seloi rodrigueslivros01.livrosgratis.com.br/cp097889.pdf · obtenção do título de mestre em...
TRANSCRIPT

Seloi Rodrigues
O gênero Potamogeton L. em rios de Bonito, Mato Grosso do Sul:
Anatomia comparada de Potamogeton gayi A. Benn. e Potamogeton
illinoensis Morong, efeitos da velocidade do fluxo da água na
morfologia, e o potencial de reprodução vegetativa de Potamogeton
illinoensis
Dissertação apresentada ao Programa de Pós-Graduação em Biologia Vegetal da Universidade Federal de Mato Grosso do Sul como um dos requisitos para a obtenção do título de Mestre em Biologia Vegetal junto ao Centro de Ciências Biológicas e da Saúde.
Orientadora: Dra. Edna Scremin-Dias
Campo Grande
2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
BANCA EXAMINADORA
_____________________________
Profa. Dra. Cleusa Bona
Universidade Federal do Paraná - UFPR
___________________________________
Prof. Dr. Etenaldo Felipe Santiago. Universidade Estadual de Mato Grosso do Sul -
UEMS
_____________________________________
Profa. Dra. Edna Scremin-Dias (Orientadora)
Universidade Federal de Mato Grosso do Sul - UFMS
Rodrigues, Seloi
O gênero Potamogeton L. em rios de Bonito, Mato Grosso do Sul: Anatomia comparada de Potamogeton gayi A. Benn. e Potamogeton illinoensis Morong, efeitos da velocidade do fluxo da água na morfologia, e o potencial de reprodução vegetativa de Potamogeton illinoensis. 61p.
Dissertação (Mestrado) – Laboratório de Botânica – Universidade Federal de Mato Grosso do Sul.
1. Potamogeton 2. Anatomia 3. Morfologia I. Laboratório de Botânica – Universidade Federal de Mato Grosso do Sul.

iii
Aos meus pais (Romaldino e Selma)
Por tudo que representam na minha vida.

iv
AGRADECIMENTOS
Agradeço a Deus por todas as coisas boas que proporciona na minha vida.
Aos meus pais por ajudarem a tornar meus sonhos possíveis
Aos meus amigos por me darem força nas horas em que pensei em desistir de tudo
Ao Edson por todo amor e carinho dedicado
Aos professores da UFMS pelo incentivo e por transmitir seus conhecimentos
A Edna Scremin-Dias pela orientação, dedicação, amizade e confiança demonstrada
durante o tempo de trabalho e pelo exemplo profissional que sempre transmitiu
Aos proprietários e funcionários das fazendas São Geraldo e Baía Bonita por permitir nossa
entrada na propriedade e pelo apoio logístico concedido, em especial ao Wellington, João
Henrique e Alex, funcionários do rio Sucuri.
A todos os estagiários e técnicos do laboratório de Anatomia vegetal: Eloise, Orlando,
Lucas, Tamires, Thales, Jane e Fabiane e em especial a Mariana e Silvana, que me
ajudaram nas coletas e preparação do material no laboratório.
Ao amigo Augusto e ao Josué pela ajuda com dicas e as análises estatísticas
Ao Prof. Fernando Paiva pelo auxílio técnico com a máquina fotográfica e o sistema de
captura de imagens.
Ao Eduardo Camargo pelas fotos 1E e 1F do capítulo II.
Aos Amigos Mathilde, Flávio e Rosana pela ajuda nas coletas e mensurações das plantas e
a Zildamara pelo auxílio na estatística final e no “socorro” com as fotografias.
A Érika Amano pela amizade e pelo auxilio concedido sempre que precisei
Ao Fernando Belan pelo auxílio nos desenhos e pranchas
Ao técnico Fábio Modesto pelo “socorro” sempre que os computadores davam “pau”

v
A Cristiane Kalife pela tradução dos resumos.
A Christiane Corrêa pela sua amizade e apoio.
A Joelma e Ana Lucia pelas sugestões no texto.
Á Profa. Dra. Maria Rosângela Sigrist, coordenadora do Projeto Gomphrena elegans, por
permitir o uso de equipamentos do projeto e pelo auxílio logístico no inicio das coletas.
A Tuka e Gisseli do Curso de Capacitação por toda a ajuda concedida.
A Propp/UFMS pelo auxilio inicial adquirindo material de consumo.
A Fundect pela Bolsa de mestrado concedida e recursos financeiros para a pesquisa.
A Rede de Sementes do Pantanal pelo empréstimo da caminhonete para as viagens de
coleta
A todos os meus colegas e professores da pós-graduação pelo apoio e incentivo.

vi
INDICE
RESUMO -----------------------------------------------------------------------------------------------1
ABSTRACT ------------------------------------------------------------------------------------------- 2
INTRODUÇÃO GERAL --------------------------------------------------------------------------- 4
CARACTERIZAÇÃO DAS ÁREAS DE ESTUDO ------------------------------------ 7
OBJETIVOS ---------------------------------------------------------------------------------- 8
REFERÊNCIAS BIBLIOGRÁFICAS ---------------------------------------------------- 8
FIGURAS ------------------------------------------------------------------------------------ 11
CAPITULO I - Anatomia comparada de Potamogeton illinoensis Morong e
Potamogeton gayi Benn. que ocorrem em rios de Bonito – MS. --------------------------- 12
INTRUÇÕES AOS AUTORES – Brazilian Archives of Biology and Tecnology - 13
ABSTRACT ----------------------------------------------------------------------------------15 INTRODUÇÃO ----------------------------------------------------------------------------- 15
MATERIAL E MÉTODOS---------------------------------------------------------------- 16
RESULTADOS ----------------------------------------------------------------------------- 16
Anatomia da folha ------------------------------------------------------------------ 16
Anatomia do pecíolo --------------------------------------------------------------- 18
Anatomia do caule e rizoma ------------------------------------------------------ 18
Anatomia da raiz ------------------------------------------------------------------- 19
DISCUSSÃO -------------------------------------------------------------------------------- 20
RESUMO ------------------------------------------------------------------------------------ 25
REFERÊNCIAS BIBLIOGRÁFICAS --------------------------------------------------- 26

vii
CAPÍTULO II - Efeito da velocidade da correnteza na morfologia de Potamogeton
illinoensis Morong e o potencial de reprodução vegetativa desta espécie, que ocorre
nos rios Sucuri e Baía Bonita – Bonito, Estado de Mato Grosso do Sul, Centro-Oeste
do Brasil. --------------------------------------------------------------------------------------------- 26
INTRUÇÕES AOS AUTORES – Aquatic Botany ------------------------------------ 27
ABSTRACT --------------------------------------------------------------------------------- 36
INTRODUÇÃO ----------------------------------------------------------------------------- 36
METODOLOGIA --------------------------------------------------------------------------- 39
RESULTADOS ----------------------------------------------------------------------------- 41
DISCUSSÃO -------------------------------------------------------------------------------- 43
REFERÊNCIAS BIBLIOGRÁFICAS --------------------------------------------------- 51
TABELA ------------------------------------------------------------------------------------- 57
FIGURAS ------------------------------------------------------------------------------------ 58
CONSIDERAÇÕES FINAIS --------------------------------------------------------------------- 59

1
Resumo
Muitas são as plantas aquáticas submersas distribuídas nos rios de Bonito-MS, entre elas
está Potamogeton illinoensis Morong, que ocorre nos rios Sucuri e Baía Bonita e
Potamogeton gayi A. Benn. somente no rio Baía Bonita. Potamogeton illinoensis é
elemento importante na paisagem subaquática destes rios, no entanto seu desenvolvimento
está em desequilíbrio no rio Baía Bonita, constituindo problema para a atividade de
ecoturismo desenvolvida no local. Além disso, pode ser prejudicial para a comunidade de
peixes que habita este rio. O capitulo I desta dissertação apresenta dados sobre a anatomia
dos órgãos vegetativos destas duas espécies, relacionando os caracteres diferenciais
consistentes para delimitá-las taxonomicamente. No capitulo II são descritas as alterações
morfológicas dos indivíduos de P. illinoensis que se desenvolvem em locais de correnteza e
remanso em ambos os rios, relacionando as adaptações desta espécie ao sucesso na
colonização em ambientes distintos. Também são descritos os tipos de estruturas
responsáveis pela reprodução vegetativa desta espécie, bem como a potencialidade de
regeneração de fragmentos de caule que consistem em propágulos vegetativos não
especializados. Os resultados contidos neste trabalho são relevantes, pois contribuem para o
entendimento da biologia de P. illinoensis, servindo como base para trabalhos futuros que
subsidiarão a realização do manejo desta espécie no rio Baía Bonita.
Palavras-chave: Potamogeton illinoensis, Potamogeton gayi, anatomia, morfologia,
regeneração, velocidade da água.

2
ABSTRACT
Submerged aquatic plants are broadly distributed in the rivers of Bonito, with Potamogeton
illinoensis Morong occuring in Sucuri and Baía Bonita rivers and Potamogeton gayi A.
Benn. only in the last one. P. illinoensis consists in an important element for the subaquatic
scenery of those rivers, however its development is in disequilibrium in Baía Bonita River,
constituting a problem for the ecotourism activity developed at this place, and it can still be
harmful to the community of fish that inhabits this river. The chapter I of this dissertation
presents data on anatomy of the vegetative organs of these two species, relating the
consistent differential characters to delimitate them taxonomically. In chapter II
morphological changes in individuals of P. illinoensis that grow in running and stagnant
water in both rivers are described, relating the adaptations of this species to the success in
the colonization of the different environments. The types of structures responsible for the
vegetative reproduction of this species are also described, as well as the potentiality of
regeneration of stem fragments that consist in non-specialized vegetative propagules. The
results presented in this work are important, because they contribute to the understanding of
the biology of P. illinoensis, serving as a base for future works that will subsidize the
management of this species in Baía Bonita River.
Key Words: Potamogeton illinoensis, Potamogeton gayi, anatomy, morphology, water
velocity, propagules.

3
________________________________________________________________________________________
INTRODUÇÃO GERAL

4
O gênero Potamogeton L. é cosmopolita (Judd et al., 2002), ocorrendo desde águas
temperadas até locais de águas tropicais (Wiegleb & Kaplan, 1998). Este gênero pertence à
Família Potamogetonaceae, possuindo outros três gêneros e aproximadamente 100
espécies. Para o Brasil são identificadas 15 espécies, em dois gêneros, sendo a maioria
encontrada na região sul (Souza & Lorenzi, 2005). Para Mato Grosso do Sul, até o
momento, foram identificadas duas espécies Potamogeton gayi A. Benn., encontrado
somente no rio Baía Bonita, e Potamogeton illinoensis Morong, encontrado em dois rios, o
Sucuri e Baía Bonita, no município de Bonito.
Estas duas espécies são exclusivamente submersas e, Potamogeton gayi apresenta
indivíduos pequenos, com órgãos vegetativos e de reprodução sexuada não atingindo a
superfície da água. Potamogeton illinoensis apresenta indivíduos com caules alongados que
atingem a superfície da água, emitindo os escapos florais e inflorescências em ambiente
aéreo, podendo ter folhas flutuantes em ambientes lênticos (Rodrigues & Irgang, 2001).
As plantas aquáticas apresentam uma série de estruturas e/ou mecanismos que
podem ser associados a sua sobrevivência e adaptação no ambiente aquático (Sculthorpe,
1967). Dentre essas características podem ser ressaltadas, alongamento de pecíolos e
entrenós logo após a subida do nível da água (Scremin-Dias, 2000), redução na área foliar
(Scremin-Dias, 1992), heterofilia, caules do tipo rizoma ou estolho, raízes adventícias,
presença de muitas lacunas aeríferas e redução dos tecidos mecânicos (Scremin-Dias et al.,
1999).
Apesar de muitas espécies aquáticas se reproduzirem sexuadamente, o que
prevalece no ambiente aquático é a reprodução vegetativa, a qual tem um papel principal na
manutenção das populações no habitat, sendo, a reprodução sexuada, importante na
colonização de novas áreas (Barrat-Segretain, 1996). Esta autora afirma que a reprodução

5
sexuada é comum em inúmeras espécies de Potamogeton, produzindo grande número de
sementes, mas a germinação nem sempre é possível. Neste aspecto, a reprodução vegetativa
contribui para a manutenção das populações nos rios (Brux et al., 1987 apud Barrat-
Segretain, 1996), sendo a principal forma de propagação para algumas espécies aquáticas.
A reprodução por fragmentos de ramos tende a ter um alto grau de regeneração em muitas
macrófitas aquáticas (Cronk & Fennessy, 2001), já que potencialmente podem produzir
muitas novas plantas num curto espaço de tempo. Esta característica comum para muitas
plantas aquáticas, deve ser levada em conta, quando há necessidade de manejo destas
plantas em algum ambiente (Barrat-Segretain, 1996).
As plantas aquáticas possuem muitas características anatômicas que podem ajudar a
minimizar os efeitos dos diversos fatores ambientais aos quais estão submetidas, sendo que
o sucesso da colonização destas espécies no ambiente pode ser determinado por estas
características. A deficiência de luminosidade, a anoxia e a baixa quantidade de nutrientes
dissolvidos podem ser evitados por células epidérmicas permeáveis, cutícula fina, muitas
raízes adventícias, folhas finas e maleáveis, cloroplastos na epiderme foliar, além de órgãos
com parênquima aerífero muito desenvolvido, que permitem sua flutuação e o
armazenamento de oxigênio (Sculthorpe, 1967).
No entanto os estudos anatômicos dos órgãos vegetativos das plantas brasileiras
ainda são escassos, mas de fundamental importância para o entendimento das adaptações
das espécies em ambientes diversos. Trabalhos realizados no Pantanal e nos rios de Bonito
por Scremin-Dias (1992), Resende (1996), Bona (1999) e Scremin-Dias (2000) se referem a
plantas anfíbias, emersas fixas, que têm seus órgãos aéreos submetidos ao alagamento em
determinada época do ano.

