teoria da coalescÊncia - fc.up.pt · a teoria da coalescência centra-se na genealogia da amostra...
TRANSCRIPT

TEORIA DA
COALESCÊNCIA
Estudo das propriedades genealógicas de linhagens

APLICABILIDADE
• É o instrumento interpretativo mais usado na análise dos
processos populacionais que estão na base da variação
de sequências de DNA.

CARACTERÍSTICAS
• Abordagem genealógica
• Modelação dos processos mutacionais no sentido
presentepassado e não presentefuturo
• Limitação da análise à amostra disponível sem que seja
necessário ter em conta toda a população
• Dissociação conceptual entre processos mutacionais e
genealógicos

AAGCTTGTTCAGAGGAGTGACAATCTGCTTACATCTTGACCTT
AAGCTTGTTCAGAGGAGTGATAATCTGCTTACATCTTGACCTT
SNP
TIPO DE DADOS

TIPO DE DADOS
Nat Rev Genet 7: 669-680 (2006)

indiv
iduals
site of variation
http://pga.gs.washington.edu/
TIPO DE DADOS

A ABORDAGEM GENEALÓGICA
ab cd
ad ac tzxy
ax ay at az
ab cd
ad ac
ax ay at az

A ABORDAGEM GENEALÓGICA
a
a a
a a a a
ab cd
ad ac
ax ay at az



A teoria da coalescência centra-se na genealogia da amostra

MODELO DE WRIGHT-FISHER
• N constante
• Acasalamento ao acaso
• Ausência subdivisões geográficas (ou outras)
• Ausência de selecção
• Gerações discretas
• O insucesso reprodutivo está distribuído ao acaso
– Para valores de N grandes, o número de descendentes por
indivíduo tem uma distribuição Poisson.

CDAB EF GH KLIJ
MODELO DE WRIGHT-FISHER

PROPRIEDADES DAS GENEALOGIAS NO
MODELO DE WRIGHT-FISHER
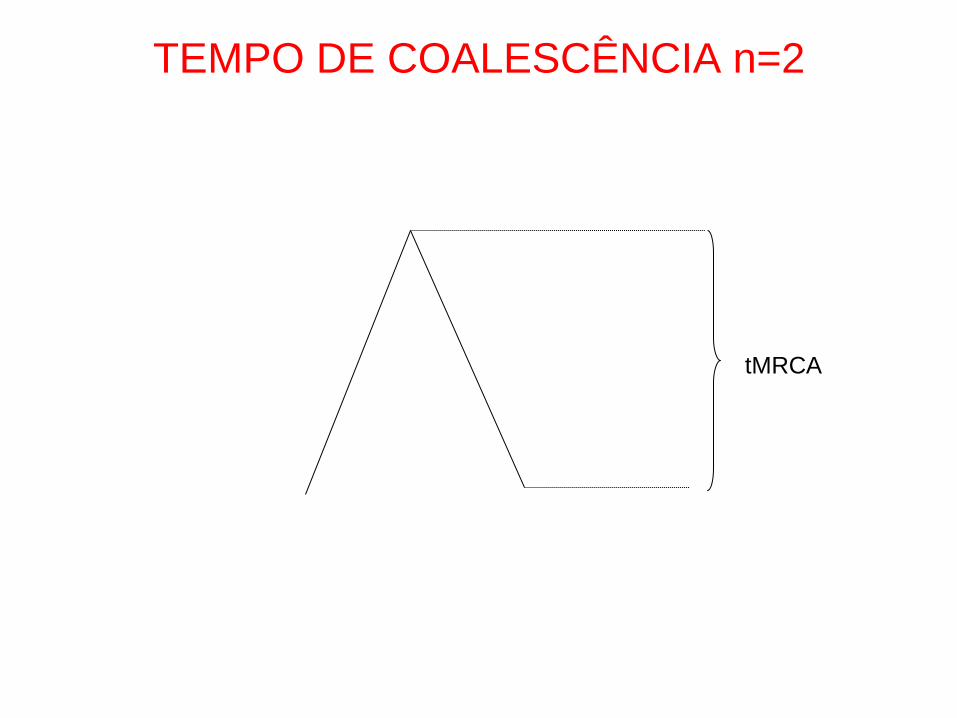
TEMPO DE COALESCÊNCIA n=2
tMRCA

