sÉrgio de araÚjo pereira - livros grátislivros01.livrosgratis.com.br/cp126494.pdf ·...
TRANSCRIPT

I�STITUTO OSWALDO CRUZ
Doutorado em Biologia Celular e Molecular
A importância de polimorfismo genético do Vírus Linfotrópico para células T
Humanas do tipo 1 (HTLV-1) na determinação da origem da epidemia e na
patogênese da mielopatia no Brasil
SÉRGIO DE ARAÚJO PEREIRA
Rio de Janeiro 2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
I�STITUTO OSWALDO CRUZ
Pós-Graduação em Biologia Celular e Molecular
SÉRGIO DE ARAÚJO PEREIRA
A importância de polimorfismo genético do Vírus Linfotrópico para células T
Humanas do tipo 1 (HTLV-1) na determinação da origem da epidemia e na
patogênese da mielopatia no Brasil
Dissertação apresentada com vistas à obtenção do título de Doutor em Ciências na área de Biologia Celular e Molecular. Trabalho realizado no Laboratório
Avançado de Saúde Pública (LASP), Centro de Pesquisas Gonçalo Moniz, FIOCRUZ/BA
Orientador (es): Prof. Dr. Bernardo Galvão Castro Filho Prof. Dr. Luis Carlos Alcântara Júnior
Rio de Janeiro 2009

iii
I�STITUTO OSWALDO CRUZ
Pós-Graduação em Biologia Celular e Molecular
SÉRGIO DE ARAÚJO PEREIRA
A importância de polimorfismo genético do Vírus Linfotrópico para células T
Humanas do tipo 1 (HTLV-1) na determinação da origem da epidemia e na
patogênese da mielopatia no Brasil
Orientador (es): Prof. Dr. Bernardo Galvão Castro Filho Prof. Dr. Luis Carlos Alcântara Júnior Aprovada em: 01 de dezembro de 2009 EXAMI�ADORES: Prof. Dra. Ana Carolina Paulo Vicente Prof. Dra. Rita Elizabeth Moreira Mascarenhas Prof. Dra. Sandra Rocha Gadelha
Rio de Janeiro, 01 de dezembro de 2009

iv
“Há um tempo em que é preciso abandonar as roupas usadas, que já têm a forma do nosso
corpo, e esquecer os nossos caminhos, que nos levam sempre aos mesmos lugares. É tempo da
travessia: e, se não ousarmos fazê-la, teremos ficado, para sempre, à margem de nós mesmos.”
(Fernando Pessoa)

v
Aos meus pais, Heldo e Lília (in memoriam), alicerces da minha formação moral e pelo
amor incondicional.

vi
Agradecimentos
A Deus, por ter me iluminado, pela força e coragem para continuar lutando apesar das
adversidades.
Aos meus pais, que sempre estiveram ao meu lado e nunca pouparam esforços para que mais esta
etapa da minha vida fosse realizada, pelo amor incondicional, por seus ensinamentos, apoio e
incentivo.
Aos meus familiares, pelo incansável apoio e incentivo para minha realização profissional e
pessoal.
A Dr. Bernardo Galvão Castro Filho uma grande admiração e muitos agradecimentos: pelo
acolhimento neste laboratório desde 1995 quando comecei iniciação científica, pela oportunidade
de continuar no LASP para desenvolver o Mestrado e Doutorado, pela orientação desta tese, e
pelos exemplos de sabedoria.
A Dr. Luis Carlos Alcântara Júnior, meus agradecimentos pela oportunidade de desenvolver o
doutorado, pela orientação desta tese e paciência durante este exercício.
A amiga Maria Eugênia Pondé, minha gratidão pelos conselhos de mãe que tanto me
incentivaram no decorrer da minha trajetória profissional, por compartilhar os momentos de
felicidades e de dificuldades e pela confiança em mim depositado.
A Dra. Fernanda Grassi, pelo carinho, incentivo e apoio.
A Dra. Flora Fernandes, por ter plantado as sementes das árvores e regado-as com muito amor,
pela amizade e carinho.

vii
A todas as pessoas da secretaria, Elisabeth Deliège, Claúdio Paulo dos Santos e Rodrigo Gaspar,
pelo companheirismo no trabalho, disponibilidade e apoio.
Aos amigos Rita Mascarenhas e Marco Mello, pela amizade e incentivo.
A Noílson Gonçalves e Jurema Carrilho, pela amizade e pelo apoio na realização deste trabalho.
Ao diretor do Centro de Pesquisas Gonçalo Moniz, Dr. Mitermayer Galvão dos Reis, pelo
incentivo, apoio e que através da competente administração conferiu-me para a realização deste
trabalho.
A todo o grupo do Centro Integrativo e Multidisciplinar do HTLV-1 e Hepatites virais da
Fundação Bahiana para o Desenvolvimento das Ciências, pela possibilidade de participar do
grupo e de realizar este trabalho.
Ao Prof. Dr. Antônio Andrade Filho, presidente da Fundação Bahiana de Neurologia e
Neurocirurgia, pelo acesso aos pacientes, obtenção das amostras para a realização deste trabalho
e contribuição na discussão do manuscrito 3.
Ao Curso de Pós-Graduação em Biologia Celular e Molecular/IOC/Instituto Oswaldo Cruz, no
qual realizei o doutorado.
A Cleide de Souza e Daniele Lobato, da secretaria do Curso da Pós-Graduação em Biologia
Celular e Molecular/IOC/Instituto Oswaldo Cruz, pelo apoio e disponibilidade.
A Ana Christina Caldeira, Adelvania de Araújo, Ana Maria Fiscina e Celeste Santana, pelo apoio
nas consultas bibliográficas e pela obtenção rápida de artigos na Biblioteca do CPqGM.

viii
A Eliana Garcia, pela amizade, pelas palavras de conforto nos momentos difíceis e apoio nas
consultas bibliográficas na Biblioteca do CPqGM.
A Nancy Azerêdo, por está sempre disposta a ajudar e pela revisão deste trabalho.
Aos funcionários do Centro de Pesquisa Gonçalo Moniz/FIOCRUZ, pelos serviços prestados.
Aos órgãos Financiadores CNPq, Capes e FAPESB.
A todos aqueles que diretamente ou indiretamente contribuíram para a realização deste trabalho.

ix
Lista de Figuras
Figura 1.1.2. Representação esquemática de um vírion HTLV-1 (adaptado de Alcântara, 2002). Detalhes sobre as proteínas do vírion são dados no texto. Figura 2.1.3. Organização genômica do HTLV-1. O genoma proviral está representado com as indicações das posições dos genes conhecidos e seus produtos proteicos. A escala representada acima corresponde ao genoma proviral total (Cann & Chen, 1996).
Figura 3.1.3. Processo de Transcrição do HTLV-1 via ativação pela proteína Tax. Figura 4.1.4. Ciclo de multiplicação viral do HTLV-1 (adaptado de Salemi, 1999).
Figura 5.1.7. Distribuição geográfica e origem do HTLV-1 (adaptado Alcântara, 2003).

x
Lista de Abreviaturas
ATK-1 Protótipo japonês do HTLV-1
ATLL Leucemia e Linfoma de células T do Adulto
ATLV Vírus da Leucemia de células T do Adulto
CA Capsídeo
CBP “CREB binding protein”
CREB “cyclic AMP response element binding protein”
DNA Ácido desoxirribonucleico
dNTPs Desoxirribonucleotideos trifosfato
dsDNA DNA dupla fita
ELISA “Enzime Linked Imuno Sorbent Assay”
enhancers Amplificadores da regulação gênica
gag “grup-specific antigen”
GLUT Transportador de glicose
GM-CSF fator estimulante de colônias de macrófago-granulócito
GP Glicoproteína
gp21 (TM) Glicoproteína 21 (Transmembranar)
gp46 (SU) Glicoproteína 46 (Superfície)
HAM Mielopatia Associada ao HTLV-1
HTLV-1 Vírus Linfotrópico de células T Humanas tipo 1
HTLV-2 Vírus Linfotrópico de células T Humanas tipo 2
HTLV-3 Vírus Linfotrópico de células T Humanas tipo 3
HTLV-4 Vírus Linfotrópico de células T Humanas tipo 4
Kb Kilobases
IL Interleucina
Linfócitos T CD4 Linfócitos T cluster differenciation 4
Linfócitos T CD8 Linfócitos T cluster differenciation 8
LTR “Long Terminal Repeats”

xi
3’LTR Região carboxi longo terminal repetitivo do ácido nucléico
5’LTR Região amino longo terminal repetitivo do ácido nucléico
LRT Razão de verossimilhança “Likelihood Ratio Test”
MA Matriz protéica
MHC-I Complexo principal de histocompatibilidade de classe I
ML Método de “Maximum Likelihood”
NC Nucleocapsídeo
NF-kB Fator de transcrição “Nuclear Factor kB”
NJ Método de “Neighbour Joining”
nt Nucleotídeos
OMS Organização Mundial de Saúde
ORF “open reading frame”
p15 (NC) Proteína 15 (Nucleocapsídeo)
p19 (MA) Proteína 19 (Matriz)
p24 (CA) Proteína 24 (Capsídeo)
pb Pares de bases
PBMC Células Mononucleares do Sangue Periférico
PCR Reação em cadeia da polimerase
pol Gene da polimerase
primers Sequência de oligonucleotídeos iniciadora da transcrição
Pro Protease
PTHrP Proteína relacionada ao hormônio da paratireóide
px Região que contém os genes reguladores tax e rex
rex Gene regulador da expressão do HTLV
Rex Proteína reguladora da expressão do HTLV
RNA Ácido ribonucléico
mRNA Ácido ribonucléico mensageiro
RT Transcriptase reversa

xii
SNP “Single nucleotide polymorphism”
ssDNA DNA fita simples
ssRNA RNA fita simples
STLV Vírus Linfotrópico de Células T de Símios
SU Glicoproteína de superfície
Taq Enzima polimerase
tax Gene de transativação
TATA box Região que antecede o sítio de transcrição
Tax Proteína de transativação
TGF-β Fator de crescimento tumoral β
TM Glicoproteína transmembrana
TNF-β Fator de necrose tumoral β
TRE Elementos conservados de resposta a Tax
TSP/HAM Paraparesia Espástica Tropical/ Mielopatia Associada ao HTLV-1
TSP Paraparesia Espástica Tropical

xiii
RESUMO
A importância de polimorfismo genético do Vírus Linfotrópico para células T
Humanas do tipo 1 (HTLV-1) na determinação da origem da epidemia e na
patogênese da mielopatia no Brasil
O Vírus Linfotrópico de Células-T Humanas do tipo 1 (HTLV-1) é o agente etiológico da Leucemia da Células T do Adulto, da Paraparesia Espástica Tropical/Mielopatia associada ao HTLV-1 (TSP/HAM) e Uveítes. No Brasil, a infecção apresenta-se em todas as regiões. Entretanto, esta infecção não possui uma distribuição uniforme e sua origem não está totalmente esclarecida. A cidade do Salvador apresenta a maior taxa de soroprevalência (aproximadamente 1,8%). Além disso, a correlação entre o subgrupo/subtipo do HTLV-1 e o TSP/HAM foi verificada somente na população japonesa. O mecanismo da transcrição proviral da sequência do HTLV-1 depende da proteína viral transativadora Tax que se liga na sequência (enhancer) de três repetições de 21- bp da região U3-LTR. O objetivo do presente trabalho foi tentar elucidar melhor a origem do HTLV-1 no Brasil e investigar se os polimorfismos na região U3-LTR poderiam estar associados com a susceptibilidade à TSP/HAM. Foram estudadas 214 sequências obtidas de pacientes infectados pelo HTLV-1 (110 assintomáticos e 104 sintomáticos), por meio da técnica da PCR e utilizando o software BioEdit. O protótipo ATK1 foi usado como sequência padrão. Neighbor-joining e análise filogenética máxima verossimilhança (ML) foram realizadas com PAUP * software. A análise filogenética da região LTR total das amostras dos doadores de sangue do Acre (2 isolados) e provenientes da cidade do Salvador (23 isolados) demonstrou que todas as amostras encontraram-se no subgroupo Transcontinental (A), subtipo Cosmopolita (a), com valores de bootstrap de 71% e 78% (p<0.001 para ML), respectivamente. A maioria das sequências oriundas de Salvador (n=18) ficou no cluster maior da América Latina (denominado beta) e 4 sequências ficaram inseridas em um cluster monofilético com outras sequências da América do Sul (denominado cluster alfa), com valor de bootstrap de 93%. Além disso, a soroprevalalência do HTLV-1 em doadores de sangue do Acre foi de 0,46% (1/219). Em relação à frequência das mutações pontuais da região U3-LTR destas cepas, nós identificamos uma inserção (C) entre as posições 8582 e 8583 que estava presente apenas no grupo assintomático (8/110, p = 0.004). Ainda, a frequência de mutações significativas C> T na posição 8583 (14/110; p = 0.001) e C> A na posição 8606 (9/110; p=0.012) foram observadas no grupo de assintomáticos. No grupo sintomático foi detectada uma inserção (G) entre as posições 8422 e 8423 (11/104, p=0.042), uma inserção (A), entre 8582 e 8583 (12/104, p=0.001) e uma deleção de um (C) na posição 8427 (17/104, p=0.002). Não foi observada correlação entre as mutações encontradas com a idade dos pacientes. Os resultados da análise filogenética sugerem que ocorreram introduções recentes, e que há uma relação mais próxima entre os isolados da América Latina com os do Sul da África do que em relação aos isolados do Oeste Africano e confirmam a introdução pós-colombiana do vírus em Salvador. Porém, não exclui uma introdução pré-colombiana. As mutações pontuais encontradas não estão associadas com os subgrupos do HTLV-1 e podem ter um impacto na transcrição viral e no desenvolvimento da TSP/HAM

xiv
ABSTRACT
The role of HTLV-1 genetic polymorphism in tracing the origin and dissemination
of the virus and in the pathogenesis of mielopathy in Brazil.
The Human T–Cell Lymphotropic Virus type 1 (HTLV-1) is the causative agent of Adult T-Cell Leukemia (ATL), Tropical Spastic Paraparesis/HTLV-associated myelopathy-1 (TSP/HAM) and uveitis. The correlation between the subgroup/subtype of HTLV-1 and TSP/HAM was observed only in the Japanese population. In Brazil, the infection is present in all regions although it is not equally distributed. The city of Salvador has the highest seroprevalence (approximately 1.8%). The origin of the virus in Brazil is not fully understood. The mechanism of transcription of HTLV-1 proviral sequence depends on Tax viral protein that transacts the three 21 bp repeated sequences in the U3-LTR region. In a further attempt to understand the possible origin of HTLV-1 infection in Brazil and the possible association of polymorphisms in the U3-LTR region with susceptibility to TSP/HAM, was studied in the sequences obtained from 214 HTLV-1 patients (110 asymptomatic and 104 symptomatic), trough PCR and using the BioEdit software. The ATK1 prototype was used as standard sequence. Neighbor-joining and maximum likelihood phylogenetic analysis were performed with PAUP* software. The phylogenetic analysis of the LTR region of all the samples of blood donors from Acre (2 isolates) and from the city of Salvador (23 isolates) showed that all samples are in the Transcontinental subgroup (A), Cosmopolitan subtype (a), with bootstrap values of 71% and 78% (p <0.001 for ML), respectively. Most sequences (n=18) from Salvador clustered within the larger Latin America cluster (named cluster beta) and 4 sequences clustered monophyletically with other sequences from South America (named cluster alpha), supported by a bootstrap value of 93%. Besides, the seroprevalence of HTLV-1 in blood donors of Acre was 0.46% (1/219). Regarding the frequency of mutations of U3-LTR region of these strains, an insertion (C) between the positions 8582 and 8582 was detected only in the asymptomatic group (8/110; p=0.004). Moreover, significant frequency of mutations C>T at position 8583 (14/110; p=0.001) and C>A at position 8606 (9/110; p=0.012) were observed in the asymptomatic group. In the symptomatic group, several insertions were detected: an insertion (G) between positions 8422 and 8423 (11/104; p=0.042), an insertion (A) between 8582 and 8583 (12/104; p=0.001) and a deletion at position 8427 (17/104; p=0.002). There was no correlation between the mutations found with age of patients. The results of phylogenetic analysis suggest that recent introductions occurred and that there is a closer relation between Latin American and South African strains than with West African HTLV-1 isolates and the post-Columbian introduction of this virus in Salvador is confirmed by the analysis though a pre-Columbian introduction is not excluded. It appears that the punctual mutations found are not associated with HTLV-1subgroup and may have an impact in the viral transcription and TSP/HAM development.

xv
SUMÁRIO
Lista de Figuras ix Lista de abreviaturas x RESUMO xiii ABSTRACT xiv 1. CAPÍTULO I 1
1.1. Introdução 1
1.2. Estrutura do HTLV-1 4
1.3. Estrutura Genômica do HTLV-I e Seus Produtos Proteicos 5
1.4. Ciclo de Multiplicação Viral do HTLV-1 9
1.5. Vias de Transmissão 11
1.6. Epidemiologia 11
1.7. Caracterização Filogenética 13
1.8. Manifestações Clínicas relacionadas ao HTLV-1 14
1.8.1. TSP/HAM 14 1.8.2. ATL 15
1.9. Patogênese da Infecção 15
1.9.1. Aspectos relacionados ao Hospedeiro 16 1.9.2. Aspectos relacionados ao vírus 16
2. RACIO�AL 18 3. OBJETIVOS 20
3.1. Geral 20
3.2. Específicos 20

xvi
4. CAPÍTULO II 21
The Close Relationship between South African and Latin American HTLV Type 1
Strains Corroborated in a Molecular Epidemiological Study of the HTLV Type 1
Isolates from a Blood Donor Cohort.
5. CAPÍTULO III 22
HTLV-1 Infection in donors from the Western Brazilian Amazon region: Seroprevalence
and Molecular study of viral isolates.
6. CAPÍTULO IV 23
Possible role of single nucleotide polymorphism in the U3-LTR region of HTLV-1 in the
pathogenesis of TSP/HAM in Brazil.
7. DISCUSSÃO 24 8. CO�CLUSÃO 33 9. REFERÊ�CIAS BIBLIOGRÁFICAS 34 10. A�EXO 50

1
1. CAPÍTULO I
1.1. Introdução
No ano de 1911, Rous e colaboradores, estudando a etiologia de neoplasias, descobriram
a existência de um vírus capaz de produzir sarcoma em galinhas. A partir dessa investigação,
inúmeros estudos experimentais foram realizados, todos concluindo pela existência de uma
numerosa quantidade de viroses causadoras de leucemias, linfomas e tumores sólidos em
animais. Posteriormente, a maioria desses vírus oncogênicos foi classificada como pertencente a
uma mesma família – os retrovírus.
Até a década de 70, não havia evidências da existência de vírus oncogênicos humanos.
Com a descoberta da transcriptase reversa (Baltimore, 1970) e o desenvolvimento de técnicas que
permitiram a obtenção de linfócitos T em cultura, utilizando a interleucina-2, vários
pesquisadores puderam estudar melhor esses vírus. Jarrett e colaboradores isolaram um retrovírus
associado a leucemias em felinos, em uma espécie de macaco (Charman et al., 1975) e em
bovinos (Ferrer et al., 1975). Finalmente, na década de 80, a partir de uma linhagem linfocitária
obtida de amostras de um paciente com linfoma cutâneo de células T, Poiesz e colaboradores
isolaram o primeiro retrovírus humano, vírus linfotrópico de células T humanas do tipo 1
(HTLV-1). Além disso, demonstraram que este vírus era o agente da leucemia, devido à presença
de anticorpos específicos existentes no soro destes pacientes, e a observação do DNA proviral
integrado ao DNA das células infectadas do hospedeiro (Poiesz et al., 1980).
Ao mesmo tempo, e de modo independente, estudos de pesquisadores japoneses
demonstraram a presença de um retrovírus do tipo C em células provenientes de um paciente com
leucemia de células T do adulto, descrita no Sudeste do Japão, conhecida como leucemia de
células T do adulto (Adult T cell Leukemia-ATL) (Uchiyama et al., 1977), e o agente infeccioso
causador desta patologia foi denominado de vírus da leucemia de células T do adulto (Vírus
Adult T Cell Leukemia-ATLV) (Hinuma et al., 1981). Posteriormente, verificou-se com análise
comparativa dos isolados ATLV e HTLV, que se tratava do mesmo vírus descrito por Poiesz
(Miwa et al., 1984).
Em 1982, o HTLV-2 foi isolado em células de um paciente com tricoleucemia, um tipo
raro de leucemia conhecida como leucemia de células T (Kalyanaraman et al., 1982). Este vírus

2
não está definitivamente associado a nenhuma doença humana até o momento, embora, cada vez
mais alterações neurológicas sejam descritas em indivíduos cronicamente infectados pelo HTLV-
2 (Jacobson et al., 1993; Peters et al., 1999; Biglione et al., 2003). Recentemente, foram isolados:
o HTLV-3 de uma paciente da África Central, da região de Camarões (Calattini et al., 2005) e o
HTLV-4 de um caçador, que relatava contato com primatas não humanos (Wolfe et al., 2005).
Entretanto, até o momento não foram associados a patologias.
Em 1983, um grupo de pesquisadores liderados por De Thé Guy realizou um inquérito
epidemiológico sobre doenças hematológicas malignas em Fort-de-France, Martinica. No curso
dos trabalhos, os pesquisadores encontraram um paciente com uma gamopatia oligoclonal e
paraparesia espástica crônica que apresentava anticorpos anti-HTLV-1, sugerindo a associação
deste vírus a esta alteração neurológica (Gessain et al., 1985). No ano seguinte, o HTLV-1 foi
associado a uma mielopatia espástica crônica frequente no Japão, que foi denominada mielopatia
associada ao HTLV-1 (HAM) (Osame et al., 1986). Vernant Jean-Claude e colaboradores (1988),
que haviam diagnosticado um grande número de pacientes com TSP ao longo de 14 anos, na
Martinica, demonstraram uma correlação entre essa última patologia e a presença de anticorpos
anti-HTLV-1 no soro desses pacientes. Esses resultados indicaram fortemente a associação do
HTLV-1 com uma doença do sistema nervoso. A relação entre TSP e HTLV-1 foi mais tarde
observada em muitas áreas do mundo (Roman and Osame, 1988). Finalmente, em 1989, um
consenso recomendou a denominação paraparesia espástica tropical/mielopatia associada ao
HTLV-1 (TSP/HAM), por se tratar de uma mesma doença (Osame, 1990). Mais tarde, foi
demonstrado que o HTLV-1 era também o agente causal de uveítes (Mochizuki et al., 1992).
Adicionalmente, diversas patologias inflamatórias como artrites, dermatite infectiva,
alveolites, polimiosites, cirrose biliar primária e síndrome de Sjögren também têm sido
relacionadas como sendo manifestações associadas à infecção viral (Morgan et al., 1989;
Mochizuki et al., 1996; La Grenade, 1996), sugerindo mecanismos patogênicos complexos
multifatoriais, envolvendo fatores genéticos do hospedeiro, fatores virais e ambientais.
Além disso, evidências clínicas sugerem que pode existir imunossupressão durante o
curso da infecção com grau distinto de morbidade e mortalidade associadas a inúmeras doenças
infecciosas como Tuberculose (Pedral-Sampaio et al., 1997; Marinho et al., 2005; Kristien et al.,
2007), hanseníase (Kashala et al., 1994), estrongiloidíase (Neva et al. 2001; Porto et al., 2002;

3
Kristien et al., 2007), e escabiose severa (Brites et al., 2002; Blas et al., 2005, Kristien et al.,
2007).
Até o momento, ainda não se sabe quais são os fatores que participam na patogênese das
doenças causadas ou associadas ao HTLV-1. Entre os fatores destacam-se aqueles relacionados
ao vírus, ao hospedeiro e com a resposta imune. Como por exemplo, subtipos virais, a carga
proviral e polimorfismo em região promotora de genes de citocinas.
Os subtipos virais se relacionam com a origem geográfica do vírus, sendo excelentes
marcadores de migração de populações humanas (Vidal et al., 1994). No entanto, não há
nenhuma associação clara, até o momento, com a patogênese das doenças (Renjifo et al., 1995).
Em relação à carga proviral, existe uma associação entre níveis elevados da mesma e
gravidade da doença, mas há relatos de indivíduos assintomáticos com carga proviral elevada.
Já em relação aos polimorfismos em determinadas regiões do vírus HTLV-1, existem
alguns trabalhos relacionando-os à patogênese da doença, mas os resultados são contraditórios.
Por exemplo, foi demonstrado através da análise da sequência do gene tax do HTLV-1 que as
mutações observadas se agrupavam em dois subgrupos: taxA e taxB. Os isolados dos subgrupos
taxA foram mais frequentes em pacientes com TSP/HAM do que em pacientes assintomáticos
(Furukawa et al., 2000). Além disso, mudanças de nucleotídeos nas sequências da região U3 do
LTR (Mukhopadhyaya & Sadaie, 1993) e troca de uma tirosina por uma citosina no gene tax
aumentam o risco do desenvolvimento da TSP/HAM (Renjifo et al., 1995). Por outro lado, em
um estudo em isolados brasileiros não foi encontrada nenhuma correlação entre a região tax A
com doença (Kashima et al., 2006).
No presente trabalho, estudamos o papel da região promotora LTR como marcador de
migrações de populações humanas, contribuindo assim, para um melhor entendimento da origem
do HTLV-1 no Brasil, principalmente em Salvador, Bahia, e Rio Branco, Acre. Além disso,
obtivemos evidências bastante sugestivas do papel de polimorfismos de único nucleotídeo (SNP-
single nucleotide polymorphism) na patogênese da TSP/HAM.

4
1.2. Estrutura do HTLV-1
O HTLV-1 está classificado na família Retroviridae, subfamília Oncovirinae, gênero
deltavirus. É um vírus que tem um nucleocapsídeo icosaédrico central de 80-100 nm de diâmetro
circundado por um envelope circular (Figura 1.1.2.). O vírion é visto como uma partícula
esférica de cerca de 100-140 nm de diâmetro por microscopia eletrônica. Um único complexo
proteico está presente na superfície do vírion, constituído de duas subunidades protéicas
glicosiladas: a proteína de superfície (SU) denominada gp46 e a proteína transmembrana (TM)
denominada gp21, que são importantes para a fusão com a célula alvo e posterior infecção. O
core interno é constituído de três proteínas: proteína do nucleocapsídeo (NC) ou p19, do capsídeo
(CA) ou p24 e da matriz (MA) ou p15. Neste core, um ácido ribonucleico (RNA) dimérico (35S
cada fita) de 8-9 kb (kilobases) está empacotado junto com outras proteínas importantes no
desenvolvimento do papel catalítico antes do ciclo de replicação viral: a transcriptase reversa, a
integrase e a protease. De acordo com o seu aspecto através da microscopia eletrônica, o HTLV-1
apresenta morfologia do tipo C: extracelular, núcleo eletro-denso central e brota da membrana
das células do hospedeiro.
Figura 1.1.2. Representação esquemática do vírion do HTLV-1 (adaptado de http://researchnews.osu.edu/archive/htlvil2pics.htm). Detalhes sobre as proteínas do vírion são dados no texto.