6
Em Bonito, as populações de Potamogeton illinoensis distribuídas ao longo do rio
Baía Bonita, o qual é utilizado para o ecoturismo de flutuação, têm aumentado visivelmente
no decorrer dos últimos anos. Como esta espécie é submersa e atinge a superfície lançando
as estruturas de reprodução sexuada, causa desconforto aos turistas por tocarem o corpo nas
partes submersas superficiais das plantas, durante a flutuação dos banhistas (Observações
de campo). Este aspecto, aliado a demasiada densidade populacional desta espécie no rio
Baia Bonita, levou os proprietários deste atrativo turístico a buscar auxílio para o manejo da
espécie, objetivando minimizar os impactos negativos de Potamogeton illinoensis na
atividade ecoturística e no ambiente.
Os estudos anatômicos com espécies de Potamogeton disponíveis na literatura
abordam aspectos taxonômicos, já que a presença de híbridos entre espécies do gênero são
comuns (Kaplan & Wolff, 2004) tornando difícil a delimitação taxonômica destas espécies.
Anatomia vegetal aplicada a taxonomia é de fundamental importância, haja vista diversos
autores afirmarem que características anatômicas auxiliam na resolução de problemas
taxonômicos, sendo tão importantes quanto os caracteres morfológicos (Tomlinson, 1982,
Metcalfe & Chalk,1983).
Trabalhos relacionados à anatomia ecológica de plantas submersas do gênero
Potamogeton, ou de espécies aquáticas submersas pertencentes a outros grupos
taxonômicos, são raros na literatura. Conhecer a anatomia de espécies aquáticas e as
adaptações destas espécies ao tipo de ambiente onde se desenvolvem, é também
fundamental para a conservação das espécies e do ambiente, bem como para o manejo
destas em locais cuja propagação vegetativa ocorre em demasia. Em Bonito, onde os cursos
d’água compõem atrativos para turistas e são responsáveis pela geração de renda para a
população, as belezas cênicas naturais devem ser conservadas. Neste aspecto, o presente

7
trabalho objetivou descrever e comparar a anatomia das espécies de Potamogeton que
ocorrem nos rios de Bonito, verificar se a velocidade da correnteza influencia na
morfologia de Potamogeton illinoensis, e o potencial para reprodução vegetativa desta
espécie.
CARACTERIZAÇÃO DA ÁREA DE ESTUDO
Este trabalho foi conduzido em dois rios da região de Bonito no Mato Grosso do Sul
(Figura 1). Este município é conhecido por suas belezas naturais que são exploradas
economicamente por meio do ecoturismo. Os dois rios estudados são utilizados no turismo
de flutuação, onde são conduzidas atividades de contemplação da beleza proporcionada
pela paisagem subaquática. Esta paisagem é caracterizada pela grande diversidade de
peixes e de macrófitas aquáticas que vicejam em águas extremamente límpidas (Figuras 2 e
3).
O rio Baía Bonita (56º26’24’’W 21º9’46’’S) (Fig. 2), também chamado Aquário
Natural, situa-se na Fazenda Baía Bonita, tem aproximadamente 1000m de extensão, possui
águas calmas com raros locais de correnteza. O rio Sucuri (Fig. 3), situado aos 56°33´32”W
21°15´56”S, faz parte da Fazenda São Geraldo, possui aproximadamente 1800m de
extensão, é caracterizado por vários locais de forte correnteza, tendo áreas de remanso que,
em geral, estão próximas aos bancos de Gomphrena elegans
A maioria dos rios da região de Bonito, possuem águas extremamente límpidas,
devido à presença de calcário muito puro dissolvido (Boggiani, 1999), que provoca a
precipitação das partículas argilosas em suspensão na água. Esta característica, de águas
transparentes, propicia o desenvolvimento de grande diversidade de macrófitas aquáticas
(Scremin-Dias et al., 1999).

8
Objetivos
• Comparar a anatomia de Potamogeton illinoensis e Potamogeton gayi que
ocorrem nos rios Sucuri e Baia Bonita - Bonito/MS;
• Verificar os efeitos da velocidade da água na morfologia de P. illinoensis;
• Descrever o potencial de reprodução vegetativa que P. illinoensis.
Referências bibliográficas
Barrat-Segretain, M. H. 1996. Strategies of reproduction, dispersion, and competition in
river plants: A review. Vegetatio. Vol. 123, p. 13-17.
Boeger, M. R. T. & Poulson, M. E. 2003. Morphological adaptations and photosynthetic
rates of amphibious Veronica anagallis-aquatica L. (Scrophulariaceae) under different flow
regimes. Aquatic Botany. 75, 123-135.
Boggiani, P. C. 1999. Por que Bonito é Bonito? In: Scremin-Dias, E., Pott, V. J. , Hora, R.
C., Souza, P. R. 1999. Nos Jardins Submersos da Bodoquena. Guia para identificação
das plantas aquáticas de Bonito e região. Campo Grande/MS. Editora UFMS. 160p.
Bona, C. 1999. Adaptações morfo-anatômicas dos órgãos vegetativos de Bacopa
salzmanii (Benth.) Wettst ex Edwall e Bacopa monnieriodoides (Cham.) Robinson
(scrophulariaceae) em ambiente terrestre e aquático. Tese de Doutorado/Instituto de
Biociências/USP, São Paulo.

9
Cronk, J. K. & Fenessy M. S. 2001. Wetland Plants: Biology and Ecology. Ed. Lewis
Publishers, 462p.
Judd, W. S., Campbell, C. S., Kellogg, E. A., Stevens, P. F., Donoghue, M. J. 2002. Plant
Systematics: A phylogenetic approach. Sinauer associates, Inc. Publishers. Sunderland,
Massachusetts, USA. Second edition. 576p.
Kaplan, Z. & Wolff, P. 2004. A morphological, anatomical and isozyme study of
Potamogeton xschrebeii: confirmation of its recent occurrence in Germany and first
documented record in France. Preslia. Vol. 76(2), p. 141-161
Metcalfe, C. R. & Chlak, L. (1983), Anatomy of de Dicotyledons: Wood structure and
conclusion of the general introduction. V.2. New York, Oxford University Press.
Resende, U. M. 1996. Análise estrutural de Neptunia plena (L.) Benth. (Mimosaceae) em
ambiente inundado e livre de inundação, no Pantanal Mato-Grossense, Município de
Corumbá, Mato Grosso do Sul. Curitiba. Dissertação de Mestrado. UFPR.
Rodrigues, R. S. & Irgang, 2001. Potamogetonaceae Dumort. no Rio Grande do Sul, Brasil.
Iheringia, Ser. Bot., Porto Alegre, n. 56, p. 3-49.

10
Scremin-Dias, E. 1992. Morfoanatomia dos órgãos vegetativos de Ludwigia sedoides
(Humb. & Bonpl.) Hara (Onagraceae) ocorrente no Pantanal Sul-Mato-Grossense.
Dissertação de Mestrado. Universidade Federal do Paraná.
Scremin-Dias, E., Pott, V. J. , Hora, R. C., Souza, P. R. 1999. Nos Jardins Submersos da
Bodoquena. Guia para identificação das plantas aquáticas de Bonito e região. Campo
Grande/MS. Editora UFMS. 160p.
Scremin-Dias, E.2000. A plasticidade fenotípica das macrófitas aquáticas em resposta a
dinâmica ambiental. In Tópicos atuais em Botânica, 51 Congresso Nacional de Botânica,
Brasília DF.
Sculthorpe, C. D. 1967. The Biology of aquatic vascular plants. Eduard Arnold Publ.
London.
Souza, V. C. & Lorenzi, H. 2005. BOTÂNICA SISTEMÁTICA: Guia ilustrado para
identificação de angiospermas da flora brasileira, baseado em APG II. Instituto
Plantarum, 640p.
Tomlinson, P. B. (1982), Anatomy of the Monocotyledons. Vol. 7: Helobiae
(Alismatidae). Editado por Metcalfe, C. R. Clarendon Press. Oxford. 560p
Wiegleb, G. & Kaplan, Z. 1998. An account of the species of Potamogeton L.
(Potamogetonaceae). Folia Geobotanica, Upssala, v. 33, 241-316.

11
Rio Baía Bonita
Rio Sucuri
Bonito
Figura 1 - Mapa indicando os dois rios estudados no município de Bonito – MS, evidenciando a Serra da Bodoquena à esquerda (área verde).
2
3
Figura 2 -Rio Baía Bonita com (seta). Figura 3 - Rio Sucuri, mostrando diversidade de espécies de macrófitas, e local onde ocorre na correnteza (no leito do rio, seta) e remanso junto ao banco de
(na parte inferior da figura, seta)
Potamogeton illinoensis
P. illionoensis Gomphrena
elegans .

12
_________________________________________________________________________CAPITULO I
Anatomia comparada de Potamogeton illinoensis Morong e Potamogeton gayi A. Benn. que ocorrem em rios de Bonito – MS

14
INSTRUCTIONS FOR AUTHORS
Submission of papers Brazilian Archives of Biology and
Technology publishes original research papers, Short notes and Review articles in English in the interdisciplinary areas of biological sciences and engineering/technology. Submission of paper implies that it has not been published or being considered for publication elsewhere. Care should be taken to prepare a compact manuscript with precision in presentation, which will help authors in its acceptance. All the papers are subjected to review by referees.
Manuscript
Three copies of the single-spaced typed
manuscript (maximum 12 pages for original and review articles and 2-4 pages for short notes) on a high grade A-4 size paper (210x297 mm), with margins (left 25, right 20, superior and inferior 30 mm) should be prepared. This should be divided under the following headings: ABSTRACT, INTRODUCTION, MATERIALS AND METHODS, RESULTS, DISCUSSION, ACKNOWLEDGEMENTS, RESUMO, REFERENCES. These headings should be
typed in bold upper case (12 font). For review articles, authors should make their own headings along with Abstract and Introduction.
Title
The title (18 font, bold) of the paper should
clearly reflect its contents. It should be followed by the name(s) of author(s) with expanded initials (12 font, bold) and the address(s) (italic, 10 font) of the institution(s) where the work has been carried out.
ABSTRACT
Each paper should be provided with an abstract (italic) of 100-150 words, describing briefly on the purpose and results of the study. It should be prepared as concisely as possible.
Key words
Authors should provide three to six key words that will be used in indexing their paper.
INTRODUCTION
This should describe the background and relevant information about the work. It should also state the objective of the work.
MATERIALS AND METHODS
Authors must take care in providing
sufficient details so that others can repeat the work. Standard procedures need not be described in detail.
RESULTS AND DISCUSSION
Results and Discussion may be presented
separately or in combined form (authors may decide easier way for them). Preliminary work or less relevant results are not to be described. The reproducibility of the results, including the number of times the experiment was conducted and the number of replicate samples should be stated clearly.
RESUMO
An abstract of the paper should also be prepared in Portuguese and placed before the list of References. Authors from other than Latin American countries can seek the help of Editor's office to prepare Portuguese resumo of their papers.
REFERENCES
References in the text should be cited at the appropriate point by the name(s) of the author(s) and year (e.g. Raimbault & Roussos, 1996; Raimbault et al., 1997). A list of references, in the alphabetic order (10 font), should appear at the end of the manuscript. All references in the list should be indicated at some point in the text and vice versa. Unpublished results should not be included in the list. Examples of references are given below.
In journals Pandey, A. (1992), Recent developments in
solid state fermentation. Process Biochem., 27, 109-117 Thesis: Chang, C. W. (1975), Effect of fluoride

15
pollution on plants and cattle. PhD Thesis, Banaras Hindu University, Varanasi, India
In books: Tengerdy, R. P. (1998), Solid substrate
fermentation for enzyme production. In-Advances in Biotechno-logy, ed. A. Pandey. Educational Publishers & Distributors, New
Delhi, pp. 13-16 Pandey, A. (1998), Threads of Life. National Institute of Science Communication, New Delhi
In conferences: Davison, A. W. (1982), Uptake, transport
and accumulation of soil and airborne fluorides by vegetation. Paper presented at 6th International Fluoride Symposium, 1-3 May, Logan, Utah Tables and Figures
Tables and figures, numbered consecutively
with Arabic numerals must be inserted at appropriate place in the text. These should be used to present only those data, which can not be described in the text Units and Abbreviations
The SI system should be used for all
experimental data. In case other units are used, these should be added in parentheses. Only standard abbreviations for the units should be used. Full stop should not be included in the abbreviation (e.g. m, not m. or rpm, not r.p.m.). Authors should use '%' and '/' in place of 'per cent' and 'per'. INSTRUCTIONS FOR AUTHORS Manuscript lay-out
It is suggested that authors consult a recent issue of
the journal for the style and layout. Except the title,
abstract and key words, entire text should

16
Anatomia comparada de Potamogeton illinoensis Morong e
Potamogeton gayi A. Benn. que ocorrem em rios de Bonito – MS.¹ Seloi Rodrigues²,³e Edna Scremin-Dias². ²Depto de Biologia, Lab. de Botânica, Universidade Federal de Mato Grosso do Sul. 3Bolsista Fundect
Caixa Postal 549, CEP: 79070-900 Campo Grande/MS, Brazil
ABSTRACT The compared anatomy of the root, rhizome, floating stem and leaves of Potamogeton illinoensis Morong and P. gayi A. Benn. that happen in Sucuri and Baía Bonita rivers in Bonito, Mato Grosso do Sul, Brazil, is described. Differential characters of these species are related; among them the presence of stele with three or four protoxylem gaps in P. gayi and eight to fourteen protoxylem gaps in P. illinoensis. This species has cortical bundles with phloem and fibers, absent in P. gayi that has groups of subepidermal fibers. Wide aeriferous gaps are distributed in the leaves, floating stems and rhizomes, with diaphragms intercepting them. The auriferous gaps are smaller in the roots that do not present epidermis in the maturity, remaining the exodermis as covering. Besides the anatomical description, this work discusses the adaptive characteristics of the tissues and organs of P. illinoensis and P. gayi relating them to their environment.
Key Words: Potamogeton gayi, Potamogeton illinoensis, anatomy, submerged plants. INTRODUÇÃO
O gênero Potamogeton L. pertence à
Família Potamogetonaceae, possui cerca de 100 espécies, exclusivamente submersas. Este gênero é cosmopolita, ocorrendo tanto em águas de regiões temperadas quanto de águas tropicais (Wiegleb & Kaplan, 1998). Algumas espécies emitem os escapos florais para o fora da água e, é descrito para outras, dentre elas P. illinoensis, folhas flutuantes em indivíduos de ambientes lênticos (Rodrigues & Irgang, 2001).
Para o Brasil são citadas cerca de 15 espécies, em dois gêneros, sendo a maioria encontrada na região sul (Souza & Lorenzi, 2005). Para o Mato Grosso do Sul, até o momento, foram identificadas duas espécies: Potamogeton gayi A. Benn., coletado no rio Baía Bonita, e Potamogeton illinoensis Morong, distribuída em dois rios, o Sucuri e o Baía Bonita, no município de Bonito/MS.
As espécies do gênero Potamogeton são caracterizadas por serem muito semelhantes, tanto na morfologia (Rodrigues & Irgang, 2001) quanto na anatomia (Tomlinson, 1982). Este autor destaca que as espécies de Potamogetonaceae são amplamente distribuídas nos diversos continentes, e dificilmente definidas, sendo, portanto difícil sua identificação taxonômica, quando as estruturas de reprodução estão ausentes. De acordo com Tomlinson (1982), os gêneros pertencentes à família Potamogetonaceae possuem anatomia simplificada, sendo a redução de tecidos bem mais acentuada em espécies totalmente submersas, que apresentam muitas lacunas de ar e ausência de tecidos mecânicos. Para este autor o sistema vascular do caule é similar em todas as espécies de Potamogetonaceae e, progressivamente, mais simplificado nas espécies de Potamogeton, devido à redução do xilema a uma lacuna ao longo de todo o caule desde o rizoma até os caules flutuantes. Tomlinson (1982) afirma que as características