CDAB EF GH KLIJ
Prob. de partilha de ancestral na geração anterior
AA = [1/12]2
BB = [1/12]2
CC = [1/12]2
DD = [1/12]2
EE = [1/12]2
FF = [1/12]2
GG = [1/12]2
HH = [1/12]2
II = [1/12]2
JJ= [1/12]2
KK = [1/12]2
LL = [1/12]2
= [1/12]2 . 12 = 1/12 = 1/2N
TEMPO DE COALESCÊNCIA n=2

TEMPO DE COALESCÊNCIA n=2
• n=2
• Prob. de partilha de ancestral na
geração anterior:
• 1/2N
• Prob. de não partilha de ancestral na
geração anterior:
• 1- (1/2N)
• Probabilidade de coalescência na
geração t+1
• 1/2N [1- (1/2N) ]t

Distribuição geométrica
0
0,005
0,01
0,015
0,02
0,025
0,03
0,035
0,04
0,045
0,05
1 8 15 22 29 36 43 50 57 64 71 78 85 92 99
t
P(M
RC
A=
t)
n=2; N=10
Distribuição geométrica
0
0,005
0,01
0,015
0,02
0,025
0,03
1 8 15 22 29 36 43 50 57 64 71 78 85 92 99
t
P(M
RC
A=
t)
Distribuição exponencial
0
0,005
0,01
0,015
0,02
0,025
0,03
1 9 17 25 33 41 49 57 65 73 81 89 97
t
P(M
RC
A=
t)P(MRCA=t)= 1/2N [1- (1/2N) ]t P(MRCA=t)= 1/2N e-t/2N
n=2; N=20
P(MRCA=t)= 1/2N [1- (1/2N) ]t
Distribuição exponencial
0
0,005
0,01
0,015
0,02
0,025
0,03
0,035
0,04
0,045
0,05
1 9 17 25 33 41 49 57 65 73 81 89 97
t
P(M
RC
A=
t)
P(MRCA=t)= 1/2N e-t/2N
• P (MRCA=t)=1/(2N) e-t/2N
• E (t)=2N gerações
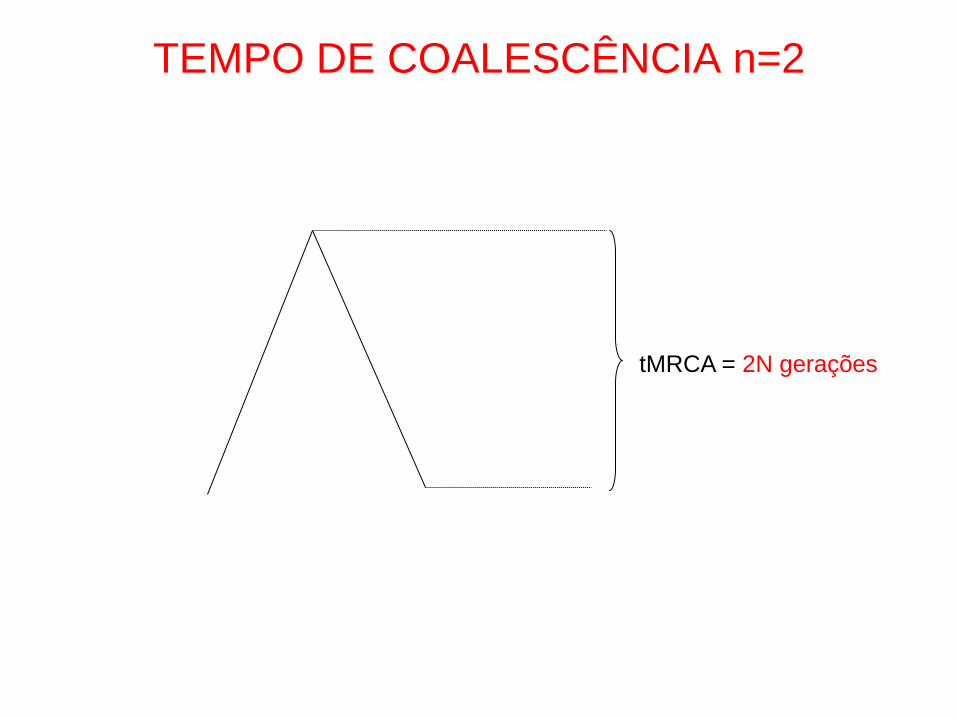
TEMPO DE COALESCÊNCIA n=2
tMRCA = 2N gerações

