universidade estadual de feira de santana … discente/mestrado... · minha mãe, minha esposa,...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA
PROGRAMA DE PÓS GRADUAÇÃO EM BIOTECNOLOGIA
CARLOS LÍVERTON DA SILVA BORGES
PURIFICAÇÃO E CARACTERIZAÇÃO BIOLÓGICAS
PARCIAIS DA PEÇONHA DA ARANHA CARANGUEJEIRA
Acanthoscurria natalensis.
Feira de Santana, BA
2008

CARLOS LÍVERTON DA SILVA BORGES
PURIFICAÇÃO E CARACTERIZAÇÃO BIOLÓGICAS
PARCIAIS DA PEÇONHA DA ARANHA CARANGUEJEIRA
Acanthoscurria natalensis.
Dissertação apresentada ao Programa de Pós-graduação em Biotecnologia, da Universidade Estadual de Feirade Santana(UEFS) como requisito parcial para obtenção do título de Mestre em Biotecnologia. Orientadora: Prof. Dr. Maria Elena de Lima Pérez Garcia Laboratório de Venenos e Toxina Animais Departamento de Bioquímca e Imunologia Universidade Federal de Minas Gerais. Co-Orientadoras: Prof. Ms. Ilka Biondi Laboratório de Animais Peçonhentos e Herpetologia Departamento de Ciência Biológicas(UEFS) Prof. Dr. Sandra Aparecida de Assis Laboratório de Enzimologia Departamento de Saúde (UEFS)
Feira de Santana, BA
2008

Este trabalho é dedicado a minha família
a qual com muito amor me apoiou nos
diversos momentos em que precisei, em especial
minha mãe, minha esposa, minha filha,
minha irmã e meus tios.
E uma homenagem póstuma a minha querida
Avó Inés

AGRADECIMENTOS
Agradeço de todo coração a todos aqueles que me apoiaram e deram aquela força para a
concretização desse trabalho.
Agradeço primeiramente a Professora Ilka Biondi, pela co-orientação(LAPH-UEFS) sem
a qual não estaria realizando esta fase da minha vida acadêmica e a todos os alunos, estagiários e
funcionários do LAPH em especial o Alexandre (O mano), a Dulce, a Rafaela e a Josemir meu
braço direito com as aranha.
À professora Sandra Aparecida de Assis pela co-orientação .
À professora Maria Elena (LVTA-UFMG), pela paciência,e orientação deste trabalho. E
a todo o pessoal do Laboratório de Venenos e Toxinas Animais –LVTA-UFMG, em especial ao
Breno Rates pela imensa colaboração, à Karla, à Marcella, ao Vitor, à Camila, ao Filipe e à
Juliana (Jú).
Aos Amigos, do Laboratório de Enzimologia e Bioquímica de Proteínas, em especial ao
Jamil pelo aprendizado com sua experiência, ao Alexandre Santos e à Kádima pelas orientações
e discussões esclarecedoras.
Ao pesquisador Dr. Antônio Brescovit (Instituto Butantã) pela confirmação da
identificação da espécie Acanthoscurria natalensis.
Ao Amigo Hugo Leonardo da eletrofisiologia pela realização dos experimentos nesta
área, e enfim, a todos os que direta ou indiretamente colaboraram com esse trabalho.
Ao proprietário do Ranário Vivendas da Rainha- Ribeirão das Neves (MG), por ceder
gentilmente as pulpas das moscas utilizadas nesse estudo.
E a Deus, meu Senhor e Criador, acima de tudo pelo sustento e graça a mim concedido.

“ Ainda que eu fale as línguas dos homens e dos anjos, se não tiver amor, serei como o
sino que ressoa ou como prato que retine. Ainda que eu tenha o dom de profecia e
saiba todos os mistérios e todo conhecimento, e tenha uma fé capaz de mover
montanhas, se não tiver amor, nada serei. (...) O amor nunca perece; mas as profecias
desaparecerão, as línguas cessarão, o conhecimento passará...”
I Corintios 13:1-8

RESUMO
O estudo biotecnológico de compostos bioquímicos de invertebrados especialmente
peçonhas de aranhas e escorpiões representa uma fonte para a formulação de novos fármacos.
Assim, o presenta trabalho tem como objetivo a purificação parcial da peçonha da Acantoscurria
natalensis. Para a purificação da peçonha utilizou-se a metodologia de cromatografia líquida de
alta performance (CLAE). A determinação das massas moleculares e o seqüenciamento parcial
das toxinas isoladas foram feitas utilizando-se espectrometria de massa e degradação de Edman,
Eletroforeses uni e bi-dimensionais foram utilizadas para auxiliar na caracterização da peçonha
e toxinas isoladas. Os testes de atividade biológica compreenderam ensaios de toxicidade em
mosca domestica (Musca domestica) e ensaios eletrofisiológicos em canais iônicos (sódio e
cálcio) utilizando-se células de gânglio dorsal de rato. O fracionamento por cromatografia de
troca catiônica resultou na separação de 32 picos que foram submetidos à cromatografia em
coluna de fase reversa, onde foram isoladas duas toxinas (AnTxI e AnTxII). Estudos
eletrofisiológicos com AnTx1, em neurônios do gânglio dorsal de rato (DRG), indicaram ação
bloqueadora em canais para sódio e para cálcio. Resultados preliminares indicam que a toxina
causa inibição da corrente de cálcio nas células DRG, em concentrações nanomolares. Em
conclusão, neste trabalho isolamos e caracterizamos parcialmente duas toxinas (AnTxI e
AnTxII) da peçonha da aranha A.natalensis e mostramos que uma delas (AnTx1) bloqueia com
alta potëncia, canais para cálcio, podendo representar uma importante ferramenta para estudos
farmacológicos destes canais.
Palavras-chave: Mygalomorphae, Acanthoscurria natalensis, toxina, canais iônicos

ABSTRACT
The biochemical and pharmacological studies from venoms especially that of invertebrates,
including spiders and scorpions, represent a source for the formulation of new medicines. Like
this, the present work has as objective the partial purification and biological characterization of
the venom of the spider Acanthoscurria. natalensis. This work was proposed considering the
lack of studies with this venom and its potential to present new molecules from biological
interest. The purification of this venom was done by using high performance liquid
chromatography (HPLC) techniques. The molecular mass and the partial sequences were
obtained by using mass spectrometry, Edman degradation and automatic sequence
determination, as techniques. Electrophoresis uni and bi-dimensional were used to complement
the characterization of the venom and of their toxins. Biological activity tests included toxicity
assays in fly (Musca domestica) and electrophysiological assays in ionic channels (sodium and
calcium) in cells from dorsal root ganglion of rat (DRG). The chromatography by cationic
exchange in HPLC originated 32 fractions that were submitted to a reverse phase column. Two
toxins (ANTx1 and ANTxII) were isolated. Electrophysiological studies showed that AnTx1, in
nanomolar doses, blocks the calcium currents in DRG neurons from rat., and in less extension
the sodium current. These results indicated that AnTx1 is a promising tool o study calcium
channels.
Key words: Mygalomorphae, Acanthoscurria natalensis, toxin, ionic channels, calcium
channels.

Lista de abreviações e siglas
Termos Técnicos ACN Acetonitrila BSA Albumina de soro bovino. Cav Canais para cálcio voltagem-dependentes Ca++ Íon cálcio Cl- Íon cloreto CLAE Cromatografia Líquida de Alta Eficiëncia CIEX “Chromatography cationic ion- exchange” cm Centímetro DRG “Dorsal Root Ganglion” -Glânglio da Raiz Dorsal DHP 1,4 dihidropiridina Da Dalton DMEM “Dulbecco's Modified Eagle Médium”- Meio de cultura Modificado de Eagle. DL50 Dose letal para morte de 50% do grupo teste ECA Enzima Conversora de angiotensina HVA “High Voltage Activated”- Ativado por alta voltagem HBSS Hank’s Balanced Salt Solution -Solução fisiológica de Hank HPLC “ High Performance Liquid Chromatography” K+ Íon potássio Kv Canal para potássio, voltagem-dependente kDa Kilodalton LVA “Low Voltage Activated”- Ativado por baixa voltagem M Molar mM Milimolar mL Mililitro mV Milivoltes µL Microlitro µΩ Microohm µM Micromolar ng Nanograma nM Nanomolar Nav Canal para sódio, voltagem-dependente Na+ Íon sódio PI Ponto isoelétrico pS picosegundo RPM Rotações por minuto RPC “Reverse phase chromatografy” - Cromatografia de fase reversa SDS Dodecil sulfato de sódio TFA Ácido trifluoracético TEMED Tetrametiletilenodiamino V Volts v/v Volume por volume
i

Lista de Tabelas
Página
Tabela 1 Lista de toxinas isoladas das peçonhas da infra-ordem das Mygalomorphae
08
Tabela 2 Nomenclatura das subunidades α1 de canais para cálcio, dependentes de voltagem clonados
13
Tabela 3 Alinhamento das toxinas homólogas aos cinco fragmentos das toxinas, AnTxI e AnTxII, comparadas pelo BLAST-p
39
ii

Lista de Figuras
Figura 1 Morfologia das aranhas caranguejeiras. 04
Figura 2 Theraphosidae – A..natalensis. 05
Figura 3 Estrutura básica dos canais iônicos. 10
Figura 4 Extração da peçonha da A. natalensis 17
Figura 5 Aplicação das amostras em diferentes concentrações da peçonha e toxinas em moscas (Musca domestica).
22
Figura 6 Perfil eletroforético do gel de poliacrilamida da peçonha ( de A. natalensis e da toxina isolada ( AnTxI ))
30
Figura 7 Perfil eletroforético do gel bidimensional da peçonha das fêmeas de A. natalensis.
31
Figura 8 Perfil cromatográfico (troca catiônica) da peçonha de Fêmea e de Macho.
33
Figura 9 Perfil cromatográfico da toxina isolada AnTxI da A. natalensis em cromatografia de fase reversa.
34
Figura 10 Perfil cromatográfico das frações de CIEX obtidas da peçonha da A. natalensis
35
Figura 11 Perfil da cromatografia em fase reversa (RPC) da toxina isolada - AnTxI.
37
Figura 12 Efeito do curso temporal da corrente após exposição à toxina AnTxI.
42
Figura 13 Traçados representativos das correntes totais carreadas pelos canais para cálcio sensíveis à voltagem no neurônio DRG.
43
iii

Indice
PÁGINA
1. INTRODUÇÃO 1
1.1 Revisão da literatura 2
1.1.1 Biologia das aranhas 2
1.2 Características gerais da peçonha das aranhas caranguejeiras
6
1.2.1 Toxinas isoladas de peçonhas de caranguejeiras
7
1.2.2 Atividade em canais iônicos de toxinas isoladas de peçonhas de caranguejeiras
7
1.3 Os canais iônicos 9
1.3.1 Canais para cálcio - breve introdução
11
1.4 Células do gânglio dorsal 14
2. OBJETIVOS 15
2.1 Objetivo geral 15
2.2 Objetivos específicos 15
3. MATERIAL E MÉTODOS 16
3.1 Coleta e manutenção A. natalensis 16
3.2 Identificação da A. natalensis 16

3.3 Extração da peçonha de A. natalensis 16
3.4 Dosagem da concentração de proteínas 18
3.5 Fracionamento da peçonha de A. natalensis 18
3.5.1 Cromatografia de troca catiônica-CIEX
18
3.5.2 Cromatografia de fase reversa (CLAE-FR)
18
3.6 Eletroforese Unidimensional em poliacrilamida - SDS (SDS-PAGE)
19
3.7 Eletroforese em gel bidimensional (2D-PAGE) 20
3.8 Bioensaios com a peçonha bruta e toxinas, em insetos
20
3.8.1 Testes de toxicidade em insetos. 20
3.8.2 Testes eletrofisiológicos da AnTxI em canais para Ca 2+
23
3.8.3 Cultura celular de células DRG do gânglio dorsal de ratos
23
3.9 Espectrometria de Massa 25
3.9.1 Análise da peçonha isolada por ESI-Q-TOF-MS
25
3.9.2 Análise da peçonha por MALDI-TOF/TOF-MS
26
3.10 Determinação da seqüência N-terminal das toxinas AnTxI e AnTxII
27
4.RESULTADOS E DISCUSSÃO 28

4.1 Eletroforese unidimensional em poliacrilamida-SDS (SDS-PAGE) e Eletroforese em gel bidimensional (2D-PAGE)
28
4.2 Cromatografia da peçonha de A. natalensis 32
4.3 Análise por Espectrometria de Massas 36
4.4 Determinação da seqüência N-terminal do peptídeo AnTxI
38
4.5 Testes biológicos 40
4.5.1 Teste de toxicidade em insetos 40
4.5.2 Testes eletrofisiológicos com a toxina AnTxI em canais para cálcio voltagem-dependentes.
41
5. CONCLUSÕES 44
6. REFERÊNCIAS BIBLIOGRÁFICAS 47
ANEXOS 55
Formulação para a eletroforese em poliacrilamida-SDS (SDS-PAGE)
55
Formulação para a solução de Hank's 56
Formulação para o experimento de “patch-clamp”
56

1 INTRODUÇÃO
A busca por princípios ativos utilizando-se da biodiversidade, tanto da fauna, como da
flora, tem se intensificado ao longo dos anos. Nessa óptica, os princípios ativos de peçonhas
animais vêm despertando interesse da indústria farmacêutica e biotecnológica. Um exemplo
ilustrativo, foi a descoberta de que um peptídeo do veneno da Jararaca (Bothops jararaca) inibe
a enzima conversora da angiotensina I (ECA) em angiotensina II (que leva ao aumento da
pressão arterial) na década de 60, pelos Doutores Sérgio Ferreira e Lewis J. Greene. Este e
outros peptídeos da peçonha são capazes de abaixar a pressão arterial. A partir dessa descoberta
a indústria formulou um produto conhecido comercialmente como captropil®, cujo uso no
tratamento de problemas hipertensivos vem salvando muitas vidas em todo o mundo.
O conhecimento dos compostos presentes nos venenos e peçonhas de animais é
importante não só para a formulação de soros no tratamento nos envenenamentos, como também
para o descobrimento de princípios ativos para sua utilização como possíveis fármacos. Assim
como os vertebrados, os invertebrados, por exemplo, aranhas e escorpiões, representam um
campo interessante para a descoberta de toxinas e de outras substâncias ativas (DE LIMA et
al,2007, ESCOUBAS et al., 2000b).
Várias toxinas já estudadas, isoladas de diferentes peçonhas de artrópodos, interferem
com estruturas e processos biológicos diversos, como canais iônicos, por atuar como
antibacterianos, anti-hipertensivos, etc (LAMPE et al, 1993; DE LIMA et al, 2007; PERREIRA
et al, 2006; GRISHIN, 1999, VERANO BRAGA et al., 2008). Um exemplo pode ser visto na
peçonha da aranha armadeira, Phoneutria nigriventer que apresenta várias toxinas com ampla
gama de ação em canais iônicos, como sódio, potássio e cálcio, possuindo atividade em
mamíferos e algumas com atividade inseticida específica (GOMEZ et al, 2002; DE LIMA et al.,
2007; FIGUEIREDO et al., 1995; FIGUEIREDO et al.,2001; DE LIMA et al., 2002;
OLIVEIRA et al., 2003; NUNES et al., 2008). Várias destas toxinas interagem com sítios
específicos do sistema nervoso, modificando funções, atuando na neurotransmissão central ou
periférica, pré ou pós-sinapticamente, na liberação de neurotrasmissores e/ou interferindo com a
transdução de sinais.
SENFE- RIBEIRON et al., 2008, faz um revisão sobre toxinas de aranhas com aplicação
biotecnológica e destaca dentre outras, a toxina de Loxosceles sp como de grande importância
para o estudo na linha de imunoterápicos, modulador de resposta inflamatória e imunopatológica
e como biopesticidas. Outro trabalho que merece destaque focaliza um peptídeo rico em glicina
1

