resistÊncia de cafeeiros À mancha- aureolada: aspectos
TRANSCRIPT

TESE
RESISTÊNCIA DE CAFEEIROS À MANCHA-
AUREOLADA: aspectos morfoanatômicos,
fitopatológicos, genéticos e moleculares
LUCAS MATEUS RIVERO RODRIGUES
Campinas, SP
2015

INSTITUTO AGRONÔMICO
CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
RESISTÊNCIA DE CAFEEIROS À MANCHA-
AUREOLADA: aspectos morfoanatômicos,
fitopatológicos, genéticos e moleculares
LUCAS MATEUS RIVERO RODRIGUES
Orientador: Oliveiro Guerreiro Filho
Campinas, SP
Dezembro de 2015
Tese submetida como requisito parcial para
obtenção do grau de Doutor em Agricultura
Tropical e Subtropical, Área de Concentração
em Genética, Melhoramento Vegetal e
Biotecnologia.

ii
FICHA ELABORADA PELA BIBLIOTECÁRIA DO NÚCLEO DE INFORMAÇÃO
E DOCUMENTAÇÃO DO INSTITUTO AGRONÔMICO - IAC

iii

iv
A meus pais Júlio e Sueli,
DEDICO.

v
AGRADECIMENTOS
Ao Dr. Oliveiro Guerreiro Filho, pela oportunidade e orientação para
realização deste estudo, além de transmitir valioso conhecimento sobre o melhoramento
de plantas, principalmente do cafeeiro.
A meu pai Dr. Júlio Rodrigues Neto, pelo grande incentivo e contribuições
dentro e fora do laboratório, que auxiliaram na realização desta tese.
Aos pesquisadores do Instituto Biológico Msc. Irene Maria Gatti da Almeida,
Dr. Luis Otávio Saggion Beriam e Dra. Flávia Rodrigues Alves Patrício, pelos
ensinamentos transmitidos, incentivo às pesquisas e amizade.
Aos pesquisadores do Instituto Agronômico Msc. Masako Toma Braghini e
Dr. Luiz Carlos Fazuoli, pela auxílio na realização desta tese e pelos valiosos
conhecimentos transmitidos.
À Dra. Rachel Benetti Queiroz-Voltan, pelo auxilo nos estudos de
histopatologia.
À Dra. Mirian Perez Maluf, pela orientação nos experimentos de biologia
molecular.
À Pós-graduação do IAC, pela oportunidade de realização do curso de
doutorado.
À CAPES, pela concessão da bolsa de estudos.
À Dra. Suzete Aparecida Lanza Destéfano, Curadora da Coleção de Culturas
de Fitobactérias do Instituto Biológico, pelo fornecimento das culturas bacterianas
utilizadas neste estudo.
Ao pesquisador do Laboratório de Ciências de Plantas Daninhas, do Instituto
Biológico, Dr. Marcus B. Matallo, por ceder parte da estufa para realização das
inoculações.
A todos os funcionários do Centro de Café Alcides Carvalho, em especial á D.
Ivone e Vânia, pela auxílio na coleta de sementes e, Anderson, pela amizade e auxílio
em diversos procedimentos da pesquisa.
Aos amigos do curso de Pós-graduação em Agricultura Tropical e Subtropical,
Bárbhara, Juliana, Vinícius, Alex, Gustavo, Cristiane e André pela amizade e
colaborações indiretas à pesquisa.
Às amigas do Laboratório de Bacteriologia Vegetal, Karen, Renata, Lucilene,
Denise e Daniele pela contribuições indiretas ao trabalho.

vi
Às colegas do laboratório de Biologia Molecular Patrícia, Juliana, Paula,
Bruna e Lígia pela amizade e auxílio nos experimentos de expressão gênica.
A meus Irmãos Tatiana e Renê e a minha namorada Mariara, pelo contínuo
incentivo para realização deste trabalho.
Aos professores e funcionários do Curso de Pós-graduação em Agricultura
Tropical e Subtropical (IAC) pela colaboração e ensinamentos;
A todos que contribuíram de alguma forma para a realização desta tese.

vii
SUMÁRIO
LISTA DE ABREVIATURAS ....................................................................................... xv
RESUMO ..................................................................................................................... xvii
ABSTRACT .................................................................................................................. xix
1. INTRODUÇÃO ............................................................................................................ 1
2. REVISÃO DE LITERATURA .................................................................................... 3
2.1. Trajetória e aspectos socioeconômicos da cultura do café ........................................ 3
2.2. Mancha-aureolada-do-cafeeiro .................................................................................. 5
2.2.1. Origem da mancha-aureolada-do-cafeeio e condições para ocorrência ................. 5
2.2.2. Sintomatologia da mancha-aureolada ..................................................................... 7
2.2.2.1. Mudas de cafeeiro ................................................................................................ 7
2.2.2.2. Plantas adultas de cafeeiro ................................................................................... 7
2.2.2.3. Confusão da mancha-aureolada com outros distúrbios ....................................... 8
2.2.3. Distribuição geográfica da mancha-aureolada do cafeeiro ..................................... 9
2.2.4. Pseudomonas syringae pv. garcae ......................................................................... 9
2.2.5. Controle da mancha-aureolada-do-cafeeiro.......................................................... 12
2.2.6. Resistência genética de cafeeiros a Pseudomonas syringae pv. garcae............... 13
2.2.6.1. Métodos para avaliação da resistência à mancha-aureolada ............................. 14
2.3. Melhoramento genético de Coffea arabica ............................................................. 14
2.3.1. Classificação botânica, origem e características de Coffea arabica ..................... 14
2.3.2. Histórico, métodos e perspectivas ........................................................................ 15
2.4. Mecanismos de resistência de plantas a patógenos ................................................. 18
3. MATERIAL E MÉTODOS ........................................................................................ 25
3.1. Alterações morfológicas em tecidos foliares de cafeeiros infectados por
Pseudomonas syringae pv. garcae ................................................................................. 25
3.1.1. Material vegetal, inoculação e condições experimentais...................................... 25
3.1.2. Análises histopatológicas ..................................................................................... 26
3.2. Natureza e agressividade de isolados de Pseudomonas syringae pv. garcae ......... 26
3.2.1. Isolados bacterianos utilizados no estudo ............................................................. 26
3.2.1.1. Reativação dos isolados de Pseudomonas syringae pv. garcae ........................ 27
3.2.3. Agressividade de isolados de Pseudomonas syringae pv. garcae........................ 28
3.2.4. Produção e sensibilidade as bacteriocinas entre isolados de Pseudomonas
syringae pv. garcae ........................................................................................................ 30
3.3. Metodologia de inoculação de Pseudomonas syringae pv. garcae para seleção de
plantas de cafeeiros resistentes à mancha-aureolada ...................................................... 31
3.4. Identificação de fontes de resistência a Pseudomonas syringae pv. garcae em
Coffea spp. ...................................................................................................................... 35
3.4.1. Obtenção das mudas de cafeeiros ......................................................................... 35

viii
3.4.2. Avaliação da resistência do germoplasma de Coffea spp. a Pseudomonas syringae
pv. garcae ....................................................................................................................... 39
3.5. Análise do efeito pleiotrópico do alelo SH1 na resistência a Pseudomonas syringae
pv. garcae ....................................................................................................................... 40
3.5.1. Material vegetal de cafeeiros ................................................................................ 40
3.5.2. Avaliação quanto à resposta de cafeeiros à infecção por Pseudomonas syringae
pv. garcae e a Hemileia vastatrix ................................................................................... 40
3.6. Análise da expressão diferencial de genes possivelmente ligados à resistência a
Pseudomonas syringae pv. garcae ................................................................................. 42
3.6.1. Extração de RNA total das folhas de cafeeiro, tratamento com RNase e síntese de
cDNA .............................................................................................................................. 42
3.6.1.1. Extração de RNA total das folhas de cafeeiro ................................................... 42
3.6.1.2. Tratamento dos RNAs com DNase ................................................................... 43
3.6.1.3. Síntese de cDNAs .............................................................................................. 43
3.6.2. Análise da expressão relativa em tempo real de genes ligados á defesa vegetal .. 44
3.6.2.1. Genes normalizadores e/ou endógenos ............................................................. 44
3.6.2.2. Genes alvo ......................................................................................................... 44
3.6.2.3. Reações de qRT-PCR ........................................................................................ 45
3.6.2.4. Análise dos resultados ....................................................................................... 45
4. RESULTADOS .......................................................................................................... 46
4.1. Alterações morfológicas em tecidos foliares de cafeeiros infectados por
Pseudomonas syringae pv. garcae ................................................................................. 46
4.2. Agressividade de isolados de Pseudomonas syringae pv. garcae........................... 51
4.2.1. Produção e sensibilidade as bacteriocinas entre isolados de Pseudomonas
syringae pv. garcae ........................................................................................................ 53
4.3. Metodologia de inoculação de Pseudomonas syringae pv. garcae para seleção de
plantas de cafeeiros resistentes à mancha-aureolada ...................................................... 54
4.4. Identificação de fontes de resistência a Pseudomonas syringae pv. garcae em
Coffea spp. ...................................................................................................................... 60
4.5. Análise do efeito pleiotrópico do alelo SH1 na resistência a Pseudomonas syringae
pv. garcae ....................................................................................................................... 73
4.6. Análise da expressão diferencial de genes possivelmente ligados à resistência a
Pseudomonas syringae pv. garcae ................................................................................. 75
5. DISCUSSÃO .............................................................................................................. 79
5.1. Alterações morfológicas em tecidos foliares de cafeeiros infectados por
Pseudomonas syringae pv. garcae ................................................................................. 79
5.2. Agressividade de isolados de Pseudomonas syringae pv. garcae........................... 81
5.2.1. Produção e sensibilidade as bacteriocinas entre isolados de Pseudomonas
syringae pv. garcae ........................................................................................................ 83
5.3. Metodologia de inoculação de Pseudomonas syringae pv. garcae para seleção de
plantas de cafeeiros resistentes à mancha-aureolada ...................................................... 85

ix
5.4. Identificação de fontes de resistência a Pseudomonas syringae pv. garcae em
Coffea spp. ...................................................................................................................... 88
5.5. Análise do efeito pleiotrópico do alelo SH1 na resistência a Pseudomonas syringae
pv. garcae ....................................................................................................................... 93
5.6. Análise da expressão diferencial de genes possivelmente ligados à resistência a
Pseudomonas syringae pv. garcae ................................................................................. 94
6. CONCLUSÕES .......................................................................................................... 98
7. REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................... 99
8. ANEXOS .................................................................................................................. 114

x
ÍNDICE DE FIGURAS Pag.
Figura 1 - Lesões provocadas por Pseudomonas syringae pv. garcae. Lesões
anasarcadas circundadas por halo clorótico, típicas da mancha-
aureolada (A). Morte de ponteiro provocada pela bactéria (B)
Superbrotamento induzido após a necrose do ápice da muda (C)............ 7
Figura 2 - Modelo da sinalização dos sinais de JOHANSSON (2015) representa o
efeito cascata ocorrido após reconhecimento do patógeno em algumas
possível vias de defesa, iniciado pela ativação dos fluxos de cálcio,
MAPKs e, pela sinalização de espécies reativos de oxigênio (ROS),
fosfolipase, óxido nítrico (ON), etileno, jasmonato (JA) e ácido
salicílico (SA)........................................................................................... 21
Figura 3 - Ilustração dos métodos de inoculação de Pseudomonas syringae pv.
garcae em folhas de cafeeiros. Inoculação por aspersão (A); inoculação
com agulhas múltiplas (B)........................................................................ 32
Figura 4 - Método de inoculação de Pseudomonas syringae pv. garcae, em folhas
de cafeeiro por abrasão. Fricção da lixa contra a superfície abaxial da
folha (A); Ferimentos ocasionados pela inoculação e deposição da
suspensão bacteriana na superfície lesionada (B)..................................... 34
Figura 5 - Cortes paradérmicos mostrando mesofilo de folhas de cafeeiro
inoculadas com solução salina (A); Células adjacentes ao ponto de
inoculação (B); detalhes do parênquima paliçádico (C); parênquima
esponjoso (D); Seta indica o tecido de cicatrização. PI = ponto de
inoculação. (Barra = 25 µm).................................................................... 48
Figura 6 - Cortes paradérmicos de folhas de cafeeiro mostrando mesofilo no local
da inoculação com Pseudomonas syringae pv. garcae, 10 dias após a
inoculação (A); parênquima esponjoso com destruição parcial de
cloroplastos, bem como massas de bactérias no interior das células e
em espaços intercelulares (B); parênquima paliçádico (C); parênquima
esponjoso com massas bacterianas (D); massas bacterianas em células
do feixe vascular (E). Secção transversal da folha com células dos
feixes vasculares bloqueadas por bactérias e células do floema
desestruturadas (F). b = bactérias; PI = ponto de inoculação, PP =
parênquima paliçádico; PE = parênquima esponjoso (Barra = 25 µm).... 49
Figura 7 - Cortes paradérmicos de folhas de cafeeiro mostrando mesofilo no local
da inoculação com Pseudomonas syringae pv. garcae, 20 dias após a
inoculação. Células do mesofilo rompidas em locais distantes do ponto
de inoculação (A); sintomas externos acompanhando as nervuras (B);
destruição dos cloroplastos de células do parênquima esponjoso (C);
secção transversal da folha com hiperplasia no parênquima esponjoso
(D); massa bacteriana presente no espaço intercelular de células do
parênquima esponjoso (E); inclusões presentes em células do
parênquima esponjoso (F). b = bactérias; PI ponto de inoculação, PP =
parênquima paliçádico; PE = parênquima esponjoso. (Barra = 25 µm)...
50
Figura 8 - Insensibilidade bacteriocinogênica dos isolados de Pseudomonas
syringae pv. garcae testados. Isolados candidatos a produção de
bacteriocinas, i – IBSBF 65, ii – IBSBF 75, iii – IBSBF 1197, iv –
IBSBF 248; isolados indicadores de atividade: v – IBSBF 1293, vi –
IBSBF 2840, vii – IBSBF 3024 e viii – IBSBF 3051 (A) Isolados
candidatos a produção de bacteriocinas: i – IBSBF 1293, ii – IBSBF
2840, iii – IBSBF 3024, iv – IBSBF 3051; isolados indicadores de

xi
atividade: v – IBSBF 65, vi – IBSBF 75, vii – IBSBF 1197 e viii –
IBSBF 248 (B)..........................................................................................
54
Figura 9 - Sintoma da mancha-aureolada na cultivar Catuaí Vermelho IAC 144,
aos 14 dias após a inoculação, resultado da inoculação por aspersão...... 57
Figura 10 - Curva da evolução da mancha-aureolada em cafeeiros de espécies de
Coffea spp., avaliados por meio de escala de zero a cinco pontos e da
Área abaixo da curva de progresso da doença (AACPD).........................
62
Figura 11 - Sintomas observados aos 21 dias após a inoculação com Pseudomonas
syringae pv. garcae, em folhas de espécies de Coffea spp. pelo método
de abrasão. C. arabica cv. IAC 125 RN (A); C. congensis (B); C.
kapakata (C); C. eugenioides (D); C. salvatrix (E); C. stenophylla (F);
C. liberica var. passipagore (G); C. anthonyi (H); C. humilis (I); C.
heterocalyx (J); C. canephora (K); C. arabica cv. Catuaí Vermelho
IAC 81 (L)................................................................................................ 64
Figura 12 - Face abaxial de folha de cafeeiro da cultivar IAC 125 RN de Coffe
arabica, inoculada com Pseudomonas syringae pv. garcae pelo
método de abrasão, presença de colonização dos tecidos foliares
adjacentes aos ferimentos, ocasionados pela inoculação por abrasão...... 65
Figura 13 - Expressão relativa de genes associados à resposta de defesa, avaliados
em amostras foliares das cultivares IAC 125 RN e IPR 102 de Coffea
arabica inoculadas com Pseudomonas syringae pv. garcae e com
solução salina (controle). As barras representam desvios padrão para as
triplicatas independentes. C. arabica cv. IAC 125 RN x Solução salina
( ); C. arabica cv. IAC 125 RN x P. syringae pv. garcae ( ); C.
arabica cv. IPR 102 x Solução salina ( ); C. arabica cv. IPR 102 x
P. syringae pv. garcae ( ).................................................................... 77

xii
ÍNDICE DE TABELAS Pag.
Tabela 1 - Identificação, local de origem, ano de isolamento e planta
hospedeira dos isolados de Pseudomonas syringae pv. garcae e
relação dos experimentos em que foram utilizados.............................. 27
Tabela 2 - Germoplasma utilizado na comparação dos métodos das agulhas
múltiplas e por abrasão, para inoculação de Pseudomonas syringae
pv. garcae, em cafeeiros....................................................................... 34
Tabela 3 - Relação de espécies, cultivares, variedades, introduções da Etiópia,
híbridos, acessos, formas botânicas e mutações do BAG de cafeeiros
do IAC avaliados para resistência aos isolados de Pseudomonas
syringae pv garcae IBSBF 75 e IBSBF 1197...................................... 36
Tabela 4 - Cafeeiros derivados do cruzamento entre Coffea arabica e C.
eugenioides avaliados em relação à resistência de suas progênies à
mancha-aureolada................................................................................. 39
Tabela 5 - Sequência dos primers utilizados na reação de qRT-PCR................... 44
Tabela 6 - Valores médios das lesões (mm) em folhas do terceiro internódio de
cafeeiro da cultivar Mundo Novo IAC 376-4 causadas por
Pseudomonas syringae pv. garcae, aos 10 e 20 dias após a
inoculação............................................................................................. 46
Tabela 7 - Comprimento médio das lesões em centímetros aos 21 dias após a
inoculação (DAI) nas cultivares Mundo Novo IAC 376-4 e Bourbon
Amarelo IAC J 30 de Coffea arabica, inoculadas por punctura com
diferentes isolados de Pseudomonas syringae pv. garcae e
combinação de isolados e períodos de incubação (PI) calculados a
partir de sintomas observados em folhas do primeiro internódio em
dias após a inoculação (DAI)............................................................... 52
Tabela 8 - Médias, intervalos de confiança (IC), desvio padrão (DP) e
coeficiente de variação (CV) das variáveis escala de pontos (EP),
área abaixo da curva de progresso da doença (AACPD) e incidência,
provocada por Pseudomonas syringae pv. garcae inoculada pelos
métodos de aspersão e agulhas múltiplas, em diferentes pares de
folhas....................................................................................................
56
Tabela 9 - Média, intervalo de confiança (IC), desvio padrão (DP) e coeficiente
de variação (CV) em porcentagem das variáveis escala de pontos
(EP) e área abaixo da curva de progresso da doença (AACPD),
provocadas por Pseudomonas syringae pv. garcae inoculada pelos
métodos de agulhas múltiplas e abrasão, em germoplasma diverso de
Coffea arabica...................................................................................... 58
Tabela 10 - Estimativas das correlações de Spearman entre os valores da
AACPD aos 7, 14, 21, 35 e 42 dias após a inoculação (DAI) de
Pseudomonas syringae pv. garcae em folhas de onze genótipos de
cafeeiros, pelos métodos de agulhas múltiplas e
abrasão..................................................................................................
59
Tabela 11 - Frequência de cafeeiros resistentes (R), moderadamente resistentes
(MR) e suscetíveis (S) segundo escala de 0 a 5 pontos em Coffea
spp. em resposta a infecção por Pseudomonas syringae pv. garcae,
avaliadas no experimento 1..................................................................
60

xiii
Tabela 12 - Frequência de cafeeios resistentes (R), moderadamente resistentes
(MR) e suscetíveis (S) segundo a avaliação pela escala de 0-5
pontos, aos 42 dias após a inoculação com Pseudomonas syringae
pv. garcae, avaliadas no experimento 4...............................................
63
Tabela 13 - Total de cafeeiros inoculados com Pseudomonas syringae pv.
garcae, frequência de plantas resistentes (R), moderadamente
resistentes (MR) e suscetíveis (S) segundo a avaliação pela escala de
0 a 5 pontos, aos 42 dias após a inoculação e severidade média da
mancha-aureolada comparada pela área abaixo da curva de
progresso do doença (AACPD), provocadas pelas inoculações
artificiais em 22 introduções de Coffea arabica oriundas da Etiópia,
avaliadas no experimento dois.............................................................
67
Tabela 14 - Total de cafeeiros inoculados com Pseudomonas syringae pv.
garcae, frequência de plantas resistentes (R), moderadamente
resistentes (MR) e suscetíveis (S) segundo a avaliação pela escala de
0 a 5 pontos, aos 42 dias após a inoculação e severidade média da
mancha-aureolada comparada pela área abaixo da curva de
progresso do doença (AACPD), provocadas pelas inoculações
artificiais em 23 introduções de Coffea arabica oriundas da Etiópia,
avaliadas no experimento três.............................................................. 68
Tabela 15 - Número total de plantas e frequência de plantas resistentes (R),
moderadamente resistentes (MR) e suscetíveis (S) de progênies de
híbridos (Coffea eugenioides x C. arabica) segundo a avaliação pela
escala de 0 a 5 pontos aos 42 dias após a inoculação com
Pseudomonas syringae pv. garcae....................................................... 70
Tabela 16 - Número total de cafeeiros inoculados, frequência de plantas
resistentes (R), moderadamente resistentes (MR) e suscetíveis (S),
segundo avaliação pela escala de 0-5 pontos aos 42 dias após a
inoculação e área abaixo da curva de progresso da doença (AACPD)
do germoplasma avaliado em relação à infecção pela bactéria
Pseudomonas syringae pv. garcae.......................................................
71
Tabela 17 - Número de cafeeiros inoculados, frequência de plantas resistentes
(R), moderadamente resistentes (MR) e suscetíveis (S), segundo
avaliação pela escala de 0 a 5 pontos e, reação de seleções de Coffea
arabica inoculadas com Pseudomonas syringae pv. garcae, pela
média da área abaixo da curva de progresso da doença (AACPD)...... 72
Tabela 18 - Reação de progênies F2 dos cafeeiros H8089-2 e H8089-7, híbridos
entre a cultivar Catuaí Vermelho IAC 24 e a seleção Geisha IAC
1137-5, à infecção pela bactéria Pseudomonas syringae pv. garcae e
pelos isolados I/2015 (v2, v5) e II/2015 (v1, v2, v5) de Hemileia
vastatrix................................................................................................ 73
Tabela 19 - Número total de cafeeiros F2 de duas progênies do híbrido H8089
resistentes e suscetíveis a Pseudomonas syringae pv. garcae e ao
isolado I/2015 de Hemileia vastatrix....................................................
74
Tabela 20 - Estimativa de parâmetros relacionados à resposta de cafeeiros F2 de
duas progênies do híbrido H8089 à infecção pela bactéria
Pseudomonas syringae pv. garcae e pelo fungo Hemileia
vastatrix................................................................................................ 74

xiv
LISTA DE ANEXOS Pag.
Anexo 1 - Representação da escala de zero a cinco pontos (EP) utilizada como
referência nas avaliações da severidade da mancha-aureolada em
cafeeiros inoculadas com Pseudomonas syringae pv. garcae, pelos
métodos de agulhas múltiplas e abrasão..............................................
114
Anexo 2 - Comprimento médio das lesões (cm) aos 21 dias após a inoculação
(DAI) originados em folhas dos quatro primeiros internódios da
cultivar Bourbon Amarelo IAC J 30 de Coffea arabica, inoculadas
por punctura com diferentes isolados de Pseudomonas syringae pv.
garcae e uma combinação de isolados e períodos de incubação (PI)
calculados a partir da observação dos sintomas inicias da doença,
provocados pelas inoculações em DAI................................................
115
Anexo 3 - Levantamento de dados (1977-1991) referentes a produção de café
cereja no período entre 1977 e 1989, ocorrência/severidade da
ferrugem em 1980, vigor e maturação dos frutos das introduções
oriundas da Etiópia avaliadas para resistência a Pseudomonas
syringae pv. garcae..............................................................................
117

xv
LISTA DE ABREVIATURAS
AACPD = Área abaixo da curva de progresso da doença.
APX = Ascorbato peroxidase.
AS = Ácido salicílico.
BAG = Banco Ativo de Germoplasma.
C = Cova.
CAT = Catalase
Ct = Cycle threshold.
cv. = Cultivar.
CTAB = Cetiltrimetil amônio brometo.
CV = Coeficiente de variação.
DAI = Dias após a inoculação.
ddH2O DEPC = Água bi-destilada, tratada com DEPC.
DEPC = Dicarbonato de dietila (diethylpyrocarbonate).
DP = Desvio padrão (s2).
EDTA = Ácido etilenodiaminotetracético.
ET = Etileto.
GLR = Glutationa redutase
HAI = Horas após a inoculação.
HT = Híbrido de Timor
IAC = Instituto Agronomico de Campinas.
IAPAR = Instituto Agronomico do Paraná.
IC = Intervalo de confiança.
ICL = Isocitrato liase.
JA = Jasmonato.
L = Lote.
ON = Óxido nítrico.
P.A. = Produto Ativo.
POX = Peroxidase.

xvi
PVP = Polivinilpirrolidona.
qRT-PCR = PCR quantitativo em tempo real.
RSA = Resistência sistêmica adquirida.
SOD = Super óxido dismutase.
UFC = Unidades formadoras de colônia.
var. = Variedade.

xvii
Resistência de cafeeiros à mancha-aureolada: aspectos morfoanatômicos,
fitopatológicos, genéticos e moleculares
RESUMO
A mancha-aureolada, causada por Pseudomonas syringae pv. garcae, afeta cafezais dos
principais estados produtores do país, sendo considerada fator limitante do cultivo em
regiões que apresentam condições favoráveis à doença. Como subsídio ao programa de
melhoramento genético (PMG) de cafeeiros do Instituto Agronômico (IAC), objetivou-
se no presente trabalho definir uma estratégia para a seleção de cafeeiros resistentes à
mancha-aureolada. Inicialmente, estudou-se as alterações anatômicas causadas pela
bactéria em folhas de cafeeiro. Posteriormente, foi avaliada a agressividade de isolados
de P. syringae pv. garcae, de forma individual ou em mistura. Em seguida, comparou-
se técnicas de inoculação em mudas, por meio de aspersão, agulhas múltiplas e abrasão,
a fim de se padronizar um método eficiente para reprodução dos sintomas da doença. A
seguir, estudou-se a resistência dos acessos do Banco Ativo de Germoplasma (BAG) de
Coffea spp. à bactéria, incluindo-se espécies do gênero, híbridos inter e intraespecíficos,
introduções, formas botânicas, variedades e mutantes de C. arabica, e, seleções de
gerações avançadas do PMG. O efeito pleiotrópico do alelo SH1, possivelmente ligado a
resistência à bactéria, foi avaliado em germoplasma com resistência a isolados de
Hemileia vastatrix, portadores ou não do alelo v1 de virulência. Finalmente, estudos
sobre a expressão relativa dos genes associados à resistência a fitopatógenos EDS1,
PAD4, ascorbato peroxidase, catalase e superóxido dismutase, peroxidase, β-1,3-
glucanase, isocitrato liase e glutationa redutase foram conduzidos por meio da técnica
de qRT-PCR, utilizando-se genótipos contrastantes para a resistência (IAC 125 RN
suscetível e IPR 102 resistente). Análises histopatológicas mostraram que, no mesofilo
de folhas jovens, os sintomas causados pela bactéria correspondem ao dobro em releção
ao visualizados na epiderme, externamente. Também, a bactéria foi detectada nos feixes
vasculares, colonizando regiões afastadas do ponto de inoculação. Os testes de
patogenicidade revelaram grande variação em agressividade, sendo os isolados IBSBF
65, IBSBF 75, IBSBF 1197, IBSBF 1293 e IBSBF 2212 os mais agressivos, e os
isolados IBSBF 248, IBSBF 1372, IBSBF 2996, IBSBF 2999, IBSBF 3005, IBSBF
3019, IBSBF 3051 e IBSBF 3065 caracterizados pela menor agressividade. Análises

xviii
comparativas entre os métodos de inoculação, determinaram que a técnica de inoculação
por abrasão resultou em maior expressão dos sintomas da mancha-aureolada nas
condições testadas, sendo a mais indicada em experimentos para se avaliar a resistência
de cafeeiros à doença. Ainda, avaliações da severidade mostraram que as folhas do
primeiro internódio foram mais propícias para os experimentos. Análises de correlação
revelaram alta similaridade no ranqueamento dos genótipos, nas avaliações realizadas
no 14° dia após a inoculação (DAI), com avaliações aos 42 DAI, validando a
proposição de seleção precoce para a característica de resistência à bactéria. Quanto aos
acessos do BAG de Coffea do IAC, detectou-se resistência nas espécies C. salvatrix, C.
eugenioides C. stenophylla e C. canephora. Dentre as 45 introduções de C. arabica de
origem etíope avaliadas, apenas duas, ETP 3 e ETP 12 apresentaram 80% ou mais de
suas progênies suscetíveis à bactéria. Entretanto, uma única introdução (ETP 22)
apresentou 100% de plantas resistentes, ou seja, homozigota para a característica.
Confirmou-se alta resistência à P. syringae pv. garcae na cultivar IPR 102, e a
variedade Pálido Viridis de C. arabica foi caracterizada como resistente. Nas seleções
do PMG do IAC avaliadas, o genótipo H 13439-8-C1736 L109 foi o mais promissor,
com resistência moderada à bactéria em sua progênie. O estudo sobre o alelo SH1
revelou que este gene não atua na resistência à P. syringae pv. garcae. Finalmente, os
resultados das análises de expressão de genes ligados a defesa sugerem a possibilidade
da via do ácido salicílico ser ativado no processo de defesa. Espécies reativas de
oxigênio parecem ser outro mecanismo com potencial ligação na resposta de defesa da
cultivar IPR 102 a P. syringae pv. garcae, contudo, estudos adicionais deverão ser
realizados para confirmar estas hipóteses.
Palavras chave: melhoramento genético; Pseudomonas syringae pv. garcae; métodos
de inoculação; qRT-PCR.

xix
Resistance to bacterial halo blight on coffee: morphoanatomics, phytopathologic,
genetic and molecular aspects.
ABSTRACT
Bacterial halo blight (Pseudomonas syringae pv. garcae) affect coffee plantations on
the major producing states in Brazil and is considered a limiting factor of cultivation in
areas with favorable conditions to the pathogen. As support to the genetic breeding
program (GBP) of coffee in the Instituto Agronômico (IAC), the aim of this study was
to outline a strategy for the selection of resistant coffee to bacterial halo blight. Initially,
were studied the anatomical changes caused by the bacteria on coffee leaves.
Afterwards, aggressiveness of bacterial strains, inoculated individually or in
combination was evaluated. Subsequently, it was compared inoculation techniques in
plants by sprinkling, multiple needles and abrasion, in order to standardize an efficient
method to reproduce disease symptoms. Later, were studied the resistance of accesses
maintained in the IAC Active Germplasm Bank (AGB) of Coffea spp. to the bacterium,
including species of the genus, hybrids inter and intraspecific, introductions, botanical
forms, varieties and mutants of C. arabica, and selections of advanced GBP
generations. Pleiotropic effect of SH1 allele, possibly linked to resistance to the
pathogen was evaluated in the germplasm with resistance to Hemileia vastatrix strains,
with or without v1 virulence allele. Finally, studies on the expression of associated
genes with resistance to phytopathogens EDS1, PAD4, ascorbate peroxidase, catalase
and superoxide dismutase, peroxidase, β-1,3-glucanase, isocitrate lyase and glutathione
reductase were conducted through of qRT-PCR, using contrasting genotypes for
resistance (IAC 125 RN susceptible and IPR 102 resistant). Histopathological analyzes
showed that, in the mesophyll of young leaves, symptoms caused by the bacteria are
two times higher than the displayed in the epidermis externally. Also, the bacterium was
detected in the vascular bundles, colonizing distant regions from the inoculation site.
Pathogenicity tests revealed great variability of aggressiveness, being strains IBSBF 65,
IBSBF 75, IBSBF 1197, IBSBF 1293 and IBSBF 2212 most aggressive, and IBSBF
248, IBSBF 1372, IBSBF 2996, IBSBF 2999, IBSBF 3005, IBSBF 3019, IBSBF 3051 e
IBSBF 3065 strains, characterized by lower aggressiveness. Comparative analysis of
inoculation methods, established that abrasion inoculation technique results in higher

xx
expression of symptoms of bacterial halo blight, in the tested conditions and it is most
appropriated in resistance experiments of coffee to the disease. Also, the severity
evaluations showed that leaves of first internode were more favorable for the
experiments. Correlation analyzes revealed high similarity in genotypes ranking of
evaluations performed at 14th day after inoculation (DAI) with assessments at 42 DAI,
validating the use of early selection for resistance to Pseudomonas syringae pv. garcae.
Regarding Coffea AGB accesses of IAC, it was detected resistance in C. salvatrix, C.
eugenioides, C. stenophylla and C. canephora species. Among the 45 C. arabica
introductions from Ethiopia evaluated, only two, ETP 3 and ETP 12, showed 80% or
more of their progeny susceptible to the pathogen. However, only the introduction ETP
22 showed 100% resistant plants, i.e., homozygous for the resistence to the bacteria. It
was confirmed high resistance to P. syringae pv. garcae in the IPR 102 cultivar and,
Pálido Viridis variety of C. arabica was determined as resistant to bacterial halo bllight.
In the IAC GBP evaluated selections, the H 13439-8-C1736 genotype was the most
promising, with moderate resistance to bacteria in their progeny. Study of SH1 allele
revealed that this gene does not act in resistance to P. syringae pv. garcae. Finally,
results of gene expression analysis of resistance related genes suggested the possibility
of defense process active salicylic acid via. Reactive oxygen species appear to be
another mechanism in connection with potential to defense response of IPR 102 cultivar
to P. syringae pv. garcae, however, additional studies are required to confirm these
hypothesis.
Keywords: genetic breeding; Pseudomonas syringae pv. garcae; inoculation methods;
qRT-PCR.

1
1. INTRODUÇÃO
O café possui alto valor econômico, sendo uma das principais commodities
agrícola do Brasil, ficando atrás apenas do petróleo. É a segunda bebida mais
consumida do mundo e, quase toda sua produção é utilizada para tal fim. Os cafés
consumidos são oriundos principalmente de duas espécies, Coffea arabica e C.
canephora. Partes menores da produção são destinadas às indústrias farmacêutica,
alimentícia e de cosméticos.
O Brasil é o maior produtor mundial, e estima-se que a área cultivada seja de
aproximadamente dois milhões de hectares. No ano de 2014, a produção atingiu 45,3
milhões de sacas de café beneficiado, das quais aproximadamente 36 milhões foram
exportadas (CONAB, 2015).
Atualmente, a produção dos cafezais tem sido afetada principalmente pelo
déficit hídrico, má condução da lavoura, além de outros fatores, com destaque para a
ocorrência de pragas e doenças.
Dentre as doenças que afetam o cafeeiro, a mancha-aureolada, moléstia de
etiologia bacteriana, vem sendo comumente detectada em viveiros de mudas e, no
campo, sua maior ocorrência está associada aos períodos chuvosos do ano e/ou em
locais principalmente de altitute elevada com alta umidade e ventos fortes (SERA,
2001).
O patógeno, a bactéria Pseudomonas syringae pv. garcae, apresenta certa
facilidade de seleção para a resistência a produtos fitossanitários, como sais de cobre e
antibióticos (MOHAN, 1977; YAMADA et al., 2014b), portanto, em regiões favoráveis
a sua ocorrência, que apresentam alto potencial de inóculo, medidas culturais de manejo
como formação de quebra ventos e utilização de mudas sadias são necessárias.
A bactéria pode estar presente como epífita no limbo foliar ROBBS (1977;
1978) e/ou confundida com outros distúrbios e não ser detectada no momento do
plantio. Uma vez instalada no cafezal, a doença pode se manifestar nas folhas, causando
lesões necróticas, irregulares, de aspecto encharcado, normalmente circundado por halo
amarelado. Nos ramos novos, infecta a epiderme e provoca seca destes ramos. Há
indícios de que a bactéria coloniza o sistema vascular e causa sintomas de secas de
ramos e eventualmente, a morte das plantas afetadas.

2
Quando instalada no cafezal, a mancha-aureolada representa uma ameaça para
toda a lavoura, pois, é de difícil controle e, em condições favoráveis, causa surtos de
intensa severidade, causando prejuízos ligados diretamente à produção. Devido ao
aumento da ocorrência dessa doença do cafeeiro, há preocupação em incorporar genes
de resistência ao patógeno em cultivares comerciais de café, visando ao aumento da
produção e com características agronômicas e sensoriais de interesse.
A utilização de cultivares resistentes é o método de controle mais indicado por
apresentar diversas vantagens aos produtores, ao consumidor final, além de favorecer o
meio ambiente. Atualmente, apenas a cultivar IPR 102 foi selecionada como resistente a
P. syringae pv. garcae (ITO et al., 2002).
O melhoramento genético de cafeeiros vem sendo aprimorado no sentido de se
aliar alta produtividade, resistência a estresses bióticos e tolerância à abióticos e ainda,
melhores qualidades tecnológicas. Entretanto, carece de longos períodos de seleção até
o lançamento de cultivares com as características desejadas.
Para a seleção de indivíduos resistentes a P. syringae pv. garcae, é indispensável
a utilização de métodos capazes de diferenciar a reação de genótipos à bactéria. Assim,
inoculações artificiais fazem parte da estratégia. Diversas metodologias de inoculação
são rotineiramente utilizadas no melhoramento de plantas a fitobacterioses, tais como
corte com lâmina, punctura por agulhas múltiplas, aspersão, infiltração, entre outros
(COSTA et al., 1957; OLIVEIRA & ROMEIRO, 1990; ITHIRU et al., 2013). Para
tanto, é necessário conhecimento prévio da eficiência dos métodos, uma vez que,
técnicas diferentes podem resultar em reações distintas entre plantas ou genótipos
avaliados.
O banco ativo de germoplasma de Coffea conservado no Instituto (IAC) é uma
das principais reservas genéticas do gênero, reunindo diversos híbridos inter e
intraespecíficos e espécies de interesse para o melhoramento, em especial para a
resistência a doenças. Portanto, cada acesso mantido em bancos de germoplasma bem
como seus híbridos resultantes, são plantas únicas e necessitam do conhecimento de
suas características para serem aproveitadas.
Neste contexto, os objetivos do presente trabalho foram: i) analisar as alterações
anatômicas em folhas de cafeeiros infectadas pela bactéria; ii) identificar diversidade

3
patogênica de isolados de P. syringae pv. garcae; iii) determinar o melhor método de
inoculação de P. syringae pv. garcae em mudas de cafeeiros para seleção de plantas
resistentes; iv) identificar fontes de resistência a P. syringae pv. garcae em cafeeiros do
banco de germoplasma de Coffea do Instituto Agronômico; v) investigar o possível
efeito pleiotrópico do alelo SH1 presente em cafeeiros resistentes à ferrugem (Hemileia
vastatrix) na expressão da resistência à bactéria e, vi) estudar a expressão diferencial de
genes potencialmente envolvidos na interação Coffea spp. - P. syringae pv. garcae.
2. REVISÃO DE LITERATURA
2.1. Trajetória e aspectos socioeconômicos da cultura do café
Do conhecido até o momento, as primeiras observações referidas sobre as
propriedades estimulantes do café são atribuídas ao pastor etíope Kaldi, na região
conhecida como Abissínia, atual Etiópia, quando as cabras de seu rebanho ingeriam as
folhas e bagas da planta apresentavam comportamento “enfeitiçado”. Após a notícia se
espalhar pela região, os próprios etíopes provavelmente experimentaram diferentes
formas de consumo, incluindo a infusão de bagas de café (qishr), hoje conhecido como
kisher (PENDERGRAST, 2010).
Com o passar do tempo, a bebida bem como plantas de café chegaram às
arábias, ficando conhecido como qahwa, palavra árabe que significa vinho (MARTINS,
2008; PENDERGRAST, 2010).
Os primeiros relatos do cultivo datam do século VI, em um monastério
localizado na região do Yêmen, por causa do valor econômico e grande demanda pela
bebida. Logo a bebida comercializada ficou conhecida como “vinho das arábias”, e até
o século XVII era cultivado apenas pelos árabes. Os peregrinos muçulmanos levaram o
café para mundo islâmico, Pérsia, Egito, Turquia e África do Norte, aumentando o valor
comercial do café. Em 1616 foi levado à Holanda, e seu cultivo experimentado pelos
holandeses em 1699 (PENDERGRAST, 2010).
Em meados de 1706, uma planta de café foi levada de Java para o Jardim
Botânico de Amsterdan e posteriormente, uma progênie desta planta foi introduzida em
Paris. Com a expansão do cultivo, sementes e mudas de café foram levadas para as

4
colônias francesas chegando à América em 1718, através do Suriname, e em seguida
São Domingos, Cuba, Porto Rico e Guianas (PENDERGRAST, 2010).
No Brasil, a primeira muda foi conseguida pelo sargento-mor Francisco Mello
Palheta, em visita a Guiana Francesa em 1727, e plantada em Belém do Pará. A partir
daí cafezais foram instalados pelo Maranhão, Bahia, Rio de Janeiro, São Paulo, Paraná e
Minas Gerais. Em 1825 a cultura se estabeleceu na região do vale do Paraíba, em São
Paulo, iniciando um novo ciclo econômico para o país (MARTINS, 2008;
PENDERGRAST, 2010).
Atualmente, o Brasil é líder na produção e na exportação de grãos de café, sendo
responsável por aproximadamente um terço da produção mundial. A estimativa de
colheita em 2015 está entre 44,1 e 46,6 milhões de sacas (60kg) de café beneficiado
(CONAB, 2015).
O café é o quinto ítem agrícola mais exportado do País, sendo que na safra de
2014, a produção foi de aproximadamente 45 milhões de sacas, das quais 34,5 milhões
foram exportadas para União Europeia, Estados Unidos e Japão (CONAB, 2015). A
demanda por café no mundo é crescente, e, no período entre os anos 2011 e 2014, o
consumo passou de 139 milhões de sacas para 149 milhões (ICO, 2015).
Para a economia do país, a indústria de café gera mais de oito milhões de
empregos ligados à cadeia produtiva, que, por sua vez, propiciam acesso à educação e à
saúde para os trabalhadores e familiares (MAPA, 2015).
O parque nacional cafeeiro é composto por cultivares do grupo arábica e robusta,
com grande predominância do café arábica. O café robusta ou conilon (C. canephora) é
cultivado principalmente nos estados do Espírito Santo, Rondônia e Bahia,
representando 27% da safra brasileira de café (CONAB, 2014). Este tipo de café possui
menor valor econômico e gera bebida de qualidade típica da espécie mas, inferior à
arábica. No entanto, apresenta boas características para produção de cafés solúveis, e
por isso vem sendo utilizado em mistura com café arábica a fim de reduzir a acidez e
atribuir “corpo” à bebida. Além disso, seu custo de produção é menor em relação ao
café arábica, em virtude da rusticidade da espécie (MATIELLO, 1998).
O cafeeiro arábica (C. arabica) constitui praticamente 70% das plantações de
café do Brasil, sendo cultivado principalmente no Estado de Minas Gerais, cuja a safra

5
em 2014 foi de 22,7 milhões de sacas, seguido pelos estados de São Paulo, Espírito
Santo, Bahia e Paraná com 4,2; 2,8; 1,2 e 0,5 milhões de sacas, respectivamente
(CONAB, 2014).
2.2. Mancha-aureolada-do-cafeeiro
2.2.1. Origem da mancha-aureolada-do-cafeeio e condições para ocorrência
A maioria das doenças em espécies de plantas introduzidas no Brasil tiveram
origem junto ao material vegetal ou, posteriormente, por agentes de disseminação. No
caso da mancha-aureolada, sua origem permanece desconhecida, pois o cafeeiro tem
origem africana e, por ter sido trazido para o Brasil seria presumível que a introdução da
doença teria ocorrido por materiais vegetais oriundos do continente africano. Embora o
patógeno apresente a capacidade de ser disseminado via sementes (BELAN, 2014), os
poucos isolados de origem brasileira e queniana, comparados até hoje, exibiram
diferenças relevantes tanto em aspectos bioquímicos (KAIRU, 1997) quanto em
diversidade genética (MACIEL 2012).
Outra hipótese é que mutantes bacterianos poderiam ter se adaptado à cultura do
café, uma vez que a bactéria P. syringae pv. garcae apresenta similaridade patogênica
com P. savastanoi pv. phaseolicola (sin P. syringae pv. phaseolicola, patogênica às
plantas de feijão), P. syringae pv. tabaci (patogênica ao fumo e inúmeras outras
culturas) COSTA et al. (1957), ou ainda P. syringae pv. coronafaciens e P. syringae pv.
striafaciens, que possuem características morfológicas semelhantes e são patogênicas à
aveia, e que em uma classificação mais atual pertencem ao grupo 4 de genomospécies,
assim como P. syringae pv. garcae (BULL & KOIKE, 2015).
A primeira constatação da doença foi realizada por AMARAL, TEIXEIRA &
PINHEIRO (1956), no final do ano de 1955, em cafezal no município de Garça, Estado
de São Paulo. O patógeno causador foi denominado Pseudomonas garcae (AMARAL
et al., 1956), sendo posteriormente reclassificado para Pseudomonas syringae patovar
garcae (AMARAL et al., 1956; YOUNG et al., 1978), classificação esta que permanece
até os dias de hoje. Seu nome popular está associado a um de seus sintomas, na forma
de halo amarelado presente ao redor das lesões necróticas.

6
Após a confirmação da etiologia da doença, a mesma foi considerada de menor
importância econômica (COSTA et al., 1957) e, por aproximadamente 17 anos, apenas
casos isolados da bacteriose foram constatados (MOHAN, 1976). Nos anos de 1973 e
1975, relatou-se a ocorrência severa da doença no viveiro de mudas do Instituto
Agronômico (IAC). Também, em lavouras do Estado do Paraná, a doença foi observada
em cafezais que se recuperavam da ocorrência de geadas (KIMURA et al., 1973;
MOHAN, 1976).
CARDOSO & MOHAN (1980) relacionaram fatores climáticos com a
incidência da mancha-aureolada no Estado do Paraná. Os autores verificaram que o
aumento da intensidade da doença estava relacionado às baixas temperaturas associadas
com períodos de maior precipitação, assim como à ocorrência de chuvas e ventos.
Resultados de ensaios conduzidos por ROBBS (1979), indicaram a
predisposição de cafeeiros da espécie C. arabica à maior infecção por P. syringae pv.
garcae, quando expostos a temperaturas mais baixas. O autor observou alta severidade
da doença em mudas inoculadas por aspersão e mantidas por 60 minutos em condições
de baixa temperatura (2 a -2° C). Com a evolução da doença, ocorreu necrose de gemas
e seca de ramos. Nos tratamentos controles, mantidos em condições normais de
temperatura, a doença muito pouco se manifestou. Segundo o autor, baixas temperaturas
podem causar macro e micro lesões, que atuam como porta de entrada para o patógeno,
entretanto, a alta severidade desenvolvida pode ser também atribuída a uma
predisposição da planta, induzida por períodos de baixas temperaturas, como notado na
interação P. syringae – pessegueiros, por WEAVER (1978). A atividade de nucleação
de gelo, característica de algumas bactérias do agrupamento P. syringae, e também de
P. syringae pv. garcae (GONÇALVES & MASSAMBANI, 2011), pode estar
relacionada com este efeito, especialmente em plantas de café, cultura sensível aos
efeitos da geada.
Atualmente a mancha-aureolada é considerada fator limitante para o cultivo do
café em regiões de clima ameno e expostas a ventos, tendo alta incidência em lavouras
em formação ou recém-podadas e em viveiros (SERA et al., 2002; SERA et al., 2004;
PATRÍCIO et al., 2010; ZOCCOLI et al., 2011; ALMEIDA et al., 2012a), ou ainda, em
áreas com alto potencial de inóculo. Nessas condições, a doença pode apresentar alta
severidade, ocasionando queda de produtividade.

7
2.2.2. Sintomatologia da mancha-aureolada
2.2.2.1. Mudas de cafeeiro
Em mudas de cafeeiro cultivadas em telados, o adensamento e
consequentemente excesso de umidade nas folhas proporcionam ambiente ideal para o
desenvolvimento da mancha-aureolada. Os sintomas iniciais observados nas folhas se
manifestam sob a forma de pequenas lesões necróticas, irregulares, anasarcadas, de
coloração marrom escuro, que aumentam em tamanho, sendo posteriormente
circundadas por halos amarelados, típicos da doença, e que deram origem à
denominação dessa moléstia bacteriana (Figura 1A)
Com o desenvolvimento das lesões, a bactéria se dissemina pela própria planta e
para mudas adjacentes, podendo colonizar também tecidos do caule e do ápice,
causando necrose das folhas novas e do ponteiro (Figura 1B), também, pode ocasionar
superbrotamento (Figura 1C) e, muitas vezes com morte das mudas.
Embora P. syringae pv. garcae seja a bactéria predominante em viveiros, outras
bacterioses também podem incidir nas mudas de cafeeiro, porém a frequência com que
são detectadas é baixa. Em mudas doentes, a sintomatologia das diferentes bacterioses é
similar, podendo causar dúvida no diagnóstico presuntivo do agente causal da doença.
Figura 1 - Lesões provocadas por Pseudomonas syringae pv. garcae. Lesões
anasarcadas circundadas por halo clorótico, típicas da mancha-aureolada (A). Morte de
ponteiro provocada pela bactéria (B) Superbrotamento induzido após a necrose do ápice
da muda (C).
2.2.2.2. Plantas adultas de cafeeiro
Em condições de campo, a mancha-aureolada incide principalmente em lavouras
de café em formação, na faixa de 3-4 anos de idade e em cafezais reformados com
podas e/ou sujeitos a ventos constantes e frios, especialmente após longos períodos de
A B C

8
umidade elevada. Os sintomas característicos são lesões foliares necróticas, inicialmente
anasarcadas, de coloração marrom escuro, circundadas por halo amarelado, que podem
coalescer formando extensas áreas necrosadas. A doença também ocorre em rosetas,
frutos (frutos novos são mais suscetíveis) e ramos do cafeeiro, causando queda
prematura de folhas e seca das hastes e ramos. Na tentativa de se recuperar, as plantas
emítem novos ramos, originando sintomas de superbrotamento. Ao colonizar as hastes
do cafeeiro, a bactéria interfere no pegamento das flores, o que prejudica a produção das
plantas no ano seguinte. O período chuvoso está associado com o período de expansão
do fruto do café. A infecção que ocorre nesta fase, especialmente com seca dos ramos,
provoca a queda dos frutos, e causa danos na produção (COSTA & SILVA, 1960).
ROBBS (1977; 1978) mostrou que a bactéria pode sobreviver como epífita entre
os períodos favoráveis à ocorrência da mancha-aureolada e que esta é uma das fontes de
inóculo primário para a epidemia.
2.2.2.3. Confusão da mancha-aureolada com outros distúrbios
Sintomas de lesões foliares de coloração parda circundada por halo amarelo
podem ser causados por outros patógenos do cafeeiro, tais como os fungos Cercospora
coffeicola e Phoma tarda, responsáveis pela cercosporiose e mancha de phoma,
respectivamente. Outros sintomas decorrentes da infecção por P. syringae pv. garcae,
como seca de ramos e desfolha, podem também gerar confusão no diagnóstico, uma vez
que P. tarda, ou ainda distúrbios nutricionais e/ou climáticos, podem causar sintomas
semelhantes, sendo que em condições de campo poderá haver ocorrência simultânea
com a bactéria (KIMATI et al., 2005).
É importante ressaltar a ocorrência de outras doenças de etiologia bacteriana em
cafeeiros. As seguintes espécies bacterianas: Pseudomonas cichorii, causadora do
“crestamento-bacteriano-do-cafeeiro”, detectada em viveiros e campo no Estado de
Minas Gerais (ROBBS et al., 1974) e em viveiro no Estado de São Paulo (ALMEIDA et
al., 2012b); Burkholderia andropogonis (sin. P. andropogonis), causadora da “mancha-
escura-bacteriana” (RODRIGUES NETO et al., 1981), constatada em viveiro de mudas
no Estado de Santa Catarina e P. syringae pv. tabaci causadora da “mancha-bacteriana”
(RODRIGUES NETO et al., 2006; DESTÉFANO et al., 2010), detectada em viveiro no
Estado de São Paulo, também ocasionam sintomas de manchas foliares em viveiro e/ou
campo.

9
Pesquisas sugerem a ocorrência de patotipo similar a P. syringae pv. garcae em
condições de campo no Estado do Paraná (PETEK et al., 2006). Isolamentos efetuados
recentemente a partir de folhas com sintomas de mancha-aureolada provenientes do
Paraná parecem indicar a presença de novo patotipo de P. syringae em cultivos naquele
estado (dados não publicados). Este fato implica diretamente nas medidas de manejo do
patógeno, uma vez que não há estudos relativos a esse patossistema.
2.2.3. Distribuição geográfica da mancha-aureolada do cafeeiro
Com exceção do Brasil, não foram encontrados na literatura relatos da
ocorrência de P. syringae pv. garcae em outro país do continente Americano. A
mancha-aureolada foi oficialmente detectada nas regiões produtoras de café dos Estados
de São Paulo (AMARAL et al., 1956), Paraná (KIMURA et al., 1976; MOHAN et al.,
1977) e Minas Gerais (KIMURA et al., 1976) sobretudo nas regiões Sul, Triângulo
Mineiro, Alto do Paranaíba e Cerrados (MOHAN, 1976; SERA et al., 2002; SERA et
al., 2004; PATRÍCIO et al., 2010; ZOCCOLI et al., 2011).
Além do Brasil, a doença foi diagnosticada no Quênia (HAYWARD, 1962;
RAMOS & SHAVDIA, 1976), Etiópia (KOROBKO & WONDIMIGEGNE 1997;
ADUGNA et al., 2013), Uganda (CHEN, 2002) e China (CHEN, 2002; XUEHUI et al.,
2013).
2.2.4. Pseudomonas syringae pv. garcae
Esta espécie pertence ao filo Proteobacteria, classe Gamma Proteobacteria,
ordem Pseudomonadales, família Pseudomonadaceae, gênero Pseudomonas, espécie P.
syringae, patovar garcae. Os isolados produzem discreta quantidade de pigmento
fluorescente em meio de cultura King B (KB) (KING et al., 1954) e, em meios de
cultura como Batata Dextrose Agar (BDA) e Nutriente Ágar (NA), produzem pigmento
marrom (AMARAL et al., 1956), citado por BARTA & WILLIS (2005), como sendo
melanina. KIMURA et al. (1973) descreveram a capacidade de um isolado bacteriano
de sintetizar fluoresceina em meio de Clara.
P. syringae pv. garcae é Gram negativa, com células em forma bastonetes retos
ou levemente curvados, móveis por meio de um a sete flagelos polares e medidas entre
0,5-1,0 x 1,5-4,0 µm. Pertence ao grupo I das bactérias fluorescentes [LOPAT + - - - +

10
(L= produção de Levan, O= atividade de oxidase, P= proctopectinase em discos de
batata, A= utilização de arginina dihidrolase, T= hipersensibilidade em folhas de fumo)]
(Lelliott et al., 1966). Utiliza L-ascorbato, meso-inositol, manitol, D-sorbitol, triacetina
e D-xylose, porém não utiliza L-histidina, DL-homoserina, DL-lactato, α-lecitina,
linolenato, L-malate, D(-) tartarato ou L(+) tartarato (BRADBURY, 1986).
De acordo com COSTA et al. (1957) e BRADBURY (1986), as hospedeiras
naturais são plantas de diversas espécies do gênero Coffea. Sintomas foram obtidos por
meio de inoculação artificial em citrus (Citrus spp.), ligustro (Ligustrum lucidum),
tomateiro (Solanum lycopersicum), oliveira (Olea europea), feijoeiro (Phaseulus
vulgaris), jurubeba (Solanum paniculatum var. acutilobum), batata (Solanum
tuberosum) (BRADBURY, 1986). No entanto, cabe ressaltar os resultados de testes de
patogenicidade obtidos por KIMURA et al. (1973), no qual os autores contestaram a
patogenicidade de P. syringae pv. garcae em diversas hospedeiras, uma vez que os
sintomas por eles observados foram devidos à reação de hipersensibilidade. Por este
motivo, KIMURA et al. (1973) concluíram que a gama de hospedeiros da bactéria é
restrita a cafeeiros.
BARTA & WILLIS (2005), também por meio de inoculações artificiais,
obtiveram sintomas de P. syringae pv. garcae em aveia (Avena sativa). Entretanto, não
se tem registro de infecção natural em todas as espécies de plantas citadas.
RIEMKE et al. (1978) demonstraram que P. syringae pv. garcae é capaz de
utilizar nitrato de sódio, uréia e sulfato de amônio, na presença de sacarose como fonte
de carbono, por isso os autores concluíram que a bactéria possui potencial para provocar
danos ao metabolismo nitrogenado do cafeeiro e que adubações nitrogenadas via foliar
podem servir como substrato para o patógeno, bem como, melhorar as condições de
sobrevivência como epífita nas folhas de cafeeiros, fato demonstrado por ROBBS
(1978).
Com relação às características morfofisiológicas da bactéria, pesquisadores do
Quênia, país onde a doença está presente em muitos cafezais, demonstraram que os
isolados bacterianos lá obtidos diferem do isolado patotipo (brasileiro) em relação a
características bioquímicas, produção de bacteriocinas e quanto à virulência em C.
arabica cv. SL 28, na qual apenas os isolados quenianos foram capazes de produzir
sintomas da doença. Outra diferença importante observada está relacionada à produção

11
de pigmentos em meio de cultura, uma vez que os isolados quenianos produziram maior
quantidade de pigmento amarelo-esverdeado em meio KB quando observados sob luz
ultra violeta, quando comparados com isolados brasileiros, que produzem menores
quantidades de pigmento azulado. No entanto, isolados brasileiros produziram pigmento
marrom difusível em meio de cultura básicos (Amaral et al., 1956; KAIRU, 1997),
indicando a grande diversidade genética e a possibilidade da existência de variabilidade
intraespecíficas entre isolados dos dois países.
Pesquisas realizada por BARTA & WILLIS (2005) revelaram alta similaridade
nos padrões moleculares, obtidos por meio digestão do DNA total com enzima de
restrição, entre linhagens de P. syringae pv. coronafaciens, agente causal da “mancha-
de-halo-amarelo” em aveia (Avena sativa), P. syringae pv. striafaciens, causadora da
“mancha-estriada” aveia e P. syringae pv. garcae. Semelhanças também foram
observadas em testes fisiológicos para utilização de fontes de carbono aplicados as
patovares acima mencionadas e por teste de patogenicidade em aveia observadas por
BARTA & WILLIS (2005). A produção de tabtoxina (BENDER et al., 1999), foi
deficiente para a patovar striafaciens, entretanto devido à proximidades do conjunto de
dados obtidos, os autores propuseram que a taxonomia entre essas patovares seria
redundante, sendo elas um só patovar.
Sobre a infecção pela bactéria, estudos apontaram diferenças na agressividade
entre folhas novas (não expandidas) e velhas (a partir do primeiro par, completamente
expandidas). OLIVEIRA & ROMEIRO (1990) provaram por meio de inoculações
artificiais na cultivar Catuaí, que folhas velhas são menos afetadas pelo patógeno, fato
esse confirmado posteriormente por ZOCOLLI et al. (2011). Ensaios subsequentes de
difusão em gel, realizados por OLIVEIRA & ROMEIRO (1991), demonstraram que
extrato obtido a partir de folhas velhas foram capazes de inibir o crescimento in vitro de
P. syringae pv. garcae. A bactéria se multiplica dez vezes mais em folhas novas, em
comparação com as folhas velhas, na qual a fase lag de crescimento é mais curta
(OLIVEIRA et al., 1991).
Estudos realizados por MACIEL (2013) analisando a diversidade genética de P.
syringae pv. garcae de origem nacional e incluindo alguns isolados quenianos, por meio
da técnica molecular e de rep-PCR, resultou na formação de dois grupos distintos, com
relação à origem dos isolados. O grupo menor foi composto pelos isolados oriundos do
Quênia. Os isolados de origem brasileira formaram um grupo maior, composto por

12
subgrupos que alocaram as linhagens por região de origem, ou seja, São Paulo e Minas
Gerais já que isolados do Estado do Paraná não foram utilizados no estudo.
É importante também, ressaltar a ocorrência de mutantes bacterianos que
apresentaram resistência a produtos fitossanitários. A primeira constatação desse fato
foi realizada por MOHAN (1977). O autor verificou em isolamentos a partir de plantas
com sintomas oriundas de viveiro com aplicações de estreptomicina, colônias capazes
de crescerem na presença de 100 ppm do produto. Testes com concentrações maiores
revelaram alta resistência ao antibiótico, que cresceu em meio de cultura com o produto
na dosagem de 10.000 ppm.
Recentemente YAMADA et al. (2014b) avaliaram a tolerância de 55 isolados de
P. syringae pv. garcae provenientes do sul do Estado de Minas Gerais ao sulfato de
cobre (CuSO4). Cinco isolados bacterianos foram capazes de tolerar a maior
concentração utilizada (0,25 mM), enquanto que menos da metade dos isolados
cresceram na concentração de 0,15 mM e três isolados apresentaram alta sensibilidade
ao sulfato de cobre, desenvolvendo-se apenas na sua ausência. Também, YAMADA et
al. (2014a), por meio de PCR com primers específicos, identificaram no DNA
plasmidial o gene de resistência ao cobre cop A em 53 dos 55 isolados de P. syringae
pv. garcae testados. Estes resultados implicam na estratégia para o manejo da mancha-
aureolada, pois, a facilidade para seleção de genes de resistência ao cobre em
populações do patógeno, resultará na busca de novas formas de controle dessa doença.
2.2.5. Controle da mancha-aureolada-do-cafeeiro
Quando a doença é constatada no cafezal, seu controle torna-se bastante
complexo. Exemplo da dificuldade do manejo está descrito em trabalhos publicados por
COSTA & SILVA (1960). Os autores relataram ligeira recuperação das plantas em
cafezal altamente afetado, onde foram aplicados produtos contendo como princípio
ativo sais de cobre, embora tenham sido necessárias 24 pulverizações com intervalo de
cinco dias.
De acordo com PARADELA et al. (2000), para o controle da mancha-aureolada
em viveiros de mudas, devem ser realizadas aplicações quinzenais de produtos contendo
como o antibiótico cloridrato de casugamicina, na dosagem de 300 mL do produto
comerial com 2% do principio ativo (P.A.), para 100 L de água. As aplicações devem

13
ser intercaladas com oxicloreto de cobre (0,3%), evitando-se a seleção e proliferação de
bactérias resistentes ao antibiótico.
Experimentos conduzidos por PATRICIO et al. (2010), avaliaram o efeito de
tratamentos químicos para o controle da mancha-aureolada em região montanhosa no
município de Caconde, SP. Houve redução da doença nas parcelas pulverizadas com
casugamicina (1,5 L/ha), casugamicina (1,5 L/ha) + hidróxido de cobre (2,5 kg/ha),
entre outros, tendo sido empregados óleos vegetais e minerais, adesivo siliconado e
acibenzolar-S-methyl. No entanto, independentemente dos produtos utilizados, os
autores observaram maior incidência da doença nas parcelas expostas à ação dos ventos,
indicando que a implementação de quebra-ventos é muito importante para o manejo da
doença.
Atualmente os produtos registrados no MAPA para controle da mancha-
aureolada se restringem ao hidróxido de cobre e também ao antibiótico casugamicina, o
qual é indicado para utilização apenas em vieiro de mudas (AGROFIT, 2015).
2.2.6. Resistência genética de cafeeiros a Pseudomonas syringae pv. garcae
Estudos realizados por MORAES et al. (1974; 1975), por meio de inoculações
artificiais com agulhas múltiplas, detectaram resistência a P. syringae pv. garcae nas
variedades exóticas de C. arabica Harar, Dilla & Alghe. S. 12 Kaffa e Geisha.
Posteriormente, MOHAN et al. (1978), em estudo semelhante, além das variedades
Semierecta e Ennarea de C. arabica, também detectaram resistência à doença nas
espécies C. congensis, C. eugenioides, C. stenophylla e C. canephora, pertencentes ao
banco de germoplasma de Coffea spp. do IAPAR (Instituto Agronômico do Paraná). A
pesquisa revelou ainda que dentre 138 introduções de C. arabica provenientes da
Etiópia, 38 apresentaram alto grau de resistência à mancha-aureolada.
Pesquisas realizadas por SERA et al. (1980), CARDOSO & SERA (1983),
PETEK et al. (2001), SERA et al. (2002), PETEK et al. (2006) e ITO et al. (2007;
2008), em cafeeiros expostos à ocorrência natural da doença, resultaram na seleção da
cultivar IPR 102, resistente a P. syringae pv. garcae.
Dentre o germoplasma avaliado por MORAES et al. (1975), as variedades
exóticas de C. arabica de origem etíope, conhecidas como Harar, Dilla & Alghe, S 12
Kaffa e Geisha, apresentaram resistência a P. syringae pv. garcae sendo todas elas
portadoras do alelo SH1.

14
Segundo CARVALHO (1988), a resistência múltipla à ferrugem e à mancha-
aureolada apresentada por essas variedades exóticas, poderia ser explicada por ligação
gênica (linkage) entre alelos de resistência a ambos os patógenos ou pelo efeito
pleiotrópico do alelo SH1. SERA (2001) e FAZUOLI et al. (2009) também atribuíram
ao alelo SH1 presente nas variedades etíopes (BETTENCOURT & CARVALHO, 1968),
avaliadas por MORAES et al. (1975), responsável pela resistência simultânea a algumas
raças de H. vastatrix e a P. syringae pv. garcae.
Segundo BETTENCOURT & CARVALHO (1968), o alelo SH1 é bastante
difundido nas principais zonas cafeeiras da Etiópia. Estudos conduzidos pelos autores
permitiram sua identificação em seleções diversas como Barbuk Sudan, BE-2 Ghembi,
BE-4 Ennarea, BE-5 Wush-Wush, BE-6 Moderalo, BE-7 Boggia, BE-8 Era, BE-14
Loulo, Dilla & Alghe, Geisha, Lejeune's, S 6 Cioiccie, S 9 Arba Gougou, S 12 Kaffa, S
17 Yrgalem, U l Dalecho, ocorrendo tanto isoladamente, como em associação com
outros genes de resistência, como SH4 (SH1 SH4), SH5 (SH1 SH5) e SH4 e SH5 (SH1 SH4
SH5) e que conferem resistência a 16 das 24 raças de H. vastatrix.
2.2.6.1. Métodos para avaliação da resistência à mancha-aureolada
A inoculação artificial de P. syringae pv. garcae tem sido efetuada a partir de
diferentes métodos como, infiltração, ferimento por agulhas múltiplas, fricção com
carborundum, aspersão (COSTA et al., 1957), corte com tesoura, infiltração,
atomização (OLIVEIRA & ROMEIRO, 1990); deposição de gota do inóculo seguido de
punctura com agulha ou com corte por lâmina e ainda, pela combinação entre eles
(ITHIRU et al., 2013). No entanto, apenas os trabalhos conduzidos por ITHIRU et al.
(2013) visaram a seleção de plantas com resistência a P. syringae pv. garcae. Os
autores verificaram variação nos resultados obtidos entre os métodos testados, embora
ambos possam ser utilizados para este fim.
2.3. Melhoramento genético de Coffea arabica
2.3.1. Classificação botânica, origem e características de Coffea arabica
Os cafeeiros têm como centro de origem a região da Abissínia, atual Etiópia
(CHEVALIER, 1947) e pertencem a família Rubiaceae, gênero Coffea, cuja

15
classificação taxonômica atual admite 124 espécies (DAVIS et al., 2011). Destas,
apenas três possuem importância econômica, C. arabica Linnaeu (café arábica), C.
canephora Pierre ex A. Froehner (café canephora, robusta ou conilon) e C. liberica
Bull. ex Hiern (café libérica ou excelsa) (DAVIS et al., 2006), com imensa
predominância das duas primeiras.
Coffea arabica, a principal espécie em importância econômica, é anfidiploide,
alotetraploide (2n = 4x = 44 cromossomos) resultado da provável hibridação natural
entre as espécies C. canephora e C. eugenioides (LASHERMES et al., 1999). Devido às
divergências genéticas existentes entre as espécies LASHERMES et al. (1999)
sugeriram que a hibridação ocorreu provavelmente na região da África Central. Os
autores demonstraram por meio de hibridação genômica in situ que, o genoma de C.
arabica é composto pela combinação de dois grupos cromossômicos e, ainda, utilizando
sondas específicas de DNA puderam identificar as espécies C. canephora e C.
eugenioides como parentais.
É conhecido que a espécie C. arabica é nativa da região entre o sudeste da
Etiópia, Sudeste do Sudão e Norte do Quênia em altitudes que variam de 1.000 a 3.000
metros (CARVALHO, 1946). Plantas desta espécie desenvolvem-se melhor em regiões
de clima ameno, com temperatura média entre 18,5 e 21,4° C, precipitação entre 1.200 e
2.000 mm e períodos de estiagem entre 3 a 4 meses (NARASIMHASWAMY, 1968).
Plantas de C. arabica são arbustos, com um ramo ortotrópico com até 4 metros
de altura, sendo sua base genética pouco diversificada devido à reprodução por
autofecundação, de modo que, as variedades Típica e Bourbon são genitoras da maioria
das cultivares atualmente utilizadas (ANTHONY et al., 2002). Apesar da baixa taxa de
fecundação cruzada (5-10%), ampla variabilidade morfológica em variedades de C.
arabica é reconhecida devido à ocorrência de hibridações naturais e mutações (KRUG
et al., 1939; CARVALHO, 1969; CARVALHO, 2007).
2.3.2. Histórico, métodos e perspectivas
Os primeiros cafeeiros trazidos para o Brasil são provavelmente originários de
uma única planta matriz, cultivada no Jardim Botânico de Amsterdam, Holanda. Por
este motivo, CARVALHO & FAZUOLI (1993) trataram os primeiros cultivos de café

16
nacionais como uma enorme progênie de apenas um cafeeiro. Essa cultivar ficou
conhecida pelos nomes de Arábica, Nacional, Típica ou Crioulo (CARVALHO, 1997).
No ano de 1870 mutações do cafeeiro Típica foram observadas no município de
Maragogipe, Estado da Bahia, resultaram na seleção da variedade Maragogipe, que
possui grãos maiores em relação à cultivar de Típica (MÔNACO, 1960). No ano
seguinte, outra mutação foi observada em Botucatu, Estado de São Paulo, onde plantas
que produziam frutos de coloração amarela, as quais foram selecionadas e resultaram na
variedade Amarelo de Botucatu. Contudo, essas seleções foram pouco cultivadas, por
apresentarem menor produtividade em relação à cultivar Típica (CARVALHO &
FAZUOLI, 1993).
Em 1930, foi encontrada no município de Pederneiras, Estado de São Paulo,
uma variedade de cafeeiro bastante promissora em produtividade, denominada Bourbon
Amarelo, originada presumivelmente do cruzamento natural entre as variedades
Bourbon Vermelho e Amarelo Botucatu (CARVALHO et al., 1957). Treze anos mais
tarde foi descrita a cultivar Mundo Novo, encontrada no município de Urupês, Estado
de São Paulo, tendo como origem provável, a hibridação natural entre Bourbon
Vermelho e Sumatra (CARVALHO et al., 1952), esta última, introduzida no Brasil no
ano de 1896 (KRUG et al., 1939).
Outras mutações do cafeeiro com elevada importância econômica, designadas de
Caturra Vermelho e Caturra Amarelo, foram originadas no Estado de Minas Gerais,
cujas sementes foram enviadas e cultivadas no Vale do Caparaó (Estado do Espírito
Santo) e, posteriormente em 1937, ao Instituto Agronômico (IAC). Esta mutação
derivou-se a partir da cultivar Bourbon Vermelho, e tem como principal característica a
redução do comprimento dos internódios, levando à redução de porte das plantas. Foi
descoberto por KRUG et al. (1949), que esta característica é governada por um par de
alelos.
O Instituto Agronômico (IAC) foi o pioneiro em estudos envolvendo seleção de
linhagens mais produtivas, e com características agronômicas desejáveis. Em 1931,
instalou-se nessa instituição um experimento com as cultivares em evidência na época,
ou seja, Típica, Amarelo Botucatu, Bourbon Amarelo, Bourbon Vermelho, Maragogipe
e Sumatra, visando seleção de variedades mais produtivas, além de outras características
como, época de maturação e tamanho de sementes (MENDES, 1951). Em paralelo,

17
diversos estudos foram conduzidos a fim de se elucidar aspectos da biologia da
reprodução, citológicos e genética de características (MENDES & KRUG, 1938;
KRUG, 1939).
Segundo MEDINA FILHO et al. (1984), dentre as estratégias aplicadas ao
melhoramento genético do cafeeiro, faz parte a introdução de plantas, seleção de plantas
individuais e posterior teste de progênies e o avanço das gerações dado pelo método
genealógico, podendo estar associado à retrocruzamentos. Também, por meio de
seleção de plantas individuais, seguida de teste de progênie é possível obter bons
resultados no melhoramento de cafeeiros. Esse método tem como pretexto a existência
de variabilidade genética entre plantas de uma mesma cultivar (BONOMO, 2002).
Em meados dos anos 40, foi instalado no IAC ensaios com a cultivar Mundo
Novo, realizando-se seleções entre e dentro das progênies, as quais foram consideradas
bastante efetivas (FAZUOLI, 1977; CARVALHO 2007), resultando em linhagens
produtivas, vigorosas e adaptadas às diversidades climáticas. Uma delas destacou-se por
apresentar características similares à Mundo Novo, porém com sementes maiores e alta
produtividade de sementes, dando origem à cultivar denominada Acaiá.
No contexto do melhoramento, um grande programa de hibridações inter e intra
específicas foi iniciado na década de 30. Os cruzamentos entre as cultivares de
C. arabica Caturra Amarelo e Mundo Novo, realizados em 1949, seguidas de diversas
seleções, viriam a gerar em 1972 as cultivares Catuaí Vermelho e Catuaí Amarelo, cujas
características incorporam rusticidade, alta produção, porte reduzido e boa qualidade de
bebida (CARVALHO & MÔNACO, 1972; CARVALHO, 2008).
Pesquisas desenvolvidas pelo pesquisador científico Dr. Alcides Carvalho foram
imprescindíveis para garantir a produção brasileira de café. Seleções de progênies do
híbrido entre C. canephora cv. Robusta (tetraploide) e C. arabica cv. Bourbon
Vermelho retrocruzado com C. arabica cv. Mundo Novo deram origem à cultivar Icatu,
que apresenta genes de resistência a algumas raças de H. vastatrix (FAZUOLI, 1991).
Dependendo da distância genética entre as plantas matrizes, deve-se realizar
retrocruzamentos sucessivos a fim de transferir a característica almejada, sem deixar
vestígios indesejáveis na cultivar comercial resultante. Essa estratégia vem sendo
utilizada na seleção de plantas resistentes ao bicho-mineiro (Leucoptera coffeella),

18
onde, híbridos interespecíficos entre C. arabica e C. racemosa estão sendo submetidos a
retrocruzamentos sucessivos com a espécie arábica. Neste caso, a seleção de plantas é
dificultada pela existência de ampla variabilidade genética nas progênies, sendo
necessária a avaliação de um grande número de plantas, para conciliar as características
de resistência ao inseto, à ferrugem, boa produtividade e qualidade de bebida ente outras
características de interesse (GUERREIRO FILHO et al., 1990; GUERREIRO FILHO,
2006; CARVALHO, 2007).
Uma das principais vantagens da utilização de acessos de C. arabica para o
programa de melhoramento genético (PMG) é a sua estreita relação genética com as
cultivares comerciais, característica maior que proporciona facilidade na obtenção de
plantas hibridas e na eliminação de plantas com características indesejáveis.
As introduções de C. arabica provenientes da Etiópia (FAO, 1968) são parte da
reserva genética da espécie, e têm grande importância para o melhoramento de plantas,
bem como, para a conservação do germoplasma da espécie. Na década de 50,
conhecendo a rápida disseminação e a devastação causada pela ferrugem-do-cafeeiro
(H. vastatrix) na Ásia e, o risco à cafeicultura brasileira, foram introduzidos no IAC
sessenta e sete acessos supostamente resistentes ao fungo. Em 1956, uma parceria foi
iniciada entre o IAC e o Centro de Investigações das Ferrugens do Cafeeiro (CIFC) –
Oeiras, Portugal, com a finalidade de manter intercâmbio de informações sobre aquele
patossistema (CARVALHO, 1988).
Essas introduções compõem importante fonte de genes de resistência a P.
syringae pv. garcae, como demostrado por MOHAN et al. (1978).
2.4. Mecanismos de resistência de plantas a patógenos
As plantas desenvolveram durante sua evolução, mecanismos para contornar as
adversidades do ambiente em que vivem, para que perpetuem sua espécie. Mecanismos
de defesa contra herbivoria, poluentes, fatores climáticos, competição com outras
plantas e ataque de patógenos, podem se manifestar de forma diversa como gomas,
espinhos, substâncias tóxicas e/ou detoxificadoras. Os mecanismos de resistência são
classificados como estruturais ou bioquímicos e, em ambas as formas podem estar
presentes antes ou após o reconhecimento do agente agressor (AGRIOS, 2005).

19
A resposta de defesa acontece com a ativação de uma complexa rede de sinais,
os quais, por efeito cascata, desencadeiam diversas vias, interligadas ou não, cujas
combinações determinam ações como restrição do fluxo de água no local da infecção
com consequente morte de células do tecido vegetal, o confinamento e a morte do
patógeno. Essas ações metabólicas podem atuar concomitantemente com a produção e o
acúmulo de compostos tóxicos aos invasores.
Um dos mecanismos de resistência de plantas a patógenos com funções
conhecidas são os genes de resistência (R). De maneira geral, genes de avirulência (avr)
e genes de patogenicidade (P) por parte do patógeno são reconhecidos por receptores
das plantas hospedeiras que dispõem de genes R e de resposta defensiva (PR) para se
defender do ataque do agente patogênico. Os genes R são aqueles relacionados com a
detecção de sinais ou elicitores do patógeno (DIXON & HARRISON, 1990).
Os genes avr e R, evoluiram reconhecidamente seguindo a teoria gene-a-gene,
proposta por Flor (BISHOP et al., 2000), por isso, é associada à resistência a patógenos
específicos. Segundo DANGL & JONES (2001), as vias de defesa desencadeadas pelos
genes R culminam num efeito cascata que resulta em uma rajada oxidativa, que por fim,
ocasionam o acúmulo de óxido nítrico (ON), superóxidos e peróxidos de hidrogênio,
induzindo reação de hipersensibilidade (HR) e/ou associações ligadas à produção de
metabólitos antimicrobianos.
Em contrapartida, os genes PR desencadeiam a expressão de moléculas que
constituem um sistema protetor nas plantas. As formas mais comumente notadas são: os
compostos fenólicos depositados na parede celular das plantas hospedeiras, conhecidos
principalmente pela ação antifúngica e antiviral; a síntese de fitoalexinas, consideradas
como biocidas ou metabólitos secundários tóxicos a fungos, bactérias, nematoides,
plantas e animais, produzidos e acumulados logo após a exposição do hospedeiro ao
patógeno DIXON & HARRISON, 1990; STANGARLIN et al., 2011) e as proteínas
relacionadas à patogênese (proteínas RP), principais compostos induzidos após a
ocorrência de reação de hipersensibilidade dos tecidos vegetais (GIANINAZZI &
MARTIN, 1970). GRAHAM & GRAHAM (1991) salientam que, estas proteínas RP
são constitutivas do tecido vegetal, estando presentes em baixas concentrações. Desta
forma, a síntese ou acúmulo de fitoalexinas e proteínas RP é resultado da indução por
patógenos e/ou agentes eliciadores. De modo geral, após o reconhecimento de elicitores

20
de patógenos, como a proteína flagelina, componente dos flagelos das bactérias, ou
ainda proteínas específicas codificadas por genes de avirulência (MARTIN et al., 2003),
são desencadeadas respostas de defesa que compreendem, entre outros processos, a
síntese de proteínas PR, que estão classificadas em 17 famílias, conforme suas funções
na resistência das plantas aos patógenos (GORJANOVIĆ, 2009).
O reconhecimento da virulência do patógeno por plantas hospedeiras acontece
mediante padrões moleculares associados aos patógenos chamados de PAMPs
(Pathogen Associated Molecular Patterns) ou MAMPs (Microbe Associated Molecular
Patterns) (BOLLER & FELIX, 2009). Proteínas presentes na membrana das células
vegetais atuam como receptoras de reconhecimento de padrões, PRRs (Pattern
Recognized Receptors), sinalizando PAMPs/MAMPs (BOLLER & FELIX, 2009).
Após a ativação do sistema de defesa, uma complexa rede de sinais resultará em
HR, produção de espécies reativas de oxigênio (EROs), síntese de proteínas RP,
compostos como fitoalexinas, suplemento da parede celular como lignificação e caloses
e, por fim, podem resultar em resistência sistêmica adiquirida (SAR) ou induzida.
Resultados obtidos por JOHANSSON (2015) a respeito da sinalização contra P.
syringae, revelaram que entre os eventos ocasionados mais precocemente estão
incluídos o fluxo de íons Ca2+, a resposta por rajada oxidativa, a ativação de proteína-
quinases ativadas por mitógeno (MAPKs), seguida pela expressão de genes que regulam
a síntese de substâncias antagonistas ao patógeno. As respostas mais tardias estão
relacionadas à uma rede complexa de sinalização de fito-hormônios, sendo os principais
o ácido salicílico (AS), o jasmonato (JA) e o etileno (ET) (GIMENEZ-IBANEZ &
RATHJEN, 2010). O modelo idealizado por JOHANSSON (2015) ilustra as respostas
basais de defesa (Figura 2).
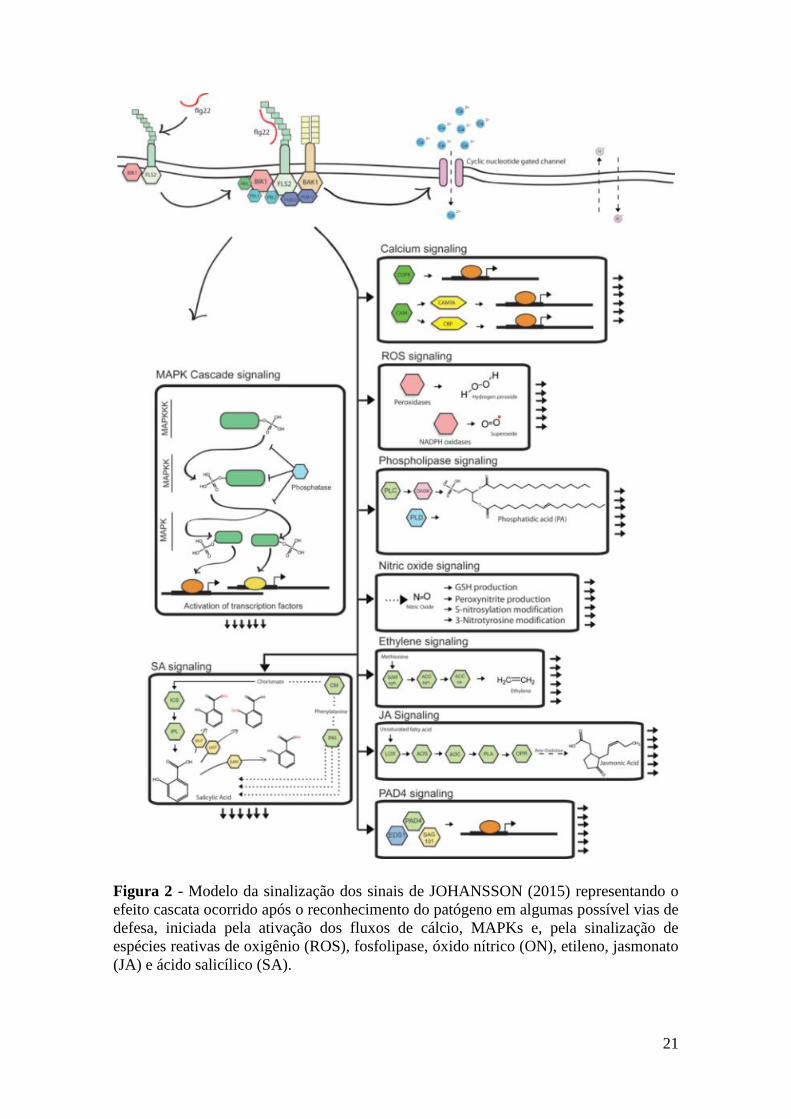
21
Figura 2 - Modelo da sinalização dos sinais de JOHANSSON (2015) representando o
efeito cascata ocorrido após o reconhecimento do patógeno em algumas possível vias de
defesa, iniciada pela ativação dos fluxos de cálcio, MAPKs e, pela sinalização de
espécies reativas de oxigênio (ROS), fosfolipase, óxido nítrico (ON), etileno, jasmonato
(JA) e ácido salicílico (SA).

22
Outro mecanismo de imunidade nas plantas é o silenciamento gênico mediado
por pequenas sequências de RNAs, que podem causar degradação dos transcritos
relacionados aos receptores do hormônio auxina sendo que esta redução da auxina
auxilia na restrição do crescimento de P. syringae em Arabdopsis (NAVARRO et al.,
2006).
A determinação da atividade de compostos associados a estresses é uma
ferramenta que auxilia no conhecimento da resposta de tolerância e resistência a fatores
abióticos e bióticos diversos.
GUZZO (2004) investigou o comportamento das enzimas quitinases e β-1,3-
glucanases em plantas de C. arabica cv. Mundo Novo tratadas com acibenzolar-S-metil
(ASM) e sua correlação com a redução da severidade da ferrugem. Os resultados
indicaram que o incremento na atividade dessas enzimas pode estar associado à
resistência local e a resistência sistêmica adquirida (RSA), induzidas pelo ASM.
A RSA é iniciada no local da infecção mas, é observada em tecidos distantes. A
ocorrência da RSA requer o acúmulo de acido salicílico (AS) e ocasiona acúmulo de
proteínas RP. O gene NPR1 (nonexpressor of PR-1) funciona como sinalizador do
acúmulo de AS por meio da interação com outros fatores de transcrição (DURRANT &
DONG, 2004).
Em berinjeleira, ZHOU et al. (2012) estudaram a relação da atividade das
enzimas catalase, polifenoloxidase, peroxidase, fenilalanina amônia-liase e superóxido
dismutase, na severidade da murcha-de-verticilium (Verticillium dahliae), em quatorze
cultivares de berinjela, com diferentes níveis de resistência. Os autores verificaram
correlações significativas entre a atividade das enzimas polifenoloxidase, peroxidase e
fenilalanina amônialiase e a resistência ao patógeno, de modo que, os níveis dessas
enzimas podiam atuar como indicadores da resistência em auxílio a programas de
melhoramento genético.
Outra abordagem sobre o assunto se relaciona ao uso de técnicas de biologia
molecular, às quais deram origem a informações importantes e mais refinadas sobre o
mecanismo de reconhecimento e defesa contra fitopatógenos, bem como, a ativação e
tradução de sinais originados pelos genes de defesa (CRAMER et al., 1993,
SILVESTRINI et al., 2005a).

23
Estudos aplicando a genética molecular em cafeeiros evoluíram rapidamente,
especialmente após inúmeros trabalhos envolvendo sequenciamento, que originaram
uma biblioteca de ESTs (Expressed Sequence Tags), das espécies C. arabica, C.
canephora e C. racemosa (disponível em: http://www.lge.ibi.unicamp.br/cafe). Com
esse estudo, foi possível identificar dentro das ESTs, 33.000 unigenes diferentes. Essa
ferramenta genômica possibilita a condução de estudos relacionados a inúmeros fatores
associados à condições de estresse, as quais os cafeeiros são submetidos constantemente
na natureza (VIEIRA et al., 2006).
Uma das vertentes que o sequenciamento do genoma funcional do cafeeiro
permitiu é que, a partir das sequências dos genes, existe a possibilidade de se
“desenhar” primers específicos e estudar a expressão das ESTs que estão associadas a
processos metabólicos específicos sob diversas condições, como é o caso do estresse
causado pela ofensiva de fitopatógenos.
A técnica de qRT-PCR (PCR quantitativo em tempo real) vem sendo utilizada
com sucesso para análises de expressão de genes que possuem importante papel na
resistência a pragas e doenças. Esse tipo de reação ocorre em termociclador com sistema
ótico, o qual permite decifrar a excitação da fluorescência emitida pelas amostras, e
desse modo, um software específico é capaz de quantificar a expressão gênica. Durante
os ciclos iniciais da reação é formada uma linha de base (baseline), útil para o gráfico
das amplificações e, em seguida, é definido o parâmetro Ct (cycle threshold),
determinando como o ciclo em que a flourescência ultrapassa a linha de base. As
análises de expressão relativa são baseadas nos valores de Ct obtidos no sistema
computacional, sendo por isso possível avaliar a expressão de um mesmo gene em
diferentes tecidos vegetais, e/ou, em situações de estresse causados por fatores bióticos
ou abióticos (APPLIED BYOSISTEMS).
VINECKY et al. (2011) estudaram a expressão gênica de 14 genes candidatos
para a tolerância à seca em quatro genótipos de C. canephora, por meio de qRT-PCR.
Por meio das análises de expressão relativa, os autores verificaram que grande parte dos
genes selecionados apresentaram expressão diferencial na condição de estresse hídrico,
e ainda, ocorreram diferenças no padrão da expressão observados entre os genótipos,
dando suporte à hipótese da existência de outros mecanismos associados à tolerância à
seca.

24
Alguns estudos demonstraram estreita ligação entre os genes EDS1 e PAD4 em
resposta à infecção contra fitopatógenos. Segundo AARTS et al. (1998), o gene EDS1 é
requerido durante a sinalização pela TIR-NBS-LRR, uma classe de proteína R, que
apresenta um sítio rico em repetições de leucina. Posteriormente, estudos indicaram a
atuação do gene PAD4, em conjunto ao EDS1, na sinalização da via do AS (FEYS et
al., 2005).
Sabe-se que, plantas de Arabidopsis mutadas para deficiência do gene EDS1
mostram aumento na suscetibilidade a Peronospora parasítica, agente causal do míldio
em brássicas (PARKER et al., 1996). Outro estudo demonstrou que os componentes
EDS1 e PAD4 apresentam importante papel na sinalização e morte celular mediada pelo
gene lsd1 (lesions simulating disease resistance response), em Arabidopsis inoculadas
com o fungo P. parasitica (RUSTÉRUCCI et al., 2001). Ainda, os autores
demonstraram diferenças na deposição de superóxido nas margens das lesões nas
plantas mutantes, ausentes do gene lsd1. Estudos mais recentes, desenvolvidos por
WIERMER et al. (2005), evidenciaram a atuação do gene SAG101 (Senescence
Associated Gene 101), em conjunto em EDS1 e PAD4 mediando a resposta de
resistência via AS.
A enzima β-1,3-glucanase possui importante atução na resistência de plantas a
fitopatógenos. ANDRADE et al. (2013), observaram que plantas de tomate tratadas
com ácido jasmônico, etileno ou acibenzolar-S-metil produziram maiores quantidades
das enzimas peroxidase, polifenoloxidase, lipoxigenase e β-1,3-glucanase, resultando
em menor expressão dos sintomas da pinta bacteriana causada por P. syringae pv.
tomato.
Com relação à expressão de genes em resposta à infecção por nematoides da
espécie Meloidogyne exigua, SILVESTRINI et al. (2005a) analisaram a amplificação de
transcritos dos genes ligados a resistência contra fitopatógenos, superóxido dismutase,
WIPK (fator de transcrição WipkA), glucanases e proteínas de resistência ResproB e
5RM1, em plantas de C. arabica cv. Icatu segregantes para resistência ao fitonematoide.
De acordo com os autores, os genes apresentaram expressão basal, mesmo nas plantas
não infectadas, ou seja, os genes testados não apresentaram variações significativas na
expressão.

25
Recentemente, FATOBENE (2014) analisou a expressão relativa de 12 genes em
resposta à infecção por Meloidogyne paranaensis em genótipos de cafeeiros
contrastantes classificados como resistentes e suscetível ao nematoide. Seis dos genes
estudados participam das funções relacionados à resistência de plantas a patógenos, três
ao metabolismo antioxidante e três associados a funções do ciclo celular. Foi relatada
ativação dos genes de defesa EDS1, PAD4, LTP e NPR1 em resposta à infecção, e por
este motivo, a autora concluiu que o mecanismo de defesa do cafeeiro a M. paranaensis
está relacionado com a via do AS, uma vez que os genes citados atuam na referida rota
metabólica.
3. MATERIAL E MÉTODOS
3.1. Alterações morfológicas em tecidos foliares de cafeeiros infectados por
Pseudomonas syringae pv. garcae
3.1.1. Material vegetal, inoculação e condições experimentais
Mudas de cafeeiros da cultivar Mundo Novo IAC 376-4, com aproximadamente
seis meses de idade, tiveram folhas do terceiro internódio inoculadas por meio de
puncturas, com alfinete entomológico #00 previamente mergulhado em suspensão
bacteriana de P. syringae pv. garcae, proveniente do isolado IBSBF 1197, na
concentração de 3x108 UFC.mL-1, em dois pontos de cada lado da nervura central. Um
tratamento controle foi estabelecido com folhas punciondas com os alfinetes
entomológicos, previamente mergulhados em solução salina.
O experimento foi conduzido em modelo inteiramente casualizado com arranjo
fatorial 2x3, sendo o período de desenvolvimento da doença (2) e o tecido infectado (3)
os efeitos procipais do modelo. As parcelas de plantas inoculadas foram representadas
por uma muda em cinco repetições e as parcelas controle, por uma planta em três
repetições.
Avaliou-se a magnitude do dano pela mensuração do maior (L1) e do menor
comprimento (L2) das lesões, externamente, na epiderme e, internamente, nos
parênquimas paliçádico e esponjoso, aos 10 e 20 dias após a inoculação (DAI).

26
3.1.2. Análises histopatológicas
Características histopatológicas da infecção de folhas de C. arabica, por P.
syringae pv. garcae, foram estudadas por meio de análises anatômicas sob microscopia
de luz. Lesões observadas em folhas aos 10 e 20 DAI foram recortadas, fixadas em
solução FAA (formaldeído-ácido acético-álcool etílico 50%) (JOHANSEN, 1940) e
submetidas à vácuo por 48 horas, sendo posteriormente desidratadas em série alcoólica-
etílica e incluídas em parafina. Foram realizadas secções transversais e paradérmicas em
micrótomo rotativo manual a 12 µm com posterior distensão e colagem com adesivo
Haupt (JOHANSEN, 1940). As lâminas foram coradas com safranina, seguido por azul
de alcian, montadas em adesivo “Permount”. As análises foram realizadas utilizando-se
de microscópio óptico Leica, modelo DMLB acoplado a uma câmera de captura de
imagens e posterior análise no programa Motic Image Plus.
Para determinar a extensão do dano provocado pela bactéria nas estruturas do
mesofilo, mensurou-se, a partir dos cortes transversais, o maior (L1) e o menor (L2)
comprimento do tecido afetado, provocado pelas bactérias, nos parênquimas paliçádico
e esponjoso, separadamente. As análises estatísticas dos dados obtidos foram realizadas
com auxílio do software Statistica 7.0. Para comparação das médias do tamanho das
lesões, utilizou-se o teste de Student-Newman-Keuls (STELL & TORRIE, 1980),
adotando-se nível de significância igual a 5%.
3.2. Natureza e agressividade de isolados de Pseudomonas syringae pv. garcae
3.2.1. Isolados bacterianos utilizados no estudo
Dezenove isolados bacterianos foram utilizados no estudo. Todos são oriundos
da Coleção de Culturas de Fitobactérias do Instituto Biológico (IBSBF
http://www.biologico.sp.gov.br/pdf/catalogo_IBSBF.pdf) (Tabela 1) e foram
selecionados pela região geográfica de origem.
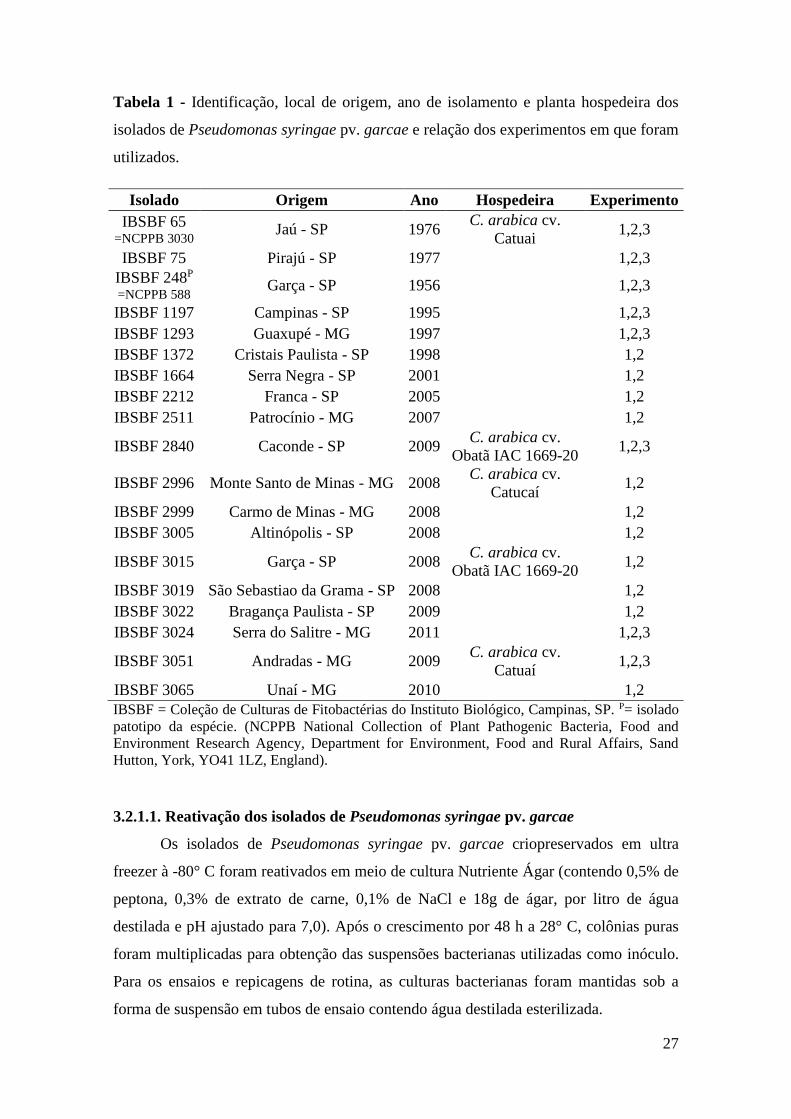
27
Tabela 1 - Identificação, local de origem, ano de isolamento e planta hospedeira dos
isolados de Pseudomonas syringae pv. garcae e relação dos experimentos em que foram
utilizados.
Isolado Origem Ano Hospedeira Experimento
IBSBF 65 =NCPPB 3030
Jaú - SP 1976 C. arabica cv.
Catuai 1,2,3
IBSBF 75 Pirajú - SP 1977 1,2,3
IBSBF 248P
=NCPPB 588 Garça - SP 1956 1,2,3
IBSBF 1197 Campinas - SP 1995 1,2,3
IBSBF 1293 Guaxupé - MG 1997 1,2,3
IBSBF 1372 Cristais Paulista - SP 1998 1,2
IBSBF 1664 Serra Negra - SP 2001 1,2
IBSBF 2212 Franca - SP 2005 1,2
IBSBF 2511 Patrocínio - MG 2007 1,2
IBSBF 2840 Caconde - SP 2009 C. arabica cv.
Obatã IAC 1669-20 1,2,3
IBSBF 2996 Monte Santo de Minas - MG 2008 C. arabica cv.
Catucaí 1,2
IBSBF 2999 Carmo de Minas - MG 2008 1,2
IBSBF 3005 Altinópolis - SP 2008 1,2
IBSBF 3015 Garça - SP 2008 C. arabica cv.
Obatã IAC 1669-20 1,2
IBSBF 3019 São Sebastiao da Grama - SP 2008 1,2
IBSBF 3022 Bragança Paulista - SP 2009 1,2
IBSBF 3024 Serra do Salitre - MG 2011 1,2,3
IBSBF 3051 Andradas - MG 2009 C. arabica cv.
Catuaí 1,2,3
IBSBF 3065 Unaí - MG 2010 1,2
IBSBF = Coleção de Culturas de Fitobactérias do Instituto Biológico, Campinas, SP. P= isolado
patotipo da espécie. (NCPPB National Collection of Plant Pathogenic Bacteria, Food and
Environment Research Agency, Department for Environment, Food and Rural Affairs, Sand
Hutton, York, YO41 1LZ, England).
3.2.1.1. Reativação dos isolados de Pseudomonas syringae pv. garcae
Os isolados de Pseudomonas syringae pv. garcae criopreservados em ultra
freezer à -80° C foram reativados em meio de cultura Nutriente Ágar (contendo 0,5% de
peptona, 0,3% de extrato de carne, 0,1% de NaCl e 18g de ágar, por litro de água
destilada e pH ajustado para 7,0). Após o crescimento por 48 h a 28° C, colônias puras
foram multiplicadas para obtenção das suspensões bacterianas utilizadas como inóculo.
Para os ensaios e repicagens de rotina, as culturas bacterianas foram mantidas sob a
forma de suspensão em tubos de ensaio contendo água destilada esterilizada.

28
Para o preparo do inóculo, colônias bacterianas na fase logarítmica de
crescimento (24h) foram suspendidas em solução salina (NaCl 0,85%). A concentração
foi padronizada em espectrofotômetro a 0,3 de absorbância (~600nm), resultando em
inóculo com aproximadamente 3x108 UFC/mL-1 (LELLIOT et al., 1966).
3.2.3. Agressividade de isolados de Pseudomonas syringae pv. garcae
A agressividade de isolados bacterianos foi avaliada em três experimentos, a
partir da manifestação dos sintomas, em folhas de diferentes estágios de maturação de
cultivares suscetíveis de C. arabica.
Os experimentos 1 e 2 foram realizados entre os meses de janeiro a março de
2012, sendo que a temperatura média no período das avaliações foi de 23,7° C. o
experimento 3 foi realizado durante o mês de junho de 2012, sendo que a temperatura
média foi de 19° C.
Experimento 1: Reação da cultivar Mundo Novo IAC 376-4 a P. syringae pv. garcae.
Mudas de C. arabica cultivar Mundo Novo IAC 376-4 com cerca de cinco
meses, contendo 4-5 pares de folhas foram inoculadas entre janeiro a março de 2012
com dezenove isolados (Tabela 1). As inoculações foram realizadas por meio de
punctura em dois pontos de cada lado do limbo foliar, nos dois primeiros pares de
folhas, a partir do ápice, utilizando-se alfinete entomológico #00, previamente
mergulhado nas suspensões bacterianas. Após as inoculações, as plantas foram mantidas
em câmara úmida por 48 h e, posteriormente, em casa-de-vegetação com irrigação
controlada e umidade relativa (UR) em torno de 63%.
As parcelas experimentais, representadas por três plantas, foram dispostas em
delineamento de blocos casualizados com quatro repetições, sendo avaliados os efeitos
dos isolados bacterianos em função do dano causado nas plantas aos 21 dias após a
inoculação (DAI), por meio da medição da maior extensão das lesões.
Para análise dos dados experimentais os valores mensurados foram submetidos à
análise de variância, sendo as médias comparadas pelo teste de Student Newman Keuls
a 5% de probabilidade, utilizando o software Statistica v. 7.0.

29
Experimento 2: Reação da cultivar Bourbon Amarelo IAC J 30 a isolados de P.
syringae pv. garcae
Folhas dos quatro primeiros internódios das plantas da cultivar de C. arabica
Bourbon Amarelo IAC J 30 com aproximadamente seis meses de idade, foram
inoculadas segundo o mesmo procedimento descrito no experimento 1, com dezenove
isolados (Tabela 1) e também, por uma mistura composta por quatro isolados (mistura I
- IBSBF 75, IBSBF 1197, IBSBF 1293, IBSBF 3024) considerados os mais agressivos
no experimento 1 realizado com a cultivar Mundo Novo IAC 376-4. Após as
inoculações, as plantas foram mantidas em câmara úmida por 48 horas e,
posteriormente, em casa-de-vegetação com irrigação controlada e UR em torno de 63%.
As parcelas experimentais, representadas por três plantas, foram dispostas em
arranjo fatorial, em delineamento de blocos casualizados com quatro repetições, sendo
avaliados os efeitos dos isolados bacterianos e da idade da folha em função do dano
causado nas plantas aos 21 dias após a inoculação (DAI), por meio da medição da maior
extensão das lesões.
Para análise dos dados experimentais, os valores mensurados foram submetidos
à análise de variância, sendo as médias comparadas pelo teste de Student Newman
Keuls a 5% de probabilidade, utilizando o software Statistica v. 7.0.
Adicionalmente, a partir das inoculações foram realizadas observações diárias
das puncturas, anotando-se o início do aparecimento dos sintomas, a fim de se calcular
o período de incubação do patógeno, sendo considerado como período de incubação o
tempo médio para aparecimento dos primeiros sintomas em todas as puncturas.
Experimento 3: Reação da cultivar Mundo Novo IAC 376-4 às misturas de isolados
bacterianos.
A agressividade dos quatro isolados IBSBF 65, IBSBF 75, IBSBF 1197 e IBSBF
1293, que constituíram a mistura I, considerados como os mais agressivos nos
experimentos anteriores, foi novamente avaliada de forma individual ou em
combinações de isolados (Mistura II: 65 e 75, Mistura III: 65 e 1197, Mistura IV: 65 e
1293, Mistura V: 75 e 1197, Mistura VI: 75 e 1293, Mistura VII: 1197 e 1293 e Mistura
VIII: 65, 75, 1197 e 1293) a partir de inoculações realizadas em folhas dos dois
primeiros internódios de mudas da cultivar Mundo Novo IAC 376-4 de C. arabica.
Quatro outros isolados foram também incluídos no estudo, sendo IBSBF 248, isolados

30
patotipo da espécie, IBSBF 2840 e 3024, pelo comportamento agressivo e IBSBF 3051
pela baixa agressividade apresentada nos experimentos anteriores.
Como nos experimentos anteriores, após as inoculações, as plantas foram
mantidas em câmara úmida por 48 horas, e posteriormente em casa-de-vegetação, com
irrigação controlada e UR em torno de 63%.
O delineamento experimental adotado foi o de blocos ao acaso com quatro
repetições e parcelas representadas por três plantas, avaliados os efeitos das
combinações entre os isolados bacterianos em função do dano causado nas plantas,
avaliado aos 21 dias após a inoculação (DAI) por meio da medição da maior extensão
das lesões, mensuradas em centímetros.
Os dados referentas aos valores mensurados foram submetidos à análise de
variância sendo as médias comparadas pelo teste de Student Newman Keuls a 5% de
probabilidade, utilizando o software Statistica v. 7.0.
3.2.4. Produção e sensibilidade as bacteriocinas entre isolados de Pseudomonas
syringae pv. garcae
A compatibilidade dos isolados IBSBF 65, IBSBF 75, IBSBF 1197, IBSBF 1293
e IBSBF 3024 utilizados de forma combinada (Experimentos 2 e 3) foi investigada por
meio de testes in vitro relacionados à produção e sensibilidade à bacteriocinas de acordo
com o método proposto por BIAGI & AZEVEDO (1992) e ROMEIRO (2001).
Além dos isolados utilizados nas inoculações de cafeeiros, avaliou-se também a
atividade em ralação à bacteriocinas dos isolados IBSBF 248, patotipo da espécie e dos
isolados IBSBF 2840 e IBSBF 3051, caracterizados pela baixa e alta agressividade,
respectivamente, apresentada nos experimentos anteriores.
Os ensaios foram realizados em triplicata, repicando-se os isolados, potenciais
produtores de bacteriocinas, por meio de estrias paralelas e equidistantes, uns aos outros
nas placas de Petri contendo meio de cultura 523 de KADO & HESKETT (1970),
contendo 1% sacarose, 0,8% caseína hidrolisada, 0,4% extrato de levedo, 0,2% fosfato
de potássio (K2HPO4), 0,03% sulfato de magnésio (MgSO4.7H2O) e 1,5% ágar e, pH
ajustado para 7,0. Após 72 h a 28° C, procedeu-se a inativação dos isolados, por meio
da adição de 5 mL de clorofórmio na superfície interna das tampas das placas de Petri,
mantidas invertidas por 20 minutos. Em seguida, os resíduos de clorofórmio foram
eliminados e uma camada de aproximadamente 1 mm do meio de cultura 523 foi vertida

31
sobre os isolados. Após a solidificação do meio, procedeu-se o semeio dos isolados
indicadores, por meio de estrias paralelas entre elas e, perpendiculares aos isolados
produtores. As placas foram novamente incubadas a 28° C por 72 h, sendo as leituras
realizadas a partir da análise de descontinuidade do crescimento dos isolados semeados
na sobre camada, em relação aos isolados produtores de bacteriocina, indicativo da
atividade bacteriocinogênica.
3.3. Metodologia de inoculação de Pseudomonas syringae pv. garcae para seleção
de plantas de cafeeiros resistentes à mancha-aureolada
O método mais adequado para a inoculação de P. syringae pv. garcae em
plantas de cafeeiro foi definido a partir da realização de dois experimentos, sendo no
primeiro, comparado os métodos de aspersão e das agulhas múltiplas e, no segundo, os
métodos das agulhas múltiplas e da inoculação por abrasão. Nos dois experimentos, o
inóculo foi proveniente da mistura dos isolados IBSBF 75 e IBSBF 1197, sendo as
suspensões preparadas de acordo com o procedimento descrito em 3.2.2.2.
Experimento 1: Comparação entre os métodos de inoculação por aspersão e agulhas
múltiplas
O material vegetal utilizado foi constituído de cultivares de C. arabica com
níveis variáveis de resistência (MOHAN et al., 1978; ITO et al., 2002), sendo Mundo
Novo IAC 376-4 e Catuaí Vermelho IAC 144 suscetíveis, IAPAR-59 classificada como
moderadamente resistente e IPR-102 como resistente.
Método de Aspersão: As inoculações por aspersão foram realizadas pulverizando-se a
suspensão bacteriana, direcionada para a face abaxial das folhas, até o ponto de
escorrimento (Figura 3A).
As avaliações da severidade da doença foram realizadas utilizando-se escala de 0
a 5 pontos, adaptada de PARADELA et al. (2000), sendo: 0 = ausência de sintomas; 1 =
início do aparecimento dos sintomas, com até 0,4% da área foliar com sintoma
característico da doença; 2 = 0,5 a 1,3% da área foliar com sintomas da doença; 3 = 1,4
a 4,0% da área foliar com sintomas; 4 = 4,1 a 12,5% da área foliar com sintomas
causados pela bactéria; 5 = 13 a 32% da área foliar apresentando sintomas da doença.
Método das Agulhas múltiplas: As inoculações foram realizadas com auxílio de um
anteparo redondo (Ø1,5 cm) contendo sete alfinetes entomológicos #00, fixados
equidistantemente a 0,4 cm, sendo seis ao redor do anteparo e um ao centro. Os

32
ferimentos foram efetuados transpassando as folhas dos cafeeiros, apoiadas em algodão
embebido com suspensão bacteriana, deixando depositado no local dos ferimentos uma
gotícula do inóculo (Figura 3B).
As avaliações da severidade da doença foram realizadas utilizando-se escala de 0
a 5 (ANEXO 1, página 114) pontos, de PARADELA FILHO et al. (1974), onde: 0 =
ausência de clorose e/ou anasarca, ou ainda, reação de hipersensibilidade ao redor das
lesões; 1 = início de amarelecimento ao redor das puncturas , até 10% de área foliar
apresentando sintomas da doença; 2 = 11-25% da área inoculada apresentando
anasarcas, podendo ou não estar circundadas por halo amarelado; 3 = 26-50% da área
inoculada anasarcada, halo amarelado pronunciado ao redor das puncturas; 4 = 51-75%
da área necrosada, halo amarelado por toda área inoculada; 5 = 100% da área inoculada
necrosada.
Figura 3 - Ilustração dos métodos de inoculação de Pseudomonas syringae pv. garcae
em folhas de cafeeiros. Inoculação por aspersão (A); inoculação com agulhas múltiplas
(B).
As plantas foram acondicionadas em câmara úmida 24 h antes e 48 h após as
inoculações, sendo transferidas em seguida para casa-de-vegetação com irrigação
controlada e UR em torno de 63%.
O experimento foi conduzido durante o mês de agosto de 2012, sendo que a
temperatura média durante o período experimental foi de 20,4° C. Adotou-se
delineamento inteiramente casualizado em arranjo fatorial com doze repetições e
parcelas de uma planta. As avalições foram realizadas aos 7, 14 e 21 DAI nos quatro
primeiros pares de folhas.
Os dados relativos à severidade da doença, obtidos por meio das escala de
pontos (EP) foram utilizados para o cálculo da área abaixo da curva de progresso da
A B

33
doença (AACPD) (CAMPBELL & MADDEN, 1990; SIMKO & PIEPHO, 2012), pela
seguinte fórmula:
∑(𝑦𝑖 + 𝑦𝑖+1
2
𝑛−1
𝑖=1
) ∗ (𝑡𝑖+1 − 𝑡𝑖)
Onde: n é o número total de observações; yi e yi+1 são os valores das escalas de
pontos relativos à severidade da doença na i-ésima observação; (ti+1 - ti) é o intervalo
entre as avaliações.
As médias dos tratamentos, relativos à EP e AACPD, foram comparadas através
do intervalo de confiança (IC), sendo a precisão dos métodos analisada pelos
respectivos desvios padrão (DP) e coeficiente de variação (CV).
Os valores de incidência foram calculados através da divisão do número folhas
inoculadas que desenvolveram qualquer sintoma da doença pelo número total de folhas
inoculadas e o valor obtido multiplicado por 100.
Experimento 2: Comparação entre os métodos de inoculação por agulhas múltiplas e
Abrasão
Em conformidade com o experimento 1, e com a finalidade de aprimorar a
eficácia do método de inoculação, uma nova técnica, por abrasão com o inóculo foi
desenvolvida. Deste modo, o métodos das agulhas múltiplas e o método abrasivo foram
comparados a partir de análises realizadas com as cultivares SL 28 considerada
resistente, K7 e Bourbon Vermelho com reação desconhecida, IAC 125 RN suscetível e
também progênies do híbrido 6839 (Catuaí Vermelho IAC 46 x BA 10 IAC 1110-8) e
4309 (Catuaí Vermelho IAC 81 x genitor desconhecido) (Tabela 2), obtidas de
polinização aberta de matrizes selecionadas pela baixa incidência da mancha-aureolada
em um campo experimental localizado em Pirajú, SP, onde a doença encontra-se
disseminada.

34
Tabela 2 - Germoplasma utilizado na comparação dos métodos das agulhas múltiplas e
por abrasão, para inoculação de Pseudomonas syringae pv. garcae, em cafeeiros.
Germoplama Acesso Natureza País
SL 28 IAC 1153-1 Col. 5 Cultivar Quênia
K7 IAC 1151-4 Col. 5 Cultivar Quênia
Bourbon Vermelho IAC 662 Col. 5 Cultivar Brasil
Icatu Vermelho IAC 5383 Material em seleção Brasil
IAC Catuaí SH3 H6839-5-7-6-4-10-8-4 Cultivar Brasil
IAC Catuaí SH3 H6839-5-7-6-4-10-8-84 Cultivar Brasil
IAC Catuaí SH3 H6839-5-7-6-4-10-8-101 Cultivar Brasil
IAC 4309 EP 575 C 34 Material em seleção Brasil
IAC 4309 EP 575 C 65 Material em seleção Brasil
IAC 4309 EP 575 C 110 Material em seleção Brasil
IAC 125 RN IAC 1669-13 Cultivar Brasil
H = híbrido; EP = Ensaio de Progênie; Col. = Coleção; C = Cova.
Métodos das Agulhas múltiplas: O procedimento de inoculação pelo método das
agulhas múltiplas encontra-se descrito em 3.3.
Técnica da Abrasão: A técnica desenvolvida nessa pesquisa consiste na fricção de uma
lixa média disposta em área circular (Ø1,5 cm) previamente embebida em suspensão
bacteriana, contra a superfície abaxial das folhas, causando ferimentos que não
transpassam as folhas, depositando-se suspensão bacteriana na superfície da área
inoculada (Figura 4).
Figura 4 - Método de inoculação de Pseudomonas syringae pv. garcae, em folhas de
cafeeiro por abrasão. Fricção da lixa contra a superfície abaxial da folha (A);
Ferimentos ocasionados pela inoculação e deposição da suspensão bacteriana na
superfície lesionada (B).
Para ambos os métodos utilizou-se a escala de notas descrita no anexo 1 (Página
114).
A B

35
O experimento foi conduzido durante os meses de janeiro a março de 2013,
sendo que a temperatura média durante o período experimental foi de 23,9 ° C.
O modelo experimental adotado foi o delineamento inteiramente casualizado,
com números diferentes de repetições por tratamento e, parcela de uma planta. Após as
inoculações as plantas foram mantidas em condições de câmara úmida durante 48 horas,
sendo transferidas posteriormente para casa-de-vegetação com irrigação controlada.
Durante o período experimental a umidade relativa média na casa-de-vegetação foi de
63% e a temperatura média de 23,5°C.
As avalições foram realizadas aos 7, 14, 21, 28, 35 e 42 DAI.
Os dados relativos à severidade da doença (EP) foram utilizados para o cálculo
da AACPD (CAMPBELL & MADDEN, 1990; SIMKO & PIEPHO, 2012) e os dados
relativos a EP e AACPD foram comparados através do IC, E a precisão dos métodos foi
analisada pelos DP e CV apresentados nos experimentos.
Com base na AACPD obtida nesse experimento, foram estimadas as correlações
classificatórias de Spearman (ρ) (STEEL et al., 1997) entre os períodos de avaliação
para cada método de inoculação, visando-se determinar o período de avaliação ideal
para a seleção de plantas resistentes à bactéria, similarmente ao descrito por PEREIRA
et al. (2008).
3.4. Identificação de fontes de resistência a Pseudomonas syringae pv. garcae em
Coffea spp.
3.4.1. Obtenção das mudas de cafeeiros
Sementes do germoplasma, listado nas tabelas 3 e 4, foram germinadas em
caixas de areia e após aproximadamente 40 dias, no estádio de palito de fósforo, foram
transplantadas para tubetes de 180 cm3 contendo substrato vegetal, adicionado de 30g/L
de basacote® (16% N; 8 % P2O5; 12% K2O; 2% MgO; 5% S; 0,4% Fe; 0,02% B; 0,02%
Zn; 0,06% Cu; 0,06% Mn; 0,015% Mo). As plantas foram mantidas em casa-de-
vegetação com irrigação controlada, sendo selecionada as plantas ausentes de doenças e
insetos praga para as inoculações. As mudas foram inoculadas com aproximadamente
seis meses de idade, entre 5 e 7 pares de folhas.
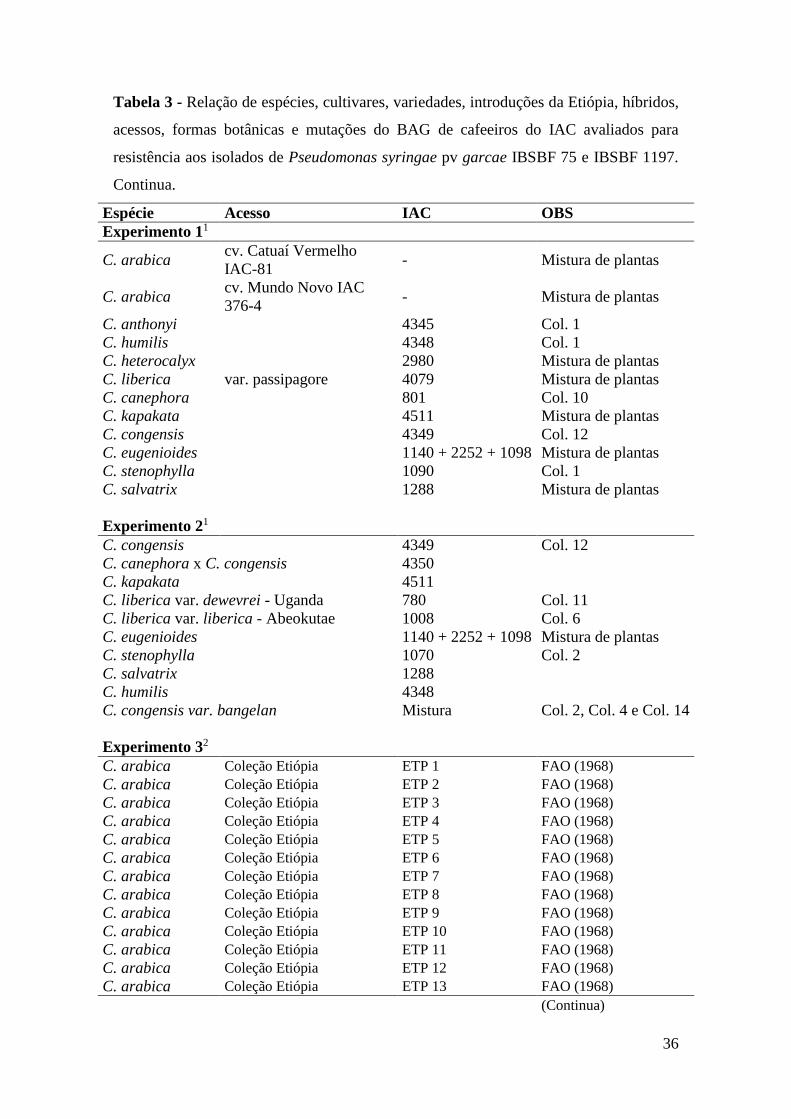
36
Tabela 3 - Relação de espécies, cultivares, variedades, introduções da Etiópia, híbridos,
acessos, formas botânicas e mutações do BAG de cafeeiros do IAC avaliados para
resistência aos isolados de Pseudomonas syringae pv garcae IBSBF 75 e IBSBF 1197.
Continua.
Espécie Acesso IAC OBS
Experimento 11
C. arabica cv. Catuaí Vermelho
IAC-81 - Mistura de plantas
C. arabica cv. Mundo Novo IAC
376-4 - Mistura de plantas
C. anthonyi 4345 Col. 1
C. humilis 4348 Col. 1
C. heterocalyx 2980 Mistura de plantas
C. liberica var. passipagore 4079 Mistura de plantas
C. canephora 801 Col. 10
C. kapakata 4511 Mistura de plantas
C. congensis 4349 Col. 12
C. eugenioides 1140 + 2252 + 1098 Mistura de plantas
C. stenophylla 1090 Col. 1
C. salvatrix 1288 Mistura de plantas
Experimento 21
C. congensis 4349 Col. 12
C. canephora x C. congensis 4350
C. kapakata 4511
C. liberica var. dewevrei - Uganda 780 Col. 11
C. liberica var. liberica - Abeokutae 1008 Col. 6
C. eugenioides 1140 + 2252 + 1098 Mistura de plantas
C. stenophylla 1070 Col. 2
C. salvatrix 1288
C. humilis 4348
C. congensis var. bangelan Mistura Col. 2, Col. 4 e Col. 14
Experimento 32
C. arabica Coleção Etiópia ETP 1 FAO (1968)
C. arabica Coleção Etiópia ETP 2 FAO (1968)
C. arabica Coleção Etiópia ETP 3 FAO (1968)
C. arabica Coleção Etiópia ETP 4 FAO (1968)
C. arabica Coleção Etiópia ETP 5 FAO (1968)
C. arabica Coleção Etiópia ETP 6 FAO (1968)
C. arabica Coleção Etiópia ETP 7 FAO (1968)
C. arabica Coleção Etiópia ETP 8 FAO (1968)
C. arabica Coleção Etiópia ETP 9 FAO (1968)
C. arabica Coleção Etiópia ETP 10 FAO (1968)
C. arabica Coleção Etiópia ETP 11 FAO (1968)
C. arabica Coleção Etiópia ETP 12 FAO (1968)
C. arabica Coleção Etiópia ETP 13 FAO (1968)
(Continua)
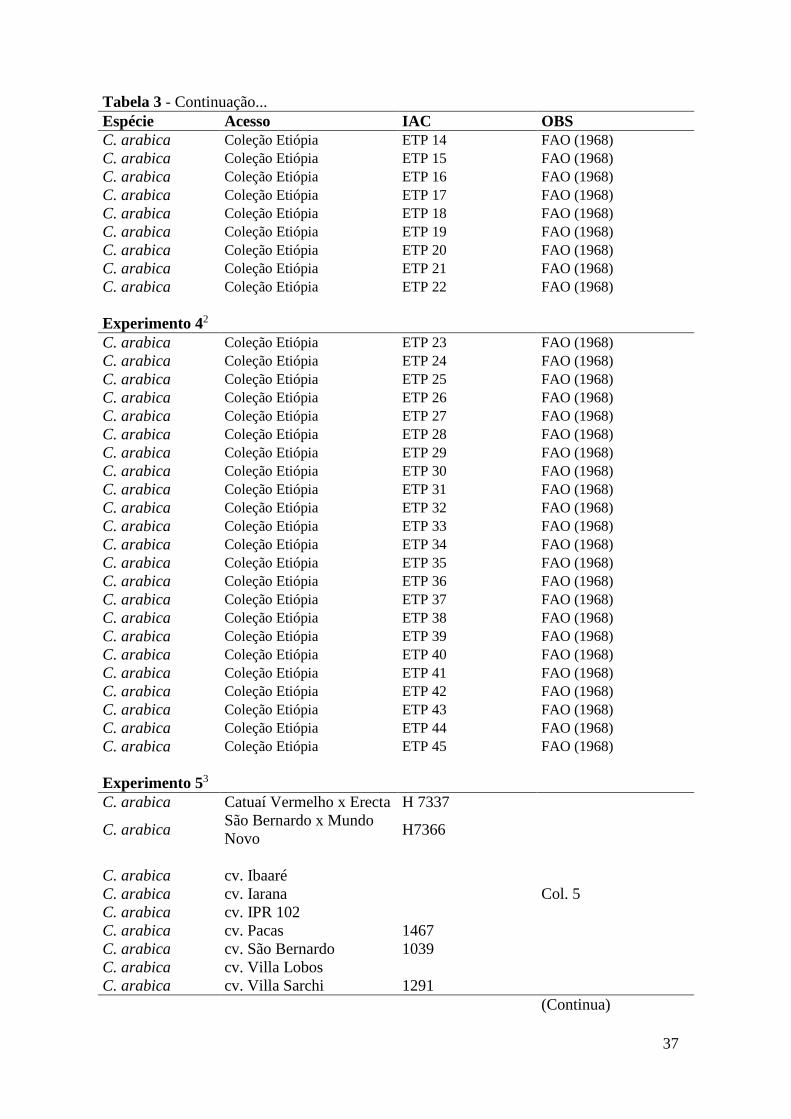
37
Tabela 3 - Continuação...
Espécie Acesso IAC OBS
C. arabica Coleção Etiópia ETP 14 FAO (1968)
C. arabica Coleção Etiópia ETP 15 FAO (1968)
C. arabica Coleção Etiópia ETP 16 FAO (1968)
C. arabica Coleção Etiópia ETP 17 FAO (1968)
C. arabica Coleção Etiópia ETP 18 FAO (1968)
C. arabica Coleção Etiópia ETP 19 FAO (1968)
C. arabica Coleção Etiópia ETP 20 FAO (1968)
C. arabica Coleção Etiópia ETP 21 FAO (1968)
C. arabica Coleção Etiópia ETP 22 FAO (1968)
Experimento 42
C. arabica Coleção Etiópia ETP 23 FAO (1968)
C. arabica Coleção Etiópia ETP 24 FAO (1968)
C. arabica Coleção Etiópia ETP 25 FAO (1968)
C. arabica Coleção Etiópia ETP 26 FAO (1968)
C. arabica Coleção Etiópia ETP 27 FAO (1968)
C. arabica Coleção Etiópia ETP 28 FAO (1968)
C. arabica Coleção Etiópia ETP 29 FAO (1968)
C. arabica Coleção Etiópia ETP 30 FAO (1968)
C. arabica Coleção Etiópia ETP 31 FAO (1968)
C. arabica Coleção Etiópia ETP 32 FAO (1968)
C. arabica Coleção Etiópia ETP 33 FAO (1968)
C. arabica Coleção Etiópia ETP 34 FAO (1968)
C. arabica Coleção Etiópia ETP 35 FAO (1968)
C. arabica Coleção Etiópia ETP 36 FAO (1968)
C. arabica Coleção Etiópia ETP 37 FAO (1968)
C. arabica Coleção Etiópia ETP 38 FAO (1968)
C. arabica Coleção Etiópia ETP 39 FAO (1968)
C. arabica Coleção Etiópia ETP 40 FAO (1968)
C. arabica Coleção Etiópia ETP 41 FAO (1968)
C. arabica Coleção Etiópia ETP 42 FAO (1968)
C. arabica Coleção Etiópia ETP 43 FAO (1968)
C. arabica Coleção Etiópia ETP 44 FAO (1968)
C. arabica Coleção Etiópia ETP 45 FAO (1968)
Experimento 53
C. arabica Catuaí Vermelho x Erecta H 7337
C. arabica São Bernardo x Mundo
Novo H7366
C. arabica cv. Ibaaré
C. arabica cv. Iarana Col. 5
C. arabica cv. IPR 102
C. arabica cv. Pacas 1467
C. arabica cv. São Bernardo 1039
C. arabica cv. Villa Lobos
C. arabica cv. Villa Sarchi 1291
(Continua)

38
Tabela 3 - Continuação
Espécie Acesso IAC OBS
C. arabica var. Abssínica Col. 8
C. arabica var. Crassinérvia
C. arabica var. Dilla & Alghe 1156-3 Col. 5
C. arabica var. Gláucia Col. 8
C. arabica var. Mokka Col. 5
C. arabica var. Pacas
C. arabica var. Palido viridis Col. 1
C. arabica var. Pêndula 81/25 Col. 2
Experimento 64
C. arabica Icatu Ct Ct 4553 EP 536 C 456
C. arabica Catuaí Vermelho IAC 81
x Geisha IAC 1137-5 H 7316 L 109 C 648 (F6)
C. arabica Catuaí Vermelho IAC 81
x Geisha IAC 1137-5 H 7316 L 109 C 1036 (F6)
C. arabica
Catuaí Vermelho x
(Catuaí Vermelho x HT
32/1)
H 13439-8 L 109 C 1736 (F6RC1)
C. arabica cv. IAC Catuaí SH3 EP565 P 89 PL5
C. arabica cv. IAC Catuaí SH3 EP565 P 99 PL 5
C. arabica cv. Mundo Novo LC EP568 P 32 PL 7
C. arabica cv. Icatu Amarelo 2944-6 EP567 P 46 PL 1
C. arabica cv. IAC Catuaí SH3 L 108 C 1621
C. arabica cv. IAC Catuaí SH3 L 108 C 1611
C. arabica cv. IAC Ourama Mococa Pl 13
C. arabica cv. IAC Ourama Mococa Pl 51 1Espécies do gênero Coffea; 2Acessos de cafeeiros oriundos da Etiópia (Coleção FAO 1968); 3Variedades, cultivares exóticas e híbridos intervarietais de C. arabica; 4Cultivares de C.
arabica.
Nos experimentos 3 e 4, avaliou-se o desempenho de 45 introduções
provenientes da Etiópia frente à infecção pela bactéria, sendo as introduções
previamente identificadas como resistentes em trabalho desenvolvido por MOHAN et
al. (1978), no Instituto Agronômico do Paraná (IAPAR), utilizando cafeeiros da coleção
de germoplasma de Coffea do IAC, recebidos do CATIE (Centro Agronómico Tropical
de Investigación y Enseñanza), Turrialba, Costa Rica.
Progênies de cafeeiros pertencentes a gerações diversas, oriundas de cruzamento
entre as espécies C. arabica e C. eugenioides, foram avaliadas em um sétimo
experimento, em relação à resistência à mancha-aureolada neste estudo, encontradas
listadas não tabela 4.

39
Tabela 4 – Cafeeiros derivados do cruzamento entre Coffea arabica e C. eugenioides
avaliados em relação à resistência de suas progênies à mancha-aureolada.
Cafeeiro Geração Genealogia
H3497-14 F1 C. arabica var. típica (10 ex. 4) x C. eugenioides (1098–6)
H3423-1 F1 C. arabica cv. Laurina (1097–18) x C. eugenioides (1098–5)
H5999-1 F1 C. eugenioides duplicado (1140-16 Dp.) x C. arabica cv Mundo
Novo IAC 376-4
H14772-1 F1 C. arabica cv. Catuaí Vermelho IAC 81 (Coleção 5) x
C. eugenioides duplicado (1140-16 Dp.)
H14503-1 RC1 C. arabica cv. Catuaí Vermelho IAC 81 (Coleção 5) x H5999-1
H = Híbrido. Dp = planta com número de cromossomos duplicado; cv. = cultivar.
3.4.2. Avaliação da resistência do germoplasma de Coffea spp. a Pseudomonas
syringae pv. garcae
O inóculo foi preparado como descrito no ítem 3.1.1. Em conformidade aos
experimentos realizados anteriormente, o inóculo selecionado para os ensaios de
resistência foi composto pelos isolados mistura V (IBSBF 75 e IBSBF 1197).
As inoculações foram realizadas pelo método de abrasão com lixa embebida no
inóculo, técnica que proporcinou os melhores resultados nos experimentos anteriores,
constituído pela mistura V (IBSBF 75 e IBSBF 1197).
Nove experimentos foram efetuados para a avaliação da resistência do
germoplasma, sendo a divisão dos experimentos realizada conforme a similaridade e
disponibilidade dos genótipos.
Os experimentos foram modelados em delineamento inteiramente casualizado,
com número variável de repetições por tratamento e parcelas de uma planta. Durante 48
h após as inoculações, as plantas foram mantidas em codição de camaa úmida. A
cultivar IAC 125 RN de C. arabica foi utilizada como padrão de suscetibilidade, em
todos os experimentos.
As avalições foram realizadas aos 7, 14, 21, 28, 35 e 42 DAI, atribuindo-se
pontos entre zero e cinco conforme escala ilustrada no anexo 1, página 114.
Os dados relativos à severidade da doença (EP) foram utilizados para o cálculo
da AACPD (Campbell & Madden, 1990; Simko & Piepho, 2012), sendo as médias da

40
variável submetidas à análise de variância não paramétrica pelo teste de Kruskal-Wallis
ao nível de 5% de probabilidade.
3.5. Análise do efeito pleiotrópico do alelo SH1 na resistência a Pseudomonas
syringae pv. garcae
Nas hipóteses de o alelo SH1 apresentar efeito pleiotrópico ou de haver ligação
gênica entre alelos que conferem resistência a H. vastatrix e a P. syringae pv. garcae,
como reportado na literatura científica por CARVALHO (1988), cafeeiros portadores
do alelo SH1 deveriam ser resistentes a ambos os patógenos.
3.5.1. Material vegetal de cafeeiros
Foi avaliada a resposta de cafeeiros quanto a infecção por P. syringae pv. garcae
e o fungo H. vastatrix, patógeno causador da ferrugem do cafeeiro.
Foram utilizadas progênies de plantas do hibrido H8089, derivados de
cruzamento entre a cultivar Catuaí Vermelho IAC 24 e o cafeeiro IAC 1137-5 da
seleção Geisha, proveniente da Etiópia e introduzida na coleção de cafeeiros do Instituto
Agronômico (IAC) em 1953, oriunda do Departamento de Agricultura dos Estados
Unidos sob o registro PI 205.928
O material avaliado foi composto pelas progênies na geração F2, dos cafeeiros
H8089-2 e H8089-7, representadas por 34 e 29 plantas respectivamente.
3.5.2. Avaliação quanto à resposta de cafeeiros à infecção por Pseudomonas
syringae pv. garcae e a Hemileia vastatrix
As progênies na geração F2, dos cafeeiros H8089-2 e H8089-7 foram inoculadas
com mistura dos isolados IBSBF 75 e IBSBF 1197 de P. syringae pv. garcae, em
concentração aproximada de 108 UFC / mL-1, pelo método de abrasão, conforme
descrito no ítem 3.3.2.2.
As avaliações foram realizadas atribuindo-se de zero e cinco pontos (descita no
item 3.3.). A escala utilizada encontra-se Ilustrada no anexo 1, página 114.
Para confirmação da presença do alelo dominante SH1, as 63 progenies doas
cafeeiros foram inoculadas com dois isolados de H. vastatrix selecionados a partir de
testes em plantas diferenciadoras, mantidas pelo Centro de Café Alcides Carvalho, do

41
IAC, sendo o isolado I/2015, portador dos alelos V2 e V5 e o isolado II/2015, portador
dos alelos de virulência V1, V2 e V5.
Discos obtidos a partir de folhas jovens dos mesmos cafeeiros foram inoculados
com suspensão de uredosporos dos isolados de H. vastarix, em concentração de 1 mg.mL-1
(1,5 x 105 uredosporos) dos isolados I/2015 e II/2015, sendo posteriormente mantidos em
câmaras úmidas, segundo método de ESKES & TOMA-BRAGHINI (1981). Avaliou-se a
densidade de lesões, através de escala de 0 a 9 pontos, sendo 0, atribuído às plantas
resistentes, sem sintomas e 9 pontos, às plantas com alta incidência de lesões grandes,
regulares e esporulação intensa. O tipo de reação das lesões foi avaliado por escala de 0 a 4
pontos, onde 0 foi atribuído às plantas imunes, sem lesões e 4 pontos, àquelas suscetíveis,
com esporulação intensa.
O possível efeito pleiotrópico do alelo SH1 para resistência simultânea a P.
syringae pv. garcae e H. vastatrix, ou a eventual ligação gênica entre alelos de
resistência aos patógenos foi investigado por meio das respostas das plantas à infecção
por P. syringae pv. garcae e H. vastatrix isolado I/2015, sendo a concordância entre os
métodos avaliada por meio dos seguintes parâmetros:
Exatidão do método (EM) = (a+d)/n: calculada a partir do somatório de plantas
resistentes à ferrugem (a) e suscetíveis à mancha-aureolada (d), dividido pelo número
total (n) de plantas.
Taxa de falso positivo (TFP) = b/(b+d): calculada pela divisão do número de plantas
resistentes à ferrugem, mas suscetíveis à mancha-aureolada (b) pelo total de plantas
suscetíveis à mancha-aureolada (b+d).
Taxa de falso negativo (TFN) = c/(a+c): calculada pela divisão do número de plantas
suscetíveis à ferrugem, mas resistentes à mancha-aureolada (c) pelo total de plantas
resistentes à mancha-aureolada (a+c).
Taxa total de erro (TE) = (b+c)/n: calculada pelo somatório do número de plantas
resistentes à ferrugem, mas suscetíveis à mancha-aureolada (b) e do número de plantas
suscetíveis à ferrugem, mas resistentes à mancha-aureolada (c), dividido pelo número
total (n) de plantas.
Qui-quadrado de McNemar com a correção de continuidade de Yates (2 McNemar) = (b-
c-0,5)2/b+c: utilizado para estimar a significância entre as duas avaliações.

42
P valor = medida quantitativa da robustez da significância calculada no teste de 2
McNemar.
Coeficiente de associação de Yule (Q): medida do grau de associação entre as classes
determinadas nas avaliações das plantas em relação à resistência à ferrugem e à
mancha-aureolada.
3.6. Análise da expressão diferencial de genes possivelmente ligados à resistência a
Pseudomonas syringae pv. garcae
A análise de expressão diferencial foi realizada avaliando-se a tendência de
expressão dos genes alvo, na cultivar IPR 102 (resistente) e IAC 125 RN (suscetível).
Folhas de cafeeiro do primeiro internódio, completamente expandidas, foram
inoculadas por suspensão bacteriana a partir da mistura dos isolados IBSBF 75 e IBSBF
1197 de P. syringae pv. garcae, em concentração aproximada de 108 UFC / mL-1, pelo
método de abrasão, conforme descrito no ítem 3.3.2.2. Utilizou-se como controle as
folhas inoculadas com solução salina. Após as inoculações, as plantas foram mantidas
em condições de câmara úmida até o momento das coletas. Amostras foliares foram
coletadas, antes das inoculações (T0), 15 minutos após as inoculações, 3, 6, 12 e 24
horas após as inoculações. Assim que coletadas as folhas foram imersas em nitrogênio
líquido e, em seguida, armazenadas em ultra-freezer (-80° C).
3.6.1. Extração de RNA total das folhas de cafeeiro, tratamento com RNase e
síntese de cDNA
3.6.1.1. Extração de RNA total das folhas de cafeeiro
O material utilizado em todo o processo de manipulação de RNA foi tratado com
DEPC (0,05%) para inativar RNases.
Procedeu-se a extração dos RNAs com base no protocolo determinado por
CHANG et al. (1993). As amostras foliares foram maceradas em nitrogênio líquido,
adicionadas de
7 mL de tampão de extração, contendo: CTAB 2%, PVP (K10) 2%, Tris HCl pH 8,0
10%, EDTA 5%, NaCl 40% e ddH2O DEPC 41% e, em seguida, incubadas em banho-
maria a 67° C, por 30 minutos.

43
Após esse período, as amostras foram centrifugadas a 10.000 rpm por 10
minutos a 10° C, recuperando-se 1.000 µL do sobrenadante em novos microtubos,
seguido pela adição de clorofórmio (1:0,80 v/v%), homogeneização e centrifugação, a
10.000 rpm por 10 minutos (10° C).
Cuidadosamente, 600 µL da fase aquosa foram recolhidos em novos microtubos,
acrescentados de 1.000 µL de TRIzol, homogeneizados e mantidos no gelo por 5
minutos. Adicionou-se 200 µL de clorofórmio e, a mistura foi centrifugada 10.800 rpm
por 15 minutos a 10° C.
O sobrenadante foi recuperado, tendo sido adicionado isoprapanol gelado (1:1),
homogeneizado por inversão durante 10 minutos e deixados durante a noite em freezer a
-20° C. Posteriormente, procedeu-se à centrifugação a 10.800 rpm por 10 minutos (10°
C). Nesta etapa o precipitado (“pellet”) formado foi lavado de duas a três vezes com
1.000 µL etanol (75%), seguido de centrifugação a 7.000 rpm por 6 minutos. Após a
lavagem, o etanol foi removido e as amostras mantidas em baixa temperatura até a
completa secagem. Finalmente, o precipitado foi eluído em 20 µL de ddH2O DEPC.
3.6.1.2. Tratamento dos RNAs com DNase
Após as extrações, as amostras foram quantificadas em espectrofotômetro
NanoDrop L-Quant 2® (Loccus Biotecnologia) e a integridade dos RNAs totais foi
verificada atravéz de eletroforese em gel de agarose 1%, corado com brometo de etídio
e visualizado sob luz ultravioleta. Posteriormente, os RNAs foram tratados com o kit
RQ1 RNase-Free DNase (Promega©), seguindo-se as recomendações do fabricante e,
em seguida, realizou-se nova quantificação como descrito anteriormente.
3.6.1.3. Síntese de cDNAs
A partir dos RNAs tratados, a síntese da primeira fita de cDNA foi através do kit
ImProm-II™ Reverse Transcription System (Promega), seguindo as especificações do
fabricante. Em seguida, as amostras de cDNA foram quantificadas em
espectrofotômetro L-Quant 2® (Loccus Biotecnologia).

44
3.6.2. Análise da expressão relativa em tempo real de genes ligados á defesa vegetal
3.6.2.1. Genes normalizadores e/ou endógenos
Para a seleção dos genes utilizados como normalizadores nas análises de qRT-
PCR, foram avaliados sete genes frequentemente empregados como controles
endógenos em estudos de microarranjos realizados no Laboratório de Biologia
Molecular do Centro de Café Alcides Carvalho do IAC. Os genes testados foram:
Actina (Act), ADH, GAPDH (gliceraldeído-3-fosfato desidrogenase) 12323, GAPDH
7234, GAPDH 2, Poli Ubiquitina A (PolyA) e 1089 ribossomal (40S). Os genes foram
ranqueados e, aqueles selecionados, foram testados em todas as amostras
separadamente. O consenso das análises definiu o gene GAPDH.2 como normalizador
para as análises subsequentes de expressão gênica dos genes alvo. A eficiência da
reação também foi analisada, sendo todos os testes realizados em triplicata utilizando-se
amostras de cDNA . Definiu-se 500ng como a quantidade mínima de cDNA inicial a ser
utilizada nas reações de PCR quantitativas em tempo real (qRT-PCR).
3.6.2.2. Genes alvo
Avaliou-se no estudo a expressão dos genes de defesa “enhanced disease
susceptibility 1” (EDS-1), “phytoalexin deficiente 4” (PAD4), ascorbato peroxidase
(APX), catalase (CAT), superóxido dismutase (SOD), peroxidase (POX), β-glucanase,
(PR-2), isocitrato liase (2669-ICL) e glutationa redutase (Glut R). As sequências e
descrições dos primers utilizados estão apresentadas na tabela 5.
Tabela 5 - Sequência dos primers utilizados na reação de qRT-PCR.
Gene Sequência do primer (5’ 3’) no. Nucleotídeos
GAPDH.2 F- TTG GGC GGT GCA AA 17
R- AAC ATG GGT GCA TCC TTG CT 20
EDS1 F- TGA AGT GCG CAA GGT AGG T 19
R- AGG TTC AAC CAA CCG CCT A 19
PAD4 F- GCC TCA ACA GTG CCA ATC TG 20
R- CCT GGC AAG CTT ATG CCT AT 20
(Continua)

45
Tabela 5 – Continuação...
Gene Sequência do primer (5’ 3’) no. Nucleotídeos
APX F- TTC CGA ATA GAA CCG TTT GG 20
R- GGA TGC GGA GTA CCT GAA AG 20
CAT F- GCC GGT TGG TAT TGA ACA AC 20
R- CAG TAG AGG GGA TGG GAT GC 20
SOD F- AGG ATG TCA GTC GGG TTT TG 20
R- CTG TGA TGT CTG GGA ACA CG 20
POX F- CTC CTT CTT TGC CTT CTC TGC 21
R- GTG TTC TTG TGG CGC TTG TA 20
PR-2 F- TAA GTG GGC GAA GAT GGT G 19
R- GCT GAG AGG CTA CGC AAA T 19
2669 - ICL F- GGC CAG GAG CAA CAG ACA TT 20
R- ATT CTC TCA CAA TCT TGA CTT TGC A 25
GLUT R. F- CAA GCG GCT ACT GTC AAA TG 20
R- GCT CAC AAA CTA CAC GAA CCC TA 23
F: sequência forward do primer; R: sequência reverse do primer.
3.6.2.3. Reações de qRT-PCR
Os ensaios de expressão em tempo real de genes relacionados com a resposta de
defesa foram efetuados em sistema de detecção mediada pelo marcador SYBR Green®.
As reações foram realizadas para o volume final de 15 µL. A mistura de reação foi
composta por 500ng de cDNA, 0,45 µL de cada primer (10 nM) e 7,5 µL de SYBR
Green®. Todas as reações foram realizadas em triplicata em placas de 96 poços e, em
sistema de detecção de sequências ABI PRISM 7300 (Applied Biosystems).
O programa padrão de amplificação das amostras consistiu de um ciclo a 50
°C/2min e um ciclo 95 °C/10min; seguidos de 40 ciclos 95 °C/15 seg; 60 °C/1min.
Curvas de dissociação foram geradas após o último ciclo de amplificação.
3.6.2.4. Análise dos resultados
Os resultados das reações de qRT-PCR referentes à expressão dos genes alvo,
foram analisados utilizando-se a quantificação relativa, calculada a partir do valor do
ciclo de detecção Ct (“Cycle Threshold”), onde a fluorescência é detectável, e seu valor

46
é inversamente proporcional ao logaritmo do número inicial de cópias. A expressão
relativa dos genes transcritos foi determinada pelo calculo do 2−∆∆𝐶𝑡, com base na
seguinte fórmula:
2−[(𝐶𝑡 𝑎𝑙𝑣𝑜−𝐶𝑡 𝑒𝑛𝑑ó𝑔𝑒𝑛𝑜)𝑎𝑚𝑜𝑠𝑡𝑟𝑎−(𝐶𝑡 𝑎𝑙𝑣𝑜−𝐶𝑡 𝑎𝑙𝑣𝑜)𝑐𝑎𝑙𝑖𝑏𝑟𝑎𝑑𝑜𝑟] = 2−∆∆𝐶𝑡
Onde: Gene alvo = gene de interesse; Gene endógeno = GAPHD2. Utilizou-se
como calibrador nos cálculos da a expressão do gene alvo na cultivar IAC 125 RN sem
contato com a bactéria.
Ajustes na concentração do “RNA template” foram realizados a fim de se
detectar alguns genes, com expressões de menor magnitude.
4. RESULTADOS
4.1. Alterações morfológicas em tecidos foliares de cafeeiros infectados por
Pseudomonas syringae pv. garcae
Observou-se rápido desenvolvimento dos sintomas da mancha-aureolada em
folhas do terceiro internódio de C. arabica cv. Mundo Novo IAC 376-4. A análise de
variância dos valores mensurados nas lesões da epiderme das folhas (Tabela 6) mostrou
diferença significativa (F = 8,57; P = 0,0021) entre o dano médio observado nas folhas
aos 10 e aos 20 DAI. Lesões aos 20 DAI (12,89 milímetros) foram caracterizadas por
uma grande área de tecido necrótico com aproximadamente, o dobro do tamanho das
lesões aos 10 DAI (6,75 mm). Aos 20 DAI as lesões apresentavam-se circundadas por
um halo amarelo pronunciado, consequência da clorose do tecido foliar.
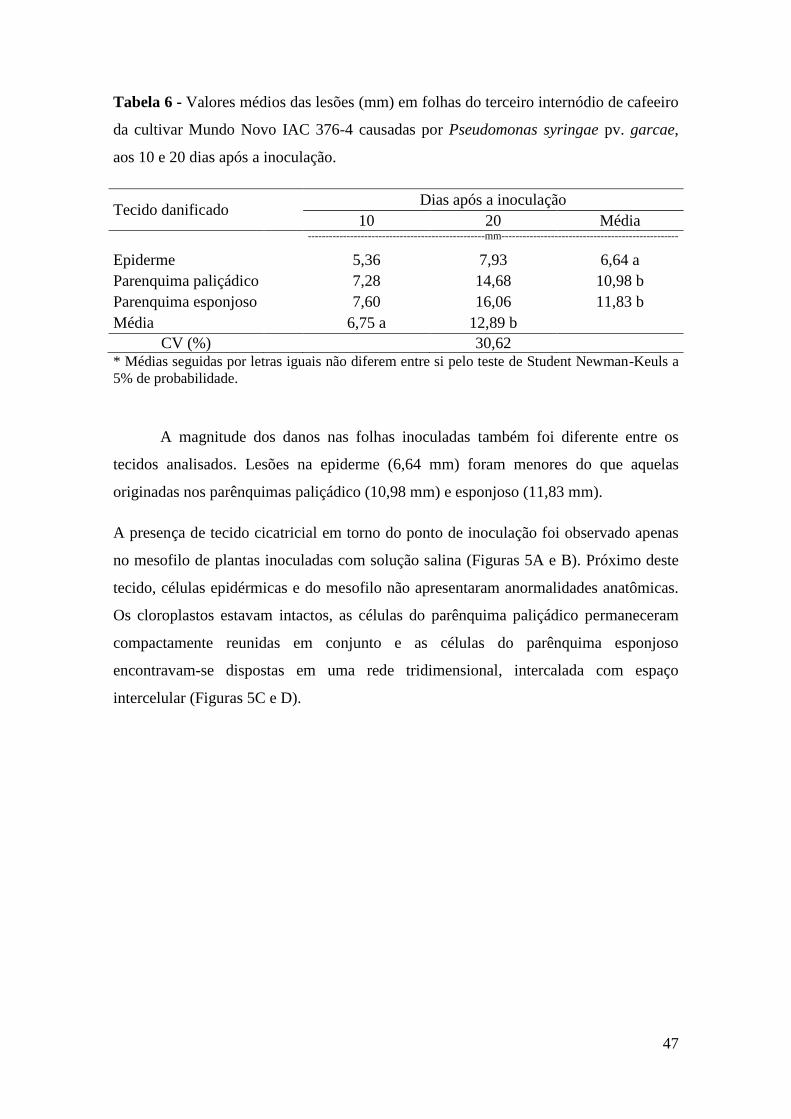
47
Tabela 6 - Valores médios das lesões (mm) em folhas do terceiro internódio de cafeeiro
da cultivar Mundo Novo IAC 376-4 causadas por Pseudomonas syringae pv. garcae,
aos 10 e 20 dias após a inoculação.
Tecido danificado Dias após a inoculação
10 20 Média
--------------------------------------------------mm--------------------------------------------------
Epiderme 5,36 7,93 6,64 a
Parenquima paliçádico 7,28 14,68 10,98 b
Parenquima esponjoso 7,60 16,06 11,83 b
Média 6,75 a 12,89 b
CV (%)
30,62 * Médias seguidas por letras iguais não diferem entre si pelo teste de Student Newman-Keuls a
5% de probabilidade.
A magnitude dos danos nas folhas inoculadas também foi diferente entre os
tecidos analisados. Lesões na epiderme (6,64 mm) foram menores do que aquelas
originadas nos parênquimas paliçádico (10,98 mm) e esponjoso (11,83 mm).
A presença de tecido cicatricial em torno do ponto de inoculação foi observado apenas
no mesofilo de plantas inoculadas com solução salina (Figuras 5A e B). Próximo deste
tecido, células epidérmicas e do mesofilo não apresentaram anormalidades anatômicas.
Os cloroplastos estavam intactos, as células do parênquima paliçádico permaneceram
compactamente reunidas em conjunto e as células do parênquima esponjoso
encontravam-se dispostas em uma rede tridimensional, intercalada com espaço
intercelular (Figuras 5C e D).
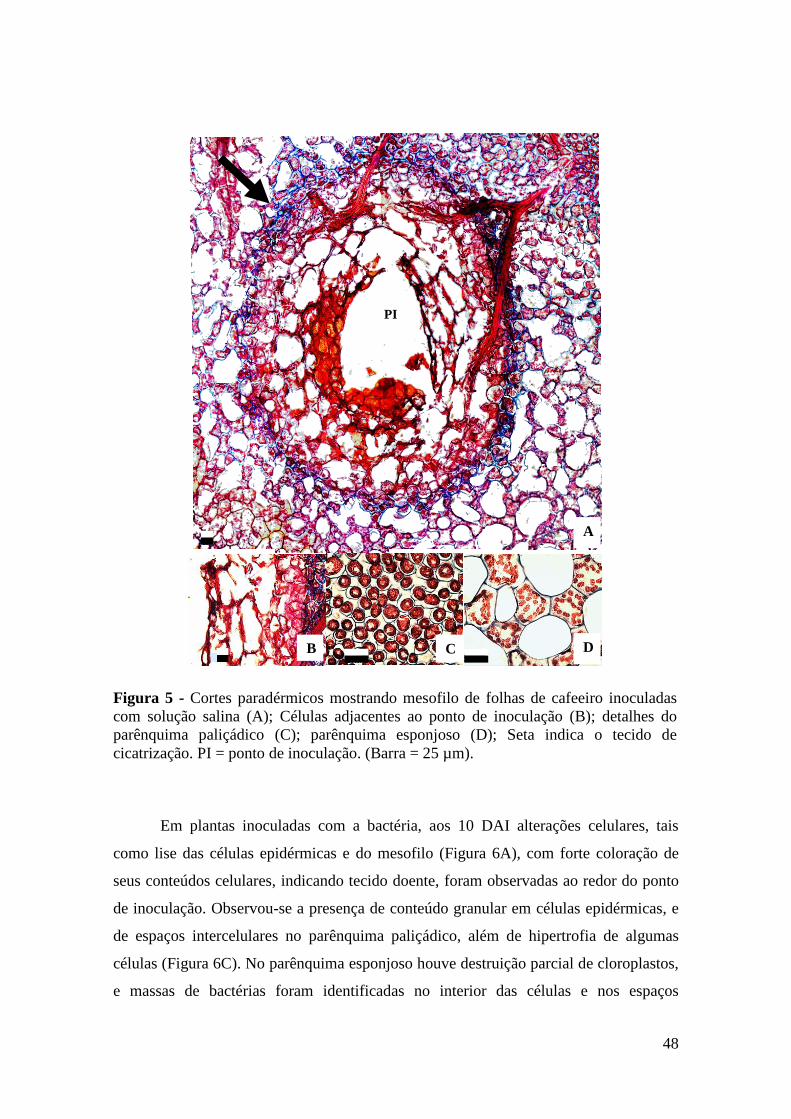
48
Figura 5 - Cortes paradérmicos mostrando mesofilo de folhas de cafeeiro inoculadas
com solução salina (A); Células adjacentes ao ponto de inoculação (B); detalhes do
parênquima paliçádico (C); parênquima esponjoso (D); Seta indica o tecido de
cicatrização. PI = ponto de inoculação. (Barra = 25 µm).
Em plantas inoculadas com a bactéria, aos 10 DAI alterações celulares, tais
como lise das células epidérmicas e do mesofilo (Figura 6A), com forte coloração de
seus conteúdos celulares, indicando tecido doente, foram observadas ao redor do ponto
de inoculação. Observou-se a presença de conteúdo granular em células epidérmicas, e
de espaços intercelulares no parênquima paliçádico, além de hipertrofia de algumas
células (Figura 6C). No parênquima esponjoso houve destruição parcial de cloroplastos,
e massas de bactérias foram identificadas no interior das células e nos espaços
D B
A
C
PI
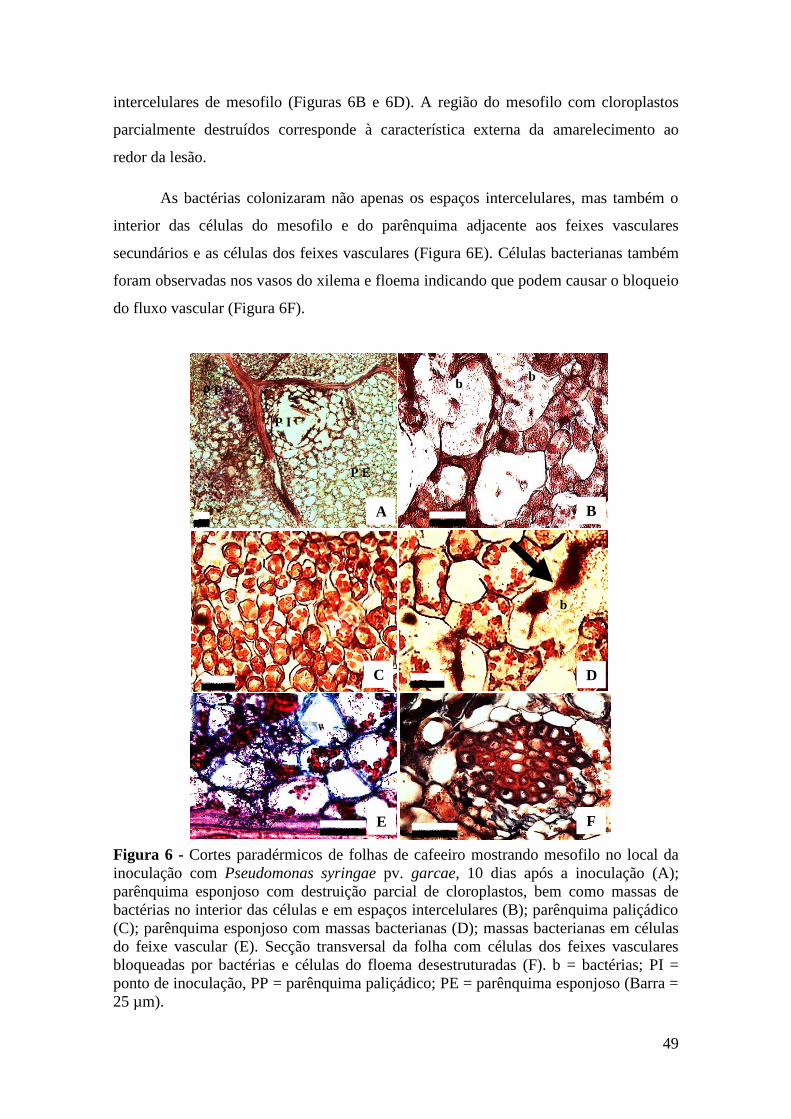
49
intercelulares de mesofilo (Figuras 6B e 6D). A região do mesofilo com cloroplastos
parcialmente destruídos corresponde à característica externa da amarelecimento ao
redor da lesão.
As bactérias colonizaram não apenas os espaços intercelulares, mas também o
interior das células do mesofilo e do parênquima adjacente aos feixes vasculares
secundários e as células dos feixes vasculares (Figura 6E). Células bacterianas também
foram observadas nos vasos do xilema e floema indicando que podem causar o bloqueio
do fluxo vascular (Figura 6F).
Figura 6 - Cortes paradérmicos de folhas de cafeeiro mostrando mesofilo no local da
inoculação com Pseudomonas syringae pv. garcae, 10 dias após a inoculação (A);
parênquima esponjoso com destruição parcial de cloroplastos, bem como massas de
bactérias no interior das células e em espaços intercelulares (B); parênquima paliçádico
(C); parênquima esponjoso com massas bacterianas (D); massas bacterianas em células
do feixe vascular (E). Secção transversal da folha com células dos feixes vasculares
bloqueadas por bactérias e células do floema desestruturadas (F). b = bactérias; PI =
ponto de inoculação, PP = parênquima paliçádico; PE = parênquima esponjoso (Barra =
25 µm).
b
B
b b
C D
E F
A
P E
P P
P I
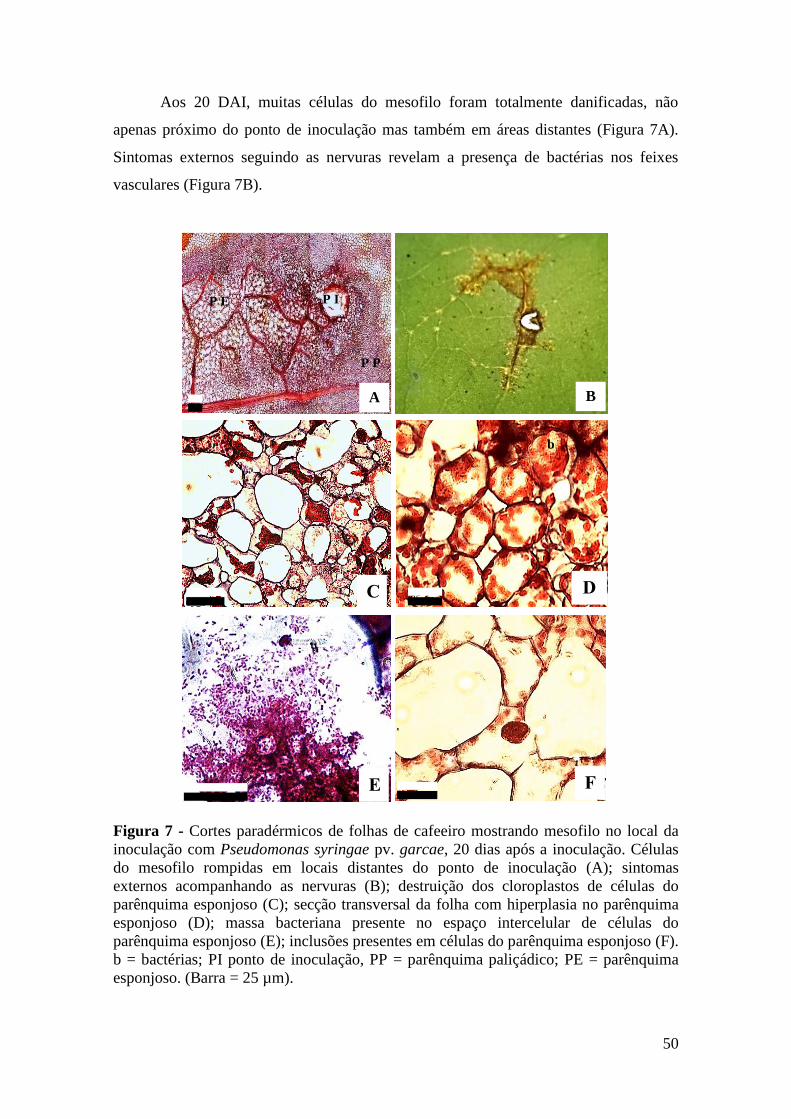
50
Aos 20 DAI, muitas células do mesofilo foram totalmente danificadas, não
apenas próximo do ponto de inoculação mas também em áreas distantes (Figura 7A).
Sintomas externos seguindo as nervuras revelam a presença de bactérias nos feixes
vasculares (Figura 7B).
Figura 7 - Cortes paradérmicos de folhas de cafeeiro mostrando mesofilo no local da
inoculação com Pseudomonas syringae pv. garcae, 20 dias após a inoculação. Células
do mesofilo rompidas em locais distantes do ponto de inoculação (A); sintomas
externos acompanhando as nervuras (B); destruição dos cloroplastos de células do
parênquima esponjoso (C); secção transversal da folha com hiperplasia no parênquima
esponjoso (D); massa bacteriana presente no espaço intercelular de células do
parênquima esponjoso (E); inclusões presentes em células do parênquima esponjoso (F).
b = bactérias; PI ponto de inoculação, PP = parênquima paliçádico; PE = parênquima
esponjoso. (Barra = 25 µm).
C
A
P E
P P
P I
B
F
D
b
E

51
Em seções transversais da lâmina foliar, a destruição dos cloroplastos no
mesofilo aos 20 DAI foi intensa, com presença de células apresentando hipertrofia e
hiperplasia, principalmente no parênquima esponjoso, que, por sua vez apresentou
decréscimo nos espaços intercelulares (Figuras 7C e D) e intensa quantidade de massa
bacteriana (Figura 7E). Inclusões também foram observados em ambos os lados da
epiderme e no mesofilo (Figura 7F).
4.2. Agressividade de isolados de Pseudomonas syringae pv. garcae
Aos 21 DAI, os sintomas nas plantas induzidos pelos isolados variaram de
pequenas áreas anasarcadas a lesões com 1,2 cm de comprimento na cultivar Mundo
Novo IAC 376-4, e até 1,0 cm na cultivar Bourbon Amarelo IAC J 30. Os resultados
dos ensaios revelaram ampla diversidade patogênica entre os dezenove isolados testados
individualmente ou de forma combinada nos três experimentos.
Os dados referentes ao comprimento médio das lesões encontram-se listados na
tabela 7.
No primeiro experimento (1) os isolados mais agressivos foram: IBSBF 65,
IBSBF 75, IBSBF 1197, IBSBF 1293, IBSBF 2212, IBSBF 1664, IBSBF 2511, IBSBF
2840 e IBSBF 3024. O isolado IBSBF 3005 foi caracterizado com agressividade
intermediária, enquanto que os isolados IBSBF 248, IBSBF 1372, IBSBF 2996, IBSBF
2999, IBSBF 3015, IBSBF 3019, IBSBF 3051 e IBSBF 3065 revelaram-se menos
agressivos, em relação à cultivar Mundo Novo IAC 376-4.
Os resultados do segundo experimento (2) evidenciaram a alta agressividade dos
isolados IBSBF 3024, IBSBF 65, IBSBF 75, IBSBF 1197, IBSBF 1293, IBSBF 1664 e
IBSBF 2840. A mistura I, composta pelos isolados IBSBF 75, IBSBF 1197, IBSBF
1293 e IBSBF 3024 caracterizados como agressivos, também foi classificada como
agressiva, por este motivo, outras diferentes misturas foram estudadas no terceiro
experimento em relação à agressividade (Experimento 3) e também à atividade
bactericinogênica dos isolados envolvidos (Item 4.2.1.). Os isolados que se revelaram
menos agressivos em relação a cultivar Bourbon Amarelo IAC J 30 no experimento 2
foram: IBSBF 248, IBSBF 1372, IBSBF 2511, IBSBF 3015, IBSBF 3022 e IBSBF
2212, IBSBF 2996, IBSBF 2999, IBSBF 3005, IBSBF 3019 e IBSBF 3051.
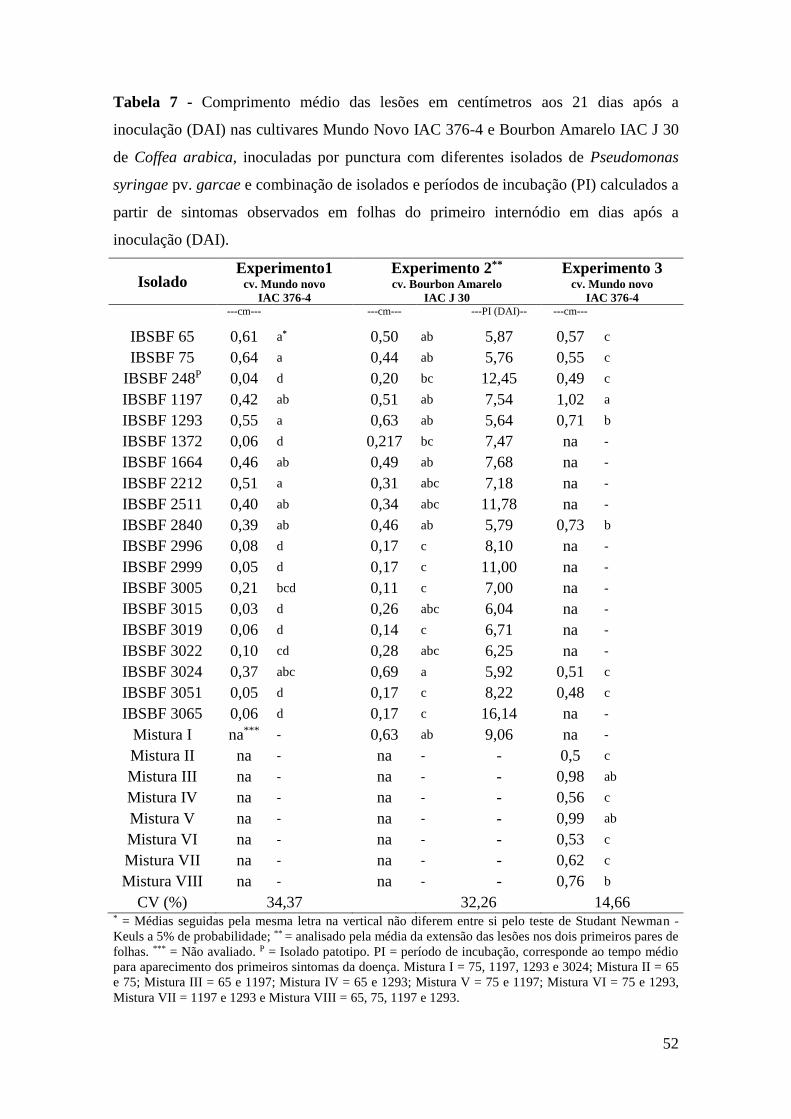
52
Tabela 7 - Comprimento médio das lesões em centímetros aos 21 dias após a
inoculação (DAI) nas cultivares Mundo Novo IAC 376-4 e Bourbon Amarelo IAC J 30
de Coffea arabica, inoculadas por punctura com diferentes isolados de Pseudomonas
syringae pv. garcae e combinação de isolados e períodos de incubação (PI) calculados a
partir de sintomas observados em folhas do primeiro internódio em dias após a
inoculação (DAI).
Isolado Experimento1
cv. Mundo novo
IAC 376-4
Experimento 2**
cv. Bourbon Amarelo
IAC J 30
Experimento 3 cv. Mundo novo
IAC 376-4
---cm--- ---cm--- ---PI (DAI)-- ---cm---
IBSBF 65 0,61 a* 0,50 ab 5,87 0,57 c
IBSBF 75 0,64 a 0,44 ab 5,76 0,55 c
IBSBF 248P 0,04 d 0,20 bc 12,45 0,49 c
IBSBF 1197 0,42 ab 0,51 ab 7,54 1,02 a
IBSBF 1293 0,55 a 0,63 ab 5,64 0,71 b
IBSBF 1372 0,06 d 0,217 bc 7,47 na -
IBSBF 1664 0,46 ab 0,49 ab 7,68 na -
IBSBF 2212 0,51 a 0,31 abc 7,18 na -
IBSBF 2511 0,40 ab 0,34 abc 11,78 na -
IBSBF 2840 0,39 ab 0,46 ab 5,79 0,73 b
IBSBF 2996 0,08 d 0,17 c 8,10 na -
IBSBF 2999 0,05 d 0,17 c 11,00 na -
IBSBF 3005 0,21 bcd 0,11 c 7,00 na -
IBSBF 3015 0,03 d 0,26 abc 6,04 na -
IBSBF 3019 0,06 d 0,14 c 6,71 na -
IBSBF 3022 0,10 cd 0,28 abc 6,25 na -
IBSBF 3024 0,37 abc 0,69 a 5,92 0,51 c
IBSBF 3051 0,05 d 0,17 c 8,22 0,48 c
IBSBF 3065 0,06 d 0,17 c 16,14 na -
Mistura I na*** - 0,63 ab 9,06 na -
Mistura II na - na - - 0,5 c
Mistura III na - na - - 0,98 ab
Mistura IV na - na - - 0,56 c
Mistura V na - na - - 0,99 ab
Mistura VI na - na - - 0,53 c
Mistura VII na - na - - 0,62 c
Mistura VIII na - na - - 0,76 b
CV (%) 34,37
32,26 14,66 * = Médias seguidas pela mesma letra na vertical não diferem entre si pelo teste de Studant Newman -
Keuls a 5% de probabilidade; ** = analisado pela média da extensão das lesões nos dois primeiros pares de
folhas. *** = Não avaliado. P = Isolado patotipo. PI = período de incubação, corresponde ao tempo médio
para aparecimento dos primeiros sintomas da doença. Mistura I = 75, 1197, 1293 e 3024; Mistura II = 65
e 75; Mistura III = 65 e 1197; Mistura IV = 65 e 1293; Mistura V = 75 e 1197; Mistura VI = 75 e 1293,
Mistura VII = 1197 e 1293 e Mistura VIII = 65, 75, 1197 e 1293.

53
Durante o experimento 2 foram realizadas observações diárias dos ferimentos
causados pelas puncturas. Os sintomas iniciais da colonização pela bactéria, foram
observados até o 17° DAI, tendo sido iniciado a partir do segundo DAI. A partir dessas
observações, puderam ser calculados o período médio de incubação de P. syringae pv.
garcae nas folhas mais afetadas, ou seja, as folhas jovens, que variou entre 5,64 e 7,68
DAI (Anexo 2, página 115) pelos isolados mais agressivos, e o período de latência
calculado entre 5 e 7 DAI (Tabela 7). Em média, as folhas jovens do primeiro
internódio apresentaram os sintomas iniciais da doença aos sete DAI, e, esse período foi
crescente conforme a idade das folhas, sendo o dobro nas folhas do quarto intenódio, a
tabela completa dos resultados é apresentados no anexo 2, página 115. Não se constatou
relação entre a agressividade dos isolados (Tabela 7) com a região de origem e/ou o ano
de isolamento das culturas bacterianas (Tabela 1) nas duas cultivares avaliadas.
No terceiro experimento, os tratamentos formados por misturas de isolados se
comportaram como medianamente agressivos, com exceção das misturas III (IBSBF 65
e IBSBF 1197) e V (IBSBF 75 e IBSBF 1197), cuja agressividade semelhante foi
aquele apresentado pelo isolado considerado agressivo IBSBF 1197 (Tabela 7). Neste
experimento, os isolados IBSBF 65, IBSBF 75, IBSBF 3024, anteriormente
classificados como agressivos, revelaram-se pouco agressivos, próximos ao isolado
IBSBF 3051.
Em geral, os isolados que se confirmaram como mais agressivos nos três
experimentos e foram: IBSBF 65, IBSBF 75, IBSBF 1197, IBSBF 1293, IBSBF 1664,
IBSBF 2212, IBSBF 2511, IBSBF 2840 e IBSBF 3024, uma vez que esses isolados
desenvolveram sintomas da doença entre 80-100% dos ferimentos provocados pela
inoculação por punctura e resultaram nas maiores lesões observadas nos experimentos.
Os isolados IBSBF 3015 e 3022 apresentaram comportamento diferencial entre os
experimentos, sendo mais agressivos à cultivar Bourbon Amarelo IAC J 30. Os isolados
IBSBF 248, IBSBF 1372, IBSBF 2996, IBSBF 2999, IBSBF 3005, IBSBF 3019,
IBSBF 3051 e IBSBF 3065 foram classificados como menos agressivos.
4.2.1. Produção e sensibilidade as bacteriocinas entre isolados de Pseudomonas
syringae pv. garcae
Com relação à produção e sensibilidade às bacteriocinas, não foi constatada
atividade bacteriocinogênica pelos isolados confrontados nos testes in vitro, com base
na metodologia descrita por BIAGI & AZEVEDO (1992) e ROMEIRO (2001) (Figura
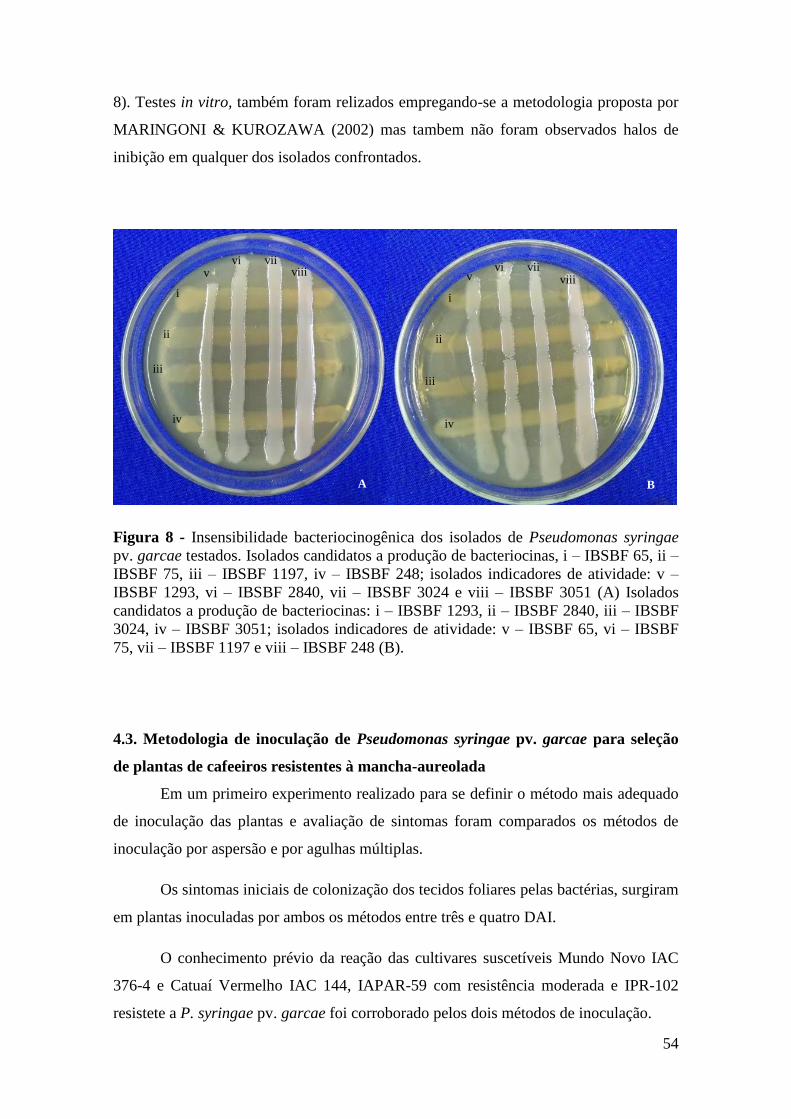
54
8). Testes in vitro, também foram relizados empregando-se a metodologia proposta por
MARINGONI & KUROZAWA (2002) mas tambem não foram observados halos de
inibição em qualquer dos isolados confrontados.
Figura 8 - Insensibilidade bacteriocinogênica dos isolados de Pseudomonas syringae
pv. garcae testados. Isolados candidatos a produção de bacteriocinas, i – IBSBF 65, ii –
IBSBF 75, iii – IBSBF 1197, iv – IBSBF 248; isolados indicadores de atividade: v –
IBSBF 1293, vi – IBSBF 2840, vii – IBSBF 3024 e viii – IBSBF 3051 (A) Isolados
candidatos a produção de bacteriocinas: i – IBSBF 1293, ii – IBSBF 2840, iii – IBSBF
3024, iv – IBSBF 3051; isolados indicadores de atividade: v – IBSBF 65, vi – IBSBF
75, vii – IBSBF 1197 e viii – IBSBF 248 (B).
4.3. Metodologia de inoculação de Pseudomonas syringae pv. garcae para seleção
de plantas de cafeeiros resistentes à mancha-aureolada
Em um primeiro experimento realizado para se definir o método mais adequado
de inoculação das plantas e avaliação de sintomas foram comparados os métodos de
inoculação por aspersão e por agulhas múltiplas.
Os sintomas iniciais de colonização dos tecidos foliares pelas bactérias, surgiram
em plantas inoculadas por ambos os métodos entre três e quatro DAI.
O conhecimento prévio da reação das cultivares suscetíveis Mundo Novo IAC
376-4 e Catuaí Vermelho IAC 144, IAPAR-59 com resistência moderada e IPR-102
resistete a P. syringae pv. garcae foi corroborado pelos dois métodos de inoculação.
i
ii
iii
iv
v vi vii
viii
i
ii
iii
iv
v vi vii
viii
B A

55
No entanto, os danos avaliados pelas variáveis EP e AACPD, foram maiores em
plantas inoculadas pelo métodos de agulhas múltiplas e em folhas jovens do primeiro
internódio, em ambos os métodos de inoculação (Tabela 8).
Os valores de IC gerados pelo método de inculação por de aspersão foram
menores nas folhas do primeiro internódio e nos pares de folhas subsequentes obteve
variação menor, mas de amplitude similar entre eles. Já os valores de IC resultates das
plantas inoculadas por agulhas múltiplas foi maior nas folhas do primeiro internódio,
em relação aos demais.
Independente do par de folhas analisado, o método de aspersão obteve os
menores valores de IC. No entanto, a severidade da doença ocasionada por este método
representada pelas notas da EP foram menores em relação a técnica de agulhas
múltiplas.
Os valores de DP apresentaram tendência semelhante aos valores de IC, no
entanto, houve variação menor entre os métodos, sendo em alguns casos menor nas
folhas inoculadas por agulhas múltiplas.
As plantas inoculadas por agulhas múltiplas resultaram em maiores CV em
quase todos os materiais e pares de folhas testados, a variável AACPD resultou em
menores valores de CV, independente da técnica de inoculação empregada.
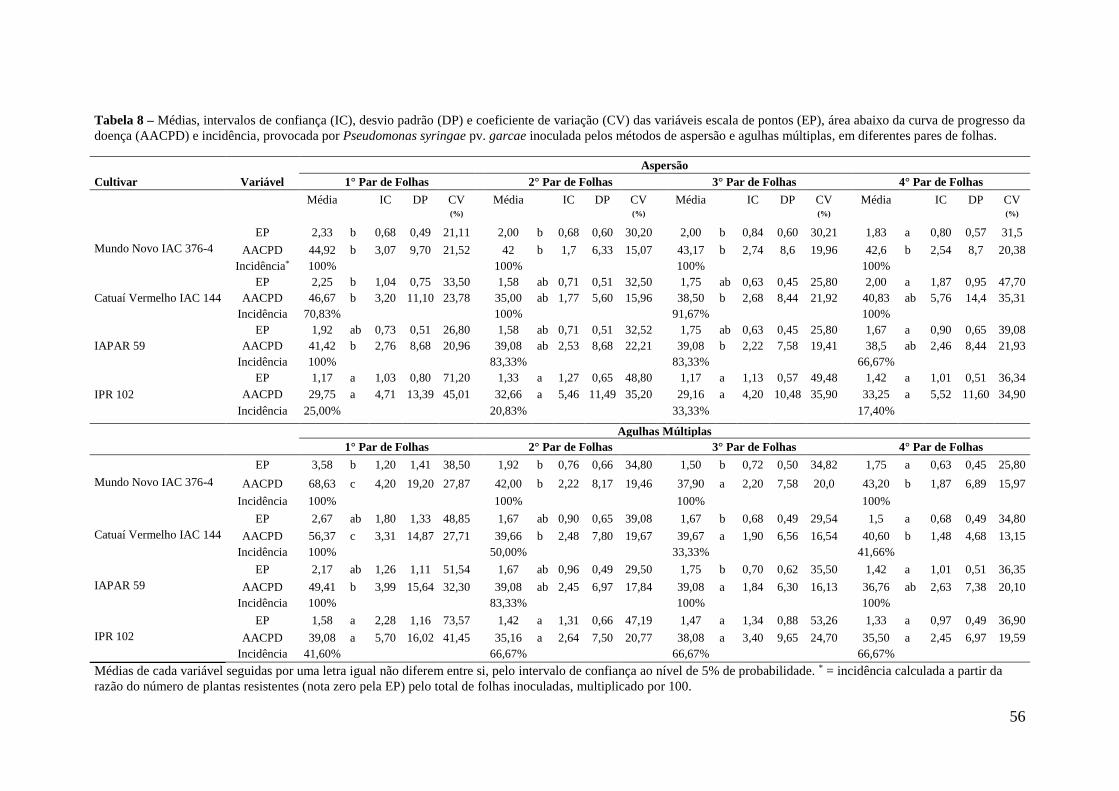
56
Tabela 8 – Médias, intervalos de confiança (IC), desvio padrão (DP) e coeficiente de variação (CV) das variáveis escala de pontos (EP), área abaixo da curva de progresso da
doença (AACPD) e incidência, provocada por Pseudomonas syringae pv. garcae inoculada pelos métodos de aspersão e agulhas múltiplas, em diferentes pares de folhas.
Aspersão
Cultivar Variável 1° Par de Folhas
2° Par de Folhas
3° Par de Folhas
4° Par de Folhas
Média
IC DP CV
Média
IC DP CV
Média
IC DP CV
Média
IC DP CV
(%) (%) (%) (%)
Mundo Novo IAC 376-4
EP 2,33 b 0,68 0,49 21,11
2,00 b 0,68 0,60 30,20
2,00 b 0,84 0,60 30,21
1,83 a 0,80 0,57 31,5
AACPD 44,92 b 3,07 9,70 21,52
42 b 1,7 6,33 15,07
43,17 b 2,74 8,6 19,96
42,6 b 2,54 8,7 20,38
Incidência* 100%
100%
100%
100%
Catuaí Vermelho IAC 144
EP 2,25 b 1,04 0,75 33,50
1,58 ab 0,71 0,51 32,50
1,75 ab 0,63 0,45 25,80
2,00 a 1,87 0,95 47,70
AACPD 46,67 b 3,20 11,10 23,78
35,00 ab 1,77 5,60 15,96
38,50 b 2,68 8,44 21,92
40,83 ab 5,76 14,4 35,31
Incidência 70,83%
100%
91,67%
100%
IAPAR 59
EP 1,92 ab 0,73 0,51 26,80
1,58 ab 0,71 0,51 32,52
1,75 ab 0,63 0,45 25,80
1,67 a 0,90 0,65 39,08
AACPD 41,42 b 2,76 8,68 20,96
39,08 ab 2,53 8,68 22,21
39,08 b 2,22 7,58 19,41
38,5 ab 2,46 8,44 21,93
Incidência 100%
83,33%
83,33%
66,67%
IPR 102
EP 1,17 a 1,03 0,80 71,20
1,33 a 1,27 0,65 48,80
1,17 a 1,13 0,57 49,48
1,42 a 1,01 0,51 36,34
AACPD 29,75 a 4,71 13,39 45,01
32,66 a 5,46 11,49 35,20
29,16 a 4,20 10,48 35,90
33,25 a 5,52 11,60 34,90
Incidência 25,00%
20,83%
33,33%
17,40%
Agulhas Múltiplas
1° Par de Folhas
2° Par de Folhas
3° Par de Folhas
4° Par de Folhas
Mundo Novo IAC 376-4
EP 3,58 b 1,20 1,41 38,50
1,92 b 0,76 0,66 34,80
1,50 b 0,72 0,50 34,82
1,75 a 0,63 0,45 25,80
AACPD 68,63 c 4,20 19,20 27,87 42,00 b 2,22 8,17 19,46 37,90 a 2,20 7,58 20,0 43,20 b 1,87 6,89 15,97
Incidência 100% 100% 100% 100%
Catuaí Vermelho IAC 144
EP 2,67 ab 1,80 1,33 48,85
1,67 ab 0,90 0,65 39,08
1,67 b 0,68 0,49 29,54
1,5 a 0,68 0,49 34,80
AACPD 56,37 c 3,31 14,87 27,71
39,66 b 2,48 7,80 19,67
39,67 a 1,90 6,56 16,54
40,60 b 1,48 4,68 13,15
Incidência 100% 50,00% 33,33% 41,66%
IAPAR 59
EP 2,17 ab 1,26 1,11 51,54
1,67 ab 0,96 0,49 29,50
1,75 b 0,70 0,62 35,50
1,42 a 1,01 0,51 36,35
AACPD 49,41 b 3,99 15,64 32,30
39,08 ab 2,45 6,97 17,84
39,08 a 1,84 6,30 16,13
36,76 ab 2,63 7,38 20,10
Incidência 100% 83,33% 100% 100%
IPR 102
EP 1,58 a 2,28 1,16 73,57
1,42 a 1,31 0,66 47,19
1,47 a 1,34 0,88 53,26
1,33 a 0,97 0,49 36,90
AACPD 39,08 a 5,70 16,02 41,45
35,16 a 2,64 7,50 20,77
38,08 a 3,40 9,65 24,70
35,50 a 2,45 6,97 19,59
Incidência 41,60% 66,67% 66,67% 66,67%
Médias de cada variável seguidas por uma letra igual não diferem entre si, pelo intervalo de confiança ao nível de 5% de probabilidade. * = incidência calculada a partir da
razão do número de plantas resistentes (nota zero pela EP) pelo total de folhas inoculadas, multiplicado por 100.

57
Os sintomas da mancha-aureolada observados nas plantas inoculadas por
aspersão em muitos os casos tiveram inicio somente a partir dos bordos foliares (Figura
9), local onde ocorreu o acúmulo de água.
Figura 9 – Sintoma da mancha-aureolada na cultivar Catuaí Vermelho IAC 144, aos 14
dias após a inoculação, resultado da inoculação por aspersão.
A incidência da doença foi baixa em cafeeiros da cultivar resistente IPR 102,
inoculados por aspersão, com valores médios variando entre 17,40% e 33,33% em
folhas dos quarto primeiros internódios. Quando inoculadas pelo método de agulhas
múltiplas, a incidência da mancha-aureolada na mesma cultivar variou entre 41,60% e
66,67% (Tabela 8).
Em geral, os valores de CV foram considerados baixos. Embora os valores
gerados por ambas as técnicas de inoculção tenham sido similares, o método de
inoculação de aspersão resultou em menores porcentagens de CV. A mesma tendência
também ocorreu para a variável DP, sendo mais acentuada em cafeeiros inoculados por
agulhas múltiplas.
Em um segundo experimento, o método de inoculação por agulhas múltiplas foi
comparado ao método de inoculação por abrasão, sendo os resultados apresentados na
tabela 9.
Os dois métodos de inoculação resultaram em 100% de incidência da doença em
folhas de germoplasma diverso da espécie C. arabica. Apenas a cultivar SL 28,
resistente à bactéria, apresentou folhas com nota zero da EP, em 50% da progênie
avaliada. Sem exceção , as médias das variáveis EP e AACPD das cultivares inoculadas
pelo métodos de abrasão foram superiores àquelas do mesmo germoplasma inoculado
pelo método de agulhas múltiplas (Tabela 9).
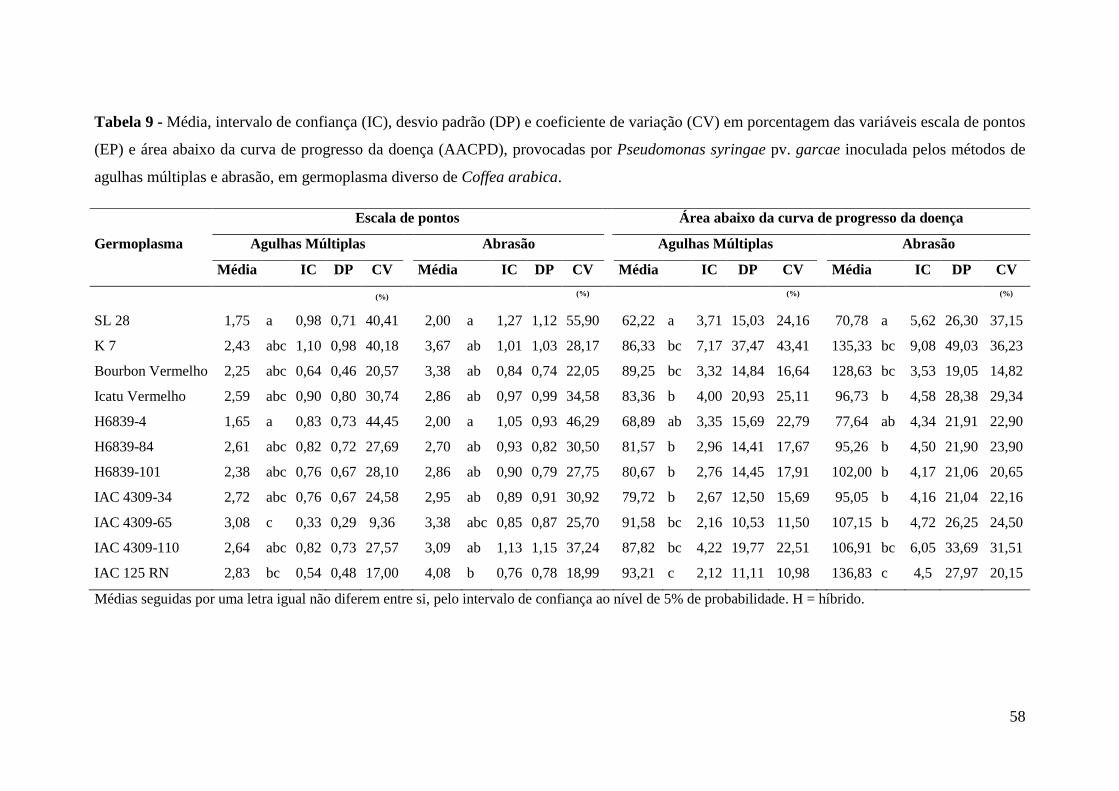
58
Tabela 9 - Média, intervalo de confiança (IC), desvio padrão (DP) e coeficiente de variação (CV) em porcentagem das variáveis escala de pontos
(EP) e área abaixo da curva de progresso da doença (AACPD), provocadas por Pseudomonas syringae pv. garcae inoculada pelos métodos de
agulhas múltiplas e abrasão, em germoplasma diverso de Coffea arabica.
Germoplasma
Escala de pontos Área abaixo da curva de progresso da doença
Agulhas Múltiplas
Abrasão Agulhas Múltiplas Abrasão
Média
IC DP CV
Média
IC DP CV Média IC DP CV Média IC DP CV
(%) (%) (%) (%)
SL 28 1,75 a 0,98 0,71 40,41
2,00 a 1,27 1,12 55,90 62,22 a 3,71 15,03 24,16 70,78 a 5,62 26,30 37,15
K 7 2,43 abc 1,10 0,98 40,18
3,67 ab 1,01 1,03 28,17 86,33 bc 7,17 37,47 43,41 135,33 bc 9,08 49,03 36,23
Bourbon Vermelho 2,25 abc 0,64 0,46 20,57
3,38 ab 0,84 0,74 22,05 89,25 bc 3,32 14,84 16,64 128,63 bc 3,53 19,05 14,82
Icatu Vermelho 2,59 abc 0,90 0,80 30,74
2,86 ab 0,97 0,99 34,58 83,36 b 4,00 20,93 25,11 96,73 b 4,58 28,38 29,34
H6839-4 1,65 a 0,83 0,73 44,45
2,00 a 1,05 0,93 46,29 68,89 ab 3,35 15,69 22,79 77,64 ab 4,34 21,91 22,90
H6839-84 2,61 abc 0,82 0,72 27,69
2,70 ab 0,93 0,82 30,50 81,57 b 2,96 14,41 17,67 95,26 b 4,50 21,90 23,90
H6839-101 2,38 abc 0,76 0,67 28,10
2,86 ab 0,90 0,79 27,75 80,67 b 2,76 14,45 17,91 102,00 b 4,17 21,06 20,65
IAC 4309-34 2,72 abc 0,76 0,67 24,58
2,95 ab 0,89 0,91 30,92 79,72 b 2,67 12,50 15,69 95,05 b 4,16 21,04 22,16
IAC 4309-65 3,08 c 0,33 0,29 9,36
3,38 abc 0,85 0,87 25,70 91,58 bc 2,16 10,53 11,50 107,15 b 4,72 26,25 24,50
IAC 4309-110 2,64 abc 0,82 0,73 27,57
3,09 ab 1,13 1,15 37,24 87,82 bc 4,22 19,77 22,51 106,91 bc 6,05 33,69 31,51
IAC 125 RN 2,83 bc 0,54 0,48 17,00
4,08 b 0,76 0,78 18,99 93,21 c 2,12 11,11 10,98 136,83 c 4,5 27,97 20,15
Médias seguidas por uma letra igual não diferem entre si, pelo intervalo de confiança ao nível de 5% de probabilidade. H = híbrido.

59
Os valores de IC obtidos nos tratamentos cujas plantas foram inoculadas por
agulhas múltiplas, foram menores em relação às plantas por abrasão, independente da
variável analisada.
Como no primeiro experimento, os níveis variáveis de resistência à mancha-
aureolada do germoplasma avaliado foi corroborado pelos dois métodos de inoculação
das plantas.
O CV das variáveis EP e AACPD do germoplasma avaliado no segundo
experimento variaram entre 9,36% e 40,41%, sendo inferiores àqueles observados no
primeiro experimento, com variação entre 13,5% e 73,7%.
Já para o DP, a técnica de inoculação por abrasão resultou em menores valores,
especialmente nas análises por meio da AACPD, sendo esses valores similares entre os
dois métodos de inoculação avaliados.
A variável AACPD foi calculada segundo a equação descrita no item 3.3, com
base em dados da escala de pontos utilizada para avaliação da severidade da doença aos
7, 14, 21, 28, 35 e 42 DAI. Dessa forma, além da AACPD total, aos 42 DAI, também
foram obtidos valores parciais da variável AACPD (14 a 35 DAI) correspondentes aos
períodos de avaliação da EP, os quais submetidos a uma análise de correlação geraram
os resultados apresentados na tabela 10.
Os resultados indicaram que a correlação da AACPD total, ou seja calculada a
partir na somatória de todos os períodos de avaliação, com as AACPD parciais são
sempre maiores em cafeeiros inoculados pelo método de abrasão. Neste método a
correlação da ACPD aos 42 DAI com os valores parciais obtidos aos 7, 14, 21, 28 e 35
foi igual ou superior a 0,90.
Tabela 10 - Estimativas das correlações de Spearman entre os valores da AACPD aos
7, 14, 21, 35 e 42 dias após a inoculação (DAI) de Pseudomonas syringae pv. garcae
em folhas de onze genótipos de cafeeiros, pelos métodos de agulhas múltiplas e abrasão.
DAI Agulhas Múltiplas (DAI) Abrasão (DAI)
14 21 28 35 42 14 21 28 35 42
7 0,96* 0,93* 0,93* 0,87* 0,77* 0,98* 0,97* 0,93* 0,95* 0,91*
14
0,98* 0,98* 0,92* 0,80*
0,96* 0,93* 0,93* 0,90*
21
1,00* 0,96* 0,88*
0,97* 0,96* 0,92*
28
0,96* 0,88*
0,99* 0,97*
35
0,95*
0,98*
* = significativo a 5% de probabilidade.
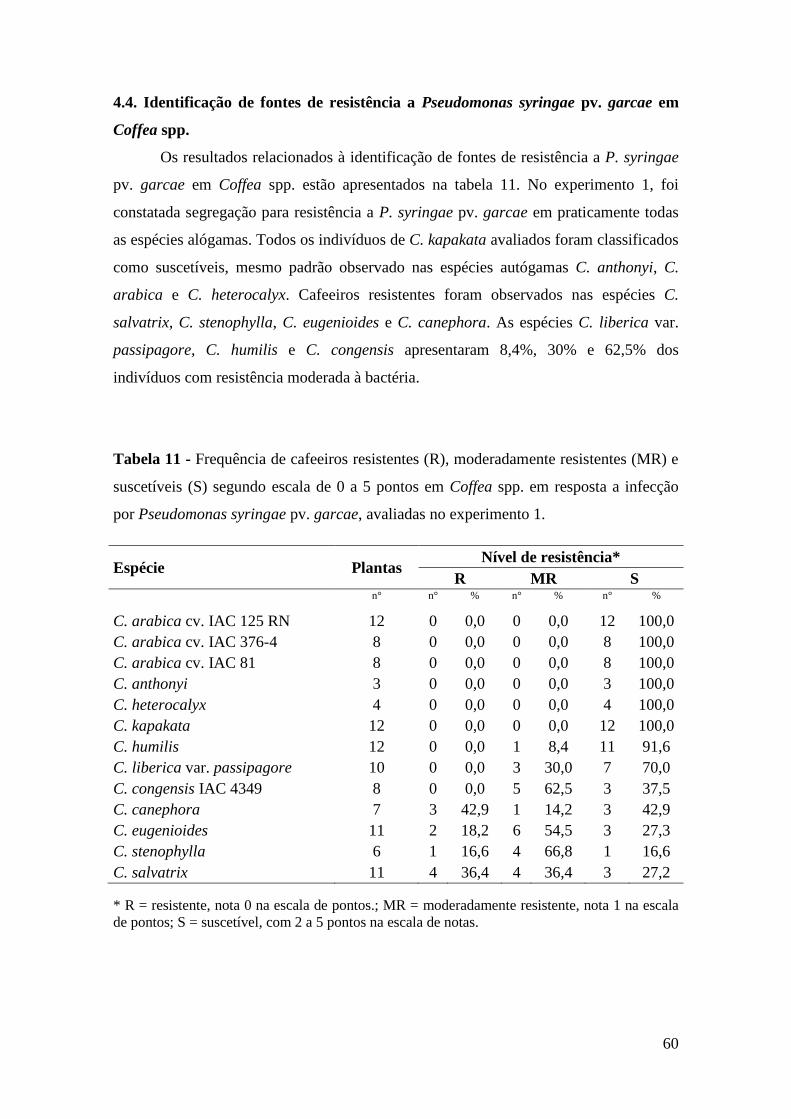
60
4.4. Identificação de fontes de resistência a Pseudomonas syringae pv. garcae em
Coffea spp.
Os resultados relacionados à identificação de fontes de resistência a P. syringae
pv. garcae em Coffea spp. estão apresentados na tabela 11. No experimento 1, foi
constatada segregação para resistência a P. syringae pv. garcae em praticamente todas
as espécies alógamas. Todos os indivíduos de C. kapakata avaliados foram classificados
como suscetíveis, mesmo padrão observado nas espécies autógamas C. anthonyi, C.
arabica e C. heterocalyx. Cafeeiros resistentes foram observados nas espécies C.
salvatrix, C. stenophylla, C. eugenioides e C. canephora. As espécies C. liberica var.
passipagore, C. humilis e C. congensis apresentaram 8,4%, 30% e 62,5% dos
indivíduos com resistência moderada à bactéria.
Tabela 11 - Frequência de cafeeiros resistentes (R), moderadamente resistentes (MR) e
suscetíveis (S) segundo escala de 0 a 5 pontos em Coffea spp. em resposta a infecção
por Pseudomonas syringae pv. garcae, avaliadas no experimento 1.
* R = resistente, nota 0 na escala de pontos.; MR = moderadamente resistente, nota 1 na escala
de pontos; S = suscetível, com 2 a 5 pontos na escala de notas.
Espécie Plantas Nível de resistência*
R MR S
n° n° % n° % n° %
C. arabica cv. IAC 125 RN 12 0 0,0 0 0,0 12 100,0
C. arabica cv. IAC 376-4 8 0 0,0 0 0,0 8 100,0
C. arabica cv. IAC 81 8 0 0,0 0 0,0 8 100,0
C. anthonyi 3 0 0,0 0 0,0 3 100,0
C. heterocalyx 4 0 0,0 0 0,0 4 100,0
C. kapakata 12 0 0,0 0 0,0 12 100,0
C. humilis 12 0 0,0 1 8,4 11 91,6
C. liberica var. passipagore 10 0 0,0 3 30,0 7 70,0
C. congensis IAC 4349 8 0 0,0 5 62,5 3 37,5
C. canephora 7 3 42,9 1 14,2 3 42,9
C. eugenioides 11 2 18,2 6 54,5 3 27,3
C. stenophylla 6 1 16,6 4 66,8 1 16,6
C. salvatrix 11 4 36,4 4 36,4 3 27,2

61
A variabilidade intra-específica presente em algumas espécies do gênero Coffea,
representada pela curva de evolução da doença e pela AACPD em plantas individuais, é
ilustrada na figura 10.
Cafeeiros das cultivares Catuaí Vermelho IAC 81 e IAC 125 RN de C. arabica
apresentaram um mesmo padrão de resposta, com evolução continuada no crescimento
das lesões do 7° ao 42 DAI do patógeno (Figura 10K e L).
Embora as espécies diploides autógamas C. heterocalyx e C. anthonyi tenham se
revelado suscetíveis em relação a todos os indivíduos avaliados, as curvas de evolução
da doença em plantas individuais se revelaram distintas umas das outras. Em C.
anthonyi, o cafeeiro 1 apresentou, aos 14 DAI, lesões correspondentes a 3 pontos na
escala de avaliação de danos, enquanto que dos outros indivíduos da espécies foram
atribuídas apenas 1 ponto, no mesmo período (Figura 10A). Situação semelhante foi
observada entre cafeeiros da espécie C. heterocalyx (Figura 10E).
Entre as espécies diploides alógamas (C. canephora, C. eugenioides, C.
stenophylla e C. salvatrix), nos segregantes para a resistência à doença, observou-se as
mesmas variações descritas para as espécies diploides autógamas.
Em C. canephora, os cafeeiros 3 e 5 (Figura 10B), ambos suscetíveis à mancha-
aureolada, foram classificados com 5 pontos, na escala de danos, aos 42 DAI. No
entanto, esta marca na escala foi alcançada pelo cafeeiro 5, apenas aos 35 DAI enquanto
o mesmo nível de danos se manifestou duas semanas antes no cafeeiro 3.
Diferenças na evolução da doença em cafeeiros altamente suscetíveis,
classificados com 5 pontos aos 42 DAI, foram também observadas em outras espécies
diploides, como C. kapakata (Figura 10G). Aos 28 DAI, o cafeeiro 11 apresentava
sintomas equivalentes a 2 pontos na escala utilizada para avaliação do dano causado
pela bactéria, enquanto que no cafeeiro 3, os danos eram de grande magnitude,
correspondentes a 5 pontos na mesma escala.
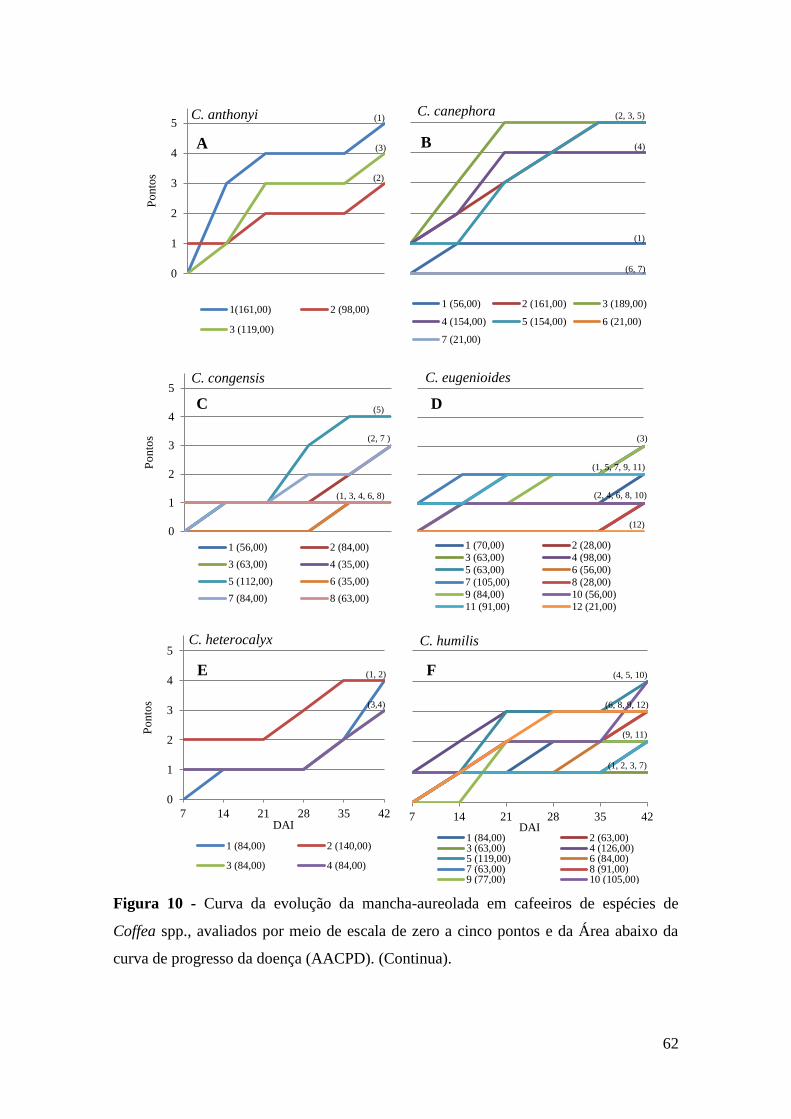
62
Figura 10 - Curva da evolução da mancha-aureolada em cafeeiros de espécies de
Coffea spp., avaliados por meio de escala de zero a cinco pontos e da Área abaixo da
curva de progresso da doença (AACPD). (Continua).
0
1
2
3
4
5
Po
nto
s
C. anthonyi
1(161,00) 2 (98,00)
3 (119,00)
C. canephora
1 (56,00) 2 (161,00) 3 (189,00)
4 (154,00) 5 (154,00) 6 (21,00)
7 (21,00)
0
1
2
3
4
5
Po
nto
s
C. congensis
1 (56,00) 2 (84,00)
3 (63,00) 4 (35,00)
5 (112,00) 6 (35,00)
7 (84,00) 8 (63,00)
C. eugenioides
1 (70,00) 2 (28,00)
3 (63,00) 4 (98,00)
5 (63,00) 6 (56,00)
7 (105,00) 8 (28,00)
9 (84,00) 10 (56,00)
11 (91,00) 12 (21,00)
0
1
2
3
4
5
7 14 21 28 35 42
Po
nto
s
DAI
C. heterocalyx
1 (84,00) 2 (140,00)
3 (84,00) 4 (84,00)
7 14 21 28 35 42DAI
C. humilis
1 (84,00) 2 (63,00)3 (63,00) 4 (126,00)5 (119,00) 6 (84,00)7 (63,00) 8 (91,00)9 (77,00) 10 (105,00)
(1)
(3)
(2, 3, 5)
(4)
(1)
(6, 7)
(2)
(5)
(2, 7 )
(1, 3, 4, 6, 8)
(3)
(1, 5, 7, 9, 11)
(2, 4, 6, 8, 10)
(12)
(1, 2)
(3,4)
(4, 5, 10)
(6, 8, 9, 12)
(9, 11)
(1, 2, 3, 7)
A
B
C
D
E
F
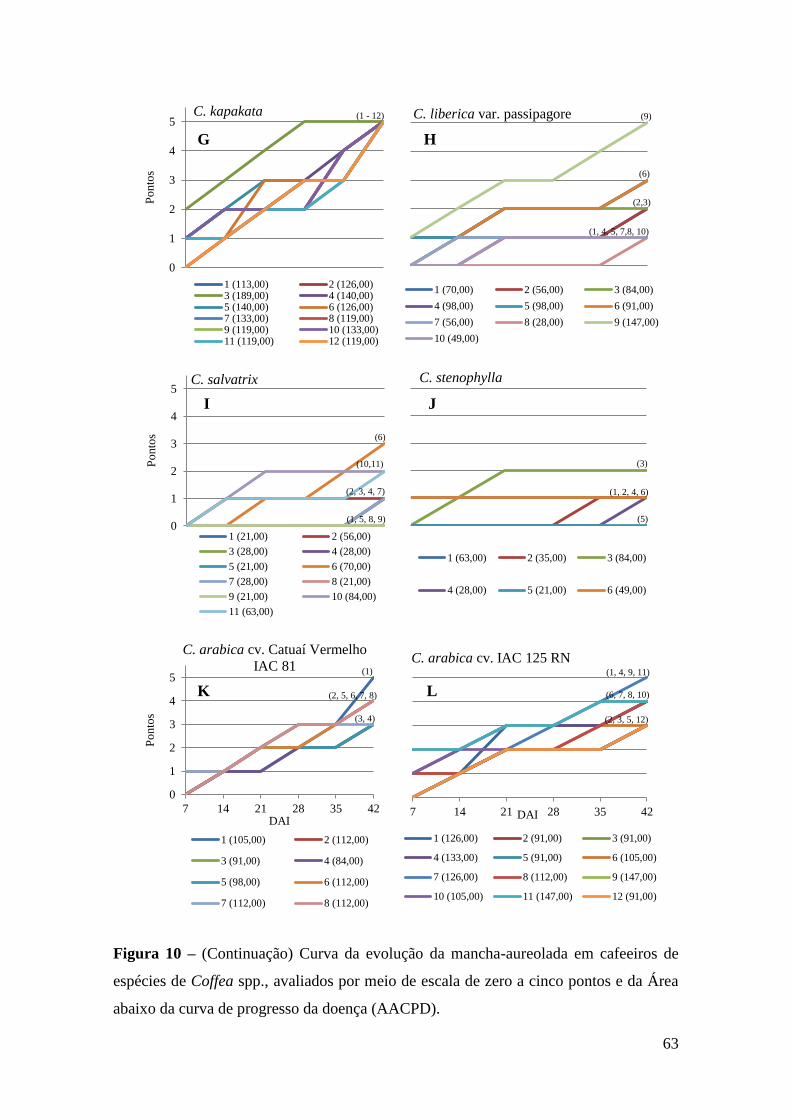
63
Figura 10 – (Continuação) Curva da evolução da mancha-aureolada em cafeeiros de
espécies de Coffea spp., avaliados por meio de escala de zero a cinco pontos e da Área
abaixo da curva de progresso da doença (AACPD).
0
1
2
3
4
5
Po
nto
s
C. kapakata
1 (113,00) 2 (126,00)3 (189,00) 4 (140,00)5 (140,00) 6 (126,00)7 (133,00) 8 (119,00)9 (119,00) 10 (133,00)11 (119,00) 12 (119,00)
C. liberica var. passipagore
1 (70,00) 2 (56,00) 3 (84,00)
4 (98,00) 5 (98,00) 6 (91,00)
7 (56,00) 8 (28,00) 9 (147,00)
10 (49,00)
0
1
2
3
4
5
Po
nto
s
C. salvatrix
1 (21,00) 2 (56,00)
3 (28,00) 4 (28,00)
5 (21,00) 6 (70,00)
7 (28,00) 8 (21,00)
9 (21,00) 10 (84,00)
11 (63,00)
C. stenophylla
1 (63,00) 2 (35,00) 3 (84,00)
4 (28,00) 5 (21,00) 6 (49,00)
0
1
2
3
4
5
7 14 21 28 35 42
Po
nto
s
DAI
C. arabica cv. Catuaí Vermelho
IAC 81
1 (105,00) 2 (112,00)
3 (91,00) 4 (84,00)
5 (98,00) 6 (112,00)
7 (112,00) 8 (112,00)
7 14 21 28 35 42DAI
C. arabica cv. IAC 125 RN
1 (126,00) 2 (91,00) 3 (91,00)
4 (133,00) 5 (91,00) 6 (105,00)
7 (126,00) 8 (112,00) 9 (147,00)
10 (105,00) 11 (147,00) 12 (91,00)
(6)
(2,3)
(9)
(1, 4, 5, 7,8, 10)
(1 - 12)
(2, 3, 4, 7)
(3)
(6)
(10,11)
(1, 2, 4, 6)
(5)
(3, 4)
(1, 4, 9, 11)
(6, 7, 8, 10)
(2, 3, 5, 12)
(1, 5, 8, 9)
(1)
G
(2, 5, 6, 7, 8)
H
I
J
A
K
L

64
Sintomas de natureza variável foram observados em folhas das espécies de
Coffea spp., inoculadas com P. syringae pv. garcae pelo método de abrasão, conforme
ilustrado na figura 11.
Figura 11 - Sintomas observados aos 21 dias após a inoculação com Pseudomonas
syringae pv. garcae, em folhas de espécies de Coffea spp. pelo método de abrasão. C.
arabica cv. IAC 125 RN (A); C. congensis (B); C. kapakata (C); C. eugenioides (D); C.
salvatrix (E); C. stenophylla (F); C. liberica var. passipagore (G); C. anthonyi (H); C.
humilis (I); C. heterocalyx (J); C. canephora (K); C. arabica cv. Catuaí Vermelho
IAC 81 (L).
Cafeeiros das espécies C. salvatrix (Figura 11E), C. canephora (Figura 11K) e
C. congensis (Figura 11B), considerados resistentes à doença, exibiram lesões com
aspecto seco, sem alterações visíveis ao redor dos ferimentos causados pela inoculação.
Em plantas resistentes das espécies C. eugenioides e C. stenophylla, observou-se
escurecimento ao redor dos pontos de inoculação e, em alguns ferimentos, notou-se a
presença de halo amarelado, indicação provável de danos ocasionados pela tabtoxina
B A D C
F E H G
J I L K

65
sintetizada pela bactéria (BENDER et al., 1999). No entanto, não foram observados
sinais visuais da colonização, como exemplificado na figura 12, e o exames de
exsudação em gota, visto ao microscópio foram negativos.
Figura 12 - Face abaxial de folha de cafeeiro da cultivar IAC 125 RN de Coffe arabica,
inoculada com Pseudomonas syringae pv. garcae pelo método de abrasão, presença de
colonização dos tecidos foliares adjacentes aos ferimentos, ocasionados pela inoculação
por abrasão.
Reação distinta foi observada em C. kapakata (Figura 11C). Aos sete DAI, os
sintomas desenvolvidos pela bactéria ultrapassavam a nota três da escala de pontos e,
aos 14 DAI, praticamente todas as plantas já apresentavam nota cinco. Entre o sétimo e
o vigésimo primeiro DAI, praticamente todas as folhas inoculadas sofreram abscisão e,
em alguns os casos, a bactéria foi capaz de colonizar outras partes das mudas.
Em um segundo experimento, realizado em 2014 com espécies de Coffea spp.,
observou-se a variabilidade similar à descrita no germoplasma avaliado no ano anterior
(Tabela 12). A maior porcentagem de cafeeiros resistentes (42,1%) foi encontrada em
progênies da espécie C. eugenioides, sendo que, aproximadamente 52,6% dos
indivíduos das outras espécies, revelaram-se suscetíveis ao patógeno.
As espécies C kapakata (21), C. humilis (2) e C. congensis Bangelan
apresentaram 100% de cafeeiros suscetíveis, assim como, a cultivar IAC 125 RN de C.
arabica.
As demais espécies segregaram em proporção variável em relação ao nível de
resistência a P. syringae pv. garcae.
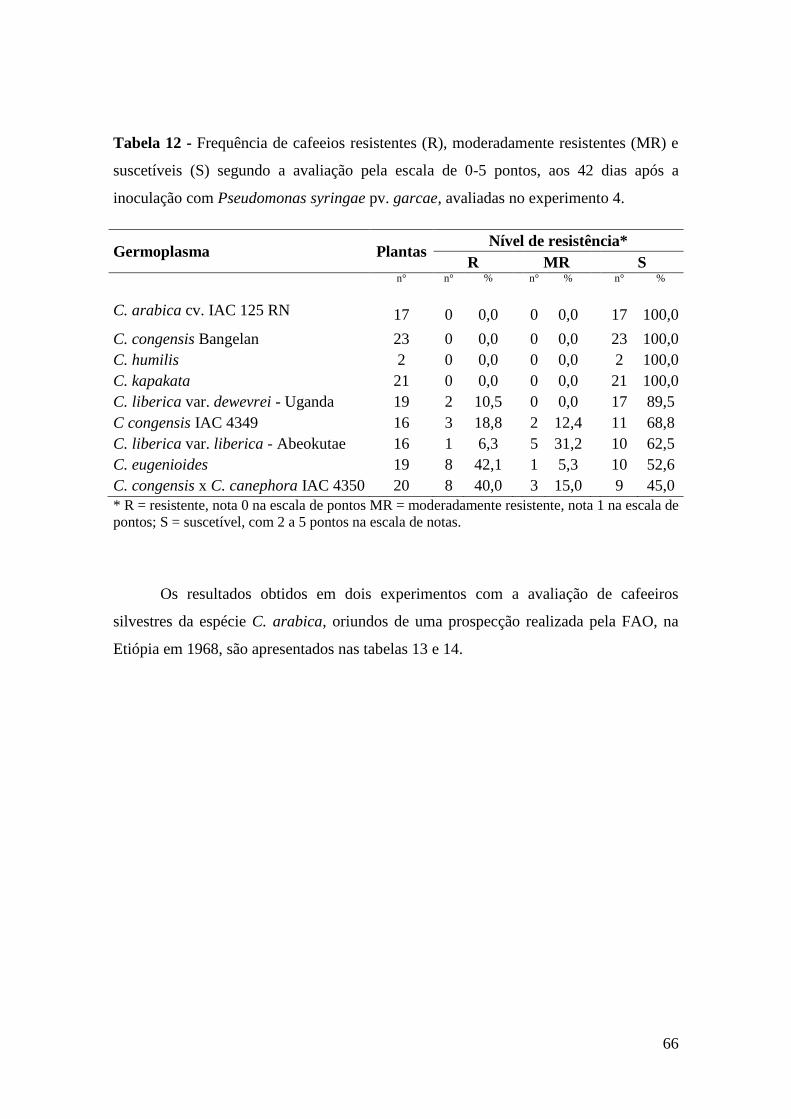
66
Tabela 12 - Frequência de cafeeios resistentes (R), moderadamente resistentes (MR) e
suscetíveis (S) segundo a avaliação pela escala de 0-5 pontos, aos 42 dias após a
inoculação com Pseudomonas syringae pv. garcae, avaliadas no experimento 4.
Germoplasma Plantas Nível de resistência*
R MR S
n° n° % n° % n° %
C. arabica cv. IAC 125 RN 17 0 0,0 0 0,0 17 100,0
C. congensis Bangelan 23 0 0,0 0 0,0 23 100,0
C. humilis 2 0 0,0 0 0,0 2 100,0
C. kapakata 21 0 0,0 0 0,0 21 100,0
C. liberica var. dewevrei - Uganda 19 2 10,5 0 0,0 17 89,5
C congensis IAC 4349 16 3 18,8 2 12,4 11 68,8
C. liberica var. liberica - Abeokutae 16 1 6,3 5 31,2 10 62,5
C. eugenioides 19 8 42,1 1 5,3 10 52,6
C. congensis x C. canephora IAC 4350 20 8 40,0 3 15,0 9 45,0
* R = resistente, nota 0 na escala de pontos MR = moderadamente resistente, nota 1 na escala de
pontos; S = suscetível, com 2 a 5 pontos na escala de notas.
Os resultados obtidos em dois experimentos com a avaliação de cafeeiros
silvestres da espécie C. arabica, oriundos de uma prospecção realizada pela FAO, na
Etiópia em 1968, são apresentados nas tabelas 13 e 14.
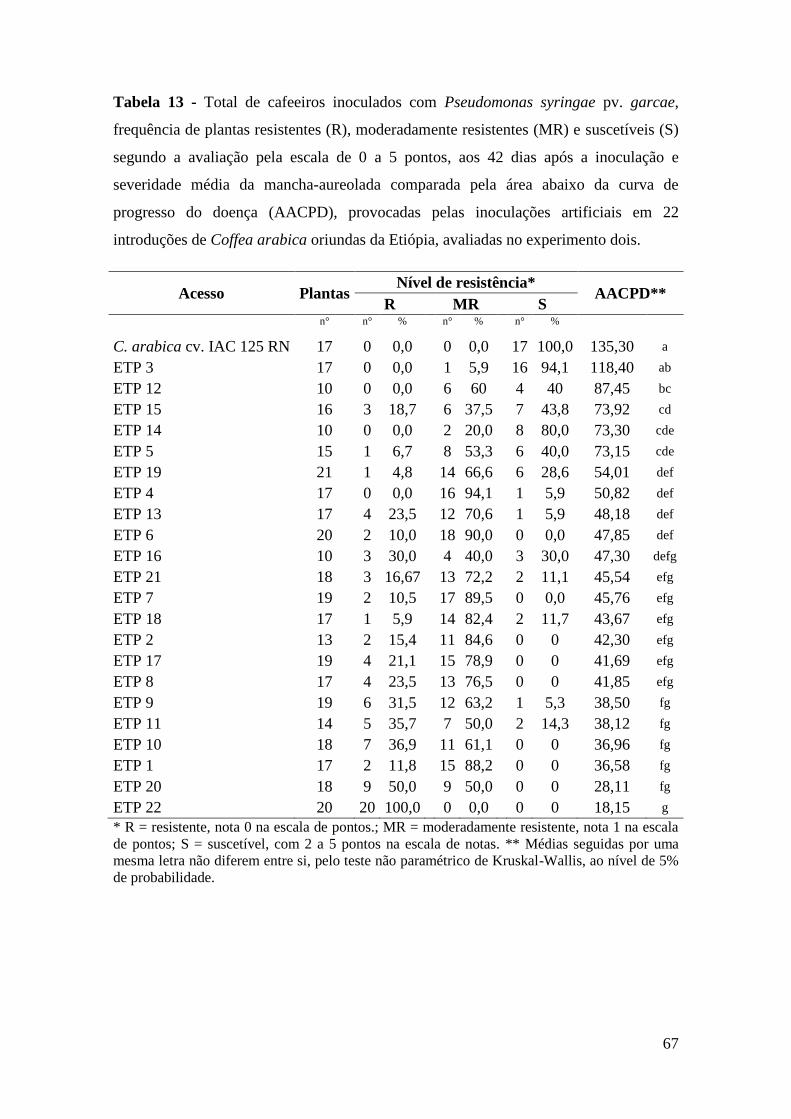
67
Tabela 13 - Total de cafeeiros inoculados com Pseudomonas syringae pv. garcae,
frequência de plantas resistentes (R), moderadamente resistentes (MR) e suscetíveis (S)
segundo a avaliação pela escala de 0 a 5 pontos, aos 42 dias após a inoculação e
severidade média da mancha-aureolada comparada pela área abaixo da curva de
progresso do doença (AACPD), provocadas pelas inoculações artificiais em 22
introduções de Coffea arabica oriundas da Etiópia, avaliadas no experimento dois.
Acesso Plantas Nível de resistência*
AACPD** R MR S
n° n° % n° % n° %
C. arabica cv. IAC 125 RN 17 0 0,0 0 0,0 17 100,0 135,30 a
ETP 3 17 0 0,0 1 5,9 16 94,1 118,40 ab
ETP 12 10 0 0,0 6 60 4 40 87,45 bc
ETP 15 16 3 18,7 6 37,5 7 43,8 73,92 cd
ETP 14 10 0 0,0 2 20,0 8 80,0 73,30 cde
ETP 5 15 1 6,7 8 53,3 6 40,0 73,15 cde
ETP 19 21 1 4,8 14 66,6 6 28,6 54,01 def
ETP 4 17 0 0,0 16 94,1 1 5,9 50,82 def
ETP 13 17 4 23,5 12 70,6 1 5,9 48,18 def
ETP 6 20 2 10,0 18 90,0 0 0,0 47,85 def
ETP 16 10 3 30,0 4 40,0 3 30,0 47,30 defg
ETP 21 18 3 16,67 13 72,2 2 11,1 45,54 efg
ETP 7 19 2 10,5 17 89,5 0 0,0 45,76 efg
ETP 18 17 1 5,9 14 82,4 2 11,7 43,67 efg
ETP 2 13 2 15,4 11 84,6 0 0 42,30 efg
ETP 17 19 4 21,1 15 78,9 0 0 41,69 efg
ETP 8 17 4 23,5 13 76,5 0 0 41,85 efg
ETP 9 19 6 31,5 12 63,2 1 5,3 38,50 fg
ETP 11 14 5 35,7 7 50,0 2 14,3 38,12 fg
ETP 10 18 7 36,9 11 61,1 0 0 36,96 fg
ETP 1 17 2 11,8 15 88,2 0 0 36,58 fg
ETP 20 18 9 50,0 9 50,0 0 0 28,11 fg
ETP 22 20 20 100,0 0 0,0 0 0 18,15 g
* R = resistente, nota 0 na escala de pontos.; MR = moderadamente resistente, nota 1 na escala
de pontos; S = suscetível, com 2 a 5 pontos na escala de notas. ** Médias seguidas por uma
mesma letra não diferem entre si, pelo teste não paramétrico de Kruskal-Wallis, ao nível de 5%
de probabilidade.
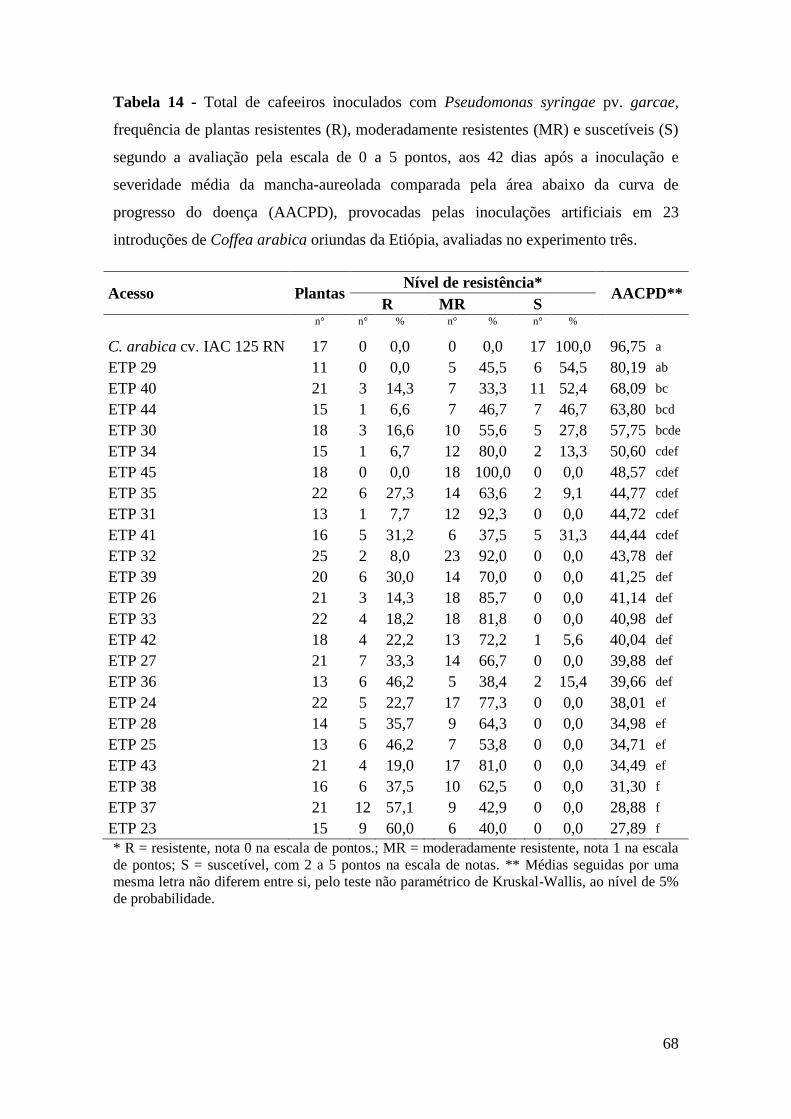
68
Tabela 14 - Total de cafeeiros inoculados com Pseudomonas syringae pv. garcae,
frequência de plantas resistentes (R), moderadamente resistentes (MR) e suscetíveis (S)
segundo a avaliação pela escala de 0 a 5 pontos, aos 42 dias após a inoculação e
severidade média da mancha-aureolada comparada pela área abaixo da curva de
progresso do doença (AACPD), provocadas pelas inoculações artificiais em 23
introduções de Coffea arabica oriundas da Etiópia, avaliadas no experimento três.
Acesso Plantas Nível de resistência*
AACPD** R MR S
n° n° % n° % n° %
C. arabica cv. IAC 125 RN 17 0 0,0 0 0,0 17 100,0 96,75 a
ETP 29 11 0 0,0 5 45,5 6 54,5 80,19 ab
ETP 40 21 3 14,3 7 33,3 11 52,4 68,09 bc
ETP 44 15 1 6,6 7 46,7 7 46,7 63,80 bcd
ETP 30 18 3 16,6 10 55,6 5 27,8 57,75 bcde
ETP 34 15 1 6,7 12 80,0 2 13,3 50,60 cdef
ETP 45 18 0 0,0 18 100,0 0 0,0 48,57 cdef
ETP 35 22 6 27,3 14 63,6 2 9,1 44,77 cdef
ETP 31 13 1 7,7 12 92,3 0 0,0 44,72 cdef
ETP 41 16 5 31,2 6 37,5 5 31,3 44,44 cdef
ETP 32 25 2 8,0 23 92,0 0 0,0 43,78 def
ETP 39 20 6 30,0 14 70,0 0 0,0 41,25 def
ETP 26 21 3 14,3 18 85,7 0 0,0 41,14 def
ETP 33 22 4 18,2 18 81,8 0 0,0 40,98 def
ETP 42 18 4 22,2 13 72,2 1 5,6 40,04 def
ETP 27 21 7 33,3 14 66,7 0 0,0 39,88 def
ETP 36 13 6 46,2 5 38,4 2 15,4 39,66 def
ETP 24 22 5 22,7 17 77,3 0 0,0 38,01 ef
ETP 28 14 5 35,7 9 64,3 0 0,0 34,98 ef
ETP 25 13 6 46,2 7 53,8 0 0,0 34,71 ef
ETP 43 21 4 19,0 17 81,0 0 0,0 34,49 ef
ETP 38 16 6 37,5 10 62,5 0 0,0 31,30 f
ETP 37 21 12 57,1 9 42,9 0 0,0 28,88 f
ETP 23 15 9 60,0 6 40,0 0 0,0 27,89 f
* R = resistente, nota 0 na escala de pontos.; MR = moderadamente resistente, nota 1 na escala
de pontos; S = suscetível, com 2 a 5 pontos na escala de notas. ** Médias seguidas por uma
mesma letra não diferem entre si, pelo teste não paramétrico de Kruskal-Wallis, ao nível de 5%
de probabilidade.

69
A cultivar IAC 125 RN, suscetível à mancha-aureolada apresentou os maiores
valores da AACPD nos dois experimentos com acesss Etíopes.
No primeiro experimento (Tabela 13) realizado com 22 acessos oriundos de
região de origem e diversificação da espécia C. arabica, a progênies ETP 22 apresentou
100% de seus cafeeiros resistentes à mancha-aureolada. As progênies ETP 1, ETP 2,
ETP 6, ETP 7, ETP 8, ETP 10, ETP 17, e ETP 20) apresentaram 100% de plantas de
suas progênies consideradas resistentes e moderadamente resistentes à doença. Apenas
duas progênies (ETP 3 e ETP 12) foram consideradas, em média, suscetíveis a P.
syringae pv. garcae, em função da elevada frequência de cafeeiros com notas de dano
igual ou superior a cinco. Apenas o acesso ETP 3 apresentou AACPD de mesma
magnitude que a cultivar testemunha IAC 125 RN.
No segundo experimento (Tabela 14), dos 23 acessos avaliados, 14 acessos
apresentaram 100% de plantas resistentes a moderadamente resistentes em suas
progênies e apenas três acessos, ETP 44, ETP 40 e ETP 29 apresentaram entre 46,7 e
54,5% de plantas suscetíveis nas progênies. A AACPD do controle suscetível, IAC 125
RN, foi semelhante apenas àquela observada no aceso ETP 29. As maiores porcentagens
de cafeeiros resistentes à doença foram encontradas nos acessos ETP 23 (60,0%) e ETP
37 (57,1%) que também apresentaram as menores médias de AACPD, 27,89 e 28,88,
respectivamente.
Em algumas progênies (ETP 5, ETP 8, ETP 9, ETP 13, ETP 17 e ETP 30) dos
dois experimentos o patógeno foi capaz de infectar tecidos foliares no local das
inoculações e, posteriormente, as lesões foram caracterizadas como reação de
hipersensibilidade localizada, com ocorrência de abscisão do tecido, semelhantes às
lesões do tipo “shot-hole”.
A resistência de progênies de quatro cafeeiros oriundos da hibridação entre C.
eugenioides (resistente), e C. arabica (suscetível) foi avaliada em dois experimentos,
sendo o primeiro constituído por progênies de polinização aberta e o segundo por
progênies de autofecundação. Os resultados relacionados à severidade da mancha-
aureolada são apresentados na tabela 15.
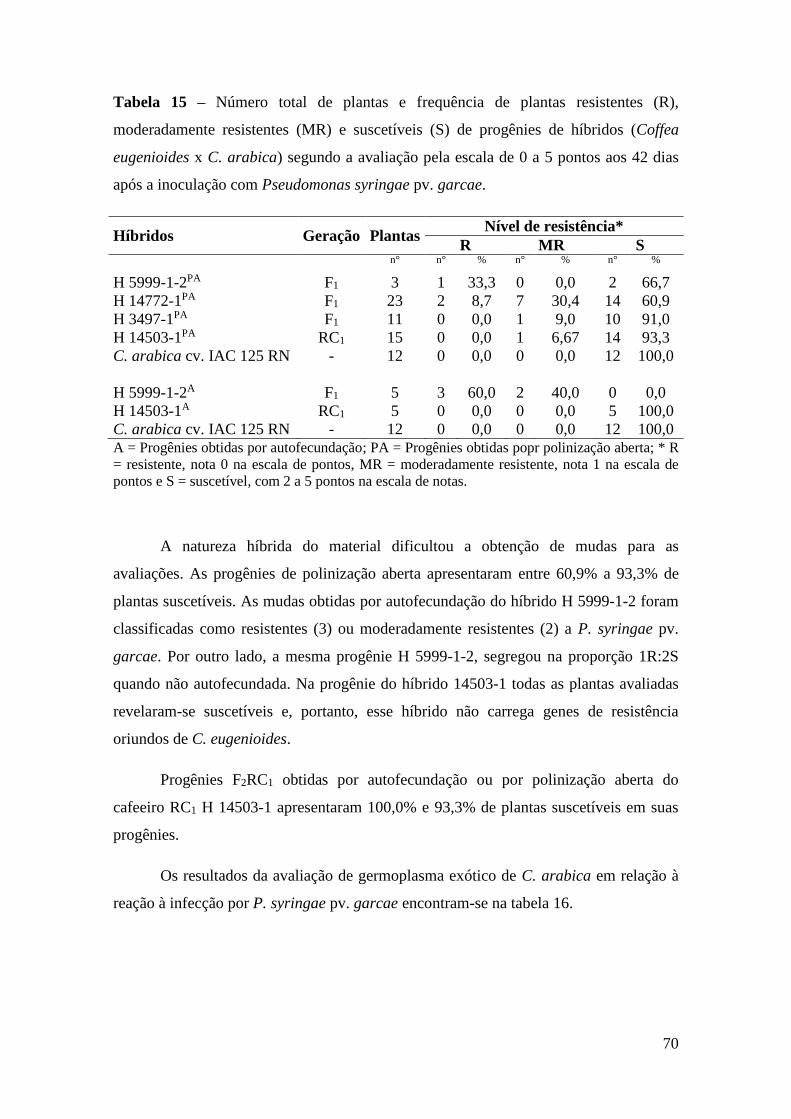
70
Tabela 15 – Número total de plantas e frequência de plantas resistentes (R),
moderadamente resistentes (MR) e suscetíveis (S) de progênies de híbridos (Coffea
eugenioides x C. arabica) segundo a avaliação pela escala de 0 a 5 pontos aos 42 dias
após a inoculação com Pseudomonas syringae pv. garcae.
Híbridos Geração Plantas Nível de resistência*
R MR S n° n° % n° % n° %
H 5999-1-2PA F1 3 1 33,3 0 0,0 2 66,7
H 14772-1PA F1 23 2 8,7 7 30,4 14 60,9
H 3497-1PA F1 11 0 0,0 1 9,0 10 91,0
H 14503-1PA RC1 15 0 0,0 1 6,67 14 93,3
C. arabica cv. IAC 125 RN - 12 0 0,0 0 0,0 12 100,0
H 5999-1-2A F1 5 3 60,0 2 40,0 0 0,0
H 14503-1A RC1 5 0 0,0 0 0,0 5 100,0
C. arabica cv. IAC 125 RN - 12 0 0,0 0 0,0 12 100,0 A = Progênies obtidas por autofecundação; PA = Progênies obtidas popr polinização aberta; * R
= resistente, nota 0 na escala de pontos, MR = moderadamente resistente, nota 1 na escala de
pontos e S = suscetível, com 2 a 5 pontos na escala de notas.
A natureza híbrida do material dificultou a obtenção de mudas para as
avaliações. As progênies de polinização aberta apresentaram entre 60,9% a 93,3% de
plantas suscetíveis. As mudas obtidas por autofecundação do híbrido H 5999-1-2 foram
classificadas como resistentes (3) ou moderadamente resistentes (2) a P. syringae pv.
garcae. Por outro lado, a mesma progênie H 5999-1-2, segregou na proporção 1R:2S
quando não autofecundada. Na progênie do híbrido 14503-1 todas as plantas avaliadas
revelaram-se suscetíveis e, portanto, esse híbrido não carrega genes de resistência
oriundos de C. eugenioides.
Progênies F2RC1 obtidas por autofecundação ou por polinização aberta do
cafeeiro RC1 H 14503-1 apresentaram 100,0% e 93,3% de plantas suscetíveis em suas
progênies.
Os resultados da avaliação de germoplasma exótico de C. arabica em relação à
reação à infecção por P. syringae pv. garcae encontram-se na tabela 16.
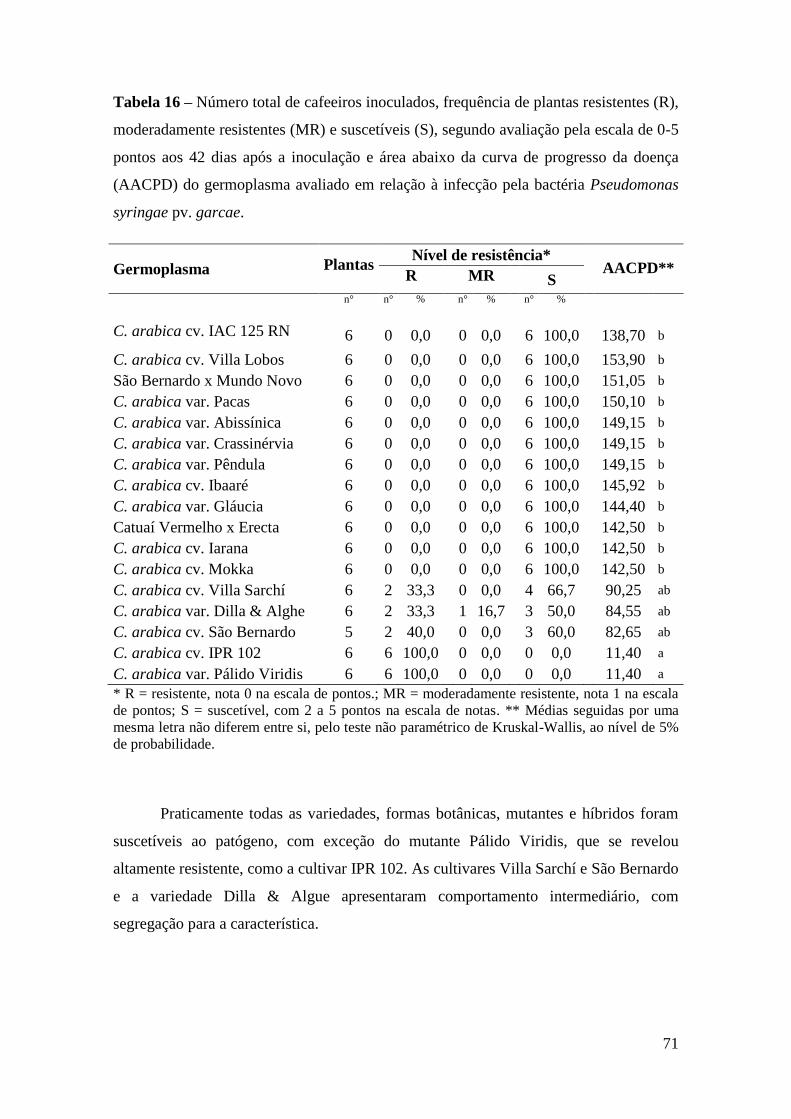
71
Tabela 16 – Número total de cafeeiros inoculados, frequência de plantas resistentes (R),
moderadamente resistentes (MR) e suscetíveis (S), segundo avaliação pela escala de 0-5
pontos aos 42 dias após a inoculação e área abaixo da curva de progresso da doença
(AACPD) do germoplasma avaliado em relação à infecção pela bactéria Pseudomonas
syringae pv. garcae.
Germoplasma Plantas Nível de resistência*
AACPD** R MR S
n° n° % n° % n° %
C. arabica cv. IAC 125 RN 6 0 0,0 0 0,0 6 100,0 138,70 b
C. arabica cv. Villa Lobos 6 0 0,0 0 0,0 6 100,0 153,90 b
São Bernardo x Mundo Novo 6 0 0,0 0 0,0 6 100,0 151,05 b
C. arabica var. Pacas 6 0 0,0 0 0,0 6 100,0 150,10 b
C. arabica var. Abissínica 6 0 0,0 0 0,0 6 100,0 149,15 b
C. arabica var. Crassinérvia 6 0 0,0 0 0,0 6 100,0 149,15 b
C. arabica var. Pêndula 6 0 0,0 0 0,0 6 100,0 149,15 b
C. arabica cv. Ibaaré 6 0 0,0 0 0,0 6 100,0 145,92 b
C. arabica var. Gláucia 6 0 0,0 0 0,0 6 100,0 144,40 b
Catuaí Vermelho x Erecta 6 0 0,0 0 0,0 6 100,0 142,50 b
C. arabica cv. Iarana 6 0 0,0 0 0,0 6 100,0 142,50 b
C. arabica cv. Mokka 6 0 0,0 0 0,0 6 100,0 142,50 b
C. arabica cv. Villa Sarchí 6 2 33,3 0 0,0 4 66,7 90,25 ab
C. arabica var. Dilla & Alghe 6 2 33,3 1 16,7 3 50,0 84,55 ab
C. arabica cv. São Bernardo 5 2 40,0 0 0,0 3 60,0 82,65 ab
C. arabica cv. IPR 102 6 6 100,0 0 0,0 0 0,0 11,40 a
C. arabica var. Pálido Viridis 6 6 100,0 0 0,0 0 0,0 11,40 a
* R = resistente, nota 0 na escala de pontos.; MR = moderadamente resistente, nota 1 na escala
de pontos; S = suscetível, com 2 a 5 pontos na escala de notas. ** Médias seguidas por uma
mesma letra não diferem entre si, pelo teste não paramétrico de Kruskal-Wallis, ao nível de 5%
de probabilidade.
Praticamente todas as variedades, formas botânicas, mutantes e híbridos foram
suscetíveis ao patógeno, com exceção do mutante Pálido Viridis, que se revelou
altamente resistente, como a cultivar IPR 102. As cultivares Villa Sarchí e São Bernardo
e a variedade Dilla & Algue apresentaram comportamento intermediário, com
segregação para a característica.
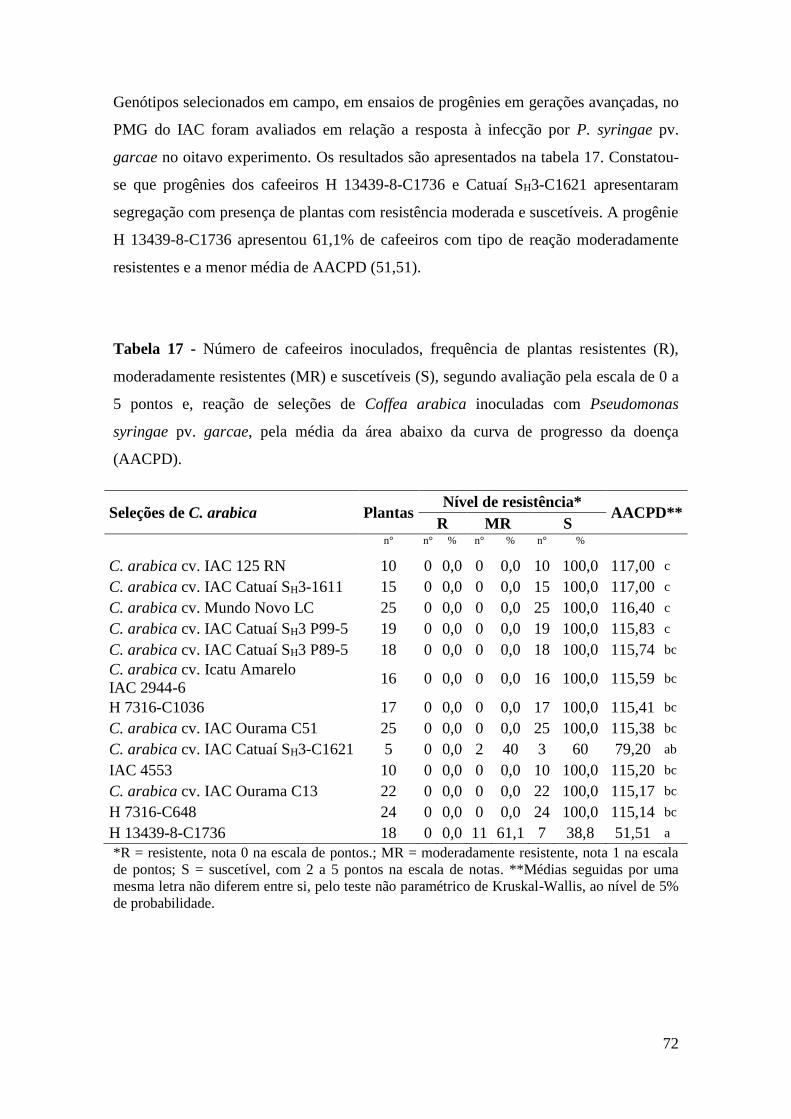
72
Genótipos selecionados em campo, em ensaios de progênies em gerações avançadas, no
PMG do IAC foram avaliados em relação a resposta à infecção por P. syringae pv.
garcae no oitavo experimento. Os resultados são apresentados na tabela 17. Constatou-
se que progênies dos cafeeiros H 13439-8-C1736 e Catuaí SH3-C1621 apresentaram
segregação com presença de plantas com resistência moderada e suscetíveis. A progênie
H 13439-8-C1736 apresentou 61,1% de cafeeiros com tipo de reação moderadamente
resistentes e a menor média de AACPD (51,51).
Tabela 17 - Número de cafeeiros inoculados, frequência de plantas resistentes (R),
moderadamente resistentes (MR) e suscetíveis (S), segundo avaliação pela escala de 0 a
5 pontos e, reação de seleções de Coffea arabica inoculadas com Pseudomonas
syringae pv. garcae, pela média da área abaixo da curva de progresso da doença
(AACPD).
Seleções de C. arabica Plantas Nível de resistência*
AACPD** R MR S
n° n° % n° % n° %
C. arabica cv. IAC 125 RN 10 0 0,0 0 0,0 10 100,0 117,00 c
C. arabica cv. IAC Catuaí SH3-1611 15 0 0,0 0 0,0 15 100,0 117,00 c
C. arabica cv. Mundo Novo LC 25 0 0,0 0 0,0 25 100,0 116,40 c
C. arabica cv. IAC Catuaí SH3 P99-5 19 0 0,0 0 0,0 19 100,0 115,83 c
C. arabica cv. IAC Catuaí SH3 P89-5 18 0 0,0 0 0,0 18 100,0 115,74 bc
C. arabica cv. Icatu Amarelo
IAC 2944-6 16 0 0,0 0 0,0 16 100,0 115,59 bc
H 7316-C1036 17 0 0,0 0 0,0 17 100,0 115,41 bc
C. arabica cv. IAC Ourama C51 25 0 0,0 0 0,0 25 100,0 115,38 bc
C. arabica cv. IAC Catuaí SH3-C1621 5 0 0,0 2 40 3 60 79,20 ab
IAC 4553 10 0 0,0 0 0,0 10 100,0 115,20 bc
C. arabica cv. IAC Ourama C13 22 0 0,0 0 0,0 22 100,0 115,17 bc
H 7316-C648 24 0 0,0 0 0,0 24 100,0 115,14 bc
H 13439-8-C1736 18 0 0,0 11 61,1 7 38,8 51,51 a
*R = resistente, nota 0 na escala de pontos.; MR = moderadamente resistente, nota 1 na escala
de pontos; S = suscetível, com 2 a 5 pontos na escala de notas. **Médias seguidas por uma
mesma letra não diferem entre si, pelo teste não paramétrico de Kruskal-Wallis, ao nível de 5%
de probabilidade.

73
4.5. Análise do efeito pleiotrópico do alelo SH1 na resistência a Pseudomonas
syringae pv. garcae
Os resultados relacionados à expressão da resistência em progênies F2 dos
cafeeiros H8089-2 e H8089-7 a P. syringae pv. garcae e aos isolados I/2015 e II/2015
de H. vastatrix, encontram-se na tabela 18.
Tabela 18 - Reação de progênies F2 dos cafeeiros H8089-2 e H8089-7, híbridos entre a
cultivar Catuaí Vermelho IAC 24 e a seleção Geisha IAC 1137-5, à infecção pela
bactéria Pseudomonas syringae pv. garcae e pelos isolados I/2015 (v2, v5) e II/2015 (v1,
v2, v5) de Hemileia vastatrix.
Parâmetro H8089-2 H8089-7 Total
Plantas (n°) 34 29 63
PHV I I/20151
(R:S) 6:28 10:19 16:47
PHV I II/20152
(R:S) 0:34 0:29 0:63
PPSG3
(R:S) 0:34 15:14 15:48 1Proporção (P) de plantas resistentes (R) e suscetíveis (S) ao isolado I/2015 de Hemileia
vastatrix.2 Proporção de plantas resistentes (R) e suscetíveis (S) ao Isolado II/2015 de Hemileia
vastatrix. 3 Proporção de plantas resistentes (R) e suscetíveis (S) a Pseudomonas syringae pv.
garcae.
De acordo com os resultados obtidos, as progênies F2 segregaram para a
resistência ao isolado I/2015 de H. vastatrix, mas revelaram-se suscetíveis ao isolado
II/2015 do fungo. Por outro lado, as 34 plantas da progênie H8089-2 se mostraram
suscetíveis à mancha-aureolada, enquanto a progênie H8089-7 segregou na proporção
aproximada de 1R:1S (Tabela 18).
Em resposta à infecção pelo isolado I/2015 de H. vastatrix, do total de progênies
analisadas, 16 plantas apresentaram reação de resistência e 47, de suscetibilidade. Para
P. syringae pv. garcae, foram identificadas 15 plantas com reação de resistência
enquanto que as 48 restantes apresentaram reação de suscetibilidade (Tabela 18).
Na tabela 19, encontram-se os dados referentes à análise comparativa da reação
das progênies F2 em relação à infecção por P. syringae pv. garcae e ao isolado I/2015
de H. vastatrix.
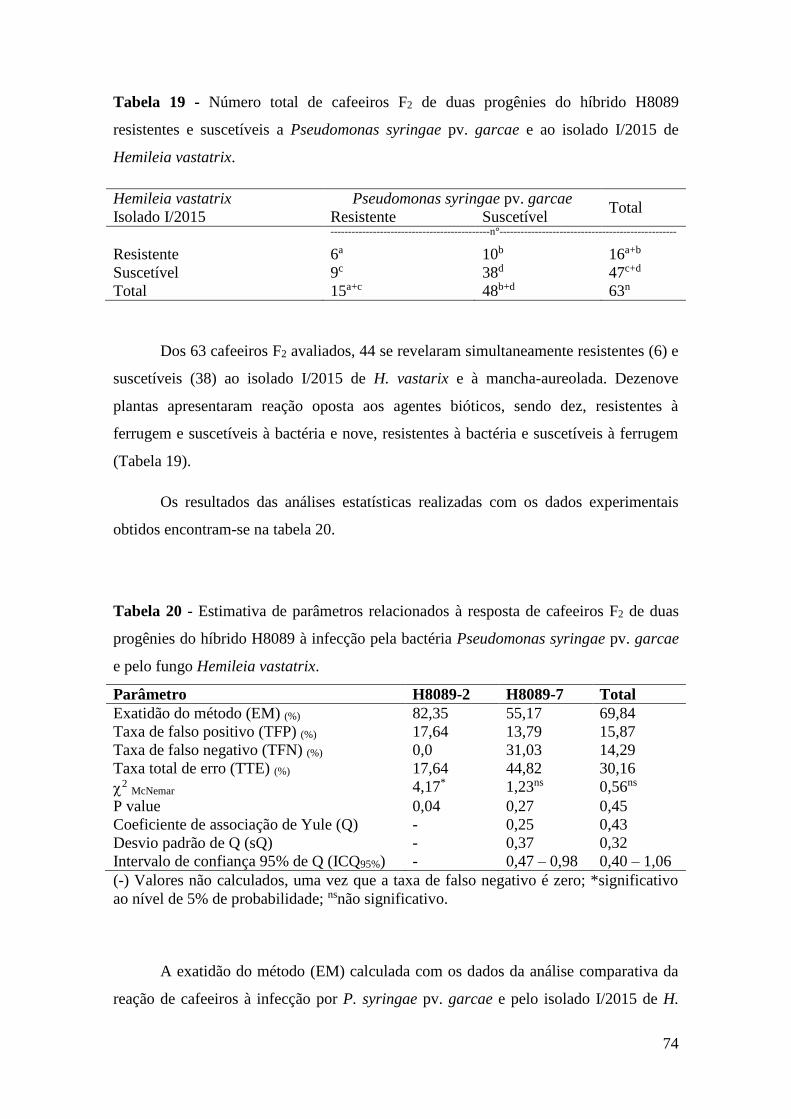
74
Tabela 19 - Número total de cafeeiros F2 de duas progênies do híbrido H8089
resistentes e suscetíveis a Pseudomonas syringae pv. garcae e ao isolado I/2015 de
Hemileia vastatrix.
Hemileia vastatrix
Isolado I/2015
Pseudomonas syringae pv. garcae Total
Resistente Suscetível
---------------------------------------------n°--------------------------------------------------
Resistente 6a 10b 16a+b
Suscetível 9c 38d 47c+d
Total 15a+c 48b+d 63n
Dos 63 cafeeiros F2 avaliados, 44 se revelaram simultaneamente resistentes (6) e
suscetíveis (38) ao isolado I/2015 de H. vastarix e à mancha-aureolada. Dezenove
plantas apresentaram reação oposta aos agentes bióticos, sendo dez, resistentes à
ferrugem e suscetíveis à bactéria e nove, resistentes à bactéria e suscetíveis à ferrugem
(Tabela 19).
Os resultados das análises estatísticas realizadas com os dados experimentais
obtidos encontram-se na tabela 20.
Tabela 20 - Estimativa de parâmetros relacionados à resposta de cafeeiros F2 de duas
progênies do híbrido H8089 à infecção pela bactéria Pseudomonas syringae pv. garcae
e pelo fungo Hemileia vastatrix.
Parâmetro H8089-2 H8089-7 Total
Exatidão do método (EM) (%) 82,35 55,17 69,84
Taxa de falso positivo (TFP) (%) 17,64 13,79 15,87
Taxa de falso negativo (TFN) (%) 0,0 31,03 14,29
Taxa total de erro (TTE) (%) 17,64 44,82 30,16
2 McNemar 4,17* 1,23ns 0,56ns
P value 0,04 0,27 0,45
Coeficiente de associação de Yule (Q) - 0,25 0,43
Desvio padrão de Q (sQ) - 0,37 0,32
Intervalo de confiança 95% de Q (ICQ95%) - 0,47 – 0,98 0,40 – 1,06
(-) Valores não calculados, uma vez que a taxa de falso negativo é zero; *significativo
ao nível de 5% de probabilidade; nsnão significativo.
A exatidão do método (EM) calculada com os dados da análise comparativa da
reação de cafeeiros à infecção por P. syringae pv. garcae e pelo isolado I/2015 de H.

75
vastatrix foi de 69,84% quando se considerou o total de plantas em F2, sendo no mesmo
grupo de plantas a taxa total de erro (TTE) igual a 30,16%.
O teste de 2 McNemar revelou-se significativo na progênie H8089-2 (4,17; p =
0,04) e não significativo na progênie H8089-7 (1,23; p = 0,27) e no grupo total de
plantas (0,56; p = 0,45). O coeficiente de associação de Yule foi igual a 0,25 para a
progênie H8089-7 e 0,43 no grupo total de plantas, não tendo sido calculado para a
progênie H8089-2, uma vez que a taxa de falso negativo, ou seja, de plantas suscetíveis
ao isolado I/2015 de H. vastatrix, mas resistentes a P. syringae pv. garcae foi igual a
zero.
4.6. Análise da expressão diferencial de genes possivelmente ligados à resistência a
Pseudomonas syringae pv. garcae
Os resultados das análises de expressão gênica estão apresentados na figura 13.
A expressão do gene EDS1 foi crescente até três horas após a inoculação (HAI) na
cultivar IPR 102, tanto em plantas inoculadas com a bactéria como em plantas controle,
tratadas com solução salina. Na cultivar IAC 125 RN houve também aumento da
expressão nas plantas inoculadas com P. syringae pv. garcae até a terceira HAI,
enquanto que, nas plantas controle houve queda na expressão aos 15 minutos após a
inoculação, seguida por pequeno aumento as três HAI. Entre três e 12 HAI, houve
queda acentuada do nível da expressão em todos os tratamentos mas, valor maior da
magnitude foram observados 24 HAI em todas as amostras.
O gene PAD4 apresentou expressão relativa semelhante ao gene EDS1. O pico
de atividade deste gene deu-se no tempo três HAI. Contudo, seis HAI todos os
tratamentos apresentaram redução da expressão, com tendência à estabilização. No
período seguinte, ou seja, 12 HAI a expressão gênica em plantas da cultivar IPR 102 foi
maior do que aquela observada na cultivar IAC 125 RN, independentemente do tipo de
inóculo das plantas. A expressão do gene PAD4 em, 24 HAI foi maior na cultivar IPR
102 inoculada com a bactéria P. syringae pv. garcae e menor nos demais tratamentos.
A expressão do gene APX esteve mais elevada aos 15 minutos após a inoculação
na cultivar IAC 125 RN inoculada com solução salina. No período entre três e seis HAI
detectou-se uma queda nível de expressão gênica que voltou a aumentar até 24 HAI,

76
especialmente em folhas da cultivar IAC 125 inoculadas com a bactéria ou mesmo com
solução salina.
O gene que codifica a enzima CAT, apresentou tendência de aumento da
expressão logo após a inoculação, seguida por queda, em todos os tratamentos até seis
HAI. A expressão voltou crescer após 12 HAI, estabilizando-se até 24 HAI, nos
tratamentos inoculados com solução salina, mas reduzindo os níveis de expressão nas
cultivares inoculadas com a bactéria.
A expressão do gene que codifica a SOD foi diferencial entre os genótipos. A
cultivar IPR 102 oscilou entre picos e queda acentuada nos níveis de expressão durante
o período de colonização avaliado. A cultivar IAC 125 RN apresentou expressão mais
estável para esta enzima, mantendo-se próximos aos níveis de expressão das plantas
tratadas com solução salina. Seis HAI, os níveis de expressão da SOD foram maiores
em cafeeiros da cultivar IPR 102, independentemente do inóculo utilizado, sendo
àquelas inoculadas com a batéria P. syringae pv. garcae mais elevada em relação
àquelas inoculadas com solução salina. Ainda, as 12 HAI a expressão mostrou-se mais
elevada nas plantas da cultivar IPR 102 inoculada com solução salina.
O gene que codifica a enzima POX apresentou expressão diferencial entre o
genótipo suscetível e resistente. Plantas da cultivar suscetível IAC 125 RN inoculada
com solução salina tiveram maior da expressão em 15 minutos após a inoculação e nos
demais períodos avaliados houve pequena variação sendo similar às plantas inoculadas
com o patógeno. Já as plantas da cultivar resistente IPR 102 apresentaram maior
expressão antes da exposição ao patógeno. Após as inoculações em ambos os
tratamentos houve queda acentuada na taxa de expressão até o tempo seis HAI, e 12
HAI ocorreu aumento na expressão sendo que as 24 HAI observou-se nova tendência de
redução da expressão.
A expressão do gene que codifica a enzima β-1,3-glucanase (PR-2) apresentou
padrão de expressão semelhante em todos os tratamentos estudados. No período que
corresponde até 15 minutos após a inoculação plantas da cultivar IAC 125 RN
inoculadas com solução salina tiveram queda no nível de expressão, sendo
discretamente menor que a tendência apresentada pelos demais tratamentos no mesmo
período analisado. Nos períodos subsequentes até 12 HAI houve decréscimo no nível de
expressão, obtendo expressão relativa similar ao período que antecedeu as inoculações.
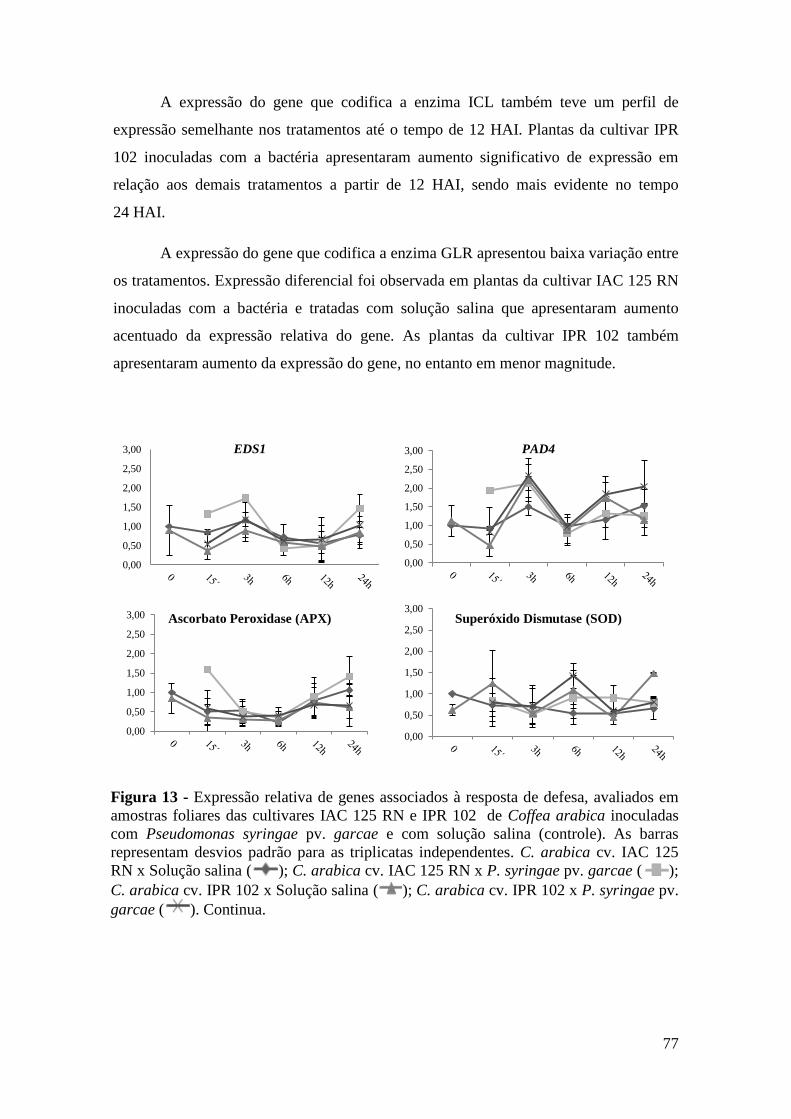
77
A expressão do gene que codifica a enzima ICL também teve um perfil de
expressão semelhante nos tratamentos até o tempo de 12 HAI. Plantas da cultivar IPR
102 inoculadas com a bactéria apresentaram aumento significativo de expressão em
relação aos demais tratamentos a partir de 12 HAI, sendo mais evidente no tempo
24 HAI.
A expressão do gene que codifica a enzima GLR apresentou baixa variação entre
os tratamentos. Expressão diferencial foi observada em plantas da cultivar IAC 125 RN
inoculadas com a bactéria e tratadas com solução salina que apresentaram aumento
acentuado da expressão relativa do gene. As plantas da cultivar IPR 102 também
apresentaram aumento da expressão do gene, no entanto em menor magnitude.
Figura 13 - Expressão relativa de genes associados à resposta de defesa, avaliados em
amostras foliares das cultivares IAC 125 RN e IPR 102 de Coffea arabica inoculadas
com Pseudomonas syringae pv. garcae e com solução salina (controle). As barras
representam desvios padrão para as triplicatas independentes. C. arabica cv. IAC 125
RN x Solução salina ( ); C. arabica cv. IAC 125 RN x P. syringae pv. garcae ( );
C. arabica cv. IPR 102 x Solução salina ( ); C. arabica cv. IPR 102 x P. syringae pv.
garcae ( ). Continua.
0,00
0,50
1,00
1,50
2,00
2,50
3,00 EDS1
0,00
0,50
1,00
1,50
2,00
2,50
3,00 PAD4
0,00
0,50
1,00
1,50
2,00
2,50
3,00 Ascorbato Peroxidase (APX)
0,00
0,50
1,00
1,50
2,00
2,50
3,00Superóxido Dismutase (SOD)
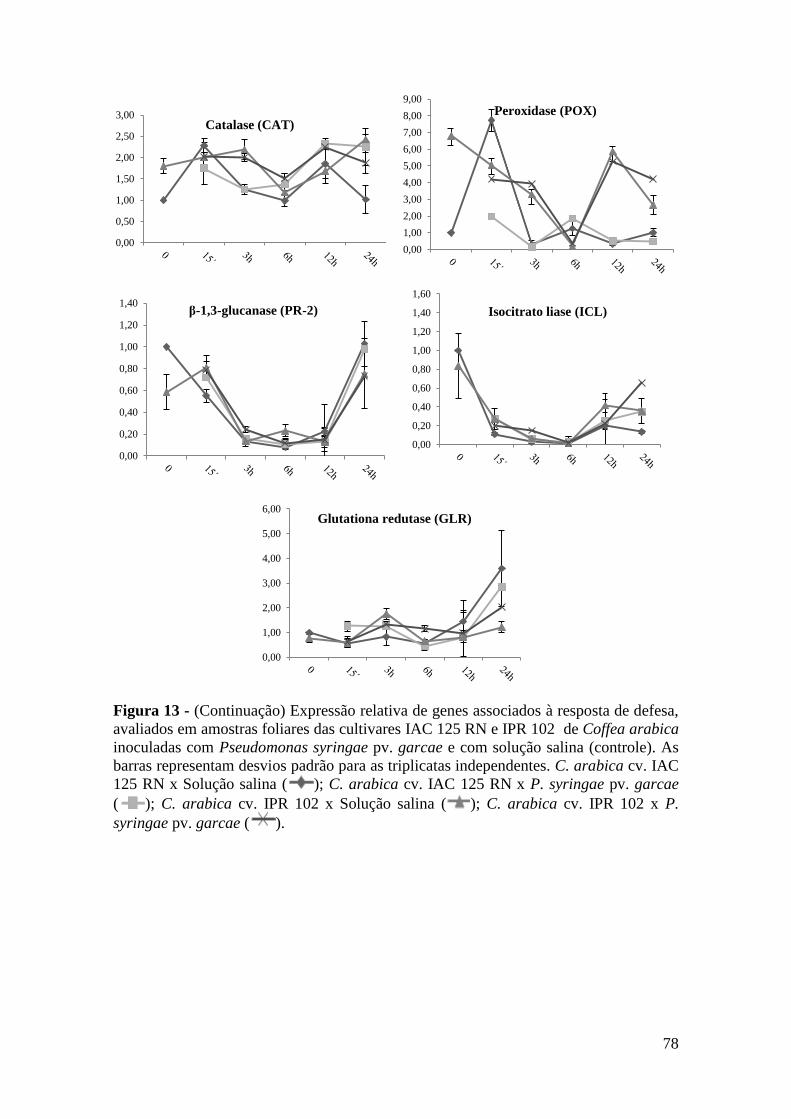
78
Figura 13 - (Continuação) Expressão relativa de genes associados à resposta de defesa,
avaliados em amostras foliares das cultivares IAC 125 RN e IPR 102 de Coffea arabica
inoculadas com Pseudomonas syringae pv. garcae e com solução salina (controle). As
barras representam desvios padrão para as triplicatas independentes. C. arabica cv. IAC
125 RN x Solução salina ( ); C. arabica cv. IAC 125 RN x P. syringae pv. garcae
( ); C. arabica cv. IPR 102 x Solução salina ( ); C. arabica cv. IPR 102 x P.
syringae pv. garcae ( ).
0,00
0,50
1,00
1,50
2,00
2,50
3,00Catalase (CAT)
0,00
1,00
2,00
3,00
4,00
5,00
6,00
7,00
8,00
9,00
Peroxidase (POX)
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40β-1,3-glucanase (PR-2)
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
Isocitrato liase (ICL)
0,00
1,00
2,00
3,00
4,00
5,00
6,00Glutationa redutase (GLR)

79
5. DISCUSSÃO
5.1. Alterações morfológicas em tecidos foliares de cafeeiros infectados por
Pseudomonas syringae pv. garcae
Os resultados obtidos podem ser úteis, tanto na compreensão do modo de
colonização por P. syringae pv. garcae no tecido foliar, bem como, da interação
bactéria-cafeeiro. Além disso, podem se transformar em importante ferramenta para
auxiliar programas de melhoramento genético de cafeeiro, visando resistência à
mancha-aureolada, principalmente na interpretação dos sintomas da doença.
Esse estudo possibilitou elucidar as alterações morfológicas e mecanismos de
infecção/colonização em folhas do terceiro internódio de café, infectadas pela bactéria
P. syringae pv. garcae. As alterações anatômicas observadas são semelhantes aos danos
induzidos por tabtoxina, com degradação da membrana tilacoide e cloroplastos,
favorecendo a colonização dos tecidos e sua utilização como substrato.
As alterações descritas por BENDER et al. (1999) foram claramente
identificadas neste estudo. Foi demonstrado que, o amarelecimento nos tecidos internos
do mesofilo, corresponde a aproximadamente o dobro dos danos que são observados
externamente na epiderme das folhas. Alterações como plasmólise do conteúdo celular
no mesofilo, bem como a hipertrofia e hiperplasia foram observadas nas folhas de
cafeeiro 20 DAI. Alterações similares foram observadas por Palmer & Bender (1995),
provocadas pela toxina coronatina em tomateiros inoculados com P. syringae pv.
tomato, como citado por BENDER et al. (1999) em revisão de literatura.
Na natureza, folhas de café são infectadas por bactérias fitopatogênicas através
de estômatos ou por ferimentos, causados principalmente por partículas de areia levadas
pelo vento (COSTA et al., 1957). Após a penetração no tecido foliar, as bactérias
ocupam os espaços inter e intracelulares, como observado na figura 6, sendo que as
bactérias são capazes de invadir o sistema vascular e de colonizar rapidamente os
tecidos, dando origem a extensas lesões, ocasionalmente em regiões mais distantes do
local de inoculação (Figura 6A).
Obstrução dos feixes vasculares foram observadas em nervuras, que estavam em
contato com a margem de colonização do tecido mesofílico colonizado, em folhas de

80
cafeiro da cultivar Mundo Novo IAC 376-4 de C. arabica (Figura 6F). Esta estratégia é
conhecida em algumas fitobactérias.
Os resultados aqui observados foram semelhantes àqueles obtidos por ROOS &
HATTINGH (1987). Os autores demostraram que P. syringae pv. morsprumorum, ao
colonizar folhas da cerejeira (Prunus avium), podia ser detectada a 3 cm, ou mais, do
ponto de inoculação, aos 10 DAI. Na interação entre P. syringae pv. syringae - tabaco
(Nicotiana benthamiana), MISAS-VILLAMIL et al. (2011) também demonstraram
forma similar de colonização de tecidos distantes do ponto de inoculação por meio de
vasos do xilema para movimentação, sendo esta uma propriedade importante desses
fitopatógenos.
ROOS & HATTINGH (1987) e HATTINGH et al. (1989) sugerem que P.
syringae pv. morsprumorum e P. syringae pv. syringae têm o mesmo comportamento
de penetração no hospedeiro, ou seja, infectam as folhas através dos estômatos, e
limitam-se inicialmente aos espaços intercelulares do mesofilo, próximos ao ponto de
inoculação, depois penetram em células do parênquima da bainha e vão para o sistema
vascular, movendo-se a partir daí para a nervura central, migrando para outras regiões
mais distantes. Esta estratégia aumenta a chance de sobrevivência, a longo prazo, do
agente patogênico, até que condições climáticas ideais proporcionem condições para
novos surtos. Os resultados do presente trabalho suportam esta hipótese, pois, a partir
de inoculações próximas ao feixe vascular, P. syringae pv. garcae colonizou outros
tecidos internos distantes do ponto de inoculação (Figuras 6E, 6F, 7A e 7B). Com base
nestas informações parece razoável supor que no cafeeiro, a morte do ápice em mudas
de café freqüentemente observadas em viveiros, causada por P. syringae pv. garcae,
pode ser potencializada pela colonização e eventual facilidade no transporte da bactéria
nos vasos do xilema e floema.
A presença de bactérias em feixes vasculares também confirma a hipótese de
movimentação e/ou de transporte para regiões distantes do local de colonização pelo
patógeno, ou seja, P. syringae pv. garcae causa a morte de folhas e ramos de cafeeiros,
movimentando-se mais rapidamente em direção ao ápice, através do xilema. Além
disso, as folhas e ramos situados abaixo do ponto de colonização também podem ser
infectados através do floema e também do xilema, podendo resultar na morte de folhas,
ramos e de plantas doentes.

81
5.2. Agressividade de isolados de Pseudomonas syringae pv. garcae
De fato, o melhoramento genético de plantas perenes visando obter plantas
resistentes às bacterioses necessita de métodos eficientes, e com boa repetibilidade para
a avaliação do germoplasma a ser pesquisado. Uma vez que é necessário testar grande
quantidade de plantas, o que requer disponibilidade de espaço e recursos. É
recomendado que a seleção para resistência a bacterioses seja realizada por meio de
inoculações artificiais, utilizando-se isolados sabidamente agressivos, para aumentar o
rigor da seleção e, com isso, reduzir a possibilidade de quebra da resistência pelo
patógeno.
Os testes de patogenicidade efetuados, evidenciaram ampla diversidade
patogênica entre os isolados de P. syringae pv. garcae oriundos dos estados de São
Paulo e Minas Gerais. Fatores como a produção de polissacarídeo extracelular,
expressão de genes de virulência (como avr) e produção de toxinas e fito-hormonios,
normalmente estão associados às diferenças em agressividade por bactérias
(PRASANNATH, 2013).
Em conjunto com outros mecanismos, esses fatores podem justificar a reação
diferencial em relação à agressividade dos isolados, quando inoculados em diferentes
genótipos, como observado por PARADELA et al. (1974). Neste caso as diferenças em
agressividade podem ser explicadas pela capacidade do patógeno em responder as vias
de defesa da planta hospedeira.
Nesse contexto, a estreita ligação dos resultados deste trabalho com fatores
observados por PARADELA et al. (1974), KAIRU (1997) e por ITHIRU et al. (2013)
indicam a possibilidade da ocorrência de variabilidade intraespecífica do patógeno,
como conhecida em outras patovares, com a ocorrência de biótipos e raças distinguíveis
da espécie P. syringae, como por exemplo os patovares P. syringae pv. phaseolicola
(patogênica ao feijoeiro), P. syringae pv. tomato (patogênica ao tomateiro), P. syringae
pv. glycinea (patogênica à soja), P. syringae pv. pisi (patogênica à ervilha) e também
em outras espécies de fitobactérias como Xanthomonas oryzae pv. oryzae (patogênica
ao arroz), X. vesicatoria (patogênica ao tomateiro), X. campestris pv. malvacearum
(patogênica ao algodoeiro), X. axonopodis pv. manihotis (patogênica à mandioca),
Ralstonia solanacearum (patogênica a diversas culturas), entre outras, como citado por
JANSE (2005).

82
Os resultados obtidos nos experimentos deste trabalho são corroborados pelas
análises de patogenicidade de isolados realizados por MACIEL (2012). O autor
caracterizou a agressividade de isolados de P. syringae pv. garcae em relação à cultivar
Mundo Novo de C. arabica, onde obteve resultados semelhantes em termos de
diversidade, sendo os isolados IBSBF 65, IBSBF 75 e IBSBF 3024 os mais agressivos e
os isolados IBSBF 248, IBSBF 2999, IBSBF 3005 e IBSBF 3065 os menos agressivos.
Além da relação entre agressividade de isolados bacterianos em genótipos de
cafeeiros, descrita por ITHIRU et al. (2013), outro aspecto relevante é a possibilidade
de interação entre isolados e a temperatura durante o período de avaliação dos sintomas.
Variação na agressividade dos isolados IBSBF 65, IBSBF 75 e IBSBF 3024 foi
observada no experimento 3, em relação ao experimento 1, onde, utilizou-se
concentração de inóculo análoga, método de inoculação e condições de umidade relativa
em casa-de-vegetação semelhantes, os isolados foram menos agressivos no experimento
3. As menores temperaturas no período de realização do terceiro experimento estão
provavelmente associadas a essas observações, demonstrando a importância de se
utilizar mais de um isolado na seleção de plantas resistentes. De fato, a severidade de
doenças bacterianas pode ser influenciada pela temperatura.
Estudos realizados por JONES et al. (1984) em outro patossistema
demonstraram que P. cichorii desenvolvem lesões de tamanho crescente em gerânio e
crisântemo, em temperaturas entre 16 a 28 °C, com forte redução do tamanho das lesões
em temperaturas mais elevadas. Estudos sobre a agressividade de diferentes isolados de
uma mesma espécie bacteriana, sob diferentes temperaturas, são escassos na literatura.
As variações em agressividade dos isolados IBSBF 65, IBSBF 75 e IBSBF
3024, nos experimentos 1 e 3, indicam que estes isolados poderão causar diferentes
reações nas plantas, dependendo das condições locais de temperatura por ocasião dos
experimentos. Além deste fator, a origem da cultivar também pode estar correlacionada
com a agressividade. Experimentos futuros poderão confirmar estas hipóteses e ainda,
estabelecer com mais rigor, os isolados que devem ser prioritariamente utilizados nas
inoculações para a seleção de plantas resistentes.
Quanto à resposta de folhas de diferentes internódios em relação à agressividade
dos isolados, demonstrou-se que folhas já expandidas do primeiro par são mais
suscetíveis, uma vez que esta tendência de reação foi detectada em praticamente todos
os dezenove isolados testados (Anexo 2, página 115).

83
Estes dados corroboram os obtidos por OLIVEIRA et al. (1991) e ZOCCOLI et
al. (2011). Ambos já haviam observados que folhas jovens são mais suscetíveis à
mancha-aureolada. Embora as diferenças médias para o aparecimento dos sintomas
iniciais tenha sido de apenas quatro DAI, a severidade da doença foi similar em folhas
do segundo, terceiro e quarto internódios.
Segundo OLIVEIRA & ROMEIRO (1990), os compostos fenólicos presentes
em folhas velhas, sadias ou produzidos após as inoculações, são capazes de inibir
desenvolvimento da doença.
Em contrapartida, folhas inoculadas do quarto internódio resultaram nos maiores
PI, sendo similares às folhas do segunda e terceiro internódio, em média 12 DAI
(Anexo 2, página 115). Isso demonstra que os compostos fenólicos são produzidos em
diferentes quantidades nas folhas com diferentes estágios de maturação, e
provavelmente diferem quanto ao tipo de composto produzido. Também, os compostos
atrasam o desevolvimento da bactéria, mas não são capazes de inibi-la. Tal efeito foi
enfatizado pelo isolado IBSBF 1372, no experimento 2, sendo mais agressivo nas folhas
do quarto internódio em relação as demais folhas inoculadas (Anexo 2, página 115).
5.2.1. Produção e sensibilidade as bacteriocinas entre isolados de Pseudomonas
syringae pv. garcae
Um dos fatores de importância fundamental para o sucesso do PMG é a
avaliação do germoplasma contra mais de um isolado bacteriano. No entanto, devido às
dificuldades a respeito do melhoramento de plantas perenes, a utilização de isolados de
forma combinada nas inoculações pode reduzir o tempo e os recursos empregados.
Contudo, é recomendado ter conhecimento sobre a atividade bacteriocinogênica dos
isolados (MARINGONI & KUROZAWA, 2002).
Os resultados aqui relatados são contrastantes em relação àqueles obtidos por
KAIRU (1997), que demonstrou que isolados brasileiros de P. syringae pv. garcae
produzem bacteriocinas capazes de inibir o crescimento de linhagens bacterianas do
Quênia, e também, em alguns casos, de isolados brasileiros, incluindo-se os isolados
IBSBF 65 e IBSBF 248 que apresentaram atividade inibitória entre si.
KAIRU (1997) utilizou isolados (possivelmente produtores de bateriocinas)
crescidos em meio NA líquido, contra os isolados desenvolvidos em meio NA
adicionado de sacarose, para a tipificação quanto à sensibilidade a bacteriocinas. O

84
desenvolvimento dos isolados em meio líquido pode ter resultado em maior taxa de
multiplicação das células bacterianas e, consequentemente, em maior quantidade de
compostos antimicrobianos sintetizados pelos isolados bacterianos, resultando na
inibição de diversos isolados do patógeno.
Segundo VIDAVIER (1983) a produção de bacteriocinas pode ser afetada por
fatores como, meio de cultura empregado no teste, temperatura de incubação e também
pela fase de crescimento da bactéria. Portanto, as divergências dos resultados
observadas nos experimentos desta tese, em relação aos resultados obtidos por KAIRU
(1997), devem ser relacionados a esses fatores, uma vez que foram utilizados meios de
cultura bem como temperatura de incubação diferentes. Cabe ressaltar os resultados
obtidos por BIAGI & AZEVEDO (1992) que demonstraram que o meio de cultura 523
(KADO & HESKETT, 1970), utilizado neste estudo, resulta em melhores condições de
análise para este tipo de ensaio in vitro.
Os resultados obtidos neste trabalho indicam que os isolados utilizados em
mistura não possuem atividade antagônica entre si (Figura 8). Experimentos adicionais
in vitro relacionados à análise da produção e da sensibilidade à bacteriocinas,
conduzidos de acordo com metodologia utilizada por MARINGONI & KUROZAWA
(2002), reforçaram a não existência de atividade bacterinogênica entre isolados de P.
syringae pv. garcae.
A ausência de evidências sobre a produção/sensibilidade à bacteriocinas indica
não haver restrições quanto ao uso de misturas para inoculações. No entanto, a
combinação contento quatro isolados resultaram em menor expressão de sintomas,
devendo ser evitada. Todavia, inoculações com mistura de dois isolados foi realizada
com sucesso em ensaios de controle químico da mancha-aureolada do cafeeiro
(PATRÍCIO et al., 2008). O mesmo foi observado em outros patossistemas, por
exemplo, testes de resistência em pessegueiros (Prunus persica) e damasco (P.
armeniaca) a Xanthomonas arboricola pv. pruni, realizados por SOCQUET-
JUGLARD et al. (2012).
A inoculação com mistura de isolados pode reduzir ou anular o efeito de reação
diferencial de plantas de um mesmo genótipo, quando inoculados por isolados distintos,
conforme mencionado por PARADELA FILHO et al. (1974) e ITHIRU et al. (2013).
Experimentos comparando-se a inoculação de isolados de forma individual ou
combinada, em genótipos contrastantes para resistência à bactéria poderão validar ou
não esta hipótese.

85
5.3. Metodologia de inoculação de Pseudomonas syringae pv. garcae para seleção
de plantas de cafeeiros resistentes à mancha-aureolada
No primeiro experimento, onde foram confrontados os métodos por aspersão e
agulhas múltiplas, os sintomas iniciais de colonização bacteriana surgiram entre três e
quatro dias após a inoculação, em ambos os métodos.
As plantas inoculadas por aspersão resultaram em baixa expressão dos sintomas
e apenas nas folhas jovens, ainda tenras, nas quais foi verificada maior severidade, em
geral, com lesões se desenvolvendo a partir dos bordos foliares. No entanto, este
método proporcionou as menores amplitudes de IC, resultado da baixa severidade da
doença e por consequência notas próximas da EP.
Considerando-se apenas os dados do primeiro par de folhas, a eficiência do
método por aspersão foi razoável na identificação de cafeeiros resistentes e suscetíveis a
P. syringae pv. garcae, entretanto, sua utilização requer atenção especial uma vez que
este método permitiu a ocorrência de escapes, resultado de incidências menores em
relação ao método de agulhas múltiplas, que proporcionou incidência de 100% nas
cultivares suscetíveis. Também inoculações por aspersão necessitam do preparo de
maiores quantidades de inóculo e a manutenção das plantas em câmara úmida por 24
horas antes das inoculações.
A cultivar resistente IPR 102 apresentou incidência considerada alta, até 66%
das folhas inoculadas. No entanto, para o cálculo desta variável foram considerados
como suscetível folhas com 1 ponto na escala, caso essa classe de plantas fossem
consideradas como resistentes a incidência passa a ser de apenas 17%, sendo esse
aspecto normal para uma cultivar nas fases finais de seleção. Nas avaliações realizadas
em experimento subsequentes com a mesma cultivar, mas com sementes oriundas de
geração mais avançada e plantas inoculadas por abrasão, 100% das plantas avaliadas
foram classificadas como resistentes (Tabela 16).
Compostos fenólicos estão envolvidos na resistência das plantas, e sua produção
pode variar devido a diversos fatores, entre eles a idade das folhas (OLIVEIRA &
ROMEIRO, 1990; OLIVEIRA et al., 1991; SALGADO et al., 2008). Deste modo, a
produção de fenóis em folhas do segundo e terceiro internódios pode estar associada às
diferenças na categorização dos genótipos. No entanto, os resultados obtidos em folhas
do quarto internódio sugerem queda na taxa de produção desses compostos, uma vez

86
que, as notas atribuídas à severidade da doença foram, em geral, similares ou maiores,
que aquelas conferidas às folhas dos segundo e especialmente do terceiro internódio.
A classificação dos genótipos pela AACPD, calculada pela evolução da doença
em folhas do quarto internódio mostrou-se ser eficiente. Contudo, seu uso apresenta
certa dificuldade operacional em relação às folhas do primeiro internódio, que se
mostraram mais indicadas para testes de resistência genética pela facilidade do
manuseio das mudas e também pela maior intensidade de danos observados nos tecidos
jovens.
Em geral o IC e CV foram semelhantes entre os dois métodos, já o DP foi mais
acentuado nos tratamentos inoculados por agulhas múltiplas. Estes resultados podem ser
atribuídos às maiores severidades ocasionadas pelo método, que resulta em maior
evidência das diferenças de suscetibilidade/resistência dentro dos tratamentos.
Os métodos testados no primeiro experimento resultaram em baixa severidade
da doença, avaliada pela escala de 0 a 5 pontos, que pode ser atribuído à baixa umidade
relativa verificada na casa-de-vegetação, onde os experimentos foram conduzidos.
Também, no segundo experimento, o método de agulhas múltiplas ocasionou baixa
severidade, mesmo com período maior de avaliação, reforçando a hipótese de
necessidade de alta umidade em experimentos que utilizam este método.
Uma vez que P. syringae pv. garcae necessita de condições de alta umidade
relativa para seu desenvolvimento, as condições de casa-de-vegetação com umidade
relativa em torno de 63% foi considerada baixa para os ensaios, fator minimizado
através das inoculações por abrasão. Esta técnica possibilitou a obtenção de resultados
promissores nos experimentos.
Uma variação dessa metodologia, utilizando fricção com carborundum, foi
testada por COSTA et al. (1957). No entanto, os autores observaram baixa taxa de
infecção, sugerindo que o estabelecimento do patógeno se dá principalmente, por
ferimentos. Possivelmente, a abrasão com carborundum resulta em maior oxidação dos
tecidos vegetais, que atrapalha a colonização da bactéria, resultando em baixa taxa de
infecção.
Deste modo, a metodologia por abrasão oferece condições ideais para a
penetração e infecção pela bactéria, devido ao maior número de ferimentos no tecido
foliar, e ainda que, devido aos ferimentos não transpassarem as folhas, aparentemente as
células bacterianas ficam protegidas da dessecação durante o processo de infecção. Um
método semelhante com bons resultados, é utilizado em inoculações com P. syringae

87
pv. tomato, para seleção de tomateiros resistentes a pinta-bacteriana, pela fricção com
gaze embebida em suspensão bacterianas (KOZIK & SOBICZEWSKI, 2007).
Com relação às respostas obtidas através das análises das variáveis EP e
AACPD, o IC foi, em geral, mais elevadas em folhas inoculadas por meio de agulhas
múltiplas em relação ao método de aspersão.
Nas respostas de folhas de diferentes internódios, o primeiro par de folhas
resultou em maiores valores do IC, em relação aos demais pares de folhas inoculados
por aspersão, enquanto que, as folhas inoculadas pela técnica de agulhas múltiplas essa
diferença foi mais pronunciada, sendo o IC maior nas folhas do primeiro internódio, no
entanto, as diferenças observadas foram de baixa magnitude.
Os valores de DP seguiram a mesma tendência do observado para os valores do
IC, sendo que, a similaridade dos resultados entre folhas de diferentes idades e ainda
menor magnitude desses parâmetros nas folhas do primeiro internódio, demonstrando
que as inoculações realizadas no primeiro par de folhas são suficientes para a
caracterização do germoplasma disponível. Estes resultados são corroborados por
valores de CV, que foram menores para as inoculações em folhas do primeiro
internódio. Em todos os casos o CV foi menor quando a análise foi realizada por meio
da variável AACPD, indicando maior confiabilidade dos resultados nos dados
transformados para essa variável.
Folhas inoculadas pela técnica das agulhas múltiplas apresentaram maior
amplitude nos valores das variáveis analisadas, por resultarem em maiores índices de
severidade, especialmente para a cultivar IPR 102, que demonstrou segregação para a
característica de resistência à mancha-aureolada.
O germoplasma avaliado por meio de inoculações por agulhas múltiplas e por
abrasão resultaram em valores de IC e DP similares, para ambas técnicas. Novamente,
os valores de CV foram menores nas análises resultantes da variável AACPD,
corroborando a eficiência dessa transformação de dados para a análise do germoplasma
disponível. Ainda, a técnica de inoculação por abrasão possibilitou os menores valores
de CV em relação às inoculações mediadas por agulhas múltiplas.
Portanto, testes de média a partir de valores da AACPD mostraram ser mais
confiáveis em relação às médias de EP, por resultar em valores de CV menores. Outra

88
observação interessante é que os valores do IC, DP e CV dos genótipos em fase de
seleção foram maiores em relação aos das cultivares, indicando possível seleção dentro
dos genótipos mais promissores, que apresentaram resistência parcial ao patógeno.
O melhoramento para resistência de plantas a bacterioses depende de
procedimentos eficazes, para avaliação de plantas mantidas em bancos de germoplasma,
sendo que inoculações artificiais fazem parte da estratégia (JANSE 2005). Os métodos
de inoculação que proporcionam fácil reprodução dos sintomas, boa repetitividade, além
de rapidez nos resultados, são considerados mais eficientes (ZAITER & COYNE,
1984), além disso, devem ser preferencialmente realizados com isolados agressivos do
patógeno, individualmente ou em misturas. Neste caso, deve-se conhecer a
compatibilidade das linhagens utilizadas por meio de testes in vitro e in vivo.
Os experimentos realizados para comparação dos métodos de inoculação,
mostraram que a técnica de abrasão desenvolvida nesta pesquisa, foi a mais consistente,
devendo ser empregada prioritariamente na seleção de genótipos de cafeeiros resistentes
a P. syringae pv. garcae.
5.4. Identificação de fontes de resistência a Pseudomonas syringae pv. garcae em
Coffea spp.
Os resultados obtidos no presente trabalho, corroboram à resistência a P.
syringae pv. garcae em genótipos estudados por MORAES et al. (1974; 1975),
MOHAN et al. (1978) e KAIRU (1997), em indivíduos presentes no BAG de Coffea do
Instituto Agronômico (IAC). Também, foram descritas novas fontes de resistência no
BAG do IAC, cujas informações poderão fornecer subsídios para programas diversos de
melhoramento, visando incorporar a resistência a P. syringae pv. garcae em cultivares
que estão sendo plantadas em larga escala no Brasil, suscetíveis ao patógeno. Fontes de
resistência foram identificadas em 53 acessos de cafeeiros do BAG de Coffea que
representam 50,5% dos genótipos avaliados.
Alguns genótipos merecem atenção especial pois, além de conter genes de
resistência à bacteriose, apresentam diversas características desejáveis para o programa
de melhoramento, tais como, tolerância à seca e resistência a pragas e outras doenças.
Dentre as espécies avaliadas, C. salvatrix apresentou os menores níveis de
doença. De acordo com a literatura, esta espécie apresenta resistência ao bicho mineiro

89
(GUERREIRO-FILHO et al. 1991), contém baixos teores de cafeína nas sementes
(0,72%) e, segundo MAZZAFERA & CARVALHO (1992) e MAZZAFERA et al.
(1998), possui alta quantidade de óleo (29%) no endosperma. Somadas, estas
características valorizam o potencial da espécie no desenvolvimento de novas cultivares
por meio de hibridações interespecífica.
Dos híbridos interespecíficos entre C. arabica e C. salvatrix, encontrados no
BAG de Coffea do IAC, duas plantas híbridas, na geração F1RC1 obtidas por
retrocruzamento para a espécie C. arabica ((C. arabica IAC 376-4 X C. salvatrix IAC
1288) x C. arabica IAC 376-4) são as mais promissoras. Essas plantas deverão ser
clonadas para a avaliação da resistência a P. syringae pv. garcae e demais
características, para melhor aproveitamento no PMG de cafeeiros do IAC, e também
retrocruzadas novamente para obtenção de características próximas às do café arábica.
Outra espécie com potencial para utilização em programas de melhoramento é
C. eugenioides. Esta espécie possui diversas características agronômicas de interesse,
sendo as principais, a resistência à broca-do-café, Hypothenemus hampei (SERA et al.,
2010) e ao bicho-mineiro, Leucoptera coffeella (GUERREIRO-FILHO et al. 1991).
Plantas desta espécie apresentam baixos teores de cafeína segundo MAZZAFERA &
CARVALHO (1992).
Seleções visando redução do teor de cafeína em C. arabica e C. canephora, a
partir de hibridações com C. eugenioides, vêm sendo realizadas por NAGAI et al.
(2008). Os autores já identificaram plantas com qualidade de bebida superior e com
menores teores de cafeína.
A respeito de C. stenophylla, outra espécie com alta resistência à bactéria, a
literatura dispõe de poucas informações sobre suas características. Sabe-se que
exemplares desta espécie são resistentes ao bicho-mineiro GUERREIRO-FILHO et al.
(1991) e apresentam teores de cafeína mais elevados, em relação à C. arabica
MAZZAFERA & CARVALHO (1992). Cafeeiros desta espécie apresentam maturação
tardia, com frutos de cor púrpura e a bebida originada de seus grãos com qualidade
sensorial bastante inferior à espécie C. arabica.
A espécie C. canephora apresenta alta resistência a P. syringae pv. garcae, e sua
utilização em programas de melhoramento é muito valiosa. Inúmeros híbridos entre C.

90
arabica e C. canephora comportaram-se como resistentes nas avaliações realizadas por
MOHAN et al. (1978), incluindo-se alguns híbridos obtidos a partir de cruzamentos
entre C. arabica e Híbrido de Timor (HT), grupo de cafeeiro que apresentam resistência
à ferrugem (H. vastatrix) e ao “Coffee Berry Disease” (CBD) (Colletotrichum
kahawae), doença não presente no Brasil, e ainda ao nematoide das galhas Meloidogyne
exigua (FAZUOLI & LORDELLO, 1978; RODRIGUES JUNIOR et al., 2004).
Híbridos entre C. arabica cv. Villa Sarchí (971/10) e HT (832/2) resultaram na seleção
da cultivar IAPAR 59, que exibe porte reduzido e resistência moderada à mancha-
aureolada (PETEK et al., 2001). Além disso, alguns genótipos de C. canephora
mostram maior tolerância à seca.
As espécies que apresentaram nível intermediário em relação à severidade da
mancha-aureolada, C. congensis e C. liberica, possuem especial utilização no
melhoramento de cafeeiros, uma vez que esses genótipos apresentam resistência ao
agente causal da ferrugem (BETTENCOURT & CARVALHO, 1968), bem como,
possuem rusticidade e certa tolerância à seca, a exemplo de C. canephora. O híbrido
natural entre C. canephora e C. congensis (IAC 4350) compreende importante
germoplasma para estudos futuros, pois carregam genes favoráveis para resistência à
mancha-aureolada e possuem potencial para aliar também a resistência à ferrugem e
tolerância à seca.
Com base nas relações e viabilidade entre cruzamentos interespecíficos em
Coffea spp., apresentadas por CARVALHO et al. (1985), pode-se deduzir que os genes
de resistência a P. syringae pv. garcae, presentes nas espécies silvestres, apresentam
maior dificuldade de serem aproveitados, sendo necessários diversos retrocruzamentos
com C. arabica a fim de serem selecionadas características agronômicas desejáveis.
Do ponto de vista aplicado, a transferecia de genes de resistência à mancha-
aureolada é facilitada pela utilização de germoplasa mais próximos geneticamente a C.
arabica. Por esse motivo, foram avaliadas diversas introduções da Etiópia, formas
botânicas, variedades e mutantes de C. arabica.
Das introduções da Etiópia avaliadas, apenas duas foram consideradas sucetíveis
ao patógeno, uma vez que, apresentaram 80% ou mais das progênies com notas entre
dois e cinco da EP. Portanto, grande parte do material avaliado mostrou comportamento
semelhante ao descrito por MOHAN et al. (1978).

91
Entretanto, o germoplasma silvestre de C. arabica provavelmente está
constituído por genes em condições heterozigóticas, pois praticamente quase todos os
acessos segregaram para a característica. Na prática, esse fator implica na necessidade
de um número maior de cruzamentos, a fim de se obter maior quantidade de plantas
com alelos favoráveis à resistência. Cem por cento das progênies avaliadas do acesso
ETP 22 comportaram-se como resistentes à mancha-aureolada, portanto a planta matriz
provavelmente encontra-se em homozigoze para esta característica. Essa condição
permitirá estudos exploratórios da heterose de híbridos F1 utilizando ETP 22 como
parental, uma vez que a característica de homozigose é um aspecto fundamental para tal
finalidade (VAN DER VOSSEN et al., 2015).
O aproveitamento da heterose em híbridos F1 tem sido utilizada de forma
associada à resistência à mancha-aureolada, (ITO et al., 2011; ANDREAZI et al.,
2015), bem como, à resistência múltipla à ferrugem, bicho-mineiro e à nematoides
(SERA et al., 2005).
Segundo SERA e colaboradores (2005), o processo de exploração do vigor
híbrido possibilita reduzir o tempo para obtenção de uma cultivar para
aproximadamente 12 anos. Além das características de resistência a fatores bióticos, é
possível a obtenção de cultivares mais produtivas em relação aos genitores, como
observado por ITO et al. (2011).
Além disso, as introduções de origem etíope compreendem importante reserva
genética da espécie, uma vez que esse germoplasma contém indivíduos com inúmeras
características de interesse. No anexo 3 (página 117), está apresentado um levantamento
dos dados de produção (1977 a 1989), ocorrência/severidade da ferrugem (1980), vigor
e tipo de maturação dos frutos (1977 a 1991) do referido germoplasma, de modo que,
esses dados poderão auxiliar na escolha dos parentais a serem utilizados nos
cruzamentos, a fim de se introduzir a resistência a P. syringae pv. garcae em cultivares
comerciais.
Além das variedades já identificadas como resistentes Villa Sarchí, Dilla &
Alghe (MORAES et al. 1975) e da cutivar IPR 102 (ITO et al., 2002), foi observado no
germoplasma de C. arabica avaliado, que compreendeu variedades, cultivares, formas
botânicas e mutantes (Tabela 16), resistência na variedade Palido Viridis (100% da
progênie) e na cultivar São Bernardo (33,3% da progênie).

92
Segundo CARVALHO et al. (1991) a cultivar São Bernardo é originaria da
Guatemala, possui porte menor e internódios mais curtos em relação ao Caturra, sendo a
característica de porte reduzido atribuída a um par de alelos dominates denominados
SbSb. Sendo assim, a utilização dessa cultivar em hibridações poderá resultar plantas de
pequeno porte e com resistência a P. syringae pv. garcae. No entanto, será necessário
elevado número de cruzamentos para obtenção de numerosas plantas híbridas, uma vez
que este material encontra-se em condições hererozigóticas, para a resistência à
mancha-aureolada que resultará na segregação para a característica. Por este motivo, a
falta de seleção para esta característica em plantas do híbrido IAC 7366 (São Bernardo x
Mundo Novo) resultou em 100% da progênie suscetíveis ao patógeno. Esta seleção
deverá ser realizada na geração F1.
A identificação de fontes de resistência no BAG do IAC possibilitou a busca por
genótipos em fase avançada de seleção, e que possuem em sua genealogia parentais
resistentes a P. syringae pv. garcae. Dessa forma, é possível realizar uma triagem
prévia do germoplasma e avaliar número maior de progênies na busca de plantas
resistentes. Essa estratégia poderá resultar na seleção de genótipos promissores, que em
poucos ciclos de seleção poderão originar uma cultivar resistente a P. syringae pv.
garcae.
O germoplasma que compreende plantas com o gene SH3 apresentam grande
valor para o melhoramento genético de Coffea, uma vez que este gene confere
resistência a diversas raças de H. vastatrix (BETTENCOURT & CARVALHO, 1968) e,
até o presente, não teve sua resistência quebrada por novas raças daquele patógeno
(BRAGHINI et al., 2014).
Dentre as seleções testadas, o material mais promissor ao PMG de cafeeiros do
IAC são os descendentes do híbrido H 13439-8-C1736, que possuem resistência
moderada à mancha-aureolada e carregam genes de resistência a diversas raças de H.
vastatrix. Esta seleção que vem sendo desenvolvida no IAC apresenta características de
porte reduzido e alta produtividade, altamente desejáveis.

93
5.5. Análise do efeito pleiotrópico do alelo SH1 na resistência a Pseudomonas
syringae pv. garcae
Os resultados obtidos a partir da inoculação das progênies F2 dos cafeeiros
H8089-2 e H8089-7 com os isolados I/2015 (v2 v5) e II/2015 (v1 v2 v5) de H. vastatrix
evidenciaram a natureza heterozigota (SH1 sH1) das matrizes F1, sendo que, conforme já
salientado por BETTENCOURT & CARVALHO (1968), os parentais, a cultivar Catuaí
Vermelho IAC 24 e a seleção Geisha IAC 1137-5, são respectivamente homozigotos
para os alelos sH1 e SH1. Ainda, de acordo com os resultados, foi confirmada a presença
do alelo dominante SH1 em um terço das plantas, sendo seis cafeeiros da progênie
H8089-2 e dez cafeeiros da progênie H8089-7. De acordo com a teoria gene-a-gene
(FLOR, 1971) estes cafeeiros, portadores de, ao menos, um alelo dominante SH1,
tiveram a resistência quebrada quando inoculados com o isolado II/2015 portador do
alelo v1 (Tabela 18).
Como reportado por CARVALHO (1988), se o alelo SH1 apresentasse efeito
pleiotrópico ou ocorrência de ligação gênica entre alelos que conferem resistência a H.
vastatrix e a P. syringae pv. garcae, cafeeiros portadores do alelo SH1 deveriam ser
resistentes a ambos os patógenos.
Assim, as progênies F2, segregantes, quando inoculadas com o isolado I/2015 de
H. vastatrix, deveriam apresentar a mesma reação quando expostas a P. syringae pv.
garcae.
No entanto, o valor significativo do teste de 2 McNemar calculado na progênie
H8089-2 revela a discordância entre as duas classificações; o número de cafeeiros
resistentes ao isolado I/2015 de H. vastatrix (6) não é o mesmo número de cafeeiros
resistentes a P. syringae pv. garcae (zero).
Por outro lado, embora o valor do 2 McNemar não tenha sido significativo, na
análise do total de plantas avaliadas, a resistência aos agentes bióticos não se revelou
simultânea, uma vez que, dez plantas resistentes ao isolado I/2015 de H. vastatrix,
portadoras do alelo SH1, revelaram-se suscetíveis à mancha-aureolada e, nove plantas
resistentes à mancha-aureolada revelaram-se suscetíveis ao isolado I/2015 de H.
vastatrix, sendo as taxas de falso positivo no primeiro e no segundo caso iguais a
15,87% e 14,29%, respectivamente, e a taxa total de erro igual a 30,16%.

94
A magnitude do coeficiente de Yule, que varia entre zero e ± 1, observado na
progênie H8089-7, individualmente, e na população F2, como um todo (H8089-2 e
H8089-7), corrobora a ausência de associação entre as classes R e S determinadas nas
avaliações das plantas em relação à resistência à ferrugem e à mancha-aureolada.
Portanto, de acordo com os dados obtidos fica evidente que não há efeito
pleiotrópico do gene SH1 ou ligação gênica entre genes que conferem resistência a
isolados homozigotos para o alelo V1 de H. vastatrix e a P. syringae pv. garcae. Por este
motivo, a busca por fontes de resistência à mancha-aureolada, para uso em programas
de melhoramento, visando ao desenvolvimento de cultivares com resistência simultânea
aos agentes bióticos, não deve ser restrita ao germoplasma de Coffea portadoras do gene
SH1.
5.6. Análise da expressão diferencial de genes possivelmente ligados à resistência a
Pseudomonas syringae pv. garcae
Do conhecido até o momento, o presente experimento relata os primeiros
estudos sobre a expressão de genes ligados à defesa de plantas, com foco nos períodos
iniciais da infecção (até 24 horas após a inoculação) e potencialmente envolvidos na
interação entre Pseudomonas syringae pv. garcae e cafeeiros. Cabe lembrar que, a
cultivar IPR 102 utilizada neste estudo tem origem a partir da hibridação artificial entre
C. canephora tetraploide e C. arabica cv. Bourbon Vermelho, realizada no IAC em
1950, e identificada como H 2460 (CARVALHO et al., 1983). Ou seja, a resistência da
cultivar IPR 102 é resultado na manifestação de genes introgredidos da espécie C.
canephora.
A respeito dos genes alvo analisados no presente estudo, na literatura
encontram-se numerosos trabalhos sobre os possíveis mecanismos de resistência
associados a diversos fatores bióticos e abióticos, incluindo-se respostas de defesa a
bactérias do agrupamento P. syringae.
Em um estudo semelhante, efetuado também por meio da expressão relativa de
diversos genes, comparou-se a atividade dos genes EDS1 e PAD4, em cafeeiros
resistente e suscetível ao nematoide M. paranaensis. Os resultados revelaram maior
expressão desses genes na cultivar resistente, sugerindo a possibilidade da ocorrência de
RSA (FATOBENE, 2014), outro mecanismo de defesa interferido pelo AS.

95
Os resultados obtidos neste trabalho evidenciaram maior expressão do gene
PAD4 24 HAI na cultivar IPR 102, mas não permitem associar a resposta de proteção
da planta a este gene, que pode desencadear a síntese de AS, como mencionado
anteriormente.
Durante a infecção por patógenos, a enzima SOD pode se associar com a CAT,
protegendo as células do dano oxidativo, mantendo os níveis de H2O2 regulados durante
a infecção (SAKAMOTO et al., 1995). NOWOGÓRSKA & PATYKOWSKI (2015)
demonstraram que SOD é crucial na resposta de defesa em feijão (P. vulgaris) cultivar
Korona a P. syringae pv. phaseolicola, uma vez que essa enzima apresentou maior
atividade nas plantas resistentes ao patógeno às 12 e 24 HAI. A indução de HR pelo
estresse oxidativo ligado a SOD, ocorre em plantas de soja (Glycine max) cultivar
Williams 82, resistente ao crestamento bacteriano (P. syringae pv. glycinea),
aproximadamente oito horas após a inoculação com o patógeno (ZOU et al., 2005).
Segundo VAN CAMP et al. (1997) e DESIKAN et al. (1996), citados por VAN
BREUSEGEM et al. (2001), durante a resposta via espécies reativas de oxigênio
(ERO), a enzima SOD é fundamental para o sistema antioxidante que regula a
concentração de peróxido de hidrogênio (H2O2), tóxico a fitopatógenos. As enzimas
CAT e POX, são os principais componentes na realocação do H2O2, atuando em
estresse oxidativo, por isso são relacionadas às respostas de defesa mediadas pela
sinalização de EROs e/ou de óxido nítrico (ON) (TRAPET et al., 2014). O efeito
causado pelo aumento da atividade de CAT, POX, bem como a APX às moléculas de
H2O2 resultam na morte celular programada no sitio de infecção (LEVINE et al., 1994;
ZOU et al., 2005).
No mesmo sentido, a inibição da enzima CAT levará ao acumulo de H2O2 que
por sua vez funcionará como sinalizador de genes PR (CHEN et al., 1993). Alguns
estudos demonstraram a ligação entre H2O2 e EROs que representa um um sistema de
defesa contra fitopatógenos (CLARK et al., 2000). Uma estreita ligação entre EROs e
AS foi sugerida por CHEN et al. (1993) e também por estudos posteriores, onde AS
reduziu a atividade de CAT e APX.
Cabe ressaltar que moléculas de AS podem desencadear o processo de RSA, em
resposta à infecção por fitopatógenos em diversas interações patógeno – hospedeiro, e
também atuam como sinalizador de efetores, que resultam na resistência local

96
(FERNADES et al., 2009). SONG & GOODMAN (2001), demonstraram que o ON é
necessário para que a RSA ocorra em plantas de tabaco (Nicotiana tabacum) cultivar
Xanthi nc, em resposta à infecção pelo vírus do mosaico do tabaco (Tobacco mosaic
virus).
Os genes associados ao estresse oxidativo por ON analisado neste trabalho, APX
e GLR, apresentaram comportamento similar nos genótipos resistente e suscetível à
bactéria. Segundo CLARK et al. (2000) ON pode atuar inibindo, de forma reversível, a
atividade da enzima APX, bem como CAT, em plantas de tabaco. Coincidentemente, a
expressão relativa destes genes neste estudo foram iguais ou menores em relação aos
tratamentos controle, mas não permitem inferir sobre o envolvimento do ON na resposta
de defesa, a participação deste composto estimula o acúmulo de AS (CHEN et al., 1993;
CLARK et al., 2000).
A enzima POX em conjunto com a enzima CAT atuam no mecanismo de defesa,
contribuindo para detoxificação do H2O2 das células, oxidação de compostos fenólicos
precursores da síntese de lignina, suberização, entre outras funções (MARGIS-
PINHEIRO et al., 1993; ALVAREZ et al., 1998). A expressão relativa do gene que
codifica a POX foi maior na cultivar resistente IPR 102 às 12 HAI, no entanto, os
resultados deste trabalho não permitem inferir a respeito da atuação desta enzima no
processo de defesa. Sabe-se que, para o acúmulo de EROs em Arabidopsis selvagem
contra P. syringae é necessária a ativação do complexo EDS1 – PAD4, o fator de
transcrição bZIP10 e também de POX e CAT da parade celular (LI et al. 2013).
A enzima β-1,3-glucanase (PR-2) possui importante papel na defesa contra
agentes fitopatogênicos, como demsntrado por ANDRADE et al. (2013) no
patossistema P. syringae pv. tomato - tomateiro. No entanto, a expressão relativa do
gene que codifica a PR-2 foi similar em todos os tratamento, portanto, essa enzima
parece não atuar na resposta de defesa contra P. syringae pv. garcae, durante o período
de ifecção avaliado.
A enzima ICL participa do ciclo do glioxilato. Segundo KUNZE et al. (2006),
esse metabolismo resulta em moléculas de carbono que podem ser reutilizadas no ciclo
de Krebs, dessa forma provendo precursores na biossíntese de carbohidratos e
aminoácidos. Portanto, pode-se pressupor que a tendência de aumento 24 HAI na
expressão relativa do gene que codifica a ICL, na cultivar resistente IPR 102, deve-se à

97
necessidade de produção de energia, para produção dos metabólitos envolvidos no
processo de defesa contra o patógeno.
Enfim, os resultados deste trabalho sugerem a possibilidade do mecanismo de
defesa a P. syringae pv. garcae ser intermediado via AS, uma vez que, a combinação
dos resultados sendo, maior expressão do gene PAD4 e do gene que codifica a SOD nas
plantas resistentes inoculadas com a bactéria e também baixa expressão dos genes que
codificam as enzimas APX, CAT e POX relacionados ao referido mecanismo de defesa,
corroboram esta hipótese.
Diversos trabalhos demonstram a biossíntese de etileno em tecidos
mecanicamente feridos (COLEMAN & HODGES, 1987). Segundo ISHIGUE et al.
(1993), as peroxidases representam um grupo de compostos associado a biossíntese de
etileno. Desse ponto de vista, as injurias causadas na inoculação da bactéria pelo
método abrasivo podem afetar diretamente a expressão de genes de defesa. em geral, a
resposta de expressão relativa das genes estudados, resultaram em tendências similares
de comportamento entre os tratamentos inoculados com P. syringae pv. garcae ou
solução salina. Vinte e quatro horas após as inoculações, houve diferenças na tendência
de expressão somente nos genes PAD4, CAT e ICL em relação ao tipo de inóculo.
Períodos mais extensos de coleta e um tratamento adicional com plantas sadias (não
inocladas) seriam úteis para demostrar os efeitos ocasionados por ocasião das
inoculações.
A enzima POX, que poderia indicar a resposta de defesa via etileno ou ácido
jasmônico, apresentou tendência de expressão similar entre os tratamentos estudados,
não sendo possível determinar sua relação na resposta de defesa.
Também o estresse oxidativo relacionado com a enzima SOD pode estar atuando
simultaneamente ao mecanismo de defesa da planta. Essa resposta pode desencadear a
deposição de compostos fenólicos, ligações proteicas ou morte celular localizada.
Estudos deverão ser elaborados utilizando períodos maiores e mais avançados de coletas
para elucidar a resposta final da resistência.

98
6. CONCLUSÕES
Pseudomonas syringae pv. garcae coloniza tecidos distantes do ponto de inoculação
em folhas de cafeeiro, por se movimentar mais facilmente nos vasos condutores;
Os isolados de P. syringae pv. garcae, oriundos de diferentes localidades dos
estados de São Paulo e Minas Gerais, apresentam diversidade em relação à
patogenicidade em cafeeiros da espécie C. arabica, sendo que, aparentemente, não
há relação entre o local de origem e/ou ano de isolamento com a agressividade dos
isolados;
O método de inoculação pela técnica de abrasão com lixa embebida em suspensão
bacteriana, é eficiente nas condições testadas e foi utilizado com sucesso na
caracterização de acessos do BAG de Coffea do IAC, bem como, na seleção de
plantas resistentes a P. syringae pv. garcae, permitindo a seleção de plantas de
cafeeiro em um curto período de tempo após as inoculações;
Fontes de resistência a P. syringae pv. garcae estão presentas em germoplasma
diverso de Coffea spp.. As seleções H6839-4, H13439-8 C1736 e IAC Catuaí SH3
C1621 do PMG do IAC apresentam potencial para o desenvolvimento de cultivares
que poderão aliar resistência moderada a P. syringae pv. garcae e resistência às
raças de Hemileia vastatrix com alta produtividade;
As introduções ETP 10, ETP 20, ETP 22, ETP 23, ETP 37 e ETP 38 apresentaram
os maiores índices de resistência, mas somente ETP 22 revelou-se homozigota para
a resistência a P. syringae pv. garcae;
O gene SH1 não confere resistência a P. syringae pv. garcae e também não possui
ligação com os genes de resistênciaa H. vastatrix na seleção Geisha (IAC 1137-5);
Os resultados da expressão relativa dos genes sugerem que o mecanismo de
resistência apresentado pela cultivar IPR 102, herdado de C. canephora,
desencadeia rotas metabólicas da via do AS e/ou biossíntense de EROs.

99
7. REFERÊNCIAS BIBLIOGRÁFICAS
AARTS, N.; METZ, M.; HOLUB, E.; STASKAWICZ, B.J.; DANIELS, M.J.;
PARKER, J.E. Different requirements for EDS1 and NDR1 by disease resistance genes
define at least two R gene-mediated signaling pathways in Arabidopsis. Proceedings of
the National Academy of Sciences USA, v. 95, p. 10306–10311, 1998.
ADUGNA, G.; JEFUKA, C.;TEFERI, D.; ABATE, S. New record and outbreaks of
bacterial blight of coffee (Pseudomonas syringae) in Southern Ethiopia: impact of
climate change scenarios. IN: INTERNATIONAL CONFERENCE ON COFFEE
SCIENCE, 24, 2012, 2012, San José. Proceedings. San José: ASIC, 2012. p.660.
AGRIOS, G.N. Plant Pathology, 5th. ed. San Diego: Elsevier Academic Press, p.922,
2005.
ALMEIDA, I.M.G.; MACIEL, K.W.; BERIAM, L.O.S.; RODRIGUES, L.M.R.;
DESTÉFANO, S.A.L.; RODRIGUES NETO, J.; PATRÍCIO, F.R.A.; Increase in
incidence of bacterial halo blight (Pseudomonas syringae pv. garcae), in coffee
producing areas in Brazil. IN: INTERNATIONAL CONFERENCE ON COFFEE
SCIENCE, 24.,.
ALMEIDA I.M.G.; RODRIGUES L.M.R.; PATRÍCIO F.R.A. Crestamento bacteriano
das folhas causado por Pseudomonas cichorii em mudas de café no Estado de São
Paulo. IN: XXXV CONGRESSO PAULISTA DE FITOPATOLOGIA. Jaguariuna –
SP, 2012. Resumos. 1CD-ROM, 2012.
ALVAREZ, M.E.; PENNELL, R.I.; MEIJER, P.J.; ISHIKAWA, A.; DIXON, R.A.;
LAMB, C. Reactive oxygen intermediates mediate a systemic signal network in the
establishment of plant immunity. Cell, v.92, p.773-84, 1998.
AMARAL, J.F. do; TEIXEIRA, C.G.; PINHEIRO, E.D. O bactério causador da
mancha-aureolada do cafeeiro. Arquivos do Instituto Biológico, v.23, p.151-155,
1956.
ANDRADE, C.C.L.; RESENDE, R.S.; RODRIGUES, F.A; SILVEIRA, P.R.; RIOS,
J.A.; OLIVEIRA, J.R.; MARIANO, R.L.R. Indutores de resistência no controle da pinta
bacteriana do tomateiro e na atividade de enzimas de defesa. Tropical Plant Pathology,
v.38, p.28-34, 2013.
ANDREAZI, E.; SERA, G.H.; FARIA, R.T.; SERA, T.; SHIGUEOKA, L.H.;
CARVALHO, F.G.; CARDUCCI, D.C. Desempenho de híbridos F1 de café arábica
com resistência simultânea a ferrugem, mancha aureolada e bicho mineiro. Coffee
Science, v. 10, p. 375 - 382, 2015.
ANTHONY, F.; COMBES, M.; ASTORGA, C.; BERTRAND, B.; GRAZIOSI, G.;
LASHERMES, P.. The origin of cultivated Coffea arabica L. varieties revealed by
AFLP and SSR markers. Theoretical and Applied Genetics v.104, p. 894 – 900, 2002.

100
BARTA, T.M. & WILLIS, D.K. Biological and molecular avidence that Pseudomonas
syringae patovars coronafaciens, striafaciens and garcae are likely the same pathovar.
Journal of Phytopathology v.153, p.492-499, 2005.
BELAN, L.L. Detecção de Pseudomonas syringae pv. garcae em sementes,
microanálise de raios X e desenvolvimento de escala diagramática para a mancha-
aureolada do cafeeiro. Tese (Doutorado) - Universidade Federal de Lavras, Lavras –
MG, 2014, 126p.
BENDER, C.L., ALARCON-CHAIDEZ, F., GROSS D.C. Pseudomonas syringae
phytotoxins: mode of action, regulation, and biosynthesis by peptide and polyketide
synthetases. Microbiology and Molecular Biology Reviews, v.63, n.2, p.266–292,
1999.
BETTENCOURT, A.J. & CARVALHO, A. Melhoramento visando à resistência do
cafeeiro à ferrugem. Bragantia, v.27, p.35-68, 1968.
BIAGI, C.M.R. & AZEVEDO, J.L. Detecção de bacteriocinas produzidas por bactérias
fitopatogênicas dos gêneros Pseudomonas e Xanthomonas. Scientia Agricola, v.49,
p.1-8, 1992.
BISHOP, J.G.; DEAN, A.M.; MITCHELL-OLDS, T. Rapid evolution in plant
chitinases: molecular targets of selection in plant–pathogen coevolution. Proceedings
of the National Academy of Sciences USA, v.97, p.5322–5327, 2000.
BOLLER, T. & FELIX, G. A renaissance of elicitors: perception of microbe-associated
molecular patterns and danger signals by pattern-recognition receptors. Annual Review
of Plant Biology, v.60, p.379-406, 2009.
BRADBURY, J.F. Guide to plant pathogenic bacteria. Farhan House: C.A.B.
International, 1986. 332 p.
BRAGHINI, M.T.; FAZUOLI, L.C.; GUERREIRO FILHO, O.; MISTRO, J.C.;
ESKES, A.B. Durable resistance to coffee leaf rust provided by the SH3 gene evaluated
at the Instituto Agronômico de Campinas, SP, Brazil. IN: INTERNATIONAL
CONFERENCE ON COFFEE SCIENCE, 25, 2014, Armenia – Colombia,
Proceedings. Colombia: ASIC, 2014, p.42-46. 1 CD ROM.
BONOMO, P. Metodologias biométricas para seleção de progênies no
melhoramento genético do cafeeiro. Tese (Doutorado) - Universidade Federal de
Viçosa, Viçosa - MG. 2002. 124p.
BULL, C.T. & KOIKE S. T. Practical Benefits of Knowing the Enemy: Modern
Molecular Tools for Diagnosing the Etiology of Bacterial Diseases and Understanding
the Taxonomy and Diversity of Plant Pathogenic Bacteria. Annual review of
phytopathology v. 53, p. 157-180.
CAMPBELL, C.L. & MADDEN, L.V. (Ed.). Introduction to plant disease
epidemiology. New York, NY: Wiley, 532p., 1990.

101
CARDOSO, R.M.L. & MOHAN, S.K. Ocorrência e intensidade do crestamento
bacteriano causado por Pseudomonas garcae em relação a alguns fatores climáticos no
Estado do Paraná. IN: CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS, 7,
1979, Araxá, MG Resumos... Rio de Janeiro: IBC/GERCA. p.65-66, 1980.
CARDOSO, R.M.L. & SERA, T. Obtenção de cultivares de Coffea arabica L. com
resistência simultânea a Hemileia vastatrix Berk. & Br. e Pseudomonas syringae pv.
garcae Amaral et al. no Estado do Paraná. In: SIMPÓSIO SOBRE FERRUGENS DO
CAFEEIRO, 1983. Oeiras. Resumos... Lisboa, CIFC/IICT, 1984, p. 417-419, 1983.
CARVALHO, A. Distribuição geográfica e classificação botânica do gênero Coffea
com referência especial à espécie arábica. V. Origem e classificação botãnica do C.
arabica L. Bol Super Serv Café, n.21, p. 174 – 180, 1946.
CARVALHO, A. Histórico do desenvolvimento do cultivo do café no Brasil.
Documentos IAC, 34, Instituto Agronômico de Campinas, Campinas - SP. 9p. 1997
CARVALHO, A. Principles and practice of coffee plant breeding for productivity and
quality factors: Coffea arabica. IN: CLARKE, R. J., MACRAE, R. (eds.). Coffee:
agronomy. Elsevier Applied Science, v.4, p.129 - 165, 1988.
CARVALHO, A; ANTUNES FILHO, H.; MENDES, J.E.T.; LAZZARINI, W; REIS,
A.J.; ALOISI SOBRINHO, J.; MORAES, M.V. de; NOGUEIRA, P.K. & ROCHA,
T.R. da. Melhoramento do cafeeiro: XIII. Café Bourbon Amarelo. Bragantia, v.16,
p.411-454, 1957.
CARVALHO, A.; FAZUOLI, L.C. Café. IN: FURLANI, A.M.C., VIÉGAS, G.P.(Eds).
O melhoramento de plantas no Instituto Agronômico, v. 1, p. 29-76, 1993.
CARVALHO, A.; FEWERDA, F.P.; FRAHM-LELIVELD, J.A.; MEDINA, D.M.;
MENDES, A.J.T.; MÔNACO, L. C. Coffee. In: FERWERDA, F. P. (Ed.). Outlines of
perennial crop breeding in the tropics. Wageningen: Veenman, 1969. p.189-241.
CARVALHO, A.; KRUG, C.A.; MENDES, J.E.T.; ANTUNES FILHO, H; MORAES,
H. de; ALOISI SOBRINHO, J.; MORAES, M.V. de & ROCHA, T.R. da.
Melhoramento do cafeeiro: IV. Café Mundo Novo. Bragantia, v.12, p.97-129, 1952.
CARVALHO, A.; MEDINA FILHO, H.P.; FAZUOLI, L.C. Evolução e melhoramento
do cafeeiro. IN: I COLÓQUIO SOBRE CITOGENÉTICA E EVOLUÇÃO DE
PLANTAS , 1984. São Paulo, SP. Tópicos de Citogenética e Evolução de Plantas.
São Paulo: Sociedade Brasileira de Genética, 215-234p., 1985
CARVALHO, A.; MÔNACO, L.C. Transferencia do fator caturra para o cultivar
Mundo Novo de Coffea arabica. Bragantia, v. 31, p. 379-399, 1972.
CARVALHO, A.; COSTA, W.M.; FAZUOLI, L.C. Auto-incompatibilidade,
produtividade, ocorrência de sementes do tipo moca e mudas anormais no café Icatu.
Bragantia, v. 42, p.157-169, 1983.

102
CARVALHO, C.H.S. Cultivares de café: origem, características e recomendações. 01.
ed. EMBRAPA CAFÉ. Brasília-DF, 2008, 332 p.
CHANG, S.; PURYEAR, J.; CAIRNEY, J. A simple and efficient method for isolating
RNA from pine trees. Plant Mol Biol Rep, v. 11, p.113-116, 1993.
CHEN, Z. Morphocultural and pathogenic comparisons between Colletotrichum
kahawae and C. gloeosporioides isolated from coffee berries. PhD Thesis -
Universidade Técnica de Lisboa. Lisboa. 2002.
CHEN, Z.; SILVA, H.; KLESSIG, D.F. Active oxygen species in the induction of plant
systemic acquired resistance by salicylic acid, Science, v.262, p.1883-1886, 1993.
CHEVALIER, A. Les caféiers du globe III, systhématique dês caféiers et fauxcaféiers,
maladies et insects nuisibles. Encyclopédie Biologique, v.281, p.356, 1947.
COLEMAN, L. W.; HODGES, C. F. Ethylene biosynthesis in Poa pratensis leaves in
response to injury or infection by Bipolaris sorokiniana. Phytopathology, v.77,
p. 1280-1283, 1987.
CONAB, Companhia Nacional de Abastecimento. Anuário Brasileiro do Café 2014,
Benno Bernardo Kist ... [et al.]. – Santa Cruz do Sul : Editora Gazeta Santa Cruz. 128
p., 2014.
CONAB, Companhia Nacional de Abastecimento. Acompanhamento da safra
brasileira. Café. Safra 2015, Primeiro levantamento, Janeiro de 2015. Brasília, p. 1-41,
2015.
COSTA, A.S.; AMARAL, J.F.; VIEGAS, A.P.; SILVA, D.M.; TEIXEIRA, C.G.;
PINHEIRO, E.D. Bacterial halo blight of coffee in Brazil. Phytopathologische
Zeitschrift, v.28, p.427-444, 1957.
COSTA, A.S., SILVA, D. M. A Mancha-aureolada do cafeeiro. Bragantia, Campinas,
v.19, p.LXII-LXVIII, 1960.
CLARK, D.; DURNER, J.; NAVARRE, D.A.; KLESSIG, D.F.Nitric oxide inhibition of
tobacco catalase and ascorbate peroxidase. Molecular Plant-Microbe Interactions,
v.13, p.1380-1384, 2000.
CRAMER, C.L.; WEISSENBORN, D.; COTTINGHAM, C.K.; DENBOW, C.J.;
EISENBACK, J.D.; RADIN, D.N. & YU, X. Regulation of defense related gene
expression during plant-pathogen interractions. Journal of Nematology, v.25, p. 507-
519, 1993.
DANGL, J.L. & JONES, J.D.G. Plant pathogens and integrated defense responses to
infection. Nature, v.411, p.826 - 833, 2001.
DAVIS, A.P. Psilanthus mannii, the type species of Psilanthus, transferred to Coffea.
Botanical Journal of the Linnean Society, Londres, v. 29, Issue 4, p. 471 - 472, 2011.

103
DAVIS, A.P.; GOVAERTS, R.; BRIDSON, D.M.; STOFFELEN, P. An annotated
taxonomic conspectus of the genus Coffea (Rubiaceae). Botanical Journal of the
Linnean Society, v. 152, n. 4, p. 465 - 512, 2006.
DESTÉFANO, S.A.L.; RODRIGUES, L.M.R.; BERIAM, L.O.S.; PATRÍCIO, F.R.A.;
THOMAZIELLO, R.A.; RODRIGUES NETO, J. Bacterial leaf spot caused by
Pseudomonas syringae pv. tabaci in Brazil. New Disease Reports, v.22, n.5, 2010.
DIXON, R.A. & HARRISON, M.J. Activation, structure and organization of genes
involved in microbial defense in plants. Adv. Genet., v.28, p.165-234, 1990.
DURRANT W.E. & DONG X. Systemic acquired resistance. Annual Review of
Phytopathology, v.42, p.185-209, 2004.
ESKES, A.B.; TOMA-BRAGHINI, M. Assessment methods for resistance to coffee
leaf rust (Hemileia vastatrix Berk. & Br.). FAO Plant Protection Bulletin, v.29, p.56-
66, 1981.
F.A.O. FAO coffee misson to Ethipia 1946 – 1965 – Report, FAO. Ed. Rome (Itália),
200p., 1968.
FATOBENE B.J.R. Seleção de cafeeiros com resistência múltipla a nematoides do
gênero Meloidogyne. Tese (Doutorado) - Instituto Agronômico de Campinas,
Campinas – SP. 2014, 71fls.
FAZUOLI, L.C. Avaliação de progênies de café “Mundo Novo” (Coffea arabica L.).
Dissertação (Mestrado em agronomia) - Escola Superior de Agricultura “Luiz de
Queiroz”, Piracicaba, SP, ESALQ, 1977.
FAZUOLI, L.C. Metodologia, critérios e resultados de seleção de progênies de café
Icatu com resistência a Hemileia vastatrix. Tese (Doutorado) - Universidade Estadual de
Campinas, UNICAMP, Campinas – SP, 1991, 322p.
FAZUOLI, L.C. & LORDELLO R.R.A. Resistência de cafeeiros Híbrido de Timor a
Meloidogyne exigua. Ciência y Cultura, v.30, 1978.
FAZUOLI, L.C.; BRAGHINI, M.T.; SILVAROLLA, M.B.; MISTRO, J.C.;
PATRÍCIO, F.R.A. Melhoramento do cafeeiro visando à resistência a doenças. Curso
de Atualização em Café, 9. Documentos IAC, v.91, p.1-16, 2009.
FERNANDES, C.F.; VIEIRA JUNIOR, J.R.; SILVA, D.S.G.; REIS, N.D.; ANTUNES
JUNIOR, H. Mecanismos de defesa de plantas contra o ataque de agentes
fitopatogênicos. Embrapa Rondônia, (Documentos, 133), 14p., 2009.
FEYS, B.J.; WIERMER, M.; BHAT, R.A.; MOISAN, L.J.; MEDINA-ESCOBAR, N.;
NEU, C.; CRUZ-CABRAL, A.; PARKER, J.E. Arabidopsis SENESCENCE-
ASSOCIATED GENE101 stabilizes and signals within an enhanced disease
susceptibility1 complex in plant innate immunity. Plant Cell, v.17, p. 2601-2613, 2005.

104
FLOR, H. H. Current status of the gene-for-gene concept. Annual Review of
Phytopathology, v.9, p.275-296, 1971.
GIANINAZZI, C.; MARTIN, J.C.V. Hypersensibilité aux virus, température et
protéines solubles chez le Nicotiana xanthi nc. Apparition de nouvelles macromolécules
lors de la répression de la synthèse virale. CR Academy of Science of Paris. v.270, p.
2383-2386, 1970.
GIMENEZ-IBANEZ, S.; RATHJEN, J.P. The case for the defense: plants versus
Pseudomonas syringae. Microbes Infect, v.12, p.428-437, 2010.
GONÇALVES, F. L. T., & MASSAMBANI O. Bacteria and fungi spores as ice nuclei
in Coffea arabica L., Revista Ciência e Natura, v.33, p.73-93, 2011.
GONÇALVES, M. M.; RODRIGUES, M.L.; MEXIA, J. N.; DAEHNHARDT, E.
Melhoramento da cafeicultura em Timor face à Hemileia vastatrix B. & Br. Garcia de
Orta, Serie de Estudos Agronómicos, v.5, p.3-10, 1978.
GORJANOVIĆ, S. A. Biological and Technological Functions of Barley Seed
Pathogenesis-Related Proteins (PRs). Journal of The Institute of Brewing, v.115,
p.334-360, 2009.
GRAHAM, T.L.; GRAHAM, M.Y. Cellular coordination of molecular responses in
plant defense. Molecular Plant-Microbe Interactions. v.4, p.415-422, 1991.
GUERREIRO FILHO, O. Coffee leaf miner resistance. Brazilian Journal of Plant
Physiology, v. 18, n. 1, p. 109-117, 2006.
GUERREIRO-FILHO, O.; MEDINA-FILHO, H.P.; CARVALHO, A. Fontes de
resistência ao bicho mineiro, Perileucoptera coffeella em Coffea spp. Bragantia, v.50,
p.45-55, 1991.
GUERREIRO FILHO, O.; MEDINA FILHO, H.P.; CARVALHO, A.; GONÇALVES,
W. Melhoramento do cafeeiro. XLIII – Seleção de cafeeiros resistentes ao bicho
mineiro (Perileucoptera coffeella). Bragantia, v.49, p. 291-304, 1990.
GUZZO S.D. Aspectos Bioquímicos e Moleculares da Resistência Sistêmica
Adquirida em Cafeeiro contra Hemileia vastatrix. Tese (Doutorado), Centro de
Energia Nuclear na Agricultura, Piracicaba – SP, 2004, 236p.
HATTINGH, M.J.; ROOS, I.M.M.; MANSVELT, E.L. Infection and systemic invasion
of decidous fruit trees by Pseudomonas syringae in South Africa. Plant Disease v.73,
n.10, p.784-789, 1989.
HAYWARD, A.C. Annual Report for 1962/1963, Coffee Res Sta Rvirv, and Coffee
Res. Serv. Kenya. (resumos em R.A.M. v.43, p.744), 1962.
INTERNATIONAL COFFEE ORGANIZATION World coffee consumption.
Disponível em: http://www.ico.org/prices/new-consumption-table.pdf. Acesso em: 28
de abril de 2015.

105
ISHIGE, F.; MORI, H.; YAMAZAKI, K.; IMASEKI, H. Identification of a basic
glycoprotein induced by ethylene in primary leaves of azuki bean as a cationic
peroxidase. Plant Physiology, v.101, p.193-199, 1993.
ITHIRU, J.M.; GICHRU, E.K.; GITONGA, P.N.; CHESEREK, J.J.; GICHIMU, B.M.
Methods for early evaluation for resistance to bacterial blight of coffee. African
Journal of Agricultural Research, v.8, p.2450-2454, 2013.
ITO, D.S.; SERA, T.; SERA, G.H.; ALEGRE, C.R.; RIBEIRO FILHO, C.; MATA,
J.S.; KANAYAMA, F.S.; AZEVEDO, J.A. Seleção de cafeeiros arábicos resistentes à
mancha-aureolada (Pseudomonas syringae pv. garcae) em condições de campo. IN:
CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS, 5, Águas de Lindóia, SP
Anais. Brasília, D.F. Embrapa Café, 2007. 1 CD-ROM.
ITO, D.S.; SERA, T., SERA, G.H.; GROSSI, L.D.; KANAYAMA, F.S. Resistance to
bacterial blight in arabica coffee cultivars. Crop Breeding and Applied
Biotechnology, v.8, p.99-103, 2008.
ITO, D.S.; SERA, T., SERA, G.H.; Shigueoka, L.H.; ANDREAZI, E.; SERA, G.H.;
GARDINO, C.G. Heterose em híbridos de café arábicos com resistência à ferrugem,
mancha aureolada e aos nematoides. IN SIMPÓSIO DE PESQUISA DOS CAFÉS DO
BRASIL, VII, Araxá – MG Resumos, Brasília – DF, EMBRAPA Café, 2011.
JANSE, J.D. Phytobacteriology: principles and practice. CABI Publishing, 2005, 360p.
JOHANSEN D.A. Plant microtechnique. McGraw-Hill, New York. 1940
JOHANSSON, O.N. Plant pathogen defense: Signalling, resistance and cell death.
Tese (Doutorado) - Göteborgs Universitet, Gotemburgo, Suécia, 2015. 54p.
JONES, J.B.; RAJU, B.C.; ENGELHARD, A.W. Effects of temperature and leaf
wetness on development of bacterial leaf spot of geraniums and chrysanthemums
incited by Pseudomonas cichorii. Plant Disease, v.68, p.248-251, 1984.
KADO, C.I. & HESKETT, M.G. Selective media for isolation of Agrobacterium,
Corynebacterium, Erwinia, Pseudomonas, and Xanthomonas. Phytopathology, v.60, p.
969-976, 1970.
KAIRU, M.G. Biochemical and pathogenic differences between Kenyan and Brazilian
isolates of Pseudomonas syringae pv. garcae. Plant Pathology, v.46, p.239-246, 1997.
KIMATI, H., AMORIM, L., REZENDE, J. A. M., BERGAMIN FILHO, A. &
CAMARGO, L. E. A. Manual de Fitopatologia; doenças das plantas cultivadas; São
Paulo: Editora Agronômica Ceres, 4ª ed., 2005, 663p.
KIMURA, O.; ROBBS, C.F.; FERRARI, J.A.R. Algumas observações relacionadas
com as bacterioses do cafeeiro. IN: CONGRESSO BRASILEIRO DE PESQUISAS
CAFEEIRAS, 4, 1976, Caxambu. Anais. Caxambu: IBC/GERCA, p.104, 1976.

106
KIMURA, O.; ROBBS, C.F.; RIBEIRO, R.L.D. Estudos sobre o agente da “Mancha-
aureolada do cafeeiro” (Pseudomonas garcae Amaral et al.) Arquivos da
Universidade Federal Rural, Rio de Janeiro, v.3, n.2, p.15-18, 1973.
KING, E.O.; WARD, M.K.; RANEY, D.E. Two simple media for the demonstration of
pyocyanin and fluorescein. Journal of Laboratory and Clinical Medicine, v.44,
p.301-307, 1954.
KOROBKO, A.; WONDINAGEGNE, E. Bacterial blight of coffee (Psedomonas
syringae pv. garcae) in Ethiopia. IN: RUDOLDH, K.; BURR, T.J.; MANSFIELD,
J.W.; STEAD, D.; VIVIAN, A.; VON KIETZELE, J. Pseudomonas syringae and
related patogens. Netherlands Springer, 1997. p. 538-541, 1997.
KOZIK, E.U. & SOBICZEWSKI, P. Assessment of inoculation techniques suitability
for determination of tomato plants resistance to bacterial speck (Pseudomonas syringae
pv. tomato). Phytopathologia polonica, v.44, p.17-25, 2007.
KRUG, C.A.; MENDES, J.E.T.; CARVALHO, A.. Taxonomia de Coffea arabica L.
Instituto Agronomico do Estado, em Campinas, Boletim Técnico 62, 57p. 1939.
KUNZE, M.; PRACHAROENWATTANA, I.; SMITH, S.M.; HARTIG, A. A central
role for the peroxisome membrane in glyoxylate cycle function. Biochimica et
Biophysica Acta, v.1763, p.1441–1452, 2006.
LASHERMES, P.; COMBES, M-C.; ROBERT, J.; TRIUSLOT, P.; D’HONT, A.;
ANTHONY, F.; CHARRIER, A. Molecular characterisation and origin of the Coffea
arabica L. genome. Molecular Genetics and Genomics, v.261, p.259-266, 1999.
LELLIOT, R.A.; BILLING, E.; HAYWARD A,C. A determinative scheme for the
fluorescent plant pathogenic Pseudomonads. Journal of Applied Bacteriology, v.29,
n.3, p.470-589, 1966.
LEVINE, A.; TENHAKEN, R.; DIXON, R.; LAMB, C. H2O2 from the oxidative burst
orchestrates the plant hypersensitive disease response. Cell, v.79, p.583-593, 1994.
LI, Y.; CHEN, L.; MU, J.; ZUO, J. Lesion simulating disease1 interacts with catalases
to regulate hypersensitive cell death in Arabidopsis. Plant physiology, v. 163, p.1059-
1070, 2013.
MACIEL, K.W. Caracterização de linhagens de Pseudomonas syringae pv. garcae,
agente causal da mancha-aureolada do cafeeiro. Dissertação (Mestrado), Instituto
Biológico, São Paulo – SP, 2013, 71p.
MARGIS-PINHEIRO, M.; MARTIN, C.; DIDIERJAN, L.; BURKARD, G. Differential
expression of bean chitinase genes by virus infection, chemical treatment and UV
irradiation. Plant Molecular Biology, v.22, p.659-668, 1993.
MARINGONI, A.C.; KUROZAWA, C. Tipificação de isolados de Curtobacterium
flaccumfaciens pv. flaccumfaciens por bacteriocinas. Pesquisa Agropecuária
Brasileira, v.37, p.1339-1355, 2002.

107
MARTIN, G.B.; BODANOVE, A.J.; SESSA, G. Understanding the functions of plant
disease resistance proteins. Annual Review in Plant Biology, v.54, p.23-61, 2003.
MARTINS, A. L. História do café. São Paulo: Contexto, 2008. 316p.
MATIELLO, J. B. Café conillon. Rio de Janeiro: MAA, SDR, PROCAFÉ, PNFC,
1998. 162 p.
MAZZAFERA, P.; & CARVALHO, A. Breeding for low seed caffeine content of
coffee (Coffea L.) by interspecific hybridization. Euphytica, v.59, p.55-60, 1992.
MAZZAFERA, P.; SOAVE, D, TEIXEIRA ZULLO, M.A.; GUERREIRO FILHO, O.
Oil content of green beans from some coffee species. Bragantia, v. 57, p.45-48, 1998.
MEDINA FILHO, H.P.; CARVALHO, A.; SONDAHL, M.R.; FAZUOLI, L.C.;
COSTA, W.M. Coffee breeding and related evolutionary aspects. In: JANICK, J. (ed.).
Plant breeding reviews. Westport: AVI Publishing Co.,1984. p.157-194.
MENDES, J.E.T. Ensaio de variedades de cafeeiros. III. Bragantia, v. 11, p. 29-43,
1951.
MENDES, J.E.T. & KRUG, C.A. O cafeeiro e sua cultura: pesquisas e trabalhos
experimentais na andamento no Instituto Agronomico do Estado de São Paulo, em
Campinas. Boletim Técnico, 54, Campinas, Instituto Agronomico, 37p., 1938.
MINISTÉRIO DA AGRICULTURA, PECUÁRIA E ABASTECIMENTO. Café.
Disponivel em: http://www.agricultura.gov.br/vegetal/culturas/cafe/ acesso em: 27 de
abril de 2015.
MISAS-VILLAMIL, J.C.; KOLODZIEJEK, I.; VAN DER HOORN, R.A.L.
Pseudomonas syringae colonizes distant tissues in Nicotiana benthamiana through
xylem vessels. The Plant Journal, v.67, p.774-782, 2011.
MOHAN, S.K. Investigações sobre Pseudomonas garcae Amaral et al. em cafeeiros.
In: CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS, 4, 1976, Caxambu.
Resumos...Rio de Janeiro: IBC, p.56, 1976.
MOHAN, S.K. Mutante de Pseudomonas garcae Amaral et al., altamente resistente a
estreptomicina. IN: CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS, 5,
1977, Guarapari - ES. Anais. IBC/GERCA, p.31, 1977.
MOHAN, S. K.; CARDOSO, R. M. L.; PAVAN, M. A. Fontes de resistência à
Pseudomonas garcae Amaral et al. em germoplasma de Coffea arabica L. e C.
canephora Pierre. In: CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS, 5,
1977, Guarapari. Resumos... Rio de Janeiro: IBC/GERCA. p. 24, 1977.
MOHAN, S.K.; CARDOSO, R.M.L.; PAIVA, M.A. Resistência em germoplasma de
Coffea ao crestamento bacteriano incitado por Pseudomonas garcae Amaral et al.
Pesquisa Agropecuária Brasileira, v.13, p.53-64, 1978.

108
MÔNACO, L.C. Efeito das lojas vazias sobre o rendimento do café Mundo Novo.
Bragantia, v.19, p.1-12, 1960.
MORAES, S.A.; SUGIMORE, M.H.; TOMAZIELLO-FILHO, M.; CARVALHO,
P.C.T. Resistência de cafeeiros à Pseudomonas garcae. IN: CONGRESSO
BRASILEIRO DE PESQUISAS CAFEEIRAS, 2, 1974, Poços de Caldas, MG.
Resumos... Rio de Janeiro: IBC. p. 183, 1974.
MORAES SA, SUGIMORI, M.H.; TOMAZELLO FILHO, M.; CARVALHO, P.C.T.
Resistência de cafeeiros a Pseudomonas garcae. Summa Phytopathologica, v.1,
p.105-110, 1975.
NAGAI, C.; RAKOTOMALALA, J.J.; KATAHIRA, R.; LI, Y.; YAMAGATA, K.;
ASHIHARA, H. Production of a new low-caffeine hybrid coffee and the biochemical
mechanism of low caffeine accumulation. Euphytica, v.164, p. 133-142, 2008.
NARASIMHASWAMY, R.L. Report on visit to coffee areas in Ethiophia 1964-65. IN:
FAO, Roma, FAO Coffee mission to Ethiophia 1946-65. Roma, p. 33-48, 1968.
NAVARRO, L.; DUNOYER, P.; JAY, F.; ARNOLD, B.; DHARMASIRI, N.; Estelle,
M.; VOINNET, O.; JONES, J. A plant miRNA contributes to antibacterial resistance by
repressing auxin signaling. Science v.312, p.436-439, 2006.
NOWOGÓRSKA A. & PATYKOWSKI J. Selected reactive oxygen species and
antioxidant enzymes in common bean after Pseudomonas syringae pv. phaseolicola and
Botrytis cinerea infection. Acta physiologiae plantarum, v.37, p.1-10, 2015.
OLIVEIRA, J.R.; ROMEIRO, R.S. Reação de folhas novas e velhas de cafeeiros a
infecção por Pseudomonas cichorii e P. syringae pv. garcae. Fitopatologia Brasileira,
v.15, p.355-356, 1990.
OLIVEIRA, J.R.; ROMEIRO, R.S. Compostos Fenólicos, idade das folhas e resistência
do cafeeiro a Pseudomonas cichorii e Pseudomonas syringae pv. garcae. Revista
Ceres, v.38, p.445-452, 1991.
OLIVEIRA, J.R.; ROMEIRO, R.S.; MUCHOVEJ, J.J. Population tendencies of
Pseudomonas cichorii and P. syringae pv. garcae in young and mature coffee leaves.
Journal of Phytopathology, v,131, p.210-214, 1991.
PARADELA, A.; ALVAREZ, J.A.A.; BATISTA, M.F; SILVA, C.L. Eficiência do
fungicida-bactericida Hokko Kasumin (kasugamicina) no controle de Mancha-aureolada
(Pseudomonas syringae pv. garcae) em mudas de café (Coffea arabica L.) IN:
CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS, 26, 2000, Marília.
Resumos... Ministério da Agricultura, PROCAFÉ, p. 247-248, 2000.
PARADELA, FILHO, O.; RIBEIRO, I.J.A.; SUGIMORI, M.H. Comportamento
diferencial em progênies de cafeeiros de 2 isolados do agente causador da Mancha-
aureolada do cafeeiro. Congresso da Sociedade Brasileira de Fitopatologia 7, 1974,
Brasilia. Resumos..., 1974.

109
PRASANNATH, K. Pathogenicity and Virulence Factors of Phytobacteria. Scholars
Academic Journal of Biosciences v.1, p. 24-33, 2013.
PARKER, J.E.; HOLUB, E.B.; FROST, L.N.; FALK, A.; GUNN, N.D.; DANIELS,
M.J. Characterization of eds1, a mutation in Arabidopsis suppressing resistance to
Peronospora parasitica specified by several different RPP genes. Plant Cell, v. 8, p.
2033–2046, 1996.
PATRÍCIO, F.R.A.; ALMEIDA, I.M.G.; BARROS, B.C.; SANTOS, A.S.; FRARE,
P.M. Effectiveness of acibenzolar-S-methyl, fungicides and antibiotics for the control of
brown eye spot, bacterial blight, brown leaf spot and coffee rust in coffee. Annals of
Applied Biology, v.152, p.29-39, 2008.
PATRÍCIO, F.R.A., BERIAM, L.O.S., ROSSI, A., MORAES, A., ALMEIDA, I.M.G.
Controle químico da mancha-aureolada em uma região montanhosa. Tropical Plant
Pathology, v. 35 (supl.), p.S119, 2010.
PEREIRA, M.J.Z.; RAMALHO, M.A.P.; ABREU, A.F.B. Estratégias para eficiência da
seleção de feijoeiro comum quanto á resistência à murcha-de-fusário. Pesquisa
Agropecuária Brasileira, v.43, p.721-728, 2008.
PETEK, M.R.; SERA, T.; ALTÉIA, M.Z.; TRILLER, C.; AZEVEDO, J.A.;
COLOMBO, L.A.; DA MATA, J.S.; SERA, G.H. Resistência à bacteriose
(Pseudomonas syringae pv. garcae) derivado do cruzamento entre ‘Iapar-59’ e Coffea
arabica SH1 x ‘Catuaí’. IN: SIMPÓSIO DE RECURSOS GENÉTICOS PARA A
AMÉRICA LATINA E CARIBE, 3, 2001, Londrina, PR. Resumos... Londrina:
IAPAR, 2001, p. 674-676.
PETEK, M.R.; SERA, T.; SERA, G.H.; FONSECA, I.C.B.; ITO, D.S. Selection of
progenies of Coffea arabica with simultaneous resistance to bacterial blight and leaf
rust. Bragantia, v.65, p.65-73, 2006.
PENDERGRAST, M. Uncommon grounds: the history of coffee and how it
transformed our world. New York: Basic Books 2010, p.
RAMOS, A.H.; SHAVDIA, L.D. A dieback of coffee in Kenya. Plant Disease
Reporter, v.60, p.831-835, 1976.
RIEMKE, E.; QUEIROZ, P.P.S.; OBERSTERN, S. Utilização de fontes nitrogenadas
pela Pseudomonas garcae Amaral et al. IN: CONGRESSO BRASILEIRO DE
PESQUISAS CAFEEIRAS, 6, 1978, Ribeirão Preto - SP. Resumos... IBC/GERCA,
p.64-66, 1978.
ROBBS, F.C. Epiphytic survival of Pseudomonas garcae on coffee. IN:
INTERNATIONAL CONFERENCE ON PLANT PATHOGENIC BACTERIA, 4,
1978, Angers, Proceeding. p.747-748, 1978.
ROBBS, F.C. Sobrevivência de Pseudomonas garcae como epífita de folhas de
cafeeiros. IN: CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS, 5, 1977,
Guarapari - ES. Anais. IBC/GERCA, p.152-153, 1977.

110
ROBBS, F.C. Predisposição de Coffea arabica a Pseudomonas garcae incitada por
temperaturas baixas. IN: CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS,
7, 1979, Araxa - MG. Resumos... IBC/GERCA, p.288, 1979.
ROBBS, C.F., KIMURA, O., RIBEIRO, R.L.D., OYADOMARI, L.C. “Crestamento
bacteriano das folhas”: nova enfermidade do cafeeiro (Coffea arabica L.) incitada por
Pseudomonas cichorii (Swingle) Stapp. Arquivos da Universidade Federal Rural,
v.4, n.2, p.1-5, 1974.
RODRIGUES JÚNIOR, C.J.; GONÇALVES, M.M.; VÁRZEA, V.M.P. Importância do
Híbrido de Timor para o território e para o melhoramento da cafeicultura mundial.
Revista de Ciências Agrárias, v.27, p.203-213, 2004.
RODRIGUES NETO, J.; FIGUEIREDO, P.; MARIOTTO, P.R.; ROBBS, C.F.
Pseudomonas andropogonis (Smith, 1911) Stapp, 1928, agente causal da “mancha
escura bacteriana” em folhas de cafeeiro (Coffea arabica L.). Arquivos do Instituto
Biológico, v.48, p.31-36, 1981.
RODRIGUES NETO, J.; SILVA, C.H.D.; BERIAM, L.O.S.; PATRÍCIO, F.R.A.;
RODRIGUES, L.M.R.; THOMAZIELO, R.A. Mancha bacteriana do cafeeiro causada
por Pseudomonas syringae pv. tabaci. Summa Phytopathologica, v.32, supl., p.85,
2006.
ROMEIRO, R. da S. Métodos em bacteriologia de plantas. Viçosa: UFV, 2001. 279p.
ROOS, I.M.M. & HATTINGH, M.J. Systemic invasion of cherry leaves and petioles by
Pseudomonas syringae pv. morsprunorum. Ecology and Epidemiology v.77, p.1246-
1252, 1987.
RUSTÉRUCCI, C; AVIV, D.H., HOLT, BF III; DANGL, J.L.; PARKER J.E. The
disease resistance signaling components EDS1 and PAD4 are essential regulators of the
cell death pathway controlled by LSD1 in Arabidopsis. Plant Cell, v.13, p.2211-2224,
2001.
SALGADO, P.R.; FAVARIN, J.L.; LEANDRO, R.A.; LIMA FILHO, O.F. Total
phenol concentrations in coffee tree leaves during fruit development. Scientia Agricola,
v.65, p.354-359, 2008.
SAKAMOTO, A.; OKUMURA, T.; KAMINAKA, H.; SUMI, K., TANAKA, K.
Structure and differential response to abscisic acid of two promoters for the cytosolic
copper/zinc-superoxide dismutase genes, SodCc1 and SodCc2, in rice protoplasts.
FEBS Lett, v.358, p.62-66, 1995.
SERA, T. Coffee Genetic Breeding at IAPAR. Crop Breeding and Applied
Biotechnology, v.1, p.179-199, 2001.
SERA, T.; ALTEIA, M.Z.; PETEK, M.R. Melhoramento do cafeeiro: variedades
melhoradas no Instituto Agronômico do Paraná (IAPAR). In: Zambolim, L (Ed.). O
estado da arte de tecnologias na produção de café. Viçosa: UFV, p.217-251, 2002.

111
SERA, T.; CARDOSO, R.M.L.; MOHAN, S.K. Resistência ao crestamento bacteriano
(Pseudomonas garcae) em materiais segregantes para resistência à ferrugem (Hemileia
vastatrix). IN: CONGRESSO BRASILEIRO DE PESQUISAS CAFEEIRAS, 7, 1979,
Araxá, MG. Resumos... Rio de Janeiro: IBC/GERCA. p.60-61, 1980.
SERA, G. H.; SERA, T.; ITO, D. S.; AZEVEDO, J. A. de; MATA, J. S.; RIBEIRO
FILHO, C.; PETEK, M. R.; COTARELLI, V. M.; DOI, D. S. Identificação de híbridos
de café com resistência múltipla à ferrugem, bicho-mineiro e nematóides. In:
SIMPÓSIO DE PESQUISA DOS CAFÉS DO BRASIL, 4, 2005, Londrina. Anais, CD-
ROM, 2005.
SERA G.H.; SERA T.; ITO D.S.; RIBEIRO FILHO C.; VILLACORTA A.;
KANAYAMA F.S. Coffee Berry Borer Resistance in Coffee Genotypes. Braz. arch.
biol. technol., v.53, p. 261-268, 2010.
SERA, G.H., SERA, T., ALTÉIA, M.Z., ANDROCIOLI FILHO, A., AZEVEDO, J.A.,
PETEK, M.R., ITO, D.S. Associação de Pseudomonas syringae pv. garcae com
algumas características agronômicas em cafeeiros F2 segregantes para o gene erecta.
Ciência e Agrotecnologia, v.28, n.5, p.974-977, 2004.
SILVESTRINI, M.; MALUF, M.P.; GUERREIRO-FILHO, O.; GONÇALVES, W.
Expressão de genes de resistência em resposta à infecção por nematóides. IN:
SIMPÓSIO DE PESQUISA DOS CAFÉS DO BRASIL, 4., 2005, Londrina. Anais...
Brasília, DF: Embrapa Café, 2005.
SILVESTRINI, M.; JUNQUEIRA, M.G.; FAVARIN, A.C.; GUERREIRO FILHO, O.;
MALUF, M.P.; SILVAROLLA, M.B.; COLOMBO, C. Diversidade genética da espécie
Coffea arabica avaliada por marcadores microssatélites. IN: Simpósio de pesquisas dos
cafés do Brasil, 4. Londrina,PR, 2005.
SIMKO, I. & PIEPHO, H.P. The area under the disease progress stairs: Calculation,
advantage, and application. Phytopathology, v.102, p.381-389, 2012.
SOCQUET-JUGLARD, D.; PATOCCHI, A.; POTHIER, J.F.; CHRISTEN, D.;
DUFFY, B. Evaluation of Xanthomonas arboricola pv. pruni inoculation techniques to
screen for bacterial spot resistance in apricot. Journal of Plant Pathology, v.94, p.
S1.91–S1.96, 2012.
SONG, F. & GOODMAN, R.M. Activity of nitric oxide is dependent on, but is partially
required for function of, salicylic acid in the signaling pathway in tobacco systemic
acquired resistance. Molecular Plant-Microbe Interactions, v. 14,
p. 1458-1462, 2001.
STANGARLIN, J.R.; KUHN, O.J.; TOLEDO, M.V.; PORTZ, R.L.; SCHWAN-
ESTRADA, K.R.F.; PASCHOLATI, S. F. A defesa vegetal contra fitopatógenos.
Scientia Agraria Paranaenis, v. 10, p.18-46, 2011.
STEEL, R.G.D. & TORRIE, J.H. Principles and procedures of statistics: a
biometrical approach. 2rd ed. Boston: McGraw-Hill Companies, 633p, 1980.

112
TRAPET, P.; KULIK, K; LAMOTTE, O.; JEANDROZ, S.; BOURQUE, S.;
NICOLAS-FRANCÈS, V.; ROSNOBLET, C.; BESSON-BARD, A.; WENDEHENNE,
D. NO signaling in plant immunity: a tale of messengers. Phytochemistry, v.112, p.72–
79, 2014.
VAN BREUSEGEM, F.; VRANOVÁ, E.; DAT, J.F.; INZÉ, D. The role of active
oxygen species in plant signal transduction. Plant Science v.161, p.405-414, 2001.
VAN DER VOSSEN, H.; BERTRAND, B.; CHARRIER A. Next generation variety
development for sustainable production of arabica coffee (Coffea arabica L.): A review.
Euphytica, v.204, p.243-256, 2015
VÁRZEA, V.M.P. & MARQUES, D.V. Population variability of Hemileia vastatrix vs.
coffee durable resistance. In: Zambolim, L.; Zambolim, E.M.; Várzea, V.M.P.
(Eds.). Durable Resistance to Coffee Leaf Rust. Viçosa: UFV, 2005. p.53–74.
VIDAVER, A. K. Bacteriocins: the lure and the reality. Plant Disease, v. 67, p. 471-
475, 1983.
VIEIRA, L.G.E.; ANDRADE, A.C.; COLOMBO, C.A.; MORAES, A.H. de A.;
METHA, Â.; OLIVEIRA, A.C. de; LABATE, C.A.; MARINO, C.L.; MONTEIRO-
VITORELLO, C. de B.; MONTE, D. de C.; GIGLIOTI, E.; KIMURA, E.T.;
ROMANO, E.; KURAMAE, E.E.; LEMOS, E.G.M.; ALMEIDA, E.R.P. de; JORGE,
É.C.; ALBUQUERQUE, É.V.S.; SILVA, F.R. da; VINECKY, F.; SAWAZAKI, H.E.;
DORRY, H.F.A.; CARRER, H.; ABREU, I.N.; BATISTA, J.A.N.; TEIXEIRA, J.B.;
KITAJIMA, J.P.; XAVIER, K.G.; LIMA, L.M. de; CAMARGO, L.E.A. de; PEREIRA,
L.F.P.; COUTINHO, L.L.; LEMOS, M.V.F.; ROMANO, M.R.; MACHADO, M.A.;
COSTA, M.M. do C.; SÁ, M.F.G. de; GOLDMAN, M.H.S.; FERRO, M.I.T.; TINOCO,
M.L.P.; OLIVEIRA, M.C.; SLUYS, M.-A.V.; SHIMIZU, M.M.; MALUF, M.P.; EIRA,
M.T.S. da; GUERREIRO FILHO, O.; ARRUDA, P.; MAZZAFERA, P.; MARIANI,
P.D.S.C.; OLIVEIRA, R.L.B.C. de; HARAKAVA, R.; BALBAO, S.F.; TSAI, S.M.;
MAURO, S.M.Z. di; SANTOS, S.N.; SIQUEIRA, W.J.; COSTA, G.G.L.;
FORMIGHIERI, E.F.; CARAZZOLLE, M.F.; PEREIRA, G.A.G. Brazilian coffee
genome project: an EST-based genomic resource. Brazilian Journal of Plant
Physiology, v.18, p.95-108, 2006.
VINECKY, F.; ALVES, G.S.C; VIEIRA, N.G.; FREIRE, L.P.; HEIMBECK, I.G.R.;
FERRÃO, R.G.; DA FONSECA, A.F.A.; FERRÃO, M.A.G.; SILVA, V.A.
MARRACCINI, P.; ANDRADE, A.C. Análises da expressão gênica por PCR
quantitativo em tempo real (qPCR) de genes candidatos para a tolerância à seca em
cafeeiro. IN: VII Simpósio de Pesquisa dos Cafés do Brasil, 2011, Araxá ‐ MG,
Resumos...1 CD-ROM.
WEAVER, D.J. Interaction of Pseudomonas syringae and freezing in bacterial canker
on excised peach twigs. Phytopathology, v.68, p.1460-1463, 1978.
WIERMER, M.; FEYS, B.J.; PARKER, J.E. Plant immunity: the EDS1 regulatory
node. Current Opinion in Plant Biology, v.8, p.383-389, 2005

113
XUEHUI, B.; LIHONG, Z.; YONGLIANG, H.; GUANGHAI, J.; JINHONG, L.;
ZHANG, H. Isolation and identification of the pathogen of coffee bacterial blight
disease. Chinese Journal of Tropical Crops v.34, p.738-742, 2013.
YAMADA, J.K.; SOUZA, R.M.; SILVA, G.M.; MATHIONI, S.M.; FIGUEIRA, A.R.
Ocorrência do gene de resistência a cobre, cop A, em isolados de Pseudomonas syringae
pv. garcae. IN: CONGRESSO BRASILEIRO DE FITOPATOLOGIA, 47, Londrina –
PR, Anais, 1 CD-ROM, 2014a.
YAMADA, J.K.; SOUZA, R.M.; SILVA, G.M.; NUNES, K.V.; PAULA, A.F.L.
Resistência in vitro de isolados de Pseudomonas syringae pv. garcae ao sulfato de
Cobre. IN: Congresso Brasileiro de Fitopatologia, 47, Londrina – PR, Anais, 1 CD-
ROM, 2014b.
YOUNG, J.M.; DYE, D.W.; BRADBURY, J.F.; PANAGOPOULOS, C.G.; ROBBS,
C.F. A proposed nomenclature and classification for plant pathogenic bacteria. New
Zealand Journal of Agricultural Research, v.21, p.153-177, 1978.
ZAITER, H.Z.; COYNE, D.P. Testing inoculation methods and sources of resistance to
the halo blight bacterium (Pseudomonas syringae pv. phaseolicola) in Phaseolus
vulgaris. Euphytica, v.33, p.133-141, 1984.
ZHOU, B.; CHEN, Z.; DU, L.; YE, X.; LI, N. Correlation betwen resistance of egplant
and defense-related enzymes and biochemical substances of leaves. African Journal of
Biotechnology, v. 1, p. 13896- 13902, 2012.
ZOCCOLI, D.M; TAKATSU, A.; UESUGI, C.H. Ocorrência de mancha-aureolada em
cafeeiros na Região do Triângulo Mineiro e Alto Paranaíba. Bragantia, v.70, p.843-
849, 2011.
ZOU, J.; RODRIGUEZ-ZAS, S.; ALDEA, M.; Li, M.; ZHU, J.; GONZALEZ, D.O.;
VODKIN, L.O.; DELUCIA, E.; CLOUGH, S.J. Expression profiling soybean response
to Pseudomonas syringae reveals new defenserelated genes and rapid HR-specific
downregulation of photosynthesis. Molecular Plant-Microbe Interactions, v.18,
p.1161-1174, 2005.

114
8. ANEXOS
Anexo 1 – Representação da escala de zero a cinco pontos (EP) utilizada como referência nas avaliações da severidade da mancha-aureolada em
cafeeiros inoculadas com Pseudomonas syringae pv. garcae, pelos métodos de agulhas múltiplas e abrasão.
EP Agulhas múltiplas Abrasão
0
1
2
EP Agulhas múltiplas Abrasão
3
4
5
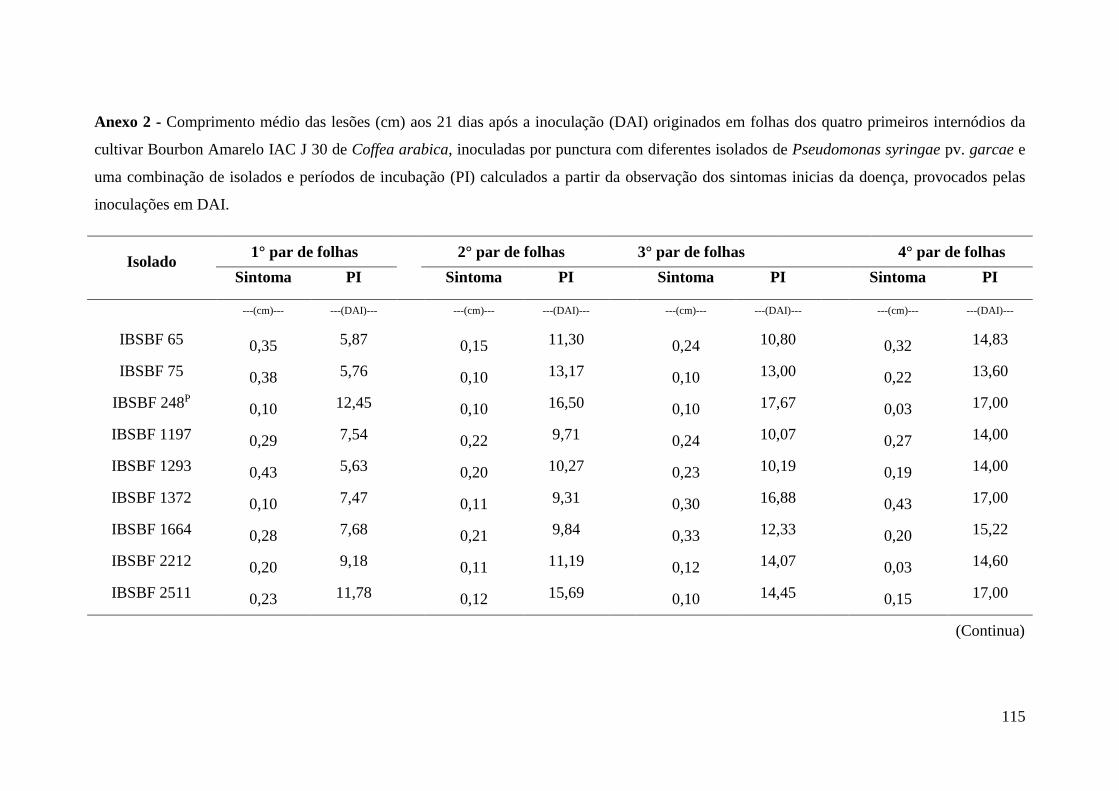
115
Anexo 2 - Comprimento médio das lesões (cm) aos 21 dias após a inoculação (DAI) originados em folhas dos quatro primeiros internódios da
cultivar Bourbon Amarelo IAC J 30 de Coffea arabica, inoculadas por punctura com diferentes isolados de Pseudomonas syringae pv. garcae e
uma combinação de isolados e períodos de incubação (PI) calculados a partir da observação dos sintomas inicias da doença, provocados pelas
inoculações em DAI.
Isolado 1° par de folhas 2° par de folhas 3° par de folhas 4° par de folhas
Sintoma PI Sintoma PI Sintoma PI Sintoma PI
---(cm)--- ---(DAI)--- ---(cm)--- ---(DAI)--- ---(cm)--- ---(DAI)--- ---(cm)--- ---(DAI)---
IBSBF 65 0,35 5,87 0,15 11,30 0,24 10,80 0,32 14,83
IBSBF 75 0,38 5,76 0,10 13,17 0,10 13,00 0,22 13,60
IBSBF 248P 0,10 12,45 0,10 16,50 0,10 17,67 0,03 17,00
IBSBF 1197 0,29 7,54 0,22 9,71 0,24 10,07 0,27 14,00
IBSBF 1293 0,43 5,63 0,20 10,27 0,23 10,19 0,19 14,00
IBSBF 1372 0,10 7,47 0,11 9,31 0,30 16,88 0,43 17,00
IBSBF 1664 0,28 7,68 0,21 9,84 0,33 12,33 0,20 15,22
IBSBF 2212 0,20 9,18 0,11 11,19 0,12 14,07 0,03 14,60
IBSBF 2511 0,23 11,78 0,12 15,69 0,10 14,45 0,15 17,00
(Continua)
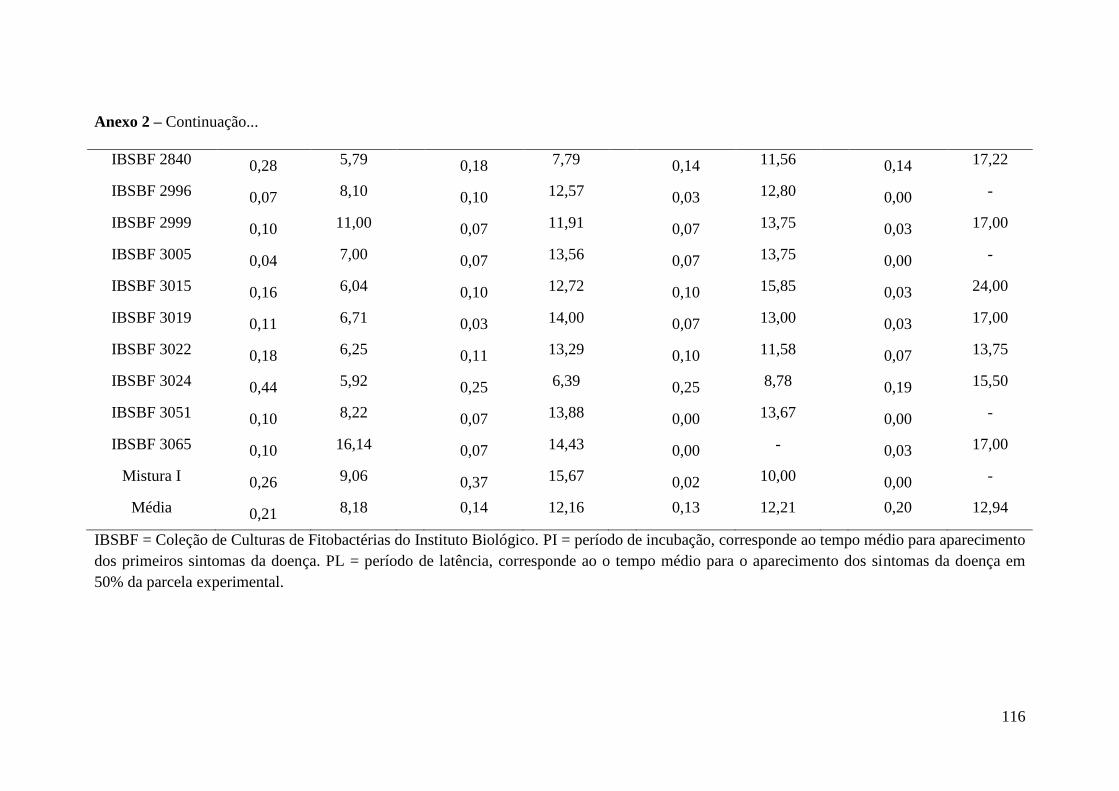
116
Anexo 2 – Continuação...
IBSBF 2840 0,28 5,79 0,18 7,79 0,14 11,56 0,14 17,22
IBSBF 2996 0,07 8,10 0,10 12,57 0,03 12,80 0,00 -
IBSBF 2999 0,10 11,00 0,07 11,91 0,07 13,75 0,03 17,00
IBSBF 3005 0,04 7,00 0,07 13,56 0,07 13,75 0,00 -
IBSBF 3015 0,16 6,04 0,10 12,72 0,10 15,85 0,03 24,00
IBSBF 3019 0,11 6,71 0,03 14,00 0,07 13,00 0,03 17,00
IBSBF 3022 0,18 6,25 0,11 13,29 0,10 11,58 0,07 13,75
IBSBF 3024 0,44 5,92 0,25 6,39 0,25 8,78 0,19 15,50
IBSBF 3051 0,10 8,22 0,07 13,88 0,00 13,67 0,00 -
IBSBF 3065 0,10 16,14 0,07 14,43 0,00 - 0,03 17,00
Mistura I 0,26 9,06 0,37 15,67 0,02 10,00 0,00 -
Média 0,21 8,18 0,14 12,16 0,13 12,21 0,20 12,94
IBSBF = Coleção de Culturas de Fitobactérias do Instituto Biológico. PI = período de incubação, corresponde ao tempo médio para aparecimento
dos primeiros sintomas da doença. PL = período de latência, corresponde ao o tempo médio para o aparecimento dos sintomas da doença em
50% da parcela experimental.
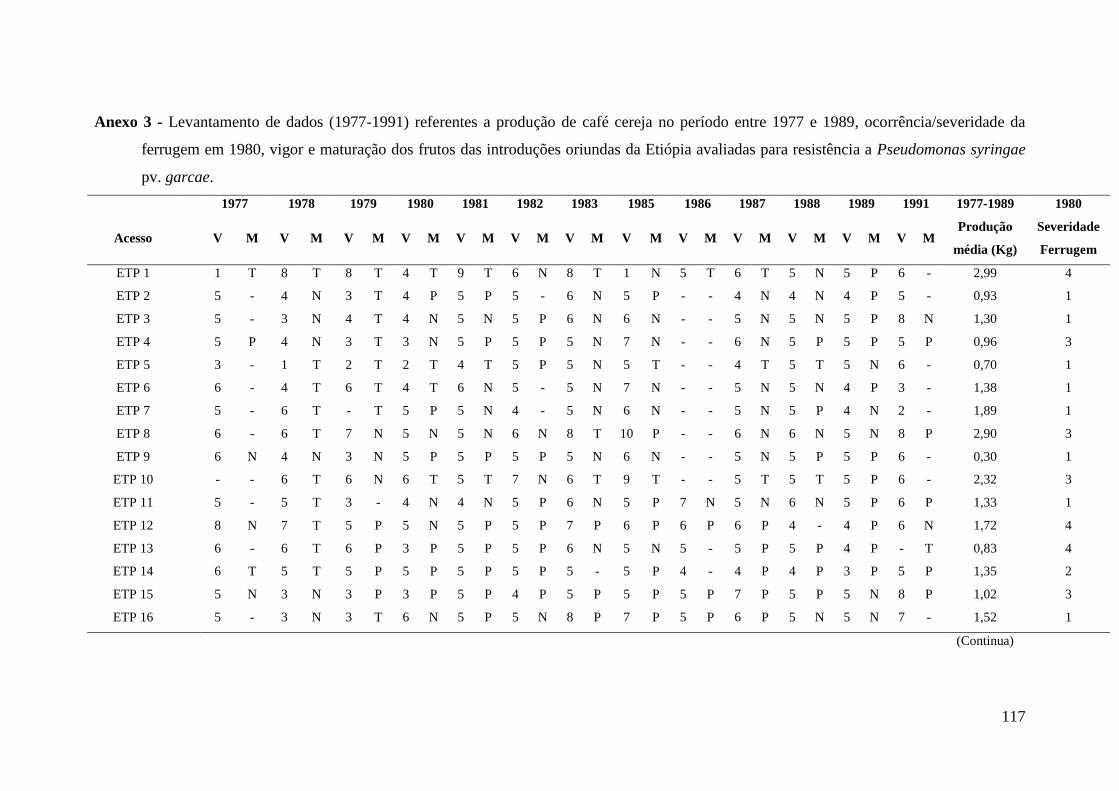
117
Anexo 3 - Levantamento de dados (1977-1991) referentes a produção de café cereja no período entre 1977 e 1989, ocorrência/severidade da
ferrugem em 1980, vigor e maturação dos frutos das introduções oriundas da Etiópia avaliadas para resistência a Pseudomonas syringae
pv. garcae.
1977 1978 1979 1980 1981 1982 1983 1985 1986 1987 1988 1989 1991 1977-1989 1980
Acesso
V M V M V M V M V M V M V M V M V M V M V M V M V M Produção
média (Kg)
Severidade
Ferrugem
ETP 1 1 T 8 T 8 T 4 T 9 T 6 N 8 T 1 N 5 T 6 T 5 N 5 P 6 - 2,99 4
ETP 2 5 - 4 N 3 T 4 P 5 P 5 - 6 N 5 P - - 4 N 4 N 4 P 5 - 0,93 1
ETP 3 5 - 3 N 4 T 4 N 5 N 5 P 6 N 6 N - - 5 N 5 N 5 P 8 N 1,30 1
ETP 4 5 P 4 N 3 T 3 N 5 P 5 P 5 N 7 N - - 6 N 5 P 5 P 5 P 0,96 3
ETP 5 3 - 1 T 2 T 2 T 4 T 5 P 5 N 5 T - - 4 T 5 T 5 N 6 - 0,70 1
ETP 6 6 - 4 T 6 T 4 T 6 N 5 - 5 N 7 N - - 5 N 5 N 4 P 3 - 1,38 1
ETP 7 5 - 6 T - T 5 P 5 N 4 - 5 N 6 N - - 5 N 5 P 4 N 2 - 1,89 1
ETP 8 6 - 6 T 7 N 5 N 5 N 6 N 8 T 10 P - - 6 N 6 N 5 N 8 P 2,90 3
ETP 9 6 N 4 N 3 N 5 P 5 P 5 P 5 N 6 N - - 5 N 5 P 5 P 6 - 0,30 1
ETP 10 - - 6 T 6 N 6 T 5 T 7 N 6 T 9 T - - 5 T 5 T 5 P 6 - 2,32 3
ETP 11 5 - 5 T 3 - 4 N 4 N 5 P 6 N 5 P 7 N 5 N 6 N 5 P 6 P 1,33 1
ETP 12 8 N 7 T 5 P 5 N 5 P 5 P 7 P 6 P 6 P 6 P 4 - 4 P 6 N 1,72 4
ETP 13 6 - 6 T 6 P 3 P 5 P 5 P 6 N 5 N 5 - 5 P 5 P 4 P - T 0,83 4
ETP 14 6 T 5 T 5 P 5 P 5 P 5 P 5 - 5 P 4 - 4 P 4 P 3 P 5 P 1,35 2
ETP 15 5 N 3 N 3 P 3 P 5 P 4 P 5 P 5 P 5 P 7 P 5 P 5 N 8 P 1,02 3
ETP 16 5 - 3 N 3 T 6 N 5 P 5 N 8 P 7 P 5 P 6 P 5 N 5 N 7 - 1,52 1
(Continua)
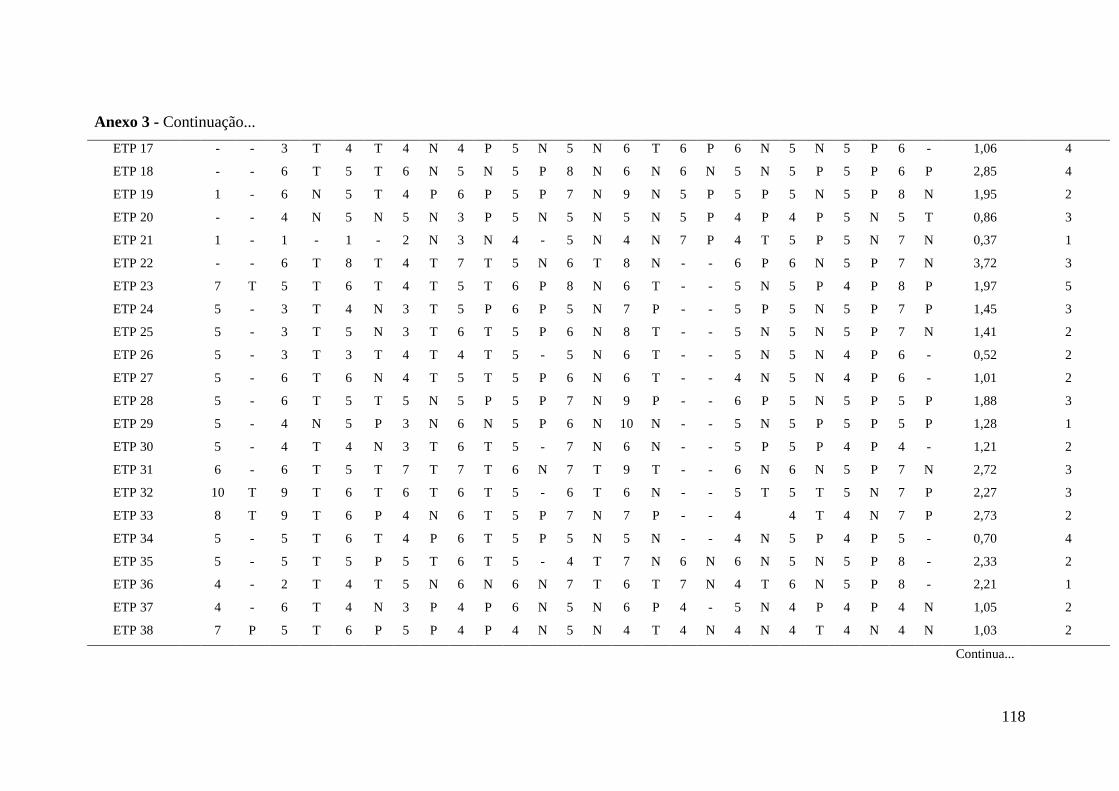
118
Anexo 3 - Continuação...
ETP 17 - - 3 T 4 T 4 N 4 P 5 N 5 N 6 T 6 P 6 N 5 N 5 P 6 - 1,06 4
ETP 18 - - 6 T 5 T 6 N 5 N 5 P 8 N 6 N 6 N 5 N 5 P 5 P 6 P 2,85 4
ETP 19 1 - 6 N 5 T 4 P 6 P 5 P 7 N 9 N 5 P 5 P 5 N 5 P 8 N 1,95 2
ETP 20 - - 4 N 5 N 5 N 3 P 5 N 5 N 5 N 5 P 4 P 4 P 5 N 5 T 0,86 3
ETP 21 1 - 1 - 1 - 2 N 3 N 4 - 5 N 4 N 7 P 4 T 5 P 5 N 7 N 0,37 1
ETP 22 - - 6 T 8 T 4 T 7 T 5 N 6 T 8 N - - 6 P 6 N 5 P 7 N 3,72 3
ETP 23 7 T 5 T 6 T 4 T 5 T 6 P 8 N 6 T - - 5 N 5 P 4 P 8 P 1,97 5
ETP 24 5 - 3 T 4 N 3 T 5 P 6 P 5 N 7 P - - 5 P 5 N 5 P 7 P 1,45 3
ETP 25 5 - 3 T 5 N 3 T 6 T 5 P 6 N 8 T - - 5 N 5 N 5 P 7 N 1,41 2
ETP 26 5 - 3 T 3 T 4 T 4 T 5 - 5 N 6 T - - 5 N 5 N 4 P 6 - 0,52 2
ETP 27 5 - 6 T 6 N 4 T 5 T 5 P 6 N 6 T - - 4 N 5 N 4 P 6 - 1,01 2
ETP 28 5 - 6 T 5 T 5 N 5 P 5 P 7 N 9 P - - 6 P 5 N 5 P 5 P 1,88 3
ETP 29 5 - 4 N 5 P 3 N 6 N 5 P 6 N 10 N - - 5 N 5 P 5 P 5 P 1,28 1
ETP 30 5 - 4 T 4 N 3 T 6 T 5 - 7 N 6 N - - 5 P 5 P 4 P 4 - 1,21 2
ETP 31 6 - 6 T 5 T 7 T 7 T 6 N 7 T 9 T - - 6 N 6 N 5 P 7 N 2,72 3
ETP 32 10 T 9 T 6 T 6 T 6 T 5 - 6 T 6 N - - 5 T 5 T 5 N 7 P 2,27 3
ETP 33 8 T 9 T 6 P 4 N 6 T 5 P 7 N 7 P - - 4
4 T 4 N 7 P 2,73 2
ETP 34 5 - 5 T 6 T 4 P 6 T 5 P 5 N 5 N - - 4 N 5 P 4 P 5 - 0,70 4
ETP 35 5 - 5 T 5 P 5 T 6 T 5 - 4 T 7 N 6 N 6 N 5 N 5 P 8 - 2,33 2
ETP 36 4 - 2 T 4 T 5 N 6 N 6 N 7 T 6 T 7 N 4 T 6 N 5 P 8 - 2,21 1
ETP 37 4 - 6 T 4 N 3 P 4 P 6 N 5 N 6 P 4 - 5 N 4 P 4 P 4 N 1,05 2
ETP 38 7 P 5 T 6 P 5 P 4 P 4 N 5 N 4 T 4 N 4 N 4 T 4 N 4 N 1,03 2
Continua...
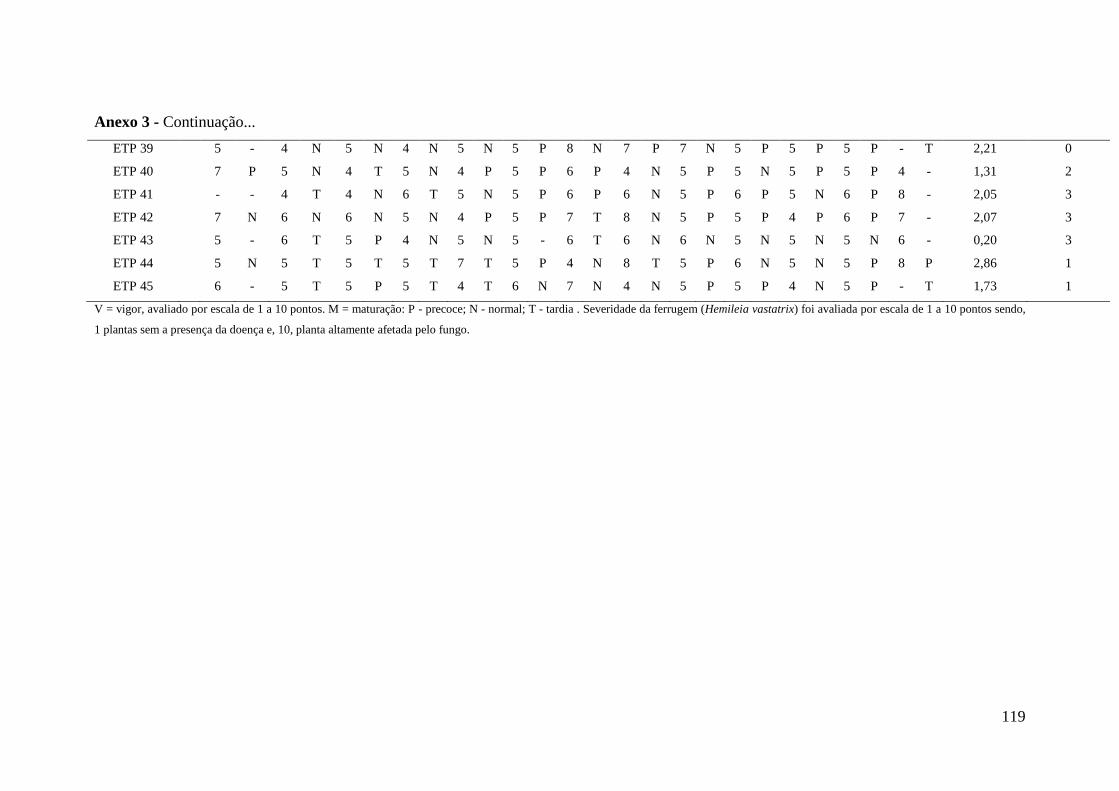
119
Anexo 3 - Continuação...
ETP 39 5 - 4 N 5 N 4 N 5 N 5 P 8 N 7 P 7 N 5 P 5 P 5 P - T 2,21 0
ETP 40 7 P 5 N 4 T 5 N 4 P 5 P 6 P 4 N 5 P 5 N 5 P 5 P 4 - 1,31 2
ETP 41 - - 4 T 4 N 6 T 5 N 5 P 6 P 6 N 5 P 6 P 5 N 6 P 8 - 2,05 3
ETP 42 7 N 6 N 6 N 5 N 4 P 5 P 7 T 8 N 5 P 5 P 4 P 6 P 7 - 2,07 3
ETP 43 5 - 6 T 5 P 4 N 5 N 5 - 6 T 6 N 6 N 5 N 5 N 5 N 6 - 0,20 3
ETP 44 5 N 5 T 5 T 5 T 7 T 5 P 4 N 8 T 5 P 6 N 5 N 5 P 8 P 2,86 1
ETP 45 6 - 5 T 5 P 5 T 4 T 6 N 7 N 4 N 5 P 5 P 4 N 5 P - T 1,73 1
V = vigor, avaliado por escala de 1 a 10 pontos. M = maturação: P - precoce; N - normal; T - tardia . Severidade da ferrugem (Hemileia vastatrix) foi avaliada por escala de 1 a 10 pontos sendo,
1 plantas sem a presença da doença e, 10, planta altamente afetada pelo fungo.

120