TEMPO DE COALESCÊNCIA n=n
tMRCA
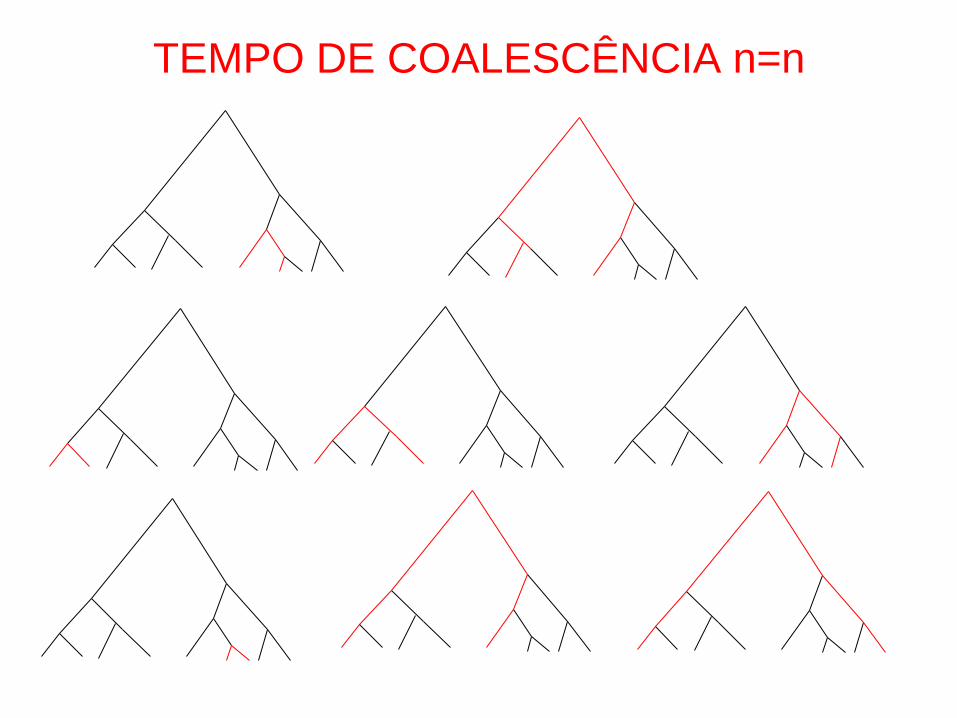
TEMPO DE COALESCÊNCIA n=n

TEMPO DE COALESCÊNCIA n=n
E (tMRCA com n=2) = 2N
Número de pares possíveis= n (n-1) / 2
Tempo médio para a primeira coalescência
numa amostra de n linhagens:
1/ [(n (n-1) / 2)] . 2N = 4N / n(n-1)

TEMPO DE COALESCÊNCIA n=n
T9
T8T7T6T5T4
T3
T2
4N / 9 (9-1) = 1/36 . 2N4N / 8 (8-1) = 1/28 . 2N4N / 7 (7-1) = 1/21 . 2N4N / 6 (6-1) = 1/15 . 2N4N / 5 (5-1) = 1/10 . 2N4N / 4 (4-1) = 1/6 . 2N4N / 3 (3-1) = 1/3 . 2N
4N / 2 (2-1) = 1 . 2N
0,78 * 2NTOTAL = E(tMRCA)=3,56 N = 4N [1- (1/n)]

TEMPO DE COALESCÊNCIA n=n
• E (tMRCA)=4N [1- (1/n)]
• n
• E (tMRCA)=4N
• Ex:
• n=2; E (t)=2N
• n=10; E (t)=3,6N
• .
• .
• .
• .
0
20
40
60
80
100
n
E(t)
4 N

TEMPO DE COALESCÊNCIA
• Propriedades médias
2N
2N
4N

TEMPO DE COALESCÊNCIA
• Aleatoridade
Stable
GrowingContracting

TEMPO DE COALESCÊNCIA
• Consequências
• Inutilidade de aumento da amostra: a estrutura da genealogia é basicamente determinada a partir da altura em que já há poucos ancestrais
• A inclusão da bifurcação mais antiga na amostra é obtida com um número surpreendentemente baixo de indivíduos:
• (n-1)/(n+1) é a probabilidade de incluir o MRCA numa amostra de tamanho n
• A informação obtida a partir de um único gene é muito limitada.
• É melhor aumentar o número de loci analisados do que o número de amostras em cada locus.

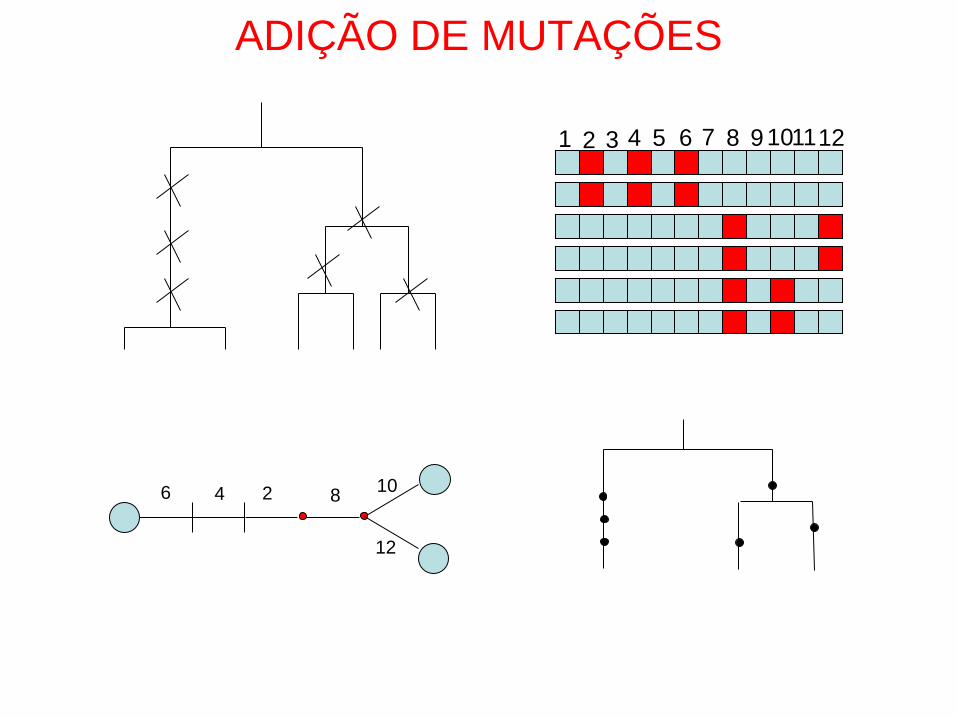
ADIÇÃO DE MUTAÇÕES

ADIÇÃO DE MUTAÇÕES
• Os padrões de diversidade genética observados
reflectem a sobreposição de mutações numa genealogia

ADIÇÃO DE MUTAÇÕES-ISM
• Cada mutação ocorre numa posição diferente de
uma sequência de DNA: ISM
t t + x t + x + y
ADIÇÃO DE MUTAÇÕES
246 810
12
1 2 3 4 5 6 7 8 9101112

ADIÇÃO DE MUTAÇÕES
Exemplo de aplicação do programa GENETREE

MEDIDAS SUMÁRIAS DE DIVERSIDADE GENÉTICA
t
2 t
E() =2 t
E(t)=2 N
=4N
ou
= 4NNº Médio de diferenças
entre um par de
sequências =
E() =2 E(t)
= taxa de
mutação por
locus por
geração

Comprimento total da árvore L = 4T4 + 3 T3 + 2 T2
Nº de mutações (posições
com segregação) numa
amostra de tamanho n = S
T4
T3
T2
MEDIDAS SUMÁRIAS DE DIVERSIDADE GENÉTICA
n
i
iiTL2
1
1222
14
1
4
)1(
4)()(
n
i
n
i
n
i
n
i
i
iN
i
N
ii
NiTiELE
1
1
1
1
114)(
n
i
n
i iiNSE
Nº esperado de mutações
1
1
1/
n
i
w
iS
Diapositivo 23

MEDIDAS SUMÁRIAS DE DIVERSIDADE GENÉTICA
Exemplo numérico
S=6
θ= 6/(1/1 + 1/2 + 1/3 + 1/4 + 1/5)
θ =2,63
1
1
1/
n
i
w
iS

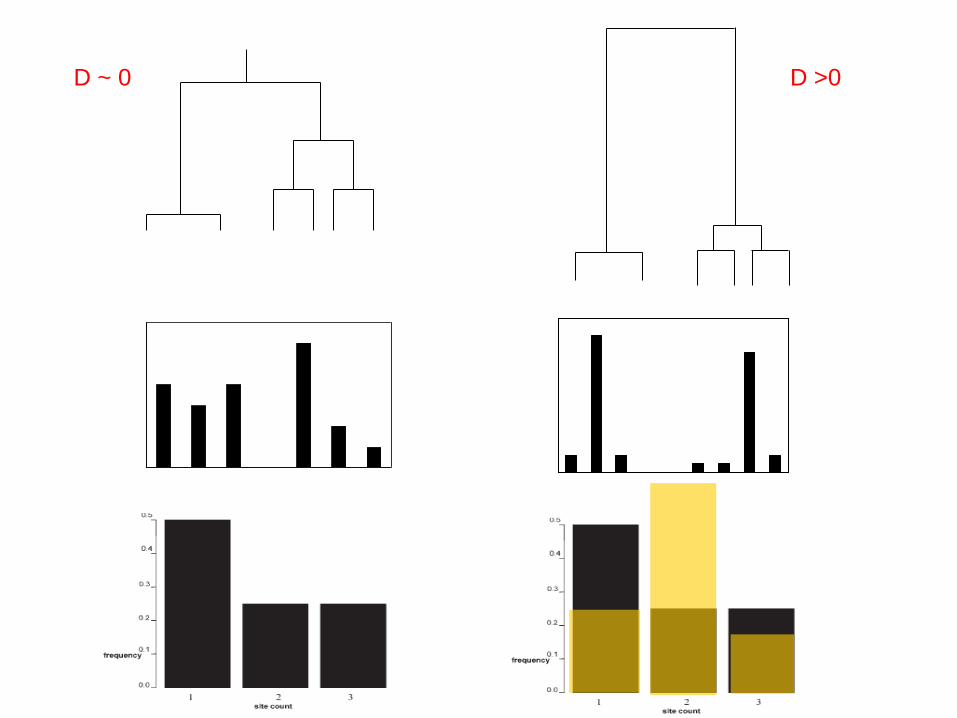
VARIAÇÃO NO TAMANHO
POPULACIONAL

Crescimento da população

N
Topologia das genealogias
t
N
t

Stable
GrowingContractingEstável
(maior aleatoridade)
Crescimento
(menor aleatoridade)
• Aleatoridade das genealogias

Stable
GrowingContracting
Distribuição de mutações

6
5
4
3
2
1
1 2 3
Espectro de frequências
Crescimento populacional

6
5
4
3
2
1
1 2 3
6
5
4
3
2
1
1 2 3
Espectro de frequências
Crescimento populacional

Crescimento populacional

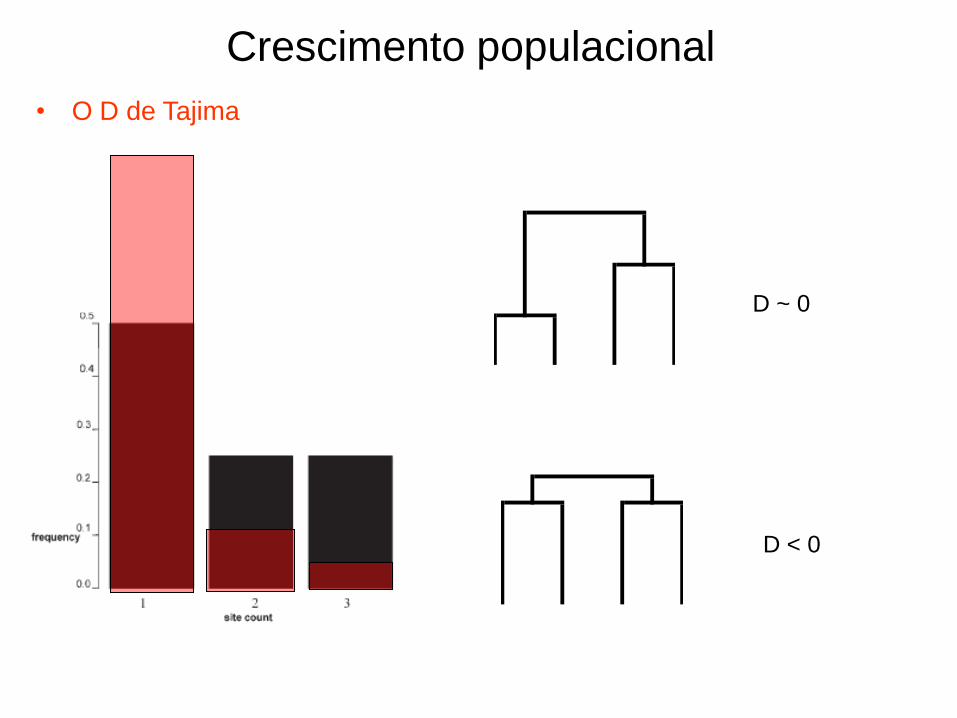
• O D de Tajima
Crescimento populacional
1
1
1/
n
i
w
iS = 2,73
1
1
1/
n
i
w
iS = 2,73
= θ = 2,25 = θ = 1,875
)(
48,0
)(
73,225,2
)(
sdsdsdD
)(
855,0
)(
73,2875,1
)(
sdsdsdD
• O D de Tajima
Crescimento populacional
D ~ 0
D < 0