5
1.3. Estrutura genômica do HTLV-1 e seus produtos proteicos
Este retrovírus humano tem um genoma diplóide composto por duas fitas simples de
RNA de aproximadamente 8.500 pares de bases (bp), com uma estrutura genética similar aos
outros retrovírus, contendo os genes estruturais: gag (group-specific antigen), pro, pol
(polimerase) e env (envelope), além de uma sequência próxima à extremidade 3’LTR conhecida
como região X, a qual contém os genes reguladores tax e rex, responsáveis pela produção das
proteínas Tax e Rex, que determinam a ativação transcricional do vírus e regulação do (splicing)
do RNA mensageiro (RNAm), respectivamente (Cann and Chen, 1996) (Figura 2.1.3.).
Figura 2.1.3. Organização genômica do HTLV-1. O genoma proviral está representado com as indicações das posições dos genes conhecidos e seus produtos proteicos. A escala representada acima corresponde ao genoma proviral total (Cann & Chen, 1996).
O DNA proviral do HTLV-1 possui 9032 pb (Seiki et al., 1983). As extremidades do
genoma são flanqueadas por duas regiões repetidas, chamadas LTR (Long Terminal Repeats), que
contêm as regiões U3, R e U5, em ambas as extremidades do DNA proviral. Essas duas regiões
LTRs idênticas, localizadas nas extremidades 5´e 3,´ são essenciais na integração do DNA
proviral dentro do DNA cromossômico do hospedeiro e, também, na regulação transcricional do
genoma do HTLV. A LTR é a região mais divergente do genoma do HTLV-1, possuindo
elementos que são importantes para expressão dos genes virais. A região U3 é constituída por três

6
elementos essenciais: três repetições de 21 pares de bases, necessárias para a transativação
mediada pela proteína Tax (Brady et al., 1987) (Figura 3.1.3.); o TATA box, que antecede o sítio
de iniciação transcricional, e o sinal de poliadenilação. As regiões flanqueadoras são
extremamente variáveis e, portanto, fonte de informação em estudos de epidemiologia molecular.
Figura 3.1.3. Processo de Transcrição do HTLV-1 via ativação pela proteína Tax.
Durante o processo de transcrição, três espécies de mRNAs são transcritos do DNA
proviral do HTLV-1: uma fita de mRNA de 8,5 kilobases (Kb), transcrito da extremidade U3-R
na porção 5' LTR até R-U5 na extremidade 3´LTR utilizada para a síntese dos produtos dos genes
gag e pol; um mRNA de 4,2 Kb subgenômico feito a partir de um único processamento (single-
spliced) e que codifica o produto do gene env, e um mRNA de 2,1Kb subgenômico processado
pela remoção de dois íntrons e que codifica para as proteínas regulatórias Tax e Rex.
Região gag. A região gag está compreendida entre os nucleotídeos 824 a 2113 no
genoma viral no protótipo ATK-1 (Número de acesso Genbank J02029) (Seiki et al., 1983), e seu
final 3’LTR sobrepõe-se ao início da ORF que codifica para a protease. Esta região é inicialmente
traduzida como um precursor poliprotéico e, a clivagem dá origem às proteínas GAG maduras: a

7
proteína da matriz de 19 kDa (p19), a proteína do capsídeo de 24 kDa (p24) e a proteína do
nucleocapsídeo de 15 kDa (p15).
Região pro. A protease (PRO) é codificada pela sequência de nucleotídeos que
compreende a parte 3’LTR da região gag e a parte 5’LTR da região pol. Assim, a síntese da
protease é feita como parte do precursor poliproteico gag, acompanhado por desvio de leitura
ribossomal. A protease é responsável pelo processamento do produto gag maduro e pela sua
própria clivagem para gerar a molécula de protease madura encontrada nas partículas virais. A
região pro é responsável por codificar a protease viral. Os genes gag e pro expressam duas
poliproteínas precursoras, a p53GAG e a p76GAG-PRO, respectivamente. A protease viral
produzida pela p76GAG-PRO cliva os precursores da proteína GAG nas proteínas maduras: MA
(p19), CA (p24) e NC (p15) (Hattori et al., 1984; Nam et al., 1988).
Região pol. A região pol está localizada entre as posições 2520 a 5207 no genoma do
protótipo ATK1. A porção 5’LTR do gene pol codifica a transcriptase reversa, e a sequência
downstream codifica a integrase. Os produtos das regiões pro e pol constituem as atividades
enzimáticas dos retrovírus e sua síntese se faz pela extensão ocasional dos produtos de gag,
resultando na presença de um número muito menor destas enzimas, quando comparado com a
síntese das proteínas estruturais.
A transcriptase reversa (RT) é a enzima responsável pela síntese do DNA viral a partir
de seu genoma RNA e é, portanto, fundamental para a fase inicial do ciclo de replicação dos
retrovírus. Além desta atividade de DNA polimerase, ela possui uma atividade de RNAse H, a
qual é responsável pela remoção da fita de RNA da molécula híbrida DNA-RNA. No genoma do
HTLV-1, ela é codificada pela região pol entre os nucleotídeos 2497 a 5187. A transcriptase
reversa está presente no core da partícula viral e sua atividade se dá no citoplasma da célula
hospedeira.
A integrase, também codificada pela região pol, é a enzima que dá prosseguimento ao
ciclo de replicação dos retrovírus após a síntese do DNA viral pela transcriptase reversa. Ela é
responsável pela integração do DNA viral no genoma da célula hospedeira, através da clivagem
do DNA da célula e sua ligação com o DNA viral.
Região env. A região env, que vai do nucleotídeo 5203 a 6669 no genoma viral ATK1
(Seiki et al., 1983), codifica as proteínas do envelope viral, responsáveis pela entrada do vírus na
célula-alvo. A proteína precursora do envelope é clivada para formar os produtos maduros: a

8
glicoproteína de superfície de 46 kDa (gp46-SU) e a proteína transmembrana de 21 kDa (p21-
TM). Como no caso de outros retrovírus, ocorre associação não covalente entre as proteínas de
superfície (SU) e a proteína TM, de modo que a mesma ancora a proteína SU no envelope da
partícula viral (Delamarre et al., 1996), permitindo a ligação do vírus com o receptor no início da
infecção, resultando na penetração do core viral no citoplasma da celular.
As glicoproteínas do envelope são altamente conservadas entre os diferentes isolados de
HTLV-1. Entretanto, vários estudos têm sido realizados com intuito de investigar as regiões
gênicas envolvidas no processo de fusão celular. Sagara e colaboradores (1996) demonstraram
que peptídeos sintéticos contendo sequências específicas de aminoácidos em gp46 e gp21 foram
capazes de inibir formação de sincícios induzidos pela infecção do HTLV-1. As glicoproteínas do
envelope, por induzirem resposta humoral e celular, seriam candidatas para reprodução de uma
possível vacina para o HTLV.
Região X. Esta região tem início logo após o gene env. O mRNA viral na região X
contém pelo menos 4 fases de leitura aberta (ORFs I, II, III e IV) e as proteínas regulatórias virais
Tax e Rex são ambas codificadas pelo mesmo mRNA policistrônico, através das ORFs IV e III,
respectivamente. Tax é uma fosfoproteína nuclear que regula a transcrição do genoma proviral
indiretamente ao interagir com diferentes proteínas regulatórias celulares. A Tax também pode
induzir indiretamente a expressão de genes celulares (Franchini, 1995; Ferreira et al., 1997). Isto
pode ter um papel importante na transformação da célula infectada pelo HTLV e, portanto, na
patogênese caracterizada por esta infecção viral. A proteína Tax é essencial para a eficiência da
expressão viral (Sodroski et al., 1984) e desenvolve um importante papel na ativação de genes
celulares, tais como genes de citocinas (Siekevitz et al., 1987; Mori et al., 1998) e
protooncogenes (Fujii et al., 1988). Além disso, a proteína Tax é um alvo antigênico de linfócitos
T citotóxicos (CTL) na resposta ao HTLV-1 (Kannagi et al., 1992; Parker et al., 1992).
Diferenças no gene pX podem influenciar na carga viral em virtude de diferenças na habilidade
de transativação da proteína Tax ou pela diferença no reconhecimento dos CTL (Niewiesk et al.,
1995). Diferenças na atividade de transativação no gene da citocina pela proteína Tax também
pode influenciar no desenvolvimento da infecção. Diversos estudos mostraram que a proteína
Tax induziu o aumento da expressão de diferentes genes relacionados com o crescimento celular,
como os protooncogenes c-fos, c-myc e erg, fatores de crescimento ou seus receptores como IL-1
(interleucina 1), IL-2, IL-3, IL-6, IL-2Rα (cadeia α do receptor para IL-2), c-sis (PDGF, fator de

9
crescimento derivado de plaqueta), GM-CSF (fator estimulante de colônias de macrófago-
granulócito), TGF-β (fator de crescimento tumoral β), PTHrP (proteína relacionada ao hormônio
da paratireoide), vimetina (proteína do citoesqueleto), MHC-I (complexo principal de
histocompatibilidade de classe I), NF-κB (fator nuclear κB) e TNF-β (fator de necrose tumoral
β). O único gene até então descrito como tendo a sua transcrição inibida pela proteína Tax é o
gene que codifica para a polimerase β, uma enzima envolvida no reparo do DNA (Franchini
1995; Ferreira et al., 1997).
Rex também é uma fosfoproteína nuclear e que atua como um regulador pós-
transcricional do genoma do HTLV-1 ao controlar o processamento (splicing) do mRNA viral.
Rex tem um efeito positivo sobre a transcrição de ambos mRNAs (unspliced), que codificam as
proteínas Gag/Pol e (single-spliced), que codificam as proteínas do envelope. Ao mesmo tempo,
Rex tem um efeito negativo sobre o processamento e transporte dos mRNAs (double-spliced) que
codificam a própria proteína Rex, Tax e outros mRNAs com processamentos alternativos.
Portanto, Rex favorece o acúmulo de proteínas estruturais em detrimento das proteínas
regulatórias e um fino balanço entre a expressão e atividade de Tax e Rex pode ditar o estado de
replicação viral nas células infectadas. Tax e Rex não apenas regulam a expressão viral, mas
também podem interferir com as funções da célula hospedeira em diferentes níveis, afetando a
transcrição e tradução de vários genes celulares. Estes efeitos de Tax e Rex são relevantes na
patogênese das doenças associadas ao HTLV-1 (Franchini, 1995; Ferreira, 1997).
1.4. Ciclo de Multiplicação Viral do HTLV-1
O ciclo de vida do HTLV-1 é similar ao dos demais retrovírus, dependendo da presença
da enzima transcriptase reversa. Inicialmente, a partícula viral se liga à superfície celular, entre o
domínio de ligação do receptor (Receptor Binding Domain-RBD), na glicoproteína de superfície
do vírus e a molécula de GLUT-1 (Glucose transporter), que é um transportador de glicose, na
membrana celular. Foi demosntrado (Coskun et al., 2005) (Figura 4.1.4.). Essa interação
provoca modificações conformacionais na glicoproteína transmembrana, permitindo a fusão entre
as membranas e consequentemente a internalização do capsídeo viral no citoplasma da célula
hospedeira. A fita simples de RNA viral é, então, transcrita reversamente pela transcriptase

10
reversa em um DNA de fita dupla. Durante a transcrição reversa, o RNA genômico é removido
pela atividade RNAse H da transcriptase reversa viral. A fita dupla de DNA linear migra para o
núcleo e integra-se no genoma do hospedeiro pela ação da integrase viral (Seiki et al., 1984).
Uma vez integrado, o provírus utiliza a maquinaria celular para a transcrição primária do RNA
genômico. Parte do RNA viral sintetizado é processado para gerar o RNAm que será traduzido
nas proteínas virais apropriadas no citoplasma. Como último passo, o core viral é montado e o
vírus é liberado da superfície celular por um processo mais ou menos simultâneo (Cann and Chen
et al., 1996).
O HTLV-1 é um vírus que se replica pouco. A replicação viral in vivo ocorre mais
devido à expansão clonal das células infectadas, via mitose, que via transcrição reversa (Wattel et
al., 1995; Cimarelli et al., 1996).
Figura 4.1.4. Ciclo de multiplicação viral do HTLV-1 (adaptado de Salemi, 1999).

11
1.5. Vias de Transmissão
A transmissão do HTLV-1 ocorre, principalmente por três vias: via horizontal (contato
sexual), sendo a infecção mais frequentemente transmitida do homem para a mulher. Presume-se
que a infecção adquirida através da atividade sexual seja consequência dos linfócitos infectados
presentes no sêmen e na secreção vaginal (Kajiyama et al., 1986a); via vertical (da mãe para o
filho) por transmissão transplacentária, durante o parto e, predominantemente, pela
amamentação. Esta última é a principal via de transmissão do HTLV-1 em áreas endêmicas; via
parenteral, através transfusão de sangue contaminado e seus produtos, bem como uso de seringas
contaminadas (Dourado Andrade et al., 1998; Carneiro-Proietti et al., 2002). A transmissão
sanguínea está associada ao desenvolvimento mais rápido para TSP/HAM (Osame et al., 1986).
A transmissão do HTLV-1 é menos eficiente que a do vírus da imunodeficiência humana
(HIV) devido à baixa carga viral do HTLV-1, cuja infecção é dependente do contato célula-célula
(Kamihira et al., 1987; Bangaham et al., 2003).
1.6. Epidemiologia
Estima-se em 10-20 milhões o número de pessoas infectadas pelo HTLV-1 no mundo.
(Edlich et al., 2000). As principais áreas endêmicas do HTLV-1 são Japão, Caribe (Levine and
Blatter et al., 1997; Muller et al., 1996), América Central e do Sul, África sub-Saara (Gessain and
De Thé, 1996), Irã e Melanésia (Yamaguchi et al., 1994; Muller et al., 1996). A prevalência do
HTLV-1 nessas áreas endêmicas não é uniforme. O sul do Japão (Kyushu, Shikoku e Okinawa) é
uma das regiões mais endêmicas do mundo (Hinuma et al., 1982; Kajiyama et al., 1986b). Nesta
região a soroprevalência pode atingir 20% da população (Muller et al., 1996). O HTLV-1 foi
também encontrado em Hokkaido (norte do Japão) em uma comunidade denominada Ainu,
descendente de uma população aborígene (Ishida et al., 1985). A existência de infecções pelo
HTLV-1 na África Equatorial (Saxinger et al., 1984; Fleming et al, 1986; Delaporte et al., 1989;
Verdier et al., 1994), juntamente com a presença do HTLV-1 entre descendentes de africanos
residentes no Caribe e na América Latina, sugere que a África seria o reservatório primário desse
vírus (Vandamme et al., 1998). As infecções pelo HTLV-1 também foram registradas entre

12
diferentes tribos de pigmeus que habitam em Camarões, na República da África Central e
nordeste do Congo (Goubau et al., 1996; Mahieux et al., 1997; Salemi et al., 1998). Os casos de
HTLV-1 relatados nos Estados Unidos ocorrem em áreas endêmicas isoladas ou foram
encontrados entre os usuários de drogas (Lee et al., 1991). O HTLV-1 foi também isolado em
índios norte-americanos (Gessain et al., 1994), em aborígenes do Alasca (Davidson et al., 1993) e
em índios canadenses que apresentaram sintomas das doenças relacionadas ao vírus (Dekaban et
al., 1994). Além disso, o virus é endêmico em diversos países da América Central e do Sul,
incluido Honduras (De Rivera et al., 1995), Panamá (Reeves et al., 1990), Colômbia (Maloney et
al., 1989; Fujiyama et al., 1993), Venezuela (Merino et al., 1984), Bolívia (Tsugane et al., 1988),
Brasil (Bouzas et al., 1990; Maloney et al., 1992; Fujiyoshi et al., 1995; Salemi et al., 1998;
Talarmin et al., 1999), Perú (Gotuzzo et al., 1996; Johnson et al., 1988), Chile (Cartier et al.,
1993) e Argentina (Bouzas et al., 1990).
No Brasil, o HTLV-1 foi primeiramente descrito por Kitagawa e colaboradores (1986),
em uma comunidade japonesa residente na cidade de Campo Grande, Estado do Mato Grosso do
Sul. Os dados epidemiológicos são relativamente escassos e em sua maioria é descrição da
prevalência em populações específicas (doadores de sangue). Atualmente, estima-se que mais de
2,5 milhões de indivíduos estejam infectados pelo HTLV-1 (Carneiro-Proietti et al., 2002;
Catalan-Soares et al., 2004). A prevalência do HTLV-1 é heterogênea, sendo as maiores
observadas em: Salvador (1.8% na população geral, ou 0.094% em doadores de sangue), São
Luís (0.10%), Belém (0.091%), Rio Branco (0,08%), Recife (0.075%) em doadores de sangue
(Colin et al., 2003; Dourado et. al., 2003; Catalan-Soares et al., 2005). Provavelmente, esta
elevada prevalência em Salvador deve-se à população dessa cidade ter características sócio-
demográficas semelhantes às cidades africanas situadas abaixo do Saara (Galvão-Castro et al.,
1997). Vale à pena ressaltar que a prevalência em algumas tribos indígenas brasileiras também é
significativa, tais como 0.62% na tribo Wayampí (Shindo et al., 2002).

13
1.7. Caracterização Filogenética
Tem sido demonstrada uma estreita associação entre o polimorfismo genético dos
retrovírus e a sua origem geográfica. Por exemplo, os primeiros isolados do HTLV-1 do Japão e
do Caribe mostraram uma variabilidade na sequência do gene env menor que 3% (Seiki et al.,
1983; Malik et al., 1988; Gray et al., 1990). O isolamento subsequente, no Congo, de uma cepa
do HTLV-1 divergente, diferindo da cepa protótipo japonesa ATK1 em torno de 4% na região
env indicou que variantes mais distintas existem em diferentes regiões geográficas (Hahn et al.,
1984; Ratner et al., 1985). Este fato foi confirmado pela identificação de variantes do HTLV-1,
muito divergentes (divergência em torno de 8,5%) na Melanésia (Gessain et al., 1993).
O HTLV-1 se diferencia do vírus linfotrópico de células-T humanas do tipo 2 (HTLV-
2), principalmente no gene pX, sendo, no entanto, homólogos em cerca de 60%. O HTLV-1 e o
HTLV-2 têm propriedades biológicas similares e tropismo para linfócitos T (Hollsberg and
Hafler, 1993; Hall, 1994a; Hall et al., 1994b), sendo que o HTLV-1 infecta preferencialmente
linfócitos T CD4+, enquanto o HTLV-2 tem tropismo para linfócitos T CD8+ com efeito
hematológico diferente do HTLV-1 (Murphy et al., 1996).
Para o estudo da origem, disseminação e evolução dos retrovírus e das migrações de
populações humanas pode-se utilizar análises dos polimorfismos genéticos do HTLV-1, devido a
sua grande estabilidade genética e análises filogenéticas do DNA mitocondrial (Black et al.,
1997). A análise filogenética baseada no gene LTR classifica o HTLV-1 em sete subtipos: Ia ou
Cosmopolita, é endêmico em diferentes regiões geográficas na Europa, sul da América do Norte
e na América do Sul, incluindo o Brasil (Miura et al., 1994; Miura et al., 1997); Ib ou da África
Central, é endêmico na África Central (Hahn et al., 1984; Vandamme et al., 1994); Ic ou
Melanésio, é endêmico na Austrália e Papua Nova Guiné (Gessain et al., 1991; Bastian et al.,
1993) e Id, descrito como um novo subtipo isolado de pigmeus de Camarões e de um indivíduo
do Gabão (Chen et al., 1995; Mahieux et al., 1997); HTLV-1e, isolado de pigmeus na República
Democrática do Gongo (Salemi et al., 1998); HTLV-1f, de um indivíduo do Gabão e HTLV-1g,
recentemente isolado como um novo subtipo em Camarões, na África Central (Wolfe et al.,
2005). O subtipo Cosmopolita consiste de cinco subgrupos, disseminado em diferentes regiões
geográficas: A - Transcontinental, B – Japonês, é endêmico no Japão e em áreas isoladas da
América do Sul; C - Oeste da África e D - Norte da África e E- Negro do Peru. (Figura 5.1.7.).

14
Figura 5 1.7. Distribuição geográfica e origem do HTLV-1 (adaptado Alcântara, 2003).
No Brasil, estudos de caracterização molecular do HTLV-1 têm demonstrado
predominância do subtipo Cosmopolita, sendo a maioria dos isolados pertencentes ao subgrupo
Transcontinental (Segurado et al., 2002; Alcantara et al., 2003), e em proporção menor o
subgrupo Japonês (Yamashita et al., 1999; Alcantara et al., 2003).
1.8. Manifestações Clínicas relacionadas ao HTLV-1
1.8.1 TSP/HAM
A TSP/HAM é uma doença inflamatória crônica que lentamente debilita o sistema
nervoso central, tendo como consequência a paraparesia dos membros inferiores (Roman &
Osame, 1988). Dificuldade progressiva à marcha associada frequentemente a dor lombar,
incontinência urinária, parestesia em extremidades, obstipação intestinal, impotência sexual, e
cãibras são apresentações clínicas comuns relacionadas à doença. O início dos sintomas
neurológicos da mielopatia associada ao HTLV-1 ocorre, habitualmente, na quarta década de
vida, sendo incomum seu aparecimento abaixo dos 20 ou acima dos 70 anos. Portanto, os
indivíduos infectados pelo HTLV-1 permanecem assintomáticos por longo período, apresentando

15
apenas os anticorpos antivirais em altos títulos (Gessain and Gout, 1992). A prevalência da
TSP/HAM é de 1 a 5% em pacientes infectados pelo HTLV-1 (Kaplan et al., 1990). Existe
predomínio de acometimento das mulheres em comparação aos homens, sendo esta relação de
3:1 (Moreno-Carvalho et al., 1992). A desmielinização local atribuída à infiltração da célula
infectada no sistema Nervoso Central (SNC), e o desencadeamento de uma resposta inflamatória
crônica são algumas implicações relacionas à patogênese da TSP/HAM.
1.8.2. ATL
A ATL é uma doença linfo-proliferativo agressiva, de caráter endêmico, associada a
lesões ósseas, hipercalcemia e rápida progressão para óbito, caracterizada pela proliferação clonal
maligna de linfócitos T multilobulados (flower cells) (Uchiyama et al., 1997), que tem o seu
curso clínico classificado em 5 estágios: agudo, linfoma, crônico/smouldering, pré-leucêmico e
assintomático (Shimoyama et al., 1991; Yamaguchi & Takatsuki et al., 1993). Na sua forma
aguda leucêmica, a doença caracteriza-se por leucocitose intensa com a presença de (flower
cells), linfadenopatia, hepatoesplenomegalia rapidamente progressivas, envolvimento cutâneo,
pulmonar, da medula óssea, do trato intestinal e do sistema nervoso central. A hipercalcemia
ocorre com ou sem lesões osteolíticas. As células neoplásticas têm características
imunofenotípicas de células T (Uchiyama et al., 1997). A forma (smoldering) caracteriza-se pela
presença de um número relativamente pequeno de células leucêmicas circulantes, além de lesões
cutâneas e, ocasionalmente, pulmonares, manifestando-se de forma indolente, podendo persistir
por vários anos.
1.9. Patogênese da Infecção
Não há evidências de correlações entre a patogenicidade e os vários subtipos de HTLV-
1. Ainda não se sabe por que a infecção pelo HTLV-1 na maioria dos indivíduos resulta em uma
infecção assintomática, enquanto poucos desenvolvem doença. Também não está claro porque
alguns indivíduos desenvolvem TSP/HAM e outros ATLL.

16
1.9.1 Aspectos relacionados ao Hospedeiro
A forma mais simples de variação de DNA entre os indivíduos é a substituição de um
único nucleotídeo por outro. Esta variação é chamada polimorfismo de nucleotídeo único (SNP),
e ele surgem devido a ocorrências de mutações aleatórias. Acredita-se que diferenças ou
variações nos genomas entre dois indivíduos selecionados aleatoriamente é de aproximadamente
0,1% (Barkur et al., 2007). Diversos estudos comparativos sobre gêmeos idênticos e fraternais
(Martin et al., 1997) sugerem que o polimorfismo no DNA é um dos fatores associados com a
susceptibilidade para muitas doenças comuns, para traços humanos, como por exemplo, o cabelo
crespo, individualidade e diferença interindividual na resposta à droga. A variação na sequência
de DNA é também considerada como responsável pela evolução do genoma. Com base nessas
observações, foi proposto que as catalogações dos polimorfismos de DNA em diferentes
populações e de espécies diferentes poderiam contribuir para: o conhecimento do genoma com
base na susceptibilidade de um indivíduo em desenvolver doenças, como por exemplo, artrite
reumatóide, esclerose sistêmica, diabetes tipo II, para produzir com mais segurança e eficácia
medicamentos para os pacientes e compreender os processos evolutivos (Barkurt et al., 2007).
1.9.2. Aspectos relacionados ao vírus
Komurian e colaboradores, em 1991, tentaram correlacionar o desenvolvimento de
TSP/HAM e ATLL com variações específicas nos genes LTR, env e pX estudando um número
pequeno de indivíduos (4 japoneses, 2 de Costa do Marfim e 2 caribenhos), porém concluíram
que as duas distintas patologias não estavam relacionadas às mutações específicas nas sequências
analisadas. Em 1999, Beby-Defaux e colaboradores analisaram a sequência das regiões LTR e pX
de 15 indivíduos com diferentes patologias associadas ao HTLV-1 (4 com TSP/HAM e síndrome
de Sicca, 2 com síndrome de Sicca, 1 com TSP/HAM, 4 com ATL e 4 indivíduos
assintomáticos), porém não correlacionaram nenhuma mutação com o desenvolvimento dessas
patologias. Posteriormente, Furukawa e colaboradores (2000), através da análise da sequência do
gene tax do HTLV-1 de 61 pacientes com TSP/HAM, 55 ATLL e 62 assintomáticos,
classificaram as mutações observadas em dois subgrupos: taxA (sequências que apresentavam 4
mutações específicas) e taxB (sequências que não apresentavam tais mutações). Comparando a
análise filogenética do gene LTR com a análise filogenética do gene tax observaram que o
subgrupo taxA era relacionado ao subtipo Cosmopolita, subgrupo Transcontinental e o subgrupo

17
taxB era relacionado ao subtipo Cosmopolita, subgrupo Japonês. Analisando todos os isolados
dos subgrupos taxA e taxB verificaram que o taxA é mais frequente em pacientes com TSP/HAM
do que em pacientes assintomáticos. Além disso, foi feita a comparação do resultado da carga
proviral com as mutações taxA e taxB, porém não foi encontrada uma diferença significativa.
Portanto, estudos das propriedades moleculares e biológicas do HTLV-1 em indivíduos
infectados poderão fornecer informações que contribuirão para o melhor entendimento da
patogenicidade deste vírus.

18
2. RACIO+AL
A infecção pelo HTLV-1 é endêmica em várias partes do mundo (Mueller 1991; Gessain
and De Thé, 1996). No Brasil, esta infecção foi detectada em todas as regiões, mas a distribuição
não é uniforme. Em um trabalho recente classificaram-se as regiões do Brasil de acordo com as
taxas de prevalências em baixa (0,0/1000 a 3,3/1000); média (3,4/1000 a 6,6/1000) e elevada (6,7
a 10/1000) (Catalan Soares et al., 2005). Alguns estados do Norte (Pará e Amapá) e no Nordeste
(Maranhão, Bahia e Pernambuco) apresentaram taxas mais elevadas do que estados do sul e
centro-oeste. Nos estados do Amazônia e Acre a prevalência foi considerada média. A população
da cidade do Rio Branco é constituída de uma elevada taxa de miscigenação indígena e há poucos
estudos de caracterização molecular do HTLV-1.
Há pelo menos duas hipóteses para explicar a origem do vírus na América do Sul. A
primeira é a introdução Pré-Colombiana, baseada na migração de populações oriundas do norte
da Ásia, através do Estreito de Bering, há cerca de 15.000-35.000 anos atrás (Neel et al., 1994;
Biggar et al., 1996). A segunda, Pós-Colombiana, especula que o HTLV-1 foi disseminado a
partir da África para o Novo Mundo durante o tráfico de escravos, no período compreendido entre
os séculos XVI a XIX (Van Dooren et al., 1998; Yamashita et al., 1999), e do Japão para o
Ocidente no início do século XX.
A associação do HTLV-1 com doença neurológica está amplamente demonstrada. No
entanto, a maioria dos indivíduos infectados pelo HTLV-1 permanece assintomático, somente 1%
a 5 % desenvolve alguma doença crônica inflamatória (Orland et al., 2003; Lemasson et al.,
2004). Entretanto, ainda não está elucidado porque alguns indivíduos desenvolvem essas
patologias. Existem evidências que as manifestações patológicas associadas à infecção causada
pelo HTLV-1 resultam de eventos multifatoriais. Entre os fatores estudados destacam-se aqueles
relacionados ao hospedeiro, como por exemplo, a resposta imune e aos fatores virais, tais como,
subtipos virais, carga proviral e polimorfismo em região promotora.
No entanto, as evidências indicam que os subtipos virais se relacionam com a origem
geográfica do HTLV-1 não havendo nenhuma associação com a patogênese de doença. Por outro
lado, há relatos de trabalhos realizados na população do Japão que mudanças de nucleotídeos na
sequência da região U3 da LTR estão associadas com a mieloneuropatia. Entretanto, outros
estudos em pacientes brasileiros não confirmaram esta associação.

19
Utilizando técnicas moleculares e a bioinformática como ferramenta, analisamos
polimorfismos na região da LTR para esclarecer melhor a origem do HTLV-1 no Brasil e
contribuir para o melhor entendimento da patogênese da mieloneuropatia.

20
3. OBJETIVOS
3.1 Geral:
� Avaliar a associação de polimorfismos na região U3-LTR proviral do HTLV-1 com a
origem geográfica e a patogênese da Paraparesia Espástica Tropical/Mielopatia associada
ao HTLV-1 nas populações das cidades do Salvador, Bahia e de Rio Branco, Acre.
3.2 Específicos:
� Determinar a prevalência do HTLV-1 em doadores de sangue atendidos no HEMOACRE
(Rio Branco-Acre);
� Analisar filogeneticamente as sequências da região U3-LTR dos isolados do HTLV-1
provenientes das cidades do Salvador, Bahia, e de Rio Branco, Acre;
� Inferir sobre a origem do HTLV-1 no Brasil, através da análise filogenética da região LTR
proviral;
� Analisar a região U3-LTR de todos os isolados do HTLV-1, provenientes de indivíduos
assintomáticos e portadores de TSP/HAM, variações polimórficas nas três primeiras
repetições de 21pb;
� Correlacionar as alterações polimórficas encontradas na região U3-LTR dos isolados do
HTLV-1 com a idade dos pacientes;

21
4. CAPÍTULO II
Artigo publicado: AIDS Research and Human Retroviruses, 2007
The Close Relationship between South African and Latin American HTLV Type 1 Strains
Corroborated in a Molecular Epidemiological Study of the HTLV Type 1 Isolates from a
Blood Donor Cohort.
Augusto Cesar Andrade Mota, Sonia Van Dooren, Flora Maria de Campos Fernandes, Sérgio de
Araújo Pereira, Artur Trancoso Lopo Queiroz, Viviana Olavarria Gallazzi, Anne-Mieke
Vandamme, Bernardo Galvão-Castro, Luiz Carlos Junior Alcântara.