17
anatômicas são tão importantes quanto as morfológicas na identificação de espécimes estéreis. Metcalfe & Chalk (1983) também ressaltam que os dados anatômicos dos órgãos vegetativos de uma espécie servem como critério adicional para resolver problemas taxonômicos. Sculthorpe (1967) já ilustrava a redução do estelo para as espécies do gênero Potamogeton, distinguindo seis tipos básicos. Tanto no trabalho publicado por Tomlinson (1982) quanto na ampla revisão taxonômica publicada por Wiegleb & Kaplan (1998) para gênero, são citadas as características anatômicas do estelo, como consistentes para delimitação de espécies.
A carência de estudos anatômicos mais detalhados para os órgãos vegetativos do gênero Potamogeton, bem como a relação das estruturas anatômicas em função das adaptações ao ambiente, objetivaram a realização do presente trabalho. Neste, são descritas a anatomia de Potamogeton illinoensis e Potamogeton gayi que ocorrem no Mato Grosso do Sul, visando contribuir com informações anatômicas que auxiliam na taxonomia e filogenia destas espécies e da família.
MATERIAL E MÉTODOS As coletas foram realizadas nos rios
Sucuri e Baía Bonita, ambos explorados pelo ecoturismo no município de Bonito em Mato Grosso do Sul. Foi feita uma coleta de indivíduos de P. illinoensis por mês durante cinco meses, em dois locais de remanso e dois de correnteza, em cada rio, e os espécimes de P. gayi foram coletados nas populações do rio Baía Bonita. Para as análises foram coletados folhas, caule, rizomas e raízes.
As amostras foram fixadas em Formalina Neutra Tamponada (Lillie, 1948 apud Clark, 1981) e armazenadas em álcool 70 (Jensen 1962). Foram confeccionadas lâminas
permanentes, de material incluído em historresina, corados com azul de toluidina (Kraus & Arduim, 1997), e lâminas semipermanentes, de material seccionado com o auxilio de lâmina de barbear e utilizando isopor como suporte, corados com azul de astra 1% e safranina 1 % (Kraus & Arduim, 1997). Os materiais coletados foram analisados em cada mês de coleta, visando verificar a existência de possíveis modificações no padrão dos tecidos e órgãos. Para a análise das folhas foi seccionada a região mediana e para o caule flutuante foi seccionado o 8º entrenó. Para detectar a presença de amido utilizou-se lugol (Jensen 1962).
Indivíduos em fase de reprodução sexuada foram prensados e desidratados em estufa, sendo as exsicatas depositadas no herbário CGMS/UFMS, sob o número 16817 (P. illinoensis) e número 16818 (P. gayi).
RESULTADOS
Limbo foliar – A epiderme, em secção transversal, da lâmina foliar de P. illinoensis possui células quadradas a retangulares, sendo menores sobre a nervura mediana e nos bordos (Figs. 1 a 5), destacando-se um tricoma em algumas regiões do bordo (Fig. 4). A parede periclinal externa das células epidérmicas é mais espessa que a periclinal interna e as anticlinais (Fig. 1). P. gayi apresenta células epidérmicas com mesmo padrão (Fig. 2) observado em P. illinoensis, tendo, em ambas espécies, cloroplastídeos distribuídos nestas células (Figs. 1 e 2).
O mesofilo nas áreas laterais da lâmina foliar de P. illinoensis constitui-se predominantemente de uma a duas camadas de células parenquimáticas que se estendem até o bordo (Fig. 4). Nos pontos onde ocorrem os feixes vasculares, pode haver mais de duas

18
ClNu
La
Ep
Cl
Fl
Fi
Px
La
Fi
Ep
FV
Fi
Ep
LaFl
Lpx
FiEp
FV
Cl
TrEp
Cl
FiFi
1 23
4 5
FV
Df
La
6
7
8 9
Fi
Px
La
FV
Figuras 1 - 10 - Secções transversais da folha de Potamogeton illinoensis e P. gayi. (1) Detalhe da epiderme de P. illinoensis com parede epidérmica espessa e cloroplastos. (2) Detalhe da nervura mediana de P. gayi, indicando cloroplastos na epiderme. (3) Mesofilo da região lateral do limbo de P. Illinoensis. (4) Bordo de P. illinoensis evidenciando fibras e tricoma. (5) Bordo foliar de P. gayi com fibras. (6) Nervura mediana de P. illinoensis. (7) Nervura mediana de P. gayi, evidenciando fibras subepidérmicas e feixe vascular com lacuna de protoxilema e floema. (8) Detalhe do diafragma com células braciformes com cloroplastos. (9) Diafragmas sem cloroplastos. (10) Feixe vascular central da nervura mediana de P. illinoensis evidenciando lacunas de protoxilema, e floema com elementos de tubo crivado evidentes. (Figuras 1, 2, 3, 4, 5, 7,8 e 9; Barra = 50 mm); (Figura 6; Barra = 100 mm). Cl, Cloroplastos; Ep, Epiderme; Fi, Fibras; Fl, Floema; FV, Feixe Vascular; La, Lacuna aerífera; Px, Lacuna de Protoxilema; Nu, Núcleo; Pr, Parênquima regular; Tr, Tricoma. *Elemento de tubo crivado.

19
camadas de células de parênquima clorofiliano, que delimitam espaços intercelulares (Fig. 3). A extensão do mesofilo de P. gayi é menor do que em P. illinoensis (Figs. 6 e 7). O mesofilo na região do bordo em ambas espécies possui predominantemente uma única camada de parênquima, com pequenos espaços intercelulares (Figs. 4 e 5). Nesta região ocorre feixe de fibras visualmente mais robustas e em maior número nos indivíduos de P. gayi (Figs. 4 e 5).
A região central do limbo o mesofilo é formada por parênquima clorofiliano de espaços intercelulares amplos, constituindo típico aerênquima em ambas as espécies (Figs. 6 e 7). Diafragmas de células braciformes interceptam transversalmente as lacunas aeríferas (Fig. 6), a maioria destes com cloroplastos (Fig. 8), podendo ocorrer alguns sem esta organela (Fig. 9). Limitando os feixes vasculares, ocorrem duas a três camadas celulares de parênquima regular (Figs. 6, 7 e 10). Calotas de fibras de esclerênquima ocorrem nos dois lados dos feixes vasculares em ambas espécies (Figs. 7 e 10).
As células que limitam os espaços aeríferos na região central do limbo em P. illinoensis são isodiamétricas, conectando-se a epiderme diretamente, ou por meio de uma camada de células parenquimáticas dispostas internamente a epiderme (Fig. 6). Na região de conexão das células parenquimáticas do mesofilo com a epiderme, ocorrem feixes de fibras de esclerênquima em P. gayi (Fig. 7). Esta característica não foi observada em P. illinoensis (Fig. 6).
Os feixes vasculares do limbo das duas espécies são formadas por número variável de células de floema, com amplos elementos de tubo crivado e número variável de células companheiras, margeados por parênquima (Fig. 10). O xilema nestas espécies restringe-se a uma grande lacuna de protoxilema, limitada por
células de parênquima, não possuindo elementos condutores funcionais (Figs. 7 e 10).
Pecíolo – P. gayi possui folhas sésseis, e P. illinoensis possui pecíolo com formato de meia lua em secção transversal, sendo levemente expandido lateralmente à medida que se aproxima do limbo (Fig. 11). A epiderme, em secção transversal, é composta por células arredondadas ou mais altas que largas (Figs. 12, 13 e 14a). A parede periclinal externa é mais espessada que a periclinal interna e as anticlinais (Figs. 12, 13 e 14a), sendo visíveis estrias epicuticulares (Fig. 14a). O parênquima clorofiliano do pecíolo restringe-se as camadas mais externas de parênquima e às células epidérmicas.
Lateralmente, o pecíolo possui parênquima regular com paredes espessadas (Fig. 12). O sistema vascular do pecíolo é constituído por um feixe vascular central e dois alinhados lateralmente a este (Fig. 11). Pode ocorrer uma bainha de fibras de esclerênquima envolvendo cada feixe vascular e, por fora destas, uma a duas camadas de células parenquimáticas (Figs. 12 e 13) podendo, a bainha de esclerênquima, estar interrompida por células parênquimáticas. Lacunas aeríferas são interrompidas transversalmente por uma camada de células braciformes com cloroplastos - os diafragmas (Fig. 14b). É comum, nas células de parênquima, a presença de amiloplastos, ocorrendo em maior densidade nas células associadas aos feixes vasculares.
Caule flutuante e rizoma – Em secção transversal o formato do caule flutuante e do rizoma de Potamogeton illinoensis é arredondado. P. gayi, possui caule flutuante levemente ovalado, e rizoma arredondado.
As células epidérmicas destes dois tipos de caules, em secção transversal, são mais altas que largas tendo parede periclinal externa espessada em P. illinoensis (figs. 15 e 16) e células levemente quadradas em P. gayi (Fig.

20
17). As paredes das células epidérmicas desta espécie são menos espessadas que P. illinoensis (Fig. 15 - 17).
Nas duas espécies, internamente a epiderme, tanto no rizoma quanto no caule flutuante, ocorrem uma ou mais camadas de células parênquimáticas com distribuição regular (Figs. 16 e 17). Em alguns pontos este parênquima pode não ocorrer, e a epiderme limitar as lacunas aeríferas do córtex caulinar (Fig. 15). Logo abaixo da epiderme ou das camadas corticais periféricas, nas duas espécies, várias camadas de células parênquimáticas delimitam amplas lacunas (Figs. 17 a 20) constituindo, o parênquima cortical, um típico aerênquima (Fig. 20).
P. illinoensis possui feixes corticais, distribuídos entre as células do parênquima aerífero (Fig. 18), formando um a dois arcos concêntricos (Fig. 20), sendo que cada feixe é envolto por fibras de esclerênquima (Fig. 18). Este tipo de feixe cortical não foi observado em P. gayi, estando presentes nesta espécie somente fibras de esclerênquima dispostas, exclusivamente, na camada subepidérmica (Fig. 17). Diafragmas estão presentes interceptando transversalmente os espaços intercelulares (Fig. 19), dos dois tipos caulinares. Nos diafragmas dos rizomas e caules flutuantes são comuns amiloplastos. E neste ultimo, há riqueza de cloroplastos nos diafragmas tanto nas áreas mais periféricas quanto mais interna do córtex.
Limitando o cilindro central do caule flutuante de P. illinoensis a endoderme apresenta espessamento em “U” (Fig. 21). P. gayi, apresenta espessamento em “O” na endoderme lignificada tanto no caule flutuante quanto no rizoma (Fig. 22), tendo células de passagem com estrias de Caspary evidentes. No rizoma de P. illinoensis o cilindro central é limitado do córtex por endoderme lignificada e com espessamento em “U” (Fig. 23), no entanto predomina neste órgão espessamento em “O”
(Fig. 24). A lignificação da endoderme é mais intensa em P. illinoensis (Fig. 24) quando comparado com P. gayi (Fig. 22). Células de passagem sem lignificação ocorrem na endoderme do rizoma de P. illinoensis (Fig. 23).
Internamente a endoderme ocorre uma camada de periciclo, que possui predominantemente células com paredes espessadas e lignificadas nos indivíduos de P. illinoensis (Figs. 23 e 24). No cilindro vascular dos rizomas e do caule flutuante de P. illinoensis ocorrem de oito a quatorze lacunas de protoxilema (Fig. 25) que podem apresentar dois ou mais anéis de espessamento (Fig. 26 e 27) e, internamente as lacunas de protoxilema, feixes de fibras (Figs. 25 e 26).
P. gayi, apresenta de três a quatro lacunas de protoxilema no cilindro vascular (Fig. 22). Cada lacuna, tanto em P. gayi, quanto em P. illinoensis é acompanhada por grupos de células de floema composto por elementos de tubo crivado de lume amplo (Figs. 22-24, 27). No parênquima associado aos feixes corticais de P. illinoensis, e nas células parenquimáticas do cilindro vascular, ocorre grande quantidade de amiloplastos (Figs. 20 e 24).
Raiz – Tanto em P. illinoensis quanto em P. gayi, a epiderme da raiz só é observada na zona de absorção, sendo visíveis pêlos radiciais (Figs. 28 e 29) nesta região. A epiderme é perdida na zona de maturação, ficando a exoderme em contato com o meio. A exoderme é formada predominantemente por duas camadas celulares, de paredes periclinais externas espessas (Fig. 30 e 31).
O córtex interno é composto por parênquima regular com espaços intercelulares reduzidos, quando comparado com o córtex externo (Figs. 28 a 31). O aerênquima da raiz é esquizógeno, podendo haver colabamento das células que delimitam as lacunas em alguns

21
Ep
La
Fv
La
Ep
Fi
Ep
La
Fi
EpLa
PxFl
Df
11
12
13
14A
14B
Df
FV
Figuras 11 –14 - Secções transversais do pecíolo de Potamogeton illinoensis. (11) Feixes vasculares alinhados com detalhe da expansão lateral do pecíolo. (12) Células parenquimáticas com parede levemente espessada e epiderme de células arredondadas. (13-14A) Feixe vascular envolto por fibras e epiderme com paredes espessadas. (14B) Em detalhe diafragma em vista frontal. Cl, Cloroplastos; Df, diafragma; Ep, Epiderme; Fi, Fibras; Fl, Floema; FV, Feixe Vascular; La, Lacuna aerífera; Px, Lacuna de Protoxilema. (Figuras 12 e 14, barra = 50 μm. Figuras 11, 13 e 15, barra = 100 μm).
pontos, ampliando os espaços intercelulares (Fig. 30 e 31). É comum nas células corticais a presença de amiloplastos (Fig. 32). A endoderme possui estrias de Caspary em P. illinoensis.
Na região proximal, no ponto de inserção da raiz ao rizoma a endoderme tem espessamento em “U” e é lignificada (Fig. 32).
O periciclo possui somente uma camada parenquimática em ambas as espécies (Figs. 31 e 32) sendo, em alguns pontos, interrompida por elemento de tubo crivado (Figs. 31 e 32). Os elementos de transporte do cilindro vascular, nas duas espécies, é formado geralmente por cinco pólos de protoxilema. Em geral, na região central do cilindro vascular ocorre um elemento de metaxilema podendo, em alguns casos, serem observados dois (Figs. 31 e 32). O floema é composto por células localizadas entre os pólos de protoxilema, na região do periciclo (Figs. 31 e 32).
DISCUSSÃO
Células epidérmicas com paredes delgadas e cutícula fina, ou por vezes cutícula ausente, são características de plantas aquáticas, principalmente daquelas com órgãos submersos, para facilitar a troca de gases entre o meio aquoso e a planta (Sculthorpe, 1967). No entanto, Potamogeton illinoensis e P. gayi revelam parede periclinal externa da epiderme acentuadamente espessa, contrariando estas premissas. Mauseth (1988) relata que quando a parede de células epidérmicas é espessa, geralmente esse espessamento é assimétrico, ocorrendo nas paredes periclinais externas ou internas estando ausente nas anticlinais, situação presente nas duas espécies de Potamogeton estudadas, e já citada por Tomlinson (1982) para o gênero Potamogeton. Bona (1999) encontrou para espécies submersas

22
20
FcLa Px
En
Ep
LaDf
En
19
Ep
Cl
Fi
La
18
Fc
Px
En
17
En CpCp
21 22
23 24
26
16
Ep
15La
PxEn
25
Px
En 27 Figuras 15 –28 - Secções do caule flutuante e do rizoma de Potamogeton illinoensis e P. gayi. (15-16) Detalhe da epiderme do caule flutuante de Potamogeton illinoensis com parede celular espessa e cloroplastos evidentes. (17) Rizoma de P. gayi evidenciando grupos de fibras subepidérmicas. (18) Detalhe dos feixes corticais do caule de P. illinoensis. (19) Corte longitudinal de caule jovem de P. illinoensis, evidenciando diafragmas. (20) Córtex do caule de P. illinoensis com aerênquima típico indicando a grande quantidade de amido (células com conteúdo escuro) ao redor das unidades vasculares e do cilindro central. (21) Detalhe do caule de P. illinoensis mostrando endoderme com espessamento em U. (22) Rizoma de P. gayi com estelo tipo quatro feixes, evidenciando endoderme pouco lignificada e com espessamento em O. (23) Rizoma de P. illinoensis com endoderme e espessamento em O, com células de passagem. (24) Rizoma de P. illinoensis com endoderme e espessamento em U. (25) Cilindro vascular do rizoma de P. illinoensis, evidenciando o estelo tipo trio. (26) detalhe de um feixe vascular com lacuna de protoxilema evidente e anéis de espessamento da traqueíde (setas). (27) Caule de P. gayi indicando as lacunas de protoxilema e elementos de tubo crivado. Cl, Cloroplastos; Cp, Células de passagem; Df, Diafragma; En, endoderme; Ep, Epiderme; Fc, Feixe cortical; Fi, Fibras; Fl, Floema; La, Lacuna aerífera; Px, Lacuna de Protoxilema; *Elemento de tubo crivado.(Barra = 50 μm. Figs. 24, barra = 100 μm).
23
Ep
Ex
Pr
30
Mx PxEn
Per
33
29Mx
En
Ex
Mx
Ep
30 31
CeCe
CiCi
En MxPerEi
Ex
32
Ce
Ci
CiCe
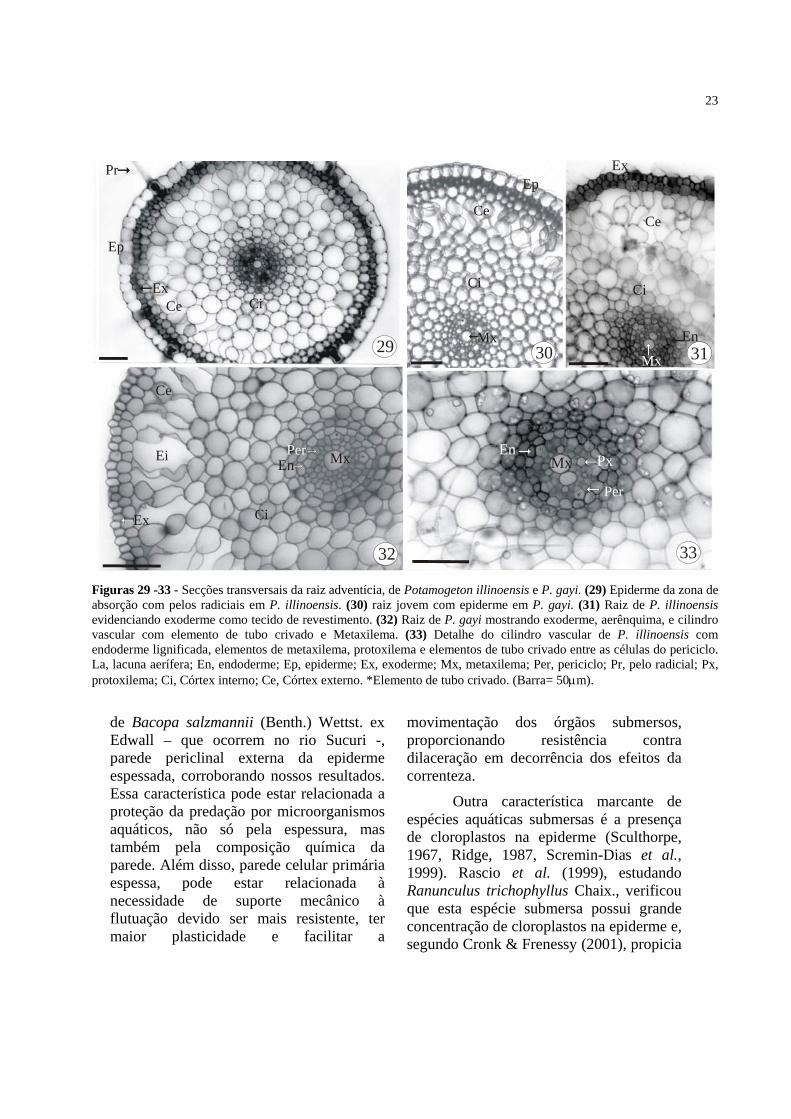
Figuras 29 -33 - Secções transversais da raiz adventícia, de Potamogeton illinoensis e P. gayi. (29) Epiderme da zona de absorção com pelos radiciais em P. illinoensis. (30) raiz jovem com epiderme em P. gayi. (31) Raiz de P. illinoensis evidenciando exoderme como tecido de revestimento. (32) Raiz de P. gayi mostrando exoderme, aerênquima, e cilindro vascular com elemento de tubo crivado e Metaxilema. (33) Detalhe do cilindro vascular de P. illinoensis com endoderme lignificada, elementos de metaxilema, protoxilema e elementos de tubo crivado entre as células do periciclo. La, lacuna aerífera; En, endoderme; Ep, epiderme; Ex, exoderme; Mx, metaxilema; Per, periciclo; Pr, pelo radicial; Px, protoxilema; Ci, Córtex interno; Ce, Córtex externo. *Elemento de tubo crivado. (Barra= 50μm).
de Bacopa salzmannii (Benth.) Wettst. ex Edwall – que ocorrem no rio Sucuri -, parede periclinal externa da epiderme espessada, corroborando nossos resultados. Essa característica pode estar relacionada a proteção da predação por microorganismos aquáticos, não só pela espessura, mas também pela composição química da parede. Além disso, parede celular primária espessa, pode estar relacionada à necessidade de suporte mecânico à flutuação devido ser mais resistente, ter maior plasticidade e facilitar a
movimentação dos órgãos submersos, proporcionando resistência contra dilaceração em decorrência dos efeitos da correnteza.
Outra característica marcante de espécies aquáticas submersas é a presença de cloroplastos na epiderme (Sculthorpe, 1967, Ridge, 1987, Scremin-Dias et al., 1999). Rascio et al. (1999), estudando Ranunculus trichophyllus Chaix., verificou que esta espécie submersa possui grande concentração de cloroplastos na epiderme e, segundo Cronk & Frenessy (2001), propicia

24
obter a maximização da captação de luz para fotossíntese, que é reduzida em ambientes aquáticos (Sculthorpe, 1967). Sendo assim, a epiderme de plantas submersas desempenha função importante no processo de fotossíntese (Rascio et al., 1999). Inclusive Fahn (1990) descreve que a epiderme de espécies submersas, dentre elas Potamogeton, possui maior concentração de cloroplastos que os tecidos internos. Além dos cloroplastos na epiderme, P. gay e P. illinoensis possuem cloroplastos nas células de parênquima que limitam as lacunas aeríferas, no entanto a densidade destas organelas diminui, em direção as áreas mais internas do pecíolo e do córtex do caule flutuante. Estas organelas também estão distribuídas nas células do diafragma, dispostas interceptando as lacunas aeríferas, no interior de caules e folhas das duas espécies estudadas. A presença destas organelas nos diafragmas das plantas aquáticas consiste em áreas extras para realização de fotossíntese (Scremin-Dias et al., 1999), nas partes mais internas dos órgãos.
Os espaços intercelulares do mesofilo e do córtex do caule flutuante, do rizoma, e da raiz são amplos nos órgãos das duas espécies de Potamogeton aqui estudadas, sendo que nas raízes os espaços são aumentados pelo colabamento das paredes celulares. Scremin-Dias (2000), encontrou para as raízes de indivíduos aquáticos de Echinodorus tenellus , células corticais colabadas. Característica igualmente encontrada por Bona (1999) para espécies de Bacopa, sendo comum nas raízes onde a hipóxia é maior.
Espaços intercelulares amplos indicam adaptação destas ao ambiente aquático, sendo o volume do aerênquima dependente do genótipo da planta e das condições ambientais a que as espécies estão sujeitas (Colmer, 2003). O aerênquima de P. illinoensis e P. gayi é
esquizógeno, sendo formado sem a ruptura das células parenquimáticas para sua formação (Evans, 2003). Sobre este aspecto, Voesenek et al. (2006), relatam que informações sobre os processos que regulam a formação do aerênquima esquizógeno ainda são ausentes.
O aerênquima tem por função estocar e transportar gases nas plantas aquáticas (Cronk & Frenessy, 2001), oferecer resistência mecânica e permitir a flutuação dos órgãos submersos (Scremin-Dias et al., 1999) e diminuir a resistência ao fluxo de oxigênio e outros gases (Vartapetian & Jackson, 1997), já que este é um dos problemas impostos pelo ambiente aquático, onde os gases se difundem mais lentamente (Levitt, 1980).
Os espaços intercelulares amplos de P. illinoensis e P. gayi, são interceptados por diafragmas de células braciformes que, de acordo com Sculthorpe, (1967) são característicos nas Nymphaeaceae e na maioria das monocotiledôneas aquáticas. Estes diafragmas são importantes para plantas aquáticas submersas ou submetidas ao alagamento, pois no caso de injúria mecânica de alguma parte submersa da planta, constituem em barreira contra a entrada de água por toda a planta (Sculthorpe, 1967, Armstrong & Armstrong, 1988, Soukup et al., 2000). No local onde a planta sofre a injúria e o diafragma está presente, pode ocorrer a formação de um “callus”, o qual, mais tardiamente pode prevenir contra ataque microbianos (Armstrong, 2000).
Como em plantas aquáticas o suporte aos órgãos submersos é oferecido pela própria água e pela flutuação, fibras em geral são ausentes. No entanto, elas podem ser observadas em algumas partes da planta, dentre elas os bordos foliares (Mauseth, 1988, Scremin-Dias, 2000), e associadas aos feixes vasculares (Scremin-

25
Dias, 2000). A presença de fibras no bordo foi observada nas duas espécies estudadas, sendo que estas células conferem certa elasticidade e oferecem resistência à dilaceração dos órgãos submersos das plantas aquáticas (Mauseth, 1988, Scremin-Dias et al., 1999). Para Usherwood et al. (1997) é mais comum observar a presença de elementos de sustentação na região central dos caules, pois assim eles podem resistir melhor as forças impostas pela corrente da água e assim resistir a quebra. Este foi o padrão encontrado para os caules das espécies objeto do presente estudo, que apresentaram o sistema vascular confinado no cilindro central. No entanto fibras também foram observadas internamente a epiderme de P. gayi e P. illinoensis e entre as células que limitam as lacunas aeríferas em P. illinoensis, em todos os órgãos, exceto nas raízes das duas espécies e nas folhas de P. illinoensis.
Tomlinson (1982) cita ser comum a todas as espécies do gênero Potamogeton a presença de fibras lignificadas nos órgãos submersos. Iida et al. (2004), relatam que a presença ou ausência destas fibras, abaixo da epiderme e/ou entre as células que limitam os espaços aeríferos, juntamente com o tipo de estelo são freqüentemente utilizadas como ferramenta para a taxonomia do gênero Potamogeton. Wiegleb & Kaplan (1998) citam, além das características anteriormente descritas, que auxiliam a diagnose das espécies, o formato das células da endoderme, além da presença, ausência ou a quantidade de camadas sub-epidérmicas. Neste aspecto, as fibras do bordo foliar bem como o espessamento das células da endoderme podem auxiliar na separação entre P. gayi e P. illinoensis.
Nas duas espécies em questão, cada unidade vascular é formada por uma lacuna de protoxilema e por elementos de tubo crivado amplos, no entanto o cilindro vascular central é formado por várias destas unidades vasculares.
Alguns autores citam que as características taxonômicas e filogenéticas da Família Potamogetonaceae, tendem a progressiva redução da complexidade do estelo (Sculthorpe, 1967, Tomlinson 1982). Estes autores apresentam alguns tipos de estelo encontrados nas espécies do gênero Potamogeton. Sculthorpe (1967) indica a ilustra estágios de fusão e redução dos tecidos vasculares em Potamogeton. Wiegleb & Kaplan (1998) reconhecem para o gênero Potamogeton cinco tipos de estelo – proto (com usualmente 10-14 feixes vasculares livres), trio (ou tipo oito feixes, com 1 ou 2 grupos de floema adicional no trio de feixe), o tipo quatro feixes e, finalmente, o tipo circular (com um feixe). Os resultados desta investigação, indicaram que P. illinoensis se enquadra no tipo proto e P. gayi no tipo quatro feixes.
Segundo Sculthorpe (1967) as unidades vasculares são dispersas nas espécies com folhas amplas, ocorrendo a fusão dos elementos do xilema em espécies com folhas lineares. Em P. gayi o cilindro vascular é mais reduzido que em P. illinoensis, isto pode ser em virtude de P. gayi possuir folhas menores ou devido os indivíduos desta espécie ocorrerem em maior profundidade na lâmina d’água. Esta espécie não atinge a superfície, ao contrário de P. illinoensis. Tomlinsom (1982) já afirmava que espécies de Potamogeton totalmente submersas apresentam maior redução dos tecidos vasculares.
Tecidos vasculares e de sustentação reduzidos são característicos em plantas aquáticas, em decorrência da homogeneidade do ambiente, do suprimento de água não consistir em problema, bem como do suporte que o meio aquático oferece aos órgãos submersos, dispensando tecidos de sustentação em quantidade (Scremin-Dias et al., 1999). Quanto ao sistema vascular das plantas submersas, tanto o floema quanto o xilema

26
são muito simplificados, sendo este último, altamente reduzido (Mauseth, 1988).
Pedersen & Sand-Jensen (1993) ressaltam que provavelmente não há pressão negativa no xilema e por isso investimentos em paredes lignificadas não são necessários. Mauseth (1988) afirma ser obvia a redução do xilema em plantas aquáticas, visto que a água e nutrientes são absorvidos diretamente pela epiderme do caule e folhas. No entanto Pedersen & Sand-Jensen (1993) demonstraram através de experimentos com diversas espécies submersas, que há fluxo de água através dos feixes vasculares, mas que não há relação nenhuma entre o tamanho do tecido vascular e o fluxo de água. Entretanto, os autores não souberam explicar se o fluxo ocorria somente no xilema ou também em outros tecidos do cilindro vascular. No entanto, a redução do xilema nas espécies de Potamogeton objeto deste estudo é muito evidente, restringindo-se a uma lacuna de protoxilema por unidade vascular. É provável que haja transporte de água e nutrientes pelas lacunas. Já o floema é bem desenvolvido, com vários elementos de tubo crivado de lume amplo dispostos entre a lacuna de protoxilema e o periciclo, sendo comum para plantas aquáticas floema mais desenvolvido que o xilema (Cutter, 1987).
Nas plantas submersas não ocorre o processo de transpiração como nas plantas terrestres, em decorrência disso há ausência de pressão negativa (Pedersen & Sand-Jensen, 1993), sendo que as plantas aquáticas devem possuir outra forma para manter o fluxo de água no sistema vascular, haja vista principalmente a necessidade da manutenção de fluxo hídrico para absorção de nutrientes. Scremin-Dias (2000) hipotetiza que substâncias osmoticamente ativas, como o amido, podem ser mobilizados para o interior dos vasos produzindo potencial osmótico nos feixes
da raiz e do caule, evitando que o fluxo hídrico seja interrompido, quando não há transpiração. Talvez seja essa a explicação para a presença de amido principalmente no parênquima localizado ao redor do cilindro vascular central e também nas células que circundam os feixes corticais das espécies de Potamogeton aqui estudadas. Pedersen & Sand-Jensen (1993) relataram que as taxas do transporte no sistema vascular de Sparganium emersum diminuíram quando esta espécie era mantida no escuro por longo período de tempo, sugerindo que a produção fotossintética de compostos orgânicos e energia é necessária para sustentar o transporte.
As raízes adventícias de P. gayi e P. illinoensis apresentam muitos pêlos radiciais na zona de absorção, e perdem a exoderme na região de maturação, semelhante ao encontrado por Bona & Morretes (2003), para Bacopa salzmanii, espécie que se desenvolve no rio Sucuri.
RESUMO
É descrita a anatomia comparada da raiz, rizoma, caule flutuante e das folhas de Potamogeton illinoensis Morong e P. gayi A. Benn. que ocorrem nos rios Sucuri e Baía Bonita em Bonito, Mato Grosso do Sul, Brasil. São relacionados caracteres diferenciais destas espécies, entre eles a presença de estelo com três ou quatro lacunas de protoxilema em P. gayi e oito a quatorze lacunas de protoxilema em P. illinoensis. Esta espécie possui feixes corticais com floema e fibras, ausentes em P. gayi que possui grupos de fibras subepidérmicas. Amplas lacunas aeríferas distribuem-se nas folhas, caule flutuante e rizomas, tendo diafragmas interceptando-as. As lacunas aeríferas são menores nas raízes, que não apresentam epiderme na maturidade, restando a exoderme como

27
revestimento. Além da descrição anatômica, este trabalho discute as características adaptativas dos tecidos e órgãos de Potamogeton illinoensis e P. gayi relacionando-as ao ambiente que habitam. Palavras-chave: Potamogeton gayi, Potamogeton illinoensis, anatomia, plantas submersas. Agradecimentos
Agradecimento especial à Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do estado de Mato Grosso do Sul (Fundect), pela concessão da bolsa de mestrado à primeira autora e pelo financiamento de projeto. Aos proprietários e funcionários das Fazendas São Geraldo (Rio Sucuri) e Baía Bonita pelo apoio logístico oferecido.
REFERÊNCIAS BIBLIOGRÁFICAS
Armstrong, J. & Armstrong, W. (1988), Phragmites australis: a preliminary study of soil-oxidizing sites and intenal gas transport pathways. New Phytologist. 108:373-382.
Armstrong, W. (2000), Diaphragms and survival. New Phytologist. 145: 1-5.
Bona, C. (1999), Adaptações morfo-anatômicas dos órgãos vegetativos de Bacopa salzmannii (Benth.) Wettst ex Edwall e Bacopa monnieriodoides (Cham.) Robinson (Scrophulariaceae) em ambiente terrestre e aquático. Tese de Doutorado/Instituto de Biociências/USP, São Paulo.
Clark, G. (1981). Staining procedure. 4º ed. Baltimore: Williams & Wikins, p. 171-215.
Colmer, T. D. (2003), Long-distance transport of gases in plants: a perspective on internal aeration and radial oxygen loss from roots. Plant, Cell and Environment. 26: 17-36.
Cronk, J. K. & Frenessy, M. S. (2001), Wetland Plants: Biology and Ecology. Lewis Publishers. 462p.
Cutter, E. G. (1987), Anatomia Vegetal: Experimento e interpretação Parte II: Órgãos. Ed. Rocca. 336p.
Evans, D. E. (2003), Aerenchyma formation. New Phytologist. 161: 35-49
Fahn, A. (1990), Plant Anatomy. Ed. 4. Oxford: Pergamon Press.
Iida, S., Kosuge, K., Kadono, Y. (2004). Molecular phylogeny of Japanese Potamogeton species in light of noncoding chloroplast sequences. Aquatic Botany, 80, 115-127.
Kraus, J. E. & Arduim, M. (1997). Manual Básico de Métodos em morfologia Vegetal. Seropédica, RJ: EDUR.
Levitt, J. (1980), Responses of Plants to Environmental Stress: Water, Radiation, Salt and other Stresses. V. II. New York, Academic Press.
Mauseth, J. D. (1988), Plant Anatomy. California: Benjamin/Cummings.
Metcalfe, C. R. & Chlak, L. (1983), Anatomy of de Dicotyledons: Wood structure and conclusion of the general introduction. V.2. New York, Oxford University Press.
Pedersen, O. & Sand-Jensen, K. (1993), Water transport in submerged macrophytes. Aquatic Botany, 44, 385-406.
Rascio, N., Cuccato, F., Dalla Vecchia, F., La Rocca, N., Larcher, W. (1999), Structural and functional features of the leaves of Ranunculus trichophhyllus Chaix., a freshwater submerged macrophyte. Plant, Cell and Environment, 22, 205-212.
Ridge, I. (1987), Ethilene and growth control in amphibious plants. In: Crawford, R. M. M. Plant life in aquatic and amphibious habitats. Oxford: Blackwell Scientific Publications, pp 53-76.
Rodrigues, R. S. & Irgang, (2001), Potamogetonaceae Dumort. no Rio Grande do Sul, Brasil. Iheringia, Ser. Bot., Porto Alegre, 56, 3-49.
Scremin-Dias, E., Pott, V. J., Hora, R. C., Souza, P. R. (1999). Nos Jardins Submersos da Bodoquena: guia para a identificação de plantas aquáticas de Bonito e região. Ed. UFMS. Campo Grande, MS.
Scremin-dias, E. (2000), Caracterização morfo-anatômica dos órgãos vegetativos de Echinodorus paniculatus Micheli e Echinodorus tenellus (Mart.) Buchenau (Família Alismataceae), durante os períodos de cheia e seca no Pantanal Sul-Mato-Grossense. Tese de Doutorado/Instituto de Biociências/USP, São Paulo.
Sculthorpe, C. D. (1967), The Biology of aquatic vascular plants. Eduard Arnold Publ. London.
Souza, V. C. & Lorenzi, H. (2005), Botânica Sistemática: Guia ilustrado para identificação de

28
angiospermas da flora brasileira, baseado em APG II. Instituto Plantarum, 640p.
Soukup, A., Votrubová, O. Cisková, H. (2000), Internal segmentation of rizhomes of Phragmites australis: protection of the internal aeration system against being flooded. New Phytologist. 145: 71-75.
Tomlinson, P. B. (1982), Anatomy of the Monocotyledons. Vol. 7: Helobiae (Alismatidae). Editado por Metcalfe, C. R. Clarendon Press. Oxford. 560p
Usherwood, J. R., Ennos, A. R., Ball, D. J. (1997), Mechanical and adaptations in terrestrial and aquatic buttercups to their respective environments. Journal of Experimental Botany. 48 (312), 1469-1475.
Vartapetian, B. B. & Jackson, M. B. (1997), Plant adaptations to anaerobic stress. Annals of Botany. 79, 3-20.
Voesenek, L. A. C., Colmer T. D., Pierik, R., Millenaar, F. F., Peeters, A. J. M. (2006), How plants cope with complete submergence. New Phytologist. 170: 213-226.
Wiegleb, G. & Kaplan, Z. (1998), An account of the species of Potamogeton L. (Potamogetonaceae). Folia Geobotanica, Upssala, 33, 241-316

29

30
CAPITULO II: Efeito da velocidade da correnteza na morfologia de Potamogeton
illinoensis Morong e o potencial de reprodução vegetativa desta espécie, que ocorre nos rios Sucuri e Baía Bonita – Bonito, Estado de
Mato Grosso do Sul, Centro-Oeste do Brasil

31
An International Scientific Journal dealing with Applied and Fundamental Research on Submerged, Floating and Emergent Plants in Marine and Freshwater Ecosystems
Guide for Authors Aquatic Botany is concerned with fundamental studies on structure, function, dynamics and classification of plant-dominated aquatic communities and ecosystems, as well as molecular, biochemical and physiological aspects of aquatic plants. It is also an outlet for papers dealing with applied research on plant-dominated aquatic systems, including the consequences of disturbance (e.g. transplantation, influence of herbicides and other chemicals, thermal pollution, biological control, grazing and disease), the use of aquatic plants, conservation of resources, and all aspects of aquatic plant production and decomposition. Types of contribution 1. Original research papers (Regular Papers) 2. Review articles 3. Short Communications 4. Technical Communications 5. Letters to the Editor 6. Book Reviews Regular papers should report the results of original research. The material should not have been previously published elsewhere, except in a preliminary form. Review articles should cover subjects falling within the scope of the journal which are of active current interest. They may be submitted or invited. A Short Communication is a concise but complete description of a limited investigation, which will not be included in a later paper. Short Communications should be as completely documented, both by reference to the literature and description of the experimental procedures employed, as a regular paper. They should not occupy more than 6 printed pages (about 12 manuscript pages, including figures, tables and references). Technical Notes should be brief descriptions of experimental procedures, technical operations or applied activities within laboratories or in the field. Letters to the Editor offering comment or appropriate critique on material published in the journal are welcomed. The decision to publish submitted letters rests purely with the Editor-in-Chief. Book Reviews are included in the journal on a range of relevant books which are not more than 2 years old. Book reviews will be solicited by the Book Review Editor. Other publications read will be listed. Unsolicited reviews will not usually be accepted, but suggestions for appropriate books for review may be sent to the Book Review Editor: T.V. Madsen University of Aarhus Department of Plant Ecology Nordlandsvej 68 D-K8240 Risskov Denmark E-mail: [email protected] Submission of manuscripts Submission of an article is understood to imply that the article is original and is not being considered for publication elsewhere. Submission also implies that all authors have approved the paper for release and are in agreement with its content. Upon acceptance of the article by the journal, the author(s) will be asked to transfer the copyright of the article to the publisher. This transfer will ensure the widest possible dissemination of information. Papers for consideration should be submitted to: http://ees.elsevier.com/aqbot Electronic manuscripts Submission to this journal proceeds totally on-line. Use the following guidelines to prepare your article. Via the

32
"Author Gateway" page of this journal ( http://authors.elsevier.com/ ) you will be guided stepwise through the creation and uploading of the various files. The system automatically converts source files to a single Adobe Acrobat PDF version of the article, which is used in the peer-review process. Please note that even though manuscript source files are converted to PDF at submission for the review process, these source files are needed for further processing after acceptance. All correspondence, including notification of the Editor's decision and requests for revision, takes place by e-mail and via the Author's homepage, removing the need for a hard-copy paper trail. Electronic format requirements for accepted articles We accept most wordprocessing formats, but Word, WordPerfect or LaTeX is preferred. Always keep a backup copyof the electronic file for reference and safety. Save your files using the default extension of the program used. Wordprocessor documents It is important that the file be saved in the native format of the wordprocessor used. The text should be in single-column format. Keep the layout of the text as simple as possible. Most formatting codes will be removed and replaced on processing the article. In particular, do not use the wordprocessor's options to justify text or to hyphenate words. However, do use bold face, italics, subscripts, superscripts etc. Do not embed 'graphically designed' equations or tables, but prepare these using the wordprocessor's facility. When preparing tables, if you are using a table grid, use only one grid for each individual table and not a grid for each row. If no grid is used, use tabs, not spaces, to align columns. The electronic text should be prepared in a way very similar to that of conventional manuscripts (see also the Author Gateway's Quickguide. Do not import the figures into the text file but, instead, indicate their approximate locations directly in the electronic text and on the manuscript. See also the section on Preparation of electronic illustrations. To avoid unnecessary errors you are strongly advised to use the 'spellchecker' function of your wordprocessor. Preparation of manuscripts 1. Manuscripts should be written in English. Authors whose native language is not English are strongly advised to have their manuscripts checked by an English-speaking colleague prior to submission. English language help service: Upon request, Elsevier will direct authors to an agent who can check and improve the English of their paper (before submission). Please contact [email protected] for further information. 2. Manuscripts should be prepared with numbered lines, with wide margins and double spacing throughout, i.e. also for abstracts, footnotes and references. Every page of the manuscript, including the title page, references, tables, etc. should be numbered. However, in the text no reference should be made to page numbers; if necessary, one may refer to sections. Underline words that should be in italics, and do not underline any other words. Avoid excessive use of italics to emphasize part of the text. 3. Manuscripts in general should be organized in the following order: Title (should be clear, descriptive and not too long) Name(s) of author(s) Complete postal address(es) of affiliations Full telephone, Fax. no. and E-mail of the corresponding author Present address(es) of author(s) if applicable Complete correspondence address to which the proofs should be sent Abstract Key words (indexing terms), normally 3-6 items Introduction Material studied, area descriptions, methods, techniques Results Discussion Conclusion Acknowledgements and any additional information concerning research grants, etc. References Tables Figure captions 4. In typing the manuscript, titles and subtitles should not be run within the text. They should be typed on a separate line, without indentation. Use lower-case lettertype. 5. Units and abbreviations In principle SI units should be used except where they conflict with current practise or are confusing. Other

33
equivalent units may be given in parentheses. Abstracts The abstract should be clear, descriptive and not longer than 400 words. Tables 1. Authors should take notice of the limitations set by the size and lay-out of the journal. Large tables should be avoided. Reversing columns and rows will often reduce the dimensions of a table. 2. If many data are to be presented, an attempt should be made to divide them over two or more tables. 3. Tables should be numbered according to their sequence in the text. The text should include references to all tables.4. Each table should be typewritten on a separate page of the manuscript. Tables should never be included in the text.5. Each table should have a brief and self-explanatory title. 6. Column headings should be brief, but sufficiently explanatory. Standard abbreviations of units of measurement should be added between parentheses. 7. Vertical lines should not be used to separate columns. Leave some extra space between the columns instead. 8. Any explanation essential to the understanding of the table should be given as a footnote at the bottom of the table. Electronic Illustrations Submitting your artwork in an electronic format helps us to produce your work to the best possible standards, ensuring accuracy, clarity and a high level of detail. 1. Make sure you use uniform lettering and sizing of your original artwork. 2. Save text in illustrations as "graphics" or enclose the font. 3. Only use the following fonts in your illustrations: Arial, Courier, Helvetica, Times, Symbol. 4. Number the illustrations according to their sequence in the text. 5. Use a logical naming convention for your artwork files, and supply a separate listing of the files and the software used. 6. Provide all illustrations as separate files. 7. Provide captions to illustrations separately. 8. Produce images near to the desired size of the printed version. A detailed guide on electronic artwork is available on our website: http://authors.elsevier.com/artwork You are urged to visit this site; some excerpts from the detailed information are given here. Formats Regardless of the application used, when your electronic artwork is finalised, please "save as" or convert the images to one of the following formats (Note the resolution requirements for line drawings, halftones, and line/halftone combinations given below.): EPS: Vector drawings. Embed the font or save the text as "graphics". TIFF: Colour or greyscale photographs (halftones): always use a minimum of 300 dpi TIFF: Bitmapped line drawings: use a minimum of 1000 dpi. TIFF: Combinations bitmapped line/half-tone (colour or greyscale): a minimum of 500 dpi is required. DOC, XLS or PPT: If your electronic artwork is created in any of these Microsoft Office applications please supply "as is". Please do not: • Supply embedded graphics in your wordprocessor (spreadsheet, presentation) document; • Supply files that are optimised for screen use (like GIF, BMP, PICT, WPG); the resolution is too low; • Supply files that are too low in resolution; • Submit graphics that are disproportionately large for the content. Colour illustrations If, together with your accepted article, you submit usable colour figures then Elsevier will ensure, at no additional charge, that these figures will appear in colour on the web (e.g., ScienceDirect and other sites) regardless of whether or not these illustrations are reproduced in colour in the printed version.

