solos e paisagem - ministério do meio ambiente 3.pdf · brasília, df. raimundo paulo barros...
TRANSCRIPT

73
Solos e paisagem
Capítulo 3Capítulo 3Capítulo 3Capítulo 3Capítulo 3
Influência da história,solo e fogo nadistribuição edinâmica das
fitofisionomias nobioma do Cerrado
Capítulo 3Capítulo 3Capítulo 3Capítulo 3Capítulo 3
Influência da história,solo e fogo nadistribuição edinâmica das
fitofisionomias nobioma do Cerrado
FOTO
: A
LDIC
IR S
CA
RIO
TE
FOTO
: A
LDIC
IR S
CA
RIO
TE
Raimundo Paulo Barros HenriquesDepartamento de Ecologia
Universidade de BrasíliaBrasília, DF.
Raimundo Paulo Barros HenriquesDepartamento de Ecologia
Universidade de BrasíliaBrasília, DF.

74
Lima & Silva

Produção hídrica
75
INTRODUÇÃO
O bioma do Cerrado é provavelmentea maior savana do mundo, ocupandoaproximadamente 2.000.000 km2 noBrasil central, mais áreas disjuntas nosoutros biomas adjacentes. Uma dasprincipais questões sobre o bioma doCerrado é a determinação dos fatoresresponsáveis pela sua distribuição e adinâmica das suas fitofisionomias.
Frost et al. (1986) indicou quatrofatores, principais responsáveis pelospadrões e processos das comunidadesde savanas: água, nutrientes, fogo eherbivoria. Para as savanas da regiãoNeotropical, foram incluídos juntamentecom os três primeiros fatores o clima eeventos históricos (Sarmiento &
Monastério, 1975). A herbivoria tempouca importância nas savanas da regiãoNeotropical, devido à pequena biomassade ungulados.
Este capítulo, propõe que aocorrência e a dinâmica dos diferentestipos de fitofisionomias no bioma doCerrado resultam principalmente dainfluência de três fatores: história, soloe fogo. Um modo de abordar estaquestão é observar quais os padrões eprocessos que ocorrem nasfitofisionomias que podem e não podemser explicados usando estes fatores.
A seguir as idéias que serãoexaminadas ao longo deste trabalho:

76
Henriques
(i) A ocorrência de áreas disjuntascom vegetação de cerrado sensulato nos biomas adjacentespode ser atribuída a uma maiordistribuição geográfica da suaárea contínua no Brasil central,no passado. Nesse caso, asatuais áreas disjuntas seriamremanescentes desta distri-buição original. Com amudança do clima para maisúmido, as áreas com cerradosficaram isoladas em outrosbiomas;
(ii) Parte das diferenças observadasentre as fitofisionomias nocerrado sensu lato pode serexplicada pela profundidade eumidade do solo. Normal-mente, a densidade e a alturada vegetação lenhosa aumen-tam proporcionalmente a essesfatores;
(iii) O terceiro fator importante é ofogo, que tem ampla ocorrênciano bioma do Cerrado, provo-cando uma série de modifica-ções na estrutura da vegetação.
A influência do fogo na dinâmicadas fitofisionomias do Cerrado é umprocesso complexo ainda poucoconhecido. Após uma perturbação pelofogo pode ocorrer uma fase de imigraçãode espécies, com crescimento no númerode indivíduos e de área basal, sendoseguida de uma fase de homeostase, comequilíbrio nas taxas de imigração eextinção, recrutamento e mortalidade,respectivamente (Hallé et al., 1978).Altas taxas de imigração de espécies, derecrutamento de indivíduos e incrementode biomassa sugerem que algumas áreascom fisionomia de cerrado sensu strictoe provavelmente, campo sujo, sãocomunidades fora do equilíbrio, estandoatualmente em uma fase inicial de
crescimento sucessional (Henriques &Hay, 2002). O tempo para umacomunidade em fase inicial decrescimento atingir a fase de equilíbrio(homeostática) na ausência do fogo vaidepender de outros fatores ecológicoscomo: disponibilidade de água enutrientes no solo e distância da fontede propágulos.
Foram propostos por Pivello &Coutinho (1996) e Meirelles et al. (1997)modelos que, sugerem a evoluçãosucessional das fisionomias abertas paraas fisionomias fechadas do cerrado sensulato, em função de vários fatoresambientais. Nesses modelos, asformações abertas (campo limpo, camposujo, etc.) são consideradas formasderivadas do cerradão e florestasestacionais, pela ação do homem (ex.fogo, pastoreio), para onde a vegetaçãoinvariavelmente converge na ausência deperturbações humanas.
Portanto, o conhecimento dahistória do solo e do fogo é fundamentalpara se conhecer a distribuição e adinâmica das fitofisionomias no biomado Cerrado. Este capítulo sintetiza oconhecimento atual sobre a influênciadesses fatores, na sua distribuição epropõe um modelo para explicar adinâmica das fisionomias do cerradosensu lato, em função do fogo e dosfatores edáficos.
TERMINOLOGIA E DEFINIÇÕESBÁSICAS
O bioma do Cerrado se distribuicomo área contínua no Brasil central ecomo áreas disjuntas em outros biomas(Figura 1), como na floresta Amazô-nica, Caatinga, floresta Atlântica,Pantanal e floresta de Pinheiro do sul doBrasil (Rizzini, 1979; Cole, 1986; Furley

77
História, solo, fogo e fitofisionomias
& Ratter, 1988; Prance, 1996). O conceitode bioma empregado aqui, refere-se aoconjunto das unidades fisionômico–estrutural da vegetação que ocorrem naregião do Cerrado, além das áreasdisjuntas em outros biomas. Esteconceito é semelhante ao usado porOliveira-Filho & Ratter (2002), mas parauma conceituação diferente, vejaCoutinho (2000). A vegetaçãopredominante do bioma do Cerrado éformada por um mosaico heterogêneode fisionomias vegetais, com asformações campestres em umaextremidade e as formações florestais emoutra extremidade, formando umgradiente de altura-densidade (Eiten,1972; 1982).
Embora existam diferenças entre osautores, usando a altura e a densidadede plantas lenhosas, podemos ordenaras fisionomias vegetais em quatro tiposprincipais (conhecidas como cerradosensu lato): campo limpo; campo sujo;
cerrado sensu stricto, cerradão (Figura2). Este gradiente forma um continuumvegetacional, não havendo limitesdefinidos entre uma fisionomia e outra,portanto, formas intermediárias podemocorrer entre elas. Apenas porconveniência, foram reconhecidos algunstipos predominantes de fitofisionomiase que serão usados ao longo dessetrabalho. Em função das característicasestruturais, foram reconhecidos quatrotipos fisionômicos do cerrado sensu lato:campo limpo é a fisionomia com a maisalta cobertura de gramínea; campo sujoapresenta uma alta cobertura degramíneas e uma baixa cobertura dearbustos; o cerrado sensu strictoapresenta uma menor cobertura degramíneas, e uma maior coberturaarbustivo-arbórea e o cerradão é umaformação florestal que apresentaausência de cobertura de gramíneas e amaior cobertura arbórea. No gradientede cerrado sensu lato, o cerradãoapresenta algumas espécies de arbustos
Figura 1
Distribuição geográficado bioma do Cerrado
no Brasil. As áreasdisjuntas nos outros
biomas adjacentes sãoindicadas.

78
Henriques
e árvores restritas a esta fisionomia,como a árvore Emmotun nitens (Furley& Ratter, 1988), sendo aqui usada comoindicadora da fisionomia de cerradão.Essa classificação dos tipos fisionômicosé aplicável, principalmente, para a regiãodo Brasil central
O campo limpo, incluído nestegradiente, é a fisionomia que ocorresobre solos Litossólicos, rasos (~30 cmde profundidade) (Eiten, 1978, 1979,1984) não se refere, portanto, ao campoúmido ao lado das matas de galeria(Goldsmith, 1974). Estes campospossuem uma flora distinta com baixaafinidade florística com a flora herbáceado cerrado sensu lato (Araújo et al.,2002).
A floresta estacional pode ocorrer emdiferentes partes do gradientefisionômico de vegetação de cerradosensu lato. Esta fisionomia apresentaestreita afinidade florística com ocerradão (Rizzini 1963; 1979; Ratter etal., 1971; 1973, 1977; 1978a; 1978b;Heringer et al., 1977; Oliveira-Filho &Ratter, 1995). A floresta estacional podeocorrer, no topo dos interflúvios sobre
solos Latossolos férteis, derivados derochas alcalinas ou nas vertentesinferiores aluviais sobre solosPodzólicos, derivados de rocha calcária,ao lado da mata de galeria (Ratter et al.,1978a; 1978b; Eiten 1978; 1984).
Existem outras formações vegetaisno bioma do Cerrado, com estruturas efisionomias similares às do gradientefisionômico do cerrado sensu lato, masdiferenciam-se pela composição florísticae determinantes edáficos (ex. camposrupestres, campos úmidos, mata degaleria; Eiten, 1982) e que não serãotratadas neste capítulo.
A ORIGEM DA VEGETAÇÃODISJUNTA DO CERRADO SENSU
LATO
A ocorrência de áreas isoladas comvegetação de cerrado sensu lato, emoutros biomas como, a florestaAmazônica, Caatinga, floresta Atlânticae floresta de Pinheiro no sul do Brasil(Figura 1), levou vários autores aproporem, que no passado houve umadistribuição mais ampla, da área
Figura 2
Diagrama de bloco dadistribuição dasfisionomias de cerradosensu lato em relação àprofundidade do solona vertente de um vale.

79
História, solo, fogo e fitofisionomias
contínua do bioma do Cerrado no Brasilcentral, (Hueck, 1957; Ab’Saber, 1963;1971; Rizzini, 1979; Cole, 1986; Filho,1993; Prance, 1996). Segundo essesautores, a distribuição mais extensa dobioma do Cerrado, seria decorrente deum clima mais seco que teria favorecidoa distribuição da sua vegetação nopassado.
A hipótese de uma distribuiçãopleistocênica para as áreas disjuntas doscerrados é baseada em dois tipos deevidências (Gottsberg & Morawetz, 1986;Prance, 1996): (1) A similaridadeflorística entre as áreas disjuntas doscerrados com a flora da sua área contínuade ocorrência no Brasil central, e (2) obaixo nível de endemismo de espéciesnas áreas disjuntas da Amazônia e dafloresta Atlântica. O teste desta hipóteserequer o registro de polens noQuaternário que indiquem a ocorrênciada flora do cerrado sensu lato, nas áreasintermediárias entre a região contínua dobioma do Cerrado do Brasil central e asáreas disjuntas nos outros biomas.Vários estudos detectaram a ocorrênciade polens de Curatella americana e deoutras espécies do cerrado sensu lato,em áreas atualmente com floresta dePinheiro e floresta Atlântica no sudestee sul do Brasil (Ledru et al., 1996; 1998;Behling, 1998; Behling & Hooghimstra,2001). Estes resultados indicam que avegetação do bioma do Cerrado do Brasilcentral se expandiu além do seu limiteatual leste, sudeste e sul. As áreasdisjuntas de cerrado sensu latoatualmente isoladas na floresta Atlânticae floresta de Pinheiro, na região Sul eSudeste, são remanescentes destadistribuição mais extensa no passado(Hueck, 1957; Behling, 1998). Aexpansão das florestas úmidas, emdireção à área central do bioma doCerrado, pode ter ocorrido aproxima-damente nos últimos 1.000 anos A. P.
no sudeste e sul do Brasil (Behling &Hooghiemstra, 2001), o que indica umisolamento recente destas áreas.
A hipótese da distribuição do biomado Cerrado, na área da florestaAmazônica durante os períodos maissecos do final do Pleistoceno e Holoceno,para explicar as ocorrências das áreasdisjuntas de cerrado sensu lato nestebioma ainda é controversa (Colinvaux,1979; 1997; Colinvaux et al., 1996). Asevidências baseadas na presença depolens, indicadores da ocorrência devegetação de cerrado sensu lato,demonstram que para as áreasatualmente com florestas úmidas nolimite sudoeste e sul da Amazônia, estavegetação esteve presente em váriosperíodos no final do Pleistoceno (65.000A.P., 49.000 A. P., 41.000 A. P., 23.000A. P., 13.000 A. P.) (Behling &Hooghiemstra, 2001; van de Hammem& Hooghiemstra, 2000).
DETERMINANTES EDÁFICOS DASFISIONOMIAS DO CERRADO SENSU
LATO
O gradiente fisionômico devegetação no cerrado sensu latoapresenta uma variação inversa docomponente lenhoso (densidade, altura)e do componente herbáceo, dominadopor gramíneas (Goodland, 1971;Goodland & Ferri, 1979). Esta variaçãofisionômica - estrutural da vegetação foicorrelacionada com a fertilidade do solo(Goodland & Pollard, 1973, Lopes & Cox,1977), ocorrendo a maior densidade ealtura de plantas lenhosas onde o soloapresentava maior fertilidade. Noentanto, vários estudos encontraramresultados que não corroboram aexistência desta correlação (Gibbs etal.,1983; Oliveira Filho et al., 1989;Moreira, 2000; Ribeiro et al., 1982;

80
Henriques
Ruggiero et al., 2002). Diferenças deescala e de metodologia podem explicar,em parte, os resultados conflitantesencontrados por esses autores.
Relativamente, espera-se maiorfertilidade onde a vegetação é mais altae densa, como no cerradão e florestaestacional, devido à maior contribuiçãoda matéria orgânica para o solo nestasfisionomias. Devido à maior capacidadeda matéria orgânica reter nutrientes, ossolos das fisionomias com maiorcobertura vegetal (cerrado, cerradão)tornam-se mais férteis do que aquelescom menor cobertura (campo limpo,campo sujo). Isto não indica queoriginalmente as áreas com formaçõesde maior cobertura possuíssem solosmais férteis.
Diferenças na fertilidade do soloentre fisionomias foram registradas paradois subtipos de cerradão (Furley &Ratter, 1988), os distróficos, de baixafertilidade e os mesotróficos, de maiorfertilidade, principalmente naconcentração de cálcio. Estascaracterísticas nutricionais estavamassociadas também a diferençasflorísticas, com as espécies do primeirosubtipo classificadas como calcífugas eas do segundo como calcífilas. Noentanto, a grande similaridade florísticados cerradões mesotróficos com asflorestas estacionais (Ratter et al., 1978;Oliveira Filho & Ratter, 1995), podeindicar que ambos pertençam ao mesmotipo de unidade florístico-fisionômica.
A floresta estacional ocorre em soloscom maior fertilidade (Ratter et al.,1978a), associada a afloramentos derochas básicas. A distribuição deste tipode fisionomia independente do gradientevegetacional do cerrado sensu lato éconsistente com a sua associação aossubstratos ricos em rochas básicas. Istopode ser verificado na comparação das
diferenças de duas característicasedáficas de 47 amostras de solos, para aregião core dos cerrados em Goiás (Krejciet al, 1982). Observa-se que, as florestasestacionais ocorrem em solos com maiorconcentração de nutrientes do que asfisionomias de cerrado sensu lato(Figura 3). A saturação média de basesem solos de floresta estacional foi maior(55,6 ± 8,7) do que em solos doscerrados (18,4 ± 14,7). Outra diferençaobservada foi na razão ki, o valor médiopara os solos do cerrado sensu lato foimenor (1,5 ± 0,8) do que para os solosda floresta estacional (2,1 ± 0,9) (Figura3). A razão ki (razão molecular do SiO
2
para Al2O
3) mede o grau de
latossolização e indica a maturidade dosolo. Quanto maior o grau delatossolização mais jovem é o solo emaior o valor de ki. Independentementeda rocha matriz do solo, baixos valoresde ki estão associados com baixos
Figura 3
Distribuição dos valoresde saturação de bases(%) e razão ki (razãomolecular de SiO
2 para
Al2O
3, veja texto), nas
áreas com cerrado sensu
lato e florestasestacionais no Brasilcentral. A caixa para95% dos valores, indicaa média (linha contínua)a mediana (linhapontilhada) e o desviopadrão.

81
História, solo, fogo e fitofisionomias
conteúdos de nutrientes. Com base nolevantamento de solos do Estado de SãoPaulo, foi proposto por Eiten (1972), queo cerrado sensu lato ocorria apenas emsolos que apresentassem um valor de kiinferior a 1,8 e que, onde o soloapresentasse baixo conteúdo denutrientes (ex. Latossolo Vermelho -Amarelo textura arenosa e Regossolo),apenas cerrado sensu lato era observado.
A Figura 3 mostra que para a regiãodo Brasil central, embora, existasobreposição nos valores de ki entre ocerrado sensu lato e a floresta estacional,o valor máximo de ki foi de 1,9 para95% dos valores, o qual foi inferior aovalor para as florestas estacionais (2,8).O valor máximo para o cerrado sensulato observado no Brasil central, foipróximo ao valor de 1,8 registrado parao cerrado sensu lato para o Estado deSão Paulo (Eiten 1972).
Todos estes resultados sugeremfortemente que as fisionomias de cerradosensu lato diferentemente da florestaestacional, estão associadas a solos degrande maturidade, e altamenteintemperizados, como indicado pelosbaixos valores de ki, o que resultou emsolos com baixo conteúdo de nutrientese, na maioria dos casos, também comalta saturação de alumínio. Comosugerido por Eiten (1972), parece que oconteúdo de nutrientes, expresso pelasoma de bases, e o valor de ki são osmelhores fatores edáficos para separaro cerrado sensu lato da florestaestacional.
O primeiro modelo explicativo dasdiferenças fisionômicas para a vegetaçãoprimária do bioma do Cerrado foirealizado por Eiten (1972). Nestemodelo, são indicados três fatores paraexplicar esta diferenciação: pro-fundidade, drenagem e fertilidade do
solo. Pelo exposto anteriormente, ficaevidente que a fertilidade não explica asdiferenças entre as fisionomias docerrado sensu lato. As evidências para ainfluência da profundidade do solo navariação das fisionomias do cerradosensu lato são baseadas nos resultadosde Eiten, (1972, 1978, 1979, 1982, 1984,1994) e Oliveira Filho et al. (1989).Devido ao baixo conteúdo de nutrientes,os aumentos da densidade e da alturada vegetação da fisionomia de cerradãosão limitados pela profundidade do solo.Apenas em uma profundidade maior, osolo possuiu um estoque de nutrientesuficiente para o desenvolvimento deuma maior biomassa da vegetação.Paralelamente, resultados de Franco(2002) e Kanegae et al. (2000) mostramque o conteúdo de água na superfíciedo solo (até 30cm) das fisionomiasabertas para as fechadas no final da secaaumenta.
Baseado nos estudos acima e nasobservações do autor no Distrito Federal,uma distribuição hipotética dos tiposfisionômicos de vegetação do cerradosensu lato é apresentada, em função daprofundidade e do conteúdo de água dosolo, no fim da estação seca (Figura 4).As fisionomias são colocadas na suaposição relativa aos dois fatoresambientais e representam o potencialmáximo de desenvolvimento davegetação para as referidas condiçõesambientais. A figura mostra também que,a fisionomia que apresenta o mínimoimpedimento edáfico para odesenvolvimento de espécies arbóreas éo cerradão. Neste caso, além disso, ascondições para o estabelecimento edesenvolvimento de uma vegetaçãoarbórea (cerradão) nas fisionomiasabertas (ex. campo limpo e campo sujo),podem ser limitadas pelo conteúdo deágua na estação seca e pelo menorestoque de nutrientes.
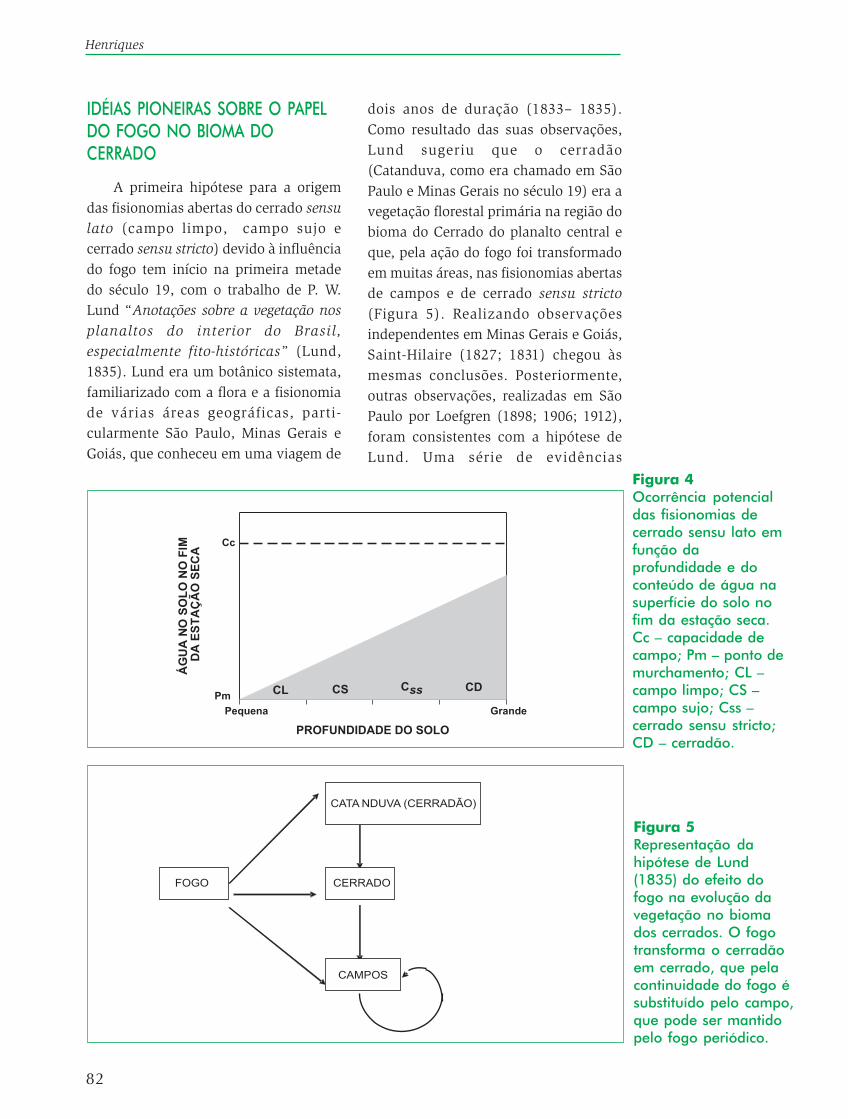
82
Henriques
IDÉIAS PIONEIRAS SOBRE O PAPELDO FOGO NO BIOMA DOCERRADO
A primeira hipótese para a origemdas fisionomias abertas do cerrado sensulato (campo limpo, campo sujo ecerrado sensu stricto) devido à influênciado fogo tem início na primeira metadedo século 19, com o trabalho de P. W.Lund “Anotações sobre a vegetação nosplanaltos do interior do Brasil,especialmente fito-históricas” (Lund,1835). Lund era um botânico sistemata,familiarizado com a flora e a fisionomiade várias áreas geográficas, parti-cularmente São Paulo, Minas Gerais eGoiás, que conheceu em uma viagem de
dois anos de duração (1833– 1835).Como resultado das suas observações,Lund sugeriu que o cerradão(Catanduva, como era chamado em SãoPaulo e Minas Gerais no século 19) era avegetação florestal primária na região dobioma do Cerrado do planalto central eque, pela ação do fogo foi transformadoem muitas áreas, nas fisionomias abertasde campos e de cerrado sensu stricto(Figura 5). Realizando observaçõesindependentes em Minas Gerais e Goiás,Saint-Hilaire (1827; 1831) chegou àsmesmas conclusões. Posteriormente,outras observações, realizadas em SãoPaulo por Loefgren (1898; 1906; 1912),foram consistentes com a hipótese deLund. Uma série de evidências
Figura 5
Representação dahipótese de Lund(1835) do efeito dofogo na evolução davegetação no biomados cerrados. O fogotransforma o cerradãoem cerrado, que pelacontinuidade do fogo ésubstituído pelo campo,que pode ser mantidopelo fogo periódico.
Figura 4
Ocorrência potencialdas fisionomias decerrado sensu lato emfunção daprofundidade e doconteúdo de água nasuperfície do solo nofim da estação seca.Cc – capacidade decampo; Pm – ponto demurchamento; CL –campo limpo; CS –campo sujo; Css –cerrado sensu stricto;CD – cerradão.

83
História, solo, fogo e fitofisionomias
observacionais, realizadas depois destesestudos, sugerem que, em muitas áreas,o cerrado se originou pela ação do fogono cerradão (Ab’Saber & Junior, 1951;Aubreville, 1959; Schnell, 1961; Eiten,1972, Rizzini, 1963, 1979).
ECONOMIA DE ÁGUA E O CARÁTERSECUNDÁRIO DAS FISIONOMIASABERTAS DO CERRADO SENSU LATO
A hipótese de Lund de que, pela açãodo fogo, o cerradão pode dar lugar àsfisionomias abertas do bioma do Cerrado(campo limpo, campo sujo, cerradosensu lato) foi aceita parcialmente porWarming (1892), que não achavapossível que este processo tivesseocorrido em tão grande extensãogeográfica. A ocorrência de fisionomiasabertas do cerrado sensu lato eraatribuída à limitação por água. Warming,que considerava o cerrado uma vege-tação adaptada à deficiência de água(xerofítica) (sensu Schimper 1903),supunha que as fisionomias abertas docerrado sensu lato, ocorriam devido aoperíodo seco e à precipitação menor queas das áreas de florestas (ex. florestaAtlântica). Esta hipótese foi refutada apartir dos resultados obtidos em umasérie de estudos por Felix Rawitscher ecolaboradores (Ferri, 1944; Rachid, 1947;Rawitscher et al., 1943; Rawitscher,1948; 1950, 1951). Os principaisresultados destes estudos mostraram: (1)que as espécies mostravam de pequenaa nenhuma adaptação fisiológica para aseca; (2) que a maioria das plantaslenhosas possuía sistemas radicularesprofundos tendo acesso às camadas desolo com água; e (3) que o solo comfisionomia de cerrado sensu strictoapresentava água disponível para avegetação o ano todo. Baseado nestas
evidências, Rawitscher et al. (1943,1948), refutam a hipótese de Warming(1892), de que a limitação por água eraa causa da ausência das florestas emáreas ocupadas com fisionomias abertasdo cerrado sensu lato, demonstrandotambém, que o conteúdo de água no solopoderia manter formações florestas.Rawitscher (1948) propõe que, em Emas,São Paulo, o cerrado sensu stricto,poderia ser uma vegetação secundáriaresultante da ação do fogo em umafisionomia florestal primária.Concluindo, considera que o solo nocerrado sensu lato, tem condições demanter formações florestais, talvez dotipo cerradão, e que as fisionomiasabertas poderiam ser formaçõessecundárias resultantes da ação do fogo(Rawitscher, 1950; 1951).
IMPACTO DO FOGO NAVEGETAÇÃO DO CERRADO SENSU
LATO
O fogo é um drástico agente deperturbação na vegetação do bioma doCerrado com grande impacto nadinâmica das populações das plantas. Ofogo causa a diminuição da altura davegetação (Hoffmann & Moreira, 2002)e, uma mortalidade de plantas lenhosasvariando de 13 a 16%, dez vezes maiorem relação às áreas protegidas, incluindoárvores de 21cm de diâmetro e 8,5m dealtura (Sato & Miranda, 1996). O fogotambém tem um grande efeito nacomposição de espécies do cerradão,eliminando espécies características destafisionomia e sensíveis ao fogo como,Emmotum nitens, Ocotea pomaderroidese Alibertia edulis (Hoffmann & Moreira,2002).
A mortalidade das plântulas pelofogo é ainda maior (33% a 100%), o

84
Henriques
mesmo ocorrendo com as rebrotas decrescimento vegetativo (7% a 47%)(Hoffmann, 1996; 1999). Em áreas comaté um ano depois de queimadas, oestabelecimento das plantas é tambémdrasticamente reduzido (Hoffmann,1996). Esse efeito é maior nas espéciescaracterísticas de cerradão, comoAlibertia macrophyla, Pera glabrata eOcotea pomaderroides.
Com maior freqüência dequeimadas, as taxas anuais de cresci-mento populacional, favorecem asformas de crescimento menores(arbustos) em detrimento das maiores(árvores) (Hoffmann, 1999; Hoffmann &Moreira, 2002). O fogo também aumentaa importância da reprodução vegetativa,em relação à da reprodução sexuada(Hoffmann, 1998; 1999). Isto ocorre pelaestimulação, pelo fogo, da reproduçãovegetativa e do seu maior valor desobrevivência em relação à das plântulas.Além disso, algumas espécies de plantaslenhosas reduzem drasticamente aprodução de sementes ao nívelpopulacional nas áreas recentementequeimadas (Hoffmann, 1998). Isto sereflete na redução do banco de sementesdestas espécies nas áreas queimadas, em
relação ao das áreas protegidas(Andrade, 2002). Nas áreas queimadastambém ocorre um aumento daabundância das gramíneas e do seubanco de sementes (Miranda, 2002;Andrade, 2002). A maior abundância degramíneas pode diminuir drasticamentea sobrevivência de plântulas de espécieslenhosas (Heringer, 1971).
Baseado nos resultados obtidosnesses estudos até o momento, a Figura6 mostra um modelo geral descrevendoos efeitos do fogo na dinâmica davegetação do cerrado sensu lato. Estemodelo mostra as complexas relaçõesentre os principais processos internos,modificando a vegetação. Algumascaracterísticas deste modelo devem serressaltadas. Os processos mostrados naFigura 6 podem ocorrer em qualquervegetação submetida ao efeito do fogo,mas alguns processos são maisimportantes na vegetação dos cerrados,como exemplo, a reprodução vegetativae o rápido aumento da abundância degramíneas. A maior espessura das setasrepresenta a sua importância relativa aosoutros processos e também aimportância da dependência entre osprocessos. A magnitude do efeito dos
Figura 6
Esquema dos efeitosdo fogo nosprocessos quedeterminam afisionomia aberta navegetação doscerrados. As setasmais grossasindicam os principaisprocessos.

85
História, solo, fogo e fitofisionomias
processos na vegetação está nadependência da freqüência com queocorre o fogo (Hoffmann, 1996; 1998;1999). Além disso, os fatores externosnão foram considerados neste modelo,como o efeito da variação deprecipitação, a ocorrência de veranicos,herbivoria, geadas, ou fertilidade dosolo, os quais podem mudar aimportância relativa dos diferentesprocessos. Todos os processosapresentados na Figura 6 enfatizam ogrande impacto causado pelo fogo namodificação das fisionomias doscerrados, de fisionomia fechada paraaberta, principalmente no que se refereà modificação de fisionomias com maiordensidade/altura de lenhosas e baixaabundância de gramíneas (ex. cerradão)para uma fisionomia com baixa altura/densidade de lenhosas e alta coberturade gramíneas (ex. cerrado sensu stricto,campo limpo, campo sujo).
Através de simulações de modelospopulacionais, foi estimado que comuma freqüência de queima maior quequatro ou cinco anos, as populações dealgumas espécies de árvores não podemse manter no cerrado sensu lato(Hoffmann, 1998; 1999). Nas condiçõestípicas de queimadas nos cerrados, deuma vez a cada dois anos (Eiten, 1972),as fisionomias mais fechadas doscerrados (ex. cerradão), podem estarsendo substituídas por fisionomias maisabertas (ex. campo limpo, campo sujo,cerrado sensu stricto), com drásticasmodificações na composição de espécies.Nestas fisionomias abertas, sãofavorecidas as plantas não sensíveis aofogo, que se reproduzem vegeta-tivamente. As alterações na composiçãode espécies que acompanham estasubstituição podem estar diminuindodrasticamente a diversidade dascomunidades vegetais do cerrado sensu
lato, como observado por Moreira(2000).
O CARÁTER SUCESSIONAL DASFISIONOMIAS ABERTAS DOSCERRADOS
Coutinho (1982, 1990) realizou umasérie de observações que mostraram queas fisionomias abertas dos cerrados,aumentavam de altura e densidade, coma proteção contra o fogo. No cerradosensu lato, a proteção contra o fogoresulta em um progressivo aumento davegetação lenhosa (Henriques & Hay,2002; Hoffmann & Moreira, 2002).Portanto, onde as fisionomias abertas docerrado (campo limpo, campo sujo,cerrado sensu stricto) não sãodeterminadas por limitação edáfica(Figura 4), mas resultantes da ação dofogo em fisionomias mais fechadas, a suaproteção contra o fogo deve permitir aevolução sucessional em direção àfisionomia primária mais fechada.
Em um gradiente fisionômicoiniciando em campo sujo e indo até ocerradão, no Brasil central, Moreira(2000) mostrou que, depois de 18 anosde proteção contra o fogo, as áreasprotegidas apresentavam aumentosignificativo no número de plantaslenhosas e na riqueza de espécies, emrelação às áreas não protegidas. Algumasespécies arbóreas do cerradão, comoBlepharocalix salicifolius e Sclerolobiumpaniculatum, apresentaram maiorabundância no cerradão protegido dofogo do que no queimado enquantoespécies características como Emmotumnitens e Ocotea pomaderroides (Furley &Ratter 1988), apenas foram encontradasno cerradão protegido do fogo.
Estudando a dinâmica daspopulações de plantas lenhosas de um

86
Henriques
cerrado protegido do fogo, Henriques &Hay (2002) encontraram fortesevidências que suportam a hipótese deque o cerrado sensu stricto pode ser umacomunidade fora do equilíbrio, tendouma natureza sucessional. Considerandoa extensa ocorrência e a alta freqüênciadas queimadas no bioma do Cerrado épossível que esta hipótese possa seraplicada para uma grande área, ocupadaatualmente com as fisionomias abertasdo cerrado sensu lato. Esta hipótese écorroborada pelos resultados deHoffmann (1999), que mostram que adiminuição da freqüência de fogo, podepermitir o crescimento líquido positivode populações de árvores, aumentandoa densidade e cobertura de lenhosas.
Se a vegetação do cerrado sensu lato,protegido do fogo, incrementa emdensidade e riqueza de espécies, quaisseriam as trajetórias sucessionais paraas fisionomias dos cerrados? Em umaanálise de agrupamento de fisionomiasqueimadas e protegidas do fogo, Moreira(2000) encontrou maior similaridadeflorística entre as áreas protegidas decampo sujo e cerrado sensu strictoqueimado, e cerrado sensu strictoprotegido com cerradão queimado. Estesresultados sugerem uma seqüênciasucessional do tipo campo sujo – cerrado– cerradão.
Usando fotografias aéreas, Duriganet al., (1987) analisaram o compor-tamento das fisionomias do cerradosensu lato após 22 anos de proteçãocontra o fogo em Assis, São Paulo. Seusresultados mostraram que a densidadee a altura da vegetação das fisionomiasabertas evoluíram para uma fisionomiaflorestal mais densa, de porte mais altoapós a proteção contra o fogo. Apresença na fisionomia mais madura dePlatypodium elegans e Machaeriumacutifolium, espécies características da
floresta estacional seca (Pennington etal., 2000; Ratter et al., 1978a,1978b),sugerem que esta fisionomia pode tersido a vegetação primária nesta área. Domesmo modo um penúltimo estágio quefoi identificado na área, apresenta asespécies: Bowdichia virgilioides eCaryocar brasiliense, características docerradão (Heringer et al., 1977; Ratter,1971, 1991). Esses resultados sugeremque, as áreas inicialmente comfisionomias abertas no estágio de camposujo, foram substituídas por cerradosensu stricto, as áreas com cerrado sensustricto por cerradão e as com cerradãopor floresta estacional, quando a áreafoi protegida do fogo.
CONSIDERAÇÕES FINAIS
Do que foi apresentado anterior-mente, podemos observar que naausência ou baixa freqüência do fogo,os diferentes tipos de vegetação nogradiente fisionômico podem serresultantes de condições edáficas. E quecada um dos diferentes tiposfisionômicos é o estágio mais maduroque a vegetação pode alcançar em cadaposição no gradiente edáfico. Nessesentido, em termos sucessionais, nocerrado existem vários estágios finais desucessão para a mesma condiçãoclimática. Esta idéia é consistente com oconceito de clímax-gradiente deWhittaker (1953), onde ocorre umacontinuidade espacial dos diferentestipos de comunidades clímax (gradientefisionômico), variando paralelamentecom o gradiente ambiental, e nãonecessariamente diferentes comunidadesclímax discretas separadas, como noconceito de policlímax. Eiten (1972)também considerou o bioma Cerradodentro do conceito clímax-gradiente deWhittaker.

87
História, solo, fogo e fitofisionomias
Assim cada um dos tipos fisio-nômicos é considerado aqui como umtipo de clímax. Na ocorrência do fogo,todos os tipos fisionômicos sofrem umprocesso de regressão para umafisionomia (estágio) mais aberta, comdesenvolvimento do estrato inferiordominado por gramíneas e diminuiçãodo componente lenhoso arbustivo –arbóreo (Figura 6). Com uma altafreqüência de queima, espécies arbóreassensíveis ao fogo não conseguem manteruma taxa positiva de crescimentopopulacional, particularmente asespécies arbóreas do cerradão (Figura 6).
Figura 7
Modelo conceitual desucessão e regressão
das fisionomias doscerrados, em funçãoda profundidade do
solo e do fogo noBrasil central.
Com a proteção contra o fogo pode seiniciar o processo de sucessão davegetação. Um modelo conceitualresumindo as seqüências de estágiossucessionais hipotéticos é apresentadona Figura 7. Segundo este modelo, asfisionomias abertas dos cerrados (campolimpo, campo sujo e cerrado sensustricto), ocorrendo em solos, profundose estando protegidas do fogo, podemapresentar o estabelecimento ecrescimento das populações de arbustose árvores (Henriques & Hay, 2002;Hoffmann & Moreira, 2002). Esteincremento na densidade é acom-

88
Henriques
panhado de aumento da cobertura ealtura da vegetação. Se não houverimpedimento edáfico (Figura 4), avegetação poderá evoluir até umafisionomia arbórea como o cerradão.
Considerando que a região do biomado Cerrado pode estar apresentando umafreqüência de fogo acima do regimenormal, devido à ação antrópica, éprovável que as fisionomias abertas, emparticular a de cerrado sensu stricto emáreas sem impedimento edáfico, estejamem diferentes estágios sucessionais apóso fogo, em uma fisionomia comvegetação mais desenvolvida.
Esta hipótese tem várias implicaçõespara estudos ecológicos de vegetação,principalmente em estudos fitosso-ciológicos comparativos entre áreas comcerrado sensu stricto. Considerando queestas áreas podem ter diferentes históriasdo fogo, e se acharem em diferentesestágios sucessionais, os resultados de
REFERÊNCIAS BIBLIOGRÁFICAS
Ab’Saber, A. 1963. Contribuição ageomorfologia dos cerrados. In Ferri, M.G. (Ed) Simpósio sobre o cerrado. p. 117-124. Editora da Universidade de SãoPaulo, São Paulo.
__________. 1971. A organização natural daspaisagens inter e subtropicaisbrasileiras. In Ferri, M. G. (Coord) IIISimpósio do cerrado. , 3. p. 1-14. EditoraEdgard Blücher & Editora daUniversidade de São Paulo, São Paulo.
__________ & M. C. Júnior, 1951.Contribuição ao estudo do sudoestegoiano. B. Geogr. 9: 123-138.
Andrade, L. A. Z. 2002. Impacto do fogo nobanco de sementes de cerrado sensustricto. Dissertação de Doutorado.Universidade de Brasília, Brasília, DF.
Araújo, G. M. de, A. A. A. Barbosa, A. A.Arantes, & A. F. Amaral, 2002.
análises de similaridade florística entreelas, podem apresentar nenhumacongruência espacial, por exemplo, comáreas próximas geograficamenteapresentando menor similaridadeflorística do que áreas mais afastadas,por se encontrarem em diferentesestágios sucessionais após o fogo.
Este capítulo apresenta a existênciade correspondência entre atributos eprocessos da vegetação em relação a trêsfatores fundamentais: história, solo efogo. Esses três fatores são consideradosos agentes que determinam a forma e aocorrência das fitofisionomias do cerradosensu lato e floresta estacional. Noentanto, embora ajudem precisamente aentender os resultados disponíveis nomomento, servindo para estabelecerfuturas prioridades de pesquisa, sãonecessárias mais informações de modoa aceitar ou rejeitar as hipóteses aquiapresentadas.
Composição florística de vereda nomunicípio de Uberlândia, MG. Rev. Bras.Bot. 4: 475-493.
Aubréville, A. 1959. As florestas do Brasil:estudo fitogeográfico e florestal. Anu.Bras. Econ. Flor. 11: 201-232.
Behling, I. 1998. Late Quaternary vegetationand climatic changes in Brazil. Rev.Palaeobot. Palynol. 99: 143-156.
__________ & H. Hooghiemstra, 2001.Neotropical savanna environments inspace and time: Late Quaternaryinterhemispheric comparison. InMarkgraf, V. (Ed) Interhemisphericclimate linkages. p. 307-323. AcademicPress, New York.
Cole, M. M. 1986. The savannas.Biogeography and geobotany. AcademicPress, Harcourt Brace JavanovichPublishers, Publishers. London, UK.