que apresenta atividade antimicrobiana, extraído da hemolinfa da aranha Acanthoscurria
gomesiana (LORENZINI, D. M. et al ,2003).
O presente estudo teve como objetivo estudar a peçonha da aranha Acanthoscurria
natalensis, do semi-ariado baiano, como possível potencial biotecnológico..
1.1 Revisão da literatura
1.1.1 Biologia das aranhas
As aranhas são animais pertencentes ao filo Arthopoda, classe Arachnida. Compreendem
cerca de 40 mil espécies já descritas (FOELIX, 1996). Esses animais são cosmopolitas, ocorrem
em regiões tropicais e subtropicais, tendo baixa freqüência de habitat nas regiões polares
(CODDINGTON e LEVI,1991). De acordo com seus hábitos, as aranhas são classificadas em
subterrâneas, terrículas, cavernículas, dendrícolas, adaptando-se também à vida subaquática
(FOELIX, 1996; RAVEN, 1985). São carnívoras caçando as suas presas vivas, através de duas
estratégias básicas : (i) a construção de teia para aprisionar as presas; (ii) saltando sobre a presa,
segurando-as com as garras e inoculando a peçonha. As aranhas das famílias Licosidae,
(tarântulas), Ctenidae (armadeiras), Salticidae (papa-moscas) e Theraphosidae (caranguejeiras),
apresentam essa segunda estratégia, e em geral, são errantes, caçadoras e solitárias.
Aranhas, exceto as da família Uloboridae, possuem um par de glândulas situadas no
cefalotórax com ductos que chegam até as quelíceras. As quelíceras são utilizadas para secretar a
peçonha e paralisar suas presas. Segundo FRIEDEL, 1989, as aranhas utilizam como função
primária a peçonha para imobilização de suas presas. Entretanto a peçonha, por apresentar
variabilidade de componentes, acaba por provocar efeito letal. Esta gama de compostos ativos,
na sua grande maioria age no sistema nervoso, sendo que a açao no sistema muscular passa a ser
um efeito secundário. O conhecimento da biologia juntamente com os efeitos causados por estas
peçonhas vem possibilitando o estudo de princípios ativos, conhecendo-se suas estruturas e
estudando-se seus modos de ação. Estima-se que exista cerca de 19.000.000 de moléculas, nas
peçonhas das duas infra ordem de aranhas descritas Araneomorphae e Mygalomorphae
demonstrado-se assim, a necessidade de estudos bioquímicos e farmacológicos (ESCOUBAS,
2006).
As Araneomorphae são conhecidas como aranhas verdadeiras e compreendem o maior
grupo de espécies, onde podemos destacar os gêneros Phoneutria, Lycosa e Loxosceles, por
2

serem responsáveis pelo maior número de acidentes em humanos no Brasil, justificando vários
estudos realizados (GOMEZ et al., 2002. FRONTALI, N., GRASSO, A, 1964, DE LIMA et al.,
2007; DE LIMA et al., 2002; FIGUEIREDO et al., 1995; FIGUEIREDO et al.,2001;
OLIVEIRA et al., 2003). As Mygalomorphae são representadas pelas caranguejeiras e são
conhecidas como falsas aranhas devido a posição anatômica das suas quelíceras. Para esta infra-
ordem estão descritas 15 famílias, 300 gêneros e aproximadamente 2.500 espécies (HEDIN &
BOND, 2006). As caranguejeiras são caracterizadas, anatomicamente, por apresentar pêlos no
corpo, agrupamento ocular do tipo cômoro ocular, e quelíceras em posição paralela, com as
glândulas localizadas na porção basal das quelíceras (BRAZIL, 1925). (FIG. 1). Dentro desta
infra-ordem está alocada a família Theraphosidae, com 112 gêneros e 906 espécies (THE
WORLD SPIDER CATALOG, 2008). 45% destes gêneros habitam as regiões neotropicais
(HEDIN & BOND, 2006).
Para o Brasil, estão descritos 32 gêneros da família Theraphosidae (THE WORLD
SPIDER CATALOG, 2008). Destes, apenas os gêneros Acanthoscurria, Grammostola,
Lasiodora e Vitalius vêm sendo mais estudados e são encontrados em todo território brasileiro
principalmente, n o Norte e no Nordeste (BARRAVIEIRA,1994; MONTANDON, 2007).
As espécies do gênero Acanthoscurria (FIG. 2) estendem-se pela América do Sul e
Central. São aranhas terrestres, alimentando-se basicamente, de insetos, pequenos lagartos, de
roedores, rãs, minhocas e outras aranhas. São principalmente escavadoras e caçadoras,
paralisando suas presas pela inoculação de peçonha (FOELIX, 1996).
3

Figura 1- Morfologia das aranhas caranguejeiras. A-Anatomia externa de aranhas caranguejeiras B- Aspecto
externo das aranhas caranguejeiras C-Anatomia Interna do cefalotórax das aranhas. Fonte: Soerensen, B., 1997;
FOELIX, S. D. Biology of spiders. Oxford University Press, inc., 1996.
A B
C
4

Figura 2 - Theraphosidae - Acanthoscurria natalensis ;
Fonte: http://www.birdspiders.com/archive/15B036F2KD0B7KAEC2K14B24A24C046092C.html
5

1.2 Características gerais da peçonha das aranhas caranguejeiras
Apesar do vasto número de espécies descritas para as aranhas, poucas delas apresentam
seu veneno bem caracterizado, como nos escorpiões, serpentes e outros animais peçonhentos
(GOMEZ et al., 2002. FRONTALI, N., GRASSO, A, 1964, DE LIMA et al., 2007; 2002;
FIGUEIREDO et al., 1995; 2001; OLIVEIRA et al., 2003).
De um modo geral, a peçonha das aranhas caranguejeiras compreende uma mistura de
sais, nucleotídeos, aminoácidos livres, neurotransmissores, poliaminas, peptídeos, proteínas e
enzimas (ESCOUBAS e RASH, 2004; ESCOUBAS et al., 2000b; SAVEL-NIEMANN, 1989;
RASH e HODGSON, 2002). Dentre esses componentes o grupo de moléculas de maior interesse
são as neurotoxinas que podem ser divididas em dois grupos: o das acilpoliaminas e o das
toxinas polipeptídicas, que são classificadas de acordo com suas características funcionais e
moleculares (GRISHIN, 1999).
As acilpoliaminas ou poliaminas amidadas são compostos orgânicos neurotóxicos
formados por uma cadeia de poliamina alifática com um grupo acil aromático final, de baixa
massa molecular (300-700Da). Este tipo de toxina foi descrito pela primeira vez na peçonha da
aranha japonesa Nephila clavada por KAWAI et al. (1982). As acilpoliaminas são compostos
que bloqueiam especificamente receptores de glutamato (HIDAI et al., 1999), sendo o glutamato
um neurotrasmissor excitatório em músculo esquelético de insetos e também o principal
neurotransmissor excitatório no sistema nervoso central de mamíferos. Assim as poliaminas são
ferramentas para estudos da neurofisiologia de vertebrados e de invertebrados, tendo potencial
como agente farmacológico ou agroquímico, que pode ser explorado (HIDAI et al., 1999; KAN
et al.,2002).
Já as toxinas peptídicas, isoladas de peçonhas de aranhas em geral, têm cadeia de 30 a 77
resíduos de aminoácidos. As neurotoxinas com baixa massa molecular geralmente interagem
com canais iônicos das membranas de células excitáveis, bloqueando a transmissão sináptica
por interferir com os potenciais de ação, enquanto que as de alto peso molécula, em geral,
ligam-se a unidades de receptores de membrana pré-sináptica, intensificando a secreção de
neurotransmissores (GRISHIN,1999).
6

1.2.1 Toxinas isoladas de peçonhas de caranguejeiras
Os trabalhos com isolamento de toxinas de peçonhas de caranguejeiras iniciaram-se no
final da década de 80, com teste de toxicidade em baratas com a peçonha da aranha Eutypelma
californicum, onde as toxinas ESTxI e ESTxII foram isoladas e testadas por Savel-Niemman e
Roth em 1989. Em aranhas chinesas do gênero Selenocosmia foram isolados vários peptídeos
com diversas funções, entre eles estão: 1) Huwentoxina I (HwTxI), 2) SHL-I, 3) Huwentoxina II
(HwTxII), 4) Huwentoxina IV (HwTxIV) e 5) Hainatoxinas IV e V entre outros peptídeos de
outras aranhas (TABELA 1). Esses peptídeos atuam em receptores nicotínicos, outros possuem
atividades hemaglutinante, inseticida, ou inibem canais para sódio (LIANG, et al 1993; LIANG
e PAN, 1995; PENG et al., 2001; SHU e LIANG, 1999; PENG et al., 2002; LIU et al. 2003;
XIAO e LIANG, 2003). Dentre essas atividades a ação em canais iônicos tem despertado
interesse especial, pois a alta seletividade para receptores e subtipos de canais iônicos ainda é
pouco conhecida. LAMPE et al, 1993, isolaram um peptídeo que bloqueia canais para cálcio e
posteriormente LAMPE, 1999 isolaram um peptídeo com ação analgésica, ambos das peçonhas
de Grammostola spatulata.
1.2.2 Atividade em canais iônicos de toxinas isoladas de peçonhas de
caranguejeiras
As toxinas ativas em canais iônicos possuem em geral, massa molecular entre 4 a 10
kDa com grande quantidade de resíduos de cisteína, variando de seis a quatorze pontes
dissulfeto. A ação das toxinas em canais iônicos leva à paralisia das presas por inibição ou
excessiva estimulação da transmissão nervosa. No primeiro caso, as toxinas inibem canais para
sódio (Nav) ou para cálcio (Cav) - voltagem-dependentes. No segundo caso, ocorre uma
excessiva estimulação de Nav ou inibição do canal para potássio (Kv)-voltagem-dependentes
(GRISHIN,1999; SWARTZ, 2007). De acordo com levantamento feito por Escoubas & Rash
(2004), os canais iônicos para sódio e para potássio são os mais susceptíveis à ação pelas toxinas
de caranguejeiras (TABELA 1).
7

Tabela 1-Lista de toxinas isoladas das peçonhas da infra-ordem das Mygalomorphae Toxinas Nome Espécies Massa Molecular Referências TxP5 Toxic protein 5 B. smithi 3959.56 ?? Kaiser et al., 1994 CvTxII Covalitoxin II Coremiocnemis validus 3406.94 ?? Balaji et al., 2000 GSAFI – G. spatulata 3707.46Analgésico Lampe, 1999 GSAFII – G. spatulata 3979.76Analgésico/ antiarritímico Lampe and Sachs, 1999 GsMTx2 Grammostola mechanotoxin 2 G. spatulata 3922.73 MSC ?? Oswald et al., 2002 GsMTx4 Grammostola mechanotoxin 4 G. spatulata 4095.90 MSC (SAC) Suchyna et al., 2000 HaTx1 Hanatoxin 1 G. spatulata 4114.73 Kv2.1 Swartz and MacKinnon, 1995 HaTx2 Hanatoxin 2 G. spatulata 4098.73 Kv2.2 Swartz and MacKinnon, 1995 HmTx1 Hetereroscodratoxin 1 H. maculata 3997.45 Kv2 / Kv4 (4.1) Escoubas et al., 2002 HmTx2 Hetereroscodratoxin 2 H. maculata 4757.55 Kv2 Escoubas et al., 2002 HnTxI Hainantoxin-I S. hainana 3607.22 ?? PDB entry 1NIX HnTxIV Hainantoxin-IV S. hainana 3987.59 Nav TTX-S Liu et al., 2003 HnTxV Hainantoxin-V S. hainana 3972.57 Nav TTX-S Xiao and Liang, 2003 HwTxI Huwentoxin-I S. huwena 3750.42 tipo N (L?) Liang et al., 1993 HwTxIV Huwentoxin-IV S. huwena 4107.82 Nav TTX-S Peng et al., 2002 PaTx1 Phrixotoxin 1 P. auratus 3548.36 Kv4.2 / 4.3 Diochot et al., 1999 PaTx2 Phrixotoxin 2 P. auratus 3921.75 Kv4.2 / 4.3 Diochot et al., 1999 PcTx1 Psalmotoxin 1 P. cambridgei 4689.45 ASIC1a Escoubas et al., 2000a ProTxI Protoxin I T. pruriens 3987.55 Nav1.2, 1.5, 1.7, 1.8 Middleton et al., 2002 ProTxII Protoxin II T. pruriens 3826.64 Nav1.2, 1.5, 1.7, 1.8 Middleton et al., 2002 ScTx1 Stromatoxin 1 S. calceata 3791.36 canal p/ K v2.1 / 2.2 / 4.2 Escoubas et al., 2002 SGTx1 – Scodra griseipesb 3776.32 cK ?? Marvin et al., 1999 SHL-1 SHLP-I S. huwenaa 3540.05 células vermelhas do sangue ?? Liang and Pan, 1995 SNX-482 – H. gigas 4495.06 Cav E classe Newcomb et al., 1998 VSTx1 Voltage sensor toxin 1 G. spatulata 3997.73 canal p/ K Ruta et al., 2003 v-GsTx SIA v-grammotoxin SIA G. spatulata 4110.74 tipo Cav N&P/Q Lampe et al., 1993 TxP1 Toxic protein 1 B. smithi 4399.38 ?? Kaiser et al., 1994 ESTx1 Eurypelma spider toxin E. californicum 4413.41 ?? Savel-Niemann, 1989 ESTx2 Eurypelma spider toxin E. californicum 4399.38 ?? Savel-Niemann, 1989 HwTxII (Gln) Huwentoxin-II S. huwenaa 4299.16 ?? Shu and Liang, 1999 HwTxII (Ileu) Huwentoxin-II S. huwenaa 4284.19 ?? Shu and Liang, 1999 LpTx1 Lasiotoxin 1 L. parahybana 5722.86 ?? Escoubas et al., 1997b LpTx2 Lasiotoxin 2 L. parahybana 5674.77 ?? Escoubas et al., 1997b
Fonte: P. Escoubas & L. Rash, 2004
8

