tesis estructura poblacional del …biblio.uabcs.mx/tesis/te3321.pdf · para confirmar la identidad...
TRANSCRIPT

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTO DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
POSGRADO EN CIENCIAS MARINAS Y COSTERAS
TESIS
ESTRUCTURA POBLACIONAL DEL RORCUAL TROPICAL (Balaenoptera edeni
brydei), EN EL GOLFO DE CALIFORNIA CON BASE EN TÉCNICAS
ACÚSTICAS Y MOLECULARES
QUE COMO REQUISITO PARA OBTENER EL GRADO DE
DOCTOR EN CIENCIAS MARINAS Y COSTERAS
CON ORIENTACIÓN EN BIOLOGÍA MARINA
PRESENTA
LORENA VILORIA GÓMORA
DIRECCIÓN:
DR. JORGE URBÁN RAMÍREZ
LA PAZ, B. C. S., JUNIO DE 2015
.

ii

iii
AGRADECIMIENTOS
Al CONACyT por la beca de manutención (165753), y al proyecto SEP-CONACyT
134251. Al Programa de Investigación de Mamíferos Marinos (PRIMMA), en la
Universidad Autónoma de Baja California Sur (UABCS), a cargo del Dr. Jorge Urbán, por
el apoyo económico para realizar salidas al campo, estancia y congresos. Al Instituto
SCRIPPS de Oceanografía de la Universidad de California, por donar hidrófono y
grabadora. A la Alianza WWF-TELCEL y a Ecology Project International por las becas y
el apoyo económico para realizar salidas al campo.
A mi comité tutorial por el apoyo recibido durante el doctorado, Dr. Jorge Urbán, Dr. Oscar
Arizpe, Dr. Ricardo Vázquez, Dr. Eduardo Romero y Dr. Luis Pastene. A Neftalí Gutiérrez
Rivera y Griselda Gallegos Simental por la asesoría técnica en el procesamiento de las
muestras en el Laboratorio de Biología Molecular del CIBNOR. Al Dr. Ricardo Vázquez
por el apoyo económico para el procesamiento de dichas muestras. Al Dr. Eduardo Romero
Vivas por la asesoría en el tema de la Bioacústica, y el apoyo económico para congresos. Al
Dr. Luis Pastene, y todos sus colaboradores, por el apoyo durante mi estancia en The
Institute of Cetacean Research, Japón.
A los amigos y compañeros del PRIMMA-UABCS, por su apoyo incondicional,
especialmente a Sergio, Hiram y Carlos por la ayuda en el mar, mi estancia en La Paz y en
todo momento. A Úrsula, Mariana, Tabata, Braulio, Anaid, Pamela, Flor y Adriana, por su
invaluable contribución en campo. A Jorge Urbán por aceptarme en el Laboratorio, y
apoyarme siempre para la realización de este proyecto.
A mi familia, a Pablo Hernández Almaraz, y a todos mis seres queridos, quienes me
apoyaron durante el camino hasta el doctorado.
El material analizado en la presente investigación se recolectó gracias a los permisos
SGPA/DGVS/08434/09–2009, SGPA/DGVS/0073411–2011 y 138 SGPA/DGVS/0099/14–
2014, otorgado por la Secretaría de Medio Ambiente y Recursos Naturales.

iv
TABLA DE CONTENIDO
LISTA DE FIGURAS ........................................................................................................................ vi
LISTA DE CUADROS ..................................................................................................................... vii
CAPÍTULO I
INTRODUCCIÓN .............................................................................................................................. 1
RESUMEN ...................................................................................................................................... 2
ABSTRACT .................................................................................................................................... 3
PRÓLOGO ...................................................................................................................................... 4
INTRODUCCIÓN GENERAL ....................................................................................................... 5
Generalidades del rorcual tropical, Balaenoptera edeni ............................................................. 5
Morfología ............................................................................................................................... 5
Alimentación ........................................................................................................................... 6
Conducta y reproducción ........................................................................................................ 6
Distribución y abundancia ....................................................................................................... 6
Distribución en el Golfo de California .................................................................................... 7
Taxonomía ............................................................................................................................... 8
Estudios poblacionales del rorcual tropical en el Golfo de California ........................................ 9
PLANTEAMIENTO DEL ESTUDIO .......................................................................................... 11
OBJETIVO GENERAL ................................................................................................................ 12
OBJETIVOS PARTICULARES ................................................................................................... 12
MÉTODOS GENERALES ........................................................................................................... 13
Área de estudio .......................................................................................................................... 13
Trabajo de mar .......................................................................................................................... 16
Registro acústico ................................................................................................................... 17
Toma de biopsias ................................................................................................................... 19
Análisis de datos........................................................................................................................ 19
RESULTADOS GENERALES..................................................................................................... 20
Trabajo de mar .......................................................................................................................... 20
DISCUSIÓN ................................................................................................................................. 23
CONCLUSIONES ........................................................................................................................ 24
LITERATURA CITADA .............................................................................................................. 25
CAPÍTULO II
IDENTIFICACIÓN DE LA FORMA TAXONÓMICA DEL RORCUAL TROPICAL DEL
GOLFO DE CALIFORNIA CON TÉCNICAS MOLECULARES, Y SU RELACIÓN CON
POBLACIONES DE LOS OCÉANOS PACÍFICO E ÍNDICO ....................................................... 31
RESUMEN .................................................................................................................................... 32
ABSTRACT .................................................................................................................................. 33
INTRODUCCIÓN ........................................................................................................................ 34
OBJETIVOS ................................................................................................................................. 35
MÉTODO ...................................................................................................................................... 36
Muestras .................................................................................................................................... 36
Extracción de ADN ............................................................................................................... 37
Análisis de datos........................................................................................................................ 38
Análisis filogenético .............................................................................................................. 38
Estructura poblacional ........................................................................................................... 41

v
Diversidad genética ........................................................................................................... 41
Diferenciación poblacional ............................................................................................... 42
RESULTADOS ............................................................................................................................. 44
Reconstrucción filogenética ...................................................................................................... 44
Estructura poblacional ............................................................................................................... 46
Diversidad genética ............................................................................................................... 49
Diferenciación poblacional ................................................................................................... 49
DISCUSIÓN ................................................................................................................................. 52
CONCLUSIONES ........................................................................................................................ 54
LITERATURA CITADA .............................................................................................................. 56
CAPÍTULO III
ESTRUCTURA GENÉTICA POBLACIONAL DEL RORCUAL TROPICAL (Balaenoptera edeni
brydei) EN EL GOLFO DE CALIFORNIA CON BASE EN ADN MITOCONDRIAL ................. 60
RESUMEN .................................................................................................................................... 61
ABSTRACT .................................................................................................................................. 62
INTRODUCCIÓN ........................................................................................................................ 63
OBJETIVOS ................................................................................................................................. 64
MÉTODO ...................................................................................................................................... 65
Análisis de datos........................................................................................................................ 65
RESULTADOS ............................................................................................................................. 66
DISCUSIÓN ................................................................................................................................. 70
CONCLUSIONES ........................................................................................................................ 72
LITERATURA CITADA .............................................................................................................. 73
CAPÍTULO IV
VARIACIÓN GEOGRÁFICA EN VOCALIZACIONES DE LAS POBLACIONES DEL
RORCUAL TROPICAL (Balaenoptera edeni brydei) EN EL GOLFO DE CALIFORNIA ........... 76
RESUMEN .................................................................................................................................... 77
ABSTRACT .................................................................................................................................. 78
INTRODUCCIÓN ........................................................................................................................ 79
OBJETIVOS ................................................................................................................................. 84
MÉTODO ...................................................................................................................................... 85
Registros acústicos .................................................................................................................... 85
Vocalizaciones: con confirmación visual .............................................................................. 85
Vocalizaciones: hidrófono fijo .............................................................................................. 85
Análisis de las vocalizaciones ................................................................................................... 87
Identificación de vocalizaciones de la región norte y sur.......................................................... 89
Análisis estadísticos .................................................................................................................. 91
Identificación de vocalizaciones de la región norte y sur...................................................... 91
RESULTADOS ............................................................................................................................. 92
Vocalizaciones con confirmación visual ................................................................................... 92
Descripción de las vocalizaciones ............................................................................................. 93
Vocalizaciones con hidrófono fijo ............................................................................................ 96
Comparación entre regiones: ................................................................................................. 96
Comparación estadística .................................................................................................... 96
Comparación morfológica ................................................................................................. 97
Comparación del gradiente de energía .............................................................................. 97
DISCUSIÓN ................................................................................................................................. 99
CONCLUSIONES ...................................................................................................................... 101

vi
LITERATURA CITADA ............................................................................................................ 102
CAPÍTULO V
TRANSMISIÓN CULTURAL Y SU IMPACTO EN LA ESTRUCTURA GENÉTICA DEL
RORCUAL TROPICAL (Balaenoptera edeni brydei) EN EL GOLFO DE CALIFORNIA ........ 106
RESUMEN .................................................................................................................................. 107
ABSTRACT ................................................................................................................................ 108
INTRODUCCIÓN ...................................................................................................................... 109
OBJETIVO .................................................................................................................................. 110
MÉTODO .................................................................................................................................... 111
Identificación de diferencias genéticas: .................................................................................. 111
Identificación de diferencias acústica: .................................................................................... 111
RESULTADOS ........................................................................................................................... 112
Identificación de diferencias genéticas.................................................................................... 112
Identificación de diferencias acústica: .................................................................................... 112
DISCUSIÓN ............................................................................................................................... 113
Teoría de la transmisión cultural ............................................................................................. 113
Balaenoptera edeni brydei en el Golfo de California ............................................................. 119
CONCLUSIONES ...................................................................................................................... 122
LITERATURA CITADA ............................................................................................................ 124
ANEXO 1 .................................................................................................................................... 131
ANEXO 2 .................................................................................................................................... 132
ANEXO 3……………………………………………………………………………………… 133
LISTA DE FIGURAS
CAPÍTULO I
Figura 1. 1. Balaenoptera edeni. ......................................................................................................... 5
Figura 1. 2. Detalle de la cabeza del rorcual tropical. ......................................................................... 6
Figura 1. 3. Cuatro stock del rorcual tropical en el Pacifico Norte. .................................................... 7
Figura 1. 4. Biopsias analizadas por Dizon et al. (1996). ................................................................... 8
Figura 1. 5. Área de estudio. ............................................................................................................. 15
Figura 1. 6. Embarcaciones medianas. .............................................................................................. 16
Figura 1.7. Embarcación menor. ....................................................................................................... 17
Figura 1. 8. Hidrófono de arrastre y grabadora Fostex. .................................................................... 18
Figura 1. 9. Esquema de hidrófono fijo HARP. ................................................................................ 18
Figura 1. 10. Flechas para biopsias de cetáceos. ............................................................................... 19
Figura 1. 11. Corte de biopsia. .......................................................................................................... 19
Figura 1. 12. Rutas navegadas en el Golfo de California durante Enero 2010–Julio 2014, en
embarcación menor. .......................................................................................................................... 21
Figura 1. 13. Distribución de los avistamientos de B. edeni brydei. ................................................. 22
CAPÍTULO II
Figura 2. 1. Ubicación de las muestras. ............................................................................................. 36
Figura 2. 2. Número de muestras por región geográfica. .................................................................. 42
Figura 2. 3. Reconstrucción filogenética. .......................................................................................... 44
CAPÍTULO III
Figura 3. 1. Distribución de las biopsias de B. edeni brydei en el Golfo de California. ................... 66
Figura 3. 2. Red de ocho haplotipos. ................................................................................................. 67
Figura 3. 3. Relación filogenética de haplotipos únicos. ................................................................... 69
CAPÍTULO IV

vii
Figura 4. 1. Ubicación del hidrófono fijo. ......................................................................................... 86
Figura 4. 2. Parámetros de las vocalizaciones. .................................................................................. 88
Figura 4. 3. Morfología de una vocalización en proyección 3D. ...................................................... 88
Figura 4. 4. Gradiente de energía en las vocalizaciones. .................................................................. 89
Figura 4. 5. Relación señal-ruido. ..................................................................................................... 90
Figura 4. 6. Puntos de medición de la duración a -24dB. .................................................................. 90
Figura 4. 7. Distribución de las vocalizaciones identificadas. .......................................................... 93
Figura 4. 8. Vocalización Be1. .......................................................................................................... 94
Figura 4. 9. Vocalización Be2. .......................................................................................................... 94
Figura 4. 10. Vocalización Be3 ......................................................................................................... 95
Figura 4. 11. Vocalización Be4. ........................................................................................................ 95
Figura 4. 12. Morfología y gradiente de energía de la vocalización Be4 del norte. .......................... 97
Figura 4. 13. Morfología y gradiente de energía de la vocalización Be4 del sur. ............................. 98
CAPÍTULO V
Figura 5. 1. Diferencias acústicas y genéticas en Balaenoptera edeni brydei del Golfo de California.
......................................................................................................................................................... 120
LISTA DE CUADROS
CAPÍTULO I
Cuadro 1. 1. Esfuerzo de navegación y número de organismos. ....................................................... 22
CAPÍTULO II
Cuadro 2. 1. Biopsias y secuencias. .................................................................................................. 38
Cuadro 2. 2. Secuencias de ADNmt usadas del GenBank. ............................................................... 40
Cuadro 2. 3. Frecuencias haplotípicas de cinco poblaciones de B. edeni brydei. ............................. 48
Cuadro 2. 4. Diversidad genética en poblaciones de B. edeni brydei. .............................................. 49
Cuadro 2. 5. Distancia neta entre poblaciones de B. edeni brydei. ................................................... 50
Cuadro 2. 6. Resultados de la prueba de heterogeneidad entre regiones. ......................................... 50
Cuadro 2. 7. Valores de Fst por región. ............................................................................................ 51
CAPÍTULO III
Cuadro 3. 1. Frecuencias haplotípicas por región. ............................................................................ 67
Cuadro 3. 2. Diversidad genética en poblaciones. ............................................................................ 68
Cuadro 3. 3. Valores de heterogeneidad entre regiones y estadístico Fst. ........................................ 68
CAPÍTULO IV
Cuadro 4. 1. Características de las vocalizaciones de B. edeni. ........................................................ 82
Cuadro 4. 2. Esfuerzo de grabación. ................................................................................................. 92
Cuadro 4. 3. Parámetros estadísticos de la duración medida a -24dB. .............................................. 96

1
CAPÍTULO I
INTRODUCCIÓN

2
RESUMEN
El rorcual tropical es una especie de misticeto en la que características biológicas y ecológicas
son poco conocidas, y cuyo status taxonómico aún no ha sido resuelto. Actualmente se
reconocen dos sub-especies Balaenoptera edeni brydei y Balaenoptera edeni edeni. Para el
Golfo de California (GC) se sugiere la presencia de B. edeni brydei; además, datos de foto-
identificación y de un estudio preliminar de genética que analizó ADN mitocondrial (ADNmt)
de cinco muestras, proponen la existencia de una población residente al norte y otra transeúnte
al sur (Capítulo I). Para confirmar la identidad de la sub-especie de rorcual tropical que se
distribuye en el GC y conocer su estructura poblacional, en el presente trabajo se efectuaron
conjuntamente análisis genéticos y acústicos. A través de un análisis filogenético que comparó
el ADNmt de 401 muestras del Pacífico Noroeste, 24 del Pacífico Suroeste (Fiji), 24 del
Pacífico Sureste (Perú), 23 del Océano Índico (Java) y 82 del GC, se logró confirmar que el
rorcual tropical del GC pertenece a la sub-especie Balaenoptera edeni brydei (Capítulo II).
Análisis de diversidad genética y diferencias poblacionales, realizados a secuencias de 299pb
de ADNmt de 82 muestras separadas a priori como representantes de las regiones norte (29) y
sur (53) del GC, indicaron que no existen diferencias significativas entre dichas regiones
(Capítulo III). Sin embargo, la comparación de elementos matemáticos, morfológicos y
gradiente de energía de 103 vocalizaciones de la región sur contra 78 de la región norte,
evidenciaron diferencias regionales norte-sur (Capítulo IV). Finalmente se relacionan las
diferencias genéticas y acústicas en el marco de la teoría de la transmisión cultural y su
impacto en la estructura genética de las poblaciones. De acuerdo con esta teoría los resultados
obtenidos en el presente trabajo pueden ser explicados por tres hipótesis alternativas, las
variaciones en las vocalizaciones del rorcual tropical del GC son una consecuencia de las
características físicas del ambiente, o que estas variaciones no intervienen en la reproducción,
o que sean variaciones recientes que aún no se expresan en la estructura genética (Capítulo V).

3
ABSTRACT
Bryde’s whale is a little known species; their taxonomic status has not been resolved yet. Two
subspecies are currently recognized, Balaenoptera edeni brydei and Balaenoptera edeni edeni.
In the Gulf of California (GC) the presence of the B. edeni brydei is suggested. Besides photo-
identification studies, and a preliminary genetic work with five samples of mitochondrial
DNA (mtDNA), suggested the presence of two populations. One resident at the north, and
another transient at the south of the GC (Chapter I). The analyses were conducted to confirm
the species identity of whales in the GC, and to identify population differences of Bryde’s
whale. The analyses to confirm the species identity included 401 samples from the western
North Pacific, 24 from the western South Pacific (Fiji), 24 from the eastern South Pacific
(Peru), 23 from the eastern Indian Ocean (Java) and 82 from GC. Phylogenetic analysis
associated the Bryde’s whales from the GC with the B. edeni brydei (Chapter II). Analyzes of
genetic (mtDNA) and acoustics data, were conducted to identify population differences in the
GC. The genetic analyzes included 82 samples, 29 from northern and 53 from southern of GC.
A total of eight haplotypes were identified, five were present in the North (one exclusive), and
seven in the South (three exclusive). The statistical analysis of heterogeneity based on Chi-
square, Kst*, Hst and Fst showed no differences between regions (Chapter III). The acoustics
analyzes included 191 Be4 calls, with high Signal to Noise Ratio (SNR > 9dB), 103 from the
North and 88 from the South. The main component were characterized using standard
parameters (maximum frequency, minimum frequency, peak frequency and duration), and
new morphological and energy gradient based criteria proposed for the present study. The
results showed differences between regions (Chapter IV). Finally, genetic and acoustics
differences were related within the framework of the theory of cultural transmission and its
impact on the genetic structure of populations. According to this theory the results obtained in
this study could be explained by three alternative hypotheses: 1) changes in the Bryde's whale
vocalizations in the GC are a consequence of the physical characteristics of the environment,
2) these variations are not involved in the reproduction, and 3) these changes are recent and
still not expressed in the genetic structure (Chapter V).

4
PRÓLOGO
El presente documento se dividió en cinco capítulos. En el primero son presentados los
fundamentos generales de la biología de la especie, así como la descripción de los métodos y
resultados relativos al trabajo de campo y obtención de muestras. Esta información da
elementos para abordar el análisis de la identidad de la forma taxonómica del rorcual tropical
que se distribuye en el GC y la relación que guarda con poblaciones del Océano Índico,
Pacífico Norte, Fiji y Perú (Capítulo II), así como la estructura genética dentro del GC
(Capítulo III). Posteriormente, permite abordar en el cuarto capítulo, la variación geográfica
de las vocalizaciones de B. edeni brydei en el GC y la descripción de vocalizaciones que no
habían sido reportadas. La información de la estructura genética y de la variación geográfica
de las vocalizaciones se reúne y se discute con base en la teoría de la transmisión cultural en el
quinto capítulo.

5
INTRODUCCIÓN GENERAL
Generalidades del rorcual tropical, Balaenoptera edeni
Morfología
El rorcual tropical se clasifica dentro del género Balaenoptera, en la familia Balaenopteridae
(Rice, 1998). Presenta un cuerpo alargado de color gris oscuro, la parte ventral es blanca
amarillenta (Reeves et al., 2002) (Figura 1.1), los límites entre el área oscura y clara es difusa.
Su aleta dorsal es extremadamente falcada y en algunos casos presenta marcas con las que se
pueden distinguir individualmente a los animales. Su rostro tiene forma de “V” y exhibe tres
cresta prominentes, las cuales se disponen en una cresta central y dos laterales (Figura 1.2);
dichas crestas representan una característica externa que distingue al rorcual tropical de los
otros miembros de la familia (Cummings, 1985).
Puede considerarse como un balenoptérido de tamaño medio, alcanza una longitud de
hasta 15m (Cummings, 1985); el balenoptérido con menor talla llega a medir 10m (ballena
minke, B. acutorostrata) y el de mayor talla alcanza 33m (ballena azul, B. musculus) (Niño et
al., 2011). El rorcual tropical presenta variaciones de talla dependientes del sexo y distribución
geográfica, siendo las hembras ligeramente más grandes que los machos (Cummings, 1985), y
los individuos del Hemisferio Sur más grandes que los del Hemisferio Norte (Kato y Perrin,
2009).
Figura 1. 1. Balaenoptera edeni (Imagen tomada de Guerrero et al., 2006).

6
Figura 1. 2. Detalle de la cabeza del rorcual tropical. Foto: L. Viloria-Gómora.
Alimentación
Es descrito como un consumidor oportunista, pues se alimenta de diversas especies de peces,
tales como sardina, macarela, arenque, anchoveta, entre otros; así como de cefalópodos y de
algunos crustáceos, entre los que se registran cangrejos rojos, eufáusidos y copépodos
(Cummings, 1985).
Conducta y reproducción
Viaja principalmente solo, aunque en ocasiones se le puede ver en grupos de dos o tres
individuos (Kato y Perrin, 2009). La madurez sexual la alcanza a los 12m. El periodo de
gestación es de 11.5 a 12 meses. Las crías al nacer miden hasta cuatro metros, pesan hasta dos
toneladas y se alimentan de leche materna por alrededor de seis meses (Cummings, 1985). Al
igual que otros balenoptéridos, el rorcual tropical emite vocalizaciones simples de baja
frecuencia (0-1000 Hz) (Cummings et al., 1986), sin embargo la información de su repertorio
acústico es escasa y algunas de las descripciones de sus vocalizaciones no están basadas en
características físicas del sonido, sino en la forma en cómo el investigador escucha o percibe la
vocalización.
Distribución y abundancia
Se distribuye esencialmente en la región tropical y subtropical, entre los 40° latitud Norte y los
40° latitud Sur, pero puede encontrarse en aguas ligeramente frías. Particularmente, se observa
en el Pacífico Occidental, desde Japón a Nueva Zelanda y en el Pacífico Oriental desde Baja
California (México) a Chile (Pastene et al., 1984, datos no publicados). En el Océano
Atlántico se ha visto en Virginia, el Golfo de México, el Mar Caribe y hacia el sur de Brasil.
También de Marruecos hasta el sur de Cabo de Buena Esperanza, en África (Cummings,

7
1985). En el Océano Índico se distribuye de norte al sur desde el Golfo Pérsico hasta el Cabo
de Buena Esperanza, y desde Burma hasta el oeste de Australia (Kato y Perrin, 2009).
Para el Océano Pacífico, la Comisión Ballenera Internacional (IWC, por sus siglas en
inglés) reconoce cuatro stock (Figura 1.3) (Reilly et al., 2008), el del Pacífico Noroeste con
una abundancia estimada en 20,500 (ca. 95% CI 10,700-39,200) (IWC, 2008), el del Pacífico
Oriental tropical con 13,000 (CV=0.202) (Wada y Gerrodette, 1993), el del Golfo de
California con 400 individuos (95% CI 435-2085) (Gerrodette y Palacios, 1994), y el de la
región del Este de China con 105 (CV=2.79) (IWC, 1996).
Figura 1. 3. Cuatro stock del rorcual tropical en el Pacifico Norte. Mapa tomado de IWC (1996), con
modificaciones según Reilly et al. (2008).
Distribución en el Golfo de California
El rorcual tropical se ha registrado de norte a sur en ambas costas del golfo, peninsular y
continental (Urbán y Flores, 1996). Se estima un tamaño poblacional total de 400 individuos
(95% CI 435-2085) (Gerrodette y Palacios, 1994), aunque estudios recientes indican
variaciones de abundancia, particularmente en la boca del golfo; hay animales que entran o

8
salen hacia aguas adyacentes en busca de alimento, según la temporada del año o evento
climatológico (v.g., Evento del Niño) (Vidal et al., 1987; Salvadeo et al., 2011). Además, con
base en datos de foto-identificación y recaptura, se propone que existen individuos residentes
en zonas norteñas, como el Canal de Ballenas (Breese y Tershy, 1987), y con datos obtenidos
de un análisis del ADNmt de cinco muestras colectadas en Bahía de Loreto, Bahía de La Paz y
La Bahía de Banderas (Figura 1.4), se propone que existen dos poblaciones, una residente del
golfo y otra transeúnte que forma parte del Pacífico Oriental tropical (Dizon et al., 1996). Sin
embargo, la identificación de estas poblaciones y su potencial distribución no han sido
corroboradas.
Taxonomía
Actualmente el rorcual tropical se asocia a la nominación Balaenoptera edeni (Kato y Perrin,
2009; IWC, 2010) y el Comité Taxonómico (Committee on Taxonomy, 2014) reconoce la
existencia de dos sub-especies, B. edeni edeni y B edeni brydei; su distinción se basa en la
talla de los especímenes (la primera sub-especie es más pequeña que la segunda) (Rice, 1998).
Figura 1. 4. Biopsias analizadas por Dizon et al. (1996).
25°N
20°N
110°W 105°W
Loreto
(1 muestra)
Bahía de Banderas
(1 muestra)
Bahía de La Paz (3 muestras)

9
Sin embargo, la identidad y número de taxones del complejo edeni/brydei aún no han sido
esclarecidos (Reilly et al., 2008).
Con base en observaciones de campo (Breese y Tershy, 1987) y datos morfológicos
obtenidos de especímenes varados (PRIMMA-UABCS, datos no publicados), se sugiere que el
rorcual tropical distribuido en el Golfo de California pudiera pertenecer a la subespecie B.
edeni brydei, sin embargo esta determinación aún no ha sido confirmada.
Estudios poblacionales del rorcual tropical en el Golfo de California
A pesar de los avances antes mencionados, la estructura poblacional del rorcual tropical en el
GC permanece pobremente conocida. Una compilación particular para el GC fue hecha por
Urbán y Flores (1996). En este trabajo los autores reúnen toda la información generada hasta
la primera mitad de la década de los 90's. Entre las citas hay pocas relacionadas con la
estructura poblacional, pero destaca una de las primeras investigaciones realizadas para la
especie en la región del golfo; en ella se obtuvo información mediante foto-identificación y
fue posible proponer la existencia de un grupo de individuos residentes en la Bahía de los
Ángeles (Breese y Tershy, 1987). También llama la atención una investigación posterior, en la
que se estimaron 235±62 individuos en la Bahía de La Paz, utilizando también la técnica de
foto-identificación (Chávez, 1995).
Técnicas recientes han aportado información de interés para el estudio poblacional de
la especie en el GC. Con el uso de Sistemas de Información Geográfica, Salvadeo et al. (2011)
analizaron la relación del rorcual tropical con la variabilidad ambiental provocada por el
Evento del Niño y la disponibilidad de presas; los datos de este trabajo proponen que los
animales se desplazan en busca de alimento, y particularmente hay una mayor abundancia en
la región sur cuando hay Eventos del Niño. Utilizando análisis de isótopos estables y ácidos
grasos, López-Montalvo (2012) caracterizó la ecología alimentaria y propone la existencia de
dos agregaciones, una en la región norte del golfo asociada a zonas costeras y otra en la región
sur asociada a zonas oceánicas. Otra investigación realizada con isótopos estables de barbas de
especímenes varados (Olavarrieta-García, 2012) encontró que el rorcual tropical presenta
cambios periódicos en el tipo de alimentación y sugiere la presencia de dos grupos, uno que se

10
alimenta preferentemente de peces y se distribuye cerca de la costa y otro oportunista que se
alimenta tanto de eufáusidos como de peces.
Aproximaciones multidisciplinarias en el que se emplean conjuntamente marcadores
poblacionales tradicionales (tales como foto-identificación) y alternativos (tales como métodos
acústicos), han sido utilizados con éxito en el análisis de otras ballenas (Castellote et al.,
2011). En el presente trabajo se hace uso de dos herramientas, la acústica y la genética, para
identificar poblaciones de manera individual, y al mismo tiempo conocer aspectos evolutivos
de la especie, usando como base la teoría de la transmisión cultural y su impacto en la
estructura genética de las poblaciones (Macdougall-Shackleton y Macdougall-Shackleton,
2001).

11
PLANTEAMIENTO DEL ESTUDIO
La discriminación de poblaciones de una misma especie se ha logrado exitosamente mediante
la identificación de variaciones geográficas de rasgos culturales. La transmisión diferencial
estable de estos rasgos, eventualmente puede conducir a una potencial disminución del flujo
génico, aún entre poblaciones adyacentes (MacDougall-Shackleton y MacDougall-Shackleton,
2001). Por lo anterior, la utilización de ambas herramientas, es decir, técnicas de identificación
de diferencias entre rasgos culturales -tales como las vocalizaciones- nos permitirá conocer
cambios recientes (decadales) (Hatch y Clark, 2004) en conjunto con un análisis de genética
de poblaciones la cual aportaría información sobre cambios ocurridos en miles de años (Hartl
y Clark, 2007). Lo anterior representa una oportunidad para interpretar aspectos biológicos,
ecológicos e historia evolutiva.
El rorcual tropical es una especie abundante en el Golfo de California, sin embargo se
desconoce la subespecie a la que pertenece. Parece estar representada por diferentes
poblaciones en simpatría parcial, según se propone en estudios de foto-identificación (Tershy
et al., 1990), genéticos (Dizon et al., 1996) y ecológicos (Salvadeo et al., 2011). La existencia
potencial de estas poblaciones podría estar apoyada por la variación geográfica de rasgos
culturales, tales como registros de ciertas vocalizaciones (Oleson et al., 2003). Para determinar
la existencia de las unidades poblacionales propuestas y saber su grado de aislamiento, el
presente trabajo plantea contrastar sonidos y análisis de ADNmt del rorcual tropical en el GC,
así como interpretar los resultados con la ayuda de la teoría de la transmisión cultural.
Considerando el status taxonómico de B. edeni, el presente parte de determinar la
subespecie a la que pertenece el rorcual tropical del GC.

12
OBJETIVO GENERAL
Determinar la estructura poblacional del rorcual tropical (Balaenoptera edeni brydei)
en el Golfo de California.
OBJETIVOS PARTICULARES
Identificar la forma taxonómica del rorcual tropical que se distribuye en el Golfo de
California.
Conocer el grado de diversidad genética con base en su ADNmt.
Determinar el grado de variación de la diversidad genética en espacio y tiempo.
Caracterizar las vocalizaciones de la especie en el Golfo de California.
Identificar variaciones de las vocalizaciones en el Golfo de California.
En el marco de la teoría de la trasmisión cultural, relacionar las diferencias acústicas y
la estructura genética de la población de rorcual tropical que se distribuye en el Golfo de
California.