89
História, solo, fogo e fitofisionomias
Colinvaux, P. A. 1979. The ice-age Amazon.Nature 278: 399-400.
__________. 1997. The ice-age Amazon andthe problem of diversity. In NWO/Huygenslezing 1997. p. 7-30. TheHague: Netherlands Organization forSicentific Research.
__________. P. E. de, Oliveira, J. E. Moreno,M. C. Miller, & M. B. Bush, 1996. A longpollen record from lowland Amazonia:forest and cooling in glacial times.Science 274: 85-87.
Coutinho, L. M.1982. Ecological Effects offire in Brasilien cerrado. In Huntley, B.J. & B. H. Walker, (eds) Ecology oftropical savannas. p. 273-291. Springer-Verlag, Berlin.
__________. 1990. Fire in the ecology of theBrazilian cerrado. In Goldmmer, J. G.(Ed) Fire in the tropical biota. p. 82-105.Springer Verlag, Berlin.
__________. 2002. O bioma do cerrado. InKlein, A. L. (Org.) Eugen Warming e ocerrado brasileiro: um século depois. p.77-91. Editora da UNESP, São Paulo.
Durigan, G.; I. R. Saraiva; L. M. A. G.Garrido; M. A. O. Garrido & A. P. Filho,1987. Fitossociologia e evolução dadensidade da vegetação do cerrado.Assis. SP. Bol. Tec. Inst. Flor. 41: 59-78.
Eiten, G. 1972. The cerrado vegetation ofBrazil. Bot. Rev. 38: 201 – 341.
__________, 1978. Delimitation of thecerrado concept. Vegetatio 36: 169-178.
__________. 1979. Formas fisionômicas docerrado. Revista Brasileira de Botânica2: 139-148.
__________. 1982. Brazilian “savanas”. InHuntley, B. J. & Walker, B. H. (Eds)Ecology of tropical savanas. P. 24 – 47.Springer Verlag, Berlim.
__________. 1984. Vegetation of Brazilia.Phytoeconologia 12: 271-292.
__________ 1994. Vegetação do cerrado. InPinto, M. N. (Ed) Cerrado:caracterização, ocupação e perspectivas.EDUNB e SEMATEC, Brasília, DF.
Ferri, M. G. 1944. Transpiração de plantaspermanentes dos cerrados. Bol. Fac. Fil.Cien. Letr. Univ. São Paulo, Botânica,4:159-224. 1993.
Filho, A. 1993. Cerrados amazônicos: fósseisvivos? Algumas reflexões. Rev. Inst.Geol. 14: 63-68.
Franco, A. C. 2002. Ecophysiology of woodyplants. In Oliveira, P. S. & Marquis, R.S. Ecology and Natural History of aNeotropical savanna: The cerrados ofBrazil. P. 178-197. The University ofColumbia Press.
Frost, P., E. Medina, J. C. Menaut, Solbrig,O., Swift, M. & Walker, B. 1986.Responses of savannas to stress anddisturbance. IUBS Special Issue no. 10:1-82.
Furley, P. A. & J. A. Ratter, 1988. Soilresources and plant communities of thecentral Brazilian cerrado and theirdevelopment. J. Biogeog. 15: 96-108.
Gibbs, P. E.; H. F. Leitão Filho; & G.Shepherd, 1983. Floristic compositionand community structure in an area ofcerrado in SE Brazil. Flora 173: 433-449.
Goldsmith, F. B. 1974. Multivariate analysisof tropical grassland communities inMato Grosso, Brazil. J. Biogeog. 1: 111-122.
Goodland, R. 1971. A physiognomic analysisof the ‘cerrado’vegetation of centralBrasil. J. Ecol. 59: 411-419.
__________ & M. G. Ferri, 1979. Ecologia docerrado. EDUSP/Livraria Itatiaia EditoraLTDA., São Paulo.
__________ & R. Pollard, 1973. The braziliancerrado vegetation: A fertility gradient.J. Ecol. 6: 219-224.
Gottsberger, G. & W. Morawetz, (1986).Florístic, structural andPhytogeographical analysis of thesavannas of Humaitá (Amazonas). Flora178: 41-71.
Hallé, F., R. A. A. Oldeman, & P. B.Tomlinson, 1978. Tropical trees andforests. Springer Verlag, Berlim.
Henriques, R. P. B. & J. D. Hay, 2002. Patternsand dynamics of plant populations. InOliveira, P. S. & R. S. Marquis, Ecologyand Natural History of a Neotropicalsavanna: The cerrados of Brazil. p. 140-178. The University of Columbia Press.
Heringer, P. E. 1971. Propagação e sucessãode espécies arbóreas do cerrado em

90
Henriques
função do fogo, do cupim, da capina edo aldrim (inseticida). In Ferri, M. G.(Coord) III Simpósio do cerrado. P. 167-174. Editora Edigard Bücher & Editorada Universidade de São Paulo, SãoPaulo.
Heringer, E. P., G. M., Barroso, J. A Rizzo &C. T. Rizzini, 1977. A flora do Cerrado.In Ferri, M. G. (Coord) Simpósio sobreo cerrado base para a utilizaçãoagropecuária. p. 211-232. Editora daUniversidade de São Paulo & EditoraItatiaia, São Paulo.
Hoffmann, W. A. 1996. The effects of fireand cover on seedling establishment ina neotropical savanna. J. Ecol. 84: 383-393.
__________. 1998. Post-burn reproduction ofwoody plants in a neotropical savanna:the relative importance of sexual andvegetative reproduction. J. Ecol. 35: 422-433.
__________. 1999. Fire and populationdynamics of woody plants in aneotropical savanna: matrix modelprojections. Ecology 80: 1354-1369.
__________ & A. Moreira, 2002. The role offire in population dynamics of woodyplants. In Oliveira, P. S. & R. S. Marquis.Ecology and Natural History of aNeotropical savanna: The cerrados ofBrazil. p. 159-177. The University ofColumbia Press.
Hooghiemstra, H. 1997. Tropical rain forestversus savanna: two sides of a preciousmedal. NWO/Huygenslezing 1997. p.31-43. The Hague: NetherlandsOrganization for Sicentific Research.
Hueck, K.1957. Sobre a origem dos camposcerrados do Brasil e algumas novasobservações no seu limite meridional.Rev. Brasil. Geog. 19: 67-81.
Kanegae, M. F.; Braz, V. S. & Franco, A. C.2000. Efeitos da seca sazonal edisponibilidade de luz na sobrevivênciae crescimento de Bowdichia virgilioidesem duas fitofisionomias típicas doscerrados do Brasil Central. Rev. Brasil.Bot. 23: 457-466.
Krejci, L. C., Fortunato, F. F. & Corréa, P. R.S. 1982. Pedologia. Levantamento
exploratório de solos. In Brasil.Ministério de Minas e Energia. ProjetoRADAMBRASIL. Folha SD.23 Brasília,Rio de Janeiro. (Levantamento deRecursos Naturais, 29).
Ledru, M.P.; P. I. S. Braga; F. Soubiès; M.Fournier; L. Martin; K. Suguio & B.Turcq, 1996. The last 50,000 years inthe Neotropics (Southern Brazil):evolution of vegetation and climate.Palaeogeogr. Palaeoclimatol. Palaeoecol.123: 239-257.
__________ Salgado-Labouriau, M. L. & M.L. Lorscheitter, 1998. Vegetationdynamics in southern and central Brazilduring the last 10,000 yr B.P. Rev.Palaeobot. Palynl. 99: 131-142.
Löefgren, A. 1898. Ensaio para umadistribuição dos vegetais nos diversosgrupos florísticos no Estado de SãoPaulo. Bol. Com. Geog. Geol. 11: 1-50.
__________. 1906. La flore de St. Paul. Rev.Centr. Sci. Let. Art. Campinas 5: 53-61.
__________. 1912. Ensaio preliminar parauma phytogeographia brasileira. Rev.Centr. Sci. Let. Art. Campinas 11: 27-47.
Lopes, A. S. & F. R. Cox, 1977. Cerradovegetation in Brazil: An edaphicgradient. Agron. J. 69: 828-831.
Lund, P. W. 1835. Bemaerkninger overvegetation paa de indre hogsletter ofBrasilien, isaer i plantehistoriskhenseende Kgl. Danske Videnskab.Selsk. Skrifter 6: 145-188.
Meirelles, M. L., C. A. Klink, & J. C. S. Silva,1997. Um modelo de estado ytransiciones para el cerrado brasileño.Ecotropicos 10: 45-50.
Miranda, M. I. 2002. Efeitos de diferentesregimes de queimadas sobre acomunidade de gramíneas do cerrado.Dissertação de Doutorado. Universidadede Brasília, Brasília, DF.
Moreira, A. G. 2000. Effects of fire protectionon savanna structure in Central Brazil.J. Biog. 27: 1021-1029.
Oliveira Filho, A. T. & J. A. Ratter, 1995. Astudy of the origin of central Brazilianforests by the analysis of plant species

91
História, solo, fogo e fitofisionomias
distribution patterns. Edinb. J. Bot. 52:141-194.
__________ & Ratter, J. A. 2002. Vegetationphysiognomies and woody flora of theCerrado biome. In Oliveira, P. S. &Marquis, R. S. Ecology and NaturalHistory of a Neotropical savanna: Thecerrados of Brazil. p. 91-120. TheUniversity of Columbia Press.
__________; Sheperd, G. J.; F. R. Martins &W. H. Stubblebine, 1989. Environmentalfactors affecting physiognomic andfloristic variation in an area of cerradoin central Brazil. J. Trop. Ecol. 5: 413-431.
Pennington, R. T.; D. E. Prado & C. A. Pendry,2000. Neotropical seasonally dry forestsand Quaternary vegetation changes. J.Biog. 27: 261-273.
Prance, G. T. 1996. Island in Amazonia.Philos. Trans. Roy. Soc. B 351: 823-833.
Pivello, V. R. & L. M. Coutinho, 1996. Aquantitative successional model to assistin the management of Braziliancerrados. For. Ecol. Manag. 87: 127-138.
Rachid, M. 1944. Transpiração e sistemasubterrâneos da vegetação de verão doscampos cerrados de Emas. Bol. Fac. Fil.Ciênc. Letr. Univ. São Paulo, Botânica5: 1-135.
Ratter, J. A. 1971. Some notes on two typesof cerradão occurring in north easternMato Grosso. In Ferri, M. G. (Coord) IIISimpósio do cerrado. p. 100-102. EditoraEdigard Bücher & Editora daUniversidade de São Paulo, São Paulo.
__________. 1991. Transitions betweencerrado and forest vegetation in Brasil.In Furley, P. A.; J. Proctor & J. A.Ratter,(Eds) Nature and Dynamics of forest-savanna boundaries. p. 417-429.Chapman & Hall, London.
__________; G. P. Askew; R. F. Montgomery& D. R. Gifford, 1977. Observaçõesadicionais sobre o cerradão de solosmesotróficos no Brasil central. In Ferri,M. G. (Coord) IV Simpósio sobre ocerrado. p. 303-316. Editora daUniversidade de São Paulo, São Paulo.
__________. & J. F. Ribeiro, 1996.Biodiversity of the flora of the cerrado.
In Pereira, R. C. & Nasser, L. C. B. (Eds.)Anais do VIII Simpósio sobre o cerrado.p. 3-5. EMBRAPA, Planaltina.
__________; Richards, P. W., Argent, G. &Gifford, D. R. 1973. Observations on thevegetation of northeastern Mato GrossoI. The woody vegetation types of theXavantina-Cachimbo Expedition area.Philos. Trans. Roy. Soc. B 266: 449-492.
__________; Askew, G. P.; Montgomery, R.F. & Gifford, D. R.1978a. Observationson forests of some mesotrophic soils incentral Brazil. Rev. bras. Bot. 1: 47-58.
Ratter, J. A.; Askew, G. P.; Montgomery, R.F. & Gifford, D. R.1978b. Observationson the vegetation of northeastern MatoGrosso II. Forests and Soils of the RioSuiá-Missu area. Proc. Roy. Soc. B 203:191-208.
__________; Bridgewater, S.; Atkinson, R. &Ribeiro, J. F. 1996. Analysis of florísticcomposition of the Brazilian cerradovegetation II: Comparison of the woodyvegetation of 98 areas. Einburg Journalof Botany 53: 153-180.
__________; Ribeiro, J. F. & Bridgewater,S.1996. The Brazilian cerrado vegetationand threats to its biodiversity. Ann. Bot.80: 223-230.
Rawitscher, F. 1948. The water economy ofthe vegetation of the çampos cerrados’in southern Brazil. J. Ecol. 36: 16-32.
__________. 1950. O problema das savanasbrasileiras e das savanas em geral. Anu.Bras. Econ. Flor. 3: 32-38.
__________. 1951. Novos ensinamentos daecologia tropical. Cienc. Cult. 3: 232-242.
__________; Ferri, M. G. & Rachid, M. 1943.Profundidade dos solos e vegetação emcampos cerrados do Brasil meridional.An. Acad. Bras. Ci 15: 267-294.
Ribeiro, J. F.; Silva, J. C. & Azevedo, L. G.1982. Estrutura e composição florísticaem tipos fisionômicos dos cerrados esua relação com alguns parâmetros dosolo. In Anais do XXXII Congr. Nac. Bot.P. 141-156. Sociedade Botânica do Brasil,Terezina.
Rizzini, C. T. 1963. A flora do cerrado. InFerri, M. G. (Ed) Simpósio sobre o

92
Henriques
cerrado. p. 127-177. Editora daUniversidade de São Paulo, São Paulo.
__________. 1979. Tratado de Fitogeografiado Brasil. Aspectos florísticos. EditoraHucitec & Editora da Universidade deSão Paulo, São Paulo.
Ruggiero, P. G. C, M. A. Batalha, V. R. Pivello& S. T. Meirelles 2002. Soil – vegetationrelationships in cerrado (Braziliansavanna) and semideciduous forest,southeastern Brazil. Plant Ecol. 160: 1-16.
Saint-Hilaire, A. 1827. Voyage aux sourcesdu Rio de S. Francisco et dans laprovince de Goyaz. Berlin Imprimeur-Librarie, Paris.
__________. 1831. Tableau de la végétationprimitive dans la province de MinasGerais. Ann. Sci. Nat. 24: 64-83.
Sarmiento, G. & M. Monastério, 1975. Acritical consideration of theenvironmental conditions associatedwith the occurrence of savannaecosystems in tropical america. InGolley, F. B. & Medina, E. (Eds) Tropicalecological systems: trends in terrestrialand aquatic research. p. 223-250.Springer-Verlag, New York.
Sato, M. N. & H. S. Miranda, 1996.Mortalidade de plantas lenhosas do
cerrado sensu stricto submetidas adiferentes regimes de queima. InMiranda, H. S., C. H. Saito, & B. F. S. deSouza Dias, (Eds) Impactos dequeimadas em áreas de cerrado erestinga. p. 102-111. Departamento deEcologia, Universidade de Brasília,Brasília, DF.
Schimper, A. F. W. 1903. Plant-geographyupon a physiological basis. CaredonPress, Oxford.
Schnell, R. 1961. Le probléme deshomologies phytogeographiques entrel’Áfrique et l’Amerique tropicales. Mem.Mus. Hist. Nat. 11: 137-241.
Van der Hammen, T. & H. Hooghiesmstra,2000. Neogene and Quaternary historyof vegetation, climate, and plantdiversity in Amazonia. Quat. Res. 19:725-742.
Warming, E. 1892. Lagoa Santa. Et. Beitragtil den biologiske plantegeografi. D. Kgl.Danske Vidensk. Selsk. Skr. 6, RaekkeVI. (Trad. Do dinamarqués por A.Loefgren, 2a ed., São Paulo, Editora daUniversidade de São Paulo e EditoraItatiaia, 1973).
Whittaker, R. H. 1953. A consideration ofclimax theory: the climax as apopulation and pattern. Ecol. Monog.23: 41-78.