universidade de lisboarepositorio.ul.pt/bitstream/10451/6211/1/ulsd062640_td_pedro_fale.pdf ·...
TRANSCRIPT

UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE QUÍMICA E BIOQUÍMICA
BIOLOGICAL ACTIVITIES OF Plectranthus barbatus
AQUEOUS EXTRACTS.
IN VITRO AND IN VIVO STUDIES OF ACTIVITY,
BIOAVAILABILITY AND METABOLISM.
Pedro Luis Vieira Falé
DOUTORAMENTO EM BIOQUÍMICA
(Bioquímica Farmacêutica e Toxicológica)
2011


UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE QUÍMICA E BIOQUÍMICA
BIOLOGICAL ACTIVITIES OF Plectranthus barbatus
AQUEOUS EXTRACTS.
IN VITRO AND IN VIVO STUDIES OF ACTIVITY,
BIOAVAILABILITY AND METABOLISM.
Pedro Luis Vieira Falé
DOUTORAMENTO EM BIOQUÍMICA
(Bioquímica Farmacêutica e Toxicológica)
Tese orientada por
Prof. Doutora Maria Luísa Mourato Oliveira Marques Serralheiro
Prof. Doutora Lia Maria Pereira de Ascensão Santos e Sousa
2011


The work presented in this thesis was performed in the
Centro de Química e Bioquímica da Faculdade de
Ciências da Universidade de Lisboa, with the financial
support of Fundação para a Ciência e a Tecnologia
(SFRH/BD/37547/2007)


v
De acordo com o disposto no Artigo nº 41 do Regulamento de Estudos Pós‐Graduados da
Universidade de Lisboa, Deliberação nº 1506/2006, publicada no Diário da República, 2ª
série—Nº 209—30 de Outubro de 2006, foram incluidos nesta dissertação resultados dos
seguintes artigos:
Falé PL, Borges C, Madeira PJA, Ascensão L, Araújo MEM, Florêncio MH, Serralheiro MLM. 2009.
Rosmarinic acid, scutellarein 4’‐methyl ether 7‐O‐glucuronide and (16S)‐coleon E are the main
compounds responsible for the antiacetylcholinesterase and antioxidant activity in herbal tea of
Plectranthus barbatus (‘‘falso boldo”). Food Chem. 114: 798–805.
Porfirio S, Falé PL, Madeira PJA, Florêncio H, Ascensão L, Serralheiro MLM. 2010.
Antiacetylcholinesterase and antioxidant activities of Plectranthus barbatus tea, after in vitro
gastrointestinal metabolism, Food Chem. 122: 798–805.
Falé PL, Madeira PJ, Florêncio MH, Ascensão L, Serralheiro MLM. 2011. Function of Plectranthus
barbatus herbal tea as neuronal acetylcholinesterase inhibitor. Food Funct. 2: 130‐136.
Falé PL, Ascensão L, Serralheiro MLM, Haris PI. 2011. Interaction between Plectranthus barbatus herbal
tea components and human serum albumin and lysozyme: binding and activity studies. Spectroscopy.
26: 79–92.
Falé PL, Ascensão L, Serralheiro MLM, Haris PI. Interaction between Plectranthus barbatus herbal tea
components and acetylcholinesterase: binding and activity studies. Submitted to Food Funct.
Falé PL, Ascensão L, Serralheiro MLM. Bioavailability of rosmarinic acid, luteolin, apigenin‐modelling
plant herbal teas through Caco‐2 cell monolayers. To be submitted to Food Chem.
Falé PL, Filipe MA, Ascensão L, Serralheiro MLM, Mira L. Activity of Plectranthus barbatus extract against
inflammatory response in human neutrophils. To be submitted to Plant Food Hum. Nutr.
No cumprimento do disposto na referida deliberação, esclarece‐se serem da minha
responsabilidade a execução das experiências que estiveram na base dos resultados
apresentados, assim como a interpretação e discussão dos mesmos.


vii
Aknowledgements
Many people have contributed, directly or indirectly, to the successful completion of
this work. First, I would like to express my deepest gratitude to Professor Maria Luísa
Serralheiro for her guidance and counseling, all the challenges, the knowledge and, most of all,
the practical (engineer-like) point of view. The elaboration of this work under her guidance
was not only a scientifically rewarding experience, but also a pleasant one.
I would like also to thank to Professor Lia Ascensão for the guidance and attention to
details the elaboration of this work. I would also like to thank her for the support, friendship,
and continuous encouragement all these years.
To Professor Parvez Haris, his group, and staff at the De Montfort University, in
Leicester, I would like to thank for the hospitality and for all the support in the studies
developed there.
To Professor Maria Helena Florêncio I would like to thank the expertise in the
compound identification by mass spectrometry. To Paulo Madeira I would also like to thank,
not only the help in mass spectrometry anlysis, but also for the lighter moments and all his
patience for listening expressions such as “Are you sure about this structure? I don’t like it!”.
I would also like to thank to Sara Porfírio, Ines Sousa Lima, Rita Carilho, Susana Santos,
Catarina Costa, Leticia Silva, Pedro Cleto, Neusa Figueiredo, Ana Margarida Rodrigues,
Francesca Amaral and Catarina Ferreira, the Master students that helped, even if only by their
enthusiasm.
Last, but not least, I am grateful to my friends from the “family of C4 library”, for
providing the encouragement and a stable (geek) environment. To Charlie, for lightening (and
enlightening) my life even in the toughest moments. And to my family, especially my Mother,
for the support through this period, but also in my dedication to such an odd area as Science,
instead of Arts.


ix
Summary
The Plectranthus barbatus herbal tea is traditionally used to treat a wide range of
health conditions, including psychiatric problems, gastrointestinal disturbances and
inflammation‐related conditions. The aim of this work was to determine if P. barbatus herbal
tea may be useful in the treatment of acetylcholinesterase or inflammation related diseases,
such as Alzheimer’s disease.
In vitro activities of the P. barbatus aqueous extract were determined, namely anti‐
acetylcholinesterase activity, antioxidant activity as radical scavenger and preventing lipid
peroxidation, and anti‐inflammatory activity by decreasing the amount of hypochlorous acid
produced by activated neutrophils. The values obtained for the in vitro activities were very
promising and related with its content in rosmarinic acid, flavonoid glucuronides and abietane
diterpenoids. After in vitro digestion with gastric and pancreatic juices, bacterial
β‐glucuronidase and metabolization by Caco‐2 cells, the anti‐acetylcholinesterase activity of
the plant extract suffered a slight decrease due to the loss of one active diterpenoid.
The bioavailability of the plant extract was determined by administering it to rats and
analyzing the rat plasma and brain. The extract components suffered glucuronidation, sulfation
and methylation by the intestine and by the liver, but the plant compounds were found in rat
brains and brain acetylcholinesterase activity showed an inhibition reaching 30%. As the
bioavailability of rosmarinic acid was not the same when in the extract and when alone, the
interference of plant phenolics on the permeability of each other was tested in Caco‐2
monolayers using rosmarinic acid and two flavonoids, apigenin and luteolin. This study showed
that the compounds have higher intestinal permeability when in a mixture due to the
inhibition of the efflux transporters that limit their bioavailability.
The compounds from the P. barbatus extract can bind to the protein structure of
acetylcholinesterase, human serum albumin and lysozyme by weak interactions such as
hydrophobic interactions and hydrogen bonds. These interactions are the cause of the
reversible inhibition of the enzymatic activity of acetylcholinesterase and lysozyme; therefore
the compounds are less susceptible to cause side effects when used therapeutically.
The P. barbatus herbal tea may be used to treat cholinesterase‐related problems such
as gastrointestinal conditions and Alzheimer’s disease as its active components may reach the
target organs, and brain acetylcholinesterase inhibition was detected. The compounds may
circulate in the plasma bound to albumin and lysozyme, and may decrease inflammation by
their radical scavenger activity, by decreasing neutrophil‐produced hypochlorous acid and by
inhibiting lysozyme activity.

x
Keywords: Acetylcholinesterase; Antioxidant; Bioavailability; Plectranthus barbatus;
Rosmarinic acid.

xi
Resumo
Infusões e decocções de Plectranthus barbatus são usadas tradicionalmente para uma
grande diversidade de fins terapeuticos, incluido o tratamento de problemas psiquiátricos,
distúrbios gastro‐intestinais e doenças relacionadas com processos inflamatórios. O objectivo
deste estudo é determinar se o extracto aquoso de P. barbatus, preparado como decocção,
poderá ser útil no tratamento de problemas relacionados com a actividade do enzima
acetilcolinesterase ou com processos inflamatórios.
Um extracto aquoso de P. barbatus foi preparado como decocção e foram
determinadas, in vitro, as actividades anti‐acetilcolinesterase, antioxidante e anti‐inflamatória.
Nestas actividades in vitro foram obtidos resultados muito promissores, com valores de IC50
baixos para a inibição da actividade do enzima acetilcolinesterase, actividade antioxidante no
sequestro de radicais livres e protegendo lipidos de peroxidação, e na actividade anti‐
inflamatória pela diminuição da quantidade de ácido hipocloroso produzido por neutrófilos
activados. Estas actividades estavam relacionadas com a composição do extracto de P.
barbatus, sendo ácido rosmarinico o composto maioritário, mas encontrando‐se também
presentes flavonóides glucuronados (apigenina 7‐O‐glucurónido, luteolina 7‐O‐glucurónido e
acacetina 7‐O‐glucurónido) e diterpenoides. Após a digestão in vitro do extracto vegetal com
sucos gástrico e pancreático artificiais, da acção de β‐glucuronidase de bactérias da microflora
intestinal e da metabolização por células Caco‐2, como modelo de células intestinais humanas,
a actividade do extracto sofreu uma diminuição devido à perda de um diterpenoide activo, no
entanto os outros compostos activos permaneceram intactos.
A biodisponibilidade da decocção de P. barbatus foi determinada por administração
intragástica e intraperitoneal a ratos, recolhendo‐se e analisando‐se o plasma e o cérebro dos
ratos por HPLC. Os componentes da decocção foram metabolisados no intestino e no fígado,
sofrendo glucuronidação, sulfatação e metilação. Encontrou‐se ácido rosmarínico no cérebro
dos ratos após a administração do extracto, e o cérebro apresentava uma inibição da
actividade da acetilcolinesterase atingindo aproximadamente 30%, sugerindo que outros
compostos ou metabolitos dos componentes da decocção pudessem estar presentes em
quantidades inferiores ao limite de detecção, mas que mesmo assim iriam influenciar a
actividade enzimática. Como se encontraram algumas alterações entre a biodisponibilidade do
ácido rosmarínico administrado no extracto ou administrado isolado, procedeu‐se ao estudo
da interferência que compostos fenólicos possam ter na permeabilidade uns dos outros
quando administrados em misturas, em membranas de células Caco‐2. Nesse estudo
recorreu‐se ao método de “central composite design” CCD para determinar se existiriam

xii
diferenças entre a permeabilidade e metabolização de ácido rosmarínico, apigenina e luteolina
em membranas de células Caco‐2, induzidas pela presença uns dos outros. Foram também co‐
administrados substractos de dois sistemas transportadores conhecidos, o transportador de
ácidos monocarbóxilicos (MCT) e a glicoproteina‐P (Pgp), e concluiu‐se que os compostos da
decocção de P. barbatus tinham maior permeabilidade por estarem em conjunto, uma vez que
uns compostos inibiam os transportadores de efluxo dos outros. A biodisponibilidade dos
substratos dos transportadores de efluxo presentes na membrana apical aumenta com a
inibição dos transportadores, pois estes transportam activamente os seus substratos para o
lúmen do intestino, limitando assim a passagem para a circulação sanguínea.
A interacção entre os compostos do extracto de P. barbatus e proteinas foi avaliado
através de técnicas de espectrometria de fluorescência e FTIR. Os componentes da decocção
de P. barbatus têm a capacidade de se ligar à estrutura proteica da acetilcolinesterase, da
albumina de soro humano e da lisozima através de interacções fracas, nomeadamente
interacções hidrofóbicas e pontes de hidrogénio. Por se tratarem de ligações fracas e não se
terem encontrado alterações da estrutura secundária das proteinas, estas interacções serão
responsáveis pela inibição reversivel da actividade enzimática da acetilcolinesterase e da
lisozima. Este tipo de interacções são as mais recomendadas para compostos a ser utilizados
para fins terapeuticos pois evitam efeitos secundários resultantes da inibição destes enzimas
por inibidores que formam complexos através de ligações mais fortes, como ligações
covalentes, e induzem alterações profundas na estrutura proteica levando à desnaturação.
Decocções de P. barbatus poderão ser utilizadas para tratar problemas relacionados
com a actividade da acetilcolinesterase, como a doença de Alzheimer, pois após a
administração os seus componentes poderão ser encontardos a a nivel do intestino, plasma e
cérebro, inibindo a actividade da acetilcolinesterase no cérebro. Os componentes da decocção
poderão circular na corrente sanguínea associados à albumina e à lisozima, e poderão reduzir
processos inflamatórios devido à sua actividade sequestradora de radicais livres, à sua
capacidade de diminuir a quantidade de àcido hipocloroso produzido por neutrófilos
activados, e por inibir a atividade enzimática da lisozima.
Palavras‐chave: Acetilcolinesterase; Ácido Rosmarínico; Antioxidante; Biodisponibilidade;
Plectranthus barbatus.

xiii
Table of Contents
Acknowledgments vii
Summary ix
Resumo xi
Table of Contents xiii
Figure List xvii
Table List xx
Abbreviations xxii
CHAPTER 1 – GENERAL INTRODUCTION 1
1. Literature Review 3
1.1. Plectranthus species and their ethnobotanical uses 3
1.2. Herbal tea components and their bioavailability 4
1.2.1. Phenolic acids 4
1.2.2. Flavonoids 6
1.3. Biological activities of herbal teas and their components 9
1.3.1. Acetylcholinesterase inhibitors 9
1.3.1.1. Acetylcholinesterase inhibitors and Alzheimer’s disease 10
1.3.1.2. Acetylcholinesterase inhibitors to treat gastrointestinal disorders 13
1.3.1.3. Finding acetylcholinesterase inhibitors ‐ from in vitro to in vivo studies 14
1.3.2. Inflammation and Antioxidants 16
2. Thesis overview 19
CHAPTER 2 – MATERIALS AND METHODS 21
1. Plant material 23
2. Animals 23
3. Chemicals 23
4. Extract preparation 24
5. Acetylcholinesterase inhibition 24
6. Determination of antioxidant activity 25
7. HPLC analysis 26
8. NMR spectroscopy 26
9. Mass spectrometry experiments 26
10. In vitro intragastric metabolism assays 27
10.1. In vitro metabolism by the gastric juice 27
10.2. In vitro metabolism by pancreatic juice 27
10.3. Glucuronidase activity 28
10.4. Metabolism by the Caco‐2 cells 28
10.5. Antiacetylcholinesterase and antioxidant activities of the digested extracts 28
11. In vitro conjugation studies for metabolites identification 29
11.1. Preparation of cell‐free extracts 29
11.2. Glucuronidation assay 29

xiv
11.3. Synthesis and identification of methyl rosmarinic acid 29
12. In vivo studies protocol 30
12.1. Intragastric and intraperitoneal administration 30
12.2. Plasma and brain sample preparation 30
12.3. Determination of glucuronidated and sulfated metabolites 30
12.4. Preparation of samples for HPLC analysis 31
12.5. Determination of acetylcholinesterase activity in brain samples 31
13. Protein fluorescence measurements 31
14. FTIR measurements 32
15. Lysozyme activity measurements 33
16. Measurement of hypochlorous acid in activated human neutrophils 33
16.1. Isolation of human neutrophils 33
16.2.Measurement of hypochlorous acid formation by human neutrophils 33
17. Caco‐2 bioavailability experiments 34
17.1. Study of the permeability and the metabolism of rosmarinic acid, luteolin and
apigenin 34
17.2. Study of the effect of transport systems (MCT and Pgp) on the permeation of
apigenin, luteolin and rosmarinic acid 35
17.3. HPLC analysis and bioavailability quantification 35
18. Statistical analysis 36
CHAPTER III ‐ SCREENING FOR ANTIACETYLCHOLINESTERASE AND
ANTIOXIDANT ACTIVITIES IN PLECTRANTHUS SPECIES. IN VITRO METABOLISM
AND ANTI‐INFLAMMATORY STUDIES 37
1. Screening for antiacetylcholinesterase and antioxidant activities in Plectranthus
species 39
1.1. Introduction 39
1.2. Materials and Methods 40
1.3. Results 40
1.3.1. General 40
1.3.2. Acetylcholinesterase inhibition 41
1.3.3. Antioxidant Activity 42
1.3.4. Identification of the main component of the Plectranthus extracts,
responsible for the enzyme inhibition activity 42
1.4. Discussion 45
1.5. Conclusion 46
2. In vitro Digestion Activities of Plectranthus barbatus Aqueous Extract and Activities
of the Digested Product 47
2.1. Introduction 47
2.2. Materials and Methods 48
2.3. Results and Discussion 48
2.3.1. Main composition of P. barbatus herbal tea 48
2.3.2. In vitro metabolism of the extract by the gastric and pancreatic juices.
Biological activity of the resulting products 50

xv
2.3.3. Metabolism of the plant extract by Caco-2 cells and biological activity of
the final products 53
2.3.4. Metabolism of the plant extract by the β-glucuronidase from E. coli,
biological activities and Caco-2 cells permeation of the final products 55
2.4. Conclusion 57
3. Activity of Plectranthus barbatus extract against inflammatory response in human
neutrophils 59
3.1. Introduction 59
3.2. Materials and Methods 60
3.3. Results and Discussion 60
3.4. Conclusions 63
4. Conclusions 64
CHAPTER IV – BIOAVAILABILITY STUDIES IN RATS AND CACO-2 CELL MONOLAYERS 65 1. Plectranthus barbatus aqueous extract bioavailability and resulting neuronal acetylcholinesterase inhibition in rats 67 1.1. Introduction 67 1.2. Materials and Methods 68 1.3. Results and Discussion 68
1.3.1. Intragastric administration of P. barbatus extract 68 1.3.2. Intraperitoneal administration of P. barbatus extract 71
1.4. Conclusions 76 2. Bioavailability of mixtures of rosmarinic acid, luteolin and apigenin through Caco-2 cell monolayers, modeling the bioavailability of plant herbal teas. 77 2.1. Introduction 77 2.2. Material and Methods 78 2.3. Results 78
2.3.1. Bioavailability of Plectranthus barbatus herbal tea 79 2.3.2. Bioavailability of a mixture of the standards rosmarinic acid, apigenin and luteolin 80 2.3.4. Effect of MCT and Pgp transporter systems on the bioavailability of the polyphenol mixture 82
2.4. Discussion 86 2.5 Conclusions 89 3. Conclusions 91
CHAPTER V – INTERACTIONS BETWEEN THE Plectranthus barbatus HERBAL TEA AND THE PROTEINS ACETYLCHOLINESTERASE, HUMAN SERUM ALBUMIN AND LYSOZYME 93 1. Interaction between the Plectranthus barbatus extract and acetylcholinesterase. Binding of herbal tea components to the protein structure and inhibition of enzymatic activity 95 1.1. Introduction 95 1.2. Materials and Methods 96 1.3. Results 97
1.3.1. Fluorescence studies on the binding of P. barbatus water extract to acetylcholinesterase 97

xvi
1.3.2. Analysis of binding equilibria of P. barbatus water extract to acetylcholinesterase
99
1.3.3. Determination of interaction forces between P.barbatus extract and AChE 101 1.3.4. Determination of protein structure changes caused by P. barbatus extract and its plasma metabolites by FTIR spectroscopy 102
1.4. Discussion 105 1.5. Conclusions 107 2. Interaction between Plectranthus barbatus herbal tea components and human serum albumin and lysozyme: binding and activity studies 109 2.1. Introduction 109 2.2. Material and Methods 110 2.3. Results 110
2.3.1 Binding of P. barbatus to albumin and lysozyme 110 2.3.2. Analysis of binding equilibria 112 2.3.3. Determination of interaction forces between P. barbatus extract metabolites and HSA and lysozyme 115 2.3.4. Determination of protein structure changes caused by P. barbatus extract and its plasma metabolites by FTIR 116 2.3.5. Effect of P. barbatus extract on lysozyme activity 118
2.4. Discussion 118 2.5. Conclusions 122 3. Conclusions 123
CHAPTER VI – GENERAL DISCUSSION AND CONCLUSIONS 125
References 133

xvii
Figure List
Figure 1.1. Plectranthus barbatus 4
Figure 1.2. Chemical structures of some common phenolic acids. 5
Figure 1.3. Basic chemical structures of the main classes of flavonoids. 7
Figure 1.4. Structures of (a) quercetin, (b) apigenin and (c) luteolin. 7
Figure 1.5. Active gorge of acetylcholinesterase (Abu‐Donia, 2003). 10
Figure 1.6. Tacrine binding to the active gorge of acetylcholinesterase, and details showing the
amino acid residues involved in the interaction between the two molecules (PDB 1ACJ, Harel
et al., 1993). 11
Figure 1.7. Biochemical pathways associated with the formation of amyloid plaques and
neurofibrillary tangles in Alzheimers’s Disease patients. (www.calbiochem.com/alzheimers). 12
Figure 1.8. Reaction associated with the Ellman assay to quantify acetylcholinesterase activity.
The final product TNB can be spectrofotometrically detected due to its absorption at a
wavelength of 405 nm. (adapted from Frasco et al., 2005) 15
Figure 1.9. Antioxidant strategies in Alzheimer’s disease. Solid arrows represent the
mechanisms of the disease and dashed arrows represent the mechanisms of antioxidant
therapy (Adapted from Dumond and Bael (2011)). 18
Figure 3.1. HPLC chromatogram of decoctions: (a) Plectranthus barbatus and (b) Plectranthus
verticillatus.
43
Figure 3.2. UV spectra obtained by HPLC‐diode array of (a) compounds with retention time
19.2 min and (b) caffeic acid. 43
Figure 3.3. NMR spectra of (a) Plectranthus barbatus extract, (b) Plectranthus verticillatus
extract and (c) rosmarinic acid standard. 44
Figure 3.4. Structure of the compounds with retention time 19.2 min, rosmarinic acid. 44
Figure 3.5. Overlay of UV spectra obtained by HPLC‐diode array of compounds with a retention
time 19.2 min (——) and the standard rosmarinic acid (___). 45
Fig. 3.6. HPLC chromatogram of Plectranthus barbatus herbal tea: 1, luteolin 7‐O‐glucuronide
(retention time 8.68 min); 2, rosmarinic acid (RT: 9.38); 3, apigenin 7‐O‐glucuronide (RT: 10.11);
4, hydrolysed abietane (RT: 12.58); 5, acacetin 7‐O‐glucuronide (RT: 13.62); 6, abietane
diterpenoid (RT: 14,15); 7, (16S)‐coleon E (RT: 18.54). 49
Figure 3.7. Chemical structure of compounds present in P. barbatus herbal tea:
1, luteolin 7‐O‐ glucuronide; 2, rosmarinic acid; 3, apigenin 7‐O‐glucuronide; 4, hydrolysed
abietane; 5, acacetin 7‐O‐glucuronide; 6, abietane diterpenoid; 7, (16S)‐coleon E. 50

xviii
Figure 3.8. HPLC chromatograms before and after the incubation of Plectranthus barbatus
extract with: (a) gastric juice, (b) pancreatic juice. *Indicates the residue of pancreatin. For the
identification of the peak numbers, refer to Figure 3.4.
51
Figure 3.9. Variations in peak areas of compounds present in herbal tea after 4 h digestion with
artificial pancreatic juice. () Luteolin 7‐O‐glucuronide; () rosmarinic acid;
() apigenin 7‐O‐glucuronide; () hydrolysed abietane; (○) acacetin 7‐O‐glucuronide; (●)
abietane diterpenoid. 53
Figure 3.10. Chromatograms of compounds inside the Caco‐2 cells (homogenates) after 6 h
contact with (a) P. barbatus extract, (b) rosmarinic acid standard. 2, rosmarinic acid; *, residual
peak from the Caco‐2 cells. 55
Figure 3.11. HPLC chromatogram after the action of β‐glucuronidase from E. coli on the herbal
tea. Compounds identified with numbers 1–7, refer to Figure 3.6. Peaks signaled with arrows: 8,
luteolin; 9, apigenin; 10, acacetin. 56
Figure 3.12. Variations in peak areas of: (a) apigenin and (b) luteolin. Each figure shows the
metabolites after 6 h in the presence of Caco‐2 cells. Peak areas are presented as percentage of
the initial area. () aglycones and () glucuronides. 57
Figure 3.13. Decrease of taurine chloration in the presence of several concentrations of
P. barbatus extract (a and b) or standard rosmarinic acid (b). The concentration of P. barbatus is
expressed in µg.mL‐1 (a) or by its content in rosmarinic acid in µM (b). 61
Figure 4.1. HPLC analysis, 30 and 60 min after intraperitoneal administration of P. barbatus
extract, of (a) plasma and (b) brain. 1: luteolin 7‐O‐glucuronide (retention time 9.6 min); 2:
rosmarinic acid (RT: 10.4); 3: apigenin 7‐O‐glucuronide (RT: 11.2); 4: abietane diterpenoid (RT:
13.8); 5: acacetin 7‐O‐glucuronide (RT: 15.1); 2m: monomethylated rosmarinic acid; 1g: luteolin
glucuronide derivative; 3’: apigenin. 72
Figure 4.2. Permeation surfaces for (a) rosmarinic acid, (b) luteolin and (c) apigenin with
different concentrations of the other two components, built with the CCD experimental plan.
The relative errors are 0.930 (a), 0.718 (b), and 0.817 (c). 81
Figure 4.3. Glucuronidation surfaces for (a) luteolin and (b) apigenin with different
concentrations of rosmarinic acid and of the other flavonoid, built with the CCD experimental
plan. The relative errors are 0.981 (a), and 0.766 (b). 82
Figure 4.4. Distribution of (a) rosmarinic acid, (b) luteolin and (c) apigenin six hours after being
placed in the apical side of a Caco‐2 cell monolayer. The effects of co‐administration of digoxin
and benzoic acid were analysed. Statistical analysis: c ‐ different from the control (P<0.05); c* ‐
different from the control (P<0.1); s – different from the other substrate (P<0.05).
83

xix
Figure 4.5. Distribution of (a) benzoic acid and (b) digoxin six hours after being placed in the
apical side of a Caco‐2 cell monolayer. The effects of co‐administration of a standard mixture
(SM) with rosmarinic acid, luteolin and apigenin, 50µM each, was analysed. Statistical analysis:
* ‐ different from the control (P<0.05).
84
Figure 4.6. Glucuronidation of (a) luteolin and (b) apigenin six hours after being placed in the
apical side of a Caco‐2 cell monolayer. The effects of co‐administration of digoxin and benzoic
acid were analysed. Statistical analysis: c ‐ different from the control (P<0.05); c* ‐ different
from the control (P<0.1); s – different from the other substrate (P<0.05). 85
Figure 5.1. Molecular structure of (a) rosmarinic acid, (b) quercetin, (c) luteolin, and (d)
apigenin. 96
Figure 5.2. Fluorescence emission spectra of acetylcholinesterase with the addition of
P. barbatus aqueous extract. Arrow points to increasing concentrations of P. barbatus plant
extract, ranging 0; 0.5; 1; 5; 10; 33; 50; 100 µg.ml‐1. 97
Figure 5.3. Stern‐Volmer plot (a) and plot of log(F0‐F)/F vs. log[Q] (b) of acetylcholinesterase
with P. barbatus aqueous extract. [Q] is the concentration of P. barbatus extract in mg.ml‐1. 99
Figure 5.4. FTIR spectra of the acetylcholinesterase alone, with P. barbatus extract, rosmarinic
acid (RA), luteolin (Lut), apigenin (Api) and quercetin (Quer). The spectra obtained are shown (a)
in the form of absorbance spectra and (b) the second derivative spectra (negative peaks). 103
Figure 5.5. Percentage of decrease of hydrogen‐deuterium exchange rate of AChE in the
presence of P. barbatus extract, rosmarinic acid, luteolin, apigenin and quercetin. 104
Figure 5.6. Fluorescence emission spectra of (a) HSA and (b) lysozyme with the addition of
P. barbatus aqueous extract. Arrow points to increasing concentrations of P. barbatus plant
extract, ranging 0; 0.5; 0.75; 1; 2.5; 5; 7.5; 100 μg.ml−1. 111
Figure 5.7. Stern–Volmer plots of HSA and lysozyme with P. barbatus aqueous extract. [Q] is the
concentration of P. barbatus in μg.ml−1. 112
Figure 5.8. Plots of log([F0 − F]/F) vs. log[Q] for HSA and lysozyme with P. barbatus aqueous
extract. [Q] is the concentration of P. barbatus in g.l−1. 114
Figure 5.9. FTIR spectra of the proteins alone, with P. barbatus extract, rosmarinic acid (RA),
luteolin (Lut) and apigenin (Api). The absorbance spectra obtained for HSA are shown in (a) and
the second derivatives are in (c). The absorbance spectra obtained for lysozyme are shown in (b)
and the second derivatives are in (d). 117
Figure 5.10. Percentage of change in protein (HSA and lysozyme) hydrogen–deuterium
exchange rate, determined from the analysis of the amide II band, in the presence of
P. barbatus extract, rosmarinic acid, luteolin or apigenin, in comparison with the
hydrogen–deuterium exchange rate of the protein alone. 118

xx
Table List
Table 2.1. Concentrations of rosmarinic acid, luteolin and apigenin used in the phenolic
mixtures to determine the bioavailability by Caco‐2 cells, according to the CCD experimental
plan. 35
Table 3.1. Amount of dry plant extract obtained. 40
Table 3.2. Inhibition of AChE activity (%), antioxidant activity and rosmarinic acid content of
water extracts of the leaves of several Plectranthus species. 41
Table 3.3. Detected ions and attribution errors (ppm) for the collected fractions corresponding
to peaks 1, 3, 4, 5, 6 and 7. 49
Table 3.4. Antiacetylcholinesterase and antioxidant activity of P. barbatus herbal tea after in
vitro gastrointestinal digestion. The action of the pancreatic juice on the inhibition activity of
rosmarinic acid (RA) is also shown. 52
Table 4.1. Concentration of rosmarinic acid, its metabolites and flavonoid glucuronide
derivatives in the plasma and in the brain, 30 and 60 min after the intragastric and
intraperitoneal administration of P. barbatus extract. 69
Table 4.2. Brain acetylcholinesterase inhibition (%) 30 and 60 min after administration
(intragastric and intraperitoneal) of rosmarinic acid and P. barbatus extract. Results significantly
different from the control are marked with * (P < 0.05) and ** (P < 0.1). Values that are not
significantly different (P < 0.05) are marked from a to d. 70
Table 4.3. Retention time of compounds from the P. barbatus extract found in plasma and their
decrease from 30 to 60 min after the extract intraperitoneal administration. 74
Table 4.4. Permeation of the P. barbatus aqueous extract constituents through the Caco‐2 cell
monolayer. 79
Table 5.1. Stern‐Volmer binding parameters (KSV, Kq), binding equilibria parameters (Kb, n) and
thermodynamic parameters (ΔHo, ΔSo, ΔGo) for the binding of P. barbatus extract, rosmarinic
acid (RA), luteolin (Lut), apigenin (Api) and quercetin (Quer) to acetylcholinesterase (R2>0.99 to
all linear regressions). Rates of Amide II/Amide I variation, reflecting the rate of hydrogen
deuterium exchange in the presence of similar amount of RA, Lut, Api and Quer or 10mg/mL of
P. barbatus extract (for AChE without ligand, ‐2.345mAU.min‐1). IC50 values for the inhibition of
acetylcholinesterase activity by P. barbatus, RA, Lut and Api. (*) and Quer for P. barbatus
extract the values are expressed in l.mg‐1, l.mg‐1s‐1, and mg.ml‐1 but the molarity was estimated
based on the content of rosmarinic acid, luteolin and apigenin (1.1122 mmol.g‐1). 100

xxi
Table 5.2. Binding parameters (KSV, Kq, Kb, n) and thermodynamic parameters (ΔHo, ΔSo, ΔGo) for
the binding of P. barbatus extract, rosmarinic acid (RA), luteolin (Lut) and apigenin (Api) to HSA
and to lysozyme (R2>0.99 to all linear regressions). Rates of Amide II/Amide I variation in the
presence of similar amount of RA, Api and Lut, or 10mg/mL of P. barbatus extract (without
ligand, ‐0.716 for HSA and ‐1.815 for lysozyme). IC50 values for the inhibition of lysozyme
activity. For P. barbatus extract the values are expressed in l.mg‐1(a), l.mg‐1s‐1(b) and mg.l‐1(c).
113

xxii
Abbreviations
ABC transporters ATP‐binding cassette transporters
AChE acetylcholinesterase
AChI acetylthiocholine
AD Alzheimer’s disease
ANOVA Analysis of Variance
Api apigenin
APP amyloid precursor protein
Asp aspartic acid
ATP adenosine triphosphate
AU absorbance unit
Aβ β‐amyloid
BCRP breast cancer resistance protein
BHT butylated hydroxytoluene
CCD central composite design
COMT catechol o‐methyl transferase
DMEM Dulbecco's Modified Eagle Medium
DMSO dimethyl sulfoxide
DMSO‐d6 deuterated dimethyl sulfoxide
DPPH di(phenyl)‐(2,4,6‐trinitrophenyl)iminoazanium
DTNB 5,5'‐dithiobis‐(2‐nitrobenzoic acid)
DTT dithiothreitol
EDTA ethylenediamine tetraacetic acid
ESI‐MS electrospray ionization mass spectrometry
FBS fetal bovine serum
FDA Food and Drug Administration
FTIR Fourier transform infrared spectroscopy
Glu glutamic acid
h hour
HBSS Hank’s balanced salt solution
HEPES 4‐(2‐hydroxyethyl)‐1‐piperazineethanesulfonic acid
His histidine

xxiii
HPLC high precision liquid chromatography
HPLC‐DAD high precision liquid chromatography coupled with a diode array
detector
HSA human serum albumine
I(%) inhibition in percentage
IC50 concentration of inhibitor causing 50% inhibition
Kb binding constant
Kq scatter collision quenching constant
KRC Krebs‐Ringer solution with calcium chloride
KSV Stern‐Volmer quenching constant
Lut luteolin
MCT monocarboxylic acid transporter
min minute
MRP multidrug resistance protein
MS mass spectrometry
MTT 3‐(4,5‐Dimethylthiazol‐2‐yl)‐2,5‐diphenyltetrazolium bromide
NADPH nicotinamide adenine dinucleotide phosphate
NMR nuclear magnetic resonance spectroscopy
PBS phosphate buffer saline
PDB Protein Data Bank
PET positron emission tomography
Pgp P‐glycoprotein
PMA phorbol myristate acetate
Quer quercetin
RA rosmarinic acid
ROS radical oxygen species
RT retention time
SAM S‐adenosyl methionine
Ser serine
TEER trans‐epithelial electric resistance
TFA trifluoroacetic acid
TMB 3,3’,5,5’‐Tetramethylbenzidine
TNB 2‐nitro‐5‐thiobenzoate
Tris tris(hydroxymethyl)aminomethane

xxiv
Trp tryptophan
Tyr tyrosine
UDPGA 5'‐diphospho‐glucuronic acid
UV ultraviolet
UV‐Vis ultraviolet‐visible
ΔGo free energy change
ΔHo enthalpy change
ΔSo entropy change

Chapter I
General Introduction


Chapter I
3
1. Literature review
1.1. Plectranthus species and their ethnobotanical uses
The genus Plectranthus L’Hér., belonging to the Mint family (Lameaceae), comprises
about 300 species widely distributed in the savannahs and forest regions of Africa, Asia and
Australia. The majority of the Plectranthus species can be described as tender shrubs or
groundcovers growing in the shade of large forests or in the partial shade of the forest edge.
A few species occur in drier regions with rocky soils and have succulent or semi-succulent leaves
and stems, which help the plants to survive in these habitats (Codd, 1975; van Jaarsveld, 2006).
The geographical distribution of Plectranthus is highly variable. Some species, as
P. laxiflorus, occur in smaller areas in the south-east of the African continent, while others, like
P. barbatus, may naturally occur in vast areas, in Africa, India and South America (Codd, 1975;
Lukhoba et al., 2006). The later species, endemic form India, was taken to Brazil probably during
the colonial period (Codd, 1975; Lukhoba et al., 2006; Lorenzi & Matos, 2002).
The value of Plectranthus species as garden plants has been long recognized. They have
ornamental leaves and flowers, are easy and fast growing plants, resistant to most pests and
diseases, and make colourful displays in autumn when other flowers are often scarce. Several
Plectranthus species were introduced in Europe during the expansion of the British Empire and
the founding of the Royal Botanic Gardens of Kew. In the nineteenth century some species were
very popular in Scandinavia and commonly planted in window boxes and hanging baskets, as
P. oertendahii, nowadays known as “Swedish ivy”, and other Plectranthus groundcover species
(P. madagascariensis, P. verticillatus, P. strigosus, etc.) Some other species are tall shrubs that
can be pruned and form colourful hedges (P. ecklonii, P. fruticosus, P. barbatus, etc.).
A great variety of ethnobotanical uses are reported for Plectranthus species. Some, due
to their aromatic nature, are used as culinary plants to flavour food, or as insect repellents
(P. ornatus). Others have tubers with high starch content and are eaten as vegetables
(P. esculentus, P. rotundifolius). However, the most common uses are to treat a wide range of
diseases (Lukhoba et al., 2006). Infusions of several Plectranthus species are used to treat
coughs and colds (P. hadiensis, P. laxiflorus), liver complaints (P. hereroensis), and ailments in
respiratory system and skin (P. madagascariensis).
Among the Plectranthus species used in traditional medicines P. barbatus is one of the
most important (Luckhoba et al., 2006). P. barbatus (Figure 1.1) is used in Hindu and Ayurveda
treataments, and in traditional medicine from Brazil, tropical Africa and China (Alabashi &
Melzig, 2010a). Extensive reviews have been written about the ethnobotanical importance of
P. barbatus (Alabashi & Melzig 2010a, 2010b); Lukhoba et al., 2006). The leaves from P. barbatus

General Introduction
4
are usually prepared as a decoction or infusion and used in the treatment of pain from different
aetiologies, inflammation, infections, colds and coughs (Lukhoba et al., 2006), which suggest
anti‐inflammatory and antioxidant properties. P. barbatus is also used to treat psychiatric
disorders in Tanzania (Lukhoba et al., 2006).
Figure 1.1. Plectranthus barbatus
Other significant uses for P. barbatus reported by Lukhoba et al. (2006) and Alabashi &
Melzig (2010a) are as food, the leaves being cooked as vegetable in Kenya and Yemen, and as
ornamental plant or garden herb, in hedges, fences or boundary markers.
1.2. Herbal tea components and their bioavailability
The main components of most medicinal teas, used traditionally by people, are phenolic
acids and flavonoids (Cai et al., 2004). The flavonoids in herbal teas are usually present in
glycosylated forms, which have higher solubility in water, although some flavonoid aglycones
may also occur in aqueous extracts (Cai et al., 2004). As herbal teas are usually taken orally,
many studies have focused on the metabolism and bioavailability of their active components.
1.2.1. Phenolic acids
Phenolic acids are a class of polyphenols commonly found in herbal medicines and plant‐
derived food (Lafay and Gil‐Izquierdo, 2008). The most commonly found phenolic acids, shown
in Figure 1.2, are gallic acid, a component of hydrolysable tannins, and hydroxycinnamic acids, as
cinnamic acid, caffeic acid (3,4‐dihydroxycinnamic acid), coumaric acid (4‐hydroxycinnamic acid),
ferulic acid (3‐methoxy‐4‐hydroxycinnamic acid) and the ester derivatives of caffeic acid, the
chlorogenic acid (3‐caffeoylquinic acid) and the rosmarinic acid.

Chapter I
5
Figure 1.2. Chemical structures of some common phenolic acids.
When phenolic acids are ingested their bioavailability is dependent on the permeability
through the intestinal barrier, and on the conjugation (glucuronidation, sulfation or methylation)
by intestinal and liver cells. Bioavailability studies using rats as model animals show that
hydroxycinnamic acids such as coumaric acid and caffeic acid can be found in plasma after
intragastric administration, in unconjugated, sulfated and/or glucuronidated forms (Konishi et
al., 2004; Konishi et al., 2005). Both coumaric and caffeic acids presented higher bioavailability
than gallic acid, which is a simpler phenolic acid (Konishi et al., 2004 & 2005). Ferulic acid, like
coumaric acid showed a higher bioavailability than caffeic acid (Konishi et al., 2006). The caffeic
acid esters rosmarinic acid and chlorogenic acid showed much lower bioavailability than simpler
hydroxycinnamic acids and similar to gallic acid (Konishi et al., 2005 & 2006). Some authors
could not detect chlorogenic acid circulating in rat plasma after its administration, but found
traces of caffeic acid and ferulic acid conjugates 6 hours after administration, suggesting that
hydrolysis of chlorogenic acid might have occurred and the resulting metabolites might be more
easily available (Azuma et al., 2000).
Many studies have focused on the bioavailability of rosmarinic acid, in the rats and
humans, as it has a number of interesting biological activities such as antioxidant, antiviral,
antibacterial and anti‐inflammatory activities that can be potentially important for public health
(Peterssen and Simmonds, 2003). When rosmarinic acid was administered to rats, intact
rosmarinic acid, monomethyl‐rosmarinic acid, and coumaric acid were found in plasma, mostly
in conjugated forms (Baba et al., 2004). In humans the same compounds plus caffeic acid and
ferulic acid were found circulating in plama, also in conjugated forms, after the intake of a
rosmarinic acid‐rich Perilla extract (Baba et al., 2005). In both models the maximum amount of

General Introduction
6
free rosmarinic acid in plasma was reached around 0.5h after administration, while conjugated
forms of rosmarinic acid had maximum peaks later, from 40 mins to 2h after administration
(Baba et al., 2004 & 2005; Konishi et al., 2005). Eighty three per cent of the ingested rosmarinic
acid was excreted between 8 and 18 hours after intake in rats (Baba et al., 2004), and 75%
within the first 6h in humans (Baba et al., 2005), suggesting that rosmarinic acid metabolites
may circulate for a relatively long time before being excreted and repeated intakes may be
favourable to the accumulation of these active compounds in the bloodstream.
The bioavailability results obtained with Caco‐2 cell monolayers are in agreement with
the in vivo results obtained in the rat model for the order of bioavailability of the phenolic acids,
which is gallic acid = rosmarinic acid = chlorogenic acid < caffeic acid < p‐coumaric acid (Konishi
et al., 2005), which suggests that the same kind of transport mechanisms for phenolic acids may
be present in both models. The high permeation of caffeic acid, p‐coumaric acid and ferulic acid
in Caco‐2 cells, and presumably in rat intestine, seems to be due to the fact that these
compounds are substrates of the monocarboxylic acid transporter (MCT) (Konishi and
Kobayashi, 2004a & 2004b). Rosmarinic acid and chlorogenic acid, which showed a very low
transepithelial permeation, do not seem to be substrates of MCT, being transported by
paracellular diffusion (Konishi and Kobayashi 2004a; Konishi and Kobayashi, 2005).
Compounds that were reported as metabolites of caffeic acid by gut microflora such as
m‐coumaric acid and m‐hydroxyphenylpropionic acid are also substrates of the MCT and showed
a higher influx than caffeic acid (Konishi et al., 2004b), suggesting that the action of gut
microflora may increase the bioavailability of these active compounds.
1.2.2. Flavonoids
Flavonoids are a group of polyphenolic compounds that share the same structural
features, namely a C6‐C3‐C6 carbon framework (Figure 1.3) or, more specifically, a
phenylbenzopyran functionality. Depending on the position of linkage of the aromatic ring to the
benzopyrano moiety, this group can be divided in three classes: flavonoids (2‐
phenylbenzopyrans) as shown in Figure 1.2; isoflavonoids (3‐phenylbenzopyrans) as the
isoflavone in Figure 1.3; and neoflavonoids (4‐phenylbenzopyrans) the rarest group of flavonoids
(not shown). In some flavonoids the C3 moiety between the two aromatic rings may not form a
heterocyclic ring (ring C) in their structure, such as the chalcones (Figure 1.3) (Marais et al.,
2006). Depending on the degree of saturation and oxidation of the C‐ring, some sub‐groups may
be considered, the most common structures can be seen in Figure 1.3 (Marais et al., 2006). The
pattern of hydroxylation of flavonoids is determinant for their identification and properties, as in

Chapter I
7
the case of quercetin, apigenin and luteolin (Figure 1.4), which are among the most commonly
occurring flavonoids.
Figure 1.3. Basic chemical structures of the main classes of flavonoids.
Figure 1.4. Structures of (a) quercetin, (b) apigenin and (c) luteolin.
Flavonoids often occur in the form of flavonoid glycosides, in which the sugar residue is
bound to the flavonoid structure by one of the hydroxyl groups. The most common
monosaccharide residues found in flavonoid glycosides are glucose, rhamnose and glucuronic
acid.
Ingested flavonoids may undergo several changes before reaching target organs where
they may have beneficial effects. Studies with quercetin, which is often used as a model of
flavonoid, showed the presence of glucuronidated, sulfated and methylated forms of quercetin
OH
O
O
OH
OH
OH
OH
OH
O
O
OH
OH
OH
OH
O
O
OH
OH
a
b
c

General Introduction
8
in rat plasma after its intragastrical administration to rats (Morand et al., 1998; da Silva et al.,
1998; Justino et al., 2004). Although the bioavailability of quercetin is low, the antioxidant
activity in rat plasma after its administration increased significantly (da Silva et al., 1998; Justin
et al., 2004). Liver sulfotransferases and glucuronyltranferases seem to be highly responsible for
the metabolization of flavonoids, as in vitro studies showed that liver enzymatic extracts are able
to conjugate quercetin into sulfated and glucuronidated metabolites (da Silva et al., 1998;
Justino et al., 2004). Studies with liver cell lines, such as HepG2, also showed that liver cells may
conjugate quercetin through sulfation, glucuronidation, and methylation reactions (O’Leary et
al., 2003), and the multidrug resistant protein 2 (MRP2) may promote the efflux of quercetin
conjugates from the cells (O’Leary et al., 2003). Although quercetin is mainly found circulating in
plasma in the conjugated form, one of the characteristic features of quercetin conjugates is a
slow elimination, with reported half‐times from 11 to 28 hours, favouring its accumulation in
plasma after repeated intakes (Manech et al., 2005).
Several studies on the bioavailability and metabolism of flavonoids by the intestine have
been performed using Caco‐2 cell monolayers as models for the intestinal barrier. Flavonoid
aglycones have shown a high permeability through these membranes, which is due to active
transport (Walgren et al., 1998; Walle et al., 1999). As flavonoids in food are present mainly in
glycosylated form, some studies suggest that the gut microflora may increase their
bioavailability by hydrolysing into flavonoid aglycones (Liu & Hu, 2002; Kobayashi et al., 2008).
Studies of bioavailability of hesperetin (aglycone) and hesperidin (hesperetin glycoside) showed
that the transepithelial transport of the aglycone is mainly active transport, while the glycoside
passes through Caco‐2 cell monolayers mainly by passive diffusion (Kobaiashi et al., 2008), which
is in good agreement with results obtained by Walgren et al. (2008) for quercetin and its
glycosides. Transporters from the MCT family seem to be related to the transepithelial transport,
across the Caco‐2 monolayers, of flavonoids such as hesperetin, naringenin, erydictiol and
epicatechins (Kobayashi & Konishi, 2008; Vaidyanathan and Walle, 2003). Several reports show
that flavonoids may be conjugated – preferentially glucuronidated, but also sulfated – by Caco‐2
cells, and transported back to the culture medium by transporters of the type multidrug
resistance pump (MRP) after conjugation (Ng et al., 2005; Walle et al., 1999).
The relationship between flavonoids and intestinal membrane transporters is highly
complex. Flavonoid aglycons may be transported to the bloodstream by some active
transporters (Walle et al., 1999). Some flavonoids may be substrate to efflux transporters such
as P‐glycoprotein (Pgp) and MRP and may return to gut lumen (Wang et al., 2009), while others
may act as inhibitors of Pgp and MRP, without being their substrates (Brand et al., 2006).

Chapter I
9
Although these complex relationships are still under study, it seems that the flavonoid
bioavailability depends on the balance of active influx transport, conjugation, efflux transport
and inhibition of the active transporters by other compounds.
1.3. Biological activities of herbal teas and their components
Many therapeutic properties have been reported for flavonoids and phenolic acids,
especially as enzyme inhibitors and antioxidants. The use of herbal teas for medicinal purposes
usually gives an indication of these activities.
The present thesis focuses on the anti‐acetylcholinesterase, anti‐inflammatory and
antioxidant activities, which can be useful in the treatment of several health disorders, such as
gastrointestinal disturbances, or the symptomatic treatment of Alzheimer’s disease.
1.3.1. Acetylcholinesterase inhibitors
Acetylcholine is usually found in the synaptic clefts in peripheral and central nervous
systems and has a neurotransmitter function. Acetylcholine is synthesized by neurons, released
to the synaptic cleft upon stimulation, and binds to acetylcholine receptors on the other end of
the synaptic cleft.
Acetylcholinesterase is an abundant enzyme in the synaptic cleft as it hydrolyses
acetylcholine into the inactive metabolites choline and acetate, clearing acetylcholine from the
synapse and ceasing the stimulation of the post‐synaptic receptors (Randall et al., 2000). The
active site of acetylcholinesterase is located in an active gorge (Figure 1.5) where some groups of
amino acid residues play an important role by positioning the acetylcholine molecule, the
peripheral binding site (Trp286, Tyr72, Tyr124 and Asp74), the choline binding site (Trp86,
Glu202, Tyr337) and the acyl pocket (Phe295, Phe297). The hydrolysis of acetylcholine is then
directly catalysed by the catalytic triad (Glu334, His447, Ser203), located in the bottom of the
active gorge (Abu‐Donia, 2003).
Acetylcholinesterase inhibitors can be used for the treatment of some dysfunctions, such
as Alzheimer’s disease and gastrointestinal disturbances, as will be discussed in the next
sections. These drugs, for instance tacrine, often bind to the active gorge with non‐covalent
bonds usually to the acetylcholine binding and positioning sites and to the catalytic site, as it is
shown in Figure 1.6 (Harel et al., 1993). Acetylcholinesterase inhibitors found by chemical
synthesis, as carbamates and organophosphates, are irreversible inhibitors binding to
acetylcholinesterase by ionic or covalent bonds, which makes them highly toxic and suitable to
be used as pesticides (Abu‐Donia, 2003; Eyer et al. 2007).

General Introduction
10
Figure 1.5. Active gorge of acetylcholinesterase (Abu‐Donia, 2003).
Galanthamine, an acetylcholinesterase inhibitor approved by the Food and Drug
Administration (FDA) to be used in the treatment of Alzheimer’s disease, is an alkaloid first
discovered in Galanthus species, and nowadays obtained for commercialization from Narcissus
spp. or synthetically (Heinrich & Teoh, 2004). The successful application of galanthamine led to
the search of new acetylcholinesterase inhibitors, more effective and with less side effects, in
plants used in traditional medicine (Houghton et al., 2004, Adsersen et al., 2006, Vinutha et al.,
2007, Ferreira et al., 2006).
1.3.1.1. Acetylcholinesterase inhibitors and Alzheimer’s disease
Alzheimer’s disease (AD) is the most common form of dementia among older people,
and its occurrence is of great concern in occidental populations where the life expectancy
increased in the 20th century.
AD is a degenerative terminal disease characterized by a progressive memory loss and
bodily functions, and symptoms generally include confusion, irritability, aggression, mood
swings, language breakdown, long term memory loss, and decline of the senses, which leads to
the withdrawal of the patients (Alzheimer’s Association, 2006).
On a biochemical point of view, AD is a protein misfolding disease, as it results from the
accumulation of abnormally folded amyloid beta peptides (Aβ). The Aβ may accumulate in the
cell within the neuronal endoplasmatic reticulum, and outside the cell forming senile plaques in
the brain of AD patients (Hashimoto et al., 2003).

Chapter I
11
Figure 1.6. Tacrine binding to the active gorge of acetylcholinesterase, and details showing the amino acid
residues involved in the interaction between the two molecules (PDB 1ACJ, Harel et al., 1993).
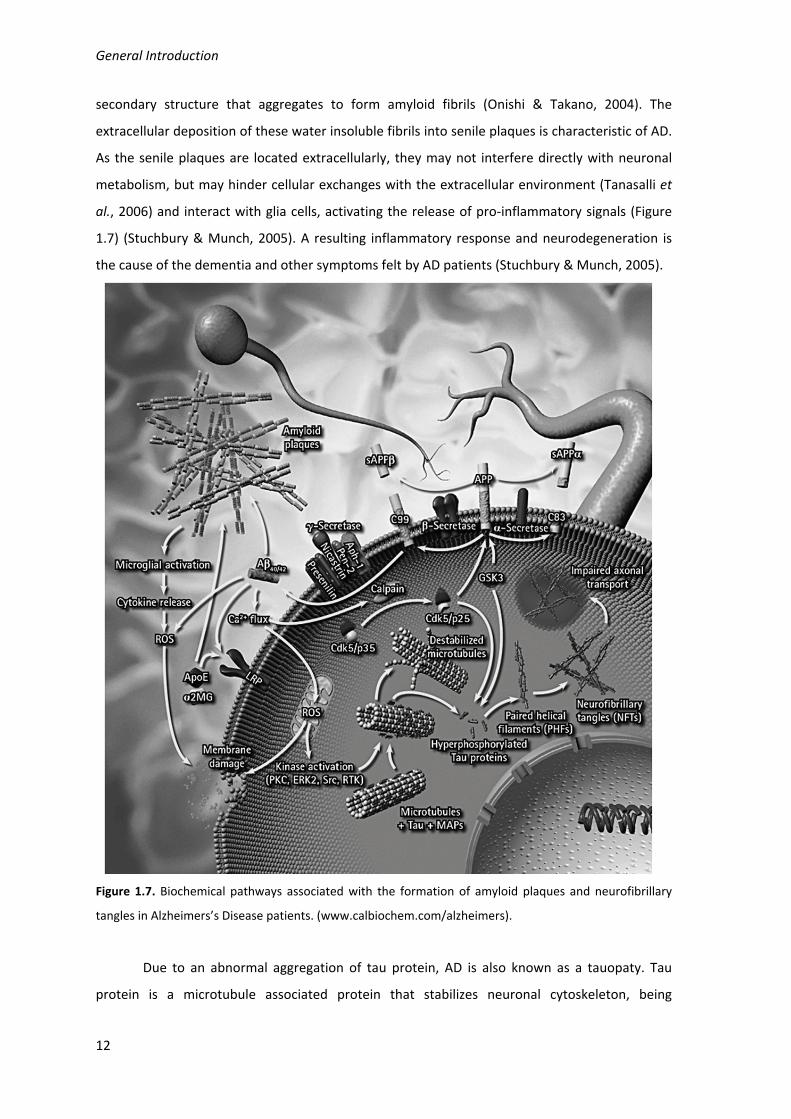
Aβ is a short peptide that results from the proteolytic cleavage by β‐secretase and
ϒ‐secretase of the amyloid precursor protein (APP) (Figure 1.7), a transmembrane protein whose
function is still unknown but seems to be involved in the early development of the nervous
system (Kerr & Small, 2005). Although Aβ are usually water soluble innocuous peptides with
short regions of β‐sheet and predominately α‐helix secondary structures in solution, they
undergo dramatic conformational changes at high concentrations to a form rich in β‐sheet
General Introduction
12
secondary structure that aggregates to form amyloid fibrils (Onishi & Takano, 2004). The
extracellular deposition of these water insoluble fibrils into senile plaques is characteristic of AD.
As the senile plaques are located extracellularly, they may not interfere directly with neuronal
metabolism, but may hinder cellular exchanges with the extracellular environment (Tanasalli et
al., 2006) and interact with glia cells, activating the release of pro‐inflammatory signals (Figure
1.7) (Stuchbury & Munch, 2005). A resulting inflammatory response and neurodegeneration is
the cause of the dementia and other symptoms felt by AD patients (Stuchbury & Munch, 2005).
Figure 1.7. Biochemical pathways associated with the formation of amyloid plaques and neurofibrillary
tangles in Alzheimers’s Disease patients. (www.calbiochem.com/alzheimers).
Due to an abnormal aggregation of tau protein, AD is also known as a tauopaty. Tau
protein is a microtubule associated protein that stabilizes neuronal cytoskeleton, being

Chapter I
13
regulated by phosphorylation. In AD patients tau protein is hyperphosphorylated and
accumulates as paired helical filaments, which aggregates into masses known as neurofibrillary
tangles inside the nerve cell bodies, also associated with the amyloid plaques (Figure 1.7)
(Stuchbury & Munch, 2005). Although some studies approach the treatment of AD by
modulating the activity of ϒ‐secretase, the enzyme mainly responsible for Aβ formation, the
most studied approach is by the administration of inhibitors of acetylcholinesterase activity
(Salawu et al., 2011).
Patients of AD show low concentration of acetylcholine in the brain as a consequence of
the cellular dysfunctions caused by the abnormal protein aggregations. The administration of
acetylcholinesterase inhibitors has proved to be effective in the symptomatic treatment of AD
patients (Rauf et al., 2002). This strategy to increase the acetylcholine levels in the brain is
presently the most commonly used, and the drugs that have been approved by the Food and
Drug Administration (FDA) to treat AD in the US are the acetylcholinesterase inhibitors tacrine,
rivastigmine, donepezil and galanthamine, which have all been successful in slowing down the
neurodegenerative process in AD patients (McGleenon et al., 1999; Heinrich & Teoh, 2004).
The major problem related with acetylcholinesterase inhibition treatments of AD is the
bioavailability of the inhibitors, as they must reach the brain passing through the blood‐brain
barrier to inhibit the brain acetylcholinesterase. If the inhibitors are too potent, or very high
concentrations are needed for the therapeutic effect, unwanted side effects may arise, such as
gastrointestinal and hepatic disturbances (McGleenon et al., 1999, Heinrich & Teoh, 2004). The
research for new acetylcholinesterase inhibitors for the treatment of AD continues, with the aim
of finding reversible inhibitors with higher specificity to brain acetylcholinesterase that may
cause less side effects than the currently used acetylcholinesterase inhibitors.
1.3.1.2. Acetylcholinesterase inhibitors to treat gastrointestinal disorders
The intestinal wall consists mainly of layers of muscle, which contract and relax in a
coordinated fashion, propelling food through the intestine to the anus. This complex pattern of
motility is coordinated by excitatory and inhibitory pathways of the enteric nervous system, to
which acetylcholine is the major excitatory neurotransmitter responsible for the peristaltic
contractions (Holzer and Maggi, 1994). Acetylcholine in gut epithelial cells is also responsible for
controlling ion transport, and therefore, water secretion for gut hydration. This process, very
important in establishing a proper aqueous environment for the enzymatic digestion and
absorption of nutrients, also provides surface lubrication to propel intestinal contents by
peristaltic movements (Hirota and McKay, 2006).

General Introduction
14
Acetylcholinesterase inhibition within the enteric nervous system prevents the
degradation of acetylcholine, increasing gastrointestinal motility (Jarvie et al., 2008). The
acetylcholinesterase inhibitor neostigmine has been used to treat conditions related to
impairment of gastrointestinal motility, such as colonic pseudo‐obstruction (Ponec et al., 1999)
and post‐operative impairment after colorectal surgery (Kreis et al., 2001). Other inhibitors have
also been used to treat gastric motility dysfunctions, such as metochlopramide and vinitidine
(Sasha et al., 1995). Conditions that may be associated with disturbances of gastrointestinal
motility and treated with acetylcholinesterase inhibitors include dysphagia, gastric stasis,
achalasia, abdominal pain, paralytic ileus, vomiting and constipation (Sasha et al., 1995).
However the side effects of acetylcholinesterase inhibitors as discussed in section 3.2. may be an
inconvenient of these therapies, being nausea and diarrhoea the most commonly associated
with neostigmine (Jarvie et al., 2008).
Some plant extracts are empirically used to treat gastrointestinal disorders, such as
extracts from Plectranthus species, and specifically, P. barbatus (Lukhoba et al., 2006). This
therapeutic activity may be related to the inhibition of acetylcholinesterase activity by the plant
extract components, which may be a potential alternative for the currently used medicines.
1.3.1.3. Finding acetylcholinesterase inhibitors ‐ from in vitro to in vivo studies
The research of acetylcholinesterase inhibitors led to the development of methods to
quantify acetylcholinesterase activity. The direct detection of acetylcholine (substrate) or choline
(product) is not easy. Therefore several indirect methods were developed, as the titration of the
acetic acid formed by the hydrolysis of acetylcholine (Jacobson et al., 1957), the use of
acetylcholine analogues as substrates, such as acetylthiocholine (Ellman et al., 1961) or
radiolabelled acetylcholine (Johnson and Rumel, 1975), or by hydrolysing the choline
enzymatically with choline oxidase, the H2O2 formed in the hydrolysis, with the addition of
luminol and peroxidase, can be detected by chemoluminescence (Birman et al., 1985).
The most commonly used in vitro assay was the one described by Ellman et al. (1961), in
which acetylthiocholine is used as substrate for acetylcholinesterase, forming acetate and
thiocholine. The thiocholine reacts with 5,5’‐dithio‐bis‐(2‐nitrobenzoic acid) (DTNB), in a very
fast chemical reaction, originating 2‐nitrobenzoate‐5‐mercaptothiocholine and
5‐thio‐2‐nitrobenzoate (TNB), which has a peak of light absorbance at 405nm (Figure 1.8)
(Ellman et al., 1961). This method is very used due to its sensitivity, fastness and accuracy, which
is mostly due to the fast reaction between thiocholine and DTNB, however it has some
limitations related with inhibitors to be tested (Sinko et al., 2007). Some compounds may react

Chapter I
15
directly with DTNB, such as thiols (Ellman, 1958), or may hydrolyse chemically acetylthiocholine,
such as oximes (Sinko et al., 2007), and in both cases there is an increase in the rate of TNB
formation that is not related with acetylcholinesterase activity.
Figure 1.8. Reaction associated with the Ellman assay to quantify acetylcholinesterase activity. The final
product TNB can be spectrofotometrically detected due to its absorption at a wavelength of 405 nm.
(adapted from Frasco et al., 2005)
In spite of its limitations, the Ellman method is very versatile and can be adapted in
order to determine the effect of acetylcholinesterase inhibitors by measuring cortical
acetylcholinesterase activity after administration (Chattipakorn et al., 2007). However, this is an
ex vivo technique that involves the sacrifice of animals, and therefore must be justified by
preliminary studies.
Acetylcholinesterase activity can also be measured in vivo by positron emission
tomography (PET), as described by Iyo and co‐workers (1997), who compared
acetylcholinesterase activity in brains of healthy controls and in brains of Alzheimer’s disease
patients. This method was also used to assess the efficiency of acetylcholinesterase inhibitors in
the brains of Alzheimer disease patients, such as donepezil (Bohnen et al., 2005) and
galanthamine (Kadir et al., 2008). However, this technique involves the administration of
Acetylthiocholine
Acetylcholinesterase
H2O
ThiocholineAcetate
5,5’‐Dithio‐bis(2‐nitrobenzoic acid)
(DTNB)
5‐thio‐2‐nitrobenzoic acid (TNB)
(detection at 405nm)

General Introduction
16
radiolabelled acetylcholine analogues to humans, and so it is considered just for potentially
active compounds that have been selected by in vitro and animal testing.
1.3.2. Inflammation and Antioxidants
Inflammation is a complex biological response to harmful stimuli involving the vascular
system, the immune system and the injured cells, in a highly coordinated process involving
multiple factors acting in a complex network as stimulators or inhibitors. Upon a stimulus
(infection) endogenous mediators are released, such as cytokines and chemokines, that
contribute to the recruitment of circulating leukocytes to the inflammation site (Gouwy et al.,
2005). These cells, highly specialized in phagocytosis, have developed mechanisms for
intracellular digestion of particles, such as pathogens and cell debris, involving the production of
radical oxygen species (ROS) and a range of hydrolytic and proteolytic enzymes.
Phagocytosis in leucocytes is followed by a sharp but transient increase in oxygen
uptake, which is used to produce O2‐ (superoxide ion) by the one‐electron reduction of oxygen, a
reaction catalysed by NADPH oxidase at the expense of NADPH. Most of the superoxide reacts
with itself forming H2O2 (hydrogen peroxide), and from these agents a large number of highly
reactive oxidants are formed, including HOCl (hypochlorous acid), which is produced by the
myeloperoxidase‐ catalyzed oxidation of Cl‐ by H2O2; OH. (hydroxyl radical), produced by the
reduction of H2O2 by Fe2+ or Cu+; ONOO‐ (peroxynitrite), formed by the reaction between O2
‐ and
NO‐; and many others. (Babior, 2000; Klebanoff, 2005) This battery of ROS not only kill the
invading particles but it also may damage nearby tissues. Although inflammation has primarily a
protective function, the destructive effects can largely surpass the gravity of the stimulus. Severe
inflammatory responses are often associated with a large number of diseases such as
emphysema, acute respiratory distress syndrome, atherosclerosis, reperfusion injury,
malignancy and rheumatoid arthritis (Babior, 2000).
Lysozyme, one of the lytic enzymes produced by the neutrophils, is a glycoside hydrolase
whose main function is to hydrolyse peptidoglycans in bacterial cell walls, especially in Gram
positive bacteria (Laible & Germaine, 1985). During inflammation lysozyme is discharged from
lysosomes of neutrophils to destroy the phagosomes, however it also may damage the animal
tissue itself, increasing the inflammation process. Therefore, excessive lysozyme activity is
known to be related to allergic conditions and violent inflammatory responses of the immune
system against pathogens (Makino et al., 2003; Ronca et al., 1998; Wu et al., 2006).
Inflammation is also involved in Alzheimer’s disease, with ROS acting as secondary
messengers (Stuchbury and Munch, 2005). Microglia are a type of glial cells that are responsible

Chapter I
17
for the immune defence in the central nervous system, acting as macrophages in the brain and
spinal chord, while astroglia, another kind of glial cells, provide biochemical support and have an
important role in repair and scarring processes of brain and spinal chord after traumatic injuries.
The β‐amyloid plaques in Alzheimer disease patients lead to the rapid activation of glial cells and
an over‐expression of glial mediators, namely free radicals (ROS) and citokines (Dickson et al.,
1993; Meda et al., 1995, 2001). The propagation of the consequent oxidative stress and
inflammation by the cytotoxic activation of glial cells plays a key‐role in the pathogenesis and
progressive degeneration characteristic of Alzheimer’s disease (Figure 1.7) (Meda et al., 2001).
The use of antioxidants to treat Alzheimer’s disease patients is still a controversial issue.
Many studies focused on the treatment of Alzheimer’s disease with antioxidant compounds that
may act through different mechanism, from free radical scavengers (citoplasmatic and
mitochondrial) and metal chelators to anti‐inflammatory agents and transcriptional activators
(Figure 1.9). Although the results obtained from animal models were usually promising, when
the compounds were tested in human clinical trials the results often did not present a significant
difference from the control groups (Dumond and Beal, 2011; Mecocci and Polidori, 2011).
Several factors were proposed to explain this disparity, being the bioavailability of the
compounds one of the most important, namely in the absorption, transport, distribution and
retention in the target area of the human body (Dumont and Bael, 2011). Other important
factors include the reaction kinetics, as the free radicals must be neutralized faster than the
damage they cause in the target tissue, and the mechanism of action itself (Mecocci and
Polidori, 2011; Viña et al., 2004; Dumont and Bael, 2011). Patients treated with vitamin E, for
instance, showed an improvement in cognitive function when a variation of antioxidant function
was detected, suggesting that the effectiveness of vitamin E over Alzheimer disease patients was
dependent on its function as antioxidant, which was highly variable among the patients (Viña et
al., 2004). Therefore it is suggested that a higher efficacy can be achieved by administering
several molecules acting with different antioxidant mechanisms, and monitoring the antioxidant
status of the patients (Mecocci and Polidori, 2011). However, many clinical trials of antioxidant
compounds are still undergoing, and most of the compounds known to be effective in in vitro
and in vivo systems are still untested in clinical trials (Dumont and Bael, 2011; Mecocci and
Polidori, 2011).

General Introduction
18
Figure 1.9. Antioxidant strategies in Alzheimer’s disease. Solid arrows represent the mechanisms of the
disease and dashed arrows represent the mechanisms of antioxidant therapy (Adapted from Dumond and
Bael (2011)).
Plant‐derived natural products have been and will continue to be extremely important to
mankind as sources of medicinal drugs. The continued interest of pharmaceutical industry in
plant‐derived drugs led to the screening of species used in traditional medicines to treat illnesses
and/or promote health. Therefore it is not surprising Cai et al. (2004) studied the antioxidant
activity of extracts of 112 plant species used in Chinese traditional medicine.

Chapter I
19
2. Thesis overview
Herbal teas are considered functional drinks, since almost all of the chemical
components of these aqueous extracts have biological functions. The ethnobotanical uses of
these drinks can often be explained through the biochemical activities found for the complete
extracts or for their isolated components. Despite of the great popularity of P. barbatus tea in
Brazil, Africa and India to treat a wide range of diseases, the present knowledge on its biological
activities is still very limited. The experimental work developed and presented in this thesis is a
contribution to the validation of the ethnobotanical medicinal uses.
The main objective of this thesis is to evaluate the potential of P. barbatus herbal tea in
therapies related with acetylcholinesterase inhibition, antioxidant and anti‐inflammatory
activities. Briefly, to approach the main objective, this thesis aimed to:
Provide information about the acetylcholinesterase inhibition, antioxidant and
anti‐inflammatory activities of P. barbatus herbal tea.
Study the bioavailability of the P. barbatus extract, and whether the active
components, or metabolites, can reach the target organs.
Analyse the interaction between the plant extract components and proteins to
provide information on some of the mechanisms involved in the extract’s
bioavailability and activities.
Contribute to the evaluation of the potential of P. barbatus herbal tea for the
symptomatic treatment of Alzheimer’s disease and gastrointestinal disorders.
This thesis is organized in six chapters.
Chapter I is a general introduction that compiles the existing information about the
ethnobotanic uses of extracts of Plectranthus species, especially the most used for medicinal
purposes, P. barbatus. The most common active components of aqueous extracts (herbal teas)
are reviewed especially regarding the current knowledge of their metabolism and bioavailability
when ingested. As the aim of this thesis is the potential of using the P. barbatus herbal tea as
acetylcholinesterase inhibitor, antioxidant and anti‐inflammatory, a brief review of the current
knowledge about these activities and of their most common therapeutic uses is presented.
Chapter II describes the general methodologies used throughout the practical work.
Chapter III reports in vitro studies of the activities of P. barbatus herbal tea. A screening
of antioxidant and acetylcholinesterase inhibition activities for several Plectranthus species is
presented. The aim of this screening was to prove that P. barbatus, the most used species for

General Introduction
20
medicinal treatments, was the most interesting in terms of its composition and activity. This
chapter also describes the in vitro digestion P. barbatus herbal tea with artificial gastric and
pancreatic juices, β‐glucuronidase from gut bacteria, and with Caco‐2 cells, modelling the
metabolism by intestinal cells, with the objective to know whether the extract components were
metabolized, and if the remaining acetylcholinesterase activity has a value that justifies carrying
out in vivo studies in rats. In this chapter the anti‐inflammatory activity of the P. barbatus extract
is also evaluated by measuring its ability to decrease the amount of hypochlorous acid produced
by activated neutrophils.
Chapter IV explores the bioavailability of the P. barbatus herbal tea in order to
investigate if, when administered to rats, the active compounds present in this aqueous extract
or their metabolites occur in the bloodstream and in the brain, and if the neuronal
acetylcholinesterase activity is affected by the administration of the P. barbatus extract. The
bioavailability of the herbal tea is also studied in Caco‐2 cell monolayers, to determine if the
extract components interfere with the permeability and metabolisation of each other.
Substrates of the transport systems MCT and Pgp are co‐administered in order to know the
involvement of these transporters in the bioavailability of the plant extract compounds.
Chapter V describes studies on the interactions between the P. barbatus herbal tea
components and proteins, using fluorescence and FTIR spectroscopic methods. The relationship
between the interactions and the inhibition of the enzymatic activity is also discussed. The
interactions between the plant extract and acetylcholinesterase are studied in order to elucidate
the mechanism by which the enzyme is inhibited by the herbal tea components. In addition, the
binding of the P. barbatus extract to the human plasma proteins albumin and lysozyme is also
analysed, to know if the extract components may be transported in the bloodstream bound to
these transport proteins, increasing their bioavailability in target organs. The influence of the
P. barbatus herbal tea on lysozyme activity is also evaluated with the objective of knowing if the
plant extract could be useful in alleviating inflammation and allergic conditions by inhibiting
lysozyme hydrolytic activity.
Chapter VI presents a global discussion and summarizes the main conclusions of this
thesis.

Chapter II
Materials and Methods


Chapter II
23
1. Plant material
The leaves of five Plectranthus species (P. barbatus, P. ecklonii, P. fruticosus,
P. lanuginosus and P. verticillatus) cultivated in the Botanic Garden of the University of Lisbon
were collected during spring (March–June) 2006 to prepare the extracts for the studies in the
section 1 of Chapter III and in September 2008 for all the other studies.
Vouchers specimens from each species have been deposited in the Herbarium of this
Botanic Garden. P. barbatus Andr. (LISU: 214625), P. ecklonii Benth. (LISU: 146895),
P. fruticosus L’Herit (LISU: 214627), P. lanuginosus (Benth.) Agnew. (LISU: 177258),
P. verticillatus (L.f.) Druce (LISU: 171088).
2. Animals
All experiments were carried out in accordance with the guidelines of the European
Communities Council Directive of 24th November 1986 (86/609/ECC). Adult male
Sprague–Dawley rats (3–4 months old) were obtained from Instituto de Investigação Cientifica
Bento da Rocha Cabral (Lisbon, Portugal). Two rats per cage were maintained in a room at 22 oC
under 12 h dark/ light cycling and ad libitum access to water and regular chow.
3. Chemicals
All chemicals were of analytical grade. Acetylcholinesterase (AChE) type VI-S, from
electric eel 349 U/mg solid, 411 U/mg protein, 5,50-dithiobis[2-nitrobenzoic acid] (DTNB),
acetylthiocholine iodide (AChI), tris[hydroxymethyl]aminomethane (Tris buffer),
dimethylsulphoxide (DMSO), 2,2-diphenyl-1-picrylhydrazyl (DPPH), linoleic acid, β-carotene,
2,6-di-tert-butyl-4-hydroxytoluene (BHT), Tween 40, β-glucuronidase type IX-A from Escherichia
coli, HEPES buffer, pancreatin, pepsin, thiazolyl tetrazolium bromide (MTT), polyethylenoglycol,
sulfatase from Helix pomatia, cathecol o-methyltransferase (COMT) from porcine liver,
S-adenosylmethionine (SAM), L-cystein non-animal source, human serum albumin (HSA),
lysozyme from human milk (EC 3.2.1.17), lyophilized Micrococcus lysodeiktcus quercetin,
luteolin, apigenin, caffeic acid and rosmarinic acid were obtained from Sigma. Trifluoroacetic
acid was acquired from Merck. DMEM (Dulbecco’s Modified Eagle Medium), HBSS (Hank’s
Balanced Salt Solution), glutamine, Pen–Strep (penicillin–streptomycin), PBS (phosphate
buffered saline), FBS (foetal bovine serum) and trypsin were bought from Lonza.

Materials and Methods
24
4. Extract preparation
Aqueous plant extracts were prepared as decoctions and infusions. Decoctions were
prepared using 10 g of grinded fresh plant material boiled for 10 min in 100 ml of distilled water.
Infusions were prepared using 10 g of grinded fresh plant material in 100 ml of freshly boiled
distilled water. Both infusions and decoctions were filtered through Whatman paper and
lyophilized.
The Plectranthus barbatus extract used in the all the work, except section 1 of Chapter III, was
the same, prepared as a decoction of 10 g of ground fresh leaves boiled for 10 min at 100oC, in
100 ml of distilled water and filtered through grade 1 Whatman paper. The extract was
lyophilised and the yield of extraction was approximately 140 mg of extract/g of plant.
5. Acetylcholinesterase inhibition
Acetylcholinesterase enzymatic activity was measured using adaptations of the method
described by Ingkaninan et al. (2003).
In microplates (in Chapter III, section 1): 90µl 50 mM Tris–HCl buffer pH 8, 30 µl sample
and 7.5 µl acetylcholinesterase solution containing 0.26 U/ml were mixed in a microplate and
left to incubate for 15 min. Subsequently, 22.5 µl of a solution of AChI (0.023 mg/ml) and 142 µl
of 3 mM DTNB were added. The absorbance at 405 nm was read when the reaction reached
equilibrium. A control reaction was carried out using water instead of extract and it was
considered 100% activity.
I(%) = 100 – (Asample/Acontrol) x 100
where I(%) is the percent inhibition of acetylcholinesterase, Asample is the absorbance of the
extract containing reaction and Acontrol the absorbance of the reaction control. Tests were carried
out in triplicate and a blank with buffer instead of enzyme solution was used.
In spectrophotometer cuvettes: 325 µl of 50mM Tris buffer (or 50 mM HEPES, when the
samples contained methanol) pH 8, 100 µl of sample and 25 µl acetylcholinesterase solution
containing 0.26 U/ml were mixed in a spectrophotometer cuvette and left to incubate for 15 min
at 25 oC. Subsequently, 75 µl of a solution of AChI (0.023 mg/ml) and 475 µl of 3 mM DTNB were
added. The absorbance at 405 nm was read during the first 5 min of the reaction and the initial
velocity was calculated. A control reaction was carried out using water, which was considered to
have 100% activity.

Chapter II
25
I(%) = 100 – (Vsample/Vcontrol) x 100
where I is the percent inhibition of acetylcholinesterase, Vsample is the initial velocity of the
extract containing reaction and Vcontrol is the initial velocity of the control reaction. Tests were
carried out in triplicate and a blank with buffer instead of enzyme solution was used. When
different volumes sample were used (maximum 200 µl), the final volume was corrected to 1ml
by adjusting the amount of buffer added in the beginning, and control reactions were carried
out in the same conditions.
6. Determination of antioxidant activity
Antioxidant activity was measured by DPPH and by a β-carotene/linoleic acid method,
both as described by Tepe et al. (2005), the latter with a slight modification.
DPPH assay: To a 2.5 ml solution of DPPH, (0.002% in methanol), was added 25 µl of
plant extract. The mixture was incubated for 30 min at room temperature. The absorbance was
measured at 517 nm against a corresponding blank. The antioxidant activity was calculated as
AA(%) = 100 x (ADPPH – Asample)/ADPPH
where AA is the antioxidant activity, ADPPH is the absorption of the DPPH solution against the
blank, Asample is the absorption of the sample against the blank. The tests were carried out in
triplicate and the extract concentration providing 50% of antioxidant activity (IC50) was obtained
by plotting the antioxidant activity against the plant extract concentration.
β-carotene–linoleic acid assay (just used in Chapter III, section 1): β-carotene (0.5 mg),
25 µl of linoleic acid and 200 mg of Tween 40 were dissolved in 1 ml of chloroform. The organic
solvent was evaporated. The residue was redissolved in 100 ml of aerated water. To an aliquot
of 2.5 ml of this solution, 300 µl of each sample were added. The test tubes were incubated in
hot water (50 oC) for 2 h, together with two blanks, one containing the standard antioxidant BHT
(as a positive control) and the other one with the same volume of distilled water instead of the
extracts. In the tube containing BHT the characteristic yellow colour of the β-carotene was
maintained. The absorbance was measured at 470 nm. The antioxidant activity (AA) of the
extracts was calculated using the following equation:
AA(%)= 100 x β-carotene content after 2 h assay/initial β-carotene content

Materials and Methods
26
The tests were carried out in triplicate and the extract concentration that gave 50% of
antioxidant activity (IC50) was obtained by plotting the percentage of antioxidant activity against
the extract concentration.
7. HPLC analysis
The HPLC analysis was carried out in a Liquid Chromatograph Finnigan™ Surveyor® Plus
Modular LC System equipped with a Purospher® STAR RP-18 column, from Merck and Xcalibur
software.
Generally, the composition of the extracts was analysed by injecting 25 µl and using a
gradient composed of solution A (0.05% trifluoroacetic acid acid), solution B (acetonitrile) and
solution C (methanol) as follows: 0 min, 90% A, 10% B and 50 min 5% A, 80% B, 15% C. Standard
compounds were run under the same conditions and the detection was carried out between 200
and 600 nm with a diode array detector. To purify the three main compounds from the
P. barbatus water extract, the HPLC system referred to above was also used. The peaks were
collected separately and this process was repeated several times.
8. NMR spectroscopy
1H-NMR spectra were recorded using a 400 MHz Bruker Advance 400. Samples were
dried, dissolved in DMSO-d6 and referenced to the residual solvent resonance at δH 2.50 ppm.
9. Mass spectrometry experiments
The mass spectrometric identification of rosmarinic acid was carried out on a
Thermoquest LCQ Duo quadrupole ion trap mass spectrometer equipped with an electrospray
interface. The flow rate of the electrospray solution to the ion source was 5 µl/min and the
instrumental parameters (sheath gas flow rate, ion spray voltage, capillary temperature,
capillary voltage, lens and octapole voltages) were optimised for maximum abundance of the
ions of interest. All mass spectrometric data were acquired in the negative ionisation mode. The
full scan mass spectra were measured using 50 ms for collection of the ions in the trap and the
three micro scans were summed. The data for mass spectra were based on 10–100 scans.
MS/MS experiments were performed in order to ascertain characteristic patterns. Helium was
used as collision gas and the collision energy was gradually increased until both the precursor
and product ions could be observed. MS/MS spectra were measured using 200 ms for collection

Chapter II
27
of the ions in the trap and the three micro scans were averaged. The data for mass spectra were
also based on 10–100 scans.
Further experiments were performed in an ApexQe FTICR Mass Spectrometer from
Bruker Daltonics (Billerica, MA, USA) equipped with an electrospray ion source and a 7 T actively
shielded superconducting magnet. The samples were introduced, by means of an infusion pump
from KD Scientific (Holliston, MA, USA), with a flow rate of 120 µl/h. The mass spectrometer was
calibrated using a 2.8 x 10-6 mol/l solution of polyethylenglycol acquired from Sigma-Aldrich
(St. Louis, MO, USA) in methanol HPLC grade acquired from Panreac (Barcelona, Spain) and
acidified with 0.1% (V/V) of acetic acid acquired from Fluka (Seelze, Germany).
The mass spectra were acquired in the positive ion broad band mode, with an
acquisition size of 512 k, in the mass range of 50-500. The nebuliser gas flow rate was set to
2.5 l/min, the dry gas flow rate was set to 4.0 l/min at a temperature of 220 oC. The capillary
voltage was set to 5000 V and the spray shield voltage was set to 4500 V. All mass spectra
presented are the average of 32 mass spectra.
10. In vitro intragastric metabolism assays
10.1. In vitro metabolism by the gastric juice
The assay was adapted from Yamamoto et al. (1999). Two and a half millilitres of gastric
juice were added to 2.5 ml of extract solution (5 mg/ml) or rosmarinic acid solution (2 mg/ml).
The mixture was left to incubate at 37 oC for 4 h. Samples (100 µl) were taken hourly, added to
900 µl of ice-cold methanol and analysed by HPLC. The gastric juice (100 ml) consisted of 320 mg
of pepsin, 200 mg NaCl, pH 1.2 (with HCl). Assays were done in triplicate.
10.2. In vitro metabolism by pancreatic juice
The assay was adapted from Yamamoto et al. (1999). Two and a half millilitres of
pancreatic juice were added to 2.5 ml of extract solution (10 mg/ml) or rosmarinic acid solution
(2 mg/ml). The mixture was left to incubate at 37 oC for 4 h. Samples (100 µl) were taken hourly,
added to 900 µl of ice-cold methanol, and centrifuged for 5 min at 5000g. The supernatant was
analysed by HPLC.
Two hundred microlitre samples were taken at the same time, centrifuged 5 min at
5000g and the supernatant was analysed for acetylcholinesterase activity, against a blank with
water instead of plant extract. The gastric juice consisted of 250 mg of pancreatin in 10 ml of
potassium-phosphate buffer 50 mM, pH 8. Assays were done in triplicate.

Materials and Methods
28
10.3. Glucuronidase activity
An adaptation of the method described by Justino et al. (2004) was used. The mixture
contained 567.3 U/ml of β-glucuronidase and 5 mg/ml aqueous extract of P. barbatus, in
potassium-phosphate buffer (50 mM, pH 7.4). The reaction was incubated for 2 h at 37 oC. After
this time the mixture was centrifuged at 5000g for 10 min and the supernatant was analysed by
HPLC. Two control reactions were analysed, one consisting of the reaction mixture without plant
extract and the other consisting of the reaction mixture without β-glucuronidase.
10.4. Metabolism by the Caco-2 cells
The assay for the metabolism of plant extracts by Caco-2 cells was adapted from Kern et
al. (2003). Caco-2 cells (ATCC#HTB-37), a colorectal adenocarcinoma epithelial cell line, were
cultured in DMEM medium supplemented with 10% FBS, 100 U/ml penicillin, 100 U/ml
streptomycin, and 2 mM L-glutamine, at 37 oC in an atmosphere with 5% CO2. The culture
medium was changed every 48–72 h.
For the assays, 2 x 104 cells were seeded in 4 cm diameter Petri dishes and grown for 10
days (to approximately 90% confluence). The medium was replaced with 2.5 ml HBSS plus
1–7 mg/ml of plant extract. For the assays with standards, the concentrations used were
1 mg/ml for rosmarinic acid and 50 µM for apigenin or luteolin. The cells were left in the
incubator and 100 µl samples of the medium were collected at 0, 1, 2, 4, and 6 h, added to 900
µl of ice-cold water, centrifuged 10 min at 5000g, and analysed by HPLC. Samples (100 µl) were
also collected at the same incubation times, centrifuged and analysed for acetylcholinesterase
inhibition, against a blank of HBSS incubated with Caco-2 cells under the same conditions. All
assays were done in triplicate.
After the 6-h assay, the cells in each Petri dish were washed with 500 µl HBSS and
incubated with 1 ml solution of 1:10 TFA (0.05% in methanol) for 30 min. The extract was
centrifuged for 10 min at 5000g, and the supernatant was analysed by HPLC.
10.5. Antiacetylcholinesterase and antioxidant activities of the digested extracts
Aliquots were withdrawn at the beginning of the experiment and after each hour. The
sample was centrifuged at 5000g to discard the bottom phase containing the enzyme, pepsin,
pancreatin or glucuronidase. The upper phase was used for the determination of
acetylcholinesterase inhibition activity as well as antioxidant activity by the DPPH method
according to the procedures described in the sections 4 and 5 of this Chapter. The aliquots from

Chapter II
29
the incubations were diluted to concentrations in the assays corresponding to the IC50 values of
the undigested extract.
11. In vitro conjugation studies for metabolites identification
11.1. Preparation of cell-free extracts
A cell-free liver extract was prepared, essentially as described by Justino and co-workers
(2004), to obtain the hepatic enzymes that metabolize flavonoids. In brief, rat livers were
collected and homogenized in ice-cold 10 mM phosphate buffer, pH 7.4, containing
10 mM 2-mercaptoethanol (1 g wet tissue/2.5 mL buffer). The homogenate was then
centrifuged for 90 min at 12 000 g at 4 oC. The protein content of the supernatant was
immediately determined by the Lowry method (Lowry et al., 1951) and the remainder
supernatant was lyophilized and stored at -20 oC until required.
11.2. Glucuronidation assay
The standard assay mixture contained 0.5 mg.mL-1, P. barbatus extract, or 40 mM
rosmarinic acid, or 50 mM flavonoid standards, 2 mM UDPGA, and cell-free extract (5 mg.mL-1)
in 10 mM potassium phosphate buffer, pH 7.4, final volume of 1 mL (Justino et al., 2004). The
reaction was initiated by addition of the cytosolic fraction extracted and the mixture was
incubated for 30 min at 37 oC without shaking. The reaction was stopped and the metabolites
were extracted as described in the HPLC analysis section below.
11.3. Synthesis and identification of methyl rosmarinic acid
The methylation of rosmarinic acid was done by the porcine liver enzyme catechol-o-
methyl-transferase (COMT), using the cofactor S-adenosylmethionine (SAM), with an adaptation
of the method described by Baba and co-workers (2004). Briefly, 20 U of COMT were dissolved
in 200 ml of deoxygenated 10 mM K-phosphate buffer, 20 mM L-cystein, 2 mM MgCl2, pH 7.4
and pre-incubated at 37 oC for 10 min under inert atmosphere. Five hundred microlitres of
7.1 mM SAM and 100 ml of rosmarinic acid, both dissolved in the reaction buffer, were added.
Two hundred microlitres of 7.1 mM were added every two hours, and the reaction was stopped
at 6 h by the process described in the section for HPLC analysis. The compounds were isolated by
HPLC, collected and analyzed by electrospray ionization mass spectrometry (ESI-MS). All
experiments were performed using a LCQ Duo ion trap mass spectrometer from Thermo
Scientific (San Jose, CA, USA) equipped with an ESI source. Samples were introduced, via a

Materials and Methods
30
syringe pump (flow rate of 5 mL.min-1), into the stainless steel capillary of the ESI source. The
applied spray voltage in the source was 4.5 kV, the capillary voltage was 10 V and the capillary
temperature was 220 oC. All the mass spectrometer parameters were adjusted in order to
optimize the signal-to-noise ratios for the ions of interest. Nitrogen was used as nebulising and
auxiliary gas in the source. All mass spectrometry data were acquired in the negative ion mode,
the full scan spectra were recorded in the range m/z 100–1000 and three micro-scans were
averaged. MS/MS experiments were performed with helium as collision gas.
12. In vivo studies protocol
12.1. Intragastric and intraperitoneal administration
P. barbatus herbal tea was administered to rats through intragastric procedure
(600 mg.kg-1, equivalent to 150 mmol RA kg-1) or intraperitoneal injection (1000 mg.kg-1,
equivalent to 250 mmol rosmarinic acid kg-1). For each experiment, twelve adult male Sprague–
Dawley rats weighing approximately 400 g and 16 h fasted were randomly divided into two
groups. Rosmarinic acid standard was administered dissolved in ethanol saline solution (20%
ethanol, 0.9% NaCl solution) in a concentration of 550 mmol kg-1. The high dose administered is
justified by the need of obtaining high levels of the different metabolites for analytical purposes.
To control animals only the ethanol saline solution was administered. Blood was withdrawn
from three rats, thirty minutes and 1 h after administration, by cardiac punction into K3EDTA
tubes and kept on ice. The brains were than collected in ice cold K-phosphate buffer 10 mM pH
7.4 10 mM mercaptoethanol and also kept on ice.
12.2. Plasma and brain sample preparation
Plasma was separated from red blood cells by centrifugation at 5000 g for 5 min at 4 oC
and samples were stored at -80 oC until further studies. Each rat brain was weighed and
homogenized in 5 ml 10 mM K-phosphate buffer pH 7.4 10 mM mercaptoethanol. The
homogenate was then centrifuged for 90 min at 12 000 g, at 4 oC and the supernatant (brain
extract) stored at -80 oC until further studies.
12.3. Determination of glucuronidated and sulfated metabolites
An adaptation of the method described by Justino and co-workers (2004) was used. To
determine the amounts of glucuronidated, sulfated and glucuronidated-sulfated compounds
sets comprised one assay with b-glucuronidase, one assay with sulfatase, one assay with both

Chapter II
31
enzymes and a control with no addition of enzymes were prepared. To 500 ml of sample were
added 1000 U of b-glucuronidase, and/or 25 U sulfatase and incubated at 37 oC for 1 h.
The samples were then processed as described in the section 12.4 for HPLC analysis.
12.4. Preparation of samples for HPLC analysis
Methanol was added to 500 ml of plasma or brain extract to 1 ml solution and the
mixture was vortexed, left to precipitate for one hour at 4 oC, and centrifuged for 10 min at
10 000 g, at 4 oC. The supernatant was recovered, left to precipitate another hour and
centrifuged in the same conditions prior to HPLC analysis. Controls were done using rosmarinic
acid and caffeic acid as internal standards added to plasma and brain extracts to determine the
losses related to this purification process. This protocol showed negligible loss of the standard
compounds (less than 10%) and the yield was calculated taking into account the concentrations.
12.5. Determination of acetylcholinesterase activity in brain samples
Acetylcholinesterase enzymatic activity was measured using an adaptation of the
methods described by Chattipakorn and coworkers (2007) and in section 5. Four hundred
microlitres of 50 mM HEPES buffer pH 8 and 50 ml brain extract were mixed in
spectrophotometer cuvette and left to incubate for 15 min at 25 oC. Subsequently, 75 ml of a
solution of AChI (0.023 mg ml-1) and 475 ml of 3 mM DTNB in HEPES 50mM pH 8 were added.
The absorbance at 405 nm was read during the first five minutes of the reaction and the initial
velocity was calculated as mAU min-1 mg-1 and converted into nmole min-1 mg-1 of tissue mass. A
control reaction was carried out using the brain extract from the control rats, and it was
considered 100% activity for calculations of enzymatic inhibition.
13. Protein fluorescence measurements
The fluorescence quenching study on HSA was performed in 3 ml solutions containing
1 × 10−8 M HSA in Tris-HCl buffer (0.20 M, pH 7.4) containing 0.10 M NaCl and the appropriate
quantities of quencher – P. barbatus extract, rosmarinic acid, luteolin or apigenin (Xiao et al.,
2008).
Lysozyme intrinsic fluorescence quenching was studied in 3 ml solutions containing
0.02 mg.ml−1 and the appropriate amount of quencher in K-phosphate buffer (0.01 M, pH 7.4).
All samples were pre-incubated for 1 h at 37◦C (Xiao et al., 2008).
The fluorescence quenching study on of acetylcholinesterase was performed according
Xiao and coworkers (2008), in 3ml solutions containing 5U of AChE in Tris-HCl buffer (50mM,

Materials and Methods
32
pH 8) and the appropriate quantities of quencher – P. barbatus extract, rosmarinic acid,
quercetin, luteolin or apigenin.
All fluorescence measurements were performed on a LS55 spectrofluorometer (Perkin-
Elmer, UK) in a 1 cm quartz cell. Excitation and emission bandwidths were set to 10 nm. Emission
spectra were recorded from 300 to 500 nm, under excitation wavelength of 280 nm. The
fluorescence intensity was measured at 335 nm under the same excitation wavelength, at
different temperatures (293, 298 and 303 K).
14. FTIR measurements
FTIR measurements were carried out at 21◦C using a Bruker Vector 22 Spectrometer.
Each spectrum was acquired by averaging between 32 and 300 scans in the spectral range of
400–4000 cm−1 at 4 cm−1 resolution.
Fifty microliters of reaction mixture were transferred into a CaF2 infrared cell, fitted with
a 50 μm path Teflon spacer and placed in the spectrometer. Spectra were acquired every 2 min
for 1 h. For each sample a spectrum was acquired after 2 h, averaging 300 scans.
The hydrogen–deuterium exchange was initiated when HSA was dissolved in Tris buffer,
50 mM pH 7.4, containing 25 mM NaCl, prepared in deuterium oxide, and the compounds in
study in a final concentration of 500 μM HSA, 500 μM of standard compounds or 10 mg/ml of P.
barbatus extract.
For the measurements with lysozyme, the hydrogen–deuterium exchange was started
when lysozyme was dissolved in K-phosphate buffer 10 mM pH 7.4, prepared in deuterium
oxide, containing lysozyme in the final concentration of 2 mM, standard compounds in the same
concentration or P. barbatus extract in the final concentration of 10 mg · ml−1.
The AChE hydrogen-deuterium exchange started when AChE was dissolved in Tris buffer,
50mM pH8, prepared in deuterium oxide. and the compounds in this study in the final
concentrations of 7080 U.mL-1 AChE, 1 mM of standard compounds or 10mg/mL of P. barbatus
extract.
The FTIR spectra recorded were analysed using the Bruker OPUS software. FTIR spectra
of the buffer were recorded under identical conditions and the OPUS software was used to
subtract the spectrum of the buffer from the spectrum of the protein in buffer using previously
described procedures (Haris et al., 1990). Subsequently, second-derivative analysis was carried
on the absorbance spectra to reveal the overlapping amide I components (Haris et al., 1990).

Chapter II
33
15. Lysozyme activity measurements
Lysozyme activity was measured with an adaptation of a method previously described
(Laible and Germaine, 1985). Five hundred microliters of M. lysodeikticus suspension in
K-phosphate buffer (0.10 M, pH 7.4) were added to 500 μl of a solution containing lysozyme
(0.4 mg) and an appropriate amount of inhibitor – P. barbatus extract, rosmarinic acid, luteolin
or apigenin – in the same buffer. The decrease in absorbance was followed at 570 nm for the
first five minutes of reaction and compared with the appropriate controls (without inhibitor and
without enzyme). All assays were done in triplicate and IC50 values were calculated.
16. Measurement of hypochlorous acid in activated human neutrophils
16.1. Isolation of human neutrophils
The isolation of human neutrophils was carried out as described by Coelho (2004). A
dextran solution (6% in phosphate buffer 0.1M, pH7.4, NaCl 0.9%) was added to the blood
collected form human donors in a proportion 5:1 (V:V), gently mixed by inversion, and left for
90 min in the dark to separate the red blood cells. A NH4Cl 0.87% (m:V) solution was added to
the supernatant in the proportion 1:2 (V:V) and left to incubate for 5 min at room temperature
for the lysis of the remaining red blood cells, and the leucocytes were collected by centrifuging
5 min at 250g. The cells were washed 3 times with phosphate buffer 16mM, pH 7.4, containing
NaCl 122 mM, KCl 4.89 mM, MgSO4 1.22 mM, D-glucose 1 mg/mL and EDTA 0.03% (m:V) and
collected by centrifuging 5 min at 250g. The cells were ressuspended in 1.5 mL ice cold KRC
solution (phosphate buffer 16mM, pH 7.4, containing NaCl 122 mM, KCl 4.89 mM, MgSO4
1.22 mM, D-glucose 1 mg/mL and CaCl2 0.6 mM), and kept on ice for no longer than 3h.
16.2.Measurement of hypochlorous acid formation by human neutrophils
The hypochlorous acid produced by human neutrophils was measured after activation
with phorbol 12-myristate 13-acetate (PMA), with an adaptation of the method described by
Dypbukt et al. (2005). The neutrophils were incubated for 30 min at 37oC in a 250 µL solution
containing 2x106 cells/mL, taurine 5 mM, PMA 100 ng/mL, and several concentrations of
rosmarinic acid or P. barbatus extract, in KRC buffer. The reaction was stopped by adding
20 µg /mL catalase and incubating on ice for 10 min. The cells were removed by centrifuging
5 min at 11000g, and the hypochlorous acid in the supernatant was measured by the taurin
chlorination method. Briefly, 40 µL of a revealing solution (3,3’,5,5’-tetrametilbenzidina (TMB)
2mM, KI 100µM, in acetate buffer 400mM pH 5.4, 30% dimethylformamide) were added to

Materials and Methods
34
160 µL of the supernatant in a 96-well plate. The absorbance was read at 655 nm after 5 min
incubation at room temperature.
17. Caco-2 bioavailability experiments
Caco-2 cells (ATCC#HTB37), a human colorectal adenocarcinoma epithelial cell line, were
cultured as described in section 10.4.
For transport and metabolism experiments, the cells were seeded at a density of
2-4x104 cells /cm2 in 12-well Transwell plate inserts with 10.5mm diameter, 0.4 µm pore size
(BD Falcon™). The monolayers were formed after 21-26 days. The integrity of the monolayers
was evaluated by measuring the permeability of phenol red or the transepithelial electrical
resistance (TEER). The membranes were considered fit when the permeability of phenol red
from apical to basolateral sides was less than 1% in one hour, or the TEER was higher than
250 Ω.cm2.
In general, to start the assays, the cells were washed with HBSS and 0.5 mL of the
solutions to be tested, in HBSS, were applied into the Transwell inserts (apical side of the cells).
1.5mL of HBSS were added to the plate well (basolateral side of the cells). After 6 hours of
incubation at 37 ºC, 5% CO2, the solutions in both sides of the cells were collected and analysed
by HPLC. The cells were washed with HBSS, and ressuspended by scraping with HBSS. The cells
were sonicated 5x10 s, centrifuged 10 min ant 5000 g, and the supernatant was analysed by
HPLC.
Before carrying out the assays the concentrations of the solutions were proved not to
affect Caco-2 cells growth by performing the MTT viability test (Mosmann, 1983). All
concentrations used showed 0% toxicity and did not affect the Caco-2 cell membrane integrity.
17.1. Study of the permeability and the metabolism of rosmarinic acid, luteolin and
apigenin
The effect of rosmarinic acid, luteolin and apigenin on the permeation and
metabolization of each other on Caco-2 cells was studied by applying several concentrations of
these compounds in the apical side of the cells according to a factorial design known as CCD
(Barker, 1985). The solutions to be used in this study were prepared in HBSS, and the
concentrations of rosmarinic acid, luteolin and apigenin were listed in Table 2.1.
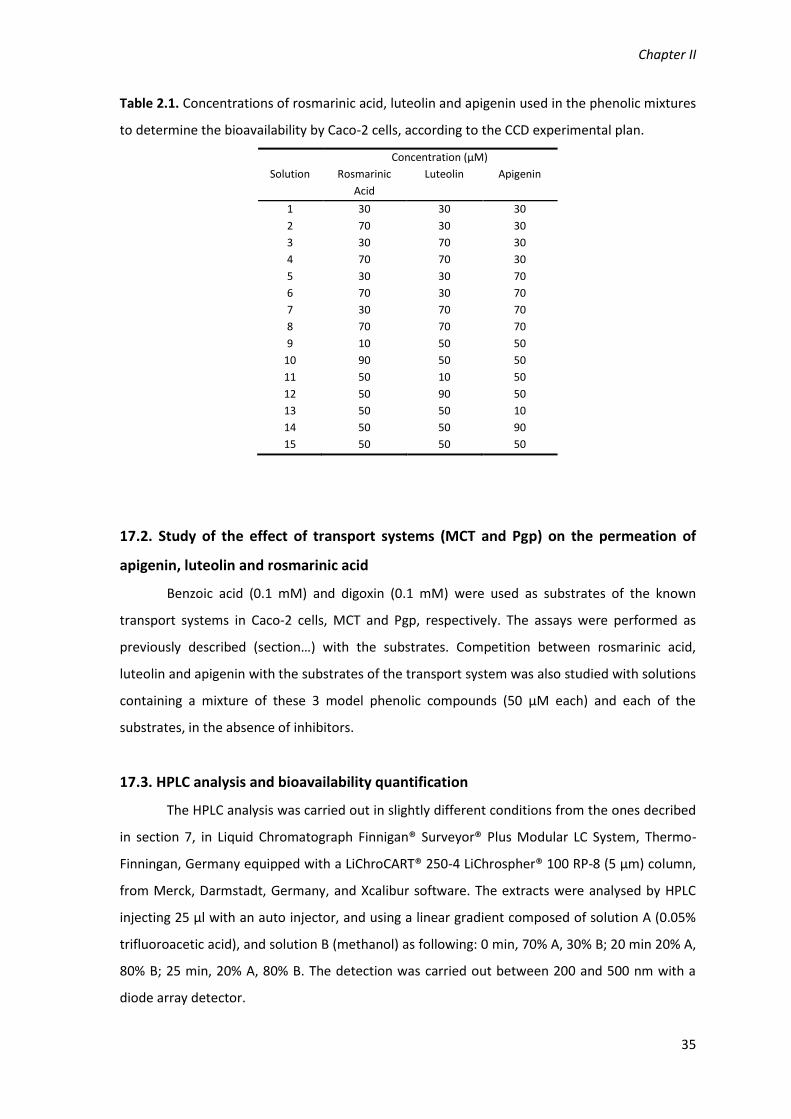
Chapter II
35
Table 2.1. Concentrations of rosmarinic acid, luteolin and apigenin used in the phenolic mixtures
to determine the bioavailability by Caco-2 cells, according to the CCD experimental plan.
Solution
Concentration (µM)
Rosmarinic
Acid
Luteolin Apigenin
1 30 30 30
2 70 30 30
3 30 70 30
4 70 70 30
5 30 30 70
6 70 30 70
7 30 70 70
8 70 70 70
9 10 50 50
10 90 50 50
11 50 10 50
12 50 90 50
13 50 50 10
14 50 50 90
15 50 50 50
17.2. Study of the effect of transport systems (MCT and Pgp) on the permeation of
apigenin, luteolin and rosmarinic acid
Benzoic acid (0.1 mM) and digoxin (0.1 mM) were used as substrates of the known
transport systems in Caco-2 cells, MCT and Pgp, respectively. The assays were performed as
previously described (section…) with the substrates. Competition between rosmarinic acid,
luteolin and apigenin with the substrates of the transport system was also studied with solutions
containing a mixture of these 3 model phenolic compounds (50 µM each) and each of the
substrates, in the absence of inhibitors.
17.3. HPLC analysis and bioavailability quantification
The HPLC analysis was carried out in slightly different conditions from the ones decribed
in section 7, in Liquid Chromatograph Finnigan® Surveyor® Plus Modular LC System, Thermo-
Finningan, Germany equipped with a LiChroCART® 250-4 LiChrospher® 100 RP-8 (5 µm) column,
from Merck, Darmstadt, Germany, and Xcalibur software. The extracts were analysed by HPLC
injecting 25 µl with an auto injector, and using a linear gradient composed of solution A (0.05%
trifluoroacetic acid), and solution B (methanol) as following: 0 min, 70% A, 30% B; 20 min 20% A,
80% B; 25 min, 20% A, 80% B. The detection was carried out between 200 and 500 nm with a
diode array detector.

Materials and Methods
36
The amount of rosmarinic acid, apigenin and luteolin in the samples were determined by
calibration curves of concentration/peak area, and the bioavailability was quantified by the
distribution of each compound in each compartment of the Caco-2 monolayer (apical,
basolateral, and intracellular).
18. Statistical analysis
All results are presented as mean ± standard deviation of three replicates (or five replicates, in
the in vitro digestions with gastric and pancreatic juices) and the software used was Microsoft
Excel®. Additionally analysis of variance (ANOVA) was performed with p = 0.05.

Chapter III
Screening for antiacetylcholinesterase and antioxidant
activities in Plectranthus species. In vitro metabolism and anti‐
inflammatory studies
Falé PL, Borges C, Madeira PJM, Ascensão L, Araújo MEM, Florêncio MH, Serralheiro MLM. 2009.
Rosmarinic acid, scutellarein 4’‐methyl ether 7‐O‐glucuronide and (16S)‐coleon E are the main
compounds responsible for the antiacetylcholinesterase and antioxidant activity in herbal tea of
Plectranthus barbatus (‘‘falso boldo”). Food Chem. 114: 798–805.
Porfirio S, Falé PLV, Madeira PJA, Florêncio H, Ascensão L, Serralheiro MLM. 2010.
Antiacetylcholinesterase and antioxidant activities of Plectranthus barbatus tea, after in vitro
gastrointestinal metabolism, Food Chem. 122: 798–805.
Falé PLV, Filipe MA, Ascensão L, Serralheiro MLM, Mira L. Activity of Plectranthus barbatus extract
against inflammatory response in human neutrophils. To be submitted to Plant Food Hum. Nutr.


Chapter III
39
1. Screening for antiacetylcholinesterase and antioxidant
activities in Plectranthus species
1.1. Introduction
The genus Plectranthus L’Hér., belonging to the Mint family (Lameaceae), comprises
about 300 species widely distributed in Africa, Asia and Australia. A great variety of
ethnobotanical uses are reported for Plectranthus species. However, the most common uses are
to treat a wide range of diseases, including digestive problems, inflammation‐related conditions
and nervous system disorders (Lukhoba et al., 2006). Infusions of several Plectranthus species
are used to treat coughs and colds, liver complaints, and ailments in respiratory system and skin.
Plectranthus barbatus (Lamiaceae) is drunk as an herbal tea to treat a wide range of
diseases, and is also used in food recipes, particularly in South America, Africa and Eastern
regions (Lukhoba et al., 2006). In order to know which were the main compounds that could be
responsible for the healing properties attributed to P. barbatus, infusions and decoctions of this
species were analysed. The inhibition capacity of the extracts on the enzyme
acetylcholinesterase (AChE) was studied. Acetylcholinesterase is the enzyme that catalyses the
hydrolysis of acetylcholine, a neurotransmitter found in the synaptic gap, and the inhibition of
this enzyme could give some explanation for the action of the herbal tea on the nervous system.
Nowadays the AChE inhibition is used in the treatment of the Alzheimer’s disease (AD) (Heinrich
& Teoh, 2004). Therefore the most successful therapy for AD, at present, consists of increasing
the levels of acetylcholine through the inhibition of acetylcholinesterase activity (Heinrich &
Teoh, 2004).
Most of the compounds isolated from the plant polar extract fraction are polyphenols
(Cai et al., 2004; Trouillas et al., 2003). These compounds also have a high antioxidant activity
(Cai et al., 2004; Djeridane et al., 2006). The antioxidant activity found in some compounds has
been connected to the capacity to scavenge the free radicals that are formed during the
inflammation processes (Gomes et al., 2008). It is becoming the objective of research to develop
or find new drugs that exhibit many simultaneous activities, as most of the diseases are not the
result of a unique mechanism, but the consequence of several biochemical processes
(Bembenek et al., 2008). This is one of the reasons why, nowadays, the herbal teas are gaining
popular acceptance. They are a mixture of several compounds, so they can act on different
diseases simultaneously. A bioguided study has been carried out where the inhibition of AChE
was looked for and the P. barbatus water extracts were analysed in order to explain some of its

In vitro Studies of Activity and Metabolism
40
traditional uses. In order to confirm the results obtained with P. barbatus in this work, four
different Plectranthus species were also analysed. P. ecklonii and P. fructicosus also have
medicinal applications but are used mostly to treat skin conditions such as infections and burns
(Lukhoba et al., 2006). P. lanuginosus herbal tea is used for digestive conditions and P.
verticillatus is only used as ornamental.
1.2. Materials and Methods
The materials and methods are described in detail in the Chapter II.
The plant aqueous extracts were prepared as infusions and decoctions. Aliquots of 1 ml
of each extract were lyophilized and used to determine the dry weight. Data present in Table 3.1
are the average of three independent measures.
Table 3.1. Amount of dry plant extract obtained.
Plant species Plant extract (mg dry extract/g fresh plant)
Infusion Decoction
P. barbatus 16 ± 1 36 ± 2
P. ecklonii 36 ± 5 48 ± 5
P. fruticosus 25 ± 3 22 ± 4
P. lanuginosus 12 ± 4 13 ± 3
P. verticillatus 3 ± 1 3 ± 1
Acetylcholinesterase inhibition was determined in microplates, as described in Chapter II
section 5, and the antioxidant activity was quantified by the DPPH and β‐carotene‐linoleic acid
methods, as described in Chapter II, section 6. The composition of the plant extracts was
analysed by HPLC as described in Chapter II section 7, and the main components, isolated by
HPLC, were identified by NMR spectroscopy (Chapter II, section 8) and mass spectrometry
(Chapter II, section 9.).
1.3. Results
1.3.1. General
In this work the aerial part of P. barbatus, P. ecklonii, P. fructicosus, P. lanuginosus and P.
verticillatus was separated into a vegetative part (leaves) and floral part (flowers), which were
analysed separately, not only for their action as AChE inhibitors but also for their antioxidant
activity. The antioxidant activity was evaluated using two tests, the DPPH and the β‐
carotene/linoleic acid bleaching test. The first gives information about the ability of the tested
compounds to scavenge free radicals. The second gives information on the ability of the tested

Chapter III
41
extracts to delay the effect of lipid peroxidation on biological compounds by reacting with chain‐
propagating peroxyl radicals faster than these radicals can react with proteins or fatty acid side‐
chains. In order to establish the results as a function of the dry weight of the extracts, 1 ml of
each extract was lyophilized and its dry weight was determined. As it was shown in Table 3.1,
P. ecklonii and P. barbatus gave the highest yield of plant extract. All the calculations carried out
in the next sections are expressed in dry weight of the extracts.
1.3.2. Acetylcholinesterase inhibition
The inhibition activity of leaves and flowers of P. barbatus demonstrated that the
extracts obtained with the leaves were more active than those obtained with the flowers (<10%
when using 0.5 mg of extract/ml). All the subsequent results refer to the leaves of the
Plecthrantus species.
All the decoctions showed higher activities than the infusions. The decoction of P.
barbatus inhibited the activity of AChE by approximately 31.5%, when using 0.5 mg of extract/ml
of test solution (Table 3.2). This result is within the range of the values found in the literature for
the inhibition of this enzyme with plant extracts (Mata et al., 2007; Orhan et al., 2004) and
corresponds to approximately one small bag of tea made from the leaves (14 mg). The values
obtained with other species of Plectranthus showed that other species also inhibit AChE (Table
3.2). P. verticillatus and P. ecklonii were the species studied that gave higher values, 59.6% and
62.8%, respectively. With the decoctions of P. lanuginosus and P. fruticosus, the inhibition values
obtained when using 0.5 mg/ml, were 10.0% and 31.3%, respectively.
Table 3.2. Inhibition of AChE activity (%), antioxidant activity and rosmarinic acid content of water
extracts of the leaves of several Plectranthus species.
Plectranthus
species
AChE inhibition (%)
with 0.5 mg/ml
dry extract
DPPH
(IC50 µg dry extract/ml)
β-Carotene
(IC50 µg dry extract/ml)
Rosmarinic
acid
(mg/mg
extract)
Infusion Decoction Infusion Decoction Infusion Decoction Decoction
P. barbatus 17.0 ± 5.4 31.5 ± 3.3 10.4 ± 0.3 45.8 ± 0.5 226.4 ± 11.5 69.8 ± 3.1 0.14
P. ecklonii 46.7 ± 0.4 62.8 ± 6.5 2.4 ± 0.1 3.8 ± 0.1 142.1 ± 8.0 109.0 ± 3.1 0.48
P. fructicosus 11.3 ± 1.8 31.3 ± 2.6 6.2 ± 0.9 4.4 ± 0.2 367.1 ± 15.6 234.6 ± 5.4 0.28
P. lanuginosus 13.7 ± 7.2 10.0 ± 1.1 5.2 ± 0.4 3.3 ± 0.2 362.9 ± 5.9 196.7 ± 2.3 0.18
P. verticillatus 47.4 ± 5.4 59.6 ± 7.6 1.5 ± 0.7 1.2 ± 0.4 37.7 ± 1.5 28.9 ± 1.5 0.44
BHTa - 15.7 ± 0.2 12.0 ± 0.7 - aMata et al. (2007)

In vitro Studies of Activity and Metabolism
42
1.3.3. Antioxidant Activity
Antioxidant activity of infusions and decoctions of the Plectranthus species was
evaluated using the DPPH method. Results are presented in Table 3.2 as IC50 (µg dry weight
extract/ml of the test solution). The antioxidant activity of these extracts, on what concerns their
ability to scavenge free radicals, determined by the DPPH, is very high, and similar to the
antioxidant activity of a known synthetic standard, BHT, previously determined, IC50 = 15.7 ± 0.2
µg/ml (Mata et al., 2007). Both extracts of P. verticillatus show the highest antioxidant activity,
IC50 = 1.5 µg/ml (infusion) and IC50 = 1.2 µg/ml (decoction). Also the extracts from P. ecklonii, P.
fructicosus and P. lanuginosus have a comparatively high antioxidant activity. These values are
lower (higher activity) than those recently reported for other Lamiaceae species from genera
Mentha and Salvia (Kanat et al., 2007; Mata et al., 2007; Parejo et al., 2004; Tepe et al., 2005)
and have the same magnitude as those reported for green tea extract, a reference extract highly
commercialised for its antioxidant activity (Atouin et al., 2005).
The capacity to prevent the lipid peroxidation, measured through the β‐carotene–
linoleic acid test, gave values higher, which means less activity, than the one obtained with the
standard BHT. An exception is P. verticillatus that gave a value of the same magnitude as BHT
(Table 3.2). P. barbatus (decoction) and P. verticillatus (infusion and decoction) were the species
showing higher antioxidant activity when measured by this test, 69.8 µg/ml, 37.7 µg/ml and 28.9
µg/ml, respectively.
1.3.4. Identification of the main component of the Plectranthus extracts, responsible for
the enzyme inhibition activity
In order to assess whether the extracts contained a large number of different
compounds, and in an attempt to identify these constituents, HPLC–DAD analysis was carried
out with the decoctions of the Plectranthus species. The decoctions were selected to identify the
main compounds because, in general, they gave higher enzyme inhibition activity than the
infusions. The chromatograms for P. barbatus and P. verticillatus are shown in Figure 3.1a and b,
respectively. The chromatograms of P. ecklonii, P. fruticosus and P. lanuginosus are similar to the
one obtained with P. verticillatus. In all the extracts there is a major peak with the retention time
at ~19.2 min. The decoctions of P. barbatus showed two additional peaks on the total absorption
of the wavelength, Figure 3.1a. In P. barbatus chromatogram the main peak (retention time of
19.2 min) represents 51% of the total area, whilst in all the other chromatograms this peak
represents ~80%. The compounds with retention times of 23.4 min and 33.0 min found in P.

Chapter III
43
barbatus chromatogram represent 6% and 20%, respectively. The amount of the compound,
with a retention time of 19.2 min in the extracts, depends on the plant analysed.
Figure 3.1. HPLC chromatogram of decoctions: (a) Plectranthus barbatus and (b) Plectranthus verticillatus.
Analysing the UV spectra of the peak with a retention time of 19 min, Figure 3.2a, a
similar spectrum to the one obtained with caffeic acid was observed, Figure 3.2b. These
compounds have characteristic absorptions at 330 nm with a shoulder between 300 and 295
nm, Figure 3.2.
Figure 3.2. UV spectra obtained by HPLC‐diode array of (a) compounds with retention time 19.2 min and
(b) caffeic acid.
The extracts of P. barbatus and P. verticillatus were subjected to NMR spectroscopy,
Figure 3.3a and b, respectively. As the P. verticillatus extract was very pure, it was used to
identify the peak with a retention time 19.2 min. 1H‐NMR spectra showed the presence of two
doublets (7.3 ppm and 6.1 ppm, J = 15.88 Hz) corresponding to the double bond of a cinnamic
derivative. The presence in the aromatic region of the spectra, 7.1–6.4 ppm, of two singlets and
four doublets, each one integrating for one proton, suggested the presence of two aromatic
rings with the same pattern of substitution.
The existence of two double doublets at 3.0 ppm and 2.7 ppm coupled with another
double doublet at 4.8 ppm indicated that these two aromatic rings should have different carbon
chain substitutions: a α,β unsaturated and a saturated one, results not shown. 13C‐NMR spectra

In vitro Studies of Activity and Metabolism
44
showed 18 signals corresponding to: two different carbon carbonyls, eight unsaturated
methines, four aromatic quaternary carbons, one deshielded methylene and one methane group
bonded to an oxygen. Compound was identified as rosmarinic acid. 1H‐NMR spectra of the
standard rosmarinic acid was identical with the one obtained with the pure compound (Figure
3.3c). The identification of rosmarinic acid was further confirmed by MS and MS/MS. For this
compound and for the standard rosmarinic acid, an ion at m/z 359, corresponding to [M–H]‐,
was found using ESI–MS (not shown). MS/MS analysis of this ion (not shown) generated a
product ion at m/z 161, which is in agreement with the observation of m/z 163 in the positive
ion mode, considered to be typical of caffeic acid esters (Grayer et al., 2003). The MS data
obtained confirmed the identification of this major compound as rosmarinic acid, which is
consistent with a previous literature report by Parejo et al. (2004). The structure of rosmarinic
acid is shown in Figure 3.4a.
Figure 3.3. NMR spectra of (a) Plectranthus barbatus extract, (b) Plectranthus verticillatus extract and (c)
rosmarinic acid standard.
Figure 3.4. Structure of the compounds with retention time 19.2 min, rosmarinic acid.

Chapter III
45
Standard rosmarinic acid was analysed by HPLC and the retention time could be
confirmed, as well as the UV spectra (Figure 3.5). In this figure a complete overlay between the
spectra of the compound with retention time 19.2 min and rosmarinic acid can be seen.
Figure 3.5. Overlay of UV spectra obtained by HPLC‐diode array of compounds with a retention time 19.2
min (——) and the standard rosmarinic acid (___).
Rosmarinic acid was quantified in plant extracts, by HPLC, using a calibration curve with
a range of 1–50 µg/ml rosmarinic acid, R2 = 0.99. The results are indicated in Table 3.2. The
rosmarinic acid content correlates with the activity found for P. ecklonii, P. fructicosus, P.
lanuginosus and P. verticillatus, R2 = 0.998. The values found with P. barbatus extract cannot
correlate with rosmarinic acid content because it contains other compounds that also have AChE
inhibition activity.
The inhibition of AChE by rosmarinic acid was determined and an IC50 of 0.44 ± 0.03
mg/ml was obtained. Orhan et al. (2008) found a value of 85.8% inhibition when using 1 mg/ml
of rosmarinic acid, a value similar to the one obtained in this study, showing a linear relationship
between the rosmarinic acid content and the IC50 values. The activity found with P. barbatus
extract could not be completely explained by the presence of rosmarinic acid, suggesting that
the other extract components may also be active.
1.4. Discussion
Rosmarinic acid was the main compound found in all the aqueous extracts of the
Plectranthus species analysed and it is known to be a rather common compound in the
Lamiaceae family (Abdel‐Mogib et al., 2002). We could only find a report of the presence of
rosmarinic acid in P. fruticosus (Pederson, 2000). Rosmarinic acid was also found to be the major
constituent in Coleus aromaticus (Kumaran & Karunakaran, 2007), a species with several
synonyms that is also referred to in the literature as P. aromaticus and P. amboinicus (Lukhoba
et al., 2006). The AChE inhibition activity of P. ecklonii, P. fructicosus, P. lanuginosus and

In vitro Studies of Activity and Metabolism
46
P. verticillatus was due only to the presence of rosmarinic acid. The activity found in the
analysed species correlated rather well with the quantity of rosmarinic acid present (R2 = 0.998).
Rosmarinic acid is the main component of polar extracts from many plants of the
Lamiaceae family, commonly used in human diet. Besides being an enzyme inhibitor, it also has
antioxidant activity, with a value of IC50 = 1.68 ± 0.21 µg/ml in the present study. The antioxidant
activity of rosmarinic acid is well studied and the presence of this compound is considered one
of the reasons for the several biological activities found in species of the Lamiaceae family.
Besides its antioxidant activity, rosmarinic acid also inhibits several enzymes, for instance, the
angiotensin‐converting enzyme (Li et al., 2008) and phospholipase A2 from snake venom (Ticli et
al., 2005). It is also known to interfere with gene expression (Lee et al., 2007) and signalling
pathways related to cancer prevention (Lee et al., 2007; Lin et al., 2007). All these biological
activities may contribute to the large uses that the people in the South Latin America and in
other parts of the world do with these Plectranthus species.
It may be speculated that the rosmarinic acid may fit into the active site gorge of AChE.
This active site is composed of tryptophan at the peripheral site, phenylalanine and tyrosine
residues down the entrance of this gorge (Silman & Sussman, 2008). These residues may interact
with the aromatic rings of rosmarinic acid, and occupy the enzyme active site due to the affinity
to the amino acid residues localized at the entrance of the gorge. This peripheral binding site
defines whether the compounds will act on the enzyme or not as described in the literature
(Eastman et al., 1995).
Rosmarinic acid is the main compound responsible for the results obtained with all the
Plectranthus species P. barbatus, P. ecklonii, P. fructicosus, P. lanuginosus and P. verticillatus
studied in this work. Although the last two species are not traditionally applied in folk medicine,
they could be a good source of rosmarinic acid for future research.
1.5. Conclusion
The inhibition of acetylcholinesterase by the water extracts of Plectranthus species
seems to correlate mainly with the concentration of rosmarinic acid present. The most active
extracts were from P. verticillatus, P. ecklonii and P. barbatus; however the activities found in
the extracts of the first two species were exclusively due to the content in rosmarinic acid. The
beneficial health activity described with the use of P. barbatus may be attributed to the
presence not only of rosmarinic acid, but to other active components. The rosmarinic acid in the
water extracts and decoctions, consumed by the population, may explain some of the health
benefits described through its traditional applications.

Chapter III
47
2. In vitro Digestion Activities of Plectranthus barbatus Aqueous
Extract and Activities of the Digested Product.
2.1. Introduction
Plectranthus barbatus, is one of the most used Plectranthus species, especially for
therapeutic applications. In Brazil, where it is known as ‘‘falso boldo”, infusions or decoctions of
the leaves are drunk to treat a wide range of diseases such as digestive and nervous system
disorders (Lukhoba et al., 2006). The same species is also used to prevent or alleviate
inflammation conditions. It is also sold as food supplement due to its healing properties. P.
barbatus is also reported to be edible in Africa where the leaves are cooked as a vegetable
(Lukhoba et al., 2006).Previous work demonstrated that this tea has antiacetylcholinesterase as
well as antioxidant activity, and rosmarinic acid was the main compound present in this herbal
tea. (Falé et al., 2009). Rosmarinic acid is widespread in Nepetoideae species of Lamiaceae
family, such as rosemary (Rosmarinus officinalis), mint (Mentha arvense), basil (Ocimum
basilicum) (Petersen & Simmonds, 2003). In opposition to the other Plectranthus species in the
previous section, rosmarinic extract was not the only constituent responsible for the activities
shown by P. barbatus extract. Also for this reason, but mainly because P. barbatus is one of the
most used Plectranthus species, the present studied continued with a decoction of P. barbatus.
Several compounds can be found in different herbal teas and it is important to
determine their metabolism after being consumed as a beverage in order to evaluate the final
biological activity. The first step in the digestion process is the contact between the beverage
and the gastrointestinal tract, since saliva does not seem to have a strong effect on the
metabolism of these compounds (Spencer 2003). Rosmarinic acid may be subject to hydrolysis
either at acidic or basic pH, as it is an ester of caffeic acid, which means that it may be degraded
under the physiological conditions of the gastrointestinal tract before reaching the intestinal
barrier. The acidic pH of the gastric juice has been reported to hydrolyze flavanol oligomers to its
monomers, (Spencer et al., 2000) and also saponins from ginseng (Kong et al., 2009), which may
facilitate their further degradation. In spite of studies focusing on the metabolism in the
gastrointestinal tract being scarce, research on this topic has demonstrated that when subjecting
polyphenols, from fruit beverages, to acidic stomach conditions followed by pancreatic medium,
a decrease in their content can be observed. The exact amount of the decrease depends on the
compound under evaluation (Cilla et al., 2009; Yoshino et al., 1999).

In vitro Studies of Activity and Metabolism
48
To simulate the metabolism occurring during the absorption process, Caco‐2 cells are
commonly used, either in studies with intact cells or in the homogenate form (Galijatovic et al.,
2000). Studies carried out with intact cells indicated that polyphenols were not metabolised by
Caco‐2 cells (Cilla et al., 2009); on the other hand, studies of the action of cell homogenates on
polyphenolic compounds, namely flavonoids, indicated that metabolites might be obtained
during the permeation process (Galijatovic et al., 2000). In addition, when rosmarinic acid was
administered orally to rats, only approximately 5% appeared in the blood stream, and most of it
in derivatized forms, such as methyl‐rosmarinic acid, caffeic acid, ferulic acid and coumaric acid
(Baba et al., 2004). Very few reports tackled the problem of the remaining biological activity
after the biodegradation process and it is important to know if metabolism increases or
diminishes the initial biological activities of the herbal teas. Therefore, the aims of this study
were to investigate if the active compounds found in the herbal tea of P. barbatus were stable
under gastrointestinal tract conditions, what metabolites could be found in contact with the
intestinal wall and whether the metabolised extract had similar antiacetylcholinesterase and
antioxidant activities as the ones previously determined (Falé et al., 2009). This study will allow
an insight into the metabolism of herbal teas containing a mixture of polyphenols and terpenoid
compounds.
2.2. Materials and Methods
The materials and methods are described in detail in the Chapter II.
The P. barbatus extract was prepared as a decoction, as described in Chapter II section 4,
and subjected to in vitro digestion by gastric juice (Chapter II, section 10.1), pancreatic juice
(Chapter II, section 10.2), β‐glucuronidase from E. coli (Chapter II, section 10.3) and Caco‐2 cells
(Chapter II, section 10.4). During the digestions the composition of the plant extract was
followed by HPLC (Chapter II, section 7), and the biological activities – antioxidant activity (DPPH
in Chapter II, section 6) and acetylcholinesterase inhibition (Chapter II, section 5). The
identification of compounds was carried out by mass spectrometry, as described in Chapter II,
section 9..
2.3. Results and Discussion
2.3.1. Main composition of P. barbatus herbal tea
The water extract of P. barbatus leaves was analysed by HPLC‐DAD (Figure 3.6). The
compounds after being separated by HPLC were identified by mass spectrometry comparatively
to standard libraries. The compounds from the that P. barbatus that had not been identified

Chapter III
49
before are shown in Table 3.3. All the compounds have already been identified in plants of
Plectranthus species (Abdel‐Mogib et al., 2002; Batista et al., 1996). The structures are shown in
Figure 3.7.
Fig. 3.6. HPLC chromatogram of Plectranthus barbatus herbal tea: 1, luteolin 7‐O‐glucuronide (retention
time 8.68 min); 2, rosmarinic acid (RT: 9.38); 3, apigenin 7‐O‐glucuronide (RT: 10.11); 4, hydrolysed
abietane (RT: 12.58); 5, acacetin 7‐O‐glucuronide (RT: 13.62); 6, abietane diterpenoid (RT: 14,15); 7, (16S)‐
coleon E (RT: 18.54).
Table 3.3. Detected ions and attribution errors (ppm) for the collected fractions corresponding to peaks 1,
3, 4, 5, 6 and 7.
Chromatogram
peak Attribution
m/z Error
(ppm) Experimental Theoretical
1 [C21H18O12 + H]+ 463.08571 463.08710 3.0
3 [C21H18O11 + H]+ 447.09064 447.09219 3.5
4 [C22H26O8 + H]+ 419.16973 419.17004 0.8
5 [C22H20O5 + H]+ 461.10754 461.10784 0.6
6 [C22H26O7 + H]+ 403.17361 403.17513 3.8
7 [C20H22O5 + H]+ 343.15341 343.15657 1.7

In vitro Studies of Activity and Metabolism
50
Figure 3.7. Chemical structure of compounds present in P. barbatus herbal tea: 1, luteolin 7‐O‐
glucuronide; 2, rosmarinic acid; 3, apigenin 7‐O‐glucuronide; 4, hydrolysed abietane; 5, acacetin 7‐O‐
glucuronide; 6, abietane diterpenoid; 7, (16S)‐coleon E.
2.3.2. In vitro metabolism of the extract by the gastric and pancreatic juices. Biological
activity of the resulting products
The digestion of the P. barbatus water extract was carried out for 4 h under stomach
acidic conditions (pepsin, pH 1). Although the digestion in the stomach may take 60–110 min
(Kong et al., 2009), it was decided to continue the process to see if the bioconversion increased
with the time. Aliquots were withdrawn during the 4 h and the HPLC of each aliquot was
analysed. None of the extract constituents was hydrolyzed under the stomach acidic conditions,
even after 4 h digestion (Figure 3.8.a). Pepsin, an enzyme that hydrolysis proteins, does not have
the capacity to hydrolyze esters, unlike other proteases, because the ester, rosmarinic acid, was
not hydrolyzed at pH 1.
The antiacetylcholinesterase and antioxidant activities of P. barbatus water extract were
analysed during the 4 h of the in vitro gastric process; the results are shown in Table 3.4. It can
be seen that after the gastric digestion the inhibition capacity of the extract showed a small
decline that is not statistically significant at the 95% level. The same was noticed for the
antioxidant activity. The standard rosmarinic acid was subject to the same in vitro conditions and
the results were similar to those obtained in the herbal tea mixture.

Chapter III
51
Figure 3.8. HPLC chromatograms before and after the incubation of Plectranthus barbatus extract with: (a)
gastric juice, (b) pancreatic juice. *Indicates the residue of pancreatin. For the identification of the peak
numbers, refer to Figure 3.4.
There is very scarce information about the gastric metabolism of phenolic acids and even
less about terpenes. Polyphenols from procyanidin oligomers were hydrolyzed by stomach acid
conditions (Spencer et al., 2000). When analysing the effect of the stomach pH on ginsenosides,
deglycosylation was observed (Kong et al., 2009). In the present study deglucuronidation was
not observed at acidic pH and the hydrolysis of rosmarinic acid, forming caffeic acid, could not
be detected.
Pancreatic juice contains a mixture of hydrolytic enzymes, pancreatin (amylase, lipase
and protease), at basic pH, which was used to simulate the behaviour of the P. barbatus extract
under small intestine conditions (Yamamoto et al., 1999). The effect of the buffer solution at pH
8, without pancreatin, on compound degradation was also analysed. The study was carried out
for 4 h with five replicates. The extracts were analysed by HPLC and the chromatograms (Figure
3.8b) allowed the establishment of the graph shown in Figure 3.9. The abietane diterpenoid
decreased, whilst its hydroxylated form increased. Rosmarinic acid showed a 25% decrease in its
concentration (Figure 3.9). Luteolin 7‐O‐glucuronide, apigenin 7‐O‐glucoronide and acacetin 7‐O‐
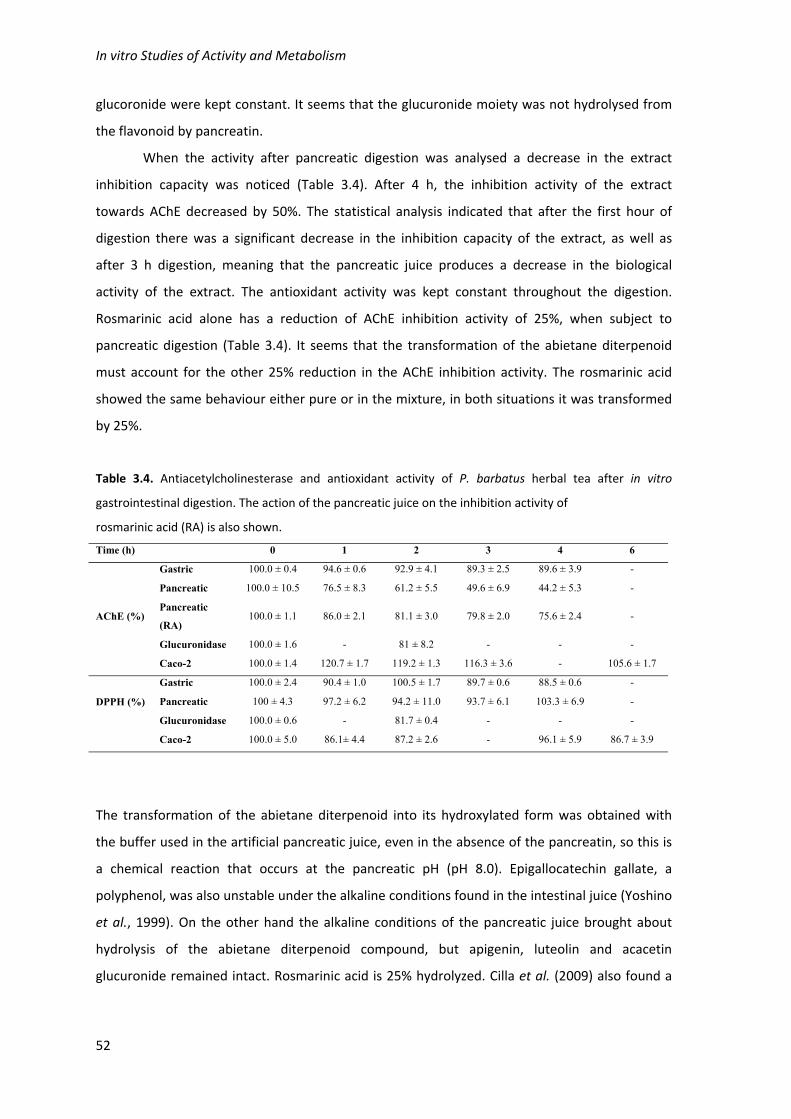
In vitro Studies of Activity and Metabolism
52
glucoronide were kept constant. It seems that the glucuronide moiety was not hydrolysed from
the flavonoid by pancreatin.
When the activity after pancreatic digestion was analysed a decrease in the extract
inhibition capacity was noticed (Table 3.4). After 4 h, the inhibition activity of the extract
towards AChE decreased by 50%. The statistical analysis indicated that after the first hour of
digestion there was a significant decrease in the inhibition capacity of the extract, as well as
after 3 h digestion, meaning that the pancreatic juice produces a decrease in the biological
activity of the extract. The antioxidant activity was kept constant throughout the digestion.
Rosmarinic acid alone has a reduction of AChE inhibition activity of 25%, when subject to
pancreatic digestion (Table 3.4). It seems that the transformation of the abietane diterpenoid
must account for the other 25% reduction in the AChE inhibition activity. The rosmarinic acid
showed the same behaviour either pure or in the mixture, in both situations it was transformed
by 25%.
Table 3.4. Antiacetylcholinesterase and antioxidant activity of P. barbatus herbal tea after in vitro
gastrointestinal digestion. The action of the pancreatic juice on the inhibition activity of
rosmarinic acid (RA) is also shown.
Time (h) 0 1 2 3 4 6
AChE (%)
Gastric 100.0 ± 0.4 94.6 ± 0.6 92.9 ± 4.1 89.3 ± 2.5 89.6 ± 3.9 -
Pancreatic 100.0 ± 10.5 76.5 ± 8.3 61.2 ± 5.5 49.6 ± 6.9 44.2 ± 5.3 -
Pancreatic
(RA) 100.0 ± 1.1 86.0 ± 2.1 81.1 ± 3.0 79.8 ± 2.0 75.6 ± 2.4 -
Glucuronidase 100.0 ± 1.6 - 81 ± 8.2 - - -
Caco-2 100.0 ± 1.4 120.7 ± 1.7 119.2 ± 1.3 116.3 ± 3.6 - 105.6 ± 1.7
DPPH (%)
Gastric 100.0 ± 2.4 90.4 ± 1.0 100.5 ± 1.7 89.7 ± 0.6 88.5 ± 0.6 -
Pancreatic 100 ± 4.3 97.2 ± 6.2 94.2 ± 11.0 93.7 ± 6.1 103.3 ± 6.9 -
Glucuronidase 100.0 ± 0.6 - 81.7 ± 0.4 - - -
Caco-2 100.0 ± 5.0 86.1± 4.4 87.2 ± 2.6 - 96.1 ± 5.9 86.7 ± 3.9
The transformation of the abietane diterpenoid into its hydroxylated form was obtained with
the buffer used in the artificial pancreatic juice, even in the absence of the pancreatin, so this is
a chemical reaction that occurs at the pancreatic pH (pH 8.0). Epigallocatechin gallate, a
polyphenol, was also unstable under the alkaline conditions found in the intestinal juice (Yoshino
et al., 1999). On the other hand the alkaline conditions of the pancreatic juice brought about
hydrolysis of the abietane diterpenoid compound, but apigenin, luteolin and acacetin
glucuronide remained intact. Rosmarinic acid is 25% hydrolyzed. Cilla et al. (2009) also found a

Chapter III
53
reduction in all the phenolic compounds of a fruit beverage after pancreatic digestion, although
the chemical composition of the juice was not specified.
Figure 3.9. Variations in peak areas of compounds present in herbal tea after 4 h digestion with artificial
pancreatic juice. () Luteolin 7‐O‐glucuronide; () rosmarinic acid; () apigenin 7‐O‐glucuronide; ()
hydrolysed abietane; (○) acacetin 7‐O‐glucuronide; (●) abietane diterpenoid.
The results found here may give an explanation the traditional use as purgative
described for P. barbatus herbal tea (El‐Kamali, 2009). It is known that inhibitors of AChE
stimulate gastrointestinal motility (Cellek et al., 2008; Jarvie et al., 2008). The results shown here
point out that the activity of the herbal tea in the stomach is not reduced by the gastric juice,
and although by passing through the pancreatic juice it reduces its inhibitory capacity to 50% of
its initial activity, it may go on with its purgative activity through the gut. The fact that P.
barbatus herbal tea can act as an inhibitor of AChE may facilitate stomach and gut motility,
explaining the purgative effect described for this herbal tea.
2.3.3. Metabolism of the plant extract by Caco‐2 cells and biological activity of the final
products
The aim of this study was to simultaneously investigate if the extract components might
disappear from the culture medium, being incorporated into the Caco‐2 cells, or if any
extracellular protein could produce a change in the chemical composition of the extract.
The permeation of phenolic acids or flavonoids is not completely established.
Polyphenols seem to be absorbed through the intestinal barrier, since after their ingestion there
is a rising in the antioxidant capacity of blood plasma (Scalbert & Williamson, 2000). Flavonoids
like glycosylated quercetin seem to permeate into the bloodstream more easily than the parent

In vitro Studies of Activity and Metabolism
54
flavonoid (Hollman & Katan, 1997). On the other hand there are very few studies pointing out
that absorption of flavonoids occurs preferentially when they are deglycosylated by the
intestinal bacteria (Liu & Hu, 2002) and that the absorption of phenolic acids depends largely on
their structure. Rosmarinic acid is very slowly absorbed, whilst coumaric or ferulic acids, for
instance, are more easily absorbed (Konishi & Kobayashi, 2005). Therefore, further studies are
necessary in order to elucidate these procedures. As the extract used in the present
investigation is a mixture of glycosylated flavonoids and phenolic acids (like rosmarinic acid), this
study was carried out in order to further elucidate the absorption process.
After having analysed that the culture medium did not produce any change to the
extract chemical composition, the herbal tea was added to the culture medium in the presence
of Caco‐2 cells. The extract was used at concentrations from 1 to 10 mg/ml in the culture
medium. After evaluating the cells’ viability by the MTT method (results not shown) 1 mg/ml of
extract was chosen in order to accomplish this study. At this concentration the P. barbatus water
extract was not toxic to the Caco‐2 cells.
The extract remained in contact with the cells for 6 h and the aliquots withdrawn and
analysed by HPLC; this analysis revealed that there were no changes in the extract composition.
The results indicated that the Caco‐2 cells did not produce extracellular enzymes capable of
metabolizing the constituents of the herbal tea, and these compounds were not able to
penetrate the cells in an appreciable quantity, since the peak areas were kept constant. At the
end of the 6 h, a homogenate of the cells was prepared and the content inside the cells was
analysed. This test had the objective of knowing if any compound could be detected inside the
cellular system. In fact, a very small quantity of rosmarinic acid could be detected inside the cells
(retention time 9.3 min; Figure 3.10a).
The permeation of rosmarinic acid standard through Caco‐2 cells was studied. It could be
seen that only 10% of its concentration disappeared from the medium. Nevertheless when the
content inside the cells was analysed, a very small quantity of rosmarinic acid was found inside
the cell homogenate (Figure 3.10b). The UV–Vis spectrum of the compound present with
retention time around 9.38 min confirms that it is rosmarinic acid. Rosmarinic acid was
previously studied regarding its permeation through Caco‐2 cell layers, and it was verified that
permeation occurred mainly by paracellular diffusion. In addition, rosmarinic acid was not
hydrolyzed by Caco‐2 esterases (Konishi & Kobayashi, 2005). When a tablet containing
rosmarinic acid was given to healthy humans, 6.3% of the ingested rosmarinic acid appeared in
the blood in several metabolized forms (Baba et al., 2004). The behaviour of rosmarinic acid was
identical in the different extracts in spite of being inside a mixture of several flavonoids.
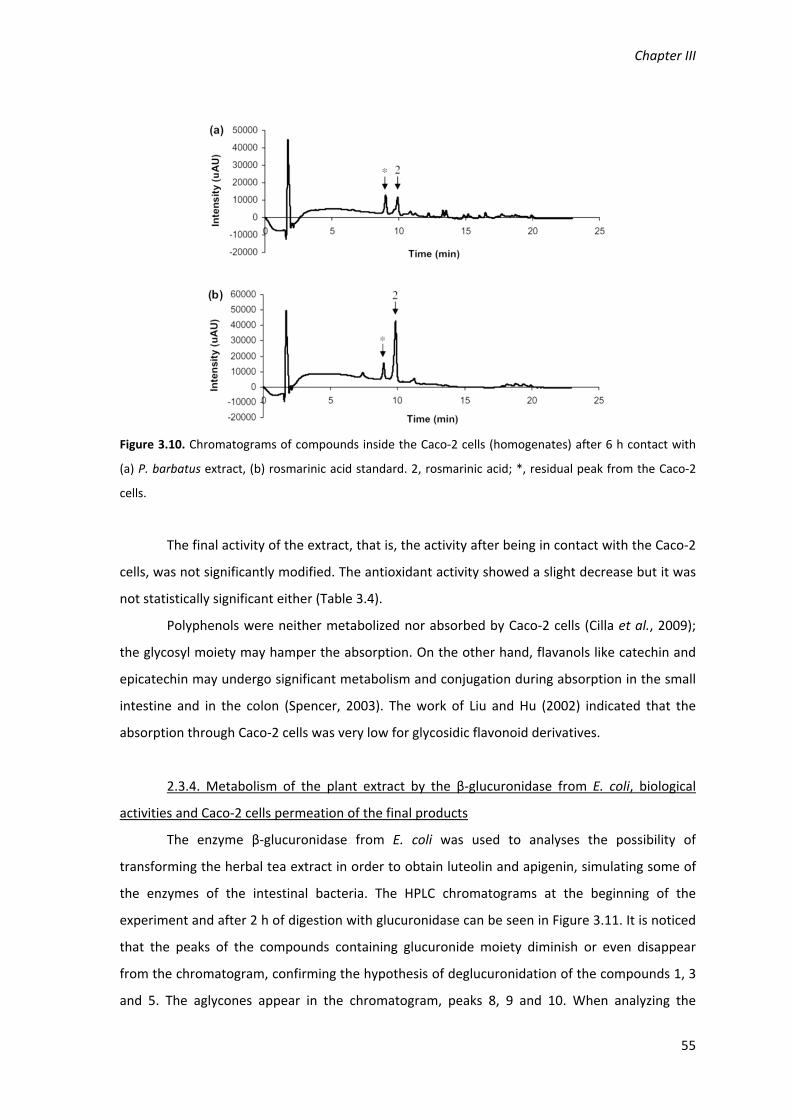
Chapter III
55
Figure 3.10. Chromatograms of compounds inside the Caco‐2 cells (homogenates) after 6 h contact with
(a) P. barbatus extract, (b) rosmarinic acid standard. 2, rosmarinic acid; *, residual peak from the Caco‐2
cells.
The final activity of the extract, that is, the activity after being in contact with the Caco‐2
cells, was not significantly modified. The antioxidant activity showed a slight decrease but it was
not statistically significant either (Table 3.4).
Polyphenols were neither metabolized nor absorbed by Caco‐2 cells (Cilla et al., 2009);
the glycosyl moiety may hamper the absorption. On the other hand, flavanols like catechin and
epicatechin may undergo significant metabolism and conjugation during absorption in the small
intestine and in the colon (Spencer, 2003). The work of Liu and Hu (2002) indicated that the
absorption through Caco‐2 cells was very low for glycosidic flavonoid derivatives.
2.3.4. Metabolism of the plant extract by the β‐glucuronidase from E. coli, biological
activities and Caco‐2 cells permeation of the final products
The enzyme β‐glucuronidase from E. coli was used to analyses the possibility of
transforming the herbal tea extract in order to obtain luteolin and apigenin, simulating some of
the enzymes of the intestinal bacteria. The HPLC chromatograms at the beginning of the
experiment and after 2 h of digestion with glucuronidase can be seen in Figure 3.11. It is noticed
that the peaks of the compounds containing glucuronide moiety diminish or even disappear
from the chromatogram, confirming the hypothesis of deglucuronidation of the compounds 1, 3
and 5. The aglycones appear in the chromatogram, peaks 8, 9 and 10. When analyzing the
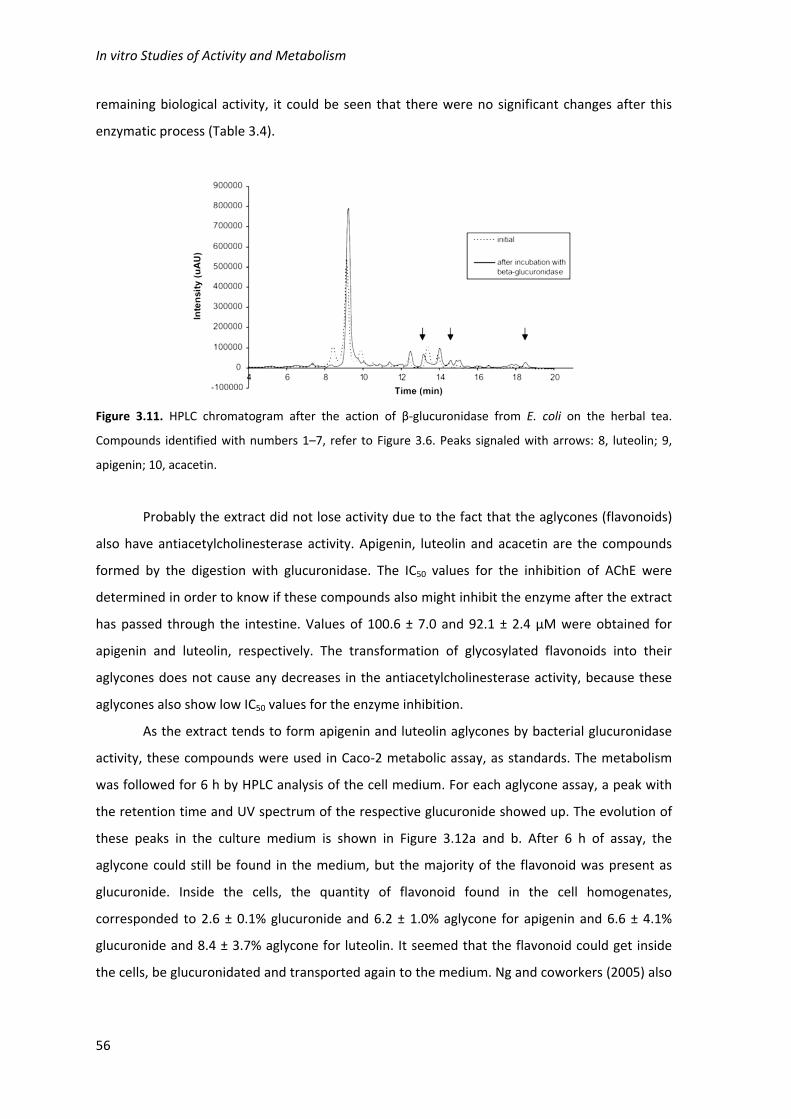
In vitro Studies of Activity and Metabolism
56
remaining biological activity, it could be seen that there were no significant changes after this
enzymatic process (Table 3.4).
Figure 3.11. HPLC chromatogram after the action of β‐glucuronidase from E. coli on the herbal tea.
Compounds identified with numbers 1–7, refer to Figure 3.6. Peaks signaled with arrows: 8, luteolin; 9,
apigenin; 10, acacetin.
Probably the extract did not lose activity due to the fact that the aglycones (flavonoids)
also have antiacetylcholinesterase activity. Apigenin, luteolin and acacetin are the compounds
formed by the digestion with glucuronidase. The IC50 values for the inhibition of AChE were
determined in order to know if these compounds also might inhibit the enzyme after the extract
has passed through the intestine. Values of 100.6 ± 7.0 and 92.1 ± 2.4 µM were obtained for
apigenin and luteolin, respectively. The transformation of glycosylated flavonoids into their
aglycones does not cause any decreases in the antiacetylcholinesterase activity, because these
aglycones also show low IC50 values for the enzyme inhibition.
As the extract tends to form apigenin and luteolin aglycones by bacterial glucuronidase
activity, these compounds were used in Caco‐2 metabolic assay, as standards. The metabolism
was followed for 6 h by HPLC analysis of the cell medium. For each aglycone assay, a peak with
the retention time and UV spectrum of the respective glucuronide showed up. The evolution of
these peaks in the culture medium is shown in Figure 3.12a and b. After 6 h of assay, the
aglycone could still be found in the medium, but the majority of the flavonoid was present as
glucuronide. Inside the cells, the quantity of flavonoid found in the cell homogenates,
corresponded to 2.6 ± 0.1% glucuronide and 6.2 ± 1.0% aglycone for apigenin and 6.6 ± 4.1%
glucuronide and 8.4 ± 3.7% aglycone for luteolin. It seemed that the flavonoid could get inside
the cells, be glucuronidated and transported again to the medium. Ng and coworkers (2005) also
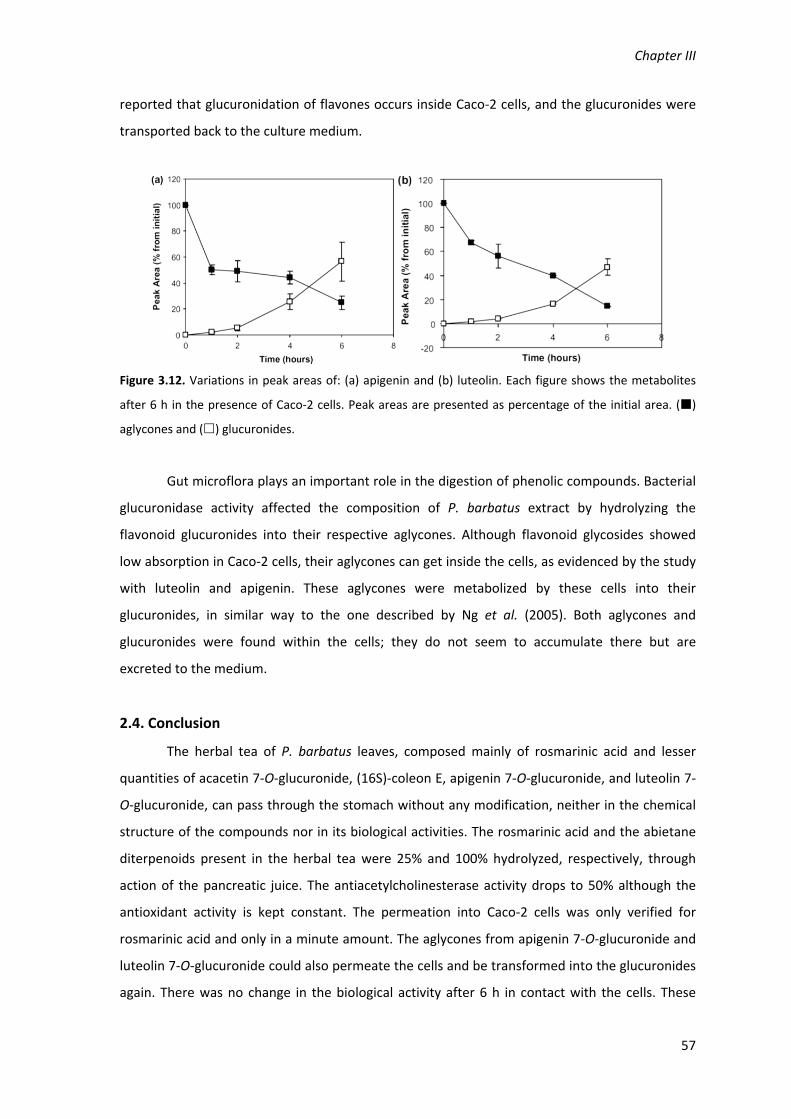
Chapter III
57
reported that glucuronidation of flavones occurs inside Caco‐2 cells, and the glucuronides were
transported back to the culture medium.
Figure 3.12. Variations in peak areas of: (a) apigenin and (b) luteolin. Each figure shows the metabolites
after 6 h in the presence of Caco‐2 cells. Peak areas are presented as percentage of the initial area. ()
aglycones and () glucuronides.
Gut microflora plays an important role in the digestion of phenolic compounds. Bacterial
glucuronidase activity affected the composition of P. barbatus extract by hydrolyzing the
flavonoid glucuronides into their respective aglycones. Although flavonoid glycosides showed
low absorption in Caco‐2 cells, their aglycones can get inside the cells, as evidenced by the study
with luteolin and apigenin. These aglycones were metabolized by these cells into their
glucuronides, in similar way to the one described by Ng et al. (2005). Both aglycones and
glucuronides were found within the cells; they do not seem to accumulate there but are
excreted to the medium.
2.4. Conclusion
The herbal tea of P. barbatus leaves, composed mainly of rosmarinic acid and lesser
quantities of acacetin 7‐O‐glucuronide, (16S)‐coleon E, apigenin 7‐O‐glucuronide, and luteolin 7‐
O‐glucuronide, can pass through the stomach without any modification, neither in the chemical
structure of the compounds nor in its biological activities. The rosmarinic acid and the abietane
diterpenoids present in the herbal tea were 25% and 100% hydrolyzed, respectively, through
action of the pancreatic juice. The antiacetylcholinesterase activity drops to 50% although the
antioxidant activity is kept constant. The permeation into Caco‐2 cells was only verified for
rosmarinic acid and only in a minute amount. The aglycones from apigenin 7‐O‐glucuronide and
luteolin 7‐O‐glucuronide could also permeate the cells and be transformed into the glucuronides
again. There was no change in the biological activity after 6 h in contact with the cells. These

In vitro Studies of Activity and Metabolism
58
results point out that P. barbatus herbal tea will reach the intestinal barrier with a reduction in
50% of its AChE inhibitory activity and without modification in its antioxidant activity. The
permeation through the intestinal barrier will be very low for the extract compounds, but may
increase due to their glucuronidase digestion by the gut microflora. The aglycones thus can be
formed and then metabolized by intestinal cells back into their glucuronides, as was shown here
for the Caco‐2 model. The P. barbatus extract showed the same activity with its flavonoids
glucuronidated or as aglycones. The AChE inhibition activity found during the gastrointestinal
digestion may explain, at least, the purgative effect found for this herbal tea.

Chapter III
59
3. Activity of Plectranthus barbatus extract against inflammatory
response in human neutrophils
3.1. Introduction
Inflammation is a complex biological response to harmful stimuli involving the vascular
system, the immune system and the injured cells. Although it has primarily a protective function,
the destructive effects can largely surpass the gravity of the stimulus. Therefore, serious
inflammatory responses are often associated with diseases, as atherosclerosis, allergies and
myopathies.
Polymorphonuclear neutrophils are white blood cells in mammals and form have an
essential part in the innate immune system. These cells, highly specialized in phagocytosis, have
developed mechanisms for intracellular digestion of particles, such as pathogens and cell debris,
involving the production of radical oxygen species (ROS) and an range of hydrolytic and
proteolytic enzymes.
Microorganisms areusually coated with opsonins (generally complement and/or
antibody) that bind to specific receptors on the surface of the phagocyte, inducing the
invagination of the cell membrane and the incorporation of microorganisms into an intracellular
phagosome. There follows a sharp but transient increase in oxygen uptake, which is used to
produce O2‐ (superoxide ion) by the one‐electron reduction of oxygen, a reaction catalysed by
NADPH oxidase at the expense of NADPH. Most of the superoxide reacts with itself forming H2O2
(hydrogen peroxide), and from these agents a large number of highly reactive oxidants are
formed, including HOCl (hypochlorous acid), which is produced by the myeloperoxidase‐
catalyzed oxidation of Cl‐ by H2O2; OH. (hydroxyl radical), produced by the reduction of H2O2 by
Fe2+ or Cu+; ONOO‐ (peroxynitrite), formed by the reaction between O2‐ and NO‐; and many
others. (Babior, 2000; Klebanoff, 2005) This battery of reactive oxidizing agents not only kills the
invading particles but also inflicts harm on nearby tissues, and is thought to be of pathogenic
significance in a large number of diseases such as emphysema, acute respiratory distress
syndrome, atherosclerosis, reperfusion injury, malignancy and rheumatoid arthritis (Babior,
2000). The myeloperoxidase‐catalysed reaction is directly related with neutrophil stimulation,
the resulting inflammatory process, tissue injury and related pathologies, and therefore serves
as a good marker for neutrophil production of reactive oxygen species (Klebanoff, 2005; Deby‐
Dupont et al., 1999; Zeraik et al., 2011).

In vitro Studies of Activity and Metabolism
60
Plectranthus barbatus Andrews (Lamiaceae), known as “falso boldo”, is used in South
Africa and South America for a wide range of therapeutic purposes. Some of the traditional uses
of P. barbatus suggest that its extract may have anti‐inflammatory activity, such as the
treatment of aches from diverse etiologies, burns, sores, insect bites, allergies, and to reduce
swelling on bruises (Lukhoba et al., 2006). Previous in vitro studies showed that a P. barbatus
extract, prepared as decoction, has promising antioxidant and antiacetylcholinesterase
properties, and the extract composition is almost unaltered during an in vitro simulated
digestion, especially in the content of its main component, rosmarinic acid (Porfirio et al., 2010).
3.2. Materials and Methods
The materials and methods are described in detail in the Chapter II.
The P. barbatus extract was prepared as a decoction, as described in Chapter II section 4.
Neutrophils were isolated from blood of human donors as described in Chapter II section 17.1.
and were activated by phorbol 12‐myristate 13‐acetate, to stimulate the production of reactive
oxygen species (Chapter II section 17.2.). The hypochlorous acid produced was measured by the
taurine chlorination method as described in section Chapter II section 17.2., in the presence and
in the absence of P. barbatus extract and rosmarinic acid.
3.3. Results and Discussion
The taurine chlorination by hypochlorous acid is a rapid chemical reaction that allows to
quantify hypochlorous acid in a sample, as the amount chlorotaurine formed (measured
spectrophotometrically) is directly proportional to the amount of hypochlorous acid (Weiss et
al., 1982). Therefore the decrease of taurin chlorination represented in Figure 3.13 shows a
decrease in hypochlorous acid production by human neutrophils relatively to a untreated control
(no antioxidants added), where 0% means that the neutrophils produced the same amount of
hypochlorous acid as the control, and 100% means that no hypochlorous acid was produced. To
evaluate the effect of the P. barbatus extract on the modulation of hipochlorous acid production
in neutrophils, we tested several concentrations of the extract. As it is shown in Figure 3.13a, the
effect is dose‐dependent, decreasing the production of hypochlorous acid with higher
concentrations of the plant extract. The value that inhibits 50% of the hypochlorous acid
production (IC50) was estimated as 10.7±3.1 µg.mL‐1. For isolated rosmarinic acid, the main
component of the P. barbatus extract, the same dose‐dependent effect was also observed
(Figure 3.13b), and the IC50 value was estimated as 3.62 ± 0.42 µM.

Chapter III
61
The amount of rosmarinic acid present in the plant extract was estimated by the peak
area in the HPLC chromatogram, comparing with a calibration curve with standard rosmarinic
acid. By plotting the amount of rosmarinic acid in neutrophil assay with P. barbatus extract
versus the decrease in taurine chlorination, together with the data with standard rosmarinic acid
(Figure 3.13b), it can be observed that the decrease in taurine chlorination is very similar. This
observation suggests that the decrease in the hypochlorous acid production by the P. barbatus
extract is mainly due to its content in rosmarinic acid.
Figure 3.13. Decrease of taurine chloration in the presence of several concentrations of P. barbatus
extract (a and b) or standard rosmarinic acid (b). The concentration of P. barbatus is expressed in µg.mL‐1
(a) or by its content in rosmarinic acid in µM (b).
Several studies have demonstrated the decrease of ROS released by activated
neutrophils in the presence of several plant extracts. The P. barbatus extract induced a decrease
in the release of hypochlorous acid in the same magnitude of Ribus nigrum water‐acetone
extract (Tabart et al., 2011), but was less potent than the ethanolic extract of Baccharis trimera
(Padua et al., 2010) and pomegranate peel extract (Bachoual et al., 1998), which showed IC50
values of approximately 0.5 µg.mL‐1 and 0.1µg.mL‐1, respectively. These values, however,

In vitro Studies of Activity and Metabolism
62
correspond to the decrease in total amount of ROS, not only hypochlorous acid as in the present
study.
To our knowledge no studies were performed on the effect of rosmarinic acid in
activated neutrophils. However, it has been shown that, when administered to rats, rosmarinic
acid reduces the inflammatory response and the myeloperoxidase activity after induction of
inflammation by 12‐tetracanoylphorbol 13‐acetate (TPA) (Osakabe et al., 2004) or by septic
shock (Jiang et al., 2009). The effect of rosmarinic acid in TPA‐induced inflammation was also
observed by the administration of Perilla frutescens extract, another Lamiaceae species, whose
main component is rosmarinic acid, but this effect was not observed with the flavonoid luteolin
(Osakabe et al., 2004).
In the present study the activity of the plant extract seems to be mainly due to the
rosmarinic acid content, however other studies showed that flavonoids may also be active.
Myricetin induced 72.7% decrease of hypochlorous acid in neutrophils with a concentration of
100 µM (Meotti et al., 2008), and isoorientin – a luteolin glucoside derivative – decreased 66% of
the release of ROS in with a concentration of 8.9 µM (Zeraik et al., 2011). These effects seemed
to be due to the inhibition of myeloperoxidase, as the same authors observed that the enzyme
activity was inhibited in the presence of those flavonoids (Meotti et al., 2008; Zeraik et al.,
2011). The hydroxylation pattern seems also to play an important role in the inhibition of
myeloperoxidase (Meotti et al., 2008), and quercetin isoquercetin and rutin showed IC50 values
in nanomolar concentrations for in vitro assays (Regasini et al., 2008). In in vivo studies a
decrease superior to 50% in myeloperoxidase activity in the lungs of rats treated with
70µmol.Kg‐1 of luteolin after acute lung injury (Kuo et al., 2011).
The decrease of ROS released by activated neutrophils in the presence of plant phenolic
compounds seems to be mainly due to the inhibition of myeloperoxidase. Bachoual and co‐
workers (1998) showed that pomegranate peel extract does not inhibit NADPH oxidase or the
formation of superoxide anion, and does not scavenge hydrogen peroxide. The same authors
observed that the inhibition of myeloperoxidase was the only biochemical effect related to ROS
production in the presence of pomegranate extract, and therefore concluded that it may be the
primary cause of the ROS decrease.
This preliminary study suggests that P. barbatus extract and rosmarinic acid may be
potentially useful in the treatment of inflammation related health conditions, especially if
P. barbatus extract components are to be found circulating in the bloodstream after the oral
administration of the water extract. The decrease of the amount of hypochlorous acid produced
by neutrophils may explain the traditional use of P. barbatus in the treatment of swellings and

Chapter III
63
pain related to inflammatory processes (Lukhoba et al., 2006). However further studies would
be useful to enlighten some aspects related to the mechanism of action of the plant extract
components, especially if the action is intracellular or extracellular. Intracellular ROS are
responsible for the destruction of phagocytised particles, while extracellular myeloperoxidase
and ROS, although playing a role in the defence mechanisms, are the main cause of damage by
inflammation (Deby‐Dupont, 1999; Klebanoff, 2005).
3.4. Conclusions
P. barbatus extract was able to scavenge hypochlorous acid produced by the activated
neutrophils in a dose‐dependent process.
The content of rosmarinic acid in the plant extract seems to be the main responsible for
the hypochlorous scavenging activity presented by the extract.
The low IC50 values shown by the plant extract and its main component for the
scavenging of hypochlorous acid, produced by human neutrophils, allow to consider that
P. barbatus herbal tea and rosmarinic acid may be useful in the treatment of inflammation
related health problems.

In vitro Studies of Activity and Metabolism
64
4. Conclusions
In summary, the in vitro tests allowed to conclude that all the Plectranthus species
tested showed antioxidant and anticholinesterase activity due to their content in rosmarinic
acid. In contrast, the activities shown by Plectranthus barbatus were not only due to rosmarinic
acid, but also to other compounds, which may explain some health benefits that are attributed
to this aqueous extracts in folk medicine.
The major component of P. barbatus aqueous extract is rosmarinic acid, but it also
contains flavonoid glucuronides (apigenin 7‐O‐glucuronide, luteolin 7‐O‐glucuronide and
acacetin 7‐O‐glucuronide), and diterpenoids. During in vitro digestion of the extract with gastric
juice and pancreatic juice all compounds are stable except one of the abietane diterpenoids,
which suffers hydroxylation in pancreatic conditions, and a consequent reduction of the
acetylcholinesterase inhibitory activity was observed. The flavonoid glucuronides were
deglucuronidated by β‐glucuronidases from gut microbiota, but the aglycones can be
glucuronidated by intestinal cells, as it was shown with the Caco‐2 cell model.
The P. barbatus aqueous extract also showed a high in vitro anti‐inflammatory activity,
by reducing the amount of hypochlorous acid formed by activated neutrophils. The content of
rosmarinic acid in the plant extract seems to be the main responsible for this activity.

Chapter IV
Bioavailability studies in rats and Caco‐2 cell monolayers
Falé PL, Madeira PJ, Florêncio MH, Ascensão L, Serralheiro ML. 2011. Function of Plectranthus barbatus
herbal tea as neuronal acetylcholinesterase inhibitor. Food Funct. 2: 130‐136.
Falé PL, Ascensão L, Serralheiro MLM. Bioavailability of rosmarinic acid, luteolin, apigenin‐modelling
plant herbal teas through Caco‐2 cell monolayers. To be submitted to Food Chem.


Chapter IV
67
1. Plectranthus barbatus aqueous extract bioavailability and
resulting neuronal acetylcholinesterase inhibition in rats
1.1. Introduction
Herbal teas may be considered as functional drinks (Percival et al., 2007), as indeed
almost all of the chemical components of these water extracts possess some biological function
in the human body. Some of the ethnobotanical uses of these herbs can be explained through
the biochemical activities found and described in the literature either for the complete extracts
or for the isolated compounds. Leaves of Plectranthus barbatus (Lamiaceae) were studied
previously concerning the antiacetylcholinesterase as well as the antioxidant activity. The
activities found in the P. barbatus herbal tea could be attributed to its main constituent,
rosmarinic acid, together with other compounds, although present in much lesser quantity,
abietane diterpenoids and flavonoid glucuronides (Falé et al., 2009; Porfirio et al., 2010). The
function of the herbal teas depends on the metabolism that the compounds present in the
extract may be subject to during the gastrointestinal digestion process. The compounds may be
transformed into metabolites with different biological activity compared to the one initially
determined. In the case of P. barbatus, some of the active compounds found were transformed
when the extract was submitted to in vitro conditions simulating the gastrointestinal tract, what
caused a small decrease in the biological activity (Porfirio et al., 2010). The fact that the herbal
tea could pass the digestive tract and keep some of its function lead to an in vivo experiment in
order to see if the compounds present in the water extract could reach the brain and still be
active there. Recently, for instance, an ethanol extract of Tabernaemontana divaricata proved to
be effective in inhibiting neuronal acetylcholinesterase when administered to rats (Chattipakorn
et al., 2007).
The quantification of the metabolites of the herbal tea components in the blood stream,
and the study of the remaining biological activity in the target organ, especially on what
concerns the acetylcholinesterase activity, are topics that are seldom referred to in scientific
papers. Therefore, the aim of this study was to investigate if the active compounds present in
the herbal tea of P. barbatus, or their derivatives, when administered to rats were found in the
blood stream and in the brain, and if the neuronal acetylcholinesterase activity was affected by
the herbal tea administration.

Bioavailability Studies
68
1.2. Materials and Methods
The materials and methods are described in detail in the Chapter II.
The P. barbatus extract was prepared as a decoction, as described in Chapter II section 4.
To know what metabolites to expect after the extract administration to rats, standards of the
metabolites were produced in vitro (Chapter II section 11). Rat liver enzymes (Chapter II section
11.1) were used to produce glucuronide derivatives of the plant extract components as
described in Chapter II section 11.2. Catechol o‐methyltransferase (COMT) from porcine liver
was used to make methylated derivatives of rosmarinic acid, as described in section Chapter II
section 11.3.
The P. barbatus extract and its main component, rosmarinic acid, were administered to
rats as described in Chapter II section 12.1. Plasma and brain were collected from the rats 30
and 60 minutes after administration, and treated as described in section II.12.2. The samples
were analyzed by HPLC‐DAD (Chapter II sections 12.4. and II.7.) and the HPLC samples were pre‐
treated as mentioned in Chapter II section 12.3 to determine the glucuronidated and sulfated
metabolites. The acetylcholinesterase activity of the rat brains was determined as described in
Chapter II section 12.5.
1.3. Results and Discussion
P. barbatus herbal tea proved to have antiacetylcholinesterase activity in previous
studies, with an IC50 of 1.02 ± 0.02 mg of dry leaves ml‐1 and this activity was kept constant after
in vitro gastric digestion and lost approximately 50% after the in vitro pancreatic studies (Falé et
al., 2009; Porfirio et al., 2010). Rosmarinic acid, luteoline 7‐O‐glucuronide, apigenine 7‐O‐
glucuronide, two abietane diterpenoid, acacetin 7‐O‐glucuronide and (16S)‐coleon E, were the
compounds identified in the water extract (Porfirio et al., 2010), being rosmarinic acid the main
component. Although all the compounds demonstrated inhibition activity relatively to AChE, an
IC50 of 0.44 mg.ml‐1 for the main component was determined (Falé et al., 2009). Inhibition
studies demonstrated that the process was reversible (unpublished studies). Due to these
previous results, the study was continued by analyzing the action of P. barbatus herbal tea in
vivo. In the present work the metabolism of the herbal tea after intragastric and intraperitoneal
administration to laboratory animals was analyzed.
1.3.1. Intragastric administration of P. barbatus extract
Plasma. After intragastric administration of P. barbatus extract, the plasma was analyzed
by HPLC and rosmarinic acid was the only compound detected. In order to confirm if this was

Chapter IV
69
indeed the only compound present in the plasma or if some metabolites could be present but in
a low amount that was beneath the detection limit of the system, β‐glucuronidase and sulfatase
were added and allowed to react under the conditions described in Chapter II, section 12.3. The
plasma was analyzed once again by HPLC, and this time the rosmarinic acid showed an increase
in its area and the aglycones from the flavonoid derivatives were also detected. The results from
this study confirmed that not only the presence of rosmarinic acid glucuronide and sulfo‐
derivatives, but also the presence of the flavonoid glucuronide derivatives in the plasma. The
concentrations of these rosmarinic acid metabolites found in the plasma are shown in Table 4.1.
It can be seen that the derivatives of rosmarinic acid in circulation in the blood stream decrease
after 30 min. This study indicated that, in fact, the flavonoid derivatives found initially in the
herbal tea could pass through the gastrointestinal barrier and appear in the plasma (Table 4.1). A
vestigial peak with retention time corresponding to acacetin was also found. The quantity of
total rosmarinic acid found in the plasma relatively to the amount of rosmarinic acid
administered in the extract can be calculated, assuming that a male Sprague‐Dawley rat contains
4.12 ml plasma per 100 g body weight (Probst et al., 2006), as 0.009% and 0.005% for 30 and 60
min, respectively.
Table 4.1. Concentration of rosmarinic acid, its metabolites and flavonoid glucuronide derivatives in the plasma and in
the brain, 30 and 60 min after the intragastric and intraperitoneal administration of P. barbatus extract.
Compound
Concentration in plasma Concentration in the brain
Intragastric administration (nM)
Intraperitoneal administration (µM)
Intraperitoneal administration (µM)
Time (min) 30 60 30 60 30 60
P. barbatus extract
Rosmarinic Acid (RA)
<0.5 10.1 1440±144 745282 24.11.1 20.40.4
RA Glucuronides
112.1 87.8 - - - -
RA Sulfates 210.4 77.3 - - - -
RA methyl - - 17.82.3 11.13.6 - -
Luteolin glucuronide
1.9 - 40.55.9 29.98.0 - -
Apigenin glucuronide
6.5 - 20.03.4 3.70.3 - -
Rosmarinic acid standard
RA 4.9 x 103 1.9 x 103 1042.3192.1 - 25.40.9 -
RA methyl - - 16.75.6 - - -

Bioavailability Studies
70
Rosmarinic acid was also intragastrically administered, in a higher quantity than that
found in the herbal extract, and analyzed under the same conditions. Rosmarinic acid was
present in the plasma 30 min and 60 min after the intragastric administration. The quantities
found (Table 4.1) corresponded to 0.036% and 0.015% of the amount of rosmarinic acid
administered to rats, 30 and 60 min after administration, respectively. The results one hour after
administration showed a decrease in the rosmarinic acid concentration in circulation.
The test with β‐glucuronidase was also carried out, but an increase in the rosmarinic acid
area in the HPLC chromatogram was not detected. This means that there were no
glucuronidated metabolites of this phenolic acid formed in the plasma. It seems that rosmarinic
acid, as pure compound or inside the herbal tea, behaves differently.
Brain AChE. When rat brains were analyzed by HPLC no compounds or metabolites of P.
barbatus extract or rosmarinic acid were found. However, when the brain acetylcholinesterase
activity was measured, a decrease of 10.0 ± 1.8% and 5.5 ± 1.7% of the enzymatic activity was
observed, 30 and 60 min after the extract administration, respectively (Table 2), from the control
value of activity of 0.149 ± 0.002 nmol min‐1 mg‐1. In the case of rosmarinic acid injection a 13.5%
decrease in AChE activity was detected after 30 min.
Table 4.2. Brain acetylcholinesterase inhibition (%) 30 and 60 min after administration (intragastric and
intraperitoneal) of rosmarinic acid and P. barbatus extract. Results significantly different from the control
are marked with * (P < 0.05) and ** (P < 0.1). Values that are not significantly different (P < 0.05) are
marked from a to d.
Brain acetylcholinesterase inhibition (%)
Intragastric
administration
Intraperitonial
administration
P. barbatus 30 minutes 10.0±1.8*a 29.0±2.3*c
60 minutes 5.5±1.7**b 24.9±3.7*d
Rosmarinic acid 30 minutes 13.5±1.7*a 10.7±5.0**ab
60 minutes 12.8±2.4**a -
When standard rosmarinic acid was administered intragastrically the concentration of
rosmarinic acid found in blood showed values in the same magnitude of those found in other
studies (Baba et al., 2004; Konishi et al., 2005). However, in the present work, when rosmarinic
acid was present within the extract, a lower quantity of this acid was found in plasma, suggesting
that the other extract components may interfere in the gut permeation of rosmarinic acid.
Previous pharmacokinetic studies also reported that the highest concentration of rosmarinic
acid was reached in less than 30 min after intragastric administration. Methylated and sulfated

Chapter IV
71
forms of rosmarinic acid were found, as well as simpler hydroxycinnamic acids generated by the
hydrolysis of rosmarinic acid (Baba et al., 2004; Konishi et al., 2005; Nakazawa & Ohsawa, 1998).
The present study also observed a decrease in the rosmarinic acid concentration after 30
min, but the methylated form of rosmarinic acid was not detected. This suggests that when
rosmarinic acid is administered alone, it can pass the intestinal barrier without undergoing
metabolisation by the intestinal cells, but it can be glucuronidated and sulfated as confirmed by
the action of β‐glucuronidase and sulfatase.
Taking into account that the amount of rosmarinic acid in the extract was approximately
one third of the rosmarinic acid standard, it is seen that the decrease in the enzyme activity is
not linearly correlated with the rosmarinic acid concentration. This is due to the fact that the
extract contains other compounds besides rosmarinic acid. These compounds also inhibit
acetylcholinesterase (Porfirio et al., 2010) and the present study indicates that they can also
reach the brain and act together with rosmarinic acid as enzyme inhibitors. This fact is
corroborated by the analysis of the rosmarinic acid concentration in the plasma (less than
0.5 nM when the herbal tea is administered and 4.9 mM when the standard is given to the
laboratory animals) and the enzyme inhibitory activity, which is similar in both situations. These
results can only be explained with the inhibitory activity of all the compounds, including the
rosmarinic acid derivatives, towards AChE.
1.3.2. Intraperitoneal administration of P. barbatus extract
Plasma. After intraperitoneal administration of P. barbatus aqueous extract, rat plasma
was analysed by HPLC (Figure 4.1a). With the exception of the diterpenoid 6, all the extract
compounds were present in rat plasma in higher amount at 30 min than 60 min after injection
(Figure 4.1a). The abietane diterpenoids 4 has a structure similar to the abietane diterpenoid 6,
and is present in the extract as a minor compound. Previous studies reported that it may be
formed from abietane diterpenoid 6 in the small intestine conditions (Porfirio et al., 2010), this
suggests that abietane diterpenoid 6 might have been completely transformed into abietane
diterpenoid 4 under the biological blood‐circulation conditions.
Rosmarinic acid, compound 2, is the major compound found in the chromatogram
(Figure 4.1a). The amount of rosmarinic acid present in the plasma was calculated to be
1439.8 ± 144.0 mM and 744.5 ± 282.3 mM for 30 and 60 min after the extract administration,
respectively (Table 4.1). The amount of rosmarinic acid found in the plasma corresponded to
23.7 ± 2.4% and 12.3 ± 4.3% of the administered amount. In the chromatogram, a peak of
methylated rosmarinic acid, 2m, could be detected. According to a standard previously

Bioavailability Studies
72
developed by using a commercial COMT from porcine liver and SAM as cofactor in a
methodology described in the Experimental section, it was possible to prove the formation of a
methylated derivative from rosmarinic acid. The amount of methyl rosmarinic acid was 17.8 ±
2.3 mM and 11.1 ± 3.6 mM for the same administration time‐points (Table 4.1). The search for
glucuronidated/sulfated metabolites was carried out by reacting β‐glucuronidase and sulfatase
with the plasma. The only increase in area was observed for flavonoid aglycons, corresponding
to the decrease in the corresponding area of the respective glucuronide peaks. The fact that no
significant amounts of glucuronidated/sulfated rosmarinic acid metabolites were found after
intraperitoneal administration could be explained by the predominance of methylation occurring
in the liver over other metabolisation reactions, such as sulfation and glucuronidation. A similar
situation was also reported by O’Leary and co‐workers (2003).
Figure 4.1. HPLC analysis, 30 and 60 min after intraperitoneal administration of P. barbatus extract, of (a)
plasma and (b) brain. 1: luteolin 7‐O‐glucuronide (retention time 9.6 min); 2: rosmarinic acid (RT: 10.4); 3:
apigenin 7‐O‐glucuronide (RT: 11.2); 4: abietane diterpenoid (RT: 13.8); 5: acacetin 7‐O‐glucuronide (RT:
15.1); 2m: monomethylated rosmarinic acid; 1g: luteolin glucuronide derivative; 3’: apigenin.
In the chromatogram shown in Figure 4.1a, a different glucuronide of luteolin than the
one initially present in the water extract (compound numbered 1g) could be detected.
Compound 1g was identified after preparing, in vitro, the glucuronidation derivatives according
to Materials and Methods. In this chromatogram the aglycone apigenin (compound numbered
3’) could also be seen. No luteolin or acacetin aglycones were detected in the plasma samples.
These results suggest that the deglucuronidation of plasma apigenin glucuronide may be the

Chapter IV
73
main reaction occurring to this compound. Studies concerning the oral administration of luteolin
suggested that the presence of the unconjugated luteolin in plasma might be due to the
permeation of the aglycone without undergoing conjugation by the intestinal cells (Shimoi et al.,
1998). However, in the present study apigenin was already administrated in its glucuronidated
form, and so the high amount of aglycon found in plasma must be due to deglucuronidation of
the apigenin glucuronide. The deglucuronidation of flavonoid glucuronides by the liver was
previously reported by O’Leary and co‐workers (2003). These authors remarked that the
conversion of the quercetin‐7‐ and quercetin‐3‐glucuronide to the mono‐sulfate conjugate
showed an intermediate step of deglucuronidation by β‐glucuronidase activity, allowing
transient contact of the free aglycone with the cellular environment.
The detection only of the apigenin aglycone in plasma may be due to a higher
deglucuronidation of the apigenin glucuronide, comparatively to the other flavonoids and/or to
a lower re‐glucuronidation of apigenin by the liver. The in vitro glucuronidation assay showed a
lower rate of glucuronidation for apigenin than for luteolin, which may explain the existence of
apigenin aglycone relatively to luteolin. This hypothesis is also supported by the appearance of a
luteolin glucuronide, which could only be produced by deglucuronidation of the extract’s
luteolin 7‐O‐glucuronide, followed by glucuronidation in a different position (Figure 4.1a). Little
is known about these de‐conjugation and re‐conjugation reactions in flavonoids, especially when
they are administered in their glucuronidated forms. The present study differs from the majority
of the in vivo flavonoid administration reports in the fact that the several flavonoids were
simultaneously administered as glucuronide derivatives, as they are commonly found in herbal
teas, instead of their aglycones.
All extract compounds showed a decrease from 30 to 60 min after extract
administration, with rates of elimination ranging from 25% to 85% (Table 4.3), the lowest value
for the luteolin glucuronide with retention time 13.8 min, numbered 1g (24.4 ± 6.9%), and the
highest value for the apigenin glucuronide, numbered 3 (84.5% ± 4.6%). Only the aglycone from
apigenin (retention time of 17.3 min, numbered 3’) (Figure 4.1a) was seen. No luteolin or
acacetin aglycones were detected in the plasma.
Brain AChE. After intraperitoneal administration of P. barbatus extract, brains from rats
were also analyzed after 30 and 60 min by HPLC (Figure 4.1b). The chromatograms showed only
the presence of unconjugated rosmarinic acid, the other extract compounds or their metabolites
were not detected (Figure 4.1b). No increase of the rosmarinic acid peak area or the aglycones
from the flavonoid derivatives were observed after the reactions with β‐glucuronidase or
sulfatase. The amount of rosmarinic acid in rat brains was quantified and a relationship with the

Bioavailability Studies
74
amount of rosmarinic acid in the plasma was calculated (Table 4.1). The amount of rosmarinic
acid found in the brain was 0.40 ± 0.02% and 0.34 ± 0.01% of the administered amount, for 30
and 60 min after administration, respectively. Although the plasma rosmarinic acid
concentration showed a decrease from 30 to 60 min (around 52%, Table 4.3) the brain
rosmarinic acid concentration decreased around 15% (14.90 ± 7.11%, Table 4.1). This difference
is reflected in the ratio of brain/plasma rosmarinic acid concentration, which is higher 60 min
after the extract administration than at 30 min (0.027 at 60 min and 0.016 at 30 min). The
brain/plasma concentration ratio reflects the permeability of the compounds from the plasma to
the brain, which may be restricted primarily by the blood brain barrier, and ranges from 0 (no
permeability) to 1 (total permeability) (Zheng et al., 2006). These ratios indicate that rosmarinic
acid has the ability to pass to the brain, but the permeability is low.
Table 4.3. Retention time of compounds from the P. barbatus extract found in plasma and their decrease
from 30 to 60 min after the extract intraperitoneal administration.
compound Retention Time (min) Decrease from 30 to 60 minutes
(%)
Luteolin 7-O-glucuronide (1) 9.7 44.91±13.48
Rosmarinic Acid (2) 10.5 51.66±15.60
Apigenin 7-O-glucuronide (3) 11.3 84.57±4.59
Monomethylated Rosmarinic Acid (2m) 12.0 41.7±14.5
Abietane Diterpenoid (4) 13.5 36.33±15.61
Luteolin glucuronide (1g) 13.8 24.36±6.88
Acacetin 7-O-glucuronide (5) 15.2 56.75±0.10
Apigenin (3’) 17.3 33.4±12.8
Rosmarinic acid diffuses slowly to the brain, but it is retained there longer than in the
plasma, where a higher decrease was observed from 30 to 60 min. Brains of rats to which
standard rosmarinic acid was administered, (results not shown) showed the same concentration
of rosmarinic acid as when rosmarinic acid was given within the extract, leading to the
conclusion that the other extract compounds do not interfere with the permeability of
rosmarinic acid in the blood‐brain barrier, in opposition to the gut barrier where a interference
of the other extract compounds was observed in the permeability of rosmarinic acid.
Thirty minutes after the extract administration the rat brain showed an inhibition in
acetylcholinesterase activity of 29.0 ± 2.3%, and after 60min an inhibition of 24.9 ± 3.7% (Table
4.2), in comparison with the control rats, where the activity was 0.145 ± 0.012 nmol.min‐1.mg‐1.
Brains of rats where standard rosmarinic acid was administered showed the same concentration

Chapter IV
75
of rosmarinic acid as that detected when the extract was administered, but demonstrated to
have less acetylcholinesterase inhibition (10.7 ± 5.0% after 30 min). This confirms the inhibition
of AChE by the other extract components, although they were not detected by HPLC.
The percentages of acetylcholinesterase inhibition achieved were of the same
magnitude of the ones reported to galanthamine in the rat model, however the amounts of
interaperitoneal injection of galanthamine were lower: 3 mg.kg‐1 for 10% inhibition (Geerts et
al., 2005) and 10 mg.kg‐1 for 28% inhibition (Chattipakorn et al., 2007). Higher inhibitions can be
obtained with lower amounts of donepezil, 39% with 3 mg.kg‐1 (Geerts et al., 2005), however,
this inhibitor may present a higher incidence of adverse effects in high amounts. The
dosage/activity in humans for galanthamine is 16–24 mg.day‐1 to 30–40% brain
acetylcholinesterase inhibition (Kadir et al., 2008), and for donepezil is 10 mg.day‐1 to 19–27%
inhibition (Bohnen et al., 2005). As there is an interest in decreasing the adverse peripheral
effects of acetylcholinesterase inhibition, mostly related to gastrointestinal and hepatic
disturbances, herbal teas may offer an alternative for mild treatments of Alzheimer’s disease. P.
barbatus water extracts are traditionally prepared as decoctions (as in the present report) and
drunk by people and those secondary effects were not reported (Lukhoba et al., 2006). In fact,
the use of this plant extract to treat gastrointestinal and hepatic conditions is widely reported
(Lukhoba et al., 2006). Rosmarinic acid when administered in low doses seems to act as an
anxiolytic‐like compound, only when administered in high doses it seem to act on the peripheral
nervous system (Pereira et al., 2005).
Several recent reports showed that flavonoids, such as quercetin and its metabolites,
have the ability to reach the brain (Huebbe et al., 2010). Coleta and co‐workers (2008)
demonstrated that intragastrically‐administered luteolin, or its metabolites, should reach the
brain, causing anxiolytic‐like effects. Gujinski and co‐workers (2009) reported that ethanolic
extracts from Melissa officinalis, a Lamiaceae species, and its main component rosmarinic acid,
when administered to rats produced dose‐related antinociception in several models of chemical
pain, through mechanisms that involved cholinergic systems. Other studies also show that
rosmarinic acid, when administered through intraperitoneal injection (2–8 mg.kg‐1), induced
neurobehavioral changes in rats (Pereira et al., 2005). Although rosmarinic acid in the brain was
not quantified in those studies, its presence in the brain was suggested by the activities it
exerted in vivo.
The bioavailability of the currently used acetylcholinesterase inhibitors is highly variable,
depending mostly to the structure of the compounds, their metabolisation and elimination, and
their ability to permeate the intestinal and blood‐brain barriers (McGleenon et al., 1999). Their

Bioavailability Studies
76
time of action also depends on the mode of acetylcholinesterase inhibition. Donepezil, for
instance, has a peak plasma level 4 h after administration and a half‐life of over 70 h, while
tacrine shows low bioavailability when taken orally and has a half‐life in plasma of 1.4 h
(McGleenon et al., 1999). However, tacrine reaches a brain concentration that is 10‐fold that of
plasma, its dosage and brain bioavailability are not proportional, and so higher doses and
multiple dosing are used to increase bioavailability and the half‐life (McGleenon et al., 1999).
In the present study rosmarinic acid and the other components of the extract also do not
seem to follow a direct relation between dose and response. First, because the intestinal
absorption and metabolisation of rosmarinic acid is influenced by the presence of the other
extract compounds. Also, although the concentration of rosmarinic acid was always much lower
in brain than in the plasma, the relationship between plasma and brain rosmarinic acid
concentrations did not seem to be linear, as a decrease of more than 50% in plasma rosmarinic
acid concentration corresponded just to a slight decrease in the brain (Table 4.1). However, the
amount of detected rosmarinic acid and the activity seem to be related, as the decrease in the
brain acethylcholinesterase inhibition was similar to the decrease in the content in rosmarinic
acid. As the phenolic compounds from the plant extract seem to act in the brain in very low
concentrations, and their amount in brain has less fluctuations than the amount in the plasma,
taking a large volume of herbal tea during a long period of time, and several times per day, as it
is usually taken, may be the correct way of increasing its effects.
Apart from the beneficial effect of increasing the acetylcholine mediated neuronal
transmission, the treatment of Alzheimer’s disease with acetylcholinesterase inhibitors also
seems to protect free radical toxicity, β‐amyloid injury and to attenuate cytokine release from
microglial cells (Tabet, 2006). Apart from this ‘‘cholinergic anti‐inflammatory pathway’’,
rosmarinic acid is also a well‐known antioxidant and free radical scavenger (Falé et al., 2009). As
far as we know, this is the first report confirming the neurologic activity of rosmarinic acid by the
quantification its amount in the brain.
1.4. Conclusions
In this study it was demonstrated that the rosmarinic acid, present in herbal teas, may
cross the intestinal barrier, as well as the blood brain barrier, and be detected in the brain,
where it inhibits the enzyme AChE. The metabolisation and bioavailability of the herbal tea
components is different from the administration of pure compounds.

Chapter IV
77
2. Bioavailability of mixtures of rosmarinic acid, luteolin and
apigenin through Caco‐2 cell monolayers, modeling the
bioavailability of plant herbal teas.
2.1. Introduction
Herbal teas are among the oldest and most common drinks in human culture and are
drunk for their pleasant flavour and/or for their health improving properties. These plant
aqueous extracts are often composed of phenolic compounds such as phenolic acids and
flavonoids, which have proved to be therapeutically active (see Komes et al., 2010 for the
composition of several taxonomically unrelated medicinal plants).
Previous studies demonstrated that the Plectranthus barbatus herbal tea has relevant in
vitro antioxidant and anti‐acetylcholinesterase activities, which are due to its main components:
rosmarinic acid, flavonoid glucuronides (apigenin, luteolin and acacetin glucuronides), and
abietane diterpenoids (Falé et al., 2009; Porfirio et al., 2010). Rosmarinic acid and the flavonoid
components remained after in vitro digestion of the P. barbatus extract with gastric and
pancreatic juices, but flavonoid glucuronides were hydrolysed into their aglycones by the
β‐glucuronidase of the microorganisms in the gut flora (Porfirio et al., 2010). When P. barbatus
extract was intragastrically administered to rats its components were found in the plasma and a
decrease in brain acetylcholinesterase activity was observed (Falé et al., 2011).
As herbal teas are complex mixtures of compounds, we aimed to determine the effect of
the presence of rosmarinic acid, luteolin and apigenin in the permeation and metabolisation of
each other in Caco‐2 cell monolayers, an in vitro model for the intestinal absorption and
metabolism. Rosmarinic acid, apigenin and luteolin are the main constituents of P. barbatus
herbal tea, but they are very commonly found together in many other herbal teas and
foodstuffs. However, little is known about the way that these compounds affect the
bioavailability of each other. Nevertheless, it is known that phenolic acids and flavonoids
interact with transport systems in intestinal cells, such as the monocarboxylic acid transporters
(MCT) (Konishi et al., 2003; Konishi et al., 2004), the ABC transporters P‐glycoprotein (Pgp) and
multidrug resistance proteins (MRP), being actively transported or inhibiting them (for review
see Brand et al., 2006). The MCT4, MCT5, MRP1, MRP3 and MRP5 transporters, located in the
basolateral membrane of the intestinal cells, promote the transport of their substrates to the
bloodstream, while Pgp and MRP2, located in the apical membrane, promote the transport of
their substrates from the intracellular compartment to the intestinal lumen (Gill et al., 2005,

Bioavailability Studies
78
Konishi et al., 2003, 2004; Brand et al., 2006). The MCT1 transporter is located in the apical
membrane, but transports its substrates from the intestinal lumen to the intracellular
compartment (Gill et al., 2005).
The aim of this study is to evaluate the permeation through Caco‐2 cell monolayers of
the aqueous extract of P. barbatus, and of mixtures of its main phenolic compounds (rosmarinic
acid, luteolin, apigenin), in different concentrations, but in a relative proportion similar to the
usually found in herbal teas. The effect of two intestinal epithelium transporters, Pgp and MCT,
on the bioavailability of the polyphenolic mixture was also investigated.
2.2. Material and Methods
The materials and methods are described in detail in the Chapter II.
The P. barbatus extract was prepared as a decoction, as described in Chapter II section 4.
The bioavailability studies were performed in Caco‐2 cell monolayers as referred in Chapter II
section 17. This methodology was used to evaluate the bioavailability of P. barbatus extract and
of mixtures of phenolic compounds (rosmarinic acid, luteolin and apigenin) according to a
factorial design known as central composite design (CCD) (Barker, 1985), as described in Chapter
II section 17.1. The concentrations used in the mixtures are listed in Table 2.1.
The involvement of the transport systems MCT and Pgp in the bioavailability of a mixture
of rosmarinic acid, luteolin and apigenin was evaluated using the substrates for these systems,
benzoic acid and digoxin, as described in Chapter II section 17.2.
The HPLC analysis was carried out as described in Chapter II section 17.3.
2.3. Results
Previous studies with Plectranthus barbatus herbal tea revealed the presence of
rosmarinic acid and flavonoid glucuronide derivatives together with two abietane diterpenoids
(Porfirio et al., 2010). Rosmarinic acid, the poplyphenol present in higher amount in the water
extract, is a characteristic secondary metabolite of species from the Lamiaceae family, where
P. barbatus is included. In the present study we evaluated the intestinal bioavailability of the
aqueous extract of P. barbatus as a model extract, and of mixtures of three of its main
components, rosmarinic acid, luteolin and apigenin. We aimed to evaluate the differences in
bioavailability of these compounds, which are very commonly found in plant infusions and
decoctions.

Chapter IV
79
2.3.1. Bioavailability of Plectranthus barbatus herbal tea
In order to study the bioavailability of the major polyphenol compounds in the herbal
tea of P. barbatus, the water extract was applied to the apical side of a Caco‐2 cell monolayer
and the polyphenols were quantified in the apical and basolateral sides after 6h of incubation.
Forty three per cent of the rosmarinic acid in the plant extract permeated the Caco‐2 membrane
to the basolateral side. The flavonoid glucuronides initially present permeated the cell
membranes 34‐35% and the abietane diterpenoids permeated the same system in a slightly
higher amount, 43‐49% (Table 4.4).
Table 4.4. Permeation of the P. barbatus aqueous extract constituents through the Caco‐2 cell monolayer.
Compound Permeation (%)
Rosmarinic acid 43.3±5.3
Luteolin 7-O-glucuronide 35.3±3.4
Apigenin 7-O-glucuronide 34.1±4.6
Acacetin 7-O-glucuronide 33.8±4.1
Abietane diterpenoid 1 43.5±4.1
Abietane diterpenois 2 49.9±1.8
Although P. barbatus abietane diterpenoids were the compounds that showed highest
bioavailability in Caco‐2 cells, previous studies showed that they may be hydroxylated in the
small intestine by the pancreatic juice into non‐active metabolites (Porfirio et al., 2010).
Furthermore, when the plant extract was administered intragastrically and intraperitonially to
rats these compounds were found in the plasma just in the non‐active form (Falé et al., 2011),
suggesting that their metabolisation is very fast processes. For this reason the abietane
diterpenoids shall not be considered in ulterior experiments.
In the digestive tract, rosmarinic acid does not undergo extensive degradation or
metabolisation and the flavonoid glucuronides may be deglucuronidated by the gut microflora,
as it was demonstrated by the in vitro digestion (Porfirio et al., 2010). Pursuing the investigation
of P. barbatus aqueous extract, the effect of rosmarinic acid, apigenin and luteolin on the
intestinal permeation and metabolisation of each other by Caco‐2 cells were analysed, as well as
the interaction of these compounds with two known transport systems to further enlighten the
mechanism by which these active compounds are made bioavailable.
In summary, when the bioavailability of the P. barbatus aqueous extract was tested with
the Caco‐2 cell system all the compounds permeated the cell membrane after 6h, including the
glucuronide derivatives.

Bioavailability Studies
80
2.3.2. Bioavailability of a mixture of the standards rosmarinic acid, apigenin and luteolin
An experimental plan involving the effect of several concentrations of rosmarinic acid,
luteolin and apigenin was set in order to evaluate the effect of the concentration of these
compounds on each other’s permeability through a Caco‐2 cell monolayer. The response surface
methodology allowed drawing the permeability curves (response surface) of each compound in
function of the concentration of the other two compounds (Figure 4.2). Using this methodology
it was possible to study interactions in the permeability by the presence of different compounds,
the real situation found in the herbal teas.
The curves show that rosmarinic acid has higher permeation in the Caco‐2 cells in the
presence of very high amounts of both flavonoids (Figure 4.2a). This means that the flavonoids
interfere with the phenolic acid permeation, probably by inhibiting the efflux transport systems.
The permeation of luteolin was facilitated by rosmarinic acid and by apigenin (Figure 4.2b), as
they may have facilitated the transport or inhibited the efflux mechanisms. The permeation of
luteolin increased with increasing concentrations of rosmarinic acid, being this effect more
pronounced than the one found with increasing concentrations of apigenin. The behaviour of
apigenin permeation is more complex (Figure 4.2c). A maximum value for apigenin permeation
can be obtained with a luteolin concentration, and this maximum can be obtained with a higher
concentration of luteolin at low concentration of rosmarinic acid, or with lower concentration of
luteolin at high concentration of rosmarinic acid. On the other hand, minimum apigenin
permeability can be seen by varying the concentration of rosmarinic acid, which can be obtained
with lower concentrations of rosmarinic acid at low concentration of luteolin, or with higher
concentration of rosmarinic acid at high concentration of luteolin. This complex permeation
behaviour suggests that these compounds may inhibit the efflux systems. When they are both
present they may also compete with apigenin for an uptake transport system, or inhibit it.
Apigenin permeation shows the lowest permeability in the presence of high concentrations of
rosmarinic acid and luteolin together.
Although only the aglycones were applied in the cell culture medium of the apical side of
the Caco‐2 system, the glucuronides derivatives could be detected on the basolateral side of the
cells. Caco‐2 cells have the ability to glucuronidate flavonoids, as was previously reported for
apigenin and luteolin (Porfirio et al., 2010). The effect of the concentration of rosmarinic acid,
luteolin and apigenin on the glucuronidation and bioavailability of these flavonoids was assessed
using the same CCD factorial design. The response – glucuronidation – surfaces curves obtained
are shown in Figure 4.3. For the glucuronidation of each flavonoid, the presence of the other
flavonoid and rosmarinic acid seemed to have a similar effect whether in presence of high or low

Chapter IV
81
amount of each other, increasing with the increase of the concentration of rosmarinic acid and
decreasing with the increase of the concentration of the other flavonoid, which suggests that
the both flavonoids are substrates of the same enzyme and may compete for the same
glucuronosyltransferases.
Figure 4.2. Permeation surfaces for (a) rosmarinic acid, (b) luteolin and (c) apigenin with different
concentrations of the other two components, built with the CCD experimental plan. The relative errors are
0.930 (a), 0.718 (b), and 0.817 (c).

Bioavailability Studies
82
Figure 4.3. Glucuronidation surfaces for (a) luteolin and (b) apigenin with different concentrations of
rosmarinic acid and of the other flavonoid, built with the CCD experimental plan. The relative errors are
0.981 (a), and 0.766 (b).
2.3.4. Effect of MCT and Pgp transporter systems on the bioavailability of the polyphenol
mixture
The monocarboxylic acid transporter system (MCT), located in the basolateral
membrane of intestinal cells, is known to act on the permeability of monocarboxylic acids and it
is also recognized that the flavonoids may interfere with these systems (Konishi et al., 2003). As
rosmarinic acid, a monocarboxylic acid, is present in a mixture containing flavonoids, the
involvement of the MCT transporter system on the mixture bioavailability was analysed. On the
other hand Pgp efflux system is known to be inhibited by flavonoids (Brand et al., 2006), so the
participation of these systems on the bioavailability of the two flavonoids together with the
monocarboxylic acid (rosmarinic acid) were also analysed. In order to evaluate the participation
of MCT and Pgp on the bioavailability of a mixture of rosmarinic acid with the two flavonoids,
the studies were conducted in the absence and in the presence of the transporter system
substrates, benzoic acid and digoxin, respectively (Tsuji et al., 1994).
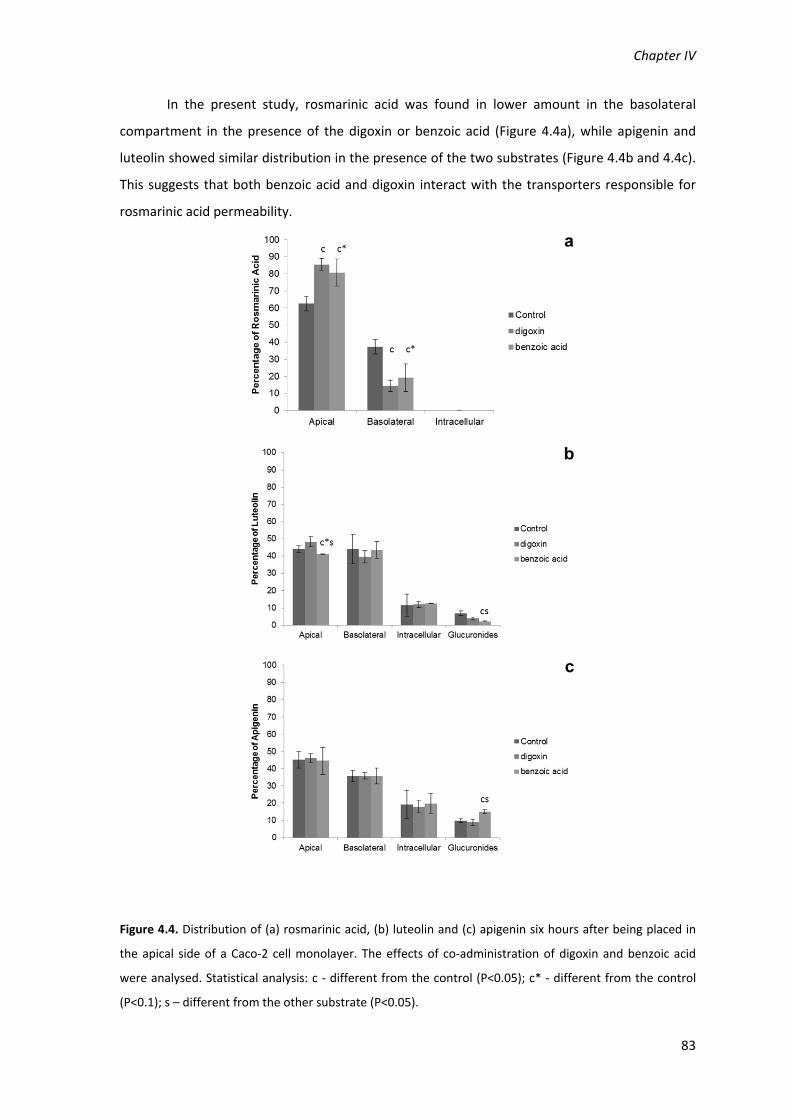
Chapter IV
83
In the present study, rosmarinic acid was found in lower amount in the basolateral
compartment in the presence of the digoxin or benzoic acid (Figure 4.4a), while apigenin and
luteolin showed similar distribution in the presence of the two substrates (Figure 4.4b and 4.4c).
This suggests that both benzoic acid and digoxin interact with the transporters responsible for
rosmarinic acid permeability.
Figure 4.4. Distribution of (a) rosmarinic acid, (b) luteolin and (c) apigenin six hours after being placed in
the apical side of a Caco‐2 cell monolayer. The effects of co‐administration of digoxin and benzoic acid
were analysed. Statistical analysis: c ‐ different from the control (P<0.05); c* ‐ different from the control
(P<0.1); s – different from the other substrate (P<0.05).

Bioavailability Studies
84
In the case of benzoic acid, transported by the system MCT (Figure 4.5a) the standard
mixture of polyphenols affected the permeation of the substrate, because it was found in higher
amounts in the basolateral side of the cells, comparatively to the amounts found in the absence
of the mixture. In the case of the Pgp system, digoxin decreased on the apical side and increased
slightly in the basolateral side when in the presence of polyphenols (Figure 4.5b). This result may
indicate that the Pgp transporter is inhibited by this system and the polyphenol mixture
(rosmarinic acid, apigenin and luteolin) may indeed alter the distribution of Pgp substrates
around the cell membranes and in the intracellular compartment (Figure 4.5b).
Figure 4.5. Distribution of (a) benzoic acid and (b) digoxin six hours after being placed in the apical side of
a Caco‐2 cell monolayer. The effects of co‐administration of a standard mixture (SM) with rosmarinic acid,
luteolin and apigenin, 50µM each, was analysed. Statistical analysis: * ‐ different from the control
(P<0.05).
On what concerns the bioavailability of the flavonoid glucuronide derivatives, studies
were also conducted in the absence and in the presence of the MCT and Pgp substrates. In this
study it is not possible to distinguish the effect of the transporter substrates on the
glucuronosyltransferase enzymatic activity and on the bioavailability of the flavonoid
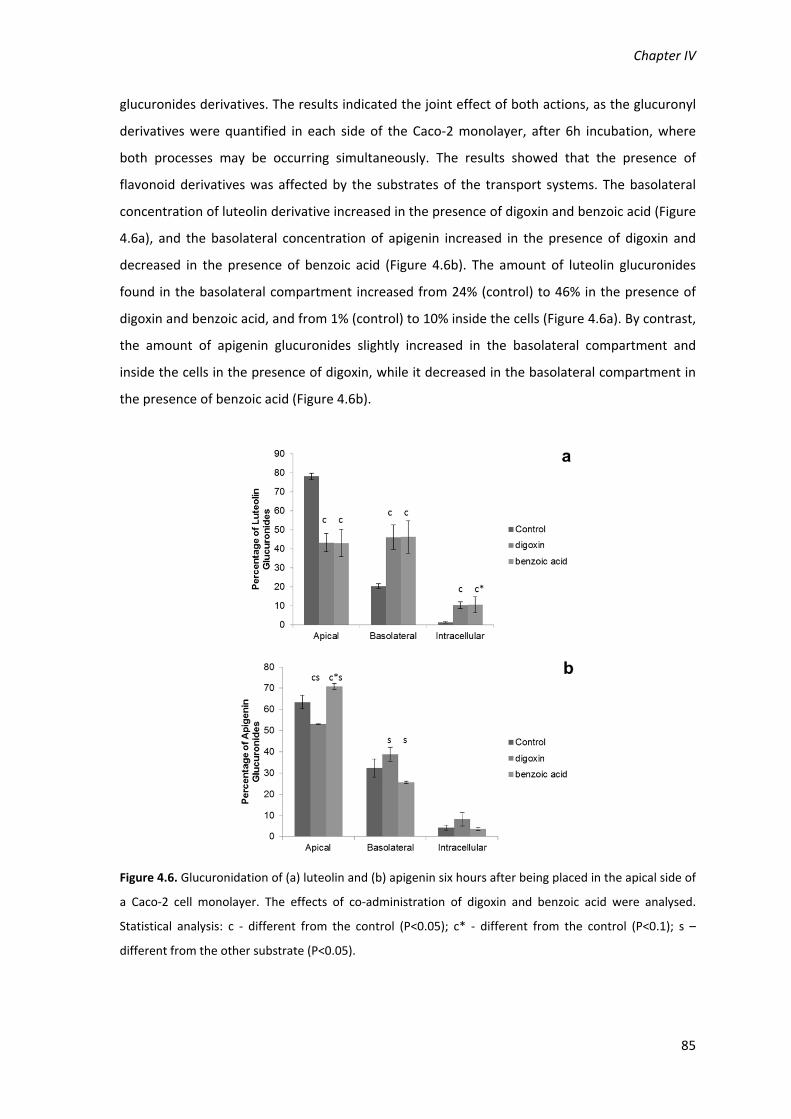
Chapter IV
85
glucuronides derivatives. The results indicated the joint effect of both actions, as the glucuronyl
derivatives were quantified in each side of the Caco‐2 monolayer, after 6h incubation, where
both processes may be occurring simultaneously. The results showed that the presence of
flavonoid derivatives was affected by the substrates of the transport systems. The basolateral
concentration of luteolin derivative increased in the presence of digoxin and benzoic acid (Figure
4.6a), and the basolateral concentration of apigenin increased in the presence of digoxin and
decreased in the presence of benzoic acid (Figure 4.6b). The amount of luteolin glucuronides
found in the basolateral compartment increased from 24% (control) to 46% in the presence of
digoxin and benzoic acid, and from 1% (control) to 10% inside the cells (Figure 4.6a). By contrast,
the amount of apigenin glucuronides slightly increased in the basolateral compartment and
inside the cells in the presence of digoxin, while it decreased in the basolateral compartment in
the presence of benzoic acid (Figure 4.6b).
Figure 4.6. Glucuronidation of (a) luteolin and (b) apigenin six hours after being placed in the apical side of
a Caco‐2 cell monolayer. The effects of co‐administration of digoxin and benzoic acid were analysed.
Statistical analysis: c ‐ different from the control (P<0.05); c* ‐ different from the control (P<0.1); s –
different from the other substrate (P<0.05).

Bioavailability Studies
86
2.4. Discussion
In order to evaluate the involvement of two membrane transporter systems on the
permeability pattern found for rosmarinic acid, luteolin and apigenin in the herbal decoction
through Caco‐2 cell monolayers, the permeability of these phenolic compounds was analysed in
the presence of substrates of known transport systems, namely benzoic acid (substrate of MCT
(Tsuji et al., 1994)) and digoxin (substrate to Pgp).
The permeability of rosmarinic acid through the Caco‐2 cell monolayer decreased in the
presence of benzoic acid (Figure 4.4a), suggesting a competition for the transport across this
membrane system indicating that it may be via one transporter of the MCT family. Previous
studies pointed out that the permeability of rosmarinic acid through MCT may be very low
because it showed lower affinity to the transporter than ferulic acid, for instance (Konishi and
Kobayashi 2005). The low affinity is also suggested by the increase of benzoic acid in the
basolateral compartment in the presence of the standard mixture (Figure 4.5a), suggesting that
the affinity of benzoic acid to MCT is much higher than rosmarinic acid.
The distribution of the rosmarinic acid was also affected by digoxin, decreasing in the
same way as benzoic acid did, from the apical to the basolateral side. Digoxin permeates the
Caco‐2 monolayer predominantly by transcellular passive diffusion, and Pgp, responsible for
digoxin efflux, is the only known transporter for this molecule (d’Souza et al., 2003). Pgp does
not seem involved in the efflux of rosmarinic acid, as the competition of digoxin with rosmarinic
acid would lead to an increase in the amount of rosmarinic acid in the basolateral compartment,
which was not observed. Therefore these results suggest that digoxin may inhibit the
transporters involved in the permeation of rosmarinic acid from the apical to the basolateral
side. Although these transporters were not studied in the present work, digoxin is known to
interact with active transport in muscle cells, as an inhibitor of Na+‐K+‐ATPase pump (Rochetti et
al., 2003).
The flavonoids luteolin and apigenin are known inhibitors of the ABC transporters
(Schutte et al., 2008). Our results suggest that luteolin and apigenin are not substrates of MCT
or Pgp transporters, since benzoic acid and digoxin did not affect the transport of these
flavonoids across the Caco‐2 cell monolayers (Figures 4.4b and 4.4c). However, the
bioavailability of apigenin is strongly affected by the presence of luteolin, this means that other
transport system beyond Pgp or MCT must be involved. Flavonoids such as quercetin,
kaempferol and isorhamnetin have been shown to be substrates of Pgp, which limits their
bioavailability (Wang et al., 2005).

Chapter IV
87
The effect of benzoic acid and digoxin on the distribution of flavonoid glucuronides
(Figure 4.6) suggests that both transporter systems seem to be involved in the permeability of
these compound because significant differences were found on the bioavailability of the
glucuronides in the presence of the substrates.
The behaviour of the flavonoids and rosmarinic acid towards the permeation of one
another seems to be a balance of two factors: competition for the transepithelial transporters
and inhibition of the efflux via ABC transporters. Previous studies showed that flavonoid
aglycones may permeate the Caco‐2 cell monolayers by active transport (Kobayashi et al., 2008)
and the co‐administration of flavonoids inhibited the efflux of hesperitin, a flavonoid aglycone,
by ABC transporters (Brand et al., 2010). The result of this balance may be seen in the
permeation surfaces of Figure 4.2. For rosmarinic acid the flavonoids caused a decrease of
permeability when a low concentration of apigenin and a high concentration of luteolin is used.
These changes may be due to the inhibition of the membrane transporters by the flavonoids. In
lower concentrations of flavonoids, the inhibition of transporters, such as MCT, may be
dominant effect, decreasing the bioavailability of rosmarinic acid. By contrast, in higher
concentrations of flavonoids may be dominant the inhibition of efflux transporters, such as the
ABC transporters. Luteolin and apigenin were previously reported to inhibit MCT transporters
(Wang and Morris, 2007) and ABC transporters (Brand et al., 2006). The permeation of
rosmarinic acid increases when the concentration of apigenin also increases, this may be due to
a higher inhibition of the efflux transporters by this flavonoid relatively to luteolin (Figure 4.2a).
The efflux transporter of rosmarinic acid that seems to be inhibited by the flavonoids may
belong to the ABC family, but it is not Pgp, since the co‐administration of digoxin did not
increase the bioavailability of rosmarinic acid (Figure 4.4a)
For luteolin the effect seems to be the inhibition of the efflux systems by rosmarinic acid
and apigenin (Figure 4.2b), while for apigenin there is a joint effect competition for the same
transporter of luteolin and inhibition of the efflux transporters by rosmarinic acid and luteolin
(Figure 4.2c). The clearly competitive effect shown by luteolin in the permeation surface of
apigenin, suggests that the transporter of both molecules has more affinity to luteolin than
apigenin. The permeation of luteolin across the Caco‐2 monolayer increases with higher
concentrations of rosmarinic acid, and the same happens with the permeation of apigenin when
the concentration of luteolin is low, which suggests that rosmarinic acid may inhibit the
transporters responsible for the efflux of both flavonoids. Although rosmarinic acid has been
reported not to have effect on Pgp or MRP1 (Kobayashi et al., 2003), it is known that it inhibits

Bioavailability Studies
88
the efflux of some molecules, like ovalbumin, involving other transporters (Nabekura et al.,
2010).
The competition of luteolin and apigenin for the same transporters is also clearly evident
in the glucuronidation surfaces of both flavonoids (Figures 4.3a and 4.3b). As rosmarinic acid
may decrease the efflux of the flavonoids in the apical membrane, their concentration in the
intracellular compartment, increasing the glucuronidation. Apigenin and luteolin may also be
glucuronidated by the same enzymes, and for this reason compete for the
glucuronosyltransferases, resulting in the decrease of glucuronidation of one of these flavonoids
in the presence of the other one, as shown in Figures 2a and 2b. Again, luteolin seems to have
higher affinity to the glucuronosyltransferases, as the glucuronidation of apigenin was more
affected by luteolin than the glucuroidnation of luteolin was affected by apigenin, suggesting
that the presence of an extra hydroxyl group may increase the affinity for the enzyme active site.
Other studies also showed this difference in affinity for the enzymes, mainly those involved in
phase II of the metabolism. For instance, a decrease of phase II metabolisation of the flavanone
hesperetin by the co‐administration of other flavonoid, especially flavonols and flavones as
luteolin and apigenin, may be also due to the higher affinity of the phase II enzymes to these
compounds (Brand et al., 2010).
To confirm the involvement of PgP and MCT in the polyphenol bioavailability, the effect
of the herbal tea on the permeation of bezoic acid and digoxin was also evaluated. The increase
of bioavailability of benzoic acid in the presence of the standard mixture (Figure 4.5a) may be
due to the inhibition of a benzoic acid efflux mechanism by the standard polyphenolic mixture.
Although the efflux mechanism of benzoic acid is not clearly elucidated, flavonoids are inhibitors
of a wide variety of efflux processes related with ABC transporters (as reviewed by Brand et al.,
2006). Therefore it can be concluded that the consumption of herbal teas containing rosmarinic
acid together with these two flavonoids may interfere with drugs based on carboxylic acid
structures.
An increase of the amount of digoxin in the basolateral compartment was observed also
in the presence of the standard mixture (Figure 4.5b), which may be assumed as due to the
inhibition of the efflux system Pgp, probably due to apigenin and luteolin that are known
inhibitors of this transporter (Brand et al., 2006). However, previous studies suggest that
rosmarinic acid is not a substrate and has no inhibitory effect on Pgp (Konishi and Kobayashi
2005; Nabekura et al., 2010). Nevertheless, the efflux of digoxin may also be partially inhibited
by the competition with the luteolin and apigenin glucuronides, which appear to be substrates
of Pgp, as discussed previously.

Chapter IV
89
The permeation of rosmarinic acid in the P. barbatus aqueous extract and in the
standard mixture, in the presence of luteolin and apigenin, was similar. However, the
permeation of rosmarinic acid after 6h was higher than that expected according to the literature
(Konishi & Kobayashi 2005), which may be due to the inhibition of the efflux mechanisms by the
flavonoids. The inhibition of the intake and efflux mechanisms points out that rosmarinic acid
may be substrate of intestinal transporters. As far as we know this is the first report suggesting
that the flux of rosmarinic acid may be mediated by transport systems, although with low
affinity. To this conclusion may contribute the larger times of incubation performed, six hours in
contrast to 40 minutes in similar studies (Konishi et al., 2005). The flavonoid glucuronides also
showed a higher bioavailability than expected, which may be due to the inhibition of the efflux
mechanisms by the co‐administration of the two flavonoids together with rosmarinic acid. A
similar increase of flavonoid glucuronides in the basolateral compartment by the co‐
administration of flavonoids has been reported (Brand et al., 2010). These authors have
demonstrated that the increase of flavonoid glucuronides was induced by the inhibition of the
efflux mechanisms exclusively in the apical membrane.
The results obtained in this study with the P. barbatus aqueous extract may be
extrapolated to other teas, since rosmarinic acid seems to be one key component in aqueous
extracts of Lamiaceae species (Janicsak et al., 1999), to which belong some of the most common
species used in herbal teas (mints, rosemary, thyme, sage, lavender and basil). The occurrence of
flavones in these species, especially luteolin and apigenin glucuronides, together with rosmarinic
acid is very common and widely described in the literature (Fecka & Turek 2007, for peppermint,
melissa and sage).
2.5 Conclusions
The compounds of P. barbatus aqueous extract have shown higher bioavailability
together than the values expected for the isolated standards, since the bioavailability of
mixtures of phenolic compounds depends on the balance between competition for the same
transporters (reducing bioavailability), and the inhibition of efflux transport systems (leading to
an increase of bioavailability).
Rosmarinic acid seems to be substrate of MCT, but not of Pgp. The flavonoids apigenin
and luteolin do not seem to be substrates of MCT or Pgp, but they may inhibit Pgp. The
glucuronide derivatives of apigenin and luteolin, however, seem to be substrates of Pgp.

Bioavailability Studies
90
The high diversity of compounds often present in medicinal plant extracts is favourable
to a high bioavailability of the compounds, as it decreases the competition for the same intake
transporters while promoting the inhibition of efflux transporters.
These results may also predict an interaction between herbal teas containing these
mixtures of polyphenols with pharmaceuticals presenting carboxylic acid groups in their
structure.

Chapter IV
91
3. Conclusions
The bioavailability studies allowed to conclude that rosmarinic acid, present in most
herbal teas, may cross the intestinal barrier (in rats and Caco‐2 monolayers), as well as the blood
brain barrier (in rats), and be detected in the brain.
The administration (intragastric and intraperitoneal) of the P. barbatus aqueous extract
and the rosmarinic acid caused the inhibition of the brain acetylcholinesterase activity.
The metabolisation and bioavailability of the herbal tea components is different from
the administration of pure compounds, as observed in the bioavailability studies with rats and
with Caco‐2 monolayers.
The compounds of P. barbatus aqueous extract have shown, in the Caco‐2 cell
monolayer model, a higher bioavailability together than the values expected for the isolated
standards. This fact is due to a predominant effect of inhibition of efflux transport systems,
leading to an increase of bioavailability.
The high diversity of compounds often present in medicinal plant extracts is favourable
to a high bioavailability of the compounds, as it decreases the competition for the same intake
transporters while promoting the inhibition of efflux transporters.


Chapter V
Interactions between the Plectranthus barbatus herbal tea
and the proteins acetylcholinesterase, human serum albumin
and lysozyme
Falé PL, Ascensão L, Serralheiro MLM, Haris PI. Interaction between Plectranthus barbatus herbal tea
components and acetylcholinesterase: binding and activity studies. Submitted to Food Funct.
Falé PL, Ascensão L, Serralheiro MLM, Haris PI. 2011. Interaction between Plectranthus barbatus herbal
tea components and human serum albumin and lysozyme: binding and activity studies. Spectroscopy. 26:
79–92.


Chapter V
95
1. Interaction between the Plectranthus barbatus extract and
acetylcholinesterase. Binding of herbal tea components to the
protein structure and inhibition of enzymatic activity
1.1. Introduction
Acetylcholinesterase (AChE) is the enzyme that catalyses the hydrolysis of acetylcholine,
a neurotransmitter found in the synaptic gap, thus terminating the synaptic transmission.
Alzheimer’s disease is a very complex ailment, and the most successful therapy to present
consists of increasing the levels of acetylcholine in the brain through the inhibition of AChE
activity by reversible inhibitors (Heinrich & Teoh, 2004). However, irreversible inhibitors cannot
be used for therapeutic purposes, as complete inactivation of AChE leads to a toxic accumulation
of acetylcholine and failure of synaptic transmission, with consequent deterioration of
neuromuscular junctions, flaccid muscle paralysis and seizures in the central nervous system
(Harel et al., 2008).
Plants have proven to be an important source of useful AChE inhibitors for the
symptomatic treatment of Alzheimer’s disease (Mukjerjee et al., 2007), as for instance,
galantamine, a commercially available compound that was first isolated from Galanthus species
(Heinrich & Teoh, 2004). The herbal tea of Plectranthus barbatus leaves, composed mainly of
rosmarinic acid and flavonoid glucuronides, has been shown to be a promising in vitro AChE
inhibitor (Falé et al., 2009), even after being subjected to an in vitro simulation of the
gastrointestinal digestion (Porfirio et al., 2010). Furthermore, when the tea was intraperitoneal
administered to rats, its components and their metabolites were found in rat plasma and brain,
and a significant decrease in brain AChE activity was observed. The flavonoid glucuronides from
the plant extract were metabolized when the extract was administered to rats, and were found
in the plasma as glucuronides and as aglycones (Falé et al., 2011).
The aim of the present work was to determine the mode of interaction of AChE with
rosmarinic acid, the main constituent of P. barbatus extracts, as well as with luteolin and
apigenin (Figure 5.1), the flavonoid aglycons found in the plasma after the extract
administration. Furthermore the interaction of AChE with quercetin was also studied, as this
flavonoid has an extra hydroxyl group in position 3 of the C ring (Figure 5.1) that makes it
interesting for a comparative study. Although quecetin is not present in P. barbatus extract, it is
very common in plants, and plant‐derived food (Erlund, 2004). These findings will allow the
determination of structural features that may be more favourable to the binding of plant

Interactions with Proteins
96
phenolic compounds to the enzyme, and which forms of interaction may be more useful to
increase the reversible inhibition of AChE. This knowledge may lead to the prediction of the
potential of a plant extract in the treatment of Alzheimer’s disease by AChE inhibition, based on
its chemical constitution.
Figure 5.1. Molecular structure of (a) rosmarinic acid, (b) quercetin, (c) luteolin, and (d) apigenin.
1.2. Materials and Methods
The materials and methods are described in detail in the Chapter II.
The P. barbatus extract was prepared as a decoction, as described in Chapter II section 4.
The interactions between the plant extract, or the standards rosmarinic acid, luteolin, apigenin
and quercetin, with acetylcholinesterase were evaluated by fluorescence spectroscopy, as
described in Chapter II section 13, and by FTIR spectroscopy, as referred in Chapter II
section 14. The acetylcholinesterase inhibition was determined as described in Chapter II
section 5.
OHOOH
OH
O
O
OH
OH
OH
O
O
OH
OH
OH
OH
OH
O
O
OH
OH
OH
OH
O
O
OH
OH
a
b
c
d

Chapter V
97
1.3. Results
1.3.1. Fluorescence studies on the binding of P. barbatus water extract to
acetylcholinesterase
The binding of P. barbatus extract to AChE was studied by fluorescence spectroscopy.
AChE, alone, showed an excitation spectrum with a maximum wavelength at 280 nm and
emission spectrum with a maximum at 335 nm, under excitation at 280 nm. Figure 5.2 shows the
emission fluorescence spectra of AChE with the addition of increasing concentrations of P.
barbatus plant extract. The intensity of the intrinsic fluorescence of the protein decreased
significantly – was quenched – with the increase of concentration of the plant extract, which
acted as a quencher.
Figure 5.2. Fluorescence emission spectra of acetylcholinesterase with the addition of P. barbatus
aqueous extract. Arrow points to increasing concentrations of P. barbatus plant extract, ranging 0; 0.5; 1;
5; 10; 33; 50; 100 µg.ml‐1.
The dominant fluorophore in proteins is the indole group of the tryptophan residues and
the fluorescence quenching by the plant extract is caused by the interaction between the plant
compounds and the protein, in the vicinity of the tryptophan residues (Lakowicz, 2006). The
emission spectrum of the indole may be shifted towards lower wavelengths (blue shifted), if the
group is buried within the native protein, or its emission may be shifted towards larger
wavelengths (red shifted), when the protein is unfolded (Lakowicz, 2006). These effects were not
observed with the plant extract in the present study (Figure 5.2), suggesting that, although the
extract components may bind to the protein in close proximity of the tryptophan residues for
the fluorescence quenching to occur, they do not change the tryptophan residue exposition by
altering the secondary or tertiary structure of the protein. The quenching of AChE intrinsic

Interactions with Proteins
98
fluorescence by the standards rosmarinic acid, apigenin, luteolin and quercetin also occurred in
a similar way, without a red or blue shift of the peak maximum.
The fluorescence data were analysed by the Stern‐Volmer equation (Equation 1), which
allows to calculate the Stern‐Volmer quenching constant (KSV) and the quenching rate constant
(Kq) of the fluorescence quenching reaction.
1 1 (Equation 1)
Where F0 and F are the steady state fluorescence in the absence and presence of
quencher, respectively, [Q] is the concentration of quencher and τ0 the average lifetime of the
protein fluorescence in the absence of quencher.
The Stern‐Volmer plot for the fluorescence quenching of AChE by P.barbatus extract can
be found in Figure 5.3a, where it is shown that the fluorescence quenching by the plant extract
follows the behaviour of single‐compound binding to the protein. As the extract is composed of
many compounds, the concentration is expressed in µg.ml‐1 and the KSV values cannot be
calculated in M‐1, just estimated in l.mg‐1. The molarity of the P. babatus extract was estimated,
based on the concentration of rosmarinic acid, luteolin glucuronide and apigenin glucuronide as
1.11 mmol per gram of plant extract, calculated by the HPLC chromatogram peak areas (Porfirio
et al., 2010). The enzyme intrinsic fluorescence quenching was also analysed for the main
component of the plant extract, rosmarinic acid, and for luteolin and apigenin, which were also
found as aglycons in the plasma of rats after the intraperitoneal administration of P. barbatus
herbal tea (Falé et al., 2011). The values for the Stern‐Volmer quenching constant (KSV) and the
quenching rate constant (Kq) for AChE in the presence of rosmarinic acid, apigenin, luteolin or
quercetin are shown in Table 5.1. The estimated KSV and Kq for the P. barbatus extract at 298K
were respectively 93598 M‐1 and 9.36x1011 M‐1s‐1, which were approximate to the values found
for the extract’s main component, rosmarinic acid. The maximum scatter collision quenching
constant (Kq) value of various quenchers with a biopolymer is reported to be 2x1010 l.M‐1s‐1.9 As
the Kq values obtained in the present work were higher (ranging from 6x1010 to 8x1011 l.M‐1s‐1)
than the Kq obtained for a scatter mechanism, it is implied that the quenching was not initiated
by dynamic collision but rather originated by the formation of a complex (Lakowicz & Weber,
1973).

Chapter V
99
Figure 5.3. Stern‐Volmer plot (a) and plot of log(F0‐F)/F vs. log[Q] (b) of acetylcholinesterase with P.
barbatus aqueous extract. [Q] is the concentration of P. barbatus extract in mg.ml‐1.
1.3.2. Analysis of binding equilibria of P. barbatus water extract to acetylcholinesterase
Fluorescence quenching data also allows one to determine the equilibrium constants
between free and bound molecules, as well as the number of binding sites on a macromolecule,
by using Equation 2 (Jin & Zhang, 2008),
log log log (Equation 2)
Where F0, F and Q have the same meaning as in Equation 1, Kb is the binding constant
and n is the number of binding sites.
As it is shown in Figure 5.3b, the binding of P. barbatus extract follows the behaviour of
a single molecule binding to a macromolecule that is modelled by Equation 2. Similarly to the
Stern‐Volmer relationship, the Kb value could not be determined in M‐1 to the plant extract, just
be expressed in terms of its concentration in mg.L‐1, and estimated based on concentration of
rosmarinic acid, luteolin glucuronide and apigenin glucuronide as 91395 M‐1. The fluorescence
quenching effects by rosmarinic acid, luteolin, apigenin and quercetin standards were analysed
independently, and the values of Kb and n can be found in Table 5.1.

Interactions with Proteins
100
Table 5.1. Stern‐Volmer binding parameters (KSV, Kq), binding equilibria parameters (Kb, n) and
thermodynamic parameters (ΔHo, ΔSo, ΔGo) for the binding of P. barbatus extract, rosmarinic acid (RA),
luteolin (Lut), apigenin (Api) and quercetin (Quer) to acetylcholinesterase (R2>0.99 to all linear
regressions). Rates of Amide II/Amide I variation, reflecting the rate of hydrogen deuterium exchange in
the presence of similar amount of RA, Lut, Api and Quer or 10mg/mL of P. barbatus extract (for AChE
without ligand, ‐2.345mAU.min‐1). IC50 values for the inhibition of acetylcholinesterase activity by
P. barbatus, RA, Lut and Api. (*) and Quer for P. barbatus extract the values are expressed in
l.mg‐1, l.mg‐1s‐1, and mg.ml‐1 but the molarity was estimated based on the content of rosmarinic acid,
luteolin and apigenin (1.1122 mmol.g‐1).
T (K) P.barbatus RA Lut Api Quer
KSV (M-1)
293 - 84696.54 51112.81 10516.96 49757.31
298 104.10 l.mg-1 (90598
M-1*) 83788.32 42921.28 7178.62 59176.54
303 - 78474.03 57734.37 6457.27 62860.43
Kq (M-1s-1)
293 - 8.47E+11 5.11E+11 1.05E+11 4.98E+11
298 1.04E+09 L.mg-1s-1 (9.36E+11 M-1s-1*)
8.38E+11 4.29E+11 7.18E+10 5.92E+11
303 - 7.85E+11 5.77E+11 6.46E+10 6.29E+11
Kb (M-1)
293 - 62517.27 2854.96 8661.63 5845208
298 101.65 l.mg-1 (91395
M-1*) 144178.3 4390.36 7314.75
1.31E+07 303 - 287342.6 5057.08 6180.16 1.47E+08
n 293 - 0.9871 0.7365 0.9911 1.4692 298 1.0371 1.0595 0.7721 1.0005 1.4298 303 - 1.1281 0.7783 0.9987 1.7651
ΔH0 (J.mol-1)
- - 112636.2 42417.34 -24915.1 237336.6
ΔS0 (J.mol-1K-1)
- - 476.4026 210.9631 -9.64146 937.2904
ΔG0 (J.mol-1)
293 - -26949.81 -19494.85 -22090.20 -37289.49 298 - -29331.83 -20549.67 -22041.99 -41975.94 303 - -31713.84 -21604.48 -21993.78 -46662.4
Rates of AAmideII/AmideI
intensity ratios (mAU.min-1)
298 0.752 0.497 1.119 1.129 0.588
IC50 (µM) 298 1.02±0.02 mg.ml-1
(1134±22 µM*) 1221±73 92.12±2.36 100.57±6.93 50.88±7.25
The Kb and n values estimated for the interaction of P. barbatus extract with AChE were
within the range of those found for the isolated extract compounds/metabolites, suggesting that
all compounds bind to the protein in just one and the same site. Quercetin however showed
higher Kb values and seems to be able to bind to two sites in the enzyme.
The values for Kb and n for rosmarinic acid, luteolin and quercetin to AChE increased
with the temperature (Table 5.1), suggesting that temperature may increase the binding of the
molecules to the AChE structure. For apigenin these values decreased with temperature,

Chapter V
101
suggesting that higher temperature may destabilise the interaction, which is in agreement with
the decrease of Kq to values closer to the ones reported to scatter mechanisms (Lakowicz &
Weber, 1973).
Apigenin, in spite of decreasing Kb with temperature, showed a higher Kb values than
luteolin, suggesting that the lack of the catechol group may increase the hydrophobic interaction
between the flavonoids and the proteins. This interaction, although it may be stronger than the
weak polar interactions made by the catechol group, is more susceptible to breakage by
increasing the energy in the system.
1.3.3. Determination of interaction forces between P.barbatus extract and AChE
The thermodynamic parameters, enthalpy change (ΔHo) and entropy change (ΔSo) of the
interaction of the plant phenolic compounds and the proteins, allow elucidating the nature of
the bond between the plant extract components and AChE macromolecules. For this purpose,
the variation of the binding constant Kb with temperature was studied at three different
temperatures: 293, 298 and 303K.
The thermodynamic parameters were determined by the Van’t Hoff equation (Equation
3), considering that the enthalpy change does not vary significantly with temperature (Jin &
Zhang, 2009). The free energy change (ΔGo) was determined by equation 4.
ln (Equation 3)
(Equation 4)
Where R is the gas constant and T is the temperature. The values of the thermodynamic
parameters of the interaction of rosmarinic acid, apigenin, luteolin and quercetin with AChE can
be found in Table 5.1.
The negative values of ΔGo indicate that the binding processes occurred spontaneously
in all studied cases. From the water structure point of view, a positive ΔSo value is a typical
evidence of hydrophobic interaction (Yang et al., 2008). In the case of rosmarinic acid, luteolin
and quercetin, both ΔHo and ΔSo values were positive (Table 5.1), suggesting that hydrophobic
association is the dominant form of interaction of these plant phenolics with AChE (Ross &
Subramanian, 1973). The values of ΔHo and ΔSo in the interaction of apigenin with AChE were
negative, suggesting that van der Waals interactions between phenolic compounds and certain
domains of AChE may originate from the higher affinity of apigenin to the hydrophobic moiety

Interactions with Proteins
102
and, consequent higher proximity between the flavonoid and the enzyme (Ross & Subramanian,
1973). In this case, the major contribution to ΔGo arises from the ΔHo term rather than from ΔSo,
which implies that the binding process is enthalpy driven.
1.3.4. Determination of protein structure changes caused by P. barbatus extract and its
plasma metabolites by FTIR spectroscopy
FTIR spectroscopy may provide additional evidence on the interaction between protein
and a ligand in aqueous media. The protein secondary structure is reflected in the amide I band,
in the region 1600‐1700 cm‐1 (mainly C=O stretch), and in the amide II band, ≈1540 cm‐1 (C‐N
stretch coupled with NH bending mode). Amide I is more sensitive to secondary structure
changes than amide II (Haris & Severcan, 1999; Xiao et al., 2008; Tantipolphan et al., 2006),
hence its analysis is more useful for the study of structural changes induced by diverse factors.
The FTIR spectra of AChE, obtained by subtracting the absorption of the buffer solution,
are shown in Figure 5.4a, in the absence and in the presence of P. barbatus extract, rosmarinic
acid, luteolin, apigenin or quercetin. The second derivative spectra are shown in Figure 5.4b. As
can be seen from the absorbance and second derivative spectra, the position and the overall
shape of the amide I band in the presence and absence of the ligands are virtually identical. As
the amide I is composed of the sum of absorbance from different secondary structures (β‐turn,
β‐sheet and α‐helix) and irregular structures (random coil) (Haris & Severcan, 1999; He et al.,
2006), it can be concluded that the presence of P. barbatus extract or its metabolites did not
interfere with the secondary structure or irregular structure of AChE. This is in good agreement
with the fluorescence results, where no shifts were observed in the peak maximum of the
emission spectra.
The amide I band maximum is located at 1938 cm‐1. Several shoulders can be observed
and features of the protein secondary structure can be attributed to them, according to the
general attributions of secondary structure features to amide I peak components for proteins in
deuterium oxide (Haris et al., 1996; Jackson et al., 1991), or the specific case of
acetylcholinesterase (Gorne‐Tschelnokow et al., 1993): β‐sheet to 1630 cm‐1; α‐helix or random
coil to 1645 cm‐1; α‐helix to 1651 cm‐1 and 1658 cm‐1 (different populations); 3(10) helices to
1665 cm‐1; β‐turns to 1675 cm‐1; and antiparallel β‐sheet structure to 1682 cm‐1, which might
also have contributed to the shoulder at 1630 cm‐1. The maximum band at 1638 cm‐1 is usually
associated with predominantly β‐sheet structures (Haris & Severcan, 1999). X‐ray
crystallography data for this protein indicate that its secondary structure is predominantly
helical (35%, 25 helices, 194 residues) (Bourne et al., 1999). Previous studies have shown that
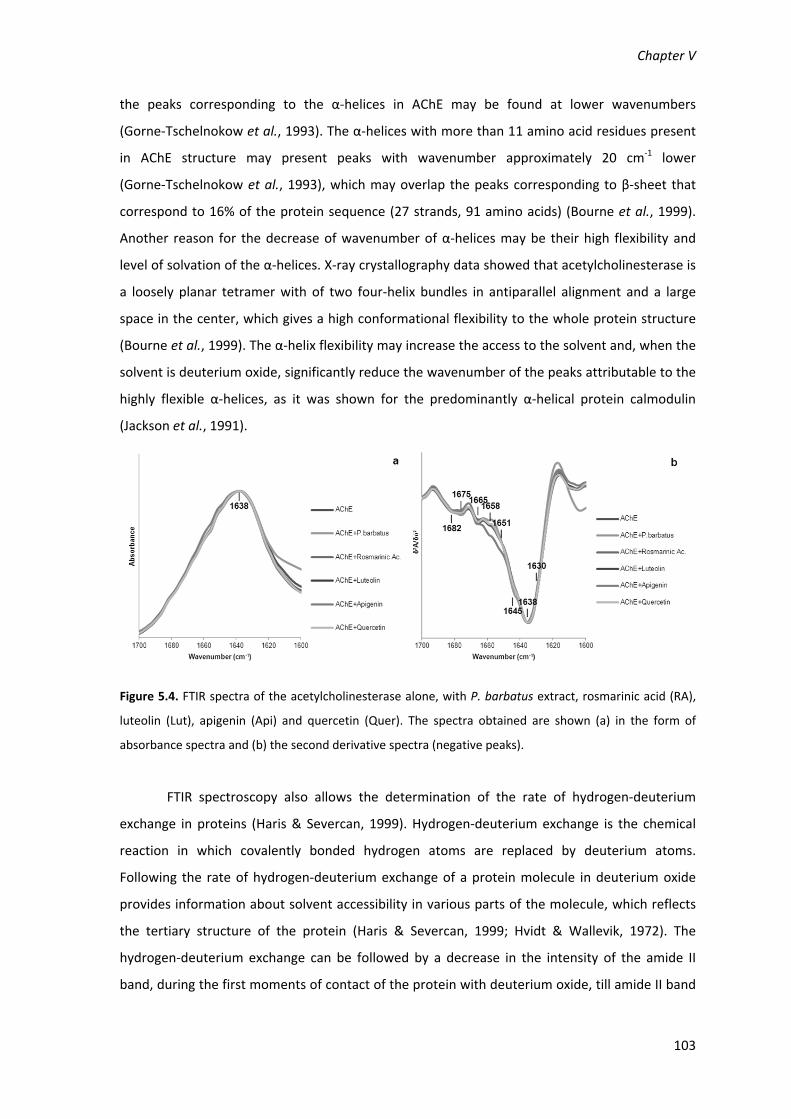
Chapter V
103
the peaks corresponding to the α‐helices in AChE may be found at lower wavenumbers
(Gorne‐Tschelnokow et al., 1993). The α‐helices with more than 11 amino acid residues present
in AChE structure may present peaks with wavenumber approximately 20 cm‐1 lower
(Gorne‐Tschelnokow et al., 1993), which may overlap the peaks corresponding to β‐sheet that
correspond to 16% of the protein sequence (27 strands, 91 amino acids) (Bourne et al., 1999).
Another reason for the decrease of wavenumber of α‐helices may be their high flexibility and
level of solvation of the α‐helices. X‐ray crystallography data showed that acetylcholinesterase is
a loosely planar tetramer with of two four‐helix bundles in antiparallel alignment and a large
space in the center, which gives a high conformational flexibility to the whole protein structure
(Bourne et al., 1999). The α‐helix flexibility may increase the access to the solvent and, when the
solvent is deuterium oxide, significantly reduce the wavenumber of the peaks attributable to the
highly flexible α‐helices, as it was shown for the predominantly α‐helical protein calmodulin
(Jackson et al., 1991).
Figure 5.4. FTIR spectra of the acetylcholinesterase alone, with P. barbatus extract, rosmarinic acid (RA),
luteolin (Lut), apigenin (Api) and quercetin (Quer). The spectra obtained are shown (a) in the form of
absorbance spectra and (b) the second derivative spectra (negative peaks).
FTIR spectroscopy also allows the determination of the rate of hydrogen‐deuterium
exchange in proteins (Haris & Severcan, 1999). Hydrogen‐deuterium exchange is the chemical
reaction in which covalently bonded hydrogen atoms are replaced by deuterium atoms.
Following the rate of hydrogen‐deuterium exchange of a protein molecule in deuterium oxide
provides information about solvent accessibility in various parts of the molecule, which reflects
the tertiary structure of the protein (Haris & Severcan, 1999; Hvidt & Wallevik, 1972). The
hydrogen‐deuterium exchange can be followed by a decrease in the intensity of the amide II
band, during the first moments of contact of the protein with deuterium oxide, till amide II band

Interactions with Proteins
104
stabilizes, when all exchangeable hydrogen’s atoms were replaced by deuterium atoms (Haris &
Severcan, 1999; Hartshorne & Stracher, 1965).
When P. barbatus extract (10 mg.mL‐1) was in contact with AChE, the rate of
hydrogen/deuterium exchange decreased by 67.93% compared to the protein in the absence of
the extract (Figure 5.5). The effects of rosmarinic acid, luteolin and apigenin were analysed
separately for each of the compounds in the same concentration of the protein. Rosmarinic acid
caused a 78.81% decrease in the rate of hydrogen‐deuterium exchange of AChE, while the
flavonoids luteolin and apigenin caused a decrease of 52.28 % and 51.86 %, respectively
(Figure 5.5). Quercetin caused a decrease in hydrogen‐deuterium exchange of 74.93%. As there
was no apparent difference in the shape of the amide I band of AChE in the presence of
P.barbatus extract, the differences found in the rate of hydrogen‐deuterium exchange are not
likely to be due to changes in the overall secondary structure of the protein (Haris et al., 1986;
Hartshorne & Stracher, 1965).
Figure 5.5. Percentage of decrease of hydrogen‐deuterium exchange rate of AChE in the presence of P.
barbatus extract, rosmarinic acid, luteolin, apigenin and quercetin.
The decrease in the rate of hydrogen‐deuterium exchange of the amide protons in AChE,
in the presence of P. barbatus extract, appears to be due to rosmarinic acid, as it is the major
component, while the flavonoids play a smaller role in the global hydrogen‐deuterium exchange.
The decrease in the protein hydrogen‐deuterium exchange in the presence of the plant extract
may be due to a blocking of the accessibility of the solvent to the peptide bonds connecting the
amino acid residues within the protein. The exchangeable hydrogens in the peptide groups may
be protected from the hydrogen‐deuterium exchange process due to the binding of the extract
components to the active gorge of the enzyme, preventing solvent accessibility. An alternative
explanation could be that the binding of the compounds results in an overall rigidification of the

Chapter V
105
structure such as through increased hydrogen‐bonding. However, as we did not detect any
major shifts in the amide I components, the former explanation is more likely.
1.4. Discussion
The in vitro acetylcholinesterase (AChE) inhibition for P. barbatus aqueous extract, used
in the present study, and its main component, rosmarinic acid, was previously determined
(Porfirio et al., 2010; Falé et al., 2011), showing IC50 values of 1.02±0.02 mg.ml‐1 (P. barbatus)
and 1221±73 µM (rosmarinic acid), as shown in Table 5.1. The standards apigenin and luteolin
also inhibit AChE activity, with IC50 values of 100.57±6.93 µM and 92.12±2.36 µM, respectively
(Porfirio et al., 2010). Quercetin is a flavonoid that, although it is not present in P. barbatus
extract, is very common in plants, and plant‐derived food, as its glycoside and aglycon forms
(Erlund, 2004). The extra hydroxyl group in position 3 of the C ring (Figure 5.1) makes this
flavonoid interesting to add to this study. Quercetin showed an IC50 for AChE inhibition of
50.88±7.25 µM, the lowest value found for the flavonoids under evaluation.
Spectroscopic analysis show that the interaction between the plant extract, or the
isolated compounds, with the AChE do not change the protein secondary structure, as no shift
was observed in either the fluorescence emission peak or on the amide I peak in the FTIR
spectrum. The fact that drastic structural changes do not occur in the protein is in agreement
with previous reports showing that the enzyme recovers its activity when the concentration of
the extract components decreases (Falé et al., 2011), which confirms that enzyme inhibition is
reversible.
The inhibition of AChE by quercetin was shown to be competitive (Khan et al., 2009) and,
several docking studies pointed out that the inhibitory activity of flavonoids to this enzyme is
related to the binding to amino acid residues in the active gorge of the enzyme (Khan et al.,
2009; Sheng et al., 2009). The active gorge of AChE is a narrow and deep gorge in the enzyme
structure, with amino acid residues that trap and position the substrate, acetylcholine, to the
active site, which is located at the base of the gorge (Harel et al., 2008). The decrease of
hydrogen‐deuterium exchange rate observed in AChE with the plant extract and its isolated
constituents indicated a decrease in the accessibility of the exchangeable amide hydrogens in
the peptide groups. This situation is probably due to the binding of the plant phenolic
compounds to the active gorge of the enzyme that may cause small changes in the protein
structure as it was already seen by X‐ray crystallography studies of AChE‐inhibitor complexes
from mouse (Bourne et al., 2010) and Torpedo (Sanson et al., 2011). These changes are usually
small orientation changes of amino acid residues, especially related to aromatic amino acids,

Interactions with Proteins
106
which may affect considerably the enzymatic activity, and occur near the four‐helix bundles of
AChE (Bourne et al., 2010; Sanson et al., 2011; Harel et al., 1993). As the peak at 1638 cm‐1 by
FTIR is not shifted to higher or lower wavenumbers, the final amount of hydrogen‐deuterium
exchange of the α‐helices may not be altered by the binding of the inhibitors. However, the
decrease of the rate of hydrogen‐deuterium exchange in AChE complexes may be due to small
changes, maybe a decrease in α‐helix‐related flexibility (Bourne et al., 1999), which may hinder
the accessibility of deuterium oxide to certain regions of AChE.
The thermodynamic data (Table 5.1) suggest that the dominant interactions in the
binding of rosmarinic acid, luteolin and quercetin to AChE are hydrophobic interactions (Ross &
Subramanian, 1981). Docking studies suggest that the hydrophobic interactions in the binding
and positioning of quercetin to the active gorge of electric eel AChE are related to the residues
Trp86, Glu202, His447 and Ser125 (Khan et al., 2009). Other authors presented a similar docking
of flavonoids to rat brain acetylcholinesterase, relating the interactions to homologous amino
acid residues (Sheng et al., 2009). Docking studies also refer the occurrence of hydrogen bonds
between the hydroxyl groups of quercetin and amino acid residues of the active site of electric
eel AChE (Khan et al., 2009). The higher Kb and lower ΔG values shown by quercetin, when
compared to luteolin and apigenin (Table 5.1), may be due to a hydrogen bond between the
Trp86 of the enzyme and the hydroxyl group in position 3 of the B ring of quercetin (Khan et al.,
2009), which is absent in the other two flavonoids. This interaction caused by this particular
hydroxyl group in quercetin seems to play an important role in stabilizing the complex flavonoid‐
protein, causing a significantly higher competitive inhibition of AChE by quercetin and, a
decrease in the accessibility of the solvent molecules to the active gorge, which is suggested by
the low hydrogen‐deuterium exchange rate presented by AChE with quercetin (Table 5.1), and
may explain the low IC50.
Although all flavonoids presented negative ΔG values for the binding to AChE, apigenin
showed the lowest for ΔHo and ΔSo, and quercetin the highest. If a plot is drawn with the
number of hydroxyl groups in the flavonoid and the values of ΔHo and ΔSo, the linear regressions
have R2 of 0.93 for ΔHo and 0.91 for ΔSo, showing that there is a direct relationship between the
number of hydroxyl groups and the thermodynamic parameters. Docking studies showed that
the hydroxyl groups in flavonoids may establish hydrogen bridges with the amino acid residues
in the active gorge, being thus positioned in a specific manner (Khan et al., 2009). The lowest the
number of hydroxyl groups, the looser is the positioning of the flavonoid in the active site, which
may explain the decrease in ΔSo with a lower the number of hydroxyl groups (Table 5.1). The
changes in ΔSo are compensated with changes in ΔHo to give negative ΔGo values in an enthalpy

Chapter V
107
driven binding. The increase in number of hydroxyl groups is also related to a decrease in the
ΔGo values (Table 5.1), suggesting that the binding is more favourable with the higher number
hydrogen bridges that may be established between the flavonoid and the amino acid residues in
the active gorge.
Rosmarinic acid presented ΔSo and ΔHo values that suggested that the dominant forces
in the rosmarinic acid‐protein complex were hydrophobic interactions (Ross & Subramanian,
1981). The enzymatic inhibition by rosmarinic acid was lower than the one presented by the
flavonoids (Table 5.1). Conversely to rosmarinic acid, the structure of flavonoids present a
hydrophobic back opposite to a hydrophilic side, with a higher number of hydroxyl groups,
allowing them to hold a clear position inside the active gorge, by establishing hydrophobic
interactions in one side and hydrogen bonds in the other with the amino acid residues (Khan et
al., 2009; Sheng et al., 2009). Also the B ring in the flavonoids molecule can rotate around the
C2‐C1’ bond, letting the hydroxyl groups to adjust the conformation, binding to the active sites of
diverse target proteins (Ji & Zhang, 2006). For these reasons, rosmarinic acid may be able to bind
to AChE through hydrophobic interactions but, may not establish such a complex interactions
with the protein as the flavonoids do. Consequently, rosmarinic acid may be a weaker
competitive inhibitor of the enzyme activity, as the 10‐fold higher IC50 seems to confirm
(Table 5.1).
To our knowledge, this is the first spectroscopic study on the interaction of plant
phenolic compounds to AChE, and relating these interactions with the enzymatic inhibition.
Although docking studies are available, spectroscopic data may present an important
complement to this knowledge, confirming docking studies and clarifying some details related to
the dominance of the binding forces, as for instance, the dominance of the hydrophobic
interactions in the binding of plant phenolics to AChE.
1.5. Conclusions
P. barbatus extract components bind to AChE forming a complex. The analysis of the
binding of the isolated standard compounds suggested that the interaction of the plant extract
components with the amino acid residues, in the active gorge of acetylcholinesterase, may be
responsible for the AChE inhibition by P. barbatus extract.
The interactions between the plant phenolics and AChE do not cause secondary
structure changes, which might be too drastic and lead to the denaturation of the enzyme. Thus
the change appears to be subtle and the drastic reduction in hydrogen‐deuterium exchange can

Interactions with Proteins
108
be explained by the binding of the phenolic compounds in the region of the active gorge
blocking solvent accessibility to a significant portion of the protein molecule.
Flavonoids seem to be better AChE inhibitors than rosmarinic acid, since this acid has a
structure less able to establish interactions with the amino acid residues in the active gorge,
while the flavonoid structure seems to fit to the active gorge of the enzyme, interacting
dominantly by hydrophobic interactions but also by hydrogen bonds established between the
hydroxyl groups of the flavonoids and the amino acid residues, which allow the flavonoid to hold
a specific position inside the gorge.
The position of the hydroxyl groups in the flavonoids seems to play an important role in
the binding to the enzyme and in the inhibitory activity, particularly in some positions, such as
the position 3 of the C ring, that is responsible for the binding of quercetin to Trp86, increasing
its inhibitory activity to the double, when compared to luteolin. Therefore, spectroscopic studies
proved to be useful in confirming and clarifying docking studies related to AChE inhibitors.

Chapter V
109
2. Interaction between Plectranthus barbatus herbal tea
components and human serum albumin and lysozyme: binding
and activity studies
2.1. Introduction
Plasma proteins play an important role in transportation and deposition of substances in
the circulatory system, such as fatty acids, hormones and medicinal drugs. Therefore, it is
important to reveal the interaction between drugs and proteins in the bloodstream, as it may
affect the bioavailability, distribution and elimination of pharmaceutical or nutraceutical active
compounds. Albumin is the major protein of plasma, and its main function is the regulation of
colloidal osmotic pressure and the binding and transport of substances in the bloodstream (Li et
al., 2007). The interaction of human serum albumin with chemically synthesized drugs used in
medicine may influence their bioavailability and effectiveness, and so many recent studies have
focused on these interactions (Varshney et al., 2010). The interaction between HSA and plant
secondary metabolites traditionally taken by people as “natural medicines” has also been
subject to many reports (Rawel et al., 2006). Lysozyme is also known to play a role in the
transportation of drugs (Jin & Zhang, 2010), although its main function is to hydrolyse
peptidoglycans, which are found in bacterial cell walls (especially in Gram positive bacteria), as
part of the innate immune system (Laible & Germaine, 1985). The most dramatic
lysozyme‐related conditions are caused by the decrease or lack of its activity (Laible & Germaine,
1985). However, its excessive activity is known to be related to allergic conditions and worsening
of inflammation in the immune response to pathogens (Makino et al., 2003; Ronca et al., 1998;
Wu et al., 2006).
Plectranthus barbatus (Lamiaceae) herbal tea has antiacetylcholinesterase as well as
antioxidant activity (Falé et al., 2009), which is related to its main components: rosmarinic acid,
flavonoid glucuronides (glucuronidated apigenin, luteolin and acacetin) and abietane
diterpenoids. After the oral administration of the plant extract to rats, its main component,
rosmarinic acid, was found in the plasma, as well as the flavonoids, in both the glucuronidated
and as aglycone forms (Falé et al., 2011).
The aim of the present study was to investigate if P. barbatus water extract compounds
and metabolites, that were previously found circulating in the plasma can interact with human
albumin and lysozyme, by binding and being transported by these proteins, or affecting
lysozyme activity.

Interactions with Proteins
110
2.2. Material and Methods
The materials and methods are described in detail in the Chapter II.
The P. barbatus extract was prepared as a decoction, as described in Chapter II section 4.
The interaction between the plant extract, or the standards rosmarinic acid, luteolin and
apigenin, with the human proteins albumin and lysozyme were evaluated by fluorescence
spectroscopy, as described in Chapter II section 13, and by FTIR spectroscopy, as referred in
Chapter II section 14. The lysozyme inhibition was determined as described in Chapter II
section 15.
2.3. Results
2.3.1 Binding of P. barbatus to albumin and lysozyme
Both lysozyme and human serum albumin (HSA) showed excitation spectra with a
maximum wavelength at 280 nm and emission spectra with a maximum at 335 nm, under
excitation at 280 nm. Figure 5.6 shows the emission fluorescence spectra of HSA and lysozyme
with the addition of increasing concentrations of P. barbatus aqueous extract. The intensity of
the intrinsic fluorescence of both proteins decreased significantly – was quenched – with the
increase of concentration of the plant extract, which acted as a quencher.
The dominant fluorophore in these proteins is the indole group of the tryptophan
residues, and the fluorescence quenching by the plant extract is caused by the interaction
between the plant compounds and the protein, in the vicinity of the tryptophan residues
(Lakowicz, 2006). The emission spectrum of the indole may be shifted towards lower
wavelengths (blue shifted) if the group is buried within the native protein, or its emission may be
shifted towards larger wavelengths (red shifted) when the protein is unfolded (Lakowicz, 2006).
These effects were not observed in the present study (Figure 5.6a and b), suggesting that,
although the flavonoids may bind to the proteins in close proximity of the tryptophan residues
for the fluorescence quenching to occur, they do not change the tryptophan residue exposition
by altering the secondary or tertiary structure of the protein.
The fluorescence data were analysed by the Stern–Volmer equation (Equation 1), which
allows to calculate the Stern–Volmer quenching constant (KSV) and the quenching rate constant
(Kq) of the fluorescence quenching reaction, as previously shown in this Chapter, section 1.2.1.

Chapter V
111
Figure 5.6. Fluorescence emission spectra of (a) HSA and (b) lysozyme with the addition of P. barbatus
aqueous extract. Arrow points to increasing concentrations of P. barbatus plant extract, ranging 0; 0.5;
0.75; 1; 2.5; 5; 7.5; 100 μg.ml−1.
The Stern–Volmer plots for the fluorescence quenching of HSA and lysozyme by
P. barbatus extract can be found in Figure 5.7, where it is shown that the fluorescence
quenching by the plant extract follows the behaviour of single‐compound binding for both
proteins. As the extract is composed of many compounds, the concentration is expressed in
μg.ml−1 and the KSV values cannot be calculated in M−1, just estimated in l.mg−1. The molarity of
the P. barbatus extract was estimated, based on the concentration of rosmarinic acid, luteolin
glucuronide and apigenin glucuronide as 1.1122 mmol per gram of plant extract, calculated by
the HPLC chromatogram peak areas (Porfirio et al., 2010). The protein intrinsic fluorescence
quenching was also analysed for the main component of the plant extract, rosmarinic acid, and
for luteolin and apigenin, which could be found in plasma as aglycones after the administration
of P. barbatus herbal tea, due to the liver β‐glucuronidase activity (Falé et al., 2011).

Interactions with Proteins
112
Figure 5.7. Stern–Volmer plots of HSA and lysozyme with P. barbatus aqueous extract. [Q] is the
concentration of P. barbatus in μg.ml−1.
The values for the Stern–Volmer quenching constant (KSV) and the quenching rate
constant (Kq) for HSA and lysozyme in the presence of rosmarinic acid, apigenin or luteolin are
shown in Table 5.2.
The estimated KSV and Kq for the P. barbatus extract were, respectively, 461,518 M−1 and
4.61 × 1013 M−1s−1 for HSA, and 68,513M−1 and 6.85×1012 M−1s−1 for lysozyme, which were
approximate to the values found for the extract’s main component, rosmarinic acid. The
maximum scatter collision quenching constant (Kq) value of various quenchers with a biopolymer
is reported to be 2 × 1010 l.M−1s−1 (Lakowicz & Weber, 1973). As the Kq values obtained in the
present work were higher, ranging from 1011 to 1013, than the Kq obtained for a scatter
mechanism, it is implied that the quenching was not initiated by dynamic collision but rather
originated by the formation of a complex.
2.3.2. Analysis of binding equilibria
Fluorescence quenching data also allows to determine the equilibrium constants
between free and bound molecules, as well as the number of binding sites on a macromolecule,
by using Equation 2 (Jin & Zhang, 2010), as shown in section 1.2.2, in this Chapter.
In Figure 5.8, the binding of P. barbatus extract follows the behaviour of a single
molecule binding to a macromolecule that is modelled by Equation 2, in both cases of HSA and
lysozyme.
Similarly to the Stern–Volmer relationship, the Kb value could not be determined in M−1
to the plant extract, just be expressed in terms of its concentration in mg.l−1, and estimated
based on concentration of rosmarinic acid, luteolin glucuronide and apigenin glucuronide as
5.84×105 and 7.92×105 M−1 for HSA and lysozyme, respectively. The fluorescence quenching

Chapter V
113
effects by rosmarinic acid, luteolin and apigenin standards were analysed independently, and
the values of Kb and n can be found in Table 5.2.
Table 5.2. Binding parameters (KSV, Kq, Kb, n) and thermodynamic parameters (ΔHo, ΔSo, ΔGo) for the
binding of P. barbatus extract, rosmarinic acid (RA), luteolin (Lut) and apigenin (Api) to HSA and to
lysozyme (R2>0.99 to all linear regressions). Rates of Amide II/Amide I variation in the presence of similar
amount of RA, Api and Lut, or 10mg/mL of P. barbatus extract (without ligand, ‐0.716 for HSA and ‐1.815
for lysozyme). IC50 values for the inhibition of lysozyme activity. For P. barbatus extract the values are
expressed in l.mg‐1(a), l.mg‐1s‐1(b) and mg.l‐1(c).
T(K) Human Serum Albumin Lysozyme
P.barbatus RA Lut Api P.barbatus RA Lut Api
KSV(M-1)
293 - 245192 194500 28029 - 87179 4191.9 5344.4
298 0.5133(a) 355856 250748 8090.5 0.0762(a) 98081 3381.4 3805.1
303 - 482757 326653 5886.5 - 143377 3533.7 3250.8
Kq(M-1s-1)
293 - 2.45E+13 1.95E+13 2.8E+12 - 8.72E+12 4.19E+11 5.34E+11
298 5.13E+7(b) 3.56E+13 2.51E+13 8.09E+11 7.62E+6(b) 9.81E+12 3.38E+11 3.81E+11
303 - 4.83E+13 3.27E+13 5.89E+11 - 1.43E+13 3.53E+11 3.25E+11
Kb(M-1)
293 - 1.32E+08 3.03E+08 3.07E+04 - 3069729 2760.57 7084.35
298 0.6493(a) 3.96E+08 1.12E+09 1.85E+06 0.8806(a) 205541.7 3498.64 642.98
303 - 7.79E+08 6.92E+09 1.61E+09 - 84781.29 4926.06 363.74
n
293 - 1.4998 1.5849 1.0101 - 1.1132 0.9551 0.9935
298 1.4643 1.5796 1.5403 1.4283 1.4667 1.0673 0.9839 0.8253
303 - 1.6267 1.6267 2.0964 - 0.9617 0.9789 0.7814
ΔH0
(kJ.mol-1) - - 131.16 230.67 96.35 - -265.66 42.69 -219.90
ΔS0
(J.mol-1K-1) - - 603.68 948.93 338.72 - -784.91 211.45 -679.24
ΔG0
(kJ.mol-1)
293 - -45.551 -47.576 -25.172 - -36.389 -19.302 -21.598
298 - -49.048 -51.619 -35.757 - -36.389 -20.218 -16.021
303 - -51.577 -57.081 -53.413 - -28.588 -21.420 -14.855
Rates of
Amide
II/Amide I
intensity
ratios
(mAU/min)
298 -0.979 -1.071 -0.071 -0.715 -1.425 -1.820 -0.086 -0.449
IC50 (µM) 298 - - - - 9.02±0.40(c) 97.2±3.9 106.5±9.6 91.0±1.4

Interactions with Proteins
114
Figure 5.8. Plots of log([F0 − F]/F) vs. log[Q] for HSA and lysozyme with P. barbatus aqueous extract. [Q] is
the concentration of P. barbatus in g.l−1.
The n value estimated for the interaction of P. barbatus extract with HSA was within the
range of the one found for the isolated compounds, while the Kb value was lower, suggesting
that the extract components may interfere with the binding of each other to HSA. For lysozyme,
the plant extract seemed to bind to more than one binding site (n = 1.5, Table 5.2) while for the
isolated compounds n is approximately 1, which may be the cause of the higher Kb value
estimated for the plant extract than for the isolated compounds (Table 5.2).
The values for Kb and n for the three compounds to HSA increased with the temperature,
suggesting that temperature may increase the binding of the molecules to two sites in the HSA
structure. The binding of luteolin showed higher Kb values than apigenin, which agrees with the
results of Xiao and coworkers (2008) for the binding of flavonols to bovine serum albumin,
where it is reported that the binding constants increased with the number of hydroxyl groups in
the B‐ring of flavonoids.
A decrease in Kb and n was observed for lysozyme with rosmarinic acid and apigenin,
suggesting that temperature may be unfavourable to the binding in these cases. For the binding
of luteolin with lysozyme, the value of n remained similar through the changes of temperature,
while the Kb value increased. This suggests that the catechol group that is present in luteolin, but
not in apigenin, may play an important role in stabilizing the complex flavonoid–protein at
higher temperatures. Apigenin, in spite of decreasing Kb with temperature, showed a higher Kb
with a lower temperature than luteolin, suggesting that the lack of the catechol group may
increase the hydrophobic interaction between flavonoids and proteins.
This interaction, although it may be stronger than the weak polar interactions made by
the catechol group, is more susceptible to be broken by increasing the energy in the system. At
higher temperatures, closer to body temperature, the binding affinities of luteolin to HSA and
lysozyme are higher than the ones for the binding of apigenin. These facts, shown by Xiao and

Chapter V
115
coworkers (2011) for total plasma proteins extracted from rat blood, demonstrate that the
hydroxyl group in position 3’ of the B ring in the flavonoid structure has great influence in
flavonoid affinity to the proteins.
2.3.3. Determination of interaction forces between P. barbatus extract metabolites and
HSA and lysozyme
The thermodynamic parameters enthalpy change (ΔHo) and entropy change (ΔSo) of the
interaction of the plant phenolic compounds and the proteins allow to elucidate the nature of
the bond between the plant extract components and the HSA/lysozyme macromolecules. For
this purpose, the variation of the binding constant Kb with temperature was studied at three
temperatures: 293, 298 and 303 K.
The thermodynamic parameters were determined by the van’t Hoff equation (Equation
3) and the free energy change (ΔGo) was determined by Equation 4, as shown in this Chapter,
section 1.2.3. The values of the thermodynamic parameters of the interaction of rosmarinic acid,
apigenin and luteolin with HSA and Lysozyme can be found in Table 5.2.
The negative values of ΔGo indicate that the binding processes occurred spontaneously
in all studied cases. From the view‐point of water structure, a positive ΔSo value is a typical
evidence of hydrophobic interaction (Yang et al., 2008). In the case of HSA, both ΔHo and ΔSo
values were positive (Table 5.2), suggesting that hydrophobic association is the dominant form
of interaction of the tested plant phenolics with HSA (Ross & Subramanian, 1981).
Previous reports on the interaction of apigenin with HSA (Yuan et al., 2007), and luteolin
with bovine serum albumin (Yang et al., 2008), also showed positive values for ΔSo. However,
Yang and coworkers (2008) presented a negative value for ΔHo, which may be due to a
strengthening of the interaction by other forces, such as van der Waals or hydrogen bonds,
introduced by hydrophobic effect in higher temperature conditions than the interval used in the
present study (Ross & Subramanian, 1981).
The values of ΔHo and ΔSo in the interaction of rosmarinic acid and apigenin with
lysozyme were negative, suggesting van der Waals interactions between phenolic compounds
and certain domains of lysozyme (Ross & Subramanian, 1981). In these cases, the major
contribution to ΔGo arises from the ΔHo term rather than from ΔSo, which implies that the
binding processes are enthalpy driven. In the interaction of luteolin and lysozyme, both ΔHo and
ΔSo values were positive, which points to hydrophobic association as the main form interaction
between the molecules. The difference in thermodynamic parameters found between the
apigenin and luteolin may have been caused by the higher polarity and solubility in water of

Interactions with Proteins
116
luteolin (Kim et al., 2008), that arises from the higher hydroxylation of the B‐ring. The higher
affinity to water of luteolin may cause a weaker hydrophobic association that did not induce the
electrostatic interactions in our interval of temperatures (Ross & Subramanian, 1981).
2.3.4. Determination of protein structure changes caused by P. barbatus extract and its
plasma metabolites by FTIR
FTIR spectroscopy may provide additional evidence on the interaction between a protein
and a ligand. The protein secondary structure is reflected in the amide I band, in the region
1600–1700 cm−1 (mainly C=O stretch), and in the amide II band, ≈1540 cm−1 (C–N stretch coupled
with NH bending mode). Amide I is more sensitive to secondary structure changes than amide II
(Haris & Severcan, 1999; Tantipolphan et al., 2007; Xiao et al., 2011), hence its analysis is more
useful for the study of structural changes induced by diverse factors.
The FTIR spectra of HSA and lysozyme, obtained by subtracting the absorption of the
buffer solution, are shown in Figure 5.9a and b, for HSA and Lysozyme, respectively, in the
absence and in the presence of P. barbatus extract, rosmarinic acid, luteolin or apigenin. The
second derivatives are shown in Figure 5.9c and d for both HSA and lysozyme, respectively. The
position of the amide I band in region 1651–1653 cm−1 indicates the presence of predominantly
α‐helical structure in these two proteins which is consistent with their known X‐ray structures.
As it can be seen in the absorbance spectra and second derivatives, the amide I band showed
the same peak and shape in the presence and absence of the ligands for both proteins. As the
amide I is composed of the sum of β‐turn, β‐sheet and α‐helix absorbance contributions (He et
al., 2006; Kim et al., 2008), it can be concluded that the presence of P. barbatus extract or its
metabolites did not interfere with the secondary structure of HSA or lysozyme.
FTIR spectroscopy also allows one to determine the rate of hydrogen–deuterium
exchange in proteins (for a review see Kim et al., 2008). Following the rate of hydrogen–
deuterium exchange of a protein molecule in deuterium oxide medium may give information
about solvent accessibility in various parts of the molecule, which reflects the tertiary structure
of the protein (Haris et al., 1990). The hydrogen–deuterium exchange can be followed by a
decrease in amide II band during the first moments of contact of the protein with deuterium
oxide (Haris & Severcan, 1999).
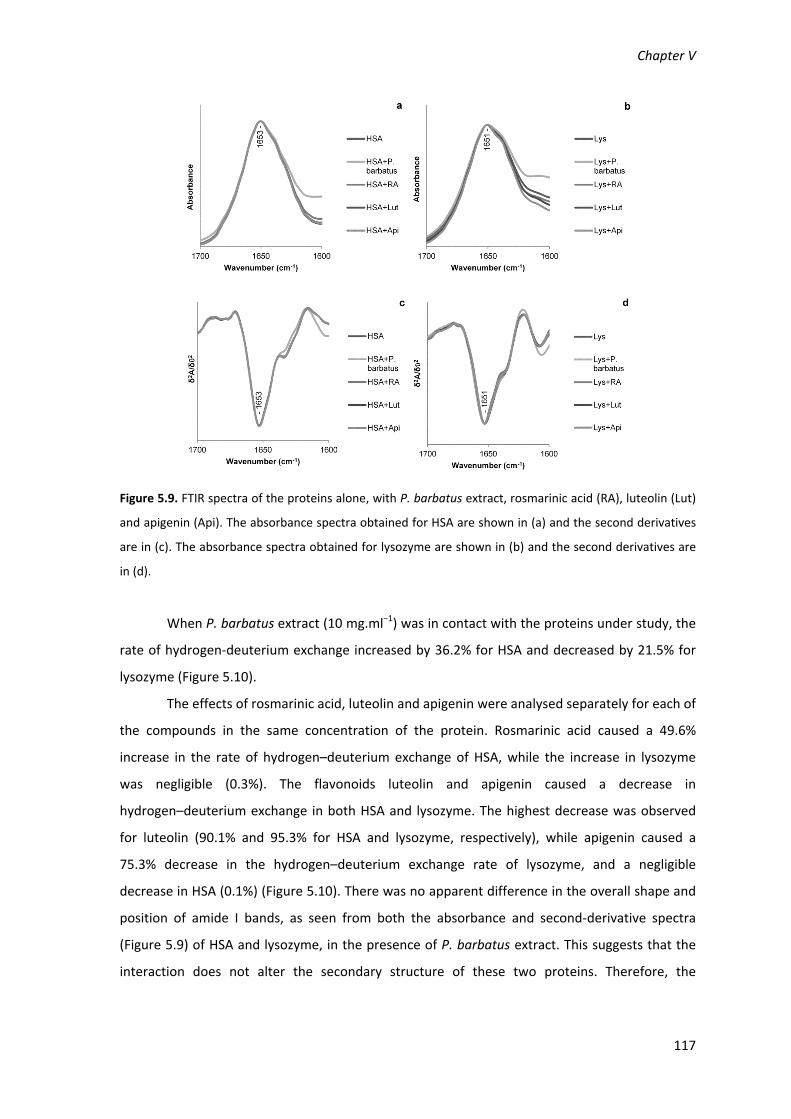
Chapter V
117
Figure 5.9. FTIR spectra of the proteins alone, with P. barbatus extract, rosmarinic acid (RA), luteolin (Lut)
and apigenin (Api). The absorbance spectra obtained for HSA are shown in (a) and the second derivatives
are in (c). The absorbance spectra obtained for lysozyme are shown in (b) and the second derivatives are
in (d).
When P. barbatus extract (10 mg.ml−1) was in contact with the proteins under study, the
rate of hydrogen‐deuterium exchange increased by 36.2% for HSA and decreased by 21.5% for
lysozyme (Figure 5.10).
The effects of rosmarinic acid, luteolin and apigenin were analysed separately for each of
the compounds in the same concentration of the protein. Rosmarinic acid caused a 49.6%
increase in the rate of hydrogen–deuterium exchange of HSA, while the increase in lysozyme
was negligible (0.3%). The flavonoids luteolin and apigenin caused a decrease in
hydrogen–deuterium exchange in both HSA and lysozyme. The highest decrease was observed
for luteolin (90.1% and 95.3% for HSA and lysozyme, respectively), while apigenin caused a
75.3% decrease in the hydrogen–deuterium exchange rate of lysozyme, and a negligible
decrease in HSA (0.1%) (Figure 5.10). There was no apparent difference in the overall shape and
position of amide I bands, as seen from both the absorbance and second‐derivative spectra
(Figure 5.9) of HSA and lysozyme, in the presence of P. barbatus extract. This suggests that the
interaction does not alter the secondary structure of these two proteins. Therefore, the

Interactions with Proteins
118
differences found in the rate of hydrogen–deuterium exchange may be due, predominantly to
changes in the tertiary structure of the proteins (Haris & Severcan, 1999).
Figure 5.10. Percentage of change in protein (HSA and lysozyme) hydrogen–deuterium exchange rate,
determined from the analysis of the amide II band, in the presence of P. barbatus extract, rosmarinic acid,
luteolin or apigenin, in comparison with the hydrogen–deuterium exchange rate of the protein alone.
The decrease of lysozyme hydrogen–deuterium exchange by P. barbatus extract may be
due to its flavonoid components, since its main component, rosmarinic acid, does not affect the
hydrogen–deuterium exchange of lysozyme. The increase in the rate of hydrogen–deuterium
exchange in HAS, when in contact with P. barbatus extract appears to be due to rosmarinic acid,
as it is the major component, while the flavonoids play a smaller role in altering the global
hydrogen–deuterium exchange.
2.3.5. Effect of P. barbatus extract on lysozyme activity
Lysozyme is a glycoside hydrolase able to hydrolyse the peptidoglycan that constitutes
the cell walls of bacteria such as Micrococcus sp. (Laible & Germaine, 1985).
The aqueous extract of P. barbatus inhibited lysozyme activity, with an IC50 value of 9.02
μg.ml−1 (Table 5.2). The IC50 values for the inhibition of lysozyme by rosmarinic acid, apigenin and
luteolin were similar and around 100 μM, as it is shown in Table 5.2.
2.4. Discussion
The structure of human serum albumin, alone and when bound to diverse molecules,
has been determined using X‐ray crystallography (for a review see Curry (2009)). In this study we

Chapter V
119
have demonstrated using fluorescence spectroscopy that flavonoids including rosmarinic acid
alter the tertiary structure of HSA. Furthermore, our FTIR spectroscopic analysis reveals
significant reduction in amide proton hydrogen– deuterium exchange rate for HSA complexed to
flavonoids. This is indicative of a decrease in the accessibility of the exchangeable hydrogens to
the solvent (deuterium oxide). However, we did not detect any changes in the secondary
structure of HSA, through the analysis of the amide I band, as a consequence of interaction with
the flavonoids. This finding is in good agreement with the results of X‐ray crystallographic
studies which have consistently shown that there are no gross changes in the secondary
structure of HSA associated with binding ligands although changes in tertiary structure such as
rigid‐body‐rotation of domains have been detected (for a review see Curry (2009)). The latter
changes in the tertiary structure may explain the differences observed in the fluorescence
spectra and hydrogen–deuterium exchange rates.
Among the different compounds tested, luteolin induced the highest decrease of
hydrogen–deuterium exchange and it also presented the highest values for the binding
constants and lowest ΔGo values, suggesting a higher stability for the HSA‐luteolin complex
compared to the HSA‐apigenin complex. The increased stability of the protein associated with
the bonding between the flavonoids and the HSA molecule makes it more difficult for the amide
protons to be substituted with deuterium. Furthermore, the interaction may result in the
shielding of certain segments of the protein from being accessible to solvent. All this explains the
significant reduction in hydrogen–deuterium exchange upon binding of the flavonoids to HSA.
Rosmarinic acid, like luteolin, had no effect on the secondary structure of HSA. However,
unlike luteolin, rosmarinic acid increased the hydrogen–deuterium exchange of HSA, suggesting
that the nature of its interaction with the transport protein is different. Our results suggest that
hydrophobic regions of the protein becomes accessible to solvent due to alterations in the
protein tertiary structure (Militello et al., 2004), without any changes in the secondary structure.
The nature of the change could be a large rigid‐body rotation of the subunits that have been
shown to occur on fatty acid binding to the main fatty acid binding sites in HSA, as was
extensively reviewed by Curry (2009). X‐ray crystallographic analysis has shown that fatty acid
binding results in a HSA molecule that is 10 Å wider than the defatted HSA. It is thus possible,
that whilst the binding may reduce hydrogen–deuterium exchange for molecules that are
interacting with rosmarinic acid, a much larger change, such as widening of the molecule, could
expose a greater number of amino acid residues to solvent resulting in an overall increase in
hydrogen–deuterium exchange. We propose that rosmarinic acid may induce a similar type of
tertiary structural change as induced by binding of fatty acids to HSA. In five of the seven known

Interactions with Proteins
120
fatty acid binding sites the lipid is anchored by the interaction of the carboxylic group with a
basic or polar group at the pocket entrance. Some non‐lipid acidic compounds are known to
interact with these binding sites, specifically by interacting with the carboxylic acid binding
residues (Curry, 2009). Rosmarinic acid and fatty acids both share a common functional group
(carboxylic acid group) which is not the case for luteolin and this may explain as to why they
behave differently in altering the structure of the HSA molecule.
Flavonoids have been reported to bind to the drug binding sites (Rawel et al., 2006; Yuan
et al., 2007), which are different from the fatty acid binding sites. The drug binding sites 1 and 2
comprise largely apolar cavities with defines polar features located in sub‐domains IIA and IIIA,
respectively. Most drugs bind to the drug binding site 1, which has preference for flat aromatic
compounds that fit between Leu238 and Ala291 in the centre of the pocket (Curry, 2009). The
planar structure of flavonoids may be responsible for their binding predominantly to the drug
site 1, as it was reported for apigenin (Yuan et al., 2007) and quercetin (Rawel et al., 2006).
Usually small adjustments in the side chains in the site 1 cavity can happen upon the binding of
the compounds. These changes are not so extensive that may lead to changes in the secondary
structure of HSA (Curry, 2009), but may be the cause of the decrease in hydrogen–deuterium
exchange rate in the binding of luteolin to HSA.
It is interesting that like with HSA, luteolin and apigenin interaction with lysozyme results
in a decrease in amide proton hydrogen–deuterium exchange rate. This can be due to one or
more of the following reasons:
(i) due to the amide protons participating in stronger H‐bonds within the secondary
structural elements as a consequence of interaction with these compounds. This will make it
more difficult to break the H‐bonds for the deuterium substitution to occur;
(ii) due to amide protons forming H‐bonds to other groups, including luteolin and
apigenin making it more difficult for the hydrogen–deuterium exchange reaction to occur;
(iii) or it could be due to movement of domains/regions of protein that results in a
reduction in solvent accessibility so that deuterium oxide is unable to penetrate into the core of
the protein to replace the amide protons with deuterium.
Similarly to HSA, luteolin induced a greater decrease in lysozyme hydrogen deuterium
exchange (Table 5.2), and presented a higher binding constant and lower ΔGo value at room
temperature, suggesting that the binding of luteolin to lysozyme is more stable at room
temperature than the binding of apigenin.

Chapter V
121
Once again, like with what we saw for HSA, the interaction of rosmarinic acid with
lysozyme is different to that observed for luteolin. Although, there is no significant increase in
hydrogen–deuterium exchange, it at least does not reduce the hydrogen–deuterium exchange.
This could be due to weaker interaction between rosmarinic acid and lysozyme or that the
nature of the binding does not alter the tertiary structure in a way that would make the amide
protons more vulnerable to hydrogen–deuterium exchange.
Although we have detected changes in hydrogen–deuterium exchange, no significant
alteration in the secondary structure was detected. However, other studies concerning the
interaction of flavonoids detected small secondary structure changes, as well as tertiary
structure changes, by the flavonoids alpinetin and cardamonin (He et al., 2006). Our findings
reveal that alterations in protein secondary structure were not found with either luteolin or
apigenin, by FTIR or fluorescence spectrometry, suggesting that the alterations in rate of
hydrogen deuterium exchange is mainly due to changes in the tertiary structure.
As the proteins and the plant phenolic compounds seemed to interact by weak forces by
fluorescence spectrometry, the bonds formed may be reversible and the compounds released
under certain conditions. Studies with total plasma protein extracted from rat blood report that
the affinity of flavonoids to the total protein may be even lower than to purified albumin, which
may be due to presence of metallic ions such as Zn2+, Cu2+, Ca2+ and Mg2+ (Xiao et al., 2011). The
weak and reversible interaction between these plant compounds and plasma proteins is in
agreement with in vivo studies that show a decrease in the concentration of plant phenolic
compounds in plasma, due to metabolism and excretion, as was observed in P. barbatus extract
components (Falé et al., 2011), rosmarinic acid (Baba et al., 2004; Falé et al., 2011) and the
flavonoids apigenin and luteolin (Shi et al., 2011). Therefore, it is important that the structural
changes in plasma proteins when bound to compounds are not so drastic that their function may
be compromised after the release of the compounds.
Lysozyme, apart from drug transport, has an important function related to the immune
response process (Laible & Germaine, 1985). When degranulation occurs, after neutrophils
reach the injured tissue by margination, adhesion and emigration, lysozyme is discharged from
lysosomes of neutrophils and destroys not only the phagosomes but also damages the animal
tissue itself, thus aggravating the response to inflammation (Ronca et al., 1998; Wu et al., 2006).
Previous reports showed that phenolic compounds from Leggera species extracts decreased the
release of lysozyme from the neutrophils to the serum (Wu et al., 2006). In the present study,
P. barbatus extract phenolic compounds inhibited directly lysozyme activity. It is long known
that some flavonoids may act as lysozyme inhibitors (Rodney et al., 1950). Other studies also

Interactions with Proteins
122
reported that plant extracts mainly composed of apigenin, luteolin and luteolin glycosides
decreased allergy symptoms caused by egg‐white lysozyme sensitization in rats (Iwaoka et al.,
2010). The same anti‐allergic effect was observed for Perilla frutescens water extract, and by its
major component, rosmarinic acid (Makino et al., 2003).
As P. barbatus components bind to lysozyme with weak interactions, our data suggests
that the enzyme does not undergo major conformational changes, the plant compounds are
eliminated from plasma in a short period of time (Falé et al., 2011), and the antibacterial activity
of lysozyme is not permanently compromised. Therefore, these compounds may be helpful in
decreasing the damage caused by a high lysozyme activity, in response to pathogens or in
allergic conditions.
P. barbatus extract has also proved to have antioxidant activity as a radical scavenger
more powerful than the commercial antioxidant BHT (Falé et al., 2009). This activity is mainly
due to its main component, rosmarinic acid. The present results suggest that P. barbatus water
extract may be useful to treat inflammatory conditions as its components and metabolites may
be transported in circulatory system binding to albumin, the most abundant transport protein,
and to lysozyme, and decrease the inflammation process by two mechanism: as free radical
scavengers, and as lysozyme inhibitors.
2.5. Conclusions
Protein intrinsic fluorescence quenching proved that the P. barbatus extract components
and its metabolites found in rat plasma are able to bind to the human transport proteins
albumin and lysozyme, allowing them to be carried in the bloodstream to organs where they can
have a beneficial activity. The spectroscopic data suggest that the interaction of the plant
phenolic compounds and the proteins in this study is made by weak interactions, causing some
changes in protein tertiary structure, but not in the secondary structure. This suggests that the
compounds may be released from the complexes they form with the proteins, without
compromising the albumin or lysozyme function in plasma.
P. barbatus extract components and metabolites also inhibit lysozyme activity, which
may be helpful in decreasing the aggravation of the inflammation caused by the immune system
in response to pathogens and in allergies. This mechanism can be added to the already well
known radical scavenger capacity of the extract components, making the anti‐inflammatory
activity of plant extract very promising.

Chapter V
123
3. Conclusions
P. barbatus extract components can bind to acetylcholinesterase, human serum albumin
and lysozyme with the formation of a complex protein‐phenolic compound.
The spectroscopic data suggest that the plant phenolic compounds bind to the proteins
through weak interactions, causing some changes in protein tertiary structure, but not in the
secondary structure. This suggests that the compounds may be released from the complexes
they form with the proteins, without compromising irreversibly the acetylcholinesterase,
albumin or lysozyme functions.
The extract components bind to the active gorge of acetylcholinesterase and the
position of the hydroxyl groups in the flavonoids seems to play an important role in the binding
to the enzyme and in the inhibitory activity.
The binding of the P. barbatus extract components to the human transport proteins
albumin and lysozyme allows them to be carried in the bloodstream to organs where they can
have a beneficial activity, and it may explain the bioavailability and brain acetylcholinesterase
inhibition in rats, shown in Chapter IV.
P. barbatus extract components and metabolites also inhibit lysozyme activity, which
may be helpful in decreasing the aggravation of the inflammation caused by the immune system
in response to pathogens and in allergies.


Chapter VI
General Discussion and Conclusions


Chapter VI
127
The use of plant aqueous extracts, prepared as infusions and decoctions, for medicinal
purposes by a wide range of cultures originates an interest in finding the scientific validation of
the activities attributed to these traditional therapies. The use of herbal teas in milder health
disorders may be an alternative therapy with fewer side effects than the use of regular
medications. Plant extracts proved to be important sources of bioactive compounds that may
modulate biochemical reactions, as galanthamine, an acetylcholinesterase inhibitor first
extracted from Galanthus species (Heinrich & Teoh, 2004).
Aqueous extracts of Plectranthus barbatus leaves are reported in the literature to be
used in the treatment of psychiatric and digestive conditions (Lukhoba et al., 2006), suggesting
that these aqueous extract may inhibit acetylcholinesterase activity. Although it’s extracts were
not the most active among the other extracts of Plectranthus species tested for
acetylcholinesterase inhibition, it was the only Plectranthus species whose activities were
associated with other compounds than rosmarinic acid, the main component of all plant extracts
in Chapter III. The IC50 values obtained for acetylcholinesterase inhibition by the components of
the P. barbatus extract ranged from 10‐3 to 10‐4 M, which are much higher (less potent) than the
IC50 value, 5.85 x 10‐7 M, obtained for galanthamine in our lab, in the same experimental
conditions. However, the efficacy of the acetylcholinesterase inhibitors in therapies depends on
their bioavailability, and on the mechanism of inhibition, as irreversible inhibitors are toxic. The
currently commercialized acetylcholinesterase inhibitors present a wide diversity of in vitro
acetylcholinesterase inhibition values, but their therapeutic value is controlled by the mode of
administration, which modulates their bioavailability (McGleenon et al., 1999).
The P. barbatus aqueous extract components also showed antioxidant activity as radical
scavengers, and anti‐inflammatory activity as lysozyme inhibitors and decreasing the amount of
hypochlorous acid in human neutrophils. These biological activities are potentially useful in the
treatment of several inflammation‐related pathologies.
The composition in phenolic compounds of the P. barbatus aqueous extracts is very
similar to extracts of other species from the Lamiaceae family, since it is composed mainly of
rosmarinic acid and flavonoid glucuronides (Abdel‐Mogib et al., 2002; Batista et al., 1996). Apart
from the phenolic compounds, the P. barbatus decoction is also composed of abietane
diterpenoids, compounds also very common in Plectranthus species. However the most active
diterpenoid was metabolized into a non‐active form during the in vitro digestion with pancreatic
juice. Although the diterpenoids showed a high permeability in Caco‐2 monolayers, when the
plant extract was intraperitonially administered to rats just the non‐active diterpenoid was
observed in plasma, suggesting that the active diterpenoid is quickly metabolized and its activity

128
lost. Diterpenes from Salvia milthiorrizha, also a species from the Lamiaceae family, have shown
a high acetylcolinesterase inhibitory activity, with IC50 values in micromolar concentrations
(Wong et al., 2010). These diterpenoids from S. milthiorrizha were also metabolized into
hydroxylated metabolites by rat liver (Li et al., 2006; Wong et al., 2010), in a similar way to the
observed to the diterpenoids of P. barbatus in Chapter III, in pancreatic juice, and in Chapter IV,
possibly by the liver after intraperitoneal administration of the aqueous extract. According to
Wong et al. (2010) the hydroxylation is followed by furan ring cleavage, oxidation, conjugation of
the resulting metabolites and excretion. Although in the present study the active diterpenoid
have a short life, and therefore a limited usefulness to the treatment of Alzheimer’s disease, a
diterpenoid isolated from P. barbatus was shown to inhibit the gastric H+K+ATPase, decreasing
this way the acid secretion in the stomach, which may be an approach in the prevention or
treatment of gastric ulcers (Schultz et al., 2007).
The flavonoids in the P. barbatus aqueous extract were found in the glucuronidated
form, which is more water soluble and therefore easier to extract from the plant material than
the aglycones. The flavonoid glucuronides were stable during the in vitro digestion with gastric
and pancreatic juices, which is in agreement with other reports in the literature for this class of
compounds (Bouayed et al., 2011; Gião et al, 2011; Laurent et al., 2007). The only flavonoid
whose degradation with gastric juice was reported in the literature was procyanidin B2 (Bouayed
et al., 2012), a dimmer of epicatechin that is hydrolysed into monomeric epicatechin in the
acidic conditions of the gastric juice. Anthocyanins were also reported to be hydrolysed to some
extent in the pancreatic juice (McDougall et al., 2005; McDougall et al., 2007).
A limitation in the bioavailability of ingested flavonoids, and phenolic compounds in
general, was reported to be due to the binding of these plant compounds to the protein in the
pancreatic juice (Laurent et al., 2007) and in protein‐rich food such as milk (Cilla et al., 2009), by
hydrophobic interactions and hydrogen bonds. These interactions between plant phenolics and
proteins were also observed in the present study for the components of the P. barbatus aqueous
extract and the proteins acetylcholinesterase, albumin and lysozyme. Although flavonoid
derivatives are generally stable in pancreatic conditions and in contact with intestinal cells
(Chapter III, section 2; Laurent et al., 2007), the glucuronide derivatives may be hydrolysed into
their aglycones by the gut flora β‐glucuronidase activity. Several other authors also reported the
same reaction for flavonoid glycosylated derivatives (Bouayed et al., 2012; Berthiller et al.,
2011), and showed that the bacterial glycosidases are the only responsible for flavonoid
deglycosylation, as flavonoid glycosides showed low affinity for human glycosidases (Berthiller et
al., 2011).

Chapter VI
129
In the present studies rosmarinic acid was stable in gastric but 25% of its amount in the
plant extract was hydrolysed in pancreatic conditions. Other authors also referred that some
degradation may occur in the passage from gastric to pancreatic conditions (Gião et al., 2011). A
similar degradation also occurs with chlorogenic acid, with formation of caffeic acid (Gião et al.,
2011; Fazzari et al., 2008; Bouayed et al., 2012). Both rosmarinic acid and chlorogenic acid are
esters of caffeic acid, and therefore may be relatively stable in acidic conditions but easily
hydrolysed in basic conditions.
When P. barbatus decoction was intragastrically administered to rats, and when it was
applied to the Caco‐2 cell monolayers for the permeability assay, low amounts of unconjugated
rosmarinic acid was found to permeate the intestinal barrier, which may be due to the low
affinity of rosmarinic acid to the MCT trasporters (Chapter IV section 2). The low affinity of
rosmarinic acid to the membrane transporters led other authors to suggest that no transporters
were involved in the permeability of rosmarinic acid. However, in the present study assays were
carried out during longer times, 6 hours instead of 40 minutes (Konishi & Kobayashi, 2005),
which allowed to detect the differences in permeability in the presence and absence of
competitors for the same transporters. Glucuronidated and sulfated metabolites of rosmarinic
acid were found in the plasma of rats after intragastric administration, while methylated
metabolites were present after intraperitonial administration. These facts suggest that
rosmarinic acid may be predominantly glucuronidated and sulphated in the intestine, and
undergo mainly methylation by the liver. In Caco‐2 cell monolayers rosmarinic acid remained
intact (Chapters III and IV), which is not in agreement with the results found in in vivo studies
with rats (Chapter IV), or with the literature (Konishi & Kobayashi, 2005). Nevertheless
rosmarinic acid in the Caco‐2 cell monolayer studies in the Chapters III and IV was co‐
administered with flavonoids that may have more affinity to phase II enzymes. Also the Caco‐2
monolayers and the rats are different bioavailability models that may present slightly different
results (Keldenish, 2009).
The flavonoid glucuronides in the plant extract permeated the intestinal barrier both in
the Caco‐2 and in the rat models. Most studies in the literature focus on flavonoid aglycones,
which can be metabolized by Caco‐2 cells in flavonoid glucuronides and sulfates (Walle et al
1999; Barrington et al., 2009). The conjugates are reported by the same authors to be effluxed
mainly to the apical side of the Caco‐2 monolayer, corresponding to the gut lumen in the
intestine, limiting their bioavailability. In chapter IV it is shown that the glucuronides can also be
transported from the apical to the basolateral side of the Caco‐2 monolayers, although the
apigenin and luteolin glucuronides may also be effluxed back to the apical side possibly with the

130
involvement of Pgp transporter. Studies regarding the bioavailability of flavonoid glycosylated
derivatives are scarce in the literature, however the quercetin derivative rutin was found to
permeate the Caco‐2 monolayers from apical to basolateral side (Gião et al 2011). The flavonoid
glycosylated derivatives may be hydrolysed into their aglycones as discussed above, or by the
liver β‐glucuronidases, as observed and discussed in Chapter IV. In the present study the
glucuronidation of flavonoid aglycones was observed in Caco‐2 cells, but sulfated metabolites
were not detected. Also, when the plant extract was intraperitonially administered to rats,
although liver β‐glucuronidase activity occurred, no sulfated metabolites of the flavonoids were
found. The sulfotransferase activity was shown to occur in the rat liver, as the sulfated
derivatives of the phenolic compounds were synthesised in vitro using rat liver protein extracts
(Chapter IV). Other authors have reported that the glucuronidation of flavonoids was
predominant, and occurred in more positions than the sulfation of the compounds (Barrington
et al., 2009). This suggests a lower affinity of the flavonoids to the sulfotransferases than to the
glucuronyltransferases.
Due to the differences found when administering rosmarinic acid in the extract or as an
isolated compound, a study was performed in Caco‐2 cell monolayers using central composite
design (CCD) to determine if the phenolic compounds interfere with the bioavailability of one
another, and if transporters were involved in the bioavailability of the compounds. It was found
that rosmarinic acid, apigenin and luteolin can permeate Caco‐2 cell monolayers through
transporters and, generally, have higher bioavailability together than when isolated. This
increase of bioavailability is mainly due to inhibition or competition for efflux transporters that
transport compounds back to the apical side of the Caco‐2 monolayers (Chapter IV section 2).
The P. barbatus extract components followed the same behaviour predicted by the model
mixture of rosmarinic acid, luteolin and apigenin.
Flavonoids are well known inhibitors of ABC transporters, such as Pgp, MRP and BCRP
(Brand et al., 2006). Some flavonoids are also substrates of these transporters, which limits their
bioavailability, such as quercetin, kaempferol, isorhametin (Wang et al., 2005), and chrysin
(Walle et al., 1999). As substrates and inhibitors of this type of efflux transporters, the
coadministration of many compounds increases their bioavailability by decreasing the transport
from basolateral to apical side, as it was described in Chapter IV. In the same way the inhibition
of effux systems may increase the bioavailability of some orally administered medicines (Brand
et al., 2006), as for instance, doxorubicin, an anticancer drug whose bioavailability is increased
by inhibiting Pgp with quercetin (Choi et al., 2011).

Chapter VI
131
As it was discussed in Chapter IV, the values for the inhibition of acetylcholinesterase
activity in rat brains after the administration of the plant extract were comparable to those
found in the literature for galanthamine, even though a lower amount of galanthamine was
administered (Geerts et al., 2005; Chattipakorn et al., 2007). Only a low amount of extract
compounds was found in the brain of rats. Although the amount of these plant compounds, or
their metabolites, was much higher, the amount found in the brain kept constant for a longer
period of time. This fact suggests that if the extract is administered several times a day, the
amount of extract components in the brain should be kept relatively constant, even though the
amounts of extract metabolites in the plasma may have high variations during the day.
Differences of plasma bioavailability, half‐life, and brain bioavailability lead to different
strategies in the administration of acetylcholinesterase inhibitors. Tacrine has high brain
bioavailability, but when taken orally, it has low plasma bioavailability and low half‐life.
Therefore the administration of tacrine follows a similar strategy to the one suggested for the
decoction of P. barbatus in this study, of several administrations during the day (McGleenon et
al., 1999).
Peripheral effects of acetylcholinesterase inhibition, such as gastrointestinal and hepatic
disturbances, are usually associated with the administration of large dosages of
acetylcholinesterase inhibitors. However, as mentioned in Chapters III (section 2) and IV (section
2), with 1mg/ml of the plant extract or 90 μM of any of the standards no toxicity was observed in
Caco‐2 cells, and 100μg/mL of plant extract or 20μM of pure compound would be the maximum
values accepted to consider the occurrence of toxicity (Cárdenas et al. 2006; Yu et al., 2007).
Furthermore, toxic effects are not reported for P. barbatus, on the contrary, the herbal tea of
this plant is usually drunk to treat gastrointestinal problems (Lukhoba et al., 2006). The effect of
the P. barbatus decoction on the gastrointestinal tract may be due to a mild inhibition of
acetylcholinesterase activity, increasing gastrointestinal motility and gut hydration (Jarvie et al.,
2008; Hirota and McKay 2006).
The binding of the plant extract components to the plasma proteins, albumin and
lysozyme, may also decrease the side‐effects of the acetylcholinesterase inhibition by reducing
the availability of the compounds to peripheral acetylcholinesterase while transporting them
through the bloodstream to the target organ, the brain. Although, as was discussed in chapter V,
the compounds seem to bind in the active gorge of acetylcholinesterase, the weak interactions
between the plant phenolics and acetylcholinesterase do not cause large conformational
changes in the enzyme and the enzymatic activity may be easily recovered after the excretion of
the compounds. Therefore, the weak interactions between the plant phenolics and

132
acetylcholinesterase may also be preferable, decreasing the side effects of excessive
acetylcholinesterase inhibition.
The P. barbatus extract showed radical scavenging activity with the DPPH and protected
fatty acids from peroxidation in the β‐carotene/linoleic acid assay, and therefore it may be
useful preventing oxidative stress‐related cell membrane disruption and apoptosis. This cell
death mechanism is similar to the one caused by the oxidative stress resultant from the
accumulation of amyloid plaques in Alzheimer’s disease patients. The plant extract and
rosmarinic acid also decreased the amount of hypochlorous acid produced by activated
neutrophils (Chapter III) and inhibited lysozyme activity (Chapter V), which can be associated
with a immune response, but also with a cytotoxic inflammatory reaction (Ronca et al., 1998;
Deby‐Dupont, 1999). The cognition status of Alzheimer’s disease patients has been correlated
with the systemic oxidative stress status (Viña et al., 2004), and therefore the plant extract may
be useful in this approach to the treatment of Alzheimer’s disease. Furthermore, the
components of the plant extract act as antioxidants through different mechanisms, which has
been proposed as the most efficient strategy in the antioxidant treatment of Alzheimer’s disease
(Macocci & Polidori, 2011).
In conclusion,
The P. barbatus decoction showed promising in vitro antioxidant, anti‐inflammatory and
anti‐acetylcholinesterase activities.
The P. barbatus herbal tea showed no toxicity in Caco‐2 cells. The low toxicity is also
supported by the weak interactions established with proteins without causing secondary
structure changes.
Some active components of the P. barbatus herbal tea undergo metabolization in the
gastrointestinal tract, but the phenolic compounds can cross the intestinal barrier and
reach the brain, where they can inhibit acetylcholinesterase activity, similarly to the
currently used acetylcholinesterase inhibitors.
The phenolic compounds can circulate in the bloodstream bound to the plasma proteins
and decrease oxidative stress‐related processes, such as inflammation, by decreasing
the amount of hypochlorous acid released by neutrophils and also by inhibiting lysozyme
activity.
The use of polyphenolic compound mixtures such as herbal teas may increase the
bioavailability of the active compounds and, as the herbal tea components show many
activities, may be useful in the treatments with multiple approaches, as it is thought to
be the most efficient for Alzheimer’s disease.

References


References
135
Abdel‐Mogib M, Albar HA, Batterjee SM. 2002. Chemistry of the genus Plectranthus.
Molecules, 7: 271–301.
Abou‐Donia MB. 2003. Organophosphorous ester‐induced chronic neurotoxicity. Arch
Environ Health. 58:484‐497.
Adsersen A, Gauguin B, Gudiksen L, Jäger AK. 2006. Screening of plants used in Danish folk
medicine to treat memory dysfunction for acetylcholinesterase inhibitory activity. J
Ethnopharmacol. 104: 418‐22.
Alasbahi RH, Melzig MF. 2010a. Plectranthus barbatus: a review of phytochemistry,
ethnobotanical uses and pharmacology ‐ Part 1. Planta Med. 76: 653‐661.
Alasbahi RH, Melzig MF. 2010b. Plectranthus barbatus: a review of phytochemistry,
ethnobotanical uses and pharmacology ‐ part 2. Planta Med. 76: 753‐765.
Albuquerque RL, Kentopff MR, Machado MIL, Silva MG, Matos FJA. 2007. Diterpenos tipo
abietano isolados de Plectranthus barbatus Andrews. Química Nova. 30: 1882–1886.
Alzheimer’s association. 2008. Alzheimer’s disease facts and figures. Alzheimer’s &
Dementia. 4: 110–133.
Atoui AK, Mansouri A, Bosku G, Kefalas P. 2005. Tea and herbal infusions: their antioxidant
activity and phenolic profile. Food Chem., 89: 27–36.
Azuma K, Ippoushi K, Nakayama M, Ito H, Higashio H, Terao J. 2000. Absorption of
Chlorogenic Acid and Caffeic Acid in Rats after Oral Administration. J. Agric. Food Chem. 48:
5496‐5500.
Baba S, Osakabe N, Natsume M, Terao J. 2004. Orally administered rosmarinic acid is present
as the conjugated and/or methylated forms in plasma, and is degraded and metabolized to
conjugated forms of caffeic acid, ferulic acid and m‐coumaric acid. Life Sci. 75: 165–178.
Baba S, Osakabe N, Natsume M, Yasuda A, Muto Y, Hiyoshi K, Takano H, Yoshikawa T, Terao
J. 2005. Absorption, metabolism, degradation and urinary excretion of rosmarinic acid after
intake of Perilla frutescens extract in humans. Eur J Nutr. 44: 1–9.
Babior BM. 2000. Phagocytes and oxidative stress. Am J Med. 109:33‐44.
Bachoual R, Talmoudi W, Boussetta T, Braut F, El‐Benna J. 2011. An aqueous pomegranate
peel extract inhibits neutrophil myeloperoxidase in vitro and attenuates lung inflammation
in mice. Food Chem Toxicol. 49:1224‐8.
Barker, T. B., in: Schilling, E. G. (Ed.), Quality by Experimental Design, Marcel Dekker, New
York 1985, pp. 310–325.
Barrington R, Williamson G, Bennett RN, Davis BD, Brodbelt JS, Kroon PA. 2009. Absorption,
conjugation and efflux of the flavonoids, kaempferol and galangin, using the intestinal Caco‐
2/TC7 Cell Model. J Funct Foods. 1:74‐87.

References
136
Batista O, Simões MF, Nascimento J, Riberio S, Duarte A, Rodríguez B, de la Torre MC. 1996.
A rearranged abietane diterpenoid from Plectranthus hereroensis. Phytochemistry. 41: 571–
573.
Bembenek SD, Keith JM, Letavic MA, Apodaca R, Barbier AJ, Dvorak L, Aluisio L, Miller KL,
Lovenberg TW, Carruthers NI. 2008. Lead identification of acetylcholinesterase inhibitors –
histamine H3 receptor antagonists from molecular modelling. Bioorg Med Chem. 16: 2968‐
2973.
Berthiller F, Krska R, Domig KJ, Kneifel W, Juge N, Schuhmacher R, Adam G. 2011. Hydrolytic
fate of deoxynivalenol‐3‐glucoside during digestion. Toxicol Lett. 206: 264‐267.
Birman S. 1985. Determination of acetylcholinesterase activity by a new chemiluminescence
assay with the natural substrate. Biochem. J. 225: 825‐828.
Bohnen NI, Kaufer DI, Hendrickson R, Ivanco LS, Lopresti BJ, Koeppe RA, Meltzer CC,
Constantine G, Davis JG, Mathis CA, Dekosky ST, Moore RY. 2005. Degree of inhibition of
cortical acetylcholinesterase activity and cognitive effects by donepezil treatment in
Alzheimer’s disease. J. Neurol., Neurosurg. Psychiatry. 76: 315–319.
Bouayed J, Deusser H, Hoffmann L, Bohn T. 2012. Bioaccessible and dialysable polyphenols
in selected apple varieties following in vitro digestion vs. their native patterns. Food Chem.
131: 1466‐1472.
Bouayed J, Hoffmann L, Bohn T. 2011. Total phenolics, flavonoids, anthocyanins and
antioxidant activity following simulated gastro‐intestinal digestion and dialysis of apple
varieties: Bioaccessibility and potential uptake. Food Chem. 128: 14‐21.
Bourne Y, Grassi J, Bougis PE, Marchot P. 1999. Conformational flexibility of the
acetylcholinesterase tetramer suggested by X‐ray crystallography. J. Biol. Chem. 274: 30370–
30376.
Bourne Y, Radić Z, Taylor P, Marchot P. 2010. Conformational remodeling of femtomolar
inhibitor − acetylcholinesterase complexes in the crystalline state. J Am Chem Soc. 132: 18292‐
18300.
Brand W, Padilla B, van Bladeren PJ, Williamson G, Rietjens IM. 2010. The effect of co‐
administered flavonoids on the metabolism of hesperetin and the disposition of its
metabolites in Caco‐2 cell monolayers. Mol Nutr Food Res. 54: 851‐60.
Brand W, Schutte ME, Williamson G, van Zanden JJ, Cnubben NH, Groten JP, van Bladeren PJ,
Rietjens IM. 2006. Flavonoid‐mediated inhibition of intestinal ABC transporters may affect
the oral bioavailability of drugs, food‐borne toxic compounds and bioactive ingredients.
Biomed Pharmacother. 60: 508‐519.
Brand W, van der Wel PA, Rein MJ, Barron D, Williamson G, van Bladeren PJ, Rietjens IM.
Metabolism and transport of the citrus flavonoid hesperetin in Caco‐2 cell monolayers. Drug
Metab Dispos. 36: 1794‐1802.

References
137
Cai Y, Luo Q, Sun M, Corke H. 2004. Antioxidant activity and phenolic compounds of 112
traditional Chinese medicinal plants associated with anticancer. Life Sci. 74: 2157‐2184.
Cárdenas M, Mardel M, Blank VC, Roguin LP. 2006. Antitumor activity of some natural
flavonoids and synthetic derivatives on various human and murine cancer cell lines. Bioorg
Med Chem. 14: 2966‐2971.
Cellek S, Thangiah R, Jarvie EM, Vivekanandan S, Lalude O, Sanger GJ. 2008. Synergy
between 5‐HT4 receptor activation and acetylcholinesterse inhibition in human colon and
rat forestomach. Neurogastroenterol Motil. 20: 539‐545.
Chattipakorn S, Pongpanparadorn A, Pratchayasakul W, Pongchaidacha A, Ingkaninan K,
Chattipakorn N. 2007. Tabernaemontana divaricata extract inhibits neuronal
acetylcholinesterase activity in rats. J. Ethnopharmacol. 110: 61–68.
Choi JS, Piao YJ, Kang KW. 2011. Effects of quercetin on the bioavailability of doxorubicin in
rats: role of CYP3A4 and P‐gp inhibition by quercetin. Arch Pharm Res. 34: 607‐13.
Cilla A, Gonzalez‐Sarrias A, Tomas‐Baeberan F, Espin JC, Barbera R. 2009. Availability of
polyphenols in fruit beverages subjected to in vitro gastrointestinal digestion and their
effects on proliferation, cell‐cycle and apoptosis in human colon cancer Caco‐2 cells. Food
Chem. 114: 813–820.
Codd LE. 1975. Plectranthus (Labiatae) and allied genera in southern Africa. Bothalia. 11:
371‐442.
Coelho M. 2004. Eixo Renina‐Angiotensina. Polimorfismos genéticos e interacções
fenotípicas. Lisboa: Universidade de Lisboa.
Coleta M, Camposa MG, Cotrim MD, Lima MCT, Cunha AP. 2008. Assessment of luteolin
(3’,4’,5,7‐tetrahydroxyflavone) neuropharmacological activity, Behav. Brain Res. 189: 75–82.
Curry S. 2009. Lessons from the crystallographic analysis of small molecule binding to human
serum albumin, Drug Metab Pharmacok. 24: 342–357.
da Silva EL, Piskula MK, Yamamoto N, Moon JH, Terao J. 1998. Quercetin metabolites inhibit
copper ion‐induced lipid peroxidation in rat plasma. FEBS Lett. 430: 405‐408.
Deby‐Dupont G, Deby C, Lamy M. 1999. Neutrophil myeloperoxidase revisited: it’s role in
health and disease. Intensivmed. 36: 500–513.
Dickson D, Lee S, Mattiace L, Yen S, Brosnan C. 1993. Microglia and cytokines in neurological
disease, with special reference to AIDS and Alzheimer’s disease. Glia. 7: 75‐83.
Djeridane A, Yousfi M, Nadjemi B, Boutassouna D, Stocker P, Vidal N. 2006. Antioxidant
activity of some Algerian medicinal plants extracts containing phenolic compounds. Food
Chem. 97: 654–660.
D'Souza VM, Shertzer HG, Menon AG, Pauletti GM. 2003. High glucose concentration in
isotonic media alters Caco‐2 cell permeability. AAPS PharmSci. 5:, E24.

References
138
Dumont M, Beal MF. 2011. Neuroprotective strategies involving ROS in Alzheimer disease.
Free Radic Biol Med. 51: 1014‐1026.
Dypbukt J, Bishop C, Brooks W, Thong B, Eriksson H,Kettle A. 2005. A sensitive and selective
assay for chloramine production by mieloperoxidase. Free Radic Biol Med. 39: 1468‐1477.
Eastman J, Wilson EJ, Cerveñansky C, Rosenberry TL. 1995. Fasciculin 2 binds to the
peripheral site on acetylcholinesterase and inhibits substrate hydrolysis by slowing a step
inhibiting proton transfer during enzyme action. J Biol Chem. 270: 19694‐19701.
Eliman GL, Courtney KD, Andres V.,Featherstone RM. 1961. A new and rapid colorimetric
determination of acetylcholinesterase activity. Biochem. Pharmacol. 7: 88‐95
El‐Kamali HH. 2009. Medicinal plants in East and Central Africa: Challenges and constraints.
Ethnobotanical Leaflets. 13: 364–369.
Ellman GL. 1958. A colorimetric method for determining low concentrations of mercaptans.
Arch Biochem Biophys. 74: 443‐450.
Erlung I. 2004. Review of the flavonoids quercetin, hesperetin, and naringenin. Dietary
sources, bioactivities, bioavailability, and epidemiology. Nutr. Res. 24: 851–874.
Eyer P, Szinicz L, Thiermann H, Worek F, Zilker T. 2007. Testing of antidotes of
organophosphorus compounds: Experimental procedures and clinical reality. Toxicol. 233:
108‐119.
Falé PL, Borges C, Madeira PJA, Ascensão L, Araújo MEM, Florêncio MH, Serralheiro MLM.
2009. Rosmarinic acid, scutellarein 4’‐methyl ether 7‐O‐glucuronide and (16S)‐coleon E are
the main compounds responsible for the antiacetylcholinesterase and antioxidant activity in
herbal tea of Plectranthus barbatus (‘‘falso boldo’’), Food Chem. 114: 798– 805.
Falé PLF, Madeira PJA, Florêncio MH, Ascensão L, Serralheiro MLM. Function of Plectranthus
barbatus herbal tea as neuronal acetylcholinesterase inhibitor. Food Funct. 2: 130‐136.
Fazzari M, Fukumoto L, Mazza G, Livrea MA, Tesoriere L, Marco LD. 2008. In vitro
bioavailability of phenolic compounds from five cultivars of frozen sweet cherries (Prunus
avium L.). J Agric Food Chem. 56: 3561‐3568.
Fecka I, Turek S. 2007. Determination of water‐soluble polyphenolic compounds in
commercial herbal teas from Lamiaceae: peppermint, melissa, and sage. J. Agric. Food
Chem. 55: 10908‐10917.
Ferreira A, Proença C, Serralheiro MLM, Araújo MEM. 2006. The in vitro screening for
acetylcholinesterase inhibition and antioxidant activity of medicinal plants from Portugal. J
Ethnopharm. 108: 31‐37.
Galijatovic A, Walle UK, Walle T. 2000. Induction of UDPglucuronosyltransferase by the
flavonoid chrysin and quercetin in Caco‐2. Pharm Res. 17: 21–26.

References
139
Geerts H, Guillaumat PO, Grantham C, Bode W, Anciaux K, Sachak S. 2005. Brain levels and
acetylcholinesterase inhibition with galantamine and donepezil in rats, mice, and rabbits,
Brain Res. 1033: 186–193.
Gião MS, Gomes S, Madureira AR, Faria A, Pestana D, Calhau C, Pintado ME, Azevedo I,
Malcata FX. 2011. Effect of in vitro digestion upon the antioxidant capacity of aqueous
extracts of Agrimonia eupatoria, Rubus idaeus, Salvia sp. and Satureja Montana. Food Chem.
doi:10.1016/j.foodchem.2011.09.030.
Gill RK, Saksena S, Alrefai WA, Sarwar Z, Goldstein JL, Carroll RE, Ramaswamy K, Dudeja PK.
2005. Expression and membrane localization of MCT isoforms along the length of the human
intestine. Am J Physiol Cell Physiol. 289: C846‐852.
Gomes A, Fernandes E, Lima JFC, Mira L, Corvo ML. 2008. Molecular mechanisms of anti‐
inflammatory activity mediated flavonoids. Curr. Med. Chem. 15: 1586–1605.
Gorne‐Tschelnokow U, Naumann D, Weise C, Hucho F. 1993. Secondary structure and
temperature behaviour of acetylcholinesterase Studies by Fourier‐transform infrared
spectroscopy. Eur J Biochem. 213: 1235‐1242.
Gouwy M, Struyf S,Proost P, Damme JV. 2005. Synergy in cytokine and chemokine networks
amplifies the inflammatory response. Cytokine Growth Factor Rev. 16: 561–580.
Grayer RJ, Eckert MR, Veitch NC, Kite GC, Marin PD, Kokubun T, Simmonds MS, Paton AJ.
2003. The chemotaxonomic significance of two bioactive caffeic acid esters, nepetoidins A
and B, in the Lamiaceae. Phytochemistry. 64: 519–528.
Grayer, R. J., Veitch, N. C., Kite, G. C., Paton, A. J., & Garnock‐Jones, P. J. (2002). Scutellarin
4’‐methyl ether glycosides as taxonomic markers in Teucridium and Tripora (Lamiaceae,
Ajugoideae). Phytochemistry, 60, 727–731.
Guginski G, Luiz AP, Silva MD, Massaro M, Martins DF, Chaves J, Mattos RW, Silveira D,
Ferreira VM, Calixto JB, Santos AR. 2009. Mechanisms involved in the antinociception caused
by ethanolic extract obtained from the leaves of Melissa officinalis (lemon balm) in mice,
Pharmacol., Biochem. Behav. 93: 10–16.
Harel M, Schalk I, Ehret‐Sabatier L, Bouet F, Goeldner M, Hirth C, Axelsen PH, Silman I,
Sussman JL. 1993. Quaternary ligand binding to aromatic residues in the active‐site gorge of
acetylcholinesterase. Proc Natl Acad Sci U S A. 90: 9031–9035.
Harel M, Sonoda LK, Silman I, Sussman JL, Rosenberry TL. 2008. Crystal structure of thioflavin
T bound to the peripheral site of Torpedo californica acetylcholinesterase reveals how
thioflavin T acts as a sensitive fluorescent reporter of ligand binding to the acylation site, J.
Am. Chem. Soc. 130: 7856–7861.
Haris PI, Chapman D, Harrison RA, Smith KF, Perkins SJ. 1990. Conformational transition
between native and reactive center cleaved forms of α1‐antitrypsin by Fourier transform
infrared spectroscopy and small‐angle neutron scattering. Biochemistry. 29: 1377–1380.

References
140
Haris PI, Lee DC, Chapman D. 1986. A Fourier transform infrared investigation of the structural
differences between ribonuclease A and ribonuclease S. Biochim Biophys Acta. 874: 255‐265.
Haris PI, Severcan F. 1999. FTIR spectroscopic characterization of protein structure in
aqueous and non‐aqueous media. J Mol Catal B Enzym. 7: 207‐221
Hartshorne DJ, Stracher A. 1965. Deuterium‐hydrogen exchange of muscle proteins,
Biochemistry. 4: 1917‐1923.
Hashimoto M, Rockenstein E, Crews L, Masliah E. 2003. Role of protein aggregation in
mitochondrial dysfunction and neurodegeneration in Alzheimer's and Parkinson's diseases.
Neuromolecular Med. 4: 21‐36.
He W, Li Y, Tang J, Luan F, Jin J, Hu Z. 2006. Comparison of the characterization on binding of
alpinetin and cardamonin to lysozyme by spectroscopic methods. Int. J. Biol. Macromol. 39:
165–173.
Heinrich M, Teoh HL. 2004. Galanthamine from snowdrop – the development of a modern
drug against Alzheimer’s disease from local Caucasian knowledge. J Ethnopharmacol. 92:
147–162.
Hirota CL, McKay DM. 2006. Cholinergic regulation of epithelial ion transport in the
mammalian intestine. Br J Pharmacol. 149: 463‐479.
Hollman PC, Katan MB. 1997. Absorption, metabolism and health effects of dietary
flavonoids in man. Biomed Pharmacother. 51: 305–310.
Holzer P, Maggi CA. 1994. Synergistic role of muscarinic acetylcholine and tachykinin NK‐2
receptors in intestinal peristalsis. Naunyn Schmiedebergs Arch Pharmacol. 349: 194‐201.
Hostettmann K, Borloz A, Urbain A, Marston A. 2006. Natural product inhibitors of
acetylcholinesterase. Curr Org Chem. 10: 825–847.
Houghton PJ, Agbedahunsi JM, Adegbulugbe A. 2004. Choline esterase inhibitory properties
of alkaloids from two Nigerian Crinum species. Phytochemistry, 65: 2893‐2896.
Huebbe P, Wagner AE, Boesch‐Saadatmandi C, Sellmer F, Wolffram S, Rimbach G. 2010.
Effect of dietary quercetin on brain quercetin levels and the expression of antioxidant and
Alzheimer’s disease relevant genes in mice. Pharmacol Res. 61: 242–246.
Hvidt A, Wallevik K. 1972. Conformational changes in human serum albumin as revealed by
hydrogen‐deuterium exchange studies. J. Biol. Chem. 247: 1530‐1535.
Ingkaninan K, Temkitthawon P, Chuenchon K, Yuyaem T, Thongnoi W. 2003. Screening for
acetylcholinesterase inhibitory activity in plants used in Thai traditional rejuvenating and
neurotonic remedies. J. Ethnopharmacol. 89: 261–264.
Iwaoka E, Oku H, Iinuma M, Ishiguro K. 2010. Allergy‐preventive effects of the flowers of
Impatiens textori. Biol Pharm Bull. 33: 714‐716.

References
141
Iyo M, Namba H, Fukushi K, Shinotoh H, Nagatsuka S, Suhara T, Sudo Y, Suzuki K, Irie T. 1997.
Measurement of acetylcholinesterase by positron emission tomography in the brains of
healthy controls and patients with Alzheimer’s disease. Lancet. 349: 1805–1809.
Jaarsveld E. 2006. The southern African Plectranthus and the art of turning shade to glade.
Fernwood Press. Simon’s Town, South Africa.
Jackson M, Haris PI, Chapman D. 1991. Fourier transform infrared spectroscopic studies of
calcium‐binding proteins. Biochemistry. 30: 9681‐9686.
Jacobsen CF, Leonis J, Linderstrom‐Lang K, Otteren M. 1957. The pH‐stat and its use in
biochemistry. Methods Biochem Anal. 4: 171‐210.
Janicsák G, Máthé I, Miklóssy‐Vári V, Blunden G. 1999. Comparative studies of the rosmarinic
and caffeic acid contents of Lamiaceae species. Biochem Syst Ecol. 27: 733‐738.
Jarvie E, Cellek S, Sanger G. 2008. Potentiation by cholinesterase inhibitors of cholinergic
activity in rat isolated stomach and colon. Pharmacol Res. 58: 297–301.
Ji H‐F, Zhang H‐Y. 2006., Theoretical evaluation of flavonoids as multipotent agents to
combat Alzheimer's disease. J Mol Struct THEOCHEM. 767: 3–9.
Jiang WL, Chen XG, Qu GW, Yue XD, Zhu HB, Tian JW, Fu FH. 2009. Rosmarinic acid protects
against experimental sepsis by inhibiting proinflammatory factor release and ameliorating
hemodynamics. Shock. 32: 608‐13.
Jin J, Zhang X. 2008. Spectrophotometric studies on the interaction between pazufloxacin
mesilate and human serum albumin or lysozyme. J Luminescence. 128: 81–86.
Johnson CD, Russell RL. 1975. A rapid, simple radiometric assay for cholinesterase, suitable
for multiple determinations. Anal Biochem. 64: 229‐238.
Justino GC, Santos MR, Canário S, Borges C, Florêncio MH, Mira L. 2004. Plasma quercetin
metabolites: structure–antioxidant activity relationships. Arch Biochem Biophys. 432: 109–
121.
Kadir A, Darreh‐Shori T, Almkvist O, Wall A, Grut M, Strandberg B, Ringheim A, B Eriksson,
Blomquist G, Langstrom B, Nordberg A. 2008. PET imaging of the in vivo brain
acetylcholinesterase activity and nicotine binding in galantaminetreated patients with AD.
Neurobiol Aging. 29: 1204–1217.
Kanatt SR, Chander R, Sharma A. 2007. Antioxidant potential of mint (Mentha spicata L.) in
radiation‐processed lamb meat. Food Chem. 100: 451–458.
Keldenich J. 2009. Measurement and prediction of oral absorption. Chem Biodivers. 6: 2000‐
2013.
Kern SM, Bennett RN, Needs PW, Mellon FA, Kroon PA, Garcia‐Conesa M‐T. 2003.
Characterization of metabolites of hydroxycinnamates in the in vitro model of human small
intestinal epithelium Caco‐2 cells. J Agric Food Chem. 51: 7884–7891.

References
142
Kerr ML, Small DH. 2005. Cytoplasmic domain of the beta‐amyloid protein precursor of
Alzheimer's disease: function, regulation of proteolysis, and implications for drug
development. J Neurosci Res. 80: 151‐159.
Khan MT, Orhan I, Senol FS, Kartal M, Sener B, Dvorská M, Smejkal K, Slapetová T. 2009.
Cholinesterase inhibitory activities of some flavonoid derivatives and chosen xanthone and
their molecular docking studies. Chem Biol Interact. 181: 383‐389.
Kim H, Kim H, Jung S. 2008. Aqueous solubility enhancement of some flavones by
complexation with cyclodextrins, Bull Korean Chem Soc. 29: 590–594.
Klebanoff SJ. 2005. Myeloperoxidase: friend and foe. J Leukoc Biol. 77: 598‐625.
Kobayashi S, Konishi Y. 2008. Transepithelial transport of flavanone in intestinal Caco‐2 cell
monolayers. Biochem. Biophys. Res. Commun. 368: 23‐29.
Kobayashi S, Tanabe S, Sugiyama M, Konishi Y. 2008. Transepithelial transport of hesperetin
and hesperidin in intestinal Caco‐2 cell monolayers. Biochim Biophys Acta. 1778: 33‐41.
Kobayashi S, Watanabe J, Fukushi E, Kawabata J, Nakajima M, Watanabe M. 2003.
Polyphenols from some foodstuffs as inhibitors of ovalbumin permeation through Caco‐2
cell monolayers. Biosci. Biotechnol. Biochem. 67: 1250‐1257.
Komes D, Belščak‐Cvitanović A, Horžić D, Rusak G, Likić S, Berendika M. 2011. Phenolic
composition and antioxidant properties of some traditionally used medicinal plants affected
by the extraction time and hydrolysis. Phytochem. Anal. 22: 172‐180.
Kong H, Wang M, Venema K, Maathuis A, van der Heijden R, van der Greef J, Xu G,
Hankemeier T. 2009. Bioconversion of red ginseng saponins in the gastro‐intestinal tract in
vitro model studied by high‐performance liquid chromatography–high resolution Fourier
transform ion cylcotron resonance mass spectrometry. J Chromatogr A. 1216: 2195–2203.
Konishi Y, Hitomi Y, Yoshioka E. 2004. Intestinal Absorption of p‐Coumaric and Gallic Acids in
Rats after Oral Administration. J. Agric. Food Chem. 52: 2527‐2532
Konishi Y, Kobayashi S, Shimizu M. 2003. Tea polyphenols inhibit the transport of dietary
phenolic acids mediated by the monocarboxylic acid transporter (MCT) in intestinal Caco‐2
cell monolayers. J Agric Food Chem. 51: 7296‐7302.
Konishi Y, Kobayashi S. 2004a. Microbial Metabolites of Ingested Caffeic Acid Are Absorbed
by the Monocarboxylic Acid Transporter (MCT) in Intestinal Caco‐2 Cell Monolayers. J. Agric.
Food Chem. 52: 6418‐6424
Konishi Y, Kobayashi S. 2004b. Transepithelial Transport of Chlorogenic Acid, Caffeic Acid,
and Their Colonic Metabolites in Intestinal Caco‐2 Cell Monolayers. J. Agric. Food Chem. 52:
2518‐2526.
Konishi Y, Kobayashi S. 2005. Transepithelial transport of rosmarinic acid in intestinal Caco‐2
monolayers. Biosci Biotechnol Biochem. 69: 583‐591.

References
143
Konishi Y, Zhao Z, Shimizu M. 2006. Phenolic Acids Are Absorbed from the Rat Stomach with
Different Absorption Rates. J. Agric. Food Chem. 54: 7539‐7543.
Konishi Y., Hitomi Y, Yoshida M, Yoshioka E. 2005. Pharmacokinetic Study of Caffeic and
Rosmarinic Acids in Rats after Oral Administration. J Agric Food Chem. 53: 4740–4746.
Kreis ME, Kasparek M, Zittel TT, Becker HD, Jehle EC. 2001. Neostigmine increases
postoperative colonic motility in patients undergoing colorectal surgery. Surgery. 130: 449‐
456.
Kumaran A, Karunakaran RJ. 2007. Activity‐guided isolation and identification of free radical‐
scavenging components from an aqueous extract of Coleus aromaticus. Food Chem., 100:
356–361.
Kuo MY, Liao MF, Chen FL, Li YC, Yang ML, Lin RH, Kuan YH. 2011. Luteolin attenuates the
pulmonary inflammatory response involves abilities of antioxidation and inhibition of MAPK
and NFκB pathways in mice with endotoxin‐induced acute lung injury. Food Chem Toxicol.
49: 2660‐2666.
Lafay S, Gil‐Izquierdo A. 2008. Bioavailability of phenolic acids. Phytochem Rev. 7: 301–311.
Laible NJ, Germaine GR. 1985. Bactericidal activity of human lysozyme, muramidase‐inactive
lysozyme, and cationic polypeptides against Streptococcus sanguis and Streptococcus
faecalis: inhibition by chitin oligosaccharides. Infect Immun. 48: 720–728.
Lakowicz JR, Weber G. 1973. Quenching of fluorescence by oxygen. Probe for structural
fluctuations in macromolecules. Biochemistry. 12: 4161‐4170.
Lakowicz JR. Principles of Fluorescence Spectroscopy, Springer, New York, 3rd edn., 2006, ch.
8, pp. 278‐292.
Laurent C, Besancon P, Caporiccio B. 2007. Flavonoids from a grape seed extract interact
with digestive secretions and intestinal cells as assessed in an in vitro digestion/Caco‐2 cell
culture model. Food Chem. 100: 1704–1712.
Lee HJ, Jeong YI, Lee TH, Jung ID, Lee JS, Lee CM, Kim JI, Joo H, Lee JD, Park YM. 2007.
Rosmarinic acid inhibits indoleamine 2,3‐dioxygenase expression in murine dendritic cells.
Biochem Pharmacol. 73: 1412‐21.
Lee J, Kim YS, Park D. 2007. Rosmarinic acid induces melanogenesis through protein kinase A
activation signaling. Biochem Pharmacol. 74: 960–968.
Li P, Wang GJ, Li J, Hao HP, Zheng CN. 2006. Identification of tanshinone IIA metabolites in
rat liver microsomes by liquid chromatography‐tandem mass spectrometry. J Chromatogr A.
1104:366‐369.
Li Q‐L, Li B‐G, Zhang Y, Gao X‐P, Li C‐Q, Zhang G‐L. 2008. The angiotensin‐converting enzyme
inhibitors from Rbdosia coetsa. Phytomedicine. 15: 386–388.

References
144
Li Y, He WY, Liu H, Yao X, Hu Z. 2007. Daidzein interaction with human serum albumin
studied using optical spectroscopy and molecular modeling methods, J Mol Struct. 831: 144–
150.
Lin C‐S, Kuo C‐L, Wang J‐P, Cheng J‐S, Huang Z‐W, Chen C‐F. 2007. Growth inhibitory and
apoptosis inducing effect of Perilla frutescens extract on human hepatoma HepG2 cells. J
Ethnopharmacol. 112: 557–567.
Liu G, Ruedi P. 1996. Phyllocladanes (13b‐kauranes) from Plectranthus ambiguus.
Phytochemistry. 41: 1563–1568.
Liu Y, Hu M. 2002. Absorption and metabolism of flavonoids in the caco‐2 cell culture model
and a perused rat intestinal model. Drug Metab Dispos. 30: 370‐377.
Lorenzi H, Matos FJA. 2002. Plantas medicinais no Brasil ‐ nativas e exóticas, 2nd Edition.
Instituto Plantarum de Estudos da Flora, Nova Odessa, Brasil.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. 1951. Protein measurement with the Folin
phenol reagent, J. Biol. Chem. 193: 265–275.
Lukhoba CW, Simmonds MSJ, Paton AJ. 2006. Plectranthus: A review of ethnobotanical uses.
J Ethnopharmacol. 103: 1‐24.
Makino T, Furuta Y, Wakushima H, Fujii H, Saito K, Kano Y. 2003. Anti‐allergic effect of Perilla
frutescens and its active constituents. Phytother Res. 17: 240‐243.
Manach C, Williamson G, Morand C, Scalbert A, Rémésy C. 2005. Bioavailability and
bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am J Clin Nutr.
81: 230S‐242S.
Marais JPJ, Deavours B, Dixon RA, Ferreira D. 2006. The stereochemistry of flavonoids in The
Science of Flavonoids, ed. Grotewold E, Springer, New York, pp 1‐46.
Mata AT, Proença C, Ferreira AR, Serralheiro MLM, Nogueira JMF, Araújo MEM. 2007.
Antioxidant and antiacetylcholinesterase activities of five plants used as Portuguese food
spices. Food Chem. 103: 778–786.
McDougall GJ, Fyffe S, Dobson P, Stewart D. 2005. Anthocyanins from red wine‐‐their
stability under simulated gastrointestinal digestion. Phytochemistry. 66: 2540‐2548.
McDougall GJ, Fyffe S, Dobson P, Stewart D. 2007. Anthocyanins from red cabbage‐‐stability
to simulated gastrointestinal digestion. Phytochemistry. 68: 1285‐94.
McGleenon BM, Dynan KB, Passmore AP. 1999. Acetylcholinesterase inhibitors in
Alzheimer’s disease. Br. J. Clin. Pharmacol. 48: 471–480.
Mecocci P, Polidori MC. 2011.Antioxidant clinical trials in mild cognitive impairment and
Alzheimer's disease. Biochim Biophys Acta. doi:10.1016/j.bbadis.2011.10.006.

References
145
Meda L, Baron P, Scarlato G. 2001. Glial Activation in Alzheimer's disease: the role of Abeta
and its associated proteins. Neurobilogy of Aging, 22: 885‐893.
Meda L, Cassatella M, Szendrei G, Otvos L, Baron P, Villalba M, Ferrari D, Rossi F. 1995.
Activation of microglial cells by β‐amyloid protein and interferron‐γ. Nature, 374: 647‐650.
Meotti FC, Senthilmohan R, Harwood DT, Missau FC, Pizzolatti MG, Kettle AJ. 2008.
Myricitrin as a substrate and inhibitor of myeloperoxidase: implications for the
pharmacological effects of flavonoids. Free Radic Biol Med. 44: 109‐20.
Militello V, Casarino C, Emanuele A, Giostra A, Pullara F, Leone M. 2004. Aggregation kinetics
of bovine serum albumin studied by FTIR spectroscopy and light scattering. Biophys Chem.
107: 175‐187
Morand C, Crespy V, Manach C, Besson C, Demigné C, Rémésy C. 1998. Plasma metabolites
of quercetin and their antioxidant properties. Am J Physiol Regulatory Integrative Comp
Physiol. 275: 212‐219.
Mosmann, T. 1983. Rapid colorimetric assay for cellular growth and survival: Application to
proliferation and cytotoxicity assays. J. Immunol. Methods. 65: 55‐63.
Mukjerjee P K, Kumar V, Mal M, Houghton PJ. 2007. Acetylcholinesterase inhibitors from
plants. Phytomedicine. 14: 289–300.
Nabekura T, Yamaki T, Hiroi T, Ueno K, Kitagawa S. 2010. Inhibition of anticancer drug efflux
transporter P‐glycoprotein by rosemary phytochemicals. Pharmacol. Res. 61: 259‐263.
Nakazawa T, Ohsawa K. 1998. Metabolism of rosmarinic acid in rats. J. Nat. Prod. 61: 993–
996.
Ng SP, Wong KY, Zhang L, Zuo Z. 2005. Evaluation of the first‐pass glucoronidation of
selected flavones in gut by Caco‐2 monolayer model. J Pharm Pharm Sci. 8: 1‐9.
Ohnishi S, Takano K. 2004. Amyloid fibrils from the viewpoint of protein folding. Cell Mol Life
Sci. 61: 511‐24.
O'Leary KA, Day AJ, Needs PW, Mellon FA, O'Brien NM, Williamson G. 2003. Metabolism of
quercetin‐7‐ and quercetin‐3‐glucuronides by an in vitro hepatic model: the role of human β‐
glucuronidase, sulfotransferase, catechol‐O‐methyltransferase and multi‐resistant protein 2
(MRP2) in flavonoid metabolism. Biochem. Pharmacol. 65: 479–491.
Orhan I, Aslant S, Kartal M, Sener B, Baser KHC. 2008. Inhibitory effect of Turkish Rosmarinus
officinalis L. on acetylcholinesterase and butyrylcholinesterase enzymes. Food Chem. 108:
663–668.
Orhan I, Sener B, Choudhary M I, Khalid A. 2004. Acetylcholinesterase and
butyrylcholinesterase inhibitory activity of some Turkish medicinal plants. J Ethnopharmacol.
91: 57–60.

References
146
Osakabe N, Yasuda A, Natsume M, Yoshikawa T. 2004. Rosmarinic acid inhibits epidermal
inflammatory responses: anticarcinogenic effect of Perilla frutescens extract in the murine
two‐stage skin model. Carcinogenesis. 25: 549‐557.
Pádua BC, Silva LD, Rossoni Júnior JV, Humberto JL, Chaves MM, Silva ME, Pedrosa ML, Costa
DC. 2010. Antioxidant properties of Baccharis trimera in the neutrophils of Fisher rats. J
Ethnopharmacol. 129: 381‐386.
Parejo I, Caprai E, Bastida J, Viladomat F, Jauregui O, Codina C. 2004. Investigation of
Lepechinia graveolens for its antioxidant activity and phenolic composition. J
Ethnopharmacol. 94: 175–184.
Pederson JA. 2000. Distribution and taxonomic implications of some phenolics in the family
Lamiaceae determined by ESR spectroscopy. Biochem. Syst Ecol. 28: 229–253.
Percival SS, Turner RE. 2007. Applications of Herbs to Functional Foods. In: Handbook of
Nutraceuticals and Functional Foods, 2nd Ed. (Edited by R. E. C. Wildman), CRC Press, pp 269–
284.
Pereira P, Tysca D, Oliveira P, Brum LFS, Picada JN, Ardenghi P. 2005. Neurobehavioral and
genotoxic aspects of rosmarinic acid, Pharmacol. Res. 52: 199–203.
Petersen M, Simmonds MSJ. 2003. Rosmarinic acid. Phytochemistry. 62: 121–125.
Ponec RJ, Saunders MD, Kimmey MB. 1999. Neostigmine for the treatment of acute colonic
pseudo‐obstruction. N Engl J Med. 341: 137‐141.
Porfírio S, Falé PL, Madeira PJA, Florêncio MH, Ascensão L, Serralheiro MLM. 2010.
Antiacetylcholinesterase and antioxidant activities of Plectranthus barbatus tea, after in vitro
gastrointestinal metabolism. Food Chem. 122: 179‐87.
Probst RJ, Lim JM, Bird DN, Pole GL, Sato AK, Claybaugh JR. 2006. Gender differences in the
blood volume of conscious Sprague‐Dawley rats. J. Am. Assoc. Lab. Anim. Sci. 45: 49–52.
Randall D, Burggren W, French K. 2000. Eckert Animal Physiology, Fourth Edition. W.H.
Freeman and Company. New York, USA.
Rawel HW, Frey SK, Meidtner K, Kroll J, Schweigert FJ. 2006. Determining the binding
affinities of phenolic compounds to proteins by quenching of the intrinsic tryptophan
fluorescence. Mol Nutr Food Res. 50:, 705–713.
Regasini LO, Vellosa JC, Silva DH, Furlan M, de Oliveira OM, Khalil NM, Brunetti IL, Young MC,
Barreiro EJ, Bolzani VS. 2008. Flavonols from Pterogyne nitens and their evaluation as
myeloperoxidase inhibitors. Phytochemistry. 69: 1739‐44.
Rocchetti M, Besana A, Mostacciuolo G, Ferrari P, Micheletti R, Zaza A. 2003. Diverse toxicity
associated with cardiac Na+/K+ pump inhibition: evaluation of electrophysiological
mechanisms. J. Pharmacol. Exp. Ther. 305: 765‐771.

References
147
Rodney G, Swanson AL, Wheeler LM, Smith GN, Worrel CS. 1950. The effect of a series of
flavonoids on hyaluronidase and some other related enzymes. J Biol Chem. 183: 739–747.
Ronca F, Palmieri L, Panicucci P, Ronca G. 1998. Anti‐inflammatory activity of chondroitin
sulphate. Osteoarthritis Cartilage. 6: 14‐21.
Ross PD, Subramanian S. 1981. Thermodynamics of protein association reactions: forces
contributing to stability, Biochemistry. 20: 3096‐3102.
Salawu FK, Umar JT, Olokoba AB. 2011. Alzheimer's disease: A review of recent
developments. Ann Afr Med. 10: 73‐79.
Sanson B, Colletier JP, Xu Y, Lang PT, Jiang H, Silman I, Sussman JL, Weik M. 2011. Backdoor
opening mechanism in acetylcholinesterase based on X‐ray crystallography and molecular
dynamics simulations. Protein Sci. 20: 1114‐1118.
Sasho S, Obase H, Ichikawa S, Kitazawa T, Nonaka H, Yoshizaki R, Ishii A, Shuto K. 1995.
Synthesis of 2‐imidazolidinylidene Propanedinitrile Derivatives as Stimulators of
Gastrointestinal Motility – III. Bioorg Med Chem. 3: 279‐287.
Scalbert A, Williamson G. 2000. Dietary intake and bioavailability of polyphenols. J Nutr. 130:
2073S–2085S.
Schultz C, Bossolani MP, Torres LM, Lima‐Landman MT, Lapa AJ, Souccar C. 2007. Inhibition
of the gastric H+,K+ ‐ATPase by plectrinone A, a diterpenoid isolated from Plectranthus
barbatus Andrews. J Ethnopharmacol. 111: 1‐7.
Schutte ME, Boersma MG, Verhallen DA, Groten JP, Rietjens IM. 2008. Effects of flavonoid
mixtures on the transport of 2‐amino‐1‐methyl‐6‐phenylimidazo[4,5‐b]pyridine (PhIP)
through Caco‐2 monolayers: an in vitro and kinetic modeling approach to predict the
combined effects on transporter inhibition. Food Chem Toxicol. 46: 557‐566.
Sheng R, Lin X, Zhang J, Chol KS, Huang W, Yang B, He Q, Hu Y. 2009. Design, synthesis and
evaluation of flavonoid derivatives as potent AChE inhibitors. Bioorg Med Chem. 17: 6692–
6698.
Shi R, Qiao S, Yu D, Shi X, Liu M, Jiang X, Wang Q, Zhang L. 2011. Simultaneous determination
of five flavonoids from Scutellaria barbata extract in rat plasma by LC‐MS/MS and its
application to pharmacokinetic study. J Chromatogr B Analyt Technol Biomed Life Sci. 879:
1625‐1632.
Shimoi K, Okada H, Furugori M, Goda T, Takase S, Suzuki M, Hara Y, Yamamoto H, Kinae N.
1998. Intestinal absorption of luteolin and luteolin 7‐O‐beta‐glucoside in rats and humans,
FEBS Lett. 438: 220–224.
Silman I, Sussman JL. 2008. Acetylcholinesterase: How is structure related to function? Chem
Biol Interact. 175: 3‐10.

References
148
Sinko G, Calić M, Bosak A, Kovarik Z. 2007. Limitations of the Ellman method: cholinesterase
activity measurement in the presence of oximes. Anal Biochem. 370: 223‐227.
Spencer JP, Chaudry F, Pannala AS, Srai SK, Debnam E, Rice‐Evans C. 2000. Decomposition of
cocoa procyanindins in the gastric milieu. Biochem Biophys Res Commun. 27: 236–241.
Spencer JPE. 2003. Metabolism of tea flavonoids in the gastrointestinal tract. J Nutr. 133:
3255S–3261S.
Stuchbury G, Münch G. 2005. Alzheimer’s associated inflammation, potential drug targets
and future therapies. J Neural Transm. 112: 429‐453.
Tabart J, Franck T, Kevers C, Pincemail J, Serteyn D, Defraigne J‐O, Dommes J. 2011.
Antioxidant and anti‐inflammatory activities of Ribes nigrum extracts. Food Chem. In Press.
doi:10.1016/j.foodchem.2011.09.076
Tabet N. 2006. Acetylcholinesterase inhibitors for Alzheimer’s disease: anti‐inflammatories
in acetylcholine clothing! Age Ageing. 35: 336–338.
Tantipolphan R, Rades T, McQuillan AJ, Medlicott NJ. 2007. Adsorption of bovine serum
albumin (BSA) onto lecithin studied by attenuated total reflectance Fourier transform
infrared (ATR‐FTIR) spectroscopy, Int. J. Pharm. 337: 40–47.
Temple MD, Perrone GG, Dawes IW. 2005. Complex cellular responses to reactive oxygen
species. Trends in Cell Biology, 15: 319‐326
Tepe B, Daferera D, Sokmen A, Sokmen M, Polissiou M. 2005. Antimicrobial and antioxidant
activities of essential oil and various extracts of Salvia tomentosa Miller (Lamiaceae). Food
Chem. 90: 333–340.
Ticli FK, Hage LI, Cambraia RS, Pereira PS, Magro AJ, Fontes MR, Stábeli RG, Giglio JR, França
SC, Soares AM, Sampaio SV. 2005. Rosmarinic acid, a new snake venom phospholipase A2
inhibitor from Cordia verbenacea (Boraginaceae): Antiserum action potentiation and
molecular interaction. Toxicon. 46: 318–327.
Trouillas P, Calliste C‐A, Allais D‐P, Simon A, Marfak A, Delage C, Duroux J‐L. 2003.
Antioxidant, anti‐inflammatory and antiproliferative properties of sixteen water plant
extracts used in Limousin countryside as herbal teas. Food Chem. 80: 399–407.
Tsuji A, Takanaga H, Tamai I, Terasaki T. 1994. Transcellular transport of benzoic acid across
Caco‐2 cells by a pH‐dependent and carrier‐mediated transport mechanism. Pharm Res. 11:
30‐37.
Vaidyanathan JB, Walle T. 2003. Cellular uptake and efflux of the tea flavonoid
(‐)epicatechin‐3‐gallate in the human intestinal cell line Caco‐2. J Pharmacol Exp Ther. 307:
745‐52.
Varshney A, Sen P, Ahmad E, Rehan M, Subbarao N, Khan RH. 2010. Ligand binding strategies
of human serum albumin: How can the cargo be utilized? Chirality 22: 77–87.

References
149
Viña J, Lloret A, Ortí R, Alonso D. 2004. Molecular bases of the treatment of Alzheimer’s
disease with antioxidants: prevention of oxidative stress. Mol Aspects Med. 25:117‐123.
Vinutha B, Prashanth D, Salma K, Sreeja SL, Pratiti D, Padmaja R, Radhika S, Amit A,
Venkateshwarlu K, Deepak M. 2006. Screening of selected Indian medicinal plants for
acetylcholinesterase inhibitory activity. J Ethnopharmacol. 109: 359‐363.
Walgren RA, Walle UK, Walle T. 1998. Transport of Quercetin and Its Glucosides across
Human Intestinal Epithelial Caco‐2 Cells. Biochem Pharmacol. 55: 1721‐1727.
Walle UK, Galijatovic A, Walle T. 1999. Transport of the flavonoid chrysin and its conjugated
metabolites by the human intestinal cell line Caco‐2. Biochem Pharmacol. 58: 431‐438.
Wang Q, Morris ME. 2007. Flavonoids modulate monocarboxylate transporter‐1‐mediated
transport of gamma‐hydroxybutyrate in vitro and in vivo. Drug Metab Dispos. 35: 201–208.
Wang Y, Cao J, Zeng SJ. 2005. Involvement of P‐glycoprotein in regulating cellular levels of
Ginkgo flavonols: quercetin, kaempferol, and isorhamnetin. J Pharm Pharmacol. 57: 751–
758.
Weiss S, Klein R, Slivka A, Wei M. 1982. Chlorination of Taurine by Human Neutrophils. J Clin
Invest. 70: 598‐607.
Wong KK, Ngo JC, Liu S, Lin HQ, Hu C, Shaw PC, Wan DC. 2010. Interaction study of two
diterpenes, cryptotanshinone and dihydrotanshinone, to human acetylcholinesterase and
butyrylcholinesterase by molecular docking and kinetic analysis. Chem Biol Interact. 187:
335‐339.
Wu Y, Zhou C, Song L, Li X, Shi S, Mo J, Chen H, Bai H, Wu X, Zhao J, Zhang R, Hao X, Sun H,
Zhao Y. 2006. Effect of total phenolics from Laggera alata on acute and chronic
inflammation models, J Ethnopharmacol 108: 243–250.
Xiao J, Cao H, Chen T, Yang F, Liu C, Xu X. 2011. Molecular property‐binding affinity
relationship of flavonoids for common rat plasma proteins in vitro. Biochimie 93: 134–140.
Xiao J, Suzuki M, Jiang X, Chen X, Yamamoto K, Ren F, Xu M. 2008. Influence of B‐ring
hydroxylation on interactions of flavonols with bovine serum albumin, J. Agric. Food Chem.
56: 2350–2356.
Yamamoto Y, Takahashi Y, Kawano M, Iizuka M, Matsumoto T, Saeki S, Yamaguchi H. 1999.
In vitro digestibility and fermentability of levan and its hypocholesterolemic effects in rats. J
Nutr Biochem. 10: 13–18.
Yang Y, Hua Q, Fan Y, Shen H. 2008. Study on the binding of luteolin to bovine serum
albumin. Spectrochim Acta A Mol Biomol Spectrosc. 69: 432–436.
Yoshino K, Suzuki M, Saski K, Miyase T, Sano M. 1999. Formation of antioxidants from (‐)‐
epigallocatechin gallate in mild alkaline fluids, such as authentic intestinal juice and mouse
plasma. J Nutr Biochem. 10: 223–229.

References
150
Yu JQ, Liao ZX, Cai XQ, Lei JC, Zou GL. 2007. Composition, antimicrobial activity and
cytotoxicity of essential oils from Aristolochia mollissima. Environ Toxicol Pharmacol. 23:
162‐167.
Yuan J, lv Z, Liu Z, Hu Z, Zou G. 2007. Study on interaction between apigenin and human
serum albumin by spectroscopy and molecular modelling, J Photochem Photobiol A Chem.
191: 104–113.
Zeraik ML, Serteyn D, Deby‐Dupont G, Wauters J‐N, Tits M, Yariwake JH, Angenot L, Franck T.
2011.Evaluation of the antioxidant activity of passion fruit (Passiflora edulis and Passiflora
alata) extracts on stimulated neutrophils and myeloperoxidase activity assays. Food Chem.
128: 259–265
Zheng GZ, Bhatia P, Kolasa T, Patel M, El Kouhen OF, Chang R, Uchic ME, Miller L, Baker S,
Lehto SG, Honore P, Wetter JM, Marsh KC, Moreland RB, Brioni JD, Stewart AO. 2006.
Correlation between brain/plasma ratios and efficacy in neuropathic pain models of
selective metabotropic glutamate receptor 1 antagonists, Bioorg. Med. Chem. Lett. 16:
4936–4940.