1.3 Os canais Iônicos
As células excitáveis dos animais são capazes de, no estado celular de equilíbrio, alterar a
polaridade da membrana, gerando sinalizações elétricas rápidas, com participação ativa dos
canais iônicos. Esta atividade é vista em neurônios, células musculares e endócrinas, onde essa
resposta rápida é essencial para o funcionamento do sistema.
Os canais iônicos (FIG.3) são constituídos de glicoproteínas transmembrânicas,
seletivas a íons e estão ancorados na membrana plasmática. Estes canais possuem um poro onde
os íons atravessam um filtro seletivo, e um mecanismo de abertura de comportas, conhecido
como “gating”, onde ocorre a abertura e o fechamento dos canais em milissegundos, como
resposta aos estímulos elétricos/químicos ou mecânicos, mudança de voltagem, alteração da
concentração de ligantes como neurotransmissores, peptídeos e hormônios (HILLE, 1992). A
Figura 4 ilustra a estrutura geral esquemática, dos canais iônicos para sódio, cálcio e potássio.
A presença dos canais iônicos representa uma economia energética para as células. Sua
eficiência no controle do gradiente celular diminui gastos com a entrada e saída de milhões de
íons por segundo. Os canais são também substratos para processos biofísicos como a
transmissão sináptica, a contração muscular, o processamento de informações intracelulares e
atuam na produção e na liberação de substâncias celulares. Os principais canais iônicos são
classificados quanto à sua permeabilidade e seletividade a íons. Eles são divididos em quatro
grupos ou famílias envolvendo os seguintes íons: Na+ , Ca++ , Cl- e K+. Estes canais podem
responder diferentemente a diferentes voltagens e a ligantes externos ou internos.
Em geral, os canais apresentam dois estados, um aberto e outro fechado, mas alguns
canais podem possuir um estado inativo no qual o poro vai estar ocluso em uma região, mesmo
estando aberto. Esta abertura e o fechamento são diretamente controlados por fatores como
voltagem, modificações covalentes ou ação de fatores exógenos como toxinas e algumas drogas
que bloqueiam a ação do canal e podem ou não ser reversíveis. Assim, o estudo de toxinas
animais e o conhecimento específico das suas ações nesses canais e nos processos
eletrofisiológicos das células representam interesse primário na busca de possíveis modelos de
fármacos e do maior conhecimento estrutural e funcional do sistema nervoso, em nível
molecular.
9

Figura 3 - Estrutura básica dos canais iônicos. Estrutura esquemática para os canais para sódio e para cálcio,
voltagem-dependentes (Nav e Cav). A subunidade alfa, principal responsável pela função dos canais iônicos, forma
quatro domínios (I a IV), com seis segmentos, que atravessam a membrana. O segmento 4 (S4-representado em
verde), é o sensor de voltagem dos canais. À direita – representação esquemática de subunidades de canais para
potássio (Kv). Os canais para sódio e para cálcio possuem sub-unidades acessorias, não representadas aqui. Fonte:
Catterall, W.A. et al., 2007
externo
interno
Subunidade α
10

1.3.1 Canais para cálcio - breve introdução.
As células nervosas e musculares têm suas funções ligadas à entrada e saída de íons.
Dentre esses íons, o cálcio desempenha papel importante na liberação de neurotransmissores, na
expressão gênica, dentre outras. Estas ações são iniciadas a partir de canais para cálcio
dependentes de voltagem, que foram descritos primeiramente em células musculares de
crustáceos por Fatt e Katz (1953).
A importância dos canais para cálcio tem sido demonstrada em testes eletrofisiológicos,
neuroquímicos e neurofarmacológicos (HILLE, 1992). Sua classificação básica está diretamente
ligada ao tipo de corrente gerada para a despolarização da membrana, podendo existir correntes
de baixa voltagem (“Low Voltage Activated”- LVA) e correntes de alta voltagem (“High
Voltage Activated” - HVA). Estas correntes apresentam, respectivamente, condutância na faixa
de 5-9 pS e 13-24 pS. A partir dessas correntes e da ação nas diferentes células, os canais para
cálcio são classificados em cinco grupos:T, L, N, P/Q e R (NOWYCKY et al, 1985.;
CARBONE & LUX, 1984; MINTZ et al, 1992; BIRNBAUMER et al, 1994; HOFMANN et al,
1994; ERTEL et al, 2000).
Os canais tipo T utilizam correntes LVA e são denominados assim por serem transitórios
e de rápida inativação. Os demais utilizam correntes tipos HVA e apresentam-se com
despolarização de longa duração. Os canais tipo L (de “long lasting”) estão presentes em células
musculares esqueléticas e cardíacas e são sensíveis a drogas como 1,4 dihidropiridina (DHP). Os
canais tipo N (de “neuron”) são semelhantes aos tipo L, mas apresentam-se em neurônios, sendo
insensíveis a drogas do tipo DHP. Em células de Purkinje ocorre uma variação da corrente, em
sua maioria, chamada de P (de “Purkinge”). Esta corrente é bloqueada seletivamente pela toxina
peptídica da peçonha da aranha Agelenopsis aperta (ω-agatoxina IVA - ‘ω-Aga IVA’) em
concentrações < de 90 nM. Em concentrações maiores, esta toxina bloqueia também um
componente chamado Q. Assim estes canais foram classificados como canais P/Q. Em alguns
casos, existe uma corrente que não é bloqueada pelos inibidores dos canais citados acima, sendo
denominada de R (de “resistent”) pela sua resistência a tais drogas.
Além disso, os canais para cálcio (FIG. 3) vão ter variações decorrentes de suas
subunidades α e β, chamadas de sub-unidades acessórias e que modulam a ação destes canais,
estando entre os canais mais diversificados e heterogêneos, dentre os canais iônicos. A
subunidade α ou α1 é constituída por um poro condutor, pelo sensor de voltagem, pelo sistema
de regulação e pelos sítios de regulação do canal. Esta subunidade é típica para os canais iônicos,
contendo quatro domínios (I-V) subdivididos em seis segmentos transmembrânicos (S1-S6) com
11

uma alça entre os segmentos S5 e S6 (FIG.3). As subunidades β são subunidades
transmembrânicas ligadas por pontes dissulfeto à subunidade α entre os domínios I e II, podendo
variar em quatro subtipos (β1 – β4). Essa interação com a subunidade β gera ainda mais variação
e diversidade de ação dos canais para cálcio (PRAGNELL et al, 1994).
Atualmente, assim como para os demais íons, a nomenclatura utiliza o símbolo químico
do íon com um subscrito (Cav ), um indicador numérico correspondente à família da subunidade
α (1-3) e outro para a ordem da descoberta da subunidade dentro daquela família (1 em diante).
Exemplo, Cav 1.1 que corresponderia a uma corrente do tipo L contendo a subunidade α1S. Os
demais tipos estão representados na TABELA 2 .
12
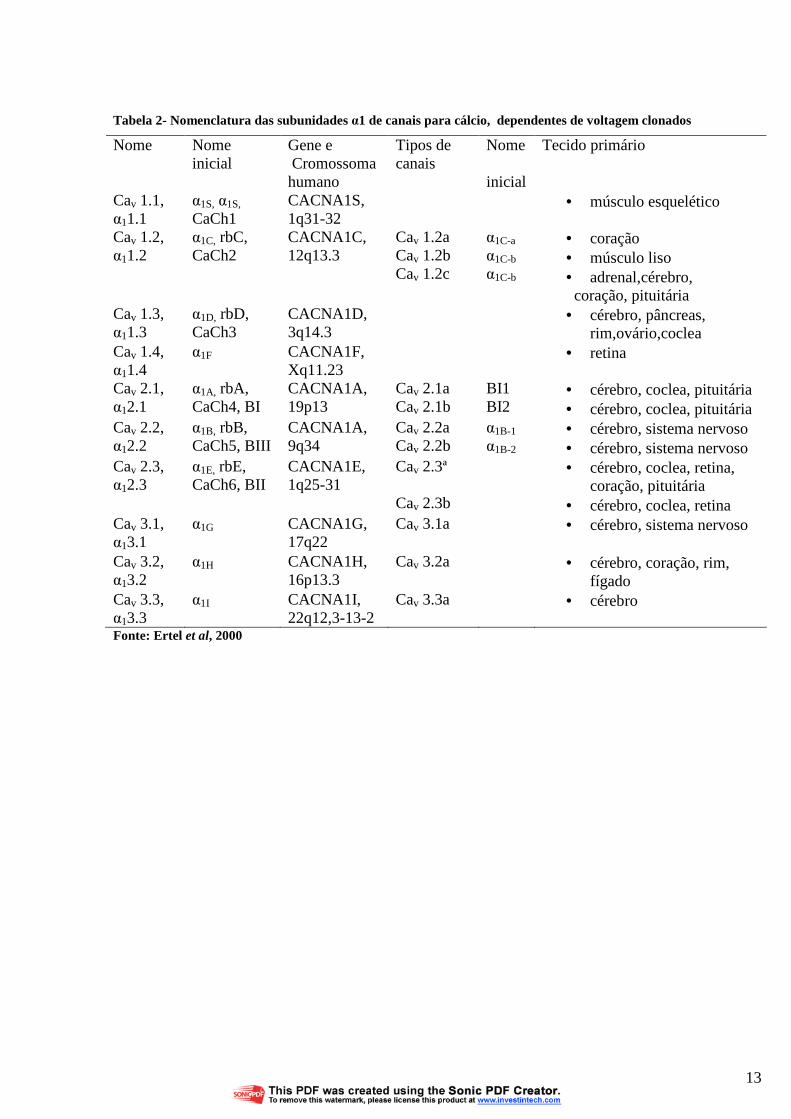
Tabela 2- Nomenclatura das subunidades α1 de canais para cálcio, dependentes de voltagem clonados
Nome Nome inicial
Gene e Cromossoma humano
Tipos de canais
Nome inicial
Tecido primário
Cav 1.1, α11.1
α1S, α1S,
CaCh1 CACNA1S, 1q31-32
• músculo esquelético
Cav 1.2, α11.2
α1C, rbC, CaCh2
CACNA1C, 12q13.3
Cav 1.2a Cav 1.2b Cav 1.2c
α1C-a
α1C-b
α1C-b
• coração • músculo liso • adrenal,cérebro,
coração, pituitária Cav 1.3, α11.3
α1D, rbD, CaCh3
CACNA1D, 3q14.3
• cérebro, pâncreas, rim,ovário,coclea
Cav 1.4, α11.4
α1F CACNA1F, Xq11.23
• retina
Cav 2.1, α12.1
α1A, rbA, CaCh4, BI
CACNA1A, 19p13
Cav 2.1a Cav 2.1b
BI1 BI2
• cérebro, coclea, pituitária • cérebro, coclea, pituitária
Cav 2.2, α12.2
α1B, rbB, CaCh5, BIII
CACNA1A, 9q34
Cav 2.2a Cav 2.2b
α1B-1
α1B-2
• cérebro, sistema nervoso • cérebro, sistema nervoso
Cav 2.3, α12.3
α1E, rbE, CaCh6, BII
CACNA1E, 1q25-31
Cav 2.3ª Cav 2.3b
• cérebro, coclea, retina, coração, pituitária
• cérebro, coclea, retina Cav 3.1, α13.1
α1G CACNA1G, 17q22
Cav 3.1a
• cérebro, sistema nervoso
Cav 3.2, α13.2
α1H CACNA1H, 16p13.3
Cav 3.2a
• cérebro, coração, rim, fígado
Cav 3.3, α13.3
α1I CACNA1I, 22q12,3-13-2
Cav 3.3a
• cérebro
Fonte: Ertel et al, 2000
13

1.4 Células do gânglio dorsal
As células DRG (“dorsal root ganglion”) são células que apresentam uma diversidade de
canais iônicos. Dentre esses, os canais para cálcio são bem representados nestas células.
Em células DRG cardíacas foram encontradas e testadas correntes do tipo L, N, P/Q e R
(ROLA, et al., 2003) o que demonstra a ampla variedade de canais para cálcio existentes nestas
células. Neurotoxinas como ω-CgTx GVIA, ω –CgTx MVII C (Omega conotoxinas de moluscos
do gênero Conus) e ω -Aga IVA (toxina da aranha Agelonopsis aperta) bloqueiam, em
vertebrados, com especificidade, correntes para cálcio do tipo N, Q, e P, respectivamente
(DUNLAP et al. 1995; MILJANICH e RAMACHANDRAN, 1995; OLIVERA et al.1994,
WICHER e PENZLIN, 1997).
Nos insetos, em células DRG de gânglios nervosos, são encontrados canais para cálcio
com correntes T, L e N, com propriedades diferentes daquelas dos vertebrados, além das
correntes P/Q (WICHER e PENZLIN, 1997).
14

2 OBJETIVOS
2.1 Objetivo geral
Este trabalho teve como objetivo iniciar a caracterização estrutural, bioquímica e
farmacológica da peçonha da aranha caranguejeira Achantoscurria natalensis.
2.2 Objetivos específicos
Purificar e caracterizar bioquímica- e farmacologicamente, pelo menos uma
toxina desta peçonha.
; (este objetivo esta sem sentido...)
Verificar o perfil protéico da peçonha A. natalensis expressanda por por técnicas
cromatográficas e de eletroforese uni- e bi-dimensional;
Testar a atividade insetotóxica da peçonha e da(s) toxina(s) isolada(s).
Analisar a atividade biológica da(s) toxina(s) isolada(s), por estudos
eletrofisiológicos.
Caracterizar estrutural e parcialmente, as massas moleculares e as seqüências N-
terminais das toxinas purificadas;
Comparar as seqüências obtidas com outras, depositadas em bancos de dados;
buscando-se possíveis homologias.
15

3 MATERIAL E MÉTODOS
3.1 Coleta e manutenção A. natalensis
As aranhas caranguejeiras A. natalensis foram coletadas na cidade de Feira de Santana e
foram mantidas em cativeiro sob controle de humidade e temperatura no Laboratório de Animais
Peçonhentos e Herpetologia (LAPH-UEFS), com alimentação realizada de 15 em 15 dias.
3.2 Identificação da A. natalensis
A identificação dos espécimes A. natalensis foi realizada por nós. Posteriormente exemplares
destes espécimes foram enviados para o Dr. Antônio Brescovit, do Instituto Butantan (IB-São
Paulo) que confirmou nossa identificação.
3.3 Extração da peçonha de A. natalensis
A peçonha da aranha foi coletada com o uso de micropipetas, após a estimulação das
quelíceras com choque elétrico (9V) (FIG.4). A peçonha foi armazenada em tubos Falcon de
50 mL contendo 30 mL de solução aquosa de TFA 0,1% (v/v). Esta suspensão foi centrifugada a
10000 RPM, durante 10 minutos (centrifuga Eppendorf 5804-R). O sobrenadante foi coletado,
liofilizado e armazenado a - 4ºC.
16

Figura 4- Extração da peçonha da Acanthoscurria natalensis utilizando-se micropipetas, após a estimulação das
quelíceras com choque elétrico (9V).
17

3.4 Dosagem da concentração de proteína
A dosagem de proteína foi realizada em um espectrofotômetro Shimadzu UV-160 A . A
absorbância foi medida nos comprimentos de onda ( λ) = 280 e 260. A partir destas leituras
utilizou-se a estimativa: x mg/mL de proteína = 1,55.A280 – 0,76.A260 (WALKER, J. M,
2002).
3.5 Fracionamento do peçonha de Acanthoscurria natalensis
3.5.1- Cromatografia de troca catiônica-CIEX
A separação parcial dos componentes da peçonha de A. natalensis (machos e fêmeas) foi
realizada no Laboratório de Enzimologia e Físicoquímica de Proteínas (UFMG) e no
Laboratório de Venenos e Toxinas Animais - LVTA (UFMG). Foram utilizados 2,0 mL da
solução de peçonha bruta, solubilizada em água deionizada.
As análises cromatográfias foram realizadas nos sistema do ÄKTA Explorer 100 HPLC
system (Amersham Biosciences, Uppsala, Sweden), controlado pelo software UNICORN 4.11 e
equipado com coletor automático de frações Frac920 (Amersham Bioscience, Uppsala, Sweden).
Alternativamente, foi utilizado o sistema SHIMADZU-10 A, com coleta manual de frações. A
eluição foi monitorada através da absorbância de luz ultravioleta (λ = 214 e 280 nm, no sistema
ÄKTA Explorer 100 e λ = 214 no sistema SHIMADZU-10 A).
A peçonha foi centrifugada a 10000 RPM, (micro-centrifuga Hettich- Mikro 200R). O
sobrenadante foi aplicado em uma coluna de troca catiônica TSK-Gel CM-SW 15 cm X 4.6 mm
(Tosoh Biosep, Montgomeryvile, USA), equilibrada com acetato de sódio (50 mmol.L-1 , pH 5)
e fluxo de 0,75 mL/min . O sistema foi eluído com um gradiente linear de NaCl 1M, pH 5 em
acetato de sódio (50 mmol.L-1 ), durante 123 minutos.
3.5.2 – Cromatografia de fase reversa
Após a cromatografia de troca catiônica eram feitos “pools” dos picos.
1 mL de cada destas frações eram eram aplicados em uma coluna de fase reversa SourceTM 15
4,6/100 (Pharmacia Biotech, Uppsala,Sweeden). A coluna era previamente equilibrada com uma
18

solução aquosa de 0.1% (v/v) de ácido trifluoroacético com um fluxo de 1,0 mL/min e com
eluição em gradiente linear (0–100%) de acetonitrila durante 50 minutos.
3.6 Eletroforese Unidimensional em poliacrilamida-SDS (SDS-PAGE) em duas
etapas:
(i) A peçonha foi solubilizada em uma solução aquosa de TFA 0,1% (v/v), 30 µL desta
solução foram acrescentados a 30 µL do tampão da amostra (2 mL de SDS à 10%, 500 µL de 2-
mercaptoetanol, 2 mL de glicerol, 400 µL de azul de bromofenol à 2%, 2,5 mL de Tris-HCl-0,5
M-pH 6,8, e 10 mL de água destilada e deionizada). Essa solução era aquecida à 80ºC por 10
minutos. O gel teve como padrão molecular o kit SIGMA M0671(“Recombinant Molecular
Weight Marker”) de 15, 25, 35, 50, 75, 100 e 150 kDa. Previamente, os géis de corrida e de
concentração foram montados como: I-Gel de corrida à 15%; II- Gel de concentração à 5% (
FORMULAÇÃO EM ANEXO). O tempo de corrida foi cerca de 2:30 à 3h, voltagem de 110V.
Após a corrida o gel foi submetido à coloração com prata, sendo este procedimento realizado
conforme descrito (WEBER & OSBORN, 1969; LAEMMLI, 1970).
(ii) A fração contendo 30 µL a toxina (AnTx1) originada na cromatografia de fase reversa,
contendo TFA-H2O (1%) e ACN (cerca de 35%) foi adicionada a 30 µL do tampão da amostra
(2 mL de SDS à 10%, 500 µL de 2-mercaptoetanol, 2 mL de glicerol, 400 µL de azul de
bromofenol à 2%, 2,5 mL de Tris-HCl-0,5 M-pH 6,8, e 10 mL de água destilada e deionizada).
Essa solução era aquecida à 80ºC por 10 minutos. O gel teve como padrão molecular o kit da
SIGMA M0671 (“Recombinant Molecular Weight Marker”) (M.W. 15, 25, 35, 50, 75, 100 e
150 kDa). Previamente, os géis de corrida e de concentração foram montados como: I-Gel de
corrida à 15%; II- Gel de concentração à 5% ( FORMULAÇÃO EM ANEXO). O tempo de
corrida foi cerca de 2:30 à 3h, voltagem de 110V. Após a corrida o gel foi submetido à
coloração com prata. Este procedimento foi realizado conforme já descrito (Weber & Osborn,
1969; Laemmli, 1970).
19

3.7 Eletroforese em gel bidimensional (2D-PAGE)
As peçonhas de fêmeas (50 microgramos de peçonha) foram diluídas com o tampão para
a primeira dimensão da isoeletro-focalização acrescidas de DTT (10 mg/mL) em um volume
total de 250 µL, com agitação por 1 hora e centrifugação por 30 minutos (10.000 RPM,
centrifuga Hettich- Mikro 200R). O sobrenadante foi aplicado no suporte da “strip” e coberto
com a “strip” de 13 cm deixando hidratar por 12 horas. Após a hidratação as “strips” foram
levadas para a eletro-focalização no Ettan IPGphor II, por 15 horas, com programação previa
para “strips” de 13 cm, de acordo com instruções do fabricante, sendo retiradas e congeladas em
seguida. Posteriormente, as “strips” foram levadas à segunda dimensão do gel passando por um
gel de poliacrilamida de 18 cm, com concentração de 12 %, com glicerol a 5 % com corrida a
150 V por 14 horas. Em seguida, o gel foi corado com Comassie Blue G-250 a 2 % por 24 horas
(O’FARRELL, 1975).
3.8 Bioensaios em insetos, com a peçonha bruta e toxinas.
3.8.1 Testes de toxicidade em insetos.
Para testar a atividade biológica da peçonha da Acanthoscurria natalensis em insetos
foram utilizadas moscas domésticas (Musca domestica). As moscas utilizadas, provenientes do
Ranário Vivendas da Rainha-Ribeirão das Neves (MG) foram mantidas no insetário do
Laboratório de Venenos e Toxinas Animais, Departamento de Bioquímica e Imunologia,
UFMG, durante várias gerações até sua utilização.
As moscas, pesando em média 20 mg cada, com 3 a 4 dias de vida, foram anestesiadas
pelo abaixamento da temperatura corporal. Elas foram colocadas em tubos Falcon de 50 mL
tampados e inseridos em um recipiente contendo banho de gelo com temperatura em torno de
4ºC, por cerca de 2-3 minutos. As injeções foram preparadas utilizando-se um controle com
solução salina (0,9 % p/v de NaCl) contendo 0,1 % (p/v) de BSA. As amostras eram diluídas na
solução controle e continham diferentes concentrações da peçonha bruta, ou da toxina. As
amostras foram injetadas via-intratoráxica (FIGUEIREDO et al. ,1995) entre os dois primeiros
pares de patas. Em cada concentração utilizaram-se dez moscas (n=10), o volume das injeções
foi de 0,5 µL/ mosca.
20

As amostras foram injetadas utilizando-se uma seringa Hamilton®contendo na
extremidade um tubo capilar de vidro (FIG.5) estirado (a quente) para o afinamento da ponta.
Após as injeções as moscas foram acondicionadas em recipientes plásticos, tampados com furos
para ventilação, limpos, contendo solução aquosa de sacarose (5 %), abrigados de outros insetos.
Foram feitas observações do comportamento (paralisia, agitação, vôo e demais atividades) nas
primeiras horas, 24 hs e 48 hs após as injeções. Este ensaio foi repetido por três vezes, pelo
menos.
21

Figura 5- Fotografia mostrando a aplicação de amostra (controle, peçonha ou toxina) em moscas (Musca
domestica) utilizando-se seringa Hamilton® com capacidade de 1mL. Observe o tubo capilar (estirado) inserido na
ponta da agulha e utilizado para inserção no abdome da mosca.
22

3.8.2 Testes eletrofisiológicos da AnTxI em canais para Ca2+
Para verificar os possíveis efeitos da toxina AnTxI em canais para cálcio foram utilizadas
células DGR (“Dorsal glanglion root”) do gânglio dorsal de ratos, provindas do Laboratório de
Membranas Excitáveis no Instituto de Ciências Biológicas (ICB-UFMG).
3.8.3 Cultura celular de células DRG do gânglio dorsal de ratos
Animais
Ratos Wistar (Rattus novergicus), machos, pesando entre 90-150 g, foram sacrificados
por decapitação, utilizando-se uma guilhotina. Para cada cultura foram utilizados dois ratos,
separadamente.
Cultura Celular
Os gânglios DRG, de onde emanam as raízes do nervo isquiático na medula espinhal:
região lombar (L4 e L5) e sacral (S1) foram dissecados, removidos cirurgicamente e transferidos
imediatamente para dois tubos Falcon de 15 mL, contendo, em separado, 4 mL de solução
fisiológica de Hank (Hank’s Balanced Salt Solution- HBSS) sem adição de Ca2+ e Mg2+ (Hank’s
Modificada), gelada, acrescida de 1% de antibiótico penicilina/estreptomicina (Sigma, St. Louis,
Mo, USA). Os gânglios foram lavados com a mesma solução de Hank Modificada, gelada, num
total de quatro repetições para cada tubo Falcon.
Os gânglios foram transferidos para tubos Falcon contendo 5 mL de meio DMEM com
alta glicose (Sigma, St. Louis, Mo, USA), acrescido de 1% de antibiótico
penicilina/estreptomicina, contendo 1 mg/mL de enzima colagenase (Tipo 1) (Sigma, St. Louis,
Mo, USA). Esta suspensão foi incubada por 75 min, a 37°C, em banho-maria. Ao final do
período, os gânglios foram lavados com meio DMEM alta glicose acrescido de 1% de
antibiótico penicilina/estreptomicina, num total de três repetições.
Posteriormente, os gânglios foram incubados com uma solução contendo 0,25% (p/v) de
tripsina (Sigma, St. Louis, Mo, USA) e 0,025% de EDTA (Ecibra, Sto Amaro/SP, Brasil) em
HBSS sem Ca2+ e Mg2+, por um período de 15 minutos, a 37°C, em banho-maria. Após o
período de incubação, os tubos foram lavados com 5 mL de meio DMEM alta glicose acrescido
de 1% de antibiótico penicilina/estreptomicina com de 10% de soro fetal bovino (SBF-Cultilab,
Brasil) – DMEM completo, para inativação enzimática, num total de três lavagens.
23

Os gânglios foram então cuidadosamente triturados com uma pipeta Pasteur de vidro em
1,5 mL de meio DMEM para induzir a dissociação mecânica do tecido. O diâmetro da ponta da
pipeta correspondia a dois terços do diâmetro dos gânglios.
As células dissociadas e em suspensão foram colocadas em lamínulas de vidro (20 x 20
mm) (Corning, México) (200 µL) pré-recobertas (período mínimo de 24 horas) com 200 µL por
lamínula poli-L-lisina (60µg/mL) (Sigma, St. Louis, Mo, USA). O excesso de poli-L-lisina foi
removido das lamínulas antes das células serem plaqueadas na placas de Petri. As lamínulas com
as células aderidas foram introduzidas em placas de Petri (35 x 10 mm) (Sarstedt, Newton,
USA), as células foram mantidas a 37 °C em uma atmosfera humidificada a 100% e aerada a
95% O2 e 5% CO2 durante um período de 120 minutos. Posteriormente, foram acrescentados 2
mL de meio DMEM em cada placa de Petri.
Experimentos de “ PATCH-CLAMP”
Os experimentos eletrofisiológicos foram realizados num período de 6 a 12 horas após o
término da cultura. Para os experimentos de “whole-cell patch-clamp”, cada lamínula foi
dividida em quatro partes, por uma caneta com a ponta de diamante. Este experimento teve o
objetivo de medir as correntes provocadas pelos íons cálcio nas células DRG. As correntes de
cálcio foram medidas por meio da técnica de “patch-clamp”, na configuração “voltage-clamp,
whole-cell” em células DRG à temperatura ambiente (25ºC). Os experimentos foram executados
em um microscópio, no qual foi acoplado um sistema de micromanipulação sob uma mesa
antivibratória no Laboratório de Membranas Excitáveis e Eletrofisiologia do Instituto de
Ciências Biológicas (ICB-UFMG). Foram utilizadas três soluções fisiológicas na preparação de
“patch-clamp” denominada solução interna de pipeta (SI), solução externa I ou solução de
Tyrode (SE1) e solução externa II ou solução externa para isolar a corrente total de Ca2+ (SE2) .
As lamínulas com as células foram lavadas com a solução externa I e as células
permaneciam incubadas nesta solução durante a etapa inicial do registro eletrofisiológico à
temperatura ambiente.
As pipetas foram confeccionadas a partir de capilares de vidro a fim de obter-se
resistências na ordem de 1,5 – 2,5 µΩ. As pipetas então eram cuidadosamente preenchidas com a
solução interna. As lamínulas foram focalizadas no microscópio e uma célula alvo, ou neurônio
do tipo fibra C (fibra C- fibra nervosa, nociceptiva, com diâmetro entre 20 à 30 µM) foi
escolhida e posicionada em frente à microperfusão.
24

O sistema de microperfusão, formado por um solenóide e um capilar de perfusão,
permitiu expor a célula a dois tipos de soluções (controle, SE2) e a experimental contendo a
toxina (100 nM) diluída na solução externa II. A perfusão manteve as células totalmente
banhadas pelas soluções com 35 µL (controle ou experimental) enquanto as leituras das
correntes eram obtidas.
Um amplificador HEKA EPC-9 acoplado a um microcomputador Mac PowerPC realizou
a monitoração das correntes iônicas em tempo real correspondentes aos protocolos aplicados,
com seqüência de pulsos de voltagem disparados em ordem temporal definida. A configuração
“whole cell” era obtida no momento em que a membrana se rompia e ocorria a difusão da S1
para dentro da célula, permitindo o registro das correntes iônicas totais para o cálcio e a sua
estabilidade com as soluções.
Na configuração “whole cell”, um pulso teste de voltagem foi aplicado para se verificar a
corrente total na célula. Esse procedimento inicialmente mantinha as células em um potencial de
-80 mV (potencial de holding), injetando em seguida um pré-pulso hiperpolarizante de -120 mV,
com duração de 1s, de modo a requisitar o maior número possível de canais de cálcio no estado
fechado. Um potencial despolarizante era então aplicado para +50 mV, com duração de 1s,
potencial no qual a probabilidade de abertura dos canais para cálcio é maior e o traçado obtido
refletia a abertura desses canais nas células DRG.
A solução contendo a toxina só foi liberada quando o pulso ou registro se estabilizou. O
traçado em tempo real visualizado não podia mostrar alterações significativas enquanto a célula
estivesse exposta à solução externa (SE1).
3.9 Espectrometria de Massa
3.9.1 Analise da peçonha isolada por ESI-Q-TOF-MS
As análises por espectrometria de massa foram realizadas em um aparelho Q-ToF Micro TM (Micromass, Grã Bretanha), pertencente ao Laboratório de Biomoléculas no ICB/UFMG. O
aparelho está equipado com fonte de ionização electrospray operada em modo positivo. A
voltagem do capilar foi estabelecida em 3-3,5kV e as voltagens sample cones foram 30-40 V. A
calibração do espectrômetro de massa foi feita com NaI, com janela de 1000 a 20000 m/z. As
amostras foram diluídas em acetonitrila 50%/TFA 0,1%/água deionisada/destilada e introduzidas
no aparelho utilizando-se uma bomba de seringa com fluxo entre 5-10µL/min. Os dados obtidos
foram analisados utilizando-se o programa MassLynx® 4.0.
25

3.9.2 Analise da peçonha por MALDI-TOF/TOF-MS
As análises de MALDI-TOF/TOF MS (matrix assisted laser desorpotion/ionization time
of flight tandem time of flight mass spectrometry) foram realizados em um aparelho Autoflex III
(Bruker Daltonics, Alemanha), pertencente ao Laboratório de Biomoléculas no ICB/UFMG,
com o controle através do programa FelxControl 2.4.30.0 (Bruker Daltonics, Alemanha). Cada
amostra foi misturada em uma solução com 5 mg ácido α-ciano-4-hidroxicinâmico, 200 µL de
água destilada, 250 µL de acetonitrila (ACN) e 50 µL de TFA a 3%(v/v) na proporção de 1 da
amostra para 3 partes da solução e inserida com micropipetas (0,5 µL) em uma placa de amostra
(MTP Anchorchip 600/384). A calibração do aparelho foi feita com padrões externos (Protein
Calibration Standard I, Bruker Daltonics, Alemanha). Os espectros foram obtidos em modo
positivo/refletido, com a freqüência do laser justada em 50 Hz. Na fragmentação de peptídeos
foram empregados CID (Collision Induced Dissociation) (N2) e LID (Laser Induced
Decomposition). Os dados obtidos foram analisados através do programa flexAnalysis 2.4
(Bruker Daltonics, Alemanha) e com o programa PepSeq (Micromass, Grã - Bretanha).
3.9.2.1 Preparação e proteólise das amostras
As amostras foram diluídas em 20 µL de tampão NH4HCO3 (0,4M) contendo uréia (8M),
posteriormente adicionaram-se 5 µL de DTT- 45 mM (Di-tio-treitol) e procedeu-se a incubação
a 50ºC, por 15 minutos. Após resfriamento adicionaram-se 5 µL de iodoacetamina (100mM) e
seguiu-se incubação durante 15 minutos, ao abrigo da luz para não degradar o iodoacetamina.
Em seguida, as amostras receberam 130 µL de água deionisada e seus conteúdos foram
aliquotados em dois grupos de 80µL. Na primeira alíquota acrescentou-se 0,8 mL de tripsina e
na outra alíquota adicionou-se 1,2 mL da protease V8. As alíquotas foram incubadas a 37ºC
durante 24 h., posteriormente foram dessalinizadas através de micropurificação em ZipTips®
C18 (Millipore-Bedford, E.U.A.). A micropurificação ocorreu com a preparação do ZipTips®
C18 passando três vezes com 8 µL de acetonitrila (ACN) desprezando-se a acetonitrila após essa
preparação. As amostras foram lavadas por três vezes com TFA 0,1% e em seguida o conteúdo
foi lavado em solução aquosa de 50% (v/v) de ACN em solução aquosa de 0,1% (v/v) de TFA
por dez vezes. Feito isso, as amostras foram submetidas à análise por MALDI-TOF/TOF.
26

3.10 Determinação da sequência n-terminal das toxinas AnTxI E AnTxII
Foi utilizado um seqüenciador automático de proteínas, Shimadzu PSSQ-21A para o
seqüenciamento através da degradação de Edman. Em uma solução de 0,1:0,5:0,5
TFA:H2O:ACN foram colocadas 50 µL da amostra, essa mistura foi aplicada sobre um disco de
fibra de vidro contendo o polímero SEQUA-BRENE (Sigma), previamente tratado com
procedimentos padronizados do equipamento PSSQ-21A. A derivação foi feita com solução de
fenilisotiocianato em n-heptano a 5% com solução de ácido trifluoroácético e trimetilamina a
12%. A derivação foi seguida de uma etapa de conversão do intermediário tiazolina em PTH-
aminoácido por solução aquosa de ácido trifluoroacético a 25%. A separação dos PTH-
aminoácidos ocorreu em uma coluna de fase reversa (WAKOSIL-PTH 25 cm x 4,6 mm, Wako,
Japão) com a eluição conduzida em condição isocrática por solução denominada de “PTH-AA
mobile Phase” (Wako, Japão). Os tempos de retenção dos PTG-aminoácidos obtidos foram
comparados com padrões.
27

4 RESULTADOS E DISCUSSÃO
4.1 Eletroforese unidimensional em poliacrilamida-SDS (SDS-PAGE) e Eletroforese
em gel bidimensional (2D-PAGE)
A análise eletroforética do gel de poliacrilamida (SDS-PAGE) da peçonha da A.
natalensis (FIG. 6) demonstra a presença de pelo menos sete bandas visualizadas nos géis. A
canelata (A) está representada pelos padrões de massa molecular. Na canaleta (B) observa-se o
perfil da peçonha com bandas de massas moleculares diversas: abaixo de 15 kDa, uma banda em
torno de 35 kDa, outras acima de 50kDa até 150 kDa, demonstrando a variedade de
componentes presentes na peçonha analisada. Chama atenção a existência de várias bandas nas
faixas de mais altas massas moleculares. De modo geral, nas peçonhas de aranhas, predominam
os peptídeos, em detrimento de proteínas com mais altas massas moleculares. Montandon (2007)
trabalhando com a peçonha de Grammmostola iheringi, aranha caranguejeira da região do Sul
do Brasil, mostrou perfil eletroforético com certas semelhanças ao encontrado na peçonha de A.
natalensis, por exemplo, uma quantidade maior de bandas moleculares abaixo de 15 kDa.
Herzig et al, 2004 e Ricardson et al, 2006, verificaram a presença de massas moleculares abaixo
de 15 kDa e acima de 50kDa para peçonha do gênero Phoneutria. Pretel et al, 2005 obtiveram
resultados semelhantes para a peçonha de Loxosceles sp. Estes dados, em certa medida
confirmam um padrão já característico de massas moleculares, encontrados nas peçonhas de
aranha.
Na canaleta (C) observa-se que a toxina AnTx1 migrou como uma grande banda
espalhada. Isto se deve, provavelmente, à quantidade relativamente grande de toxina aplicada ,
cerca de 30 microgramas.. Há poucos trabalhos relacionados com peçonhas de aranhas da
família Theraphosidae. Como exemplo podemos citar os trabalhos realizados por Kalapothakis
et al, 2003, com a peçonha de Lasiodora sp., onde verificou-se os efeitos desta em corações
isolados de ratos.
Apesar de que, relativamente poucas bandas terem sido encontradas, deve-se considerar
que em muitos casos a eletroforese uni-dimensional não separa moléculas diferentes mas que
têm a mesma massa molecular. Buscando-se esclarecer esta questão utilizou-se também a
eletroforese bi-dimensional.
As peçonhas de A. natalensis (fêmeas e machos) quando resolvidas em um gel (2D-
PAGE) não demonstraram deferenças aparentes entre os sexos. Um experimento representativo
destes resultados é observado para o gel 2D com a peçonha de fêmeas da A. natalensis (FIG.7)
28

Este gel foi escolhido por mostrar melhor resolução no momento de coloração. Neste gel
verifica-se uma gama de componentes protéicos de diferentes PI´s. Isto comprova que no gel
unidimensional uma mesma banda pode congregar diferentes componentes, que são separados
quando submetidos ao gel bidimensional. Embora não tivéssemos a oportunidade de explorar
mais os diferentes “spots”, este experimento foi importante no sentido de mostrar claramente, a
diversidade de componentes da peçonha. Ricardson et al, 2006 analisaram três peçonhas de
espécies do gênero Phoneutria por 2D-PAGE, determinando uma média de 80 spots,
caracterizados como proteínas de massa molecular abaixo de 12 kDa e alguns peptídeos com
massa em torno de 4kDa.
Apesar do presente experimento (gel 2D-PAGE) ter sido realizado uma única vez para cada
peçonha (macho e fêmea) e apesar dos padrões de massas moleculares apresentarem-se
irregulares na migração, dificultando a avaliação das massas moleculares que constituem a
peçonha, nossos resultados mostram um maior agrupamento de compostos com PIs
intermediários, e com massas molecular acima de 55kDa para as peçonhas de ambos os sexos de
A. natalensis. Infelizmente, no presente estágio deste trabalho não foi possível avaliar-se as
massas dos “spots”, por espectrometria de massas. O que se faz é a digestão enzimática dos
“spots” no gel, posterior coleta dos fragmentos protéicos e análise das massas por
espectrometria de massas.
29

Figura 6 – Perfil eletroforético do gel de poliacrilamida 15%(SDS-PAGE), da peçonha de A. natalensis e da toxina
isolada (AnTxI –Massa molecular de 5374 Da.). (A) Padrão de massas moleculares - kit da SIGMA “Recombinant
Molecular Weight Marker”SIGMA M0671 -(M.W. 15, 25, 35, 50, 75, 100 e 150 kDa- ”); (B) Peçonha da aranha
A.natalensis ; (C) Toxina (AnTxI). Cada amostra aplicada continha 30 µg da peçonha e da toxina.
PadrãoMassa Molecular
15 kDa
25 kDa
35 kDa
50 kDa
75 kDa100 kDa150 kDa
BA
30

Figura 7- Perfil eletroforético do gel bidimensional da peçonha de fêmeas de A. natalensis (50 microgramas). Os padrões moleculares foram: miosina (200 kDa), beta-galactosidase (116,3 kDa), Fosfolipase B (97,4 kDa), BSA (66,3 kDa), glutamato desidrogenase (55,4 kDa), lactato desidrogenase (36,5kDa), amidrase carbônica (31,0 kDa),inibidor de tripsina (21,5 kDa), lisozima (14,4 kDa), aprotinina (6,0 kDa), cadeia β da insulina (3,5 kDa), cadeia α da insulina ( 2,5 kDa)
PI
MW
Padrão
31

4.2 Cromatografia da peçonha de A. natalensis
A cromatografia foi realizada em duas etapas: cromatografia em troca catiônica e em fase
reversa foram escolhidas visando-se a obtenção da melhor resolução dos componentes da
peçonha de machos e de fêmeas de A. natalensis. A análise das duas peçonhas (machos e
fêmeas) por cromatografia de troca catiônica (CIEX) resultou na coleta de 32 picos para ambos
os sexos. O perfil cromatográfico demonstra que as peçonhas dos machos apresentaram uma
maior área e maior altura nas frações compreendidas entre os picos 04 e 12 (FIG.8). Todas as
frações coletadas (machos e fêmeas) foram re-fracionadas na segunda fase através de
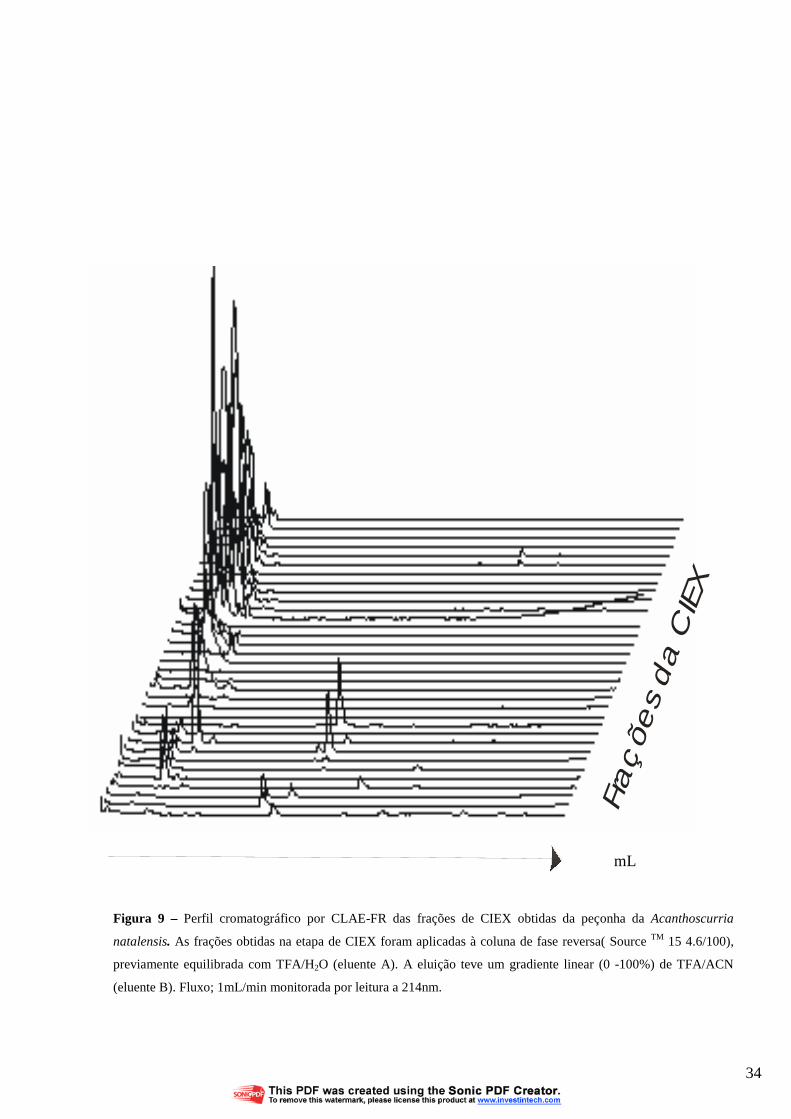
cromatografia de fase reversa (CLAE-FR). A grande maioria das frações resultantes da fase
reversa das duas peçonhas de A. natalensis (machos e fêmeas) demonstram baixa
hidrofobicidade, eluindo-se no inicio da corrida, entre 9 a 10ml do tampão de eluição (FIG. 9).
Em ambas as peçonhas (de machos e fêmeas) dois picos eluíram com 60ml de tampão de corrida
(FIG.9). Estes picos foram eluídos com maior grau de pureza (FIG.10). Como pode-se
perceber, apesar das poucas bandas apresentadas na eletroforese unidimensional a peçonha
apresenta uma grande quantidade de compostos, o que concorda com os resultados da
eletroforese 2D.
32

A
B
0 40 80 120
ABS
214
0
ABS
214
0
100
100
mL
%
%
Figura 8 – A CIEX foi desenvolvida em uma coluna TSK-Gel CM-SW 15cm x 4.6 mm(Tosoh Biosep) equilibrada
com 50mmol.L-1 de acetato de sódio pH 5, e fluxo de 0,75mL/min e gradiente linear de 0 a 1.0 mol/L de NaCl (1M-
pH 5) durante 123 minutos. Monitorado pela absorbância à 214 e 280nm, apresentando 32 picos demonstrando a
semelhança entre as Peçonhas. A linha vermelha representa a leitura da absorbância a 280 nm, a linha azul a
absorbância a 214 nm, a linha verde a concentração de NaCl e a marrom a condutância.
33
Fra
çõe
s d
a C
IEX
Figura 9 – Perfil cromatográfico por CLAE-FR das frações de CIEX obtidas da peçonha da Acanthoscurria
natalensis. As frações obtidas na etapa de CIEX foram aplicadas à coluna de fase reversa( Source TM 15 4.6/100),
previamente equilibrada com TFA/H2O (eluente A). A eluição teve um gradiente linear (0 -100%) de TFA/ACN
(eluente B). Fluxo; 1mL/min monitorada por leitura a 214nm.
mL
34

14 16 18 20 22 24 26 28 30 32 34 360
100
200
300
400
500
600
700
Eluição (mL)
Abs
214
nm
(m
AU
)
10
15
20
25
30
35
40
45
50
Gra
dien
te (
%B
)
Figura 10 - Perfil cromatográfico da toxina isolada AnTxI da A. natalensis em CLAE-FR. Aplicou-se 1mL da
fração 12 (vide figura anterior) onde foi encontrada a toxina, em uma coluna Source TM 15 4.6/100 (Pharmacia
Biotech, Uppsala, Sweeden) equilibrada com TFA 0,1%(solução A), eluída em um gradiente linear com
acetonitrila/TFA 0,1% (solução B) durante 50 minutos, e monitorado pela absorbância à A214.
35

4.3 Análise por Espectrometria de Massas
As frações re-purificadas da peçonha de A. natalensis (machos e fêmeas) pela
cromatografia de fase reversa, foram submetidas à análise por espectrometria de massa
(MALDI-TOF/TOF MS). Para todos os picos que apresentaram baixa hidrofobicidade não foi
possível detectar as massas moleculares correspondentes, por espectrometria de massa. Por outro
lado, para os dois picos eluídos mais tardiamente, as massas moleculares detectadas foram de
5374 Da e 6763 Da, para as duas peçonhas (machos e fêmeas). O grau de pureza das duas
toxinas foi verificado utilizando-se (MALDI-TOF/TOF MS) (FIG. 11), e pelo electrospray –
(ESI-Q-ToF) (dados não mostrados). As toxinas foram denominamos de AnTxI e AnTxII,
respectivamente, fazendo-se referência ao nome da espécie Acanthoscurria natalensis.
Entendemos que se faz necessário averiguar posteriormente, a razão pela qual os vários
componentes que apresentaram baixa hidrofobicidade aparentemente não foram ionizados
durante a análise de massas, não sendo assim detectados. Escoubas et al, 2006, trabalhando com
aranha Atrax rubustos, ordem Mygalomorphae que habitam a região da Austrália, obtiveram
toxinas com massa moleculares próximas às encontradas na peçonha de A. Natalensis. Estes
dados vêm reforçar mais uma vez, que o perfil da massa molecular das peçonhas de aranhas se
mantém com grande semelhança entre as Mygalomorphae.
36

