universidade de lisboa faculdade de ciÊncias ... -...
TRANSCRIPT

1
UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
Aplicação do método de captura-recaptura de larvas fotoidentificadas para avaliação de parâmetros demográficos:
comparação entre anura e caudata
Joana Cristina Cardoso Teixeira Ribeiro
MESTRADO EM BIOLOGIA DA CONSERVAÇÃO
2009

2
UNIVERSIDADE DE LISBOA
FACULDADE DE CIÊNCIAS
DEPARTAMENTO DE BIOLOGIA ANIMAL
Aplicação do método de captura-recaptura de larvas fotoidentificadas para avaliação de parâmetros demográficos:
comparação entre anura e caudata
Joana Cristina Cardoso Teixeira Ribeiro
Dissertação orientada por:
Professor Doutor Rui Rebelo
Mestrado em Biologia da Conservação
2009

3
RESUMO
A conservação dos anfíbios passa impreterivelmente pela melhor
compreensão da sua biologia, demografia e ecologia. Frequentemente o estudo da
distribuição e dinâmica das populações de anfíbios envolve a utilização de métodos
de marcação individual artificial tanto em adultos como nas formas larvares.
Contudo, as técnicas mais utilizadas podem ter consequências negativas sobre os
indivíduos e, por conseguinte, sobre as populações - razão que leva à crescente
popularização de técnicas não-invasivas, como a fotoidentificação.
Neste trabalho recorreu-se pela primeira vez à fotoidentificação de larvas de
anura (Alytes cisternasii) e caudata (Salamandra salamandra) distribuídas ao longo
de um sistema de pegos de ribeira estimando-se com sucesso vários parâmetros
demográficos. De acordo com os dados obtidos, a probabilidade de sobrevivência
depende da quantidade de manta morta e da profundidade média do pego, atingindo
valores inesperadamente elevados. A probabilidade de sobrevivência das larvas de S.
salamandra variou mais entre pegos e mostrou-se dependente do grau de
ensombramento e da vegetação aquática do pego. Adicionalmente verificou-se que as
elevadas densidades não afectaram a probabilidade de sobrevivência e que as larvas
conseguem desenvolver-se em habitats lotados. Os resultados obtidos sugerem que as
larvas de A. cisternasii são muito resistentes a ataques predatórios, ao contrário das
larvas de S. salamandra, cuja morfologia as torna mais susceptível a sofrer ataques
letais.
A captura-recaptura de larvas de anfíbios fotoidentificadas assume-se como
um método viável e preferível para obter estimativas fiáveis de parâmetros
demográficos quando os indivíduos apresentam padrões distintos.
Palavras-chave: Alytes cisternasii, captura-recaptura, fotoidentificação, probabilidade
de sobrevivência, Salamandra salamandra, taxa de crescimento.

4
SUMMARY
Amphibian conservation is highly dependent on a better understanding of
these animals’ biology, demography and ecology. Often, studies on distribution and
population dynamics of amphibians require the use of artificial marking methods on
individuals. However, even the most popular marking techniques are nowadays
known for having negative effects on individuals and, consequently, on amphibian
populations – leading to the increasing search for non-invasive marking techniques,
such as photoidentification.
We successfully applied a mark-recapture method on Alytes cisternasii and
Salamandra salamandra larvae living along a system of stream pools and obtained
reliable estimates of various demographic parameters. Our results suggest that A.
cisternasii larvae’s survival is unexpectedly high and influenced by the pond’s
amount of leaf litter and average depth. S. salamandra larvae’s survival is less stable
and influenced by the pond’s shading and amount of aquatic vegetation. We also
proved that high densities do not seem to influence survival for either species,
showing that larvae can develop on crowded ponds. In addition, A. cisternasii larvae
are highly resilient to predator attacks, unlike S. salamandra larvae that, as a result of
their morphology, are more likely to die if preyed upon.
Mark-recapture of photoidentified amphibian larvae should be the chosen
method to obtain reliable estimates of demographic parameters when the individuals
display distinctive, unique patterns.
Keywords: Alytes cisternasii, growth rate, larvae, mark-recapture,
photoidentification, Salamandra salamandra, survival.

5
AGRADECIMENTOS
Agradeço à minha família sem a qual não seria ninguém, em especial à minha mãe e à minha avó que são as mulheres mais incríveis deste planeta.
Claro, ao Prof. Rui Rebelo, mastermind da ideia central deste trabalho.
Aos amigos do costume, ao Jójó, que tanto me ajudou e ajuda, ao Daniel, que está sempre lá para o melhor e para o pior, ao Porquinho, à Silvie e ao resto da “galera do mal”, pelo companheirismo, pela galhofa e toda a alegria!

6
ÍNDICE 1. Introdução
1.1. Importância ecológica dos anfíbios……………………………………….7 1.2. Estudos de captura-recaptura e a conservação de populações selvagens....7 1.2.1. O modelo Cormack-Jolly-Seber……….………………………..............9 1.2.2. O modelo Jolly-Seber.............................................................................10
1.3. O Programa MARK..............................................................................................................12
1.4. Marcação individual através de fotoidentificação...............................................................................................13 1.5. Factores……………………………………..…....……....…..…..............14
2. Materiais e métodos 2.1. Espécies em estudo……………………………………………......…......17 2.1.1. O Sapo-parteiro-ibérico……………………………….........….............17 2.1.2. A Salamandra-comum………………………………………................18 2.2. Área de estudo…….......……………………………………………........19 2.3. Datas e métodos de amostragem………………………………….….......20 2.4. Caracterização dos pegos……………………………………..…….........20 2.5. Fotoidentificação das larvas………………………………………….......21 2.6. Validação do método de identificação com o I3S……………………......22 2.7. Determinação das taxas de crescimento das larvas…………………........23 2.8. Avaliação dos indícios de predação……….…………………………......23
2.9. Determinação dos estádios de desenvolvimento larvar….……………......23 2.10. Modelação CR………..……………………………….………………....23 2.11. Outras análises estatísticas…..………………………….……………......26
3. Resultados 3.1. Caracterização dos pegos………………………………………….…......27 3.2. Capturas…………………………………………………………….........29 3.3. Validação do método de identificação com o I3S…………………..........30 3.4. Recapturas……………………………………………………………......32 3.5. Modelação CR…………………………………………………………....33
3.5.1. Alytes cisternasii………………………………………...…….…...33 3.5.1.1. Validação dos pressupostos dos modelos CR…………...…34 3.5.1.2. Modelos………………………………………………...…..34 3.5.1.3. Estimativas dos parâmetros demográficos……………...…..35
3.5.1.3.1. Probabilidades de sobrevivência………………….......35 3.5.1.3.2. Probabilidade de captura………………………….......38 3.5.1.3.3. Probabilidades de entrada e “nascimentos”……….......39 3.5.1.3.4. Abundâncias e densidades (total e por ocasião)…....…41
3.5.1.4. Indícios de predação…………………………………...…....44 3.5.1.5. Taxas de crescimento…………………………………..........46 3.5.1.6. Estádios de desenvolvimento…………………………..........46
3.5.2. Salamandra salamandra……………………………………….....49 3.5.2.1. Validação dos pressupostos dos modelos CR……………49 3.5.2.2. Modelos………………………………………………….50
3.5.2.3. Estimativas dos parâmetros demográficos……………………….51 3.5.2.3.1. Probabilidades de sobrevivência…………………………..51 3.5.2.3.2. Probabilidade de captura…………………………………..53 3.5.2.3.3. Probabilidades de entrada e “nascimentos”………………..54

7
3.5.2.3.4. Abundâncias e densidades (total e por ocasião)…………...55 3.5.2.4. Indícios de predação……………………………………………...57 3.5.2.5. Taxas de crescimento…………………………………………….58
3.6. Movimentos entre pegos……………………………………………………58 4. Discussão….……………………………………………………………………..60 5. Referências Bibliográficas……………………………………………………….60

8
1. NTRODUÇÃO
1.1. Importância ecológica dos anfíbios
Foi em 1989, no I Congresso Mundial de Herpetologia, que a comunidade
científica reconheceu que as populações de anfíbios estavam em declínio, a nível
global [1]. Desde então, têm sido avançadas várias causas para este fenómeno,
nomeadamente o aumento da radiação ultravioleta [2], as alterações climáticas [3], a
degradação de habitats [4], as epidemias, a poluição [4], e mais frequentemente, a
interacção entre alguns destes factores [1].
A importância do declínio dos anfíbios não se prende apenas com questões
éticas relacionadas com o valor intrínseco deste grupo taxonómico; prende-se
sobretudo com o papel importantíssimo que desempenham na dinâmica trófica dos
ecossistemas e com o seu valor como bioindicadores de stress ambiental [1]. Assim,
a urgência da conservação destes vertebrados passa pela promoção de estudos que
esclareçam a importância das variáveis bióticas e abióticas na dinâmica das
populações naturais [5].
Uma vez que estes estudos envolvem frequentemente a marcação de formas
larvares e que esta pode ser particularmente complexa, as investigações nesta área
são geralmente limitadas a ambientes laboratoriais cuja simplicidade intrínseca não
permite que reproduzam fielmente as tendências das populações nos meios naturais
[6].
Para travar o declínio dos anfíbios há que procurar estudar as populações in
situ, desenvolvendo e aperfeiçoando os métodos e delineamentos experimentais
necessários.
1.2. Estudos de captura-recaptura (CR) e a conservação de populações
selvagens
A monitorização de populações biológicas tem recebido cada vez mais atenção
em muitos países [7]. O recurso a indivíduos marcados e o desenvolvimento dos
métodos de delineamento e modelação de dados provenientes de estudos de captura-
recaptura (CR) constituem ferramentas muito importantes neste processo. A
obtenção de estimativas de abundância fiáveis é de extrema relevância para o estudo,

9
monitorização e conservação de populações selvagens em geral e de populações de
anfíbios em particular [8].
Apesar da compreensão da dinâmica de populações animais depender
frequentemente da análise de parâmetros biológicos, é virtualmente impossível obter
estimativas para a sobrevivência ou abundância directamente através do seguimento
de indivíduos até à sua morte [9]. Assim, a análise quantitativa de parâmetros
populacionais passa pelo delineamento de estudos CR, instrumentos-chave na
estimação de abundâncias e de diversos parâmetros demográficos como a
sobrevivência, recrutamento e dispersão [10; 11].
Durante a última década, o principal objectivo da análise de dados CR passou
da estimação da dimensão da população para a estimação da sobrevivência, por duas
razões: (1) a sobrevivência varia com características individuais (idade, sexo, massa
corporal, etc) e em função de variáveis bióticas e abióticas, logo, os testes de
hipóteses relacionados com a estimação de taxas de sobrevivência revelaram-se mais
importantes para o conhecimento da dinâmica das populações do que a estimação da
abundância [11]; e (2) os estimadores da sobrevivência são substancialmente mais
robustos a pequenos desvios ao cumprimento dos pressupostos do que os estimadores
da abundância [11].
A maior diferença entre as técnicas de CR e as técnicas tradicionais de
amostragem reside no facto dos animais serem amostrados diversas vezes, sendo
inicialmente capturados e marcados para posterior identificação em eventos de
amostragem subsequentes (recapturas) [9; 10].
Os modelos CR são frequentemente classificados em dois grupos: modelos
para populações “abertas” e modelos para populações “fechadas” [10; 11]. Os
modelos para populações fechadas consideram a não ocorrência de nascimentos,
mortes, emigração ou imigração [12; 13]. O principal objectivo da utilização destes
modelos é a estimação da dimensão da população. Os pressupostos associados a estes
modelos incluem a presunção de que durante todo o período de amostragem a
população se encontra demográfica e geograficamente fechada [10]. Assim, não é
permitida a inclusão (nascimentos ou emigração) ou remoção (morte ou imigração)

10
de qualquer indivíduo da população em estudo. Estes modelos admitem que a
probabilidade de sobrevivência é máxima e constante ao longo dos curtos intervalos
definidos no estudo [13; 14].
Os modelos para populações “abertas” admitem a ocorrência de nascimentos, mortes,
emigrações ou imigrações [12; 13]. Os animais são capturados em k ocasiões e
marcados de forma única. Após a ocasião inicial, tanto animais marcados como não
marcados são capturados. Nas sessões subsequentes, os indivíduos não marcados
capturados são nesse momento marcados e posteriormente libertados.
Os primeiros métodos de análise de dados CR baseavam-se em modelos ad hoc
para a sua interpretação. No entanto, no final da década de 1960 o método da
máxima verosimilhança (Maximum Likelihood), conhecido por produzir estimativas
com boas propriedades para uma vasta gama de condições [15], serviu como base
para o cálculo de estimadores de parâmetros e da variância e covariância de
delineamentos CR [16]. Este método tem duas etapas: (1) construção do modelo que
dá a probabilidade de observar os dados considerados em função dos parâmetros
desconhecidos de interesse - função da máxima verosimilhança; e (2) obtenção das
estimativas dos parâmetros desconhecidos que maximizam a função verosimilhança
[13].
Neste trabalho optou-se por recorrer a modelos CR “abertos”, nomeadamente
ao modelo Cormack-Jolly-Seber (CJS) e ao modelo Jolly–Seber (JS) por se
adaptarem à biologia das espécies em estudo e facultarem estimativas dos parâmetros
demográficos pretendidos.
1.2.1. O Modelo Cormack-Jolly-Seber
O modelo Cormack-Jolly-Seber (CJS), inicialmente desenvolvido por Cormack
[17] e posteriormente por Jolly [18] e Seber [19], constitui um dos modelos mais
usados em estudos CR [20]. Este modelo permite o cálculo das probabilidades de
captura (pi) e sobrevivência aparente (Φi) condicionais às libertações feitas ao longo
das ocasiões de marcação [13]
Como em todos os modelos CR “abertos”, o modelo CJS estima apenas a
sobrevivência aparente, i.e., 1- Φi representa quer animais que morreram quer

11
animais que deixaram a população (emigração) e em geral, Φi ≤ S, sendo S a
verdadeira probabilidade de sobrevivência [11; 14].
Tal como qualquer modelo CR, o modelo CJS exige o cumprimento de
alguns pressupostos para assegurar a validade das suas estimativas – muitos dos
quais são comuns a todos os modelos CR [13; 14; 16]:
1. Qualquer animal marcado presente na população no tempo de amostragem i
tem a mesma probabilidade pi de ser recapturado ou re-identificado;
2. Qualquer animal marcado presente na população imediatamente a seguir à
ocasião de amostragem i tem a mesma probabilidade Φi de sobreviver até à
ocasião de amostragem i+1;
3. As marcas não são perdidas nem se tornam indetectáveis e não afectam o
comportamento ou o destino dos animais marcados;
4. Os períodos de amostragem são instantâneos, ou seja, durante o processo de
amostragem não ocorrem nascimentos, mortes, imigração ou emigração, e os
animais recapturados são libertados imediatamente;
5. Toda a emigração da área é permanente;
6. O destino de cada animal no que diz respeito às probabilidades de captura e
sobrevivência é independente do destino de qualquer outro animal; e
7. Os parâmetros estimados para a população marcada podem ser aplicados à
população não marcada no estudo.
A realização de testes de ajustamento do modelo geral é de grande utilidade
na verificação do cumprimento de alguns destes pressupostos.
De todos os modelos para populações “abertas”, este é considerado o mais
robusto à presença de heterogeneidade nos dados [10].
1.2.2. O Modelo Jolly-Seber
Ao contrário do modelo CJS, no modelo JS [18; 19] os animais não marcados
capturados na ocasião i são considerados para o cálculo dos parâmetros. Os
parâmetros estimados incluem para além da sobrevivência aparente (Φi) e da
probabilidade de captura (pi), a dimensão da população (N) e o número de entradas
em cada ocasião (B) [14] Apesar de se tratar de um modelo biologicamente mais

12
apelativo, o facto de ter muitos a parâmetros a estimar, pode resultar em estimativas
pouco satisfatórias se os dados forem escassos [14].
À semelhança do modelo CJS, o modelo JS também exige o cumprimento de
alguns pressupostos para assegurar a validade da sua aplicação. Estes pressupostos
são semelhantes aos do modelo CJS à excepção do 2º - no modelo JS a assumpção da
homogeneidade de probabilidade de sobrevivência estende-se aos indivíduos não
marcados, possibilitando assim a estimação do parâmetro para todos os indivíduos
pertencentes à população [10; 14].
Todos os estimadores do modelo JS se baseiam na estimação da dimensão da
população marcada, Mi. Como estes modelos consideram a ocorrência de saídas da
população (morte e emigração), Mi tem obrigatoriamente de ser estimado. Assim,
equacionam-se as probabilidades de captura futuras de dois grupos distintos de
animais presentes na população na ocasião i: (1) os que já foram marcados mas não
foram capturados em i (Mj - mj); e (2) os que são capturados e libertados em i+1 (Ri).
Assumindo a homogeneidade de captura, as probabilidades de captura futuras destes
dois grupos devem ser equivalentes. Assim, sendo zi e ri os membros de Mi - mi e Ri
que são recapturados posteriormente, então
���� − �� ≈ ����
e
�� = �� + ������
para i = 2,…,k-1.
A estimativa para Φi obtém-se a partir da razão entre o número de indivíduos
marcados presentes em i+1 e os presentes em i,
�� = ������ + �� − ��
em que i = 1,…,k-2 e o termo Ri-mi representa o número de indivíduos recém-
marcados libertados em i. Assim, os indivíduos que eventualmente entrem na

13
população no intervalo referido e morram antes da sessão de captura i+1 são
excluídos. No entanto, este estimador não consegue discernir as origens das entradas;
considera conjuntamente entradas por nascimentos in-situ e por imigração.
A probabilidade de captura, pi, é estimada através da razão entre o número de
animais marcados capturados em i (mi) e o número de indivíduos marcados presentes
na população em i (Mi),
�̂� = ����
para i = 2,3,…,k-1.
A dimensão populacional na ocasião i pode-se determinar através das
proporções entre indivíduos marcados e não marcados e a totalidade de animais,
���� ≈ ����
ou seja,
�� = ������
ou
�� = ���̂�
em que ni representa o número de animais capturados em cada sessão de
amostragem, mi dos quais já se encontravam marcados.
A estimação dos nascimentos, Bi, é obtida através da diferença entre a
dimensão da população em i + 1 e i, considerando as perdas por morte natural (1-Φi)
e a probabilidade de captura (ni - Ri). Assim,
��� = ���� − ��[�� − (�� − ��)]

14
para i = 2,…,k-2. Os valores de Bi traduzem o valor líquido de entradas para a
população entre as sessões de amostragem i e i+1.
1.2.3. Programa MARK
O desenvolvimento e acessibilidade a software eficaz na análise de dados de
CR permitiram a proliferação de estudos que exploram esta metodologia [21].
Para a estimação das taxas de sobrevivência, a utilização dos modelos
lineares generalizados (GLM) como suporte ao estudo de modelos de CR para
populações abertas e a disponibilização de software flexível como os programas
SURGE [22] e MARK [13] popularizou esta poderosa metodologia [21]. O programa
MARK oferece ao utilizador um software para Windows de fácil compreensão,
permitindo a análise de uma vasta gama de modelos [12]. Actualmente, este
programa oferece 47 modelos CR, permitindo, através de um grande número de
análises, a obtenção das estimativas dos parâmetros e da sua precisão [23]. A análise
destes modelos inclui a construção de gráficos das estimativas dos parâmetros, a
construção de modelos a partir de matrizes de delineamento para estimar o conjunto
dos parâmetros de interesse biológico, a aplicação de medidas de correcção para a
dispersão dos dados, o uso de metodologias para avaliar o ajustamento do modelo e
as ferramentas necessárias para a selecção do melhor modelo [13].
Pelas razões supramencionadas optou-se por utilizar o programa MARK para
a análise dos dados de CR relativos às formas larvares de anfíbios.
1.3. Marcação individual através de fotoidentificação
Nos modelos CR as estimativas da probabilidade de sobrevivência são
relativamente robustas à heterogeneidade de captura. No entanto, se a forma de
marcação influenciar a sobrevivência ou se as marcas desaparecerem ou forem
difíceis de detectar, as estimativas de Φi podem ficar severamente subestimadas e a
abundância sobrestimada [24]. Assim, a escolha do método de marcação utilizado no
estudo CR assume-se como uma etapa muito importante, determinando
frequentemente a validade das estimativas produzidas [10; 13].

15
A utilização bem sucedida de características corporais naturais na
identificação de animais em ambientes terrestres e aquáticos para uma série de
espécies tornou a fotoidentificação uma técnica relativamente popular,
principalmente com o advento da fotografia digital [25]. Esta técnica envolve o
recurso a marcas físicas, padrões ou colorações para distinguir indivíduos
conspecíficos de forma não invasiva, duradoura, prática e económica [26]. As
principais condições para que este método seja fiável prendem-se com a qualidade,
resolução, nitidez, estabilidade do ângulo e luminosidade da fotografia [27].
Apesar de ser muito utilizado em várias espécies animais [26], a
fotoidentificação não é um método comummente usado em anfíbios. Os poucos
estudos CR com indivíduos fotoidentificados concentraram-se na fase juvenil/adulta
[28; 29] e não existe qualquer estudo que tenha aplicado a fotoidentificação a formas
larvares. A marcação de girinos passa tradicionalmente pela aplicação de técnicas de
marcação individual artificial como o fin clipping (Turner, 1960 in [9]), a imersão em
soluções corantes (Guttman & Creasey, 1973 in [9]) e a injecção de corantes
orgânicos nos girinos [30]. Infelizmente estes métodos de marcação são
frequentemente pouco praticáveis por razões económicas, logísticas, éticas ou ainda
por poderem afectar a probabilidade de recaptura ou sobrevivência dos indivíduos,
violando os pressupostos dos métodos CR [27]. Não existem estudos bem sucedidos
da ecologia das formas larvares de anfíbios que recorram à fotoidentificação porque
o tamanho das larvas, associado à ocorrência de alterações nos padrões
identificativos, reduz a aplicabilidade do método. No entanto, esta técnica pode vir a
assumir-se como uma alternativa viável e preferível para espécies cujos girinos
possuam marcas individuais e de fácil identificação.
A identificação manual de fotografias pode ser difícil, morosa e propensa a
erros de identificação [27], pelo que as metodologias mais fiáveis até ao momento
recorrem a programas de análise de imagem como o I3S. Este programa foi
desenvolvido inicialmente para identificar indivíduos de Carcharias taurus (tubarão-
touro) [31], recorrendo ao padrão de manchas que esta espécie apresenta e baseia-se
na análise de padrões de pontos. O I3S depende do utilizador para construir as bases a
partir das quais faz as comparações. No início, o utilizador aponta as características
mais distintas na foto do indivíduo desconhecido, procurando distinguir as marcas de
outro “ruído” que exista na fotografia (reflexos, sombras, partículas em suspensão).

16
De seguida, o software compara automaticamente a imagem em questão com todas
as outras imagens que se encontram na base de dados e faculta uma lista ordenada de
imagens. Por fim, o utilizador é responsável por verificar a validade das
correspondências entre a imagem desconhecida e uma imagem da base de dados,
inspeccionando visualmente as potenciais correspondências [32]. Esta abordagem
aumenta a precisão, reduz o tempo de processamento e permite analisar bases de
dados de dimensões consideráveis [31].
1.4. Factores que influenciam a sobrevivência e robustez das larvas de
anfíbios
A dinâmica das populações larvares de anfíbios é influenciada por variáveis
abióticas e bióticas que actuam em sinergia, produzindo as condições a que os
animais se encontram expostos [33]. A performance das formas larvares pode
influenciar os efectivos da espécie, pelo que a gestão e conservação dos anfíbios tem
de passar pelo estudo das fases iniciais do seu complexo desenvolvimento [34; 35;
36].
O hidroperíodo determina o tempo que as larvas dispõem para se desenvolver
até poderem subsistir em meio terrestre, bem como a densidade e natureza dos
competidores e predadores a que estão expostas [34; 37]. Peixes, insectos aquáticos e
tritões são os predadores de larvas e ovos de anfíbios mais importantes [38]. De uma
forma geral, o número de espécies predadoras e a abundância total de predadores
aumenta com o hidroperíodo porque, à excepção dos insectos aquáticos, que são
ubíquos, os predadores evitam meios aquáticos efémeros [33; 38].
Em zonas húmidas florestadas, a distribuição de vários invertebrados [39] e
anfíbios [35] está em geral relacionada com o coberto vegetal. A vegetação ripícola
influencia a disponibilidade alimentar, a quantidade de oxigénio dissolvido na água,
a incidência de luz solar e a temperatura da água [40]. De uma forma geral, zonas
húmidas ensombradas apresentam temperaturas baixas, baixas concentrações de
oxigénio dissolvido e menor produtividade primária do que zonas com menor grau
de ensombramento [41]. O coberto vegetal influencia também a entrada de material
vegetal no corpo de água, podendo alterar dramaticamente as cadeias tróficas

17
aquáticas. De uma forma geral, as zonas húmidas ensombradas recebem material
alóctone principalmente sob a forma de matéria vegetal senescente. Em zonas
húmidas menos ensombradas, os maiores níveis de energia solar permitem o
crescimento de vegetação herbácea, incluindo macrófitos submersos e emergentes
que dominam os substratos bênticos [41]. A natureza do material vegetal senescente
que entra no corpo de água pode igualmente influenciar as cadeias tróficas aquáticas
[42]. Pegos com manta morta de origem herbácea podem poduzir uma produtividade
primária mais elevada e metamórficos mais precoces e robustos do que pegos com
manta morta lenhosa, tendência que resulta da maior quantidade e/ou qualidade de
recursos alimentares para as larvas, tendência que resulta da maior quantidade e/ou
qualidade de recursos alimentares para as larvas [40; 41]. Material herbáceo
decompõe-se a um ritmo mais rápido do que material lenhoso em sistemas aquáticos
[43], aumentando a disponibilidade de nutrientes e facilitando o crescimento
microbiano. Por outro lado, os ácidos húmicos e taninos libertados na água por
matéria lenhosa senescente atenuam a radiação fotossinteticamente activa e afectam
a produtividade do sistema e consequentemente da performance larvar [40; 41]. A
vegetação aquática aumenta a complexidade estrutural do habitat e reduz a
frequência de contactos entre predadores e presas, podendo fornecer alguma
protecção aos girinos (Luckinbill, 1973 in [44]).
As dimensões e forma da massa de água são também importantes para o
sucesso reprodutor dos anfíbios porque influenciam o hidroperíodo, os micro-
habitats disponíveis para as larvas e ainda a complexidade do habitat [38; 45].
A densidade larvar pode ter uma influência indirecta na sobrevivência das
larvas através da diminuição da disponibilidade de recursos alimentares do aumento
da pressão canibalista em algumas espécies, podendo resultar na diminuição da taxa
de desenvolvimento larvar, diminuindo também a probabilidade de completarem a
metamorfose [37; 46; 47].
A sobrevivência e robustez dos indivíduos recém-metamorfoseados pode ter
consequências para a população adulta. Vários estudos [36; 48; 49] indicam que a
sobrevivência dos juvenis é maior para indivíduos que metamorfoseiam mais cedo ou
maiores. Este efeito pode simultaneamente reduzir directamente a mortalidade no
primeiro ano de vida e antecipar a idade da primeira reprodução [48].

18
As alterações na sobrevivência e performance de larvas de anfíbios têm
potencial para induzir cascadas de alterações nas comunidades das zonas húmidas.
Os anfíbios são consumidores abundantes nestes ecossistemas e, no caso das
salamandras, são predadores de topo [50]. Os recursos consumidos pelas larvas dos
anfíbios em zonas húmidas são transferidos para os habitats terrestres depois da
metamorfose [51]. Assim, diferenças na sobrevivência e robustez das larvas afectam
directamente a biomassa que entra nos ecossistemas terrestres já que regulam o
recrutamento de juvenis.
Este trabalho pretende estimar a qualidade de vários tipos de pegos de uma
ribeira temporária para a reprodução do sapo-parteiro-ibérico (Alytes cisternasii) e da
salamandra-comum (Salamandra salamandra). Serão estudados os efeitos das
principais características dos pegos na abundância, sobrevivência e desenvolvimento
das formas larvares destas espécies. Este tipo de estudo é frequente para
comunidades piscícolas [52], mas não para larvas de anfíbios devido às dificuldades
inerentes à obtenção de estimativas fiáveis para os parâmetros demográficos. Como
as larvas de anfíbios podem ser muito abundantes, sendo difícil estimar as suas
densidades e sobrevivência até à metamorfose, aplicar-se-á pela primeira vez o
método de fotoidentificação para as seguir individualmente.
Para cada pego considerado pretende-se estimar:
� Densidade (indivíduos/m2);
� Taxas de crescimento individuais e
� Taxa de sobrevivência até à metamorfose.
Por fim, relacionar-se-ão os resultados obtidos com as características de cada
corpo de água para analisar quais favorecem a produção de metamórficos destas
espécies.

19
2. MATERIAIS E MÉTODOS
2.1. Espécies em estudo
2.1.1. O sapo-parteiro-ibérico (Alytes cisternasii Boscá, 1879)
O sapo-parteiro-ibérico é um endemismo da Península Ibérica e em Portugal
encontra-se maioritariamente a Sul do Tejo, estendendo-se para Norte ao longo da
fronteira até ao extremo Este do Parque Natural de Montesinho [53]. Esta espécie é
comummente encontrada em climas áridos e quentes, associada a habitats
mediterrânicos de florestas abertas de Quercus ilex e Q. suber [54].
É um anfíbio com hábitos nocturnos e fossadores, podendo passar por vários
períodos de inactividade durante os meses com temperaturas mais extremas [55].
Este anuro reproduz-se no Outono e na Primavera [54], períodos durante os quais os
machos vocalizam durante a noite, principalmente depois do pôr-do-sol. Depois da
fecundação, os machos enrolam os cordões de ovos nos membros posteriores,
podendo transportar as posturas de várias fêmeas, até um máximo de 180 ovos [53].
Após três semanas, os machos deslocam-se até uma massa de água, onde os ovos
eclodem sincronizadamente, independentemente da altura em que as posturas foram
recolhidas [54].
As larvas podem alcançar os 70 mm de comprimento e possuem manchas
características concentradas na parte superior e inferior da cauda. O período larvar
pode durar 3 - 5 meses e os indivíduos recém-metamorfoseados são muito
semelhantes aos adultos [53]. As larvas alimentam-se principalmente de matéria
vegetal e de invertebrados aquáticos. Os principais predadores das larvas de A.
cisternasii são as cobras de água, os escaravelhos aquáticos (adultos e larvas), as
ninfas de libélula e as larvas de salamandra [53].
2.1.2. A salamandra-comum (Salamandra salamandra Linnaeus, 1758))
A salamandra-comum é um urodelo de ampla distribuição geográfica,
ocupando grande parte do continente europeu. Ocorre em todo o território nacional
continental, à excepção das zonas agrícolas do Baixo Alentejo [53].
A sua actividade é quase exclusivamente nocturna e está muito dependente de
condições ambientais favoráveis, nomeadamente humidade relativa elevada,
temperaturas amenas e ausência de vento [53]. A sua actividade anual está

20
concentrada nos meses mais húmidos, normalmente entre Setembro e Maio, período
em que ocorre a reprodução [54]. Trata-se de uma espécie ovovivípara ou vivípara,
podendo as fêmeas depositar entre 20 e 120 larvas recém-eclodidas na água [53].
Reproduzem-se preferencialmente em meios com águas limpas e correntes, tais como
ribeiros, fontes e minas [54].
As larvas têm aspecto robusto, cabeça grande e larga com brânquias externas
plumosas bem desenvolvidas, e a cauda mais curta do que o corpo, com terminação
arredondada [53]. As larvas recém-depositadas na água têm 25 - 38 mm de
comprimento e possuem já os quatro membros bem desenvolvidos. Inicialmente são
acinzentadas ou acastanhadas com um ponteado escuro mas à medida que se vão
desenvolvendo as manchas escuras tornam-se mais evidentes e na altura da
metamorfose tornam-se negras e as suas manchas brancas ficam amarelas. Em geral
as larvas atingem a metamorfose 2-6 meses após o nascimento [53].
Os adultos alimentam-se de invertebrados terrestres e as larvas são
particularmente vorazes e alimentam-se principalmente de artrópodes aquáticos [53].
Este caudata é predado por cobras-de-água, víboras e, ocasionalmente,
algumas aves. As formas larvares são frequentemente predadas por trutas, cobras-de-
água, insectos aquáticos, aves aquáticas e larvas da própria espécie [55].
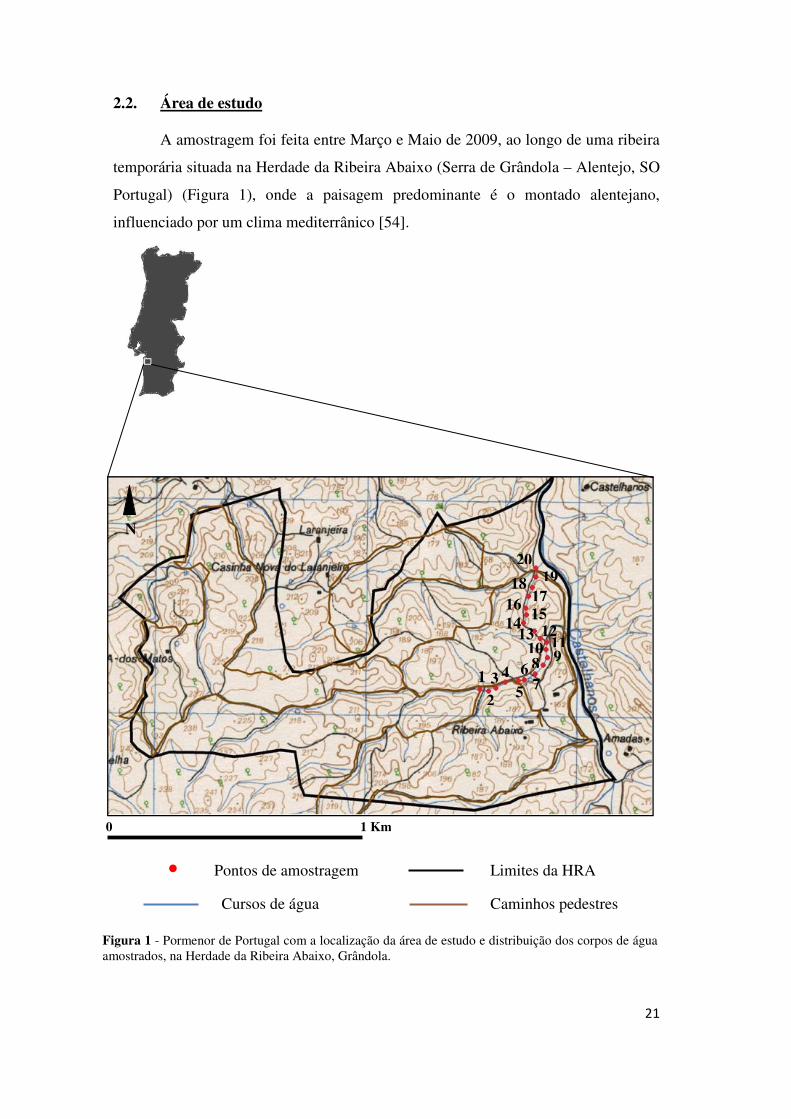
21
2.2. Área de estudo
A amostragem foi feita entre Março e Maio de 2009, ao longo de uma ribeira
temporária situada na Herdade da Ribeira Abaixo (Serra de Grândola – Alentejo, SO
Portugal) (Figura 1), onde a paisagem predominante é o montado alentejano,
influenciado por um clima mediterrânico [54].
• Pontos de amostragem
Cursos de água
Limites da HRA
Caminhos pedestres
N
Figura 1 - Pormenor de Portugal com a localização da área de estudo e distribuição dos corpos de água amostrados, na Herdade da Ribeira Abaixo, Grândola.
0 1 Km
1 7
13 15
19
2 3 4
5
6 8 9 10 11 12 14
16 17 18
20

22
2.3. Datas e métodos de amostragem
Amostraram-se 20 pegos entre 9 de Março e 6 de Maio de 2009, com 6
sessões de amostragem de larvas de A. cisternasii e 5 de larvas de S. salamandra.
A amostragem foi feita em média durante 20 minutos por pego, usando um
camaroeiro de rede de cor verde com 1 mm de malha. Depois de capturadas, todas as
larvas foram fotografadas no local. Cada larva foi colocada numa caixa de acrílico (C
6,5 cm x A 5,5 cm x P 1 cm) com escala milimétrica onde o lado direito do seu corpo
foi fotografado pelo menos 2 vezes com uma máquina fotográfica Canon EOS
1000D, objectiva Canon EFS 18-55mm.
No final de cada sessão, as fotografias foram transferidas para o computador,
renomeadas com um código individual, editadas e organizadas num álbum digital.
2.4. Caracterização dos pegos
A caracterização dos pegos foi feita dia 21 de Abril de 2009, sensivelmente a
meio do período total de amostragem. Foram medidas as dimensões de cada pego
(largura, comprimento e profundidade máxima). Foi também estimado o grau de
ensombramento, a percentagem de vegetação aquática, e a composição do substrato
de cada pego utilizando o método de quadrados pontuais [56]. Para tal utilizou-se
uma estrutura semelhante a um pente, com 1m de comprimento e 10 “pregos”,
separados por intervalos de 10 cm. Após colocar o “pente” dentro de água, foi
registado quantos “pregos” entravam em contacto com vegetação aquática, com
cascalho, gravilha, areia, argila e manta morta e ainda quantos se encontravam
ensombrados. Este procedimento foi repetido várias vezes ao longo de cada pego, em
pontos distanciados por 1 m. No final calculou-se a percentagem de cobertura destas
variáveis a partir da frequência de contacto com os “pregos”, para a totalidade dos
pontos de cada pego.
Procedeu-se ainda à medição do pH, da condutividade, da salinidade, da
quantidade de oxigénio dissolvido e da temperatura utilizando um medidor
multiparamétrico das características físico-químicas da água (YSI, modelo 556
MPS).

23
2.5. Fotoidentificação das larvas
A comparação fotográfica foi realizada através do programa I3S (Interactive
Individual Identification System), v. 2.0.
Antes de proceder a comparações e correspondências, o I3S requer que o
utilizador defina um sistema de referência composto por três pontos comuns a todas
as imagens. Para além de definir a área de referência para a marcação das manchas
corporais que permitirão a identificação individual, o sistema de referência
possibilita ainda operações que maximizam o processo de correspondência como
transformações de escala, rotação e correcção de perspectiva [31; 32].
De seguida, o utilizador escolhe as manchas mais distintas do indivíduo,
formando o fingerprint que é guardado num ficheiro com a extensão .fgp [31; 32] na
mesma directoria onde se encontra a fotografia. Posteriormente, a imagem poderá ser
inserida na base de dados e/ou ser comparada com as imagens existentes.
Ao iniciar um processo de comparação, o fingerprint da imagem
desconhecida é comparado com todos os fingerprints das imagens que se encontram
na base de dados. Deste processo resulta uma lista de 50 possíveis correspondências
ordenadas por ordem crescente de um índice linear, sendo a correspondência mais
provável listada em primeiro lugar.
O índice linear que está na base das correspondências é calculado através da
razão da soma das distâncias entre cada par de pontos pelo quadrado do número de
pares de pontos. Assim, um par de pontos é aceite como um bom par se o candidato
mais próximo estiver, pelo menos, ao dobro da distância do par actual.
O sistema de referência utilizado para as larvas de A. cisternasii foi composto
pela intersecção ânus-cauda, pelo início da membrana caudal e pelo ponto em que a
membrana caudal atinge a altura máxima e foi testado previamente [57] (Figura 2). O
sistema de referência usado para as larvas de S. salamandra foi composto pela
intersecção do membro posterior com o corpo, pelo ponto paralelo a este, no dorso
da larva e pelo ponto que assinala o meio da cauda (Figura 3). Em ambas as espécies
assinalaram-se 24 pintas para a construção dos fingerprints [57].

Para cada indivíduo foram processadas duas fotos
cada correspondência listada até encontrar uma correspondência verdadeira ou até à
50ª posição.
2.6. Validação do método de
Determinou-se o índice linear, o
correspondências), o tempo levado a identificar
identificações nos tops 1, 10, 20, 30, 40 e 50
Verificou-se igualmente se o
desenvolvimento do indivíduo
temporalmente.
Figura 3 – Girino de A. cisternasii
assinalados.
Figura 2 - Larva de S. salamandra
assinalados.
uo foram processadas duas fotos, verificando-se
cada correspondência listada até encontrar uma correspondência verdadeira ou até à
Validação do método de fotoidentificação com o I3S
se o índice linear, o ranking (posição na listagem de 50
o tempo levado a identificar cada foto e a percentagem
identificações nos tops 1, 10, 20, 30, 40 e 50 da lista de 50 correspondências
se o ranking médio das re-identificações se alterou
desenvolvimento do indivíduo por comparação entre recapturas próximas e afastadas
A. cisternasii com as pintas e o sistema de referência para a fotoidentificação
S. salamandra com as pintas e o sistema de referência para a fotoidentificação
24
se visualmente
cada correspondência listada até encontrar uma correspondência verdadeira ou até à
(posição na listagem de 50
a percentagem de
da lista de 50 correspondências.
identificações se alterou com o
por comparação entre recapturas próximas e afastadas
para a fotoidentificação
com as pintas e o sistema de referência para a fotoidentificação

25
2.7. Determinação das taxas de crescimento das larvas
Registou-se o comprimento total das larvas de A. cisternasii e S. salamandra,
a partir das fotografias, recorrendo ao programa Sigma Scan Pro, v.5. As taxas de
crescimento (r) foram calculadas através de
� = �� − ���
onde cf corresponde ao comprimento final, ci ao inicial e t ao tempo decorrido (em
dias).
2.8. Avaliação dos indícios de predação;
Registaram-se as frequências de ocorrência de indícios de predação (rasgões
e/ou excisões na parte anterior do corpo das larvas) para ambas as espécies (ANEXO
III).
2.9. Determinação dos estádios de desenvolvimento larvar
Determinou-se o estádio larvar de todas as larvas de A. cisternasii capturadas
de acordo com Gosner (1960) [58]. Para facilitar posteriores análises, os estádios de
desenvolvimento dos girinos de A. cisternasii foram agrupados por classes: Classe I
(estádio 25 ao 28); Classe II (estádio 29 ao 33); Classe III (estádio 33 ao 38); Classe
IV (estádio 39 ao 43).
2.10. Modelação CR
Os métodos estatísticos para a estimação dos parâmetros desconhecidos e
para a realização de testes de hipóteses são baseados nos historiais de captura. Estes
historiais são matrizes constituídas por séries de 1’s (captura ou recaptura) e 0’s (não
captura ou recaptura) [13]. Assim, existindo k ocasiões de amostragem, o historial de
captura consiste numa série de k 0’s e 1’s [16] (ANEXO IV)
No cálculo das estimativas dos parâmetros Φi e pi foram usados os historiais
de captura de 893 girinos de A. cisternasii e de 833 larvas de S. salamandra nos 20
pegos.

26
De um ponto de vista biológico e prático, é admissível que os parâmetros Φi e
pi sejam função de variáveis externas como variáveis ambientais ou medidas de
esforço de captura. Existem vantagens em expressar estes constrangimentos num tipo
de estrutura usada nos modelos lineares generalizados para relacionar variáveis
resposta e variáveis preditoras externas [11; 13]. Neste trabalho considerou-se que as
variáveis ambientais poderiam influenciar a probabilidade de sobrevivência das
larvas nos diferentes pegos considerados, pelo que se procedeu ao ajustamento de
modelos com essas variáveis através da manipulação da matriz de delineamento1 do
MARK.
Para proceder à análise dos dados CR foi seguido o procedimento geral
aconselhado neste tipo de estudos [11; 16]:
1. Ajustar o modelo como totalmente dependente do tempo e do grupo
(diferentes pegos), o que é compatível com a biologia das espécies e com o
delineamento do estudo e verificar o seu ajustamento aos dados;
2. Testar as questões biológicas mais importantes pela comparação deste modelo
com modelos vizinhos;
3. Seleccionar o modelo mais parcimonioso usando o Critério de Informação de
Akaike (AIC);
4. Obter estimativas dos parâmetros do modelo através do Método de Máxima
Verosimilhança (MLE).
Para os dados relativos às larvas de A. cisternasii, optou-se por ajustar
modelos Jolly-Seber, com a formulação POPAN. Trata-se de uma modificação do
modelo JS original que considera a existência de uma super-população composta por
todos os animais que entrarão para a população e o parâmetro bi, que representa a
probabilidade de um animal desta hipotética super-população entrar na população no
intervalo i � i+1 [59]. Optou-se por utilizar esta formulação porque das cinco
formulações para o modelo JS disponíveis no MARK, a POPAN é a única que, para
além das probabilidades de sobrevivência, de captura e de entrada, estima ainda os
1 Matriz utilizada pelo MARK para descrever o delineamento, ligando os parâmetros de interesse
biológico às variáveis preditoras.

27
“nascimentos” líquidos, a abundância total (N) e por cada ocasião de captura (Ni).
Para os dados relativos às larvas de S. salamandra optou-se por ajustar
modelos CJS para obter estimativas de Φi e pi e modelos JS para estimar os restantes
parâmetros de interesse. Os dados CR das larvas de S. salamandra são mais escassos
que os de A. cisternasii e como o modelo JS estima muitos parâmetros com a mesma
função verosimilhança, a estimação de cada parâmetro em particular acaba por ser
preterida pela estimação de todos os parâmetros da forma mais verosímil, o que pode
resultar em estimativas pouco plausíveis [10; 11; 13]. Assim, a probabilidade de
sobrevivência (Φi) acaba por não ser tão bem estimada com o modelo JS como com o
modelo CJS, pelo que dos modelos JS considerou-se apenas as abundâncias e os
“nascimentos”.
Para testar a qualidade do ajustamento do modelo geral (todos os parâmetros
dependentes do tempo e, quando aplicável, dos grupos) aos dados das larvas de A.
cisternasii foram realizados os Testes 2 e 3 de ajustamento do modelo [16]
desenvolvidos especificamente para dados CR. Estes testes constituem um
procedimento diagnóstico para testar os pressupostos 1 e 2 do modelo CJS e JS
(homogeneidade dos parâmetros), admitindo que os restantes são cumpridos. O
cálculo dos Testes 2 e 3 é realizado pelo software RELEASE [16], actualmente
integrado no programa MARK [13].
O ajustamento do modelo geral para as larvas de S. salamandra não foi feito
através dos testes RELEASE porque os dados foram escassos. Recorreu-se então à
análise dos resíduos do desvio para o modelo CJS e a um teste de sensibilidade
baseado no factor de inflação da variância (variance inflaction factor), �̂ [17] para o
modelo JS. Os resíduos do desvio (rh) permitem verificar se os valores esperados
para cada historial de captura são muito ou pouco diferentes dos obtidos [17]. Assim,
�� = ��� !("� − #�)$2(#� − "� + "�log ("�#�)

28
onde Sinal corresponde ao sinal (+/-) do valor O-E, #� ao número de indivíduos
esperados com o historial de captura h e "� ao número de indivíduos observados
com o historial de captura h. Num modelo bem ajustado aos dados a diferença entre
os valores esperados e os observados será mínima e, portanto os resíduos serão quase
nulos. A estimação de �̂ permite verificar se as probabilidades de sobrevivência ou de
recaptura são homogéneas dentro de cada grupo e pode ser obtido a partir de uma
estatística de teste χ2 e dos graus de liberdade (gl) associados à distribuição
assimptótica desta estatística de teste [47], com
�̂ = *+,-
Assim, a um ajustamento “perfeito” do modelo aos dados corresponde �̂ = 1.
Quando os dados não permitem a utilização dos testes de ajustamento tradicionais
em modelação CR é frequente recorrer a um teste de sensibilidade a �̂ [9]. Se o
ranking dos modelos ajustados se alterar à medida que se vai variando gradualmente
o valor de �̂, pode-se estar perante um fraco ajustamento do modelo aos dados, o que
compromete a validade das inferências resultantes. Neste caso o valor de �̂ foi
alterado de 1 a 7, verificando-se se o modelo que melhor se ajustava aos dados se
mantinha [13].
2.11. Outras análises estatísticas
Para avaliar se a taxa de crescimento das larvas de A. cisternasii e S.
salamandra variaram significativamente entre pegos recorreu-se à ANOVA
univariada. O pressuposto da distribuição normal da variável dependente (taxa de
crescimento) nos diferentes grupos foi avaliado através do teste de Kolmogorov-
Smnirnov e o pressuposto da homogeneidade de variância foi verificado pelo teste de
Levene. Em caso de não cumprimento dos pressupostos, recorreu-se a um teste não-
paramétrico, o Kruskal-Wallis. Posteriormente utilizou-se a Regressão Linear
Múltipla para verificar se alguma das variáveis externas medidas estaria relacionada
com as taxas de crescimento de cada pego. Analisaram-se os pressupostos do
modelo, nomeadamente o da distribuição normal, homogeneidade e independência

29
dos erros. Os dois primeiros pressupostos foram validados graficamente e o
pressuposto da independência foi validado com a estatística de Durbin-Watson.
O teste Qui-quadrado de independência foi utilizado nos seguintes testes:
independência dos indícios de predação entre pegos; independência do estádio de
desenvolvimento (classe modal) entre pegos; igualdade nas frequências de
movimentos entre pegos entre as duas espécies. Quando se registaram células com
valores esperados inferiores a 5 o teste do χ2 não pôde ser aplicado e recorreu-se à
técnica de simulação de Monte-Carlo. A simulação de Monte-Carlo é um método
estatístico que procura determinar a probabilidade de ocorrência de uma determinada
situação experimental através de um conjunto elevado de simulações, baseado na
geração aleatória de amostras a partir do conhecimento empírico da população sob
estudo.
As análises estatísticas descritivas, gráficas e inferenciais foram executadas
com o software SPSS (v. 17.0, SPSS Inc., Chicago, IL). Consideraram-se
estatisticamente significativos os efeitos cujo p-value foi inferior ou igual a 0.05.
3. RESULTADOS
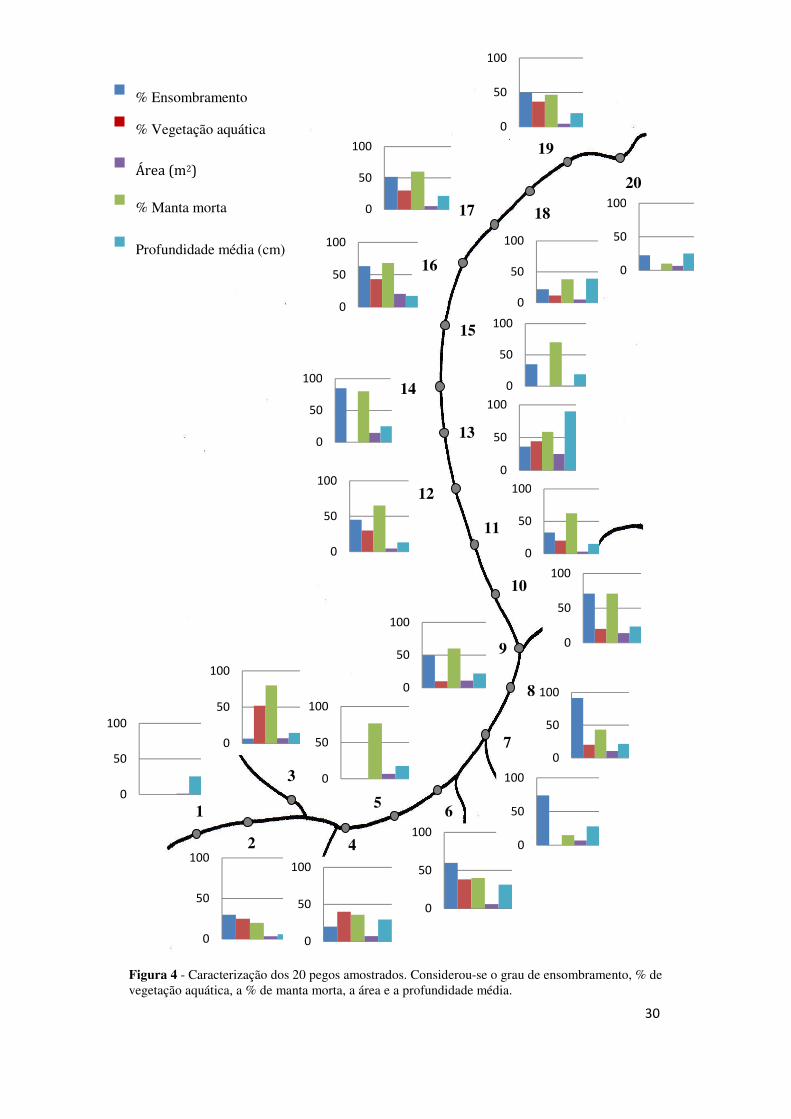
3.1. Caracterização dos pegos
Como se pode ver pela Figura 4, os pegos não são muito distintos no que toca
às variáveis medidas. Podem-se distinguir 4 pegos com graus de ensombramento
elevados: o pego 7, o 8, o 10 e o 14. Os pegos 3, 4, 6, 13, 16 e 19 destacam-se com as
maiores percentagens de vegetação aquática. Os pegos com área ≥ 10 m2 são o 10, o
13, o 14 e o 16. Os pegos em média mais profundos são o 4, o 8, o 13 e o 18.
Distinguem-se dois grupos de pegos semelhantes e próximos espacialmente:
(1) pegos 2, 4, 6, 8 e 9; e (2) pegos 16, 17 e 19. Há pegos que se destacam dos
restantes, caso dos pegos 1, 5, 7 e 20.
Não existem evidências de alterações graduais das características dos pegos
ao longo da ribeira.
30
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100
0
50
1000
50
100
0
50
1000
50
100
0
50
100
0
50
100
0
50
100
0
50
100
0
50
100
Figura 4 - Caracterização dos 20 pegos amostrados. Considerou-se o grau de ensombramento, % de vegetação aquática, a % de manta morta, a área e a profundidade média.
1
2
3
4
5 6
7
8
9
10
11
12
13
14
15
16
17 18
19
20
� % Manta morta
� % Vegetação aquática
� Área (m2)
� Profundidade média (cm)
� % Ensombramento

31
0
50
100
150
200
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Nº d
e in
diví
duos
Pegos
A. cisternasii S. salamandra
Para além das variáveis referidas foi também medida a temperatura,
condutividade, salinidade, % de oxigénio dissolvido (DO) e pH de cada pego.
Nenhum destes parâmetros apresentou grande variabilidade ao longo da ribeira: a
temperatura de água variou entre 14 e 19.24 ºC; a condutividade variou entre 16.54 e
os 19.58 mS/cm; a salinidade variou entre 12.25 e 14.60 mg/L; a percentagem de
oxigénio dissolvido foi 0.1 para todos os pegos à excepção de um para o qual foi 0.2;
e o pH variou entre 7.69 e 8.39, sem agrupamento espacial de pegos semelhantes.
3.2. Capturas
Pela Figura 5 podemos verificar que houve um predomínio de larvas de A.
cisternasii nos pegos mais a montante (1 a 5) e nos pegos mais a jusante (17 a 20),
sendo que os pegos intermédios da ribeira pareceram dominados pelas larvas de S.
salamandra. Pode ainda verificar-se que o pego 8 foi o único onde só foram
encontradas larvas de S. salamandra.
Figura 5 - Total das capturas de larvas de A. cisternasii e S. salamandra para cada pego amostrado.
32
0102030405060708090
100
Id no Top 1
Id no Top 10
Id no Top 20
Id no Top 30
Id no Top 40
Id no Top 50
NID
%
A. cisternasii S. salamandra
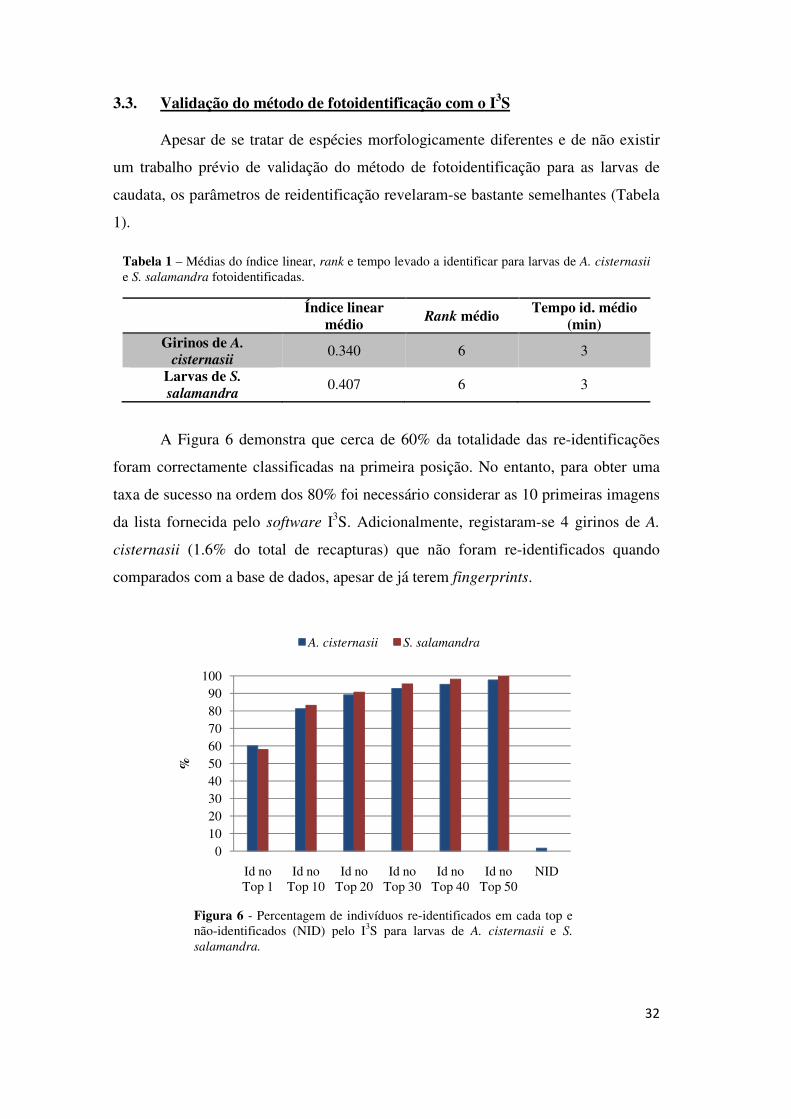
3.3. Validação do método de fotoidentificação com o I3S
Apesar de se tratar de espécies morfologicamente diferentes e de não existir
um trabalho prévio de validação do método de fotoidentificação para as larvas de
caudata, os parâmetros de reidentificação revelaram-se bastante semelhantes (Tabela
1).
Tabela 1 – Médias do índice linear, rank e tempo levado a identificar para larvas de A. cisternasii e S. salamandra fotoidentificadas.
Índice linear
médio Rank médio
Tempo id. médio (min)
Girinos de A.
cisternasii 0.340 6 3
Larvas de S.
salamandra 0.407 6 3
A Figura 6 demonstra que cerca de 60% da totalidade das re-identificações
foram correctamente classificadas na primeira posição. No entanto, para obter uma
taxa de sucesso na ordem dos 80% foi necessário considerar as 10 primeiras imagens
da lista fornecida pelo software I3S. Adicionalmente, registaram-se 4 girinos de A.
cisternasii (1.6% do total de recapturas) que não foram re-identificados quando
comparados com a base de dados, apesar de já terem fingerprints.
Figura 6 - Percentagem de indivíduos re-identificados em cada top e não-identificados (NID) pelo I3S para larvas de A. cisternasii e S.
salamandra.

33
0
20
40
60
80
100
top 1 top 5 top 15
%
Sessão 2 Sessão 3 Sessão 4 Sessão 5 Sessão 6
0
20
40
60
80
100
top 1 top 5 top 15
%
Sessão 2 Sessão 3 Sessão 4 Sessão 5
Figura 8 - Percentagem de larvas de S. salamandra capturadas na sessão 1 e re-identificadas em sessões posteriores no top 1, 5 e 15.
Como é ilustrado pela Figura 7, a percentagem de girinos re-identificados no
top 1 não dependeu do tempo passado entre a captura e a recaptura, i.e., o
desenvolvimento da larva aparentemente não prejudicou o processo de re-
identificação por parte do I3S. A diferença máxima registada entre uma sessão de
captura e uma recaptura foi de 56 dias (indivíduo capturado na sessão 1 e
recapturado na sessão 6), com a transição do estádio 25 (3.88 cm de comprimento
total) para o estádio 37 (4.83 cm).
O desenvolvimento das larvas de S. salamandra também parece não ter
influenciado o processo de re-identificação do I3S porque não se verificou uma
tendência para uma maior percentagem de re-identificações nas posições mais
cimeiras da lista entre sessões mais próximas (Figura 8).
Figura 7 - Percentagem de larvas de A. cisternasii capturadas na sessão 1 e re-identificadas em sessões posteriores

34
A diferença máxima entre a sessão de captura e a de recaptura registada foi de
38 dias (indivíduo capturado na sessão 1 e recapturado na sessão 5), com um
comprimento total inicial de 3.36 cm e final de 3.98 cm.
Contrariando o estudo de validação do método [45], registou-se alguma
dificuldade de processamento dos dados por parte do I3S à medida que a base de
dados aumentava até às 1786 fingerprints de A. cisternasii e 1666 de S. salamandra.
Houve um aumento de ca. de 70% no tempo de processamento entre a altura em que
base de dados possuía < 200 fingerprints e a altura em que possuía > 600
fingerprints. O aumento da dimensão da base de dados provocou um aumento
considerável do tempo de processamento e da ocorrência de problemas de software.
3.4. Recapturas
Nos 20 pegos registaram-se 893 capturas de larvas de A. cisternasii (6
sessões), incluindo 249 episódios de recaptura, e 833 capturas de larvas de S.
salamandra (5 sessões), incluindo 108 episódios de recaptura (Tabela 2).
Tabela 2 – Número de indivíduos amostrados, capturados e recapturados, para A. cisternasii e S.
salamandra.
Girinos A. cisternasii Larvas S. salamandra Amostragem Total 893 833
Totalidade indivíduos capturados
646 725
Totalidade de recapturas 249 108 Totalidade de migrações 32 11
A percentagem de recapturas de larvas de A. cisternasii por sessão manteve-
se relativamente semelhante, enquanto que a das larvas de S. salamandra sofreu uma
queda de quase 50% na percentagem de recaptura entre a segunda sessão e as
restantes (Tabela 3).

35
Tabela 3 - Percentagem de recapturas por sessão, para A. cisternasii e S. salamandra.
Girinos A. cisternasii Larvas de S. salamandra Sessão 2 15 39 Sessão 3 21 23 Sessão 4 20 16 Sessão 5 29 22 Sessão 6 11 --
Registaram-se consideravelmente mais recapturas de girinos do que de larvas,
sendo que 15 girinos foram recapturados mais do que duas vezes, algo que nunca
aconteceu com as larvas (Tabela 4).
Tabela 4 - Número de indivíduos recapturados 1, 2, 3 e 4 vezes, para A. cisternasii e S. salamandra.
Girinos A. cisternasii Larvas S. salamandra
Indivíduos recapturados 1x 106 94 Indivíduos recapturados 2x 48 7 Indivíduos recapturados 3x 13 0 Indivíduos recapturados 4x 2 0
3.5. Modelação CR
Antes de proceder à modelação CR há que verificar se os pressupostos de
homogeneidade de captura e de sobrevivência foram respeitados através da
realização de testes de ajustamento do modelo geral (todos os parâmetros
dependentes do tempo e do grupo, quando aplicável). Se estas condições são
verificadas, procede-se ao ajustamento de modelos mais simples, que podem
envolver variáveis externas na estimação dos diferentes parâmetros, sendo a escolha
final do modelo feita através do Critério de Informação de Akaike (AIC).
Seguidamente apresenta-se o procedimento da modelação e as estimativas dos
parâmetros, separadamente para cada espécie.
3.5.1. Alytes cisternasii
Uma vez que todos os 20 pegos amostrados apresentaram taxas de recaptura
válidas (> 10%) foram todos considerados na modelação CR [11].

36
3.5.1.1. Validação dos pressupostos dos modelos CR
O ajustamento do modelo geral foi aferido pelos Testes 2 e 3 do RELEASE
(ANEXO V). Foi registada heterogeneidade na probabilidade de captura e de
sobrevivência em 6 pegos, o que constitui uma violação dos pressupostos dos
modelos CR. Assim, esses pegos foram excluídos da modelação e,
consequentemente, os parâmetros demográficos foram estimados para 14 pegos.
3.5.1.2. Modelos
De seguida procedeu-se ao ajustamento de modelos mais simples,
considerando a influência de algumas variáveis ambientais como o grau de
ensombramento ou percentagem de vegetação aquática no pego sobre a
probabilidade de sobrevivência dos girinos.
Tabela 5 - Lista do conjuntos dos modelos JS com a formulação POPAN que melhor se ajustou aos dados das larvas de A. cisternasii. Representa-se a probabilidade de sobrevivência (Φi) dependente do pego (g) e do tempo (t). A probabilidade de captura (pi) depende apenas de cada pego (g) e a probabilidade de entradas (penti) depende apenas do efeito do tempo (t).
Modelo AICc ∆AICc Nº de
parâmetros
Φ(p(mantamorta)*t)p(g)pent(t) 1619.40 0.00 89
Φ(p(profmedia)*t)p(g)pent(t) 1621.52 2.12 89
Φ(p(area)*t)p(g)pent(t) 1621.81 2.41 89
Como se pode observar na Tabela 5, o modelo JS que melhor se ajustou aos
dados considera Φi dependente da percentagem de manta morta presente no leito do
pego, pi dependente apenas do pego e pent i dependente do tempo. No entanto, o
valor do ∆AICc indicia alguma importância da variável profundidade média para a
probabilidade de sobrevivência.
37
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 1
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 2
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 3
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 4
3.5.1.3. Estimativas dos parâmetros demográficos
3.5.1.3.1. Probabilidades de sobrevivência
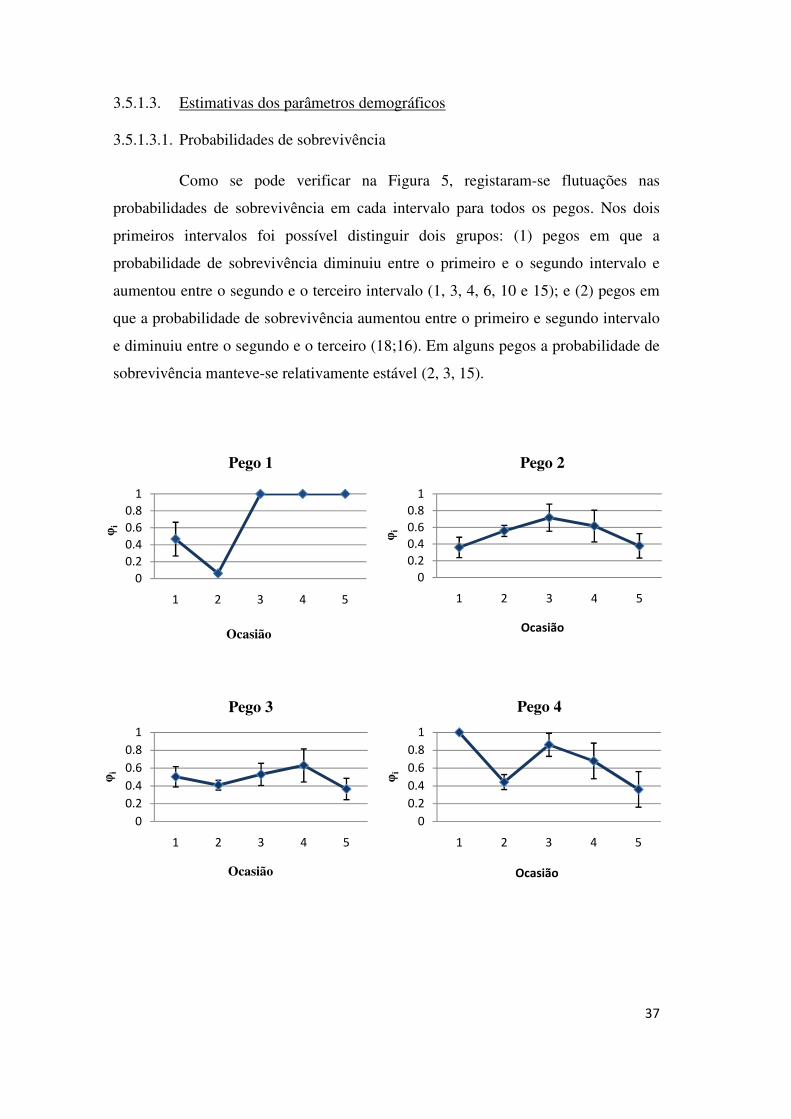
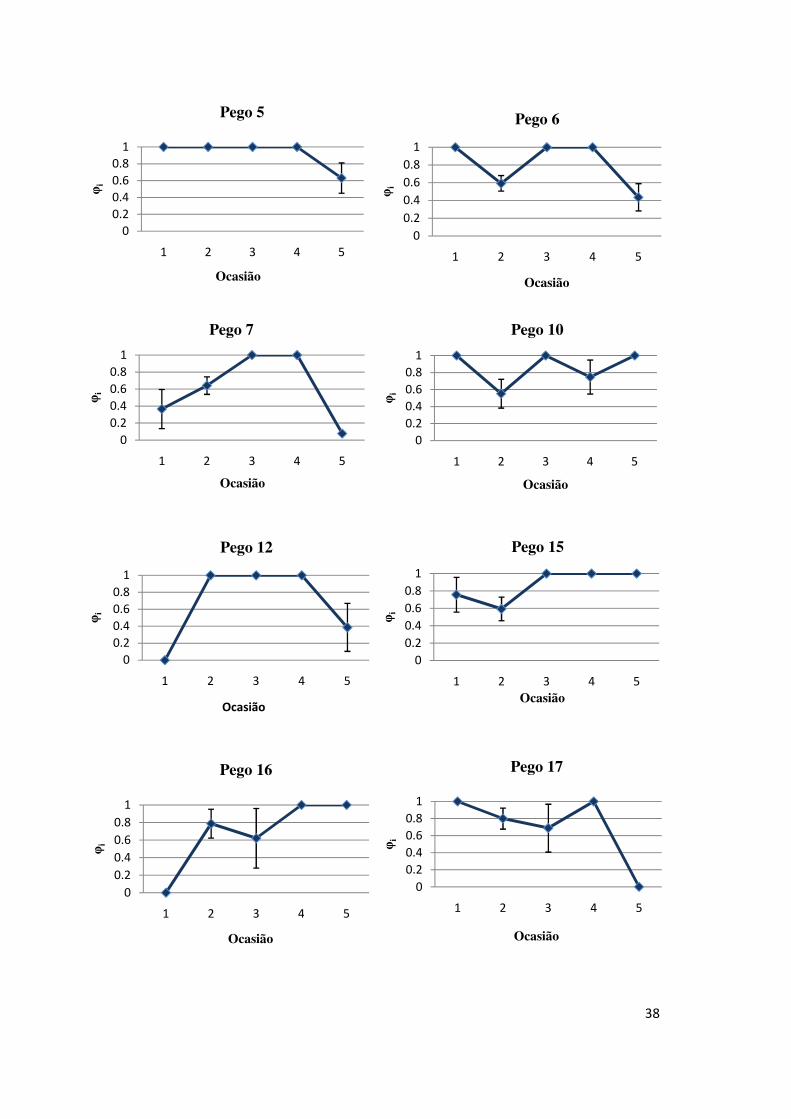
Como se pode verificar na Figura 5, registaram-se flutuações nas
probabilidades de sobrevivência em cada intervalo para todos os pegos. Nos dois
primeiros intervalos foi possível distinguir dois grupos: (1) pegos em que a
probabilidade de sobrevivência diminuiu entre o primeiro e o segundo intervalo e
aumentou entre o segundo e o terceiro intervalo (1, 3, 4, 6, 10 e 15); e (2) pegos em
que a probabilidade de sobrevivência aumentou entre o primeiro e segundo intervalo
e diminuiu entre o segundo e o terceiro (18;16). Em alguns pegos a probabilidade de
sobrevivência manteve-se relativamente estável (2, 3, 15).
38
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 5
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 7
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 6
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 10
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 12
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 15
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 16
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 17
39
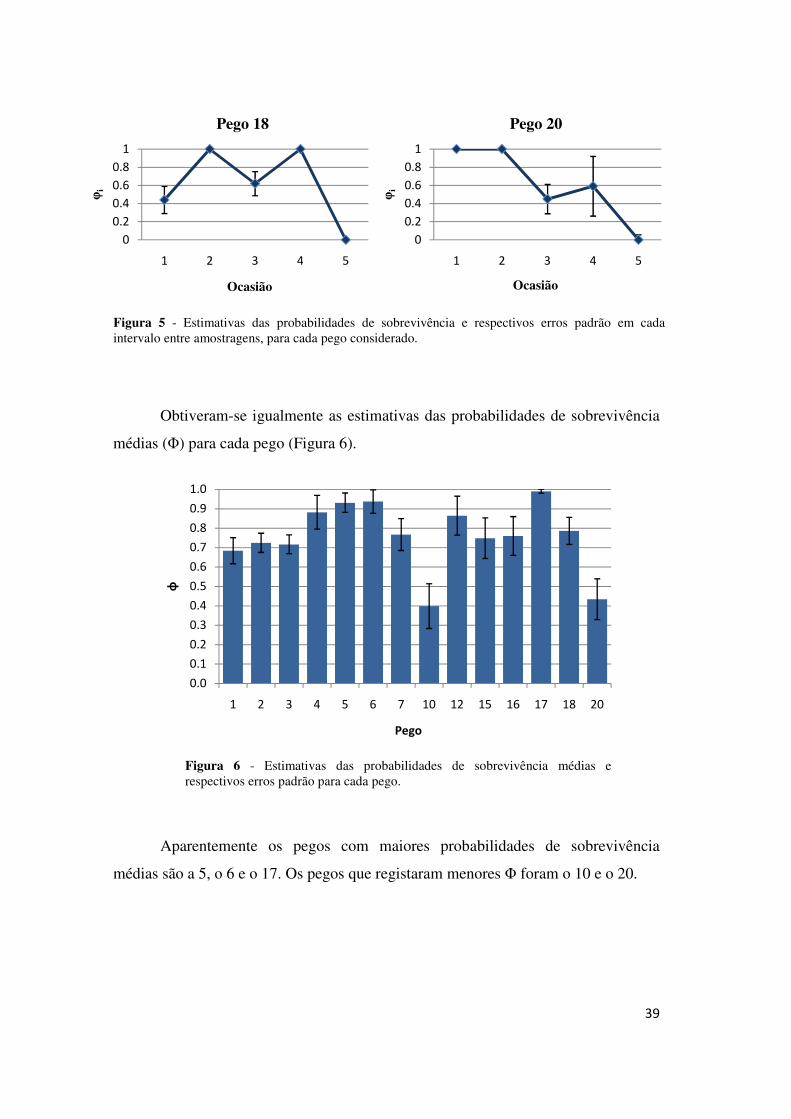
Figura 5 - Estimativas das probabilidades de sobrevivência e respectivos erros padrão em cada intervalo entre amostragens, para cada pego considerado.
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 18
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5
φi
Ocasião
Pego 20
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
1 2 3 4 5 6 7 10 12 15 16 17 18 20
φ
Pego
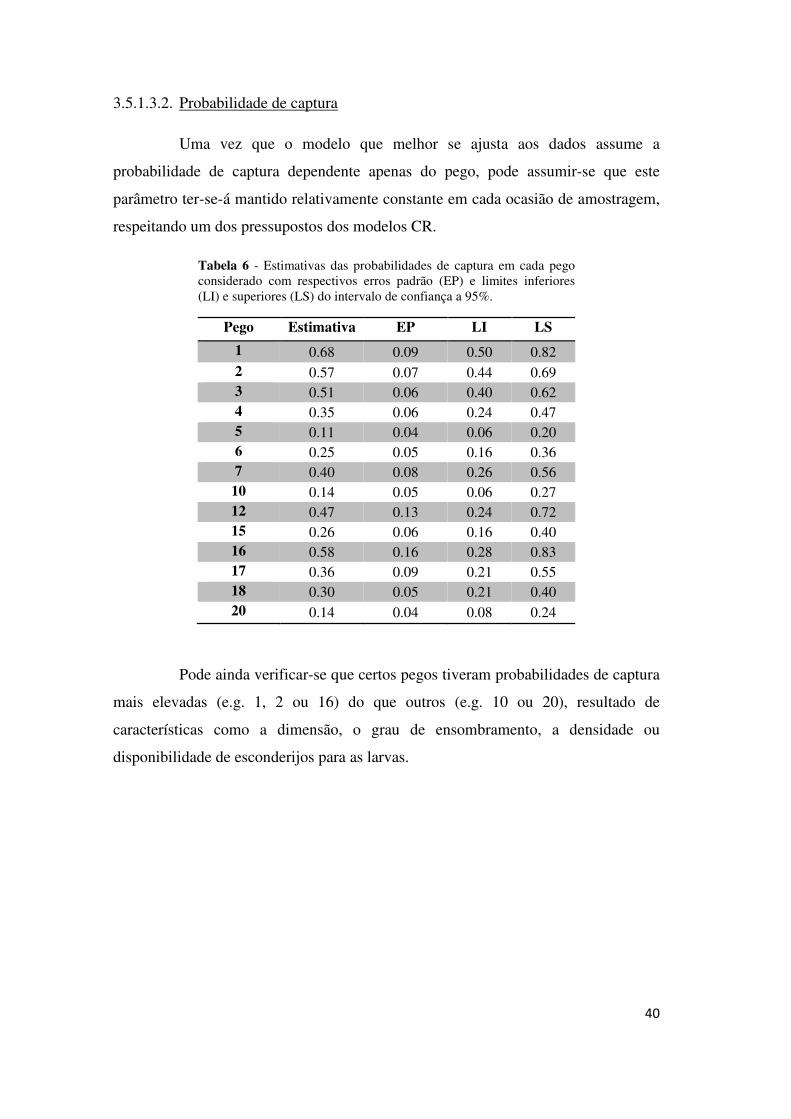
Obtiveram-se igualmente as estimativas das probabilidades de sobrevivência
médias (Φ) para cada pego (Figura 6).
Aparentemente os pegos com maiores probabilidades de sobrevivência
médias são a 5, o 6 e o 17. Os pegos que registaram menores Φ foram o 10 e o 20.
Figura 6 - Estimativas das probabilidades de sobrevivência médias e respectivos erros padrão para cada pego.
40
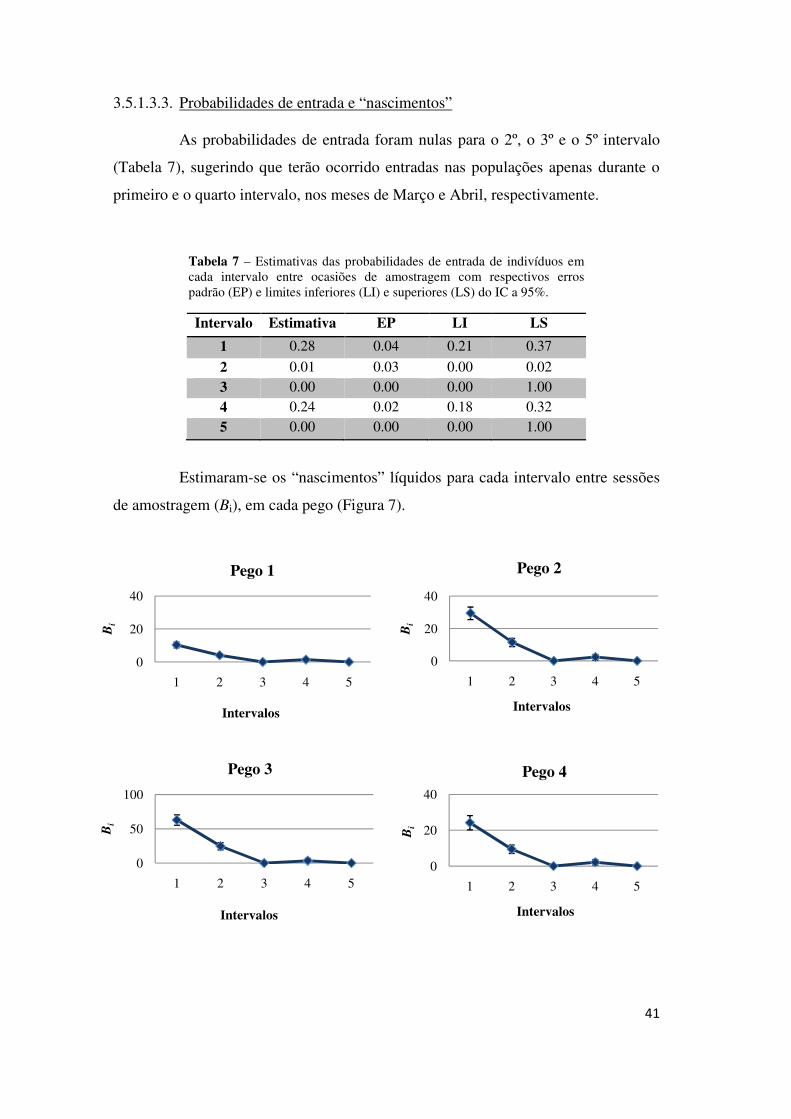
3.5.1.3.2. Probabilidade de captura
Uma vez que o modelo que melhor se ajusta aos dados assume a
probabilidade de captura dependente apenas do pego, pode assumir-se que este
parâmetro ter-se-á mantido relativamente constante em cada ocasião de amostragem,
respeitando um dos pressupostos dos modelos CR.
Tabela 6 - Estimativas das probabilidades de captura em cada pego considerado com respectivos erros padrão (EP) e limites inferiores (LI) e superiores (LS) do intervalo de confiança a 95%.
Pego Estimativa EP LI LS
1 0.68 0.09 0.50 0.82 2 0.57 0.07 0.44 0.69 3 0.51 0.06 0.40 0.62 4 0.35 0.06 0.24 0.47 5 0.11 0.04 0.06 0.20 6 0.25 0.05 0.16 0.36 7 0.40 0.08 0.26 0.56
10 0.14 0.05 0.06 0.27 12 0.47 0.13 0.24 0.72 15 0.26 0.06 0.16 0.40 16 0.58 0.16 0.28 0.83 17 0.36 0.09 0.21 0.55 18 0.30 0.05 0.21 0.40 20 0.14 0.04 0.08 0.24
Pode ainda verificar-se que certos pegos tiveram probabilidades de captura
mais elevadas (e.g. 1, 2 ou 16) do que outros (e.g. 10 ou 20), resultado de
características como a dimensão, o grau de ensombramento, a densidade ou
disponibilidade de esconderijos para as larvas.
41
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 2
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 1
0
50
100
1 2 3 4 5
Bi
Intervalos
Pego 3
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 4
3.5.1.3.3. Probabilidades de entrada e “nascimentos”
As probabilidades de entrada foram nulas para o 2º, o 3º e o 5º intervalo
(Tabela 7), sugerindo que terão ocorrido entradas nas populações apenas durante o
primeiro e o quarto intervalo, nos meses de Março e Abril, respectivamente.
Tabela 7 – Estimativas das probabilidades de entrada de indivíduos em cada intervalo entre ocasiões de amostragem com respectivos erros padrão (EP) e limites inferiores (LI) e superiores (LS) do IC a 95%.
Intervalo Estimativa EP LI LS
1 0.28 0.04 0.21 0.37
2 0.01 0.03 0.00 0.02 3 0.00 0.00 0.00 1.00 4 0.24 0.02 0.18 0.32 5 0.00 0.00 0.00 1.00
Estimaram-se os “nascimentos” líquidos para cada intervalo entre sessões
de amostragem (Bi), em cada pego (Figura 7).
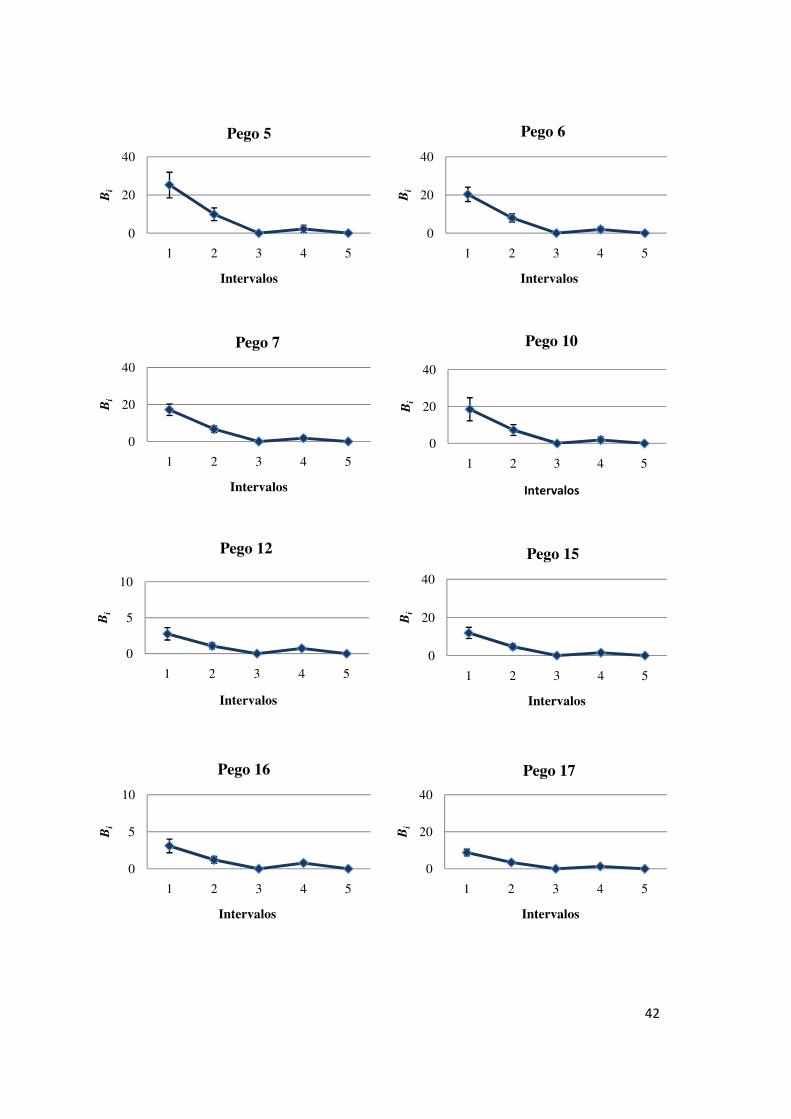
42
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 5
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 6
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 7
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 10
0
5
10
1 2 3 4 5
Bi
Intervalos
Pego 12
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 15
0
5
10
1 2 3 4 5
Bi
Intervalos
Pego 16
0
20
40
1 2 3 4 5
Bi
Intervalos
Pego 17
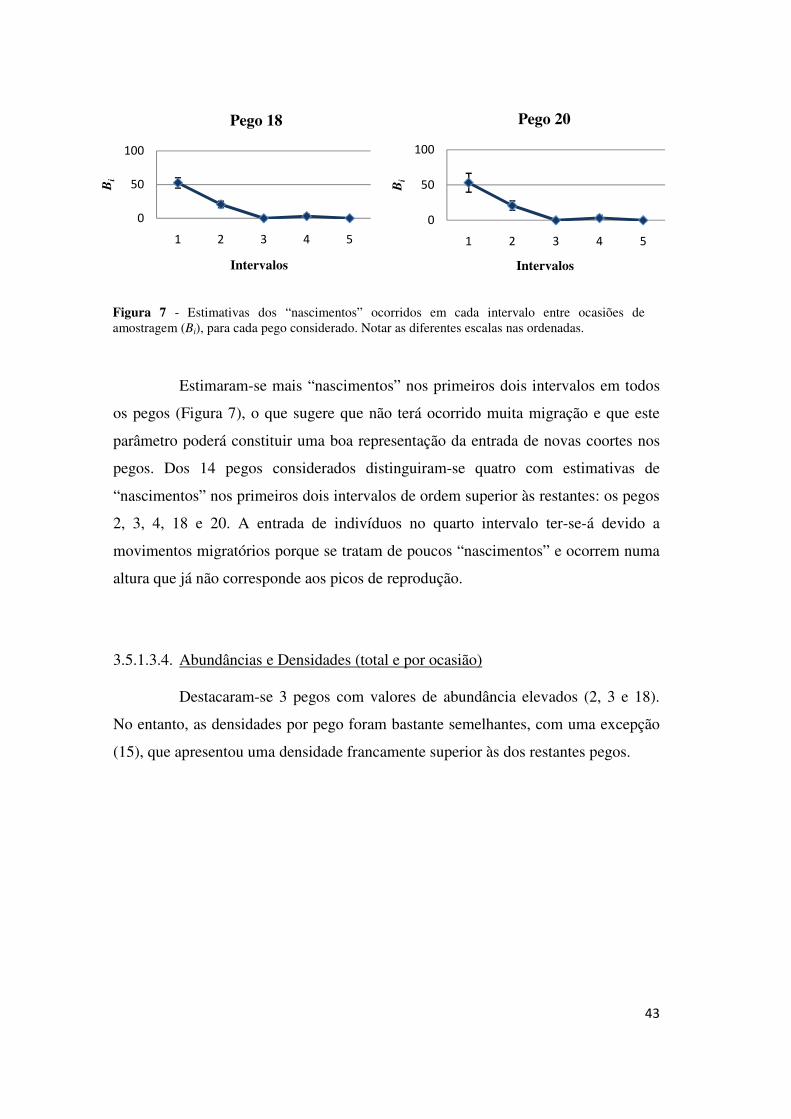
43
0
50
100
1 2 3 4 5
Bi
Intervalos
Pego 18
0
50
100
1 2 3 4 5
Bi
Intervalos
Pego 20
Estimaram-se mais “nascimentos” nos primeiros dois intervalos em todos
os pegos (Figura 7), o que sugere que não terá ocorrido muita migração e que este
parâmetro poderá constituir uma boa representação da entrada de novas coortes nos
pegos. Dos 14 pegos considerados distinguiram-se quatro com estimativas de
“nascimentos” nos primeiros dois intervalos de ordem superior às restantes: os pegos
2, 3, 4, 18 e 20. A entrada de indivíduos no quarto intervalo ter-se-á devido a
movimentos migratórios porque se tratam de poucos “nascimentos” e ocorrem numa
altura que já não corresponde aos picos de reprodução.
3.5.1.3.4. Abundâncias e Densidades (total e por ocasião)
Destacaram-se 3 pegos com valores de abundância elevados (2, 3 e 18).
No entanto, as densidades por pego foram bastante semelhantes, com uma excepção
(15), que apresentou uma densidade francamente superior às dos restantes pegos.
Figura 7 - Estimativas dos “nascimentos” ocorridos em cada intervalo entre ocasiões de amostragem (Bi), para cada pego considerado. Notar as diferentes escalas nas ordenadas.
44
0
10
20
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 1
0
40
80
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 2
0
100
200
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 3
0
40
80
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 4
0
100
200
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 5
0
40
80
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 6
0
40
80
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 7
0
40
80
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 10
� Abundância � Densidade
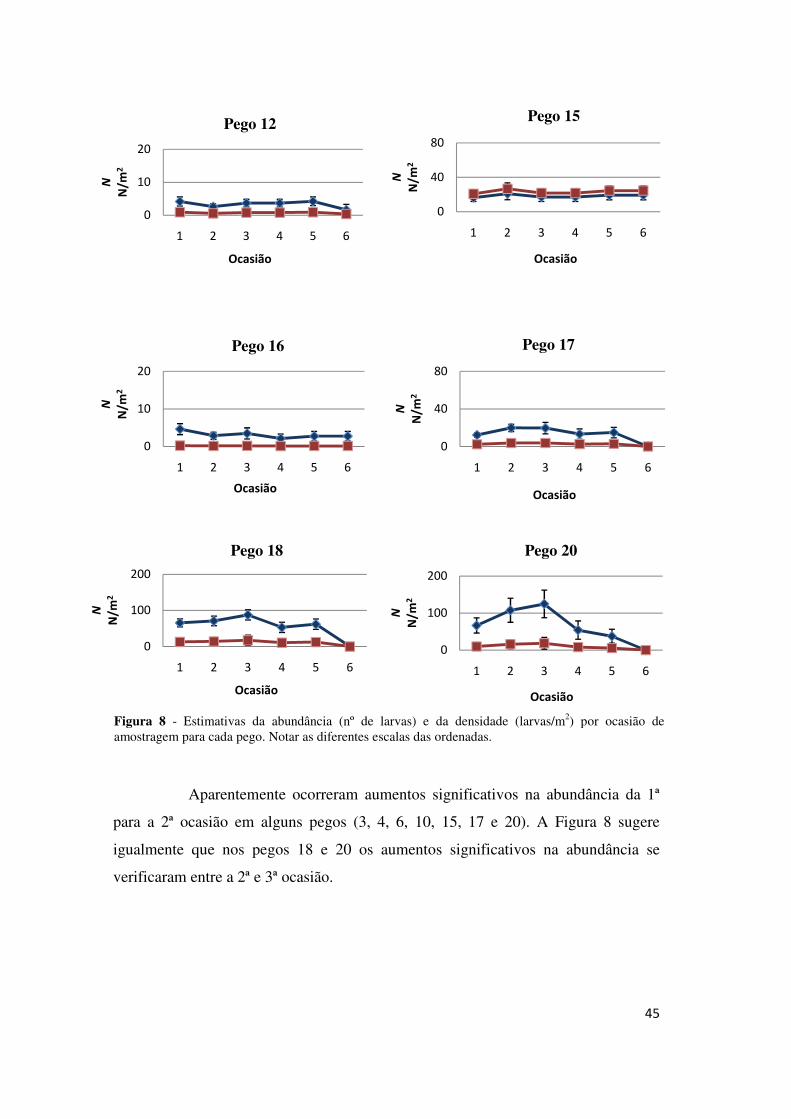
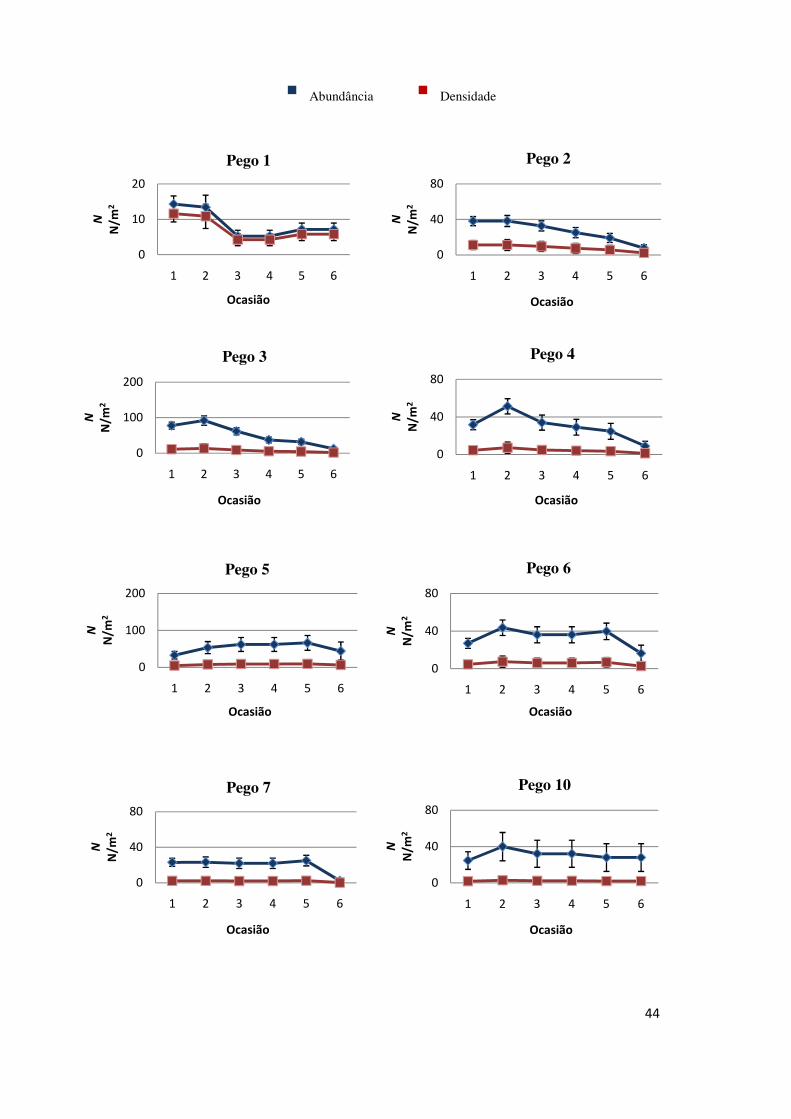
45
0
40
80
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 15
0
10
20
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 12
0
100
200
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 18
0
100
200
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 20
0
10
20
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 16
0
40
80
1 2 3 4 5 6
N
N/m
2
Ocasião
Pego 17
Aparentemente ocorreram aumentos significativos na abundância da 1ª
para a 2ª ocasião em alguns pegos (3, 4, 6, 10, 15, 17 e 20). A Figura 8 sugere
igualmente que nos pegos 18 e 20 os aumentos significativos na abundância se
verificaram entre a 2ª e 3ª ocasião.
Figura 8 - Estimativas da abundância (nº de larvas) e da densidade (larvas/m2) por ocasião de amostragem para cada pego. Notar as diferentes escalas das ordenadas.
46
52 %
48 %
Ausentes Presentes
0
50
100
150
200
250
1 2 3 4 5 6 7 10 12 15 16 17 18 20
N
N/m
2
Pegos
Abundância Densidade
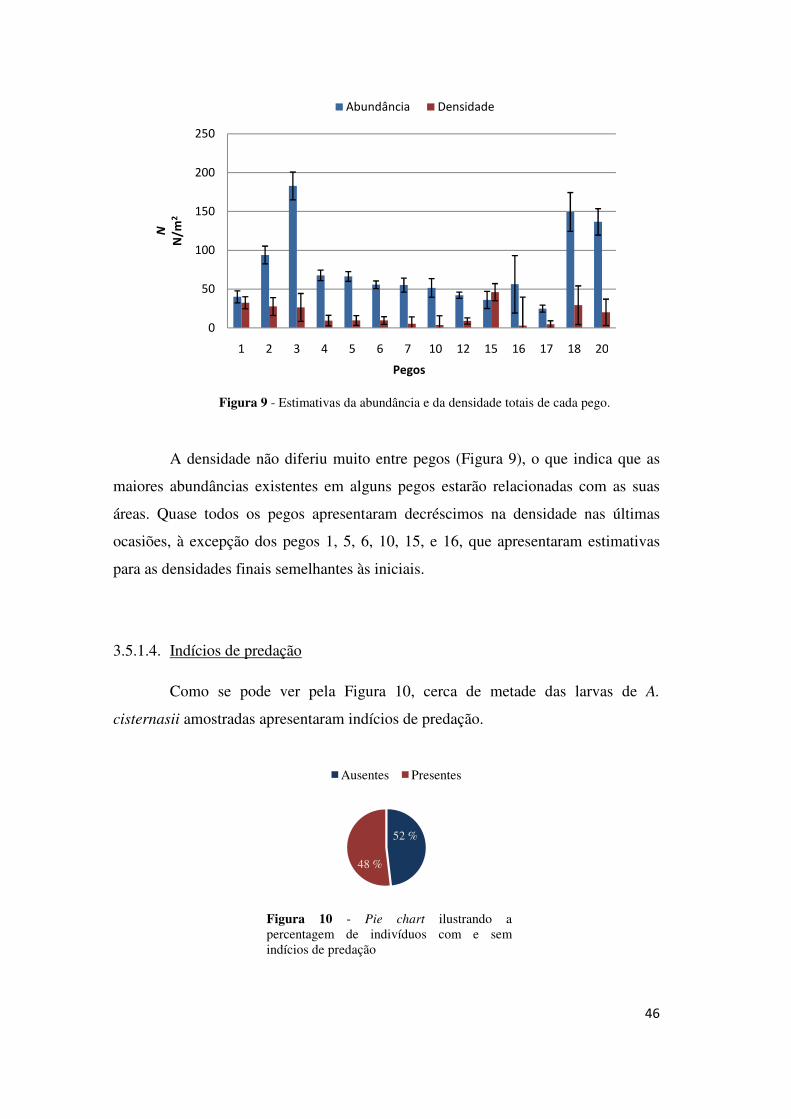
A densidade não diferiu muito entre pegos (Figura 9), o que indica que as
maiores abundâncias existentes em alguns pegos estarão relacionadas com as suas
áreas. Quase todos os pegos apresentaram decréscimos na densidade nas últimas
ocasiões, à excepção dos pegos 1, 5, 6, 10, 15, e 16, que apresentaram estimativas
para as densidades finais semelhantes às iniciais.
3.5.1.4. Indícios de predação
Como se pode ver pela Figura 10, cerca de metade das larvas de A.
cisternasii amostradas apresentaram indícios de predação.
Figura 9 - Estimativas da abundância e da densidade totais de cada pego.
Figura 10 - Pie chart ilustrando a percentagem de indivíduos com e sem indícios de predação
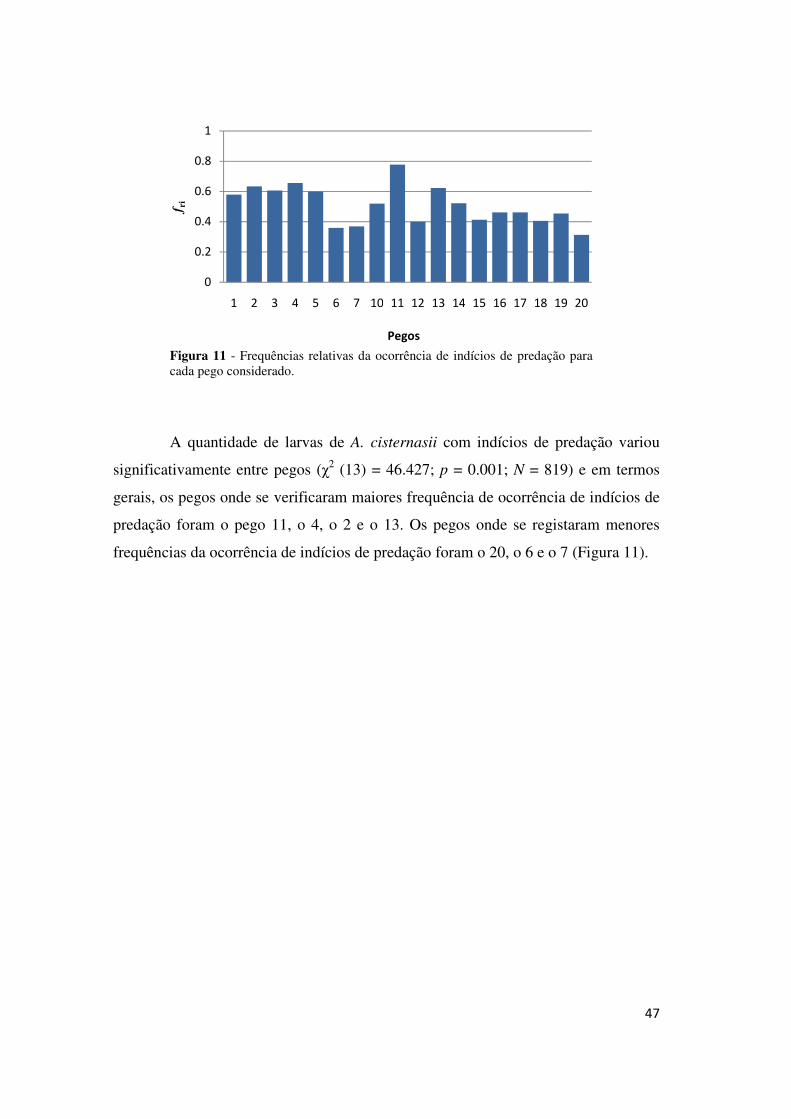
47
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6 7 10 11 12 13 14 15 16 17 18 19 20
f ri
Pegos
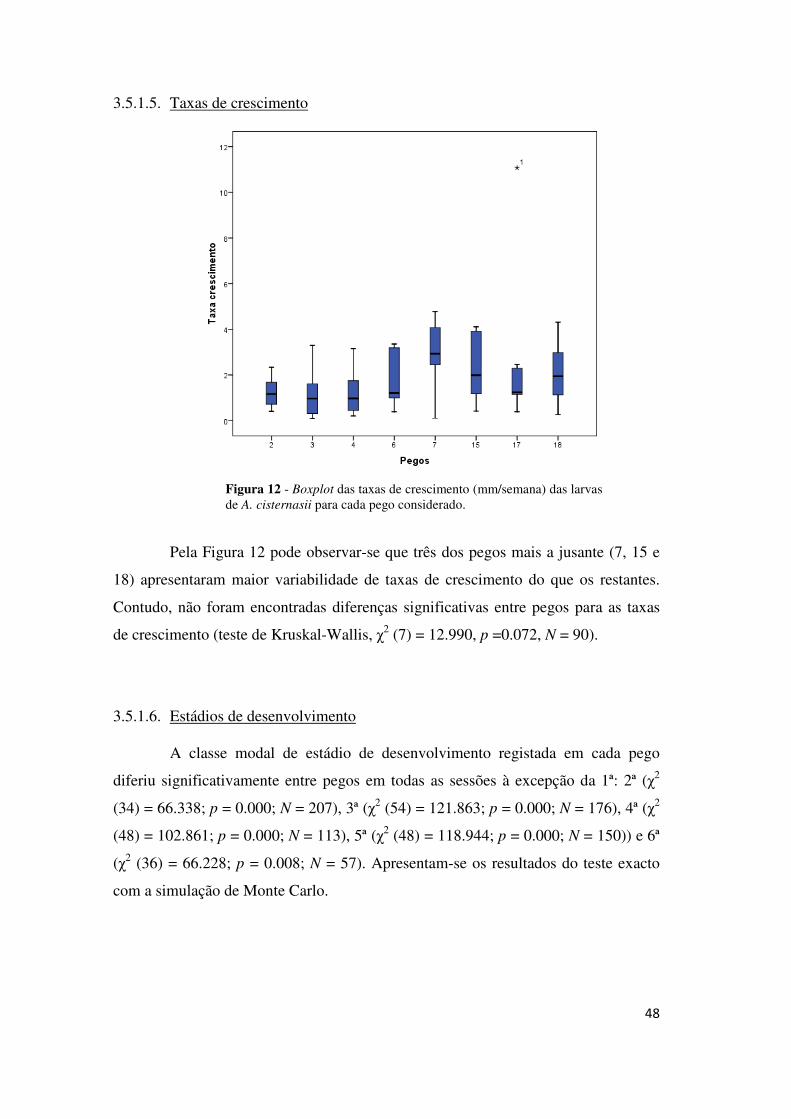
A quantidade de larvas de A. cisternasii com indícios de predação variou
significativamente entre pegos (χ2 (13) = 46.427; p = 0.001; N = 819) e em termos
gerais, os pegos onde se verificaram maiores frequência de ocorrência de indícios de
predação foram o pego 11, o 4, o 2 e o 13. Os pegos onde se registaram menores
frequências da ocorrência de indícios de predação foram o 20, o 6 e o 7 (Figura 11).
Figura 11 - Frequências relativas da ocorrência de indícios de predação para cada pego considerado.
48
3.5.1.5. Taxas de crescimento
Pela Figura 12 pode observar-se que três dos pegos mais a jusante (7, 15 e
18) apresentaram maior variabilidade de taxas de crescimento do que os restantes.
Contudo, não foram encontradas diferenças significativas entre pegos para as taxas
de crescimento (teste de Kruskal-Wallis, χ2 (7) = 12.990, p =0.072, N = 90).
3.5.1.6. Estádios de desenvolvimento
A classe modal de estádio de desenvolvimento registada em cada pego
diferiu significativamente entre pegos em todas as sessões à excepção da 1ª: 2ª (χ2
(34) = 66.338; p = 0.000; N = 207), 3ª (χ2 (54) = 121.863; p = 0.000; N = 176), 4ª (χ2
(48) = 102.861; p = 0.000; N = 113), 5ª (χ2 (48) = 118.944; p = 0.000; N = 150)) e 6ª
(χ2 (36) = 66.228; p = 0.008; N = 57). Apresentam-se os resultados do teste exacto
com a simulação de Monte Carlo.
Figura 12 - Boxplot das taxas de crescimento (mm/semana) das larvas de A. cisternasii para cada pego considerado.

49
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
f ri
Pegos
Sessão 1
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
f ri
Pegos
Sessão 2
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
f ri
Pegos
Sessão 3
� Classe I � Classe II � Classe III � Classe IV
50
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
f ri
Pegos
Sessão 4
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
f ri
Pegos
Sessão 5
0
0.2
0.4
0.6
0.8
1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
f ri
Pegos
Sessão 6
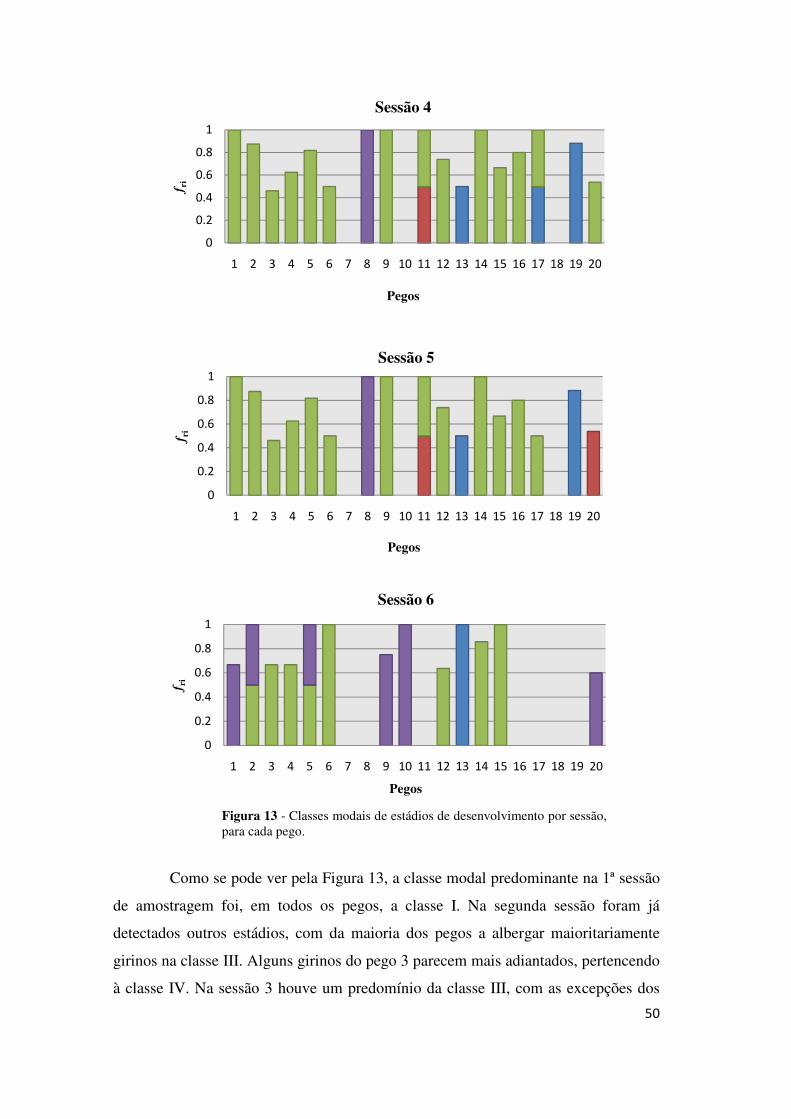
Como se pode ver pela Figura 13, a classe modal predominante na 1ª sessão
de amostragem foi, em todos os pegos, a classe I. Na segunda sessão foram já
detectados outros estádios, com da maioria dos pegos a albergar maioritariamente
girinos na classe III. Alguns girinos do pego 3 parecem mais adiantados, pertencendo
à classe IV. Na sessão 3 houve um predomínio da classe III, com as excepções dos
Figura 13 - Classes modais de estádios de desenvolvimento por sessão, para cada pego.

51
pegos 1, 10 e 20 com a maioria dos girinos na classe IV. Na sessão 4 continuou a
predominar a classe III, com a excepção do pego 8, dominado por girinos na classe
IV. Na sessão 5 verificou-se novamente o predomínio da classe III, à excepção do
pego 8 que apresentou mais indivíduos na classe IV. Pode igualmente verificar-se
que o pego 11 é particularmente heterogéneo. Os pegos 13 e 19 constituíram também
excepções, com predomínio da classe I. Aquando da sessão 6, muitos pegos (6) já se
encontravam secos mas para os restantes continuou a predominar a classe III. Os
girinos dos pegos 1, 9, 10 e 20 encontravam-se maioritariamente na classe IV de
desenvolvimento. Adicionalmente, verificou-se que os pegos 2 e 5 foram os mais
heterogéneos e que no pego 13 só se encontraram indivíduos na classe I.
3.5.2. Salamandra salamandra
Apesar de se ter amostrado 20 pegos, apenas os dados relativos aos pegos
com taxas de recaptura válidas (> 10%) foram considerados na modelação CR [11].
3.5.2.1. Validação dos pressupostos dos modelos CR
Uma vez que os dados CR relativos às larvas de S. salamandra foram
escassos, o ajustamento do modelo geral CJS foi testado através da análise dos
resíduos do desvio. De uma forma geral os resíduos parecem estar distribuídos
aleatoriamente (ANEXO VI). A maioria dos resíduos encontra-se próxima da linha
do zero e estão todos na área do gráfico definida por ± 2.0, sugerindo que não existe
muita dispersão e que os resíduos têm uma distribuição normal. Assim, pode
assumir-se que os pressupostos de homogeneidade de captura e sobrevivência foram
respeitados. O ajustamento do modelo geral JS foi aferido através do teste de
sensibilidade a �̂. Como a ordem dos modelos ajustados não se alterou à medida que
se alterava o valor de �̂, admitiu-se que o modelo estava razoavelmente bem ajustado
aos dados.
3.5.2.2. Modelos
Depois de testar o ajustamento do modelo geral, procedeu-se ao ajustamento
de modelos CJS e JS mais simples, examinando a influência de variáveis externas.
52
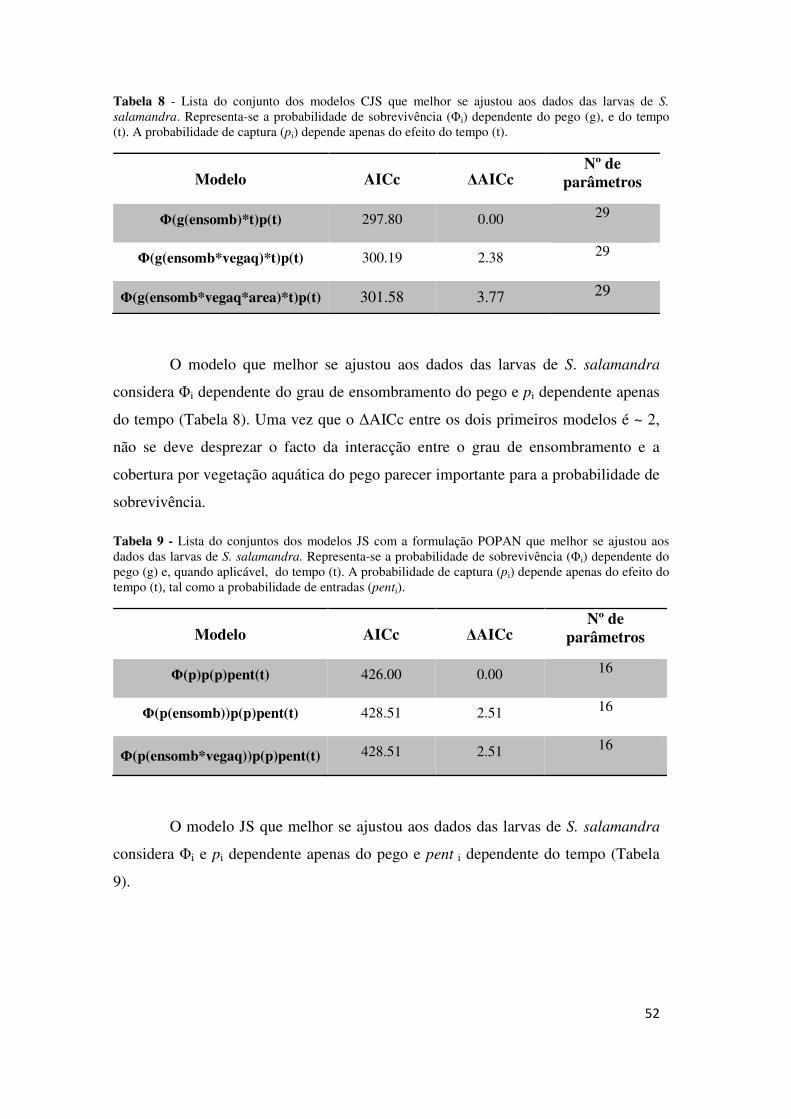
Tabela 8 - Lista do conjunto dos modelos CJS que melhor se ajustou aos dados das larvas de S.
salamandra. Representa-se a probabilidade de sobrevivência (Φi) dependente do pego (g), e do tempo (t). A probabilidade de captura (pi) depende apenas do efeito do tempo (t).
Modelo AICc ∆AICc Nº de
parâmetros
Φ(g(ensomb)*t)p(t) 297.80 0.00 29
Φ(g(ensomb*vegaq)*t)p(t) 300.19 2.38 29
Φ(g(ensomb*vegaq*area)*t)p(t) 301.58 3.77 29
O modelo que melhor se ajustou aos dados das larvas de S. salamandra
considera Φi dependente do grau de ensombramento do pego e pi dependente apenas
do tempo (Tabela 8). Uma vez que o ∆AICc entre os dois primeiros modelos é ~ 2,
não se deve desprezar o facto da interacção entre o grau de ensombramento e a
cobertura por vegetação aquática do pego parecer importante para a probabilidade de
sobrevivência.
Tabela 9 - Lista do conjuntos dos modelos JS com a formulação POPAN que melhor se ajustou aos dados das larvas de S. salamandra. Representa-se a probabilidade de sobrevivência (Φi) dependente do pego (g) e, quando aplicável, do tempo (t). A probabilidade de captura (pi) depende apenas do efeito do tempo (t), tal como a probabilidade de entradas (penti).
Modelo AICc ∆AICc Nº de
parâmetros
Φ(p)p(p)pent(t) 426.00 0.00 16
Φ(p(ensomb))p(p)pent(t) 428.51 2.51 16
Φ(p(ensomb*vegaq))p(p)pent(t) 428.51 2.51 16
O modelo JS que melhor se ajustou aos dados das larvas de S. salamandra
considera Φi e pi dependente apenas do pego e pent i dependente do tempo (Tabela
9).
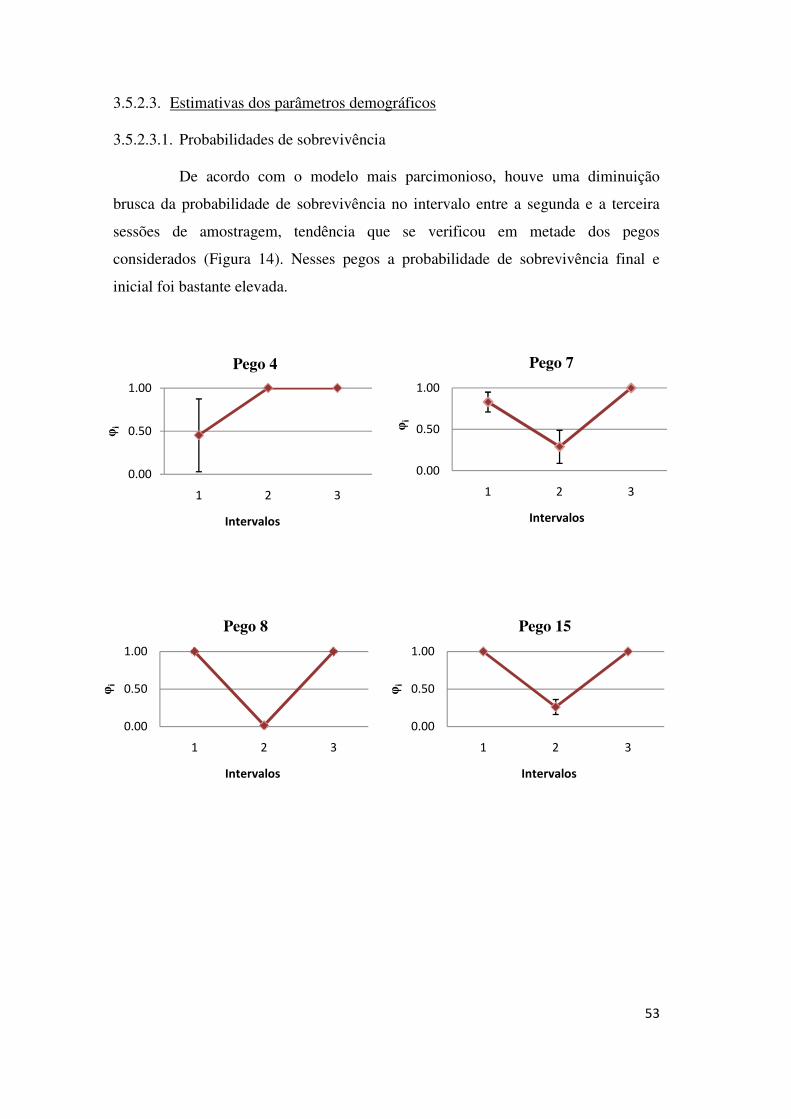
53
0.00
0.50
1.00
1 2 3
φi
Intervalos
Pego 4
0.00
0.50
1.00
1 2 3
φi
Intervalos
Pego 7
0.00
0.50
1.00
1 2 3
φi
Intervalos
Pego 8
0.00
0.50
1.00
1 2 3
φi
Intervalos
Pego 15
3.5.2.3. Estimativas dos parâmetros demográficos
3.5.2.3.1. Probabilidades de sobrevivência
De acordo com o modelo mais parcimonioso, houve uma diminuição
brusca da probabilidade de sobrevivência no intervalo entre a segunda e a terceira
sessões de amostragem, tendência que se verificou em metade dos pegos
considerados (Figura 14). Nesses pegos a probabilidade de sobrevivência final e
inicial foi bastante elevada.
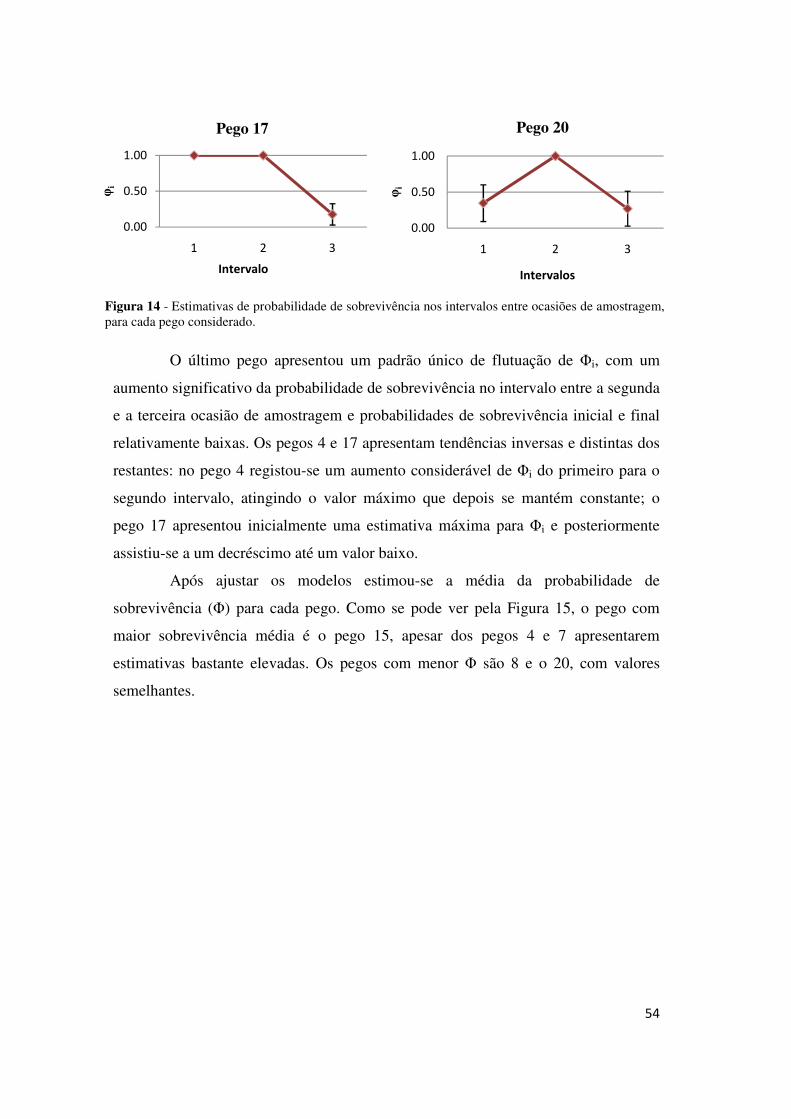
54
0.00
0.50
1.00
1 2 3
φi
Intervalo
Pego 17
0.00
0.50
1.00
1 2 3
φi
Intervalos
Pego 20
O último pego apresentou um padrão único de flutuação de Φi, com um
aumento significativo da probabilidade de sobrevivência no intervalo entre a segunda
e a terceira ocasião de amostragem e probabilidades de sobrevivência inicial e final
relativamente baixas. Os pegos 4 e 17 apresentam tendências inversas e distintas dos
restantes: no pego 4 registou-se um aumento considerável de Φi do primeiro para o
segundo intervalo, atingindo o valor máximo que depois se mantém constante; o
pego 17 apresentou inicialmente uma estimativa máxima para Φi e posteriormente
assistiu-se a um decréscimo até um valor baixo.
Após ajustar os modelos estimou-se a média da probabilidade de
sobrevivência (Φ) para cada pego. Como se pode ver pela Figura 15, o pego com
maior sobrevivência média é o pego 15, apesar dos pegos 4 e 7 apresentarem
estimativas bastante elevadas. Os pegos com menor Φ são 8 e o 20, com valores
semelhantes.
Figura 14 - Estimativas de probabilidade de sobrevivência nos intervalos entre ocasiões de amostragem, para cada pego considerado.

55
0.00
0.10
0.20
0.30
0.40
1 2 3 4
pi
Ocasião
3.5.2.3.2. Probabilidades de captura
A probabilidade de captura manteve-se relativamente constante ao longo
das 4 sessões de amostragem estimáveis (Figura 16), assegurando o cumprimento de
um dos pressupostos mais importantes dos estudos de CR [11; 13].
Figura 16 - Probabilidades de captura de larvas de S. salamandra
estimadas para cada ocasião de amostragem
Figura 15 - Probabilidades de sobrevivência médias (com erro padrão) das larvas de S. salamandra para cada pego.
0.0
0.2
0.4
0.6
0.8
1.0
4 7 8 15 17 20
Φ
Pegos
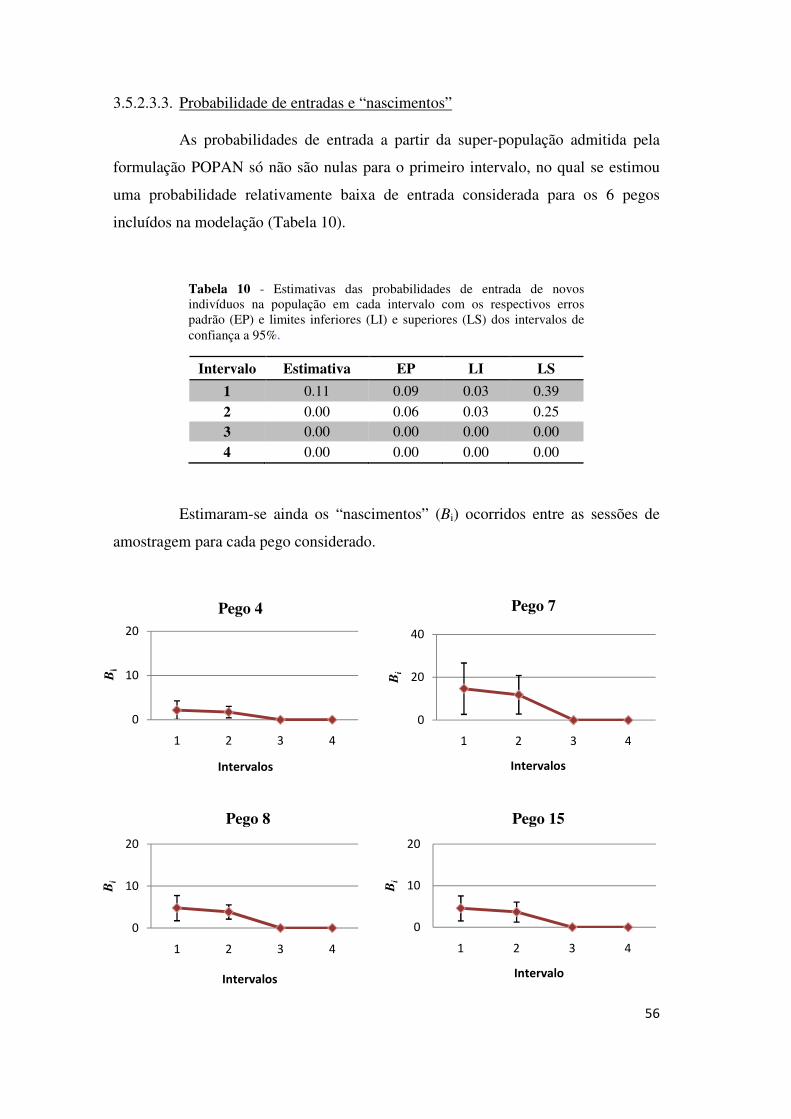
56
0
10
20
1 2 3 4
Bi
Intervalos
Pego 4
0
20
40
1 2 3 4
Bi
Intervalos
Pego 7
0
10
20
1 2 3 4
Bi
Intervalos
Pego 8
0
10
20
1 2 3 4
Bi
Intervalo
Pego 15
3.5.2.3.3. Probabilidade de entradas e “nascimentos”
As probabilidades de entrada a partir da super-população admitida pela
formulação POPAN só não são nulas para o primeiro intervalo, no qual se estimou
uma probabilidade relativamente baixa de entrada considerada para os 6 pegos
incluídos na modelação (Tabela 10).
Tabela 10 - Estimativas das probabilidades de entrada de novos indivíduos na população em cada intervalo com os respectivos erros padrão (EP) e limites inferiores (LI) e superiores (LS) dos intervalos de confiança a 95%.
Intervalo Estimativa EP LI LS
1 0.11 0.09 0.03 0.39
2 0.00 0.06 0.03 0.25 3 0.00 0.00 0.00 0.00
4 0.00 0.00 0.00 0.00
Estimaram-se ainda os “nascimentos” (Bi) ocorridos entre as sessões de
amostragem para cada pego considerado.
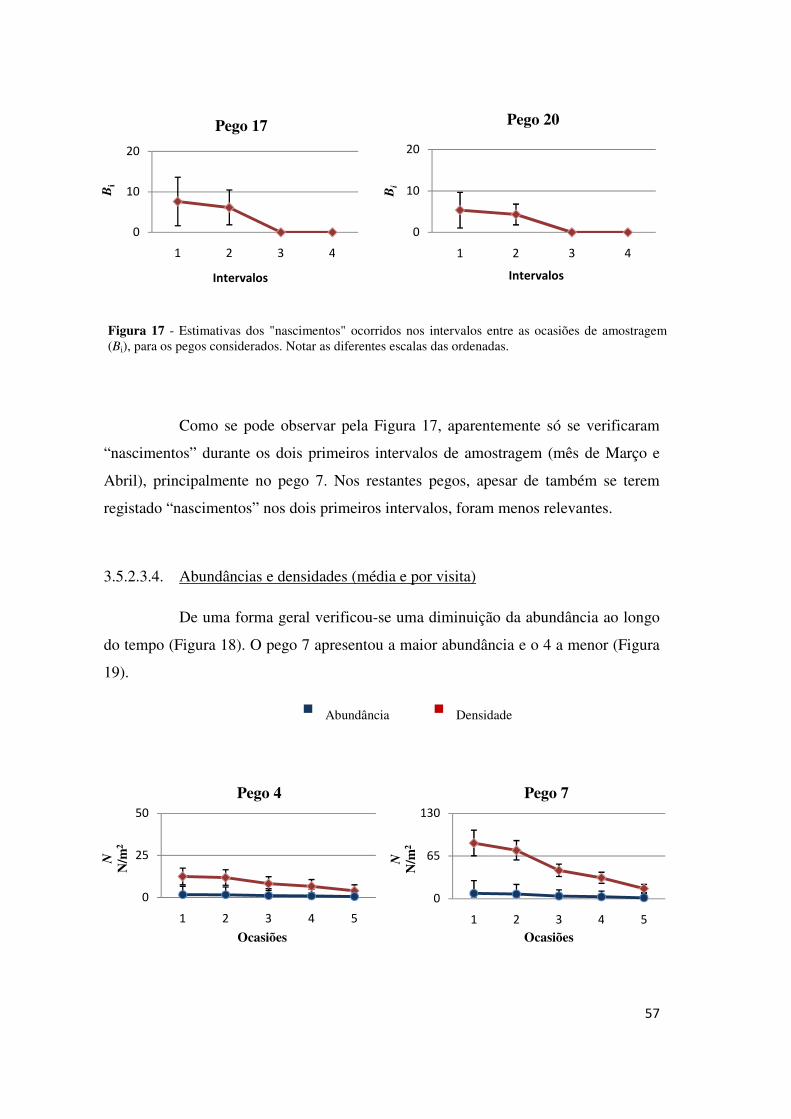
57
0
25
50
1 2 3 4 5
N
N/m
2
Ocasiões
Pego 4
0
65
130
1 2 3 4 5
N
N/m
2
Ocasiões
Pego 7
0
10
20
1 2 3 4
Bi
Intervalos
Pego 17
0
10
20
1 2 3 4
Bi
Intervalos
Pego 20
Como se pode observar pela Figura 17, aparentemente só se verificaram
“nascimentos” durante os dois primeiros intervalos de amostragem (mês de Março e
Abril), principalmente no pego 7. Nos restantes pegos, apesar de também se terem
registado “nascimentos” nos dois primeiros intervalos, foram menos relevantes.
3.5.2.3.4. Abundâncias e densidades (média e por visita)
De uma forma geral verificou-se uma diminuição da abundância ao longo
do tempo (Figura 18). O pego 7 apresentou a maior abundância e o 4 a menor (Figura
19).
Figura 17 - Estimativas dos "nascimentos" ocorridos nos intervalos entre as ocasiões de amostragem(Bi), para os pegos considerados. Notar as diferentes escalas das ordenadas.
� Abundância � Densidade
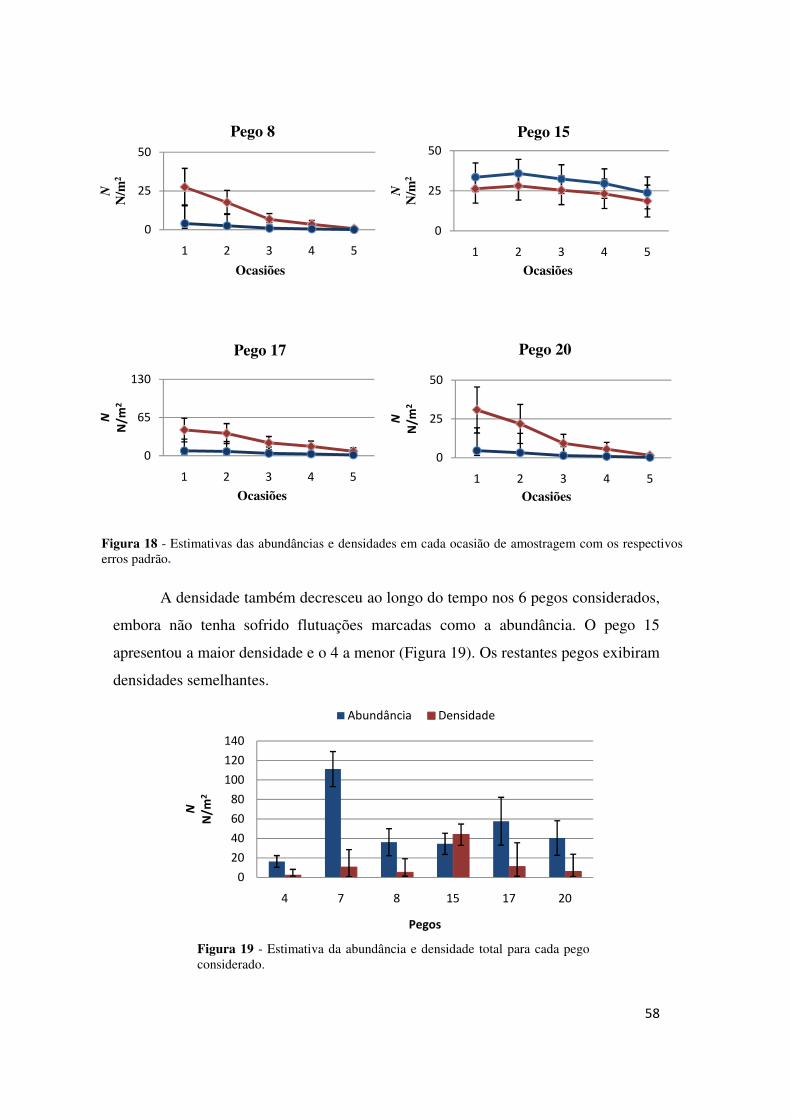
58
0
25
50
1 2 3 4 5
N
N/m
2
Ocasiões
Pego 15
0
65
130
1 2 3 4 5
N
N/m
2
Ocasiões
Pego 17
0
25
50
1 2 3 4 5
N
N/m
2
Ocasiões
Pego 20
0
25
50
1 2 3 4 5
N
N/m
2
Ocasiões
Pego 8
0
20
40
60
80
100
120
140
4 7 8 15 17 20
N
N/m
2
Pegos
Abundância Densidade
A densidade também decresceu ao longo do tempo nos 6 pegos considerados,
embora não tenha sofrido flutuações marcadas como a abundância. O pego 15
apresentou a maior densidade e o 4 a menor (Figura 19). Os restantes pegos exibiram
densidades semelhantes.
Figura 19 - Estimativa da abundância e densidade total para cada pego considerado.
Figura 18 - Estimativas das abundâncias e densidades em cada ocasião de amostragem com os respectivos erros padrão.
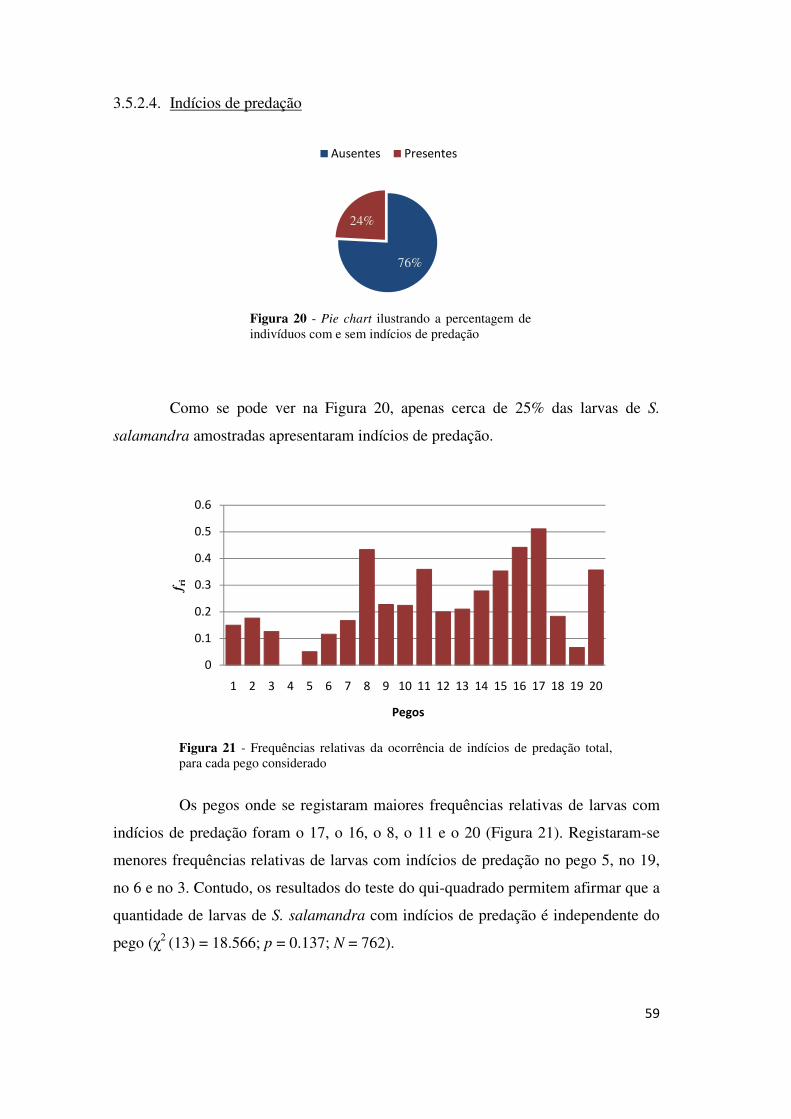
59
76%
24%
Ausentes Presentes
3.5.2.4. Indícios de predação
Como se pode ver na Figura 20, apenas cerca de 25% das larvas de S.
salamandra amostradas apresentaram indícios de predação.
Os pegos onde se registaram maiores frequências relativas de larvas com
indícios de predação foram o 17, o 16, o 8, o 11 e o 20 (Figura 21). Registaram-se
menores frequências relativas de larvas com indícios de predação no pego 5, no 19,
no 6 e no 3. Contudo, os resultados do teste do qui-quadrado permitem afirmar que a
quantidade de larvas de S. salamandra com indícios de predação é independente do
pego (χ2 (13) = 18.566; p = 0.137; N = 762).
Figura 20 - Pie chart ilustrando a percentagem de indivíduos com e sem indícios de predação
Figura 21 - Frequências relativas da ocorrência de indícios de predação total, para cada pego considerado
0
0.1
0.2
0.3
0.4
0.5
0.6
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
f ri
Pegos

60
3.5.2.5. Taxas de crescimento
Como o teste de Kruskal-Wallis indicou (χ2 (7) =15.376, p = 0.031, N = 63),
pela Figura 22 pode observar-se que parece existir alguma heterogeneidade entre as
taxas de crescimento das larvas de S. salamandra de cada pego considerado.
A regressão linear múltipla não identificou nenhuma das variáveis externas
consideradas como preditores significativos da taxa de crescimento (F (2, 57) =
1.560; p = 0.219; R2=0.019). De uma forma geral os pegos com maior variabilidade
de taxas de crescimento são o 5, o 12 e o 13 e os com menor variabilidade são o 6, o
14 e o 16.
3.6. Movimentos entre pegos
Através da recaptura de indivíduos fotoidentificados registaram-se 32
movimentos de larvas de A. cisternasii e 7 de larvas de S. salamandra. A frequência
de movimentos foi significativamente diferente entre as duas espécies estudadas (χ2 =
13.278; p = 0.000; N = 1371).
Figura 22 - Boxplot das taxas de crescimento de larvas de S.
salamandra nos pegos considerados
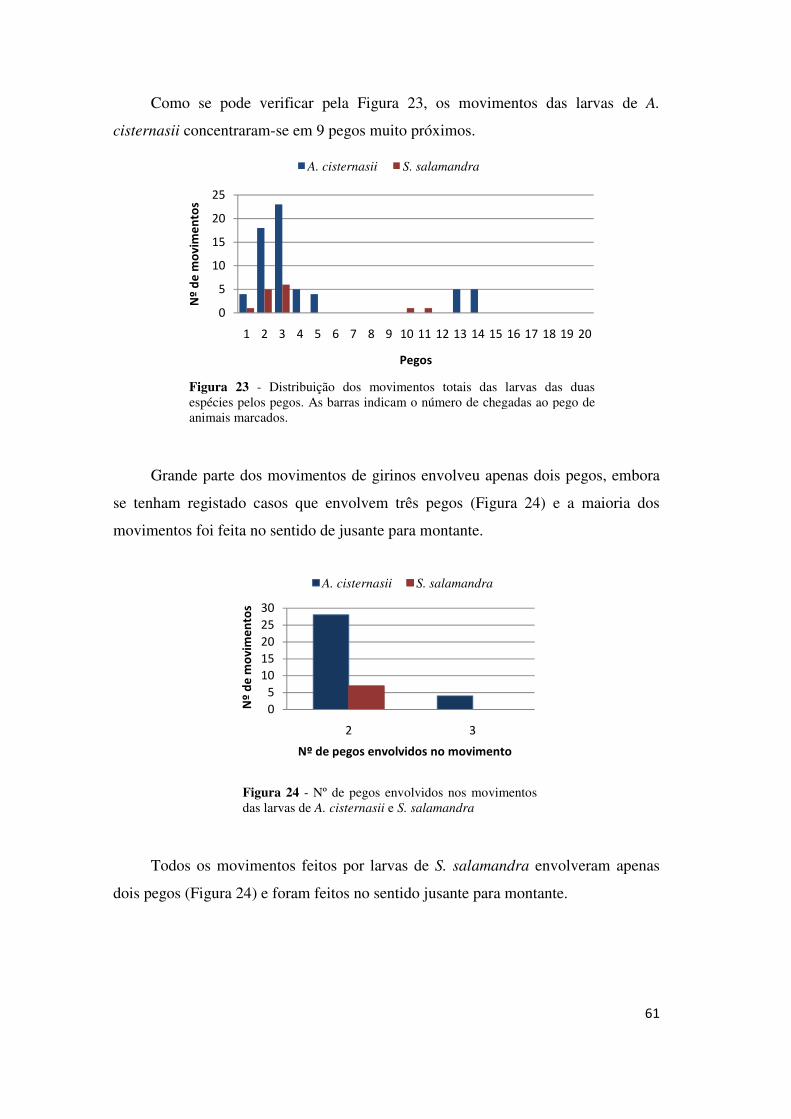
61
0
5
10
15
20
25
30
2 3
Nº
de
mo
vim
en
tos
Nº de pegos envolvidos no movimento
A. cisternasii S. salamandra
0
5
10
15
20
25
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Nº
de
mo
vim
en
tos
Pegos
A. cisternasii S. salamandra
Como se pode verificar pela Figura 23, os movimentos das larvas de A.
cisternasii concentraram-se em 9 pegos muito próximos.
Grande parte dos movimentos de girinos envolveu apenas dois pegos, embora
se tenham registado casos que envolvem três pegos (Figura 24) e a maioria dos
movimentos foi feita no sentido de jusante para montante.
Todos os movimentos feitos por larvas de S. salamandra envolveram apenas
dois pegos (Figura 24) e foram feitos no sentido jusante para montante.
Figura 24 - Nº de pegos envolvidos nos movimentos das larvas de A. cisternasii e S. salamandra
Figura 23 - Distribuição dos movimentos totais das larvas das duas espécies pelos pegos. As barras indicam o número de chegadas ao pego de animais marcados.

62
4. Discussão
Com este trabalho foi pela primeira vez possível obter estimativas da
sobrevivência, abundância e densidade de larvas de anura e caudata, utilizando o
método CR com indivíduos fotoidentificados. O estudo da dinâmica das
comunidades de formas larvares de anfíbios não está muito desenvolvido porque os
métodos de marcação necessários para os estudos CR dificilmente são aplicáveis a
larvas [6]. A fotoidentificação tem sido usada em alguns estudos para marcar
anfíbios adultos mas nunca larvas, devido à dimensão dos indivíduos e à ausência ou
inconstância de padrões individuais.
Este trabalho provou que a fotoidentificação de larvas de anura e de caudata é
possível e produz resultados válidos. A utilização do I3S revelou-se eficaz e
imprescindível para evitar que o processo de re-identificação se tornasse
incomportavelmente moroso e propenso a erros. Provou-se igualmente que é possível
re-identificar indivíduos ao longo do seu desenvolvimento quase até ao final da
metamorfose, apesar de no caso das larvas de A. cisternasii a absorção da cauda
poder dificultar o processo e, no caso das larvas de S. salamandra, a alteração do
padrão de coloração poder inviabilizar a re-identificação nas fases finais da
metamorfose. A grande limitação associada à fotoidentificação prendeu-se com a
fraca capacidade do I3S para lidar com bases de dados de grandes dimensões.
A fotoidentificação ainda não permite acompanhar o indivíduo até à forma
juvenil e adulta porque o padrão utilizado se altera. Esta possibilidade de
acompanhamento seria de grande utilidade para o seguimento de coortes, permitindo
estudos populacionais a médio e longo prazo. Apesar de não acreditarmos ser
possível acompanhar o desenvolvimento do juvenil de A. cisternasii até à idade
adulta, talvez seja admissível para as larvas de S. salamandra porque quando
começam a desenvolver as características manchas amarelas ainda se encontram na
água, sendo facilmente capturáveis. Contudo, como o padrão de cada indivíduo só
estabiliza quando iniciam a reprodução - por volta dos 4 anos [55] -, ter-se-ia de
estudar a evolução do padrão de pigmentação até essa idade para verificar se é
estável e individual.
As larvas de A. cisternasii são herbívoras, com grande capacidade natatória e sem
tendência para apresentarem comportamentos gregários [53].

63
Verificou-se que as probabilidades de sobrevivência médias desta espécie foram
elevadas para grande parte dos pegos considerados, apenas dois pegos a registarem
valores consideravelmente inferiores aos dos restantes. Valores de sobrevivência de
larvas de anura em cursos de água temporários da ordem dos 40% são considerados
normais e da ordem dos 80% elevados [29]. Assim e como a comunidade de
predadores potenciais das larvas de A. cisternasii é bastante diversa [53], as
probabilidades de sobrevivência foram inesperadamente elevadas. As flutuações de
Φi em cada pego não resultaram de alterações significativas no habitat e terão
resultado de factores estocásticos como a presença ocasional de predadores vorazes.
Ao contrário do verificado em muitos estudos laboratoriais [60; 61], as
probabilidades de sobrevivência não foram influenciadas pela densidade total por
pego ou por ocasião. A maioria dos pegos considerados apresentou densidades ~ 10
larvas/m2 havendo, havendo uma excepção (pego 15), com uma densidade total de
40 larvas/ m2 e uma das probabilidades de sobrevivência médias mais elevada.
No pego com maior densidade foi registada uma elevada heterogeneidade das
taxas de crescimento, apesar de não se terem estimado “nascimentos” significativos e
da classe de estádios de desenvolvimento dominante a partir da 2ª sessão ser a III,
sugerindo que o crescimento e desenvolvimento de algumas larvas pode ter sido mais
lento. Nesse caso, a elevada competição intra-específica nos pegos com maiores
densidades afectará provavelmente o tamanho à metamorfose e/ou o período larvar,
mas não a sobrevivência. Os girinos do pego 15 apresentaram também uma
velocidade de desenvolvimento aparentemente inferior às dos restantes pegos, sendo
frequente que se registem desenvolvimentos mais lentos em locais maiores
densidades devido à competição intra-específica [46; 48].
As estimativas de abundância por ocasião indicaram aumentos na segunda sessão
para 5 pegos e na 3ª para 4 pegos. Estas tendências vão de encontro às estimativas de
nascimentos por intervalo, em cada pego. Assim, estes aumentos na abundância não
terão resultado de migração, mas sim do surgimento de novas coortes. Verificou-se
também um aumento da abundância na 5ª sessão para alguns (3) pegos, algo que não
é confirmado pelos nascimentos estimados nem pela classe modal dos estádios de
desenvolvimento. Contudo, surgiu uma nova coorte na 4ª sessão em dois desses
pegos, composta por larvas que, graças às suas dimensões, só terão podido ser

64
amostradas na sessão seguinte. No outro pego não se registou um aumento da
proporção de larvas na classe I de estádios de desenvolvimento, pelo que neste caso
talvez possamos estar perante um fenómeno migratório.
Não foram registadas diferenças significativas entre as taxas de crescimento de
cada pego; contudo, três dos pegos mais a jusante apresentaram maior variabilidade
nessas taxas. A análise de alguns dos parâmetros estudados sugere que dois desses
pegos receberam novas coortes durante o decorrer do estudo. Como as larvas mais
novas apresentam tipicamente taxas de crescimento superiores às das larvas mais
desenvolvidas, pode assumir-se que a elevada variabilidade de taxas de crescimento
terá resultado da mistura de muitos indivíduos recém-eclodidos com outros nas fases
mais tardias do desenvolvimento.
Registaram-se diferenças significativas entre as classes modais de estádio de
desenvolvimento de cada pego. Na sessão 1 todos os pegos foram dominados por
larvas na classe I mas na 2ª sessão muitos pegos albergavam já maioritariamente
larvas na classe III e IV. Na 4ª sessão só se encontraram representadas as classe III e
I, pelo que os indivíduos que nas sessões anteriores se encontravam na classe IV
terão sofrido uma elevada mortalidade ou terão completado a metamorfose e
abandonado o meio aquático.
Neste trabalho verificou-se que cerca de metade das larvas amostradas
apresentou indícios de predação e que a frequência de ocorrência destes variou com
os pegos. Destacaram-se os pegos 11, 4, 2, 13 e 3 com fri > 0.6, particularmente o
pego 11, onde 80% das larvas apresentou indícios de predação. Uma vez que as
probabilidades de sobrevivência médias para os pegos considerados foram elevadas e
que as frequências de ocorrência dos indícios de predação em cada sessão não
coincidiram com as flutuações na probabilidade de sobrevivência em cada intervalo,
pode deduzir-se que a elevada ocorrência de indícios de predação será o resultado de
uma boa capacidade de fuga a tentativas de predação e não de uma elevada
susceptibilidade a ataques predatórios. O corpo dos girinos de A. cisternasii é
composto maioritariamente por uma cauda grande e musculada [53] que proporciona
uma boa capacidade natatória, constituindo simultaneamente um alvo para potenciais
predadores que, atacando apenas a cauda, acabam por não capturar o girino,
potenciando a sobrevivência a ataques.

65
O modelo CR que melhor se ajustou aos dados considera que a percentagem de
manta morta presente no leito do pego é importante para a probabilidade de
sobrevivência dos girinos. Contudo, a profundidade média do pego também se
revelou de alguma importância.
Os pegos com maior percentagem de manta morta no leito foram os que
apresentaram maior estabilidade nas flutuações de Φi. Verificou-se igualmente que
os pegos com maiores probabilidades de sobrevivência médias tinham manta morta
de natureza herbácea, o que pode estar relacionado com uma maior libertação de
nutrientes e produtividade do sistema, aumentando os recursos alimentares das larvas
[40].
O pego 18 apresentou a maior profundidade média, revelando ser o mais instável
no que diz respeito às estimativas de Φi. Entre os 3 pegos mais profundos, os
restantes dois estão entre os que apresentam maiores probabilidades de sobrevivência
médias. Estes são igualmente os que, apresentando maiores profundidades médias,
não sofreram grandes alterações de profundidade ao longo da estação, ao contrário
do pego 18. As menores profundidades estão frequentemente associadas a maior
abundância de predadores, nomeadamente odonatas [38], tendência não confirmada
neste estudo. No entanto, talvez exista menos pressão predatória em pegos em média
mais profundos ou talvez as maiores profundidades proporcionem aos girinos uma
maior possibilidade de fuga a predadores.
As larvas de S. salamandra são predadoras vorazes e caracteristicamente
sedentárias [53].
As probabilidades de sobrevivência médias para cada pego foram muito elevadas
para metade dos pegos (4, 7 e 15) e mais baixas para os restantes (8, 17, 20). Ao
comparar as probabilidades de sobrevivência estimadas para cada intervalo com as
frequências de ocorrência de indícios de predação verificou-se que coincidiam: em
grande parte dos pegos, um aumento na frequência de ocorrência de indícios de
predação numa sessão correspondeu a uma diminuição da probabilidade de
sobrevivência no intervalo imediatamente anterior a essa sessão. Adicionalmente, os
pegos com maiores probabilidades de sobrevivência médias foram os que registaram
menores frequências relativas de indícios de predação enquanto que os pegos com
menores probabilidades de sobrevivência médias foram os que apresentaram as

66
maiores frequências de indícios de predação. Estas tendências, aliadas ao facto da
grande maioria dos indivíduos amostrados não apresentar qualquer indício de
predação, sugerem que as larvas de S. salamandra são muito susceptíveis a ataques
por predadores e possuem pouca capacidade de fuga.
A maioria dos pegos apresentou densidades por ocasião relativamente estáveis -
apesar das flutuações consideráveis da abundância - e densidades totais da ordem das
15 larvas/m2. O pego 15 é novamente o que se destaca com uma densidade bem
superior às dos restantes (ca. de 40 larvas/m2). Apesar disto, os “nascimentos”
estimados para o pego 15 não foram relevantes, nem nos primeiros intervalos. É de
salientar que a elevada densidade parece não ter afectado a probabilidade de
sobrevivência já que os valores para este parâmetro, bem como as suas flutuações,
são semelhantes às de outros pegos.
As taxas de crescimento diferiram significativamente entre pegos, com os pegos
3, 6, 14 e 16 a apresentar menor variabilidade de taxas e os pegos 5, 12 e 13 maior
variabilidade. Os pegos 7 e 15 apresentaram as menores variabilidades e as medianas
mais baixas, o que se pode dever às suas elevadas densidades. No entanto, apesar de
aparentemente os indivíduos crescerem pouco e de forma semelhante, registou-se
uma elevada probabilidade de sobrevivência. Dentro dos pegos considerados para
estimar as taxas de crescimento, o 15 e o 16 apresentaram a menor heterogeneidade,
a mediana mais baixa e a maior frequência de ocorrência de indícios de predação.
Assim, talvez a pressão predatória tenha conduzido à aceleração do desenvolvimento
em detrimento do crescimento, já que a probabilidade de sobrevivência média no
pego 15 foi muito elevada.
De acordo com os modelos CR, o ensombramento e, com menor importância, a
vegetação aquática, influenciam as probabilidades de sobrevivência. Os 2 pegos com
maiores probabilidades de sobrevivência médias têm ambos baixos a médios graus
de ensombramento, enquanto que os 2 pegos com menores probabilidades de
sobrevivência foram o 37 e o 51, ambos sem vegetação aquática. Estes factores
poderão estar relacionados principalmente com a disponibilidade alimentar para as
larvas. Estas consomem principalmente ninfas de Ephemeroptera [62] que se
alimentam de detritos e de matéria vegetal. Assim, quanto maior for a incidência de
radiação solar e mais vegetação aquática houver no pego, mais ninfas de

67
Ephemeroptera existirão e, consequentemente, maior disponibilidade de recursos
alimentares para as larvas. Adicionalmente, mais vegetação aquática pode aumentar
a complexidade estrutural do habitat e a disponibilidade de refúgios [44].
Apesar de ser admissível que estas variáveis exerçam maior influência sobre a
sobrevivência larvar, os meios naturais são intrinsecamente complexos e os factores
que influenciam a performance larvar exercem efeitos sinergéticos difíceis de
detectar [33; 35]. As flutuações estimadas para a probabilidade de sobrevivência ao
longo do período de amostragem não são totalmente explicáveis pelo grau de
ensombramento ou pela vegetação aquática, uma vez que estes se mantiveram
relativamente constantes. Contudo, poderão explicar algumas das diferenças entre
pegos. Por exemplo, os pegos 15 e 7 apresentaram condições que podem ter
favorecido a produção de larvas mais robustas; o pego 15 apresenta um grau de
ensombramento mediano e o 36, apesar de ser muito ensombrado em geral, tinha
muitos locais totalmente expostos à radiação, daí apresentar alguma vegetação
aquática. Por outro lado, o pego 8 é o que apresenta a flutuação mais marcada, com a
queda da probabilidade de sobrevivência até zero, tratando-se de um pego com
elevado grau de ensombramento e nenhuma vegetação aquática. Desta forma, as
diferentes características dos pegos podem possibilitar o desenvolvimento de larvas
maiores, que tenham maior probabilidade de escapar a ataques de predadores.
Apesar de carnívoras e potencialmente canibais [53] não se registaram indícios
evidentes de ocorrência de canibalismo entre larvas de S. salamandra já que os pegos
com maiores densidades foram os que registaram maiores probabilidades de
sobrevivência.
As larvas de A. cisternasii concentraram-se mais nos pegos a montante e a
jusante enquanto que as de S. salamandra se concentraram nos pegos da secção
intermédia da ribeira. Esta tendência poderá estar relacionada com a densidade de
indivíduos adultos das duas espécies nas regiões referidas e com o facto destes não se
deslocarem muito para depositar as suas larvas. Geralmente as salamandras-comuns
adultas encontram-se em locais com muita cobertura arbórea, sob a manta morta [53;
54] e a secção intermédia da ribeira em questão era sem dúvida a que possuía a
galeria ribeirinha com maior cobertura arbórea. Contudo, de acordo com Rebelo,

68
2000 (com.Pess.) esta secção é onde se encontram menos adultos de S. salamandra,
sendo mais abundantes na secção da ribeira mais a montante.
De acordo com os dados obtidos, os sapos-parteiros-ibéricos depositam mais
posturas, de maior dimensão e durante um período mais longo do que as salamandras
comuns uma vez que os “nascimentos” registados para A. cisternasii são de maior
magnitude e mais frequentes dos que os registados para S. salamandra.
As sobrevivências médias de S. salamandra foram bastante diferentes entre
pegos, com a separação em dois grupos bastante distintos. As probabilidades de
sobrevivência médias de larvas de A. cisternasii não variaram muito entre pegos,
tendo sido em geral elevadas. Isto talvez se deva à maior capacidade de adaptação
das larvas de A. cisternasii a muitos microhabitats ou talvez estes valores estejam a
ser enviesados pelo facto do desenvolvimento larvas das S. salamandra ser mais
acelerado e a probabilidade de sobrevivência considerar tanto mortes como saídas da
área de amostragem.
A análise dos parâmetros estimados pelos modelos CR para as larvas de caudata
e anura e das frequências de ocorrência de indícios de predação permite afirmar que
as larvas de S. salamandra serão menos capazes de sobreviver a um episódio de
predação do que as larvas de A. cisternasii. O facto de, apesar da grande maioria dos
indivíduos amostrados não apresentarem qualquer indício de predação, quase todas
as sessões onde se registaram mais indivíduos com indícios de predação
corresponderem a diminuições na probabilidade de sobrevivência, sugere que a
morfologia destas larvas não lhes permite escapar frequentemente a ataques
predatórios. Pelo contrário, as larvas de A. cisternasii apresentam morfologicamente
todas as características que potenciam a sobrevivência a um ataque predatório.
Uma vez que os pegos onde se registaram maiores frequências de ocorrência de
indícios de predação em larvas de A. cisternasii não coincidiram com os pegos com
maiores densidades de larvas de S. salamandra pode assumir-se que estas não terão
sido os principais predadores dos girinos de A. cisternasii. Apesar das larvas de S.
salamandra serem predadores vorazes, não atacarão muitas larvas de A. cisternasii,
provavelmente por constrições da cavidade bucal e pela baixa probabilidade de
sucesso na captura de presas com uma grande capacidade natatória.

69
As larvas de A. cisternasii aparentam ser muito mais móveis do que as larvas de
S. salamandra. Os movimentos parecem ter-se concentrado nos 5 pegos mais a
montante e em 4 pegos na secção intermédia da ribeira, de uma forma geral
relativamente pequenos, muito próximos e muito pouco ensombrados. Os
movimentos por parte de larvas de S. salamandra foram bem inferiores aos
registados noutro estudo que indicou uma taxa de movimentação de 19 - 41% [63]. O
mesmo estudo concluiu que as larvas que se movimentavam eram as recém-eclodidas
e as menos nutridas, sendo que a movimentação das primeiras era involuntária e a
das outras comportamental. Neste trabalho as larvas que se movimentaram não eram
recém-eclodidas, e o pego de origem não foi incluído na modelação por falta de
dados para 4 sessões; contudo, era um pego com grande abundância de salamandras.
Assim, talvez a movimentação por parte destas larvas tenha sido motivada pela falta
de recursos alimentares no pego original. Uma vez que a grande maioria dos
movimentos registados foi feito de jusante para montante, pode excluir-se a migração
involuntária, ou seja, as larvas não terão sido arrastadas. De uma forma geral, os
movimentos concentraram-se entre as 3 primeiras sessões, coincidindo com grandes
frequências de ocorrência de indícios de predação em alguns pegos, o que sugere que
os movimentos podem ser motivados por ataques predatórios, constituindo
mecanismos de fuga. Como já foi referido, as larvas de A. cisternasii estão
morfologicamente preparadas para fugir, ao contrário das de S. salamandra e talvez
por isso se tenham verificado diferenças tão marcadas na quantidade de movimentos
efectuados pelas larvas das duas espécies.
As larvas de A. cisternasii apresentaram maiores densidades do que as registadas
para S. salamandra, com mais pegos a atingir a fasquia das 20 larvas/m2. O pego 15
tem elevada densidade de larvas de ambas as espécies.
Registaram-se menos diferenças entre as taxas de crescimento de larvas de A.
cisternasii entre pegos. Como a mediana das taxas de crescimento relativas às larvas
de anura nunca ultrapassa os 2 mm/semana e as relativas às larvas de caudata
ultrapassam essa fasquia em quase todos os pegos pode sugerir-se que as larvas de S.
salamandra crescem mais rapidamente, o que estará relacionado com a velocidade
de desenvolvimento.
Com este trabalho foi possível verificar que as larvas de A. cisternasii têm
elevadas taxas de sobrevivência, grande capacidade de fuga a ataques por parte de

70
predadores e que a sobrevivência não é prejudicada pelo aumento da densidade. Foi
também demonstrado que ambas as espécies conseguem atingir taxas de
sobrevivência elevadas em meios com elevadas densidades, ao contrário do que
alguns estudos laboratoriais sugerem [60; 61]. Apesar da influência de densidades
elevadas sobre os indivíduos ser incontestável, não é frequente que em meios
naturais as densidades atinjam o limite a partir do qual passam registar-se efeitos
individuais negativos [34; 46]. Os factores que mais influenciaram a sobrevivência
das larvas de A. cisternasii foram a quantidade e eventualmente o tipo de manta
morta e a profundidade média do pego, acentuando a importância dos recursos
alimentares e da existência de microhabitats para evitar predadores. Já a
sobrevivência das larvas de S. salamandra parecer ser mais influenciada pelo grau de
ensombramento e quantidade de vegetação aquática do pego, ambos relacionados
com a disponibilidade das presas preferenciais e, eventualmente com a defesa contra
predadores.
A identificação de factores reguladores tem fortes implicações sobre a gestão e
conservação de populações selvagens. No caso de animais com ciclos de vida
complexos como os anfíbios os diferentes estágios são geralmente dependentes de
factores distintos, tornando a investigação e conservação destas populações bem mais
difícil. Como neste trabalho não se verificou efeito algum da densidade larvar de
ambas as espécies sobre a sobrevivência, futuras acções de gestão ou conservação
deverão incidir preferencialmente sobre o habitat terrestre dos juvenis e adultos.
O presente trabalho vem suportar a afirmação da fotoidentificação como um
método de marcação aplicável a larvas de anfíbios. Face à presente situação
ecologicamente instável destes vertebrados é urgente desenvolver estudos de
dinâmica populacional que não prejudiquem a capacidade de sobrevivência,
crescimento e/ou desenvolvimento das formas larvares.
Actualmente está na fase final de desenvolvimento um software de análise de
imagem desenvolvido para estudos CR com fotoidentificação da autoria de uma
equipa do Massachusetts Institute of Technology. Este programa terá vantagens sobre
o I3S relacionadas principalmente com a capacidade de manipulação de ângulos e
eliminação de “ruído”, potenciando a eficácia da fotoidentificação.

71
REFERÊNCIAS BIBLIOGRÁFICAS2
1. Blaustein, A.R. & Wake, D.B. (1990) Declining amphibian populations: a global
phenomenon? Trends in Ecology and Evolution, 5, 203-204.
2. Blaustein, A.R., Kiesecker, J.M., Chivers, D.P., Hokit, D.G., Marco, A., Belden,
L.K. & Hatch, A. (1998) Effects of ultraviolet radiation on amphibians: Field
experiments. American Zoologist, 38, 799-812.
3. Carey, C., Heyer, W.R., Wilkinson, J., Alford, R.A., Arntzen, J.W., Halliday, T.,
Hungerford, L., Lips, K.R., Middleton, E.M., Orchard, S.A. & Rand, S.A. (2001)
Amphibian declines and environmental change: Use of remote sensing data to
identify environmental correlates. Conservation Biology, 15, 903-913.
4. Kiesecker, J.M., Blaustein, A.R. & Belden, L.K. (2001) Complex causes of
amphibian population declines. Nature, 410, 681–684.
5. Alford, R.A. & Richards, S. J. (1999) Global Amphibian Declines: A Problem in
Applied Ecology. Annual Review of Ecology and Systematics, 30, 133-165.
6. Donnelly, M. A., Guyer, G., Juterbock, J. E. & Alford, R. A. (1994) Techniques
for marking amphibians. In: Measuring and Monitoring Biological Diversity.
Standard Methods for Amphibians (Eds. Heyer, W. R., Donnelly, M. A.,
McDiarmid, R. W., Hayek, L. C. & Foster, M. S.). Smithsonian Institution Press,
Washington. pp. 277-284.
7. Likens, G. E. (Ed.) (1989) Long-term studies in ecology. Springer-Verlag. Berlim.
214 pp.
8. Whiles, M.R., Lips, K.R., Pringle, C.M., Kilham, S.S., Bixby, R.J., Brenes, R.,
Connelly, S., Colon-Gaud, J.C., Hunte-Brown, M., Huryn, A.D., Montgomery, C.,
& Peterson, S. (2006) The effects of amphibian population declines on the
structure and function of Neo-tropical stream ecosystems. Frontiers in Ecology
and the Environment, 4, 27-34.
9. Pollock, K. H., J. D. Nichols, C. Brownie, & Hines, J. E. (1990) Statistical
inference for capture-recapture experiments. Wildlife Monographs, 107, 97 pp.
10. Amstrup, S.C., McDonald, T. L. & Manly, B.F.J. (Eds.) (2005) Handbook of
Capture-Recapture Analysis. Princeton University Press, New Jersey. 313 pp.
2 De acordo com as regras do Journal of Animal Ecology.

72
11. Lebreton, J.D., Burnham, K.P., Clobert, J. & Anderson, D.R. (1992) Modeling
survival and testing biological hypothesis using marked animals: a unified
approach with case studies. Ecological Monographs, 62, 67-118.
12. Pine, W.E., Pollock, K.H., Hightower, J.E., Kwak, T.J. & Rice, J.A. (2003) A
review of tagging methods for estimating fish population size and components of
mortality. Fisheries, 28, 10-23.
13. White, G.C. & Burnham, K.P. (1999) Program MARK: Survival estimation from
populations of marked animals. Bird Study, 46, 120-138.
14. Lindberg, M. & Rexstad, E. (2002) Capture-recapture sampling designs. In El-
Shaarawi, A.H. e Piegorsch, W.W. (Eds). Encyclopedia of Environmetrics, 1, pp
251-262. John-Wiley & Sons Ltd, Chichester.
15. Murteira, B., Ribeiro, C.S., Silva, J.A. & Pimenta, C. (2002) Introdução à
Estatística. McGraw-Hill. Lisboa. 647 pp.
16. Burnham, K.P., Anderson, D.R., White, G.C., Brownie, C. & Pollock P.H. (1987)
Design and analysis methods for fish survival experiments based on release–
recapture. American Fisheries Society Monograph, 5.
17. Cormack, R.M. (1964) Estimates of survival from sightings of marked animals.
Biometrika, 51, 429-438.
18. Jolly, G. M. (1965) Explicit estimates from capture-recapture data with both death
and immigration: stochastic model. Biometrika, 52, 225-247.
19. Seber, G.A.F. (1965) A note on multiple recapture census. Biometrika, 52, 249-
259.
20. Link, W.A. & Barker, R.J. (2005) Modeling association among demographic
parameters in analysis of open population capture-recapture data. Biometrics, 61,
46-54.
21. Choquet, R., Reboulet, A.M., Pradel, R., Gimenez, O. & Lebreton, J.D. (2004)
M–SURGE: an integrated software for multi-state recapture models. Animal
Biodiversity and Conservation, 27, 207–215.
22. Reboulet, A.M., Viallefont, A., Pradel, R. & Lebreton, J.D. (1998) Selection of
survival and recruitment models with Surge 5.0. Bird Study, 46, 148–156.
23. Barker, R.J. & White, G.G. (2004) Towards the mother-of-all-models: customized
construction of the mark recapture likelihood function. Animal Biodiversity and
conservation, 27, 177-185.

73
24. Bonesi, L. & MacDonald, D.W. (2003) Evaluation of sign surveys as a way to
estimate the relative abundance of American mink (Mustela vison). Journal of
Zoology, 262, 65-72.
25. Begon, M., Townsend, C.R. & Harper, J.L. (2007) Ecologia: de indivíduos e
ecossistemas. Artmed. Porto Alegre. 752 pp.
26. Frisch, A.J. & Hobbs, J.P.A. (2007) Photographic identification based on unique,
polymorphic colour patterns: A novel method for tracking a marine crustacean.
Journal of Experimental Marine Biology and Ecology, 351, 294-299.
27. Sagar, J.P., Olson, D.H. & Schmitz, R.A. (2007) Survival and growth of larval
Coastal Giant Salamanders (Dicamptodon tenebrosus) in streams of the Oregon
Coast Range. Copeia, 123-130.
28. Rebelo, R. & Leclair, M.H. (2003) Site tenacity in the terrestrial salamandrid
Salamandra salamandra. Journal of Herpetology, 37, 440-445.
29. Bradfield, K.S. (2004) Photographic identification of individual Archey’s frog,
Leiopelma archeyi, from natural markings. DOC Science Internal Series 191.
Department of Conservation. Wellington. 36pp.
30. Cecil, S. G. & Just, J.J. (1978) Use of acrylic polymers for marking of tadpoles
(Amphibia, Anura). Journal of Herpetology, 12, 95-96.
31. Van Tienhoven, A.M., Den Hartog, J.E., Reijns, R.A. & Peddemors, V.M. (2007)
A computer-aided program for pattern-matching of natural marks on the spotted
raggedtooth shark Carcharias Taurus. Journal of Applied Ecology, 44, 273-280.
32. Hartog, J. & Reijns, R. (2007) Interactive Individual Identification System (I3S).
Reijns Free Software Foundation Inc.. Boston. 27pp.
33. Van Buskirk, J. (2005) Local and landscape influence on amphibian occurrence
and abundance. Ecology, 86, 1936-1947.
34. Manenti, R., Ficetola, G.F. & Bernardi, F. (2009) Water, stream morphology and
landscape: complex habitat determinants for the fire salamander Salamandra
salamandra. Amphibia-Reptilia, 30, 7-15.
35. Skelly, D.K., Werner, E.E. & Cortwright, S.A. (1999) Long-term distributional
dynamics of a Michigan amphibian assemblage. Ecology, 80, 2326–2337.
36. Block, M & Stoks, R. (2005) Fitness effects from egg to reproduction: bridging
the life history transition. Ecology, 86, 185-197.

74
37. Lauck, B., Swain, R. & Barmuta, L. (2005) Breeding site characteristics
regulating life history traits of the brown tree frog, Litoria ewingii. Hydrobiologia,
537,135-146.
38. Pearman, P. (1995) Effects of pond size and consequent predator density on two
species of tadpoles. Oecologia, 102, 1-8.
39. Corbet, P.S. (1999) Dragonflies: Behavior and ecology of Odonata. Comstock
Press. Cornell University. Ithaca. 829 pp.
40. Williams, B.K., Rittenhouse, T.A. & Semlitsch, R. (2008) Leaf litter input
mediates tadpole performance across forest canopy treatments. Oecologia, 155,
377-384.
41. Schiesari, L. (2006) Pond canopy cover: a resource gradient for anuran larvae.
Freshwater Biology, 51, 412–423.
42. Rubbo, M.J., Mirza, R.S., Belden, L.K., Falkenbach V, J.J., Storrs, S. I. &
Kiesecker, J.M. (2006) Evaluating a predator-prey interaction in the field: the
interaction between beetle larvae (predator) and tadpoles (prey). Journal of
Zoology 269, 1-5.
43. Webster, J.R. & Benweld, E.F. (1986) Vascular plant breakdown in freshwater
ecosystems. Annual Review of Ecology and Systematics, 17, 567–594.
44. Sredl, M.J. & Collins, J.P. (1992) The interaction of predation, competition and
habitat complexity in structuring an amphibian community. Copeia, 607-614.
45. Marsh, D.M., Fegraus, E.H. & Harrison, S. (1999) Effects of breeding pond
isolation on the spatial dynamics of pond use by the tungara frog Physalaemus
pustulosus. Journal of Animal Ecology, 68, 804-814.
46. Loman, J. (2001) Intraspecific competition in tadpoles of Rana arvalis: does it
matter in nature? A field experiment. Population Ecology, 43, 253-263.
47. Nowakowski, A. & Maerz, J. (2009) Estimation of larval stream salamander
densities in three proximate streams in Georgia Piedmont. Journal of
Herpetology, 43, 503-509.
48. Morey, S. & Reznick, D. (2001) Effects of larval density on postmetamorphic
spadefoot toads (Spea hammondii). Ecology, 82, 510-522.
49. Binckley, C.A. & Resetarits, W.J. (2007) Effects of forest canopy on habitat
selection in treefrogs and aquatic insects: implications for communities and
metacommunities. Oecologia, 153, 951–958.

75
50. Wissinger, S.A., Bohonak, A.J., Whiteman, H.H. & Brown, W.S. (1999)
Subalpine wetlands in Colorado: habitat permanence, salamander predation and
invertebrate communities. Pp 757–790. In Batzer, D.P., Rader, R.B. & Wissinger,
S. A. (Eds.) Invertebrates in Freshwater Wetlands of North America: Ecology and
Management. John Wiley and Sons. New York. USA. 1120 pp.
51. Gibbons, J.W., Winne, C.T., Scott, D.E., Willson, J.D., Glaudas, X., Andrews,
K.M., Todd, B.D., Fedewa, L.D., Wilkinson, L., Tsaliagos, R.N., Harper, S.J.,
Greene, J.L., Tuberville, T.D., Metts, B.S., Dorcas, M.E., Nestor, J.P., Young,
C.A., Akre, T., Reed, R.N., Buhlmann, K.A., Norman, J., Croshaw, D.A., Hagen,
C. & Rothermel, B.B. (2006) Remarkable amphibian biomass and abundance in
an isolated wetland: implications for wetland conservation. Conservation Biology,
20, 1457–1465.
52. Mesquita, N., Coelho, M.M. & Magalhães, M.F. (2006) Spatial variation in fish
assemblages across small Mediterranean drainages: effects of habitat and
landscape context. Environmental Biology of Fishes, 77, 105-120.
53. Almeida, N.F., Almeida, P.F., Gonçalves, H., Sequeira, F., Teixeira, J. &
Almeida, F.F. (2001) Guia FAPAS Anfíbios e Répteis de Portugal. FAPAS. Porto.
249pp.
54. Rebelo, R. & Crespo, E.G. (1999) Anfíbios. Pp 177-188. In M. Santos-Reis & A.I.
Correia, (Eds). Caracterização da flora e fauna do montado da Herdade da
Ribeiro Abaixo. CBA. Lisboa. 262 pp.
55. Barbadilho, E. & Javier, L. (1987) La guia de incafo de los anfíbios y reptiles de
la Peninsula Iberica, islas baleares y Canárias. Incafo, S.A. 694 pp.
56. Mueller-Dombois, D. & Ellenberg, H. (1974) Aims and methods of vegetation
ecology. John-Wiley & Sons. New York. 547 pp.
57. Guerreiro, C. (2007) Métodos de fotoidentificação individual de girinos Alytes
cisternasii (Boscá, 1879). Projecto de Tese de Mestrado em Biologia da
Conservação. Faculdade de Ciências da Universidade de Lisboa.
58. Gosner, K.L. (1960) A simplified table for staging anuran embryos and larvae
with notes on identification. Herpetologica, 16, 183-190.
59. Arnason, A. N. & Schwarz, C. J. (1995) POPAN-4: enhancements to a system for
the analysis of mark–recapture data from open populations. Journal of Applied
Statistics, 22, 785–800.

76
60. Alvarez, R. & Herraez, M.P. (1996) Growth and metamorphosis of Rana perezi
larvae in culture: Effects of larval density. Aquaculture, 142, 163-170.
61. Browne, R.K., Pomering, M. & Hamer, A.J. High density effects on the growth,
development and survival of Litoria aurea tadpoles. Aquaculture, 215, 109-121.
62. Ferreira, G. (2000) Haverá polimorfismo trófico em larvas de Salamandra
salamandra? Relatório de estágio para obtenção da Licenciatura em Biologia
Aplicada aos Recursos Animais – Variante Terrestres. Faculdade de Ciências da
Universidade de Lisboa.
63. Thiesmeier, B. & Schuhmacher, H. (1990) Causes of larval drift of the fire
salamander Salamandra salamandra terrestris and its effects on population
dynamics. Oecologia, 82, 259-263.

77
ANEXOS

Larva de Alytes cisternasii
Figura 25 – Larva de 2009.
Figura 26 - Larva de A. cisternasii
2009.
Figura 27 - Larva de A. cisternasii
2009.
Figura 28 - Larva de A. cisternasii
ANEXO I
recapturado 3 vezes ao longo do período de amostragem.
de A. cisternasii capturada pela primeira vez dia 12 de Março de
A. cisternasii recapturada pela primeira vez dia 20 de Março de
A. cisternasii recapturada pela segunda vez dia 10 de Abril de
A. cisternasii recapturada pela terceira vez dia 20 de Abril de 2009.
78
recapturado 3 vezes ao longo do período de amostragem.
capturada pela primeira vez dia 12 de Março de
20 de Março de
recapturada pela segunda vez dia 10 de Abril de
recapturada pela terceira vez dia 20 de Abril de 2009.

79
ANEXO II
Larva de Salamandra salamandra recapturada 3 vezes ao longo do período de
amostragem.
Figura 1 - Larva de S. salamandra capturada pela primeira vez dia 9 de Abril de 2009.
Figura 2 - Larva de S. salamandra recapturada pela primeira vez dia 15 de Abril de 2009.
Figura 3 - Larva de S. salamandra recapturada pela segunda vez dia 19 de Abril de 2009.
ANEXO III

80
Exemplos de indícios de predação em larvas de A. cisternasii
Exemplos de indícios de predação em larvas de S. salamandra.
ANEXO IV

81
Excerto de um ficheiro com os historiais de captura de larvas de A. cisternasii em 7
pegos. A primeira coluna corresponde ao historial de captura dos indivíduos, as
restantes colunas contabilizam o número de indivíduos com o historial de captura
respectivo, para cada grupo/pego (7 colunas correspondentes a 7 pegos).
111100 0 1 0 0 0 0 0 1;
100000 7 17 27 6 4 5 2 13;
110000 1 2 8 3 0 0 1 1;
111010 0 1 1 0 0 1 1 0;
101000 0 1 1 1 1 0 0 2;
110100 0 1 0 0 0 0 1 0;
111101 0 1 0 0 0 0 0 0;
100001 1 0 0 0 0 0 0 0;
111011 0 0 1 0 0 0 0 0;
111000 0 0 4 0 0 0 0 1;
100110 0 0 1 0 0 0 1 0;
100101 0 0 0 1 0 0 0 0;
110010 0 0 0 1 0 1 0 1;
100100 0 0 0 1 1 0 0 0;
110110 0 0 0 1 1 0 0 0;
100010 0 0 0 0 1 0 2 1;
010000 7 10 25 9 7 4 2 5;
011010 0 2 1 0 2 0 0 1;
011000 0 3 4 2 2 0 1 1;
011100 0 2 1 1 0 0 0 0;
010100 0 1 4 1 0 0 0 0;
011001 0 1 2 0 0 0 0 0;
010001 0 0 1 0 0 0 0 0;
010110 0 0 1 0 0 1 0 1;
010010 0 0 0 1 2 1 1 0;
011110 0 0 0 0 0 0 0 1;
ANEXO V
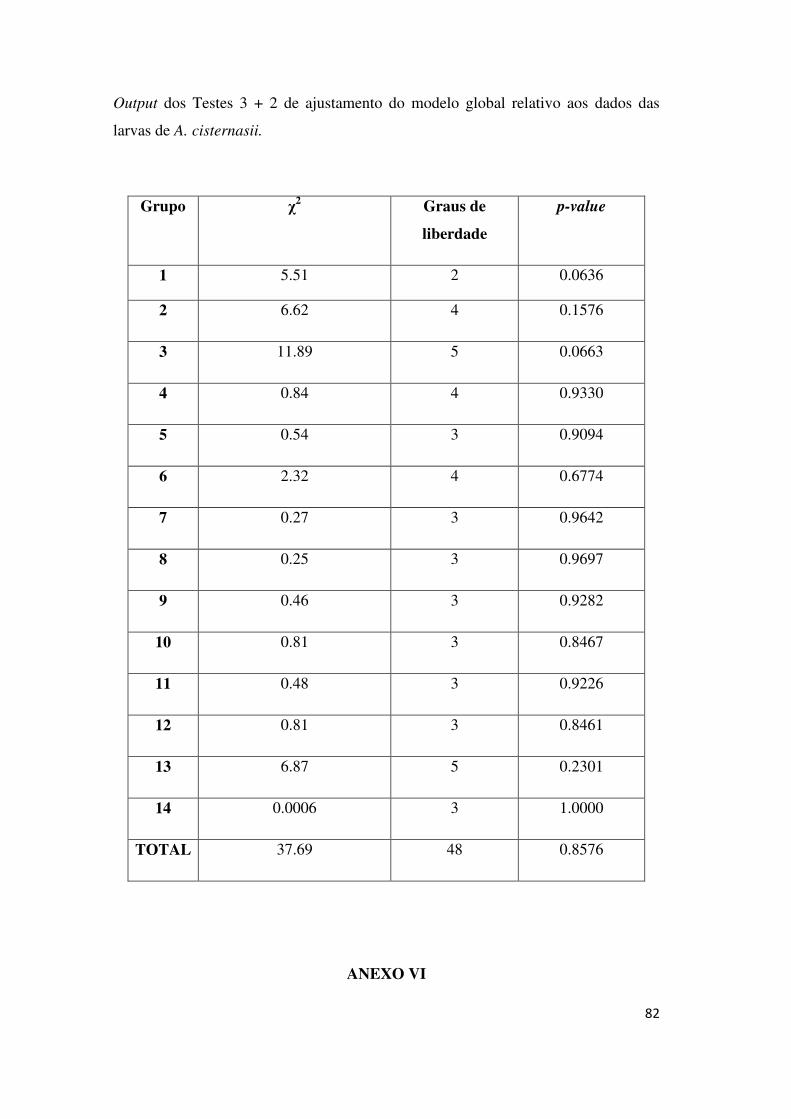
82
Output dos Testes 3 + 2 de ajustamento do modelo global relativo aos dados das
larvas de A. cisternasii.
ANEXO VI
Grupo χ2 Graus de
liberdade
p-value
1 5.51 2 0.0636
2 6.62 4 0.1576
3 11.89 5 0.0663
4 0.84 4 0.9330
5 0.54 3 0.9094
6 2.32 4 0.6774
7 0.27 3 0.9642
8 0.25 3 0.9697
9 0.46 3 0.9282
10 0.81 3 0.8467
11 0.48 3 0.9226
12 0.81 3 0.8461
13 6.87 5 0.2301
14 0.0006 3 1.0000
TOTAL 37.69 48 0.8576

Resíduos do modelo geral CJS para os dados de CR das larvas de
geral CJS para os dados de CR das larvas de S. salamandra
ANEXO VII
83
S. salamandra
84
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão I
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão II
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão III
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão IV
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão V
Frequências relativas de ocorrência de indícios de predação em larvas de S.
salamandra, por sessão em cada pego.
ANEXO VIII

85
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão 1
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão 2
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão 3
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão 4
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão 5
0
0.5
1
1 3 5 7 9 11 13 15 17 19
f ri
Pegos
Sessão 6
Frequências relativas de ocorrência de indícios de predação em larvas de A.
cisternasii para cada pego, por ocasião.