6
5
4
3
2
1
1 2 3
Teste de D
,, S Simulações/WF,, , S
D obs
D esp
Crescimento populacional

Número de diferenças entre pares de sequências comparadas (mismatch distributions).
Média=
Crescimento populacional

Crescimento populacional

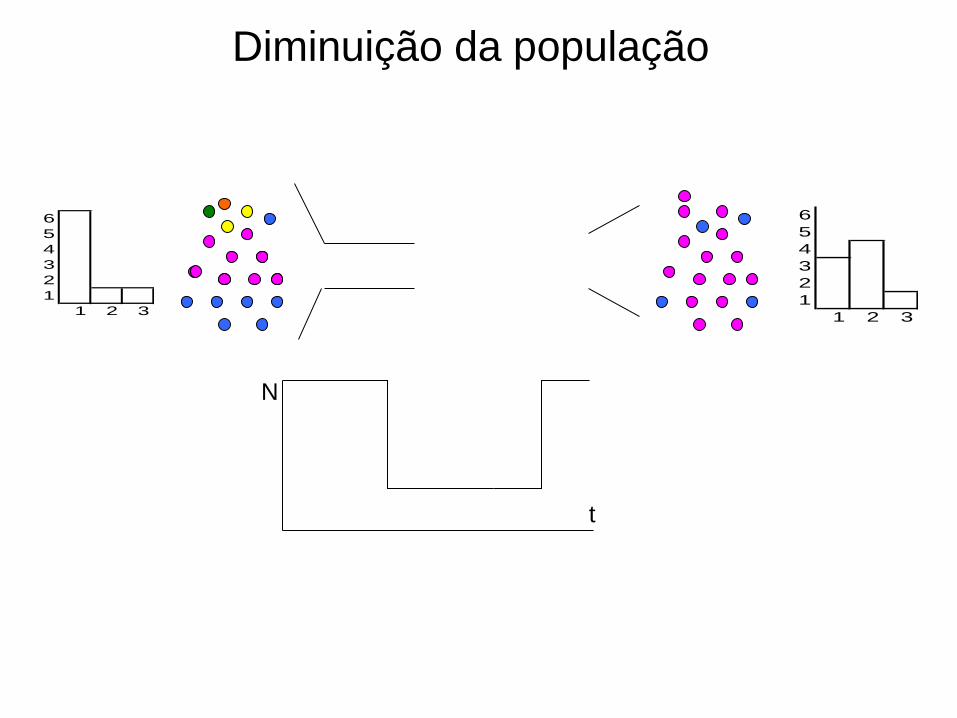
Diminuição da população

N
Topologia das genealogias
N
tt
t
N
6
5
4
3
2
1
1 2 3
6
5
4
3
2
1
1 2 3
Diminuição da população

Alterações no tamanho

Subdivisão populacional

Subdivisão da população


Fst
Subdivisão da população

PANMIXIASUBESTRUTURAÇÃO
Subdivisão populacional
D ~ 0 D >0

Alterações no tamanho e subdivisão