AIDS RESEARCH AND HUMAN RETROVIRUSES
Volume 23, Number 4, 2007, pp. 503–507
© Mary Ann Liebert, Inc.
DOI: 10.1089/aid.2006.0203
The Close Relationship between South African and Latin American HTLV Type 1 Strains Corroborated in aMolecular Epidemiological Study of the HTLV Type 1
Isolates from a Blood Donor Cohort
AUGUSTO CESAR ANDRADE MOTA,1 SONIA VAN DOOREN,2
FLORA MARIA DE CAMPOS FERNANDES,1,3 SERGIO ARAUJO PEREIRA,3
ARTUR TRANCOSO LOPO QUEIROZ,1 VIVIANA OLAVARRIA GALLAZZI,3
ANNE-MIEKE VANDAMME,2 BERNARDO GALVÃO-CASTRO,1,3
and LUIZ CARLOS JUNIOR ALCANTARA1,3
ABSTRACT
It has been difficult to explain why all HTLV-1 sequences in Salvador, a city in the northeast of Brazil, be-
long to the Transcontinental (A) subgroup of the Cosmopolitan (a) subtype, since according to historical data
the vast majority of slaves brought to Brazil (through Salvador) came from west Africa, where only the west-
ern African subgroup (C) has been found. To shed more light on this subject we conducted a phylogenetic
analysis of 23 isolates from blood donors of Salvador. DNA was extracted and submitted to a nested PCR for
amplification of the entire LTR region. The PCR products were purified and sequenced on an automated se-
quencer. Neighbor-joining and maximum likelihood phylogenetic analyses were performed. None of the new
sequences from Salvador clustered within the West-African subgroup C. Confirming previous results, all se-
quences belonged to the Transcontinental subgroup (A) of the Cosmopolitan subtype, and clustered in two
Latin American clusters. In addition we showed sequences from southern Africa clustering in both Latin
American clusters. One of the new sequences is ancestral to the larger Latin American cluster beta due to a
duplication of a 12-bp long fragment, a finding that has not been previously described. These findings sup-
port the hypothesis that HTLV-1 isolates circulating in Latin America have a closer relationship to South
African compared to West-African HTLV-1 strains. The 12-bp-long duplications in one of the sequences has
no obvious clinical or biological implications yet.
503
INTRODUCTION
MOLECULAR EPIDEMIOLOGICAL STUDIES have demonstratedthat the HTLV-1 genome has a high genetic stability.1 This
stability can be explained mainly by the fact that proviral repli-cation depends on the clonal expansion of HTLV-1-infectedcells.2 This low HTLV-1 variability has been used to give sup-port to theories about the origin, dissemination, and evolution ofthis retrovirus, and also about the migrations of the ancestor hu-man population across the world.1 Seven different genetic sub-
types of HTLV-1 have been proposed based on phylogeneticanalyses: a, or Cosmopolitan, which is distributed worldwide;3
b, from Central Africa;4 c, highly divergent Melanesian strainfrom Papua New Guinea and Australia;5 d, isolated from Cen-tral African Republic (CAR) pygmies, and from two patients inCameroon and Gabon;6,7 e, isolated in a sample from an Efepygmy in Democratic Republic of Congo (DRC) and f, from aGabonese individual;8 and g, recently described as a new sub-type in Cameroon.9 The Cosmopolitan subtype is divided intofive subgroups based on geographical distribution: Transconti-
1Bahia School of Medicine and Public Health/Foundation for Scientific Development, Salvador, Bahia, Brazil.2Rega Institute for Medical Research, Katholieke Universiteit Leuven, Leven, Belgium.3Advanced Public Health Laboratory, Gonçalo Moniz Research Center, Oswaldo Cruz Foundation (LASP/CPqGM/FIOCRUZ), Salvador,
Bahia, Brazil.

nental (A), Japanese (B), West African/Caribbean(C), NorthAfrican (D), and Black Peruvian (E).3,10–12
The HTLV-1 infection is endemic in Japan, the Caribbeanbasin, and some regions of Africa and South America.13 Sal-vador, a city located in the northeast of Brazil, has the highestseroprevalence of HTLV-1 in the country.14 The origin and mo-lecular epidemiology of the HTLV-1 infection in Salvador havebeen enigmatic and difficult to trace. According to historicaldata, most of the Africans brought to this city came from west-ern Africa,15,16 where only the C subgroup of the HTLV-1a hasbeen found.3 In contrast, HTLV-1 isolates found in Salvadorare exclusively of the A subgroup.17,18
Previous studies, comparing both the molecular phylogeniesof the HTLV-1 sequences and the genetic background of theinfected hosts from Salvador with those from South Africa, havehypothesized that the virus was introduced in Salvador by in-fected individuals from the southern region of Africa, duringthe post-Columbian slave trade.17,18 The phylogenetic analysisrevealed that South African sequences cluster inside the LatinAmerican group of the Transcontinental subgroup.18
In a further attempt to understand the possible origin ofHTLV-1 infection in Salvador, we conducted a phylogeneticanalysis with sequences from infected blood donors from Sal-vador who have participated in a case–control study exploringthe risk factors for this infection.19 In this paper we report theresults of this new phylogenetic analysis, indicating a closer re-lation between Latin American and South African compared toWest African HTLV-1 strains.
MATERIALS AND METHODS
DNA was extracted from peripheral blood mononuclear cellsof 23 positive samples using the QIAamp Blood kit (QiagenInc., Chatsworth, CA). Nested polymerase chain reaction (PCR)was performed and long terminal repeat (LTR) regions wereamplified as two overlapping fragments: an LTR-gag segmentof 473 bp and a tax-LTR segment of 479 bp, as previously de-scribed.12,17 All amplification products were electrophoresed on1% agarose gel stained with ethidium bromide, and visualizedwith ultraviolet light.
Ninety-one HTLV-1-infected blood donors residing in the cityof Salvador were enrolled in a case–control study of risk factorsfor HTLV-1 infection, as previously described.19 In brief, blooddonors were routinely screened for bloodborne infections (includ-ing HLTV-1) as recommended by the Brazilian Ministry of Health.Those who had a positive screening test by the ELISA methodhad new blood samples reanalyzed in duplicate, and if persistentlypositive were subsequently confirmed by Western Blot analysis.DNA was extracted from peripheral blood mononuclear cells of23 participants from whom blood samples were available, usingthe QIAamp Blood kit (Qiagen Inc., Chatsworth, CA). Nested PCRwas performed and LTR regions were amplified as two overlap-ping fragments: an LTR-gag segment of 473 bp and a tax-LTRsegment of 479 bp, as previously described.18 All amplificationproducts were electrophoresed on 1% agarose gel stained withethidium bromide, and visualized with ultraviolet light.
The LTR amplification products were purified using theQIAquick Gel Extraction kit (Qiagen) and sequenced directly onan automated 3100 genetic analyzer (Applied Biosystems Inc.,Foster City, CA). Sequencing reactions were carried out using the
inner PCR primers, and a Taq FS Dye terminator cycle sequenc-ing kit (Applied Biosystems). The new LTR sequences and se-lected references strains (including sequences from different eth-nic groups and different geographic regions, and all African LTRsequences of the Transcontinental subgroup) were downloadedfrom the GenBank/EMBL database, aligned with ClustalX andDambe,20 and manually edited using the GeneDoc programs.21
Phylogenetic trees were generated using the neighbor-joining (NJ)and maximum-likelihood (ML) methods of PAUP*, version4.0b10.22 The Tamura–Nei evolutionary model (which takes intoaccount different substitution rates for transversions and transi-tions, as well as intersite substitution rate heterogeneity, using ag-distribution) was selected as the best model for the phyloge-netic analysis (alpha parameter 5 0.5124). The likelihood ratiotest was employed to select the model as well as to test the mo-lecular clock hypothesis. The NJ tree was constructed with an op-timized nucleotide substitution rate matrix and a g-shaped pa-rameter, using empirical base frequencies. The reliability of theNJ trees was assessed by analyzing 1000 bootstrap replicates. ForML trees, a heuristic search was performed with a subtree-prun-ing-regrafting branch swapping algorithm using the NJ tree as thestarting material, including its optimized parameters. The likeli-hood ratio test (RT) method was used to calculate statistical sup-port (expressed in p-values) for the branches. Trees were drawnwith TreeView, version 1.4.
Accession numbers: the GenBank accession numbers of thenew HTLV-1 fragments included in our phylogenetic studywere as follows: HB3167, DQ471187; HB3104, DQ471188;HB3133, DQ471189; HB2966, DQ471190; HB3168, DQ471191;HB3171, DQ471192; HB3170, DQ471193; HB3311, DQ471194;HB3229, DQ471195; HB3114, DQ471196; HB2562, DQ471197;HB3205, DQ471198; HB3135, DQ471199; HB3166, DQ471200;HB3160, DQ471201; HB3119, DQ471202; HB3230, DQ471203;HB3102, DQ471204; HB3203, DQ471205; HB3120, DQ471206;HB3134, DQ471207; HB3202, DQ471208; HB3194, DQ471209.The other HTLV-1 strains were: pyg19, L76310; ITIS, Z32527;MEL5, L02534; HS35; D00294; GH78, D23693; CH26,D23690; Bo, U12804; OD, U12805; Pr52, U12806; MOMJ,Z31659 Pr144, U12807; GB233, D23692; RKI4, AF054627;MT4, Z31661; HTLV24, DQ005565; HTLV04, DQ005557;FNN100, DQ005547; Ni1.Peru, Y16484; Bl1.Peru, Y16481;Me1.Peru, Y16478; Bl3.Peru, Y16483; Bl2.Peru, Y16482;Qu1.Peru, Y16475; Me3.Peru, Y16480; Qu3.Peru, Y16477;Me2.Peru, Y16479 and Ni2.Peru, Y16487; ATM, J02030;ATK1, J02029; MT2, L03562; CR1, K02722; TBH2 to -4,L76025, L76034 and L76028; TBH6 and -7, L76030 andL76029; BOI, L36905; AMA, CMC, FCR and MAQS,X88871–X88873 and X88876; MWMG, Z31662; StDen,L76306; PH236, L76307; GAB7, L76311; 12503A, L76309;H24, L76308; Ni3.Peru, Y16485; HKN, X88874; TBH5,L76027; Cam, AF 063819; Nar, AF063820; H5, M37299;AINU, D23694; Qu2.Peru, Y16476.
RESULTS
The phylogenetic analysis of the total LTR region showedthat all 23 sequences belonged to the Transcontinental (A) sub-group of the Cosmopolitan (a) subtype, a finding that was sup-ported by a bootstrap value of 78% (p , 0.001 for ML analy-ses) (Fig. 1). The majority of our new sequences (n 5 18)
MOTA ET AL.504

clustered within the larger Latin American cluster (named clus-ter beta in Fig. 1). One of these sequences formed a mono-phyletic group with two Peruvian sequences (Me1 and Me2),yet this cluster was supported by neither bootstrap nor ML. Four
of the other new sequences clustered monophyletically withother sequences from South America, forming a smaller LatinAmerican cluster (named cluster alpha in Fig. 1), supported bya bootstrap value of 93% and by ML analysis.
MOLECULAR STUDY OF HTLV-1 ISOLATES FROM BRAZIL 505
FIG. 1. Rooted neighbor-joining tree of 80 HTLV-1 strains based on a 718-bp fragment of the LTR region. The bootstrap val-ues (.60% and using 1000 bootstrap samples) on the branches represent the percentage of trees for which the sequences locatedat the right end of the branch form a monophyletic group. Mel5 was used as the out-group. Geographic origin is given in paren-theses. Newly sequenced LTRs included in this analysis are in bold. **The ML method was highly statistically significant (p ,
0.001). The rectangle represents part of the sequenced LTR region showing the 12-bp duplication of the isolate HB3166 (the up-per line represents the reference sequence ATK1).

Two HTLV-1 prototypes from South Africa (HTLV24 andHTLV04) belonged to each of the Latin American clusters. Thesequence HTLV24 clustered into the smaller Latin Americancluster a (alpha) (93% bootstrap; p , 0.001 for ML), andHTLV04 into the larger Latin American cluster b (beta) (65%bootstrap; p , 0.001 for ML). We have tested the molecularclock hypothesis for all branches intending to date our data. Inall cases the null hypothesis was rejected.
One of our new sequences (HB3166) clusters ancestral to thelarger Latin American cluster b (beta), yet with no bootstrap orML support. This HB3166 sequence has a 12-bp duplicationwithin its LTR region, corresponding to position 9018 to 9029of the ATK1 prototype. This position does not overlap tax andis distal from the functional regions in the LTR.
DISCUSSION
In line with previous reports,17,18 our phylogenetic analysishas shown that all sequences belonged to the Cosmopolitan sub-type, Transcontinental subgroup. Moreover, this study demon-strates that sequences from southern Africa cluster into both al-pha and beta Latin American groups. These findings indicate acloser relation between Latin American and South Africanstrains compared to West African HTLV-1 isolates. In addition,with our previously published reports we present the hypothe-sis that infected individuals from the southern region of Africamight have introduced HTLV-1 into Latin America.12,17,18
However, in the current phylogenetic analysis there is no con-clusive evidence on the direction of migration, either fromSouthern Africa to Latin America or from Latin America tosouthern Africa.
The fact that sequences clustered into two different clusterssuggests that there were multiple introductions of HTLV-1 intoSalvador. Nonetheless, the observation that the sequences clus-tered homogeneously within each Latin American group sug-gests that the introduction of HTLV-1 among people in Sal-vador was relatively recent, most likely occurring during theslave trade that took place during the seventeenth, eighteenth,and nineteenth centuries, as previously reported.12,17,18,23,24
Since the molecular clock was rejected, we could not date theinterior nodes.
However, these findings appear to contradict historical data,according to which the vast majority of slaves brought to thisregion originally came from western Africa,15,16 where HTLV-1aC predominates.2 A possible explanation for this discrepancyis that individuals originally from the Central Africa continent(and infected with HTLV-1aA) migrated to southern Africa assuggested by the presence of the bA-globin CAR haplotype insouthern Africa.18 The CAR haplotype is prevalent in CentralAfrica, South Africa, Mozambique, and other regions of south-ern Africa. The source of the CAR haplotype in South Africais not known, but may reflect the migration of the Bantu pop-ulation from the Central to South African continent during thelast 3000 years. A plausible alternative explanation is that rel-atively few isolates from all these regions have been analyzed,hampering definitive conclusions.
The only sequence that clustered ancestral to the Latin Amer-ican cluster beta had a 12-bp duplication in its LTR region (se-quencing was performed three times for confirmation purposes).
As far as we know this is the first time that such a duplicationhas been reported. The isolate is from an asymptomatic 37-year-old male blood donor. It is not known at this point whether thisinsertion could have any clinical or biological implication. Weintend to follow him up for clinical symptoms and proviral loadand to investigate his relatives.
ACKNOWLEDGMENTS
This paper is part of Augusto Cesar Andrade Mota’s Ph.D.thesis, Escola Bahiana de Medicina e Saúde Publica, Postgrad-uate Course in Medicine and Human Health. This study waspartially supported by FAPESB (Grants 303/03) and by theBrazilian Ministry of Health (306/04 and 307/04). The authorsare grateful to Mr. Noilson Lazaro and Mrs. Elisabeth Deliège,respectively, for their technical and editing assistance.
REFERENCES
1. Gessain A, Gallo R, and Franchini G: Low degree of human T-cellleukemia/lymphoma virus type I genetic drift in vivo as a meansof monitoring viral transmission and movement of ancient humanpopulations. J Virol 1992;66:2288–2295.
2. Wattel E, Vartanian J-P, Pannetier C, et al.: Clonal expansion ofhuman T-cells leukemia virus type I infected cells in asymptomaticsymptomatic carriers without malignancy. J Virol 1995;69:2863–2868.
3. Miura T, Fukunaga T, Igarashi T, et al.: Phylogenetic subtypes ofhuman T-lymphotropic virus type I and their relations to the an-thropological background. Proc Natl Acad Sci USA 1994;91:1124–1127.
4. Hahn BH, Shaw GM, Popovic M, et al.: Molecular cloning andanalysis of a new variant of human T-cell leukemia virus (HTLV-Ib) from an African patient with adult T-cell leukemia-lymphoma.Int J Cancer 1984;34:613–618.
5. Gessain A, Boeri E, Yanagihara R, et al.: Complete nucleotide se-quence of a highly divergent human T-cell leukemia (lympho-tropic) virus type I (HTLV-I) variant from Melanesia: Genetic andphylogenetic relationship to HTLV-I strains from other geograph-ical regions. J Virol 1993;67:1015–1023.
6. Mahieux R, Chappey C, Georges-Courbot MC, et al.: Simian T-cell lymphotropic virus type I from Mandrillus sphinx as a simiancounterpart of human T-cell lymphotropic virus type I subtype D.J Virol 1998;72:10316–10322.
7. Mboudjeka I, Zekeng L, Yamashita M, et al.: Prevalence and phy-logenetic analysis of HTLV-I isolates in Cameroon, including thoseof the Baka Pygmy. Jpn J Cancer Res 1997;88:619–624.
8. Salemi M, Van Dooren S, Audenaert E, et al.: Two new human T-lymphotropic virus type I phylogenetic subtypes in seroindetermi-nates, a Mbuti pygmy and a Gabonese, have close relatives amongAfrican STLV-I strains. Virology 1998;246:277–287.
9. Wolfe ND, Heneine W, Carr JK, et al.: Emergence of unique pri-mate T-lymphotropic viruses among central African bush meathunters. Proc Natl Acad Sci USA 2005;102:7994–7999.
10. Gasmi M, Farouqi B, d’Incan M, et al.: Long terminal repeat se-quence analysis of HTLV type I molecular variants identified infour North African patients. AIDS Res Hum Retrovirol 1994;10:1313–1315.
11. Vidal AU, Gessain A, Yoshida M, et al.: Phylogenetic classifica-tion of human T cell leukaemia/lymphoma virus type I genotypesin five major molecular and geographical subtypes. J Gen Virol1994;75:3655–3666.
MOTA ET AL.506

12. Van Dooren S, Gotuzzo E, Salemi M, et al.: Evidence for a post-Columbian introduction of HTLV-I in Latin America. J Gen Virol1998;79:2695–2708.
13. Proietti FA, Carneiro-Proietti ABF, Catalan-Soares BC, et al.:Global epidemiology of HTLV-I infection and associated diseases.Oncogene 2005;24:6058–6068.
14. Dourado I, Alcantara LCJ, Barreto ML, et al.: HTLV-I in the gen-eral population of Salvador, Brazil: A city with African ethnic andsociodemographic characteristics. J Acquir Immune Defic Syndr2003;34:527–531.
15. Curtin PD: The Slave Atlantic Trade: A Census. The University ofWisconsin Press, Milwaukee, 1969.
16. Verger P: Flux et reflux de la traite des nègres entre le Golfe de
Benin et Bahia de Todos os Santos. Mouton, Paris, 1968.17. Alcantara LC Jr, Van Dooren S, Gonçalves MS, et al.: Globin hap-
lotypes of human T-cell lymphotropic virus type I-infected indi-viduals in Salvador, Bahia, Brazil, suggest a post-ColumbianAfrican origin of this virus. J Acquir Immune Defic Syndr 2003;33:536–542.
18. Alcantara LCJ, Oliveira T, Gordon M, et al.: Tracing the origin ofBrazilian HTLV-1 as determined by analysis of host and viralgenes. AIDS 2006;20:780–782.
19. Mota A, Nunes C, Mello A, et al.: A case-control study of HTLV-1 infection among blood donors in Salvador, Bahia, Brazil—As-sociated risk factors and trends towards declining prevalence.Brazil J Hematol Hemother 2006;28(2):120–126.
20. Xia X: DAMBE: Data analysis in molecular biology and evolu-tion. Department of Ecology and Biodiversity, University of HongKong, Hong Kong, 2000.
21. Nicholas KB, Nicholas HBJ, and Deerfield DW: GeneDoc: Analysisand visualization of genetic variation. EMBNEW NEWS 1997:30.
22. Swofford DL, Olsen GJ, Waddell PJ, et al.: Phylogenetic infer-ence. In: Molecular Systematics, 2nd ed. (Hillis DM et al., eds.).Sinauer, Sunderland, MA, 1996, pp. 407–514.
23. Yamashita M, Veronesi R, Menna-Barreto M, et al.: Molecular epi-demiology of human T-cell leukemia virus type I (HTLV-1) Brazil:The predominant HTLV-1s in South America differ from HTLV-Is of Japan and Africa, as well as those of Japanese immigrantsand their relatives in Brazil. Virology 1999;261:59–69.
24. Van Dooren S, Pybus OG, Salemi M, et al.: The low evolutionaryrate of human T-cell lymphotropic virus type-1 confirmed by anal-ysis of vertical transmission chains. Mol Biol Evol 2004;21:603–611.
Address reprint requests to:Luiz Carlos Junior Alcantara
LASP/CPqGM/FIOCRUZ
Rua Waldemar Falcão 121
Brotas, Salvador
Bahia, Brazil 40295-001
E-mail: [email protected]
MOLECULAR STUDY OF HTLV-1 ISOLATES FROM BRAZIL 507

22
4. CAPÍTULO III
Artigo Publicado: J medical Virology, 80 (11): 1966-1971, 2008.
HTLV-1 infection in blood donors from Western Brazilian Amazon region: seroprevalence
and molecular study of viral isolates.
Aline Cristina Mota-Miranda, Sérgio de Araújo Pereira, Denise Duizit Colin, Simone Kashima,
Dimas Tadeu Covas, José Tavares-Neto, Bernardo Galvão-Castro, Luiz Carlos Júnior Alcântara.








23
4. CAPÍTULO IV
Artigo submetido: J medical Virology
Possible role of single nucleotide polymorphism in the U3-LTR region of HTLV-1 in the
pathogenesis of TSP/HAM .in Brazil.
Authors: Sérgio Araújo Pereira±, Luiz Carlos Júnior Alcântara
± ¥, Joana Paixão Monteiro
±, Antônio Andrade Filho
€,
Simone Kashima§, and Bernardo Galvão-Castro
±¥.
Author's affiliation: ±
Laboratório Avançado de Saúde Pública, Centro de Pesquisa Gonçalo Muniz, Fundação
Oswaldo Cruz, Salvador, Bahia, Brasil; €Fundação de (eurologia e (eurocirurgia da Bahia, Salvador, Ba, Brazil
,
§Hemocentro de Ribeirão Preto, São Paulo, Brasil,
¥ Escola Bahiana de Medicina e Saúde Pública, Fundação
Bahiana para o Desenvolvimento das Ciências, Salvador, Bahia, Brasil.
Institution where the analysis were performed:
Laboratório Avançado de Saúde Pública, Centro de Pesquisa Gonçalo Muniz, Fundação Oswaldo Cruz, Salvador,
Bahia, Brasil

1
Possible role of single nucleotide polymorphism in the U3-LTR region of HTLV-1 in
the pathogenesis of TSP/HAM in Brazil.
Authors: Sérgio Araújo Pereira±, Luiz Carlos Júnior Alcântara
± ¥, Joana Paixão Monteiro
±, Antônio Andrade
Filho€, Simone Kashima
§, and Bernardo Galvão-Castro
±¥.
Author's affiliation: ±
Laboratório Avançado de Saúde Pública, Centro de Pesquisa Gonçalo Muniz, Fundação
Oswaldo Cruz, Salvador, Bahia, Brasil; €Fundação de (eurologia e (eurocirurgia da Bahia, Salvador, Ba,
Brazil,, §Hemocentro de Ribeirão Preto, São Paulo, Brasil,
¥ Escola Bahiana de Medicina e Saúde Pública,
Fundação Bahiana para o Desenvolvimento das Ciências, Salvador, Bahia, Brasil.
Institution where the analysis were performed:
Laboratório Avançado de Saúde Pública, Centro de Pesquisa Gonçalo Muniz, Fundação Oswaldo Cruz,
Salvador, Bahia, Brasil
Corresponding author:
Prof. Bernardo Galvão Castro, MD, PhD.
Laboratório Avançado de Saúde Pública, Centro de Pesquisa Gonçalo Moniz, Fundação Oswaldo Cruz.
Rua Waldemar Falcão 121, Candeal, Salvador, Bahia, Brasil – 40296-610.
Telephone # 55 71 31762255
Fax # 55 71 31762300
E-mail: [email protected]
Shortened title: Mutations in the LTR region of HTLV-1 patients associated with
TSP/HAM

2
The transcription of HTLV-1 proviral sequence depends of Tax viral protein that transacts
the three 21 bp repeated sequences in the U3-LTR region. To further investigate if
polymorphisms in the U3 region could be associated with susceptibility to TSP/HAM, the
frequency of U3 punctual mutations was studied in the sequences obtained from 214 HTLV-
1 patients (110 asymptomatic and 104 symptomatic), using the BioEdit software. The ATK1
prototype was used as standard sequence. Neighbor-joining and maximum likelihood
phylogenetic analysis were performed with PAUP* software. It was detected an insertion
(C) between the positions 8582+ that were only present in the asymptomatic group (8/110;
p=0.004). Besides, significant frequency of mutations C>T at position 8583 (14/110;
p=0.001) and C>A at position 8606 (9/110; p=0.012) were observed of the asymptomatic
group. In the symptomatic group, were detected an insertion (G) between positions 8422+
(11/104; p=0.042), an insertion (A) between 8582+ (12/104; p=0.001) and an deletion at
position 8427 (17/104; p=0.002). When we correlated the phylogenetic analysis with the
punctual mutations, we identified that all new sequences belonged to the Transcontinental
group of the Cosmopolitan subtype, and clustered in two Latin American clusters. The
phylogenetic results confirm the post-Columbian introduction of this virus in Salvador, as
described in previous analysis and suggest that punctual mutations are not associated with
HTLV-1 subgroup. Furthermore, the mutations detected in the U3 region may have an
impact in the viral transcription and TSP/HAM development.
Key-words: HTLV-1, Polymorphisms, U3-LTR region, TSP/HAM.

3
I(TRODUCTIO(
The Human T-cell lymphotropic virus type 1 (HTLV-1) is the etiologic agent of adult T-cell
Leukemia lymphoma (ATLL) [Poiesz et al., 1980], tropical spastic paraparesis/HTLV-1
associated myelopathy (HAM/TSP) [Gessain et al., 1985; Osame et al., 1986] and HT V-1
associated uveitis (HAU) [Mochizuki et al, 1992].
The HTLV-1 infection is endemic in several parts of the world. 2.5 million people are
infected with HTLV-1[Carneiro-Proietti et al., 2002]. This infection has high prevalence
rates in Southern Japan, in the Caribbean basin, and some African regions, and to a lesser
rate in Latin American [Proietti et al., 2005]. In Brazil, according to epidemiological data,
the prevalence of HTLV-1 infection is not uniform, in the Southeast (0.18%-0.41%, São
Paulo) and the North (0.08%, Manaus) regions [Carneiro-Proietti et al., 2002; Mota-
Miranda et al., 2008]. The Salvador city, located in northeast of Brazil, has a highly mixed
population, mostly of African origin, and the highest seroprevalence (around 1.85%) of
HTLV-1 in the country [Dourado et al., 2003; Mota et al., 2006].
According to the virus long terminal repeat (LTR) region analysis, seven HTLV-1 genetic
subtypes have been proposed [Miura et al., 1994]: subtype a, Cosmopolitan, which is
distributed worldwide; subtype b, from Central Africa; subtype c, circulating in Melanesia;
subtype d, isolated from Central African pygmies, Cameroon and Gabon; subtype e, isolated
from Democratic Republic of Congo pygmies; subtype f, isolated initially from an infected
individual from Democratic Republic of Congo; and subtype g, also described in Cameroon.
The Cosmopolitan subtype is further divided in five subgroups: A-Transcontinental, B-
Japanese, C-West African/Caribbean, D-North African and E-Black Peruvian [Miura et al.,
1994; Gasmi et al., 1994; Van Dooren et al., 1998].

4
While the majority of HTLV-1 infected individuals remains asymptomatic, 1-3% develop a
disabling chronic inflammatory disease [Bangham et al., 1999; Jeffery et al., 1999],
indicating that viral, host and environmental factors are involved [Orland et al., 2003;
Lemasson et al., 2004]. For instance, HLA-A*02 type was shown to decrease the risk of
TSP/HAM [Jeffery et al., 1999], suggesting that HLA subtype may influence the outcome of
HTLV-I infection. However, the mechanisms by which HTLV-1 causes disease are not fully
elucidated. The gene expression of retroviruses is regulated by cis elements in the LTRs of
the proviral genome [Dan et al., 1999]. HTLV-1 transcription is activated by the interaction
of the viral Tax protein directly with the unphosphorylated cyclic AMP responsive element
binding protein (CREB). This binds to another transcription factor, CREB binding to
phosphorylated protein (CBP), forming CREB-Tax-CBP complex that finally binds to the
21-base pairs (bp) sequence located within the U3-LTR region (Tax-responsive element-
TRE). This process is repeated three times in LTR [Dan et al., 1999; Lemasson et al., 2004].
Previous studies have demonstrated that several nucleotide substitutions in the U3-LTR
region seem to be common among isolates from patients with HAM/TSP [Shimotohno et
al., 1986; Mukhopadhyaya et al., 1993] and that the presence of the nucleotide substitution
T→C in tax gene of the isolates from asymptomatic HTLV-1 infected individuals was
associated with high risk of TSP/HAM development [Renjifo et al., 1995]. In addition,
studies have revealed that only two direct repeats of the 21-bp sequence are sufficient for
the efficient activation by Tax [Fujisawa et al., 1986; Tanimura et al., 1998; Dan et al.,
1999] and that deletions in the sequence TRE2S decrease significantly HTLV-1 Tax-
mediated transcriptional activity [Brady et al., 1987; Tanimura et al., 1993; Tanimura et
al.,1998]. In this context, the U3-LTR region of HTLV-1 isolates from asymptomatic and

5
symptomatic (TSP/HAM) patients were analyzed, particularly, variations in the first three
21-pb repetitions were investigated in attempt to associate polymorphisms in the U3-LTR
region of the HTLV-1 with the development of TSP/HAM. The results suggest that
mutations in this region could be important to the understanding of HTLV-1 pathogenesis.

6
METHODS
Study Subjects and Serology
A total of 214 infected individuals were studied. Out of these 214 infected individuals were
included in this study (40 asymptomatic carriers, 23 oligosymptomatic, and 36 TSP/HAM
patients were recruited from the HTLV reference center located at Bahia Foundation School
of Medicine and Public Health/Bahia Foundation for the Scientific Development (EBMSP)
between June 2005 and July 2006. Additionally 12 asymptomatic carriers and 30 TSP/HAM
patients from the Foundation of Neurology and Neurosurgery (FNN) [Alcantara et al.,
2003], 3 asymptomatic carriers from the general population of Salvador, Bahia, Brazil
[Dourado et al., 2003], 3 asymptomatic carries blood donors from the (FUNDACRE) and 67
sequences from GenBank database (29 asymptomatic and 38 TSP/HAM).
Serum samples repeatedly positive in duplicate were screened for antibodies against HTLV-
1 and 2 by ELISA (HTLV-1; HTLV-2 Ab-Capture ELISA Test System, Ortho, Clinical
Diagnostic Inc., Raritan, NJ), according to the algorithm recommended by the Brazilian
Ministry of Health, and were confirmed and discriminated between HTLV-1 and HTLV-2
using Western Blot (HTLV Blot 2.4, Genelabs Diagnostics, Science Park Drive, Singapore).
The TSP/HAM diagnosis was done according to the World Health Organization diagnostic
criteria. All subjects provided written consent. The infected individuals classified as
oligosymptomatic were clinically different from TSP/HAM patients. They presented some
neurological symptoms, such as disabilities in gait and sphincter control areas, weakness,
bladder, and erectile dysfunctions, but they did not fulfill the TSP/HAM criteria. The study
received official institutional and ethical approval of the EBMSP/FBDC ethic committee.