Figura 11- Espectrometria de massas (MALDI-TOF/TOF MS) do peptídeos AnTxI e AnTxII isolados da
peçonha da A. natalensis .
ANTxI
ANTxII
5374 Da
6763 Da
37

4.4 Determinação da seqüência N-terminal do peptídio AnTxI
O peptídeo AnTxI foi clivado com tripsina e com V8 e os produtos resultantes foram
submetidos ao seqüenciamento pelo MALDI-TOF/TOF MS. O N-terminal de AnTxI foi
também seqüenciado pela técnica de degradação de Edman (TABELA 3 ). Após a determinação
da massa molecular (5374 Da), por espectrometria de massas, o peptídeo AnTxI foi seqüenciado
parcialmente, e apresentou os seguintes resíduos de aminoácidos:
IIECFFSCEIEKDGKSKEGKPCKPKG .
Este peptídeo teve sua seqüência comparada com outras depositadas em um banco de
dados (BLAST-P). Os resultados estão expressos na tabela 3, onde verifica-se que as
huwentoxinas (LIANG et al., 1993), toxinas encontradas na peçonha de certas aranhas chinesas,
Selenocosmia huwena foram os peptídeos que apresentaram similaridade de seqüência com a
AnTxI. . A toxina AnTxI teve sua sequência N-terminal similar com neurotoxinas como a
Huwentoxina-2 (HWTxII), ESTx1, ao TxP1 e o precursor das Toxina LTx2, HwTx II, HwTx-
VII . A toxina AnTxII apresentou sequência com similaridade às toxinas magi-15, a OcF32-8, e
à Phospholipase A2 (DIAO et al 2003; SHU & LIANG 1999; SAVEL-NIEMANN 1989;
VIEIRA et al. 2004; KAISER et al 1994; TSAI et al,2000; SATAKEA et al, 2004).
As toxinas Huwentoxina-2 e magi-15 apresentam ação paralisante reversível 6h após a
sua aplicação em insetos. A toxina LTx2, presente na peçonha das Lasiodoras sp (VIEIRA,A.L.,
et al. 2004), apresenta ação inibitória nos canais para cálcio e modulação dos canais para sódio.
Estas propriedades também foram observadas para a toxina estudada no presente trabalho (ver
adiante).
Pela comparação citada acima, as possíveis ações para AnTxI poderiam ser : i) inseticida;
ii) ação bloqueadora em canais para cálcio e/ou sódio, iii) aglutinação de lectina, iv) inibidor de
tripsina e v) paralisante, devido a ações no sistema nervoso central. Com tais informações,
iniciaram-se os testes biológicos “in vivo” em animais e `in vitro`em células, buscando-se
verificar as ações da peçonha e das toxinas isoladas.
38

Tabela 3- Alinhamento dos (cinco) fragmentos das toxinas isoladas (AnTxI e AnTxII) com toxinas
homólogas, depositadas em banco de dados ( BLAST-p)
proteína alinhamento Ação referência
AnTxI-Fragmento tripsina LLECFFSCELEK
Huwentoxin-2 form 1 precursor (HwTx-II) LFECSFSCEIEK Diao,J., et al,2003 Huwentoxin-7 precursor (HwTx-VII)
LFECSFSCEIEK
Diao,J., et al,2003 Huwentoxin-2 forma 2(HwTx-II)
LFECSFSCEQEK
Shu,Q. and Liang,S.P.,1999
AnTxI-N-terminal
IIECFFSCEIEKDGKSKEGKPCKPKG
Neurotoxina ESTx1
IFECVFSCDIE -----KEGKPCKPKG Savel-Niemann,A.1989 Toxina LTx2 precursor
ECTFECDI----K -KEGKPCKPKG
Vieira,A.L., et al. 2004 Veneno protein 1 (TXP1) IFECVFSCDIE -----KEGKPCKPKG Kaiser,I.I., et al1994 AnTxII -Fragmento 1
NLYQFKNMIQCTNTR
phospholipase A2
NLYQFKNMI KCTNTR
Tsai,H.Y., et al,2005
AnTxII-Fragmento 2
AE_ICATVYVGR
Neurotoxina magi-15
ATVICGTIYVGGKEEN Satakea, H., et al, 2004
AnTxII-Fragmento 3
CNNHCDCE
Neurotoxina Oc F32-8
39

4.5 Testes biológicos
4.5.1 Teste de toxicidade em insetos
Os testes biológicos foram realizados com as duas toxinas (AnTxI e AnTxII) entretanto
optamos por apresentar os dados referentes aos ensaios com a toxina AnTxI, por ter sido
possível fazer os mesmos em triplicadas devido a maior quantidade disponível desta toxina
purificada. Os testes de toxicidade em insetos foram realizados com a peçonha e com a toxina
purificada..
A peçonha foi diluída em salina contendo albumina (0,1%). A solução foi injetada por
via intratoráxica em moscas (Musca domestica) em diferentes doses (0,25 µg; 0,5 µg; 1 µg; e 2
µg/mosca). Observou-se ação paralisante da peçonha sobre os insetos com doses a partir de 1,0
µg/mosca. A paralisia foi reversível nas primeiras horas após a injeção da dose de 1,0 µg/mosca
e após 12h para a dose de 2,0 µg/mosca. Além da ação paralisante, as moscas apresentavam
espasmos musculares, com movimentação das patas de forma não coordenada, sugerindo ação
no sistema neuro-muscular.
A toxina AnTxI foi diluída na mesma solução (salina + albumina) e quando injetada em
moscas (Musca domestica) causou excitação, seguida de efeito paralisante reversível após as
primeiras horas da injeção. O efeito era notado a partir de 35 ng/mosca. 48 horas após a injeção
da maior dose (70 ng/mosca) ocorria letalidade acentuada. Neste caso observa-se um efeito
inseticida desta toxina, embora em doses bastante superiores a outras já conhecidas como, por
exemplo, a toxina AaH-IT do escorpião africano Androctonus australis Hector, cuja toxicidade
(DL50) é em torno de 2 ng (DE DIANOUS, et al,1987; LEGROS, et al 2005). Há que se
considerar, entretanto, que os testes utilizados para a AaH-IT referem-se à ação paralisante desta
toxina em larvas de moscas varejeiras (Spodopera frugiperda). Teste semelhante ao do presente
trabalho, com moscas adultas (M.domestica), foi utilizado para ensair toxinas obtidas do veneno
da aranha “armadeira” Phoneutia nigriventer. Neste caso, a toxicidade (DL50) da toxina TX4(6-
1) que mostrou propriedades inseticidas, foi de 3,8 ± 2 ng/mosca (FIGUEIREDO et al, 1995).
Esta toxina apresentou uma taxa de mortalidade alta com efeitos excitatórios semelhantes aos de
toxinas peptídicas presentes em peçonhas de escorpiões e de outras aranhas (ZLOTKIN, 1983;
ZLOTKIN et al., 1985; ADAMS et al., 1989). Comparando-se a toxina AnTxI com as toxinas
inseticidas de escorpião e da P. nigriventer, estas últimas apresentam-se mais potentes para
insetos. Como dito acima, efeitos letais da toxina em estudo, ocorreram com dose de 70
ng/mosca contra 3,8 ng para TX4(6.1) da aranha armadeira.
40

4.5.2 Testes eletrofisiológicos com a toxina AnTxI em canais para cálcio
voltagem-dependentes (Cavs)
Os ensaios com a toxina AnTxI foram realizados com neurônios do gânglio da raiz dorsal
(DRG) de ratos. Testes preliminares evidenciaram ação inibitória da toxina sobre as correntes de
sódio e de cálcio. A inibição da corrente de sódio foi baixa e não foi explorada neste trabalho.
Contudo, a inibição da corrente de cálcio foi bastante intensa com a perfusão de 100nM da
toxina (FIG. 12). Estes experimentos de isolamento da corrente para o cálcio foram realizados
com o devido cuidado, evitando-se a interferência do íon sódio nos registros.
A baixas doses da toxina (100nM) observou-se inibição imediata na corrente total
carreada pelos canais para cálcio, sensíveis à voltagem, indicando alta afinidade deste peptídeo
por estes canais. Nas figuras 12 e 13 verifica-se que apesar da reversão, após lavagem da
preparação, parte da corrente manteve-se inibida, indicando que a AnTxI mantém interações
fortes com parte dos sítios dos canais (responsáveis pelo seus comportamentos cinéticos), ou
com alguns canais específicos que compõem a corrente total, que não se desfez após a lavagem.
Conclui-se com estes resultados, que a toxina AnTxI causou inibição da corrente dos
canais para cálcio. Esse resultado corrobora com outros obtidos com a toxina LTx2, da peçonha
da aranha caranguejeira Lasiodoras sp, que apresenta ação inibitória nos canais para cálcio e
modulação nos canais para sódio (VIEIRA,A.L., et al. 2004). Essa atuação nos canais para
cálcio pode concorrer para explicar os espasmos (excitação) e a paralisia, apresentadas pelas
moscas que foram injetadas com esta toxina. Sabe-se que os canais para cálcio estão diretamente
envolvidos na contração de células musculares (WITHERS, 1992).
Embora as células (DRG) utilizadas nos estudos eletrofisiológicos tenham sido oriundas
de rato, Rattus norvegicus, outros estudos (em andamento por grupo colaborador) indicam ações
desta toxina em canais para cálcio e para sódio, em células de inseto (De Lima, comunicação
pessoal). Com os ensaios realizados nas células nervosas (DRG) de rato ainda não se pode
afirmar qual (quais) o(s) tipo(s) de canais para cálcio que são os alvos da toxina.
41

Tempo (s)
0 100 200 300 400
Cor
rent
e (p
A)
-1e-8
-8e-9
-6e-9
-4e-9
-2e-9
0
AnTxI (100nM) Lavagem
Figura 12 – Efeito no curso temporal da corrente (em pico Amperes, pA) após exposição à toxina AnTxI, na
concentração de 100 nM. O pico da corrente total carreada pelos canais para cálcio sensíveis à voltagem no
neurônio DRG, no período de estabilização do registro é representado por círculos pretos, por círculos vermelhos no
instante de exposição à toxina e por círculos azuis após lavagem, (quando a solução contendo a toxina era trocada
pela solução sem a toxina). Observa-se a reversão da maior parte da inibição da corrente causada pela toxina.
42

Figura 13 - Traçados representativos das correntes totais carreadas pelos canais para cálcio sensíveis à voltagem no
neurônio DRG. A linha de base, no período de estabilização da corrente (linha preta), o pico de inibição pela AnTxI
na concentração de 100nM (linha vermelha) e a corrente após lavagem (linha azul).
Tempo (ms)
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Cor
rent
e (A
)
-8e-9
-6e-9
-4e-9
-2e-9
0
2e-9
ControleAnTxI (100nM)Lavagem
(pA)
43

5 CONCLUSÕES
A peçonha da aranha caranguejeira A.natalensis representa uma fonte de substâncias,
com atividades biológicas, a grande maioria a ser explorada. A complexidade da peçonha é
evidenciada quando a mesma foi submetida à cromatografia e à eletroforese bidimensional, que
possibilitam melhor separação dos componentes protéicos demonstrando a presença de vários
picos e `spots`, rescpetivamente. Pela cromatografia de troca iônica observam-se trinta e dois
picos, tanto em peçonha de espécimens machos como de fêmeas, mostrando que não existem
diferenças aparentes do ponto de vista qualitativo, quanto ao gênero. Entretanto, diferenças
quantitativas foram evidentes em algumas regiões, por exemplo, naquela dos picos de 4 a 12. É
possível ainda, que existam outras diferenças que não foram evidenciadas por esta metodologia.
A peçonha, quando injetada em mosca doméstica (Musca domestica), mostrou atividade
de excitação seguida de paralisia nestes insetos. Em doses mais baixas, como 0,25 µg e 0,5 µg, o
efeito da peçonha foi reversível. Doses maiores, como 2 µg, foram letais.
Os experimentos envolvendo cromatografia de troca iônica seguida de cromatografia de
fase reversa mostraram-se eficientes para a separação de toxinas da peçonha da A. natalensis. Na
cromatografia de fase reversa a quase totalidade dos componentes é eluída no início do processo,
mostrando a baixa hidrofobicidade dos mesmos. Com essa metodologia foram isoladas duas
toxinas de baixa massa molecular, a ANTxI com 5374Da. e a ANTxII com 6763 Da. Por
espectrometria de massas não foram detectados outros componentes, o que deverá ainda ser
averiguado. Ambas as toxinas ANTxI e a ANTxII foram testadas em moscas e apresentaram
toxicidade na ordem de nanogramos. A dose de 70ng por mosca foi a maior dose aplicada e os
efeitos observados foram idênticos aos obtidos quando da aplicação de doses maiores de
peçonha. Verificou-se portanto, que tanto a peçonha como as duas toxinas isoladas apresentam
efeitos tóxicos em moscas, sendo as toxinas mais potentes.
A toxina AnTxI teve sua seqüência N-terminal determinada
(IIECFFSCEIEKDGKSKEGKPCKPKG ) e, quando esta foi comparada a outras
sequências de toxinas depositadas em um banco de dados, mostrou homologia com toxinas
ativas em canais para cálcio e para sódio. Ensaios eletrofisiológicos (patch clamp) em neurônios
de gânglio dorsal de rato mostraram que a toxina bloqueou a corrente para cálcio destas células,
em doses nanomolares, portanto apresentando alta afinidade. Este processo é parcialmente
44

reversível com a lavagem da preparação. A toxina também apresentou bloqueio sobre a corrente
para sódio tanto nas células DRG de rato, como em céluas de inseto (De Lima, com.pessoal).
Em conclusão, a toxina obtida em estado puro neste trabalho, mostra ação mais seletiva
em canais para cálcio, o que poderia relacionar-se à sua atividade excitatória e paralisante em
insetos. A ação em canais para sódio parece menos potente e deve ainda ser investigada.
Acreditamos que esta toxina possa vir a ser uma ferramenta farmacológica importante
para o estudo de canais para cálcio voltagem dependentes. Estes estudos, quando concluídos,
contribuirão para avaliar o real potencial desta toxina em possíveis aplicações em saúde e ou
biotecnologia. Por outro lado, os demais componentes da peçonha também representam um
valioso material a ser estudado.
45

Perspectivas futuras
Este trabalho representa uma primeira avaliação da peçonha de A. natalensis, que ocorre
na região do semi-árido baiano. Entendemos que pontos fundamentais para se conhecer melhor
os componentes desta peçonha necessitam de continuidade, como: .
(i) concluir o seqüênciamento das toxinas isoladas;
(ii) concluir os estudos eletrofisiológicos com as toxinas isoladas, verificando seus
possíveis efeitos em diferentes sub-tipos de canais para cálcio e s;
(iii) avaliar a ação das toxinas isoladas em canais para sódio;
(iv) separar e analisar (bioquímica e farmacologicamente) os demais componentes desta
peçonha;
(v) verificar quais os compostos e/ou toxinas que podem ser utilizados como um provável
produto biotecnológico, para a realização de ensaios específicos.
46

6 REFERÊNCIAS BIBLIOGRÁFICAS
____, http://research.amnh.org/entomology/spiders/catalog/COUNTS.html The World Spider Catalog, Version 8.5 by Norman I. Platnick , © Copyright 2000 — 2008 American Museum of Natural History. All Rights Reserved, Acesso data- 23/02/2008 _____, www.birdspiders.com/archive/15B036F2KD0B7KAEC2K14B24A24C046092C.html
___, Fonte: http://www.socanestesia.cl/rev_anestesia/0406/images/im03.jpg
ADAMS, M. E., HEROLD, E. E. AND VENENA, V. J. Two classes of channel-specific toxins from funnel web spider venom. J. camp. Physiol. n. A164, p. 333-342, 1989. BALAJI, R.A., SASAKI, T., GOPALAKRISHNAKONE, P., SATO, K., KINI, R.M., BAY, B.H.,. Purification, structure determination and synthesis of covalitoxin-II, a short insect-specific neurotoxic peptide from the venom of the Coremiocnemis validus (Singapore tarantula). FEBS Lett . v. 474, p. 208–212. 2000 BARRAVIEIRA, B. ,Venenos Animais: Uma visão integrada. Rio de Janeiro; Ed. De Publicações Científicas, p.441,1994 BIRNBAUMER, L.;CAMPBELL, K.P.; CATTERALL, W.A.; HARPOLD, M.M.; HOFMANN, F.; HORNE, W.A.; MORI, Y.; SCHWARTZ, A.; SNUTCH, T.P.; TANABE, T., The naming of voltage-gated calcium channels. Neuron. v.13, n.3, p.505-506, 1994. BRASIL, V.; VELLARD, J. Contribuição ao estudo do veneno das aranhas-aranhas inimigas das serpents-gênero Grammostola.Rio de Janeiro: Brazil-Medico, v. 39,n.1, p. 47-51. 1925 BU¨CHERL, W.; SPIDERS. IN: BU¨CHERL, W., BUCKLEY, E.E. (Eds.), Venomous Animals and their Venoms, vol. III. Academic Press,New York, pp. 197–277. 1971. CARBONE, E. & LUX, H. D., A low voltagem-activated fully incectivatins Ca+2 channel in vertebrate sensory neurons. Nature, v. 310, p. 501-502, 1984. CATTERALL, W. A.; CESTÈ LE S.; YAROV-YAROVOY, V.; YU, F. H.; KONOKI, K.; SCHEUER , T. Voltage-gated ion channels and gating modifier toxins. Toxicon v.49, p. 124–141; 2007 CHIEN, A.J.; ZHAO, X.; SHIROKOV, R.E.; PURI, T.S.; CHANG, C.F.; SUN, D.; RIOS, E.; HOSEY, M.M., Roles of a membrane-localized beta subunit in the formation and targeting of functional L-type Ca2+ channels. J Biol Chem. v.270, n.50, p.30036-30044, 1995. CODDINGTON, J. A. & LEVI, H. W.. Systematics and evolution of spiders(Araneae). Annu. Rev. Ecol. Syst. V.22, p.565-592. 1991.
47

DE DIANOUS, S.,HOARAU, F.,ROCHAT,H. Re-examination of the specificity of the scorpion Androctonus australis Hector insect toxin towards arthropods. Toxicon V.25, n.4, p.411-417, 1987.
DE LIMA,M.E.; MARTIN,M.F.; DINIZ,C.R.; ROCHAT,H. Tityus serrulatus toxin VII bears pharmacological properties of both beta-toxin and insect toxin from scorpion venoms. Biochem.Biophys.Res.Commun. v. 139, p. 296-302.1986. DE LIMA,M.E.; COURAUD,F.; LAPIED,B.; PELHATE,M.; RIBEIRO,DINIZ C.; ROCHAT,H. Photoaffinity labeling of scorpion toxin receptors associated with insect synaptosomal Na+ channels. Biochem.Biophys.Res.Commun. v. 151, p. 187-192. 1988. DE LIMA,M.E.; STANKIEWICZ,M.; HAMON,A.; DE FIGUEIREDO,S.G.; CORDEIRO,M.N.; DINIZ,C.R.; MARTIN-EAUCLAIRE,M.; PELHATE,M., The toxin Tx4(6-1) from the spider Phoneutria nigriventer slows down Na(+) current inactivation in insect CNS via binding to receptor site 3. J.Insect Physiol. v.48, p. 53-61, 2002 DE LIMA,M.E.; FIGUEIREDO,S.G.; PIMENTA,A.M.; SANTOS,D.M.; BORGES,M.H.; CORDEIRO,M.N.; RICHARDSON,M.; OLIVEIRA,L.C.; STANKIEWICZ,M.; PELHATE,M. Peptides of arachnid venoms with insecticidal activity targeting sodium channels. Comp Biochem.Physiol C.Toxicol.Pharmacol. v.146, p. 264-279, 2007. DIAO,J., LIN,Y., TANG,J. AND LIANG,S. cDNA sequence analysis of seven peptide toxins from the spider Selenocosmia huwena. Toxicon v.42,n.7,p. 715-723,2003. DIOCHOT, S., DRICI, M.D., MOINIER, D., FINK, M., LAZDUNSKI, M., Effects of phrixotoxins on the Kv4 family of potassium channels and implications for the role of Ito1 in cardiac electrogenesis. Br. J. Pharmacol. v. 126, p . 251–263. 1999. DUNLAP, K., LUEBKE, J. I., AND TURNER, T. Exocytotic Ca 2+ channels in mammalian neurons. Trends Neurosci. v.18, p. 89–98, 1995. ESCOUBAS, P., CÉLÉRIER, M.L., NAKAJIMA, T.,. High-performance liquid chromatography matrix-assisted laser desorption/ ionization time-of-flight mass spectrometry peptide fingerprinting of tarantula venoms in the genus Brachypelma: chemotaxonomic and biochemical applications. Rapid Commun. Mass Spectrom. v. 11, p. 1891–1899. 1997a ESCOUBAS, P., CÉLÉRIER, M.L., ROMI-LEBRUN, R., NAKAJIMA, T., Two novel peptide neurotoxins from the venom of the tarantula Lasiodora parahybana. Toxicon. v. 35, p . 805–806. 1997b. ESCOUBAS, P., CHAMOT-ROOKE, J., STO¨CKLIN, R., WHITELEY, B.J., CORZO, G., GENET, R., NAKAJIMA, T.,. A comparison of matrix-assisted laser desorption/ionization time-of-flight and liquid chromatography electrospray ionization mass spectrometry methods for the analysis of crude tarantula venoms in the Pterinochilus group. Rapid Commun. Mass Spectrom. v. 13, p. 1861–1868. 1999. ESCOUBAS, P., DE WEILLE, J.R., LECOQ, A., DIOCHOT, S., WALDMANN, R., CHAMPIGNY, G., MOINIER, D., ME´NEZ, A., LAZDUNSKI, M.,. Isolation of a tarantula toxin specific for a class of proton-gated Naþ channels. J. Biol. Chem. v. 275, p. 25116–25121. 2000a
48

ESCOUBAS, P., DIOCHOT, S., CORZO, G.,. Structure and pharmacology of spider venom neurotoxins. Biochimie. v. 82, p. 893–907. 2000b ESCOUBAS, P., CORZO, G., WHITELEY, B.J., CÉLÉRIER, M.L., NAKAJIMA, T., Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry and high-performance liquid chromatography study of quantitative and qualitative variation in tarantula spider venoms. Rapid Commun. Mass Spectrom. v. 16, p .403–413. 2002a. ESCOUBAS, P., DIOCHOT, S., CÉLÉRIER, M.L., NAKAJIMA, T., LAZDUNSKI, M.,. Novel tarantula toxins for subtypes of voltagedependent potassium channels in the Kv2 and Kv4 subfamilies. Mol. Pharmacol. v. 62, p. 48–57. 2002b ESCOUBAS, P., BERNARD, C., LAMBEAU, G., LAZDUNSKI, M., DARBON, H., Recombinant production and solution structure of PcTx1, the specific peptide inhibitor of ASIC1a proton-gated channels. Protein Sci. v. 12, p. 1332–1343. 2003. ESCOUBAS, P., RASH, L. Tarantulas: eight-legged pharmacists and combinatorial chemists. Toxicon, v. 43, p. 555–574. 2004. ESCOUBAS, P. Molecular diversification in spider venoms: A web of combinatorial peptide libraries. Springer, V.10, P.545-554, 2006. ERTEL, E.A; CAMPBELL, K.P.; HARPOLD, M.M.; HOFMANN, F.; MORI, Y.; PEREZ-REYES, E.; SCHWARTZ, A.; SNUTCH, T.P.; TANABE, T.; BIRNBAUMER, L.; TSIEN, R.W.; CATTERALL, W.A. Nomenclature of voltage-gated calcium channels. Neuron. v.25, n.3, p.533-535, 2000 FATT, P. & KATZ, B. The electrical properties of crustacean muscle fibres. J. Physiol, V. 120, p. 171-204, 1953. FIGUEIREDO, S. G.; GARCIA, M. E.; VALENTIM, A. C., CORSEIRO, M. N., DINIZ, C. R.; RICHARDSON, M., Purification and amino acid sequence of the insecticidal neurotoxin Tx4 (6-1) from the venom of the “armed” spider Phoneutria nigriventer (Keys). Toxicon, v. 33, p. 83-93, 1995. FIGUEIREDO, S.G.; DE LIMA M.E.; NASCIMENTO CORDEIRO. M.; DINIZ, C.R.; PATTEN, D.; HALLIWELL, R.F.; GILROY, J.; RICHARDSON, M., Purification and amino acid sequence of a highly insecticidal toxin from the venom of the brazilian spider Phoneutria nigriventer which inhibits NMDA-evoked currents in rat hippocampal neurones. Toxicon. v.39, n. 2-3, p.309-317, 2001 FOELIX, S. D. 1996. Biology of spiders. Oxford University Press, inc. FRONTALI, N., GRASSO, A., Separation of three toxicologically different protein components from the venom of the spider Latrodectus tredecimguttatus. Arch. Biochem. Biophys. V.106, p. 213–218, 1964. FRIEDEL,T., NENTWIG, W. Immobilizing and lethal effects of spider venoms on the cockroach and commom mealbeetle. Toxicon, v.27p. 305-316
49

GOMEZ. M.V., KALAPOTHAKIS E., GUATIMOSIM C., AND PRADO, M.A. Phoneutria nigriventer venom: a cocktail of toxins that affect ion channels. Cell Mol Neurobiol V.22, p.579–588, 2002. GOUVÊA DOS SANTOS,R., Estudo do mecanismo de ação de ω-phonetoxina IIA da peçonha de aranha Phoneutria nigriventes, Tese de doutorado, Departamento de Imunologia e Bioquímica, -ICB-UFMG, 2002. GRISHIN, E. Polypeptide neurotoxins from spider venoms. European Journal of Biochemistry. v. 264, p. 276-280, 1999. HEDIN, M., BOND, J. E. Molecular phylogenetics of the spider infraorder Mygalomorphae using nuclear rRNA genes( 18S and 28S) ;Conflict and agreement with the current system of classification. Molecular Phylogenetics and evolution, v. 41, p. 454-471, 2006 HERZING, V. , WARD, R. J. SANTOS, W. F. Ontogenetic changes in Phoneutria nigriventer(Araneae, Ctenidae) spider venom. Toxicon, v. 44, p. 635-640, 2004 HIDAI, Y., KAN, T., FUHUYAMA, T. Total synthesis of polyamine toxin HO-416b utilizing the 2-nitrobenzenesulfonamide protecting group. Tetrahedron Letters, v. 40, p. 4711-4714. 1999. HILLE, B. Ion channels of excitable cells. 3. Ed, Sinauer Associates: Sunderland, Massachusetts, p.83-114, 1992 HOFMANN, F.; ANAGLI, J.; CARAFOLI, E.; VORHERR, T., Phosphorylation of the calmodulin binding domain of the plasma membrane Ca2+ pump by protein kinase C reduces its interaction with calmodulin and with its pump receptor site. J Biol Chem. v.269, n.39, p.24298-24303,1994. KAISER, I.I., GRIFFIN, P.R., AIRD, S.D., HUDIBURG, S., SHABANOWITZ, J., FRANCIS, B., JOHN, T.R., HUNT, D.F., ODELL, G.V., Primary structures of two proteins from the venom of the Mexican red knee tarantula (Brachypelma smithii). Toxicon. v. 32, p. 1083–1093. 1994. KALAPOTHAKIS, E., KUSHMERICK, C., GUSMÃO, D.R., FAVARON, G.O., FERREIRA, A.J., GOMEZ, M.V., PINTO DE ALMEIDA, A. Effects of the venom of a Mygalomorph spider (Lasiodora sp.) on the isolated rat heart. Toxicon. v.41, n.1, p. 23-28,2003. KAN, T. et al. Efficient macrocyclization by means of 2-nitrobenzenesulfonamine and total synthesis of lipogrammistin-A. Tetrahedron Letters, v. 58, p. 6267-6276. 2002. KAWAI, N., MIWA, A., ABE, T.,. Effect of a spider venom toxin on glutaminergic synapses in mammalian brain. Biomed. Res. v.3, p. 353–355. 1982. KUSHMERICK, C., MESQUITA DE CARVALHO, F., DE MARIA, M., MASSENSIN,I A.R., ROMANO-SILVA, M.A., GOMEZ, M.V., KALAPOTHAKIS E., PRADO M.A., Effects of a Lasiodora spider venom on Ca2+ and Na+ channels. Toxicon. v.39, n.7., p.991-1002. 2001
50

LAEMMLI, U. K. , Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4, Nature v.227, p. 680 - 685 ,1970
LAMPE, R.A., DEFEO, P.A., DAVISON, M.D., YOUNG, J., HERMAN, J.L., SPREEN, R.C., HORN, M.B., MANGANO, T.J., KEITH, R.A.,.Isolation and pharmacological characterization of omegagrammotoxin SIA, a novel peptide inhibitor of neuronal voltage-sensitive calcium channel responses. Mol. Pharmacol. v. 44, p. 451–460. 1993. LAMPE, R.A. Analgesic peptides from venom of Grammostola spatulata and use thereof. US Patent 5,877,026. 1999. LEGROS, C.; CÉARD, B. ; VACHER, H.; MARCHOT, P.;BOUGIS, P.E.; MARTIN-EAUCLAIRE, M., Expression of the standard scorpion alpha-toxin AaH II and AaH II mutants leading to the identification of some key bioactive elements, Biochimica et Biophysica Acta . v.1723, p. 91– 99, 2005. LIANG, S.P., ZHANG, D.Y., PAN, X., CHEN, Q., ZHOU, P.A., Properties and amino acid sequence of huwentoxin-I, a neurotoxin purified from the venom of the Chinese bird spider Selenocosmia huwena. Toxicon. v. 31, p. 969–978. 1993. LIANG, S.P., PAN, X. A lectin-like peptide isolated from the venom of the Chinese bird spider Selenocosmia huwena. Toxicon. v. 33, p. 875–882. 1995. LIU, Z., DAI, J., CHEN, Z., HU, W., XIAO, Y., LIANG, S., Isolation and characterization of hainantoxin-IV, a novel antagonist of tetrodotoxin-sensitive sodium channels from the Chinese bird spider Selenocosmia hainana. Cell. Mol. Life Sci. v. 60, p. 972–978. 2003. LORENZINI,D. M., SILVA Jr, P. I., FOGAÇA, A. C., BULET,P. DAFFRE, S. ,Achantoscurrin: a novel glycine-rich ntimicrobial peptide constitutively expressed in the hemocytes of the spider Acanthoscurria gomesiana. Developmental and Comparative Immunology v.27, p.781-791,2003 MARVIN, L., DE, E., COSETTE, P., GAGNON, J., MOLLE, G., LANGE, C., Isolation, amino acid sequence and functional assays of SGTx1. The first toxin purified from the venom of the spider Scodra griseipes. Eur. J. Biochem. v. 265, p. 572–579. 1999. MIDDLETON, R.E., WARREN, V.A., KRAUS, R.L., HWANG, J.C., LIU, C.J., DAI, G., BROCHU, R.M., KOHLER, M.G., GAO, Y.D., GARSKY, V.M., BOGUSKY, M.J., MEHL, J.T., COHEN, C.J., SMITH, M.M., Two tarantula peptides inhibit activation of multiple sodium channels. Biochemistry v. 41, p. 14734–14747. 2002. MILJANICH, G. P. & RAMACHANDRAN, J. Antagonists of neuronal calcium channels: structure, function and therapeutic implications. Annu. Rev. Pharmacol. Toxicol. 35: 707–734, 1995. MINTZ, I.M.; ADAMS, M.E.; BEAN, B.P., P-type calcium channels in rat central and peripheral neurons. Neuron. v.9, n.1,p.85-95,1992. NEWCOMB, R., SZOKE, B., PALMA, A., WANG, G., CHEN, X., HOPKINS, W., CONG, R., MILLER, J., URGE, L., TARCZY-HORNOCH, K., LOO,J.A., DOOLEY, D.J., NADASDI, L.,
51

TSIEN, R.W., LEMOS, J., MILJANICH, G., Selective peptide antagonist of the class E calcium channel from the venom of the tarantula Hysterocrates gigas. Biochemistry v. 37, p. 15353–15362. 1998. NOWYCKY, M.C.; FOX, A.P.; TSIEN, R.W., Three types of neuronal calcium channel with different calcium agonist sensitivity. Nature. v.316, n.6027, p.440-443., 1985 NUNES, E.A, BONATTO, S.J, DE OLIVEIRA. H.H., RIVERA N.L., MAIORKA, A., KRABBE, E.L., TANHOFFER, R.A., FERNANDES, L.C., The effect of dietary supplementation with 9-cis:12-trans and 10-trans:12-cis conjugated linoleic acid (CLA) for nine months on serum cholesterol, lymphocyte proliferation and polymorphonuclear cells function in Beagle dogs. Res Vet Sci. v. 84, n.1, p.62-67, 2008. O’FARRELL, P. H. High resolution two-dimensional electrophoresis of proteins. J. Biol. Chem. v. 250, p. 4007–4021 ,1975. OLIVERA, B. M., MILJANICH, G. P., RAMACHANDRAN, J., & ADAMS, M. E. Calcium channel diversity and neurotransmitter release: the v-conotoxins and v-agatoxins. Annu. Rev. Biochem. 63: 823–867, 1994. OLIVEIRA, L.C., DE LIMA, M.E., PIMENTA, A.M., MANSUELLE, P., ROCHAT, H., CORDEIRO, M.N., RICHARDSON, M., FIGUEIREDO, S.G. PnTx4-3, a new insect toxin from Phoneutria nigriventer venom elicits the glutamate uptake inhibition exhibited by PhTx4 toxic fraction. Toxicon. v. 42, p. 793–800. 2003. OSWALD, R.E., SUCHYNA, T.M., MCFEETERS, R., GOTTLIEB, P., SACHS, F., Solution structure of peptide toxins that block mechanosensitive ion channels. J. Biol. Chem. v. 277, p. 34443–34450. 2002. PENG K.; CHEN X.D.; LIANG S.P. The effect of Huwentoxin-I on Ca2+ channels in differentiated NG108-15 cells, a patch-clamp study. Toxicon. V. 39, n. 4, p. 491-498(8). 2001 . PENG, K., SHU, Q., LIU, Z., LIANG, S., Function and solution structure of Huwentoxin-IV, a potent neuronal tetrodotoxin (TTX)-sensitive sodium channel antagonist from Chinese bird spider Selenocosmia huwena. J. Biol. Chem. v. 277, p. 47564–47571. 2002. PEREIRA, L. S., SILVA JR. P. I. , MIRANDA, T. M. M. , ALMEIDA, I.C. ,NAOKI, H. , KONNO, K. , DAFFRE, S., Structural and biological characterization of one antibacterial acylpolyamine isolated from the hemocytes of the spider Acanthocurria gomesiana . Biochemical and Biophysical Research Communications v.352, p. 953–959, 2007. PIMENTA, A.M., STO¨CKLIN, R., FAVREAU, P., BOUGIS, P.E., MARTIN-EAUCLAIRE, M.F. Moving pieces in a proteomic puzzle: mass fingerprinting of toxic fractions from the venom of Tityus serrulatus (Scorpiones, Buthidae). Rapid Commun. Mass Spectrom. v. 15, p. 1562–1572. 2001. PRAGNELLE, M.; De WAARD, M.; MORE, Y.; TANABE, T.; SNUTCH, T. D.; CAMPBELL, K. P. , Calcium channel β-subunit binds to a conserved motif in the I-II cytoplasmic linker of the α1-subunit. Nature . v.368, n.6466, p. 67-70,1994.
52

PRETEL, F., GONÇALVES-DE-ANDRADE, R. M., MAGNOLI, F. C., da SILVA, M. E. R., FERREIRA Jr., J. M. C., VAN DEN BERG, C. W., TAMBOURGI, D. V., Analysis of the toxic potential of venom from Loxosceles adelaida, a Brazilian brown from karstic areas. Toxicon, n. 45, p. 449-458,2005 RASH, L.D., HODGSON, W.C.,. Pharmacology and biochemistry of spider venoms. Toxicon. v. 40, p. 225–254. 2002. RAVEN, R., The spider infraorder Mygalomorphae (Araneae). Bull. Am. Mus. Nat. Hist. v. 182, p. 1–180. 1985. RICHARDSON, M., PIMENTA, A.M.C., BEMQUERER, M.P., SANTORO, M.M., BEIRÃO, P.S.L., LIMA, M. E., FIGUEIREDO, S. G., BLOCH Jr, C., VASCONCELOS, E.A.R., CAMPOS, F.A.P., GOMES, P.C., CORDEIRO, M.N., Comparison of the partial proteomeos of venoms of Brazilian spiders of the genus Phoneutria, Comparative Biochemistry and Physiology, parte C, n.142, p. 173-187, 2006. RUTA, V., JIANG, Y., LEE, A., CHEN, J., MACKINNON, R.,.Functional analysis of an archaebacterial voltage-dependent Kþ channel. Nature v. 422, p. 180–185. 2003. SATAKE, H., VILLEGAS,E., OSHIRO,N., TERADA, K., SHINADA, T., CORZO,G.,. Rapid and efficient identification of cysteine-rich peptides by random screening of a venom gland cDNA library from the hexathelid spider Macrothele gigas. Toxicon , v.44, p. 149–156, 2004. SAVEL-NIEMANN, A., Tarantula (Eurypelma californicum) venom, a multicomponent system. Biol. Chem. Hoppe Seyler . v. 370, p. 485–498. 1989. SENFF-RIBEIRO, A. , SILVA, P. H., CHAIM, O. M., GREMSKI, L. H., PALUDO, K. S., SILVEIRA, R. B., GREMSKI, W., MANGILI, O. C., VEIGA, S. S. , Biotecnological applications of brown spider (Loxosceles genus) venom toxins. Biotechnology Advences, n. 26, p. 210-218. 2008 SOERENSEN, B. Acidentes com Animais Peçonhentos, Editora Atheneu, São Paulo, 1997
SHU, Q., LIANG, S.P. Purification and characterization of huwentoxin-II, a neurotoxic peptide from the venom of the Chinese bird spider Selenocosmia huwena. J. Pept. Res. v. 53, p. 486–491. 1999. SHU, Q., HUANG, R., LIANG, S., Assignment of the disulfide bonds of huwentoxin-II by Edman degradation sequencing and stepwise thiol modification. Eur. J. Biochem. v. 268, p.2301–2307. 2001. SHU, Q., LU, S.Y., GU, X.C., LIANG, S.P., The structure of spider toxin huwentoxin-II with unique disulfide linkage: evidence for structural evolution. Protein Sci. v. 11, p. 245–252. 2002. SUCHYNA, T.M., JOHNSON, J.H., HAMER, K., LEYKAM, J.F., GAGE, D.A., CLEMO, H.F., BAUMGARTEN, C.M., SACHS, F., Identification of a peptide toxin from Grammostola spatulata spider venom that blocks cation-selective stretch-activated channels. J. Gen. Physiol. v.115, p. 583–598. 2000.
53

SWARTZ, K. J. Tarantula toxins interacting wint voltage sensors in potassium channels. Toxicon, v. 49, p. 213-312. 2004. VERANO-BRAGA, T., ROCHA-RESENDE, C., SILVA, D.M., IANZER, D., MARTIN-EAUCLAIRE, M.F., BOUGIS, P.E., DE LIMA, M.E., SANTOS, R.A., PIMENTA, A.M., Tityus serrulatus Hypotensins: a new family of peptides from scorpion venom. Biochem Biophys Res Commun. v.371, n. 3 , p.515-520, 2008 VIEIRA,A.L.,MOURA,M.B.,BABA,E.H.,CHAVEZ-OLORTEGUI,C., KALAPOTHAKIS,E. & CASTRO,I.M. Molecular cloning of toxins expressed by the venom gland of Lasiodora sp. Toxicon v.44 , n.8, p. 949-952, 2004. TAREILUS, E.; ROUX, M.; QIN, N.; OLCESE, R.; ZHOU, J.; STEFANI, E.; BIRNBAUMER, L., A Xenopus oocyte β subunit: Evidence for a role in the assembly/expression of voltage-gated calcium channels that is separate from its role as aregulatorysubunit. Proc Natl Acad Sci U S A. v.94, n.5,p.1703-1708,1997 TSAI,H.Y., TSAI,I.H. & WANG,Y.M., Characterization and sequencing of venom proteins of Micrurus f.fulvius subimetido ao Biological Chemistry , Academia Sinica, P.O. Box 23-106, Taipei, Taiwan 10798, Republic of China em 21-maio-2005 XIAO, Y.C., LIANG, S.P., Purification and characterization of Hainantoxin-V, a tetrodotoxin-sensitive sodium channel inhibitor from the venom of the spider Selenocosmia hainana. Toxicon. v. 41, p. 643–650. 2003. WALKER, J. M., The Protein Protocols Handbook, University of Hertfordshire, Hatfield, UK, 2ª Ed.,Editora Humana Press Totowa, New Jersey, 2002 WEBER, K., & OSBORN, M., J. Biol. Chem., v.244, p. 4406 ,1969. WICHER, D. & PENZLIN, H., Ca2+ Currents in Central Insect Neurons: Electrophysiological and Pharmacological Properties. J Neurophysiol, v. 77, p. 186-199, 1997; ZLOTKIN, E. Insect selective toxins derived from scorpion venoms: an approach to insect neuropharmacology. Insect Biochem. n.13, p.219-236, 1983. ZLOTKIN, E., KADOURI, D., GORDON, D., PELHATE, M., MARTIN, M. F. AND ROCHAT, H. An excitatory and a depressant insect toxin from scorpion venom both affect sodium conductance and possess a common binding site. Arch. Biochem. Biophys. n.240, p. 877-887,1985.
54

ANEXOS
FORMULAÇÃO PARA A ELETROFORESE EM
POLIACRILAMIDA-SDS (SDS-PAGE)
tampão da amostra : 2 mL de SDS à 10%, 500 µL de 2 –mercaptoetanol, 2 mL de
glicerol, 400 µL de Azul de bromofenol à 2%, 2,5 mL de Tris-HCl(0,5 M-pH 6,8), e 10
mL de água destilada e deionizada.
I-Gel de corrida à 15%: 12,5mL de acrilamida 30%; 6,25mL de TRIS-HCl -1,5M ,pH-
8,8; 250µL de dodecil sulfato de sódio à 10%; 5,85 mL de água tipo II(deionizada e
destilada) ; 125 µL de persulfato de amônio; 25 µL de TEMED,
II- Gel de concentração à 5%: 0,834 mL de acrilamida 30%; 1,25mL de tris-HCl -
1,5M ,Ph - 6,8; 50µL de Dodecil Sulfato de Sódio à 10%; 2,83 mL de água ; 25 µL de
persulfato de amônio; 10 µL de TEMED.
Solução de prata amoniacal: solução A- 15mL de NaOH e 0,5 mL de NH4OH e
Solução B- 0,3 g de AgNO3 e 5mL de água deionizada completando a solução com
água deionizada até completar 50 mL.
Pré-fixado por 1 hora no pré-fixador I contendo uma solução de 50% de etanol e 7,5%
de ácido acético.
Pré-fixador II com uma solução de etanol 5% e ácido acético 7,5% .
Procedimento: Após a corrida o gel era submetido à coloração com prata sendo pré-
fixado por 1 hora no pré-fixador I, posteriormente foi pré-fixado por mais 1 hora no
pré-fixador II, feita está prefixação foram realizadas lavagens no gel para a fixação
com o glutaraldeido a 10% durante 30 minutos. Depois da fixação o gel foi novamente
lavado até a extração do excesso do glutaraldeído e incubado com a solução de prata
amoniacal. Em seguida o gel foi lavado com uma solução de 50 µL de ácido cítrico
(2,3M), 200 µL Formaldeído ( 38% em 200 mL de água deionizada) e inativado com
ácido acético (1% v/v).
55

FORMULAÇÃO PARA A SOLUÇÃO DE HANK'S
A composição de HBSS é em mM: NaCl 137,93 (J.J. Baker, México); KH2PO4 0,44
(Merck, Darmstadt, Alemanha); KCl 5,33 (Sigma, St. Louis, Mo, USA); NaHCO3 4 mM
(Sigma, St. Louis, Mo, USA); Na2HPO4 0,3 (Sigma, St. Louis, Mo, USA); Glucose 5,6 (Sigma,
St. Louis, Mo, USA); CaCl2 1,8 mM (Sigma, St. Louis, Mo, USA); MgCl2 1 mM (Labsynth,
Diadema / SP, Brasil) ajustado a pH 7,4 com NaOH.
FORMULAÇÃO PARA O EXPERIMENTO DE “PATCH-CLAMP”
solução interna de pipeta (SI-20 mM de TEA-Cl, 100 mM de CsCl, 2 mM de MgCl2,10
mM de HEPES, 11 mM de EGTA, 1 mM de CaCl2 ,4 mM de ATP-Mg, 100 mM de
GTP-Li, pH= 7,2, ajustado com CsOH ),
solução externa I ou solução de Tyrode (SE1- 130 mM de NaCl, 5 mM de KCl, 2 mM de
CaCl2, 1 mM de MgCl2 ,10 mM de HEPES e 10 mM de Glicose, pH=7,4 ajustado com
NaOH )
solução externa II ou solução externa para isolar a corrente total de Ca2+ (SE2-35 mM de
TEA-Cl, 100 mM de cholina-Cl, 10 mM de CsCl, 0,8 mM de MgCl2 , 10mM de HEPES,
2 mM de CaCl2 , 10 mM de Glicose, ph 7,4, ajustado com CsOH) .
56