13
MÉTODOS GENERALES
Área de estudio
El Golfo de California se ubica entre los 20° y 32° de latitud Norte, y entre los -105° y los
-115° de longitud Oeste (Figura 1.5). Es una cuenca alargada y estrecha de ca. 1400 km de
largo y 150 km de ancho (Roden y Groves, 1959), con alta productividad (Álvarez-Borrero et
al., 1978; Álvarez-Borrego y Gaxiola-Castro, 1988).
La dinámica del Golfo de California está afectada por varios factores como el viento
(Parés-Sierra et al., 2003), las mareas (Álvarez-Borrego y Gaxiola-Castro, 1988) y la radiación
solar (evaporación; Lavín y Organista, 1988). La circulación de los vientos ocurren
principalmente a los largo del eje del golfo debido a la topografía alta a ambos lados, la Sierra
Madre Occidental de Sonora y Sinaloa, y la Sierra de San Pedro Mártir al norte y La Giganta
al sur de la Península de Baja California, esto genera una cuenca semicerrada y un efecto de
embudo (Parés-Sierra et al., 2003). Estos vientos inducen una circulación superficial y a gran
escala, conocidos como giros ciclónicos en verano (junio-noviembre) y anticiclónicos en
invierno (noviembre-mayo; Lavín et al., 1997; Marinone, 2003).
El golfo tienen una compleja circulación desde y hacia el Pacífico (Mascarenhas et al.,
2004). Este intercambio es variable espacial y temporal, tienen un componente horizontal con
flujo hacia dentro, principalmente en el centro y lado occidental del golfo, aunque algunas
veces tiene núcleos de flujo de adentro hacia afuera que se alternan (Roden, 1972, Castro et al.
2006). También existe un intercambio vertical entre el golfo y el Pacífico, en el que agua
superficial y subsuperficial menos densa, más caliente y salada, y pobre en nutrientes se
mueve hacia afuera del golfo, y agua más profunda más densa y fría, con salinidad menor y
rica en nutrientes fluye hacia dentro del golfo; este intercambio tienen una implicación
ecológica importante, pues es un mecanismo natural de fertilización (Álvarez-Borrego, 2012).
Los fenómenos estacionales antes mencionados influyen en la distribución y
abundancia de los recursos marinos, a partir de los cuales se pueden reconocer cinco regiones:
la Norte, la de las Grandes Islas, la del Canal de Ballenas, la Central y la Sur (Santamaría-del-
Ángel et al., 1994). La región Norte es definida como una zona poco profunda que se
enriquece con las corrientes de marea, gran turbidez, sedimentos gruesos, temperaturas

14
extremas, alta evaporación y alta salinidad (Santamaría-del Ángel et al., 1994), con
circulación ciclónica en verano y anticiclónica en la fría (Marrione, 2003). La región de las
Grandes Islas, con topografía particular, se caracteriza por aguas superficiales relativamente
frías, asociadas a niveles de alta producción primaria (Santamaría-del-Ángel et al., 1994). El
Canal de Ballenas se caracteriza por ser una zona muy profunda (1500m), en la que se
producen corrientes de marea fuertes, las cuales provocan una temperatura superficial baja y
alta productividad durante todo el año (López, 2005). La región Central se caracteriza por la
presencia de subcuencas, baja amplitud de mareas, considerables surgencias estacionales
(Maluf, 1983) y diferencias de temperatura entre invierno y verano (Soto-Mardones et al.,
1999). La región Sur tiene características más oceánicas por la influencia directa del Pacífico,
en ella, se identifican tres masas de agua superficiales, Agua de la Corriente de California (fría
y de baja salinidad), Agua del Golfo de California (cálida y de mayor salinidad; Álvarez-
Borrego et al., 1978; Lavin y Marinone, 2003) y Agua Superficial tropical (cálida y de
salinidad intermedia; Álvarez- Borrego, 1983).

15
Figura 1. 5. Área de estudio. Imagen modificada de Lluch-Cota et al. (2007).

16
Trabajo de mar
De enero 2010 a julio 2014 se realizaron salidas al mar en dos tipos de embarcación, mediana
y menor, ambas con motor fuera de borda. El área monitoreada incluyó un recorrido iniciado
en San José del Cabo en el extremo sur de la Península de Baja California, hasta Bahía de los
Ángeles en la costa peninsular y Bahía Kino en la costa continental, en el extremo norte. Los
recorridos entre bahías se realizaron en embarcación mediana, 14-16m de largo (Figura 1.6), y
los recorridos dentro de cada bahía se realizaron en embarcación menor, 7m de largo (Figura
1.7). En estos últimos se navegó sin una trayectoria predefinida. Los objetivos de todas las
salidas fueron identificar, registrar y tomar biopsias del rorcual tropical.
A
B
Figura 1. 6. Embarcaciones medianas de 14 y 16m de largo, A y B respectivamente. Fotos:
PRIMMA-UABCS.
En todos los avistamientos de rorcual tropical se registró la fecha, posición geográfica,
número de individuos, presencia de crías, fotografías de la aleta dorsal para su posterior
identificación, así como algunos datos ambientales (temperatura superficial del agua,
nubosidad, estado del mar en la escala de Beaufort). Todas las navegaciones se hicieron en
horas de luz (0700 – 1900 hrs) y en estado del mar máximo de tres Beaufort.

17
Figura 1.7. Embarcación menor de 7m de largo. Foto: PRIMMA-UABCS.
Registro acústico
Los datos acústicos se obtuvieron de dos fuentes, a partir de grabaciones con confirmación
visual, utilizando un hidrófono de arrastre y una grabadora Fostex (Figura 1.8) y a partir de un
paquete autónomo de grabación pasiva o hidrófono fijo, denominado HARP (High-Frequency
Acoustics Recording Package) diseñado por el Instituto SCRIPPS de Oceanografía de la
Universidad de California (Figura 1.9). El método y el análisis de estos datos se describen de
manera explícita en el Capítulo IV.

18
A
B
Figura 1. 8. Hidrófono de arrastre (A) y grabadora Fostex (B). Fotos: PRIMMA-UABCS.
Figura 1. 9. Esquema de hidrófono fijo HARP. Imagen modificada de Wiggins y Hildebrand (2007).

19
Toma de biopsias
Las biopsias se tomaron con una ballesta (150lb), flechas con flotador y puntas de acero
inoxidable (2.5cm de largo/0.5cm de diámetro) diseñadas específicamente para toma de
muestras de cetáceos (Figura 1.10).
Figura 1. 10. Flechas para biopsias de cetáceos. Fotos: PRIMMA-UABCS.
Todas las biopsias colectadas se cortaron en tres secciones, una de grasa y dos
fragmentos de piel (Figura 1.11). Uno de los fragmentos de piel se conservó en alcohol grado
molecular para su posterior procesamiento en laboratorio (ver Capítulo II). El otro fragmento
se conservó en seco y a 4°C y la grasa se conservó en nitrógeno líquido.
Figura 1. 11. Corte de biopsia. Foto: PRIMMA-UABCS.
Análisis de datos
Los análisis genéticos se explican en los capítulos II y III y los de acústica en el capítulo IV.

20
RESULTADOS GENERALES
Trabajo de mar
De enero 2010 a julio 2014 se realizaron 35 salidas en embarcaciones medianas con
trayectorias que llegaron a San José del Cabo, Bahía de Loreto y Bahía de los Ángeles; en
todos los casos el punto de partida y llegada fue Bahía de La Paz (Figura 1.12). También se
efectuaron 81 salidas en embarcación menor dentro de las tres Bahías (Figura 1.13).
El esfuerzo de navegación, observación, toma de biopsias y registro acústico se resume
en el Cuadro 1.1. La frecuencias de individuos de la especie (número de animales registrados
entre el número de horas navegadas) indican que se observaron 0.08 rorcuales tropicales por
hora de esfuerzo. Durante el periodo de muestreo se tuvo un éxito 23% en la toma de biopsias
y el 42% en las grabaciones, esto si tomamos en cuenta al número de individuos (198) como el
100% de éxito (Cuadro 1.1).

21
Figura 1. 12. Rutas navegadas en el Golfo de California durante Enero 2010–Julio 2014, en embarcación menor. Localidades: Bahía de los
Ángeles (arriba izquierda), Bahía de la Paz (abajo), y Bahía Kino (arriba derecha).

22
Cuadro 1. 1. Esfuerzo de navegación (en horas) y número de organismos de rorcual tropical durante
Enero 2010–Julio 2014.
Año Horas Organismos No Biopsias No. Grabaciones
2010 611.4 42 7 31
2011 498.9 52 8 11
2012 578.0 64 27 33
2013 545.95 28 3 6
2014 281.67 12 1 3
Total 2515.92 198 46 84
Promedio anual 503.184 39.6 9.2 16.8
Frecuencia de organismos 0.08
Éxito de biopsias 23%
Éxito de grabaciones 42%
Los lugares con mayor número de avistamientos fueron: corredor Bahía de los Ángeles-Canal
de Ballenas (143) y el corredor Loreto-La Paz (33). No se registraron avistamientos durante
los recorridos efectuados en Bahía Kino (Figura 1.13).
Figura 1. 13. Distribución de los avistamientos de B. edeni brydei durante Enero 2010–Julio
2014.

23
DISCUSIÓN
Durante las diferentes rutas navegadas se registró un mayor número de avistamientos en las
zonas Bahía de la Paz-Loreto y Bahía de los Ángeles-Canal de Ballenas. Este patrón de
distribución de avistamientos parece reflejar el esfuerzo de navegación, se navegaron 1074.02
hrs totales en el recorrido Loreto-La Paz y 311.22 hrs totales en Bahía de los Ángeles-Canal
de Ballenas. El hecho de no registrar avistamientos en Bahía Kino puede ser atribuido a un
limitado esfuerzo de muestreo o a la capacidad de desplazamiento que presenta el rorcual
tropical dentro del golfo, como lo han reportado otros autores. Flores et al. (1997) y Salvadeo
et al. (2011), reportan un mayor número de ballenas en el Canal de Ballenas y una
disminución en la Bahía de La Paz durante eventos de El Niño, y un comportamiento contrario
durante eventos de La Niña. Esto sugiere que los movimientos del rorcual tropical en el GC
están influenciados por la variabilidad interanual, y probablemente mediados por la
disponibilidad de alimento (Salvadeo et al., 2011), al respecto se sabe que en el GC la sardina
del Pacífico (Sardinops sagax caeruleus) es una de las presas principales del rorcual tropical
(Urbán y Flores-Ramírez, 1996), y presenta cambios de abundancia y distribución
relacionadas con eventos de El Niño (Lluch-Belda et al., 1986).
La frecuencia de organismos registrada en este trabajo (0.08 organismos/hora) es
similar a la reportada en el trabajo de Salvadeo et al. (2011). Estos autores analizaron datos de
la especie de 1998 a 2006 de la región suroeste del golfo y reportan 0.09 organismos por hora.
Esto sugiere que la frecuencia de observación del rorcual tropical es, en promedio, constante a
lo largo de los años y en la región de Bahía de los Ángeles y el corredor Loreto-La Paz.
El porcentaje de biopsias que se obtuvo del rorcual tropical (23%) es bajo comparado
con el que obtuvo en el rorcual común (47%) durante el mismo periodo (2010-2014). Esto
puede deberse a dos factores, la conducta propia de la especie y a las condiciones ambientales.
El primer factor hace referencia a la característica evasiva de los animales ante la presencia de
embarcaciones, a que se observa una ruta indefinida al navegar (“navegación errática”) y a que
al hacer inmersión expone una fracción pequeña del dorso, dificultando su avistamiento, todas
estas conductas también observadas por otros autores (Kato y Perrin, 2009; Kerosky et al.,
2012). El segundo factor se refiere a que para la toma de biopsias es necesario que las

24
condiciones del mar se mantengan entre 0 y 2 en escala de Beaufort, de lo contrario aumenta
la dificultad de seguir y observar el dorso del organismo, durante los avistamientos efectuados
en el presente trabajo se registró un Beaufort de 0-3. A pesar de las limitaciones provocadas
por la conducta y las condiciones del mar el número de muestras obtenidas es comparable con
el de otros trabajos (Kershaw et al., 2013).
El éxito de grabación con confirmación visual fue de 21.5% (23 de 109 grabaciones
tuvieron vocalizaciones; Cuadro 1.2). Este éxito de grabación puede considerarse bajo si,
además del número de grabaciones, tomamos en cuenta que de 2515.92 horas de grabación a lo
largo de cuatro años, sólo se registraron ca. 23 minutos de vocalizaciones. En un estudio de la
población y señales acústicas de esta especie en la Bahía de Tosa, Japón, también se obtuvo un
bajo éxito de grabación; en 159 días de grabación sistemática durante un período anual
(número de horas no precisadas) sólo se registraron dos vocalizaciones (Chiu, 2009). El éxito
bajo de grabación pudiera estar relacionado con un número bajo de emisión de vocalizaciones,
lo cual pudiera estar relacionado con conductas propias de la especie, pues los individuos del
rorcual tropical generalmente viajan solos y no se han registrado períodos de reproducción ni
sonidos asociados al cortejo (Kato y Perrin, 2009), en contraste con especies como la ballena
azul (Bradbury y Vehrencamp, 1998), el rorcual común (Croll et al., 2002) y la ballena
jorobada (Darling y Bérubé, 2001), en las que se presenta una marcada temporalidad en la
reproducción y diversos patrones de vocalización específicos para el cortejo.
CONCLUSIONES
La distribución y abundancia de los avistamientos responde a las regiones con mayor esfuerzo:
Bahía de los Ángeles y corredor Loreto-La Paz y pudiera estar relacionada a la disponibilidad
de alimento. El número de organismos registrados en los avistamientos sugiere que la
frecuencia de observación del rorcual tropical es constante durante todo el año en las zonas
donde se realizó el mayor esfuerzo. La toma de biopsias y registro de vocalizaciones de esta
especie es difícil si la comparamos con otras especies (v.g., B. physalus o B. musculus), debido
a su comportamiento evasivo, a que no forma agregaciones y a que no tiene temporadas de
reproducción marcada.

25
LITERATURA CITADA
Álvarez-Borrego, S. 2012. New phytoplankton production as a tool to estimate the vertical
component of water exchange between the Gulf of California and the Pacific. Ciencias
Marinas. 38: 89–99.
Álvarez-Borrego, S. 1983. Gulf of California, 427–450. En: Ketchum, B. K. (Ed.) Ecosystems
of the world 26. Estuaries and enclosed areas. Elsevier, Amsterdam.
Álvarez-Borrego, S., Alberto-Rivera, J. A., Gaxiola-Castro, G., Acosta-Ruiz, M. J. y
Schwartzlose, R. A. 1978. Nutrientes en el Golfo de California: Ciencias Marinas. 5:
53–71.
Álvarez-Borrego, S. y Gaxiola-Castro, G. 1988. Photosynthetic parameters of northern Gulf of
California phytoplankton: Continental Shelf Research. 8: 37–47.
Bradbury, J. W. y Vehrencamp, S. L. 1998. Principles of animal communication. Sinauer
Associates, Sunderland, MA. EE. UU. 882pp.
Breese, D. y Tershy, B. R. 1987. Residency patterns in female Bryde’s whales in the Gulf of
California, Mexico. Abstract: Seven Biennial Conference on the Biology of Marine
Mammals. Miami, FL. EE. UU.
Castellote, M., Clark, W. C. y Lammers, O. M. 2011. Fin whale (Balaenoptera physalus)
population identity in the western Mediterranean Sea. Marine Mammal Science. 28(2),
325–344
Castro, R., Durazo, R., Mascarenhas, A., Collins, C. A. y Trasviña, A. 2006. Thermohaline
variability and geostrophic circulation in the southern portion of the Gulf of California.
Deep-Sea Res. I 53:188–200.
Chávez, R. S. 1995. Estimación poblacional del rorcual tropical Balaenoptera edeni
(Anderson, 1878) en la Bahía de la Paz, B. C. S. México. Tesis de Maestría.
CICIMAR. 55p.
Chiu, P. H. 2009. Studies on the population and acoustic signals of Bryde’s whales
(Balaenoptera edeni) in Tosa Bay, Japan. Tesis de Maestría, Institute of Marine
Biology, National Sun Yat-sen University, Kaohsiung, Taiwan. 75p.
Committee on Taxonomy. 2014. List of marine mammal species and subspecies. Society for
Marine Mammalogy, www.marinemammalscience.org, consultado el 22 dic 2014.

26
Croll, D. A., Clark, W. C., Acevedo, A., Tershy, B., Flores, S., Gedamke, J. y Urban, J. 2002.
Only male fin whales sing loud songs. Nature. 417: 809.
Cummings, W. C. 1985. Bryde’s whale Balaenoptera edeni Anderson, 1878. Vol 3: 137–154.
En: Ridgway, S, H. y Harrison, R. (eds.) 1985. Handbook of Marine Mammals.
Academia Press, London.
Cummings, W. C., Thompson, P. O. y Samuel, J. H. 1986. Sounds from Bryde, Balaenoptera
edeni, and Finback, B. physalus, whales in the Gulf of California. Fishery Bulletin.
84(2): 359–370.
Darling, J. D. y Bérubé, M. 2001. Interactions of singing humpback whales with other males.
Marine Mammal Science. 17: 570–584.
Dizon, A., Lux, C. A., LeDuc, R. G., Urbán, R, J., Henshaw, M. y Brownell, R. Jr. 1996. An
interin phylogenetic analysis of sei and Bryde’s whale mitochondrial DNA control
region sequences. SC/47/NP23 presented to the IWC Scientific Committee. 12p.
Flores, R. S., Urbán, R. J., Delgado. G. O. y Vargas, S. J. 1997. Avistamientos del rorcual
tropical, Balaenoptera edeni (Cetacea: Balaenopteride) y temperatura del agua, en Baja
California Sur, México. Rev. Biol. Trop. 45(3): 1209–1214.
Gerrodette, T. y Palacios, D. N. 1994. Estimates of cetacean abundance in EEZ waters of the
Eastern Tropical Pacific. Southwest Fisheries and Science Center. Administrative
Report LJ-96–10. 28 p.
Guerrero, R. M., Urbán, J. R. y Rojas, B. L. 2006. Las ballenas del Golfo de California.
SEMARNAT, INE. 537p.
Hartl, D. L., y Clark, A. G. 2007. Principles of population genetics. Sunderland: Sinauer
associates. 652p.
Hatch, L. T. y Clark, C. W. 2004. Acoustic differentiation between fin whales in both the
North Atlantic and North Pacific Oceans, and integration with genetic estimates of
divergence. SC/56/SD6 IWC Scientific Committee (unpublished). 37p.
IWC, 1996. Report of the subcommittee on North Pacific Bryde’s whales, Annex G, Report of
the Scientific Committee. Rep. Int. Whal. Commn. 47: 163–168.
IWC, 2008. North Pacific Bryde’s whales. J. Cetacean Res. Manage. Editorial. 10(2): iii–xii.
IWC, 2010. Classification of the order cetacean. J. Cetacean Res. Manage. Appendix 3.11(1):
xi–xii.

27
Kato, H. y Perrin, W. F. 2009. Bryde’s Whales Balaenoptera edeni/brydei. En: William, F. P.,
Würsig, B. y Thewissen, J. G. M. 2009. Encyclopedia Marine Mammals 2a ed.
Elservier. 157–162 pp.
Kershaw, F., Leslie, M. T., Collins, T., Mansur, R. M., Smith, B. D., Mington, G., Baldiwn,
R., Leduc, R. G., Anderson, R. C., Brownell, R. L. JR. y Rosenbaum, H. C. 2013.
Population Differentiation of 2 Forms of Bryde’s Whales in the Indian and Pacific
Oceans. Journal of Heredity. 104.6: 755–764.
Kerosky, M. S., Širović, A., Roche, L. K., Baumann-Pickering, B., Wiggins, S. M. y
Hildebrand, A. J. 2012. Bryde’s whale seasonal range expansion and increasing
presence in the Southern California Bight from 2000 to 2010. Deep Sea Research.
1(65):125–132.
Lavín, M. F. y Organista, S. 1988. Surface heat flux in the Northern Gulf of California,
Journal of Geophysical Research. 93: 14033–14038.
Lavín, M. F., Beier. E. y Badan, A. 1997. Estructura hidrográfica y circulación del Golfo de
California: Escalas estacional e interanual. En: Lavín, M. F. (ed.) Contribuciones a la
Oceanografía Física en México. Monografía No. 3, Unión Geofísica Mexicana,
Ensenada, Baja California. 141–171 pp.
Lavín, M. F. y Marinone, S. G. 2003. An overview of the physical oceanography of the Gulf
of California. En: Nonlinear Processes in Geophysical Fluid Dynamics. O. U. Velasco
Fuentes et al. (Eds). Kluwer Academic Publishers. Países Bajos. 173–204 pp.
López, J. M. 2005. Variabilidad anual e interanual de la clorofila (SeaWiFS) y el viento
superficial (QuikSCAT) en el Alto Golfo de California: su circulación y asociación.
Tesis de Maestria. Universidad Autónoma de Baja California. 62 pp.
López-Montalvo, C. 2012. Caracterización de la ecología alimentaria del rorcual de Bryde,
Balaenoptera edeni (Anderson, 1879), en el Golfo de California, con base en análisis
de isótopos estables de nitrógeno, carbón y ácidos grasos. Tesis de maestría. Instituto
del Mar y Limnología, UNAM. 76 pp.
Lluch-Belda, D., Magallón, B. F. J. y Schwartzlose, R. A. 1986. Large fluctuations in the
sardine fishery in the Gulf of California: Possible causes. CalCOFI Rep. 27: 136–140.
Lluch-Cota, S. E., Aragon-Noriega, E. A., Arreguín-Sánchez, F., Aurioles-Gamboa, D.,
Bautista-Romero, J. J., Brusca, R. C. y Sierra-Beltrán, A. P. 2007. The Gulf of

28
California: review of ecosystem status and sustainability challenges. Progress in
Oceanography. 73(1): 1–26.
Macdougall-Shackleton, E. A. y Macdougall-Shackleton, S. A. 2001. Cultural and genetic
evolution in mountain white-crowned sparrows: song dialects are associated with
population structure. Evolution. 55(12): 2568–2575.
Maluf, L. Y. 1983. The Physical Oceanography, 26-45. En: Case, T. J. y M. L. Cody (Eds.)
Island Biogeography in the Sea of Cortez. University of California Press, Berkeley.
Marinone, S. G. 2003. A three-dimensional model of the mean and seasonal circulation of the
Gulf of California. J. Geophys. Res. 108(C10): 3325.
Mascarenhas, Jr A. S., Castro, R., Collins, C. A. y Durazo, R. 2004. Seasonal variation of
geostrophic velocity and heat flux at the entrance to the Gulf of California, Mexico. J.
Geophys. Res. 109: C07008.
Niño-Torres, C. A., Urbán-Ramírez, J. y Vidal, O. 2011. Mamíferos Marinos del Golfo de
California: Guía ilustrada. Publicación Especial No. 2, Alianza WWF México-Telcel.
192 pp.
Olavarrieta-García, T. 2012. Hábitos alimenticios del rorcual tropical (Balaenoptera edeni) en
el Golfo de California. Tesis de Licenciatura. Universidad Autónoma de Baja
California Sur. 38 pp.
Oleson, E. M., Barlow, J., Gordon, J., Rankin, S. y Hildebrand, J. A. 2003. Low frequency
calls of Bryde’s whales. Mar. Mamm. Sci. 19(2): 160–172.
Parés-Sierra, A., Mascarenhas, A., Marinone, S. G., Castro, R., 2003. Temporal and spatial
variation of the surface winds in the Gulf of California: Geophysical Research Letters,
30: 1–4.
Pastene, A. L., Acevedo, M y Gallardo, V. A. 1984. A note on Chilean Bryde’s whales.
SC/35/Ba4. Datos no publicados.
Reeves, R. R., Steward, B., Clapham, P. J. y Powell, J. A. 2002. Guide to Marine Mammals or
the World. National Audubon Society. U.S.A. 527 pp.
Reilly, S. B., Bannister, J. L., Best, P. B., Brown, M., Brownell Jr., R. L., Butterworth, D.S.,
Clapham, P. J., Cooke, J., Donovan, G. P., Urbán, J. y Zerbini, A. N. 2008.
Balaenoptera edeni. The IUCN Red List of Threatened Species. Version 2014.3.
www.iucnredlist.org, consultado el 12 Mayo 2015.

29
Rice, W. D. 1998. Marine Mammals of the World. Systematic and distribution. The Society
for marine Mammalogy. Special Publication. 4. 231 pp.
Roden, G. I. y Groves, G. W. 1959. Recent oceanographic observations in the Gulf of
California. J. Mar. Res. 18: 10–35.
Roden, G. I. 1972. Thermohaline structure and baroclinic flow across the Gulf of California
entrance and in the Revillagigedo Islands region. J. Phys. Oceanogr. 2: 177–183.
Salvadeo, C., Flores-Ramírez, S., Gómez-Gallardo, A., MacLeod, C., Llunch-Belda, D.,
Jaime-Schinkel, S. y Urbán, R. 2011. Bryde’s whale (Balaenoptera edeni) in the
southwestern Gulf of California: Relationship with ENSO variability and prey
availability. Ciencias Marinas. 37(2): 215–225.
Santamaría del Ángel, E. Álvarez-Borrego, S. y Muller-Karger, F. 1994. Gulf of California
biogeographic regions based on coastal zone color scanner imagery. Journal of
Geophisical Research. 99(C4): 7411–7421.
Soto-Mardones, L., Marinone ,S. G. y Parés-Sierra, A. 1999. Variabilidad espacio-temporal de
la temperatura superficial del mar en el Golfo de California. Ciencias Marinas. 25(1):
1–30.
Tershy, B. R., Breese, D. y Strong, C. S. 1990. Abundance, seasonal distribution and
population composition of balaenopterid whales in the Canal de Ballenas, Gulf of
California, Mexico. Rep. Int. Whal. Commn. Special Issue. 12: 369–375.
Urbán, R. J. y Flores, S. R. 1996. A note on Bryde’s Whales (Balaenoptera edeni) in the Gulf
of California, Mexico. Rep. Int. Whal. Commn. 46: 453–457.
Vidal, O. Aguayo, A., Findley, L., Robles, A., Bourillón, L., Vomend, I., Turk, P., Gárate, K.,
Maroñas, L. y Rosas, J. 1987. Avistamientos de mamíferos marinos durante el crucero
“Guaymas I” en la región superior del Golfo de California, primavera de 1984. En:
Memorias de la X Reunión Internacional sobre Mamíferos Marinos. La Paz, B.C.S.,
México. 7–35 pp.
Wada, P. R. y Gerrodette, T. 1993. Estimates of cetacean abundance and distribution in the
eastern tropical Pacific. Rep. Int. Whal. Commn. 43: 477–493.
Wiggins, S. M. y Hildebrand, J. A. 2007. High-frequency Acoustic Recording Package
(HARP) for broad-band, long-term marine mammal monitoring. En: Underwater

30
Technology and Workshop on Scientific Use of Submarine Cables and Related
Technologies. 551–557 pp.

31
CAPÍTULO II
IDENTIFICACIÓN DE LA FORMA TAXONÓMICA DEL RORCUAL
TROPICAL DEL GOLFO DE CALIFORNIA CON TÉCNICAS
MOLECULARES, Y SU RELACIÓN CON POBLACIONES DE LOS
OCÉANOS PACÍFICO E ÍNDICO

32
RESUMEN
El rorcual tropical es una especie poco conocida, su status taxonómico no se ha resuelto.
Actualmente se reconocen dos sub-especies: una pequeña (~12m) Balaenoptera edeni edeni y
la normal (~14m) Balaenoptera edeni brydei. En el Golfo de California (GC) varios autores
reconocen al rorcual tropical como de la sub-especie Balaenoptera edeni brydei, sin embargo,
no existen evidencias genéticas para tal designación. Con el objetivo de confirmar la identidad
de la especie, así como la relación genética de la población del GC con la de otras regiones se
realizó una comparación de secuencias 299pb de la región control del ADN mitocondrial
(ADNmt) de individuos del GC con datos publicados de los océanos Pacífico e Índico. El
análisis incluyó 401 muestras del Pacífico Noroeste, 24 del Pacífico Suroeste (Fiji), 24 del
Pacífico Sureste (Perú), 23 del Océano Índico (Java) y 82 del GC. Los resultados del análisis
filogenético identificaron al rorcual tropical del GC como de la sub-especie B. edeni brydei.
De los 56 haplotipos identificados, ocho se encontraron en el GC, de los cuales seis son
específicos para esta región y dos son compartidos con Perú. El análisis estadístico de
heterogeneidad basado en Chi-cuadrado, Hst, Kst* y Fst mostró diferencias significativas
entre los rorcuales del GC y los de cada región analizada. Datos de distancia genética
mostraron que la población de Perú es la más cercana a la del GC. Con esta información se
concluye que el rorcual tropical del GC pertenece a la sub-especie B. edeni brydei y que
conforman una población diferente de otras regiones de los océanos Pacífico e Índico.

33
ABSTRACT
Currently there are two recognized sub-species of Bryde’s whale: Balaenoptera edeni edeni
(~12m) and Balaenoptera edeni brydei (~14m). In Mexico, several authors have associated the
Bryde’s whales distributed in Mexican Pacific with the sub-specie Balaenoptera edeni brydei;
however, there is no genetic information to support this association. Here we present results of
the comparison of mitochondrial DNA (mtDNA) control region sequences of Bryde’s whales
from the Gulf of California (GC) and published sequences of whales from the Pacific and
Indian Oceans. The analyses were conducted to confirm the species identity of whales in the
GC and to elucidate the genetic relationship of this population with populations from other
oceanic regions. The analyses included 401 samples from the western North Pacific, 24 from
the western South Pacific (Fiji), 24 from the eastern South Pacific (Peru), 23 from the eastern
Indian Ocean (Java) and 82 from GC. Phylogenetic analysis associated the Bryde’s whales
from the GC with the Balaenoptera edeni brydei. A total of 56 haplotypes were identified in
the total sample, of these 8 were found in the GC 6 specific to this locality and 2 shared with
the Peruvian population. The statistical analysis of heterogeneity based on Chi-square, Hst,
Kst* and Fst showed significant differences between the GC and each of the other regions.
The smallest genetic distance resulted from the comparison between the GC and Peruvian
populations. We concluded that the Bryde’s whales of the GC belong to sub-specie B. edeni
brydei, and that this population is genetically differentiated from other populations in the
Pacific and Indian Oceans.

34
INTRODUCCIÓN
Balaenoptera edeni fue descrita por Anderson (1878/9, en Rice, 1998) como un animal
pequeño, con una longitud de alrededor de 11.5m y la llamó B. edeni. Olsen (1913) describió
el esqueleto de otro espécimen con una longitud mayor (alrededor de 14.5m) y le asignó el
nombre de B. brydei. Posteriormente Junge (1950) realizó una revisión detallada de los
esqueletos de B. edeni y B. brydei, y concluyó que las dos especies eran la misma, de modo
que permaneció en uso el nombre de B. edeni. Después de varios estudios Omura (1959) y
Best (1960) soportaron la conclusión de Junge (1950). Wada et al. (2003), con base en análisis
de morfología externa, osteología y ADNmt, describen una nueva especie, Balaenoptera
omurai, y proponen que B. edeni y B. brydei son especies separadas. Sasaki et al. (2006), con
análisis de la secuencia completa del ADNmt y de elementos cortos repetitivos intercalados
(SINE, por sus siglas en inglés), confirman los resultados de Wada et al. (2003).
Posteriormente Kanda et al. (2007) compararon 508 secuencias parciales de ADNmt de la
forma común (Balaenoptera brydei) con la forma pigmea (B. edeni), y confirmaron la
separación de estas dos en especies. Sin embargo, para algunos investigadores, estas
diferencias sólo serían suficientes para considerarlas a nivel de subespecies (Kato y Perrin,
2009), como concluyen Kershaw et al. (2013) quienes con base en análisis del ADNmt
concluyen que las diferencias que ellos encuentran son a nivel de sub-especie, donde B. edeni
edeni es la forma pequeña con distribución restringida y B. edeni brydei es la forma grande
con amplia distribución.
Sin embargo el problema taxonómico no se ha resuelto. Según Kato y Perrin (2009) y
IWC (2010), todos los tipos de rorcual tropical se consideran provisionalmente como una sola
especie, Balaenoptera edeni. En el presente estudio se adopta la clasificación del Comité
Taxonómico (Committee on Taxonomy, 2014), que emplea la división sub-específica B. edeni
edeni y B. edeni brydei.
Con base en observaciones (Tershy et al., 1990) y datos de longitud tomados en
varamientos (PRIMMA-UABCS, datos no publicados) se sugiere que el rorcual tropical que
se distribuye en el Golfo de California pertenece a B. edeni brydei, pero no existe ningún
estudio genético y/o morfológico que corrobore esta propuesta. Por lo anterior, el presente
trabajo busca determinar la sub-especie del rorcual tropical que se distribuye en el GC y

35
conocer su relación genética con algunas poblaciones de los océanos Pacífico e Índico, a
través del análisis de su ADNmt.
OBJETIVOS
• Identificar la sub-especie de rorcual tropical que se distribuye en el Golfo de
California.
• Determinar la relación genética entre las poblaciones del rorcual tropical del Golfo de
California y océanos Pacífico e Índico.

36
MÉTODO
Muestras
Durante 2010-2013 se tomaron 50 biopsias del rorcual tropical en diferentes regiones del
Golfo de California (Figura 2.1). A esta recolecta se sumaron 44 biopsias tomadas en años
anteriores (1994-2009) por integrantes del Proyecto de Investigación Mamíferos Marinos
(PRIMMA) de la Universidad Autónoma de Baja California Sur (UABCS). A un total de 94
muestras totales se les extrajo ADN. Para evitar duplicar individuos en los análisis, se realizó
una revisión fotográfica de cada individuo con biopsia y se omitieron las muestras repetidas.
Figura 2. 1. Ubicación de las muestras tomadas durante 2010-2013 en el Golfo de California.

37
Extracción de ADN
Se extrajo el ADN genómico a 94 muestras de individuos diferentes, con el protocolo
modificado de Miller et al. (1988). El procedimiento consistió en una digestión con proteinasa
K (20mg/ml) en 400µl de buffer de lisis (NaCl a 100mM, Tris pH 8 a 50mM, EDTA pH 8 a
100mM, SDS a 1% y agua desionizada), la muestra se incubó por una hora a 65°C, se le
agregó NaCl (6M) saturado, se agitó y se incubó en hielo, se centrifugó a 10,000 rpm durante
10 minutos, el sobrenadante se trasfirió a un tubo con 1ml de etanol absoluto. Como el ADN
no precipitó, se centrifugó a 13,000 rpm 5 minutos, se decantó y se lavó con etanol al 70%, se
centrifugó (13,000 rpm 5 minutos) y se re-suspendió en agua.
Para amplificar la región control de 299pb de ADNmt se utilizaron primers que no
fueron diseñados específicamente para B. edeni brydei, pese a lo cual se tuvo éxito en la
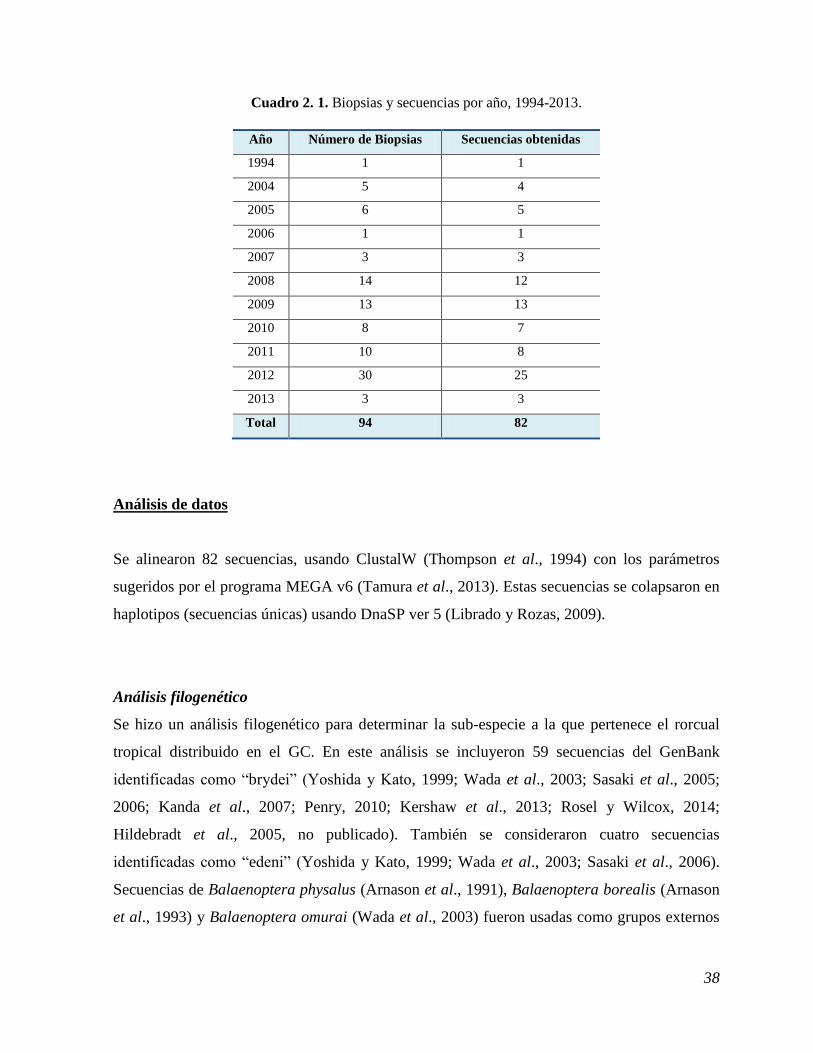
amplificación. De 94 muestras procesadas, se obtuvieron 82 secuencias de buena calidad
(Cuadro 2.1). Los cromatogramas obtenidos presentaron picos únicos, bien separados entre sí
y con escasa sobre-posición en las señales de las bases nitrogenadas alternas (“ruido de
fondo”). La utilización de primers no específicos para taxones de este grupo de rorcuales
también se ha llevado a cabo en otros estudios con resultados confiables (Yoshida y Kato,
1999; Kanda et al., 2007).
La amplificación vía reacción en cadena de la polimerasa (PCR) se hizo en una
solución de agua destilada, MgCl2 (25mM), dNTPs (2.5mM), solución buffer con colorante
(5x), taq polimerasa (5u/µl), y primers (250µg/ml) t-PRO 5’-
CCTCCCTAAGACTCAAGGAA-3’ (Arnason et al., 1993) y el 5’-
GAAGAGGGATCCCTGCCAAGCGG-3’ (Hori et al., 1994, en Yoshida y Kato 1999). De
esta mezcla, se agregó 11.5 µl + 1 µl de ADN 100ng/µl. Las muestras se llevaron a un
termociclador (GenAmp® PCRSystem 9700) que se programó: 1min a 96°C más 42 ciclos de
1min a 94°C, 1min a 60°C y 1 min a 72°C. Los productos de la PCR se corrieron en geles de
agarosa al 2%. Cada uno se revisó con luz ultravioleta. Los fragmentos se purificaron y fueron
secuenciados en laboratorios comerciales.
38
Cuadro 2. 1. Biopsias y secuencias por año, 1994-2013.
Año Número de Biopsias Secuencias obtenidas
1994 1 1
2004 5 4
2005 6 5
2006 1 1
2007 3 3
2008 14 12
2009 13 13
2010 8 7
2011 10 8
2012 30 25
2013 3 3
Total 94 82
Análisis de datos
Se alinearon 82 secuencias, usando ClustalW (Thompson et al., 1994) con los parámetros
sugeridos por el programa MEGA v6 (Tamura et al., 2013). Estas secuencias se colapsaron en
haplotipos (secuencias únicas) usando DnaSP ver 5 (Librado y Rozas, 2009).
Análisis filogenético
Se hizo un análisis filogenético para determinar la sub-especie a la que pertenece el rorcual
tropical distribuido en el GC. En este análisis se incluyeron 59 secuencias del GenBank
identificadas como “brydei” (Yoshida y Kato, 1999; Wada et al., 2003; Sasaki et al., 2005;
2006; Kanda et al., 2007; Penry, 2010; Kershaw et al., 2013; Rosel y Wilcox, 2014;
Hildebradt et al., 2005, no publicado). También se consideraron cuatro secuencias
identificadas como “edeni” (Yoshida y Kato, 1999; Wada et al., 2003; Sasaki et al., 2006).
Secuencias de Balaenoptera physalus (Arnason et al., 1991), Balaenoptera borealis (Arnason
et al., 1993) y Balaenoptera omurai (Wada et al., 2003) fueron usadas como grupos externos

39
(Ver cuadro 2.2). Las 74 secuencias resultantes (66 del GenBank y 8 del GC) se alinearon en
el MEGA v6 (Tamura et al., 2013).
Para la reconstrucción filogenética se calculó el mejor modelo de sustitución
nucleotídica, se tomaron en cuenta los criterios de información de Akaike (AC) y Bayesiano
(BIC) usando el MEGA v6 (Tamura et al., 2013). Posteriormente se construyeron arboles
filogenéticos con el programa MrBayesv3.2.1 (Huelsenbeck y Ronquist, 2001). El análisis fue
realizado con cuatro iteraciones de Monte Carlo de Cadenas de Markov (MCMC), con
5,000,000 de generaciones y los valores propuestos en el programa MrBayesv3.2.1
(Huelsenbeck y Ronquist, 2001). El Burn-in se fijó en 25%. Se hizo una inspección visual de
los valores del parámetro estimado de MCMC con el programa TRACER 1.5 (Rambaut y
Drummond, 2009), el árbol consenso fue generado con el programa FigTreev1.4.0 (Rambaut,
2012).
A las secuencias identificadas como “brydei” y “edeni” se les aplicó el análisis de
Atributos de Caracter (CAs) (Davis y Nixon, 1992; Sarkar et al., 2002; Lowenstein et al.,
2009), considerando que un estado de carácter está presente en un clado pero no en su grupo
hermano (Sarkar et al., 2002), es decir es único para la especie (Lowenstein et al., 2009). Se
tomaron en cuenta únicamente caracteres “puros” (Sarkar et al., 2002), es decir, exclusivos
para cada taxón. Siguiendo a Kershaw et al. (2013), se tomaron nueve sitios diagnósticos para
clasificar a los rorcuales tropicales del Golfo de California dentro de las formas conocidas.
Para este análisis se utilizaron los mismos grupos externos antes mencionados.
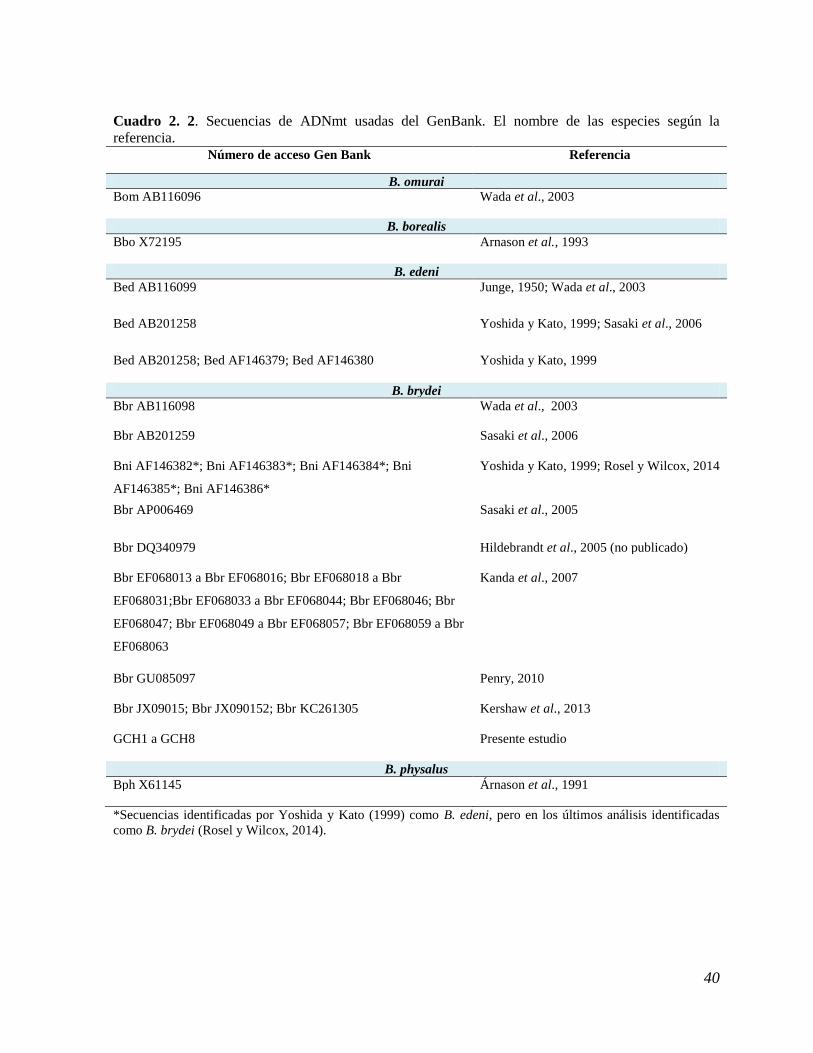
40
Cuadro 2. 2. Secuencias de ADNmt usadas del GenBank. El nombre de las especies según la
referencia. Número de acceso Gen Bank Referencia
B. omurai
Bom AB116096 Wada et al., 2003
B. borealis
Bbo X72195 Arnason et al., 1993
B. edeni
Bed AB116099 Junge, 1950; Wada et al., 2003
Bed AB201258 Yoshida y Kato, 1999; Sasaki et al., 2006
Bed AB201258; Bed AF146379; Bed AF146380 Yoshida y Kato, 1999
B. brydei
Bbr AB116098 Wada et al., 2003 Bbr AB201259 Sasaki et al., 2006
Bni AF146382*; Bni AF146383*; Bni AF146384*; Bni
AF146385*; Bni AF146386*
Yoshida y Kato, 1999; Rosel y Wilcox, 2014
Bbr AP006469 Sasaki et al., 2005
Bbr DQ340979 Hildebrandt et al., 2005 (no publicado)
Bbr EF068013 a Bbr EF068016; Bbr EF068018 a Bbr
EF068031;Bbr EF068033 a Bbr EF068044; Bbr EF068046; Bbr
EF068047; Bbr EF068049 a Bbr EF068057; Bbr EF068059 a Bbr
EF068063
Kanda et al., 2007
Bbr GU085097 Penry, 2010
Bbr JX09015; Bbr JX090152; Bbr KC261305 Kershaw et al., 2013
GCH1 a GCH8 Presente estudio
B. physalus
Bph X61145 Árnason et al., 1991
*Secuencias identificadas por Yoshida y Kato (1999) como B. edeni, pero en los últimos análisis identificadas
como B. brydei (Rosel y Wilcox, 2014).

41
Estructura poblacional
Posterior a la identificación de la sub-especie de rorcual tropical, se comparó la
población del GC con poblaciones de los océanos Pacífico Norte, Pacífico Sur, e Índico. Para
este análisis se incluyeron 82 secuencias del Golfo de California (Figura 2.2) y 473 secuencias
tomadas del análisis Kanda et al. (2007) de las cuales, 401 son del Pacífico norte occidental,
23 de Java, 24 de Fiji y 25 de las costas de Perú. Todas las secuencias fueron colapsadas en
haplotipos.
Diversidad genética
Las 555 muestras fueron agrupadas según la región geográfica a la que pertenecían y se
calculó el número de haplotipos, diversidad haplotípica y diversidad nucleotídica para cada
área, según Nei y Li (1979) con la corrección de Jukes-Cantor y la diferencia en el número
nucleotídica promedio, también se consideraron las inserciones y supresiones. Todos los
cálculos se realizaron con el programa DnaSP (Librado y Rozas, 2009).

42
Figura 2. 2. Número de muestras por región geográfica, incluidas en el análisis de estructura
poblacional.
Diferenciación poblacional
Se calculó la distancia genética entre secuencias únicas (haplotipos) usando el método 2
parámetros de Kimura (Kimura, 1980).
Se calculó la heterogeneidad entre regiones, según Hudson et al. (1992) con tres
estadísticos, Chi-cuadrada, Hst y Kst*.
a) Chi-cuadrada (x2)
Donde L es el número de localidades; K, es el número de haplotipos totales en la muestra; ni,
es el tamaño de muestra de cada localidad; nij, es el número de copias observadas por
haplotipo por localidad; p, es la frecuencia haplotípica de la muestra total.

43
b) Diversidad haplotípica HST
HST=1-(HS/HT)
Donde Hs es el promedio ponderado de la estimación de la diversidad haplotípica en las
subpoblaciones; HT, es la estimación de la diversidad haplotípica de la población total.
c) Kst*
KST*=1-(KS/KT);
Donde KS es el promedio ponderado de Ki; KT, es el número promedio de diferencias entre
dos secuencias sin tomar en cuenta a las localidades.
Para KST*, Ki*
Donde dij,lk es el número de diferencias entre cada secuencia de la localidad (i) y cada
secuencia de la localidad (l). La función logarítmica en Ki* tiene como objetivo disminuir el
valor de dij,ik en caso de este sea grande.
El nivel de significancia se estimó a partir de 100,000 simulaciones de Monte Carlo.
Adicionalmente se calculó el estadístico Fst en el programa ARLEQUINv3.11 (Excoffier et
al., 2005), su significancia fue probada con 10,000 permutaciones.

44
RESULTADOS
De las 82 secuencias analizadas se observaron 11 sitios segregantes (sitio donde las secuencias
difieren; Castillo-Cobián, 2007). Todos considerados transiciones (sustitución de una base
púrica a otra base púrica, o de una pirimidica a otra pirimidica; Hartl y Clark, 2007) que
discriminaron en ocho haplotipos. A los haplotipos se le denominó GCH1, GCH2, hasta
GCH8. Los ocho haplotipos se incluyeron en la reconstrucción filogenética (Figura 2.3).
Reconstrucción filogenética
El mejor modelo de sustitución nucleotídica calculado el programa fue el tres parámetros de
Tamura T92+G. El árbol filogenético obtenido tras el análisis bayesiano muestra que los
haplotipos de la población del GC se agrupan con los haplotipos conocidos de B edeni brydei,
y todos forman un clado separado de B. edeni edeni, B. omurai, B. physalus y B. borealis
(Figura 2.3). Estos los grupos fueron soportados por valores mayores de 0.9 de probabilidad
posterior.
Con el análisis de atributos característicos (CAs) (Davis y Nixon, 1992; Sarkar et al.,
2002, Lowenstein et al., 2009) se identificaron 10 CAs puros que distinguen a los haplotipos
de B. edeni brydei de los de B. edeni edeni publicados en GenBank. Las secuencias obtenidas
en el presente trabajo coincidieron con B. edeni brydei, y compartieron los 10 CAs de este
grupo (Figura 2.3).
De los 10 CAs identificados en el presente estudio, 9 coincidieron con los CAs
descritos por Kershaw et al. (2013; posiciones 15563, 15592, 15609, 15614, 15616, 15681,
15722, 15726 y 15769) y uno por Rosel y Wilcox (2014; CA de la posición 15682) (Figura
2.3).

45
Figura 2.3: Reconstrucción filogenética (izquierda) de haplotipos de la región control de DNAmt de
individuos del Golfo de California (GC-indicados con flechas) y con individuos identificados como B.
edeni y B. brydei. 10 atributos diagnósticos característicos (CAs) (derecha), usados para la
identificación de grupos. La posición nucleotídica corresponde a la secuencia de B. brydei del genoma
mitocondrial en la posición 15477-16410 (ACCN:AB201259; Sasaki et al., 2006). Para ambos casos
grupos B. omurai, B. borealis y B. physalus fueron grupos externos (subrayados).

46
Estructura poblacional
De las 555 secuencias de B. edeni brydei analizadas, se observaron 37 sitios segregantes que
discriminaron a 57 haplotipos; 51 publicados previamente por Kanda et al. (2007), y seis
exclusivos para el Golfo de California. Siguiendo la nomenclatura de Kanda et al. (2007),
tenemos que GCH1 a GCH6 es igual a H52 a H57 (Figura 2.4; Cuadro 2.3). Y dos de los
haplotipos identificados para el GC fueron compartidos con la población de Perú, GCH7 y
GCH8 = H41 y H45, respectivamente (Figura 2.4; Cuadro 2.3). Estos resultados están
apoyados por valores de bootstrap mayores a 50%.

47
Figura 2. 4. Relación filogenética entre los 8 haplotipos identificados en el Golfo de California (GC) y
los 51 haplotipos de B. edeni brydei publicados por Kanda et al. (2007). En gris haplotipos exclusivos
para el GC y en negritas haplotipos compartidos con Perú.

48
Cuadro 2. 3. Frecuencias haplotípicas de cinco poblaciones de B. edeni brydei. En gris oscuro se
indican haplotipos exclusivos al Golfo de California y en gris claro los que comparte con la población
de Perú.
HAPLOTIPOS JAPÓN FIJI PERÚ GC JAVA TOTAL
1 50 15 1 0 0 66
2 2 0 0 0 0 2
3 35 0 1 0 0 36
4 3 0 0 0 0 3
5 156 3 1 0 0 160
6 8 0 0 0 0 8
7 14 0 0 0 0 14
8 14 0 1 0 0 15
9 3 0 0 0 0 3
10 6 0 0 0 0 6
11 5 0 0 0 0 5
12 4 0 0 0 0 4
13 5 0 0 0 0 5
14 1 0 0 0 0 1
15 8 0 0 0 0 8
16 6 0 0 0 0 6
17 2 0 1 0 0 3
18 14 0 0 0 0 14
19 8 0 0 0 0 8
20 4 0 0 0 0 4
21 3 0 0 0 0 3
22 5 0 0 0 0 5
23 2 0 0 0 0 2
24 2 0 0 0 0 2
25 7 0 0 0 0 7
26 7 0 0 0 0 7
27 1 0 0 0 0 1
28 6 0 0 0 0 6
29 8 0 0 0 0 8
30 2 0 0 0 0 2
31 1 0 0 0 0 1
32 1 0 0 0 0 1
33 3 0 0 0 0 3
34 2 0 0 0 0 2
35 1 0 0 0 0 1
36 2 3 1 0 0 6
37 0 2 0 0 0 2
38 0 1 0 0 0 1
39 0 0 1 0 0 1
40 0 0 1 0 0 1
41 0 0 1 2 0 3
42 0 0 5 0 0 5
43 0 0 1 0 0 1
44 0 0 1 0 0 1
45 0 0 3 25 0 28
46 0 0 5 0 0 5
47 0 0 1 0 0 1
48 0 0 0 0 19 19
49 0 0 0 0 2 2
50 0 0 0 0 1 1

49
51 0 0 0 0 1 1
52 0 0 0 31 0 31
53 0 0 0 13 0 13
54 0 0 0 5 0 5
55 0 0 0 3 0 3
56 0 0 0 1 0 1
57 0 0 0 2 0 2
Total 401 24 25 82 23 555
Diversidad genética
Los resultados de diversidad genética muestran que la población del Golfo de California es
más diversa que la de Java y Fiji, y ligeramente menor que la de las poblaciones de Perú y
Japón (Cuadro 2.4).
Cuadro 2. 4. Diversidad genética en poblaciones de B. edeni brydei, por región geográfica. N: número
de secuencias.
REGIÓN N Sitios
segregantes Haplotipos
Diversidad
haplotípica
Diversidad
Nucleotídica
π(JC)
JAPÓN 401 33 36 0.820
(DE 0.017) 0.0103
FIJI 24 7 5 0.594
(DE 0.105) 0.00733
PERÚ 25 19 14 0.840
(DE 0.070) 0.0111
JAVA 23 11 4 0.320
(DE 0.121) 0.00652
GC 82 14 8 0.729
(DE 0.027) 0.0111
Diferenciación poblacional
Los valores de distancia neta entre poblaciones, según el método 2 parámetros de Kimura,
indican que las poblaciones más lejanas genéticamente son en Golfo de California y Fiji
(0.01497) y las más cercanas son Japón y Fiji (0.00122). Se observa que la población más
cercana al GC es la de Perú (0.00369) seguida de Japón (0.01160) y Java (0.01185) (Cuadro
2.5).

50
Cuadro 2. 5. Distancia neta entre poblaciones de B. edeni brydei.
Japón
N=401
Fiji
N=24
Perú
N=25
Java
N=23
GC
N=82
Japón 0000 0.00122 0.00477 0.01145 0.01160
Fiji
0000 0.00685 0.01304 0.01497
Perú
00000 0.00888 0.00369
Java
0000 0.01185
GC
00000
Los valores de los tres estadísticos de heterogeneidad (Hst, Kst* y x2) entre regiones
geográficas muestra que la población del Golfo de California es diferente significativamente
de las otras poblaciones (Cuadro 2.6), y a su vez, las otras poblaciones analizadas (Japón, Fiji,
Java y Perú) también son diferentes entre sí.
Cuadro 2. 6. Resultados de la prueba de heterogeneidad entre regiones geográficas de B. edeni brydei
(GC= Golfo de California).
Hst Kst* x2
Japón-Fiji 0.0215 (P=0.0000) 0.0166 (P=0.0000) P=0.0000
Japón -Perú 0.0140 (P=0.0000) 0.0377 (P=0.0000) P=0.0000
Japón -GC 0.0273 (P=0.0000) 0.0867 (P=0.0000) P=0.0000
Japón -Java 0.0528 (P=0.0000) 0.0885 (P=0.0000) P=0.0000
Fiji-Perú 0.1366 (P=0.0000) 0.2347 (P=0.0000) P=0.0000
Fiji-GC 0.1640 (P=0.0000) 0.3299 (P=0.0000) P=0.0000
Fiji-Java 0.3760 (P=0.0000) 0.4722 (P=0.0000) P=0.0000
Peru-GC 0.0511 (P=0.0000) 0.1296 (P= 0.0000) P= 0.0000
Perú-Java 0.2377 (P=0.0000) 0.3336 (P=0.0000) P=0.0000
GC-Java 0.2550 (P=0.0000) 0.3268 (P=0.0000) P=0.0000

51
Los valores de Fst también indican que la población del Golfo de California es
diferente a todas las otras poblaciones analizadas y que todas las poblaciones son diferentes
entre sí (Cuadro 2.7).
Cuadro 2. 7. Valores de Fst por región de B. edeni brydei (P<0.05).
Japón Fiji Perú Java GC
Japón 0000
Fiji 0.098 0000
Perú 0.373 0.396 0000
Java 0.500 0.576 0.482 0000
GC 0.525 0.536 0.198 0.472 00000

52
DISCUSIÓN
El rorcual tropical que se distribuye en el Golfo de California pertenece a Balaenoptera edeni
brydei. Esta población es genéticamente distinta de las poblaciones de Japón, Fiji, Java y Perú
(Kanda et al., 2007). Estos resultados coinciden con lo propuesto por la IWC (1997), donde
reconocen a la población del rorcual tropical del GC como un stock separado del Pacífico
Norte.
Tershy et al. (1990) propusieron que Balaenoptera edeni brydei es la sub-especie que
se distribuye en el Golfo de California. Esta propuesta se corrobora con los haplotipos
identificados en el presente estudio. La relación filogenética que guardan éstos haplotipos con
los haplotipos de otras poblaciones B. edeni brydei identificados por otros autores (Yoshida y
Kato, 1999; Wada et al., 2003; Sasaki et al., 2005 y 2006; Kanda et al., 2007; Penry, 2010;
Kershaw et al., 2013; Hildebrandt et al., 2005, no publicado), como puede verse en el árbol de
reconstrucción calculado en el presente trabajo (Figura 2.3). En este árbol filogenético,
además es posible apreciar que todas secuencias de B. edeni brydei pertenecen a un grupo
diferente, claramente distinto de B. edeni edeni, como se ha propuesto en otros estudios (Wada
et al., 2003; Sasaki et al., 2006; Kanda et al., 2007; Penry, 2010; Kershaw et al., 2013; Rosel
y Wilcox, 2014). La comparación de un fragmento de 302pb de la región control del ADNmt
de 64 secuencias del GenBank y la reconstrucción filogenética bayesiana apoyan esta
distinción entre taxones. La ubicación de la población del rorcual tropical del Golfo de
California dentro de B. edeni brydei, y la separación de B. edeni brydei y B. edeni edeni, es
confirmado con la identificación de atributos característicos puros (CAs). Esta técnica ha sido
de utilidad para distinguir especies (Davis y Nixon, 1992; Sarkar et al., 2002; Lowenstein et
al., 2009; Kershaw et al., 2013; Rosel y Wilcox, 2014). La identificación de 10 CAs (Figura
2.3) y su coincidencia con los registrados por Kershaw et al. (2013) y por Rosel y Wilcox
(2014) muestra que estos sitios de la secuencia son diagnósticos y de utilidad para la distinción
de los grupos.
La comparación de las secuencias del Golfo de California muestran que la diversidad
genética de esta población (Hd= 0.729; π(JC)= 0.0111) es tan alta como las poblaciones que se
distribuyen en las regiones de Perú y Japón (Hd= 0.840; π(JC)= 0.0111 y Hd= 0.820; π(JC)=
0.0103, respectivamente). Una diversidad genética alta se relaciona con un mayor éxito

53
reproductivo y capacidad de adaptación a las condiciones ambientales, entre otros, y permite
la persistencia de la especie en tiempo y espacio (Watson, 2006; Piñero et al., 2008). El origen
de la magnitud de esta diversidad llama la atención porque es comparable con la diversidad
genética de ballenas con poblaciones numerosas, como la ballena jorobada cuyo tamaño
poblacional es de ca. 20 000 individuos, Hd= 0.766 y π(JC)= 0.009 (Baker et al., 1994; Piñero
et al., 2008).
Los resultados de distancia genética calculados en el presente trabajo (Cuadro 2.5) se
añaden a los registros obtenidos por Kanda et al. (2007) para poblaciones del oeste del Océano
Pacífico Norte (Japón), oeste y este del Pacífico Sur (Fiji y Perú), y este del Océano Índico
(Java). Estos valores indican que la distancia más grande entre poblaciones se presenta entre
las poblaciones del Golfo de California y Fiji (0.01497), mientras que las poblaciones más
cercanas son las del Golfo de California y Perú (0.00369), seguida de las distancias con Japón
(0.01160) y Java (0.01185). Estas distancia genética son bajas comparadas con las registradas
en otras ballenas, tales como ballena jorobada (GST= 0.3), esto puede deberse a una separación
más reciente en tiempo evolutivo y/o a que la distribución del rorcual tropical se restringe a
zonas tropicales y subtropicales; la propuesta es que una distribución restringida se relaciona
con un flujo génico más efectivo en distancias cortas, en contraste con especies que tienen una
migración latitudinal mayor, como la ballena jorobada (Kanda et al., 2007).
La heterogeneidad entre regiones geográficas a través los índices Hst, Kst*, x2 y Fst
(Cuadros 2.6 y 2.7), reitera que la población del Golfo de California es diferente de las
poblaciones comparadas. Estos índices han sido empleados con éxito para comparar
poblaciones de otras especies de mamíferos marinos (Baker et al., 1994; Bérubé et al., 2002;
Ruíz-García et al., 2008). Los valores calculados al comparar la población del Golfo de
California con las de Japón, Fiji, Perú y Java (Cuadro 2.7), implican migración de individuos
entre poblaciones (Kanda et al., 2007). Considerando que los valores de Fst indican que las
diferencias son pocas (0 – 0.05), moderadas (0.05 – 0.15), grandes (0.15 – 0.25) o que todo es
diferente (>0.25; Hartl y Clark, 2007), se encontró que todas las poblaciones comparadas son
diferentes entre sí. Además, estos valores también sugieren intercambio genético reciente;
pues mientras mayor es el Fst, menor es el número de migrantes entre poblaciones (Hartl y
Clark, 2007). Sin embargo para determinar si el flujo genético continua o si el hecho de que
las poblaciones sean diferentes pero sin una separación filogenética, se debe a una

54
clasificación de linaje incompleto o a un polimorfismo ancestral residual (en el cual no hay
migración), es necesario aplicar métodos estadísticos como el de Nielsen y Wakeley (2001)
que ayuden a resolver la interrogante.
El análisis filogenético del ADNmt muestra que la población del Golfo de California
tiene dos haplotipos compartidos con la población de Perú (H41 y H45) y seis haplotipos
privados exclusivos (H52-57; Figura 4). Los haplotipos compartidos indican un flujo génico
entre dichas poblaciones. Se estima que el tiempo de divergencia de Balaenoptera edeni fue
hace 6.3±2.0 millones de años, y la divergencia entre subespecies B. edeni brydei y B. edeni
edeni fue hace 0.7±0.6 millón de años (Sasaki et al., 2006). La teoría de la coalescencia
propone que el haplotipo más frecuente corresponde al linaje más antiguo (Posada y Crandall,
2001). A partir de este postulado es posible inferir un evento de flujo génico en el pasado,
desde el Golfo de California, pues los haplotipos 45 y 41 son más frecuente en el golfo (25 y
2, respectivamente) con respecto a Perú (3 y 1, respectivamente). Esto parece ser confirmado
por la baja distancia genética entre estas poblaciones (0.00369). La población de Perú pudiera
ser un punto importante de intercambio génico en un tiempo determinado entre poblaciones
del Océano Pacífico porque comparte haplotipos con Japón, Fiji y el Golfo de California, y las
distancias genéticas son cortas entre dichas poblaciones (0.00477 con Japón, y 0.00685 con
Fiji). Los haplotipos exclusivos parecen indicar una acumulación de diferenciación genética
debido a una alta fidelidad del sitio y a la filopatría sexo específica, entre otros factores
(Kanda et al., 2007).
CONCLUSIONES
El rorcual tropical que se distribuye en el Golfo de California pertenece a la subespecie B.
edeni brydei según la clasificación del Comité Taxonómico (Committee on Taxonomy, 2014).
La población de B. edeni brydei del GC es diferente genéticamente de otras poblaciones de los
océanos Pacífico e Índico. Existe una relación inversa entre las distancia genética y la
distancia geográfica, es decir entre más cercanas geográficamente estén las poblaciones,
menores los niveles de diferenciación genética.

55
Este es el primer estudio donde se identifica genéticamente la sub-especie del rorcual
tropical que se distribuye el Golfo de California y que aporta datos que amplían la información
sobre la distribución de Balaenoptera edeni brydei.

56
LITERATURA CITADA
Anderson, J. 1878/9. Comprising an account of the zoological results of the two expeditions to
western Yunan in 1868 and 1875; and a mono-graph of the two cetacean genera. En:
Anatomical and Zoological Researches. B. Quaritch, London. 1: 551–564.
Árnason, Ú., Gullberg, A. y Widegren, B. 1991. The complete nucleotide sequence of the
mitochondrial DNA of the fin whale, Balaenoptera physalus. J Mol Evol. 33: 556–568.
Árnason, Ú., Gullberg, A. y Widegren, B. 1993. Cetacean mitochondrial DNA control region
sequences of all extant baleen whales and two sperm whale species. Mol Biol Evol. 10:
960–970.
Baker, C. S., Slade, R. W., Bannister, J. L., Abernethy, R. B., Weinrich, M. T., Lien, J.,
Palumbi, S. R. 1994. Hierarchical structure of mitochondrial DNA gene flow among
humpback whales Megaptera novaeangliae, worldwide. Molecular Ecology. 3(4):
313–327.
Bérubé, M., Urbán, J., Dizon, A. E., Brownell, R. L. y Palsbøll, P. J. 2002. Genetic
identification of a small and highly isolated population of fin whales (Balaenoptera
physalus) in the Sea of Cortez, Mexico. Conservation Genetics. 3(2): 183–190.
Best, P. B. 1960. Further information on Bryde’s whale (Balaenoptera edeni Anderson) from
Saldanha Bay, South Africa. Nor. Hval.-tid. 49: 201–215.
Castillo-Cobián, A. 2007. La selección natural a nivel molecular. Capítulo 1. En Eguiarte, L.
E., Sousa, V. y Aguirre X., (Compiladores). Ecología Molecular. Secretaría de Medio
Ambiente y Recursos Naturales, Instituto Nacional Ecología, Universidad Nacional
Autónoma de México y Comisión Nacional para el Conocimiento y Uso de la
Biodiversidad. 25–48 pp.
Committee on Taxonomy. 2014. List of marine mammal species and subspecies. Society for
Marine Mammalogy, www.marinemammalscience.org, consultado el 22 dic 2014.
Davis, J. I., y Nixon, K. C. 1992. Populations, genetic variation, and the delimitation of
phylogenetic species. Systematic Biology. 41: 421–435.

57
Excoffier, L., Laval, G. y Schneider, L. 2005. Arlequin ver. 3.0: An integrated software
package for population genetics data analysis. Evolutionary Bioinformatics Online. 1:
47–50.
Hartl, D. L. y Clark, A. G. 2007. Principles of population genetics. Sunderland: Sinauer
associates. 652 pp.
Hildebrandt, S., Lopez Jurado, L. F. y Lopez, J. M. A. 2005. First genetic description of the
Bryde's whale (Balaenoptera brydei) in the North Atlantic Ocean.
Hori, H., Bessho, Y., Kawabata, R., Watanabe, I., Koga, A. y Pastene, L. A. 1994. World-
wide population structure of minke whales deduced from mitochondrial DNA
control region sequences. Unpublished document SC/46/SH14, 46th IWC Scientific
Committee. 11pp. Available from Division of Biological Sciences, Graduate School
of Science, Nagoya University, Chikusa, Nagoya. 464–8602.
Hudson, R. R., Boos, B. B. y Kaplan, N. L. 1992. A statistical test for detecting geographic
subdivision. Molecular Biology and Evolution. 9: 138–151.
Huelsenbeck, J. P. y Ronquist, F. 2001. MRBAYES: Bayesian inference of phylogenetic trees.
Bioinformatics. 17: 754–755.
IWC, 1997. Report of the subcommittee on North Pacific Bryde’s whales, Annex G, Report of
the Scientific Committee. Rep. Int. Whal. Commn. 47: 163–168.
Junge, G. C. 1950. On a specimen of the rare fin whale, Balaenoptera edeni Anderson,
stranded on Pulu Sugi near Singapore. Zoologische Verhandelingen. 9: 1–26.
Kanda, N., Goto, M., Kato, H., McPhee, M. F. y Pastene, L. A. 2007. Population genetic
structure of Bryde’s whales (Balaenoptera brydei) at the inter oceanic and trans
equatorial levels. Conserv. Genet. 8: 853–864.
Kato, H. y Perrin, W. F. 2009. Bryde’s Whales Balaenoptera edeni/brydei. En: William, F. P.,
Würsig, B. y Thewissen, J. G. M. 2009. Encyclopedia Marine Mammals 2a ed.
Elservier. 157–162 pp.
Kershaw, F., Leslie, M. T., Collins, T., Mansur, R. M., Smith, B. D., Mington, G., Baldiwn,
R., Leduc, R.G., Anderson, R. C., Brownell JR, R. L. y Rosenbaum, H. C. 2013.
Population Differentiation of 2 Forms of Bryde’s Whales in the Indian and Pacific
Oceans. Journal of Heredity. 104.6: 755–764.

58
Kimura, M. 1980. A simple method for estimating evolutionary rates of base substitutions
through comparative studies of nucleotide sequences. J Mol Evol. 16: 111–120.
Lowenstein, J. H., Amato. G. y Kolokotronis, S-O. 2009. The Real maccoyii: Identifying Tuna
Sushi with DNA Barcodes – Contrasting Characteristic Attributes and Genetic
Distances. PloS One. 4e7866.
Librado, P. y Rozas, J. 2009. DnaSPv5: A software for comprehensive analysis of DNA
polymorphism data. Bioinformatics. 25: 1451–1452.
Miller, S. A., Dykes, D. D. y Polesky, H. F. 1988. A simple salting out procedure for
extracting DNA from human nucleated cells. Nucleic Acids Research. 16(3): 1225.
Nielsen, R. y Wakeley, J. 2001. Distinguishing Migration From Isolation: A Markov Chain
Monte Carlo Approach. Genetics. 158: 885–896.
Nei, M. y Li, W. H. 1979. Mathematical Model for Studying Genetic Variation in Terms of
Restriction Endonucleases. PNAS. 76(10): 5269–5273.
Olsen, Ö. 1913. On the external characters and biology of Bryde’s whale (Balaenoptera
brydei), a new rorquals from the coast of South Africa. Proc. Zool. Soc. Lond. 1073–
1090.
Omura, H. 1959. Bryde’s whale from the coast of Japan. Sci. Rep. Whales Res. Inst., Tokyo.
14: 1–33.
Penry, G. S. 2010. The biology of South African Bryde's whales. PhD dissertation, University
of St. Andrews.
Piñero, D., et al., 2008. La diversidad genética como instrumento para la conservación y el
aprovechamiento de la biodiversidad: estudios en especies mexicanas, en Capital
natural de México, vol. I: Conocimiento actual de la biodiversidad. Conabio, México.
437–494 pp.
Posada, D. y Crandall, K. 2001. Intraspecific gene genealogies: trees grafting into networks.
TRENDS in Ecology and Evolution. 16(1): 37–45.
Rambaut, A. y Drummond, A. 2009. MCM Trace Analysis Tool. Version v1.5.0. Institute of
Evolutionary Biology, University of Edinburgh. http://beas.bio.ed.ac.uk/.
Rambaut, A. 2012. Tree Figure Drawing Tool. Version 1.4.0. Institute of Evolutionary
Biology, University of Edinburgh. http://tree.ed.ac.uk/

59
Rice, W. D. 1998. Marine Mammals of the World. Systematic and distribution. The Society
for marine Mammalogy. Special Publication. 4. 231 pp.
Rosel, P. E. y Wilcox, L. A. 2014. Genetic evidence reveals a unique lineage of Bryde’s
whales in the northern Gulf of Mexico. Endangered Species Research. 25: 19–34.
Ruíz-García, M., Caballero, S., Martínez-Agüero, M. y Shostell, J. M. 2008. Molecular
differentiation among Inia geoffrensis and Inia boliviensis (Iniidae, Cetacea) by means
of nuclear intron sequences. Population genetics research progress. 177–223.
Sarkar, I. N., Thornton, J. W., Planet, P. J., Figurski, D. H., Schierwater, B. y DeSalle, R.
2002. An automated phylogentic key for classiyng homoboxes. Mol. Phylogeneet Evol.
24:388–399.
Sasaki, T., Nikaido, M., Hamilton, H., Goto, M. Kato, H., Kanda, N., Pastene, L., Cao, Y.,
Fordyce, R., Hasegawa, M. y Okada, N. 2005. Mitochondrial phylogenetics and
evolution of mysticete whales. Syst Biol 54: 77–90.
Sasaki, T., Nikaido, M., Wada, S., Yamada, T. K., Cao, Y., Hasegawa, M. y Okada, N. 2006.
Balaenoptera omurai is a newly discovered baleen whale that represents an ancient
evolutionary lineage. Mol. Phylogenetics Evol. 41: 40–52.
Tamura, K., Stecher, G., Peterson, D., Filipski, A. y Kumar, S. 2013. MEGA6: Molecular
Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution 30:
2725–2729.
Tershy, B. R., Breese, D. y Strong, C. S. 1990. Abundance, seasonal distribution and
population composition of balaenopterid whales in the Canal de Ballenas, Gulf of
California, Mexico. Rep. Int. Whal. Commn. Special Issue. 12: 369–375.
Thompson, J., Higgins, D. G., y Gibson, T. 1994. ClustalW. Nucleic Acids Res. 22: 4673–
4680.
Wada, S., Oishi, M. y Yamada, T. K. 2003. A newly discovered species of living baleen
whale. Nature. 426(6964): 278–281.
Watson, J. D. 2006. Biología Molecular del Gen. 5ª ed. Editorial Médica Panamericana. 776
pp.
Yoshida, H. y Kato, H. 1999. Phylogenetic relationships of Bryde’s whale in the western
North Pacific and adjacent waters inferred from mitochondrial DNA sequences. Mar.
Mamm. Sci. 15: 1269–1286.

60
CAPÍTULO III
ESTRUCTURA GENÉTICA POBLACIONAL DEL RORCUAL TROPICAL
(Balaenoptera edeni brydei) EN EL GOLFO DE CALIFORNIA CON BASE
EN ADN MITOCONDRIAL

61
RESUMEN
El conocimiento que se tiene sobre la estructura poblacional del rorcual tropical en el Golfo de
California (GC) es limitado. Estudios de foto-identificación sugieren la existencia de animales
residentes en la región norte y análisis de movimientos estacionales sugieren la presencia de
animales transeúntes en la región sur. Datos de isótopos estables, de contenido de ácidos
grasos y estudios preliminares sobre la estructura genética, son consistentes con esta
propuesta. Sin embargo, hasta ahora no se ha hecho un análisis particular para probar dicha
hipótesis. En el presente trabajo se compararon secuencias de 299pb de la región control del
ADNmt de 82 muestras separadas a priori como representantes del norte (29) y sur (53), a fin
de conocer el grado de diversidad y variación genética espacial de Balaenoptera edeni brydei.
Los análisis permitieron discriminar ocho haplotipos a partir de once sitios segregantes; cuatro
de los haplotipos son compartidos por ambas regiones, mientras que tres son exclusivos para
la región sur y uno para la región norte. Un análisis de la diversidad genética y diferencias
poblacionales, estimadas por el método 2 parámetros de Kimura y los estadísticos Chi-
cuadrada, Hst, Kst* y Fst, indican que no existe diferencia significativa entre las regiones
norte y sur. Es posible que tal semejanza genética entre regiones sea el reflejo de un flujo
génico facilitado por la capacidad de desplazamiento propio de la especie, y también una
evidencia de que los rorcuales tropicales forman una sola población en el Golfo de California.
Sin embargo, será necesario utilizar muestras del Pacífico adyacente o analizar marcadores
nucleares para confirmar los resultados presentados en este trabajo.

62
ABSTRACT
The knowledge we have on the population structure of Bryde's whale in the Gulf of California
(GC) is limited. Photo-identification studies suggest the existence of resident animals in the
northern region and analysis of seasonal movements suggest transient animals in the south.
Stable isotope data, fatty acid, and preliminary studies on the genetic structure are consistent
with this proposal. However, there is not specific analysis to test this hypothesis. Therefore, in
this work we want to identify population differences of Bryde’s whale in the Gulf of
California, based on their haplotype diversity of mitochondrial DNA (mtDNA). The analyses
included 82 samples, 29 from northern and 53 from southern of GC. A total of eight
haplotypes were identified, five were present in the north (one exclusive), and seven in the
south (three exclusive). The statistical analysis of heterogeneity based on Chi-square, Kst*,
Hst and Fst showed no differences between samples from north and south of GC. These results
suggest that Bryde's whales are a population in the Gulf of California. However, samples from
adjacent Pacific or nuclear markers are necessary to confirm.

63
INTRODUCCIÓN
El rorcual tropical se distribuye de norte a sur en el Golfo de California, y en ambas costas,
peninsular y continental (Urbán y Flores, 1996). Su abundancia se estima en 400 individuos
(Gerrodette y Palacios, 1994; Chávez, 1995; Urbán y Flores, 1996), los cuales son
considerados como un stock particular con respecto a otras poblaciones del mundo (IWC,
1997) (Figura 1.3).
El conocimiento sobre la estructura poblacional del rorcual tropical en el golfo es
limitado. Estudios de foto-identificación sugieren la existencia de animales residentes en la
zona norteña (Breese y Tershy, 1987), y análisis de los movimientos estacionales de la especie
sugieren la presencia de animales del Pacífico adyacente en el suroeste (Vidal et al., 1987;
Salvadeo et al., 2011). Estudios de isótopos estables (López-Montalvo, 2012; Olavarrieta-
García, 2012) y contenido de ácidos grasos (López-Montalvo, 2012) sugieren la presencia de
dos poblaciones, una que se alimenta en zonas con características más costeras y la otra se
alimenta en zonas con características más oceánicas. Además, el único trabajo que se tienen
sobre la estructura genética del rorcual tropical, propone la existencia de dos poblaciones, una
residente y otra transeúnte (Dizon et al., 1996), sin embargo, esta propuesta está basada en un
número reducido de muestras. Estos antecedentes dejan ver que el problema no ha quedado
resuelto y que es necesario hacer más estudios, al respecto, es necesario considerar que para
lograr conclusiones más robustas sobre la estructura poblacional de la especie, es
recomendable analizar un mayor número de muestras y tratar de que éstas representen
diferentes regiones geográficas del GC.
Con base en lo anterior, el presente trabajo plantea conocer el grado de diversidad y la
variación genética espacial del rorcual tropical (Balaenoptera edeni brydei) en el Golfo de
California. Se propone el análisis de ADNmt como se ha hecho en poblaciones distribuidas en
las costas de Japón, Perú, Océano Índico, Pacifico Noroeste entre otras (Pastene et al., 1997;
Yoshida y Kato, 1999; Kanda et al., 2007; Kershaw et al., 2011).

64
OBJETIVOS
Conocer el grado de diversidad genética del rorcual tropical que se distribuye en el
Golfo de California con base en su ADNmt.
Determinar el grado de variación de la diversidad genética en espacio y tiempo.

65
MÉTODO
Obtención de muestras y extracción de ADN, ver detalles en Capítulo II.
Análisis de datos
Las secuencias se editaron, alinearon (MEGA v6; Tamura et al., 2013) y se separaron
por región geográfica (norte y sur). Se calculó el número de haplotipos, número de sitios
polimórficos, diversidad nucleotídica y haplotípica (DnaSPv.5.10.01; Librado y Rozas, 2009).
Para visualizar las frecuencias haplotípicas y la relación entre haplotipos, se construyó una red
de haplotipos usando el método Median-joining network (NETWORKv4.2.0.1; Bandelt et al.,
1999).
Para conocer el grado de diversidad genética e identificar diferencias poblacionales, se
calcularon los siguientes estimadores:
1) Distancia genética usando el método: 2 parámetros de Kimura (Kimura, 1980).
2) Heterogeneidad entre regiones, según Hudson et al. (1992) con tres estadísticos: Chi-
cuadrada, Hst y Kst*.
3) Se calculó el estadístico Fst (Excoffier et al., 2005).
Para detalles de los estimadores, ver Capítulo II.
Se construyeron árboles filogenéticos usando el método de Máxima Verosimilitud
(MEGA v6; Tamura et al., 2013). Se tomaron secuencias de B. omurai y B. edeni del
GenBank (Núm. Acceso AB116096 y AB201258, respectivamente), y se determinó el mejor
modelo de sustituciones nucleónicas (MEGA v6; Tamura et al., 2013). Para la estimación de
los intervalos de confianza, se corrieron 10000 simulaciones de bootstraps.

66
RESULTADOS
Durante 1994-2013 se colectaron 94 biopsias, de las cuales 39 fueron tomadas en la región
norte y 55 en la sur del GC. Del total de muestras se logró obtener 82 secuencias de buena
calidad y de animales diferentes. Como parte de las muestras del sur, se incluyó una muestra
del Pacífico Oriental tropical (Figura 3.1).
Figura 3. 1. Distribución de las biopsias de B. edeni brydei en el Golfo de California.
De las secuencias 82 analizadas se identificaron 11 sitios segregantes que
discriminaron ocho haplotipos. De los cuales, cuatro son compartidos entre la región sur y
norte (H1, H2, H4 y H8), entre ellos están los tres más frecuentes, H1 con 38%, H2 con 16% y
H8 con 30%. En la región sur se encontraron tres haplotipos exclusivos (H3, H5 y H7) y en la

67
región norte sólo uno, H6 (Cuadro 3.1, Figura 3.2). El mayor número de diferencias (6) entre
haplotipos se observan entre los haplotipos H1-H2, y H5-H8. Existen cuatro pares de
haplotipos que presentan un cambio en la secuencia, H2-H6, H6-H8, H8-H3 y H3-H4 (Figura
3.2). La muestra tomada en el Pacífico Oriental tropical se ubicó en el haplotipo 8.
Cuadro 3. 1. Frecuencias haplotípicas por región.
Haplotipos Norte Sur Total
H1 12 19 31
H2 5 8 13
H3 0 5 5
H4 2 1 3
H5 1 0 1
H6 0 2 2
H7 0 2 2
H8 9 16 25
Total 29 53 82
Figura 3. 2. Red de ocho haplotipos únicos identificados en individuos B. edeni brydei del Golfo de
California. El tamaño del círculo es proporcional a la frecuencia haplotípica por región según el color.
La posición y cantidad de mutaciones entre cada haplotipo están indicadas con números sobre las líneas.

68
El análisis de diversidad genética muestra que las regiones norte y sur presentan una
diversidad haplotípica y diversidad nucleotídica similares. Los sitios segregantes y el número
de haplotipos fueron mayores en el sur con respecto al norte (Cuadro 3.2).
Cuadro 3. 2. Diversidad genética en poblaciones de B. edeni brydei, por región geográfica. N: número
de secuencias.
REGIÓN N Sitios
segregantes
No.
Haplotipos
Diversidad
haplotípica
Diversidad
Nucleotídica
π(JC)
Norte 29 8 5 0.722
(DE 0.049) 0.0111
Sur 53 11 7 0.760
(DE 0.033) 0.0116
El valor de la distancia genética entre poblaciones calculado con el Método dos
parámetros de Kimura (Kimura, 1980), fue de -0.01974. Los resultados de los tres estadísticos
sobre la heterogeneidad entre los individuos de la región norte y sur, demostraron que no
existe diferencia significativa entre dichas regiones. El resultado del estadístico Fst confirma
que los rorcuales del Golfo de California pertenecen a una sola población (Cuadro 3.3).
Cuadro 3. 3. Valores de heterogeneidad entre regiones B. edeni brydei, y estadístico Fst.
Regiones Hst Kst* x2 Fst
Norte-Sur 0.00668
(P<0.05)
-0.00861
(P<0.05)
0.312
(P<0.05)
-0.01412
(P<0.01)
La relación filogenética calculada para los ocho haplotipos coloca en la base del árbol
filogenético al haplotipo H1 y lo separa del resto, esta separación es apoyada por el 90% de
bootstrap. Los otros siete haplotipos forman dos grupos, uno formado por H2 y H6, y el
segundo grupo constituido por los otros cinco haplotipos (Figura 3.3).

69
Figura 3. 3. Relación filogenética de haplotipos únicos de la región control del rorcual tropical, usando
el método Máxima Verosimilitud, basado en el modelo tres parámetros de Tamura (T92) y secuencias
de Balaenoptera edeni edeni (Bed) y Balaenoptera omurai (Bom) como grupos externos. Se muestran
los valores de bootstrap (basados en 10000 iteraciones), mayores al 50%; la barra equivale a 0.005
sustitución/sitio.

70
DISCUSIÓN
El análisis de la región control del ADNmt de B. edeni brydei mostró que no hay
diferenciación genética estadísticamente significativa entre individuos del norte y sur,
sugiriendo la ocurrencia de una sola población.
La presencia y abundancia de los haplotipos compartidos indican flujo génico en un
tiempo no definido. Este flujo podría estar favorecido por la capacidad de desplazamiento de
la especie, ya que los individuos pueden movilizarse a lo largo del golfo y ser avistados
durante todo el año, se han registrado desplazamientos estacionales en función de la
disponibilidad de alimento y como una respuesta a eventos climáticos propios de la región y
eventuales fenómenos como la Niña y el Niño (Urbán y Flores, 1996; Salvadeo et al., 2011).
Este desplazamiento es factible gracias a que el rorcual tropical no se enfrenta a barreras
geográficas obvias a lo largo del Golfo de California. Este libre tránsito y el flujo génico entre
regiones norte y sur también se observa en animales con alta movilidad como el tiburón
ballena (Ramírez-Macías et al., 2007). Avise (1992) señala que uno de los factores que
también influye en el flujo génico, además de la capacidad de dispersión, es la historia de vida
de la especie.
Adicionalmente, el haplotipo H1, siendo el más frecuente (representa el 37% de la
población) y más ampliamente distribuido en el Golfo de California, podría ser considerado el
linaje mitocondrial más antiguo, como lo describe la teoría de la coalescencia para haplotipos
frecuentes y abundantes (Posada y Crandall, 2001). Esta condición es apoyada por el resultado
del análisis filogenético, en el que el H1 se ubica en la posición basal (Figura 3.3). Esta
posición basal implica que la población del rorcual tropical del Golfo de California fue
fundada por una población pequeña con el haplotipo H1 y tuvo una expansión rápida de la que
surgieron el resto de los haplotipos. Este mecanismo se conoce como efecto fundador (Bush,
1975) y se ha propuesto para otras poblaciones en el Golfo de California, como el tiburón
ballena y vaquita marina (Rosel y Rojas-Bracho, 1999; Ramírez-Macías et al., 2007).
La diversidad haplotípica calculada en el presente trabajo fue muy parecida entre el
norte y sur (0.722 y 0.759, respectivamente), esto se debe a que cuatro de los cinco haplotipos
son compartidos entre norte y sur (H1, H2, H4 y H8). Este nivel de diversidad haplotípica es
alto si consideramos que el intervalo de valores para diversidad se expresa en una escala de 0 a

71
1, donde 0 significa que no hay diversidad haplotípica en la población y 1 indica que la
población es diversa (Hartl y Clark, 2007). Un ejemplo de baja diversidad haplotípica se
observa en Balaenoptera physalus, para la que se calcula una diversidad haplotípica de 0.007
en la población del Golfo de California, siendo un reflejo de su aislamiento y ausencia de flujo
génico con otras poblaciones de la especie (Bérubé et al., 2002).
El valor de la diversidad nucleotídica registrado (Cuadro 3.2) indica pocas diferencias
en las secuencias del Golfo de California, ya que este análisis compara el parecido
nucleotídico (Hartl y Clark, 2007). Dicha semejanza se puede observar en la longitud de las
ramas del árbol filogenético, donde los haplotipos más parecidos poseen las ramas más cortas
(Figura 3.3).
El valor de la distancia genética (-0.01974), los resultados de los tres estadísticos de
heterogeneidad entre regiones (Hst, Kst* y x2) y los valores de Fst (Cuadro 3.3) coinciden en
que no hay diferencia poblacional entre regiones norte y sur. Esto resultados apoyan las
deducciones resultantes de la red de haplotipos y los valores de divergencia genética (la
diversidad haplotípica y nucleotídica) en los que se plantea que en el Golfo de California hay
sólo una población de Balaenoptera edeni brydei.
Los resultados del presente estudio contrastan con trabajos previos. La existencia de
una población residente al norte y otra población transeúnte al sur del GC, no coincide con los
resultados del presente trabajo. Las 82 muestras analizadas no reflejan las dos poblaciones
propuesta por Dizon et al. (1996) a parir de cinco muestras. La única muestra analizada en el
presente trabajo para Pacífico Oriental tropical, corresponde con el haplotipo 8 (haplotipo 45
según clasificación de Kanda et al., 2007), el cual es el segundo haplotipo más abundante
dentro del golfo y se comparte con la población de Perú (Capitulo II). Al respecto, Dizon et al.
(1996) pudieron haber encontrado un haplotipo diferente para la zona, sin embargo, las
secuencias no son disponibles y no pudieron ser comparadas. Los trabajos donde se utilizan
ácidos grasos (López-Montalvo, 2012) e isótopos estables (López-Montalvo, 2012;
Olavarrieta-García, 2012) encuentran diferencias espaciales y estacionales en la dieta, lo que
pudiera reflejar el movimientos de los animales en función de la disponibilidad de alimento
(Vidal et al., 1987; Salvadeo et al., 2011), y no la existencia de dos poblaciones. Además, los
resultados de estos marcadores ecológicos reflejan cambios de meses (López-Montalvo,
2012), o en el mejor de los casos cambios en términos de años (2.7 años; Olavarrieta-García,

72
2012), por lo que pudieran corresponder a diferencias en las estrategias alimenticias, y no una
separación poblacional. Adicionalmente, el movimiento de individuos registrado con foto-
identificación (Chávez, 1995) pudiera ser el reflejo de la capacidad de movilidad de la especie
y su potencial interacción a través del GC.
El número de muestras y el tiempo analizado en el presente trabajo le confiere robustez
a los resultados mostrados, sin embargo es necesario realizar estudios que incluyan mayor
número de muestras del Pacífico adyacente y/o análisis con marcadores nucleares o
fragmentos más grandes del ADNmt, a fin de corroborar las conclusiones expuestas en este
trabajo.
CONCLUSIONES
Se identificaron ocho haplotipos en el Golfo de California, uno fue exclusivo a la región norte
y tres a la región sur, el resto fueron compartidos entre ambas regiones. Los resultados de
diversidad, distancia y las pruebas de heterogeneidad (chi cuadrada, Hst y Kst* y Fst.)
genética indicaron que B. edeni brydei del Golfo de California no presenta diferenciación
geográfica, es decir pertenece a una población. Sin embargo es necesario realizar estudios con
muestras el Pacífico adyacente o con marcadores nucleares para corroborar las conclusiones
expuesta.

73
LITERATURA CITADA
Avise, J. C. 1992. Molecular population structure and the biogeographic history of a regional
fauna: a case history with lessons for conservation biology. Oikos. 62–76.
Bandelt, H. J., Forster, P. y Rol, A. 1999. Median-joining networks for inferring intraspecific
phylogenies. Mol. Biol. Evol. 16: 37–48.
Bérubé, M., Urbán, J., Dizon, A. E., Brownell, R. L. y Palsbøll, P. J. 2002. Genetic
identification of a small and highly isolated population of fin whales (Balaenoptera
physalus) in the Sea of Cortez, Mexico. Conservation Genetics. 3(2): 183–190.
Breese, D. y Tershy, B. R. 1987. Residency patterns in female Bryde’s whales in the Gulf of
California, Mexico. Abstract: Seven Biennial Conference on the Biology of Marine
Mammals. Miami, FL. EE. UU.
Bush, G. L. 1975. Modes of animal speciation. Annual Review of Ecology and Systematics.
339–364.
Chávez, R, S. 1995. Estimación poblacional del rorcual tropical Balaenoptera edeni
(Anderson, 1878) en la Bahía de la Paz, B. C. S. México. Tesis de Maestría.
CICIMAR. 55 pp.
Dizon, A., Lux, C. A., LeDuc, R. G., Urbán, R, J., Henshaw, M. y Brownell, R. Jr. 1996. An
interin phylogenetic analysis of sei and Bryde’s whale mitochondrial DNA control
region sequences. SC/47/NP23 presented to the IWC Scientific Committee. 12 pp.
Excoffier, L., Laval, G. y Schneider, L. 2005. Arlequin ver. 3.0: An integrated software
package for population genetics data analysis. Evolutionary Bioinformatics Online. 1:
47–50.
Gerrodette, T. y Palacios, D. N. 1994. Estimates of cetacean abundance in EEZ waters of the
eastern Tropical Pacific. Southwest Fisheries and Science Center. Administrative
Report LJ-96-10. 28 pp.
Hartl, D. L. y Clark, A. G. 2007. Principles of population genetics. Sunderland: Sinauer
associates. 652 pp.
Hudson, R. R., Boos, B. B. y Kaplan, N. L. 1992. A statistical test for detecting geographic
subdivision. Molecular Biology and Evolution. 9: 138–151.

74
IWC, 1997. Report of the subcommittee on North Pacific Bryde’s whales, Annex G, Report of
the Scientific Committee. Rep. Int. Whal. Commn. 47: 163–168.
Kanda, N., Goto, M., Kato, H., McPhee, M. F. y Pastene, L. A. 2007. Population genetic
structure of Bryde’s whales (Balaenoptera brydei) at the inter oceanic and trans-
equatorial levels. Conserv. Genet. 8: 853–864.
Kershaw, F. Leslie, S. M., Collins, T., Mansur, M. R., Smith, D. B., Minton G., Baldwin, R.,
LeDuc, G. L., Anderson, C., Brownell, L. Jr. y Rosenbaum, C. H. 2013. Population
differentiation of 2 forms of Bryde’s Whales in the Indian and Pacific Oceans. Journal
of Heredity. 104(6): 755–764.
Kimura, M. 1980. A simple method for estimating evolutionary rates of base substitutions
through comparative studies of nucleotide sequences. J Mol Evol. 16: 111–120.
Librado, P. y Rozas, J. 2009. DnaSPv5: A software for comprehensive analysis of DNA
polymorphism data. Bioinformatics. 25: 1451–1452.
López-Montalvo, C. 2012. Caracterización de la ecología alimentaria del rorcual tropical
(Balaenoptera edeni), en el Golfo de California con base en análisis de isotopos
estables N y C, y Ácidos Grasos. Tesis Maestría. Instituto de Ciencias del Mar y
Limnología. UNAM. 76pp
Olavarrieta-García, T. 2012. Hábitos alimenticios del rorcual tropical (Balaenoptera edeni) en
el Golfo de California. Tesis Licenciatura, Universidad Autónoma de Baja California
Sur. 43pp
Pastene, L. A., Goto, M., Itoh, S., Wada, S. y Kato, H. 1997. Intra-and inter-oceanic patterns
of mitocondrial DNA variation in the bryde´s whale, Balaenoptera edeni. Rep. Int.
Whal. Commn. (SC/48/NP15). 47: 569–574.
Posada, D. y Crandall, K. 2001. Intraspecific gene genealogies: trees grafting into networks.
TRENDS in Ecology and evolution. 16(1): 37–45.
Ramírez-Macías, D., Vázquez-Juárez, R., Galván-Magaña, F. y Munguía-Vega, A. 2007.
Variations of the mitochondrial control region sequence in whale sharks (Rhincodon
typus) from the Gulf of California, Mexico. Fisheries Research. 84(1): 87–95.
Rosel, P. E. y Rojas‐Bracho, L. 1999. Mitochondrial DNA variation in the critically
endangered vaquita Phocoena sinus Norris and Macfarland, 19581. Mar. Mamm. Sci.
15(4): 990–1003.

75
Salvadeo, C., Flores-Ramírez, S., Gómez-Gallardo, A., MacLeod, C., Llunch-Belda, D.,
Jaume-Schinkel, S. y Urbán, R. 2011. Bryde’s whale (Balaenoptera edeni) in the
southwestern Gulf of California Relationship with ENSO variability and prey
availability. Ciencias Marinas. 37(2): 215–225.
Tamura, K., Stecher, G., Peterson, D., Filipski, A. y Kumar, S. 2013. MEGA6: Molecular
Evolutionary Genetics Analysis version 6.0. Mol. Biol. and Evol. 30: 2725–2729.
Urbán, R, J. y Flores, R, S. 1996. A note on Bryde’s Whales (Balaenoptera edeni) in the Gulf
of California, Mexico. Rep. Int. Whal. Commn. 46: 453–457.
Vidal, O., Aguayo, A., Findley, L., Robles, A., Bourillón, L., Vomend, I., Turk, P., Gárate, K.,
Maroñas, L. y Rosas, J. 1987. Avistamientos de mamíferos marinos durante el crucero
“Guaymas I” en la región superior del Golfo de California, primavera de 1984. En:
Memorias de la X Reunión Internacional sobre Mamíferos Marinos. La Paz, B.C.S.,
México. 7–35 pp.
Yoshida, H. y Kato, H. 1999. Phylogenetic relationships of Bryde’s whale in the western
North Pacific and adjacent waters inferred from mitochondrial DNA sequences. Mar.
Mamm. Sci. 15: 1269–1286.

76
CAPÍTULO IV
VARIACIÓN GEOGRÁFICA EN VOCALIZACIONES DEL RORCUAL
TROPICAL (Balaenoptera edeni brydei) EN EL GOLFO DE CALIFORNIA

77
RESUMEN
De un total de 109 grabaciones hechas con confirmación visual a individuos adultos de
Balaenoptera edeni brydei, representantes de las regiones norte y sur del Golfo de California
(GC), se identificaron cuatro diferentes tipos de vocalizaciones, una de las cuales (Be4) ha
sido registrada en el Pacífico Oriental tropical y en el sur de California. Las tres
vocalizaciones restantes no habían sido registradas previamente para la especie. Estas últimas
vocalizaciones, denominadas Be10, Be11 y Be12, fueron grabadas durante avistamientos de
individuos solos, mientras que la vocalización Be4 siempre fue grabada en avistamientos con
al menos dos individuos. Los pocos registros acústicos disponibles para Balaenoptera edeni
han sido una limitante para el uso de técnicas de monitorización acústica pasiva, por lo que los
registros efectuados con confirmación visual realizados en el presente trabajo, son pertinentes
para la monitorización y estudio poblacional de la especie. Las vocalizaciones Be4 obtenidas
con un Paquete de Grabación de Acústica de Alta Frecuencia (HARP por sus siglas en inglés)
fueron analizadas para probar la hipótesis de la presencia de dos poblaciones en el GC, una
residente al norte y otra transeúnte al sur. Las vocalizaciones grabadas con confirmación
visual se clasificaron según su Proporción Señal-Ruido (SNR). Sólo los componentes
principales de 191 vocalizaciones con SNR alta (> 9dB) fueron caracterizados para cada una
de las zonas; su caracterización consideró elementos matemáticos (frecuencia máxima,
frecuencia mínima, frecuencia pico y duración), morfológicos y de gradiente de energía. En el
análisis morfológico se seleccionó un umbral a -24 dB para resaltar las diferencias de duración
no visibles a un rango dinámico completo; una prueba de t de Student mostró diferencias
significativas entre vocalizaciones del norte (n=88) y sur (n=103), para este parámetro. Se
observaron diferencias morfológicas en los espectrogramas tridimensionales de las
vocalizaciones de cada una de las regiones y las representaciones de la distribución de la
energía confirmaron concentración alta en las vocalizaciones del norte. Tales diferencias
sugieren que hay variación geográfica en las vocalizaciones de la especie en el GC.

78
ABSTRACT
A total of 109 recordings were collected during sightings of adult of Bryde´s whale. Four call
types were identified. Three were different to previously documented calls for the species,
while the fourth call type matched the Be4 call type reported for the Eastern Tropical Pacific
and south of California. While the three new call types were recorded during sightings of
individual Bryde’s whales; there were always at least two whales in sight during recordings of
the Be4 call type. Previous acoustic recordings of Bryde’s whale are few and several have had
no accompanying visual confirmation of the species or number of individuals. The visual
documentation provided by this study makes a significant contribution to further acoustic
monitoring of this species. Analysis of Be4 type calls obtained from a High-frequency
Acoustic Recording Package (HARP) were conducted to test whether acoustic data supports
the hypothesis of the presence of two distinct populations, one resident in the north and
another transient in the south of the Gulf of California (GC). Calls confirmed with recordings
of the species in the wild were classified by their Signal to Noise Ratio into Low SNR (<
5.9dB), Medium SNR (6-8.9 dB) and high SNR (> 9dB). The main component of 191 calls
with high SNR from North (103) and south (88) were characterized using standard parameters
(maximum frequency, minimum frequency, peak frequency and duration), and new
morphological and energy gradient based criteria proposed for the present study. Through
morphological analysis, a threshold at -24 dB was selected to highlight the duration
differences not seen at full dynamic range. A t-Student test showed significant differences
between the calls of the north and the south, for this parameter. Morphological differences in
the tridimensional spectrograms of calls from each region were observed, and energy
distribution representations confirmed higher concentration in north calls. These results
suggest that there is a geographic variation in the vocalizations of B. edeni in the GC, which
coincides with records of geographic variation in the eastern tropical Pacific.

79
INTRODUCCIÓN
El monitoreo acústico en mamíferos marinos se ha incrementado en las últimas décadas
debido a que es una herramienta efectiva, precisa y no invasiva que permite conocer varios
aspectos de las especies tales como la presencia, distribución, abundancia e identidad de
poblaciones o individuos, entre otras características biológicas (Au y Hasting, 2009).
Particularmente el uso más común del monitoreo acústico es el análisis de la estructura
poblacional, ya que es un método idóneo que permite obtener muestras de lugares donde
logísticamente es casi imposible (Mellinger y Barlow, 2003).
A diferencia de los marcadores tradicionales, la variación registrada con las
vocalizaciones (sonidos producidos por las cuerdas vocales de la laringe; Au y Hasting, 2009)
refleja cambios recientes y rápidos en las poblaciones, tales como movimientos o patrones de
asociación (McDonald et al., 2006). Incluso algunos autores sugieren el uso de estas
vocalizaciones como una herramienta más efectiva en la identificación de unidades de manejo
(Mellinger y Barlow, 2003; Delarue et al., 2009; Rendell et al., 2011).
Diversos estudios efectuados en cetáceos utilizan los sonidos emitidos por estos
animales para identificar variaciones geográficas. La primera especie en la que se utilizaron
vocalizaciones como indicadores de stocks fue en la ballena Jorobada (Megaptera
novaeangliae) (Payne, 1979). En tiempo recientes, se han logrado identificar poblaciones con
base en patrones de variación acústica, como en el rorcual común (Balaenoptera physalus)
(Hatch y Clark, 2004) y la ballena azul (Balaenoptera musculus) (McDonald et al., 2006). En
esta última especie ha sido posible identificar hasta nueve tipos canciones (patrones fijos de
sonidos o vocalizaciones que se repiten en ciclos y con duración de varios minutos, Payne y
McVay, 1971) y asociar cada una de las canciones a una región geográfica particular
(McDonald et al., 2006). Además, estudios en odontocetos demuestran que es posible
distinguir grupos de Orcas (Orcinus orca) que viven en simpatría, y asignar conductas, tipo de
vocalización, genética y hábitos alimenticios de manera particular para cada grupo (Baird,
2000). En el cachalote (Physeter macrocephalus) también es posible reconocer grupos
mediante patrones cortos de series de clicks (sonidos de alta frecuencia, 3-31 KHz y de muy
corta duración; Au y Hasting, 2008), llamados “codas (Rendell y Whitehead, 2003).

80
Para el rorcual tropical hay poca información sobre su repertorio acústico, además
algunas descripciones no están basadas en características físicas del sonido, sino en la forma
en cómo el investigador escucha o percibe la vocalización, dicha situación complica la
comparación del repertorio. En el Cuadro 4.1 se reúne la información de los siete trabajos que
publican descripciones de vocalizaciones de la especie. La primera descripción (Cummings et
al., 1986) se realizó en Bahía de Loreto, Golfo de California, a partir de dos sesiones de
grabación. La primera sesión se hizo con confirmación visual y la segunda sin confirmación.
En esta última sesión se asumió que eran vocalizaciones de la especie por el parecido con el
primer registro, el cual fue realizado en animales adultos y con hidrófono portátil. La segunda
descripción (Edds et al., 1993) corresponde a grabaciones oportunistas con confirmación
visual hechas en Bahía de los Ángeles, en el Golfo de California, y a registros de
vocalizaciones de un animal joven en cautiverio en Sea Word Florida, Estados Unidos,
después de que dicho individuo se varó. La tercera descripción (Oleson et al., 2003) fue hecha
en vocalizaciones registrados en el Pacífico Oriental tropical (incluida la costa suroccidental
de la Península de Baja California), Mar Caribe y costas de Japón; todos estos registros fueron
obtenidos de organismos adultos usando un hidrófono de arrastre. La cuarta descripción
(Heimlich et al., 2005) se realizó en vocalizaciones obtenidas con un hidrófono fijo en el
Pacífico Oriental tropical, tales sonidos son atribuidos a B. edeni por la similitud con la
descripción de Oleson et al. (2003). Dichas similitudes incluyen “la naturaleza del sonido
(tonal, con frecuencias ligeramente hacia arriba o hacia abajo y frecuencia modulada), así
como la duración, la banda de frecuencia aproximada, el tiempo y la frecuencia” (Oleson et
al., 2003). En ambos trabajos, el de Oleson et al. (2003) y de Heimlich et al. (2005), se
identificaron relaciones entre los tipos de vocalización y la distribución geográfica. En la
quinta descripción, hecha por Širović et al. (2014) a partir de grabaciones con confirmación
visual y acústica pasiva, se describe una vocalización en el Golfo de México, dicha
vocalización se caracteriza por tener una frecuencia entre 78-110 Hz y estar asociada a grupos
de adultos. En la misma región del Golfo de México, Rice et al. (2104) hicieron la sexta
descripción a partir de tres vocalizaciones obtenidas mediante acústica pasiva, los autores
mencionan que para validar las grabaciones es necesario realizar registros con seguimiento
local. Finalmente, Figueiredo y Simão (2014) hacen la séptima descripción en tres

81
vocalizaciones registradas con confirmación visual en las costas de Brasil, una de estas
vocalizaciones se asocia a grupos de Madre con cría.
Estudios de foto-identificación (Breese y Tershy, 1987), genéticos (Dizon et al., 1996),
de isótopos estables (López-Montalvo, 2012; Olavarrieta-García, 2012) y contenido de ácidos
grasos (López-Montalvo, 2012) sugieren que la distribución poblacional del rorcual tropical se
encuentra dividida en stock discretos dentro del Golfo de California. Se propone que pueden
distinguirse dos poblaciones (Dizon et al., 1996), una con presencia en la zona norte,
conformada por animales residentes (Breese y Tershy, 1987); y otra con distribución en la
zona sur, conformada por animales transeúntes provenientes del Océano Pacífico (Vidal et al.,
1987; Salvadeo et al., 2011). De existir tal distribución poblacional, métodos acústico podrían
aportar información para discriminar stock, como se ha observado en otros cetáceos (Hatch y
Clark, 2004; McDonald et al., 2006) y odontocetos (Baird, 2000; Rendell et al., 2011).
Con base en lo anterior, el presente trabajo propone caracterizar vocalizaciones de
Balaenoptera edeni brydei en el Golfo de California, ampliar el repertorio acústico e
identificar variación geográfica en sonidos obtenidos con hidrófono fijo y a partir de
grabaciones con confirmación visual usando un hidrófono portátil.

82
Cuadro 4. 1. Características de las vocalizaciones registrados en Balaenoptera edeni.
Método de
Identificación
Tipo de
vocalización
Presencia de
Harmónicos
Intervalo
de
frecuencia
Intervalo
de
duración
Conducta
asociada
Tipo de
grupo Lugar Autor
Confirmación
Visual Simple No 70–245 0.2–1.5 Ninguna Par
Golfo de
California
Cummings
et al., 1986
Confirmación
Visual Simple No
500–700 25–40 Ninguna Cría sola Golfo de
California
Edds et al.,
1993 100–500 0.7–1.4 Ninguna Cría y adulto
90–180 0.1–0.6 Ninguna Adulto con cría
Acústica
pasiva y
Confirmación
Visual
Simple
Be1 No 20–23 1.1–6.7 Ninguna
Más de un animal
en el área de
registro
ETP
Oleson et
al., 2003
Be2 Yes 35.7–38.2 0.5–3.0 Ninguna Single ETP
Be3 Yes 2.44–26.9 1.0–4.0 Ninguna Animal solo o en
par ETP
Be4 No 59.5–60.2 0.8–3.0 Ninguna Animal solo o en
par ETP
Be5 Yes 26.0–26.8 1.3–2.2 Ninguna Single ETP
Be6 No 57.1–
232.7 0.7–4.9
Viajando a una
velocidad
constante y
dirección
definida
Animal solo o en
grupo ETP
Acústica
pasiva Simple Be7 Si 43.7–48.7 0.8–2.5 Ninguna
Ninguna Caribe
Sistema de
hidrófonos y
cámara de
video
Simple
Be8a Si 43.0–48
0.25–
0.60 Ninguna Ninguna Japón
Be8b Si 137–192
0.30–
0.41 Ninguna Ninguna Japón
Acústica
pasiva Frase
Tonal de
barrido
alterno
No 33.5–39.0
14.8–26.0
0.3–3.0
0.4–3.2
0.3–5.6
Ninguna Ninguna
ETP Heimlich
et al., 2005 Tonal de
no barrido No
24.6–32.3
13.5–20.7
0.30–3.4
0.3–3.2 Ninguna Ninguna

83
alterno 37.1-53.9 0.3–5.6
Alta
ráfaga-
tonal
Frecuencias no
relacionadas
harmónicamente
39.9–46.6
24.8–37.9
47.5–56.4
0.3–2.5
0.4–1.8
0.3–1.0
0.3–2.8
Ninguna Ninguna
Baja
ráfaga–
tonal
Frecuencias no
relacionadas
harmónicamente
16.8–26.7
24.1–36.1
62.6–78.8
0.4–2.8
0.3–2.5
0.4–2.0
0.4–4.0
Ninguna Ninguna
Tono
harmónico Si
20.7–25.8
75.1–79.6
46.8–66.3
0.5–3.2
0.4–2.3
0.5–2.2
Ninguna Ninguna
Acústica
pasiva y
Confirmación
Visual
Simple Be9 No 110–78
143–85 0.4–0.7 Ninguna Grupo de adultos
Golfo de
México Širović et
al., 2014
Acústica
pasiva
Frase Bajo-
barrido No
96.84–
97.68a
0.2–0.2b
Ninguna Ninguna
Golfo de
México
Rice et al.,
2014 Simple
Gemido
largo
Si (En algunas
señales)
106.6–
108.1a
11.1–
11.5b
Ninguna Ninguna
Frase Tonal No 99.4–
107.4 1.9–2.7
b Ninguna Ninguna
Confirmación
Visual
Simple PS1 No 225–555 0.788 Ninguna Madre con cría
Cabo Frio
en las
costas de
Brasil
Figueiredo
et al.,
2014.
Simple LFT Si 8.5–19 1.413 Ninguna Solo
Simple FMT No 416–671 1.531 Ninguna Solo
Simple TM1 Si 85.7–
123.6 0.876
Ballena muy
cerca de la
embarcación
Solo
Simple TM2 No 49.5–
105.5 1.156
Individuo
alejándose de la
embarcación
Solo
ETP: Pacífico Oriental tropical.

84
OBJETIVOS
Caracterizar vocalizaciones de Balaenoptera edeni brydei en el Golfo de California.
Identificar variaciones de las vocalizaciones en el Golfo de California.

85
MÉTODO
Registros acústicos
Para este estudio se utilizaron registros acústicos de dos fuentes: grabaciones con
confirmación visual con hidrófono portátil y grabaciones obtenidas de un paquete autónomo
de grabación pasiva o hidrófono fijo.
Vocalizaciones: con confirmación visual
En cada salida al mar, una vez avistado e identificado un organismo de la especie, se apagó el
motor y se sumergió un hidrófono portátil [Transductores AQ-1s & ITC-1042 (10 Hz -100
KHz)] a una profundidad aproximada de 10 metros. Con una grabadora FOSTEX FR2 se
hicieron grabaciones de diez minutos en formato de audio digital sin compresión de datos (con
extensión .wav). La frecuencia de muestreo (FS1) en un inicio fue de 192600
2, resolución (R)
de 24 bits, posteriormente se modificó a FS 2050 y R de 16bits. Para evitar saturación en la
grabación se modificó la ganancia según las condiciones de grabación. Se registró el nombre
del archivo de grabación en una hoja asociada al resto de los datos del avistamiento, fecha,
hora, condiciones ambientales, número de organismos, presencia de crías, conducta y
fotografías.
Vocalizaciones: hidrófono fijo
El hidrófono fijo, denominado HARP (High-frequency Acoustics Recording Package), fue
diseñado por el Instituto SCRIPPS de Oceanografía de la Universidad de California. Estuvo
constituido por un conjunto de pilas eléctricas, un hidrófono [Transductores AQ-1s & ITC-
1042 (10 Hz -100 KHz)] y discos de memoria, todos contenidos en cilindros de plástico PVC.
La FS fue de 192600 y se programó para registrar sonidos de 0 a 2000 Hz, a intervalos de
1 FS: Frequency Sample, es el número de muestras por unidad de tiempo, tomadas de una señal continúa para
producir una señal discreta, esto durante el proceso para convertir una señal analógica a digital (Au y Hasting,
2009).
2 Según el teorema de Nyquist, la frecuencia de muestreo deberá ser al menos del doble que la frecuencia de
interés (Nyquist, 1928). En este trabajo se utilizaron frecuencias mayores, con la finalidad de obtener una forma
de onda con mayor resolución temporal, y una mejor representación de las frecuencias superiores de las
vocalizaciones.

86
cinco minutos de grabación por diez minutos de descanso (ocho horas por día), por
aproximadamente seis meses (duración de las pilas y capacidad de memoria; Ethan Roth, 2009
comn. pers).
Se colocó a 700 metros de profundidad en tres localidades del Golfo de California: 1)
Punta Pescadero (23° 48.448’ Lat N y -109° 37.687’ Long W), del 08 de diciembre 2006 a
junio de 2007 (siete meses de registro); 2) Isla Tiburón (28° 36.536’ Lat N y -112° 30.930’
Long W), de julio 2007 a diciembre 2007 (seis meses de registro); y 3) en Bahía de los
Ángeles (29° 01.750’ Lat N y -113° 22.474’ Long W), de agosto a diciembre de 2008 y de
mayo a noviembre 2009 (12 meses de registro) (Figura 4.1).
Figura 4. 1. Ubicación del hidrófono fijo en Punta pescadero (1), Isla Tiburón (2) y Bahía de los
Ángeles (3).

87
Los archivos de sonido (formato .wav) se revisaron con el programa de cómputo Triton
(v 1.64.20070724), desarrollado por el Instituto SCRIPPS para plataforma Matlab® (Matlab
Signal Processing Toolbox v 6.2).
Análisis de las vocalizaciones
La vocalizaciones se visualizaron y pre-procesaron en el programa Adobe Audition® (v 3.1)
(filtro Butterworth, pasa bajas, con una frecuencia de corte 500 Hz y normalización 90%).
Posteriormente en el programa Matlab® (v 2010b), se hizo la descripción del componente
principal de las vocalizaciones a partir de los elementos: Matemático, Morfológico y
Gradiente de energía (Figuras 4.2, 4.3 y 4.4).
El Elemento Matemático consistió en medir la Frecuencia máxima (Fa), Frecuencia
mínima (Fi), Pico de frecuencia (Pf; medido en Hertz, Hz) y Duración (Td; medida en
segundos, s) (Figura 4.2). La medición del Elemento Morfológico se hizo a partir de una
revisión visual de las vocalizaciones proyectadas en imágenes en tercera dimensión (3D;
Figura 4.3). La medición del Elemento Gradiente de energía consistió en revisar imágenes en
3D a las que se les aplicó la función de gradiente. El resultado de esta aplicación, fue
identificar el gradiente de energía de cada vocalización en función de la intensidad de color y
el valor de referencia (Figura 4.4).
Las vocalizaciones con confirmación visual caracterizadas se compararon con registros
previos (Cuadro 4.1), esto con el objetivo de saber si estaban o no descritas previamente.
Finalmente, las vocalizaciones con confirmación visual registradas durante el presente estudio
fueron el molde para encontrar vocalizaciones en los registros del hidrófono fijo.

88
Figura 4. 2. Parámetros de las vocalizaciones. Fa=Frecuencia máxima, Fi=Frecuencia mínima;
Pf=Pico de frecuencia; Td=duración.
Figura 4. 3. Morfología de una vocalización en proyección 3D, con parámetros señalados.
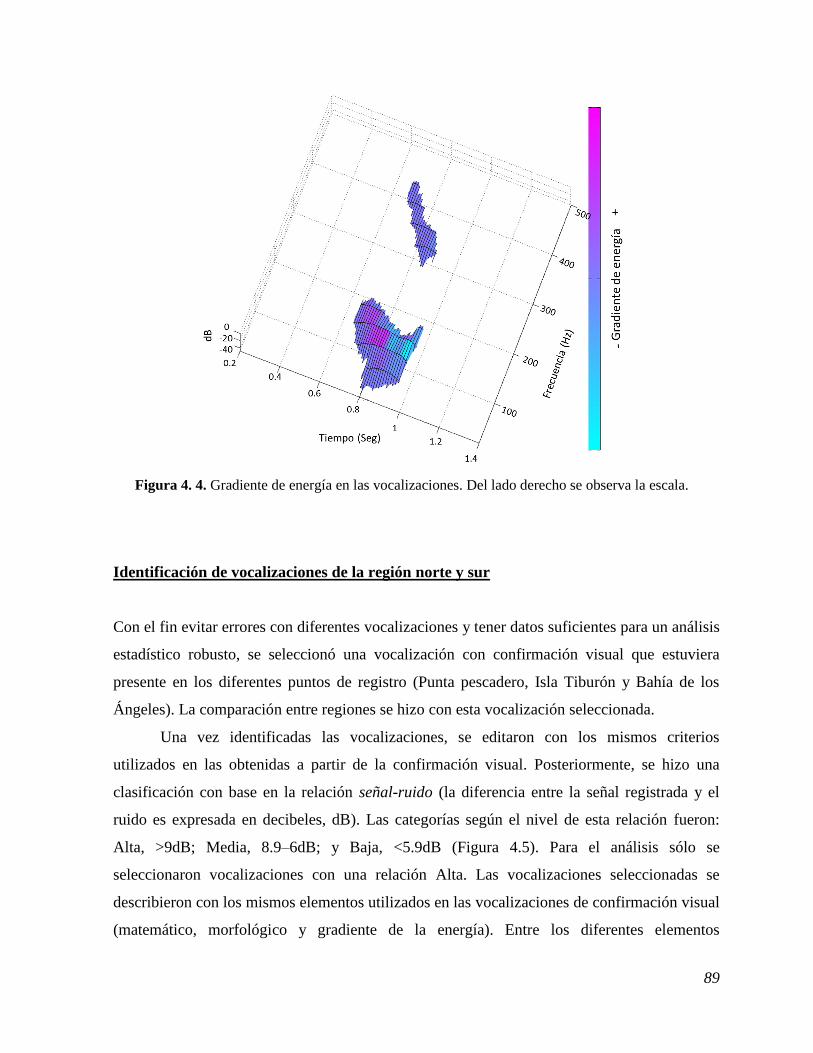
89
Identificación de vocalizaciones de la región norte y sur
Con el fin evitar errores con diferentes vocalizaciones y tener datos suficientes para un análisis
estadístico robusto, se seleccionó una vocalización con confirmación visual que estuviera
presente en los diferentes puntos de registro (Punta pescadero, Isla Tiburón y Bahía de los
Ángeles). La comparación entre regiones se hizo con esta vocalización seleccionada.
Una vez identificadas las vocalizaciones, se editaron con los mismos criterios
utilizados en las obtenidas a partir de la confirmación visual. Posteriormente, se hizo una
clasificación con base en la relación señal-ruido (la diferencia entre la señal registrada y el
ruido es expresada en decibeles, dB). Las categorías según el nivel de esta relación fueron:
Alta, >9dB; Media, 8.9–6dB; y Baja, <5.9dB (Figura 4.5). Para el análisis sólo se
seleccionaron vocalizaciones con una relación Alta. Las vocalizaciones seleccionadas se
describieron con los mismos elementos utilizados en las vocalizaciones de confirmación visual
(matemático, morfológico y gradiente de la energía). Entre los diferentes elementos
Figura 4. 4. Gradiente de energía en las vocalizaciones. Del lado derecho se observa la escala.

90
descriptivos se buscó uno que fuera informativo para diferencias regionales. El parámetro
Duración (elemento matemático) fue el seleccionado como elemento informativo. Para la
medición de este parámetro se buscó un punto donde las vocalizaciones fueran comparables,
en el que se observaran los componentes de la vocalización y se eliminara el ruido. Este punto
fue a -24dB. La medición se hizo en el componente principal, en imágenes en 3D
estandarizadas (resolución en tiempo y frecuencia constante; Figura 4.6). Estas características
permitieron realizar mediciones bajo las mismas condiciones y evitar errores de percepción.
Finalmente, se hizo una comparación de la duración, morfología y gradiente de la energía de
las vocalizaciones de cada región.
Figura 4. 5. Relación señal-ruido.
Figura 4. 6. Puntos de medición de la duración a -24dB en el componente principal de las
vocalizaciones del hidrófono fijo.

91
Análisis estadísticos
Para todas las vocalizaciones se calculó el promedio y desviación estándar de los cuatro
parámetros (Fa, Fi, Pf, y Td).
Identificación de vocalizaciones de la región norte y sur
Se obtuvieron datos de estadística descriptiva (promedio, mediana, desviación estándar y
varianza) de las vocalizaciones seleccionadas de buena calidad, y se hizo la prueba de
Kolmogorov-Smirnov para comprobar normalidad. Para detectar diferencias entre zonas se
usó la prueba de t de Student (Zar, 1999)
92
RESULTADOS
Vocalizaciones con confirmación visual
Durante el periodo de muestreo se realizaron 109 grabaciones, sólo en 23 se registraron
vocalizaciones (Cuadro 4.2; Figura 4.7), es decir, se obtuvo un 25.3 % de éxito en registro de
vocalizaciones. Al comparar las 23 vocalizaciones se lograron identificar cuatro
vocalizaciones diferentes, tres se registraron en la región sur (Be1, 4.2 y 4.3; Figuras 4.8 a
4.10) y una en la región norte (Be4; Figura 4.11).
Cuadro 4. 2. Esfuerzo de grabación durante 2010-2014.
Año Horas Organismos Grabaciones Vocalizaciones
2010 611.4 42 31 10
2011 498.9 52 11 1
2012 578.0 64 33 11
2013 545.95 28 6 1
2014 281.67 12 3 0
Total 2515.92 198 84 23
Las cuatro vocalizaciones registradas se cotejaron con trabajos anteriores (Cummings
et al., 1986; Edds et al., 1997; Oleson et al., 2003; Heimlich et al., 2005) y el resultado fue
que tres (Be1, 2 y 3) no se habían reportado previamente para la especie (Viloria et al., 2012;
Anexo 1); y la cuarta vocalización (Be4), ya se había reportado en la costa suroccidental de la
Península de Baja California (Oleson et al., 2003) y al sur de la región conocida como
California Bight (Kerosky et al., 2012).

93
Descripción de las vocalizaciones
La vocalización denominada Be1 se presentó en dos individuos adultos, solos. En uno de ellos
se registró cinco veces y en el otro individuo se registró solamente una vez. No se identificó
ninguna conducta asociada. Esta vocalización presentó una Fa promedio del componente
principal de 144.8±14.6 Hz, Fi de 82±7.4 Hz, Pf a 99.6±7.4 Hz y un Td de 0.31±0.1 s (Figura
4.8).
Figura 4. 7. Distribución de las vocalizaciones identificadas.

94
Figura 4. 8. Vocalización Be1.
La vocalización denominada Be2 (Figura 4.9) se registró dos veces en un individuo
adulto, solo. No se identificó conducta asociada. Esta vocalización presentó una Fa de
276.6±47.7 Hz, Fi de 114.6±10 Hz y un Pf 161.1±20.8 Hz. Los tres parámetros fueron los más
altos registrados entre todas las vocalizaciones. Además, la vocalización Be2 presentó un Td
de 0.4±0.1 s, similar a la vocalización Be1.
Figura 4. 9. Vocalización Be2.
La vocalización Be3 se registró una vez en un individuo adulto, solo. No se asoció
ninguna conducta. Esta vocalización presentó una Fa de 154.2 Hz, Fi de 80.5 Hz y un Pf de
105.5 Hz. A diferencia de las anteriores, la duración de esta vocalización fue superior a un
segundo, 1.2 s (Figura 4.10).

95
Figura 4. 10. Vocalización Be3
La vocalización Be4 se registró ocho veces en seis animales diferentes, todos adultos.
Esta vocalización se destacó de las otras por registrarse siempre que había más de un
individuo en la zona de grabación. Otra diferencia que la destaca de las otras vocalizaciones es
que la Fi 50.4±1.0 Hz, Fa de 63.3±1.4 Hz y Pf es más baja 56.7±0.79 Hz. El Td fue de 1.8 ±
0.5 s (Figura 4.11).
En las vocalizaciones de confirmación visual no se registraron sonidos de hembras con
cría, a pesar de que se hicieron grabaciones durante este tipo de avistamiento.
Figura 4. 11. Vocalización Be4.

96
Vocalizaciones con hidrófono fijo
En total se revisaron 1237 horas de grabación, de las cuales 527 fueron de Punta Pescadero,
305 de Isla Tiburón y 405 de Bahía de los Ángeles. La vocalización Be4 estuvo presente de
manera abundante en los tres puntos de registro, por lo que fue seleccionada para buscar
diferencias entre la región norte y sur del golfo. En Punta pescadero se identificaron 444
vocalizaciones del tipo Be4, de las cuales 103 vocalizaciones fueron de buena calidad
(relación señal/ruido>8); en Bahía de los Ángeles se identificaron 517, de las cuales 78 fueron
de buena calidad; y en Isla Tiburón se identificaron 45, de las cuales 10 fueron de buena
calidad. La diferencia entre el número de horas revisadas y el número de vocalizaciones de
buena calidad identificadas para cada punto del HARP estuvo relacionada con los niveles de
ruido de cada zona. Isla Tiburón fue el sitio con mayor número de horas con ruido, seguido de
Bahía de los Ángeles y finalmente Punta Pescadero. El origen del ruido pudiera ser ambiental
y no antropogénico; pues en los sitios de grabación ubicados al norte (Isla Tiburón y Bahía de
los Ángeles), las actividades humanas son menores que en la región del sur (Punta Pescadero).
Comparación entre regiones:
Comparación estadística
Los parámetros de estadística descriptiva de las 88 vocalizaciones del norte y 103 de la sur,
calculados para la duración medida a -24dB, se muestran en el Cuadro 4.3.
Cuadro 4. 3. Parámetros estadísticos de la duración medida a -24dB para vocalizaciones de cada
región.
REGIÓN SUR
n=103
NORTE
n=88
Media 2.46 1.93
Mediana 2.46 1.89
Desviación estándar 0.43 0.36
Varianza 0.18 0.13

97
La prueba de Kolmogorov-Smirnov (d=0.22, p>0.20), indicó una distribución normal de los
datos. La prueba de t de Student aplicada al parámetro Duración de vocalizaciones, mostró
diferencias estadísticamente significativas entre las regiones norte y sur (t= 9.13, valor crítico=
1.65, p<0.05).
Comparación morfológica
Se observaron diferencias entre las vocalizaciones de la región norte y sur, después de
comparar las imágenes 3D. Las vocalizaciones del norte se caracterizaron por presentar un
solo montículo bien definido (Figura 4.12), mientras que las del sur presentaron dos
montículos un grande principal y uno pequeño asociado a éste (Figura 4.13). Además la
vocalización del norte presentó una base más angosta con respecto a la del sur; esta
característica se relaciona con la duración de las vocalizaciones.
Comparación del gradiente de energía
La gráfica 3D permitió ver la energía concentrada en las vocalizaciones. Esta energía
concentrada fue observada como un montículo con pendientes pronunciadas y tonalidad rosa,
según el gradiente de energía (Figura 4.12 y 4.13).
Figura 4. 12. Morfología y gradiente de energía de la vocalización Be4 del norte.

98
Figura 4. 13. Morfología y gradiente de energía de la vocalización Be4 del sur.

99
DISCUSIÓN
En el presente estudio se identificaron cuatro vocalizaciones, Be1, Be2, Be3 y Be4. Las tres
primeras son nuevos registros para la especie y no fue posible su comparación entre regiones.
La cuarta vocalización, Be4, ha sido registrada en trabajos anteriores y pudieron identificarse
diferencias geográficas entre individuos del Golfo de California, siendo una vocalización más
corta en el norte que en el sur (Viloria et al., 2013; Anexo 2).
Las cuatro vocalizaciones comparten características con descripciones previas hechas
para la especie (Cummings et al., 1986; Edds et al., 1993; Oleson et al., 2003; Heimlich et al.,
2005). Son vocalizaciones simples (Clark, 1990), que no forman frases, ni tampoco se repiten
para formar canciones (terminología de Payne y McVay, 1971). Además todas son de baja
frecuencia (< 1000 Hz), según la descripción de Au y Hasting (2009). Las vocalizaciones Be1
y Be4 se caracterizan por presentar un componente principal y dos a tres armónicos, a
diferencia de las Be2 y Be3 que sólo tienen el componente principal. Esta variación en el
número de componentes se observa en otras vocalizaciones de la especie y su duración (0.3-
1.8 s) coincide con el intervalo reportado por otros autores (0.2-2.83 s; Cummings et al., 1986;
Edds et al., 1993; Oleson et al., 2003; Heimlich et al., 2005).
La vocalización con la frecuencia mínima más baja fue la Be4, a 60 Hz. Esta
vocalización fue grabada en avistamientos con más de dos adultos en la zona. Su frecuencia
mínima baja difiere de otras vocalizaciones de la especie grabadas con acústica pasiva en el
Océano Pacífico oriental, 13.5 Hz (Heimlich et al., 2005) y 20 Hz (Oleson et al., 2003), y con
grabaciones con confirmación visual en el golfo de California, 70 Hz (Cummings et al., 1986)
y 90 Hz (Edds et al., 1993). La frecuencia máxima registrada en la vocalización Be2, 276 Hz,
fue similar a lo reportado por Cummings et al. (1986) en el golfo de California, 245 Hz, pero
diferente de la frecuencia máxima registrada por otros autores (79.6 Hz, Heimlich et al., 2005;
192 Hz, Oleson et al., 2003; 500 Hz, Edds et al., 1993). Debido a que todas las vocalizaciones
que se compararon en el presente trabajo eran de animales adultos, se descarta que la variación
de las frecuencias registradas se relaciona con la edad de los individuos (Oleson et al., 2003),
por lo que debería considerarse como una ampliación en el repertorio de vocalizaciones para la
especie.

100
Las vocalizaciones Be1, Be2 y Be3 se registraron en animales solos, en algunos casos
se registraron cuando había grupos de delfines comunes de rostro largo y lobos marinos
alimentándose en bancos de peces. Como en otras especies de ballenas, este tipo de
vocalizaciones podrían estar asociada a conductas de alimentación (Oleson et al., 2007). La
vocalización Be4 se registró siempre que había al menos dos individuos adultos en el área, y
en la mayoría de los avistamientos en presencia de alimento, i.e., sardina (Tershy et al., 1993).
Kato y Perrin (2009) también describieron que los individuos de esta especie sólo se agrupan
cuando se alimentan, si la reunión representan oportunidades de socialización y uno de los
eventos más importante de socialización para las especies es la reproducción (Tyack, 2009), es
posible que esta vocalización esté asociada para tales fines. Sin embargo, es necesario
confirmar dicha asociación.
La identificación de diferencias geográficas con registros de hidrófono fijo se efectuó
con la vocalización Be4. Esta vocalización se eligió porque estuvo presente de manera
abundante en ambas regiones, 88 vocalizaciones en el norte y 109 en el sur. Tras la
comparación entre regiones se identificaron diferencias estadísticamente significativas. La
característica distintiva fue la duración, las vocalización del norte fueron más cortas
(1.8±0.29s) con respecto a las del sur (2.5±0.44s). Estos sugiere una variación geográfica en
las vocalizaciones de B. edeni brydei en el Golfo de California, coincidiendo con registros de
variación geográfica en el Pacífico Oriental tropical (Oleson et al., 2003; Heimlich et al.,
2005). Las variaciones geográficas entre vocalización se ha registrado en otras especies de
ballenas como la ballena azul (McDonald et al., 2006), la ballena jorobada (Payne, 1979) y el
rorcual común (Hatch y Clark, 2004), también en odontocetos tales como la orca (Baird, 2000)
y el cachalote (Rendell et al., 2011). Las diferencias en las vocalizaciones de estos grupos, van
desde variaciones en la frecuencia máxima, hasta vocalización particulares por localidad. En el
rorcual común se presentan variaciones geográficas en la frecuencia del componente del pulso
de 20 Hz (Hatch y Clark, 2004, Širović et al., 2009; Castellote et al., 2012); la magnitud de
variación entre localidades llega a ser de 12 Hz (Gedamke, 2009). En la ballena azul se
identifican hasta nueve poblaciones en el mundo a partir de cambios en la morfología y
características físicas de las vocalización (McDonald et al., 2006).
La forma de la Be4 del norte registrada para el rorcual tropical en el presente trabajo
coincidió casi totalmente con los registros previos del Pacífico Oriental tropical (Oleson et al.,

101
2003; Heimlich et al., 2005). Solamente se encontraron diferencias en la duración. Al
comparar los espectrogramas se observó que la vocalización de la región norte del golfo
parece ser más corta, por alrededor de un segundo, con respecto a los registros previos. Esto
implicaría una variación asociada a la región geográfica. Sin embargo, no podemos concluir al
respecto pues la información disponible no permite una comparación precisa de los
parámetros, además la calidad de las imágenes publicadas no ayuda en la interpretación.
Finalmente, la vocalización Be4 obtenida en la región sur en el presente trabajo, es idéntica a
la Be4 descrita por Oleson et al. (2003), y más recientemente a la registrada por Kerosky et al.
(2012) en el sur de California. Esto sugiere la entrada de individuos del Pacífico adyacente al
Golfo de California, como lo proponen Salvadeo et al. (2011).
CONCLUSIONES
En el presente trabajo se describen tres vocalizaciones previamente no reportadas para la
especie (Be1, Be2, Be3), y una cuarta (Be4) que se describió en trabajos anteriores en la costa
sur occidental de la Península de Baja California y al sur de California Bight. La vocalización
Be4 se registró tanto en la región norte como en la sur del GC y de acuerdo con los resultados
de la comparación de la morfología, el gradiente de energía y los parámetros matemáticos, se
identificaron diferencias entre las vocalizaciones Be4. La característica distintiva fue la
duración, las vocalizaciones del sur son más largas que las del norte. Esto sugiere una
variación geográfica de las vocalizaciones del rorcual tropical en el Golfo de California.

102
LITERATURA CITADA
Au, W. y Hasting, M. 2009. Principles of marine bioacoustics. Springer. 686 pp.
Baird, R. W. 2000. Status of killer whales, Orcinus orca, in Canada. Canadian field-naturalist.
115(4): 676–701.
Breese, D. y Tershy, B. R. 1987. Residency patterns in female Bryde’s whales in the Gulf of
California, Mexico. Abstract: Seven Biennial Conference on the Biology of Marine
Mammals. Miami, FL. EE. UU.
Castellote, M., Clark, C. W. y Lammers, M. O. 2012. Fin whale (Balaenoptera physalus)
population identity in the western Mediterranean Sea. Mar. Mamm. Sci.28(2): 325–
344.
Clark, C. W. 1990. Acoustic behavior of mysticete whales, in sensory abilities of cetaceans.
Thomas J y R Kastelein, (eds.) Plenum, New York. 571–583 pp.
Cummings, W. C., Thompson, P. O. y Samuel, J. H. 1986. Sounds from Bryde, Balaenoptera
edeni, and Finback, B. physalus, whales in the Gulf of California. Fishery Bulletin.
84(2): 359–370.
Delarue, J., Todd, S. K., Van Parijs, S. M. y Di Lorio, L. 2009. Geographic variation in
Northwest Atlantic fin whale (Balaenoptera physalus) song: Implications for stock
structure assessment. J. Acoust. Soc. Am. 125(3): 1778–1782.
Dizon, A., Lux, C. A., LeDuc, R. G., Urbán, R, J., Henshaw, M. y Brownell, R. Jr. 1996. An
interin phylogenetic analysis of sei and Bryde’s whale mitochondrial DNA control
region sequences. SC/47/NP23 presented to the IWC Scientific Committee. 12 pp.
Edds, P., Odell, P. K. y Tershy, B. R. 1993. Calls of a captive juvenile and free-ranging adult-
calf pairs of Bryde’s whales, Balaenoptera edeni. Mar. Mamm. Sci. 9: 269–284.
Figueiredo, L. D. y Simão, S. M. 2014. Bryde´s Whales (Balaenoptera edeni) vocalizations
from Southeast Brazil. Aquatic Mammals. 40(3): 225–231.
Gedamke, J. 2009. Geographic variation in Southern Ocean fin whale song. Paper
SC/61/SH16 presented to the 2009 IWC Scientific Committee (unpublished).
Hatch, L. T. y Clark, C. W. 2004. Acoustic differentiation between fin whales in both the
North Atlantic and North Pacific Oceans, and integration with genetic estimates of
divergence. SC/56/SD6 IWC Scientific Committee (unpublished). 37 pp.

103
Heimlinch, S. L., Mellinger, D. K., Nieukirk, S. L. y Fox, G. C. 2005. Types, distribution, and
seasonal occurrence of sounds attributed to Bryde’s whales (Balaenoptera edeni)
recorded in the Eastern Tropical Pacific, 1999–2001. Jour. Acoust. Soc. Am. 118 (3):
1830–1837.
Kato, H. y Perrin, W. F. 2009. Bryde’s Whales Balaenoptera edeni/brydei. En: William, F. P.,
Würsig, B. y Thewissen, J. G. M. 2009. Encyclopedia Marine Mammals 2a ed.
Elservier. 157–162 pp.
Kerosky, M. S., Širović, A., Roche, L. K., Baumann-Pickering, B., Wiggins, S. M. y
Hildebrand, A. J. 2012. Bryde’s whale seasonal range expansion and increasing
presence in the Southern California Bight from 2000 to 2010. Deep Sea Research.
1(65): 125–132.
López-Montalvo, C. 2012. Caracterización de la ecología alimentaria del rorcual tropical
(Balaenoptera edeni), en el Golfo de California con base en análisis de isotopos
estables N y C, y Ácidos Grasos. Tesis Maestría. Instituto de Ciencias del Mar y
Limnología. UNAM. 76pp.
McDonald, M., Mesnick, S. L. y Hildebrand, J. 2006. Biogeographic characterization of blue
whale song worldwide: using song to identify populations. J. Cetacean Res. Manage.
8(1): 55–65.
Mellinger, D. K. y Barlow, J. 2003. Future directions for acoustic marine mammal surveys:
stock assessment and habitat use. NOAA OAR Special Report, NOAA/PMEL
Contribution 2557. 37 pp.
Nyquist, H. 1928. Certain topics in telegraph transmission theory. American Institute of
Electrical Engineers, Transactions of the A.I.E.E. 47(2): 617-644.
Olavarrieta-García, T. 2012. Hábitos alimenticios del rorcual tropical (Balaenoptera edeni) en
el Golfo de California. Tesis Licenciatura, Universidad Autónoma de Baja California
Sur. 43pp.
Oleson, E. M., Barlow, J., Gordon, J., Rankin, S. y Hildebrand, J. A. 2003. Low frequency
calls of Bryde’s whales. Mar. Mamm. Sci. 19(2): 160–172.
Oleson, E. M., Wiggins, S. M. y Hildebrand, J. A. 2007. Temporal separation of blue whale
call types on a southern California feeding ground. Animal Behaviour. 74(4): 881–
894.

104
Payne, R. S. 1979. Humpback whale songs as an indicator of "stocks". Third Biennial
Conference on the Biology of Marine Mammals. Seattle, WA. p.46.
Payne, R. S. y McVay, S. 1971. Songs of Humpback Whales. Science. 173(3997): 85–597.
Rice, A. R., Palmer, K. J., Tielens, J. T. y Muirhead, C. A. 2014. Potential Bryde’s whales
(Balaenoptera edeni) calls recorded in the northern Gulf of Mexico. J. Acoust. Soc.
Am. 135: 3066–3076.
Rendell, L. y Whitehead, H. 2003. Vocal clans in sperm whale Physeter macrocephalus. Proc.
R. Soc. Lond. B. 270: 225–231.
Rendell, L., Mesnick, S. L., Dalebout, M. L., Burtenshaw, J. y Whitehead, H. 2011. Can
Genetic Differences Explain Vocal Dialect Variation in Sperm Whales, Physeter
macrocephalus? Behav. Genet. 42(2), 332-343.
Salvadeo, C., Flores-Ramírez, S., Gómez-Gallardo, A., MacLeod, C., Llunch-Belda, D.,
Jaume-Schinkel, S. y Urbán, R. 2011. Bryde’s whale (Balaenoptera edeni) in the
southwestern Gulf of California Relationship with ENSO variability and prey
availability. Ciencias Marinas. 37(2): 215–225.
Širović, A., Hildebrand, J. A., Wiggins, S. M. y Thiele, D. 2009. Blue and fin whale acoustic
presence around Antarctica during 2003 and 2004. Mar. Mamm. Sci. 25(1): 125–136.
Širović, A., Bassett, H. R., Johnson, S. C., Wiggins, S. M. y Hildebrand, J. A. 2014. Bryde's
whale calls recorded in the Gulf of Mexico. Mar. Mamm. Sci. 30: 399–409.
Tershy, B. R., Acevedo, G. A., Breese, D. y Strong, C. S. 1993. Diet and feeding behavior of
fin and Bryde's whales in the central Gulf of California, Mexico. Rev Inv Cient 1: 31–
38.
Tyack, P. 2009. Behavior, overview. En: William, F. P., Würsig, B. y Thewissen, J. G. M.
2009. Encyclopedia Marine Mammals 2a ed. Elservier. 102–108 pp.
Vidal, O., Aguayo, A., Findley, L., Robles, A., Bourillón, L., Vomend, I., Turk, P., Gárate, K.,
Maroñas, L. y Rosas, J. 1987. Avistamientos de mamíferos marinos durante el crucero
“Guaymas I” en la región superior del Golfo de California, primavera de 1984. En:
Memorias de la X Reunión Internacional sobre Mamíferos Marinos. La Paz, B.C.S.,
México. 7–35 pp.
Viloria, L., Romero-Vivas, E. y Urbán, J. 2012. Caracterización del repertorio de
vocalizaciones del rorcual tropical (Balaenoptera edeni) en el Golfo de California a

105
partir de registros acústicos con confirmación visual. Memorias de la 15ª Reunión de
trabajo de expertos en mamíferos acuáticos de América del Sur. 9º Congreso de
Sociedad Latinoamericana de especialistas de Mamíferos Acuáticos, SOLAMAC.
Puerto Madryn, Argentina.
Viloria, L., Romero-Vivas, E., Hildebrand, J., A. Kerosky, S. y Urbán, J. 2013. Geographic
variation of Bryde's whale (Balaenoptera edeni) Be4 type call in the Gulf of
California. 20th
Biennial Conference on the Biology of Marine Mammals. Dunedin,
New Zealand.
Zar, J. H. 1999. Biostatistical analysis. Pearson Education India. 662 pp.

106
CAPÍTULO V
TRANSMISIÓN CULTURAL Y SU IMPACTO EN LA ESTRUCTURA
GENÉTICA DE LAS POBLACIONES DEL RORCUAL TROPICAL
(Balaenoptera edeni brydei) EN EL GOLFO DE CALIFORNIA

107
RESUMEN
En muchas especies de animales las vocalizaciones son el principal medio de comunicación
entre individuos. Variaciones regionales de estas vocalizaciones a nivel intra-específico,
pueden ser un mecanismo de aislamiento. La teoría de la transmisión de rasgos culturales y su
impacto en la estructura genética de las poblaciones supone que la herencia genera variaciones
dadas por efectos de filopatría y copia imperfecta, originando aislamiento geográfico a través
de las generaciones. El presente trabajo plantea conocer la existencia de diferentes poblaciones
del rorcual tropical en el Golfo de California (GC) con base en sus vocalizaciones y diversidad
haplotípica del ADN mitocondrial (ADNmt), así como conocer aspectos evolutivos de la
especie. El análisis genético incluyó 82 muestras, 29 correspondieron al norte y 53 al sur del
GC. Un total de ocho haplotipos fueron identificados, cinco estuvieron presentes en el norte
(uno de ellos exclusivo) y siete en el sur (tres de ellos exclusivos). El análisis estadístico de
heterogeneidad basado en Chi-cuadrado, Hst y Kst, no mostró diferencias entre los rorcuales
del norte y sur del GC. Para la caracterización de las vocalizaciones se revisaron registros con
confirmación visual y se encontraron cinco diferentes. Una de ellas (Be4), se eligió como
referencia para encontrar llamadas del rorcual en los registros del HARP (base de datos de
sonidos). Se analizó el componente principal de la vocalización Be4 103 región norte y 88
región sur, este análisis mostró diferencias significativas entre regiones. Los resultados
obtenidos en el presente trabajo pueden ser explicados con base en la teoría de la transmisión
cultural; de acuerdo a sus cinco postulados se proponen tres escenarios para el rorcual tropical
del GC, las variaciones en las vocalizaciones son una consecuencia de las características
físicas del ambiente, o que estas variaciones no intervienen en la reproducción, o que sean
variaciones recientes que aún no se expresan en la estructura genética. Estudios que
identifiquen el sexo del emisor y la conducta asociada al rasgo cultural, son necesarios para
ampliar el conocimiento sobre la transmisión cultural en B. edeni brydei del Golfo de
California.

108
ABSTRACT
Vocalizations are the most important way of communication among individuals. Regional
variations of this vocalizations intraspecific level can be a mechanism of isolation. The theory
of transmission of cultural traits and their impact on the genetic structure assumes that the
inheritance generates variations. This variation is given by the combined effects of philopatry
and imperfect copy, causing geographic isolation through the generations. Therefore, in this
work we want to identify populations differences of Bryde’s whale in the Gulf of California
(GC) based on their vocalizations and haplotype diversity of mitochondrial DNA (mtDNA),
and to know evolutionary aspects of Bryde’s whale. The genetic analyses included 82
samples, 29 from the northern region and 53 from southern region of GC. A total of eight
haplotypes were identified, five were present in the north (one exclusive) and seven in the
south (three exclusive). The statistical analysis of heterogeneity based on Chi-square, Kst*,
Hst and Fst showed no differences between samples from the north and south of the GC. For
the characterizations of the vocalizations, we analyzed recordings with visual confirmation
founding five different. One of them (Be4) was chosen as a reference to find Bryde’s calls in
the HARP (database of sounds). The main component of the vocalization Be4 was analyzed,
103 north region and 88 south regions, this analysis showed significant differences between
regions. The results obtained in this study can be explained based on the theory of cultural
transmission and according to its five postulates three scenarios for the GC Bryde´s whale are
proposed. According to this theory the results obtained in this study could be explained by
three alternative hypotheses: 1) changes in the Bryde's whale vocalizations in the GC are a
consequence of the physical characteristics of the environment, 2) these variations are not
involved in the reproduction, and 3) these changes are recent and still not expressed in the
genetic structure. Studies to identify the sex of the transmitter or the behavior associated with
cultural trait are needed to increase knowledge of cultural transmission in B. edeni brydei the
Gulf of California.

109
INTRODUCCIÓN
Los sonidos emitidos por ciertas especies de animales, se encuentran entre los rasgos que son
aprendidos y transmitidos de una generación a otra. Esta transmisión cultural es de amplio
interés teórico por su posible impacto en la evolución biológica. Los sonidos de vertebrados,
tales como aves y mamíferos, han permitido analizar el postulado de que la transmisión
cultural de los cantos se relaciona con la estructura genética de las poblaciones (MacDougall-
Shackleton y MacDougall-Shackleton, 2001).
Las vocalizaciones de diversas especies de animales son el principal medio de
comunicación entre individuos. A nivel de especie, las variaciones regionales de
vocalizaciones están bien documentadas, y se sabe que pueden ser parte de un mecanismo de
aislamiento entre taxones (Marler, 1957 en MacDonald et al., 2006; West-Eberhard, 1983).
Con base en este conocimiento, la teoría de la transmisión de rasgos culturales3 y su impacto
en la estructura genética de las poblaciones ha tomado fuerza en las últimas décadas
(Macdougall-Shackleton y Macdougall-Shackleton, 2001); en gran medida esto se deba al
aumento de estudios de acústica en los diferentes grupos de animales, y a la relación de la
acústica y la genética de poblaciones. La teoría de transmisión supone que los rasgos
culturales, en este caso los cantos, son transmitidos por individuos adultos, donde la
consecución de esta transmisión–aprendizaje genera variaciones geográficas, las cuales son
provocadas por efectos combinados de la filopatría y la copia imperfecta de las canciones
(Baker y Cunningham, 1985). Debido a que una de las principales funciones del canto en aves
y cetáceos es el encuentro de pareja, se considera que ciertas diferencias en los cantos pueden
llegar a inhibir el flujo genético entre poblaciones de conspecíficos, impactando así, la
estructura genética de la población a través de la transmisión cultural (Marler y Tamura, 1962;
Nottebohm, 1969).
El rorcual tropical, Balaenoptera edeni brydei, es una especie poco estudiada, a pesar
de lo cual se han registrado variaciones geográficas en sus patrones de vocalización
(Heimlinch et al., 2005). En el Golfo de California (GC) se ha postulado la existencia de dos
3 Cultura: es la información o conducta adquirida entre individuos de la misma especie, a través de algún
tipo un aprendizaje social (Boyd y Richerson, 1996).

110
unidades poblacionales con base en datos de foto–identificación (Tershy et al., 1990) y
estudios preliminares de genética (Dizon et al., 1996). Se plantea que una de estas poblaciones
es residente y se distribuye en la región centro–norte del golfo; mientras que la otra se integra
por individuos que entran y salen del golfo, distribuyéndose principalmente al sur y se
relacionan con la población de B. edeni del Pacífico Oriental tropical (Tershy et al., 1990;
Dizon et al., 1996).
Diferencias regionales de vocalizaciones del rorcual tropical y la posible existencia de
dos poblaciones en el GC, plantean interrogantes relacionadas con la transmisión de las
vocalizaciones como rasgos culturales y sobre el estado de separación cultural–genético en el
que se encuentra la especie en esta región. Por lo que el presente trabajo propone relacionar las
diferencias acústicas y la estructura genética de la población de rorcual tropical que se
distribuye en el GC, utilizando la teoría de la transmisión cultural como una herramienta para
explicar el fenómeno.
OBJETIVO
En el marco de la teoría de la trasmisión cultural, relacionar las diferencias acústicas y
la estructura genética de la población de rorcual tropical que se distribuye en el Golfo de
California.

111
MÉTODO
Identificación de diferencias genéticas:
Se analizaron 82 secuencias de 299pb de ADNmt, obtenidas de diferentes individuos del B.
edeni brydei, en Golfo de California.
Para identificar diferencias poblacionales se calculó la distancia genética con el
Método dos parámetros de Kimura (Kimura, 1980), tres estadísticos de heterogeneidad entre
regiones (Chi-cuadrada, Hst y Kst*; Hudson et al., 1992) y Fst (Excoffier et al., 2005).
Los árboles filogénicos se construyeron con el método de Máxima verosimilitud en el
programa MEGA v6 (Tamura et al., 2013). Secuencias de B. omurai y B. edeni edeni se
usaron como grupos externos.
Identificación de diferencias acústica:
Se analizaron 1237 horas de grabaciones de tres hidrófonos fijos, dos representantes de
la región norte del GC (Isla Tiburón y Bahía de los Ángeles), y uno de la región sur (Punta
Pescadero).
Se identificó una vocalización que estuviera de manera abundante y presente en los tres
puntos de grabación. Esta vocalización se caracterizó a través de tres elementos: el elemento
Matemático (Frecuencia máxima, Frecuencia mínima, Pico de frecuencia y Duración (Td;
medida en segundos), elemento Morfológico con imágenes de la vocalización en 3D y el
elemento Gradiente de energía.
Al elemento Matemático se le calculó estadística descriptiva, y para identificar
diferencias entre regiones se usó la prueba de t de Student.

112
RESULTADOS
Identificación de diferencias genéticas
De las 82 secuencias analizadas, se identificaron ocho haplotipos (H1-H8), de los cuales
cuatro son compartidos entre la región sur y norte (entre estos están los más frecuentes, H1,
H2 y H8), tres son exclusivos para el sur y uno para el norte.
La diversidad nucleotídica (0.722 N, 0.759 S) y haplotípica (0.111 N y 0.0116 S) es
similar en ambas regiones. Los tres estadísticos de heterogeneidad entre regiones norte y sur, y
el Fst demostraron que no existe diferencia significativa.
Identificación de diferencias acústica:
La vocalización Be4 estuvo presente en los tres puntos de registro y de manera abundante. El
análisis de los tres elementos de 103 vocalizaciones de la región sur y 88 del norte indicaron
diferencias significativas entre regiones. La variación se observó en la duración, la morfología
y el gradiente de energía. La Be4 del norte es de menor duración, su forma está caracterizada
por presentar un montículo con la energía está concentrada en este y con pendientes
pronunciadas.

113
DISCUSIÓN
Teoría de la transmisión cultural
Esta teoría formula que los rasgos culturales son transmitidos por individuos adultos, y que
durante la consecución de esta transmisión–aprendizaje se pueden generar variaciones por
efectos combinados de la filopatría y una copia imperfecta (Baker y Cunningham, 1985). Si
alguna de estas variaciones se da en detrimento del apareamiento, se puede llegar a inhibir el
flujo genético entre poblaciones de conspecíficos e impactar la estructura genética de la
población (Marler y Tamura, 1962; Nottebohm, 1969). Para fundamentar el efecto de las
variaciones de rasgos culturales en la estructura poblacional, la teoría asume cinco supuestos:
1) Existe una trasmisión cultural, es decir, los rasgos culturales son trasmitidos de generación
en generación (v.g., estrategias de forrajeo, rutas de migración, cantos, vocalizaciones, etc.;
Laland y Janik, 2006). 2) Se generan variaciones por la copia imperfecta del rasgo y la
filopatría. 3) Una de las funciones del rasgo es el encuentro de pareja. 4) La transmisión
cultura del rasgo es estable. 5) Las diferencias a largo plazo pueden inhibir el flujo genético
entre grupos de individuos (MacDougall-Shackleton y MacDougall-Shackleton, 2001) y
promover la especiación (Podos y Warren, 2007).
1) Existe trasmisión cultural.
Aunque la mayor evidencia sobre el aprendizaje social o transmisión cultural proviene de
estudios sobre patrones de vocalización en aves (Catchpole y Slater, 1995; Slabbekoor y
Smith, 2002), hay evidencias de transmisión de otros rasgos culturales en mamíferos tan
disímiles como primates y mamíferos marinos (Laland y Janik, 2006). Por ejemplo, en los
chimpancés (Pan troglodites) es conocido un repertorio conductual complejo que incluye el
uso de herramientas para conseguir alimento, este repertorio es compartido (aprendido) y es
variable entre diferentes grupos de individuos (Whiten et al., 1999). En mamíferos marinos, la
transmisión cultural puede ser observada en el aprendizaje de rutas migratorias o en estrategias
de forrajeo (Laland y Janik, 2006). Las toninas (Tursiops truncatus) aprenden el uso de
esponjas como herramienta para conseguir alimento (Smolker et al., 1997; Krützen et al.,
2005), las orcas (Orcinus orca) aprenden a vararse intencionalmente para la captura de

114
cachorros de elefantes marinos (Guinet y Bouvier, 1995), y con fines pedagógicos, las
hembras adultas de delfines moteados del atlántico (Stenella frontalis) cambian la estrategia de
forrajeo en presencia de sus crías (Bender et al., 2009). En todos los ejemplos el aprendizaje
se transmite de madres a crías (Smolker et al., 1997; Krützen et al., 2005).
Teóricamente se acepta que el conocimiento sobre los patrones de vocalización en aves
puede ser tomados como ilustración en la diversificación conductual en los vertebrados (Podos
et al., 2004), y que la comunicación a través de señales acústicas sólo puede ser aprendida de
otro animal (Laland y Janik, 2006). Gran parte de las investigaciones que analizan la
trasmisión cultural en mamífero marinos, se ha enfocado en el dominio de las vocalizaciones
en especies como las toninas y la ballena jorobada (Megaptera novaeangliae) (Laland y Janik,
2006). La capacidad de imitar silbidos (whistles) o acciones a partir de otros conespecíficos ha
sido ampliamente registrada en toninas mantenidas en cautiverio (Richards et al., 1984; Reiss
y McCowan, 1993) y en delfines de vida libre (Janik, 1997 y 2000). En las ballenas jorobadas
se ha demostrado que todos los machos de una población pueden compartir el mismo patrón
de vocalización (canción) en espacio (Guinee et al., 1983; Cerchio et al., 2001) y tiempo
determinados (Payne et al., 1983; Noad et al., 2000), pero a través de una temporada esa
canción cambia gradualmente (Guinee et al., 1983), haciendo evidente la trasmisión cultural
(Laland y Janik, 2006).
Antes del presente trabajo no se había registrado ni analizado la transmisión cultural de
ningún rasgo de B. edeni brydei. La grabación de la vocalización Be4 en diferentes localidades
del GC (Punta Pescadero, Bahía de los Ángeles y Bahía de La Paz), su seguimiento por varios
años (2006-2014) y el número de registros (444 vocalizaciones en Punta Pescadero en 2006-
2007 y 517 en Bahía de los Ángeles en 2007-2008), son evidencias que describen a un
conjunto de individuos que comparten un patrón de vocalización en espacio y tiempo
determinados, y que este rasgo está siendo transmitido.
2) Se generan variaciones por la copia imperfecta de las canciones y filopatría.
Una de las evidencias sobre la transmisión cultural es la variación geográfica en los patrones
de vocalización. En este fenómeno, animales vecinos o de un grupo social particular
comparten sonidos, pero difieren en su patrón con animales distantes o de otros grupos; este
fenómeno es más ilustrativo cuando se presenta en grupos que no están aislados

115
geográficamente (Janik y Slater, 1997). Las variaciones regionales en los patrones de
vocalización está bien documentada en aves (Marler y Tamura, 1962; Slabbekoorn y Smith,
2002), particularmente en el gorrión corona blanca (Zonotrichia leucophrys) (Baker et al.,
1981; MacDougall-Shackleton y MacDougall-Shackleton, 2001), y el loro tropical (Amazona
auropalliata) (Wright, 1996).
La variación geográfica de vocalizaciones ha sido descrita en diferentes grupos
taxonómicos de mamíferos marinos. En pinnípedos se han identificado diferencias regionales
en características acústicas tales como duración, forma de la onda y el contorno de la
frecuencia en vocalizaciones de la foca de Wedell (Leptonychotes weddelli) (Thomas y
Stirling, 1983; Terhune et al., 2008). También se han registrado en la foca leopardo (Hydrurga
leptonix) (Thomas y Golladay, 1995), entre otras especies. La combinación entre fidelidad al
sitio de nacimiento y el aprendizaje se cuentan entre las razones que causan las diferencias
(Thomas y Stirling, 1983; Terhune et al., 2008).
En odoncetos, se han registrado variaciones geográficas en las vocalizaciones del
cachalote (Physeter macrocephalus) (Whitehead et al., 1998), y las orcas (Orcinus orca)
(Baird, 2000). En ambas especies se han identificados dialectos, es decir, variaciones en las
vocalizaciones con límites geográficos bien definidos (Marler y Tamura, 1962). También se
han registrado variaciones geográficas en los silbidos de la tonina del Indo-Pacífico (Tursiops
aduncus) (Morisaka et al., 2005), y del delfín tornillo (Stenella longirostris) (Bazúa-Duran y
Au, 2001). Estas variaciones se originan por factores macro y microgeográficos (Bazúa-Duran
y Au, 2001; Morisaka et al., 2005). Los factores macrogeográficos se asocian a la distancia
geográfica; y los microgeográficos sugieren que las diferencias son el resultado de la
acumulación de cambios en el proceso de aprendizaje de los silbidos (Morisaka et al., 2005), y
que las diferencias generados por este factor pueden ser usadas para identificar grupos sociales
o poblaciones de delfines (Bazúa-Duran y Au, 2001).
En misticetos, las variaciones regionales de los “cantos” han permitido reconocer
nueve poblaciones de la ballena azul en el mundo (Balaenoptera musculus) (McDonald et al.,
2006); y patrones de vocalización del rorcual común (Balaenoptera physalus) distinguen a la
población del GC, de la población de las costas de California (Hatch y Clark, 2004).
En el presente trabajo se identificaron diferencias geográficas en la vocalización Be4,
emitida por individuos de B. edeni brydei que se distribuyen en el GC. Como en todas las

116
especies antes mencionadas, la identificación de diferencias se logró utilizando elementos
matemáticos (Frecuencia máxima, Frecuencia mínima, Pico de frecuencia y Duración), pero
para su descripción se emplearon adicionalmente elementos morfológicos y de gradiente de
energía. Esta información mostró que la vocalización Be4 del norte es de menor duración con
respecto a la emitida en el sur de golfo, que la morfología en el norte consiste de una sola
cresta o montículo mientras que en la región sur presenta dos crestas, y que el gradiente de
energía indica una mayor concentración de energía en un área más angosta en la Be4 del norte
con respecto a la del sur (ver Capítulo IV). Esta variación pudiera explicarse por factores
macrogeográficos asociados a la distancia geográfica, como se describió para odontocetos
(Morisaka et al., 2005), y a una combinación entre fidelidad al sitio de nacimiento y al
aprendizaje de copias imperfectas, como se ha descrito para otras especies (Thomas y Stirling,
1983; Terhune et al., 2008).
3) Una de las funciones del “canto” es el encuentro de pareja
A los cantos se les atribuyen diversas funciones, entre ellas el estar involucradas en el proceso
de apareamiento. Estudios detallados en aves muestran que los cantos sirven en la elección de
pareja (Albrecht y Oring, 1995). En ballena jorobada (Winn y Winn, 1978; Darling, 1983) y
rorcual común (Croll et al., 2002), las canciones son producidas por machos durante la
temporada de reproducción, por lo que cantos particulares son asociados a este proceso (Payne
y Mcvay, 1971; Winn y Winn, 1978). Sin embargo, se desconoce si la función es exclusiva
para la atracción de pareja, como sucede en aves (Albrecht y Oring, 1995). Se sugiere que el
canto de los machos de ballena jorobada también pudieran ser emitidos como una señal
intersexual para mantener un territorio acústico (Tyack, 1981), es decir que los machos buscan
mantenerse alejados unos de otros para evitar peleas (Frankel et al., 1995), como se ha
demostrado en la ballena minke (Balaenoptera acuturostrata) (Gedamke et al., 2003).
El conocimiento que se dispone de la biología del rorcual tropical no permite asociar la
emisión de cantos con conductas relativas al apareamiento. Se desconoce si existe una
temporada y una región particular para las actividades de reproducción de esta especie, como
se ha observado en otros misticetos que se distribuyen en el GC (v.g., ballena jorobada;
Aguayo et al., 1993; Ladrón de Guevara, 1995). De las vocalizaciones registradas y analizadas
en el presente trabajo, la vocalización Be4 es la más abundante. Las conductas observadas

117
durante su grabación y el avistamiento de los individuos, y los registros de otros autores
(Cummings et al., 1986; Edds et al., 1993; Oleson et al., 2003; Heimlich et al., 2005; Širović
et al., 2014; Rice et al., 2014; Figueiredo y Simão, 2014), no hacen referencia a conductas
reproductivas, no obstante no debe descartarse su emisión durante este tipo de actividades.
Dicha vocalización se registró siempre que había al menos dos individuos adultos en el área, y
en la mayoría de los avistamientos en presencia de alimento, i.e., sardina (Tershy et al., 1993).
Kato y Perrin (2009) señalan que los individuos de esta especie sólo se agrupan cuando se
alimentan; si la reunión representan oportunidades de socialización, y uno de los eventos más
importante de socialización para las especies es la reproducción (Tyack, 2009), es posible que
esta vocalización esté asociada para tales fines, de manera similar a lo que sucede con el
rorcual común (Croll et al., 2002).
Estudios que pudieran confirmar la emisión de la vocalización Be4 en actividades de
apareamiento, tendrían que considerar la identificación del género (hembra, macho) y los
niveles de las hormonas de reproducción (estrógenos y testosterona) del individuo emisor. O
alternativamente, tendrían que registrarse eventos directos de atracción a la hembra o de
marcado de territorio posteriores a la emisión.
4) Transmisión cultura estable.
Registros acústicos prolongados indican que los rasgos de algunos de los caracteres pueden
mantenerse a través de varias generaciones. Por ejemplo, los rasgos de la canción propia de la
población del Pacífico Sureste de la ballena azul se ha mantenido al menos por 27 años, los de
la canción de la población del Pacífico Suroeste por más de 33 años, y los de la canción del
Pacífico Noreste por lo menos durante 40 años (McDonalds et al., 2006). En otras ballenas
como minke se observó que los rasgos de una vocalización asociada a la reproducción son
transmitidos de adultos a jóvenes y permanecen estables por lo menos durante tres años (1997-
1999; Gedamke et al., 2001). También algunas de las vocalizaciones de orcas no muestran
cambios en su estructura hasta por 13 años (Deecke et al., 2000).
Los rasgos de las vocalizaciones registradas en el presente trabajo para B. edeni brydei
en el GC, también parecen ser estables a través de la transmisión cultural. En las
vocalizaciones de esta especie se observa que los rasgos de los elementos matemáticos
(Frecuencia máxima, Frecuencia mínima, Frecuencia pico y Duración), así como la

118
morfología y el gradiente de energía, han permanecido iguales durante varios años; al menos
cuatro años si consideramos las grabaciones registradas en el HARP (2006-2009), y nueve
años si comparamos el primer registro obtenido por el HARP (2006) y la última grabación
obtenida por el PRIMMA del Laboratorio de Mamíferos Marinos de la UABCS (2014).
5) Las diferencias a largo plazo pueden inhibir el flujo genético entre grupos de individuos
La estabilidad en la transmisión cultural es de utilidad en el momento de comparar
poblaciones, pues da certeza en la definición. Además explica parcialmente ciertos procesos
de especiación. En aves se postula que los jóvenes aprenden a producir y reconocer sonidos
mientras que están en su región natal, ya de adultos usan estos sonidos transmitidos de una a
otra generación como una clave específica para la elección de pareja; si estas condiciones son
mantenidas a través del tiempo, los dialectos pueden inhibir el flujo genético en conespecíficos
hasta cierto grado (Slabbekoorn y Smith, 2002). Este escenario, conocido como hipótesis de
adaptación genética (Marler y Tamura, 1962 y 1964; Nottebohm, 1969), lleva a sugerir que los
dialectos influyen en la estructuración de las poblaciones y, eventualmente, aceleran la tasa
evolutiva (MacDougall-Shackleton y MacDougall-Shackleton, 2001), como se ha visto en
diversos grupos taxonómicos (Foster, 1999; Maan y Seehasuen, 2011; Lackey y Boughman,
2014).
Sin embargo la variación de rasgos culturales no siempre genera especiación. Algunos
rasgos neutros pueden ser observados en diversas especies como variaciones geográficas
asociadas a adaptaciones ambientales (Schluter, 2000) o variaciones netamente culturales en
las que el ambiente no interviene (Whiten et al., 1999); en ninguno de los casos las variaciones
corresponden con modificaciones en el genotipo, por lo que no intervienen en procesos de
especiación (Podos y Warren, 2007). La hipótesis de adaptación social aplicada a los cantos de
las aves, propone que los dialectos pudieran tener efectos en las interacciones sociales y
sexuales, pero señala que la variación de estos rasgos no logra el aislamiento genético. Por
ejemplo, machos adultos de algunas aves pueden modificar su canto para imitar a sus vecinos
y lograr reproducirse con hembras de dialectos diferentes (Payne, 1981). Durante el cortejo de
otras aves, también se ha demostrado que las hembras pueden elegir a machos con un dialecto
diferente (Slabbekoorn y Smith, 2002).

119
Un fenómeno particular en el que se presentan variaciones culturales y genéticas entre
poblaciones, siendo el origen de estas diferencias la distancia geográfica (MacDougall-
Shackleton y MacDougall-Shackleton, 2001), se explica por la hipótesis epifenoménica
(Andrew, 1962). Este postulado propone que los dialectos son meros subproductos de
aprendizaje vocal y no son conductualmente sobresalientes; y considera que las regiones
dialécticas pudieran ser genéticamente aisladas, principalmente por barreras geográficas,
evento fundador o filopatría estricta, más que por el apareamiento selectivo basado en
canciones (MacDougall-Shackleton y MacDougall-Shackleton, 2001). Por ejemplo, en
poblaciones de B. physalus, distribuidas en el GC y Océano Pacífico, se presentan diferencias
en sus vocalizaciones (Thompson et al., 1992) y características genéticas (Bérubé et al., 2002),
esta diferenciación podrían ser el reflejo de una población antigua aislada y con limitado flujo
genético; o el de un evento fundador, ocurrido tras la última máxima glaciación, y un flujo
genético restringido (Nigenda-Morales et al., 2008).
Balaenoptera edeni brydei en el Golfo de California
El análisis de la estructura genética de B. edeni brydei, realizado en el presente trabajo
(Capítulo III), muestra que no hay diferenciación entre regiones norte y sur en el GC. El
número de haplotipos compartidos (H1, H2, H4 y H8), las frecuencias haplotípicas para ambas
regiones (Figura 5.1), la similitud entre la diversidad genética (Norte 0.722, 0.0111; Sur 0.759,
0.0116), el bajo valor de la distancia genética (-0.0197) y los resultados de heterogeneidad
entre regiones (Hst= 0.00668; Kst*=-0.00861; X2= 0.312, P<0.05; y Fst=-0.01412, P<0.01),
son indicadores de la existencia de una sola población con flujo genético. En contraste, el
análisis de características acústicas en sus vocalizaciones (Capítulo IV), muestra la existencia
de diferencias regionales. Las características morfológicas de la vocalización Be4
(vocalización con un montículo y base angosta en el norte; con dos montículos y base ancha
en el sur) y su duración (1.8±0.29s al norte, 2.5±0.44s al sur; t= 9.13, valor crítico= 1.65,
p<0.05), son indicadores de una variación norte-sur en este rasgo cultural (Figura 5.1).

120
Figura 5. 1. Diferencias acústicas y genéticas en Balaenoptera edeni brydei del Golfo de California.

121
El hecho de que B. edeni brydei no presente diferenciación genética entre regiones
norte y sur en el GC, y que sí existan diferencias regionales en sus vocalizaciones, sugiere
tres escenarios posibles:
1) El ambiente generó las diferencias, pero éstas no representan variaciones en el genotipo
(hipótesis de adaptación social; Schluter, 2000). Las diferencias norte-sur en la
vocalización Be4, pudieron ser generadas por características físicas del ambiente, sin
promover la especiación. Este proceso de diferenciación pudiera ser un fenómeno de
selección hábitat-dependiente, donde el ambiente juega un papel importante en las
características de las vocalizaciones. Se ha observado que en hábitats diferentes, es posible
predecir características acústicas particulares (Slabbekoorn y Smith, 2002). En aves, se ha
demostrado que la presión de selección para las señales acústicas en sitios con una densidad
de vegetación alta es diferente a aquellos sitios con densidad menor (Slabbekoorn y Smith,
2002). Las bajas frecuencias se transmiten mejor en vegetación densa, y son emitidas por
especies que viven en bosques, a diferencia de las aves que se encuentran en hábitats
abiertos y emiten cantos con frecuencias más altas (Morton, 1975; Ryan y Brenowitz,
1985). En odontocetos, las marsopas comunes (Phocoena phocoena) que viven en zonas
someras y el delfín del río amazonas (Inia geoffrensis) que también vive en aguas someras
pero particularmente turbias, emiten sonidos con una frecuencia máxima mayor (160 KHz
y 200 KHz, respectivamente) a las emitidas por las toninas (Tursiops truncatus; 150 KHz)
que viven en zonas más profundas y de menor turbidez (Ketten, 1994). También, algunas
características del hábitat generan ciertos fenómenos físicos en la señal acústica (v.g,
reverberación; Richards y Wiley, 1980; Holland et al., 1998). Se ha visto que los niveles de
ruido ambiental y sus características espectrales pueden exhibir diferencias consistentes
entre hábitats (Slabbekoorn y Smith, 2002). En consecuencia estas diferencias en niveles de
ruido conducen a la elección divergente hábitat-dependiente (Ryan y Brenowitz, 1985;
Slabbekoorn y Smith, 2002).
Las diferencias encontradas en el presente trabajo entre las vocalizaciones Be4 del
norte y sur del rorcual tropical (1.93±0.36s y 2.46±0.43s, respectivamente), podrían ser
explicadas por la presencia de ruido ambiental de origen natural, asociado a características
ambientales propios de cada sitio. Durante la revisión de los archivos de sonido, al menos

122
el 60% de los archivos del HARP de Bahía de los Ángeles mostraron ruido en el intervalo
de frecuencias analizado (desde 1 a 1000 Hz), mientras que en más del 90% de los archivos
del HARP del sur en Punta Pescadero dicho ruido no fue observado.
2) Las variaciones no están involucradas en la selección de pareja, no interviene en
la separación genética (Whiten et al., 1999). La vocalización Be4 pudiera ser un rasgo
neutro cultural que no interviene en un proceso de especiación, como explica la hipótesis de
adaptación social a los rasgos neutros (Schluter, 2000); y/o que este rasgo cultural no esté
involucrado en la reproducción.
3) Existe un aislamiento genético incompleto. El rorcual tropical pudiera presentar
una diferenciación acústica con aislamiento genético incompleto, debido a que las
vocalizaciones representarían barreras parciales contra el flujo genético. Al mismo tiempo,
la distancia geográfica pudiera no representar una barrera lo suficientemente fuerte para
aislar a poblaciones y generar diversidad genética, siendo un reflejo de la capacidad de
desplazamiento de la especie. Se ha demostrado que el flujo de genes en especies altamente
móviles es factible aun entre poblaciones con diferentes dialectos (MacDougall-Shackleton
y MacDougall-Shackleton, 2001). En relación a esto, la movilidad del rorcual tropical a
través del GC, se ha evidenciado por estudios de foto-identificación que muestran
individuos recapturados en la Bahía de los Ángeles, al norte, y en la Bahía de La Paz, al sur
del GC (Chávez, 1995).
CONCLUSIONES
El flujo genético registrado en individuos de B. edeni brydei distribuidos en el GC, así
como las variaciones regionales norte-sur en sus vocalizaciones, pueden explicarse con los
postulados de la teoría de la transmisión cultural. Las vocalizaciones como rasgos
culturales, se transmiten a través de las generaciones, durante este aprendizaje se generan
variaciones estables en el tiempo, por lo que se puede postular que las variaciones acústicas
son una consecuencia de las características físicas del ambiente, o que estas variaciones no

123
intervengan en la reproducción, o que sean variaciones recientes que aún no se expresan en
la estructura genética.
Estudios donde se identifique el sexo del emisor y la conducta asociada a la emisión
acústica de la vocalización Be4, serán necesarios para ampliar el conocimiento sobre la
transmisión cultural en B. edeni brydei del Golfo de California, y de esta manera confirmar
alguna de las hipótesis expuestas.

124
LITERATURA CITADA
Aguayo, L. A., Findley, L., Rojas-B. y Vidal, O. 1983. The population of fin whales
(Balaenoptera physalus) in the Gulf of California, México. Abstracts: Fifth Biennial
Conference on the Biology of Marine Mammals. Boston, MA. EE.UU.
Albrecht, D. J. y Oring, L. W. 1995. Song in chipping sparrows, Spizella passerina:
Structure and function. Animal Behaviour. 50(5): 1233–1241.
Andrew, R. J. 1962. Evolution of intelligence and vocal mimicking. Science. 137: 585–89.
Baird, R. W. 2000. Status of killer whales, Orcinus orca, in Canada. Canadian field-
naturalist. 115(4): 676–701.
Baker, M. C. y Cunningham, M. A. 1985. The biology of bird-song dialects. Behavioral
and Brain Sciences. 8: 85–133.
Baker, M. C., Spitler-Nabors, K. J. y Bradley, D. C. 1981. Early experience determines
song dialect responsiveness of female sparrows. Science. 214: 819–821.
Bazúa-Duran, C. y Au, W. W. L. 2001. Geographic variations in the whistles of spinner
dolphins (Stenella longirostris). 14th
Biennial Conference on the Biology of Marine
Mammals. Vancouver, Canada. Pág. 19.
Bender, C. E., Herzing, D. L. y Bjorklund, D. F. 2009. Evidence of teaching in Atlantic
spotted dolphins (Stenella frontalis) by mother dolphins foraging in the presence of
their calves. Animal Cognition. 12(1): 43–53.
Bérubé, M., Urbán, R. J., Dizon, A. E., Brownell, R. L. y Palsbøll, P. J. 2002. Genetic
identification of a small and highly isolated population of fin whales (Balaenoptera
physalus) in the Sea of Cortez, México. Conservation Genetics. 3: 183–190.
Boyd, R. y Richerson, P. J. 1996. Why culture is common but cultural evolution is rare.
Proceedings of the British Academy. 88: 77–93.
Catchpole, C. K. y Slater, P. J. B. 1995 Birdsong: Biological Themes and Variations,
Cambridge University Press.
Cerchio, S., Jacobsen, J. K. y Norris, T. F. 2001. Temporal and geographical variation in
songs of humpback whales, Megaptera novaeangliae: synchronous change in
Hawaiian and Mexican breeding assemblages. Animal Behavior. 62(2): 313–329.

125
Chávez, R, S. 1995. Estimación poblacional del rorcual tropical Balaenoptera edeni
(Anderson, 1878) en la Bahía de la Paz, B. C. S. México. Tesis de Maestría.
CICIMAR. 55 pp.
Croll, D. A., Clark, W. C., Acevedo, A., Tershy, B., Flores, S., Gedamke, J. y Urbán, J.
2002 Only male fin whales sing loud songs. Nature. 417: 809.
Cummings, W. C., Thompson, P. O. y Samuel, J. H. 1986. Sounds from Bryde,
Balaenoptera edeni, and Finback, B. physalus, whales in the Gulf of California.
Fishery Bulletin. 84(2): 359–370.
Darling, J. D. 1983. Migrations, abundance and behavior of Hawaiian humpback whales,
Megaptera novaeangliae (Borowski). Ph.D. dissertation, University of California,
Santa Cruz, CA. 147 pp.
Deecke, V. B.; Ford, J. K. B. y Spong, P. 2000. Dialect change in resident killer whales:
Implications for vocal learning and cultural transmission. (Orcinus orca). Animal
Behaviour. 60(5): 629–638.
Dizon, A., Lux, C. A., LeDuc, R. G., Urbán, R, J., Henshaw, M. y Brownell, R. Jr. 1996.
An interin phylogenetic analysis of sei and Bryde’s whale mitochondrial DNA
control region sequences. SC/47/NP23 presented to the IWC Scientific Committee.
12 pp.
Edds, P., Odell, P. K. y Tershy, B. R. 1993. Calls of a captive juvenile and free-ranging
adult-calf pairs of Bryde’s whales, Balaenoptera edeni. Mar. Mamm. Sci. 9: 269–
284.
Excoffier, L., Laval, G. y Schneider, L. 2005. Arlequin ver. 3.0: An integrated software
package for population genetics data analysis. Evolutionary Bioinformatics. 1: 47–
50.
Figueiredo, L. D. y Simão, S. M. 2014. Bryde’s Whales (Balaenoptera edeni) vocalizations
from Southeast Brazil. Aquatic Mammals. 40(3): 225–231.
Foster, S. A. 1999. The geography of behaviour: an evolutionary perspective. Trends in
Ecology & Evolution. 14(5): 190–195.
Frankel, A. S., Clark, W. C., Herman, L. M. y Gabriele, C. M. 1995. Spatial distribution,
habitat utilization, and social interactions of humpback whales, Megaptera

126
novaeangliae, off Hawaii, determined using acoustic and visual techniques.
Canadian Journal of Zoology. 73(6): 1134–1146.
Gedamke, J., Costa, D. P. y Dunstan, A. 2001. Localization and visual verification of a
complex minke whale vocalization. Journal of the Acoustical society of America.
109(6): 3038–3047.
Gedamke, J., Costa, D., Clark; W. C., Mellinger, K. D. y O'neill, F. L. 2003. The role of
song in maintaining spacing between singing dwarf minke whales (Balaenoptera
acutorostrata): Preliminary results from playback experiments. (Abstract).
Environmental Consequences of Underwater Sound (ECOUS) Symposium. San
Antonio, Texas.
Guinee, L. N., Chu, K. y Dorsey, E. 1983. Changes over time in the songs of known
individual humpback whales (Megaptera novaeangliae). Communication and
behavior of whales. 76– 59-80.
Guinet, C. y Bouvier, J. 1995. Development of intentional stranding hunting techniques in
killer whale (Orcinus orca) calves at Crozet Archipelago. Can. J. Zool. 73: 27–33.
Hatch, L. T. y Clark, C. W. 2004. Acoustic differentiation between fin whales in both the
North Atlantic and North Pacific Oceans, and integration with genetic estimates of
divergence. SC/56/SD6 IWC Scientific Committee (unpublished). 37 pp.
Heimlinch, S. L., Mellinger, D. K., Nieukirk, S. L. y Fox, G. C. 2005. Types, distribution,
and seasonal occurrence of sounds attributed to Bryde’s whales (Balaenoptera
edeni) recorded in the eastern tropical Pacific, 1999–2001. Jour. Acoust. Soc. Am.
118 (3): 1830–1837.
Holland, J., Dabelsteen, T. y Pedersen, S. B. 1998. Degradation of wren Troglodytes
troglodytes song: implications for information transfer and ranging. J. Acoust. Soc.
Am. 103: 2154–2166.
Janik V. M. y Slater, J. V. 1997. Vocal Learning in Mammals. Advances in the study of
behavior. 26: 59–62.
Janik, V. M. 1997. Whistle matching in wild bottlenose dolphins. J. Acoust. Soc. Am. 101:
3136.
Janik, V. M. 2000. Whistle matching in wild bottlenose dolphins. Science 289: 1355–57.

127
Kato, H. y Perrin, W. F. 2009. Bryde’s Whales Balaenoptera edeni/brydei. En: William, F.
P., Würsig, B. y Thewissen, J. G. M. 2009. Encyclopedia Marine Mammals 2a ed.
Elservier. 157–162 pp.
Ketten, R. D. 1997. Structure and function in whale ears. Bioacoustics. The International
Journal of Animal sound and its Recording. 8: 103–135.
Kimura, M. 1980. A simple method for estimating evolutionary rates of base substitutions
through comparative studies of nucleotide sequences. J Mol Evol. 16: 111–120.
Krützen, M., Mann, J., Heithaus, R. M., Connor, C. R., Bejder, L. y Sherwin, B. W. 2005.
Cultural transmission of tool use in bottlenose dolphins. PNAS. 25: 8939–8943.
Lackey, A. C. R. y Boughman, J. W. 2014. Female discrimination against heterospecific
mates does not depend on mating habitat. Behavioral Ecology. 1–12.
Ladrón de Guevara P., P. 1995. La ballena jorobada Megaptera novaeangliae (Borowski,
1781) en la Bahía de Banderas, Nayarit-Jalisco, Mexico. (Cetacea:
Balaenopteridae). Tesis Profesional. Facultad de Ciencias, UNAM, México. 155 pp.
Laland, K. N. y Janik, V. M. 2006. The animal cultures debate. TRENDS in Ecology and
Evolution. 21(10): 542–547.
Maan, M. E. y Seehausen, O. 2011. Ecology, sexual selection and speciation. Ecology
Letters. 14(6), 591-602.
MacDougall-Shackleton, E. A. y MacDougall-Shackleton, S. A. 2001. Cultural and genetic
evolution in mountain white-crowned sparrows: song dialects are associated with
population structure. Evolution. 55(12): 2568–2575.
Marler, P. 1957. Species distinctiveness in the communication signals of birds. Behaviour.
11(1): 13–38.
Marler, P. y Tamura, M. 1962. Song "dialects" in three populations of white-crowned
sparrows. Condor. 64: 368–377.
Marler, P. y Tamura, M. 1964. Culturally transmitted patterns of vocal behavior in
sparrows. Science. 146: 1483–86
McDonald, M., Mesnick, S. L. y Hildebrand, J. 2006. Biogeographic characterization of
blue whale song worldwide: using song to identify populations. J. Cetacean Res.
Manage. 8(1): 55–65.

128
Morisaka, T., Shinohara, M., Nakahara, F. y Akamatsu, T. 2005. Geographic variations in
the whistles among three Indo-Pacific bottlenose dolphin Tursiops aduncus
populations in Japan. Fisheries Science. 71: 568–576.
Morton, E. S. 1975. Ecological sources of selection on avian sounds. Am. Nat. 109: 17–34.
Nigenda-Morales, S., Flores-Ramírez, S., Urbán-R., S. y Vázquez-Juárez, R. 2008. MHC
DQB-1 Polymorphism in the Gulf of California Fin Whale (Balaenoptera physalus)
Population. Journal of Heredity 99(1): 14–21.
Noad, M. J., Cato, D. H., Bryden, M. M., Jenner, M. N. y Jenner, K. C. S. 2000. Cultural
revolution in whale songs. Nature 408(6812): 537–537.
Nottebohm, F. 1969. The song of the chingolo, Zonotrichia capensis, in Argentina:
description and evaluation of a system of dialects. Condor 71: 299–315.
Oleson, E. M., Barlow, J., Gordon, J., Rankin, S. y Hildebrand, J. A. 2003. Low frequency
calls of Bryde’s whales. Mar. Mamm. Sci. 19(2): 160–172.
Payne, K., Tyack, P. y Payne, R. 1983. Progressive changes in the songs of humpback
whales (Megaptera novaeangliae): a detailed analysis of two seasons in Hawaii.
Communication and behavior of whales. 9-57.
Payne, R. B. 1981. Population structure and social behavior: models for testing the
ecological significance of song dialects in birds. En: R. D. Alexander and D. W.
Tinkle. (eds). Natural selection and social behavior: recent research and new theory.
Chiron Press, New York. 108–119 pp.
Payne, R. S. y McVay, S. 1971. Songs of Humpback Whales. Science 173(3997): 85–597.
Podos, J. S., Huber, K. y Taft, B. 2004. Bird song: The Interface of Evolution and
Mechanism. Annu. Rev. Ecol. Evol. Syst. 35: 55–87
Podos, J. y Warren, S. P. 2007. The Evolution of Geographic Variation in Birdsong.
Advances in the study of behavior. (37): 403–458.
Reiss, D. y McCowan, B. 1993 Spontaneous vocal mimicry and production by bottlenose
dolphins (Tursiops truncatus): evidence for vocal learning. J. Comp. Psych. 107:
301–312.
Rice, A. R., Palmer, K. J., Tielens, J. T. y Muirhead, C. A. 2014. Potential Bryde’s whales
(Balaenoptera edeni) calls recorded in the northern Gulf of Mexico. J. Acoust. Soc.
Am. 135: 3066–3076.

129
Richards, D. G. y Wiley, R. H. 1980. Reverberations and amplitude fluctuations in the
propagation of sound in a forest: implications for animal communication. Am. Nat.
115: 381–399.
Richards, D. G., Wolz, J. P. y Herman, L. M. 1984 Vocal mimicry of computer-generated
sounds and vocal labeling of objects by a bottlenose dolphin, Tursiops truncatus. J.
Comp. Psych. 98: 10–28.
Ryan, M. J. y Brenowitz, E. A. 1985. The role of body size, phylogeny, and ambient noise
in the evolution of bird song. Am. Nat. 126: 87–100.
Schluter, D. 2000. The Ecology of Adaptive Radiation. Oxford University Press, Oxford.
Širović, A., Bassett, H. R., Johnson, S. C., Wiggins, S. M. y Hildebrand, J. A. 2014.
Bryde's whale calls recorded in the Gulf of Mexico. Mar. Mamm. Sci. 30: 399–409.
Slabbekoorn, H. y Smith, B. T. 2002. Bird song, ecology and speciation. Phil. Trans. R.
Soc. Lond. B. 357: 493–503.
Smolker, R. A., Richards, A., Connor, R. C., Mann, J. y Berggren, P. 1997. Sponge
carrying by dolphins (Delphinidae, Tursiops sp): A foraging specialization
involving tool use? Ethology. 103: 454–465.
Tamura, K., Stecher, G., Peterson, D., Filipski, A. y Kumar, S. 2013. MEGA6: Molecular
Evolutionary Genetics Analysis version 6.0. Molecular Biology and Evolution. 30:
2725–2729.
Terhune, J. M., Quin, D., Dell’Apa, A., Mirhaj, M., Plötz, J.,·Kindermann, L. y
Bornemann, H. 2008. Geographic variations in underwater male Weddell seals
Trills suggest breeding area fidelity. Polar Biol. 31: 671–680.
Tershy, B. R., Acevedo, G. A., Breese, D. y Strong, C. S. 1993. Diet and feeding behavior
of fin and Bryde's whales in the central Gulf of California, Mexico. Rev Inv Cient.
1: 31–38.
Tershy, B. R., Breese, D. y Strong, C. S. 1990. Abundance, seasonal distribution and
population composition of balaenopterid whales in the Canal de Ballenas, Gulf of
California, Mexico. Rep. Int. Whal. Commn. Special Issue. 12: 369–375.
Thomas, J. A. y Stirling, I. 1983. Geographic variation in the underwater vocalizations of
Weddell seals (Leptonychotes weddelli) from Palmer Peninsula and McMurdo
Sound, Antarctica. Can. J. Zool. 61: 2203–2212.

130
Thomas, J. A. y Golladay, C. L. 1995. Geographic variation in leopard seal (Hydrurga
leptonynx) underwater vocalizations. En: Kastelein R. A, Thomas JA, Nachtigall PE
(eds) Sensory systems of aquatic mammals. De Spil Publishers, Woerden. 201–221
pp.
Thompson, O. P., Findley, T. L. y Vidal, O. 1992. 20-Hz pulses and other vocalizations of
fin whales, Balaenoptera physalus, in the Gulf of California, Mexico. J. Acoust.
Soc. Am. 92(6): 3051–3057.
Tyack, P. 1981. Interactions between singing hawaiian humpback whales and conspecifics
nearby. Behavioral Ecology Sociobiology. 8(2): 105–116.
Tyack, P. 2009. Behavior, overview. 101-108. En: William, F. P., Würsig, B. y Thewissen,
J. G. M. 2009. Encyclopedia Marine Mammals 2a ed. Elservier. 157-162 pp.
West-Eberhard, M. J. 1983. Sexual selection, social competition and speciation. Quarterly
Review of Biology. 58: 155–83.
Whitehead, H., Dillon, M., Dufault, S., Weilgart, L. y Wright, J. 1998. Non-geographically
based population structure of south Pacific sperm whales: dialects, fluke-markings
and genetics. J. Anim. Ecol. 67: 253-262.
Whiten, A., Goodall, J., McGrew, W.C., Nishida, T., Reynolds, V., Sugiyama, Y. Tutin, C.
E. G., Wrangham, R. W. y Boesch, C. 1999. Cultures in Chimpanzees. Nature. 399:
682–685.
Winn, H. E. y Winn, L. K. 1978. The song of the humpback whale Megaptera
novaeangliae in the West Indies. Marine Biology. 47(2): 97–114.
Wright, T. F. 1996. Regional dialects in the contact call of a parrot. Proc. R. Soc. Lond. B
263: 867–872.

131
ANEXO 1
132
ANEXO 2

133
ANEXO 3

Calls of Bryde’s whale (Balaenoptera edeni) recorded in the Gulf of California 1
2
Lorena Viloria Gómora. 3
Programa de Investigación de Mamíferos marinos. Departamento de Biología Marina, Universidad 4
Autónoma de Baja California Sur, La Paz, México. 5
Eduardo Romero-Vivas. 6
Centro de Investigaciones Biológicas del Noroeste, La Paz, B.C.S., Mexico. 7
Jorge Urbán-Ramíreza. 8
Programa de Investigación de Mamíferos marinos. Departamento de Biología Marina, Universidad 9
Autónoma de Baja California Sur, La Paz, México. 10
11
Running title: New calls of Bryde’s whale 12
13
14
aCorresponding author: [email protected] 15
16

Abstract: Field trips from January 2010 to June 2014 provided opportunities to identify calls of 17
Bryde’s whale recorded during sighting of adults. From a total of 109 recordings, three of the four 18
types of calls found are different to previously documented calls from the specie for the species 19
(labeled Be10, Be11, and Be12); the remaining call type was reported for the Eastern Tropical 20
Pacific and south of California (Be4). The calls were recorded during sightings of individual 21
Bryde’s whales; for Be4, there were always at least two whales in sight. The few previous acoustic 22
recordings of Bryde’s whale have the limitations of the use of passive acoustic monitoring technics, 23
Be10, Be11, and Be12 calls, made together with visual confirmation of the species, contribute to 24
monitoring capabilities. 25
26
PACS numbers: 43.80.Ka, 43.30.Sf 27
43.80Ka, Sound production by animals: mechanism, characteristics, populations, biosonar. 28
43.30Sf, Acoustical detection of marine life; passive and active. 29
30
I. INTRODUCTION 31
32
The use of passive acoustics monitoring (PAM) techniques for the study of marine 33
mammals has increased in the last decade. PAM is an effective, accurate, and non-invasive method 34
for studying different aspects of the marine mammals, such as presence, distribution, abundance, 35
differences among populations or individuals, and other characteristics (Au and Hasting, 2008). 36
However, PAM relies on previous knowledge of the acoustic repertoire for each species, since it is 37
necessary to identify the calls by comparison to a template that has been obtained from records with 38
visual confirmation of the species (Mellinger et al., 2007). For species with sparse data, PAM 39
techniques become limited. 40
Bryde’s whale is one of these species that is poorly studied globally. The whale can be 41
found year-round in temperate and tropical waters (Kato and Perrin, 2009). There is no distinctive 42
migration pattern or a specific reproduction season (Leatherwood et al., 1982), although a seasonal 43
expansion of their habitat has been reported recently (Kereosky et al., 2012). Single individuals 44
crossing are common, but this whale has occasionally been seen in groups of two or three (Kato and 45
Perrin, 2009). This species is one of the most common 46
baleen whale in the Gulf of California (Leatherwood et al., 1982; Urbán and Flores, 1996). 47
The vocal repertoire of this whale is quite limited. Currently, there are only seven studies 48
describing its calls. In three of them, the whales were seen during the recordings (Cummings et al., 49
1986; Edds et al., 1993; Figueiredo and Simão, 2014). In later studies, automated equipment and 50

PAM techniques were used (Heimlich et al., 2005; Rice et al., 2014) and the calls were associated 51
with the whale by their spectral and timing characteristics. A combination of 54 both methods has 52
also being used (Oleson et al., 2003; Širović et al., 2014), although some authors acknowledge that 53
it is necessary to validate the 56 recordings with local follow-up (Rice et al., 2014). 54
Table 1 summarizes the information available for the Bryde’s whale. It is important to notice that 55
the Be4 call at 60 Hz with no harmonics, recorded on two occasions along the southwest coast of 56
the Baja California Peninsula, reported by Oleson et al. (2003), is later reported by Kerosky et al. 57
(2012) in the Southern California Bight but with harmonics, nevertheless no distinction is made, 58
and both have been equally identified as Be4. Here, we report and describe three previously 59
undocumented calls from Bryde’s whales and the presence of the Be4 call recorded during sightings 60
in the Gulf of California. 61
62
II. METHODS 63
64
A. Recordings 65
66
From January 2010 to June 2014, 116 field trips were conducted to study the Bryde’s whale in 67
the Gulf of California (Fig. 1). Protocols were followed to minimize disturbance, including slow 68
approaches and keeping a minimum distance of 30 m. This research was sanctioned under a permit 69
from the Mexican government for studies of wild animals. 70
The whales were identified through pictures of the head, which has three distinct prominent 71
longitudinal ridges on the whale’s rostrum in front of the blowhole (Leatherwood et al., 1982). 72
After confirmation, the engines were shut off. The 10 min recordings were made with a portable 73
hydrophone made of AQ-1s & ITC–1042 transducers (10 Hz–100 kHz frequency response) 74
deployed at a depth of 10 m. Signals were digitalized with a FOSTEX FR2 solid state recorder 75
(22.05 kHz sampling frequency, 24 bit resolution). During recordings, the distance from the ship to 76
the whale was over 150 m, and no other species of whale were found in the area. Each location was 77
geo-referenced. 78
79
B. Signal pre-processing 80
81
Recording were filtered with a 10th order Butterworth band-pass filter (30–600 Hz) and 82
normalized with Adobe Audition 3.1. The resulting waveforms were analyzed with Matlab 2010b. 83
From the fft spectrograms (Hanning window, Fs = 196200, NFFT = 5845, Overlap = 80%), four 84

standard parameters were obtained for the main component of the call: maximum frequency, 85
minimum frequency, peak frequency, and duration. For signals in more than one recording, average 86
values were computed. 87
The recorded calls were compared with previous reports for Bryde’s whales. New call 88
names were assigned following the convention used by Oleson et al. (2003), thus starting with 89
Be10, Be11, and so forth. 90
91
III. RESULTS 92
93
During 48 days of cruises, 109 recordings, amounting to 11.8 h, were obtained, of which 94
3.46 hours (29.3%) of vocalizations were detected. All vocalizations corresponded to sightings of 95
individual adult whales. Recordings of mothers with calves were made, but no recording for this 96
kind of grouping were recorded. 97
Four different calls were detected. Three of them were new for the species and were labeled 98
Be10, Be11 and Be12 (Figs. 2, 3, and 4, respectively). The fourth call (Fig. 5) was similar to Be4 99
first reported by Oleson et al. (2003) for the tropical Eastern Pacific, and by Kerosky et al. (2012) 100
for the southern California Bight. 101
A summary of the main standard parameters of the calls and information related to the local 102
ambient conditions shown in Table 2. 103
104
IV. DISCUSSION 105
106
Three new calls of Bryde’s whales, with single whales in sight, were recorded in the Gulf of 107
California, presenting similar frequency characteristics, as previously reported for this species. 108
Remarkably, one individual (labeled A, Table 2) produced all three calls within a 10 min recording, 109
Be10 (six times), Be11 (one time), and Be12 (one time). During this event, long-beaked common 110
dolphins, California sea lions and the Bryde’s whale were probably feeding in the area. Since the 111
observation of specific behavior simultaneously to the emission of calls is a rare event among 112
baleen whales (Calambokidis et al., 2007; Oleson et al., 2007), the sighting and recording of Be10 113
and Be11 calls acquires relevance in the study of biology of B. edeni, because suggests a potential 114
relationship to feeding behaviors. 115
The Be4 call was not recorded during sightings of individual whales. These calls were 116
always recorded when there were at least two whales present in the area, which suggests that it is a 117

socializing call. However, since there was food present in the area, it might be also associated with 118
feeding behavior. 119
Bryde’s whales are difficult to study. They have a morphology similar to Sei’s whales 120
(Balaenoptera borealis), which makes visual identification challenging (Kerosky et al., 2012). 121
Their avoidance behaviour, little activity at the surface, and absence of gregariousness (Kato and 122
Perrin, 2009) complicates common methods of study, such as call recordings on sightings and 123
collecting biopsies for determining gender. Unlike other species, Bryde’s whales do not exhibit a 124
reproductive period or a feeding season (Breese and Tershy, 1987). This makes behavioural 125
categories difficult to resolve. For these reasons, recording of three new calls for these whales is 126
considered to contribute solidly to the knowledge of the species and advancing future studies with 127
the PAM techniques. 128
129
ACKNOWLEDGMENTS 130
131
We thank PRIMMA-UABCS members who participated in data collection. Ira Fogel of 132
CIBNOR provided editorial services. Research was carried out under scientific research permits 133
(SGPA/DGVS/08434/09–2009, SGPA/DGVS/0073411–2011, and SGPA/DGVS/0099/14–2014) 134
from the Secretaria de Medio Ambiente y Recursos Naturales of Mexico. Funding was provided by 135
Consejo Nacional de Ciencia y Tecnologia (CONACYT project 134251-Sep-CONACYT) and 136
WWF-TELCEL. L.V.G. is a recipient of a fellowship grant (CONACYT 165753). 137
138
REFERENCES 139
140
Au, W. W. L., and Hastings, M. C. (2008). Principles of Marine Bioacoustics (Springer, New 141
York). 142
Breese, D., and Tershy, B. R. (1987). “Residency patterns in female Bryde’s whales in the Gulf of 143
California, Mexico”. Abstracts, Seven Biennial Conference on the Biology of Marine 144
Mammals, 5–7 December, Miami, FL. 145
Calambokidis, J., Schorr, G. S., Steiger, G. H., Francis, J., Bakhtiari, M., Marshall, G., and 146
Robertson, K. (2007). “Insights into the underwater diving, feeding, and calling behavior of 147
blue whales from a suction-cup-attached video-imaging tag (CRITTERCAM),” Mar. 148
Technol. Soc. J. 41, 19–29. 149
Cummings, W. C., Thompson, P. O., and Samuel, J. H. (1986). “Sounds from Bryde, Balaenoptera 150
edeni, and finback, B. physalus, whales in the Gulf of California”. Fish. Bull. 84: 359–370. 151

Edds, P., Odell, P. K., and Tershy, B. R. (1993). “Calls of a captive juvenile and free-ranging adult-152
calf pairs of Bryde’s whales, Balaenoptera edeni”. Mar. Mamm. Sci. 9:269–284. 153
Figueiredo, L. D. and Simão, S. M. (2014). “Bryde’s Whales (Balaenoptera edeni) vocalizations 154
from Southeast Brazil,” Aquatic Mammals 40, 225–231. 155
Heimlich, S. L., Mellinger, D. K., Nieukirk, S. L., and Fox, G. C. (2005). “Types, distribution, and 156
seasonal occurrence of sounds attributed to Bryde’s whales (Balaenoptera edeni) recorded 157
in the eastern tropical Pacific, 1999–2001”. J. Acoust. Soc. Am. 118: 830–1837. 158
Kato, H., and Perrin, W. F. (2009). “Bryde’s Whales Balaenoptera edeni/brydei,” in Encyclopedia 159
of Marine Mammals, edited by F. P. William, B. Würsig and J. G. M. Thewissen (Elsevier), 160
pp. 157–162. 161
Kerosky, M. S., Širović, A., Roche, L. K., Baumann-Pickering, B., Wiggins, S. M., and Hildebrand, 162
A. J. (2012). “Bryde’s whale seasonal range expansion and increasing presence in the 163
Southern California Bight from 2000 to 2010”. Deep Sea Res. 1(65):125–132. 164
Leatherwood, S., Reeves, R. R., Perrin, W. F., and Evans, W. E. (1982). Whales, dolphins and 165
porpoises of the eastern North Pacific and adjacent Arctic waters: A guide to their 166
identification. NOAA Technical Report, NMFS. Circular 444. 167
Mellinger, D. K., Stafford, K. M., Moore, S. E., Dziak, R. P., and Matsumoto H. (2007). “An 168
overview of fixed passive acoustic observation methods for cetaceans”. Oceanography. 169
20(4):36–45. 170
Oleson, E. M., Barlow, J, Gordon, J., Rankin, S., and Hildebrand, J. A. (2003). “Low frequency 171
calls of Bryde’s whales.” Mar. Mamm. Sci. 19:160–172. 172
Oleson, E. M., Calambokidis, J., Burgess, W. C., McDonald, M. A., LeDuc, C. A., and Hildebrand, 173
J. A. (2007). “Behavioral context of call production by eastern North Pacific blue whales,” 174
Mar. Ecol.–Prog. Ser. 330. 175
Rice, A. R., Palmer, K. J., Tielens, J. T., and Muirhead, C. A. (2014). “Potential Bryde’s whales 176
(Balaenoptera edeni) calls recorded in the northern Gulf of Mexico”. J. Acoust. Soc. Am. 177
135: 3066–3076. 178
Širović, A., Bassett, H. R., Johnson, S. C., Wiggins, S. M., and Hildebrand, J. A. (2014). “Bryde's 179
whale calls recorded in the Gulf of Mexico”. Mar. Mamm. Sci. 30, 399–409. 180
Urbán, R. J., and Flores, R. S. (1996). “A note on Bryde’s Whales (Balaenoptera edeni) in the Gulf 181
of California, Mexico”. Rep. Int. Whal. Commn. 46, 453–457. 182
183

Figure legends 184
185
Fig. 1. Locations in the Gulf of California where Bryde’s whale vocalizations were recorded. 186
187
Fig. 2. Spectrogram and time series of a Be10 call. Spectrogram parameters: Hanning window, Fs = 188
196200, NFFT = 15845, Overlap = 80%; Butterworth filter/band pass = 60–1000 Hz. 189
190
Fig. 3. Spectrogram and time series of a Be11 call. Spectrogram parameters: Hanning window, Fs = 191
196200, NFFT = 9845, Overlap = 80%; Butterworth filter/band pass = 50–1000 Hz. 192
193
Fig. 4. Spectrogram and time series of a Be12 call. Spectrogram parameters: Hanning window, Fs = 194
196200, NFFT = 15845, Overlap = 80%; Butterworth filter/band pass = 60–500 Hz. 195
196
Fig. 5. Spectrogram and time series of a Be4 call. Spectrogram parameters: Hanning window, Fs = 197
196200, NFFT = 15845, Overlap = 80%; Butterworth filter/band pass = 60–500Hz. 198
199
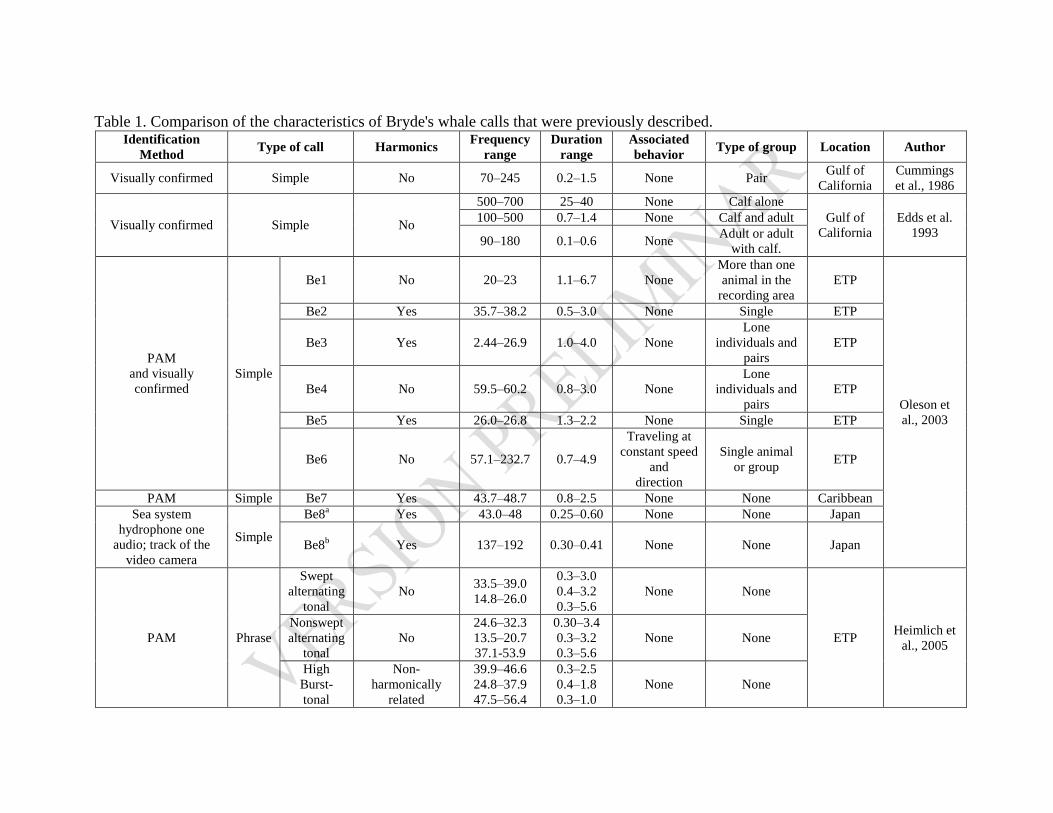
Table 1. Comparison of the characteristics of Bryde's whale calls that were previously described. Identification
Method Type of call Harmonics
Frequency
range
Duration
range
Associated
behavior Type of group Location Author
Visually confirmed Simple No 70–245 0.2–1.5 None Pair Gulf of
California
Cummings
et al., 1986
Visually confirmed Simple No
500–700 25–40 None Calf alone
Gulf of
California
Edds et al.
1993
100–500 0.7–1.4 None Calf and adult
90–180 0.1–0.6 None Adult or adult
with calf.
PAM
and visually
confirmed
Simple
Be1 No 20–23 1.1–6.7 None
More than one
animal in the
recording area
ETP
Oleson et
al., 2003
Be2 Yes 35.7–38.2 0.5–3.0 None Single ETP
Be3 Yes 2.44–26.9 1.0–4.0 None
Lone
individuals and
pairs
ETP
Be4 No 59.5–60.2 0.8–3.0 None
Lone
individuals and
pairs
ETP
Be5 Yes 26.0–26.8 1.3–2.2 None Single ETP
Be6 No 57.1–232.7 0.7–4.9
Traveling at
constant speed
and
direction
Single animal
or group ETP
PAM Simple Be7 Yes 43.7–48.7 0.8–2.5 None None Caribbean
Sea system
hydrophone one
audio; track of the
video camera
Simple
Be8a Yes 43.0–48 0.25–0.60 None None Japan
Be8b Yes 137–192 0.30–0.41 None None Japan
PAM Phrase
Swept
alternating
tonal
No 33.5–39.0
14.8–26.0
0.3–3.0
0.4–3.2
0.3–5.6
None None
ETP Heimlich et
al., 2005
Nonswept
alternating
tonal
No
24.6–32.3
13.5–20.7
37.1-53.9
0.30–3.4
0.3–3.2
0.3–5.6
None None
High
Burst-
tonal
Non-
harmonically
related
39.9–46.6
24.8–37.9
47.5–56.4
0.3–2.5
0.4–1.8
0.3–1.0
None None

frequencies 0.3–2.8
Low
burst–
tonal
Non-
harmonically
related
frequencies
16.8–26.7
24.1–36.1
62.6–78.8
0.4–2.8
0.3–2.5
0.4–2.0
0.4–4.0
None None
Harmonic
tone Yes
20.7–25.8
75.1–79.6
46.8–66.3
0.5–3.2
0.4–2.3
0.5–2.2
None None
PAM
and visually
confirmed
Simple Be9 No 110–78
143–85 0.4–0.7 None Group of adults
Gulf of
Mexico
Sirovic et
al., 2014
PAM
Phrase Down-
sweep No
96.84–
97.68a
0.2–0.2b
None None
Gulf of
Mexico
Rice et al.,
2014 Simple Long-
moan
Yes
(In some signals)
106.6–
108.1a
11.1–11.5b None None
Phrase Tonal No 99.4–107.4 1.9–2.7b None None
Visually
confirmed
Simple PS1 No 225–555 0.788 None Mother whit
calf
Cabo Frio
coast,
Brazil
Figeiredo,
et al.,
2014.
Simple LFT Yes 8.5–19 1.413 None Single
Simple FMT No 416–671 1.531 None Single
Simple TM1 Yes 85.7–123.6 0.876
Whale was
very close
to de boat
(~3m)
Single
Simple TM2 No 49.5–105.5 1.156
Whale
moved
away from
the boat
Single
aCenter frequency,
bCenter time.
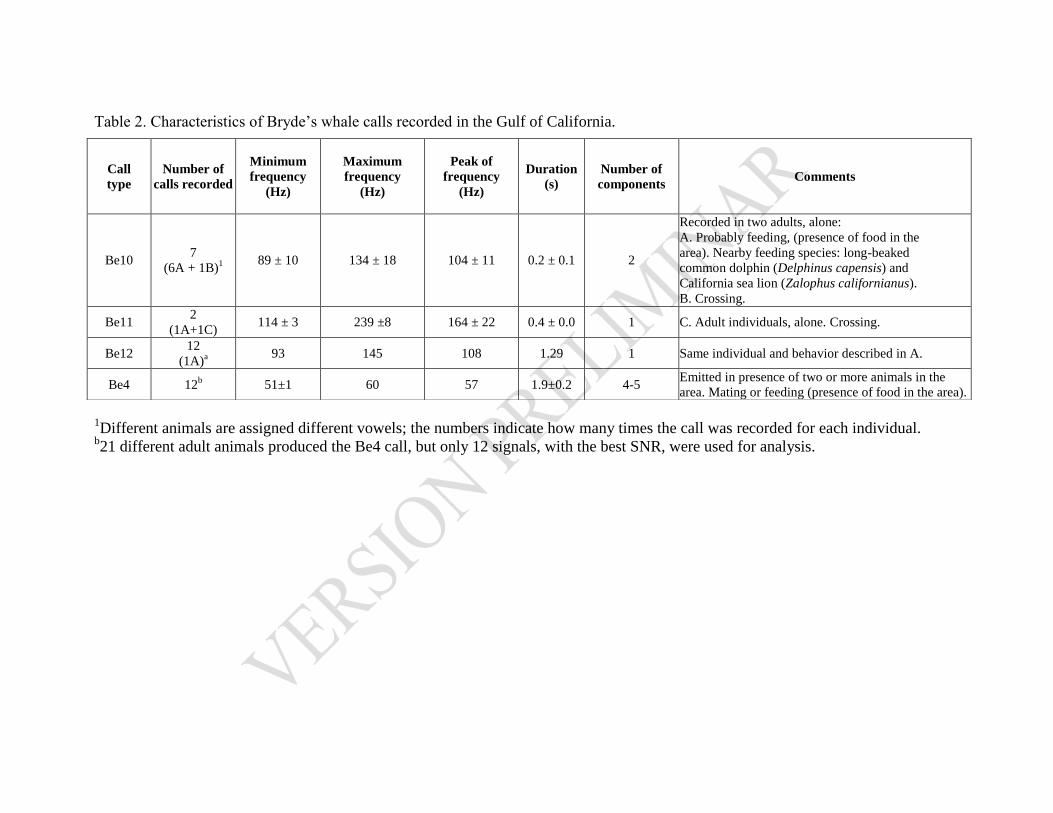
Table 2. Characteristics of Bryde’s whale calls recorded in the Gulf of California.
1Different animals are assigned different vowels; the numbers indicate how many times the call was recorded for each individual.
b21 different adult animals produced the Be4 call, but only 12 signals, with the best SNR, were used for analysis.
Call
type
Number of
calls recorded
Minimum
frequency
(Hz)
Maximum
frequency
(Hz)
Peak of
frequency
(Hz)
Duration
(s)
Number of
components Comments
Be10 7
(6A + 1B)1
89 ± 10 134 ± 18 104 ± 11 0.2 ± 0.1 2
Recorded in two adults, alone:
A. Probably feeding, (presence of food in the
area). Nearby feeding species: long-beaked
common dolphin (Delphinus capensis) and
California sea lion (Zalophus californianus).
B. Crossing.
Be11 2
(1A+1C) 114 ± 3 239 ±8 164 ± 22 0.4 ± 0.0 1 C. Adult individuals, alone. Crossing.
Be12 12
(1A)a
93 145 108 1.29 1 Same individual and behavior described in A.
Be4 12b 51±1 60 57 1.9±0.2 4-5
Emitted in presence of two or more animals in the
area. Mating or feeding (presence of food in the area).