7
D(A Extraction and Polymorphism Detection
Genomic DNA was extracted from peripheral blood mononuclear cells (PBMC) using the
QIAamp Blood Kit (Qiagen Inc., Chatsworth, CA). The U3-LTR region was amplified by
nested polymerase chain reaction (PCR), as one overlapping fragment: a U3-LTR segment
of 329 bp, as previously described [Alcantara et al., 2006]. PCR products were purified
using Qiagen columns (Qiagen Inc,Valencia, CA) and sequenced in an ABI Prism 3100
Genetic Analyzer (Applied Biosystems Inc., Foster City, CA) using the Big Dye terminator
Kit (Applied Biosystems Inc., Foster City, CA). The same inner PCR primers were used in
the sequencing reactions. Sequences were assembled using SeqMan software (DNAStar,
Madison, WI) and checked for contamination by BLAST search against the HTLV-1
sequences from the Bank/EMBL database. A multiple sequence alignment of the obtained
sequences with previously characterized sequences from the database was performed with
the Clustal algorithm [Thompson et al., 1997], and further edited using the GeneDoc
software [Nicholas et al., 1997]. The ATK1 prototype Japanese was used as standard
sequence. 53 HTLV-1 positives samples (FNN22, FNN29, FNN28, FNN64, FNN31,
FNN18, FNN39, FNN73, FNN61, FNN89, FNN87, FNN26, FNN82, FNN9, FNN35,
FNN19, FNN83, FNN32, FNN78, FNN68, FNN60, FNN54) [Alcantara et al., 2003],
(FNN155, FNN159, FNN156, FNN100, FNN158, FNN153) [Alcantara et al.,2006], (AC42,
AC129) [Miranda-Mota et al., 2008] and (HB3102, HB3166, HB3230, HB3194, HB2562,
HB3203, HB3134, HB3202, HB3120, HB3119, HB3170, HB3160, HB3135, HB3167,
HB3168, HB3205, HB3104, HB3311, HB3229, HB2966, HB3114, HB3133, HB3171)

8
[Mota et al., 2007] from previous study were included in the analysis. In addition, 67
samples from the Genbank database were included.
Statistical Methods
The frequencies (%) of mutations in the U3-LTR region (329pb) were compared between
the asymptomatic and the symptomatic groups, and between asymptomatic patients with
more (>45) or less (<45) than 45 years old. The frequencies were tested by X2 test followed
by adjusted X2 residual analysis or with Fisher’s exact test using the PEPI package, version
4.0 [Abramson and Gahlinger, 2001]. A p-value <0.05 was considered statistically
significant.
Phylogenetic Analysis
The phylogenetic analysis, based on the partial U3-LTR region, included reference strains
ITIS-Z32527, pyg19-AF076264, Bo-U12804, RKI4.Peru-AF054627, BCI2.1-U32557,
CH26-D23690, RKI1.Iran-AF3010, NM2015-AF076264, HKN-X88874, JCP-X88875,
CMC-X88872, MT2-L03562, CH-M69044, AMA-X88871, MAQS-X88876, Afs911-
L72212, ALT2-95086, Neu7-EU622618, FCR-X88873, MASU-X88877, PW1-EU622624,
BD12-EU622592, BD16-EU622596, IDU1-EU622598 from the NCBI/Nucleotide Sequence
Database (GenBank). The Mel5-L02534 reference sequence was used as outgroup. The
Neighbor-joining (NJ) and maximum-likelihood (ML) phylogenetic analysis were
performed with the PAUP* software, version 4.0.2a [Swofford, 1999], using the HKY+G
nucleotide substitution model including substitution rate heterogeneity, as determined by the
Modeltest software [Posada and Krandall, 1998]. The reliability of the NJ trees was
evaluated by analyzing 1,000 bootstrap replicates. For the ML tree reconstruction, a

9
heuristic search with the subtree-pruning-regrafting branch swapping algorithm was
performed using the NJ tree as starting tree, including its optimized parameters. The
likelihood ratio test (RT) method was used to calculate statistical support for the branches: p
< 0.001 (highly significant **) and p < 0.005 (significant*). Bootstrap and ML supports
were added to NJ tree that was drawn with TreeView 1.4 software [Page, 1996].
(ucleotide Sequence Accession (umbers
The GenBank accession numbers of the new HTLV-1 fragments were as follows: FNN169-
GQ354521, FNN164-GQ354520, FNN171-GQ354518, FNN170-GQ354522, FNN47-
GQ354519, FNN40-GQ354524, FNN174-GQ354523, FNN150-GQ354516, FNN172-
GQ354515, FNN165-GQ354517, FNN94-GQ354451, AC204-GQ354479, BA115000-
GQ354454, BA123070-GQ354452, BA155630-GQ354453, BABR3417-GQ354488,
BABR2095-GQ354467, BABR1764-GQ354466, BABR2091-GQ354465, BABR2151-
GQ354468, BABR664-GQ354506, BABR2819-GQ354489, BABR1709-GQ354487,
BABR1603-GQ354486, BABR1438-GQ354485, BABR290-GQ354442, BABR531-
GQ354443, BABR0136-GQ354439, BABR2224-GQ354438, BABR991-GQ354437,
BABR72-GQ354436, BABR0982-GQ354435, BABR1792-GQ354458, BABR394-
GQ354440, BABR643-GQ354463, BABR1910-GQ354472, BABR1790-GQ354441,
BABR3591-GQ354498, BABR3510-GQ354493, BABR2674-GQ354492, BABR3517-
GQ354484, BABR275-GQ354477, BABR1909-GQ354469, BABR2142-GQ354474,
BABR1237-GQ354444, BABR1669-GQ354445, BABR1982-GQ354473, BABR3637-
GQ354496, BABR1921-GQ354507, BABR2986-GQ354462, BABR721-GQ354513,

10
BABR945-GQ354514, BABR3140-GQ354455, BABR1260-GQ354475, BABR2707-
GQ354495, BABR3488-GQ354483, BABR2441-GQ354459, BABR2294-GQ354490,
BABR3511-GQ354480, BABR2946-GQ354509, BABR619-GQ354501, BABR3698-
GQ354497, BABR3518-GQ354494, BABR620-GQ354481, BABR1599-GQ354460,
BABR3132-GQ354434, BABR1513-GQ354446, BABR1504-GQ354500, BABR1480-
GQ354511, BABR2470-GQ354510, BABR2713-GQ354461, BABR1911-GQ354502,
BABR1759-GQ354471, BABR3632-GQ354479, BABR944-GQ354512, BABR1741-
GQ354470, BABR885-GQ354505, BABR423-GQ354447, BABR2041-GQ354449,
BABR1389-GQ354482, BABR3714-354499, BABR870-GQ354503, BABR1359-
GQ354448, BABR2177-GQ354476, BABR911-GQ354504, BABR3115-GQ354456,
BABR3173-GQ354457, BABR2672-GQ354491, BABR526-GQ354450, BABR1851-
GQ354464, BABR965-GQ354508. The other HTLV-1 strains were: BD15-EU622595,
ATL1-EU622582, Neu11-EU622621, Neu5-EU622616, Neu4-EU622615, Neu8-
EU622619, Neu12-EU622622, Neu10-EU622620, Neu13-EU622623, Neu6-EU622617,
Neu3-EU622614, Neu2-EU622613, Neu1-EU622612, FCR-X88873, ATM-J02030,
RKI2Rom-AF003012, KUW3-L42225, TSP1-M86840, H5-M37299, BOI-L36905, KUW1-
L42253, TBH5-L76027, TBH7-L76029, TBH3-L76034, TBH1-L76026, TBH2-L76025,
TBH4-L76028, TBH6-L76030, TP50-AY124607 TP67-AY124617, TP52-AY124609,
TP61-AY124614, MONJ-Z31659, KUW2-L42255, AINU-D23694, BD1-EU622584,
FSW1-EU622600, TP51-AY124608, TP62-AY125467, PW2-EU622625, BD2-EU622585,
CR1-K02722, TP39-AY124602, BD3-EU622586, BCI1.2-U32552, TP65-AY124616,
TP54-AY124610, TP44-AY124606, TP64-AY124615.

11
RESULTS
In this study we sought to correlate polymorphisms in the U3-LTR HTLV-I region isolated
from asymptomatic and TSP/HAM infected individuals from Salvador, Bahia, Brazil with
the risk of development of TSP/HAM through the inspection of nucleotide signatures in
their sequences. The sequences were stratified according to the patient symptomatic status
and compared to the ATK1 reference strain in a multiple sequence alignment created in the
Clustal software [Thompson et al., 1997]. A total of 110 (51.4%) sequences comprised the
asymptomatic group and 104 (48.6%) sequences comprised the symptomatic group. In this
analysis, 10 nucleotide substitutions were detected and six specific signatures showed a P-
value statistically significant (Table I). It was observed an insertion (C) between the
positions 8582+ in eight of 110 patients (7.27%) (OR=∞; 95%CI=1.68-∞; x
2=7.821;
p=0.004) was identified exclusively in the asymptomatic group. Therefore, significant
frequency of substitution C>T at position 8583 was present in fourteen patients (12.73%)
(OR=15.02; 95%CI=2.19-641.15; x2=11.301; p=0.001) of the asymptomatic group and in
one patient (0.96%) of the symptomatic group, and a substitution C>A at position 8606 was
found in nine of the 110 asymptomatic patients (8.18%) (OR=9.18; 95%CI=1.22-406.1;
x2=6.227; p=0.012) were observed of the asymptomatic group. In the symptomatic group, an
insertion (G) between positions 8422+ were detected in eleven patients (10.58%) (OR=3.13;
95%CI=0.07-1.13; x2=3.932; p=0.042), an insertion (A) between the positions 8582
+ in
twelve patients (11.54%) (OR=14.22; 95%CI=0.00-0.50; x2=10.536; p= 0.001) and a

12
deletion at 8427 was observed in seventeen patients of 104 symptomatic patients (16.35%)
(OR=5.18; 95%CI=0.05-0.63; x2=9.712; p=0.002).
In addition, the nucleotide substitutions frequencies (%) in the U3-LTR region (329pb) was
further compared between two groups of asymptomatic individuals: patients with more
(n=56; 69.1%) or less (n=25; 30.9%) than 45 years old (Table II). This comparative analysis
did not indicate any mutation with significant difference in frequency between the two
groups as determined by the p-value.
The phylogenetic analysis of the partial U3-LTR region demonstrated that all 147 sequences
belong to the Cosmopolitan (a) subtype, Transcontinental (A) subgroup, a finding that was
supported by a bootstrap value of 50% (p<0.001 for ML analysis) (fig. 1). The majority of
our new sequences (n=91) clustered within a large Latin American group (named Larger
Latin American cluster in fig. 1). Nine of the new sequences clustered into two
monophyletic groups, forming a smaller Latin American Cluster (named Smaller Latin
American cluster in fig. 3), one group supported by a bootstrap value of 72% and another
group, yet with no bootstrap or ML support.

13
TABLE I. Comparison of mutations frequencies (%) in the U3-LTR region (329pb) between asymptomatic
and symptomatic groups.
* The p-value was calculated by the x2 test. A multiple sequence alignment was created using the CLUSTAL
software [Thompson et al., 1997] and the studied sequences were compared to ATK1 Japanese prototype. The
following mutations were detected: 8427 (-) deletion (16.35%; p=0.002), 8422+(G) insertion (10.58%; p=
0.042), 8582+(A) insertion (11.54%; p=0.001), 8582
+ (C) insertion (7.27%; p=0.004), 8583 C>T mutations
(12.73%; p= 0.001) and 8606 C>A mutations (8.18%; p= 0.012).
Position
Status
95% CI
Mutation Hcs TSP/HAM or ATLL Odds
ATK1 n % n % P Ratio
8391
G 89 80.91 89 85.58
1.233 0.177 1.52 0.68 -3.44A 21 19.09 14 13.46
C 0 1 0.96
110 104
(-) 86 78.18 81 77.88
G 4 3.64 11 10.58 3.932 0.042 3.13 0.07-1.13
C 20 18.18 12 11.54 1.847 0.121 1.70 0.74-4.05
110 104
*8427
G 106 96.36 87 83.65
9.712 0.002 5.18 0.05-0.63(-) 4 3.64 17 16.35
110 104
8452
G 73 66.36 77 74.04
0.360 0.325 1.19 0.64-2.22A 37 33.64 31 29.81
110 104
8462
T 104 94. 55 95 91.35
0.836 0.259 1.64 0.17-2C 6 5.45 9 8.65
110 104
8546
T 7 6.36 8 7.69
0.144 0.455 1.23 0.37-4.13C 103 93.64 96 92.31
110 104
(-) 101 91.82 92 88.46
A 1 0.91 12 11.54 10.536 0.001 14.22 0.00-0.50
C 8 7.27 0 7.821 0.004 ∞ 1.68-∞
110 104
*8583
C 96 87.27 103 99.04
11.301 0.001 15.02 2.19-641.15T 14 12.73 1 0.96
110 104
8595
C 104 94.55 103 99.04
3.395 0.069 5.94 0.70-275.93T 6 5.45 1 0.96
110 104
*8606
C 3 2.72 6 5.77
G 98 89.10 97 93.27 1.148 0.203 1.70 0.19-1.71
A 9 8.18 1 0.96 6.227 0.012 9.18 1.22-406.1
110 104
x2
* 8422a
*8582a
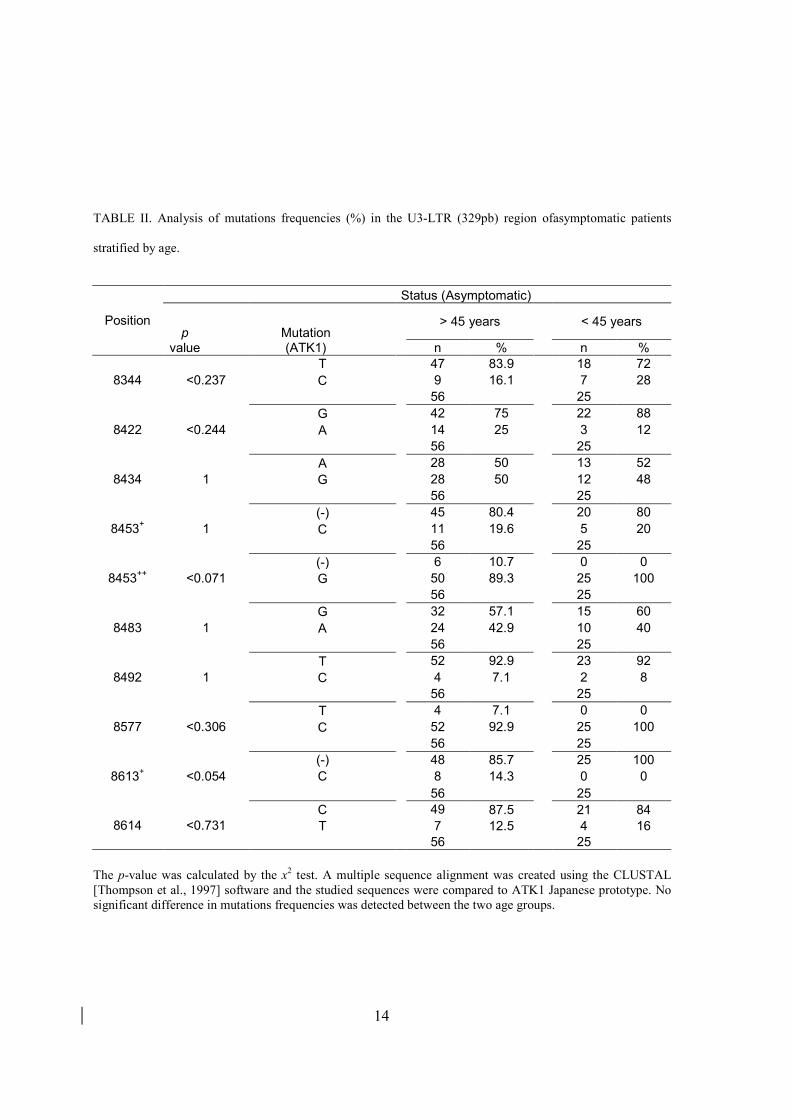
14
TABLE II. Analysis of mutations frequencies (%) in the U3-LTR (329pb) region ofasymptomatic patients
stratified by age.
Position
Status (Asymptomatic)
p value
Mutation (ATK1)
> 45 years
< 45 years
n % n %
8344 <0.237
T
47 83.9
18 72
C 9 16.1 7 28
56 25
8422 <0.244
G 42 75 22 88
A 14 25 3 12
56 25
8434 1
A 28 50 13 52
G 28 50 12 48
56 25
8453+
1
(-) 45 80.4 20 80
C 11 19.6 5 20
56 25
8453++
<0.071
(-) 6 10.7 0 0
G 50 89.3 25 100
56 25
8483 1
G 32 57.1 15 60
A 24 42.9 10 40
56 25
8492 1
T 52 92.9 23 92
C 4 7.1 2 8
56 25
8577 <0.306
T 4 7.1 0 0
C 52 92.9 25 100
56 25
8613+
<0.054
(-) 48 85.7 25 100
C 8 14.3 0 0
56 25
8614 <0.731
C 49 87.5 21 84
T 7 12.5 4 16
56 25
The p-value was calculated by the x
2 test. A multiple sequence alignment was created using the CLUSTAL
[Thompson et al., 1997] software and the studied sequences were compared to ATK1 Japanese prototype. No
significant difference in mutations frequencies was detected between the two age groups.

15

16
AC42 (Brazil)
FNN18 (Brazil)
HB3120 (Brazil)
BABR1237 (Brazil) ¥
FNN39 (Brazil) ¥
BABR1982 (Brazil) ¥ +
BABR3119 (Brazil)
BABR965 (Brazil)
FNN26 (Brazil) ¥ +
BABR3140 (Brazil)
FNN73 (Brazil) +
FNN61 (Brazil) ¥
FNN89 (Brazil) ¥ +
Neu7 (Argentina)
ATL2 (Argentina)
BD16 (Peru)
MASU (Brazil) ¤
IDU1 (Argentina)
FCR (Brazil) ¤
BABR2707 (Brazil)
FNN156 (Brazil)
BA115000 (Brazil)
BABR885 (Brazil)
BABR2470 (Brazil) +
BABR2713 (BRAZIL)
FNN54 (Brazil)
BABR1480 (Brazil) +
FNN153 (Brazil)
FNN158 (Brazil)
BABR1504 (Brazil) +
MAQS (Brazil) ¤
BABR2177 (Brazil)
BABR870 (Brazil)
BABR944 (Brazil)
BABR1851 (Brazil)
BABR526 (Brazil)
FNN57 (Brazil) +
FNN60 (Brazil)
HB3171 (Brazil)
FNN150 (Brazil)
BABR2946 (Brazil)
BABR3167 (Brazil)
BABR945 (Brazil)
BABR1513 (Brazil) ¥
BABR1909 (Brazil) ¥ +
FNN94 ¥
HB3203 (Brazil)
BABR3637 (Brazil)
BABR1790 (Brazil)
FNN148 (Brazil)
FNN31 (Brazil) +
CMC (Taiwan) ¤
FNN 47 (Brazil) +
FNN64 (Brazil)
BABR3417 (Brazil)
BABR3102 (Brazil)
AMA (Brazil) ¤
CH26 (Chile)
RKI1Iran
Afs911 (South African) ¤
CH (North American)
MT2 (Japan)
BABR72 (Brazil)
BABR1764 (Brazil) +
BABR2091 (Brazil) +
BABR2151 (Brazil) +
BABR2095 (Brazil) +
BCI2.1 (Japan)
HKN (Japan)
RKI4Peru (Peru) E- Black Peruvian
BO (Algeria) D- North African
NM2015 (French Guiana) C- West-African/ Caribbean
pyg19 HTLV-1 d
ITIS (Zaire) ¥ HTLV-1 b
MEL5 HTLV-1c
0.01
B- Japanese
A -Transcontinental
HTLV-1a
Smaller Latin American cluster
Larger Latin American cluster
*
*
**
*
*
*
*
*
*
*
* *
**
**
55
58
86
63
95
54
54
50
50

17
Fig. 1. Rooted NJ tree of 172 HTLV-1 strains based upon a 365bp fragment of the U3-LTR region contain the
three 21-base- pair (bp) enhancer element. The percent values (above 50% and using 1000 bootstrap samples)
on the branches represent the percentage of trees for which the sequences at the right end of the branch form a
monophyletic group. Mel5 is used as out-group. Geographic origin is given in parentheses. The** means that
the ML method was highly significant with a p value <0.001 or * significant with a p value <0.005. Newly
sequenced LTRs included in this analysis are in bold. The asymptomatic isolates are represented in blue and
the symptomatic isolates are in red. Symbols indicate the nucleotide substitutions: ¥ 8583 C>T, + 8582+ (A)
insertion and ¤ 8427 (-) deletion.
18
Table III. Identical sequences
References Identical sequences of the references
Similar sequences
of total (not
incluing the
reference)
CMC
BABR1709, BABR664, BABR2674, BABR3510,
BABR1438, BABR2819, BABR3591, HB3194,
BABR0136, BABR394, BABR290, BABR531,
HB3230, BA123070, FNN169, FNN164, FNN29,
FNN170, FNN171, FNN22, BABR643, JCR,
BABR2142, BABR1910, BABR275, FNN28,
BABR1603, BABR3517, HB3166, FNN47
30
BABR965 FNN174, FNN159, FNN40, BABR721 4
BABR945 BABR3518, BABR3698, BABR619, BABR620,
BABR3132, FNN9, FNN100, FNN35, FNN32,
BABR1599, FNN19, FNN83
12
MASU PW1, BD12, BABR2441, HB3160, BABR1260,
BABR3488, FNN165, FNN82 , HB3170, FCR 10
FNN26 FNN87 1
BABR1982 BABR1669 1
BH3120 FNN155, HB3202, HB3134, BABR2986,
BABR1921, AC204 6
FNN54 HB3229, BABR1911 2
BABR 1909 HB2562 1
BABR1764 BABR2091, BABR2095 2
BABR944 BABR3632 1
FNN150 FNN149 1
BABR1504 BABR911, BABR3714, BABR3511, BABR2672,
BABR3173, BABR3115, BABR1389,
BABR1480, BABR2294, BABR2041,
BABR1359, BABR423, HB3114, HB3205,
HB2966, HB3135, AC129, FNN78, BA155630,
FNN172, FNN68, HB3311, HB3133, HB3104,
BABR1759, HB3168, BABR1741
27
BABR72 BABR2224, BABR991, BABR1792, BABR0982 4

19
DISCUSSIO(
The findings of this study reveal, for the first time an association between the variation in
the U3-LTR region of HTLV-1 and the susceptibility to develop TSP/HAM among isolates
from Salvador, Bahia, Brazil.
Thus, the nucleotide substitutions found in positions 8606 (C>A), 8583 (C>T) and 8582+
insertion (A) were considered nine (OR=9.18), fifteen (OR=15.02) and infinite (OR=∞)
times, respectively, more likely to occur in the asymptomatic group than in the symptomatic
group, and could be representing a protective factor against the development of TSP/HAM.
These three nucleotide substitutions are not inside the three 21 pb repeated region but
between a TRE3 and TATA region, which may interfere somehow in the process of
transcription. On the contrary, the nucleotide substitutions observed in positions 8422+
insertion (G), 8582+ (insertion (A) and 8427 (deletion) were three (OR=3.13), fourteen
(OR=14.22) and five (5.18), times, respectively, more frequent in the symptomatic group,
being related to an increased risk of developing TSP/HAM. Interestingly, two of the three
mutations were located within the three 21 pb repeated region: 8422+ (G) is at the end of the
TRE1, 8427 (deletion) is between the TRE1 and TRE2 and the third mutation, 8582+ (A), is
located between the TRE3 and region TATA and therefore may affect the transcription
process.
The TSP/HAM is developing in up to 4% of infected individuals (Orland et al., 2003), and
as usually is expressed in the fourth decade of life (Carneiro-Proietti et al., 2006). The
nucleotide substitutions frequencies (%) in the U3-LTR region between asymptomatic
patients with more (>45) or less (<45) than 45 years old were performed. This comparative
analysis did not indicate any mutation with significant difference in frequency between the

20
two groups. The sample size and age range may be responsible for this result. With a greater
number of asymptomatic individuals the p-value could have been more significant.
Until now, few previous studies have investigated the U3-LTR region polymorphism. A
large variety of nucleotide was observed in the U3 region which contains specific sequences
that optimize transcriptional activity via tax. Among these sequences, repeated three 21-bp,
revealing the Tax-responsive elements named TRE1, TRE2 and TRE3, were located in the
center of each repeat and reported in the cyclic AMP-responsive element [Marriot et al.
1990; Fujisawa et al., 1989; Rosen et al., 1987].
There are probably more mechanisms by which Tax could influence the risk of TSP/HAM
development. Differences in the tax gene might influence the viral load by the difference in
transactivation ability of Tax protein or by the difference in recognition by the CTL
response [Parker et al., 1992; Niewiesk et al., 1995]. Nevertheless, it is possible that Tax
protein via transactivating activity on inflammatory cytokine genes such as interleukin IL-2,
tumor growth factor- β1, tumor necrosis factor-β, IL-1 α, and IL-8 could also influence the
outcome of the HTLV-1 infection [Kannagi et al., 1992]. Recently, Gadelha et al.,
demonstrated that the G allele at -174 and C allele at -634 IL-6 contribute to TSP/HAM
development. In addition, other studies showed that HLA-A*02 type [Jeffery et al., 1999]
and the HTLV-1 tax gene could be classified into two distinct subgroups, tax A and tax B,
and that tax A, based on phylogenetic analysis, probably was associated with the occurrence
of TSP/HAM [Furukawa et al., 2000].
The HTLV-1 genotypes show strong correlation with the geographic origin of the virus
carriers [Miura et al., 1994]. When comparing HTLV-1 subgroups from different countries,
other factors, such as environment or customs may influence the outcome of HTLV-1

21
infection. This creates difficulties in studying the association between HTLV-1 subgroup
and disease outcome. The phylogenetic analysis performed in this study demonstrated that
all sequences belong to the Cosmopolitan subtype, Transcontinental subgroup, and clustered
within two major Latin American clusters (Fig. 1). This suggest that multiple introductions
occurred relatively recently in Salvador, during the post-Columbian introduction, through
the slave trade, as previously reported [Alcantara et al., 2003; Alcantara et al., 2006; Mota et
al, 2007, Miranda et al., 2008]. Moreover, the results suggest that punctual mutations are
associated with HTLV-1 subgroup.
We have not concluded yet that polymorphisms in the U3-LTR region of HTLV-1 is
responsible for TSP/HAM development, but they are associated with a higher risk of
TSP/HAM development. It is also possible that other parts of the HTLV-1 genome that are
firmly associated with subgroup, could be responsible for the outcome of HTLV-1 infection.

22
ACK(OWLEDGME(T
This work was partially supported by (FAPESB) and PN-DST/AIDS, Brazilian Ministry of
Health. The authors are grateful to Elisabeth Deliege for technical assistance.
REFERE(CES
Abramson JH, Gahlinger PM. 2001. Computer program for epidemiologists: PEPI. Version
4.0. Salt Lake City, UT: Sagebrush Press.
Alcantara LC, de Oliveira T, Gordon M, Pybus O, Mascarenhas RE, Seixas MO, Gonçalves
M, Hlela C, Cassol S, Galvão-Castro B. 2006. Tracing the origin of Brazilian HTLV-1 as
determined by analysis of host and viral genes. AIDS 20(5):780-782.
Alcantara LCJ, Van Dooren S, Gonçalves MS, Kashima S, Costa MCR, Santos FLN,
Bittencourt AL, Dourado I, Andrade Filho A, Covas DT, Vandamme A-M, Galvão-Castro
B. 2003. Globin haplotypes of human T-cell lymphotropic virus type I (HTLV-I) infected
individuals in Salvador, Bahia, Brazil suggest a Post-Columbian African origin of this virus.
J Acquir Immune Defic Syndr Hum Retrovirol 33:536-542.
Bangham CRM, Hall SE, Jeffery KJM, Vine AM, Witkover A, Nowak M A, Wodarz D,
Ususku K, Osame M. 1999. Genetic control and dynamics of the cellular immune response
to the human T-cell leukaemia virus, HTLV-1. Phil Trans R Soc B 354:691-700.
Beby-Defaux A, Frugier F, Bourgoin A, Moynet D, Hajjar C, Sainte-Foie S, Guillemain B,
Agius G. 1999. Nucleotide sequence analysis of human T- cell lymphotropic virus type I pX
and LTR regions from patients with Sicca syndrome. J Med Virol 59:245-255.

23
Brady J, Jeang, Duvall J, Khoury G. 1987. Identification of p40x-responsive regulatory
sequenses within the human T-cell leukemia virus type I long terminal repeat. J Virol 61(7):
2175-2181.
Cartharius K, Frech K, Grote K, Klocke B, Haltmeier M, Klingenhoff A, Frisch M,
Bayerlein M, Werner T. 2005. MatInspector and beyond: promoter analysis based on
transcription factor binding sites. Bioinformatics 21(13):2933-2942.
Colin DD, Alcantara LCJ, Santos FLN, Uchôa R, Tavares-Neto J. 2003. Seroprevalence of
human T cell lymphotropic virus infection and associated factors of risk in blood donors of
Rio Branco city, AC, Brazil (1998-2001). Rev Soc Bras Med Trop 36(6):677-683.
Dan S, Tanimura A, Yoshida M. 1999. Interaction of Gli2 with CREB protein on DNA
elements in the long terminal repeat of human T-cell leukemia virus type 1 is responsible
for transcriptional activation by tax protein. J Virol 73:3258-63.
Dourado I, Alcantara JLC, Barreto ML, Teixeira MG, Galvão-Castro B. 2003. HTLV-1 in
the general population of Salvador, Brazil: a city with African ethnic and sociodemographic
characteristics. J Acquir Immune Defic Syndr Hum Retrovirol 34:527-531.
Fujisawa J, Seiki M, Sato M, Yoshida M. 1986. A transcriptional enhancer sequence of
HTLV-I is responsible for trans-activation mediated by p40 chi HTLV-I. EMBO J 5(4):713-
8.
Fujisawa J I, Toita M, Yoshida M, 1989. A unique enhancer element for the trans activator
(p40tax) of human T-cell leukemia virus type I that is distinct from cyclic AMP-and 12-O-
tetradecanoylphorbol-13-acetate-responsive elements. J Virol 63(8):3234-3239.

24
Furukawa Y, Yamashita M, Usuku K, Izumo S, Nakagawa M, Osame M. 2000.
Phylogenetic subgroups of human T cell lymphotropic virus (HTLV) type I in the tax gene
and their association with different risks for HTLV-I-associated myelopathy/tropical spastic
paraparesis. J Infect Dis 182(5):1343-1349.
Gadelha SR, Alcantara LCJ, Costa GC, Acosta AX, Rios D, Kashima S, Covas DT, Castro
BG. 2008. Correlation between Polymorphisms at Interleukin-6 but not at Interleukin-10
promoter and the risk of human T lymphotropic virus type 1- associated myelopathy/tropical
spastic paraparesis in Brazilian individuals. J Med Virol 80(12):2141-2146.
Gasmi M, Farouqi B, d’Incan M, Desgranges C. 1994. Long terminal repeat sequence
analysis HTLV type I molecular variants identified in four North African patients. AIDS Res
Hum Retroviruses 10(10): 1313-1315.
Gessain A, Barin F, Vernant JC, Gout O, Maurs L, Calender A, de The G. 1985. Antibodies
to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet 2:
407-410.
Iñiguez AM, Otsuki K, Gastaldello R, Gallego S, Vicente ACP. 2007. HTLV-1a tax Gene
and Long Terminal Repeat sequences from Argentinean Strains Reveal Disagreement with
tax Restriction Fragment Length Polymorphism Subtyping. AIDS Res Hum Retroviruses
23(9):1127-1130.
Jeanmougin F, Thompson JD, Goy M, Higgins DG, Gibson TJ. 1998. Multiple sequence
alignment with clustalX. Trends Biochem Sci 23:403-405.
Jeffery KJ, Usuku K, Hall SE, , Matsumoto W, Taylor GP, Procter J, Bunce M, Ogg GS,
Welsh KI, Weber JN, Lloyd AL, Nowak MA, Nagai M, Kodama D, Izumo S, Osame M,

25
Bangham CR. 1999. HLA alleles determine human T-lymphotropic virus-1 (HTLV-1)
proviral load and the risk of HTLV-1 associated myelopathy. Proc Natl Acad Sci USA
96(7):3848-3853.
Kannagi M, Shida H, Igarashi H, Kuruma K,Murai H, Aono Y,Maruyama I,Osame M,
Hattori T, Inoko H.1992. Target epitope in the tax protein of human T-cell leukemia virus
type 1 recognized by class major histocompatibility complex restricted cytotoxic T cells. J
Virol 66:2928-2933.
Kashima S, Alcantara LJ, Takayanagui OM, Cunha MAV, Castro BG, Oliveira MSB, Zago
MA, Covas DT. 2006. Distribution of human T cell lymphotropic virus type 1 (HTLV-1)
subtypes in Brazil: genetic characterization of LTR and Tax region. AIDS Res Hum
Retroviruses 22:953-959.
Lemasson I, Polakowski NJ, Laybourn PJ, Nyborg JK. 2004. Transcription regulatory
complexes bind the human T-cell leukemia virus 5´ and 3´ long terminal repeats to control
gene expression. Mol Cell Biol 24(14): 6117-6126.
Marriott SJ, Lindholm PF, Brown KM, Gitlin SD, Duvall JF, Radonovich MF, Brady JN.
1990. A 36-Kilodalton cellular transcription factor mediates an indirect interaction of human
T-cell leukemia/lymphoma virus type I Tax1 with a responsive element in the viral long
terminal repeat. Mol Cell Biol 10(8):4192-4201.
Miranda ACM, Pereira SA, Dias JP, Colin DD, Kashima S, Covas DT, Neto JT, Castro BG,
Alcantara LCJ. 2008. HTLV-1 infection in blood donors from the Western Brazilian
Amazon region: seroprevalence and molecular study of viral isolates. J Med Virol 80:1966-
1971.

26
Miura T, Fukunaga T, Igarashi T. 1994. Phylogenetic subtypes of human T-lymphotropic
virus type I and their relations to the anthropological background. Proc Natl Acad Sci USA
91(3):1124-1127.
Mochizuki M, Watanabe T, Yamaguchi K, Yoshimura K, Nakashima S, Shirao M, Araki S,
Takatsuki K, Mori S, Miyata M. Uveitis.associated with human T-cell lymphotropic virus
type I. AmericanJournal Ophthalmology 114: 123-129, 1992.
Mota ACA, Van Dooren S, Fernandes FMC, Pereira SA, Queiroz ATL, Gallazzi VO,
Vandamme AM, Castro BG, Alcantara LCJ. 2007. The close relationship between South
African and Latin American HTLV-1 type 1 strains corroborated in a molecular
epidemiological study of the HTLV type 1 isolates from a blood donors cohort. AIDS Res
Hum Retroviruses 23:503-507.
Mukhopadhyaya R, Sadaie M.R. 1993. Nucleotide sequence analysis of HTLV-I isolated
from cerebrospinal fluid of a patient with TSP/HAM: comparison to other HTLV-I isolates.
AIDS Res Hum Retroviruses 9(2):109-114.
Nicholas KB, Nicholas HBJ, Deerfield DW. 1997. GeneDoc: Analysis and visualization of
genetic variation. Embnew News 4:14.
Niewiesk S, Daenke S, Parker CE, Taylor G, Weber J, Nightingale S, Bangham CR. 1995.
Naturally occurring variants of human T-cell leukemia virus type 1 tax protein impair its
recognition by cytotoxic T lymphocytes and the transactivation function of Tax. J Virol
69:2649-2653.
Orland JR, Engstrom J, Fridey J, Sacher RA, Smith JW, Nass C, Garratty G, Newman B,
Smith D, Wang B, Loughlin K, Murphy EEL. 2003 Prevalence and clinical features of
HTLV-1 neurologic disease in the HTLV-1. Outcomes study. Neurology 61(11):1588-1594.

27
Osame M, Usuku K, Izumo S,Ijichi N,Amitani H,Igata A, Matsumoto M, Tara M. 1986.
HTLV-I associated myelopathy, a new clinical entity. Lancet 1(8488): 1031-1032.
Page RDM. 1996. TreeView: An application to display phylogenetic trees on personal
computers. Cabios Applications Note 12:357-358.
Parker CE, Daenke S, Nightingale S, Bangham CR. 1992. Activated HTLV-1-specific
cytotoxic T-lymphocytes are found in the healthy seropositives as well as in patients with
tropical spastic paraparesis. Virology 188(2):628-636.
Poiesz BJ, Ruscetti FW, Gazdar AF, Bunn PA, Minna JD, Gallo RC. 1980. Detection and
isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with
cutaneous T-cell lymphoma. Proc Natl Acad Sci USA 77:7415–7419.
Posada D, Crandall KA. 1998. MODELTEST: testing the model of DNA substitution.
Bioinformatics 14(9): 817-818
Renjifo B, Borrero I, Essex M. 1995. Tax mutation associated with tropical spastic
paraparesis/human T-cell leukemia virus type I-associated myelopathy. J Virol 69: 2611-
2616.
Rosen CA, Park R, Sodroski JG, Haseltine WA. 1987. Multiple sequence elements are
required for regulation of human T-cell leukemia virus gene expression. Proc Natl Acad Sci
USA 84(14):4919-4923.
Sabouri AH, Saito M, Usuku K, Bajestan SN, Mahmoudi M, Forughipour M, Sabouri Z,
Abbaspour Z, Goharjoo ME, Khayami E, Hasani A, Izumo S, Arimura K, Farid R, Osame
M. 2005. Differences in viral and host genetic risk factors for development of human T-cell

28
lymphotropic vírus type 1 (HTLV-1) associated myelopathy/tropical spastic paraparesis
between Iranian and Japanese HTLV-1 infected individuals. J Gen Virol. 86:773-781.
Shimotohno K, Takano M, Teruuchi T, Miwa M.1986. Requirement of multiple copies of a
21-nucleotide sequence in the U3 regions of human T-cell leukemia virus type I and type II
long terminal repeats for trans-acting activation of transcription. Proc Natl Acad Sci USA
83(21):8112-8116.
Swofford D. PAUP 4.0: 1999. Phylogenetic analysis using parsimony (and other methods),
4.0b2a. Sunderland, MA, USA: Sinauer Associates, Inc.
Tanimura A, Teshima H, Fujisawa J, Yoshida M. 1993. A new regulatory element that
augments the Tax-dependent enhancer of human T-cell leukemia virus type 1 and cloning of
cDNAs encoding its binding proteins. J Virol 67:5375-5382.
Tanimura A, Dan S, Yoshida M. 1998. Cloning of novel isoforms of the human Gli2
oncogene and their activities to enhance tax-dependent transcription of the human T-cell
leukemia virus type 1 genome. J Virol 72:3958-64.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins D. 1997. The CLUSTAL_X
Windows interface: flexible strategies for multiple sequence alignment aided by quality
analysis tools. Nucleic Acids Res 25(24):4876-4882.
Van Dooren S, Gotuzo E, Salemi M, Watts D Audenaert E, Duwe S, Ellerbrok H,
Grassmann R, Hagelberg E, Desmyter J, Vandamme AM. 1998. Evidence for a post-
Columbian introduction of human T-cell lymphotropic virus in Latin America. J Gen Virol
79:2695-2708.

29

24
7. DISCUSSÃO
A população brasileira é uma das mais heterogêneas do mundo, resultado de cerca de
500 anos de miscigenação, principalmente entre ameríndios, europeus e africanos. Mais
recentemente, ocorreu, no início do século XX, uma onda importante de migração de asiáticos,
predominantemente de japoneses. Desde o descobrimento do Brasil, no início do século XVI, até
o início da oitava década do século XX, há relatos que a população brasileira era constituída por
um pouco mais da metade (58%) de descendentes de europeus, cerca de 40% afrodescendente e
2% de descendentes asiáticos, predominantemente de japoneses (Callegari-Jacques and Salzano,
1999). Porém, esta miscigenação não é homogênea no território nacional, variando nas regiões
geográficas. Estima-se que as regiões Norte (68,97%) e Nordeste (65,8%) tenham o maior
contingente de afrodescendente (pretos + pardos) e o segundo maior percentual de negros (7,7%).
A maior concentração de indivíduos brancos é observada nas regiões Sudeste (62,4%) e Sul
(83,6%). Por fim, a maior concentração de indígenas, somente (1,65%), encontra-se na região
Norte.
A Cidade do Salvador, capital do Estado da Bahia, foi fundada como São Salvador da
Bahia de Todos os Santos, em 29 de março de 1549. Foi a primeira capital do Brasil e
permaneceu nesta condição durante 214 anos, de 1549 a 1763. Construída numa posição
estratégica no alto de uma escarpa, entre a Baía de Todos os Santos e os morros, possibilitou
ligações entre Portugal, Brasil, África e Ásia. Salvador, desde a sua fundação, desempenhou
papeis importantes tais como porto de apoio às rotas do Oriente e grande centro de exportação de
açúcar e de pau-brasil. Os africanos foram trazidos como escravos, em grande quantidade para o
cultivo da cana. A extração de pau-brasil era a maior de todo território e o porto do recôncavo,
além de mais próximo da Europa, era o principal responsável pelo escoamento de matérias-
primas. Esses fatos contribuíram para a formação de uma população mestiça de portugueses e de
africanos.
Atualmente, a população do Estado da Bahia é composta por 77,5% de afrodescendente,
o que corresponde a 10.095.282 indivíduos. Salvador tem três milhões e cerca de 80% dos
indivíduos se autodenominam como negro-pardos (Censo Demográfico-IBGE, 2000). No
entanto, existe uma distribuição heterogênea dos grupos étnicos em diferentes regiões. Observa-
se que nas regiões que tiveram importância econômica há uma predominância de afrodescente.

25
Por exemplo, as regiões litorâneas e sul e a Chapada Diamantina, onde está localizada a cidade de
Lençóis, que respectivamente, desenvolveram o cultivo da cana-de-açúcar, do cacau e a
exploração de minérios (Abes-Sandes et al., 2004).
Estima-se que a Cidade do Salvador recebeu o maior contingente dos africanos trazidos
para o Brasil durante o tráfico negreiro e que a maioria deles foi proveniente do Benin e da
Nigéria, países localizados no oeste da África (Viana Filho, 1988; Verger, 1976). Entre os anos
1678-1810, final do século XVII e início do século XIX, não existe nenhuma informação sobre a
rota dos navios negreiros vindos da África para a Bahia. Uma possibilidade é que, neste período,
os africanos poderiam ter sido trazidos de outras regiões do continente sul africano, como o sul de
Angola, Madagascar e Moçambique.
No entanto, em relação aos subtipos e subgrupos do HTLV-1 observou-se que a maioria
dos isolados do HTLV-1, provenientes de indivíduos nascidos nesta cidade, foi classificado como
do subtipo Cosmopolita (a), subgrupo Transcontinental (A) (Alcantara et al., 2003; Alcantara et
al., 2006). Um único isolado, oriundo de uma gestante, foi classificado como subgrupo Japonês.
Chama a atenção o fato de não se ter encontrado o subtipo do oeste da África e de a maioria dos
isolados se agruparem com amostras da África do Sul.
Tem sido difícil explicar porque todos os isolados provenientes de soteropolitanos
pertencem ao subtipo Cosmopolita (a), subgrupo Transcontinental (A) e não ao subgrupo (C) do
oeste da África, contradizendo dados históricos. Para explicar esta contradição tem sido sugerido
que os africanos que vieram para o Brasil durante o tráfico negreiro tenham se originado de
outras regiões da África. Analisando os ciclos de migrações no período do tráfico negreiro,
observamos que durante o primeiro ciclo, conhecido como da Guiné, os navios negreiros partiam
da costa oeste da África, região onde hoje está situado o Senegal, Gâmbia e Guiné Bissau e que
naquela época correspondia à área compreendida entre a embocadura do rio Senegal, limite da
região desértica entre Senegal e Mauritânia, até o rio Orange, no atual Gabão e era denominada
de Guiné. Todos os africanos trazidos para o Brasil, nesta época, eram de forma genérica,
considerados como oriundos da Guiné, não havendo precisão da sua proveniência (Pinsky, 1988).
No período compreendido entre 1641-1648, Angola estava sob o domínio dos holandeses e o
fluxo de navios negreiros predominava da África para Pernambuco, mas em torno de 1624,
período de dominação holandesa no Estado da Bahia, foi relatado que chegaram ao porto da
Cidade do Salvador, 7 navios negreiros sendo 6 provenientes de Angola, com 1.440 africanos, e

26
um único da Guiné com 28 africanos (Viana Filho, 1988). Esse ciclo foi pouco representativo
para a Bahia e estima-se que sete mil africanos, em sua maioria pertencente às etnias sudanesas,
tenham vindo para este estado neste período. Este fato poderia explicar a ausência do subgrupo
do oeste africano na Bahia. O segundo ciclo, denominado de Ciclo de Angola e do Congo,
compreendeu o final do século XVI, tendo início por volta de 1580 e chegou no seu final no
século XVII. Sabe-se que neste período a maioria dos africanos foi proveniente da África Austral,
banhada pelo Oceano Índico na sua costa oriental e pelo Atlântico na costa ocidental. Não se sabe
exatamente o número de africanos que vieram desta região. Além de angolanos e congos, vieram,
também, moçambicanos, a maioria deles da etnia banto tendo grande parte deles se instalado na
região do Recôncavo. O terceiro ciclo, denominado de Ciclo da Costa da Mina, ocorreu durante
os dois terços iniciais do século XVIII e contribuiu com maior número de africanos trazidos para
o Brasil. Neste período, que durou até meados do século XVIII, aproximadamente até 1775, estes
africanos, denominados negros minas, eram oriundos de quatro portos que ficavam no leste da
África, ao longo da costa do Daomé, situado onde atualmente é o Benin, região conhecida como
Costa da Mina. A maioria dos africanos era iorubá, chegando à Bahia com a denominação de
nagôs. Entre 1678 e 1814, de acordo com Verger, apenas 39 dos 1.770 navios que levavam
tabaco da Bahia para a África tinham como destino os portos congo-angolanos, sendo os outros
destinados aos portos da Costa da Mina (Florentino, 1997).
O ciclo da Baía de Benin ocorreu entre 1770 e 1850, incluindo o tráfico clandestino,
uma vez que a Bahia acusou a limitação ao tráfico ao norte do Equador em 1815, fato que explica
a forte ligação da Bahia com a África Ocidental, particularmente com a Baía de Benin
(Florentino, 1997). Nesse período, a forte presença de etnias iorubá passa a ser representativa,
visto que, desde o ciclo anterior, são encontrados daomeanos, tratados no Brasil pela
denominação de jejes, haussás e, sobretudo os nagôs-iorubás. Pelo exposto nos relatos históricos
há uma mistura da origem geográfica com as diversas etnias não havendo grande precisão da
proveniência dos africanos para a Bahia (Pinsky, 1988).
Com o objetivo de esclarecer melhor a origem do HTLV-1 em Salvador, analisou-se a
distribuição de haplótipos ligados ao grupo de genes da globina β na população estudada
(Alcantara et al., 2003; Alcantara et al., 2006). Estes haplótipos compreendem grupo de
polimorfismos genéticos associados à presença ou ausência de sítios para endonucleases de
restrição e estão relacionados a diferentes etnias e regiões geográficas, sendo utilizados como

27
marcadores de migrações populacionais em estudos antropológicos. Foi observado que 45,2%
dos haplótipos do gene da globina β eram do tipo benin; 28,9% do tipo banto e 25,9% do tipo
senegal. A prevalência encontrada do haplótipo tipo benin está de acordo com os relatos
históricos, pois a maioria dos africanos que vieram para a Bahia foi do oeste da África
principalmente no século XVIII (Viana Filho, 1988). O haplótipo tipo senegal, característico da
África Atlântica, já foi demonstrado anteriormente em Salvador (Gonçalves et al., 2002).
Portanto, a não detecção do subtipo do oeste da África do HTLV-1 continua sendo uma
incógnita. A demonstração de 29,4% de haplótipo tipo banto poderia explicar porque a maioria
dos isolados de HTLV-1 se agrupa com isolados do sul da África. Sabe-se que africanos da etnia
banto foram trazidos para a Bahia entre 1678-1810 (Verger, 1976). De fato foram trazidos para a
Bahia cerca de 2.400 africanos bantos, sendo 100 procedentes de Angola e 2.300 de Madagascar
no período compreendido entre os anos de 1817-1843 (Curtin, 1969). Há também evidências que
muitas vezes os portos de partida dos navios negreiros na África não estavam necessariamente
relacionados com as origens étnicas e geográficas dos Africanos transportados. Sabe-se que no
período de colonização da África do Sul pelos ingleses, nos séculos XVII e XVIII, muitos
africanos migravam para outras regiões vizinhas, atualmente conhecidas como Angola,
Madagascar e Moçambique, onde eram capturados e transportados para Salvador. Há também
evidências de que os bantos povoaram a Bahia, principalmente a Cidade do Salvador. Nesta
cidade, ainda persistem, sobremaneira, os marcos da cultura banto que influenciou a língua, a
religião, a música e o folclore baiano principalmente o soteropolitano.
Para obter um melhor entendimento da origem do HTLV-1 em Salvador, realizamos
uma análise filogenética com as sequências de isolados provenientes de doadores de sangue desta
cidade (Mota et al., 2007) (manuscrito1). O resultado da análise filogenética revelou que todos os
isolados pertenciam ao subtipo Cosmopolita, Subgrupo Transcontinental. Além disso,
observamos que as sequências de agrupamento do Sudeste da África estavam inseridas nos
grupos alfa e beta do grupo latino-americano. Estes resultados indicam uma relação mais próxima
entre os isolados do HTLV-1 da América Latina e do Sul-Africano do que em relação aos
isolados do HTLV-1 do Oeste Africano, corroborando com resultados anteriores (Van Dooren et
al., 1998; Alcantara et al., 2003; Alcantara et al., 2006). Além do mais, observamos que as
sequências se segregavam em dois diferentes grupos, mais de forma homogênea dentro de cada
grupo latino-americano sugerindo que houve múltiplas introduções recentes do HTLV-1, no

28
período pós-colombiano, durante os séculos XVII, XVIII e XIX, na Cidade do Salvador, como já
foi sugerido anteriormente (Van Dooren et al., 1998; Alcantara et al., 2003; Alcantara et al.,
2006; Yamashita et al., 1999; Van Dooren et al., 2004). Entretanto, não foi possível prever as
datas anteriores uma vez que o relógio molecular foi rejeitado. No entanto, estes resultados
parecem contradizer os dados históricos, segundo a qual a maioria dos escravos trazidos para
Salvador veio originalmente da África ocidental (Curtin, 1969; Verger,1976), onde o HTLV-1
AC é predominante (Wattel et al., 1995). Por outro lado, corrobora a hipótese de que os
indivíduos originários da África Central, onde predomina o haplótipo CAR, infectados pelo
HTLV-1, poderiam ter emigrado para o Sul da África. Este haplótipo é prevalente na África
Central, África do Sul, Moçambique e outras regiões do Sudeste da África. Sua origem no
Sudeste da África não é conhecida, mas poderia refletir a migração da população banto da região
Central para o Sudeste do continente africano durante os últimos 3.000 anos. A existência de
poucas sequências de isolados provenientes do continente africano dificulta uma conclusão
definitiva.
No intuito de se conhecer melhor a soro prevalência e a origem do HTLV-1 no Brasil,
estudamos isolados de doadores de sangue de Rio Branco, Acre (Miranda et al., 2008)
(manuscrito 2). Determinamos uma prevalência da infecção pelo HTLV-1 de 0,46%. Este
resultado corrobora dados anteriores (Catalan-Soares et al., 2005), mas é maior que o encontrado
(0,11%), entre 1998 e 2001 por (Colin et al., 2003).
Este foi o primeiro estudo que avaliou as características genéticas e a epidemiologia
molecular da população do Rio Branco-Acre, caracterizada por ter um componente indígena
considerável. Estudo anterior demonstrou que a frequência de características fenotípicas do grupo
misto ameríndio racial da população do Acre é de 33,3% (Colin et al., 2003).
Sabe-se também que alguns estados da região amazônica apresentam as maiores
prevalências do HTLV-1 no país (Ishak et al., 2002; Catalan-Soares et al., 2005). A análise
filogenética realizada neste estudo identificou dois isolados que foram classificados como do
subgrupo Transcontinental (A), subtipo Cosmopolita (a). Este achado está de acordo com outros
estudos que indicaram a presença de um grande numero de isolados do Brasil neste subgrupo
(Queiroz et al., 2007). Resultados das análises filogenéticas deste estudo sugerem pelo menos
duas hipóteses para a origem deste subgrupo (A) do HTLV-1, subtipo Cosmopolita (a) no Novo
Mundo. A primeira hipótese é que a introdução do vírus ocorreu durante a migração através do

29
Estreito de Bering em torno de 40.000 a 10.000 anos atrás, corroborando dados anteriores (Miura
et al., 1994). A segunda hipótese sugere que o vírus foi trazido durante o período pós-colombiano
através do tráfico de escravos africanos, cerca de 400 anos atrás (Van Dooren et al., 1998). O fato
de que os isolados do HTLV-1 provenientes do Rio Branco foram agrupados em dois clusters
distintos e bem definidos, e que estes dois clusters demonstram uma baixa divergência,
poderíamos sugerir uma propagação recente e separada do subtipo Cosmopolita do HTLV-1
nesta população latino-americana. Uma conclusão similar foi considerada em um estudo
molecular recente de isolados peruanos do HTLV-1, quando três dos dez novos isolados do
HTLV-1 ficaram inseridos no principal grupo Sul-Africano, enquanto que sete deles se
agruparam em dois clusters da América Latina (Zehender et al., 2007). Comparando-se com
outras cepas brasileiras, a maior diversidade genética entre as cepas do Acre foi de (2,7%), dando
suporte à hipótese da migração da população através do Estreito de Bering para Continente
Americano, e depois para o Norte do Brasil.
Por outro lado, a baixa diversidade genética entre as cepas do Nordeste brasileiro
(Salvador-1.2% e Fortaleza- 0.7%) poderia sugerir que o vírus também foi trazido para o Brasil,
durante o período pós-colombiano, através do tráfico de escravos. Outra possibilidade é que tenha
sido introduzido mais recentemente por usuários de drogas injetáveis (Alcantara et al., 2003;
Shindo et al., 2002). Além disto, a presença do subtipo japonês já demonstrado no território
nacional indica a introdução do HTLV-1 através de migrações de asiáticos, principalmente
japoneses, que ocorreu no século XX (Alcântara et al., 2003; Kashima et al., 2006)
Portanto, poderíamos concluir que ocorreram múltiplas introduções do HTLV-1 no
Brasil predominantemente no período pós-colombiano, mas não poderíamos descartar que houve
introdução na era pré-colombiana. De fato, observamos uma alta diversidade (2.7%) entre os
isolados do Acre e as do Nordeste do Brasil (Salvador e Fortaleza). Será importante analisamos
um maior número de amostras provenientes da população indígena brasileira.
Se é amplamente confirmado que subtipos do HTLV-1 estão correlacionados com a
origem geográfica, não existe até o momento evidência de associação destes subtipos com a
patogênese da infecção causada pelo HTLV-1. No entanto, dados da epidemiologia molecular do
HTLV-1 são extremamente relevantes, pois nos permitem compreender melhor não só a origem e
disseminação deste vírus, mas as rotas migratórias das populações humanas em tempos remotos.

30
A associação do vírus com as distintas patologias clínicas promove uma série de
incertezas relacionadas ao padrão observado nas características do vírus e do hospedeiro. A
proliferação espontânea das células T infectadas e a alta carga pro viral são efeitos observados em
pacientes com TSP/HAM quando comparados com pacientes com ATL (Gessain et al., 1990;
Tendler et al., 1990; Yoshida et al., 1989).
A expressão dos genes na maioria dos retrovírus é regulada pelos elementos cis
localizados nas regiões LTRs do genoma pro viral (Dan et al., 1999). Entretanto, o HTLV-1
apresenta um único sistema regulatório para aumentar a sua própria expressão de genes. A
transcrição do HTLV-1 é ativada pela interação da proteína viral transativadora, Tax e, os fatores
de transcrição celular, a proteína CREB (cyclic AMP response element binding protein) e, a
proteína fosforilada CBP (CREB binding protein), formando um complexo transcricional ativo
que se liga na sequência (enhancer) de três 21-bp da região LTR 5`do HTLV-1 (Tanimura et al.,
1998; Dan et al., 1999).
Estudos moleculares na região do gene tax têm sido intensamente realizados já que se
trata de uma região importante que codifica a proteína regulatória Tax. Esta proteína poderia ser
um dos fatores que promovem a patogenicidade nas células infectadas, sendo de crucial
importância para o ciclo biológico do vírus, pois aumenta significantemente a eficácia da
transcrição gênica e a multiplicação viral. Além disso, já foi evidenciada a sua capacidade de
desregular vários genes celulares, facilitando a proliferação descontrolada da célula infectada.
O HTLV-1 é caracterizado pela sua alta estabilidade genômica (Salemi et al., 1999). No
entanto, a região da LTR é a mais variável (Miura et al., 1994). Há poucas investigações que têm
determinado correlações genotípicas entre a patogenicidade e os vários subtipos de HTLV-1.
Estudos das propriedades moleculares e biológicas deste vírus em indivíduos infectados poderão
fornecer informações importantes sobre sua patogenicidade. Existem cada vez mais evidências de
que polimorfismos de único nucleotídeo (SNP) estão associados com doenças (Barkur, 2007).
Em relação à patogênese da TSP/HAM existem resultados controversos.
Komurian e colaboradores (1991) não observaram associação entre o desenvolvimento
de TSP/HAM e ATLL com variações específicas nos genes LTR, env e pX estudando um número
pequeno de indivíduos (4 japoneses, 2 de Costa do Marfim e 2 caribenhos). Em 1999, Beby-
Defaux e colaboradores analisaram a sequência das regiões LTR e pX de 15 indivíduos com
diferentes patologias associadas ao HTLV-1 (4 com TSP/HAM e síndrome de Sicca, 2 com

31
síndrome de Sicca, 1 com TSP/HAM, 4 com ATL e 4 indivíduos assintomáticos), porém não
encontraram correlação entre mutações gênicas e o desenvolvimento dessas patologias. Em 2000,
Furukawa e colaboradores, através da análise da sequência do gene tax do HTLV-1 de 61
pacientes com TSP/HAM, 55 ATLL e 62 assintomáticos, classificaram as mutações observadas
em dois subgrupos: taxA (sequências que apresentavam 4 mutações específicas) e taxB
(sequências que não apresentavam tais mutações). Comparando a análise filogenética do gene
LTR com a análise filogenética do gene tax, observaram que o subgrupo taxA era relacionado ao
subtipo Cosmopolita, subgrupo Transcontinental e o subgrupo taxB era relacionado ao subtipo
Cosmopolita, subgrupo Japonês (B). Analisando-se todos os isolados dos subgrupos taxA e taxB,
observaram que o taxA é mais frequente em pacientes com TSP/HAM do que em pacientes
assintomáticos. Mas não observaram nenhuma correlação entre níveis de carga proviral no
sangue periférico com as mutações taxA e taxB.
Várias mudanças de nucleotídeos na região U3 da LTR parecem ser comuns entre
isolados de pacientes com TSP/HAM (Mukhopadhyaya et al., 1993). Observou-se que isolados
de indivíduos assintomáticos com a mutação do nucleotídeo T7959→C no gene tax apresentam alto
risco de desenvolver TSP/HAM (Renjifo et al., 1995). Muitas deleções na região U3 da LTR do
HTLV-1 resultam em uma menor ativação da transativação transcricional (Shimotohno et al.,
1986). Vários trabalhos relatam que algumas deleções encontradas na região da LTR em isolados
do HTLV-1 e que apresentaram somente uma cópia da sequência de 21 pb (enhancer), mas que
possuíam a sequência adjacente TRE2S, foram ativados pela proteína Tax (Brady et al, 1987;
Fujisawa et al., 1986; Tanimura et al., 1998). A deleção da sequência TRE2S diminuiu
drasticamente a atividade na resposta da ativação pela proteína Tax (Brady et al., 1987; Tanimura
et al., 1993; Tanimura et al., 1998).
Neste trabalho (manuscrito 3), analisando isolados provenientes da Cidade do Salvador e
de Rio Branco, demonstramos que as substituições de nucleotídeos encontradas nas posições
8606 (C> A), 8583 (C> T) e entre as posições 8582 e 8583 uma inserção (A) foram consideradas
nove (OR = 9,18), quinze (OR = 15,02) e infinito (OR = ∞) vezes mais provável de ocorrer no
grupo assintomático do que no grupo sintomático, o que representa um possível fator de proteção
ao desenvolvimento da TSP/HAM. Estas três substituições de nucleotídeos não estão dentro da
região de 21 pb repetido três, mas entre a TRE3 e a região TATA, o que poderia interferir
também de alguma forma no processo de transcrição. Por outro lado, as substituições de

32
nucleotídeos observadas nas posições entre 8422 e 8423 uma inserção (G), 8582 e 8583 uma
inserção (A) e na posição 8427 uma deleção foram três (OR = 3,13), quatorze (OR = 14,22) e
cinco (5,18) vezes mais frequentes no grupo sintomático do que no grupo do assintomático,
respectivamente, estando relacionadas a um maior risco de desenvolvimento de TSP/HAM.
Curiosamente, duas das três mutações estão localizadas entre as três sequências de 21 pb
repetidas: entre as posições 8422 e 8423 (G) está no final da TRE1, 8427 (deleção) está entre as
TRE1 e TRE2, e a mutação (A) entre as posições 8582 e 8583 está localizada entre a TRE3 e a
região TATA e, portanto, também possível de afetar de alguma forma o processo de transcrição.
A análise comparativa das frequências (%) de substituições de nucleotídeos na região U3-LTR
entre pacientes assintomáticos com idade superior ou inferior a 45 anos não revelou diferença
significativa na frequência entre os dois grupos. O tamanho da amostra estudada entre estes
grupos pode ter sido responsável por este resultado. Com um número maior de indivíduos do
grupo assintomáticos, o valor do p poderá apresentar um valor mais significativo. Além disso, os
resultados indicam que as mutações pontuais que tiveram o valor de p significantes não estão
associadas com o subgrupo do HTLV-1, corroborando com resultados anteriores (Kashima et al.,
2006).
Estudos funcionais e de seguimento destes pacientes serão fundamentais para esclarecer
o real papel destas mutações na proteção ou patogênese da TSP/HAM.
Dados da epidemiologia molecular do HTLV-1 são extremamente relevantes, pois nos
permitem compreender melhor não só a origem e disseminação deste vírus, mas as rotas
migratórias das populações humanas em tempos remotos em torno do mundo.

33
8. CO�CLUSÃO
� A soroprevalalência do HTLV-1 em doadores de sangue do Acre foi de 0,46%.
� A análise filogenética dos fragmentos da região LTR dos isolados estudados
classifica-os como pertencentes ao subgrupo Transcontinental do subtipo
Cosmopolita, confirmando a presença majoritária deste subgrupo na população
brasileira.
� Os resultados da análise filogenética sugerem que ocorreram introduções recentes e
que há uma relação mais próxima entre isolados da América Latina com os do Sul da
África do que em relação aos isolados do Oeste Africano.
� Os resultados da análise filogenética confirmam a introdução pós-colombiana do
vírus em Salvador. Porém, não exclui uma introdução pré-colombiana.
� As mutações encontradas na região U3-LTR podem estar associadas com o
desenvolvimento da TSP/HAM.
� Os resultados sugerem que as mutações pontuais com valor de p significantes não
estão associadas com o subgrupo do HTLV-1.
� Não foi observada correlação entre as mutações encontradas com a idade dos
pacientes.

34
9. REFERÊ�CIAS BIBLIOGRÁFICAS
Alcantara L. C., Van Dooren S., Goncalves M. S., Kashima S., Costa M. C., Santos F. L.,
Bittencourt A. L., Dourado I., Filho A. A., Covas D. T., Vandamme A. M., Galvao-Castro
B.,: Globin Haplotypes of Human T-Cell Lymphotropic Virus Type 1 Infected Individuals
in Salvador, Bahia, Brazil, Suggest a Post-Columbian African Origin of this Virus. J.
Acquir. Immune. Defic Syndr. 2003; 33: 536-542.
Alcantara L. C., de Oliveira T., Gordon M., Pybus O., Mascarenhas R. E., Seixas M. O.,
Gonçalves M., Hlela C., Cassol S., Galvão-Castro B.: Tracing the origin of Brazilian
HTLV-1 as determined by analysis of host and viral genes. AIDS. 2006; 21: 20(5):780-2.
Abé-Sandes K., Silva W. A. Jr., Zago M. A.: Heterogeneity of the Y chromosome in
AfroBrazilian populations. Hum Biol. 2004; 76(1): 77-86.
Baltimore D.: RNA polymerase in virions of RNA tumor viruses. &ature. 1970; 226:1209-11.
Bangham C. R.: The immune control and cell-to-cell spread of human T-lymphotropic virus
type 1. J Gen Virol. 2003; 84(Pt 12): 3177-89.
Barkur S. S.: SNPs in disease gene mapping, medicinal drug development and evolution. J. Hum
Gent. 2007; 52: 871-880.
Bastian I., Weeb D. and Gander I.: Isolation of human T-lymphotropic virus type I strain from
Australian aborigenals. J. Virol. 1993; 67:842-851.
Beby-Defaux A., Frugier F., Bourgoin A., Moynet D., Hajjar C., Sainte-Foie S., Guillemain B.
and Agius G.: Nucleotide sequence analysis of human T-cell lymphotropic virus type I pX
and LTR regions from patients with Sicca Syndrome. J. med Virol. 1999; 59: 245-255.
Biglione M. M., Pizarro M., Salomón H. E., Berría M. I.: A possible case of myelopathy/tropical
spastic paraparesis in an Argentinian woman with human T lymphocyte virus type II. Clin.
Infect. Dis. 2003; 37: 456-8.
Biggar R. J., M. E. Taylor, J. V. Neel, B. Hjelle, P. H. Levine, F. L. Black, G. M. Shaw, P. M.
Sharp and B. H. Hahn: Genetic variants of human T-lymphotropic virus type II in
American indian groups. Virol. 1996; 216:165-173.

35
Black F. L.: Tracing prehistoric migrations by the viruses they carry: Human T-cell
lymphotropic viruses as markers of ethnic relationships. Hum. Biology. 1997; 69: 467-
482.
Blas M., F. Bravo, et al.: Norwegian scabies in Peru: the impact of human T cell lymphotropic
virus type I infection. Am J Trop Med Hyg. 2005; 72(6): 855-7.
Bouzas M. B., Picchio G., Muchinik G., Campestri R., Cahn P., Perez H., Gallo R., and Hanson
C. V.: HTLV-I in Argentina. J. Acquir. Immune Def. Syndr. Hum. Retrovol. 1990; 3:741-
742.
Brady J., K.T. Jeang, J. Duvall and G. Khoury: Identification of p40x-responsive regulatory
sequenses within the human T-cell leukemia virus type I long terminal repeat. J. Virol.
1987; 61: 2175-2181.
Brites C., Weyll M., Pedroso C., Badaró R.: Severe and Norwegian scabies are strongly
associated with retroviral (HIV-1/HTLV-1) infection in Bahia, Brazil. Aids. 2002; 16(9):
1292-3.
Calattini S., Chevalier S. A., Duprez R., Bassot S., Froment A., Mahieux R., Gessain A.:
Discovery of a new human T-cell lymphotropic virus (HTLV-3) in Central Africa.
Retrovirol. 2005; (2): 30-38.
Callegari-Jacques S. M. and Salzano F. M.: Brazilian Indian/non-Indian interactions and their
effects. Ciênc Cult. 1999; 51: 166-174.
Cann A. J. and Chen I. S. Y.: Human T-cell leukemia virus type I and II. In: Fields BN, Knipe
DM, Howley PM et al (Ed). Fields Virology. 3rd edition. Philadelphia: Raven Publishers.
1996; V. 2, p. 1849-1879.
Carneiro-Proietti A. B., J. G. Ribas, et al.: Infection and disease caused by the human T cell
lymphotropic viruses type I and II in Brazil. Rev Soc Bras Med Trop. 2002; 35(5): 499-508.
Cartier L., Araya F., Castilho J. L., Zaninovic V., Hayami M., Miura T., Imai J., Sonoda S.,
Shiraki H., Miyamoto K., and Tajima K.: Southernmost carriers of HTLV-I/II in the world.
Japanese J. Cancer Research. 1993; 84: 1-3.
Catalan-Soares B., Carneiro-Proietti A. B., Proietti F. A.: Grupo Interdisciplinar de Pesquisas
em HTLV. Human T-cell lymphotropic virus in family members of seropositive blood
donors: silent dissemination Rev Panam Salud Publica. 2004; 16(6):387-394.

36
Catalan-Soares B., Carneiro-Proietti A. B., Proietti F. A.: Interdisciplinary HTLV Research
Group. Heterogeneous geographic distribution of human T-cell lymphotropic viruses I and
II (HTLV-I/II): serological screening prevalence rates in blood donors from large urban
areas in Brazil. Cad Saude Publica. 2005; 21(3): 926-931.
Censo Demográfico. Rio de Janeiro: Fundacão IBGE; 2000.
Charman H. P., N. Kim, et al.: Natural and experimentally induced antibodies to defined
mammalian type-C virus proteins in primates. J &atl Cancer Inst. 1975; 55(6): 1419-24.
Chen J., Zekeng L., Yamashita M., Takeisha J., Miura T., Ido E., Mboudjeka I., Tsague J. M.,
Hayami M. and Kaaptue L.: HTLV isolated from a Pygmy in Cameroon is related but
distinct from the known Central African type. AIDS Res. Hum. Retroviruses. 1995; 11:
1529-1531.
Cimarelli A., C. A. Duclos, A. Gessain, C. Casoli and U. Bertazzoni.: Clonal expansion of
human T-cell leukemia virus type II in patients with high proviral load. Virol. 1996; 223:
362-364.
Colin D. D., Alcântara Júnior L. C., Santos F. L., Uchôa R., Tavares-Neto J.: Seroprevalence of
human T cell lymphotropic virus infection and associated factors of risk in blood donors of
Rio Branco city, AC, Brazil (1998-2001). Rev Soc. Bras. Med. Trop. 2003; 36(6): 667-683.
Coskun A. K., Sutton R. E.: Expression of glucose transporter 1 confers susceptibility to human
T-cell leukemia virus envelope-mediated fusion. J. Virol. 2005; 79: 4150-4158.
Curtin P. D.: The slave Atlantic trade: a census. Milwaukee: The University of Wisconsin Press. 1969.
Dan S., Tanimura A., Yoshida M.: Interaction of Gli2 with CREB protein on DNA elements in
the long terminal repeat of human T-cell leukemia virus type 1 is responsible for
transcriptional activation by tax protein. J. Virol. 1999; 73: 3258-63.
Davidson M., Braziel R. M., Lairmore M. D., Jacobson S., Sprott J. M., Tucker S. B., Levine P.
H., and Kaplan E.: Atypical human T-cell lymphotropic virus type-I-associated T-cell
lymphoma in a low-prevalence Alaska Native population; inplications for disease
survellance. Cancer. 1993; 71: 4072-4076.
De Rivera I., Amador L., Moura S., Li Z., and Rasheed S.: Geographical clusttering of human T-
cell lymphotropic virus type I infection in Honduras. J. Clin. Microb. 1995; 33: 2999-3003.
Dekaban G. A., Oger J. J. F., Foti D., King E. E., Waters D. J., Picard F. J., Arp J., Werker D.,
and Rice G. P. A.: HTLV-I infection associated with disease in aboriginal Indians from

37
Brazil Columbia: a serological and PCR analysis. Clinical Diagnostic. Virol. 1994; 2: 67-
68.
Delamarre L., Rosemberg A. R., Pique C., Pham D., Callebaut I. and Dokhélar M.: The HTLV-I
envelope glycoproteins: structure and functions. J. Acquir. Immune Defic. Syndr. Hum.
Retrovirol. 1996; 13 (suppl. 1): S85-S91.
Delaporte E., Peeters M., Durand J. P., Dupont A., Schrijvers D., Bedjabaga L. et al.:
Seroepidemiological survey of HTLV-I infection among randomized populations of
western central African countries. J. Acquir. Immune. Defic. Syndr. 1989; (4): 410-3.
De Thé G., McLane M. F., Monplaisir N., Lee T. H., Essex M.: Presence of anti-HTLV (human
T-cell leukemia virus) antibodies in blood donors from Martinique. C R Seances Acad Sci
III . 1983; 297: 195-7.
Dourado I., T. Andrade and Galvao-Castro B.: HTLV-I in Northeast Brazil: differences for male
and female injecting drug users. J. Acquir. Immune. Defic. Syndr. Hum Retrovirol. 1998;
19: 426-9.
Dourado I., Alcantara J. L. C., Barreto M. L., Teixeira M. G., Galvão-Castro B.: HTLV-1 in the
general population of Salvador, Brazil: a city with African ethnic and sociodemographic
characteristics. J. Acquir. Immune. Defic. Syndr. Hum. Retrovirol. 2003; 34: 527-531.
Edlich R. F., J. A. Arnette, et al.: Global epidemic of human T-cell lymphotropic virus type-I
(HTLV-I). J Emerg Med. 2000; 18(1): 109-19.
Ferreira Jr. O. C., Planelles V., Rosenblatt J. D.: Human T-cell leukemia viruses: epidemiology,
biology and pathogenesis. Blood Reviews. 1997; 11: 91-104.
Ferrer J. F., D. M. Bhatt, et al.: Further studies on the antigenic properties and distribution n of
the putative bovine leukemia virus. Bibl Haematol. 1975; (40): 59-66.
Fleming A.F., Maharajan R., Abraham M., Kulkarni A. G., Bhusnurmath S. R., Okpara R. A. et
al.: Antibodies to HTLV-I in Nigerian blood-donors, their relatives and patients with
leukaemias, lymphomas and other diseases. Int J Cancer. 1986; 38(6): 809-13.
Florentino M.. Em Costas Negras. Ed Companhia das Letras, São Paulo. 1997; pp 305
Franchini G.: Molecular mechanisms of human T-cell leukemia/lymphotropic virus type I
infection. Blood. 1995; 86: 1619-1639.
Fujii M., Sassone-Corsi P., Verma I. M.: c-fos promoter trans-activation by the tax1 protein of
human T-cell leukemia virus type I. Proc &atl Acad Sci U S A. 1988; 85(22): 8526-30.

38
Fujisawa J., Seiki M., Sato M., Yoshida M.: A transcriptional enhancer sequence of HTLV-I is
responsible for trans-activation mediated by p40 chi HTLV-I. EMBO J. 1986; 5(4):713-8.
Fujiyama Y., Fujiyoshi T., Miura T., Yashiki S., Matsumoto D., Zaninovic V., Blanco O.,
Harrington W., Byrnes J. J., and Hayami M.: A new endemic focus of human T-cell
lymphotropic type II carriers among Orinico natives in Colombia (Letter). J. Infectious
Diseases. 1993; 168: 1075-1077.
Fujiyoshi T., Yashiki S., Fujiyama C., Kuwayama M., Miyashita H., Omishi H., Blank M.,
Sonoda S.: Ethinic segregation of HTLV-I and HTLV-II carries among South American
native indians. Int J. Cancer. 1995; 63: 510-515.
Furukawa Y., Yamashita M., Usuku K., Izumo S., Nakagawa M., and Osame M.: Phylogenetic
subgroups of human T-cell lymphotropic virus type I in the tax gene and their association
with different risks for HTLV-I-associated myelophaty/tropical spastic paraparesis. J.
Infectious Diseases. 2000; 182: 000-000.
Galvão-Castro B., Loures L., Rodrigues L. G. M., Sereno A., Ferreira O. C., Franco Jr. L. G. P.,
et al.: Geographic distribution of human T-lymphotropic virus type-I among blood donors:
A Brazilian nationwide study. Transfusion. 1997; 37: 242.
Gessain A., Barin E., Vernant J. C., Gout O., Maurs L., Calender A. and de-The G.: Antibodies
to human T-lymphotropic virus type I in patients with tropical spastic paraparesis. Lancet.
1985; 2: 407-410.
Gessain A., F.Saal, O. Gout, M.T. Daniel, G. Flandrin, G. de Thé, J. Peries and F. Sigaux.: High
human T-cell lymphotropic virus type I proviral DNA load with policlonal integration in
peripheral blood mononuclear cells of French West Indian, Guianese and African patients
with tropical spastic paraparesis. Blood. 1990; 75: 428-433.
Gessain A., Yanagihara R., Franchini G., Garruto R. M., Jenkins C. L., Ajdukiewicz A. B., Gallo
R. C. and Gajdusek D. C.: Highly divergent molecular variants of human T-lymphotropic
virus type I from isolated population in Papua New Guinea and Solomon Islands. Pro. &atl.
Sci. 1991; 88: 7694-7698.
Gessain A. and O. Gout.: Chronic myelopathy associated with human T-lymphotropic virus type
I (HTLV-I). Annals of Internal Medicine. 1992; 117: 933-946.

39
Gessain A.,Boeri E., Yanahigara R., Gallo R. C., Franchini G.: Complete nucleotide sequence of
a highly divergent HTLV-1 variant from Melanesia. Genetic and phylogenetic relationship
to HTLV-1 strains from other geographical regions. J. Virol. 1993; 67: 1015-23.
Gessain A., Koralnik I. J., Fullen J., Boeri E., Mora C., Blank A., Salazar-Grueso E. F., Kaplan
J., Saxinger W. C., and Davidson M.: Phylogenetic study of ten new HTLV-I strains from
the Americas. AIDS Research and Hum. Retroviruses. 1994; 10:103-106.
Gessain A. and De The G.: Geographic and molecular epidemiology of primate T lymphotropic
retroviruses: HTLV-I, HTLV-II, STLV-I, STLV-PP, and PTLV-L. Adv. Virus. Res. 1996;
47: 377-426.
Gonçalves, M.S., G.C. Bomfim, E. Maciel, I. Cerqueira, I.L. Queiroz, G. Bomfim, E. Adorno, A.
Albuquerque, A. Pontes, R. Muniz, M.F. Dupuit, G.B. Fernandes and M.G. Reis.: βs-
haplotypes in sickle cell anemia patients from Salvador-Bahia-Brazil. Brazilian J. Med.
Biol. Res. 2002
Gottuzo E., Yamamoto M., Kanna M., Chauca G., and Watts D. M.: Human T-cell lymphotropic
virus type I infection among Japanese immigrants in Peru. Intern J. Infec. Dis. 1996; 1:75-
77.
Goubau P., Vandamme A.-M., Beuselinck, and Desmyter J.: Proviral HTLV-I and HTLV-II in
the Efe pygmies of northeastern Zaire. J. Acquir. Immune Defic. Syndr Hum. Retrovirol.
1996; 12: 208-210.
Gray G. S., White M., Bartman T., Mann D.: Envelope gene sequence of HTLV-1 isolates MT-2
and its comparison with other HTLV-1 isolates. Virol. 1990; 177: 391-5.
Hall W. W.: Human T cell lymphotropic virus type I and cutaneous T cell leukemia/lymphoma.
J. Exp. Med. 1994 a; 180: 1581-5.
Hall W. W., Kubo T., Ljichi S., Takahashi T. and Zhu S. W.: Human T-cell leukemia/lymphoma
virus, type II (HTLV-II): emergence of an important newly recognised pathogen. Semin.
virol. 1994b; 5: 165-78.
Hahn B., Shaw G. M., Popovic M., Lo Monico A., Gallo R. C., and Wong-Staal F.: Molecular
cloning and analysis of a new variant of human T-cell leukemia virus (HTLV-Ib) from an
African patient with adult T-cell leukemia. Int. J. Cancer. 1984; 34: 613-618.

40
Hattori S., Kiyokawa T., Imagawa K., Shimizu F., Hashimura E., Seiki M., Yoshida M.:
Identificationof gag and env gene products of human T-cell Leukemia virus (HTLV). Virol.
1984; 136(2): 338-347.
Hinuma Y., K. Nagata, et al.: Adult T-cell leukemia: antigen in an ATL cell line and detection of
antibodies to the antigen in human sera. Proc &atl Acad Sci USA. 1981; 78(10): 6476-80.
Hinuma Y., Komoda H., Chosa T., Kondo T. and Kohakura M.: Antibodies to adult T-cell
leukemia-virus-associated antigen (ATLA) in sera from patients with ATL and controls in
Japan: a nation-wide seroepidemiologic study. Int. J. Cancer. 1982; 29: 631-635.
Hollsberg P. and Hafler D. A.: Pathogenesis of diseases induced by human lymphotropic virus
type I infection. &. Engl. J. Med. 1993; 328: 1173-82.
Ishak R., Cavalcante F., Vallinoto A. C., Azevedo V. N., Ishak M. O.: HTLV-I associated
myelopathy in the northern region of Brazil (Belém-Pará): serological and clinical features
of three cases. Rev Soc Bras Med Trop. 2002 ;35(3): 243-6.
Ishida T., Yamamoto K., Omoto K., Iwanafa M., Osato T. and Hinuma Y.: Prevalence of a
human retrovirus in native Japanese: evidence for a possible ancient origin. J. Infectol.
1985; 2: 153-157.
Jacobson S., T. Lehky, et al.: Isolation of HTLV-II from a patient with chronic, progressive
neurological disease clinically indistinguishable from HTLV-I-associated
myelopathy/tropical spastic paraparesis. Ann &eurol. 1993; 33(4): 392-6.
Johnson R. T., Griffin D. E., Arregui A., Mora C., Gibbs C. J., Cuba J. M., Trelies L., and
Vaisberg A.: Spastic paraparesis and HTLV-I infection in Peru. Annals &eurol. 1988; 23:
S151-S155.
Kajiyama W., Kashiwagi S., Ikematsu H., Hayashi J. Nomura H., and Okochi K.: Intrafamilial
transmission of adult T-cell leukemia virus. J. Infect Dis. 1986 a; 154: 851-7.
Kajiyama W., Kashiwagi S., Nomura H., Ikematsu H., Hayashi J. and Ikematsu W.:
Seroepidemiologic study of antibody to adult T-cell leukemia virus in Okinawa, Japan. Am.
J. Epidemiol. 1986b; 123: 41-47.
Kalyanaraman V. S., M. G. Sarngadharan, M. Robert-Guroff, I. Myioshi, D. Golde and R. C.
Gallo. A new subtype of human T-cell leukemia virus (HTLV-II) associated with a T-cell
variant of hairy cell leukemia. Science. 1982; 218: 571-573.

41
Kamihira S., Nakasima S. and Oyakawa Y.: Transmission of human T-cell lymphotropic virus
type I by blood transfusion before and after mass screening of sera from seropositive
donors. Vox Sang. 1987; 52: 43-44.
Kannagi M., Shida H., Igarashi H., et al:. Target epitope in the tax protein of human T-cell
leukemia virus type I recognized by class I major histocompatibility complex-restricted
cytotoxic T cells. J Virol 1992; 66: 2928-33.
Kaplan J.E., M. Osame and H. Kubota: The risk of development of HTLV-I associated
myelopathy/tropical spastic paraparesis among persons infected with HTLV-I. J. Acquir.
Immune. Defic. Syndr. 1990; 3: 1096-101.
Kashala O., R. Marlink, et al.: Infection with human immunodeficiency virus type 1 (HIV-1) and
human T cell lymphotropic viruses among leprosy patients and contacts: correlation
between HIV-1 cross-reactivity and antibodies to lipoarabinomannan. J. Infect. Dis. 1994;
169(2): 296-304.
Kashima S., Alcantara L. C., Takayanagui O. M., Cunha M. A., Castro B. G., Pombo-de-
Oliveira M. S., Zago M. A., Covas D. T.: Distribution of human T cell lymphotropic virus
type 1 (HTLV-1) subtypes in Brazil: genetic characterization of LTR and tax region. AIDS
Res. Hum. Retroviruses. 2006; 22(10): 953-9.
Kitagawa T., Fujishita M., Tagushi H., Miyoshi I. and Tadokoro H.: Antibodies to HTLV-I in
Japanese immigrants in Brazil. JAMA. 1986; 256: 2342.
Komurian F., Pelloquin F. and De Thé G.: In vivo genomic variability of human T-cell leukemia
virus type I depends more upon geography than upon pathologies. J. Virol. 1991; 65(7):
3770-8.
Kristien V., Elsa González, Sonia Van Dooren, Anne-Mieke Vandamme, Guido Vanham,
Eduardo Gotuzzo. : Human T-lymphotropic virus: recent knowledge about an ancient
infection. Lancet. infect. Dis.2007; 7: 266-281.
La Grenade L.: HTLV-1 associated infective dermatitis; past, present and future. J. Acquir.
Immune Defic. Syndr Hum. Retrovirol. 1996; 13: 846-849.
Lee H., Swanson P., Rosenblatt J. D., Chen I. S. Y., Sherwood W. C., Smith D. E., Tegtmeier G.
E., Fernando L. P., Fang C. T., Osame M., and Kleinman S. H.: Relative prevalence and

42
risk factors of HTLV-I and HTLV-II infection in U.S. blood donors. Lancet. 1991;
337:1435-1439.
Lemasson I., Polakowski N. J., Laybourn P. J., Nyborg J. K.: Transcription regulatory
complexes bind the human T-cell leukemia virus 5´ and 3´ long terminal repeats to control
gene expression. Mol. Cell Biol. 2004; 24(14): 6117-6126.
Levine P. H. and Blattner W. A. The epidemiology of diseases associated with HTLV-I and
HTLV-II. Infect Dis Clin &orth Am. 1987; (3): 501-10.
Mahieux R., Ibrahim F., Mauclere P., Herve V., Michel P., Tekaia F., Chappey C., Garin B.,
Van der Ryst E., Guillemain B., Ledru E., Delaport E., De The G. and Gessain A.:
Molecular epidemiology of 58 new African Human T-cell leukemia virus type-I (HTLV-I)
strains: Identification of a new distinct HTLV-I molecular subtype in Central Africa and in
Pygmys. J. Virol. 1997; 71: 1317-1333.
Malik. K. T., Even J., Karpas A.: Molecular cloning and complete nucleotide sequence of an
adult T cell leukemia virus/human T cell leukemia virus type I (ATLV/HTLV-I) isolates of
Caribbean origin: Relationship to other members of the ATLV/HTLV-1 subgroup. J. Gen
Virol.1988; 69: 1695-710.
Maloney E. M., Ramirez H., Levin A. And Blattner W. A.: A survey of human T-cell
lymphotropic virus type I (HTLV-I) in south-western Colombia. Int. J. Cancer. 1989;
44:419-423.
Maloney E. M., Biggar R. J., Neel J. V. et al.: Endemic human T cell lymphotropic virus type II
infection among isolated Brazilian Amerindians. J. Infect. Dis. 1992; 166: 100-7.
Marinho J., Galvao-Castro B., et al.: Increased risk of tuberculosis with human T-lymphotropic
virus-1 infection: a case-control study. J Acquir Immune Defic Syndr. 2005; 40(5): 625-8.
Martin �., Boomsma D., Machin G..: A twin pronged attacks on complex traits. &at Gent. 1997;
17: 387-392
Merino F., Robert-Guroff M., Clarck J., Biondo-Brancho M., Blattner W. A. and Gallo R. C.:
Natural antibodies to human T-cell leukemia/lymphoma virus in healthy Venezuelan
populations. Int. J. Câncer. 1984; 34: 501-506.
Miranda A. C. M., Pereira S. A., Dias J. P., Colin D .D., Kashima S., Covas D. T., Neto J. T.,
Castro B. G., Alcantara L. C. J.: HTLV-1 infection in blood donors from the Western

43
Brazilian Amazon region: seroprevalence and molecular study of viral isolates. J Med
Virol. 2008; 80: 1966-1971.
Miura T., Fukunaga T., Igarashi T. et al.: Phylogenetic subtypes of human T-lymphotropic virus
type I and their relations to anthropological backgroud. Proc. &atl. Acad. Sci. USA 1994;
91:1124-7.
Miura T., Yamashita M., Zaninovic V., Catrier L., Takehisa J., Igarashi T., Ido E., Fujiyoshi T.,
Sonoda S., Tajima K. and Hayami M.: Molecular phylogeny of human T-cell leukemia
virus type-I and II of Amerindians in Columbia and in Chile. J. Mol. Evol. 1997; 44
(Suppl.1): S76-S82.
Miwa M., Shimotohno K., Hoshino H., Fujino M., Sugimura T.: Detection of pX proteins in
human T-cell leukemia virus (HTLV)-infected cells by using antibody against peptide
deduced from sequences of X-IV DNA of HTLV-I and Xc DNA of HTLV-II proviruses.
Gann. 1984; 75: 752-5.
Mochizuki M., Watanabe T., Yamaguchi K., Yoshimura K., Nakashima S., Shirao M.., Araki S.,
Takatsuki K., Mori S., Miyata M.: Uveitis.associated with human T-cell lymphotropic virus
type I. American J. Ophthalmol. .1992; 114: 123-129.
Mochizuki M., A. Ono, E. Ikeda, N. Hikita, T. Watanabe, K. Yamaguchi, K. Sagawa and K. Ito:
HTLV-I uveitis. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol.1996; 13: S50-S56.
Moreno-Carvalho O. A., Santos J. I., Credico G. and Galvão-Castro B.: Evidence of preferential
female prevalence of HTLV-I associated tropical spastic paraparesis in Bahia-Brazil. Arq.
&euro-Psiquiat. (São Paulo). 1992; 50(2): 183-188.
Mori �., Mukaida N., Ballard D. W., Matsushima K., Yamamoto N.: Human T-cell leukemia
virus type I tax transactivates human interleukin-8 gene through acting concurrently on AP-
1 and nuclear factor-κB-like sites. Cancer Res. 1998; 58: 3993-4000.
Morgan O. S., P. Rodgers-Johnson, C. Mora and G. Char.: HTLV-I and polymyositis in
Jamaica. Lancet. 1989; ii: 1184-87.
Mota A. C. A., Van Dooren S., Fernandes F. M. C., Pereira S. A., Queiroz A. T. L., Gallazzi V.
O., Vandamme A. M., Castro B. G, Alcantara L. C. J.: The close relationship between
South African and Latin American HTLV-1 type 1 strains corroborated in a molecular
epidemiological study of the HTLV type 1 isolates from a blood donors cohort. AIDS Res
Hum Retroviruses. 2007; 23: 503-507.

44
Mueller �.: The epidemiology of HTLV-I infection. Cancer Causes Control. 1991; 2(1): 37-52.
Mueller �., Okayama A., S. Stuver and N. Tachibana: Findings from the Miyazaki Cohort
Study. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1996; 13: S2-S7.
Mukhopadhyaya R. and Sadaie M. R.: Nucleotide sequence analysis of HTLV-I isolated from
cerebrospinal fluid of a patient with TSP/HAM: comparison to other HTLV-I isolates. AIDS
Res Hum Retroviruses. 1993; 9(2): 109-114.
Murphy E. L.: The clinical epidemiology of human T-lymphotropic virus type II (HTLV-II). J.
AIDS Hum. Retrovirol. 1996; 13(Suppl 1): S215-S219.
�am S. H., M. Kidokoro, H. Shida and M. Hatanaka.: Processing of gag precursor polyprotein of
human T-cell leukemia virus type I by virus-encoded preotease. J. Virol. 1988; 62: 3718-28.
�eel J. V., R. J. Biggar and R. I. Sukernik: Virologic and genetic studies related Amerindian
origins to the indigenous people of the Mongolia/ Manchuria/ southeastern Siberia region.
Proc &atl. Acad Sci. USA. 1994; 91: 10737-10741.
�eva F. A., Gam A. A., Maxwell C., Pelletier L. L.: Skin test antigens for immediate
hypersensitivity prepared from infective larvae of Strongyloides stercoralis. Am. J. Trop.
Med. Hyq. 2001; 65: 567-72.
�iewiesk S., Daenke S., Parker C. E., et al.: Naturally occurring variants of human T-cell
leukemia virus type I tax protein impair its recognition by cytotoxic T lymphocytes and the
transactivation function of tax. J Virol. 1995; 69: 2649-53.
Orland J. R., Engstrom J., Fridey J., Sacher R. A., Smith J. W., Nass C., Garratty G., Newman
B., Smith D., Wang B., Loughlin K., Murphy E. E. L.: Prevalence and clinical features of
HTLV-1 neurologic disease in the HTLV-1. Outcomes study &eurol.. 2003; 61(11): 1588-
1594.
Osame M., Usuku K., Izumo S., Ijichi N. Amitani H.and Igata A: HTLV-I associated
myelopathy, a new clinical antity. Lancet. 1986; 1: 1031-32.
Osame M.: Review of WHO Kagoshima Meeting and Diagnostic Guidelines for HAM/TSP:
Human Retrovirology: HTLV. 1990; A. B. William. New York, Raven Press, Ltd: 191-197.
Parker C. E., Daenke S., Nightingale S., Bangham C. R.: Activated, HTLV-I-specific cytotoxic
T-lymphocytes are found in healthy seropositives as well as in patientes with tropical
spastic paraparesis. Virol. 1992; 188: 628-36.

45
Pedral-Sampaio D. B., Martins Netto E., Pedrosa C., Brites C., Duarte M., Harrington W. Jr.
Co-Infection of Tuberculosis and HIV/HTLV Retroviruses: Frequency and Prognosis
Among Patients Admitted in a Brazilian Hospital: Braz J. Infect. Dis.1997; 1; 31-35.
Peters A. A., Oger J. J., Coulthart M. B., Waters D. J., Cummings H. J., Dekaban G. A. An
apparent case of human T-cell lymphotropic virus type II (HTLV-II)-associated
neurological disease: a clinical, molecular, and phylogenetic characterisation. J. Clin. Virol.
1999;14: 37-50.
Pinsky J.: O escravo negro. In Pinsky, J., A escravidão no Brasil. Ed Contexto, São Paulo. 1988; pp.24.
Poiesz B. J., Ruscetti F. W., Gazdar A. F., Bunn P. A., Minna J. D. and Gallo R.: Detection and
isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient
with cutaneous T-cell lymphoma. Proc. &atl. Acad. Sci. USA 1980; 77: 7415-19.
Porto M. A., Muniz A., Oliveira Júnior J., Carvalho E. M.: Clinical and immunological
consequences of the association between HTLV-1 and strongyloidiasis. Rev Soc Bras Med
Trop. 2002; 35(6): 641-9.
Queiroz A.T., Mota-Miranda A. C., de Oliveira T., Moreau D. R., Urpia C.de C., Carvalho C.M.,
Galvão-Castro B., Alcantara L. C.: Re-mapping the molecular features of the human
immunodeficiency virus type 1 and human T-cell lymphotropic virus type 1 Brazilian
sequences using a bioinformatics unit established in Salvador, Bahia, Brazil, to give
support to the viral epidemiology studies. Mem Inst Oswaldo Cruz. 2007; 102(2): 133-9.
Ratner L., Josephs S., Starcich B., et al..: Nucleotide sequence analysis of a variant human T cell
leukemia virus (HTLV-1b) provirus with a deletion in px- I. J. Virol. 1985; 54: 781-90.
Reeves W. C., Levine P., Cuevas M., Quiroz E., Maloney E., and Saxinger W. C.:
Seroepidemiology of human T-cell lymphotropic virus type I (HTLV-I) in the Republic of
Panama. American J. Trop Medic Hyg. 1990; 42: 374-379.
Renjifo B., Borrero I., Essex M.: Tax mutation associated with tropical spastic
paraparesis/human T-cell leukemia virus type I-associated myelopathy. J. Virol. 1995; 69:
2611-2616.
Roman C. G., M. and Osame: Identify of HTLV-I-associated tropical spastic paraparesis and
HTLV-I-associated myelopathy. Lancet. 1988; i: 651.

46
Rous P.: A sarcoma of the fowl transmissible by an agent separable from the tumor cells. J. Exp.
Med. 1911; 13: 397.
Sagara Y., Inoue Y., Shiraki H., Jinno A., Hoshino H., Maeda Y.: Identification and mapping of
functional domains on human T-cell lymphotropic virus type 1 envelope proteins by using
synthetic peptides. J. Virol. 1996; 70(3): 1564-9.
Salemi M. :Molecular investigations of the origin and genetic stability of the human T-cell
lymphotropic viruses. PHD tesis in Katholieke Universiteit Leuven. Rega Institut. 1999.
Salemi M., Van Dooren S., Audenaert E., Delaport E., Goubau P., Desmyter J. and Vandamme
A-M.: Two new Human T-lymphotropic virus type-I phylogenetic subtypes in
seroindeterminates, a Mbuti Pygmy and a Gabonese, have relatives among African HTLV-I
strains. Virol. 1998; 246: 277-278.
Saxinger W., Blattner W. A., Levine P. H., Clark J, Biggar R., Hoh M., et al.: Human T-cell
leukemia virus (HTLV-I) antibodies in Africa. Sci. 1984; 225(4669): 1473-6.
Segurado A. A., Biasutti C., Zeigler R., Rodrigues C., Damas C. D., Jorge M. L., Marchiori P.
E.: Identification of human T-lymphotropic virus type I (HTLV-I) subtypes using restricted
fragment length polymorphism in a cohort of asymptomatic carriers and patients with
HTLV-I-associated myelopathy/tropical spastic paraparesis from São Paulo, Brazil. Mem
Inst Oswaldo Cruz. 2002; 97(3): 329-33.
Seiki M., Hattori S., Hirayama Y. and Yoshida M.: Human adult T-cell leukemia virus: complete
nucleotide sequence of the provirus genome integrated in leukemia cell DNA. Proc &at
Acad. Sci. USA. 1983; 80: 3618-3622.
Seiki M., R. Eddy, T. B. Shows and M. Yoshida: Nonspecific integration of the HTLV provirus
genome into Adult t-cell leukemia cells. &ature. 1984; 309: 640-642.
Shindo �., Alcantara L. C., Van Dooren S., Salemi M., Costa M., Kashima S., Covas D. T., Teva
A., Pellegrini M., Brito I., Vandamme A. M., Galvao-Castro B.: Human retroviruses (HIV
and HTLV) in Brazilian Indians: seroepidemiological study and molecular epidemiology of
HTLV type 2 isolates. AIDS Res. Hum. Retroviruses.2002; 18: 71-77.

47
Shimotohno K.., Takano M., Teruuchi T., Miwa M.: Requirement of multiple copies of a 21-
nucleotide sequence in the U3 regions of human T-cell leukemia virus type I and type II
long terminal repeats for trans-acting activation of transcription. Proc &atl Acad Sci USA.
1986; 83(21): 8112-8116.
Shimoyama M.: Diagnostic criteria and classification of clinical subtypes of adult T-cell
leukaemia-lymphoma. A report from the lymphoma study group. British J. Haematol. 1991;
79: 428-437.
Siekevitz M., Feinberg M. B., Holbrook N., Wong Staal F., Greene W. C.: Activation of
interleukin-2 and interleukin-2- receptor (Tac) promoter expression by the trans-activator
(tat) gene product of human T-cell leukemia virus, type I. Proc &atl Acad Sci USA. 1987;
84: 5389-93.
Sodroski J. G., Rosen C. A., Haseltine W. A.: Trans-acting transcriptional activation of the long
terminal repeat of human T lymphotropic viruses in infected cells. Sci. 1984; 225: 381-5.
Tanimura A., Teshima H., Fujisawa J., Yoshida M.: A new regulatory element that augments
the Tax-dependent enhancer of human T-cell leukemia virus type 1 and cloning of cDNAs
encoding its binding proteins. J Virol. 1993; 67: 5375-5382.
Tanimura A., Dan S., Yoshida M.: Cloning of novel isoforms of the human Gli2 oncogene and
their activities to enhance tax-dependent transcription of the human T-cell leukemia virus
type 1 genome. J Virol. 1998; 72: 3958-64.
Talarmin A., Vion B., Ureta-Vidal A., Du Fou G., Marty C., and Kazanji M.: Frist
seroepidemiological study and phylogenetic of human T-cell lymphotropic virus type I and
II infection among Amerindians in French Guiana. J. Gen. Virol. 1999; 80: 3083-3088.
Tendler C. L., Greenberg S. J., Blattner W. A., Manns A., Murphy E., Fleisher T., Hanchard B.,
Morgan O., Burton J. D., Nelson D. L.: Transactivation of interleukin 2 and its receptor
induces immune activation in human T-cell lymphotropic virus type I-associated
myelopathy:pathogenic implications and a rationale for immunotherapy. Proc &atl Acad
Sci USA. 1990; 87(13): 5218-5222.
Tsugane S., Watanabe S., Sugumura H., Otsu T., Tobinai K., Shimoyama M., Nanri S., and Ishii
H.: Infectious state of human T-cell lymphotropic virus type I and hepatitis B virus among
Japanese immigrants in the Republic of Bolivia. American J. Epidemiol. 1988; 128: 1153-
1161.

48
Uchiyama T., J. Yodoi, K. Sagawa, K. Takatsuki and H. Uchino: Adult T-cell leukemia: clinical
and hematologic features of 16 cases. Blood. 1977; 50: 481-92.
Van Dooren S., Gottuzo E., Salemi M., Watts D., Audenaert E., Duwe S., Ellerbrok H.,
Grassmann R., Hagelberg J., and Vandame A-M.: Evidence for a post-Columbian
introduction of human T-cell lymphotropic virus in Latin America. J. General Virol. 1998;
79: 2695-2708.
Van Dooren S., Pybus O. G., Salemi M., Liu H. F., Goubau P., Remondegui C., Talarmin A.,
Gotuzzo E., Alcantara L. C., Galvão-Castro B., Vandamme A.M.: The low evolutionary
rate of human T-cell lymphotropic virus type-1 confirmed by analysis of vertical
transmission chains. Mol Biol Evol. 2004; (3): 603-11. Epub 2004.
Vandamme A.-M., Liu H., Goubou P. and Desmyter J.: Primate T-Lymphotropic virus type I
LTR sequence variation and its phylogenetic analysis: compatibility with an African origin
of PTLV-I. Virol. 1994; 202: 212-23.
Vandamme A-M., Salemi M., Desmyter J.: The simian origens of the pathogenic human T-cell
lynphotropic virus type I. Trends in Microbilogy. 1998; 6(12): 477-483.
Verdier M., Bonis J., and Denis F. A.: The prevalence and incidence of HTLVs in Africa. 1994.
In M. Essex (ed), AIDS in Africa, vol 9, p173-193. Raven Press, New York.
Verger, P. Mouvement des navires. P.12. In Verger, P., Trade relations between the Bight of
Benin and Bahia, 17th to 19th century, trans. Evelyn Crawford, Ibadan, Ibadan University
Press. 1976; pp.24-26.
Vernant J. C., Buisson G., Magdeleine, J., Thore J., Jouannelle A., Monplaisir N.: T-lymphocyte
alveolites, tropical spastic paraparesis and Sjogren’s syndrome. Lancet. 1988; 1: 177.
Viana Filho L.:. O negro na Bahia. 31-224. Nova Fronteira (3a ed), Rio de Janeiro, Brasil. 1988.
Vidal A.U., Gessain A., Yoshida M., Tekaia F., Garin B., Guillemain B., Schulz T., Farid R., De
Thé G.: Phylogenetic classification of human T cell leukaemia/lymphoma virus type I
genotypes in five major molecular and geographical subtypes. J. Gen. Virol. 1994; 75:
3655-3366.

49
Yamaguchi K., Kiyokawa T., Watanabe T., Ideta T., Asayama K., Mochizuki M., Blank A.,
Takatsuki K.,: Increased serum levels of C- terminal parathyroid hormone-related protein in
different diseases associated with HTLV-1 infection. Leukemia. 1994; 8: 1708-1711.
Yamaguchi K. & Takatsuki K., Adult T cell leukaemia-Lymphoma. Baillieres Clin. Haematol.
1993; 6: 899-915.
Yamashita M., R. Veronesi, M. Menna-Barreto, W. J. Jr. Harrington, C. Sampio, C. Brites, R.
Badaro, A. S. Andrade-Filho, S. Okhura, T. Igarashi, T. Miura, D. Chamone, O. Bianchini,
C. Jardim, S. Sonoda and M. Hayami: Molecular epidemiology of human T-cell leukemia
virus type I (HTLV-1) Brazil: the predominant HTLV-1s in South America differ from
HTLV-Is of Japan and Africa, as well as those of Japanese immigrants and their relatives in
Brazil. Virol. 1999; 261: 59-69.
Yoshida M., J. Inoue, et al.: Molecular mechanisms of regulation of HTLV-1 gene expression
and its association with leukemogenesis. Genome. 1989; 31(2): 662-7.
Zehender G., Ebranati E., Bernini F., De Maddalena C., Giambelli C., Collins J., Valverde A.,
Montin Z., Galli M.: Phylogeny of human T cell lymphotropic virus type 1 in Peru: a high degree
of evolutionary relatedness with South African isolates. AIDS Res Hum Retroviruses. 2007;
23(9): 1146-9.
Wattel E., J.-P. Vartanian, C. Pannetier and S. Wain-Hobson: Clonal Expansion of Human T-
Cell Leukemia Virus Type I- Infected cells in asymptomatic and symptomatic carriers
without malignancy. J. Virol. 1995; 69: 2863-2868.
Wolfe �.D., Heneine W., Carr J. K., Garcia A. D., Shanmugam V., Tamoufe U., Torimiro J. N,
Prosser A. T., Lebreton M., Mpoudi-Ngole E., McCutchan F. E., Birx D. L., Folks T. M.,
Burke D. S., Switzer W. M.: Emergence of unique primate T-lymphotropic viruses among
central African bushmeat hunters. Proc. &atl. Acad. Sci. USA. 2005; 7994-9.

50
10. A�EXO
Artigo aceito para publicação

3Gaz. méd. Bahia 2009;79:1(Jan-Dez):3-10 Epidemiologia e Origem do HTLV-1 em Salvador, Bahia, Brasil
www.gmbahia.ufba.br
EPIDEMIOLOGIA E ORIGEM DO HTLEPIDEMIOLOGIA E ORIGEM DO HTLEPIDEMIOLOGIA E ORIGEM DO HTLEPIDEMIOLOGIA E ORIGEM DO HTLEPIDEMIOLOGIA E ORIGEM DO HTLVVVVV-1 EM SAL-1 EM SAL-1 EM SAL-1 EM SAL-1 EM SALVVVVVADOR ESTADOR ESTADOR ESTADOR ESTADOR ESTADO DA BAHIA: A CIDADE COM A MAISADO DA BAHIA: A CIDADE COM A MAISADO DA BAHIA: A CIDADE COM A MAISADO DA BAHIA: A CIDADE COM A MAISADO DA BAHIA: A CIDADE COM A MAISELEVELEVELEVELEVELEVADA PREVADA PREVADA PREVADA PREVADA PREVALÊNCIA DESTALÊNCIA DESTALÊNCIA DESTALÊNCIA DESTALÊNCIA DESTA INFECÇÃO NO BRASILA INFECÇÃO NO BRASILA INFECÇÃO NO BRASILA INFECÇÃO NO BRASILA INFECÇÃO NO BRASILHTLV-1 EPIDEMIOLOGY AND ORIGIN IN SALVADOR, STATE OF BAHIA: THE CITY WITH THE HIGHESTPREVALENCE OF THIS INFECTION IN BRAZIL
Bernardo Galvão-Castro1,2, Luiz C. Junior Alcântara1,2, Maria Fernanda R. Grassi1,2, Aline C.A. Mota-Miranda1, ArturT.L. de Queiroz1, Filipe F.A. Rego1, Augusto Cesar A. Mota3, Sérgio A. Pereira1, Temístocles Magalhães2, JoséTavares-Neto4, Marilda Gonçalves1,5, Inês Dourado61Fundação Oswaldo Cruz/Centro de Pesquisas Gonçalo Moniz, Salvador-BA; 2Escola Bahiana de Medicina e Saúde Pública(EBMSP); 3Hospital São Rafael; 4Faculdade de Medicina da Bahia, Universidade Federal da Bahia; 5Escola de Farmácia daUniversidade Federal da Bahia; 6Instituto de Saúde Coletiva, Universidade Federal da Bahia; Salvador, Bahia, BrasilA cidade de Salvador, Bahia apresenta a mais elevada prevalência de HTLV-1 no Brasil. Esta prevalência é maior emmulheres, aumenta com a idade consideravelmente nos indivíduos com mais de 51 anos (8,4%) e é maior naquelescom baixa condição socioeconômica. Salvador tem uma população de 3 milhões de habitantes sendo que 80% são afrodescendentes. A maioria dos africanos que veio para esta cidade, durante o tráfico de escravos, foi originária do Oesteda África (Benin, Nigéria, Norte de Angola).A análise filogenética da região LTR demonstrou que as seqüênciasestudadas eram do subtipo Cosmopolita (a), subgrupo Transcontimental (A). Estes resultados são discordantes emrelação aos dados históricos que indicam que a maioria dos africanos que veio para Salvador foi originária do oeste daÁfrica onde somente circula o subgrupo Oeste da África (C). Algumas seqüências do HTLV-1 se agrupavam em umgrupo da America Latina que continha seqüências da África do Sul. Este grupo da America Latina era segregado apartir de um mesmo ancestral do outro grupo que continha uma seqüência da África Central. Esta relação deancestralidade sugere que este subgrupo foi inicialmente introduzido na África do Sul devido a migração de umapopulação Banto da África Central para o Sul da África há cerca de 3000 anos e após para o Brasil durante o tráfico deescravos no período compreendido entre os séculos XVI e XVII. Nossos dados corroboram a hipótese de que ocorrerammúltiplas introduções do HTLV-1 em Salvador na era pós-colombiana.Palavras-chave: HTLV-1, origem, LTR, haplótipos da globina βββββ, Banto, Salvador-Brazil, Sul da África.
The city of Salvador, in the Bahia state, has the highest prevalence of HTLV-1 in Brazil. This prevalence is higher in female
and it is associated with age, increasing substantially among those older than 51 years (8.4%), and is greater among those
with lower income, less education and worse living conditions. The population of Salvador is estimated at 3 million and
approximately 80% of which have African ethnic ancestry. Most Africans brought to Salvador; Bahia during the slave trade
came from West Africa (Benin, Nigeria, and northern Angola). Phylogenetic analysis of the LTR region demonstrated that
all sequences analyzed belong to the Transcontinental (A) subgroup of the Cosmopolitan (a) subtype. These results are not
in agreement to historical data that affirms the vast majority of African brought to Salvador came from West Africa, where
only the western African subgroup (C) has been found. Some HTLV- 1 sequences formed a well-supported clade within one
of the Latin American clusters that contain a South African sequence. This Latin American cluster that segregated from the
same ancestor as the other clade contained a Central African sequence. This ancestral relationship suggests that this
subgroup was first introduced into South Africa as a result of the migration of the Bantu population from Central Africa to
Southern Africa over the past 3000 years, and afterward to Brazil during the slave trade between the sixteenth and nineteenth
centuries. Our data support the hypothesis of multiple post-Columbian introductions of African HTLV-1 strains in Salvador,
Bahia.
Key words: HTLV-1, origin, LTR, βββββ-globin haplotypes, Bantu, Salvador, Brazil, Southern Africa.
Recebido em 02/06/2009 Aceito em 15/06/2009Endereço para correspondência: Prof. Bernardo Galvão-Castro. CentroIntegrativo e Multidisciplinar de HTLV e Hepatites Virais – EscolaBahiana de Medicina e Saúde Pública, Av. Dom João VI, n0 274, Brotas,Salvador, BA, 40.290-000. Tel.: 55 71 32768281; Fax: 5571 32768290.E-mail [email protected].
Gazeta Médica da Bahia 2009;79:1(Jan-Dez):3-10© 2009 Gazeta Médica da Bahia. Todos os direitos reservados.
A história dos retrovírus começa em 1908 com a induçãode leucose aviária por agentes ultrafiltráveis realizada porEllermann e Bang. Peyton Rous, em 1911, induziu neoplasiasem galinhas injetando filtrados livres de células obtidos de
sarcomas aviários. Os vírus isolados destes sarcomas foramdenominados Vírus do Sarcoma de Rous. Estes experimentosforam confirmados em 1914 pelos pesquisadores japonesesFujinani e Inamoto. A importância dessa descoberta só foireconhecida 55 depois, com a outorga do Prêmio Nobel deFisiologia e Medicina (PNFM) a Peyton Rous em 1966. A enzimatranscriptase reversa, necessária para a replicação destes vírus,foi descoberta por Howard Temin e David Baltimore em 1970 epor isto receberam o PNFM em 1975. Michel Bishop e HaroldVarmus identificaram, isolaram e determinaram a seqüêncianucleotídica completa de oncogenes nesses tumores em 1980e foram agraciados com PNFM em 1989. (31 45)

4 Gaz. méd. Bahia 2009;79:1(Jan-Dez):3-10Bernardo Galvão-Castro, et al.
www.gmbahia.ufba.br
Esses fatos contribuíram para a identificação dos primeirosretrovírus humanos, os vírus linfotrópicos de células Thumanas tipo1 (HTLV-1) e do tipo 2 (HTLV-2), em 1980 e 1982respectivamente (46 67) . Dois novos tipos de HTLV, 3 e 4, foramidentificados em populações da África Central em 2005 (16 88).
A Síndrome da Imunodeficiência Adquirida (AIDS) foiidentificada em 1981 (39) e o seu agente causal, o vírus daimunodeficiência humana (HIV), em 1983 (7). Os cientistas,Françoise Barré-Sinoussi e Luc Montagnier, do InstitutoPasteur de Paris, que isolaram pela primeira vez o HIV foramagraciados com o PNFM em 2008 (31).
O HTLV-1 e HIV-1 são da mesma família, mas apresentamalgumas características distintas. O primeiro está disseminadopelo mundo e a maioria dos indivíduos infectados por eleadoece. O segundo é mais freqüente em determinadas regiõesgeográficas, mas somente 3-5% dos infectados pelo HTLV-1virão adoecer.
Pressupõe-se que 20 milhões de pessoas estejaminfectadas pelo HTLV-1 no mundo (26). No entanto, aprevalência varia de acordo com a região geográfica, ospadrões sócio-comportamentais e étnicos das populações.Esta infecção caracteriza-se, também, por ser mais freqüenteem mulheres e aumentar com a idade (68).
Este vírus se transmite por três vias: a) sexual, com maioreficiência de transmissão de homem para mulher. Estima-seque esta eficiência seja de cerca de 60%, sendo de 4% nosentido inverso; b) sangüínea, através de transfusão desangue ou pelo uso de agulhas e/ou seringas contaminadas ec) perinatal, da mãe para o filho, principalmente peloaleitamento materno. Para que ocorra a transmissão há anecessidade da presença de células infectadas (68).
As principais áreas endêmicas da infecção compreendemÁfrica, América do Sul e Central, Caribe, Japão Melanésia eOriente Médio. Na América do Sul e Central se destacamArgentina, Bolívia, Brasil, Chile Colômbia, Honduras, Panamá,Peru, e Venezuela (68). A província equatorial do Congoapresenta a prevalência mais elevada desta infecção naÁfrica(40). Foram detectadas infecções em tribos de pigmeusque vivem isoladas em áreas remotas da República Centro-Africana, Camarões e nordeste do Congo (74). Baseado naexistência destas infecções na África Equatorial e emdescendentes de africanos no Caribe e na América Latinasugere-se que a África seria a fonte primária deste vírus (83).As doenças associadas a resposta imune ao HTLV-1 estãodescritas em (Grassi et al., 2008, neste volume).
O HTLV-1 foi detectado pela primeira vez no Brasil, emimigrantes japoneses na cidade de Campo Grande, MatoGrosso do Sul em 1986 (47). Posteriormente foi constatado emoutros grupos populacionais (20). A maioria dos dadosepidemiológicos da infecção no Brasil refere-se à prevalênciaem populações específicas. Em doadores de sangue observa-se um gradiente de indivíduos infectados, com taxas maiselevadas nas regiões norte/nordeste e as mais baixas na regiãosul (21). Salvador e São Luiz são as cidades que apresentam asmaiores prevalências em doadores de sangue no território
nacional (21 30), Tem-se observado uma diminuição daprevalência em doadores de sangue no Brasil (21). Em Salvadoresta prevalência diminuiu de 1,35% para 0,48% em uma década(59).
Não existe ainda um estudo epidemiológico, commetodologia adequada, que tenha determinado a prevalênciana população geral do Brasil. No entanto, baseados nos dadosde prevalência em doadores de sangue estima-se que no Brasilexistam 2,5 milhões de portadores de HTLV-1 (17 18).
O único estudo de base populacional no Brasil foi realizadoem Salvador, Bahia e demonstrou 1,8% de prevalência napopulação geral, sendo que 2% em mulheres e 1,2% emhomens, mas, aumentando em indivíduos com mais de 50 anospara 9 e 6,3% nos sexos masculino e femininorespectivamente(26).
Em gestantes a prevalência nacional foi de 0,28 % (13) e naBahia de 0,9 e 1,0% respectivamente, em Salvador e Cruz dasAlmas, no Recôncavo Baiano (10 52 75). Na população de usuáriosde drogas injetáveis em Salvador detectaram-se prevalênciasde 25,5% e 8,8% para HTLV-1 e HTLV-2 respectivamente (25).As prevalências de co-infecções foram para HTLV-1/ HIV-1;HTLV-2 / HIV-1 e HTLV-1/ HTLV-2 /HIV-1 de 21,6%; 10,6% e0,9% respectivamente (3). Observou-se, dois anos depois, umaredução das prevalências de infecções pelo HIV-1 e HTLV-1/2 de 49.5% e 35.2% em 1998 para 7.1% e 5.0% em 2000 (4). Estasdiminuições poderiam ter sido decorrentes do programa dediminuição de danos desde que se tem observado umadiminuição da prevalência para HIV, HBV e HCV em usuáriosde drogas injetáveis após a introdução deste programa emvarias cidades brasileiras (43 56 78). No entanto, não se podedescartar um viés de amostragem e deve ser tambémconsiderado o curto espaço de tempo em que estasprevalências foram obtidas.
As patologias associadas à infecção pelo HTLV-1 foramdemonstradas em Salvador. Por exemplo, PET/MAH (37 6 65 28 36
71). Têm sido descritos casos graves de escabiose (“sarnanorueguesa”) e estrongiloidíase em portadores de HTLV-1 (14
19 51). Os pacientes infectados por este vírus apresentam maiorprevalência e curso mais insidioso de tuberculose (55 58 65).Como já foi observado em outras regiões, esta infecção emSalvador atinge mais freqüentemente, os indivíduos menosfavorecidos socialmente, particularmente as mulheres, commenor grau de escolaridade (26 62) .
Baseado na prevalência da infecção na população geralestima-se que 40 mil pessoas estejam infectadas em SalvadorDourado (26).Tem sido, também, demonstrando a presença doHTLV-1 em outras cidades do estado da Bahia (15 52 72).
Associação de Subtipos do HTLV-1 e a Origem GeográficaOs HTLV-1 e 2 infectam populações humanas há milhares
de anos e, por terem estabilidade genômica elevada, sãoexcelentes marcadores para traçar as migrações destaspopulações (34). A estabilidade genomica elevada dos HTLVspossibilitou o estabelecimento da associação entre opolimorfismo genético viral e a origem geográfica (34 48 49 80 87).

5Gaz. méd. Bahia 2009;79:1(Jan-Dez):3-10 Epidemiologia e Origem do HTLV-1 em Salvador, Bahia, Brasil
www.gmbahia.ufba.br
Análises da variabilidade na seqüência do gene env e o LTRmostraram variantes distintas obtidas de diferentes regiõesgeográficas. A identificação de variantes divergentes doHTLV-1 provenientes de indivíduos do Japão, Congo eMelanésia apresentavam respectivamente variabilidade naseqüência do gene env em torno de 3%, 4% e 8,5% (35 41 44 54 70
76).Atualmente, baseada nas análises filogenéticas das regiões
LTR e env de cepas provenientes de todas as regiõesgeográficas do mundo, o HTLV-1 foi classificado em setesubtipos: 1) a ou Cosmopolita disseminado por tudo o mundo(57). 2) b ou subtipo Centro-africano de isolados provenientesde populações da África Central (44 81). 3) c ,ou subtipo daMelanésia caracterizado a partir de isolados provenientes deindivíduos de Papua Nova Guiné e de aborígenes australianos(8 33),4) d, isolados de pigmeus da Republica Central Africanae de dois pacientes de Camarões e Gabão (23 53) ; e os subtipos:e, e f isolados respectivamente de um pigmeu Efe Mbuti doCongo e de um indivíduo do Gabão (74) e 6) g proveniente daRepublica Central Africana (88). O subtipo Cosmopolita édividido em cinco subgrupos: 1) A ou Transcontinental; 2) Bou Japonês; 3) C ou Norte - africano; 4) D ou Oeste-africano(32 57 86) e 5 ) E isolado proveniente de negros do Peru (79).
Origem do HTLV- 1 no Estado da BahiaPara o estudo da origem, disseminação e evolução dos
retrovírus e das migrações de populações humanas utilizam-se análises do polimorfismo genético dos HTLV e análisesfilogenéticas do DNA mitocondrial, como também, marcadoresgênicos de ancestralidade dos seres humano (11).
É possível que o HTLV-1 tenha se originado de infecçãointerespecies, pela transmissão do STLV-1 de primatas nãohumanos (PNH) para a espécie humana. Este evento poderiater acontecido, há milhares de anos antes da separação daspopulações africanas e não africanas que provavelmenteocorreu há cerca de 75.000-287.000 anos tendo o fluxo gênicoocorrido dos pigmeus para as populações vizinhas (22). Aexistência de Pigmeus Efe Mbuti,isolados e infectados peloHTLV-1 na Republica do Congo, considerados descendentesdos povos proto-africanos corrobora esta hipótese. A primeirapovoação da America ocorreu, no leste e no sul do continenteamericano no período pré-colombiano, há 15.000-35.000 anos,decorrente de uma ou mais ondas de migrações de populaçõesasiáticas através do Estreito de Bering (12 42).
A outra possibilidade é que esta introdução tenha ocorridono período pós-colombiano decorrente de migrações depopulações africanas (tráfico de africanos) e/ou de populaçõesasiáticas (filipinos, chineses, japoneses e indianos) nosséculos XVI, XVII e XX (57 79 89). As migrações das populaçõesafricanas ocorreram principalmente do oeste e centro da Áfricapara as Américas, sendo que 40% dos quase 10 milhões deafricanos desembarcaram em portos brasileiros, no períodocompreendido entre os séculos XVI e XIX, abrangendo osseguintes ciclos: a) Ciclo da Guiné, durante a 2a metade doséculo XVI; b) Ciclo de Angola e do Congo no século XVII; c)
Ciclo da Costa da Mina, durante os três primeiros quartos doséculo XVIII e d) Ciclo da baía de Benin nos séculos XVIII eXIX. O ciclo da Costa da Mina contribuiu com maior númerode africanos trazidos para o Brasil. Porem há evidências queafricanos foram, também, trazidos, de regiões do sul da África,hoje conhecidas como sul de Angola, África do Sul eMoçambique (73).
Com o objetivo de esclarecer a origem do HTLV-1 no Brasilanalisamos 243 seqüências da região LTR de isoladosprovenientes de brasileiros descendentes de diversas etniase de diferentes regiões geográficas do país, depositadas emum banco de dados (Genbank database) até 05 de outubrode 2006 (69). Todas foram classificadas como do subtipoCosmopolita, sendo que a maioria (235 das 243; 96.7%) era dosubgrupo A, Transcontinental e somente oito (3.3%) eram dosubgrupo B, Japonês. Destas, cinco eram provenientes daregião sudoeste, dois da norte e um da nordeste. Três destascinco foram provenientes de imigrantes japoneses, queresidiam em São Paulo e uma de uma gestante da regiãonordeste, cujo cônjuge viveu alguns anos em São Paulo. Estesresultados confirmam dados anteriores e indicam que existeuma predominância do subtipo Cosmopolita, subgroupo A,transcontimental circulando no território nacional (1 2 52 60 72 77
89). Posteriormente, verificamos que isolados de HTLV-1provenientes de doadores de sangue do Acre foramclassificados como subtipo Cosmopolita, subgrupo A,transcontinental, corroborando a presença deste subtipo noBrasil (62). A grande variabilidade genética e a ampla distribuiçãodeste subgrupo no território nacional sugerem que aintrodução do HTLV-1 ocorreu por meio da migração (tráfico)dos povos africanos no período compreendido entre osséculos XVI e XIX. Por outro lado, a baixa variação dasseqüências observadas no subgrupo Japonês sugere que elesforam introduzidos recentemente no Brasil. Sabe-se queocorreu uma intensa migração japonesa para o Brasil,principalmente para São Paulo, no inicio do século XIX, o queexplicaria a presença deste subgrupo no Brasil.
Em relação a Salvador, observamos que a maioria dosisolados do HTLV-1, provenientes de indivíduos nascidosnesta cidade, foi classificado como do subtipo Cosmopolita,subgrupo A, Transcontinental (1 2 60). Um único isolado,oriundo de uma gestante, e já referido anteriormente, foiclassificado como subgrupo Japonês. Chama a atenção osfatos de não termos encontrados o subtipo do oeste da Áfricae da maioria dos isolados se agruparem com amostras da Áfricado Sul.
Estima-se que a Bahia recebeu o maior contingente dosafricanos trazidos para o Brasil durante o trafico negreiro eque a maioria foi proveniente do Benin e da Nigéria, paíseslocalizados no oeste da África (83 84 85). Salvador é uma cidadecom cerca de 80% da população constituída por afro-descendentes (5).
No primeiro ciclo de migração, o ciclo da Guiné, os naviosnegreiros partiam da costa oeste da África, região onde hojeestá situado o Senegal, Gâmbia e Guiné Bissau e que naquela

6 Gaz. méd. Bahia 2009;79:1(Jan-Dez):3-10Bernardo Galvão-Castro, et al.
www.gmbahia.ufba.br
época correspondia a área compreendida entre a embocadurado rio Senegal, limite da região desértica entre Senegal eMauritânia, até o do rio Orange, no atual Gabão e eradenominada de Guiné. Todos os africanos trazidos para oBrasil, nesta época, eram de forma genérica, consideradoscomo oriundos da Guiné não havendo precisão daproveniência dos mesmos (66). No período compreendido entre1641-1648, Angola estava sob o domínio dos holandeses e ofluxo de navios negreiros predominava da África paraPernambuco, mas em torno de 1624, período de dominaçãoholandesa na Bahia, foi relatado que havia no porto da cidadede Salvador, 7 navios negreiros sendo 6 proveniente de Angolacom 1440 africanos e um único da Guiné com 28 africanos (84
85) .Esse ciclo foi pouco representativo para a Bahia e estima-se que sete mil africanos, em sua maioria, pertencentes àsetnias sudanesas, tenham vindo para este estado nesteperíodo. Este fato poderia explicar a ausência do subgrupo dooeste africano na Bahia. O segundo ciclo denominado de Ciclode Angola e do Congo compreendeu o final do século XVI,tendo inicio em 1580 e o final do XVII. Neste período a maioriados africanos foi proveniente da África Austral, regiãolocalizada no sul da África banhada pelo Oceano Índico nasua costa oriental e pelo Atlântico na costa ocidental. Não sesabe exatamente o número de africanos que vieram desta região.Além de angolas e congos, vieram, também, moçambicanos, amaioria da etnia banto, sendo que grande parte deles seinstalaram na região do Recôncavo. O terceiro ciclodenominado de Ciclo da Costa da Mina ocorreu durante osdois terços iniciais do século XVIII e contribuiu com maiornúmero de africanos trazidos para o Brasil. Neste período,que durou até meados de 1775, no século XVIII, os africanoseram denominado negros minas e oriundos de quatro portosque ficavam no leste da África, ao longo da costa do Daomésituado onde atualmente é o Benin, região conhecida comoCosta da Mina. A maioria dos africanos era ioruba, chegandoà Bahia com a denominação de nagôs. Entre 1678 e 1814,apenas 39 dos 1770 navios que levavam tabaco da Bahia paraa África tinham como destino os portos congo-angolanos,sendo os outros destinados aos portos da Costa da Mina (84)..O ciclo da Baía de Benin ocorreu entre 1770 e 1850, incluindoo tráfico clandestino, uma vez que a Bahia acusou a limitaçãoao tráfico ao norte do Equador em 1815, fato que explica aforte ligação da Bahia com a África Ocidental, particularmentecom a Baía de Benin (29). Nesse período, a forte presença deetnias iorubá passa a ser representativa, onde desde o cicloanterior, são encontrados daomeanos, tratados no Brasil peladenominação de jejes, haussás e, sobretudo os nagôs-iorubás.Pelo exposto nos relatos históricos há uma mistura da origemgeográfica com as diversas etnias não havendo grandeprecisão da proveniência dos africanos para a Bahia (66).
Com o objetivo de esclarecer melhor a origem do HTLV-1em Salvador analisamos a distribuição de haplótipos ligadosao grupo de genes da globina β na população estudada (1 2).Estes haplótipos compreendem grupo de polimorfismosgenéticos associados a presença ou ausência de sítios para
endonucleases de restrição e estão relacionados a diferentesetnias e regiões geográficas, sendo utilizados comomarcadores de migrações populacionais em estudosantropológico (63)., Observamos que 45,2% dos haplótipos dogene da globina β eram do tipo Benin; 28,9% do tipo Banto e25,9% do tipo Senegal. A prevalência encontrada do haplótipotipo Benin está de acordo com os relatos históricos, pois amaioria dos africanos que vieram para a Bahia foi do oeste daÁfrica principalmente no século XVIII (85). O haplótipo tipoSenegal, característico da África Atlântica, já foi demonstradoanteriormente em Salvador (38). Portanto, a não detecção dosubtipo do oeste da África do HTLV-1 continua sendo umaincógnita. No entanto, a demonstração de 29,4% de haplótipotipo Banto poderia explicar porque a maioria dos isolados deHTLV-1 se agrupa com isolados do sul da África. Sabe-se queafricanos da etnia banto foram trazidos para Bahia entre 1678-1810 (84). Por exemplo, foram trazidos para a Bahia cerca de2400 africanos Bantos sendo 100 procedentes de Angola e2300 de Madagascar no período compreendidos entre os anosde 1817-1843 (24). Há, também, evidências de que muitas vezesos portos de partida dos navios negreiros na África nãoestavam necessariamente relacionados, com as origens étnicase geográficas dos Africanos transportados. Sabe-se que noperíodo de colonização da África do Sul pelos ingleses, nosséculos XVII e XVIII, muitos africanos migravam para regiõesvizinhas, atualmente conhecidas como Angola, Madagascare Moçambique onde eram capturados e transportados paraSalvador. Portanto, há evidencias de que os Bantos povoarama Bahia, principalmente Salvador. Nesta cidade, aindapersistem, sobremaneira, os marcos da cultura banto queinfluenciou a língua, a religião, a musica e o folclore baianosprincipalmente o soteropolitano.
Recentemente, verificamos que isolados obtidos de outrascidades do estado da Bahia foram classificados com subtipo,cosmopolita, subgrupo A, transcontinental com seqüênciasque se agrupam com as da África do Sul, corroborando osdados anteriores (72).Neste trabalho observamos, também, queestas seqüências apresentavam um ancestral comum com umisolado da África Central e da África do Sul, sugerindo que oHTLV-1 poderia ter se originado na África Central e sedisseminado para a África do Sul pela migração dos bantos(Figura 1). Além disso, a presença de isolados da Bahia emdiferentes grupamentos sugere a introdução múltipla do HTLV-1 no período pós-colombiano neste estado.
Devido ao pequeno número de amostras analisadas seriatemerário concluir que não existe o subgrupo Oeste Africanoem Salvador. Outro fator limitante é a escassez de informaçõessobre amostras de HTLV-1 do continente africano. Estudosestão em andamento com o objetivo de se obter um númeromaior de seqüências africanas do HTLV- 1, provenientes tantodo sul como do oeste da África para que possamos melhorcompará-las com as amostras de Salvador.
Baseado nos nossos resultados e considerando a extensãocontinental e a grande variedade de grupos étnicos no Brasil,sugerimos que múltiplas introduções do HTLV-1 ocorreram

7Gaz. méd. Bahia 2009;79:1(Jan-Dez):3-10 Epidemiologia e Origem do HTLV-1 em Salvador, Bahia, Brasil
www.gmbahia.ufba.br
Figura 1. Hipótese provável da origem do HTLV na Bahia.

8 Gaz. méd. Bahia 2009;79:1(Jan-Dez):3-10Bernardo Galvão-Castro, et al.
www.gmbahia.ufba.br
neste país e na Bahia na época pós-colombiana (1 2 52 59 61 72);Estudos contemplando isolados do HTLV-1 de diferentesregiões e provenientes de indivíduos de diferentesancestralidades serão de extrema importância para esclarecermelhor as origens deste vírus no Brasil.
Referências1. Alcantara LC, Van Dooren S, Gonçalves MS, Kashima S, Costa
MC, Santos FL, Bittencourt AL, Dourado I, Filho AA, Covas DT,Vandamme AM, Galvão-Castro B Globin haplotypes of human T-cell lymphotropic virus type I-infected individuals in Salvador,Bahia, Brazil, suggest a post-Columbian African origin of this virus.J Acquir Immune Defic Syndr 1:536-542, 2003.
2. Alcantara LCJ, De Oliveira T, Gordon M, Pybus O, MascarenhasRE, Seixas MO, Gonçalves M, Hlela C, Cassol S, Galvão-Castro B.Tracing the origin of Brazilian HTLV-1 as determined by analysisof host and viral genes. AIDS 20: 780-782, 2006.
3. Andrade TM,. Dourado I, Galvao-Castro B. Associations amongHTLV-I, HTLV-II, and HIV in injecting drug users in Salvador,Brazil. J Acquir Immune Defic Syndr Hum Retrovirol 18:186-187,1998.
4. Andrade T, Farias A , Dourado I, Galvão-Castro B.. Changing patternsof HIV-1 and HTLV-I/II seroprevalence among IDUs from differentdistricts of Salvador: Efficiency of prevention strategies. AIDS ResHum Retroviruses. 17:S35, 2001.
5. Azevedo ES, Fortuna CM, Silva KM, Sousa MG, Machado MA,Lima AM, Aguiar ME, Abe K, Eulalio MC, Conceicao MM, SantosMG . Spread and diversity of human populations in Bahia, Brazil.Hum Biol 54:329–341, 1982.
6. Barbosa HS, Bittencourt AL, Barreto de Araújo I, Pereira Filho CS,Furlan R, Pedrosa C, Lessa G, Harrington W Junior, Galvão CastroB. Adult T-cell leukemia/lymphoma in northeastern Brazil: aclinical, histopathologic, and molecular study. J Acquir ImmuneDefic Syndr 1:65-71, 1999.
7. Barré-Sinoussi F, Chermann JC, Rey F, Nugeyre MT, Chamaret S,Gruest J, Dauguet C, Axler-Blin C, Vezinet-Brun F, Rouzioux C,Rozenbaum W, Montagnier L. Isolation of a T-lymphotropicretrovirus from a patient at risk for acquired immune deficiencysyndrome (AIDS). Science 220:868-871, 1983.
8. Bastian I, Gardner J ,Webb D , Gardner I. Isolation of a human T-lymphotropic virus type I strain from Australian aboriginals. JVirol. 67:843-51, 1993.
9. Biggar RJ, Taylor ME, Neel JV, Hjelle B, Levine PH, Black FL,Shaw GM, Sharp PM, Hahn BH. Genetic variants of human T-lymphotropic virus type II in American indian groups. Virology216:165-173, 1996.
10. Bittencourt AL, Dourado I, Filho PB, Santos M, Valadão E,Alcantara LC, Galvão-Castro B. Human T-cell lymphotropic virustype 1 infection among pregnant women in northeastern Brazil. JAcquir Immune Defic Syndr 26:490-4, 2001.
11. Black FL. Tracing prehistoric migrations by the viruses they carry:Human T-cell lymphotropic viruses as markers of ethnicrelationships. Human Biology 69: 467-482, 1997.
12. Bonatto SL, Salzano FM. A single and early migration for thepeopling of the Americas supported by mitochondrial DNA sequencedata. Proc Natl Acad Sci U S A. 94:1866-1871, 1997.
13. Brasil. Ministério da Saúde. Programa Nacional de DoençasSexualmente Transmissíveis e AIDS Projeto sentinela: gestantes /Sentinel Project- Pregnant Women Brasília; Brasil. Ministério daSaúde; 12 p. 1997.
14. Brites C, Weyll M, Pedroso C, Badaró R. Severe and Norwegianscabies are strongly associated with retroviral (HIV-1/HTLV-1)infection in Bahia, Brazil. AIDS. 16:1292-3, 2002.
15. Britto, APCR, Galvão-Castro B, Straatmann A, Santos-Torres S,Tavares-Neto J. Infecção pelo HTLV-I/II no Estado da Bahia. RevSoc Bras Med Trop 31: 35-41, 1998.
16. Calattini S, Chevalier SA, Duprez R, Bassot S, Froment A, MahieuxR, Gessain A. Discovery of a new human T-cell lymphotropic virus(HTLV-3) in Central Africa. Retrovirology 2:30, 2005.
17. Carneiro-Proietti AB, Ribas JG, Catalan-Soares BC, Martins ML,Brito-Melo GE, Martins-Filho OA, Pinheiro SR, Araújo Ade Q,Galvão-Castro B, de Oliveira MS, Guedes AC, Proietti FA. Infectionand disease caused by the human T cell lymphot ropic viruses typeI and II in Brazil. Rev Soc Bras Med Trop 35:499-508, 2002.
18. Carneiro-Proietti AB, Catalan-Soares BC, Castro-Costa CM, MurphyEL, Sabino EC, Hisada M, Galvão-Castro B, Alcantara LC, RemondeguiC, Verdonck K, Proietti FA. HTLV in the Americas: challenges andperspectives. Rev Panam Salud Publica 19:44-53, 2006.
19. Carvalho EM, Da Fonseca Porto A. Epidemiological and clinicalinteraction between HTLV-1 and Strongyloides stercoralis. ParasiteImmunol. 26:487-97, 2004.
20. Catalan-Soares BC, Proietti FA, Carneiro-Proietti AB Os víruslinfotrópicos de células T humanos (HTLV) na última década (1990-2000) Aspectos epidemiológicos Rev Bras Epidemiol 4:81-95, 2001.
21. Catalan-Soares B, Carneiro-Proietti AB, Proietti FA.Interdisciplinary HTLV Research Group. Heterogeneous geographicdistribution of human T-cell lymphotropic viruses I and II (HTLV-I/II): serological screening prevalence rates in blood donors fromlarge urban areas in Brazil. Cad Saude Publica 21:926-931, 2005.
22. Cavalli-Sforza LL, Menozzi P, Piazza A. The history and geographyof human genes. New Jersey, USA: Princeton University Press,1994.
23. Chen J, Zekeng L,Yamashita M, Takeisha J, Miura T,Ido E,Mboudjeka I,Tsague JM, Hayami M, Kaptue L. HTLV isolatedfrom a Pygmy in Cameroon is related but distinct from the knownCentral African type. AIDS Res Hum Retroviruses 11:1529-1531,1995.
24. Curtin PD. The slave Atlantic trade: a census. Milwaukee: TheUniversity of Wisconsin Press, 1969.
25. Dourado I, Andrade T, Carpenter CL, Galvão-Castro B. Risk factorsfor human T cell lymphotropic virus type I among injecting drugusers in northeast Brazil: possibly greater efficiency of male tofemale transmission. Mem Inst Oswaldo Cruz. 94:13-8, 1999.
26. Dourado I, Alcantara LCJ, Barreto ML, Teixeira MG, B. Galvao-Castro. HTLV-I in the general population of Salvador, Brazil\: acity with African ethnic and sociodemographic characteristics. JAcquir Immune Defic Syndr 34: 527–531, 2003.
27. Edlich RF, Arnette JA, Williams FM. Global epidemic of human T-cell lymphotropic virus type-I (HTLV-I). J Emerg Med 18:109-119, 2000.
28. Ferraz-Chaoui AK, Atta AM, Atta ML, Galvão-Castro B, SantiagoMB. Study of autoantibodies in patients with keratoconjunctivitissicca infected by the human T cell lymphotropic virus type 1.Rheumatol Int. Jul 29. 2009 [Epub ahead of print].
29. Florentino M. Em Costas Negras. Ed Companhia das Letras, SãoPaulo. pp 305 1997.
30. Galvão-Castro B, Loures L, Rodrigues LG, Sereno A, Ferreira JuniorOC, Franco LG, Muller M, Sampaio DA, Santana A, Passos LM,Proietti F. Distribution of human T-lymphotropic virus type Iamong blood donors: a nationwide Brazilian study. Transfusion37:242-243, 1997.
31. Galvão-Castro B, BongertzV, Morgado MG. “Importance del’isolement du VIH dans le programme brésilien de contrôle duVIH/SIDA (A importância do isolamento do HIV no ProgramaBrasileiro de Controle do HIV/AIDS) Cendotec, 2008.
32. Gasmi M, Farouqi B, d’Incan M, Desgranges C. Long terminalrepeat sequence analysis of HTLV type I molecular variantsidentified in four North African patients. AIDS Res Hum Retroviruses10:1313-1315, 1994.
33. Gessain A, Yanagihara R, Francini G, Garruto RM, Jenkins CL,Ajdukiewicz AB, R.C. Gallo and D.C.Gajdusek. Highly divergentmolecular variants of human T-lymphotropic virus type fromisolated populations in Papua New Guinea and the Solomon Islands.Proc Natl Acad Sci USA 88:7694-7698, 1991.

9Gaz. méd. Bahia 2009;79:1(Jan-Dez):3-10 Epidemiologia e Origem do HTLV-1 em Salvador, Bahia, Brasil
www.gmbahia.ufba.br
34. Gessain A, Gallo RC, Franchini G. Low degree of human T-cellleukemia/ lymphoma virus type I genetic drift in vivo as a meansof monitoring viral transmission and movement of ancient humanpopulations. J Virol 66:2288-2295, 1992.
35. Gessain, A., E. Boeri, R. Yanagihara, R.C. Gallo and G. Francini.Complete nucleotide sequence of a highly divergent human T-cell leukemia (lymphotropic) virus type I (HTLV-I) variantfrom Melanesia: genetic and phylogenetic relationship toHTLV-I strains from other geographical regions. J Virol67:1015-1023, 1993.
36. Giozza SP, Santos SB, Martinelli M, Porto MA, Muniz AL,Carvalho. Salivary and lacrymal gland disorders and HTLV-1infection Rev Stomatol Chir Maxillofac. 109:153-7. Epub2008 Mar 28.
37. Gomes I, Melo A, Proietti FA , Moreno-Carvalho O , Loures L.A.M,Daza M.C, Said G,Larouzé B, Galvão-Castro B . Human Tlymphotropic vírus type I (HTLV-I) infection in neurologicalpatients in Salvador, Bahia, Brazil. J Neurol Sci 165:84-89, 1999.
38. Gonçalves MS, Bomfim GC, Maciel E, Cerqueira I, Lyra I, ZanetteA, Bomfim G, Adorno EV, Albuquerque AL, Pontes A, Dupuit MF,Fernandes GB, Reis MG. bs-haplotypes in sickle cell anemia patientsfrom Salvador-Bahia-Brazil. Braz J Med Biol Res 36:1283-1288,2003.
39. Gottlieb MS, Schroff R, Schanker HM, Weisman JD, Fan P.T, WolfRA, . Saxon A. Pneumocystis carinii pneumonia and mucosalcandidiasis in previously healthy homosexual men: evidence of anew acquired cellular immunodeficiency. N Engl J Med 305:1425-1431, 1981.
40. Goubau P, Carton H, Kazadi K, Muya K.W, Desmyter J. HTLVseroepidemiology in a central African population with highincidence of tropical spastic paraparesis. Trans R Soc Trop MedHyg 84:577-579, 1990.
41. Gray GS, White M, Bartman T, Mann D. Envelope gene sequenceof HTLV-1 isolate MT-2 and its comparison with other HTLV-1isolates.Virology. 177:391-395, 1990.
42. Greenberg J, Turner C Zegura S. The settlement of the Americas:a comparison of the linguistic, dental and genetical evidence. CurrAnthropol 27:477-497, 1986.
43. Hacker MA., Friedman SR, Telles PR. The role of “long-term” and“new” injectors in a declining HIV/AIDS epidemic in Rio de Janeiro,Brazil. Subst Use Misuse 4 :99-123, 2005.
44. Hahn BH, Shaw GM, Popovic M, Lo-Monico A, Gallo RC, Staal-Wong F. Molecular cloning and analysis of a new variant of humanT-cell leukemia virus (HTLV-Ib) from an African patient with adultT-cell Leukemia-lymphoma. Int J Cancer 34:613-618, 1984.
45. Hehlmann R, Erfle V.. Introduction to retroviruses. In: Belshe RB(ed), Textbook of human virology. 2a.edição, St. Louis: Mosby, p242-273, 1991.
46. Kalyanaraman VS, Sarngadharan M.G, Robert-Guroff M, MyioshiI, Golde D, Gallo RC. A new subtype of human T-cell leukemia virus(HTLV-II) associated with a T-cell variant of hairy cell leukemia.Science 218:571-573, 1982.
47. Kitagawa T, Fujishita M, Tacagushi H, Miyoshi I, Tadokoro H.Antibodies to HTLV-I in Japanese immigrants in Brazil. JAMA256:2342, 1986.
48. Kourian F, Pelloquin F de Thé G. In vivo genomic variability ofhuman T-cell leukemia virus type I depends more upon geographythan upon pathologies. J Virol 65:3770-3778, 1991.
49. Liu HF, Vandamme AM, Kazadi K, Carton H, Desmyter J, GoubauP. Familial transmission and minimal sequence variability of humanT-lymphotropic virus type-I (HTLV-I) in Zaire. AIDS Res HumRetroviruses 10:1135-1142, 1994.
50. Liu HF,.Goubau P, Van Brussel M, Van Laethem K, Chen YC,Desmyter J, Vandamme, AM. The three human T-lymphotropicvirus type I subtypes arose from three geographically distinct simianreservoirs. J Gen Virol. 77:359-368, 1996.
51. Maciel E, Espinheira L, Brites C. Strongyloidiasis as an opportunisticinfection in a HAM/TSP Patient. Braz J Infect Dis 3:23-27, 1999.
52. Magalhães T, Mota-Miranda AC, Alcântara LCJ, Olavarria V,Galvao-Castro B, Rios-Grassi MF. Phylogenetic and MolecularAnalysis of HTLV-1 Isolates From a Medium Sized Town in Northernof Brazil: Tracing a Common Origin of the Virus From the MostEndemic City in the Country. J Med Virol 80: 2040-2045, 2008.
53. MahieuxR, Ibrahim F, Mauclere P, Herve V, Michel P, Telaoa F,Chappey C ,Garin B, Van Der Ryst E, Guillemain B, Ledru E,Delaporte E,de Thé G, Gessain A. Molecular epidemiology of 58new African Human T-cell Leukemia Virus Type I (HTLV-I) strains:identification of a new and distinct HTLV-1 molecular subtype inCentral Africa in Pygmies. J Virol. 71:1317-1333, 1997.
54. Malik KTA, Even J Karpas A. Molecular cloning and completenucleotide sequence of an adult T-cell leukemia virus/ human T-cellleukemia virus type I (ATLV/ HTLV-I) isolate of Caribbean origin:Relationship to other members of the ATLV/ HTLV-I subgroup. JGen Virol. 69:1695-1710, 1988.
55. Marinho J, Galvao-Castro B, Rodrigues LC, Barreto ML. Increasedrisk of tuberculosis with human T-lymphotropic virus-1 infection: acase-control study. J Acquir Immune Defic Syndr 15:625-628, 2005.
56. Mesquita F, Kral A, Reingold A. Trends of HIV infection amonginjection drug users in Brazil in the 1990s: the impact of changes inpatterns of drug use. J Acquir Immune Defic Syndr 28: 208 – 302,2001.
57. Miura T, Fukunaga T, Igarashi T. Phylogenetic subtypes of humanT-lymphotropic virus type I and their relations to theanthropological background. Proc Natl Acad Sci U S A. 91:1124-1127, 1994.
58. Moreira JrED, Ribeiro TT, Swanson P,Sampaio-Filho C, Melo A, BritesC, Badaró R, Toedter G, Lee H, Harrington Jr W. Seroepidemiology ofhuman T-cell lymphotropic vírus type I/II in Northeastern Brazil. J ofAcquir Immune Defic Syndr 6:959-963, 1993.
59. Mota ACA, NunesC; Melo A, Romeo M, Boa-Sorte N; Dourado I,Alcantara LCJ, Galvão-Castro B. A case control study of HTLV-infection among blood donors in Salvador, Bahia, Brazil: Associatedrisk factors and trend towards declining prevalence. Rev BrasHematol Hemoter 28:120-126, 2006.
60. Mota AC, Van Dooren S, Fernandes FM, Pereira SA, Queiroz AT,Gallazzi VO, Vandamme AM, Galvão-Castro B, Alcantara LC. Theclose relationship between South Africa and Latin American HTLV-1 strains corroborated in a molecular epidemiological study of theHTLV-1 isolates from a blood donor cohort. AIDS Res HumRetroviruses23:503–507, 2007.
61. Mota-Miranda AC, Araújo SP, Dias JP, Colin DD, Kashima S, CovasDT, Tavares-Neto J, Galvão-Castro B, Alcantara LC. HTLV-1infection in blood donors from the Western Brazilian Amazonregion: seroprevalence and molecular study of viral isolates. J MedVirol 80:1966-1971, 2008.
62. Moxoto I, Boa-Sorte I, Nunes C, Mota AC, Dumas A, Dourado I,Galvão-Castro B. Perfil Sócio-Demográfico Epidemiológico eComportamental de Mulheres infectadas pelo HTLV-1 em Salvador-Bahia, uma área endêmica para o HTLV. Rev Soc Bras Med Trop42:37-41, 2007.
63. Nagel RL, Ranney HM.. Genetic epidemiology of structuralmutations of the b-globin gene. Semin Hematol 27:342-343, 1990.
64. Oliveira M de F, Brites C, Ferraz N, Magalhaes P, Almeida F,Bittencourt AL. Infective dermatitis associated with the human Tcell lymphotropic virus type I in Salvador, Bahia, Brazil. ClinInfect Dis 40:90-96, 2005.
66. Pedral-Sampaio DB, Martins Netto E, Pedrosa C, Brites C, DuarteM, Harrington W. Co-infection of tuberculosis and HIV/HTLVretroviruses: frequency and prognosis among patients admitted in aBrazilian Hospital Brazil. Clin Infect Dis1:31-35, 1997.
66. PinskyJ.. O escravo negro: In Pinsky J, A escravidão no Brasil. SãoPaulo: Contexto, . pp. 24, 1988.
67. Poiesz BJ, Ruscetti FW, Gazdar AF , Bunn PA, Minna JA, . GalloR.C. Detection and isolation of type C retrovirus particles fromfresh and cultured lymphocytes of a patient with cutaneous T-celllymphoma.. Proc Natl Acad Sci U S A 77:7415-7419, 1980.

10 Gaz. méd. Bahia 2009;79:1(Jan-Dez):3-10Bernardo Galvão-Castro, et al.
www.gmbahia.ufba.br
68. Proietti FA, Carneiro-Proietti ABF, Catalan-Soares BC , MurphyEL. Global epidemiology of HTLV-1 infection and associateddiseases. Oncogene 24:6058-6068, 2005.
69. Queiroz AT, Mota-Miranda AC, de Oliveira T, Moreau DR, Urpia C,Carvalho CM, Galvão-Castro B, Alcantara LC. Re-mapping the molecularfeatures of the human immunodeficiency virus type 1 and human T-celllymphotropic virus type 1 Brazilian sequences using a bioinformaticsunit established in Salvador, Bahia, Brazil, to give support to the viralepidemiology studies. Mem Inst Oswaldo Cruz 102:133-9, 2007.
70. Ratner L, Josephs SF, Starcich, B. Hahn, Shaw G.M,. Gallo RC,.Wong-Staal, F.. Nucleotide sequence analysis of a variant human T-cell leukemia virus (HTLV-Ib) provirus with a deletion in pX-I. JVirol. 54:781-790, 1985.
71. Rathsam-Pinheiro RH, Boa-Sorte N, Pinheiro CA , Castro–Lima-Vargens C, Castro-Lima H, Galvão-Castro B. Ocular lesions inHTLV-1 infected patients from Salvador (Bahia), the city with thehighest prevalence of this infection in Brazil., Rev Soc Brasileirade Medicina Trop (no prelo).
72. Rego FF, Alcantara LC, Moura Neto JP, Miranda AC, Pereira Ode S,Goncalves Mde S, et al. HTLV type 1 molecular study in Brazilianvillages with African characteristics giving support to the post-Columbian introduction hypothesis. AIDS Res Hum Retroviruses24:673-677, 2008.
73. Rodrigues N. Os africanos no Brasil. (5a ed), São Paulo, CompanhiaEditora Nacional, 1977.
74. Salemi M, Van Dooren S, Audenaert E, Delaporte E, Goubau P,Desmyter J, Vandamme AM. Two new human T-lymphotropicvirus type I phylogenetic subtypes in seroindeterminates, a Mbutipygmy and a Gabonese, have closest relatives among African STLV-I strains Virology. 246 :277-287, 1998.
75 Santos JI, Lopes MAA, Deliége-Vasconcelos E, Couto-FernandezJC, Patel BN, Barreto M.L, .Ferreira Jr OC, Galvão-Castro B. .Seroprevalence of HIV, HTLV-I/II and otheperinatally-transmittedpathogens in Salvador, Bahia. Rev. Inst. Med. Trop. S. Paulo37:343-348, 1995.
76. Seiki M, Hattory S, Hyraiama Y Yoshida M. Human adult T-cellleukemia virus: completed nucleotide sequence of the provirusgenome integrated in leukemia cell DNA. Proc Natl Acad Sci U S A80:3618-3622, 1983.
77. Souza LA, Lopes IG, Maia EL, Azevedo VN, Machado LF, Ishak MO,Ishak R, Vallinoto AC. Molecular characterization of HTLV-1 amongpatients with tropical spastic paraparesis/HTLV-1 associated myelopathyin Belém, Pará. Rev Soc Bras Med Trop. 39(5):504-6, 2006.
78. Teixeira SLM, Bastos FI, Telles PR. HIV-1 infection Sanchez-Albornoz, N., R. Morse, P. Bakewell, E. Florescano, M. Morner,M.J. Macleod, J. Lockhart and A. Lavrin. . Colonial Latin Americain The Cambridges History of Latin America volume 2,Cambridge:Cambridge University Press, , pp.15-335 1984.
79. Van Dooren S, Gotuzzo E, Salemi M , Watts , E. Audenaert, S.Duwe, H. Ellerbrok, R. Grassmann, J. Desmyter and A.-M.Vandamme.. Evidence for a Post-Colombian introduction of humanT-cell lymphotropic virus in Latin America. J Gen Virol 79:2695-2708, 1998.
80. Van Dooren S, Pybus OG, Salemi M, Liu HF, Goubau P,Remondegui C, Talarmin A, Gotuzzo E, Alcantara LC, Galvão-Castro B, Vandamme AM. The low evolut ionary rate ofhuman T-ce l l lymphot rop ic v i rus type-1 conf i rmed byanalysis of vert ical transmission chains. Mol Biol Evol.21:603-611, 2004.
81. Vandamme A.M., Liu HF, Goubau P ,Desmyter J. Primate T-lymphotropic virus type I LTR sequence variation and itsphylogenetic analysis: compatibility with an African origin of PTLV-I. Virology. 202:212-223, 1994.
82. Verdier M, Bonis J, Denis FA. The prevalence and incidence ofHTLV in Africa.In. Essex M (ed.), AIDS in Africa, vol. 9. NewYork: Raven Press,. p. 173-193, 1994.
83. Verger, P. Flux et reflux de la traite des nègres entre le Golfe deBenin et Bahia de Todos os Santos. Mouton, 1968.
84. Verger, P. Mouvement des navires. In Verger, P. Trade relationsbetween the Bight of Benin and Bahia, 17th to 19th century, trans.Evelyn Crawford, Ibadan, Ibadan University Press. pp.24-26, 1976.
85. Viana Filho L. O negro na Bahia. 3a edição. Rio de Janeiro: NovaFronteira, p. 31-224, 1988.
86. Vidal AU, Gessain A , Yoshida M, Tekaia F, Garin B, GuillermainB, Schuls T, Farid R, De The G. Phylogenetic classification ofhuman T cell leukaemia/lymphoma virus type I genotypes infive major molecular and geographical subtypes. J Gen Virol75:3655, 1994.
87. Wattel E, Vartanian JP, Pannetier C, Wain-Hobson S. ClonalExpansion of Human T-Cell Leukemia Virus Type I- Infected cellsin asymptomatic and symptomatic carriers without malignancy. JGen Virol. 69:2863-2868, 1995.
88. Wolfe N, Heneine W, Carr JK, Garcia A, Shanmugam V, TamoufeU,Torimiro J, Prosser A, LeBreton M, Mpoudi-Ngole E,Mccutchan F, Birx DL, Folks T, Burke DS, Switzer WM.Emergence of unique primate of T-lymphotropic viruses amongcentral África bushmeat hunters. Proc Natl Acad Sci USA; 102:7994-7999, 2005.
89. Yamashita M, Veronesi R, Menna-Barreto M, Harrington WJ Jr,Sampaio C, Brites C, Badaro R, Andrade-Filho AS, Okhura S,Igarashi T, Miura T, Chamone D, Bianchini O, Jardim C, SonodaS, Hayami M. Molecular epidemiology of human T-cell leukemiavirus type I (HTLV-I) in Brazil: the predominant HTLV-Is inSouth America differ from HTLV-Is of Japan and Africa, as wellas those of Japanese immigrants and their relatives in Brazil.Virology 261:59-69, 1999.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo