regenerao natural em uma floresta ombrfila densa … · extinção*** no estado do espírito santo...
TRANSCRIPT

REGENERAÇÃO NATURAL EM UMA FLORESTA OMBRÓFILA
DENSA ALUVIAL SOB DIFERENTES USOS DO SOLO NO DELTA DO
RIO DOCE
JOSÉ MANOEL LÚCIO GOMES
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE-UENF
CAMPOS DOS GOYTACAZES-RJ
MARÇO-2006

II

REGENERAÇÃO NATURAL EM UMA FLORESTA OMBRÓFILA
DENSA ALUVIAL SOB DIFERENTES USOS DO SOLO NO DELTA DO
RIO DOCE
JOSÉ MANOEL LÚCIO GOMES
Tese apresentada ao Centro de Biociências e Biotecnologia da Universidade Estadual do Norte Fluminense, como parte das exigências para obtenção do título de Doutor em Biociências e Biotecnologia na área de concentração em Ciências Ambientais.
Orientador: Prof. Dr. Marcelo Trindade Nascimento
CAMPOS DOS GOYTACAZES-RJ
MARÇO-2006
III

REGENERAÇÃO NATURAL EM UMA FLORESTA OMBRÓFILA
DENSA ALUVIAL SOB DIFERENTES USOS DO SOLO NO DELTA DO
RIO DOCE
JOSÉ MANOEL LÚCIO GOMES
Tese apresentada ao Centro de Biociências e Biotecnologia da Universidade Estadual do Norte Fluminense, como parte das exigências para obtenção do título de Doutor em Biociências e Biotecnologia na área de concentração em Ciências Ambientais.
Aprovada em 15 de março de 2006
Comissão examinadora:
__________________________________________________________
Prof. Dr. Haroldo Cavalcante de Lima (Doutor em Ciências – Ecologia) - JBRJ
____________________________________________________________
Profa. Dra. Maria Cristina Gaglianone (Doutora em Ciências – Entomologia) - UENF
____________________________________________________
Profa. Dra. Dora Maria Villela (Doutora em Ciências – Ecologia) - UENF
____________________________________________________
Prof. Dr. Carlos Eduardo de Rezende (Doutor em Ciências – Ecologia) - UENF
_________________________________________________________________
Prof. Dr. Marcelo Trindade Nascimento (Doutor em Ciências – Ecologia) -UENF
Orientador
IV

AGRADECIMENTOS
- Aos Professores do Setor de Botânica, Departamento de Biologia e à Reitoria da
Universidade Federal do Espírito Santo, pela autorização de afastamento para
realização desta Tese.
- Ao Laboratório de Ciências Ambientais (LCA) do Centro de Biociências e
Biotecnologia (CBB) da Universidade Estadual do Norte Fluminense (UENF), pela
oportunidade de realizar o curso de Doutorado.
- À minha família (Vera Lúcia Santos Gomes, Vivian Santos Gomes, Breno Lúcio
Gomes e Caio Lúcio Andrade Gomes), pela paciência nestes últimos meses.
- Aos meus pais Alda dos Santos Gomes e José Lúcio Gomes, aos quais serei
eternamente grato.
- Ao orientador Marcelo Trindade Nascimento, pela amizade, confiança e
profissionalismo, dedicados durante estes quatro anos de convivência.
- Aos componentes da banca examinadora, pela revisão da tese e valiosas
sugestões.
- Aos amigos, que durante estes quatros anos pude compartilhar algumas
sugestões de trabalhos (Fabrício A. de Carvalho, Ana Paula da Silva, Josival
Santos Souza, Andreia Vita Reis Mendonça, João Marcelo Braga, dentre outros) e
aos demais professores, colegas e funcionários do LCA.
- Ao Engo Florestal Renato Moraes de Jesus, pelo acesso ao Herbário (CVRD) da
Floresta Natural da Companhia Vale do Rio Doce, Linhares/ES.
- Ao Eng.° Florestal Vinícius Catane Vitor, pelo auxílio na digitação e cálculos.
- Ao Eng.° Agrônomo Lênio Bandeira, pelo auxílio na classificação do solo.
- Aos funcionários aposentados do INCAPER, Agnaldo Lorenzutti, Vitalino Pereira
e Amadeu, pelas preciosas informações sobre o histórico da área de estudo.
- Ao Biólogo Oberdan José Pereira, pelo incentivo para realização deste trabalho.
- Ao Eng.° Florestal Jorge Fernandes, pela ajuda em todos os momentos difíceis.
V

SUMÁRIO
Resumo..............................................................................................................
XVIII
Abstract..............................................................................................................
XIX
Capítulo 1: Introdução Geral.............................................................................
01
1.1. A Floresta Atlântica - o uso da terra e fragmentação.................................
01
1.2. Regeneração de florestas tropicais.............................................................
05
1.3. Florestas de baixada...................................................................................
06
1.4. Objetivos gerais..........................................................................................
08
1.5. Área de estudo............................................................................................
09
1.5.1. Localização........................................................................................
09
1.5.2. Histórico e uso do solo......................................................................
10
1.5.3. Solo e geomorfologia.........................................................................
12
1.5.4. Clima.................................................................................................
13
VI

Capítulo 2: Estrutura e composição florística do estrato arbóreo de um
remanescente de Floresta Atlântica Aluvial no Delta do Rio Doce, Norte
do Estado do Espírito Santo
2.1. Introdução...................................................................................................
15
2.2. Material e métodos......................................................................................
17
2.2.1. Área de estudo – vide capítulo 1....................................................... 17
2.2.2. Levantamento florístico e estrutural.................................................. 18
2.2.3. Análise dos dados.............................................................................
20
2.3. Resultados.................................................................................................. 22
2.3.1. Estrutura da floresta..........................................................................
22
2.3.2. Aspectos florísticos e fitossociológicos.............................................
24
2.3.3. Diversidade, riqueza e similaridade de espécies..............................
28
2.4. Discussão....................................................................................................
31
2.4.1. Classificação da vegetação...............................................................
31
2.4.2. Estrutura da floresta..........................................................................
32
2.4.3. Aspectos florísticos e fitossociológicos.............................................
34
2.4.4. Diversidade, riqueza e similaridade de espécies.............................. 35
2.5. Conclusões.................................................................................................
37
VII

Capítulo 3: Regeneração de espécies arbustivo-arbóreas da Floresta
Atlântica sob vários usos do solo no norte do estado do Espírito Santo
3.1. Introdução...................................................................................................
38
3.2. Material e métodos...................................................................................... 42
3.2.1. Área de estudo – vide capítulo 1....................................................... 42
3.2.2. Instalação das parcelas e coleta dos dados..................................... 44
3.2.3. Aspectos florísticos e fitossociológicos............................................. 45
3.2.4. Aspectos florísticos e estruturais da mata........................................ 46
3.2.5. Determinação da diversidade e similaridade de espécies................ 46
3.2.6. Análise estatística............................................................................. 47
3.3. Resultados.................................................................................................. 48
3.3.1. Comparações estruturais.................................................................. 48
3.3.1.1. Alturas e diâmetros................................................................. 48
3.3.1.2. Densidade e área basal.......................................................... 52
3.3.1.3. Árvores perfilhadas e densidade de lianas............................. 54
3.3.2. Comparações florísticas e fitossociológicas...................................... 56
3.3.2.1. Riqueza e diversidade de espécies........................................ 56
VIII

3.3.2.2. Comparações florísticas entre os sítios.................................. 62
3.4. Discussão.................................................................................................... 70
3.4.1. Comparações estruturais.................................................................. 70
3.4.2. Comparações florísticas.................................................................... 73
3.5. Conclusões................................................................................................. 76
Capítulo 4: Considerações Finais.....................................................................
78
Referências bibliográficas..................................................................................
81
IX

Lista de Figuras
Figura 1.5.1-1. Localização do Estado do Espírito Santo, do Município de
Linhares e da FLONA de Goytacazes (área de estudo) em relação ao Brasil.
10
Figura 1.5.2-1. Fotografia área da FLONA de Goytacazes (centro) e seu
entorno em 2000, Linhares/ES. Fonte: MAPLAN.
11
Figura 1.5.3-1. Área de estudo, Rio Doce e sua foz, Linhares/ES. Fonte:
Martin et al. (1997).
12
Figura 1.5.4-1. Precipitação média mensal entre 1975-2004 (30 anos) e entre
1995-2004 (10 anos), estação pluviométrica de povoação (Linhares/ES),
código – 01939002, Agência Nacional das Águas.
14
Figura 2.2.1-1. Fotografia aérea da FLONA de Goytacazes (centro) em 2000,
com os dois sítios (S1-sítio1 e S2-sítio2), Linhares/ES. Fonte: MAPLAN.
18
Figura 2.3.1-1. Distribuição do número de indivíduos por classe diamétrica,
nos sítios1 e 2, FLONA de Goytacazes, Linhares/ES.
23
Figura 2.3.1-2. Distribuição da porcentagem de lianas por parcela, na floresta
da FLONA de Goytacazes, Linhares/ES.
24
Figura 2.3.2-1. Distribuição das dez famílias com maior Valor de Cobertura
nos sítios1 e 2 estudados, FLONA de Goytacazes, Linhares/ES.
28
Figura 2.3.3-1 Dendograma de similaridade (Morisita modificado de Horn
1966 – Brower e Zar, 1977) de espécies e indivíduos entre as parcelas
amostradas no sítio1 (parcelas-P: 1, 2, 3, 4, 5) e sítio2 (parcelas-P: 6, 7, 8, 9,
10), nível de fusão (NF), FLONA de Goytacazes, Linhares/ES.
29
Figura 2.3.3-2 Dendograma de similaridade (Jaccard) de espécies entre
amostragens realizadas em florestas no Espírito Santo, Rio de Janeiro e
31
X

Pernambuco. (Tipo Florestal: ESTB= Estacional Semidecidual das Terras
Baixas, ESA= Estacional Semidecidual Aluvial, ODTB= Ombrófila Densa das
Terras Baixas, ODA= Ombrófila Densa Aluvial, ODS= Ombrófila Densa
Submontana, ODM= Ombrófila Densa Montana, R= Restinga, M=
Mussununga).
Figura 3.2.1-1. Fotografia área da FLONA de Goytacazes em 1970, com as
seis clareiras (A) e em 2000 (B), com os quatro sítios (1-capoeira, 2-cabruca,
3-floresta, 4-capoeirinha), Linhares/ES. Fonte: MAPLAN.
44
Figura 3.3.1.1-1. Comparação entre as curvas da distribuição das alturas (A,
B, C, D) e da distribuição relativa das alturas (E, F, G, H), das árvores,
arvoretas, varas e jovens, respectivamente, entre os sítios na FLONA de
Goytacazes, Linhares/ES.
49
Figura 3.3.1.1-2. Comparação entre as curvas da distribuição das classes
diamétricas (A, B, C, D) e distribuição relativa dos diâmetros (E, F, G, H), das
árvores, arvoretas, varas e jovens, respectivamente, entre os sítios na
FLONA de Goytacazes, Linhares/ES.
51
Figura 3.3.1.2-1. Comparação das densidades (A, B, C, D, E) e das áreas
basais (F, G, H, I, J), das árvores, arvoretas, varas, jovens e plântulas,
respectivamente, entre os sítios na FLONA de Goytacazes, Linhares/ES.
53
Figura 3.3.1.3-1. Comparação relativa dos indivíduos com perfilhos (A, B, C)
e da densidade de lianas (D, E, F), entre os sítios na FLONA de Goytacazes,
Linhares/ES.
55
Figura 3.3.2.1-1. Comparação das riquezas de espécies de árvores (A),
arvoretas (B), varas (C) e dos jovens (D) e diversidade de árvores (E),
arvoretas (F), varas (G) e dos jovens (H), entre os sítios na FLONA de
Goytacazes, Linhares/ES.
58
Figura 3.3.2.1-2. Curva de rarefação da diversidade de espécies (H’) com 61
XI

valores médios ± intervalo de confiança para árvores e arvoretas na floresta
(A, E), capoeira (B, F), cabruca (C, G) e capoeirinha (D, H), FLONA de
Goytacazes, Linhares/ES.
Figura 3.3.2.1-3. Curva de rarefação da diversidade de espécies (H’) com
valores médios ± intervalo de confiança para varas e jovens na floresta (A,
E), capoeira (B, F), cabruca (C, G) e capoeirinha (D, H), FLONA de
Goytacazes, Linhares/ES.
62
Figura 3.3.2.2-1. As coordenadas dos sítios nos dois eixos da DCA
(detrended correspondence analysis) formaram quatro grupos (G1, G2, G3,
G4), calculados com as espécies e sua abundância em cada estrato de cada
sítio [floresta (árvores - Fl 10, arvoretas - Fl 5, varas - Fl 2,5 e jovens - Fl
0,5); capoeira (árvores - Cap 10, arvoretas - Cap 5, varas - Cap 2,5 e jovens
- Cap 0,5); Cabruca (árvores - Cab 10, arvoretas - Cab 5, varas - Cab 2,5 e
jovens - Cab 0,5); Capoeirinha (árvores - Ca 10, arvoretas - Ca 5, varas - Ca
2,5 e jovens - Ca 0,5)], FLONA de Goytacazes, Linhares/ES.
64
Figura 3.3.2.2-2. Dendograma de similaridade (Sorensen) de espécies entre
os estratos dos sítios realizadas na FLONA de Goytacazes, Linhares/ES.
[floresta (árvores - Fl 10, arvoretas - Fl 5, varas - Fl 2,5 e jovens - Fl 0,5);
capoeira (árvores - Cap 10, arvoretas - Cap 5, varas - Cap 2,5 e jovens - Cap
0,5); Cabruca (árvores - Cab 10, arvoretas - Cab 5, varas - Cab 2,5 e jovens
- Cab 0,5); Capoeirinha (árvores - Ca 10, arvoretas - Ca 5, varas - Ca 2,5 e
jovens - Ca 0,5)].
65
Figura 3.5-1. Modelo de sucessão secundária em uma floresta Ombrófila
Densa Aluvial após corte e queima para uso agrícola na FLONA de
Goytacazes, Linhares/ES. Dados baseados no estrato arbóreo (DAP ≥ 10
cm). AB – área basal; N – número de indivíduos; H’- diversidade de
Shannon; N.spp. – número de espécies.
77
XII

Lista de Tabelas
Tabela 1.1-1. Área ocupada e participação na produção das diversas
atividades agrícolas na década de 1990 no Estado do Espírito Santo (Fonte:
Espírito Santo, 1999).
04
Tabela 2.3-1. Número de indivíduos (No Ind.), espécies (N.spp.), número de
indivíduos mortos, área basal por hectare (AB.ha-1), perfilhadas (P), altura
média (Alt. M.), diversidade (H’) e equidade (J’), por parcela amostrada nos
sítios1 e 2, FLONA de Goytacazes, Linhares/ES. Não houve diferença
estatística entre os valores médios das matas (teste t, P < 0,05) para um
mesmo parâmetro.
22
Tabela 2.3.2-1. Distribuição das espécies e famílias amostradas nos dois
sítios (S1 e S2) com o número de parcelas que ocorreram, caducifolia - C (D
– decídua, BD - brevidecíduas; SV – sempre verde e ameaçadas de
extinção*** no Estado do Espírito Santo e no Brasil (Decreto Estadual n0
1.499-R de 14/06/05 e Portaria n0 37-N/IBAMA, de 3/04/02), FLONA de
Goytacazes, Linhares/ES.
25
Tabela 2.4.2-1. Dados estruturais obtidos em alguns trabalhos realizados em
Floresta Atlântica (Tipo Florestal: ODTB = Ombrófila Densa de Tabuleiro,
ODS = Ombrófila Densa Submontana, ODM = Ombrófila Densa Montana,
ODA = Ombrófila Densa Aluvial, FV= Floresta de Várzea, ESA = Estacional
Aluvial, ESTB = Estacional das terras baixas, R = Restinga, M =
Mussununga); Área = área amostral em ha; DAP = diâmetro à altura do peito
em cm; H’ = índice de Shannon; N.spp. = número de espécies; Den =
densidade por hectare; AB = área basal por hectare em m2).
33
Tabela 3.3.1.2-1. Comparação da densidade de indivíduos por hectare entre
os sítios através de cálculos estatísticos pelo método de Mann-Whitney (U)
ao nível de 5% de probabilidade (P), na FLONA de Goytacazes,
Linhares/ES.
54
XIII

Tabela 3.3.1.2-2. Taxa de perda de indivíduos entre os estratos
subseqüentes, FLONA da Goytacazes, Linhares/ES.
54
Tabela 3.3.1.2-3. Comparação da área basal por hectare entre os sítios
através de cálculos estatísticos pelo método de Mann-Whitney (U) ao nível
de 5% de probabilidade (P), na FLONA de Goytacazes, Linhares/ES.
54
Tabela 3.3.1.3-1. Comparação dos indivíduos perfilhados entre os sítios
através de cálculos estatísticos pelo método de Mann-Whitney (U) ao nível
de 5% de probabilidade (P), na FLONA de Goytacazes, Linhares/ES.
56
Tabela 3.3.1.3-2. Comparação da densidade de lianas entre os sítios
através de cálculos estatísticos pelo método de Mann-Whitney (U) ao nível
de 5% de probabilidade (P), na FLONA de Goytacazes, Linhares/ES.
56
Tabela 3.3.1.3-3. Proporção de perda de lianas por faixa de diâmetro à
altura do solo (DAS), FLONA da Goytacazes, Linhares/ES.
56
Tabela 3.3.2.1-1. Riqueza (R), eqüidade (J’) e diversidade (H’) de espécies
para os sítios e estratos, FLONA de Goytacazes, Linhares/ES.
59
Tabela 3.3.2.1-2. Comparação da riqueza de espécies entre os sítios
através de cálculos estatísticos pelo método de Mann-Whitney (U) em nível
de 5% de probabilidade (P), na FLONA de Goytacazes, Linhares/ES.
59
Tabela 3.3.2.1-3. Proporção do número de espécies por estrato (N.spp.),
FLONA da Goytacazes, Linhares/ES.
59
Tabela 3.3.2.1-4. Proporção do número de espécies (N.spp.) em comum
com a floresta, FLONA de Goytacazes, Linhares/ES.
59
Tabela 3.3.2.1-5. Comparação da diversidade de espécies entre os sítios
através de cálculos estatísticos pelo método de Mann-Whitney (U) ao nível
de 5% de probabilidade (P), na FLONA de Goytacazes, Linhares/ES.
60
XIV

Tabela 3.3.2.2-1. As 113 espécies com os maiores valores de cobertura,
amostradas nos estratos e sítios, FLONA de Goytacazes, Linhares/ES.
(DAP - diâmetro à altura do peito; Alt. - altura; Flo - floresta; Cap - capoeira;
Cab - cabruca e Ca - capoeirinha).
66
XV

Anexo do capítulo 2
Anexo 2-1. Parâmetros fitossociológicos para as espécies amostradas na
floresta (total de 1 ha), com inclusão dos indivíduos com diâmetro à altura do
peito ≥ 10 cm, FLONA de Goytacazes, Linhares/ES. (NI - número de
indivíduos, DR - densidade relativa, FR - freqüência relativa, DoR -
dominância relativa, VC - valor de cobertura, VI - valor de importância).
95
Anexo do capítulo 3
Anexo 3-1. Espécies amostradas com presença (1) e ausência (0), por
estrato nos respectivos sítios, FLONA de Goytacazes, Linhares/ES. (DAP -
diâmetro à altura do peito; Alt. – altura; Flo - floresta; Cap - capoeira; Cab -
cabruca e Ca - capoeirinha).
99
XVI

Resumo
Este trabalho teve por objetivos: 1) investigar a composição florística, estrutura
arbórea e classificar uma Floresta Ombrófila Densa Aluvial no delta do Rio Doce
(FLONA de Goytacazes), além de compará-la com outras florestas da região, e 2)
avaliar a regeneração natural em três trechos desmatados para uso agropecuário e
diferente tempo de abandono. Um total de cinco parcelas de 20 x 50 m (n=5) foram
estabelecidas em dois sítios (1 e 2) de floresta e nos sítios de regeneração natural
(capoeirinha, capoeira e cabruca). Todos os indivíduos arbóreos (DAP ≥ 10 cm) e
lianas com DAS ≥ 10 cm (com exceção do sítio2), foram medidos e plaqueados.
Estas parcelas foram subdivididas em três grupos de subparcelas (10 x 25 m; 5 x 5
m e 2,5 x 1 m). No primeiro grupo (subparcelas de 10 x 25 m, n=5/sítio) foram
medidos os indivíduos com 5 ≤ DAP < 10 cm (arvoretas), os com 2,5 ≤ DAP < 5 cm
(varas) e as lianas com 2,5 ≤ DAS < 10 cm. Nas parcelas de 5 x 5 m (n=25/sítio)
foram amostrados os indivíduos com DAP < 2,5 cm e altura ≥ 0,5 m (jovens). Para
os indivíduos < 0,5 m de altura (plântulas), foram medidas as alturas e os DAS dos
amostrados em subparcelas de 2,5 m x 1 m (n=25/sítio). A comunidade arbórea da
floresta apresentou alta riqueza (157 espécies/ha) e diversidade (H’= 4,38), com
área basal média de 30 m2.ha-1. As espécies com maior valor de importância (VI)
foram Polyandrococos caudescens, Eschweilera ovata e Hydrogaster trinervis. Os
resultados indicaram que a floresta da FLONA de Goytacazes é madura e embora
tenha sofrido impactos de origem antrópica ainda possui uma alta riqueza e
diversidade florística, com ocorrência de espécies raras e ameaçadas. Em geral, os
parâmetros estruturais (altura, DAP, área basal, densidade total, riqueza e
diversidade de espécies) ocorreram na seguinte seqüência: floresta > capoeira >
cabruca > capoeirinha (indivíduos com DAP ≥ 10 cm). Entretanto, quando
consideradas as outras classes de tamanho (arvoretas, varas, jovens e plântulas)
foi observado um padrão geral (floresta = capoeira > cabruca > capoeirinha). Foi
encontrada uma alta similaridade de espécies no estrato de jovens entre a floresta
e os sítios secundários de capoeira (62%) e cabruca (30%), indicando com isso
que as espécies da floresta estão colonizando estas áreas. A capoeirinha com
nove anos e a capoeira com 33 anos de abandono da atividade agrícola,
encontravam-se em estágios de macega e médio de regeneração da Floresta
Atlântica, respectivamente. A alta diversidade e riqueza de espécies encontrada
para o mosaico de vegetação (sítios secundários e floresta madura) na FLONA de
XVII

Goytacazes, indicam a importância da manutenção e conservação dessa Unidade
de Conservação para a Floresta Atlântica do Estado do Espírito Santo.
XVIII

Abstract
The aims of this work were 1) to assess the floristic composition and the structure of
tree community in an evergreen Atlantic forest in the delta of Rio Doce (FLONA de
Goytacazes) and to compare the results to others from the region, and 2) to survey
the natural regeneration in three deforested sites for agriculture and with different
time of abandonment. A total of five plots (20 m x 50 m) were set up in each
deforested site and in two forest sites (control areas). All tree individuals (diameter
at breast height (DBH) ≥ 10 cm) and lianas [Diameter at soil height (DSH) ≥ 10 cm]
were tagged and measured. These plots were divided into three groups of sub-plots
(10 x 25 m; 5 x 5 m e 2,5 x 1 m). At the first one (subplots of 10 m x 25 m, n=5/site)
were measured all individuals of small trees (5 cm ≤ DBH < 10 cm), sapling (2.5 ≤
DBH < 5 cm) and lianas (2,5 ≤ DSH < 10 cm). In plots of 5 m x 5 m (n=25/site) were
sampled all individuals with DBH < 2,5 cm and height ≥ 0,5 m (young). For
individuals < 0,5 m height were sampled and measured for DSH and height in
subplots of 2,5 m x 1 m (n=25/site). In the forest sites the tree community showed a
high species richness (157 spp.ha-1) and diversity index (H’= 4,38), with a total
basal area of 30 m2.ha-1. The most important species (IV) were Polyandrococos
caudescens, Eschweilera ovata e Hydrogaster trinervis. These results suggest that
the forest of the FLONA de Goytacazes is mature and although had been disturbed
in the past, it still has a high species richness and floristic diversity, with occurrence
of rare and endangered species. In general, the structural parameters (height, DBH,
total basal area, total density, species richness and diversity) occurred in the
following sequence: forest>capoeira>cabruca>capoeirinha for individuals with DBH
> 10 cm. However, when considered the other size classes (small trees, saplings,
young and seedlings) a general pattern (forest=capoeira>cabruca>capoeirinha).
The capoeirinha with nine years and the capoeira with 33 years of abandonment of
agricultural activity meeting in stage of macega and half of regeneration of Atlantic
Forest, respectivement. A high species similarity in the young class was found
between forest and the secondary areas (capoeira, 62% and cabruca, 30%),
suggesting that forest species area colonizing these areas. The high values of
species diversity and richness found for the vegetation mosaic (secondary areas
and mature forest) of the FLONA de Goytacazes suggest the importance of this
conservation unit for the maintenance and conservation of the Atlantic forest in the
Espírito Santo State.
XIX

1
CAPÍTULO 1
INTRODUÇÃO GERAL
1.1. A Floresta Atlântica - o uso da terra e fragmentação
As florestas tropicais constituem um dos biomas terrestres de maior
biodiversidade. Nelas habitam cerca de 70% das plantas e animais do globo
terrestre, representadas por mais de 13 milhões de espécies, onde existem cerca de
70% das plantas vasculares, 30% das espécies de aves e 90% de invertebrados
(vide Roper e Roberts, 1999).
As áreas de florestas tropicais estão sendo desflorestadas nas últimas
décadas a passos largos, principalmente para ampliação das fronteiras agrícolas
(Tilman, 1999). Segundo estimativas da FAO (1997), nos países em
desenvolvimento vêm sendo desmatados cerca de 15,5 milhões de hectares por
ano, entre 1980 e 1990 e 13,7 milhões entre 1990 e 1995, totalizando
aproximadamente 200 milhões de hectares entre 1980 e 1995.
Segundo MMA (2002), o Brasil possui a maior cobertura de florestas tropicais
do planeta, embora em 1995 encontrava-se em primeiro lugar no ranking dos países
com maior perda anual de florestas tropicais (2.550.000 ha) seguido pela Indonésia
com menos da metade (FAO, 1997). Dentre as florestas tropicais brasileiras
destacam-se as florestas Amazônica e Atlântica.
A Floresta Atlântica que no início do século passado recobria 16 Estados ao
longo da costa brasileira, representando cerca de 1.100.000 Km2 (SOS Mata
Atlântica,1993) e que se encontra distribuída em faixa paralela ao oceano Atlântico
com 120 a 160 km de largura, desde o Rio Grande do Sul ao Rio Grande do Norte,
apresenta redução drástica de sua área original, sendo considerado o ecossistema
brasileiro mais impactado por ações antrópicas (Bigarella et al., 1975; Rizzini, 1979;
Myers et al., 2000; Morellato e Haddad, 2000).
Na década de 1990 a Floresta Atlântica ocupava apenas 95.641 Km2, sendo
composta por florestas de planície e de altitude, matas costeiras e de interior, ilhas
oceânicas, encraves e brejos interioranos, incluindo os ecossistemas associados
como Restingas, Manguezais e Campos de Altitude. Correspondendo a 8,8% da sua
área original (MMA, 2002), seus principais remanescentes concentram-se nos

2
Estados das regiões Sul e Sudeste, recobrindo parte das Serras do Mar e da
Mantiqueira, onde o processo de ocupação foi dificultado pelo relevo acidentado e
pouca infra-estrutura de transporte (Capobianco e Lima, 1997). Sendo os
remanescentes florestais do Sudeste referidos pela literatura como tendo uma das
mais altas biodiversidades conhecidas, no que tange a flora fanerogâmica (Giullieti,
1992), além de deter um grande endemismo (Mori et al., 1981) e considerados como
de alta prioridade para conservação (Myers et al., 2000).
A Floresta Atlântica no Sudeste do Brasil devido a repetidos e sucessivos
distúrbios antrópicos, gerados pela expansão humana tais como: atividades
agropecuárias, urbanização, industrialização, dentre outras, teve uma ocupação
desordenada, principalmente a partir da década de 1950, onde os distúrbios se
intensificaram produzindo uma paisagem consideravelmente fragmentada (Dean,
2000; SOS Mata Atlântica, 2002).
Segundo Ruschi (1950), dos 45.597 Km2 da área do território do Estado do
Espírito Santo, 37.800 Km2 eram ocupados pela Floresta Atlântica (Floresta das
Terras Baixas - Tabuleiros, Floresta Montanas e Florestas Altimontanas - Encosta),
já em 1950, restavam 16.760 Km2, correspondendo a 55% da área ocupada. Este
autor cita que as áreas desmatadas até aquela época eram cultivadas
principalmente por plantas agrícolas como: café (Coffea sp.), milho (Zea sp.), feijão
(Phaseolus sp.), arroz (Oryza sp.), cana-de-açúcar (Saccharum sp.), banana (Musa
sp.), laranja (Citrus sp.), cacau (Theobroma cacao L.), mandioca (Manihot sp.) e
pastagens compostas por diversas espécies de Gramineae como: colonião (Panicum
maximum Hochst. ex A.Rich.) e braquiária (Brachiaria spp.).
Relata ainda o crescente volume da madeira serrada em toras e dormentes
para utilização em indústrias de móveis, esquadrias e engradamento de telhados,
madeira para energia como lenha e carvão vegetal, este último destinado para
siderúrgicas. Constatou também que, com a crescente expansão demográfica
espírito-santense, o alto preço da madeira e a facilidade de acesso às florestas, a
tendência foi de aumento desse consumo e de destruição das florestas.
Mais tarde, em 1968, a área de floresta no Estado do Espírito Santo estava
reduzida 3.650 Km2, correspondendo a 9,7 % de seu território (Mori et al., 1981).
Posteriormente, segundo dados do SOS Mata Atlântica (1993), em 1990 restavam
4.023,92 Km2, representando 10,64 % da cobertura de Floresta Atlântica neste
Estado.

3
Em 2002 foi lançado um Atlas da cobertura vegetal da Floresta Atlântica (SOS
Mata Atlântica, 2002), indicando que no Estado do Espírito Santo há 7.656,18 Km2,
correspondendo a 20% de remanescentes de florestas em relação à área ocupada
originalmente pela Floresta Atlântica, incluídas as áreas com no mínimo 10 ha. Já
Thomas et al. (1998), indicaram que em 1990 no Estado do Espírito Santo havia
2,62 % da área de Floresta Atlântica madura e 13,56 % perturbada.
As diferenças de área de Floresta Atlântica remanescente para o Estado do
Espírito Santo, podem ter ocorrido devido à baixa precisão das ferramentas e da
escala utilizadas, bem como da possibilidade de inclusão de matas secundárias em
estágios inicial e médio na avaliação.
Estes dados confirmam a previsão de Ruschi (1950) sobre a ampliação da
fragmentação e de destruição, com aumento principalmente das fronteiras
agropecuárias, resultando numa situação agrária com predominância de pastagens
(54,7%), em sua maioria mal cuidadas e com processos erosivos instalados. Apesar
da situação agrária atual no Estado do Espírito Santo estar representada por
predominância de pequenas propriedades (90% menor que 100 ha), que ocupam
3.327.417 ha, equivalente a 73% da área Estadual (Espírito Santo, 1999), a
ocupação foi totalmente desordenada e sem levar em conta a conservação a
ambiental (tabela 1.1-1).
Em relação à região norte do Espírito Santo, Egler (1951) a considerou
complicada devido à relação de ocupação pelos índios Botocudos, a ponto de em
1819 Saint-Hilaire ser desaconselhado a realizar a viagem ao Rio Doce. Portanto, a
expansão do povoamento da cidade de Linhares passou a ocorrer a partir da
segunda metade do século XIX.
Em 1815 o príncipe Wied-Neuweid descreve a cidade de Linhares, como uma
das mais importantes da região norte do Estado, composta por casebres cobertos
por folhas de palmeiras, com a ocupação da porção sul do Rio Doce tendo início
somente após a expulsão dos índios Botocudos. Esta ação levou a instalação da
fazenda denominada Bom Jardim, nas proximidades da área onde hoje se encontra
a Floresta Nacional (FLONA) de Goytacazes, iniciando assim o desmatamento e a
implantação de cultivos, principalmente de cana-de-açúcar e arroz (Wied, 1989).
Até o início da década de 1930 a maior parte da região norte do Estado era
recoberta pela Floresta Atlântica. Este cenário começou a ser alterado com a

4
construção da ponte do Rio Doce em 1928, fato que facilitou o acesso às áreas
florestadas da região. A partir daí, a região passou a ser responsável por um longo
tempo como grande produtora e exportadora de madeira no Estado. Com a
exaustão do estoque de madeira nativa, os exploradores voltaram as explorações
para o sul da Bahia e mais recentemente para a Amazônia (Garcia, 2002).
Tabela 1.1-1. Área ocupada e participação na produção das diversas atividades agrícolas na década de 1990 no Estado do Espírito Santo (Fonte: Espírito Santo, 1999).
Área Atividade ha % Café 550.000 16,5 Pastagem/Produção Animal 1.821.069 54,7 Fruticultura 65.000 1,9 Olericultura 9.530 0,3 Culturas Alimentares Temporárias 118.600 3,6 Outras Culturas 68.666 2,1 Florestas Naturais 371.862 11,2 Florestas Plantadas 172.735 5,2 Terras não Aproveitadas 149.955 4,5
Total 3.327.417 100,0
Após a exploração madeireira em florestas maduras, as áreas desflorestadas
foram sendo substituídas principalmente por pastagem e agricultura, com a
vegetação remanescente altamente fragmentada e distribuída na sua maioria em
fragmentos com áreas em geral inferiores a 1.000 ha, localizadas, principalmente,
em propriedades privadas (SOS Mata Atlântica, 1998; 2002). Estes fragmentos de
matas secundarias, no presente, podem ser classificados, segundo critérios
previstos na legislação (CONAMA, 1994; IDAF, 1999) produzida a partir reuniões de
trabalho com especialistas com as seguintes denominações: macega (capoeirinha –
Veloso et al., 1991), estágios inicial (capoeirinha – Veloso et al., 1991), médio
(capoeira – Veloso et al., 1991) e avançado (capoeirão – Veloso et al., 1991) de
regeneração da Floresta Atlântica, além de floresta.
Dentre os municípios do Estado do Espírito Santo, Marataízes se destaca
com zero de remanescentes da Floresta Atlântica e Marechal Floriano com a maior
porcentagem equivalente a 53% (SOS Mata Atlântica, 2002). Linhares se encontra
com 648,87 Km2 de florestas, destes, cerca de 450 Km2 se encontravam localizados
em área contínua composta pelas Reservas Natural da Vale do Rio Doce e Biológica
de Sooretama (SOS Mata Atlântica, 2002) e cerca de 200 Km2 no delta do Rio Doce

5
na sua maioria em cabruca, constituída por áreas compostas por árvores
remanescentes da floresta madura raleada e com cultivo de cacau sob a sombra
(Rolim e Chiarello, 2004; Rolim et al., no prelo).
1.2. Regeneração de florestas tropicais
Segundo Grime (1979), a regeneração natural em áreas fortemente
degradadas depende na primeira fase do estabelecimento da vegetação e das
respostas por parte das espécies vegetais às restrições ambientais. Assim, as
espécies colonizadoras possuem, em geral, características relacionadas ao vigor
vegetativo a um ciclo de vida curto e uma alta produção de sementes. Já em fase
mais avançada de regeneração, o sucesso no estabelecimento está mais
relacionado ao ciclo de vida mais longo das espécies e na produtividade do habitat,
proporcionando uma base que pode explicar as diversas seqüências existentes na
sucessão secundária.
A ampliação do conhecimento acerca do comportamento das espécies
vegetais em diferentes ambientes e com idades distintas de regeneração nas zonas
tropicais do planeta é desejável, para que possamos entender as tendências
evolutivas da biota, permitindo e facilitando o estabelecimento de manejo adequado,
principalmente com relação à recuperação deste tipo de vegetação (Gómez-Pompa
e Vázquez-Yanes, 1976). Este fato está diretamente ligado à importância da função
da floresta, das espécies e do fornecimento de benefícios oriundos de seus recursos
florestais (Saito, 1992).
A recuperação da vegetação em áreas alteradas pela ação antrópica pode ser
realizada pela interferência direta, com a utilização de técnicas silviculturais e
conseqüente redução do prazo necessário à reconstituição natural ou sem a
intervenção humana. Por outro lado, segundo Kageyama e Castro (1989), para que
ocorra a regeneração sem intervenção é necessário haver disponibilidade de
sementes de espécies pioneiras no solo, baixa intensidade e freqüência de
distúrbios, além de condições ambientais favoráveis.
Segundo Seitz (1994), na maioria dos casos de degradação de áreas de
florestas no Brasil podem ser recuperadas por dinâmica natural do ambiente.
Durigan (2003) sugere técnicas e parâmetros para avaliar o potencial da capacidade
de regeneração natural em áreas de cerrado e de floresta, sem plantio e com plantio
e recuperação de solo. Estes autores apontam para a necessidade urgente e

6
crescente de informações sobre como ocorrem os processos de regeneração da
Floresta Atlântica, uma vez que, dependendo do grau de degradação sofrido pelo
sítio, a regeneração natural da vegetação é o procedimento mais viável para sua
recuperação.
A velocidade e a qualidade da regeneração está diretamente ligada à
proximidade de fragmentos para fornecimento de propágulos bem como da
intensidade das intervenções realizadas, principalmente no solo do sítio
(Buschbacher, 1986; Aide et al.,1996; Uhl et al.,1988; Durigan, 2003)
1.3. Florestas de baixada
O desenvolvimento da vegetação e sua fitofisionomia estão condicionados a
vários fatores do ambiente como disponibilidade de água, de nutrientes do solo,
relevo (Resende et al.2002) e a composição florística é influenciada diretamente por
estes fatores e pode tornar-se mais dissimilar com o distanciamento (Oliveira-Filho,
et al, 2005). Estas variações do ambiente irão condicionar também as variações na
fitofisionomia e na flora em comunidades próximas ou distantes entre si, criando um
mosaico de formações vegetais característico das florestas tropicais.
Ruschi (1950) classificou a vegetação do Estado do Espírito Santo em várias
formações e, segundo sua classificação, parte da área do delta do Rio Doce
enquadra-se no domínio da Floresta dos Tabuleiros, também indicada pelo projeto
Radambrasil (IBGE, 1987), pelo Mapa de Vegetação do Brasil (IBGE, 1993) e por
Veloso et al. (1991) como Floresta Ombrófila Densa das Terras Baixas. Apesar
destas classificações, toda a área do delta encontra-se sobre sedimentos aluviais
holocênicos (Martin et al., 1997).
A floresta aluvial é uma formação ribeirinha ou floresta ciliar, que ocorre ao
longo dos cursos de água ocupando os terraços das planícies quaternárias, com
muitas palmeiras no estrato inferior (Veloso et al., 1991). Sua composição florística é
variável em função dos aspectos físicos, como níveis de umidade e constituição do
solo (Rodrigues e Shepherd, 1999) e também em função do gradiente latitudinal,
como por exemplo, a da bacia do Rio Doce é diferente daquela da bacia do Paraíba
do Sul e nestas duas é diversa da que ocorre na bacia do rio Itajaí (Veloso et al.,
1991).
Rodrigues e Shhepherd (2001) definem dois tipos básicos de floresta ciliar: (1)
aquelas que ocorrem com interferência direta e atual do rio ou do lençol freático,

7
podendo ser faixa marginal periodicamente inundada pela elevação do rio ou do
lençol freático e permanentemente inundado pelo afloramento do lençol freático e (2)
floresta ciliar sem interferência direta e atual do rio ou do lençol freático, sobre
aluvião ou não.
No caso da FLONA de Goytacazes, ocorrem áreas permanentemente
alagadas, periodicamente alagáveis e não alagáveis, todas sobre solos aluviais.
Sendo que a área de estudo encontra-se sobre solo auvial sem alagamento.
Mais de 30 levantamentos florísticos e fitossociológicos foram desenvolvidos
em áreas de floresta ciliar na região extra-amazônica (Rodrigues e Nave, 2001).
Todavia, há carência de informações sobre os aspectos florísticos e estruturais e
sobre o processo de regeneração das florestas em planícies quaternárias aluviais
sem inundação sob diferentes usos do solo, com apenas dois trabalhos publicados
para o Estado do Espírito Santo (Rolim e Chiarelo, 2004; Rolim et al., no prelo), além
de vários outros publicados em áreas ciliares extra-amazônicas (Rodrigues e Nave,
2001). O primeiro (Rolim e Chiarelo, 2004) sobre florística e fitossociologia em áreas
sob “cabruca” com cultivo de cacau e o segundo (Rolim et al., no prelo) sobre a flora
arbórea da floresta aluvial no vale do Rio Doce.
Entretanto, várias pesquisas vêm sendo desenvolvidas nas comunidades
arbóreas próximas da FLONA de Goytacazes e no Espírito Santo, no norte
Fluminense, no Sul da Bahia e em Pernambuco.
Destacam-se na planície Quaternária, oriunda da regressão marítima,
levantamentos florísticos, fitossociológicos e descrições de Matas Secas de
Restinga, produzidos por Pereira (1990), Pereira e Gomes (1994), Fabris e Cesar
(1996), Pereira et al. (1998), Pereira e Assis (2000), Pereira e Zambom (1998),
Pereira e Araújo (2000), Pereira et al. (2000) e Assunpção e Nascimento (2001).
Na Mussununga (Simonelli, 1998), localizada na região dos Tabuleiros
Costeiros, com solo composto principalmente por areia, em platôs com cerca de 30-
40 m de altitude e constituída por vegetação com ocorrência predominante na Mata
Seca de Restinga.
Na Floresta das Terras Baixas, localizadas principalmente no Terciário, foram
realizados trabalhos semelhantes tais como: Heinsdijk et al. (1965), Peixoto (1981),
Peixoto (1982), Peixoto e Gentry (1990), Jesus e Garcia (1992), Rizzini (2000),
Souza et al. (1998), Silva e Nascimento (2001), Martini (2002), Andrade e Rodal
(2004), Jesus e Rolim (2005) e Villela et al. (2006).

8
Na Floresta Montana, no pré – cambriano, (Thomaz, 1996; Moreno et al,
2003).
Com relação à dinâmica de populações vegetais e dispersão de sementes
são mais escassos ainda, como na Floresta das Terras Baixas no Espírito Santo,
principalmente Costa et al. (1992), Nunes (1996), Rolim (1997), Jesus (2001) e no
norte do Estado do Rio de Janeiro (Souza, 2005).
Podemos citar ainda listas das espécies ocorrentes no Sul da Bahia tais
como: na Floresta da Esperança, Parque Nacional Monte Pascoal, Reserva Bilógica
de Una, Parque Estadual Serra do Condurú, Floresta da Serra Grande e Serra do
Teimoso (vide Thomas e Amorim, 2005a,b,c,d,e,f), além de Mori et al. (1983).
Todas estas formações possuem peculiaridades que podem influenciar na
sua composição florística, podendo serem similares ou dissimilares (Oliveira-Filho et
al.,2005).
Segundo Martin et al (1997) o delta do Rio Doce foi formado a partir do
continente em direção ao oceano, com a vegetação se estabelecendo logo após a
deposição dos sedimentos aluviais.
A Floresta das Terras Baixas existente no entorno Floresta Aluvial da FLONA
de Goytacazes sem alagamento, provavelmente, foi quem contribuiu mais com sua
composição florística (Jesus e Rolim, 2005; Rodrigues e Nave, 2001).
As Matas Secas de Restinga encontram-se sobre sedimentos arenosos
oriundos da regressão marítima e fluviomarinha (Martim et al., 1997) e a
Mussununga também em solo arenoso e fitofisionomia e composição florística muito
parecidos com os da Restinga (Simonelli, 1998). As Florestas Montanas encontram-
se em altitudes mais elevadas que as anteriores.
1.4. Objetivos gerais
Esta pesquisa tem como objetivos: (1) descrever a estrutura e a composição
florística da Floresta Aluvial da FLONA de Goytacazes, realizar sua classificação
quanto à mudança foliar e compará-la com outras do norte do estado do Rio de
Janeiro, Espírito Santo e Pernambuco (parte do Sudeste e Nordeste), (2) avaliar
através do levantamento florístico, da análise estrutural e da regeneração natural de
áreas de Floresta Aluvial sem inundação com distintos níveis de perturbações, usos
do solo e tempo de abandono, o estabelecimento de espécies vegetais, fornecendo

9
base para projetos de conservação, manejo e recuperação deste tipo de vegetação
e (3) testar as seguintes hipóteses:
1) A composição florística e estrutural do estrato arbóreo da Floresta Aluvial
madura e sem inundação (em bom estado de conservação) da FLONA de
Goytacazes são similares às existentes na Floresta das Terras Baixas e
dissimilares da Floresta de Mussununga, da Floresta Seca de Restinga e da
Floresta Atlântica Montana, abrangendo o norte do estado do Rio de Janeiro,
Espírito Santo e Pernambuco (parte do Sudeste e Nordeste).
2) A estrutura e a composição florística da regeneração (espécies arbustivas-
arbóreas) natural em três trechos desmatados para cultivo agrícola adjacentes
ao remanescente de Floresta Aluvial madura sem inundação (FLONA de
Goytacazes), com diferentes idades e usos do solo, difere entre si e da floresta
adjacente (controle).
Devido aos diversos temas abordados nesta tese, a organização estrutural foi
estabelecida em capítulos. O primeiro, introdutório e com a descrição da área de
estudo e os objetivos gerais. O segundo, descritivo, onde se compara a estrutura e a
composição florística da floresta estudada com a de parte do Sudeste e Nordeste. Já
o terceiro aborda a regeneração de uma floresta Aluvial Atlântica madura
comparando-a com outras áreas em regeneração após supressão da floresta e usos
agrícolas. Por último, o capítulo quatro, onde se encontram as considerações finais.
1.5. Área de estudo
1.5.1. Localização
A área estudada está localizada na FLONA de Goytacazes, com cerca de
1.350 ha, nas coordenadas 4005’W e 19025’S (figura 1.5.1-1) e integra o complexo
vegetacional do delta do Rio Doce, distando cerca de um quilômetro ao Sul da
cidade de Linhares e 400 m das margens do Rio Doce.
No entorno da FLONA de Goytacazes há regiões de domínio da Floresta
Ombrófila Densa das Terras Baixas (IBGE, 1987; 1993) em terrenos do Terciário, de
áreas de influência marinha ou Restinga (IBGE, 1987; 1993; Bellini et al., 1990) e
Mussununga (Simonelli, 1998), protegidos na Reserva Natural da Vale do Rio Doce
e Reserva Biológica de Sooretama.

10
Além destas, há outras unidades de conservação de dimensões bem menores
como a Reserva Biológica Córrego do Veado, FLONA do Rio Preto e FLONA de
Goytacazes. Destas, apenas a FLONA de Goytacazes encontra-se totalmente em
área do Quaternário Holocênico de influência fluvial (Martin et al., 1997).
O Rio Doce atravessa o Estado do Espírito Santo de Leste a Oeste e é um
divisor entre o Norte e o centro-sul do Estado, com sua foz no Oceano Atlântico a
cerca de 36 km da FLONA de Goytacazes.
Figura 1.5.1-1. Localização do Estado do Espírito Santo, do Município de Linhares e
da FLONA de Goytacazes (área de estudo) em relação ao Brasil.
1.5.2. Histórico e uso do solo
A área estudada pertencia ao Instituto de Pesquisa Agropecuária do Centro
Sul do Ministério da Agricultura (IPEACS), sendo transferida para a Empresa
Brasileira de Pesquisa Agropecuária em 1973 que posteriormente a transferiu em
regime de comodato para a Empresa Capixaba de Pesquisa Agropecuária
(EMCAPA), hoje denominado de Instituto de Pesquisa Agropecuária e Extensão
Rural do Espírito Santo (INCAPER). Em dezembro de 2002 a área foi transferida
para o Instituto Brasileiro de Meio Ambiente e Recursos Naturais (IBAMA), sendo
então criada a FLONA de Goytacazes, com cerca de 1.350 ha.
Uma parte da área foi utilizada para experimentos agrícolas e pecuários até
1994. Dentre as atividades agropecuárias se destacaram os experimentos com
cacau (Theobroma cacao L.), forrageiras (capim elefante - Pennisetum sp.), pasto
(colonião - Panicum maximum Hochst. ex A.Rich.) e cereais (milho - Zea sp; feijão -
Flona de Goytacazes

11
Phaseolus sp.), (comunicação pessoal de Agnaldo Lorenzutti, 2002). Atualmente a
área possui cerca de 90% de seu solo coberto por floresta com coleta de madeira
com várias intensidades e o restante com cultivos abandonados de cereais, pasto e
de cacau. Assim, áreas outrora cultivadas hoje se encontram com vegetação
secundária em vários estágios de regeneração.
A matriz da paisagem no entorno da FLONA encontra-se composta por
atividades agropecuárias como pastagem, principalmente de Brachiaria spp.,
produção de milho, mamão (Carica sp.) e de cacau através do cultivo em sistema de
cabruca (raleamento da floresta madura para cultivo de cacau sob sombra) e/ou
utilizando exóticas para seu sombreamento e fragmentos em vários estágios de
regeneração da Floresta Atlântica (figura 1.5.2-1).
Segundo Embrapa (1978), na região com ocorrência de Neossolo Flúvico
eutrófico (solo Aluvial) no delta do Rio Doce a cultura predominante era de cacau,
além de banana (Musa sp.), milho, feijão e pasto. Atualmente, esta área (delta do
Rio Doce) é considerada prioritária para a conservação da biodiversidade da
Floresta Atlântica da região (MMA, 2002).
Figura 1.5.2-1. Fotografia aérea da FLONA de Goytacazes (centro) e seu entorno
em 2000, Linhares/ES. Fonte: MAPLAN.
Br101
Rio Doce
FLONA de Goytacazes
Cabruca
Agricultura
Linhares
Pastagem
Cabruca
N↑
750 m

12
1.5.3. Solo e geomorfologia
A floresta estudada encontra-se sobre terrenos do Quaternário (Holoceno),
com sedimentos de origem continentais aluviais e coluviais, havendo também
sedimentos arenosos de paleocanais, conforme descrição e mapeamento realizados
por Martin et al. (1997).
O gradiente topográfico observado nas margens do Rio Doce é muito
pequeno, constituindo uma extensa planície, com alagados que podem propiciar
aumento de umidade nos períodos chuvosos, principalmente devido à influência do
lençol freático, ou até mesmo de lagoas temporárias. Depois do município de
Linhares, ele corre por cerca de 36 km, com um declive menor que 2 m/km, até
desembocar no oceano (Rolim et al., no prelo), (figura 1.5.3-1).
As margens do Rio Doce e a área de estudo são ocupadas por Neossolos
Flúvicos, podendo ser eutróficos (solos Aluviais) e, em sua maior parte em sistema
de cabruca com cultivo de cacau e nas porções mais distantes das margens do Rio
Doce ocorrem os Neossolos Flúvicos distróficos (solos Aluviais), (Lani, 1998;
Embrapa, 1978). Feitoza et al. (1999) classificaram como de terras quentes, planas,
transição chuvosa/seca, férteis, enxutas e de textura fina.
Figura 1.5.3-1. Área de estudo, Rio Doce e sua foz, Linhares/ES. Fonte: Martin et al.
(1997).
Área de estudo
Foz do Rio DocePaleocanais Paleolagoas
Terciário(─)
Quaternário (─)
Terciário(─)
N↑
5 km
Oceano Atlântico

13
Pela classificação da Embrapa (1999), os solos Aluviais passaram a integrar
os Neossolos, compreendendo solos constituídos por material mineral ou por
material orgânico pouco espesso, com baixa intensidade de atuação dos processos
pedogenéticos, resultando em pequena expressão dos processos pedogenéticos,
que não conduziram, ainda, às modificações expressivas do material originário, pela
sua resistência ao intemperismo ou devido ao relevo plano, que poderá impedir ou
limitar a evolução desses solos.
Os Neossolos Flúvicos (solos Aluviais) são solos derivados de sedimentos
aluviais com horizonte A sobre horizonte C, perfil constituído de camadas
estratificadas e presença de carbono orgânico.
1.5.4. Clima
Nos últimos trinta anos a precipitação média anual na estação meteorológica
de Povoação foi de 1.206 mm, sendo os meses de junho e agosto com menos de 60
mm. Nos últimos 10 anos a precipitação média foi de 1.121 mm, com os meses de
junho e agosto apresentando menos de 60 mm, acompanhando as médias dos
últimos trinta anos. Os meses de maior precipitação (mais de 100 mm) foram:
janeiro, março, abril, outubro, novembro e dezembro (figura 1.5.4-1). Não houve
diferença estatística (p < 0,05) entre os dois períodos de precipitação analisados
(1975 a 2003 e 1995 a 2004).
Segundo Embrapa (1978), o clima da região é do tipo AW da classificação de
Köppen, apresentando inverno com estiagem bem menos pronunciada. Feitoza et al.
(1999) cita que na região os períodos úmido e seco abrangem dois meses cada,
com excedente hídrico de 71 mm e déficit de 281 mm. Temperatura média na região
de 23,3oC com média mínima de 14,8 oC e máxima de 34,2 oC (Jesus e Rolim,
2005).

14
Figura 1.5.4-1. Precipitação pluviométrica média mensal (mm) entre 1975-2004 (30
anos) e entre 1995-2004 (10 anos), estação pluviométrica de povoação
(Linhares/ES), código – 01939002, Agência Nacional das Águas.
125
74
123112
84
55
82
46
71
104
173159
76
51
112
135
72
50
81
28
77 73
217
151
0
50
100
150
200
250
Jane
iro
Favere
iro
Março
Abril
MaioJu
nho
Julho
Agosto
Setembro
Outubro
Novembro
Dezembro
Meses
Pre
cipi
taçã
o m
édia
1975-20041995-2004

15
CAPÍTULO 2
ESTRUTURA E COMPOSIÇÃO FLORÍSTICA DO ESTRATO
ARBÓREO DE UM REMANESCENTE DE FLORESTA ATLÂNTICA
ALUVIAL NO DELTA DO RIO DOCE, NORTE DO ESTADO DO
ESPÍRITO SANTO 2.1. Introdução
A Floresta atlântica dentro de uma visão global é considerada prioritária para
conservação (Myers et al., 2000; MMA, 2002) e a maior parte de seus
remanescentes encontra-se sob o domínio de propriedades privadas (Viana et al.,
1997).
Estes remanescentes encontram-se restritos a fragmentos, muitas vezes nos
altos dos morros e entremeados por monoculturas, o que em alguns casos, têm
levado à extinção de grande número de espécies (Giulietti e Forero, 1990).
As pressões e distúrbios de origem antropogênica continuam acontecendo
(Oliveira et al. 1994), criando mosaicos de vegetação, que após o abandono da
exploração e do controle sobre a regeneração, tendem a evoluir para estágios mais
avançados de sucessão da floresta madura (Tabarelli et al., 1999).
A ocupação do território no Estado do Espírito Santo, inicialmente se deu de
forma lenta e, posteriormente, passou a ser acelerada. Até meados do século XVIII a
colonização se limitava à zona litorânea e o Estado possuía 80% da cobertura
florestal.
Em 1888 apenas 15,4% do território encontravam-se explorado. Isto devido
ao interesse de se manter uma barreira natural protetora para zonas de mineração
de Minas Gerais (Espírito Santo, 1989).
Com o ciclo do café (Coffea spp.), cacau (Theobroma cacao L.), exploração
de madeira, abertura de espaços para agropecuária e urbanização, ocorridos com
maior intensidade a partir do século XIX, com intensificação da fragmentação das
florestas no Estado, restando uma cobertura de floresta madura inferior a 10% (SOS
Mata Atlântica, 1998; 2002).
A exuberância da Floresta Atlântica sobre a planície costeira do Estado do
Espírito Santo, principalmente na região norte, foi descrita por pesquisadores que

16
aqui empreenderam expedições científicas no passado, como Saint-Hilaire em 1819
(Saint-Hilaire, 1974). Atualmente os maiores remanescentes vegetais dessa região
estão restritos às Reserva Natural da Vale do Rio Doce e Reserva Biológica de
Sooretama, que juntas chegam próximo dos 45.000 ha de floresta contínua,
constituindo um rico patrimônio genético.
Dentre as comunidades vegetais que recobriam o território do Estado do
Espírito Santo, a floresta aluvial é uma das menos representativas em termos de
área, localizada principalmente ao longo das margens do Rio Doce, integrando o seu
delta, considerado como área prioritária para conservação (MMA, 2002).
Sobre solos aluviais e relevo plano, com ocorrência de algumas espécies de
ampla distribuição geográfica, como Virola surinamensis Warb., Tapirira guianensis
Aubl., Garcinia brasiliensis e no sub-bosque com palmeiras dos Gêneros Mauritia e
Euterpe.
Muitas destas áreas são adequadas ao cultivo de cacau, que foi plantado em
cerca de 17.000 ha, na maioria dos casos sob o dossel raleado da floresta aluvial
“cabruca”. Prática esta, que segundo Rolim e Chiarello (2004), alterou a estrutura da
comunidade, ocasionando diminuição da diversidade e da riqueza de espécies, além
disso, no processo de manutenção do cultivo do cacaueiro é realizado o controle da
regeneração impedindo que a floresta se recupere.
Pouco se conhece sobre a composição florística e estrutura das florestas em
solos aluviais no Estado do Espírito Santo (Rolim e Chiarelo, 2004; Rolim et al., no
prelo). Outras comunidades vêm sendo pesquisadas com mais intensidade com
relação à florística e estrutura que se encontram principalmente no entorno da
FLONA de Goytacazes, como as produzidas principalmente no Estado do Espírito
Santo, norte Fluminense, sul da Bahia e Pernambuco (vide capítulo 1).
A devastação acentuada nos ecossistemas naturais da região Sudeste e,
principalmente no Estado do Espírito Santo, é também um fato a ser considerado,
sendo nesse caso necessário estudo da composição de sua vegetação, fornecendo
base para conservação de áreas com remanescentes vegetais e com potencial para
estabelecimento de corredores ecológicos, assim como para recomposição de
ambientes degradados.
Este trabalho teve por objetivos: descrever a estrutura e a composição
florística da Floresta Aluvial da FLONA de Goytacazes, realizar sua classificação
quanto à mudança foliar e compará-la com outras do norte do estado do Rio de

17
Janeiro, Espírito Santo e Pernambuco (parte do Sudeste e Nordeste) e testar a
seguinte hipótese: a composição florística e estrutural do estrato arbóreo da
Floresta Aluvial madura e sem inundação (em bom estado de conservação) da
FLONA de Goytacazes são similares às existentes na Floresta das Terras Baixas e
dissimilares da Floresta de Mussununga, da Floresta Seca de Restinga e da
Floresta Atlântica Montana, abrangendo o norte do estado do Rio de Janeiro,
Espírito Santo e Pernambuco (parte do Sudeste e Nordeste).
2.2. Material e métodos
2.2.1. Área de estudo – vide capíulo 1
Sítios 1 e 2
As duas áreas com 50 ha foram selecionadas em função das características
próximas de fitofisionomia, solo, profundidade do lençol freático e coleta seletiva de
madeira. Utilizaram-se duas áreas na mesma floresta com o intuito de compará-las e
aferir as semelhanças ou não de estrutura e florística.
Estes sítios (figura 2.2.1-1), encontram-se recobertos por floresta madura,
formando um único fragmento florestal. Inseridos em região de coleta seletiva de
madeira (Dalbergia nigra Allem. ex Benth. – jacarandá; Manilkara bella Monachino -
parajú e Melanoxilon brauna Schott - braúna) ocorrida até o final da década de 1970
e, de extração de palmito (Polyandrococos caudescens (Mart.) Barb. Rodr.) –
palmito de brejaúba (Astrocarium aculeatissimum (Scott) Burret), para alimentação
humana até 2002. Entretanto, nas áreas (sítios) de instalação das parcelas não foi
observada a presença de tocos remanescentes (com exceção de palmito amargoso
e brejaúba), apresentando as características de floresta madura, como presença de
árvores emergentes com mais de 40 m de altura, cipós com diâmetro a altura do
solo superior a 10 cm, sub-bosque bem definido (estratificado) e serrapilheira
espessa e contínua.

18
Figura 2.2.1-1. Fotografia aérea da FLONA de Goytacazes (centro) em 2000, com os
dois sítios (S1 – sítio1 e S2 – sítio2), Linhares/ES. Fonte: MAPLAN.
2.2.2. Levantamento florístico e estrutural
A FLONA de Goytacazes foi mapeada através de fotografias aéreas e nesta
foram delimitadas duas áreas de 50 ha na floresta madura, distantes 2 km entre si.
Em cada área denominadas de sítio1 (S1) e sítio2 (S2) (figura 2.2.1-1), foram
sorteadas cinco parcelas de 20 x 50 m. Para amostragem das árvores, em abril de
2004 foram medidos os diâmetros à altura do peito (DAP) dos indivíduos com 10 cm
ou mais e sua altura. Foram contabilizadas as lianas com diâmetro mínimo de 0,5
cm que se apoiavam nas árvores a 1,30 m do solo.
As coletas botânicas foram efetuadas durante dois anos, ao longo das áreas
das parcelas e em seu entorno. O material botânico foi coletado com cerca de cinco
ramos férteis e/ou estéreis de cada espécie, retirados com auxílio de podão de
ramos altos e escalada de árvores. Todo material botânico foi depositado no
Herbário UENF do Centro de Biociências e Biotecnologia (CBB) da Universidade
Estadual do Norte Fluminense (UENF) e duplicata dos férteis enviados para o
Herbário (VIES) da Universidade Federal do Espírito Santo.
S1
S2
Br101
Rio Doce
Alagado
FLONA de Goytacazes
N↑
750 m

19
A identificação do material botânico coletado foi realizada por meio de
literatura especializada, por especialistas nos grupos e por comparação e consulta
em Herbários (Companhia Vale do Rio Doce em Linhares/ES, CVRD; Museu de
Biologia Professor Mello Leitão em Santa Teresa/ES, MBML; Universidade Federal
do Espírito Santo em Vitória/ES, VIES; e no da Universidade Estadual do Norte
Fluminense em Campos dos Goytacazes/RJ, UENF).
Foi adotado para famílias o sistema de Cronquist (1968), com exceção das
Leguminosae, que seguiu Polhill et al. (1981), separando as espécies nas
subfamílias, mas na análise dos dados considerou-se a família Leguminosae. Para
a categorização das espécies ameaçadas de extinção, seguiu-se Espírito Santo
(2005) e IBAMA (1992).
As espécies quanto às mudanças foliar (caducifolia) foram classificadas
conforme observações pessoais de José Manoel Lúcio Gomes e Domingos Folli em
observações ao longo de três anos por ocasião das coletas quinzenais registrando a
presença ou ausência de folhas.
Utilizou-se também os resultados obtidos por Engel (2001), que realizou uma
pesquisa na Floresta das Terras Baixas na Reserva Natural da Vale do Rio Doce
distante cerca de 35 km da FLONA de Goytacazes. Esta pesquisa versou sobre a
fenodinâmica com 41 espécies arbóreas por período de 10 anos (1982-1992), com
frequência quinzenal, observando presença ou ausência de folhas nos indivíduos.
Assim, as espécies foram classificadas de acordo com os seguintes critérios:
espécies decíduas – perdem as folhas no fim da estação seca e brotam na chuvosa;
brevidecíduas – a queda das folhas ocorre ao mesmo tempo em que o rebento das
gemas e emissão de folhas novas, ficando totalmente desfolhada por no máximo
uma semana, ou parcialmente desfolhada por um período maior; sempre verdes – a
queda das folhas ocorre bem após a brotação, ou então a formação e perda das
folhas são contínuas ao longo do ano. Além de informações obtidas na literatura
como (Lorenzi, 1992; 1998).
A profundidade do lençol freático na região da FLONA foi estimada através de
observações de poços existentes e afloramentos do lençol freático na área de
estudo e em seu entorno.

20
2.2.3. Análise dos dados
A análise dos dados fitossociológicos foi baseada em (Mueller-Dombois e
Ellenberg, 1974) através do programa Fitopac (Shepherd, 1994) e Floresta Nativa
(Cientec, 2005), considerando a Freqüência Absoluta (FAi = (Pi / P) . 100),
Freqüência Relativa [FR = (FAi / ∑ FA) . 100], Densidade Absoluta (DA = ni/A),
Densidade Relativa [DR = (ni / N) . 100], Dominância Absoluta (DoA = ABi / A),
Dominância Relativa (DoR = ABi / AB), Valor de Importância (VI = DR + FR + DoR),
Valor de Cobertura (DR + DoR). Onde,
Pi = número de parcelas onde ocorre uma determinada espécie
P = número total de parcelas da amostragem
FAi = Freqüência absoluta de uma determinada espécie
ni = Número de indivíduos de uma determinada espécie
A = Área amostrada em m2
N = Número de indivíduos de todas as espécies presentes na amostragem
ABi = Área basal por hectare, considerando a fórmula para cálculo área basal de um
círculo (AB = D2. π / 4)
AB = Área basal total
D = Diâmetro do tronco de cada indivíduo, medido à altura do solo
Para o cálculo da diversidade (H’) utilizou-se o índice de Shannon, foi
calculada também a diversidade máxima (H’máx) e eqüidade ou uniformidade de
Pielou (J’), (Brower e Zar, 1977; Gotelli e Entsminger, 2001).
H’= - ∑ pi Ln pi, onde:
pi = ni/N
H’máx = Ln S
J’= H’/ H’máx
Onde:
S = Número total de espécies da amostra
N = Número total de indivíduos ou cobertura total
ni = Número de indivíduos da espécie i ou valor de cobertura da espécie i
Ln = Logaritmo natural
Para comparação com outros estudos foi realizada a análise qualitativa da
similaridade de espécies utilizando o índice de Jaccard e para comparação entre as
parcelas foi utilizada a análise de similaridade quantitativa de Morisita (Im) modificado

21
de Horn 1966 (Brower e Zar, 1977). E para comparação dos dados estruturais entre
os dois sítios utilizou-se o teste t (Zar, 1996).
Jaccard:
IJ= 2C/A+B+C
Onde:
C= número de espécies comuns às comunidades A e B;
A= número total de espécies na comunidade A;
B= número total de espécies na comunidade B.
Morisita (mod. Horn):
IM= 2*∑Xi*Yi/(A+B)NxNy
Onde:
A= Xi(Xi-1)/Nx(Nx-1);
B= Yi(Yi-1)/Ny(Ny-1);
Xi= número de indivíduos da espécie i na comunidade X;
Yi= número de indivíduos da espécie i na comunidade Y;
Nx= número total de indivíduos na comunidade X;
Ny= número total de indivíduos na comunidade Y.

22
2.3. Resultados
As duas áreas quando comparadas quanto às espécies, apresentaram
índices de similaridade de 0,39 (Jaccard) e 0,56 (Morisita modificado de Horn, 1966).
Quanto ao número de indivíduos vivos, número de espécies, número de árvores
mortas em pé, área basal por hectare, número de indivíduos perfilhados, índice de
diversidade de Shannon e eqüidade de Pielou, não houve diferença estatística entre
os valores médios das matas (teste t, P < 0,05) para um mesmo parâmetro (tabela
2.3-1).
Tabela 2.3-1. Número de indivíduos (No Ind.), espécies (N.spp.), número de
indivíduos mortos, área basal por hectare (AB.ha-1), perfilhadas (P), altura média
(Alt. M.), diversidade (H’) e eqüidade (J’), por parcela amostrada no sítio1 e sítio2,
FLONA de Goytacazes, Linhares/ES. Não houve diferença estatística entre os
valores médios dos sítios da floresta (teste t, P < 0,05) para um mesmo parâmetro. Parcelas No Ind. N.spp. Mortos AB.ha-1(m2) P Alt. M.(m) H’ J’
1 45 26 2 20,65 1 17 3,05 0,94
2 67 41 3 35,58 2 19 3,51 0,95
3 62 40 2 33,62 0 18 3,50 0,95
4 62 43 0 30,31 0 18 3,67 0,98
5 58 40 3 28,09 0 17 3,58 0,97
Média ± DP 58,8±8,3 38±7 2±1 29,6±5,8 0,6±0,9 18±0,8 3,46±0,2 0,96±0,02
Total– sítio1 294 109 10 _ 3 - 4,30 0,92
6 56 25 3 29,03 0 15 2,79 0,87
7 54 34 4 34,33 1 14 3,12 0,88
8 61 38 6 33,95 0 15 3,26 0,90
9 59 29 6 26,37 1 14 2,82 0,84
10 57 31 0 26,94 0 13 2,96 0,86
Média ± DP 57,4±2,7 31±5 4±2 30,1±3,8 0,4±0,5 14±0,9 3±0,19 0,9±0,02
Total– sítio2 287 109 19 _ 2 - 3,83 0,82
2.3.1. Estrutura da floresta
O número de indivíduos amostrados por parcela variou de 45 a 67, a área
basal por hectare de 20,65 a 35,58 m2 (média de 30) e a altura média por parcela de
13,26 a 18,80 m (tabela 2.3-1). A maior densidade de árvores ocorreu na classe
diamétrica de 10-15 cm (figura 2.3.1-1). Foi amostrado um baixo número de árvores
perfilhadas (n = 5), perfazendo 0,9 % do total de indivíduos (tabela 2.3-1).

23
A distribuição dos indivíduos em classes diamétricas incluindo todas as
árvores amostradas (figura 2.3.1-1), apresentou um padrão de distribuição do tipo “J”
reverso, indicando ocorrência de regeneração.
Foi encontrado um total de 29 indivíduos mortos e em pé nas 10 parcelas (1
ha), correspondendo a 5 % da densidade total (tabela 2.3-1).
020406080
100120140160180
12,5 17,5 22,5 27,5 32,5 37,5 42,5 47,5 52,5 > 57,5
Centros de classes diamétricas
Núm
ero
de in
diví
duos Sítio1
Sítio2
Figura 2.3.1-1. Distribuição do número de indivíduos por classe diamétrica (cm), nos
sítios1 e 2, FLONA de Goytacazes, Linhares/ES.
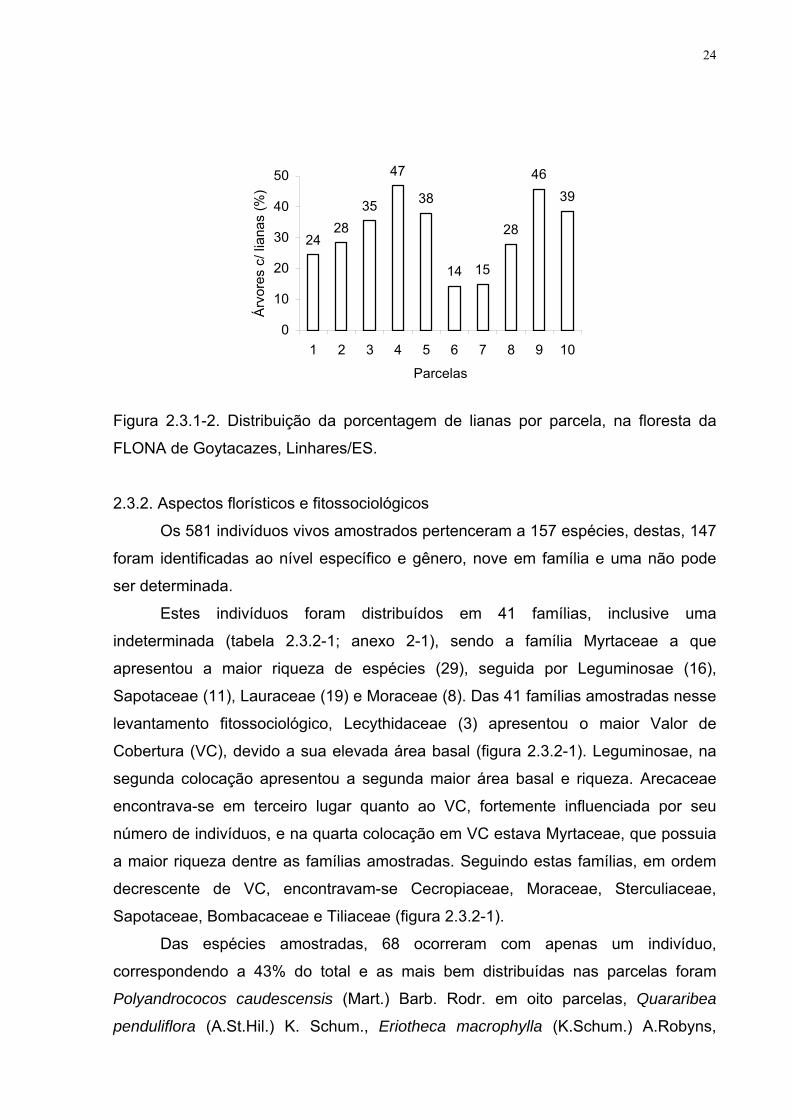
A porcentagem de árvores com lianas por parcela, variou de 14 a 47% (figura
2.3.1-2), média de 32%. Dentre as espécies com maior número de lianas,
destacaram-se Licania salzmannii (Hook. f.) Fritsch com 25, Ficus mariae C.C. Berg,
Emydio e Carauta e Glycydendron amazonicum Ducke com 18, além de Lecythis
pisonis Cambess. com 15. Todas estas árvores adultas, com DAP de 55, 25, 40 e 35
cm e altura de 28, 25, 15 e 35 m, respectivamente.
Considerando a existência de semelhança entre os dois sítios de uma mesma
floresta, eles foram tratados como uma única área para os parâmetros estruturais e
florísticos.
24
2428
35
47
38
14 15
28
4639
0
10
20
30
40
50
1 2 3 4 5 6 7 8 9 10
Parcelas
Árv
ores
c/ l
iana
s (%
)
Figura 2.3.1-2. Distribuição da porcentagem de lianas por parcela, na floresta da
FLONA de Goytacazes, Linhares/ES.
2.3.2. Aspectos florísticos e fitossociológicos
Os 581 indivíduos vivos amostrados pertenceram a 157 espécies, destas, 147
foram identificadas ao nível específico e gênero, nove em família e uma não pode
ser determinada.
Estes indivíduos foram distribuídos em 41 famílias, inclusive uma
indeterminada (tabela 2.3.2-1; anexo 2-1), sendo a família Myrtaceae a que
apresentou a maior riqueza de espécies (29), seguida por Leguminosae (16),
Sapotaceae (11), Lauraceae (19) e Moraceae (8). Das 41 famílias amostradas nesse
levantamento fitossociológico, Lecythidaceae (3) apresentou o maior Valor de
Cobertura (VC), devido a sua elevada área basal (figura 2.3.2-1). Leguminosae, na
segunda colocação apresentou a segunda maior área basal e riqueza. Arecaceae
encontrava-se em terceiro lugar quanto ao VC, fortemente influenciada por seu
número de indivíduos, e na quarta colocação em VC estava Myrtaceae, que possuia
a maior riqueza dentre as famílias amostradas. Seguindo estas famílias, em ordem
decrescente de VC, encontravam-se Cecropiaceae, Moraceae, Sterculiaceae,
Sapotaceae, Bombacaceae e Tiliaceae (figura 2.3.2-1).
Das espécies amostradas, 68 ocorreram com apenas um indivíduo,
correspondendo a 43% do total e as mais bem distribuídas nas parcelas foram
Polyandrococos caudescensis (Mart.) Barb. Rodr. em oito parcelas, Quararibea
penduliflora (A.St.Hil.) K. Schum., Eriotheca macrophylla (K.Schum.) A.Robyns,

25
Eschweilera ovata (Cambess.) Miers., (amostradas em sete parcelas) e Hydrogaster
trinervis Kuhlm., Cecropia glaziovi Snethl., Joannesia princips Vell., Virola gardneri
(A.DC.) Warb., Eugenia ubensis Camb. e Schoepfia oblongifolia Turez, ocorreram
em seis parcelas e o restante das espécies em cinco ou menos (tabela 2.3.2-1).
Entre as espécies, Polyandrococos caudescens ocorreu com o maior Valor de
Importância (VI), influenciado pelo elevado número de indivíduos, seguido por
Eschweilera ovata e Hidrogaster trinervis (anexo 2-1). As 25 primeiras espécies em
VI somaram 146 e as 132 restantes 154.
Nas amostragens foram detectadas ainda oito espécies ameaçadas de
extinção. Destas, seis foram registradas ocorrendo com apenas um indivíduo
amostrado (Terminalia kuhlmannii - em perigo, Beilschmiedia linharensis - em perigo,
Ocotea confertiflora - vulnerável, Moldenhawera papillanthera - em perigo, Dalbergia
nigra - vulnerável, Mollinedia marquetiana - vulnerável, e duas espécies
(Campomanesia espiritosantensis – em perigo e Myrcia folli - vulnerável), ocorrendo
com dois e oito indivíduos, respectivamente (tabela 2.3.2-1).
Dentre as 157 espécies amostradas seis foram consideradas brevidecíduas
com 32 indivíduos e nove foram decíduas com 16 indivíduos. Totalizando 15
espécies e 48 indivíduos dos 581 amostrados, as outras 142 espécies e 533
indivíduos foram sempre-verdes (tabela 2.3.2.1).
Tabela 2.3.2-1. Distribuição das espécies e famílias amostradas nos dois sítios (S1 e
S2) com o número de parcelas que ocorreram, caducifolia - C (D – decídua, BD –
brevidecídua; SV – sempre verde e ameaçadas de extinção***, (Decreto Estadial no
1.499-R de 14/06/05 e Portaria no 37-N/IBAMA, de 3/04/02), FLONA de Goytacazes,
Linhares/ES.
Família/Espécie C S1 S2 Família/Espécie C S1 S2ANACARDIACEAE MALPIGHIACEAE Astronium concinnum (Engl.) Schott BD 3 1 Byrsonima stipulacea (Juss.) Nied. SV 1 - Astronium graveolens Jacq. D - 1 MELASTOMATACEAE Spondias venulosa Mart. Ex Engl. SV - 1 Miconia hypoleuca (Benth.) Triana SV 1 - Tapirira guianensis Aubl. SV 2 1 MELIACEAE ANNONACEAE Guarea guidonia (L.) Sleumer SV 3 - Duguetia flagellaris Huber. SV 3 1 Trichilia casaretti C.DC. SV 1 - Oxandra nitida R.E.Fries SV - 1 Trichilia lepidota Mart. SV 1 1 Oxandra sp.1 SV 4 1 Trichilia quadrijuga H.B. & K. SV 3 1 Rollinia laurifolia Schltdl. SV 2 1 Trichilia aff. surinamensis C.DC. SV - 1 Xylopia laevigata (Mart.) R. E. Fries SV 2 - MONIMIACEAE APOCYNACEAE ***Mollinedia marquetiana Peixoto SV - 1

26
Tabela 2.3.2-1 (continuação1) Família/Espécie C S1 S2 Família/Espécie C S1 S2
Aspidosperma illustre (Vell.) Kuhlm. e Piraja SV - 1 MORACEAE
Aspidosperma pyricollum Muell. Arg. SV 1 1 Brosimum glaucum Taub. SV 2 -
ARECACEAE Brosimum guianense (Aubl.) Huber SV - 2
Astrocaryum aculeatissimum (Schott) Burret SV - 1 Brosimum lactescens (S. Moore)
C.C. Berg SV 1 2
Polyandrococos caudescens (Mart.) Barb. Rodr. SV 3 5 Brosimum sp.1 SV 2 2
BIGNONIACEAE Ficus mariae C.C. Berg, Emydio e Carauta D - 2
Tabebuia heptaphylla (Vell.) Toledo D 1 1 Maclura tinctoria (L.) Don. ex Steudel SV 1 -
Tabebuia roseo-alba (Ridley) Sandwith D - 1 Naucleopsis oblongifolia (Kuhlm.)
Carauta SV 1 1
BOMBACACEAE Sorocea guilleminiana Gaudich. SV 2 3
Chorisia glaziovii (Kuntze) E.Santos D - 1 MYRISTICACEAE Eriotheca macrophylla (K. Schum.) A. Robyns BD 3 4 Virola gardneri (A.DC.) Warb. SV 3 3
Pseudobombax grandiflorum (Cav.) A. Robyns D 1 2 Virola oleifera (Schott) A. C. Smith SV 2 -
Quararibea penduliflora (A.St.Hil.) K. Schum. SV 5 2 MYRTACEAE
BORAGINACEAE Calyptranthes sp.2 SV - 1
Cordia sellowiana Cham. SV 1 - ***Campomanesia espiritosantensis Landrum SV 1 1
BURSERACEAE Campomanesia guazumifolia (Cambess.) O.Berg SV 1 1
Protium heptaphyllum (Aubl) March. SV 1 1 Eugenia brasiliensis Lam. SV 1 2
CAPPARACEAE Eugenia excelsa Berg SV - 1
Capparis flexuosa (L.) L. SV - 1 Eugenia itapemirinensis Camb. SV - 1
CECROPIACEAE Eugenia microcarpa Berg SV 3 -
Cecropia glaziovi Snethl. SV 4 2 Eugenia ubensis Camb. SV 3 3
Coussapoa curranii Blake SV 1 - Eugenia sp.1 SV - 2
Pourouma velutina Miq. SV 3 - Eugenia sp.2 SV - 1
CELASTRACEAE Eugenia sp.3 SV 1 1
Maytenus sp.1 SV - 2 Marlierea grandifolia Berg SV 2 1
CHRYSOBALANACEAE Marlierea obversa Legrand. SV 1 -
Hirtella burchellii Britton SV 3 1 Marlierea regeliana Berg SV - 1
Hirtella sp.1 SV - 1 Marlierea sylvatica (Gardner) Kiaersk. SV - 1
Licania salzmannii (Hook. f.) Fritsch SV 3 - Marlierea sp.1 SV 1 -
Licania sp.1 SV - 1 Myrcia falax (Richard) DC. SV 1 -
Chrysobalanaceae sp.1 SV 1 1 ***Myrcia follii Barroso et Peixoto SV 3 2
CLUSIACEAE Myrcia cf. subrugosa M.Sobral SV - 1
Garcinia brasiliensis Mart. SV - 1 Myrcia sp.1 SV 1 - Garcinia gardneriana (Triana & Planch.) D.C. Zappi SV 1 - Myrciaria jaboticaba (Vell.) Berg SV 1 1
Garcinia sp.1 SV 2 - Plinia aff. grandiflora (Matos) Sobral SV - 1
COMBRETACEAE Myrtaceae sp.1 SV 1 -

27
Tabela 2.3.2-1 (continuação2) Família/Espécie C S1 S2 Família/Espécie C S1 S2
***Terminalia kuhlmannii Alwan & Stace D 1 - Plinia aff. grandiflora (Mattos)
Sobral SV 1 1
ELAEOCARPACEAE Myrtaceae sp.5 SV - 1
Sloanea eichleri K.Schum. SV 1 - Myrtaceae sp.6 SV - 1
ERYTHROXYLACEAE Plinia involucrata (Berg) McVaugh. SV 1 -
Erythroxylum columbinum Mart. SV - 1 Plinia rivularis (Cambess.) Rotman SV 2 - Erythroxylum plowmanianum A. Cogollo P. e J.J. Pipoly SV - 1 Psidium sp.1 SV - 1
Erythroxylum pulchrum A. St. Hil. SV 1 1 NYCTAGINACEAE
Erythroxylum sp.2 SV - 1 Andradaea floribunda Allemao SV 2 1
EUPHORBIACEAE Guapira opposita (Vell.) Reitz SV 1 2
Actinostemon estrellensis Pax SV 1 - Guapira subferruginosa (Mart. Ex Schum.) Lundell SV - 2
Glycydendron amazonicum Ducke SV 1 1 OLACACEAE
Joannesia princeps Vell. BD 3 3 Schoepfia oblongifolia Turez SV 5 1
Paradrypetes ilicifolia Kuhlm. SV - 1 PHYTOLACCACEAE Pausandra morisiana (Casar.) Radlk. SV - 1 Gallesia integrifolia (Spreng.)
Harms. SV 2 -
Pera leandri Baill. SV 1 - RHAMNACEAE SV
Senefeldera multiflora Mart. SV - 1 Ziziphus glaviovii Warm. SV 1 1
FLACOURTIACEAE RUBIACEAE Carpotroche brasiliensis (Raddi.) A. Gray SV - 1 Alseis floribunda Schott. D* 1 -
Casearia sp.1 SV 1 1 Simira sampaioana (Standl.) Steyerm SV - 2
INDETERMINADA Rubiaceae sp.1 SV - 1
Indeterminada sp.1 SV 1 1 LAURACEAE RUTACEAE
Aniba sp.1 SV - 1 Neoraputia alba (Nees e Mart.) Emmerich SV - 2
***Beilschmiedia linharensis Nishida SV 1 - SAPINDACEAE
Cryptocarya aschersoniana Mez SV 1 - Allophylus petiolulatus Radlk. SV 1 1
Licaria bahiana Kutz SV 1 - Cupania scrobiculata L.C. Rich. SV 2 3
Ocotea aciphylla (Nees) Mez SV 3 - Cupania zanthoxyloides Camb. SV - 1
Ocotea cernua Mez SV 1 - Talisia intermedia Radlk. SV 2 2
Ocotea conferta Coe Teixeira SV 2 1 SAPOTACEAE
***Ocotea confertiflora (Meisn.) Mez SV 1 - Chrysophyllum lucentifolium Cronquist SV 1 -
Lauraceae sp.1 SV 1 1 Chrysophyllum splendens Spreng. SV 3 1
LECYTHIDACEAE Chrysophyllum sp.1 SV 1 -
Cariniana legalis (Mart.) Kuntze. BD 3 2 Ecclinusa ramiflora Mart. SV 1 1 Eschweilera ovata (Cambess.) Miers. SV 5 2 Micropholis crassipedicellata
(Mart. & Eichler.) Pierre SV 1 -
Lecythis pisonis Cambess. BD 1 2 Micropholis sp.1 SV 1 -
LEG. CAESALPINIOIDEAE Pouteria bangii (Rusby) T.D.Pennington SV 3 -
Copaifera langsdorffii Desv. SV 1 2 Pouteria bullata (S.Moore) Baehni SV 1 -
Dialium guianense (Aubl.) Sandwith SV 1 4 Pouteria coelomatica Rizzini SV 3 1
Hymenaea rubiflora Ducke SV 1 1 Sapotaceae sp.1 SV 4 -

28
Tabela 2.3.2-1 (continuação3) Família/Espécie C S1 S2 Família/Espécie C S1 S2
***Moldenhawera papillanthera L.P.Queiroz, G.P.Lewis & R.Allkim SV 1 - Sapotaceae sp.2 SV 1 -
Tachigalia paratyensis (Vell.) H.C.Lima SV 1 - SIMAROUBACEAE
LEG. FABOIDEAE Simaruba amara Aubl. SV 2 1
Andira fraxinifolia Benth. SV 1 1 Simaba subcymosa A. St. Hil. Til. SV - 4 ***Dalbergia nigra (Vell.) Allemao ex Benth. BD - 1 STERCULIACEAE
Exostyles venusta Schott ex Spreng. SV - 1 Guazuma crinita Mart. SV - 1
Pterocarpus rohrii Vahl. SV 3 1 Pterygota brasiliensis Fr. All. SV 1 2 Vatairea heteroptera (Allem.) Ducke ex de Assis Iglesias D - 2 Sterculia elata Ducke SV 1 2
Zollernia latifolia Benth. SV 1 - TILIACEAE
Zollernia sp.1 SV - 1 Hidrogaster trinervis Kuhlm. SV 3 3
LEG. MIMOSOIDEAE ULMACEAE
Inga capitata Desv. SV 2 1 Amphelocera glabra Kuhlm. SV - 2
Inga flagelliformis (Vell.) Mart. SV 1 -
Inga striata Benth. SV 1 -
Inga thibaudiana DC. SV 1 -
0 5 10 15 20 25 30 35
Lecythidaceae
Leguminosae
Arecaceae
Myrtaceae
Cecropiaceae
Moraceae
Sterculiaceae
Sapotaceae
Bombacaceae
Tiliaceae
Outras (n=31)
Fam
ílias
Valor de cobertura (%)
Figura 2.3.2-1. Distribuição das dez famílias com maior Valor de Cobertura nos
sítios1 e 2 estudados, FLONA de Goytacazes, Linhares/ES.
2.3.3. Diversidade, riqueza e similaridade de espécies
O índice de diversidade de Shannon encontrado para a amostragem geral,
incluindo as dez parcelas (1 ha), foi de 4,38 nats/indivíduo e valor de eqüidade de
0,87 nats/indivíduo. Entre as parcelas, os valores de diversidade de espécies
variaram de 2,82 a 3,67 nats/indivíduo, com os valores do índice de similaridade de

29
espécies arbóreas entre parcelas (Morisita modificado de Horn, 1966) variando entre
0,18 a 0,42 (figura 2.3.3-1).
O dendrograma obtido do agrupamento florístico entre 16 sítios de Floresta
Atlântica, como florestas montana, das terras baixas, Restinga e Mussununga nos
Estados do Espírito Santo, Rio de Janeiro e Pernambuco, indicou de uma maneira
geral baixa similaridade entre as áreas (figura 2.3.3-2), com formação de grupos de
acordo com o tipo de ambiente: um primeiro grupo com os dois sítios da Reserva
Natural da Vale do Rio Doce (Jesus, 2001; Rolim, 1997), juntamente com os dois
sítios da FLONA (presente estudo e Paquele, dados não publicados); um grande
grupo com as florestas montana do Espírito Santo e Rio de Janeiro (Guedes – Bruni,
1998; Rodrigues, 2004; Thomaz,1996); um grupo formado pelas duas áreas de
Restinga e Mussununga (Fabris, 1995; Assis et al., 2004; Simonelli, 1998) e por fim
um grupo com as florestas estacionais no Rio de Janeiro e Pernambuco (Carvalho -
dados não publicados; Silva e Nascimento, 2001; Andrade e Rodal, 2004). Os dados
obtidos para a floresta em estudo na FLONA de Goytacazes foi mais similar ao
grupo da Reserva Natural da Vale do Rio Doce – 24% (Jesus, 2001; Rolim, 1997) do
que uma outra área na da própria FLONA – 12% (Paquele, dados não publicados).
NF P
Figura 2.3.3-1 Dendrograma de similaridade (Morisita modificado de Horn, 1966 –
Brower e Zar, 1977) de espécies e indivíduos entre as parcelas amostradas no sítio1
(parcelas – P: 1,2,3,4,5) e sítio2 parcelas – P: 6,7,8,9,10), nível de fusão (NF),
FLONA de Goytacazes, Linhares/ES.

30
Localidade
Fonte
Tipo Florestal
Nível de Fusão
Linhares/ES
Jesus (2001)
ODTB 0,82
Linhares/ES Rolim (1997)
ODTB 0,24
Linhares/ES Presente estudo
ODA 0,12
Linhares/ES Paquele (dados não publicados)
ODA 0,09
Poço das Antas/RJ
Guedes–Bruni (1998)
ODTB 0,24
União/RJ Rodrigues (2004)
ODS 0,16
Paraiso/RJ Guedes–Bruni (1998)
ODS 0,10
Santa Teresa/ES Thomaz (1996) ODM 0,07
Macaé de cima/RJ Guedes–Bruni (1998)
ODM 0,12
Itatiaia/RJ Guedes–Bruni (1998)
ODM 0,05
Guarapari/ES Fabris (1995) R 0,38
Guarapari/ES Assis et al. (2004)
R 0,14
Linhares/ES Simonelli (1998) M 0,04
Campos dos Goytacazes/RJ
Carvalho (dados não publicados)
ESA 0,08
Campos dos Goytacazes/RJ
Silva e Nascimento (2001)
ESTB 0,06
São Lourenço da Mata/PE
Andrade e Rodal (2004)
ESTB 0,04
Figura 2.3.3-2 Dendrograma de similaridade (Jaccard) de espécies entre
amostragens realizadas em florestas no Espírito Santo, Rio de Janeiro e
Pernambuco. (Tipo Florestal: ESTB= Estacional Semidecidual das Terras Baixas,
ESA= Estacional Semidecidual Aluvial, ODTB= Ombrófila Densa das Terras Baixas,
ODA= Ombrófila Densa Aluvial, ODS= Ombrófila Densa Submontana, ODM=
Ombrófila Densa Montana, R= Restinga, M= Mussununga).

31
2.4. Discussão
2.4.1. Classificação da vegetação
No mapeamento da vegetação brasileira (IBGE, 1987; 1993), a vegetação no
delta do Rio Doce e inclusive da FLONA de Goytacazes, foi classificada como
Floresta Ombrófila Densa, apesar de terem sido observadas algumas espécies
decíduas. Além destes mapas, Thomas e Amorim (2005a), Oliveira-Filho e Fontes
(2000) e Oliveira-Filho et al. (2005), também as consideraram Ombrófila Densa.
Por outro lado, Rolim et al. (no prelo) consideraram a vegetação do delta
como Estacional Semidecidual. Engel (2001) analisou as fenofases de 41 espécies
arbóreas na Floresta das Terras Baixas da Reserva Natural da Vale do Rio Doce e a
classificou como Estacional Perenifólia, considerada um tipo transicional entre a
floresta Ombrófila Densa e Estacional Semidecídua (Veloso et al., 1991). Contudo,
ela não considerou a fitossociologia e a quantificação dos indivíduos com caducifolia
na comunidade.
Segundo Rizzini (1979), são considerados como meses secos do ponto de
vista ecológico o mês que apresentar menos de 60 mm de chuva, porém se um
determinado mês com menos de 60 mm for precedido de outro com mais de 100 mm
será contado como úmido. Neste sentido, são consideradas áreas úmidas, aquelas
com 0-3 meses secos por ano e estacionalmente secas com 4-5 meses secos. Já
Veloso et al. (1991), consideraram como estacional semidecidual florestas em
regiões com mais de dois meses com menos de 60 mm cada e caducifólia entre 20 e
50% dos indivíduos e para IBGE (1993) período seco de 90-120 dias e caducifólia
entre 20 e 50%.
As médias de precipitação pluviométrica dos últimos 30 anos, coletadas no
delta do Rio Doce (Estação de Povoação), indicaram dois meses com menos de 60
mm (figura 1.5.4-1) e nos últimos dez anos, fevereiro também aparece com menos
de 60 mm. Uma análise quali-quantitativa dos indivíduos caducifólios na amostragem
realizada (tabela 2.3.2-1), indicou que 9,5% das espécies perdem as folhas (3,8%
brevidecíduas e 5,7% decíduas) e 7% dos indivíduos perdem as folhas (5,5%
brevidecíduos e 2,7% decíduos), bem aquém do mínimo que deve ser acima de
20%.
Além disto, a floresta na área de estudo encontra-se sobre solo com lençol
freático com variação entre 1 e 3m de profundidade. O que facilita o acesso à água
por parte das raízes, além da percolação por capilaridade da água no perfil do solo

32
em direção à superfície (Molchanov, 1963), aumentando sua disponibilidade e
conseqüente absorção pelos vegetais e suprimento dos tecidos com maior teor de
água. Esta disponibilidade de água subterrânea pode estar influenciando também no
regime de troca de folhas.
Segundo Oliveira-Filho et al. (2005), a flora arbórea das florestas
semidecíduas é em boa parte um subconjunto da flora das florestas ombrófilas
selecionando espécies com maior capacidade de resistir à seca. Detectou também
que as duas fisionomias (florestas semideciduais e ombrófilas) encontram-se
próximas florísticamente dentro de uma mesma faixa latitudinal. Isto nos leva a crer
que a maior disponibilidade de água no solo, pode condicionar se a floresta é
semidecidual ou ombrófila.
Com base nestes resultados e no sistema de classificação de Veloso et al.
(1991), pode-se afirmar que a vegetação da FLONA de Goytacazes se encaixa em
Floresta Ombrófila Densa Aluvial.
2.4.2. Estrutura da floresta
Quando comparamos os valores de densidade total (581 ind.ha-1) e área
basal (30 m2.ha-1) encontrados na floresta aluvial da FLONA de Goytacazes com
outras florestas dos Estados do Espírito Santo, Rio de Janeiro e Minas Gerais
(tabela 2.4.2-1), observou-se grande variação nesses parâmetros, em função não
apenas da diferença entre as tipologias florestais mas também da metodologia
empregada em cada estudo. Os dados indicam uma tendência de maior densidade
de indivíduos com a elevação da altitude (Rodrigues, 2004; Moreno et al., 2003).
Considerando estudos realizados com a mesma metodologia (tamanho da
amostragem e critério de inclusão), observa-se que a FLONA de Goytacazes
apresenta densidade e área basal menor apenas que as áreas do Imbé e União, no
Rio de Janeiro (tabela 2.4.2-1). Os valores desses parâmetros na floresta estudada
ficaram muito próximos aos encontrados na Reserva Natural da Vale do Rio Doce,
representada pela Floresta Ombrófila Densa das Terras Baixas, apesar das
diferenças edáficas e geomorfológica entre as áreas.
A baixa pressão antrópica nos locais estudados foi evidenciada pelo pequeno
número de árvores perfilhadas (1% e 0,7% do total, nos sítios1 e 2,
respectivamente). Valores semelhantes foram encontrados para áreas de matas
maduras (Rodrigues, 2004 – 1,6%) e bem menores que os encontrados para áreas

33
secundárias (Rodrigues, 2004 - 16,9% em bordas; Carvalho, 2005 - 6,1 a 11,5%;
Pessoa, 2003 - 3 a 11,3%), neste último as ramificações surgiram após incêndio.
Tabela 2.4.2-1. Dados estruturais obtidos em alguns trabalhos realizados em
Floresta Atlântica (Tipo Florestal: ODTB = Ombrófila Densa das Terras Baixas, ODS
= Ombrófila Densa Submontana, ODM = Ombrófila Densa Montana, ODA =
Ombrófila Densa Aluvial, FV= Floresta de Várzea, ESA = Estacional Aluvial, ESTB =
Estacional das Terras Baixas, R = Restinga, M = Mussununga); Área = área
amostral em ha; DAP = diâmetro à altura do peito em cm; H’ = índice de Shannon;
N.spp. = número de espécies; Den = densidade por hectare; AB = área basal por
hectare em m2).
Localidade Fonte Tipo Florestal Área DAP H’ N.spp. Den AB
Guarapari/ES Fabris (1995) R 0,50 ≥ 4,8 3,70 96 3082 32,1Guarapari/ES Assis et al. (2004) R 1,00 ≥ 4,8 3,73 92 2106 27,5Imbé/RJ Moreno et al. (2003) ODS 0,60 ≥ 10,0 4,30 145 792 34,6Imbé/RJ Moreno et al. (2003) ODS 0,60 ≥ 10,0 4,21 125 767 41,9Itatiaia/RJ Guedes-Bruni (1998) ODM 1,00 ≥ 5,0 3,86 150 569 29,2Linhares/ES Simonelli (1998) M 0,93 ≥ 5,0 3,37 79 1643 23,1Linhares/ES Presente estudo ODA 1,00 ≥ 10,0 4,38 157 581 30,0Linhares/ES Rizzini (2000) FV 0,37 ≥ 6,3 4,15 139 1019 - Linhares/ES Rizzini (2000) ODTB 0,37 ≥ 6,3 4,26 146 1147 - Linhares/ES Jesus (2001) ODTB 2,50 ≥ 10,0 4,82 243 527,6 28,0Linhares/ES Rolim e Chiarelo (2004) ODA 4,80 ≥ 5,0 3,99 110 104 23,8
Linhares/ES Paquele (dados não publicados) ODA 1,20 ≥ 5,0 4,44 168 1116 36,6
Linhares/ES Paquele (dados não publicados) ODA 1,20 ≥ 10,0 - 150 541 34,4
Macaé de Cima/RJ Guedes-Bruni (1998) ODM 1,00 ≥ 5,0 4,08 119 510 26,3Madre Teresa de Deus/MG Vilela et al. (2000) ESA 0,63 ≥ 5,0 0,93 8 322 22,9Mata do Carvão/RJ Silva e Nascimento (2001) ESTB 1,00 ≥ 10,0 3,21 84 564 15,0Mergulhão/RJ Nascimento (não publicado) ODA 0,60 ≥ 10,0 3,81 69 465 18,2Paraíso/RJ Guedes-Bruni (1998) ODS 1,00 ≥ 5,0 4,43 130 438 48,8Poço das Antas/RJ Guedes-Bruni (1998) ODTB 1,00 ≥ 5,0 3,99 97 486 23,8Poço das Antas/RJ Guedes-Bruni (1998) ODS 1,00 ≥ 5,0 4,55 169 564 25,6Santa Teresa/ES Thomaz (1996) ODM 1,02 ≥ 6,3 5,50 476 2130 47,5União/RJ Rodrigues (2004) ODS 1,20 ≥ 10,0 4,90 208 735 32,0
A proporção de árvores mortas (5%) está pouco acima da indicada para
florestas maduras (ca. 3%) (Moreno et al., 2003) e abaixo de florestas secundárias
(9,75%; 11,1-32,7%) no Estado do Rio de Janeiro (Silva e Nascimento, 2001;
Carvalho, 2005) e esta mortalidade estaria relacionada à fatores como senescência,
doenças e queda por chuva e vento.
A porcentagem de árvores com lianas (DAP ≥ 0,5 cm) se apoiando em seu
tronco ou na copa (32%), foi mais alto que o encontrado em florestas maduras (13%
na Reserva Biológica União – Carvalho, 2005) e bem abaixo de florestas fortemente
fragmentadas no Norte Fluminense (79%; 84%; 64% e 94% - Carvalho, 2005),

34
indicando uma maior concentração nas áreas com vegetação perturbada. As lianas
são componentes importantes nas florestas tropicais, contribuindo para a
biodiversidade (Schnitzer e Bongers, 2002), em processos como no sequestro de
carbono (Schnitzer e Bongers, 2002) e alimento para a fauna, principalmente nas
épocas mais secas, produzindo flores e frutos comestíveis (Morellato, 1991). Por
outro lado, lianas em florestas fragmentadas podem competir por luz, provocar
danos nas copas, promover sombreamento em plântulas e devido ao excesso de
peso nas copas e às vezes com auxílio de ventanias provocar a queda e acelerar a
mortalidade de árvores (Putz, 1984; Schnitzer e Bongers, 2002). Segundo Souza et
al. (2002), o corte de cipós em uma Floresta Ombrófila Densa secundária favoreceu
a dinâmica da sucessão secundária, promovendo aumento da área basal e grande
elevação de ingresso de novos indivíduos no estrato arbóreo.
A distribuição dos indivíduos em classes diamétricas em forma de “J” reverso
(figura 2.3-1) segue o padrão encontrado em outras florestas tropicais (Assis et al.,
2004; Rodrigues, 2004; Jesus, 2001; Guedes-Bruni, 1998; Simonelli, 1998; Thomaz,
1996; Silva, 1980), com poucos indivíduos de grande porte diamétrico. A altura
média das árvores foi menor no sítio2, influenciada pelo grande número de
indivíduos de Polyandrococos caudescens.
2.4.3. Aspectos florísticos e fitossociológicos
As famílias com maior número de espécies na FLONA de Goytacazes
(Myrtaceae e Leguminosae) seguem o padrão de riqueza para a costa brasileira
mencionado por Mori et al. (1983) e Peixoto e Gentry (1990). Sapotaceae é indicada
por Gentry (1988) como uma das famílias de maior riqueza de espécies para
florestas neotropicais, o que foi observado em estudos no Estado do Espírito Santo,
em diferentes tipos de ambientes florestais (Assis et al., 2004; Peixoto e Gentry,
1990; Thomaz e Monteiro, 1997; Simonelli, 1998), que também indicaram
Sapotaceae dentre as famílias de maior riqueza, o que pode ser um padrão para a
Floresta Atlântica do Sudeste.
Segundo Peixoto e Gentry (1990), Lecythidaceae é característica da floresta
amazônica, e vem sendo demonstrada sua importância também na Floresta
Ombrófila Densa das Terras Baixas do Espírito Santo (Peixoto e Gentry, 1990;
Jesus, 2001), assim como nesse trabalho, comprovando mais uma vez a ampla
ocorrência de algumas famílias nas florestas tropicais.

35
Arecaceae é uma família comum nas florestas tropicais de solo úmido
(Emmons e Gentry, 1983) e destacou-se neste estudo pela elevada densidade de
Polyandrococos caudescens, espécie que ocorre em trechos da Floresta Atlântica de
baixada (Rizzini, 2000; Simonelli, 1998) e montana (Thomaz, 1996) e na Restinga
(Pereira e Araujo, 2000) do Estado do Espírito Santo, mas não é citada no
levantamento de Rodrigues e Nave (2001) para 43 áreas de matas ciliares extra-
amazônicas.
Da mesma forma Eschweilera ovata (Cambess.) Miers., a 2ª colocada em
valor de importância (VI) também não consta no levantamento de Rodrigues e Nave
(2001), ocorrendo na região amazônica e Floresta Atlântica de Pernambuco ao
Espírito Santo e Rio de Janeiro (Lorenzi, 1998). No Espírito Santo essa espécie
ocorre tanto em áreas de Floresta (tabuleiro) Ombrófila Densa das Terras Baixas
(Jesus, 2001) quanto em Mussununga (Simonelli, 1998) e Restinga (Pereira e
Araujo, 2000). Segundo Lorenzi (1992), tanto Polyandrococos caudescens quanto
Hidrogaster trinervis ocorrem na Floresta Atlântica da Bahia, Espírito Santo e Minas
Gerais e no delta do Rio Doce, mas Lopes et al. (2002) não as encontraram em seu
estudo no Parque Estadual do Rio Doce (MG), embora ocorram na região de
Linhares tanto na Floresta Ombrófila Densa das Terras Baixas (Jesus, 2001; Rizzini,
2000) como na Floresta Ombrófila Densa Aluvial na foz do Rio Doce (Rolim e
Chiarelo, 2004).
Nenhuma das dez principais espécies em VI neste estudo (Anexo 2-1) consta
na lista das mais freqüentes em florestas ciliares extra-amazônicas do levantamento
de Rodrigues e Nave (2001), demonstrando a peculiaridade florística deste tipo de
formação aluvial no Estado do Espírito Santo.
2.4.4. Diversidade, riqueza e similaridade de espécies
A Floresta Aluvial da FLONA de Goytacazes apresentou um dos maiores
índices de diversidade e riqueza de espécies quando comparada à estudos em
outros trechos de florestas da costa brasileira que utilizaram o mesmo critério de
inclusão (DAP ≥ 10 cm), ficando atrás apenas da Floresta Ombrófila Densa
Submontana da REBIO União no Rio de Janeiro (Rodrigues, 2004) e da Floresta
Ombrófila Densa das Terras Baixas (tabuleiro) da Reserva Natural da CVRD (Jesus,
2001), (tabela 2.4.2-1). Isto realça a importância de sua conservação, sobretudo pelo
tipo de ambiente em que está inserida, um dos poucos remanescentes de Floresta

36
Aluvial no vale do Rio Doce no Estado do Espírito Santo. A alta diversidade na
FLONA pode estar relacionada a proximidade com Floresta Ombrófila Densa das
Terras Baixas e as variações de ambientes existentes na área de estudo.
A formação dos grupos no dendrograma (figura 2.3.3-2) parece refletir
condições semelhantes de geomorfologia, solo e clima, como a união entre as áreas
da Floresta Ombrófila Densa Submontana e as da Restinga com a Mussununga.
A maior similaridade florística da floresta estudada na FLONA de Goytacazes
com a Floresta Ombrófila Densa das Terras Baixas da Reserva Natural da Vale do
Rio Doce (Jesus, 2001; Rizzini, 2000) do que com áreas da Floresta Ombrófila
Densa Aluvial próxima (Paquele, dados não publicados; Rolim e Chiarelo, 2004),
pode ser reflexo das variações entre a profundidade do lençol freático e sua
influência sobre a vegetação. Na FLONA de Goytacazes foi realizada extração
seletiva de madeira no passado, principalmente de Dalbergia nigra (Vell.) Allemao ex
Benth. (jacarandá da Bahia) para produção de móveis, Manilkara bella Monachino
(parajú) muito usada no engradamento de telhados e Melanoxylon brauna Schott.
(braúna) para utilização como postes de cerca nas fazendas. Estas duas últimas
espécies não foram amostradas, mas foram observados indivíduos adultos fora das
parcelas. M. brauna é considerada ameaçada de extinção, na categoria de
criticamente em perigo, no Estado do Espírito Santo (Decreto Estadual n0 1.499-R
de 14/06/05).
Outro aspecto observado na FLONA é a extração de palmito. Este fato foi
detectado tanto nas parcelas, quanto fora delas através de vestígios recentes de
corte de Polyandrococos caudescens (palmito amargoso) e de Astrocarium
aculeatissimum (Scott) Burret (brejaúba), para extração, comercialização e consumo
do palmito.
Também não foi detectada na amostragem nem observada fora das parcelas
amostradas a presença de Euterpe edulis Mart. (palmito doce), embora seja
normalmente comum em florestas tropicais, principalmente relacionadas a solos
úmidos e presente na Floresta Ombrófila Densa das Terras Baixas (tabuleiro) em
registros antigos para as margens do Rio Doce (Ruschi, 1950; Wied, 1989). Rolim et
al. (no prelo) em levantamento florístico em fragmentos da Floresta Ombrófila Densa
Aluvial e em cultivos de cacau pelo sistema de cabruca no delta do Rio Doce
também não encontraram esta espécie. A sua ausência pode estar relacionada à
grande coleta seletiva para consumo do palmito.

37
Os resultados indicam que a Floresta Ombrófila Densa Aluvial estudada da
FLONA de Goytacazes, embora tenha sofrido impactos de origem antrópica, ainda
detém uma alta riqueza e diversidade florística, com ocorrência de espécies raras e
ameaçadas de extinção. Carvalho et al. (2004), observaram que embora pequenos
fragmentos tenham perdido uma boa parte da diversidade de espécies arbóreas
antes existente, o conjunto de pequenos fragmentos em áreas com paisagem
fragmentada de Floresta Atlântica no norte fluminense mantêm ao final uma
importante diversidade e assim estes fragmentos devem ser conservados. A alta
diversidade e riqueza de espécies observadas para a FLONA de Goytacazes,
indicam a importância da manutenção e conservação dessas áreas florestadas
também no Estado do Espírito Santo.
Dada a baixa representatividade de Florestas Ombrófilas Densas Aluviais
como unidade de conservação restrita no Espírito Santo e a importância florística já
demonstrada para a FLONA de Goytacazes, sugere-se a mudança desta categoria
de unidade de conservação para uma categoria de uso mais restritivo como a de
Reserva Biológica.
2.5. Conclusões
Apesar de ter sofrido impactos antrópicos, a vegetação arbórea da FLONA de
Goytacazes (Floresta Ombrófila Densa Aluvial) é uma das mais ricas e diversas (H’)
da costa brasileira (Floresta Atlântica), além de exibir biomassa expressiva e conter
várias espécies vegetais ameaçadas de extinção. Foi confirmada a hipótese de que
composição florística e estrutural da vegetação da FLONA são similares às
existentes na Floresta das Terras Baixas e dissimilares da Floresta de Mussununga,
da Floresta Seca de Restinga e da Floresta Atlântica Montana.

38
CAPÍTULO 3
REGENERAÇÃO DE ESPÉCIES ARBUSTIVO-ARBÓREAS DA
FLORESTA ATLÂNTICA SOB VÁRIOS USOS DO SOLO NO NORTE
DO ESTADO DO ESPÍRITO SANTO 3.1. Introdução
A Floresta Atlântica dentro de uma visão global é considerada prioritária para
conservação (MMA, 2002; Myers et al., 2000) e a maior parte de seus
remanescentes encontram-se sob o domínio de propriedades privadas (Viana et al.,
1997), sendo que as pressões e distúrbios de origem antropogênica continuam
acontecendo (Oliveira et al., 1994), criando mosaicos de vegetação, que após o
abandono da exploração e do controle sobre a regeneração, tendem a evoluir para
estágios mais avançados de sucessão da floresta madura (Tabarelli et al., 1999).
A extração do pau-brasil foi a primeira forma de degradação sofrida pela
Floresta Atlântica, seguida de outros grandes ciclos econômicos como o da cana-de-
açúcar e o do café, além da mineração (Dean, 2000). Outro fator que potencializou a
devastação da Floresta Atlântica foi a instalação e respectivo crescimento das
cidades na sua faixa de ocupação, ocorrendo a supressão da vegetação em função
do inevitável avanço da malha urbana e dos núcleos industriais.
De acordo com Martins (2001), o processo de ocupação do Brasil
caracterizou-se pela falta de planejamento e conseqüente destruição de boa parte
dos recursos naturais, particularmente as florestas. As matas ciliares e aluviais não
escaparam, sendo até hoje alvo de ações antrópicas, em função principalmente das
atividades agropecuárias e do surgimento de muitas cidades às margens dos rios,
onde a vegetação deu lugar às edificações residenciais e industriais, pagando-se um
alto preço por isto, através das inundações constantes. Mesmo assim, apesar da
reconhecida importância ecológica, ainda mais evidente nesta virada de século, em
que a água é considerada o recurso mais importante para a humanidade, as
florestas ciliares e aluviais continuam sendo eliminadas ou sofrendo algum tipo de
agressão pelo homem.
Paiva e Gonçalves (2002) citam que nos últimos anos a visão ecológica
mudou muito a maneira de o homem perceber o meio ambiente. Desde uma atitude

39
denunciativa até uma atitude politicamente correta, o caminho trilhado vem
favorecendo positivamente as questões ligadas ao meio ambiente. Assim, a
paisagem passa a ser vista como uma interação de fatores envolvendo valores
ecológicos para uma melhor qualidade de vida, tendo o homem como elemento
principal. Neste contexto, a vegetação constitui um dos principais elementos na
melhoria da paisagem.
As florestas secundárias exercem em média três diferentes efeitos, que são
extraordinariamente importantes para o desenvolvimento da vegetação no processo
sucessional, o primeiro diz respeito à transferência dos nutrientes do solo para a
comunidade biótica, reduzindo sua perda; o segundo indica a melhoria da estrutura
edáfica pela produção de grande quantidade de matéria orgânica em forma de
serrapilheira e o terceiro tem haver com a modificação do microclima, que reduz a
flutuação térmica e aumenta a umidade relativa da atmosfera local. Estas mudanças
permitem o estabelecimento de espécies de etapas posteriores que substituem as
pioneiras nas comunidades (Vazquez-Yanes, 1980).
A regeneração em florestas na maioria das vezes está relacionada com a
ocupação das clareiras, causadas por algum tipo de perturbação, ocorrendo a partir
daí o processo de sucessão secundária (Gómez-Pompa e Burley, 1991). Assim, o
estrato regenerativo de uma floresta é composto das fases juvenis das espécies
(Passos, 1998). Neste sentido, em florestas tropicais Africanas, Swaine e Hall (1988)
consideraram que os juvenis encontravam-se com o diâmetro à altura do peito (DAP)
< 30 cm, porém, Fimbel e Fimbel (1996) também na África (Uganda), detectaram
que a maior concentração de indivíduos regenerantes se encontravam nos estratos
inferiores a 5 cm de DAP.
Vários autores como Buschbacher (1986), Aide et al. (1996), e Uhl et al.
(1988), verificaram que em áreas de pastagens abandonadas, nos primeiros dez
anos, a densidade de plantas lenhosas, área basal, número de espécies e a
diversidade foram baixos, a partir daí aumentaram rapidamente. Dentre os fatores
que podem influenciar a regeneração, destacam-se a distância da fonte de
sementes, a idade e a altitude, estes dois últimos influenciam diretamente a
composição de espécies. Após 40 anos de idade, os bosques em regeneração não
podem ser distinguidos de sítios inalterados em termos de densidade, área basal,
número de espécies e diversidade e parece que há necessidade de centenas de
anos para que estes sítios regressem a uma composição de espécies similar a área

40
original (Aide et al, 1996). Alguns autores, indicam que são necessários entre 140 a
200 anos para que as florestas tropicais desenvolvidas em áreas de cultivos
abandonados adquiram a fisionomia de floresta madura (Uhl e Murphy, 1981; Uhl e
Saldarriaga, 1986).
Áreas de floresta madura (ilhas de diversidade de árvores) próximas às áreas
a serem restauradas, funcionam como áreas fontes, doadoras de propágulos que
vão se estabelecendo conforme as condições físicas da área, às vezes dispensando
o plantio. Estas também podem ser pequenas áreas remanescentes ou plantadas,
com diferentes densidades e diversidades de espécies arbóreas. São úteis para
atrair dispersores de sementes das espécies presentes, assim como para trazer
propágulos de outras espécies de áreas florestais remanescentes que estejam
próximas às áreas que se quer restaurar, possibilitando a recolonização por diversas
espécies e o restabelecimento do fluxo gênico entre as populações arbóreas,
aumentando assim a biodiversidade, até mesmo a restauração da conectividade dos
fragmentos florestais (Kageyama et al., 2003). A importância de ilhas de vegetação
natural, como elementos para aumentar a heterogeneidade da paisagem em áreas
degradadas em regeneração, também foram ressaltados por Guevara et al., (1998).
Desta forma pode-se concluir que a recuperação de fragmentos florestais em
diferentes estágios de sucessão pode ser operada, ampliando a sua diversidade de
espécies vegetais, através da utilização de ilhas de vegetação – remanescentes ou
plantadas. Em locais onde existe a presença de fragmentos remanescentes de
floresta madura e em seu entorno após a desativação de uso agrícola, durante o
processo de regeneração pode ocorrer a migração das espécies existentes nas ilhas
para os vários estágios de sucessão em andamento.
Budowiski (1965) estudando florestas tropicais separou as espécies vegetais
quanto à sucessão em pioneiras, secundárias iniciais, secundárias tardias e
clímaxes. Propôs que as espécies pioneiras são sombreadoras, as clímaxes
sombreadas e as secundárias iniciais com papel de tutorar as secundárias tardias.
Assim, as pioneiras e secundárias iniciais são espécies de pleno sol, sendo as
primeiras com longevidade entre 5-15 anos e copa densa (sombreadora), as últimas
com vida mais longa de 30-50 anos e copa estreita. As secundárias tardias
desenvolvem-se à sombra parcial e as clímaxes estão adaptadas à sombra do
dossel. Posteriormente, Swaine e Whitmore (1988) classificaram as espécies
arbóreas quanto à sucessão em dois grupos ecológicos básicos: as pioneiras, que

41
surgem após perturbações que expõem o solo à luz, e as clímaxes que, embora
também tenham este comportamento, vivem mais que as pioneiras e
freqüentemente tornam-se grandes árvores emergentes. Dividiram ainda, o último
grupo em espécies clímaxes exigentes de luz e tolerantes à sombra.
Kageyama e Castro (1989) consideraram ainda que, os conceitos relativos à
tolerância a luz-sombra são básicos para as atividades de manejo. Nos plantios
mistos julgaram necessário delinear sistemas em que as árvores de diferentes
classes de tolerância tenham oportunidades iguais, cada uma em seu nicho
apropriado. Concluem que o reflorestamento misto deve ser composto por espécies
de diferentes estágios de sucessão, assemelhando-se à floresta madura. No caso de
algumas espécies não apresentarem comportamento esperado, pode estar
relacionado não apenas com a aplicação de conceitos teóricos de sucessão vegetal,
ainda não confirmados para determinadas espécies, como pode ser reflexo de um
comportamento diferenciado das mesmas, ou seja, de resultados mascarados por
fatores ligados a efeitos alelopáticos ou de competição interespecífica, estabelecidos
nos modelos impostos nos plantios de diferentes espécies (Barbosa et al., 1992).
A dispersão, dormência, sobrevivência de sementes e crescimento de
plântulas têm sido considerados essenciais para o sucesso do estabelecimento de
uma espécie vegetal (Piña-Rodrigues et al., 1990; Santos, 1991).
A germinação é considerada um dos pontos críticos das fases de crescimento
e desenvolvimento do indivíduo, sendo fortemente influenciada por fatores abióticos,
como luz, temperatura e umidade. O recrutamento via banco de sementes, de
plântulas e da rebrota de partes das plantas, estão relacionados à regeneração da
floresta (Mack et al., 1999; Martinez-Ramos, 1994).
Neste sentido, torna-se necessária a pesquisa do comportamento das
espécies ocorrentes em determinada região, procurando obter indicações que
poderão ser utilizadas na proposta de modelos relativos principalmente à
composição e proporção entre as espécies que serão utilizadas, como o estudo de
parâmetros como: distribuição vertical dos indivíduos (Svensson e Jeglum, 2001),
mudanças na composição e na riqueza de espécies ao longo do tempo (Garbutt e
Sparks, 2001).
A devastação acentuada nos ecossistemas naturais da região sudeste e,
principalmente no Estado do Espírito Santo, é fato, sendo nesse caso, necessários
estudos dos componentes de sua vegetação, fornecendo bases para conservação

42
de áreas com remanescentes vegetais e com potencial para estabelecimento de
corredores ecológicos, assim como para recomposição de ambientes degradados.
Esta pesquisa servirá como um registro da diversidade dos componentes
arbóreos da floresta madura e das espécies que participam dos processos de
sucessão natural em áreas cultivadas no passado, fornecendo base para ações de
manejo florestal na flora do Espírito Santo.
Este trabalho, teve por objetivo avaliar através do levantamento florístico, da
análise estrutural e da regeneração natural em áreas de floresta com distintos níveis
de perturbações, usos do solo e tempo de abandono, o estabelecimento de espécies
vegetais, fornecendo base para projetos de conservação, manejo e recuperação
deste tipo de vegetação. Partimos da hipótese de que a estrutura e a composição
florística da regeneração natural em três trechos desmatados para cultivo agrícola,
adjacentes ao remanescente de Floresta Atlântica (FLONA de Goytacazes), com
diferentes idades e uso do solo, difere entre si e da floresta adjacente.
3.2. Material e Métodos
3.2.1. Área de estudo – vide capítulo 1
Três áreas com diferentes usos do solo e tempo de abandono foram
selecionadas na FLONA de Goytacazes e denominadas da seguinte forma: capoeira
(sítio 1), cabruca (sítio 2) e capoeirinha (sítio 4), (figura 3.2.1-1B). Como área
controle foi selecionado um trecho com 50 ha de floresta madura (sítio 3) adjacente
a estes sítios.
Sítio 1. Capoeira
No início da década de 1970 foi desmatada uma área de floresta madura para
plantio de Theobroma cacao L. (cacau), sendo a madeira retirada e o restante do
material vegetal queimado no local. Com a desistência do plantio de cacau, a área
com cerca de 10 ha denominada de sítio 1 (figura 3.2.1-1A e B), foi abandonada na
mesma época (1971) e surgiu espontaneamente Panicum maximum Hochst. ex
A.Rich. (capim colonião), sendo utilizada por vários anos para pastoreio por gado
bovino, mas sem qualquer tipo de intervenção direta, principalmente o controle da
sucessão (comunicação pessoal de Vitalino Pereira, 2002), então a regeneração
natural de espécies arbustivo-arbóreas evoluiu para a capoeira.
Sítio 2. Cabruca

43
No início da década de 1980 foi aberta outra clareira deixando árvores
remanescentes e esparsas (cabruca), onde foram plantadas 1.300 mudas de cacau
da variedade Parazinho, abrangendo cerca de 2 ha, denominada de sítio 2 (figura
3.2.1-1A e B). Em 1994 os tratos culturais deixaram de ser efetuados, como por
exemplo, o corte das plantas em regeneração, estando a área em processo de
regeneração natural (comunicação pessoal de Agnaldo Lorenzutti, 2002).
Sítio 3. Floresta
O sítio 3 (figura 3.2.1-1B) encontra-se recoberto por floresta madura, inserida
em região de coleta seletiva de madeira (Dalbergia nigra Allem. ex Benth. –
jacarandá; Manilkara bella Monachino - parajú e Melanoxilon brauna Schott - braúna)
até o final da década de 1970, e de palmito de Polyandrococos caudescens (Mart.)
Barb. Rodr. - palmito amargoso e Astrocarium aculeatissimum (Scott) Burret –
brejaúba, para alimentação humana até 2002, mas na área de instalação das
parcelas não foi observada a presença de tocos remanescentes (com exceção de
palmito amargoso e brejaúba), apresentando as características de floresta madura,
como presença de árvores emergentes com mais de 40 m de altura, cipós com
diâmetro a altura do solo superior a 10 cm, sub-bosque bem definido e serrapilheira
espessa e contínua.
Sítio 4. Capoeirinha
Em 1950 foram abertas seis clareiras através do corte raso (figura 3.2.1-1A),
destocamento, aração e queima, com área variando entre 2 e 5 ha cada no interior
da floresta, onde cultivaram Triticum sp. (trigo) por cerca de dois anos.
Posteriormente este cultivo foi abandonado e a partir daí os processos de
regeneração natural passaram a não serem mais controlados. Em 1968, as
capoeiras formadas foram desmatadas para plantio de Zea sp. (milho), Phaseolus
sp. (feijão) e Gossypium sp. (algodão).
Em uma delas, denominada de sítio 4 (figura 3.2.1-1A e B), que em 1975 era
recoberta predominantemente por duas gramíneas, conhecidas por Pennisetum sp.
(capim elefante) e Brachiaria sp. (angola), foram retiradas para cultivo de milho. Com
o fim do cultivo, surgiu espontaneamente o colonião e o capim elefante, então a área
passou a ser utilizada para pastoreio pelo gado bovino até 1994, ficando a partir daí
sem utilização direta (comunicação pessoal de Agnaldo Lorenzutti, 2002).

44
Figura 3.2.1-1. Fotografia aérea da FLONA de Goytacazes em 1970, com as seis
clareiras (A) e em 2000 (B), com os quatro sítios (1- capoeira, 2- cabruca, 3- floresta,
4- capoeirinha), Linhares/ES. Fonte: MAPLAN.
3.2.2. Instalação das parcelas e coleta dos dados
Em 2002 foram estabelecidas aleatoriamente cinco parcelas na área com
floresta madura (floresta) e áreas em regeneração natural, compostas por
capoeirinha, capoeira e por cultivo de cacau (através do sistema de cabruca). Estas
áreas foram mapeadas através de fotografias aéreas, subdivididas em parcelas de
20 x 50 m e estas subdivididas em três grupos de subparcelas (10 x 25 m; 5 x 5m e
2,5 x 1m).
Os indivíduos arbustivo-arbóreos com diâmetro à do peito (DAP) ≥ 10 cm a
1,30 m do solo e as lianas com diâmetro à altura do solo (DAS) ≥ 10 cm foram
amostrados no ano de 2004 em parcelas de 20 x 50 m (n= 5) por tratamento e
marcados com plaquetas de alumínio numeradas e fixadas com auxílio de prego
galvanizado 10 cm acima do local de medição. As alturas dos indivíduos com mais
de 2 m foram estimadas com auxílio do cabo do podão de ramos com extensão total
de oito metros.
Para a amostragem das arvoretas e varas foram medidos os indivíduos com
2,5 cm ≤ DAP < 10 cm e as lianas com 2,5 cm ≤ DAS < 10 cm, em subparcelas de
10 x 25 m, sorteadas dentre quatro por parcela de 20 x 50 m, totalizando cinco por
Alagado
A
2 3
4
1
B
12
3
4 Br101
Rio Doce
Alagado
FLONA de Goytacazes FLONA de Goytacazes
N↑ N↑
600 m 650 m

45
sítio. Os indivíduos amostrados foram marcados com plaquetas de alumínio
numeradas e fixadas com auxílio de prego galvanizado 10 cm acima do local de
medição. As alturas dos indivíduos arbustivo-arbóreos com mais de 2 m foram
estimadas com auxílio do cabo do podão de ramos com extensão total de oito
metros. Para os cálculos, os indivíduos foram separados em duas categorias, sendo
uma delas com 5 cm ≤ DAP < 10 cm e outro grupo com 2,5 cm ≤ DAP < 5 cm. Os
arbustivo-arbóreos com 5 cm ≤ DAP < 10 cm foram denominados de arvoretas e
aqueles com 2,5 cm ≤ DAP < 5 cm de varas.
Na amostragem dos indivíduos jovens, foram computados aqueles de
espécies arbustivo-arbóreas com DAP menor que 2,5 cm e com meio metro ou mais
de altura, em uma subparcela de 5 x 5 m sorteada dentre as dez possíveis em cada
quadrante de 10 x 25 m e uma sorteada dentre os quatro quadrantes no centro da
parcela de 20 X 50 m, totalizando vinte e cinco por sítio. Foi medida a altura e o
diâmetro coletado à altura do peito (DAP) naqueles indivíduos com altura superior a
1,30 m do solo e o diâmetro à altura do solo (DAS) em todos os indivíduos.
Para os indivíduos das espécies arbustivo-arbóreas com até 0,5 m de altura,
foram medidas as alturas e os DAS de todos os indivíduos amostrados em uma
subparcela de 2,5 x 1 m sorteada dentre as doze possíveis por subparcela de 5 x 5
m, totalizando vinte e cinco por sítio.
3.2.3. Aspectos florísticos e fitossociológicos
Amostras de material botânico fértil ou vegetativo, com cinco ramos para cada
espécie, foram coletadas com o auxílio do podão de ramos altos e escalador de
árvores e foi depositado no Herbário (UENF) do Centro de Biociências e
Biotecnologia (CBB) da Universidade Estadual do Norte Fluminense (UENF) e
duplicatas do material fértil foram depositadas no Herbário (VIES) da Universidade
Federal do Espírito Santo (UFES).
A identificação do material botânico coletado foi realizada por meio de
literatura especializada, por especialistas nos grupos e por comparação e consulta
em Herbários nacionais (Companhia Vale do Rio Doce em Linhares/ES, CVRD;
Museu de Biologia Professor Mello Leitão em Santa Teresa/ES, MBML;
Universidade Federal do Espírito Santo em Vitória/ES, VIES; e no da Universidade
Estadual do Norte Fluminense em Campos dos Goytacazes, UENF), adotando para
famílias o sistema de Cronquist (1968), com exceção das Leguminosae, que seguiu

46
Polhill et al. (1981), separando as espécies nas subfamílias, mas na análise dos
dados considerou-se a família Leguminosae. As espécies tiveram seus binômios
confirmados e atualizados através do “software” índice de espécies do Royal
Botanical Garden (IK, 1997), ou por meio de literatura especializada.
Para os indivíduos com menos de 2,5 cm de DAP foi realizada a identificação
no campo com auxílio de mateiro experiente.
3.2.4. Aspectos florísticos e estruturais da mata
A análise dos dados fitossociológicos foi baseada em (Mueller-Dombois e
Ellenberg, 1974) através do programa Fitopac (Shepherd, 1994) e Floresta Nativa
(Cientec, 2005), considerando a Freqüência Absoluta (FAi = (Pi / P) . 100),
Freqüência Relativa [FR = (FAi / ∑ FA) . 100], Densidade Absoluta (DA = ni/A),
Densidade Relativa [DR = (ni / N) . 100], Dominância Absoluta (DoA = ABi / A),
Dominância Relativa (DoR = ABi / AB), Valor de Importância (VI = DR + FR + DoR),
Valor de Cobertura (DR + DoR). Onde,
Pi = número de parcelas onde ocorre uma determinada espécie
P = número total de parcelas da amostragem
FAi = Freqüência absoluta de uma determinada espécie
ni = Número de indivíduos de uma determinada espécie
A = Área amostrada em m2
N = Número de indivíduos de todas as espécies presentes na amostragem
ABi = Área basal por hectare, considerando a fórmula para cálculo da área basal de
um círculo (AB = D2. π / 4)
AB = Área basal total
D = Diâmetro do tronco de cada indivíduo, medido à altura do solo
3.2.5. Determinação da diversidade e similaridade de espécies
Para o cálculo da diversidade (H’) utilizou-se o índice de Shannon, foi
calculada também a diversidade máxima (H’máx) e eqüidade ou uniformidade de
Pielou (J’), (Brower e Zar, 1977; Gotelli e Entsminger, 2001).
H’= - ∑ pi Ln pi, onde:
pi = ni/N
H’máx = Ln S
J’= H’/ H’máx

47
Onde:
S = Número total de espécies da amostra
N = Número total de indivíduos ou cobertura total
ni = Número de indivíduos da espécie i ou valor de cobertura da espécie i
Ln = Logaritmo natural
Para o cálculo da similaridade de espécies foi utilizado o método de Sorensen
- IS (Brower e Zar, 1977).
IS= (2w/A+B)100
Onde:
w= total das espécies das áreas;
A= total das espécies da área A;
B= total das espécies da área B.
A curva de rarefação da diversidade (Shannon) de espécies e número de
indivíduos foi elaborada segundo Gotelli e Entsminger (2001).
3.2.6. Análise estatística
Os dados foram avaliados quanto à normalidade pelo teste de Kolmogorov-
Smirnov e homocedasticidade através do teste de Bartlett (Zar, 1996).
A distribuição das classes diamétricas e das alturas foram testadas pelo
método de Kolmogorov (K-S).
Os quatro sítios e os estratos da vegetação foram comparados quanto à
densidade dos indivíduos arbustivo-arbóreos e de lianas nos respectivos estratos,
área basal, riqueza de espécies, diversidade e número de árvores perfilhadas, pelo
teste de Kruskal-Wallis (H) e Mann-Whitney (Zar, 1996).
A análise multivariada de ordenação foi realizada pelo método Decorama
(Detrended Correspondence Analysis - DCA).

48
3.3. Resultados
3.3.1. Comparações estruturais
3.3.1.1. Alturas e diâmetros
Em 2004 o estrato arbóreo da capoeira com 33 anos em regeneração
apresentou uma curva de distribuição das classes de altura semelhante a da floresta
madura (figura 3.3.1.1-1 A). Fato não observado para a cabruca e a capoeirinha
após nove anos de abandono do controle da sucessão que ainda diferiram da
floresta. A floresta apresentou altura média com 18 m, a capoeira 14 m, a cabruca
17 m e a capoeirinha 8 m. Em relação ao estrato das arvoretas, na capoeira foi
semelhante e na capoeirinha apresentou tendência de ser diferente da floresta
madura e na cabruca foi diferente (figura 3.3.1.1-1 B). As curvas de varas e de
jovens na capoeira e na cabruca foram semelhantes a da floresta madura e na
capoeirinha houve uma tendência de ser diferente (figura 3.3.1.1-1 C, D).
As árvores na capoeirinha apresentaram ainda 80% dos indivíduos na classe
de 5 a 10 m de altura, havendo uma redução drástica nas classes superiores. A
capoeira apresentou indivíduos concentrados entre 5 e 25 m e a cabruca destacou-
se pela presença de indivíduos na classe de 30 m ou mais, muito semelhante a da
floresta madura (figura 3.3.1.1-1 E). A maior concentração de indivíduos de
arvoretas ocorreu entre 4 e 10 m de altura, sendo que na classe de 4 a 6 m
destacaram-se pelo número relativo de indivíduos a capoeirinha e a cabruca, na
penúltima classe a floresta e a capoeira, e na última a floresta e a cabruca (figura
3.3.1.1-1 F). Todos os sítios apresentaram os indivíduos de varas concentrados
entre dois e oito metros de altura (figura 3.3.1.1-1 G). Quanto aos jovens,
apresentaram concentração na primeira classe de altura em todos os sítios
estudados, principalmente na floresta, foi reduzindo gradativamente em direção à
última (figura 3.3.1.1-1 H).

49
0
50
100
150
200
250
Classes de altura
Den
sida
de (n
/ha)
Floresta n= 588±83 Capoeira n= 576±106Cabruca n= 182±100 Capoeirinha n= 34±39
K-S= 0,57 ns
K-S= 1,73*
K-S= 1,73*
5 - 10 10 - 15 - 20 20 - 25 25 - 30 ≥ 300
102030405060708090
5 - 10 10 - 15 15 - 20 20 - 25 25 - 30 ≥ 30Classes de altura
Den
sida
de (%
)
Floresta n= 588±83 Capoeira n= 576±106Cabruca n= 182±100 Capoeirinha n= 34±39
0
50
100
150
200
250
300
Classes de altura
Den
sida
de (n
/ha)
Floresta n= 832±310 Capoeira n= 760±150Cabruca n= 312±209 Capoeirinha n= 304±205
K-S= 0,57 ns
K-S= 1,44*
K-S= 0,86 ns
2 - 4 4 - 6 6 - 8 8 - 10 10 - 12 ≥1205
101520253035404550
2 - 4 4 - 6 6 - 8 8 - 10 10 - 12 ≥ 12Classes de altura
Den
sida
de (%
)
Floresta n= 832±310 Capoeira n= 760±150Cabruca n= 312±209 Capoeirinha n= 304±205
0100200300400500600700800
Classes de altura
Den
sida
de (n
/ha)
Floresta n= 1200±486 Capoeira n= 1360±445Cabruca n= 560±397 Capoeirinha n= 488±433
K-S= 0,31 ns
K-S= 0,63 ns
K-S= 0,94 ns
< 2 2 - 4 4 - 6 6 - 8 ≥ 8
0
10
20
30
40
50
60
< 2 2 - 4 4 - 6 6 - 8 ≥ 8 Classes de altura
Den
sida
de (%
)
Floresta n= 1200±486 Capoeira n= 1360±445Cabruca n= 560±397 Capoeirinha n= 488±433
0
1000
2000
3000
4000
5000
6000
7000
Classes de altura
Den
sida
de (n
/ha)
Floresta n= 11360±7903 Capoeira n= 9248±3097Cabruca n= 4369±2365 Capoeirinha n= 1808±1793
K-S= 0,26 ns
K-S= 0,8 ns
K-S= 1,06 ns
< 1 1 - 2 2 - 3 3 - 4 4 - 5 5 - 6 ≥ 6
0
10
20
30
40
50
60
< 1 1 - 2 2 - 3 3 - 4 4 - 5 5 - 6 ≥ 6Classes da altura
Den
sida
de (%
)
Floresta n= 11360±7903 Capoeira n= 9248±3097Cabruca n= 4369±2365 Capoeirinha n= 1808±1793
Figura 3.3.1.1-1. Comparação entre as curvas da distribuição das alturas - classes
em metros - (A, B, C, D) e da distribuição relativa das alturas - classes em metros-
(E, F, G, H), das árvores, arvoretas, varas e jovens, respectivamente, entre os sítios
na FLONA de Goytacazes, Linhares/ES.
A - Árvores E - Árvores
B - Arvoretas F - Arvoretas
D - Jovens H - Jovens
C - Varas G - Varas

50
A comparação das curvas da distribuição diamétrica do estrato arbóreo,
revelou que o sítio mais semelhante foi a capoeira (figura 3.3.1.1-2 A). Os estratos
das arvoretas, varas e jovens apresentaram-se semelhantes em todos os sítios à
curva da floresta, com exceção da capoeirinha que mostrou uma tendência da curva
ser diferente daquela da floresta (figura 3.3.1.1-2 B, C, D).
Quanto à distribuição relativa dos diâmetros das árvores, a capoeirinha possui
cerca de 80% dos indivíduos concentrados na primeira classe, seguida da capoeira
com cerca de 55% (figura 3.3.1.1-1 E). Com relação às arvoretas, a capoeirinha
possui a maior concentração de todos os sítios, cerca de 45% dos indivíduos na
primeira classe (figura 3.3.1.1-1 F).
Quanto às varas, todos os sítios possuem uma distribuição muito parecida em
todas as classes (figura 3.3.1.1-1 F). Já os diâmetros dos jovens estavam
concentrados principalmente nas primeiras classes diamétricas, principalmente na
floresta, com espécimes em todos os sítios, em maior concentração entre um e três
centímetros de diâmetro à altura do solo, com tendência de ser diferente da floresta
(figura 3.3.1.1-2 H).

51
0
50
100
150
200
250
300
350
Classes de diâmetro
Den
sida
de (n
/ha)
Floresta n= 588±83 Capoeira n= 576±106Cabruca n= 182±100 Capoeirinha n= 34±39
10 - 15 15 - 20 20 - 25 25 - 30 30 - 35 ≥ 35
K-S= 0,89 ns
K-S= 1,15 ns
K-S= 1,73*0
102030405060708090
10 - 15 15 - 20 20 - 25 25 - 30 30 - 35 ≥ 35
Classes de diâmetro
Den
sida
de (%
)
Floresta n= 588±83 Capoeira n= 576±106Cabruca n= 182±100 Capoeirinha n= 34±39
0
50
100
150
200
250
300
Classes de diâmetro
Den
sida
de (n
/ha)
Floresta n= 832±310 Capoeira n= 760±150Cabruca n= 312±209 Capoeirinha n= 304±205
5 - 6 6 - 7 7 - 8 8 - 9 ≥ 9
K-S = 0,63 ns
K-S = 1,26 ns
K-S = 1,26 ns05
1015202530354045
5 - 6 6 - 7 7 - 8 8 - 9 ≥ 10Classes de diâmetro
Den
sida
de (%
)
Floresta n= 832±310 Capoeira n= 760±150Cabruca n= 312±209 Capoeirinha n= 304±205
0100200300400500600700800
Classes de diâmetro
Den
sida
de (n
/ha)
Floresta n= 1200±486 Capoeira n= 1360±445Cabruca n= 560±397 Capoeirinha n= 488±433
2,5 - 3 3 - 4 ≥ 4
K-S = 0,41 ns
K-S = 1,22 ns
K-S = 1,22 ns
-
0
10
20
30
40
50
60
2,5 - 3 3 - 4 ≥ 4Classes de diâmetro
Den
sida
de (%
)
Floresta n= 1200±486 Capoeira n= 1360±445
Cabruca n= 560±397 Capoeirinha n= 488±433
0
5000
10000
15000
20000
25000
30000
35000
Classes de diâmetro
Den
sida
de (n
/ha)
Floresta n= 11360±7903 Capoeira n= 9248±3097Cabruca n= 4369±2365 Capoeirinha n= 1808±1793
< 1 1 - 2 2 - 3 3 - 4 4 - 5 ≥ 5
K-S = 0,28 ns
K-S = 0,57 ns
K-S = 0,86 ns0
10
20
30
40
50
60
<1 1 - 2 2 - 3 3 - 4 4 - 5 ≥ 5
Classes de diâmetro
Den
sida
de (%
)
Floresta n= 11360±7903 Capoeira n= 9248±3097Cabruca n= 4369±2365 Capoeirinha n= 1808±1793
Figura 3.3.1.1-2. Comparação entre as curvas da distribuição das classes
diamétricas - classes em centímetros - (A, B, C, D) e distribuição relativa dos
diâmetros - classes em centímetros - (E, F, G, H), das árvores, arvoretas, varas e
jovens, respectivamente, entre os sítios na FLONA de Goytacazes, Linhares/ES.
A - Árvores E - Árvores
B - Arvoretas F - Arvoretas
C - Varas G - Varas
D - Jovens H - Jovens

52
3.3.1.2. Densidade e área basal
A densidade de árvores por hectare nos diferentes estratos foi sempre maior
na floresta e na capoeira seguindo seqüência em valor decrescente: floresta =
capoeira > cabruca > capoeirinha (figura 3.3.1.2-1 A, B, C, D, E; tabela 3.3.1.2-1). A
densidade de indivíduos arbóreos, arvoretas, varas, jovens e plântulas, na capoeira
com 33 anos em regeneração representaram respectivamente 97%, 91%, 113%,
81% e 36% da densidade da floresta. A cabruca apresentou uma densidade relativa
à floresta de 31%, 37%, 47%, 38% e 38%, respectivamente, para árvores, arvoretas,
varas, jovens e plântulas. Já para a capoeirinha esses estratos encontravam-se com
6%, 37%, 37%, 20% e 5% dos indivíduos encontrados na floresta. A
proporcionalidade de indivíduos entre os estratos foi sendo reduzida
progressivamente a partir de plântulas em direção ao arbóreo, com perdas de
indivíduos de até 92% de plântulas para jovens, para a floresta (tabela 3.3.1.2-2).
Em relação à área basal, foi observado um padrão para o estrato regenerante
(floresta = capoeira > cabruca = capoeirinha), com exceção do estrato arbóreo que
teve a cabruca semelhante a capoeira e a floresta (figura 3.3.1.2-1. F, G, H, I, J e
tabela 3.3.1.2-3).
±Std. Dev.
±Std. Err.
Mean
Sítio
N/H
a
-100
0
100
200
300
400
500
600
700
800
Floresta Capoeira Cabruca Capoeirrinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Ab/H
a
-5
0
5
10
15
20
25
30
35
40
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
N/H
a
0
200
400
600
800
1000
1200
1400
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Ab/H
a
0
1
2
3
4
5
Floresta Capoeira Cabruca Capoeirinha
832±310
760±150
312±209 304±205
3,2±1,2
3,3±0,7
1,4±0,9 1,3±0,7
B G
H= 15,98 P= 0,001
H= 15,29 P= 0,001
29,7±5,8
12,8 ±2,9 21,7±9,6
0,6±0,8
H= 15,98 P= 0,001
588±83 576±106
182±100 34±39
H= 15,28 P= 0,001
A F Árvores
Arvoretas

53
±Std. Dev.
±Std. Err.
Mean
Sítio
N/H
a
-200
200
600
1000
1400
1800
2200
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Ab/
Ha
-0,2
0,2
0,6
1,0
1,4
1,8
2,2
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
N/H
a
-2000
2000
6000
10000
14000
18000
22000
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Ab/H
a
-0,5
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
N/H
a
-40000
20000
80000
140000
200000
260000
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Ab/H
a
-0,2
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Floresta Capoeira Cabruca Capoeirinha
Figura 3.3.1.2-1. Comparação das densidades (A, B, C, D, E) e das áreas basais (F,
G, H, I, J), das árvores, arvoretas, varas, jovens e plântulas, respectivamente, entre
os sítios na FLONA de Goytacazes, Linhares/ES.
1200±486 1360±445
560±397 488±433
1,3±0,5
1,5±0,4
0,7±0,4 0,6±0,5
C H
H= 9,78 P= 0,02 H= 9,78
P= 0,02
11360±7903
9200±3097
4368±2365
1808±17932,3±0,6
2,6±0,9
1,3±0,8 0,6±0,6
D I
H= 14,15 P= 0,02
H= 10,17 P= 0,01
Varas
Jovens
146560±83118
46400±31179
17600±11071
6560±11834 0,7±0,3 0,5±0,2
0,2±0,1 0,03±0,07
E J
H= 14,93 P= 0,02
H= 14,24 P= 0,02
Plântulas

54
Tabela 3.3.1.2-1. Comparação da densidade de indivíduos por hectare entre os
sítios através de cálculos estatísticos pelo método de Mann-Whitney (U) ao nível de
5% de probabilidade (P), na FLONA de Goytacazes, Linhares/ES. Árvores Arvoretas Varas Jovens Plântulas Sítio U P U P U P U P U P
Floresta x capoeira 12 1,00 11 0,84 11 0,84 12 1,00 2 0,03* Floresta x cabruca 0 0,007* 1 0,01* 3 0,05 0 0,007* 0 0,007* Floresta x capoeirinha 0 0,007* 1 0,01* 3 0,05 0 0,007* 0 0,007* Capoeira x cabruca 0 0,007* 1 0,01* 2 0,02* 2 0,03* 0 0,05 Capoeira x capoeirinha 0 0,007* 1 0,01* 1 0,01* 0 0,007* 1 0,001* Cabruca x capoeirinha 0,5 0,007* 12,5 1 10,5 0,69 3 0,005* 4 0,09 * Significativo ao nível de 5%.
Tabela 3.3.1.2-2. Taxa de perda de indivíduos entre os estratos subseqüentes,
FLONA da Goytacazes, Linhares/ES. Sítio / taxa de perda (%)
Estratos Floresta Capoeira Cabruca Capoeirinha
Plântulas para jovens 92 80 75 74
Jovens para varas 89 85 87 73
Varas para arvoretas 31 44 44 38
Arvoretas para árvores 29 24 42 89
Tabela 3.3.1.2-3. Comparação da área basal por hectare entre os sítios através de
cálculos estatísticos pelo método de Mann-Whitney (U) ao nível de 5% de
probabilidade (P), na FLONA de Goytacazes, Linhares/ES. Árvores Arvoretas Varas Jovens Plântulas Sítio U P U P U P U P U P
Floresta x capoeira 0 0,007* 9 0,540 7 0,310 10 0,690 7 0,310 Floresta x cabruca 5 0,150 2 0,030* 4 0,090 6 0,220 0 0,007* Floresta x capoeirinha 0 0,007* 2 0,030* 3 0,050 0 0,007* 0 0,007* Capoeira x cabruca 0 0,150 1 0,010* 1 0,010* 4 0,090 4 0,090 Capoeira x capoeirinha 0 0,007* 1 0,010* 2 0,030* 1 0,010* 1 0,010* Cabruca x capoeirinha 0 0,007* 12 1 10 0,690 6 0,220 1 0,010* * Significativo ao nível de 5%.
3.3.1.3. Árvores perfilhadas e densidade de lianas
O número de indivíduos arbustivo-arbóreos perfilhados em geral foi menor
para floresta em todos os sítios amostrados, indiferente do estrato (figura 3.3.1.3-1
A, B, C; tabela 3.3.1.3-1).
A densidade de lianas com diâmetro à altura do solo (DAS ≥ 10 cm) dentre os
quatro sítios mostrou-se maior na floresta e na capoeira (figura 3.3.1.3-1 D; tabela
3.3.1.3-2). Já a densidade de lianas com 5 ≤ DAS < 10 cm foi semelhante entre as
áreas, mas com tendência de ser mais elevada na floresta e na capoeira (figura

55
3.3.1.3-1 E; tabela 3.3.1.3-2). O número de lianas com 2,5 ≤ DAS < 5 cm foi maior
na capoeira (figura 3.3.1.3-1 F; tabela 3.3.1.3-2).
Houve uma redução progressiva no número de lianas a partir de varas para
arvoretas e de arvoretas para árvores em todos os sítios (tabela 3.3.1.3-3)
.
±Std. Dev.
±Std. Err.
Mean
Sítio
%
-30
-10
10
30
50
70
90
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
SítioD
ensi
dade
(N/H
a)
-2
2
6
10
14
18
22
26
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
%
-5
5
15
25
35
45
55
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Den
sida
de (N
/Ha)
-40
0
40
80
120
160
200
240
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
%
-4
0
4
8
12
16
20
24
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Den
sida
de (N
/Ha)
-200
0
200
400
600
800
1000
1200
1400
Floresta Capoeira Cabruca Capoeirinha
Figura 3.3.1.3-1. Comparação relativa dos indivíduos com perfilhos (A, B, C) e da
densidade de lianas (D, E, F), entre os sítios na FLONA de Goytacazes,
Linhares/ES.
1,0±1,4
8,7±6,7
15,1±10,1 24,4±43
16±5,5
8±4,5 0 ± 0 0 ± 0
A
D
H= 8,14 P= 0,04
H= 16,08 P= 0,001
6,5±7,6
7,5±2,628,3±18,2
24,5±16,7
80±98
120±102
22±40 8±18
B
E
H= 8,39 P= 0,03
H= 7 P= 0,07
3,8±2,3
13,9±5,5 10,4±7,1
8,2±9,8
248±184
872±434
80±40 56±78
C
F
H= 5,1 P= 0,16
H= 13,26 P= 0,004
Árvoretas
Varas
Árvores Lianas DAS≥10cm
Lianas 5≤DAS<10cm
Lianas 2,5≤DAS<5cm

56
Tabela 3.3.1.3-1. Comparação dos indivíduos perfilhados entre os sítios através de
cálculos estatísticos pelo método de Mann-Whitney (U) ao nível de 5% de
probabilidade (P), na FLONA de Goytacazes, Linhares/ES. Árvores Arvoretas Sítio U P U P
Floresta x capoeira 0 0,007* 9 0,540 Floresta x cabruca 0 0,007* 4 0,090 Floresta x capoeirinha 10,5 0,690 3 0,050 Capoeira x cabruca 4 0,090 5 0,150 Capoeira x capoeirinha 4 0,690 0 0,007* Cabruca x capoeirinha 9 0,540 10,5 0,690 * Significativo ao nível de 5%.
Tabela 3.3.1.3-2. Comparação da densidade de lianas entre os sítios através de
cálculos estatísticos pelo método de Mann-Whitney (U) ao nível de 5% de
probabilidade (P), na FLONA de Goytacazes, Linhares/ES. DAS≥10cm 2,5≤DAS<5cm Sítio U P U P
Floresta x capoeira 4 0,090 0,5 0,007* Floresta x cabruca 0 0,007* 5 0,150 Floresta x capoeirinha 0 0,007* 3,5 0,050 Capoeira x cabruca 2,5 0,030* 0 0,007* Capoeira x capoeirinha 2,5 0,030* 0 0,007* Cabruca x capoeirinha 12,5 - 9 0,540 * Significativo ao nível de 5%.
Tabela 3.3.1.3-3. Proporção da perda de lianas por faixa de diâmetro à altura do solo
(DAS), FLONA da Goytacazes, Linhares/ES. Sítio / taxa de perda (%)
Dimensões Floresta Capoeira Cabruca Capoeirinha
2,5 cm ≤ DAS < 5 cm para 5 cm ≤ DAS < 10 cm 68 86 73 86
5 cm ≤ DAS < 10 cm para DAS ≥ 10 cm 80 93 100 100
3.3.2. Comparações florísticas e fitossociológicas
3.3.2.1. Riqueza e diversidade de espécies
A riqueza de espécies de árvores (média entre as parcelas) foi diferente entre
todos os sítios (figura 3.3.2.1-1. A; tabelas 3.3.2.1-1 e 3.3.2.1-2). Com relação às
arvoretas, varas e jovens foram sempre semelhantes entre floresta e capoeira que
apresentaram maior riqueza que a cabruca e a capoeirinha (figura 3.3.2.1-1B, C, D,
E, F, G, H; tabelas 3.3.2.1-1 e 3.3.2.1-2).
A porcentagem de espécies por estrato variou de 100% na floresta a 8% em
árvores para a capoeirinha e de 94% de varas para a capoeira (tabela 3.3.2.1-3). E o
número de espécies por estrato comuns à floresta, variou de duas na capoeirinha

57
(2%) para o arbóreo a 60 (48%) na capoeira para jovens e, entre os estratos nos
respectivos sítios, variou de 16 (24%) para arvoretas a 69 (55%) para jovens (tabela
3.3.2.1-4). Das 109, 68, 80 e 125 espécies ocorrentes nos estratos de árvores,
arvoretas, varas e jovens na floresta, 43, 16, 28 e 69, ocorreram nos respectivos
estratos regenerantes.
Quanto à diversidade de espécies, ela sempre foi maior na floresta em todos
os estratos, com exceção do sítio capoeira que foi semelhante à floresta para
arvoretas, varas e jovens (tabelas 3.3.2.1-1, 3.3.2.1-5 e figuras 3.3.2.1-2 e 3).
Na floresta a curva da diversidade das árvores possui um incremento grande
de até 100 indivíduos, mas ainda continuou crescendo, não apresentando
estabilidade, já na capoeira o maior incremento foi até 50 indivíduos e a partir dos
150 ela ficou praticamente estabilizada, indicando que a diversidade máxima na
capoeira, ao contrário da floresta, foi captada pela amostragem. Na cabruca a
amostragem foi suficiente para detectar o incremento mais expressivo da
diversidade, mas insuficiente para captar a diversidade máxima da área e na
capoeirinha a amostragem foi insuficiente para detectar o incremento inicial mais
expressivo da diversidade (figura 3.3.2.1-2).
O número de indivíduos amostrados no estrato de arvoretas, varas e jovens
foi insuficiente para captar a diversidade máxima na cabruca e na capoeirinha,
havendo uma redução gradativa entre a floresta, capoeira, cabruca e capoeirinha
(figura 3.3.2.1-2 e 3).
Os valores de eqüidade dos sítios e respectivos estratos variaram de 0,74
(jovens) a 0,95 (arvoretas), mostrando que houve maior dominância de algumas
espécies no estrato de jovens em todos os sítios, com exceção de capoeira que
apresentou uma maior dominância para árvores (tabela 3.3.2.1-1).

58
±Std. Dev.
±Std. Err.
Mean
Sítio
Riq
ueza
de
espé
cies
-5
5
15
25
35
45
55
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Div
ersi
dade
(H')
-1
0
1
2
3
4
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Riq
ueza
de
espé
cies
0
4
8
12
16
20
24
28
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Div
ersi
dade
(H')
0,4
1,0
1,6
2,2
2,8
3,4
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Riq
ueza
de
espé
cies
-2
4
10
16
22
28
34
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Div
ersi
dade
(H')
0,4
0,8
1,2
1,6
2,0
2,4
2,8
3,2
3,6
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Riq
ueza
de
espé
cies
-5
5
15
25
35
45
55
65
Floresta Capoeira Cabruca Capoeirinha
±Std. Dev.
±Std. Err.
Mean
Sítio
Div
ersi
dade
(H')
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
Floresta Capoeira Cabruca Capoeirinha
Figura 3.3.2.1-1. Comparação das riquezas de espécies de árvores (A), arvoretas
(B), varas (C) e dos jovens (D) e diversidade de árvores (E), arvoretas (F), varas (G)
e dos jovens (H), entre os sítios na FLONA de Goytacazes, Linhares/ES.
2,6±2,7
3,5±0,2
2,5±0,1 2,2±0,4
0,7±0,9
E
H= 17,01 P= 0,007
H= 15,29 P= 0,001
A
11,8±5,4
21,6±3,6
38±6,8
2,7±0,4 2,4±0,3
1,6±0,5
17,6±6,5
13,2±2,9
6±2,7
3,6±1,7
0,9±0,3
B F
H= 15,43 P= 0,001
H= 15,25 P= 0,001
23,8±7,2 24±5,1
8±4
4±2,3
3±0,2 2,9±0,2
1,8±0,4
1,1±0,5
H= 15,06 P= 0,001
H= 15,5 P= 0,001
C G
54±5
43±9
18±7 5±3
3,4±0,7
3,4±0,2
2,2±0,2
1,2±0,5
D H
H= 16,63 P= 0,0008
H= 15,73 P= 0,001
Árvores
Arvoretas
Varas
Jovens
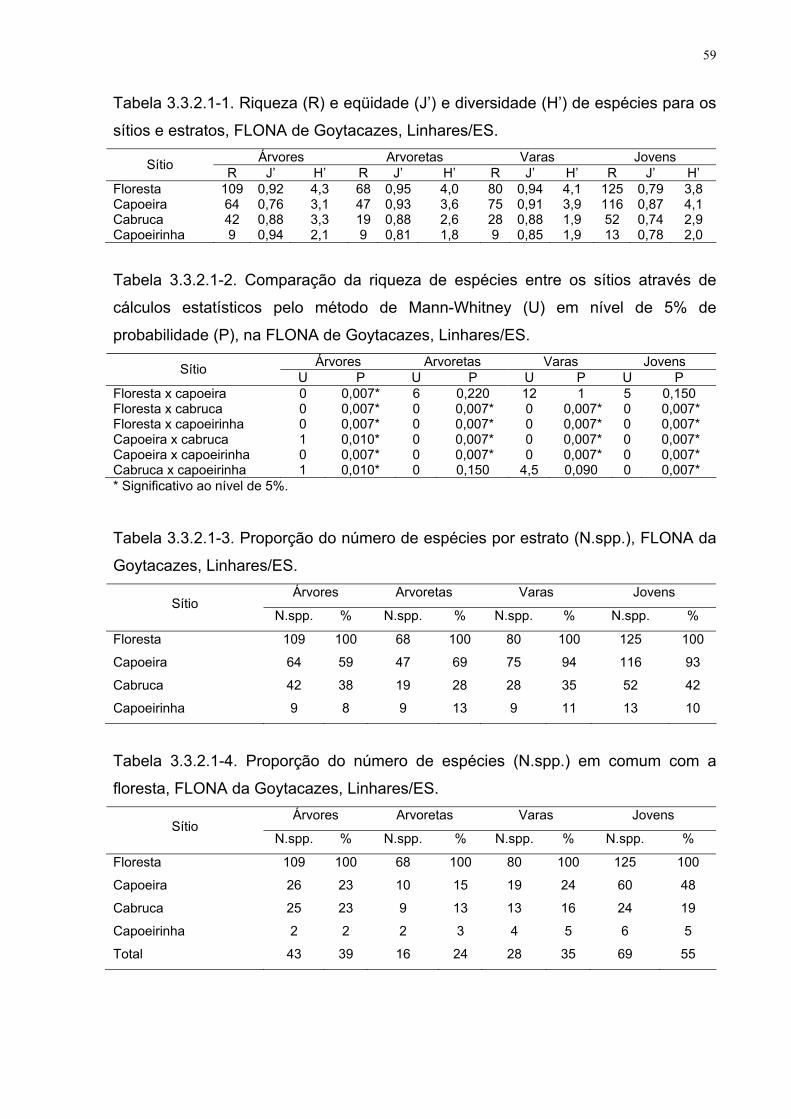
59
Tabela 3.3.2.1-1. Riqueza (R) e eqüidade (J’) e diversidade (H’) de espécies para os
sítios e estratos, FLONA de Goytacazes, Linhares/ES. Árvores Arvoretas Varas Jovens Sítio R J’ H’ R J’ H’ R J’ H’ R J’ H’
Floresta 109 0,92 4,3 68 0,95 4,0 80 0,94 4,1 125 0,79 3,8 Capoeira 64 0,76 3,1 47 0,93 3,6 75 0,91 3,9 116 0,87 4,1 Cabruca 42 0,88 3,3 19 0,88 2,6 28 0,88 1,9 52 0,74 2,9 Capoeirinha 9 0,94 2,1 9 0,81 1,8 9 0,85 1,9 13 0,78 2,0
Tabela 3.3.2.1-2. Comparação da riqueza de espécies entre os sítios através de
cálculos estatísticos pelo método de Mann-Whitney (U) em nível de 5% de
probabilidade (P), na FLONA de Goytacazes, Linhares/ES. Árvores Arvoretas Varas Jovens Sítio U P U P U P U P
Floresta x capoeira 0 0,007* 6 0,220 12 1 5 0,150 Floresta x cabruca 0 0,007* 0 0,007* 0 0,007* 0 0,007* Floresta x capoeirinha 0 0,007* 0 0,007* 0 0,007* 0 0,007* Capoeira x cabruca 1 0,010* 0 0,007* 0 0,007* 0 0,007* Capoeira x capoeirinha 0 0,007* 0 0,007* 0 0,007* 0 0,007* Cabruca x capoeirinha 1 0,010* 0 0,150 4,5 0,090 0 0,007* * Significativo ao nível de 5%.
Tabela 3.3.2.1-3. Proporção do número de espécies por estrato (N.spp.), FLONA da
Goytacazes, Linhares/ES. Árvores Arvoretas Varas Jovens
Sítio N.spp. % N.spp. % N.spp. % N.spp. %
Floresta 109 100 68 100 80 100 125 100
Capoeira 64 59 47 69 75 94 116 93
Cabruca 42 38 19 28 28 35 52 42
Capoeirinha 9 8 9 13 9 11 13 10
Tabela 3.3.2.1-4. Proporção do número de espécies (N.spp.) em comum com a
floresta, FLONA da Goytacazes, Linhares/ES. Árvores Arvoretas Varas Jovens
Sítio N.spp. % N.spp. % N.spp. % N.spp. %
Floresta 109 100 68 100 80 100 125 100
Capoeira 26 23 10 15 19 24 60 48
Cabruca 25 23 9 13 13 16 24 19
Capoeirinha 2 2 2 3 4 5 6 5
Total 43 39 16 24 28 35 69 55

60
Tabela 3.3.2.1-5. Comparação da diversidade de espécies entre os sítios através de
cálculos estatísticos pelo método de Mann-Whitney (U) ao nível de 5% de
probabilidade (P), na FLONA de Goytacazes, Linhares/ES. Árvores Arvoretas Varas Jovens Sítio U P U P U P U P
Floresta x capoeira 0 0,007* 6 0,220 12 0,840 5 0,150 Floresta x cabruca 0 0,007* 0 0,007* 0 0,007* 2 0,030* Floresta x capoeirinha 0 0,007* 0 0,007* 0 0,007* 0 0,007* Capoeira x cabruca 6,5 0,220 1,5 0,010* 0 0,007* 0 0,007* Capoeira x capoeirinha 0 0,007* 0 0,007* 0 0,007* 0 0,007* Cabruca x capoeirinha 1 0,010* 3 0,050 2,5 0,030* 0 0,007* * Significativo ao nível de 5%.

61
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150 200 250 300Número de indivíduos
Div
ersi
dade
(H')
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150 200Número de indivíduos
Div
ersi
dade
(H')
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150 200 250 300Número de indivíduos
Div
ersi
dade
(H')
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150 200Número de indivíduos
Div
ersi
dade
(H')
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150 200 250 300Número de indivíduos
Div
ersi
dade
(H')
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150 200Número de indivíduos
Div
ersi
dade
(H')
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150 200 250 300Número de indivíduos
Div
ersi
dade
(H')
00,5
11,5
22,5
33,5
44,5
5
0 50 100 150 200Número de indivíduos
Div
ersi
dade
(H')
Figura 3.3.2.1-2. Curva de rarefação da diversidade de espécies (H’) com
valores médios ± intervalo de confiança para árvores e arvoretas na floresta
(A, E), capoeira (B, F), cabruca (C, G) e capoeirinha (D, H), FLONA de
Goytacazes, Linhares/ES.
A
B
C
D
E
F
G
H
Árvores
Árvores
Árvores
Árvores
Arvoretas
Arvoretas
Arvoretas
Arvoretas
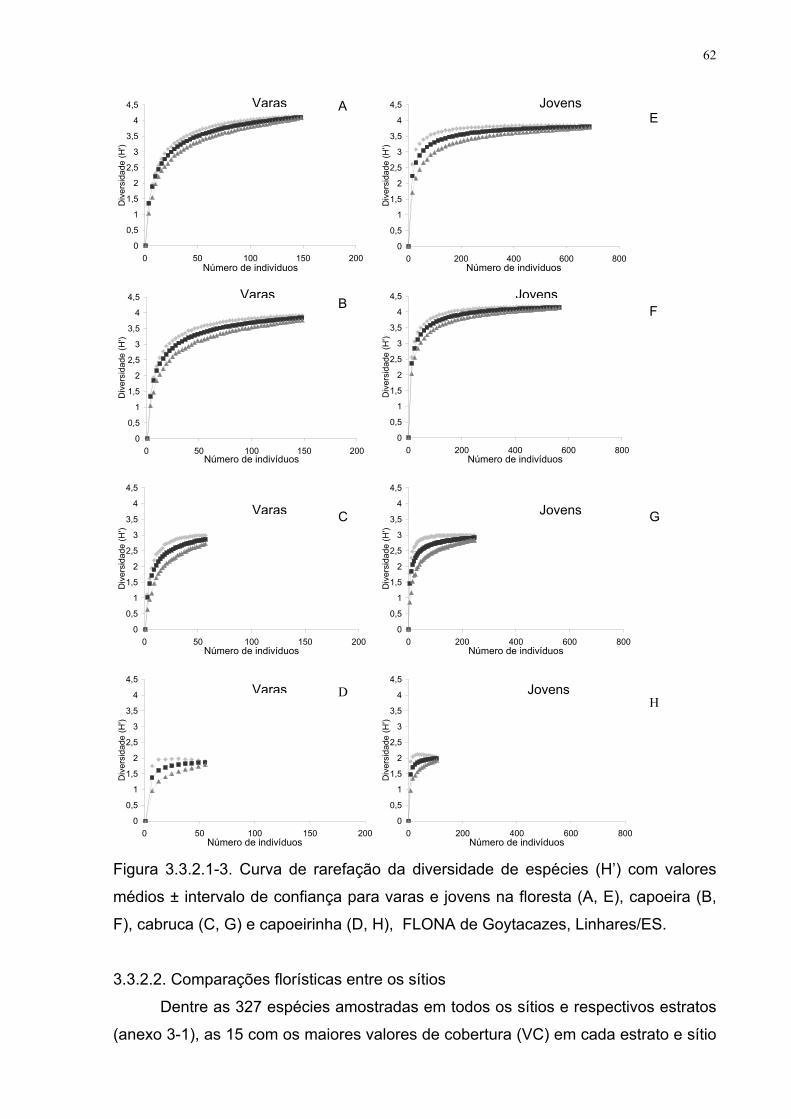
62
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 50 100 150 200Número de indivíduos
Div
ersi
dade
(H')
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 200 400 600 800Número de indivíduos
Div
ersi
dade
(H')
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 50 100 150 200Número de indivíduos
Div
ersi
dade
(H')
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 200 400 600 800Número de indivíduos
Div
ersi
dade
(H')
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 50 100 150 200Número de indivíduos
Div
ersi
dade
(H')
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 200 400 600 800Número de indivíduos
Div
ersi
dade
(H')
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 50 100 150 200Número de indivíduos
Div
ersi
dade
(H')
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 200 400 600 800Número de indivíduos
Div
ersi
dade
(H')
Figura 3.3.2.1-3. Curva de rarefação da diversidade de espécies (H’) com valores
médios ± intervalo de confiança para varas e jovens na floresta (A, E), capoeira (B,
F), cabruca (C, G) e capoeirinha (D, H), FLONA de Goytacazes, Linhares/ES.
3.3.2.2. Comparações florísticas entre os sítios
Dentre as 327 espécies amostradas em todos os sítios e respectivos estratos
(anexo 3-1), as 15 com os maiores valores de cobertura (VC) em cada estrato e sítio
A
B
D
E
F
G
H
C
Varas
Varas
Varas
Varas
Jovens
Jovens
Jovens
Jovens

63
analisados (com exceção da capoeirinha que teve um total de espécies menor)
estão na tabela 3.3.2.2-1. Dentre as 15 espécies de maior VC na floresta, 10
ocorrem em pelo menos um outro sítio e estrato, destacando-se Astronium
concinnum (Engl.) Schott (gonçalo alves) e Pterocarpus rohrii Vahl. (pau sangue),
presentes na maioria das estratos e sítios (tabela 3.3.2.2-1). Das espécies com os
maiores valores de cobertura, apenas Gallesia integrifolia (Spreng.) Harms. (pau
d’alho) e Tapirira guianensis Aubl. (cupuba) na cabruca e Psidium guajava Linn.
(goiaba) e Dalbergia nigra (jacarandá) na capoeirinha, ocorreram no estrato de
árvores e nos estratos regenerantes.
A ordenação através da DCA para o número de espécies e respectivo número
de indivíduos, por sítio e estrato, produziu autovalores intermediários,
respectivamente 0,586 e 0,380 para os eixos de ordenação 1 e 2, indicando a
existência de gradiente, ou seja, parte das espécies distribuiu-se por todo o
gradiente e parte delas foi exclusiva. Produziu quatro grupos pelo eixo1, e o eixo2
influenciou principalmente o subgrupo pertencente ao grupo três - G3B (figura
3.3.2.2-1). O primeiro grupo foi formado por todos os estratos da floresta (árvores,
arvoretas, varas e jovens), o segundo grupo foi aglomerado pelos estratos de varas
e jovens, pertencentes à capoeira. O terceiro grupo foi composto por três estratos da
capoeirinha (arvoretas, varas e jovens), um da capoeira (arvoretas) e da cabruca
(jovens), o quarto grupo foi formado pelo estrato de árvores da capoeirinha e da
capoeira. O eixo2 separou principalmente o grupo3 em dois subgrupos: 1 - G3A com
cabruca (árvores, arvoretas e varas); 2 - G3B com capoeirinha (arvoretas, varas e
jovens), capoeira (arvoretas) e cabruca (jovens). Muitas espécies foram exclusivas e
influenciaram o eixo1 como Hirtella buchellii Britton, Simaba subcymosa A. St. Hil.
Til., Lecythis pisonis Cambess., dentre outras, principalmente para o G1. O outro
extremo do eixo1 foi influenciado por Psidium guajava, Psidium guineense SW.,
Dalbergia nigra(Vell.) Allemao ex Benth., dentre outras (G4) e o eixo2 por Spondias
nombim L., Trema micrantha (L.) Blume e Tapirira guianensis (G3B).

64
Figura 3.3.2.2-1. As coordenadas dos sítios nos dois eixos da DCA (detrended
correspondence analysis) formaram quatro grupos (G1, G2, G3, G4), calculados com
as espécies e sua abundância em cada estrato de cada sítio [floresta (árvores - Fl
10, arvoretas - Fl 5, varas - Fl 2,5 e jovens - Fl 0,5); capoeira (árvores - Cap 10,
arvoretas - Cap 5, varas - Cap 2,5 e jovens - Cap 0,5); cabruca (árvores - Cab 10,
arvoretas - Cab 5, varas - Cab 2,5 e jovens - Cab 0,5); capoeirinha (árvores – Ca 10,
arvoretas - Ca 5, varas - Ca 2,5 e jovens - Ca 0,5)], FLONA de Goytacazes,
Linhares/ES.
Os grupos formados pelo método de Sorensen foram os seguintes: o primeiro
constituído por três estratos da capoeira (árvores, arvoretas e varas), o segundo por
todos os estratos da floresta e jovens da capoeira, o terceiro por todos os estratos da
cabruca e o quarto por todos os estratos da capoeirinha (figura 3.3.2.2-2).
Os resultados da similaridade pelo método de Sorensen (figura 3.3.2.2-2)
corroboraram em grande parte a ordenação formada pela DCA, onde os estratos e
os sítios formaram também quatro grandes grupos (figura 3.3.2.2-1).
G1 G2
G3
G4
G3B
G3A

65
Figura 3.3.2.2-2. Dendrograma de similaridade (Sorensen) de espécies entre os
estratos dos sítios realizadas na FLONA de Goytacazes, Linhares/ES. [floresta
(árvores - Fl 10, arvoretas - Fl 5, varas - Fl 2,5 e jovens - Fl 0,5); capoeira (árvores -
Cap 10, arvoretas - Cap 5, varas - Cap 2,5 e jovens - Cap 0,5); cabruca (árvores -
Cab 10, arvoretas - Cab 5, varas - Cab 2,5 e jovens - Cab 0,5); capoeirinha (árvores
- Ca 10, arvoretas - Ca 5, varas - Ca 2,5 e jovens - Ca 0,5)].
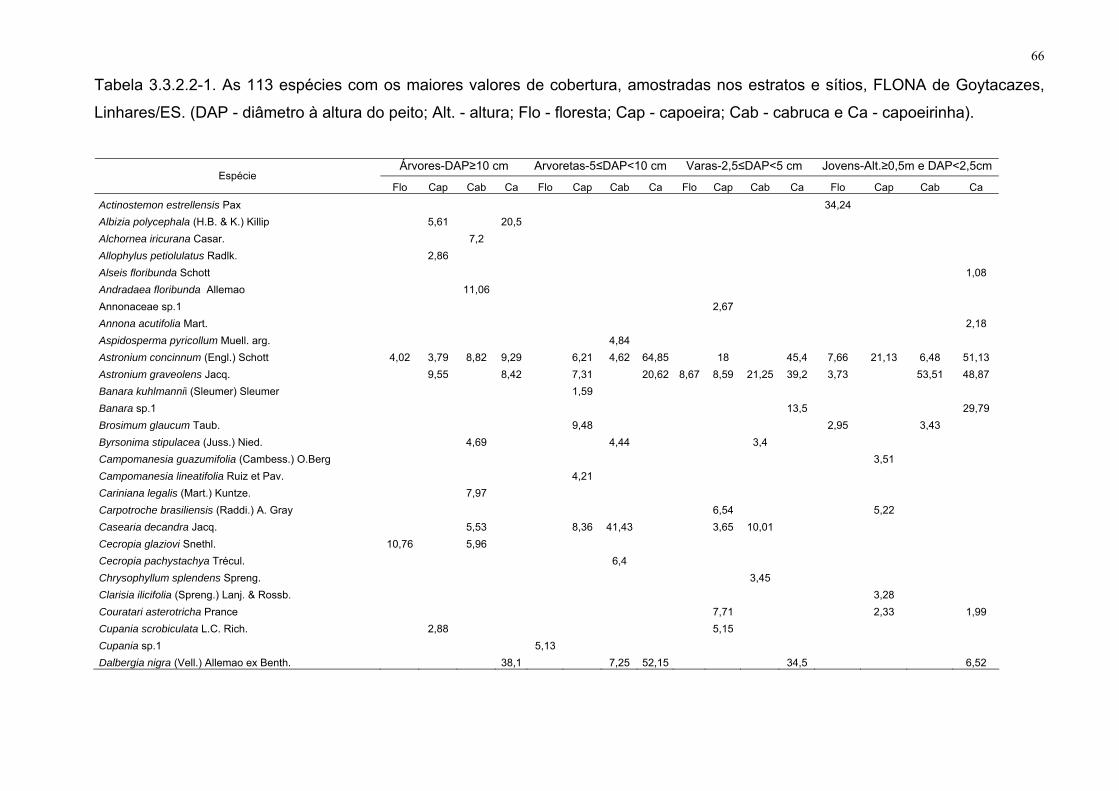
66
Tabela 3.3.2.2-1. As 113 espécies com os maiores valores de cobertura, amostradas nos estratos e sítios, FLONA de Goytacazes,
Linhares/ES. (DAP - diâmetro à altura do peito; Alt. - altura; Flo - floresta; Cap - capoeira; Cab - cabruca e Ca - capoeirinha).
Árvores-DAP≥10 cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5 cm Jovens-Alt.≥0,5m e DAP<2,5cm
Espécie Flo Cap Cab Ca Flo Cap Cab Ca Flo Cap Cab Ca Flo Cap Cab Ca
Actinostemon estrellensis Pax 34,24 Albizia polycephala (H.B. & K.) Killip 5,61 20,5 Alchornea iricurana Casar. 7,2 Allophylus petiolulatus Radlk. 2,86 Alseis floribunda Schott 1,08 Andradaea floribunda Allemao 11,06 Annonaceae sp.1 2,67 Annona acutifolia Mart. 2,18 Aspidosperma pyricollum Muell. arg. 4,84 Astronium concinnum (Engl.) Schott 4,02 3,79 8,82 9,29 6,21 4,62 64,85 18 45,4 7,66 21,13 6,48 51,13 Astronium graveolens Jacq. 9,55 8,42 7,31 20,62 8,67 8,59 21,25 39,2 3,73 53,51 48,87 Banara kuhlmannii (Sleumer) Sleumer 1,59 Banara sp.1 13,5 29,79 Brosimum glaucum Taub. 9,48 2,95 3,43 Byrsonima stipulacea (Juss.) Nied. 4,69 4,44 3,4 Campomanesia guazumifolia (Cambess.) O.Berg 3,51 Campomanesia lineatifolia Ruiz et Pav. 4,21 Cariniana legalis (Mart.) Kuntze. 7,97 Carpotroche brasiliensis (Raddi.) A. Gray 6,54 5,22 Casearia decandra Jacq. 5,53 8,36 41,43 3,65 10,01 Cecropia glaziovi Snethl. 10,76 5,96 Cecropia pachystachya Trécul. 6,4 Chrysophyllum splendens Spreng. 3,45 Clarisia ilicifolia (Spreng.) Lanj. & Rossb. 3,28 Couratari asterotricha Prance 7,71 2,33 1,99 Cupania scrobiculata L.C. Rich. 2,88 5,15 Cupania sp.1 5,13 Dalbergia nigra (Vell.) Allemao ex Benth. 38,1 7,25 52,15 34,5 6,52
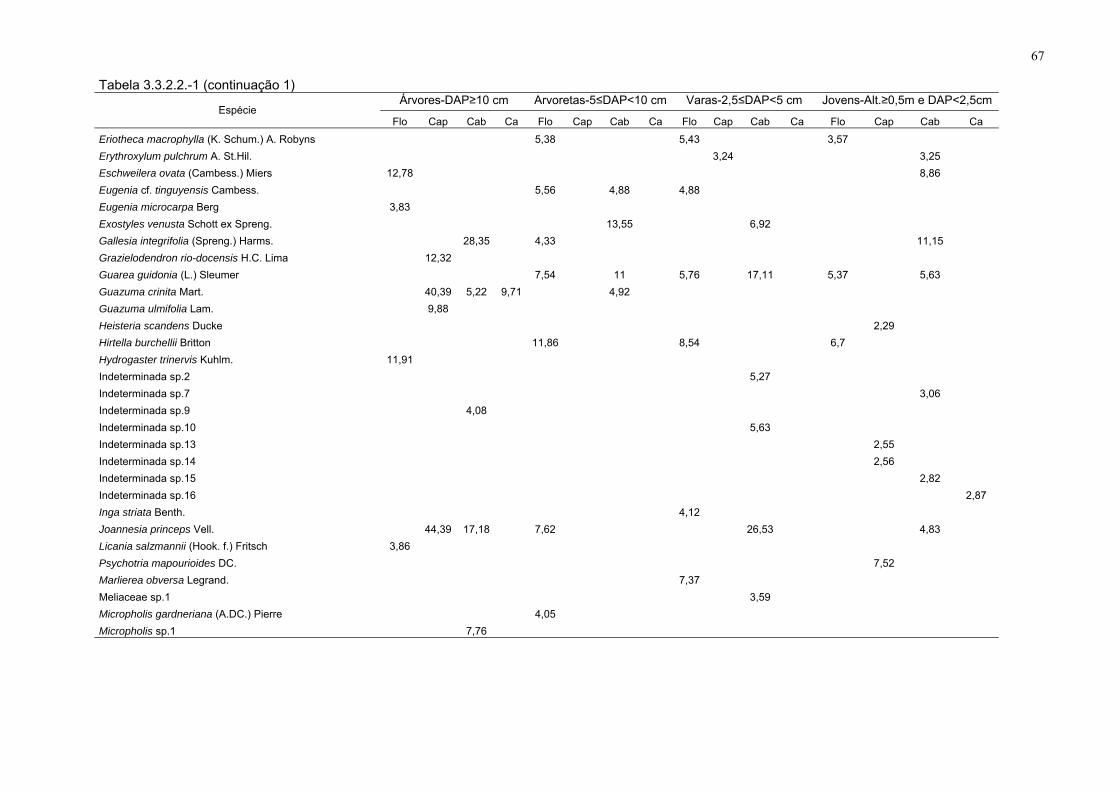
67
Tabela 3.3.2.2.-1 (continuação 1) Árvores-DAP≥10 cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5 cm Jovens-Alt.≥0,5m e DAP<2,5cm
Espécie Flo Cap Cab Ca Flo Cap Cab Ca Flo Cap Cab Ca Flo Cap Cab Ca
Eriotheca macrophylla (K. Schum.) A. Robyns 5,38 5,43 3,57 Erythroxylum pulchrum A. St.Hil. 3,24 3,25 Eschweilera ovata (Cambess.) Miers 12,78 8,86 Eugenia cf. tinguyensis Cambess. 5,56 4,88 4,88 Eugenia microcarpa Berg 3,83 Exostyles venusta Schott ex Spreng. 13,55 6,92 Gallesia integrifolia (Spreng.) Harms. 28,35 4,33 11,15 Grazielodendron rio-docensis H.C. Lima 12,32 Guarea guidonia (L.) Sleumer 7,54 11 5,76 17,11 5,37 5,63 Guazuma crinita Mart. 40,39 5,22 9,71 4,92 Guazuma ulmifolia Lam. 9,88 Heisteria scandens Ducke 2,29 Hirtella burchellii Britton 11,86 8,54 6,7 Hydrogaster trinervis Kuhlm. 11,91 Indeterminada sp.2 5,27 Indeterminada sp.7 3,06 Indeterminada sp.9 4,08 Indeterminada sp.10 5,63 Indeterminada sp.13 2,55 Indeterminada sp.14 2,56 Indeterminada sp.15 2,82 Indeterminada sp.16 2,87 Inga striata Benth. 4,12 Joannesia princeps Vell. 44,39 17,18 7,62 26,53 4,83 Licania salzmannii (Hook. f.) Fritsch 3,86 Psychotria mapourioides DC. 7,52 Marlierea obversa Legrand. 7,37 Meliaceae sp.1 3,59 Micropholis gardneriana (A.DC.) Pierre 4,05 Micropholis sp.1 7,76

68
Tabela 3.3.2.2.-1 (continuação 2) Árvores-DAP≥10 cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5 cm Jovens-Alt.≥0,5m e DAP<2,5cm
Espécie Flo Cap Cab Ca Flo Cap Cab Ca Flo Cap Cab Ca Flo Cap Cab Ca
Myrcia follii Barroso et Peixoto 6,6 6,34 Myrcia riodocensis G.M. Barroso et Peixoto 7,42 Myrciaria floribunda (West. ex Willd.) Berg 4,22 Myrtaceae sp.1 3,33 9 Myrtaceae sp.13 5,39 Ocotea aciphylla (Nees) Mez 17,52 16,82 Ocotea confertiflora (Meisn.) Mez 5,76 Ocotea divaricata (Poir.) Mez 7,88 5,47 Ocotea elegans Mez 2,56 Oxandra sp.1 3,34 5,5 Parapiptadenia pterosperma (Benth.) Brenan 30,8 3,85 38,5 42,88 Piptadenia gonoacantha (Mart.) Macbr. 4,5 Piptadenia paniculata Benth. 3,65 Platymiscium floribundum Vogel 2,57 Plinia involucrata (Berg) McVaugh. 4,14 Pourouma velutina Miq. 7,54 Pouteria bangii (Rusby) T.D.Pennington 4,25 3,18 Protium heptaphyllum (Aubl.) Marchand. 3,15 4,68 2,89 Pseudima frutescens (Aubl.) Radlk. 7,51 Pseudobombax grandiflorum (Cav.) A. Robyns 2,73 11,76 Pseudopiptadenia contorta (DC.) G.P.Lewis & M.P.Lima 1,23 Psidium guajava Linn. 42,4 14,41 Psidium guianeense Sw. 22,73 9,47 4,11 Psidium sp.1 2,13 Pterocarpus rohrii Vahl. 9,28 2,54 14,21 6,37 6,05 12,8 3,68 Quararibea penduliflora (A.St.Hil.) K. Schum. 3,52 6,61 Rauwolfia sp.1 4,27 Rollinia laurifolia Schltdl. 4,3 Sapium glandulatum (Vell.) Pax. 4,1 Sapotacea sp. 4 5,67

69
Tabela 3.3.2.2.-1 (continuação 3) Árvores-DAP≥10 cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5 cm Jovens-Alt.≥0,5m e DAP<2,5cm
Espécie Flo Cap Cab Ca Flo Cap Cab Ca Flo Cap Cab Ca Flo Cap Cab Ca
Sapotaceae sp.1 4,87 2,24 Sapotaceae sp.2 8,51 3,02 Schoepfia oblongifolia Turez 5,91 5,2 8,3 4,84 Simaruba amara Aubl. 4,41 6,44 Simira grazielae A. L. Peixoto 10,4 3,41 Sorocea guilleminiana Gaudich. 3,78 Spondias mombin L. 6,51 Sterculia elata Ducke 4,62 Swartzia simplex SW. Spreng var. ochnacea (DC.) R.S.Cowan 1,59 Tabebuia serratifolia (Vahl) Nichols. 3,23 5,09 Tachigalia paratyensis (Vell.) H.C. Lima 3,73 Talisia intermedia Radlk. 2,27 Tapirira guianensis Aubl. 23,02 22,5 36,8 19,82 13,57 Theobroma cacao L. 4,62 Tovomita brevistaminea Engl. 8,78 8,56 5,31 Trichilia casaretti C.DC. 4,49 Trichilia sp.1 22,32 10,8 5,25 Vatairea heteroptera (Fr. All.) Ducke ex de Assis Iglesias 18,3 10,79 14,1 Vernonanthura phosphorica (Vell. Conc.) H. Rob. 6,18 8,28 Virola gardneri (A.DC.) Warb. 5,58 3,7 Vitex orinocensis HBK 3,28 Xylopia sericea A. St. Hil. 2,85 Zanthoxylon cf. juriperinum Poeppig & Endl. 8,45 Zanthoxylon rhoifolium Lam. var. petiolatum Engl. 5,11 4,58 Zollernia sp.1 29,09

70
3.4. Discussão
3.4.1. Comparações estruturais
Trabalhos realizados em áreas em regeneração com idades variando entre 20
e 30 anos, mostraram que a altura do dossel nestas áreas encontrava-se em torno
de 75% das alturas do dossel das florestas maduras (Brown e Lugo, 1990; Kappelle
et al., 1996). A observação de que a capoeira com 33 anos em regeneração
alcançou 80% da altura do dossel da floresta, corrobora esta afirmação. Entretanto,
a cabruca com apenas 9 anos em regeneração também apresentou altura média
próxima da floresta, todavia, a curva da distribuição das alturas diferiu da floresta. No
caso da capoeira, o período de 33 anos de abandono foi suficiente para que as
espécies que se instalaram no início da regeneração fossem responsáveis pela
proximidade, ao contrário da cabruca, visto que a técnica de manejo da área
(cabruca) deixa presença de árvores de grande porte remanescentes da floresta
madura, para fornecerem sombra aos indivíduos de cacau que foram plantados e as
menores foram eliminadas. As poucas árvores de grande porte remanescentes
foram responsáveis pela elevada média das alturas, porém, o fato da distribuição
das alturas diferir da floresta, indica que a altura média não foi o parâmetro mais
indicado para descrever o estrato arbóreo da cabruca. Este método (cabruca) tem
sido considerado um tipo de exploração agrícola (agrofloresta) com potencial para
conservação da biodiversidade (Rice e Greenberg, 2000; Sambuichi, 2002). Porém,
Rolim e Chiarello (2004) sugerem que a conservação de árvores no sistema cabruca
encontra-se comprometida, dentre outros fatores, pelo sistema de manutenção
dessas áreas, que elimina o estrato regenerante, prejudicando a dinâmica natural da
vegetação e a substituição do estrato arbóreo.
Os resultados para as alturas dos estratos regenerantes (arvoretas, varas e
jovens) foram muito próximos da floresta, indicando que a regeneração,
principalmente na capoeira e na cabruca, foi muito semelhante quanto ao tamanho
dos indivíduos ao observado na floresta, confirmando indicações de outros autores
(Nunes, 1996).
O fato da densidade na capoeira ter alcançado valores próximos aos da
floresta (ca. 90%) nos diferentes estratos amostrados, sugere que o tempo de
abandono e o uso do solo influenciam na taxa de regeneração. Visto que esta alta
densidade atingida pela capoeira nos primeiros quatro estratos, pode estar
relacionada à elevada resiliência da área da capoeira e o tempo de abandono, que

71
possibilitaram a aceleração do crescimento das espécies que foram se instalando na
área ainda com a presença do Panicum maximum (colonião) e do pastoreio pelo
gado. Já a densidade menor na capoeira que na floresta no estrato de plântulas
(36%) pode estar ligada a maior lentidão da entrada de indivíduos de espécies
secundárias tardias e restrições ambientais para as secundárias iniciais, devido,
principalmente, à cobertura em função da alta densidade de indivíduos no estrato
arbóreo. Padrão também detectado por Nunes (1996), comparando floresta e
capoeira com 30 anos em regeneração, no domínio da Floresta Ombrófila Densa
das Terras Baixas.
Apesar de possuírem o mesmo tempo de abandono do controle da sucessão
pelas atividades de manutenção, a cabruca em relação à capoeirinha apresentou
uma densidade de indivíduos e desenvolvimento mais rápido e uniforme entre os
estratos, este fato parece estar relacionado ao tipo de uso do solo, que foi distinto
para as duas áreas, principalmente, devido a grande dominância e cobertura do
Pennisetum sp. (capim elefante) na capoeirinha, que parece estar dificultando o
estabelecimento de indivíduos oriundos de propágulos vindos da floresta adjacente.
A redução progressiva na proporcionalidade de indivíduos entre os estratos
de plântulas para árvores, encontradas para todos os sítios e estratos, está de
acordo com o observado por Nunes (1996). Esta redução se deve à grande
mortalidade nos primeiros estágios de vida dos indivíduos, principalmente de
plântulas e jovens.
Em relação à área basal, ficou evidente que no estrato arbóreo, a observação
de indivíduos de grande porte no sistema de cabruca (área basal 27% menor que
para a floresta), foi o fator responsável pela ocorrência de valor semelhante com o
encontrado na floresta, como já ressaltado anteriormente, todavia, a densidade de
árvores foi diferente entre os dois sítios. Este padrão não foi observado para a
capoeira que, embora com cerca de 33 anos de abandono agrícola, possui valor de
área basal por hectare de apenas 43% do observado para a floresta.
A ocorrência de valores semelhantes de área basal por hectare na capoeira e
na floresta, quando comparamos apenas os estratos regenerantes, sugere que a
área basal do estrato arbóreo (DAP ≥ 10 cm) parece ser um dos indicadores para a
classificação do estado de regeneração de Floresta Ombrófila Densa Aluvial, do que
o uso das áreas basais dos estratos inferiores. Entretanto, como salientado por
outros autores (ex. Clark e Clark, 1992) o dossel de uma floresta madura em geral é

72
composto por indivíduos com DAP ≥ 30 cm, assim o uso da área basal de indivíduos
com DAP superiores a este valor seria também indicado. Entretanto, a resolução
Conama (1994) e IDAF (1999), sugerem a utilização da área basal com inclusão dos
indivíduos com DAP ≥ 10 cm como um dos parâmetros para que se possam
classificar os estágios de regeneração da Floresta Atlântica no Estado do Espírito
Santo. Segundo esta resolução, e com base na área basal, a capoeira estudada se
encaixa no estágio médio de regeneração da Floresta Atlântica, a cabruca no
avançado e a capoeirinha em macega. Com relação a cabruca, devido a presença
de algumas árvores de grande porte remanescente e isoladas houve provocou ruido
na classificação, pois quando as excluimos, a área passa a se encaixar em estágio
médio, mais condizente com sua fitofisionomia. O uso do parâmetro área basal para
situações como o da cabruca deve ser utilizado com cautela devido a manutenção
de grandes árvores, responsáveis por grande parte da área basal total, para
sombreamento do cacau.
A expressiva taxa de árvores com perfilhos detectadas nas áreas secundárias
(capoeira - 870%, cabruca - 1510% e capoeirinha - 2440%) em relação à floresta,
está condizente com o padrão encontrado por (Carvalho, 2005) em fragmentos
degradados no Norte Fluminense. Nos estratos inferiores ocorrem valores um pouco
mais elevados na floresta, porém, de maneira geral se mantêm maiores nas áreas
secundárias.
Segundo Grime (1979), algumas espécies apresentam mecanismos de
rebrota pós-injúria ou perturbação. A rebrota funciona como estratégia de
sobrevivência importante para algumas espécies florestais, pois o sistema radicular
já se encontra estabelecido, economizando assim a energia necessária para a
formação das raízes a partir da semente, além de ser um mecanismo útil para
reocupar espaço e promover a cicatrização de áreas abertas, como vem sendo
verificado para algumas florestas do Estado do Rio de Janeiro (Assunpção e
Nascimento, 2001; Oliveira, 2002; Pessoa, 2003).
As lianas são abundantes nas florestas tropicais maduras (Gentry, 1991;
Carsten, 2002; Burnham, 2002) e em áreas perturbadas são mais numerosas ainda,
devido principalmente ao fato de serem heliófilas (Putz, 1984). A porcentagem de
lianas em relação ao número de indivíduos amostrados em todos os estratos da
floresta foi de 24%, estando compatível com o detectado por Gentry (1991). Para as
áreas secundárias variou de 120% para a capoeira, 62% cabruca e 237% na

73
capoeirinha. Na cabruca a porcentagem de lianas obtida foi bem menor, pois quando
da manutenção do cultivo de cacau boa parte delas foi suprimida e parte das árvores
não, alterando a proporcionalidade. Segundo Dewalt et al. (2000), em áreas de
florestas secundárias no Panamá (20 a 40 anos de idade) as lianas são mais
abundantes que em florestas mais antigas (70 anos ou mais), coerente com os
resultados obtidos neste trabalho (floresta, 38.520 lianas/ha; capoeira, 70.072
lianas/ha).
Segundo Nunes (1996), ocorre uma grande redução no número de lianas com
menores diâmetros em relação a aqueles com diâmetros mais elevados. Que pode
ser atribuído à elevada mortalidade dos indivíduos mais novos. Fato detectado
também no presente trabalho, onde houve grande redução no número de indivíduos
a partir de lianas com 2,5 cm ≤ DAS < 5 cm em relação aos diâmetros maiores (5 cm
≤ DAS < 10 cm e DAS ≥ 10 cm)
3.4.2. Comparações florísticas
A riqueza de espécies vasculares na Floresta Atlântica é uma das maiores do
planeta (Giullieti, 1992). Esta pode estar relacionada ao elevado número de
indivíduos por área (Martini, 2002), ao grande número de espécies com apenas um
indivíduo por hectare (Martins, 1991; Rolim e Nascimento, 1997; Rizzini, 2000) e a
sua dinâmica (Phillips e Gentry, 1994), principalmente nas florestas maduras. Os
dados de riqueza e diversidade de espécies observados para a floresta estudada
corroboram esta afirmação e podem ser considerados altos e na faixa superior do
observado para outras áreas de Floresta Atlântica (Thomaz e Monteiro, 1997;
Moreno et al., 2003; Rodrigues, 2004; Jesus e Rolim, 2005). Entretanto, essa alta
riqueza de espécies, em geral, não ocorre nas áreas secundárias com menor tempo
de abandono (capoeirinha) e infestada por Gramineae (Penisetum sp.).
A velocidade do processo de regeneração em determinada área degradada
no domínio de florestas depende de vários fatores tais como: disponibilidade de
sementes, presença de dispersores bióticos e abióticos, proximidade ou não de
remanescentes, do nível de degradação sofrida pela vegetação e pelo solo da área
(Uhl et al., 1988; Kageyama e Castro, 1989) em função da influência do uso agrícola
sofrido.
A cabruca é um bom exemplo, visto que o processo de cultivo do cacau
(Theobroma cação) deixou árvores remanescentes da floresta e o cacau não tem

74
demonstrado ser uma espécie agressiva e competitiva a ponto de dificultar a
proliferação das nativas do entorno. Por outro lado, áreas densamente cobertas por
gramíneas, podem afetar a sobrevivência e o crescimento de plântulas, devido à
alteração da qualidade e quantidade luminosa que chega ao solo, competição por
água, nutrientes, alelopatia ou alterando a atividade e a abundância de herbívoros
(Holl, 2002).
O Pennisetum sp. (capim elefante), parece estar competindo fortemente e
dificultando o estabelecimento de espécies nativas na área da capoeirinha. Se
compararmos o número de indivíduos de espécies arbustivo-arbóreas que se
encontra em processo de estabelecimento na cabruca e na capoeirinha, com mesmo
tempo de suspensão da manutenção da atividade agrícola (ca. de nove anos) as
varas e os jovens na área da cabruca são no mínimo três vezes maiores. Corrobora
com resultados obtidos por Sambuichi (2002) e Guevara et al. (2004). O primeiro
ressalta a importância biológica das áreas de cabruca no sul da Bahia e o segundo
detectou estabelecimento de bosque denso com 4-5 m de altura sob copa de Ficus
spp. num período de três anos.
As áreas com cabruca já instalada, realmente podem possuir alta resiliência
quanto à regeneração se as atividades de controle da sucessão forem suspensas,
mas, segundo Rolim e Chiarello (2004), essas áreas em longo prazo tendem a ter
sua riqueza e diversidade reduzidas, devido o impedimento da renovação das
espécies adultas que forem morrendo, em função da manutenção do cultivo de
cacau, sendo na atualidade, inconcebível o raleamento de floresta madura para
plantio de cacau no sistema de cabruca na Floresta Atlântica.
Em áreas secundárias em regeneração, tem sido observado que sua riqueza
e diversidade de espécies aumentam em função do período de tempo decorrido da
última perturbação (Saldarriaga et al., 1988; Brown e Lugo, 1990; Tabarelli e
Mantovani, 1999). Confirmando as tendências de sucessão entre áreas secundárias,
a diversidade de espécies foi maior na capoeira, que apresentou valores
semelhantes à floresta para arvoretas, varas e jovens.
Nas florestas tropicais maduras muitas espécies estão adaptadas à sombra
do dossel ou às pequenas clareiras oriundas da queda de árvores de grande porte,
não ocorrendo predomínio absoluto de determinada espécie (Uhl e Murphy, 1981;
Clark e Clark, 1987; 1992). A presença de espécies como Eschweilera ovata e
Hydrogaster trinervis com os maiores valores de cobertura para árvores na floresta,

75
com baixa ocorrência nos estratos regenerantes da floresta, está de acordo com a
dinâmica natural das florestas tropicais. Por outro lado, Cecropia glaziovi,
considerada pioneira (Lorenzi, 1998), aparece tanto no estrato arbóreo como no de
arvoretas da floresta, indicando que surgiu, provavelmente, após queda de árvore de
grande porte ou corte seletivo de madeira.
Em florestas secundárias, geralmente ocorre dominância de algumas
espécies que se instalam logo após a perturbação e vão sendo substituídas a longo
do tempo (Kleim, 1980; Uhl et al., 1988; Tabarelli et al., 1993). Os resultados indicam
que isto vem ocorrendo com a Joannesia princeps e Guazuma crinita, que na
capoeira predominam no estrato arbóreo e aparecem em baixa densidade nos
estratos regenerantes, com nítidas características de secundárias iniciais. Ainda
neste sentido, das 109, 68, 80 e 125 espécies que ocorreram na floresta nos
estratos de árvores, arvoretas, varas e jovens, respectivamente, 43, 16, 28 e 69
espécies encontravam-se com ocorrência nos estratos (árvores, arvoretas, varas e
jovens) das áreas secundárias, indicando com isso que indivíduos destas espécies,
provavelmente, oriundos de sementes da floresta estão se estabelecendo nestas
áreas.
O predomínio de Dalbergia nigra (jacarandá da Bahia) na capoeirinha e a
Tapirira guianensis na cabruca, ocorrendo em todos os estratos destes sítios e em
alguns estratos regenerantes de outros sítios, indica que estas espécies possuem
preferência por clareiras grandes, embora possam ser encontradas na floresta
madura. O resultado para a Dalbergia nigra está de acordo com Lorenzi (1992), já o
da Tapirira guianensis corrobora em parte, visto que embora esta espécie atue como
pioneira ou secundária inicial, ela foi detectada no estrato regenerante da floresta e
da capoeira.
Trabalhos realizados em florestas maduras no Norte Fluminense, Espírito
Santo e sul da Bahia, detectaram que nestas áreas há uma grande riqueza e
diversidade de espécies vegetais (Thomaz e Monteiro, 1997; Moreno et al., 2003;
Rodrigues, 2004; Jesus e Rolim, 2005; Thomas e Amorim, 2005a,b,c,d,f;).
Tendências semelhantes, também foram observadas para florestas com fortes
impactos antrópicos, principalmente, coleta de madeira e efeito e borda (Silva e
Nascimento, 2001; Rodrigues, 2004; Carvalho, 2005; Villela et al., 2006; Rolim et al.,
no prelo). Neste sentido, os resultados obtidos na FLONA de Goytacazes,
confirmaram estas observações e, indicam que a regeneração da uma floresta

76
suprimida e com 33 anos de abandono vem ocorrendo com sucesso (62% de
similaridade do estrato de jovens com aquele da floresta madura), apesar de não ter
sofrido qualquer intervenção para auxiliar na sua recuperação.
Os resultados indicam que a análise de riqueza, diversidade e composição
florística são de fundamental importância para a classificação da vegetação em
regeneração, conforme indicado de forma subjetiva pela resolução Conama (1994) e
IDAF (1999), devendo no futuro serem fixados faixas de valores para estes
parâmetros (riqueza, diversidade), tornando a classificação mais objetiva.
3.5. Conclusões
Os resultados obtidos a partir da comparação de áreas com diferentes usos
do solo confirmam a hipótese de que a regeneração natural em três trechos
desmatados e com infestação de Gramineae e cultivo cacau (Theobroma cacao L.),
adjacentes ao remanescente de Floresta Atlântica (Floresta Ombrófila Densa Aluvial)
diferem pelo tipo de uso do solo, como por exemplo a cabruca (estágio médio) e
capoeirinha (mecega), que com o mesmo tempo de abandono apresentaram
diferenças significativas quanto à regeneração.
A capoeira com 33 anos em regeneração após infestação com Panicum
maximum (colonião), apresentou florística e estrutura do estrato regenerante
(arvoretas, varas, jovens, plântulas) próxima da floresta madura, entretanto para o
estrato arbóreo foi expressivamente inferior, encaixando na categoria de estágio
médio de regeneração da Floresta Atlântica. A cabruca com nove anos em
regeneração contém biomassa aérea próxima da floresta madura, mas a diversidade
e o estrato regenerante encontram-se bem distintos da floresta madura, podendo ser
classificada como em estágio médio de regeneração da Floresta Atlântica. Já a
capoeirinha, também com nove anos de abandono, porém com infestação de
Gramineae (Penisetum sp.), encontra-se muito mais dissimilar da floresta madura,
tanto para parâmetros estruturais quanto florísticos, podendo ser classificada como
em estágio de macega.
Estudos de sucessão ecológica em áreas abandonadas após distúrbios
(desmatamento, queimadas, etc.) de Floresta Atlântica, com uso leve do solo e
adjacentes à floresta, são escassos o que dificulta estabelecar prognósticos sobre o
tempo de recuperação dessas áreas, todavia, a partir dos resultados obtidos neste
trabalho pode-se prever que o tempo para atingir os estágios de macega (ca. nove

77
anos) e médio de regeneração da Floresta Atlântica (ca. 33 anos) e para atingir o
estágio maduro, baseado em informações da literatura para outras florestas (Uhl e
Murphy, 1981; Uhl e Saldarriaga, 1986) podem ser estimados em cerca de 200 anos
(figura 3.5-1). A cabruca não foi incluída no fluxograma da referida figura por ter
sofrido um tipo de interferência muito distinta da capoeira e da capoeirinha.
A floresta madura bem como as áreas em regeneração (estágios inicial,
médio e avançado de regeneração da Floresta Atlântica), constitui um mosaico rico
em espécies e com diversidade alta, riqueza e fluxo de espécies (regeneração) para
o complexo de vegetação na FLONA de Goytacazes e indicam a importância da
manutenção e conservação dessas áreas florestadas no Estado do Espírito Santo.
Figura 3.5-1. Modelo de sucessão secundária em uma floresta Ombrófila Densa Aluvial após corte e queima para uso agrícola na FLONA de Goytacazes, Linhares/ES. Dados baseados no estrato arbóreo (DAP ≥ 10 cm). AB – área basal; N – número de indivíduos; Nspp. – número de espécies; H’- diversidade de Shannon.
Floresta Madura
Uso Agrícola e Pastoreio
Macega - Capoeirinha
Estágio Médio - Capoeira
Floresta Madura
Desmatamento e queima
Abandono por 9 anos
Abandono por mais 24 anos
??? 140 - 200 Anos
AB.ha -1 = 30 m2 N.ha -1 = 588
Nspp.0,5ha -1 = 109 H’ = 4,3
AB.ha -1 = 0,6 m2 N.ha -1 = 34
Nspp. 0,5.ha -1 = 9 H’ = 2,1
AB.ha -1 = 12,8 m2 N.ha -1 = 576
Nspp. 0,5ha -1 = 64 H’ = 3,1
AB. ha -1 ≥ 30 m2. N.ha -1 ≥ 500
Nspp.0,5ha -1 ≥ 110 H’ ≥ 4
Eschweilera ovata; Hidrogaster trinervis; Pterocarpus rohrii; Polyandrococos caudescens.
Penisetum sp.; Panicum maximum; Dalbergia nigra; Psidium guajava; Parapiptadenia pterosperma.
Eschweilera ovata; Hidrogaster trinervis; Pterocarpus rohrii; Polyandrococos caudescens.
Joannesia princeps; Guazuma crinita; Grazielodendron rio-docensis; Guazuma ulmifolia.

78
CAPÍTULO 4
CONSIDERAÇÕES FINAIS
Embora as florestas tropicais detenham a maior biodiversidade do planeta,
elas vêm sendo desflorestadas a passos largos nas últimas décadas. Fato
facilmente comprovado pela avaliação da cobertura de remanescentes de Floresta
Atlântica, com menos de 10% de sua área original. Este conjunto encontra-se
composto por fragmentos, que em sua maioria, sofreu perturbações de origem
antrópica como coleta seletiva de madeira.
No Estado do Espírito Santo, este padrão também é fato, sendo necessários
estudos que nos forneçam base para estabelecimento de estratégias de
conservação de áreas com remanescentes vegetais e com potencial para criação de
corredores ecológicos, assim como para recomposição de ambientes degradados.
Muitas áreas cujo solo outrora foi utilizado para produção agrícola, hoje se
encontram abandonadas e em estágio de regeneração natural. A partir de uma
fiscalização mais rígida, na última década, áreas em estágios médios e avançados
de regeneração vêm sendo mantidas, mas pouco se sabe sobre a estrutura e
florística desses remanescentes.
A velocidade da regeneração de áreas florestadas após uso agrícola depende
de uma série de fatores, dentre estes, se destaca o tempo de abandono, o uso
anterior do solo, crescimento do banco de sementes e proximidade de área fonte de
propágulos (floresta suporte). Esses remanescentes florestais funcionam como
áreas com alta diversidade e são doadoras de propágulos. Desta forma, pode-se
concluir que o processo de recuperação de fragmentos florestais ou de áreas
degradadas, com o aumento da diversidade de espécies vegetais é em muito
favorecido se existe uma floresta no entorno da respectiva área. Assim, a
conservação de remanescentes florestais do Bioma Mata Atlântica é fundamental
para manutenção de sua alta biodiversidade e de viabilizar a recuperação
(regeneração) de áreas degradadas em seu entorno.
Neste contexto, a Floresta Ombrófila Densa Aluvial da FLONA de Goytacazes
é um remanescente existente sobre sedimentos aluviais do holoceno (quaternário),
no delta do Rio Doce e sendo a única unidade de conservação com toda sua área

79
localizada em planície de sedimentos aluviais, tornando-se única e de relevância
ímpar quanto à conservação e possibilitando estudos, cujos resultados tornam-se de
suma importância para o entendimento dos processos, tanto na floresta com
razoável nível de conservação, de áreas degradadas no entorno de florestas, bem
como em relação à recuperação de áreas degradadas em geral.
De certa forma, os resultados obtidos no presente trabalho, convergem para
um quadro positivo para a recuperação de áreas degradadas, cujo solo foi utilizado
para atividades agrícolas e com períodos variados em fase de regeneração natural e
que se encontram adjacentes a floresta madura.
A vegetação da FLONA de Goytacazes sofreu no passado várias pressões
como desmatamento para pesquisa agrícola, coleta seletiva de madeira, coleta de
palmeiras para consumo do palmito, incêndios na sua bordadura e caça, devido
principalmente ao fato de estar localizada próxima a área de alta densidade urbana,
cerca de 3 km da cidade de Linhares.
Apesar de todas as pressões sofridas, a floresta madura da FLONA contem
uma das maiores diversidades vegetais para a costa brasileira, com presença
inclusive de oito espécies da flora capixaba e brasileira ameaçadas de extinção
(Terminalia kuhlmannii, Beilschmiedia linharensis, Ocotea confertiflora,
Moldenhawera papillanthera, Dalbergia nigra, Mollinedia marquetiana,
Campomanesia espiritosantensis e Myrcia folli).
Este fragmento é fundamental para o estabelecimento de grandes corredores
ecológicos, funcionando como importante elo para a conectividade, entre fragmentos
ao norte e sul do Rio Doce, servindo como refúgio para a fauna, facilitando fluxos
biológicos, de sementes e grãos de pólen e consequentemente o processo de
regeneração natural das comunidades florestais no seu entorno. Assim a FLONA de
Goytacazes assume fundamental papel como banco genético de espécies vegetais,
para programas de florestamento e recuperação ambiental. Os resultados podem
auxiliar também na recomendação de espécies vegetais e serem utilizadas em
projetos de revegetação ou de enriquecimento de áreas em varios estágios de
sucessão.
Existem poucos estudos no domínio da Floresta Atlântica e até mesmo das
florestas tropicais com relação à regeneração de florestas a partir de áreas que
foram degradadas para uso agrícola. Especialmente no Brasil, muitas das
informações acerca da regeneração são generalizadas a partir de estudos

80
desenvolvidos na América Central e na Amazônia. Na Amazônia, foram realizados
alguns trabalhos a partir da década de 1980, que trazem informações preciosas
sobre os processos que envolvem a regeneração.
Para a Floresta Atlântica, na última década foram realizados trabalhos
abordando este tema, também trazendo informações importantes. Entretanto, para o
domínio da Floresta Atlântica, ainda é muito pouco o que tem sido feito em termos
de pesquisas. As decisões têm sido tomadas utilizando informações extrapoladas e
a partir de poucas observações empíricas.
Com mais de 90% da Floresta Atlântica desflorestada, a sua recuperação
passa por plantios de pequenas áreas com níveis de degradação fortes e que sejam
irrecuperáveis ao longo do tempo sem uma intervenção através do plantio e manejo
do solo. Contudo, a recuperação da diversidade vegetal máxima de determinada
área só é viável através dos processos regenerativos (mesmo após a revegetação),
uma vez que, o conhecimento sobre as propriedades e exigências das espécies
ainda estão longe de serem desvendados, devido a enorme riqueza existente nas
áreas florestadas do domínio da Floresta Atlântica e os custos financeiros
necessários.
No manejo destas áreas sugere-se como, por exemplo, o controle do
Pennisetum sp. na macega, permitindo um maior incremento no processo de
regeneração e acelerando o tempo necessário para o retorno da floresta madura.
Sugere-se também que seja dada continuidade na coleta e análise de dados
relativos à dinâmica nas diferentes áreas para que se possa entender melhor a
substituição das espécies ao longo do tempo.

81
REFERÊNCIAS BIBLIOGRÁFICAS
Aide, T. M., Zimmerman, J. K., Rosario, M., Marcano, H. (1996) Forest recovery in
abandoned cattle pastures along an elevational gradient in northeastern Puerto
Rico. Biotropica. 28 (4):537-548.
Andrade, K.V.S.A., Rodal, M.J.N. (2004) Fisionomia e estrutura de um remanescente
de floresta estacional semidecidual de terras baixas no nordeste do Brasil.
Revista Brasileira de Botânica 27 (3):463-474.
Assis, A.M., Thomaz, L.D., Pereira, O. J. (2004) Florística de um trecho de floresta
de restinga no munícipio de Guarapari, Espírito Santo, Brasil. Acta Botanica
Brasilica 18 (1):191-201.
Assunpção, J., Nascimento, M.T. (2001) Estrutura e composição florística de quatro
formações vegetais de restinga no complexo lagunar Grussaí/Iquipari, São João
da Barra, RJ, Brasil. Acta Botanica Brasilica 14 (3):301-315.
Barbosa, L. M., Asperti, L. M., Bedinelli, C., Barbosa, J. M., Belasque, E. F. , Pirré, E.
(1992) Informações básicas para modelos de recuperação de áreas degradadas
de Mata Ciliares. Revista do Instituto Florestal 4:640-644.
Bellini, C., Martins Filho, S., Thomé, J.C., Moreira, L.M.P., Sá, S.S. (1990)
Caracterização ambiental e mapeamento das interferências antrópicas na região
do ecossistema Rio-Lagoa Monsarás, Povoação, ES. Anais do Simpósio de
Ecossistemas da Costa Sul e Sudeste Brasileira, Estrutura, Função e Manejo, 2.
Águas de Lindóia: ACIESP-SP 71 (2):73-85.
Bigarella, J.J., Andrade-Lima, D., Riehs, P.J. (1975) Considerações a respeito das
mudanças paleoambientais na distribuição de algumas espécies vegetais e
animais no Brasil. Anais Academia Brasileira de Ciências, v.47, p.411-464.
Brower, J.E., Zar, J.H. (1977) Field and laboratory methods for general ecology.
Iwoa: W. C. Brown Company Publishers.
Brown, S., Lugo, A. (1990) Tropical secondary forests. Journal of Tropical Ecology
6:1-32.
Budowski, G. (1965) Distribution of Tropical American Rain Forrest Species in the
Light of Successional Processes. Turrialba 15 (1):40-42.
Burnham J.B. (2002) Dominance, diversity and distribution of lianas in Yasuní,
Ecuador: who is on top?. Journal of Tropical Ecology 18:845-864.

82
Buschbacher, R. (1986) Tropical deforestation and pasture development. Bioscience
36:23-27.
Capobianco, J.P.R., Lima, A.A. (1997) A evolução da proteção legal da Floresta
Atlântica. Documento do Instituto Socioambiental (ISA) no 4.
Carsten, L.D., Juolat, F.A., Male, T.D., Cherry, S. (2002) Host associations of lianas
in a south-east Qeensland rain forest. Journal of Tropical Ecology 18:107-120.
Carvalho, F.A. (2005) Efeitos da fragmentação florestal na florística e estrutura da
Floresta Atlântica submontanha da região de Imbaú, município da Silva Jardim,
RJ. Dissertação (Mestrado em Biociências e Biotecnologia) - Campos dos
Goytacazes- RJ, Universidade Estadual de Norte Fluminense – UENF, 124p.
Carvalho, F.A., Nascimento, M.T., Oliveira, P.P., Rambaldi, D.M., Fernandes, R.V.
(2004) A importância dos remanescentes florestais da Mata Atlântica da baixada
costeira fluminense para a conservação da biodiversidade na APA da bacia do
rio São João/Mico-Leão-Dourado/IBAMA – RJ. Anais do Congresso Brasileiro de
Unidades de Conservação, Curitiba, v.1, p.106-113.
Cientec (2005) Sistema para análise fitossociológica e elaboração de planos de
manejo de florestas nativas, CD-ROM, Viçosa, MG.
Clark, D. A., Clark, D. B. (1987) Análisis de la regeneración de árboles del dosel en
bosque muy húmedo tropical: aspectos teóricos y prácticos. Revista de
BiologiaTropical 35 (Supl. 1):41-54.
Clark, D.A., Clark, D.B. (1992) Life history diversity of canopy and emergent trees in
a neotropical rain forest. Ecologycal monographs 62 (3):315-344.
Conama (1994) Resolução do Conselho Nacional de Meio Ambiente n0 29 de
setembro de 1994. Brasília: MMA.
Costa, L.G.S., Piña-Rodrigues, F.C.M., Jesus, R.M. (1992) Grupos ecológicos e a
dispersão de sementes de espécies arbóreas em trecho da florestal tropical na
Reserva Florestal de Linhares (ES). Revista do Instituto Florestal 4:303-305.
Cronquist, A. (1968) The evolution and classification of the flowers plants. Nova York:
Willian C. Steere, 395p.
Dean, W. (2000) A ferro e a fogo: a história e a devastação da Floresta Atlântica.
São Paulo: Schwarcz, 484p.
Dewalt, S.J., Shnitzer, S.A., Denslow, J.S. (2000) Density and diversity of lianas
along a chronosequence in a Panamaniam lowland forest. Journal of Tropical
Ecology 16:1-19.

83
Durigan, G. (2003) Bases e diretrizes para a restauração da vegetação do cerrado.
In: Kageyama, P.Y., Oliveira, R.E. de, Moraes, L.F.D de, Engel, V.L., Gandara,
F.B (orgs.). Restauração ecológica de ecossistemas naturais. Botucatú: FEPAG,
240p.
Egler, W.A. (1951) A zona pioneira ao norte do Rio Doce. Revista Brasileira de
Geografia 13 (2):223-264.
Embrapa (1978) Serviço nacional de levantamento e conservação de solos.
Levantamento e reconhecimento dos solos do Estado do Espírito Santo. Rio de
Janeiro. Boletim Técnico 5, 461p.
Embrapa (1999) Sistema brasileiro de classificação de solos. Brasília: Serviço de
produção de informação, 412p.
Emmons, L.H., Gentry, A.H. (1983) Tropical forest structure and the distribution of
gliding and prehensile-tailed vertebrates. The Americam Naturalist 121:513-524.
Engel, V.L. (2001) Estudo fenológico de espécies arbóreas de uma floresta tropical
em Linhares, ES. Tese (Doutorado em Ecologia) - Campinas - SP, Universidade
Estadual de Campinas – UNICAMP, 137p.
Espírito Santo (Governo do Estado do Espírito Santo) (2005) Lista de espécies
ameaçadas de extinção no Espírito Santo, Decreto Estadual n0 1.499-R de 14 de
junho de 2005.
Espírito Santo (Governo do Estado do Espírito Santo) (1989) Programa de
desenvolvimento florestal do Espírito Santo, V.1, p. 109.
Espírito Santo (Secretaria de Estado da Agricultura – SEAG) (1999) Rumos da
agricultura capixaba: Plano de ação 2002-2003. Vitória, ES, 72p.
Fabris, L.C. (1995) Composição florística e fitossociológica de uma faixa de floresta
arenosa litorânea do Parque Estadual de Setiba, município de Guaraparí, ES.
Dissertação (Mestrado em Biologia Vegetal) - Rio Claro - SP, Universidade
Estadual Paulista, 195p.
Fabris, L.C., Cesar, O. (1996) Estudos florísticos em uma Floresta litorânea no sul do
Estado do Espírito Santo, Brasil. Boletim do Museu de Biologia Mello Leitão
5:15-46.
FAO (1997) State of the World’s Forests. Rome: Food and Agriculture Organization
of the United Nations. Rome, Italy, 200p.
Feitoza, L.R., Castro, L.L.F., Resende, M., Zangrane, M.B., Stocking, M., Borel,
R.M.A., Cerqueira, A.F., Salgado - Filho, J.S., Feitosa, H.N., Fullin, E.A., Stock,

84
L.A., Dessaune – Filho, N., Mank, A.M., Feringa, W. e Martinez, J.A. (1999)
Mapa das unidades naturais do Espírito Santo. Vitória: EMCAPA, ES.
Fimbel, R.A., Fimbel, C.C. (1996) The role of exotic conifer plantations in
rehabilitating degraded tropical forest lands: A case study from the Kibale Forest
in Uganda. Forest Ecology and Management 81: 215-226.
Garbutt, R.A., Sparks, T.H. (2001) Changes in the botanical diversity of uma species
rich ancient hedgerow between two surveys (1971–1998). Biological
conservation 106:273-278.
Garcia, A. (2002) O fomento florestal como vetor para o desenvolvimento rural do
Estado do Espírito Santo. In: Programa Nacional de Florestas – PNF/MMA.
Vitória: Projeto FAO UTF/BRA/047, p.1-29.
Gentry, A.H. (1988) Changes in plant community diversity and floristic composition
on envirommental and geographical gradientes. Annals of the Missouri Botanical
Gardem 75 (1):1-34.
Gentry, A.H. (1991) The distribution and avolution of climbing plants. In: Putz, F.E.,
Mooney, H.A. (eds.) The Biology of vines. Cambridge University Press, p.3-39.
Giulietti, A.M. (1992) Biodiversidade da região sudeste. Revista do Intituto Florestal
4(especial), p.125-130.
Giulietti, A.M., Forero, E. (1990) Workshop: diversidade taxonômica e padrões de
distribuição das angiospermas brasileiras. Introdução. Acta Botanica Brasilica 4
(1):3-9.
Gómez-Pompa, A., Burley, Y.F.W. (1991) The management of natural tropical
forests. In: Gomez-Pompa, A., Whitmore, T.C., Hadley, M. (eds.). Rain Forest:
regeneration and manegement. New York: Parthenon Publishing, p.3-17.
Gómez-Pompa, A., Vázques-Yanes, C. (1976) Estudios sobre sucesion secundaria
en los tropicos calidos-humedos: el ciclo de vida de las especies secundarias. In:
Gómez-Pompa, A., Vázques-Yanes, C., Delamo, S., Butando, A. (eds.)
Regeneracion de selvas. México: Cia editorial Continental, p.579-591.
Gotelli, N.J., Entsminger, G.L. (2001) EcoSim: Null models softwere for ecology.
Verson 7.0. Acquired intelligence Inc., Kesey Bear. Disponível em
http://homepages.together.net~gentsmin/ecosim.htm. Acesso em 15 de
setembro de 2005.
Grime, J.P. (1979) Estratégias de adaptación de las plantas y procesos que
controlan la vegetación. México: Noriega Editores, 291p.

85
Guedes-Bruni, R.R.B. (1998) Composição, Estrutura e Similaridade Florística de
Dossel em Seis Unidades Fisionômicas de Floresta Atlântica no Rio de Janeiro.
Tese (Doutorado em Ecologia) - São Paulo, Universidade de São Paulo - USP,
231p.
Guevara, S., Laborde, J., Sanchez, G. (1998) Are isolated remnant trees in pastures
a fragmented canopy? Selbyana 19 (1):34-43.
Guevara, S., Laborde, J., Sanchez-Rios, G. (2004) Rain forest regeneration beneath
the canopy of Fig trees isolated in pastures of Los Tuxtlas, Mexico. Biotropica 36
(1):99-108.
Heinsdijk, D., Macedo, J.G., Angel, S., Ascoly, R.B. (1965) A floresta do norte do
Espírito Santo: dados e conclusões de um inventário florestal piloto.
Departamento de Recursos Naturais Renováveis 9 (7):1-69.
Holl, K.D. 2002 Efect of shrubs on tree seedling establishment in an abandoned
tropical pasture. Journal of Ecology 90:179-187.
IBAMA (Instituto Brasileiro de Meio Ambiente e dos Recursos Renováveis) 1992
Lista das espécies da flora brasileira ameaçada de extinção, Portaria Nº 37-
N/IBAMA, de 3 de abril de 1992.
IBGE (Instituto Brasileiro de Geografia e Estatística) (1987) Levantamento de
recursos naturais, folha SE 24 Rio Doce. Rio de Janeiro, 544p.
IBGE (Instituto Brasileiro de Geografia e Estatística) (1993) Mapa de vegetação do
Brasil. Diretoria de Goeciências.
IDAF (Instituto de defesa agropecuária e florestal do Espírito Santo) (1999)
Legislação florestal do Espírito Santo, Lei 5.361. Vitória-ES, 112p.
IK (1997) Index kewensis 2.0, CD-ROM, Royal Botanic Gardens, Kew.
Jesus, R.M. (2001) Manejo Florestal: Impactos ecológicos de diferentes níveis de
remoção e os impactos de sua sustentabilidade. Tese (Doutorado em Ecologia) -
Campinas - SP, Universidade Estadual de Campinas - UNICAMP, 244p.
Jesus, R.M., Garcia, A. (1992) Index seminum - Reserva Florestal de Linhares,
Espírito Santo - Brasil. Revista do Instituto Florestal 4:306 – 317.
Jesus, R.M., Rolim, G.S. (2005) Fitossociologia da Mata Atlântica de Tabuleiro.
Boletim Técnico SIF 19, 136p.
Kageyama, P.Y., Castro, C.F. de A. (1989) Sucessão secundária, estrutura genética
e plantações de espécies arbóreas nativas. IPEF, Piracicaba - SP, (41/42):83-

86
93.
Kageyama, P.Y., Oliveira, R. E., Moraes, L.F. de (2003) Restauração ecológica de
ecossistemas naturais. Botucatu-SP: Fundação de estudos e pesquisas
agrícolas e florestais – FEPAF, 340p.
Kappelle, M., Geuze, T., Leal, M.E., Cleef, A.M. (1996) Successional age and forest
structure in a Costa Rican upper montane Quercus forest. Journal of Tropical
Ecology 12:681-698.
Lani, J.L. (1998) Delta dos rios Doce e Itapemirim: solos, com ênfase nos
tiomórficos, água e impacto ambiental do uso. Tese - Viçosa - MG, Universidade
Federal de Viçosa – UFV, 169p.
Lopes, W.P., Silva, A.F., Neto, J.A.A.M. (2002) Estrutura fisiológica de um trecho de
vegetação arbórea no Parque Estadual do Rio Doce (Minas Gerais, Brasil). Acta
Botanica Brasilica 16(4): 443-456.
Lorenzi, H. (1992) Árvores brasileiras: Manual de identificação e cultivo de plantas
arbóreas nativas do Brasil. Nova Odessa, SP: Editora Plantarum, v.1, 373p.
Lorenzi, H. (1998) Árvores brasileiras: Manual de identificação e cultivo de plantas
arbóreas nativas do Brasil. 2. ed. Nova Odessa, SP: Editora Plantarum, v.2,
352p.
Mack, A.L., Ickes, k., Jessen, J.H., Kennedy, B., Sinclair, J.R. (1999) Ecology of
Aglaia mackiana, seedlings in a New Guinea rain forest. Biotropica 31 (1):111-
120.
Molchanov, A.A. (1963) Hidrologia florestal. Lisboa: Ed. Fundação Calouste
Gulbenkian, 419p.
Martin, L., Suguio, K., Dominguez, J.M.L., Flexor, J-M. (1997) Geologia do
Quaternário costeiro do litoral norte do Rio de Janeiro e do Espírito Santo. Belo
Horizonte: CPRM, 112 p.
Martinez-Ramos, M. (1994) Regeneracíon natural y diversidad de especies arbóreas
en selvas húmidas. Boletín de la Sociedad Botánica de México 54:179-224.
Martini, A.M.Z. (2002) Estrutura e composição da vegetação e chuva de sementes
em sub-bosque, clareiras naturais e área perturbada por fogo em Floresta
Tropical no Sul da Bahia. Tese (Doutorado em Ecologia) - Campinas-SP,
Universidade Estadual de Campinas - Unicamp, 125p.
Martins, F. R. (1991) Estrutura de uma floresta mesófila. Campinas: Editora da
Unicamp, 246p

87
Martins, S.V. (2001) Recuperação de matas ciliares. UFV. Viçosa: Aprenda Fácil,
146p.
MMA (2002) Biodiversidade brasileira: avaliação e identificação de áreas e ações
prioritárias para conservação, utilização sustentável e repartição dos benefícios
da biodiversidade nos biomas brasileiros. Brasília: Ministério do Meio Ambiente,
Secretaria de biodiversidade e florestas. 404p.
Morellato, L.P.C. (1991) Estudos da fenologia de árvores, arbustos e lianas de uma
floresta semidecídua no sudeste do Brasil. Tese (Doutorado em Ecologia) -
Campinas-SP, Universidade Estadual de Campinas - Unicamp, 176p.
Morellato, L.P.C., Haddad, C.F.B. (2000) Introduction: The Brazilian Atlantic Forest.
Biotropica 32 (4b):786-792.
Moreno, M.R., Nascimento, M.T., Kurtz, B.C. (2003) Estrutura e composição florística
do estrato arbóreo em duas zonas altitudinais na Floresta Atlântica de Encosta
da região do Imbé – RJ. Acta Botanica Brasilica 17 (3):371-386.
Mori, S.A., Boom, B.M., Carvalho, A.M., Talmón, S. S. (1983) Southern Bahian Moist
Forest. The New York Botanical Garden, New York, 49 (2):150-204.
Mori, S.A., Boom, B.M., Prance, G.T. (1981) Distribuition of eastern Brazilian costal
forest tree species. Brittonia 33 (2):233-245.
Mueller-Dombois, D., Ellenberg, H. (1974) Aims and methods of vegetation ecology.
New York: John Wiley & Sons, 525p.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., Fonseca, G.A.B., Kent, J. (2000)
Biodiversity hotspots for conservation priorities. Nature 403:845-853.
Nunes, M.F.S.Q.C. (1996) Estudo do potencial de regeneração das espécies de uma
floresta tropical de tabuleiros - Linhares, E.S. Tese (Doutorado em Ecologia) -
São Carlos – MG, Universidade Federal de São Carlos v.1, p.212., v.2, p.103.
Oliveira, R.R. (2002) Ação antrópica e resultantes sobre a estrutura e composição da
Floresta Atlântica na Ilha Grande, RJ. Rodriguésia 53:33-58.
Oliveira, R.R., Sampaio, P.D., Silva, R.F., Toffoli, D.G. (1994) Roça caicara, um
sistema “primitivo” auto sustentável. Ciência Hoje 18:44-51.
Oliveira-Filho, A.T., Fontes, M.A.L (2000) Paterns of floristic differentiation among
Atlantic forests in south-eastern Brazil, and the influence of climate. Biotropica 32
(4b):793-810.
Oliveira-Filho, A.T., Tameirão-Neto, E., Carvalho, W.A.C., Werneck, M., Brina, A.E.,
Vidal, C.V., Rezende, S.C., Pereira, J.A.A. (2005) Análise florística do

88
comportamento arbóreo de áreas de floresta Atlântica sensu lato na região das
Bacias do leste (Bahia, Minas Gerais, Espírito Santo e Rio de Janeiro).
Rodriguésia 56 (87):85-235.
Paiva, H.N., Gonçalves, W. (2002) Florestas urbanas: Planejamento para melhoria
da qualidade de vida. Viçosa: Aprenda Fácil, 180p.
Passos, M.J. (1998) Estrutura da vegetação arbórea e regeneração natural em
remanescentes de Floresta ciliar do rio Mogi Guaçú – SP. Dissertação -
Piracicaba – SP, Escola Superior de Agricultura Luiz de Queiroz-ESALQ.
Peixoto, A.L. (1981) Novas espécies de Simira Aublet no norte do Espírito Santo.
Boletim do Museu Municipal, Curitiba 44:1-7.
Peixoto, A.L. (1982) Considerações preliminares sobre a flora e a vegetação da
Reserva Florestal da Companhia Vale do Rio Doce (Linhares - Espírito Santo).
Caderno de Pesquisa - Série Botânica 1:41-48.
Peixoto, A.L., Gentry, A.H. (1990) Diversidade e composição florística da Floresta
dos Tabuleiros na Reserva Natural da Vale do Rio Doce (Espírito Santo, Brasil).
Revista Brasileira Botânica 13:19-25.
Pereira, O.J. (1990) Caracterização fitofisionômica da restinga de Setiba-Guarapari-
Espírito Santo. Anais Simpósio de Ecossistemas da costa sul e sudeste
brasileira: Estrutura, função e manejo, 2. Águas de Lindóia: ACIESP, v.3, p.207-
219.
Pereira, O.J., Araújo, D.S. D. (2000) Análise florística das restingas dos Estados do
Espírito Santo e Rio de Janeiro. In: Esteves, F.A., Lacerda, L.D. (orgs.)
Oecologia Brasiliensis: Ecologia de Restingas e lagoas costeiras. Macaé – RJ:
NUPEM/UFRJ, p.25-63.
Pereira, O.J., Assis, A.M. (2000) Florística da restinga de Camburi, Vitória, ES. Acta
Botanica Brasilica 14 (1):99-111.
Pereira, O.J., Assis, A.M., Souza, R.L.D. (1998) Vegetação da restinga de Pontal do
Ipiranga, Município de Linhares (ES). Anais do Simpósio de Ecossistemas
Brasileiros, 4. São Paulo: ACIESP, V.3, p. 117-128
Pereira, O.J., Borgo, J.H., Rodrigues, I.D., Assis, A.M. (2000) Composição florística
de uma floresta de restinga no município de Serra-ES. Anais do Simpósio de
Ecossistemas Brasileiros: Conservação, 5. Vitória: ACIESP, p. 74-83.
Pereira, O.J., Gomes, J.M.L. (1994) Levantamento florístico das comunidades
vegetais de restinga no município de Conceição da Barra, ES. Anais do

89
Simpósio de Ecossistemas da costa brasileira, 3. Serra Negra, SP: ACIESP, v.3,
p. 67-78.
Pereira, O.J., Zambom, O. (1998) Composição florísitca da restinga de Interlagos,
Vilha Velha (ES). Anais do Simpósio de Ecossistemas Brasileiros, 4. São Paulo:
ACIESP, v.3, p. 129-139.
Pessoa, S.V.A. (2003) Aspectos da fragmentação em remanescentes florestais da
planície costeira do estado do Rio de Janeiro. Dissertação - Seropédica- RJ,
Universidade Federal Rural do Rio de Janeiro - UFRRJ, 96p.
Phillips, O.L., Gentry, A.H. (1994) Increasing turnover through time in tropical forests.
Science 263:954-957.
Piña-Rodrigues, F.C.M., Costa, L.S., Reis, A. (1990) Estratégias reprodutivas de
espécies arbóreas e o manejo de florestas tropicais. Congresso Florestal
Brasileiro, 6. Campos do Jordão, SP. Sociedade Brasileira de Silvicultura, v.3, p.
672-690.
Polhill, R.M., Raven, P.H., Stirton, C.H. (1981) Evolution and systematics of the
Fabaceae. In: Polhill, R.M., Raven, P.H. (eds). Advances in legume Systematics,
Kew : Royal Botanic Gardens. v.1, p.1-26.
Putz, F.E. (1984) The natural history of lianas on Barro Colorado Island, Panama.
Ecology 65:1713-1724.
Resende, M., Curi, N., Rezende, S.B., Corrêa, G.F. (2002) Pedologia: base para
distinção de ambientes. Viçosa: NEPUT. 338p.
Rice, R.A.; Greenberg, R. (2000) Cacao cultivation and the conservation of biological
diversity. Ambio 29 (3):167-173.
Rizzini, C.M. (2000) Diversidade funcional do estrato arbóreo como indicador do
status da biodiversidade em floresta atlântica de tabuleiros (Linhares - ES). Tese
(Doutorado em Geografia) - Rio de Janeiro, Universidade Federal do Rio de
Janeiro - UFRJ. v.1, 150p.
Rizzini, C.T. (1979) Tratado de fitogeografia do Brasil: Aspectos sociológicos e
florísticos. 2.ed. São Paulo: HUCITEC, v.2. 374 p.
Rodrigues, P.J.F.P. (2004) A vegetação da Reserva Biológica União e os efeitos de
borda na Floresta Atlântica fragmentada. Tese (Doutorado em Biociências e
Biotecnologia) - Campos dos Goytacazes - RJ, Universidade Estadual do Norte
Fluminense - UENF, 153p.

90
Rodrigues, R.R. (2001) Florestas ciliares? Uma discussão nomenclatural das
formações ciliares. In: Rodrigues, R.R., Leitão Filho, H.F. (orgs.) Matas Ciliares:
Conservação e Recuperação. São Paulo: Edusp/Fapesp, p.91-99.
Rodrigues, R.R., Nave, A.G. (2001) Heterogeneidade florística de matas ciliares. In:
Rodrigues, R.R., Leitão Filho, H.F. (orgs.) Matas Ciliares: Conservação e
Recuperação. São Paulo: Edusp/Fapesp, p.91-99.
Rodrigues, R.R., Shepherd, G.J. (2001) Fatores condicionantes da vgetação ciliar.
In: Rodrigues, R.R., Leitão Filho, H.F. (orgs.) Matas Ciliares: Conservação e
Recuperação. São Paulo: Edusp/Fapesp, p.101-108.
Rolim, S.G. (1997) Dinâmica da Floresta Atlântica em Linhares(ES). Dissertação
(Mestrado em Ciências Florestais) - Piracicaba - SP, Universidade de São Paulo
- USP, 87p.
Rolim, S.G., Chiarello, A.G. (2004) Slow death of Atlantica forest trees in cocoa
agroforestry in southeastern Brazil. Biodiversity and Conservation 13:2679-2694.
Rolim, S.G., Ivanauskas, N.M., Rodrigues, R.R., Nascimento, M.T.N., Gomes, J.M.L.
(no prelo) Composição florística do estrato arbóreo da floresta atlântica sobre a
planície aluvial do rio Doce (ES). Acta Botanica Brasilica.
Rolim, S.G., Nascimento, H.E.M. (1997) Análise da riqueza, diversidade e relação
espécie-abundância de uma comunidade arbórea tropical em diferentes
intensidades amostrais. Scientia Forestalis 52:7-16.
Roper, J., Roberts, R.W. (1999) Deforestation: tropical forests in decline. Disponível
em: http//WWW.rcfa-cfan.org. Acesso em 15 de dezembro de 2005.
Ruschi, A. (1950) Fitogeografia no Espírito Santo. Boletim do Museu de Biologia
Mello Leitão, Série Botânica 1:349.
Saint-Hilaire, A. (1974) Viagem ao Espírito Santo e Rio Doce. São Paulo: Ed.
Itatiaia,121 p.
Saito, M. (1992) As três funções da floresta e a importância das espécies indígenas.
Revista do Instituto Florestal 4:1029 - 1034.
Saldarriaga, J.G., West, D.C., Tharp, M. L., Uhl, C. (1988) Long-term
chronosequence in the upper Rio Negro of Colombia and Venezuela. Journal of
Ecology 76:938-958.

91
Sambuichi, R.H.R. (2002) Fitossociologia e diversidade de espécies arbóreas em
cabruca (Mata Atlântica raleada sobre plantação de cacau) na região sul da
Bahia, Brasil. Acta Botânica Brasilica 16 (1):89-101.
Santos, F.A.M. (1991) Padrão espacial de jovens em relação a adultos de espécies
arbóreas de cerrado que ocorrem no Estado de São Paulo. Tese - Campinas -
SP, Universidade Estadual de Campinas.
Schnitzer, S.A., Bongers, F. (2002) The ecology of lianas and their role in forests.
Trends in Ecology & Evolution 17 (5):223-230.
Seitz, R.A. (1994) A regeneração natural na recuperação de áreas degradadas.
Anais do I Simpósio Sul-Americano e II Nacional de Recuperação de Áreas
Degradadas. Foz do Iguaçú, p. 103-111.
Shepherd, G.J. (1994) FITOPAC 1: Manual de usuário. Departamento de Botânica,
Campinas: Universidade Estadual de Campinas, 93p.
Silva, A.F. (1980) Composição florística e estrutura de um trecho de Floresta
Atlântica de encosta no município de Ubatuba, SP. Dissertação - Campinas - SP,
Universidade Estadual de Campinas - Unicamp, 152p.
Silva, G.C. da, Nascimento, M.T. (2001) Fitossociologia de um remanescente de
Floresta sobre tabuleiros no norte do estado do Rio de Janeiro (Floresta do
Carvão). Revta Brasileira de Botânica, São Paulo, 24 (1):51-62.
Simonelli, M. (1998) Composição florística e estrutura do estrato arbóreo de uma
Mussununga no Reserva Florestal de Linhares, Espírito Santo. Dissertação
(Mestrado em Botânica) - Viçosa - MG, Universidade Federal de Viçosa - UFV,
101p.
SOS Mata Atlântica (1993) Atlas da evolução dos remanescentes florestais e
ecossistemas associados no domínio da Floresta Atlântica no período de 1985-
1990. São Paulo: Fundação SOS Mata Atlântica/INPE/ISA.
SOS Mata Atlântica (1998) Atlas da evolução dos remanescentes florestais e
ecossistemas associados no domínio da Floresta Atlântica no período de 1990-
1995. São Paulo: Fundação SOS Mata Atlântica/INPE/ISA.
SOS Mata Atlântica (2002) Atlas da evolução dos remanescentes florestais e
ecossistemas associados no domínio da Floresta Atlântica no período de 1995-
2000. São Paulo: Fundação SOS Mata Atlântica/INPE/ISA.
Souza, A.L., Schetino, S., Jesus, R.M., Vale, A.B. (2002) Dinâmica da regeneração
natural em uma Floresta Ombrófila Densa secundária, após corte de cipós,

92
Reserva Natural da Companhia Vale do Rio Doce S.A., Estado do Espírito
Santo, Brasil. Revista Árvore 26 (4):411-419.
Souza, A.L., Meira Neto, J.A.A., Schettino, S. (1998) Avaliação florística,
fitossociológica e paramétrica de um fragmento de floresta atlântica secundária,
município de Pedro Canário, Espírito Santo. Documento SIF, Viçosa, 18:117.
Souza, J.S. (2005) Efeito do corte seletivo de madeira na dinâmica de uma Floresta
Atlântica de Tabuleiro no Norte Fluminense. Tese (Doutorado em Biociências e
Biotecnologia) – Campos dos Goytacazes – RJ, Centro de Biociências e
Biotecnologia, Universidade Estadual do Norte Fluminense – UENF, 177p.
Svensson, S.J., Jeglum, J.K. (2001) Structure and dynamics of an undisturbed old-
growth Norway spruce forest on the rising Bothnian coastline. Forest ecology and
Management 151:67-79.
Swaine, M.D., Hall, J.B. (1988) The mosaic theory of forest regeneration and the
determination of forest composition in Ghana. Jornal Tropical Ecology 4:253-269.
Swaine, M.D., Whitmore, T.C. (1988) On the definition of ecological species grups in
tropical rain forests. Vegetatio 75:81-86.
Tabarelli, M., Mantovani, W. (1999) A regeneração de uma floresta tropical montana
após corte e queima (São Paulo - Brasil). Revista Brasileira de Biologia 59 (2):
239-250.
Tabarelli, M., Mantovani, W., Peres, C.A. (1999) Effects of habitat fragmentation on
plant guild structure in the montane Atlantic Forest of southeastern Brazil.
Biological Conservation 91:119-127.
Tabarelli, M., Villani. P.J., Mantovani, W. (1993) Aspectos da sucessão secundária
em trecho da Floresta Atlântica no Parque estadual da Serra do Mar, SP.
Revista Instituto Florestal, São Paulo, 5 (1):99-112.
Thomas, W.W., Amorim, A.M. (2005a) The Atlantic Coastal Forest Project of
Northeastern Brazil. New York, New York Botanical Garden. Disponível em:
http//www.nybg.org/bsci. Acesso em 16 de novembro de 2005.
Thomas, W.W., Amorim, A.M. (2005b) Preliminary checklist of the plants of the
Floresta da Esperança. New York, New York Botanical Garden. Disponível em:
http//www.nybg.org/bsci/res/bahia/Mata-Esp.html. Acesso em 16 de novembro
de 2005.
Thomas, W.W., Amorim, A.M. (2005c) Preliminary checklist of the plants of Monte

93
Paschoal National Park. New York, New York Botanical Garden. Disponível em:
http//www.nybg.org/bsci/res/bahia/MPNP.html. Acesso em 16 novembro de
2005.
Thomas, W.W., Amorim, A.M. (2005d) Preliminary checklist of the plants of the Una
Biological Reserve. New York, New York Botanical Garden. Disponível em:
http//www.nybg.org/bsci/res/bahia/Una.html. Acesso em 16 de novembro de
2005.
Thomas, W.W., Amorim, A.M. (2005e) Preliminary checklist of the plants of The
Serra Grande Floresta and Serra do Condurú State Park. New York, New York
Botanical Garden. Disponível em: http//www.nybg.org/bsci/res/bahia/SG.html.
Acesso em 16 de novembro de 2005.
Thomas, W.W., Amorim, A.M. (2005f) Preliminary checklist of the plants of The Serra
do Teimoso Natural Reserve. New York, New York Botanical Garden . Disponível
em: http//www.nybg.org/bsci/res/bahia/Teimoso-Reserve.html. Acesso em 18 de
novembro de 2005.
Thomas, W.W., Carvalho, A.M.V., Amorim, A.M.A., Garrison, J., Arbeláez, A.L.
(1998) Plant endemism in two forest in southern Bahia, Brazil. Biodiversity and
Conservation 7:311-322.
Thomaz, L.D. (1996) Florística e fitossociologia da Floresta Atlântica na Estação
Biológica de Santa Lúcia, Santa Teresa/ES. Tese (Doutorado em Biologia
Vegetal) - Rio Claro-SP, Universidade Estadual Paulista - UNESP, 322 p.
Thomaz, L.D., Monteiro, R. (1997) Composição florística da Floresta Atlântica de
encosta da estação Biológica de Santa Lúcia, município de Santa Teresa-ES.
Boletim do Museu de Biologia Mello Leitão (nova série) 7:3-48.
Tilman, D. (1999) Global environmental impacts of agricultural expansion: The need
for sustainable and efficient practices. Proceedings of the National Academy of
Sciences USA 96:5995-6000.
Uhl, C., Murphy, P.G. (1981) Composition, structure and regeneration of a tierra
firme forest in the Amazon Bazin of Venezuela. Tropical Ecology 22 (2):219-237.
Uhl, C., Buschbacher, R., Serrão, E.A.S. (1988) Abandoned pastures in eastern
Amazonia. l. Petterns of plant sucession. Journal of Ecology 76:663-681.
Vasquez-Yanes, C. (1980) Notas sobre la autoecologia de los arboles pioneros de
rapido crecimento de la selva tropical lluviosa. Tropical Ecology 21 (1):103-112.

94
Veloso, H.P., Rangel-Filho, A.L.R.R., Lima, J.C.A. (1991) Classificação da
Vegetação Brasileira, Adaptada a um Sistema Universal. IBGE (Instituto
Brasileiro de Geografia e Estatística). Rio de Janeiro: IBGE, Departamento de
Recursos Naturais e Estudos Ambientais, 124p.
Viana, V.M., Tabanez, A.A. J., Batista, A.J. (1997) Dynamics and restaration of forest
fragments in the Brazilian Atlantic moist forest. In: Bierregaard Jr., R. O (org.),
Tropical Forest Remnants, Management and Conservation. Chicago: The
University of Chicago Press, p. 351-365.
Vilela, E.A., Oliveira-Filho, T., Carvalho, D.A., Guilherme, F.A.G., Appolinário, V.
(2000) Caracterização Estrutural de Floresta ripária do Alto rio Grande, em
Madre de Deus de Minas, MG. Cerne 6 (2):41-54.
Villela, D.M., Nascimento, M.T.N., Aragão, L.E.O.C., Gama, D.M. (2006) Effect of
selective logging on forest structure and nutrient cycling in a seasonally dry
Brazilian Atlantic forest. Journal of Biogeography.
Wied, M.P. von (1989) Viagem ao Brasil. São Paulo: Editora da Universidade de São
Paulo, 662p.
Zar, J.H. (1996) Biostatistical analysis. 3 ed. New Jersey: Prentice Hall, 662p.
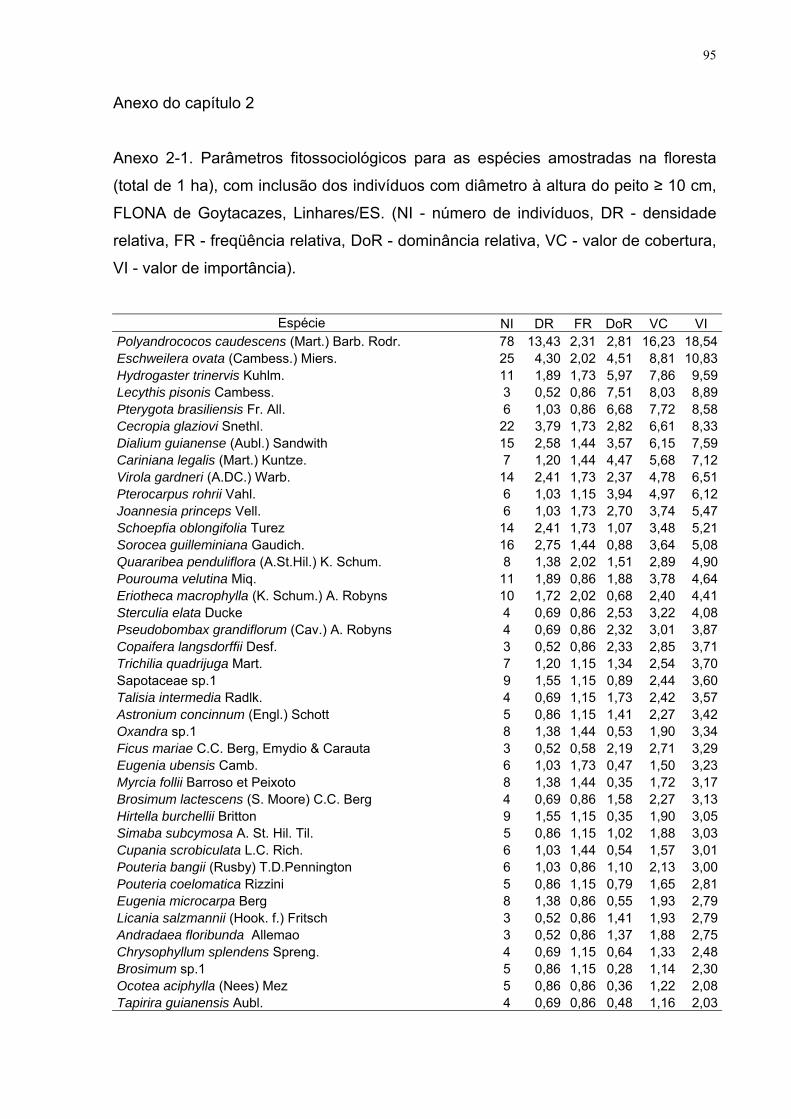
95
Anexo do capítulo 2
Anexo 2-1. Parâmetros fitossociológicos para as espécies amostradas na floresta
(total de 1 ha), com inclusão dos indivíduos com diâmetro à altura do peito ≥ 10 cm,
FLONA de Goytacazes, Linhares/ES. (NI - número de indivíduos, DR - densidade
relativa, FR - freqüência relativa, DoR - dominância relativa, VC - valor de cobertura,
VI - valor de importância).
Espécie NI DR FR DoR VC VI
Polyandrococos caudescens (Mart.) Barb. Rodr. 78 13,43 2,31 2,81 16,23 18,54Eschweilera ovata (Cambess.) Miers. 25 4,30 2,02 4,51 8,81 10,83Hydrogaster trinervis Kuhlm. 11 1,89 1,73 5,97 7,86 9,59Lecythis pisonis Cambess. 3 0,52 0,86 7,51 8,03 8,89Pterygota brasiliensis Fr. All. 6 1,03 0,86 6,68 7,72 8,58Cecropia glaziovi Snethl. 22 3,79 1,73 2,82 6,61 8,33Dialium guianense (Aubl.) Sandwith 15 2,58 1,44 3,57 6,15 7,59Cariniana legalis (Mart.) Kuntze. 7 1,20 1,44 4,47 5,68 7,12Virola gardneri (A.DC.) Warb. 14 2,41 1,73 2,37 4,78 6,51Pterocarpus rohrii Vahl. 6 1,03 1,15 3,94 4,97 6,12Joannesia princeps Vell. 6 1,03 1,73 2,70 3,74 5,47Schoepfia oblongifolia Turez 14 2,41 1,73 1,07 3,48 5,21Sorocea guilleminiana Gaudich. 16 2,75 1,44 0,88 3,64 5,08Quararibea penduliflora (A.St.Hil.) K. Schum. 8 1,38 2,02 1,51 2,89 4,90Pourouma velutina Miq. 11 1,89 0,86 1,88 3,78 4,64Eriotheca macrophylla (K. Schum.) A. Robyns 10 1,72 2,02 0,68 2,40 4,41Sterculia elata Ducke 4 0,69 0,86 2,53 3,22 4,08Pseudobombax grandiflorum (Cav.) A. Robyns 4 0,69 0,86 2,32 3,01 3,87Copaifera langsdorffii Desf. 3 0,52 0,86 2,33 2,85 3,71Trichilia quadrijuga Mart. 7 1,20 1,15 1,34 2,54 3,70Sapotaceae sp.1 9 1,55 1,15 0,89 2,44 3,60Talisia intermedia Radlk. 4 0,69 1,15 1,73 2,42 3,57Astronium concinnum (Engl.) Schott 5 0,86 1,15 1,41 2,27 3,42Oxandra sp.1 8 1,38 1,44 0,53 1,90 3,34Ficus mariae C.C. Berg, Emydio & Carauta 3 0,52 0,58 2,19 2,71 3,29Eugenia ubensis Camb. 6 1,03 1,73 0,47 1,50 3,23Myrcia follii Barroso et Peixoto 8 1,38 1,44 0,35 1,72 3,17Brosimum lactescens (S. Moore) C.C. Berg 4 0,69 0,86 1,58 2,27 3,13Hirtella burchellii Britton 9 1,55 1,15 0,35 1,90 3,05Simaba subcymosa A. St. Hil. Til. 5 0,86 1,15 1,02 1,88 3,03Cupania scrobiculata L.C. Rich. 6 1,03 1,44 0,54 1,57 3,01Pouteria bangii (Rusby) T.D.Pennington 6 1,03 0,86 1,10 2,13 3,00Pouteria coelomatica Rizzini 5 0,86 1,15 0,79 1,65 2,81Eugenia microcarpa Berg 8 1,38 0,86 0,55 1,93 2,79Licania salzmannii (Hook. f.) Fritsch 3 0,52 0,86 1,41 1,93 2,79Andradaea floribunda Allemao 3 0,52 0,86 1,37 1,88 2,75Chrysophyllum splendens Spreng. 4 0,69 1,15 0,64 1,33 2,48Brosimum sp.1 5 0,86 1,15 0,28 1,14 2,30Ocotea aciphylla (Nees) Mez 5 0,86 0,86 0,36 1,22 2,08Tapirira guianensis Aubl. 4 0,69 0,86 0,48 1,16 2,03
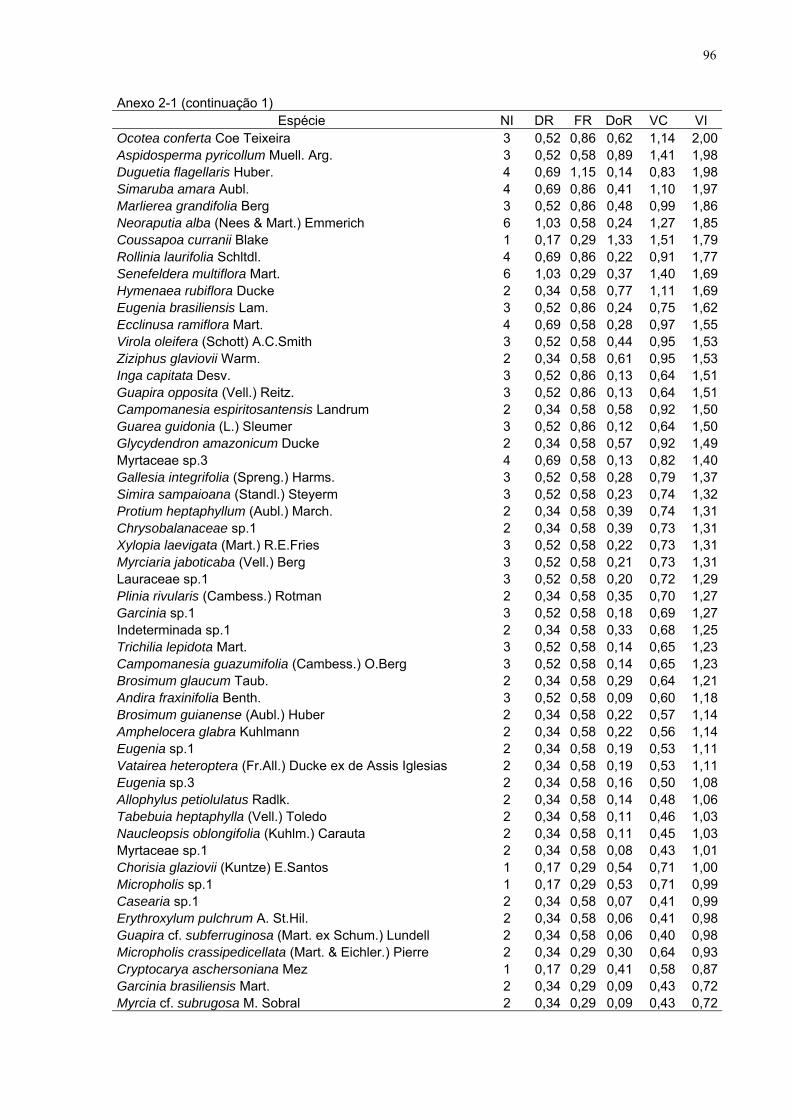
96
Anexo 2-1 (continuação 1) Espécie NI DR FR DoR VC VI
Ocotea conferta Coe Teixeira 3 0,52 0,86 0,62 1,14 2,00Aspidosperma pyricollum Muell. Arg. 3 0,52 0,58 0,89 1,41 1,98Duguetia flagellaris Huber. 4 0,69 1,15 0,14 0,83 1,98Simaruba amara Aubl. 4 0,69 0,86 0,41 1,10 1,97Marlierea grandifolia Berg 3 0,52 0,86 0,48 0,99 1,86Neoraputia alba (Nees & Mart.) Emmerich 6 1,03 0,58 0,24 1,27 1,85Coussapoa curranii Blake 1 0,17 0,29 1,33 1,51 1,79Rollinia laurifolia Schltdl. 4 0,69 0,86 0,22 0,91 1,77Senefeldera multiflora Mart. 6 1,03 0,29 0,37 1,40 1,69Hymenaea rubiflora Ducke 2 0,34 0,58 0,77 1,11 1,69Eugenia brasiliensis Lam. 3 0,52 0,86 0,24 0,75 1,62Ecclinusa ramiflora Mart. 4 0,69 0,58 0,28 0,97 1,55Virola oleifera (Schott) A.C.Smith 3 0,52 0,58 0,44 0,95 1,53Ziziphus glaviovii Warm. 2 0,34 0,58 0,61 0,95 1,53Inga capitata Desv. 3 0,52 0,86 0,13 0,64 1,51Guapira opposita (Vell.) Reitz. 3 0,52 0,86 0,13 0,64 1,51Campomanesia espiritosantensis Landrum 2 0,34 0,58 0,58 0,92 1,50Guarea guidonia (L.) Sleumer 3 0,52 0,86 0,12 0,64 1,50Glycydendron amazonicum Ducke 2 0,34 0,58 0,57 0,92 1,49Myrtaceae sp.3 4 0,69 0,58 0,13 0,82 1,40Gallesia integrifolia (Spreng.) Harms. 3 0,52 0,58 0,28 0,79 1,37Simira sampaioana (Standl.) Steyerm 3 0,52 0,58 0,23 0,74 1,32Protium heptaphyllum (Aubl.) March. 2 0,34 0,58 0,39 0,74 1,31Chrysobalanaceae sp.1 2 0,34 0,58 0,39 0,73 1,31Xylopia laevigata (Mart.) R.E.Fries 3 0,52 0,58 0,22 0,73 1,31Myrciaria jaboticaba (Vell.) Berg 3 0,52 0,58 0,21 0,73 1,31Lauraceae sp.1 3 0,52 0,58 0,20 0,72 1,29Plinia rivularis (Cambess.) Rotman 2 0,34 0,58 0,35 0,70 1,27Garcinia sp.1 3 0,52 0,58 0,18 0,69 1,27Indeterminada sp.1 2 0,34 0,58 0,33 0,68 1,25Trichilia lepidota Mart. 3 0,52 0,58 0,14 0,65 1,23Campomanesia guazumifolia (Cambess.) O.Berg 3 0,52 0,58 0,14 0,65 1,23Brosimum glaucum Taub. 2 0,34 0,58 0,29 0,64 1,21Andira fraxinifolia Benth. 3 0,52 0,58 0,09 0,60 1,18Brosimum guianense (Aubl.) Huber 2 0,34 0,58 0,22 0,57 1,14Amphelocera glabra Kuhlmann 2 0,34 0,58 0,22 0,56 1,14Eugenia sp.1 2 0,34 0,58 0,19 0,53 1,11Vatairea heteroptera (Fr.All.) Ducke ex de Assis Iglesias 2 0,34 0,58 0,19 0,53 1,11Eugenia sp.3 2 0,34 0,58 0,16 0,50 1,08Allophylus petiolulatus Radlk. 2 0,34 0,58 0,14 0,48 1,06Tabebuia heptaphylla (Vell.) Toledo 2 0,34 0,58 0,11 0,46 1,03Naucleopsis oblongifolia (Kuhlm.) Carauta 2 0,34 0,58 0,11 0,45 1,03Myrtaceae sp.1 2 0,34 0,58 0,08 0,43 1,01Chorisia glaziovii (Kuntze) E.Santos 1 0,17 0,29 0,54 0,71 1,00Micropholis sp.1 1 0,17 0,29 0,53 0,71 0,99Casearia sp.1 2 0,34 0,58 0,07 0,41 0,99Erythroxylum pulchrum A. St.Hil. 2 0,34 0,58 0,06 0,41 0,98Guapira cf. subferruginosa (Mart. ex Schum.) Lundell 2 0,34 0,58 0,06 0,40 0,98Micropholis crassipedicellata (Mart. & Eichler.) Pierre 2 0,34 0,29 0,30 0,64 0,93Cryptocarya aschersoniana Mez 1 0,17 0,29 0,41 0,58 0,87Garcinia brasiliensis Mart. 2 0,34 0,29 0,09 0,43 0,72Myrcia cf. subrugosa M. Sobral 2 0,34 0,29 0,09 0,43 0,72

97
Anexo 2-1 (continuação 2) Espécie NI DR FR DoR VC VI
Marlierea regeliana Berg 1 0,17 0,29 0,25 0,42 0,71Actinostemon estrellensis Pax 2 0,34 0,29 0,06 0,40 0,69Sapotaceae sp.2 1 0,17 0,29 0,20 0,37 0,66Licania sp.1 1 0,17 0,29 0,16 0,34 0,62Guazuma crinita Mart. 1 0,17 0,29 0,15 0,33 0,61Spondias venulosa Mart. ex Engl. 1 0,17 0,29 0,15 0,32 0,61Sloanea eichleri K.Schum. 1 0,17 0,29 0,13 0,30 0,59Paradrypetes ilicifolia Kuhlm. 1 0,17 0,29 0,12 0,29 0,58Aspidosperma illustre (Vell.) Kuhlm. & Piraja 1 0,17 0,29 0,11 0,28 0,57Pouteria bullata (S.Moore) Baehni 1 0,17 0,29 0,11 0,28 0,57Trichilia aff. surumuensis C.DC. 1 0,17 0,29 0,11 0,28 0,57Inga thibaudiana DC. 1 0,17 0,29 0,11 0,28 0,57Astronium graveolens Jacq. 1 0,17 0,29 0,10 0,28 0,56Dalbergia nigra (Vell.) Fr. All. ex Benth. 1 0,17 0,29 0,10 0,27 0,56Maytenus sp.1 1 0,17 0,29 0,10 0,27 0,56Chrysophyllum sp.1 1 0,17 0,29 0,09 0,27 0,55Hirtella sp.1 1 0,17 0,29 0,09 0,26 0,55Exostyles venusta Schott 1 0,17 0,29 0,09 0,26 0,55Rubiaceae sp.1 1 0,17 0,29 0,08 0,26 0,54Terminalia kuhlmannii Alwan & Stace 1 0,17 0,29 0,08 0,25 0,54Erythroxylum sp.2 1 0,17 0,29 0,08 0,25 0,54Psidium sp.1 1 0,17 0,29 0,08 0,25 0,54Ocotea cernua (Nees) Mez Vell. 1 0,17 0,29 0,08 0,25 0,54Tachigalia paratyensis (Vell.) H.C.Lima 1 0,17 0,29 0,07 0,24 0,53Inga flagelliformis (Vell.) Mart. 1 0,17 0,29 0,07 0,24 0,53Carpotroche brasiliensis (Raddi.) A. Gray 1 0,17 0,29 0,07 0,24 0,53Marlierea sylvatica (Gardner) Kiaersk. 1 0,17 0,29 0,06 0,23 0,52Pera leandri Baill. 1 0,17 0,29 0,06 0,23 0,52Moldenhawera papillanthera L.P.Queiroz, G.P.Lewis & R.Allkin 1 0,17 0,29 0,06 0,23 0,52Calyptrantes sp.2 1 0,17 0,29 0,06 0,23 0,52Miconia hypoleuca (Benth.) Triana 1 0,17 0,29 0,06 0,23 0,52Oxandra nitida R.E.Fries 1 0,17 0,29 0,06 0,23 0,52Myrtaceae sp.5 1 0,17 0,29 0,05 0,23 0,51Inga striata Benth. 1 0,17 0,29 0,05 0,22 0,51Aniba sp.1 1 0,17 0,29 0,05 0,22 0,51Myrcia falax (Richard) DC. 1 0,17 0,29 0,05 0,22 0,51Eugenia sp.2 1 0,17 0,29 0,05 0,22 0,51Garcinia gardneriana (Triana & Planch.) D.C.Zappi 1 0,17 0,29 0,04 0,22 0,50Byrsonima stipulacea (Juss.) Nied. 1 0,17 0,29 0,04 0,22 0,50Plinia involucrata (Berg) McVaugh. 1 0,17 0,29 0,04 0,22 0,50Eugenia itapemirinensis Cambess. 1 0,17 0,29 0,04 0,21 0,50Maclura tinctoria (L.) Don ex Steudel 1 0,17 0,29 0,04 0,21 0,50Licaria bahiana Kutz 1 0,17 0,29 0,04 0,21 0,50Cordia sellowiana Cham. 1 0,17 0,29 0,04 0,21 0,50Erythroxylum plowmanii Amaral Jr. 1 0,17 0,29 0,04 0,21 0,50Ocotea confertiflora (Meisn.) Mez 1 0,17 0,29 0,04 0,21 0,50Myrcia sp.1 1 0,17 0,29 0,04 0,21 0,50Marlierea obversa Legrand. = 464 1 0,17 0,29 0,04 0,21 0,50Alseis floribunda Schott 1 0,17 0,29 0,04 0,21 0,50Capparis flexuosa (L.) L. 1 0,17 0,29 0,04 0,21 0,50Eugenia excelsa Berg 1 0,17 0,29 0,04 0,21 0,50Tabebuia roseo-alba (Ridley) Sandwith 1 0,17 0,29 0,03 0,21 0,50

98
Anexo 2-1 (continuação 3) Espécie NI DR FR DoR VC VI
Zollernia latifolia Benth. 1 0,17 0,29 0,03 0,21 0,50Plinia aff. grandifolia (Mattos) Sobral 1 0,17 0,29 0,03 0,21 0,49Chrysophyllum lucentifolium Cronquist 1 0,17 0,29 0,03 0,21 0,49Erythroxylum columbinum Mart. 1 0,17 0,29 0,03 0,21 0,49Trichilia casaretti C.DC. 1 0,17 0,29 0,03 0,20 0,49Zollernia sp.1 1 0,17 0,29 0,03 0,20 0,49Cupania zanthoxyloides Camb. 1 0,17 0,29 0,03 0,20 0,49Mollinedia marquetiana Peixoto 1 0,17 0,29 0,03 0,20 0,49Marlierea sp.1 1 0,17 0,29 0,03 0,20 0,49Beilschmiedia linharensis Nishida 1 0,17 0,29 0,03 0,20 0,49Pausandra morisiana (Casar.) Radlk. 1 0,17 0,29 0,03 0,20 0,49Astrocaryum aculeatissimum (Schott) Burret 1 0,17 0,29 0,03 0,20 0,49Myrtaceae sp.6 1 0,17 0,29 0,03 0,20 0,49
Total 581 100 100 100 200 300

99
Anexo do capítulo 3
Anexo 3-1. Espécies amostradas com presença (1) e ausência (0), por estrato nos respectivos sítios, FLONA de Goytacazes, Linhares/ES. (DAP - diâmetro à altura do peito; Alt. – altura; Flo - floresta; Cap - capoeira; Cab - cabruca e Ca- capoeirinha).
Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca
ANACARDIACEAE Astronium concinnum (Engl.) Schott 1 1 1 1 1 1 1 1 1 0 1 1 1 1 1 1 Astronium graveolens Jacq. 1 1 0 1 1 0 1 1 1 1 1 1 1 1 1 1 Spondias mombin L. 0 1 0 0 0 1 0 1 0 0 0 0 0 0 0 0 Tapirira guianensis Aubl. 0 1 1 1 0 1 1 0 0 1 1 0 1 1 1 0 ANNONACEAE Annonaceae sp.1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Annona acutiflora Mart. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 1 Duguetia flagellaris Huber. 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 Oxandra nitida R.E.Fries 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 Oxandra sp.1 0 0 1 0 0 0 1 0 1 0 1 0 1 0 1 0 Oxandra sp.3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Rollinia laurifolia Schltdl. 1 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 Xylopia laevigata (Mart.) R.E.Fries 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 Xylopia sericea A. St. Hil. 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 APOCYNACEAE Aspidosperma illustre (Vell.) Kuhlm. & Piraja 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Aspidosperma pyricollum Muell. Arg. 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 Rauwolfia sp.1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Tabernaemontana salzmanni A. DC. 0 0 0 0 0 0 0 0 1 0 0 0 1 0 1 0 ARECACEAE Polyandrococos caudescens (Mart.) Barb. Rodr. 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 ASTERACEAE Vernonanthura phosphorica (Vell. Conc.) H. Rob. 0 0 0 0 0 1 0 0 0 1 0 0 0 1 0 0 BIGNONIACEAE Jacaranda puberula Cham. 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Tabebuia heptaphylla (Vell.) Toledo 1 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 Tabebuia riodocensis A. Gentry 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Tabebuia roseo-alba (Ridley) Sandwith 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0

100
Anexo 3-1 (continuação 1)
Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca
Tabebuia serratifolia (Vahl) Nichols. 1 0 0 0 1 0 0 0 1 0 0 0 1 0 1 1 BOMBACACEAE Bombacopsis stenopetala (Casar.) A. Robyns 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Eriotheca macrophylla (K. Schum.) A. Robyns 0 0 1 0 0 0 1 0 0 0 1 0 0 0 1 0 Pseudobombax grandiflorum (Cav.) A. Robyns 1 1 1 0 0 0 0 0 0 0 0 0 0 1 0 0 Quararibea penduliflora (A.St.Hil.) K. Schum. 1 0 1 0 0 0 1 0 1 0 1 0 1 0 1 0 BORAGINACEAE Boraginaceae sp.1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Cordia ecalyculata Vell. 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 Cordia sellowiana Cham. 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Cordia taguahyensis Vell 0 0 0 0 0 0 0 0 0 0 1 0 1 0 1 0 BURSERACEAE Protium aff. warmingianum March. 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 0 Protium heptaphyllum (Aubl.) March. 1 0 1 0 0 0 1 0 1 1 1 0 1 0 1 0 CAPPARACEAE Capparis brasiliensis DC. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Capparis flexuosa (L.) L. 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 Crataeva tapia L. 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 CECROPIACEAE Cecropia glaziovi Snethl. 0 1 1 0 0 0 1 0 0 0 0 0 0 0 0 0 Cecropia pachystachya Trécul. 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 Coussapoa curranii Blake 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Pourouma guianensis Aubl. 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Pourouma velutina Miq. 1 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 CELASTRACEAE Maytenus cestrifolia Reiss. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Maytenus sp.1 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 CHRYSOBALANACEAE Chrysobalanaceae sp.1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Couepia schottii Fritsch 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Hirtella burchellii Britton 0 0 1 0 0 0 1 0 0 0 1 0 0 0 1 0 Licania salzmannii (Hook. f.) Fritsch 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 CLUSIACEAE Garcinia gardneriana (Triana & Planch.) D.C.Zappi 0 0 1 0 0 0 0 0 1 0 0 0 0 0 1 0

101
Anexo 3-1 (continuação 2) Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie
Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Garcinia sp.1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Tovomita brevistaminea Engl. 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 0 COMBRETACEAE Terminalia kuhlmannii Alwan & Stace 1 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 DICHAPETALACEAE Stephanopodium blanchetianum Baill. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 ELAEOCARPACEAE Sloanea eichleri K.Schum. 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 ERYTHROXYLACEAE Erythroxylum columbinum Mart. 0 0 0 0 0 0 0 0 1 0 0 0 1 0 1 0 Erythroxylum pulchrum A. St.Hil. 0 0 1 0 0 0 0 0 1 1 1 0 1 1 1 0 EUPHORBIACEAE Actinostemon estrellensis Pax 0 0 1 0 0 0 0 0 0 0 1 0 1 0 1 0 Alchornea iricurana Casar. 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Glycydendron amazonicum Ducke 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 Joannesia princeps Vell. 1 1 1 0 1 1 1 0 0 1 1 0 0 1 0 0 Margaritaria nobilis Linn.f. 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Pera leandri Baill. 1 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 Pera sp.1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Sapium glandulatum (Vell.) Pax. 1 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 Sebastiania brasiliensis Spreng. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 FLACOURTIACEAE Banara kuhlmannii (Sleumer) Sleumer 0 0 0 0 1 0 0 0 0 0 0 0 1 0 1 0 Banara sp.1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 Carpotroche brasiliensis (Raddi.) A. Gray 1 0 0 0 1 0 0 0 1 0 0 0 1 0 1 0 Casearia decandra Jacq. 1 1 0 0 1 1 0 0 1 1 0 0 1 0 0 0 Casearia sp.1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Casearia sp.2 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Flacourtiaceae sp.1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Flacourtiaceae sp.2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 INDETERMINADA Indeterminada sp.1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Indeterminada sp.2 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 Indeterminada sp.3 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0

102
Anexo 3-1 (continuação 3) Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie
Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Indeterminada sp.4 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Indeterminada sp.5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Indeterminada sp.6 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Indeterminada sp.7 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Indeterminada sp.8 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 Indeterminada sp.9 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Indeterminada sp.10 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 Indeterminada sp.11 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Indeterminada sp.12 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Indeterminada sp.13 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 Indeterminada sp.14 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Indeterminada sp.15 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Indeterminada sp.16 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 Indeterminada sp.17 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 LAURACEAE Aniba canelillla (HBK.) Mez 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Aniba firmula (Nees & C. Mart.) Mez 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Beilschmiedia linharensis Nishida 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Cryptocarya aschersoniana Mez 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Cryptocarya saligna Mez. 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 0 Lauraceae sp.1 0 0 1 0 0 0 1 0 0 0 0 0 0 1 1 0 Lauraceae sp.2 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 Licaria bahiana Kutz 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Ocotea aciphylla (Nees) Mez 0 1 1 0 0 1 0 0 0 1 0 0 0 0 0 0 Ocotea aff. teleiandra Mez 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Ocotea cernua Mez 0 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 Ocotea cf. nitida (Meissn.) J.G.Rohwer 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Ocotea conferta Coe Teixeira 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 Ocotea confertiflora (Meisn.) Mez 0 0 1 0 0 0 1 0 0 0 1 0 0 1 1 0 Ocotea divaricata (Poir.) Mez 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 0 Ocotea elegans Mez 0 0 0 0 1 0 0 0 1 1 1 0 1 0 0 0 Ocotea longifolia HBK. 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 LECYTHIDACEAE Cariniana estrellensis (Raddi.) Kuntze. 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0
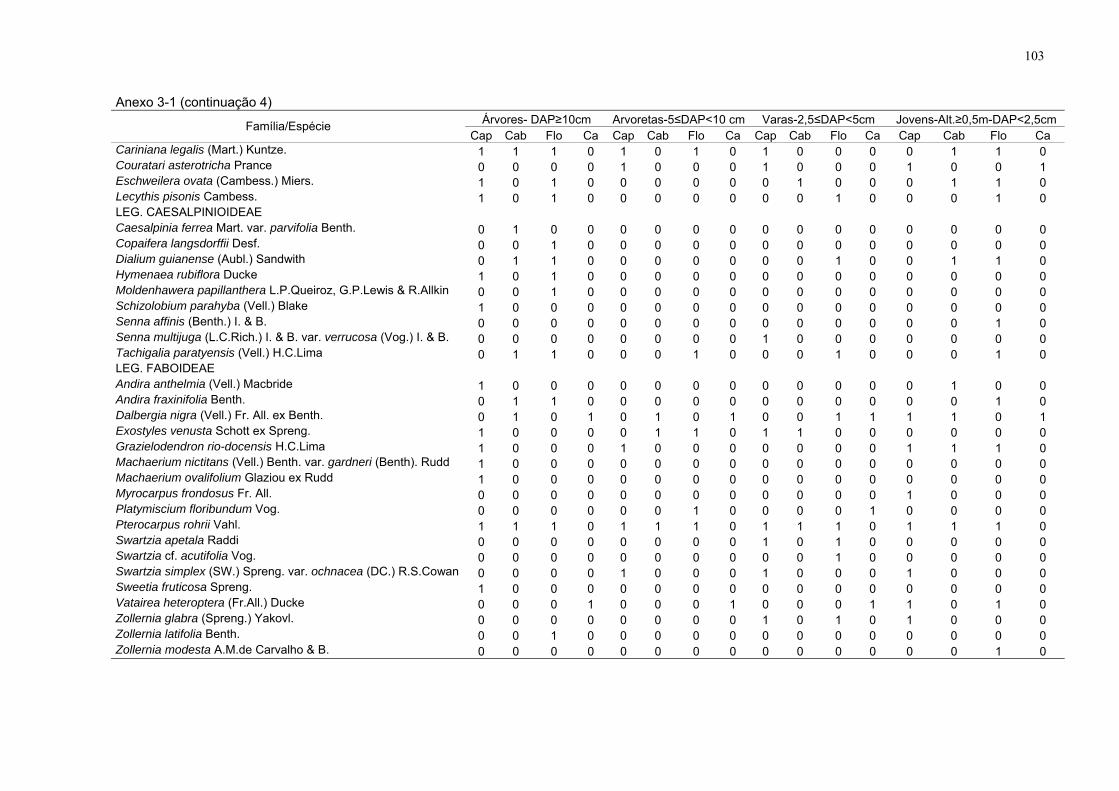
103
Anexo 3-1 (continuação 4) Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie
Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cariniana legalis (Mart.) Kuntze. 1 1 1 0 1 0 1 0 1 0 0 0 0 1 1 0 Couratari asterotricha Prance 0 0 0 0 1 0 0 0 1 0 0 0 1 0 0 1 Eschweilera ovata (Cambess.) Miers. 1 0 1 0 0 0 0 0 0 1 0 0 0 1 1 0 Lecythis pisonis Cambess. 1 0 1 0 0 0 0 0 0 0 1 0 0 0 1 0 LEG. CAESALPINIOIDEAE Caesalpinia ferrea Mart. var. parvifolia Benth. 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Copaifera langsdorffii Desf. 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Dialium guianense (Aubl.) Sandwith 0 1 1 0 0 0 0 0 0 0 1 0 0 1 1 0 Hymenaea rubiflora Ducke 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Moldenhawera papillanthera L.P.Queiroz, G.P.Lewis & R.Allkin 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Schizolobium parahyba (Vell.) Blake 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Senna affinis (Benth.) I. & B. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Senna multijuga (L.C.Rich.) I. & B. var. verrucosa (Vog.) I. & B. 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Tachigalia paratyensis (Vell.) H.C.Lima 0 1 1 0 0 0 1 0 0 0 1 0 0 0 1 0 LEG. FABOIDEAE Andira anthelmia (Vell.) Macbride 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Andira fraxinifolia Benth. 0 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 Dalbergia nigra (Vell.) Fr. All. ex Benth. 0 1 0 1 0 1 0 1 0 0 1 1 1 1 0 1 Exostyles venusta Schott ex Spreng. 1 0 0 0 0 1 1 0 1 1 0 0 0 0 0 0 Grazielodendron rio-docensis H.C.Lima 1 0 0 0 1 0 0 0 0 0 0 0 1 1 1 0 Machaerium nictitans (Vell.) Benth. var. gardneri (Benth). Rudd 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Machaerium ovalifolium Glaziou ex Rudd 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Myrocarpus frondosus Fr. All. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Platymiscium floribundum Vog. 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 Pterocarpus rohrii Vahl. 1 1 1 0 1 1 1 0 1 1 1 0 1 1 1 0 Swartzia apetala Raddi 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 Swartzia cf. acutifolia Vog. 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Swartzia simplex (SW.) Spreng. var. ochnacea (DC.) R.S.Cowan 0 0 0 0 1 0 0 0 1 0 0 0 1 0 0 0 Sweetia fruticosa Spreng. 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Vatairea heteroptera (Fr.All.) Ducke 0 0 0 1 0 0 0 1 0 0 0 1 1 0 1 0 Zollernia glabra (Spreng.) Yakovl. 0 0 0 0 0 0 0 0 1 0 1 0 1 0 0 0 Zollernia latifolia Benth. 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Zollernia modesta A.M.de Carvalho & B. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0

104
Anexo 3-1 (continuação 5) Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie
Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Zollernia sp.1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 LEG. MIMOSOIDEAE Albizia polycephala (Benth.) Killip 1 0 0 1 1 0 0 0 1 0 0 0 0 0 0 0 Inga cabelo T.D. Penn. 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Inga capitata Desv. 0 0 1 0 0 0 1 0 0 0 1 0 1 0 0 0 Inga flagelliformis (Vell.) Mart. 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 Inga striata Benth. 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 Inga thibaudiana DC. 0 1 1 0 0 0 0 0 0 0 0 0 0 1 1 0 Parapiptadenia pterosperma (Benth.) Brenan 1 1 0 1 0 0 0 1 1 0 1 1 1 1 1 1 Piptadenia gonoacantha (Mart.) Macbr. 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Piptadenia paniculata Benth. 1 0 0 0 0 1 1 0 0 0 1 0 0 0 0 0 Pithecellobium pedicellare (DC.) Benth. 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Pseudopiptadenia contorta (DC.) G.P.Lewis & M.P.Lima 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 LOGANIACEAE Strychnos cf. hirsuta Spruce ex Benth. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 MALPIGHIACEAE Byrsonima cacaophila W.R. Anderson 1 0 0 0 1 0 1 0 0 0 1 0 0 0 0 0 Byrsonima stipulacea (Juss.) Nied. 0 1 1 0 0 1 0 0 0 1 1 0 0 0 0 0 MELASTOMATACEAE Miconia cf. cinnamomifolia (DC.) Naudin 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Miconia hypoleuca (Benth.) Triana 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 MELIACEAE Cedrela odorata Linn. 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Guarea guidonia (L.) Sleumer 0 1 1 0 0 1 1 0 0 1 1 0 0 1 1 0 Meliaceae sp.1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 Trichilia casaretti C.DC. 1 0 1 0 1 0 0 0 1 0 1 0 1 0 1 0 Trichilia lepdota Mart. subsp. schumanniana (Harms) T.D. Pennington 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 Trichilia lepidota Mart. 1 0 1 0 1 0 0 0 0 0 1 0 0 0 1 0 Trichilia pallens C. DC. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Trichilia quadrijuga H.B. & K. 1 0 1 0 0 0 0 0 0 0 1 0 1 0 0 0 Trichilia sp.1 1 0 0 0 1 0 0 0 1 0 0 0 1 0 0 0 MONIMIACEAE Monimiaceae sp.1 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0
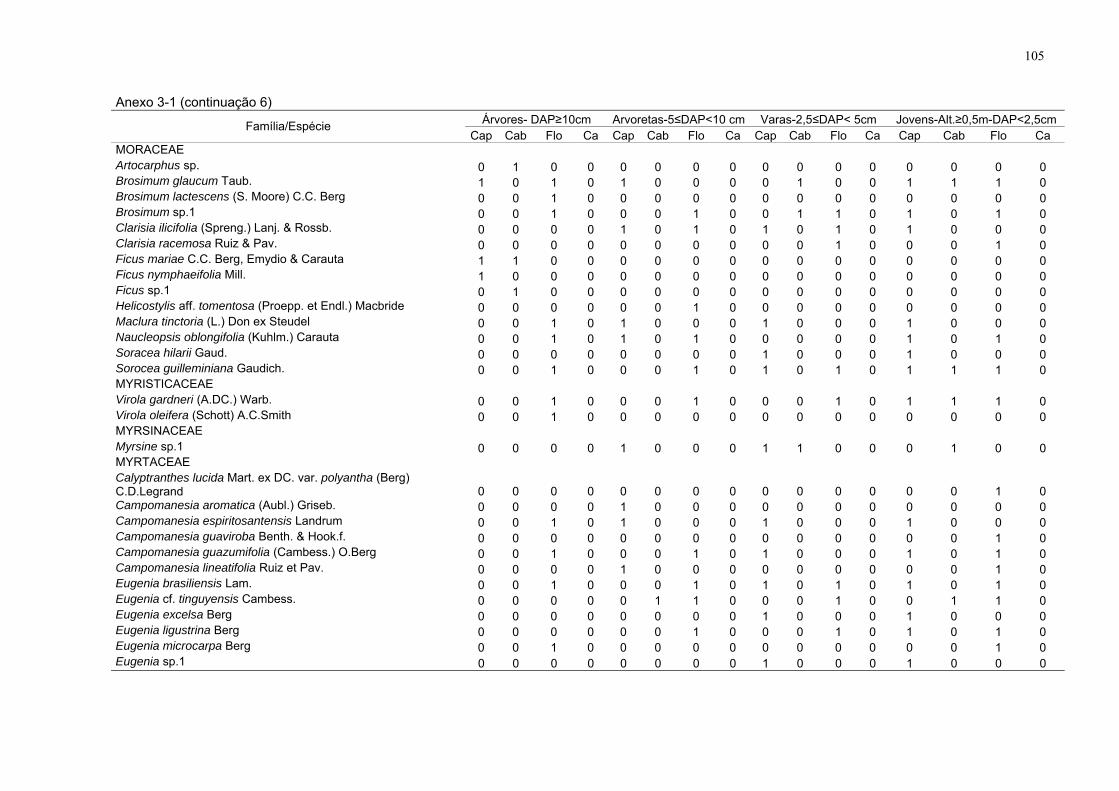
105
Anexo 3-1 (continuação 6) Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP< 5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie
Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca MORACEAE Artocarphus sp. 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Brosimum glaucum Taub. 1 0 1 0 1 0 0 0 0 1 0 0 1 1 1 0 Brosimum lactescens (S. Moore) C.C. Berg 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Brosimum sp.1 0 0 1 0 0 0 1 0 0 1 1 0 1 0 1 0 Clarisia ilicifolia (Spreng.) Lanj. & Rossb. 0 0 0 0 1 0 1 0 1 0 1 0 1 0 0 0 Clarisia racemosa Ruiz & Pav. 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 Ficus mariae C.C. Berg, Emydio & Carauta 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Ficus nymphaeifolia Mill. 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Ficus sp.1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Helicostylis aff. tomentosa (Proepp. et Endl.) Macbride 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Maclura tinctoria (L.) Don ex Steudel 0 0 1 0 1 0 0 0 1 0 0 0 1 0 0 0 Naucleopsis oblongifolia (Kuhlm.) Carauta 0 0 1 0 1 0 1 0 0 0 0 0 1 0 1 0 Soracea hilarii Gaud. 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 Sorocea guilleminiana Gaudich. 0 0 1 0 0 0 1 0 1 0 1 0 1 1 1 0 MYRISTICACEAE Virola gardneri (A.DC.) Warb. 0 0 1 0 0 0 1 0 0 0 1 0 1 1 1 0 Virola oleifera (Schott) A.C.Smith 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 MYRSINACEAE Myrsine sp.1 0 0 0 0 1 0 0 0 1 1 0 0 0 1 0 0 MYRTACEAE Calyptranthes lucida Mart. ex DC. var. polyantha (Berg) C.D.Legrand 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Campomanesia aromatica (Aubl.) Griseb. 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 Campomanesia espiritosantensis Landrum 0 0 1 0 1 0 0 0 1 0 0 0 1 0 0 0 Campomanesia guaviroba Benth. & Hook.f. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Campomanesia guazumifolia (Cambess.) O.Berg 0 0 1 0 0 0 1 0 1 0 0 0 1 0 1 0 Campomanesia lineatifolia Ruiz et Pav. 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 Eugenia brasiliensis Lam. 0 0 1 0 0 0 1 0 1 0 1 0 1 0 1 0 Eugenia cf. tinguyensis Cambess. 0 0 0 0 0 1 1 0 0 0 1 0 0 1 1 0 Eugenia excelsa Berg 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 Eugenia ligustrina Berg 0 0 0 0 0 0 1 0 0 0 1 0 1 0 1 0 Eugenia microcarpa Berg 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 Eugenia sp.1 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0
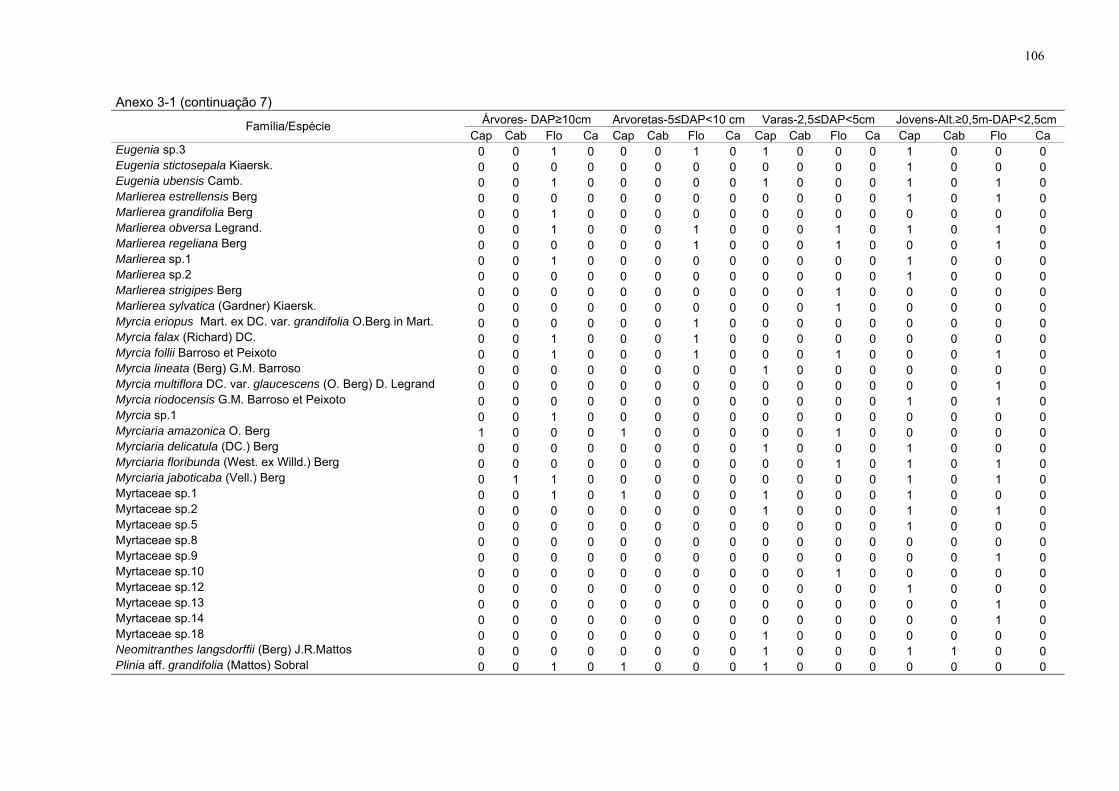
106
Anexo 3-1 (continuação 7) Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie
Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Eugenia sp.3 0 0 1 0 0 0 1 0 1 0 0 0 1 0 0 0 Eugenia stictosepala Kiaersk. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Eugenia ubensis Camb. 0 0 1 0 0 0 0 0 1 0 0 0 1 0 1 0 Marlierea estrellensis Berg 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Marlierea grandifolia Berg 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Marlierea obversa Legrand. 0 0 1 0 0 0 1 0 0 0 1 0 1 0 1 0 Marlierea regeliana Berg 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 0 Marlierea sp.1 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 Marlierea sp.2 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Marlierea strigipes Berg 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Marlierea sylvatica (Gardner) Kiaersk. 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Myrcia eriopus Mart. ex DC. var. grandifolia O.Berg in Mart. 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Myrcia falax (Richard) DC. 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 Myrcia follii Barroso et Peixoto 0 0 1 0 0 0 1 0 0 0 1 0 0 0 1 0 Myrcia lineata (Berg) G.M. Barroso 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Myrcia multiflora DC. var. glaucescens (O. Berg) D. Legrand 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Myrcia riodocensis G.M. Barroso et Peixoto 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Myrcia sp.1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Myrciaria amazonica O. Berg 1 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 Myrciaria delicatula (DC.) Berg 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 Myrciaria floribunda (West. ex Willd.) Berg 0 0 0 0 0 0 0 0 0 0 1 0 1 0 1 0 Myrciaria jaboticaba (Vell.) Berg 0 1 1 0 0 0 0 0 0 0 0 0 1 0 1 0 Myrtaceae sp.1 0 0 1 0 1 0 0 0 1 0 0 0 1 0 0 0 Myrtaceae sp.2 0 0 0 0 0 0 0 0 1 0 0 0 1 0 1 0 Myrtaceae sp.5 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Myrtaceae sp.8 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Myrtaceae sp.9 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Myrtaceae sp.10 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Myrtaceae sp.12 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Myrtaceae sp.13 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Myrtaceae sp.14 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Myrtaceae sp.18 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Neomitranthes langsdorffii (Berg) J.R.Mattos 0 0 0 0 0 0 0 0 1 0 0 0 1 1 0 0 Plinia aff. grandifolia (Mattos) Sobral 0 0 1 0 1 0 0 0 1 0 0 0 0 0 0 0

107
Anexo 3-1 (continuação 8) Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie
Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Plinia involucrata (Berg) McVaugh. 0 0 1 0 0 0 1 0 0 0 0 0 1 0 1 0 Plinia rivularis (Cambess.) Rotman 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Plinia stictophylla Barroso & Peixoto 0 0 0 0 0 0 0 0 0 0 1 0 1 1 0 0 Plinia strigipes (Berg) Sobral 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Psidium aff. macrospermum Berg 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Psidium guajava Linn. 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 Psidium guianeense Sw. 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 Psidium sartorianum (Berg) Nied. 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Psidium sp.1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 NYCTAGINACEAE Andradaea floribunda Allemao 1 1 1 0 0 0 1 0 0 0 0 0 0 0 1 0 Guapira cf. subferruginosa (Mart. ex Schum.) Lundell 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Guapira noxia (Netto) Lundell 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Guapira opposita (Vell.) Reitz. 1 0 1 0 1 0 0 0 1 0 1 0 1 1 1 0 OCHNACEAE Ouratea cuspidata (A.St.Hil.) Engl. 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 OLACACEAE Heisteria cf. ovata Benth. 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Heisteria periantomega (Vellozo) H.O.Sleumer 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Heisteria scandens Ducke 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Heisteria sp.1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Liriosma sp.1 0 0 0 0 0 0 0 0 0 0 1 0 1 0 1 0 Olacaceae sp.1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Schoepfia oblongifolia Turez 0 0 1 0 0 0 1 0 0 0 1 0 0 0 1 0 PHYTOLACACEAE Gallesia integrifolia (Spreng.) Harms. 0 1 1 0 0 0 1 0 1 0 0 0 1 1 1 0 POLYGONACEAE Coccoloba sp.1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Coccoloba tenuiflora Lindau 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Coccoloba warmingii Meisn 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Ruprechtia laurifolia C.A.Mey. 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 RHAMNACEAE Rhamnidium glabrum Reissek 1 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 Ziziphus glaviovii Warm. 0 1 1 0 0 0 1 0 0 0 1 0 0 0 0 0

108
Anexo 3-1 (continuação 9) Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie
Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca RUBIACEAE Alseis floribunda Schott 0 0 1 0 1 0 0 0 1 0 0 0 1 0 0 1 Amaioua intermedia Mart. var. brasiliana (A.Rich.) Steyerm. 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Anisomeris pubescens (A. Rich.) Standl. 0 0 0 0 0 0 0 0 0 0 1 0 1 0 0 0 Melanopsidium nigrum Colla 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Posoqueria latifolia (Rudge) Roem & Schult. 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Psychotria carthagenensis Jacq. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Psychotria mapourioides DC. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Psychotria myriantha Muell. Arg. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Randia armata D.C. 0 0 0 0 0 0 0 0 1 0 0 0 1 0 1 0 Randia sp.1 0 0 0 0 0 0 0 0 1 0 1 0 0 0 0 0 Rubiaceae sp.1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Rubiaceae sp.2 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Rubiaceae sp.3 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Rubiaceae sp.4 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 Rubiaceae sp.5 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Simira grazielae A. L. Peixoto 0 0 0 0 1 0 0 0 1 0 1 0 1 1 1 0 Simira sampaioana (Standl.) Steyerm 0 1 0 0 0 0 1 0 0 0 0 0 1 0 1 0 RUTACEAE Erythrochiton brasiliensis Nees & Mart. 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 Neoraputia alba (Nees & Mart.) Emmerich 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Pilocarpus grandiflora Engl. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Rutaceae sp.1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Zanthoxylon cf. juniperinum Poepp.& Endl. 1 0 0 0 1 0 0 0 1 0 0 0 1 0 0 0 Zanthoxylum rhoifolium Lam. var. petiolatum Engl. 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 SAPINDACEAE Allophylus petiolulatus Radlk. 1 1 1 0 0 0 0 0 0 0 0 0 1 0 1 0 Cupania bracteosa RadlK. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Cupania rugosa Radlk. 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 Cupania scrobiculata L.C. Rich. 1 0 1 0 1 0 0 0 1 0 0 0 1 0 1 0 Cupania sp.1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Pseudima frutescens (Aubl.) Radlk. 0 0 0 0 1 0 1 0 0 0 0 0 1 0 1 0 Sapindaceae sp.1 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 Talisia intermedia Radlk. 1 0 1 0 0 0 1 0 1 0 1 0 1 0 1 1 Toulicia patentinervis Radlk. 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 0

109
Anexo 3-1 (continuação 10)
Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca
SAPOTACEAE Chrysophyllum lucentifolium Cronquist 0 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 Chrysophyllum sp.1 1 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 Chrysophyllum splendens Spreng. 0 1 1 0 1 0 0 0 0 1 1 0 1 0 1 0 Ecclinusa ramiflora Mart. 0 0 1 0 0 0 0 0 0 0 1 0 0 0 1 0 Manilkara bella Monach. 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Manilkara salzmannii (A.DC.) H.J.Lam 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 Micropholis crassipedicellata (Mart. & Eichler.) Pierre 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Micropholis gardneriana (A.DC.) Pierre 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 Micropholis sp.1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 Pouteria aff. hispida Eyma 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 Pouteria bangii (Rusby) T.D.Pennington 0 0 1 0 0 0 0 0 1 0 0 0 1 1 1 0 Pouteria bullata (S.Moore) Baehni 0 0 1 0 0 0 0 0 0 0 0 0 0 1 0 0 Pouteria coelomatica Rizzini 0 0 1 0 1 0 1 0 0 0 0 0 0 0 1 0 Pouteria psammophila (Mart.) Radlk. 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Pouteria reticulata (Engl.) Eyma 0 0 0 0 0 0 0 0 1 0 0 0 1 0 0 0 Pradosia lactescens (Vellozo) Radlk. 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 Sapotaceae sp.1 0 0 1 0 0 0 0 0 0 0 0 0 1 0 1 0 Sapotaceae sp.2 0 0 1 0 0 0 1 0 0 0 1 0 0 0 1 0 Sapotaceae sp.3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Sapotaceae sp.4 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 SIMAROUBACEAE Simaba cedron Planchon 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0 Simaba subcymosa A. St. Hil. Til. 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 Simaruba amara Aubl. 0 1 1 0 0 1 1 0 0 0 1 0 0 0 0 0 SOLANACEAE Cyphomandra divaricata (Mart.) Scndth. 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 Solanaceae sp.1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 Solanum inaequale Vell. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 Solanum sp.1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 STERCULIACEAE Guazuma crinita Mart. 1 1 0 1 1 1 0 0 1 1 0 0 1 0 0 0 Guazuma ulmifolia Lam. 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Pterygota brasiliensis Fr. All. 1 0 1 0 0 0 0 0 0 0 0 0 1 0 1 0

110
Anexo 3-1 (continuação 11)
Árvores- DAP≥10cm Arvoretas-5≤DAP<10 cm Varas-2,5≤DAP<5cm Jovens-Alt.≥0,5m-DAP<2,5cm Família/Espécie Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca Cap Cab Flo Ca
Sterculia elata Ducke 0 1 1 0 0 0 0 0 0 0 0 0 0 0 1 0 Theobroma cacao L. 0 0 0 0 0 0 0 0 0 1 1 0 0 1 1 0 THEOPHRASTACEAE Clavija caloneura Mart. Miq. 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0 TILIACEAE Hydrogaster trinervis Kuhlm. 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 ULMACEAE Amphelocera glabra Kuhlmann 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 Trema micrantha (L.) Blume 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 VERBENACEAE Vitex orinocensis HBK 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 VIOLACEAE Rinorea bahiensis (Moric.) Kuntze 0 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0
Total 64 42 109 9 47 19 68 9 75 28 80 9 116 52 125 13