34
For colour reproduction in print, you will receive information regarding the costs from Elsevier after receipt of your accepted article. For further information on the preparation of electronic artwork, please see http://authors.elsevier.com/artwork. Please note: Because of technical complications which can arise by converting colour figures to grey scale (for the printed version should you opt to not pay for colour in print) please submit in addition usable black and white prints corresponding to all the colour illustrations. As only one figure caption may be used for both colour and black and white versions of figures, please ensure that the figure captions are meaningful for both versions, if applicable. References 1. All publications cited in the text should be presented in a list of references following the text of the manuscript. The manuscript should be carefully checked to ensure that the spelling of author's names and dates are exactly the same in the text as in the reference list. 2. In the text refer to the author's name (without initial) and year of publication, followed if necessary by a short reference to appropriate pages. Examples: "Since Peterson (1988) has shown that..." "This is in agreement with results obtained later (Kramer, 1989, pp. 12-16)". 3. If reference is made in the text to a publication written by more than two authors the name of the first author should be used followed by "et al.". This indication, however, should never be used in the list of references. In this list names of first author and co-authors should be mentioned. 4. References cited together in the text should be arranged chronologically. The list of references should be arranged alphabetically on authors' names, and chronologically per author. If an author's name in the list is also mentioned with co-authors the following order should be used: publications of the single author, arranged according to publication dates - publications of the same author with one co-author - publications of the author with more than one co-author. Publications by the same author(s) in the same year should be listed as 1994a, 1994b, etc. 5. Use the following system for arranging your references: a. For periodicals Stewart, D.A., Agnew, D., Boyd, R., Briggs, R., Toland, P., 1993. The derivation of changes in Nephrops per unit effort values for the Northern Ireland fishing fleet. Fish. Res. 17, 273-292. b. For edited symposia, special issues, etc. published in a periodical Roberts, R.J., 1993. Ulcerative dermal necrosis (UDN) in wild salmonids. In: Bruno, D.W. (Ed.), Pathological conditions of wild salmonids. Fish. Res. 17, 3 14. c. For books Gaugh, Jr., H.G., 1992. Statistical Analysis of Regional Yield Trials. Elsevier, Amsterdam. d. For multi-author books Bucke, D., 1989. Histology. In: Austin, B., Austin, D.A. (Eds.), Methods for the Microbiological Examination of Fish and Shellfish. Wiley, New York, pp. 69-97. 6. Abbreviate the titles of periodicals mentioned in the list of references according to the International List of Periodical Title Word Abbreviations. 7. In the case of publications in any language other than English, the original title is to be retained. However, the titles of publications in non-Latin alphabets should be transliterated, and a notation such as "(in Russian)" or "(in Greek, with English abstract)" should be added. 8. Work accepted for publication but not yet published should be referred to as "in press". 9. References concerning unpublished data and "personal communications" should not be cited in the reference list but may be mentioned in the text. Formulae 1. Formulae should be typewritten, if possible. Leave ample space around the formulae. 2. Subscripts and superscripts should be clear. 3. Greek letters and other non-Latin symbols should be explained in the margin where they are first used. Take special care to show clearly the difference between zero (0) and the letter O, and between one (1) and the letter l. 4. Give the meaning of all symbols immediately after the equation in which they are first used. 5. For simple fractions use the solidus (/) instead of a horizontal line. 6. Equations should be numbered serially at the right-hand side in parentheses. In general only equations explicitly referred to in the text need be numbered. 7. The use of fractional powers instead of root signs is recommended. Also powers of e are often more conveniently denoted by exp. 8. Levels of statistical significance which can be mentioned without further explanation are *P<0.05, **P<0.01 and ***P<0.001.

35
9. In chemical formulae, valence of ions should be given as, e.g., Ca2+ and not as Ca++. 10. Isotope numbers should precede the symbols, e.g., 18O. 11. The repeated writing of chemical formulae in the text is to be avoided where reasonably possible; instead, the name of the compound should be given in full. Exceptions may be made in the case of a very long name occurring very frequently or in the case of a compound being described as the end product of a gravimetric determination (e.g., phosphate as P2O5). Footnotes 1. Footnotes should be used only if absolutely essential. In most cases it should be possible to incorporate the information in normal text. 2. If used, they should be numbered in the text, indicated by superscript numbers, and kept as short as possible. Nomenclature 1. Authors and editors are, by general agreement, obliged to accept the rules governing biological nomenclature, as laid down in the International Code of Botanical Nomenclature, the International Code of Nomenclature of Bacteria, and the International Code of Zoological Nomenclature. 2. All biotica (crops, plants, insects, birds, mammals, etc.) should be identified by their scientific names when the English term is first used, with the exception of common domestic animals. 3. All biocides and other organic compounds must be identified by their Geneva names when first used in the text. Active ingredients of all formulations should be likewise identified. 4. For chemical nomenclature, the conventions of the International Union of Pure and Applied Chemistry and the official recommendations of the IUPAC IUB Combined Commission on Biochemical Nomenclature should be followed. Preparation of supplementary data Elsevier now accepts electronic supplementary material to support and enhance your scientific research. Supplementary files offer the author additional possibilities to publish supporting applications, movies, animation sequences, high-resolution images, background datasets, sound clips and more. Supplementary files supplied will be published online alongside the electronic version of your article in Elsevier web products, including ScienceDirect:
http://www.sciencedirect.com. In order to ensure that your submitted material is directly usable, please ensure that data is provided in one of our recommended file formats. Authors should submit the material in electronic format together with the article and supply a concise and descriptive caption for each file. For more detailed instructions please visit our Author Gateway at http://authors.elsevier.com. Copyright 1. An author, when quoting from someone else's work or when considering reproducing an illustration or table from a book or journal article, should make sure that he is not infringing a copyright. 2. Although in general an author may quote from other published works, he should obtain permission from the holder of the copyright if he wishes to make substantial extracts or to reproduce tables, plates, or other illustrations. If the copyright-holder is not the author of the quoted or reproduced material, it is recommended that the permission of the author should also be sought. 3. Material in unpublished letters and manuscripts is also protected and must not be published unless permission has been obtained. 4. A suitable acknowledgement of any borrowed material must always be made. Proofs One set of proofs will be sent to the corresponding author as given on the title page of the manuscript. Only typesetter's errors may be corrected; no changes in, or additions to, the edited manuscript will be allowed. Elsevier will do everything possible to get your article corrected and published as quickly and accurately as possible. Therefore, it is important to ensure that all of your corrections are sent back to us in one communication. Subsequent corrections will not be possible, so please ensure your first sending is complete. Reprints The corresponding author, at no cost, will be provided with a PDF file of the article via e-mail or, alternatively, 25 free paper offprints. The PDF file is a watermarked version of the published article and includes a cover sheet with

36
the journal cover image and a disclaimer outlining the terms and conditions of use. Additional offprints can be ordered on are print order form, which is included with the proofs. UNESCO coupons are acceptable in payment of extra offprints. GenBank/DNA sequence linking Many Elsevier journals cite "gene accession numbers" in their running text and footnotes. Gene accession numbers refer to genes or DNA sequences about which further information can be found in the databases at the National Center for Biotechnical Information (NCBI) at the National Library of Medicine. Elsevier authors wishing to enable other scientists to use the accession numbers cited in their papers via links to these sources, should type this information in the following manner: For each and every accession number cited in an article, authors should type the accession number in bold, underlined text. Letters in the accession number should always be capitalised. This combination of letters and format will enable Elsevier's typesetters to recognize the relevant texts as accession numbers and add the required link to GenBank's sequences. Example: "GenBank accession nos. AI631510, AI631511, AI632198, and BF223228". Please check accession numbers used very carefully as an error in a letter or number can result in a dead link. Author Services Authors can also keep a track on the progress of their accepted article, and set up e-mail alerts informing them of changes to their manuscript's status, by using the "Track a Paper" feature of Elsevier's Author Gateway .

37
Efeito da velocidade da correnteza na morfologia de Potamogeton illinoensis Morong e o
potencial de reprodução vegetativa desta espécie, que ocorre nos rios Sucuri e Baía
Bonita – Bonito, Estado de Mato Grosso do Sul, Centro-Oeste do Brasil¹.
Seloi Rodrigues ² & Edna Scremin-Dias²
¹Parte da dissertação de mestrado da primeira autora.
²Laboratório de Botânica, Departamento de Biologia, Universidade Federal de Mato
Grosso do Sul, Caixa Postal 549, Campo Grande, 79070-900, Brazil
Autor para correspondência. Tel. +55-67-3345-7399; fax: +55-67-3345-7306.
E-mail: [email protected] [email protected]
Abstract
The present study focuses the morphology of Potamogeton illinoensis Morong that occurs in
running and stagnant water, emphasizing the methods of vegetative reproduction that this
species presents in the studied rivers. The materials were collected in Sucuri and Baía Bonita
Rivers in Bonito, Mato Grosso do Sul, Brazil. For the morphologic analysis total length of
individuals, length of the petiole and the leaf blade and the width of the leaf blade were
measured. It was verified that this species does not present significant differences in the
length of individuals as well as in the length of leaves developed in different environments,
however, the width of the leaves of the individuals that grow in running water of Baía Bonita
River, differed significantly of the leaves of individuals from the other places. In spite of the
differences in the total length of the individual do not present significant value, the
individuals from the stagnant water were always smaller when compared with those of
running water. The population patches that occur in the two rivers are different, and in Baía
Bonita River the populations are densely distributed, being characterized as invader in this

38
environment. This species spreads vegetatively by the emission of rhizomes, colonizing areas
adjacent to the patches by peripherical propagation. Floating stems have the capacity of
regenerating itself after fragmentation, which can happen in a natural way or by human
action. The human interference is common in the studied places, as they are used in the
ecotourism. Our results indicate that it is necessary a great care in management of those
rivers, because they are fragile environments and ecologically and economically important.
Key Words: Potamogeton illinoensis, morphology, water velocity, regenerating itself,
vegetative fragment
Introdução
Plantas aquáticas estão sujeitas às diferentes condições dos ambientes aquáticos, como
a exposição às ondas, às distintas velocidades do fluxo de água, além da deficiência no
suprimento de nutrientes, oxigênio, dióxido de carbono e restrição aos processos de
reprodução sexuada (Santamaría, 2002). Dentre estes fatores, o movimento da água é um
importante fator regulador do crescimento e da distribuição de macrófitas aquáticas
submersas. O aumento da velocidade da água pode, em alguns casos, contribuir para a
diminuição da biomassa e densidade dos ramos em plantas aquáticas (Chambers et al., 1991).
A velocidade do fluxo da água também pode influenciar indiretamente no crescimento destas
plantas, pois pode determinar o tamanho das partículas do substrato (Madsen et al., 2001) e,
conseqüentemente, influenciar na disponibilidade de nutrientes. No entanto, a força
hidrodinâmica aplicada sobre os organismos, pode depender do tamanho dos organismos, ou
seja, da área de contato entre o corpo da planta e o fluxo de água (Puijalon & Bornette, 2004).

39
Em experimentos com duas espécies de gêneros distintos Mentha aquática L. e Berula
erecta (Huds.) Coville, Puijalon & Bornette (2004) verificaram a espécie B. erecta que cresce
verticalmente formando rosetas sofreu diminuição no tamanho dos ramos e na densidade da
população, a medida que houve aumento da velocidade da água. Já M. aquatica com parte dos
ramos crescendo paralelos ao substrato e parte eretos, teve aumento na biomassa e no
crescimento dos ramos, à medida que a velocidade do fluxo da água aumentou.
Alguns trabalhos demonstram que na maioria das vezes, o aumento de profundidade e
da velocidade da corrente de água provoca a diminuição da biomassa da planta aquática
(Barrat-Segretain, 2001; Strand & Weisner, 2001), no entanto este pode não ser o padrão para
todas as espécies (Puijalon & Bornette, 2004). Também pode ocorrer alocação de biomassa
para ramos ou partes subterrâneas naquelas espécies expostas às ondas, e a diferentes
profundidades (Barrat-Segretain, 2001; Strand & Weisner, 2001; Leeflang et al., 1998).
Dall’Armellina et al. (1996a), amostrando a diversidade e biomassa das espécies de
macrófitas que ocorrem em canais de irrigação no Vale do Rio Negro – Argentina,
demonstrou que as espécies de Potamogeton se desenvolvem fácil e rapidamente em
ambientes com características diversas, indicando a presença de Potamogeton illinoensis em
100% das amostras retiradas dos experimentos, sendo que a biomassa representou mais de
93% da biomassa total do canal.
Mesmo com as barreiras impostas pelo ambiente aquático, algumas espécies se
desenvolvem em demasia e se tornam invasoras, como é o caso do gênero Potamogeton, que é
referido como espécie invasora na Argentina (Dall’Armellina et al., 1996a). Esta espécie
nativa nos rios Sucuri e Baía Bonita, no município de Bonito em Mato Grosso do Sul, vem
crescendo em demasia no rio Baia Bonita, tornando-se dominante naquele ambiente
(Observação pessoal). Abundância de crescimento de uma determinada espécie pode
restringir ou impedir o crescimento de outras espécies, igualmente importantes para

40
manutenção da integridade e diversidade ambiental. Ambientes onde há predominância de
uma espécie de macrófita, constituem habitats menos adequados a aves aquáticas e peixes, no
que diz respeito a fonte de alimento e abrigo (Herb & Stefan, 2006), e o crescimento
excessivo de macrófitas interfere negativamente nestes ambientes, inclusive quando usados
para recreação (Chamber et al., 1991, Herb & Stefan, 2006).
As populações de muitas espécies de macrófitas aquáticas são mantidas no ambiente
aquático principalmente devido a eficiência no processo de reprodução vegetativa. As
estratégias desta reprodução podem ser por meio da emissão de rizomas, estolhos, botões de
inverno e por fragmentos não especializados, como caules desprendidos da planta mãe
(Barrat-Segretain, 1996), que podem funcionar como propágulos.
Em vários rios da região de Bonito, em Mato Grosso do Sul, devido a alta
disponibilidade de calcário dissolvido, ocorre a precipitação das partículas argilosas em
suspensão, resultando em corpos d’água de extrema limpidez e grande diversidade biológica,
explorada pelo turismo contemplativo de flutuação (Scremin-Dias et al., 1999). No entanto, a
prática inadequada de manejo de ambientes aquáticos, e a degradação do ambiente, podem
provocar o aumento da biomassa de espécies oportunistas nas comunidades de macrófitas,
alterando o equilíbrio e ocorrendo em alta densidade (Chambers et al., 1999). O crescimento
em demasia da população de Potamogeton illinoensis no rio Baia Bonita tem provocado
desconforto aos turistas que praticam ecoturismo naquele ambiente, devido as características
de seu hábito – densas populações, com caules atingindo a superfície. Em outro rio da região
– o rio Sucuri – a população desta espécie encontra-se em equilíbrio, estando os indivíduos
distribuídos na calha do rio, onde a água apresenta grande velocidade, e também em remanso,
local em que a velocidade da água é quase imperceptível.
Entender a biologia de uma espécie, bem como detectar a existência de estruturas
especializadas na propagação vegetativa, consiste no primeiro passo para propor estratégias

41
para manutenção do equilíbrio das populações no ambiente. Este trabalho objetivou estudar
indivíduos de Potamogeton illinoensis que se desenvolvem em locais com diferentes
velocidades de correnteza em dois rios de Bonito, explorados economicamente pelo
ecoturismo. Além disso, são apresentados dados sobre a potencialidade de regeneração dos
caules, resultantes da fragmentação dos indivíduos, causada por diversos fatores.
Material e Métodos
Local de estudo e população de Potamogeton illinoensis
O trabalho foi desenvolvido em dois rios do município de Bonito, no Estado de Mato
Grosso do Sul, onde são desenvolvidas atividades de turismo de contemplação subaquática,
através da flutuação, e geralmente se utiliza barco a remo como apoio ao turistas. Devido a
região ter formação calcária, os rios Sucuri (56°33’32’’W 21°15’56’’S) e Baía Bonita
(56º26’24’’W 21º9’46’’S) possuem águas extremamente límpidas, e flora e fauna
subaquáticas exuberantes.
Foram selecionados agrupamentos de indivíduos de P. illinoensis em três locais de
remanso e de correnteza, e realizadas uma coleta mensal durante cinco meses, na população
distribuída ao longo dos rios Sucuri e Baía Bonita. Em cada ponto de coleta foi anotada a
profundidade da lâmina d’água e a velocidade do fluxo de água com auxílio de Fluxímetro
Global Water Fp 201. A velocidade do fluxo de água no rio Sucuri teve uma média de
0,34m/s nas regiões de correnteza e média de 0,04m/s nas áreas de remanso. No rio Baía
Bonita a velocidade média foi de 0,24m/s e 0,06m/s, para correnteza e remanso
respectivamente. O rio Baía Bonita teve profundidade média nos locais de correnteza 1,22m e
nos locais de remanso 1,11m. No rio Sucuri as médias foram 1,25m para correnteza e 1,03m
para remanso.

42
Dos indivíduos coletados em cada mês foram feitas mensurações com o auxílio de
régua graduada, obtendo o comprimento e a largura do limbo foliar e comprimento dos
pecíolos, de folhas do 4º e 5º nós e comprimento total do caule.
Análise dos dados
Das mensurações obtidas de 20 indivíduos em média, por local – remanso e correnteza
– e por coleta, foram obtidas as médias das amostras mensais. Foi utilizada a análise de
variância para comparar os valores médios das medições obtidas nos grupos independentes e
aplicado teste de Tukey para verificar se a diferença entre as amostras era significante.
Regeneração de fragmentos de caule
Visando testar se havia a potencialidade de regeneração dos fragmentos dos caules
flutuantes e subterrâneos foi estabelecido experimento no rio Sucuri, com 10 fragmentos de
rizomas e 10 de caule flutuante – do 9º ao 10º entrenós -, amarrados com corda de nylon®, e
fixados entre duas estacas. Destes 10 fragmentos, cinco foram depositados junto ao substrato
e cinco na superfície da água. Após um mês de permanência dos fragmentos no rio, estes
foram retirados e analisados morfologicamente. Também foram coletados e analisados
fragmentos de caule acumulados na superfície da água nas populações de P. illinoensis,
presentes em remansos ao longo do rio Baía Bonita.
Resultados
Potamogeton illinoensis é uma planta aquática totalmente submersa, rizomatosa, tendo
folhas que podem alcançar a superfície da água, mas raramente a ultrapassa, e escapos florais
emergentes. Os indivíduos se desenvolvem em correnteza forte e em locais de águas mais
lentas. Nos locais de maior correnteza, os indivíduos de P. illinoensis ficam prostrados em

43
direção a corrente e assim quase nunca encostam suas folhas na superfície da água, e
raramente emitem estruturas de reprodução sexuada.
As mensurações obtidas do comprimento total dos indivíduos de P. illinoensis
mostraram que indivíduos desenvolvidos em remanso foram maiores que os de correnteza, em
ambos os rios. Todavia as diferenças de valores obtidos para este parâmetro não foram
significativas (Tabela 1).
Quando comparado o comprimento do limbo foliar dos indivíduos de remanso com
aqueles desenvolvidos em correnteza, apesar dos indivíduos de remanso apresentarem folhas
maiores, nenhum valor obtido foi significativamente diferente (Tabela 1). Para as medidas da
largura do limbo foliar, os indivíduos presentes em correnteza no rio Sucuri tiveram folhas
mais estreitas que aqueles de remanso, no entanto a diferença não foi significativa. O mesmo
padrão de largura de limbo foi obtido para os indivíduos do rio Baía Bonita, no entanto as
diferenças das medições obtidas foram significativas quando comparados os indivíduos de
correnteza, com os de remanso neste rio (Tabela 1).
As medições do comprimento dos pecíolos, tanto dos indivíduos do rio Sucuri quanto
daqueles distribuídos no rio Baía Bonita, mostraram comprimento médio menor para os
indivíduos de correnteza em relação aos de remanso. Essa diferença não é significativa
quando comparados remanso e correnteza dentro do mesmo rio (Tabela 1).
P. illinoensis tem o rápido crescimento e desenvolvimento dos indivíduos facilitado
pela formação dos rizomas (Fig. 1A e 1B), o que possibilita a colonização de vastas áreas nos
dois rios. Além dos rizomas, que são emitidos pela planta mãe, fragmentos desprendidos do
caule flutuante tem capacidade de formar novas plantas. Do experimento estabelecido para
testar a potencialidade de regeneração do caule desta espécie, houve rebrota em todos os
fragmentos de caule (Fig. 1C) após o período de quatro semanas. No caso dos rizomas, não
houve surgimento de novos indivíduos nos nós, no entanto os fragmentos permaneceram

44
vivos durante o período do experimento. Dos fragmentos de caule que se desprenderam das
plantas mães naturalmente ou pela ação dos remos, coletados sobre as populações de P.
illinoensis no rio Baía Bonita (Fig. 1D), vários possuíam brotações (Fig. 1E). Em alguns dos
fragmentos coletados nesta situação, já havia desenvolvido raízes adventícias (Fig. 1F). Estes
fragmentos podem se acumular em determinadas partes do rio (Fig. 1G), devido o rio Baía
Bonita possuir águas calmas. Dos fragmentos coletados nesta situação foi observado que,
mesmo apresentando sinais de deterioração, alguns possuíam brotos, com raízes emitidos a
partir dos nós da planta mãe (Fig. 1F), demonstrando potencialidade para o estabelecimento
de novas plantas.
Discussão
As espécies de Potamogeton podem ter grande plasticidade fenotípica, devido a fatores
químicos e físicos do ambiente onde se desenvolvem (Sculthorpe, 1967). Para P. illinoensis a
velocidade da água influencia no tamanho dos indivíduos, sendo aqueles que ocorrem em
maior velocidade de correnteza sempre de menor tamanho, quando comparados aos que se
desenvolvem em velocidades menores. Estes dados corroboram os descritos por vários
autores sobre o efeito mecânico do fluxo da água sobre o crescimento de macrófitas aquáticas
(Chambers et al., 1991, Puijalon & Bornette, 2004). Resposta semelhante já foi descrita para
Potamogeton pectinatus L. quando os ramos foram expostos ao efeito de ondas e vento,
produzindo indivíduos menores nestas condições, comparados aqueles desenvolvidos em
locais mais calmos, sugerindo que o menor tamanho dos indivíduos, diminui o risco de
desenraizamento (Idestam-Almquist & Kautsky, 1995), pela ação da correnteza. Para Power
(1996) a velocidade da correnteza pode ser responsável pela definição da estrutura da
comunidade de macrófitas submersas.

45
Outros fatores, além da velocidade da água, poderiam estar influenciando o
desenvolvimento de macrófitas ao longo dos rios como disponibilidade de nutrientes, luz,
temperatura e herbivoria, no entanto Chambers et al. (1991), demonstraram que para
Potamogeton pectinatus a velocidade do fluxo de água afeta diretamente o crescimento dos
indivíduos, independente dos efeitos indiretos do tipo de sedimento, pois influencia
amplamente na densidade e biomassa dos ramos, sendo que em velocidades maiores ocorre a
menor densidade de macrófitas e, em velocidades maiores que 1m/s, macrófitas são raras.
Algumas plantas, como é o caso de P. illinoensis, tendem a desenvolver ramos eretos,
em velocidades de fluxo da água menores, e ramos alinhados à correnteza quando a
velocidade do fluxo da água é maior, diminuindo assim a superfície exposta diretamente ao
fluxo, e reduzindo o estresse em velocidades de corrente maiores (Puijalon & Bornette, 2004).
O menor comprimento dos caules pode refletir a necessidade de maior flexibilidade das
plantas em locais de água mais corrente (Boeger & Poulson, 2003), visando diminuir a
resistência ao fluxo (Schutten & Davy, 2000) e minimizar os danos mecânicos. Redução no
tamanho do indivíduo com aumento da velocidade da corrente é resposta comum para
determinadas espécies que são submetidas ao estresse mecânico (Puijalon et al., 2005).
Também, plantas aquáticas submetidas a maior correnteza, tendem a ter menor rigidez
nos caules que plantas que se desenvolvem em águas mais lentas (Usherwood et al., 1997),
diminuindo assim a resistência dos indivíduos ao fluxo. Macrófitas submersas são mais
sensíveis a altas velocidades do fluxo da água, sendo que locais de águas mais lentas,
possuem manchas de vegetação submersas bem maiores que nos locais de correnteza (Gantes
& Caro, 2001), característica corroborada pelos dados obtidos no presente estudo com P.
illinoensis.
A força aplicada pelo fluxo da água sobre as plantas pode causar quebra e
desenraizamento da planta, sendo que a força aplicada pela corrente nas espécies muito

46
pequenas, pode não ser significante (Schutten & Davy, 2000), para provocar fragmentação
dos indivíduos. Por essa razão, plantas pequenas encontram menores forças hidrodinâmicas
(Puijalon & Bornette, 2004), quando comparadas com as maiores.
Apesar dos indivíduos de P. illinoensis distribuídos em áreas de maior correnteza,
possuírem tamanho menor e ficarem prostradas perto do substrato, alinhados em direção ao
fluxo da água, estas características não são suficientes para impedir a fragmentação dos caules
flutuantes pois, de acordo com nossos resultados, a fragmentação pode ocorrer pela ação da
correnteza. Também a ação antrópica, resultante das atividades desenvolvidas no local resulta
na formação de fragmentos, que constituem-se em propágulos vegetativos não especializados,
que podem ser carreados pela correnteza ou permanecerem depositados sobre a superfície da
água em áreas de remanso e, potencialmente, podem gerar novos indivíduos.
Sculthorpe (1967) já enfatizava que, como muitas plantas não possuem método
especializado para reprodução vegetativa, qualquer fragmento de ramo desprendido pode ser
hábil para rebrotar e formar novos indivíduos, visto que estes fragmentos vegetativos podem
se tornar propágulos com alto poder de regeneração (Barrat-Segretain 1996, Cronk &
Frenessy, 2001). Algumas espécies podem apresentar diferenças na habilidade de regeneração
e recolonização de áreas perturbadas dependendo da época do ano em que a fragmentação
ocorre (por ex. outono ou primavera), mesmo assim esta característica consiste num excelente
meio para a colonização de áreas abertas (Barrat-Segretain & Bornette, 2000).
Em algumas espécies de plantas aquáticas, os fragmentos flutuam por um longo
período de tempo podendo, depois disso, afundar no sedimento e desenvolver novas plantas.
No entanto para outras, os fragmentos só se estabelecem quando as águas são rasas, sendo a
propagação periférica, por meio de rizomas, a melhor forma de colonizar novas áreas (Barrat-
Segretain et al., 1998). Os fragmentos de P. illinoensis flutuantes na superfície da água,
observados no rio Baía Bonita, possuíam brotos na região dos nós e, saindo da base desses

47
brotos, raízes, mesmo nos caule apresentando sinais de deterioração. Estes fatos evidenciam
que ramos desprendidos ou quebrados podem se regenerar e dar origem a um novo individuo,
contribuindo para a rápida colonização de novas áreas por esta espécie. A habilidade dos
fragmentos de flutuar pode contribuir para a dispersão e estabelecimento dos propágulos
longe da planta-mãe, aumentando a probabilidade de encontrar um local favorável para o
estabelecimento de novos indivíduos (Barrat-Segretain et al., 1998).
Barrat-Segretain et al. (1998), estudando regeneração de diversas espécies submersas,
verificaram que fragmentos da espécie Potamogeton pusillus L. não desenvolveram
propágulos ou raízes, no entanto fragmentos desta espécie já foram descritos como sendo
hábeis para rebrotar (Kadono, 1984 apud Barrat-Segretain et al., 1998). Portanto, os autores
enfatizam que provavelmente os fragmentos utilizados no experimento não tinham maturação
suficiente para formar novos brotos, visto que a espécie foi coletada no início da estação de
crescimento. Estas informações poderiam explicar a maior potencialidade de regeneração nos
fragmentos de 9º e 10º entrenó de P. illinoenses, que apresentaram 100% de rebrota,
contrastando com a ausência de regeneração nos ramos próximos ao meristema apical,
corroborando o descrito por Barrat-Segretain et al. (1998).
O tipo de sedimento também pode influenciar no estabelecimento dos fragmentos de
plantas em novas áreas, sendo demonstrado que indivíduos de Potamogeton crispus L.
apresentaram crescimento mais rápido e plasticidade fenotípica em locais com sedimentos
mais ricos em nutrientes como argila (Zhou et al., 2006). No rio Sucuri, onde há maior
velocidade de correnteza e o sedimento predominante são areia e seixos calcários, a
distribuição da população de P. illinoensis é mais esparsa, contrastando com o rio Baía Bonita
que possui como principal tipo de sedimento a argila, rica em nutrientes. Neste rio a
população de P. illinoensis é muito mais densa, sendo espécie dominante neste ambiente.

48
É provável que o desequilíbrio na densidade populacional de P. illinoensis tenha sido
facilitada pela riqueza de nutrientes do substrato, no entanto o manejo inadequado daquele
local pode constituir o principal fator, haja vista as constantes podas – naturais ou artificiais –
as quais a população é submetida.
Além disso, P. illinoensis possui acentuado potencial de reprodução vegetativa por
meio da emissão de rizomas, característica mencionada por vários autores como sendo
comum para as plantas aquáticas em geral, além de outras estruturas vegetativas como os
estolhos, tubérculos e botões de inverno (Barrat-Segretain, 1996, Cronk & Frenessy, 2001),
presentes em várias espécies aquáticas. Propagação por rizomas é relacionada para quase
todas as espécies do gênero Potamogeton (Wiegleb & Kaplan, 1998), sendo de particular
importância para P. illinoensis (Bezic et al., 1996). Espécies do gênero Potamogeton podem
potencialmente produzir sementes e rizomas (Combroux & Bornette, 2004), além de outras
formas de reprodução. Estas formas de reprodução estão presentes em P. illinoensis, no
entanto alguns autores sugerem que o ambiente aquático inibe a reprodução sexual,
favorecendo a dependência da reprodução vegetativa (Sculthorpe, 1967), e afirmam que os
métodos de reprodução clonal presente em espécies aquáticas são altamente efetivos (Grace,
1993). Contudo, a emissão de grande quantidade de escapos florais é comum nos indivíduos
estudados, excetuando naqueles de correnteza, cujo meristema apical encontra-se totalmente
submerso.
Considerando as condições ecológicas dos ambientes aquáticos, o crescimento clonal e
a fragmentação parecem ser métodos bem adequados de reprodução, visto que mantêm uma
forma de crescimento altamente competitivo e de rápida propagação das plantas aquáticas,
dentro da população (Wijesinghe & Handel, 1994, Santamaría, 2002). Além disso, este tipo
de reprodução é muito eficiente, consistindo em adaptação importante para as espécies que
habitam locais onde as alterações de habitats são imprevisíveis. Espécies com amplitude

49
ecológica estreita tendem a promover dispersão de propágulos aumentando a oportunidade de
se estabelecer em locais favoráveis e, aquelas mais tolerantes às alterações das condições
ecológicas, ancoram mais eficientemente em locais mais próximos (Barrat-Segretain, 1998).
No entanto a sobrevivência, regeneração e colonização dependem do tipo de
fragmento e da época em que a fragmentação ocorre (Barrat-Segretain & Bornette, 2000). Os
fragmentos vegetativos de macrófitas aquáticas podem ter sucesso na colonização de
ambientes de maneira aleatória, se estabelecendo nas proximidades das manchas, ou pela
propagação lateral de manchas para áreas abertas adjacentes, ou ainda por tipos intermediários
(Campbell, 2003). Devido as condições do ambiente a colonização ocorre pela propagação
periférica (Nielsen et al., 2006), denominada de efeito de borda (Barrat-Segretain, 1998),
apesar de muitas macrófitas aquáticas possuírem propágulos vegetativos hábeis para se
estabelecer e originar novas plantas (Nielsen et al., 2006). Em P. illinoensis foi observado que
os rizomas potencialmente colonizam áreas periféricas da população e, apesar de ser citada
para esta espécie a presença de estruturas especializadas que potencialmente produzem novos
rizomas após alterações ambientais (Dall Armellina et al., 1996a; Bezic et al., 1996), estas
estruturas não foram observadas nos indivíduos presentes nos rios estudados, provavelmente
devido à uniformidade do ambiente e a ausência, durante o período de estudo, de qualquer
evento sazonal que impusesse restrições ambientais às populações.
Pode-se afirmar que P. illinoensis está adaptado a distintas condições de velocidade da
corrente e natureza do substrato estabelecendo indivíduos saudáveis em locais de menor ou
maior velocidade da corrente, em substrato arenoso ou argiloso. No entanto, indivíduos desta
espécie podem ocorrer de maneira esparsa em rios com substrato arenoso e velocidade de
fluxo maiores, como é o caso do rio Sucuri, ou em grandes adensamentos como no rio Baía
Bonita, que possui águas mais calmas e substrato argiloso.

50
Potamogeton illinoensis é uma espécie invasora de canais de irrigação da Argentina
(Dall’Armellina et al., 1996a) e outros ambientes aquáticos. Considerando que esta espécie é
tolerante a locais com altas perturbações, associados ao controle mecânico regular, é possível
que o manejo mantenha a dominância desta espécie nos canais de irrigação da Argentina
(Sabbatini et al., 1998). Neste sentido, a ação antrópica resultante da exploração econômica
por meio do ecoturismo no rio Baía Bonita, bem como os reflexos da atividade agropecuária
desenvolvida nas áreas de entorno, sem práticas adequadas de manejo de solo e água, podem
estar propiciando o desenvolvimento extensivo de Potamogeton illinoensis.
O entendimento da biologia das plantas aquáticas é importante para propor a resolução
de problemas relacionados ao manejo de plantas invasoras de ecossistemas aquáticos
(Nichols, 1991), sendo evidente que a redução da biomassa é mais efetiva quando o controle
da população é feito após a estação de crescimento da planta (Dall’Armellina et al., 1996b).
No entanto, em ambientes aquáticos naturais extremamente frágeis, como é o caso do rio Baía
Bonita, os métodos para controle de plantas aquáticas devem ser muito bem analisados, e
todos os possíveis impactos devem ser mensurados. A principal meta ao se propor o manejo
deve ser produzir comunidades de plantas aquáticas estáveis, diversa e com alta porcentagem
de espécies desejáveis (Nichols, 1991).
O potencial invasor apresentado por P. illinoensis em outros ambientes da América do
Sul e por outras espécies do gênero, invasoras de ambientes aquáticos, bem como as
características relacionadas à densidade de indivíduos da população presente no rio Baia
Bonita despertam preocupação e o controle de crescimento desta espécie será necessário.
Qualquer alteração na integridade daquele ambiente colocará em risco a biodiversidade e,
conseqüentemente, a atividade de ecoturismo desenvolvida no local.
Os resultados apresentados demonstraram que esta espécie tem alto potencial de
reprodução vegetativa o que facilita o rápido desenvolvimento e colonização de novas áreas,

51
influenciando nos usos da água para recreação, sendo que em alguns trechos do rio Baia
Bonita, é dominante, impedindo o desenvolvimento de outras espécies vegetais. Até o
desenvolvimento deste trabalho, não se tinha conhecimento da potencialidade reprodutiva dos
propágulos não especializados, resultante das podas naturais ou pela ação antrópica. Sendo
assim, nossos resultados apontam para o cuidado necessário quanto ao manejo desta espécie,
bem como para a necessidade de estudos complementares quanto aos períodos de maior
crescimento dos indivíduos.
Agradecimentos
Agradecimento especial à Fundação de Apoio ao Desenvolvimento do Ensino, Ciência
e Tecnologia do estado de Mato Grosso do Sul (Fundect), pela concessão da bolsa de
mestrado à primeira autora e pelo financiamento de projeto. Aos proprietários e funcionários
das Fazendas São Geraldo (Rio Sucuri) e Baía Bonita pelo apoio logístico oferecido. E a Dra.
Maria Rosangela Sigrist, coordenadora do Projeto Gomphrena elegans pelo apoio logístico na
fase inicial deste trabalho.
Referências
Barrat-Segretain, M. H. 1996. Strategies of reproduction, dispersion, and competition in river
plants: A review. Vegetatio. 123, 13-17.
Barrat-Segretain, M. H., Bornette, G., Hering-Vilas-Boas, A. 1998. Comparative abilities of
vegetative regeneration among aquatic plants growing in disturbed habitats. Aquatic Botany.
60, 201-211.

52
Barrat-Segretain, M. H. & Bornette, G. 2000. Regeneration and colonization abilities of
aquatic plant fragments: effect of disturbance seasonality. Hydrobiologia 421: 31-39p.
Barrat-Segretain, M. H. 2001. Biomass allocation in three macrophyte species in relation to
the disturbance level of their habitat. Freshwater Biology. 41, 935-945.
Bezic, C., Dall’Armellina, A., Gajardo, O. 1996. Studies on vegetative production of
Potamogeton illinoensis Morong in southern Argentina. Hydrobiologia. 340, 7-10.
Boeger, M. R. T. & Poulson, M. E. 2003. Morphological adaptations and photosynthetic rates
of amphibious Veronica anagallis-aquatica L. (Scrophulariaceae) under different flow
regimes. Aquatic Botany. 75, 123-135.
Campbell, M. L. 2003. Recruitment and colonisation of vegetative fragments of Posidonia
australis and Posidonia coriacea. Aquatic Botany. 76, 175-184.
Chambers, P. A.; Prepas, E. E.; Hamilton, H. /r.; Bothwell, M. L. 1991. Current velocity and
its effect on aquatic macrophytes in flowing waters. Ecological Applications. 1, 249-257.
Chambers, P. A.; De Wreede, R. E.; Irlandi, E. A.; Vandermenhen, H. 1999. Manegement
issues in aquatic macrophyte ecology: a Canadian perspective. Can. J. Bot., 77, 471-487.
Combroux, I. C. S. & Bornette, G. 2004. Propagule banks and regenerative strategies of
aquatic plants. Journal of Vegetation Science. 15, 13-20.

53
Cronk, J. K. & Fenessy M. S. 2001. Wetland Plants: Biology and Ecology. Ed. Lewis
Publishers, 462p.
Dall” Armellina, A., Bezic, C. R., Gajardo, O. A. 1996a. Propagation and Mechanical Control
of Potamogeton illinoensis Morong in Irrigation Canals in Argentina. J. Aquat. Planta
Manage. 34, 12-16.
Dall” Armellina, A., Gajardo, A., Bezic, C., Luna, E., Britto, A., Dall’Armellina, V. 1996b.
mechanical aquatic weed management in the lower valley of the Río Negro, Argentina.
Hydrobiologia. 340, 225-228.
Gantes, H. P. & Caro, A. S. 2001. Environmental heterogeneity and spatial distribution of
macrophytes in plain streams. Aquatic botany. 70, 225-236.
Grace, J. B. 1993. The adaptative significance of clonal reproduction in angiosperms: an
aquatic perspective. Aquatic Botany. 44, 159-180.
Herb, W. R. & Stefan, H. G. 2006. Seasonal growth of submersed macrophytes in lakes: The
effects of biomass density and light competition. Ecological Modelling. 193: 560-574.
Idestam-Almquist & Kautsky, L. 1995. Plastic responses in morphology of Potamogeton
pectinatus L. to sediment and above-sediment conditions at two sites in the northern Baltic
proper. Aquatic Botany. 52, 205-216.

54
Leeflang, L., During, H. J., Werger, M. J. A. 1998. The role of petioles in light acquisition by
Hydrocotyle vulgaris L. in a vertical light gradient. Oecologia. 117, 235-238.
Madsen, J. D., Chambers P. A., James, W. F., Koch, E. W., Westlake, D. F. 2001. The
interaction between water movement, sediment dynamics and submersed macrophytes.
Hydrobiologia. 444, 71-84.
Nichols, S. A. 1991. The interaction between biology and the management of aquatic
macrophytes. Aquatic Botany. 41, 225-252.
Nielsen, U. N., Riis, T., Brix, H. 2006. The importance of vegetative and sexual dispersal of
Luronium natans. Aquatic Botany. 84, 165-170.
Power, P. 1996. Effects of current velocity and substrate composition on growth of Texas
wildrice (Zizania texana). Aquatic Botany. 55, 199-204.
Puijalon, S. & Bornette, G. 2004. Morphological variation of two taxonomically distant plant
species along a natural flow velocity gradient. New Phytologist. 163, 651-660.
Puijalon, S.; Bornette, G.; Sagnes, P. 2005. Adaptations to increasing hydraulic stress:
morphology, hydrodynamics and fitness of two higher aquatic plant species. Journal of
Experimental Botany. 56, 777-786.
Sabbatini, M. R., Murphy, K. J., Irigoyen, J. H. 1998. Vegetation-environment relationships
in irrigation Channel systems of southern Argentina. Aquatic Botany. 60, 119-133.

55
Santamaría, L. 2002. Why are most aquatic plants widely distributed? Dispersal, clonal
growth and small-scale heterogeneity in a stressful environment. Acta Oecologica. 23, 137-
154.
Schutten, J. & Davy, A. J. 2000. Predicting the hydraulic forces on submerged macrophytes
from current velocity, biomass and morphology. Oecologia. 123, 445-452.
Scremin-Dias, E., Pott, V. J. , Hora, R. C., Souza, P. R. 1999. Nos Jardins Submersos da
Bodoquena. Guia para identificação das plantas aquáticas de Bonito e região. Campo
Grande/MS. Editora UFMS. 160p.
Sculthorpe, C. D. 1967. The Biology of aquatic vascular plants. Eduard Arnold Publ. London.
Strand, J. A. & Weisner, S. E. B. 2001. Morphological plastic responses to wather depth and
wave exposure in na aquatic plant ( Myriophyllum spicatum). Journal of ecology. 89, 166-
175.
Usherwood, J. R., Ennos, A. R., Ball, D. J. 1997. Mechanical and anatomical adaptations in
terrestrial and aquatic buttercups to their respective environments. Journal of Experimental
Botany. 48, 1469-1475.
Wiegleb, G. & Kaplan, Z. 1998. An Account of the species of Potamogeton L.
(Potamogetonaceae). Folia Geobotanica, Upssala. 33, 241-216.

56
Wijesinghe, D. K. & Handel, S. N. 1994. Advantages of clonal growth in heterogeneous
habitats: An experiment with Potentilla simplex. Journal of Ecology. 82, 495-502.
Zhou, C., An, S., Jiang, J., Yin, D., Wang, Z., Fang, C., Sun Z., Qian, C. 2006. An in vitro
propagation protocol of two submerged macrophytes for lake revegetation in east China.
Aquatic Botany. 85, 44-52.

57
Tabela 1. Valores médios para a biometria de P. illinoensis. Os valores seguidos de letras iguais não diferem significativamente entre si em linhas. Tukey < 0,05. Rio Baía Bonita Rio Sucuri
Correnteza
Remanso
Correnteza
Remanso
Comprimento da folha 9,54 a 9,63 a 9,24 a 10,35 a
Largura da folha 1,5 b 1,8 a 2,06 a 2,27 a
Comprimento do pecíolo 0,87 c 0,93 c,b 1,49 b,a 1,93 a
Comprimento total do indivíduo
53,92 a 89,93 a 78,92 a 87,78 a

58
Ra
F
C
D E
GFigura 1. Detalhe dos indivíduos de . izoma com raízes adventícias e, desenvolvendo-se a partir dos nós ntrenós de caules flutuantes provenientes do experimento, indicando brotações nos nós. amos coletados sobre a superfície da água no rio Baía Bonita, mostra brotações dos fragmentos (setas). Aspecto geral do fragmento de caule flutuante de , ainda com ápice caulinar, indicando as brotações nos nós basais. Detalhe do fragmento do caule flutuante, com regeneração mesmo apresentando sinais de deterioração, evidenciando a presença de raiz adventícia na base do caule . Aspecto geral do rio Baía Bonita com população de ramos desprendidos (setas) na superfície da água.
Potamogeton illinoensis
P. illinoensis
P. illinoensis
Fig. 1A e 1B - Rcaules verticais flutuantes . Fig. 1C - E
Fig. 1D - Rndo
Fig. 1E - Figura 1F -
(seta) Fig. 1G - , e
A B

59
_______________________________________________________________ CONSIDERAÇÕES FINAIS

60
Duas espécies de Potamogeton illinoensis Morong foram encontradas nos rios de
Bonito, Mato Grosso do Sul – P. illinoensis ocorrendo nos rios Sucuri e Baía Bonita, e
Potamogeton gayi A. Benn. ocorrendo somente neste último. P. illinoensis, é uma espécie
bem mais agressiva que P. gayi podendo formar densa vegetação submersa nestes rios, sendo
espécie dominante no rio Baia Bonita, contrastando com P. gayi, que distribui-se em
pequenas manchas de plantas, vicejando em equilíbrio.
As duas espécies possuem anatomia semelhante, bastante simplificada, podendo ser
relacionado alguns dados consistentes para diferenciá-las anatomicamente:
• Distinta quantidade de fibras no bordo foliar, visivelmente maior em P. gayi e
presença de feixes corticais com floema e fibras somente em P. illinoensis.
• Presença de grupos de fibras subepidérmicas nas folhas, rizoma e caule
flutuante somente em P. gayi.
• Endoderme dos caules de parede celular pouco espessada e lignificada em P.
gayi, contrastando com espessamento parietal e lignificação maiores em P.
illinoensis.
• Estelo com até quatro lacunas de protoxilema em P. gayi e de oito a quatorze
lacunas em P. illinoensis.
O ambiente em que P. illinoensis se desenvolve – remanso ou correnteza -, influencia
o desenvolvimento e densidade dos indivíduos, mesmo que para alguns fatores amostrados, os
dados biométricos não foram significativamente diferentes, como esperado, os indivíduos de
correnteza são sempre menores em relação aos de remanso.
Verificou-se de maneira geral, em relação inversamente proporcional, entre a
velocidade do fluxo da corrente e a densidade dos indivíduos submersos, que quanto maior a
correnteza, mais esparsas são as manchas de vegetação submersa tendo, o rio Baía Bonita,

61
velocidades menores de correnteza, e a população de P. illinoensis mais densamente
distribuída. Por esta espécie se reproduzir vegetativamente emitindo rizomas, há grande
facilidade para a colonização de áreas adjacentes a população, e a expansão rápida da mancha
de plantas. Esta característica, aliada a riqueza do substrato presente naquele rio, são as
prováveis causas do aumento da densidade de indivíduos e da extensão da população.
Além disso, muitos propágulos vegetativos não especializados são produzidos por
meio da fragmentação dos caules flutuantes – naturalmente ou pela ação antrópica -, que são
hábeis para regenerar e formar novas plantas. Aparentemente, grande parte dos caules
depositados sobre a superfície da água, no rio Baia Bonita, foram retirados com objeto
cortante, haja vista a homogeneidade do tamanho dos fragmentos. A confirmação parcial
desta informação foi dada por guias que atuam no local. Caso esta forma de manejo esteja
sendo efetuada, não houve utilização de estudos prévios sobre a biologia desta espécie, nem
do comportamento das populações de P. illinoensis em relação a sazonalidade.
O uso inconsciente do ecossistema, sem o manejo adequado, tanto do rio quanto das
áreas de entorno, favoreceu o crescimento exagerado desta planta no rio Baía Bonita,
tornando esta espécie um problema, tanto para a diversidade da flora quanto para a
comunidade de peixes que habitam o local. Isto pode refletir na atividade econômica, o
ecoturismo, desenvolvido naquele rio.
Este trabalho corrobora os desenvolvidos com outras espécies de plantas aquáticas,
quanto a necessidade do conhecimento da biologia da espécie, bem como do comportamento
destas às variações ambientais, quando se pretende realizar o manejo de uma espécie. Os
estudos sobre o ciclo de vida de P. illinoensis, detectando o período de maior crescimento
vegetativo, deverá ser o próximo passo para as pesquisas que subsidiarão o manejo da
população do rio Baia Bonita, sem causar maiores impactos para o ambiente.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo