Encefalite Caprina
CAEV – Vírus da Artrite Encefalite Caprina
CAEV-Cork – Cepa padrão de CAEV
CE – Ceará
c-fos – Fator celular presente nos macrófagos que participa da ativação viral
c-jun – Fator celular presente nos macrófagos que participa da ativação viral
cELISA – ELISA competitivo
cm2 –
centímetro quadrado
CNPC – Centro Nacional de Pesquisa em Caprinos/ Embrapa Caprinos e Ovinos
CO – Monóxido de Carbono
CO2 – Dióxido de Carbono
CPE – sigla do inglês para Efeito Citopático Característico
DNA – Ácido desoxirribonucléico
dNTPs – Desoxinucleotídeo trifosfato
dUTPase - Enzima codificada pelo gene pol

20
e - ativador de expressão
ECP – Efeito citopático
EDTA – Ethylenediaminetetraacetic acid
EIAV – sigla do inglês para anemia infecciosa equina
ELISA – Enzyme-linked immunosorbent assay
env - Gene que codifica as proteínas do envelope viral
ENV - proteínas do envelope viral
EUA – Estados Unidos da América
EV-1 – cepa britânica do vírus MV
FAVET – Faculdade de Veterinária da UECE
FBS – sigla do inglês para soro fetal bovino
Fig – Figura
FIV – Vírus da Imunodeficiência Felina
FMVZ – Faculdade de Medicina Veterinária e Zootecnia da USP
Fos - produto de c-fos
FUNCAP – Fundação Cearense de Apoio à Pesquisa
g – Unidade da força centrífuga relativa
gag - Gene viral que codifica as proteínas interna dos vírus
GAG – proteínas interna dos vírus
GenBank – Banco de dados genéticos
gp – Glicoproteína
GSM – Membrana Sinovial Caprina
HCl – Ácido Clorídrico
HIV – Vírus da Imunodeficiência Humana
IDGA – Imunodifusão em gel de Agar
IgG – Imunoglobulina G
K1514 – cepa Islandesa do vírus MV
kb – kilobases
KCl – Cloreto de Potássio
kDa – Kilodaltons
KV1772 – cepa Islandesa do vírus MV
LABMAS – Laboratório Aplicado à Biologia Molecular e Sorologia - USP
LABOVIR - Laboratório de Virologia - UECE
LSP - Leucócitos do sangue periférico

21
LTR – Seqüências longas repetidas
LVPRs – Lentivírus de pequenos ruminantes
M – molar
MA – Proteína da Matriz
MEM – Meio Essencial Mínimo
mg – miligramas
Mg2+
– Magnésio
MHC I ou II – Complexo Principal de Histocompatibilidade
min – minutos
mL - Mililitros
mM – mili-molar
MSC – Membrana sinovial caprina
MV – Maedi-visna
MVV – Vírus Maedi-Visna
NC – Proteína do nucleocapsídeo
ng – nanograma
NJ - neighbor joining
nm – nanômetro
nt – nucleotídeo
NUBIS – Núcleo de Biotecnologia de Sobral
ºC – Graus Celsius
OIE – Organização Internacional de Epizootias
ORFs – Pequena região do genoma viral
P1OLV – cepa portuguesa do vírus MV
p28 – protein do capsídeo
pb – Pares de bases
PBL – sigla do inglês para Leucócitos do sangue periférico
PBS – Solução de tampão de fosfato
PCR – Reação em cadeia da Polimerase
pH – potencial hidrogeniônico
pmol – picomol
pol - Gene que codifica as enzimas virais
POL – proteína codificada a partir do gene pol
PPGCV – Programa de Pós-graduação em Ciências Veterinárias

22
PPO – Pneumonia Progressiva Ovina
RNA – Ácido ribonucléico
RNAm – Ácido Rribonucléico mensageiro
rpm – rotação por minuto
SA-OMVV – cepa Sul-Africana do vírus MV
SIV – Vírus da imunodeficiência símia
SMF - sistema monocítico fagocitário
SNC – Sistema Nervoso Central
SRLVs – sigla em inglês para Lentivírus de pequenos ruminantes
SU – Glicoproteína de superfície
Taq – DNA Taq Polimerase
TBE - Tris-Borato; 1 mM EDTA
TE - Tris 10 mM/ EDTA 1 mM pH 8,0
TM – proteína transmembranica
Tris – Trisma
U – Unidade
U3 – upstrem 3´
U5 – upstrem 5´
UECE – Universidade Estadual do Ceará
UFC – Universidade Federal do Ceará
USA – United States of America
USP – Universidade de São Paulo
UVA – Universidade Estadual Vale do Acarú
vif - Gene de regulação viral
VISNA – Doença causada pelo vírus Maedi-visna
vpr –gene envolvido na regulação da síntese e processamento do RNA viral e outras
funções na replicação
WB - Western Blotting
ZP – Zona pelúcida
L – microlitro
M – micromol
- sinal de encapsidação

23
SUMÁRIO
1 INTRODUÇÃO.......................................................................... 25
2 REVISÃO DE LITERATURA................................................. 27
2.1 Histórico....................................................................................... 27
2.2 Taxonomia dos Retrovírus........................................................... 28
2.3 Agente Etiológico e Classificação Viral..................................... 28
2.4 Estrutura e Genoma dos vírus...................................................... 29
2.5 Replicação e Transcrição............................................................. 31
2.6 Heterogeneidade Genética........................................................... 34
2.7 Tropismo celular.......................................................................... 39
2.8 Epidemiologia e Transmissão...................................................... 43
2.9 Sinais Clínicos............................................................................. 46
2.10 Diagnóstico clínico...................................................................... 46
2.11 Diagnóstico laboratorial............................................................... 46
2.12 Ensaio imunoenzimático (ELISA) indireto................................ 47
2.13 Isolamento viral............................................................................ 47
2.14 Reação em cadeia de polimerase (PCR) ..................................... 48
2.15 Imunodifusão em gel de agarose (IDGA) ................................... 49
2.16 Western blotting ou imunoblotting.............................................. 50
2.17 Prevenção e controle.................................................................... 50
3 JUSTIFICATIVA....................................................................... 52
4 HIPÓTESES CIENTÍFICAS.................................................... 53
5 OBJETIVOS............................................................................... 54
5.1 Objetivos Gerais........................................................................... 54
5.2 Objetivos Específicos................................................................... 54
6 CAPÍTULO I.............................................................................. 55

24
7 CAPÍTULO II............................................................................ 69
8 CAPÍTULO III........................................................................... 83
9 CAPÍTULO IV........................................................................... 101
10 CONCLUSÕES GERAIS.......................................................... 138
11 PERSPECTIVAS....................................................................... 140
12 REFERÊNCIAS BIBLIOGRÁFICAS..................................... 141
13 ANEXOS..................................................................................... 162

25
INTRODUÇÃO
Os Lentivírus de Pequenos Ruminantes (LVPRs) estão amplamente distribuídos
entre caprinos e ovinos no mundo, causando grandes perdas econômicas. Diferentes
formas clínicas da doença têm sido descritas em pequenos ruminantes infectados com
LVPRs no Brasil, como artrite, pneumonia, mastite infectando caprinos e também
recentemente a forma nervosa da doença em ovinos (BENAVIDES et al., 2006). O Vírus
da Artrite Encefalite Caprina (CAEV) e o Vírus Maedi-Visna (MVV) têm sido
considerados como patógenos geneticamente distintos, mas antigenicamente
relacionados entre si. Estes vírus estão constantes e facilmente transpondo a barreira
entre espécies de caprinos e ovinos (VALAS et al., 2000; KARR et al., 1996; SHAH et al.,
2004b).
Em meados dos anos 90, as primeiras sequências genômicas tornaram-se
disponíveis. LVPRs de regiões geográficas distintas foram agrupados de acordo com as
espécies que foram isoladas. Dois grupos distintos, mas taxonomicamente relacionados
mostraram-se evidentes, um grupo evoluindo a partir de sequências de ovinos e os
outros evoluindo a partir sequências caprinas. A partir desse período, como mais
sequências de LVPRs foram caracterizadas, uma distribuição mais complexa
tornou-se evidentes sobre reconstruções filogenéticas, sugerindo que não os LVPRs
não podiam mais ser agrupados exclusivamente com base nas espécies que foram
obtidas. Numerosos estudos filogenéticos subsequentes trouxeram evidências adicionais
desde fato. Essa descoberta modificou a percepção sobre a relação entre os LVPRs e
seus hospedeiros e levou ao conceito de transmissão cruzada entre
espécies. Atualmente, MVV e CAEV não são mais vistos como duas espécies virais
infectantes distintas exclusivamente de ovinos e caprinos, respectivamente, e sim
como quasispecies virais que podem ser transmitidos entre pequenos
ruminantes sob condições favoráveis (LʼHOMME, et al., 2011).
Dados recentes baseados em estudos filogenéticos ajudaram a classificar os
LVPRs de acordo com seus hospedeiros (CASTRO et al., 1999; LEROUX et al., 1997;
CHEBLOUNE et al., 1996). Estes estão classificados dentro de cinco grupos relativamente
equidistantes (SHAH et al., 2004a). A alta heterogeneidade das sequências de
nucleotídeos e aminoácidos em lentivírus pode determinar a sua antigenicidade,

26
virulência e crescimento e pode ainda afetar sua persistência e o seu mecanismo de
evasão.
A alta heterogeneidade das sequências de nucleotídeos e aminoácidos em
lentivírus pode determinar a sua antigenicidade, virulência e crescimento e pode ainda
afetar sua persistência e o seu mecanismo de evasão.
A caracterização molecular desses LVPRs pode ajudar a entender suas relações
genéticas, protéicas e antigênicas, sua patogênese, epidemiologia, relações filogenéticas
e sua inclusão nos grupos recentemente estabelecidos (SHAH et al., 2004a). Isto também
pode ser relevante para o desenvolvimento de testes de diagnósticos mais sensíveis e
específicos.
A análise filogenética de sequências de CAEV e MVV e consequentemente a
aquisição do conhecimento sobre a heterogeneidade genética dos LVPRs é de imensa
importância para a análise epidemiológica de infecções por lentivírus de pequenos
ruminantes e também para estudar as diferenças de virulência entre linhagens de LVPRs
circulantes.

27
REVISÃO DE LITERATURA
Histórico
A síndrome Artrite-encefalite caprina (CAE) foi inicialmente descrita nos
Estados Unidos da América, sob a forma clínica de leucoencefalomielite em cabritos e o
vírus foi isolado em 1980, por CRAWFORD e colaboradores em explants de membrana
sinovial de um caprino adulto com artrite.
A Maedi-Visna (MV) ou a equivalente norte-americana Pneumonia Progressiva
Ovina (PPO) foi uma das primeiras doenças denominadas como infecção lenta
(SIGURDSSON, 1954). Os dois nomes são de origem islandesa: Maedi, que significa
dispnéia e é caracterizada por pneumonia intersticial progressiva, e Visna, que significa
“desorientação”, caracterizada por leucoencefalomielite (DAWSON, 1980). Inicialmente,
Visna/Maedi foi reconhecida como duas entidades distintas.
O primeiro isolado de MVV foi feito por SIGURDSSON et al. (1960) a partir do
Sistema Nervoso Central (SNC) de ovinos. Quando as primeiras amostras dos vírus
foram isoladas de ovinos infectados com Visna e Maedi (SIGURDARDÓTTIR E THOMAR,
1964), foi observada similaridade entre esses vírus. Posteriormente estudos revelaram
que tanto Maedi quanto Visna eram doenças causadas pelo mesmo agente, originando
assim a denominação Maedi-Visna ou MVV. Os vírus foram introduzidos através da
importação, de ovinos Karakul da Alemanha em 1933, para melhorar rebanhos da
Islândia, mas esta raça também introduziu a adenomatose pulmonar, paratuberculose e
visna.
Os primeiros relatos da doença com sinais clínicos semelhantes à Maedi-Visna
ocorreram na África do Sul em 1915 e, posteriormente, nos Estados Unidos em 1923.
A doença foi observada, mais tarde, em outras regiões do mundo, sendo a síndrome
respiratória de maior prevalência. Na Holanda esta enfermidade recebeu o nome de
Zwoegerziekte (DE BÔER, 1975; HOUWERS, 1985), Na África do Sul, recebeu o nome de
Jaagsiekte (VERWOERD et al., 1983) e nos Estados Unidos, de Pneunonia Progressiva
Ovina (PPO) e Doença Pulmonar da Montanha (CUTLIP E LAIRD, 1976).

28
Taxonomia dos Retrovírus
A família Retroviridae inclui a subfamília Orthoretrovirinae a qual contém o
gênero Lentivirus. Deste fazem parte as nove espécies: Vírus da artrite encefalite
caprina (CAEV), Vírus Maedi-Visna (MVV), Vírus da Anemia Infecciosa
Equina (AIE), Vírus da imunodeficiência felina (FIV), Vírus da imonodeficiência
humana (HIV tipo-1), Vírus da imonodeficiência humana (HIV tipo-2), Lentivírus de
Puma, Vírus da Imunodeficiência Símia (SIV) e Vírus imunodeficiência bovina (BIV)
(ICTV, 2011).
Agente Etiológico e Classificação Viral
O vírus Maedi-visna (MVV) e o vírus da artrite encefalite caprina (CAEV)
afetam ovinos e caprinos, respectivamente, pertencem à família Retroviridae e ao
gênero Lentivirus (PASICK, 1998). São vírus complexos e não oncogênicos. Os
lentivírus geralmente limitam a infecção a um simples hospedeiro, levando a sérios
problemas ou à morte de caprinos jovens e de ovinos; multiplicam-se também em
células em repouso (não ativadas); infectam monócitos, macrófagos e/ou linfócitos,
causando infecção persistente e multissistêmica (AIEV, MVV e CAEV), associada à
síndrome de imunodeficiência (SIV, HIV e FIV); com altas taxas de mutação, e
consequente diversidade genotípica, fenotípica e antigênica (CLEMENTS E PAYNE, 1994;
GONDA, 1994; REISCHAK, 2000; PINHEIRO, 2001).
Os LVPRs podem ser classificados em cinco grupos filogenéticos: O genótipo
A, que compreende o grupo Maedi-visna (SHAH et al., 2004), este grupo pode ser
subdividido em nove subtipos A1-A9, onde o subtipo A1 é identificado por MVV por
ser genética e geograficamente heterogêneos. Recentemente, foram encontrados dois
novos subtipos do Grupo A, A8 (isolado de caprino) e A9 (isolado tanto de caprino
quanto de ovino), na Itália (GREGO et al., 2007).
O genótipo B, inluindo CAEV como isolados (PISONI et al., 2005), compreende
apenas dois subtipos distintos, B1 e B2, que estão associados principalmente com artrite
em caprinos (PISONI et al., 2005) e ovinos (GLARIA et al., 2009; ROSATI et al., 2004),
respectivamente.

29
O genótipo C, afeta pequenos ruminantes norueguêses (GJERSET et al., 2007). O
genótipo D foi caracterizado através do gene pol em isolados suíços e espanhóis (REINA
et al., 2006; SHAH et al., 2004) e o genótipo E inclui os isolados caprinos italianos
(GREGO et al., 2007; REINA et al., 2009a). O último grupo foi identificado pela primeira
vez em rebanhos assintomático da raça local Roccaverano, daí o nome dado à cepa. Até
o momento o genótipo E compreende dois subtipos: E1, cepa Roccaverano, descrita
como de baixa patogenicidade por não levar à sintomas clínicos em caprinos (REINA et
al., 2011) e E2, cepa Seui.
Estrutura e Genoma dos vírus
MVV e CAEV, como os demais lentivírus apresentam-se como vírions
envelopados, com diâmetro de 80 a 100 nm, núcleo cônico e denso, contendo duas
moléculas idênticas de RNA fita simples, não complementares, unidas por pontes de
hidrogênio, de polaridade positiva. Neste caso, o RNAm serve como molde para a
transcriptase reversa. O genoma tem cerca de 9.3 kb, uma molécula de transcriptase
reversa dependente de Mg2+
e proteínas do nucleocapsídeo (GONDA et al., 1986). O
envelope está associado covalentemente com glicoproteínas transmembranárias (TM) e
de superfície (SU). A matriz (MA) também é outra estrutura presente na partícula viral e
esta está situada entre o capsídeo e o envelope (PÉPIN et al., 1998). São pleomórficos,
esferóides e têm um coeficiente de sedimentação de 1,14 a 1,18 g/mL (CLEMENTS et al.,
1980), apresentando uma grande quantidade de ácido N-acetilneurâminico (ácido
siálico) em sua superfície (HUSO et al., 1988) (Figura 1).
O genoma é constituído por genes que codificam as proteínas estruturais (gag),
um precursor que é posteriormente clivado em três proteínas principais: matriz (MA),
capsídeo (CA) e nucleocapsídeo (NC) (PÉPIN et al., 1998), e o gene (env), que codifica
as glicoproteínas de superfície (SU) e transmembranária (TM) do envelope viral. O
gene (pol) que codifica as proteínas de atividade enzimática: transcriptase reversa,
protease, integrase e dUTPase, além de pequenos ORFs, que codificam proteínas que
regulam a expressão gênica como os genes acessórios (tat, vif), (ou Q, no HIV) e (rev),
codificantes de proteínas de regulação da expressão viral (CLEMENTS E PAYNE 1994).

30
Estas proteínas não se encontram nos vírus e somente são traduzidas durante a
replicação viral, na qual participam ativamente.
Figura 1. Estrutura dos Vírus da Artrite Encefalite Caprina (Coffin, 1996).
Adaptado por Rizaldo (2001).
Os tamanhos das proteínas codificadas são: MA ~17kDa; CA~24 kDa; NC ~7-
11 kDa; PR ~14 kDa; RT ~66 kDa; DU (em todos, exceto nos lentivírus de primatas),
IN ~32 kDa; SU ~120 kDa; TM ~41 kDa. Além dos genes estruturais gag, pro, pol, env,
existem genes adicionais, dependendo do vírus (p. e., vírus da imunodeficiência
humana 1, HIV-1): vif, vpr, vpu, tat, rev, nef em que os produtos estão envolvidos na
regulação da síntese e processamento do RNA viral e outras funções na replicação. A
LTR tem cerca de 600 nt de comprimento, dos quais 450 nt compõem a região U3, 100
nt na região R e aproximadamente 80 nt na região U5. Estes vírus estão associados com
uma variedade de doenças, incluindo imunodeficiencias, desordens neurológicas,
artrites entre outros (FAUQUET et al., 2005).

31
Os genes gag e pol são os mais conservados, enquanto o gene env é altamente
heterogêneo (NARAYAN E CLEMENTS, 1989) e são típicos da família Retroviridae. Estes
genes, únicos de lentivírus, apresentam um nucleotídeo ou aminoácido homólogo
diferenciando as várias lentiviroses, no entanto, suas funções são preservadas
(CLEMENTS E PAYNE, 1994).
Replicação e Transcrição
O ciclo de replicação dos retrovírus é constituído por uma série de passos que,
após transferência de informação genética do RNA para as moléculas de DNA, leva ao
estabelecimento de uma infecção persistente após a integração do DNA proviral nas
células do hospedeiro (Figura 2). Para iniciar a infecção, os retrovírus interagem com
receptores específicos na superfície das células-alvo por meio das proteínas do envelope
presentes do lado de fora da membrana viral.
Após a ligação inicial, as proteínas de envelope estão sujeitas à mudanças
conformacionais que levam à fusão viral com as membranas celulares e posterior
liberação do núcleo viral para o citoplasma.
Os eventos iniciais após a penetração são, até à data, pouco compreendidos.
Enquanto em alguns casos, propõe-se que o desencapsulamento do núcleo viral ocorre
após a internalização, existem outras indicações sugerindo que este capsídeo pode
permanecer intacto, permitindo que a transcrição reversa ocorra neste ambiente
fechado. Após a penetração do capsídeo viral no citoplasma da célula-alvo, a transcrição
reversa começa e segue para a geração de uma fita dupla de DNA a partir do genoma
RNA de fita simples. Este passo característico tem dado o nome a esta família de vírus.
A molécula de DNA deve então entrar no núcleo, a fim de integrar no genoma
do hospedeiro, dando origem a uma "provírus", que será permanentemente estabelecido
no genoma do hospedeiro. A fase final do ciclo de replicação, que geralmente depende
do maquinário celular, ocorre após esta etapa de integração. O RNA viral é expresso a
partir de seu promotor, localizado na região U3, e a regulação da transcrição é
controlado por fatores virais, bem como, fatores de transcrição do hospedeiro.

32
Durante a fase inicial da expressão do DNA proviral, a transcrição é processada
para dar origem a uma série de forma sub-genômica que serão usadas para
tradução. Mais tarde o RNA genômico será empacotado em partículas nascentes virais.
Figura 2 adaptada do Livro Viral Genome Replication (2009). Um esboço
do ciclo infeccioso de retrovírus. Painel A: Primeiros passos da primeira fase do ciclo
de replicação. Fixação do vírus ao se ligar aos receptores de membrana presentes nas
células hospedeiras suscetíveis ocorre através das proteínas virais Env. Esta
interação leva a alterações conformacionais nas proteínas Env que facilitam a fusão das
membranas celular e viral, entregando o capsídeo viral para o citoplasma. Painel B: A
transcrição reversa ocorre no citoplasma e produz uma molécula de DNA de fita
dupla que se integra no genoma do hospedeiro, gerando um provírus. Últimos
estágios do ciclo de vida incluem a expressão do RNA viral a partir do provírus. Alguns
desses RNAs são emendados e exportados para o citoplasma juntamente com RNAs não
processado. O RNA ainda não processado serve tanto como RNA genômico como para
a síntese das poliproteínas Gag e Gag-Pol. Formas unidas são usadas para fazer
proteínas de Env e complexo de proteínas reguladoras do retrovírus.
Painel C: Montagem das proteínas virais e encapsidação dos RNAs genômicos são
representados, levando à formação da progênie viral.

33
Uma vez que o produto dupla-fita final da transcrição reversa é gerado, o DNA
pré-proviral deve ser integrado ao genoma da célula infectada, um processo mediado
pela integrase retroviral. Para que o DNA pré-proviral possa ter acesso aos
cromossomos celulares, certos retrovírus requerem a desintegração da membrana
nuclear
ou progressão do ciclo celular de suas células-alvo. No caso dos lentivírus, os
complexos pré-integração podem entrar no núcleo das células em não divisão através do
transporte ativo do DNA viral pela membrana nuclear intacta, provavelmente mediada
por fatores celulares e virais. Ambas as extremidades das moléculas de DNA geradas
após transcrição reversa são constituídas pela LTRs, que contêm as sequências
reconhecidas pela integrase. A integração ocorre em duas etapas: numa primeira
instância, a remoção de dois nucleotídeos a partir de extremidades 3' do DNA pré-
proviral ocorre, enquanto na segunda etapa, os grupamento 3'OH
ataca o DNA-alvo e, através de um processo de transferência de cadeia, essas
são ligadas ao DNA do hospedeiro. Este processo gera uma duplicação de sequências
curtas que flanqueiam o DNA proviral, cujo tamanho dependerá da distância em que o
ataque foi feito pela integrase no DNA do hospedeiro. Esta distância varia de 4 a
6 nt dependendo do retrovírus. A integração no DNA da célula infectada não ocorre em
sequências específicas. No entanto, as preferências para a integração em regiões
ativamente transcritas foram relatadas (SCHRODER et al., 2002; WU et al., 2003). Vírus
diferentes marcam preferências por diferentes sequências (MITCHELL et al., 2004).
Após a integração há formação do provírus; síntese do RNA viral pela RNA
polimerase II celular, utilizando o provírus como molde; transcrição do genoma em
RNA-mensageiros (RNAm); síntese das proteínas virais; montagem; construção do
capsídeo e brotamento do vírus (COFFIN, 1996; PERTURSON et al., 1992).
O provírus, uma vez integrado, é estável, não existindo evidência de qualquer
mecanismo para remoção dos provírus do DNA celular, transposição direta dele para
outro sítio ou replicação independente (COFFIN, 1996). A integração do DNA proviral
ao DNA celular não é essencial à replicação dos lentivírus, mas é fundamental para a
persistência da infecção (HAASE, 1986).
A transcrição e o genoma viral iniciam um papel crucial no ciclo de vida dos
vírus, como prover o molde para a síntese de proteínas estruturais e regulatórias,

34
resultando na geração de novas cópias de RNA viral. O DNA do provírus resultante da
retrotranscrição apresenta no centro do genoma viral, as sequências trans (GAG, ENV e
POL) que codificam as proteínas estruturais e enzimas. Essas sequências são
enquadradas por duas sequências cis, necessárias para sua expressão, como o enhancer
(e, ativador de expressão) e ( sinal de encapsidação) (Figura 3). A repetição das
sequências de transcrição forma as repetições terminais longas (LTR, na sigla em
inglês), necessárias à integração do provírus ao genoma da célula (PAGÈS E PIVER,
2008).
Figura 3. DNA províral resultante da retrotranscrição. (Revista American
Science, 2008).
A transcrição é regulada pela ligação de várias proteínas celulares à sequência de
DNA. AP-1, que é um ativador de proteína, atua com um dos fatores de transcrição,
sendo este um produto de c-jun, que interage com Fos (produto de c-fos) para formar
um dímero. Apresentam dois domínios funcionais, um responsável pela ligação ao DNA
e um para ligação à Fos, e inclui o zíper de leucina. AP2 interage com elementos virais
e celulares estimulando a transcrição de genes selecionados. Vários estudos têm
demonstrado que AP-1 e AP-4 ligam-se a sítios, que estão localizados na região
promotora U3 da LTR, tendo um papel central na regulação e transcrição viral (BARROS
et al., 2004, 2005; CAMPBELL E AVERY, 1996; GABUZDA et al., 1989; GDOVIN E
CLEMENTS, 1992; HESS et al., 1986; SUTTON et al., 1997).
Heterogeneidade Genética
A enorme diversidade dos lentivírus é devida às altas taxas de mutação, de 0,5 a
1 genoma por ciclo de replicação, ou seja, por dia cerca de 10 bilhões de partículas

35
virais são produzidas in vivo, todas essas partículas têm pelo menos uma mutação em
seu código genético, que quando são acumuladas, auxiliam à rápida evolução desse
grupo de vírus (CASTRO, 1998) e são atribuídas ao fato de que esses vírus de RNA em
geral, especialmente os lentivírus, têm altas taxas de erro provocadas pela RNA
polimerase e pela falta do mecanismo de correção de bases (STEINHAUER E HOLLAND,
1987). Alguns autores relataram que estas mutações decorrem, geralmente, de erros nas
polimerizações, durante o ciclo de replicação dos lentivírus com substituições de bases,
deslocamento de janelas, rearranjo genético, recombinação e hipermutações (COFFIN,
1996; PRESTON E DOUGHERTY, 1996).
A transcriptase reversa é a principal responsável por erros e, ao contrário das
polimerases celulares, apresenta baixa fidelidade, devido à ausência da ação da enzima
3' - 5' exonuclease, necessária às correções de erros surgidos durante a polimerização.
Porém algumas mutações podem ser causadas durante a replicação do DNA proviral,
por erros da DNA polimerase celular, e da RNA polimerase celular II, quando da
síntese de RNA a partir do DNA proviral (PRESTON E DOUGHERTY, 1996).
Essa alta variabilidade genética dos lentivírus (CLEMENTS e ZINK,1996; LEROUX
et al.,1997), exemplificada pela variação observada nas regiões de LTR do genoma
viral, foi associada com a alta capacidade dos LVPRs serem ativados até o nível
transcricional em muitos ambientes celulares diferentes, podendo favorecer a seleção de
vírus que podem replicar em células e órgãos particulares, portanto afetando
diretamente o tropismo celular e a patogenicidade dos vírus (CLEMENTS e PAYNE, 1994;
AGNARSDOTTIR et al., 2000; ANGELOPOULOU et al., 2006a).
Mundialmente são conhecidas apenas as seguintes sequências genômicas
completas de LVPRs, as estirpes Islandesas em K1514 e KV1772 (ANDRÉSSON et al.,
1993; SONIGO et al., 1985), CAEV Cork (SALTARELLI et al., 1990), os vírus Sul
Africano em (SA-OMVV) (QUERAT et al., 1990) e Britânico (EV-1) (SARGAN et al.,
1991), todos eles do tipo rápido/alto. Em 2004, FEVEREIRO et al., através de análise
filogenética baseada nos alinhamentos das sequências nucleotídicas dos genes pol e env,
revelaram uma nova cepa de MVV em Portugal, denominada P1OLV. Até o momento,
este é o único lentivírus ovino com padrão de replicação do tipo lento/baixo cujo
genoma foi totalmente sequenciado. Além das estirpes recentemente publicadas
VOLTERRA e FONNI (Bertolotti et al., 2011); SEUI (Reina et al., 2010) e FESC-752
(Ramirez et al., 2011, dados não publicado). Os LVPRs podem ser classificados como

36
rápido/alto ou baixo/lento de acordo com sua replicação in vitro. A linhagem do tipo
rápida/alta replica rapidamente, induzindo a lise celular e alcançando altos títulos,
enquanto o tipo baixo/lento cresce lentamente e apresenta baixos títulos (BARROS et al.,
2004; LAIRMORE et al., 1987; QUERAT et al., 1984; WOODWARD et al., 1995).
LEROUX et al. (1997) sugerem que as cepas K1514, SA-OMVV e EV1 tenham
uma origem comum, talvez a Alemanha. As sequências nucleotídicas desses lentivírus
encontram-se registradas em bancos de dados genéticos internacionais (Genbank)
(MARCHESIN et al., 1997; FEITOSA et al., 2010).
Estudos subsequentes, baseados em sequências parciais dos genes gag, pol e
env, mostraram que estes vírus de ovinos e caprinos estão entremeando na árvore
filogenética independente do hospedeiro, sugerindo que as infecções interespécies têm
ocorrido entre os animais hospedeiros (CHEBLOUNE et al., 1996; LEROUX et al., 1997;
ZANONI, 1998; CASTRO et al., 1999; GREGO et al., 2002; ROLLAND et al., 2002; SHAH et
al., 2004a;).
O genoma completo da linhagem de CAEV-Cork dos Estados Unidos,
considerado o protótipo de lentivirus caprino, foi clonado e a sequência de DNA
proviral, determinada (SALTARELLI et al., 1990). Sequências nucleotídicas completas e
parciais de muitos outros lentivírus têm sido recentemente determinadas a partir de
isolados de caprinos e ovinos como as do Brasil, Irlanda, Estados Unidos, Suíça, Grécia,
França, Espanha, Itália, Finlândia, Polônia, Noruega e recentemente no Canadá
(MARCHESIN, 1997; RAVAZZOLO et al., 2001; ROLLAND et al., 2002; ABELSON E
SCHOBORG, 2003; HERRMANN et al., 2004; SHAH et al., 2004a; ANGELOPOULOU et al.,
2005; PISONI et al., 2005,2006; GERMAIN E VALAS E GERMAIN, 2006; REINA et al., 2006;
CONTRERAS et al., 2006; LAAMANEN et al., 2007; KUZMAK et al., 2007; GJERSET et al.,
2007; FEITOSA et al., 2010, L'HOMME et al., 2011).
Análises filogenéticas utilizando as sequências gag/pol revelaram a presença dos
subtipos de LVPRs do grupo A em diferentes países da Europa, incluindo França, Itália,
Portugal, Espanha e Suíça (BARROS et al., 2004; GREGO et al., 2002; LEROUX et al.,
1997; SHAH et al., 2004a; GREGO et al., 2007).
Embora a classificação filogenética de LVPRs esteja correta quando algumas
sequências de isolados são avaliadas para mais de um gene (ZANONI, 1998), tais como a
caracterização das sequências gag e pol, nenhuma caracterização para as sequências env

37
está completa. Este gene codifica glicoproteínas do envelope que iniciam um papel
importante no hospedeiro, na infectividade e na progressão da doença. Vários estudos
demonstraram que as linhagens de lentivírus podem diferir em suas propriedades
biológicas, incluindo efeito citopático e tropismo in vitro (BARROS et al., 2004; BRUETT
e CLEMENTS, 2001; HÖTZEL e CHEEVERS, 2001; LAIRMORE et al., 1987; QUERAT et al.,
1984).
No Brasil, os primeiros estudos de caracterização molecular e isolamento de
parte do gene gag foram realizados em isolados nos Estados de Minas Gerais e Rio
Grande do Sul. Em Minas Gerais, cinco amostras de CAEV isoladas de animais
soropositivos foram comparadas com outros lentivírus existentes no GenBank e
revelaram-se únicas e distintas das demais amostras de LVPRs com maior identidade
com amostras de CAEV do que com a de MVV. Entretanto, no Rio Grande do Sul,
outros cinco isolados de CAEV, de animais infectados foram comparados com as
sequências de seis lentivírus de pequenos ruminantes, permitindo separar as amostras
em três grupos distintos (MARCHESIN, 1997).
CASTRO (1998) amplificou e sequenciou parte do gene pol e o gene tat de
isolados de CAEV dos estados de Minas Gerais e Pernambuco. A análise filogenética
das sequências destes genes indicou que o isolado de Pernambuco foi estreitamente
relacionado ao isolado islandês MVV K1514, enquanto os isolados de Minas Gerais
foram mais relacionados ao CAEV.
REISCHAK (2000), estudando três isolados caprinos e um isolado ovino do Rio
Grande do Sul, verificou que dois destes apresentavam características líticas nos
cultivos celulares secundários e de linhagem, indicando diversidade quanto ao ciclo de
replicação viral in vitro destes isolados.
No Ceará, foi realizado pela primeira vez um estudo filogenético de lentivírus de
pequenos ruminantes. FEITOSA et al (2010) amplificou e sequenciou o gene gag a partir
de PCR de quatro isolados de CAEV no Estado do Ceará. Os alinhamentos dessas
sequências cearenses quando comparadas com as sequências padrões e outros isolados
nacionais, já disponíveis em bancos de dados genéticos, revelaram que essas linhagens
estão relacionadas com o subtipo B1 de CAEV.
Porém ao comparar as sequências dos isolados do Ceará com noventa sequências
do gene gag disponíveis em bancos de dados genéticos, a árvore filogenética gerada
mostrou as sequências Cearenses agrupadas em um grupo distinto, demostrando uma

38
forte relação filogenética com as sequências dos isolados da França, Noruega e uma do
Brasil, mais especificamente do Rio Grande do Sul.
Recentemente GLARIA e colaboradores (2011), observaram pela primeira vez na
Espanha um ovino apresentando a forma artrítica da doença. Ao estudarem a sequência
completa do vírus isolado, observaram que este pertencia ao subgrupo B2 de CAEV de
acordo com a nova nomenclatura, sugerindo assim, uma adaptação do vírus caprino ao
novo hospedeiro ovino, adquirindo propriedades genéticas que conferem o fenótipo do
tipo rápido/alto.
Ao longo do tempo, estudos de epidemiologia molecular têm mostrado que
CAEV e MVV estão relacionados, mas são vírus geneticamente distintos e que
pertencem a dois conjuntos filogenéticos principais (ROLLAND et al., 2002; SHAH et al.,
2004a; ZANONI, 1998). Os pequenos ruminantes são reservatórios de lentivírus com
uma linha filogenética própria e os dendogramas inferidos a partir das sequências dos
genes gag, pol e env de vários lentivírus ovinos e caprinos, sugerem que o CAEV teve
origem no MVV (VALAS et al., 1997). À medida que um maior número de sequências
de lentivírus for incorporado ao Genbank, será possível avaliar se os perfis observados
estão mais relacionados com CAEV ou com MVV.
Trabalhos baseados em estudos filogenéticos têm ajudado na classificação de
lentivirus caprino e ovino de acordo com seus hospedeiros. Foi demonstrado que estes
vírus estão constantes e facilmente transgredindo a barreira entre caprinos e ovinos
(SHAH et al., 2004b; BANKS et al., 1983), devendo isto ser levado em consideração nos
estudos epidemiológicos e nos planos de erradicação das infecções por LVPRs. Dados
de análise filogenética mostraram também que sequências de lentivírus caprino/ovino
estão frequentemente dispersos dentro de árvores filogenéticas, indicando assim várias
transmissões entre caprinos e ovinos e vice-versa, e que o gene env pode ser mais
informativo que as regiões gag e pol, pois este último gene, por ser uma região do
genoma viral bastante conservado, retém menos sinais filogenéticos (FEITOSA et al.,
2010; ROLLAND et al., 2002).

39
Tropismo celular
Os lentivírus de pequenos ruminantes têm tropismo por as células da linhagem
monocito-fagocitária. Nestas células, a expressão viral está estreitamente associada à
diferenciação e maturação dos monócitos em macrófagos (GENDELMAN et al., 1986;
NARAYAN et al., 1983).
Os lentivírus infectam células da linhagem dos macrófagos, incluindo microglia,
astrócitos, e oligodendrócitos, assim como os tipos I e II de pneumócitos, fibroblastos,
fibroblastos imortalizados e células epideliais e endoletiais (TEIXEIRA et al., 1997;
SANNA et al., 1999; CARROZZA et al., 2003; CHI et al., 2000).
Células epiteliais do leite de caprinos têm demonstrado susceptibilidade à
infecção com CAEV tanto in vitro quanto in vivo (MSELLI-LAKHAL et al., 1999). Estes
vírus podem ser detectados por hibridização in situ e/ou por métodos de
imunohistoquímica em drenagem de nódulos linfáticos, porém a frequência de células
infectadas é baixa, sugerindo que apenas a minoria das células são infectadas
(BLACKLAWS et al., 1995; CARROZZA et al., 2003; GROSSI et al., 2005; LECHNER et al.,
1997; STORSET et al., 1997).
In vivo, infectam principalmente células do sistema monocitos-fagocitário, sendo
os macrófagos a grande maioria das células infectadas (NARAYAN E CLEMENTS, 1989;
LUJÁN et al., 1994; BRODIE et al., 1995). A infecção das células depende da presença de
receptores para os vírus. Poucos monócitos têm estes receptores, entretanto estes
aumentam após sua maturação (GENDELMAN et al., 1986). Linfócitos também são
infectados, entretanto sem multiplicação viral (ZINK E JOHNSON, 1994). Ainda foi
detectado o RNA viral em células endoteliais, epiteliais, fibroblásticas, várias células do
sistema nervoso (plexo coróide, células microgliais, astrócitos, oligodentrócitos e
neurônios) e da glândula mamária (ZINK et al., 1990; BRODIE et al., 1995; SANNA et al.,
1999, MSELLI-LAKHAL et al., 1999; LERONDELLE et al., 1999). Os vírus também já
foram detectados na medula óssea (fibrócitos, células endotelias e adipócitos) de
caprinos positivos para CAEV através de imunohistoquimica, porém células
hematopoéticas foram negativas (GROSSI et al., 2005).

40
Nos tecidos-alvos, tais como sinóvias, interstício pulmonar, plexo coróide e
úbere, a ativação da replicação viral, juntamente com a maturação dos macrófagos,
induzem à formação das lesões. Segundo ZINK et al. (1990), outras células podem
também servir como áreas de replicação viral, como as células da membrana sinovial,
oligodentrócitos (PERK, 1990), células epiteliais intestinais, células tubulares renais, e
células glandulares da paratireóide, adrenais e da tireóide. Contudo, é o tropismo do
vírus por células do sistema imune, particularmente monócitos e macrófagos, o
principal fator responsável pela habilidade dos lentivírus em causar infecções crônicas,
que persistem por toda a vida do animal.
A forma nervosa da doença em ovinos jovens doentes sugere a existência de
linhagens de MVV neurotrópicas e altamente neurovirulentas (BENAVIDES et al., 2006).
Em outros casos de encefalites lentivirais (O´NEIL et al., 2004) foi afirmado que a
progressão da doença está associada com a carga viral e resposta inicial do hospedeira à
infecção. Isto também ocorre na infecção por MVV (DAWSON, 1980; HOUWERS, 1990)
quando animais jovens infectados, sendo incapazes de uma resposta defensiva,
permetiriam que os vírus alcançassem o encéfalo (BENAVIDES et al., 2006).
A detecção de RNA viral e DNA proviral em secções de tecidos caprinos e
ovinos infectados, revelou que os LVPRs poderiam entrar em uma gama de tipos
celulares como células dendríticas, linfócitos, plasmócitos, células endoteliais e
fibroblastos, adipócitos, células microgliais e pericitos, assim como em células epiteliais
dos brônquios, alvéolos, glândulas mamárias e plexo coróide, intestino delgado e
túbulos renais e terceira pálpebra (GEORGSSON et al., 1989; ZINK et al., 1990; STASKUS
et al., 1991; RYAN et al., 2000; CAPUCCHIO et al., 2003; CARROZZA et al., 2003;
PREZIUSO et al., 2004; BIESCAS et al., 2005; ANGELOPOULOU et al., 2006b; BOLEA et al.,
2006).
Ainda células endoteliais caprinas podem ser infectadas in vitro por
transmigração de leucócitos infectados com CAEV (LECHAT et al., 2005). Entretanto a
infecção produtiva é restrita às células da linhagem dos macrófagos (DE LA CONCHA-
BERMEJILLO, 1995) e da glândula mamária (MSELLI-LAKHAL et al., 1999; LERONDELLE
et al., 1999).

41
Um estudo para determinar a carga viral em áreas selecionadas e a expressão de
RNAm de CAEV em caprinos, oito anos após a infecção, demonstrou que os nódulos
linfáticos são um importante reservatório viral. A glândula mamária e células do leite
são os sítios preferidos para a replicação do vírus. A carga viral variou
significantemente entre os animais, demostrando a importância da carga genética frente
à infecção. Foi encontrada uma associação clara entre a ocorrência de lesões
histopatológicas e carga viral em sítios específicos. As análises da expressão de RNAm
de várias citocinas não revelam diferenças significantes entre animais o que poderiam
esclarecer uma considerável variação individual na carga viral observada (RAVAZZOLO
et al., 2006).
Recentemente, utilizando a técnica de PCR e imunohistoquímica, MVV foram
indiretamente detectados no fígado e no coração de seis ovinos naturalmente infectados.
Além disso, estes testes revelaram antígenos no citoplasma de hepatócitos e
cardiomiócitos. Porém exames histopatológicos mostraram níveis de leve a moderado
de colangi-hepatite e miocardite linfocítica crônica, mas não a presença de pequenos
agregados linfóides, as lesões linfoproliferativas típicas de MVV nestes órgãos
(BRELLOU et al., 2007).
A evidência de células infectadas com CAEV no útero e no oviduto coborroram
com o achado destas células em secreções genitais pós-parto. Recentemente CAEV
proviral foi detectado em tecidos do trato genital: útero, oviduto e ovários. Células da
granulosa (LAMARA et al., 2001) e células epiteliais do oviduto (LAMARA et al., 2002a)
são completamente susceptíveis à infecção por CAEV in vitro.
O DNA proviral de CAEV foi detectado em células do cumulus oophorus, mas
não em oócitos de cabras naturalmente infectadas. Um estudo mostrou que vinte e dois
por cento dos oócitos com células do cumulus intactas foram positivos para a presença
de DNA proviral e nenhum dos oócitos em que as células do cumulus foram removidas
mecanicamente foram positivos para o CAEV através de RT-PCR (ALI AL AHMAD et
al., 2005). Este resultado contradiz estudos feitos por FIENI et al. (2002, 2003) em que
sugerem a presença de CAEV em células infectadas no meio de lavagem do oviduto de
cabras infectadas doadoras de embriões e uma potencial transmissão de CAEV para o
embrião ou feto. Este estudo demonstra que oócitos livres de CAEV podem ser obtidos

42
de cabras infectadas. O DNA proviral do CAEV foi detectado pela primeira vez em
folículos pré-antrais de cabras sorologicamente positivas (SILVA, 2006).
Estudos in vitro mostraram que a zona pelúcida intacta é uma forte barreira que
protege o embrião caprino na infecção por CAEV, mas embriões sem zona pelúcida
quando incubados com CAEV e lavados extensivamente, poderiam transmitir a infecção
para células MSC (LAMARA et al., 2002b). Células embrionárias prematuras caprinas
são susceptíveis à infecção por CAEV e a infecção com estes vírus são produtivas,
porém, a única barreira natural que pode prevenir a infecção por CAEV de blastômeros
caprinos é a presença de uma zona pelúcida intacta (ZP) (ALI AL AHMAD et al., 2006).
In vitro os LVPRs infectam e replicam em cultivo celulares de macrófagos
(NARAYAN et al., 1983), membrana sinovial (NARAYAN et al., 1980; QUÉRAT et al.,
1984), glândula mamária (MSELLI-LAKHAL et al., 1999; LERONDELLE et al., 1999),
células fetais do plexo coróide (SIGURDSSON et al., 1960), músculo cardíaco (LEROUX et
al., 1995), baço, timo, linfonodo escapular (NARAYAN et al., 1980), células da linhagem
fibroblástica imortalizadas (TEIXEIRA et al., 1997) e tecido epitelial (LEE et al., 1996). E
produzem efeito citopático, que consiste na formação de células gigantes
multinucleadas ou sincícios e lise celular (CAREY E DALZIEL, 1983).
JUGANARU et al., (2010) demostraram ser fraca a associação entre patologia
induzida por lentivírus e atividade promotora de LTR, pelo menos nos lentivírus do
genótipo E, de baixa patogenicidade, fornecendo evidências indiretas de que o tropismo
celular pode ser mediado por sítio de ligação à receptores ao invés de fatores de
transcrição.
Embora o mecanismo de tropismo tecidual e patogênese da doença por CAEV
permanceça indeterminado, várias hipóteses possíveis são previstas: (i) a região do
envelope (env) é a porção menos conservada do genoma viral e códigos para ligantes do
receptor viral (proteínas de superfície-SU e transmembrânica-TM), onde a variabilidade
na sequência de nucleotídeos no gene env poderá afetar o tropismo tecidual, direta ou
indiretamente. Tem sido ainda proposto que as alterações no tropismo tecidual podem
ser responsáveis, pelo menos em parte, por conduzir mudanças na região variável de
env. (ii) cofatores poderiam estar atuando em sinergia com o vírus para influenciar o
tropismo e (iii) fatores relacionados ao hospedeiro poderiam determinar ou afetar o

43
tropismo tecidual (em caprinos MHC I ou II) ou (iv) o tropismo tecidual de CAEV pode
vir a culminar com eventos estocáticos aleatórios que ocorrem no início da infecção
(MURPHY, 2010).
Epidemiologia e Transmissão
A menor soroprevalência de infecção por CAEV através de cELISA encontrada
foi no México, 0,4% (TORRES-ACOSTA et al., 2003); TURQUIA 1,9% (BURGU et al.,
1994); Suíça, 2% (KRIEG and PETERHANS, 1990); Itália, 4,2% (GUFLER e
BAUMGARTNER, 2007) e Reino Unido, 4,3% (DAWSON E WILESMITH, 1985). Ao
contrário, os países com maior soroprevalência foram BRASIL, 36,5%; Noruega, 49,5%
(NORD et al., 1998); EUA, 73-81% (CUTLIP et al., 1992; ADAMS et al. 1984) e Austrália,
82% (GREWAL et al., 1986).
Por meio de isolamento a partir de células infectadas de ovinos soropositivos,
que foram submetidas à microscopia eletrônica, foi concluído o primeiro isolamento
deste vírus no país (MOOJEN et al., 1986) nos Estados do Rio Grande do Sul, Rio de
Janeiro, Minas Gerais, e por ABREU e colaboradores no ano de 1998 em Pernambuco.
No Brasil a infecção por LVPRs está amplamente distribuída. Achados nacionais
revelam que 36,5% dos rebanhos caprinos e ovinos estão infectados com vírus
(Bandeira et al., 2009).
No Ceará, 31,61% dos ovinos avaliados foram positivos para o Maedi-visna,
através de levantamento soro epidemiológico utilizando a técnica de imunodifusão em
gel de agarose (IDGA) feito por ALMEIDA e TEIXEIRA (1999). Em 2004, ARAÚJO e
colaboradores (2004) verificaram que 4,96 % dos 222 ovinos predestinados ao abate
foram positivos para MVV na região metropolitana de Fortaleza. Em 2006, em estudo
epidemiológico da região Norte do Ceará, PRIMO e colaboradores (2006) confirmaram a
presença de ovinos positivos, com prevalência de 0,5% dos 415 animais testados pelo
IDGA. CAMINHA e colaboradores (2008) realizaram o levantamento sorológico por
IDGA, de CAEV, na região metropolitana de Fortaleza e encontraram 16 (12,8%)
positivos entre os 125 caprinos testados.

44
Nos outros estados da região Nordeste, a prevalência encontrada para ovino e/ou
caprino foi de 13,40%; 0,73% e 26,1% na Bahia (ALMEIDA et al., 2001; OLIVEIRA et al.,
2006; TIGRE et al., 2006), de 4,4% e 2,5% no Piauí (PINHEIRO et al., 1996; BATISTA et
al., 2004), de 2,2% e 8,2% na Paraíba (CASTRO et al., 2002; BANDEIRA et al., 2009), de
11,0% no Rio Grande do Norte (SILVA et al., 2005), de 50,6% no Maranhão (ALVES e
PINHEIRO, 1997), de 17,7% e 3,9% em Pernambuco (CASTRO et al., 1994, 2002) e de
4,25% em Sergipe (MELO et al., 2003). SILVA e colaboradores (2005) analisaram
amostras de soro caprino no Estado do Rio Grande do Norte, verificando na ocasião
que, do total dos 184 animais testados, 2,71% foram positivos. Em Pernambuco,
OLIVEIRA et al. (2006), analisando amostras de soro caprino, provenientes de
abatedouros, notificaram o fato de que 3,8% eram positivos a CAEV.
Nos estados da região Sudeste do Brasil, a ocorrência de animais com CAE
observada em São Paulo foi de 43,01% e 34,93% (LEITE et al., 2004; MADUREIRA e
GOMES, 2007), em Minas Gerais de 33,3% e 23,6% (ASSIS e GOUVEIA, 1994; GOUVEIA
et al., 1998), Espírito Santo de 47,5% (GOUVEIA et al., 1998) e no Rio de Janeiro de
29,7%; 21,0% e 10,6% (ASSIS e GOUVEIA, 1994; CUNHA e NASCIMENTO, 1995;
GOUVEIA et al., 1998).
Na região Sul do Brasil a prevalência encontrada no Paraná foi de 28,2%
(SOTOMAIORe MILCZEWSKI, 1997). No Rio Grande do Sul, de 6,0% (MOOJEN et al.,
1986) e em Santa Catarina de 6,72% (SELL, 2000). Em estados de outras regiões
brasileiras como, Goiás, a prevalência relatada foi de 10,0% (GOUVEIA et al., 1998) e no
Pará de 40,0% (RAMOS et al., 1996).
Evidências diretas de transmissão natural de ovinos para caprinos e vice-versa,
particularmente dos subtipos A4 e B1, dos vírus têm sido relatadas (SHAH et al., 2004b;
PISONI et al., 2005). Até agora, os subtipos B1 e B2 (VALAS E GERMAIN, 2006; GREGO
et al., 2007), assim como quatro de sete subtipos do grupo A (A1, A3, A4 e A6), foram
isolados de ambos, caprinos e ovinos, sugerindo que os subtipos são particularmente
propensos a atravessar a barreira entre espécies (VALAS E GERMAIN, 2006).
O fato de alguns lentivírus terem sido isolados de caprinos e ovinos indica que a
transmissão interespécie deve ter ocorrido pelo menos uma vez em cada um dos
subtipos, embora a frequência e direção da transmissão permaneçam desconhecidas
(LEROUX et al., 1997; RAVAZZOLO et al., 2001; ROLLAND et al., 2002; ZANONI, 1998).

45
A principal forma de infecção de rebanhos livres se dá por meio da introdução
de animais portadores oriundos de rebanhos contaminados (ZINK E JOHNSON, 1994). A
infecção pelo leite/colostro é a principal via de transmissão em cabritos seguida da
transmissão horizontal através de secreções contaminadas (PETERHANS et al., 2004;
BLACKLAWS et al., 2004; HERRMANN-HOESING, 2007).
A via digestiva, geralmente ocorre no período neonatal, através do leite e/ou
colostro de mães infectadas (ROWE et al., 1992), onde os vírus podem se encontrar tanto
livre como em células somáticas.
Estudos recentes sugerem que a depuração de células associadas aos lentivírus
na circulação após a transferência passiva de anticorpos do colostro pode ocorrer após
32 semanas em resposta ao provírus e aos anticorpos anti-LVPRs e estes permanecem
não detectáveis até os quatro anos de idade (HERRMANN-HOESING, 2007).
GUEDES (1999) verificou que em ovinos, em regime de criação intensivo,
existem evidências da transmissão do MVV via secreção respiratória ou aerossóis
(HOUWERS et al., 1987).
Em relação ao potencial risco de transmissão sexual, estudos recentes
verificaram a presença de RNAm viral e DNA proviral de CAEV no sêmen (plasma
seminal e células não espermáticas, respectivamente) e em tecidos do trato genital de
machos naturalmente infectados (ALI AL AHMAD et al., 2008). Recentemente, RICARTE e
colaboradores (2010), através de análise de imunohistoquímica e ultraestrutural
detectaram a presença do vírus em espermatozóides de caprinos infectados com CAEV.
Outros estudos anteriores mostraram que a infecção recorrente ou inflamação
nos testículos induziram a liberação do MVV e CAEV no sêmen (LAMARA et al.,
2002b; ANDRIOLI et al., 2006). Porém, carneiros infectados com MVV utilizados para
procriação com ovelhas não infectadas, e alojadas separadamente, não causou
soroconversão nas mesmas (LAMARA et al., 2002b).
Experimentos recentes, usando inoculações de MVV intranasal e intratraqueal
em ovinos, têm mostrado que a rota intratraqueal, em infecções experimentais, é mais
eficiente que a rota de transmissão intranasal (TORSTEINSDOTTIR et al., 2003). Além das
vias de infecção, devem-se levar em conta os fatores que afetam o risco de transmissão
como estresse, imunossupressão, dose do vírus e rota de infecção, cepa do vírus, idade,
alta densidade e tempo de exposição aos vírus (BLACKLAWS et al., 2004).

46
Sinais Clínicos
O período de incubação da CAE é variável, podendo se estender por vários anos.
O aparecimento da manifestação clínica da doença e a produção de patologia
significativa induzida pelo vírus podem ocorrer meses ou anos após a infecção
(NARAYAN e CLEMENTS, 1989). A manifestação sintomatológica da CAE pode ser
dividida em quatro aspectos clínicos principais, podendo ocorrer de forma isolada ou
simultânea, incluindo artrite, encefalite, mamite e pneumonia (FRANKE, 1998). A CAE
também compromete o estado geral dos animais infectados. A infecção tem longa
incubação e a doença evolução crônica, com instalação progressiva das lesões, sendo,
na maioria dos casos, manifestada em animais adultos (CASTRO, 1998).
Diagnóstico clínico
Observam-se as manifestações da sintomatologia como pneumonia, artrite,
mamite ou encefalite, dados epidemiológicos e manejo dos animais, porém, o
diagnóstico de CAE só é confirmado com testes laboratoriais (PINHEIRO, 2001).
Diagnóstico laboratorial
Muitos fatores influenciam no diagnóstico sorológico de caprinos infectados
com CAEV, incluindo a variação no tempo relativo ao início da infecção, especificidade
da proteína viral dos anticorpos do soro e sua quantidade, efeitos da cepa viral e
diferenças baseadas nos hospedeiros com relação à resposta imunológica para infecção
(DEMARTINI et al., 1999). As características da relação hospedeiro-lentivírus que
afetam a detecção em caprinos infectados incluem: heterogeneidade viral, mutação em
animais infectados, bem como baixos níveis de replicação viral in vivo.
Os caprinos infectados podem ser detectados com base na presença de
anticorpos antivirais no soro, pela detecção de proteínas ou do ácido nucléico, além do
isolamento viral (DEMARTINI et al., 1999). Essa enfermidade pode ser diagnosticada por
vários métodos, sendo o principal a imunodifusão em gel de agarose (IDGA), que é o
teste recomendado pela Organização Mundial de Saúde Animal (OIE). Existe também o
teste de ELISA, Western Blot, imunofluorescência indireta, isolamento do vírus por

47
cultivo celular, reação em cadeia de polimerase (PCR), imunohistoquímica e
microscopia eletrônica, porém esses procedimentos são mais empregados nas pesquisas
científicas (DANTAS, et al., 2008).
Ensaio imunoenzimático (ELISA) indireto
Nesse teste utilizam-se duas IgG, uma para reconhecer o antígeno e outra (anti-
IgG) produzida em diferentes espécies de animal que reconhece a primeira IgG, com a
qual se ligará (ALMEIDA; LIMA, 2001). É um ensaio amplamente utilizado para a
detecção e/ou quantificação de anticorpos em amostras de soro, com destaque em
estudos soroepidemiológicos. A especificidade dessa prova é garantida principalmente
pela qualidade do antígeno adsorvido à placa (MADRUGA; ARAÚJO; SOARES, 2001).
Vários métodos de ELISA indiretos usando vírus inteiro, proteínas do capsídeo CAEV
ou recombinante têm sido descritos (REYBURN et al., 1992; CAREY; ROY; DALZIEL,1993;
DANTAS et al., 2008). Esse teste permite a detecção de estirpes de um mesmo vírus que,
embora sejam sorologicamente relacionadas, apresentam maior variação entre si
(ALMEIDA; LIMA, 2001).
Isolamento viral
O isolamento viral em cultivo celular é a principal técnica utilizada para o
diagnóstico da maioria das infecções virais. Baseia-se na capacidade de demonstrar a
propagação dos vírus em cultura de células, mediante observação de alterações
morfofisiológicas apresentadas pela monocamada celular ou efeito citopático (CPE).
Essa técnica é aplicável ao diagnóstico de quase todas as viroses de interesse veterinário
inclusive CAE, capaz de detectar quantidades mínimas de vírus, sendo considerado um
teste padrão de diagnóstico em virologia. Existem, no entanto, algumas restrições, uma
vez que é demorada, dispendiosa, necessita da implantação de cultivos celulares
especiais e, além disso, é incapaz de detectar vírus que não causam efeito citopático
(KNOWLES, 1997). Geralmente, o CAEV pode ser isolado de animais vivos pelo
cocultivo de células, como leucócitos do sangue periférico, células somáticas de leite ou
células de membrana sinovial caprina (MSC) (PINHEIRO, 2001).

48
Reação em cadeia de polimerase (PCR)
A detecção de DNA proviral pode ser feita também através da PCR. Esta é
sensível na detecção de pequenas quantidades de ácidos nucléicos virais e é utilizada
para demonstrar a presença de DNA proviral tanto in vivo quanto in vitro.
Vários trabalhos têm demonstrado, com sucesso, o uso da técnica de PCR na
detecção do DNA proviral dos LVPRs. A PCR permite por amplificação direta de parte
do ácido nucléico viral específica de fluidos e tecidos de um animal infectado (ZANONI
et al.,1990; REDDY et al., 1993; RIMSTAD et al., 1993; WAGTER et al., 1998). O DNA
proviral pode ser detectado em células mononucleares e granulócitos presentes no soro
de leite, inferindo que o soro lácteo pode ser infectante, difundindo, consequentemente,
os vírus (RUSSO et al., 1997).
As variações de PCR aplicadas são: PCR quantitativa em tempo real que foi
desenvolvida e caracterizada in vitro (GUDMUNDSSON et al., 2003; ZHANG et al., 2000) e
semi-nested PCR utilizando primers degenerados, desenvolvida por ELTAHIR et al.
(2006).
REDDY et al. (1993) verificaram também que três animais negativos por IDGA
foram positivos por PCR, enquanto dois animais positivos por IDGA apresentaram
resultado negativo por PCR.
Devido à alta variabilidade genômica dos lentivírus, a escolha dos primers em
genes relativamente conservados como pol, gag e da LTR, combinado a com as
condições de reação para a máxima sensibilidade é decisiva para detecção de um grande
espectro de cepas de campo de ambos CAEV e MVV (ZANONI, 1998).
WAGTER e colaboradores (1998) através de PCR, utilizando um mix de seis
primers de três cepas de lentivírus ovino e um de caprino da região conservada do gene
gag, encontraram resultados discordantes com quantidades diferentes de amostras de
sangue (monócitos) demonstrando que a sensibilidade do teste é dependende da carga
viral, e que o volume da amostra é diretamente proporcional a possibilidade de detecção
do vírus . Verificaram que quando amostraram 1 mL de sangue de seis animais nenhum
reagiu positivamente, entretanto quando aumentaram a alíquota para 5 mL, cinco dos
seis animais foram positivos e quando utilizaram 10 mL de sangue todos os animais
foram positivos. De 60 animais soropositivos pela IDGA, o PCR do sangue detectou
somente 53. Observaram, também, que a resposta sorológica pode ser bem lenta até 18

49
meses após a detecção pelo PCR ou até mesmo não soroconverter e que a PCR é mais
eficiente em detectar DNA no sangue nos estágios iniciais da doença. A detecção de
infecção por LVPR através de PCR é indicativa de uma infecção persistente e é
dependente da quantidade amplificada da sequência alvo e da especificidade do primer.
Entretanto esta técnica poderá ser utilizada em programas de erradicação,
quando estiver disponível rotineiramente, para identificar os animais não diagnosticados
por sorologia, inclusive antes da soroconversão. É recomendado que esta técnica seja
empregada para esclarecer resultados sorológicos indeterminados ou negativos, devido
ao alto custo e aos resultados discordantes entre testes sorológicos e PCR (KNOWLES,
1997; RIET-CORREA, 2001).
Imunodifusão em gel de agarose (IDGA)
Recomendado pela OIE, por ser de fácil aplicabilidade e não exigir
equipamentos nem instalações sofisticadas, o IDGA é a forma diagnóstica mais
utilizada em todo o mundo, principalmente em programas de controle da doença
(MOOJEN et al., 1986). O IDGA tem alta especificidade, característica essa que o
credencia a ser empregado como padrão no diagnóstico de triagem (VAREA et al., 2001).
Segundo VITU e colaboradores (1982), o teste de IDGA é capaz de identificar animais
experimentalmente infectados com CAEV cerca de quatro a cinco meses após a
infecção. Os autores indicam ainda que esse teste se mostrou mais sensível que o teste
de fixação do complemento e, ao ser comparado com o ELISA, mostrou resultados
bastante correlacionados. Vale salientar que, ao se empregar o ELISA, a infecção pode
ser identificada de modo mais precoce, até sete semanas após o período de incubação.
Esse fato foi confirmado por LARSEN et al., (1982), ao avaliarem caprinos
experimentalmente infectados, afirmando a presença de anticorpos precipitantes, os
quais podiam ser detectados de maneira mais precoce e com maior intensidade do que
na fixação do complemento.
O teste IDGA pode ser utilizado para a detecção de anticorpos anti-CAEV tanto
no soro sanguíneo quanto no colostro de animais infectados e a presença de anticorpos
no colostro pode ser utilizada na detecção da infecção no rebanho (ALKAN e TAN,
1998).

50
Western blotting ou imunoblotting
É uma técnica imunoenzimática de detecção de proteínas. Foi largamente
empregado em estudos bioquímicos e imunológicos e com o advento das técnicas de
Biologia Molecular, tal método é aplicado como ferramenta valiosa nos estudos de
expressão gênica. As análises de Western blotting são deveras úteis na identificação e
quantificação de proteínas específicas, em uma mistura complexa de proteínas, com a
vantagem de não se empregar qualquer marcação radioativa (REGITANO e COUTINHO,
2001).
Prevenção e controle
Até o presente, não existe tratamento nem vacina contra CAEV. Com isso, o
controle dessa lentivirose é muito difícil, devendo-se impedir a entrada de animais
infectados no plantel e fazer exames periódicos do rebanho, para que os animais doentes
possam ser identificados e separados dos sadios ou mesmo eliminados do rebanho
(CASTRO, 1998). Quando a prevalência for maior do que 30% sugere-se a formação de
um rebanho de animais negativos separado dos positivos, de modo a estabelecer o
saneamento da propriedade mediante um rigoroso manejo sanitário de ambos os grupos
e da gradativa eliminação do grupo dos animais infectados (FRANKE, 1998). Nos
plantéis suspeitos ou sabidamente positivos, devem-se separar os cabritos
imediatamente após o nascimento, evitando ao máximo seu contato com as secreções da
mãe; administrar colostro de cabras soronegativas por IDGA tratado a 56 ºC por 60 min,
ou de vaca; alimentar os cabritos com leite de vaca ou leite de cabras pasteurizado;
testar os cabritos a cada seis meses e separar os positivos (CASTRO, 1998).
Apesar da inexistência de tratamento, ARAÚJO (2008), estudando a atividade
antiviral de inibidores da enzima transcriptase reversa (lamivudina, estavudina,
zidovudina e efavirens), observou que estes inibiram in vitro a replicação dos LVPRs,
enquanto os inibidores da enzima protease não impediram a multiplicação do agente.
Nesse mesmo trabalho, o autor avaliou a atividade antiviral dos extratos, tanto do
cinamomo (Melia azedarach L.) quanto do nim (Azadirachta indica A. Juss) e verificou
que estes inibiram in vitro a replicação dos vírus da artrite encefalite caprina e do Vírus
Maedi-visna.

51
O desenvolvimento de uma vacina é um processo longo e dispendioso. Várias
vacinas já foram e estão sendo testadas contra os LVPRs, porém nenhuma apresentou
uma proteção eficaz, embora algumas tenham apresentado produção de anticorpos
precipitantes e neutralizantes. Pesquisadores utilizando um vírus inativado por calor,
formalina com ou sem adjuvante incompleto de Freund e adjuvante de hidróxido de
alumínio para vacinar ovinos, observaram que os ovinos produziram anticorpos
precipitantes contra o vírus, os quais não protegeram contra a infecção quando
desafiado com o MVV in vivo. Não obstante, experimentos de vacinação com o vírus
da imunodeficiência símia (SIV) atenuado têm mostrado reduzir a carga viral e atrasar a
progressão da infecção para a doença clínica (WYAND et al., 1996). Resultados similares
são relatados com vírus da imunodeficiência felina (FIV) (PISTELLO et al., 2003).
PERTUSSON et al., (2005) utilizando clone do vírus MVV atenuado, por via
intratraqueal, observaram em dois dos ovinos vacinados que os anticorpos
neutralizantes foram específicos para o vírus vacinado, mas não neutralizaram o vírus
do desafio, apesar da grande similaridade genética de ambos os vírus. Caprinos Saanen
foram imunizados com glicoproteína de superfície gp 135 purificada de isolado 79-63
(CAEV-63), e avaliados para a produção de anticorpos neutralizantes homólogos e a
ocorrência de reação cruzada. Os anticorpos neutralizantes CAEV-63 foram detectados
em todos os animais após sete imunizações com gp 135 em adjuvante Quil A. Além
disso, o soro de três dos quatro testados neutralizou um isolado do CAEVco; todavia, o
soro de um dos caprinos não manifestou neutralização para o CAEVco heterólogo,
porém inibiu um CAEVco neutralizado por outro soro. Em suma, o estudo demonstrou
que a vacina de subunidades da glicoproteína desse lentivírus pode induzir resposta
humoral neutralizante cruzada em caprinos (KEMP et al., 2000).

52
JUSTIFICATIVA
A natureza de uma doença, especialmente sua epidemiologia e impacto
econômico potencial sobre populações de animais são fatores de elevada importância
para autoridades veterinárias nacionais, quando forem ponderar as ameaças a países
importadores, regiões ou rebanhos (GARNER et al., 1995).
No Brasil, importações de animais de alta linhagem procedentes de países com
alta prevalência para a lentivirose caprina e ovina têm ocorrido no decorrer de vários
anos. Relatos comprovaram que a introdução e disseminação do CAEV e MVV no
rebanho nacional ocorreram pelas importações de raças leiteiras, precedentes de
distintos países, sem adequada supervisão, acarretando graves problemas sanitários
(ASSIS E GOUVEIA, 1994).
A introdução de novas cepas merece atenção especial, pois estas podem se
apresentar mais virulentas que as nativas, além disso, muitos agentes como os vírus de
RNA, que não apresentam a função de leitura e correção genética, podem ter altas taxas
de mutação, de aproximadamente um milhão de vezes mais que a de animais e plantas.
Consequentemente há uma enorme diversidade de populações de vírus de RNA que
podem se tornar mais patogênicos e causarem novas doenças quando recém-
introduzidos em novos ambientes (ANDRIOLI, 2001).
Dessa forma, a caracterização molecular dessas amostras é importante para
evidenciar a diversidade de propriedades biológicas dos vírus que podem redundar em
diferenças de citopatogenicidade tanto in vivo quanto in vitro, e de manifestações
clínicas da doença, além de auxiliar o desenvolvimento de métodos de diagnósticos
mais sensíveis.
O isolamento e a caracterização de novas amostras virais provenientes de
regiões geográficas distintas possibilitarão um estudo mais detalhado da
heterogeneidade genética entre os LVPRs, fundamental tanto para os estudos
epidemiológicos relacionados à transmissão intra e interespecífica dos vírus, quanto
para esclarecer a origem e a evolução desse grupo viral.
O conhecimento dos sorotipos virais existentes servirá de base para a produção
de antígenos para testes de diagnósticos mais eficientes e possíveis antígenos vacinais.

53
HIPÓTESE CIENTÍFICA
Devido à alta variabilidade genética dos lentivírus, as sequências dos genes gag,
env e pol de isolados de animais naturalmente infectados de vários estados brasileiros
são diferentes das sequências dos genes gag, env e pol obtidas das cepas padrões
(CAEV-Cork e K1514).

54
OBJETIVOS
Objetivos Gerais
Isolar cepas virais circulantes, de animais naturalmente infectados, em vários
Estados do Brasil, tais como: Ceará, Piauí, Rio Grande do Norte, Bahia e Minas
Gerais.
Caracterizar molecularmente as sequências parciais dos genes gag, env e pol,
obtidas de amostras virais isoladas.
Implementar uma biblioteca genômica de LVPRs isolados no Brasil.
Objetivos Específicos
Classificar as amostras virais isoladas nos tipo rápido/alto ou lento/baixo por
isolamento viral;
Amplificar os genes de interesse através da PCR.
Sequenciar parcialmente os genes gag, env e pol das amostras virais isoladas;
Comparar as sequências parciais de gag, env e pol com outras sequências
disponíveis no GenBank através da construção de uma árvore filogenética.

55
CAPÍTULO I
Primeiro Isolamento de Lentivírus de Pequenos Ruminantes em Caprino
Naturalmente Infectado em Rebanho do Rio Grande do Norte, Brasil
First Isolation of Small Ruminant Lentiviruses from Naturally Infected
Goat in Flock of Rio Grande do Norte, Brazil.
Periódico: Arquivos do Instituto Biológico , São Paulo, v.78, n.4, p.501-505, out./dez.,
2011

56
Resumo
O Vírus da Artrite Encefalite Caprina (CAEV) e Vírus Maedi-visna (MVV)
pertencem ao gênero Lentivírus da família Retroviridae. São considerados
geneticamente distintos, mas antigenicamente relacionados. O objetivo desde trabalho
foi isolar o vírus da CAE de um animal oriundo de um rebanho do Rio Grande do
Norte,positivo pelo teste de Imunodifusão em Gel de Agarose (IDGA), através do
cocultivo de leucócitos infectados do sangue periférico em Membrana Sinovial Caprina
(MSC). Dezesseis caprinos da raça Saanen, com suspeitas clínicas para CAE foi testado
por IDGA e Western Blotting. Para o isolamento viral, os leucócitos do sangue
periférico foram isolados por cocultivo em MSC. Monócitos/Macrófagos coletados
foram inoculados em monocamadas pré-formadas em garrafas A25. O resultado do
IDGA foi positivo para um animal, confirmado por Western Blotting. Após 50 dias de
cocultivo, foi realizada a coloração da monocamada com cristal de violeta a 0,1% para
visualização do ECP e realizada Nested-PCR do sobrenadante do cocultivo, com
confirmação do efeito citopático viral. A cepa isolada, denominada BrRN-CNPC.G1 foi
considerada o primeiro isolamento do CAEV no Estado do Rio Grande do Norte. Esse
estudo permitirá em breve, realizar a caracterização molecular do genoma do vírus
isolado, através da análise de seus diferentes genes estruturais e comparar com outras
sequencias virais isoladas para identificar a provável origem da infecção desse animal e
estabelecer as possíveis divergências entre cepas padrões de Lentivírus e cepas
regionais circulantes.
Palavras-chave: Cocultivo, Membrana Sinovial Caprina, CAEV.

57
Abstract
Caprine Arthritis Encephalitis virus (CAEV) and Visna/maedi virus (VISNA) belong to
the genus Lentivirus family Retroviridae. The aim of the present study was to isolate the
CAE goat from positive goats when tested by agarose gel immunodifusion (AGID) by
cocultivation techniques of infected peripheral blood leukocytes in goat synovial
membrane (GSM). In this study, one flock of 16 goats, obtained from a flock of Rio
Grande do Norte with suspected clinics for CAEV, was screened using Agar-gel
Immunodiffusion (AGID) and Western Blotting. The result was positive for an animal,
confirmed by Western Blotting. For virus isolation, peripheral blood leukocytes (PBL)
were isolated from the blood for the co-culture on goat sinovial membrane (GSM) cells.
Monocytes/macrophages collected were inoculated in monolayer of 90% semiconfluent
cells in the A25 culture bottles. After 50 days co-culture was performed staining of the
monolayer with crystal violet 0.1% for viewing viral cytopathic effects (CPE)
characteristic of viruses and nested-PCR performed with the supernatant of co-culture
for confirmation of CPE. The isolate, named BrRN-CNPC.G1 was considered the first
isolation of Small Ruminant Lentiviruses from Naturally Infected Goat in Flock of Rio
Grande do Norte, Brazil. This study will soon perform the characterization molecular
isolation of the virus genome, by analysis of their different structural genes and
compare with other viral sequences isolated to identify the likely source of infection of
that animal and establish the possible differences between strains of lentiviruses and
regional strains circulating.
Key-words: Co-culture, Goat sinovial membrane, CAEV.

58
Primeiro Isolamento de Lentivírus de Pequenos Ruminantes em Caprino
Naturalmente Infectado em Rebanho do Rio Grande do Norte, Brasil.
1Aryana Lushese Vasconcelos Lima Feitosa;
2Maria Fátima da Silva Teixeira;
3Raymundo Rizaldo Pinheiro;
4Dalva Alana Aragão de Azevedo;
4Samilly Mesquita
Alves, 3Alice Andrioli Pinheiro
1Autor correspondente: Programa de Pós-Graduação em Ciências Veterinárias;
Laboratório de Virologia, Universidade Estadual do Ceará – UECE, Av. Paranjana
1700, 60.740-093, Fortaleza, CE, Brasil, Tel.: +55 85 96332990; Fax: +55 85
31019849. [email protected]
2Programa de Pós-Graduação em Ciências Veterinárias; Laboratório de Virologia,
Universidade Estadual do Ceará – UECE,
3Embrapa Caprinos e Ovinos, Laboratório de Virologia;
4Iniciação Científica CNPq/PIBIC e FUNCAP, Universidade Estadual Vale do Acaraú –
UVA.

59
Introdução
Os Lentivírus de Pequenos Ruminantes (LVPRs) estão amplamente distribuídos
entre caprinos e ovinos. Diferentes formas clínicas da doença foram descritas em
pequenos ruminantes infectados com o vírus, tais como artrite progressiva crônica,
pneumonia e mastite em animais adultos e, mais raramente, leucoencefalomielite em
animais de dois a quatro anos de idade (CRAWFORD E ADAMS, 1981). O Vírus da
Artrite Encefalite Caprina (CAEV) e Vírus Maedi-visna (VISNA) pertencem ao gênero
Lentivírus da família Retroviridae. São considerados geneticamente distintos, mas
antigenicamente relacionados. De acordo com a nova nomenclatura, os LVPRs são
classificados em cinco grupos: A-D proposto por SHAH et al. (2004) e o grupo E,
altamente divergente, proposto por REINA et al. (2009). Segundo CORREL et al. 1992,
CAEV possui tropismo por células do sistema imune da linhagem dos
monócitos/macrófagos, na qual incorpora seu genoma, escapando dos mecanismos de
defesa do hospedeiro.
No Brasil, por meio de isolamento em células infectadas de ovinos
soropositivos, que foram submetidas à microscopia eletrônica, foi concluído o primeiro
isolamento deste vírus no país (MOOJEN, et al., 1986) nos Estados do Rio Grande do
Sul, Rio de Janeiro, em Minas Gerais, por ABREU e colaboradores no ano de 1998 em
Pernambuco, TIGRE et al. (2006) na Bahia e por FEITOSA et al. (2010) no Estado do
Ceará. O isolamento de cepas regionais contribui para um melhor conhecimento da
diversidade genética circulantes desses vírus e pode ainda contribuir para melhorar o
controle da doença. O objetivo desde trabalho foi isolar o vírus da CAE de um animal
oriundo de um rebanho do Rio Grande do Norte, positivo pelo teste de Imunodifusão

60
em Gel de Agarose (IDGA), através do cocultivo de leucócitos infectados do sangue
periférico em Membrana Sinovial Caprina (MSC).
Material e Métodos
IDGA e Western Blotting
Dezesseis caprinos da raça Saanen, obtido de um rebanho do Rio Grande do
Norte com suspeitas clínicas para CAE foi testado sorologicamente para a presença de
anticorpos contra a proteína Gag do capsídeo (p28) usando o kit IDGA produzido pela
Embrapa Caprinos e Ovinos, este sendo previamente testado contra o controle positivo
do kit Americano (Caprine Arthritis-Encephalitis/Ovine Progressive Pneumonia
Antibody Test Kit. Veterinary Diagnostic Technology, Inc® - USA). O rebanho foi
posteriormente testado para a CAE por Western Blotting, segundo a metodologia
descrita por PINHEIRO (2001).
Isolamento dos Leucócitos do Sangue Periférico
Foram coletados 10 ml de sangue por venipuntura da jugular em tubo com
anticoagulante (EDTA) para obtenção dos leucócitos do sangue periférico. Estes foram
isolados por centrifugação a 380 g por 10 minutos. Os eritrócitos foram hemolisados
com 8 ml de cloreto de amônio (0,84%), por duas vezes. O pellet foi lavado em 10 ml
de solução salina fosfato PBS a 380 g por 5 min e finalmente ressuspendido em 1 ml de
PBS para uso no cocultivo em células MSC.

61
Cultivo de Células de Membrana Sinovial Caprina (MSC)
Explantes das membranas sinoviais caprinas das articulações intercarpais de
membros anteriores de cabritos livres de CAEV foram cultivados em Meio Essencial
Mínimo (MEM-G), acrescido de 10% de Soro Fetal Bovino (SFB), incubadas a 37 oC
em atmosfera de 5% de CO2 (ABREU et al., 1998) em garrafas de cultura celular A25.
O meio foi trocado até a monocamada obter 90% de confluência.
Cocultivo de Monócito/Macrófagos com MSC
Monócitos/Macrófagos (200 L) coletados foram inoculados em monocamadas
pré-formadas em garrafas A25 (capacidade 25 cm2) e incubados durante 60 minutos em
estufa 37 oC a 5% de CO2 acrescidos de 1 ml de meio essencial mínimo (MEM) sem
soro fetal bovino (SFB) (Sigma®; BDV Free). Após o período de incubação foram
adicionados 3,5 ml de MEM com 10% de soro fetal bovino. O meio foi trocado a cada
sete dias e a passagem foi realizada a cada 21 dias, com adição de 200 L de células da
MSC a cada troca de meio ou passagem. A monocamada foi examinada diariamente em
microscópio invertido para verificação de efeito citopático viral (ECP), formação de
sincícios, fusão nuclear ou vacuolização. Após 50 dias de cocultivo, foi realizada a
coloração da monocamada com cristal de violeta a 0,1% para visualização do ECP.
Extração de DNA proviral a partir de sobrenadante de cocultivo
O sobrenadante de cocultivo coletado a cada sete dias foi utilizado para extração
do DNA proviral. Uma proporção de 1:1 de PBS 1X foi utilizada para centrifugar 5 ml

62
de sobrenadante de cocultivo a 250 g por 5 minutos. Após a centrifugação a fase
superior foi descartada e o pellet novamente lavado em PBS nas mesmas condições
anteriores. Em seguida o pellet foi ressuspendido em 2 ml de tampão hipertônico,
durante 2 minutos a temperatura ambiente, para lise do citoplasma celular, seguido de
centrifugação a 1100 g por 10 minutos. O pellet foi ressuspendido em 2 ml de PBS e
submetido a centrifugação a 1100 g por 10 minutos. Por fim, o sedimento foi
novamente ressuspendido em 250 L de tampão de PCR e homogeneizado. O material
foi, então, tratado com proteinase K (0,1g/L), durante 60 minutos, em banho-maria
a 56 oC. A proteinase K foi inativada termicamente em água fervente a 100
oC por 10
minutos e acondicionadas em freezer a -20 oC até a sua utilização.
Nested-PCR
Duas rodadas de PCR (Nested-PCR) foram realizados para amplificar um
fragmento de 187 pb de DNA proviral, correspondente ao gene gag de CAEV. Todos os
primers foram desenhados baseados na sequencia padrão de CAEV-Cork publicadas por
SALTARELLI et al., (1990). Foram utilizados dois pares de primers. Os
oligonucleotídeos externos GEX5 (5’-CAAGCAGCAGGAGGGAGAAGCTG-3’) e
GEX3 (5’-TCCTACCCCCATAATTTGATCCAC-3’) foram utilizados na primeira
amplificação, correspondendo as bases 953-975 e o complemento 1249-1226,
respectivamente. Os oligonucleotídeos internos GIN5 (5’-
GTTCCAGCAACTGCAAACAGTAGCAATG-3’) e GIN3 (5’-
ACCTTTCTGCTTCTTCATTTAATTTCCC- 3’) foram usados na segunda
amplificação, correspondendo as bases 997-1024 e o complemento 1181-1154,
respectivamente. Para cada round de amplificação, foi utilizado um controle positivo

63
referente ao CAEV-Cork e um controle negativo sem amostra de DNA em paralelo com
todas as amostras. O mix de PCR constituiu de 34,6 L de água sem DNA, 5 L de
tampão sem MgCl2 (Tris HCl 10 mM, KCl 50 mM,) 3,5 L de MgCl2 (1,5 mM), 1L
de cada dNTPs (100 M) (Invitrogen®), 2 L de cada primers (20 pmol), 0,4 L de
Taq (2U) e 5 L de amostra de DNA, tanto para o primeiro como para o segundo round.
As amplificações foram realizadas usando termociclador da BioRad em 35 ciclos sob as
seguintes condições: desnaturação a 94 °C por cinco minutos, 1 minuto a 94 °C,
anelamento a 56 °C por um minuto e extensão a 72 °C por 45 segundos, com extensão
final por 7 minutos 72 °C no último ciclo. Os produtos obtidos foram analisados por
eletroforese em gel de agarose a 1 %, corado com brometo de etídio e visualizados em
foto documentador de luz ultravioleta com densidade ótica de 260 nm.
Resultados
IDGA e Western Blotting
Um caprino naturalmente infectado foi detectado do teste de IDGA e confirmado
por Western Blotting.
Isolamento dos Leucócitos do Sangue Periférico e Cocultivo
A garrafa de cultura celular foi corada no dia 50. O cocultivo de
Monócito/Macrófagos com MSC apresentou formação de poucos sincícios na
monocamada, a partir do 27º dia de cocultivo, por observação em microscópio
invertido. A cepa isolada, denominada BrRN-CNPC.G1 apresentou fenotipagem
altamente lítica, causando a destruição quase total da monocamada desde os primeiros

64
dias até o final do cocultivo sendo necessário a adição de mais células a garrafa A25. A
presença do efeito citopático viral na monocamada de MSC foi observada, com células
gigantes, multinucleadas, variando de 4 a 8 núcleos por célula, confirmando a presença
do vírus em células do sistema monocítico fagocitário (SMF).
Nested-PCR
A confirmação do isolamento foi feita pela amplificação do DNA proviral de
CAEV, através de Nested-PCR que detectou a presença fragmento de 187 bp do gene
gag no sobrenadante de cultivo celular em todas as 6 coletas de sobrenadante do
cocultivo.
Discussão
A partir de um caprino da raça Saanen, foi isolada uma cepa de LVPR,
denominada BrRB-CNPC.G1. Através da técnica de PCR, foi confirmada a presença do
DNA proviral no sobrenadante do cocultivo. Os achados desse isolamento viral foram
compatíveis com outros relacionados a amostras líticas de LVPR. Os Vírus da Artrite
Encefalite Caprina (CAEV) e os Vírus Maedi-visna (MVV) têm sido considerados
como patógenos geneticamente distintos, mas antigenicamente relacionados entre si.
Estes vírus estão constantes e facilmente transpondo a barreira entre espécies de
caprinos e ovinos (VALAS et al., 2000; KARR et al., 1996; SHAH et al., 2004). Os
LVPRs estão classificados dentro de cinco grupos relativamente equidistante,
denominados de A-E (SHAH et al., 2004; REINA et al., 2009). A alta heterogeneidade
de sequências de nucleotídeos e aminoácidos em lentivírus pode determinar a sua

65
antigenicidade, virulência e crescimento e pode afetar sua persistência e o seu
mecanismo de evasão. Apesar das diferenças no tropismo celular, órgãos alvo e
patogenicidade, comumente encontrados entre CAEV e MVV, ambos os vírus foram
recentemente atribuído a um único grupo, os LVPRs. A introdução de novas cepas
merece atenção especial, pois podem se apresentar mais virulentas que as nativas, além
do que muitos agentes como os vírus de RNA, que não apresentam a função de leitura e
correção genética, podem ter altas taxas de mutação, de aproximadamente um milhão de
vezes mais que a de animais e plantas. Consequentemente há uma enorme diversidade
de populações de vírus de RNA que podem se tornar mais patogênicos e causarem
novas doenças quando recém-introduzidos em ambientes (ANDRIOLI, 2001). Somente
um pequeno número de LVPRs foi isolado no Brasil. Até agora foi concluído o
isolamento deste vírus no país nos Estados do Rio Grande do Sul, Rio de Janeiro, Minas
Gerais, Pernambuco, Bahia e no Ceará. O isolamento de cepas regionais contribui para
um melhor conhecimento da diversidade genética desses vírus (FEITOSA et al., 2010).
O conhecimento da variabilidade genética pode ajudar a entender melhor os
mecanismos de evolução molecular e heterogeneidade epidemiológica da infecção. A
variabilidade genotípica e fenotípica das cepas circulantes pode ser útil para melhorar as
ferramentas de diagnóstico, importantes para o controle e erradicação da doença.
Futuros estudos de caracterização molecular das amostras virais isoladas poderão ajudar
a desenvolver testes de diagnósticos mais sensíveis. Esse trabalho permitirá em breve,
estudar a caracterização molecular do genoma do vírus isolado, através da análise de
seus diferentes genes estruturais, tais como gag, env e pol e comparar com outras
sequencias virais isoladas para identificar a provável origem da infecção desse animal e
estabelecer as possíveis divergências entre cepas padrões de Lentivírus e cepas
regionais circulantes.

66
Conclusão
Foi realizado o primeiro isolamento do CAEV no Estado do Rio Grande do
Norte a partir de monócitos/macrófagos co-cultivados em monocamadas de membrana
sinovial caprina. A cepa foi caracterizada fenotipicamente como lítica, do tipo
rápida/alta.
Agradecimentos
UECE, EMBRAPA Caprinos e Ovinos, FUNCAP, CNPq e Banco do Nordeste
pelo apoio financeiro para realização deste projeto.
Referências Bibliográficas
ABREU, S. R. O., CASTRO, R. S., NASCIMENTO, S. A., 1998. Produção de antígeno
nucleoprotéico dos vírus da artrite encefalite caprina e comparação com os vírus
Maedivisna para imunodifusão em ágar-gel. Pesq. Vet. Bras. 18, 57-60.
ANDRIOLI, A. Vírus da Artrite Encefalite Caprina: PCR e Isolamento Viral em
amostras de sêmen, fluido uterino e embriões. Belo Horizonte – MG: Escola de
Veterinária – UFMG, 2001. Tese (Doutorado).
CORREL, M. D.; BRANDON, M. R.; SHEFFER D. et al., 1992. Ovine lentivirus is
macrophagetropic and does not replicate productively in Tlymphocytes. J. Virol. 66,
2679-2688.

67
CRAWFORD, T. B.; ADAMS, D. S., 1981. Caprine arthritis-encephalitis: clinical
features and presence of antibody in selected populations. J. Am. Vet. Med. Assoc., v.
178, 713-719.
FEITOSA, A.L.V.L., TEIXEIRA, M. F. S., PINHEIRO, R. R., et al., 2010.
Phylogenetic analysis of small ruminant lentiviruses from Northern Brazil. Small
Ruminant Res., 94(1-3), 205-209.
KARR BM, CHEBLOUNE Y, LEUNG K, NARAYAN O., 1996. Genetic
characterization of two phenotipically distinct North American ovine lentiviruses and
their possible origin from caprine arthritisencephalitis virus. Virology. 225(1),1–10.
MOOJEN, V., SOARES, H.C., RAVAZZOLO, A.P., PIZZOL, M., GOMES, M., 1986.
Evidência de infecção pelo lentivirus (Maedi-visna/artrite encefalite caprina) em
caprinos no Rio Grande do Sul, Brasil. Arq. Fac. Med. Vet. UFRGS. 14, 77-78.
PINHEIRO, R.R., Vírus da artrite encefalite caprina: Desenvolvimento e padronização
de ensaios imunoenzimáticos (ELISA e Dot-Blot) e estudo epidemiológico no Estado
do Ceará. Belo Horizonte: UFMG, 2001. 115p. Tese (Doutorado) – Escola de
Veterinária, Universidade Federal de Minas Gerais.
REINA, R., GREGO, E., PROFITI, M., GLARIA, I., ROBINO, P., QUASSO, A.,
AMORENA, B., ROSATI, S., 2009. Development of specific diagnostic test for small
ruminant lentivirus genotype E. Vet. Microbiol. 138, 251– 257.
SALTARELLI, M., QUERAT, G., KONINGS, D.A., VIGNE, R., CLEMENTS, J.E.,
1990. Nucleotide sequence and transcriptional analysis of molecular clones of CAEV
which generate infectious virus. Virology 179, 347– 364.
SHAH, C., BONI, J., HUDER, J.B., VOGT, H.R., MUHLHERR, J., ZANONI,
R.,MISEREZ, R., LUTZ, H., SCHUPBACH, J., 2004. Phylogenetic analysis and
reclassification of caprine and ovine lentiviruses based on 104 new isolates: evidence

68
for regular sheep-to-goat transmission and worldwide propagation through livestock
trade. Virology. 319 (1), 12–26.
TIGRE, D. M., CAMPOS, G. S., SILVIA, I. S., 2006. Isolamento e identificação do
vírus da Artrite encefalite caprina, a partir do cocultivo de células mononucleares do
sangue com células de membrana sinovial de cabra. R. Ci. Méd., biol. 5 (2), 124-131.
VALAS S, BENOIT C, BAUDRY C, PERRIN G, MAMOUN RZ., 2000. Variability
and immunogenicity of caprine arthritis-encephalitis virus surface glycoprotein. J Virol.
74(13),6178–85.

69
CAPÍTULO II
Isolamento do Vírus da Artrite Encefalite Caprina (CAEV) realizado por
cocultivo de leucócitos de caprinos naturalmente infectados provenientes
de estados do Nordeste e Sudeste (CE, PI, BA e MG) do Brasil
Isolation of Caprine arthritis-encephalitis virus (CAEV) through by
coculture of leukocytes from naturally infected goats from Brazil
Periódico: artigo a ser submetido.

70
Resumo
O Vírus da Artrite Encefalite Caprina (CAEV) e Vírus Maedi-visna (MVV)
pertencentes ao gênero Lentivírus da família Retroviridae, são considerados
geneticamente distintos, mas antigenicamente relacionados. O objetivo desde trabalho
foi isolar o vírus da CAE de caprinos positivos, de diferentes raças, idades e sexo,
oriundos dos rebanhos de quatro Estados do Brasil (Ceará e Piauí, Bahia e Minas
Gerais) que foram positivos pelo teste de Imunodifusão em Gel de Agarose (IDGA). Os
resultados positivos para o IDGA foram confirmados por Western Blotting. Para o
isolamento viral, leucócitos do sangue periférico foram destinados ao cocultivo em
Membrana Sinovial Caprina (MSC). Os leucócitos coletados foram inoculados em
monocamadas pré-formadas em garrafas A25. Após o cocultivo, foi realizada a
coloração da monocamada com cristal de violeta a 0,1% e a visualização do ECP
(Efeito Citopático). Os resultados demonstraram que os vírus isolados foram
identificados como sendo pertencente ao gênero lentivírus pela observação de células
gigantes multinucleadas (sincícios) em Membrana Sinovial Caprina (MSC),
característico para esse tipo de vírus em isolamento viral. Uma Nested-PCR realizada a
partir do sobrenadante do cocultivo confirmou a infecção por CAEV por amplificar
parte do gene gag. As cepas isoladas mostraram-se fenotipicamente líticas, do tipo
rápido/alto.
Palavras-chave: Cocultivo. Membrana Sinovial Caprina. CAEV.

71
Isolamento do Vírus da Artrite Encefalite Caprina através de cocultivo de
macrófagos no Brasil
1Aryana Lushese Vasconcelos Lima Feitosa;
2Maria Fátima da Silva Teixeira;
3Raymundo Rizaldo Pinheiro;
4Dalva Alana Aragão de Azevedo;
5Ronaldo Pereira
Dias, 3Alice Andrioli Pinheiro
1. Doutoranda do Programa de Pós-Graduação em Ciências Veterinárias; Laboratório de
Virologia, Universidade Estadual do Ceará – UECE,
2. Professora Orientadora do Programa de Pós-Graduação em Ciências Veterinárias;
Laboratório de Virologia, Universidade Estadual do Ceará – UECE; Bolsista de
Produtividade em Pesquisa do CNPq.,
3. Pesquisador Embrapa Caprinos e Ovinos, Laboratório de Virologia;
4. Bolsista de Iniciação Científica CNPq/PIBIC e FUNCAP, Universidade Estadual
Vale do Acaraú – UVA.
5. Mestrando do Programa de Pós-Graduação em Ciências Veterinárias; Laboratório de
Virologia, Universidade Estadual do Ceará – UECE.
* Corresponding author at: Laboratory of Virology, State University of
Ceará, Av. Paranjana, 1700, 60.740-903, Fortaleza, CE, Brazil.
Phone.: +55 8596332990; Fax: +55 8531019849.

72
Abstract
Caprine arthritis encephalitis virus and Visna/Maedi virus belong to Lentivirus genus of
Retroviridae family. They are considered genetically distinct but antigenically related.
The aim of the present study was to isolate the CAE virus from positive goats of
different breeds, ages and sex, which were positive when tested by agarose gel
immunodifusion (AGID). Cocultivation techniques of infected peripheral blood
leukocytes in goat synovial membrane (GSM) were performed in animals from the
herds of the Ceará, Piauí, Bahia and Minas Gerais states, Brazil. The positives results
were confirmed by Western Blotting. For virus isolation, peripheral blood leukocytes
were isolated for the cocultivation on GSM. The collected monocytes/macrophages
were inoculated into preformed monolayers in bottles A25. After cocultivation, the
monolayer was stained with crystal violet 0,1% followed by the observation of the CPE
(cytopathic effect). The lentivirus was identified as CAEV by observation of
multinucleated giant cells (syncytium) in GSM. A nested-PCR performed from the
supernatant of the cocultivation confirmed infection by CAEV amplifies a portion of
gag gene. The isolated strains were shown to be phenotypically like rapid/high.
Key-words: Cocultivation, goat synovial membrane (GSM), CAEV.

73
1.Introdução
A CAE é uma doença causada pelo Vírus da Artrite Encefalite Caprina (CAEV),
um retrovírus pertencente à família Retroviridae, subfamília Lentivirinae, gênero
Lentivírus e acomete caprinos de todas as raças, idades e sexos (CORK et al., 1974). A
sintomatologia nervosa, ou seja, a encefalite ocorre em cabritos, o que leva a quadros de
ataxia secundária e paresia. A forma articular é a mais comum e se manifesta
essencialmente nos adultos (RUSSO, 1984), bem como a forma pulmonar e mamária.
A doença acarreta grandes perdas econômicas para os caprinocultores, tais como
diminuição da produção láctea e do período de lactação, assim como perda de peso
ocasionada pela dificuldade de locomoção, redução do peso ao nascer e da taxa de
crescimento e morte (PINHEIRO et al., 2003), sendo recomendado o abate dos animais
infectados.
O isolamento e a análise filogenética de cepas regionais circulantes aumentam o
conhecimento da diversidade genética desses vírus e contribuem para o controle da
doença. O objetivo desde trabalho foi isolar o vírus da CAE, de animais positivos pelo
teste de Imunodifusão em Gel de Agarose (IDGA) e Western Blotting, por meio
cocultivo de leucócitos infectados do sangue periférico em Membrana Sinovial Caprina
(MSC), oriundos dos rebanhos dos Estados da Bahia, Ceará, Piauí e Minas Gerais.
Material e Métodos
IDGA e Western Blotting
Mil e duzentos caprinos e ovinos de diversas idades, raças e sexo dos estados da
Bahia, Ceará, Piauí e Minas Gerais, com suspeitas clínicas para CAE, foram testados

74
sorologicamente para a presença de anticorpos contra a proteína Gag do capsídeo (p28)
usando o kit IDGA produzido pela Embrapa Caprinos e Ovinos, este sendo previamente
testado contra o controle positivo do kit Americano (Caprine Arthritis-
Encephalitis/Ovine Progressive Pneumonia Antibody Test Kit. Veterinary Diagnostic
Technology, Inc® - USA). Os animais positivos no IDGA para a CAE foram
confirmados por Western Blotting, segundo a medodologia descrita por Pinheiro (2001).
Leucócitos do Sangue Periférico e Cultivo de MSC
Foram escolhidos 37 animais positivos por ambos os testes, IDGA e Western
blotting. Destes foram coletados 10 mL de sangue por venipuntura da jugular em tubo
com anticoagulante (EDTA) para obtenção dos leucócitos do sangue periférico, isolados
por centrifugação a 380 g por 10 minutos. Os eritrócitos foram hemolisados com 8 mL
de cloreto de amônio (0,84%), por duas vezes. O pellet foi lavado em 10 mL de solução
salina fosfato (PBS), centrifugado a 380 g por 5 min e, finalmente, ressuspendido em 1
mL de PBS para uso no cocultivo em células MSC.
Explantes das membranas sinoviais caprinas das articulações intercarpais de
membros anteriores de cabritos livres de CAEV foram cultivados em Meio Essencial
Mínimo (MEM-G), acrescido de 10% de Soro Fetal Bovino (SFB), incubadas a 37 oC
em atmosfera de 5% de CO2 (ABREU et al., 1998) em garrafas de cultura celular A25.
O meio foi trocado a cada 48-72 horas até a monocamada obter 90% de confluência.

75
Cocultivo de Leucócitos com MSC
Leucócitos (200 L) coletados foram inoculados em monocamadas pré-
formadas em garrafas A25 (capacidade 25 cm2) e incubados durante 60 minutos em
estufa 37 oC a 5% de CO2 acrescidos de 1 mL de meio essencial mínimo (MEM) sem
soro fetal bovino (SFB) (Sigma®; BDV Free). Após o período de incubação foram
adicionados 3,5 mL de MEM com 10% de soro fetal bovino. O meio foi trocado a cada
sete dias e a passagem foi realizada a cada 21 dias, com adição de 200 L de células da
MSC a cada troca de meio ou passagem. A monocamada foi examinada diariamente em
microscópio invertido para verificação de efeito citopático viral (ECP), formação de
sincícios, fusão nuclear ou vacuolização. Após 50 dias de cocultivo, foi realizada a
coloração da monocamada com cristal de violeta a 0,1% para visualização do ECP.
Extração de DNA proviral
O sobrenadante de cocultivo coletado a cada sete dias foi utilizado para extração
do DNA proviral. Uma proporção de 1:1 de PBS 1X foi utilizada para centrifugar 5 mL
de sobrenadante de cocultivo a 250 g por 5 minutos. Após a centrifugação, a fase
superior foi descartada e o pellet novamente lavado em PBS nas mesmas condições
anteriores. Em seguida, o pellet foi ressuspendido em 2 mL de tampão hipertônico,
durante 2 minutos a temperatura ambiente, para lise do citoplasma celular, seguido de
centrifugação a 1100 g por 10 minutos. O pellet foi ressuspendido em 2 mL de PBS e
submetido a centrifugação a 1100 g por 10 minutos. Por fim, o sedimento foi
novamente ressuspendido em 250 L de PCR buffer e homogeneizado. O material foi,
então, tratado com proteinase K (0,1 mg/mL), durante 60 minutos, em banho-maria a 56

76
oC. A proteinase K foi inativada termicamente em água fervente a 100
oC por 10
minutos e acondicionadas em freezer a -20 oC até a sua utilização.
Nested-PCR
Dois rounds de PCR (Nested-PCR) foram realizadas para amplificar um
fragmento de 187 pb de DNA proviral, correspondente ao gene gag de CAEV. Todos os
primers foram desenhados baseados na sequência padrão de CAEV-Cork publicadas
por Saltarelli et al., (1990). Tabela 1.
Tabela 1 - Sequências de primers utilizados nas reações de Nested-PCR e seus
respectivos tamanhos de fragmentos amplificados
Gene Primers Sequências Tamanho dos
fragmentos (bp)
gag
1º.
round
gag1
(senso) 5’-CAAGCAGCAGGAGGGAGAAGCTG-3’ 296
gag2
(antissenso) 5’-TCCTACCCCCATAATTTGATCCAC-3’
gag
2º.
round
gag3
(senso) 5’-GTTCCAGCAACTGCAAACAGTAGCAATG-3’ 187
gag4
(antissenso) 5’-ACCTTTCTGCTTCTTCATTTAATTTCCC- 3’
As reações de PCR foram realização em um volume final de 50 L. Estas
consistiam de 20 pmol para cada primer, 100 M de dNTPs, 1,5 mM de MgCl2, 1 X de
PCR buffer, 2 U de DNA polimerase Taq (Invitrogen, Carlsbad, CA) e 5 L ou 1 L de
DNA extraído para o primeiro ou segundo round, respectivamente.
As condições utilizadas na amplificação pela PCR foram de 94 oC por 5 min, 35
ciclos de 94 oC por 1min, 56
oC por 1 minuto s e 72 oC por 45 segundos, seguidos de

77
uma extensão final de 72 oC por 7 min, usando termocliclador BioRad®.
Para cada round de amplificação, foi utilizado um controle positivo referente ao
CAEV-Cork e um controle negativo sem amostra de DNA em paralelo com todas as
amostras. Os produtos obtidos foram analisados por eletroforese em gel de agarose a
1%, corado com brometo de etídio e visualizados em transiluminador de luz ultravioleta
com densidade óptica de 260 nm.
Resultados e Discussão
IDGA e Western Blotting
Dos 1.200 animais testados, 37 caprinos foram positivos pelo teste de IDGA e
confirmados por Western Blotting. Estes foram escolhidos para o isolamento viral por
apresentarem forte reação no IDGA e WB. Nenhum Ovino foi diagnosticado como
positivo para ambos os testes.
Cocultivo de leucócitos com MSC
O cocultivo de leucócitos com MSC apresentou formação de poucos sincícios na
monocamada, a partir do 27º dia de cocultivo, seguido por observação em microscópio
invertido. As cepas isoladas apresentaram fenotipagem altamente lítica, causando a
destruição quase total da monocamada desde os primeiros dias até o final do cocultivo,
sendo necessária a adição de mais células nas garrafas A25. A presença do efeito
citopático viral na monocamada de MSC foi observada, com presença de células
gigantes, multinucleadas, variando de 4 a 8 núcleos por célula, confirmando a presença
de vírus em células do sistema monocítico fagocitário (SMF) nos 37 isolados (Figura 1).

78
Nested-PCR
A Nested-PCR realizada a partir do sobrenadante do cocultivo confirmou a
infecção por CAEV por amplificar um fragmento de 187 pb do gene gag em todas as
coletas de sobrenadante do cocultivo (Tabela 2) em trinta e sete cepas de LVPR isoladas
dos Estados do Ceará, Piauí, Bahia e Minas Gerais.
Figura 1. Isolamento a partir de cocultivo de monócitos/macrófagos em MSC
apresentando efeito citopático característico do CAEV com presença de
células gigantes multinucleadas.

79
Os achados desses isolamentos virais foram compatíveis com outros
relacionados a amostras líticas de LVPR. A introdução de novas cepas merece atenção
especial, pois podem se apresentar mais virulentas que as nativas, além do que, muitos
agentes como os vírus de RNA, que não apresentam a função de leitura e correção
genética, podem ter altas taxas de mutação, de aproximadamente um milhão de vezes
mais que a de animais e plantas. Consequentemente há uma enorme diversidade de
populações de vírus de RNA que podem se tornar mais patogênicos e causarem novas
doenças quando recém-introduzidos em ambientes (ANDRIOLI, 2001).
Somente um pequeno número de LVPRs foi isolado no Brasil. O isolamento de
cepas regionais contribui para um melhor conhecimento da diversidade genética desses
vírus (FEITOSA et al., 2010). O conhecimento da variabilidade genética pode ajudar a
entender melhor os mecanismos de evolução molecular e heterogeneidade
epidemiológica da infecção. A variabilidade genotípica e fenotípica das cepas
circulantes pode ser útil para melhorar as ferramentas de diagnóstico, importantes para o
controle e erradicação da doença.
Dia do experimento Coleta de Sobrenadante Resultado Nested-PCR
Dia 0 Início do cocultivo -
Dia 07 1ª. Troca de Meio Positivo
Dia 14 2ª. Troca de Meio Positivo
Dia 21 1a. Passagem Positivo
Dia 28 4ª. Troca de Meio Positivo
Dia 36 5ª. Troca de Meio Positivo
Dia 43 2ª. Passagem Positivo
Dia 50 Coloração da monocamada -
Tabela 2. Correlação: dia do experimento, coleta de sobrenadante e resultado de PCR

80
É importante que a partir desse estudo seja realizada uma caracterização
molecular do genoma desses vírus isolados, mediante análise de seus diferentes genes
estruturais, bem como compará-los com outras sequências virais isoladas para
identificar a provável origem da infecção dos animais.
Conclusão
O vírus da Artrite Encefalite Caprina está presente nos Estados do Ceará, Piauí,
Bahia e Minas Gerais. As cepas isoladas revelaram-se fenotipicamente líticas, do tipo
rápido/alto. O cocultivo de monócitos/macrófagos em monocamadas de membrana
sinovial caprina demostrou ser uma técnica eficiente no isolamento de lentivírus de
pequenos ruminantes.
Apoio:
UECE, EMBRAPA Caprinos e Ovinos, FUNCAP, CNPq e Banco do Nordeste pelo
apoio financeiro para realização deste projeto.

81
Referências Bibliográficas
ABREU, S. R. O., CASTRO, R. S., NASCIMENTO, S. A. Produção de antígeno
nucleoprotéico dos vírus da artrite encefalite caprina e comparação com os vírus
Maedivisna para imunodifusão em ágar-gel. Pesquisa Veterinária Brasileira. v. 18, p.
57-60. 1998
ANDRIOLI, A. Vírus da Artrite Encefalite Caprina: PCR e Isolamento Viral em
amostras de sêmen, fluido uterino e embriões. Belo Horizonte – MG: Escola de
Veterinária – UFMG. Tese de Doutorado. 2001.
CORK, L. C.; HADLOW, W. J.; CRAWFORD, T. B.; GORHAM, J. R.; PIPER, R. C.
Infectious leukoencephalomyelitis of young goats. Journal Infeccio Disease, v. 129,
n.2, p.134-141, 1974.
FEITOSA, A.L.V.L., TEIXEIRA, M. F. S., PINHEIRO, R. R., et al. Phylogenetic
analysis of small ruminant lentiviruses from Northern Brazil. Small Ruminant Research.
v. 94, n. 1-3, p. 205-209. 2010.
KARR BM, CHEBLOUNE Y, LEUNG K, NARAYAN O. Genetic characterization of
two phenotipically distinct North American ovine lentiviruses and their possible origin
from caprine arthritisencephalitis virus. Virology. v. 225, n. 1, p. 1–10. 1996.
PINHEIRO, R. R.; CHAGAS, A. C. S.; ANDRIOLI, A.; ALVES, F. S. F. Viroses de
pequenos ruminantes. Sobral, CE: Embrapa Caprinos. 30 p. (Série Documentos,
46). 2003.

82
PINHEIRO, R.R., Vírus da artrite encefalite caprina: Desenvolvimento e padronização
de ensaios imunoenzimáticos (ELISA e Dot-Blot) e estudo epidemiológico no Estado
do Ceará. Belo Horizonte: UFM. 115p. Tese de Doutorado – Escola de Veterinária,
Universidade Federal de Minas Gerais. 2001.
REINA, R., GREGO, E., PROFITI, M., GLARIA, I., ROBINO, P., QUASSO, A.,
AMORENA, B., ROSATI, S. Development of specific diagnostic test for small
ruminant lentivirus genotype E. Veterinary Microbiology. v.138, p. 251– 257. 2009.
RUSSO, P. Virus de l’arthrite-encéphalite caprine (CAEV). Bréve Revue. Annales de
Recherches Vétérinaires, Paris, v. 15, n. 1, p. 3 – 6, 1984.
SALTARELLI, M., QUERAT, G., KONINGS, D.A., VIGNE, R., CLEMENTS, J.E.
Nucleotide sequence and transcriptional analysis of molecular clones of CAEV which
generate infectious virus. Virology. v. 179, p. 347– 364. 1990.
SHAH, C., BONI, J., HUDER, J.B., VOGT, H.R., MUHLHERR, J., ZANONI,
R.,MISEREZ, R., LUTZ, H., SCHUPBACH, J. Phylogenetic analysis and
reclassification of caprine and ovine lentiviruses based on 104 new isolates: evidence
for regular sheep-to-goat transmission and worldwide propagation through livestock
trade. Virology. v. 319, n. 1, p. 12–26.2004.
VALAS S, BENOIT C, BAUDRY C, PERRIN G, MAMOUN RZ.. Variability and
immunogenicity of caprine arthritis-encephalitis virus surface glycoprotein. The Journal
of Virology. v. 74, n. 1), p.6178–85.2000.

83
CAPÍTULO III
Análise Filogenética de Lentivírus de Pequenos Ruminantes no
Nordeste do Brasil
Phylogenetic analysis of small ruminant lentiviruses from
Northern Brazil
Periódico: Small Ruminant Research. 94: 205–209, 2010

84
Resumo
O Vírus da artrite encefalite caprina (CAEV) e o Vírus Maedi-visna (MVV) são
considerados geneticamente distintos, mas antigenicamente relacionados. Neste estudo,
um rebanho de 250 cabras foi testado para o IDGA, sendo observada uma
soroprevalência de 4.4% (11 animais soropositivos). Quatro caprinos positivos para o
IDGA foram testados para a Reação em Cadeia da Polimerase (PCR), que amplifica
parte do gene gag. Todos os animais foram positivos para o teste biomolecular. As
sequências nucleotídicas do gene gag foram determinadas a partir de quatro lentivírus
BR/CNPC isolados de caprinos naturalmente infectados. As quatro sequências do gene
gag BR/CNPC de LVPRs foram comparadas com sequências do GenBank e os
resultados demonstraram que estas estão mais relacionadas com linhagens caprinas que
ovinas. A análise filogenética de parte do gene gag mostrou que estes isolados
pertencem ao subtipo B1 do grupo de CAEV. A análise também indicou que estes vírus
são geneticamente estáveis.

85
Phylogenetic analysis of small ruminant lentiviruses from Northern Brazil
Aryana Lushese Vasconcelos Lima Feitosa a,*, Maria Fátima da Silva Teixeira a,
Raymundo Rizaldo Pinheiro b,**, Rodrigo Maranguape Silva da Cunha c, João Paulo
Matos Santos Lima d, Alice Andrioli b, Tânia Valeska Medeiros Dantas a, Valeska
Shelda Pessoa de Melo a, Diana Célia Sousa Nunes Pinheiro a
a Laboratory of Virology and Immunology – Veterinary Faculty UECE – State
University of Ceará, Fortaleza, Brazil
b National Goat Research Center, Embrapa-CNPC, Brazil
c Center of Biotechnology of Sobral, NUBIS, Brazil
d Laboratory of Bioinformatics – Universidade Federal do Ceará – UFC, Brazil
* Corresponding author at: Laboratory of Virology, State University of
Ceará, Av. Paranjana, 1700, 60.740-903, Fortaleza, CE, Brazil.
Tel.: +55 8596332990; fax: +55 8531019849.
** Corresponding author at: Embrapa Goats, Fazenda Três Lagoas,
Estrada Sobral-Groaíras Km 4, P.O. Box 145, 62011-970, Sobral, CE, Brazil.
Tel.: +55 8596332990; fax: +55 8531019849.
E-mail addresses: [email protected] (A.L.V.L. Feitosa),
[email protected] (R.R. Pinheiro).
0921-4488/$ – see front matter © 2010 Elsevier B.V. All rights reserved.
doi:10.1016/j.smallrumres.2010.07.020

86
Abstract
Caprine Arthritis Encephalitis virus (CAEV) and Visna/maedi virus (VISNA), are
considered to be genetically distinct but antigenically related pathogens of goats and
sheep. In this study, one flock of 250 goats was screened using Agar-gel
Immunodiffusion (AGID), and the level of seroprevalence observed was 11 animals
(4.4%) of positive tested serum. Four goats positive to AGID were analyzed using a
Polymerase Chain Reaction (PCR), which amplifies part of the gag gene. All the
animals were found to be positive. The nucleotide sequences of gag gene were
determined from four BR/CNPC lentivirus isolates from naturally infected goats. The
four gag gene sequences BR/CNPC small ruminant lentivirus (SRLV) were compared
to GenBank, and the results demonstrated that these sequences were more related to the
caprine rather than the ovine strains. Further phylogenetic analysis of the proviral gag
sequences showed that they constituted subtype B1 of the CAEV group. The analyses of
the sequences also showed that the viruses were genetically stable.
Keywords: Small ruminant lentiviruses, CAEV, VISNA, Phylogeny, Gag

87
1. Introduction
Small ruminant lentivirus (SRLVs) is distributed worldwide among sheep and
goats. Different clinical forms of the disease have been described in Brazil among
SRLV-infected small ruminants. CAEV and VISNA are considered to be genetically
distinct, but antigenically related pathogens of goats and sheep. According to a recently
proposed nomenclature based on the 1.8 kb gag–pol and 1.2 kb pol gene sequences, the
SRLV are classified into four equidistantly related groups, A–D (Shah et al., 2004). An
SRLV belonging to the highly divergent genotype E has recently been identified in an
Italian goat breed (Reina et al., 2009). The high level of heterogeneity of nucleotide and
amino acid sequences in lentiviruses may determine their antigenicity, virulence and
growth, as well as their persistence and escape from immunesystem. Despite differences
in the target organs, cell tropism and pathogenicity commonly found between the
CAEV and the ovine VISNA, both viruses have recently been assigned to small
ruminant lentiviruses (SRLV). A genetic analysis of these SRLVs may provide
additional insights into the genetic, protein and antigenic makeup of these viruses, their
pathogenesis, epidemiology, and phylogenetic relationships, and thus, their position in
the recently established SRLV groups (Shah et al., 2004). Genetic studies mayalso be
relevant for the development of sensitive and local-breed diagnostic tests. The main aim
of this work was to establish an SRLV phylogeny as a basis for future studies of the
molecular epidemiology of lentiviral infections in Northeast Brazil dairy goat herds,
with particular interest in monitoring the effects of ongoing and future eradication
programs on the distribution of SRLV strains.

88
2. Materials and methods
2.1. Animals and AGID
Two hundred and fifty naturally infected goats of the Saanen and Anglo-nubiana
breeds were selected from a dairy flock belonging the National Goat Research Center
(CNPC), where, according to a previous serological survey, there was a known chronic
infection of goats with CAEV. The flock was located in Northeast Brazil. The
seroprevalence was determined using directed antibodies against the major Gag capsid
protein of both VISNA and CAEV using the AGID detection Kit (Caprine Arthritis-
Encephalitis/Ovine Progressive Pneumonia Antibody Test Kit, Veterinary Diagnostic
Technology, Inc.®, USA). Sensitivity was 65.6% and specificity was 94.1%.
2.2. Isolation of peripheral blood leukocytes
Peripheral blood leukocytes (PBL) were isolated from the blood by
centrifugation at 1500 rpm for 10 min. The erythrocytes were hemolyzed by osmotic
shock, the cell pellets were used for co-culture with goat sinovial membrane (GSM)
cells.
2.3. Co-culture
Cells derived from the GSM were used to propagate monolayer cultures of cells
in six-well microtiter plates. The medium was changed until there was a monolayer of
90% semiconfluent cells. The leukocytes were isolated from each AGID-positive animal
and were placed in each well, added with 500 l of MEM without fetal bovine serum

89
(FBS) (Sigma, BDV Free). The culture plate remained in the CO2 incubator for 1 h,
added by 1 mL of MEM with 5% FBS was then changed to the wells and the cells were
incubated until there were cytopathic effects (CPE) characteristic of viruses.
2.4. DNA preparation
The culture plate containing the virus-infected cell culture was frozen at -80 oC
and thawed at room temperature three times. The purified DNA stocks were stored at -
20 oC until use.
2.5. Nested-PCR and DNA quantification
Two rounds of PCR amplification (nested-PCR) were used to detect the 187 bp
proviral DNA fragment, corresponding to the leader gag sequence of the CAEV
genome. All of the primers were selected on the basis of the published sequence of the
CAEV-Co strain (Saltarelli et al., 1990). The primers GEX5 (5´-
CAAGCAGCAGGAGGGAGAAGCTG-3´) and GEX3 (5´-
TCCTACCCCCATAATTTGATCCAC-3´) were used for the first amplification,
corresponding to bases 953–975 and the complement of 1249–1226, respectively.
Primers GIN5 (5´-GTTCCAGCAACTGCAAACAGTAGCAATG-3´) and GIN3 (5´-
ACCTTTCTGCTTCTTCATTTAATTTCCC-3´-) were used for the second
amplification, corresponding to bases 997–1024 and the complementof 1181–1154,
respectively. For each amplification, a positive control of CAEV-Co DNA and a no-
template negative control were run in parallel with all of the specimens.

90
2.6. DNA sequencing and sequence analysis
The products of the PCR amplification from the first round were sequenced
using Mega Base 7.0. The sequencing was performed using the external primers GEX5
(5´-CAAGCAGCAGGAGGGAGAAGCTG-3´) and GEX3 (5´-
TCCTACCCCCATAATTTGATCCAC-3´). The bases were called using Phred version
0.020425 (Ewing and Green, 1998; Ewing et al., 1998) and subsequently, the sequence
reads were aligned and a single consensus sequence was assembled with Phrap version
0.990329 (Phil Green, University of Washington).
All of the positions with ambiguous codes or alignment gaps were excluded
from the analyses. The editor program Consed version 16.0 (Gordon et al., 1998) was
used to prepare the final version of the consensus sequences. The pairwise genetic
distances were calculated by using MEGA version 4.0 (Tamura et al., 2007) with the
Tamura–Nei (NJ) substitution model. The phylogenetic tree was determined using the
neighbor joining (NJ) method (Saitou and Nei, 1987) implemented in MEGA, with the
Tamura–Nei gamma distance (Tamura and Nei, 1993). The SRLV strains were
compared to the K1514-VMV (Sonigo et al., 1985), EV-1 VMV (Sargan et al., 1991),
SAOVMV (Querat et al., 1990), P1OLV (Barros et al., 2004), CAEV (Saltarelli et al.,
1990), BR/MG (Drumond and Resende, 2001) and BR/UFRGS (Ravazzolo et al., 2001)
strains (GenBank accession numbers: M60609, S51392, M31646, AF479638, M33677,
AF402668 and AJ305040, respectively).

91
3. Results
3.1. Serology
The results of the AGID test showed that 11 (4.4%) of the 250 goats were
AGID-positive for SRLV. All these animals AGID-positive were descending of animals
of the French flock.
3.2. Co-culture
Between 14 and 21 days after infection, the characteristic CPE, syncytium was
formed, confirming the presence of SRLV.
3.3. Nested-PCR
The presence of SRLV in the goat synovial membrane cells was confirmed by
the detection of a gag fragment by nested-PCR. Four to 11 samples were amplified of
the naturally infected animals. A 187-nucleotide region of the gag SRLV gene (997–
1181) was detected by an electrophoretic analysis of nested-PCR products. The
amplification products were sequenced and denoted as BR/CNPC-G1, BR/CNPC-G2,
BR/CNPC-G3, BR/CNPC-G4 (BR, Brazil; CNPC, National Goat Research Center; G,
goat).

92
3.4. DNA sequencing and phylogenetic analysis
The phylogenetic analysis revealed that the BR/CNPC viruses formed a cluster
of identical or highly related sequences within the genetically heterogeneous CAEV-Co
group. A phylogenetic tree (Fig. 1) was obtained using the NJ method with the gag
nucleotide sequences, and this tree showed the relationship between BR/CNPC and the
other known SRLV strains. The bootstrap values derived from the 100 bootstrap
replicates indicating the confidence of the branching pattern are shown above the
respective branches. All of the analysis parameters were standard. The analyses also
showed a close relationship between the prototype CAEV-Co strain and the two
Brazilian strains, CAEVBrUFRGS and the CAEVBR-MG. All of the BR/CNPC SRLV
strains had a similar TTG trinucleotide at positions 19–21 of the gag gene, which would
result in a leucine in the gag protein from the SRLV BR/CNPC stains, compared to a
leucine in the same position in the CAEV-Co protein (Fig. 2). This nucleotide change
would result in the translation of a different amino acid, isoleucine in CAEVCo and
leucine in the four BR/CNPC SRLV strains (Fig. 2). The pairwise distance calculations
were obtained by comparing the nucleotide and amino acids sequences among the four
BR/CNPC strains and known SRLV strains. The values closest to zero represent the
strains that are the most similar.

93
Fig. 1. In the phylogenetic tree a group identified by the prototype VISNA was clearly
distinguished from the CAEVs with a high bootstrap value of 100.
3.5. Nucleotide sequence accession number
All of the new sequences were entered in the GenBank database and are
available under check numbers EU300976, EU300977, EU300978 and EU300979.
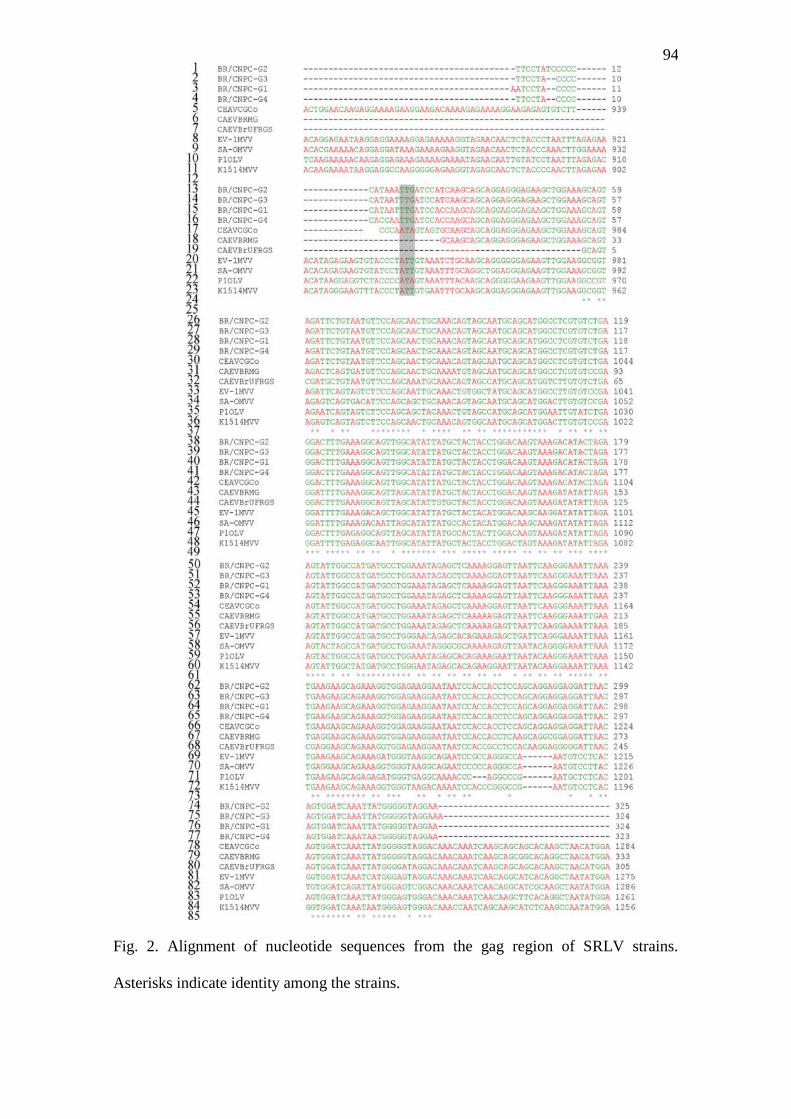
94
Fig. 2. Alignment of nucleotide sequences from the gag region of SRLV strains.
Asterisks indicate identity among the strains.

95
Fig. 3. Alignment of deduced amino acid sequences from the capsid protein of SRLV
strains. Asterisks indicate identity between the strains.
4. Discussion
The four gag gene sequences from the BR/CNPC SRLV strains were compared
to available GenBank strains, and the results of this comparison demonstrated that these
sequences were more related to the caprine strains than to the ovine strains. Further
phylogenetic analyses of the proviral gag sequences confirmed the initial classification
and showed that these sequences were of the B1 subtype of the CAEV group. The
analyses of the sequences also showed viruses were genetically stable, which was
previously observed by Laamanen et al. (2007). Although some authors indicate that
future studies should concentrate on the env regions, which are sufficiently divergent,
rather than the gag region, because the gag region is highly conserved and retains less
phylogenetic signal (Rolland et al., 2002), but this study cannot infer the results only
with the sequence gag, being necessary more comparative studies with other sequences
of viruses.

96
All of the four SRLV BR/CNPC strains had the same TTG trinucleotide at
positions 19–21 of the capsid peptide, which was different from the sequence in CAEV-
Co. This nucleotide change would result in the translation of a different amino acid,
isoleucine in CAEV-Co and leucine in the four BR/CNPC SRLV strains (Figs. 2 and 3).
There was an absence of transversions, but transitions were observed. The high level of
similarity among the four sequences from Northeast Brazil and the CAEV-Co strain
maybe explained by vertical transmission through infected females over the generations.
The negative results from these tests suggest that the animals were already
infected prior to the tests and had not produced antibodies that could be detected in the
test that was used (data unpublished).
This gag region of the genome may be used to differentiate the viral strains
present in infected animals and to develop more specific diagnostic tests for local
breeds. It may also be informative for the establishment of eradication programs
according to the distribution of different lineages of SRLV and the development of
diagnostic reagents with greater specificity and sensitivity in local flocks (Germain and
Valas, 2006), different of the sequences gag gene CAEV-Cork researched by Saltarelli
et al. (1990). The results indicate that more studies should be conducted on viral
genomic area obtained from animals of Northeast Brazil.
5. Conclusion
Eleven animals were AGID-Positive. Four goats positive to AGID were
analyzed using a PCR, which amplifies part of the gag gene. The nucleotide sequences
of gag gene were determined from four BR/CNPC lentivirus isolates from naturally

97
infected goats. The four gag gene sequences BR/CNPC small ruminant lentivirus
(SRLV) were compared to GenBank, and the results demonstrated that these sequences
were more related to the caprine rather than the ovine strains. Further phylogenetic
analysis of the proviral gag sequences showed that they constituted subtype B1 of the
CAEV group. The alignment of deduced amino acid sequences from the capsid protein
of SRLV strains, indicates a divergence between the strains in this study and the strain
CAEV-Co. This divergence is important for a better understanding of the local
sequences previously described.
Acknowledgements
Laboratory of Virology and Laboratory of Immunology – Veterinary Faculty
UECE – State University of Ceará; Laboratory of Bioinformatics – Federal University
of Ceará (UFC), Fortaleza, Brazil; National Goat Research Center (CNPCEmbrapa);
Center of Biotechnology of Sobral (NUBIS); CAPES, FUNCAP and Banco do
Nordeste.
References
Barros, S.C., Ramos, F., Duarte, M., Fagulha, T., Cruz, B., Fevereiro, M., 2004.
Genomic characterization of a slow/low Maedi visna virus. Virus Genes (29), 199–210.
Drumond, B.P., Resende, M., 2001. Phylogenetic analysis of small ruminant
lentiviruses in naturally infected goats, based on sequence of capsid protein gene. In: 4◦.

98
Encontro de Virologia do Mercosul e XII Encontro Nacional de Virologia. Caldas
Novas – GO. Virus Rev. Res. 6, 172.
Ewing, B., Green, P., 1998. Base calling of automated sequencer traces using phred. II.
Error probabilities. Genome Res. (8), 186–194.
Ewing, B., Hiller, L., Wendl, M.C., e Green, P., 1998. Base-calling of automated
sequencer traces using phred. I. Accuracy assessment. Genome Res. (8), 175–185.
Gordon, D., Abajian, C., Green, P., 1998. Consed: a graphical tool for sequence
finishing. Genome Res. (8), 195–202.
Germain, K., Valas, S., 2006. Distribution and heterogeneity of small ruminant
lentivirus envelope subtypes in naturally infected French sheep. Virus Res. (120), 156–
162.
Laamanen, I., Jakava-Viljanen, M., Sihvonen, L., 2007. Genetic characterization of
maedi-visna virus (VMV) detected in Finland. Vet. Microbiol. (122), 357–365.
Querat, G., Audoly, G., Sonigo, P., Vigne, R., 1990. Nucleotide sequence analysis of
SA-OVMV, a visna-related ovine lentivirus: phylogenetic history of lentiviruses.
Virology (175), 434–447.
Ravazzolo, A.P., Reischak, D., Peterhans, E., Zanoni, R., 2001. Phylogenetic analysis of
small ruminant lentiviruses from Southern Brazil. Virus Res. (79), 117–123.

99
Reina, R., Grego, E., Profiti, M., Glaria, I., Robino, P., Quasso, A., Amorena, B.,
Rosati, S., 2009. Development of specific diagnostic test for small ruminant lentivirus
genotype E. Vet. Microbiol. 138, 251–257.
Rolland, M., Mooney, J., Valas, S., Perrin, G., Mamoun, R.Z., 2002. Characterisation of
an Irish caprine lentivirus strain-SRLV phylogeny revisited. Virus Res. (85), 29–39.
Sargan, D.R., Bennet, I.D., Cousens, C., Roy, D.J., Blackaws, B.A., Dalziel, R.G., Watt,
N.J., Mcconnell, I., 1991. Nucleotide sequence of EV1, a British isolate of Maedi-visna
virus. J. Gen. Virol. 72, 1893–1903.
Saitou, N., Nei, M., 1987. The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Saltarelli, M., Querat, G., Konings, D.A., Vigne, R., Clements, J.E., 1990. Nucleotide
sequence and transcriptional analysis of molecular clones of CAEV which generate
infectious virus. Virology 179, 347–364.
Shah, C., Boni, J., Huder, J.B., Vogt, H.R., Muhlherr, J., Zanoni, R., Miserez, R., Lutz,
H., Schupbach, J., 2004. Phylogenetic analysis and reclassification of caprine and ovine
lentiviruses based on 104 new isolates: evidence for regular sheep-to-goat transmission
and worldwide propagation through livestock trade. Virology (319), 12–26.

100
Sonigo, P., Alizon, M., Staskus, K., Klatzmann, D., Cole, S., Danos, O., Retzel, E.,
Tiollais, P., Haase, A., Wain-Hobson, S., 1985. Nucleotide sequence of the visna
lentivirus: relationship to the AIDS virus. Cell (42), 369–382.
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: Molecular Evolutionary
Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24, 1596–1599,
www.megasoftware.net.
Tamura, K., Nei, M., 1993. Estimation of the number of nucleotide substitutions in the
control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol.
(10), 512–526.

101
CAPÍTULO IV
Caracterização Molecular de Lentivírus de Pequenos Ruminantes Isolados
de Caprinos no Brasil baseados na análise das sequências dos genes gag e
pol
Molecular characterization of Small Ruminant Lentiviruses isoleted from
Brazil goats based on gag and pol gene sequence analysis
Periódico: artigo a ser submetido

102
Resumo
A CAE é uma doença causada pelo Vírus da Artrite Encefalite Caprina (CAEV),
um retrovírus pertencente à família Retroviridae, subfamília Lentivirinae, gênero
Lentivírus e acometem caprinos, de todas as raças, idades e sexos. A sintomatologia
nervosa, ou seja, a encefalite ocorre em cabritos, o que leva a quadros de ataxia
secundária e paresia. A sintomatologia articular é a mais comum e se manifesta
essencialmente nos adultos, bem como a forma pulmonar e mamária. O objetivo desde
trabalho foi isolar e sequenciar cepas circulantes do vírus da CAE. Trinta e oito isolados
de lentivírus caprino foram selecionados para o isolamento viral através de cocultivo de
leucócitos. Destes, foram sequenciadas amostras nos estados do Ceará, Rio Grande do
Norte e Minas Gerais. Estas foram comparadas com outras já conhecidas depositadas no
Genbank. As amostras circulantes apresentam elevada conservação nucleotídica, sendo
99,3% para o gene gag quando comparado com a cepa padrão CAEV-Cork e
relativamente conservado, 70,1% para o gene pol, apresentando, portanto uma
diversidade intragenotípica. Pela genealogia proposta com base nas sequências
nucleotídicas dos genes gag e pol, a sua topologia mostrou-se compatível com os
genótipos do subtipo B de CAEV. O alinhamento das sequências de aminoácidos da
região gag, que correspondem à proteína p28 do capsídeo, indicou divergências entre as
linhagens deste estudo com a cepa padrão CAEV-Cork.
Palavras-chave: Filogenia, CAEV.

103
Caracterização Molecular de Lentivírus de Pequenos Ruminantes
Isolados de Caprinos no Brasil baseados na análise das sequências dos
genes gag e pol
1Aryana Lushese Vasconcelos Lima Feitosa;
2Maria Fátima da Silva Teixeira;
3Raymundo Rizaldo Pinheiro;
3Alice Andrioli Pinheiro,
4Dalva Alana Aragão de
Azevedo; 5Paulo Eduardo Brandão,
5Fábio Gregori,
5Alessandra Marnie Martins
Gomes de Castro, 5Sheila Oliveira de Souza Silva
1Autor correspondente: Programa de Pós-Graduação em Ciências Veterinárias;
Laboratório de Virologia, Universidade Estadual do Ceará – UECE, Av. Paranjana
1700, 60.740-093, Fortaleza, CE, Brasil, Tel.: +55 85 96332990; Fax: +55 85
31019849. [email protected]
2Programa de Pós-Graduação em Ciências Veterinárias; Laboratório de Virologia,
Universidade Estadual do Ceará – UECE, Pesquisador de Produtividade em Pesquisa
CNPq,
3Embrapa Caprinos e Ovinos, Laboratório de Virologia;
4Iniciação Científica CNPq/PIBIC e FUNCAP, Universidade Estadual Vale do Acaraú –
UVA.
5Laboratório Aplicado à Biologia Molecular e Sorologia – LABMAS, Faculdade de
Medicina Veterinária e Zootecnia – FMVZ-VPS, Universidade de São Paulo - USP

104
Abstract
CAE is a disease caused by caprine arthritis encephalitis virus (CAEV), a retrovirus
belonging to the family Retroviridae, subfamily Lentivirinae, and gender Lentivirus. It
affects lentiviruses and goats of all breeds, ages and sex. The neurological symptoms,
ie, encephalitis occurs in young goats, which leads to secondary ataxia and paresis. The
articular symptomatology is the most common and it affects adults mainly, as well as
pulmonary and mammary forms. The disease causes huge economic losses, such as
decreased milk production and lactation, weight loss caused by limited mobility,
reduced birth weight, growth rate and death, leading to the slaughter of infected
animals. The aim of the present study was to isolate and sequence circulating strains of
CAEV. Thirty-eight isolates of goat lentivirus were selected for virus isolation by
cocultivation of leukocytes. These samples were sequenced in the states of Ceara, Rio
Grande do Norte and Minas Gerais. These were compared with other known sequences
deposited in Genbank. The samples show high circulating nucleotide conservation of
99.3% for the gag gene as compared to the standard strain CAEV-Cork and is relatively
conservative, 70.1%, for the pol gene, showing an intragenotype diversity. Through the
proposed genealogy based on the nucleotide sequences of gag and pol genes, their
topology was compatible with the genotypes of subtype B of CAEV. Alignment of the
amino acid sequences of the gag region, corresponding to the p28 capsid protein,
indicated differences between strains of this study with the standard strain CAEV-Cork.
This divergence of amino acids was even more pronounced when compared to the
amino acid sequences of the pol gene, which codifies for dUTPase and integrase
proteins. These differences are important for a better understanding of the condition and
diversity of these described sequences.
Key-words: Phylogeny, CAEV, gene pol, gene gag

105
1. Introdução
O virus da artrite encefalite caprina (CAEV) e o virus Maedi-visna (MVV) são
membros do grupo de Lentivírus de Pequenos Ruminantes (LVPRs), pertencem à
família Retroviridae, infectando caprinos e ovinos mundialmente (Pasick, J. 1998;
Pepin et al., 1998). Os Lentivírus de espécies de animais diferentes têm em comum sua
organização genética, a indução de doenças de forma progressiva e lenta e um amplo
espectro de órgãos-alvo, sinais, sintomas e a capacidade de persistir em seus
hospedeiros apesar da forte resposta imunológica.
A transmissão dos LVPRs ocorre principalmente pela ingestão de leite ou
colostro infectado, mas, pelo menos em ovinos, a transmissão horizontal pode ter um
papel importante (Rowe J. e East N., 1997; Narayan O. et al., 1985). Os sinais clínicos
causados pela infecção por LVPRs incluem desordens neurológicas, dispnéia,
emaciação, mastite e artrite (Pepin M. et al., 1998; Narayan O. et al., 1989; Alvarez V.
et al., 2006).
A organização genômica de LVPRs é típica de lentivírus: dímeros de RNA de
senso positivo com aproximadamente 9 kb de tamanho que consiste de duas LTRs
(sequências longas repetidas) nas extremidades e flanqueando genes gag, que codificam
as proteínas estruturais e são responsáveis por um grupo de antígenos específicos (Pépin
et al., 1998): pol (polimerase) que codifica as proteínas de atividade enzimática; env
(envelope), que codifica as glicoproteínas de superfície (SU) e proteínas estruturais
transmembranárias (TM) do envelope viral, além de pequenos ORFs, que codificam
proteínas que regulam a expressão gênica (Clements e Payne 1994). Os genes gag e pol
são relativamente bem conservados entre os LVPRs e por isso estes genes são alvos
ideais para desenhos de primers para PCRs (Pépin M., el al., 1998).

106
Originalmente, CAEV e MVV são linhagens protótipas e foram vistas como
espécies virais distintas restritas aos seus respectivos hospedeiros, ou seja, vírus
isolados de ovinos estão relacionados ao MVV e vírus isolados de caprinos relacionados
com CAEV. Porém nas últimas duas décadas, com mais sequências disponíveis em
bancos de dados genéticos, tornou-se evidente que os lentivírus de pequenos ruminantes
podem atravessar a barreira entre espécies com vírus caprino infectando ovino e vice-
versa, uma vez que algumas cepas de caprino e ovino compartilham o mesmo cluster
nas árvores filogenéticas.
Apesar do fato dos lentivírus circularem e causarem doenças em caprinos e
ovinos no Brasil há mais de quatro décadas, poucas sequências para caracterização
molecular das linhagens de vírus circulantes no país foram obtidas. O objetivo deste
estudo foi caracterizar molecularmente sequências parcias do gene gag e pol isolados de
caprinos naturalmente infectados em três estados brasileiros.
Material e Métodos
Animais e IDGA
Caprinos infectados naturalmente, de diferentes raças, idades e sexo, dos estados
de Minas Gerais, Rio Grande do Norte e Ceará, Brasil, foram testados pelo IDGA para
o vírus da artrite encefalite caprina utilizando o kit de detecção de anticorpos contra a
proteína Gag do capsídeo (Caprine Arthritis-Encephalitis/Ovine Progressive Pneumonia
Antibody Test Kit. Veterinary Diagnostic Technology,Inc® - USA). A sensibilidade do
teste foi de 65.6% e a especificidade foi de 94.1%. Os animais positivos no IDGA para
CAEV foram confirmados por Western Blotting (WB), segundo a metodologia de
Pinheiro (2001).

107
Isolamento de leucócitos do Sangue Periférico
Foram escolhidos 38 animais positivos para ambos os testes, IDGA e WB.
Leucócitos do sangue periférico (LSP) foram isolados do sangue por centrifugação a
1500 rpm por 10 min. Os eritrócitos foram hemolizados por choque osmótico e o pellet,
contendo as células, foram usados no cocultivo de células da membrana sinovial caprina
(MSC).
Cocultivo
Explantes das membranas sinoviais caprinas das articulações intercarpais de
membros anteriores de cabritos livres de CAEV foram cultivados em Meio Essencial
Mínimo (MEM-G), acrescido de 10% de Soro Fetal Bovino (SFB), incubadas a 37 oC
em atmosfera de 5% de CO2 (Abreu et al., 1998) em garrafas de cultura celular A25. O
meio foi trocado a cada 48-72 horas até a monocamada obter 90% de confluência.
Monócitos/Macrófagos (200 mL) coletados foram inoculados em monocamadas pré-
formadas em garrafas A25 (capacidade 25 cm2) e incubados durante 60 minutos em
estufa 37 oC a 5% de CO2 acrescidos de 1 mL de meio essencial mínimo (MEM) sem
soro fetal bovino (SFB) (Sigma®; BDV Free).
Após o período de incubação foram adicionados 3,5 mL de MEM com 10% de
soro fetal bovino. O meio foi trocado a cada sete dias e a passagem foi realizada a cada
21 dias, com adição de 200 L de células da MSC a cada troca de meio ou passagem. A
monocamada foi examinada diariamente em microscópio invertido para verificação de
efeito citopático viral (ECP), formação de sincícios, fusão nuclear ou vacuolização.
Após 50 dias de cocultivo, foi realizada a coloração da monocamada com cristal de

108
violeta a 0,1% para visualização do ECP. As garrafas de cultura contendo as células
infectadas pelo vírus foram congeladas a -80 oC.
Extração de DNA
A extração de DNA foi realizada a partir de sobrenadante de cocultivo de
macrófagos/monócitos em MSC (200 L) utilizando 600 uL de tiocianato de guanidina
acrescido de fenol pH 7,5. A suspensão foi homogeneizada em vórtex por 15 segundos
e adicionado 100 L de clorofórmio, seguido de homogeneização em vórtex e
centrifugação a 12.000 g por 5 minutos a 4 oC. O sobrenadante (400 L) foi transferido
para um outro tudo eppendorf contendo 600 L de propanol, novamente
homogeneizado em vórtex por 15 segundos e acondicionada em freezer a -20 oC por
duas horas.
Após essa etapa, a suspensão foi centrifugada a 12000 g por minutos a 4 oC e
descartado o sobrenadante. Ao pellet foi adicionado 500 L de etanol a 70% e
homogeneizado em vórtex por 15 segundos e centrifugação sob as mesmas condições
supracitadas. O sobrenadante foi descartado e o pellet foi seco por 15 minutos em
termobloco. Este último foi ressuspendido em 30 L de TE (Tris 10 mM/ EDTA 1 mM
pH 8,0), incubado a 56 oC por 15 minutos, seguidos de homogeneização e spin
(Adaptado de Chomczynski, 1993).

109
Reação em Cadeia da Polimerase (PCR)
A reação de amplificação (PCR) foi realizada para detectar um fragmento de 620
pb do DNA proviral, correspondente ao fragmento parcial do gene gag, e de
aproximadamente 472 pb do gene pol do genoma de CAEV. Os primers utilizados na
PCR para ambos os genes estão expostos na Tabela 1.
Tabela 1 - Sequências de primers utilizados nas reações de PCR e seus respectivos
tamanhos de fragmentos amplificados
Gene Primers Sequências Tamanhos dos
fragmentos (bp)
Referências
gag
Tvet-1
(senso) 5´AGC TGG AAA GCA GTA GAT TC 3´
620 Dantas et al.,
(2009)
Tvet-2
(antissenso) 5´GAA CCC TTC TGA TCC CAC ATC 3´
pol
Pol 1
(senso) 5’ ACA GAA GAG AAA TTA AAAGG-3’
472
Leroux et al.,
(1997)
Pol 2
(antissenso) 5’ CAT CCA TAT ATA TGC CAA ATT G-3’
As reações de PCR foram realização em um volume final de 50 L. Estas
consistiram de 20 pmol para cada primer, 100 M de dNTPs, 1,5 mM de MgCl2, 1X de
PCR buffer, 2 U de DNA polimerase Platinum Taq (Invitrogen, Carlsbad, CA) e 10 L
de DNA extraído.
As condições utilizadas na amplificação pela PCR foram de 94 oC por 5 min, 40
ciclos de 94 oC por 1min, 53
oC por 45 s e 72 oC por 1min, seguidos de uma extensão
final de 72 oC por 8 min.

110
Detecção do amplicon
Os fragmentos amplificados pela PCR foram analisados em eletroforese em cuba
horizontal. Uma alíquota de 10 L de cada amostra foi disposta em gel de agarose a
1,5% previamente imerso em tampão TBE (0,045 M Tris-Borato; 1 mM EDTA). Após
a corrida eletroforética, o gel foi corado em solução de brometo de etídio (0,5 g/ mL)
por 20 minutos. As bandas foram visualizadas em transiluminador com luz ultravioleta
de 260 nm. Os tamanhos dos fragmentos amplificados foram comparados ao padrão de
peso molecular de 100 pb (ladder).
Purificação e Quantificação do amplicon
As bandas de interesse foram cortadas e purificadas utilizando o kit GFXTM
(Amersham Biosciences) de acordo com as indicações do fabricante. No caso de bandas
únicas, foi utilizado o reagente de purificação Exo SAP-IT TM
(GE Healthcare Life
Sciences) também de acordo com as instruções do fabricante.
A concentração do DNA amplificado presente nas amostras purificadas foi
determinada por densidade óptica em espectrofotômetro (Thermo Scientific Nanodrop®
2000) utilizando 2 L do purificado.
Reação de Sequenciamento
Para a reação de sequenciamento empregou-se o kit BigDye Terminator Cycle
Sequencing Ready Reaction (Perkin Elmer). As quantidades de reagentes foram
determinadas a partir da concentração de DNA purificado. Para uma reação de 10 L de

111
volume final, foram utilizados 2 L de BigDyeTM
v. 3.1. (Applied Biosystems), 2 L de
tampão Save Money 5X (200 mM Tris-HCl; 5 mM MgCl2; pH 9,0), 10 pmoles de
primer (senso ou antissenso) e 28 ng de DNA purificado. Em amostras com
concentração abaixo de 12 ng/L, o volume final da reação foi dobrado, assim como as
concentrações dos reagentes, exceto a do BigDye e do primer utilizado.
Precipitação do produto amplificado
Após a reação de sequenciamento, os produtos foram purificados por
precipitação em álcool. A cada amostra foi utilizado 32,5 L ou 65 L do mix (Etanol
100%; EDTA 125 mM) para as reações de volumes finais de 10 L e 20 L,
respectivamente. Após homogeneização, as amostras foram mantidas ao abrigo da luz
por 30 min, e em seguida, centrifugadas a 12.000 g por 30 min a 4 oC. Descartou-se o
sobrenadante por inversão de tubos e o sedimento foi lavado com 30 L ou 60 L de
etanol a 70%. Procedeu-se nova centrifugação a 12.000 g por 20 min. O excesso de
etanol foi desprezado por inversão total dos tubos, sendo sua total remoção conduzida
em termobloco a 95 oC por 10 min. As amostras foram mantidas a -20
oC até sua
utilização no sequenciamento.
Sequenciamento
As amostras foram homogeneizadas em 3,4 L de formamida e Blue Dextran-
EDTA (Applied Biosystems, USA) na porção de 5:1, desnaturadas a 95 oC por 3 min e
colocadas no gelo por 2 minutos. Posteriormente, foram submetidas à eletroforese em
sequenciador automático modelo ABI PrismTM
377 DNA Sequencers (Applied

112
Biosystems, USA).
As condições da reação de sequenciamento consistiram de desnaturação inicial a
96 oC por 1 min e 40 ciclos de 96
oC por 15 s, 50
oC por 15 s e 60
oC por 4 min.
Edição e Análise das sequências
As sequências de nucleotídeos foram analisadas e editadas com o auxílio do
programa Phred online (http://asparagin.cenargen.embrapa.br/phph/). O programa
BioEdit Sequence Alignment Editor (Hall, 1999) foi usado para preparar a versão final
das sequências consensos. Todas as posições com códigos ambíguos ou gaps nos
alinhamentos foram excluídas nas análises. O pareamento das distâncias genéticas foi
calculado pelo programa MEGA 5.03 (Tamura et al., 2011) com o modelo de
substituição Tamura-Nei (NJ).
Filogenia
A árvore filogenética foi determinada usando o método de Neighbor Joining
(NJ) (Saitou e Nei, 1987) implementado no programa MEGA 5.03 com análise das
distâncias Tamura-Nei (Tamura-Nei, 1993). As sequências resultantes foram
comparadas com sequências homólogas disponíveis no GenBank (Tabela 2).
Tabela 2 - Sequências de nucleotídeos analisados neste estudo.
Linhagem do vírus Número de acesso no
Genbank
Referência
CAEV-Cork M33677 Saltarelli et al., 1990
FESC-752 HM210570 Ramirez et al., não publicado
Volterra JF502417 Bertolotti et al., 2011
Fonni JF502416 Bertolotti et al., 2011
Seui GQ381130 Reina et al., 2010
MVV - K1514 M60609 Sonigo et al., 1985

113
MVVEV-1 S51392 Sargan et al., 1991
SA-OMVV M31646 Querat et al., 1990
P1OLV AF479638 Barros et al., 2004
Demais amostras - Deste estudo
Resultados
Trinta e oito animais de três estados brasileiros, de diferentes rebanhos, positivos
para os testes de IDGA e WB foram escolhidos para o isolamento viral através de
cocultivo em MSC, até o aparecimento do efeito citopático (ECP). Um fragmento de
aproximadamente 620 e 472 nucleotídeos foi amplificado para o gene gag e pol,
respectivamente.
Amplificação dos genes gag e pol através da PCR
Das 38 amostras de LVPRs isoladas pelo cocultivo, foi possível amplificar o
fragmento parcial de 620 nt para o gene gag em 18 delas (18/38 ou 47,4%) e para o
gene pol foi possível detectar um fragmento de 472 nt em 17 das 38 amostras,
correspondendo a 44,7%.
Reação de sequenciamento nucleotídico para os genes gag e pol
A partir das 18 amostras amplificadas do gene gag, submetidas à reação de
sequenciamento de nucleotídeos, foi possível definir a sequência parcial de 10
amplificados para o gene gag (69LVCE, 137CE, 229CE, 2361CE, 3596CE, 6900CE,
Animal02CE, Pan5CE, Pan7CE, Pan12CE) (anexo 1). Das 17 amostras amplificadas do

114
gene pol, foi possível determinar a sequência parcial de nucleotídeos através de
sequenciamento de 16 amplificados, (69LVCE, 137CE, 229CE, 2361CE, 3596CE,
237CE, 6900CE, Animal02CE, Pan5CE, Pan7CE, 1604MG, 1625MG, 2239CE,
SQSBCE, 31RN, 32CE) (anexo 2). A identificação das amostras e tamanho do
fragmento sequenciado encontra-se na Tabela 3.
Tabela 3 – Relação da origem das amostras e tamanho do fragmento dos respectivos
genes sequenciados.
Amostra Origem Fragmento
Sequenciado
Gene gag
Fragmento
Sequenciado
Gene pol
69LVCE Ceará 620 nt 472 nt
137CE Ceará 620 nt 472 nt
229CE Ceará 620 nt 472 nt
2361CE Ceará 620 nt 472 nt
3596CE Ceará 620 nt 472 nt
237CE Ceará - 472 nt
6900CE Ceará 620 nt 472 nt
Animal02CE Ceará 620 nt 472 nt
Pan5CE Ceará 620 nt 472 nt
Pan7CE Ceará 620 nt 472 nt
Pan12CE Ceará 620 nt -
1604MG Minas Gerais - 472 nt
1625MG Minas Gerais - 472 nt
2239CE Ceará - 472 nt
SQSBCE Ceará - 472 nt
31RN Rio Grande do Norte - 472 nt
32CE Ceará - 472 nt
Gene gag
Alinhamento das sequências geradas
Todas as linhagens deste estudo apresentaram transição de uma guanina por uma
adenina na posição 328, de uma citosina para uma timina na posição 401 e 410 e uma
transversão de uma timina para uma adenina na posição 420, resultando na mudança de
aminoácidos: valina (apolar) para isoleucina (apolar); treonina (polar neutro) para

115
metionina (apolar); prolina (apolar) para serina (polar neutro); ácido aspártico (polar
ácido) para ácido glutâmico (polar ácido), respectivamente. Além dessas mudanças, na
cepa 229CE ocorreu a troca de alanina (apolar) para um ácido aspártico (polar ácido) na
posição 164. E na cepa Pan12CE houve mudança de uma prolina (apolar) para uma
histidina (polar básico) na posição 157 e de uma glicina (apolar) para uma arginina
(polar básico) na posição 180, tomando como referência a cepa padrão CAEV-Cork
(Figuras 2 e 3). A matriz de identidade (Tabela 4), com os cálulos das distâncias dos
pareamentos, foi obtida comparando as sequências de nucleotídeos das cepas deste
estudo com outras já conhecidas. Os valores próximos a zero revelam maior
similaridade entre as sequências.

116
Figura 2. Alinhamento do fragmento parcial de 620 nucleotídeos do gene codificador da
proteína p28 do capsídeo gerados no presente estudo comparadas com sequências
padrões depositadas no Genbank.

117
Figura 3. Alinhamento das sequências de aminoácidos do gene gag correspondente à
proteína p28 do capsídeo.
Tabela 4. Matriz de identidade com os valores de similaridade nucleotídica do
fragmento parcial do gene gag comparadas com as demais cepas depositadas do
Genbank.

118
Inferêcia filogenética a partir das sequências nucleotídicas – Gene gag
A análise filogenética revelou que as cepas deste estudo formaram um grupo
altamente relacionado com o grupo de CAEV-Cork. A árvore filogenética (Figura 4) foi
obtida usando o método de NJ com sequências nucleotídicas do gene gag e mostrou a
relação das cepas deste estudo com outras já conhecidas. Os valores de bootstrap
derivados de 100 repetições indicam confiança no padrão de ramificação dos
respectivos ramos da árvore. Todos os parâmetros das análises foram padrões. MVV
está claramente separado do grupo de CAEV.
Figura 4. Árvore filogenética do gene gag diferenciando os grupos de CAEV e MVV.

119
Gene pol
Alinhamento das sequências de aminoácidos – Gene pol
Todas as linhagens deste estudo apresentaram transição e/ou tranversão em
diferentes posições das sequências, resultando na mudança de aminoácidos para outros
de grupos diferentes, tomando como referência a cepa padrão CAEV-Cork (Figura 5 e
6). A matriz de identidade (Tabela 5), com os cálculos das distâncias dos pareamentos,
foi obtida comparando as sequências de nucleotídeos das cepas deste estudo com outras
já conhecidas. Os valores próximos a um revelam maior similaridade entre as
sequências.
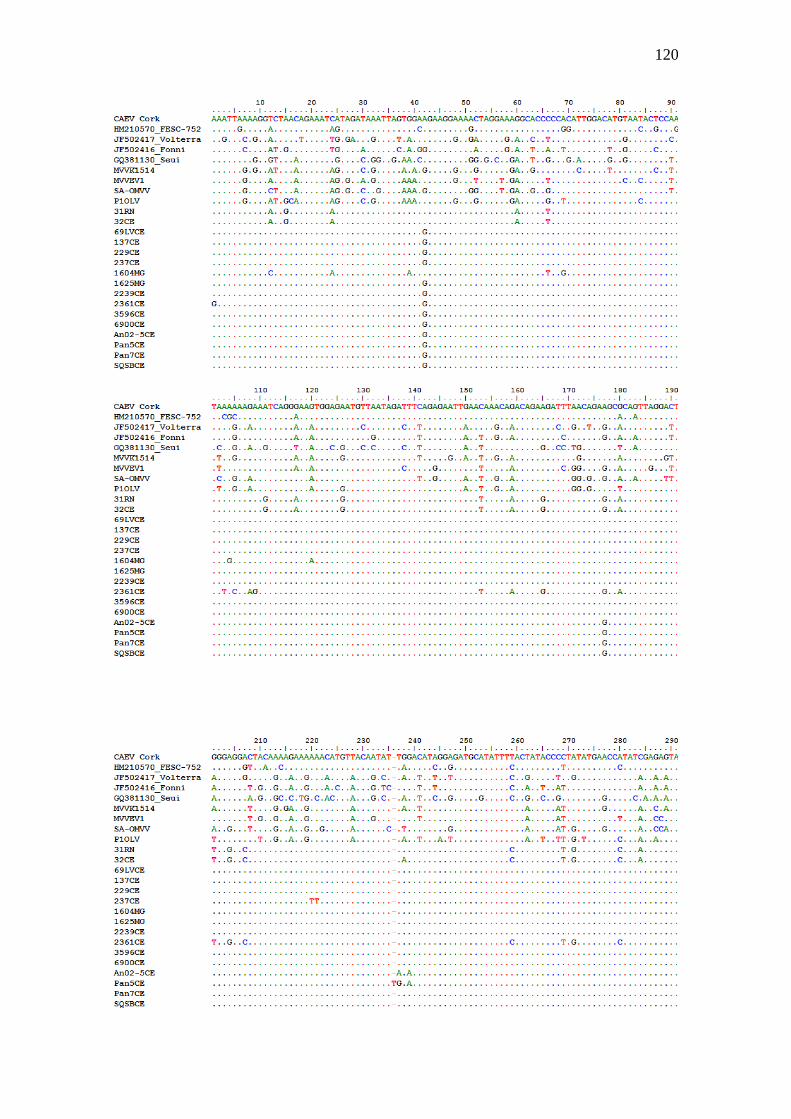
120

121
Figura 5 - Alinhamento do fragmento parcial de 472 nucleotídeos do gene pol gerados
no presente estudo comparadas com sequências padrões depositadas no Genbank.

122
Figura 6. Alinhamento das sequências de aminoácidos do gene pol correspondente a
160 aminoácidos referente à região intergênica de dUTPase e integrase.
Tabela 5. Matriz de identidade com os valores de similaridade nucleotídica do
fragmento parcial do gene pol comparadas com as demais cepas depositadas do
Genbank.

123
Inferêcia filogenética a partir das sequências nucleotídicas – Gene pol
A análise filogenética revelou que as cepas deste estudo formaram um grupo
estreitamente relacionado com o grupo de CAEV-Cork. A árvore filogenética (Figura 7)
foi obtida usando o método de NJ com sequências nucleotídicas do gene pol e mostrou a
relação das cepas deste estudo com outras já conhecidas. Os valotes de bootstrap
derivados de 100 repetições indicam confiança no padrão de ramificação dos
respectivos ramos da árvore. Todos os parâmetros das análises foram padrões. Observa-
se que as cepas de MVV estam claramente separados das demais de CAEV. Porém as
cepas 31RN e 32CE apresentaram ramificações distintas divergindo para um cluster
separado das demais amostras deste estudo.

124
Figura 7. Árvore filogenética do gene pol diferenciando os grupos de CAEV e MVV.

125
Discussão
Trinta e oito animais infectados, de diferentes estados brasileiros, foram
escolhidos para o isolamento viral por cocultivo de leucócitos. Destes, apenas 18
amostras amplificaram para o gene gag e 17 para o gene pol.
Deve-se considerar que as amostras possuem diferentes concentrações virais e
que a reação de PCR não contempla uma etapa de reamplificação (“nested” PCR). Estes
fatos em conjunto podem ter conferido um maior limiar de detecção, levando a uma
perda de sensibilidade diagnóstica.
Finalmente, devido aos mecanismos de variabilidade genética dos lentivírus
(shifts, drifts, rearranjos e recombinações) é possível que tenha havido falhas de
hibridização dos primers. Nesse sentido, julga-se necessário o desenho de novos
primers que contemplem a diversidade genética dos lentivírus.
As amplificações foram submetidas ao sequenciamento de nucleotídeos. Foram
obtidas 10 sequências viáveis para o gene gag e 16 sequências para o gene pol. Atribui-
se esta perda a fatores relacionados principalmente à quantidade de DNA presente na
amostra e à purificação insatisfatória dos produtos submetidos à reação de
sequenciamento. Outra hipótese para a perda de sequências seria a condição isotérmica
na reação de sequenciamento, não favorendo o anelamento dos primers.
Ao analizar o gene gag, foi observado, por eletroferograma, a presença de
polimorfismo nas amostras 229CE e Pan12CE, pela visualização da sobreposição de
bases (adenina e citosina) tanto na sequência senso quanto na antissenso. Da mesma
forma, nas sequências do gene pol, foi observado a presença de polimorfismo na
amostra Pan7CE, pela da visualização da sobreposição de bases (adenina e guanina)
tanto na sequência senso quanto na antissenso, sugerindo que um mesmo animal pode

126
ter sido infectado por um vírus que sofreu mutações dentro do hospedeiro, corroborando
com os achados de Ellis e colaboradores (1987) em que afirma que ambas as viroses
têm a capacidade de apresentar variação antigênica significativa e que dentro do mesmo
animal infectado pode existir mais de uma cepa viral ao mesmo tempo. Uma alternativa
para este problema seria clonar e sequenciar novamente estas cepas para garantir a
presença desse achado.
Além disso, para o gene gag, foi observada em todas as cepas isoladas a
presença de mutações pontuais de nucleotídeos, com a substituição de purina por outra
purina e de purina por pirimidina e vice-versa, o que provocou a mudança de
aminoácidos por outro do mesmo grupo ou por aminoácidos de grupos diferentes. O
fragmento sequenciado corresponde à proteína do capsídeo p28 que tem influência na
antigenicidade, podendo este fato, estar diretamente relacionado com a resposta imune
do hospedeiro, embora esta seja ineficaz para prevenir a infecção viral, seu papel é de
enorme importância no controle da intensidade da infecção até que o organismo comece
a produzir elementos específicos de defesa, o que demanda entre cinco e sete dias. Os
principais mecanismos da resposta imune natural consistem no bloqueio à infecção de
novas células, inibição da replicação do vírus dentro das células e eliminação de células
infectadas (MADRUGA et al., 2001). Sendo assim, essa proteína apresenta características
que são importantes tanto para antigenicidade quanto para a patogenicidade desdes
lentivírus (SHAH et al., 2004a), podendo ainda afetar sua persistência e o seu mecanismo
de evasão. A heterogeneidade das sequências de nucleotídeos e aminoácidos da proteína
p28 de lentivírus pode auxiliar no desenvolvimento de vacinas no combate ao agente.
Além disso, poderá ajudar a compreender sua epidemiologia e relações filogenéticas.
Para o gene pol foi também foi observado em todas as cepas isoladas a presença
de mutações pontuais de nucleotídeos, com a presença de trasições e/ou transversões,

127
resultando mudança de aminoácidos por outro do mesmo grupo ou por aminoácidos de
grupos diferentes. O fragmento sequenciado do gene pol corresponde às proteínas de
atividade enzimática, e abrange uma região intergênica de dUTPase e integrase. Estudos
recentes relataram que deleções de nucleotídeos na porção que codifica para a dUTPase
afetam na fidelidade da transcriptase reversa em células em não-divisão (REINA et al.,
2010). Portando, mutações na região que codificam para essas proteínas podem exercer
um papel fundamental na regulação da replicação e transcrição viral.
Entre as sequências geradas neste estudo frente às demais selecionadas no
Genbank, mediante visualização do alinhamento nucleotídico da matriz de identidade,
constatou-se que as amostras 69LV, 137CE, 2361CE, 3596CE, 6900CE, Animal02-
5CE, Pan5CE e Pàn7CE tiveram 99,3% de semelhança com o CAEV enquanto a maior
diferença observada foi de 70,1% entre as amostras Pan12CE e GQ381130 para o gene
gag. E que as amostras 69LV, 137CE, 229CE, 2239CE, 3596CE, 6900CE tiveram
99,7% de semelhança com CAEV-Cork enquanto a maior diferença observada foi de
77,4 entre as amostras Animal02-5CE e P1OLV para o gene pol.
A árvore filogenética foi construída para cada gene separadamente. De forma
geral, as árvores propostas guardaram correspondência com suas respectivas matriz de
identidade e com as cepas de CAEV já publicadas. Observou-se que as amostras do
gene gag agruparam-se no clado correspondente ao de CAEV, amparado com elevado
valor de bootstrap (Figura 4). A árvore construída a partir de sequências pol mostrou
que as amostras desse gene agruparam no mesmo clado de CAEV, porém as amostras
31RN e 32CE segreraram para um cluster isolado (Figura 7). Sendo assim, conclui-se
que tanto pela topologia quanto pelos valores de identidade nucleotídica, as amostras
geradas neste estudo pertencem ao subtipo B do grupo de CAEV.
Através do estudo proteômico dos lentivírus, os testes de diagnóstico podem

128
tornar-se específicos para cada região. Devido à ampla distribuição destes vírus em
certas regiões e às perdas econômicas resultantes, vários países têm adotado programas
de erradicação com base na segregação dos vírus de caprinos e ovinos (GREENWOOD et
al., 1995; HOUWERS et al., 1987; PERETZ et al., 1994; ROWE E EAST, 1997; SCHEER-
CZECHOWSKI et al., 2000).
Estudos na Finlândia, comparando o protótipo de CAEV com MVV,
demonstraram uma inserção de sete aminoácidos (181–187) na linhagem K1514 de
MVV na região C-terminal da MA e uma deleção de dois aminoácidos (entre 283 e 284)
na região N-terminal do CA. Uma comparação das sequências de lentivírus com
algumas sequências encontradas em banco de dados genéticos revelou que em contraste
com o CAEV, todos os protótipos de MVV contêm uma deleção de seis nucleotídeos na
região do gene gag resultando na deleção de dois resíduos de aminoácidos na região
central do capsídeo. Esta deleção pode ser utilizada como marcador na análise de
LVPRs, especialmente quando se considerar uma possível transmissão de lentivírus
entre caprinos e ovinos.
Os autores verificaram que a similaridade entre as sequências de lentivírus
caprino e ovino é maior na região N-terminal que na região C-terminal das proteínas
analisadas do capsídeo. É interessante observar que a maior região de homologia é
menos conservada que a região N-terminal do capsídeo, demonstrando diferenças entre
LVPRs de ovinos e caprinos (RAVAZZOLO et al., 2001). Embora o domínio C-terminal
das proteínas do capsídeo de lentivírus tenha sido crucial para a montagem dos vírus
(FREED, 1998), dados indicam que a região N-terminal também pode ser importante
para a montagem e liberação dos vírus (LI et al., 1997; WANG et al., 1998).
O modelo baseado na estrutura de cristalografia da p26 (capsídeo) de lentivírus
equino propõe que o domínio N-terminal inicia um papel essencial na maturação do

129
core (JIN et al., 1999). Um peptídeo de cinco aminoácidos LYPNL (Motif III),
encontrado em todas as estirpes de MVV isoladas na Polônia, mas não nas estirpes
padrão, pode ser utilizado como um marcador para distinguir estirpes de vírus (PISONI et
al., 2006). Um segundo peptídeo KNLTPEESNKQDFMSL (Motif II), foi
especificamente relacionado com a estirpe de CAEV. Em contraste, 6/16 mudanças de
aminoácidos (Q - - - - - - TS - RE - A - -) foram específicas para todas as estirpes de
MVV (KUZMAK et al., 2007).
As mudanças similares de aminoácidos foram observadas em dois isolados
italianos de ovinos, geograficamente distantes, anteriormente classificados no grupo de
ovinos e genotipicamente como CAEV (GREGO et al., 2005). Esses peptídeos foram
considerados como uma das principais regiões imunodominantes da matriz. Assim,
ambos os peptídeos (Motif II e III), podem ser utilizados em testes de diagnóstico para
distinguir infecção com os vírus MVV e CAEV. Isto seria importante para análise de
lentivírus de ovinos e caprinos que vivem em rebanhos mistos e aqueles em contato
direto, o que pode elucidar os acontecimentos da transmissão cruzada destes vírus após
o salto da barreira entre espécies (KUZMAK, 2007).
Em relação ao gene pol, A Uracila no DNA aparece como resultado da
incorporação defeituosa ou desaminação da citosina (HARRIS et al., 2003). A enzima
dUTPase age diretamente sobre o conjunto de nucleotídeos prevenindo a incorporação
defeituosa de resíduos de uracila no DNA viral após o processo de retrotranscrição,
mantendo uma relação baixa de dUTP: sTTP (PRIET et al., 2003). A dUTPase é
codificada pelo gene pol e é uma proteína associada ao vírion presente em lentivírus de
não-primatas como (EIAV e CAEV) (CHEN et al., 2002), não sendo encontrado em
nenhum dos lentivírus de primatas (HIV, SIV). A expressão de dUTPase celular é rica
em células indiferenciadas em divisão e pobre em células diferenciadas em não-divisão

130
(MILLER et al., 2000). Além disso, a replicação de dUTPase-negativas de CAEV em
células em não-divisão (p.e. macrófagos) leva a uma redução da carga viral 10 a 100
vezes mais em comparação com o vírus do tipo selvagem. Experimentos in vivo
mostraram que cepas dUTPase-negativas produzem lesões menos graves (TURELLI et
al., 1997). A deleção artificial dos genes dUTPase em CAEV-Co resultou na replicação
viral tardia, acumulação de mutações pontuais de G por A e decréscimo da carga viral
(HARMACHE et al., 1996; TURELLI et al., 1997).
Conclusão
Trinta e oito animais de amostras circulantes de lentivírus caprino foram
selecionados para o isolamento viral por cocultivo de leucócitos. Destas, 18 amostras
amplificaram para o gene gag e 17 para o gene pol. A partir das 18 amostras
amplificadas do gene gag, submetidas à reação de sequenciamento de nucleotídeos, foi
possível definir a sequência parcial de 10 amplificados. Das 17 amostras amplificadas
do gene pol, foi possível determinar a sequência parcial de nucleotídeos através de
sequenciamento de 16 amplificados. Foram sequenciadas amostras nos estados do
Ceará, Rio Grande do Norte e Minas Gerais. Estas foram comparadas com outras já
conhecidas depositadas no Genbank. As amostras circulantes apresentam elevada
conservação nucleotídica sendo 99,3% para o gene gag quando comprado com a cepa
padrão CAEV-Cork e relativamente conservado, 70,1% para o gene pol, apresentando,
portanto uma diversidade intragenotípica. Pela genealogia proposta com base nas
sequências nucleotídicas dos genes gag e pol, a sua topologia mostrou-se compatível
com os genótipos do subtipo B de CAEV.

131
Agradecimentos
UECE, EMBRAPA Caprinos e Ovinos, FUNCAP, CNPq e Banco do Nordeste pelo
apoio financeiro para realização deste projeto.

132
REFERÊNCIAS BIBLIOGRÁFICAS
Almeida, M. G. A. R.; Anunciação, A. V. M.; Figueredo, A.; Martinez, T. C. N.;
Laborda, S. S. Dados sorológicos sobre a presença e distribuição da Artrite - Encefalite
Caprina (CAE) no estado da Bahia, Brasil. Revista Brasileira de Saúde e Produção
Animal, v. 1, n. 3, p. 78 – 83, 2001.
Alvarez, V., Daltabuit-Test, M., Arranz, J., Leginagoikoa, I., Juste, R.A.,
Amorena, B., De Andres, D., Lujan, L.L., Badiola, J.J., Barriatua, E. PCR detection of
colostrum-associated Maedi–Visna virus (MVV) infection and relationship with
ELISA-antibody status in lambs. Research in Veterinary Science. v. 80, p. 226–234,
2006.
Alves, F.S.F. & Pinheiro, R.R.. Presença de artrite-encefalite caprina a vírus
(CAEV) no Estado do Maranhão. In: XXV Congresso Brasileiro de Medicina
Veterinária, Gramado, p.278. (Resumo). 1997.
Assis, A. P. M. V.; Gouveia, A. M. G. Evidência sorológica de Lentivírus
(Maedi Visna/Atrite-encefalite caprina) em rebanhos nos Estados de MG, RJ, BA e CE.
In: 94 Congresso Brasileiro de Medicina Veterinária, XXIII, 1994, Recife. Anais.
SPMV. p. 104. 1994.
Bandeira, D. A., De Castro, R. S., Azevedo, E. O., De Souza Seixas Melo, L. &
De Melo, C. B. Seroprevalence of caprine arthritis-encephalitis virus in goats in the
Cariri region, Paraiba state, Brazil. Veterinary Journal v. 180, p. 399-401. 2009.
Barquero, N., Arjona, a, Domenech, a, Toural, C., de las Heras, a, Fernández-
Garayzabal, J. F., Ruiz-Santa Quiteria, J. a, et al. Diagnostic performance of PCR and
ELISA on blood and milk samples and serological survey for small ruminant
lentiviruses in central Spain. The Veterinary record, 168(1), 20. doi:10.1136/vr.c4951.
2011.
Barros, S.C., Ramos, F., Duarte, M., Fagulha, T., Cruz, B., Fevereiro. Genomic
characterization of a slow/low Maedi visna virus. Vírus Genes 29, 199–210. 2004.
Batista, M. C. S.; De Castro, R. S.; Carvalho, F. A. A.; Cruz, M. S. P.; Silva, S.
M. M. S.; Rego, E. W.; Lopes, J. B. Anticorpos anti-Lentivírus de pequenos ruminantes
em caprinos integrantes de nove municípios Piauienses. Ciência Veterinária nos
Trópicos, v.7, n 2 e 3, p. 75 – 81, 2004.
Batista, M. C. S.; De Castro, R. S.; Carvalho, F. A. A.; Cruz, M. S. P.; Silva, S.
M. M. S.; Rego, E. W.; Lopes, J. B. Anticorpos anti-Lentivírus de pequenos ruminantes
em caprinos integrantes de nove municípios Piauienses. Ciência Brasileira de Medicina
Veterinária, v. 17, n. 2, p. 72 - 75, 1995.
Bertolotti L, Mazzei M, Puggioni G, Carrozza M. L, Dei Giudici S, Muz
D, Juganaru M, Patta C, Tolari F, Rosati S. Characterization of new small ruminant
lentivirus subtype B3 suggests animal trade within the Mediterranean Basin. Journal of
General Virology. v. 92, n. 8, p. 1923-9. 2011.

133
Castro, R. S.; Azevedo, E. O.; Tabosa, I.; Nascimento, S. A.; Oliveira, M. M. M.
Anticorpos para o Vírus da Artrite-Encefalite Caprina em animais sem raça definida
(SRD) de abatedouros dos estados de Pernambuco e Paraíba. Ciência Veterinária nos
Trópicos, v. 5, n. 2 – 3, p. 121 – 123, 2002.
Castro, R. S.; Nascimento, S. A.; Abreu, S. R. O. Evidência sorológica de
infecção pelo vírus da artrite-encefalite caprina em caprinos leiteiros do Estado de
Pernambuco. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.46, n. 5, p.
571-572, 1994.
Chen, R., Wang, H., Mansky, L.M. Roles of uracil-DNA glycosylase and
dUTPase in virus replication. Journal of General Virology. v. 83, Pt 10, p. 2339–2345.
2002.
Chomczynski, P. A. A reagent for the single-step simultaneous isolation of
RNA, DNA and protein from the cell and tissues samples. Biotechniques, v. 15, n. 3, p.
532-537, 1993.
Clements, J. E. And Payne, S. L. Molecular basis of the pathobiology of
lentiviruses. Virus Research, 32, 97-109. 1994.
Cunha, R. G.; Nascimento, M. D. Ocorrência de anticorpos para o vírus da
artrite-encefalite caprina em soros de caprinos do estado do Rio de Janeiro. Revista
Brasileira de Medicina Veterinária, v. 17, n. 2, p. 72 - 75, 1995.
Dantas, T.V.M.; Proteína Recombinante Do Vírus Da Artrite Encefalite Caprina
Com Potencial Antigênico Fortaleza: UECE. 120p. Tese de Doutorado. 2009.
Ellis, T. M.; Wilcox, G.E.; Robinson, W.F. Antigenic variation of caprine
arthritis encephalitis virus during persistent infection of goats. Journal of General
Virology, v.63, n.12, p. 3145-3152, 1987.
Freed, E.O. HIV-1 Gag proteins: diverse functions in the virus life cycle.
Virology. v. 251, p. 1–15, 1998
Gouveia, A. M. G.; Coura, M. A.; Brandão, H. M.; Atanásio, C. Distribuição
sorológica do lentivírus caprino em amostragem por demanda. In: Encontro De
Pesquisa da Escola de Veterinária da UFMG, Belo Horizonte. Anais... Belo Horizonte.
p. 116. 1998.
Greenwood, P. L. Effects of Caprine Arthritis-Encephalitis Virus on productivity
and health of dairy goats in New South Wales, Australia. Preventive Veterinary
Medicine, v. 1-2, n. 22, p. 71-87, 1995.
Grego, E., Bertolotti, L., Carrozza, M.L., Profiti, M., Mazzei, M., Tolari, F.,
Rosati, S. Genetic and antigenic characterization of the matrix protein of two genetically
distinct ovine lentiviruses. Veterinary microbiology. v. 106, p. 179–185, 2005.

134
Hall, T.A. BioEdit: a user-friendly biological sequence alignment editor and
analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series. v. 41, p.
95-98. 1999.
Harmache, A. et al. Requirement of caprine arthritis encephalitis virus vif gene
for in vivo replication. Virology, 224, 246–255. 1996.
Harris, R.S., Bishop, K.N., Sheehy, A.M., Craig, H.M., Petersen-Mahrt, S.K.,
Watt, I.N., Neuberger, M.S., Malim, M.H. DNA deamination mediates innate immunity
to retroviral infection. Cell. v. 113, n. 6, p. 803–809. 2003.
Houwers, D.J., Konig, C.D., Bakker, J., De Boer, M.J., Pekelder, J.J., Sol, J.,
Vellema, P., de VRIES, G. Maedi– visna control in sheep: III. Results and evaluation of
a voluntary control program in The Netherlands over a period of four years. The
Veterinary quarterly. v. 9, p. 29S–36S, 1987.
Jin, Z., Jin, L., Peterson, D.L., Lawson, C.L. Model for lentivirus capsid core
assembly based on crystal dimers of EIAV p26. Journal of molecular biology. v. 286, p.
83–93, 1999.
Kuzmak J, et al. Molecular characterization of lentiviruses from goats from
Poland based on gag gene sequence analysis. Comparative Immunology, Microbiology
and Infectious Diseases. v. 30, p. 211-223, 2007.
Leite, B. L. S.; Modolo, J. R.; Padovani, C. R.; Stachissini, A. V. M.; Castro, R.
S.; Simões, L. B. Avaliação da taxa comparativa de ocorrência da Artrite-Encefalite
Caprina a Vírus pelas regionais de defesa agropecuária do estado de São Paulo, Brasil, e
seu mapeamento por meio de sistema de informações geográficas. Arquivos do Instituto
Biológico, v. 71, n. 1, p. 21 -26, 2004.
Leroux, C., Lerondelle C., Chastang, J., Mornex J. F. RT-PCR detection of
lentiviruses in milk or mammary secretions of sheep or goats from infected flocks.
Veterinary Research. v. 28, p. 115-121. 1997.
Li, X., Yuan, B., Goff, S.P. Genetic analysis of interactions between Gag
proteins of Rous Sarcoma virus. Journal of virology. v. 71, p. 5624–5630, 1997.
Madruga, C. R.; Araújo, F. R.; Soares, C. O. Princípios, padronização e
validação de provas sorológicas. In: Madruga, C. R.; Araújo, F. R.; Soares, C. O.
Imunodiagnóstico em medicina Veterinária. Campo Grande: EMBRAPA gado de corte
(Campo Grande- Mato Grosso do Sul- Brasil), p.145-175. 2001.
Madureira, K. M.; Gomes, V. Prevalência da Artrite-Encefalite Caprina (CAE)
em propriedades leiteiras do estado de São Paulo. Disponível em:
<http://www.unifian.edu.br/programasinst/Revistas/revistas2007/veterinaria/Prevalencia
.2007
Melo, C. B.; De Castro, R. S.; Oliveira, A. A.; Fontes, L. B.; Callado, A. K. C.;
Nascimento, S. A. Estudo preliminar sobre a infecção por lentivírus de pequenos

135
ruminantes em ovinos e caprinos de Sergipe. In: Congresso Latino Americano De
Buiatria, 5°, Salvador. Anais... Salvador: SBB, 2003.
Miller, R.J., Cairns, J.S., Bridges, S., Sarver, N. Human immunodeficiency virus
and AIDS: insights from animal lentiviruses. Journal of Virology. v. 74, n,16, p.7187–
7195. 2000.
Narayan, O., Clements, J.E., Biology and pathogenesis of lentiviruses.
Journal of General Virology. v. 70, p. 1617–1639. 1989.
Narayan O, Cork LC: Lentiviral diseases of sheep and goats: chronic pneumonia
leukoencephalomyelitis and arthritis. Reviews of infectious diseases. v. 7, n. 1, p. 89-98.
1985.
Oliveira, M. M. M.; Castro, R. S.; Carneiro, K. L.; Nascimento, S. A.; Callado,
A. K. C.; Alencar, C. S. A.; Costa L. S. P. Anticorpos contra lentivírus de pequenos
ruminantes em caprinos e ovinos em abatedouros do estado de Pernambuco. Arquivo
Brasileiro de Medicina Veterinária e Zootecnia, v. 58, n.5, p. 947-949, 2006.
Pasick J: Maedi-visna virus and caprine arthritis-encephalitis virus: distinct
species or quasispecies and its implications for laboratory diagnosis. The Canadian
Journal of Veterinary Research. v. 62, n. 4, p. 241-4. 1998.
Pepin M, Vitu C, Russo P, Mornex JF, Peterhans E: Maedi-visna virus infection
in sheep: a review. Veterinary Research. v. 29, n. 3-4, p. 341-67, 1998.
Peretz, G., Bugnard, F., Calavas, D. Study of a prevention programme for
caprine arthritis– encephalitis. Veterinary research. v. 25, p. 322– 326, 1994.
Pinheiro, R. R.; Gouveia, A. M. G.; Alves, F. S. F. Prevalência da infecção pelo
Vírus da Artrite-Encefalite Caprina no estado do Ceará, Brasil. Ciência Rural, v. 31, n.
3, p. 449 – 454, 2001.
Pinheiro, R. R.; Alves, F. S. F.; Girão, E. S.; Medeiros, L. P. A.; Girão, R. N.
Presença da Artrite-Encefalite Caprina a Vírus (CAEV), em Teresina – Piauí. In:
Congresso Brasileiro de Medicina Veterinária, 24°, Goiânia. Anais...Goiânia. p. 161.
1996.
Pisoni G, Bertoni G, Boettcher P, Ponti W, Moroni P. Phylogenetic analysis of
the gag region encoding the matrix protein of small ruminant lentiviruses: comparative
analysis and molecular epidemiological applications. Virus research. v. 116, n. 1–2, p.
159–67, 2006.
Priet, S., Navarro, J.M., Gros, N., Querat, G., Sire, J. Differential incorporation
of uracil DNA glycosylase UNG2 into HIV-1, HIV-2, and SIV(MAC) viral particles.
Virology v. 307, n. 2, p. 283–289.2003.

136
Querat, G., Audoly, G., Sonigo, P., Vigne, R. Nucleotide sequence analysis of
SA-OMVV, a visna-related ovine lentivirus: phylogenetic history of lentiviruses.
Virology. v. 175, p. 434–447. 1990.
Ravazzolo, A.P., Reischak, D., Peterhans, E., Zanoni, R. Phylogenetic analysis
of small ruminant lentiviruses from Southern Brazil. Virus research. v. 79, n. 1-2, p.
117– 123, 2001.
Reina R, Bertolotti L, Dei Giudici S, Puggioni G, Ponti N, Profiti M, Patta
C, Rosati S. Small ruminant lentivirus genotype E is widespread in Sarda goat.
Veterinary Microbiology. v. 144, n. 1-2, p. 24-31. 2010.
Rowe JD, East NE: Risk factors for transmission and methods for control of
caprine arthritis-encephalitis virus infection. Veterinary Clinics of North America: Food
Animal Practice v. 13, n.1, p.35-53, 1997.
Saitou, N., Nei, M., The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Molecular Biology and Evolution. v. 4, p. 406–425.
1987.
Saltarelli, M., Querat, G., Konings, D.A., Vigne, R., Clements, J.E. Nucleotide
sequence and transcriptional analysis of molecular clones of CAEV which generate
infectious virus. Virology 179, 347–364. 1990.
Sargan, D.R., Bennet, I.D., Cousens, C., Roy, D.J., Blacklaws, B.A., Dalziel,
R.G., Watt, N.J., Mcconnell, I. Nucleotide sequence of EV1, a British isolate of Maedi-
visna virus. Journal of General Virology. v. 72, p. 1893–1903. 1991.
Scheer-Czechowski, P., Vogt, H.R., Tontis, A., Peterhans, E., Zanoni, R. Pilot
project for eradicating Maedi – visna in Walliser blacknose sheep. Schweizer Archiv fur
Tierheilkunde. v. 142, p. 155– 164, 2000.
Silva, J. S.; Castro, R. S.; Melo, C. B.; Feijó, F. M. C.. Infecção pelo vírus da
artrite encefalite caprina no Rio Grande do Norte. Arquivo Brasileiro de Medicina
Veterinária e Zootecnia, v. 57, n.6, p. 727-731, 2005.
Sonigo, P., Alizon, M., Staskus, K., Klatzmann, D., Cole, S., Danos, O., Retzel,
E., Tiollais, P., Haase, A., Wain-Hobson, S. Nucleotide sequence of the visna lentivirus:
relationship to the AIDS virus. Cell. n. 42, p. 369–382. 1985.
Tamura K, Dudley J, Nei M E Kumar S Mega 5.03: Molecular Evolutionary
Genetics Analysis (MEGA) software version 5.03. Molecular Biology and Evolution v.
24, p. 1596-1599. (Publication PDF at http://www.kumarlab.net/publications). 2011.
Tamura, K., Nei, M., Estimation of the number of nucleotide substitutions in the
control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology
and Evolution . v. 10, p. 512– 526. 1993.
Tigre, D. M.; Campos, G. S.; Sardi, S. I. Isolamento e identificação do Vírus da
Artrite Encefalite Caprina, a partir do cocultivo de células mononucleares do sangue
com células de membrana sinovial de cabra. Revista de Ciências Médicas e Biológicas,
v. 5, n. 2, p. 124 – 131, maio/ agosto, 2006.

137
Turelli, P., Guiguen, F., Mornex, J.F., Vigne, R., Querat, G. dUTPase-minus
caprine arthritis-encephalitis virus is attenuated for pathogenesis and accumulates G-to-
A substitutions. Journal of virology. v. 71, n. 6, p. 4522–4530.1997.
Wang, C., Lai, H., Li, J. Analysis of minimal human immunodeficiency virus
type 1 gag coding sequences capable of virus-like particle assembly and release.
Journal of virology. v. 72, p. 7950–7959, 1998.

138
CONCLUSÕES GERAIS
Foram isoladas cepas virais circulantes de animais naturalmente infectados nos
estados do Rio Grande do Norte, Ceará, Piauí, Bahia e Minas Gerais.
A cepa isolada, denominada BrRN-CNPC.G1 foi considerada o primeiro
isolamento do CAEV no Estado do Rio Grande do Norte.
Todas as cepas isoladas mostraram-se ser fenotipicamente líticas, do tipo
rápido/alto.
Foram amplificadas e sequenciadas cepas virais isoladas do Ceará e Minas
Gerais e Rio Grande do Norte, tanto para o gene gag quanto para o gene pol.
Não foi possível obter amplificação por PCR dos isolados do Piauí e Bahia com
a finalidade de sequenciar.
Os alinhamentos das sequências – Capítulo III (EU300976, EU300977,
EU300978 e EU300979) com outras já disponíveis em bancos de dados
genéticos revelaram que estas linhagens estão mais relacionadas com CAEV que
MVV. A classificação mostrou que estas pertecem ao subtipo B1 do grupo de
CAEV.
As demais amostras circulantes (Capítulo IV) apresentaram elevada conservação
nucleotídica sendo 99,3% para o gene gag quando comprado com a cepa padrão
CAEV-Cork e relativamente conservado, 70,1% para o gene pol, apresentando,
portanto uma diversidade intragenotípica.
Pela genealogia proposta com base nas sequências nucleotídicas dos genes gag e
pol, a sua topologia mostrou-se compatível com os genótipos do subtipo B de
CAEV.
O alinhamento das sequências de aminoácidos da região gag indicaram

139
divergências entre as linhagens deste estudo com a cepa padrão CAEV-Cork.
Essa divergência de aminoácidos foi ainda mais acentuada quando comparas
sequências de aminoácidos do gene pol.
Não foram obtidas sequências viavéis em nenhuma das amostras isoladas para o
gene env.

140
PERSPECTIVAS DA TESE
Estabelecer programas de erradicação de acordo com a distribuição das
linhagens de LVPR poderá fundamentar o desenvolvimento de reagentes para
diagnóstico de maior especificidade e sensibilidade do que os atualmente disponíveis no
mercado em rebanhos locais.
A aplicação da análise filogenética para sequências de CAEV e MVV é de
imensa importância para este tipo de análise epidemiológica de infecções por lentivírus
de pequenos ruminantes e também para estudar as potenciais diferenças de virulência
entre linhagens de LVPR circulantes, ajudando a entender as propriedades genéticas,
protéicas e antigênicas desses vírus, sua patogênese, epidemiologia e relacionamento
filogenético e poderão ainda ser estudados possíveis antígenos vacinais.

141
REFERÊNCIAS BIBLIOGRÁFICA
ABELSON, M. L. and SCHOBORG, R. V. Characterization of the caprine arthritis
encephalitis virus (CAEV) rev N-terminal elements required for efficient interaction
with the RRE. Virus Research, v. 92, p.23-35, 2003.
ABREU, S. R. O., CASTRO, R. S., NASCIMENTO, S. A., (1998). Produção de
antígeno nucleoprotéico dos vírus da artrite encefalite caprina e comparação com os
Visna/Maedi Vírus para imunodifusão em ágar-gel. Pesquisa veterinária brasileira. v.
18, p. 57-60, 1998.
ADAMS, D.S., OLIVER, R.E., AMEGHINI, E., DEMARTINI, J.C., VERWOERD,
D.W., HOUWERS,D.J.,WAGHELA,S.,GORHAM,J.R., HYLLSETH,
B.,DAWSON,M., TRIGO, F.J., MCGUIRE, C. Global survey of serological evidence
of caprine arthritis-encephalitis virus infection. Veterinary Record. v.15, p. 493–495.
1984.
AGNARSDOTTIR, G., THORSTEINSDOTTIR, H., OSKARSSON, T.,
MATTHIASDOTTIR, S., St HAFLIDADOTTIR, B., ANDRESSON, O. S. and
ANDRESDOTTIR,V. The long terminal repeat is a determinant of cell tropism of
Maedi-visna virus. Journal of General Virology. v. 81, p.1901-1905, 2000.
ALI AL AHMAD MZ, FIENI F, PELLERIN, J.L., GUIGUEN, F., CHEREL, Y.,
CHATAGNON, G., BOUZAR, A.B., CHEBLOUNE, Y. Detection of viral genomes of
caprine arthritis-encephalitis vírus (CAEV) in semen and in genital tract tissues of male
goat. Theriogenology. v. 69, p. 473–480, 2008.
ALI AL AHMAD, M.Z., FIENI, F., GUIGUEN, F., LARRAT, M., PELLERIN, J.L.,
ROUX, C., CHEBLOUNE, Y. Cultured early goat embryos and cells are susceptible to
infection with caprine encephalitis vírus. Virology. v. 353, p. 307–315. 2006.
ALI AL AHMAD, M.Z., FIENI, F., MARTIGNAT, L., CHATAGNON, G., BARIL,
G., BOUVIER, F., CHEBLOUNE, Y. Proviral DNA of caprine arthritis encephalitis
vírus (CAEV) is detected in cumulus oophorus cells but not in oocytes from naturally
infected goats. Theriogenology. v. 64, p. 1656–1666, 2005.
ALKAN, F.; TAN, M. T. A. A comparative study on the diagnosis of Maedi-Visna
infection in serum and colostrum samples using agar gel immunodiffusion (AGID)
technique. Deutsche Tierarztliche Wochenschrift, v. 105, n.7, p.276-278, 1998.
ALMEIDA, A. M. R.; LIMA, J. A. A. Princípios e técnicas de diagnose em
fitovirologia. Brasília/Fortaleza: Publicação SBF. 186p. 2011.
ALMEIDA, M. G. A. R.; ANUNCIAÇÃO, A. V. M.; FIGUEREDO, A.; MARTINEZ,
T. C. N.; LABORDA, S. S. Dados sorológicos sobre a presença e distribuição da Artrite
- Encefalite Caprina (CAE) no estado da Bahia, Brasil. Revista Brasileira de Saúde e
Produção Animal, v. 1, n. 3, p. 78 – 83, 2001.

142
ALMEIDA, N. C.; TEIXEIRA, M. F. S. Levantamento sorológico de lentivírus ovino
em animais destinados ao abate na região metropolitana de Fortaleza. Anais da IV
Semana Universitária da UECE, Fortaleza: Universidade Estadual do Ceará. v.1, p. 300-
300, 1999.
ALVAREZ, V., ARRANZ, J., DALTABUIT, M., LEGINAGOIKOA, I., JUSTE, R.A.,
AMORENA, B., de ANDRES, D., LUJAN, L.L., BADIOLA, J.J., BARRIATUA, E.
Relative contribution of colostrum from Maedi–visna virus (MVV) infected ewes to
MVV-seroprevalence in lambs. Research in veterinary science. v. 78, p. 237–243, 2005.
ALVES, F.S.F. & PINHEIRO, R.R. Presença de artrite-encefalite caprina a vírus
(CAEV) no Estado do Maranhão. In: XXV Congr. Bras. Med. Vet., Gramado, p.278.
(Resumo). 1997.
ANDRÉSSON OS, ELSER JE, TOBIN GJ, GREENWOOD JD, GONDA MA,
GEORGSSON G, ANDRESDOTTIR V, BENEDIKTSDOTTIR E, CARLSDOTTIR
HM, MANTYLA EO, RAFNAR B, PALSSON PA, CASEY JM, PETURSSON P.
Nucleotide sequence and biological properties of a pathogenic proviral molecular clone
of neurovirulent visna virus. Virology. v. 193, p. 89±105. 1993.
ANDRIOLI, A., GOUVEIA, A. M. G., MARTINS, A. S., PINHEIRO, R. R., SANTOS,
D. O. Fatores de risco na transmissão do lentivírus caprino pelo sêmen. Pesquisa
Agropecuária Brasileira. v.41 n.8, p.1313-1319, 2006.
ANDRIOLI, A. Vírus da Artrite Encefalite Caprina: PCR e Isolamento Viral em
amostras de sêmen, fluido uterino e embriões. Belo Horizonte – MG: Escola de
Veterinária – UFMG. Tese (Doutorado), 2001.
ANGELOPOULOU K, KARANIKOLAOU K, PANASTASOPOULOU M,
KOUMPATI-ARTOPIOU M, VLEMMAS I, PAPADOPOULOS O, et al. First partial
characterization of small ruminant lentiviruses from Greece. Veterinary microbiology.
v. 109, p. 1–9, 2005.
ANGELOPOULOU, K., BRELLOU, G. D., GREENLAND,T. and VLEMMAS, I. A
novel deletion in the LTR region of a Greek small ruminant lentivirus may be
associated with low pathogenicity.Virus Research, v. 118, p. 178-184, 2006a.
ANGELOPOULOU, K., BRELLOU, G. D. and VLEMMAS, I. Detection of Maedi-
visna virus in the kidneys of naturally infected sheep. Journal of Comparative
Pathology, v. 134, p. 329-335. 2006b.
ARAÚJO, S. A. C. Avaliação in vitro da atividade antiviral de produtos sintéticos e
naturais contra lentivírus de pequenos ruminantes. 120p. Tese (Doutorado em Ciências
Veterinárias) – Universidade Estadual do Ceará, Fortaleza, 2008.
ARAÚJO, S.A.C. et al. Identificação do Maedi-visna vírus em pulmão de ovinos
infectados naturalmente. Arquivos do Instituto Biológico, v. 71 n.4, p. 431-436, 2004.
ASSIS, A. P. M., GOUVEIA, A.M.G., Evidência sorológica de lentivírus (Maedi-visna/
Artrite Encefalite Caprina) em rebanhos nos estados de Minas Gerais, Rio de Janeiro,

143
Bahia, Ceará. In Congresso Brasileiro de Medicina Veterinária. Anais. Recife, v. 23, p.
104, 1994.
BANDEIRA, D. A. B.; DE CASTRO, R. S.; AZEVEDO, E. O.; MELO, L. S. S.;
MELO, C. B. Seroprevalence of Caprine Arthritis–Encephalitis Virus in goats in the
Cariri region, Paraiba state, Brazil. The Veterinary Journal, v. 180, n. 3, p. 399-401,
2009.
BANKS, K. L.; ADAMS, D. L.; McGUIRE, T. C.; JAM, N; CARLSON, B. S.
Experimental infection of sheep by caprine arthritis-encephalitis virus and gotas by
progressive pneumonia virus. American Journal Veterinary Research, v. 44. n. 2, p.
2307-2310, 1983.
BARROS, S.C., ANDRESDOTTIR, V., FEVEREIRO, M. Cellular specificity and
replication rate of Maedi visna virus in vitro can be controlled by LTR sequences.
Archives of virology. v. 150, p. 201–213, 2005.
BARROS, S.C., RAMOS, F., DUARTE, M., FAGULHA, T., CRUZ, B., FEVEREIRO,
M. Genomic characterization of a slow/low Maedi visna virus. Vírus Genes. v. 29, p.
199–210, 2004.
BATISTA, M. C. S.; DE CASTRO, R. S.; CARVALHO, F. A. A.; CRUZ, M. S. P.;
SILVA, S. M. M. S.; REGO, E. W.; LOPES, J. B. Anticorpos anti-Lentivírus de
pequenos ruminantes em caprinos integrantes de nove municípios Piauienses. Ciência
Veterinária nos Trópicos, v.7, n 2 e 3, p. 75 – 81, 2004.
BENAVIDES, J., GOMEZ, N., GELMETTI, D., FERRERAS, M.C., GARCIA-
PARIENTE, C., FUERTES, M., GARCIA-MARIN, J.F., PEREZ, V. Diagnosis of the
nervous form of Maedi-visna infection with a high frequency in sheep in Castilla y
Leon, Spain. The Veterinary Record. v. 158, n. 7, p. 230–235, 2006.
BERTOLOTTI L, MAZZEI M, PUGGIONI G, CARROZZA M. L, DEI GIUDICI
S, MUZ D, JUGANARU M, PATTA C, TOLARI F, ROSATI S. Characterization of
new small ruminant lentivirus subtype B3 suggests animal trade within the
Mediterranean Basin. Journal of General Virology. v. 92, n. 8, p. 1923-9. 2011.
BIESCAS, E., PREZIUSO, S., BULGIN, M. and DEMARTINI, J. C. Ovine lentivirus-
associated leucomyelitis in naturally infected North American sheep. Journal of
Comparative Pathology, v. 132, p. 107-116. 2005.
BLACKLAWS, B.A., BERRIATUA, E., TORSTEINSDOTTIR, S., WATT, N.J., de
ANDRES, D., KLEIN, D., HARKISS, G.D. Transmission of small ruminant
lentiviruses. Veterinary microbiology. v. 101, p. 199–208, 2004.
BLACKLAWS, B.A., BIRD, P., ALLEN, D., ROY, D.J., MACLENNAN, I.C.M.,
HOPKINS, J., SARGAN, D.R., MCCONNELL, I. Initial lentivirus-host interactions
within lymph nodes: a study of Maedi-Visna virus infection in sheep. Journal of
Virology. v. 69, p. 1400–1407. 1995.

144
BOLEA, R., MONLEON, E., CARRASCO, L.,VARGAS, A., de ANDREC, D.,
AMORENA, B., BADIOLA, J. J. and LUJAN, L. Maedi-visna virus infection of ovine
mammary epithelial cells. Veterinary Research, v. 37, p. 133-144. 2006.
BRELLOU, G.D.; ANGELOPOULOU, K; POUTAHIDIS, T.; VLEMMAS, T.
Detection of Maedi-Visna Virus in the Liver and Heart of Naturally Infected Sheep.
Journal of Comparative Pathology. v.136, p. 27-35. 2007.
BRODIE, S.; PEARSON, L.; ZINK, M. Ovine lentivirus expression and disease. Virus
replication, bu no entry, is restricted to macrophages of especific tissues. American
Journal of Pathology, v.146, n.1, p. 250-263, 1995.
BRUETT, L., CLEMENTS, J.E. Functional murine leukaemia virus vectors
pseudotyped with the visna virus envelope show expanded visna vírus cell tropism.
Journal of virology. v. 75, p. 11464–11473, 2001.
BURGU, I., AKCA, Y., ALKAN, F., ÖZKUL, A., KARAGLU, T., C¸ ABALAR, M.,
Anti- body prevalence of caprine arthritis encephalitis virus (CAEV) in goats in Turkey.
Deutsche Tierarztliche Wochenschrift. v. 10, p. 390–391.1994.
CAMINHA, C. M. M.; TEIXEIRA, M. F. S.; FEITOSA, A. L. V. L.; MELO, V. S. P.;
MARTINS, G. R.; DANTAS, T. V. M. ; PINHEIRO, R. R. Levantamento Sorológico
Do Vírus Da Artrite Encefalite Caprina E Do Visna/Maedi Vírus Na Região Norte E
Nordeste Do 96 Ceará. In: XIII SEMANA UNIVERSITÁRIA DA UECE, 2008,
Fortaleza; V FEIRA DE CIÊNCIA, CULTURA, TECNOLOGIA E INOVAÇÃO DO
ESTADO DO CEARÁ, 2008,Fortaleza. Anais...Fortaleza, 2008.
CAMPBELL, B.J., AVERY, R.J. Sequence analysis and transcriptional activity of the
LTR of OLV-CU1, a North American ovine lentivirus. The Journal of general virology.
v. 77, p. 2999–3004, 1996.
CAPUCCHIO, M.T., SANNA, E., SANNA, M. P., FARIGU, S., MINELLI, R. and
GUARDA, F. Maedi-visna virus detection in ovine third eyelids. Journal of
Comparative Pathology, v. 129, p. 37-43. 2003.
CAREY, N.; ROY, D. J.; DALZIEL, R. G. Use of recombinant gp 135 to study
pitopespecific antibody responses to Maedi-Visna virus. Journal of Virological
Methods, v. 43, n.2, p. 221-323, 1993.
CARROZZA, M.L., MAZZEI, M., BANDECCHI, P., ARISPICI, M., TOLARI, F. In
situ PCR-associated immunohistochemistry identifies cell types harbouring the
MaediVisna virus genome in tissue sections of sheep infected naturally. J. Virol.
Methods Journal of Virological Methods. v. 107, p. 121–127. 2003.
CASTRO, R. S.; AZEVEDO, E. O.; TABOSA, I.; NASCIMENTO, S. A.; OLIVEIRA,
M. M. M. Anticorpos para o Vírus da Artrite-Encefalite Caprina em animais sem raça
definida (SRD) de abatedouros dos estados de Pernambuco e Paraíba. Ciência
Veterinária nos Trópicos, v. 5, n. 2 – 3, p. 121 – 123, 2002.

145
CASTRO, R.S., GREENLAND, T., LEITE, R.C., GOUVEIA, A., MORNEX, J.F.,
CORDIER, G. Conserved sequence motifs involving the tat reading frame of Brazilian
caprine lentiviruses indicate affiliations to both caprine arthritisencephalitis virus and
visna-Maedi virus. The Journal of general virology. v. 80, p. 1583–1589, 1999.
CASTRO, R. S. Lentivírus de pequenos ruminantes: ensaios imunoenzimáticos, Perfil
sorológico e inferências filogenéticas. Belo Horizonte. MG: UFMG – Escola de
Veterinária. 132p. Tese (Doutorado), 1998.
CASTRO, R. S.; NASCIMENTO, S. A.; ABREU, S. R. O. Evidência sorológica de
infecção pelo vírus da artrite-encefalite caprina em caprinos leiteiros do Estado de
Pernambuco. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v.46, n. 5, p.
571-572, 1994.
CHEBLOUNE, Y., KARR, B., SHEFFER, D., LEUNG, K., NARAYAN, O. Variation
in lentiviral gene expression in monocyte-derived macrophages from naturally infected
sheep. The Journal of general virology. v.77, p.2037-2051, 1996.
CHEN, R., WANG, H., MANSKY, L.M. Roles of uracil-DNA glycosylase and
dUTPase in virus replication. Journal of General Virology. v. 83, Pt 10, p. 2339–2345.
2002.
CHI, D., HENRY, J., KELLEY, J., THORPE, R., SMITH, J.K., KRISHNASWARNY,
G. The effects of HIV infection on endothelial functions. Endothelium. v. 7, p. 223–242.
2000.
CLEMENTS, J. E. and ZINK, M. C. Molecular biology and pathogenesis of animal
lentivirus infections. Clinical Microbiology Reviews. v. 9, p.100-117, 1996.
CLEMENTS, J. E. and PAYNE, S. L. Molecular basis of the pathobiology of
lentiviruses. Virus Research. v. 32, p. 97-109, 1994.
CLEMENTS, J.E., NARAYAN, O., CORK, L.C. Biochemical characterization of the
virus causing leucoencephalitis and arthritis in goats. The Journal of general virology.
v. 50, p. 423-427, 1980.
COFFIN, J. M. Retroviridae: the virus their of replication. In: FIELDS, B. N,; KNIPE,
D. M.; HOWLEY, P. M.; CNANOCK, R. M.; MELNICK, J. L.; MONATH, T. P.;
ROIZMAN, B.; STRAUS, S. E. 3 ed. Fields Virology. Philadelphia, Lippincoh-Raven,
p. 1767-1847, 1996.
CONTRERAS, A. et al. Molecular characterization and phylogenetic study of Maedi
visna and Caprine Arthritis Encephalitis viral sequences in sheep and goats from Spain.
Virus Research. v. 121, p. 189–198, 2006.
CRAWFORD, T.B., ADAMS, D.S., CHEEVERS, W.P., CORK, L.C. Chronic arthritis
in goats caused by a retrovirus. Science. v. 207, p. 997–999, 1980.

146
CUNHA, R. G.; NASCIMENTO, M. D. Ocorrência de anticorpos para o vírus da
artrite-encefalite caprina em soros de caprinos do estado do Rio de Janeiro. Revista
Brasileira de Medicina Veterinária, v. 17, n. 2, p. 72 - 75, 1995.
CUTLIP, R. C.; LEHMKUHL, H. D.; SACKS, J. M.; WEAVER, A. L. Prevalence of
antibody to caprine arthritis-encephalitis virus in goat in the United States. Journal of
American Veterinary Medical Association, v. 200, p. 802 -805, 1992.
CUTLIP, R. C., LAIRD, G. A. Isolation and characterization of a virus associated with
progressive pneumonia (Maedi) of sheep. American Journal Veterinary Research, v. 37,
p. 1377-1382, 1976.
DANTAS, T. V. M.; ARAÚJO, S. A. C.; PINHEIRO, R. R.; ARAGÃO, M. A. C.;
SILVA, J. B. A.; RICARTE, A. R. F.; RIBEIRO, A. L.; TEIXEIRA, M. F. S.
Desenvolvimento e padronização de um ELISA indireto para Diagnóstico de Maedi
Visna em ovinos. Ciência Animal Brasileira, v. 9, n. 1, p. 181-187, 2008.
DAWSON, M. Maedi/visna: a review. Veterinary Record. v. 106, p. 212–216, 1980.
DAWSON, M., WILESMITH, J.W. Serological survey of lentivirus (maedi-
visna/caprine arthritis encephalitis) infection in British goat herds. Veterinary Record. v. 117, p. 86–89.1985.
DE BÔER, G. F. Zwoergerziekte virus, the causative agent for progressive interstitial
pneumonia (Maedi) and meningo-leucoencephalitis (visna) in sheep. Research
Veterinary Science, v. 18, p. 15-25, 1975.
DE LA CONCHA – BERMEJILLO, A., BRODIE, S. J., MAGNUS – CORRAL, S.,
BOWEN, R. A., DEMARTINI, J. C. Pathologic and serological responses of isogeneic
twin lambs to phenotypicalty distinct lentiviruses . J. Acquir. immune defice. syndr.
human retrovirol Journal of Acquired Immune Deficiency Syndromes & Human
Retrovirology. 8: 116-123, 1995.
DeMARTINI, J. C.; HALSEY, W.; BOSHOFF, C.; DENNIS, Y.; HOWELL, M. D.
Comparison of a Maedi- Visna virus CA-TM fusion protein ELISA with other assays
for detecting sheep infected with North American ovine lentivírus strains. Veterinary
Immunology and Immunopathology, v. 71, n., 1, p. 29-40, 1999.
ELTAHIR, Y. M., DOVAS, C.I., PAPANASTASSOPOULOU, M., KOUMBATI, M.,
GIADINIS, N., VERGHESE-NIKOLAKAKI, S., KOPTOPOULOS, G. Development
of a semi-nested PCR using degenerate primers for the generic detection of small
ruminant lentivirus proviral DNA. Journal of Virological Methods. v. 135, p. 240–246.
2006.
FAUQUET, C. M., MAYO, M. A., MANILOFF, J., DESSELBERGER, U., BALL, L.
A. Virus Taxonomy. 2ed., San Diego, California: Elsevier, 2005, p. 421.
FEITOSA, A. L. V. L. Análise filogenética de Lentivírus de pequenos ruminantes
isolados do Ceará. Fortaleza, CE: UECE – Programa de Pós-graduação em Ciências
Veterinárias, 87p. Dissertação (Mestrado), 2007.

147
FEITOSA, A.L.V.L., TEIXEIRA, M. F. S., PINHEIRO, R. R., et al., 2010.
Phylogenetic analysis of small ruminant lentiviruses from Northern Brazil. Small
Ruminant Res., 94(1-3), 205-209.
FIENI, F., ROWE, J., VAN HOOSEAR, K., BURUCOA, C., OPPENHEIM, S.,
ANDERSON, G., MURRAY, J., BONDURAN, R. Presence of caprine
arthritisencephalitis virus (CAEV) proviral DNA in genital tract tissues of
superovulated dairy goat does. Thcriogenology. v. 59, p. 1515-1523. 2003.
FIENI, F., ROWE, J., VAN HOOSEAR, K., BURUCOA, C., OPPENHEIM, S.,
ANDERSON, G., MURRAY, J., BONDURANT, R. Presence of caprine
arthritisencephalitis virus (CAEV) infected cells in flushing media following oviductal-
stage embryo collection. Theriogenology. v. 57, p. 931–940. 2002.
FRANKE, C. R. Controle sanitário da artrite-encefalite caprina . Salvador: EDUFBA,
70p. 1998.
FREED, E.O. HIV-1 Gag proteins: diverse functions in the virus life cycle. Virology. v.
251, p. 1–15, 1998.
GABUZDA, D.H., HESS, J.L., SMALL, J.A., CLEMENTS, J. Regulation of the visna
virus long terminal repeat in macrophages involves cellular factors that bind sequences
containing AP-1 sites. Molecular and cellular biology. v. 9, p. 2728–2733, 1989.
GARNER, M. G., LACK, M. B. Modelling the potential impact of exotic disease on
regional Australia. Australian veterinary journal. v. 72, p. 81-87, 1995.
GDOVIN, S., CLEMENTS, J. Molecular mechanisms of visna virus tat: identification
of the targets for transcriptional activation and evidence for a post-transcriptional effect.
Virology. v. 188, p. 438–450, 1992.
GENDELMAN, H. E., NARAYAN, O., KENNEDY-STOSKPF, S., KENNEDY, P. G.
E., GHOTBI, Z., CLEMENTS, J. E., STANLEY, J., PEZESHKPOUR, G. Tropism of
sheep lentivirus for monocytes: susceptibility to infection and virus gene expression
increase during maturation of monocytes to macrophages. Journal Virology. n. 58, v. 1,
p. 67-74, 1986.
GEORGSSON, G., HOUWERS, D. J., PALSSON, P. A. and PETURSSON, G.
Expression of viral-antigens in the central nervous system of visna-infected sheep: an
immunohistochemical study on experimental visna induced by vírus strains of increased
neurovirulence. Acta Neuropathologica, v. 77, p. 299-306. 1989.
GERMAIN, K.; VALAS, S. Distribution and heterogeneity of small ruminant lentivirus
envelope subtypes in naturally infected French sheep. Virus Research. v. 120, p. 156–
162, 2006.
GJERSET, B., JONASSEN, C.M., RIMSTAD, E. Natural transmission and
comparative analysis of small ruminant lentiviruses in the Norwegian sheep and goat
populations. Virus Research. v. 125, p. 153–161, 2007.

148
GJERSET, B., STORSET, A.K., RIMSTAD, E. Genetic diversity of small-ruminant
lentiviruses: characterization of Norwegian isolates of Caprine arthritis encephalitis
virus. The Journal of general virology. v. 87, p. 573–580, 2006.
GLARIA, I., REINA, R., CRESPO, H., DE ANDRÉS, X, RAMÍREZ, H., BIESCAS,
E., PÉREZ, M M., BADIOLA, J., LUJÁN, L., AMORENA, B., DE ANDRÉS, D.
Phylogenetic analysis of SRLV sequences from an arthritic sheep outbreak
demonstrates the introduction of CAEV-like viruses among Spanish sheep. Veterinary
microbiology. v. 138, p. 156-62, 2011.
GLARIA, I., REINA, R., CRESPO, H., DE ANDRÉS, X., RAMÍREZ, H., BIESCAS,
E., PÉREZ, M. M., et al. Phylogenetic analysis of SRLV sequences from an arthritic
sheep outbreak demonstrates the introduction of CAEV-like viruses among Spanish
sheep. Veterinary microbiology, v. 138, n.1-2, p.156-62. 2009.
GONDA, M. A. Molecular biology and virus-host interactions of lentiviruses. Annals of
the New York Academy of Sciences. v. 724, p.22-42, 1994.
GONDA, M. A.; BRAUM, M. J.; CLEMENTS, J. E.; PYPER, J. M.; WONGSTAAL,
F.; GALLO, R. C.; GILDEN, R. V. Human T-cell lymphotropic virus type III shares
sequence homology with a family of pathogenic lentiviruses. Proceedings National
Academy Science. USA, v. 83, p. 4007-4011, 1986.
GOUVEIA, A. M. G.; COURA, M. A.; BRANDÃO, H. M.; ATANÁSIO, C.
Distribuição sorológica do lentivírus caprino em amostragem por demanda. In:
ENCONTRO DE PESQUISA DA ESCOLA DE VETERINÁRIA DA UFMG, Belo
Horizonte. Anais... Belo Horizonte. p. 116. 1998.
GREENWOOD, P. L. Effects of Caprine Arthritis-Encephalitis Virus on productivity
and health of dairy goats in New South Wales, Australia. Preventive Veterinary
Medicine, v. 1-2, n. 22, p. 71-87, 1995.
GREENWOOD, P.L., NORTH, R.N., KIRKLAND, P.D. Prevalence, spread and
control of caprine arthritis –encephalitis virus in dairy goat herds in New South Wales.
Australian veterinary journal. v. 72, p. 341– 345, 1995.
GREGO E, BERTOLOTTI L, QUASSO A, PROFITI M, LACERENZA D, MUZ D,
ROSATI S. Genetic characterization of small ruminant lentivirus in Italian mixed
flocks: evidence for a novel genotype circulating in a local goat population. The Journal
of general virology, v.88 (Pt 12), p.3423-7, Dec., 2007.
GREGO, E., BERTOLOTTI, L., CARROZZA, M.L., PROFITI, M., MAZZEI, M.,
TOLARI, F., ROSATI, S. Genetic and antigenic characterization of the matrix protein
of two genetically distinct ovine lentiviruses. Veterinary microbiology. v. 106, p. 179–
185, 2005.
GREGO, E., PROFITI, M., GIAMMARIOLI, M., GIANNINO, L., RUTILI, D.,
WOODALL, C., ROSATI, S. Genetic heterogeneity of small ruminant lentiviruses
involves immunodominant epitope of capsid antigen and affects sensitivity of single-

149
strain-based immunoassay. Clinical and diagnostic laboratory immunology. v. 9, p.
828–832, 2002.
GREWAL, A.S., GREENWOOD, P.E., BURTON, R.W., SMITH, J.E., BATTY, E.M.,
NORTH, R. Caprine retrovirus infection in New South Wales: virus isolations, clinical
and histopathological findings and prevalence of antibody. Australian Veterinary
Journal. v. 63, p. 245–248.1986.
GROSSI, P., GIUDICE, C., BERTOLETTI, I., CIOCCARELLI, G., BROCCHI, E.,
CAMMARATA, G., GELMETTI, D. Immunohistochemical detection of the p27 capsid
protein of caprine arthritis–encephalitis virus (CAEV) in bone-marrow cells of
seropositive goats. Journal of Comparative Pathology. v. 133, p. 197–200. 2005.
GUDMUNDSSON, B., BJARNADOTTIR, H., KRISTJANSDOTTIR, S., JONSSON,
J.J. Quantitative assays for Maedi-visna virus genetic sequences and RNAm’s based on
RT-PCR with real-time FRET measurements. Virology. v. 307, p. 135–142. 2003.
GUEDES, M.I.M.C. Infecção experimental pelos vírus da artrite encefalite caprina em
cabritos de nove a vinte e sete dias de idade. Belo Horizonte, MG: UFMG - Escola de
Veterinária. 59p. Dissertação (Mestrado), 1999.
GUFLER, H., BAUMGARTNER, W. Overview of herd and CAEV status in dwarf
goats in South Tyrol, Italy. Veterinary Questions and Answers Archive. v.29, p. 68–
70.2007.
HAASE, A. T., Pathogenesis lentivirus infection. Nature, v. 322, n. 10, p. 130-136.
1986.
HARMACHE, A., VITU, C., GUIGUEN, F., RUSSO, P., BERTONI, G., PEPIN, M.,
VIGNE, R., SUZAN, M. Priming with tat-deleted caprine arthritis encephalitis virus
(CAEV) proviral DNA or live virus protects goats from challenge with pathogenic
CAEV. Journal of Virology. v. 72, n. 8, p. 6796–6804. 1998.
HARMACHE, A. et al. Requirement of caprine arthritis encephalitis virus vif gene for
in vivo replication. Virology, 224, 246–255. 1996.
HARMACHE, A., VITU, C., RUSSO, P., BOUYAC, M., HIEBLOT, C., PEVERI, P.,
VIGNE, R., SUZAN, M. The caprine arthritis encephalitis virus tat gene is dispensable
for efficient viral replication in vitro and in vivo. Journal of Virology. v. 69 n. 9, p.
5445–5454. 1995.
HARRIS, R.S., BISHOP, K.N., SHEEHY, A.M., CRAIG, H.M., PETERSEN-MAHRT,
S.K., WATT, I.N., NEUBERGER, M.S., MALIM, M.H. DNA deamination mediates
innate immunity to retroviral infection. Cell. v. 113, n. 6, p. 803–809. 2003.
HERRMANN-HOESING, L. M., PALMER, G. H., KNOWLES, D. P. Evidence of
proviral clearance following postpartum transmission of an ovine lentivirus. Virology. v.
362, p. 226–234, 2007.

150
HERRMANN, L. M., HÖTZEL, I., CHEEVERS, W.P., TOP, K.P.O., LEWIS, G.S.,
KNOWLES, D.P. Seven new ovine progressive pneumonia virus (OPPV) field isolates
from Dubois Idaho sheep comprise part of OPPV clade II based on surface envelope
glycoprotein (SU) sequences . Virus Research. v. 102, p. 215–220, 2004.
HESS, J.L., PYPER, J.M., CLEMENTS, J.E. Nucleotide sequence and transcriptional
activity of the caprine arthritis encephalitis virus long terminal repeat. Journal of
virology. v. 60, p. 385–393, 1986.
HÖTZEL, I., KUMPULA-MCWHIRTER, N., CHEEVERS, W. P. Rapid evolution of
two discrete regions of the caprine arthritis-encephalitis virus envelope surface
glycoprotein during persistent infection. Virus Research. v. 84, p. 17–25, 2002.
HÖTZEL, I., CHEEVERS, W.P. Host range of small ruminant lentivirus cytopathic
variants determined with a selectable caprine arthritis-encephalitis virus pseudotype
system. Journal of virology. v. 75, p. 7384–7391, 2001.
HOUWERS, D.J. Economic importance, epidemiology and control. In: Pétursson, G.,
Hoff-Jørgensen, R. (Eds.), Maedi-Visna and Related Diseases. Kluwer Academic
Publishers, Boston, pp. 83–117. 1990.
HOUWERS, D.J., KONIG, C.D., BAKKER, J., de BOER, M.J., PEKELDER, J.J.,
SOL, J., VELLEMA, P., de VRIES, G. Maedi– visna control in sheep: III. Results and
evaluation of a voluntary control program in The Netherlands over a period of four
years. The Veterinary quarterly. v. 9, p. 29S–36S, 1987.
HOUWERS, D. J. Experimental Maedi-visna control in the Netherlands. In Sharp J. M.
Holf-Jorgensen T (eds). Slow virus in sheep, goats and cattle, Luxembourg.
Commission of the European Communities, p. 115 – 121, 1985.
HUSO, L.D., NARAYAN, O., HART, W.G. Sialic acids on the surface of caprine
arthritis-encephalitis virus define the biological properties of the virus. Journal of
virology. v. 62, p.1974-1980, 1988.
ICTV - International Committee on Taxonomy of Viruses. Release 2009.
http://www.ictvonline.org/virusTaxonomy.asp. 2011.
JIN, Z., JIN, L., PETERSON, D.L., LAWSON, C.L. Model for lentivirus capsid core
assembly based on crystal dimers of EIAV p26. Journal of molecular biology. v. 286, p.
83–93, 1999.
JUGANARU, M., REINA, R., GREGO, E., PROFITI, M., & ROSATI, S. LTR
promoter activity of SRLV genotype E, strain Roccaverano. Veterinary research
communications, 34 Suppl 1, S47-51. 2010.
KARR, BM, CHEBLOUNE Y, LEUNG K, NARAYAN O. Genetic characterization of
two phenotipically distinct North American ovine lentiviruses and their possible origin
from caprine arthritisencephalitis virus. Virology. v. 225, n. 1, p. 1–10, 1996.

151
KEMP, R. K. ; KNOWLES, D. P.; PERRY, L. L.; McGUIRE, T. C.; CHEEVERS, W.
Crossreactive neutralizing antibodies induced by immunization with caprine arthritis
encephalitis virus surface glycoprotein. Vaccine, v.18, n.13, p.1282-1287, 2000.
KNOWLES, D. P. Laboratory diagnostic test for Retrovirus infections of small
ruminants. Veterinary Clinics of North American: Food Animal Practice, v. 13, n.1, p.
1-11, 1997
KNOWLES, D.P., CHEEVERS,W.P., MCGUIRE, T.C., BRASSFIELD, A.L.,
HARWOOD,W.G., STEM, T.A. Structure and genetic variability of envelope
glycoproteins of two antigenic variants of caprine arthritis-encephalitis lentivirus.
Journal of virology. v. 65, p. 5744–5750, 1991.
KRIEG, A., PETERHANS, E. Caprine arthritis-encephalitis in Switzer- land:
epidemiological and clinical studies. Schweiz. Arch. Tierheilkd. v. 132, p. 345–352.
1990.
KUZMAK J, et al. Molecular characterization of lentiviruses from goats from Poland
based on gag gene sequence analysis. Comparative Immunology, Microbiology and
Infectious Diseases. v. 30, p. 211-223, 2007.
L’HOMME, Y., OUARDANI, M., LÉVESQUE, V., BERTONI, G., SIMARD, C., &
PISONI, G. Molecular characterization and phylogenetic analysis of small ruminant
lentiviruses isolated from Canadian sheep and goats. Virology journal, v. 8, n.1, p.271.
2011
LAAMANEN, I. et al., Genetic characterization of Maedi-visna virus (MVV) detected
in Finland. Veterinary microbiology. v. 122, p. 357-365, 2007.
LACERENZA, D., GIAMMARIOLI, M., GREGO, E., MARINI, C., PROFITI, M.,
RUTILI, D., ROSATI, S. Antibody response in sheep experimentally infected with
different small ruminant lentivirus genotypes. Veterinary
Immunology and Immunopathology. v. 112, p. 264–271. 2006.
LAIRMORE, D.P., AKITA, G.Y., RUSSELL, H.I., DEMARTINI, J.C. Replication and
cytopathic effects of ovine lentivirus strains in alveolar macrophages correlate with in
vivo pathogenicity. Journal of virology. v. 61, p. 4038–4042, 1987.
LAMARA, A., FIENI, F., MSELLI-LAKHAL, L., TAINTURIER, D., CHEBLOUNE,
Y. Epithelial cells from goat oviduct are highly permissive for productive infection with
caprine arthritis-encephalitis virus (CAEV). Virus research. v. 87, p. 69–77, 2002a.
LAMARA, A., FIENI, F., MSELLI-LAKHAL, L., CHATAGNON, G., BRUYAS, J.F.,
TAINTURIER, D., BATTUT, I., FORNAZERO, C., CHEBLOUNE, Y. Early
embryonic cells from in vivo-produced goat embryos transmit the caprine arthritis-
encephalitis virus (CAEV). Theriogenology. v. 58, n. 6, p. 1153–1163, 2002b.
LAMARA, A., FIENI, F., MSELLI-LAKHAL, L., TAINTURIER, D., CHEBLOUNE,
Y. Efficient replication of caprine arthritis-encephalitis virus in goat granulosa cells.
Virus research. v. 79, n. 165–172, 2001.

152
LARSEN, H. J.; HYLLSETH, B.; KROGSRUD, J. Experimental maedi virus infection
in sheep: cellular and humoral immune response during three years following intranasal
inoculation. American Journal of Veterinary Research, v. 43, n. 3, p. 384-389, 1982.
LECHAT, E., MILHAU, N., BRUN, P., BELLATON., C., GREENLAND, T,
MORNEX, J. F., LE JAN, C. Goat endothelial cells may be infected in vitro by
transmigration of caprine arthritis-encephalitis virus-infected leucocytes. Veterinary
Immunology and Immunopathology. v. 104, p. 257–263. 2005.
LEE, W. C.; McCONNEL, I., BLACKLAWS, B. A. Electron microscope studies of the
replication of a British isolate of Maedi-visna virus in macrophages and skin cell lines.
Veterinary Microbiology, v. 43, p. 93-104, 1996.
LEITE, B. L. S.; MODOLO, J. R.; PADOVANI, C. R.; STACHISSINI, A. V. M.;
CASTRO, R. S.; SIMÕES, L. B. Avaliação da taxa comparativa de ocorrência da
Artrite-Encefalite Caprina a Vírus pelas regionais de defesa agropecuária do estado de
São Paulo, Brasil, e seu mapeamento por meio de sistema de informações geográficas.
Arquivos do Instituto Biológico, v. 71, n. 1, p. 21 -26, 2004.
LERONDELLE, C., GODET, M., MORNEX, JF. Infection of primary cultures of
mammary epithelial cells by small ruminant lentiviruses. Veterinary Reshearch. n.30,
v.5, p.467-474, 1999.
LEROUX, C., CHASTANG, J., GREENLAND, T. and MORNEX, J. F. Genomic
heterogeneity of small ruminant lentiviruses: existence of heterogeneous populations in
sheep and of the same lentiviral genotypes in sheep and goats. Archives of Virology. v.
142, p.1125-1137, 1997.
LEROUX, C., VUILLERMOZ, S., MORNEX, J.F. et al. Genomic heterogeneity in
the poolregion of ovine lentivirus obtained from bronchoalveolar cells of infected sheep
from France. Journal of General Virology. v.76, p.1533-1537, 1995.
LI, X., YUAN, B., GOFF, S.P. Genetic analysis of interactions between Gag proteins of
Rous Sarcoma virus. Journal of virology. v. 71, p. 5624–5630, 1997.
LUJÁN, L., BEGARA, I., COLLIE, D.D.S., WATT, N.J. Ovine lentivirus (Maedi-visna
virus) protein expression in sheep alveolar macrophages. Veterinary Pathology. v.31 ,
p.695-703, 1994.
MADRUGA, C. R.; ARAÚJO, F. R.; SOARES, C. O. Princípios, padronização e
validação de provas sorológicas. In: _____ MADRUGA, C. R.; ARAÚJO, F. R.;
SOARES, C. O. Imunodiagnóstico em medicina Veterinária. Campo Grande:
EMBRAPA gado de corte (Campo Grande- Mato Grosso do Sul- Brasil), p.145-175.
2001.
MADUREIRA, K. M.; GOMES, V. Prevalência da Artrite-Encefalite Caprina (CAE)
em propriedades leiteiras do estado de São Paulo. 2007. Disponível em:
http://sare.unianhanguera.edu.br/index.php/rensc/article/viewFile/370/371. Acesso em:
07 out 2011.

153
MARCHESIN, D.M. Caracterização molecular de parte do gene gag dos lentivírus
artrite-encefalite caprina (CAEV) e Maedi-visna dos ovinos (MVV), isolados de animais
naturalmente infectados do Rio Grande do Sul, Brasil. Porto Alegre, RS: UFRGS -
Faculdade de Veterinária. 111p. Dissertação (Mestrado - Ciências Veterinárias), 1997.
MELO, C. B.; DE CASTRO, R. S.; OLIVEIRA, A. A.; FONTES, L. B.; CALLADO,
A. K. C.; NASCIMENTO, S. A. Estudo preliminar sobre a infecção por lentivírus de
pequenos ruminantes em ovinos e caprinos de Sergipe. In:
CONGRESSOLATINOAMERICANO DE BUIATRIA, 5°, Salvador. Anais...
Salvador: SBB, p.47 – 48. 2003.
MILLER, R.J., CAIRNS, J.S., BRIDGES, S., SARVER, N. Human immunodeficiency
virus and AIDS: insights from animal lentiviruses. Journal of Virology. v. 74, n,16,
p.7187–7195. 2000.
MITCHELL RS, BEITZEL BF, SCHRODER ARW, SHINN P, CHEN H, ET AL.
Retroviral DNA Integration: ASLV, HIV, and MLV Show Distinct Target Site
Preferences. PLoS Biology. v. 2, n. 8, p. 234. 2004.
MOOJEN, V., SOARES, H.C., RAVAZZOLO, A.P., PIZZOL, M., GOMES, M.
Evidência de infecção pelo lentivirus (Maedi-visna/artrite encefalite caprina) em
caprinos no Rio Grande do Sul, Brasil. Arquivos da Faculdade de Veterinária UFRGS.
v. 14, p.77-78, 1986.
MSELLI-LAKHAL, L., GUIGUEN, F., FORNAZERO, C., DU, J., FAVIER, C.,
DURAND, J., et al. Goat milk epithelial cells are highly permissive to CAEV infection
in vitro. Virology. v. 259, p. 67–73. 1999.
MURPHY, B., MCELLIOTT, V., VAPNIARSKY, N., OLIVER, A, & ROWE, J.
Tissue tropism and promoter sequence variation in caprine arthritis encephalitis virus
infected goats. Virus research, v. 151, n. 2, p. 177-84. 2010.
MURPHY, B., JASMER, D.P., WHITE, S.N., KNOWLES, D. Localization of a TNF-
activated transcription site and interactions with the gamma activated site within the
CAEV U3 70 base pair repeat. Virology. v. 364, n.1, p. 196–207. 2007.
NARAYAN, O., CLEMENTS, J.E. Biology and pathogenesis of lentiviruses. The
Journal of general virology. v.70, p.1617-1639, 1989.
NARAYAN, O., KENNEDY-STOSKOPF, S., SHEFFER, D., GRIFFIN, D.E.,
CLEMENTS, J.E. Activation of caprine arthritis-encephalitis virus expression during
maturation of monocytes to macrophages. Infection and Immunity. v. 41, n. 1, p. 67-73,
1983.
NARAYAN, O., CLEMENTS, J. E., STRANDBERG, J. D., CORK, L. C., AND
GRIFFIN, D. E. Biological characterization of the virus causing leukoencephalitis and
arthritis in goats. Journal of General Virology. v. 50, p. 69– 79. 1980.

154
NORD, K., LOKEN, T., ORTEN, A. Control of caprine arthritis-encephalitis virus
infection in three Norwegian goat herds. Small Ruminant Research. v. 28, p. 109–114.
1998.
O’NEIL, S.P., SUWYN, C., ANDERSON, D.C., NIEDZIELA, G., BRADLEY, J.,
NOVEMBRE, F.J., HERNDON, J.G., MCCLURE, H.M. Correlation of acute humoral
response with brain virus burden and survival time in pig-tailed macaques infected with
the neurovirulent simian immunodeficiency virus SIVsmmFGb. American Journal of
Pathology. v. 164, p. 1157–1172. 2004.
OLIVEIRA, M. M. M.; CASTRO, R. S.; CARNEIRO, K. L.; NASCIMENTO, S. A.;
CALLADO, A. K. C.; ALENCAR, C. S. A.; COSTA L. S. P. Anticorpos contra
lentivírus de pequenos ruminantes em caprinos e ovinos em abatedouros do estado de
Pernambuco. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 58, n.5, p.
947-949, 2006.
OSKARSSON, T., HREGGVIDSDOTTIR, H.S., AGNARSDOTTIR, G.,
MATTHIASDOTTIR, S., OGMUNDSDOTTIR, M.H., JONSSON, S.R.,
GEORGSSON, G., INGVARSSON, S., ANDRESSON, O.S., ANDRESDOTTIR, V.
Duplicated sequence motif in the long terminal repeat of maedi-visna virus extends cell
tropismandisassociatedwithneurovirulence. Journal of Virology. v. 81, n.8, p.4052–
4057. 2007.
PAGÈS, J-C; PIVER, E. Vírus, vetores de moléculas terapêuticas. Scientific American:
Vírus, Inimigos Úteis, Brasil, p.44-49, out. 2008.
PASICK, J. Maedi-visna virus and caprine Arthritis-Encephalitis virus: Distinct species
or quasispecies and its implications for laboratory diagnoses Canada Journal
Veterinary. v. 62, p. 241- 244, 1998.
PÉPIN, M., VITU, C., RUSSO, P., MORNEX, J.F., PETERHANS, E. Maedi-visna
virus infection in sheep: a review. Veterinary research. v. 29, 341–367, 1998.
PERETZ, G., BUGNARD, F., CALAVAS, D. Study of a prevention programme for
caprine arthritis– encephalitis. Veterinary research. v. 25, p. 322– 326, 1994.
PERK, K. Presence of viral particles in neural cells of goats with caprine arthritis
encephalitis. Research in Veterinary Science. v. 49, n.3, p. 367-369, 1990.
PERTURSON, G.; ANDRÉSDÓTIR, O. S.; ANDRÉSSON, G. Lentivírus disease of
sheep and goat: Maedi Visna an caprine Arthritis encephalitis. In: ___ SPEEDY, A. W.
Progress in sheep and Goat Research, Keldur: Hardcover. p. 107-129. 1992.
PERTUSSON, G.; MTTHIASDOTTIR, S.; SVANSSON, V.; ANDRESDOTTIR, V.;
GEORGSSON, G.; MARTIN, A. H.; AGNARSDOTTIR, G.; GISLADÓTTIR, E.;
ÁRNADÓTTIR, S.; HOGNADÓTTIR, S.; JÓHNSSON, S. R.; ANDRÉSSON, O. S.;
TORSTEINSDÓTTIR, S. Mucosal vaccination with an attenuated maedi-visna virus
clone. Vaccine, v.23, n.24 3223-3228, 2005.
PETERHANS, E., GREENLAND, T., BADIOLA, J., HARKISS, G., BERTONI, G.,
AMORENA, B., ELIASZEWICZ, M., JUSTE, R.A., KRASSNIG, R., LAFONT, J.P.,

155
LENIHAN, P., PETURSSON, G., PRITCHARD, G., THORLEY, J., Vitu, C.,
MORNEX, J.F., PÉPIN, M. Routes of transmission and consequences of small ruminant
lentiviruses (SRLVs) infection and eradication schemes. Veterinary research. v. 35, p.
257–274, 2004.
PINHEIRO, R. R.; GOUVEIA, A. M. G.; ALVES, F. S. F. Prevalência da infecção pelo
Vírus da Artrite-Encefalite Caprina no estado do Ceará, Brasil. Ciência Rural, v. 31, n.
3, p. 449 – 454, 2001.
PINHEIRO, R.R., Vírus da artrite encefalite caprina: Desenvolvimento e padronização
de ensaios imunoenzimáticos (ELISA e Dot-Blot) e estudo epidemiológico no Estado do
Ceará. Belo Horizonte: UFMG. 115p. Tese (Doutorado) – Escola de Veterinária,
Universidade Federal de Minas Gerais, 2001.
PINHEIRO, R. R.; ALVES, F. S. F.; GIRÃO, E. S.; MEDEIROS, L. P. A.; GIRÃO, R.
N. Presença da Artrite-Encefalite Caprina a Vírus (CAEV), em Teresina – Piauí. In:
CONGRESSO BRASILEIRO DE MEDICINA VETERINÁRIA, 24°, Goiânia. Anais...
Goiânia. p. 161. 1996.
PISONI G, BERTONI G, BOETTCHER P, PONTI W, MORONI P. Phylogenetic
analysis of the gag region encoding the matrix protein of small ruminant lentiviruses:
comparative analysis and molecular epidemiological applications. Virus research. v.
116, n. 1–2, p. 159–67, 2006.
PISONI, G., QUASSO, A., MORONI, P. Phylogenetic analysis of small-ruminant
lentivirus subtype B1 in mixed flocks: Evidence for natural transmission from goats to
sheep. Virology. v. 339, p. 147–152, 2005.
PISTELLO, M.; MATTEUCCI, D. ; DONCI, F; ISOLA, P.; MAZZETTI, P.;
ZACCARO, L.; MERICO, A.; MAURO, D. D.; FLYNN, N. ; BENDINELLI, M.
AIDS vaccination studies using an vivo feline immunodeficiency virus model:
protection fro an intrclade challenge administered systemally or mucosally by an
attenuated vaccine. Journal of Virology, v.77, n.20, p.10740-10750. 2003.
PRESTON, B., DOUGHERTY, J. P. Mechanisms of retrovial mutation. Trends in
microbiology. v. 4, p. 16-21, 1996.
PREZIUSO, S., RENZONI, G., ALLEN,T. E.,TACCINI, E., ROSSI, G., DEMARTINI,
J. C. and BRACA, G. Colostral transmission of Maedi visna virus: sites of viral entry in
lambs born from experimentally infected ewes.Veterinary Microbiology, v. 104, p. 157-
164. 2004.
PRIET, S., NAVARRO, J.M., GROS, N., QUERAT, G., SIRE, J. Differential
incorporation of uracil DNA glycosylase UNG2 into HIV-1, HIV-2, and SIV(MAC)
viral particles. Virology v. 307, n. 2, p. 283–289.2003.
PRIMO, T.S. Enquete soroepidemiológica da Maedi Visna no estado do Ceará. In:
SEMANA DA CAPRINOCULTURA E OVINOCULTURA BRASILEIRAS. Campo
Grande, Anais. Campo Grande, CD-ROM, 2006.

156
QUÉRAT, G., AUDOLY, G., SONICO, P., VIGNE, R. Nucleotide sequence analysis of
SA-OMW, a visna-related ovine lentivirus: phylogenetic history of lentiviroses.
Virology. v. 175, p. 434-447, 1990.
QUERAT, G., BARBAN, V., SAUZE, N., FILIPPI, P., VIGNE, R., RUSSO, P., VITU,
C. Highly lytic and persistent lentiviruses naturally present in sheep with progressive
pneumonia are genetically distinct. Journal of virology. v. 52, p. 672–679, 1984.
RAMOS, O. S.; SILVA, A. C. S; MONTENEGRO, A. J. D.; FREITAS, J. A.;
WATANABE, N. A. Anticorpos para o Vírus da Artrite Encefálica no município de
Castanhal – Pará. Revista de Ciências Agrárias (Belém), v. 26, p. 107 – 111, 1996.
RAVAZZOLO, A. P.; NENCI, C.; VOGT, H. R.; WALDVOGEL, A.; OBEXER- UFF,
G.; PETERHANS, E.; BERTONI, G. Viral load, organ distribution, histopathological
lesions, and cytokine RNAm expression in goats infected with a molecular clone of the
caprine arthritis encephalitis vírus. Virology. v.350, p.116–127. 2006.
RAVAZZOLO, A.P., REISCHAK, D., PETERHANS, E., ZANONI, R. Phylogenetic
analysis of small ruminant lentiviruses from Southern Brazil. Virus research. v. 79, n.
1-2, p. 117– 123, 2001.
REDDY, P.G., SAPP, W.J., HENEINE, W. Detection of caprine arthritis-encephalitis
virus by polymerase chain reaction. Journal of Clinical Microbiology. v. 31, n. 11, p.
3042-3043, 1993.
REGITANO, L. C. A.; COUTINHO, L. L. Biologia molecular aplicada à produção
animal. Brasília: Embrapa. Informação tecnológica. 215p. 2001.
REINA, R., JUGANARU, M. M., PROFITI, M., CASCIO, P., CERRUTI, F.,
BERTOLOTTI, L., DE MENEGHI, D., et al. Immunological parameters in goats
experimentally infected with SRLV genotype E, strain Roccaverano. Veterinary
immunology and immunopathology, v. 139, n. 2-4, p. 237-44. 2011.
REINA, R., BERTOLOTTI, L., DEI GIUDICI, S., PUGGIONI, G., PONTI, N.,
PROFITI, M., PATTA, C., et al. Small ruminant lentivirus genotype E is widespread in
Sarda goat. Veterinary microbiology, v. 144, n.1-2, p. 24-31. 2010.
REINA, R., GREGO, E., BERTOLOTTI, L., DE MENEGHI, D., ROSATI, S. Genome
analysis of small-ruminant lentivirus genotype E: a caprine lentivirus with natural
deletions of the dUTPase subunit, vpr-like accessory gene, and 70-base-pair repeat of
the U3 region. Journal of Virology. v. 83, n. 2, p. 1152–1155. 2009a.
REINA R, GREGO E, BERTOLOTTI L, DE MENEGHI D, ROSATI S. Genome
analysis of small-ruminant lentivirus genotype E: a caprine lentivirus with natural
deletions of the dUTPase subunit, vpr-like accessory gene, and 70-base-pair repeat of
the U3 region. Journal of Virology v. 83, n.2, p.1152-5. 2009b.
REINA, R., MORA, M.I., GLARIA, I., GARCÍA, I., SOLANO, C., LUJÁN, L.,
BADIOLA, J.J., CONTRERAS, A., BERRIATUA, E., JUSTE, R., MAMOUN, R.Z.,
ROLLAND, M., AMORENA, B., DE ANDRÉS, D. Molecular characterization and

157
phylogenetic study of Maedi visna and Caprine Arthritis Encephalitis viral sequences in
sheep and goats from Spain. Virus Research. v. 121, p. 189–198, 2006.
REISCHAK, D. Lentivirus de pequenos ruminantes imunofluorescência utilizando
isolados brasileiros para diagnosticar sorológico de infecção em ovinos e caprinos.
Dissertação de Mestrado - Universidade Federal do Rio Grande do Sul. Porto Alegre,
132p, 2000.
REYBURN, H. T.; ROY, D. J.; BLACKLAWS, B. A.; SARGAN, D. R.; McCONELL,
I. Expression of Maedi-visna virus major core protein p. 25: development of a sensitive
p 25 antigen detection assay. Journal of Virological Methods, v. 37, n. 3, p. 305-320,
1992.
RICARTE, A. R. F., ANDRIOLI, A., PINHEIRO, R. R., BÁO, S. N., SILVA, J. S.,
BRAZ, S. V, NAME, K. P. O., LIMA-VERDE, I. B., BRITO, F. F., DIAS, R. P.,
FREITAS AGUIAR, T. D., DANTAS, T. V. M., ARAÚJO, S. A., CAVALCANTI, D.
M. L. P., PAULA, N. R. O., TEIXEIRA, M. F. S. Avaliação Imunohistoquímica e
Ultraestrutural de Gametas e Embriões Caprinos Infectados com o CAEV. Arquivos do
Instituto Biológico. v.77, n.2, p.217-223, 2010.
RIET-CORRÊA, F. Doenças de Ruminantes e Equinos. v.1, São Paulo: Varela. p.998.
2001.
RIMSTAD, E., EAST, N.E., TORTEN, M., HIGGINS, B.S., DEROCK, E.,
PEDERSEN, N.C. Delayed seroconversion following naturally acquired caprine
arthritis-encephalitis virus infection in goats. American Journal of Veterinary Research.
v. 54, p. 1858–1862.1993.
ROLLAND, M., MOONEY, J., VALAS, S., PERRIN, G., MAMOUN, R.Z.
Characterisation of an Irish caprine lentivirus strain-SRLV phylogeny revisited. Virus
research. v. 85, p. 29–39, 2002.
ROSATI, S., PROFITI, M., GREGO, E., CARROZZA, M.L., MAZZEI, M.,
BANDECCHI, P. Antigenic variability of ovine lentivirus isolated in Italy. Veterinary
Research Communications. v. 28 (Suppl. 1), p. 319–322. 2004.
ROWE, J.D., EAST, N.E. Risk factors for transmission and methods for control of
caprine arthritis– encephalitis virus infection. The Veterinary clinics of North America.
Food animal practice. v. 13, p. 35–53, 1997.
ROWE, J.D., EAST, N. E., THUNNOND, M.C., FRANTI, C. E., PEDERSEN, N.C.
Cohort study of natural transmission and two methods for control of caprine arthritis-
encephalitis virus infection in goats on a California dairy. American journal of
veterinary research. v. 53, p. 2386-95, 1992.
RUSSO, P., VITU, C., BOURGOGNE, A., VIGNONI, M., ABADIE, G., DAVID, V.,
PÉPIN, M. Caprine arthritis-encephalitis virus: detection of proviral DNA in lactoserum
cells. Veterinary Record. v. 140, n. 18, p. 483-484, 1997.

158
RYAN, S.,TILEY, L., MCCONNELL, I. and BLACKLAWS, B. Infection of dendritic
cells by the Maedi-visna lentivirus. Journal of Virology, v. 74, p. 10096-10103. 2000.
SALTARELLI, M., QUERAT, G., KONINGS, D.A., VIGNE, R., CLEMENTS, J.E.
Nucleotide sequence and transcriptional analysis of molecular clones of CAEV which
generate infectious virus. Virology. v. 179, p. 347–364, 1990.
SANNA, E., SANNA, M.P., VITALI, C.G., RENZONI, G., SANNA, L., SPANO, S.,
ROSSI, G., LEONI, A. Proviral DNA in the brains of goats infected with caprine
arthritis-encephalitis virus. Journal of Comparative Pathology. v. 121, p. 271–276.
1999.
SARGAN, D.R., BENNET, I.D., COUSENS, C., ROY, D.J., BLACKLAWS, B.A.,
DALZIEL, R.G., WATT, N.J., McCONNELL, I. Nucleotide sequence of EV1, a British
isolate of Maedi-visna virus. The Journal of general virology. v. 72, p. 1893-1903,
1991.
SCHEER-CZECHOWSKI, P., VOGT, H.R., TONTIS, A., PETERHANS, E.,
ZANONI, R. Pilot project for eradicating Maedi – visna in Walliser blacknose sheep.
Schweizer Archiv fur Tierheilkunde. v. 142, p. 155– 164, 2000.
SCHRODER AR, SHINN P, CHEN H, et al. HIV-1 integration in the human genome
favors active genes and local hotspots. Cell. v. 110, n.4, p. 521–9. 2002.
SELL, B. E. Prevalência de anticorpos para o vírus da Artrite-Encefalite Caprina em
soros de caprinos no estado de Santa Catarina. 2000. 23 f. Monografia (Especialização
em Sanidade Animal) – Universidade do Estado de Santa Catarina, Lages, 2000.
SHAH, C., BONI, J., HUDER, J.B., VOGT, H.R., MUHLHERR, J., ZANONI, R.,
MISEREZ, R., LUTZ, H., SCHUPBACH, J. Phylogenetic analysis and reclassification
of caprine and ovine lentiviruses based on 104 newisolates: evidence for regular sheep-
to-goat transmission and worldwide propagation through livestock trade. Virology. v.
319, n.1, p. 12–26, 2004a.
SHAH, C., HUDER, J.B., BONI, J., SCHONMANN, M., MUHLHERR, J., LUTZ, H.,
SCHUPBACH, J. Direct evidence for natural transmission of small-ruminant
lentiviruses of subtype A4 from goats to sheep and vice versa. Journal of virology. v.
78, n. 74, p. 7518–7522, 2004b.
SIGURDARDÓTTIR, B., THORMAR, H. Isolation of a viral agent from the Iungs of
sheep affected with Maedi. J. Infec. Dis., v. 114, n. 1, p. 55-60, 1964.
SIGURDSSON, B.; THORMAR, H.; PALSSON, P. A. Cultivation of visna virus in
tissue culture. Archives Gesante Virusforsch, v. 10, p. 368-381, 1960.
SIGURDSSON, B. Maedi, a slow progressive pneumonie of sheep: an epizooiogical
and pathological study. The British veterinary journal. v.110, p.225-270, 1954.

159
SILVA, J. S.; CASTRO, R. S.; MELO, C. B.; FEIJÓ, F. M. C.. Infecção pelo vírus da
artrite encefalite caprina no Rio Grande do Norte. Arquivo Brasileiro de Medicina
Veterinária e Zootecnia, v. 57, n.6, p. 727-731, 2005.
SILVA, J., Vírus da Artrite Encefalite Caprina (CAEV) em folículos pré-antrais de cabras
naturalmente infectadas/ Fortaleza: UECE. 148p. Tese (Doutorado) – Programade Pos-
Graduação em Ciências Veterinárias, Universidade Estadual do Ceará, 2006.
SKRABAN, R., MATTHÍASDÓTTIR, S., TORSTEINSDÓTTIR, S.,
AGNARSDÓTTIR, G., GUDMUNDSSON, B., GEORGSSON, G., MELOEN, R.H.,
ANDRÉSSON, O.S., STASKUS, K.A., THORMAR, H., ANDRÉSDÓTTIR, V.
Naturally occurring mutations within 39 amino acids in the envelope glycoprotein of
Maedi-visna virus alter the neutralization phenotype. Journal of virology. v. 73, p.
8064–8072, 1999.
SONIGO, P., ALIZON, M., STASKUS, K., KLATZMANN, D., COLE, S., DANOS,
O., RETZEL, E., TIOLLAIS, P., HAASE, A., WAIN-HOBSON, S. Nucleotide
sequence of the visna lentivirus: relationship to the AIDS virus. Cell. v. 42, p. 369–382,
1985.
SOTOMAIOR C.; MILCZEWSKI V. Relato de um rebanho ovino infectado pelo vírus
maedi-visna no Estado do Paraná. In: In: CONGRESSO BRASILEIRO.DE
MEDICINA VETERINÁRIA,25, 1997, Gramado. Anais...Gramado. p. 179.1997.
STASKUS, K.A., COUCH, L., BITTERMAN, P., RETZEL, E.F., ZUPANCIC, M.,
LIST, J., HAASE, A.T. In situ amplification of visna virus DNA in tissue sections
reveals a reservoir of latently infected cells. Microbial Pathogenesis. v. 11, p. 67–76.
1991.
STEINHAUER, D.A., HOLLAND, J.J. Rapid evolution of RNA viruses. Annual review
of microbiology. v. 41, p. 409-433, 1987.
STORSET, A.K., EVENSEN, O., RIMSTAD, E. Immunohistochemical identification
of caprine arthritis–encephalitis virus in paraffin-embedded specimens from naturally
infected goats. Veterinary Pathology. v. 34, p.180–188. 1997.
SUTTON, K., LIN, C.-T., HARKISS, G., MCCONNELL, I., SARGAN, D. Regulation
of the long terminal repeat in visna virus by a transcription factor related to the
AML/PEBP2/CBF superfamily. Virology. v. 229, p. 240–250, 1997.
TEIXEIRA, M. F. S.; LAMBERT, V.; MSELLI-LAKAHL, L.; CHETTAB, A.;
CHEBLOUNE, Y.; MORNEX, J. F.Immortalization of caprine fibroblasts permissive
for replication of small ruminant lentiviruses. American Journal of Veterinary
Research, v.58, n.6, p.579-584, 1997.
TIGRE, D. M.; CAMPOS, G. S.; SARDI, S. I. Isolamento e identificação do Vírus da
Artrite Encefalite Caprina, a partir do cocultivo de células mononucleares do sangue
com células de membrana sinovial de cabra. Revista de Ciências Médicas e Biológicas,
v. 5, n. 2, p. 124 – 131, 2006.

160
TORRES-ACOSTA, J.F.J., GUTIERREZ-RUIZ, E.J., BUTLER, V., SCHMIDT, A.,
EVANS, J., BABINGTON, J., BEARMAN, K., FORDHAM, T., BROWNLIE, T.,
SCHROER, S., CÁMARA, G.E., BUTLER, V. Serological survey of caprine arthritis-
encephalitis virus in 83 goat herds of Yucatan, Mexico. Small Ruminant Research. v.
49, p. 207–211. 2003.
TORSTEINSDOTTIR, S., MATTHIASDOTTIR, S., VIDARSDOTTIR, N.,
SUASSON, V., PÉTURSSON, S. Intratracheal inoculation as na efficient croute of
experimental infection with Maedi visna virus. Research Veterinary Science. v.75,
p.245-247, 2003.
TURELLI, P., GUIGUEN, F., MORNEX, J.F., VIGNE, R., QUERAT, G. dUTPase-
minus caprine arthritis-encephalitis virus is attenuated for pathogenesis and accumulates
G-to-A substitutions. Journal of virology. v. 71, n. 6, p. 4522–4530.1997.
VALAS, S., GERMAIN, K. Distribution and heterogeneity of small ruminant lentivirus
envelope subtypes in naturally infected French sheep. Virus Research. v. 120, p. 156–
162, 2006.
VALAS, S., BENOIT, C., BAUDRY, C., PERRIN, G., MAMOUN, R.Z. Variability
and immunogenicity of caprine arthritis-encephalitis virus surface glycoprotein. Journal
of virology. v. 74, p. 6178–6185, 2000.
VALAS, S., BENOIT, C., GUIONAUD, C., PERRIN, G., MAMOUN, R.Z. North
American and French caprine arthritis-encephalitis viruses emerge from ovine Maedi-
visna viruses. Virology. v. 237, p. 307–318, 1997.
VAREA, R.; MONLEON, E.; PACHECO, C.; LUJAN, L.; BOLEA, R.; VARGAS, M.
A.; VAN EYNDE, G.; SAMAN, E.; DICKSON, L.; HARKISS, G.; AMORENA, B.;
BADIOLA, J. J. Early detection of maedi-visna (ovine progressive pneumonia) virus
seroconversion in field sheep samples. Journal of Veterinary Diagnostic Investigation,
v. 13, n.4, p. 301-307, 2001.
VERWOERD, D. W.; PAYNE, A. L.; YORK D. F.; MYER, M. S. Isolation and
preliminary characterization of the Jaagsiekte retrovirus (JSRV). Onderstepoort Journal
Veterinary Research, v.50, p. 309-316, 1983.
VITU, C.; RUSSO, P.; VIGNE, R.; QUERAT, G.; GIAUFFRET, A. An ELISA Test
For Detection Of Maedi- Visna antibodies. Comparative study with Gel
Immunodiffusion na complement fixation test. Comparative Immunology and
Microbiology Infectious Disease. v.4, n.5, p .469-481, 1982.
WAGTER, L.H.A., JANSEN, A., BLEUMINK-PLUYM, J.A., LENSTRA, J.A.,
HOUWERS, D.J. PCR detection of lentiviral gag segment DNA in the white blood cells
of sheep and goats. Veterinary Research Communications., v.22, p.355-262, 1998.
WANG, C., LAI, H., LI, J. Analysis of minimal human immunodeficiency virus type 1
gag coding sequences capable of virus-like particle assembly and release. Journal of
virology. v. 72, p. 7950–7959, 1998.

161
WOODWARD, T.M., CARLSON, J.O., DE LA CONCHA-BERMEJILLO, A.,
DEMARTINI, J.C. Biological and genetic changes in ovine lentivirus strains following
passage in isogeneic twin lambs. Journal of acquired immune deficiency syndromes and
human retrovirology. v. 8, p. 124–133, 1995.
WU X, LI Y, CRISE B, et al. Transcription start regions in the human genome are
favored targets for MLV integration. Science. v. 300, n. 5626, p. 1749–51. 2003.
WYAND, M. S.; MANSON, K. H.; GARCIA-MOL, M.; MONTEFIORI, D.;
DESROSIERS , R. C. Vaccine protection by a triple deletion mutant of simian
immunodeficiency virus. Journal of Virology, v.70, n.6, p.3724-3733, 1996.
ZANONI, R.G. Phylogenetic analysis of small ruminant lentiviruses. The Journal of
general virology. v. 79, p. 1951–1961, 1998.
ZANONI, R., PAULI, LT., PETERHANS, E. Detection of caprine arthritis-encephalitis
and Maedi-visna viruses using polymerase chain reaction. Experientia, v. 46, p. 3l6-
319, 1990.
ZHANG, G.; LEUNG, C.; MURDIN, L.; ROVINSKI, B.; WHITE, K. A. In planta
expression of HIV-1 p24 protein using an RNA plant virusbased expression vector.
Molecular Biotechnoogyl, v.14, n.2, p.99-107, 2000.
ZINK, M.C., NARAYAN, O., KENNEDY, P.G.E., CLEMENTS, J.E. Pathogenesis of
visna-Maedi and caprine arthritis-encephalitis: new leads on the mechanism of restricted
virus replication and persistent inflammation. Veterinary
Immunology and Immunopathology. v.15, p.1671-80, 1987.
ZINK, M.C., JOHNSON, L.K. Pathobiology of lentivirus infections of sheep and goats.
Virus Research. v.32, p. 139-154, 1994.
ZINK, M. C.; YAGER, J. A.; MYERS, J. D. Pathogenesis of caprine arthritis
encephalites virus cellular localization of viral transcripts in tissue of infected goats.
American Journal Pathology, v. 136, n. 4, p. 843-854, 1990.

162
ANEXOS
Capítulo I

163
Capítulo III
Anexo 2. Primeira página do artigo publicado – Capítulo III

164
Capítulo IV
Anexo 3. As sequências nucleotídicas parciais do gene gag de LVPRs geradas no
presente estudo encontram-se relacionadas abaixo:
>69LVCE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGCTAACACTATCTTATACAAA
TGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGAGT
ACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGGAT
CAGAAGGGT
>137CE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGCTAACACTATCTTATACAAA
TGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGAGT

165
ACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGGAT
CAGAAGGGT
>229CE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGATAACACTATCTTATACAA
ATGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGA
GTACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGG
ATCAGAAGGGT
>2361CE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGCTAACACTATCTTATACAAA
TGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGAGT
ACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGGAT
CAGAAGGGT

166
>3596CE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGCTAACACTATCTTATACAAA
TGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGAGT
ACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGGAT
CAGAAGGGT
>6900CE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGCTAACACTATCTTATACAAA
TGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGAGT
ACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGGAT
CAGAAGGGT
>Animal02CE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA

167
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGCTAACACTATCTTATACAAA
TGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGAGT
ACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGGAT
CAGAAGGGT
>Pan5CE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGCTAACACTATCTTATACAAA
TGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGAGT
ACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGGAT
CAGAAGGGT
>Pan7CE
GATTCTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC

168
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCCTATAAAAGATTATCTAAAGCTAACACTATCTTATACAAA
TGCATCAGCAGATTGTCAGAAGCAAATGGATAGAACACTAGGACAAAGAGT
ACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGGAT
CAGAAGGGT
>Pan12CE
GATTGTGTAATGTTCCAGCAACTGCAAACAGTAGCAATGCAGCATGGCCTC
GTGTCTGAGGACTTTGAAAGGCAGTTGGCATATTATGCTACTACCTGGACAA
GTAAAGACATACTAGAAGTATTGGCCATGATGCCTGGAAATAGAGCTCAAA
AGGAGTTAATTCAAGGGAAATTAAATGAAGAAGCAGAAAGGTGGAGAAGG
AATAATCCACCACCTCCAGCAGGAGGAGGATTAACAGTGGATCAAATTATG
GGGGTAGGACAAACAAATCAAGCAGCAGCACAAGCTAACATGGATCAGGC
AAGGCAAATATGCCTGCAATGGATAATAAATGCATTAAGAGCAGTAAGACA
TATGGCGCACAGGCCAGGGAATCCAATGCTAGTAAAGCAAAAAATGAATG
AGTCATATGAAGAATTTGCAGCAAGACTGCTAGAAGCAATAGATGCAGAGC
CAGTTACACAGCATATAAAAGATTATCTAAAGCTAACACTATCTTATACAA
ATGCATCAGCAGATTGTCAGAAGCAAATCGATAGAACACTAGGACAAAGA
GTACAACAAGCTAGTGTAGAAGAAAAAATGCAAGCATGTAGAGATGTGGG
ATCAGAAGGGT

169
Anexo 4. As sequências nucleotídicas parciais do gene pol de LVPRs geradas no
presente estudo encontram-se relacionadas abaixo:
>31RN
AAATTAAAAGGACTGACAGAAATAATAGATAAATTAGTGGAAGAAGGAAA
ACTAGGAAAAGCACCTCCACATTGGACATGTAATACTCCAATATTTTGCATA
AAAAAGAAGTCAGGAAAGTGGAGGATGTTAATAGATTTCAGAGAATTGAAT
AAACAAACAGAGGATTTAACAGAGGCACAGTTAGGACTTCCACATCCTGGG
GGCCTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATAT
TTCACTATACCCTTGTATGAACCCTATAGAGAGTACACATGTTTTACTCTATT
AAGTCCTAATAATTTGGGACCATGTAAAAGATATTATTGGAAAGTGCTGCC
ACAAGGGTGGAAGCTGAGTCCTTCTGTATATCAATTTACTATGCAAGAAATT
TTAGAGGATTGGATACAGCAACATCCAGAAATTCAATTTGGCATATATATG
GATG
>32CE
AAATTAAAAGGACTGACAGAAATAATAGATAAATTAGTGGAAGAAGGAAA
ACTAGGAAAAGCACCTCCACATTGGACATGTAATACTCCAATATTTTGCATA
AAAAAGAAGTCAGGAAAGTGGAGGATGTTAATAGATTTCAGAGAATTGAAT
AAACAAACAGAGGATTTAACAGAGGCACAGTTAGGACTTCCACATCCTGGG
GGCCTACAAAAGAAAAAACATGTTACAATATTAGACATAGGAGATGCATAT
TTCACTATACCCTTGTATGAACCCTATAGAGAGTACACATGTTTTACTCTATT
AAGTCCTAATAATTTGGGACCATGTAAAAGATATTATTGGAAAGTGCTGCC
ACAAGGGTGGAAGCTGAGTCCTTCTGTATATCAATTTACTATGCAAGAAATT
TTAGAGGATTGGATACAGCAACATCCAGAAATTCAATTTGGCATATATATG
GATG
>69LVCE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA

170
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG
>137CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG
>229CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG

171
>237CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAATTACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG
>1604MG
AAATTAAAAGGCCTAACAGAAATAATAGATAAATTAGTAGAAGAAGGAAA
ACTAGGAAAGGCACCTCCGCATTGGACATGTAATACTCCAATCTTTTGCATA
AGAAAGAAATCAGGGAAATGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCAAATAACTTGGGACCATGTAAAAGATACTTTTGGAAAGTTTTGCC
TCAGGGTTGGAAACTGAGTCCATCTGTATATCAATTCACCATGCAGGAAATT
TTGGAAGATTGGATAAAACAACATCCAGAAATTCAATTTGGCATATATATG
GATG
>1625MG
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT

172
CGTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG
>2239CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG
>2361CE
GAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
TACAAAGAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAAT
AAACAAACAGAGGATTTAACAGAGGCACAGTTAGGACTTCCACATCCTGGG
GGCCTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATAT
TTCACTATACCCTTGTATGAACCCTATCGAGAGTACACATGTTTTACTCTATT
AAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGCC
ACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGATC
TTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATATG
GATG
>3596CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT

173
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG
>6900CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAAGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG
>Animal02CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAGGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATAGAACATAGGAGATGCAT
ATTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCT
ATTAAGGCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAATGCT
GCCACAAGGCTGGAAAGTGAGTCCATCGGCATATCAATTTACTATGCAGGA
GATCTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATA
TATGGATG
>Pan5CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAGGCGCAGTTAGGACTCCCGCATCCGGG

174
AGGACTACAAAAGAAAAAACATGTTACAATATTGGAACATAGGAGATGCAT
ATTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCT
ATTAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGAT
GCCACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAG
ATCTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATAT
ATGGATG
>Pan7CE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAGGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG
>SQSBCE
AAATTAAAAGGTCTAACAGAAATCATAGATAAATTAGTGGAGGAAGGAAA
ACTAGGAAAGGCACCCCCACATTGGACATGTAATACTCCAATCTTTTGCATA
AAAAAGAAATCAGGGAAGTGGAGAATGTTAATAGATTTCAGAGAATTGAA
CAAACAGACAGAAGATTTAACAGAGGCGCAGTTAGGACTCCCGCATCCGGG
AGGACTACAAAAGAAAAAACATGTTACAATATTGGACATAGGAGATGCATA
TTTTACTATACCCCTATATGAACCATATCGAGAGTACACATGTTTTACTCTAT
TAAGTCCTAATAATCTAGGACCATGTAAAAGATACTATTGGAAAGTGCTGC
CACAAGGTTGGAAATTGAGTCCATCTGTATATCAATTTACTATGCAGGAGAT
CTTAGAGGATTGGATACAGCAGCATCCAGAAATTCAATTTGGCATATATAT
GGATG

175
Anexo 5.Certificado de Bioética referente ao Capítulo I